
スフィンゴミエリナーゼ含有組成物の製造方法
【課題】スフィンゴミエリナーゼ、及び純度が高く安定性の高いスフィンゴミエリナーゼ含有組成物を提供する。
【解決手段】以下の工程(a)及び(b):(a)以下の(i)又は(ii)のアミノ酸配列:(i)特定アミノ酸配列、又は(ii)特定のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして形質転換体を得る工程;及び(b)工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程;を含む。
【解決手段】以下の工程(a)及び(b):(a)以下の(i)又は(ii)のアミノ酸配列:(i)特定アミノ酸配列、又は(ii)特定のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして形質転換体を得る工程;及び(b)工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程;を含む。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はスフィンゴミエリナーゼの製造方法、及び純度が高く安定性の高いスフィンゴミエリナーゼ含有組成物及びその製造方法に関する。
【背景技術】
【0002】
スフィンゴミエリナーゼ(Sphingomyelinase)はスフィンゴミエリン(Sphingomyelin)をセラミド(N−アシルスフィンゴシン(N−acylsphingosine))とフォスフォリルコリン(Phosphorylcholine)に加水分解する酵素である(EC 3.1.4.12)。
【0003】
スフィンゴミエリナーゼとしては、バチルス セレウス(Bacillus cereus)(非特許文献1)、レプトスピラ インターロガンス(Leptospira interrogans serovars)(非特許文献2)、リステリア イバノビ(Listeria ivanovii)(非特許文献3)、マウス(非特許文献4)、ラット(非特許文献5)、ヒト(非特許文献6)、及び、ストレプトマイセス グリセオカルネウス(Streptomyces griseocarneus)(非特許文献7)由来のもの等が知られている。非特許文献2及び3は、血清型レプトスピラ インターロガンス及びリステリア イバノビの、スフィンゴミエリナーゼと予想される遺伝子を開示する。非特許文献4は、マウス由来のスフィンゴミエリナーゼの機能を開示する。
【0004】
スフィンゴミエリナーゼは、例えば、動脈硬化の診断に重要な小粒子高比重LDL(small,denseLDL)中のコレステロールの定量等に利用されている(特許文献1)。
【0005】
スフィンゴミエリナーゼの製造法は、例えば、特許文献2及び特許文献3等に記載されている。特許文献2は、Streptomyces sp.A9107菌株を培養した培養物からスフィンゴミエリナーゼを採取することを特徴とする、スフィンゴミエリナーゼの製造法を開示する。特許文献3は、スフィンゴミエリナーゼ遺伝子を含むDNAで形質転換したバチルス・ブレビス47/pNUSMを培養し、培養物からスフィンゴミエリナーゼを採取することを特徴とする該酵素の製造法を開示する。
【0006】
さらに、スフィンゴミエリナーゼの製造法として、非特許文献1は、バチルス セレウス由来のスフィンゴミエリナーゼ遺伝子をエシェリキア コリ(Escherichia coli)HB10に導入することによりスフィンゴミエリナーゼ含有培養物を取得する方法を開示する。非特許文献5は、ラット由来のスフィンゴミエリナーゼ遺伝子をEscherichia coli JM109に導入することを含む、スフィンゴミエリナーゼ含有培養物を取得する方法を開示する。非特許文献6はヒト由来のスフィンゴミエリナーゼ遺伝子をアフリカミドリザル由来の培養細胞株COS−1細胞に導入したことを開示する。非特許文献7はストレプトマイセス グリセオカルネウス由来のスフィンゴミエリナーゼ遺伝子をストレプトマイセス リビダンス(Stpreotomyces lividans)に導入することを含む、スフィンゴミエリナーゼ含有培養物を取得する方法を開示する。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】WO2009/048143
【特許文献2】特開昭58−43786号公報
【特許文献3】特開平5−328970号公報
【非特許文献】
【0008】
【非特許文献1】Johansen T.ら、Gene,65,293−304(1988)
【非特許文献2】Nascimento A.L.T.O.ら,Bacteriol.,186,2164−2172(2004)
【非特許文献3】Gonzalez−Zornら,Mol.Microbiol.,33,510−523(1999)
【非特許文献4】Stoffel W.ら、J.Proc.Natl.Acad.Sci. U.S.A.,102,4554−4559(2005)
【非特許文献5】Mizutani Y.ら、Biochim.Biophys.Acta,1485,236−246(2000)
【非特許文献6】Schuchman E.H.ら、J.Biol.Chem.,266,8531−8539(1991)
【非特許文献7】Sugimori D., Tomita Y.ら、Biotechnol.Lett.,2010 Nov 30.
【発明の概要】
【発明が解決しようとする課題】
【0009】
特許文献2に記載の方法により得られるスフィンゴミエリナーゼ含有培養物は、純度が低く安定性が悪いという問題がある。特許文献3に記載の方法により得られるスフィンゴミエリナーゼ含有培養物は、特異性が低いという問題がある。非特許文献1に記載の方法により得られるスフィンゴミエリナーゼ含有培養物は、フォスフォリパーゼ(phospholipase)C活性を併せ持ち特異性が低いという問題がある。非特許文献5に開示された方法で得られるスフィンゴミエリナーゼ含有培養物は、リゾ血小板活性化因子を加水分解する活性を併せ持ち特異性が低いという問題があり、また、非特許文献5はスフィンゴミエリナーゼ含有培養物中のスフィンゴミエリナーゼの純度に関する記載はない。非特許文献6に開示された方法は、工業的生産に適していないという問題があり、また、非特許文献6はスフィンゴミエリナーゼ含有培養物中のスフィンゴミエリナーゼの純度を開示していない。非特許文献7に開示された方法で得られるスフィンゴミエリナーゼ含有培養物は、フォスフォリパーゼC活性を併せ持ち、特異性が低いという問題がある。
【0010】
このように従来知られているスフィンゴミエリナーゼの製造法はいずれも実用性が低く、また、微生物の培養によりスフィンゴミエリナーゼ含有培養物を得る場合、得られる培養物中のスフィンゴミエリナーゼの純度が低いという問題があり、満足のいくものではないという現状を本発明者らは問題点として強く認識した。このような背景のもと、本発明が解決しようとする課題は、スフィンゴミエリナーゼの製造方法、並びに純度が高く安定性の高いスフィンゴミエリナーゼ含有組成物及びその製造方法を提供することにある。
【課題を解決するための手段】
【0011】
本発明者らは、上記の課題を解決すべく鋭意研究を重ねた結果、驚くべきことにロドコッカス(Rhodococcus)属細菌を宿主として、当該宿主とは属が異なるストレストマイセス・エスピー(Streptomyces sp.)由来のスフィンゴミエリナーゼをコードする遺伝子を導入し、得られた形質転換体を培養することで、スフィンゴミエリナーゼを容易に製造することができ、また、得られた培養物は純度が高く安定性の高いスフィンゴミエリナーゼ含有組成物であることを見出し、本発明を完成するに至った。
【0012】
すなわち、本発明は以下のものに関する。
〔1〕
スフィンゴミエリナーゼの製造方法であって、以下の工程(a)及び(b):
(a)以下の(i)又は(ii)のアミノ酸配列:
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして形質転換体を得る工程;及び
(b)工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程;
を含む方法。
〔1−2〕
さらに以下の工程(c):
(c)工程(b)で得られた培養物から形質転換体を分離してスフィンゴミエリナーゼを得る工程
を含む、前記〔1〕に記載の製造方法。
〔1−3〕
ロドコッカス属細菌が、ロドコッカス エリスロポリスである、前記〔1〕又は〔1−2〕に記載の製造方法。
〔1−4〕
ロドコッカス エリスロポリスが、ロドコッカス エリスロポリス L88株である、前記〔1−3〕に記載の製造方法。
【0013】
〔2〕
工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、前記〔1〕〜〔1−4〕のいずれかに記載の製造方法。
〔2−2〕
工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、
(i)又は(ii)のアミノ酸配列のN末端と、
(iii)又は(iv)のアミノ酸配列のC末端とが、直接結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、前記〔2〕に記載の製造方法。
〔2−3〕
工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、
(iii)のアミノ酸配列コードする塩基配列を含む遺伝子である、前記〔2〕又は〔2−2〕に記載の製造方法。
【0014】
〔3〕
前記工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、(i)のアミノ酸配列をコードする塩基配列を含む、前記〔1〕〜〔2−3〕のいずれかに記載の製造方法。
【0015】
〔4〕
以下の工程(a)〜(c):
(a)以下の(i)又は(ii)のアミノ酸配列:
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして形質転換体を得る工程;
(b)工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程;及び
(c)工程(b)で得た培養物から形質転換体を分離してスフィンゴミエリナーゼ含有組成物を得る工程;
を含む方法により得られる、スフィンゴミエリナーゼ含有組成物。
〔4−2〕
以下の工程(a)及び(b):
(a)以下の(i)又は(ii)のアミノ酸配列:
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして形質転換体を得る工程;及び
(b)工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程;
を含む方法により得られる、形質転換体及びスフィンゴミエリナーゼ含有培養物。
〔4−3〕
以下の工程(a)及び(b):
(a)以下の(i)又は(ii)のアミノ酸配列:
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして形質転換体を得る工程;及び
(b)工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程;
を含む方法により得られる培養物の培養上清であって、スフィンゴミエリナーゼを含有する培養上清。
〔4−4〕
ロドコッカス属細菌が、ロドコッカス エリスロポリスである、前記〔4〕〜〔4−3〕のいずれかに記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔4−5〕
ロドコッカス エリスロポリスが、ロドコッカス エリスロポリス L88株である、前記〔4−4〕に記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
【0016】
〔5〕
工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、前記〔4〕〜〔4−5〕のいずれかに記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔5−2〕
工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、
(i)又は(ii)のアミノ酸配列のN末端と、
(iii)又は(iv)のアミノ酸配列のC末端とが、直接結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、前記〔5〕に記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔5−3〕
工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、
(iii)のアミノ酸配列をコードする塩基配列を含む遺伝子である、前記〔5〕又は〔5−2〕に記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
【0017】
〔6〕
前記スフィンゴミエリナーゼ含有組成物、培養物又は培養上清に水又は緩衝液を添加してスフィンゴミエリナーゼ濃度が4mg/mLの中性又は酸性の水溶液を調製し、30℃で96時間静置保存した場合、保存前と比較して50%以上のスフィンゴミエリナーゼ活性を保持する、前記〔4〕〜〔5−3〕のいずれかに記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔6−2〕
前記水溶液を30℃で96時間静置保存した場合、保存前と比較して65%以上(好ましくは70%以上、より好ましくは75%以上、さらに好ましくは78%以上)のスフィンゴミエリナーゼ活性を保持する、前記〔6〕に記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔6−3〕
前記スフィンゴミエリナーゼ含有組成物、培養物又は培養上清に水又は緩衝液を添加してスフィンゴミエリナーゼ濃度が4mg/mLのpH6.8の水溶液を調製し、30℃で96時間静置保存した場合、保存前と比較して50%以上のスフィンゴミエリナーゼ活性を保持する、前記〔4〕〜〔5−3〕のいずれかに記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔6−4〕
前記水溶液を30℃で96時間静置保存した場合、保存前と比較して65%以上(好ましくは70%以上、より好ましくは75%以上、さらに好ましくは78%以上)のスフィンゴミエリナーゼ活性を保持する、前記〔6−3〕に記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔6−5〕
前記スフィンゴミエリナーゼ含有組成物、培養物又は培養上清に水又は緩衝液を添加してスフィンゴミエリナーゼ濃度が4mg/mLのpH5.5の水溶液を調製し、30℃で96時間静置保存した場合、保存前と比較して50%以上のスフィンゴミエリナーゼ活性を保持する、前記〔4〕〜〔5−3〕のいずれかに記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔6−6〕
前記水溶液を30℃で96時間静置保存した場合、保存前と比較して75%以上(好ましくは78%以上、より好ましくは90%以上、さらに好ましくは97%以上)のスフィンゴミエリナーゼ活性を保持する、前記〔6−5〕に記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔6−7〕
得られるスフィンゴミエリナーゼが、ストレプトマイセス由来のスフィンゴミエリナーゼと同一のアミノ酸配列及び/又は特徴を有する、前記〔1〕〜〔3〕のいずれかに記載の製造方法。
【0018】
なお、上記〔1〕〜〔3〕のように引用する項番号が範囲で示され、その範囲内に〔2−2〕等の枝番号を有する項が配置されている場合には、〔2−2〕等の枝番号を有する項も引用されることを意味する。以下においても同様である。
【0019】
〔6−8〕
得られるスフィンゴミエリナーゼが、ストレプトマイセス・エスピー由来のスフィンゴミエリナーゼと同一のアミノ酸配列及び/又は特徴を有する、前記〔1〕〜〔3〕のいずれかに記載の製造方法。
〔6−9〕
得られるスフィンゴミエリナーゼが、ストレプトマイセス・エスピー A9107株由来のスフィンゴミエリナーゼと同一のアミノ酸配列及び/又は特徴を有する、前記〔1〕〜〔3〕のいずれかに記載の製造方法。
〔6−10〕
前記製造方法で得られるスフィンゴミエリナーゼあるいは前記組成物、培養物又は培養上清に含まれるスフィンゴミエリナーゼが、以下の(1)の作用を有し、さらに以下の(2)〜(5)のいずれか1つ以上の特徴を有するスフィンゴミエリナーゼである、前記〔1〕〜〔6−9〕のいずれかに記載の製造方法、組成物、培養物又は培養上清:
(1)作用
1分子の水の存在下、スフィンゴミエリン1分子を分解してセラミド1分子とフォスフォリルコリン1分子を生成する。
(2)基質特異性
スフィンゴミエリンに作用し、かつ、レシチン、リゾレシチン、フォスファチジルエタノールアミン、フォスファチジルセリン及びフォスファチジルイノシトールのいずれにも実質的に作用しない。
(3)Km
スフィンゴミエリンに対するKm値が約0.45mMである。
(4)至適pH
pH7.0〜8.0。
(5)分子量
ドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動法による分子量が約38kDaである。
〔6−11〕
前記〔1〕〜〔3〕のいずれかに記載の製造方法により得られるスフィンゴミエリナーゼ。
【0020】
〔7〕
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子を含有する組換えベクター。
〔7−2〕
前記遺伝子を、以下の群:pTip QC1、pTip QC2、pTip QT1、pTip QT2、pTip RC1、pTip RC2、pTip RT1、pTip RT2、pNit QC1、pNit QC2、pNit QT1、pNit QT2、pNit RC1、pNit RC2、pNit RT1及びpNit RT2;から選択される発現ベクターに挿入して得られる、前記〔7〕に記載の組換えベクター。
〔7−3〕
前記遺伝子が、(i)のアミノ酸配列をコードする塩基配列を含む、前記〔7〕又は〔7−2〕に記載の組換えベクター。
【0021】
〔8〕
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子を含有する組換えベクター。
〔8−2〕
前記遺伝子を、以下の群:pTip QC1、pTip QC2、pTip QT1、pTip QT2、pTip RC1、pTip RC2、pTip RT1、pTip RT2、pNit QC1、pNit QC2、pNit QT1、pNit QT2、pNit RC1、pNit RC2、pNit RT1及びpNit RT2;から選択される発現ベクターに挿入して得られる、前記〔8〕に記載の組換えベクター。
〔8−3〕
前記遺伝子が、
(i)又は(ii)のアミノ酸配列のN末端と、
(iii)又は(iv)のアミノ酸配列のC末端とが、直接結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、前記〔8〕又は〔8−2〕に記載の組換えベクター。
〔8−4〕
前記遺伝子が、(i)のアミノ酸配列をコードする塩基配列を含む、前記〔8〕〜〔8−3〕のいずれかに記載の組換えベクター。
〔8−5〕
前記遺伝子が、(iii)のアミノ酸配列をコードする塩基配列を含む、前記〔8〕〜〔8−4〕のいずれかに記載の組換えベクター。
【0022】
〔9〕
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子を有し、スフィンゴミエリナーゼを培養物中に分泌することを特徴とする、ロドコッカス属細菌形質転換体。
〔9−2〕
ロドコッカス属細菌が、ロドコッカス エリスロポリスである、前記〔9〕に記載の形質転換体。
〔9−3〕
ロドコッカス エリスロポリスが、ロドコッカス エリスロポリス L88株である、前記〔9−2〕に記載の製造方法。
【0023】
〔10〕
前記遺伝子が、
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、前記〔9〕〜〔9−3〕のいずれかに記載の形質転換体。
〔10−2〕
前記遺伝子が、
(i)又は(ii)のアミノ酸配列のN末端と、
(iii)又は(iv)のアミノ酸配列のC末端とが、直接結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、前記〔10〕に記載の形質転換体。
〔10−3〕
前記遺伝子が、(i)のアミノ酸配列をコードする塩基配列を含む、前記〔9〕〜〔10−2〕のいずれかに記載の形質転換体。
〔10−4〕
前記遺伝子が、(iii)のアミノ酸配列をコードする塩基配列を含む、前記〔10〕〜〔10−3〕のいずれかに記載の形質転換体。
【0024】
〔11〕
前記〔7〕〜〔8−5〕のいずれかに記載の組換えベクターでトランスフェクトされたロドコッカス属細菌。
〔11−2〕
ロドコッカス エリスロポリスである、前記〔11〕に記載のロドコッカス属細菌。
〔11−3〕
ロドコッカス エリスロポリス L88株である前記〔11−2〕に記載のロドコッカス属細菌。
【0025】
〔12〕
前記〔1〕〜〔3〕のいずれかに記載の製造方法で得られるスフィンゴミエリナーゼ、あるいは前記〔4〕〜〔6−10〕のいずれかに記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清を用いたスフィンゴミエリナーゼの基質の測定方法。
〔13〕
前記〔1〕〜〔3〕のいずれかに記載の製造方法で得られるスフィンゴミエリナーゼ、あるいは前記〔4〕〜〔6−10〕のいずれかに記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清を用いたスフィンゴミエリナーゼの基質の消去方法。
〔14〕
スフィンゴミエリナーゼ又はスフィンゴミエリナーゼ含有組成物の製造における前記〔7〕〜〔8−5〕のいずれかに記載の組換えベクターの使用。
〔15〕
スフィンゴミエリナーゼ又はスフィンゴミエリナーゼ含有組成物の製造における前記〔11〕〜〔11−3〕に記載のロドコッカス属細菌の使用。
【発明の効果】
【0026】
本発明により、スフィンゴミエリナーゼ、及び純度が高く、安定性の高いスフィンゴミエリナーゼ含有組成物を効率的に製造することができる。したがって、純度が高く、安定性の高いスフィンゴミエリナーゼ含有組成物を取得することができ、この組成物から容易にスフィンゴミエリナーゼを取得することもできる。
【図面の簡単な説明】
【0027】
【図1】比較例2において得られた配列番号20の塩基配列の構造と、PCR反応に用いたプライマーの位置を示す模式図である。
【図2】「実施例1 10.」記載の精製酵素と「比較例1 2.」記載の比較試験用酵素を、それぞれ、30℃で(A)72時間又は(B)96時間静置保存した際のスフィンゴミエリナーゼ活性の残存率を示す図である。
【図3】「実施例1 10.」記載の精製酵素と「比較例1 2.」記載の比較試験用酵素を純水又は酢酸ナトリウム緩衝液に溶解し、30℃又は−30℃で96時間静置保存したものを用いてSDS−PAGEを行った結果を示す図である。
【発明を実施するための形態】
【0028】
以下、本発明を実施するための形態(以下、「本実施の形態」と略すことがある)について具体的に説明する。
【0029】
本実施の形態のスフィンゴミエリナーゼの製造方法は、以下の工程(a)及び(b)を含む。
(a)以下の(i)又は(ii)のアミノ酸配列:
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして形質転換体を得る工程;及び
(b)工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程。
【0030】
工程(a)
工程(a)は、ロドコッカス属細菌に遺伝子をトランスフェクトして形質転換体を得る工程である。トランスフェクトする遺伝子は、得られた形質転換体を培養した場合にスフィンゴミエリナーゼを含む培養物が得られる限り特に限定されないが、好ましくは以下の(i)又は(ii)のアミノ酸配列をコードする塩基配列を含む遺伝子である。
【0031】
(i)配列番号1に記載のアミノ酸配列
上記工程(a)における(i)のアミノ酸配列からなるタンパク質は、ストレプトマイセス・エスピー A9107に由来するスフィンゴミエリナーゼである。
【0032】
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
上記工程(a)における(ii)のアミノ酸配列としては、例えば、配列番号1に記載のアミノ酸配列において、酵素作用に関与しない一部のアミノ酸を変異させたアミノ酸配列が例示される。配列番号3に記載のアミノ酸配列(配列番号1に記載のアミノ酸配列の50〜56番目の配列に対応)及び配列番号4に記載のアミノ酸配列(配列番号1に記載のアミノ酸配列の199〜204番目の配列に対応)(但し、配列番号3及び4中、Xaaは任意のアミノ酸を示す)は、いずれもスフィンゴミエリナーゼ活性に必須の配列として知られる(Amy E.A.O.ら、J.Biol.Chem.,280,35011−35017(2005)、Ago H.ら、J.Biol.Chem.,281,16157−16167(2006)等)。したがって、上記工程(a)における(ii)のアミノ酸配列としては、例えば、配列番号1に記載のアミノ酸配列において、配列番号3及び4に対応する配列以外の部分のアミノ酸を変異させたアミノ酸配列が例示される。性質の似たアミノ酸への置換等であれば、変異後のアミノ酸配列からなるタンパク質のスフィンゴミエリナーゼ活性に影響しないであろう。例えば、上記工程(a)における(ii)のアミノ酸配列としては、DVVX1X2X3E(式中、X1はV又はIを、X2はL又はFを、X3はD、N、Q又はEを、それぞれ表す)、及びX4X5GDX6N(式中、X4はV、I又はLを、X5はA、C又はGを、X6はF、M又はLを、それぞれ表す)の配列を含むアミノ酸配列が好ましい。
【0033】
上記工程(a)における(ii)のアミノ酸配列としては、また、配列番号1に記載のアミノ酸配列に各種のアミノ酸残基が付加したアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列も例示される。そのようなアミノ酸配列又は該アミノ酸配列からなるタンパク質としては、例えば、配列番号1に記載のアミノ酸配列のC末端側にチオレドキシンタンパク質等機能性タンパク質を付加した融合タンパク質、配列番号1に記載のアミノ酸配列に、タンパク質の精製や確認等をせしめることのできるタグと呼ばれる部分を付加した融合タンパク質等が挙げられる。タグの付加は、その全部又は一部であってもよい。例えば、本実施の形態における製造方法において、培養物からスフィンゴミエリナーゼを効率的に精製するために、配列番号1に記載のアミノ酸配列に5〜10個のHisを付加してもよい。また、配列番号1に記載のアミノ酸配列と、付加アミノ酸(配列)との間等に、数個のプロテアーゼ認識アミノ酸配列を配置して付加することもできる。更に、配列番号1のアミノ酸配列のC末端側に付加していてもよいアミノ酸残基(又はポリペプチド残基)としては、TEE配列、Sタグ、Hisタグ等が挙げられる。
【0034】
上記工程(a)における(ii)のアミノ酸配列としては、また、配列番号1に記載のアミノ酸配列において、各種のアミノ酸が欠失したアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列も例示される。そのような欠失の例としては、例えば:スフィンゴミエリナーゼの本質的な機能とは無関係の数個のアミノ酸からなるドメインの欠失;配列番号1に記載のアミノ酸配列中の複数個のアミノ酸からなるギャップの欠失;配列番号1に記載のアミノ酸配列のC末端側のArgから順に削除する欠失;配列番号1のアミノ酸配列のN末端側のAlaの欠失;それらの欠失の組み合わせ等が挙げられる。
【0035】
変異後に得られるアミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有する限り、上記の欠失、置換又は付加等の変異は、適宜組み合わせることも可能である。任意のアミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するか否かは、当業者に公知の手法により確認することができる。例えば、該アミノ酸配列をコードする塩基配列を含む遺伝子をトランスフェクトして得た形質転換体を培養し、培養物のスフィンゴミエリナーゼ活性を測定することで確認することができる。培養物が、後述する理化学的性質<1>〜<5>を有するスフィンゴミエリナーゼを含むことが好ましい。
【0036】
一態様において、工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子は、上記(i)及び(ii)のアミノ酸配列のうち、(i)のアミノ酸配列をコードする塩基配列を含むことが好ましい。別の態様として(ii)のアミノ酸配列をコードする塩基配列を含むことが好ましい場合もある。
【0037】
上記(i)又は(ii)のアミノ酸配列をコードする塩基配列は、目的とするアミノ酸配列をもとに、容易に得ることができる。配列番号1のアミノ酸配列をコードする塩基配列としては、特に限定されないが、例えば、配列番号5の塩基配列や、配列番号5の塩基配列を宿主のコドン使用頻度に合わせて変更した塩基配列が例示されるが、ロドコッカス属細菌のコドン使用頻度に合わせて変更した塩基配列が好ましく、ロドコッカス エリスロポリスのコドン使用頻度に合わせて変更した塩基配列がより好ましい。そのようなコドン使用頻度は、 http://www.kazusa.or.jp/codon/ などで容易に検索することができる。塩基配列の調製には、通常用いられる公知の遺伝子操作を利用することができる。塩基配列が所望のアミノ酸配列をコードする限り、例えば、部位特異的変異法や、目的遺伝子の特定塩基の断片を人工変異塩基で置換する等の種々の方法を用いて塩基配列に変異を加えてもよい。
【0038】
微生物等を用いて酵素等のタンパク質を製造する場合、生産性を高めること等を目的として、シグナルペプチドと呼ばれるアミノ酸配列を、目的とする酵素等のタンパク質に付加することがある。本実施の形態におけるスフィンゴミエリナーゼの製造方法においても、スフィンゴミエリナーゼのアミノ酸配列のN末端側にシグナルペプチドを付加することができる。そのようなシグナルペプチドは、本実施の形態における製造方法においてシグナルペプチドとして機能するものであれば特に限定されない。
【0039】
本実施の形態において、あるペプチドが、「シグナルペプチドとして機能する」とは、該ペプチドと、上記(i)又は(ii)に記載のアミノ酸配列とが直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして得られた形質転換体を培養した場合に、スフィンゴミエリナーゼを含む培養物を得ることができることを言う。純度が高く、安定性の高いスフィンゴミエリナーゼを含む培養物を効率的に得ることができるシグナルペプチドが好ましい。例えば、上記培養した場合に、スフィンゴミエリナーゼが培養液中に形成される(形質転換体内で生産されたスフィンゴミエリナーゼがシグナルペプチドにより培養液中に輸送される)シグナルペプチド(アミノ酸配列)が好ましい。また、シグナルペプチドとしては、得られるスフィンゴミエリナーゼの活性を低下させないものが好ましい。特に、純度が高く、安定性の高いスフィンゴミエリナーゼを含む培養物を効率的に得るという観点からは、シグナルペプチドとしてはストレプトマイセス・エスピー A9107株由来のシグナルペプチド(配列番号2)が好ましい。また、シグナルペプチドとしては、上記シグナルペプチドのアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、シグナルペプチドとして機能するアミノ酸配列を用いることもできる。
【0040】
具体的には、本実施の形態におけるスフィンゴミエリナーゼの製造方法の工程(a)において、
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子を、ロドコッカス属細菌にトランスフェクトすることで、シグナルペプチドをスフィンゴミエリナーゼに付加することができる。
【0041】
上記(iv)のアミノ酸配列としては、例えば、配列番号2に記載のアミノ酸配列のN末端側にチオレドキシンタンパク質等機能性タンパク質やHisタグ等のその他のアミノ酸配列からなる部分を付加したアミノ酸配列が例示され、融合タンパク質とすることも好ましい。場合によっては、上記の付加アミノ酸配列は、その全部又は一部であってもよい。上述の付加の例と同様に、シグナルペプチドとして機能するアミノ酸配列が得られる限り、常法によってアミノ酸の欠失又は置換を行うことができ、例えば、該アミノ酸配列中に複数個のアミノ酸からなるギャップが存在する場合、それらの欠失を組み合わせることもできる。また、上記の欠失、置換又は付加を適宜組み合わせることも可能である。
【0042】
上記(iii)又は(iv)のアミノ酸配列をコードする塩基配列は、目的とするアミノ酸配列をもとに、容易に得ることができる。配列番号2のアミノ酸配列をコードする塩基配列としては、特に限定されないが、例えば、配列番号6の塩基配列が挙げられる。このような塩基配列は上述のように宿主のコドン使用頻度にあわせて変更することができる。
【0043】
上記遺伝子に含まれる塩基配列は、
上記(i)又は(ii)のアミノ酸配列のN末端と、
上記(iii)又は(iv)のアミノ酸配列のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする。
【0044】
ここで、上記(i)又は(ii)のアミノ酸配列のN末端と、上記(iii)又は(iv)のアミノ酸配列のC末端とが、1若しくは数個のアミノ酸を介して結合する場合、該結合は、結合したアミノ酸配列をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして得られた形質転換体を培養した場合に、スフィンゴミエリナーゼを含む培養物を得ることができる限り特に限定されない。該結合は、プロテアーゼの切断部位であることが望ましく、代表的なプロテアーゼとそれに対応する配列としては、例えば、以下の配列が挙げられる(配列中、↓が切断部位を表す)。
トロンビン:LVPR↓GS
エンテロキナーゼ:DDDDK↓
第X因子(Factor Xa):IEGR↓
【0045】
好ましい態様において、上記遺伝子に含まれる塩基配列は、上記(i)又は(ii)のアミノ酸配列のN末端と、上記(iii)又は(iv)のアミノ酸配列のC末端とが、直接結合したアミノ酸配列をコードする。
【0046】
塩基配列の調製には、通常用いられる公知の遺伝子操作を利用することができる。DNA合成により調製してもよい。塩基配列が所望のアミノ酸配列をコードする限り、例えば、部位特異的変異法や、目的遺伝子の特定塩基の断片を人工変異塩基で置換する等の種々の方法を用いて塩基配列に変異を加えてもよい。
【0047】
上記の塩基配列を含む遺伝子を、常法を用いてロドコッカス属細菌にトランスフェクトして形質転換体を得ることができる。より詳細には、形質転換体において導入遺伝子を発現させるための発現ベクターに、上記の塩基配列を含む遺伝子を挿入して得られる組換えベクターを使用することが出来る。このような発現ベクターとしては、ロドコッカス属細菌を宿主としてトランスフェクトし、得られた形質転換体を培養した場合に、スフィンゴミエリナーゼを含む培養物が得られるベクターであれば特に限定されないが、純度が高く、安定性の高いスフィンゴミエリナーゼを含む培養物を効率的に得られる発現ベクターが好ましく、宿主微生物体内で自律的に増殖し得るファージ又はプラスミドのうち、遺伝子組換え用として構築されたものが好ましい。
【0048】
そのような発現ベクターとしては、例えば、λgt・λC、λgt・λB、pET−3a、pET−11a、pET−32aなどのpETベクター(Novagen社製)並びにpBR322、pBR325、pACYC184、pUC12、pUC13、pUC18、pUC19、pUC118、pIN I、BluescriptKS+、pWH1520、pUB110、pKH300PLK、pIJ680、pIJ702、YRp7、pYC1、YEp13、pTip QC1、pTip QC2、pTip QT1、pTip QT2、pTip RC1、pTip RC2、pTip RT1、pTip RT2、pNit QC1、pNit QC2、pNit QT1、pNit QT2、pNit RC1、pNit RC2、pNit RT1及びpNit RT2などが使用できる。好ましいベクターとしては、pTip QC1、pTip QC2、pTip QT1、pTip QT2、pTip RC1、pTip RC2、pTip RT1、pTip RT2、pNit QC1、pNit QC2、pNit QT1、pNit QT2、pNit RC1、pNit RC2、pNit RT1及びpNit RT2が挙げられ、pTip QC1及びpTip QC2がより好ましい。本実施の形態においては、ロドコッカス属細菌を宿主とする発現ベクターが好ましい。より好ましくは大腸菌とのシャトルベクターとして機能する発現ベクターが挙げられる。
【0049】
プロモーターは宿主中で発現出来るものであれば特に限定されないが、好ましくはpTip QC1、pTip QC2、pTip QT1、pTip QT2、pTip RC1、pTip RC2、pTip RT1、pTip RT2、pNit QC1、pNit QC2、pNit QT1、pNit QT2、pNit RC1、pNit RC2、pNit RT1及びpNit RT2のプロモーターが挙げられ、さらに好ましくはpTip QC1、pTip QC2、pTip QT1及びpTip QT2のプロモーターが挙げられる。
【0050】
発現ベクターへの遺伝子の挿入は、常法により行うことができる。まず、挿入しようとする塩基配列を制限酵素により切断して生成する塩基配列の末端と同じ末端を生成する制限酵素により上記発現ベクターを切断し、発現ベクター断片を作製する。挿入しようとする塩基配列を該制限酵素で切断することにより作製した塩基配列の断片と、発現ベクター断片とを、DNAリガーゼにより常法に従って結合し、遺伝子を発現ベクターに挿入することができる。得られたベクターに目的の遺伝子が挿入されていることは、例えばDNAシーケンスにより確認することができる。得られた組換えベクターは、本実施の形態におけるスフィンゴミエリナーゼの製造等に有用である。
【0051】
本実施の形態において、トランスフェクト(形質転換)とは、遺伝子を細胞(菌体)に導入し、形質転換体を得ることを意味し、その方法は当業者が一般に行うことができるものであれば特に限定されない。例えばコンピテントセルを用いる方法、マイクロインジェクション法、リポフェクション法が例示される。好ましくは、ロドコッカス属細菌に効率的に遺伝子を導入することができる方法であり、形質転換効率がよいという観点から、より好ましくはエレクトロポレーション法である。
【0052】
工程(a)において、遺伝子をトランスフェクトする微生物は、ロドコッカス属細菌であれば特に限定されないが、ロドコッカス エリスロポリス(Rhodococcus erythropolis)であることが好ましい。ロドコッカス エリスロポリスとしては、得られた形質転換体を培養して、スフィンゴミエリナーゼを含む培養物が得られる限り特に限定されないが、純度が高く、安定性の高いスフィンゴミエリナーゼを含む培養物が効率的に得られるものが好ましい。例えば、ロドコッカス エリスロポリス ATCC15591、ロドコッカス エリスロポリス ATCC25544、ロドコッカス エリスロポリス ATCC17041、ロドコッカス エリスロポリス NCIB8147、ロドコッカス エリスロポリス JCM3201、ロドコッカス エリスロポリス JCM3201の変異株及びロドコッカス エリスロポリス L88株(FERM BP−08444)が例示される。好ましくはロドコッカス エリスロポリス JCM3201が挙げられ、より好ましくは特開2004−073116号公報で開示されたロドコッカス エリスロポリス JCM3201の変異株が挙げられ、さらに好ましくはロドコッカス エリスロポリス L88株が挙げられる。任意の微生物が、ロドコッカス属細菌(又はロドコッカス・エリスロポリス)であるかの判断は、バージーズ・マニュアル・オブ・システマティック・バクテリオロジー(Bergey's Manual of Systematic Bacteriology)に従って行うことができる。rRNA塩基配列解析によって行ってもよい。また、微生物同定検査会社に依頼して判別してもよい。
【0053】
目的の遺伝子がトランスフェクトされた形質転換体を選択する際には、導入した発現ベクターに存在する抗生物質耐性遺伝子などの選択マーカーを利用すればよい。例えば、pTip QC1、pTip QC2、pTip RC1、pTip RC2、pNit QC1、pNit QC2、pNit RC1及びpNit RC2を発現ベクターとして用いる場合には、クロラムフェニコールに対する耐性により選択する方法が例示される。また、pTip QT1、pTip QT2、pTip RT1、pTip RT2、pNit QT1、pNit QT2、pNit RT1及びpNit RT2を発現ベクターとして用いる場合にはテトラサイクリンに対する耐性により選択する方法が例示される。得られた形質転換体は、本実施の形態におけるスフィンゴミエリナーゼの製造等に有用である。
【0054】
工程(b)
工程(b)は、上記工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程である。
【0055】
形質転換体の培養条件は、ロドコッカス属細菌の生理学的性質を考慮して選択すればよい。培養は、通常液体培養で行うが、工業的には深部通気撹拌培養を行うことが有利である。
【0056】
培養に用いる培地は、通常ロドコッカス属細菌の培養に用いられるものであれば特に限定されず、例えば、肉汁培地、肉汁ゼラチン培地、牛乳培地、コーンミール培地、トマトジュース培地、TSI培地、SIM培地、グルコース-ブイヨン培地、YM培地、Lennox培地、LB培地、Davis培地、トリス-グルコース培地、EMB糖指示培地、Spizizen最少培地、YPAD培地、Vogel培地、ツァペック-ドッグス培地等を用いることができる。さらに、例えば http://www.dsmz.de/ 等を参考にして、適切な培地を選択することもできる。
【0057】
培養温度はロドコッカス属細菌が発育し、スフィンゴミエリナーゼを含む培養物が得られる範囲で適宜変更し得る。例えば、通常は下限が4℃以上、好ましくは10℃以上、より好ましくは20℃以上、上限が50℃以下、好ましくは42℃以下、より好ましくは37℃以下が例示される。
【0058】
培養時間は、選択する培養条件によって多少異なるが、得られる培養物中のスフィンゴミエリナーゼが最大量に達する時期を見計らって適当な時期に培養を終了すればよい。例えば、後述の実施例1に記載の培養条件で培養した場合、通常は下限が10時間以上、好ましくは12時間以上、より好ましくは17時間以上、上限が100時間以下、好ましくは90時間以下、より好ましくは80時間以下が例示される。
【0059】
培地pHはロドコッカス属細菌が発育し、スフィンゴミエリナーゼを含む培養物が得られる範囲で適宜変更し得る。例えば、下限としてpH 5.8以上、好ましくはpH 6.2以上、上限としてpH 8.5以下、好ましくはpH 7.5以下が例示される。
【0060】
上記の手法で形質転換体を培養して得られる、スフィンゴミエリナーゼを含む培養物は、形質転換体内、培養液中又はその両方にスフィンゴミエリナーゼを含むことができるが、精製の容易性等の観点から、好ましくは、培養液中にスフィンゴミエリナーゼを含む。
【0061】
本実施の形態は、上記工程(a)及び(b)を含む方法により得られる、形質転換体及びスフィンゴミエリナーゼ含有培養物並びに該培養物の培養上清であって、スフィンゴミエリナーゼを含有する培養上清にも関する。
【0062】
本実施の形態は、また、上記工程(a)及び(b)に続いて、以下の工程(c)を行うことにより得られる、スフィンゴミエリナーゼ含有組成物にも関する。
【0063】
工程(c)
工程(c)は、工程(b)で得た培養物から形質転換体を分離してスフィンゴミエリナーゼ含有組成物を得る工程である。工程(b)で得た培養物は、上述の通り、好ましくは培養液中にスフィンゴミエリナーゼを含む培養物である。この場合、工程(b)で得られた培養物から、濾過又は遠心分離などの通常の手段により形質転換体を分離し、培養上清としてスフィンゴミエリナーゼ含有組成物を得ることができる。
【0064】
形質転換体の分離後、さらに、マイクローザ(旭化成ケミカルズ社製)やフィルトロン(富士フィルター社製)などを用いた限外濾過濃縮法などにより濃縮した形で、本実施の形態におけるスフィンゴミエリナーゼ含有組成物を得ることもできる。
【0065】
本実施の形態において、上記工程(c)で得られたスフィンゴミエリナーゼ含有組成物は、純度が高く、保存安定性が高いため、さらなる精製を行うことなく保存が可能であるという利点を有するが、所望により、安定剤として各種の塩類、糖類、タンパク質、脂質、界面活性剤等を加え、あるいは加えることなく、限外濾過濃縮、凍結乾燥等の方法により、液状又は固形のものとすることができる。例えば凍結乾燥を行う場合、安定剤としてサッカロース、マンニトール、食塩、アルブミン等を0.5%〜10%程度添加してもよい。このようにして得られた濃縮物や凍結乾燥物も、本実施の形態におけるスフィンゴミエリナーゼ含有組成物に含まれる。
【0066】
本実施の形態におけるスフィンゴミエリナーゼ含有組成物は、水又は緩衝液を添加してスフィンゴミエリナーゼ濃度が4mg/mLの中性又は酸性の水溶液を調製し、30℃で96時間静置保存して保存試験を行った場合、保存前と比較して50%以上のスフィンゴミエリナーゼ活性を保持することが好ましい。
【0067】
上記の保存試験における「中性又は酸性」とは、pHが中性から酸性であることを意味し、pHの下限は4.0であり、好ましくは5.0であり、より好ましくは5.5である。pHの上限は7.5であり、好ましくは7.0であり、より好ましくは6.8である。
【0068】
上記の保存試験における「緩衝液」は、スフィンゴミエリナーゼ溶液として中性又は酸性の水溶液が得られる限り特に限定されないが、例えば、弱酸の酢酸、クエン酸、リン酸、ホウ酸、又は酒石酸と、これらのナトリウム塩等を用いて調製される緩衝液であり、好ましくは酢酸緩衝液(酢酸+酢酸ナトリウム)である。
【0069】
上記の保存試験における温度の上限は、スフィンゴミエリナーゼ活性が残存する温度であればよく、例えば50℃であり、好ましくは40℃であり、より好ましくは30℃である。静置保存する場合の温度の下限は、例えば4℃であり、好ましくは10℃であり、より好ましくは20℃である。特に好ましい温度は30℃以下である。
【0070】
上記の保存試験におけるスフィンゴミエリナーゼの水溶液の濃度は、特に制限されないが、下限は、例えば1mg/mLであり、好ましくは2mg/mLであり、より好ましくは3mg/mLである。上限は、例えば30mg/mLであり、好ましくは20mg/mLであり、より好ましくは10mg/mLである。例えば、4mg/mLのスフィンゴミエリナーゼ水溶液を作成して、上記の30℃で96時間の静置保存をすることができる。
【0071】
上記の条件で保存した本実施の形態の組成物中のスフィンゴミエリナーゼ活性は、保存前と比較して、少なくとも40%より高い、例えば50%以上、好ましくは65%以上、より好ましくは70%以上、さらに好ましくは75%以上、さらにより好ましくは78%以上のスフィンゴミエリナーゼ活性を保持し、特に好ましくは80%以上のスフィンゴミエリナーゼ活性を保持する。特に、上記の水溶液のpHが酸性(例えばpH5.5)の場合、上記の条件で保存した本実施の形態の組成物中のスフィンゴミエリナーゼ活性は、保存前と比較して、例えば75%以上、好ましくは78%以上、より好ましくは90%以上、さらに好ましくは97%以上のスフィンゴミエリナーゼ活性を保持する。
【0072】
本実施の形態において、上記工程(c)で得られたスフィンゴミエリナーゼ含有組成物は、高い純度(タンパク質量あたりのスフィンゴミエリナーゼ活性)を有する。例えば、本実施の形態におけるスフィンゴミエリナーゼ含有組成物は、タンパク質量あたりのスフィンゴミエリナーゼ活性として少なくとも10U/mg以上、好ましくは50U/mg以上、より好ましくは100U/mg以上の純度を有する。
【0073】
純度は、後述のスフィンゴミエリナーゼ活性測定法と共に、タンパク質濃度の測定法を用いて決定することができる。タンパク質濃度の測定法としては、当該業者に公知の測定方法を用いればよく、特に限定されるものではない。例えば、280nmの吸光度を測定する方法(紫外吸収法)、クーマシーブルーを用いる方法(Bradford法)、フェノール試薬を用いる方法(Lowry法)、ビシンコニン酸を用いる方法(BCA法)が例示される。標準試料として用いるタンパク質も、当該業者が一般的に用いるタンパク質を用いればよく、例えばウシ血清アルブミン、オボアルブミン、ガンマグロブリン、ヒト血清アルブミン及びクリスタリンが例示され、ウシ血清アルブミン及びオボアルブミンが汎用される。
【0074】
本実施の形態において、上記工程(c)で得られたスフィンゴミエリナーゼ含有組成物は、高い保存安定性を有し、例えば70時間以上、好ましくは80時間以上、より好ましくは90時間以上、例えば96時間程度、静置保存しても、保存前と比較して、例えば50%以上、好ましくは65%以上、より好ましくは70%以上、さらに好ましくは75%以上、特に好ましくは78%以上のスフィンゴミエリナーゼ活性を保持する。保存時の温度は、上記の保存試験に関する記載も参照して適宜決定することができるが、保存温度は低いことが好ましい。
【0075】
本実施の形態において、上記工程(c)で得られたスフィンゴミエリナーゼ含有組成物の高い保存安定性は、例えば保存前後のサンプルの電気泳動によっても確認することができる。例えば、本実施の形態におけるスフィンゴミエリナーゼ含有組成物は、水又は緩衝液を添加してスフィンゴミエリナーゼ濃度が4mg/mLである中性又は酸性の水溶液を調製し、30℃又は−30℃で96時間静置保存後、ドデシル硫酸ナトリウム―ポリアクリルアミドゲル電気泳動法で、電気泳動させた場合、例えばデンシトメーターで測定した場合、保存前と比較して、スフィンゴミエリナーゼのポリペプチドを示す帯が少なくとも70%以上、好ましくは80%以上、より好ましくは90%以上残存する。
【0076】
上記の工程(b)で得た培養物又は上記の工程(c)で得た組成物から、以下に記載の通り、常法によりスフィンゴミエリナーゼ(精製物)を得ることができる。
【0077】
上記の工程(b)で得た培養物において、スフィンゴミエリナーゼが形質転換体内に含まれる場合、例えば、まず培養物を固液分離し、得られた湿潤菌体(形質転換体)をリン酸緩衝液やトリス−塩酸緩衝液などの溶液に分散し、リゾチーム処理、超音波処理、フレンチプレス処理、ダイノミル処理などの菌体破砕手段を適宜選択し組み合わせて、形質転換体内のスフィンゴミエリナーゼを抽出することができる。
【0078】
このようにして得た抽出物や、上記の工程(c)で得た組成物から、さらに純度の高いスフィンゴミエリナーゼ(精製物)を得る工程としては、例えば、上記抽出物や組成物に、アセトン、メタノール、エタノールなどの有機溶媒を添加することによる分別沈殿法、硫酸アンモニウム、食塩などによる塩析法などを適用してスフィンゴミエリナーゼを沈殿させ回収する工程が挙げられる。さらに、この沈殿物に対し、必要に応じて透析、等電点沈殿を行った後、電気泳動法などで単一の帯を示すまで、イオン交換体、ゲル濾過剤、吸着体などを用いるカラムクロマトグラフィーなどにより精製し、スフィンゴミエリナーゼ(精製物)を得ることができる。これらの工程を組み合わせることにより目的酵素の精製度が上がる場合は適宜組み合わせて行うことができる。
【0079】
上記の工程(c)で得た組成物は、純度が高いため、非常に簡便な精製法によりスフィンゴミエリナーゼ(精製物)を得ることができるという利点を有する。
【0080】
本実施の形態における製造方法で得られるスフィンゴミエリナーゼ(Sphingomyelinase)または本実施の形態におけるスフィンゴミエリナーゼ含有組成物(培養物、培養上清であってもよい)に含まれるスフィンゴミエリナーゼ(単に「本実施の形態のスフィンゴミエリナーゼ」ともいう)とは、EC 3.1.4.12に分類されるタンパク質であり、好ましくはストレプトマイセス由来、より好ましくはストレプトマイセス・エスピー由来、さらに好ましくはストレプトマイセス・エスピー A9107株スフィンゴミエリナーゼと同一のアミノ酸配列及び/又は特徴を有する。該アミノ酸配列としては、配列番号1に記載のアミノ酸配列が例示される。また、該特徴としては、以下の(2)〜(5)に記載の特徴が例示される。
【0081】
本実施の形態のスフィンゴミエリナーゼは、少なくとも以下の(1)に記載の作用を有するものであれば特に限定されないが、さらに以下の(2)〜(5)のいずれかに記載の1つ以上の性質を有することが好ましい。例えば、一態様において、スフィンゴミエリナーゼは、以下の(1)に記載の作用に加えて以下の(2)に記載の基質特異性を有することが好ましい。また、スフィンゴミエリナーゼは、以下の(1)に記載の作用に加えて以下の(2)〜(5)に記載の性質のうち、任意の2つ以上を備えていることも好ましい。特に好ましいスフィンゴミエリナーゼとしては、以下の(1)に記載の作用に加えて以下の(2)〜(5)の全てに記載の性質を有するものが挙げられる。
【0082】
(1)作用
本実施の形態におけるスフィンゴミエリナーゼの作用の一例は以下の(式1)に示す通りであり、1分子の水の存在下、スフィンゴミエリン(Shingomyelin)1分子を分解(加水分解)してセラミド(N−アシルスフィンゴシン(N−acylsphingosin))1分子とフォスフォリルコリン(Phosphorylcholine)1分子を生成する。該スフィンゴミエリンの作用は、少なくとも2価の陽イオンの存在下で行われることが好ましく、マグネシウムイオン及び/又はマンガンイオンの存在下で行われることがより好ましい。
【0083】
スフィンゴミエリン + H2O ―>
N−アシルスフィンゴシン + フォスフォリルコリン (式1)
【0084】
本実施の形態において、上記のスフィンゴミエリン(Sphingomyelin)とは、以下の一般式:
【化1】

で表されるスフィンゴ脂質の一種であれば、由来や純度は限定されず、その塩、誘導体等であってもよい。上記一般式中、Rは飽和または不飽和炭化水素を表し、Xはコリン、エタノールアミン、セリン、イノシトール等を表し、好ましくは生体中に多く存在するコリンを表す。本実施の形態において、スフィンゴミエリンは、好ましくは生体内に存在するスフィンゴミエリンであり、特に好ましくは本実施の形態の製造方法で得られるスフィンゴミエリナーゼにより加水分解されるスフィンゴミエリンであり、例えば、N−ヘキサデカノイル−スフィンゲニン−1−フォスフォリルコリン(N−(hexadecanoyl)−sphing−4−enine−1−phosphorylcholine)、N−アセチル−スフィンゲニン−1−フォスフォリルコリン(N−(acetyl)−sphing−4−enine−1−phosphorylcholine)、N−ヘキサノイル−スフィンゲニン−1−フォスフォリルコリン(N−(hexanoyl)−sphing−4−enine−1−phosphorylcholine)、N−ドデカノイル−スフィンゲニン−1−フォスフォリルコリン(N−(dodecanoyl)−sphing−4−enine−1−phosphocholine)、N−ドデカノイル−スフィンガニン−1−フォスフォリルコリン(N−(dodecanoyl)−sphinganine−1−phosphocholine)、N−ヘプタデカノイル−スフィンゲニン−1−フォスフォリルコリン(N−(heptadecanoyl)−sphing−4−enine−1−phosphorylcholine)、N−オクタデカノイル−スフィンゲニン−1−フォスフォリルコリン(N−(octadecanoyl)−sphing−4−enine−1−phosphorylcholine)、N−オクタデカノイル−スフィンゲニン−1−フォスフォリルコリン(N−(9Z−octadecenoyl)−sphing−4−enine−1−phosphorylcholine)、N−テトラコサノイル−スフィンゲニン−1−フォスフォリルコリン(N−(tetracosanoyl)−sphing−4−enine−1−phosphorylcholine)、N−テトラコセノイル−スフィンゲニン−1−フォスフォリルコリン(N−(15Z−tetracosenoyl)−sphing−4−enine−1−phosphocholine)などが例示される。本実施の形態のスフィンゴミエリナーゼは、複数のスフィンゴミエリンに同時に作用するものであってもよい。
【0085】
本実施の形態において、上記スフィンゴミエリナーゼの作用はスフィンゴミエリナーゼ活性として測定できる。スフィンゴミエリナーゼ活性とは、上記の(式1)の反応を触媒する活性であり、スフィンゴミエリナーゼ活性1単位(ユニット)は、37℃で1分間に1マイクロモルのスフィンゴミエリンを分解する酵素量とする。スフィンゴミエリナーゼ活性の測定方法を以下に例示するが、測定方法はこれに限定されるものではない。
【0086】
<スフィンゴミエリナーゼ活性測定法>
(反応試薬混合液組成)
1M Tris/HCl pH 8.0 150μL
10mM スフィンゴミエリン溶液 300μL
0.3% 4−アミノアンチピリン溶液 300μL
0.2% TODB溶液 300μL
1M 塩化マグネシウム水溶液 6μL
1M 塩化ナトリウム水溶液 30μL
10% トリトンX−100溶液 30μL
500U/mL アルカリフォスファターゼ溶液 60μL
600U/mL コリンオキシダーゼ溶液 50μL
500U/mL ペルオキシダーゼ溶液 120μL
精製水 1654μL
【0087】
小試験管に上記組成の反応試薬混合液を0.8mLとり、37℃で予備加温する。5分後、適当に希釈したスフィンゴミエリナーゼ溶液(被験液)を0.03mL加えて混和し、37℃で反応を開始し、5分間インキュベートして反応を行う。続いてこの反応液に1% ドデシル硫酸ナトリウム溶液を1.6mL加えて反応を停止する。反応停止後546nmにおける吸光度を測定し、これをAsとする。スフィンゴミエリナーゼ溶液の代わりに精製水を用いた盲検をAbとする。スフィンゴミエリナーゼ活性1単位を、37℃で1分間に1マイクロモルのスフィンゴミエリンを分解する酵素量として、上記反応液中におけるTODBのミリモル吸光係数を16として、得られたAs及びAbの値に基づき以下の(式2)を用いて算出する。
【0088】
スフィンゴミエリナーゼ活性(単位)
=[{(As−Ab)/16}/5]×(2430/30) (式2)
【0089】
(2)基質特異性
本実施の形態のスフィンゴミエリナーゼ及びスフィンゴミエリナーゼ含有組成物は、上記のスフィンゴミエリンに対する作用を有するものであれば特に限定されないが、好ましくはスフィンゴミエリンに作用し、かつ、レシチン、リゾレシチン、フォスファチジルエタノールアミン、フォスファチジルセリン及びフォスファチジルイノシトール(以下、「レシチン等」ともいう)のいずれにも実質的に作用しない。ここで、「実質的に作用しない」とは、通常、上記のスフィンゴミエリナーゼの活性測定法における条件下でレシチン等に反応しないことをいう。スフィンゴミエリナーゼ活性測定法における条件(酵素の希釈の程度、反応試薬混合液中の各組成物の濃度等)を変更してもレシチン等に反応しないことが好ましい。
【0090】
(3)至適pH
本実施の形態のスフィンゴミエリナーゼの至適pHは、上記スフィンゴミエリナーゼ活性が測定できるpHであれば特に限定されないが、上記スフィンゴミエリナーゼ活性が最大となるpHが好ましい。pHの下限としては、例えばpH4.0以上であり、好ましくはpH5.0以上であり、より好ましくはpH7.0以上である。またpHの上限としては、例えばpH10.0以下であり、好ましくはpH9.0以下であり、より好ましくはpH8.0以下である。
【0091】
(4)Km
本実施の形態のスフィンゴミエリナーゼのスフィンゴミエリンに対するKmは、温度や気圧などの測定の条件や使用機器の精度等によりその値は変化し得るが、例えば2mM以下であり、好ましくは1mM以下であり、より好ましくは約0.45mMである。
【0092】
(5)分子量
本実施の形態のスフィンゴミエリナーゼの分子量は、温度や気圧などの測定の条件や使用機器の精度等によりその値は変化し得るが、例えば30kDa〜50kDaの範囲にあり、好ましくは35kDa〜40kDaの範囲にあり、より好ましくはドデシル硫酸ナトリウム―ポリアクリルアミドゲル電気泳動法で約38kDaである。
【0093】
なお、本実施の形態におけるスフィンゴミエリナーゼの分子量は、工程(a)においてトランスフェクトする遺伝子の変異や、工程(b)における培養時の翻訳後修飾、工程(b)や(c)の後の化学修飾等によって変化する場合があり、例えば、トランスフェクトする際のベクターとしてpTip QC1やpTipQC2(特許第3793812号公報、特開2004−321013号公報、特開2004−073116号公報参照)を利用してC末端にHisタグを付加する場合には、上記の分子量が約1,000程度大きくなることがある。
【0094】
このように、翻訳後修飾(N末端側がアシル基やアルキル基等による修飾を受ける等)、化学修飾(無水コハク酸やPEG等)等を受けたスフィンゴミエリナーゼも、本実施の形態のスフィンゴミエリナーゼに含まれる。化学修飾等により、スフィンゴミエリナーゼの至適pHや安定性等の性質を、利用しやすいように変化させることが可能である。
【0095】
本実施の形態におけるスフィンゴミエリナーゼ又はスフィンゴミエリナーゼ含有組成物を用いれば、スフィンゴミエリナーゼを定性的、半定量的又は定量的に測定することができる。その方法としては、例えばスフィンゴミエリンに対するスフィンゴミエリナーゼの作用で生成したフォスフォリルコリンにさらにグリセロホスフォコリンホスフォジエステラーゼ(EC 3.1.4.2)を作用させて生成したコリンをコリンオキシダーゼ(EC 1.1.3.17)で酸化し、生成した過酸化水素を4−アミノアンチピリンとフェノールなどと縮合して発色させる方法など、公知の方法が挙げられる。
【0096】
また、本実施の形態におけるスフィンゴミエリナーゼ又はスフィンゴミエリナーゼ含有組成物を用いれば、上記(式1)の反応により、スフィンゴミエリナーゼの基質を消去することもできる。
【0097】
本実施の形態におけるスフィンゴミエリナーゼ又はスフィンゴミエリナーゼ含有組成物は、そのまま、あるいは精製、濃縮、凍結乾燥等の後、各種製品、試薬、キット、センサー、原薬、製剤、化粧品、食品等に利用することができる。
【実施例】
【0098】
以下、本発明を実施例等に基づいて説明するが、本発明の範囲は以下の実施例等に限定して解釈されるものではない。尚、記述した技術は、市販の各種酵素、キット類に添付された手順書等に従えば実施できるものである。「常法に従い」と記述した技術は、例えばマニアティスらの方法(Maniatis,T.,et al.Molecular Cloning.Cold Spring Harbor Laboratory 1982年、1989年)や蛋白質・酵素の基礎実験法(改訂第2版、堀尾武一、1994年、南光堂)、等に従えば実施できるものである。又、以下に示した測定値等は測定の条件や使用機器の精度等によりその値は変化し得る。
【0099】
なお、以下の実施例で使用する試薬類は、特に断らない限り、和光純薬工業株式会社製、シグマアルドリッチ社製、タカラバイオ株式会社製等であり、市販で容易に入手できるものを使用することができる。試薬のメーカーや純度等は特に限定されない。
【0100】
〔実施例1〕
1.DNAの抽出
配列番号1のアミノ酸配列(スフィンゴミエリナーゼ)を含むタンパク質を発現するStreptomyces sp. A9107(FERM P−22058)の菌体を50mM Tris/HCl(pH8.0)、50mM EDTA、15%シュークロースを含む1mg/mLリゾチーム溶液で37℃、10分処理した後、SDSを最終濃度0.25%になるように添加して菌体を溶解した。さらに等量のフェノール/クロロホルムの1:1混合液を加え、30分撹拌した後、遠心分離(12,000rpm、15分間)して水層を回収した。回収した水層に10分の1量の3Mの酢酸ナトリウム(pH5.5)を混合後、2倍量のエタノールを静かに重層し、ゲノムDNAをガラス棒に巻き付かせて分離した。分離したゲノムDNAを、10mM Tris/HCl(pH8.0)、1mM EDTA水溶液(以下「TE」と記載する場合もある)に溶解し、適量のRNaseAを加え、37℃で1時間保温し、混在しているRNAを分解した。次いで、等量のフェノール/クロロホルムの1:1混合液を加え、前記と同様に処理して、水層を分取した。分取した水層に10分の1量の3Mの酢酸ナトリウム(pH5.5)と2倍量のエタノールを加えて前記の方法でもう一度ゲノムDNAを分離した。このゲノムDNA(染色体)をTEに溶解し、TE飽和のフェノールとクロロホルムの1:1混合液を加え、全体を懸濁した後、同様の遠心分離を繰り返し、上層を再び別の容器に移した。この分離した上層に3Mの酢酸ナトリウム(pH5.5)とエタノールを加え、撹拌後、−70℃で5分間冷却した後、遠心分離(2,000G、4℃、15分)し、沈殿したゲノムDNAを75%エタノールで洗い、減圧乾燥した。以上の操作によりStreptomyces sp.のDNA標品を得た。
【0101】
2.精製酵素の取得
容量500mLの三角フラスコに、前培養用培地として培地A(3% 脱脂大豆ニッカミルキー(日華油脂社製)、2% コーンスターチ、0.3% 塩化カリウム、0.4% 炭酸カルシウム、pH7.0)100mLを入れて、蒸気加圧滅菌した後、Streptomyces sp. A9107の菌体を白金耳で植菌した。次いで25℃で72時間振とう培養することにより、前培養液を得た。容量30Lのジャーに、培地Aを20L入れ、121℃で20分間蒸気加圧滅菌した後、上記の前培養液を20mL植菌し、25℃、125rpmで96時間通気培養を行った。培養後、遠心分離して培養上清を得た。この培養上清を20mM 酢酸ナトリウム緩衝液を用いてpH5.0に調製し、20mM 酢酸ナトリウム緩衝液(pH5.0)で平衡化したGEヘルスケア社製のCM Sepharose Fast Flowを用いたカラムに吸着させた。20mM 酢酸ナトリウム緩衝液(pH5.5)で充分に洗浄した後、20mM 酢酸ナトリウム緩衝液(pH5.5)と100mM 酢酸ナトリウム緩衝液(pH5.5)を用いてベッド体積の10倍量のリニアグラジエントで溶出を行った。活性画分を回収し、100倍量の10mM 酢酸ナトリウム緩衝液(pH5.5)で透析し、スフィンゴミエリナーゼ(SPC)の精製酵素を得た。
【0102】
3.酵素の部分アミノ酸配列決定
「実施例1 2.」で取得した精製酵素の一部を、Laemmliの方法で5〜20%のポリアクリルアミドグラジエントゲルを用いてドデシル硫酸ナトリウム・ポリアクリルアミド電気泳動(SDS−PAGE)を行った後、ポリフッ化ビニリデン(PVDF)膜に転写した。これを用いてN末端アミノ酸配列解析を行った結果、配列番号7のアミノ酸配列を得た。また、「実施例1 2.」で取得した精製酵素の一部を消化酵素トリプシンにより処理し、上記と同様に操作を行い、N末端アミノ酸配列解析を行った結果、配列番号8のアミノ酸配列を得た。
【0103】
4.酵素遺伝子の部分塩基配列決定
「実施例1 3.」で得られた配列番号7及び8のアミノ酸配列を元に配列番号9及び10の塩基配列を有するPCR用DNAプライマーを設計した。このDNAプライマーを用い、「実施例1 1.」で調製したStreptomyces sp.のDNA標品を鋳型として常法に従ってPCR反応を行い、約450bpのDNA断片を取得した。このDNA断片をDNAシーケンスして部分塩基配列を決定した。
【0104】
5.酵素遺伝子の塩基配列決定
「実施例1 4.」で決定した部分塩基配列を元に、配列番号11及び12の塩基配列を有するPCR用DNAプライマーを設計した。「実施例1 1.」で調製したStreptomyces sp.のDNA標品の一部をSalIにより制限酵素処理した。制限酵素処理したDNA断片を常法に従ってライゲーションし、環状DNAを得た。配列番号9及び10の塩基配列を有するDNAプライマーを用いてSalI切断環状DNAを鋳型としてPCR反応を行い、約850bpのDNA断片を取得した。このDNA断片をDNAシーケンスして塩基配列を決定した。「実施例1 1.」で調製したStreptomyces sp.のDNA標品の一部をBamHIにより制限酵素処理した。制限酵素処理したDNA断片をライゲーションし、環状DNAを得た。配列番号11及び12の塩基配列を有するDNAプライマーを用いてBamHI切断環状DNAを鋳型として常法に従ってPCR反応を行い、約800bpのDNA断片を取得した。このDNA断片を常法に従ってDNAシーケンスして塩基配列を決定した。決定した塩基配列は、配列番号6(配列番号2のアミノ酸配列をコードする遺伝子配列)と、配列番号5(配列番号1のアミノ酸配列をコードする遺伝子配列)とが順に結合した塩基配列を含む塩基配列であった。
【0105】
6.PCR法による酵素遺伝子の増幅
「実施例1 5.」で決定した塩基配列を元に、配列番号13及び14の塩基配列を有するPCR用DNAプライマーを設計した。このDNAプライマーを用い、「実施例1 1.」で調製したStreptomyces sp.のDNA標品を鋳型として常法に従ってPCR反応を行った。得られた約1200bpのPCR産物は、例えばQIAGEN QIAquick PCR Purification Kitを用い精製した。
【0106】
7.発現ベクターとのライゲーション
「実施例1 6.」で取得したPCR産物を、NdeI及びHindIIIにより制限酵素処理した。このDNA断片は、例えばQIAGEN QIAquick PCR Purification Kitを用いる等、常法に従って精製した。精製したDNA断片は、NdeI及びHindIIIにより制限酵素処理し精製したpTip QC2と常法に従ってライゲーションし、配列番号15の塩基配列を有するpTip QC2/SPCを作製した。作製したpTip QC2/SPCはDNAシーケンスして挿入した塩基配列が正しいことを確認した。
【0107】
8.Rhodococcus erythropolis L88株の形質転換
Rhodococcus erythropolis L88株(FERM BP−08444)の菌体を、「実施例1 7.」で作製したpTip QC2/SPCと混合し、氷上で30分静置した後、バイオ・ラッド社のGene Pulser IIを用いてエレクトロポレーション法(2.5kV、25μF、400Ω)により形質転換した。形質転換したRhodococcus erythropolis L88株をSOC培地(2% トリプトン、0.5% 酵母エキス、10mM 塩化ナトリウム、25mM 塩化カリウム、10mM 塩化マグネシウム、10mM 硫酸マグネシウム、20mM グルコース)に懸濁した後、30℃で1時間培養し、34μg/mLのクロラムフェニコールを含むLB寒天培地(1% トリプトン、0.5% 酵母エキス、1% 塩化ナトリウム、1.5% アガー)に塗布して30℃で72時間培養し、pTip QC2/SPCにより形質転換したRhodococcus erythropolis L88株(L88/SPC)を得た。
【0108】
9.L88/SPCの培養
容量500mLの三角フラスコに、前培養用培地としてLB液体培地(1% トリプトン、0.5% 酵母エキス、1% 塩化ナトリウム)100mLを入れて、蒸気加圧滅菌した後、「実施例1 8.」で形質転換したRhodococcus erythropolis L88株(L88/SPC)を白金耳で植菌した。次いで30℃で72時間振とう培養することにより、前培養液を得た。容量2Lのジャーに、LB液体培地を1.6L入れ、121℃で20分間蒸気加圧滅菌した後、34mg/mLのクロラムフェニコールを1.6mL、40mg/mLのチオストレプトンを80μL添加し、上記の前培養液を1.6mL植菌し、30℃、300rpmで72時間通気培養を行った。培養後、遠心分離して培養上清を得た。3基培養を行ったところ、スフィンゴミエリナーゼの培養力価は50〜52U/mLであった。培養力価は、上述の<スフィンゴミエリナーゼ活性測定法>に記載の方法を用いて、培養上清を被験液として(式2)を用いて算出した。
【0109】
10.精製
「実施例1 9.」で得られた培養上清の一部を20mM 酢酸ナトリウム緩衝液を用いてpH5.0に調製し、20mM 酢酸ナトリウム緩衝液(pH5.0)で平衡化したGEヘルスケア社製のCM Sepharose Fast Flowを用いたカラムに吸着させた。20mM 酢酸ナトリウム緩衝液(pH5.5)で充分に洗浄した後、20mM 酢酸ナトリウム緩衝液(pH5.5)と100mM 酢酸ナトリウム緩衝液(pH5.5)を用いてベッド体積の10倍量のリニアグラジエントで溶出を行った。活性画分を回収し、100倍量の10mM 酢酸ナトリウム緩衝液(pH5.5)で透析した後、凍結乾燥して粉末状の精製酵素(スフィンゴミエリナーゼ)約80mgを得た。
【0110】
〔比較例1〕
1.Streptomyces sp.の培養
容量500mLの三角フラスコに、前培養用培地として培地A(実施例1と同様)100mLを入れて、蒸気加圧滅菌した後、「実施例1 1.」に記載のStreptomyces sp.の菌体を白金耳で植菌した。次いで25℃で72時間振とう培養することにより、前培養液を得た。容量30Lのジャーに、培地Aを20L入れ、121℃で20分間蒸気加圧滅菌した後、上記の前培養液を20mL植菌し、25℃、125rpmで96時間通気培養を行った。培養後、遠心分離して培養上清を得た。2基培養を行ったところ、スフィンゴミエリナーゼの培養力価は62〜67U/mLであった。
【0111】
2.精製
「比較例1 1.」で取得した培養上清を20mM 酢酸ナトリウム緩衝液を用いてpH5.0に調製し、20mM 酢酸ナトリウム緩衝液(pH5.0)で平衡化したGEヘルスケア社製のCM Sepharose Fast Flowを用いたカラムに吸着させた。100mM 酢酸ナトリウム緩衝液(pH5.5)を用いて溶出し、活性画分を回収した。活性画分をゲル濾過により脱塩し、その一部にマンニトールを重量パーセント濃度で1%となるように添加した後、凍結乾燥して粉末状の比較試験用酵素(野生型スフィンゴミエリナーゼ)約950mgを得た。
【0112】
〔比較例2〕
1.挿入遺伝子の調製
配列番号16の塩基配列を有するDNAプライマーを設計した。このDNAプライマーと配列番号13の塩基配列を有するDNAプライマーとを用いてpTip QC2/SPCを鋳型として常法に従ってPCR反応を行い、DNA断片A(配列番号6の塩基配列(配列番号2のアミノ酸配列をコードする遺伝子配列)を含む)を得た。また、配列番号17及び18の塩基配列を有するDNAプライマーを設計した。このDNAプライマーを用いて配列番号19の塩基配列を有するDNA(カンジダ シリンドラセ(Candida cylindracea)(ATCC14830)由来のコレステロールエステラーゼ(CEBP)をコードする塩基配列)を鋳型として常法に従ってPCR反応を行い、DNA断片Bを得た。配列番号13及び18の塩基配列を有するDNAプライマーを用いてDNA断片AとDNA断片Bを混合したものを鋳型として常法に従ってPCR反応を行い、配列番号20の塩基配列を有するDNA断片Cを得た。DNA断片Cを得るまでのPCR反応の模式図を図1に示す。得られたPCR産物は、例えばQIAGEN QIAquick PCR Purification Kitを用い精製した。
【0113】
2.発現ベクターとのライゲーション
「比較例1 1.」で取得したPCR産物をNdeI及びHindIIIにより制限酵素処理した。このDNA断片は、例えばQIAGEN QIAquick PCR Purification Kitを用いて精製した。精製したDNA断片は、NdeI及びHindIIIにより制限酵素処理し精製したpTip QC2とライゲーションし、配列番号21の塩基配列を有するpTip QC2/CEBPを作製した。作製したpTip QC2/CEBPは常法に従ってDNAシーケンスして挿入した塩基配列が正しいことを確認した。
【0114】
3.Rhodococcus erythropolis L88株の形質転換
「実施例1 7.」と同様の方法で、「比較例2 2.」で作製したpTip QC2/CEBPによりRhodococcus erythropolis L88株を形質転換し、pTip QC2/CEBPにより形質転換したRhodococcus erythropolis L88株(L88/CEBP)を得た。
【0115】
4.L88/CEBPの培養
容量500mLの三角フラスコに、前培養用培地としてLB液体培地100mLを入れて、蒸気加圧滅菌した後、「比較例2 3.」で形質転換したRhodococcus erythropolis L88株(L88/CEBP)を白金耳で植菌した。次いで30℃で72時間振とう培養することにより、前培養液を得た。容量2Lのジャーに、LB液体培地を1.6L入れ、121℃で20分間蒸気加圧滅菌した後、34mg/mLのクロラムフェニコールを1.6mL、40mg/mLのチオストレプトンを80μL添加し、上記の前培養液を1.6mL植菌し、30℃、300rpmで72時間通気培養を行った。培養後、遠心分離して培養上清を得た。しかしながら、配列番号19にコードされるCandida cylindracea由来のコレステロールエステラーゼの発現は確認できなかった。なお、コレステロールエステラーゼの発現は、コレステロールエステラーゼの活性として、ASAHI KASEI Diagnostic Enzymes(旭化成ファーマ株式会社2008年7月131ページの方法に従い確認した。
【0116】
〔比較例3〕
1.挿入遺伝子の調製
配列番号22の塩基配列を有するDNAプライマーを設計した。このDNAプライマーと配列番号13の塩基配列を有するDNAプライマーとを用いてpTip QC2/SPCを鋳型として常法に従ってPCR反応を行い、DNA断片A’(配列番号6の塩基配列(配列番号2のアミノ酸配列をコードする遺伝子配列)を含む)を得た。また、配列番号23及び24の塩基配列を有するDNAプライマーを設計した。このDNAプライマーを用いて配列番号25の塩基配列を有するDNA(バークホルデリア スタビリス(Burkholderia stabilis)(FERM P−21014)(特開2008−086277)由来のコレステロールエステラーゼ(CEN)及びその補助タンパク質をコードする塩基配列)を鋳型として常法に従ってPCR反応を行い、DNA断片B’を得た。配列番号13及び24の塩基配列を有するDNAプライマーを用いてDNA断片A’とDNA断片B’を混合したものを鋳型として常法に従ってPCR反応を行い、配列番号26の塩基配列を有するDNA断片C’を得た。得られたPCR産物は、例えばQIAGEN QIAquick PCR Purification Kitを用い精製した。
【0117】
2.発現ベクターとのライゲーション
「比較例3 1.」で取得したPCR産物に対して「比較例2 2.」と同様の操作を行い、配列番号27の塩基配列を有するpTip QC2/CENを作製した。作製したpTip QC2/CENは常法に従ってDNAシーケンスして挿入した塩基配列が正しいことを確認した。
【0118】
3.Rhodococcus erythropolis L88株の形質転換
「実施例1 7.」と同様の方法で、「比較例3 2.」で作製したpTip QC2/CENによりRhodococcus erythropolis L88株を形質転換し、pTip QC2/CENにより形質転換したRhodococcus erythropolis L88株(L88/CEN)を得た。
【0119】
4.L88/CEBPの培養
「比較例3 3.」で得たL88/CENに対して「比較例2 4.」と同様の操作を行い、培養上清を得た。しかしながら、配列番号25にコードされるBurkholderia stabilis由来のコレステロールエステラーゼの発現は確認できなかった。なお、コレステロールエステラーゼの発現は、コレステロールエステラーゼの活性として、ASAHI KASEI Diagnostic Enzymes (旭化成ファーマ株式会社2008年7月48ページの方法に従い確認した。
【0120】
〔比較例4〕
1.挿入遺伝子の調製
配列番号28の塩基配列を有するDNAプライマーを設計した。このDNAプライマーと配列番号13の塩基配列を有するDNAプライマーとを用いてpTip QC2/SPCを鋳型として常法に従ってPCR反応を行い、DNA断片A”(配列番号6の塩基配列(配列番号2のアミノ酸配列をコードする遺伝子配列)を含む)を得た。また、配列番号29及び30の塩基配列を有するDNAプライマーを設計した。このDNAプライマーを用いて配列番号31の塩基配列を有するDNA(クロモバクテリウム ビスコスム(Chromobacterium viscosum)(工発研菌寄第137号)(特公昭46−29787)由来のリパーゼ(LP)及びその補助タンパク質をコードする塩基配列)を鋳型として常法に従ってPCR反応を行い、DNA断片B”を得た。配列番号13及び30の塩基配列を有するDNAプライマーを用いてDNA断片A”とDNA断片B”を混合したものを鋳型として常法に従ってPCR反応を行い、配列番号32の塩基配列を有するDNA断片C”を得た。得られたPCR産物は、例えばQIAGEN QIAquick PCR Purification Kitを用い精製した。
【0121】
2.発現ベクターとのライゲーション
「比較例4 1.」で取得したPCR産物に対して「比較例2 2.」と同様の操作を行い、配列番号33の塩基配列を有するpTip QC2/LPを作製した。作製したpTip QC2/LPは常法に従ってDNAシーケンスして挿入した塩基配列が正しいことを確認した。
【0122】
3.Rhodococcus erythropolis L88株の形質転換
「実施例1 7.」と同様の方法で、「比較例4 2.」で作製したpTip QC2/LPによりRhodococcus erythropolis L88株を形質転換し、pTip QC2/LPにより形質転換したRhodococcus erythropolis L88株(L88/LP)を得た。
【0123】
4.L88/LPの培養
「比較例4 3.」で得たL88/LPに対して「比較例2 4.」と同様の操作を行い、培養上清を得た。しかしながら、配列番号30にコードされるChromobacterium viscosum由来のリパーゼの発現は確認できなかった。なお、リパーゼの発現は、リパーゼの活性として、ASAHI KASEI Diagnostic Enzymes(旭化成ファーマ株式会社2008年7月15ページ)に記載の方法に従い確認した。
【0124】
〔試験例1〕<比活性試験>
「実施例1 9.」で得た培養上清と、「比較例1 1.」で得た培養上清のタンパク質量あたりのスフィンゴミエリナーゼ活性を計算したところ、「実施例1 9.」で得た培養上清は142.7U/mg、「比較例1 1.」で得た培養上清は2.1U/mgであり、「実施例1 9.」で得た培養上清は、「比較例1 1.」で得た培養上清よりも比活性が70倍以上高かった。なお、培養上清中のタンパク質量は、ビシンコニン酸を用いる方法(BCA法)により測定した。スフィンゴミエリナーゼ活性は、上述の<スフィンゴミエリナーゼ活性測定法>に記載の方法を用いて、培養上清を被験液として(式2)を用いて算出した。これらの測定から、タンパク質量あたりのスフィンゴミエリナーゼ活性を計算した。
【0125】
〔試験例2〕<安定性試験1>
「実施例1 10.」で得た精製酵素、及び「比較例1 2.」で得た比較試験用酵素を、4mg/mLになるように純水(pH6.8)又は50mM 酢酸ナトリウム緩衝液(pH5.5)に溶解し、それぞれ−30℃又は30℃で静置保存した。保存した酵素溶液について、保存開始から72時間後及び96時間後に残存活性を測定した(保存前の活性を100%とする)。その結果、図2に示すように、30℃で静置保存した場合、「比較例1 2.」で得た比較試験用酵素は、72時間後及び96時間後の残存活性がいずれも40%以下であるのに対して、「実施例1 10.」で得られた精製酵素は、72時間後及び96時間後のいずれにおいても78%以上の活性を保持しており、高い保存安定性を示した。特に、pH5.5の緩衝液に溶解した場合、72時間後及び96時間後のいずれにおいても97.6%以上の活性を保持しており、非常に高い保存安定性を示した。なお、−30℃で静置保存した場合も同様に、「実施例1 10.」で得られた精製酵素は高い保存安定性を示した(図示せず)。
【0126】
〔試験例3〕<安定性試験2>
試験例2で得られた、−30℃又は30℃で96時間保存した酵素溶液の一部を、Laemmliの方法で5〜20%のポリアクリルアミドグラジエントゲルを用いてSDS−PAGEを行った後、常法に従ってクーマシーブルー(CBB)で染色した。結果を図3に示す。図3中(A)〜(H)のバンドは、それぞれ以下の処理を行った酵素溶液に対応する。
(A)比較例1の酵素を純水(pH6.8)に溶解し、−30℃で保存。
(B)比較例1の酵素を純水(pH6.8)に溶解し、30℃で保存。
(C)比較例1の酵素を酢酸ナトリウム緩衝液(pH5.5)に溶解し、−30℃で保存。
(D)比較例1の酵素を酢酸ナトリウム緩衝液(pH5.5)に溶解し、30℃で保存。
(E)実施例1の酵素を純水(pH6.8)に溶解し、−30℃で保存。
(F)実施例1の酵素を純水(pH6.8)に溶解し、30℃で保存。
(G)実施例1の酵素を酢酸ナトリウム緩衝液(pH5.5)に溶解し、−30℃で保存。
(H)実施例1の酵素を酢酸ナトリウム緩衝液(pH5.5)に溶解し、30℃で保存。
【0127】
図3に示すように、比較例1で得た比較試験用酵素は、30℃で96時間保存した場合((B)及び(D))には、−30℃で保存した場合((A)及び(C))と比較して明らかにスフィンゴミエリナーゼを示すバンドが薄くなり、分解されていた。一方、「実施例1 10.」で得た精製酵素は、30℃で96時間保存した場合((F)及び(H))も、−30℃で保存した場合((E)及び(G))と同程度のバンドが確認され、分解されていなかった。
【産業上の利用可能性】
【0128】
本発明は、スフィンゴミエリナーゼ、及び純度が高く安定性の高いスフィンゴミエリナーゼ含有組成物の効率的な製造に利用することができる。本発明は、特に、試薬、医薬品、化粧品等の分野において、産業上の利用可能性を有する。
【技術分野】
【0001】
本発明はスフィンゴミエリナーゼの製造方法、及び純度が高く安定性の高いスフィンゴミエリナーゼ含有組成物及びその製造方法に関する。
【背景技術】
【0002】
スフィンゴミエリナーゼ(Sphingomyelinase)はスフィンゴミエリン(Sphingomyelin)をセラミド(N−アシルスフィンゴシン(N−acylsphingosine))とフォスフォリルコリン(Phosphorylcholine)に加水分解する酵素である(EC 3.1.4.12)。
【0003】
スフィンゴミエリナーゼとしては、バチルス セレウス(Bacillus cereus)(非特許文献1)、レプトスピラ インターロガンス(Leptospira interrogans serovars)(非特許文献2)、リステリア イバノビ(Listeria ivanovii)(非特許文献3)、マウス(非特許文献4)、ラット(非特許文献5)、ヒト(非特許文献6)、及び、ストレプトマイセス グリセオカルネウス(Streptomyces griseocarneus)(非特許文献7)由来のもの等が知られている。非特許文献2及び3は、血清型レプトスピラ インターロガンス及びリステリア イバノビの、スフィンゴミエリナーゼと予想される遺伝子を開示する。非特許文献4は、マウス由来のスフィンゴミエリナーゼの機能を開示する。
【0004】
スフィンゴミエリナーゼは、例えば、動脈硬化の診断に重要な小粒子高比重LDL(small,denseLDL)中のコレステロールの定量等に利用されている(特許文献1)。
【0005】
スフィンゴミエリナーゼの製造法は、例えば、特許文献2及び特許文献3等に記載されている。特許文献2は、Streptomyces sp.A9107菌株を培養した培養物からスフィンゴミエリナーゼを採取することを特徴とする、スフィンゴミエリナーゼの製造法を開示する。特許文献3は、スフィンゴミエリナーゼ遺伝子を含むDNAで形質転換したバチルス・ブレビス47/pNUSMを培養し、培養物からスフィンゴミエリナーゼを採取することを特徴とする該酵素の製造法を開示する。
【0006】
さらに、スフィンゴミエリナーゼの製造法として、非特許文献1は、バチルス セレウス由来のスフィンゴミエリナーゼ遺伝子をエシェリキア コリ(Escherichia coli)HB10に導入することによりスフィンゴミエリナーゼ含有培養物を取得する方法を開示する。非特許文献5は、ラット由来のスフィンゴミエリナーゼ遺伝子をEscherichia coli JM109に導入することを含む、スフィンゴミエリナーゼ含有培養物を取得する方法を開示する。非特許文献6はヒト由来のスフィンゴミエリナーゼ遺伝子をアフリカミドリザル由来の培養細胞株COS−1細胞に導入したことを開示する。非特許文献7はストレプトマイセス グリセオカルネウス由来のスフィンゴミエリナーゼ遺伝子をストレプトマイセス リビダンス(Stpreotomyces lividans)に導入することを含む、スフィンゴミエリナーゼ含有培養物を取得する方法を開示する。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】WO2009/048143
【特許文献2】特開昭58−43786号公報
【特許文献3】特開平5−328970号公報
【非特許文献】
【0008】
【非特許文献1】Johansen T.ら、Gene,65,293−304(1988)
【非特許文献2】Nascimento A.L.T.O.ら,Bacteriol.,186,2164−2172(2004)
【非特許文献3】Gonzalez−Zornら,Mol.Microbiol.,33,510−523(1999)
【非特許文献4】Stoffel W.ら、J.Proc.Natl.Acad.Sci. U.S.A.,102,4554−4559(2005)
【非特許文献5】Mizutani Y.ら、Biochim.Biophys.Acta,1485,236−246(2000)
【非特許文献6】Schuchman E.H.ら、J.Biol.Chem.,266,8531−8539(1991)
【非特許文献7】Sugimori D., Tomita Y.ら、Biotechnol.Lett.,2010 Nov 30.
【発明の概要】
【発明が解決しようとする課題】
【0009】
特許文献2に記載の方法により得られるスフィンゴミエリナーゼ含有培養物は、純度が低く安定性が悪いという問題がある。特許文献3に記載の方法により得られるスフィンゴミエリナーゼ含有培養物は、特異性が低いという問題がある。非特許文献1に記載の方法により得られるスフィンゴミエリナーゼ含有培養物は、フォスフォリパーゼ(phospholipase)C活性を併せ持ち特異性が低いという問題がある。非特許文献5に開示された方法で得られるスフィンゴミエリナーゼ含有培養物は、リゾ血小板活性化因子を加水分解する活性を併せ持ち特異性が低いという問題があり、また、非特許文献5はスフィンゴミエリナーゼ含有培養物中のスフィンゴミエリナーゼの純度に関する記載はない。非特許文献6に開示された方法は、工業的生産に適していないという問題があり、また、非特許文献6はスフィンゴミエリナーゼ含有培養物中のスフィンゴミエリナーゼの純度を開示していない。非特許文献7に開示された方法で得られるスフィンゴミエリナーゼ含有培養物は、フォスフォリパーゼC活性を併せ持ち、特異性が低いという問題がある。
【0010】
このように従来知られているスフィンゴミエリナーゼの製造法はいずれも実用性が低く、また、微生物の培養によりスフィンゴミエリナーゼ含有培養物を得る場合、得られる培養物中のスフィンゴミエリナーゼの純度が低いという問題があり、満足のいくものではないという現状を本発明者らは問題点として強く認識した。このような背景のもと、本発明が解決しようとする課題は、スフィンゴミエリナーゼの製造方法、並びに純度が高く安定性の高いスフィンゴミエリナーゼ含有組成物及びその製造方法を提供することにある。
【課題を解決するための手段】
【0011】
本発明者らは、上記の課題を解決すべく鋭意研究を重ねた結果、驚くべきことにロドコッカス(Rhodococcus)属細菌を宿主として、当該宿主とは属が異なるストレストマイセス・エスピー(Streptomyces sp.)由来のスフィンゴミエリナーゼをコードする遺伝子を導入し、得られた形質転換体を培養することで、スフィンゴミエリナーゼを容易に製造することができ、また、得られた培養物は純度が高く安定性の高いスフィンゴミエリナーゼ含有組成物であることを見出し、本発明を完成するに至った。
【0012】
すなわち、本発明は以下のものに関する。
〔1〕
スフィンゴミエリナーゼの製造方法であって、以下の工程(a)及び(b):
(a)以下の(i)又は(ii)のアミノ酸配列:
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして形質転換体を得る工程;及び
(b)工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程;
を含む方法。
〔1−2〕
さらに以下の工程(c):
(c)工程(b)で得られた培養物から形質転換体を分離してスフィンゴミエリナーゼを得る工程
を含む、前記〔1〕に記載の製造方法。
〔1−3〕
ロドコッカス属細菌が、ロドコッカス エリスロポリスである、前記〔1〕又は〔1−2〕に記載の製造方法。
〔1−4〕
ロドコッカス エリスロポリスが、ロドコッカス エリスロポリス L88株である、前記〔1−3〕に記載の製造方法。
【0013】
〔2〕
工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、前記〔1〕〜〔1−4〕のいずれかに記載の製造方法。
〔2−2〕
工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、
(i)又は(ii)のアミノ酸配列のN末端と、
(iii)又は(iv)のアミノ酸配列のC末端とが、直接結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、前記〔2〕に記載の製造方法。
〔2−3〕
工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、
(iii)のアミノ酸配列コードする塩基配列を含む遺伝子である、前記〔2〕又は〔2−2〕に記載の製造方法。
【0014】
〔3〕
前記工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、(i)のアミノ酸配列をコードする塩基配列を含む、前記〔1〕〜〔2−3〕のいずれかに記載の製造方法。
【0015】
〔4〕
以下の工程(a)〜(c):
(a)以下の(i)又は(ii)のアミノ酸配列:
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして形質転換体を得る工程;
(b)工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程;及び
(c)工程(b)で得た培養物から形質転換体を分離してスフィンゴミエリナーゼ含有組成物を得る工程;
を含む方法により得られる、スフィンゴミエリナーゼ含有組成物。
〔4−2〕
以下の工程(a)及び(b):
(a)以下の(i)又は(ii)のアミノ酸配列:
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして形質転換体を得る工程;及び
(b)工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程;
を含む方法により得られる、形質転換体及びスフィンゴミエリナーゼ含有培養物。
〔4−3〕
以下の工程(a)及び(b):
(a)以下の(i)又は(ii)のアミノ酸配列:
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして形質転換体を得る工程;及び
(b)工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程;
を含む方法により得られる培養物の培養上清であって、スフィンゴミエリナーゼを含有する培養上清。
〔4−4〕
ロドコッカス属細菌が、ロドコッカス エリスロポリスである、前記〔4〕〜〔4−3〕のいずれかに記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔4−5〕
ロドコッカス エリスロポリスが、ロドコッカス エリスロポリス L88株である、前記〔4−4〕に記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
【0016】
〔5〕
工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、前記〔4〕〜〔4−5〕のいずれかに記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔5−2〕
工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、
(i)又は(ii)のアミノ酸配列のN末端と、
(iii)又は(iv)のアミノ酸配列のC末端とが、直接結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、前記〔5〕に記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔5−3〕
工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、
(iii)のアミノ酸配列をコードする塩基配列を含む遺伝子である、前記〔5〕又は〔5−2〕に記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
【0017】
〔6〕
前記スフィンゴミエリナーゼ含有組成物、培養物又は培養上清に水又は緩衝液を添加してスフィンゴミエリナーゼ濃度が4mg/mLの中性又は酸性の水溶液を調製し、30℃で96時間静置保存した場合、保存前と比較して50%以上のスフィンゴミエリナーゼ活性を保持する、前記〔4〕〜〔5−3〕のいずれかに記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔6−2〕
前記水溶液を30℃で96時間静置保存した場合、保存前と比較して65%以上(好ましくは70%以上、より好ましくは75%以上、さらに好ましくは78%以上)のスフィンゴミエリナーゼ活性を保持する、前記〔6〕に記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔6−3〕
前記スフィンゴミエリナーゼ含有組成物、培養物又は培養上清に水又は緩衝液を添加してスフィンゴミエリナーゼ濃度が4mg/mLのpH6.8の水溶液を調製し、30℃で96時間静置保存した場合、保存前と比較して50%以上のスフィンゴミエリナーゼ活性を保持する、前記〔4〕〜〔5−3〕のいずれかに記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔6−4〕
前記水溶液を30℃で96時間静置保存した場合、保存前と比較して65%以上(好ましくは70%以上、より好ましくは75%以上、さらに好ましくは78%以上)のスフィンゴミエリナーゼ活性を保持する、前記〔6−3〕に記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔6−5〕
前記スフィンゴミエリナーゼ含有組成物、培養物又は培養上清に水又は緩衝液を添加してスフィンゴミエリナーゼ濃度が4mg/mLのpH5.5の水溶液を調製し、30℃で96時間静置保存した場合、保存前と比較して50%以上のスフィンゴミエリナーゼ活性を保持する、前記〔4〕〜〔5−3〕のいずれかに記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔6−6〕
前記水溶液を30℃で96時間静置保存した場合、保存前と比較して75%以上(好ましくは78%以上、より好ましくは90%以上、さらに好ましくは97%以上)のスフィンゴミエリナーゼ活性を保持する、前記〔6−5〕に記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清。
〔6−7〕
得られるスフィンゴミエリナーゼが、ストレプトマイセス由来のスフィンゴミエリナーゼと同一のアミノ酸配列及び/又は特徴を有する、前記〔1〕〜〔3〕のいずれかに記載の製造方法。
【0018】
なお、上記〔1〕〜〔3〕のように引用する項番号が範囲で示され、その範囲内に〔2−2〕等の枝番号を有する項が配置されている場合には、〔2−2〕等の枝番号を有する項も引用されることを意味する。以下においても同様である。
【0019】
〔6−8〕
得られるスフィンゴミエリナーゼが、ストレプトマイセス・エスピー由来のスフィンゴミエリナーゼと同一のアミノ酸配列及び/又は特徴を有する、前記〔1〕〜〔3〕のいずれかに記載の製造方法。
〔6−9〕
得られるスフィンゴミエリナーゼが、ストレプトマイセス・エスピー A9107株由来のスフィンゴミエリナーゼと同一のアミノ酸配列及び/又は特徴を有する、前記〔1〕〜〔3〕のいずれかに記載の製造方法。
〔6−10〕
前記製造方法で得られるスフィンゴミエリナーゼあるいは前記組成物、培養物又は培養上清に含まれるスフィンゴミエリナーゼが、以下の(1)の作用を有し、さらに以下の(2)〜(5)のいずれか1つ以上の特徴を有するスフィンゴミエリナーゼである、前記〔1〕〜〔6−9〕のいずれかに記載の製造方法、組成物、培養物又は培養上清:
(1)作用
1分子の水の存在下、スフィンゴミエリン1分子を分解してセラミド1分子とフォスフォリルコリン1分子を生成する。
(2)基質特異性
スフィンゴミエリンに作用し、かつ、レシチン、リゾレシチン、フォスファチジルエタノールアミン、フォスファチジルセリン及びフォスファチジルイノシトールのいずれにも実質的に作用しない。
(3)Km
スフィンゴミエリンに対するKm値が約0.45mMである。
(4)至適pH
pH7.0〜8.0。
(5)分子量
ドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動法による分子量が約38kDaである。
〔6−11〕
前記〔1〕〜〔3〕のいずれかに記載の製造方法により得られるスフィンゴミエリナーゼ。
【0020】
〔7〕
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子を含有する組換えベクター。
〔7−2〕
前記遺伝子を、以下の群:pTip QC1、pTip QC2、pTip QT1、pTip QT2、pTip RC1、pTip RC2、pTip RT1、pTip RT2、pNit QC1、pNit QC2、pNit QT1、pNit QT2、pNit RC1、pNit RC2、pNit RT1及びpNit RT2;から選択される発現ベクターに挿入して得られる、前記〔7〕に記載の組換えベクター。
〔7−3〕
前記遺伝子が、(i)のアミノ酸配列をコードする塩基配列を含む、前記〔7〕又は〔7−2〕に記載の組換えベクター。
【0021】
〔8〕
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子を含有する組換えベクター。
〔8−2〕
前記遺伝子を、以下の群:pTip QC1、pTip QC2、pTip QT1、pTip QT2、pTip RC1、pTip RC2、pTip RT1、pTip RT2、pNit QC1、pNit QC2、pNit QT1、pNit QT2、pNit RC1、pNit RC2、pNit RT1及びpNit RT2;から選択される発現ベクターに挿入して得られる、前記〔8〕に記載の組換えベクター。
〔8−3〕
前記遺伝子が、
(i)又は(ii)のアミノ酸配列のN末端と、
(iii)又は(iv)のアミノ酸配列のC末端とが、直接結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、前記〔8〕又は〔8−2〕に記載の組換えベクター。
〔8−4〕
前記遺伝子が、(i)のアミノ酸配列をコードする塩基配列を含む、前記〔8〕〜〔8−3〕のいずれかに記載の組換えベクター。
〔8−5〕
前記遺伝子が、(iii)のアミノ酸配列をコードする塩基配列を含む、前記〔8〕〜〔8−4〕のいずれかに記載の組換えベクター。
【0022】
〔9〕
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子を有し、スフィンゴミエリナーゼを培養物中に分泌することを特徴とする、ロドコッカス属細菌形質転換体。
〔9−2〕
ロドコッカス属細菌が、ロドコッカス エリスロポリスである、前記〔9〕に記載の形質転換体。
〔9−3〕
ロドコッカス エリスロポリスが、ロドコッカス エリスロポリス L88株である、前記〔9−2〕に記載の製造方法。
【0023】
〔10〕
前記遺伝子が、
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、前記〔9〕〜〔9−3〕のいずれかに記載の形質転換体。
〔10−2〕
前記遺伝子が、
(i)又は(ii)のアミノ酸配列のN末端と、
(iii)又は(iv)のアミノ酸配列のC末端とが、直接結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、前記〔10〕に記載の形質転換体。
〔10−3〕
前記遺伝子が、(i)のアミノ酸配列をコードする塩基配列を含む、前記〔9〕〜〔10−2〕のいずれかに記載の形質転換体。
〔10−4〕
前記遺伝子が、(iii)のアミノ酸配列をコードする塩基配列を含む、前記〔10〕〜〔10−3〕のいずれかに記載の形質転換体。
【0024】
〔11〕
前記〔7〕〜〔8−5〕のいずれかに記載の組換えベクターでトランスフェクトされたロドコッカス属細菌。
〔11−2〕
ロドコッカス エリスロポリスである、前記〔11〕に記載のロドコッカス属細菌。
〔11−3〕
ロドコッカス エリスロポリス L88株である前記〔11−2〕に記載のロドコッカス属細菌。
【0025】
〔12〕
前記〔1〕〜〔3〕のいずれかに記載の製造方法で得られるスフィンゴミエリナーゼ、あるいは前記〔4〕〜〔6−10〕のいずれかに記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清を用いたスフィンゴミエリナーゼの基質の測定方法。
〔13〕
前記〔1〕〜〔3〕のいずれかに記載の製造方法で得られるスフィンゴミエリナーゼ、あるいは前記〔4〕〜〔6−10〕のいずれかに記載のスフィンゴミエリナーゼ含有組成物、培養物又は培養上清を用いたスフィンゴミエリナーゼの基質の消去方法。
〔14〕
スフィンゴミエリナーゼ又はスフィンゴミエリナーゼ含有組成物の製造における前記〔7〕〜〔8−5〕のいずれかに記載の組換えベクターの使用。
〔15〕
スフィンゴミエリナーゼ又はスフィンゴミエリナーゼ含有組成物の製造における前記〔11〕〜〔11−3〕に記載のロドコッカス属細菌の使用。
【発明の効果】
【0026】
本発明により、スフィンゴミエリナーゼ、及び純度が高く、安定性の高いスフィンゴミエリナーゼ含有組成物を効率的に製造することができる。したがって、純度が高く、安定性の高いスフィンゴミエリナーゼ含有組成物を取得することができ、この組成物から容易にスフィンゴミエリナーゼを取得することもできる。
【図面の簡単な説明】
【0027】
【図1】比較例2において得られた配列番号20の塩基配列の構造と、PCR反応に用いたプライマーの位置を示す模式図である。
【図2】「実施例1 10.」記載の精製酵素と「比較例1 2.」記載の比較試験用酵素を、それぞれ、30℃で(A)72時間又は(B)96時間静置保存した際のスフィンゴミエリナーゼ活性の残存率を示す図である。
【図3】「実施例1 10.」記載の精製酵素と「比較例1 2.」記載の比較試験用酵素を純水又は酢酸ナトリウム緩衝液に溶解し、30℃又は−30℃で96時間静置保存したものを用いてSDS−PAGEを行った結果を示す図である。
【発明を実施するための形態】
【0028】
以下、本発明を実施するための形態(以下、「本実施の形態」と略すことがある)について具体的に説明する。
【0029】
本実施の形態のスフィンゴミエリナーゼの製造方法は、以下の工程(a)及び(b)を含む。
(a)以下の(i)又は(ii)のアミノ酸配列:
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして形質転換体を得る工程;及び
(b)工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程。
【0030】
工程(a)
工程(a)は、ロドコッカス属細菌に遺伝子をトランスフェクトして形質転換体を得る工程である。トランスフェクトする遺伝子は、得られた形質転換体を培養した場合にスフィンゴミエリナーゼを含む培養物が得られる限り特に限定されないが、好ましくは以下の(i)又は(ii)のアミノ酸配列をコードする塩基配列を含む遺伝子である。
【0031】
(i)配列番号1に記載のアミノ酸配列
上記工程(a)における(i)のアミノ酸配列からなるタンパク質は、ストレプトマイセス・エスピー A9107に由来するスフィンゴミエリナーゼである。
【0032】
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
上記工程(a)における(ii)のアミノ酸配列としては、例えば、配列番号1に記載のアミノ酸配列において、酵素作用に関与しない一部のアミノ酸を変異させたアミノ酸配列が例示される。配列番号3に記載のアミノ酸配列(配列番号1に記載のアミノ酸配列の50〜56番目の配列に対応)及び配列番号4に記載のアミノ酸配列(配列番号1に記載のアミノ酸配列の199〜204番目の配列に対応)(但し、配列番号3及び4中、Xaaは任意のアミノ酸を示す)は、いずれもスフィンゴミエリナーゼ活性に必須の配列として知られる(Amy E.A.O.ら、J.Biol.Chem.,280,35011−35017(2005)、Ago H.ら、J.Biol.Chem.,281,16157−16167(2006)等)。したがって、上記工程(a)における(ii)のアミノ酸配列としては、例えば、配列番号1に記載のアミノ酸配列において、配列番号3及び4に対応する配列以外の部分のアミノ酸を変異させたアミノ酸配列が例示される。性質の似たアミノ酸への置換等であれば、変異後のアミノ酸配列からなるタンパク質のスフィンゴミエリナーゼ活性に影響しないであろう。例えば、上記工程(a)における(ii)のアミノ酸配列としては、DVVX1X2X3E(式中、X1はV又はIを、X2はL又はFを、X3はD、N、Q又はEを、それぞれ表す)、及びX4X5GDX6N(式中、X4はV、I又はLを、X5はA、C又はGを、X6はF、M又はLを、それぞれ表す)の配列を含むアミノ酸配列が好ましい。
【0033】
上記工程(a)における(ii)のアミノ酸配列としては、また、配列番号1に記載のアミノ酸配列に各種のアミノ酸残基が付加したアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列も例示される。そのようなアミノ酸配列又は該アミノ酸配列からなるタンパク質としては、例えば、配列番号1に記載のアミノ酸配列のC末端側にチオレドキシンタンパク質等機能性タンパク質を付加した融合タンパク質、配列番号1に記載のアミノ酸配列に、タンパク質の精製や確認等をせしめることのできるタグと呼ばれる部分を付加した融合タンパク質等が挙げられる。タグの付加は、その全部又は一部であってもよい。例えば、本実施の形態における製造方法において、培養物からスフィンゴミエリナーゼを効率的に精製するために、配列番号1に記載のアミノ酸配列に5〜10個のHisを付加してもよい。また、配列番号1に記載のアミノ酸配列と、付加アミノ酸(配列)との間等に、数個のプロテアーゼ認識アミノ酸配列を配置して付加することもできる。更に、配列番号1のアミノ酸配列のC末端側に付加していてもよいアミノ酸残基(又はポリペプチド残基)としては、TEE配列、Sタグ、Hisタグ等が挙げられる。
【0034】
上記工程(a)における(ii)のアミノ酸配列としては、また、配列番号1に記載のアミノ酸配列において、各種のアミノ酸が欠失したアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列も例示される。そのような欠失の例としては、例えば:スフィンゴミエリナーゼの本質的な機能とは無関係の数個のアミノ酸からなるドメインの欠失;配列番号1に記載のアミノ酸配列中の複数個のアミノ酸からなるギャップの欠失;配列番号1に記載のアミノ酸配列のC末端側のArgから順に削除する欠失;配列番号1のアミノ酸配列のN末端側のAlaの欠失;それらの欠失の組み合わせ等が挙げられる。
【0035】
変異後に得られるアミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有する限り、上記の欠失、置換又は付加等の変異は、適宜組み合わせることも可能である。任意のアミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するか否かは、当業者に公知の手法により確認することができる。例えば、該アミノ酸配列をコードする塩基配列を含む遺伝子をトランスフェクトして得た形質転換体を培養し、培養物のスフィンゴミエリナーゼ活性を測定することで確認することができる。培養物が、後述する理化学的性質<1>〜<5>を有するスフィンゴミエリナーゼを含むことが好ましい。
【0036】
一態様において、工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子は、上記(i)及び(ii)のアミノ酸配列のうち、(i)のアミノ酸配列をコードする塩基配列を含むことが好ましい。別の態様として(ii)のアミノ酸配列をコードする塩基配列を含むことが好ましい場合もある。
【0037】
上記(i)又は(ii)のアミノ酸配列をコードする塩基配列は、目的とするアミノ酸配列をもとに、容易に得ることができる。配列番号1のアミノ酸配列をコードする塩基配列としては、特に限定されないが、例えば、配列番号5の塩基配列や、配列番号5の塩基配列を宿主のコドン使用頻度に合わせて変更した塩基配列が例示されるが、ロドコッカス属細菌のコドン使用頻度に合わせて変更した塩基配列が好ましく、ロドコッカス エリスロポリスのコドン使用頻度に合わせて変更した塩基配列がより好ましい。そのようなコドン使用頻度は、 http://www.kazusa.or.jp/codon/ などで容易に検索することができる。塩基配列の調製には、通常用いられる公知の遺伝子操作を利用することができる。塩基配列が所望のアミノ酸配列をコードする限り、例えば、部位特異的変異法や、目的遺伝子の特定塩基の断片を人工変異塩基で置換する等の種々の方法を用いて塩基配列に変異を加えてもよい。
【0038】
微生物等を用いて酵素等のタンパク質を製造する場合、生産性を高めること等を目的として、シグナルペプチドと呼ばれるアミノ酸配列を、目的とする酵素等のタンパク質に付加することがある。本実施の形態におけるスフィンゴミエリナーゼの製造方法においても、スフィンゴミエリナーゼのアミノ酸配列のN末端側にシグナルペプチドを付加することができる。そのようなシグナルペプチドは、本実施の形態における製造方法においてシグナルペプチドとして機能するものであれば特に限定されない。
【0039】
本実施の形態において、あるペプチドが、「シグナルペプチドとして機能する」とは、該ペプチドと、上記(i)又は(ii)に記載のアミノ酸配列とが直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして得られた形質転換体を培養した場合に、スフィンゴミエリナーゼを含む培養物を得ることができることを言う。純度が高く、安定性の高いスフィンゴミエリナーゼを含む培養物を効率的に得ることができるシグナルペプチドが好ましい。例えば、上記培養した場合に、スフィンゴミエリナーゼが培養液中に形成される(形質転換体内で生産されたスフィンゴミエリナーゼがシグナルペプチドにより培養液中に輸送される)シグナルペプチド(アミノ酸配列)が好ましい。また、シグナルペプチドとしては、得られるスフィンゴミエリナーゼの活性を低下させないものが好ましい。特に、純度が高く、安定性の高いスフィンゴミエリナーゼを含む培養物を効率的に得るという観点からは、シグナルペプチドとしてはストレプトマイセス・エスピー A9107株由来のシグナルペプチド(配列番号2)が好ましい。また、シグナルペプチドとしては、上記シグナルペプチドのアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、シグナルペプチドとして機能するアミノ酸配列を用いることもできる。
【0040】
具体的には、本実施の形態におけるスフィンゴミエリナーゼの製造方法の工程(a)において、
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子を、ロドコッカス属細菌にトランスフェクトすることで、シグナルペプチドをスフィンゴミエリナーゼに付加することができる。
【0041】
上記(iv)のアミノ酸配列としては、例えば、配列番号2に記載のアミノ酸配列のN末端側にチオレドキシンタンパク質等機能性タンパク質やHisタグ等のその他のアミノ酸配列からなる部分を付加したアミノ酸配列が例示され、融合タンパク質とすることも好ましい。場合によっては、上記の付加アミノ酸配列は、その全部又は一部であってもよい。上述の付加の例と同様に、シグナルペプチドとして機能するアミノ酸配列が得られる限り、常法によってアミノ酸の欠失又は置換を行うことができ、例えば、該アミノ酸配列中に複数個のアミノ酸からなるギャップが存在する場合、それらの欠失を組み合わせることもできる。また、上記の欠失、置換又は付加を適宜組み合わせることも可能である。
【0042】
上記(iii)又は(iv)のアミノ酸配列をコードする塩基配列は、目的とするアミノ酸配列をもとに、容易に得ることができる。配列番号2のアミノ酸配列をコードする塩基配列としては、特に限定されないが、例えば、配列番号6の塩基配列が挙げられる。このような塩基配列は上述のように宿主のコドン使用頻度にあわせて変更することができる。
【0043】
上記遺伝子に含まれる塩基配列は、
上記(i)又は(ii)のアミノ酸配列のN末端と、
上記(iii)又は(iv)のアミノ酸配列のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする。
【0044】
ここで、上記(i)又は(ii)のアミノ酸配列のN末端と、上記(iii)又は(iv)のアミノ酸配列のC末端とが、1若しくは数個のアミノ酸を介して結合する場合、該結合は、結合したアミノ酸配列をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして得られた形質転換体を培養した場合に、スフィンゴミエリナーゼを含む培養物を得ることができる限り特に限定されない。該結合は、プロテアーゼの切断部位であることが望ましく、代表的なプロテアーゼとそれに対応する配列としては、例えば、以下の配列が挙げられる(配列中、↓が切断部位を表す)。
トロンビン:LVPR↓GS
エンテロキナーゼ:DDDDK↓
第X因子(Factor Xa):IEGR↓
【0045】
好ましい態様において、上記遺伝子に含まれる塩基配列は、上記(i)又は(ii)のアミノ酸配列のN末端と、上記(iii)又は(iv)のアミノ酸配列のC末端とが、直接結合したアミノ酸配列をコードする。
【0046】
塩基配列の調製には、通常用いられる公知の遺伝子操作を利用することができる。DNA合成により調製してもよい。塩基配列が所望のアミノ酸配列をコードする限り、例えば、部位特異的変異法や、目的遺伝子の特定塩基の断片を人工変異塩基で置換する等の種々の方法を用いて塩基配列に変異を加えてもよい。
【0047】
上記の塩基配列を含む遺伝子を、常法を用いてロドコッカス属細菌にトランスフェクトして形質転換体を得ることができる。より詳細には、形質転換体において導入遺伝子を発現させるための発現ベクターに、上記の塩基配列を含む遺伝子を挿入して得られる組換えベクターを使用することが出来る。このような発現ベクターとしては、ロドコッカス属細菌を宿主としてトランスフェクトし、得られた形質転換体を培養した場合に、スフィンゴミエリナーゼを含む培養物が得られるベクターであれば特に限定されないが、純度が高く、安定性の高いスフィンゴミエリナーゼを含む培養物を効率的に得られる発現ベクターが好ましく、宿主微生物体内で自律的に増殖し得るファージ又はプラスミドのうち、遺伝子組換え用として構築されたものが好ましい。
【0048】
そのような発現ベクターとしては、例えば、λgt・λC、λgt・λB、pET−3a、pET−11a、pET−32aなどのpETベクター(Novagen社製)並びにpBR322、pBR325、pACYC184、pUC12、pUC13、pUC18、pUC19、pUC118、pIN I、BluescriptKS+、pWH1520、pUB110、pKH300PLK、pIJ680、pIJ702、YRp7、pYC1、YEp13、pTip QC1、pTip QC2、pTip QT1、pTip QT2、pTip RC1、pTip RC2、pTip RT1、pTip RT2、pNit QC1、pNit QC2、pNit QT1、pNit QT2、pNit RC1、pNit RC2、pNit RT1及びpNit RT2などが使用できる。好ましいベクターとしては、pTip QC1、pTip QC2、pTip QT1、pTip QT2、pTip RC1、pTip RC2、pTip RT1、pTip RT2、pNit QC1、pNit QC2、pNit QT1、pNit QT2、pNit RC1、pNit RC2、pNit RT1及びpNit RT2が挙げられ、pTip QC1及びpTip QC2がより好ましい。本実施の形態においては、ロドコッカス属細菌を宿主とする発現ベクターが好ましい。より好ましくは大腸菌とのシャトルベクターとして機能する発現ベクターが挙げられる。
【0049】
プロモーターは宿主中で発現出来るものであれば特に限定されないが、好ましくはpTip QC1、pTip QC2、pTip QT1、pTip QT2、pTip RC1、pTip RC2、pTip RT1、pTip RT2、pNit QC1、pNit QC2、pNit QT1、pNit QT2、pNit RC1、pNit RC2、pNit RT1及びpNit RT2のプロモーターが挙げられ、さらに好ましくはpTip QC1、pTip QC2、pTip QT1及びpTip QT2のプロモーターが挙げられる。
【0050】
発現ベクターへの遺伝子の挿入は、常法により行うことができる。まず、挿入しようとする塩基配列を制限酵素により切断して生成する塩基配列の末端と同じ末端を生成する制限酵素により上記発現ベクターを切断し、発現ベクター断片を作製する。挿入しようとする塩基配列を該制限酵素で切断することにより作製した塩基配列の断片と、発現ベクター断片とを、DNAリガーゼにより常法に従って結合し、遺伝子を発現ベクターに挿入することができる。得られたベクターに目的の遺伝子が挿入されていることは、例えばDNAシーケンスにより確認することができる。得られた組換えベクターは、本実施の形態におけるスフィンゴミエリナーゼの製造等に有用である。
【0051】
本実施の形態において、トランスフェクト(形質転換)とは、遺伝子を細胞(菌体)に導入し、形質転換体を得ることを意味し、その方法は当業者が一般に行うことができるものであれば特に限定されない。例えばコンピテントセルを用いる方法、マイクロインジェクション法、リポフェクション法が例示される。好ましくは、ロドコッカス属細菌に効率的に遺伝子を導入することができる方法であり、形質転換効率がよいという観点から、より好ましくはエレクトロポレーション法である。
【0052】
工程(a)において、遺伝子をトランスフェクトする微生物は、ロドコッカス属細菌であれば特に限定されないが、ロドコッカス エリスロポリス(Rhodococcus erythropolis)であることが好ましい。ロドコッカス エリスロポリスとしては、得られた形質転換体を培養して、スフィンゴミエリナーゼを含む培養物が得られる限り特に限定されないが、純度が高く、安定性の高いスフィンゴミエリナーゼを含む培養物が効率的に得られるものが好ましい。例えば、ロドコッカス エリスロポリス ATCC15591、ロドコッカス エリスロポリス ATCC25544、ロドコッカス エリスロポリス ATCC17041、ロドコッカス エリスロポリス NCIB8147、ロドコッカス エリスロポリス JCM3201、ロドコッカス エリスロポリス JCM3201の変異株及びロドコッカス エリスロポリス L88株(FERM BP−08444)が例示される。好ましくはロドコッカス エリスロポリス JCM3201が挙げられ、より好ましくは特開2004−073116号公報で開示されたロドコッカス エリスロポリス JCM3201の変異株が挙げられ、さらに好ましくはロドコッカス エリスロポリス L88株が挙げられる。任意の微生物が、ロドコッカス属細菌(又はロドコッカス・エリスロポリス)であるかの判断は、バージーズ・マニュアル・オブ・システマティック・バクテリオロジー(Bergey's Manual of Systematic Bacteriology)に従って行うことができる。rRNA塩基配列解析によって行ってもよい。また、微生物同定検査会社に依頼して判別してもよい。
【0053】
目的の遺伝子がトランスフェクトされた形質転換体を選択する際には、導入した発現ベクターに存在する抗生物質耐性遺伝子などの選択マーカーを利用すればよい。例えば、pTip QC1、pTip QC2、pTip RC1、pTip RC2、pNit QC1、pNit QC2、pNit RC1及びpNit RC2を発現ベクターとして用いる場合には、クロラムフェニコールに対する耐性により選択する方法が例示される。また、pTip QT1、pTip QT2、pTip RT1、pTip RT2、pNit QT1、pNit QT2、pNit RT1及びpNit RT2を発現ベクターとして用いる場合にはテトラサイクリンに対する耐性により選択する方法が例示される。得られた形質転換体は、本実施の形態におけるスフィンゴミエリナーゼの製造等に有用である。
【0054】
工程(b)
工程(b)は、上記工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程である。
【0055】
形質転換体の培養条件は、ロドコッカス属細菌の生理学的性質を考慮して選択すればよい。培養は、通常液体培養で行うが、工業的には深部通気撹拌培養を行うことが有利である。
【0056】
培養に用いる培地は、通常ロドコッカス属細菌の培養に用いられるものであれば特に限定されず、例えば、肉汁培地、肉汁ゼラチン培地、牛乳培地、コーンミール培地、トマトジュース培地、TSI培地、SIM培地、グルコース-ブイヨン培地、YM培地、Lennox培地、LB培地、Davis培地、トリス-グルコース培地、EMB糖指示培地、Spizizen最少培地、YPAD培地、Vogel培地、ツァペック-ドッグス培地等を用いることができる。さらに、例えば http://www.dsmz.de/ 等を参考にして、適切な培地を選択することもできる。
【0057】
培養温度はロドコッカス属細菌が発育し、スフィンゴミエリナーゼを含む培養物が得られる範囲で適宜変更し得る。例えば、通常は下限が4℃以上、好ましくは10℃以上、より好ましくは20℃以上、上限が50℃以下、好ましくは42℃以下、より好ましくは37℃以下が例示される。
【0058】
培養時間は、選択する培養条件によって多少異なるが、得られる培養物中のスフィンゴミエリナーゼが最大量に達する時期を見計らって適当な時期に培養を終了すればよい。例えば、後述の実施例1に記載の培養条件で培養した場合、通常は下限が10時間以上、好ましくは12時間以上、より好ましくは17時間以上、上限が100時間以下、好ましくは90時間以下、より好ましくは80時間以下が例示される。
【0059】
培地pHはロドコッカス属細菌が発育し、スフィンゴミエリナーゼを含む培養物が得られる範囲で適宜変更し得る。例えば、下限としてpH 5.8以上、好ましくはpH 6.2以上、上限としてpH 8.5以下、好ましくはpH 7.5以下が例示される。
【0060】
上記の手法で形質転換体を培養して得られる、スフィンゴミエリナーゼを含む培養物は、形質転換体内、培養液中又はその両方にスフィンゴミエリナーゼを含むことができるが、精製の容易性等の観点から、好ましくは、培養液中にスフィンゴミエリナーゼを含む。
【0061】
本実施の形態は、上記工程(a)及び(b)を含む方法により得られる、形質転換体及びスフィンゴミエリナーゼ含有培養物並びに該培養物の培養上清であって、スフィンゴミエリナーゼを含有する培養上清にも関する。
【0062】
本実施の形態は、また、上記工程(a)及び(b)に続いて、以下の工程(c)を行うことにより得られる、スフィンゴミエリナーゼ含有組成物にも関する。
【0063】
工程(c)
工程(c)は、工程(b)で得た培養物から形質転換体を分離してスフィンゴミエリナーゼ含有組成物を得る工程である。工程(b)で得た培養物は、上述の通り、好ましくは培養液中にスフィンゴミエリナーゼを含む培養物である。この場合、工程(b)で得られた培養物から、濾過又は遠心分離などの通常の手段により形質転換体を分離し、培養上清としてスフィンゴミエリナーゼ含有組成物を得ることができる。
【0064】
形質転換体の分離後、さらに、マイクローザ(旭化成ケミカルズ社製)やフィルトロン(富士フィルター社製)などを用いた限外濾過濃縮法などにより濃縮した形で、本実施の形態におけるスフィンゴミエリナーゼ含有組成物を得ることもできる。
【0065】
本実施の形態において、上記工程(c)で得られたスフィンゴミエリナーゼ含有組成物は、純度が高く、保存安定性が高いため、さらなる精製を行うことなく保存が可能であるという利点を有するが、所望により、安定剤として各種の塩類、糖類、タンパク質、脂質、界面活性剤等を加え、あるいは加えることなく、限外濾過濃縮、凍結乾燥等の方法により、液状又は固形のものとすることができる。例えば凍結乾燥を行う場合、安定剤としてサッカロース、マンニトール、食塩、アルブミン等を0.5%〜10%程度添加してもよい。このようにして得られた濃縮物や凍結乾燥物も、本実施の形態におけるスフィンゴミエリナーゼ含有組成物に含まれる。
【0066】
本実施の形態におけるスフィンゴミエリナーゼ含有組成物は、水又は緩衝液を添加してスフィンゴミエリナーゼ濃度が4mg/mLの中性又は酸性の水溶液を調製し、30℃で96時間静置保存して保存試験を行った場合、保存前と比較して50%以上のスフィンゴミエリナーゼ活性を保持することが好ましい。
【0067】
上記の保存試験における「中性又は酸性」とは、pHが中性から酸性であることを意味し、pHの下限は4.0であり、好ましくは5.0であり、より好ましくは5.5である。pHの上限は7.5であり、好ましくは7.0であり、より好ましくは6.8である。
【0068】
上記の保存試験における「緩衝液」は、スフィンゴミエリナーゼ溶液として中性又は酸性の水溶液が得られる限り特に限定されないが、例えば、弱酸の酢酸、クエン酸、リン酸、ホウ酸、又は酒石酸と、これらのナトリウム塩等を用いて調製される緩衝液であり、好ましくは酢酸緩衝液(酢酸+酢酸ナトリウム)である。
【0069】
上記の保存試験における温度の上限は、スフィンゴミエリナーゼ活性が残存する温度であればよく、例えば50℃であり、好ましくは40℃であり、より好ましくは30℃である。静置保存する場合の温度の下限は、例えば4℃であり、好ましくは10℃であり、より好ましくは20℃である。特に好ましい温度は30℃以下である。
【0070】
上記の保存試験におけるスフィンゴミエリナーゼの水溶液の濃度は、特に制限されないが、下限は、例えば1mg/mLであり、好ましくは2mg/mLであり、より好ましくは3mg/mLである。上限は、例えば30mg/mLであり、好ましくは20mg/mLであり、より好ましくは10mg/mLである。例えば、4mg/mLのスフィンゴミエリナーゼ水溶液を作成して、上記の30℃で96時間の静置保存をすることができる。
【0071】
上記の条件で保存した本実施の形態の組成物中のスフィンゴミエリナーゼ活性は、保存前と比較して、少なくとも40%より高い、例えば50%以上、好ましくは65%以上、より好ましくは70%以上、さらに好ましくは75%以上、さらにより好ましくは78%以上のスフィンゴミエリナーゼ活性を保持し、特に好ましくは80%以上のスフィンゴミエリナーゼ活性を保持する。特に、上記の水溶液のpHが酸性(例えばpH5.5)の場合、上記の条件で保存した本実施の形態の組成物中のスフィンゴミエリナーゼ活性は、保存前と比較して、例えば75%以上、好ましくは78%以上、より好ましくは90%以上、さらに好ましくは97%以上のスフィンゴミエリナーゼ活性を保持する。
【0072】
本実施の形態において、上記工程(c)で得られたスフィンゴミエリナーゼ含有組成物は、高い純度(タンパク質量あたりのスフィンゴミエリナーゼ活性)を有する。例えば、本実施の形態におけるスフィンゴミエリナーゼ含有組成物は、タンパク質量あたりのスフィンゴミエリナーゼ活性として少なくとも10U/mg以上、好ましくは50U/mg以上、より好ましくは100U/mg以上の純度を有する。
【0073】
純度は、後述のスフィンゴミエリナーゼ活性測定法と共に、タンパク質濃度の測定法を用いて決定することができる。タンパク質濃度の測定法としては、当該業者に公知の測定方法を用いればよく、特に限定されるものではない。例えば、280nmの吸光度を測定する方法(紫外吸収法)、クーマシーブルーを用いる方法(Bradford法)、フェノール試薬を用いる方法(Lowry法)、ビシンコニン酸を用いる方法(BCA法)が例示される。標準試料として用いるタンパク質も、当該業者が一般的に用いるタンパク質を用いればよく、例えばウシ血清アルブミン、オボアルブミン、ガンマグロブリン、ヒト血清アルブミン及びクリスタリンが例示され、ウシ血清アルブミン及びオボアルブミンが汎用される。
【0074】
本実施の形態において、上記工程(c)で得られたスフィンゴミエリナーゼ含有組成物は、高い保存安定性を有し、例えば70時間以上、好ましくは80時間以上、より好ましくは90時間以上、例えば96時間程度、静置保存しても、保存前と比較して、例えば50%以上、好ましくは65%以上、より好ましくは70%以上、さらに好ましくは75%以上、特に好ましくは78%以上のスフィンゴミエリナーゼ活性を保持する。保存時の温度は、上記の保存試験に関する記載も参照して適宜決定することができるが、保存温度は低いことが好ましい。
【0075】
本実施の形態において、上記工程(c)で得られたスフィンゴミエリナーゼ含有組成物の高い保存安定性は、例えば保存前後のサンプルの電気泳動によっても確認することができる。例えば、本実施の形態におけるスフィンゴミエリナーゼ含有組成物は、水又は緩衝液を添加してスフィンゴミエリナーゼ濃度が4mg/mLである中性又は酸性の水溶液を調製し、30℃又は−30℃で96時間静置保存後、ドデシル硫酸ナトリウム―ポリアクリルアミドゲル電気泳動法で、電気泳動させた場合、例えばデンシトメーターで測定した場合、保存前と比較して、スフィンゴミエリナーゼのポリペプチドを示す帯が少なくとも70%以上、好ましくは80%以上、より好ましくは90%以上残存する。
【0076】
上記の工程(b)で得た培養物又は上記の工程(c)で得た組成物から、以下に記載の通り、常法によりスフィンゴミエリナーゼ(精製物)を得ることができる。
【0077】
上記の工程(b)で得た培養物において、スフィンゴミエリナーゼが形質転換体内に含まれる場合、例えば、まず培養物を固液分離し、得られた湿潤菌体(形質転換体)をリン酸緩衝液やトリス−塩酸緩衝液などの溶液に分散し、リゾチーム処理、超音波処理、フレンチプレス処理、ダイノミル処理などの菌体破砕手段を適宜選択し組み合わせて、形質転換体内のスフィンゴミエリナーゼを抽出することができる。
【0078】
このようにして得た抽出物や、上記の工程(c)で得た組成物から、さらに純度の高いスフィンゴミエリナーゼ(精製物)を得る工程としては、例えば、上記抽出物や組成物に、アセトン、メタノール、エタノールなどの有機溶媒を添加することによる分別沈殿法、硫酸アンモニウム、食塩などによる塩析法などを適用してスフィンゴミエリナーゼを沈殿させ回収する工程が挙げられる。さらに、この沈殿物に対し、必要に応じて透析、等電点沈殿を行った後、電気泳動法などで単一の帯を示すまで、イオン交換体、ゲル濾過剤、吸着体などを用いるカラムクロマトグラフィーなどにより精製し、スフィンゴミエリナーゼ(精製物)を得ることができる。これらの工程を組み合わせることにより目的酵素の精製度が上がる場合は適宜組み合わせて行うことができる。
【0079】
上記の工程(c)で得た組成物は、純度が高いため、非常に簡便な精製法によりスフィンゴミエリナーゼ(精製物)を得ることができるという利点を有する。
【0080】
本実施の形態における製造方法で得られるスフィンゴミエリナーゼ(Sphingomyelinase)または本実施の形態におけるスフィンゴミエリナーゼ含有組成物(培養物、培養上清であってもよい)に含まれるスフィンゴミエリナーゼ(単に「本実施の形態のスフィンゴミエリナーゼ」ともいう)とは、EC 3.1.4.12に分類されるタンパク質であり、好ましくはストレプトマイセス由来、より好ましくはストレプトマイセス・エスピー由来、さらに好ましくはストレプトマイセス・エスピー A9107株スフィンゴミエリナーゼと同一のアミノ酸配列及び/又は特徴を有する。該アミノ酸配列としては、配列番号1に記載のアミノ酸配列が例示される。また、該特徴としては、以下の(2)〜(5)に記載の特徴が例示される。
【0081】
本実施の形態のスフィンゴミエリナーゼは、少なくとも以下の(1)に記載の作用を有するものであれば特に限定されないが、さらに以下の(2)〜(5)のいずれかに記載の1つ以上の性質を有することが好ましい。例えば、一態様において、スフィンゴミエリナーゼは、以下の(1)に記載の作用に加えて以下の(2)に記載の基質特異性を有することが好ましい。また、スフィンゴミエリナーゼは、以下の(1)に記載の作用に加えて以下の(2)〜(5)に記載の性質のうち、任意の2つ以上を備えていることも好ましい。特に好ましいスフィンゴミエリナーゼとしては、以下の(1)に記載の作用に加えて以下の(2)〜(5)の全てに記載の性質を有するものが挙げられる。
【0082】
(1)作用
本実施の形態におけるスフィンゴミエリナーゼの作用の一例は以下の(式1)に示す通りであり、1分子の水の存在下、スフィンゴミエリン(Shingomyelin)1分子を分解(加水分解)してセラミド(N−アシルスフィンゴシン(N−acylsphingosin))1分子とフォスフォリルコリン(Phosphorylcholine)1分子を生成する。該スフィンゴミエリンの作用は、少なくとも2価の陽イオンの存在下で行われることが好ましく、マグネシウムイオン及び/又はマンガンイオンの存在下で行われることがより好ましい。
【0083】
スフィンゴミエリン + H2O ―>
N−アシルスフィンゴシン + フォスフォリルコリン (式1)
【0084】
本実施の形態において、上記のスフィンゴミエリン(Sphingomyelin)とは、以下の一般式:
【化1】
で表されるスフィンゴ脂質の一種であれば、由来や純度は限定されず、その塩、誘導体等であってもよい。上記一般式中、Rは飽和または不飽和炭化水素を表し、Xはコリン、エタノールアミン、セリン、イノシトール等を表し、好ましくは生体中に多く存在するコリンを表す。本実施の形態において、スフィンゴミエリンは、好ましくは生体内に存在するスフィンゴミエリンであり、特に好ましくは本実施の形態の製造方法で得られるスフィンゴミエリナーゼにより加水分解されるスフィンゴミエリンであり、例えば、N−ヘキサデカノイル−スフィンゲニン−1−フォスフォリルコリン(N−(hexadecanoyl)−sphing−4−enine−1−phosphorylcholine)、N−アセチル−スフィンゲニン−1−フォスフォリルコリン(N−(acetyl)−sphing−4−enine−1−phosphorylcholine)、N−ヘキサノイル−スフィンゲニン−1−フォスフォリルコリン(N−(hexanoyl)−sphing−4−enine−1−phosphorylcholine)、N−ドデカノイル−スフィンゲニン−1−フォスフォリルコリン(N−(dodecanoyl)−sphing−4−enine−1−phosphocholine)、N−ドデカノイル−スフィンガニン−1−フォスフォリルコリン(N−(dodecanoyl)−sphinganine−1−phosphocholine)、N−ヘプタデカノイル−スフィンゲニン−1−フォスフォリルコリン(N−(heptadecanoyl)−sphing−4−enine−1−phosphorylcholine)、N−オクタデカノイル−スフィンゲニン−1−フォスフォリルコリン(N−(octadecanoyl)−sphing−4−enine−1−phosphorylcholine)、N−オクタデカノイル−スフィンゲニン−1−フォスフォリルコリン(N−(9Z−octadecenoyl)−sphing−4−enine−1−phosphorylcholine)、N−テトラコサノイル−スフィンゲニン−1−フォスフォリルコリン(N−(tetracosanoyl)−sphing−4−enine−1−phosphorylcholine)、N−テトラコセノイル−スフィンゲニン−1−フォスフォリルコリン(N−(15Z−tetracosenoyl)−sphing−4−enine−1−phosphocholine)などが例示される。本実施の形態のスフィンゴミエリナーゼは、複数のスフィンゴミエリンに同時に作用するものであってもよい。
【0085】
本実施の形態において、上記スフィンゴミエリナーゼの作用はスフィンゴミエリナーゼ活性として測定できる。スフィンゴミエリナーゼ活性とは、上記の(式1)の反応を触媒する活性であり、スフィンゴミエリナーゼ活性1単位(ユニット)は、37℃で1分間に1マイクロモルのスフィンゴミエリンを分解する酵素量とする。スフィンゴミエリナーゼ活性の測定方法を以下に例示するが、測定方法はこれに限定されるものではない。
【0086】
<スフィンゴミエリナーゼ活性測定法>
(反応試薬混合液組成)
1M Tris/HCl pH 8.0 150μL
10mM スフィンゴミエリン溶液 300μL
0.3% 4−アミノアンチピリン溶液 300μL
0.2% TODB溶液 300μL
1M 塩化マグネシウム水溶液 6μL
1M 塩化ナトリウム水溶液 30μL
10% トリトンX−100溶液 30μL
500U/mL アルカリフォスファターゼ溶液 60μL
600U/mL コリンオキシダーゼ溶液 50μL
500U/mL ペルオキシダーゼ溶液 120μL
精製水 1654μL
【0087】
小試験管に上記組成の反応試薬混合液を0.8mLとり、37℃で予備加温する。5分後、適当に希釈したスフィンゴミエリナーゼ溶液(被験液)を0.03mL加えて混和し、37℃で反応を開始し、5分間インキュベートして反応を行う。続いてこの反応液に1% ドデシル硫酸ナトリウム溶液を1.6mL加えて反応を停止する。反応停止後546nmにおける吸光度を測定し、これをAsとする。スフィンゴミエリナーゼ溶液の代わりに精製水を用いた盲検をAbとする。スフィンゴミエリナーゼ活性1単位を、37℃で1分間に1マイクロモルのスフィンゴミエリンを分解する酵素量として、上記反応液中におけるTODBのミリモル吸光係数を16として、得られたAs及びAbの値に基づき以下の(式2)を用いて算出する。
【0088】
スフィンゴミエリナーゼ活性(単位)
=[{(As−Ab)/16}/5]×(2430/30) (式2)
【0089】
(2)基質特異性
本実施の形態のスフィンゴミエリナーゼ及びスフィンゴミエリナーゼ含有組成物は、上記のスフィンゴミエリンに対する作用を有するものであれば特に限定されないが、好ましくはスフィンゴミエリンに作用し、かつ、レシチン、リゾレシチン、フォスファチジルエタノールアミン、フォスファチジルセリン及びフォスファチジルイノシトール(以下、「レシチン等」ともいう)のいずれにも実質的に作用しない。ここで、「実質的に作用しない」とは、通常、上記のスフィンゴミエリナーゼの活性測定法における条件下でレシチン等に反応しないことをいう。スフィンゴミエリナーゼ活性測定法における条件(酵素の希釈の程度、反応試薬混合液中の各組成物の濃度等)を変更してもレシチン等に反応しないことが好ましい。
【0090】
(3)至適pH
本実施の形態のスフィンゴミエリナーゼの至適pHは、上記スフィンゴミエリナーゼ活性が測定できるpHであれば特に限定されないが、上記スフィンゴミエリナーゼ活性が最大となるpHが好ましい。pHの下限としては、例えばpH4.0以上であり、好ましくはpH5.0以上であり、より好ましくはpH7.0以上である。またpHの上限としては、例えばpH10.0以下であり、好ましくはpH9.0以下であり、より好ましくはpH8.0以下である。
【0091】
(4)Km
本実施の形態のスフィンゴミエリナーゼのスフィンゴミエリンに対するKmは、温度や気圧などの測定の条件や使用機器の精度等によりその値は変化し得るが、例えば2mM以下であり、好ましくは1mM以下であり、より好ましくは約0.45mMである。
【0092】
(5)分子量
本実施の形態のスフィンゴミエリナーゼの分子量は、温度や気圧などの測定の条件や使用機器の精度等によりその値は変化し得るが、例えば30kDa〜50kDaの範囲にあり、好ましくは35kDa〜40kDaの範囲にあり、より好ましくはドデシル硫酸ナトリウム―ポリアクリルアミドゲル電気泳動法で約38kDaである。
【0093】
なお、本実施の形態におけるスフィンゴミエリナーゼの分子量は、工程(a)においてトランスフェクトする遺伝子の変異や、工程(b)における培養時の翻訳後修飾、工程(b)や(c)の後の化学修飾等によって変化する場合があり、例えば、トランスフェクトする際のベクターとしてpTip QC1やpTipQC2(特許第3793812号公報、特開2004−321013号公報、特開2004−073116号公報参照)を利用してC末端にHisタグを付加する場合には、上記の分子量が約1,000程度大きくなることがある。
【0094】
このように、翻訳後修飾(N末端側がアシル基やアルキル基等による修飾を受ける等)、化学修飾(無水コハク酸やPEG等)等を受けたスフィンゴミエリナーゼも、本実施の形態のスフィンゴミエリナーゼに含まれる。化学修飾等により、スフィンゴミエリナーゼの至適pHや安定性等の性質を、利用しやすいように変化させることが可能である。
【0095】
本実施の形態におけるスフィンゴミエリナーゼ又はスフィンゴミエリナーゼ含有組成物を用いれば、スフィンゴミエリナーゼを定性的、半定量的又は定量的に測定することができる。その方法としては、例えばスフィンゴミエリンに対するスフィンゴミエリナーゼの作用で生成したフォスフォリルコリンにさらにグリセロホスフォコリンホスフォジエステラーゼ(EC 3.1.4.2)を作用させて生成したコリンをコリンオキシダーゼ(EC 1.1.3.17)で酸化し、生成した過酸化水素を4−アミノアンチピリンとフェノールなどと縮合して発色させる方法など、公知の方法が挙げられる。
【0096】
また、本実施の形態におけるスフィンゴミエリナーゼ又はスフィンゴミエリナーゼ含有組成物を用いれば、上記(式1)の反応により、スフィンゴミエリナーゼの基質を消去することもできる。
【0097】
本実施の形態におけるスフィンゴミエリナーゼ又はスフィンゴミエリナーゼ含有組成物は、そのまま、あるいは精製、濃縮、凍結乾燥等の後、各種製品、試薬、キット、センサー、原薬、製剤、化粧品、食品等に利用することができる。
【実施例】
【0098】
以下、本発明を実施例等に基づいて説明するが、本発明の範囲は以下の実施例等に限定して解釈されるものではない。尚、記述した技術は、市販の各種酵素、キット類に添付された手順書等に従えば実施できるものである。「常法に従い」と記述した技術は、例えばマニアティスらの方法(Maniatis,T.,et al.Molecular Cloning.Cold Spring Harbor Laboratory 1982年、1989年)や蛋白質・酵素の基礎実験法(改訂第2版、堀尾武一、1994年、南光堂)、等に従えば実施できるものである。又、以下に示した測定値等は測定の条件や使用機器の精度等によりその値は変化し得る。
【0099】
なお、以下の実施例で使用する試薬類は、特に断らない限り、和光純薬工業株式会社製、シグマアルドリッチ社製、タカラバイオ株式会社製等であり、市販で容易に入手できるものを使用することができる。試薬のメーカーや純度等は特に限定されない。
【0100】
〔実施例1〕
1.DNAの抽出
配列番号1のアミノ酸配列(スフィンゴミエリナーゼ)を含むタンパク質を発現するStreptomyces sp. A9107(FERM P−22058)の菌体を50mM Tris/HCl(pH8.0)、50mM EDTA、15%シュークロースを含む1mg/mLリゾチーム溶液で37℃、10分処理した後、SDSを最終濃度0.25%になるように添加して菌体を溶解した。さらに等量のフェノール/クロロホルムの1:1混合液を加え、30分撹拌した後、遠心分離(12,000rpm、15分間)して水層を回収した。回収した水層に10分の1量の3Mの酢酸ナトリウム(pH5.5)を混合後、2倍量のエタノールを静かに重層し、ゲノムDNAをガラス棒に巻き付かせて分離した。分離したゲノムDNAを、10mM Tris/HCl(pH8.0)、1mM EDTA水溶液(以下「TE」と記載する場合もある)に溶解し、適量のRNaseAを加え、37℃で1時間保温し、混在しているRNAを分解した。次いで、等量のフェノール/クロロホルムの1:1混合液を加え、前記と同様に処理して、水層を分取した。分取した水層に10分の1量の3Mの酢酸ナトリウム(pH5.5)と2倍量のエタノールを加えて前記の方法でもう一度ゲノムDNAを分離した。このゲノムDNA(染色体)をTEに溶解し、TE飽和のフェノールとクロロホルムの1:1混合液を加え、全体を懸濁した後、同様の遠心分離を繰り返し、上層を再び別の容器に移した。この分離した上層に3Mの酢酸ナトリウム(pH5.5)とエタノールを加え、撹拌後、−70℃で5分間冷却した後、遠心分離(2,000G、4℃、15分)し、沈殿したゲノムDNAを75%エタノールで洗い、減圧乾燥した。以上の操作によりStreptomyces sp.のDNA標品を得た。
【0101】
2.精製酵素の取得
容量500mLの三角フラスコに、前培養用培地として培地A(3% 脱脂大豆ニッカミルキー(日華油脂社製)、2% コーンスターチ、0.3% 塩化カリウム、0.4% 炭酸カルシウム、pH7.0)100mLを入れて、蒸気加圧滅菌した後、Streptomyces sp. A9107の菌体を白金耳で植菌した。次いで25℃で72時間振とう培養することにより、前培養液を得た。容量30Lのジャーに、培地Aを20L入れ、121℃で20分間蒸気加圧滅菌した後、上記の前培養液を20mL植菌し、25℃、125rpmで96時間通気培養を行った。培養後、遠心分離して培養上清を得た。この培養上清を20mM 酢酸ナトリウム緩衝液を用いてpH5.0に調製し、20mM 酢酸ナトリウム緩衝液(pH5.0)で平衡化したGEヘルスケア社製のCM Sepharose Fast Flowを用いたカラムに吸着させた。20mM 酢酸ナトリウム緩衝液(pH5.5)で充分に洗浄した後、20mM 酢酸ナトリウム緩衝液(pH5.5)と100mM 酢酸ナトリウム緩衝液(pH5.5)を用いてベッド体積の10倍量のリニアグラジエントで溶出を行った。活性画分を回収し、100倍量の10mM 酢酸ナトリウム緩衝液(pH5.5)で透析し、スフィンゴミエリナーゼ(SPC)の精製酵素を得た。
【0102】
3.酵素の部分アミノ酸配列決定
「実施例1 2.」で取得した精製酵素の一部を、Laemmliの方法で5〜20%のポリアクリルアミドグラジエントゲルを用いてドデシル硫酸ナトリウム・ポリアクリルアミド電気泳動(SDS−PAGE)を行った後、ポリフッ化ビニリデン(PVDF)膜に転写した。これを用いてN末端アミノ酸配列解析を行った結果、配列番号7のアミノ酸配列を得た。また、「実施例1 2.」で取得した精製酵素の一部を消化酵素トリプシンにより処理し、上記と同様に操作を行い、N末端アミノ酸配列解析を行った結果、配列番号8のアミノ酸配列を得た。
【0103】
4.酵素遺伝子の部分塩基配列決定
「実施例1 3.」で得られた配列番号7及び8のアミノ酸配列を元に配列番号9及び10の塩基配列を有するPCR用DNAプライマーを設計した。このDNAプライマーを用い、「実施例1 1.」で調製したStreptomyces sp.のDNA標品を鋳型として常法に従ってPCR反応を行い、約450bpのDNA断片を取得した。このDNA断片をDNAシーケンスして部分塩基配列を決定した。
【0104】
5.酵素遺伝子の塩基配列決定
「実施例1 4.」で決定した部分塩基配列を元に、配列番号11及び12の塩基配列を有するPCR用DNAプライマーを設計した。「実施例1 1.」で調製したStreptomyces sp.のDNA標品の一部をSalIにより制限酵素処理した。制限酵素処理したDNA断片を常法に従ってライゲーションし、環状DNAを得た。配列番号9及び10の塩基配列を有するDNAプライマーを用いてSalI切断環状DNAを鋳型としてPCR反応を行い、約850bpのDNA断片を取得した。このDNA断片をDNAシーケンスして塩基配列を決定した。「実施例1 1.」で調製したStreptomyces sp.のDNA標品の一部をBamHIにより制限酵素処理した。制限酵素処理したDNA断片をライゲーションし、環状DNAを得た。配列番号11及び12の塩基配列を有するDNAプライマーを用いてBamHI切断環状DNAを鋳型として常法に従ってPCR反応を行い、約800bpのDNA断片を取得した。このDNA断片を常法に従ってDNAシーケンスして塩基配列を決定した。決定した塩基配列は、配列番号6(配列番号2のアミノ酸配列をコードする遺伝子配列)と、配列番号5(配列番号1のアミノ酸配列をコードする遺伝子配列)とが順に結合した塩基配列を含む塩基配列であった。
【0105】
6.PCR法による酵素遺伝子の増幅
「実施例1 5.」で決定した塩基配列を元に、配列番号13及び14の塩基配列を有するPCR用DNAプライマーを設計した。このDNAプライマーを用い、「実施例1 1.」で調製したStreptomyces sp.のDNA標品を鋳型として常法に従ってPCR反応を行った。得られた約1200bpのPCR産物は、例えばQIAGEN QIAquick PCR Purification Kitを用い精製した。
【0106】
7.発現ベクターとのライゲーション
「実施例1 6.」で取得したPCR産物を、NdeI及びHindIIIにより制限酵素処理した。このDNA断片は、例えばQIAGEN QIAquick PCR Purification Kitを用いる等、常法に従って精製した。精製したDNA断片は、NdeI及びHindIIIにより制限酵素処理し精製したpTip QC2と常法に従ってライゲーションし、配列番号15の塩基配列を有するpTip QC2/SPCを作製した。作製したpTip QC2/SPCはDNAシーケンスして挿入した塩基配列が正しいことを確認した。
【0107】
8.Rhodococcus erythropolis L88株の形質転換
Rhodococcus erythropolis L88株(FERM BP−08444)の菌体を、「実施例1 7.」で作製したpTip QC2/SPCと混合し、氷上で30分静置した後、バイオ・ラッド社のGene Pulser IIを用いてエレクトロポレーション法(2.5kV、25μF、400Ω)により形質転換した。形質転換したRhodococcus erythropolis L88株をSOC培地(2% トリプトン、0.5% 酵母エキス、10mM 塩化ナトリウム、25mM 塩化カリウム、10mM 塩化マグネシウム、10mM 硫酸マグネシウム、20mM グルコース)に懸濁した後、30℃で1時間培養し、34μg/mLのクロラムフェニコールを含むLB寒天培地(1% トリプトン、0.5% 酵母エキス、1% 塩化ナトリウム、1.5% アガー)に塗布して30℃で72時間培養し、pTip QC2/SPCにより形質転換したRhodococcus erythropolis L88株(L88/SPC)を得た。
【0108】
9.L88/SPCの培養
容量500mLの三角フラスコに、前培養用培地としてLB液体培地(1% トリプトン、0.5% 酵母エキス、1% 塩化ナトリウム)100mLを入れて、蒸気加圧滅菌した後、「実施例1 8.」で形質転換したRhodococcus erythropolis L88株(L88/SPC)を白金耳で植菌した。次いで30℃で72時間振とう培養することにより、前培養液を得た。容量2Lのジャーに、LB液体培地を1.6L入れ、121℃で20分間蒸気加圧滅菌した後、34mg/mLのクロラムフェニコールを1.6mL、40mg/mLのチオストレプトンを80μL添加し、上記の前培養液を1.6mL植菌し、30℃、300rpmで72時間通気培養を行った。培養後、遠心分離して培養上清を得た。3基培養を行ったところ、スフィンゴミエリナーゼの培養力価は50〜52U/mLであった。培養力価は、上述の<スフィンゴミエリナーゼ活性測定法>に記載の方法を用いて、培養上清を被験液として(式2)を用いて算出した。
【0109】
10.精製
「実施例1 9.」で得られた培養上清の一部を20mM 酢酸ナトリウム緩衝液を用いてpH5.0に調製し、20mM 酢酸ナトリウム緩衝液(pH5.0)で平衡化したGEヘルスケア社製のCM Sepharose Fast Flowを用いたカラムに吸着させた。20mM 酢酸ナトリウム緩衝液(pH5.5)で充分に洗浄した後、20mM 酢酸ナトリウム緩衝液(pH5.5)と100mM 酢酸ナトリウム緩衝液(pH5.5)を用いてベッド体積の10倍量のリニアグラジエントで溶出を行った。活性画分を回収し、100倍量の10mM 酢酸ナトリウム緩衝液(pH5.5)で透析した後、凍結乾燥して粉末状の精製酵素(スフィンゴミエリナーゼ)約80mgを得た。
【0110】
〔比較例1〕
1.Streptomyces sp.の培養
容量500mLの三角フラスコに、前培養用培地として培地A(実施例1と同様)100mLを入れて、蒸気加圧滅菌した後、「実施例1 1.」に記載のStreptomyces sp.の菌体を白金耳で植菌した。次いで25℃で72時間振とう培養することにより、前培養液を得た。容量30Lのジャーに、培地Aを20L入れ、121℃で20分間蒸気加圧滅菌した後、上記の前培養液を20mL植菌し、25℃、125rpmで96時間通気培養を行った。培養後、遠心分離して培養上清を得た。2基培養を行ったところ、スフィンゴミエリナーゼの培養力価は62〜67U/mLであった。
【0111】
2.精製
「比較例1 1.」で取得した培養上清を20mM 酢酸ナトリウム緩衝液を用いてpH5.0に調製し、20mM 酢酸ナトリウム緩衝液(pH5.0)で平衡化したGEヘルスケア社製のCM Sepharose Fast Flowを用いたカラムに吸着させた。100mM 酢酸ナトリウム緩衝液(pH5.5)を用いて溶出し、活性画分を回収した。活性画分をゲル濾過により脱塩し、その一部にマンニトールを重量パーセント濃度で1%となるように添加した後、凍結乾燥して粉末状の比較試験用酵素(野生型スフィンゴミエリナーゼ)約950mgを得た。
【0112】
〔比較例2〕
1.挿入遺伝子の調製
配列番号16の塩基配列を有するDNAプライマーを設計した。このDNAプライマーと配列番号13の塩基配列を有するDNAプライマーとを用いてpTip QC2/SPCを鋳型として常法に従ってPCR反応を行い、DNA断片A(配列番号6の塩基配列(配列番号2のアミノ酸配列をコードする遺伝子配列)を含む)を得た。また、配列番号17及び18の塩基配列を有するDNAプライマーを設計した。このDNAプライマーを用いて配列番号19の塩基配列を有するDNA(カンジダ シリンドラセ(Candida cylindracea)(ATCC14830)由来のコレステロールエステラーゼ(CEBP)をコードする塩基配列)を鋳型として常法に従ってPCR反応を行い、DNA断片Bを得た。配列番号13及び18の塩基配列を有するDNAプライマーを用いてDNA断片AとDNA断片Bを混合したものを鋳型として常法に従ってPCR反応を行い、配列番号20の塩基配列を有するDNA断片Cを得た。DNA断片Cを得るまでのPCR反応の模式図を図1に示す。得られたPCR産物は、例えばQIAGEN QIAquick PCR Purification Kitを用い精製した。
【0113】
2.発現ベクターとのライゲーション
「比較例1 1.」で取得したPCR産物をNdeI及びHindIIIにより制限酵素処理した。このDNA断片は、例えばQIAGEN QIAquick PCR Purification Kitを用いて精製した。精製したDNA断片は、NdeI及びHindIIIにより制限酵素処理し精製したpTip QC2とライゲーションし、配列番号21の塩基配列を有するpTip QC2/CEBPを作製した。作製したpTip QC2/CEBPは常法に従ってDNAシーケンスして挿入した塩基配列が正しいことを確認した。
【0114】
3.Rhodococcus erythropolis L88株の形質転換
「実施例1 7.」と同様の方法で、「比較例2 2.」で作製したpTip QC2/CEBPによりRhodococcus erythropolis L88株を形質転換し、pTip QC2/CEBPにより形質転換したRhodococcus erythropolis L88株(L88/CEBP)を得た。
【0115】
4.L88/CEBPの培養
容量500mLの三角フラスコに、前培養用培地としてLB液体培地100mLを入れて、蒸気加圧滅菌した後、「比較例2 3.」で形質転換したRhodococcus erythropolis L88株(L88/CEBP)を白金耳で植菌した。次いで30℃で72時間振とう培養することにより、前培養液を得た。容量2Lのジャーに、LB液体培地を1.6L入れ、121℃で20分間蒸気加圧滅菌した後、34mg/mLのクロラムフェニコールを1.6mL、40mg/mLのチオストレプトンを80μL添加し、上記の前培養液を1.6mL植菌し、30℃、300rpmで72時間通気培養を行った。培養後、遠心分離して培養上清を得た。しかしながら、配列番号19にコードされるCandida cylindracea由来のコレステロールエステラーゼの発現は確認できなかった。なお、コレステロールエステラーゼの発現は、コレステロールエステラーゼの活性として、ASAHI KASEI Diagnostic Enzymes(旭化成ファーマ株式会社2008年7月131ページの方法に従い確認した。
【0116】
〔比較例3〕
1.挿入遺伝子の調製
配列番号22の塩基配列を有するDNAプライマーを設計した。このDNAプライマーと配列番号13の塩基配列を有するDNAプライマーとを用いてpTip QC2/SPCを鋳型として常法に従ってPCR反応を行い、DNA断片A’(配列番号6の塩基配列(配列番号2のアミノ酸配列をコードする遺伝子配列)を含む)を得た。また、配列番号23及び24の塩基配列を有するDNAプライマーを設計した。このDNAプライマーを用いて配列番号25の塩基配列を有するDNA(バークホルデリア スタビリス(Burkholderia stabilis)(FERM P−21014)(特開2008−086277)由来のコレステロールエステラーゼ(CEN)及びその補助タンパク質をコードする塩基配列)を鋳型として常法に従ってPCR反応を行い、DNA断片B’を得た。配列番号13及び24の塩基配列を有するDNAプライマーを用いてDNA断片A’とDNA断片B’を混合したものを鋳型として常法に従ってPCR反応を行い、配列番号26の塩基配列を有するDNA断片C’を得た。得られたPCR産物は、例えばQIAGEN QIAquick PCR Purification Kitを用い精製した。
【0117】
2.発現ベクターとのライゲーション
「比較例3 1.」で取得したPCR産物に対して「比較例2 2.」と同様の操作を行い、配列番号27の塩基配列を有するpTip QC2/CENを作製した。作製したpTip QC2/CENは常法に従ってDNAシーケンスして挿入した塩基配列が正しいことを確認した。
【0118】
3.Rhodococcus erythropolis L88株の形質転換
「実施例1 7.」と同様の方法で、「比較例3 2.」で作製したpTip QC2/CENによりRhodococcus erythropolis L88株を形質転換し、pTip QC2/CENにより形質転換したRhodococcus erythropolis L88株(L88/CEN)を得た。
【0119】
4.L88/CEBPの培養
「比較例3 3.」で得たL88/CENに対して「比較例2 4.」と同様の操作を行い、培養上清を得た。しかしながら、配列番号25にコードされるBurkholderia stabilis由来のコレステロールエステラーゼの発現は確認できなかった。なお、コレステロールエステラーゼの発現は、コレステロールエステラーゼの活性として、ASAHI KASEI Diagnostic Enzymes (旭化成ファーマ株式会社2008年7月48ページの方法に従い確認した。
【0120】
〔比較例4〕
1.挿入遺伝子の調製
配列番号28の塩基配列を有するDNAプライマーを設計した。このDNAプライマーと配列番号13の塩基配列を有するDNAプライマーとを用いてpTip QC2/SPCを鋳型として常法に従ってPCR反応を行い、DNA断片A”(配列番号6の塩基配列(配列番号2のアミノ酸配列をコードする遺伝子配列)を含む)を得た。また、配列番号29及び30の塩基配列を有するDNAプライマーを設計した。このDNAプライマーを用いて配列番号31の塩基配列を有するDNA(クロモバクテリウム ビスコスム(Chromobacterium viscosum)(工発研菌寄第137号)(特公昭46−29787)由来のリパーゼ(LP)及びその補助タンパク質をコードする塩基配列)を鋳型として常法に従ってPCR反応を行い、DNA断片B”を得た。配列番号13及び30の塩基配列を有するDNAプライマーを用いてDNA断片A”とDNA断片B”を混合したものを鋳型として常法に従ってPCR反応を行い、配列番号32の塩基配列を有するDNA断片C”を得た。得られたPCR産物は、例えばQIAGEN QIAquick PCR Purification Kitを用い精製した。
【0121】
2.発現ベクターとのライゲーション
「比較例4 1.」で取得したPCR産物に対して「比較例2 2.」と同様の操作を行い、配列番号33の塩基配列を有するpTip QC2/LPを作製した。作製したpTip QC2/LPは常法に従ってDNAシーケンスして挿入した塩基配列が正しいことを確認した。
【0122】
3.Rhodococcus erythropolis L88株の形質転換
「実施例1 7.」と同様の方法で、「比較例4 2.」で作製したpTip QC2/LPによりRhodococcus erythropolis L88株を形質転換し、pTip QC2/LPにより形質転換したRhodococcus erythropolis L88株(L88/LP)を得た。
【0123】
4.L88/LPの培養
「比較例4 3.」で得たL88/LPに対して「比較例2 4.」と同様の操作を行い、培養上清を得た。しかしながら、配列番号30にコードされるChromobacterium viscosum由来のリパーゼの発現は確認できなかった。なお、リパーゼの発現は、リパーゼの活性として、ASAHI KASEI Diagnostic Enzymes(旭化成ファーマ株式会社2008年7月15ページ)に記載の方法に従い確認した。
【0124】
〔試験例1〕<比活性試験>
「実施例1 9.」で得た培養上清と、「比較例1 1.」で得た培養上清のタンパク質量あたりのスフィンゴミエリナーゼ活性を計算したところ、「実施例1 9.」で得た培養上清は142.7U/mg、「比較例1 1.」で得た培養上清は2.1U/mgであり、「実施例1 9.」で得た培養上清は、「比較例1 1.」で得た培養上清よりも比活性が70倍以上高かった。なお、培養上清中のタンパク質量は、ビシンコニン酸を用いる方法(BCA法)により測定した。スフィンゴミエリナーゼ活性は、上述の<スフィンゴミエリナーゼ活性測定法>に記載の方法を用いて、培養上清を被験液として(式2)を用いて算出した。これらの測定から、タンパク質量あたりのスフィンゴミエリナーゼ活性を計算した。
【0125】
〔試験例2〕<安定性試験1>
「実施例1 10.」で得た精製酵素、及び「比較例1 2.」で得た比較試験用酵素を、4mg/mLになるように純水(pH6.8)又は50mM 酢酸ナトリウム緩衝液(pH5.5)に溶解し、それぞれ−30℃又は30℃で静置保存した。保存した酵素溶液について、保存開始から72時間後及び96時間後に残存活性を測定した(保存前の活性を100%とする)。その結果、図2に示すように、30℃で静置保存した場合、「比較例1 2.」で得た比較試験用酵素は、72時間後及び96時間後の残存活性がいずれも40%以下であるのに対して、「実施例1 10.」で得られた精製酵素は、72時間後及び96時間後のいずれにおいても78%以上の活性を保持しており、高い保存安定性を示した。特に、pH5.5の緩衝液に溶解した場合、72時間後及び96時間後のいずれにおいても97.6%以上の活性を保持しており、非常に高い保存安定性を示した。なお、−30℃で静置保存した場合も同様に、「実施例1 10.」で得られた精製酵素は高い保存安定性を示した(図示せず)。
【0126】
〔試験例3〕<安定性試験2>
試験例2で得られた、−30℃又は30℃で96時間保存した酵素溶液の一部を、Laemmliの方法で5〜20%のポリアクリルアミドグラジエントゲルを用いてSDS−PAGEを行った後、常法に従ってクーマシーブルー(CBB)で染色した。結果を図3に示す。図3中(A)〜(H)のバンドは、それぞれ以下の処理を行った酵素溶液に対応する。
(A)比較例1の酵素を純水(pH6.8)に溶解し、−30℃で保存。
(B)比較例1の酵素を純水(pH6.8)に溶解し、30℃で保存。
(C)比較例1の酵素を酢酸ナトリウム緩衝液(pH5.5)に溶解し、−30℃で保存。
(D)比較例1の酵素を酢酸ナトリウム緩衝液(pH5.5)に溶解し、30℃で保存。
(E)実施例1の酵素を純水(pH6.8)に溶解し、−30℃で保存。
(F)実施例1の酵素を純水(pH6.8)に溶解し、30℃で保存。
(G)実施例1の酵素を酢酸ナトリウム緩衝液(pH5.5)に溶解し、−30℃で保存。
(H)実施例1の酵素を酢酸ナトリウム緩衝液(pH5.5)に溶解し、30℃で保存。
【0127】
図3に示すように、比較例1で得た比較試験用酵素は、30℃で96時間保存した場合((B)及び(D))には、−30℃で保存した場合((A)及び(C))と比較して明らかにスフィンゴミエリナーゼを示すバンドが薄くなり、分解されていた。一方、「実施例1 10.」で得た精製酵素は、30℃で96時間保存した場合((F)及び(H))も、−30℃で保存した場合((E)及び(G))と同程度のバンドが確認され、分解されていなかった。
【産業上の利用可能性】
【0128】
本発明は、スフィンゴミエリナーゼ、及び純度が高く安定性の高いスフィンゴミエリナーゼ含有組成物の効率的な製造に利用することができる。本発明は、特に、試薬、医薬品、化粧品等の分野において、産業上の利用可能性を有する。
【特許請求の範囲】
【請求項1】
スフィンゴミエリナーゼの製造方法であって、以下の工程(a)及び(b):
(a)以下の(i)又は(ii)のアミノ酸配列:
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして形質転換体を得る工程;及び
(b)工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程;
を含む方法。
【請求項2】
工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、請求項1に記載の製造方法。
【請求項3】
前記工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、(i)のアミノ酸配列をコードする塩基配列を含む、請求項1又は2に記載の製造方法。
【請求項4】
以下の工程(a)〜(c):
(a)以下の(i)又は(ii)のアミノ酸配列:
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして形質転換体を得る工程;
(b)工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程;及び
(c)工程(b)で得た培養物から形質転換体を分離してスフィンゴミエリナーゼ含有組成物を得る工程;
を含む方法により得られる、スフィンゴミエリナーゼ含有組成物。
【請求項5】
工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、請求項4に記載のスフィンゴミエリナーゼ含有組成物。
【請求項6】
前記スフィンゴミエリナーゼ含有組成物に水又は緩衝液を添加してスフィンゴミエリナーゼ濃度が4mg/mLの中性又は酸性の水溶液を調製し、30℃で96時間静置保存した場合、保存前と比較して50%以上のスフィンゴミエリナーゼ活性を保持する、請求項4又は5に記載のスフィンゴミエリナーゼ含有組成物。
【請求項7】
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子を含有する組換えベクター。
【請求項8】
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子を含有する組換えベクター。
【請求項9】
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子を有し、スフィンゴミエリナーゼを培養物中に分泌することを特徴とする、ロドコッカス属細菌形質転換体。
【請求項10】
前記遺伝子が、
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、請求項9に記載の形質転換体。
【請求項1】
スフィンゴミエリナーゼの製造方法であって、以下の工程(a)及び(b):
(a)以下の(i)又は(ii)のアミノ酸配列:
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして形質転換体を得る工程;及び
(b)工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程;
を含む方法。
【請求項2】
工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、請求項1に記載の製造方法。
【請求項3】
前記工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、(i)のアミノ酸配列をコードする塩基配列を含む、請求項1又は2に記載の製造方法。
【請求項4】
以下の工程(a)〜(c):
(a)以下の(i)又は(ii)のアミノ酸配列:
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子をロドコッカス属細菌にトランスフェクトして形質転換体を得る工程;
(b)工程(a)で得た形質転換体を培養し、スフィンゴミエリナーゼを含む培養物を得る工程;及び
(c)工程(b)で得た培養物から形質転換体を分離してスフィンゴミエリナーゼ含有組成物を得る工程;
を含む方法により得られる、スフィンゴミエリナーゼ含有組成物。
【請求項5】
工程(a)において、ロドコッカス属細菌にトランスフェクトする遺伝子が、
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、請求項4に記載のスフィンゴミエリナーゼ含有組成物。
【請求項6】
前記スフィンゴミエリナーゼ含有組成物に水又は緩衝液を添加してスフィンゴミエリナーゼ濃度が4mg/mLの中性又は酸性の水溶液を調製し、30℃で96時間静置保存した場合、保存前と比較して50%以上のスフィンゴミエリナーゼ活性を保持する、請求項4又は5に記載のスフィンゴミエリナーゼ含有組成物。
【請求項7】
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子を含有する組換えベクター。
【請求項8】
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子を含有する組換えベクター。
【請求項9】
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
をコードする塩基配列を含む遺伝子を有し、スフィンゴミエリナーゼを培養物中に分泌することを特徴とする、ロドコッカス属細菌形質転換体。
【請求項10】
前記遺伝子が、
(i)配列番号1に記載のアミノ酸配列、又は
(ii)配列番号1に記載のアミノ酸配列において、1若しくは複数のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつ該アミノ酸配列からなるタンパク質がスフィンゴミエリナーゼ活性を有するアミノ酸配列
のN末端と、
(iii)配列番号2に記載のアミノ酸配列(シグナルペプチド)、又は
(iv)配列番号2のアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列であって、かつシグナルペプチドとして機能するアミノ酸配列
のC末端とが、直接又は1若しくは数個のアミノ酸を介して結合したアミノ酸配列をコードする塩基配列を含む遺伝子である、請求項9に記載の形質転換体。
【図1】

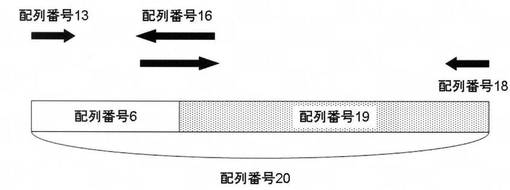
【図2】
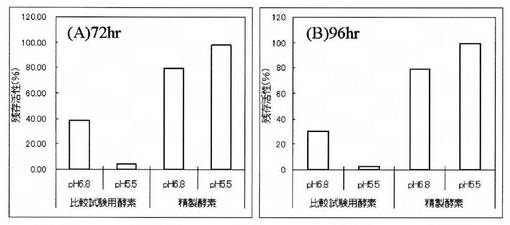
【図3】
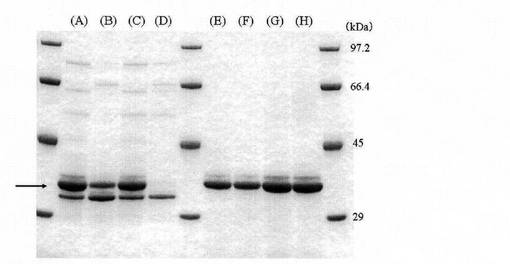
【図2】
【図3】
【公開番号】特開2012−231720(P2012−231720A)
【公開日】平成24年11月29日(2012.11.29)
【国際特許分類】
【出願番号】特願2011−101810(P2011−101810)
【出願日】平成23年4月28日(2011.4.28)
【出願人】(303046299)旭化成ファーマ株式会社 (105)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成24年11月29日(2012.11.29)
【国際特許分類】
【出願日】平成23年4月28日(2011.4.28)
【出願人】(303046299)旭化成ファーマ株式会社 (105)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]