
イヌのライム病のワクチン
【課題】本明細書に記載のワクチン組成物を提供すること。
【解決手段】上記ワクチン組成物は、免疫学的有効量の、ボレリア遺伝種の第1又は単一の株の生物を含み、ここで、該第1又は該単一の株の生物は、標準的なボレリア遺伝種の増殖条件下で増殖させる場合、B.burgdorferi ss50772(ATCC番号PTA−439)を注射した動物において誘導されるOspC特異的抗体の存在下に死滅する。一局面において、上記免疫学的有効量の生物が不活化されている。他の局面において、上記第1又は上記単一の株がその細胞表面にOspC抗原を提示し、及びイヌが上記ワクチン組成物で免疫化されると、OspC殺ボレリア菌抗体が該イヌにおいて誘導される。
【解決手段】上記ワクチン組成物は、免疫学的有効量の、ボレリア遺伝種の第1又は単一の株の生物を含み、ここで、該第1又は該単一の株の生物は、標準的なボレリア遺伝種の増殖条件下で増殖させる場合、B.burgdorferi ss50772(ATCC番号PTA−439)を注射した動物において誘導されるOspC特異的抗体の存在下に死滅する。一局面において、上記免疫学的有効量の生物が不活化されている。他の局面において、上記第1又は上記単一の株がその細胞表面にOspC抗原を提示し、及びイヌが上記ワクチン組成物で免疫化されると、OspC殺ボレリア菌抗体が該イヌにおいて誘導される。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願への相互参照
この出願は、米国特許法§119(e)の下、2006年11月3日に出願された米国仮出願第60/864,258号(この内容は、その全体が参考として本明細書に援用される)の優先権を主張する本出願である。
【0002】
本発明は、イヌのライム病に対するワクチンに関する。このワクチンを作製する方法及びこのワクチンを単独又は他の防御因子との併用で使用する方法も提供する。
【背景技術】
【0003】
イヌのライム病は、主に米国における狭義のB.burgdorferi(B.burgdorferi sensu stricto(ss))並びに欧州におけるB.burgdorferi ss、B.garinii及びB.afzeliiを含むBorrelia属(Borrelia spp.)スピロヘータによる感染が原因である(非特許文献1;非特許文献2)。これらのスピロヘータは感染したマダニ属のダニが血液を吸った時に伝染し、イヌに生じた感染は潜在性滑膜炎から急性関節炎及び関節痛に及ぶ臨床的症状をもたらす(非特許文献3;非特許文献4)。重要なことであるが、イヌのライム病の発生率は、ヒトの症例数の増加と一致して毎年増加し続けている(非特許文献5)。
【0004】
ボレリア属による感染に対して産生される抗体には異なる2つの機能があるが、従来、いずれの反応も哺乳類の宿主から隔絶されたスピロヘータを除去するという点で無効であった。この自然感染に対する正常な免疫反応の欠如に関して、抗原変異(Schwan、Biochem.Soc.Trans.2003年、31:p.108−112;Tokarzほか、Infect.Immun.2004年、72:p.5419−5432)、宿主模倣(Barbourほか、Microbiol.Rev.1986、50:p.381−400)及び細胞内局在性(Maほか、Infect.Immun.1991年、59:p.671−678)を含むいくつかの解釈が想定されている。
【0005】
最も一般的な液性免疫反応は、食細胞による貪食のためにスピロヘータに「印を付ける」非特異的結合/オプソニン化(被覆)抗体の産生である。残念ながら、オプソニン化抗体は他の微生物に共通にみられる数種の蛋白質(即ち、細菌鞭毛を構成する41kDa蛋白質)によって誘導され、このため、ワクチン接種誘導液性免疫におけるその価値はどうひいき目にみても疑わしいものとなる。
【0006】
別の一般的な免疫反応は殺ボレリア菌(致死的)抗体の産生である。オプソニン化抗体とは対照的に、殺ボレリア菌抗体は僅か数種のボレリア属菌蛋白質のエピトープを認識する。殺ボレリア菌抗体は、スピロヘータ表面の特定の標的に結合した後、最も一般的には補体を誘導して、食細胞による除去を必要とすることなくこの菌を殺す細胞膜傷害複合体を形成する。
【0007】
現在ワクチンとして用いられているイヌのライム病バクテリンが開発されて、感染されたダニが吸血する時にこの寄生虫中のOspA発現スピロヘータを殺すOspA殺ボレリア菌抗体(Hsien−Chuほか、JAVMA 1992、201:p.403−411;Maほか、Vaccine 1996年、14:p.1366−1374;Wikleほか、Intern.J.Appl.Res.Vet.Med.2006年、4:p.23−28;Straubingerほか、Vaccine 2001年、20:p.181−193)を誘導することによる防御が可能となった(非特許文献6)。Straubingerら(Vaccine 2002, 20:181−193)は、全細胞ワクチンの方が組換えOspAよりも有意に高い殺ボレリア菌抗体価を誘導したと報告している。このようなワクチンはほどほどに上出来であったが、ワクチン接種の失敗例が報告されている(Levyほか、JAVMA 1993年、202:p.1834−1838;Maほか、Vaccine 1996年、14:p.1366−1374;Schutzerほか、N.Engl.J.Med.1997年、337:p.794−795)。
【0008】
生じたOspA殺ボレリア菌抗体はOspAを発現しているB.burgdorferi ssを認識するのみであり(Jobeほか、J.Clin.Microbiol.1994、32:p.618−622;Lovrichほか、Infect.Immun.1995、63:p.2113−2119)、通常、上記ダニはOspAを発現していないB.burgdorferi ssスピロヘータに感染する(Fikrigほか、Infect.Immun.1995、63:p.1658−1662;Ohnishiほか、 Proc.Natl.Acad.Sci.2001年、98:p.670− 675)ので、現在、こうした抗体は吸血している上記ダニを殺菌できない場合が多いと理解されている。さらに、上記ダニは、一般にB.afzelii及びB.gariniiを含む他の病原性ボレリア属菌にも感染する(Omsteinほか、J.Clin.Microbiol.2001年、39:p.1294−1298)が、OspA抗体は遺伝種特異的である(Lovrichほか、Infect.Immun.1995年、63:p.2113−2119)。さらに、ダニの中腸への付着を媒介する(Palほか、J.Clin.Invest.2000年、106:p.561−569)OspAの発現は感染されたダニが吸血を始めた後まもなく下方制御される(Schwanほか、Proc.Natl.Acad.Sci.USA 1995年、92:p.2909−2913)ので、OspAボレリア抗体による防御の「絶好の機会」はスピロヘータが感受性の場合でも限定される。
【0009】
B.burgdorferi ssのOspCは殺ボレリア菌液性免疫の別の標的と考えられる(Rousselleほか、J.Infect.Dis.1998年、178:p.733−741)。この蛋白質は、殺ボレリア菌抗体の誘導に関与し、病原性ボレリア属菌類間で保存されているエピトープを有すると考えられる(Lovrichほか、Clin.Diagn.Lab.Immunol.2005年、12:p.746−751)。OspC蛋白質の具体的な機能は依然として不明であるが、OspCの発現は哺乳動物の感染には必要であるが、ダニの感染には必要とされないことが示唆されている(Grimmほか、2004年、Proc.Natl.Acad.Sci.101(9):p.3142−3147)。いずれにしても、ライム病スピロヘータはダニが吸血を開始した後まもなくOspCを発現する(Schwanほか,Proc.Natl.Acad.Sci.USA 1995,92:2909−2913)が、哺乳動物において感染が成立するためにはOspCを発現し続ける必要がある(Stewartほか、Infect Immun.2006年、74:p.3547−3553,Tillyほか、Infect.Immun.2006年、74:p.3554−3564)。従って、OspC殺ボレリア菌抗体の「有効性のウインドウ(window of effectiveness)」はOspA殺ボレリア菌抗体に比し、著しく増大する。
【0010】
OspC蛋白質が防御性殺ボレリア菌抗体を誘導することができることは分かっている(Rousselleほか、J.Infect.Dis.1998年、178:p.733−741;Ikushimaほか、FEMS Immunol.Med.Microbiol.2000年、29:p.15−21)が、これまでの一部の「マッピング」研究では上記エピトープはこの蛋白質の高度に不均一な領域に限局されている(Bucklesほか、Clin.Vacc.Immunol.2006年、13:p.1162−1165)。従って、こうした領域に対する殺ボレリア菌OspC抗体は少数種のボレリア属分離株に対する液性免疫をもたらすに過ぎないことになる。Lovrichら(Clin.Diagn.Lab.Immunol.2005年、12:p.746−751)は、上記蛋白質のC末端7アミノ酸(OspC7)内にOspC殺ボレリア菌抗体エピトープをつきとめた。最も注目に値すべきなのは、このエピトープが上記病原性ボレリア菌類の間で保存されていることである。しかしながら、OspAを発現する(即ち、ospA/ospBオペロンを含有する)通常の実験室B.burgdorferi ss分離株は、OspA殺ボレリア菌抗体誘導能を大きく損なうことなく有意なレベルのOspC殺ボレリア菌抗体をも誘導するように実験室で操作することができない。さらに、OspAを発現する通常の実験室B.burgdorferi ss分離株の死菌体を用いてワクチン接種しても殺ボレリア菌OspC抗体は誘導されない(Schwanほか、1995年、Proc.Natl.Acad.Sci USA、92:p.2909−2913;Obonyoほか、1999年、J.Clin.Microbiol.、37:p.2137−2141)。
【0011】
Callisterら(本明細書に引用により組み込まれている特許文献1及び特許文献2)は、OspCの免疫原性ポリペプチド断片を単独又はOspAポリペプチドとの併用で用いることによりライム病に対してヒト及び他の哺乳動物を防御するワクチンを調製することを提唱している。また、Liveyら(本明細書に引用により組み込まれている特許文献3)は、組換えOspA、OspB及びOspC蛋白質の組合せから調製された、ライム病に対して免疫化するワクチンを提案した。しかしながら、現在に至るまで、組換え蛋白質ワクチンで、現在市販されているワクチンより改良されたものであることが示されたものはない。従って、当該分野では長年にわたり、ライム病から哺乳動物、特にイヌを防御するワクチンの改良が求められ続けている。
【0012】
本明細書におけるいかなる文献の引用も、そのような文献が本願の「先行技術」として利用可能であるということを認めたものと解するべきではない。
【先行技術文献】
【特許文献】
【0013】
【特許文献1】米国特許第6,210,676号明細書
【特許文献2】米国特許第6,464,985号明細書
【特許文献3】米国特許第6,872,550号明細書
【非特許文献】
【0014】
【非特許文献1】Barantonほか、Int.J.Sys.Bacteriol.1992年、42:p.378−383
【非特許文献2】Hoviusほか、J.Clin.Microbiol.2000年、38:p.2611−2621
【非特許文献3】Jacobsonほか、Semin.Vet.Med.Surg.1996年、11:p.172−182
【非特許文献4】Summersほか、J.Comp.Path.2005年、133:p.1−13
【非特許文献5】Haninkovaほか、Emerg.Infect.Dis.2006年、12:p.604−610
【非特許文献6】Fikrigほか、Proc.Natl.Acad.Sci.USA 1992年、89:p.5418−5421
【発明の概要】
【課題を解決するための手段】
【0015】
従って、本発明はワクチンに用いることができる新規免疫原性組成物を提供する。本発明の一態様において、ワクチンはライム病を防御する。この種の具体的な実施態様では、ワクチンのレシピエントはイヌである。別の実施態様では、ワクチンのレシピエントは飼いネコである。ウマ及び/又はウシなどの他の家畜哺乳類も本発明のワクチン及び/又は方法によって防御することができる。本発明はさらに、ライム病及び他の疾患、例えばイヌの他の感染症に対する防御免疫を誘導する混合ワクチンを提供する。本発明のワクチンの作製方法及び使用方法も提供する。
【0016】
本発明のワクチンは、免疫学的有効量の、OspC抗原を発現する(任意選択的に不活化された)第1又は単一の株の生物を含む。この株は、標準的な培養条件で増殖させる場合、(任意選択的に補体を必要とする)OspC特異的殺ボレリア菌抗体の存在下に死滅する株である。このような抗体としては、B.burgdorferi ss50772(ATCC番号PTA−439)によってワクチン接種した動物に誘導される、保存エピトープOspC7に対する殺ボレリア抗体が挙げられる。この種の具体的な実施態様では、ボレリア遺伝種(genospecie)の上記第1又は単一の株は上記OspC抗原を構成的に発現する。より具体的な実施態様では、この第1の株はB.burgdorferi ss50772(ATCC番号PTA−439)である。
【0017】
本発明のワクチン組成物は、免疫学的有効量の、病原性ボレリア遺伝種からの1種以上の別の株(これらを総称して第2の株と名付ける場合がある)からの不活化生物をさらに含むことができる。具体的な実施態様では、この第2の株はOspA及びOspB抗原を提示する。
【0018】
適切な第2の株の例としては、以下のうちの1種以上が挙げられる:B.burgdorferi ssS−1−10(ATCC番号:PTA−1680)、B.burgdorferi ssB−31(ATCC番号35210)、B.afzelii(例えば、ATCC番号51567として入手可能)及びB.garinii(例えば、ATCC番号51383及び同51991として入手可能)、B.burgdorferi ssDK7、B.burgdorferi ss61BV3、B.burgdorferi ssZS7、B.burgdorferi ssPka、B.burgdorferi ssIP1、IP2、IP3、B.burgdorferi ssHII、B.burgdorferi ssP1F、B.burgdorferi ssMil、B.burgdorferi ss20006、B.burgdorferi ss212、B.burgdorferi ssESP1、B.burgdorferi ssNe−56、B.burgdorferi ssZ136、B.burgdorferi ss ia及び/又はこれらの任意の組合せ。
【0019】
上記ワクチン組成物は、概して、各それぞれの株の生物を1ミリリットル当たり約1×104乃至約1×1010個含む。特定の実施態様では、このワクチンは各それぞれの株の生物を1ミリリットル当たり約1×106乃至約5×109個含む。別の実施態様では、このワクチンは上記(又は各)第2の株の生物を1ミリリットル当たり約1.0×108乃至約5×108個及び上記第1の株の生物を1ミリリットル当たり約5.0×108乃至約5×109個含む。
【0020】
また、上記ワクチン組成物には、例えば、アルミニウム化合物(例えば、リン酸アルミニウム、水酸化アルミニウム)、代謝性及び非代謝性油、ブロックポリマー、免疫刺激複合体、ビタミン、ミネラル、CARBOPOL(登録商標)(例えば、CARBOPOL941)などの医薬用として許容可能なアジュバントを含めることもできる。特定の実施態様では、この医薬用として許容可能なアジュバントは、(例えば、Emulsigen(登録商標)として販売されているような)水中にミクロン径の油滴を均一分散させた乳剤を含む。
【0021】
任意選択的に、上記ワクチン組成物は医薬用として許容可能な免疫刺激剤、例えば、サイトカイン、成長因子、ケモカイン、リンパ球、単球もしくはリンパ器官由来細胞の細胞培養液の上清、植物、細菌もしくは寄生虫からの細胞調製物及び/又は抽出物又はマイトジェンをも含む。
【0022】
本発明はさらに、病原性ボレリア属菌、具体的にはB.burgdorferi ssに対してイヌ又は他の哺乳動物を免疫化する方法であって、免疫学的有効量の上記本発明ワクチンをイヌに注射することを含む方法を提供する。例えば、このようなワクチンには各それぞれの株の生物を約1×108乃至3×109個を含ませることができる。ワクチンは、例えば、筋肉内注射、皮下注射、静脈内注射、皮内注射、経口投与、鼻腔内投与及びこれらの組合せなどの経路によって投与することができる。特定の実施態様では、ワクチン接種後、免疫化されたイヌは殺ボレリア抗体を産生する。
【0023】
本発明はさらに、B.burgdorferi ss OspCに結合する殺ボレリア菌抗体を含有する、ワクチン接種動物から得られる血清を提供する。同様に、本発明はOspCに結合する精製抗体を提供する。特定の実施態様では、この血清はかなりの割合のOspC7特異的殺ボレリア菌抗体を含有する。
【0024】
本発明はさらに、本発明のボレリア遺伝種の1種以上の株を1種以上の他のイヌ病原体及び/又は免疫原との組合せで含む混合ワクチンを提供する。この免疫原としては、例えば、イヌジステンパーウイルス、イヌアデノウイルス、イヌパルボウイルス、イヌパラインフルエンザウイルス、イヌコロナウイルス、イヌインフルエンザウイルス及び/又はLeptospira血清型(例えば、Leptospira kirschneri血清型grippotyphosa、Leptospira interrogans血清型canicola、Leptospira interrogans血清型icterohaemorrhagiae及び/又はLeptospira interrogans血清型pomana)に対する免疫を誘導する免疫原が挙げられる。本発明の混合ワクチンに添加することができる別のイヌ病原体としては、Leishmania major、Leishmania infantumなどのリーシュマニア属生物、Bordetella bronchiseptica、マイコプラズマ属菌(例えば、Mycoplasma cynos)、狂犬病ウイルス及びanaplasma phagocytophilum、anaplasma platysなどのanaplasma属生物及びEhrlichia canisが挙げられる。
【0025】
本発明のこれらの態様及びそれ以外の態様については添付の図面及び詳細な説明によって一層よく理解されよう。
例えば、本発明は、以下の項目を提供する:
(項目1)
免疫学的有効量の、ボレリア遺伝種の第1又は単一の株の生物を含むワクチン組成物であって、
該第1又は該単一の株の生物は、標準的なボレリア遺伝種の増殖条件下で増殖させる場合、B.burgdorferi ss50772(ATCC番号PTA−439)を注射した動物において誘導されるOspC特異的抗体の存在下に死滅する、ワクチン組成物。
(項目2)
上記免疫学的有効量の生物が不活化されている項目1に記載のワクチン。
(項目3)
上記第1又は上記単一の株がその細胞表面にOspC抗原を提示し、及び
イヌが上記ワクチン組成物で免疫化されると、OspC殺ボレリア菌抗体が該イヌにおいて誘導される項目1に記載のワクチン。
(項目4)
かなりの割合の上記OspC特異的殺ボレリア菌抗体がOspC7に特異的である項目3に記載のワクチン組成物。
(項目5)
上記第1又は上記単一の株がB.burgdorferi ss50772(ATCC番号PTA−439)である項目3に記載のワクチン組成物。
(項目6)
免疫学的有効量の、ボレリア遺伝種由来の第2の株の不活化生物をさらに含み、該第2の株がその細胞表面にOspA抗原、OspB抗原を提示し、又はOspA抗原及びOspB抗原の両者を提示する項目1に記載のワクチン組成物。
(項目7)
イヌが上記ワクチン組成物で免疫化されると、OspC殺ボレリア菌抗体及びOspA殺ボレリア菌抗体が該イヌにおいて誘導される項目6に記載のワクチン組成物。
(項目8)
上記第2の株がB.burgdorferi ssS−1−10株(ATCC番号PTA−1680)である項目6に記載のワクチン組成物。
(項目9)
上記第1の株の生物を1ミリリットル当たり約1×104乃至約1×1010個及び上記第2の株の生物を1ミリリットル当たり約1×104乃至約1×1010個含む、項目6に記載のワクチン組成物。
(項目10)
上記第1の株の生物を1ミリリットル当たり約5.0×108乃至約5×109個及び上記第2の株の生物を1ミリリットル当たり約1.0×108乃至約5×108個含む、項目9に記載のワクチン組成物。
(項目11)
医薬用として許容可能なアジュバントをさらに含む項目1に記載のワクチン組成物。
(項目12)
医薬用として許容可能な免疫刺激剤をさらに含む項目1に記載のワクチン組成物。
(項目13)
非ボレリア病原体に対する防御免疫を誘導するための少なくとも1種の非ボレリア免疫原をさらに含む項目1に記載のワクチン組成物。
(項目14)
上記非ボレリア免疫原がイヌジステンパーウイルス、イヌアデノウイルス、イヌパルボウイルス、イヌパラインフルエンザウイルス、イヌコロナウイルス、イヌインフルエンザウイルス、Leptospira血清型、Leishmania属生物、Bordetella bronchiseptica、Mycoplasma属菌、狂犬病ウイルス、Ehrlichia canis、anaplasma属生物及びこれらの組合せからなる群から選ばれる項目13に記載のワクチン組成物。
(項目15)
上記Leptospira血清型がLeptospira kirschneri血清型grippotyphosa、Leptospira interrogans血清型canicola、Leptospira interrogans血清型icterohaemorrhagiae、Leptospira interrogans血清型pomana及びこれらの組合せからなる群から選ばれる項目14に記載のワクチン組成物。
(項目16)
上記Mycoplasma属菌がMycoplasma cynosを含む項目14に記載のワクチン組成物。
(項目17)
病原性ボレリア遺伝種に対してイヌを免疫化する方法であって、免疫学的有効量の項目1に記載のワクチンを該イヌに注射することを含む方法。
(項目18)
病原性ボレリア遺伝種に対してイヌを免疫化する方法であって、免疫学的有効量の項目6に記載のワクチンを該イヌに注射することを含む方法。
(項目19)
上記第2の株が狭義のB.burgdorferiである項目18に記載の方法。
(項目20)
上記第2の株がB.burgdorferi ssS−1−10(ATCC番号:PTA−1680)、B.burgdorferi ssB−31(ATCC番号35210)、B.afzelii(ATCC番号51567)及びB.garinii(ATCC番号51383又は同51991)、B.burgdorferi ssDK7、B.burgdorferi ss61BV3、B.burgdorferi ssZS7、B.burgdorferi ssPka、B.burgdorferi ssIP1、IP2、IP3、B.burgdorferi ssHII、B.burgdorferi
ssP1F、B.burgdorferi ssMil、B.burgdorferi ss20006、B.burgdorferi ss212、B.burgdorferi ssESP1、B.burgdorferi ssNe−56、B.burgdorferi ssZ136、B.burgdorferi ss ia及び/又はこれらの組合せからなる群から選ばれる項目18に記載の方法。
(項目21)
上記免疫化されたイヌが殺ボレリア菌抗体を産生する項目18に記載の方法。
(項目22)
上記イヌがボレリア属の各それぞれの株の生物を約5×107乃至約5×109個注射される項目18に記載の方法。
(項目23)
項目21に記載の方法によって免疫化されたイヌから得られるB.burgdorferi OspCに結合する殺ボレリア菌抗体を含む血清。
(項目24)
項目21に記載の方法によって免疫化されたイヌから得られるB.burgdorferi ss OspCに結合する殺ボレリア菌抗体。
(項目25)
OspC7特異的抗体を含む項目24に記載の殺ボレリア菌抗体。
【図面の簡単な説明】
【0026】
【図1】正常イヌ対照血清(NS)、試験品(TP:test product)を用いたワクチン接種(試験43日目)後のイヌからの血清又はB.burgdorferi ss感染ダニ(NI)による感染(試験134日目)後のイヌからの血清のウエスタンブロットの写真である。分子量約20kDaの蛋白質に対する感染特異的抗体の存在に注目されたい。
【図2A】正常イヌ対照血清(NS)又はプラセボ接種及びB.burgdorferi ss感染ダニによる感染(試験134日目)後の個々のイヌの個別のコホートからの血清(1番から15番まで番号が付けられている)のウエスタンブロットの写真である。
【図2B】正常イヌ対照血清(NS)又は試験品を用いたワクチン接種後の個々のイヌの個別のコホートからの血清(1番から15番まで番号が付けられている)のウエスタンブロットの写真である。プラセボ接種した14頭(93%)のイヌでは感染特異的20kDa抗体が存在し(図2A)、試験品を用いてワクチン接種したイヌでは同抗体が存在しなかった(p<0.0001)(図2B)ことに注目されたい。
【図3A】プラセボ接種してB.burgdorferi ssに感染させたイヌの関節における組織病理学的変化の代表的な1例を示す写真である。長方形内の領域はイヌのライム病関節炎に特有の好中球及び単核細胞の著しい浸潤を示している。
【図3B】試験品を用いてワクチン接種し、B.burgdorferi ssに感染させたイヌの関節において組織病理学的変化がみられない代表的な1例を示す写真である。長円形内の領域には好中球及び単核細胞の浸潤はみられない。
【図4】ライム症状もしくはライム関連症状の(カルテ審査による)既往歴がない個体(n=36)からの正常血清(N)、献血者(BD、n=100)もしくはコレステロール検診を受けた個体(CS、n=100)からの特性化されていない血清、又は通常B.burgdorferi ss抗原と交差反応する血液因子もしくは疾患、例えば、抗核抗体(ANA、n=20)、リウマチ因子(RF、n=20)、単核球症(EBV、n=10)、サイトメガロウイルス(CMV、n=10)、梅毒(SYPH、n=13)もしくはロッキー山熱(RMSF、n=4)を有するボランティアからの血清を用いたOspC7ELISA反応性の測定結果を示した図である。実線は、正常血清及び交差反応する可能性のある血清の平均吸光度値よりも上の3標準偏差値分を示す。この線よりも上にくる値は陽性に相当する確率が99%となる(バックグランド値 1%)。
【図5】定型ライム病病変を有する患者である、遊走性紅斑を示すほぼ確かなライム病の患者(n=86)、「非定型病巣」を有する、確からしいライム病の患者(n=22)及び全身症状(初発臨床症状)を有する、ライム病の可能性のある患者(n=49)からの血清を用いたOspC7ELISA反応性を示す図である。実線は、正常血清及び交差反応する可能性のある血清の平均吸光度値よりも上の3標準偏差値分を示す。
【発明を実施するための形態】
【0027】
本発明は、レシピエントのワクチン接種動物に効果的な殺ボレリア菌液性免疫を誘導するボレリア遺伝種の1種以上の株からの免疫学的有効量の生物を含むワクチン組成物を提供する。このような株の生物は、標準的なボレリア遺伝種の増殖条件下で増殖させる場合、OspC特異的抗体、例えば、B.burgdorferi ss50772(ATCC番号PTA−439)を用いてワクチン接種した動物において誘導される一種の抗体又は複数種の抗体の存在下に死滅する。
【0028】
本発明の一態様において、本発明のワクチンにより誘導される抗体の有効性のウインドウは、単にOspA殺ボレリア抗体に基づいた従来のワクチンに比し、著しく増大する。別の態様では、本発明のワクチンでは、例えばインビボでのOspCの発現によるなどの有益な免疫記憶反応を促進する確率が改善され、及び/又はボレリア属菌の複数種の病原性株に対する防御が追加及び/又は増強される。
【0029】
本発明の一実施態様において、そのような株の生物は、標準的なボレリア遺伝種増殖条件下で増殖させると、OspC抗原を構成的に発現する。この種の特定の実施態様では、そのような株の生物は、標準的なボレリア遺伝種増殖条件下で増殖させると、補体特異的反応によって死滅する。さらに別の実施態様では、そのような株の生物を含むワクチンにより誘導される血清中のかなりの割合のOspC特異的殺ボレリア菌抗体は、保存エピトープOspC7に特異的であることにより、複数の病原性ボレリア遺伝種(例えば、B.burgdorferi ss、B.afzelii及び/又はB.garinii)に対する防御を可能にする。さらに別の実施態様では、そのような株の生物はOspA抗原もOspB抗原も提示しない。さらに別の実施態様では、そのような株の生物はこれらの性質のうちの任意の2つ以上を有する。さらに別の実施態様では、そのような株の生物はこれらの性質のうちの任意の3つ以上を有する。さらに別の実施態様では、そのような株の生物はこれらの性質のうちの任意の4つ以上を有する。特定の実施態様では、そのような株の生物はこれらの性質の全てを有する。そのような特定の1種の株はB.burgdorferi ss50772(ATCC番号PTA−439)である。
【0030】
別の実施態様において、本発明のワクチンは、上記のような第1の株及び有効量の少なくとも第2の株の生物を含む組成物である。この第2の株は、本発明のワクチンの一部として投与された場合に殺ボレリア菌OspA抗体を誘導する病原性ボレリア遺伝種からのものであることが好ましい。また、上記組成物には1種以上の別の適合性ワクチン遺伝種を含ませることもできる。さらに、本発明のワクチンには1種以上の他の哺乳動物(例えば、イヌ)病原体及び/又は免疫原を含ませることができる。
【0031】
本発明の理解をさらに十分に進めるために、以下の定義を設ける。
【0032】
原文の英文では、記載上の便宜のために単数形の用語を用いているが、これは決してそのように限定的なものではない。すなわち、例えば、「ポリペプチド(a polypeptide)」を含む組成物に言及する場合、1種以上のそのようなポリペプチドに言及することを含む。さらに、生物(organism)に言及する場合、特に示さない限り、複数種のそのような生物(organisms)に言及することを含む。
【0033】
本明細書で使用される「ほぼ(approximately)」という用語は「約(about)」と同じ意味で用いており、値がその示された値の50%以内にあること、すなわち、1ミリリットル当たり「ほぼ」1×1010個の菌を含む組成物は1ミリリットル当たり5×109乃至5×1010個の菌を含むことを意味する。
【0034】
「遺伝種」という用語は、G.Barantonら(1992年、International J.of Systematic Bacteriology 42:p.378−383)によって最初に用いられ、定義されたが、本明細書でも、非ボレリア生物の分類を記載する際に「種」という用語を用いるのと同じ意味で用いている。
【0035】
ボレリア遺伝種を培養するための「標準的な増殖条件」では、BSK(Barbour−Stoenner−Kelly)培地中約33℃乃至約35℃の範囲の温度で増殖させることが必要とされる。本明細書に記載したBSK培地は、Callisterら[参考としてその全体が本明細書に援用されるDetection of Borreliacidal Antibodies by Flow Cytometry、セクション11.5.1 − 11.5.12、Current Protocols in Cytometry、John Wiley and Sons,Inc.Supplement26、(2003年)]に従って調製した。(BSK培地は、例えば、Sigma、St.Louis、ミズーリ州から市販されてもいる。)
本明細書で使用される「OspC7」とは、既知の病原性ボレリア属間で完全に保存されている、Callisterら(参考としてその全体が本明細書に援用される米国特許第6,210,676B1号及び同6,464,985B1号)によって開示されているOspCのC末端側50アミノ酸内の7アミノ酸領域(参考としてその全体が本明細書に援用されるLovrichほか、2005年、Clin.Diagn.Lab.Immunol.、12:p.746−751)にある免疫優勢OspC殺ボレリア菌抗体エピトープである。この保存については、Lovrichらによって報告された7アミノ酸セグメントをコードしているコドンセグメントのBLAST探索により容易に確認できる。2006年10月9日に行ったこのような探索では、上記のOspC 7−マーエピトープコードセグメントを含有する100種のボレリア種の結果リストが得られた。
【0036】
「OspC特異的殺ボレリア菌抗体」とは、例えばB.burgdorferi ss50772(ATTC番号PTA−439)でワクチン接種した動物の血清中に存在するものであり、OspC抗原の任意のエピトープに選択的に結合してスピロヘータを補体依存性又は非依存性に死滅させるものである。「OspC7特異的殺ボレリア菌抗体」とは、例えばB.burgdorferi ss50772(ATTC番号PTA−439)でワクチン接種した動物の血清中に存在するものであり、Lovrichら[参考としてその全体が本明細書に援用されるLovrichほか、Clin.Diagn.Lab.Immunol.、12:p.746−751、(2005年)]によって報告されたOspCの7個のC末端アミノ酸に選択的に結合してスピロヘータを(一般に補体媒介性細胞膜傷害複合体を誘導することにより)死滅させるものである。OspC殺ボレリア菌抗体の特異性は十分立証されている。例えば、OspC殺ボレリア菌抗体は、殺ボレリア菌抗体試験においてB.burgdorferi ss50772の感受性を測定することによって通常ライム病血清中に検出される。類縁疾患を有するヒト患者からの血清には、株50772をも死滅させる交差反応性抗体はまれに(2%)しか含まれない(Callisterほか、1996年、Clinical and Diagnostic Laboratory Immunology 3(4):p.399−402に詳細に報告されている)。さらに、OspC7殺ボレリア菌エピトープを用いるペプチドELISA法ではライム病の血清中の殺ボレリア菌抗体が正確に捕捉されるが、他の類縁疾患を有する患者からの血清にはOspC7ペプチドにも結合する交差反応性抗体はまれに(<2%)しか含まれない(図4及び5に示した)。
【0037】
ワクチンにより誘導される血清中の「かなりの割合の」OspC特異的殺ボレリア菌抗体が保存エピトープOspC7に特異的である場合、これは、OspC7によるその血清の吸収後に血清中OspC特異的殺ボレリア菌抗体が無視できないほど減少することを意味する。これは、好ましくはB.burgdorferi ss50772を用いて検出される血清の殺ボレリア菌抗体価の少なくとも2分の1、より好ましくはOspC7によるその血清の吸収後の血清の殺ボレリア菌抗体価の2分の1乃至4分の1又はこれ以下への低下と定義される。
【0038】
「補体特異的反応」とは、ボレリア属の生物(類)を殺ボレリア菌抗体によって死滅させるために血清補体の存在を必要とする抗体反応である。
【0039】
本発明では、「不活化」Borrelia burgdorferi ss生物とは、動物において免疫反応を誘導することができるが、その動物に感染することができない生物である。Borrelia burgdorferi ss分離株は、二成分エチレンイミン、ホルマリン、ベータ−プロピオラクトン、チメロサール又は熱からなる群から選ばれる作用因子によって不活化することができる。特定の実施態様では、このBorrelia burgdorferi ss分離株は二成分エチレンイミンによって不活化する。
【0040】
「アジュバント」及び「免疫刺激剤」という用語は、本明細書では同じ意味で用いており、免疫系の亢進をもたらす1種以上の物質と定義される。これに関連して、アジュバントは1種以上のワクチン抗原/分離株に対する免疫反応を増強するために使用する。アジュバントは、ワクチンの投与の前、又はこの投与と組み合わせて、又はこの投与の後に対象動物に投与することができる。本発明のアジュバントは、例えば、天然供給源、組換え供給源を含むいくつかの供給源のいずれかから入手し、及び/又は化学的に合成することができる。アジュバントとして用いる化合物の例としては、アルミニウム化合物、代謝性及び非代謝性油、ブロックポリマー、ISCOM(免疫刺激複合体)、ビタミン及びミネラル(ビタミンE、ビタミンA、セレン、ビタミンB12が挙げられるが、これらに限定されない)、キル(Quil)A(サポニン)及びポリアルケニルエーテル又はジビニルグリコールで架橋したアクリル酸のポリマー、例えば、CARBOPOL(登録商標)が挙げられるが、これらに限定されるものではない。特に免疫刺激剤と呼ばれることがあるアジュバントの別の例としては、細菌及び真菌細胞壁成分(例えば、リポ多糖、リポ蛋白質、糖蛋白質、ムラミルペプチド、ベータ−1,3/1,6−グルカン)、植物由来の各種複合糖質(例えば、グリカン、アセマンナン(acemannan))、動物由来の各種蛋白質及びペプチド(例えば、ホルモン、サイトカイン、共刺激因子)並びにウイルス及び他の供給源由来の新規核酸(例えば、二本鎖RNA、CpG)が挙げられる。さらに、上記の物質のいくつもの組合せがアジュバント効果をもたらすことができ、従って、これらは本発明のアジュバントを形成することができる。好ましいアジュバントの1つはEmulsigen(登録商標)である。
【0041】
米国特許第6,210,676号に記載されているB.burgdorferi ss50772(ATCC番号PTA−439)及び米国特許第6,316,005号に記載されているB.burgdorferi ssS−1−10(ATCC番号PTA−1680)は、それぞれ1999年7月30日及び2000年4月11日にAmerican Type Culture Collection、10801 Boulevard大学、Manassas(VA)20110に寄託された。本発明の共同権利者らは、個々に上記2特許に対する権利を保有している。
【0042】
また、本発明が本明細書に開示した特定の構造、工程段階及び材料に限定されないことは、このような構造、工程段階及び材料はある程度変えることができるので、言うまでもないことである。また、本明細書に用いた用語は、単に特定の実施態様を説明するために用いたものであり、本発明の範囲が添付の特許請求の範囲及びその均等物によってのみ限定されることになるので、限定的なものではないことも言うまでもない。
【0043】
他のワクチン株
・他のOspC株
一態様において、本発明は、単独又は他のボレリア遺伝種株との併用でライム病を防御するワクチンにおけるB.burgdorferi ss50772(ATCC番号PTA−439)の使用を提供する。特に、B.burgdorferi ss50772は、当初、ワクチンに有用な候補としては受け入れなかった。何故なら、この株はOspCを発現しないと間違って報告されていた(Andersonほか、1996年、J.Clin.Microbiol.、34:p.524−529)からである。
【0044】
本発明はさらに、本発明のワクチン用の他のそのような有用分離株を選択/特定する独自の特異的な基準を提供する。特に、OspC特異的抗体により死滅させられやすく、好ましくは、以下の属性のうちの1つ以上又は全てをも有する分離株を選択/特定することができる。即ち、その属性とは、(i)補体特異的反応を介してOspC特異的抗体により死滅させられやすいこと、(ii)この分離株でワクチン接種したレシピエントにおいてOspC殺ボレリア菌抗体を誘導することができること、(iii)そのように誘導されたかなりの割合のOspC殺ボレリア菌抗体がC末端7アミノ酸(Osp7)内の保存エピトープに特異的であること、(iv)OspA及びOspB発現能を欠くこと、及び/又は(v)インビトロで構成的にOspCを発現することができること、である。
【0045】
A.培養条件によるOspA/B発現の抑制及びOspC発現の増強
通常の病原性ボレリア属菌株(B.burgdorferi ssの株JMNT及びN40)をダニ細胞株(Ixodes scapularisISE6)と共培養することにより5日間にわたって、OspA発現を下方制御することができ、OspC発現を上方制御することができることをObonyoら(J.Clin.Microbiol.、1999年、37:p.2137−2141)は明らかにしている。最も著しい効果は37℃で培養することで認められた。従って、この方法によって1種以上の通常の病原性ボレリア菌株が本発明のワクチンに有用な形態に変換されることになる。
【0046】
B.突然変異によるOspA/B発現の抑制及びOspC発現の増強
別の実施態様において、元々OspA及びOspBを発現するボレリア属菌株に対して遺伝子操作を行ってOspA及びOspBの発現をコードする遺伝子を下方制御又は削除することによりOspC発現を上方制御する。当該分野で既知の全ての遺伝子操作方法をこの目的に使用することを想定している。例えば、入手できるボレリア属菌株の複数の菌体にOspA/OspB遺伝子の不活性化アナログを、抗生物質選択マーカを用いて調製したボレリア属菌適合性プラスミドの形で導入する(例えば、OspC不活性化及び相補プラスミドをB.burgdorferiB31−A3株に挿入することによりボレリア属菌におけるOspCの発現をブロックすることができると報告しているGrimmほか、2004年、Proc.Nat’l Acad.Sci 101(9):p.3142−3147を参照されたい)。この導入プラスミドとOspA/OspBを含む天然プラスミドとの間に組換え現象がおこることによって突然変異OspA/OspBプラスミドを含む一定数の組換えがうまくいったボレリア属菌が得られる。選択は、例えば、対応抗生物質の存在下における関連する抗生物質耐性と増殖とによることを前提とすることができる。OspA/OspBが選択的に発現されなくなったボレリア属菌はOspCの発現を上方制御することが想定されている。
【0047】
別の任意選択的な実施態様では、OspA、OspB及び/又は1種以上の別のOsp抗原を発現する1種以上の別のB.burgdorferi ss又はボレリア属生物を別の生物として前記ワクチン組成物に含ませることができる。
【0048】
各それぞれの不活化生物は約1×104乃至約1×1010個/mLの範囲の濃度で上記ワクチンに存在させることが好ましい。特に、このワクチンは、1.0×108個/mL以上のB.burgdorferi ssS−1−10及び5.0×108個/mL以上のB.burgdorferi ss50772の用量でイヌに投与することが好ましい。
【0049】
・他のOspA株
OspA抗原を有する第2の株をB.burgdorferi ssB−31(ATCC番号35210)などの通常の病原性実験室B.burgdorferi ss分離株(Barbourほか、1985年、J.Clin.Microbiol.52:p.478−484)とすることができる。特定の第2の生物は上記に例示したB.burgdorferi ssS−1−10株(ATCC番号PTA−1680)である。北米以外の地域に対して最適化されたワクチン組成物の第2の生物として用いるのに適した別の株としては、例えば、B.burgdorferi ssB−31(ATCC番号35210)、B.afzelii(例えば、ATCC番号51567として入手可能)及びB.garinii(例えば、ATCC番号51383及び同51991として入手可能)並びに下記の表1に示した株が挙げられる。
【0050】
【表1】

前記ワクチン組成物には医薬用として許容可能なアジュバントを含ませることができる。「アジュバント」とは、特定の抗原に対する免疫反応を非特異的に増大させることにより所与のワクチンにおいて必要な抗原量及び/又は対象抗原に対して十分な免疫反応を生じるのに必要な注射頻度を減少させる作用物質である。動物のワクチン接種に好適なアジュバントとしては、(ピーナッツ油、モノオレイン酸マンノイド及びモノステアリン酸アルミニウムを含有する)アジュバント65;完全又は不完全フロイントアジュバント;水酸化アルミニウム、リン酸アルミニウム、ミョウバンなどのミネラルジェル;ヘキサデシルアミン、オクタデシルアミン、リゾレシチン、臭化ジメチルジオクタデシルアンモニウム、N,N−ジオクタデシル−N’,N7−ビス(2−ヒドロキシメチル)プロパンジアミン、メトキシヘキサデシルグリセロール、プルロニックポリオールなどの界面活性剤;ピラン、硫酸デキストラン、ポリIC、ポリアクリル酸、CARBOPOL(登録商標)(例えば、CARBOPOL941)などのポリアニオン;ムラミルジペプチド、ジメチルグリシン、タフトシンなどのペプチド;及び油乳濁液が挙げられるが、これらに限定されるものではない。アジュバントに関する情報は、例えば、引用により本明細書に組み込まれているP.Tijssen によるシリーズ[Practice and Theory of Enzyme Immunoassays、第3版1987年、Elsevier、New York]に開示されている。
【0051】
任意選択的に、上記ワクチン組成物には、細菌及び真菌細胞壁成分(例えば、リポ多糖、リポ蛋白質、糖蛋白質、ムラミルペプチド)、植物由来の各種複合糖質(例えば、グリカン、アセマンナン)、動物由来の各種蛋白質及びペプチド(例えば、ホルモン、サイトカイン、共刺激因子)、ウイルス及び/又は他の供給源由来の新規核酸(例えば、二本鎖RNA、CpG)などの医薬用として許容可能な免疫刺激剤をさらに含ませることができる。
【0052】
上記ワクチン組成物は、静脈内、筋肉内、皮下、経口、鼻腔内、皮内及び/又は腹腔内ワクチン接種を含む任意の標準的経路によって容易に投与することができる。このワクチン組成物がレシピエント動物及び投与経路の各種類に対して適切に製剤化されることが好ましいことは当業者なら理解するであろう。
【0053】
従って、本発明はB.burgdorferi ss及び他のボレリア属菌に対してイヌを免疫化する方法をも提供する。そのような方法の1つは、イヌに免疫学的有効量の本発明ワクチンを注射することによりイヌに適切なOspA及びOspC殺ボレリア菌抗体を産生させることを含む。
【0054】
一実施態様において、本発明の皮下投与によってOspC7に特異的な殺ボレリア菌抗体を含む高濃度のOspA及びOspC殺ボレリア菌抗体が産生される。別の実施態様では、本発明は、有害なボレリア属菌に対して有効な各B.burgdorferi ss株を特定の最小限の量で含むワクチンを提供する。別の実施態様では、本発明のワクチンは、イヌにおいてB.burgdorferi ss及び他の病原性ボレリア属感染を防御するのに有効である。特定の実施態様では、上記ワクチンは、本発明の2種のB.burgdorferi ss株と医薬用として許容可能なアジュバントとの安全で免疫学的に有効な組合せを含む。
【0055】
本発明は、2種のB.burgdorferi ss分離株を特定の最低限の量で含むワクチンがダニ感染後のボレリア属菌からイヌを防御することを開示する。本発明はさらに、ワクチン接種したイヌにおいて特定の最小限の量のB.burgdorferi ss特異的OspA及びボレリア属菌特異的OspC殺ボレリア菌抗体を誘導するワクチンを開示する。また、本発明のワクチンは、許容可能な免疫刺激剤及び/又アジュバントと共に投与することもできる。
【実施例】
【0056】
以下の実施例は、本発明の理解をさらに進めるのに役立つものであるが、本発明の有効範囲を何ら限定するものではない。
【0057】
(実施例1)
組換えOspAを用いたワクチン接種
A.材料及び方法
・動物
Charles River Breeding Laboratories,Inc.Wilmington、マサチューセッツ州)から5乃至10週齢のLVGハムスターを入手した。ハムスターは21℃の室温でケージ当たり3乃至4匹を収容し、餌と水を自由に与えた。
【0058】
・ハムスターのワクチン接種及び血清の採取
ハムスターの頸背部皮下に市販組換えOspA(rOspA)ワクチンRECOMBITEK(登録商標)ライム(Lyme)(Merial Limited、Duluth、ジョージア州)を0.5mL接種し、この初回ワクチン接種の3週後に追加免疫を行った。初回ワクチン接種の3、7、9及び15週後に、1群5匹の各ハムスター群をノーズ・アンド・マウス・カップに含ませたエーテルの吸入により軽く麻酔し、心臓内穿刺により放血させた。この血液を凝固させ、血清を分離して、使用するまで−70℃で貯蔵した。さらに、ワクチン接種していないハムスター3匹からのハムスター血清をプールして対照正常血清として用いた。別の対照として、20匹のハムスターの両大腿後部に、B.burgdorferi ssS−1−10を含有する市販のイヌライム病全細胞ワクチン(Galaxy、Solvay Animal Health,Inc.、Mendota Heights、ミネソタ州、現Schering−Plough Animal Health、Elkhorn、ネブラスカ州)を0.25mL接種した。これらのハムスターに追加免疫した後、上述のようにして血液を採取した。
【0059】
・BSK培地の調製
B.burgdorferiのインビトロ培養のためにBarbour−Stoenner−Kelly(BSK)肉汁培地を用い、これを殺ボレリア菌抗体試験及び本発明全体にわたって記載した方法に用いる一次培地(primary substrate)とした。BSK培地は、Callisterら[参考としてその全体が本明細書に援用されるDetection of Borreliacidal Antibodies by Flow Cytometry、セクション11.5.1 − 11.5.12、Current Protocols in Cytometry、John Wiley
and Sons,Inc.Supplement26、(2003年)]により報告されたものと同様にして調製した。つまり、以下の方法によってBSK培地を調製した。
【0060】
材料:
HEPES(Sigma)
ネオペプトン(Neopeptone)(Difco)
クエン酸ナトリウム(Sigma)
グルコース(Sigma)
炭酸水素ナトリウム(Sigma)
TCイーストレート(yeastolate)(Difco)
ピルビン酸(Sigma)
N−アセチルグルコサミン(Sigma)
ウシ血清アルブミン(Sigma)
ゼラチン(微生物学用グレード;Difco)
5N NaOH 。
【0061】
L−グルタミンを含有するが、炭酸水素ナトリウムを含有しないIOxコンノート・メディカル・リサーチ・ラボラトリーズ(CMRL)1066(MP Biomedicals)又はローズウェル・パーク・メモリアル・インスティテュート(RPMI)1640培地(Sigma)
ウサギ血清(Life Technologies)、56℃45分間加熱不活化
56℃水浴
陽圧ポンプ
ミリポアフィルターマニフォールド(Millipore filter manifold)
プレフィルター(prefilter)(124mm)
0.2−、0.45−及び0.8−μmフィルター(142−mm径)
0.2−μmベル・フィルター(bell filter)
滅菌100−ml容器
暗視野顕微鏡 。
【0062】
BSK培地の調製
1.以下のものを2−リットルフラスコ中で混合し、2乃至4時間混和した。900mLミニQ二重濾過又は脱イオン蒸留水:
6.0g HEPES
5.0g ネオペプトン
0.7g クエン酸ナトリウム
5.0g グルコース
2.2g 炭酸水素ナトリウム
2.5g TCイーストレート
0.8g ピルビン酸
0.4g N−アセチルグルコサミン
50g ウシ血清アルブミン(フラクションV) 。
【0063】
2.これらの成分は攪拌板を最も低速の設定にしてゆっくりと混和した。何故なら、激しく攪拌すると、B.burgdorferiに毒性のある分解産物が生じる場合があるからである。
【0064】
3.上記BSK成分を混和している間に、ゼラチン溶液を以下の通り調製する。即ち、500−mLフラスコ中で、ミリQ二重濾過又は脱水蒸溜水200mLとゼラチン14gとを混合し、中程度の設定値で加熱し、攪拌して溶解させた。次いで、この溶液を121℃で15分間加圧滅菌した後、56℃の水浴に入れた。
【0065】
4.BSK成分が完全に溶解したとき、この溶液を5N NaOHでpH7.5に調整した。
【0066】
5.次に、上記ゼラチン溶液200mL、10xCMRL1066培地100mL及び(56℃45分間)加熱不活化ウサギ血清64mLをその他のBSK成分と混合し、よく混和した。
【0067】
BSK培地の滅菌
6.前記陽圧ポンプを用いて、124−mmプレフィルター及び最小孔径(底部)から最大孔径(最上部)まで積み重ねられた142−mm0.2−、0.45−及び0.8−μm孔径フィルターを装着したミリポアフィルターマニフォールドを通してBSK培地を送り出した。[このBSK培地は加圧滅菌によって滅菌することができない。前濾過は、滅菌濾過の前に大きな粒子を除去しておくのに必要であった。]
7.次いで、前記陽圧ポンプ及び滅菌0.2−μmベル・フィルターを用いて上記BSK培地を滅菌容器中へ滅菌濾過した。
【0068】
滅菌性の確認
8.滅菌BSKの1−mL分量を無菌的に取り出し、滅菌した1.5−mL微小遠心管に移して35℃で一夜インキュベートした。残る滅菌濾過したBSKは4℃で貯蔵した。
【0069】
9.インキュベーション後、暗視野顕微鏡を用いてこの滅菌BSKを調べ、滅菌性を確認した。
【0070】
10.次に、この滅菌BSKを滅菌貯蔵容器中に移した。[滅菌BSK培地の酸化を低減させるためにこの貯蔵容器に充填する際、その上部にできる空間を最小限に保った。]
Barbour−Stoenner−Kelly(BSK)培地の品質管理
BSK培地を使用する前に、BSKが少数のボレリア属菌を増殖させるのに有利であること、及び生育可能なスピロヘータが凝集せずに増殖することを確認することは重要である。BSK培地の極めて変化しやすい成分はウシ血清アルブミン(BSA)である。本明細書で用いている品質管理プロトコルは、Callisterほか、2003年、上記文献に詳細に記載されているものと同様に、得られた培養物をインキュベートして検査する試験培養を利用したものである。
【0071】
・OspA殺ボレリア菌抗体の検出
OspA殺ボレリア菌抗体は、フローサイトメトリー法及びB.burgdorferi ssS−1−10を用いて検出した(Callisterほか、Arch.Intern.Med.1994年、154:p.1625−1632)。スピロヘータの新鮮培養液を新鮮なBSKでほぼ5×106個/mLのスピロヘータ濃度に希釈した。同時に、血清試料を新鮮BSKで40倍希釈し、0.2ミクロン(μm)孔径微小遠心フィルターを通して滅菌した。次いで、200μLの分量を滅菌された1.5mLねじ口微小遠心管に移し、BSKで80倍から20,480倍まで連続希釈した。血清試料を56℃で10分間加熱不活化した後、上記スピロヘータ懸濁液の100μL分量(スピロヘータ:5×105個)及び滅菌モルモット補体(Sigma)10μLを添加した。こうした分析試料をよく混和し、35℃で16乃至24時間インキュベートした。
【0072】
インキュベーション後、PBS400μL及びアクリジンオレンジ1μg/mLを入れたポリプロピレン管に各分析試料懸濁液100μLを移した。次いで、FACScanフローサイトメータ(Becton Dickinson lmmunocytometry Systems)、San Jose、カリフォルニア州)を用いて殺ボレリア菌活性を検出した。スピロヘータは、ゲート処理(gating)(CellQuestソフトウェア、Becton Dickinson)によって単離し、流速を低く設定して1乃至2分間分析した。OspA殺ボレリア菌抗体は、アクリジンオレンジが水疱状の生育不能スピロヘータ中に入り込む時に生じる蛍光強度の増大を観察することによって間接的に検出した。対照正常血清との比較における平均蛍光強度の13%以上のシフトを陽性と見なした(Callisterほか、Clin.Diagn.Lab.Immunol.2002年、9:p.908−912)。次に、暗視野顕微鏡によって水疱状の運動能力のないB.burgdorferiの存在を確認した。各分析試料に対して陽性対照を含め、ラン毎の反応性を同一(+/−1希釈)にするには分析法間の変動をできるだけ少なくすることが必要であった。さらに、各イヌから採取した血液試料を同時に分析した。
【0073】
B.結果
rOspAワクチンを用いてワクチン接種すると、7週後にごくわずかな量(平均力価:<61)のOspA殺ボレリア菌抗体が誘導されたが、15週後では検出されなかった(表2)。これに対して、通常の(Barbourほか、J.Infect.Dis.1985年、152:p.478−484)B.burgdorferi ss株(S−1−10)を含有するイヌのワクチンを用いてハムスターにワクチン接種すると、高レベルの殺ボレリア菌OspA抗体が誘導され、7週目にピーク(力価:>2560)に達し、実験期間中、高値を維持した。
【0074】
表2: rOspA又はGALAXY(商標)(S−1−10)イヌライム病ワクチンを用いたワクチン接種後のOspA殺ボレリア菌抗体bの平均力価a
【0075】
【表2】
従って、イヌの組換えOspAワクチンではハムスターにおいて有意な力価のOspA殺ボレリア菌抗体を誘導できない。
【0076】
(実施例2)
組換えOspCを用いたワクチン接種
A.材料及び方法
・動物
Charles River Breeding Laboratories,Inc.(Wilmington、マサチューセッツ州)から5乃至10週齢のLVGハムスターを入手した。ハムスターは21℃の室温でケージ当たり3乃至4匹を収容し、餌と水を自由に与えた。
【0077】
・組換え(「r:recombinant」)OspCワクチンの調製
rOspCは、以前に報告されたもの(Rousselleほか、J.Infect.Dis.1998年、178:p.733−741)と同様にして、pX3−22を含有するE.coli JM109から回収した。つまり、このE.coliをアンピシリンを含有する2xTYブロス中37℃で培養し、これにイソプロピル−β−d−ガラクトピラノシド(IPTG)(0.1mM)を指数増殖期の間に添加した。次いで、細胞を遠心分離によりペレット化し、リン酸緩衝食塩水(PBS)に再懸濁して超音波処理により溶解させた。次に、トリトンX−100(1%容積/容積)を添加した後、溶解物を10,000×gで5分間遠心した。次いで、この超音波処理したE.coliを遠心によりペレット化し、上清を、アミノ末端のビオチン化精製タグを介してOspCに結合したSoftLink樹脂(Promega)を含有するカラム上を通した。次に、5mMビオチン(Sigma)をも含有する精製緩衝液でこの結合OspCを溶出させた。
【0078】
・ハムスターのワクチン接種及び血清の採取
ハムスターの頸背部皮下に、75μgのrOspCを含有するフロイント完全アジュバントを0.1mL接種し、この初回ワクチン接種の3週後に75μgのrOspCを含有するフロイント不完全アジュバントで追加免疫を行った。初回ワクチン接種の5週後に、各ハムスターをノーズ・アンド・マウス・カップに含ませたエーテルの吸入により軽く麻酔し、心臓内穿刺により放血させた。この血液を凝固させ、血清を分離して、使用するまで−70℃で貯蔵した。さらに、正常なハムスター3匹からのハムスター血清をプールして対照正常血清として用いた。
【0079】
・OspC抗体の検出
rOspCをコーティング緩衝液(0.015M Na2CO3、0.035M NaHCO3、pH9.6)で1000ng/mLとなるように希釈し、各100μL量を個々の平底アミン結合マイクロタイター・ウェル(Costar、Cambridge、マサチューセッツ州)に添加した。マイクロタイター・プレートを4℃で一夜インキュベートした。インキュベーション後、プレートをPBS(pH7.2)で3回洗浄し、0.05%TWEEN20(Sigma)及び1%ウシ血清アルブミン(Sigma)を含有するPBSで振盪しながら室温で1時間ブロックした。ブロックの後、プレートを再びPBSで洗浄した。次いで、PBS/Tweenで80倍から20,480倍まで連続希釈したハムスター血清各100μL量を個々のウェルに添加し、これらのプレートを室温で1時間インキュベートした。インキュベーション後、プレートをPBSで3回洗浄し、PBS/Tweenで3,000倍希釈した抗ハムスターIgGホースラディッシュペルオキシダーゼ結合体(Organon Teknika Cappel)100μLを各ウェルに添加し、これらのプレートを室温で1時間再度インキュベートした。次に、プレートをPBSで3回洗浄し、リン酸o−フェニレンジアミン(0.4mg/ml;Sigma)100μLを各ウェルに添加して室温で30分間インキュベートさせた。1N H2SO4を100μL添加して反応を止め、直ちに490nm(モデルEL311;Bio−Tek Inc.、Winooski、バーモント州)での吸光度を測定した。
【0080】
・OspC殺ボレリア菌抗体の検出
B.burgdorferi ss50772を用いた以外は前述(実施例1)と同様にして、OspC殺ボレリア菌抗体を検出した。
【0081】
B.結果
rOspCワクチンを用いてワクチン接種すると、ELISAにより検出される高レベルのOspC抗体が誘導されたが(以下の表3)、殺ボレリア菌活性は低レベル(力価:80)で存在するに過ぎなかった。従って、rOspCを用いてワクチン接種すると、高濃度のOspC抗体が誘導されたが、その反応は、感染に対する防御をもたらさない抗体(例えば、オプソニン化抗体)でほとんど構成された。このことは、B.burgdorferi ss50772ワクチン中の蛋白質の75μg濃度が多数の別の非OspC蛋白質の存在を計上しないで算出されることを考慮すると、特に重要であった。従って、全体的な結果(実施例1及び2)から、組換えOspでは完全な状態のB.burgdorferi ssよりも殺ボレリア菌抗体誘導の量が相当に少ないというこれまでの知見(Straubingerほか、Vaccine 2003年、20:p.181−193)が裏付けられた。
【0082】
表3: rOspCによるワクチン接種後のOspCELISA又はOspC殺ボレリア菌抗体試験bによって検出された抗体の平均力価a
【0083】
【表3】
従って、イヌの組換えOspCワクチンではハムスターにおいて有意な力価のOspC殺ボレリア菌抗体を誘導できない。
【0084】
(実施例3)
B.burgdorferi ss分離株S−1−10を用いたワクチン接種
A.材料及び方法
・生物
B.burgdorferi ss分離株S−1−10は、通常B.burgdorferi ssB31と区別できない代表的な病原性株である(Barbourほか、J.Infect.Dis., 1985年、152:p.478−484)。この分離株は元々、ウィスコンシン州La Crosse付近から1998年2月に捕獲されたシロアシネズミPeromyscus leucopusの腎臓からウィスコンシン州La CrosseのGundersen Lutheran Medical FoundationのSteven M.Callister博士によって回収され、イヌのライム病ワクチンに組み入れるためにSolvay Animal Healthによりライセンス供与されたものである。その後の市販品(GALAXY(商標)ライム(Lyme))は1997年4月17日にSchering−Plough Animal Healthによって獲得された。この菌はOspA、OspB及びOspCを発現する。しかしながら、感染性B.burgdorferi ss分離株に特有であるが、スピロヘータを実験室BSK培地で培養すると、有意により高濃度のOspA及びOspBが産生される。OspCの発現は35℃でのインキュベーションによって最大化することができることが分かっている(Schwan、Biochem.Soc.Trans.2003年、31:p.108−112)。
【0085】
・動物
8週齢のビーグル幼犬(Ridglan Farms、Mount Horeb、ウィスコンシン州)を共同飼育し、餌と水を自由に与えた。実験はSchering Plough Animal Health動物管理使用委員会(IACUC:Animal Care and Use Committee)によって審査され、承認された。
【0086】
・B.burgdorferi ssS−1−10ワクチンの調製
35℃のBSKにおけるインキュベーションにより対数増殖期に達したB.burgdorferi ssS−1−10の新鮮培養物を、二成分エチレンイミン(BEI:binary ethylenimine)を最終濃度10mMとなるように添加してさらに48時間インキュベートすることによって不活化した。不活化後、滅菌チオ硫酸ナトリウムを添加して35℃で6乃至12時間インキュベートすることによりBEIを中和した。次いで、スピロヘータを遠心によりペレット化し、≦30μg/mLのゲンタマイシン及び≦30単位/mLのナイスタチンを含有する滅菌平衡塩類溶液に再懸濁した。次に、このスピロヘータに5%Emulsigen(登録商標)(MVP Laboratories,Inc.)及び1%HEPESを混合することによって1.0mL用量が≧2.5×107個のスピロヘータを含有するようにした。
【0087】
・ワクチン接種及び血清の採取
イヌの頚部皮下に1mL用量の上記S−1−10ワクチンを接種し、21日後にさらに1mL用量を追加免疫した。追加免疫ワクチン接種(試験21日目)の直前並びに試験28、35、47、83及び113日目に頚静脈穿刺によって全血を採取した。次いで、遠心により血清を分離し、これを試験するまで−20℃で貯蔵した。
【0088】
・殺ボレリア菌抗体の検出
上記の実施例1及び2と同様に、B.burgdorferi ssS−1−10及び50772を用いてフローサイトメトリーによりOspA及びOspC殺ボレリア菌抗体を検出した。
【0089】
B.結果
B.burgdorferi ssS−1−10を用いてワクチン接種することにより高レベルの殺ボレリア菌OspA抗体が確実に誘導された。この抗体は試験21日目には検出可能であり、試験28日目にピークに達し、試験113日目でも依然として検出可能であった(以下の表4)。これに対して、OspC殺ボレリア菌抗体は検出されなかった(力価:1:80未満)。すなわち、通常のB.burgdorferi ss株では、スピロヘータを35℃でインキュベートすることによりOspC発現を最大化したにもかかわらず、OspC殺ボレリア菌抗体の産生を誘導できなかった。
【0090】
表4: B.burgdorferi ssS−1−10によるワクチン接種後のOspA又はOspC殺ボレリア菌抗体の平均力価a(=8)
【0091】
【表4】
従って、B.burgdorferi ss分離株S−1−10を用いたワクチンでは有意な力価のOspC殺ボレリア菌抗体を誘導できなかった。
【0092】
(実施例4)
B.burgdorferi ss分離株50772を用いたワクチン接種
A.材料及び方法
・菌
B.burgdorferi ss50772は元々Andersonほか、J.of Clin.Microbiol.,1996年、34:p.524−529に報告されたユニークなospA−/ospB−株であり、著者らがOspC抗原を発現するものとみなすことができなかった株である。この50772株はOspC抗原を発現するものとして米国特許第6,464,985号に開示されており、これは引用により本明細書に組み込まれているが、この株はPTA−439の番号でATCCに寄託されているとこの特許に記載されている。
【0093】
・動物
8週齢のビーグル幼犬(Ridglan Farms、Mount Horeb、ウィスコンシン州)を共同飼育し、餌と水を自由に与えた。実験はSchering Plough Animal Health動物管理使用委員会(IACUC:Animal Care and Use Committee)によって審査され、承認された。
【0094】
・B.burgdorferi ss50772ワクチンの調製
上記の実施例3で説明したものと同様にして、B.burgdorferi ss50772ワクチンを調製した。
【0095】
・ワクチン接種及び血清の採取
イヌの頚部皮下に1mL用量の上記50772ワクチンを接種し、21日後にさらに1mL用量を追加免疫した。追加免疫ワクチン接種(試験21日目)の直前並びに試験28、35、47、83及び113日目に頚静脈穿刺によって全血を採取した。次いで、遠心により血清を分離し、これを試験するまで−20℃で貯蔵した。
【0096】
・殺ボレリア菌抗体の検出
上記の実施例1及び2と同様に、B.burgdorferi ss50772を用いてフローサイトメトリーによりOspC殺ボレリア菌抗体を検出した。
【0097】
B.結果
B.burgdorferi ss50772を用いてワクチン接種することにより高レベルの殺ボレリア菌OspC抗体が確実に誘導された。この抗体は試験35日目にピークに達し、試験113日目でも依然として検出可能であった(以下の表5)。すなわち、rOspC又はB.burgdorferi ss分離株(S−1−10)によるワクチン接種とは対照的に、上記のユニークなB.burgdorferi ss株50772を用いてワクチン接種すると、有意なレベルのOspC殺ボレリア菌抗体が誘導された。
表5: B.burgdorferi ss50772によるワクチン接種後のOspC殺ボレリア菌抗体の平均力価a(=8)
【表5】
【0098】
従って、B.burgdorferi ss分離株50772を含むワクチンは高濃度のOspC殺ボレリア菌抗体を誘導する。
【0099】
(実施例5)
B.burgdorferi ss分離株S−1−10及び50772を含むワクチンの調製
A.材料及び方法
・増殖
B.burgdorferi ss分離株S−1−10及び50772の凍結保存培養物を標準的な条件下で培養した。すなわち、これらの保存培養物を、5%CO2を混入させた空気の雰囲気下で[本明細書にその全体が引用により組み込まれているCallisterほか、1990年、J.of Clinical Microbiology 28:p.363−365に準じて調製した]BSK(Barbour−Stoenner−Kelly)培地5乃至17mLを含有する個々のねじ口培養管中、それぞれ33±2℃及び35±2℃で増殖させた。対数増殖に達した後、これらの培養物を用いて150乃至200mLの新鮮BSK培地に植菌し、次に、これを培養物が対数増殖に達するまでインキュベートした。次いで、得られた懸濁液を用いて、10乃至20リットルジャグに入れた5乃至10リットルの新鮮BSKに植菌し(生産培養)た後、さらに、上記のようにしてインキュベートした。
【0100】
・不活化
生産培養物中のスピロヘータを、BEIを10mMの濃度となるように添加して24乃至48時間ゆっくりと攪拌しながらインキュベートすることによって不活化した。スピロヘータの不活化後、10.6mLの滅菌済3.015Mチオ硫酸ナトリウムを各1リットルのBSL/スピロヘータ懸濁液に添加して6乃至12時間ゆっくりと攪拌しながらインキュベートすることによりBEIを中和した。
【0101】
・濃縮及び混合
不活化スピロヘータを遠心によりペレット化し、30.0μg/mLのゲンタマイシン及び30単位/mLのナイスタチンを含有する滅菌平衡塩類溶液に再懸濁した。この試験ワクチンに5%Emulsigen(登録商標)(MVP Laboratories,Inc.)を混合し、2.0mLガラスバイアルに充填して各用量(1.0mL)に≧2.5×107個/mLのB.burgdorferi ssS−1−10、≧5.0×108個/mLのB.burgdorferi ss50772、1%HEPES、29μ/mLゲンタマイシン及び29単位/mLナイスタチンを含有させた。
【0102】
(実施例6)
B.burgdorferi ss分離株S−1−10及び50772を含むワクチンの安全性及び有効性
A.材料及び方法
・動物
8週齢のビーグル幼犬(Ridglan Farms、Mount Horeb、ウィスコンシン州)を個別又は共同飼育し、餌と水を自由に与えた。実験はSchering Plough Animal Health動物管理使用委員会(IACUC)によって審査され、承認された。
【0103】
・ワクチン接種及び血清の採取
イヌの頚部皮下に1mL用量の上記ワクチンを接種し、21日後にさらに1mL用量を追加免疫した。初回(試験−3日目)及び追加免疫ワクチン接種(試験21日目)の前並びに試験28、35、43、78、106、134、162及び197日目にも頚静脈穿刺によって全血を採取した。次いで、遠心により血清を分離し、これを試験するまで−20℃で貯蔵した。
【0104】
・ワクチン接種後の観察
各ワクチン接種後、注射部位を反応が感じられなくなるまで毎日触診し、直腸温をワクチン接種後4乃至6時間及びワクチン接種後1乃至3日目に記録した。
【0105】
・ダニ感染
2回目のワクチン接種の3週後に、イヌを胸腔の右側で剃毛し、B.burgdorferi ss感染I.scapularis tickの雌10匹及び雄10匹を入れたラバーカップを剃毛部にテープ及び包帯ラップ(bandage wrap)で固定した。これらのダニに7日間吸血させた。
【0106】
・ダニにおけるB.burgdorferi ssの検出
ダニの口器をメスで除去し、中腸を掻き裂き、スライドガラスになすりつけて室温で一夜乾燥させた。乾燥後、このスライドをアセトンで8乃至10分間固定し、風乾した。種特異的マウスOspAモノクロナール抗体H5332をPBS(pH7.2)で40倍希釈し、PBSで500倍希釈したヒツジ抗マウスフルオレセインイソチオシアネート標識免疫グロブリンG抗体を重層した。インキュベーション後、スライドをPBSですすぎ、風乾して蛍光顕微鏡により調べた。各スライドは経験豊かな二名の微生物学者により独立に調べた。
【0107】
・血液試料
試験78、106、134、162及び197日目に全血を血清分離輸送(SST)管に採取し、血清を分離して試験するまで−20℃で貯蔵した。
【0108】
・OspA抗体の除去
以前に報告されたもの(Callisterほか、J.Infect.Dis.1993年、167:p.158−164)と同様にして、OspA−グルタチオン−S−トランスフェラーゼ融合蛋白質を発現するEscherichia coli(「E.coli」)DH5αから組換え(r)OspAを回収した。つまり、この大腸菌を、アンピシリンを含有する2xTYブロス中37℃で培養し、指数増殖期中にイソプロピル−β−ガラクトピラノシド(「IPTG」)(0.1mM)を添加した。次いで、細胞を遠心によりペレット化し、リン酸緩衝食塩水(「PBS」)に再懸濁して超音波処理により溶解させた。次に、トリトンX−100(1%容積/容積)を添加した後、溶解液を10,000×gで5分(min.)間遠心した。次に、上記融合蛋白質を含有する上清をグルタチオン−セファロース4Bカラム(Pharmacia)に通し、50mMトリス−Cl(pH8.0)プラス5mM還元グルタチオンで組換えOspAを溶出し、これを精製緩衝液(50mMトリス[pH8]、50mM NaCl、2mM EDTA、0.1%トリトンX−100)に再懸濁した。
【0109】
次に、rOspA融合蛋白質を臭化シアン(CNBr)活性化によりセファロース4Bに結合させた。具体的には、0.8gのCNBr活性化セファロース4B(Pharmacia)をカップリング緩衝液(0.1M NaHCO3−0.5M NaCl、pH8.3)で洗浄し、カップリング緩衝中の2mg量のOspAをこのゲルに添加し、この混合液を室温で2時間静かに振盪した。ゲルをカップリング緩衝液で2回洗浄した後、4mLのエタノールアミン(pH9)を添加し、ゲルを2時間インキュベートして未結合部位をブロックした。次いで、ゲルを50mLの0.1M酢酸ナトリウム−0.5M NaCl(pH4.0)で3回洗浄した後、PBS中で平衡化した。PBSで10倍希釈した免疫血清の試料1mLを4回上記カラムに通した。
【0110】
・OspC抗体の除去
以前に報告された(Rousselleほか、J.Infect.Dis.1998年、178:p.733−741)ようにして、pX3−22を含有するE.coli JM109からrOspCを回収した。この組換え蛋白質の産生は上記のrOspAで述べたものと同様にして誘導された。次いで、超音波処理したE.coli細胞を遠心によりペレット化し、上清を、OspCとそのアミノ末端のビオチン化精製タグを介して結合したSoftLink樹脂(Promega)を含有するカラム上を通した。次に、5mMビオチン(Sigma)をも含有する精製緩衝液でこの結合OspCを溶出させた。
【0111】
次に、1mL容量のテトラリンク四量体アビジン樹脂(Promega)を洗浄し、40mLのPBSに懸濁し、10mm×70mmのポリエチレンカラムにロードした。1mL容量のPBS中の0.5mg量の透析rOspCを上記カラムに通し、タンパク質測定(Bio−Rad)により結合を確認した。次いで、PBSで10倍希釈した1mL容量の免疫血清を4回このカラムに通した。
【0112】
・OspC7特異的抗体の除去
以前に報告された(本明細書にその全体が引用により組み込まれているLovrichほか、Clin.Diagn.Lab.Immunol.2005年、12:p.746−751)ようにして、pXT7を含有するE.coli JM109から、OspCの7個のC末端アミノ酸(OspC7)を含有する融合蛋白質を回収した。この組換え蛋白質の産生は上記のrOspAで述べたものと同様にして誘導された。次いで、超音波処理したE.coli細胞を遠心によりペレット化し、上清を、OspC7とそのアミノ末端のビオチン化精製タグを介して結合したSoftLink樹脂(Promega)を含有するカラムに通した。次に、5mMビオチン(Sigma)をも含有する精製緩衝液でこの結合OspC7を溶出させた。次に、1mL容量のテトラリンク四量体アビジン樹脂(プロメガ社)を洗浄し、40mLのPBSに懸濁し、10mm×70mmのポリエチレンカラムにロードした。1mL容量のPBS中の0.5mg量の透析rOspCを上記カラムに通し、タンパク質測定(Bio−Rad)により結合を確認した。次いで、PBSで10倍希釈した1mL容量の免疫血清を4回このカラムに通した。
【0113】
・殺ボレリア菌抗体の検出
上記の実施例1及び2と同様に、B.burgdorferi ssS−1−10及び50772を用いてフローサイトメトリーによりOspA及びOspC殺ボレリア菌抗体を検出した。
【0114】
・皮膚生検
試験78、106、134、162及び197日目にダニ付着の近傍部位から使い捨ての4mm穿刺器具を用いて皮膚生検材料を採取した。これらの生検材料は、9mlのゼラチンを補ったBSK(Callisterほか、J.Clin.Microbiol.1990年、28:p.363−365)(BSK+G)、40乃至50マイクログラム(μg)/mLリファンピン及び8μg/mLカナマイシンを含有する別々の管に入れた。培養物は35±2℃で3週間インキュベートし、暗視野顕微鏡検査法により週1回スピロヘータを調べた。
【0115】
・免疫抑制
イヌにおいて、感染後13日から始めて5日間デキサメタゾン(0.4mg/lb体重)の連日投与により免疫を抑制した。
【0116】
・臨床的観察
イヌを、こわばり(足に全体重をかけたがらないこと)、跛行(歩行又は走行中足をかばうこと)又は歩行困難(免荷(non−weight bearing))を含む肢/関節障害について毎日観察した。採点は少なくとも2名の観察者の合意によって行い、イヌは、他の外傷を全て除外するために担当獣医によって観察した。
【0117】
・剖検
少なくとも3連続観察期間の間、肢/関節障害を有したイヌは安楽死させて剖検した。肘、手根、膝、足根、心臓、脾臓、膀胱及び両腎臓からの組織試料を採取し、培養によるB.burgdorferiの単離のために処理した。関節組織は9mLのBSK+Gを含有する個々の管中で培養した。他の組織(心臓、脾臓、膀胱及び腎臓)は9mLのBSK+Gと混合し、ストマッチャーでよくホモジナイズし、1mL量を9mLの新鮮BSK+Gを含有する別の管に移した。培養物は35±2℃でインキュベートとした。
【0118】
残りのイヌは試験終了時に安楽死させて剖検した。これらのイヌからの組織試料は肘、手根、膝及び足根の関節包及び腱から採取し、組織病理検査のために10%緩衝ホルマリン中に貯蔵した。さらに、肘、手根、膝及び足根の関節包及び腱からの試料を9mLのBSK+G又は抗生物質含有BSK+Gを入れた個々の管に入れ、35±2℃で3週間インキュベートした。
【0119】
・ウエスタンブロッティング
ウエスタンブロッティングは標準的な技術を用いて実施した。B.burgdorferi ss297を試料緩衝液中で5分間煮沸し、150μgの全蛋白質を0.1%SDS−12%ポリアクリルアミドゲル(櫛を含まない4%ポリアクリルアミド濃縮用ゲル)上にロードした。蛋白質濃度は蛋白質測定キットで測定し、電気泳動装置において2つのゲルを同時に泳動させた。電気泳動後、蛋白質をニトロセルロースに移し、このニトロセルロースを細片に切ってPBS−0.3%TWEEN20で22℃30分間ブロックした。次いで、細片を100倍希釈したイヌ血清と共に22℃で1時間インキュベートした後、PBS−0.3%TWEEN20で3回洗浄した。ホースラディッシュペルオキシダーゼ標識抗イヌIgG(Kirkegaard & Perry Laboratories、Gaithersburg、メリーランド州)を添加し、これらの細片を22℃で30分間インキュベートした後、TMB膜ペルオキシダーゼ基質システム(Kirkegaard & Perry Laboratories、Gaithersburg、メリーランド州)で発色させた
B.結果
・ワクチンの安全性
イヌは、ワクチン接種後、直腸温の上昇がないなど、臨床的に正常な状態を維持した。しかしながら、ワクチンを接種されたイヌは注射部位に軽度の腫脹を生じ、これは8日以内に消散した。その最大の反応は(ワクチン接種後24時間の)1×1×0.5cmの大きさであった。
【0120】
・ワクチン接種後の血清
プラセボ接種では、B.burgdorferi ssに特異的な抗体を誘導することができなかった。これに対し、上記ワクチンの接種では、B.burgdorferi ss分離株S−1−10又は50772を用いることで検出される殺ボレリア菌抗体が有意なレベルで誘導された。S−1−10を用いて検出される殺ボレリア菌抗体の平均力価は追加免疫ワクチン接種(試験28日目)の1週後にピークに達し、試験197日目でも依然として検出可能であった(下記表6)。同様に、分離株50772に特異的な高濃度の殺ボレリア菌抗体が試験28日目までに存在し、試験43日目ではなお高く、試験197日目でも依然として検出可能であった(下記表7)。
【0121】
表6: ワクチン接種後の殺ボレリア菌活性bの平均力価a(n=15)
【0122】
【表6】
表7: ワクチン接種後の殺ボレリア菌活性bの平均力価a(n=15)
【0123】
【表7】
・OspA及びOspC殺ボレリア菌抗体の確認
上記殺ボレリア菌活性がOspA及びOspC殺ボレリア菌抗体によるものであることを確認するために、上記ワクチンを用いてワクチン接種した5頭のイヌからの免疫血清をrOspA又はrOspCを含有する別々のカラムに通し、殺ボレリア菌活性に及ぼす影響を調べた。これらの組換え蛋白質で上記血清を吸収させることによりOspA特異的又はOspC特異的抗体を除去すると、それぞれB.burgdorferi ssS−1−10又は50772分離株を用いて検出される殺ボレリア菌抗体が著しく(4分の1以下に)減少した。従って、こうした全体的な知見から、上記ワクチンが有意なレベルのOspA及びOspC殺ボレリア菌抗体を誘導し、B.burgdorferi ssS−1−10又は50772により検出される殺ボレリア菌活性が、それぞれOspA又はOspC殺ボレリア菌抗体にほとんど限定されたものであることが確認された。下記の表8を参照されたい。
【0124】
表8:ワクチンを用いてワクチン接種したイヌ(n=5)からの血清における殺ボレリア菌活性に及ぼすOspA又はOspC抗体除去の影響
【0125】
【表8】
・OspC7特異的殺ボレリア菌抗体の確認
上記OspC殺ボレリア菌抗体がかなりの割合のOspC7特異的殺ボレリア菌抗体を含有することを確認するために、上記ワクチンを用いてワクチン接種した5頭のイヌからの免疫血清をrOspC7含有カラムに通し、殺ボレリア菌活性に及ぼす影響を調べた。この組換えOspC7蛋白質で上記血清を吸収させることによりOspC7特異的抗体を除去すると、B.burgdorferi ss50772分離株を用いて検出される殺ボレリア菌抗体が著しく(2乃至4分の1に)減少した。従って、これらの知見から、上記ワクチンにより、保存OspC7エピトープに特異的なOspC殺ボレリア菌抗体が有意なレベルで誘導されることが確認された。下記の表9を参照されたい。
【0126】
表9: ワクチンを用いてワクチン接種したイヌ(n=5)からの血清における殺ボレリア菌活性に及ぼすOspC7抗体除去の影響
【0127】
【表9】
・感染ダニに対するOspA及びOspC殺ボレリア菌抗体の滅菌能
ワクチン接種又は対照イヌにおいて吸血したダニの中腸を調べることによって、ワクチンにより誘導されたOspA及びOspC殺ボレリア菌抗体がダニを滅菌したことが確認された。プラセボ接種したイヌ15頭中13頭において吸血した雌性ダニ106匹中34匹(32%)からのダニ塗抹においてB.burgdorferi ssが検出された(下記の表10)。これに対して、ワクチン接種したイヌにおいて吸血した雌性ダニからの中腸ではスピロヘータは検出されなかった(99匹中0)(p<0.0001)。
【0128】
表10: ワクチン接種又は対照イヌから取り出した雌性ダニにおけるB.burgdorferi ssの検出
【0129】
【表10】
・皮膚からB.burgdorferi ssが回収されないようにする上記ワクチンの能力
上記ワクチンにより誘導された殺ボレリア菌抗体はスピロヘータが皮膚にコロニーを形成するのも防止した。ダニ感染後1ヶ月おきにプラセボ接種イヌから採取した71個の生検材料のうち56個(79%)からB.burgdorferi ssが回収され(下記の表11)、これら15頭のイヌのうちの14頭(93%)の少なくとも1個の皮膚生検材料からスピロヘータが回収された。これに対して、上記ワクチンを接種したイヌから採取したどの皮膚生検材料からもB.burgdorferi ssは回収されなかった(p<0.0001)。
【0130】
表11: ワクチン接種又は非ワクチン接種対照イヌの皮膚からのB.burgdorferi ssの単離
【0131】
【表11】
・感染の血清学的証拠をもたらさないようにする上記ワクチンの能力
上記ワクチンはライム病特有の抗体の発生をも阻止した。これまでの研究(SPAHC Report B01−184−01 R)によって、B.burgdorferi ssによる感染はほぼ20kDaの蛋白質に結合するイヌ抗体を確実に誘導することが確認されたが、この反応は上記ワクチンにより誘導されなかった(図1)。今回の試験では、感染特異的20kDa蛋白質に結合する抗体は15頭のプラセボ接種対照イヌのうちの14頭(93%)からの免疫血清中に容易に検出することができた(図2A)。さらに、20kDa蛋白質の抗体を含有しない血清が、皮膚からスピロヘータも得られない対照イヌから採取された。これに対して、上記ワクチンを接種したイヌのいずれによっても20kDa蛋白質の抗体は産生されなかった(p<0.0001)(図2B)。
【0132】
さらに、ライム病を有するイヌは、B.burgdorferi ss50772分離株を用いても検出できる非OspC殺ボレリア菌抗体を産生する(Callisterほか、J.Clin.Microbiol.2000年、38:p.3670−3674)。この反応の明確な標的は不明であるが、こうした抗体によって感染の高度に特異的な血清診断的確認が可能になる。上記ワクチンを接種したイヌは既にOspC殺ボレリア菌抗体を産生していたが、ダニ感染後B.burgdorferi ss50772特異的抗体のレベルの著しい(≧4倍の)上昇を生じたイヌはいなかった。従って、これらのワクチン接種イヌがスピロヘータに感染することは全くありそうになかった。これに対して、殺ボレリア菌抗体はダニ感染前のプラセボ接種イヌにより産生されなかったが、ダニに吸血させた後では15頭中10頭(67%;p=0.0002)が、B.burgdorferi ss50772を用いて検出される殺ボレリア菌抗体を顕著なレベル(力価:≧1:640)で生じた(下記の表12)。
【0133】
表12: ダニ感染後における殺ボレリア菌抗体bの平均力価a
【0134】
【表12】
・明白な肢/関節障害に対する上記ワクチンの防止能
以前の研究(Summersほか、J.Comp.Path.2005年、133:p.1−13、Wikleほか、J.Appl.Res.Vet.Med.2006年、4:p.23−28)によって、B.burgdorferi ssのみの感染では明白な肢/関節障害が生じることはまれであることが明らかにされたが、我々の知見でもこのことが裏付けられた。1頭(8%)のプラセボ接種対照イヌのみが関節異常を生じた。ダニ感染の9週後に左前肢がこわばり、B.burgdorferi ssスピロヘータが肘から回収された。しかしながら、こうした知見は重要ではなかった。何故なら、上記ワクチンを接種したイヌ2頭も1肢以上にこわばりを生じたが、プラゼボ接種イヌとは対照的に、どの組織からもスピロヘータは回収されなかったからである。
【0135】
明白な症状の発症を増幅させるために、残りのイヌに対して免疫を抑制した。その後、プラセボ接種対照イヌ3頭が歩行困難になり、これらのイヌのうち2頭からB.burgdorferi ssが回収された。これに対して、上記ワクチンを接種した免疫抑制イヌは歩行困難を生じなかった。従って、これらの知見全体から、上記ワクチンはライム関節炎の発症を防止することが示唆されたが、こうした結果は有意ではなかった(p=0.0996)。しかしながら、以下に示し、説明した別のデータを参照されたい。
【0136】
・B.burgdorferi ss感染に伴うびらん性変化に対する上記ワクチンの防止能
しかしながら、上述のイヌの関節を顕微鏡で調べたときの所見では、上記ワクチンの効果が明らかに示された。上記ワクチンを接種した残りのイヌ(n=13)からの関節包は正常であった。これに対して、プラセボ接種した残りの対照イヌ11頭(上記の表7)のうちの6頭(p=0.0034)からの1個以上の関節組織は、好中球及び単核細胞の浸潤を特徴とする著しい炎症を示した(図3)。さらに、上記ワクチンを接種したイヌからのどの組織からもB.burgdorferi ssは回収されなかったが、びらん性変化を有するプラセボ接種対照イヌ6頭のうちの5頭(83%)の関節組織からスピロヘータが回収された。従って、これらの知見全体から、上記ワクチンがイヌライム病関節炎を防止することが確認され、この症状が、ほとんどの場合、潜在性滑膜炎を特徴とするという以前の報告(Summersほか、J.Comp.Path.2005年、133:p.1−13;Wikleほか、J.Appl.Res.Vet.Med.2006年、4:p.23−28)も裏付けられた。
【0137】
(実施例7)
OspC7エピトープに対する天然殺ボレリア菌抗体の特異性
OspC7に対して哺乳動物に誘導される殺ボレリア菌抗体が特異的であることを裏付けるデータは、以下の通り、ライム病の病歴又はライム関連症状の有無とは無関係に個体(ヒト)の血清を分析することによって得られた。
【0138】
A.材料及び方法
1.血清
ライム病の既往歴(カルテ審査)もしくはライム関連症状を有しない個体(n=36)からの正常血清、献血者(n=100)もしくはコレステロール検診を受けた個体(n=100)からの特性化されていない血清又は通常B.burgdorferi ss抗原と交差反応する血液因子もしくは疾患、例えば、抗核抗体(n=20)、リウマチ因子(n=20)、単核球症(n=10)、サイトメガロウイルス(n=10)、梅毒(n=13)もしくはロッキー山熱(n=4)を有するボランティアからの血清は20℃で貯蔵された保管試料からのものとした。ライム病血清は2003年又は2004年にGundersen Lutheran Medical Centerで診断された患者から採取した。CDCサーベイランス基準(米国疾病対策予防センターMorb.Mort.Wkly.Rep.1990、39:p.19−21)を満たす遊走性紅斑(EM)を有する患者からの血清(n=86)を確からしいライム病と分類し、何度もダニに暴露されて非定型皮膚病巣を有する患者からの血清(n=22)をほぼ確かなライム病(n=49)と分類し、ダニに暴露されて主に頭痛、発熱、筋肉痛及び関節痛を含む全身症状を有する患者からの血清を可能なライム病と分類した。これらの血清は初診時に採取し、−20℃で貯蔵し、試験前に盲検化(blinded)した。
【0139】
OspC7ペプチド。7−aaOspC7ペプチド(AESPKKP;配列番号1)は、University of Wisconsin Biotechnology Center(Madison、ウィスコンシン州)で自動合成器(Protein Technologies)及びFmoc法(Fieldsほか、Peptide Res.1991年、4:p.95−101)を用いて合成された。合成後、このペプチドのアミノ末端をHBTU活性化により手動でビオチン化し、高圧液体クロマトグラフィーによって精製した。組成は、マトリックス支援レーザー脱離イオン化飛行時間(MALDI−TOF:matrix−assisted laser desorption ionization−time of flight)質量分析法(予測質量:1095.4;実測質量:1095.8)を用いて確認した。
【0140】
OspC7 ELISA。マイクロタイター・プレート(イムノロン(Immunolon)2HB、(Thermo Labsystems、Franklin、マサチューセッツ州)の個々のウェルを、ストレプトアビジン(Pierce、Rockland、イリノイ州)を炭酸塩緩衝液(90mM NaHCO3、60mM Na2CO3;pH9.6)に含有させた4μg/ml懸濁液100μLで被覆し、4℃で一夜インキュベートした。インキュベーション後、0.05%Tween20を含有するトリス緩衝食塩水(TBS−T;13mMトリスHCl、3mMトリス塩基、140mM NaCl、2.7mM KCl;pH7.4)で5回洗浄した。洗浄後、1μg/mLのビオチン化OspC7ペプチドを含有するブロッキング緩衝液(15mM NaCl、10mMトリスHCl、3%ウシ胎仔血清、0.05%Tween20)200μLを各ウェルに添加し、室温で1時間回転(150rpm)させながらインキュベートした。次いで、プレートをTBS−Tで3回洗浄し、ブロッキング緩衝液で200倍希釈した100μL量の血清と室温で1時間反応させた。第2の抗体は、ブロッキング緩衝液で15,000倍希釈したペルオキシダーゼ結合ヤギ抗ヒトIgM及びIgG(Kirkegaad Perry Laboratories、Gaithersburg、メリーランド州)とした。1時間のインキュベーション後、結合された上記第2の抗体をo−フェニレンジアミン及び過酸化水素のクエン酸緩衝液(Sigma、St.Louis、ミズーリ州、)溶液を添加し、490nmでのODを測定(スペクトラマックス(SpectraMax)250、Molecular Devices、Sunnyvale、カリフォルニア州)して定量した。
【0141】
B.結果
以前の研究(Jobeほか、Clin.Diagn.Lab.Immuno.2003年、10:p.573−578、Lovrichほか、Clin.Diagn.Lab.Immunol.2005年、12:p746−751)では、高度に保存された免疫優勢OspC殺ボレリア菌抗体エピトープはそのC末端に最も近い7アミノ酸(OspC7)内の領域に限局していた。このエピトープがボレリア属菌による感染に特異的な抗体を誘導することを確認するために、ライム病患者からの血清(図5)及び正常対象者又はボレリア属菌蛋白質とも結合することができる抗体を産生する可能性の高い他の疾患を有する患者からの血清(図4)を用いたELISAの反応性を比較した。図4及び5では、吸光度1.25の線は正常血清及び交差反応する可能性のある血清の平均吸光度の3標準偏差分だけ上の値を設定したものである。従って、この線よりも上にくる結果はいずれもその反応性が有意である確率が99%である(真陽性)。有意な反応性は正常血清又は交差反応する可能性のある血清ではまれにのみ検出された(図4)。陽性結果の大部分はライム病患者由来のものであったかも知れない血清を用いて得られたことにも注目されたい。何故なら、これらの血清(CS)は種々の病気に関してGundersen Lutheran Medical Centerで診察されている100名の患者のグループから得られたものであるからである。この地方はライム病の高度流行地であり、患者の個人情報及び病歴が不明である。これに対して、ウィスコンシン州の非流行地であるミルウォーキー(Milwaukee)の献血者(BD)から無作為に採取された血清試料(n=100)では陽性血清は検出されなかった。
【0142】
さらに、一般にライム病患者からの血清では陽性の結果(非反応性であるべき血清の平均値の3標準偏差分だけ上の値−図4)が検出され、吸光度の値は高かった(図5)。従って、これらの知見全体から、OspC7エピトープに対する上記抗体の高度の特異性及びヒトライム病におけるこの反応の免疫優勢が裏付けられた。
【0143】
上記に例示したデータをまとめると、8週齢の幼犬のグループ群(15匹)に5.0×108個のB.burgdorferi ss50772、2.5×107個のB.burgdorferi S−1−10 SS及び5%Emulsigen(登録商標)を含有するワクチン又は5%Emulsigen(登録商標)のみを含有するプラセボを接種及び追加接種を行った。このワクチンにより注射部位に軽度の腫張が生じたが、速やかに消散した。より注目に値すべきなのは、このワクチンが高濃度の殺ボレリア菌OspA及びOspC抗体を誘導し、これらの抗体の濃度が追加免疫ワクチン接種後1週間でピークに達し、試験期間の間検出可能なレベルを維持したことである。さらに、かなりの割合のOspC殺ボレリア菌抗体がOspC7内の高度に保存されたエピトープに特異的であった。
【0144】
次に、B.burgdorferi ss感染雌性シカダニ(I.scapularis tick)にワクチン接種したイヌに対して吸血させ、ダニの中腸を調べることによって吸われた血液中のOspA及びOspC殺ボレリア菌抗体がB.burgdorferi ssを完全に除去することが確認された。具体的には、B.burgdorferi ssは、プラセボを接種した対照イヌから採取した34匹(32%)のダニで検出されたが、上記ワクチンを接種したイヌに対して吸血したダニ(n=99)では検出されなかった(p<0.0001)。さらに、ワクチンのレシピエントは、感染の確認に通常用いられるいくつかの間接的及び直接的方法でライム病に対して陰性の状態を維持した。これに対して、対照イヌでは、10頭(67%)が「感染特有の」20kDaB.burgdorferi ss蛋白質に対する抗体を産生し、8頭(53%)が「感染特異的」殺ボレリア菌抗体を産生した。さらに、4頭(27%)のイヌが持続性の歩行困難を生じ、これら4頭の歩行困難なイヌのうちの3頭(75%)の関節からB.burgdorferi ssが回収された。さらに、調べたプラセボ接種イヌ11頭のうちの6頭(55%)の関節包に炎症性浸潤が生じた。より説得力あることには、14頭(93%)及び8頭(53%)の対照イヌにおいて、それぞれ皮膚及び関節からB.burgdorferi ssが回収されたのに対し、上記試験品を接種したイヌからスピロヘータは回収されなかった。すなわち、本発明のライム病ワクチンは生じた副作用がごく僅かであり、B.burgdorferi ssによる感染に対して完全に防御した。
【0145】
(実施例8)
B.burgdorferi ss分離株S−1−10及び50772を含むワクチンの免疫性持続期間
A.材料及び方法
・動物
材料及び方法は上記の実施例6で述べた通りである。
【0146】
・ワクチン接種及び血清の採取
イヌの頚部皮下に1mL用量の上記ワクチンを接種し、21日後にさらに1mL用量を追加免疫した。初回(試験−4日目)及び追加免疫ワクチン接種(試験21日目)の前並びに試験28、35、49、79、114、142、175、210、238、266、302、322、357及び394日目にも頚静脈穿刺によって全血を採取した。次いで、遠心により血清を分離し、これを試験するまで−20℃で貯蔵した。
【0147】
・ワクチン接種後の観察
イヌは毎日観察した。
【0148】
・ダニ感染
上記2回目のワクチン接種の1年後に、イヌを胸腔の右側で剃毛し、B.burgdorferi感染シカダニ(I.scapularis tick)の雌10匹及び雄10匹を入れたラバーカップを剃毛部にテープ及び包帯ラップで固定した。これらのダニに7日間吸血させた。
【0149】
・ダニにおけるB.burgdorferi ssの検出
検出は上記の実施例6に記載の方法で実施した。
【0150】
・血液試料
感染後(PC)試験43日目、PC試験77日目、PC試験112日目、PC試験147目、PC試験174目及びPC試験241日目に全血を血清分離輸送管に採取し、血清を分離して試験するまで−20℃で貯蔵した。
【0151】
・殺ボレリア菌抗体の検出
上記の実施例1及び2に記載したものと同様にして、フローサイトメトリーによりOspA及びOspC殺ボレリア菌抗体を検出した。
【0152】
・皮膚生検
感染後(PC)試験43日目、PC試験77日目、PC試験112日目、PC試験147目、PC試験174目及びPC試験241日目にダニ付着の近傍部位から使い捨ての4mm穿刺器具を用いて皮膚生検材料を採取した。これらの生検材料は、上記の実施例6に記載したものと同様に処理した。
【0153】
・免疫抑制
イヌに対して、感染のほぼ20週後から始めて5日間デキサメタゾン(0.4mg/lb体重)の連日投与により免疫を抑制した。
【0154】
・臨床的観察
臨床的観察は上記の実施例6と同様にして実施した。
【0155】
・剖検
剖検は上記の実施例6と同様にして実施した。
【0156】
B.結果
・ワクチン接種後の血清
プラセボ接種では、B.burgdorferi ssに特異的な抗体を誘導することができなかった。これに対し、上記ワクチンの接種では、B.burgdorferi ss分離株S−1−10及び50772を用いることで検出される殺ボレリア菌抗体が有意なレベルで誘導された。S−1−10に対する平均殺ボレリア菌抗体価は追加免疫ワクチン接種の1週後(試験28日目)にピークに達し、この反応は試験394日目まで依然として検出可能であった(表13)。同様に、分離株50772に特異的な高濃度の殺ボレリア菌抗体が試験28日目に検出され、力価は試験49日目でも依然として高く、79日後でも低レベルではあるが、依然として検出可能であった(表14)。
【0157】
表13: 上記ワクチン接種後の殺ボレリア菌活性bの平均力価a(n=15)
【0158】
【表13】
表14: 上記ワクチン接種後の殺ボレリア菌活性bの平均力価a(n=15)
【0159】
【表14】
・感染ダニに対するOspA及びOspC殺ボレリア菌抗体の滅菌能
ワクチン接種又は対照イヌにおいて吸血したダニの中腸を調べることによって、上記ワクチンにより誘導されたOspA及びOspC殺ボレリア菌抗体がダニを滅菌したことが確認された。プラセボ接種したイヌ15頭中12頭において吸血した雌性ダニ95匹中15匹(16%)からのダニ塗抹においてB.burgdorferi ssが検出された(表15)。これに対して、ワクチン接種イヌ15頭のうちの2頭において吸血した75匹の雌性ダニのうちのわずか2匹(3%)からのダニ塗抹でB.burgdorferi
ssが検出された(p=0.0003)。
【0160】
表15: ワクチン接種又は対照イヌから取り出した雌性ダニにおけるB.burgdorferi ssの検出
【0161】
【表15】
・皮膚からB.burgdorferi ssが回収されないようにする上記ワクチンの能力
上記ワクチンにより誘導された殺ボレリア菌抗体は皮膚におけるB.burgdorferi菌の持続感染を防止した。ダニ感染後1ヶ月おきにプラセボ接種イヌから採取した75個の生検材料のうち33個(44%)からB.burgdorferi ssが回収された(表16)。これに対して、上記ワクチンを接種したイヌから採取した75個の生検材料のうちわずか6個(8%)からB.burgdorferi ssが回収された(p<0.0001)。スピロヘータは、ワクチン接種イヌ15頭中6頭(40%)に対し、対照イヌ15頭中10頭(67%)から少なくとも1個の皮膚生検材料から回収された。しかしながら、ワクチン接種イヌからの単離はダニ感染後最初の1ヶ月に限定されたのに対して、培養陽性対照イヌ10頭中8頭(80%)はその後の試験月数全体を通して皮膚生検材料培養物がB.burgdorferi陽性であった。
【0162】
表16: ワクチン接種又は非ワクチン接種対照イヌの皮膚からのB.burgdorferi ssの単離
【0163】
【表16】
・感染の血清学的証拠をもたらさないようにするワクチンの能力
上記ワクチンはライム病特有の抗体の発生をも阻止した。ライム病を有するイヌは、(実施例6で述べたように)B.burgdorferi ss50772分離株を用いても検出できる非OspC殺ボレリア菌抗体を産生する。15頭のプラセボ接種対照イヌのうちの5頭(33%)でダニ感染後に50772株に対する殺ボレリア菌抗体のレベルの著しい上昇(≧4倍の上昇)が認められたのに対し、上記試験ワクチンを接種したイヌでは全く認められなかった。
【0164】
表17: ダニ感染後の殺ボレリア菌抗体bの平均力価a
【0165】
【表17】
・明白な肢/関節障害及びB.burgdorferi ss感染に伴うびらん性変化に対する上記ワクチンの防止能
以前の研究によって、B.burgdorferi ssのみの感染では明白な肢/関節障害(実施例6)が生じることはまれであることが明らかにされ、ダニ感染モデルはイヌの年齢が高いほど効果が低いことが分かっている。明白な症状の発症を増幅させるために、ダニ感染のほぼ3ヶ月後にデキサメタゾンによりイヌの免疫を抑制した。4頭のプラセボ接種対照イヌが歩行困難になるか、関節組織にびらん性病変を生じた。これに対して、本発明のワクチンを接種したイヌはいずれも肢/関節障害の臨床的症状や関節組織によくみられるびらん性病変を生じなかった。
【0166】
本発明は、本明細書に記載した特定の実施態様によって範囲が限定されるものではない。事実、本明細書に記載したものの他に本発明の様々な変形例があり得ることは、前記の説明から当業者には明らかであろう。このような変形例は添付の特許請求の範囲に包含されるものである。
【0167】
さらに、核酸又はポリペプチドに関して示した全ての塩基の大きさ又はアミノ酸の大きさ及び分子量値は近似値であり、説明のために示したものであることは理解されるべきである。
【0168】
本明細書では種々の刊行物が引用されているが、これらの開示内容はその全体が引用により本明細書に組み込まれている。
【技術分野】
【0001】
関連出願への相互参照
この出願は、米国特許法§119(e)の下、2006年11月3日に出願された米国仮出願第60/864,258号(この内容は、その全体が参考として本明細書に援用される)の優先権を主張する本出願である。
【0002】
本発明は、イヌのライム病に対するワクチンに関する。このワクチンを作製する方法及びこのワクチンを単独又は他の防御因子との併用で使用する方法も提供する。
【背景技術】
【0003】
イヌのライム病は、主に米国における狭義のB.burgdorferi(B.burgdorferi sensu stricto(ss))並びに欧州におけるB.burgdorferi ss、B.garinii及びB.afzeliiを含むBorrelia属(Borrelia spp.)スピロヘータによる感染が原因である(非特許文献1;非特許文献2)。これらのスピロヘータは感染したマダニ属のダニが血液を吸った時に伝染し、イヌに生じた感染は潜在性滑膜炎から急性関節炎及び関節痛に及ぶ臨床的症状をもたらす(非特許文献3;非特許文献4)。重要なことであるが、イヌのライム病の発生率は、ヒトの症例数の増加と一致して毎年増加し続けている(非特許文献5)。
【0004】
ボレリア属による感染に対して産生される抗体には異なる2つの機能があるが、従来、いずれの反応も哺乳類の宿主から隔絶されたスピロヘータを除去するという点で無効であった。この自然感染に対する正常な免疫反応の欠如に関して、抗原変異(Schwan、Biochem.Soc.Trans.2003年、31:p.108−112;Tokarzほか、Infect.Immun.2004年、72:p.5419−5432)、宿主模倣(Barbourほか、Microbiol.Rev.1986、50:p.381−400)及び細胞内局在性(Maほか、Infect.Immun.1991年、59:p.671−678)を含むいくつかの解釈が想定されている。
【0005】
最も一般的な液性免疫反応は、食細胞による貪食のためにスピロヘータに「印を付ける」非特異的結合/オプソニン化(被覆)抗体の産生である。残念ながら、オプソニン化抗体は他の微生物に共通にみられる数種の蛋白質(即ち、細菌鞭毛を構成する41kDa蛋白質)によって誘導され、このため、ワクチン接種誘導液性免疫におけるその価値はどうひいき目にみても疑わしいものとなる。
【0006】
別の一般的な免疫反応は殺ボレリア菌(致死的)抗体の産生である。オプソニン化抗体とは対照的に、殺ボレリア菌抗体は僅か数種のボレリア属菌蛋白質のエピトープを認識する。殺ボレリア菌抗体は、スピロヘータ表面の特定の標的に結合した後、最も一般的には補体を誘導して、食細胞による除去を必要とすることなくこの菌を殺す細胞膜傷害複合体を形成する。
【0007】
現在ワクチンとして用いられているイヌのライム病バクテリンが開発されて、感染されたダニが吸血する時にこの寄生虫中のOspA発現スピロヘータを殺すOspA殺ボレリア菌抗体(Hsien−Chuほか、JAVMA 1992、201:p.403−411;Maほか、Vaccine 1996年、14:p.1366−1374;Wikleほか、Intern.J.Appl.Res.Vet.Med.2006年、4:p.23−28;Straubingerほか、Vaccine 2001年、20:p.181−193)を誘導することによる防御が可能となった(非特許文献6)。Straubingerら(Vaccine 2002, 20:181−193)は、全細胞ワクチンの方が組換えOspAよりも有意に高い殺ボレリア菌抗体価を誘導したと報告している。このようなワクチンはほどほどに上出来であったが、ワクチン接種の失敗例が報告されている(Levyほか、JAVMA 1993年、202:p.1834−1838;Maほか、Vaccine 1996年、14:p.1366−1374;Schutzerほか、N.Engl.J.Med.1997年、337:p.794−795)。
【0008】
生じたOspA殺ボレリア菌抗体はOspAを発現しているB.burgdorferi ssを認識するのみであり(Jobeほか、J.Clin.Microbiol.1994、32:p.618−622;Lovrichほか、Infect.Immun.1995、63:p.2113−2119)、通常、上記ダニはOspAを発現していないB.burgdorferi ssスピロヘータに感染する(Fikrigほか、Infect.Immun.1995、63:p.1658−1662;Ohnishiほか、 Proc.Natl.Acad.Sci.2001年、98:p.670− 675)ので、現在、こうした抗体は吸血している上記ダニを殺菌できない場合が多いと理解されている。さらに、上記ダニは、一般にB.afzelii及びB.gariniiを含む他の病原性ボレリア属菌にも感染する(Omsteinほか、J.Clin.Microbiol.2001年、39:p.1294−1298)が、OspA抗体は遺伝種特異的である(Lovrichほか、Infect.Immun.1995年、63:p.2113−2119)。さらに、ダニの中腸への付着を媒介する(Palほか、J.Clin.Invest.2000年、106:p.561−569)OspAの発現は感染されたダニが吸血を始めた後まもなく下方制御される(Schwanほか、Proc.Natl.Acad.Sci.USA 1995年、92:p.2909−2913)ので、OspAボレリア抗体による防御の「絶好の機会」はスピロヘータが感受性の場合でも限定される。
【0009】
B.burgdorferi ssのOspCは殺ボレリア菌液性免疫の別の標的と考えられる(Rousselleほか、J.Infect.Dis.1998年、178:p.733−741)。この蛋白質は、殺ボレリア菌抗体の誘導に関与し、病原性ボレリア属菌類間で保存されているエピトープを有すると考えられる(Lovrichほか、Clin.Diagn.Lab.Immunol.2005年、12:p.746−751)。OspC蛋白質の具体的な機能は依然として不明であるが、OspCの発現は哺乳動物の感染には必要であるが、ダニの感染には必要とされないことが示唆されている(Grimmほか、2004年、Proc.Natl.Acad.Sci.101(9):p.3142−3147)。いずれにしても、ライム病スピロヘータはダニが吸血を開始した後まもなくOspCを発現する(Schwanほか,Proc.Natl.Acad.Sci.USA 1995,92:2909−2913)が、哺乳動物において感染が成立するためにはOspCを発現し続ける必要がある(Stewartほか、Infect Immun.2006年、74:p.3547−3553,Tillyほか、Infect.Immun.2006年、74:p.3554−3564)。従って、OspC殺ボレリア菌抗体の「有効性のウインドウ(window of effectiveness)」はOspA殺ボレリア菌抗体に比し、著しく増大する。
【0010】
OspC蛋白質が防御性殺ボレリア菌抗体を誘導することができることは分かっている(Rousselleほか、J.Infect.Dis.1998年、178:p.733−741;Ikushimaほか、FEMS Immunol.Med.Microbiol.2000年、29:p.15−21)が、これまでの一部の「マッピング」研究では上記エピトープはこの蛋白質の高度に不均一な領域に限局されている(Bucklesほか、Clin.Vacc.Immunol.2006年、13:p.1162−1165)。従って、こうした領域に対する殺ボレリア菌OspC抗体は少数種のボレリア属分離株に対する液性免疫をもたらすに過ぎないことになる。Lovrichら(Clin.Diagn.Lab.Immunol.2005年、12:p.746−751)は、上記蛋白質のC末端7アミノ酸(OspC7)内にOspC殺ボレリア菌抗体エピトープをつきとめた。最も注目に値すべきなのは、このエピトープが上記病原性ボレリア菌類の間で保存されていることである。しかしながら、OspAを発現する(即ち、ospA/ospBオペロンを含有する)通常の実験室B.burgdorferi ss分離株は、OspA殺ボレリア菌抗体誘導能を大きく損なうことなく有意なレベルのOspC殺ボレリア菌抗体をも誘導するように実験室で操作することができない。さらに、OspAを発現する通常の実験室B.burgdorferi ss分離株の死菌体を用いてワクチン接種しても殺ボレリア菌OspC抗体は誘導されない(Schwanほか、1995年、Proc.Natl.Acad.Sci USA、92:p.2909−2913;Obonyoほか、1999年、J.Clin.Microbiol.、37:p.2137−2141)。
【0011】
Callisterら(本明細書に引用により組み込まれている特許文献1及び特許文献2)は、OspCの免疫原性ポリペプチド断片を単独又はOspAポリペプチドとの併用で用いることによりライム病に対してヒト及び他の哺乳動物を防御するワクチンを調製することを提唱している。また、Liveyら(本明細書に引用により組み込まれている特許文献3)は、組換えOspA、OspB及びOspC蛋白質の組合せから調製された、ライム病に対して免疫化するワクチンを提案した。しかしながら、現在に至るまで、組換え蛋白質ワクチンで、現在市販されているワクチンより改良されたものであることが示されたものはない。従って、当該分野では長年にわたり、ライム病から哺乳動物、特にイヌを防御するワクチンの改良が求められ続けている。
【0012】
本明細書におけるいかなる文献の引用も、そのような文献が本願の「先行技術」として利用可能であるということを認めたものと解するべきではない。
【先行技術文献】
【特許文献】
【0013】
【特許文献1】米国特許第6,210,676号明細書
【特許文献2】米国特許第6,464,985号明細書
【特許文献3】米国特許第6,872,550号明細書
【非特許文献】
【0014】
【非特許文献1】Barantonほか、Int.J.Sys.Bacteriol.1992年、42:p.378−383
【非特許文献2】Hoviusほか、J.Clin.Microbiol.2000年、38:p.2611−2621
【非特許文献3】Jacobsonほか、Semin.Vet.Med.Surg.1996年、11:p.172−182
【非特許文献4】Summersほか、J.Comp.Path.2005年、133:p.1−13
【非特許文献5】Haninkovaほか、Emerg.Infect.Dis.2006年、12:p.604−610
【非特許文献6】Fikrigほか、Proc.Natl.Acad.Sci.USA 1992年、89:p.5418−5421
【発明の概要】
【課題を解決するための手段】
【0015】
従って、本発明はワクチンに用いることができる新規免疫原性組成物を提供する。本発明の一態様において、ワクチンはライム病を防御する。この種の具体的な実施態様では、ワクチンのレシピエントはイヌである。別の実施態様では、ワクチンのレシピエントは飼いネコである。ウマ及び/又はウシなどの他の家畜哺乳類も本発明のワクチン及び/又は方法によって防御することができる。本発明はさらに、ライム病及び他の疾患、例えばイヌの他の感染症に対する防御免疫を誘導する混合ワクチンを提供する。本発明のワクチンの作製方法及び使用方法も提供する。
【0016】
本発明のワクチンは、免疫学的有効量の、OspC抗原を発現する(任意選択的に不活化された)第1又は単一の株の生物を含む。この株は、標準的な培養条件で増殖させる場合、(任意選択的に補体を必要とする)OspC特異的殺ボレリア菌抗体の存在下に死滅する株である。このような抗体としては、B.burgdorferi ss50772(ATCC番号PTA−439)によってワクチン接種した動物に誘導される、保存エピトープOspC7に対する殺ボレリア抗体が挙げられる。この種の具体的な実施態様では、ボレリア遺伝種(genospecie)の上記第1又は単一の株は上記OspC抗原を構成的に発現する。より具体的な実施態様では、この第1の株はB.burgdorferi ss50772(ATCC番号PTA−439)である。
【0017】
本発明のワクチン組成物は、免疫学的有効量の、病原性ボレリア遺伝種からの1種以上の別の株(これらを総称して第2の株と名付ける場合がある)からの不活化生物をさらに含むことができる。具体的な実施態様では、この第2の株はOspA及びOspB抗原を提示する。
【0018】
適切な第2の株の例としては、以下のうちの1種以上が挙げられる:B.burgdorferi ssS−1−10(ATCC番号:PTA−1680)、B.burgdorferi ssB−31(ATCC番号35210)、B.afzelii(例えば、ATCC番号51567として入手可能)及びB.garinii(例えば、ATCC番号51383及び同51991として入手可能)、B.burgdorferi ssDK7、B.burgdorferi ss61BV3、B.burgdorferi ssZS7、B.burgdorferi ssPka、B.burgdorferi ssIP1、IP2、IP3、B.burgdorferi ssHII、B.burgdorferi ssP1F、B.burgdorferi ssMil、B.burgdorferi ss20006、B.burgdorferi ss212、B.burgdorferi ssESP1、B.burgdorferi ssNe−56、B.burgdorferi ssZ136、B.burgdorferi ss ia及び/又はこれらの任意の組合せ。
【0019】
上記ワクチン組成物は、概して、各それぞれの株の生物を1ミリリットル当たり約1×104乃至約1×1010個含む。特定の実施態様では、このワクチンは各それぞれの株の生物を1ミリリットル当たり約1×106乃至約5×109個含む。別の実施態様では、このワクチンは上記(又は各)第2の株の生物を1ミリリットル当たり約1.0×108乃至約5×108個及び上記第1の株の生物を1ミリリットル当たり約5.0×108乃至約5×109個含む。
【0020】
また、上記ワクチン組成物には、例えば、アルミニウム化合物(例えば、リン酸アルミニウム、水酸化アルミニウム)、代謝性及び非代謝性油、ブロックポリマー、免疫刺激複合体、ビタミン、ミネラル、CARBOPOL(登録商標)(例えば、CARBOPOL941)などの医薬用として許容可能なアジュバントを含めることもできる。特定の実施態様では、この医薬用として許容可能なアジュバントは、(例えば、Emulsigen(登録商標)として販売されているような)水中にミクロン径の油滴を均一分散させた乳剤を含む。
【0021】
任意選択的に、上記ワクチン組成物は医薬用として許容可能な免疫刺激剤、例えば、サイトカイン、成長因子、ケモカイン、リンパ球、単球もしくはリンパ器官由来細胞の細胞培養液の上清、植物、細菌もしくは寄生虫からの細胞調製物及び/又は抽出物又はマイトジェンをも含む。
【0022】
本発明はさらに、病原性ボレリア属菌、具体的にはB.burgdorferi ssに対してイヌ又は他の哺乳動物を免疫化する方法であって、免疫学的有効量の上記本発明ワクチンをイヌに注射することを含む方法を提供する。例えば、このようなワクチンには各それぞれの株の生物を約1×108乃至3×109個を含ませることができる。ワクチンは、例えば、筋肉内注射、皮下注射、静脈内注射、皮内注射、経口投与、鼻腔内投与及びこれらの組合せなどの経路によって投与することができる。特定の実施態様では、ワクチン接種後、免疫化されたイヌは殺ボレリア抗体を産生する。
【0023】
本発明はさらに、B.burgdorferi ss OspCに結合する殺ボレリア菌抗体を含有する、ワクチン接種動物から得られる血清を提供する。同様に、本発明はOspCに結合する精製抗体を提供する。特定の実施態様では、この血清はかなりの割合のOspC7特異的殺ボレリア菌抗体を含有する。
【0024】
本発明はさらに、本発明のボレリア遺伝種の1種以上の株を1種以上の他のイヌ病原体及び/又は免疫原との組合せで含む混合ワクチンを提供する。この免疫原としては、例えば、イヌジステンパーウイルス、イヌアデノウイルス、イヌパルボウイルス、イヌパラインフルエンザウイルス、イヌコロナウイルス、イヌインフルエンザウイルス及び/又はLeptospira血清型(例えば、Leptospira kirschneri血清型grippotyphosa、Leptospira interrogans血清型canicola、Leptospira interrogans血清型icterohaemorrhagiae及び/又はLeptospira interrogans血清型pomana)に対する免疫を誘導する免疫原が挙げられる。本発明の混合ワクチンに添加することができる別のイヌ病原体としては、Leishmania major、Leishmania infantumなどのリーシュマニア属生物、Bordetella bronchiseptica、マイコプラズマ属菌(例えば、Mycoplasma cynos)、狂犬病ウイルス及びanaplasma phagocytophilum、anaplasma platysなどのanaplasma属生物及びEhrlichia canisが挙げられる。
【0025】
本発明のこれらの態様及びそれ以外の態様については添付の図面及び詳細な説明によって一層よく理解されよう。
例えば、本発明は、以下の項目を提供する:
(項目1)
免疫学的有効量の、ボレリア遺伝種の第1又は単一の株の生物を含むワクチン組成物であって、
該第1又は該単一の株の生物は、標準的なボレリア遺伝種の増殖条件下で増殖させる場合、B.burgdorferi ss50772(ATCC番号PTA−439)を注射した動物において誘導されるOspC特異的抗体の存在下に死滅する、ワクチン組成物。
(項目2)
上記免疫学的有効量の生物が不活化されている項目1に記載のワクチン。
(項目3)
上記第1又は上記単一の株がその細胞表面にOspC抗原を提示し、及び
イヌが上記ワクチン組成物で免疫化されると、OspC殺ボレリア菌抗体が該イヌにおいて誘導される項目1に記載のワクチン。
(項目4)
かなりの割合の上記OspC特異的殺ボレリア菌抗体がOspC7に特異的である項目3に記載のワクチン組成物。
(項目5)
上記第1又は上記単一の株がB.burgdorferi ss50772(ATCC番号PTA−439)である項目3に記載のワクチン組成物。
(項目6)
免疫学的有効量の、ボレリア遺伝種由来の第2の株の不活化生物をさらに含み、該第2の株がその細胞表面にOspA抗原、OspB抗原を提示し、又はOspA抗原及びOspB抗原の両者を提示する項目1に記載のワクチン組成物。
(項目7)
イヌが上記ワクチン組成物で免疫化されると、OspC殺ボレリア菌抗体及びOspA殺ボレリア菌抗体が該イヌにおいて誘導される項目6に記載のワクチン組成物。
(項目8)
上記第2の株がB.burgdorferi ssS−1−10株(ATCC番号PTA−1680)である項目6に記載のワクチン組成物。
(項目9)
上記第1の株の生物を1ミリリットル当たり約1×104乃至約1×1010個及び上記第2の株の生物を1ミリリットル当たり約1×104乃至約1×1010個含む、項目6に記載のワクチン組成物。
(項目10)
上記第1の株の生物を1ミリリットル当たり約5.0×108乃至約5×109個及び上記第2の株の生物を1ミリリットル当たり約1.0×108乃至約5×108個含む、項目9に記載のワクチン組成物。
(項目11)
医薬用として許容可能なアジュバントをさらに含む項目1に記載のワクチン組成物。
(項目12)
医薬用として許容可能な免疫刺激剤をさらに含む項目1に記載のワクチン組成物。
(項目13)
非ボレリア病原体に対する防御免疫を誘導するための少なくとも1種の非ボレリア免疫原をさらに含む項目1に記載のワクチン組成物。
(項目14)
上記非ボレリア免疫原がイヌジステンパーウイルス、イヌアデノウイルス、イヌパルボウイルス、イヌパラインフルエンザウイルス、イヌコロナウイルス、イヌインフルエンザウイルス、Leptospira血清型、Leishmania属生物、Bordetella bronchiseptica、Mycoplasma属菌、狂犬病ウイルス、Ehrlichia canis、anaplasma属生物及びこれらの組合せからなる群から選ばれる項目13に記載のワクチン組成物。
(項目15)
上記Leptospira血清型がLeptospira kirschneri血清型grippotyphosa、Leptospira interrogans血清型canicola、Leptospira interrogans血清型icterohaemorrhagiae、Leptospira interrogans血清型pomana及びこれらの組合せからなる群から選ばれる項目14に記載のワクチン組成物。
(項目16)
上記Mycoplasma属菌がMycoplasma cynosを含む項目14に記載のワクチン組成物。
(項目17)
病原性ボレリア遺伝種に対してイヌを免疫化する方法であって、免疫学的有効量の項目1に記載のワクチンを該イヌに注射することを含む方法。
(項目18)
病原性ボレリア遺伝種に対してイヌを免疫化する方法であって、免疫学的有効量の項目6に記載のワクチンを該イヌに注射することを含む方法。
(項目19)
上記第2の株が狭義のB.burgdorferiである項目18に記載の方法。
(項目20)
上記第2の株がB.burgdorferi ssS−1−10(ATCC番号:PTA−1680)、B.burgdorferi ssB−31(ATCC番号35210)、B.afzelii(ATCC番号51567)及びB.garinii(ATCC番号51383又は同51991)、B.burgdorferi ssDK7、B.burgdorferi ss61BV3、B.burgdorferi ssZS7、B.burgdorferi ssPka、B.burgdorferi ssIP1、IP2、IP3、B.burgdorferi ssHII、B.burgdorferi
ssP1F、B.burgdorferi ssMil、B.burgdorferi ss20006、B.burgdorferi ss212、B.burgdorferi ssESP1、B.burgdorferi ssNe−56、B.burgdorferi ssZ136、B.burgdorferi ss ia及び/又はこれらの組合せからなる群から選ばれる項目18に記載の方法。
(項目21)
上記免疫化されたイヌが殺ボレリア菌抗体を産生する項目18に記載の方法。
(項目22)
上記イヌがボレリア属の各それぞれの株の生物を約5×107乃至約5×109個注射される項目18に記載の方法。
(項目23)
項目21に記載の方法によって免疫化されたイヌから得られるB.burgdorferi OspCに結合する殺ボレリア菌抗体を含む血清。
(項目24)
項目21に記載の方法によって免疫化されたイヌから得られるB.burgdorferi ss OspCに結合する殺ボレリア菌抗体。
(項目25)
OspC7特異的抗体を含む項目24に記載の殺ボレリア菌抗体。
【図面の簡単な説明】
【0026】
【図1】正常イヌ対照血清(NS)、試験品(TP:test product)を用いたワクチン接種(試験43日目)後のイヌからの血清又はB.burgdorferi ss感染ダニ(NI)による感染(試験134日目)後のイヌからの血清のウエスタンブロットの写真である。分子量約20kDaの蛋白質に対する感染特異的抗体の存在に注目されたい。
【図2A】正常イヌ対照血清(NS)又はプラセボ接種及びB.burgdorferi ss感染ダニによる感染(試験134日目)後の個々のイヌの個別のコホートからの血清(1番から15番まで番号が付けられている)のウエスタンブロットの写真である。
【図2B】正常イヌ対照血清(NS)又は試験品を用いたワクチン接種後の個々のイヌの個別のコホートからの血清(1番から15番まで番号が付けられている)のウエスタンブロットの写真である。プラセボ接種した14頭(93%)のイヌでは感染特異的20kDa抗体が存在し(図2A)、試験品を用いてワクチン接種したイヌでは同抗体が存在しなかった(p<0.0001)(図2B)ことに注目されたい。
【図3A】プラセボ接種してB.burgdorferi ssに感染させたイヌの関節における組織病理学的変化の代表的な1例を示す写真である。長方形内の領域はイヌのライム病関節炎に特有の好中球及び単核細胞の著しい浸潤を示している。
【図3B】試験品を用いてワクチン接種し、B.burgdorferi ssに感染させたイヌの関節において組織病理学的変化がみられない代表的な1例を示す写真である。長円形内の領域には好中球及び単核細胞の浸潤はみられない。
【図4】ライム症状もしくはライム関連症状の(カルテ審査による)既往歴がない個体(n=36)からの正常血清(N)、献血者(BD、n=100)もしくはコレステロール検診を受けた個体(CS、n=100)からの特性化されていない血清、又は通常B.burgdorferi ss抗原と交差反応する血液因子もしくは疾患、例えば、抗核抗体(ANA、n=20)、リウマチ因子(RF、n=20)、単核球症(EBV、n=10)、サイトメガロウイルス(CMV、n=10)、梅毒(SYPH、n=13)もしくはロッキー山熱(RMSF、n=4)を有するボランティアからの血清を用いたOspC7ELISA反応性の測定結果を示した図である。実線は、正常血清及び交差反応する可能性のある血清の平均吸光度値よりも上の3標準偏差値分を示す。この線よりも上にくる値は陽性に相当する確率が99%となる(バックグランド値 1%)。
【図5】定型ライム病病変を有する患者である、遊走性紅斑を示すほぼ確かなライム病の患者(n=86)、「非定型病巣」を有する、確からしいライム病の患者(n=22)及び全身症状(初発臨床症状)を有する、ライム病の可能性のある患者(n=49)からの血清を用いたOspC7ELISA反応性を示す図である。実線は、正常血清及び交差反応する可能性のある血清の平均吸光度値よりも上の3標準偏差値分を示す。
【発明を実施するための形態】
【0027】
本発明は、レシピエントのワクチン接種動物に効果的な殺ボレリア菌液性免疫を誘導するボレリア遺伝種の1種以上の株からの免疫学的有効量の生物を含むワクチン組成物を提供する。このような株の生物は、標準的なボレリア遺伝種の増殖条件下で増殖させる場合、OspC特異的抗体、例えば、B.burgdorferi ss50772(ATCC番号PTA−439)を用いてワクチン接種した動物において誘導される一種の抗体又は複数種の抗体の存在下に死滅する。
【0028】
本発明の一態様において、本発明のワクチンにより誘導される抗体の有効性のウインドウは、単にOspA殺ボレリア抗体に基づいた従来のワクチンに比し、著しく増大する。別の態様では、本発明のワクチンでは、例えばインビボでのOspCの発現によるなどの有益な免疫記憶反応を促進する確率が改善され、及び/又はボレリア属菌の複数種の病原性株に対する防御が追加及び/又は増強される。
【0029】
本発明の一実施態様において、そのような株の生物は、標準的なボレリア遺伝種増殖条件下で増殖させると、OspC抗原を構成的に発現する。この種の特定の実施態様では、そのような株の生物は、標準的なボレリア遺伝種増殖条件下で増殖させると、補体特異的反応によって死滅する。さらに別の実施態様では、そのような株の生物を含むワクチンにより誘導される血清中のかなりの割合のOspC特異的殺ボレリア菌抗体は、保存エピトープOspC7に特異的であることにより、複数の病原性ボレリア遺伝種(例えば、B.burgdorferi ss、B.afzelii及び/又はB.garinii)に対する防御を可能にする。さらに別の実施態様では、そのような株の生物はOspA抗原もOspB抗原も提示しない。さらに別の実施態様では、そのような株の生物はこれらの性質のうちの任意の2つ以上を有する。さらに別の実施態様では、そのような株の生物はこれらの性質のうちの任意の3つ以上を有する。さらに別の実施態様では、そのような株の生物はこれらの性質のうちの任意の4つ以上を有する。特定の実施態様では、そのような株の生物はこれらの性質の全てを有する。そのような特定の1種の株はB.burgdorferi ss50772(ATCC番号PTA−439)である。
【0030】
別の実施態様において、本発明のワクチンは、上記のような第1の株及び有効量の少なくとも第2の株の生物を含む組成物である。この第2の株は、本発明のワクチンの一部として投与された場合に殺ボレリア菌OspA抗体を誘導する病原性ボレリア遺伝種からのものであることが好ましい。また、上記組成物には1種以上の別の適合性ワクチン遺伝種を含ませることもできる。さらに、本発明のワクチンには1種以上の他の哺乳動物(例えば、イヌ)病原体及び/又は免疫原を含ませることができる。
【0031】
本発明の理解をさらに十分に進めるために、以下の定義を設ける。
【0032】
原文の英文では、記載上の便宜のために単数形の用語を用いているが、これは決してそのように限定的なものではない。すなわち、例えば、「ポリペプチド(a polypeptide)」を含む組成物に言及する場合、1種以上のそのようなポリペプチドに言及することを含む。さらに、生物(organism)に言及する場合、特に示さない限り、複数種のそのような生物(organisms)に言及することを含む。
【0033】
本明細書で使用される「ほぼ(approximately)」という用語は「約(about)」と同じ意味で用いており、値がその示された値の50%以内にあること、すなわち、1ミリリットル当たり「ほぼ」1×1010個の菌を含む組成物は1ミリリットル当たり5×109乃至5×1010個の菌を含むことを意味する。
【0034】
「遺伝種」という用語は、G.Barantonら(1992年、International J.of Systematic Bacteriology 42:p.378−383)によって最初に用いられ、定義されたが、本明細書でも、非ボレリア生物の分類を記載する際に「種」という用語を用いるのと同じ意味で用いている。
【0035】
ボレリア遺伝種を培養するための「標準的な増殖条件」では、BSK(Barbour−Stoenner−Kelly)培地中約33℃乃至約35℃の範囲の温度で増殖させることが必要とされる。本明細書に記載したBSK培地は、Callisterら[参考としてその全体が本明細書に援用されるDetection of Borreliacidal Antibodies by Flow Cytometry、セクション11.5.1 − 11.5.12、Current Protocols in Cytometry、John Wiley and Sons,Inc.Supplement26、(2003年)]に従って調製した。(BSK培地は、例えば、Sigma、St.Louis、ミズーリ州から市販されてもいる。)
本明細書で使用される「OspC7」とは、既知の病原性ボレリア属間で完全に保存されている、Callisterら(参考としてその全体が本明細書に援用される米国特許第6,210,676B1号及び同6,464,985B1号)によって開示されているOspCのC末端側50アミノ酸内の7アミノ酸領域(参考としてその全体が本明細書に援用されるLovrichほか、2005年、Clin.Diagn.Lab.Immunol.、12:p.746−751)にある免疫優勢OspC殺ボレリア菌抗体エピトープである。この保存については、Lovrichらによって報告された7アミノ酸セグメントをコードしているコドンセグメントのBLAST探索により容易に確認できる。2006年10月9日に行ったこのような探索では、上記のOspC 7−マーエピトープコードセグメントを含有する100種のボレリア種の結果リストが得られた。
【0036】
「OspC特異的殺ボレリア菌抗体」とは、例えばB.burgdorferi ss50772(ATTC番号PTA−439)でワクチン接種した動物の血清中に存在するものであり、OspC抗原の任意のエピトープに選択的に結合してスピロヘータを補体依存性又は非依存性に死滅させるものである。「OspC7特異的殺ボレリア菌抗体」とは、例えばB.burgdorferi ss50772(ATTC番号PTA−439)でワクチン接種した動物の血清中に存在するものであり、Lovrichら[参考としてその全体が本明細書に援用されるLovrichほか、Clin.Diagn.Lab.Immunol.、12:p.746−751、(2005年)]によって報告されたOspCの7個のC末端アミノ酸に選択的に結合してスピロヘータを(一般に補体媒介性細胞膜傷害複合体を誘導することにより)死滅させるものである。OspC殺ボレリア菌抗体の特異性は十分立証されている。例えば、OspC殺ボレリア菌抗体は、殺ボレリア菌抗体試験においてB.burgdorferi ss50772の感受性を測定することによって通常ライム病血清中に検出される。類縁疾患を有するヒト患者からの血清には、株50772をも死滅させる交差反応性抗体はまれに(2%)しか含まれない(Callisterほか、1996年、Clinical and Diagnostic Laboratory Immunology 3(4):p.399−402に詳細に報告されている)。さらに、OspC7殺ボレリア菌エピトープを用いるペプチドELISA法ではライム病の血清中の殺ボレリア菌抗体が正確に捕捉されるが、他の類縁疾患を有する患者からの血清にはOspC7ペプチドにも結合する交差反応性抗体はまれに(<2%)しか含まれない(図4及び5に示した)。
【0037】
ワクチンにより誘導される血清中の「かなりの割合の」OspC特異的殺ボレリア菌抗体が保存エピトープOspC7に特異的である場合、これは、OspC7によるその血清の吸収後に血清中OspC特異的殺ボレリア菌抗体が無視できないほど減少することを意味する。これは、好ましくはB.burgdorferi ss50772を用いて検出される血清の殺ボレリア菌抗体価の少なくとも2分の1、より好ましくはOspC7によるその血清の吸収後の血清の殺ボレリア菌抗体価の2分の1乃至4分の1又はこれ以下への低下と定義される。
【0038】
「補体特異的反応」とは、ボレリア属の生物(類)を殺ボレリア菌抗体によって死滅させるために血清補体の存在を必要とする抗体反応である。
【0039】
本発明では、「不活化」Borrelia burgdorferi ss生物とは、動物において免疫反応を誘導することができるが、その動物に感染することができない生物である。Borrelia burgdorferi ss分離株は、二成分エチレンイミン、ホルマリン、ベータ−プロピオラクトン、チメロサール又は熱からなる群から選ばれる作用因子によって不活化することができる。特定の実施態様では、このBorrelia burgdorferi ss分離株は二成分エチレンイミンによって不活化する。
【0040】
「アジュバント」及び「免疫刺激剤」という用語は、本明細書では同じ意味で用いており、免疫系の亢進をもたらす1種以上の物質と定義される。これに関連して、アジュバントは1種以上のワクチン抗原/分離株に対する免疫反応を増強するために使用する。アジュバントは、ワクチンの投与の前、又はこの投与と組み合わせて、又はこの投与の後に対象動物に投与することができる。本発明のアジュバントは、例えば、天然供給源、組換え供給源を含むいくつかの供給源のいずれかから入手し、及び/又は化学的に合成することができる。アジュバントとして用いる化合物の例としては、アルミニウム化合物、代謝性及び非代謝性油、ブロックポリマー、ISCOM(免疫刺激複合体)、ビタミン及びミネラル(ビタミンE、ビタミンA、セレン、ビタミンB12が挙げられるが、これらに限定されない)、キル(Quil)A(サポニン)及びポリアルケニルエーテル又はジビニルグリコールで架橋したアクリル酸のポリマー、例えば、CARBOPOL(登録商標)が挙げられるが、これらに限定されるものではない。特に免疫刺激剤と呼ばれることがあるアジュバントの別の例としては、細菌及び真菌細胞壁成分(例えば、リポ多糖、リポ蛋白質、糖蛋白質、ムラミルペプチド、ベータ−1,3/1,6−グルカン)、植物由来の各種複合糖質(例えば、グリカン、アセマンナン(acemannan))、動物由来の各種蛋白質及びペプチド(例えば、ホルモン、サイトカイン、共刺激因子)並びにウイルス及び他の供給源由来の新規核酸(例えば、二本鎖RNA、CpG)が挙げられる。さらに、上記の物質のいくつもの組合せがアジュバント効果をもたらすことができ、従って、これらは本発明のアジュバントを形成することができる。好ましいアジュバントの1つはEmulsigen(登録商標)である。
【0041】
米国特許第6,210,676号に記載されているB.burgdorferi ss50772(ATCC番号PTA−439)及び米国特許第6,316,005号に記載されているB.burgdorferi ssS−1−10(ATCC番号PTA−1680)は、それぞれ1999年7月30日及び2000年4月11日にAmerican Type Culture Collection、10801 Boulevard大学、Manassas(VA)20110に寄託された。本発明の共同権利者らは、個々に上記2特許に対する権利を保有している。
【0042】
また、本発明が本明細書に開示した特定の構造、工程段階及び材料に限定されないことは、このような構造、工程段階及び材料はある程度変えることができるので、言うまでもないことである。また、本明細書に用いた用語は、単に特定の実施態様を説明するために用いたものであり、本発明の範囲が添付の特許請求の範囲及びその均等物によってのみ限定されることになるので、限定的なものではないことも言うまでもない。
【0043】
他のワクチン株
・他のOspC株
一態様において、本発明は、単独又は他のボレリア遺伝種株との併用でライム病を防御するワクチンにおけるB.burgdorferi ss50772(ATCC番号PTA−439)の使用を提供する。特に、B.burgdorferi ss50772は、当初、ワクチンに有用な候補としては受け入れなかった。何故なら、この株はOspCを発現しないと間違って報告されていた(Andersonほか、1996年、J.Clin.Microbiol.、34:p.524−529)からである。
【0044】
本発明はさらに、本発明のワクチン用の他のそのような有用分離株を選択/特定する独自の特異的な基準を提供する。特に、OspC特異的抗体により死滅させられやすく、好ましくは、以下の属性のうちの1つ以上又は全てをも有する分離株を選択/特定することができる。即ち、その属性とは、(i)補体特異的反応を介してOspC特異的抗体により死滅させられやすいこと、(ii)この分離株でワクチン接種したレシピエントにおいてOspC殺ボレリア菌抗体を誘導することができること、(iii)そのように誘導されたかなりの割合のOspC殺ボレリア菌抗体がC末端7アミノ酸(Osp7)内の保存エピトープに特異的であること、(iv)OspA及びOspB発現能を欠くこと、及び/又は(v)インビトロで構成的にOspCを発現することができること、である。
【0045】
A.培養条件によるOspA/B発現の抑制及びOspC発現の増強
通常の病原性ボレリア属菌株(B.burgdorferi ssの株JMNT及びN40)をダニ細胞株(Ixodes scapularisISE6)と共培養することにより5日間にわたって、OspA発現を下方制御することができ、OspC発現を上方制御することができることをObonyoら(J.Clin.Microbiol.、1999年、37:p.2137−2141)は明らかにしている。最も著しい効果は37℃で培養することで認められた。従って、この方法によって1種以上の通常の病原性ボレリア菌株が本発明のワクチンに有用な形態に変換されることになる。
【0046】
B.突然変異によるOspA/B発現の抑制及びOspC発現の増強
別の実施態様において、元々OspA及びOspBを発現するボレリア属菌株に対して遺伝子操作を行ってOspA及びOspBの発現をコードする遺伝子を下方制御又は削除することによりOspC発現を上方制御する。当該分野で既知の全ての遺伝子操作方法をこの目的に使用することを想定している。例えば、入手できるボレリア属菌株の複数の菌体にOspA/OspB遺伝子の不活性化アナログを、抗生物質選択マーカを用いて調製したボレリア属菌適合性プラスミドの形で導入する(例えば、OspC不活性化及び相補プラスミドをB.burgdorferiB31−A3株に挿入することによりボレリア属菌におけるOspCの発現をブロックすることができると報告しているGrimmほか、2004年、Proc.Nat’l Acad.Sci 101(9):p.3142−3147を参照されたい)。この導入プラスミドとOspA/OspBを含む天然プラスミドとの間に組換え現象がおこることによって突然変異OspA/OspBプラスミドを含む一定数の組換えがうまくいったボレリア属菌が得られる。選択は、例えば、対応抗生物質の存在下における関連する抗生物質耐性と増殖とによることを前提とすることができる。OspA/OspBが選択的に発現されなくなったボレリア属菌はOspCの発現を上方制御することが想定されている。
【0047】
別の任意選択的な実施態様では、OspA、OspB及び/又は1種以上の別のOsp抗原を発現する1種以上の別のB.burgdorferi ss又はボレリア属生物を別の生物として前記ワクチン組成物に含ませることができる。
【0048】
各それぞれの不活化生物は約1×104乃至約1×1010個/mLの範囲の濃度で上記ワクチンに存在させることが好ましい。特に、このワクチンは、1.0×108個/mL以上のB.burgdorferi ssS−1−10及び5.0×108個/mL以上のB.burgdorferi ss50772の用量でイヌに投与することが好ましい。
【0049】
・他のOspA株
OspA抗原を有する第2の株をB.burgdorferi ssB−31(ATCC番号35210)などの通常の病原性実験室B.burgdorferi ss分離株(Barbourほか、1985年、J.Clin.Microbiol.52:p.478−484)とすることができる。特定の第2の生物は上記に例示したB.burgdorferi ssS−1−10株(ATCC番号PTA−1680)である。北米以外の地域に対して最適化されたワクチン組成物の第2の生物として用いるのに適した別の株としては、例えば、B.burgdorferi ssB−31(ATCC番号35210)、B.afzelii(例えば、ATCC番号51567として入手可能)及びB.garinii(例えば、ATCC番号51383及び同51991として入手可能)並びに下記の表1に示した株が挙げられる。
【0050】
【表1】
前記ワクチン組成物には医薬用として許容可能なアジュバントを含ませることができる。「アジュバント」とは、特定の抗原に対する免疫反応を非特異的に増大させることにより所与のワクチンにおいて必要な抗原量及び/又は対象抗原に対して十分な免疫反応を生じるのに必要な注射頻度を減少させる作用物質である。動物のワクチン接種に好適なアジュバントとしては、(ピーナッツ油、モノオレイン酸マンノイド及びモノステアリン酸アルミニウムを含有する)アジュバント65;完全又は不完全フロイントアジュバント;水酸化アルミニウム、リン酸アルミニウム、ミョウバンなどのミネラルジェル;ヘキサデシルアミン、オクタデシルアミン、リゾレシチン、臭化ジメチルジオクタデシルアンモニウム、N,N−ジオクタデシル−N’,N7−ビス(2−ヒドロキシメチル)プロパンジアミン、メトキシヘキサデシルグリセロール、プルロニックポリオールなどの界面活性剤;ピラン、硫酸デキストラン、ポリIC、ポリアクリル酸、CARBOPOL(登録商標)(例えば、CARBOPOL941)などのポリアニオン;ムラミルジペプチド、ジメチルグリシン、タフトシンなどのペプチド;及び油乳濁液が挙げられるが、これらに限定されるものではない。アジュバントに関する情報は、例えば、引用により本明細書に組み込まれているP.Tijssen によるシリーズ[Practice and Theory of Enzyme Immunoassays、第3版1987年、Elsevier、New York]に開示されている。
【0051】
任意選択的に、上記ワクチン組成物には、細菌及び真菌細胞壁成分(例えば、リポ多糖、リポ蛋白質、糖蛋白質、ムラミルペプチド)、植物由来の各種複合糖質(例えば、グリカン、アセマンナン)、動物由来の各種蛋白質及びペプチド(例えば、ホルモン、サイトカイン、共刺激因子)、ウイルス及び/又は他の供給源由来の新規核酸(例えば、二本鎖RNA、CpG)などの医薬用として許容可能な免疫刺激剤をさらに含ませることができる。
【0052】
上記ワクチン組成物は、静脈内、筋肉内、皮下、経口、鼻腔内、皮内及び/又は腹腔内ワクチン接種を含む任意の標準的経路によって容易に投与することができる。このワクチン組成物がレシピエント動物及び投与経路の各種類に対して適切に製剤化されることが好ましいことは当業者なら理解するであろう。
【0053】
従って、本発明はB.burgdorferi ss及び他のボレリア属菌に対してイヌを免疫化する方法をも提供する。そのような方法の1つは、イヌに免疫学的有効量の本発明ワクチンを注射することによりイヌに適切なOspA及びOspC殺ボレリア菌抗体を産生させることを含む。
【0054】
一実施態様において、本発明の皮下投与によってOspC7に特異的な殺ボレリア菌抗体を含む高濃度のOspA及びOspC殺ボレリア菌抗体が産生される。別の実施態様では、本発明は、有害なボレリア属菌に対して有効な各B.burgdorferi ss株を特定の最小限の量で含むワクチンを提供する。別の実施態様では、本発明のワクチンは、イヌにおいてB.burgdorferi ss及び他の病原性ボレリア属感染を防御するのに有効である。特定の実施態様では、上記ワクチンは、本発明の2種のB.burgdorferi ss株と医薬用として許容可能なアジュバントとの安全で免疫学的に有効な組合せを含む。
【0055】
本発明は、2種のB.burgdorferi ss分離株を特定の最低限の量で含むワクチンがダニ感染後のボレリア属菌からイヌを防御することを開示する。本発明はさらに、ワクチン接種したイヌにおいて特定の最小限の量のB.burgdorferi ss特異的OspA及びボレリア属菌特異的OspC殺ボレリア菌抗体を誘導するワクチンを開示する。また、本発明のワクチンは、許容可能な免疫刺激剤及び/又アジュバントと共に投与することもできる。
【実施例】
【0056】
以下の実施例は、本発明の理解をさらに進めるのに役立つものであるが、本発明の有効範囲を何ら限定するものではない。
【0057】
(実施例1)
組換えOspAを用いたワクチン接種
A.材料及び方法
・動物
Charles River Breeding Laboratories,Inc.Wilmington、マサチューセッツ州)から5乃至10週齢のLVGハムスターを入手した。ハムスターは21℃の室温でケージ当たり3乃至4匹を収容し、餌と水を自由に与えた。
【0058】
・ハムスターのワクチン接種及び血清の採取
ハムスターの頸背部皮下に市販組換えOspA(rOspA)ワクチンRECOMBITEK(登録商標)ライム(Lyme)(Merial Limited、Duluth、ジョージア州)を0.5mL接種し、この初回ワクチン接種の3週後に追加免疫を行った。初回ワクチン接種の3、7、9及び15週後に、1群5匹の各ハムスター群をノーズ・アンド・マウス・カップに含ませたエーテルの吸入により軽く麻酔し、心臓内穿刺により放血させた。この血液を凝固させ、血清を分離して、使用するまで−70℃で貯蔵した。さらに、ワクチン接種していないハムスター3匹からのハムスター血清をプールして対照正常血清として用いた。別の対照として、20匹のハムスターの両大腿後部に、B.burgdorferi ssS−1−10を含有する市販のイヌライム病全細胞ワクチン(Galaxy、Solvay Animal Health,Inc.、Mendota Heights、ミネソタ州、現Schering−Plough Animal Health、Elkhorn、ネブラスカ州)を0.25mL接種した。これらのハムスターに追加免疫した後、上述のようにして血液を採取した。
【0059】
・BSK培地の調製
B.burgdorferiのインビトロ培養のためにBarbour−Stoenner−Kelly(BSK)肉汁培地を用い、これを殺ボレリア菌抗体試験及び本発明全体にわたって記載した方法に用いる一次培地(primary substrate)とした。BSK培地は、Callisterら[参考としてその全体が本明細書に援用されるDetection of Borreliacidal Antibodies by Flow Cytometry、セクション11.5.1 − 11.5.12、Current Protocols in Cytometry、John Wiley
and Sons,Inc.Supplement26、(2003年)]により報告されたものと同様にして調製した。つまり、以下の方法によってBSK培地を調製した。
【0060】
材料:
HEPES(Sigma)
ネオペプトン(Neopeptone)(Difco)
クエン酸ナトリウム(Sigma)
グルコース(Sigma)
炭酸水素ナトリウム(Sigma)
TCイーストレート(yeastolate)(Difco)
ピルビン酸(Sigma)
N−アセチルグルコサミン(Sigma)
ウシ血清アルブミン(Sigma)
ゼラチン(微生物学用グレード;Difco)
5N NaOH 。
【0061】
L−グルタミンを含有するが、炭酸水素ナトリウムを含有しないIOxコンノート・メディカル・リサーチ・ラボラトリーズ(CMRL)1066(MP Biomedicals)又はローズウェル・パーク・メモリアル・インスティテュート(RPMI)1640培地(Sigma)
ウサギ血清(Life Technologies)、56℃45分間加熱不活化
56℃水浴
陽圧ポンプ
ミリポアフィルターマニフォールド(Millipore filter manifold)
プレフィルター(prefilter)(124mm)
0.2−、0.45−及び0.8−μmフィルター(142−mm径)
0.2−μmベル・フィルター(bell filter)
滅菌100−ml容器
暗視野顕微鏡 。
【0062】
BSK培地の調製
1.以下のものを2−リットルフラスコ中で混合し、2乃至4時間混和した。900mLミニQ二重濾過又は脱イオン蒸留水:
6.0g HEPES
5.0g ネオペプトン
0.7g クエン酸ナトリウム
5.0g グルコース
2.2g 炭酸水素ナトリウム
2.5g TCイーストレート
0.8g ピルビン酸
0.4g N−アセチルグルコサミン
50g ウシ血清アルブミン(フラクションV) 。
【0063】
2.これらの成分は攪拌板を最も低速の設定にしてゆっくりと混和した。何故なら、激しく攪拌すると、B.burgdorferiに毒性のある分解産物が生じる場合があるからである。
【0064】
3.上記BSK成分を混和している間に、ゼラチン溶液を以下の通り調製する。即ち、500−mLフラスコ中で、ミリQ二重濾過又は脱水蒸溜水200mLとゼラチン14gとを混合し、中程度の設定値で加熱し、攪拌して溶解させた。次いで、この溶液を121℃で15分間加圧滅菌した後、56℃の水浴に入れた。
【0065】
4.BSK成分が完全に溶解したとき、この溶液を5N NaOHでpH7.5に調整した。
【0066】
5.次に、上記ゼラチン溶液200mL、10xCMRL1066培地100mL及び(56℃45分間)加熱不活化ウサギ血清64mLをその他のBSK成分と混合し、よく混和した。
【0067】
BSK培地の滅菌
6.前記陽圧ポンプを用いて、124−mmプレフィルター及び最小孔径(底部)から最大孔径(最上部)まで積み重ねられた142−mm0.2−、0.45−及び0.8−μm孔径フィルターを装着したミリポアフィルターマニフォールドを通してBSK培地を送り出した。[このBSK培地は加圧滅菌によって滅菌することができない。前濾過は、滅菌濾過の前に大きな粒子を除去しておくのに必要であった。]
7.次いで、前記陽圧ポンプ及び滅菌0.2−μmベル・フィルターを用いて上記BSK培地を滅菌容器中へ滅菌濾過した。
【0068】
滅菌性の確認
8.滅菌BSKの1−mL分量を無菌的に取り出し、滅菌した1.5−mL微小遠心管に移して35℃で一夜インキュベートした。残る滅菌濾過したBSKは4℃で貯蔵した。
【0069】
9.インキュベーション後、暗視野顕微鏡を用いてこの滅菌BSKを調べ、滅菌性を確認した。
【0070】
10.次に、この滅菌BSKを滅菌貯蔵容器中に移した。[滅菌BSK培地の酸化を低減させるためにこの貯蔵容器に充填する際、その上部にできる空間を最小限に保った。]
Barbour−Stoenner−Kelly(BSK)培地の品質管理
BSK培地を使用する前に、BSKが少数のボレリア属菌を増殖させるのに有利であること、及び生育可能なスピロヘータが凝集せずに増殖することを確認することは重要である。BSK培地の極めて変化しやすい成分はウシ血清アルブミン(BSA)である。本明細書で用いている品質管理プロトコルは、Callisterほか、2003年、上記文献に詳細に記載されているものと同様に、得られた培養物をインキュベートして検査する試験培養を利用したものである。
【0071】
・OspA殺ボレリア菌抗体の検出
OspA殺ボレリア菌抗体は、フローサイトメトリー法及びB.burgdorferi ssS−1−10を用いて検出した(Callisterほか、Arch.Intern.Med.1994年、154:p.1625−1632)。スピロヘータの新鮮培養液を新鮮なBSKでほぼ5×106個/mLのスピロヘータ濃度に希釈した。同時に、血清試料を新鮮BSKで40倍希釈し、0.2ミクロン(μm)孔径微小遠心フィルターを通して滅菌した。次いで、200μLの分量を滅菌された1.5mLねじ口微小遠心管に移し、BSKで80倍から20,480倍まで連続希釈した。血清試料を56℃で10分間加熱不活化した後、上記スピロヘータ懸濁液の100μL分量(スピロヘータ:5×105個)及び滅菌モルモット補体(Sigma)10μLを添加した。こうした分析試料をよく混和し、35℃で16乃至24時間インキュベートした。
【0072】
インキュベーション後、PBS400μL及びアクリジンオレンジ1μg/mLを入れたポリプロピレン管に各分析試料懸濁液100μLを移した。次いで、FACScanフローサイトメータ(Becton Dickinson lmmunocytometry Systems)、San Jose、カリフォルニア州)を用いて殺ボレリア菌活性を検出した。スピロヘータは、ゲート処理(gating)(CellQuestソフトウェア、Becton Dickinson)によって単離し、流速を低く設定して1乃至2分間分析した。OspA殺ボレリア菌抗体は、アクリジンオレンジが水疱状の生育不能スピロヘータ中に入り込む時に生じる蛍光強度の増大を観察することによって間接的に検出した。対照正常血清との比較における平均蛍光強度の13%以上のシフトを陽性と見なした(Callisterほか、Clin.Diagn.Lab.Immunol.2002年、9:p.908−912)。次に、暗視野顕微鏡によって水疱状の運動能力のないB.burgdorferiの存在を確認した。各分析試料に対して陽性対照を含め、ラン毎の反応性を同一(+/−1希釈)にするには分析法間の変動をできるだけ少なくすることが必要であった。さらに、各イヌから採取した血液試料を同時に分析した。
【0073】
B.結果
rOspAワクチンを用いてワクチン接種すると、7週後にごくわずかな量(平均力価:<61)のOspA殺ボレリア菌抗体が誘導されたが、15週後では検出されなかった(表2)。これに対して、通常の(Barbourほか、J.Infect.Dis.1985年、152:p.478−484)B.burgdorferi ss株(S−1−10)を含有するイヌのワクチンを用いてハムスターにワクチン接種すると、高レベルの殺ボレリア菌OspA抗体が誘導され、7週目にピーク(力価:>2560)に達し、実験期間中、高値を維持した。
【0074】
表2: rOspA又はGALAXY(商標)(S−1−10)イヌライム病ワクチンを用いたワクチン接種後のOspA殺ボレリア菌抗体bの平均力価a
【0075】
【表2】
従って、イヌの組換えOspAワクチンではハムスターにおいて有意な力価のOspA殺ボレリア菌抗体を誘導できない。
【0076】
(実施例2)
組換えOspCを用いたワクチン接種
A.材料及び方法
・動物
Charles River Breeding Laboratories,Inc.(Wilmington、マサチューセッツ州)から5乃至10週齢のLVGハムスターを入手した。ハムスターは21℃の室温でケージ当たり3乃至4匹を収容し、餌と水を自由に与えた。
【0077】
・組換え(「r:recombinant」)OspCワクチンの調製
rOspCは、以前に報告されたもの(Rousselleほか、J.Infect.Dis.1998年、178:p.733−741)と同様にして、pX3−22を含有するE.coli JM109から回収した。つまり、このE.coliをアンピシリンを含有する2xTYブロス中37℃で培養し、これにイソプロピル−β−d−ガラクトピラノシド(IPTG)(0.1mM)を指数増殖期の間に添加した。次いで、細胞を遠心分離によりペレット化し、リン酸緩衝食塩水(PBS)に再懸濁して超音波処理により溶解させた。次に、トリトンX−100(1%容積/容積)を添加した後、溶解物を10,000×gで5分間遠心した。次いで、この超音波処理したE.coliを遠心によりペレット化し、上清を、アミノ末端のビオチン化精製タグを介してOspCに結合したSoftLink樹脂(Promega)を含有するカラム上を通した。次に、5mMビオチン(Sigma)をも含有する精製緩衝液でこの結合OspCを溶出させた。
【0078】
・ハムスターのワクチン接種及び血清の採取
ハムスターの頸背部皮下に、75μgのrOspCを含有するフロイント完全アジュバントを0.1mL接種し、この初回ワクチン接種の3週後に75μgのrOspCを含有するフロイント不完全アジュバントで追加免疫を行った。初回ワクチン接種の5週後に、各ハムスターをノーズ・アンド・マウス・カップに含ませたエーテルの吸入により軽く麻酔し、心臓内穿刺により放血させた。この血液を凝固させ、血清を分離して、使用するまで−70℃で貯蔵した。さらに、正常なハムスター3匹からのハムスター血清をプールして対照正常血清として用いた。
【0079】
・OspC抗体の検出
rOspCをコーティング緩衝液(0.015M Na2CO3、0.035M NaHCO3、pH9.6)で1000ng/mLとなるように希釈し、各100μL量を個々の平底アミン結合マイクロタイター・ウェル(Costar、Cambridge、マサチューセッツ州)に添加した。マイクロタイター・プレートを4℃で一夜インキュベートした。インキュベーション後、プレートをPBS(pH7.2)で3回洗浄し、0.05%TWEEN20(Sigma)及び1%ウシ血清アルブミン(Sigma)を含有するPBSで振盪しながら室温で1時間ブロックした。ブロックの後、プレートを再びPBSで洗浄した。次いで、PBS/Tweenで80倍から20,480倍まで連続希釈したハムスター血清各100μL量を個々のウェルに添加し、これらのプレートを室温で1時間インキュベートした。インキュベーション後、プレートをPBSで3回洗浄し、PBS/Tweenで3,000倍希釈した抗ハムスターIgGホースラディッシュペルオキシダーゼ結合体(Organon Teknika Cappel)100μLを各ウェルに添加し、これらのプレートを室温で1時間再度インキュベートした。次に、プレートをPBSで3回洗浄し、リン酸o−フェニレンジアミン(0.4mg/ml;Sigma)100μLを各ウェルに添加して室温で30分間インキュベートさせた。1N H2SO4を100μL添加して反応を止め、直ちに490nm(モデルEL311;Bio−Tek Inc.、Winooski、バーモント州)での吸光度を測定した。
【0080】
・OspC殺ボレリア菌抗体の検出
B.burgdorferi ss50772を用いた以外は前述(実施例1)と同様にして、OspC殺ボレリア菌抗体を検出した。
【0081】
B.結果
rOspCワクチンを用いてワクチン接種すると、ELISAにより検出される高レベルのOspC抗体が誘導されたが(以下の表3)、殺ボレリア菌活性は低レベル(力価:80)で存在するに過ぎなかった。従って、rOspCを用いてワクチン接種すると、高濃度のOspC抗体が誘導されたが、その反応は、感染に対する防御をもたらさない抗体(例えば、オプソニン化抗体)でほとんど構成された。このことは、B.burgdorferi ss50772ワクチン中の蛋白質の75μg濃度が多数の別の非OspC蛋白質の存在を計上しないで算出されることを考慮すると、特に重要であった。従って、全体的な結果(実施例1及び2)から、組換えOspでは完全な状態のB.burgdorferi ssよりも殺ボレリア菌抗体誘導の量が相当に少ないというこれまでの知見(Straubingerほか、Vaccine 2003年、20:p.181−193)が裏付けられた。
【0082】
表3: rOspCによるワクチン接種後のOspCELISA又はOspC殺ボレリア菌抗体試験bによって検出された抗体の平均力価a
【0083】
【表3】
従って、イヌの組換えOspCワクチンではハムスターにおいて有意な力価のOspC殺ボレリア菌抗体を誘導できない。
【0084】
(実施例3)
B.burgdorferi ss分離株S−1−10を用いたワクチン接種
A.材料及び方法
・生物
B.burgdorferi ss分離株S−1−10は、通常B.burgdorferi ssB31と区別できない代表的な病原性株である(Barbourほか、J.Infect.Dis., 1985年、152:p.478−484)。この分離株は元々、ウィスコンシン州La Crosse付近から1998年2月に捕獲されたシロアシネズミPeromyscus leucopusの腎臓からウィスコンシン州La CrosseのGundersen Lutheran Medical FoundationのSteven M.Callister博士によって回収され、イヌのライム病ワクチンに組み入れるためにSolvay Animal Healthによりライセンス供与されたものである。その後の市販品(GALAXY(商標)ライム(Lyme))は1997年4月17日にSchering−Plough Animal Healthによって獲得された。この菌はOspA、OspB及びOspCを発現する。しかしながら、感染性B.burgdorferi ss分離株に特有であるが、スピロヘータを実験室BSK培地で培養すると、有意により高濃度のOspA及びOspBが産生される。OspCの発現は35℃でのインキュベーションによって最大化することができることが分かっている(Schwan、Biochem.Soc.Trans.2003年、31:p.108−112)。
【0085】
・動物
8週齢のビーグル幼犬(Ridglan Farms、Mount Horeb、ウィスコンシン州)を共同飼育し、餌と水を自由に与えた。実験はSchering Plough Animal Health動物管理使用委員会(IACUC:Animal Care and Use Committee)によって審査され、承認された。
【0086】
・B.burgdorferi ssS−1−10ワクチンの調製
35℃のBSKにおけるインキュベーションにより対数増殖期に達したB.burgdorferi ssS−1−10の新鮮培養物を、二成分エチレンイミン(BEI:binary ethylenimine)を最終濃度10mMとなるように添加してさらに48時間インキュベートすることによって不活化した。不活化後、滅菌チオ硫酸ナトリウムを添加して35℃で6乃至12時間インキュベートすることによりBEIを中和した。次いで、スピロヘータを遠心によりペレット化し、≦30μg/mLのゲンタマイシン及び≦30単位/mLのナイスタチンを含有する滅菌平衡塩類溶液に再懸濁した。次に、このスピロヘータに5%Emulsigen(登録商標)(MVP Laboratories,Inc.)及び1%HEPESを混合することによって1.0mL用量が≧2.5×107個のスピロヘータを含有するようにした。
【0087】
・ワクチン接種及び血清の採取
イヌの頚部皮下に1mL用量の上記S−1−10ワクチンを接種し、21日後にさらに1mL用量を追加免疫した。追加免疫ワクチン接種(試験21日目)の直前並びに試験28、35、47、83及び113日目に頚静脈穿刺によって全血を採取した。次いで、遠心により血清を分離し、これを試験するまで−20℃で貯蔵した。
【0088】
・殺ボレリア菌抗体の検出
上記の実施例1及び2と同様に、B.burgdorferi ssS−1−10及び50772を用いてフローサイトメトリーによりOspA及びOspC殺ボレリア菌抗体を検出した。
【0089】
B.結果
B.burgdorferi ssS−1−10を用いてワクチン接種することにより高レベルの殺ボレリア菌OspA抗体が確実に誘導された。この抗体は試験21日目には検出可能であり、試験28日目にピークに達し、試験113日目でも依然として検出可能であった(以下の表4)。これに対して、OspC殺ボレリア菌抗体は検出されなかった(力価:1:80未満)。すなわち、通常のB.burgdorferi ss株では、スピロヘータを35℃でインキュベートすることによりOspC発現を最大化したにもかかわらず、OspC殺ボレリア菌抗体の産生を誘導できなかった。
【0090】
表4: B.burgdorferi ssS−1−10によるワクチン接種後のOspA又はOspC殺ボレリア菌抗体の平均力価a(=8)
【0091】
【表4】
従って、B.burgdorferi ss分離株S−1−10を用いたワクチンでは有意な力価のOspC殺ボレリア菌抗体を誘導できなかった。
【0092】
(実施例4)
B.burgdorferi ss分離株50772を用いたワクチン接種
A.材料及び方法
・菌
B.burgdorferi ss50772は元々Andersonほか、J.of Clin.Microbiol.,1996年、34:p.524−529に報告されたユニークなospA−/ospB−株であり、著者らがOspC抗原を発現するものとみなすことができなかった株である。この50772株はOspC抗原を発現するものとして米国特許第6,464,985号に開示されており、これは引用により本明細書に組み込まれているが、この株はPTA−439の番号でATCCに寄託されているとこの特許に記載されている。
【0093】
・動物
8週齢のビーグル幼犬(Ridglan Farms、Mount Horeb、ウィスコンシン州)を共同飼育し、餌と水を自由に与えた。実験はSchering Plough Animal Health動物管理使用委員会(IACUC:Animal Care and Use Committee)によって審査され、承認された。
【0094】
・B.burgdorferi ss50772ワクチンの調製
上記の実施例3で説明したものと同様にして、B.burgdorferi ss50772ワクチンを調製した。
【0095】
・ワクチン接種及び血清の採取
イヌの頚部皮下に1mL用量の上記50772ワクチンを接種し、21日後にさらに1mL用量を追加免疫した。追加免疫ワクチン接種(試験21日目)の直前並びに試験28、35、47、83及び113日目に頚静脈穿刺によって全血を採取した。次いで、遠心により血清を分離し、これを試験するまで−20℃で貯蔵した。
【0096】
・殺ボレリア菌抗体の検出
上記の実施例1及び2と同様に、B.burgdorferi ss50772を用いてフローサイトメトリーによりOspC殺ボレリア菌抗体を検出した。
【0097】
B.結果
B.burgdorferi ss50772を用いてワクチン接種することにより高レベルの殺ボレリア菌OspC抗体が確実に誘導された。この抗体は試験35日目にピークに達し、試験113日目でも依然として検出可能であった(以下の表5)。すなわち、rOspC又はB.burgdorferi ss分離株(S−1−10)によるワクチン接種とは対照的に、上記のユニークなB.burgdorferi ss株50772を用いてワクチン接種すると、有意なレベルのOspC殺ボレリア菌抗体が誘導された。
表5: B.burgdorferi ss50772によるワクチン接種後のOspC殺ボレリア菌抗体の平均力価a(=8)
【表5】
【0098】
従って、B.burgdorferi ss分離株50772を含むワクチンは高濃度のOspC殺ボレリア菌抗体を誘導する。
【0099】
(実施例5)
B.burgdorferi ss分離株S−1−10及び50772を含むワクチンの調製
A.材料及び方法
・増殖
B.burgdorferi ss分離株S−1−10及び50772の凍結保存培養物を標準的な条件下で培養した。すなわち、これらの保存培養物を、5%CO2を混入させた空気の雰囲気下で[本明細書にその全体が引用により組み込まれているCallisterほか、1990年、J.of Clinical Microbiology 28:p.363−365に準じて調製した]BSK(Barbour−Stoenner−Kelly)培地5乃至17mLを含有する個々のねじ口培養管中、それぞれ33±2℃及び35±2℃で増殖させた。対数増殖に達した後、これらの培養物を用いて150乃至200mLの新鮮BSK培地に植菌し、次に、これを培養物が対数増殖に達するまでインキュベートした。次いで、得られた懸濁液を用いて、10乃至20リットルジャグに入れた5乃至10リットルの新鮮BSKに植菌し(生産培養)た後、さらに、上記のようにしてインキュベートした。
【0100】
・不活化
生産培養物中のスピロヘータを、BEIを10mMの濃度となるように添加して24乃至48時間ゆっくりと攪拌しながらインキュベートすることによって不活化した。スピロヘータの不活化後、10.6mLの滅菌済3.015Mチオ硫酸ナトリウムを各1リットルのBSL/スピロヘータ懸濁液に添加して6乃至12時間ゆっくりと攪拌しながらインキュベートすることによりBEIを中和した。
【0101】
・濃縮及び混合
不活化スピロヘータを遠心によりペレット化し、30.0μg/mLのゲンタマイシン及び30単位/mLのナイスタチンを含有する滅菌平衡塩類溶液に再懸濁した。この試験ワクチンに5%Emulsigen(登録商標)(MVP Laboratories,Inc.)を混合し、2.0mLガラスバイアルに充填して各用量(1.0mL)に≧2.5×107個/mLのB.burgdorferi ssS−1−10、≧5.0×108個/mLのB.burgdorferi ss50772、1%HEPES、29μ/mLゲンタマイシン及び29単位/mLナイスタチンを含有させた。
【0102】
(実施例6)
B.burgdorferi ss分離株S−1−10及び50772を含むワクチンの安全性及び有効性
A.材料及び方法
・動物
8週齢のビーグル幼犬(Ridglan Farms、Mount Horeb、ウィスコンシン州)を個別又は共同飼育し、餌と水を自由に与えた。実験はSchering Plough Animal Health動物管理使用委員会(IACUC)によって審査され、承認された。
【0103】
・ワクチン接種及び血清の採取
イヌの頚部皮下に1mL用量の上記ワクチンを接種し、21日後にさらに1mL用量を追加免疫した。初回(試験−3日目)及び追加免疫ワクチン接種(試験21日目)の前並びに試験28、35、43、78、106、134、162及び197日目にも頚静脈穿刺によって全血を採取した。次いで、遠心により血清を分離し、これを試験するまで−20℃で貯蔵した。
【0104】
・ワクチン接種後の観察
各ワクチン接種後、注射部位を反応が感じられなくなるまで毎日触診し、直腸温をワクチン接種後4乃至6時間及びワクチン接種後1乃至3日目に記録した。
【0105】
・ダニ感染
2回目のワクチン接種の3週後に、イヌを胸腔の右側で剃毛し、B.burgdorferi ss感染I.scapularis tickの雌10匹及び雄10匹を入れたラバーカップを剃毛部にテープ及び包帯ラップ(bandage wrap)で固定した。これらのダニに7日間吸血させた。
【0106】
・ダニにおけるB.burgdorferi ssの検出
ダニの口器をメスで除去し、中腸を掻き裂き、スライドガラスになすりつけて室温で一夜乾燥させた。乾燥後、このスライドをアセトンで8乃至10分間固定し、風乾した。種特異的マウスOspAモノクロナール抗体H5332をPBS(pH7.2)で40倍希釈し、PBSで500倍希釈したヒツジ抗マウスフルオレセインイソチオシアネート標識免疫グロブリンG抗体を重層した。インキュベーション後、スライドをPBSですすぎ、風乾して蛍光顕微鏡により調べた。各スライドは経験豊かな二名の微生物学者により独立に調べた。
【0107】
・血液試料
試験78、106、134、162及び197日目に全血を血清分離輸送(SST)管に採取し、血清を分離して試験するまで−20℃で貯蔵した。
【0108】
・OspA抗体の除去
以前に報告されたもの(Callisterほか、J.Infect.Dis.1993年、167:p.158−164)と同様にして、OspA−グルタチオン−S−トランスフェラーゼ融合蛋白質を発現するEscherichia coli(「E.coli」)DH5αから組換え(r)OspAを回収した。つまり、この大腸菌を、アンピシリンを含有する2xTYブロス中37℃で培養し、指数増殖期中にイソプロピル−β−ガラクトピラノシド(「IPTG」)(0.1mM)を添加した。次いで、細胞を遠心によりペレット化し、リン酸緩衝食塩水(「PBS」)に再懸濁して超音波処理により溶解させた。次に、トリトンX−100(1%容積/容積)を添加した後、溶解液を10,000×gで5分(min.)間遠心した。次に、上記融合蛋白質を含有する上清をグルタチオン−セファロース4Bカラム(Pharmacia)に通し、50mMトリス−Cl(pH8.0)プラス5mM還元グルタチオンで組換えOspAを溶出し、これを精製緩衝液(50mMトリス[pH8]、50mM NaCl、2mM EDTA、0.1%トリトンX−100)に再懸濁した。
【0109】
次に、rOspA融合蛋白質を臭化シアン(CNBr)活性化によりセファロース4Bに結合させた。具体的には、0.8gのCNBr活性化セファロース4B(Pharmacia)をカップリング緩衝液(0.1M NaHCO3−0.5M NaCl、pH8.3)で洗浄し、カップリング緩衝中の2mg量のOspAをこのゲルに添加し、この混合液を室温で2時間静かに振盪した。ゲルをカップリング緩衝液で2回洗浄した後、4mLのエタノールアミン(pH9)を添加し、ゲルを2時間インキュベートして未結合部位をブロックした。次いで、ゲルを50mLの0.1M酢酸ナトリウム−0.5M NaCl(pH4.0)で3回洗浄した後、PBS中で平衡化した。PBSで10倍希釈した免疫血清の試料1mLを4回上記カラムに通した。
【0110】
・OspC抗体の除去
以前に報告された(Rousselleほか、J.Infect.Dis.1998年、178:p.733−741)ようにして、pX3−22を含有するE.coli JM109からrOspCを回収した。この組換え蛋白質の産生は上記のrOspAで述べたものと同様にして誘導された。次いで、超音波処理したE.coli細胞を遠心によりペレット化し、上清を、OspCとそのアミノ末端のビオチン化精製タグを介して結合したSoftLink樹脂(Promega)を含有するカラム上を通した。次に、5mMビオチン(Sigma)をも含有する精製緩衝液でこの結合OspCを溶出させた。
【0111】
次に、1mL容量のテトラリンク四量体アビジン樹脂(Promega)を洗浄し、40mLのPBSに懸濁し、10mm×70mmのポリエチレンカラムにロードした。1mL容量のPBS中の0.5mg量の透析rOspCを上記カラムに通し、タンパク質測定(Bio−Rad)により結合を確認した。次いで、PBSで10倍希釈した1mL容量の免疫血清を4回このカラムに通した。
【0112】
・OspC7特異的抗体の除去
以前に報告された(本明細書にその全体が引用により組み込まれているLovrichほか、Clin.Diagn.Lab.Immunol.2005年、12:p.746−751)ようにして、pXT7を含有するE.coli JM109から、OspCの7個のC末端アミノ酸(OspC7)を含有する融合蛋白質を回収した。この組換え蛋白質の産生は上記のrOspAで述べたものと同様にして誘導された。次いで、超音波処理したE.coli細胞を遠心によりペレット化し、上清を、OspC7とそのアミノ末端のビオチン化精製タグを介して結合したSoftLink樹脂(Promega)を含有するカラムに通した。次に、5mMビオチン(Sigma)をも含有する精製緩衝液でこの結合OspC7を溶出させた。次に、1mL容量のテトラリンク四量体アビジン樹脂(プロメガ社)を洗浄し、40mLのPBSに懸濁し、10mm×70mmのポリエチレンカラムにロードした。1mL容量のPBS中の0.5mg量の透析rOspCを上記カラムに通し、タンパク質測定(Bio−Rad)により結合を確認した。次いで、PBSで10倍希釈した1mL容量の免疫血清を4回このカラムに通した。
【0113】
・殺ボレリア菌抗体の検出
上記の実施例1及び2と同様に、B.burgdorferi ssS−1−10及び50772を用いてフローサイトメトリーによりOspA及びOspC殺ボレリア菌抗体を検出した。
【0114】
・皮膚生検
試験78、106、134、162及び197日目にダニ付着の近傍部位から使い捨ての4mm穿刺器具を用いて皮膚生検材料を採取した。これらの生検材料は、9mlのゼラチンを補ったBSK(Callisterほか、J.Clin.Microbiol.1990年、28:p.363−365)(BSK+G)、40乃至50マイクログラム(μg)/mLリファンピン及び8μg/mLカナマイシンを含有する別々の管に入れた。培養物は35±2℃で3週間インキュベートし、暗視野顕微鏡検査法により週1回スピロヘータを調べた。
【0115】
・免疫抑制
イヌにおいて、感染後13日から始めて5日間デキサメタゾン(0.4mg/lb体重)の連日投与により免疫を抑制した。
【0116】
・臨床的観察
イヌを、こわばり(足に全体重をかけたがらないこと)、跛行(歩行又は走行中足をかばうこと)又は歩行困難(免荷(non−weight bearing))を含む肢/関節障害について毎日観察した。採点は少なくとも2名の観察者の合意によって行い、イヌは、他の外傷を全て除外するために担当獣医によって観察した。
【0117】
・剖検
少なくとも3連続観察期間の間、肢/関節障害を有したイヌは安楽死させて剖検した。肘、手根、膝、足根、心臓、脾臓、膀胱及び両腎臓からの組織試料を採取し、培養によるB.burgdorferiの単離のために処理した。関節組織は9mLのBSK+Gを含有する個々の管中で培養した。他の組織(心臓、脾臓、膀胱及び腎臓)は9mLのBSK+Gと混合し、ストマッチャーでよくホモジナイズし、1mL量を9mLの新鮮BSK+Gを含有する別の管に移した。培養物は35±2℃でインキュベートとした。
【0118】
残りのイヌは試験終了時に安楽死させて剖検した。これらのイヌからの組織試料は肘、手根、膝及び足根の関節包及び腱から採取し、組織病理検査のために10%緩衝ホルマリン中に貯蔵した。さらに、肘、手根、膝及び足根の関節包及び腱からの試料を9mLのBSK+G又は抗生物質含有BSK+Gを入れた個々の管に入れ、35±2℃で3週間インキュベートした。
【0119】
・ウエスタンブロッティング
ウエスタンブロッティングは標準的な技術を用いて実施した。B.burgdorferi ss297を試料緩衝液中で5分間煮沸し、150μgの全蛋白質を0.1%SDS−12%ポリアクリルアミドゲル(櫛を含まない4%ポリアクリルアミド濃縮用ゲル)上にロードした。蛋白質濃度は蛋白質測定キットで測定し、電気泳動装置において2つのゲルを同時に泳動させた。電気泳動後、蛋白質をニトロセルロースに移し、このニトロセルロースを細片に切ってPBS−0.3%TWEEN20で22℃30分間ブロックした。次いで、細片を100倍希釈したイヌ血清と共に22℃で1時間インキュベートした後、PBS−0.3%TWEEN20で3回洗浄した。ホースラディッシュペルオキシダーゼ標識抗イヌIgG(Kirkegaard & Perry Laboratories、Gaithersburg、メリーランド州)を添加し、これらの細片を22℃で30分間インキュベートした後、TMB膜ペルオキシダーゼ基質システム(Kirkegaard & Perry Laboratories、Gaithersburg、メリーランド州)で発色させた
B.結果
・ワクチンの安全性
イヌは、ワクチン接種後、直腸温の上昇がないなど、臨床的に正常な状態を維持した。しかしながら、ワクチンを接種されたイヌは注射部位に軽度の腫脹を生じ、これは8日以内に消散した。その最大の反応は(ワクチン接種後24時間の)1×1×0.5cmの大きさであった。
【0120】
・ワクチン接種後の血清
プラセボ接種では、B.burgdorferi ssに特異的な抗体を誘導することができなかった。これに対し、上記ワクチンの接種では、B.burgdorferi ss分離株S−1−10又は50772を用いることで検出される殺ボレリア菌抗体が有意なレベルで誘導された。S−1−10を用いて検出される殺ボレリア菌抗体の平均力価は追加免疫ワクチン接種(試験28日目)の1週後にピークに達し、試験197日目でも依然として検出可能であった(下記表6)。同様に、分離株50772に特異的な高濃度の殺ボレリア菌抗体が試験28日目までに存在し、試験43日目ではなお高く、試験197日目でも依然として検出可能であった(下記表7)。
【0121】
表6: ワクチン接種後の殺ボレリア菌活性bの平均力価a(n=15)
【0122】
【表6】
表7: ワクチン接種後の殺ボレリア菌活性bの平均力価a(n=15)
【0123】
【表7】
・OspA及びOspC殺ボレリア菌抗体の確認
上記殺ボレリア菌活性がOspA及びOspC殺ボレリア菌抗体によるものであることを確認するために、上記ワクチンを用いてワクチン接種した5頭のイヌからの免疫血清をrOspA又はrOspCを含有する別々のカラムに通し、殺ボレリア菌活性に及ぼす影響を調べた。これらの組換え蛋白質で上記血清を吸収させることによりOspA特異的又はOspC特異的抗体を除去すると、それぞれB.burgdorferi ssS−1−10又は50772分離株を用いて検出される殺ボレリア菌抗体が著しく(4分の1以下に)減少した。従って、こうした全体的な知見から、上記ワクチンが有意なレベルのOspA及びOspC殺ボレリア菌抗体を誘導し、B.burgdorferi ssS−1−10又は50772により検出される殺ボレリア菌活性が、それぞれOspA又はOspC殺ボレリア菌抗体にほとんど限定されたものであることが確認された。下記の表8を参照されたい。
【0124】
表8:ワクチンを用いてワクチン接種したイヌ(n=5)からの血清における殺ボレリア菌活性に及ぼすOspA又はOspC抗体除去の影響
【0125】
【表8】
・OspC7特異的殺ボレリア菌抗体の確認
上記OspC殺ボレリア菌抗体がかなりの割合のOspC7特異的殺ボレリア菌抗体を含有することを確認するために、上記ワクチンを用いてワクチン接種した5頭のイヌからの免疫血清をrOspC7含有カラムに通し、殺ボレリア菌活性に及ぼす影響を調べた。この組換えOspC7蛋白質で上記血清を吸収させることによりOspC7特異的抗体を除去すると、B.burgdorferi ss50772分離株を用いて検出される殺ボレリア菌抗体が著しく(2乃至4分の1に)減少した。従って、これらの知見から、上記ワクチンにより、保存OspC7エピトープに特異的なOspC殺ボレリア菌抗体が有意なレベルで誘導されることが確認された。下記の表9を参照されたい。
【0126】
表9: ワクチンを用いてワクチン接種したイヌ(n=5)からの血清における殺ボレリア菌活性に及ぼすOspC7抗体除去の影響
【0127】
【表9】
・感染ダニに対するOspA及びOspC殺ボレリア菌抗体の滅菌能
ワクチン接種又は対照イヌにおいて吸血したダニの中腸を調べることによって、ワクチンにより誘導されたOspA及びOspC殺ボレリア菌抗体がダニを滅菌したことが確認された。プラセボ接種したイヌ15頭中13頭において吸血した雌性ダニ106匹中34匹(32%)からのダニ塗抹においてB.burgdorferi ssが検出された(下記の表10)。これに対して、ワクチン接種したイヌにおいて吸血した雌性ダニからの中腸ではスピロヘータは検出されなかった(99匹中0)(p<0.0001)。
【0128】
表10: ワクチン接種又は対照イヌから取り出した雌性ダニにおけるB.burgdorferi ssの検出
【0129】
【表10】
・皮膚からB.burgdorferi ssが回収されないようにする上記ワクチンの能力
上記ワクチンにより誘導された殺ボレリア菌抗体はスピロヘータが皮膚にコロニーを形成するのも防止した。ダニ感染後1ヶ月おきにプラセボ接種イヌから採取した71個の生検材料のうち56個(79%)からB.burgdorferi ssが回収され(下記の表11)、これら15頭のイヌのうちの14頭(93%)の少なくとも1個の皮膚生検材料からスピロヘータが回収された。これに対して、上記ワクチンを接種したイヌから採取したどの皮膚生検材料からもB.burgdorferi ssは回収されなかった(p<0.0001)。
【0130】
表11: ワクチン接種又は非ワクチン接種対照イヌの皮膚からのB.burgdorferi ssの単離
【0131】
【表11】
・感染の血清学的証拠をもたらさないようにする上記ワクチンの能力
上記ワクチンはライム病特有の抗体の発生をも阻止した。これまでの研究(SPAHC Report B01−184−01 R)によって、B.burgdorferi ssによる感染はほぼ20kDaの蛋白質に結合するイヌ抗体を確実に誘導することが確認されたが、この反応は上記ワクチンにより誘導されなかった(図1)。今回の試験では、感染特異的20kDa蛋白質に結合する抗体は15頭のプラセボ接種対照イヌのうちの14頭(93%)からの免疫血清中に容易に検出することができた(図2A)。さらに、20kDa蛋白質の抗体を含有しない血清が、皮膚からスピロヘータも得られない対照イヌから採取された。これに対して、上記ワクチンを接種したイヌのいずれによっても20kDa蛋白質の抗体は産生されなかった(p<0.0001)(図2B)。
【0132】
さらに、ライム病を有するイヌは、B.burgdorferi ss50772分離株を用いても検出できる非OspC殺ボレリア菌抗体を産生する(Callisterほか、J.Clin.Microbiol.2000年、38:p.3670−3674)。この反応の明確な標的は不明であるが、こうした抗体によって感染の高度に特異的な血清診断的確認が可能になる。上記ワクチンを接種したイヌは既にOspC殺ボレリア菌抗体を産生していたが、ダニ感染後B.burgdorferi ss50772特異的抗体のレベルの著しい(≧4倍の)上昇を生じたイヌはいなかった。従って、これらのワクチン接種イヌがスピロヘータに感染することは全くありそうになかった。これに対して、殺ボレリア菌抗体はダニ感染前のプラセボ接種イヌにより産生されなかったが、ダニに吸血させた後では15頭中10頭(67%;p=0.0002)が、B.burgdorferi ss50772を用いて検出される殺ボレリア菌抗体を顕著なレベル(力価:≧1:640)で生じた(下記の表12)。
【0133】
表12: ダニ感染後における殺ボレリア菌抗体bの平均力価a
【0134】
【表12】
・明白な肢/関節障害に対する上記ワクチンの防止能
以前の研究(Summersほか、J.Comp.Path.2005年、133:p.1−13、Wikleほか、J.Appl.Res.Vet.Med.2006年、4:p.23−28)によって、B.burgdorferi ssのみの感染では明白な肢/関節障害が生じることはまれであることが明らかにされたが、我々の知見でもこのことが裏付けられた。1頭(8%)のプラセボ接種対照イヌのみが関節異常を生じた。ダニ感染の9週後に左前肢がこわばり、B.burgdorferi ssスピロヘータが肘から回収された。しかしながら、こうした知見は重要ではなかった。何故なら、上記ワクチンを接種したイヌ2頭も1肢以上にこわばりを生じたが、プラゼボ接種イヌとは対照的に、どの組織からもスピロヘータは回収されなかったからである。
【0135】
明白な症状の発症を増幅させるために、残りのイヌに対して免疫を抑制した。その後、プラセボ接種対照イヌ3頭が歩行困難になり、これらのイヌのうち2頭からB.burgdorferi ssが回収された。これに対して、上記ワクチンを接種した免疫抑制イヌは歩行困難を生じなかった。従って、これらの知見全体から、上記ワクチンはライム関節炎の発症を防止することが示唆されたが、こうした結果は有意ではなかった(p=0.0996)。しかしながら、以下に示し、説明した別のデータを参照されたい。
【0136】
・B.burgdorferi ss感染に伴うびらん性変化に対する上記ワクチンの防止能
しかしながら、上述のイヌの関節を顕微鏡で調べたときの所見では、上記ワクチンの効果が明らかに示された。上記ワクチンを接種した残りのイヌ(n=13)からの関節包は正常であった。これに対して、プラセボ接種した残りの対照イヌ11頭(上記の表7)のうちの6頭(p=0.0034)からの1個以上の関節組織は、好中球及び単核細胞の浸潤を特徴とする著しい炎症を示した(図3)。さらに、上記ワクチンを接種したイヌからのどの組織からもB.burgdorferi ssは回収されなかったが、びらん性変化を有するプラセボ接種対照イヌ6頭のうちの5頭(83%)の関節組織からスピロヘータが回収された。従って、これらの知見全体から、上記ワクチンがイヌライム病関節炎を防止することが確認され、この症状が、ほとんどの場合、潜在性滑膜炎を特徴とするという以前の報告(Summersほか、J.Comp.Path.2005年、133:p.1−13;Wikleほか、J.Appl.Res.Vet.Med.2006年、4:p.23−28)も裏付けられた。
【0137】
(実施例7)
OspC7エピトープに対する天然殺ボレリア菌抗体の特異性
OspC7に対して哺乳動物に誘導される殺ボレリア菌抗体が特異的であることを裏付けるデータは、以下の通り、ライム病の病歴又はライム関連症状の有無とは無関係に個体(ヒト)の血清を分析することによって得られた。
【0138】
A.材料及び方法
1.血清
ライム病の既往歴(カルテ審査)もしくはライム関連症状を有しない個体(n=36)からの正常血清、献血者(n=100)もしくはコレステロール検診を受けた個体(n=100)からの特性化されていない血清又は通常B.burgdorferi ss抗原と交差反応する血液因子もしくは疾患、例えば、抗核抗体(n=20)、リウマチ因子(n=20)、単核球症(n=10)、サイトメガロウイルス(n=10)、梅毒(n=13)もしくはロッキー山熱(n=4)を有するボランティアからの血清は20℃で貯蔵された保管試料からのものとした。ライム病血清は2003年又は2004年にGundersen Lutheran Medical Centerで診断された患者から採取した。CDCサーベイランス基準(米国疾病対策予防センターMorb.Mort.Wkly.Rep.1990、39:p.19−21)を満たす遊走性紅斑(EM)を有する患者からの血清(n=86)を確からしいライム病と分類し、何度もダニに暴露されて非定型皮膚病巣を有する患者からの血清(n=22)をほぼ確かなライム病(n=49)と分類し、ダニに暴露されて主に頭痛、発熱、筋肉痛及び関節痛を含む全身症状を有する患者からの血清を可能なライム病と分類した。これらの血清は初診時に採取し、−20℃で貯蔵し、試験前に盲検化(blinded)した。
【0139】
OspC7ペプチド。7−aaOspC7ペプチド(AESPKKP;配列番号1)は、University of Wisconsin Biotechnology Center(Madison、ウィスコンシン州)で自動合成器(Protein Technologies)及びFmoc法(Fieldsほか、Peptide Res.1991年、4:p.95−101)を用いて合成された。合成後、このペプチドのアミノ末端をHBTU活性化により手動でビオチン化し、高圧液体クロマトグラフィーによって精製した。組成は、マトリックス支援レーザー脱離イオン化飛行時間(MALDI−TOF:matrix−assisted laser desorption ionization−time of flight)質量分析法(予測質量:1095.4;実測質量:1095.8)を用いて確認した。
【0140】
OspC7 ELISA。マイクロタイター・プレート(イムノロン(Immunolon)2HB、(Thermo Labsystems、Franklin、マサチューセッツ州)の個々のウェルを、ストレプトアビジン(Pierce、Rockland、イリノイ州)を炭酸塩緩衝液(90mM NaHCO3、60mM Na2CO3;pH9.6)に含有させた4μg/ml懸濁液100μLで被覆し、4℃で一夜インキュベートした。インキュベーション後、0.05%Tween20を含有するトリス緩衝食塩水(TBS−T;13mMトリスHCl、3mMトリス塩基、140mM NaCl、2.7mM KCl;pH7.4)で5回洗浄した。洗浄後、1μg/mLのビオチン化OspC7ペプチドを含有するブロッキング緩衝液(15mM NaCl、10mMトリスHCl、3%ウシ胎仔血清、0.05%Tween20)200μLを各ウェルに添加し、室温で1時間回転(150rpm)させながらインキュベートした。次いで、プレートをTBS−Tで3回洗浄し、ブロッキング緩衝液で200倍希釈した100μL量の血清と室温で1時間反応させた。第2の抗体は、ブロッキング緩衝液で15,000倍希釈したペルオキシダーゼ結合ヤギ抗ヒトIgM及びIgG(Kirkegaad Perry Laboratories、Gaithersburg、メリーランド州)とした。1時間のインキュベーション後、結合された上記第2の抗体をo−フェニレンジアミン及び過酸化水素のクエン酸緩衝液(Sigma、St.Louis、ミズーリ州、)溶液を添加し、490nmでのODを測定(スペクトラマックス(SpectraMax)250、Molecular Devices、Sunnyvale、カリフォルニア州)して定量した。
【0141】
B.結果
以前の研究(Jobeほか、Clin.Diagn.Lab.Immuno.2003年、10:p.573−578、Lovrichほか、Clin.Diagn.Lab.Immunol.2005年、12:p746−751)では、高度に保存された免疫優勢OspC殺ボレリア菌抗体エピトープはそのC末端に最も近い7アミノ酸(OspC7)内の領域に限局していた。このエピトープがボレリア属菌による感染に特異的な抗体を誘導することを確認するために、ライム病患者からの血清(図5)及び正常対象者又はボレリア属菌蛋白質とも結合することができる抗体を産生する可能性の高い他の疾患を有する患者からの血清(図4)を用いたELISAの反応性を比較した。図4及び5では、吸光度1.25の線は正常血清及び交差反応する可能性のある血清の平均吸光度の3標準偏差分だけ上の値を設定したものである。従って、この線よりも上にくる結果はいずれもその反応性が有意である確率が99%である(真陽性)。有意な反応性は正常血清又は交差反応する可能性のある血清ではまれにのみ検出された(図4)。陽性結果の大部分はライム病患者由来のものであったかも知れない血清を用いて得られたことにも注目されたい。何故なら、これらの血清(CS)は種々の病気に関してGundersen Lutheran Medical Centerで診察されている100名の患者のグループから得られたものであるからである。この地方はライム病の高度流行地であり、患者の個人情報及び病歴が不明である。これに対して、ウィスコンシン州の非流行地であるミルウォーキー(Milwaukee)の献血者(BD)から無作為に採取された血清試料(n=100)では陽性血清は検出されなかった。
【0142】
さらに、一般にライム病患者からの血清では陽性の結果(非反応性であるべき血清の平均値の3標準偏差分だけ上の値−図4)が検出され、吸光度の値は高かった(図5)。従って、これらの知見全体から、OspC7エピトープに対する上記抗体の高度の特異性及びヒトライム病におけるこの反応の免疫優勢が裏付けられた。
【0143】
上記に例示したデータをまとめると、8週齢の幼犬のグループ群(15匹)に5.0×108個のB.burgdorferi ss50772、2.5×107個のB.burgdorferi S−1−10 SS及び5%Emulsigen(登録商標)を含有するワクチン又は5%Emulsigen(登録商標)のみを含有するプラセボを接種及び追加接種を行った。このワクチンにより注射部位に軽度の腫張が生じたが、速やかに消散した。より注目に値すべきなのは、このワクチンが高濃度の殺ボレリア菌OspA及びOspC抗体を誘導し、これらの抗体の濃度が追加免疫ワクチン接種後1週間でピークに達し、試験期間の間検出可能なレベルを維持したことである。さらに、かなりの割合のOspC殺ボレリア菌抗体がOspC7内の高度に保存されたエピトープに特異的であった。
【0144】
次に、B.burgdorferi ss感染雌性シカダニ(I.scapularis tick)にワクチン接種したイヌに対して吸血させ、ダニの中腸を調べることによって吸われた血液中のOspA及びOspC殺ボレリア菌抗体がB.burgdorferi ssを完全に除去することが確認された。具体的には、B.burgdorferi ssは、プラセボを接種した対照イヌから採取した34匹(32%)のダニで検出されたが、上記ワクチンを接種したイヌに対して吸血したダニ(n=99)では検出されなかった(p<0.0001)。さらに、ワクチンのレシピエントは、感染の確認に通常用いられるいくつかの間接的及び直接的方法でライム病に対して陰性の状態を維持した。これに対して、対照イヌでは、10頭(67%)が「感染特有の」20kDaB.burgdorferi ss蛋白質に対する抗体を産生し、8頭(53%)が「感染特異的」殺ボレリア菌抗体を産生した。さらに、4頭(27%)のイヌが持続性の歩行困難を生じ、これら4頭の歩行困難なイヌのうちの3頭(75%)の関節からB.burgdorferi ssが回収された。さらに、調べたプラセボ接種イヌ11頭のうちの6頭(55%)の関節包に炎症性浸潤が生じた。より説得力あることには、14頭(93%)及び8頭(53%)の対照イヌにおいて、それぞれ皮膚及び関節からB.burgdorferi ssが回収されたのに対し、上記試験品を接種したイヌからスピロヘータは回収されなかった。すなわち、本発明のライム病ワクチンは生じた副作用がごく僅かであり、B.burgdorferi ssによる感染に対して完全に防御した。
【0145】
(実施例8)
B.burgdorferi ss分離株S−1−10及び50772を含むワクチンの免疫性持続期間
A.材料及び方法
・動物
材料及び方法は上記の実施例6で述べた通りである。
【0146】
・ワクチン接種及び血清の採取
イヌの頚部皮下に1mL用量の上記ワクチンを接種し、21日後にさらに1mL用量を追加免疫した。初回(試験−4日目)及び追加免疫ワクチン接種(試験21日目)の前並びに試験28、35、49、79、114、142、175、210、238、266、302、322、357及び394日目にも頚静脈穿刺によって全血を採取した。次いで、遠心により血清を分離し、これを試験するまで−20℃で貯蔵した。
【0147】
・ワクチン接種後の観察
イヌは毎日観察した。
【0148】
・ダニ感染
上記2回目のワクチン接種の1年後に、イヌを胸腔の右側で剃毛し、B.burgdorferi感染シカダニ(I.scapularis tick)の雌10匹及び雄10匹を入れたラバーカップを剃毛部にテープ及び包帯ラップで固定した。これらのダニに7日間吸血させた。
【0149】
・ダニにおけるB.burgdorferi ssの検出
検出は上記の実施例6に記載の方法で実施した。
【0150】
・血液試料
感染後(PC)試験43日目、PC試験77日目、PC試験112日目、PC試験147目、PC試験174目及びPC試験241日目に全血を血清分離輸送管に採取し、血清を分離して試験するまで−20℃で貯蔵した。
【0151】
・殺ボレリア菌抗体の検出
上記の実施例1及び2に記載したものと同様にして、フローサイトメトリーによりOspA及びOspC殺ボレリア菌抗体を検出した。
【0152】
・皮膚生検
感染後(PC)試験43日目、PC試験77日目、PC試験112日目、PC試験147目、PC試験174目及びPC試験241日目にダニ付着の近傍部位から使い捨ての4mm穿刺器具を用いて皮膚生検材料を採取した。これらの生検材料は、上記の実施例6に記載したものと同様に処理した。
【0153】
・免疫抑制
イヌに対して、感染のほぼ20週後から始めて5日間デキサメタゾン(0.4mg/lb体重)の連日投与により免疫を抑制した。
【0154】
・臨床的観察
臨床的観察は上記の実施例6と同様にして実施した。
【0155】
・剖検
剖検は上記の実施例6と同様にして実施した。
【0156】
B.結果
・ワクチン接種後の血清
プラセボ接種では、B.burgdorferi ssに特異的な抗体を誘導することができなかった。これに対し、上記ワクチンの接種では、B.burgdorferi ss分離株S−1−10及び50772を用いることで検出される殺ボレリア菌抗体が有意なレベルで誘導された。S−1−10に対する平均殺ボレリア菌抗体価は追加免疫ワクチン接種の1週後(試験28日目)にピークに達し、この反応は試験394日目まで依然として検出可能であった(表13)。同様に、分離株50772に特異的な高濃度の殺ボレリア菌抗体が試験28日目に検出され、力価は試験49日目でも依然として高く、79日後でも低レベルではあるが、依然として検出可能であった(表14)。
【0157】
表13: 上記ワクチン接種後の殺ボレリア菌活性bの平均力価a(n=15)
【0158】
【表13】
表14: 上記ワクチン接種後の殺ボレリア菌活性bの平均力価a(n=15)
【0159】
【表14】
・感染ダニに対するOspA及びOspC殺ボレリア菌抗体の滅菌能
ワクチン接種又は対照イヌにおいて吸血したダニの中腸を調べることによって、上記ワクチンにより誘導されたOspA及びOspC殺ボレリア菌抗体がダニを滅菌したことが確認された。プラセボ接種したイヌ15頭中12頭において吸血した雌性ダニ95匹中15匹(16%)からのダニ塗抹においてB.burgdorferi ssが検出された(表15)。これに対して、ワクチン接種イヌ15頭のうちの2頭において吸血した75匹の雌性ダニのうちのわずか2匹(3%)からのダニ塗抹でB.burgdorferi
ssが検出された(p=0.0003)。
【0160】
表15: ワクチン接種又は対照イヌから取り出した雌性ダニにおけるB.burgdorferi ssの検出
【0161】
【表15】
・皮膚からB.burgdorferi ssが回収されないようにする上記ワクチンの能力
上記ワクチンにより誘導された殺ボレリア菌抗体は皮膚におけるB.burgdorferi菌の持続感染を防止した。ダニ感染後1ヶ月おきにプラセボ接種イヌから採取した75個の生検材料のうち33個(44%)からB.burgdorferi ssが回収された(表16)。これに対して、上記ワクチンを接種したイヌから採取した75個の生検材料のうちわずか6個(8%)からB.burgdorferi ssが回収された(p<0.0001)。スピロヘータは、ワクチン接種イヌ15頭中6頭(40%)に対し、対照イヌ15頭中10頭(67%)から少なくとも1個の皮膚生検材料から回収された。しかしながら、ワクチン接種イヌからの単離はダニ感染後最初の1ヶ月に限定されたのに対して、培養陽性対照イヌ10頭中8頭(80%)はその後の試験月数全体を通して皮膚生検材料培養物がB.burgdorferi陽性であった。
【0162】
表16: ワクチン接種又は非ワクチン接種対照イヌの皮膚からのB.burgdorferi ssの単離
【0163】
【表16】
・感染の血清学的証拠をもたらさないようにするワクチンの能力
上記ワクチンはライム病特有の抗体の発生をも阻止した。ライム病を有するイヌは、(実施例6で述べたように)B.burgdorferi ss50772分離株を用いても検出できる非OspC殺ボレリア菌抗体を産生する。15頭のプラセボ接種対照イヌのうちの5頭(33%)でダニ感染後に50772株に対する殺ボレリア菌抗体のレベルの著しい上昇(≧4倍の上昇)が認められたのに対し、上記試験ワクチンを接種したイヌでは全く認められなかった。
【0164】
表17: ダニ感染後の殺ボレリア菌抗体bの平均力価a
【0165】
【表17】
・明白な肢/関節障害及びB.burgdorferi ss感染に伴うびらん性変化に対する上記ワクチンの防止能
以前の研究によって、B.burgdorferi ssのみの感染では明白な肢/関節障害(実施例6)が生じることはまれであることが明らかにされ、ダニ感染モデルはイヌの年齢が高いほど効果が低いことが分かっている。明白な症状の発症を増幅させるために、ダニ感染のほぼ3ヶ月後にデキサメタゾンによりイヌの免疫を抑制した。4頭のプラセボ接種対照イヌが歩行困難になるか、関節組織にびらん性病変を生じた。これに対して、本発明のワクチンを接種したイヌはいずれも肢/関節障害の臨床的症状や関節組織によくみられるびらん性病変を生じなかった。
【0166】
本発明は、本明細書に記載した特定の実施態様によって範囲が限定されるものではない。事実、本明細書に記載したものの他に本発明の様々な変形例があり得ることは、前記の説明から当業者には明らかであろう。このような変形例は添付の特許請求の範囲に包含されるものである。
【0167】
さらに、核酸又はポリペプチドに関して示した全ての塩基の大きさ又はアミノ酸の大きさ及び分子量値は近似値であり、説明のために示したものであることは理解されるべきである。
【0168】
本明細書では種々の刊行物が引用されているが、これらの開示内容はその全体が引用により本明細書に組み込まれている。
【特許請求の範囲】
【請求項1】
本明細書に記載のワクチン組成物。
【請求項1】
本明細書に記載のワクチン組成物。
【図1】

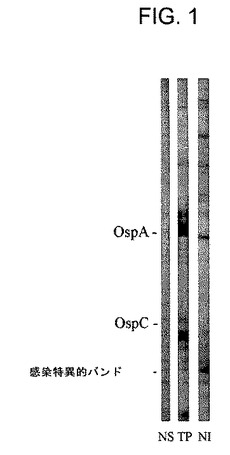
【図2A】
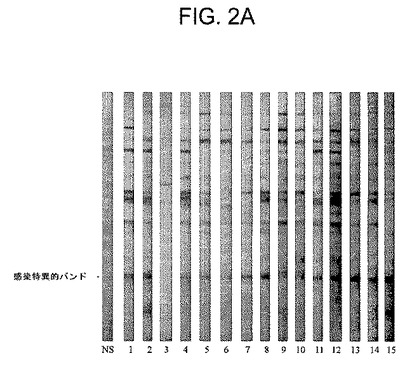
【図2B】
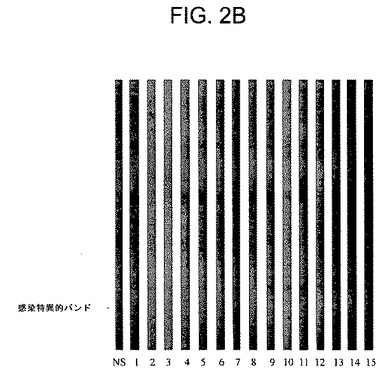
【図4】
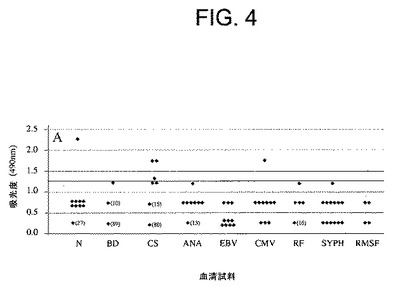
【図5】
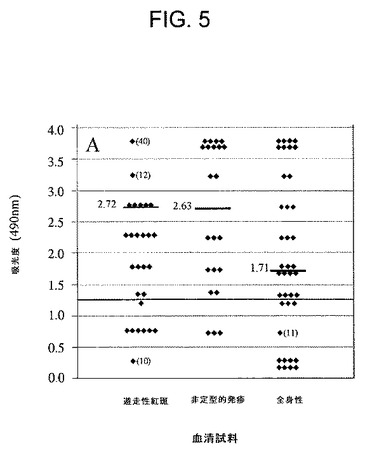
【図3A】

【図3B】

【図2A】
【図2B】
【図4】
【図5】
【図3A】
【図3B】
【公開番号】特開2012−211205(P2012−211205A)
【公開日】平成24年11月1日(2012.11.1)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−175212(P2012−175212)
【出願日】平成24年8月7日(2012.8.7)
【分割の表示】特願2009−535321(P2009−535321)の分割
【原出願日】平成19年11月1日(2007.11.1)
【出願人】(503442097)シェーリング−プラウ・リミテッド (47)
【住所又は居所原語表記】Weystrasse20,Lucerne6,CH−6000,Switzerland
【Fターム(参考)】
【公開日】平成24年11月1日(2012.11.1)
【国際特許分類】
【出願番号】特願2012−175212(P2012−175212)
【出願日】平成24年8月7日(2012.8.7)
【分割の表示】特願2009−535321(P2009−535321)の分割
【原出願日】平成19年11月1日(2007.11.1)
【出願人】(503442097)シェーリング−プラウ・リミテッド (47)
【住所又は居所原語表記】Weystrasse20,Lucerne6,CH−6000,Switzerland
【Fターム(参考)】
[ Back to top ]