
イヌ(canine)L−PBE配列
本発明は、イヌL−PBEと称される、従来知られていなかったポリペプチドをコードするcDNA;該遺伝子にコードされるイヌL−PBEポリペプチド;該ポリペプチドに対する抗体;および前記すべてを作成しそして用いる方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、イヌ(canine)L−PBEポリペプチドを含む単離ポリペプチド、および該ポリペプチドをコードするポリヌクレオチドを提供する。本発明はまた、毒性化合物または分子の指標となる代謝反応のための化合物および療法剤をスクリーニングするアッセイも提供する。
【背景技術】
【0002】
発明の背景
ペルオキシソームは、古典的なL−ヒドロキシ特異的ペルオキシソーム増殖因子誘導性ベータ酸化系を含有する。ヒト・ペルオキシソームのエノイル−CoA−ヒドラターゼ:3−ヒドロキシアシル−CoAデヒドロゲナーゼ二機能性酵素(L−PBE)は、Hoeflerら(1)によって最初に同定された。cDNA配列は、3,779ヌクレオチドに渡り、2,169ヌクレオチドのオープンリーディングフレームを有する。
【0003】
Qiらは、ノックアウトマウスを生成することによって、L−PBEの機能を検討した(2)。突然変異体マウスは生存可能であり、そして繁殖性であり、そして検出可能な著しい表現型上の欠陥を示さなかった。L−PBEノックアウトマウスは、ペルオキシソーム増殖因子に曝露されると、同様に処置された野生型マウスと比較して、ペルオキシソーム増殖反応が認識可能に鈍化していたことを除き、PPARαに制御される肝臓mRNAおよびタンパク質レベルの増加を示した。ペルオキシソーム増殖反応のこの鈍化は、L−PBEノックアウトマウス肝臓中にL−PBEタンパク質が存在しなかったことに起因すると考えられる。
【0004】
Watkinsらは、新生児副腎白質ジストロフィー(NALD)の臨床的特徴を持つ男性乳児において、ペルオキシソーム機能を評価した(3)。ペルオキシソームのベータ−酸化酵素のイムノブロット研究によって、死後の肝臓試料において、L−PBE(エノイル−CoAヒドラターゼ/3−ヒドロキシアシル−CoAデヒドロゲナーゼ)が欠損していることが示された。Wandersらもまた、ペルオキシソーム二機能性酵素欠損を記載しており、これはNALDのものより、より重度の表現型と関連しているようである(4)。
【0005】
イヌは、薬剤の毒性学を研究するための重要な種であるため、イヌL−PBE遺伝子に特異的な核酸プローブおよびイヌL−PBEタンパク質に特異的な抗体は、推定上の毒性化合物に対する代謝反応を示す手段として、産業毒性学において、大きな価値を有する。本発明はこの必要性に対処する。
【0006】
参考文献
【0007】
【化1】

【発明の開示】
【発明が解決しようとする課題】
【0008】
発明の概要
本発明は、上に同定される必要性に対処し、イヌL−PBEタンパク質をコードする単離核酸分子、該単離核酸分子を取り込んだ構築物および組換え宿主細胞;該単離核酸分子によってコードされるイヌL−PBEポリペプチド;イヌL−PBEポリペプチドに対する抗体を提供する。
【0009】
1つの態様において、本発明は、配列番号2に示すアミノ酸配列を含む、単離イヌL−PBEポリペプチドを提供する。配列番号2のポリペプチドは、いくつかのポリペプチド種を生じる、特定のタンパク質分解プロセシング事象にさらされることも可能であると理解される。
【0010】
さらに、本発明は、イヌL−PBEポリペプチドのエピトープを含む断片を提供する。「特異的なエピトープ」によって、以下に詳細に定義するように、イヌL−PBEポリペプチドに特異的な抗体によって認識可能であるイヌL−PBEポリペプチドの部分を意味する。別の態様は、配列番号2に示す完全アミノ酸配列を含む単離ポリペプチドを含む。
【0011】
イヌL−PBEのコードcDNA配列および予測されるアミノ酸配列を以下に複製し、そしてまた、それぞれ、配列番号1および配列番号2に示す。
【0012】
【化2】
【0013】
配列番号1および配列番号2は、特定のイヌ・ポリヌクレオチド配列およびポリペプチド配列を提供するが、本発明は、その範囲内に他のイヌ・アレル変異体を含むことが意図される。
【0014】
別の態様において、本発明は、本発明のポリペプチドのアミノ酸配列をコードするヌクレオチド配列を含む、単離ポリヌクレオチド(例えば一本鎖または二本鎖のcDNA、ゲノムDNA、合成DNA、RNA、またはその組み合わせ)を提供する。こうしたポリヌクレオチドは、細胞において、該酵素を組換え的に発現するのに有用であり、そしてまた、該酵素の発現を検出するのにも有用である(例えばノーザン・ハイブリダイゼーションおよびin situハイブリダイゼーションアッセイを用いる)。こうしたポリヌクレオチドはまた、培養細胞もしくは組織において、または動物において、イヌL−PBEの発現を抑制するアンチセンスおよび他の分子を設計するため、療法目的のため、あるいは異常なイヌL−PBE発現に特徴付けられる疾患のモデルを提供するために、有用である。本発明のポリヌクレオチドの定義から明確に排除されるのは、ポリヌクレオチドが元来得られた天然宿主細胞由来の完全単離染色体である。配列番号1に示すポリヌクレオチドは、天然存在イヌL−PBE配列に対応する。普遍的遺伝暗号の周知の縮重のため、やはり配列番号2のイヌL−PBEをコードする多くの他の配列が存在することが認識されるであろう。別の態様において、本発明は、配列番号1に示す配列以外の縮重イヌL−PBEコード配列のすべてに関する。
【0015】
本発明はまた、ヌクレオチド配列を含む単離ポリヌクレオチドであって、以下のハイブリダイゼーション条件下で、配列番号1に示すヌクレオチド配列または該配列に相補的な非コード鎖に特異的にハイブリダイズする、前記単離ポリヌクレオチドも提供する:
(a)50%ホルムアミド、1%SDS、1M NaCl、10%デキストラン硫酸を含むハイブリダイゼーション溶液中、42℃16時間のハイブリダイゼーション;および
(b)0.1%SSC、1%SDSを含む洗浄溶液中、60℃30分間2回の洗浄。
【0016】
本発明の1つのポリヌクレオチドは、イヌL−PBEコードDNA配列を含む配列番号1に示す配列、またはそのユニークな断片を含む。
関連する態様において、本発明は、本発明のポリヌクレオチドを含むベクターを提供する。こうしたベクターは、例えば宿主細胞において、ポリヌクレオチドを増幅し、有用な量を生じるのに有用である。他の態様において、ベクターは発現ベクターであり、該ベクター中で、本発明のポリヌクレオチドが、発現調節配列を含むポリヌクレオチドに機能可能であるように連結されている。こうしたベクターは、本発明のポリペプチドの組換え体産生に有用である。
【0017】
別の関連する態様において、本発明は、本発明のポリヌクレオチドまたは本発明のベクターで(安定にまたは一過性に)形質転換またはトランスフェクションされた宿主細胞を提供する。上述のように、こうした宿主細胞は、該ポリヌクレオチドを増幅するのに有用であり、そしてまた、該ポリヌクレオチドがコードする、イヌL−PBEポリペプチドまたはその断片を発現するのにも有用である。
【0018】
さらに別の関連する態様において、本発明は、イヌL−PBEポリペプチド(またはその断片)を産生する方法であって、栄養培地中で本発明の宿主細胞を増殖させ、そして細胞または培地から該ポリペプチドまたはその変異体を単離する工程を含む、前記方法を提供する。
【0019】
さらに別の態様において、本発明は、本発明のイヌL−PBEに特異的な抗体を提供する。抗体特異性を以下により詳細に記載する。しかし、文献に先に記載されたポリペプチドから生成可能であり、そしてイヌL−PBEと偶然に交差反応する(例えば両ポリペプチド中に類似のエピトープが偶然存在するため)ことが可能な抗体は、「交差反応性」抗体と見なされることを強調しなければならない。こうした交差反応性抗体は、イヌL−PBEに「特異的」な抗体ではない。抗体がイヌL−PBEに特異的であるか、または別の既知の酵素と交差反応性であるかの決定は、ウェスタンブロッティングアッセイまたは文献で周知のいくつかの他のアッセイを用いて行われる。イヌL−PBEを発現する細胞を同定するため、そしてまた、イヌL−PBE活性を調節するため、イヌL−PBEの活性部位に特異的に結合する抗体は、特に有用であるが、もちろん、他のエピトープに結合する抗体もまた、本発明の一部であることが意図される。
【0020】
1つの変型において、本発明はモノクローナル抗体を提供する。こうした抗体を産生するハイブリドーマもまた、本発明の側面と意図される。
別の変型において、本発明は、ポリクローナル抗体を含む細胞不含組成物を提供し、ここで少なくとも1つの抗体は、イヌL−PBEに特異的な本発明の抗体である。動物から単離される抗血清が典型的な組成物であり、水または別の希釈剤、賦形剤、もしくはキャリアーに再懸濁されている抗血清の抗体分画を含む組成物も典型的である。
【0021】
本発明はまた、本発明の抗体を用いる方法も提供する。例えば、本発明は、細胞抽出物内に存在するイヌL−PBEの量を決定する方法であって、イヌL−PBEポリペプチドとイヌL−PBEポリペプチドに特異的な抗体を、抗体がイヌL−PBEポリペプチドに結合する条件下で接触させる工程を含む、前記方法を提供する。
【0022】
本発明はまた、試料内に存在するイヌL−PBEポリヌクレオチドの量を決定する方法であって:配列番号1若しくはその断片、又はそれらの相補体を含む核酸分子と試料を、1以上の特異的ハイブリダイゼーション複合体形成のための条件下で接触させることを含む、前記方法も提供し、ここで該断片は、配列番号1の少なくとも12の連続ヌクレオチドを含むポリヌクレオチドである。
【0023】
本発明はまた、イヌにおいて、試験剤に対する代謝反応を測定する方法であって:試験剤で処置したイヌ由来の核酸を含有する試料を提供し;そして前記試料中の配列番号1若しくはその断片、又はそれらの相補体を含むポリヌクレオチドの量を決定することを含む、前記方法も提供し、ここで未処置のイヌ由来のポリヌクレオチドの量と比較した際の、処置したイヌ由来のポリヌクレオチドの量の変化が、試験剤に対する代謝反応の指標となる。
【0024】
1つの態様において、決定はハイブリダイゼーションを介して達成される。ハイブリダイゼーションは、核酸分子と試料核酸分子を、核酸分子および試料核酸分子間でハイブリダイゼーション複合体を形成するのに有効な条件下で接触させ;そしてハイブリダイゼーション複合体の存在または非存在を検出することを含む方法によって達成されることも可能である。1つの側面において、配列番号1若しくはその断片、又は配列番号1の相補体は、固体支持体アレイまたは他の固体支持体上の複数の核酸とともに存在することも可能である。
【0025】
本発明はさらに、試料内に存在するイヌL−PBEポリペプチドの量を決定する方法であって:
イヌL−PBEポリペプチドとイヌL−PBEポリペプチドに特異的な抗体を、抗体がイヌL−PBEポリペプチドに結合する条件下で接触させる
ことを含む、前記方法を提供する。所望によって、L−PBEポリペプチドまたは抗体は、固体支持体に付着していることも可能である。
【0026】
本発明はまた、イヌにおいて、試験剤に対する代謝反応を測定する方法であって:試験剤で処置したイヌ由来の試料を提供し;そして前記試料中の配列番号2を含むポリペプチド、または前記ポリペプチドに特異的なエピトープを含む該ポリペプチドの断片の量を決定することを含む、前記方法も提供し、ここで未処置のイヌ由来のポリペプチドの量と比較した際の、処置したイヌ由来のポリペプチドの量の変化が、試験剤に対する代謝反応の指標となる。
【課題を解決するための手段】
【0027】
配列表の簡単な説明
配列番号1――イヌL−PBEをコードするcDNA配列
配列番号2――イヌL−PBEの予測されるアミノ酸配列
配列番号3〜10――クローニング用プライマーおよび配列決定用プライマー
配列番号11 ヒトL−PBEをコードするcDNA配列
配列番号12 ヒトL−PBEの予測されるアミノ酸配列
配列番号13 マウスL−PBEをコードするcDNA配列
配列番号14 マウスL−PBEの予測されるアミノ酸配列
配列番号15 ラットL−PBEをコードするcDNA配列
配列番号16 ラットL−PBEの予測されるアミノ酸配列
【0028】
発明の詳細な説明
一般的な定義
本明細書において、これ以降、「ポリヌクレオチド」は、一般的に、非修飾RNAもしくはDNA、または修飾RNAもしくはDNAであることも可能な、ポリリボヌクレオチドまたはポリデオキシリボヌクレオチドいずれかを指す。「ポリヌクレオチド」には、限定なしに、一本鎖DNAおよび二本鎖DNA、一本鎖領域および二本鎖領域の混合物であるDNA、一本鎖RNAおよび二本鎖RNA、並びに一本鎖領域および二本鎖領域の混合物であるRNA、一本鎖、またはより典型的には二本鎖、あるいは一本鎖および二本鎖領域の混合物であることも可能なDNAおよびRNAを含むハイブリッド分子が含まれる。さらに、「ポリヌクレオチド」は、RNAもしくはDNAまたはRNAおよびDNA両方を含む三重鎖領域を指す。用語「ポリヌクレオチド」はまた、1以上の修飾塩基を含有するDNAまたはRNA、および安定性または他の理由のために修飾された骨格を持つDNAまたはRNAも含む。「修飾」塩基には、例えば、トリチル化塩基およびイノシンなどの普通でない塩基が含まれる。DNAおよびRNAに多様な修飾を行うことも可能であり;したがって、「ポリヌクレオチド」は、天然に典型的に見られるポリヌクレオチドの化学的、酵素的、または代謝的修飾型とともに、ウイルスおよび細胞に特徴的なDNAおよびRNAの化学型も含む。「ポリヌクレオチド」はまた、しばしばオリゴヌクレオチドと称される、比較的短いポリヌクレオチドも含む。
【0029】
本明細書において、これ以降、「ポリペプチド」は、ペプチド結合または修飾ペプチド結合によって互いに連結されたアミノ酸を含むペプチドまたはタンパク質いずれかを指す。「ポリペプチド」は、一般的にペプチド、オリゴペプチドまたはオリゴマーと称される短い鎖、および一般的にタンパク質と称される、より長い鎖両方を指す。ポリペプチドは、遺伝子にコードされる20のアミノ酸以外のアミノ酸を含有することも可能である。「ポリペプチド」には、翻訳後プロセシングなどの天然プロセスによるか、または当該技術分野に周知の化学的修飾技術によるか、いずれかによって修飾されたアミノ酸配列が含まれる。こうした修飾は、基本的な教科書、そしてより詳細なモノグラフ、さらに、膨大な研究文献によく記載されている。修飾は、ペプチド骨格、アミノ酸側鎖およびアミノ末端またはカルボキシル末端を含めて、ポリペプチドのどこに存在することも可能である。同種の修飾が、既定のポリペプチドのいくつかの部位で、同じ度合いで、または異なる度合いで存在することも可能であることが認識されるであろう。また、既定のポリペプチドが多くの種類の修飾を含有することも可能である。ポリペプチドは、ユビキチン化の結果として分枝していることも可能であるし、そして分枝を伴い、または伴わずに、環状であることも可能である。環状、分枝および分枝環状ポリペプチドは、翻訳後天然プロセスから生じることも可能であるし、または合成法によって作成されることも可能である。修飾または修飾型には、アセチル化、アシル化、ADP−リボシル化、アミド化、フラビンの共有結合、ヘム部分の共有結合、ヌクレオチドまたはヌクレオチド誘導体の共有結合、脂質または脂質誘導体の共有結合、ホスファチジルイノシトール(phosphotidylinositol)の共有結合、架橋、環状化、ジスルフィド結合形成、脱メチル化、共有架橋の形成、シスチンの形成、ピログルタミン酸の形成、ホルミル化、ガンマ−カルボキシル化、グリコシル化、GPIアンカー形成、ヒドロキシル化、ヨウ素化、メチル化、ミリストイル化、酸化、タンパク質分解プロセシング、リン酸化、プレニル化、ラセミ化、セレノイル化(selenoylation)、硫酸化、アルギニル化などのトランスファーRNAが仲介するタンパク質へのアミノ酸の付加、およびユビキチン化が含まれる(例えば、Proteins―Structure and Molecular Properties, 第2版, T.E. Creighton監修, W.H. Freeman and Company, ニューヨーク, 1993;Wold, F., Post−translational Protein Modifications:Perspectives and Prospects, 1−12ページ, Posttranslational Covalent Modification of Proteins中, B.C. Johnson監修, Academic Press, ニューヨーク, 1983;Seifterら, “Analysis for protein modifications and nonprotein cofactors”, Meth Enzymol(1990)182:626−646およびRattanら, “Protein Synthesis: Post−translational Modifications and Aging”, Ann NY Acad Sci(1992)663:4842を参照されたい)。
【0030】
「合成」は、本明細書において、そして当該技術分野に理解されるように、酵素的方法と対照的に、純粋に化学的方法によって産生されるポリヌクレオチドを指す。したがって、「完全に」合成されたDNA配列は、完全に化学的手段によって産生され、そして「部分的に」合成されたDNAは、生じるDNAのある部分のみが化学的手段によって産生されたものを含む。
【0031】
本明細書において、これ以降、「単離された」は、人の手によって天然状態から改変されたことを意味する。「単離された」組成物または物質が天然に存在する場合、その元来の環境から変化しているかまたは取り除かれているか、あるいはその両方である。例えば、生存動物に天然に存在するポリヌクレオチドまたはポリペプチドは、「単離されて」いないが、天然状態で共存する物質から分離された、同じポリヌクレオチドまたはポリペプチドは、該用語を本明細書で使用する際、「単離されて」いる。「単離された」は、本明細書において、そして当該技術分野に理解されるように、「単離」ポリヌクレオチドまたはポリペプチドいずれを指すのであっても、該ポリペプチドまたは核酸が通常見られる元来の細胞環境から分離されたことを意味するように解釈される。したがって、本明細書において、一例として、本発明のポリヌクレオチドで構築したトランスジェニック動物または組換え細胞株は、「単離」核酸を利用する。
【0032】
本明細書において、用語「イヌL−PBEポリペプチド」は、イヌL−PBE遺伝子がコードするタンパク質を意味し、保存的変化または非保存的変化を含有するアレル変異体を含む。1つのイヌL−PBEタンパク質配列を配列番号2として開示する。
【0033】
イヌL−PBEポリペプチドを、組換え細胞または生物によって産生することも可能であるし、天然組織または細胞株から実質的に精製することも可能であるし、あるいは化学的または酵素的に合成することも可能である。したがって、用語「イヌL−PBEポリペプチド」は、グリコシル化型、部分的グリコシル化型、または非グリコシル化型のタンパク質とともに、リン酸化型、部分的リン酸化型、非リン酸化型、硫酸化型、部分的硫酸化型、または非硫酸化型のタンパク質を含むことが意図される。該用語にはまた、アレル変異体、生物学的に活性であるタンパク質分解断片または他の断片を含む、PS2アミノ酸配列の他の機能的同等物、並びにイヌL−PBEポリペプチドの生理学的および病的タンパク質分解切断産物が含まれる。
【0034】
本明細書において、用語「試験剤」は、同定可能な化学薬品または分子いずれかを意味し、これには、限定されるわけではないが、小分子、ペプチド、タンパク質、糖、ヌクレオチド、または核酸が含まれる。こうした試験剤は、天然または合成であることも可能である。
【0035】
本明細書において、用語「接触」は、直接または間接的いずれかで、化合物を本発明のポリペプチドまたはポリヌクレオチドと物理的に近接させて、合わせることを意味する。該ポリペプチドまたはポリヌクレオチドは、いかなる数の緩衝剤、塩、溶液などの中に存在することも可能である。接触には、例えば、イオンチャネルポリペプチドもしくはその断片、またはイオンチャネルもしくはその断片をコードする核酸分子いずれかを含有する、ビーカー、マイクロタイタープレート、細胞培養フラスコ、または遺伝子チップ等のマイクロアレイに化合物を入れることが含まれる。
【0036】
本発明の核酸
本発明は、本明細書において、イヌL−PBEと称されるイヌL−PBEポリペプチドをコードする単離ポリヌクレオチド(例えば、センス鎖および相補的アンチセンス鎖両方の、一本鎖および二本鎖両方のDNA配列およびRNA転写物、そのスプライス変異体を含む)を提供する。本発明のDNAポリヌクレオチドには、ゲノムDNA、cDNA、および全体または部分が化学的に合成されているDNAが含まれる。
【0037】
本発明のゲノムDNAは、本発明のポリペプチドのタンパク質コード領域を含み、そしてまたそのアレル変異体を含むことも意図される。多くの遺伝子に関して、ゲノムDNAがRNA転写物に転写され、これが1以上のスプライシング事象を経て、転写物のイントロン(すなわち非コード領域)が除去されるか、または「スプライシング−アウト」されることが広く理解されている。選択的(alternative)機構によってスプライシングされることも可能であり、そしてしたがって異なるRNA配列の除去にさらされることも可能であるが、なおイヌL−PBEポリペプチドをコードするRNA転写物は、当該技術分野において、スプライス変異体と称され、本発明に含まれる。したがって、本発明に含まれるスプライス変異体は、同一の元来のゲノムDNA配列にコードされるが、別個のmRNA転写物から生じる。アレル変異体は、野生型遺伝子配列の修飾型であり、修飾は、染色体分離中の組換えから、または遺伝子突然変異を生じさせる条件への曝露から生じる。アレル変異体は、野生型遺伝子同様、天然存在配列である(in vitro操作から生じる非天然存在変異体とは対照的である)。
【0038】
本発明はまた、イヌL−PBEをコードするRNAポリヌクレオチドの逆転写を通じて得られるcDNAも含む(慣用的には、相補鎖の第二の鎖合成が続き、二本鎖DNAを提供する)。
【0039】
イヌL−PBEポリペプチドをコードするDNA配列を配列番号1に示す。当業者は、本発明のDNAが二本鎖分子を含み、例えば、DNAのワトソン−クリック塩基対形成規則にしたがって配列番号1の配列から推定可能な配列を有する相補分子(「非コード鎖」または「相補体」)を伴う、配列番号1に示す配列を有する分子を含むことを容易に認識するであろう。本発明にやはり意図されるのは、普遍的遺伝暗号の周知の縮重のため、配列番号1のポリヌクレオチドと配列が異なる、配列番号2のイヌL−PBEポリペプチドをコードする他のポリヌクレオチドである。
【0040】
当該技術分野に周知であるように、遺伝暗号の縮重のため、前述の配列番号1のポリペプチドにコードされるのと同一のポリペプチドをコードすることが可能な、多くの他のDNA分子およびRNA分子がある。したがって、本発明は、発現に際して、配列番号2のポリペプチドをコードする、他のDNA分子およびRNA分子を意図する。イヌL−PBEポリペプチドをコードするアミノ酸残基配列が同定されたら、そして各特定のアミノ酸残基をコードするすべての3つ組コドンを知っていれば、こうしたコードRNA配列およびDNA配列をすべて記載することが可能である。したがって、特定のアミノ酸に関して、単純にコドン中の変化によって特徴付けられる、本明細書に具体的に開示するもの以外のDNA分子およびRNA分子が、本発明の範囲内である。
【0041】
アミノ酸およびそれを表す略語、記号およびコドンの表を、以下の表1に示す。
表1
【0042】
【表1】
【0043】
当該技術分野に周知であるように、コドンは、mRNAおよびその対応するcDNA分子において、ヌクレオチドの3つ組配列を構成する。コドンは、mRNA分子に存在する場合、塩基ウラシル(U)によって特徴付けられるが、DNAに存在する場合、塩基チミジン(T)によって特徴付けられる。ポリヌクレオチド内の同一アミノ酸残基のコドンにおける単純な変化は、コードされるポリペプチドの配列または構造を変化させないであろう。句が、特定の3ヌクレオチド配列が特定のアミノ酸いずれかを「コードする」ことを述べる場合、一般の当業者は、上の表が問題の特定のヌクレオチドを同定する手段を提供するのを認識するであろうことが明らかである。例として、特定の3ヌクレオチド配列が、スレオニンをコードする場合、上の表は、ありうる3つ組配列がACA、ACG、ACCおよびACU(DNAの場合ACT)であることを開示する。
【0044】
本発明は、イヌL−PBE DNAの種相同体(homolog)、好ましくは哺乳動物相同体をさらに含む。種相同体は、ときに「オルソログ」と称され、本発明の配列番号1と、少なくとも89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%の相同性を共有する。本発明のポリヌクレオチドに関する配列「相同性」パーセントは、本明細書において、配列を並列させ、そして必要であればギャップを導入して最大の配列同一性パーセントを達成した後、配列番号1に示すイヌL−PBE配列のヌクレオチドと同一である、候補配列中のヌクレオチド塩基の割合として定義される。例えば、SmithおよびWaterman(Adv. Appl. Math. 2:482−489(1981))のアルゴリズムを用いる、Gapプログラム(ウィスコンシン配列解析パッケージ、バージョン8、Unix用、遺伝学コンピュータグループ、University Research Park, Madison, Wisconsin)などの、この目的に一般的に使用されるコンピュータプログラムいずれかを用いて、2つの配列を比較することによって、天然および変異体イヌL−PBE配列間の配列の割合を決定することもまた可能である。
【0045】
本発明に提供されるポリヌクレオチド配列情報は、当該技術分野に周知であり、そして日常的に実施される技術によって、コードされるポリペプチドの大規模発現を可能にする。本発明のポリヌクレオチドはまた、サザン・ハイブリダイゼーションおよび/またはノーザン・ハイブリダイゼーション、およびポリメラーゼ連鎖反応(PCR)を含む周知の技術によって、ヒト・アレル変異体および種相同体などの、関連するイヌL−PBEポリペプチドをコードするポリヌクレオチドの同定および単離も可能にする。関連するポリヌクレオチドの例には、アレル変異体を含むヒトおよび非ヒトゲノム配列、並びに、イヌL−PBEに相同なポリペプチド、およびイヌL−PBEの1以上の生物学的、免疫学的、および/または物理的特性を共有する、構造的に関連するポリペプチドをコードするポリヌクレオチドが含まれる。イヌL−PBE DNAの配列を知ることで、サザン・ハイブリダイゼーションまたはポリメラーゼ連鎖反応(PCR)の使用を通じて、プロモーター、オペレーター、エンハンサー、リプレッサー等のイヌL−PBE発現調節制御配列をコードするゲノムDNA配列の同定もまた可能になる。本発明のポリヌクレオチドはまた、細胞がイヌL−PBEを発現する能力を検出するか、またはイヌL−PBE発現レベルを測定する、ハイブリダイゼーションアッセイにおいても有用である。本発明のポリヌクレオチドはまた、単数または複数の疾患状態の根底にあるイヌL−PBE遺伝子座における遺伝子改変(単数または複数)を同定するのに有用な診断法の基礎となることも可能であり、こうした情報は、診断にも、そして療法戦略の選択にも有用である。
【0046】
イヌL−PBEポリペプチドをコードする全長ポリヌクレオチドの本明細書における開示は、一般の当業者が、全長ポリヌクレオチドのすべてのありうる断片を入手するのを容易に可能にする。したがって、本発明は、イヌL−PBEをコードするポリヌクレオチドの少なくとも12から2562(この間の1つ1つの整数値を含む)の連続ヌクレオチドを含む、イヌL−PBEコードポリヌクレオチドの断片を提供する。本発明のポリヌクレオチド(断片を含む)は、しばしば、イヌL−PBEをコードするポリヌクレオチド配列にユニークな配列を含み、そしてしたがって、非常にストリンジェントな条件下、または中程度にストリンジェントな条件下で、イヌL−PBE(またはその断片)をコードするポリヌクレオチドにのみ(すなわち「特異的に」)ハイブリダイズするであろう。本発明のゲノム配列のポリヌクレオチド断片は、コード領域にユニークな配列だけでなく、イントロン、制御領域、および/または他の非翻訳配列に由来する全長配列の断片も含む。本発明のポリヌクレオチドにユニークな配列は、他の既知のポリヌクレオチドへの配列比較を通じて認識可能であり、そして当該技術分野で日常的に利用される並列プログラム、例えば公共の配列データベースで入手可能なものの使用を通じて同定可能である。こうした配列はまた、ポリヌクレオチドがハイブリダイズするゲノムDNA断片の数を決定するサザン・ハイブリダイゼーション解析からも認識可能である。本発明のポリヌクレオチドは、放射能標識、蛍光標識、および酵素標識を含めて、その検出を可能にする方式で標識されることも可能である。
【0047】
断片ポリヌクレオチドは、全長または他の断片イヌL−PBEポリヌクレオチドの検出用のプローブとして特に有用である。1以上の断片ポリヌクレオチドは、イヌL−PBEをコードするポリヌクレオチドの存在を検出するのに用いられるか、またはイヌL−PBEをコードするポリヌクレオチド配列における変動を検出するのに用いられる、キットに含まれることも可能である。
【0048】
本発明はまた、イヌL−PBEポリペプチドをコードするDNAであって、配列番号1のポリヌクレオチドの非コード鎖または相補体に、高ストリンジェンシーハイブリダイゼーションまたは洗浄条件下でハイブリダイズする、前記DNAもまた含む。
【0049】
高ストリンジェンシーハイブリダイゼーションの概念は、以下の「本発明のアッセイ」を詳述する項で論じる。
【実施例】
【0050】
(実施例1)
イヌL−PBEのクローニング
PCRのプライマーを設計するため、DNASTARソフトウェア(DNASTAR、ウィスコンシン州)を用いて、ヒト、ラットおよびマウスL−PBEの各遺伝子のオープンリーディングフレーム(ORF)配列を並列した。各遺伝子について3つのプライマーセットを選択した。その中で、2つのセットは、それぞれ、ヒトおよびラットのオルソログの開始配列領域および停止配列領域に対応し;残りのセットは、ヒト、ラットおよびマウスのオルソログ間の保存される配列領域にしたがって設計された。プライマーは、一般的に、長さ30ヌクレオチドである。
【0051】
ヒトPSDデータベース(Proteome Inc.)およびNCBI ESTデータベース検索を通じて(相同ESTがクローニングされた組織を同定することによって)、選択したクローニング候補遺伝子の組織発現パターンを同定した。5つの組織、肝臓、腎臓、結腸、脾臓および肺が、選択した12の遺伝子すべてを含むことが同定された。
【0052】
RNeasyキット(Qiagen、カリフォルニア州)を用いて、イヌ肝臓試料からRNAを調製した。RNAをcDNAに逆転写するため、総体積12.5μl中、0.5μgのRNAを5.77μlの32.5μMランダム六量体(Amersham Biosciences、ニュージャージー州)および2.5μlの10mM dNTPと混合した。混合物を65℃で5分間インキュベーションし、そして氷上で冷却した。2μlのRNアーゼ不含水、5μlの5x第一鎖緩衝液(Invitrogen、カリフォルニア州)、0.5μlの100mM DTT、2.5μlの40U/μl RNaseOUT(Invitrogen、カリフォルニア州)および2.5μlの200U/μl Superscript II逆転写酵素(Invitrogen、カリフォルニア州)を含有する12.5μlのマスター混合物を添加した。最終反応混合物を25℃で10分間、42℃で60分間、そして70℃で15分間、連続してインキュベーションした。合成したcDNAを20倍に希釈し(最終濃度ほぼ1ng/μl)、そしてPCR反応のテンプレートとして直接使用するために、−20℃フリーザーで保存した。
【0053】
RACE(cDNA端の迅速増幅)反応から適応させた条件を用いた修飾PCR反応を用いて、イヌ遺伝子または遺伝子断片をクローニングした。50μlのPCR反応について、5μlの逆転写cDNA、5μlの10xPCR緩衝液(Clontech、カリフォルニア州)、50xdNTP混合物の1μlのdNTP(各10mM、最終濃度各0.2mM、Clontech、カリフォルニア州)、各20μMの1μlの5’プライマーおよび3’プライマーであった。用いたプライマー配列は:
【0054】
【化3】
【0055】
であった。1μlのClontech Advantage 2 Taqポリメラーゼ(50x)および36μlのPCR水(Clontech、カリフォルニア州)を混合した。混合物を94℃で1分間インキュベーションし、そして94℃15秒間そして次いで72℃4分間が5周期、94℃15秒間そして次いで70℃4分間が5周期、94℃15秒間そして次いで68℃4分間が25周期のタッチダウンPCRプロトコルを実行した。PCR反応の第一周期からクローニングされたのではない遺伝子に関しては、よりストリンジェントでないPCR条件は、94℃15秒間そして次いで68℃4分間が5周期、94℃15秒間そして次いで65℃4分間が5周期、94℃15秒間そして次いで62℃4分間が25周期である。製造者の使用説明書にしたがって、Agilent 2100 Bioanalyzer上、DNA7500チップ(Agilent、カリフォルニア州)を用いて、PCR断片の長さおよび濃度を調べる。
【0056】
DNA調製および配列決定
蛍光に基づくABI377配列決定装置(Perkin Elmer/Applied Biosystems Division、PE/ABD、カリフォルニア州フォスターシティ)およびABI PRISMTM Ready Dye−Deoxy TerminatorキットとTaq FSTMポリメラーゼを以下のプライマー:
【0057】
【化4】
【0058】
とともに用いて、PCR産物を配列決定した。
各ABI周期配列決定反応は、約0.5μgのプラスミドDNAを含有する。98℃1分間の最初の変性、次いで50周期の:98℃30秒間の変性、50℃30秒間のアニーリング、および60℃4分間の伸長を用いて、周期配列決定を行う。温度周期および時間は、Perkin−Elmer 9600サーモサイクラーによって調節される。CentriflexTMゲルろ過カートリッジ(Advanced Genetic Technologies Corp.、メリーランド州ガイザーズバーグ)を用いて、伸長産物を単離する。各反応産物をピペットによってカラム上に装填し、これを次いで1500xg、室温で4分間、スウィンギングバケット遠心分離装置(SorvallモデルRT6000B卓上遠心分離装置)中で遠心分離する。カラムで精製した試料を、真空下で約40分間乾燥させ、そして次いで、5μlのDNA装填溶液(83%脱イオン化ホルムアミド、8.3mM EDTA、および1.6mg/mlブルーデキストラン)に溶解する。次いで試料を90℃に3分間加熱し、そしてABI377配列決定装置による配列解析のため、ゲル試料ウェルに装填する。ABI377ファイルをDNASTAR(DNASTAR、ウィスコンシン州マディソン)プログラムにインポートすることによって、配列解析を行った。一般的に、700bpの配列読み取りが得られた。DNA鎖両方から配列情報を得ることによって、そして配列決定のあいまいさがすべて取り除かれるまで、異なる位置のプライマーを用いて、困難な領域を再配列決定することによって、潜在的な配列エラーを最小限にした。配列番号1は、産物の配列に相当する。
【0059】
ヒト遺伝子オルソログと配列決定データを並列し、そして配列追跡データにしたがって、手動で管理した(curated)。第一周期の配列決定後、完全には終了していないクローニング遺伝子または遺伝子断片に関しては、全プロジェクトが完了するまで、管理配列データに基づいて、さらなる配列決定プライマーを注文した。イヌ・コード配列は、それぞれ、ヒト、ラットおよびマウス相同体に86%、75%および76%同一であるタンパク質をコードする。イヌDNAコード配列は、ヒト、ラットおよびマウス・コードDNA配列に、それぞれ、88%、78.4%、78.8%同一である(比較は、ヒトmRNAに関してはGenbank NM_001966、マウスmRNAに関してはBC016899、そしてラットmRNAに関してはNM_133606に、そしてヒトタンパク質に関してはNP_001957、マウスタンパク質に関してはAAH16899、そしてラットタンパク質に関してはNP_598290に基づいた)。新規発見イヌ配列と他の3つの相同体の並列を図1および2に示す。
【0060】
配列番号1の配列を得るこの方法は例示であり、そして配列番号1を開示することによって、当業者には、配列番号1の全配列を得る多数の方法が提供されることを認識しなければならない。例えば、配列番号1に開示する配列からプローブを生成し、そしてイヌcDNAまたはゲノムライブラリーをスクリーニングして、そしてそれによって配列番号1全体またはそのゲノム同等物を得ることが可能であろう。Sambrookら(監修), Molecular Cloning:A Laboratory Manual, Cold Spring Harbor Laboratory Press:ニューヨーク州コールドスプリングハーバー(1989)。やはり例として、当業者は、配列番号1に開示する配列が与えられれば、それに次いで、配列番号1に代表される全配列を得るためのPCR増幅に適したプライマーを生成可能であることを直ちに認識するであろう(例えば、PCR Technology, H.A. Erlich監修, Stockton Press, ニューヨーク, 1989;PCR Protocols:A Guide to Methods and Applications, M.A. Innis, David H. Gelfand, John J. Sninsky, およびThomas J. White監修, Academic Press, Inc., ニューヨークを参照されたい)。
【0061】
本発明の宿主細胞およびベクター
本発明のポリヌクレオチドを取り込んだプラスミドおよびウイルスDNAベクターなどの自律複製組換え発現構築物もまた提供する。イヌL−PBEコードポリヌクレオチドが、内因性または外因性発現調節DNA配列および転写ターミネーターに、機能可能であるように連結されている発現構築物もまた、提供する。発現調節DNA配列には、プロモーター、エンハンサー、およびオペレーターが含まれ、そして一般的に、発現構築物を利用しようとする発現系に基づいて選択される。プロモーターおよびエンハンサー配列は、一般的に、遺伝子発現を増加させる能力に関して選択され、一方、オペレーター配列は、一般的に、遺伝子発現を制御する能力に関して選択される。本発明の発現構築物は、構築物を所持する宿主細胞の同定を可能にする1以上の選択可能マーカーをコードする配列も含むことも可能である。発現構築物はまた、宿主細胞における相同組換えを容易にし、そして好ましくは促進する配列も含むことも可能である。本発明の構築物はまた、宿主細胞における複製に必要な配列も含む。
【0062】
発現構築物は、好ましくは、コードされるタンパク質の産生のために利用されるが、イヌL−PBEをコードするポリヌクレオチド配列を単純に増幅するために利用することもまた可能である。
【0063】
本発明の別の側面にしたがって、コードされるイヌL−PBEポリペプチドの発現を可能にする方式で、本発明のポリヌクレオチド(または本発明のベクター)を含む、原核細胞および真核細胞を含む、宿主細胞を提供する。本発明のポリヌクレオチドを、環状プラスミドの一部として、または単離タンパク質コード領域を含む直鎖DNAとして、あるいはウイルスベクターとして、宿主細胞に導入することも可能である。当該技術分野で周知であり、そして日常的に実施されている、宿主細胞にDNAを導入する方法には、形質転換、トランスフェクション、エレクトロポレーション、核注入、またはリポソーム、ミセル、ゴースト細胞およびプロトプラストなどのキャリアーとの融合が含まれる。本発明の発現系には、細菌、酵母、真菌、植物、昆虫、無脊椎動物、および哺乳動物細胞系が含まれる。イヌL−PBEポリペプチドの発現に適した宿主細胞には、原核生物、酵母、およびより高次の真核細胞が含まれる。イヌL−PBEポリペプチドの発現に用いられるのに適した原核宿主には、限定されるわけではないが、エシェリキア属(Escherichia)、バチルス属(Bacillus)、およびサルモネラ属(Salmonella)の細菌とともに、シュードモナス属(Pseudomaonas)、ストレプトミセス属(Streptomyces)、およびブドウ球菌属(Staphylococcus)のメンバーが含まれる。
【0064】
本発明の単離核酸分子は、好ましくは、原核細胞での発現用に設計されたベクターよりも、真核細胞での発現用に設計されたベクターにクローニングされる。より高次の真核生物から得られる遺伝子の発現には、ときに真核細胞が好ましく、これはこれらのタンパク質の合成、プロセシング、および分泌のシグナルが、真核細胞では通常認識されるが、原核宿主では、しばしば、認識されないためである(Ausubelら監修, Short Protocols in Molecular Biology, 第2版, John Wiley & Sons, publishers, 16−49ページ, 1992)。イヌL−PBEの場合、N−連結グリコシル化のための2つのコンセンサス配列があり、そしてプロテインキナーゼCリン酸化およびO−グリコシル化のための、翻訳後修飾の他の部位が予測可能である。真核宿主には、限定されるわけではないが、以下の:昆虫細胞、アフリカミドリザル(African green monkey)腎臓細胞(COS細胞)、チャイニーズハムスター卵巣細胞(CHO細胞)、ヒト293細胞、およびネズミ3T3線維芽細胞が含まれることも可能である。
【0065】
原核宿主で使用する発現ベクターは、一般的に、1以上の表現型的選択可能マーカー遺伝子を含む。こうした遺伝子は、一般的に、例えば抗生物質耐性を与えるか、または栄養要求必要条件を供給するタンパク質をコードする。非常に多様なこうしたベクターが商業的供給源から容易に入手可能である。例には、pSPORTベクター、pGEMベクター(Promega)、pPROEXベクター(LTI、メリーランド州ベセスダ)、Bluescriptベクター(Stratagene)、およびpQEベクター(Qiagen)が含まれる。
【0066】
イヌL−PBEはまた、サッカロミセス属(Saccharomyces)、ピキア属(Pichia)、およびクルイベロミセス属(Kluveromyces)を含む属由来の酵母宿主細胞で発現されることも可能である。酵母宿主には、S.セレビシエ(S. cerevisiae)およびP.パストリス(P. pastoris)が含まれる。酵母ベクターはしばしば、2ミクロン酵母プラスミド由来の複製起点配列、自律複製配列(ARS)、プロモーター領域、ポリアデニル化のための配列、転写終結のための配列、および選択可能マーカー遺伝子を含有するであろう。酵母および大腸菌(E. coli)両方で複製可能なベクター(シャトルベクターと称する)もまた、使用可能である。酵母ベクターの上述の特徴に加えて、シャトルベクターはまた、大腸菌における複製および選択のための配列も含むであろう。イヌL−PBEをコードするヌクレオチド配列の5’端に酵母因子リーダー配列をコードするヌクレオチド配列を含むことによって、酵母宿主で発現されるイヌL−PBEポリペプチドの直接分泌を達成することも可能である。
【0067】
イヌL−PBEポリペプチドの発現には、昆虫宿主細胞培養系もまた、使用可能である。別の態様において、バキュロウイルス発現系を用いて、本発明のイヌL−PBEポリペプチドを発現する。昆虫細胞において異種タンパク質を発現するためのバキュロウイルス系の使用に関するさらなる情報は、LuckowおよびSummers, Bio/Technology 6:47(1988)に概説されている。
【0068】
別の態様において、イヌL−PBEポリペプチドを哺乳動物宿主細胞で発現させる。適切な哺乳動物細胞株の限定されない例には、サル腎臓細胞のCOS−7株(Gluzmanら, Cell 23:175(1981))、チャイニーズハムスター卵巣(CHO)細胞、およびヒト293細胞が含まれる。
【0069】
本発明のイヌL−PBEポリペプチドの発現に適した発現ベクターの選択は、もちろん、使用しようとする特定の宿主細胞に応じ、そして一般の当業者の技術の範囲内である。適切な発現ベクターの例には、pcDNA3(Invitrogen)およびpSVL(Pharmacia Biotech)が含まれる。哺乳動物宿主細胞で使用する発現ベクターには、ウイルスゲノム由来の転写調節配列および翻訳調節配列が含まれることも可能である。本発明で使用可能な、一般的に用いられるプロモーター配列および修飾因子配列には、限定されるわけではないが、ヒト・サイトメガロウイルス(CMV)、アデノウイルス2、ポリオーマウイルス、およびサルウイルス40(SV40)由来のものが含まれる。哺乳動物発現ベクターを構築する方法は、例えばOkayamaおよびBerg(Mol. Cell. Biol. 3:280(1983));Cosmanら(Mol. Immunol. 23:935(1986));Cosmanら(Nature 312:768(1984));EP−A−0367566;およびWO 91/18982に開示される。
【0070】
(実施例2)
真核宿主細胞におけるイヌL−PBEの発現
イヌL−PBEタンパク質を産生するため、適切な発現ベクターを用い、標準的遺伝子操作技術を用いて、適切な宿主細胞中で、イヌL−PBEをコードするポリヌクレオチドを発現させる。例えば、実施例1に記載するイヌL−PBEコード配列を商業的発現ベクターpzeoSV2(Invitrogen、カリフォルニア州サンディエゴ)にサブクローニングし、そしてトランスフェクション試薬fuGENE6(Boehringer−Mannheim)および製品挿入物に提供されるトランスフェクションプロトコルを用いて、チャイニーズハムスター卵巣(CHO)細胞にトランスフェクションする。ヒト胚性腎臓HEK293細胞およびCOS細胞を含む、他の真核細胞株もまた、適切である。100μg/mlゼオシン(Stratagene、カリフォルニア州ラホヤ)の存在下での増殖によって、イヌL−PBEを安定に発現している細胞を選択する。所望により、標準的クロマトグラフィー技術を用いて、細胞からイヌL−PBEを単離する。精製を容易にするため、イヌL−PBEアミノ酸配列の部分に対応する1以上の合成ペプチド配列に対して抗血清を作成することも可能であり、そしてこの抗血清を用いて、イヌL−PBEをアフィニティー精製する。イヌL−PBEをタグ配列(例えばポリヒスチジン、赤血球凝集素、FLAG)とインフレームで発現させて、精製を容易にすることもまた可能である。
【0071】
本発明の宿主細胞は、イヌL−PBEと特異的に免疫反応性である抗体の発展のための、免疫原の価値ある供給源である。本発明の宿主細胞はまた、イヌL−PBEポリペプチドの大規模産生の方法においても有用であり、ここで細胞を適切な培地中で増殖させ、そして当該技術分野に知られる精製法、例えば免疫アフィニティークロマトグラフィー、疎水性相互作用クロマトグラフィー、レクチンアフィニティークロマトグラフィー、サイズ排除ろ過、陽イオンまたは陰イオン交換クロマトグラフィー、高圧液体クロマトグラフィー(HPLC)、逆相HPLC等を含む慣用的クロマトグラフィー法によって、細胞から、または細胞を増殖させた培地から、望ましいポリペプチド産物を単離する。さらに他の精製法には、特異的結合パートナーまたは剤に認識される特異的タグ、標識、またはキレート部分を有する融合タンパク質として、望ましいタンパク質を発現し、そして単離するものが含まれる。単離されたタンパク質を切断して、望ましいタンパク質を得ることも可能であるし、または損なわれていない(intact)融合タンパク質としてそのままにしておくことも可能である。融合構成要素の切断は、切断プロセスの結果として、さらなるアミノ酸残基を有する望ましいタンパク質の型を生じることも可能である。
【0072】
イヌL−PBE DNA配列を知ることで、内因性イヌL−PBEの発現を可能にするか、または増加させるように、細胞を修飾することが可能になる。天然存在イヌL−PBEプロモーターと異種プロモーターのすべてまたは一部を交換して、細胞がより高いレベルでイヌL−PBEを発現するようにすることによって、細胞を修飾して(例えば相同組換えによって)、発現増加を提供することも可能である。内因性イヌL−PBEコード配列に機能可能であるように連結される方式で、異種プロモーターを挿入する[例えばPCT国際公報第WO 94/12650号、PCT国際公報第WO 92/20808号、およびPCT国際公報第WO 91/09955号を参照されたい]。異種プロモーターDNAに加えて、増幅可能マーカーDNA(例えばada、dhfr、並びにカルバミルリン酸シンターゼ、アスパラギン酸トランスカルバミラーゼ、およびジヒドロオロターゼをコードする多機能CAD遺伝子)および/またはイントロンDNAを、異種プロモーターDNAとともに挿入可能であることもまた、意図される。イヌL−PBEコード配列に連結した場合、標準的選択法によってマーカーDNAを増幅すると、細胞において、イヌL−PBEコード配列が同時に増幅される。
【0073】
本発明に提供するDNA配列情報はまた、例えば相同組換えまたは「ノックアウト」戦略[Capecchi, Science 244:1288−1292(1989)]を通じて、機能するイヌL−PBEを発現不能な動物またはイヌL−PBEの変異体を発現する動物の発展も可能にする。こうした動物(特にラット、ウサギ、およびマウスなどの実験小動物)は、イヌL−PBEのin vivo活性およびイヌL−PBEの調節因子を研究するモデルとして有用である。
【0074】
本発明によりやはり利用可能になるのは、イヌL−PBEをコードするポリヌクレオチドを認識し、そして該ポリヌクレオチドにハイブリダイズする、アンチセンスポリヌクレオチドである。全長および断片のアンチセンスポリヌクレオチドを提供する。本発明の断片アンチセンス分子には、(i)イヌL−PBEを特異的に認識し、そしてこれにハイブリダイズするもの(イヌL−PBEをコードするDNAと他の既知の分子をコードするDNAの配列比較によって決定されるようなもの)が含まれる。イヌL−PBEがコードするポリヌクレオチドにユニークな配列の同定は、公的に利用可能な配列データベースいずれかの使用を通じて、そして/または商業的に入手可能な配列比較プログラムの使用を通じて、推定可能である。全ゲノムにおける選択された配列のユニークさは、ハイブリダイゼーション解析によってさらに検証可能である。望ましい配列を同定した後、制限消化を通じて単離するか、または当該技術分野に周知の多様なポリメラーゼ連鎖反応技術いずれかを用いて増幅することも可能である。アンチセンスポリヌクレオチドは、イヌL−PBE mRNAを発現する細胞によるイヌL−PBEの発現を制御するのに特に適している。
【0075】
イヌL−PBE発現調節配列またはイヌL−PBE RNAに特異的に結合可能なアンチセンス核酸(好ましくは10〜20塩基対オリゴヌクレオチド)を(例えばウイルスベクター、またはリポソームなどのコロイド性分散系によって)細胞に導入する。アンチセンス核酸は、細胞において、イヌL−PBE標的ヌクレオチド配列に結合し、そして標的配列の転写または翻訳を妨げる。ホスホロチオエートおよびメチルホスホネート・アンチセンスオリゴヌクレオチドは、本発明による療法的使用に明確に意図される。アンチセンスオリゴヌクレオチドを、5’端で、ポリ−L−リジン、トランスフェリン、ポリリジン、またはコレステロール部分によってさらに修飾することも可能である。転写レベルまたは翻訳レベルいずれかでのイヌL−PBE発現の抑制は、異常なイヌL−PBE発現に特徴付けられる疾患の細胞モデルまたは動物モデルを生成するために、あるいは療法様式として、有用である。
【0076】
本明細書に解説されるイヌL−PBE配列は、天然細胞および動物、並びにイヌL−PBEポリヌクレオチドで形質転換またはトランスフェクションされた細胞における、イヌL−PBE発現を調節する新規転写因子の設計を容易にする。例えば、ジンクフィンガードメインを介してDNAに結合する、Cys2−His2ジンクフィンガータンパク質は、異なる標的配列の認識につながる構造変化を受けやすいことが示されてきている。これらの人工的ジンクフィンガータンパク質は、高い親和性定数および低い解離定数の特異的標的部位を認識し、そして遺伝子発現を調節する遺伝子スイッチとして作用することが可能である。本発明の特定のイヌL−PBE標的配列を知ることで、構造に基づくモデリングおよびファージディスプレーライブラリーのスクリーニングの組み合わせなどの、既知の方法を用いて、標的配列に特異的なジンクフィンガータンパク質の操作が容易になる[Segalら(1999)Proc Natl Acad Sci USA 96:2758−2763;Liuら(1997)Proc Natl Acad Sci USA 94:5525−30;GreismanおよびPabo(1997)Science 275:657−61;Chooら(1997)J Mol Biol 273:525−32]。各ジンクフィンガードメインは、通常、3以上の塩基対を認識する。一般的に、18塩基対の認識配列は、既知のゲノムいずれにおいても、該配列をユニークにするのに十分な長さであるため、ジンクフィンガーの6つのタンデムリピートからなるジンクフィンガータンパク質は、特定の配列に対する特異性を確実にすると予期されるであろう[Segalら(1999)Proc Natl Acad Sci USA 96:2758−2763]。イヌL−PBE配列に基づいて設計した、人工的なジンクフィンガーリピートを、活性化ドメインまたは抑制ドメインに融合させて、イヌL−PBE発現を促進するかまたは抑制する[Liuら(1997)Proc Natl Acad Sci USA 94:5525−30]。あるいは、ジンクフィンガーペプチドおよびTATAボックス結合因子(TBP)間のリンカー領域の長さを多様にしながら、ジンクフィンガードメインをTBPに融合させて、転写活性化因子またはリプレッサーいずれかを生成することも可能である[Kimら(1997)Proc Natl Acad Sci USA 94:3616−3620]。こうしたタンパク質、および該タンパク質をコードするポリヌクレオチドは、天然細胞、動物およびヒト;および/またはイヌL−PBEコード配列でトランスフェクションされた細胞両方のin vivoでのイヌL−PBE発現を調節するのに有用性を有する。転写因子を発現する構築物をトランスフェクションすること(遺伝子療法)によって、または該タンパク質を導入することによって、新規転写因子を標的細胞に搬送することも可能である。RNA配列に結合するように、操作ジンクフィンガータンパク質を設計して、アンチセンスまたは触媒RNA法の代替法として療法において使用することもまた可能である[McCollら(1999)Proc Natl Acad Sci USA 96:9521−6;Wuら(1995)Proc Natl Acad Sci USA 92:344−348]。本発明は、(天然の、または形質転換された)細胞において、イヌL−PBE発現を調節するのに有用な、本発明の遺伝子配列に基づく転写因子とともに、カスタマイズしたジンクフィンガータンパク質などの設計法を意図し、細胞の遺伝的相補体にはこれらの配列が含まれる。本発明はまた、本発明のポリヌクレオチドにコードされる単離イヌL−PBEポリペプチドも提供する。
【0077】
本発明のポリペプチド
イヌL−PBEポリペプチドのアミノ酸配列を配列番号2に示す。配列番号2に例示されるような本発明のアミノ酸配列は、いくつかの興味深い特徴を有する。
【0078】
本発明はまた、配列番号2に示すポリペプチドに、少なくとも87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%の同一性および/または相同性を有するポリペプチドも含む。配列番号2のポリペプチドに関するアミノ酸配列「同一性」パーセントは、本明細書において、両配列を並列させ、そして必要であればギャップを導入して最大の配列同一性パーセントを達成した後、そしていかなる保存的置換も配列同一性の一部と見なさずに、イヌL−PBE配列の残基と同一である、候補配列中のアミノ酸残基の割合として定義される。配列番号2のポリペプチドに関するアミノ酸配列「相同性」パーセントは、本明細書において、配列を並列させ、そして必要であればギャップを導入して最大の配列同一性パーセントを達成した後、そしてまたいかなる保存的置換も配列同一性の一部と見なして、イヌL−PBE配列の残基と同一である、候補配列中のアミノ酸残基の割合として定義される。1つの側面において、相同性パーセントは、長さ100アミノ酸中、4つのギャップを導入して並列を最大にすることが可能である際には、2つの配列のうち、より小さい配列の、比較しようとする配列中の同一アミノ酸残基と並列する、アミノ酸残基の割合として計算される[本明細書に援用されるDayhoff, Atlas of Protein Sequence and Structure中, Vol.5 p.124, National Biochemical Reserch Foundation, ワシントンD.C.(1972)]。
【0079】
本発明のポリペプチドは、天然細胞供給源から単離されることも可能であるし、または化学的に合成されることも可能であるが、好ましくは、本発明の宿主細胞を伴う組換え法によって産生される。哺乳動物宿主細胞を使用すると、本発明の組換え発現産物に最適な生物学的活性を与えるために必要である可能性があるような、こうした翻訳後修飾(例えばグリコシル化、一部切除(truncation)、脂質化、およびリン酸化)が提供されると予期される。イヌL−PBEポリペプチドのグリコシル化型および非グリコシル化型が含まれる。
【0080】
上述のような真核宿主および原核宿主における過剰発現は、イヌL−PBEポリペプチドの単離を容易にする。したがって、本発明には、配列番号2に示すような単離イヌL−PBEポリペプチド、並びに変異体、および保存的アミノ酸置換が含まれ、標識化ポリペプチドおよびタグ化ポリペプチドが含まれる。本発明には、「標識された」イヌL−PBEポリペプチドが含まれる。用語「標識された」は、本明細書において、酵素(例えば西洋ワサビ(horseradish)ペルオキシダーゼ、ベータ−グルクロニダーゼ、アルカリホスファターゼ、およびベータ−D−ガラクトシダーゼ)、蛍光標識(例えばフルオレセイン、ルシフェラーゼ)、および放射標識(例えば14C、125I、3H、32P、および35S)を含む適切な検出可能基いずれかを、標識しようとする化合物にコンジュゲート化するか、または共有結合することを指す。タンパク質、ペプチド、および抗体を含む、多様な化合物を標識する技術が周知である。例えば、Morrison, Methods in Enzymology 32b, 103(1974);Syvanenら, J. Biol. Chem. 284, 3762(1973);BoltonおよびHunter, Biochem. J. 133, 529(1973)を参照されたい。用語、標識された、はまた、以下に論じるように、アミノ酸タグに共有結合したポリペプチドも含むことも可能である。
【0081】
さらに、本発明のイヌL−PBEポリペプチドを、間接的に標識することも可能である。これは、部分(a moiety)をポリペプチドに共有的に付加し、そしてそれに続いて、付加された部分への特異的結合を示す標識または標識化合物に、付加された部分をカップリングすることを伴う。間接的な標識の可能性には、ペプチドをビオチン化し、それに続いて、上記標識基の1つにカップリングしたアビジンに結合させることが含まれる。別の例は、ポリヒスチジン・タグを含むイヌL−PBEポリペプチドと、ヒスチジン・タグに特異的な放射標識抗体をインキュベーションすることであろう。抗体がタグにかなりの親和性を有するため、ポリペプチドに対する放射能抗体の結合が、正味の効果である。
【0082】
本発明はまた、イヌL−PBEタンパク質の変異体(または類似体)も含む。1つの例において、1以上のアミノ酸残基がイヌL−PBEアミノ酸配列に付加されている、挿入変異体を提供する。挿入は、タンパク質のいずれかの末端または両方の末端に位置することも可能であるし、あるいはイヌL−PBEタンパク質アミノ酸配列の内部領域内に位置することも可能である。いずれかの末端または両方の末端にさらなる残基を有する挿入変異体には、例えば、融合タンパク質、およびアミノ酸タグまたは標識を含むタンパク質が含まれることも可能である。挿入変異体には、イヌL−PBE酸配列、またはその生物学的活性断片に、1以上のアミノ酸残基が付加されているイヌL−PBEポリペプチドが含まれる。
【0083】
したがって、挿入変異体はまた、イヌL−PBEのアミノ末端および/またはカルボキシ末端が別のポリペプチドに融合している融合タンパク質を含むことも可能である。多様なタグ・ポリペプチドおよびその対応する抗体が当該技術分野に周知である。例には、ポリ−ヒスチジン(ポリ−his)またはポリ−ヒスチジン−グリシン(ポリ−his−gly)タグ;インフルエンザHAタグ・ポリペプチドおよびその抗体12CA5[Fieldら, Mol. Cell. Biol., 8:2159−2165(1988)];c−mycタグ、並びに該タグに対する8F9、3C7、6E10、G4、B7および9E10抗体[Evanら, Molecular and Cellular Biology, 5:3610−3616(1985)];並びに単純疱疹ウイルス糖タンパク質D(gD)タグおよびその抗体[Paborskyら, Protein Engineering, 3(6):547−553(1990)]が含まれる。他のタグ・ポリペプチドには、Flag−ペプチド[Hoppら, BioTechnology, 6:1204−1210(1988)];KT3エピトープペプチド[Martinら, Science, 255:192−194(1992)];アルファ−チューブリン・エピトープペプチド[Skinnerら, J. Biol. Chem., 266:15163−15166(1991)];およびT7遺伝子10タンパク質ペプチドタグ[Lutz−Freyermuthら, Proc. Natl. Acad. Sci. USA, 87:6393−6397(1990)]が含まれる。さらに、イヌL−PBEポリペプチドを、ペルオキシダーゼおよびアルカリホスファターゼなどの酵素タンパク質でタグ化することも可能である。
【0084】
別の側面において、本発明は、イヌL−PBEポリペプチド中の1以上のアミノ酸残基が除去されている欠失変異体を提供する。欠失は、イヌL−PBEポリペプチドの一方の末端または両方の末端で、あるいはイヌL−PBEアミノ酸配列内の1以上の残基の除去を伴って、達成されることも可能である。したがって、欠失変異体には、イヌL−PBEポリペプチドのすべての断片が含まれる。
【0085】
本発明はまた、断片が生物学的活性(例えばリガンド結合活性またはDNA結合活性および/または他の生物学的活性)を維持する、配列番号2に示す配列のポリペプチド断片も示す。配列番号2の少なくとも10から853(この間の1つ1つの整数値を含む)の連続アミノ酸を含む断片が本発明に含まれる。当該技術分野に周知であり、そして日常的に実施される方法いずれかによって、望ましい生物学的特性を有する本発明の断片を調製することも可能である。
【0086】
本発明にはまた、残基が別の類似の特性を持つ別の残基で置換される、保存的アミノ酸置換によって、参照配列とは異なる、前述のポリペプチドの変異体も含まれる。変異体ポリペプチドには、保存的置換が、本発明のポリペプチドをコードするポリヌクレオチドの修飾によって導入されているものが含まれる。アミノ酸は、物理的特性、並びに二次および三次タンパク質構造への寄与にしたがって、分類可能である。保存的置換は、当該技術分野において、1つのアミノ酸に対して、類似の特性を有する別のアミノ酸配列を置換するものと認識される。典型的な保存的置換を、すぐ下の表2に示す(WO 97/09433、10ページ、1997年3月13日公表(PCT/GB96/02197、9/6/96出願)から)。
【0087】
表2
保存的置換I
【0088】
【表2】
【0089】
あるいは、保存的アミノ酸は、すぐ下の表3に示すように、Lehninger[Biochemistry, 第2版;Worth Publishers, Inc., ニューヨーク州ニューヨーク(1975), pp.71−77]に記載されるように分類可能である。
【0090】
表3
保存的置換II
【0091】
【表3】
【0092】
さらに別の代替物として、典型的な保存的置換をすぐ下の表4に示す。
表4
保存的置換III
【0093】
【表4】
【0094】
一般的に、イヌL−PBEポリペプチドは、主に細胞内に見られることが予期され、細胞内物質は、当業者に知られる標準的技術いずれかを用いて、宿主細胞から抽出可能である。例えば、宿主細胞を溶解して、ホモジナイズ、および/または超音波処理、その後の遠心分離によって、細胞質の内容物を遊離させることも可能である。イヌL−PBEポリペプチドは、細胞ホモジネートの遠心分離後、主に上清に見られ、そして以下の方法のいずれかによる限定されない例によって、イヌL−PBEポリペプチドを単離することも可能である。
【0095】
イヌL−PBEポリペプチドを部分的にまたは完全に単離することが好ましい状況において、当業者に周知の標準法を用いて、精製を達成することも可能である。こうした方法には、限定されるわけではないが、電気泳動による分離、その後、電気溶出、多様な種類のクロマトグラフィー(免疫アフィニティー、分子ふるい、および/またはイオン交換)、および/または高圧液体クロマトグラフィーが含まれる。いくつかの場合、完全に精製するため、これらの方法の1より多くを用いることが好ましい可能性もある。
【0096】
多様な技術を用いて、イヌL−PBEポリペプチドの精製を達成することも可能である。ポリペプチドが、そのカルボキシル末端またはアミノ末端いずれかに、ヘキサヒスチジン(イヌL−PBE/ヘキサHis)、またはFLAG(Eastman Kodak Co.、コネチカット州ニューヘブン)またはmyc(Invitrogen、カリフォルニア州カールスバッド)などの他の小ペプチドなどのタグを含有するように合成されている場合、カラムマトリックスがタグに高い親和性を有するか、またはポリペプチドに直接、高い親和性を有する(すなわち、イヌL−PBEを特異的に認識するモノクローナル抗体)アフィニティーカラムに、溶液を通過させることによって、1工程プロセスで本質的に精製可能である。例えば、ポリヒスチジンは、高い親和性および特異性でニッケルに結合し、したがってニッケルの親和性カラム(Qiagen Registered TMニッケルカラムなど)をイヌL−PBE/ポリHisの精製に用いることも可能である(例えば、Ausubelら監修, Current Protocols in Molecular Biology, セクション10.11.8, John Wiley & Sons, ニューヨーク[1993]を参照されたい)。
【0097】
精製を容易にする標識またはタグなしに、イヌL−PBEポリペプチドを調製する場合であっても、本発明のイヌL−PBEを免疫アフィニティークロマトグラフィーで精製することも可能である。これを達成するためには、当該技術分野に周知の手段によって、イヌL−PBEポリペプチドに特異的な抗体を調製しなければならない。本発明のイヌL−PBEポリペプチドまたはエピトープを所持する断片、類似体(analogue)または細胞を、動物、好ましくは非ヒトに、日常的プロトコルを用いて投与することによって、本発明のイヌL−PBEポリペプチドに対して生成された抗体を得ることも可能である。モノクローナル抗体を調製するには、連続細胞株培養によって産生される抗体を提供する、当該技術分野に知られる技術いずれを使用することも可能である。例には、Kohler, G.およびMilstein, C., Nature 256:495−497(1975);Kozborら, Immunology Today 4:72(1983);Coleら, 77−96ページ Monoclonal Antibodies and Cancer Therapy中, Alan R. Liss, Inc.(1985)のものなどの多様な技術が含まれる。
【0098】
付着するタグなしにイヌL−PBEポリペプチドを調製する場合、そして抗体がまったく入手可能でない場合、他の周知の精製法が使用可能である。こうした方法には、限定なしに、イオン交換クロマトグラフィー、分子ふるいクロマトグラフィー、HPLC、ゲル溶出と組み合わせた未変性ゲル電気泳動、および分取用等電点電気泳動(「Isoprime」装置/技術、Hoefer Scientific)が含まれる。いくつかの場合、2以上のこれらの技術を組み合わせて、純度増加を達成することも可能である。代表的な精製スキームを以下に詳述する。
【0099】
天然イヌL−PBEの阻害特性を示し、そしてより高いレベルで発現される変異体、および恒常的に活性であるイヌL−PBEポリペプチドを提供する変異体は、本発明のアッセイにおいて特に有用であり、そしてまた、異常なイヌL−PBE発現活性によって特徴付けられる疾患の細胞モデルまたは動物モデルにおいても有用である。
【0100】
本発明のポリペプチドの定義は、アミノ酸残基の挿入、欠失、または置換以外の修飾を所持するポリペプチドを含むことが意図されることを理解しなければならない。例えば、修飾は、共有性であることも可能であるし、そして例えば、ポリマー、脂質、他の有機部分、および無機部分との化学結合が含まれる。
【0101】
本発明にやはり含まれるのは、イヌL−PBEまたはその断片に特異的な抗体(例えばモノクローナル抗体およびポリクローナル抗体、一本鎖抗体、キメラ抗体、二官能性/二重特異性抗体、ヒト化抗体、ヒト抗体、および本発明のポリペプチドを特異的に認識するCDR配列を含む化合物を含む相補性決定領域(CDR)移植抗体)である。本発明の抗体には、本明細書に完全に援用される、WO93/11236、1993年6月20日公表に記載される方法にしたがって産生され、そして同定されたヒト抗体が含まれる。Fab、Fab’、F(ab’)2、およびFvを含む抗体断片もまた本発明に提供される。用語「特異的」は、本発明の抗体を記載するのに使用する場合、本発明の抗体の可変領域が、もっぱらイヌL−PBEポリペプチドを認識し、そしてこれに結合する(すなわち、イヌL−PBEおよび他の既知のポリペプチド間に局所配列同一性、相同性、または類似性が存在しうるにもかかわらず、結合親和性が測定可能に異なるため、イヌL−PBEポリペプチドをこうした他のポリペプチドから区別可能である)ことを示す。特異的抗体がまた、抗体の可変領域外の配列、そして特に分子の定常領域との相互作用を通じて、他のタンパク質(例えば黄色ブドウ球菌(S. aureus)プロテインA、またはELISA技術における他の抗体)とも相互作用可能であることが理解されるであろう。本発明の抗体の結合特異性を決定するスクリーニングアッセイが当該技術分野において周知であり、そして日常的に実施される。こうしたアッセイの包括的議論に関しては、Harlowら(監修), Antibodies A Laboratory Manual;Cold Spring Harbor Laboratory;ニューヨーク州コールドスプリングハーバー(1988), 第6章を参照されたい。本発明のイヌL−PBEポリペプチドの断片を認識し、そしてこれに結合する抗体もまた含まれるが、該抗体が、何よりもまず、イヌL−PBEポリペプチドに特異的であることが条件である。当該技術分野において周知であり、そして日常的に実施される方法いずれかを用いて、本発明の抗体を産生することも可能である。当該技術分野に知られる方法いずれかによって、非ヒト抗体をヒト化することも可能である。1つの方法では、非ヒトCDRをヒト抗体またはコンセンサス抗体フレームワーク配列に挿入する。次いで、抗体フレームワークにさらなる変化を導入して、アフィニティーまたは免疫原性を調節することも可能である。
【0102】
本発明の抗体は、イヌL−PBEを検出するかまたは定量化する診断目的に有用であるとともに、イヌL−PBEを精製するのにも有用である。本明細書に記載する目的いずれかのための、本発明の抗体を含むキットもまた、含まれる。一般的に、本発明のキットにはまた、抗体が免疫特異的である対照抗原も含まれる。
【0103】
(実施例3)
イヌL−PBEに対する抗体の生成
標準的技術を使用して、イヌL−PBE酵素に対するポリクローナル抗体またはモノクローナル抗体を生成し、そして「ヒト化」変異体を含む、その有用な抗原結合断片または変異体を生成する。こうしたプロトコルは、例えばSambrookら, Molecular Cloning:a Laboratory Manual 第2版, ニューヨーク州コールドスプリングハーバー:Cold Spring Harbor Laboratory(1989);Harlowら(監修), Antibodies A Laboratory Manual;Cold Spring Harbor Laboratory;ニューヨーク州コールドスプリングハーバー(1988);および以下に引用する他の文書に見出すことも可能である。1つの態様において、組換えイヌL−PBEポリペプチド(またはこうしたポリペプチドを含有する細胞もしくは細胞膜)を抗原として用いて、抗体を生成する。別の態様において、イヌL−PBEの免疫原部分に対応するアミノ酸配列(例えば6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、またはそれより多いアミノ酸)を有する1以上のペプチドを抗原として用いる。小さい合成ペプチドを用いて、タンパク質エピトープを模倣するため、親水性、表面配向性で、そして柔軟である配列を選択することが重要である。これは、生理学的溶液中に見られる天然存在タンパク質の大部分は、表面に親水性残基を有し、そして疎水性残基は包埋されているためである。抗体は、一般的に、天然存在タンパク質の表面上のエピトープに結合する。いくつかの既知のエピトープは、高い度合いの移動度を有する。タンパク質のN末端およびC末端は、荷電基、すなわちNH3+およびCOO−を含有するため、一般的に表面配向性である。これらは、末端に位置するため、しばしば、高い度合いの移動度も有する。これらの末端は、3つの特性すべてを所持するため、しばしば、合成候補として選択される。イヌL−PBEの表面残基に対応するペプチド、特に親水性部分が意図される。やはり意図されるのは、イヌL−PBEのアミノ末端およびカルボキシ末端に位置するペプチドである。
【0104】
当業者は、既定のタンパク質配列内の各アミノ酸残基に、親水性、表面アクセス可能性、および柔軟性の値を割り当てるために、アルゴリズムが開発されていることを認識する。各残基に抗原性指数を割り当てて、特定の配列内の残基がどの程度抗原性であるか、指標を与えるため、やはりアルゴリズムが開発されている。HoppおよびWoods, Mol. Immunol, 1983 20(4):p.483−9, HoppおよびWoods, Proc. Natl Acad. Sci USA 1981, 78(6)p.3824−8。しかし、天然抗原に結合するのに有用な抗ペプチド抗体を生成するには、親水性セグメントが広く選択されてきている。しかし抗体とは異なり、T細胞受容体は、切断およびアンフォールディング後のタンパク質抗原の比較的小さいセグメントを認識する。T細胞抗原性部位もまた、予測コンピュータモデルによって対処されてきている。Margalit, H.ら, J. Immunol. 1987 138(7):2213−29ページ。
【0105】
エピトープの予測に有用なコンピュータプログラムが商業的に入手可能である。例えばMacVector(登録商標)(Oxford Molecular、英国オックスフォード)およびProtean(登録商標)(DNAStar Madison, WI53715)。ペプチド抗原を選択し、そして合成したら、抗原をアジュバントと混合するか、またはハプテンに連結して、抗体産生を増加することも可能である。
【0106】
ポリクローナル抗体およびモノクローナル抗体
1つの典型的なプロトコルとして、モノクローナル抗体を生成するため、組換えイヌL−PBEまたはその合成断片を用いて、マウスを免疫する(またはポリクローナル抗体の場合は、ウサギなどのより大きい哺乳動物)。抗原性を増加させるため、製造者の推奨にしたがって、ペプチドをキーホール・リンペット(Keyhole Lympet)ヘモシアニン(Pierce)にコンジュゲート化する。最初の注射では、抗原をフロイントの完全アジュバントで乳化し、そして皮下注射する。2〜3週間間隔で、イヌL−PBE抗原のさらなるアリコットをフロイントの不完全アジュバントで乳化し、そして皮下注射する。最後の追加免疫注射前に、免疫したマウスから血清試料を採取し、そしてウェスタンブロットによってアッセイして、イヌL−PBEと免疫反応する抗体の存在を確認する。免疫した動物由来の血清をポリクローナル抗血清として用いるか、またはイヌL−PBEを認識するポリクローナル抗体を単離するのに用いる。あるいは、モノクローナル抗体を生成するため、マウスを屠殺して、そして脾臓を取り除く。
【0107】
モノクローナル抗体を生成するため、脾臓を10ml血清不含RPMI1640に入れ、そして2mM L−グルタミン、1mMピルビン酸ナトリウム、100単位/mlペニシリン、および100μg/mlストレプトマイシン(RPMI)(Gibco、カナダ)を補った血清不含RPMI1640中で脾臓をすりつぶすことによって、単細胞懸濁物を形成する。細胞懸濁物をろ過し、そして遠心分離によって洗浄し、そして血清不含RPMIに再懸濁する。3匹の未処置(naive)Balb/cマウスから採取した胸腺細胞を同様の方式で調製し、そしてフィーダー層として使用する。融合前に3日間、10%ウシ胎児血清(FBS)(Hyclone Laboratories, Inc.、ユタ州ローガン)を補ったRPMI中、対数期に維持した、NS−1骨髄腫細胞もまた、遠心分離し、そして洗浄する。
【0108】
ハイブリドーマ融合体を産生するため、免疫したマウス由来の脾臓細胞をNS−1細胞と合わせて、そして遠心分離し、そして上清を吸引する。試験管を軽く叩くことによって、細胞ペレットを外し、そして2mlの37℃ PEG1500(75mM Hepes、pH8.0中50%)(Boehringer Mannheim)を攪拌しながらペレットに入れ、その後、血清不含RPMIを添加する。その後、細胞を遠心分離し、そして15%FBS、100μMヒポキサンチンナトリウム、0.4μMアミノプテリン、16μMチミジン(HAT)(Gibco)、25単位/ml IL−6(Boehringer Mannheim)および1.5x106胸腺細胞/mlを含有するRPMIに再懸濁し、そして10のCorning平底96ウェル組織培養プレートに蒔く(Corning、ニューヨーク州コーニング)。
【0109】
融合後、第2日、第4日および第6日、100μlの培地を融合プレートのウェルから取り除き、そして新鮮な培地と交換する。第8日、イヌL−PBEに結合するマウスIgGの存在に関して試験するELISAによって融合体をスクリーニングする。抗イヌL−PBE抗体を産生するモノクローナル培養物が得られるまで、希釈によって、選択された融合体ウェルをさらにクローニングする。
【0110】
ファージディスプレー由来のイヌL−PBEに向けられる抗体
すべて本明細書に援用される、Aujameら, Human Antibodies, 8(4):155−168(1997);Hoogenboom, TIBTECH, 15:62−70(1997);およびRaderら, Curr. Opin. Biotechnol., 8:503−508(1997)に記載されるものなどのファージディスプレー技術によって、イヌL−PBE抗体を生成する。例えば、Fab断片の形の、または連結された一本鎖Fv断片の形の抗体可変領域を、繊維状ファージの小コートタンパク質pIIIのアミノ末端に融合させる。融合タンパク質の発現および成熟ファージコートへのその取り込みによって、表面上に抗体を提示し、そして抗体をコードする遺伝物質を含有する、ファージ粒子が生じる。こうした構築物を含むファージライブラリーを細菌中で発現させ、そして抗原−プローブとして、標識イヌL−PBEまたは固定イヌL−PBEを用いて、イヌL−PBEに特異的なファージ抗体をパニング(スクリーニング)する。
【0111】
トランスジェニックマウス由来のイヌL−PBEに向けられる抗体
BruggemannおよびNeuberger, Immunol. Today, 17(8):391−97(1996)、並びにBruggemannおよびTaussig, Curr. Opin. Biotechnol., 8:455−58(1997)に本質的に記載されるように、トランスジェニックマウスにおいて、イヌL−PBE抗体を生成する。生殖系列配置中、ヒトV遺伝子セグメントを所持し、そしてそのリンパ組織でこれらの導入遺伝子を発現するトランスジェニックマウスを、慣用的免疫プロトコルを用いて、イヌL−PBE組成物で免疫する。慣用的プロトコルを用い、免疫したマウス由来のB細胞を用いてハイブリドーマを生成し、そしてスクリーニングして、抗イヌL−PBE抗体を分泌するハイブリドーマを同定する(例えば上述のとおり)。
【0112】
本発明のアッセイ
本発明は、イヌにおいて、試験剤の影響を検出するかまたは診断する方法を提供する。既定の時間経過に渡って、好ましくは亜慢性用量(subchronic dose)の、1以上の既知のまたは推定上の毒性化合物で、イヌを処置する。次いで、L−PBEを発現する組織試料を得る。こうした処置した生物学的試料から得たイヌL−PBE遺伝子発現のレベルを、未処置の生物学的試料から得た遺伝子発現パターンと比較して、化合物に反応して、イヌL−PBEを上方制御するかまたは下方制御する化合物を同定することも可能である。試料は、試料核酸分子またはタンパク質を含み、そしてL−PBEを発現する体性組織いずれか(脳、腎臓、肝臓など)、培養細胞、生検、または他の組織調製物から得た、いかなる試料であることも可能である。
【0113】
発現レベルを、産生されるメッセンジャーRNAまたはタンパク質のレベルいずれかまたは両方で評価することも可能である。
【0114】
核酸に基づくアッセイ
イヌL−PBEを生じる核酸は、溶液中または固体支持体上にあることも可能である。いくつかの態様において、これらをマイクロアレイ中の単独のアレイ要素として、または他のアレイ要素分子と組み合わせたアレイ要素として、使用することも可能である。こうしたマイクロアレイは、毒性であることが知られるかまたは推定される試験剤と関連して、ハイブリダイゼーションによって、遺伝子発現パターンを検出し、そして性質決定するのに特に有用である。しかし、アレイは、膜に基づくハイブリダイゼーション系、または溶液ハイブリダイゼーションアッセイであっても、または実際、特異的ハイブリダイゼーションを決定可能な方法いずれによっても置き換えることも可能であることが認識される。次いで、こうした遺伝子発現パターンを比較のために用いて、やはり毒性学的反応を誘発する他の化合物を同定することも可能である。
【0115】
核酸に基づく方法は、一般的に、試料からのDNAまたはRNAの単離、およびそれに続く、配列番号1由来の特異的プライマーを用いたハイブリダイゼーションまたはPCR増幅を必要とする。DNAまたはRNAは、当業者に周知のいくつかの方法いずれにしたがって、試料から単離することも可能である。例えば、核酸精製法が、Tijssen, P.(1993)Laboratory Techniques in Biochemistry and Molecular Biology:Hybridization With Nucleic Acid Probes, パートI. Theory and Nucleic Acid Preparation, Elsevier, ニューヨーク州ニューヨークに記載される。1つの好ましい態様において、TRIZOL総RNA単離試薬(Life Technologies, Inc.、メリーランド州ガイザーズバーグ)を用いて総RNAを単離し、そしてオリゴd(T)カラムクロマトグラフィーまたはガラスビーズを用いて、mRNAを単離する。試料核酸分子を増幅する場合、試料核酸分子を増幅し、そして低存在量転写物を含めて、元来の試料の相対存在量を維持することが望ましい。RNAはin vitro、in situ、またはin vivoで増幅することも可能である(Eberwine米国特許第5,514,545号を参照されたい)。
【0116】
増幅法および標識法が、試料中の核酸分子の真の分布を変化させなかったことを確実にするため、試料内に対照を含むこともまた、好適である。この目的のため、相補アレイ化核酸分子へのハイブリダイゼーションに際して検出可能であることがあらかじめ決定された、ある量の対照核酸分子が試料に混入され(spiked)、そして核酸分子の組成物には、対照アレイ化核酸分子と特異的にハイブリダイズする参照核酸分子が含まれる。ハイブリダイゼーションおよびプロセシング後、得られるハイブリダイゼーションシグナルは、試料に添加された対照アレイ化核酸分子の量を正確に反映しなければならない。
【0117】
ハイブリダイゼーション前に、試料核酸分子を断片にすることが望ましい可能性もある。断片化は、二次構造、および試料中の他の試料核酸分子または非相補的核酸分子への交差ハイブリダイゼーションを最小限にすることによって、ハイブリダイゼーションを改善する。断片化は、機械的手段または化学的手段によって実行可能である。
【0118】
標識
試料核酸分子を1以上の標識部分で標識して、ハイブリダイズしたアレイ化/試料核酸分子複合体の検出を可能にすることも可能である。標識部分は、分光的手段、光化学的手段、生化学的手段、生体電子工学的手段、免疫化学的手段、電気的手段、視覚的手段または化学的手段によって検出することも可能な組成物を含むことも可能である。標識部分には、(32)P、(33)Pまたは(35)Sなどの放射性同位体、化学発光化合物、標識結合タンパク質、重金属原子、蛍光マーカーおよび色素などの分光マーカー、磁気標識、連結酵素、質量分析タグ、スピン標識、電子伝達供与体および受容体等が含まれる。好ましい蛍光マーカーには、Cy3およびCy5フルオロフォア(Amersham Pharmacia Biotech、ニュージャージー州ピスカタウェイ)が含まれる。
【0119】
ハイブリダイゼーション
配列番号1およびその断片の核酸分子配列を、多様な目的のため、多様なハイブリダイゼーション技術で用いることも可能である。配列番号1から、ハイブリダイゼーションプローブを設計するか、または得ることも可能である。こうしたプローブを、非常に特異的な領域から、または保存されたモチーフから作成し、そしてL−PBEメッセージ、アレル変異体、または関連配列を定量化するプロトコルで用いることも可能である。本発明のハイブリダイゼーションプローブは、DNAまたはRNAであることも可能であり、そして配列番号1の配列から、または哺乳動物遺伝子のプロモーター、エンハンサー、およびイントロンを含むゲノム配列から得ることも可能である。標識ヌクレオチドの存在下、オリゴ標識、ニックトランスレーション、末端標識、またはPCR増幅を用いて、ハイブリダイゼーションプローブまたはPCRプローブを産生することも可能である。核酸配列を含有するベクターを用いて、RNAポリメラーゼおよび標識核酸分子を添加することによって、in vitroでmRNAプローブを産生することも可能である。Amersham Pharmacia Biotechによって提供されるものなどの商業的に入手可能なキットを用いて、これらの方法を行うことも可能である。ハイブリダイゼーションのストリンジェンシーは、プローブのG+C含量、塩濃度、および温度によって、決定される。特に、塩濃度を減少させるか、またはハイブリダイゼーション温度を上昇させることによって、ストリンジェンシーを増加させることも可能である。膜に基づく、いくつかのハイブリダイゼーションに用いる溶液においては、ホルムアミドなどの有機溶媒を添加すると、反応がより低い温度で起こることが可能になる。1%ドデシル硫酸ナトリウム(SDS)を含む5xSSC、60℃などの緩衝液を用いて、低ストリンジェンシーでハイブリダイゼーションを行って、いくつかのミスマッチを含有するヌクレオチド配列間のハイブリダイゼーション複合体の形成を可能にすることも可能である。続いて、0.1%SDSを含む0.2xSSCなどの緩衝液を、45℃(中程度のストリンジェンシー)または68℃(高ストリンジェンシー)いずれかで用いて、より高いストリンジェンシーで洗浄を行う。高ストリンジェンシーでは、核酸配列がほぼ完全に相補的である箇所でのみ、ハイブリダイゼーション複合体が安定であり続けるであろう。膜に基づく、いくつかのハイブリダイゼーションにおいて、好ましくは35%または最も好ましくは50%のホルムアミドをハイブリダイゼーション溶液に添加し、ハイブリダイゼーションを行う温度を下げることも可能であるし、そしてザルコシルまたはTritonX−100などの他の界面活性剤およびサケ精子DNAなどのブロッキング剤の使用によって、バックグラウンドシグナルを減少させることも可能である。ハイブリダイゼーションのための構成要素および条件の選択は、当業者に周知であり、そしてAusubel(上記)およびSambrookら(1989)Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Press, ニューヨーク州プレーンビューに概説されている。
【0120】
典型的な高ストリンジェンシー・ハイブリダイゼーション条件は、以下のとおりである:50%ホルムアミド、1%SDS、1M NaCl、10%デキストラン硫酸を含むハイブリダイゼーション溶液中、42℃でのハイブリダイゼーション、そして0.1xSSCおよび1%SDSを含む洗浄溶液中、60℃30分間2回の洗浄。Ausubelら(監修), Protocols in Molecular Biology, John Wiley & Sons(1994), pp.6.0.3〜6.4.10に記載されるように、温度および緩衝剤、または塩濃度の変動を通じて、同等のストリンジェンシーの条件を達成可能であることが、当該技術分野において理解される。ハイブリダイゼーション条件の修飾を、経験的に決定することも可能であるし、またはプローブの長さおよびグアノシン/シトシン(GC)塩基対形成の割合に基づいて、正確に計算することも可能である。Sambrookら(監修), Molecular Cloning:A Laboratory Manual, Cold Spring Harbor Laboratory Press:ニューヨーク州コールドスプリングハーバー(1989), pp.9.47〜9.51に記載されるように、ハイブリダイゼーション条件を計算することも可能である。
【0121】
試料に既知の量で添加した特異性対照試料核酸分子に対する、特異性対照核酸分子のハイブリダイゼーションを比較することによって、ハイブリダイゼーション特異性を評価することも可能である。特異性対照アレイ化核酸分子は、対応するアレイ化核酸分子と比較して、1以上の配列ミスマッチを有することも可能である。この方式で、相補的アレイ化核酸分子のみが試料核酸分子にハイブリダイズするのか、またはミスマッチハイブリッド二重鎖が形成されるのかを決定することも可能である。
【0122】
絶対的または示差的ハイブリダイゼーション形式で、ハイブリダイゼーション反応を行うことも可能である。絶対的ハイブリダイゼーション形式では、1つの試料由来の核酸分子を、マイクロアレイ形式で分子にハイブリダイズさせ、そしてハイブリダイゼーション複合体形成後に検出されるシグナルが、試料中の核酸分子レベルに相関する。示差的ハイブリダイゼーション形式では、2つの生物学的試料中の遺伝子セットの示差的発現を解析する。示差的ハイブリダイゼーションでは、両方の生物学的試料から核酸分子を調製し、そして異なる標識部分で標識する。2つの標識核酸分子の混合物を、マイクロアレイに添加する。次いで、2つの異なる標識からの発光を個々に検出可能である条件下で、マイクロアレイを調べる。両方の生物学的試料由来の実質的に同等の数の核酸分子にハイブリダイズする、マイクロアレイ中の分子は、区別可能な組み合わされた蛍光を生じる(ShalonらPCT公報WO95/35505)。好ましい態様において、標識は、Cy3およびCy5フルオロフォアなどの、区別可能な発光スペクトルを持つ蛍光マーカーである。
【0123】
ハイブリダイゼーション後、マイクロアレイを洗浄し、ハイブリダイズしていない核酸分子を取り除き、そしてハイブリダイズ可能なアレイ要素および核酸分子間の複合体形成を検出する。複合体形成を検出する方法は当業者に周知である。好ましい態様において、核酸分子を蛍光標識で標識し、そして複合体形成を示す蛍光のレベルおよびパターンの測定を蛍光顕微鏡、好ましくは共焦点蛍光顕微鏡によって達成する。
【0124】
示差的ハイブリダイゼーション実験において、2以上の異なる生物学的試料由来の核酸分子を、異なる発光波長を持つ2以上の異なる蛍光標識で標識する。特定の波長を検出する異なる光電子増倍管セットを用いて、蛍光シグナルを別個に検出する。2以上の試料中の核酸分子の相対的存在量/発現レベルを得る。
【0125】
典型的には、同様の試験条件下で、1より多いマイクロアレイを用いる場合、マイクロアレイ蛍光強度を規準化して、ハイブリダイゼーション強度の変動を考慮に入れることも可能である。好ましい態様において、各マイクロアレイ上に含有される内部規準化対照由来の強度を用いて、個々のアレイ化試料核酸分子複合体ハイブリダイゼーション強度を規準化する。
【0126】
ポリペプチドに基づくアッセイ
本発明は、イヌL−PBEポリペプチドを検出し、そして定量化するための方法および試薬を提供する。これらの方法には、電気泳動、質量分析、クロマトグラフィー法などの解析的生化学的方法、またはラジオイムノアッセイ(RIA)、酵素連結免疫吸着アッセイ(ELISA)、免疫蛍光アッセイ、ウェスタンブロッティングなどの多様な免疫学的方法、アフィニティー捕捉質量分析、生物学的活性、および以下に記載し、そして本開示を再検討した際に当業者には明らかである他のアッセイが含まれる。
【0127】
イムノアッセイ
本発明はまた、1以上の抗イヌL−PBE抗体試薬を使用して、イヌL−PBEポリペプチドを検出する方法(すなわちイムノアッセイ)も提供する。本明細書において、イムノアッセイは、イヌL−PBEポリペプチドまたはエピトープに特異的に結合する抗体(本明細書に広く定義されるとおりであり、そして具体的には断片、キメラおよび他の結合剤が含まれる)を利用するアッセイである。
【0128】
本発明の実施に適した、いくつかのよく確立された免疫学的結合アッセイ形式が知られる(例えば米国特許第4,366,241号;第4,376,110号;第4,517,288号;および第4,837,168号を参照されたい)。例えばMethods in Cell Biology Volume 37:Antibodies in Cell Biology, Asai監修, Academic Press, Inc. ニューヨーク(1993);Basic and Clinical Immunology 第7版, Stites & Terr監修(1991);HarlowおよびLane、上記[例えば第14章]、並びにAusubelら、上記[例えば第11章]を参照されたい。典型的には、免疫学的結合アッセイ(またはイムノアッセイ)は、解析物に特異的に結合し、そしてしばしば解析物を固相に固定する、「捕捉剤」を利用する。1つの態様において、捕捉剤は、イヌL−PBEポリペプチドまたは下位配列に特異的に結合する部分、例えば抗イヌL−PBE抗体である。
【0129】
通常、アッセイしようとするイヌL−PBE遺伝子産物を直接検出するか、または検出可能標識を用いて、間接的に検出する。アッセイで用いる特定の標識または検出可能基は、アッセイに用いる単数または複数の抗体の特異的結合に有意に干渉しない限り、通常、本発明の決定的な側面ではない。標識は、捕捉剤(例えば抗イヌL−PBE抗体)に共有的に付着していることも可能であるし、またはイヌL−PBEポリペプチドに特異的に結合する別の抗体などの第三の部分に付着していることも可能である。
【0130】
本発明は、イヌL−PBEポリペプチドを検出する競合的および非競合的イムノアッセイの方法および試薬を提供する。非競合的イムノアッセイは、捕捉された解析物(この場合、イヌL−PBE)の量を直接測定するアッセイである。1つのこうしたアッセイは、イヌL−PBEタンパク質上の2つの干渉しないエピトープに反応性であるモノクローナル抗体を利用する、モノクローナル抗体に基づく2部位イムノアッセイである。背景情報に関しては、例えばMaddoxら, 1983, J. Exp. Med., 158:1211を参照されたい。1つの「サンドイッチ」アッセイにおいて、捕捉剤(例えば抗イヌL−PBE抗体)を固体支持体に直接結合させて、固定する。次いで、これらの固定された抗体が、試験試料中に存在するいかなるイヌL−PBEタンパク質も捕捉する。次いで、こうして固定されたイヌL−PBEを、すなわち標識を所持する第二のイヌL−PBE抗体に結合させることによって、標識することも可能である。あるいは、第二のイヌL−PBE抗体は標識を欠いているが、第二の抗体が得られた種の抗体に特異的な、標識された第三の抗体に結合されていることも可能である。あるいは、第二の抗体をビオチンなどの検出可能部分で修飾し、これに酵素標識ストレプトアビジンなどの、第三の標識分子が特異的に結合することも可能である。
【0131】
競合的アッセイにおいて、試料中に存在するイヌL−PBEタンパク質によって、捕捉剤(例えばイヌL−PBE抗体)から置換された(または競合により離れた(competed away))、添加された(外因性)イヌL−PBEの量を測定することによって、試料に存在するイヌL−PBEタンパク質の量を間接的に測定する。ハプテン阻害アッセイは、競合アッセイの別の例である。このアッセイにおいて、イヌL−PBEタンパク質を固体支持体上に固定する。既知の量のイヌL−PBE抗体を試料に添加し、そして次いで、試料を固定イヌL−PBEタンパク質と接触させる。この場合、固定イヌL−PBEタンパク質に結合した抗イヌL−PBE抗体の量は、試料に存在するイヌL−PBEタンパク質の量に反比例する。抗体の固定された割合または溶液中に残存する抗体の割合いずれかを検出することによって、固定された抗体の量を検出することも可能である。この側面において、抗体が標識されている場合、検出は直接であることも可能であるし、または上述のように抗体に特異的に結合する分子に標識が結合している場合、間接的であることも可能である。
【0132】
抗体に基づく他のアッセイ形式
本発明はまた、イムノブロット(ウェスタンブロット)形式を用いることによって、試料中のイヌL−PBEポリペプチドの存在を検出し、そして定量化するための試薬および方法も提供する。別のイムノアッセイは、いわゆる「側方流動クロマトグラフィー」である。側方流動クロマトグラフィーの非競合型では、試料は、例えばキャピラリー作動によって、支持体を横切って移動し、そして可動標識抗体と出会い、該抗体が解析物に結合してコンジュゲートを形成する。次いで、コンジュゲートが支持体を横切り、そして固体された第二の抗体と出会い、この抗体が解析物と結合する。したがって、標識抗体を検出することによって、固定された解析物が検出される。側方流動クロマトグラフィーの競合型では、解析物の標識型が、キャリアーを横切って移動し、そして固定された抗体との結合に関して、非標識解析物と競合する。試料中により多量の解析物があれば、標識解析物による結合はより少なくなり、そしてしたがって、シグナルはより弱くなる。例えばMayら、米国特許第5,622,871号およびRosenstein、米国特許第5,591,645号を参照されたい。
【0133】
アッセイに応じて、抗原、標的抗体、または抗カテプシンS抗体を含む、多様な構成要素を、固体表面または支持体(例えば担体(substrate)、膜、またはろ紙)に結合させることも可能である。多様な固体表面に生体分子を固定する多くの方法が当該技術分野に知られる。例えば、固体表面は、膜(例えばニトロセルロース)、マイクロタイタープレート(例えばPVC、ポリプロピレン、またはポリスチレン)、試験管(ガラスまたはプラスチック)、ディップスティック(例えばガラス、PVC、ポリプロピレン、ポリスチレン、ラテックス等)、微量遠心分離試験管、またはガラスもしくはプラスチックビーズであることも可能である。望ましい構成要素を共有結合させることも可能であるし、または非特異的結合を介して、非共有的に付着させることも可能である。
【0134】
天然および合成両方の、非常に多様な有機ポリマーおよび無機ポリマーを、固体表面の材料として使用することも可能である。実例となるポリマーには、ポリエチレン、ポリプロピレン、ポリ(4−メチルブテン)、ポリスチレン、ポリメタクリレート、ポリ(エチレンテレフタレート)、レーヨン、ナイロン、ポリ(酪酸ビニル)、ポリビニリデンジフルオリド(PVDF)、シリコーン類、ポリホルムアルデヒド、セルロース、酢酸セルロース、ニトロセルロース等が含まれる。使用可能な他の物質には、紙、ガラス、セラミックス、金属、メタロイド、半導体物質、セメント等が含まれる。さらに、タンパク質(例えばゼラチン)、リポ多糖、ケイ酸塩、アガロースおよびポリアクリルアミドなどの、ゲルを形成する物質もまた、使用可能である。いくつかの水性相を形成するポリマー、例えばデキストラン、ポリアルキレングリコールまたは界面活性剤、例えばリン脂質、長鎖(12〜24の炭素原子)アルキルアンモニウム塩等もまた適切である。固体表面が多孔である場合、系の性質に応じて、多様な孔サイズを使用することも可能である。
【0135】
質量分析
分子の質量(mass)は、しばしば、分子の同定因子として使用することも可能である。したがって、質量分析法を用いて、タンパク質解析物を同定することも可能である。質量分析装置は、イオン化解析物が飛行管を通過して、そしてイオン検出装置に検出されるのに必要な時間を測定することによって、質量を測定可能である。タンパク質のための質量分析の1つの方法は、マトリックス支援レーザー脱離イオン化質量分析(「MALDI」)である。MALDIにおいて、レーザーの波長のエネルギーを吸収するエネルギー吸収マトリックス物質と解析物を混合し、そしてプローブ表面に置く。マトリックスをレーザーで打つと、解析物がプローブ表面から脱離し、イオン化され、そしてイオン検出装置に検出される。例えばHillenkampら、米国特許第5,118,937号を参照されたい。
【0136】
タンパク質のための質量分析の他の方法が、HutchensおよびYip、米国特許第5,719,060号に記載される。1つのこうした方法は、アフィニティー捕捉のための増進表面(「SEAC」)と称され、この方法において、解析物に特異的にまたは非特異的に結合する固相アフィニティー試薬、例えば抗体または金属イオンを用いて、解析物を試料中の他の物質から分離する。次いで、捕捉された解析物を、例えばレーザーエネルギーによって固相から脱離し、イオン化し、そして検出装置によって検出する。
【0137】
本発明のさらなる特徴および変型は、詳細な説明を含めた本出願全体から、当業者に明らかであろうし、そしてこうした特徴はすべて、本発明の側面と意図される。同様に、本明細書に記載する本発明の特徴を再度組み合わせて、特徴の組み合わせが、本発明の側面または態様として、具体的に上述されているかいないかにかかわりなく、やはり本発明の側面であることも意図されるさらなる態様にすることも可能である。また、本発明に決定的であると本明細書に記載した限定のみが、限定として見なされるべきであり;決定的であると本明細書に記載されていない、限定されない本発明の変型は、本発明の側面であると意図される。
【0138】
前述の説明および実施例に特に記載されるものとは別の方式で、本発明を実施可能であることが明らかであろう。
上記の解説を考慮すると、本発明の多くの修飾および変型が可能であり、そしてしたがって、これらは本発明の側面内にある。
【0139】
本明細書に引用したすべての刊行物のすべての開示は、本明細書に援用される。
【図面の簡単な説明】
【0140】
【図1A】イヌ(配列番号1)、ヒト(配列番号11)、マウス(配列番号13)およびラット(配列番号15)L−PBEポリペプチド配列の並列。
【図1B】イヌ(配列番号1)、ヒト(配列番号11)、マウス(配列番号13)およびラット(配列番号15)L−PBEポリペプチド配列の並列。
【図2A】イヌ(配列番号2)、ヒト(配列番号12)、マウス(配列番号14)およびラット(配列番号16)L−PBEポリヌクレオチド配列の並列。
【図2B】イヌ(配列番号2)、ヒト(配列番号12)、マウス(配列番号14)およびラット(配列番号16)L−PBEポリヌクレオチド配列の並列。
【図2C】イヌ(配列番号2)、ヒト(配列番号12)、マウス(配列番号14)およびラット(配列番号16)L−PBEポリヌクレオチド配列の並列。
【図2D】イヌ(配列番号2)、ヒト(配列番号12)、マウス(配列番号14)およびラット(配列番号16)L−PBEポリヌクレオチド配列の並列。
【配列表】
【技術分野】
【0001】
発明の分野
本発明は、イヌ(canine)L−PBEポリペプチドを含む単離ポリペプチド、および該ポリペプチドをコードするポリヌクレオチドを提供する。本発明はまた、毒性化合物または分子の指標となる代謝反応のための化合物および療法剤をスクリーニングするアッセイも提供する。
【背景技術】
【0002】
発明の背景
ペルオキシソームは、古典的なL−ヒドロキシ特異的ペルオキシソーム増殖因子誘導性ベータ酸化系を含有する。ヒト・ペルオキシソームのエノイル−CoA−ヒドラターゼ:3−ヒドロキシアシル−CoAデヒドロゲナーゼ二機能性酵素(L−PBE)は、Hoeflerら(1)によって最初に同定された。cDNA配列は、3,779ヌクレオチドに渡り、2,169ヌクレオチドのオープンリーディングフレームを有する。
【0003】
Qiらは、ノックアウトマウスを生成することによって、L−PBEの機能を検討した(2)。突然変異体マウスは生存可能であり、そして繁殖性であり、そして検出可能な著しい表現型上の欠陥を示さなかった。L−PBEノックアウトマウスは、ペルオキシソーム増殖因子に曝露されると、同様に処置された野生型マウスと比較して、ペルオキシソーム増殖反応が認識可能に鈍化していたことを除き、PPARαに制御される肝臓mRNAおよびタンパク質レベルの増加を示した。ペルオキシソーム増殖反応のこの鈍化は、L−PBEノックアウトマウス肝臓中にL−PBEタンパク質が存在しなかったことに起因すると考えられる。
【0004】
Watkinsらは、新生児副腎白質ジストロフィー(NALD)の臨床的特徴を持つ男性乳児において、ペルオキシソーム機能を評価した(3)。ペルオキシソームのベータ−酸化酵素のイムノブロット研究によって、死後の肝臓試料において、L−PBE(エノイル−CoAヒドラターゼ/3−ヒドロキシアシル−CoAデヒドロゲナーゼ)が欠損していることが示された。Wandersらもまた、ペルオキシソーム二機能性酵素欠損を記載しており、これはNALDのものより、より重度の表現型と関連しているようである(4)。
【0005】
イヌは、薬剤の毒性学を研究するための重要な種であるため、イヌL−PBE遺伝子に特異的な核酸プローブおよびイヌL−PBEタンパク質に特異的な抗体は、推定上の毒性化合物に対する代謝反応を示す手段として、産業毒性学において、大きな価値を有する。本発明はこの必要性に対処する。
【0006】
参考文献
【0007】
【化1】
【発明の開示】
【発明が解決しようとする課題】
【0008】
発明の概要
本発明は、上に同定される必要性に対処し、イヌL−PBEタンパク質をコードする単離核酸分子、該単離核酸分子を取り込んだ構築物および組換え宿主細胞;該単離核酸分子によってコードされるイヌL−PBEポリペプチド;イヌL−PBEポリペプチドに対する抗体を提供する。
【0009】
1つの態様において、本発明は、配列番号2に示すアミノ酸配列を含む、単離イヌL−PBEポリペプチドを提供する。配列番号2のポリペプチドは、いくつかのポリペプチド種を生じる、特定のタンパク質分解プロセシング事象にさらされることも可能であると理解される。
【0010】
さらに、本発明は、イヌL−PBEポリペプチドのエピトープを含む断片を提供する。「特異的なエピトープ」によって、以下に詳細に定義するように、イヌL−PBEポリペプチドに特異的な抗体によって認識可能であるイヌL−PBEポリペプチドの部分を意味する。別の態様は、配列番号2に示す完全アミノ酸配列を含む単離ポリペプチドを含む。
【0011】
イヌL−PBEのコードcDNA配列および予測されるアミノ酸配列を以下に複製し、そしてまた、それぞれ、配列番号1および配列番号2に示す。
【0012】
【化2】
【0013】
配列番号1および配列番号2は、特定のイヌ・ポリヌクレオチド配列およびポリペプチド配列を提供するが、本発明は、その範囲内に他のイヌ・アレル変異体を含むことが意図される。
【0014】
別の態様において、本発明は、本発明のポリペプチドのアミノ酸配列をコードするヌクレオチド配列を含む、単離ポリヌクレオチド(例えば一本鎖または二本鎖のcDNA、ゲノムDNA、合成DNA、RNA、またはその組み合わせ)を提供する。こうしたポリヌクレオチドは、細胞において、該酵素を組換え的に発現するのに有用であり、そしてまた、該酵素の発現を検出するのにも有用である(例えばノーザン・ハイブリダイゼーションおよびin situハイブリダイゼーションアッセイを用いる)。こうしたポリヌクレオチドはまた、培養細胞もしくは組織において、または動物において、イヌL−PBEの発現を抑制するアンチセンスおよび他の分子を設計するため、療法目的のため、あるいは異常なイヌL−PBE発現に特徴付けられる疾患のモデルを提供するために、有用である。本発明のポリヌクレオチドの定義から明確に排除されるのは、ポリヌクレオチドが元来得られた天然宿主細胞由来の完全単離染色体である。配列番号1に示すポリヌクレオチドは、天然存在イヌL−PBE配列に対応する。普遍的遺伝暗号の周知の縮重のため、やはり配列番号2のイヌL−PBEをコードする多くの他の配列が存在することが認識されるであろう。別の態様において、本発明は、配列番号1に示す配列以外の縮重イヌL−PBEコード配列のすべてに関する。
【0015】
本発明はまた、ヌクレオチド配列を含む単離ポリヌクレオチドであって、以下のハイブリダイゼーション条件下で、配列番号1に示すヌクレオチド配列または該配列に相補的な非コード鎖に特異的にハイブリダイズする、前記単離ポリヌクレオチドも提供する:
(a)50%ホルムアミド、1%SDS、1M NaCl、10%デキストラン硫酸を含むハイブリダイゼーション溶液中、42℃16時間のハイブリダイゼーション;および
(b)0.1%SSC、1%SDSを含む洗浄溶液中、60℃30分間2回の洗浄。
【0016】
本発明の1つのポリヌクレオチドは、イヌL−PBEコードDNA配列を含む配列番号1に示す配列、またはそのユニークな断片を含む。
関連する態様において、本発明は、本発明のポリヌクレオチドを含むベクターを提供する。こうしたベクターは、例えば宿主細胞において、ポリヌクレオチドを増幅し、有用な量を生じるのに有用である。他の態様において、ベクターは発現ベクターであり、該ベクター中で、本発明のポリヌクレオチドが、発現調節配列を含むポリヌクレオチドに機能可能であるように連結されている。こうしたベクターは、本発明のポリペプチドの組換え体産生に有用である。
【0017】
別の関連する態様において、本発明は、本発明のポリヌクレオチドまたは本発明のベクターで(安定にまたは一過性に)形質転換またはトランスフェクションされた宿主細胞を提供する。上述のように、こうした宿主細胞は、該ポリヌクレオチドを増幅するのに有用であり、そしてまた、該ポリヌクレオチドがコードする、イヌL−PBEポリペプチドまたはその断片を発現するのにも有用である。
【0018】
さらに別の関連する態様において、本発明は、イヌL−PBEポリペプチド(またはその断片)を産生する方法であって、栄養培地中で本発明の宿主細胞を増殖させ、そして細胞または培地から該ポリペプチドまたはその変異体を単離する工程を含む、前記方法を提供する。
【0019】
さらに別の態様において、本発明は、本発明のイヌL−PBEに特異的な抗体を提供する。抗体特異性を以下により詳細に記載する。しかし、文献に先に記載されたポリペプチドから生成可能であり、そしてイヌL−PBEと偶然に交差反応する(例えば両ポリペプチド中に類似のエピトープが偶然存在するため)ことが可能な抗体は、「交差反応性」抗体と見なされることを強調しなければならない。こうした交差反応性抗体は、イヌL−PBEに「特異的」な抗体ではない。抗体がイヌL−PBEに特異的であるか、または別の既知の酵素と交差反応性であるかの決定は、ウェスタンブロッティングアッセイまたは文献で周知のいくつかの他のアッセイを用いて行われる。イヌL−PBEを発現する細胞を同定するため、そしてまた、イヌL−PBE活性を調節するため、イヌL−PBEの活性部位に特異的に結合する抗体は、特に有用であるが、もちろん、他のエピトープに結合する抗体もまた、本発明の一部であることが意図される。
【0020】
1つの変型において、本発明はモノクローナル抗体を提供する。こうした抗体を産生するハイブリドーマもまた、本発明の側面と意図される。
別の変型において、本発明は、ポリクローナル抗体を含む細胞不含組成物を提供し、ここで少なくとも1つの抗体は、イヌL−PBEに特異的な本発明の抗体である。動物から単離される抗血清が典型的な組成物であり、水または別の希釈剤、賦形剤、もしくはキャリアーに再懸濁されている抗血清の抗体分画を含む組成物も典型的である。
【0021】
本発明はまた、本発明の抗体を用いる方法も提供する。例えば、本発明は、細胞抽出物内に存在するイヌL−PBEの量を決定する方法であって、イヌL−PBEポリペプチドとイヌL−PBEポリペプチドに特異的な抗体を、抗体がイヌL−PBEポリペプチドに結合する条件下で接触させる工程を含む、前記方法を提供する。
【0022】
本発明はまた、試料内に存在するイヌL−PBEポリヌクレオチドの量を決定する方法であって:配列番号1若しくはその断片、又はそれらの相補体を含む核酸分子と試料を、1以上の特異的ハイブリダイゼーション複合体形成のための条件下で接触させることを含む、前記方法も提供し、ここで該断片は、配列番号1の少なくとも12の連続ヌクレオチドを含むポリヌクレオチドである。
【0023】
本発明はまた、イヌにおいて、試験剤に対する代謝反応を測定する方法であって:試験剤で処置したイヌ由来の核酸を含有する試料を提供し;そして前記試料中の配列番号1若しくはその断片、又はそれらの相補体を含むポリヌクレオチドの量を決定することを含む、前記方法も提供し、ここで未処置のイヌ由来のポリヌクレオチドの量と比較した際の、処置したイヌ由来のポリヌクレオチドの量の変化が、試験剤に対する代謝反応の指標となる。
【0024】
1つの態様において、決定はハイブリダイゼーションを介して達成される。ハイブリダイゼーションは、核酸分子と試料核酸分子を、核酸分子および試料核酸分子間でハイブリダイゼーション複合体を形成するのに有効な条件下で接触させ;そしてハイブリダイゼーション複合体の存在または非存在を検出することを含む方法によって達成されることも可能である。1つの側面において、配列番号1若しくはその断片、又は配列番号1の相補体は、固体支持体アレイまたは他の固体支持体上の複数の核酸とともに存在することも可能である。
【0025】
本発明はさらに、試料内に存在するイヌL−PBEポリペプチドの量を決定する方法であって:
イヌL−PBEポリペプチドとイヌL−PBEポリペプチドに特異的な抗体を、抗体がイヌL−PBEポリペプチドに結合する条件下で接触させる
ことを含む、前記方法を提供する。所望によって、L−PBEポリペプチドまたは抗体は、固体支持体に付着していることも可能である。
【0026】
本発明はまた、イヌにおいて、試験剤に対する代謝反応を測定する方法であって:試験剤で処置したイヌ由来の試料を提供し;そして前記試料中の配列番号2を含むポリペプチド、または前記ポリペプチドに特異的なエピトープを含む該ポリペプチドの断片の量を決定することを含む、前記方法も提供し、ここで未処置のイヌ由来のポリペプチドの量と比較した際の、処置したイヌ由来のポリペプチドの量の変化が、試験剤に対する代謝反応の指標となる。
【課題を解決するための手段】
【0027】
配列表の簡単な説明
配列番号1――イヌL−PBEをコードするcDNA配列
配列番号2――イヌL−PBEの予測されるアミノ酸配列
配列番号3〜10――クローニング用プライマーおよび配列決定用プライマー
配列番号11 ヒトL−PBEをコードするcDNA配列
配列番号12 ヒトL−PBEの予測されるアミノ酸配列
配列番号13 マウスL−PBEをコードするcDNA配列
配列番号14 マウスL−PBEの予測されるアミノ酸配列
配列番号15 ラットL−PBEをコードするcDNA配列
配列番号16 ラットL−PBEの予測されるアミノ酸配列
【0028】
発明の詳細な説明
一般的な定義
本明細書において、これ以降、「ポリヌクレオチド」は、一般的に、非修飾RNAもしくはDNA、または修飾RNAもしくはDNAであることも可能な、ポリリボヌクレオチドまたはポリデオキシリボヌクレオチドいずれかを指す。「ポリヌクレオチド」には、限定なしに、一本鎖DNAおよび二本鎖DNA、一本鎖領域および二本鎖領域の混合物であるDNA、一本鎖RNAおよび二本鎖RNA、並びに一本鎖領域および二本鎖領域の混合物であるRNA、一本鎖、またはより典型的には二本鎖、あるいは一本鎖および二本鎖領域の混合物であることも可能なDNAおよびRNAを含むハイブリッド分子が含まれる。さらに、「ポリヌクレオチド」は、RNAもしくはDNAまたはRNAおよびDNA両方を含む三重鎖領域を指す。用語「ポリヌクレオチド」はまた、1以上の修飾塩基を含有するDNAまたはRNA、および安定性または他の理由のために修飾された骨格を持つDNAまたはRNAも含む。「修飾」塩基には、例えば、トリチル化塩基およびイノシンなどの普通でない塩基が含まれる。DNAおよびRNAに多様な修飾を行うことも可能であり;したがって、「ポリヌクレオチド」は、天然に典型的に見られるポリヌクレオチドの化学的、酵素的、または代謝的修飾型とともに、ウイルスおよび細胞に特徴的なDNAおよびRNAの化学型も含む。「ポリヌクレオチド」はまた、しばしばオリゴヌクレオチドと称される、比較的短いポリヌクレオチドも含む。
【0029】
本明細書において、これ以降、「ポリペプチド」は、ペプチド結合または修飾ペプチド結合によって互いに連結されたアミノ酸を含むペプチドまたはタンパク質いずれかを指す。「ポリペプチド」は、一般的にペプチド、オリゴペプチドまたはオリゴマーと称される短い鎖、および一般的にタンパク質と称される、より長い鎖両方を指す。ポリペプチドは、遺伝子にコードされる20のアミノ酸以外のアミノ酸を含有することも可能である。「ポリペプチド」には、翻訳後プロセシングなどの天然プロセスによるか、または当該技術分野に周知の化学的修飾技術によるか、いずれかによって修飾されたアミノ酸配列が含まれる。こうした修飾は、基本的な教科書、そしてより詳細なモノグラフ、さらに、膨大な研究文献によく記載されている。修飾は、ペプチド骨格、アミノ酸側鎖およびアミノ末端またはカルボキシル末端を含めて、ポリペプチドのどこに存在することも可能である。同種の修飾が、既定のポリペプチドのいくつかの部位で、同じ度合いで、または異なる度合いで存在することも可能であることが認識されるであろう。また、既定のポリペプチドが多くの種類の修飾を含有することも可能である。ポリペプチドは、ユビキチン化の結果として分枝していることも可能であるし、そして分枝を伴い、または伴わずに、環状であることも可能である。環状、分枝および分枝環状ポリペプチドは、翻訳後天然プロセスから生じることも可能であるし、または合成法によって作成されることも可能である。修飾または修飾型には、アセチル化、アシル化、ADP−リボシル化、アミド化、フラビンの共有結合、ヘム部分の共有結合、ヌクレオチドまたはヌクレオチド誘導体の共有結合、脂質または脂質誘導体の共有結合、ホスファチジルイノシトール(phosphotidylinositol)の共有結合、架橋、環状化、ジスルフィド結合形成、脱メチル化、共有架橋の形成、シスチンの形成、ピログルタミン酸の形成、ホルミル化、ガンマ−カルボキシル化、グリコシル化、GPIアンカー形成、ヒドロキシル化、ヨウ素化、メチル化、ミリストイル化、酸化、タンパク質分解プロセシング、リン酸化、プレニル化、ラセミ化、セレノイル化(selenoylation)、硫酸化、アルギニル化などのトランスファーRNAが仲介するタンパク質へのアミノ酸の付加、およびユビキチン化が含まれる(例えば、Proteins―Structure and Molecular Properties, 第2版, T.E. Creighton監修, W.H. Freeman and Company, ニューヨーク, 1993;Wold, F., Post−translational Protein Modifications:Perspectives and Prospects, 1−12ページ, Posttranslational Covalent Modification of Proteins中, B.C. Johnson監修, Academic Press, ニューヨーク, 1983;Seifterら, “Analysis for protein modifications and nonprotein cofactors”, Meth Enzymol(1990)182:626−646およびRattanら, “Protein Synthesis: Post−translational Modifications and Aging”, Ann NY Acad Sci(1992)663:4842を参照されたい)。
【0030】
「合成」は、本明細書において、そして当該技術分野に理解されるように、酵素的方法と対照的に、純粋に化学的方法によって産生されるポリヌクレオチドを指す。したがって、「完全に」合成されたDNA配列は、完全に化学的手段によって産生され、そして「部分的に」合成されたDNAは、生じるDNAのある部分のみが化学的手段によって産生されたものを含む。
【0031】
本明細書において、これ以降、「単離された」は、人の手によって天然状態から改変されたことを意味する。「単離された」組成物または物質が天然に存在する場合、その元来の環境から変化しているかまたは取り除かれているか、あるいはその両方である。例えば、生存動物に天然に存在するポリヌクレオチドまたはポリペプチドは、「単離されて」いないが、天然状態で共存する物質から分離された、同じポリヌクレオチドまたはポリペプチドは、該用語を本明細書で使用する際、「単離されて」いる。「単離された」は、本明細書において、そして当該技術分野に理解されるように、「単離」ポリヌクレオチドまたはポリペプチドいずれを指すのであっても、該ポリペプチドまたは核酸が通常見られる元来の細胞環境から分離されたことを意味するように解釈される。したがって、本明細書において、一例として、本発明のポリヌクレオチドで構築したトランスジェニック動物または組換え細胞株は、「単離」核酸を利用する。
【0032】
本明細書において、用語「イヌL−PBEポリペプチド」は、イヌL−PBE遺伝子がコードするタンパク質を意味し、保存的変化または非保存的変化を含有するアレル変異体を含む。1つのイヌL−PBEタンパク質配列を配列番号2として開示する。
【0033】
イヌL−PBEポリペプチドを、組換え細胞または生物によって産生することも可能であるし、天然組織または細胞株から実質的に精製することも可能であるし、あるいは化学的または酵素的に合成することも可能である。したがって、用語「イヌL−PBEポリペプチド」は、グリコシル化型、部分的グリコシル化型、または非グリコシル化型のタンパク質とともに、リン酸化型、部分的リン酸化型、非リン酸化型、硫酸化型、部分的硫酸化型、または非硫酸化型のタンパク質を含むことが意図される。該用語にはまた、アレル変異体、生物学的に活性であるタンパク質分解断片または他の断片を含む、PS2アミノ酸配列の他の機能的同等物、並びにイヌL−PBEポリペプチドの生理学的および病的タンパク質分解切断産物が含まれる。
【0034】
本明細書において、用語「試験剤」は、同定可能な化学薬品または分子いずれかを意味し、これには、限定されるわけではないが、小分子、ペプチド、タンパク質、糖、ヌクレオチド、または核酸が含まれる。こうした試験剤は、天然または合成であることも可能である。
【0035】
本明細書において、用語「接触」は、直接または間接的いずれかで、化合物を本発明のポリペプチドまたはポリヌクレオチドと物理的に近接させて、合わせることを意味する。該ポリペプチドまたはポリヌクレオチドは、いかなる数の緩衝剤、塩、溶液などの中に存在することも可能である。接触には、例えば、イオンチャネルポリペプチドもしくはその断片、またはイオンチャネルもしくはその断片をコードする核酸分子いずれかを含有する、ビーカー、マイクロタイタープレート、細胞培養フラスコ、または遺伝子チップ等のマイクロアレイに化合物を入れることが含まれる。
【0036】
本発明の核酸
本発明は、本明細書において、イヌL−PBEと称されるイヌL−PBEポリペプチドをコードする単離ポリヌクレオチド(例えば、センス鎖および相補的アンチセンス鎖両方の、一本鎖および二本鎖両方のDNA配列およびRNA転写物、そのスプライス変異体を含む)を提供する。本発明のDNAポリヌクレオチドには、ゲノムDNA、cDNA、および全体または部分が化学的に合成されているDNAが含まれる。
【0037】
本発明のゲノムDNAは、本発明のポリペプチドのタンパク質コード領域を含み、そしてまたそのアレル変異体を含むことも意図される。多くの遺伝子に関して、ゲノムDNAがRNA転写物に転写され、これが1以上のスプライシング事象を経て、転写物のイントロン(すなわち非コード領域)が除去されるか、または「スプライシング−アウト」されることが広く理解されている。選択的(alternative)機構によってスプライシングされることも可能であり、そしてしたがって異なるRNA配列の除去にさらされることも可能であるが、なおイヌL−PBEポリペプチドをコードするRNA転写物は、当該技術分野において、スプライス変異体と称され、本発明に含まれる。したがって、本発明に含まれるスプライス変異体は、同一の元来のゲノムDNA配列にコードされるが、別個のmRNA転写物から生じる。アレル変異体は、野生型遺伝子配列の修飾型であり、修飾は、染色体分離中の組換えから、または遺伝子突然変異を生じさせる条件への曝露から生じる。アレル変異体は、野生型遺伝子同様、天然存在配列である(in vitro操作から生じる非天然存在変異体とは対照的である)。
【0038】
本発明はまた、イヌL−PBEをコードするRNAポリヌクレオチドの逆転写を通じて得られるcDNAも含む(慣用的には、相補鎖の第二の鎖合成が続き、二本鎖DNAを提供する)。
【0039】
イヌL−PBEポリペプチドをコードするDNA配列を配列番号1に示す。当業者は、本発明のDNAが二本鎖分子を含み、例えば、DNAのワトソン−クリック塩基対形成規則にしたがって配列番号1の配列から推定可能な配列を有する相補分子(「非コード鎖」または「相補体」)を伴う、配列番号1に示す配列を有する分子を含むことを容易に認識するであろう。本発明にやはり意図されるのは、普遍的遺伝暗号の周知の縮重のため、配列番号1のポリヌクレオチドと配列が異なる、配列番号2のイヌL−PBEポリペプチドをコードする他のポリヌクレオチドである。
【0040】
当該技術分野に周知であるように、遺伝暗号の縮重のため、前述の配列番号1のポリペプチドにコードされるのと同一のポリペプチドをコードすることが可能な、多くの他のDNA分子およびRNA分子がある。したがって、本発明は、発現に際して、配列番号2のポリペプチドをコードする、他のDNA分子およびRNA分子を意図する。イヌL−PBEポリペプチドをコードするアミノ酸残基配列が同定されたら、そして各特定のアミノ酸残基をコードするすべての3つ組コドンを知っていれば、こうしたコードRNA配列およびDNA配列をすべて記載することが可能である。したがって、特定のアミノ酸に関して、単純にコドン中の変化によって特徴付けられる、本明細書に具体的に開示するもの以外のDNA分子およびRNA分子が、本発明の範囲内である。
【0041】
アミノ酸およびそれを表す略語、記号およびコドンの表を、以下の表1に示す。
表1
【0042】
【表1】
【0043】
当該技術分野に周知であるように、コドンは、mRNAおよびその対応するcDNA分子において、ヌクレオチドの3つ組配列を構成する。コドンは、mRNA分子に存在する場合、塩基ウラシル(U)によって特徴付けられるが、DNAに存在する場合、塩基チミジン(T)によって特徴付けられる。ポリヌクレオチド内の同一アミノ酸残基のコドンにおける単純な変化は、コードされるポリペプチドの配列または構造を変化させないであろう。句が、特定の3ヌクレオチド配列が特定のアミノ酸いずれかを「コードする」ことを述べる場合、一般の当業者は、上の表が問題の特定のヌクレオチドを同定する手段を提供するのを認識するであろうことが明らかである。例として、特定の3ヌクレオチド配列が、スレオニンをコードする場合、上の表は、ありうる3つ組配列がACA、ACG、ACCおよびACU(DNAの場合ACT)であることを開示する。
【0044】
本発明は、イヌL−PBE DNAの種相同体(homolog)、好ましくは哺乳動物相同体をさらに含む。種相同体は、ときに「オルソログ」と称され、本発明の配列番号1と、少なくとも89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%の相同性を共有する。本発明のポリヌクレオチドに関する配列「相同性」パーセントは、本明細書において、配列を並列させ、そして必要であればギャップを導入して最大の配列同一性パーセントを達成した後、配列番号1に示すイヌL−PBE配列のヌクレオチドと同一である、候補配列中のヌクレオチド塩基の割合として定義される。例えば、SmithおよびWaterman(Adv. Appl. Math. 2:482−489(1981))のアルゴリズムを用いる、Gapプログラム(ウィスコンシン配列解析パッケージ、バージョン8、Unix用、遺伝学コンピュータグループ、University Research Park, Madison, Wisconsin)などの、この目的に一般的に使用されるコンピュータプログラムいずれかを用いて、2つの配列を比較することによって、天然および変異体イヌL−PBE配列間の配列の割合を決定することもまた可能である。
【0045】
本発明に提供されるポリヌクレオチド配列情報は、当該技術分野に周知であり、そして日常的に実施される技術によって、コードされるポリペプチドの大規模発現を可能にする。本発明のポリヌクレオチドはまた、サザン・ハイブリダイゼーションおよび/またはノーザン・ハイブリダイゼーション、およびポリメラーゼ連鎖反応(PCR)を含む周知の技術によって、ヒト・アレル変異体および種相同体などの、関連するイヌL−PBEポリペプチドをコードするポリヌクレオチドの同定および単離も可能にする。関連するポリヌクレオチドの例には、アレル変異体を含むヒトおよび非ヒトゲノム配列、並びに、イヌL−PBEに相同なポリペプチド、およびイヌL−PBEの1以上の生物学的、免疫学的、および/または物理的特性を共有する、構造的に関連するポリペプチドをコードするポリヌクレオチドが含まれる。イヌL−PBE DNAの配列を知ることで、サザン・ハイブリダイゼーションまたはポリメラーゼ連鎖反応(PCR)の使用を通じて、プロモーター、オペレーター、エンハンサー、リプレッサー等のイヌL−PBE発現調節制御配列をコードするゲノムDNA配列の同定もまた可能になる。本発明のポリヌクレオチドはまた、細胞がイヌL−PBEを発現する能力を検出するか、またはイヌL−PBE発現レベルを測定する、ハイブリダイゼーションアッセイにおいても有用である。本発明のポリヌクレオチドはまた、単数または複数の疾患状態の根底にあるイヌL−PBE遺伝子座における遺伝子改変(単数または複数)を同定するのに有用な診断法の基礎となることも可能であり、こうした情報は、診断にも、そして療法戦略の選択にも有用である。
【0046】
イヌL−PBEポリペプチドをコードする全長ポリヌクレオチドの本明細書における開示は、一般の当業者が、全長ポリヌクレオチドのすべてのありうる断片を入手するのを容易に可能にする。したがって、本発明は、イヌL−PBEをコードするポリヌクレオチドの少なくとも12から2562(この間の1つ1つの整数値を含む)の連続ヌクレオチドを含む、イヌL−PBEコードポリヌクレオチドの断片を提供する。本発明のポリヌクレオチド(断片を含む)は、しばしば、イヌL−PBEをコードするポリヌクレオチド配列にユニークな配列を含み、そしてしたがって、非常にストリンジェントな条件下、または中程度にストリンジェントな条件下で、イヌL−PBE(またはその断片)をコードするポリヌクレオチドにのみ(すなわち「特異的に」)ハイブリダイズするであろう。本発明のゲノム配列のポリヌクレオチド断片は、コード領域にユニークな配列だけでなく、イントロン、制御領域、および/または他の非翻訳配列に由来する全長配列の断片も含む。本発明のポリヌクレオチドにユニークな配列は、他の既知のポリヌクレオチドへの配列比較を通じて認識可能であり、そして当該技術分野で日常的に利用される並列プログラム、例えば公共の配列データベースで入手可能なものの使用を通じて同定可能である。こうした配列はまた、ポリヌクレオチドがハイブリダイズするゲノムDNA断片の数を決定するサザン・ハイブリダイゼーション解析からも認識可能である。本発明のポリヌクレオチドは、放射能標識、蛍光標識、および酵素標識を含めて、その検出を可能にする方式で標識されることも可能である。
【0047】
断片ポリヌクレオチドは、全長または他の断片イヌL−PBEポリヌクレオチドの検出用のプローブとして特に有用である。1以上の断片ポリヌクレオチドは、イヌL−PBEをコードするポリヌクレオチドの存在を検出するのに用いられるか、またはイヌL−PBEをコードするポリヌクレオチド配列における変動を検出するのに用いられる、キットに含まれることも可能である。
【0048】
本発明はまた、イヌL−PBEポリペプチドをコードするDNAであって、配列番号1のポリヌクレオチドの非コード鎖または相補体に、高ストリンジェンシーハイブリダイゼーションまたは洗浄条件下でハイブリダイズする、前記DNAもまた含む。
【0049】
高ストリンジェンシーハイブリダイゼーションの概念は、以下の「本発明のアッセイ」を詳述する項で論じる。
【実施例】
【0050】
(実施例1)
イヌL−PBEのクローニング
PCRのプライマーを設計するため、DNASTARソフトウェア(DNASTAR、ウィスコンシン州)を用いて、ヒト、ラットおよびマウスL−PBEの各遺伝子のオープンリーディングフレーム(ORF)配列を並列した。各遺伝子について3つのプライマーセットを選択した。その中で、2つのセットは、それぞれ、ヒトおよびラットのオルソログの開始配列領域および停止配列領域に対応し;残りのセットは、ヒト、ラットおよびマウスのオルソログ間の保存される配列領域にしたがって設計された。プライマーは、一般的に、長さ30ヌクレオチドである。
【0051】
ヒトPSDデータベース(Proteome Inc.)およびNCBI ESTデータベース検索を通じて(相同ESTがクローニングされた組織を同定することによって)、選択したクローニング候補遺伝子の組織発現パターンを同定した。5つの組織、肝臓、腎臓、結腸、脾臓および肺が、選択した12の遺伝子すべてを含むことが同定された。
【0052】
RNeasyキット(Qiagen、カリフォルニア州)を用いて、イヌ肝臓試料からRNAを調製した。RNAをcDNAに逆転写するため、総体積12.5μl中、0.5μgのRNAを5.77μlの32.5μMランダム六量体(Amersham Biosciences、ニュージャージー州)および2.5μlの10mM dNTPと混合した。混合物を65℃で5分間インキュベーションし、そして氷上で冷却した。2μlのRNアーゼ不含水、5μlの5x第一鎖緩衝液(Invitrogen、カリフォルニア州)、0.5μlの100mM DTT、2.5μlの40U/μl RNaseOUT(Invitrogen、カリフォルニア州)および2.5μlの200U/μl Superscript II逆転写酵素(Invitrogen、カリフォルニア州)を含有する12.5μlのマスター混合物を添加した。最終反応混合物を25℃で10分間、42℃で60分間、そして70℃で15分間、連続してインキュベーションした。合成したcDNAを20倍に希釈し(最終濃度ほぼ1ng/μl)、そしてPCR反応のテンプレートとして直接使用するために、−20℃フリーザーで保存した。
【0053】
RACE(cDNA端の迅速増幅)反応から適応させた条件を用いた修飾PCR反応を用いて、イヌ遺伝子または遺伝子断片をクローニングした。50μlのPCR反応について、5μlの逆転写cDNA、5μlの10xPCR緩衝液(Clontech、カリフォルニア州)、50xdNTP混合物の1μlのdNTP(各10mM、最終濃度各0.2mM、Clontech、カリフォルニア州)、各20μMの1μlの5’プライマーおよび3’プライマーであった。用いたプライマー配列は:
【0054】
【化3】
【0055】
であった。1μlのClontech Advantage 2 Taqポリメラーゼ(50x)および36μlのPCR水(Clontech、カリフォルニア州)を混合した。混合物を94℃で1分間インキュベーションし、そして94℃15秒間そして次いで72℃4分間が5周期、94℃15秒間そして次いで70℃4分間が5周期、94℃15秒間そして次いで68℃4分間が25周期のタッチダウンPCRプロトコルを実行した。PCR反応の第一周期からクローニングされたのではない遺伝子に関しては、よりストリンジェントでないPCR条件は、94℃15秒間そして次いで68℃4分間が5周期、94℃15秒間そして次いで65℃4分間が5周期、94℃15秒間そして次いで62℃4分間が25周期である。製造者の使用説明書にしたがって、Agilent 2100 Bioanalyzer上、DNA7500チップ(Agilent、カリフォルニア州)を用いて、PCR断片の長さおよび濃度を調べる。
【0056】
DNA調製および配列決定
蛍光に基づくABI377配列決定装置(Perkin Elmer/Applied Biosystems Division、PE/ABD、カリフォルニア州フォスターシティ)およびABI PRISMTM Ready Dye−Deoxy TerminatorキットとTaq FSTMポリメラーゼを以下のプライマー:
【0057】
【化4】
【0058】
とともに用いて、PCR産物を配列決定した。
各ABI周期配列決定反応は、約0.5μgのプラスミドDNAを含有する。98℃1分間の最初の変性、次いで50周期の:98℃30秒間の変性、50℃30秒間のアニーリング、および60℃4分間の伸長を用いて、周期配列決定を行う。温度周期および時間は、Perkin−Elmer 9600サーモサイクラーによって調節される。CentriflexTMゲルろ過カートリッジ(Advanced Genetic Technologies Corp.、メリーランド州ガイザーズバーグ)を用いて、伸長産物を単離する。各反応産物をピペットによってカラム上に装填し、これを次いで1500xg、室温で4分間、スウィンギングバケット遠心分離装置(SorvallモデルRT6000B卓上遠心分離装置)中で遠心分離する。カラムで精製した試料を、真空下で約40分間乾燥させ、そして次いで、5μlのDNA装填溶液(83%脱イオン化ホルムアミド、8.3mM EDTA、および1.6mg/mlブルーデキストラン)に溶解する。次いで試料を90℃に3分間加熱し、そしてABI377配列決定装置による配列解析のため、ゲル試料ウェルに装填する。ABI377ファイルをDNASTAR(DNASTAR、ウィスコンシン州マディソン)プログラムにインポートすることによって、配列解析を行った。一般的に、700bpの配列読み取りが得られた。DNA鎖両方から配列情報を得ることによって、そして配列決定のあいまいさがすべて取り除かれるまで、異なる位置のプライマーを用いて、困難な領域を再配列決定することによって、潜在的な配列エラーを最小限にした。配列番号1は、産物の配列に相当する。
【0059】
ヒト遺伝子オルソログと配列決定データを並列し、そして配列追跡データにしたがって、手動で管理した(curated)。第一周期の配列決定後、完全には終了していないクローニング遺伝子または遺伝子断片に関しては、全プロジェクトが完了するまで、管理配列データに基づいて、さらなる配列決定プライマーを注文した。イヌ・コード配列は、それぞれ、ヒト、ラットおよびマウス相同体に86%、75%および76%同一であるタンパク質をコードする。イヌDNAコード配列は、ヒト、ラットおよびマウス・コードDNA配列に、それぞれ、88%、78.4%、78.8%同一である(比較は、ヒトmRNAに関してはGenbank NM_001966、マウスmRNAに関してはBC016899、そしてラットmRNAに関してはNM_133606に、そしてヒトタンパク質に関してはNP_001957、マウスタンパク質に関してはAAH16899、そしてラットタンパク質に関してはNP_598290に基づいた)。新規発見イヌ配列と他の3つの相同体の並列を図1および2に示す。
【0060】
配列番号1の配列を得るこの方法は例示であり、そして配列番号1を開示することによって、当業者には、配列番号1の全配列を得る多数の方法が提供されることを認識しなければならない。例えば、配列番号1に開示する配列からプローブを生成し、そしてイヌcDNAまたはゲノムライブラリーをスクリーニングして、そしてそれによって配列番号1全体またはそのゲノム同等物を得ることが可能であろう。Sambrookら(監修), Molecular Cloning:A Laboratory Manual, Cold Spring Harbor Laboratory Press:ニューヨーク州コールドスプリングハーバー(1989)。やはり例として、当業者は、配列番号1に開示する配列が与えられれば、それに次いで、配列番号1に代表される全配列を得るためのPCR増幅に適したプライマーを生成可能であることを直ちに認識するであろう(例えば、PCR Technology, H.A. Erlich監修, Stockton Press, ニューヨーク, 1989;PCR Protocols:A Guide to Methods and Applications, M.A. Innis, David H. Gelfand, John J. Sninsky, およびThomas J. White監修, Academic Press, Inc., ニューヨークを参照されたい)。
【0061】
本発明の宿主細胞およびベクター
本発明のポリヌクレオチドを取り込んだプラスミドおよびウイルスDNAベクターなどの自律複製組換え発現構築物もまた提供する。イヌL−PBEコードポリヌクレオチドが、内因性または外因性発現調節DNA配列および転写ターミネーターに、機能可能であるように連結されている発現構築物もまた、提供する。発現調節DNA配列には、プロモーター、エンハンサー、およびオペレーターが含まれ、そして一般的に、発現構築物を利用しようとする発現系に基づいて選択される。プロモーターおよびエンハンサー配列は、一般的に、遺伝子発現を増加させる能力に関して選択され、一方、オペレーター配列は、一般的に、遺伝子発現を制御する能力に関して選択される。本発明の発現構築物は、構築物を所持する宿主細胞の同定を可能にする1以上の選択可能マーカーをコードする配列も含むことも可能である。発現構築物はまた、宿主細胞における相同組換えを容易にし、そして好ましくは促進する配列も含むことも可能である。本発明の構築物はまた、宿主細胞における複製に必要な配列も含む。
【0062】
発現構築物は、好ましくは、コードされるタンパク質の産生のために利用されるが、イヌL−PBEをコードするポリヌクレオチド配列を単純に増幅するために利用することもまた可能である。
【0063】
本発明の別の側面にしたがって、コードされるイヌL−PBEポリペプチドの発現を可能にする方式で、本発明のポリヌクレオチド(または本発明のベクター)を含む、原核細胞および真核細胞を含む、宿主細胞を提供する。本発明のポリヌクレオチドを、環状プラスミドの一部として、または単離タンパク質コード領域を含む直鎖DNAとして、あるいはウイルスベクターとして、宿主細胞に導入することも可能である。当該技術分野で周知であり、そして日常的に実施されている、宿主細胞にDNAを導入する方法には、形質転換、トランスフェクション、エレクトロポレーション、核注入、またはリポソーム、ミセル、ゴースト細胞およびプロトプラストなどのキャリアーとの融合が含まれる。本発明の発現系には、細菌、酵母、真菌、植物、昆虫、無脊椎動物、および哺乳動物細胞系が含まれる。イヌL−PBEポリペプチドの発現に適した宿主細胞には、原核生物、酵母、およびより高次の真核細胞が含まれる。イヌL−PBEポリペプチドの発現に用いられるのに適した原核宿主には、限定されるわけではないが、エシェリキア属(Escherichia)、バチルス属(Bacillus)、およびサルモネラ属(Salmonella)の細菌とともに、シュードモナス属(Pseudomaonas)、ストレプトミセス属(Streptomyces)、およびブドウ球菌属(Staphylococcus)のメンバーが含まれる。
【0064】
本発明の単離核酸分子は、好ましくは、原核細胞での発現用に設計されたベクターよりも、真核細胞での発現用に設計されたベクターにクローニングされる。より高次の真核生物から得られる遺伝子の発現には、ときに真核細胞が好ましく、これはこれらのタンパク質の合成、プロセシング、および分泌のシグナルが、真核細胞では通常認識されるが、原核宿主では、しばしば、認識されないためである(Ausubelら監修, Short Protocols in Molecular Biology, 第2版, John Wiley & Sons, publishers, 16−49ページ, 1992)。イヌL−PBEの場合、N−連結グリコシル化のための2つのコンセンサス配列があり、そしてプロテインキナーゼCリン酸化およびO−グリコシル化のための、翻訳後修飾の他の部位が予測可能である。真核宿主には、限定されるわけではないが、以下の:昆虫細胞、アフリカミドリザル(African green monkey)腎臓細胞(COS細胞)、チャイニーズハムスター卵巣細胞(CHO細胞)、ヒト293細胞、およびネズミ3T3線維芽細胞が含まれることも可能である。
【0065】
原核宿主で使用する発現ベクターは、一般的に、1以上の表現型的選択可能マーカー遺伝子を含む。こうした遺伝子は、一般的に、例えば抗生物質耐性を与えるか、または栄養要求必要条件を供給するタンパク質をコードする。非常に多様なこうしたベクターが商業的供給源から容易に入手可能である。例には、pSPORTベクター、pGEMベクター(Promega)、pPROEXベクター(LTI、メリーランド州ベセスダ)、Bluescriptベクター(Stratagene)、およびpQEベクター(Qiagen)が含まれる。
【0066】
イヌL−PBEはまた、サッカロミセス属(Saccharomyces)、ピキア属(Pichia)、およびクルイベロミセス属(Kluveromyces)を含む属由来の酵母宿主細胞で発現されることも可能である。酵母宿主には、S.セレビシエ(S. cerevisiae)およびP.パストリス(P. pastoris)が含まれる。酵母ベクターはしばしば、2ミクロン酵母プラスミド由来の複製起点配列、自律複製配列(ARS)、プロモーター領域、ポリアデニル化のための配列、転写終結のための配列、および選択可能マーカー遺伝子を含有するであろう。酵母および大腸菌(E. coli)両方で複製可能なベクター(シャトルベクターと称する)もまた、使用可能である。酵母ベクターの上述の特徴に加えて、シャトルベクターはまた、大腸菌における複製および選択のための配列も含むであろう。イヌL−PBEをコードするヌクレオチド配列の5’端に酵母因子リーダー配列をコードするヌクレオチド配列を含むことによって、酵母宿主で発現されるイヌL−PBEポリペプチドの直接分泌を達成することも可能である。
【0067】
イヌL−PBEポリペプチドの発現には、昆虫宿主細胞培養系もまた、使用可能である。別の態様において、バキュロウイルス発現系を用いて、本発明のイヌL−PBEポリペプチドを発現する。昆虫細胞において異種タンパク質を発現するためのバキュロウイルス系の使用に関するさらなる情報は、LuckowおよびSummers, Bio/Technology 6:47(1988)に概説されている。
【0068】
別の態様において、イヌL−PBEポリペプチドを哺乳動物宿主細胞で発現させる。適切な哺乳動物細胞株の限定されない例には、サル腎臓細胞のCOS−7株(Gluzmanら, Cell 23:175(1981))、チャイニーズハムスター卵巣(CHO)細胞、およびヒト293細胞が含まれる。
【0069】
本発明のイヌL−PBEポリペプチドの発現に適した発現ベクターの選択は、もちろん、使用しようとする特定の宿主細胞に応じ、そして一般の当業者の技術の範囲内である。適切な発現ベクターの例には、pcDNA3(Invitrogen)およびpSVL(Pharmacia Biotech)が含まれる。哺乳動物宿主細胞で使用する発現ベクターには、ウイルスゲノム由来の転写調節配列および翻訳調節配列が含まれることも可能である。本発明で使用可能な、一般的に用いられるプロモーター配列および修飾因子配列には、限定されるわけではないが、ヒト・サイトメガロウイルス(CMV)、アデノウイルス2、ポリオーマウイルス、およびサルウイルス40(SV40)由来のものが含まれる。哺乳動物発現ベクターを構築する方法は、例えばOkayamaおよびBerg(Mol. Cell. Biol. 3:280(1983));Cosmanら(Mol. Immunol. 23:935(1986));Cosmanら(Nature 312:768(1984));EP−A−0367566;およびWO 91/18982に開示される。
【0070】
(実施例2)
真核宿主細胞におけるイヌL−PBEの発現
イヌL−PBEタンパク質を産生するため、適切な発現ベクターを用い、標準的遺伝子操作技術を用いて、適切な宿主細胞中で、イヌL−PBEをコードするポリヌクレオチドを発現させる。例えば、実施例1に記載するイヌL−PBEコード配列を商業的発現ベクターpzeoSV2(Invitrogen、カリフォルニア州サンディエゴ)にサブクローニングし、そしてトランスフェクション試薬fuGENE6(Boehringer−Mannheim)および製品挿入物に提供されるトランスフェクションプロトコルを用いて、チャイニーズハムスター卵巣(CHO)細胞にトランスフェクションする。ヒト胚性腎臓HEK293細胞およびCOS細胞を含む、他の真核細胞株もまた、適切である。100μg/mlゼオシン(Stratagene、カリフォルニア州ラホヤ)の存在下での増殖によって、イヌL−PBEを安定に発現している細胞を選択する。所望により、標準的クロマトグラフィー技術を用いて、細胞からイヌL−PBEを単離する。精製を容易にするため、イヌL−PBEアミノ酸配列の部分に対応する1以上の合成ペプチド配列に対して抗血清を作成することも可能であり、そしてこの抗血清を用いて、イヌL−PBEをアフィニティー精製する。イヌL−PBEをタグ配列(例えばポリヒスチジン、赤血球凝集素、FLAG)とインフレームで発現させて、精製を容易にすることもまた可能である。
【0071】
本発明の宿主細胞は、イヌL−PBEと特異的に免疫反応性である抗体の発展のための、免疫原の価値ある供給源である。本発明の宿主細胞はまた、イヌL−PBEポリペプチドの大規模産生の方法においても有用であり、ここで細胞を適切な培地中で増殖させ、そして当該技術分野に知られる精製法、例えば免疫アフィニティークロマトグラフィー、疎水性相互作用クロマトグラフィー、レクチンアフィニティークロマトグラフィー、サイズ排除ろ過、陽イオンまたは陰イオン交換クロマトグラフィー、高圧液体クロマトグラフィー(HPLC)、逆相HPLC等を含む慣用的クロマトグラフィー法によって、細胞から、または細胞を増殖させた培地から、望ましいポリペプチド産物を単離する。さらに他の精製法には、特異的結合パートナーまたは剤に認識される特異的タグ、標識、またはキレート部分を有する融合タンパク質として、望ましいタンパク質を発現し、そして単離するものが含まれる。単離されたタンパク質を切断して、望ましいタンパク質を得ることも可能であるし、または損なわれていない(intact)融合タンパク質としてそのままにしておくことも可能である。融合構成要素の切断は、切断プロセスの結果として、さらなるアミノ酸残基を有する望ましいタンパク質の型を生じることも可能である。
【0072】
イヌL−PBE DNA配列を知ることで、内因性イヌL−PBEの発現を可能にするか、または増加させるように、細胞を修飾することが可能になる。天然存在イヌL−PBEプロモーターと異種プロモーターのすべてまたは一部を交換して、細胞がより高いレベルでイヌL−PBEを発現するようにすることによって、細胞を修飾して(例えば相同組換えによって)、発現増加を提供することも可能である。内因性イヌL−PBEコード配列に機能可能であるように連結される方式で、異種プロモーターを挿入する[例えばPCT国際公報第WO 94/12650号、PCT国際公報第WO 92/20808号、およびPCT国際公報第WO 91/09955号を参照されたい]。異種プロモーターDNAに加えて、増幅可能マーカーDNA(例えばada、dhfr、並びにカルバミルリン酸シンターゼ、アスパラギン酸トランスカルバミラーゼ、およびジヒドロオロターゼをコードする多機能CAD遺伝子)および/またはイントロンDNAを、異種プロモーターDNAとともに挿入可能であることもまた、意図される。イヌL−PBEコード配列に連結した場合、標準的選択法によってマーカーDNAを増幅すると、細胞において、イヌL−PBEコード配列が同時に増幅される。
【0073】
本発明に提供するDNA配列情報はまた、例えば相同組換えまたは「ノックアウト」戦略[Capecchi, Science 244:1288−1292(1989)]を通じて、機能するイヌL−PBEを発現不能な動物またはイヌL−PBEの変異体を発現する動物の発展も可能にする。こうした動物(特にラット、ウサギ、およびマウスなどの実験小動物)は、イヌL−PBEのin vivo活性およびイヌL−PBEの調節因子を研究するモデルとして有用である。
【0074】
本発明によりやはり利用可能になるのは、イヌL−PBEをコードするポリヌクレオチドを認識し、そして該ポリヌクレオチドにハイブリダイズする、アンチセンスポリヌクレオチドである。全長および断片のアンチセンスポリヌクレオチドを提供する。本発明の断片アンチセンス分子には、(i)イヌL−PBEを特異的に認識し、そしてこれにハイブリダイズするもの(イヌL−PBEをコードするDNAと他の既知の分子をコードするDNAの配列比較によって決定されるようなもの)が含まれる。イヌL−PBEがコードするポリヌクレオチドにユニークな配列の同定は、公的に利用可能な配列データベースいずれかの使用を通じて、そして/または商業的に入手可能な配列比較プログラムの使用を通じて、推定可能である。全ゲノムにおける選択された配列のユニークさは、ハイブリダイゼーション解析によってさらに検証可能である。望ましい配列を同定した後、制限消化を通じて単離するか、または当該技術分野に周知の多様なポリメラーゼ連鎖反応技術いずれかを用いて増幅することも可能である。アンチセンスポリヌクレオチドは、イヌL−PBE mRNAを発現する細胞によるイヌL−PBEの発現を制御するのに特に適している。
【0075】
イヌL−PBE発現調節配列またはイヌL−PBE RNAに特異的に結合可能なアンチセンス核酸(好ましくは10〜20塩基対オリゴヌクレオチド)を(例えばウイルスベクター、またはリポソームなどのコロイド性分散系によって)細胞に導入する。アンチセンス核酸は、細胞において、イヌL−PBE標的ヌクレオチド配列に結合し、そして標的配列の転写または翻訳を妨げる。ホスホロチオエートおよびメチルホスホネート・アンチセンスオリゴヌクレオチドは、本発明による療法的使用に明確に意図される。アンチセンスオリゴヌクレオチドを、5’端で、ポリ−L−リジン、トランスフェリン、ポリリジン、またはコレステロール部分によってさらに修飾することも可能である。転写レベルまたは翻訳レベルいずれかでのイヌL−PBE発現の抑制は、異常なイヌL−PBE発現に特徴付けられる疾患の細胞モデルまたは動物モデルを生成するために、あるいは療法様式として、有用である。
【0076】
本明細書に解説されるイヌL−PBE配列は、天然細胞および動物、並びにイヌL−PBEポリヌクレオチドで形質転換またはトランスフェクションされた細胞における、イヌL−PBE発現を調節する新規転写因子の設計を容易にする。例えば、ジンクフィンガードメインを介してDNAに結合する、Cys2−His2ジンクフィンガータンパク質は、異なる標的配列の認識につながる構造変化を受けやすいことが示されてきている。これらの人工的ジンクフィンガータンパク質は、高い親和性定数および低い解離定数の特異的標的部位を認識し、そして遺伝子発現を調節する遺伝子スイッチとして作用することが可能である。本発明の特定のイヌL−PBE標的配列を知ることで、構造に基づくモデリングおよびファージディスプレーライブラリーのスクリーニングの組み合わせなどの、既知の方法を用いて、標的配列に特異的なジンクフィンガータンパク質の操作が容易になる[Segalら(1999)Proc Natl Acad Sci USA 96:2758−2763;Liuら(1997)Proc Natl Acad Sci USA 94:5525−30;GreismanおよびPabo(1997)Science 275:657−61;Chooら(1997)J Mol Biol 273:525−32]。各ジンクフィンガードメインは、通常、3以上の塩基対を認識する。一般的に、18塩基対の認識配列は、既知のゲノムいずれにおいても、該配列をユニークにするのに十分な長さであるため、ジンクフィンガーの6つのタンデムリピートからなるジンクフィンガータンパク質は、特定の配列に対する特異性を確実にすると予期されるであろう[Segalら(1999)Proc Natl Acad Sci USA 96:2758−2763]。イヌL−PBE配列に基づいて設計した、人工的なジンクフィンガーリピートを、活性化ドメインまたは抑制ドメインに融合させて、イヌL−PBE発現を促進するかまたは抑制する[Liuら(1997)Proc Natl Acad Sci USA 94:5525−30]。あるいは、ジンクフィンガーペプチドおよびTATAボックス結合因子(TBP)間のリンカー領域の長さを多様にしながら、ジンクフィンガードメインをTBPに融合させて、転写活性化因子またはリプレッサーいずれかを生成することも可能である[Kimら(1997)Proc Natl Acad Sci USA 94:3616−3620]。こうしたタンパク質、および該タンパク質をコードするポリヌクレオチドは、天然細胞、動物およびヒト;および/またはイヌL−PBEコード配列でトランスフェクションされた細胞両方のin vivoでのイヌL−PBE発現を調節するのに有用性を有する。転写因子を発現する構築物をトランスフェクションすること(遺伝子療法)によって、または該タンパク質を導入することによって、新規転写因子を標的細胞に搬送することも可能である。RNA配列に結合するように、操作ジンクフィンガータンパク質を設計して、アンチセンスまたは触媒RNA法の代替法として療法において使用することもまた可能である[McCollら(1999)Proc Natl Acad Sci USA 96:9521−6;Wuら(1995)Proc Natl Acad Sci USA 92:344−348]。本発明は、(天然の、または形質転換された)細胞において、イヌL−PBE発現を調節するのに有用な、本発明の遺伝子配列に基づく転写因子とともに、カスタマイズしたジンクフィンガータンパク質などの設計法を意図し、細胞の遺伝的相補体にはこれらの配列が含まれる。本発明はまた、本発明のポリヌクレオチドにコードされる単離イヌL−PBEポリペプチドも提供する。
【0077】
本発明のポリペプチド
イヌL−PBEポリペプチドのアミノ酸配列を配列番号2に示す。配列番号2に例示されるような本発明のアミノ酸配列は、いくつかの興味深い特徴を有する。
【0078】
本発明はまた、配列番号2に示すポリペプチドに、少なくとも87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%の同一性および/または相同性を有するポリペプチドも含む。配列番号2のポリペプチドに関するアミノ酸配列「同一性」パーセントは、本明細書において、両配列を並列させ、そして必要であればギャップを導入して最大の配列同一性パーセントを達成した後、そしていかなる保存的置換も配列同一性の一部と見なさずに、イヌL−PBE配列の残基と同一である、候補配列中のアミノ酸残基の割合として定義される。配列番号2のポリペプチドに関するアミノ酸配列「相同性」パーセントは、本明細書において、配列を並列させ、そして必要であればギャップを導入して最大の配列同一性パーセントを達成した後、そしてまたいかなる保存的置換も配列同一性の一部と見なして、イヌL−PBE配列の残基と同一である、候補配列中のアミノ酸残基の割合として定義される。1つの側面において、相同性パーセントは、長さ100アミノ酸中、4つのギャップを導入して並列を最大にすることが可能である際には、2つの配列のうち、より小さい配列の、比較しようとする配列中の同一アミノ酸残基と並列する、アミノ酸残基の割合として計算される[本明細書に援用されるDayhoff, Atlas of Protein Sequence and Structure中, Vol.5 p.124, National Biochemical Reserch Foundation, ワシントンD.C.(1972)]。
【0079】
本発明のポリペプチドは、天然細胞供給源から単離されることも可能であるし、または化学的に合成されることも可能であるが、好ましくは、本発明の宿主細胞を伴う組換え法によって産生される。哺乳動物宿主細胞を使用すると、本発明の組換え発現産物に最適な生物学的活性を与えるために必要である可能性があるような、こうした翻訳後修飾(例えばグリコシル化、一部切除(truncation)、脂質化、およびリン酸化)が提供されると予期される。イヌL−PBEポリペプチドのグリコシル化型および非グリコシル化型が含まれる。
【0080】
上述のような真核宿主および原核宿主における過剰発現は、イヌL−PBEポリペプチドの単離を容易にする。したがって、本発明には、配列番号2に示すような単離イヌL−PBEポリペプチド、並びに変異体、および保存的アミノ酸置換が含まれ、標識化ポリペプチドおよびタグ化ポリペプチドが含まれる。本発明には、「標識された」イヌL−PBEポリペプチドが含まれる。用語「標識された」は、本明細書において、酵素(例えば西洋ワサビ(horseradish)ペルオキシダーゼ、ベータ−グルクロニダーゼ、アルカリホスファターゼ、およびベータ−D−ガラクトシダーゼ)、蛍光標識(例えばフルオレセイン、ルシフェラーゼ)、および放射標識(例えば14C、125I、3H、32P、および35S)を含む適切な検出可能基いずれかを、標識しようとする化合物にコンジュゲート化するか、または共有結合することを指す。タンパク質、ペプチド、および抗体を含む、多様な化合物を標識する技術が周知である。例えば、Morrison, Methods in Enzymology 32b, 103(1974);Syvanenら, J. Biol. Chem. 284, 3762(1973);BoltonおよびHunter, Biochem. J. 133, 529(1973)を参照されたい。用語、標識された、はまた、以下に論じるように、アミノ酸タグに共有結合したポリペプチドも含むことも可能である。
【0081】
さらに、本発明のイヌL−PBEポリペプチドを、間接的に標識することも可能である。これは、部分(a moiety)をポリペプチドに共有的に付加し、そしてそれに続いて、付加された部分への特異的結合を示す標識または標識化合物に、付加された部分をカップリングすることを伴う。間接的な標識の可能性には、ペプチドをビオチン化し、それに続いて、上記標識基の1つにカップリングしたアビジンに結合させることが含まれる。別の例は、ポリヒスチジン・タグを含むイヌL−PBEポリペプチドと、ヒスチジン・タグに特異的な放射標識抗体をインキュベーションすることであろう。抗体がタグにかなりの親和性を有するため、ポリペプチドに対する放射能抗体の結合が、正味の効果である。
【0082】
本発明はまた、イヌL−PBEタンパク質の変異体(または類似体)も含む。1つの例において、1以上のアミノ酸残基がイヌL−PBEアミノ酸配列に付加されている、挿入変異体を提供する。挿入は、タンパク質のいずれかの末端または両方の末端に位置することも可能であるし、あるいはイヌL−PBEタンパク質アミノ酸配列の内部領域内に位置することも可能である。いずれかの末端または両方の末端にさらなる残基を有する挿入変異体には、例えば、融合タンパク質、およびアミノ酸タグまたは標識を含むタンパク質が含まれることも可能である。挿入変異体には、イヌL−PBE酸配列、またはその生物学的活性断片に、1以上のアミノ酸残基が付加されているイヌL−PBEポリペプチドが含まれる。
【0083】
したがって、挿入変異体はまた、イヌL−PBEのアミノ末端および/またはカルボキシ末端が別のポリペプチドに融合している融合タンパク質を含むことも可能である。多様なタグ・ポリペプチドおよびその対応する抗体が当該技術分野に周知である。例には、ポリ−ヒスチジン(ポリ−his)またはポリ−ヒスチジン−グリシン(ポリ−his−gly)タグ;インフルエンザHAタグ・ポリペプチドおよびその抗体12CA5[Fieldら, Mol. Cell. Biol., 8:2159−2165(1988)];c−mycタグ、並びに該タグに対する8F9、3C7、6E10、G4、B7および9E10抗体[Evanら, Molecular and Cellular Biology, 5:3610−3616(1985)];並びに単純疱疹ウイルス糖タンパク質D(gD)タグおよびその抗体[Paborskyら, Protein Engineering, 3(6):547−553(1990)]が含まれる。他のタグ・ポリペプチドには、Flag−ペプチド[Hoppら, BioTechnology, 6:1204−1210(1988)];KT3エピトープペプチド[Martinら, Science, 255:192−194(1992)];アルファ−チューブリン・エピトープペプチド[Skinnerら, J. Biol. Chem., 266:15163−15166(1991)];およびT7遺伝子10タンパク質ペプチドタグ[Lutz−Freyermuthら, Proc. Natl. Acad. Sci. USA, 87:6393−6397(1990)]が含まれる。さらに、イヌL−PBEポリペプチドを、ペルオキシダーゼおよびアルカリホスファターゼなどの酵素タンパク質でタグ化することも可能である。
【0084】
別の側面において、本発明は、イヌL−PBEポリペプチド中の1以上のアミノ酸残基が除去されている欠失変異体を提供する。欠失は、イヌL−PBEポリペプチドの一方の末端または両方の末端で、あるいはイヌL−PBEアミノ酸配列内の1以上の残基の除去を伴って、達成されることも可能である。したがって、欠失変異体には、イヌL−PBEポリペプチドのすべての断片が含まれる。
【0085】
本発明はまた、断片が生物学的活性(例えばリガンド結合活性またはDNA結合活性および/または他の生物学的活性)を維持する、配列番号2に示す配列のポリペプチド断片も示す。配列番号2の少なくとも10から853(この間の1つ1つの整数値を含む)の連続アミノ酸を含む断片が本発明に含まれる。当該技術分野に周知であり、そして日常的に実施される方法いずれかによって、望ましい生物学的特性を有する本発明の断片を調製することも可能である。
【0086】
本発明にはまた、残基が別の類似の特性を持つ別の残基で置換される、保存的アミノ酸置換によって、参照配列とは異なる、前述のポリペプチドの変異体も含まれる。変異体ポリペプチドには、保存的置換が、本発明のポリペプチドをコードするポリヌクレオチドの修飾によって導入されているものが含まれる。アミノ酸は、物理的特性、並びに二次および三次タンパク質構造への寄与にしたがって、分類可能である。保存的置換は、当該技術分野において、1つのアミノ酸に対して、類似の特性を有する別のアミノ酸配列を置換するものと認識される。典型的な保存的置換を、すぐ下の表2に示す(WO 97/09433、10ページ、1997年3月13日公表(PCT/GB96/02197、9/6/96出願)から)。
【0087】
表2
保存的置換I
【0088】
【表2】
【0089】
あるいは、保存的アミノ酸は、すぐ下の表3に示すように、Lehninger[Biochemistry, 第2版;Worth Publishers, Inc., ニューヨーク州ニューヨーク(1975), pp.71−77]に記載されるように分類可能である。
【0090】
表3
保存的置換II
【0091】
【表3】
【0092】
さらに別の代替物として、典型的な保存的置換をすぐ下の表4に示す。
表4
保存的置換III
【0093】
【表4】
【0094】
一般的に、イヌL−PBEポリペプチドは、主に細胞内に見られることが予期され、細胞内物質は、当業者に知られる標準的技術いずれかを用いて、宿主細胞から抽出可能である。例えば、宿主細胞を溶解して、ホモジナイズ、および/または超音波処理、その後の遠心分離によって、細胞質の内容物を遊離させることも可能である。イヌL−PBEポリペプチドは、細胞ホモジネートの遠心分離後、主に上清に見られ、そして以下の方法のいずれかによる限定されない例によって、イヌL−PBEポリペプチドを単離することも可能である。
【0095】
イヌL−PBEポリペプチドを部分的にまたは完全に単離することが好ましい状況において、当業者に周知の標準法を用いて、精製を達成することも可能である。こうした方法には、限定されるわけではないが、電気泳動による分離、その後、電気溶出、多様な種類のクロマトグラフィー(免疫アフィニティー、分子ふるい、および/またはイオン交換)、および/または高圧液体クロマトグラフィーが含まれる。いくつかの場合、完全に精製するため、これらの方法の1より多くを用いることが好ましい可能性もある。
【0096】
多様な技術を用いて、イヌL−PBEポリペプチドの精製を達成することも可能である。ポリペプチドが、そのカルボキシル末端またはアミノ末端いずれかに、ヘキサヒスチジン(イヌL−PBE/ヘキサHis)、またはFLAG(Eastman Kodak Co.、コネチカット州ニューヘブン)またはmyc(Invitrogen、カリフォルニア州カールスバッド)などの他の小ペプチドなどのタグを含有するように合成されている場合、カラムマトリックスがタグに高い親和性を有するか、またはポリペプチドに直接、高い親和性を有する(すなわち、イヌL−PBEを特異的に認識するモノクローナル抗体)アフィニティーカラムに、溶液を通過させることによって、1工程プロセスで本質的に精製可能である。例えば、ポリヒスチジンは、高い親和性および特異性でニッケルに結合し、したがってニッケルの親和性カラム(Qiagen Registered TMニッケルカラムなど)をイヌL−PBE/ポリHisの精製に用いることも可能である(例えば、Ausubelら監修, Current Protocols in Molecular Biology, セクション10.11.8, John Wiley & Sons, ニューヨーク[1993]を参照されたい)。
【0097】
精製を容易にする標識またはタグなしに、イヌL−PBEポリペプチドを調製する場合であっても、本発明のイヌL−PBEを免疫アフィニティークロマトグラフィーで精製することも可能である。これを達成するためには、当該技術分野に周知の手段によって、イヌL−PBEポリペプチドに特異的な抗体を調製しなければならない。本発明のイヌL−PBEポリペプチドまたはエピトープを所持する断片、類似体(analogue)または細胞を、動物、好ましくは非ヒトに、日常的プロトコルを用いて投与することによって、本発明のイヌL−PBEポリペプチドに対して生成された抗体を得ることも可能である。モノクローナル抗体を調製するには、連続細胞株培養によって産生される抗体を提供する、当該技術分野に知られる技術いずれを使用することも可能である。例には、Kohler, G.およびMilstein, C., Nature 256:495−497(1975);Kozborら, Immunology Today 4:72(1983);Coleら, 77−96ページ Monoclonal Antibodies and Cancer Therapy中, Alan R. Liss, Inc.(1985)のものなどの多様な技術が含まれる。
【0098】
付着するタグなしにイヌL−PBEポリペプチドを調製する場合、そして抗体がまったく入手可能でない場合、他の周知の精製法が使用可能である。こうした方法には、限定なしに、イオン交換クロマトグラフィー、分子ふるいクロマトグラフィー、HPLC、ゲル溶出と組み合わせた未変性ゲル電気泳動、および分取用等電点電気泳動(「Isoprime」装置/技術、Hoefer Scientific)が含まれる。いくつかの場合、2以上のこれらの技術を組み合わせて、純度増加を達成することも可能である。代表的な精製スキームを以下に詳述する。
【0099】
天然イヌL−PBEの阻害特性を示し、そしてより高いレベルで発現される変異体、および恒常的に活性であるイヌL−PBEポリペプチドを提供する変異体は、本発明のアッセイにおいて特に有用であり、そしてまた、異常なイヌL−PBE発現活性によって特徴付けられる疾患の細胞モデルまたは動物モデルにおいても有用である。
【0100】
本発明のポリペプチドの定義は、アミノ酸残基の挿入、欠失、または置換以外の修飾を所持するポリペプチドを含むことが意図されることを理解しなければならない。例えば、修飾は、共有性であることも可能であるし、そして例えば、ポリマー、脂質、他の有機部分、および無機部分との化学結合が含まれる。
【0101】
本発明にやはり含まれるのは、イヌL−PBEまたはその断片に特異的な抗体(例えばモノクローナル抗体およびポリクローナル抗体、一本鎖抗体、キメラ抗体、二官能性/二重特異性抗体、ヒト化抗体、ヒト抗体、および本発明のポリペプチドを特異的に認識するCDR配列を含む化合物を含む相補性決定領域(CDR)移植抗体)である。本発明の抗体には、本明細書に完全に援用される、WO93/11236、1993年6月20日公表に記載される方法にしたがって産生され、そして同定されたヒト抗体が含まれる。Fab、Fab’、F(ab’)2、およびFvを含む抗体断片もまた本発明に提供される。用語「特異的」は、本発明の抗体を記載するのに使用する場合、本発明の抗体の可変領域が、もっぱらイヌL−PBEポリペプチドを認識し、そしてこれに結合する(すなわち、イヌL−PBEおよび他の既知のポリペプチド間に局所配列同一性、相同性、または類似性が存在しうるにもかかわらず、結合親和性が測定可能に異なるため、イヌL−PBEポリペプチドをこうした他のポリペプチドから区別可能である)ことを示す。特異的抗体がまた、抗体の可変領域外の配列、そして特に分子の定常領域との相互作用を通じて、他のタンパク質(例えば黄色ブドウ球菌(S. aureus)プロテインA、またはELISA技術における他の抗体)とも相互作用可能であることが理解されるであろう。本発明の抗体の結合特異性を決定するスクリーニングアッセイが当該技術分野において周知であり、そして日常的に実施される。こうしたアッセイの包括的議論に関しては、Harlowら(監修), Antibodies A Laboratory Manual;Cold Spring Harbor Laboratory;ニューヨーク州コールドスプリングハーバー(1988), 第6章を参照されたい。本発明のイヌL−PBEポリペプチドの断片を認識し、そしてこれに結合する抗体もまた含まれるが、該抗体が、何よりもまず、イヌL−PBEポリペプチドに特異的であることが条件である。当該技術分野において周知であり、そして日常的に実施される方法いずれかを用いて、本発明の抗体を産生することも可能である。当該技術分野に知られる方法いずれかによって、非ヒト抗体をヒト化することも可能である。1つの方法では、非ヒトCDRをヒト抗体またはコンセンサス抗体フレームワーク配列に挿入する。次いで、抗体フレームワークにさらなる変化を導入して、アフィニティーまたは免疫原性を調節することも可能である。
【0102】
本発明の抗体は、イヌL−PBEを検出するかまたは定量化する診断目的に有用であるとともに、イヌL−PBEを精製するのにも有用である。本明細書に記載する目的いずれかのための、本発明の抗体を含むキットもまた、含まれる。一般的に、本発明のキットにはまた、抗体が免疫特異的である対照抗原も含まれる。
【0103】
(実施例3)
イヌL−PBEに対する抗体の生成
標準的技術を使用して、イヌL−PBE酵素に対するポリクローナル抗体またはモノクローナル抗体を生成し、そして「ヒト化」変異体を含む、その有用な抗原結合断片または変異体を生成する。こうしたプロトコルは、例えばSambrookら, Molecular Cloning:a Laboratory Manual 第2版, ニューヨーク州コールドスプリングハーバー:Cold Spring Harbor Laboratory(1989);Harlowら(監修), Antibodies A Laboratory Manual;Cold Spring Harbor Laboratory;ニューヨーク州コールドスプリングハーバー(1988);および以下に引用する他の文書に見出すことも可能である。1つの態様において、組換えイヌL−PBEポリペプチド(またはこうしたポリペプチドを含有する細胞もしくは細胞膜)を抗原として用いて、抗体を生成する。別の態様において、イヌL−PBEの免疫原部分に対応するアミノ酸配列(例えば6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、またはそれより多いアミノ酸)を有する1以上のペプチドを抗原として用いる。小さい合成ペプチドを用いて、タンパク質エピトープを模倣するため、親水性、表面配向性で、そして柔軟である配列を選択することが重要である。これは、生理学的溶液中に見られる天然存在タンパク質の大部分は、表面に親水性残基を有し、そして疎水性残基は包埋されているためである。抗体は、一般的に、天然存在タンパク質の表面上のエピトープに結合する。いくつかの既知のエピトープは、高い度合いの移動度を有する。タンパク質のN末端およびC末端は、荷電基、すなわちNH3+およびCOO−を含有するため、一般的に表面配向性である。これらは、末端に位置するため、しばしば、高い度合いの移動度も有する。これらの末端は、3つの特性すべてを所持するため、しばしば、合成候補として選択される。イヌL−PBEの表面残基に対応するペプチド、特に親水性部分が意図される。やはり意図されるのは、イヌL−PBEのアミノ末端およびカルボキシ末端に位置するペプチドである。
【0104】
当業者は、既定のタンパク質配列内の各アミノ酸残基に、親水性、表面アクセス可能性、および柔軟性の値を割り当てるために、アルゴリズムが開発されていることを認識する。各残基に抗原性指数を割り当てて、特定の配列内の残基がどの程度抗原性であるか、指標を与えるため、やはりアルゴリズムが開発されている。HoppおよびWoods, Mol. Immunol, 1983 20(4):p.483−9, HoppおよびWoods, Proc. Natl Acad. Sci USA 1981, 78(6)p.3824−8。しかし、天然抗原に結合するのに有用な抗ペプチド抗体を生成するには、親水性セグメントが広く選択されてきている。しかし抗体とは異なり、T細胞受容体は、切断およびアンフォールディング後のタンパク質抗原の比較的小さいセグメントを認識する。T細胞抗原性部位もまた、予測コンピュータモデルによって対処されてきている。Margalit, H.ら, J. Immunol. 1987 138(7):2213−29ページ。
【0105】
エピトープの予測に有用なコンピュータプログラムが商業的に入手可能である。例えばMacVector(登録商標)(Oxford Molecular、英国オックスフォード)およびProtean(登録商標)(DNAStar Madison, WI53715)。ペプチド抗原を選択し、そして合成したら、抗原をアジュバントと混合するか、またはハプテンに連結して、抗体産生を増加することも可能である。
【0106】
ポリクローナル抗体およびモノクローナル抗体
1つの典型的なプロトコルとして、モノクローナル抗体を生成するため、組換えイヌL−PBEまたはその合成断片を用いて、マウスを免疫する(またはポリクローナル抗体の場合は、ウサギなどのより大きい哺乳動物)。抗原性を増加させるため、製造者の推奨にしたがって、ペプチドをキーホール・リンペット(Keyhole Lympet)ヘモシアニン(Pierce)にコンジュゲート化する。最初の注射では、抗原をフロイントの完全アジュバントで乳化し、そして皮下注射する。2〜3週間間隔で、イヌL−PBE抗原のさらなるアリコットをフロイントの不完全アジュバントで乳化し、そして皮下注射する。最後の追加免疫注射前に、免疫したマウスから血清試料を採取し、そしてウェスタンブロットによってアッセイして、イヌL−PBEと免疫反応する抗体の存在を確認する。免疫した動物由来の血清をポリクローナル抗血清として用いるか、またはイヌL−PBEを認識するポリクローナル抗体を単離するのに用いる。あるいは、モノクローナル抗体を生成するため、マウスを屠殺して、そして脾臓を取り除く。
【0107】
モノクローナル抗体を生成するため、脾臓を10ml血清不含RPMI1640に入れ、そして2mM L−グルタミン、1mMピルビン酸ナトリウム、100単位/mlペニシリン、および100μg/mlストレプトマイシン(RPMI)(Gibco、カナダ)を補った血清不含RPMI1640中で脾臓をすりつぶすことによって、単細胞懸濁物を形成する。細胞懸濁物をろ過し、そして遠心分離によって洗浄し、そして血清不含RPMIに再懸濁する。3匹の未処置(naive)Balb/cマウスから採取した胸腺細胞を同様の方式で調製し、そしてフィーダー層として使用する。融合前に3日間、10%ウシ胎児血清(FBS)(Hyclone Laboratories, Inc.、ユタ州ローガン)を補ったRPMI中、対数期に維持した、NS−1骨髄腫細胞もまた、遠心分離し、そして洗浄する。
【0108】
ハイブリドーマ融合体を産生するため、免疫したマウス由来の脾臓細胞をNS−1細胞と合わせて、そして遠心分離し、そして上清を吸引する。試験管を軽く叩くことによって、細胞ペレットを外し、そして2mlの37℃ PEG1500(75mM Hepes、pH8.0中50%)(Boehringer Mannheim)を攪拌しながらペレットに入れ、その後、血清不含RPMIを添加する。その後、細胞を遠心分離し、そして15%FBS、100μMヒポキサンチンナトリウム、0.4μMアミノプテリン、16μMチミジン(HAT)(Gibco)、25単位/ml IL−6(Boehringer Mannheim)および1.5x106胸腺細胞/mlを含有するRPMIに再懸濁し、そして10のCorning平底96ウェル組織培養プレートに蒔く(Corning、ニューヨーク州コーニング)。
【0109】
融合後、第2日、第4日および第6日、100μlの培地を融合プレートのウェルから取り除き、そして新鮮な培地と交換する。第8日、イヌL−PBEに結合するマウスIgGの存在に関して試験するELISAによって融合体をスクリーニングする。抗イヌL−PBE抗体を産生するモノクローナル培養物が得られるまで、希釈によって、選択された融合体ウェルをさらにクローニングする。
【0110】
ファージディスプレー由来のイヌL−PBEに向けられる抗体
すべて本明細書に援用される、Aujameら, Human Antibodies, 8(4):155−168(1997);Hoogenboom, TIBTECH, 15:62−70(1997);およびRaderら, Curr. Opin. Biotechnol., 8:503−508(1997)に記載されるものなどのファージディスプレー技術によって、イヌL−PBE抗体を生成する。例えば、Fab断片の形の、または連結された一本鎖Fv断片の形の抗体可変領域を、繊維状ファージの小コートタンパク質pIIIのアミノ末端に融合させる。融合タンパク質の発現および成熟ファージコートへのその取り込みによって、表面上に抗体を提示し、そして抗体をコードする遺伝物質を含有する、ファージ粒子が生じる。こうした構築物を含むファージライブラリーを細菌中で発現させ、そして抗原−プローブとして、標識イヌL−PBEまたは固定イヌL−PBEを用いて、イヌL−PBEに特異的なファージ抗体をパニング(スクリーニング)する。
【0111】
トランスジェニックマウス由来のイヌL−PBEに向けられる抗体
BruggemannおよびNeuberger, Immunol. Today, 17(8):391−97(1996)、並びにBruggemannおよびTaussig, Curr. Opin. Biotechnol., 8:455−58(1997)に本質的に記載されるように、トランスジェニックマウスにおいて、イヌL−PBE抗体を生成する。生殖系列配置中、ヒトV遺伝子セグメントを所持し、そしてそのリンパ組織でこれらの導入遺伝子を発現するトランスジェニックマウスを、慣用的免疫プロトコルを用いて、イヌL−PBE組成物で免疫する。慣用的プロトコルを用い、免疫したマウス由来のB細胞を用いてハイブリドーマを生成し、そしてスクリーニングして、抗イヌL−PBE抗体を分泌するハイブリドーマを同定する(例えば上述のとおり)。
【0112】
本発明のアッセイ
本発明は、イヌにおいて、試験剤の影響を検出するかまたは診断する方法を提供する。既定の時間経過に渡って、好ましくは亜慢性用量(subchronic dose)の、1以上の既知のまたは推定上の毒性化合物で、イヌを処置する。次いで、L−PBEを発現する組織試料を得る。こうした処置した生物学的試料から得たイヌL−PBE遺伝子発現のレベルを、未処置の生物学的試料から得た遺伝子発現パターンと比較して、化合物に反応して、イヌL−PBEを上方制御するかまたは下方制御する化合物を同定することも可能である。試料は、試料核酸分子またはタンパク質を含み、そしてL−PBEを発現する体性組織いずれか(脳、腎臓、肝臓など)、培養細胞、生検、または他の組織調製物から得た、いかなる試料であることも可能である。
【0113】
発現レベルを、産生されるメッセンジャーRNAまたはタンパク質のレベルいずれかまたは両方で評価することも可能である。
【0114】
核酸に基づくアッセイ
イヌL−PBEを生じる核酸は、溶液中または固体支持体上にあることも可能である。いくつかの態様において、これらをマイクロアレイ中の単独のアレイ要素として、または他のアレイ要素分子と組み合わせたアレイ要素として、使用することも可能である。こうしたマイクロアレイは、毒性であることが知られるかまたは推定される試験剤と関連して、ハイブリダイゼーションによって、遺伝子発現パターンを検出し、そして性質決定するのに特に有用である。しかし、アレイは、膜に基づくハイブリダイゼーション系、または溶液ハイブリダイゼーションアッセイであっても、または実際、特異的ハイブリダイゼーションを決定可能な方法いずれによっても置き換えることも可能であることが認識される。次いで、こうした遺伝子発現パターンを比較のために用いて、やはり毒性学的反応を誘発する他の化合物を同定することも可能である。
【0115】
核酸に基づく方法は、一般的に、試料からのDNAまたはRNAの単離、およびそれに続く、配列番号1由来の特異的プライマーを用いたハイブリダイゼーションまたはPCR増幅を必要とする。DNAまたはRNAは、当業者に周知のいくつかの方法いずれにしたがって、試料から単離することも可能である。例えば、核酸精製法が、Tijssen, P.(1993)Laboratory Techniques in Biochemistry and Molecular Biology:Hybridization With Nucleic Acid Probes, パートI. Theory and Nucleic Acid Preparation, Elsevier, ニューヨーク州ニューヨークに記載される。1つの好ましい態様において、TRIZOL総RNA単離試薬(Life Technologies, Inc.、メリーランド州ガイザーズバーグ)を用いて総RNAを単離し、そしてオリゴd(T)カラムクロマトグラフィーまたはガラスビーズを用いて、mRNAを単離する。試料核酸分子を増幅する場合、試料核酸分子を増幅し、そして低存在量転写物を含めて、元来の試料の相対存在量を維持することが望ましい。RNAはin vitro、in situ、またはin vivoで増幅することも可能である(Eberwine米国特許第5,514,545号を参照されたい)。
【0116】
増幅法および標識法が、試料中の核酸分子の真の分布を変化させなかったことを確実にするため、試料内に対照を含むこともまた、好適である。この目的のため、相補アレイ化核酸分子へのハイブリダイゼーションに際して検出可能であることがあらかじめ決定された、ある量の対照核酸分子が試料に混入され(spiked)、そして核酸分子の組成物には、対照アレイ化核酸分子と特異的にハイブリダイズする参照核酸分子が含まれる。ハイブリダイゼーションおよびプロセシング後、得られるハイブリダイゼーションシグナルは、試料に添加された対照アレイ化核酸分子の量を正確に反映しなければならない。
【0117】
ハイブリダイゼーション前に、試料核酸分子を断片にすることが望ましい可能性もある。断片化は、二次構造、および試料中の他の試料核酸分子または非相補的核酸分子への交差ハイブリダイゼーションを最小限にすることによって、ハイブリダイゼーションを改善する。断片化は、機械的手段または化学的手段によって実行可能である。
【0118】
標識
試料核酸分子を1以上の標識部分で標識して、ハイブリダイズしたアレイ化/試料核酸分子複合体の検出を可能にすることも可能である。標識部分は、分光的手段、光化学的手段、生化学的手段、生体電子工学的手段、免疫化学的手段、電気的手段、視覚的手段または化学的手段によって検出することも可能な組成物を含むことも可能である。標識部分には、(32)P、(33)Pまたは(35)Sなどの放射性同位体、化学発光化合物、標識結合タンパク質、重金属原子、蛍光マーカーおよび色素などの分光マーカー、磁気標識、連結酵素、質量分析タグ、スピン標識、電子伝達供与体および受容体等が含まれる。好ましい蛍光マーカーには、Cy3およびCy5フルオロフォア(Amersham Pharmacia Biotech、ニュージャージー州ピスカタウェイ)が含まれる。
【0119】
ハイブリダイゼーション
配列番号1およびその断片の核酸分子配列を、多様な目的のため、多様なハイブリダイゼーション技術で用いることも可能である。配列番号1から、ハイブリダイゼーションプローブを設計するか、または得ることも可能である。こうしたプローブを、非常に特異的な領域から、または保存されたモチーフから作成し、そしてL−PBEメッセージ、アレル変異体、または関連配列を定量化するプロトコルで用いることも可能である。本発明のハイブリダイゼーションプローブは、DNAまたはRNAであることも可能であり、そして配列番号1の配列から、または哺乳動物遺伝子のプロモーター、エンハンサー、およびイントロンを含むゲノム配列から得ることも可能である。標識ヌクレオチドの存在下、オリゴ標識、ニックトランスレーション、末端標識、またはPCR増幅を用いて、ハイブリダイゼーションプローブまたはPCRプローブを産生することも可能である。核酸配列を含有するベクターを用いて、RNAポリメラーゼおよび標識核酸分子を添加することによって、in vitroでmRNAプローブを産生することも可能である。Amersham Pharmacia Biotechによって提供されるものなどの商業的に入手可能なキットを用いて、これらの方法を行うことも可能である。ハイブリダイゼーションのストリンジェンシーは、プローブのG+C含量、塩濃度、および温度によって、決定される。特に、塩濃度を減少させるか、またはハイブリダイゼーション温度を上昇させることによって、ストリンジェンシーを増加させることも可能である。膜に基づく、いくつかのハイブリダイゼーションに用いる溶液においては、ホルムアミドなどの有機溶媒を添加すると、反応がより低い温度で起こることが可能になる。1%ドデシル硫酸ナトリウム(SDS)を含む5xSSC、60℃などの緩衝液を用いて、低ストリンジェンシーでハイブリダイゼーションを行って、いくつかのミスマッチを含有するヌクレオチド配列間のハイブリダイゼーション複合体の形成を可能にすることも可能である。続いて、0.1%SDSを含む0.2xSSCなどの緩衝液を、45℃(中程度のストリンジェンシー)または68℃(高ストリンジェンシー)いずれかで用いて、より高いストリンジェンシーで洗浄を行う。高ストリンジェンシーでは、核酸配列がほぼ完全に相補的である箇所でのみ、ハイブリダイゼーション複合体が安定であり続けるであろう。膜に基づく、いくつかのハイブリダイゼーションにおいて、好ましくは35%または最も好ましくは50%のホルムアミドをハイブリダイゼーション溶液に添加し、ハイブリダイゼーションを行う温度を下げることも可能であるし、そしてザルコシルまたはTritonX−100などの他の界面活性剤およびサケ精子DNAなどのブロッキング剤の使用によって、バックグラウンドシグナルを減少させることも可能である。ハイブリダイゼーションのための構成要素および条件の選択は、当業者に周知であり、そしてAusubel(上記)およびSambrookら(1989)Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Press, ニューヨーク州プレーンビューに概説されている。
【0120】
典型的な高ストリンジェンシー・ハイブリダイゼーション条件は、以下のとおりである:50%ホルムアミド、1%SDS、1M NaCl、10%デキストラン硫酸を含むハイブリダイゼーション溶液中、42℃でのハイブリダイゼーション、そして0.1xSSCおよび1%SDSを含む洗浄溶液中、60℃30分間2回の洗浄。Ausubelら(監修), Protocols in Molecular Biology, John Wiley & Sons(1994), pp.6.0.3〜6.4.10に記載されるように、温度および緩衝剤、または塩濃度の変動を通じて、同等のストリンジェンシーの条件を達成可能であることが、当該技術分野において理解される。ハイブリダイゼーション条件の修飾を、経験的に決定することも可能であるし、またはプローブの長さおよびグアノシン/シトシン(GC)塩基対形成の割合に基づいて、正確に計算することも可能である。Sambrookら(監修), Molecular Cloning:A Laboratory Manual, Cold Spring Harbor Laboratory Press:ニューヨーク州コールドスプリングハーバー(1989), pp.9.47〜9.51に記載されるように、ハイブリダイゼーション条件を計算することも可能である。
【0121】
試料に既知の量で添加した特異性対照試料核酸分子に対する、特異性対照核酸分子のハイブリダイゼーションを比較することによって、ハイブリダイゼーション特異性を評価することも可能である。特異性対照アレイ化核酸分子は、対応するアレイ化核酸分子と比較して、1以上の配列ミスマッチを有することも可能である。この方式で、相補的アレイ化核酸分子のみが試料核酸分子にハイブリダイズするのか、またはミスマッチハイブリッド二重鎖が形成されるのかを決定することも可能である。
【0122】
絶対的または示差的ハイブリダイゼーション形式で、ハイブリダイゼーション反応を行うことも可能である。絶対的ハイブリダイゼーション形式では、1つの試料由来の核酸分子を、マイクロアレイ形式で分子にハイブリダイズさせ、そしてハイブリダイゼーション複合体形成後に検出されるシグナルが、試料中の核酸分子レベルに相関する。示差的ハイブリダイゼーション形式では、2つの生物学的試料中の遺伝子セットの示差的発現を解析する。示差的ハイブリダイゼーションでは、両方の生物学的試料から核酸分子を調製し、そして異なる標識部分で標識する。2つの標識核酸分子の混合物を、マイクロアレイに添加する。次いで、2つの異なる標識からの発光を個々に検出可能である条件下で、マイクロアレイを調べる。両方の生物学的試料由来の実質的に同等の数の核酸分子にハイブリダイズする、マイクロアレイ中の分子は、区別可能な組み合わされた蛍光を生じる(ShalonらPCT公報WO95/35505)。好ましい態様において、標識は、Cy3およびCy5フルオロフォアなどの、区別可能な発光スペクトルを持つ蛍光マーカーである。
【0123】
ハイブリダイゼーション後、マイクロアレイを洗浄し、ハイブリダイズしていない核酸分子を取り除き、そしてハイブリダイズ可能なアレイ要素および核酸分子間の複合体形成を検出する。複合体形成を検出する方法は当業者に周知である。好ましい態様において、核酸分子を蛍光標識で標識し、そして複合体形成を示す蛍光のレベルおよびパターンの測定を蛍光顕微鏡、好ましくは共焦点蛍光顕微鏡によって達成する。
【0124】
示差的ハイブリダイゼーション実験において、2以上の異なる生物学的試料由来の核酸分子を、異なる発光波長を持つ2以上の異なる蛍光標識で標識する。特定の波長を検出する異なる光電子増倍管セットを用いて、蛍光シグナルを別個に検出する。2以上の試料中の核酸分子の相対的存在量/発現レベルを得る。
【0125】
典型的には、同様の試験条件下で、1より多いマイクロアレイを用いる場合、マイクロアレイ蛍光強度を規準化して、ハイブリダイゼーション強度の変動を考慮に入れることも可能である。好ましい態様において、各マイクロアレイ上に含有される内部規準化対照由来の強度を用いて、個々のアレイ化試料核酸分子複合体ハイブリダイゼーション強度を規準化する。
【0126】
ポリペプチドに基づくアッセイ
本発明は、イヌL−PBEポリペプチドを検出し、そして定量化するための方法および試薬を提供する。これらの方法には、電気泳動、質量分析、クロマトグラフィー法などの解析的生化学的方法、またはラジオイムノアッセイ(RIA)、酵素連結免疫吸着アッセイ(ELISA)、免疫蛍光アッセイ、ウェスタンブロッティングなどの多様な免疫学的方法、アフィニティー捕捉質量分析、生物学的活性、および以下に記載し、そして本開示を再検討した際に当業者には明らかである他のアッセイが含まれる。
【0127】
イムノアッセイ
本発明はまた、1以上の抗イヌL−PBE抗体試薬を使用して、イヌL−PBEポリペプチドを検出する方法(すなわちイムノアッセイ)も提供する。本明細書において、イムノアッセイは、イヌL−PBEポリペプチドまたはエピトープに特異的に結合する抗体(本明細書に広く定義されるとおりであり、そして具体的には断片、キメラおよび他の結合剤が含まれる)を利用するアッセイである。
【0128】
本発明の実施に適した、いくつかのよく確立された免疫学的結合アッセイ形式が知られる(例えば米国特許第4,366,241号;第4,376,110号;第4,517,288号;および第4,837,168号を参照されたい)。例えばMethods in Cell Biology Volume 37:Antibodies in Cell Biology, Asai監修, Academic Press, Inc. ニューヨーク(1993);Basic and Clinical Immunology 第7版, Stites & Terr監修(1991);HarlowおよびLane、上記[例えば第14章]、並びにAusubelら、上記[例えば第11章]を参照されたい。典型的には、免疫学的結合アッセイ(またはイムノアッセイ)は、解析物に特異的に結合し、そしてしばしば解析物を固相に固定する、「捕捉剤」を利用する。1つの態様において、捕捉剤は、イヌL−PBEポリペプチドまたは下位配列に特異的に結合する部分、例えば抗イヌL−PBE抗体である。
【0129】
通常、アッセイしようとするイヌL−PBE遺伝子産物を直接検出するか、または検出可能標識を用いて、間接的に検出する。アッセイで用いる特定の標識または検出可能基は、アッセイに用いる単数または複数の抗体の特異的結合に有意に干渉しない限り、通常、本発明の決定的な側面ではない。標識は、捕捉剤(例えば抗イヌL−PBE抗体)に共有的に付着していることも可能であるし、またはイヌL−PBEポリペプチドに特異的に結合する別の抗体などの第三の部分に付着していることも可能である。
【0130】
本発明は、イヌL−PBEポリペプチドを検出する競合的および非競合的イムノアッセイの方法および試薬を提供する。非競合的イムノアッセイは、捕捉された解析物(この場合、イヌL−PBE)の量を直接測定するアッセイである。1つのこうしたアッセイは、イヌL−PBEタンパク質上の2つの干渉しないエピトープに反応性であるモノクローナル抗体を利用する、モノクローナル抗体に基づく2部位イムノアッセイである。背景情報に関しては、例えばMaddoxら, 1983, J. Exp. Med., 158:1211を参照されたい。1つの「サンドイッチ」アッセイにおいて、捕捉剤(例えば抗イヌL−PBE抗体)を固体支持体に直接結合させて、固定する。次いで、これらの固定された抗体が、試験試料中に存在するいかなるイヌL−PBEタンパク質も捕捉する。次いで、こうして固定されたイヌL−PBEを、すなわち標識を所持する第二のイヌL−PBE抗体に結合させることによって、標識することも可能である。あるいは、第二のイヌL−PBE抗体は標識を欠いているが、第二の抗体が得られた種の抗体に特異的な、標識された第三の抗体に結合されていることも可能である。あるいは、第二の抗体をビオチンなどの検出可能部分で修飾し、これに酵素標識ストレプトアビジンなどの、第三の標識分子が特異的に結合することも可能である。
【0131】
競合的アッセイにおいて、試料中に存在するイヌL−PBEタンパク質によって、捕捉剤(例えばイヌL−PBE抗体)から置換された(または競合により離れた(competed away))、添加された(外因性)イヌL−PBEの量を測定することによって、試料に存在するイヌL−PBEタンパク質の量を間接的に測定する。ハプテン阻害アッセイは、競合アッセイの別の例である。このアッセイにおいて、イヌL−PBEタンパク質を固体支持体上に固定する。既知の量のイヌL−PBE抗体を試料に添加し、そして次いで、試料を固定イヌL−PBEタンパク質と接触させる。この場合、固定イヌL−PBEタンパク質に結合した抗イヌL−PBE抗体の量は、試料に存在するイヌL−PBEタンパク質の量に反比例する。抗体の固定された割合または溶液中に残存する抗体の割合いずれかを検出することによって、固定された抗体の量を検出することも可能である。この側面において、抗体が標識されている場合、検出は直接であることも可能であるし、または上述のように抗体に特異的に結合する分子に標識が結合している場合、間接的であることも可能である。
【0132】
抗体に基づく他のアッセイ形式
本発明はまた、イムノブロット(ウェスタンブロット)形式を用いることによって、試料中のイヌL−PBEポリペプチドの存在を検出し、そして定量化するための試薬および方法も提供する。別のイムノアッセイは、いわゆる「側方流動クロマトグラフィー」である。側方流動クロマトグラフィーの非競合型では、試料は、例えばキャピラリー作動によって、支持体を横切って移動し、そして可動標識抗体と出会い、該抗体が解析物に結合してコンジュゲートを形成する。次いで、コンジュゲートが支持体を横切り、そして固体された第二の抗体と出会い、この抗体が解析物と結合する。したがって、標識抗体を検出することによって、固定された解析物が検出される。側方流動クロマトグラフィーの競合型では、解析物の標識型が、キャリアーを横切って移動し、そして固定された抗体との結合に関して、非標識解析物と競合する。試料中により多量の解析物があれば、標識解析物による結合はより少なくなり、そしてしたがって、シグナルはより弱くなる。例えばMayら、米国特許第5,622,871号およびRosenstein、米国特許第5,591,645号を参照されたい。
【0133】
アッセイに応じて、抗原、標的抗体、または抗カテプシンS抗体を含む、多様な構成要素を、固体表面または支持体(例えば担体(substrate)、膜、またはろ紙)に結合させることも可能である。多様な固体表面に生体分子を固定する多くの方法が当該技術分野に知られる。例えば、固体表面は、膜(例えばニトロセルロース)、マイクロタイタープレート(例えばPVC、ポリプロピレン、またはポリスチレン)、試験管(ガラスまたはプラスチック)、ディップスティック(例えばガラス、PVC、ポリプロピレン、ポリスチレン、ラテックス等)、微量遠心分離試験管、またはガラスもしくはプラスチックビーズであることも可能である。望ましい構成要素を共有結合させることも可能であるし、または非特異的結合を介して、非共有的に付着させることも可能である。
【0134】
天然および合成両方の、非常に多様な有機ポリマーおよび無機ポリマーを、固体表面の材料として使用することも可能である。実例となるポリマーには、ポリエチレン、ポリプロピレン、ポリ(4−メチルブテン)、ポリスチレン、ポリメタクリレート、ポリ(エチレンテレフタレート)、レーヨン、ナイロン、ポリ(酪酸ビニル)、ポリビニリデンジフルオリド(PVDF)、シリコーン類、ポリホルムアルデヒド、セルロース、酢酸セルロース、ニトロセルロース等が含まれる。使用可能な他の物質には、紙、ガラス、セラミックス、金属、メタロイド、半導体物質、セメント等が含まれる。さらに、タンパク質(例えばゼラチン)、リポ多糖、ケイ酸塩、アガロースおよびポリアクリルアミドなどの、ゲルを形成する物質もまた、使用可能である。いくつかの水性相を形成するポリマー、例えばデキストラン、ポリアルキレングリコールまたは界面活性剤、例えばリン脂質、長鎖(12〜24の炭素原子)アルキルアンモニウム塩等もまた適切である。固体表面が多孔である場合、系の性質に応じて、多様な孔サイズを使用することも可能である。
【0135】
質量分析
分子の質量(mass)は、しばしば、分子の同定因子として使用することも可能である。したがって、質量分析法を用いて、タンパク質解析物を同定することも可能である。質量分析装置は、イオン化解析物が飛行管を通過して、そしてイオン検出装置に検出されるのに必要な時間を測定することによって、質量を測定可能である。タンパク質のための質量分析の1つの方法は、マトリックス支援レーザー脱離イオン化質量分析(「MALDI」)である。MALDIにおいて、レーザーの波長のエネルギーを吸収するエネルギー吸収マトリックス物質と解析物を混合し、そしてプローブ表面に置く。マトリックスをレーザーで打つと、解析物がプローブ表面から脱離し、イオン化され、そしてイオン検出装置に検出される。例えばHillenkampら、米国特許第5,118,937号を参照されたい。
【0136】
タンパク質のための質量分析の他の方法が、HutchensおよびYip、米国特許第5,719,060号に記載される。1つのこうした方法は、アフィニティー捕捉のための増進表面(「SEAC」)と称され、この方法において、解析物に特異的にまたは非特異的に結合する固相アフィニティー試薬、例えば抗体または金属イオンを用いて、解析物を試料中の他の物質から分離する。次いで、捕捉された解析物を、例えばレーザーエネルギーによって固相から脱離し、イオン化し、そして検出装置によって検出する。
【0137】
本発明のさらなる特徴および変型は、詳細な説明を含めた本出願全体から、当業者に明らかであろうし、そしてこうした特徴はすべて、本発明の側面と意図される。同様に、本明細書に記載する本発明の特徴を再度組み合わせて、特徴の組み合わせが、本発明の側面または態様として、具体的に上述されているかいないかにかかわりなく、やはり本発明の側面であることも意図されるさらなる態様にすることも可能である。また、本発明に決定的であると本明細書に記載した限定のみが、限定として見なされるべきであり;決定的であると本明細書に記載されていない、限定されない本発明の変型は、本発明の側面であると意図される。
【0138】
前述の説明および実施例に特に記載されるものとは別の方式で、本発明を実施可能であることが明らかであろう。
上記の解説を考慮すると、本発明の多くの修飾および変型が可能であり、そしてしたがって、これらは本発明の側面内にある。
【0139】
本明細書に引用したすべての刊行物のすべての開示は、本明細書に援用される。
【図面の簡単な説明】
【0140】
【図1A】イヌ(配列番号1)、ヒト(配列番号11)、マウス(配列番号13)およびラット(配列番号15)L−PBEポリペプチド配列の並列。
【図1B】イヌ(配列番号1)、ヒト(配列番号11)、マウス(配列番号13)およびラット(配列番号15)L−PBEポリペプチド配列の並列。
【図2A】イヌ(配列番号2)、ヒト(配列番号12)、マウス(配列番号14)およびラット(配列番号16)L−PBEポリヌクレオチド配列の並列。
【図2B】イヌ(配列番号2)、ヒト(配列番号12)、マウス(配列番号14)およびラット(配列番号16)L−PBEポリヌクレオチド配列の並列。
【図2C】イヌ(配列番号2)、ヒト(配列番号12)、マウス(配列番号14)およびラット(配列番号16)L−PBEポリヌクレオチド配列の並列。
【図2D】イヌ(配列番号2)、ヒト(配列番号12)、マウス(配列番号14)およびラット(配列番号16)L−PBEポリヌクレオチド配列の並列。
【配列表】
【特許請求の範囲】
【請求項1】
アミノ酸配列が配列番号2を含む単離イヌ(canine)L−PBEポリペプチド、または前記ポリペプチドに特異的なエピトープを含む該ポリペプチドの断片。
【請求項2】
請求項1のイヌL−PBEポリペプチドに特異的な抗体。
【請求項3】
配列番号1、または配列番号1の少なくとも12の連続ヌクレオチドを含むその断片、もしくはその相補的な非コード鎖を含む、単離ポリヌクレオチドであって、前記断片が、配列番号11、配列番号13、配列番号15に示す配列のいかなる部分とも、そしてその相補鎖とも、少なくとも1ヌクレオチド異なるヌクレオチド配列を含むという規定付きの前記ポリヌクレオチド。
【請求項4】
核酸が一本鎖である、請求項3の核酸。
【請求項5】
配列番号1の相補体の少なくとも12の連続ヌクレオチドを含む、請求項3の単離核酸。
【請求項6】
固体支持体に付着した核酸分子のアレイであって、請求項3のポリヌクレオチドを含む前記アレイ。
【請求項7】
配列番号2に少なくとも95%相同であるアミノ酸配列を含むポリペプチドをコードし、そしてL−PBE活性を有するポリペプチドをコードする、ヌクレオチド配列を含む、単離ポリヌクレオチド。
【請求項8】
配列番号2のポリペプチドをコードする、請求項7の単離ポリヌクレオチド。
【請求項9】
イヌ由来の試料内に存在するイヌL−PBEポリヌクレオチドの量を決定する方法であって:
配列番号1若しくはその断片、又はそれらの相補体を含む核酸分子と試料を、1以上の特異的ハイブリダイゼーション複合体形成のための条件下で接触させる、ここで該断片は、配列番号1の少なくとも12の連続ヌクレオチドを含むポリヌクレオチドである
ことを含む、前記方法。
【請求項10】
イヌにおいて、試験剤に対する代謝反応を測定する方法であって:
a)試験剤で処置したイヌ由来の核酸を含有する試料を提供し;そして
b)前記試料中の配列番号1若しくはその断片、又はそれらの相補体を含むポリヌクレオチドの量を決定する、ここで未処置のイヌ由来のポリヌクレオチドの量と比較した際の、処置したイヌ由来のポリヌクレオチドの量の変化が、試験剤に対する代謝反応の指標となる
ことを含む、前記方法。
【請求項11】
決定がハイブリダイゼーションを介して達成される、請求項10の方法。
【請求項12】
ハイブリダイゼーションが:
a)配列番号1若しくはその断片、又はそれらの相補体を含む核酸分子と試料を、1以上の特異的ハイブリダイゼーション複合体形成のための条件下で接触させ、ここで該断片は、配列番号1の少なくとも12の連続ヌクレオチドを含むポリヌクレオチドである;そして
b)ハイブリダイゼーション複合体を検出する、ここで未処置のイヌ由来の核酸分子から形成されるハイブリダイゼーション複合体の量と比較した際の、処置したイヌ由来の核酸分子から形成されるハイブリダイゼーション複合体の量の変化が、試験剤に対する代謝反応の指標となる
ことによって達成される、請求項11の方法。
【請求項13】
試料内に存在するイヌL−PBEポリペプチドの量を決定する方法であって:
イヌL−PBEポリペプチドとイヌL−PBEポリペプチドに特異的な抗体を、抗体がイヌL−PBEポリペプチドに結合する条件下で接触させる
ことを含む、前記方法。
【請求項14】
イヌにおいて、試験剤に対する代謝反応を測定する方法であって:
a)試験剤で処置したイヌ由来の試料を提供し;そして
b)前記試料中の配列番号2を含むポリペプチド、または前記ポリペプチドに特異的なエピトープを含む該ポリペプチドの断片の量を決定する、ここで未処置のイヌ由来のポリペプチドの量と比較した際の、処置したイヌ由来のポリペプチドの量の変化が、試験剤に対する代謝反応の指標となる
ことを含む、前記方法。
【請求項15】
決定が、イヌL−PBEポリペプチドとイヌL−PBEポリペプチドに特異的な抗体を、抗体がイヌL−PBEポリペプチドに結合する条件下で接触させることによって達成される、請求項14の方法。
【請求項1】
アミノ酸配列が配列番号2を含む単離イヌ(canine)L−PBEポリペプチド、または前記ポリペプチドに特異的なエピトープを含む該ポリペプチドの断片。
【請求項2】
請求項1のイヌL−PBEポリペプチドに特異的な抗体。
【請求項3】
配列番号1、または配列番号1の少なくとも12の連続ヌクレオチドを含むその断片、もしくはその相補的な非コード鎖を含む、単離ポリヌクレオチドであって、前記断片が、配列番号11、配列番号13、配列番号15に示す配列のいかなる部分とも、そしてその相補鎖とも、少なくとも1ヌクレオチド異なるヌクレオチド配列を含むという規定付きの前記ポリヌクレオチド。
【請求項4】
核酸が一本鎖である、請求項3の核酸。
【請求項5】
配列番号1の相補体の少なくとも12の連続ヌクレオチドを含む、請求項3の単離核酸。
【請求項6】
固体支持体に付着した核酸分子のアレイであって、請求項3のポリヌクレオチドを含む前記アレイ。
【請求項7】
配列番号2に少なくとも95%相同であるアミノ酸配列を含むポリペプチドをコードし、そしてL−PBE活性を有するポリペプチドをコードする、ヌクレオチド配列を含む、単離ポリヌクレオチド。
【請求項8】
配列番号2のポリペプチドをコードする、請求項7の単離ポリヌクレオチド。
【請求項9】
イヌ由来の試料内に存在するイヌL−PBEポリヌクレオチドの量を決定する方法であって:
配列番号1若しくはその断片、又はそれらの相補体を含む核酸分子と試料を、1以上の特異的ハイブリダイゼーション複合体形成のための条件下で接触させる、ここで該断片は、配列番号1の少なくとも12の連続ヌクレオチドを含むポリヌクレオチドである
ことを含む、前記方法。
【請求項10】
イヌにおいて、試験剤に対する代謝反応を測定する方法であって:
a)試験剤で処置したイヌ由来の核酸を含有する試料を提供し;そして
b)前記試料中の配列番号1若しくはその断片、又はそれらの相補体を含むポリヌクレオチドの量を決定する、ここで未処置のイヌ由来のポリヌクレオチドの量と比較した際の、処置したイヌ由来のポリヌクレオチドの量の変化が、試験剤に対する代謝反応の指標となる
ことを含む、前記方法。
【請求項11】
決定がハイブリダイゼーションを介して達成される、請求項10の方法。
【請求項12】
ハイブリダイゼーションが:
a)配列番号1若しくはその断片、又はそれらの相補体を含む核酸分子と試料を、1以上の特異的ハイブリダイゼーション複合体形成のための条件下で接触させ、ここで該断片は、配列番号1の少なくとも12の連続ヌクレオチドを含むポリヌクレオチドである;そして
b)ハイブリダイゼーション複合体を検出する、ここで未処置のイヌ由来の核酸分子から形成されるハイブリダイゼーション複合体の量と比較した際の、処置したイヌ由来の核酸分子から形成されるハイブリダイゼーション複合体の量の変化が、試験剤に対する代謝反応の指標となる
ことによって達成される、請求項11の方法。
【請求項13】
試料内に存在するイヌL−PBEポリペプチドの量を決定する方法であって:
イヌL−PBEポリペプチドとイヌL−PBEポリペプチドに特異的な抗体を、抗体がイヌL−PBEポリペプチドに結合する条件下で接触させる
ことを含む、前記方法。
【請求項14】
イヌにおいて、試験剤に対する代謝反応を測定する方法であって:
a)試験剤で処置したイヌ由来の試料を提供し;そして
b)前記試料中の配列番号2を含むポリペプチド、または前記ポリペプチドに特異的なエピトープを含む該ポリペプチドの断片の量を決定する、ここで未処置のイヌ由来のポリペプチドの量と比較した際の、処置したイヌ由来のポリペプチドの量の変化が、試験剤に対する代謝反応の指標となる
ことを含む、前記方法。
【請求項15】
決定が、イヌL−PBEポリペプチドとイヌL−PBEポリペプチドに特異的な抗体を、抗体がイヌL−PBEポリペプチドに結合する条件下で接触させることによって達成される、請求項14の方法。
【図1A】

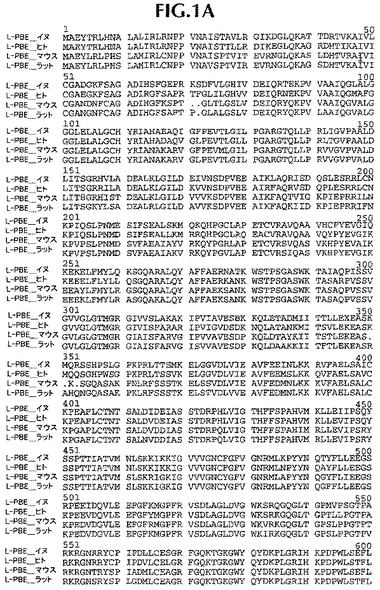
【図1B】

【図2A】
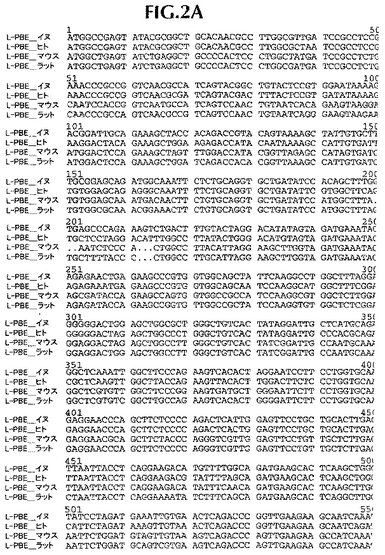
【図2B】
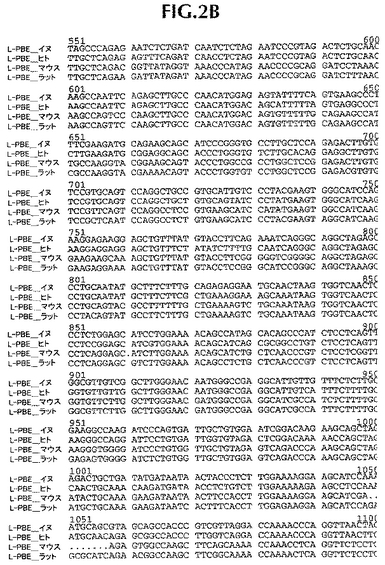
【図2C】

【図2D】
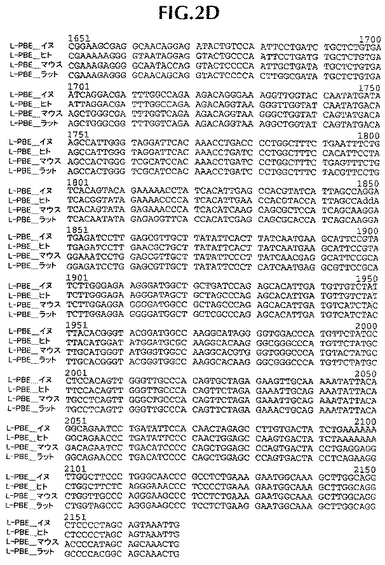
【図1B】
【図2A】
【図2B】
【図2C】
【図2D】
【公表番号】特表2006−512076(P2006−512076A)
【公表日】平成18年4月13日(2006.4.13)
【国際特許分類】
【出願番号】特願2004−563518(P2004−563518)
【出願日】平成15年12月19日(2003.12.19)
【国際出願番号】PCT/IB2003/006280
【国際公開番号】WO2004/058938
【国際公開日】平成16年7月15日(2004.7.15)
【出願人】(504396379)ファルマシア・アンド・アップジョン・カンパニー・エルエルシー (130)
【Fターム(参考)】
【公表日】平成18年4月13日(2006.4.13)
【国際特許分類】
【出願日】平成15年12月19日(2003.12.19)
【国際出願番号】PCT/IB2003/006280
【国際公開番号】WO2004/058938
【国際公開日】平成16年7月15日(2004.7.15)
【出願人】(504396379)ファルマシア・アンド・アップジョン・カンパニー・エルエルシー (130)
【Fターム(参考)】
[ Back to top ]