
ウイルス感染症の予防・治療
【課題】急性及び慢性のウイルス感染症、あるいは癌の新規免疫療法およびそのための薬剤の提供。
【解決手段】生体にとって望ましくない異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメインと、ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部とを含むキメラレセプターを発現する細胞傷害性T細胞を含有してなる、該異常細胞の除去剤、急性及び慢性ウイルス感染症、癌などの予防・治療剤。上記細胞傷害性T細胞を投与することを含む異常細胞の除去方法。
【解決手段】生体にとって望ましくない異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメインと、ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部とを含むキメラレセプターを発現する細胞傷害性T細胞を含有してなる、該異常細胞の除去剤、急性及び慢性ウイルス感染症、癌などの予防・治療剤。上記細胞傷害性T細胞を投与することを含む異常細胞の除去方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、キメラレセプターを発現する細胞傷害性T細胞を含む新規な異常細胞の除去剤、このような細胞傷害性T細胞を投与することを含む異常細胞の除去方法に関する。より具体的には、本発明は、ウイルス感染細胞や癌細胞等の異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメインと、ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部とを含むキメラレセプターを発現する細胞傷害性T細胞を利用する、急性及び慢性のウイルス感染症や癌の予防・治療に関する。
【背景技術】
【0002】
ウイルス感染に対する宿主防御は、2つの異なる免疫応答のセット(自然免疫及び獲得免疫)によって媒介される。自然免疫系の最初の防御ラインである樹状細胞(DC)は、パターン認識レセプターであるToll様レセプター(TLR)を使用し、このレセプターは、種々の病原体成分(タンパク質、DNA、RNA及び多糖を含む)と相互作用する。DCはサイトカインを迅速に分泌して、他の自然免疫細胞(例えば、ナチュラルキラー(NK)細胞)及び獲得免疫細胞(T細胞及びB細胞)の応答を誘発する。両方の免疫系の成分が、病原体からの宿主の有効な保護のために必要である。自然免疫の欠如は病原体の迅速な複製を導き、感染後数日以内に死をもたらす。対照的に、獲得免疫系の非存在下では病原体は獲得免疫監視から逃れ、ヒト免疫不全患者において報告されているように、慢性及び再発の感染に対して宿主を感受性にする。
【0003】
持続感染を引き起こすウイルスの多くは、細胞傷害性T細胞による抗原認識を回避するために宿主細胞でのMHCクラスI抗原の発現を抑制する。しかし、NK細胞はMHCクラスI抗原を認識する抑制レセプターを用いて該抗原を発現している細胞を自己とみなし、非発現細胞を非自己と認識して傷害するので、そのようなウイルス感染細胞はNK細胞の抑制レセプターに認識されなくなり、該細胞により傷害される。NK細胞はまた、細胞外領域において抑制レセプターと非常に高い相同性を示すものの、該レセプターと相反する機能を有する活性化レセプターを有している。活性化レセプターは、ウイルス感染細胞上で特異的に発現するリガンドを認識して非自己とみなし、NK細胞のキラー活性を促進する。しかしながら、多くの活性化レセプターのリガンドは不明であり、その機能は明らかになっていないものが多い。
【0004】
サイトメガロウイルス(CMV)は、免疫無防備宿主(例えば臓器移植患者)における主要な日和見感染病原体である。マウスサイトメガロウイルス(MCMV)感染は、宿主−病原体相互作用及びその免疫を研究するための有用なモデルを提供する(非特許文献1、2)。最近、MCMV感染細胞の認識に関わるNK細胞レセプター及びそのリガンドのペアとして、MCMV耐性マウス株において見出されたLy49H活性化レセプター及びMCMVゲノムにコードされるMHCクラスI様ホールドタンパク質m157が同定された(非特許文献3、4)。二次構造をもとにした立体構造上の相同性検索によると、m157はCD1やH−2M3等の非古典的MHCクラスI分子と相同性を示す。
【0005】
NK細胞の活性化は活性化レセプターの状態のみによって決定されるのではなく、抑制レセプターのシグナルと活性化レセプターのシグナルとの間のバランスによって決定され、このことは、「missing-self」仮説によって支持される(非特許文献5)。興味深いことに、m157は、MCMV感受性マウス株129/Jにおいて、抑制レセプターであるLy49I129に結合することが示されている(非特許文献3)。一方、MCMV耐性のC57BL/6マウスのLy49IB6は宿主クラスI分子を認識するにもかかわらず、m157を認識しない。
【0006】
Ly49Iは、マウスの原型的な抑制レセプターであるLy49Aと同様に、その細胞質ドメイン中に免疫レセプターチロシンベースの阻害モチーフ(immuno-receptor tyrosine-based inhibitory motifs(ITIM))を含む。対照的に、Ly49Hは、その細胞質ドメイン中にシグナル伝達モチーフを有さない。その代わりに、Ly49Hは、その表面発現及びLy49H膜貫通ドメイン中の正に荷電した残基を介して、免疫レセプターチロシンベースの活性化モチーフ(immuno-receptor tyrosine-based activation motifs(ITAM))を含むシグナル伝達アダプター分子DAP12/KARAP(非特許文献6、7)と会合する。T細胞レセプター(TCR)複合体中のCD3ζと同様に、DAP12は、リガンド認識に際し、Syk又はZap70のようなSykキナーゼファミリーのメンバーをそのITAMへとリクルートする。
【0007】
リガンド同定のために、マウスT細胞株において発現させた、オーファンNK細胞レセプターの細胞外ドメインとCD3ζの細胞質ドメインとのキメラレセプターのレパートリーが以前に使用されている(非特許文献8、9)。この系によれば、レセプターの本来の性質(抑制であるか活性化であるか)にかかわらず、リガンドを認識するとNFAT結合プロモーターの制御下でlacZ発現を誘導するので、リガンド発現細胞を検出することができる(非特許文献9)。
【非特許文献1】Tortorella, D., B.E. Gewurz, M.H. Furman, D.J. Schust, and H.L. Ploegh. 2000. Viral subversion of the immune system. Annu Rev Immunol 18:861-926.
【非特許文献2】Webb, J.R., S.H. Lee, and S.M. Vidal. 2002. Genetic control of innate immune responses against cytomegalovirus: MCMV meets its match. Genes Immun 3:250-262.
【非特許文献3】Arase, H., and L.L. Lanier. 2004. Specific recognition of virus-infected cells by paired NK receptors. Rev Med Virol 14:83-93.
【非特許文献4】Smith, H.R., J.W. Heusel, I.K. Mehta, S. Kim, B.G. Dorner, O.V. Naidenko, K. Iizuka, H. Furukawa, D.L. Beckman, J.T. Pingel, A.A. Scalzo, D.H. Fremont, and W.M. Yokoyama. 2002. Recognition of a virus-encoded ligand by a natural killer cell activation receptor. Proc Natl Acad Sci U S A 99:8826-8831.
【非特許文献5】Lanier, L.L. 2005. NK cell recognition. Annu Rev Immunol 23:225-274.
【非特許文献6】Bakker, A.B., R.M. Hoek, A. Cerwenka, B. Blom, L. Lucian, T. McNeil, R. Murray, L.H. Phillips, J.D. Sedgwick, and L.L. Lanier. 2000. DAP12-deficient mice fail to develop autoimmunity due to impaired antigen priming. Immunity 13:345-353.
【非特許文献7】Tomasello, E., P.-O. Desmoulins, K. Chemin, S. Guia, H. Cremer, J. Ortaldo, P. Love, D. Kaiserlian, and E. Vivier. 2000. Combined natural killer cell and dendritic cell functional deficiency in KARAP/DAP12 loss-of-function mutant mice. Immunity 13:355-364.
【非特許文献8】Furukawa, H., K. Iizuka, J. Poursine-Laurent, N. Shastri, and W.M. Yokoyama. 2002. A ligand for the murine NK activation receptor Ly-49D: activation of tolerized NK cells from beta(2)-microglobulin-deficient mice. Journal of Immunology 169:126-136.
【非特許文献9】Iizuka, K., O.V. Naidenko, B.F. Plougastel, D.H. Fremont, and W.M. Yokoyama. 2003. Genetically linked C-type lectin-related ligands for the NKRP1 family of natural killer cell receptors. Nat Immunol 4:801-807.
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明の目的は、急性期のみならず慢性のウイルス感染症の予防・治療にも有用な新規免疫療法およびそのための薬剤を提供することである。また、本発明の別の目的は、癌の新規免疫療法およびそのための薬剤を提供することである。
【課題を解決するための手段】
【0009】
本発明者は、上記の目的を達成するために鋭意研究を重ねた結果、ウイルス感染細胞や癌細胞等の異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメインと、ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部とを含むキメラレセプターを発現させた細胞傷害性T細胞を用いることにより、標的異常細胞上の該リガンドを認識して当該細胞を殺傷し得ることを見出した。すなわち、本発明者は、Ly49Hレセプターの細胞外ドメインとCD3ζの細胞質ドメインとを含むキメラレセプターをコードする核酸を細胞傷害性T細胞中に導入し、これをMCMV感受性マウス株に投与したところ、該マウスが急性相のMCMV感染による死亡から逃れ、さらに獲得免疫による長期保護が確立されたことを見出し、本発明を完成するに至った。
【0010】
すなわち、本発明は以下の通りである:
[1] 生体にとって望ましくない異常細胞の除去剤であって、
(a)該異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメインと、
(b)ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部と、
を含むキメラレセプターを発現する細胞傷害性T細胞を含有してなる剤。
[2] NK細胞レセプターが活性化レセプターである、上記[1]に記載の剤。
[3] 異常細胞がウイルス感染細胞である、上記[1]又は[2]に記載の剤。
[4] ウイルスがサイトメガロウイルスである、上記[3]に記載の剤。
[5] サイトメガロウイルスがヒトサイトメガロウイルスである、上記[4]に記載の剤。
[6] NK細胞レセプターがヒト由来である、上記[1]〜[5]のいずれかに記載の剤。
[7] NK細胞レセプターがNKRP1A、NKRP1C、NKRP1F、KIR2DS、KIR3DS、NKG2C、NKG2DおよびNKG2Eからなる群より選択される、上記[6]に記載の剤。
[8] サイトメガロウイルスがマウスサイトメガロウイルスである、上記[4]に記載の剤。
[9] NK細胞レセプターがマウス由来である、上記[1]〜[5]及び[8]のいずれかに記載の剤。
[10] NK細胞レセプターがLy49H、Ly49D、NKRP1A、NKRP1C、NKRP1F、NKG2C、NKG2DおよびNKG2Eからなる群より選択される、上記[9]に記載の剤。
[11] NK細胞レセプターがLy49Hである、上記[10]に記載の剤。
[12] シグナル伝達アダプター分子がCD3ζ、DAP12/KARAPおよびFcRγからなる群より選択される、上記[1]〜[11]のいずれかに記載の剤。
[13] シグナル伝達アダプター分子がCD3ζである、上記[12]に記載の剤。
[14] キメラレセプターの膜貫通ドメインがNK細胞活性化レセプター由来でない、上記[1]〜[13]のいずれかに記載の剤。
[15] 急性ウイルス感染症の予防・治療用である、上記[1]〜[14]のいずれかに記載の剤。
[16] 慢性ウイルス感染症の予防・治療用である、上記[1]〜[14]のいずれかに記載の剤。
[17] 生体にとって望ましくない異常細胞の除去方法であって、
(a)該異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメインと、
(b)ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部と、
を含むキメラレセプターを発現する細胞傷害性T細胞の有効量を、該異常細胞を有する生体に投与することを含む方法。
[18]
(a)ウイルス感染ヒト細胞で特異的に発現するリガンドを認識するヒトNK細胞レセプターの細胞外ドメインと、
(b)ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部と、
を含むキメラレセプタータンパク質。
[19] ウイルスがヒトサイトメガロウイルスである、上記[18]に記載のタンパク質。
[20] NK細胞レセプターが活性化レセプターである、上記[18]又は[19]に記載のタンパク質。
[21] NK細胞レセプターがNKRP1A、NKRP1C、NKRP1F、KIR2DS、KIR3DS、NKG2C、NKG2DおよびNKG2Eからなる群より選択される、上記[18]〜[20]のいずれかに記載のタンパク質。
[22] シグナル伝達アダプター分子がCD3ζ、DAP12/KARAPおよびFcRγからなる群より選択される、上記[18]〜[21]のいずれかに記載のタンパク質。
[23] シグナル伝達アダプター分子がCD3ζである、上記[22]に記載のタンパク質。
[24] 前記キメラレセプターの膜貫通ドメインがNK細胞活性化レセプター由来でない、上記[18]〜[23]のいずれかに記載のタンパク質。
[25] 上記[18]〜[24]のいずれかに記載のタンパク質をコードする核酸。
[26] 上記[25]記載の核酸を含む、ヒト細胞傷害性T細胞で機能し得る発現ベクター。
[27] 上記[26]記載の発現ベクターでトランスフェクトされたヒト細胞傷害性T細胞。
【発明の効果】
【0011】
本発明のキメラレセプターを発現する細胞傷害性T細胞は、NK細胞レセプターを利用した迅速な免疫応答により急性ウイルス感染症に対する予防・治療効果を発揮するだけでなく、T細胞による免疫監視により慢性のウイルス感染症に対しても予防・治療効果を奏する。
【発明を実施するための最良の形態】
【0012】
以下に、本発明を詳細に説明する。
本発明は、生体にとって望ましくない異常細胞の除去剤(以下、単に「本発明の異常細胞除去剤」ともいう)を提供する。「生体にとって望ましくない異常細胞」は、任意の疾患の病原となり得る細胞であって、正常細胞が全くあるいはほとんど発現していない分子を細胞表面に特異的に、あるいは高発現する細胞であれば、特に制限はないが、好ましくは、ウイルス感染細胞(例えば、CMV、単純ヘルペスウイルス、EBウイルス、ヒト免疫不全ウイルス、インフルエンザウイルス、SARSウイルス等に感染した細胞)、癌細胞など、より好ましくはウイルス感染細胞、いっそう好ましくはCMV感染細胞、特に好ましくはヒトサイトメガロウイルス(HCMV)もしくはMCMV感染細胞である。本発明の異常細胞除去剤は、細胞傷害性T細胞が異常細胞を傷害して除去することにより、該異常細胞を病原とする疾患の予防・治療に有用である(本明細書中で「予防・治療」とは、「予防及び/又は治療」を意味する)。したがって、異常細胞がウイルス感染細胞の場合、本発明の異常細胞除去剤はウイルス感染症の予防・治療剤として用いることができ、異常細胞が癌細胞の場合、本発明の異常細胞除去剤は癌の予防・治療剤として用いることができる。
【0013】
本発明の異常細胞除去剤は、(a)異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメインと、(b)ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部とを含むキメラレセプターを発現する細胞傷害性T細胞を含有する。
【0014】
NK細胞は、自然免疫系(innate immune system)の細胞傷害性細胞であり、腫瘍細胞やウイルス感染細胞などを殺傷する(MHC非拘束)。NK細胞は、構造的に類似した2つのタイプのレセプター:「NK活性化レセプター(本明細書中で、単に「活性化レセプター」ともいう)」及び「NK抑制レセプター(本明細書中で、単に「抑制レセプター」ともいう)」をその表面に発現し、異常細胞を自己の細胞から識別している。活性化レセプターは、膜貫通領域に存在する正荷電のアミノ酸残基を介して、膜貫通領域に負荷電のアミノ酸残基を有するシグナル伝達アダプター分子と会合する。シグナル伝達アダプター分子はITAM配列[Y-x-x-L-xもしくはY-x-x-x-L](ここで、xは任意のアミノ酸である)を有し、SykやZap70などのキナーゼ群をリクルートすることにより、活性化シグナルを伝達する。一方、抑制レセプターは細胞質ドメインにITIM配列[(I/V/L/S) -x-Y-x-x- (L/V)](ここで、xは任意のアミノ酸である)を有し、SHP−1などのホスファターゼ群をリクルートすることにより、活性化レセプターからのシグナルを遮断する。NK細胞の細胞傷害活性は、これらのレセプターからのシグナルのバランスによって制御される。
【0015】
NK細胞で発現するNK細胞レセプターの遺伝子は、主としてLRC(Leukocyte Receptor Complex;ヒト19番染色体、マウス7番染色体上)及びNKC(NK receptor Complex;ヒト12番染色体、マウス6番染色体上)の2つの遺伝子座にクラスターを形成して存在する。ペア型(活性化/抑制)レセプターとしては、例えば、ヒトLRCにコードされるKIR(Killer cell Immunoglobulin-like Receptor;KIR2DS、3DSが活性化レセプター、KIR2DL、3DLが抑制レセプター)、ヒト及びマウスで共通にNKCにコードされるNKG2(NKG2C、D、Eが活性化レセプター、NKG2Aが抑制レセプター;NKG2A、C、EはCD94とヘテロダイマーを形成し、NKG2Dはホモダイマーを形成する)及びNKRP1(NKRP1A、C、Fが活性化レセプター、NKRP1Dが抑制レセプター)、マウスNKCにコードされるLy49(Ly49D、Hが活性化レセプター、Ly49A、C、Iが抑制レセプター)が挙げられるが、これらに限定されない。本発明のキメラレセプターに使用されるNK細胞レセプターは、異常細胞(例えば、ウイルス感染細胞、癌細胞など)の細胞表面上に特異的に発現する分子(リガンド)を認識するものであれば、活性化レセプターであっても抑制レセプターであってもよい。活性化レセプターの多くはリガンド未知のオーファンレセプターであるが、異常細胞で発現するいずれかの分子をリガンドとして認識してNK細胞のキラー活性を促進するので、本発明のキメラレセプターに好ましく使用することができる。一方、抑制レセプターは、本来的には宿主クラスI分子を認識するが、ウイルスはNK細胞による攻撃を回避するために、抑制レセプターに認識され得るMHCクラスI様分子を獲得したものがあるので(例えば、CMVのm157は抑制レセプターLy49I129に認識される)、そのような抑制レセプターもまた、本発明において使用することができる。
尚、ここで「異常細胞で特異的に発現する」とは、特定の異常細胞のみで発現し、他の細胞(特に正常細胞)で全く発現しないことを意味する他、特定の異常細胞での発現頻度及び/又は発現レベルに比べて、他の細胞での発現頻度及び/又は発現レベルが無視し得る程度に低い場合をも含む意味で用いられる。
【0016】
本発明のキメラレセプターに用いられるNK細胞レセプターは、好ましくは活性化レセプターである。活性化レセプターは、標的異常細胞で特異的に発現するリガンドを認識するものであれば特に制限はなく、例えば、標的異常細胞がヒト細胞の場合、NKRP1A、NKRP1C、NKRP1F、KIR2DS、KIR3DS、NKG2C、NKG2D、NKG2E等から、また、標的異常細胞がマウス細胞の場合は、Ly49H、Ly49D、NKRP1A、NKRP1C、NKRP1F、NKG2C、NKG2D、NKG2E等から、異常細胞の種類に応じて適宜選択される。例えば、ヒトNKG2DはMICやULBP等、マウスNKG2DはRAE−1、H60、Mult−1等のウイルス感染や細胞癌化により誘導されるクラスI様分子を認識する。また、MCMV感染細胞で特異的に発現するm157を認識する活性化レセプターとしては、Ly49Hが挙げられる。
【0017】
NK細胞レセプターの細胞外ドメインは、上記した種々のNK細胞レセプターの細胞外領域のうち、標的異常細胞で特異的に発現するリガンドを認識するのに必要な部分を含むものであれば、特に制限されず、例えば、細胞外領域全体であってよい。例えば、ヒトNK細胞活性化レセプターについては、GenBank Accession No. NP_002249として登録されているNKRP1Aのアミノ酸配列の細胞外領域、NKRP1Cの細胞外領域、NKRP1Fの細胞外領域、GenBank Accession No. NP_055327として登録されているKIR2DSのアミノ酸配列の細胞外領域、GenBank Accession No. AAB95317として登録されているKIR3DSのアミノ酸配列の細胞外領域、GenBank Accession No. NP_002251として登録されているNKG2Cのアミノ酸配列の細胞外領域、NKG2Dの細胞外領域、NKG2Eの細胞外領域が、それぞれ細胞外領域として挙げられる。また、マウスNK細胞活性化レセプターについては、GenBank Accession No. AAR03586として登録されているLy49Hのアミノ酸配列の細胞外領域が挙げられる。
【0018】
「ITAM配列を含むシグナル伝達アダプター分子」は、その細胞質ドメインに上記ITAM配列を含み、細胞傷害性T細胞を活性化するシグナルを伝達して標的細胞の殺傷をもたらす分子である限り、特に限定されないが、例えば、CD3ζ、DAP12/KARAP、FcRγ等の、膜貫通領域に負に荷電したアミノ酸残基を有し活性化レセプターと会合することが知られているアダプタータンパク質などが含まれる。より好ましくはCD3ζである。
【0019】
本発明のキメラレセプターは、上記シグナル伝達アダプター分子の全体を含んでもよいし、T細胞のキラー活性を活性化するシグナルを伝達し得る限りその一部のみを含んでいてもよい。「ITAM配列を含むシグナル伝達アダプター分子の一部」としては、例えば、ITAM配列を含む細胞質領域の一部、細胞質領域全体、さらに膜貫通領域を含んだ部分などが挙げられる。より具体的には、ヒトCD3ζの細胞質領域、GenBank Accession No. AAD09436として登録されているヒトDAP12/KARAPのアミノ酸配列の細胞質領域、ヒトFcRγの細胞質領域などが挙げられる。マウスCD3ζの細胞質領域もまた挙げられる。
【0020】
本発明のキメラレセプターを構成する上記タンパク質には、その同族体(ホモログやスプライスバリアント)、変異体、誘導体、成熟体及びアミノ酸修飾体などが包含される。ここでホモログとしては、当該タンパク質に対応する他生物種のタンパク質が例示でき、これらは、既に同定された遺伝子の塩基配列から同定することができる。また変異体には、天然に存在する対立遺伝子変異体、天然に存在しない変異体、及び人為的に欠失、置換、付加又は挿入されることによって改変されたアミノ酸配列を有する変異体が包含される。なお、前記変異体としては、野生型タンパク質と、少なくとも70%、好ましくは80%、より好ましくは90%、さらにより好ましくは95%相同なタンパク質を挙げることができる。またアミノ酸修飾体には、天然に存在するアミノ酸修飾体、天然に存在しないアミノ酸修飾体が包含される。本明細書におけるアミノ酸配列の相同性は、相同性計算アルゴリズムNCBI BLAST(National Center for Biotechnology Information Basic Local Alignment Search Tool)を用い、以下の条件(期待値=10;ギャップを許す;マトリクス=BLOSUM62;フィルタリング=OFF)にて計算することができる。アミノ酸配列の相同性を決定するための他のアルゴリズムとしては、例えば、Karlinら, Proc. Natl. Acad. Sci. USA, 90: 5873-5877 (1993)に記載のアルゴリズム[該アルゴリズムはNBLASTおよびXBLASTプログラム(version 2.0)に組み込まれている(Altschulら, Nucleic Acids Res., 25: 3389-3402 (1997))]、Needlemanら, J. Mol. Biol., 48: 444-453 (1970)に記載のアルゴリズム[該アルゴリズムはGCGソフトウェアパッケージ中のGAPプログラムに組み込まれている]、MyersおよびMiller, CABIOS, 4: 11-17 (1988)に記載のアルゴリズム[該アルゴリズムはCGC配列アラインメントソフトウェアパッケージの一部であるALIGNプログラム(version 2.0)に組み込まれている]、Pearsonら, Proc. Natl. Acad. Sci. USA, 85: 2444-2448 (1988)に記載のアルゴリズム[該アルゴリズムはGCGソフトウェアパッケージ中のFASTAプログラムに組み込まれている]等が挙げられ、それらも同様に好ましく用いられ得る。
【0021】
本発明のキメラレセプターは、1回膜貫通型レセプターであり、細胞傷害性T細胞上で異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメイン部分を細胞表面に提示し、ITAM配列を含むシグナル伝達アダプター分子部分を細胞質中に含む。該キメラレセプターの膜貫通領域は、NK細胞レセプターの膜貫通領域の全部もしくは一部及び/又はシグナル伝達アダプター分子の膜貫通領域の全部もしくは一部であってもよいし、他の膜タンパク質の膜貫通領域や人為的にデザインされたアミノ酸配列を用いてもよい。NK細胞活性化レセプター及びシグナル伝達アダプター分子の膜貫通領域は、それぞれ陽性および陰性荷電のアミノ酸残基を有し、T細胞に内在のシグナル伝達アダプター分子や他の内在性タンパク質と不都合な相互作用を生じるおそれがあるので、他の膜タンパク質の膜貫通領域、好ましくは、用いられる活性化レセプターに対応する抑制レセプターの膜貫通領域をキメラレセプター中に含めることが好ましい。
【0022】
本発明のキメラレセプターを発現する細胞傷害性T細胞は、該レセプターをコードする核酸を適当なプロモーターの制御下においた発現ベクターで、細胞傷害性T細胞をトランスフェクトすることにより調製することができる。本発明のキメラレセプターをコードする核酸はDNAであってもRNAであってもよく、あるいはDNA/RNAキメラであってもよい。また、該核酸は二本鎖であっても、一本鎖であってもよい。二本鎖の場合は、二本鎖DNA、二本鎖RNAまたはDNA:RNAのハイブリッドでもよい。好ましくは、本発明のキメラレセプターをコードする核酸は二本鎖DNAである。キメラレセプターをコードするDNAは、例えば、該キメラレセプターを構成するNK細胞レセプターの細胞外ドメインを含む領域、ITAM配列を含むシグナル伝達アダプター分子の該配列を含む領域(例えば、細胞質領域)、および必要に応じて他の膜タンパク質の膜貫通領域を含む領域をコードするcDNAを、それらを発現している哺乳動物細胞(例えば、ヒト、マウスなどのNK細胞、T細胞など)もしくはそれらの細胞が存在する組織(例えば、脾臓、リンパ節、リンパ液、血液など)より調製した全RNAもしくはmRNA画分を鋳型として用い、Reverse Transcriptase-Polymerase Chain Reaction(RT−PCR)法によって直接増幅することにより調製した後、通常の遺伝子工学的手法を用いて、それらを直接もしくは適当なリンカーを介して連結することによって製造することができるが、これに限定されない。
【0023】
例えば、ヒトNK細胞活性化レセプターであるNKRP1Aの細胞外領域は、GenBank Accession No. NM_002258として登録されている塩基配列情報に基づいて、該塩基配列中の細胞外領域コード領域を増幅し得るプライマーペアをデザインし、PCRを実施することにより調製することができる。同様に、NKRP1Cの細胞外領域コード領域、NKRP1Fの細胞外領域コード領域、GenBank Accession No. NM_014512として登録されているKIR2DS塩基配列中の細胞外領域コード領域、GenBank Accession No. AF022044として登録されているKIR3DS塩基配列中の細胞外領域コード領域、GenBank Accession No. NM_002260として登録されているNKG2Cの塩基配列中の細胞外領域コード領域、NKG2Dの細胞外領域コード領域、NKG2Eの細胞外領域コード領域を、各遺伝子の塩基配列情報に基づいて作製したプライマーペアを用いて増幅することにより、各タンパク質の細胞外領域を調製することができる。また、マウスNK細胞活性化レセプターであるLy49Hの細胞外領域は、GenBank Accession No. AY338977として登録されている塩基配列情報に基づいて、該塩基配列中の細胞外領域コード領域を増幅し得るプライマーペアをデザインし、PCRを実施することにより調製することができる。
【0024】
例えば、ヒトシグナル伝達アダプター分子であるCD3ζの細胞質領域は、細胞質領域コード領域を増幅し得るプライマーペアをデザインし、PCRを実施することにより調製することができる。同様に、GenBank Accession No. AF019562として登録されているヒトDAP12/KARAPの塩基配列中の細胞質領域コード領域、ヒトFcRγの細胞質領域コード領域を、各遺伝子の塩基配列情報に基づいて作製したプライマーペアを用いて増幅することにより調製することができる。また、マウスCD3ζの細胞質領域は、細胞質領域コード領域を増幅し得るプライマーペアをデザインし、PCRを実施することにより調製することができる。
【0025】
さらに、キメラレセプターの膜貫通領域として、活性化レセプターに対応する抑制レセプターの膜貫通領域を用いる場合、例えば、用いられるNK細胞活性化レセプターがKIR2DSであれば、対応するKIR2DLの膜貫通領域は、GenBank Accession No. NM_014218として塩基配列情報に基づいて、膜貫通領域コード領域を増幅し得るプライマーペアをデザインし、PCRを実施することにより調製することができる。
【0026】
ここで使用するプライマーペアは、各増幅断片の末端に、増幅断片同士および増幅断片とベクターとを連結するのを容易にするために、適当な制限酵素認識サイトを導入するようにデザインすることもできる。
【0027】
本発明のキメラレセプターをコードする核酸分子を含む発現ベクターは、例えば、本発明のキメラレセプターを構成するドメインをコードする目的の断片を調製し、該断片を連結し、適切な発現ベクター中のプロモーターの下流に連結することにより作製することができる。
【0028】
発現ベクターの骨格は、例えば、プラスミド又はウイルスベクター(例、アデノウイルス、レトロウイルス、アデノ随伴ウイルス、ヘルペスウイルス、ワクシニアウイルス、ポックスウイルス、ポリオウイルス、シンドビスウイルス、センダイウイルス、レンチウイルスなどのウイルス由来ベクター)であり得る。発現ベクターとして、例えば、IRESバイシストロン性発現ベクター、pA1−11、pXT1、pRc/CMV、pRc/RSV、pcDNAI/Neo、pME18Sなどが使用され得る。発現ベクターとしては、目的遺伝子が選択マーカー又はレポーターと共に同一mRNAから発現される、IRESバイシストロン性発現ベクターが好ましい。IRESバイシストロン性発現ベクターとしては、例えば、pMX−IRES−GFP(Kitamuraら、Int. J. Hematol., 67, 351-9 (1998))、及びClontech社により市販されているベクターなどが挙げられる。
【0029】
発現ベクターで使用されるプロモーターは、細胞傷害性T細胞で機能し得るものであれば特に制限されず、例えば、SV40由来初期プロモーター、サイトメガロウイルスLTR、ラウス肉腫ウイルスLTR、MoMuLV由来LTR、アデノウイルス由来初期プロモーター、LCK近位プロモーター、SRαプロモーター、HSV−TKプロモーターなどのウイルスプロモーター、ならびにβ−アクチン遺伝子プロモーター、PGK遺伝子プロモーター、トランスフェリン遺伝子プロモーターなどの哺乳動物の構成タンパク質遺伝子プロモーターなどが挙げられる。また、プロモーターとして、細胞傷害性T細胞に特異的なプロモーターを用いてもよい。このようなプロモーターは、細胞傷害性T細胞に特異的に発現している任意の遺伝子のプロモーターであり得る。
【0030】
発現ベクターは、必要に応じて、エンハンサー、スプライシングシグナル、ポリA付加シグナル、選択マーカー又はレポーター遺伝子、SV40複製起点(SV40ori)などを含み得る。
【0031】
選択マーカー及びレポーター遺伝子としては、例えば、緑色蛍光タンパク質(GFP)遺伝子、ルシフェラーゼ遺伝子、β−ガラクトシダーゼ遺伝子、ジヒドロ葉酸還元酵素遺伝子、アンピシリン耐性遺伝子、ネオマイシン耐性遺伝子が挙げられる。
【0032】
該発現ベクターの導入宿主となる細胞傷害性T細胞は、細胞表面の自己のMHCクラスI分子を非自己抗原と共に認識して標的細胞を殺傷する(MHC拘束)、獲得免疫系(adaptive immune system)において機能する細胞である。このT細胞はCD8陽性であるので、CD8 T細胞ともいう。細胞傷害性T細胞は、TCR(T cell Receptor)を介してリガンドに結合すると、TCR複合体中のCD3ζのITAM配列を介して活性化シグナルを伝達し、標的細胞を殺傷する。
【0033】
本発明において使用される細胞傷害性T細胞は、任意の動物種由来のものであり得る。このような動物種としては、ヒト、ヒトを除く哺乳動物(例えば、マウス、ラット、ハムスター、モルモットなどのげっ歯類やウサギなどの実験動物、ブタ、ウシ、ヤギ、ウマ、ヒツジなどの家畜、イヌ、ネコなどのペット、サル、オランウータン、チンパンジーなどの霊長類)が挙げられるがこれらに限定されない。本発明においてはヒト及びマウスの細胞傷害性T細胞が好ましく、臨床応用の観点から、ヒトの細胞傷害性T細胞がより好ましい。
【0034】
細胞傷害性T細胞の遺伝子型は特に限定されないが、異常細胞を除去すべき投与対象の動物と同種同系、同種異系又は異種異系であり、好ましくは同種同系又は同種異系である。投与時の拒絶反応を回避するために、この細胞傷害性T細胞は、投与対象と同種同系であることが好ましく、投与対象由来(即ち、自家細胞)であることが最も好ましい。例えば、臓器移植患者などの場合、免疫抑制剤投与前に予め該患者から細胞傷害性T細胞を採取しておき、これに本発明のキメラレセプターを導入し、臓器移植後に該キメラレセプターを発現する細胞傷害性T細胞を該患者に戻すなどの態様が、好ましく例示される。
【0035】
本発明において使用される細胞傷害性T細胞は、自体公知の方法によって、上記動物種の組織(例えば、脾臓、リンパ節、リンパ液、血液など)から単離及び/又は精製することが可能である。例えば、細胞傷害性T細胞上に特異的に発現する細胞表面マーカーに対する抗体を用いて、セルソーター、パニング、抗体磁気ビーズ法などにより単離することができる。
【0036】
本発明において使用される細胞傷害性T細胞は、上記動物種より採取された細胞(例えば、初代培養細胞)又は細胞株であり得る。この細胞傷害性T細胞はポリクローナルであり得る。細胞株は既存の細胞株又は新たに作製される細胞株であり得る。細胞株は自体公知の方法により作製できる。
【0037】
発現ベクターをT細胞に導入(形質導入)する方法は、細胞の種類により適宜選択することができる。例えば、リポフェクション法、マイクロインジェクション法、リン酸カルシウム共沈殿法、PEG法、エレクトロポレーション法、ウイルス感染などが挙げられるが、これらに限定されない。
【0038】
発現ベクターが導入されたT細胞は、上記選択マーカー及びレポーター遺伝子などを利用して、セルソーターなどにより選択することができる。例えば、GFPの発現に基づいてフローサイトメーターによってソートすることができる。
【0039】
選択されたT細胞の培養は、動物細胞の培養に通常用いられる培地を基礎培地として調製した培地中で行うことができる。基礎培地としては、例えば、MEM培地、DMEM培地、αMEM培地、ハム培地、RPMI1640培地、Fischer’s培地、及びこれらの混合培地が挙げられる。培養培地は、例えば、血清(例えば、FCS)、血清代替物(例えば、knockout Serum Replacement(KSR))、脂肪酸又は脂質、アミノ酸、ビタミン、増殖因子、サイトカイン、抗酸化剤、2−メルカプトエタノール、ピルビン酸、緩衝剤、無機塩類などを含むことができる。培養温度、CO2濃度などの他の培養条件は適宜設定できる。培養温度は、特に限定されるものではないが、例えば約30〜40℃、好ましくは約37℃である。また、CO2濃度は、例えば約1〜10%、好ましくは約5%である。培地のpHは通常約6〜8であり、培養時間は通常4〜14日間、好ましくは6〜14日間である。培養における細胞数、各種因子の濃度などのその他の条件は、自体公知の方法により適宜設定できる。
【0040】
上記のようにして調製される本発明のキメラレセプターを発現する細胞傷害性T細胞(以下、「本発明の細胞傷害性T細胞」ともいう)は、該キメラレセプターの細胞外領域に存在するNK細胞レセプターの細胞外ドメインを用いて、異常細胞で特異的に発現するリガンドを認識し、該キメラレセプターの細胞質領域に存在するシグナル伝達アダプター分子部分を介した活性化シグナルによって活性化され、該異常細胞を傷害するので、本発明の細胞傷害性T細胞は、該キメラレセプターが認識し得るリガンドを発現する異常細胞の除去に有用である。したがって、本発明の細胞傷害性T細胞は、該異常細胞を病原とする各種疾患の予防・治療に用いることができる。
本発明に従って予防・治療され得る疾患としては、ウイルス感染症、癌などが挙げられる。本発明の細胞傷害性T細胞は、ウイルス感染症の予防・治療に特に適している。具体的には、本発明の細胞傷害性T細胞により予防・治療されるウイルス感染症としては、CMV(例えば、HCMVまたはMCMVなど)、単純ヘルペスウイルス、EBウイルス、ヒト免疫不全ウイルス、インフルエンザウイルス、SARSウイルスなどの感染症が挙げられるがこれらに限定されない。
【0041】
本明細書中で使用する場合、「急性ウイルス感染症(本明細書中で「急性相感染」ともいう)」とは、あるウイルスによる最初の感染によって引き起こされる疾患をいう。本明細書中で使用する場合、「慢性ウイルス感染症」は、慢性化したウイルス感染症及び再発のウイルス感染症を含み、最初に感染したウイルスが排除されないまま体内に残ること、同じウイルスの再感染などによって引き起こされる。急性ウイルス感染症に対して一次免疫応答が生じ、慢性ウイルス感染症に対して二次免疫応答が生じる。この二次免疫応答には免疫記憶応答が関与している。本明細書中で使用する場合、「長期」とは、免疫記憶の確立によってもたらされる、慢性ウイルス感染症からの保護などをいう。本発明の細胞傷害性T細胞は、急性及び慢性の両方のウイルス感染症を予防・処置することができる。
【0042】
本発明の細胞傷害性T細胞は、単独で、あるいは他の治療のための任意の有効成分との混合物として、薬理学的に許容される一種もしくはそれ以上の担体とともに製剤化することができる。
【0043】
本発明の異常細胞除去剤は、通常は、注射剤、点滴剤などの剤型で提供される。本発明の異常細胞除去剤は、好ましくは、受容者の体液(血液など)と等張な滅菌水性担体中に、本発明のキメラレセプターを発現する細胞傷害性T細胞が懸濁された懸濁液である。水性担体としては、生理食塩水、PBSなどを挙げることができる。これらの水性担体には、更に、必要に応じて溶解補助剤、緩衝剤、等張化剤、無痛化剤、保存剤、安定化剤などを添加することもできる。
【0044】
本発明の剤中に含まれる、本発明のキメラレセプターを発現する細胞傷害性T細胞の濃度は、通常、約1×105〜1×1010個/ml、好ましくは約2×105〜1×109個/mlの範囲であるが、特に限定されない。細胞濃度が低すぎると、投与に時間がかかるため患者への負担が増大し、細胞濃度が高すぎると、細胞同士が凝集してしまう可能性がある。
【0045】
本発明の剤の投与量は、本発明のキメラレセプターの発現量、投与様式、疾患の種類、疾患の重篤度、投与対象となる動物種、投与対象の受容性、体重、年齢などによって異なるが、所望の免疫活性を達成するような細胞数が適宜投与され得、通常、1回の投与につき、本発明のキメラレセプターを発現する細胞傷害性T細胞の数として、通常1×106〜1×109個/m2、好ましくは1×107〜1×109個/m2の用量で投与される。これらの投与量は、前述の種々の条件により変動する。
【実施例】
【0046】
以下の実施例は本発明を具体的に説明するが、本発明を限定するものとして解釈すべきではない。
【0047】
材料及び方法
マウス及び細胞株:
BALB/cマウス及びB6マウスは、Charles River Japanから購入した。B6バックグラウンドを有するRAG−2欠損マウスは、Taconic(Hudson,NY)から購入した。6〜8週齢のマウスを実験に使用した。DBA/2由来の肥満細胞株P815、B6由来の胸腺腫細胞株RMA、Ly49Aに対するmAb(A1)のためのハイブリドーマ及び北村敏雄博士(東京大学)によって提供されたレトロウイルスパッケージング細胞株PLAT−Eをin vitroで維持した。Ly49Hに対するmAbのための3D10ハイブリドーマは、W.M. Yokoyama博士(Washington University,St.Louis)により提供された。
【0048】
レトロウイルスベクター及び形質導入:
Ly49H−CD3ζキメラレセプター、細胞質欠失Ly49H、Ly49A−CD3ζキメラレセプター及び細胞質欠失Ly49Aのコンストラクトを、以前に記載したように(非特許文献8)PCRによって生成した。Ly49H−CD3ζキメラレセプター及び細胞質欠失Ly49Hのコンストラクトについて、Ly49HのC末端細胞外ドメインを、それぞれCD3ζの細胞質ドメインあり又はなしでLy49Aの膜貫通ドメインと融合させて、別のシグナル伝達分子との相互作用を回避した。これらのコンストラクトをpMX−IRES−GFPベクターの第1シストロン中に挿入した。W.Yokoyamaにより提供されたm157のcDNA及びH2DdのcDNAを、それぞれpMX−IRES−GFPベクター又はpMX−IRES−hCD4ベクター中に挿入した(非特許文献8)。上記細胞株へのこれらcDNAの形質導入を、以前に記載された通りに実施した(非特許文献8)。GFP又はhCD4ポジティブな標的細胞を、MoFlowフローサイトメーター(Cytomation)によってソートした。
【0049】
キメラレセプターを発現するポリクローナルCD8 T細胞の生成:
脾臓CD8 T細胞を磁気ビーズ(Miltenyi Biotec,CA)によって精製した。ソートした細胞を、0日目に、200U/mlのrIL−2を含む培地中、5μg/mlのプレート結合抗CD3 Ab及び2μg/mlのモノクローナル抗CD28 Abで刺激した。細胞を1日目及び2日目に感染させた。細胞を3日目に洗浄し、200U/mlのrIL−2と共にもう24時間増殖させた。約50〜60%のCD8 T細胞が4日目にGFPポジティブであった。これらの細胞をin vivoのウイルス保護アッセイに使用した。in vitroの殺傷アッセイのために、Ly49H又はLy49Aの表面発現についてGFPポジティブ細胞をソートすることによって、細胞を精製した。
【0050】
マウスサイトメガロウイルスの調製及び感染:
Smith株のMCMV(VR194)を、American Tissue Culture Collection(Manassas,VA)から得た。ストック溶液を、MCMVに感染したBALB/cマウス由来の唾液腺から、以前に記載されたとおりに公知技術に従って調製した。このMCMVストック懸濁物のウイルス濃度は、以前に報告されたとおりに公知技術に従ってサブコンフルエントの3T3/Swissアルビノ細胞を使用してプラークアッセイによって決定したところ、2.0×107PFU/mlであった。マウスに、ウイルスストックの腹腔内注射によってウイルスを感染させた。
【0051】
ウイルスm157のPCR増幅及び配列:
ゲノムDNAを、一般に使用されている市販のキットを使用して抽出した。ウイルスm157遺伝子を、French et.al.(French, A.R., and W.M. Yokoyama. 2003. Natural killer cells and viral infections. Curr Opin Immunol 15:45-51.)に記載されたプライマーを用いてPCR増幅した。増幅したPCR産物を、TAクローニング(Invitrogen)中にサブクローニングし、配列決定した。m157配列のPCR分析のために、図4B中に示される実験における死にかけのマウスを屠殺し、唾液腺を使用した。
【0052】
実施例1
ポリクローナルCD8 T細胞上でのキメラレセプターの発現:
初代細胞におけるキメラレセプターの機能を試験するために、バイシストロン性レトロウイルスベクターpMX−IRES−GFP(pIG)中にLy49H細胞外ドメイン、Ly49A膜貫通ドメイン及びCD3ζ細胞質ドメインを挿入したLy49H−CD3ζキメラレセプターコンストラクト(Ly49HZ)を調製した。Ly49H発現の困難性及び他のシグナル伝達分子との会合を回避するために、原型的な阻害レセプターLy49A由来の膜貫通ドメインでその膜貫通ドメインを置換した。コントロールとして、Ly49H細胞外ドメイン及びLy49A膜貫通ドメインを有するがCD3ζ細胞質ドメインを有さない、細胞質欠失Ly49Hコンストラクト(Ly49Hcyto−del)もまた調製した。レトロウイルスの系を使用して、これらのキメラレセプターをMCMV感受性BLAB/c株(H−2dハプロタイプ)由来のCD8 T細胞中に形質導入した。形質導入後に約50〜60%のGFPポジティブ細胞が得られ、これらのGFPポジティブ細胞の殆どがLy49H特異的mAbによってポジティブに染色された。GFPポジティブ細胞をエフェクター細胞としてソートしたところ、それらのうち90%より多くがLy49Hポジティブであった(図1A)。標的細胞としてDBA2由来のP815(H−2d)を準備した。P815中にpMX−m157−IRES−GFP及びpMX−IRES−GFP(コントロールとして)を形質導入し、それぞれP815−m157及びP815−vectorとした(図1B)。これらのエフェクター細胞及び標的細胞を使用して、in vitroの51Cr放出アッセイを実施した。Ly49HZレセプターを形質導入したCD8 T細胞は、P185−m157を殺傷する能力を獲得したが、Ly49Hcyto−delを形質導入したCD8 T細胞はこの能力を獲得できなかった。このことは、ポジティブなシグナルが、Ly49Hレセプターとm157との結合によって促進される他のシグナル伝達分子を介したものではなく、実際にキメラレセプターのCD3ζ部分を介したものであることを示している(図1C)。形質導入したキメラレセプターの特異性のさらなる支持として、抗Ly49H特異的mAbは、P185−m157の殺傷をブロックしたが、コントロールAbはブロックしなかった(図1C及びデータ示さず)。特に、これらの形質導入されたCD8 T細胞は、親P185細胞及びP185−vector形質導入コントロール細胞に対する寛容を維持した。これらのデータは、Ly49H−CD3ζキメラレセプターを形質導入したポリクローナルCD8 T細胞がm157を特異的に認識して標的細胞を殺傷することを示す。
【0053】
実施例2
ネイティブのレセプター機能の変換:
m157タンパク質は、129/Jマウスにおいて発現されるLy49ファミリーのメンバーである阻害NK細胞レセプターLy49I129に結合することが示されている(Arase, H., E.S. Mocarski, A.E. Campbell, A.B. Hill, and L.L. Lanier. 2002. Direct recognition of cytomegalovirus by activating and inhibitory NK cell receptors. Science296:1323-1326.)。C57BL/6由来のLy49AB6(Ly49A)は、このファミリーメンバー中の原型的なNK細胞阻害レセプターである。Ly49AはMHCクラスIのH−2Ddを認識し、その特異性は、マウスNK細胞レセプターの中で最も特徴付けられている(Karlhofer, F.M., R.K. Ribaudo, and W.M. Yokoyama. 1992. MHC class I alloantigen specificity of Ly-49+ IL-2-activated natural killer cells. Nature 358:66-70.、Tormo, J., K. Natarajan, D.H. Margulies, and R.A. Mariuzza. 1999. Crystal structure of a lectin-like natural killer cell receptor bound to its MHC class I ligand. Nature402:623-631.)。Ly49H−CD3ζキメラレセプターを用いてポリクローナルCD8 T細胞の特異性を変化させたので、次に、in vitroの51Cr放出アッセイにおいて、Ly49AとCD3ζとのキメラレセプターを操作することによって初代CD8 T細胞においてネイティブのレセプター機能を変換できるか否かを試験した。Ly49Aの細胞外ドメインと膜貫通ドメインとをCD3ζ細胞質ドメインと融合させて作製したLy49A−CD3ζキメラレセプターコンストラクト(Ly49AZ)を調製した。コントロールとして、Ly49Aの細胞外ドメイン及び膜貫通ドメインのみで作製した細胞質欠失Ly49A(Ly49Acyto−del)もまた調製した。標的細胞として、C57BL6(B6)由来のT細胞株であるRMAを準備した。RMA中にH−2Dd−IRES−GFP及びIRES−GFP(形質導入コントロールとして)を形質導入し、それぞれ、RMA−H−2Dd及びRMA−GFPとした。Ly49Aの内因性リガンドを発現しないC57BL6マウス由来のCD8 T細胞中に、Ly49Aキメラレセプターを形質導入した(図2A)。細胞質欠失Ly49Aレセプターを形質導入したCD8 T細胞はRMA−H2Ddを殺傷することができなかった。一方で、Ly49AZレセプターを形質導入したCD8 T細胞はRMA−H2Ddを有効に殺傷したが、RMAもRMA−GFPも殺傷しなかった(図2B)。さらに、RMA−H2Ddの殺傷は、Ly49A特異的mAbであるA1によってブロックされた(図2C)。これらのデータは、Ly49AZレセプターを形質導入したポリクローナルCD8 T細胞がそのリガンドを特異的に認識して標的細胞を殺傷することを示す。合わせて考えると、これらのデータは、細胞質CD3ζと融合したNK細胞レセプターを形質導入したCD8 T細胞が、そのレセプターのネイティブの性質(阻害又は活性化)に関わらず、ネイティブのNK細胞レセプターの特異性を獲得して、標的細胞を殺傷できることを示す。
【0054】
実施例3
Ly49Hキメラレセプターを発現するCD8 T細胞によってMCMV感染からレスキューされたBALB/cマウス
Ly49H−CD3ζキメラレセプターを形質導入したポリクローナルCD8 T細胞によって、m157発現細胞のin vitroでの首尾よい殺傷がもたらされたので、本発明者は、これらの細胞がin vivoでMCMV感染を制御する能力を評価した。Ly49Hの発現を欠くBALB/cマウスはMCMV感染に対して感受性であり、1×104pfuのMCMVでの接種後2週間以内に死亡した(図3A)。BALB/cマウス由来のCD8 T細胞を準備し、Ly49HZ又はLy49Hcyto−delのいずれかを含むレトロウイルスをこの細胞に形質導入した。MCMV感染の当日、Ly49HZレセプターを発現するCD8 T細胞1×107を注入し、次いで、致死用量のMCMVをマウスに感染させた。細胞質欠失Ly49Hを発現するコントロールCD8 T細胞を注入したマウス及び細胞を注入していないマウスは10日以内に死亡したが、Ly49HZレセプターを発現するCD8 T細胞を注入したマウスは致死用量のMCMV感染の急性相を生存した(図3A)。このことと一致して、予備研究(データ示さず)において、注入したキメラレセプター発現細胞の数と生存との間の正の相関が観察された。生存マウスは、最初の感染の1ヵ月後に健康なままであった。
【0055】
実施例4
ウイルス感染の有効な制御は、自然免疫及び獲得免疫の両方を必要とする。従って、最初のチャレンジと同じウイルス用量のMCMVでマウスを再チャレンジすることによって、これらの生存したマウスが獲得免疫を発達させたか否かをさらに試験した。最初の感染の40日後に、CD8 T細胞をさらに注入することなくマウスを再チャレンジし、生存を観察した。ナイーブなBALB/cマウスは10日以内に死亡したが、再チャレンジしたマウスは全て生存し、このことは、これらのマウスにおける有効な記憶応答の確立を示す(図3B)。記憶応答における注入されたCD8 T細胞の寄与を評価するために、再チャレンジの10日後に生存マウスの脾臓中のCD8 T細胞を評価した。興味深いことに、CD8集団中にGFPポジティブ細胞もLy49Hポジティブ細胞も検出されなかった(図3C)。再チャレンジの7日後に分析した脾細胞からは、GFPポジティブ細胞は観察されなかった(データ示さず)。
【0056】
実施例5
RAG−2欠損B6マウスの一過的なレスキュー:
上記実験において、移入した全てのCD8 T細胞を、形質導入前にα−CD3 mAb及びα−CD28 mAbで刺激したところ、注入したCD8 T細胞の約40〜50%はGFP発現についてネガティブであった。内因性T細胞又は注入されたT細胞のいずれが抗CMV記憶応答に寄与しているかは明らかではなく、この記憶の形成が生存に如何に寄与したかも明らかではなかった。これらの問題点に取り組むため、B6 RAG2欠損(B6−RAG2)マウスを使用した。B6−RAG2マウスはT細胞及びB細胞を欠くので、内因性T細胞からCMVクリアランスに必要な獲得記憶応答を形成することができない。Ly49HZを形質導入した後に、B6マウス由来のCD8 T細胞をB6−RAG2マウス中に移入した。B6−RAG2マウスはLy49Hを発現するNK細胞を有するのでMCMVに対して耐性であるため、より高い用量のMCMV(マウス1匹当たり1×107pfu)でチャレンジした。コントロールとして、CD8 T細胞中にLy49Hcyto−delを形質導入し、これらをRAG2欠損B6マウス中に注入した。BALB/cモデルと同様に、全てのRAG2欠損B6マウス、及びコントロールの細胞質欠失Ly49Hを発現するCD8 T細胞を注入したRAG2欠損B6マウスの殆どは10日以内に死亡したが、Ly49HZを発現するCD8 T細胞を移入したRAG2欠損B6マウスは生存した(図4A)。これらのデータは、急性相感染が、移入されたCD8 T細胞によって実際に制御されたことを示す。第2の実験において、第1の実験と同じ短期生存パターンが観察された。しかし、感染の1ヵ月後、全ての生存マウスが死亡した(図4B)。これらのデータは、注入されたCD8 T細胞自体が長期記憶を形成することができたわけではないことを示唆する。この遅延相の死亡の機構に取り組むために、死にかけのRAG2欠損B6マウス(n=1)から脾臓及び唾液腺を単離した。平均ウイルス力価は、脾臓、肝臓、唾液腺及び肺においてそれぞれ4.4×106、2.0×107、3.6×108及び5.7×107であり、このことは、遅延相の死亡がMCMVの再活性化に起因するものであることを示唆する。
【0057】
本明細書において、致死用量のMCMVでチャレンジしたマウスをレスキューするための、首尾よい細胞免疫学的ストラテジーを記載してきた。内因性のNK細胞が致死的なウイルスチャレンジからB6−RAG−2マウスを保護できない条件において、ウイルスタンパク質に特異的なキメラレセプターを発現するCD8 T細胞の注入により、この致死的なチャレンジが克服された。Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞によって生存したBALB/cマウスは、高用量のウイルス再チャレンジを明らかに制御し、このことは、記憶細胞の有効な増殖及び細胞傷害活性を示唆する。従って、本明細書中に提供されるデータは、キメラ活性化レセプター遺伝子を移入したポリクローナルCD8 T細胞が、NK細胞の機能を模倣することによって、自然免疫応答の重要な役割を果たし得ることを示している。さらに、in vitroデータは、ネイティブの阻害レセプターが、その細胞質ドメインをCD3ζの細胞質ドメインと交換することによって、初代CD8 T細胞において活性化レセプターに変換され得ることもまた示唆している。
【0058】
ペプチド特異的CD8 T細胞の生成が所望され、免疫無防備の移植患者において疾患を引き起こすいくつかのウイルスについて確立されている(Riddell, S.R., and P.D. Greenberg. 1995. Principles for adoptive T cell therapy of human viral diseases. Annual Review of Immunology 13:545-586.)。ヒト同種移植の設定において所定のMHC拘束様式であつらえたペプチド特異的CD8 T細胞の生成は、リガンドの認識についてMHC拘束を必要とするが、CD3ζの細胞質ドメインを有するウイルスタンパク質特異的NK細胞キメラレセプターは、リガンドの認識についてMHC拘束を必要としない。ヒト及びマウスの両方において、末梢中の細胞の最初の割合が異なるので、多数のT細胞を得ることはNK細胞よりも容易である。
移入されたCD8 T細胞は急性相感染を有効に制御し、これは、有効な長期獲得免疫を導く。本明細書中のデータは、CMV感染に対する獲得免疫応答が内因性T細胞によって媒介されることを示唆する。
【産業上の利用可能性】
【0059】
本発明のキメラレセプターを発現する細胞傷害性T細胞は、NK細胞によるMHC非拘束的なリガンド認識により、ウイルス感染症や癌などにおける異常細胞(即ち、ウイルス感染細胞や癌細胞)を認識してこれを傷害することができ、なおかつウイルス感染や癌からの長期的な保護をもたらし得るので、急性及び慢性のウイルス感染症、あるいは癌およびその再発を予防・治療するために有用である。
【図面の簡単な説明】
【0060】
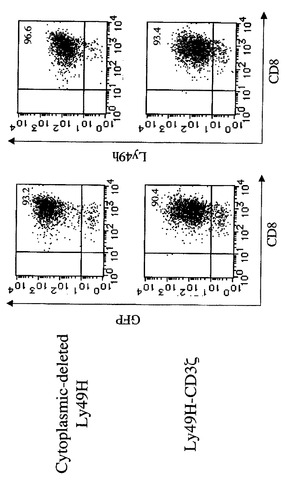
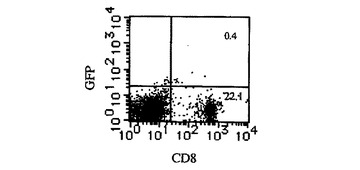
【図1A】Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞によるm157ポジティブ標的細胞の特異的殺傷を示す図である。BALB/c CD8 T細胞に、細胞質欠失Ly49H又はLy49H−CD3ζキメラレセプターのいずれかを含むレトロウイルスを感染させた。細胞を、そのGFP発現によってエフェクター細胞としてソートした。ソートした細胞上のCD8、GFP及びLy49Hの発現が示される。
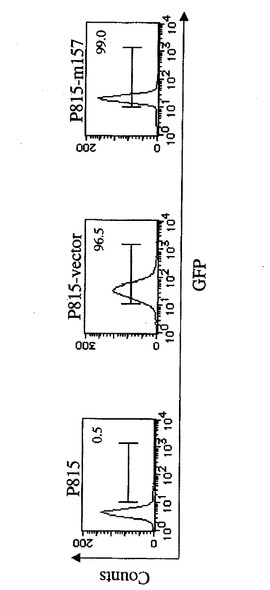
【図1B】Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞によるm157ポジティブ標的細胞の特異的殺傷を示す図である。P815細胞に、m157−IRES−GFP又はIRES−GFPのいずれかを形質導入し、そのGFP発現に基いてソートした。各形質導入体中のGFP発現が示される。
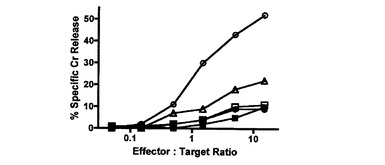
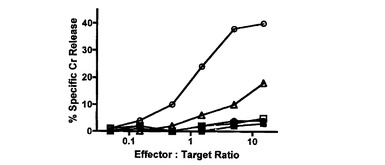
【図1C】Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞によるm157ポジティブ標的細胞の特異的殺傷を示す図である。51Cr放出アッセイを、m157−IRES−GFPを発現するP815細胞(丸)又はIRES−GFPを発現するP815細胞(四角)を標的細胞として用い、Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞(白抜き記号)又は細胞質欠失Ly49Hを発現するCD8 T細胞(黒塗り記号)をエフェクター細胞として用いて実施した。Ly49H Ab(培養物上清の1/20希釈)の存在下での細胞傷害活性を白抜きの三角で示した。4回の独立した実験からの代表的なデータが示される。別個の実験において、標的及びエフェクターの各組み合わせについて、コントロールmAbの存在下では効果は観察されなかった(データ示さず)。
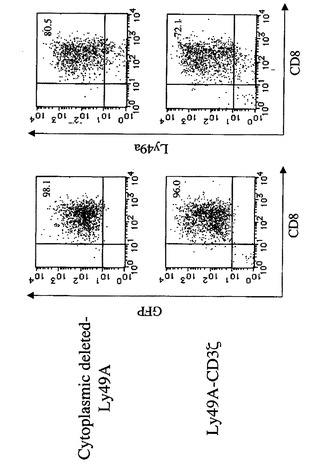
【図2A】CD3ζキメラレセプターとの融合による、ネイティブの阻害レセプター機能の活性化レセプターへの変換を示す図である。B6 CD8 T細胞に、細胞質欠失Ly49A又はLy49A−CD3ζキメラレセプターのいずれかを含むレトロウイルスを感染させた。細胞を、そのGFP発現によってエフェクター細胞としてソートし、Ly49A表面発現について分析した。
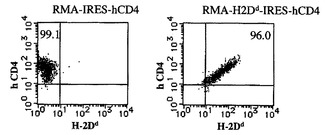
【図2B】CD3ζキメラレセプターとの融合による、ネイティブの阻害レセプター機能の活性化レセプターへの変換を示す図である。RMA細胞に、コントロールとしてのIRES−ヒトCD4(hCD4)又はH2Dd−IRES−hCD4のいずれかを形質導入し、hCD4発現によってソートした。これらの細胞のH−2Dd及びhCD4の発現を、細胞溶解アッセイの前に決定した。
【図2C】CD3ζキメラレセプターとの融合による、ネイティブの阻害レセプター機能の活性化レセプターへの変換を示す図である。51Cr放出アッセイを、IRES−hCD4を形質導入したRMA細胞(四角)又はH2Dd−IRES−hCD4を形質導入したRMA細胞(丸及び三角)を標的細胞として用い、Ly49A−CD3ζキメラを発現するCD8 T細胞(白抜き記号)又は細胞質欠失Ly49Aを発現するCD8 T細胞(黒塗り記号)をエフェクター細胞として用いて実施した。Ly49A Ab(培養物上清の1/20希釈)の存在下での細胞傷害活性を三角で示す。3回の独立した実験からの代表的なデータが示される。別個の実験において、標的及びエフェクターの各組み合わせについて、アイソタイプが一致したコントロールmAbの存在下では効果は観察されなかった(データ示さず)。
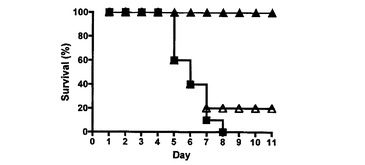
【図3A】Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞による致死的MCMV感染からのBALB/cマウスの保護、及び記憶応答の確立を示す図である。Ly49H−CD3ζキメラ(黒塗り三角)を発現するか、細胞質欠失Ly49H(白抜き三角)を発現するか、又は何も発現しない(黒塗り四角)5×106のCD8 T細胞を、BALB/cマウス(1群当たり10匹)中に注射した。次いで、マウスにMCMV(マウス1匹当たり104PFU)を感染させ、生存についてモニタリングした。
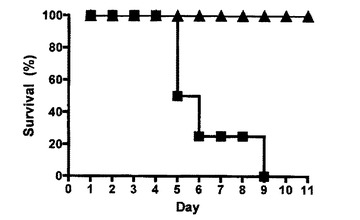
【図3B】Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞による致死的MCMV感染からのBALB/cマウスの保護、及び記憶応答の確立を示す図である。Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞によって最初のMCMV感染から保護された10匹のBALB/cマウス(黒塗り三角)を、最初の感染の40日後に、CD8 T細胞をさらに注入することなくMCMVで再チャレンジした。コントロールとして、ナイーブな未感染BALB/cマウス(黒塗り四角)(1群当たり5匹のマウス)に、同量のウイルスを感染させた。2回の実験からの代表的なデータが示される。
【図3C】Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞による致死的MCMV感染からのBALB/cマウスの保護、及び記憶応答の確立を示す図である。図3Bと同様に処理したマウス由来の脾細胞(黒塗り三角の群)を、再チャレンジの10日後にGFPポジティブなCD8 T細胞の存在について分析した。3匹のマウスからの代表的なデータが示される。
【図4A】Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞による、致死的なMCMV感染からの免疫不全マウスの保護を示す図である。Ly49H−CD3ζキメラ(黒塗り三角)を発現するか、細胞質欠失Ly49H(白抜き三角)を発現するか、又は何も発現しない(黒塗り四角)5×106のCD8 T細胞を、B6−RAG2欠損マウス(1群当たり5匹)中に注射した。次いで、マウスにMCMV(マウス1匹当たり107PFU)を感染させ、生存についてモニタリングした。
【図4B】Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞による、致死的なMCMV感染からの免疫不全マウスの保護を示す図である。マウス(1群当たり5匹)を、図4Aと同様に処理し、延長された期間にわたってモニタリングした。
【技術分野】
【0001】
本発明は、キメラレセプターを発現する細胞傷害性T細胞を含む新規な異常細胞の除去剤、このような細胞傷害性T細胞を投与することを含む異常細胞の除去方法に関する。より具体的には、本発明は、ウイルス感染細胞や癌細胞等の異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメインと、ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部とを含むキメラレセプターを発現する細胞傷害性T細胞を利用する、急性及び慢性のウイルス感染症や癌の予防・治療に関する。
【背景技術】
【0002】
ウイルス感染に対する宿主防御は、2つの異なる免疫応答のセット(自然免疫及び獲得免疫)によって媒介される。自然免疫系の最初の防御ラインである樹状細胞(DC)は、パターン認識レセプターであるToll様レセプター(TLR)を使用し、このレセプターは、種々の病原体成分(タンパク質、DNA、RNA及び多糖を含む)と相互作用する。DCはサイトカインを迅速に分泌して、他の自然免疫細胞(例えば、ナチュラルキラー(NK)細胞)及び獲得免疫細胞(T細胞及びB細胞)の応答を誘発する。両方の免疫系の成分が、病原体からの宿主の有効な保護のために必要である。自然免疫の欠如は病原体の迅速な複製を導き、感染後数日以内に死をもたらす。対照的に、獲得免疫系の非存在下では病原体は獲得免疫監視から逃れ、ヒト免疫不全患者において報告されているように、慢性及び再発の感染に対して宿主を感受性にする。
【0003】
持続感染を引き起こすウイルスの多くは、細胞傷害性T細胞による抗原認識を回避するために宿主細胞でのMHCクラスI抗原の発現を抑制する。しかし、NK細胞はMHCクラスI抗原を認識する抑制レセプターを用いて該抗原を発現している細胞を自己とみなし、非発現細胞を非自己と認識して傷害するので、そのようなウイルス感染細胞はNK細胞の抑制レセプターに認識されなくなり、該細胞により傷害される。NK細胞はまた、細胞外領域において抑制レセプターと非常に高い相同性を示すものの、該レセプターと相反する機能を有する活性化レセプターを有している。活性化レセプターは、ウイルス感染細胞上で特異的に発現するリガンドを認識して非自己とみなし、NK細胞のキラー活性を促進する。しかしながら、多くの活性化レセプターのリガンドは不明であり、その機能は明らかになっていないものが多い。
【0004】
サイトメガロウイルス(CMV)は、免疫無防備宿主(例えば臓器移植患者)における主要な日和見感染病原体である。マウスサイトメガロウイルス(MCMV)感染は、宿主−病原体相互作用及びその免疫を研究するための有用なモデルを提供する(非特許文献1、2)。最近、MCMV感染細胞の認識に関わるNK細胞レセプター及びそのリガンドのペアとして、MCMV耐性マウス株において見出されたLy49H活性化レセプター及びMCMVゲノムにコードされるMHCクラスI様ホールドタンパク質m157が同定された(非特許文献3、4)。二次構造をもとにした立体構造上の相同性検索によると、m157はCD1やH−2M3等の非古典的MHCクラスI分子と相同性を示す。
【0005】
NK細胞の活性化は活性化レセプターの状態のみによって決定されるのではなく、抑制レセプターのシグナルと活性化レセプターのシグナルとの間のバランスによって決定され、このことは、「missing-self」仮説によって支持される(非特許文献5)。興味深いことに、m157は、MCMV感受性マウス株129/Jにおいて、抑制レセプターであるLy49I129に結合することが示されている(非特許文献3)。一方、MCMV耐性のC57BL/6マウスのLy49IB6は宿主クラスI分子を認識するにもかかわらず、m157を認識しない。
【0006】
Ly49Iは、マウスの原型的な抑制レセプターであるLy49Aと同様に、その細胞質ドメイン中に免疫レセプターチロシンベースの阻害モチーフ(immuno-receptor tyrosine-based inhibitory motifs(ITIM))を含む。対照的に、Ly49Hは、その細胞質ドメイン中にシグナル伝達モチーフを有さない。その代わりに、Ly49Hは、その表面発現及びLy49H膜貫通ドメイン中の正に荷電した残基を介して、免疫レセプターチロシンベースの活性化モチーフ(immuno-receptor tyrosine-based activation motifs(ITAM))を含むシグナル伝達アダプター分子DAP12/KARAP(非特許文献6、7)と会合する。T細胞レセプター(TCR)複合体中のCD3ζと同様に、DAP12は、リガンド認識に際し、Syk又はZap70のようなSykキナーゼファミリーのメンバーをそのITAMへとリクルートする。
【0007】
リガンド同定のために、マウスT細胞株において発現させた、オーファンNK細胞レセプターの細胞外ドメインとCD3ζの細胞質ドメインとのキメラレセプターのレパートリーが以前に使用されている(非特許文献8、9)。この系によれば、レセプターの本来の性質(抑制であるか活性化であるか)にかかわらず、リガンドを認識するとNFAT結合プロモーターの制御下でlacZ発現を誘導するので、リガンド発現細胞を検出することができる(非特許文献9)。
【非特許文献1】Tortorella, D., B.E. Gewurz, M.H. Furman, D.J. Schust, and H.L. Ploegh. 2000. Viral subversion of the immune system. Annu Rev Immunol 18:861-926.
【非特許文献2】Webb, J.R., S.H. Lee, and S.M. Vidal. 2002. Genetic control of innate immune responses against cytomegalovirus: MCMV meets its match. Genes Immun 3:250-262.
【非特許文献3】Arase, H., and L.L. Lanier. 2004. Specific recognition of virus-infected cells by paired NK receptors. Rev Med Virol 14:83-93.
【非特許文献4】Smith, H.R., J.W. Heusel, I.K. Mehta, S. Kim, B.G. Dorner, O.V. Naidenko, K. Iizuka, H. Furukawa, D.L. Beckman, J.T. Pingel, A.A. Scalzo, D.H. Fremont, and W.M. Yokoyama. 2002. Recognition of a virus-encoded ligand by a natural killer cell activation receptor. Proc Natl Acad Sci U S A 99:8826-8831.
【非特許文献5】Lanier, L.L. 2005. NK cell recognition. Annu Rev Immunol 23:225-274.
【非特許文献6】Bakker, A.B., R.M. Hoek, A. Cerwenka, B. Blom, L. Lucian, T. McNeil, R. Murray, L.H. Phillips, J.D. Sedgwick, and L.L. Lanier. 2000. DAP12-deficient mice fail to develop autoimmunity due to impaired antigen priming. Immunity 13:345-353.
【非特許文献7】Tomasello, E., P.-O. Desmoulins, K. Chemin, S. Guia, H. Cremer, J. Ortaldo, P. Love, D. Kaiserlian, and E. Vivier. 2000. Combined natural killer cell and dendritic cell functional deficiency in KARAP/DAP12 loss-of-function mutant mice. Immunity 13:355-364.
【非特許文献8】Furukawa, H., K. Iizuka, J. Poursine-Laurent, N. Shastri, and W.M. Yokoyama. 2002. A ligand for the murine NK activation receptor Ly-49D: activation of tolerized NK cells from beta(2)-microglobulin-deficient mice. Journal of Immunology 169:126-136.
【非特許文献9】Iizuka, K., O.V. Naidenko, B.F. Plougastel, D.H. Fremont, and W.M. Yokoyama. 2003. Genetically linked C-type lectin-related ligands for the NKRP1 family of natural killer cell receptors. Nat Immunol 4:801-807.
【発明の開示】
【発明が解決しようとする課題】
【0008】
本発明の目的は、急性期のみならず慢性のウイルス感染症の予防・治療にも有用な新規免疫療法およびそのための薬剤を提供することである。また、本発明の別の目的は、癌の新規免疫療法およびそのための薬剤を提供することである。
【課題を解決するための手段】
【0009】
本発明者は、上記の目的を達成するために鋭意研究を重ねた結果、ウイルス感染細胞や癌細胞等の異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメインと、ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部とを含むキメラレセプターを発現させた細胞傷害性T細胞を用いることにより、標的異常細胞上の該リガンドを認識して当該細胞を殺傷し得ることを見出した。すなわち、本発明者は、Ly49Hレセプターの細胞外ドメインとCD3ζの細胞質ドメインとを含むキメラレセプターをコードする核酸を細胞傷害性T細胞中に導入し、これをMCMV感受性マウス株に投与したところ、該マウスが急性相のMCMV感染による死亡から逃れ、さらに獲得免疫による長期保護が確立されたことを見出し、本発明を完成するに至った。
【0010】
すなわち、本発明は以下の通りである:
[1] 生体にとって望ましくない異常細胞の除去剤であって、
(a)該異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメインと、
(b)ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部と、
を含むキメラレセプターを発現する細胞傷害性T細胞を含有してなる剤。
[2] NK細胞レセプターが活性化レセプターである、上記[1]に記載の剤。
[3] 異常細胞がウイルス感染細胞である、上記[1]又は[2]に記載の剤。
[4] ウイルスがサイトメガロウイルスである、上記[3]に記載の剤。
[5] サイトメガロウイルスがヒトサイトメガロウイルスである、上記[4]に記載の剤。
[6] NK細胞レセプターがヒト由来である、上記[1]〜[5]のいずれかに記載の剤。
[7] NK細胞レセプターがNKRP1A、NKRP1C、NKRP1F、KIR2DS、KIR3DS、NKG2C、NKG2DおよびNKG2Eからなる群より選択される、上記[6]に記載の剤。
[8] サイトメガロウイルスがマウスサイトメガロウイルスである、上記[4]に記載の剤。
[9] NK細胞レセプターがマウス由来である、上記[1]〜[5]及び[8]のいずれかに記載の剤。
[10] NK細胞レセプターがLy49H、Ly49D、NKRP1A、NKRP1C、NKRP1F、NKG2C、NKG2DおよびNKG2Eからなる群より選択される、上記[9]に記載の剤。
[11] NK細胞レセプターがLy49Hである、上記[10]に記載の剤。
[12] シグナル伝達アダプター分子がCD3ζ、DAP12/KARAPおよびFcRγからなる群より選択される、上記[1]〜[11]のいずれかに記載の剤。
[13] シグナル伝達アダプター分子がCD3ζである、上記[12]に記載の剤。
[14] キメラレセプターの膜貫通ドメインがNK細胞活性化レセプター由来でない、上記[1]〜[13]のいずれかに記載の剤。
[15] 急性ウイルス感染症の予防・治療用である、上記[1]〜[14]のいずれかに記載の剤。
[16] 慢性ウイルス感染症の予防・治療用である、上記[1]〜[14]のいずれかに記載の剤。
[17] 生体にとって望ましくない異常細胞の除去方法であって、
(a)該異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメインと、
(b)ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部と、
を含むキメラレセプターを発現する細胞傷害性T細胞の有効量を、該異常細胞を有する生体に投与することを含む方法。
[18]
(a)ウイルス感染ヒト細胞で特異的に発現するリガンドを認識するヒトNK細胞レセプターの細胞外ドメインと、
(b)ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部と、
を含むキメラレセプタータンパク質。
[19] ウイルスがヒトサイトメガロウイルスである、上記[18]に記載のタンパク質。
[20] NK細胞レセプターが活性化レセプターである、上記[18]又は[19]に記載のタンパク質。
[21] NK細胞レセプターがNKRP1A、NKRP1C、NKRP1F、KIR2DS、KIR3DS、NKG2C、NKG2DおよびNKG2Eからなる群より選択される、上記[18]〜[20]のいずれかに記載のタンパク質。
[22] シグナル伝達アダプター分子がCD3ζ、DAP12/KARAPおよびFcRγからなる群より選択される、上記[18]〜[21]のいずれかに記載のタンパク質。
[23] シグナル伝達アダプター分子がCD3ζである、上記[22]に記載のタンパク質。
[24] 前記キメラレセプターの膜貫通ドメインがNK細胞活性化レセプター由来でない、上記[18]〜[23]のいずれかに記載のタンパク質。
[25] 上記[18]〜[24]のいずれかに記載のタンパク質をコードする核酸。
[26] 上記[25]記載の核酸を含む、ヒト細胞傷害性T細胞で機能し得る発現ベクター。
[27] 上記[26]記載の発現ベクターでトランスフェクトされたヒト細胞傷害性T細胞。
【発明の効果】
【0011】
本発明のキメラレセプターを発現する細胞傷害性T細胞は、NK細胞レセプターを利用した迅速な免疫応答により急性ウイルス感染症に対する予防・治療効果を発揮するだけでなく、T細胞による免疫監視により慢性のウイルス感染症に対しても予防・治療効果を奏する。
【発明を実施するための最良の形態】
【0012】
以下に、本発明を詳細に説明する。
本発明は、生体にとって望ましくない異常細胞の除去剤(以下、単に「本発明の異常細胞除去剤」ともいう)を提供する。「生体にとって望ましくない異常細胞」は、任意の疾患の病原となり得る細胞であって、正常細胞が全くあるいはほとんど発現していない分子を細胞表面に特異的に、あるいは高発現する細胞であれば、特に制限はないが、好ましくは、ウイルス感染細胞(例えば、CMV、単純ヘルペスウイルス、EBウイルス、ヒト免疫不全ウイルス、インフルエンザウイルス、SARSウイルス等に感染した細胞)、癌細胞など、より好ましくはウイルス感染細胞、いっそう好ましくはCMV感染細胞、特に好ましくはヒトサイトメガロウイルス(HCMV)もしくはMCMV感染細胞である。本発明の異常細胞除去剤は、細胞傷害性T細胞が異常細胞を傷害して除去することにより、該異常細胞を病原とする疾患の予防・治療に有用である(本明細書中で「予防・治療」とは、「予防及び/又は治療」を意味する)。したがって、異常細胞がウイルス感染細胞の場合、本発明の異常細胞除去剤はウイルス感染症の予防・治療剤として用いることができ、異常細胞が癌細胞の場合、本発明の異常細胞除去剤は癌の予防・治療剤として用いることができる。
【0013】
本発明の異常細胞除去剤は、(a)異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメインと、(b)ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部とを含むキメラレセプターを発現する細胞傷害性T細胞を含有する。
【0014】
NK細胞は、自然免疫系(innate immune system)の細胞傷害性細胞であり、腫瘍細胞やウイルス感染細胞などを殺傷する(MHC非拘束)。NK細胞は、構造的に類似した2つのタイプのレセプター:「NK活性化レセプター(本明細書中で、単に「活性化レセプター」ともいう)」及び「NK抑制レセプター(本明細書中で、単に「抑制レセプター」ともいう)」をその表面に発現し、異常細胞を自己の細胞から識別している。活性化レセプターは、膜貫通領域に存在する正荷電のアミノ酸残基を介して、膜貫通領域に負荷電のアミノ酸残基を有するシグナル伝達アダプター分子と会合する。シグナル伝達アダプター分子はITAM配列[Y-x-x-L-xもしくはY-x-x-x-L](ここで、xは任意のアミノ酸である)を有し、SykやZap70などのキナーゼ群をリクルートすることにより、活性化シグナルを伝達する。一方、抑制レセプターは細胞質ドメインにITIM配列[(I/V/L/S) -x-Y-x-x- (L/V)](ここで、xは任意のアミノ酸である)を有し、SHP−1などのホスファターゼ群をリクルートすることにより、活性化レセプターからのシグナルを遮断する。NK細胞の細胞傷害活性は、これらのレセプターからのシグナルのバランスによって制御される。
【0015】
NK細胞で発現するNK細胞レセプターの遺伝子は、主としてLRC(Leukocyte Receptor Complex;ヒト19番染色体、マウス7番染色体上)及びNKC(NK receptor Complex;ヒト12番染色体、マウス6番染色体上)の2つの遺伝子座にクラスターを形成して存在する。ペア型(活性化/抑制)レセプターとしては、例えば、ヒトLRCにコードされるKIR(Killer cell Immunoglobulin-like Receptor;KIR2DS、3DSが活性化レセプター、KIR2DL、3DLが抑制レセプター)、ヒト及びマウスで共通にNKCにコードされるNKG2(NKG2C、D、Eが活性化レセプター、NKG2Aが抑制レセプター;NKG2A、C、EはCD94とヘテロダイマーを形成し、NKG2Dはホモダイマーを形成する)及びNKRP1(NKRP1A、C、Fが活性化レセプター、NKRP1Dが抑制レセプター)、マウスNKCにコードされるLy49(Ly49D、Hが活性化レセプター、Ly49A、C、Iが抑制レセプター)が挙げられるが、これらに限定されない。本発明のキメラレセプターに使用されるNK細胞レセプターは、異常細胞(例えば、ウイルス感染細胞、癌細胞など)の細胞表面上に特異的に発現する分子(リガンド)を認識するものであれば、活性化レセプターであっても抑制レセプターであってもよい。活性化レセプターの多くはリガンド未知のオーファンレセプターであるが、異常細胞で発現するいずれかの分子をリガンドとして認識してNK細胞のキラー活性を促進するので、本発明のキメラレセプターに好ましく使用することができる。一方、抑制レセプターは、本来的には宿主クラスI分子を認識するが、ウイルスはNK細胞による攻撃を回避するために、抑制レセプターに認識され得るMHCクラスI様分子を獲得したものがあるので(例えば、CMVのm157は抑制レセプターLy49I129に認識される)、そのような抑制レセプターもまた、本発明において使用することができる。
尚、ここで「異常細胞で特異的に発現する」とは、特定の異常細胞のみで発現し、他の細胞(特に正常細胞)で全く発現しないことを意味する他、特定の異常細胞での発現頻度及び/又は発現レベルに比べて、他の細胞での発現頻度及び/又は発現レベルが無視し得る程度に低い場合をも含む意味で用いられる。
【0016】
本発明のキメラレセプターに用いられるNK細胞レセプターは、好ましくは活性化レセプターである。活性化レセプターは、標的異常細胞で特異的に発現するリガンドを認識するものであれば特に制限はなく、例えば、標的異常細胞がヒト細胞の場合、NKRP1A、NKRP1C、NKRP1F、KIR2DS、KIR3DS、NKG2C、NKG2D、NKG2E等から、また、標的異常細胞がマウス細胞の場合は、Ly49H、Ly49D、NKRP1A、NKRP1C、NKRP1F、NKG2C、NKG2D、NKG2E等から、異常細胞の種類に応じて適宜選択される。例えば、ヒトNKG2DはMICやULBP等、マウスNKG2DはRAE−1、H60、Mult−1等のウイルス感染や細胞癌化により誘導されるクラスI様分子を認識する。また、MCMV感染細胞で特異的に発現するm157を認識する活性化レセプターとしては、Ly49Hが挙げられる。
【0017】
NK細胞レセプターの細胞外ドメインは、上記した種々のNK細胞レセプターの細胞外領域のうち、標的異常細胞で特異的に発現するリガンドを認識するのに必要な部分を含むものであれば、特に制限されず、例えば、細胞外領域全体であってよい。例えば、ヒトNK細胞活性化レセプターについては、GenBank Accession No. NP_002249として登録されているNKRP1Aのアミノ酸配列の細胞外領域、NKRP1Cの細胞外領域、NKRP1Fの細胞外領域、GenBank Accession No. NP_055327として登録されているKIR2DSのアミノ酸配列の細胞外領域、GenBank Accession No. AAB95317として登録されているKIR3DSのアミノ酸配列の細胞外領域、GenBank Accession No. NP_002251として登録されているNKG2Cのアミノ酸配列の細胞外領域、NKG2Dの細胞外領域、NKG2Eの細胞外領域が、それぞれ細胞外領域として挙げられる。また、マウスNK細胞活性化レセプターについては、GenBank Accession No. AAR03586として登録されているLy49Hのアミノ酸配列の細胞外領域が挙げられる。
【0018】
「ITAM配列を含むシグナル伝達アダプター分子」は、その細胞質ドメインに上記ITAM配列を含み、細胞傷害性T細胞を活性化するシグナルを伝達して標的細胞の殺傷をもたらす分子である限り、特に限定されないが、例えば、CD3ζ、DAP12/KARAP、FcRγ等の、膜貫通領域に負に荷電したアミノ酸残基を有し活性化レセプターと会合することが知られているアダプタータンパク質などが含まれる。より好ましくはCD3ζである。
【0019】
本発明のキメラレセプターは、上記シグナル伝達アダプター分子の全体を含んでもよいし、T細胞のキラー活性を活性化するシグナルを伝達し得る限りその一部のみを含んでいてもよい。「ITAM配列を含むシグナル伝達アダプター分子の一部」としては、例えば、ITAM配列を含む細胞質領域の一部、細胞質領域全体、さらに膜貫通領域を含んだ部分などが挙げられる。より具体的には、ヒトCD3ζの細胞質領域、GenBank Accession No. AAD09436として登録されているヒトDAP12/KARAPのアミノ酸配列の細胞質領域、ヒトFcRγの細胞質領域などが挙げられる。マウスCD3ζの細胞質領域もまた挙げられる。
【0020】
本発明のキメラレセプターを構成する上記タンパク質には、その同族体(ホモログやスプライスバリアント)、変異体、誘導体、成熟体及びアミノ酸修飾体などが包含される。ここでホモログとしては、当該タンパク質に対応する他生物種のタンパク質が例示でき、これらは、既に同定された遺伝子の塩基配列から同定することができる。また変異体には、天然に存在する対立遺伝子変異体、天然に存在しない変異体、及び人為的に欠失、置換、付加又は挿入されることによって改変されたアミノ酸配列を有する変異体が包含される。なお、前記変異体としては、野生型タンパク質と、少なくとも70%、好ましくは80%、より好ましくは90%、さらにより好ましくは95%相同なタンパク質を挙げることができる。またアミノ酸修飾体には、天然に存在するアミノ酸修飾体、天然に存在しないアミノ酸修飾体が包含される。本明細書におけるアミノ酸配列の相同性は、相同性計算アルゴリズムNCBI BLAST(National Center for Biotechnology Information Basic Local Alignment Search Tool)を用い、以下の条件(期待値=10;ギャップを許す;マトリクス=BLOSUM62;フィルタリング=OFF)にて計算することができる。アミノ酸配列の相同性を決定するための他のアルゴリズムとしては、例えば、Karlinら, Proc. Natl. Acad. Sci. USA, 90: 5873-5877 (1993)に記載のアルゴリズム[該アルゴリズムはNBLASTおよびXBLASTプログラム(version 2.0)に組み込まれている(Altschulら, Nucleic Acids Res., 25: 3389-3402 (1997))]、Needlemanら, J. Mol. Biol., 48: 444-453 (1970)に記載のアルゴリズム[該アルゴリズムはGCGソフトウェアパッケージ中のGAPプログラムに組み込まれている]、MyersおよびMiller, CABIOS, 4: 11-17 (1988)に記載のアルゴリズム[該アルゴリズムはCGC配列アラインメントソフトウェアパッケージの一部であるALIGNプログラム(version 2.0)に組み込まれている]、Pearsonら, Proc. Natl. Acad. Sci. USA, 85: 2444-2448 (1988)に記載のアルゴリズム[該アルゴリズムはGCGソフトウェアパッケージ中のFASTAプログラムに組み込まれている]等が挙げられ、それらも同様に好ましく用いられ得る。
【0021】
本発明のキメラレセプターは、1回膜貫通型レセプターであり、細胞傷害性T細胞上で異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメイン部分を細胞表面に提示し、ITAM配列を含むシグナル伝達アダプター分子部分を細胞質中に含む。該キメラレセプターの膜貫通領域は、NK細胞レセプターの膜貫通領域の全部もしくは一部及び/又はシグナル伝達アダプター分子の膜貫通領域の全部もしくは一部であってもよいし、他の膜タンパク質の膜貫通領域や人為的にデザインされたアミノ酸配列を用いてもよい。NK細胞活性化レセプター及びシグナル伝達アダプター分子の膜貫通領域は、それぞれ陽性および陰性荷電のアミノ酸残基を有し、T細胞に内在のシグナル伝達アダプター分子や他の内在性タンパク質と不都合な相互作用を生じるおそれがあるので、他の膜タンパク質の膜貫通領域、好ましくは、用いられる活性化レセプターに対応する抑制レセプターの膜貫通領域をキメラレセプター中に含めることが好ましい。
【0022】
本発明のキメラレセプターを発現する細胞傷害性T細胞は、該レセプターをコードする核酸を適当なプロモーターの制御下においた発現ベクターで、細胞傷害性T細胞をトランスフェクトすることにより調製することができる。本発明のキメラレセプターをコードする核酸はDNAであってもRNAであってもよく、あるいはDNA/RNAキメラであってもよい。また、該核酸は二本鎖であっても、一本鎖であってもよい。二本鎖の場合は、二本鎖DNA、二本鎖RNAまたはDNA:RNAのハイブリッドでもよい。好ましくは、本発明のキメラレセプターをコードする核酸は二本鎖DNAである。キメラレセプターをコードするDNAは、例えば、該キメラレセプターを構成するNK細胞レセプターの細胞外ドメインを含む領域、ITAM配列を含むシグナル伝達アダプター分子の該配列を含む領域(例えば、細胞質領域)、および必要に応じて他の膜タンパク質の膜貫通領域を含む領域をコードするcDNAを、それらを発現している哺乳動物細胞(例えば、ヒト、マウスなどのNK細胞、T細胞など)もしくはそれらの細胞が存在する組織(例えば、脾臓、リンパ節、リンパ液、血液など)より調製した全RNAもしくはmRNA画分を鋳型として用い、Reverse Transcriptase-Polymerase Chain Reaction(RT−PCR)法によって直接増幅することにより調製した後、通常の遺伝子工学的手法を用いて、それらを直接もしくは適当なリンカーを介して連結することによって製造することができるが、これに限定されない。
【0023】
例えば、ヒトNK細胞活性化レセプターであるNKRP1Aの細胞外領域は、GenBank Accession No. NM_002258として登録されている塩基配列情報に基づいて、該塩基配列中の細胞外領域コード領域を増幅し得るプライマーペアをデザインし、PCRを実施することにより調製することができる。同様に、NKRP1Cの細胞外領域コード領域、NKRP1Fの細胞外領域コード領域、GenBank Accession No. NM_014512として登録されているKIR2DS塩基配列中の細胞外領域コード領域、GenBank Accession No. AF022044として登録されているKIR3DS塩基配列中の細胞外領域コード領域、GenBank Accession No. NM_002260として登録されているNKG2Cの塩基配列中の細胞外領域コード領域、NKG2Dの細胞外領域コード領域、NKG2Eの細胞外領域コード領域を、各遺伝子の塩基配列情報に基づいて作製したプライマーペアを用いて増幅することにより、各タンパク質の細胞外領域を調製することができる。また、マウスNK細胞活性化レセプターであるLy49Hの細胞外領域は、GenBank Accession No. AY338977として登録されている塩基配列情報に基づいて、該塩基配列中の細胞外領域コード領域を増幅し得るプライマーペアをデザインし、PCRを実施することにより調製することができる。
【0024】
例えば、ヒトシグナル伝達アダプター分子であるCD3ζの細胞質領域は、細胞質領域コード領域を増幅し得るプライマーペアをデザインし、PCRを実施することにより調製することができる。同様に、GenBank Accession No. AF019562として登録されているヒトDAP12/KARAPの塩基配列中の細胞質領域コード領域、ヒトFcRγの細胞質領域コード領域を、各遺伝子の塩基配列情報に基づいて作製したプライマーペアを用いて増幅することにより調製することができる。また、マウスCD3ζの細胞質領域は、細胞質領域コード領域を増幅し得るプライマーペアをデザインし、PCRを実施することにより調製することができる。
【0025】
さらに、キメラレセプターの膜貫通領域として、活性化レセプターに対応する抑制レセプターの膜貫通領域を用いる場合、例えば、用いられるNK細胞活性化レセプターがKIR2DSであれば、対応するKIR2DLの膜貫通領域は、GenBank Accession No. NM_014218として塩基配列情報に基づいて、膜貫通領域コード領域を増幅し得るプライマーペアをデザインし、PCRを実施することにより調製することができる。
【0026】
ここで使用するプライマーペアは、各増幅断片の末端に、増幅断片同士および増幅断片とベクターとを連結するのを容易にするために、適当な制限酵素認識サイトを導入するようにデザインすることもできる。
【0027】
本発明のキメラレセプターをコードする核酸分子を含む発現ベクターは、例えば、本発明のキメラレセプターを構成するドメインをコードする目的の断片を調製し、該断片を連結し、適切な発現ベクター中のプロモーターの下流に連結することにより作製することができる。
【0028】
発現ベクターの骨格は、例えば、プラスミド又はウイルスベクター(例、アデノウイルス、レトロウイルス、アデノ随伴ウイルス、ヘルペスウイルス、ワクシニアウイルス、ポックスウイルス、ポリオウイルス、シンドビスウイルス、センダイウイルス、レンチウイルスなどのウイルス由来ベクター)であり得る。発現ベクターとして、例えば、IRESバイシストロン性発現ベクター、pA1−11、pXT1、pRc/CMV、pRc/RSV、pcDNAI/Neo、pME18Sなどが使用され得る。発現ベクターとしては、目的遺伝子が選択マーカー又はレポーターと共に同一mRNAから発現される、IRESバイシストロン性発現ベクターが好ましい。IRESバイシストロン性発現ベクターとしては、例えば、pMX−IRES−GFP(Kitamuraら、Int. J. Hematol., 67, 351-9 (1998))、及びClontech社により市販されているベクターなどが挙げられる。
【0029】
発現ベクターで使用されるプロモーターは、細胞傷害性T細胞で機能し得るものであれば特に制限されず、例えば、SV40由来初期プロモーター、サイトメガロウイルスLTR、ラウス肉腫ウイルスLTR、MoMuLV由来LTR、アデノウイルス由来初期プロモーター、LCK近位プロモーター、SRαプロモーター、HSV−TKプロモーターなどのウイルスプロモーター、ならびにβ−アクチン遺伝子プロモーター、PGK遺伝子プロモーター、トランスフェリン遺伝子プロモーターなどの哺乳動物の構成タンパク質遺伝子プロモーターなどが挙げられる。また、プロモーターとして、細胞傷害性T細胞に特異的なプロモーターを用いてもよい。このようなプロモーターは、細胞傷害性T細胞に特異的に発現している任意の遺伝子のプロモーターであり得る。
【0030】
発現ベクターは、必要に応じて、エンハンサー、スプライシングシグナル、ポリA付加シグナル、選択マーカー又はレポーター遺伝子、SV40複製起点(SV40ori)などを含み得る。
【0031】
選択マーカー及びレポーター遺伝子としては、例えば、緑色蛍光タンパク質(GFP)遺伝子、ルシフェラーゼ遺伝子、β−ガラクトシダーゼ遺伝子、ジヒドロ葉酸還元酵素遺伝子、アンピシリン耐性遺伝子、ネオマイシン耐性遺伝子が挙げられる。
【0032】
該発現ベクターの導入宿主となる細胞傷害性T細胞は、細胞表面の自己のMHCクラスI分子を非自己抗原と共に認識して標的細胞を殺傷する(MHC拘束)、獲得免疫系(adaptive immune system)において機能する細胞である。このT細胞はCD8陽性であるので、CD8 T細胞ともいう。細胞傷害性T細胞は、TCR(T cell Receptor)を介してリガンドに結合すると、TCR複合体中のCD3ζのITAM配列を介して活性化シグナルを伝達し、標的細胞を殺傷する。
【0033】
本発明において使用される細胞傷害性T細胞は、任意の動物種由来のものであり得る。このような動物種としては、ヒト、ヒトを除く哺乳動物(例えば、マウス、ラット、ハムスター、モルモットなどのげっ歯類やウサギなどの実験動物、ブタ、ウシ、ヤギ、ウマ、ヒツジなどの家畜、イヌ、ネコなどのペット、サル、オランウータン、チンパンジーなどの霊長類)が挙げられるがこれらに限定されない。本発明においてはヒト及びマウスの細胞傷害性T細胞が好ましく、臨床応用の観点から、ヒトの細胞傷害性T細胞がより好ましい。
【0034】
細胞傷害性T細胞の遺伝子型は特に限定されないが、異常細胞を除去すべき投与対象の動物と同種同系、同種異系又は異種異系であり、好ましくは同種同系又は同種異系である。投与時の拒絶反応を回避するために、この細胞傷害性T細胞は、投与対象と同種同系であることが好ましく、投与対象由来(即ち、自家細胞)であることが最も好ましい。例えば、臓器移植患者などの場合、免疫抑制剤投与前に予め該患者から細胞傷害性T細胞を採取しておき、これに本発明のキメラレセプターを導入し、臓器移植後に該キメラレセプターを発現する細胞傷害性T細胞を該患者に戻すなどの態様が、好ましく例示される。
【0035】
本発明において使用される細胞傷害性T細胞は、自体公知の方法によって、上記動物種の組織(例えば、脾臓、リンパ節、リンパ液、血液など)から単離及び/又は精製することが可能である。例えば、細胞傷害性T細胞上に特異的に発現する細胞表面マーカーに対する抗体を用いて、セルソーター、パニング、抗体磁気ビーズ法などにより単離することができる。
【0036】
本発明において使用される細胞傷害性T細胞は、上記動物種より採取された細胞(例えば、初代培養細胞)又は細胞株であり得る。この細胞傷害性T細胞はポリクローナルであり得る。細胞株は既存の細胞株又は新たに作製される細胞株であり得る。細胞株は自体公知の方法により作製できる。
【0037】
発現ベクターをT細胞に導入(形質導入)する方法は、細胞の種類により適宜選択することができる。例えば、リポフェクション法、マイクロインジェクション法、リン酸カルシウム共沈殿法、PEG法、エレクトロポレーション法、ウイルス感染などが挙げられるが、これらに限定されない。
【0038】
発現ベクターが導入されたT細胞は、上記選択マーカー及びレポーター遺伝子などを利用して、セルソーターなどにより選択することができる。例えば、GFPの発現に基づいてフローサイトメーターによってソートすることができる。
【0039】
選択されたT細胞の培養は、動物細胞の培養に通常用いられる培地を基礎培地として調製した培地中で行うことができる。基礎培地としては、例えば、MEM培地、DMEM培地、αMEM培地、ハム培地、RPMI1640培地、Fischer’s培地、及びこれらの混合培地が挙げられる。培養培地は、例えば、血清(例えば、FCS)、血清代替物(例えば、knockout Serum Replacement(KSR))、脂肪酸又は脂質、アミノ酸、ビタミン、増殖因子、サイトカイン、抗酸化剤、2−メルカプトエタノール、ピルビン酸、緩衝剤、無機塩類などを含むことができる。培養温度、CO2濃度などの他の培養条件は適宜設定できる。培養温度は、特に限定されるものではないが、例えば約30〜40℃、好ましくは約37℃である。また、CO2濃度は、例えば約1〜10%、好ましくは約5%である。培地のpHは通常約6〜8であり、培養時間は通常4〜14日間、好ましくは6〜14日間である。培養における細胞数、各種因子の濃度などのその他の条件は、自体公知の方法により適宜設定できる。
【0040】
上記のようにして調製される本発明のキメラレセプターを発現する細胞傷害性T細胞(以下、「本発明の細胞傷害性T細胞」ともいう)は、該キメラレセプターの細胞外領域に存在するNK細胞レセプターの細胞外ドメインを用いて、異常細胞で特異的に発現するリガンドを認識し、該キメラレセプターの細胞質領域に存在するシグナル伝達アダプター分子部分を介した活性化シグナルによって活性化され、該異常細胞を傷害するので、本発明の細胞傷害性T細胞は、該キメラレセプターが認識し得るリガンドを発現する異常細胞の除去に有用である。したがって、本発明の細胞傷害性T細胞は、該異常細胞を病原とする各種疾患の予防・治療に用いることができる。
本発明に従って予防・治療され得る疾患としては、ウイルス感染症、癌などが挙げられる。本発明の細胞傷害性T細胞は、ウイルス感染症の予防・治療に特に適している。具体的には、本発明の細胞傷害性T細胞により予防・治療されるウイルス感染症としては、CMV(例えば、HCMVまたはMCMVなど)、単純ヘルペスウイルス、EBウイルス、ヒト免疫不全ウイルス、インフルエンザウイルス、SARSウイルスなどの感染症が挙げられるがこれらに限定されない。
【0041】
本明細書中で使用する場合、「急性ウイルス感染症(本明細書中で「急性相感染」ともいう)」とは、あるウイルスによる最初の感染によって引き起こされる疾患をいう。本明細書中で使用する場合、「慢性ウイルス感染症」は、慢性化したウイルス感染症及び再発のウイルス感染症を含み、最初に感染したウイルスが排除されないまま体内に残ること、同じウイルスの再感染などによって引き起こされる。急性ウイルス感染症に対して一次免疫応答が生じ、慢性ウイルス感染症に対して二次免疫応答が生じる。この二次免疫応答には免疫記憶応答が関与している。本明細書中で使用する場合、「長期」とは、免疫記憶の確立によってもたらされる、慢性ウイルス感染症からの保護などをいう。本発明の細胞傷害性T細胞は、急性及び慢性の両方のウイルス感染症を予防・処置することができる。
【0042】
本発明の細胞傷害性T細胞は、単独で、あるいは他の治療のための任意の有効成分との混合物として、薬理学的に許容される一種もしくはそれ以上の担体とともに製剤化することができる。
【0043】
本発明の異常細胞除去剤は、通常は、注射剤、点滴剤などの剤型で提供される。本発明の異常細胞除去剤は、好ましくは、受容者の体液(血液など)と等張な滅菌水性担体中に、本発明のキメラレセプターを発現する細胞傷害性T細胞が懸濁された懸濁液である。水性担体としては、生理食塩水、PBSなどを挙げることができる。これらの水性担体には、更に、必要に応じて溶解補助剤、緩衝剤、等張化剤、無痛化剤、保存剤、安定化剤などを添加することもできる。
【0044】
本発明の剤中に含まれる、本発明のキメラレセプターを発現する細胞傷害性T細胞の濃度は、通常、約1×105〜1×1010個/ml、好ましくは約2×105〜1×109個/mlの範囲であるが、特に限定されない。細胞濃度が低すぎると、投与に時間がかかるため患者への負担が増大し、細胞濃度が高すぎると、細胞同士が凝集してしまう可能性がある。
【0045】
本発明の剤の投与量は、本発明のキメラレセプターの発現量、投与様式、疾患の種類、疾患の重篤度、投与対象となる動物種、投与対象の受容性、体重、年齢などによって異なるが、所望の免疫活性を達成するような細胞数が適宜投与され得、通常、1回の投与につき、本発明のキメラレセプターを発現する細胞傷害性T細胞の数として、通常1×106〜1×109個/m2、好ましくは1×107〜1×109個/m2の用量で投与される。これらの投与量は、前述の種々の条件により変動する。
【実施例】
【0046】
以下の実施例は本発明を具体的に説明するが、本発明を限定するものとして解釈すべきではない。
【0047】
材料及び方法
マウス及び細胞株:
BALB/cマウス及びB6マウスは、Charles River Japanから購入した。B6バックグラウンドを有するRAG−2欠損マウスは、Taconic(Hudson,NY)から購入した。6〜8週齢のマウスを実験に使用した。DBA/2由来の肥満細胞株P815、B6由来の胸腺腫細胞株RMA、Ly49Aに対するmAb(A1)のためのハイブリドーマ及び北村敏雄博士(東京大学)によって提供されたレトロウイルスパッケージング細胞株PLAT−Eをin vitroで維持した。Ly49Hに対するmAbのための3D10ハイブリドーマは、W.M. Yokoyama博士(Washington University,St.Louis)により提供された。
【0048】
レトロウイルスベクター及び形質導入:
Ly49H−CD3ζキメラレセプター、細胞質欠失Ly49H、Ly49A−CD3ζキメラレセプター及び細胞質欠失Ly49Aのコンストラクトを、以前に記載したように(非特許文献8)PCRによって生成した。Ly49H−CD3ζキメラレセプター及び細胞質欠失Ly49Hのコンストラクトについて、Ly49HのC末端細胞外ドメインを、それぞれCD3ζの細胞質ドメインあり又はなしでLy49Aの膜貫通ドメインと融合させて、別のシグナル伝達分子との相互作用を回避した。これらのコンストラクトをpMX−IRES−GFPベクターの第1シストロン中に挿入した。W.Yokoyamaにより提供されたm157のcDNA及びH2DdのcDNAを、それぞれpMX−IRES−GFPベクター又はpMX−IRES−hCD4ベクター中に挿入した(非特許文献8)。上記細胞株へのこれらcDNAの形質導入を、以前に記載された通りに実施した(非特許文献8)。GFP又はhCD4ポジティブな標的細胞を、MoFlowフローサイトメーター(Cytomation)によってソートした。
【0049】
キメラレセプターを発現するポリクローナルCD8 T細胞の生成:
脾臓CD8 T細胞を磁気ビーズ(Miltenyi Biotec,CA)によって精製した。ソートした細胞を、0日目に、200U/mlのrIL−2を含む培地中、5μg/mlのプレート結合抗CD3 Ab及び2μg/mlのモノクローナル抗CD28 Abで刺激した。細胞を1日目及び2日目に感染させた。細胞を3日目に洗浄し、200U/mlのrIL−2と共にもう24時間増殖させた。約50〜60%のCD8 T細胞が4日目にGFPポジティブであった。これらの細胞をin vivoのウイルス保護アッセイに使用した。in vitroの殺傷アッセイのために、Ly49H又はLy49Aの表面発現についてGFPポジティブ細胞をソートすることによって、細胞を精製した。
【0050】
マウスサイトメガロウイルスの調製及び感染:
Smith株のMCMV(VR194)を、American Tissue Culture Collection(Manassas,VA)から得た。ストック溶液を、MCMVに感染したBALB/cマウス由来の唾液腺から、以前に記載されたとおりに公知技術に従って調製した。このMCMVストック懸濁物のウイルス濃度は、以前に報告されたとおりに公知技術に従ってサブコンフルエントの3T3/Swissアルビノ細胞を使用してプラークアッセイによって決定したところ、2.0×107PFU/mlであった。マウスに、ウイルスストックの腹腔内注射によってウイルスを感染させた。
【0051】
ウイルスm157のPCR増幅及び配列:
ゲノムDNAを、一般に使用されている市販のキットを使用して抽出した。ウイルスm157遺伝子を、French et.al.(French, A.R., and W.M. Yokoyama. 2003. Natural killer cells and viral infections. Curr Opin Immunol 15:45-51.)に記載されたプライマーを用いてPCR増幅した。増幅したPCR産物を、TAクローニング(Invitrogen)中にサブクローニングし、配列決定した。m157配列のPCR分析のために、図4B中に示される実験における死にかけのマウスを屠殺し、唾液腺を使用した。
【0052】
実施例1
ポリクローナルCD8 T細胞上でのキメラレセプターの発現:
初代細胞におけるキメラレセプターの機能を試験するために、バイシストロン性レトロウイルスベクターpMX−IRES−GFP(pIG)中にLy49H細胞外ドメイン、Ly49A膜貫通ドメイン及びCD3ζ細胞質ドメインを挿入したLy49H−CD3ζキメラレセプターコンストラクト(Ly49HZ)を調製した。Ly49H発現の困難性及び他のシグナル伝達分子との会合を回避するために、原型的な阻害レセプターLy49A由来の膜貫通ドメインでその膜貫通ドメインを置換した。コントロールとして、Ly49H細胞外ドメイン及びLy49A膜貫通ドメインを有するがCD3ζ細胞質ドメインを有さない、細胞質欠失Ly49Hコンストラクト(Ly49Hcyto−del)もまた調製した。レトロウイルスの系を使用して、これらのキメラレセプターをMCMV感受性BLAB/c株(H−2dハプロタイプ)由来のCD8 T細胞中に形質導入した。形質導入後に約50〜60%のGFPポジティブ細胞が得られ、これらのGFPポジティブ細胞の殆どがLy49H特異的mAbによってポジティブに染色された。GFPポジティブ細胞をエフェクター細胞としてソートしたところ、それらのうち90%より多くがLy49Hポジティブであった(図1A)。標的細胞としてDBA2由来のP815(H−2d)を準備した。P815中にpMX−m157−IRES−GFP及びpMX−IRES−GFP(コントロールとして)を形質導入し、それぞれP815−m157及びP815−vectorとした(図1B)。これらのエフェクター細胞及び標的細胞を使用して、in vitroの51Cr放出アッセイを実施した。Ly49HZレセプターを形質導入したCD8 T細胞は、P185−m157を殺傷する能力を獲得したが、Ly49Hcyto−delを形質導入したCD8 T細胞はこの能力を獲得できなかった。このことは、ポジティブなシグナルが、Ly49Hレセプターとm157との結合によって促進される他のシグナル伝達分子を介したものではなく、実際にキメラレセプターのCD3ζ部分を介したものであることを示している(図1C)。形質導入したキメラレセプターの特異性のさらなる支持として、抗Ly49H特異的mAbは、P185−m157の殺傷をブロックしたが、コントロールAbはブロックしなかった(図1C及びデータ示さず)。特に、これらの形質導入されたCD8 T細胞は、親P185細胞及びP185−vector形質導入コントロール細胞に対する寛容を維持した。これらのデータは、Ly49H−CD3ζキメラレセプターを形質導入したポリクローナルCD8 T細胞がm157を特異的に認識して標的細胞を殺傷することを示す。
【0053】
実施例2
ネイティブのレセプター機能の変換:
m157タンパク質は、129/Jマウスにおいて発現されるLy49ファミリーのメンバーである阻害NK細胞レセプターLy49I129に結合することが示されている(Arase, H., E.S. Mocarski, A.E. Campbell, A.B. Hill, and L.L. Lanier. 2002. Direct recognition of cytomegalovirus by activating and inhibitory NK cell receptors. Science296:1323-1326.)。C57BL/6由来のLy49AB6(Ly49A)は、このファミリーメンバー中の原型的なNK細胞阻害レセプターである。Ly49AはMHCクラスIのH−2Ddを認識し、その特異性は、マウスNK細胞レセプターの中で最も特徴付けられている(Karlhofer, F.M., R.K. Ribaudo, and W.M. Yokoyama. 1992. MHC class I alloantigen specificity of Ly-49+ IL-2-activated natural killer cells. Nature 358:66-70.、Tormo, J., K. Natarajan, D.H. Margulies, and R.A. Mariuzza. 1999. Crystal structure of a lectin-like natural killer cell receptor bound to its MHC class I ligand. Nature402:623-631.)。Ly49H−CD3ζキメラレセプターを用いてポリクローナルCD8 T細胞の特異性を変化させたので、次に、in vitroの51Cr放出アッセイにおいて、Ly49AとCD3ζとのキメラレセプターを操作することによって初代CD8 T細胞においてネイティブのレセプター機能を変換できるか否かを試験した。Ly49Aの細胞外ドメインと膜貫通ドメインとをCD3ζ細胞質ドメインと融合させて作製したLy49A−CD3ζキメラレセプターコンストラクト(Ly49AZ)を調製した。コントロールとして、Ly49Aの細胞外ドメイン及び膜貫通ドメインのみで作製した細胞質欠失Ly49A(Ly49Acyto−del)もまた調製した。標的細胞として、C57BL6(B6)由来のT細胞株であるRMAを準備した。RMA中にH−2Dd−IRES−GFP及びIRES−GFP(形質導入コントロールとして)を形質導入し、それぞれ、RMA−H−2Dd及びRMA−GFPとした。Ly49Aの内因性リガンドを発現しないC57BL6マウス由来のCD8 T細胞中に、Ly49Aキメラレセプターを形質導入した(図2A)。細胞質欠失Ly49Aレセプターを形質導入したCD8 T細胞はRMA−H2Ddを殺傷することができなかった。一方で、Ly49AZレセプターを形質導入したCD8 T細胞はRMA−H2Ddを有効に殺傷したが、RMAもRMA−GFPも殺傷しなかった(図2B)。さらに、RMA−H2Ddの殺傷は、Ly49A特異的mAbであるA1によってブロックされた(図2C)。これらのデータは、Ly49AZレセプターを形質導入したポリクローナルCD8 T細胞がそのリガンドを特異的に認識して標的細胞を殺傷することを示す。合わせて考えると、これらのデータは、細胞質CD3ζと融合したNK細胞レセプターを形質導入したCD8 T細胞が、そのレセプターのネイティブの性質(阻害又は活性化)に関わらず、ネイティブのNK細胞レセプターの特異性を獲得して、標的細胞を殺傷できることを示す。
【0054】
実施例3
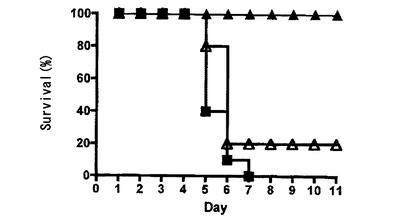
Ly49Hキメラレセプターを発現するCD8 T細胞によってMCMV感染からレスキューされたBALB/cマウス
Ly49H−CD3ζキメラレセプターを形質導入したポリクローナルCD8 T細胞によって、m157発現細胞のin vitroでの首尾よい殺傷がもたらされたので、本発明者は、これらの細胞がin vivoでMCMV感染を制御する能力を評価した。Ly49Hの発現を欠くBALB/cマウスはMCMV感染に対して感受性であり、1×104pfuのMCMVでの接種後2週間以内に死亡した(図3A)。BALB/cマウス由来のCD8 T細胞を準備し、Ly49HZ又はLy49Hcyto−delのいずれかを含むレトロウイルスをこの細胞に形質導入した。MCMV感染の当日、Ly49HZレセプターを発現するCD8 T細胞1×107を注入し、次いで、致死用量のMCMVをマウスに感染させた。細胞質欠失Ly49Hを発現するコントロールCD8 T細胞を注入したマウス及び細胞を注入していないマウスは10日以内に死亡したが、Ly49HZレセプターを発現するCD8 T細胞を注入したマウスは致死用量のMCMV感染の急性相を生存した(図3A)。このことと一致して、予備研究(データ示さず)において、注入したキメラレセプター発現細胞の数と生存との間の正の相関が観察された。生存マウスは、最初の感染の1ヵ月後に健康なままであった。
【0055】
実施例4
ウイルス感染の有効な制御は、自然免疫及び獲得免疫の両方を必要とする。従って、最初のチャレンジと同じウイルス用量のMCMVでマウスを再チャレンジすることによって、これらの生存したマウスが獲得免疫を発達させたか否かをさらに試験した。最初の感染の40日後に、CD8 T細胞をさらに注入することなくマウスを再チャレンジし、生存を観察した。ナイーブなBALB/cマウスは10日以内に死亡したが、再チャレンジしたマウスは全て生存し、このことは、これらのマウスにおける有効な記憶応答の確立を示す(図3B)。記憶応答における注入されたCD8 T細胞の寄与を評価するために、再チャレンジの10日後に生存マウスの脾臓中のCD8 T細胞を評価した。興味深いことに、CD8集団中にGFPポジティブ細胞もLy49Hポジティブ細胞も検出されなかった(図3C)。再チャレンジの7日後に分析した脾細胞からは、GFPポジティブ細胞は観察されなかった(データ示さず)。
【0056】
実施例5
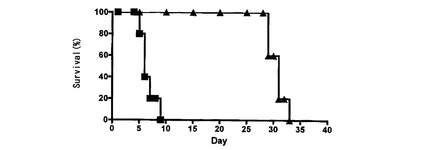
RAG−2欠損B6マウスの一過的なレスキュー:
上記実験において、移入した全てのCD8 T細胞を、形質導入前にα−CD3 mAb及びα−CD28 mAbで刺激したところ、注入したCD8 T細胞の約40〜50%はGFP発現についてネガティブであった。内因性T細胞又は注入されたT細胞のいずれが抗CMV記憶応答に寄与しているかは明らかではなく、この記憶の形成が生存に如何に寄与したかも明らかではなかった。これらの問題点に取り組むため、B6 RAG2欠損(B6−RAG2)マウスを使用した。B6−RAG2マウスはT細胞及びB細胞を欠くので、内因性T細胞からCMVクリアランスに必要な獲得記憶応答を形成することができない。Ly49HZを形質導入した後に、B6マウス由来のCD8 T細胞をB6−RAG2マウス中に移入した。B6−RAG2マウスはLy49Hを発現するNK細胞を有するのでMCMVに対して耐性であるため、より高い用量のMCMV(マウス1匹当たり1×107pfu)でチャレンジした。コントロールとして、CD8 T細胞中にLy49Hcyto−delを形質導入し、これらをRAG2欠損B6マウス中に注入した。BALB/cモデルと同様に、全てのRAG2欠損B6マウス、及びコントロールの細胞質欠失Ly49Hを発現するCD8 T細胞を注入したRAG2欠損B6マウスの殆どは10日以内に死亡したが、Ly49HZを発現するCD8 T細胞を移入したRAG2欠損B6マウスは生存した(図4A)。これらのデータは、急性相感染が、移入されたCD8 T細胞によって実際に制御されたことを示す。第2の実験において、第1の実験と同じ短期生存パターンが観察された。しかし、感染の1ヵ月後、全ての生存マウスが死亡した(図4B)。これらのデータは、注入されたCD8 T細胞自体が長期記憶を形成することができたわけではないことを示唆する。この遅延相の死亡の機構に取り組むために、死にかけのRAG2欠損B6マウス(n=1)から脾臓及び唾液腺を単離した。平均ウイルス力価は、脾臓、肝臓、唾液腺及び肺においてそれぞれ4.4×106、2.0×107、3.6×108及び5.7×107であり、このことは、遅延相の死亡がMCMVの再活性化に起因するものであることを示唆する。
【0057】
本明細書において、致死用量のMCMVでチャレンジしたマウスをレスキューするための、首尾よい細胞免疫学的ストラテジーを記載してきた。内因性のNK細胞が致死的なウイルスチャレンジからB6−RAG−2マウスを保護できない条件において、ウイルスタンパク質に特異的なキメラレセプターを発現するCD8 T細胞の注入により、この致死的なチャレンジが克服された。Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞によって生存したBALB/cマウスは、高用量のウイルス再チャレンジを明らかに制御し、このことは、記憶細胞の有効な増殖及び細胞傷害活性を示唆する。従って、本明細書中に提供されるデータは、キメラ活性化レセプター遺伝子を移入したポリクローナルCD8 T細胞が、NK細胞の機能を模倣することによって、自然免疫応答の重要な役割を果たし得ることを示している。さらに、in vitroデータは、ネイティブの阻害レセプターが、その細胞質ドメインをCD3ζの細胞質ドメインと交換することによって、初代CD8 T細胞において活性化レセプターに変換され得ることもまた示唆している。
【0058】
ペプチド特異的CD8 T細胞の生成が所望され、免疫無防備の移植患者において疾患を引き起こすいくつかのウイルスについて確立されている(Riddell, S.R., and P.D. Greenberg. 1995. Principles for adoptive T cell therapy of human viral diseases. Annual Review of Immunology 13:545-586.)。ヒト同種移植の設定において所定のMHC拘束様式であつらえたペプチド特異的CD8 T細胞の生成は、リガンドの認識についてMHC拘束を必要とするが、CD3ζの細胞質ドメインを有するウイルスタンパク質特異的NK細胞キメラレセプターは、リガンドの認識についてMHC拘束を必要としない。ヒト及びマウスの両方において、末梢中の細胞の最初の割合が異なるので、多数のT細胞を得ることはNK細胞よりも容易である。
移入されたCD8 T細胞は急性相感染を有効に制御し、これは、有効な長期獲得免疫を導く。本明細書中のデータは、CMV感染に対する獲得免疫応答が内因性T細胞によって媒介されることを示唆する。
【産業上の利用可能性】
【0059】
本発明のキメラレセプターを発現する細胞傷害性T細胞は、NK細胞によるMHC非拘束的なリガンド認識により、ウイルス感染症や癌などにおける異常細胞(即ち、ウイルス感染細胞や癌細胞)を認識してこれを傷害することができ、なおかつウイルス感染や癌からの長期的な保護をもたらし得るので、急性及び慢性のウイルス感染症、あるいは癌およびその再発を予防・治療するために有用である。
【図面の簡単な説明】
【0060】
【図1A】Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞によるm157ポジティブ標的細胞の特異的殺傷を示す図である。BALB/c CD8 T細胞に、細胞質欠失Ly49H又はLy49H−CD3ζキメラレセプターのいずれかを含むレトロウイルスを感染させた。細胞を、そのGFP発現によってエフェクター細胞としてソートした。ソートした細胞上のCD8、GFP及びLy49Hの発現が示される。
【図1B】Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞によるm157ポジティブ標的細胞の特異的殺傷を示す図である。P815細胞に、m157−IRES−GFP又はIRES−GFPのいずれかを形質導入し、そのGFP発現に基いてソートした。各形質導入体中のGFP発現が示される。
【図1C】Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞によるm157ポジティブ標的細胞の特異的殺傷を示す図である。51Cr放出アッセイを、m157−IRES−GFPを発現するP815細胞(丸)又はIRES−GFPを発現するP815細胞(四角)を標的細胞として用い、Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞(白抜き記号)又は細胞質欠失Ly49Hを発現するCD8 T細胞(黒塗り記号)をエフェクター細胞として用いて実施した。Ly49H Ab(培養物上清の1/20希釈)の存在下での細胞傷害活性を白抜きの三角で示した。4回の独立した実験からの代表的なデータが示される。別個の実験において、標的及びエフェクターの各組み合わせについて、コントロールmAbの存在下では効果は観察されなかった(データ示さず)。
【図2A】CD3ζキメラレセプターとの融合による、ネイティブの阻害レセプター機能の活性化レセプターへの変換を示す図である。B6 CD8 T細胞に、細胞質欠失Ly49A又はLy49A−CD3ζキメラレセプターのいずれかを含むレトロウイルスを感染させた。細胞を、そのGFP発現によってエフェクター細胞としてソートし、Ly49A表面発現について分析した。
【図2B】CD3ζキメラレセプターとの融合による、ネイティブの阻害レセプター機能の活性化レセプターへの変換を示す図である。RMA細胞に、コントロールとしてのIRES−ヒトCD4(hCD4)又はH2Dd−IRES−hCD4のいずれかを形質導入し、hCD4発現によってソートした。これらの細胞のH−2Dd及びhCD4の発現を、細胞溶解アッセイの前に決定した。
【図2C】CD3ζキメラレセプターとの融合による、ネイティブの阻害レセプター機能の活性化レセプターへの変換を示す図である。51Cr放出アッセイを、IRES−hCD4を形質導入したRMA細胞(四角)又はH2Dd−IRES−hCD4を形質導入したRMA細胞(丸及び三角)を標的細胞として用い、Ly49A−CD3ζキメラを発現するCD8 T細胞(白抜き記号)又は細胞質欠失Ly49Aを発現するCD8 T細胞(黒塗り記号)をエフェクター細胞として用いて実施した。Ly49A Ab(培養物上清の1/20希釈)の存在下での細胞傷害活性を三角で示す。3回の独立した実験からの代表的なデータが示される。別個の実験において、標的及びエフェクターの各組み合わせについて、アイソタイプが一致したコントロールmAbの存在下では効果は観察されなかった(データ示さず)。
【図3A】Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞による致死的MCMV感染からのBALB/cマウスの保護、及び記憶応答の確立を示す図である。Ly49H−CD3ζキメラ(黒塗り三角)を発現するか、細胞質欠失Ly49H(白抜き三角)を発現するか、又は何も発現しない(黒塗り四角)5×106のCD8 T細胞を、BALB/cマウス(1群当たり10匹)中に注射した。次いで、マウスにMCMV(マウス1匹当たり104PFU)を感染させ、生存についてモニタリングした。
【図3B】Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞による致死的MCMV感染からのBALB/cマウスの保護、及び記憶応答の確立を示す図である。Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞によって最初のMCMV感染から保護された10匹のBALB/cマウス(黒塗り三角)を、最初の感染の40日後に、CD8 T細胞をさらに注入することなくMCMVで再チャレンジした。コントロールとして、ナイーブな未感染BALB/cマウス(黒塗り四角)(1群当たり5匹のマウス)に、同量のウイルスを感染させた。2回の実験からの代表的なデータが示される。
【図3C】Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞による致死的MCMV感染からのBALB/cマウスの保護、及び記憶応答の確立を示す図である。図3Bと同様に処理したマウス由来の脾細胞(黒塗り三角の群)を、再チャレンジの10日後にGFPポジティブなCD8 T細胞の存在について分析した。3匹のマウスからの代表的なデータが示される。
【図4A】Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞による、致死的なMCMV感染からの免疫不全マウスの保護を示す図である。Ly49H−CD3ζキメラ(黒塗り三角)を発現するか、細胞質欠失Ly49H(白抜き三角)を発現するか、又は何も発現しない(黒塗り四角)5×106のCD8 T細胞を、B6−RAG2欠損マウス(1群当たり5匹)中に注射した。次いで、マウスにMCMV(マウス1匹当たり107PFU)を感染させ、生存についてモニタリングした。
【図4B】Ly49H−CD3ζキメラレセプターを発現するCD8 T細胞による、致死的なMCMV感染からの免疫不全マウスの保護を示す図である。マウス(1群当たり5匹)を、図4Aと同様に処理し、延長された期間にわたってモニタリングした。
【特許請求の範囲】
【請求項1】
生体にとって望ましくない異常細胞の除去剤であって、
(a)該異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメインと、
(b)ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部と、
を含むキメラレセプターを発現する細胞傷害性T細胞を含有してなる剤。
【請求項2】
NK細胞レセプターが活性化レセプターである、請求項1に記載の剤。
【請求項3】
異常細胞がウイルス感染細胞である、請求項1又は2に記載の剤。
【請求項4】
ウイルスがサイトメガロウイルスである、請求項3に記載の剤。
【請求項5】
サイトメガロウイルスがヒトサイトメガロウイルスである、請求項4に記載の剤。
【請求項6】
NK細胞レセプターがヒト由来である、請求項1〜5のいずれかに記載の剤。
【請求項7】
NK細胞レセプターがNKRP1A、NKRP1C、NKRP1F、KIR2DS、KIR3DS、NKG2C、NKG2DおよびNKG2Eからなる群より選択される、請求項6に記載の剤。
【請求項8】
サイトメガロウイルスがマウスサイトメガロウイルスである、請求項4に記載の剤。
【請求項9】
NK細胞レセプターがマウス由来である、請求項1〜5及び8のいずれかに記載の剤。
【請求項10】
NK細胞レセプターがLy49H、Ly49D、NKRP1A、NKRP1C、NKRP1F、NKG2C、NKG2DおよびNKG2Eからなる群より選択される、請求項9に記載の剤。
【請求項11】
NK細胞レセプターがLy49Hである、請求項10に記載の剤。
【請求項12】
シグナル伝達アダプター分子がCD3ζ、DAP12/KARAPおよびFcRγからなる群より選択される、請求項1〜11のいずれかに記載の剤。
【請求項13】
シグナル伝達アダプター分子がCD3ζである、請求項12に記載の剤。
【請求項14】
キメラレセプターの膜貫通ドメインがNK細胞活性化レセプター由来でない、請求項1〜13のいずれかに記載の剤。
【請求項15】
急性ウイルス感染症の予防・治療用である、請求項1〜14のいずれかに記載の剤。
【請求項16】
慢性ウイルス感染症の予防・治療用である、請求項1〜14のいずれかに記載の剤。
【請求項17】
生体にとって望ましくない異常細胞の除去方法であって、
(a)該異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメインと、
(b)ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部と、
を含むキメラレセプターを発現する細胞傷害性T細胞の有効量を、該異常細胞を有する生体に投与することを含む方法。
【請求項18】
(a)ウイルス感染ヒト細胞で特異的に発現するリガンドを認識するヒトNK細胞レセプターの細胞外ドメインと、
(b)ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部と、
を含むキメラレセプタータンパク質。
【請求項19】
ウイルスがヒトサイトメガロウイルスである、請求項18に記載のタンパク質。
【請求項20】
NK細胞レセプターが活性化レセプターである、請求項18又は19に記載のタンパク質。
【請求項21】
NK細胞レセプターがNKRP1A、NKRP1C、NKRP1F、KIR2DS、KIR3DS、NKG2C、NKG2DおよびNKG2Eからなる群より選択される、請求項18〜20のいずれかに記載のタンパク質。
【請求項22】
シグナル伝達アダプター分子がCD3ζ、DAP12/KARAPおよびFcRγからなる群より選択される、請求項18〜21のいずれかに記載のタンパク質。
【請求項23】
シグナル伝達アダプター分子がCD3ζである、請求項22に記載のタンパク質。
【請求項24】
前記キメラレセプターの膜貫通ドメインがNK細胞活性化レセプター由来でない、請求項18〜23のいずれかに記載のタンパク質。
【請求項25】
請求項18〜24のいずれかに記載のタンパク質をコードする核酸。
【請求項26】
請求項25記載の核酸を含む、ヒト細胞傷害性T細胞で機能し得る発現ベクター。
【請求項27】
請求項26記載の発現ベクターでトランスフェクトされたヒト細胞傷害性T細胞。
【請求項1】
生体にとって望ましくない異常細胞の除去剤であって、
(a)該異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメインと、
(b)ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部と、
を含むキメラレセプターを発現する細胞傷害性T細胞を含有してなる剤。
【請求項2】
NK細胞レセプターが活性化レセプターである、請求項1に記載の剤。
【請求項3】
異常細胞がウイルス感染細胞である、請求項1又は2に記載の剤。
【請求項4】
ウイルスがサイトメガロウイルスである、請求項3に記載の剤。
【請求項5】
サイトメガロウイルスがヒトサイトメガロウイルスである、請求項4に記載の剤。
【請求項6】
NK細胞レセプターがヒト由来である、請求項1〜5のいずれかに記載の剤。
【請求項7】
NK細胞レセプターがNKRP1A、NKRP1C、NKRP1F、KIR2DS、KIR3DS、NKG2C、NKG2DおよびNKG2Eからなる群より選択される、請求項6に記載の剤。
【請求項8】
サイトメガロウイルスがマウスサイトメガロウイルスである、請求項4に記載の剤。
【請求項9】
NK細胞レセプターがマウス由来である、請求項1〜5及び8のいずれかに記載の剤。
【請求項10】
NK細胞レセプターがLy49H、Ly49D、NKRP1A、NKRP1C、NKRP1F、NKG2C、NKG2DおよびNKG2Eからなる群より選択される、請求項9に記載の剤。
【請求項11】
NK細胞レセプターがLy49Hである、請求項10に記載の剤。
【請求項12】
シグナル伝達アダプター分子がCD3ζ、DAP12/KARAPおよびFcRγからなる群より選択される、請求項1〜11のいずれかに記載の剤。
【請求項13】
シグナル伝達アダプター分子がCD3ζである、請求項12に記載の剤。
【請求項14】
キメラレセプターの膜貫通ドメインがNK細胞活性化レセプター由来でない、請求項1〜13のいずれかに記載の剤。
【請求項15】
急性ウイルス感染症の予防・治療用である、請求項1〜14のいずれかに記載の剤。
【請求項16】
慢性ウイルス感染症の予防・治療用である、請求項1〜14のいずれかに記載の剤。
【請求項17】
生体にとって望ましくない異常細胞の除去方法であって、
(a)該異常細胞で特異的に発現するリガンドを認識するNK細胞レセプターの細胞外ドメインと、
(b)ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部と、
を含むキメラレセプターを発現する細胞傷害性T細胞の有効量を、該異常細胞を有する生体に投与することを含む方法。
【請求項18】
(a)ウイルス感染ヒト細胞で特異的に発現するリガンドを認識するヒトNK細胞レセプターの細胞外ドメインと、
(b)ITAM配列を含むシグナル伝達アダプター分子の全部もしくは一部と、
を含むキメラレセプタータンパク質。
【請求項19】
ウイルスがヒトサイトメガロウイルスである、請求項18に記載のタンパク質。
【請求項20】
NK細胞レセプターが活性化レセプターである、請求項18又は19に記載のタンパク質。
【請求項21】
NK細胞レセプターがNKRP1A、NKRP1C、NKRP1F、KIR2DS、KIR3DS、NKG2C、NKG2DおよびNKG2Eからなる群より選択される、請求項18〜20のいずれかに記載のタンパク質。
【請求項22】
シグナル伝達アダプター分子がCD3ζ、DAP12/KARAPおよびFcRγからなる群より選択される、請求項18〜21のいずれかに記載のタンパク質。
【請求項23】
シグナル伝達アダプター分子がCD3ζである、請求項22に記載のタンパク質。
【請求項24】
前記キメラレセプターの膜貫通ドメインがNK細胞活性化レセプター由来でない、請求項18〜23のいずれかに記載のタンパク質。
【請求項25】
請求項18〜24のいずれかに記載のタンパク質をコードする核酸。
【請求項26】
請求項25記載の核酸を含む、ヒト細胞傷害性T細胞で機能し得る発現ベクター。
【請求項27】
請求項26記載の発現ベクターでトランスフェクトされたヒト細胞傷害性T細胞。
【図1A】


【図1B】
【図1C】
【図2A】
【図2B】
【図2C】
【図3A】
【図3B】
【図3C】
【図4A】
【図4B】
【図1B】
【図1C】
【図2A】
【図2B】
【図2C】
【図3A】
【図3B】
【図3C】
【図4A】
【図4B】
【公開番号】特開2008−5722(P2008−5722A)
【公開日】平成20年1月17日(2008.1.17)
【国際特許分類】
【出願番号】特願2006−177195(P2006−177195)
【出願日】平成18年6月27日(2006.6.27)
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【Fターム(参考)】
【公開日】平成20年1月17日(2008.1.17)
【国際特許分類】
【出願日】平成18年6月27日(2006.6.27)
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【Fターム(参考)】
[ Back to top ]