
キャンディダ・ユティリスを用いた有用物質の製造法
【課題】キャンディダ・ユティリスの培養において、培養液のpHを調節するための方法の提供。
【解決手段】キャンディダ・ユティリスの培養中に培養液のpHを上方に調節するための方法であって、培養液中に硝酸塩を添加することを含んでなる方法。
【解決手段】キャンディダ・ユティリスの培養中に培養液のpHを上方に調節するための方法であって、培養液中に硝酸塩を添加することを含んでなる方法。
【発明の詳細な説明】
【発明の背景】
【0001】
技術分野
本発明は、クラブトゥリー陰性酵母であるキャンディダ・ユティリス(Candida utilis)を宿主とした有用物質の製造方法に関する。
【0002】
背景技術
将来的に化石資源が枯渇するリスクに鑑み、近年は、化成品およびその素材を、植物バイオマスを原料に用いて発酵法で生産する取り組みに注目が集められている。プラスチックもその1つであり、その素材として生物の代謝物である有機酸を利用することができる。有機酸を素材とするプラスチックの例としては、例えば、乳酸の重合体であるポリ乳酸がある。
【0003】
乳酸にはL−体とD−体がある。高い融点を有するポリ乳酸を製造するためには、光学純度の高いL−体またはD−体の乳酸を重合させる必要がある。また、ポリL乳酸とポリD乳酸を等量混ぜてステレオコンプレックスとすることにより、融点をさらに高めることが可能である。このため、光学純度の高いL−乳酸とD−乳酸をそれぞれ独立に生産することが求められている。すなわち、D−乳酸の発酵生産系がステレオコンプレックス型の乳酸製造の鍵である。
【0004】
乳酸の発酵生産においては、産物である乳酸の蓄積に伴ってpHが極端に酸性域に偏ることにより、菌体の活性が低下し、あるいは菌が死滅してしまうという問題がある。したがって、乳酸を高濃度で生産するためには、菌は、高い発酵能を有するだけでなく、高濃度の乳酸に対して耐性を有することが求められる。
【0005】
工業的な乳酸の発酵生産には、pHの極端な低下を避けるために、炭酸カルシウムを始めとする中和剤が一般的に用いられている。しかしながら、化学プロセスを経て合成される中和剤の使用は、環境負荷の低減というバイオマス・プラスチックの生産コンセプトに必ずしも合致しない。これは、ピルビン酸[非特許文献1:Appl Environ Microbiol Vol.70: p159-66 (2004)]、リンゴ酸やコハク酸[非特許文献2:Appl Environ Microbiol Vol.74: p.2766-77 (2008)]といった乳酸以外の各種有機酸を発酵生産する場合にも共通する課題である。
【0006】
酵母のうち、今日まで最も良く研究され、遺伝学的知見が蓄積しているものにはサッカロマイセス属酵母があり、この酵母は種々の物質生産の宿主として検討がなされている。
また、近年、サッカロマイセス属酵母以外の酵母としてピキア属酵母、ハンセヌラ属酵母、クルイベロマイセス属酵母、キャンディダ属酵母などのいくつかの種について、それらを形質転換する手法が開発され、有用物質生産の宿主として検討されている。このうち、キャンディダ属酵母は、特に、炭素資化域が広いなど、サッカロマイセス属酵母にない特性を有している。
【0007】
キャンディダ属酵母の中でも、キャンディダ・ユティリスは、キシロースをはじめとするペントースに対する優れた資化性を示す。また、サッカロマイセス酵母と異なり、好気的条件下での培養でエタノールを製造せず、それによる増殖阻害も受けないことから、高密度での連続培養による効率的な菌体製造が可能である。従って、キャンディダ・ユティリスは、かつて蛋白質源として注目され、ペントースを多く含む広葉樹の糖化液や亜硫酸パルプ廃液を糖源とした菌体の工業生産が行われていた。また、この酵母は米国FDA(Food and Drug Administration)により、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)、サッカロマイセス・フラジリス(Saccharomyces fragilis)とともに、食品添加物として安全に使用できる酵母として認められている。実際に、キャンディダ・ユティリスは、ドイツをはじめとして、米国や台湾、ブラジルなど世界各国で製造され、食飼料として使用されている。また、このような微生物蛋白質としての利用以外にも、キャンディダ・ユティリスは、ペントースやキシロースの発酵株、エチルアセテート、L−グルタミン、グルタチオン、インベルターゼ等の製造株として広く産業界で利用されてきた。
【0008】
【非特許文献1】Appl Environ Microbiol Vol.70: p159-66 (2004)
【非特許文献2】Appl Environ Microbiol Vol.74: p.2766-77 (2008)]
【発明の概要】
【0009】
本発明者らは、キャンディダ・ユティリスを培養する際に、培養液中に硝酸塩を添加することにより、培養液のpHが経時的に上昇することを見出した。さらに、本発明者らは、キャンディダ・ユティリスの培養によって有機酸を製造する際に、培養液中に硝酸塩を添加することにより、有機酸の生成に伴う培養液のpHの低下が抑制されることを見出した。さらに、本発明者らは、遺伝子操作により、D−乳酸を生産するキャンディダ・ユティリス酵母菌株を開発することに成功した。本発明は、これらの知見に基づくものである。
【0010】
従って、本発明の目的は、キャンディダ・ユティリスの培養において、培養液のpHを調節するための方法およびpH調節剤、キャンディダ・ユティリスの培養によって有機酸を製造する方法、ならびにD−乳酸を高い効率で製造する酵母菌株を提供することにある。
【0011】
本発明によるpH調節法は、キャンディダ・ユティリスの培養中に培養液のpHを上方に調節するための方法であって、培養液中に硝酸塩を添加することを含んでなる方法である。
【0012】
本発明による有機酸の製造法は、キャンディダ・ユティリスを培養することにより有機酸を製造する方法であって、培養液中に硝酸塩を添加することを含んでなる方法である。
【0013】
本発明によるpH調節剤は、硝酸塩を含んでなる、キャンディダ・ユティリスの培養中に培養液のpHを上方に調節するためのpH調節剤である。
【0014】
本発明による酵母菌株は、D−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子の発現を可能にするプロモーター配列が機能的に結合した該遺伝子の少なくとも1コピーにより形質転換されてなり、かつ、ピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードしている内因性遺伝子が破壊されている、キャンディダ・ユティリスの酵母菌株である。
【0015】
本発明によれば、キャンディダ・ユティリスの培養において培養液のpHを上方に調節することができる。このような効果は、例えば、キャンディダ・ユティリスの培養によって有機酸を製造する場合などに、有機酸の生成に伴うpHの低下によってもたらされる悪影響、例えば菌体の死滅、を防止するのに有利である。また、このようなpHの低下を防止するために中和剤を必要としない点も、本発明の利点である。さらに、本発明による酵母菌株のような、D−乳酸を高濃度で生産するキャンディダ・ユティリス株の培養において、上記の効果は特に有利である。
【発明の具体的説明】
【0016】
本発明によるpH調節法は、キャンディダ・ユティリスの培養中に培養液のpHを上方に調節するための方法である。該方法は、培養液中に硝酸塩を添加することを含んでなる。
【0017】
本明細書において、「pHを上方に調節する」という用語およびその派生語は、培養液に硝酸塩を添加しない場合と比較して、培養液のpHをアルカリ側に調節することを意味する。すなわち、この用語は、培養開始から経時的にpHが上昇することを意味するものではなく、pHの経時変化を示すグラフの曲線が、硝酸塩を添加しない場合と比べてアルカリ側にあることを意味する。従って、培養開始から経時的にpHが酸性側に低下したとしても、硝酸塩を添加しない場合と比較してアルカリ側にあれば、それは「pHを上方に調節する」ことに該当する。
【0018】
本発明に用いられる酵母は、クラブトゥリー陰性酵母であるキャンディダ・ユティリスである。キャンディダ・ユティリスの菌株は当技術分野において公知の様々な株、例えば、NBRC0626株、NBRC0639株、NBRC0988株、NBRC1086株等であってよい。
【0019】
本発明に用いられる硝酸塩は、キャンディダ・ユティリスが資化しうるものであればよく、特に制限されない。このような硝酸塩は当技術分野において周知であるが、好ましくは硝酸ナトリウム、硝酸カリウムもしくは硝酸カルシウム、またはこれらの任意の組み合わせとすることができ、より好ましくは硝酸ナトリウムとされる。
【0020】
硝酸塩の培養液への添加量は、キャンディダ・ユティリスの培養に用いられる通常の窒素源の量であればよく、特に制限されない。よって、硝酸塩の添加量は、当業者であれば適宜決定することができるが、好ましくは、硝酸塩に由来する硝酸イオンの濃度として1〜80mM、より好ましくは20〜80mM、最も好ましくは約50mMとすることができる。
【0021】
硝酸塩の培養液への添加の時期は、培養液のpHを上方調節したい時など、当業者であれば適切に選択することができるため、特に制限されない。例えば、硝酸塩は、培養開始時に既に添加されていてもよく、培養開始後の培養の途中で添加されてもよい。また、硝酸塩は、培養開始時に添加しておき、培養の途中でさらに添加してもよい。
【0022】
培養液は、硝酸塩以外の他の窒素源を含んでいてもよく、これにより硝酸塩によるpH調節効果が損なわれることはない。しかし、高いpH調節効果を得るためには、培養液中の硝酸塩以外の窒素源はできるだけ少ない方が好ましい。例えば、硝酸塩以外の窒素源のモル濃度は、硝酸塩に由来する硝酸イオンのモル濃度と同じか、または少ないことが好ましい。本発明の好ましい実施態様によれば、添加される硝酸塩は、キャンディダ・ユティリスによって資化される主要な窒素源として培養液に存在する。ここで、「主要な窒素源」との用語は、資化される窒素の50%以上が硝酸塩に由来することを意味する。本発明の特に好ましい実施態様によれば、添加される硝酸塩は、キャンディダ・ユティリスによって資化される実質的に唯一の窒素源として培養液に存在する。ここで、「実質的に」との用語は、硝酸塩以外の窒素源の量がキャンディダ・ユティリスの生育に寄与しうる程度の量に満たないこと、すなわち、その培養液から硝酸塩を除いた場合にキャンディダ・ユティリスが生育できないことを意味する。
【0023】
培養液の窒素源以外の組成は、キャンディダ・ユティリスの培養に用いられる通常の培地に倣って決定することができる。例えば、培養液に含まれる炭素源としては、例えば、グルコースのほかにもキシロースやスクロースでも資化できれば用いることができる。また、培養液に含まれる栄養源としては、例えば、酵母エキス、ペプトン、ホエーなどが用いられる。さらに、培養液は、無機物栄養源として、リン酸カリウム、硫酸マグネシウムやFe(鉄)、Mn(マンガン)化合物などを含んでもよい。また、培養液はSC培地などの完全合成培地であってもよいが、炭素源として廃糖蜜および/または亜硫酸パルプ廃液を含み、窒素源としてペプトン類を含む天然培地であってもよい。
【0024】
上述したように、pHの低下を防止するための中和剤を必要としない点も、本発明の利点である。よって、本発明の好ましい実施態様によれば、用いられる培養液は、中和剤などのpH調節剤を含まないものとされる。
【0025】
培養温度は、用いるキャンディダ・ユティリス菌株の生育可能な範囲で選択することができる。培養温度は、例えば、約15℃〜45℃とすることができ、より好ましくは20〜40℃、さらに好ましくは25〜35℃、最も好ましくは約30℃とする。
【0026】
培養時間は特に限定されず、本発明の効果が認められる限り任意の反応時間で実施される。これらの条件の最適化は、当業者であれば容易に行うことができる。
【0027】
本発明において、キャンディダ・ユティリスの培養は、キャンディダ・ユティリスの発酵による物質の製造を目的として行うことができる。特に、製造の目的とする物質が有機酸などの酸性を示す物質である場合には、その物質の生産に伴って下降するはずのpHが硝酸塩の効果によって上方へ調節されるため、キャンディダ・ユティリスの培養に適したpHを維持する上で有利である。
【0028】
従って、本発明の他の態様によれば、キャンディダ・ユティリスを培養することにより有機酸を製造する方法が提供され、該方法は、培養液中に硝酸塩を添加することを含んでなる。
【0029】
本発明において製造される有機酸はキャンディダ・ユティリスの野生株または変異株の培養によって製造しうる有機酸であればよく、特に制限されるものではない。このような有機酸としては、限定されるものではないが、例えば、乳酸、ピルビン酸、リンゴ酸、コハク酸などが挙げられる。本発明の好ましい実施態様によれば、本発明により製造される有機酸はL−乳酸またはD−乳酸とされ、より好ましくはD−乳酸とされる。
【0030】
上記の有機酸の製造には、キャンディダ・ユティリスの野生株を遺伝的に改変して得られる変異株を用いることが有利である。よって、本発明の好ましい実施態様によれば、前記キャンディダ・ユティリスは遺伝的に改変された菌株とされる。酵母の遺伝子操作については当技術分野においてよく知られており、よって、当業者であれば、所望の有機酸を生産するようにキャンディダ・ユティリスを遺伝的に改変することができる。例えば、L−乳酸を生産するキャンディダ・ユティリス菌株は、国際公開第2010/095751号パンフレットに記載されている。
【0031】
本発明の好ましい実施態様によれば、本発明において用いられるキャンディダ・ユティリス菌株はD−乳酸を生産するものとされ、このような菌株は、例えば、キャンディダ・ユティリスの野生株に対して、(i)D−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子の発現を可能にするプロモーター配列が機能的に結合した該遺伝子の少なくとも1コピーにより形質転換されており、かつ、(ii)ピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードしている内因性遺伝子が破壊されている、ものとされる。この(i)および(ii)の遺伝子改変がなされたキャンディダ・ユティリス菌株は、特に優れたD−乳酸生産能を有しており、よって、このキャンディダ・ユティリス菌株は本発明の一つの態様をなす。このようなキャンディダ・ユティリス菌株を培養すると、そのD−乳酸の高効率生産によってpHが急激に低下するため、硝酸塩の存在下で培養することによりそのpHを上方に調節することが好適である。
【0032】
本発明による酵母菌株は、D−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子(D−LDH遺伝子)を保持している。酵母は元来乳酸製造能を持たないので、本発明による酵母菌株が有する乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子(D−LDH)は外来性である。本発明において使用するD−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子としては、天然由来のD−LDHの他、化学合成的または遺伝子工学的な手法により人工合成されたD−LDHも包含している。D−LDHをもつ生物としては、乳酸菌等の原核生物、カビ等の真核生物、植物や動物並びに昆虫等の高等真核生物などが挙げられる。本発明において使用するD−LDHとして好ましいのは乳酸菌由来のものであり、特にラクトバチルス・ヘルベチカス(Lactobacillus helveticus)由来のものが適している。
【0033】
本発明の好ましい実施態様によれば、D−乳酸脱水素酵素の活性を有するポリペプチドは、配列番号2で表されるアミノ酸配列を含むポリペプチドとされる。また、D−乳酸脱水素酵素の活性を有するポリペプチドは、配列番号2で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつD−乳酸脱水素酵素の活性を有するポリペプチドであってもよい。
【0034】
ここで、アミノ酸の欠失、置換、付加、又は挿入は、上記ポリペプチドをコードする遺伝子を、当技術分野で公知の手法によって改変することによって行うことができる。遺伝子に変異を導入するには、Kunkel法又はGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キット、例えばMutant−K(タカラバイオ社)やMutant−G(タカラバイオ社)などを用いて、あるいは、タカラバイオ社のLA PCR in vitro Mutagenesisシリーズキット、KOD−Plus−Mutagenesis Kit(TOYOBO)などを用いて変異を導入することができる。D−乳酸脱水素酵素の活性は、当技術分野において公知の手法により確認することができる。
【0035】
さらに、宿主に導入するD−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子は、配列番号2に記載したラクトバチルス・ヘルベチカス(Lactobacillus helveticus)由来の酵素のアミノ酸配列をコードするヌクレオチド配列を、キャンディダ・ユティリスのコドン使用頻度を考慮して人工的に合成したものが好ましい。このような人工的な合成は当業者であれば適切に行うことができるが、特に好ましいヌクレオチド配列は、配列番号1で表されるヌクレオチド配列である。
【0036】
本発明の好ましい実施態様によれば、D−乳酸脱水素酵素の活性を有するポリペプチドをコードしている遺伝子は、配列番号1で表されるヌクレオチド配列を含む遺伝子、またはその同等物とされる。この同等物は、配列番号1で表されるヌクレオチド配列を含む遺伝子と同等の機能を有することを条件に、一部のヌクレオチド残基が異なる遺伝子を意味する。このような同等物としては、配列番号1で表されるヌクレオチド配列と70%以上、好ましくは80%以上、より好ましくは85%以上、さらに好ましくは90%以上、最も好ましくは95%以上の相同性があり、かつD−乳酸脱水素酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。前記同等物としてはさらに、配列番号1で表されるヌクレオチド配列もしくはその相補配列とストリンジェントな条件下でハイブリダイズし、かつD−乳酸脱水素酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。前記同等物としてはさらに、配列番号1で表されるヌクレオチド配列において1もしくは数個のヌクレオチド残基が欠失、置換、付加、または挿入された配列を含み、かつD−乳酸脱水素酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。本発明の特に好ましい実施態様によれば、D−乳酸脱水素酵素の活性を有するポリペプチドをコードしている遺伝子は、配列番号1で表されるヌクレオチド配列を含む遺伝子とされる。
【0037】
ここで、ヌクレオチド残基の欠失、置換、付加、又は挿入は、上記配列を含む遺伝子を、当技術分野で公知の手法によって改変することによって行うことができる。遺伝子に変異を導入するには、Kunkel法又はGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キットを用いればよい。例えばMutant−K(タカラバイオ社)やMutant−G(タカラバイオ社)などを用いて、あるいは、タカラバイオ社のLA PCR in vitro Mutagenesisシリーズキット、KOD−Plus−Mutagenesis Kit(TOYOBO)などを用いて変異を導入することができる。D−乳酸脱水素酵素の活性は、当技術分野において公知の手法により確認することができる。
【0038】
相同性を示す数値(%)は、塩基配列比較用プログラム:例えばGENETYX−WIN7.0.0を用いて、デフォルト(初期設定)のパラメーターにより算出されるものである。すなわち、酵母染色体上の各遺伝子が、同一ではないが同等の機能、すなわち各活性を有するポリペプチドをコードする遺伝子によって相同組換え等を介して置換されていてもよい。D−乳酸脱水素酵素の活性は、当技術分野において公知の手法により確認することができる。
【0039】
ストリンジェントな条件とは、例えば、Rapid−Hyb Buffer(GEヘルスケアバイオサイエンス社製)を用い、温度条件を好ましくは40〜70℃、より好ましくは60℃として、その他は添付のプロトコールに従って行うハイブリダイゼーション条件である。その後、例えば当業者の一般的な方法を用い、2×SSCと0.1%(w/v)SDSから成り立つ溶液での5分間の洗浄、続いて1×SSCと0.1%(w/v)SDSから成り立つ溶液での10分間の洗浄、さらに0.1×SSCと0.1%(w/v)SDSから成り立つ溶液での10分間の洗浄を行うことを指す。ただしハイブリダイゼーション時の温度条件や、その後のメンブレンの洗浄に用いる溶液の塩濃度等の条件を適宜設定することにより、ある一定(70%、80%、85%、90%、95%のいずれか)以上の相同性を有する塩基配列を含むDNAをクローニングできる。そのようにして得られる遺伝子が、配列上は同一ではないが同等の機能、すなわち各活性を有するポリペプチドをコードする遺伝子によって相同組換え等を介して置換されていてもよい。D−乳酸脱水素酵素の活性は、当技術分野において公知の手法により確認することができる。
【0040】
D−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子は、強力なプロモーター活性を有するプロモーターの制御下で発現可能に備えられていることが好ましい。例えば、キャンディダ・ユティリスでは、キャンディダ・ユティリスのグリセロアルデヒド−3−リン酸脱水素酵素の活性を有するポリペプチドをコードするGAP遺伝子のプロモーター、ホスホグリセリン酸キナーゼの活性を有するポリペプチドをコードするPGK遺伝子のプロモーター、原形質膜プロトンATPaseの活性を有するポリペプチドをコードするPMA遺伝子のプロモーター(以上、特開2003−144185号公報)等が例示されるが、さらに好ましくはピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードする遺伝子1(CuPDC1遺伝子)のプロモーターである。
【0041】
D−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子は、酵母染色体上のCuPDC1遺伝子プロモーターの制御下で発現可能に備えられていることが好ましい。本発明による酵母菌株の宿主として用いるキャンディダ・ユティリスは少なくとも1種類のPDC遺伝子を有していると推定される(CuPDC1遺伝子)。このCuPDC1遺伝子プロモーターによって制御されるCuPDC1遺伝子が破壊されて乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子が代わりに発現されることで、効果的にピルビン酸脱炭酸酵素活性の低下と乳酸脱水素酵素活性の発現とを同時に実現できる。
【0042】
本発明による酵母菌株においては、ピルビン酸脱炭酸酵素(PDC)活性がないか、または低下している。この酵素は、アルコール発酵経路においてピルビン酸をアセトアルデヒドに変換する酵素であり、アルコール発酵を行う酵母はピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードする遺伝子を染色体上に本来的に有している。サッカロマイセス・セレビシエにはピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードする遺伝子が3種類(ScPDC1、ScPDC5およびScPDC6)存在し、これらはいわゆるオートレギュレーション機構により機能している。また、各遺伝子のヌクレオチドレベルでの相同性も70%以上と高い。これらの遺伝子がコードするタンパク質はN末端側のTPP結合領域とC末端側のPDC活性領域から構成されている。PDCをコードする遺伝子は他の酵母でも存在しており、例えば、クルイベロマイセス・ラクティスのKlPDC1遺伝子はScPDC1遺伝子との高い相同性を有する。一方、キャンディダ・ユティリスにはピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードする1種類の遺伝子(CuPDC1)が存在し、他にも同様の遺伝子が存在する可能性があるが、少なくともCuPDC1遺伝子を破壊することによりアルコール発酵はほぼ全く行われなくなる。
【0043】
ここで「PDC活性がないか、または低下している」とは、PDC活性が全くないか、または野生型よりも低い活性の該酵素が生産されているか、あるいは該酵素の生産量が野生型よりも少ないことを意味する。PDC活性がないか、または低下している酵母菌株は、人工的な操作により得られたものであっても、あるいはスクリーニングによって見出されたものであってもよい。このような酵素活性の消滅または低下のための人工的操作は、RNAiを利用する方法、選択マーカーの全部または一部の配列などの他の遺伝子と入れ替える方法、無意味な配列を遺伝子内部に挿入する方法など、当技術分野において周知の方法で行うことができる。この中でも、当該酵素の活性を有するポリペプチドをコードする遺伝子を破壊(ノックアウト)することが好ましく、このような方法として、上記の各手法のうち、選択マーカーの全部または一部の配列などの他の遺伝子とPDCをコードする遺伝子とを入れ替える方法が挙げられる。
【0044】
破壊対象のピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードする遺伝子は、元々キャンディダ・ユティリスに存在し、例えば、NBRC0988株に存在するCuPDC1遺伝子のアレルのうちの1つのヌクレオチド配列は配列番号7で表され、コードされるアミノ酸配列は配列番号8で表される。キャンディダ・ユティリスの他の株、例えばNBRC0626株、NBRC0639株、NBRC1086株等を用いる場合には、仮に当該配列と相違していても、同等の機能、すなわち活性を有するものが存在していればそれを破壊対象とすることができる。
【0045】
本発明の好ましい実施態様によれば、破壊対象となるピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードする内因性遺伝子は、配列番号8で表されるアミノ酸配列を含むポリペプチドをコードする遺伝子、より好ましくは配列番号7で表されるヌクレオチド配列を含む遺伝子とされる。
【0046】
本発明による酵母菌株の分子育種は、宿主酵母に対してD−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子を発現可能な状態で導入することによって行うことができる。その際に、宿主酵母に対してPDCをコードする遺伝子の破壊を伴っていることが好ましい。
【0047】
PDC遺伝子の破壊とL−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子の導入については、国際公開第2010/095751号パンフレットに記載されている。よって、国際公開第2010/095751号パンフレットの記載に従って、L−乳酸脱水素酵素の代わりに上述のD−乳酸脱水素酵素を用いることにより、本発明による酵母菌株を得ることが可能である。また、キャンディダ・ユティリスのNBRC0988株において、ピルビン酸脱炭酸酵素をコードするCuPDC1遺伝子が完全に欠損した株:Cupdc1delta4株が、Ikushimaらによって構築されている[Biosci Biotechnol Biochem. Vol.73: p1818-24 (2009)]。よって、本発明による酵母菌株の育種にあたっては、このCupdc1delta4株を利用することが有利である。
【0048】
D−乳酸脱水素酵素の活性を有するポリペプチドをコードしている遺伝子が酵母ゲノムに組込まれる染色体上の位置は特に制限されるものではないが、ピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードする遺伝子座とすることが有利である。これにより、乳酸脱水素酵素の活性を有するポリペプチドをコードしている遺伝子を、PDC遺伝子の全長プロモーターの制御下におくことができ、よって、高い発現効率を得ることができる。
【0049】
D−乳酸およびL−乳酸以外にも、ピルビン酸、リンゴ酸、コハク酸などの有機酸を生産するキャンディダ・ユティリス菌株を構築することが可能である。例えば、PDC遺伝子が完全に欠損したサッカロミセス・セレビシエ(S. cerevisiae)を連続培養することにより、高濃度のグルコースの存在下でも増殖できる株が選抜され、この株を、一定のpHの下、10%グルコースを糖源とする条件下でこの株を好気培養することにより、高濃度かつ高効率でピルビン酸を生産できることが報告されている[Appl. Environ. Microbiol. 70 (1), p159-66 (2004)]。さらに、このグルコース耐性株に次の改変を加えることにより、リンゴ酸高生産株を構築できることが報告されている:ピルビン酸カルボキシラーゼ酵素(PYC2)遺伝子の過剰発現、細胞質で機能するように組換えられたリンゴ酸脱水素酵素(MDH3)遺伝子の高発現、および出芽酵母Schizosaccharomyces pombe由来のリンゴ酸トランスポーター(SpMAE1)遺伝子の機能的な発現[Appl. Environ. Microbiol. 74 (9), p.2766-77 (2008)]。また、このリンゴ酸高生産株は、コハク酸を生産することも確認されている[Appl. Environ. Microbiol. 74 (9), p.2766-77 (2008)]。これらの手法をキャンディダ・ユティリスに適用することにより、ピルビン酸、リンゴ酸、またはコハク酸を生産する酵母菌株を構築することが可能である。
【実施例】
【0050】
以下、実施例によって本発明をより具体例に説明するが、これら実施例は本発明の技術的範囲を制限するものではない。
【0051】
実施例1:D−LDH遺伝子を導入したキャンディダ・ユティリス株の構築
トルラ酵母キャンディダ・ユティリス(Candida utilis)のNBRC0988株において、ピルビン酸脱炭酸酵素をコードするCuPDC1遺伝子が完全に欠損した株:Cupdc1delta4株が既にIkushimaらによって構築されている[Biosci Biotechnol Biochem. Vol.73: p1818-24 (2009)]。この株に、乳酸菌Lactobacillus helveticus由来のD−乳酸脱水素酵素(D−LDH)遺伝子を導入した。ここで用いたDNA断片は、アミノ酸の置換がないようにコドン使用頻度をキャンディダ・ユティリスに適合させて人工合成したものである(配列番号1)。その断片を含むプラスミドをpDLDHとした。以下、導入に用いたプラスミドの構築手順を示す。
【0052】
まず、次の3種類のポリメラーゼ連鎖反応(PCR)を行った:(1)C. utilis NBRC0988株のゲノムDNAを鋳型として、IM-341(配列番号3:actcgcggccgctctagaCACCAACTTTGAAGATAGGG)とIM-678(配列番号4:gtaggcgaagaccttggtcatGGTATCGATTGTTTTAGTTTTGTTTGTTTGTTGTGTATAACGGG)をプライマーとしてPCRを行い(伸長反応2分)、CuPDC1遺伝子のプロモーター領域を増幅した;(2)pDLDHを鋳型とし、IM-671(配列番号5:cccgttatacacaacaaacaaacaaaactaaaacaatcgataccATGACCAAGGTCTTCGCCTAC)とIM-672(配列番号6:atactcagatctTCATCAGAACTTGTTCTTGTTC)をプライマーとしてPCRを行い(伸長反応1分)、D−LDH構造遺伝子を増幅した;(3)鋳型として(1)および(2)で増幅したDNA断片を用い、プライマーとしてIM-341とIM-672を用いてPCRを行った(伸長反応3分)。次に、(3)で増幅したDNA断片をNotIとBglIIで消化し、得られたDNA断片を、NotIとBamHIで切断したpCU675(別名:pPGtHPt)に連結した。ここで、pCU675は、Ikushimaらによって既に構築されているプラスミドである[Biosci Biotechnol Biochem. Vol.73: p1818-24 (2009)]。
【0053】
上述の工程を経て得られたプラスミドpCU871は、pBluescriptIISK(+)のBssHII部位に、順に、CuPDC1遺伝子プロモーター領域、D−LDH構造遺伝子、PGK(3−ホスホグリセロキナーゼ)遺伝子ターミネーター、loxP、PGK遺伝子プロモーター、HPT遺伝子(ハイグロマイシンB耐性遺伝子)、GAP(グリセルアルデヒド3リン酸脱水素酵素)遺伝子ターミネーター、loxP、CuPDC1遺伝子の下流領域からなるDNA断片が挿入されている。2つのBssHII認識配列の挿入DNA断片側には、当該認識配列の直後にBglII認識配列が備わっている。そこで、pCU871をBglIIで消化することにより、上述のCuPDC1遺伝子プロモーターからCuPDC1遺伝子の下流領域からなるDNA断片を取得し、これを用いて電気パルス法でcupdc1delta4の形質転換を行った。ここで、パルス条件は、Ikushimaらの方法に従った[Biosci Biotechnol Biochem. Vol.73: p879-84 (2009)]。陽性クローンとしては、600μg/mlのハイグロマイシンBを含むYPD培地(1%酵母エキス、2%ペプトン、2%グルコース)で生育した株を選抜した。これをCDLA01株と命名した。
【0054】
こうして得られたCDLA01株を、500mlスケールの坂口フラスコを用い、100mlのYPD液体培地(1%酵母エキス、2%ペプトン、2%グルコース)中、30℃で好気的に24時間振とう培養した。この株を、100ml三角フラスコに入った15mlの富栄養培地(1%酵母エキス、2%ペプトン、90.7g/lグルコース)に、OD600が0.3となるように接種した。なお、この培地には、45g/lの炭酸カルシウムを中和剤として添加した。培養開始24時間後には、全ての糖が消費され、代わりに、光学純度が99.9%のD−乳酸が86.8g/lの濃度で生産された。培地中に含まれる糖から理論的に最大でD−乳酸が作られる場合を収率100%とすると、今回の収率は95%を越えている。
【0055】
次に、CDLA01株を、300mLスケールの坂口フラスコ中において、YPD2液体培地(1%酵母エキス、2%ペプトン、2%グルコース)中、30℃で好気的に振盪培養した。この株を、5Lスケールのジャーファーメンターにおいて、2.0Lの糖蜜培地(18%廃糖蜜、2%ペプトン)に初期OD600が0.1となるように接種し、35℃で攪拌培養した。通気量は1.0vvm、撹拌速度は300rpmとし、10N NaOHおよび10N HClを中和剤として用いることでpHを5.0に維持した。培養開始から72時間が経過した時点で、99.5%を上回る光学純度のD−乳酸が160g/Lの濃度で生産された(図1)。これは、当該時点で消費された糖に対する変換効率で100%に達する値である。
【0056】
実施例2:硝酸塩を唯一の窒素源とする培地におけるキャンディダ・ユティリスの培養
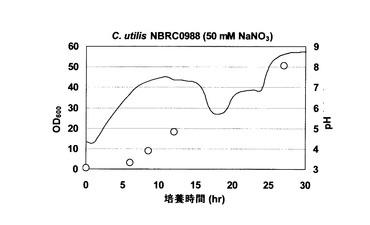
C. utilis NBRC 0988野生株を30℃で好気的に振盪培養した。この株を、5Lスケールのジャーファーメンターにおいて、2.0Lの合成培地(5%グルコース、0.17% Yeast Nitrogen Base、50mM 硝酸ナトリウム)に初期OD600が0.5となるように接種し、30℃で撹拌培養した。通気量は2.0L/min、撹拌速度は300rpmとした。その結果、培養液のpHは経時的に上昇し、培養開始27時間後にはpH8.6にまで達した(図2)。
【0057】
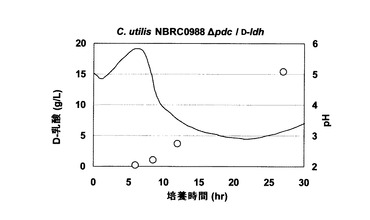
次に、CDLA01株について、上述の合成培地を用いて同様の試験を行ったところ、中和剤を添加すること無く、培養液のpHは3程度で安定し、乳酸が蓄積しても極端な酸性域に偏ることはなかった。この効果によって、pHの低下による酵母の死滅を回避することができた。この結果、培養開始27時間後には15.4g/LのD−乳酸が生産された(図3)。
【0058】
実施例3:硝酸塩と他の窒素含有物質を窒素源とする培地におけるキャンディダ・ユティリスの培養
窒素源として25mM硝酸ナトリウムおよび25mM塩化アンモニウムの組み合わせを含む合成培地において、実施例2と同様にC. utilis NBRC 0988野生株の培養を行った。塩化アンモニウムを単一窒素源とした場合には、通常、pHの急激な低下とともに、培養24時間程度で生菌数の大幅な低下が生じる。これに対して、塩化アンモニウムと同じモル濃度の硝酸ナトリウムを混合した場合には、pHの低下が抑制され、40時間以上にわたって培養を継続できることが明らかとなった(図4)。
【0059】
この結果から、硝酸塩を窒素源として含む培地におけるキャンディダ・ユティリスの培養において、他の窒素源が共存したとしても、硝酸塩によるpH上昇効果が阻害されないことが示される。
【0060】
実施例4:様々な硝酸塩を唯一の窒素源とする培地におけるキャンディダ・ユティリスの培養
50mM硝酸ナトリウムに代えて、50mM硝酸カリウムまたは25mM硝酸カルシウムを単一窒素源として加えた培地を用いて、実施例2と同様にC. utilis NBRC 0988野生株の培養を行った。その結果、いずれの場合においても、硝酸ナトリウムを用いたときと同様の経時的なpHの上昇を示した(図5)。
【0061】
この結果から、硝酸塩を窒素源として含む培地におけるキャンディダ・ユティリスの培養において、硝酸塩によるpH上昇効果は硝酸塩のカチオンの種類に影響されないことが示される。
【0062】
実施例5:硝酸塩を唯一の窒素源とする培地におけるキャンディダ・ユティリスの様々な株の培養
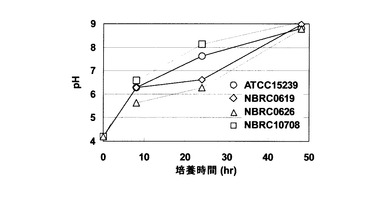
キャンディダ・ユティリスのATCC 15239株、NBRC 0619株、NBRC 0626株、およびNBRC 10708株を用いて、培養液のpH上昇効果を試験した。具体的には、200mLスケールの三角フラスコにおいて、各株を、実施例2に記載の硝酸培地50mLに初期OD600が0.5となるように接種し、30℃で振盪培養した。その結果、いずれの株においても、NBRC 0988株と同様に経時的な培養液のpH上昇が認められた(図6)。
【0063】
この結果から、硝酸塩を窒素源として含む培地におけるキャンディダ・ユティリスの培養において、硝酸塩によるpH上昇効果は、株の違いに影響されず、キャンディダ・ユティリス全般に認められることが示される。
【0064】
実施例6:硝酸塩を唯一の窒素源とする培地におけるキャンディダ・ユティリス以外の酵母の培養
硝酸塩を単一窒素源として生育できるキャンディダ・ユティリス以外の酵母として、ピキア・アングスタ(Pichia angusta)、パキソレン・タンノフィルス(Pachysolen tannophilus)、およびハンセヌラ・ポリモルファ(Hansenula polymorpha)の3種を用いて、実施例5と同様の培養試験を行った。その結果を表1に示す。
【0065】
【表1】

【0066】
表1から明らかなように、3種の酵母のいずれを培養した場合にも、キャンディダ・ユティリスに見られたような急激なpHの上昇は認められなかった。
【図面の簡単な説明】
【0067】
【図1】図1は、CDLA01株を、5Lスケールのジャーファーメンターにおいて、2.0Lの糖蜜培地(18%廃糖蜜、2%ペプトン)に初期OD600が0.1となるように接種し、35℃で撹拌培養した結果を示す。この培養では、通気量を1.0vvm、撹拌速度を300rpmとし、10N NaOHおよび10N HClを中和剤として用いることによりpHを5.0に維持した。図中の白丸および白三角は培養混合物中のD−乳酸の濃度を示し、黒丸および黒三角は培養混合物中のグルコースの濃度を示す。
【図2】図2は、C. utilis NBRC 0988野生株を、硝酸ナトリウムを唯一の窒素源として含む培地において培養した場合のpHの変化を示す。図中の実線は培養液のpHを示し、白丸(ドット)はOD600の値を示す。
【図3】図3は、CDLA01株を、硝酸ナトリウムを唯一の窒素源として含む培地において培養した場合のpHの変化およびD−乳酸の生成を示す。図中の実線は培養液のpHを示し、白丸(ドット)はD−乳酸の濃度(g/l)を示す。
【図4】図4は、C. utilis NBRC 0988野生株を、硝酸ナトリウムと塩化アンモニウムの組み合わせを窒素源として含む培地において培養した場合のpHの変化を示す。図中の実線は培養液のpHを示し、白丸(ドット)はOD600の値を示す。
【図5】図5は、C. utilis NBRC 0988野生株を、硝酸ナトリウム(白丸)、硝酸カリウム(白菱形)および硝酸カルシウム(白三角)のいずれか一つを唯一の窒素源として含む培地において培養した場合のpHの変化を示す。
【図6】図6は、キャンディダ・ユティリスATCC 15239株(白丸)、NBRC 0619株(白菱形)、NBRC 0626株(白三角)、およびNBRC 10708株(白四角)のそれぞれを、硝酸ナトリウムを唯一の窒素源として含む培地において培養した場合のpHの変化を示す。
【発明の背景】
【0001】
技術分野
本発明は、クラブトゥリー陰性酵母であるキャンディダ・ユティリス(Candida utilis)を宿主とした有用物質の製造方法に関する。
【0002】
背景技術
将来的に化石資源が枯渇するリスクに鑑み、近年は、化成品およびその素材を、植物バイオマスを原料に用いて発酵法で生産する取り組みに注目が集められている。プラスチックもその1つであり、その素材として生物の代謝物である有機酸を利用することができる。有機酸を素材とするプラスチックの例としては、例えば、乳酸の重合体であるポリ乳酸がある。
【0003】
乳酸にはL−体とD−体がある。高い融点を有するポリ乳酸を製造するためには、光学純度の高いL−体またはD−体の乳酸を重合させる必要がある。また、ポリL乳酸とポリD乳酸を等量混ぜてステレオコンプレックスとすることにより、融点をさらに高めることが可能である。このため、光学純度の高いL−乳酸とD−乳酸をそれぞれ独立に生産することが求められている。すなわち、D−乳酸の発酵生産系がステレオコンプレックス型の乳酸製造の鍵である。
【0004】
乳酸の発酵生産においては、産物である乳酸の蓄積に伴ってpHが極端に酸性域に偏ることにより、菌体の活性が低下し、あるいは菌が死滅してしまうという問題がある。したがって、乳酸を高濃度で生産するためには、菌は、高い発酵能を有するだけでなく、高濃度の乳酸に対して耐性を有することが求められる。
【0005】
工業的な乳酸の発酵生産には、pHの極端な低下を避けるために、炭酸カルシウムを始めとする中和剤が一般的に用いられている。しかしながら、化学プロセスを経て合成される中和剤の使用は、環境負荷の低減というバイオマス・プラスチックの生産コンセプトに必ずしも合致しない。これは、ピルビン酸[非特許文献1:Appl Environ Microbiol Vol.70: p159-66 (2004)]、リンゴ酸やコハク酸[非特許文献2:Appl Environ Microbiol Vol.74: p.2766-77 (2008)]といった乳酸以外の各種有機酸を発酵生産する場合にも共通する課題である。
【0006】
酵母のうち、今日まで最も良く研究され、遺伝学的知見が蓄積しているものにはサッカロマイセス属酵母があり、この酵母は種々の物質生産の宿主として検討がなされている。
また、近年、サッカロマイセス属酵母以外の酵母としてピキア属酵母、ハンセヌラ属酵母、クルイベロマイセス属酵母、キャンディダ属酵母などのいくつかの種について、それらを形質転換する手法が開発され、有用物質生産の宿主として検討されている。このうち、キャンディダ属酵母は、特に、炭素資化域が広いなど、サッカロマイセス属酵母にない特性を有している。
【0007】
キャンディダ属酵母の中でも、キャンディダ・ユティリスは、キシロースをはじめとするペントースに対する優れた資化性を示す。また、サッカロマイセス酵母と異なり、好気的条件下での培養でエタノールを製造せず、それによる増殖阻害も受けないことから、高密度での連続培養による効率的な菌体製造が可能である。従って、キャンディダ・ユティリスは、かつて蛋白質源として注目され、ペントースを多く含む広葉樹の糖化液や亜硫酸パルプ廃液を糖源とした菌体の工業生産が行われていた。また、この酵母は米国FDA(Food and Drug Administration)により、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)、サッカロマイセス・フラジリス(Saccharomyces fragilis)とともに、食品添加物として安全に使用できる酵母として認められている。実際に、キャンディダ・ユティリスは、ドイツをはじめとして、米国や台湾、ブラジルなど世界各国で製造され、食飼料として使用されている。また、このような微生物蛋白質としての利用以外にも、キャンディダ・ユティリスは、ペントースやキシロースの発酵株、エチルアセテート、L−グルタミン、グルタチオン、インベルターゼ等の製造株として広く産業界で利用されてきた。
【0008】
【非特許文献1】Appl Environ Microbiol Vol.70: p159-66 (2004)
【非特許文献2】Appl Environ Microbiol Vol.74: p.2766-77 (2008)]
【発明の概要】
【0009】
本発明者らは、キャンディダ・ユティリスを培養する際に、培養液中に硝酸塩を添加することにより、培養液のpHが経時的に上昇することを見出した。さらに、本発明者らは、キャンディダ・ユティリスの培養によって有機酸を製造する際に、培養液中に硝酸塩を添加することにより、有機酸の生成に伴う培養液のpHの低下が抑制されることを見出した。さらに、本発明者らは、遺伝子操作により、D−乳酸を生産するキャンディダ・ユティリス酵母菌株を開発することに成功した。本発明は、これらの知見に基づくものである。
【0010】
従って、本発明の目的は、キャンディダ・ユティリスの培養において、培養液のpHを調節するための方法およびpH調節剤、キャンディダ・ユティリスの培養によって有機酸を製造する方法、ならびにD−乳酸を高い効率で製造する酵母菌株を提供することにある。
【0011】
本発明によるpH調節法は、キャンディダ・ユティリスの培養中に培養液のpHを上方に調節するための方法であって、培養液中に硝酸塩を添加することを含んでなる方法である。
【0012】
本発明による有機酸の製造法は、キャンディダ・ユティリスを培養することにより有機酸を製造する方法であって、培養液中に硝酸塩を添加することを含んでなる方法である。
【0013】
本発明によるpH調節剤は、硝酸塩を含んでなる、キャンディダ・ユティリスの培養中に培養液のpHを上方に調節するためのpH調節剤である。
【0014】
本発明による酵母菌株は、D−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子の発現を可能にするプロモーター配列が機能的に結合した該遺伝子の少なくとも1コピーにより形質転換されてなり、かつ、ピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードしている内因性遺伝子が破壊されている、キャンディダ・ユティリスの酵母菌株である。
【0015】
本発明によれば、キャンディダ・ユティリスの培養において培養液のpHを上方に調節することができる。このような効果は、例えば、キャンディダ・ユティリスの培養によって有機酸を製造する場合などに、有機酸の生成に伴うpHの低下によってもたらされる悪影響、例えば菌体の死滅、を防止するのに有利である。また、このようなpHの低下を防止するために中和剤を必要としない点も、本発明の利点である。さらに、本発明による酵母菌株のような、D−乳酸を高濃度で生産するキャンディダ・ユティリス株の培養において、上記の効果は特に有利である。
【発明の具体的説明】
【0016】
本発明によるpH調節法は、キャンディダ・ユティリスの培養中に培養液のpHを上方に調節するための方法である。該方法は、培養液中に硝酸塩を添加することを含んでなる。
【0017】
本明細書において、「pHを上方に調節する」という用語およびその派生語は、培養液に硝酸塩を添加しない場合と比較して、培養液のpHをアルカリ側に調節することを意味する。すなわち、この用語は、培養開始から経時的にpHが上昇することを意味するものではなく、pHの経時変化を示すグラフの曲線が、硝酸塩を添加しない場合と比べてアルカリ側にあることを意味する。従って、培養開始から経時的にpHが酸性側に低下したとしても、硝酸塩を添加しない場合と比較してアルカリ側にあれば、それは「pHを上方に調節する」ことに該当する。
【0018】
本発明に用いられる酵母は、クラブトゥリー陰性酵母であるキャンディダ・ユティリスである。キャンディダ・ユティリスの菌株は当技術分野において公知の様々な株、例えば、NBRC0626株、NBRC0639株、NBRC0988株、NBRC1086株等であってよい。
【0019】
本発明に用いられる硝酸塩は、キャンディダ・ユティリスが資化しうるものであればよく、特に制限されない。このような硝酸塩は当技術分野において周知であるが、好ましくは硝酸ナトリウム、硝酸カリウムもしくは硝酸カルシウム、またはこれらの任意の組み合わせとすることができ、より好ましくは硝酸ナトリウムとされる。
【0020】
硝酸塩の培養液への添加量は、キャンディダ・ユティリスの培養に用いられる通常の窒素源の量であればよく、特に制限されない。よって、硝酸塩の添加量は、当業者であれば適宜決定することができるが、好ましくは、硝酸塩に由来する硝酸イオンの濃度として1〜80mM、より好ましくは20〜80mM、最も好ましくは約50mMとすることができる。
【0021】
硝酸塩の培養液への添加の時期は、培養液のpHを上方調節したい時など、当業者であれば適切に選択することができるため、特に制限されない。例えば、硝酸塩は、培養開始時に既に添加されていてもよく、培養開始後の培養の途中で添加されてもよい。また、硝酸塩は、培養開始時に添加しておき、培養の途中でさらに添加してもよい。
【0022】
培養液は、硝酸塩以外の他の窒素源を含んでいてもよく、これにより硝酸塩によるpH調節効果が損なわれることはない。しかし、高いpH調節効果を得るためには、培養液中の硝酸塩以外の窒素源はできるだけ少ない方が好ましい。例えば、硝酸塩以外の窒素源のモル濃度は、硝酸塩に由来する硝酸イオンのモル濃度と同じか、または少ないことが好ましい。本発明の好ましい実施態様によれば、添加される硝酸塩は、キャンディダ・ユティリスによって資化される主要な窒素源として培養液に存在する。ここで、「主要な窒素源」との用語は、資化される窒素の50%以上が硝酸塩に由来することを意味する。本発明の特に好ましい実施態様によれば、添加される硝酸塩は、キャンディダ・ユティリスによって資化される実質的に唯一の窒素源として培養液に存在する。ここで、「実質的に」との用語は、硝酸塩以外の窒素源の量がキャンディダ・ユティリスの生育に寄与しうる程度の量に満たないこと、すなわち、その培養液から硝酸塩を除いた場合にキャンディダ・ユティリスが生育できないことを意味する。
【0023】
培養液の窒素源以外の組成は、キャンディダ・ユティリスの培養に用いられる通常の培地に倣って決定することができる。例えば、培養液に含まれる炭素源としては、例えば、グルコースのほかにもキシロースやスクロースでも資化できれば用いることができる。また、培養液に含まれる栄養源としては、例えば、酵母エキス、ペプトン、ホエーなどが用いられる。さらに、培養液は、無機物栄養源として、リン酸カリウム、硫酸マグネシウムやFe(鉄)、Mn(マンガン)化合物などを含んでもよい。また、培養液はSC培地などの完全合成培地であってもよいが、炭素源として廃糖蜜および/または亜硫酸パルプ廃液を含み、窒素源としてペプトン類を含む天然培地であってもよい。
【0024】
上述したように、pHの低下を防止するための中和剤を必要としない点も、本発明の利点である。よって、本発明の好ましい実施態様によれば、用いられる培養液は、中和剤などのpH調節剤を含まないものとされる。
【0025】
培養温度は、用いるキャンディダ・ユティリス菌株の生育可能な範囲で選択することができる。培養温度は、例えば、約15℃〜45℃とすることができ、より好ましくは20〜40℃、さらに好ましくは25〜35℃、最も好ましくは約30℃とする。
【0026】
培養時間は特に限定されず、本発明の効果が認められる限り任意の反応時間で実施される。これらの条件の最適化は、当業者であれば容易に行うことができる。
【0027】
本発明において、キャンディダ・ユティリスの培養は、キャンディダ・ユティリスの発酵による物質の製造を目的として行うことができる。特に、製造の目的とする物質が有機酸などの酸性を示す物質である場合には、その物質の生産に伴って下降するはずのpHが硝酸塩の効果によって上方へ調節されるため、キャンディダ・ユティリスの培養に適したpHを維持する上で有利である。
【0028】
従って、本発明の他の態様によれば、キャンディダ・ユティリスを培養することにより有機酸を製造する方法が提供され、該方法は、培養液中に硝酸塩を添加することを含んでなる。
【0029】
本発明において製造される有機酸はキャンディダ・ユティリスの野生株または変異株の培養によって製造しうる有機酸であればよく、特に制限されるものではない。このような有機酸としては、限定されるものではないが、例えば、乳酸、ピルビン酸、リンゴ酸、コハク酸などが挙げられる。本発明の好ましい実施態様によれば、本発明により製造される有機酸はL−乳酸またはD−乳酸とされ、より好ましくはD−乳酸とされる。
【0030】
上記の有機酸の製造には、キャンディダ・ユティリスの野生株を遺伝的に改変して得られる変異株を用いることが有利である。よって、本発明の好ましい実施態様によれば、前記キャンディダ・ユティリスは遺伝的に改変された菌株とされる。酵母の遺伝子操作については当技術分野においてよく知られており、よって、当業者であれば、所望の有機酸を生産するようにキャンディダ・ユティリスを遺伝的に改変することができる。例えば、L−乳酸を生産するキャンディダ・ユティリス菌株は、国際公開第2010/095751号パンフレットに記載されている。
【0031】
本発明の好ましい実施態様によれば、本発明において用いられるキャンディダ・ユティリス菌株はD−乳酸を生産するものとされ、このような菌株は、例えば、キャンディダ・ユティリスの野生株に対して、(i)D−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子の発現を可能にするプロモーター配列が機能的に結合した該遺伝子の少なくとも1コピーにより形質転換されており、かつ、(ii)ピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードしている内因性遺伝子が破壊されている、ものとされる。この(i)および(ii)の遺伝子改変がなされたキャンディダ・ユティリス菌株は、特に優れたD−乳酸生産能を有しており、よって、このキャンディダ・ユティリス菌株は本発明の一つの態様をなす。このようなキャンディダ・ユティリス菌株を培養すると、そのD−乳酸の高効率生産によってpHが急激に低下するため、硝酸塩の存在下で培養することによりそのpHを上方に調節することが好適である。
【0032】
本発明による酵母菌株は、D−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子(D−LDH遺伝子)を保持している。酵母は元来乳酸製造能を持たないので、本発明による酵母菌株が有する乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子(D−LDH)は外来性である。本発明において使用するD−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子としては、天然由来のD−LDHの他、化学合成的または遺伝子工学的な手法により人工合成されたD−LDHも包含している。D−LDHをもつ生物としては、乳酸菌等の原核生物、カビ等の真核生物、植物や動物並びに昆虫等の高等真核生物などが挙げられる。本発明において使用するD−LDHとして好ましいのは乳酸菌由来のものであり、特にラクトバチルス・ヘルベチカス(Lactobacillus helveticus)由来のものが適している。
【0033】
本発明の好ましい実施態様によれば、D−乳酸脱水素酵素の活性を有するポリペプチドは、配列番号2で表されるアミノ酸配列を含むポリペプチドとされる。また、D−乳酸脱水素酵素の活性を有するポリペプチドは、配列番号2で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつD−乳酸脱水素酵素の活性を有するポリペプチドであってもよい。
【0034】
ここで、アミノ酸の欠失、置換、付加、又は挿入は、上記ポリペプチドをコードする遺伝子を、当技術分野で公知の手法によって改変することによって行うことができる。遺伝子に変異を導入するには、Kunkel法又はGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キット、例えばMutant−K(タカラバイオ社)やMutant−G(タカラバイオ社)などを用いて、あるいは、タカラバイオ社のLA PCR in vitro Mutagenesisシリーズキット、KOD−Plus−Mutagenesis Kit(TOYOBO)などを用いて変異を導入することができる。D−乳酸脱水素酵素の活性は、当技術分野において公知の手法により確認することができる。
【0035】
さらに、宿主に導入するD−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子は、配列番号2に記載したラクトバチルス・ヘルベチカス(Lactobacillus helveticus)由来の酵素のアミノ酸配列をコードするヌクレオチド配列を、キャンディダ・ユティリスのコドン使用頻度を考慮して人工的に合成したものが好ましい。このような人工的な合成は当業者であれば適切に行うことができるが、特に好ましいヌクレオチド配列は、配列番号1で表されるヌクレオチド配列である。
【0036】
本発明の好ましい実施態様によれば、D−乳酸脱水素酵素の活性を有するポリペプチドをコードしている遺伝子は、配列番号1で表されるヌクレオチド配列を含む遺伝子、またはその同等物とされる。この同等物は、配列番号1で表されるヌクレオチド配列を含む遺伝子と同等の機能を有することを条件に、一部のヌクレオチド残基が異なる遺伝子を意味する。このような同等物としては、配列番号1で表されるヌクレオチド配列と70%以上、好ましくは80%以上、より好ましくは85%以上、さらに好ましくは90%以上、最も好ましくは95%以上の相同性があり、かつD−乳酸脱水素酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。前記同等物としてはさらに、配列番号1で表されるヌクレオチド配列もしくはその相補配列とストリンジェントな条件下でハイブリダイズし、かつD−乳酸脱水素酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。前記同等物としてはさらに、配列番号1で表されるヌクレオチド配列において1もしくは数個のヌクレオチド残基が欠失、置換、付加、または挿入された配列を含み、かつD−乳酸脱水素酵素の活性を有するポリペプチドをコードするヌクレオチド配列を含む遺伝子が挙げられる。本発明の特に好ましい実施態様によれば、D−乳酸脱水素酵素の活性を有するポリペプチドをコードしている遺伝子は、配列番号1で表されるヌクレオチド配列を含む遺伝子とされる。
【0037】
ここで、ヌクレオチド残基の欠失、置換、付加、又は挿入は、上記配列を含む遺伝子を、当技術分野で公知の手法によって改変することによって行うことができる。遺伝子に変異を導入するには、Kunkel法又はGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キットを用いればよい。例えばMutant−K(タカラバイオ社)やMutant−G(タカラバイオ社)などを用いて、あるいは、タカラバイオ社のLA PCR in vitro Mutagenesisシリーズキット、KOD−Plus−Mutagenesis Kit(TOYOBO)などを用いて変異を導入することができる。D−乳酸脱水素酵素の活性は、当技術分野において公知の手法により確認することができる。
【0038】
相同性を示す数値(%)は、塩基配列比較用プログラム:例えばGENETYX−WIN7.0.0を用いて、デフォルト(初期設定)のパラメーターにより算出されるものである。すなわち、酵母染色体上の各遺伝子が、同一ではないが同等の機能、すなわち各活性を有するポリペプチドをコードする遺伝子によって相同組換え等を介して置換されていてもよい。D−乳酸脱水素酵素の活性は、当技術分野において公知の手法により確認することができる。
【0039】
ストリンジェントな条件とは、例えば、Rapid−Hyb Buffer(GEヘルスケアバイオサイエンス社製)を用い、温度条件を好ましくは40〜70℃、より好ましくは60℃として、その他は添付のプロトコールに従って行うハイブリダイゼーション条件である。その後、例えば当業者の一般的な方法を用い、2×SSCと0.1%(w/v)SDSから成り立つ溶液での5分間の洗浄、続いて1×SSCと0.1%(w/v)SDSから成り立つ溶液での10分間の洗浄、さらに0.1×SSCと0.1%(w/v)SDSから成り立つ溶液での10分間の洗浄を行うことを指す。ただしハイブリダイゼーション時の温度条件や、その後のメンブレンの洗浄に用いる溶液の塩濃度等の条件を適宜設定することにより、ある一定(70%、80%、85%、90%、95%のいずれか)以上の相同性を有する塩基配列を含むDNAをクローニングできる。そのようにして得られる遺伝子が、配列上は同一ではないが同等の機能、すなわち各活性を有するポリペプチドをコードする遺伝子によって相同組換え等を介して置換されていてもよい。D−乳酸脱水素酵素の活性は、当技術分野において公知の手法により確認することができる。
【0040】
D−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子は、強力なプロモーター活性を有するプロモーターの制御下で発現可能に備えられていることが好ましい。例えば、キャンディダ・ユティリスでは、キャンディダ・ユティリスのグリセロアルデヒド−3−リン酸脱水素酵素の活性を有するポリペプチドをコードするGAP遺伝子のプロモーター、ホスホグリセリン酸キナーゼの活性を有するポリペプチドをコードするPGK遺伝子のプロモーター、原形質膜プロトンATPaseの活性を有するポリペプチドをコードするPMA遺伝子のプロモーター(以上、特開2003−144185号公報)等が例示されるが、さらに好ましくはピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードする遺伝子1(CuPDC1遺伝子)のプロモーターである。
【0041】
D−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子は、酵母染色体上のCuPDC1遺伝子プロモーターの制御下で発現可能に備えられていることが好ましい。本発明による酵母菌株の宿主として用いるキャンディダ・ユティリスは少なくとも1種類のPDC遺伝子を有していると推定される(CuPDC1遺伝子)。このCuPDC1遺伝子プロモーターによって制御されるCuPDC1遺伝子が破壊されて乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子が代わりに発現されることで、効果的にピルビン酸脱炭酸酵素活性の低下と乳酸脱水素酵素活性の発現とを同時に実現できる。
【0042】
本発明による酵母菌株においては、ピルビン酸脱炭酸酵素(PDC)活性がないか、または低下している。この酵素は、アルコール発酵経路においてピルビン酸をアセトアルデヒドに変換する酵素であり、アルコール発酵を行う酵母はピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードする遺伝子を染色体上に本来的に有している。サッカロマイセス・セレビシエにはピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードする遺伝子が3種類(ScPDC1、ScPDC5およびScPDC6)存在し、これらはいわゆるオートレギュレーション機構により機能している。また、各遺伝子のヌクレオチドレベルでの相同性も70%以上と高い。これらの遺伝子がコードするタンパク質はN末端側のTPP結合領域とC末端側のPDC活性領域から構成されている。PDCをコードする遺伝子は他の酵母でも存在しており、例えば、クルイベロマイセス・ラクティスのKlPDC1遺伝子はScPDC1遺伝子との高い相同性を有する。一方、キャンディダ・ユティリスにはピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードする1種類の遺伝子(CuPDC1)が存在し、他にも同様の遺伝子が存在する可能性があるが、少なくともCuPDC1遺伝子を破壊することによりアルコール発酵はほぼ全く行われなくなる。
【0043】
ここで「PDC活性がないか、または低下している」とは、PDC活性が全くないか、または野生型よりも低い活性の該酵素が生産されているか、あるいは該酵素の生産量が野生型よりも少ないことを意味する。PDC活性がないか、または低下している酵母菌株は、人工的な操作により得られたものであっても、あるいはスクリーニングによって見出されたものであってもよい。このような酵素活性の消滅または低下のための人工的操作は、RNAiを利用する方法、選択マーカーの全部または一部の配列などの他の遺伝子と入れ替える方法、無意味な配列を遺伝子内部に挿入する方法など、当技術分野において周知の方法で行うことができる。この中でも、当該酵素の活性を有するポリペプチドをコードする遺伝子を破壊(ノックアウト)することが好ましく、このような方法として、上記の各手法のうち、選択マーカーの全部または一部の配列などの他の遺伝子とPDCをコードする遺伝子とを入れ替える方法が挙げられる。
【0044】
破壊対象のピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードする遺伝子は、元々キャンディダ・ユティリスに存在し、例えば、NBRC0988株に存在するCuPDC1遺伝子のアレルのうちの1つのヌクレオチド配列は配列番号7で表され、コードされるアミノ酸配列は配列番号8で表される。キャンディダ・ユティリスの他の株、例えばNBRC0626株、NBRC0639株、NBRC1086株等を用いる場合には、仮に当該配列と相違していても、同等の機能、すなわち活性を有するものが存在していればそれを破壊対象とすることができる。
【0045】
本発明の好ましい実施態様によれば、破壊対象となるピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードする内因性遺伝子は、配列番号8で表されるアミノ酸配列を含むポリペプチドをコードする遺伝子、より好ましくは配列番号7で表されるヌクレオチド配列を含む遺伝子とされる。
【0046】
本発明による酵母菌株の分子育種は、宿主酵母に対してD−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子を発現可能な状態で導入することによって行うことができる。その際に、宿主酵母に対してPDCをコードする遺伝子の破壊を伴っていることが好ましい。
【0047】
PDC遺伝子の破壊とL−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子の導入については、国際公開第2010/095751号パンフレットに記載されている。よって、国際公開第2010/095751号パンフレットの記載に従って、L−乳酸脱水素酵素の代わりに上述のD−乳酸脱水素酵素を用いることにより、本発明による酵母菌株を得ることが可能である。また、キャンディダ・ユティリスのNBRC0988株において、ピルビン酸脱炭酸酵素をコードするCuPDC1遺伝子が完全に欠損した株:Cupdc1delta4株が、Ikushimaらによって構築されている[Biosci Biotechnol Biochem. Vol.73: p1818-24 (2009)]。よって、本発明による酵母菌株の育種にあたっては、このCupdc1delta4株を利用することが有利である。
【0048】
D−乳酸脱水素酵素の活性を有するポリペプチドをコードしている遺伝子が酵母ゲノムに組込まれる染色体上の位置は特に制限されるものではないが、ピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードする遺伝子座とすることが有利である。これにより、乳酸脱水素酵素の活性を有するポリペプチドをコードしている遺伝子を、PDC遺伝子の全長プロモーターの制御下におくことができ、よって、高い発現効率を得ることができる。
【0049】
D−乳酸およびL−乳酸以外にも、ピルビン酸、リンゴ酸、コハク酸などの有機酸を生産するキャンディダ・ユティリス菌株を構築することが可能である。例えば、PDC遺伝子が完全に欠損したサッカロミセス・セレビシエ(S. cerevisiae)を連続培養することにより、高濃度のグルコースの存在下でも増殖できる株が選抜され、この株を、一定のpHの下、10%グルコースを糖源とする条件下でこの株を好気培養することにより、高濃度かつ高効率でピルビン酸を生産できることが報告されている[Appl. Environ. Microbiol. 70 (1), p159-66 (2004)]。さらに、このグルコース耐性株に次の改変を加えることにより、リンゴ酸高生産株を構築できることが報告されている:ピルビン酸カルボキシラーゼ酵素(PYC2)遺伝子の過剰発現、細胞質で機能するように組換えられたリンゴ酸脱水素酵素(MDH3)遺伝子の高発現、および出芽酵母Schizosaccharomyces pombe由来のリンゴ酸トランスポーター(SpMAE1)遺伝子の機能的な発現[Appl. Environ. Microbiol. 74 (9), p.2766-77 (2008)]。また、このリンゴ酸高生産株は、コハク酸を生産することも確認されている[Appl. Environ. Microbiol. 74 (9), p.2766-77 (2008)]。これらの手法をキャンディダ・ユティリスに適用することにより、ピルビン酸、リンゴ酸、またはコハク酸を生産する酵母菌株を構築することが可能である。
【実施例】
【0050】
以下、実施例によって本発明をより具体例に説明するが、これら実施例は本発明の技術的範囲を制限するものではない。
【0051】
実施例1:D−LDH遺伝子を導入したキャンディダ・ユティリス株の構築
トルラ酵母キャンディダ・ユティリス(Candida utilis)のNBRC0988株において、ピルビン酸脱炭酸酵素をコードするCuPDC1遺伝子が完全に欠損した株:Cupdc1delta4株が既にIkushimaらによって構築されている[Biosci Biotechnol Biochem. Vol.73: p1818-24 (2009)]。この株に、乳酸菌Lactobacillus helveticus由来のD−乳酸脱水素酵素(D−LDH)遺伝子を導入した。ここで用いたDNA断片は、アミノ酸の置換がないようにコドン使用頻度をキャンディダ・ユティリスに適合させて人工合成したものである(配列番号1)。その断片を含むプラスミドをpDLDHとした。以下、導入に用いたプラスミドの構築手順を示す。
【0052】
まず、次の3種類のポリメラーゼ連鎖反応(PCR)を行った:(1)C. utilis NBRC0988株のゲノムDNAを鋳型として、IM-341(配列番号3:actcgcggccgctctagaCACCAACTTTGAAGATAGGG)とIM-678(配列番号4:gtaggcgaagaccttggtcatGGTATCGATTGTTTTAGTTTTGTTTGTTTGTTGTGTATAACGGG)をプライマーとしてPCRを行い(伸長反応2分)、CuPDC1遺伝子のプロモーター領域を増幅した;(2)pDLDHを鋳型とし、IM-671(配列番号5:cccgttatacacaacaaacaaacaaaactaaaacaatcgataccATGACCAAGGTCTTCGCCTAC)とIM-672(配列番号6:atactcagatctTCATCAGAACTTGTTCTTGTTC)をプライマーとしてPCRを行い(伸長反応1分)、D−LDH構造遺伝子を増幅した;(3)鋳型として(1)および(2)で増幅したDNA断片を用い、プライマーとしてIM-341とIM-672を用いてPCRを行った(伸長反応3分)。次に、(3)で増幅したDNA断片をNotIとBglIIで消化し、得られたDNA断片を、NotIとBamHIで切断したpCU675(別名:pPGtHPt)に連結した。ここで、pCU675は、Ikushimaらによって既に構築されているプラスミドである[Biosci Biotechnol Biochem. Vol.73: p1818-24 (2009)]。
【0053】
上述の工程を経て得られたプラスミドpCU871は、pBluescriptIISK(+)のBssHII部位に、順に、CuPDC1遺伝子プロモーター領域、D−LDH構造遺伝子、PGK(3−ホスホグリセロキナーゼ)遺伝子ターミネーター、loxP、PGK遺伝子プロモーター、HPT遺伝子(ハイグロマイシンB耐性遺伝子)、GAP(グリセルアルデヒド3リン酸脱水素酵素)遺伝子ターミネーター、loxP、CuPDC1遺伝子の下流領域からなるDNA断片が挿入されている。2つのBssHII認識配列の挿入DNA断片側には、当該認識配列の直後にBglII認識配列が備わっている。そこで、pCU871をBglIIで消化することにより、上述のCuPDC1遺伝子プロモーターからCuPDC1遺伝子の下流領域からなるDNA断片を取得し、これを用いて電気パルス法でcupdc1delta4の形質転換を行った。ここで、パルス条件は、Ikushimaらの方法に従った[Biosci Biotechnol Biochem. Vol.73: p879-84 (2009)]。陽性クローンとしては、600μg/mlのハイグロマイシンBを含むYPD培地(1%酵母エキス、2%ペプトン、2%グルコース)で生育した株を選抜した。これをCDLA01株と命名した。
【0054】
こうして得られたCDLA01株を、500mlスケールの坂口フラスコを用い、100mlのYPD液体培地(1%酵母エキス、2%ペプトン、2%グルコース)中、30℃で好気的に24時間振とう培養した。この株を、100ml三角フラスコに入った15mlの富栄養培地(1%酵母エキス、2%ペプトン、90.7g/lグルコース)に、OD600が0.3となるように接種した。なお、この培地には、45g/lの炭酸カルシウムを中和剤として添加した。培養開始24時間後には、全ての糖が消費され、代わりに、光学純度が99.9%のD−乳酸が86.8g/lの濃度で生産された。培地中に含まれる糖から理論的に最大でD−乳酸が作られる場合を収率100%とすると、今回の収率は95%を越えている。
【0055】
次に、CDLA01株を、300mLスケールの坂口フラスコ中において、YPD2液体培地(1%酵母エキス、2%ペプトン、2%グルコース)中、30℃で好気的に振盪培養した。この株を、5Lスケールのジャーファーメンターにおいて、2.0Lの糖蜜培地(18%廃糖蜜、2%ペプトン)に初期OD600が0.1となるように接種し、35℃で攪拌培養した。通気量は1.0vvm、撹拌速度は300rpmとし、10N NaOHおよび10N HClを中和剤として用いることでpHを5.0に維持した。培養開始から72時間が経過した時点で、99.5%を上回る光学純度のD−乳酸が160g/Lの濃度で生産された(図1)。これは、当該時点で消費された糖に対する変換効率で100%に達する値である。
【0056】
実施例2:硝酸塩を唯一の窒素源とする培地におけるキャンディダ・ユティリスの培養
C. utilis NBRC 0988野生株を30℃で好気的に振盪培養した。この株を、5Lスケールのジャーファーメンターにおいて、2.0Lの合成培地(5%グルコース、0.17% Yeast Nitrogen Base、50mM 硝酸ナトリウム)に初期OD600が0.5となるように接種し、30℃で撹拌培養した。通気量は2.0L/min、撹拌速度は300rpmとした。その結果、培養液のpHは経時的に上昇し、培養開始27時間後にはpH8.6にまで達した(図2)。
【0057】
次に、CDLA01株について、上述の合成培地を用いて同様の試験を行ったところ、中和剤を添加すること無く、培養液のpHは3程度で安定し、乳酸が蓄積しても極端な酸性域に偏ることはなかった。この効果によって、pHの低下による酵母の死滅を回避することができた。この結果、培養開始27時間後には15.4g/LのD−乳酸が生産された(図3)。
【0058】
実施例3:硝酸塩と他の窒素含有物質を窒素源とする培地におけるキャンディダ・ユティリスの培養
窒素源として25mM硝酸ナトリウムおよび25mM塩化アンモニウムの組み合わせを含む合成培地において、実施例2と同様にC. utilis NBRC 0988野生株の培養を行った。塩化アンモニウムを単一窒素源とした場合には、通常、pHの急激な低下とともに、培養24時間程度で生菌数の大幅な低下が生じる。これに対して、塩化アンモニウムと同じモル濃度の硝酸ナトリウムを混合した場合には、pHの低下が抑制され、40時間以上にわたって培養を継続できることが明らかとなった(図4)。
【0059】
この結果から、硝酸塩を窒素源として含む培地におけるキャンディダ・ユティリスの培養において、他の窒素源が共存したとしても、硝酸塩によるpH上昇効果が阻害されないことが示される。
【0060】
実施例4:様々な硝酸塩を唯一の窒素源とする培地におけるキャンディダ・ユティリスの培養
50mM硝酸ナトリウムに代えて、50mM硝酸カリウムまたは25mM硝酸カルシウムを単一窒素源として加えた培地を用いて、実施例2と同様にC. utilis NBRC 0988野生株の培養を行った。その結果、いずれの場合においても、硝酸ナトリウムを用いたときと同様の経時的なpHの上昇を示した(図5)。
【0061】
この結果から、硝酸塩を窒素源として含む培地におけるキャンディダ・ユティリスの培養において、硝酸塩によるpH上昇効果は硝酸塩のカチオンの種類に影響されないことが示される。
【0062】
実施例5:硝酸塩を唯一の窒素源とする培地におけるキャンディダ・ユティリスの様々な株の培養
キャンディダ・ユティリスのATCC 15239株、NBRC 0619株、NBRC 0626株、およびNBRC 10708株を用いて、培養液のpH上昇効果を試験した。具体的には、200mLスケールの三角フラスコにおいて、各株を、実施例2に記載の硝酸培地50mLに初期OD600が0.5となるように接種し、30℃で振盪培養した。その結果、いずれの株においても、NBRC 0988株と同様に経時的な培養液のpH上昇が認められた(図6)。
【0063】
この結果から、硝酸塩を窒素源として含む培地におけるキャンディダ・ユティリスの培養において、硝酸塩によるpH上昇効果は、株の違いに影響されず、キャンディダ・ユティリス全般に認められることが示される。
【0064】
実施例6:硝酸塩を唯一の窒素源とする培地におけるキャンディダ・ユティリス以外の酵母の培養
硝酸塩を単一窒素源として生育できるキャンディダ・ユティリス以外の酵母として、ピキア・アングスタ(Pichia angusta)、パキソレン・タンノフィルス(Pachysolen tannophilus)、およびハンセヌラ・ポリモルファ(Hansenula polymorpha)の3種を用いて、実施例5と同様の培養試験を行った。その結果を表1に示す。
【0065】
【表1】
【0066】
表1から明らかなように、3種の酵母のいずれを培養した場合にも、キャンディダ・ユティリスに見られたような急激なpHの上昇は認められなかった。
【図面の簡単な説明】
【0067】
【図1】図1は、CDLA01株を、5Lスケールのジャーファーメンターにおいて、2.0Lの糖蜜培地(18%廃糖蜜、2%ペプトン)に初期OD600が0.1となるように接種し、35℃で撹拌培養した結果を示す。この培養では、通気量を1.0vvm、撹拌速度を300rpmとし、10N NaOHおよび10N HClを中和剤として用いることによりpHを5.0に維持した。図中の白丸および白三角は培養混合物中のD−乳酸の濃度を示し、黒丸および黒三角は培養混合物中のグルコースの濃度を示す。
【図2】図2は、C. utilis NBRC 0988野生株を、硝酸ナトリウムを唯一の窒素源として含む培地において培養した場合のpHの変化を示す。図中の実線は培養液のpHを示し、白丸(ドット)はOD600の値を示す。
【図3】図3は、CDLA01株を、硝酸ナトリウムを唯一の窒素源として含む培地において培養した場合のpHの変化およびD−乳酸の生成を示す。図中の実線は培養液のpHを示し、白丸(ドット)はD−乳酸の濃度(g/l)を示す。
【図4】図4は、C. utilis NBRC 0988野生株を、硝酸ナトリウムと塩化アンモニウムの組み合わせを窒素源として含む培地において培養した場合のpHの変化を示す。図中の実線は培養液のpHを示し、白丸(ドット)はOD600の値を示す。
【図5】図5は、C. utilis NBRC 0988野生株を、硝酸ナトリウム(白丸)、硝酸カリウム(白菱形)および硝酸カルシウム(白三角)のいずれか一つを唯一の窒素源として含む培地において培養した場合のpHの変化を示す。
【図6】図6は、キャンディダ・ユティリスATCC 15239株(白丸)、NBRC 0619株(白菱形)、NBRC 0626株(白三角)、およびNBRC 10708株(白四角)のそれぞれを、硝酸ナトリウムを唯一の窒素源として含む培地において培養した場合のpHの変化を示す。
【特許請求の範囲】
【請求項1】
キャンディダ・ユティリスの培養中に培養液のpHを上方に調節するための方法であって、培養液中に硝酸塩を添加することを含んでなる、方法。
【請求項2】
添加される硝酸塩が、キャンディダ・ユティリスによって資化される主要な窒素源として培養液に存在する、請求項1に記載の方法。
【請求項3】
キャンディダ・ユティリスを培養することにより有機酸を製造する方法であって、培養液中に硝酸塩を添加することを含んでなる、方法。
【請求項4】
添加される硝酸塩が、キャンディダ・ユティリスによって資化される主要な窒素源として培養液に存在する、請求項3に記載の方法。
【請求項5】
前記有機酸がL−乳酸またはD−乳酸である、請求項3または4に記載の方法。
【請求項6】
キャンディダ・ユティリスが遺伝的に改変された菌株である、請求項3〜5のいずれか一項に記載の方法。
【請求項7】
前記菌株が、キャンディダ・ユティリスの野生株に対して、
D−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子の発現を可能にするプロモーター配列が機能的に結合した該遺伝子の少なくとも1コピーにより形質転換されており、かつ、
ピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードしている内因性遺伝子が破壊されているものである、請求項6に記載の方法。
【請求項8】
硝酸塩を含んでなる、キャンディダ・ユティリスの培養中に培養液のpHを上方に調節するためのpH調節剤。
【請求項9】
D−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子の発現を可能にするプロモーター配列が機能的に結合した該遺伝子の少なくとも1コピーにより形質転換されてなり、かつ、ピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードしている内因性遺伝子が破壊されている、キャンディダ・ユティリスの酵母菌株。
【請求項10】
D−乳酸脱水素酵素の活性を有するポリペプチドが、
(a)配列番号2で表されるアミノ酸配列を含むポリペプチド、または
(b)配列番号2で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつD−乳酸脱水素酵素の活性を有するポリペプチド
である、請求項9に記載の酵母菌株。
【請求項11】
D−乳酸脱水素酵素の活性を有するポリペプチドをコードしている遺伝子が、
(a)配列番号1で表されるヌクレオチド配列、
(b)配列番号1で表されるヌクレオチド配列と85%以上の相同性があり、かつD−乳酸脱水素酵素の活性を有するポリペプチドをコードするヌクレオチド配列、または
(c)配列番号1で表されるヌクレオチド配列もしくはその相補配列とストリンジェントな条件下でハイブリダイズし、かつD−乳酸脱水素酵素の活性を有するポリペプチドをコードするヌクレオチド配列
を含むものである、請求項9または10に記載の酵母菌株。
【請求項12】
ピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードしている内因性遺伝子が、配列番号8で表されるアミノ酸配列をコードするヌクレオチド配列、または配列番号7で表されるヌクレオチド配列を含むものである、請求項9〜11のいずれか一項に記載の酵母菌株。
【請求項1】
キャンディダ・ユティリスの培養中に培養液のpHを上方に調節するための方法であって、培養液中に硝酸塩を添加することを含んでなる、方法。
【請求項2】
添加される硝酸塩が、キャンディダ・ユティリスによって資化される主要な窒素源として培養液に存在する、請求項1に記載の方法。
【請求項3】
キャンディダ・ユティリスを培養することにより有機酸を製造する方法であって、培養液中に硝酸塩を添加することを含んでなる、方法。
【請求項4】
添加される硝酸塩が、キャンディダ・ユティリスによって資化される主要な窒素源として培養液に存在する、請求項3に記載の方法。
【請求項5】
前記有機酸がL−乳酸またはD−乳酸である、請求項3または4に記載の方法。
【請求項6】
キャンディダ・ユティリスが遺伝的に改変された菌株である、請求項3〜5のいずれか一項に記載の方法。
【請求項7】
前記菌株が、キャンディダ・ユティリスの野生株に対して、
D−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子の発現を可能にするプロモーター配列が機能的に結合した該遺伝子の少なくとも1コピーにより形質転換されており、かつ、
ピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードしている内因性遺伝子が破壊されているものである、請求項6に記載の方法。
【請求項8】
硝酸塩を含んでなる、キャンディダ・ユティリスの培養中に培養液のpHを上方に調節するためのpH調節剤。
【請求項9】
D−乳酸脱水素酵素の活性を有するポリペプチドをコードする遺伝子の発現を可能にするプロモーター配列が機能的に結合した該遺伝子の少なくとも1コピーにより形質転換されてなり、かつ、ピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードしている内因性遺伝子が破壊されている、キャンディダ・ユティリスの酵母菌株。
【請求項10】
D−乳酸脱水素酵素の活性を有するポリペプチドが、
(a)配列番号2で表されるアミノ酸配列を含むポリペプチド、または
(b)配列番号2で表されるアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換、付加もしくは挿入されたアミノ酸配列を含み、かつD−乳酸脱水素酵素の活性を有するポリペプチド
である、請求項9に記載の酵母菌株。
【請求項11】
D−乳酸脱水素酵素の活性を有するポリペプチドをコードしている遺伝子が、
(a)配列番号1で表されるヌクレオチド配列、
(b)配列番号1で表されるヌクレオチド配列と85%以上の相同性があり、かつD−乳酸脱水素酵素の活性を有するポリペプチドをコードするヌクレオチド配列、または
(c)配列番号1で表されるヌクレオチド配列もしくはその相補配列とストリンジェントな条件下でハイブリダイズし、かつD−乳酸脱水素酵素の活性を有するポリペプチドをコードするヌクレオチド配列
を含むものである、請求項9または10に記載の酵母菌株。
【請求項12】
ピルビン酸脱炭酸酵素の活性を有するポリペプチドをコードしている内因性遺伝子が、配列番号8で表されるアミノ酸配列をコードするヌクレオチド配列、または配列番号7で表されるヌクレオチド配列を含むものである、請求項9〜11のいずれか一項に記載の酵母菌株。
【図1】


【図2】
【図3】
【図4】

【図5】

【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2012−235714(P2012−235714A)
【公開日】平成24年12月6日(2012.12.6)
【国際特許分類】
【出願番号】特願2011−105417(P2011−105417)
【出願日】平成23年5月10日(2011.5.10)
【出願人】(000253503)キリンホールディングス株式会社 (247)
【Fターム(参考)】
【公開日】平成24年12月6日(2012.12.6)
【国際特許分類】
【出願日】平成23年5月10日(2011.5.10)
【出願人】(000253503)キリンホールディングス株式会社 (247)
【Fターム(参考)】
[ Back to top ]