
ヒト化免疫グロブリン遺伝子座
【課題】ヒト化抗体を発現するトランスジェニック非ヒト動物を作製することにおいて有用な、新規の免疫グロブリン重鎖および免疫グロブリン軽鎖の構築物、組み換えおよびトランスジェニックベクター、トランスジェニック動物、ならびにヒト化免疫グロブリン調製物を提供する。
【解決手段】本発明は、複数のヌクレオチド配列が隣接するヒト免疫グロブリン遺伝子セグメントを含む単離された核酸分子に関する。
【解決手段】本発明は、複数のヌクレオチド配列が隣接するヒト免疫グロブリン遺伝子セグメントを含む単離された核酸分子に関する。
【発明の詳細な説明】
【技術分野】
【0001】
特許協力条約の実施細則の801(a)(i)およびPCT規則5.2に従い、本出願は、2004年7月15日に1.489KBのサイズで作成された、ファイル39691−0007PCTとして保存された配列表を含むコンパクトディスクと、配列表の紙コピーに代わる2部のコピーによって行われる。その内容全体が、引用により本明細書中に明確に組み込まれる。
【0002】
(発明の分野)
本発明は、非ヒトトランスジェニック動物からヒト化抗体を生産させるための方法および手段に関する。本発明は、具体的には、新規の免疫グロブリン重鎖および軽鎖構築物、ヒト化抗体を発現する非ヒトトランスジェニック動物を作成することにおいて有用である組み換えのトランスジェニックベクター、トランスジェニック動物、ならびにヒト化免疫グロブリン調製物に関する。トランスジェニックベクターには、ヒト化された免疫グロブリン遺伝子座が含まれる。これには、非ヒト動物内因性の調節機構および抗体生産機構を本質的には完全なまま残し、多様化させられたヒト化抗体を生じるように、非ヒトトランスジェニック動物中で、遺伝子再構成、遺伝子変換、および超変異を受けさせることができる。得られるヒト化抗体は、ヒトに対する最少の免疫原性を有しており、ヒト患者の治療的処置における使用に適している。
【背景技術】
【0003】
(発明の背景)
抗体は、薬学的産物の1つの重要なクラスである。これは、ガン、アレルギー性疾患のような種々のヒトの疾患および症状の処置、移植による拒絶反応、宿主対移植片反応の予防において良好に使用されている。
【0004】
動物から得られる抗体調製物の重要な問題点は、ヒト患者において本来備わっている、非ヒト免疫グロブリンについての免疫原性である。非ヒト抗体の免疫原性を低下させるためには、動物の可変(V)領域のエキソンをヒト定常(C)領域のエキソンと融合させることにより、キメラ抗体遺伝子を得ることができることが示されている。
【0005】
ヒト化モノクローナル抗体もまた開発されており、これは、臨床的に使用されている。ヒト化モノクローナル抗体は、通常は、ヒト抗体であり、その中のいくつかのCDR残基と、場合によってはいくつかのFR残基が、非ヒト動物、例えば、齧歯類の抗体の中に類似している部位に由来する残基で置換されている。ヒト化は、本質的には、Winterとその共同研究者らの方法に従って(Jones et al.,Nature,321:522(1986);Riechmann et al.,Nature,332:323(1988);Verhoeyen et al.,Science,239:1534(1988))、非ヒト動物、例えば、齧歯類のCDRまたはCDR配列を、ヒトのモノクローナル抗体の対応する配列で置換することによって、行うことができる。
【0006】
キメラの生殖細胞系列変異体マウスにおける抗体重鎖−結合領域(JH)遺伝子のホモ接合体欠失により、内因性抗体の生産の完全な阻害が生じることが記載されている。ヒトの生殖細胞系列の免疫グロブリン遺伝子配列を、そのような生殖細胞系列変異体マウスに導入することにより、抗原でのチャレンジの際にヒト抗体の生産が生じる。例えば、Jakobovits et al.,Proc.Natl.Acad.Sci.USA,90:2551(1993);Jakobovits et al.,Nature,362:255(1993);Bruggemann et al.,Year in Immunol.,7:33(1993)を参照のこと。この遺伝子操作によるアプローチによって、遺伝子操作されたマウス中でヒト免疫グロブリンポリペプチドの発現が生じるが、ヒト免疫グロブリンの発現レベルは低い。これは、免疫グロブリンの効率的な発現に不可欠である免疫グロブリン遺伝子座の中の種特異的調節エレメントが原因である。トランスフェクトされた細胞株において示されるように、ヒトの免疫グロブリン遺伝子中に存在する調節エレメントは、非ヒト動物においては適正には機能しない場合がある。
【0007】
実際に、免疫グロブリン遺伝子中のいくつかの調節エレメントが記載されている。特に注目すべきは、重鎖定常領域の下流(3’)のエンハンサーと、軽鎖遺伝子中にあるイントロンのエンハンサーである。さらに、他の、すでに同定されている制御エレメントもまた、免疫グロブリン遺伝子中に存在し得る。マウスでの研究では、免疫グロブリン分子の膜と、膜形態の細胞質尾部が、ヒトCγ1遺伝子についてホモ接合型であるマウスの血清中でのヒト−マウスキメラ抗体の発現レベルにおいて重要な役割を果たしていることが示されている。したがって、動物中でのヘテロ接合型免疫グロブリン遺伝子の発現については、エンハンサーエレメントと、膜貫通部分(M1エキソン)および細胞質尾部(M2エキソン)をコードするエキソンを含む配列を、動物において通常見ることができる配列と、同様の位置で、置き換えることが望まれる。
【0008】
非ヒト抗体の潜在的な免疫原性によって生じる問題に加えて、モノクローナル抗体の使用は、通常は、キメラ抗体、ヒト化抗体、またはヒト抗体を問わず、ガンのような破壊的な疾患、および伝染力の強い病原体での感染が、その複雑さ、様々な原因によるとされる病因論、および適応性にあることが原因で1つの標的を攻撃することによって処置することは困難であるという事実によって、さらに限定される。著しく定義された標的に対して指向させられたモノクローナル抗体は、その標的が変化する、進化する、および突然変異すると、うまくいかない。したがって、悪性腫瘍は、標準的なモノクローナル抗体での治療に対して耐性を獲得する場合がある。この問題の解決方法は、ポリクローナル抗体の使用である。ポリクローナル抗体は、複雑な疾患に関係している複数の進化しつつある標的を標的化して、攻撃する能力を有している。ポリクローナル抗体は、また、細菌やウイルスの毒素を中和する能力も有しており、病原体を死滅させ、排除するための免疫反応を指示する。
【0009】
したがって、高力価の、親和性の高いヒト化ポリクローナル抗体およびモノクローナル抗体の大量生産に適している新しいアプローチが、臨床的に非常に必要とされている。
【0010】
マウスのゲノム中にヒト免疫グロブリン遺伝子を導入することにより、遺伝子操作されたマウスにおいて多様化させられたヒト抗体のレパートリーの発現が生じる。マウスとヒトのいずれにおいても、主な抗体の多様性は、遺伝子再構成によって作成される。このプロセスにより、種々の抗原結合部位を有している多数の抗体分子をコードする多数の異なる組み換えV(D)Jセグメントが作成される。しかし、ウサギ、ブタ、ウシ、および鳥類のような他の動物においては、主な抗体の多様性は、実質的に異なる機構、すなわち、鋳型を用いる突然変異、または遺伝子変換、および鋳型を用いない突然変異、または超変異によって生じる。例えば、ウサギとニワトリにおいては、VDJ再構成が極めて限られており(免疫グロブリンのおよそ90%が3’近傍にVH1エレメントを伴って生じる)、抗体の多様性は、遺伝子変換と超変異によって生じる。対照的に、マウスとヒトの遺伝子変換は、仮にあったとしても極めて稀に生じる。したがって、遺伝子変換および超変異によってそれらの主な抗体のレパートリーを多様化させる動物においては、遺伝子の再構成に基づく遺伝子操作アプローチにより、低い抗体力価と限られた抗体多様性を有している動物が得られる。したがって、ヒトの治療のための非免疫原性の抗体調製物を生産するための大動物の遺伝子操作には、別の遺伝子操作ストラテジーが必要である。
【0011】
非ヒトトランスジェニック動物中でのヒト化抗体の生産は、2002年2月14日に公開されたPCT公開番号WO02/12437に記載されている。この開示は、その全体を参照することにより明確に本明細書中に組み入れられる。WO02/12437には、1つ以上のヒト化免疫グロブリン遺伝子座を含む、遺伝子操作された非ヒト動物が記載されている。ヒト化免疫グロブリン遺伝子座は、非ヒトトランスジェニック動物中で遺伝子再構成と遺伝子変換を受けることができる。非ヒトトランスジェニック動物には、その中での抗体多様性が主に遺伝子変換によって生じて、多様化されたヒト化抗体を生じる動物が含まれる。得られたヒト化抗体は、ヒトに対して最少の免疫原性を有しており、ヒト患者の治療的処置における使用に適している。これには、さらに、種々の非ヒト哺乳動物、例えば、ニワトリ、ウシ、ヒツジ、およびウサギの免疫グロブリン重鎖定常領域セグメントの5’および3’隣接領域に由来する新規のヌクレオチド配列が記載されている。ヒト免疫グロブリン重鎖遺伝子セグメントに、そのような5’および3’配列に相同な配列が隣接している組み換えベクターは、非ヒト動物の免疫グロブリン重鎖遺伝子セグメントを、対応するヒト免疫グロブリン重鎖遺伝子セグメントで置き換えるために有用であることが示されている。
【発明の概要】
【課題を解決するための手段】
【0012】
(発明の要旨)
1つの局面においては、本発明は、複数のヌクレオチド配列が隣接するヒト免疫グロブリン遺伝子セグメントを含む単離された核酸分子に関する。ここで、隣接配列は、同じである場合も、異なる場合もあり、これには、遺伝子変換および/または超変異によって主に抗体多様性を生じる動物の免疫グロブリン重鎖もしくは軽鎖遺伝子に由来するスペーサー配列の、あるいは、2つ以上のスペーサー配列のコンセンサス配列に由来する、少なくとも約20個の連続しているヌクレオチドが含まれる。
【0013】
別の局面においては、本発明は、複数のヌクレオチド配列が隣接している、ヒト免疫グロブリン重鎖または軽鎖定常領域(C)遺伝子セグメントを含む単離された核酸分子に関する。ここで、隣接配列は、同じである場合も、異なる場合もあり、これには、非霊長類動物の免疫グロブリン重鎖もしくは軽鎖遺伝子に由来するスペーサー配列の、あるいは、2つ以上のスペーサー配列のコンセンサス配列に由来する、少なくとも約20個の連続しているヌクレオチドが含まれる。
【0014】
さらなる局面においては、本発明は、複数のヌクレオチド配列が隣接するヒト免疫グロブリン重鎖または軽鎖遺伝子セグメントを含む単離された核酸分子に関する。ここで、隣接配列は、同じである場合も、異なる場合もあり、これには、配列番号1から185(表1)からなる群より選択されるスペーサー配列の、あるいは、2つ以上のスペーサー配列のコンセンサス配列に由来する、少なくとも約20個の連続しているヌクレオチドが含まれる。
【0015】
1つの実施形態においては、隣接配列には、スペーサー配列の少なくとも約50個の連続しているヌクレオチドが含まれる。
【0016】
別の実施形態においては、ヒト免疫グロブリン遺伝子セグメントは、重鎖のV、D、またはJセグメントであり、Vセグメントは、例えば、VH3、VH1、VH5、またはVH4ファミリーのメンバーであり得る。
【0017】
さらなる実施形態においては、ヒト免疫グロブリン遺伝子セグメントは、軽鎖VまたはJセグメントであり、Vセグメントは、例えば、κ軽鎖遺伝子セグメント、例えば、Vκ1、Vκ3、またはVκ4である場合も、また、λ軽鎖セグメント、例えば、Vλ1、Vλ2、またはVλ3である場合もある。
【0018】
さらなる実施形態においては、主に遺伝子変換および/または体細胞超変異により抗体多様性を生じる非霊長類動物とは、例えば、ウサギ、ブタ、鳥類(例えば、ニワトリ、シチメンチョウ、アヒル、あるいはガチョウ)、ヒツジ、ヤギ、ウシ、ウマ、またはロバであるが、霊長類ではない他の動物、例えば、齧歯類もまた、本発明の範囲に具体的に含まれる。
【0019】
別の局面においては、本発明は、上記の核酸分子のいずれかを含む組み換えベクターに関する。
【0020】
さらに別の局面においては、本発明は、ヒト化免疫グロブリン(Ig)遺伝子座を含むトランスジェニックベクターに関する。ここで、ヒト化Ig遺伝子座は、非ヒト動物のIg遺伝子座またはIg遺伝子座の一部に由来するものであり、これには、複数のIgセグメントが含まれる。ここで、
(a)少なくとも1つの遺伝子セグメントが、スペーサー配列に由来するか、または2つ以上のそのようなスペーサー配列のコンセンサス配列に由来する少なくとも20個の連続しているヌクレオチドを含むヌクレオチド配列が隣接しているヒトIg遺伝子セグメントであり;
(b)複数の遺伝子セグメントが、再構成されていない立体配置、部分的に再構成された立体配置、または完全に再構成された立体配置において並列しており;そして
(c)非ヒト動物が遺伝子を変換する動物であり、非ヒト動物中でヒト化された免疫グロブリンのレパートリーを生産する場合には、ヒト化Ig遺伝子座が遺伝子再構成を受けることができ、必要に応じて、さらに遺伝子変換および/または超変異を受けることができる。
【0021】
さらなる実施形態においては、トランスジェニックベクター中に存在するヒト化Ig重鎖遺伝子座は、約5から100個のV遺伝子セグメントを含み、これには少なくとも1つのヒトV遺伝子セグメントが含まれる。特異的な実施形態においては、ヒト化Ig重鎖遺伝子座には、1つ以上のヒトV遺伝子セグメントが含まれる。
【0022】
別の実施形態においては、トランスジェニックベクター中に存在するヒト化Ig重鎖遺伝子座には、約5から25個のDの遺伝子セグメントが含まれる。特異的な実施形態においては、ヒト化Ig重鎖遺伝子座には、1つまたはいくつかのヒトD遺伝子セグメントが含まれる。
【0023】
さらに別の実施形態においては、トランスジェニックベクター中に存在するヒト化Ig重鎖遺伝子座には、約1から10個のJ遺伝子セグメントが含まれ、これには少なくとも1つのヒトJ遺伝子セグメントが含まれる。特異的な実施形態においては、ヒト化Ig重鎖遺伝子座には、1つ以上のヒトJ遺伝子セグメントが含まれる。
【0024】
別の実施形態においては、トランスジェニックベクター中に存在するヒト化Ig重鎖遺伝子座には、約1から25個のC領域セグメントが含まれ、これには少なくとも1つのヒトC領域セグメントが含まれる。特異的な実施形態においては、トランスジェニックベクター中に存在するヒト化Ig重鎖遺伝子座には、1つ以上のヒトC遺伝子セグメントが含まれる。
【0025】
なおさらに別の実施形態においては、トランスジェニックベクター中に存在するヒト化Ig遺伝子座は、非ヒト動物の軽鎖遺伝子座であり、これには、少なくとも1つのV遺伝子セグメントと、少なくとも1つのJ遺伝子セグメントと、少なくとも1つの定常(C)領域遺伝子セグメントが含まれる。ここで、少なくとも1つの遺伝子セグメントは、ヒト軽鎖VおよびJセグメント、ならびにヒト軽鎖C領域セグメントの群より選択される。特異的な実施形態においては、定常領域遺伝子セグメントは、ヒト軽鎖定常領域遺伝子セグメントであり、これは、例えば、CλまたはCκ遺伝子セグメントであり得る。別の実施形態においては、ヒト化Ig軽鎖遺伝子座には、ヒトVおよびJセグメント、ならびにヒトC領域セグメントより選択される2つ以上のセグメントが含まれる。さらなる実施形態においては、ヒト化Ig軽鎖遺伝子座には、少なくとも1つのヒトVセグメントと、少なくとも1つのヒトJセグメントと、少なくとも1つのヒトC領域セグメントが含まれる。
【0026】
さらなる実施形態においては、トランスジェニックベクター中に存在するヒト化Ig軽鎖遺伝子座には、約5から100個のV遺伝子セグメントが含まれ、これには少なくとも1つのヒトV遺伝子セグメントが含まれる。ここで、ヒトV遺伝子セグメントは、非ヒト動物の5から100個のV遺伝子セグメントに対して下流に存在する。特異的な実施形態においては、ヒトV遺伝子セグメントは、再構成された立体配置においてJ遺伝子セグメントのすぐ5’側に存在する。別の実施形態においては、トランスジェニックベクター中に存在するヒト化Ig軽鎖遺伝子座には、1つ以上のヒトV遺伝子セグメントが含まれる。
【0027】
なお別の実施形態においては、トランスジェニックベクター中に存在するヒト化Ig軽鎖遺伝子座には、約1から10個のJ遺伝子セグメントが含まれ、これには少なくとも1つのヒトJ遺伝子セグメントが含まれる。特異的な実施形態においては、トランスジェニックベクター中に存在するヒト化Ig軽鎖遺伝子座には、1つ以上のヒトJ遺伝子セグメントが含まれる。
【0028】
別の実施形態においては、トランスジェニックベクター中に存在するヒト化Ig軽鎖遺伝子座には、約1から25個のC領域セグメントが含まれ、これには少なくとも1つのヒトC領域セグメントが含まれる。特異的な実施形態においては、トランスジェニックベクター中に存在するヒト化Ig軽鎖遺伝子座には、1つ以上のヒトC遺伝子セグメントが含まれる。
【0029】
なおさらなる実施形態においては、トランスジェニックベクター中に存在するヒト化Ig遺伝子座は、非ヒト動物の軽鎖遺伝子座であり、これには、少なくとも1つのV遺伝子セグメントと、少なくとも1つのJ遺伝子セグメントと、少なくとも1つの定常(C)領域遺伝子セグメントが含まれる。ここで、少なくとも1つの遺伝子セグメントは、ヒト軽鎖VおよびJセグメント、ならびにヒト軽鎖C領域セグメントの群より選択される。特異的な実施形態においては、定常領域遺伝子セグメントは、ヒト軽鎖定常領域遺伝子セグメントであり、これは、例えば、CλまたはCκ遺伝子セグメントであり得る。別の実施形態においては、ヒト化Ig軽鎖遺伝子座には、ヒトVおよびJセグメント、ならびにヒトC領域セグメントより選択される2つ以上のセグメントが含まれる。さらなる実施形態においては、ヒト化Ig軽鎖遺伝子座には、少なくとも1つのヒトVセグメントと、少なくとも1つのヒトJセグメントと、少なくとも1つのヒトC領域セグメントが含まれる。
【0030】
異なる局面においては、本発明は、5’から3’の方向で、5’ヌクレオチド配列、ヒト免疫グロブリン配列、そして3’ヌクレオチド配列からなる2つ以上のユニットを含む核酸分子に関する。ここで、5’ヌクレオチド配列と3’ヌクレオチド配列は同じである場合も、異なる場合もあり、これには、非霊長類動物の免疫グロブリン重鎖もしくは軽鎖遺伝子中のコード領域の間隔を隔てるスペーサー配列に由来するか、あるいは、2つ以上のスペーサー配列のコンセンサス配列に由来する、少なくとも約20個の連続しているヌクレオチドが含まれる。特異的な実施形態においては、スペーサー配列は、配列番号1から185(表1)の中から選択される。別の特定の実施形態においては、核酸分子の全ての反復ユニット中の5’ヌクレオチド配列および/または3’ヌクレオチド配列は同じである。別の特定の実施形態においては、核酸分子の反復ユニットには、少なくとも2つの異なる5’配列および/または3’配列が含まれる。さらなる実施形態においては、5’ヌクレオチド配列と3’ヌクレオチド配列は互いに異なるが、全ての5’ヌクレオチド配列は同一であり、全ての3’配列も同一である。
【0031】
さらなる局面においては、本発明は、非ヒト動物中でヒト化抗体の機能的なレパートリーを生産することができるヒト化免疫グロブリン(Ig)遺伝子座を含むトランスジェニックベクターを作成するための方法に関する。この方法には:
(a)少なくとも1つのV遺伝子セグメントと、少なくとも1つのJ遺伝子セグメントと、少なくとも1つの定常領域遺伝子セグメントを含む、非ヒト動物に由来するIg遺伝子座またはその一部を含むDNAフラグメントを得る工程;ならびに、
(b)ヒト化Ig遺伝子座を生産するように、工程(a)のDNAフラグメント中に少なくとも1つのヒトIg遺伝子セグメントを組み込む工程であって;ここで、ヒトIg遺伝子セグメントには、非霊長類動物の免疫グロブリン重鎖もしくは軽鎖遺伝子中のコード領域の間隔を隔てるスペーサー配列に由来するスペーサー配列か、あるいは、2つ以上のそのようなスペーサー配列のコンセンサス配列に由来する、少なくとも約20個の連続しているヌクレオチドを含むヌクレオチド配列が隣接しており;ここで、(i)複数の遺伝子セグメントが、再構成されていない立体配置、部分的に再構成された立体配置、または完全に再構成された立体配置において並列しており;そして(ii)ヒト化Ig遺伝子座が遺伝子再構成を受けることができ、そして、必要に応じて、非ヒト動物中でヒト化免疫グロブリンのレパートリーを生産することができる、工程
が含まれる。
【0032】
ヒト化Ig遺伝子座は、ヒト化Ig重鎖または軽鎖遺伝子座であり得る。ヒト化Ig重鎖遺伝子座の場合には、工程(a)で得られたDNAフラグメントには、さらに、少なくとも1つのD遺伝子セグメントが含まれる。
【0033】
別の局面においては、本発明は、上記のヒト化免疫グロブリン遺伝子座を含むトランスジェニック動物と、そのようなトランスジェニック動物を作成するための方法に関する。1つの実施形態においては、トランスジェニック動物には、ヒト化免疫グロブリン重鎖遺伝子座と、ヒト化免疫グロブリン軽鎖遺伝子座の両方が含まれる。別の実施形態においては、トランスジェニック動物中に存在する重鎖遺伝子座と軽鎖遺伝子座のうち1つだけが、ヒト化されている。別の実施局面においては、動物の免疫グロブリン遺伝子座の少なくとも1つのV、D、J、およびC領域の全てがヒト化されている。さらに別の実施形態においては、トランスジェニック動物の内因性免疫グロブリン遺伝子座のV、D、J、およびC領域の全てがヒト化されている。
【0034】
さらなる局面においては、本発明は、本発明にしたがって生産されたトランスジェニック動物に由来するB細胞に関する。
【0035】
なおさらなる局面においては、本発明は、本発明のトランスジェニック動物を使用して生産されるヒト化免疫グロブリンと、ヒト化免疫グロブリンを含む抗体調製物または薬学的組成物に関する。
【図面の簡単な説明】
【0036】
【図1】図1は、ウサギ免疫グロブリン遺伝子の重鎖遺伝子座の模式図である。
【図2】図2は、ウサギ重鎖スペーサー配列の比較を示す。
【図3】図3は、ウサギ重鎖スペーサー配列の比較を示す。
【図4】図4は、ウサギ重鎖スペーサー配列の比較を示す。
【図5】図5は、ウサギ免疫グロブリン軽鎖遺伝子座の模式図である。
【図6】図6は、ヒトVHエレメントとウサギのスペーサーエレメントを用いた免疫グロブリン遺伝子V遺伝子座の組み立てを説明する。
【図7】図7は、redεβγ−システムを用いた相同組み換えによる2つのカセットの挿入である。上のカセットには、ゲンタマイシンカセットに隣接している2つの制限部位(FseIとAscI)が含まれている。下のカセットには、反転させられ(i)変異させられた(71)loxP部位と、FRT部位と、MluI制限部位が含まれている。修飾後、BACはFseIとAscIで消化される。
【図8】図8は、ウサギK1軽鎖遺伝子座をベースとするヒト化ウサギ軽鎖遺伝子座(rLC3−B)を示す。ウサギCκ1をヒトCκで置き換えた。ヒトの再構成ヒトVκJκが挿入されている。合成のヒトVκJκは、ウサギのVκエレメントと80%を超える配列相同性を共有する。
【図9a】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−1】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−2】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−3】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−4】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−5】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−6】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−7】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−8】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−9】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−10】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図10】図10は、ニワトリの免疫グロブリンスペーサー配列とヒトVエレメントを用いた、ヒト化免疫グロブリン遺伝子座の構築を示す概要である。
【図11】図11は、マウスの免疫グロブリンスペーサー配列またはウサギの免疫グロブリンスペーサー配列と、ヒトVエレメントを用いた、ヒト化免疫グロブリン遺伝子座の構築を示す概要である。
【図12a−1】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図12a−2】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図12a−3】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図12a−4】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図12b−1】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図12b−2】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図12b−3】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図12b−4】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図12c】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図13a−1】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13a−2】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13b−1】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13b−2】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13b−3】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13b−4】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13b−5】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13c−1】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13c−2】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13c−3】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13c−4】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13d−1】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13d−2】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13d−3】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13d−4】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13d−5】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13e】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【0037】
(発明の詳細な説明)
(A.定義)
「抗体」(Ab)と「免疫グロブリン」(Ig)は、同じ構造特性を有している糖タンパク質である。抗体は特異的な抗原に対して結合特異性を示すが、免疫グロブリンには、抗体と、抗原特異性を有していない他の抗体様分子の両方が含まれる。後者のポリペプチドは、例えば、リンパ系によっては低レベルで、そして骨髄腫によっては高レベルで生産される。
【0038】
「自然界に存在している抗体および免疫グロブリン」は、通常、約150,000ダルトンのヘテロ四量体糖タンパク質であり、これは、2つの同じ軽(L)鎖と2つの同じ重(H)鎖から構成されている。個々の軽鎖は、ジスルフィド共有結合(単数または複数)によって重鎖に連結されているが、ジスルフィド結合の数は、それぞれの免疫グロブリンイソ型の重鎖に応じて変化する。重鎖と軽鎖のそれぞれは、規則的な間隔の鎖内ジスルフィド結合もまた有する。個々の重鎖は、一方の末端に可変ドメイン(VH)を、その後に、多数の定常ドメインを有している。個々の軽鎖は、一方の末端に可変ドメイン(VL)を有しており、その他方の末端には定常ドメインを有している;軽鎖の定常ドメインは、重鎖の第1の定常ドメインとアラインメントされ、軽鎖の可変ドメインは、重鎖の可変ドメインとアラインメントされる。特定のアミノ酸残基が、軽鎖可変ドメインと重鎖可変ドメインとの間の界面を形成すると考えられている(Clothia et al.,J.Mol.Biol.186:651(1985);Novotny and Haber,Proc.Natl.Acad.Sci.U.S.A.82:4592(1985))。
【0039】
用語「可変」とは、可変ドメインの特定の部分が抗体の間で配列に関して幅広く異なり、これがその特定の抗原についてそれぞれの特定の抗体の結合と特異性において使用されるという事実を意味する。しかし、可変性は、抗体の可変ドメイン全体にわたって均一に分布しているわけではない。これは、軽鎖および重鎖のいずれの可変ドメインにおいても、相補性決定領域(CDR)または超可変領域と呼ばれる3つのセグメントに集中している。可変ドメインのより高度に保存されている部分は、フレームワーク(FR)と呼ばれる。自然界に存在している重鎖と軽鎖の可変ドメインのそれぞれには、3個のCDRによって繋がれている4個のFR領域が含まれる。それぞれの鎖のCDRはFR領域によって互いに近い位置に保たれ、他の鎖のCDRとともに、抗体の抗原結合部位の形成に関与している(Kabat et al.,Sequences of Proteins of Immunological Interest,Fifth Edition,National Institute of Health,Bethesda,MD(1991)を参照のこと)。定常ドメインは、抗原に対する抗原の結合に直接は関与していないが、抗体依存性細胞傷害性における抗体の関与のような、種々のエフェクター機能を示す。
【0040】
用語「モノクローナル抗体」は、B細胞の単一のクローンによって合成される抗体分子を意味するように使用される。
【0041】
用語「ポリクローナル抗体」は、多数のB細胞クローンによって合成される抗体分子の集団を意味するように使用される。特異的な実施形態においては、ポリクローナル抗体はいくつかのエピトープを認識する。
【0042】
用語「ヒト化抗体」と「ヒト化免疫グロブリン」は、本明細書中で使用される場合は、ヒト免疫グロブリンポリペプチド配列(または、ヒト免疫グロブリン遺伝子セグメントによってコードされるポリペプチド配列)の少なくとも一部を含む免疫グロブリン分子を意味する。本発明のヒト化免疫グロブリン分子は、ヒト化免疫グロブリン分子を生じるように遺伝子操作された非ヒトトランスジェニック動物から単離することができる。このようなヒト化免疫グロブリン分子は、動物から調製されるか、または動物に由来する細胞から調製される非ヒト化免疫グロブリン分子と比較して、霊長類、特に、ヒトに対しては免疫原性が低い。
【0043】
用語「非ヒト動物」には、本明細書中で使用される場合は、哺乳動物が含まれるがこれに限定されない。「非ヒト動物」としては、例えば、非ヒト霊長類、ウサギ、ブタ、鳥類(例えば、ニワトリ、シチメンチョウ、アヒル、ガチョウなど)、ヒツジ、ヤギ、ウシ、ウマ、および齧歯類(例えば、マウスおよびラット)が挙げられる。好ましい非ヒト動物は、遺伝子変換および/または体細胞超変異によって実質的に抗体多様性を生じる動物、例えば、ウサギ、ブタ、鳥類(例えば、ニワトリ、シチメンチョウ、アヒル、ガチョウなど)、ヒツジ、ヤギ、およびウシが挙げられる。特に好ましい非ヒト動物は、ウサギとニワトリである。
【0044】
用語「非霊長類動物」としては、本明細書中で使用される場合には、霊長類ではない哺乳動物が挙げられるがこれに限定されず、これには上記に列挙した動物が含まれる。
【0045】
語句「遺伝子変換および/または超変異によって実質的に抗体多様性を生じる動物」は、抗体多様化の主な機構が、遺伝子再構成とは対照的な遺伝子変換および/または超変異である動物を意味するように使用される。
【0046】
用語「Ig遺伝子セグメント」は、本明細書中で使用される場合は、動物およびヒトの生殖細胞系列に存在し、B細胞中で互いに集まって再構成されたIg遺伝子を形成する、Ig分子の種々の部分をコードするDNAのセグメントを意味する。したがって、本明細書中で使用される場合Ig遺伝子セグメントにはV遺伝子セグメント、D遺伝子セグメント、J遺伝子セグメント、C領域遺伝子セグメントが含まれる。
【0047】
用語「ヒトIg遺伝子セグメント」には、本明細書中で使用される場合は、ヒトIg遺伝子セグメントの自然界に存在している配列と、ヒトIgセグメントの自然界に存在している配列の縮重形態と、さらには、ヒトIg遺伝子セグメントの自然界に存在している配列によってコードされるポリペプチドと実質的に同一であるポリペプチド配列をコードする合成の配列のいずれもが含まれる。「実質的に」によって、アミノ酸配列同一性の程度が少なくとも約85%〜95%であることが意味される。特定の実施形態においては、ヒトIg遺伝子セグメントは、免疫グロブリン分子をヒトにおいて非免疫原性にする。
【0048】
本発明の特異的なヒト化免疫グロブリン分子には、ヒト重鎖可変領域ポリペプチド配列または軽鎖可変領域ポリペプチド配列の少なくとも一部が含まれる。別の特異的な免疫グロブリン分子には、ヒト重鎖もしくは軽鎖可変領域ポリペプチド配列の少なくとも一部と、ヒト定常ドメインポリペプチド配列の少なくとも一部が含まれる。
【0049】
「ヒト化抗体の調製物」または「ヒト化抗体調製物」により、非ヒトトランスジェニック動物から(例えば、動物の血清、乳汁、または卵黄から)、あるいは、非ヒトトランスジェニック動物に由来する細胞から(例えば、B細胞またはハイブリドーマ細胞から)調製された、単離された抗体産物または精製された抗体産物が意味される。
【0050】
ヒト化抗体調製物は、ヒト化免疫グロブリン分子のレパートリーを含むポリクローナル抗体の調製物であり得る。ヒト化抗体調製物は、また、モノクローナル抗体の調製物でもあり得る。
【0051】
用語「抗体多様性」および「抗体のレパートリー」は、ほぼ同じ意味で使用され、生物体が発現できる全抗体特異性の全てを意味する。
【0052】
用語「スペーサー配列」は、免疫グロブリン重鎖または軽鎖遺伝子中に存在する任意の非コードヌクレオチド配列を意味するように、本明細書中で使用される。したがって、この用語には、具体的に、免疫グロブリン重鎖遺伝子中のイントロン配列と、V、D、Jセグメント、およびC領域セグメント中の複数のコード領域の間隔を隔てる任意の他の非コード配列、免疫グロブリン軽鎖遺伝子中のイントロン配列と、VおよびJセグメント、ならびにC領域セグメント中の複数のコード領域の間隔を隔てる任意の他の非コード配列、さらには、免疫グロブリン重鎖もしくは軽鎖遺伝子中の非コード配列である隣接している調節エレメント(例えば、エンハンサー)が含まれる。さらに、膜貫通ヘリックス部分をコードするエキソンと、重鎖および軽鎖のエンハンサーとの間に、非コード配列が具体的に含まれる。
【0053】
遺伝子再構成および遺伝子変換を受ける能力を有しているIg遺伝子座は、また、本明細書中では「機能的な」Ig遺伝子座とも呼ばれ、機能的なIg遺伝子座によって生じる多様性を有している抗体は、また、本明細書中では、「機能的」抗体または抗体の「機能的な」レパートリーとも呼ばれる。
【0054】
(B.関連する文献)
免疫グロブリン遺伝子中の調節エレメントは、Bradley et al.(1999),Transcriptional enhancers and the evolution of the IgH locus;Lauster,R.et al.,Embo J 12:4615−23(1993);Volgina et al.,J Immunol 165:6400(2000);Hole et al.,J Immunol 146:4377(1991)によって記載されている。
【0055】
ニワトリおよびウサギの中での遺伝子変換による抗体多様化は、Bucchini et al.,Nature 326:409−11(1987);Knight et al.,Advances in Immunology 56:179−218(1994);Langman et al.,Res Immunol 144:422−46(1993)によって記載されている。ヒト−マウスキメラ抗体を発現するマウスの作成は、Pluschke et al.,Journal of Immunological Methods 215:27−37(1998)によって記載されている。マウスに由来する膜と細胞質テールを有しているヒト−マウスキメラ抗体を発現するマウスの作成は、Zou et al.Science 262:1271−1274(1993);Zou et al.,Curr Biol 4:1099−1103によって記載されている。ヒト免疫グロブリンポリペプチドを発現するマウスの作成は、Bruggemann et al.Curr Opin Biotechnol 8(4):455−8(1997);Lonberg et al.Int Rev Immunol 13(1):65−93(1995);Neuberger et al.,Nature 338:350−2(1989)によって記載されている。BACクローンを使用するトランスジェニックマウスの作成は、Yang et al.,Nat Biotechnol 15:859−65(1997)によって記載されている。ヒト抗体を発現するウシの作成は、Kuroiwa et al.Nature Biotech 20(9):889−894(2002)によって記載されている。
【0056】
トランスジェニックウサギの作成は、Fan,J.et al.,Pathol Int 49:583−94(1999);Brem et al.,Mol Reprod
Dev 44:56−62(1996)によって記載されている。免疫グロブリンの発現を欠損させたウサギは、McCartney−Francis et al.,Mol
Immunol 24:357−64(1987);Allegrucci,et al.,Eur J Immunol 21:411−7(1991)によって記載されている。
【0057】
トランスジェニックニワトリの作成は、Sherman et al.Nature Biotech 16:1050−1053(1998);Etches et al.,Methods in Molecular Biology 62:433−450;Pain et al.,Cells Tissues Organs 165(3−4):212−9(1999);Sang,H.,“Transgenic chickens−−methods and potential applications”,Trends Biotechnol 12:415(1994);および、WO2004003157,“Gene regulation in transgenic animals using a transposon based vector”;ならびに、WO200075300,“Introducing a nucleic
acid into an avian genome,useful for transfecting avian blastodermal cells for producing transgenic avian animals with the desired genes,by directly introducing the nucleic acid into the germinal disc of the egg.”によって記載されている。
【0058】
γグロブリン血症のニワトリは、Frommel et al.,J Immunol
105(1):1−6(1970);Benedict et al.,Adv Exp Med Biol 1977;88(2):197−205によって記載されている。
【0059】
細胞からの動物のクローニングは、T.Wakayama et al.,Nature 1998;394:369−374;J.B.Cibelli et al.,Science 280:1256−1258(1998);J.B.Cibelli et
al.,Nature Biotechnology 1998;16:642−646;A.E.Schnieke et al.,Science 278:2130−2133(1997);K.H.Campbell et al.,Nature 380:64−66(1996);Kuroiwa et al.,Nature Genetics 2004年6月6日によって記載されている。ウサギの核導入クローニングは、Stice et al.,Biology of Reproduction 39:657−664(1988)、およびChallah−Jacques et al.,Cloning and Stem Cells 8(4):295−299(2003)によって記載されている。
【0060】
トランスジェニック動物からの抗体の産生は、米国特許第5,814,318号、同第5,545,807号、および同第5,570,429号に記載されている。キメラ哺乳動物宿主についての相同組み換えは、米国特許第5,416,260号に説明されている。胚にDNAを導入するための方法は、米国特許第5,567,607号に記載されている。胚性幹細胞の維持および拡大は、米国特許第5,453,357号に記載されている。
【0061】
ブタ、ヒツジ、およびウシにおける抗体のレパートリーの多様化に関係している機構は、Butler,J.E.(1998)“Immunoglobulin diversity,B−cell and antibody repertoire development in large farm animals.”,Rev Sci Tech 17:43に概説されている。ヒツジでの抗体の多様化は、Reynaud,C.A.,C.Garcia,W.R.Heim,and J.C.Weill(1995),“Hypermutation generating the sheep immunoglobulin repertoire is an antigen−independent process”,Cell 80:115;およびDufour,V.,S.Malinge,and F.Nau.(1996),“The Sheep Ig variable region repertoire consists
of a single VH family,”J Immunol 156:2163に記載されている。
【0062】
(C.詳細な説明)
免疫グロブリン重鎖および軽鎖遺伝子には、個々の遺伝子によってコードされ、イントロン配列によって間隔を隔てられているいくつかのセグメントが含まれる。したがって、ヒト免疫グロブリン重鎖についての複数の遺伝子が、14番染色体上で見られる。重鎖(VH)の可変領域には3つの遺伝子セグメント:V、D、およびJセグメントが含まれ、その後ろに、C領域をコードする複数の遺伝子が続く。V領域は、大きなスペーサーによってC領域から隔てられおり、V、D、およびJセグメントをコードする個々の遺伝子もまた、スペーサーによって隔てられている。
【0063】
免疫グロブリンの軽鎖には2つのタイプ:κおよびλが存在する。ヒトκ軽鎖の遺伝子は2番染色体上に見られ、ヒトλ軽鎖の遺伝子は22番染色体上に見られる。抗体軽鎖の可変領域には、VセグメントとJセグメントが含まれ、これらは別々の遺伝子セグメントによってコードされている。κ軽鎖遺伝子の生殖細胞系列での立体配置においては、直線的な配置でおよそ100〜200個のV領域遺伝子が存在しており、それぞれの遺伝子がそれぞれのリーダー配列を有しており、その後ろにおよそ5個のJ遺伝子セグメントと、c領域遺伝子セグメントが続く。全てのV領域は、イントロンによって隔てられており、さらに、V、J、およびC領域遺伝子セグメントを隔てるイントロンが存在する。
【0064】
感染から防御する免疫系の能力は、抗体の多様なレパートリーを作成するように特殊化された遺伝的機構に原因がある。B細胞中では、抗体をコードする遺伝子は、可変(V)領域中に無数の結合部位の組み合わせを生じるような様式で組み立てられる。1012種類を超える可能な結合構造が、そのような機構によって生じると推定される。ヒトを含む全ての動物において、抗体を作成するプロセスは、免疫グロブリン(Ig)遺伝子座の可変(V)セグメント、多様化(D)セグメント、結合(J)セグメントを組換えることによって開始される。この工程の後、動物の種に応じて、2つの一般的な機構が、抗体の多様な結合構造を生じさせるために使用される。
【0065】
ヒトやマウスのようないくつかの動物においては、免疫グロブリン重鎖遺伝子座上に、V、D、およびJ遺伝子セグメントの複数のコピーが、免疫グロブリン軽鎖遺伝子座上にVおよびJ遺伝子セグメントの複数のコピーが存在する。これらの動物における抗体の多様性は、遺伝子再構成、すなわち、再構成された重鎖可変領域と軽鎖可変領域を形成するような遺伝子セグメントの異なる組み合わせによって、主に生じる。しかし、他の動物(例えば、ウサギ、鳥類(例えば、ニワトリ、ガチョウ、およびアヒル)、ヒツジ、ヤギ、およびウシ)においては、遺伝子再構成は、抗体の多様性を生じることにおいてはより小さい役割しか果たしていない。例えば、ウサギにおいては、ごく限られた数のV遺伝子セグメント、最も多くの場合には、V領域の3’末端のV遺伝子セグメントだけが、連続するVDJセグメントを形成するように遺伝子再構成において使用される。ニワトリにおいては、1つのV遺伝子セグメント(D領域に隣接しているもの、すなわち、「3’近接V遺伝子セグメント」)と、1つのDセグメントと、1つのJセグメントだけが、重鎖の再構成に使用される:また、1つのV遺伝子セグメント(3’近接Vセグメント)と、1つのJセグメントだけが、軽鎖の再構成に使用される。したがって、これらの動物においては、接合部の多様化によって生じる最初に再構成された可変領域配列の中には少ししか多様性は存在しない。再構成されたIg遺伝子のさらなる多様化は、遺伝子変換によって行われる。遺伝子変換は、上流のV遺伝子セグメントに由来する短い配列が再構成されたIg遺伝子中のV遺伝子セグメント内の短い配列を置き換えるプロセスである。抗体配列のさらなる多様化は、超変異によって生じる場合もある。
【0066】
免疫グロブリン(抗体)は、5つのクラス(IgG、IgM、IgA、IgE、およびIgD)に属し、これらのそれぞれは、免疫防御において異なる生物学的役割を有している。血液中で最も豊富に存在し、感染に応答する能力を有しているものは、IgGクラスである。ヒトIgGクラスの中には、Fcドメインを含む重鎖定常領域の構造によって決定される4つのサブクラス(IgG1、IgG2、IgG3、およびIgG4イソ型)が存在している。抗体のF(ab)ドメインは、抗原上の特異的な配列(エピトープ)に結合するが、抗体のFcドメインは、抗原を排除するための免疫系の他の成分を動員し、活性化する。
【0067】
抗体は、動物から得られたポリクローナル抗血清によってヒトの生命にかかわる感染症を処置することができることが見出された1890年代から、治療薬としてうまく使用されている。抗体の研究における目覚しい進歩は、抗体の組み換え生産のための方法の開発、それに続く、抗体ヒト化技術の開発と、非ヒト動物において完全なヒトモノクローナル抗体を作成するための方法の開発にともなって生じた。
【0068】
結果として、キメラモノクローナル抗体、ヒト化モノクローナル抗体、およびヒトモノクローナル抗体は、薬学的産物の重要なクラスとして、最近、浮上してきた。モノクローナル抗体をベースとする薬剤は、疾患を処置することにおいて、特定の標的(例えば、受容体またはリガンド)をブロックする場合に極めて有効であるが、ガンおよび感染性病原体への感染のような特定の破壊的疾患は、それらの複雑さ、様々な原因によるとされる病因論、および適応性が原因で、処置することが難しい場合がある。モノクローナル抗体は、集団の全体へ、または個体の中へと疾患が広がる間に、変化し、進化し、そして成熟する、明確に定義された標的に対して向けられたものである。このような適応性の進化は、単特異的薬剤(例えば、モノクローナル抗体)にとっては致命的であり、単特異的薬剤は、耐性株によって迅速に回避される。高力価の抗生物質に耐性である細菌およびウイルスの多く、ならびに、悪性であるガンは、モノクローナル抗体治療のような標準的な抗ガン剤に対する耐性を生じる。
【0069】
対照的に、ポリクローナル抗体は、複雑な疾患に関係している複数の進化しつつある標的に結合し、それらを排除する能力を有している。複数の抗原に結合することにより、ポリクローナル抗体は、抗原の成熟の事象においてもなお、標的を飽和させて活性を保つ。この後、情報伝達のカスケードの間に、ポリクローナル抗体は、強い免疫反応を誘導して、標的抗原、病原体、または細胞を排除する。これらの特性により、ポリクローナル抗体は感染性疾患やガンを処置するために理想的な抗体である。
【0070】
これまでのところ、ポリクローナル抗体の使用は、供給の問題、または非ヒトタンパク質に対する望ましくない反応によって、厳しく制限されてきた。
【0071】
本発明により、免疫グロブリン(Ig)トランス遺伝子座の免疫グロブリンをコードするエレメントの選択的なヒト化に基づく、新しいヒト化アプローチが提供される。このようなヒト−動物トランス遺伝子座の作成により、多様化された親和性の高いヒト化(ポリクローナル)抗体を高い収量で発現するトランスジェニック動物を作成することができる。
【0072】
第1段階として、ヒト以外(霊長類以外を含む)の免疫グロブリン重鎖および軽鎖のゲノム遺伝子座が同定され、配列決定される。例えば、本発明の一部として、ウサギおよびニワトリの免疫グロブリン重鎖および軽鎖のゲノム配列が決定された。これらは、図1、5、および9に示される。
【0073】
ウサギのIg重鎖ゲノム遺伝子座の分析により、免疫グロブリン重鎖可変領域(Vh)には、多数の遺伝子が含まれており、これには、機能的な遺伝子と機能的ではない偽遺伝子が含まれていることが明らかにされた。18個のVh遺伝子のアラインメントにより、ウサギの重鎖可変領域の遺伝子配列(Vh1〜Vh18)の間に高度(80〜90%)な配列同一性があることが明らかになった。ウサギの重鎖可変領域の遺伝子は、ヒトの重鎖可変領域の遺伝子のVh3グループと最も高い相同性を共有している。具体的には、ウサギのVh1−a2遺伝子のヒトVh3−23配列との配列比較により、72.8%の配列同一性が明らかになった。
【0074】
さらに、複数のウサギの重鎖可変領域の遺伝子配列を隔てる非コード(例えば、イントロン)配列を分析した。図2〜4は、ウサギの重鎖イントロン配列の比較を示す。このようなイントロン配列は2つのグループに別れ、これらが高度に保存されていることが見出された。グループ1のイントロンの特別なメンバーは、驚くほど高い(80〜90%)の配列同一性を示す。
【0075】
同様の観察を、ウサギの免疫グロブリン軽鎖可変領域のゲノム配列に分析によって行った。具体的には、ウサギの免疫グロブリン軽鎖遺伝子座の分析により、軽鎖可変領域(V1)領域に多数の遺伝子セグメントが含まれており、これらが高度な(80〜90%)配列同一性を示すことが明らかにされた。さらに、ウサギの軽鎖可変領域(Vκ)は、ヒトの軽鎖可変領域の遺伝子配列のVκ1グループと高い相同性を示すことが明らかにされた。ほとんどのVκ配列は、機能的であり、高度に保存されていることが見出された。ウサギの重鎖可変領域の遺伝子とは異なり、ウサギの軽鎖可変領域の遺伝子中では、イントロン配列は異なることが見出されている。
【0076】
ニワトリの免疫グロブリン重鎖および軽鎖のゲノム配列を用いた同様の研究によっても、同様の結果が得られた。
【0077】
1つの局面においては、本発明により、スペーサー配列が提供される。スペーサー配列は、非霊長類動物の重鎖または軽鎖の遺伝子中のコード領域の間隔を隔てる。1つの実施形態においては、本発明により、遺伝子変換によって実質的に抗体多様性を生じる動物(例えば、ウサギおよびニワトリを含む)の重鎖および軽鎖の遺伝子に由来するスペーサー配列が提供される。このようなスペーサー配列は、その後、ヒト化免疫グロブリン遺伝子座を作成するプロセスにおいて使用されるヒト免疫グロブリン重鎖または軽鎖遺伝子セグメントに隣接させるように使用される。
【0078】
スペーサー配列には、通常は、少なくとも20個のヌクレオチド、または少なくとも約30個のヌクレオチド、または少なくとも約40個のヌクレオチド、または少なくとも約50個のヌクレオチドが含まれ、そして通常は、約20個から約10000個のヌクレオチドの長さである。スペーサー配列には、非ヒト動物(例えば、霊長類以外)中の自然界に存在しているイントロン配列に由来する適切な長さのヌクレオチドの連続するストレッチが含まれる場合があり、また、人工的な配列が含まれる場合もある。人工的な配列は、例えば、2つ以上の自然界に存在しているイントロン配列のコンセンサス配列であり得る。
【0079】
スペーサー配列には、配列番号1から185(表1)からなる群より選択される配列に由来するか、あるいは、2つ以上のそのような配列のコンセンサス配列に由来する、少なくとも約20(30、40、50などであって、10ヌクレオチドずつ増大して1000まで)個の連続しているヌクレオチドが含まれる場合がある。必ずしもそうである必要はないが、その免疫グロブリンがヒト化させられるヒト以外(霊長類以外)の動物のゲノム配列中の対応する領域の間隔を隔てるスペーサー配列により、ヒト化に使用されるヒト重鎖または軽鎖の配列(例えば、V、D、J、C領域の配列)の間隔を隔てることもできる。
【0080】
一般的には、非ヒト動物の免疫グロブリン(Ig)遺伝子座のヒト化には、ヒト化免疫グロブリン遺伝子座を作成するために動物のゲノム中に1つ以上のヒトIg遺伝子セグメントを組み込むことが含まれる。したがって、ヒト化Ig重鎖遺伝子座の作成には、1つ以上のVおよび/またはDおよび/またはJセグメント、ならびに/あるいはC領域セグメントの動物のゲノムへの組み込みが含まれる。同様に、ヒト化Ig軽鎖遺伝子座の作成には、1つ以上のVおよび/またはJセグメント、ならびに/あるいはC領域セグメントの動物のゲノムへの組み込みが含まれる。
【0081】
使用されるアプローチに応じて、ヒトIg遺伝子セグメント(単数または複数)を、動物の内因性のIg遺伝子座が通常存在する染色体位置に、あるいは、動物の異なる遺伝子座に組み込むことができる。染色体の位置にはかかわらず、本発明のヒト化Ig遺伝子座は、非ヒト動物中で、遺伝子再構成、および遺伝子変換、および超変異を受ける能力を有しており、それによってヒト化Ig分子の多様化されたレパートリーを生じる。遺伝子再構成および遺伝子変換を受ける能力を有しているIg遺伝子座はまた、「機能的な」Ig遺伝子座とも呼ばれ、機能的なIg遺伝子座によって作成される多様性を有している抗体はまた、「機能的な」抗体または抗体分子の「機能的な」レパートリーとも呼ばれる。
【0082】
さらなる局面においては、本発明により、非霊長類動物のIg重鎖または軽鎖遺伝子中の複数のコード領域の間隔を隔てるスペーサー配列に由来するか、あるいは、2つ以上のそのようなスペーサー配列のコンセンサス配列に由来する、少なくとも約20個の連続しているヌクレオチドを含むヌクレオチド配列が隣接している、ヒトIg遺伝子セグメントを含む核酸分子が提供される。隣接配列中のスペーサー配列に由来するセクションとしての隣接配列は、同一であっても、異なっていてもよい。スペーサー配列に由来するか、あるいは2つ以上のスペーサー配列のコンセンサス配列に由来する、連続しているヌクレオチドは、ヒトIg遺伝子セグメントに対して直接融合させることができる。あるいは、ヒトIg遺伝子セグメントと、少なくとも1つのスペーサーを起源とするヌクレオチド配列との間の介在配列である場合もある。したがって、例えば、ヒトV遺伝子セグメントの5’末端の隣接配列にはプロモーター領域が含まれ、これは、ヒトV遺伝子セグメントに対して直接連結させられ、少なくとも20ヌクレオチドのスペーサー配列に由来するヌクレオチドのストレッチからのその間隔が隔てられる。
【0083】
さらに別の局面においては、本発明は、ヒト重鎖V、D、および/またはJ遺伝子セグメント、ならびに/あるいはC領域セグメントが、もとの非ヒト動物の免疫グロブリン遺伝子中と同じ立体配置で存在し、非霊長類動物のIg重鎖または軽鎖遺伝子中のコード領域の間隔を隔てるイントロン配列に由来する少なくとも約20個の連続しているヌクレオチドを含む配列によって間隔を隔てられている、ヒト化Ig重鎖遺伝子座に関する。別の実施形態においては、本発明により、ヒト軽鎖C領域セグメント、および/またはJ遺伝子セグメント、および/またはV領域セグメントが、もとの非ヒト動物の免疫グロブリン遺伝子と同じ立体配置で非ヒト動物(例えば、霊長類以外)のイントロン配列によって間隔を隔てられている、ヒト化軽鎖遺伝子座が提供される。特定の実施形態においては、スペーサー配列は、ヒト以外(霊長類以外)の動物の非コード配列(例えば、イントロン配列)に基づいて設計される。1つの実施形態においては、スペーサーは、ヒト以外(霊長類以外)の動物に由来する適切な非コード配列を保持し得る。あるいは、構築を簡単にするために、高度に相同である非コード(イントロン)配列に基づいて設計されるコンセンサス配列が設計され、複数のヒト重鎖または軽鎖の遺伝子セグメントの調製のための同じ型のスペーサー配列として使用される。
【0084】
本発明により、具体的には、ヒト化免疫グロブリン遺伝子座の構築に有用な、単離された核酸配列およびベクターが提供される。
【0085】
1つの実施形態においては、ヒト化されるIg遺伝子座を含むDNAフラグメントは、遺伝子変換により抗体多様性を生じる動物(例えば、ウサギおよびニワトリ)から単離される。このような大きなDNAフラグメントは、ヒト以外、例えば、非霊長類動物のゲノムDNAから調製することができる、プラスミド、コスミド、酵母人工染色体(YAC)、または細菌の人工染色体(BAC)などのライブラリーをスクリーニングすることによって単離することができる。動物のC領域の全体を、1つのプラスミドまたはコスミドクローンに含めることができ、これは続いて、ヒト化される。YACクローンには、2メガ塩基までのDNAフラグメントを持たせることができ、したがって、動物の重鎖遺伝子座全体、またはその大きな部分を1つのYACクローン中に単離することができ、また、1つのYACクローン中に含ませるように再構築することもできる。BACクローンは、より小さい大きさ(約150〜450kb)のDNAフラグメントを持つことができる。しかし、Ig遺伝子座の重複するフラグメントを含む複数のBACクローンを別々にヒト化して、その後、動物のレシピエント細胞に一緒に注入することができる。ここで、重複しているフラグメントは、レシピエント動物の細胞中で組み換わって連続しているIg遺伝子座を生じる。
【0086】
ヒトIg遺伝子セグメントは、DNAフラグメントの連結、または相同組み換えによるDNAフラグメントの挿入を含む種々の方法によって、ベクター(例えば、BACクローン)上のIg遺伝子座に組み込むことができる。ヒトIg遺伝子セグメントの組み込みは、ヒトIg遺伝子セグメントがトランス遺伝子中で宿主動物配列に動作可能であるように連結されて、機能的なヒト化Ig遺伝子座、すなわち、遺伝子再構成、および遺伝子変換、および超変異が可能であるIg遺伝子座を生じ、これによってヒト化抗体の多様化されたレパートリーの生産を導くように、行われる。
【0087】
1つの実施形態においては、ヒトIg遺伝子セグメントは、相同組み換えによってIg遺伝子座に組み込むことができる。相同組み換えは、細菌、酵母、および他の細胞中で、高頻度の相同組み換え事象で行うことができる。例えば、酵母細胞は、動物のIg遺伝子座またはその大きな部分を含むYACで形質転換される。続いて、そのような酵母細胞は、さらに、本明細書中で上記に記載されるような組み換えベクターで形質転換される。組み換えベクターは、5’隣接配列と3’隣接配列に連結されたヒトIg遺伝子セグメントを有している。組み換えベクター中の5’隣接配列と3’隣接配列は、YAC上の動物のIg遺伝子セグメントの隣接配列と相同である。相同組み換えの結果、YAC上の動物のIg遺伝子セグメントは、ヒトIg遺伝子セグメントで置き換えられる。あるいは、大腸菌(E.coli)のような細菌細胞は、動物のIg遺伝子座またはその大きな部分を含むBACで形質転換される。そのような細菌細胞は、さらに、5’隣接配列と3’隣接配列に連結されているヒトIg遺伝子セグメントを有している組み換えベクターで形質転換される。組み換えベクター中の5’隣接配列と3’隣接配列は、相同組み換えを媒介し、組み換えベクター上のヒトIg遺伝子セグメントと、BAC上の動物のIg遺伝子セグメントとを交換する。ヒト化YACとBACは、細胞から容易に単離することができ、トランスジェニック動物の作成に使用することができる。
【0088】
本発明のさらなる局面においては、ヒト化免疫グロブリンを生産することができるトランスジェニック動物を作成する方法が提供される。
【0089】
本発明にしたがって、ヒト化免疫グロブリンを作成することができるトランスジェニック動物が、レシピエント細胞または動物の細胞中に、ヒト化Ig遺伝子座を有している本明細書中の上記に記載されている1つ以上のトランスジェニックベクターを導入し、遺伝子修飾されたレシピエント細胞(単数または複数)から動物へと導くことによって、作成することができる。
【0090】
レシピエント細胞は、例えば、非ヒト動物に由来するものであり得る。非ヒト動物は、遺伝子変換、および/または超変異によって抗体多様性を生じる動物、例えば、鳥類(例えば、ニワトリ)、ウサギ、ウシなどである。このような動物においては、3’近接V遺伝子セグメントは、免疫グロブリンの生産のために優先的に使用される。動物の3’近接V遺伝子セグメントの置き換えによるか、または3’近接V遺伝子セグメントの極近くに置くことによるかのいずれかによる、トランスジェニックベクター上のIg遺伝子座へのヒトV遺伝子セグメントの組み込みにより、免疫グロブリンの大部分において、ヒトV領域ポリペプチド配列の発現が生じる。あるいは、再構成されたヒトV(D)Jセグメントを、トランスジェニックベクター上の免疫グロブリン遺伝子座のJ遺伝子座中に挿入することもできる。
【0091】
ヒト化Ig遺伝子座を含むトランスジェニックベクターは、レシピエント細胞(単数または複数)に導入され、その後、ランダムな組み込みによって、または標的化された組み込みによって、レシピエント細胞(単数または複数)のゲノム中に組み込まれる。
【0092】
ランダムな組み込みのために、ヒト化Ig遺伝子座を含むトランスジェニックベクターを、標準的なトランスジェニック技術によって動物のレシピエント細胞中に導入することができる。例えば、トランスジェニックベクターを、受精卵の前核に直接注入することができる。トランスジェニックベクターは、また、卵母細胞の受精の前に、精子をトランスジェニックベクターとともにインキュベーションすることによって導入することもできる。トランスジェニック動物は、受精卵から発生させることができる。トランスジェニックベクターを導入することに代わる別の方法は、胚性幹細胞をトランスフェクトし、その後、遺伝子修飾された胚性幹細胞を胚発生させることによる。あるいは、トランスジェニックベクター(裸のもの、または促進試薬と組み合わせたもの)を、発生しつつある胚に直接注入することができる。最終的には、キメラであるトランスジェニック動物が、トランスジェニック動物の少なくともいくつかの体細胞のゲノム中に組み込まれたヒト化Igトランス遺伝子を含む胚から作成される。
【0093】
特定の実施形態においては、ヒト化Ig遺伝子座を含むトランス遺伝子が、内因性の免疫グロブリン遺伝子の発現が欠損している動物株に由来するレシピエント細胞(例えば、受精卵、または発生しつつある胚)のゲノム中にランダムに組み込まれる。このような動物株の使用により、ヒト化トランスジェニックIg遺伝子座からの免疫グロブリン分子の優先的な発現が可能になる。このような動物の例としては、AliciaおよびBasileaウサギ株、さらには、ガンマグロブリン血症(Agammaglobinemic)ニワトリ株、さらには、免疫グロブリンノックアウトマウスが挙げられる。あるいは、ヒト化免疫グロブリントランス遺伝子または遺伝子座を有しているトランスジェニック動物を、内因性免疫グロブリンの発現が欠損している動物株と交配させることができる。内因性Ig遺伝子座の欠損と、ヒト化トランスジェニックIg遺伝子座についてホモ接合型である子孫を得ることができる。
【0094】
標的化された組み換えのためには、トランスジェニックベクターを、胚性幹細胞、またはすでに分化した体細胞のような、適切な動物のレシピエント細胞中に導入することができる。以後、トランス遺伝子が動物のゲノムに組み込まれており、相同組み換えによって対応する内因性のIg遺伝子座が置き換えられている細胞を、標準的な方法によって選択することができる。例えば、Kuroiwa et al,Nature Genetics 2004年6月6日を参照のこと。選択された細胞は、その後、徐核された核導入ユニット細胞、例えば、卵母細胞または胚性幹細胞、全能性のきちんとした機能を有している新生児を形成することができる細胞と融合させられる。融合は、十分に確立されている従来技術にしたがって行われる。卵母細胞の徐核と、核の導入は、また、注入用ピペットを使用する顕微手術によって行うこともできる。(例えば、Wakayama et al.,Nature(1998)394:369を参照のこと)。得られた卵細胞は、その後、適切な培地中で培養されて、トランスジェニック動物を作成するために同調させたレシピエントに導入される。あるいは、選択された遺伝子修飾された細胞を、発生しつつある胚に注入して、その後、キメラ動物中へと発生させることもできる。
【0095】
さらに、本発明にしたがって、ヒト化免疫グロブリンを生産することができるトランスジェニック動物を、また、レシピエント細胞(単数または複数)中に、内因性のIg遺伝子セグメントの隣接配列に相同である5’隣接配列と3’隣接配列に連結されたヒトIg遺伝子セグメントを有している、本明細書中で上記に記載されている1つ以上の組み換えベクターを導入し、内因性のIg遺伝子セグメントが相同組み換えによってヒトIg遺伝子セグメントによって置き換えられている細胞を選択し、そして選択された遺伝子修飾されたレシピエント細胞(単数または複数)から動物へと誘導することによって、作成することもできる。
【0096】
トランスジェニックベクターの標的挿入と同様に、このアプローチにおいてレシピエントとしての使用に適切な細胞としては、胚性幹細胞、またはすでに分化した体細胞が挙げられる。ヒトIg遺伝子セグメントを有している組み換えベクターを、例えば、トランスフェクションのような、任意の適切な手段によってそのようなレシピエント細胞中に導入することができる。以後、ヒトIg遺伝子セグメントが相同組み換えによって対応する内因性Ig遺伝子セグメントと置き換わっている細胞を、標準的な方法によって選択することができる。これらの遺伝子修飾された細胞を、トランスジェニック動物のクローニングのための核導入手順において、核ドナー細胞とすることができる。あるいは、選択された遺伝子修飾された胚性幹細胞を、発生しつつある胚に注入することができる。これは続いて、キメラ動物へと発達させることができる。
【0097】
上記の方法のいずれかによって作成されたトランスジェニック動物は、本発明の別の実施形態を形成する。トランスジェニック動物は、少なくとも1つ、すなわち、1つ以上の、ヒト化Ig遺伝子座をゲノム中に有しており、この遺伝子座から、ヒト化抗体の機能的なレパートリーが生産される。
【0098】
特異的な実施形態においては、本発明により、ゲノム中に1つ以上のヒト化Ig遺伝子座を有しているトランスジェニックウサギが提供される。本発明のトランスジェニックウサギは、ヒト化Ig遺伝子座を再構成および遺伝子変換することができ、ヒト化された交代の機能的なレパートリーを発現することができる。
【0099】
別の特異的な実施形態においては、本発明により、1つ以上のヒト化Ig遺伝子座をゲノム中に有しているトランスジェニックニワトリが提供される。本発明のトランスジェニックニワトリは、ヒト化Ig遺伝子座を再構成および遺伝子変換することができ、ヒト化抗体の機能的なレパートリーを発現することができる。別の特異的な実施形態においては、本発明により、1つ以上のヒト化V領域をゲノム中に有しているトランスジェニックマウスが提供される。ヒト化V領域には、非ヒトスペーサー配列が隣接している少なくとも2つのヒトV遺伝子セグメントが含まれる。トランスジェニックマウスは、ヒトVエレメントを再構成することができ、抗体の機能的なレパートリーを発現することができる。
【0100】
一旦、多様化させられたヒト化免疫グロブリン分子を生産することができる非ヒトトランスジェニック動物が作成されると、抗原に対するヒト化免疫グロブリンおよびヒト化抗体調製物を、抗原で動物を免疫化することによって容易に得ることができる。種々の抗原を使用して、トランスジェニックである宿主動物を免疫化することができる。このような抗原としては、微生物、例えば、ウイルス、および単細胞微生物(例えば、細菌および真菌)、生存している微生物、弱毒化された微生物、または死滅させた微生物、微生物のフラグメント、あるいは、微生物から単離された抗原性分子が挙げられるが、これらに限定されない。
【0101】
動物の免疫化に使用される例示的な細菌抗原としては、莢膜多糖類であるタイプ5および8のような、黄色ブドウ球菌(Staphylococcus aureus)から精製された抗原、α毒素、付着因子結合タンパク質、コラーゲン結合タンパク質、およびフィブロネクチン結合タンパク質のような感染性因子の組み換えバージョンが挙げられる。例示的な細菌抗原としては、また、黄色ブドウ球菌(S.aureus)、シュードモナス・アエルギノーサ(Pseudomonas aeruginosa)、腸球菌(enterococcus)、腸内細菌(enterobacter)、および肺炎桿菌(Klebsiella pneumoniae)の弱毒化されたバージョン、あるいは、これらの細菌細胞に由来する培養上清が挙げられる。免疫化に使用することができる他の細菌抗原としては、精製されたリポ多糖(LPS)、莢膜抗原、莢膜多糖類、および/または外膜タンパク質の組み換えバージョン、フィブロネクチン結合タンパク質、ならびに、シュードモナス・アエルギノーサ(Psuedomonas aeruginosa)、超球菌(enterococcus)、腸内細菌(enterobacter)、および肺炎桿菌(Klebsiella pneumoniae)由来の内毒素および外毒素が挙げられる。
【0102】
真菌に対する抗体の作成のための例示的な抗原としては、真菌の弱毒化されたバージョン、またはその外膜タンパク質が挙げられる。真菌としては、カンジダ・アルビカンス(Candida albicans)、カンジダ・パラシロシ(Candida parapsilosis)、カンジタ・トロピカリス(Candida tropicalis)、およびクリプトコッカス・ネオホルマンス(Cryptococcus neoformans)が挙げられるが、これらに限定されない。
【0103】
ウイルスに対する抗体を作成するための免疫化に使用される例示的な抗原としては、ウイルスの外膜タンパク質および弱毒化させられたバージョンが挙げられる。ウイルスとして、呼吸器合胞体ウイルス(respiratory syncytial virus)(RSV)(特に、F−タンパク質)、C型肝炎ウイルス(HCV)、B型肝炎ウイルス(HBV)、サイトメガロウイルス(CMV)、EBV、およびHSVが含まれるが、これらに限定されない。
【0104】
治療用抗体を、単離された腫瘍細胞または腫瘍細胞株;腫瘍関連抗原(これには、Her−2−neu抗原(それに対する抗体が乳ガンの処置に有用である);CD19、CD20、CD22、およびCD53抗原(それに対する抗体がB細胞リンパ腫の処置に有用である);(3)前立腺特異的膜抗原(PMSA)(それに対する抗体が前立腺ガンの処置に有用である)、および17−1A分子(それに対する抗体が結腸ガンの処置に有用である)が含まれるが、これらに限定されない)でトランスジェニック動物を免疫化することによって、ガンの処置のために作成することができる。
【0105】
抗原は、任意の従来の様式でトランスジェニック宿主動物に、アジュバントとともに、またはアジュバントなしで投与することができ、予め決定されたスケジュールに従って投与することができる。
【0106】
免疫化後、免疫化されたトランスジェニック動物に由来する血清または乳汁を、抗原に特異的な薬学的等級のポリクローナル抗体を精製するために分画することができる。トランスジェニックである鳥類の場合には、抗体は、また、卵黄を分画することによっても得ることができる。濃縮され、精製された免疫グロブリン画分は、クロマトグラフィー(アフィニティー、イオン交換、ゲル濾過など)、硫酸アンモニウムのような塩、エタノールのような有機溶媒、またはポリエチレングリコールのような高分子を用いる選択的な沈降によって、得ることができる。
【0107】
モノクローナル抗体を作成するために、脾細胞が、免疫化されたトランスジェニック動物から単離され、これは、ハイブリドーマの生産のための形質転換細胞株との細胞融合において使用されるか、または、抗体をコードするcDNAが、標準的な分子生物学的技術によってクローニングされ、トランスフェクトされた細胞中で発現させられるかのいずれかが行われる。モノクローナル抗体を作成するための手順は、当該分野で十分に確立されている。例えば、欧州特許第0 583 980 A1号(“Method For Generating Monoclonal Antibodies From Rabbits”)、米国特許第4,977,081号(“Stable Rabbit−Mouse Hybridomas And Secretion Products Thereof”)、WO97/16537(“Stable Chicken B−cell Line And Method of Use Thereof”)、およびEP
0 491 057 B1(“Hybridoma Which Produces Avian Specific Immunoglobulin G”)を参照のこと。これらの開示は、引用により本明細書中に組み入れられる。クローニングされたcDNA分子からのモノクローナル抗体の生体外での生産は、Andris−Widhopf et al.,“Methods for the generation of chicken monoclonal antibody fragments by phage display”、J Immunol Methods 242:159(2000)によって、そして、Burton,D.R.,“Phage display”Immunotechnology 1:87(1995)によって記載されている。それらの開示は、引用により本明細書中に組み入れられる。
【0108】
B細胞のような、本発明のトランスジェニック動物に由来する細胞、または抗原に対して免疫化されたトランスジェニック動物から確立された細胞株もまた、本発明の一部である。
【0109】
本発明のさらなる局面においては、疾患の処置に望ましい、精製されたヒト化抗体組成物、好ましくは、ヒト化ポリクローナル抗体組成物を投与することにより、霊長類、具体的には、ヒト患者の疾患を処置するための方法が提供される。
【0110】
本発明の別の局面においては、精製されたモノクローナル抗体またはポリクローナル抗体が、霊長類、特に、ヒトへの投与に適している適切な薬学的担体と混合させられて、薬学的組成物が提供される。本発明の薬学的組成物において使用することができる薬学的に許容される担体は、任意の全ての溶媒、分散媒体、等張化剤などであり得る。任意の従来の媒体、試薬、希釈剤、または担体は、レシピエントに対して、またはその中に含まれる抗体の治療有効性に対して不利益をもたらさない限りは、本発明の薬学的組成物中でのその使用は適切である。担体は、液体、半固体(例えば、ペースト状)、または固体の担体であり得る。担体の例としては、油、水、生理食塩溶液、アルコール、糖、ゲル、脂質、リポソーム、樹脂、多孔性基質、結合剤、増量剤、コーティング剤、保存料など、あるいはそれらの組み合わせが挙げられる。
【0111】
投与に使用されるヒト化ポリクローナル抗体組成物は、通常、0.1から100mg/mlまでの免疫グロブリン濃度、より一般的には、1から10mg/mlまでの濃度のポリクローナル抗体の集団を含むことを特徴とする。抗体組成物には、種々のイソ型の免疫グロブリンが含まれる場合がある。あるいは、抗体組成物には、1つにイソ型の抗体のみが含まれる場合も、また、複数の選択されたイソ型の抗体が含まれる場合もある。
【0112】
ほとんどの場合においては、抗体組成物は、修飾されていない免疫グロブリン、すなわち、さらに例えば、化学的もしくは酵素的に、修飾されることなく動物から、調製されたヒト化抗体から構成される。あるいは、免疫グロブリン画分に、酵素消化(例えば、ペプシン、パパイン、プラスミン、グリコシダーゼ、ヌクレアーゼなどで)、加熱などのような処理が行われる場合もあり、そして/また、さらに分画される場合もある。
【0113】
抗体組成物は、通常は、血管系に投与され、適切な静脈に入れられたカテーテルを通じて、注射または注入によって静脈内に投与することが好都合である。抗体組成物は、適切な速度で、通常は、約10分から約24時間の範囲で、さらに一般的には、約30分から約6時間の範囲で、患者が順応できる液体の速度によって投与される。有効投与量の投与は、1回の注入の中に存在させることができ、また、一連の注入の中に存在させることもできる。反復注入は、抗体調製物の半減期および臨床的な適応症に応じて、1日に1回、1週間に1回、1ヶ月に1回、または3ヶ月ごとに1回投与することができる。上皮表面への塗布については、抗体組成物は、意図される最終的な結果をもたらすために十分な量で処置が必要な表面に塗布され、そして必要な場合には繰り返すことができる。さらに、抗体は、例えば、筋肉内のボーラス注射として投与することができ、これには、その後、例えば、注入による持続的な投与が続く場合があるが、これは必ずしも必要ではない。
【0114】
抗体組成物は、疾患の原因となるか、あるいは、望ましくないかまたは異常な免疫反応を誘発する、ヒトの体組織中の抗原性物質に結合し、それを中和するために使用することができる。「抗原性物質」は、本明細書中では、タンパク質を含む任意の可溶性または細胞表面結合分子、さらには、細胞、または、疾患を引き起こす微生物、あるいは、少なくとも抗体に結合することができ、好ましくは、免疫反応を刺激することもできる因子を含むように定義される。
【0115】
単剤療法として、または化学療法と組み合わせた、感染性因子に対する抗体組成物の投与により、感染性粒子の排除がもたらされる。抗体の単独での投与により、感染性粒子の数は、10倍から100倍、より一般的には、1000倍以上減少する。同様に、単剤療法として、または化学療法との組み合わせにおいて使用された、悪性疾患を有している患者における抗体療法により、悪性細胞の数は、通常、10倍から100倍、あるいは、さらに1000倍減少する。治療は、感染性粒子、悪性細胞などの完全な排除が確実となるまでの期間、繰り返すことができる。いくつかの症例においては、抗体調製物での治療が、検出できる量の感染性粒子または望ましくない細胞が存在しなくなるまでの期間、続けられる。同様に、免疫反応の調節のための抗体療法の使用は、1回の治療用抗体の投与から構成される場合も、また、複数回の投与から構成される場合もある。治療は、任意の疾患の兆候がなくなるまでの期間、続けられ得る。
【0116】
この処置は、感染性疾患または悪性腫瘍を阻害するために十分な投与量で、化学療法と組み合わせて使用することができる。自己免疫疾患の患者または移植のレシピエントにおいては、抗体療法を、免疫反応を阻害するために十分な投与量の免疫抑制療法と組み合わせて使用することもできる。本発明はさらに、以下の実施例によって説明されるが、実施例は決して限定ではない。
【実施例】
【0117】
(実施例1)
(ウサギ免疫グロブリン遺伝子座を含むBACクローンの単離および配列決定)
高分子量のDNAをa2b5雄ウサギから単離した。ウサギを安楽死させ、脾臓と腎臓を取り出し、氷冷したPBS中でリンスした。脂肪と結合組織を除去し、別々に処理した。臓器をフラグメントになるように切り、予め冷却しておいたDounceホモジナイザーにおいてホモジナイズした。上清を冷却した50mlのファルコンチューブに移し、冷却したPBSと混合し、大きな組織の破片を2分間の間、底に沈ませた。上清中の細胞を、200gで4℃で10分間かけてペレット状にし、PBSで1回洗浄し、1mlのPBS中に再度懸濁させ、数えた。5×106、5×107、および5×108個の細胞のセットを、CHEF哺乳動物ゲノムDNAプラグキット(CHEF Mammalian Genomic DNA Plug Kit)(BIORAD)を使用してアガロースプラグに包埋した。HindIIIでの部分的消化の条件を最適化するために、プラグを5個の同じフラグメントに切断し、1から10単位のHindIIIで種々の時間の間、種々の温度で消化した。最良の結果は、4℃で3時間、または37℃で25分間の、2単位のHindIIIを用いた場合に得られた。消化したDNAを、以下のパラメーターを使用してパルスフィールドゲル電気泳動(Pulse Field Gel Electrophoresis)(PFGE)装置上でダブルサイズ分画した:6時間の逆行、15秒間のスイッチ時間;6時間前方へ進める、15秒間のスイッチ時間;20時間前方へ進める、90秒間のスイッチ時間;200V、14℃。所望される大きさの部分的に消化されたDNAを有しているゲルの領域を切り取り、DNAをゲレース(gelase)を使用して単離した。11ngの挿入フラグメントを、1ngのHindIIIで消化したpBELOBAC11と連結し、DH10B細胞中にエレクトロポレーションした。得られたコロニーの1%についてNotIを使用して大きさを測定し、平均の挿入フラグメントの大きさが124kbであることを明らかにした。1×105個のクローンをナイロンフィルター(Nylon filter)上にスポットし、特異的プローブとのハイブリダイゼーションによってスクリーニングした。
【0118】
スクリーニングのためのプローブを、ウサギ由来のゲノムDNAを使用してPCRによって増幅し、pBlueScriptにクローニング氏、配列決定によって確認した。プライマー対(配列番号193〜208、表2)を、公開されている配列にしたがって設計した。ウサギ重鎖および軽鎖免疫グロブリン遺伝子座を提示するいくつかのBACを単離し、マップした(図1および5)。BACs 219D23 219D23(GenBank Acc.No.AY386695)、225P18(GenBank Acc.No.AY386697)、27N5(GenBank Acc.No.AY386696)、38A2(GenBank Acc.No.AY386694)、179L1(GenBank Acc.No.AY495827)、215M22(GenBank Acc.No.AY495826)、19(GenBank Acc.No.AY495828)、およびフォスミドFos15B(GenBank Acc.No.AY3866968)を配列決定した。配列決定のためのショットガンライブラリーを、pCR−Blunt中に、1.5〜2kbの大きさの挿入フラグメントを含めて構築した。配列分析には、STADENパッケージ(Roger Staden,Cambridge,UK)を使用した。ソフトウェアによってプレギャップを調節し、gap4を組み立てとギャップを閉じるために使用した。配列の特質クリッピング(quality clipping)のために、PHRED(Washington University)とSTADENパッケージを組み合わせた。
【0119】
(実施例2)
(ヒト化ウサギ免疫グロブリン重鎖遺伝子座の構築)
ウサギの免疫グロブリン重鎖遺伝子座の配列を含むBACおよびフォスミドクローンを、定常領域、可変領域、および結合遺伝子セグメント、あるいは、3’エンハンサー領域に特異的なプローブを使用してゲノムDNAライブラリーから単離した。単離したBAC(図1)27N5(GenBank Acc.No.AY386696)、219D23(GenBank Acc.No.AY386695)、225P18(GenBank
Acc.No.AY386697)、38A2(GenBank Acc.No.AY386694)、フォスミドFos15B(GenBank Acc.No.AY3866968)を配列決定した(Ros et al.,Gene 330,49−59)。
【0120】
選択した免疫グロブリンコード配列を、ETクローニングによる大腸菌(E.coli)中での相同組み換えにより、対応するヒトの部分と交換した(E−Chiang Lee et al.,Genomics 73,56−65(2001);Daiguan
Yu et al.,PNAS 97,5978−5983(2000);Muyrers et al.,Nucleic Acids Research 27,1555−1557(1999);Zhang et al.,Nature Biotechnology 18,1314−1317(2000))。
【0121】
あるいは、DNAフラグメントを、生体外での連結によって組み換え、その後、大腸菌(E.coli)を形質転換した。BACおよび/またはFos15B、あるいはその一部を、生体外での連結および形質転換、ETクローニングによって、あるいはCreリコンビナーゼによって媒介される組み込みによって結合させた。
【0122】
ETクローニングのために、標的配列を含むベクターを、誘導性のλファージ組み換え酵素Redα、Redβおよびγを含むストレプトマイシン耐性大腸菌(E.coli)株に形質転換した。これらの組み換えタンパク質を、同時トランスフェクトさせたプラスミドから(プラスミドpSC101を有しているDH10B大腸菌(E.coli)細胞)、またはゲノムに組み込まれたλプロファージから(DY380大腸菌(E.coli)株)のいずれかから発現させた。ETクローニング手順には、2回の相同組み換え工程を含めた。
【0123】
最初の工程では、標的遺伝子座が、選択−対抗選択(selection−counter selection)カセット(例えば、neo−rpsL、これは、ネオマイシンに対する耐性(neo)と、ストレプトマイシンに対する感受性(rpsL)を与える)によって置き換えた。neo−耐性コロニーの単離後、相同組み換えによる選択カセットの挿入を、制限酵素分析と部分的な配列決定によって確認した。
【0124】
第2の工程では、rpsL−neo選択カセットを、新しい配列と交換した。ストレプトマイシン耐性クローンを制限分析と配列決定によって分析した。ETクローニング手順に使用したフラグメントには、20から50bpの長さの隣接配列が含まれており、これは、標的配列と同じであった。連結に使用した配列は、その3’末端と5’末端に適切な制限酵素部位を有していた。これらの部位は、もともと備わっている部位であるか、またはそれらは、適切な部位を含むプライマーを使用してPCRによって導入されたものであるかのいずれかであった。
【0125】
あるいは、配列を合成によって作成した。
【0126】
ヒト化重鎖を、BAC 219D23中のウサギJH、Cμ、およびBAC 27N5中のCγの、対応するヒト部分での、ETクローニングにより置き換えによって構築した。ETクローニング手順に使用したヒト配列は、ヒトゲノムDNAからPCRによって増幅した。
【0127】
ヒトCμ、Cγ、およびJH遺伝子セグメントを、ウサギの標的配列に相同である50bpのプライマー(配列番号209〜214、表2)を使用して増幅した。
【0128】
BACクローン225P18のクローン219D23との連結、およびBAC 27N5のフォスミド15Bとの連結の後、連結させた構築物を大腸菌(E.coli)に形質転換し、Creリコンビナーゼによって媒介される挿入によって繋いだ。これによって、18個のウサギの可変遺伝子、ウサギD領域、ヒトJ領域、ヒトCμ、ヒトCγ、ウサギCε、ウサギCα4、および3’エンハンサーエレメントから構成される機能的な遺伝子座が得られた。
【0129】
トランスジェニック動物の作成のために、ヒト化BACクローンを、3個の重複しているBAC(225P18と219D23とBAC27N5)、または2つの重複している結合させたBAC(225P18−229D23と、BAC27N5−フォスミド15B)を同時にか、または別々のいずれかで、あるいは、1つのBACとして(225P18−219D23−27N5−フォスミド15B)注入した。トランス遺伝子を有している創始動物をPCRによって同定した。
【0130】
(実施例3)
(合成のフラグメントを用いたヒト化免疫グロブリン重鎖遺伝子座の構築)
ヒトV配列とウサギのスペーサーを有している、ユニット1、ユニット2、ユニット3、およびユニット4と示される4個のフラグメント(図13、配列番号189〜192)を化学的に合成した。それぞれのフラグメントには、5’に、AscI制限エンドヌクレアーゼ認識部位が、3’にはlox71 Creリコンビナーゼ認識配列と、その後ろにFseIとMluI制限酵素認識配列が隣接している。ユニット1は、ウサギのスペーサーI29−30、I3−4、I2−3、およびIl−2の3’側半分(I1−2B)によって間隔を隔てられているヒトVH3−49、VH3−11、VH3−7、およびVH3−15可変遺伝子から構成されている。ユニット2は、ウサギのスペーサーI1−2A、(I1−2の5’側半分)、I7−8、I6−7、およびI4−5の3’側半分(I4−5B)によって間隔を隔てられているヒトVH3−48、VH3−43、およびVH3−64から構成されている。ユニット3は、ウサギのスペーサー配列I4−5B、I26−27、I11−12、およびI17−18によって間隔を隔てられているヒトVH3−74、VH3−30、およびVH3−9から構成されている。
【0131】
ユニット3は、上記に加えて、上流のFlpリコンビナーゼ認識標的(FRT)配列の隣接と、その後ろに、上記のAscI部位の前にSglfI制限エンドヌクレアーゼ認識配列を有している。
【0132】
ユニット4は、ウサギのスペーサーI1−2が5’側に隣接しているヒトVH3−23遺伝子、lox66 Creリコンビナーゼ標的配列と、AscIエンドヌクレアーゼ認識配列を有しており、3’には、IV−C(5’側半分)ウサギスペーサー配列が隣接しており、その後ろにMluIエンドヌクレアーゼ認識配列が続く。
【0133】
ゲンタマイシン選択カセットを、AscIとFseI部位を含むプライマー配列番号215および216(表2)を使用してPCR増幅し、AscI、FseI、およびMluIエンドヌクレアーゼ認識部位を含む修飾されたクローニング部位を有しているpGEMベクター(配列番号217および218(表2)を使用してPCRによって修飾したpGEM.Genta)に連結した。
【0134】
ユニット1、2、および3を、pGEM.Genta(Promega)ベクターにクローニングした。
【0135】
ユニット4を、HindIIIで直鎖状にしたカスタマイズしたpBELOBAC11(NEB)ベクターにサブクローニングし、PCR増幅した。正方向プライマー(配列番号219、表2)は、HindIII、PacI、およびAatIIについての制限部位を有しており、逆方向プライマー(配列番号220)は、BamHI、MluI、およびAscIについての制限部位を有している。プライマーは、pBELOBAC11クロラムフェニコール選択カセットを欠失させるように設計した。さらに、ネオマイシン選択カセットを、BamHIとHindIII制限部位を有しているプライマー配列番号221および222(表2)を用いてPCR増幅し、修飾したpBELOBAC11ベクター(pBB11.1)に連結した。
【0136】
ユニット1〜4を、記載されているようにcreによって媒介される組み換えによって組み立てた(Mejia et al.,Genomics 70(2)165−70(2000))。最初に、ユニット1をカスタマイズしたpGEM.Gentaベクターにクローニングし、FseIで消化し、その後、連結によって再度環化させた。このベクターではない(vectorless)構築物で、pBB11.1.Unit4とp706−Creプラスミドを有している大腸菌(E.coli)を形質転換した。ユニット1とPBB11.1.unit4との組み換えの後、ポジティブクローン(ユニット4/1)を、カナマイシンとゲンタマイシンを含む培地上で選択した。クローンを、種々の酵素を用いる制限分析によって特徴付けた。
【0137】
ユニット2の組み換えのために、ユニット4/1挿入フラグメントを、AscIとPacIでの二重消化によって切り出し、pBELOBAC11中に修飾したリンカーとともにクローニングした(pBB11.2:プライマー配列番号223および224(表2)を使用してPCRによって増幅した)。
【0138】
pBELOBAC11を、HindIIIで直鎖状にし、PacIおよびAatIIエンドヌクレアーゼ認識部位をコードする正方向プライマーと、MluIおよびNotIエンドヌクレアーゼ認識部位、ならびに、lox66 Creリコンビナーゼ標的部位をコードする逆方向プライマーを用いて、PCR増幅した。ユニット1/4の連結のために、pBB11.2ベクターをMluIとPacIで開環させた。
【0139】
pGEM.Genta.Unit2を、pGEM.Genta.Unit1について記載したように、環状のベクターではない構築物へと変換し、pBB11.2.Unit4/1と、生体内でのCreによって媒介される組み換えによって繋いだ。続いて、得られた構築物pBB11.2.Unit4/1/2を、ETクローニングによって野生型のloxp部位をlox66標的部位で置き換えることによる、ユニット3とのCreによって媒介される組み換えのために調製した(Muyrers et al.,Nucleic Acids Research 27,1555−1557(1999);Muyers et al.,Trends Biochem.Sci.26(5):325−31(2001))。クロラムフェニコール選択カセットを、BAC標的配列に相同である50bpの配列を含むプライマー(配列番号225および226、表2)を用いてPCRによって増幅した。逆方向プライマーには、lox66部位を含めた。ゲルから精製したPCR産物を、標的BACと、さらに相同組み換えに必要なpSC101プラスミドを有している細胞に形質転換した。ポジティブクローンを、クロラムフェニコールで選択し、制限分析と配列決定によって確認した。pGEM.Genta.Unit3を、ユニット1および2について上記に記載したように生体内での組み換えのために調製し、受容体BAC、さらには、p706−Creプラスミドを有している細胞に形質転換した。ポジティブクローンpBB11.2.Unit4/1/2/3を、ゲンタマイシンを用いて選択し、制限分析によって確認した。pBB11.2.Unit4/1/2/3を、さらにETクローニングによって、lox71標的部位を生じるように修飾した。その後、pBB11.2.Unit4/1/2/3を、BAC 219D23、27N5、およびFos15Bに由来するフラグメントに繋いだ。
【0140】
(実施例4)
(PCR増幅したフラグメントを用いるヒト化免疫グロブリン重鎖遺伝子座の構築)
ヒトVHエレメントを、ゲノムDNA(ClonTech)とプライマー配列番号227〜248(表2)を使用して増幅した。PCR産物を、ゲル電気泳動によって分析し、GENECLEANキット(Q−Biogen)を使用してゲル精製した。その後、増幅産物を、Zero−Blunt TOPO(登録商標)(Invitrogen)に、製造業者の説明書にしたがってサブクローニングした。全ての増幅したVエレメントの配列を確認した。ヒト化V領域の構築のために、Vエレメントを、鋳型としてのプラスミドDNAと、プライマー配列番号249〜270(表2)を使用して増幅した。正方向プライマーには、AscI部位、その後ろに、ウサギスプライシング部位を含めた。逆方向プライマーには、ウサギの組み換えシグナル配列(RSS)と、MluI制限部位を含めた。PCR産物を、GENECLEANキットを使用してゲル精製した。
【0141】
ヒトVHs、V3−33、V3−74、V3−49、V3−21、V3−48、V3−73、V3−7、およびV3−Dは、PCRによっては単離できず、これらは、化学合成した(BlueHeron,Bothel,WA)。制限部位とウサギの調節配列を、合成の間に付加した。
【0142】
ウサギのスペーサー配列を、鋳型としてのBAC 38A2および225P18と、プライマー配列番号271〜288(表2)を使用して増幅した。BAC 225P18をNheIで二重消化し、41kbのフラグメントをゲル精製した。このフラグメントを、スペーサーV1−2、V2−3、V3−4、V4−5、およびV5−6の増幅のための鋳型とした。
【0143】
BAC 225P18を、BstBIで消化し、スペーサーV6−7とV7−8の鋳型をゲル精製した。BAC 38A2のPacIとRsrIIでの二重消化により、スペーサーV21−22とV22−23の鋳型のゲル精製ができた。
【0144】
増幅したスペーサー配列をゲル精製し、XL−PCR−TOPO(登録商標)(Invitrogen)に、製造業者の説明書にしたがってサブクローニングした。
【0145】
VHエレメントとウサギのスペーサー配列を、修飾したpGEM(Promega)とpBS(Strategene)ベクターにサブクローニングした。pGEMベクターをNotIとHindIIIで切断し、FseI、AscI、およびMluI部位を含む化学合成したオリゴヌクレオチド配列(オリゴ1:配列番号289;オリゴ2:配列番号290;表2)に連結させた。ベクターpBSをSacIとKpnIで切断し、制限部位FseI、AscI、およびMluIを含む化学合成したオリゴヌクレオチド配列(オリゴ1:配列番号291;オリゴ2:配列番号292;表2)と連結させた。
【0146】
ゲンタマイシンとネオマイシン選択カセットを、FseIまたはAscI部位を有するプライマー(配列番号293〜296、表2)を使用して増幅し、修飾したpGEMとpBSベクターに連結した。
【0147】
最終的な構築物を、修飾したpBeloBACIIベクター中で組み立てた。pBeloBACIIベクターを、BamHIとHindIIIで開環し、クローニング部位を、FseI、AscI、MluI部位を含むように、化学合成したオリゴヌクレオチド配列(オリゴ1:配列番号297;オリゴ2:配列番号298;表2)を使用して修飾した。
【0148】
BAC 219D23を、ETクローニングを使用して制限部位を導入することによって修飾した(Muyrers et al.,Nucleic Acids Research 27,1555−1557(1999);Muyers et al.,Trends Biochem.Sci.26(5):325−31(2001))。ネオマイシン選択カセットを、プライマー配列番号299および配列番号300(表2)を用いて増幅した。正方向プライマーにはFseI部位を含め、逆方向プライマーにはAscI部位を含めた。
【0149】
精製したPCR産物で、BAC 219D23と、相同組み換えに必要なプラスミドpSC101を有している大腸菌(E.coli)を形質転換した。カセットとBAC中の標的部位の相同組み換えの後、導入した制限部位を制限分析によって確認した。その後、修飾したBAC 219D23をFseIとMluIで消化し、得られた17kbのフラグメント(FseI−Neo−AscIカセット)を、PFGEによって分離し、電気的溶出によって精製した。この精製したフラグメントを、FseIとMluIで開環させた修飾したpBeloBAC IIベクターと連結させた。
【0150】
ヒトVHエレメントをコードする精製したDNAフラグメントを、AscIとMluIで開環させた修飾したpGEM.neoベクターと連結させた。同様に、スペーサー配列を、修飾したpGEM.gentaベクターにサブクローニングした。その後、スペーサー配列を有しているpGEM.gentaベクターをFseIとMluIで切断し、挿入フラグメントを、FseIとAscIで開環させたpGEM.neo.VHベクターと連結させた。この工程を数回繰り返して、いくつかのスペーサーとVHセグメントから構成されるフラグメントを組み立てた。このようなフラグメントをFseIとMluIで切り出し、FseIとAscIで直鎖状にした修飾したpBeloBACIIベクターと連結させた。これらのプロセスを、大きな免疫グロブリンV領域を組み立てるために繰り返した(図6)。ヒト化重鎖遺伝子座を、トランスジェニック動物の作成に使用した。
【0151】
(実施例5)
(ヒト化Cκとヒト化再構成VJを含む、ヒト化ウサギ軽鎖遺伝子座の構築)
ウサギのゲノムBACライブラリーのスクリーニングにより、ウサギの軽鎖κ1遺伝子セグメントを含む3個のBAC(215M22、179L1、および196O2;それぞれ、Gene Bank Acc.No.AY495826、AY495827、およびAY495828)が得られた。ウサギのCκ1を、記載されている(E−Chiang
Lee et al.,Genomics 73,56−65(2001);Daiguan Yu et al.,PNAS 97,5978−5983(2000);Muyrers et al.,Nucleic Acids Research 27,1555−1557(1999);Zhang et al.,Nature Biotechnology 18,1314−1317(2000))ようなETクローニングによって、ヒトCκアロタイプKm3と交換した。ヒトCκ(アロタイプKm3)を、標的配列に相同である50bpの配列を含むプライマー(配列番号301および302、表2)を用いてPCRによって増幅した。相同アームを、ウサギの生殖細胞系列κ(b5;GenBank Accession No.K01363)の公開されている配列に基づいて設計し、Cκのイントロン−エキソン境界と適合させた。BAC 179L1中でのウサギのCκのヒトCκへの交換を、配列決定によって確認した。
【0152】
BAC 179L1−huCκを2回のETクローニングによって修飾した。ネオマイシン選択カセットを、BAC 179L1に相同である50bpの配列を含むプライマー(配列番号303および304、表2)を用いて増幅した。正方向プライマーには、さらに、i−CeuIメガヌクレアーゼ部位を含めた。PCR産物をETクローニングに使用した。ポジティブクローンをネオマイシンで選択し、制限酵素消化と配列決定によって正確であることをチェックした。ゼオシン選択カセットを、BAC 179L1に相同である50bpの配列を含むプライマー(配列番号305および306、表2)を用いて増幅した。正方向プライマーには、さらに、i−SceIメガヌクレアーゼ部位を含めた。PCR産物をETクローニングに使用した。ポジティブクローンをゼオシンで選択し、制限酵素消化と配列決定によって正確であることをチェックした。
【0153】
BAC 215M22を1回のETクローニングによって修飾した。ゲンタマイシン耐性遺伝子を、BAC 215M22に相同である50bpの配列を含むプライマー(配列番号307および308、表2)を用いて増幅した。正方向プライマーには、さらに、i−CeuIメガヌクレアーゼ部位を含め、逆方向プライマーには、i−SceIメガヌクレアーゼ部位を含めた。PCR産物をETクローニングに使用した。得られたクローンをゲンタマイシンで選択し、制限酵素消化と配列決定によって分析した。
【0154】
修飾したBAC 179L1および225M22を、i−CeuIとi−SceIで切断した。98kbのフラグメントと、132kbのフラグメントを精製し、連結させた。得られたクローンを、カナマイシンとクロラムフェニコールで選択し、制限酵素消化、i−SceIとi−CeuI制限部位を含む領域のPCR、および配列決定によって正確であることをチェックした。得られたBACを、179−215−huCκと呼んだ。
【0155】
BAC 179−215−huCκのウサギのJκ1とJκ2を、ETクローニングによって、合成のヒトの再構成したVκJκ遺伝子と置き換えた。ウサギのプロモーター、ウサギのリーダー、ウサギのイントロンと、ヒトVκJκ遺伝子を有しているDNAフラグメントを化学合成した。合成のヒトVJのコドン使用は、ウサギのVκ遺伝子に最も高く相同であるDNA配列となるように最適化した。
【0156】
合成のヒトVJを、BAC 179L1に相同である50bpの配列を含む正方向プライマー(配列番号309、表2)と、ゲンタマイシン耐性遺伝子に相同である配列およびFRT部位を含む逆方向プライマー(配列番号310、表2)を用いてPCR増幅した。ゲンタマイシン耐性遺伝子を、FRT部位を含む正方向プライマー(配列番号311、表2)と、BAC 179L1に相同である50bpおよびFRT部位を有している逆方向プライマー(配列312、表2)を用いて増幅した。合成のヒトVJとゲンタマイシン耐性遺伝子とを、合成のヒトVJ遺伝子についての正方向プライマーと、ゲンタマイシン耐性遺伝子についての逆方向プライマーを使用する重複伸張(overlap extension)PCRによって結合させた。得られたフラグメントをETクローニングに使用した。ポジティブクローンをゲンタマイシンで選択し、制限酵素消化と配列決定によって正確であることを確認した。
【0157】
ゲンタマイシン耐性遺伝子を、Flpリコンビナーゼの発現を通じて、部位特異的組み換えによって除去した。組み換え後、1つのFRTが残った。このFRT部位をETクローニングによって欠失させた。合成のヒトVJに由来する232bpのフラグメントを、(プライマー配列番号313および314(表2)を使用して)PCRによって増幅し、ETクローニングに使用した。得られたコロニーを、FRT部位が存在しないことについて(プライマー配列番号315および316(表2)を使用して)PCRによってスクリーニングし、配列決定によって確認した。
【0158】
BAC 179−215−huCκのネオマイシン耐性遺伝子をETクローニングによって置き換えた。ゲンタマイシン耐性(pRep−Genta;Genebridges)遺伝子を、BAC 179−215−huCκに相同である50bpの配列を含むプライマー(配列番号317および318、表2)を用いてPCRによって増幅した。正方向プライマーには、さらに、loxP部位、attB部位、およびPvuI制限部位を持たせておいた。得られたクローンをゲンタマイシンで選択し、制限酵素消化と配列決定によって正確であることをチェックした。
【0159】
得られたBAC(rLC3−B、図8)を、トランスジェニック動物の作成に使用した。
【0160】
(実施例6)
(複数のヒトVκエレメント、ニワトリのスペーサーエレメント、および再構成されたヒトVJを含む、ヒト化ウサギ軽鎖遺伝子座の構築)
ウサギのゲノムBACライブラリーのスクリーニングにより、ウサギの軽鎖κ1遺伝子セグメントを含む3個のBAC(215M22、179L1、および196O2;それぞれ、Gene Bank Acc.No.AY495826、AY495827、およびAY495828)が得られた。ウサギのCκ1を、記載されている(E−Chiang
Lee et al.,Genomics 73,56−65(2001);Daiguan Yu et al.,PNAS 97,5978−5983(2000);Muyrers et al.,Nucleic Acids Research 27,1555−1557(1999);Zhang et al.,Nature Biotechnology 18,1314−1317(2000))ようなETクローニングによって、ヒトCκアロタイプKm3と交換した。ヒトCκ(アロタイプKm3)を、標的配列に相同である50bpの配列を含むプライマー(配列番号301および302、表2)を用いてPCRによって増幅した。相同アームを、ウサギの生殖細胞系列κ(b5;GenBank Accession No.K01363)の公開されている配列に基づいて設計し、Cκのイントロン−エキソン境界と適合させた。BAC 179L1中でのウサギのCκのヒトCκへの交換を、配列決定によって確認した。
【0161】
2つのDNAフラグメント、17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含むユニット1(12,235bp、図12a、配列番号187)と、リーダーを伴う機能的な再構成されたヒトκVJ遺伝子フラグメント、11個のヒトV偽遺伝子、12個のニワトリのスペーサー配列、イントロン1、およびイントロン2の一部を含むユニット2(13,283bp、図12b、配列番号188)を化学合成し、ベクターpBR322にクローニングした。
【0162】
ユニット1およびユニット2を、制限酵素NgoMIVとAsiSI、またはNgoMIVとAscIでそれぞれ消化し、3個のフラグメントの連結により修飾されたリンカーを有しているpBELOBAC11に連結させた。修飾されたリンカーには、BsiWI制限部位、FRT5部位、rpsL−Neoカセット、AscI部位、およびAsiSI部位が含まれている。リンカーフラグメントを、高品質のポリメラーゼ(Roche)、プライマーCE_1_001_012904(配列番号319、表2)、および鋳型としてのプラスミドpRpsL−Neo(Genebridges)を用いて増幅した。その後、増幅した産物を、pBELOBAC11のBamHIとHindIII部位に連結した。ユニット1とユニット2の連結のために、修飾されたpBELOBAC11をAsiSIとAscIを用いて開環させた。ポジティブクローン(pBELOBAC11 Unit1/2)を制限酵素消化によってチェックした。
【0163】
BAC 179L1(GENBANK Acc.No.AY495827)を、ETクローニングによる2つの修飾された選択カセットの挿入によって修飾した。カセット1は、BAC 179L1に相同である50bpの配列とAscI部位を逆方向プライマー中に含むプライマー(配列番号321および322、表2)を用いて増幅したゲンタマイシン耐性遺伝子である。カセット2は、BAC 179L1に相同である50bpの配列、attB部位、FRT5部位、およびBsiWI部位を正方向プライマー中に含むプライマー(配列番号323および324、表2)を用いて増幅したrps1−Neo選択カセットである。
【0164】
精製したPCR産物で、BACと、相同組み換えに必要なプラスミドpSC101を有している大腸菌(E.coli)を形質転換した。相同組み換えの後、BACの修飾がうまくできたことを、制限消化分析、サザンブロット、および配列決定によって確認した。
【0165】
修飾したBAC 179L1を、制限酵素AscIとBsiWIで切り出した。ヒトCκを含むフラグメントを精製し、同じ制限酵素で開環させたpBELOBAC11 Unit1/2と連結させた。ポジティブクローンを、制限酵素消化によってチェックした。最終的な構築物(図12c)を、トランスジェニック動物の作成のために使用した。
【0166】
(実施例7)
(複数のヒトVκエレメント、ニワトリのスペーサー、および再構成されていないヒトJκ遺伝子座を含む、ヒト化ウサギ軽鎖遺伝子座の構築)
実施例6に記載した構築物を、以下のようなETクローニングによって修飾した。再構成された機能的なVJ配列を、機能的な組み換えシグナル配列(RSS)が隣接している機能的なV1と交換した。RSSを、pBELOBAC11 Unit1/2のV1に相同である50bpの配列を含む正方向プライマー(配列番号325、表2)、および、AscI制限酵素部位を含み、ゲンタマイシン耐性遺伝子に対する相同性を有している逆方向プライマー(配列番号326、表2)を用いて、BAC 179L1からPCR増幅した。ゲンタマイシン耐性遺伝子を、RSSの増幅に使用した逆方向プライマーに相同である配列を含む正方向プライマー(配列番号327、表2)、およびpBELOBAC11
Unit1/2に相同である50bpの配列と、BsiWI制限酵素部位を含む逆方向プライマー(配列番号328、表2)を用いて増幅した。
【0167】
RSSとゲンタマイシン耐性遺伝子を、RSSの増幅のための正方向プライマーと、ゲンタマイシン耐性遺伝子の増幅のための逆方向プライマーを使用する、重複伸張PCRによって結合させた。得られたフラグメントを、ETクローニングによりpBELOBAC11 Unit1/2を修飾するために使用した。ポジティブクローンをゲンタマイシンで選択し、制限酵素消化と配列決定によって分析した。
【0168】
ヒトのCκを有しているBAC 179L1を、ETクローニングによってさらに修飾した。カナマイシン選択カセットを、BAC 179L1に相同である50bpの配列を含むプライマー(配列番号329および330、表2)を用いて増幅した。逆方向プライマーにはまた、AscI制限酵素部位とFRT部位を含めておいた。PCR産物をETクローニングに使用した。アンピシリン選択カセットを、BAC 179L1に相同である50bpの配列を含むプライマー(配列番号331および332、表2)を用いて増幅した。正方向プライマーには、また、attB部位、AsiSI制限酵素部位、およびFRT5部位も含めておいた。逆方向プライマーには、BsiWI制限酵素部位とFRT部位を含めておいた。PCR産物をETクローニングに使用した。ヒトJ領域を、BAC 179L1に相同である50bpの配列を含むプライマー(配列番号333および334、表2)を用いて、ヒトのゲノムDNAから増幅した。PCR産物をETクローニングに使用した。得られたクローンを、制限酵素消化と配列決定によって分析した。
【0169】
ポジティブクローンをAscIとBsiWIで切断した。得られたフラグメントを精製し、同じ制限酵素で切断した修飾したpBELOBAC11 Unit1/2に連結した。ポジティブクローンをアンピシリンで選択し、制限酵素消化と配列決定によって分析した。正しいクローンを、トランスジェニック動物を作成するために使用した。
【0170】
(実施例8)
(複数のヒトVκエレメントを含むヒト化ウサギ軽鎖遺伝子座の構築)
ウサギのゲノムBACライブラリーのスクリーニングにより、ウサギの軽鎖κ1遺伝子セグメントを含む3個のBAC(215M22、179L1、および196O2;それぞれ、Gene Bank Acc.No.AY495826、AY495827、およびAY495828)が得られた。ウサギのCκ1を、記載されている(E−Chiang
Lee et al.,Genomics 73,56−65(2001);Daiguan Yu et al.,PNAS 97,5978−5983(2000);Muyrers et al.,Nucleic Acids Research 27,1555−1557(1999);Zhang et al.,Nature Biotechnology 18,1314−1317(2000))ようなETクローニングによって、ヒトCκアロタイプKm3と交換した。ヒトCκ(アロタイプKm3)を、標的配列に相同である50bpの配列を含むプライマー(配列番号301および302、表2)を用いてPCRによって増幅した。相同アームを、ウサギの生殖細胞系列κ(b5;GenBank Accession No.K01363)の公開されている配列に基づいて設計し、Cκのイントロン−エキソン境界と適合させた。BAC 179L1中でのウサギのCκのヒトCκへの交換を、配列決定によって確認した。
【0171】
Vκ1ファミリー(O2、L8、L4、A30、L11、L1、L5、L15、O8、L19、L12、A20、O4、L14、L23、L9、A4、L24、O6、L22、A9、A25、A15、O9)のヒトVκエレメントを、プライマー(配列番号335〜382、表2)、および鋳型としてのヒトゲノムDNAを使用してPCRによって増幅した。
【0172】
増幅産物をゲル電気泳動によって分析し、GENECLEAN(Q−Biogen)を使用してゲル精製し、Zero−Blunt TOPO(登録商標)ベクター(Invitrogen)にサブクローニングし、配列決定した。再構成されたヒトVκ(O2)Jκ(J4)エレメントが、PCR増幅によって生じ、これをサブクローニングし、配列決定した。ヒトVκエレメントをウサギのスペーサーと結合させるために、ヒトVκエレメントを、プラスミドDNAを鋳型として使用して、プライマー(配列番号383〜430、表2)を用いて、PCRによって増幅した。プライマーには、AscI部位またはMluI部位を含めた。
【0173】
ウサギのスペーサー配列を、プライマー配列番号431〜450(表2)を使用してPCRによって増幅した。BAC 179L1および215M22を、SpeI、NheI、AclI、SfoI、MluI、およびSalI/XhoIで消化した。フラグメントをゲル精製し、増幅鋳型として使用した。
【0174】
5’末端に存在するスペーサー配列を、FRTとattB部位を含む上流オリゴヌクレオチドによって増幅した。PCR産物を、GENECLEANキットを使用してゲル精製し、XL−PCR−TOPO(登録商標)(Invitrogen)に、製造業者の説明書にしたがってサブクローニングした。
【0175】
ヒトVκエレメントとウサギのスペーサー配列を、実施例4に記載したように修飾したpGEM(Promega)にクローニングした。
【0176】
ヒトVκと修飾したpGEM.gentaベクターを、AscIとMluIで消化し、これらを連結させた。同様に、ウサギのスペーサー配列をpGEM.neoにクローニングした。その後、pGEM.neoVκをFseIとAscIで切断し、FseIとMluIで切り出したpGem.genta.spacerの精製した挿入フラグメントと連結させた。AscIとMluIの相補末端の連結により、それらの制限酵素部位は破壊され、いくつかのVκとスペーサーエレメントを含むVκ遺伝子座の構築のために、AscIとMluIを繰り返し使用することができるようになる。ヒト化BAC 179L1および215M22のフラグメントと、ヒト化Vκ領域から構成される最終的な構築物を、pBeloBACの中で組み立てた。BAC 179L1および215M22を修飾し、結合させた。その後、BAC 179L1−215M22−huCκを、ETクローニングによってさらに修飾した。制限酵素部位、選択マーカー、およびさらなる機能的部位を含む2つのカセットを、図7に示すようにETクローニングによってベクター中に挿入した。カセットの増幅のために使用したプライマー(配列番号451〜454)を、表2に列挙する。
【0177】
最終的な構築物を組み立てるために、ヒトVエレメント、ウサギのスペーサーエレメント、および耐性マーカーから構成されるユニットを、pGEMからFseIとMluIで切り出し、FseIとAscIで消化したBAC 179L1−215M22と連結させた。その後、耐性マーカーを、ヒトVエレメント、ウサギのスペーサーエレメント、および別の耐性マーカーから構成される新しい挿入フラグメントと置き換えた。数回繰り返した後、最終的な構築物は、ウサギのスペーサー配列によって隔てられた多くのVκセグメント(L8、L4、A30、L11、L1、L5、L15、O8、L19、L12、A20、O4、L14、L23、L9、A4、L24、O6、L22、A9、A25、A15、O9)から構成される。ヒト化軽鎖遺伝子座を、トランスジェニック動物の作成のために使用する。
【0178】
(実施例9)
(ニワトリ重鎖遺伝子座スペーサー配列を有するヒト化重鎖遺伝子座の構築)
ニワトリのスペーサー配列によって隔てられている、ウサギD領域、ヒトJ領域、ヒトCμ、ヒトCγ、ウサギCα4、ウサギ3’αエンハンサー、およびヒトVHエレメント(プロモーター、および7マー/9マーの配列を含む)を含む、合成のヒト化重鎖遺伝子座を構築する。
【0179】
修飾したウサギBAC 27N5(「実施例2」を参照のこと)を、ETクローニングによってさらに修飾した。この構築物には、ヒト化CμおよびCγと、2つの1つずつしか存在しないBsiWIおよびAsiSI制限部位が、α−4膜エキソンの下流に含まれている。DNAを、オリゴヌクレオチド配列番号455および456(表2)を用いて増幅した。
【0180】
フォスミドFos15BをNheIで消化し、3’αエンハンサーを含む得られた13kbのフラグメントを、挿入フラグメントがBsiWIおよびAsiSI部位と隣接するように、クローニングベクター中にサブクローニングした。その後、挿入フラグメントを、BsiWIおよびAsiSIで切断し、修飾したBAC 27N5と連結させてBAC
27N5Fosを形成させた。
【0181】
BAC 219D23中のウサギJ領域を、ETクローニングによって対応するヒトJ領域と交換した。ヒトJ領域を、プライマー配列番号457および458(表2)を使用してPCRによって増幅した。
【0182】
1つずつしか存在しない制限酵素部位を、BAC 219D23のD領域の上流(A)とCμの上流(B)に挿入した。BAC 27N5Fos中では、制限部位Aを、リンカー領域の上流に挿入し、Bを、BAC 219D23に相同である配列の中に挿入した。酵素AおよびBでの消化後、ヒトJ領域とウサギD領域を含むフラグメントを単離し、BAC 27N5Fosと連結させて、BAC 219D23/27N5Fosを作成した。
【0183】
ニワトリの重鎖スペーサー配列を、ニワトリ重鎖V偽遺伝子(Mansikka et
al.,J Immunol 145(11),3601−3609(1990)、Reynaud et al.,Cell 59(1),171−183(1989))に特異的なプライマー(配列番号459および460、表2)を使用して、PCRによってニワトリのゲノムDNAから増幅した。あるいは、スペーサー配列を化学合成した。
【0184】
PCR産物をゲル精製し、pTOPO(Invitrogen)にクローニングし、配列決定した。
【0185】
ヒト重鎖可変エレメントを、GENBANKに公開されている配列(例えば、Acc.No.NG_001019)にしたがって設計したプライマーを用いてPCRによって増幅したか、あるいは化学合成した。ヒトVエレメントには、ヒトプロモーター領域、ヒトリーダー配列、リーダーとVをコードする領域との間のヒトのイントロン、ヒトVをコードする領域、およびヒトの組み換えシグナル配列が含まれている。増幅したか、または合成したフラグメントには、特異的な制限エンドヌクレアーゼ認識部位が隣接している。ニワトリのスペーサー配列とヒトのVエレメントを、ヒト化免疫グロブリン遺伝子座を含む1つまたはいくつかの大きなDNAフラグメントとなるように結合させた。構築物を、トランスジェニック動物を作成するために使用した。
【0186】
(実施例10)
(ヒトVエレメントおよび非ヒトスペーサー配列を含む(プロモーター領域とRSSは含まない)ヒト化免疫グロブリン遺伝子座の構築)
非ヒトゲノムDNAを用いて作成したBACライブラリーを、免疫グロブリンに特異的なプローブを用いてスクリーニングし、重鎖および軽鎖免疫グロブリンC、J、およびD領域を含むBACクローンを同定した。BACクローンを修飾して、制限酵素部位を含ませた。ヒト重鎖および軽鎖可変エレメントを、GenBank(例えば、Acc.No.NG_001019)に公開されている配列にしたがって設計したプライマーを使用してPCRによって増幅した。配列は、ゲノムDNAから増幅したか、または化学合成した。ヒトVエレメントには、ヒトのプロモーター領域、ヒトのリーダー配列、リーダーとVをコードする領域との間のヒトのイントロン、ヒトVをコードする領域、およびヒトの組み換えシグナル配列(RSS)が含まれている。増幅したか、または合成したフラグメントは、末端に特異的な制限エンドヌクレアーゼ認識部位を有している。非ヒトスペーサー配列は、PCR増幅したか、または化学合成した。非ヒトスペーサー配列とヒトのVエレメントを、ヒト化免疫グロブリン遺伝子座を含む1つまたはいくつかの大きなDNAフラグメントとなるように結合させた。構築物を、トランスジェニック動物を作成するために使用した。ニワトリのスペーサー配列を用いたヒト化V領域の構築の例を、図10に示す。
【0187】
(実施例11)
(ヒトVエレメントおよび非ヒトスペーサー配列を含むヒト化免疫グロブリン遺伝子座の構築)
非ヒトゲノムDNAを用いて作成したBACライブラリーを、免疫グロブリンに特異的なプローブを用いてスクリーニングし、重鎖および軽鎖免疫グロブリンC、J、およびD領域を含むBACクローンを同定した。BACクローンを修飾して、制限酵素部位を含ませた。ヒト重鎖および軽鎖可変エレメントを、GenBank(例えば、Acc.No.NG_001019)に公開されている配列にしたがって設計したプライマーを使用してPCRによって増幅した。配列は、ゲノムDNAから増幅したか、または化学合成した。ヒトVエレメントには、ヒトVをコードする領域が含まれている。非ヒトスペーサー配列は、PCRによって増幅したか、または化学合成した。これには、組み換えシグナル配列、スペーサー配列、プロモーター領域、リーダー配列、およびリーダーとVをコードする領域との間のイントロンが含まれている。このような非ヒトスペーサー配列を、ヒトのVエレメントと、1つまたはいくつかの大きなDNAフラグメントとなるように結合させ、トランスジェニック動物を作成するために使用した。マウスまたはウサギのスペーサー配列を用いたヒト化V領域の構築の例を、図11に示す。
【0188】
(実施例12)
(ヒト化免疫グロブリンを発現するトランスジェニックウサギ)
トランスジェニックウサギを、Fan et al.(Pathol.Int.49:583−594,1999)に記載されているように作成した。簡単に説明すると、雌のウサギを標準的な方法を使用して過剰排卵させ、雄のウサギと交配させた。前核段階の受精卵を卵管から回収し、20%のウシ胎児血清を補充したダルベッコのリン酸緩衝化生理食塩水のような適切な培養液に入れた。ヒト化免疫グロブリン遺伝子座を含むBACを、1対のマニピュレーターの力を借りて雄の前核にマイクロインジェクションした。形態学的に生存している受精卵を、偽妊娠させたウサギの卵管に移した。偽妊娠は、ヒトの絨毛膜性腺刺激ホルモン(hCG)の注射によって誘導した。ヒト化軽鎖構築物を受精卵の4645個の前核に注入した後、4043個の卵母細胞を132匹のレシピエントに移した。全部で253匹の生存性の子孫が産まれ、そのうちの11匹がトランスジェニックであった。ヒトκ軽鎖の発現を、ヒトκ軽鎖特異的試薬(例えば、マウスの抗ヒトκ、Southern Biotech,9220−01;ヤギの抗ヒトκ、Southern Biotech 2063−08)を使用してELISAによって検出した。
【0189】
ヒト化重鎖構築物を、4083個の受精卵の前核に注入した。3485個の卵母細胞を119匹のレシピエントに移し、433匹の子孫が生まれた。PCRおよびFISHによる分析により、これらの動物のうちの20匹がトランスジェニックであることが明らかになった。創始動物の血液中のヒト化重鎖を、ヒトIgM/IgGに特異的な抗体(例えば、ウサギの抗ヒトIgM、Rockland 609−4131;ウサギの抗ヒトIgM、Rockland 609−4631;ウサギの抗ヒトIgG、Pierce 31142、ウサギの抗ヒトIgG、Southern Biotech 6145−08;ウサギの抗ヒトIgG、Pierce 31784)を使用してELISAによって検出した。
【0190】
ヒト化κ、μ、およびγ鎖を検出するサンドイッチ型ELISAを、標準的な手順を使用して行った。簡単に説明すると、マイクロタイタープレートを捕捉抗体でコーティングし、希釈した血清試料とともにインキュベートした。結合したヒト免疫グロブリンを、二次標識抗体と、ペルオキシダーゼ−ストレプトアビジン結合体(Sigma、S2438)を使用して検出した。
【0191】
ヒト化重鎖免疫グロブリン鎖とヒト化軽鎖免疫グロブリン鎖の両方を発現する二重トランスジェニックである動物を、創始動物を交配させることによって作成した。
【0192】
(実施例13)
(ヒト化免疫グロブリンを発現するトランスジェニックマウス)
トランスジェニックマウスを、Nagy et al.,(Manipulating
the Mouse Embryo:A Laboratory Manual;Cold Spring Harbor Laboratory Press,New York,2003)に記載されているように作成した。簡単に説明すると、雌のマウスを標準的な方法を使用して過剰排卵させ、雄のマウスと交配させた。前核段階の受精卵を卵管から回収し、M2培地のような適切な培養液に入れた。ヒト化免疫グロブリン遺伝子座を含むBACを、1対のマニピュレーターの力を借りて雄の前核にマイクロインジェクションした。形態学的に生存している受精卵を、偽妊娠させた雌のマウスの卵管に移した。偽妊娠は無精子症の雄との交配によって誘導した。ヒト化軽鎖構築物を受精卵の1325個の受精卵の前核への注入の後、787個の卵母細胞を29匹のレシピエントに移した。全部で55匹の生存性の子孫が産まれ、そのうちの11匹がトランスジェニックであった。
【0193】
ヒト化重鎖構築物を、1050個の受精卵の前核に注入した。650個の卵母細胞を25匹のレシピエントに移し、64匹の生存している子孫が生まれた。PCRによる分析から、これらの動物のうちの19匹がトランスジェニックであることが明らかになった。
【0194】
ヒト化重鎖免疫グロブリン鎖とヒト化軽鎖免疫グロブリン鎖の両方を発現する二重トランスジェニックである動物を、創始動物を交配させることによって作成した。ヒト化κ、μ、およびγ鎖の発現を、標準的な手順を使用してELISAによって検出した。簡単に説明すると、マイクロタイタープレートを捕捉抗体でコーティングし、希釈した血清試料とともにインキュベートした。結合したヒト免疫グロブリンを、二次標識抗体と、ペルオキシダーゼ−ストレプトアビジン結合体(Sigma、S2438)を使用して検出した。
【0195】
明細書を通じて引用された全ての参考文献は、引用により本明細書中に明確に組み入れられる。本発明は、特定の実施例を参照することによって説明されるが、それらに限定されない。当業者であれば、種々の変更はバリエーションが、本発明の本質から逸脱することなく可能であることを認識するであろう。全てのこのような変更とバリエーションが、本明細書の範囲に具体的に含まれる。
【0196】
【表1−1】

【0197】
【表1−2】
【0198】
【表1−3】
【0199】
【表1−4】
【0200】
【表1−5】
【0201】
【表2−1】
【0202】
【表2−2】
【0203】
【表2−3】
【0204】
【表2−4】
【0205】
【表2−5】
【0206】
【表2−6】
【0207】
【表2−7】
【0208】
【表2−8】
【0209】
【表2−9】
【技術分野】
【0001】
特許協力条約の実施細則の801(a)(i)およびPCT規則5.2に従い、本出願は、2004年7月15日に1.489KBのサイズで作成された、ファイル39691−0007PCTとして保存された配列表を含むコンパクトディスクと、配列表の紙コピーに代わる2部のコピーによって行われる。その内容全体が、引用により本明細書中に明確に組み込まれる。
【0002】
(発明の分野)
本発明は、非ヒトトランスジェニック動物からヒト化抗体を生産させるための方法および手段に関する。本発明は、具体的には、新規の免疫グロブリン重鎖および軽鎖構築物、ヒト化抗体を発現する非ヒトトランスジェニック動物を作成することにおいて有用である組み換えのトランスジェニックベクター、トランスジェニック動物、ならびにヒト化免疫グロブリン調製物に関する。トランスジェニックベクターには、ヒト化された免疫グロブリン遺伝子座が含まれる。これには、非ヒト動物内因性の調節機構および抗体生産機構を本質的には完全なまま残し、多様化させられたヒト化抗体を生じるように、非ヒトトランスジェニック動物中で、遺伝子再構成、遺伝子変換、および超変異を受けさせることができる。得られるヒト化抗体は、ヒトに対する最少の免疫原性を有しており、ヒト患者の治療的処置における使用に適している。
【背景技術】
【0003】
(発明の背景)
抗体は、薬学的産物の1つの重要なクラスである。これは、ガン、アレルギー性疾患のような種々のヒトの疾患および症状の処置、移植による拒絶反応、宿主対移植片反応の予防において良好に使用されている。
【0004】
動物から得られる抗体調製物の重要な問題点は、ヒト患者において本来備わっている、非ヒト免疫グロブリンについての免疫原性である。非ヒト抗体の免疫原性を低下させるためには、動物の可変(V)領域のエキソンをヒト定常(C)領域のエキソンと融合させることにより、キメラ抗体遺伝子を得ることができることが示されている。
【0005】
ヒト化モノクローナル抗体もまた開発されており、これは、臨床的に使用されている。ヒト化モノクローナル抗体は、通常は、ヒト抗体であり、その中のいくつかのCDR残基と、場合によってはいくつかのFR残基が、非ヒト動物、例えば、齧歯類の抗体の中に類似している部位に由来する残基で置換されている。ヒト化は、本質的には、Winterとその共同研究者らの方法に従って(Jones et al.,Nature,321:522(1986);Riechmann et al.,Nature,332:323(1988);Verhoeyen et al.,Science,239:1534(1988))、非ヒト動物、例えば、齧歯類のCDRまたはCDR配列を、ヒトのモノクローナル抗体の対応する配列で置換することによって、行うことができる。
【0006】
キメラの生殖細胞系列変異体マウスにおける抗体重鎖−結合領域(JH)遺伝子のホモ接合体欠失により、内因性抗体の生産の完全な阻害が生じることが記載されている。ヒトの生殖細胞系列の免疫グロブリン遺伝子配列を、そのような生殖細胞系列変異体マウスに導入することにより、抗原でのチャレンジの際にヒト抗体の生産が生じる。例えば、Jakobovits et al.,Proc.Natl.Acad.Sci.USA,90:2551(1993);Jakobovits et al.,Nature,362:255(1993);Bruggemann et al.,Year in Immunol.,7:33(1993)を参照のこと。この遺伝子操作によるアプローチによって、遺伝子操作されたマウス中でヒト免疫グロブリンポリペプチドの発現が生じるが、ヒト免疫グロブリンの発現レベルは低い。これは、免疫グロブリンの効率的な発現に不可欠である免疫グロブリン遺伝子座の中の種特異的調節エレメントが原因である。トランスフェクトされた細胞株において示されるように、ヒトの免疫グロブリン遺伝子中に存在する調節エレメントは、非ヒト動物においては適正には機能しない場合がある。
【0007】
実際に、免疫グロブリン遺伝子中のいくつかの調節エレメントが記載されている。特に注目すべきは、重鎖定常領域の下流(3’)のエンハンサーと、軽鎖遺伝子中にあるイントロンのエンハンサーである。さらに、他の、すでに同定されている制御エレメントもまた、免疫グロブリン遺伝子中に存在し得る。マウスでの研究では、免疫グロブリン分子の膜と、膜形態の細胞質尾部が、ヒトCγ1遺伝子についてホモ接合型であるマウスの血清中でのヒト−マウスキメラ抗体の発現レベルにおいて重要な役割を果たしていることが示されている。したがって、動物中でのヘテロ接合型免疫グロブリン遺伝子の発現については、エンハンサーエレメントと、膜貫通部分(M1エキソン)および細胞質尾部(M2エキソン)をコードするエキソンを含む配列を、動物において通常見ることができる配列と、同様の位置で、置き換えることが望まれる。
【0008】
非ヒト抗体の潜在的な免疫原性によって生じる問題に加えて、モノクローナル抗体の使用は、通常は、キメラ抗体、ヒト化抗体、またはヒト抗体を問わず、ガンのような破壊的な疾患、および伝染力の強い病原体での感染が、その複雑さ、様々な原因によるとされる病因論、および適応性にあることが原因で1つの標的を攻撃することによって処置することは困難であるという事実によって、さらに限定される。著しく定義された標的に対して指向させられたモノクローナル抗体は、その標的が変化する、進化する、および突然変異すると、うまくいかない。したがって、悪性腫瘍は、標準的なモノクローナル抗体での治療に対して耐性を獲得する場合がある。この問題の解決方法は、ポリクローナル抗体の使用である。ポリクローナル抗体は、複雑な疾患に関係している複数の進化しつつある標的を標的化して、攻撃する能力を有している。ポリクローナル抗体は、また、細菌やウイルスの毒素を中和する能力も有しており、病原体を死滅させ、排除するための免疫反応を指示する。
【0009】
したがって、高力価の、親和性の高いヒト化ポリクローナル抗体およびモノクローナル抗体の大量生産に適している新しいアプローチが、臨床的に非常に必要とされている。
【0010】
マウスのゲノム中にヒト免疫グロブリン遺伝子を導入することにより、遺伝子操作されたマウスにおいて多様化させられたヒト抗体のレパートリーの発現が生じる。マウスとヒトのいずれにおいても、主な抗体の多様性は、遺伝子再構成によって作成される。このプロセスにより、種々の抗原結合部位を有している多数の抗体分子をコードする多数の異なる組み換えV(D)Jセグメントが作成される。しかし、ウサギ、ブタ、ウシ、および鳥類のような他の動物においては、主な抗体の多様性は、実質的に異なる機構、すなわち、鋳型を用いる突然変異、または遺伝子変換、および鋳型を用いない突然変異、または超変異によって生じる。例えば、ウサギとニワトリにおいては、VDJ再構成が極めて限られており(免疫グロブリンのおよそ90%が3’近傍にVH1エレメントを伴って生じる)、抗体の多様性は、遺伝子変換と超変異によって生じる。対照的に、マウスとヒトの遺伝子変換は、仮にあったとしても極めて稀に生じる。したがって、遺伝子変換および超変異によってそれらの主な抗体のレパートリーを多様化させる動物においては、遺伝子の再構成に基づく遺伝子操作アプローチにより、低い抗体力価と限られた抗体多様性を有している動物が得られる。したがって、ヒトの治療のための非免疫原性の抗体調製物を生産するための大動物の遺伝子操作には、別の遺伝子操作ストラテジーが必要である。
【0011】
非ヒトトランスジェニック動物中でのヒト化抗体の生産は、2002年2月14日に公開されたPCT公開番号WO02/12437に記載されている。この開示は、その全体を参照することにより明確に本明細書中に組み入れられる。WO02/12437には、1つ以上のヒト化免疫グロブリン遺伝子座を含む、遺伝子操作された非ヒト動物が記載されている。ヒト化免疫グロブリン遺伝子座は、非ヒトトランスジェニック動物中で遺伝子再構成と遺伝子変換を受けることができる。非ヒトトランスジェニック動物には、その中での抗体多様性が主に遺伝子変換によって生じて、多様化されたヒト化抗体を生じる動物が含まれる。得られたヒト化抗体は、ヒトに対して最少の免疫原性を有しており、ヒト患者の治療的処置における使用に適している。これには、さらに、種々の非ヒト哺乳動物、例えば、ニワトリ、ウシ、ヒツジ、およびウサギの免疫グロブリン重鎖定常領域セグメントの5’および3’隣接領域に由来する新規のヌクレオチド配列が記載されている。ヒト免疫グロブリン重鎖遺伝子セグメントに、そのような5’および3’配列に相同な配列が隣接している組み換えベクターは、非ヒト動物の免疫グロブリン重鎖遺伝子セグメントを、対応するヒト免疫グロブリン重鎖遺伝子セグメントで置き換えるために有用であることが示されている。
【発明の概要】
【課題を解決するための手段】
【0012】
(発明の要旨)
1つの局面においては、本発明は、複数のヌクレオチド配列が隣接するヒト免疫グロブリン遺伝子セグメントを含む単離された核酸分子に関する。ここで、隣接配列は、同じである場合も、異なる場合もあり、これには、遺伝子変換および/または超変異によって主に抗体多様性を生じる動物の免疫グロブリン重鎖もしくは軽鎖遺伝子に由来するスペーサー配列の、あるいは、2つ以上のスペーサー配列のコンセンサス配列に由来する、少なくとも約20個の連続しているヌクレオチドが含まれる。
【0013】
別の局面においては、本発明は、複数のヌクレオチド配列が隣接している、ヒト免疫グロブリン重鎖または軽鎖定常領域(C)遺伝子セグメントを含む単離された核酸分子に関する。ここで、隣接配列は、同じである場合も、異なる場合もあり、これには、非霊長類動物の免疫グロブリン重鎖もしくは軽鎖遺伝子に由来するスペーサー配列の、あるいは、2つ以上のスペーサー配列のコンセンサス配列に由来する、少なくとも約20個の連続しているヌクレオチドが含まれる。
【0014】
さらなる局面においては、本発明は、複数のヌクレオチド配列が隣接するヒト免疫グロブリン重鎖または軽鎖遺伝子セグメントを含む単離された核酸分子に関する。ここで、隣接配列は、同じである場合も、異なる場合もあり、これには、配列番号1から185(表1)からなる群より選択されるスペーサー配列の、あるいは、2つ以上のスペーサー配列のコンセンサス配列に由来する、少なくとも約20個の連続しているヌクレオチドが含まれる。
【0015】
1つの実施形態においては、隣接配列には、スペーサー配列の少なくとも約50個の連続しているヌクレオチドが含まれる。
【0016】
別の実施形態においては、ヒト免疫グロブリン遺伝子セグメントは、重鎖のV、D、またはJセグメントであり、Vセグメントは、例えば、VH3、VH1、VH5、またはVH4ファミリーのメンバーであり得る。
【0017】
さらなる実施形態においては、ヒト免疫グロブリン遺伝子セグメントは、軽鎖VまたはJセグメントであり、Vセグメントは、例えば、κ軽鎖遺伝子セグメント、例えば、Vκ1、Vκ3、またはVκ4である場合も、また、λ軽鎖セグメント、例えば、Vλ1、Vλ2、またはVλ3である場合もある。
【0018】
さらなる実施形態においては、主に遺伝子変換および/または体細胞超変異により抗体多様性を生じる非霊長類動物とは、例えば、ウサギ、ブタ、鳥類(例えば、ニワトリ、シチメンチョウ、アヒル、あるいはガチョウ)、ヒツジ、ヤギ、ウシ、ウマ、またはロバであるが、霊長類ではない他の動物、例えば、齧歯類もまた、本発明の範囲に具体的に含まれる。
【0019】
別の局面においては、本発明は、上記の核酸分子のいずれかを含む組み換えベクターに関する。
【0020】
さらに別の局面においては、本発明は、ヒト化免疫グロブリン(Ig)遺伝子座を含むトランスジェニックベクターに関する。ここで、ヒト化Ig遺伝子座は、非ヒト動物のIg遺伝子座またはIg遺伝子座の一部に由来するものであり、これには、複数のIgセグメントが含まれる。ここで、
(a)少なくとも1つの遺伝子セグメントが、スペーサー配列に由来するか、または2つ以上のそのようなスペーサー配列のコンセンサス配列に由来する少なくとも20個の連続しているヌクレオチドを含むヌクレオチド配列が隣接しているヒトIg遺伝子セグメントであり;
(b)複数の遺伝子セグメントが、再構成されていない立体配置、部分的に再構成された立体配置、または完全に再構成された立体配置において並列しており;そして
(c)非ヒト動物が遺伝子を変換する動物であり、非ヒト動物中でヒト化された免疫グロブリンのレパートリーを生産する場合には、ヒト化Ig遺伝子座が遺伝子再構成を受けることができ、必要に応じて、さらに遺伝子変換および/または超変異を受けることができる。
【0021】
さらなる実施形態においては、トランスジェニックベクター中に存在するヒト化Ig重鎖遺伝子座は、約5から100個のV遺伝子セグメントを含み、これには少なくとも1つのヒトV遺伝子セグメントが含まれる。特異的な実施形態においては、ヒト化Ig重鎖遺伝子座には、1つ以上のヒトV遺伝子セグメントが含まれる。
【0022】
別の実施形態においては、トランスジェニックベクター中に存在するヒト化Ig重鎖遺伝子座には、約5から25個のDの遺伝子セグメントが含まれる。特異的な実施形態においては、ヒト化Ig重鎖遺伝子座には、1つまたはいくつかのヒトD遺伝子セグメントが含まれる。
【0023】
さらに別の実施形態においては、トランスジェニックベクター中に存在するヒト化Ig重鎖遺伝子座には、約1から10個のJ遺伝子セグメントが含まれ、これには少なくとも1つのヒトJ遺伝子セグメントが含まれる。特異的な実施形態においては、ヒト化Ig重鎖遺伝子座には、1つ以上のヒトJ遺伝子セグメントが含まれる。
【0024】
別の実施形態においては、トランスジェニックベクター中に存在するヒト化Ig重鎖遺伝子座には、約1から25個のC領域セグメントが含まれ、これには少なくとも1つのヒトC領域セグメントが含まれる。特異的な実施形態においては、トランスジェニックベクター中に存在するヒト化Ig重鎖遺伝子座には、1つ以上のヒトC遺伝子セグメントが含まれる。
【0025】
なおさらに別の実施形態においては、トランスジェニックベクター中に存在するヒト化Ig遺伝子座は、非ヒト動物の軽鎖遺伝子座であり、これには、少なくとも1つのV遺伝子セグメントと、少なくとも1つのJ遺伝子セグメントと、少なくとも1つの定常(C)領域遺伝子セグメントが含まれる。ここで、少なくとも1つの遺伝子セグメントは、ヒト軽鎖VおよびJセグメント、ならびにヒト軽鎖C領域セグメントの群より選択される。特異的な実施形態においては、定常領域遺伝子セグメントは、ヒト軽鎖定常領域遺伝子セグメントであり、これは、例えば、CλまたはCκ遺伝子セグメントであり得る。別の実施形態においては、ヒト化Ig軽鎖遺伝子座には、ヒトVおよびJセグメント、ならびにヒトC領域セグメントより選択される2つ以上のセグメントが含まれる。さらなる実施形態においては、ヒト化Ig軽鎖遺伝子座には、少なくとも1つのヒトVセグメントと、少なくとも1つのヒトJセグメントと、少なくとも1つのヒトC領域セグメントが含まれる。
【0026】
さらなる実施形態においては、トランスジェニックベクター中に存在するヒト化Ig軽鎖遺伝子座には、約5から100個のV遺伝子セグメントが含まれ、これには少なくとも1つのヒトV遺伝子セグメントが含まれる。ここで、ヒトV遺伝子セグメントは、非ヒト動物の5から100個のV遺伝子セグメントに対して下流に存在する。特異的な実施形態においては、ヒトV遺伝子セグメントは、再構成された立体配置においてJ遺伝子セグメントのすぐ5’側に存在する。別の実施形態においては、トランスジェニックベクター中に存在するヒト化Ig軽鎖遺伝子座には、1つ以上のヒトV遺伝子セグメントが含まれる。
【0027】
なお別の実施形態においては、トランスジェニックベクター中に存在するヒト化Ig軽鎖遺伝子座には、約1から10個のJ遺伝子セグメントが含まれ、これには少なくとも1つのヒトJ遺伝子セグメントが含まれる。特異的な実施形態においては、トランスジェニックベクター中に存在するヒト化Ig軽鎖遺伝子座には、1つ以上のヒトJ遺伝子セグメントが含まれる。
【0028】
別の実施形態においては、トランスジェニックベクター中に存在するヒト化Ig軽鎖遺伝子座には、約1から25個のC領域セグメントが含まれ、これには少なくとも1つのヒトC領域セグメントが含まれる。特異的な実施形態においては、トランスジェニックベクター中に存在するヒト化Ig軽鎖遺伝子座には、1つ以上のヒトC遺伝子セグメントが含まれる。
【0029】
なおさらなる実施形態においては、トランスジェニックベクター中に存在するヒト化Ig遺伝子座は、非ヒト動物の軽鎖遺伝子座であり、これには、少なくとも1つのV遺伝子セグメントと、少なくとも1つのJ遺伝子セグメントと、少なくとも1つの定常(C)領域遺伝子セグメントが含まれる。ここで、少なくとも1つの遺伝子セグメントは、ヒト軽鎖VおよびJセグメント、ならびにヒト軽鎖C領域セグメントの群より選択される。特異的な実施形態においては、定常領域遺伝子セグメントは、ヒト軽鎖定常領域遺伝子セグメントであり、これは、例えば、CλまたはCκ遺伝子セグメントであり得る。別の実施形態においては、ヒト化Ig軽鎖遺伝子座には、ヒトVおよびJセグメント、ならびにヒトC領域セグメントより選択される2つ以上のセグメントが含まれる。さらなる実施形態においては、ヒト化Ig軽鎖遺伝子座には、少なくとも1つのヒトVセグメントと、少なくとも1つのヒトJセグメントと、少なくとも1つのヒトC領域セグメントが含まれる。
【0030】
異なる局面においては、本発明は、5’から3’の方向で、5’ヌクレオチド配列、ヒト免疫グロブリン配列、そして3’ヌクレオチド配列からなる2つ以上のユニットを含む核酸分子に関する。ここで、5’ヌクレオチド配列と3’ヌクレオチド配列は同じである場合も、異なる場合もあり、これには、非霊長類動物の免疫グロブリン重鎖もしくは軽鎖遺伝子中のコード領域の間隔を隔てるスペーサー配列に由来するか、あるいは、2つ以上のスペーサー配列のコンセンサス配列に由来する、少なくとも約20個の連続しているヌクレオチドが含まれる。特異的な実施形態においては、スペーサー配列は、配列番号1から185(表1)の中から選択される。別の特定の実施形態においては、核酸分子の全ての反復ユニット中の5’ヌクレオチド配列および/または3’ヌクレオチド配列は同じである。別の特定の実施形態においては、核酸分子の反復ユニットには、少なくとも2つの異なる5’配列および/または3’配列が含まれる。さらなる実施形態においては、5’ヌクレオチド配列と3’ヌクレオチド配列は互いに異なるが、全ての5’ヌクレオチド配列は同一であり、全ての3’配列も同一である。
【0031】
さらなる局面においては、本発明は、非ヒト動物中でヒト化抗体の機能的なレパートリーを生産することができるヒト化免疫グロブリン(Ig)遺伝子座を含むトランスジェニックベクターを作成するための方法に関する。この方法には:
(a)少なくとも1つのV遺伝子セグメントと、少なくとも1つのJ遺伝子セグメントと、少なくとも1つの定常領域遺伝子セグメントを含む、非ヒト動物に由来するIg遺伝子座またはその一部を含むDNAフラグメントを得る工程;ならびに、
(b)ヒト化Ig遺伝子座を生産するように、工程(a)のDNAフラグメント中に少なくとも1つのヒトIg遺伝子セグメントを組み込む工程であって;ここで、ヒトIg遺伝子セグメントには、非霊長類動物の免疫グロブリン重鎖もしくは軽鎖遺伝子中のコード領域の間隔を隔てるスペーサー配列に由来するスペーサー配列か、あるいは、2つ以上のそのようなスペーサー配列のコンセンサス配列に由来する、少なくとも約20個の連続しているヌクレオチドを含むヌクレオチド配列が隣接しており;ここで、(i)複数の遺伝子セグメントが、再構成されていない立体配置、部分的に再構成された立体配置、または完全に再構成された立体配置において並列しており;そして(ii)ヒト化Ig遺伝子座が遺伝子再構成を受けることができ、そして、必要に応じて、非ヒト動物中でヒト化免疫グロブリンのレパートリーを生産することができる、工程
が含まれる。
【0032】
ヒト化Ig遺伝子座は、ヒト化Ig重鎖または軽鎖遺伝子座であり得る。ヒト化Ig重鎖遺伝子座の場合には、工程(a)で得られたDNAフラグメントには、さらに、少なくとも1つのD遺伝子セグメントが含まれる。
【0033】
別の局面においては、本発明は、上記のヒト化免疫グロブリン遺伝子座を含むトランスジェニック動物と、そのようなトランスジェニック動物を作成するための方法に関する。1つの実施形態においては、トランスジェニック動物には、ヒト化免疫グロブリン重鎖遺伝子座と、ヒト化免疫グロブリン軽鎖遺伝子座の両方が含まれる。別の実施形態においては、トランスジェニック動物中に存在する重鎖遺伝子座と軽鎖遺伝子座のうち1つだけが、ヒト化されている。別の実施局面においては、動物の免疫グロブリン遺伝子座の少なくとも1つのV、D、J、およびC領域の全てがヒト化されている。さらに別の実施形態においては、トランスジェニック動物の内因性免疫グロブリン遺伝子座のV、D、J、およびC領域の全てがヒト化されている。
【0034】
さらなる局面においては、本発明は、本発明にしたがって生産されたトランスジェニック動物に由来するB細胞に関する。
【0035】
なおさらなる局面においては、本発明は、本発明のトランスジェニック動物を使用して生産されるヒト化免疫グロブリンと、ヒト化免疫グロブリンを含む抗体調製物または薬学的組成物に関する。
【図面の簡単な説明】
【0036】
【図1】図1は、ウサギ免疫グロブリン遺伝子の重鎖遺伝子座の模式図である。
【図2】図2は、ウサギ重鎖スペーサー配列の比較を示す。
【図3】図3は、ウサギ重鎖スペーサー配列の比較を示す。
【図4】図4は、ウサギ重鎖スペーサー配列の比較を示す。
【図5】図5は、ウサギ免疫グロブリン軽鎖遺伝子座の模式図である。
【図6】図6は、ヒトVHエレメントとウサギのスペーサーエレメントを用いた免疫グロブリン遺伝子V遺伝子座の組み立てを説明する。
【図7】図7は、redεβγ−システムを用いた相同組み換えによる2つのカセットの挿入である。上のカセットには、ゲンタマイシンカセットに隣接している2つの制限部位(FseIとAscI)が含まれている。下のカセットには、反転させられ(i)変異させられた(71)loxP部位と、FRT部位と、MluI制限部位が含まれている。修飾後、BACはFseIとAscIで消化される。
【図8】図8は、ウサギK1軽鎖遺伝子座をベースとするヒト化ウサギ軽鎖遺伝子座(rLC3−B)を示す。ウサギCκ1をヒトCκで置き換えた。ヒトの再構成ヒトVκJκが挿入されている。合成のヒトVκJκは、ウサギのVκエレメントと80%を超える配列相同性を共有する。
【図9a】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−1】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−2】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−3】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−4】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−5】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−6】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−7】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−8】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−9】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図9b−10】図9aは、ニワトリの軽鎖ゲノム遺伝子座を含むBACクローンの配列を示す。ニワトリの軽鎖ゲノム遺伝子座のヌクレオチド配列を図9bに示す(配列番号186)。
【図10】図10は、ニワトリの免疫グロブリンスペーサー配列とヒトVエレメントを用いた、ヒト化免疫グロブリン遺伝子座の構築を示す概要である。
【図11】図11は、マウスの免疫グロブリンスペーサー配列またはウサギの免疫グロブリンスペーサー配列と、ヒトVエレメントを用いた、ヒト化免疫グロブリン遺伝子座の構築を示す概要である。
【図12a−1】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図12a−2】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図12a−3】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図12a−4】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図12b−1】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図12b−2】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図12b−3】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図12b−4】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図12c】図12aから12cは、ヒト化軽鎖遺伝子座を示す。17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含む合成の配列(図12a、ユニット1、12,235bp、配列番号187)を(a)に示す。機能的な再構成されたヒトVκJκ遺伝子フラグメントと、11個のヒトV偽遺伝子と、12個のニワトリのスペーサー配列と、2つのイントロンを含む、第2の合成の配列(図12b、ユニット2、13,283bp、配列番号188)を(b)に示す。ユニット1とユニット2を、ヒトCκと、ウサギのイントロンと、スペーサー配列(図12c)を含むBAC179L1に由来するフラグメントを用いて結合させた。
【図13a−1】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13a−2】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13b−1】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13b−2】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13b−3】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13b−4】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13b−5】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13c−1】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13c−2】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13c−3】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13c−4】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13d−1】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13d−2】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13d−3】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13d−4】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13d−5】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【図13e】図13a〜eは、ヒト化重鎖遺伝子座を示す。ヒトVH3遺伝子フラグメントと、ウサギのスペーサーと、イントロン配列からなる4個の合成のDNAフラグメント(ユニット1〜4、図13a〜d、配列番号189、190、191、192)を、(図13e)に示すように、BAC 219D23、27N5、およびFos15Bの一部と結合させた。
【0037】
(発明の詳細な説明)
(A.定義)
「抗体」(Ab)と「免疫グロブリン」(Ig)は、同じ構造特性を有している糖タンパク質である。抗体は特異的な抗原に対して結合特異性を示すが、免疫グロブリンには、抗体と、抗原特異性を有していない他の抗体様分子の両方が含まれる。後者のポリペプチドは、例えば、リンパ系によっては低レベルで、そして骨髄腫によっては高レベルで生産される。
【0038】
「自然界に存在している抗体および免疫グロブリン」は、通常、約150,000ダルトンのヘテロ四量体糖タンパク質であり、これは、2つの同じ軽(L)鎖と2つの同じ重(H)鎖から構成されている。個々の軽鎖は、ジスルフィド共有結合(単数または複数)によって重鎖に連結されているが、ジスルフィド結合の数は、それぞれの免疫グロブリンイソ型の重鎖に応じて変化する。重鎖と軽鎖のそれぞれは、規則的な間隔の鎖内ジスルフィド結合もまた有する。個々の重鎖は、一方の末端に可変ドメイン(VH)を、その後に、多数の定常ドメインを有している。個々の軽鎖は、一方の末端に可変ドメイン(VL)を有しており、その他方の末端には定常ドメインを有している;軽鎖の定常ドメインは、重鎖の第1の定常ドメインとアラインメントされ、軽鎖の可変ドメインは、重鎖の可変ドメインとアラインメントされる。特定のアミノ酸残基が、軽鎖可変ドメインと重鎖可変ドメインとの間の界面を形成すると考えられている(Clothia et al.,J.Mol.Biol.186:651(1985);Novotny and Haber,Proc.Natl.Acad.Sci.U.S.A.82:4592(1985))。
【0039】
用語「可変」とは、可変ドメインの特定の部分が抗体の間で配列に関して幅広く異なり、これがその特定の抗原についてそれぞれの特定の抗体の結合と特異性において使用されるという事実を意味する。しかし、可変性は、抗体の可変ドメイン全体にわたって均一に分布しているわけではない。これは、軽鎖および重鎖のいずれの可変ドメインにおいても、相補性決定領域(CDR)または超可変領域と呼ばれる3つのセグメントに集中している。可変ドメインのより高度に保存されている部分は、フレームワーク(FR)と呼ばれる。自然界に存在している重鎖と軽鎖の可変ドメインのそれぞれには、3個のCDRによって繋がれている4個のFR領域が含まれる。それぞれの鎖のCDRはFR領域によって互いに近い位置に保たれ、他の鎖のCDRとともに、抗体の抗原結合部位の形成に関与している(Kabat et al.,Sequences of Proteins of Immunological Interest,Fifth Edition,National Institute of Health,Bethesda,MD(1991)を参照のこと)。定常ドメインは、抗原に対する抗原の結合に直接は関与していないが、抗体依存性細胞傷害性における抗体の関与のような、種々のエフェクター機能を示す。
【0040】
用語「モノクローナル抗体」は、B細胞の単一のクローンによって合成される抗体分子を意味するように使用される。
【0041】
用語「ポリクローナル抗体」は、多数のB細胞クローンによって合成される抗体分子の集団を意味するように使用される。特異的な実施形態においては、ポリクローナル抗体はいくつかのエピトープを認識する。
【0042】
用語「ヒト化抗体」と「ヒト化免疫グロブリン」は、本明細書中で使用される場合は、ヒト免疫グロブリンポリペプチド配列(または、ヒト免疫グロブリン遺伝子セグメントによってコードされるポリペプチド配列)の少なくとも一部を含む免疫グロブリン分子を意味する。本発明のヒト化免疫グロブリン分子は、ヒト化免疫グロブリン分子を生じるように遺伝子操作された非ヒトトランスジェニック動物から単離することができる。このようなヒト化免疫グロブリン分子は、動物から調製されるか、または動物に由来する細胞から調製される非ヒト化免疫グロブリン分子と比較して、霊長類、特に、ヒトに対しては免疫原性が低い。
【0043】
用語「非ヒト動物」には、本明細書中で使用される場合は、哺乳動物が含まれるがこれに限定されない。「非ヒト動物」としては、例えば、非ヒト霊長類、ウサギ、ブタ、鳥類(例えば、ニワトリ、シチメンチョウ、アヒル、ガチョウなど)、ヒツジ、ヤギ、ウシ、ウマ、および齧歯類(例えば、マウスおよびラット)が挙げられる。好ましい非ヒト動物は、遺伝子変換および/または体細胞超変異によって実質的に抗体多様性を生じる動物、例えば、ウサギ、ブタ、鳥類(例えば、ニワトリ、シチメンチョウ、アヒル、ガチョウなど)、ヒツジ、ヤギ、およびウシが挙げられる。特に好ましい非ヒト動物は、ウサギとニワトリである。
【0044】
用語「非霊長類動物」としては、本明細書中で使用される場合には、霊長類ではない哺乳動物が挙げられるがこれに限定されず、これには上記に列挙した動物が含まれる。
【0045】
語句「遺伝子変換および/または超変異によって実質的に抗体多様性を生じる動物」は、抗体多様化の主な機構が、遺伝子再構成とは対照的な遺伝子変換および/または超変異である動物を意味するように使用される。
【0046】
用語「Ig遺伝子セグメント」は、本明細書中で使用される場合は、動物およびヒトの生殖細胞系列に存在し、B細胞中で互いに集まって再構成されたIg遺伝子を形成する、Ig分子の種々の部分をコードするDNAのセグメントを意味する。したがって、本明細書中で使用される場合Ig遺伝子セグメントにはV遺伝子セグメント、D遺伝子セグメント、J遺伝子セグメント、C領域遺伝子セグメントが含まれる。
【0047】
用語「ヒトIg遺伝子セグメント」には、本明細書中で使用される場合は、ヒトIg遺伝子セグメントの自然界に存在している配列と、ヒトIgセグメントの自然界に存在している配列の縮重形態と、さらには、ヒトIg遺伝子セグメントの自然界に存在している配列によってコードされるポリペプチドと実質的に同一であるポリペプチド配列をコードする合成の配列のいずれもが含まれる。「実質的に」によって、アミノ酸配列同一性の程度が少なくとも約85%〜95%であることが意味される。特定の実施形態においては、ヒトIg遺伝子セグメントは、免疫グロブリン分子をヒトにおいて非免疫原性にする。
【0048】
本発明の特異的なヒト化免疫グロブリン分子には、ヒト重鎖可変領域ポリペプチド配列または軽鎖可変領域ポリペプチド配列の少なくとも一部が含まれる。別の特異的な免疫グロブリン分子には、ヒト重鎖もしくは軽鎖可変領域ポリペプチド配列の少なくとも一部と、ヒト定常ドメインポリペプチド配列の少なくとも一部が含まれる。
【0049】
「ヒト化抗体の調製物」または「ヒト化抗体調製物」により、非ヒトトランスジェニック動物から(例えば、動物の血清、乳汁、または卵黄から)、あるいは、非ヒトトランスジェニック動物に由来する細胞から(例えば、B細胞またはハイブリドーマ細胞から)調製された、単離された抗体産物または精製された抗体産物が意味される。
【0050】
ヒト化抗体調製物は、ヒト化免疫グロブリン分子のレパートリーを含むポリクローナル抗体の調製物であり得る。ヒト化抗体調製物は、また、モノクローナル抗体の調製物でもあり得る。
【0051】
用語「抗体多様性」および「抗体のレパートリー」は、ほぼ同じ意味で使用され、生物体が発現できる全抗体特異性の全てを意味する。
【0052】
用語「スペーサー配列」は、免疫グロブリン重鎖または軽鎖遺伝子中に存在する任意の非コードヌクレオチド配列を意味するように、本明細書中で使用される。したがって、この用語には、具体的に、免疫グロブリン重鎖遺伝子中のイントロン配列と、V、D、Jセグメント、およびC領域セグメント中の複数のコード領域の間隔を隔てる任意の他の非コード配列、免疫グロブリン軽鎖遺伝子中のイントロン配列と、VおよびJセグメント、ならびにC領域セグメント中の複数のコード領域の間隔を隔てる任意の他の非コード配列、さらには、免疫グロブリン重鎖もしくは軽鎖遺伝子中の非コード配列である隣接している調節エレメント(例えば、エンハンサー)が含まれる。さらに、膜貫通ヘリックス部分をコードするエキソンと、重鎖および軽鎖のエンハンサーとの間に、非コード配列が具体的に含まれる。
【0053】
遺伝子再構成および遺伝子変換を受ける能力を有しているIg遺伝子座は、また、本明細書中では「機能的な」Ig遺伝子座とも呼ばれ、機能的なIg遺伝子座によって生じる多様性を有している抗体は、また、本明細書中では、「機能的」抗体または抗体の「機能的な」レパートリーとも呼ばれる。
【0054】
(B.関連する文献)
免疫グロブリン遺伝子中の調節エレメントは、Bradley et al.(1999),Transcriptional enhancers and the evolution of the IgH locus;Lauster,R.et al.,Embo J 12:4615−23(1993);Volgina et al.,J Immunol 165:6400(2000);Hole et al.,J Immunol 146:4377(1991)によって記載されている。
【0055】
ニワトリおよびウサギの中での遺伝子変換による抗体多様化は、Bucchini et al.,Nature 326:409−11(1987);Knight et al.,Advances in Immunology 56:179−218(1994);Langman et al.,Res Immunol 144:422−46(1993)によって記載されている。ヒト−マウスキメラ抗体を発現するマウスの作成は、Pluschke et al.,Journal of Immunological Methods 215:27−37(1998)によって記載されている。マウスに由来する膜と細胞質テールを有しているヒト−マウスキメラ抗体を発現するマウスの作成は、Zou et al.Science 262:1271−1274(1993);Zou et al.,Curr Biol 4:1099−1103によって記載されている。ヒト免疫グロブリンポリペプチドを発現するマウスの作成は、Bruggemann et al.Curr Opin Biotechnol 8(4):455−8(1997);Lonberg et al.Int Rev Immunol 13(1):65−93(1995);Neuberger et al.,Nature 338:350−2(1989)によって記載されている。BACクローンを使用するトランスジェニックマウスの作成は、Yang et al.,Nat Biotechnol 15:859−65(1997)によって記載されている。ヒト抗体を発現するウシの作成は、Kuroiwa et al.Nature Biotech 20(9):889−894(2002)によって記載されている。
【0056】
トランスジェニックウサギの作成は、Fan,J.et al.,Pathol Int 49:583−94(1999);Brem et al.,Mol Reprod
Dev 44:56−62(1996)によって記載されている。免疫グロブリンの発現を欠損させたウサギは、McCartney−Francis et al.,Mol
Immunol 24:357−64(1987);Allegrucci,et al.,Eur J Immunol 21:411−7(1991)によって記載されている。
【0057】
トランスジェニックニワトリの作成は、Sherman et al.Nature Biotech 16:1050−1053(1998);Etches et al.,Methods in Molecular Biology 62:433−450;Pain et al.,Cells Tissues Organs 165(3−4):212−9(1999);Sang,H.,“Transgenic chickens−−methods and potential applications”,Trends Biotechnol 12:415(1994);および、WO2004003157,“Gene regulation in transgenic animals using a transposon based vector”;ならびに、WO200075300,“Introducing a nucleic
acid into an avian genome,useful for transfecting avian blastodermal cells for producing transgenic avian animals with the desired genes,by directly introducing the nucleic acid into the germinal disc of the egg.”によって記載されている。
【0058】
γグロブリン血症のニワトリは、Frommel et al.,J Immunol
105(1):1−6(1970);Benedict et al.,Adv Exp Med Biol 1977;88(2):197−205によって記載されている。
【0059】
細胞からの動物のクローニングは、T.Wakayama et al.,Nature 1998;394:369−374;J.B.Cibelli et al.,Science 280:1256−1258(1998);J.B.Cibelli et
al.,Nature Biotechnology 1998;16:642−646;A.E.Schnieke et al.,Science 278:2130−2133(1997);K.H.Campbell et al.,Nature 380:64−66(1996);Kuroiwa et al.,Nature Genetics 2004年6月6日によって記載されている。ウサギの核導入クローニングは、Stice et al.,Biology of Reproduction 39:657−664(1988)、およびChallah−Jacques et al.,Cloning and Stem Cells 8(4):295−299(2003)によって記載されている。
【0060】
トランスジェニック動物からの抗体の産生は、米国特許第5,814,318号、同第5,545,807号、および同第5,570,429号に記載されている。キメラ哺乳動物宿主についての相同組み換えは、米国特許第5,416,260号に説明されている。胚にDNAを導入するための方法は、米国特許第5,567,607号に記載されている。胚性幹細胞の維持および拡大は、米国特許第5,453,357号に記載されている。
【0061】
ブタ、ヒツジ、およびウシにおける抗体のレパートリーの多様化に関係している機構は、Butler,J.E.(1998)“Immunoglobulin diversity,B−cell and antibody repertoire development in large farm animals.”,Rev Sci Tech 17:43に概説されている。ヒツジでの抗体の多様化は、Reynaud,C.A.,C.Garcia,W.R.Heim,and J.C.Weill(1995),“Hypermutation generating the sheep immunoglobulin repertoire is an antigen−independent process”,Cell 80:115;およびDufour,V.,S.Malinge,and F.Nau.(1996),“The Sheep Ig variable region repertoire consists
of a single VH family,”J Immunol 156:2163に記載されている。
【0062】
(C.詳細な説明)
免疫グロブリン重鎖および軽鎖遺伝子には、個々の遺伝子によってコードされ、イントロン配列によって間隔を隔てられているいくつかのセグメントが含まれる。したがって、ヒト免疫グロブリン重鎖についての複数の遺伝子が、14番染色体上で見られる。重鎖(VH)の可変領域には3つの遺伝子セグメント:V、D、およびJセグメントが含まれ、その後ろに、C領域をコードする複数の遺伝子が続く。V領域は、大きなスペーサーによってC領域から隔てられおり、V、D、およびJセグメントをコードする個々の遺伝子もまた、スペーサーによって隔てられている。
【0063】
免疫グロブリンの軽鎖には2つのタイプ:κおよびλが存在する。ヒトκ軽鎖の遺伝子は2番染色体上に見られ、ヒトλ軽鎖の遺伝子は22番染色体上に見られる。抗体軽鎖の可変領域には、VセグメントとJセグメントが含まれ、これらは別々の遺伝子セグメントによってコードされている。κ軽鎖遺伝子の生殖細胞系列での立体配置においては、直線的な配置でおよそ100〜200個のV領域遺伝子が存在しており、それぞれの遺伝子がそれぞれのリーダー配列を有しており、その後ろにおよそ5個のJ遺伝子セグメントと、c領域遺伝子セグメントが続く。全てのV領域は、イントロンによって隔てられており、さらに、V、J、およびC領域遺伝子セグメントを隔てるイントロンが存在する。
【0064】
感染から防御する免疫系の能力は、抗体の多様なレパートリーを作成するように特殊化された遺伝的機構に原因がある。B細胞中では、抗体をコードする遺伝子は、可変(V)領域中に無数の結合部位の組み合わせを生じるような様式で組み立てられる。1012種類を超える可能な結合構造が、そのような機構によって生じると推定される。ヒトを含む全ての動物において、抗体を作成するプロセスは、免疫グロブリン(Ig)遺伝子座の可変(V)セグメント、多様化(D)セグメント、結合(J)セグメントを組換えることによって開始される。この工程の後、動物の種に応じて、2つの一般的な機構が、抗体の多様な結合構造を生じさせるために使用される。
【0065】
ヒトやマウスのようないくつかの動物においては、免疫グロブリン重鎖遺伝子座上に、V、D、およびJ遺伝子セグメントの複数のコピーが、免疫グロブリン軽鎖遺伝子座上にVおよびJ遺伝子セグメントの複数のコピーが存在する。これらの動物における抗体の多様性は、遺伝子再構成、すなわち、再構成された重鎖可変領域と軽鎖可変領域を形成するような遺伝子セグメントの異なる組み合わせによって、主に生じる。しかし、他の動物(例えば、ウサギ、鳥類(例えば、ニワトリ、ガチョウ、およびアヒル)、ヒツジ、ヤギ、およびウシ)においては、遺伝子再構成は、抗体の多様性を生じることにおいてはより小さい役割しか果たしていない。例えば、ウサギにおいては、ごく限られた数のV遺伝子セグメント、最も多くの場合には、V領域の3’末端のV遺伝子セグメントだけが、連続するVDJセグメントを形成するように遺伝子再構成において使用される。ニワトリにおいては、1つのV遺伝子セグメント(D領域に隣接しているもの、すなわち、「3’近接V遺伝子セグメント」)と、1つのDセグメントと、1つのJセグメントだけが、重鎖の再構成に使用される:また、1つのV遺伝子セグメント(3’近接Vセグメント)と、1つのJセグメントだけが、軽鎖の再構成に使用される。したがって、これらの動物においては、接合部の多様化によって生じる最初に再構成された可変領域配列の中には少ししか多様性は存在しない。再構成されたIg遺伝子のさらなる多様化は、遺伝子変換によって行われる。遺伝子変換は、上流のV遺伝子セグメントに由来する短い配列が再構成されたIg遺伝子中のV遺伝子セグメント内の短い配列を置き換えるプロセスである。抗体配列のさらなる多様化は、超変異によって生じる場合もある。
【0066】
免疫グロブリン(抗体)は、5つのクラス(IgG、IgM、IgA、IgE、およびIgD)に属し、これらのそれぞれは、免疫防御において異なる生物学的役割を有している。血液中で最も豊富に存在し、感染に応答する能力を有しているものは、IgGクラスである。ヒトIgGクラスの中には、Fcドメインを含む重鎖定常領域の構造によって決定される4つのサブクラス(IgG1、IgG2、IgG3、およびIgG4イソ型)が存在している。抗体のF(ab)ドメインは、抗原上の特異的な配列(エピトープ)に結合するが、抗体のFcドメインは、抗原を排除するための免疫系の他の成分を動員し、活性化する。
【0067】
抗体は、動物から得られたポリクローナル抗血清によってヒトの生命にかかわる感染症を処置することができることが見出された1890年代から、治療薬としてうまく使用されている。抗体の研究における目覚しい進歩は、抗体の組み換え生産のための方法の開発、それに続く、抗体ヒト化技術の開発と、非ヒト動物において完全なヒトモノクローナル抗体を作成するための方法の開発にともなって生じた。
【0068】
結果として、キメラモノクローナル抗体、ヒト化モノクローナル抗体、およびヒトモノクローナル抗体は、薬学的産物の重要なクラスとして、最近、浮上してきた。モノクローナル抗体をベースとする薬剤は、疾患を処置することにおいて、特定の標的(例えば、受容体またはリガンド)をブロックする場合に極めて有効であるが、ガンおよび感染性病原体への感染のような特定の破壊的疾患は、それらの複雑さ、様々な原因によるとされる病因論、および適応性が原因で、処置することが難しい場合がある。モノクローナル抗体は、集団の全体へ、または個体の中へと疾患が広がる間に、変化し、進化し、そして成熟する、明確に定義された標的に対して向けられたものである。このような適応性の進化は、単特異的薬剤(例えば、モノクローナル抗体)にとっては致命的であり、単特異的薬剤は、耐性株によって迅速に回避される。高力価の抗生物質に耐性である細菌およびウイルスの多く、ならびに、悪性であるガンは、モノクローナル抗体治療のような標準的な抗ガン剤に対する耐性を生じる。
【0069】
対照的に、ポリクローナル抗体は、複雑な疾患に関係している複数の進化しつつある標的に結合し、それらを排除する能力を有している。複数の抗原に結合することにより、ポリクローナル抗体は、抗原の成熟の事象においてもなお、標的を飽和させて活性を保つ。この後、情報伝達のカスケードの間に、ポリクローナル抗体は、強い免疫反応を誘導して、標的抗原、病原体、または細胞を排除する。これらの特性により、ポリクローナル抗体は感染性疾患やガンを処置するために理想的な抗体である。
【0070】
これまでのところ、ポリクローナル抗体の使用は、供給の問題、または非ヒトタンパク質に対する望ましくない反応によって、厳しく制限されてきた。
【0071】
本発明により、免疫グロブリン(Ig)トランス遺伝子座の免疫グロブリンをコードするエレメントの選択的なヒト化に基づく、新しいヒト化アプローチが提供される。このようなヒト−動物トランス遺伝子座の作成により、多様化された親和性の高いヒト化(ポリクローナル)抗体を高い収量で発現するトランスジェニック動物を作成することができる。
【0072】
第1段階として、ヒト以外(霊長類以外を含む)の免疫グロブリン重鎖および軽鎖のゲノム遺伝子座が同定され、配列決定される。例えば、本発明の一部として、ウサギおよびニワトリの免疫グロブリン重鎖および軽鎖のゲノム配列が決定された。これらは、図1、5、および9に示される。
【0073】
ウサギのIg重鎖ゲノム遺伝子座の分析により、免疫グロブリン重鎖可変領域(Vh)には、多数の遺伝子が含まれており、これには、機能的な遺伝子と機能的ではない偽遺伝子が含まれていることが明らかにされた。18個のVh遺伝子のアラインメントにより、ウサギの重鎖可変領域の遺伝子配列(Vh1〜Vh18)の間に高度(80〜90%)な配列同一性があることが明らかになった。ウサギの重鎖可変領域の遺伝子は、ヒトの重鎖可変領域の遺伝子のVh3グループと最も高い相同性を共有している。具体的には、ウサギのVh1−a2遺伝子のヒトVh3−23配列との配列比較により、72.8%の配列同一性が明らかになった。
【0074】
さらに、複数のウサギの重鎖可変領域の遺伝子配列を隔てる非コード(例えば、イントロン)配列を分析した。図2〜4は、ウサギの重鎖イントロン配列の比較を示す。このようなイントロン配列は2つのグループに別れ、これらが高度に保存されていることが見出された。グループ1のイントロンの特別なメンバーは、驚くほど高い(80〜90%)の配列同一性を示す。
【0075】
同様の観察を、ウサギの免疫グロブリン軽鎖可変領域のゲノム配列に分析によって行った。具体的には、ウサギの免疫グロブリン軽鎖遺伝子座の分析により、軽鎖可変領域(V1)領域に多数の遺伝子セグメントが含まれており、これらが高度な(80〜90%)配列同一性を示すことが明らかにされた。さらに、ウサギの軽鎖可変領域(Vκ)は、ヒトの軽鎖可変領域の遺伝子配列のVκ1グループと高い相同性を示すことが明らかにされた。ほとんどのVκ配列は、機能的であり、高度に保存されていることが見出された。ウサギの重鎖可変領域の遺伝子とは異なり、ウサギの軽鎖可変領域の遺伝子中では、イントロン配列は異なることが見出されている。
【0076】
ニワトリの免疫グロブリン重鎖および軽鎖のゲノム配列を用いた同様の研究によっても、同様の結果が得られた。
【0077】
1つの局面においては、本発明により、スペーサー配列が提供される。スペーサー配列は、非霊長類動物の重鎖または軽鎖の遺伝子中のコード領域の間隔を隔てる。1つの実施形態においては、本発明により、遺伝子変換によって実質的に抗体多様性を生じる動物(例えば、ウサギおよびニワトリを含む)の重鎖および軽鎖の遺伝子に由来するスペーサー配列が提供される。このようなスペーサー配列は、その後、ヒト化免疫グロブリン遺伝子座を作成するプロセスにおいて使用されるヒト免疫グロブリン重鎖または軽鎖遺伝子セグメントに隣接させるように使用される。
【0078】
スペーサー配列には、通常は、少なくとも20個のヌクレオチド、または少なくとも約30個のヌクレオチド、または少なくとも約40個のヌクレオチド、または少なくとも約50個のヌクレオチドが含まれ、そして通常は、約20個から約10000個のヌクレオチドの長さである。スペーサー配列には、非ヒト動物(例えば、霊長類以外)中の自然界に存在しているイントロン配列に由来する適切な長さのヌクレオチドの連続するストレッチが含まれる場合があり、また、人工的な配列が含まれる場合もある。人工的な配列は、例えば、2つ以上の自然界に存在しているイントロン配列のコンセンサス配列であり得る。
【0079】
スペーサー配列には、配列番号1から185(表1)からなる群より選択される配列に由来するか、あるいは、2つ以上のそのような配列のコンセンサス配列に由来する、少なくとも約20(30、40、50などであって、10ヌクレオチドずつ増大して1000まで)個の連続しているヌクレオチドが含まれる場合がある。必ずしもそうである必要はないが、その免疫グロブリンがヒト化させられるヒト以外(霊長類以外)の動物のゲノム配列中の対応する領域の間隔を隔てるスペーサー配列により、ヒト化に使用されるヒト重鎖または軽鎖の配列(例えば、V、D、J、C領域の配列)の間隔を隔てることもできる。
【0080】
一般的には、非ヒト動物の免疫グロブリン(Ig)遺伝子座のヒト化には、ヒト化免疫グロブリン遺伝子座を作成するために動物のゲノム中に1つ以上のヒトIg遺伝子セグメントを組み込むことが含まれる。したがって、ヒト化Ig重鎖遺伝子座の作成には、1つ以上のVおよび/またはDおよび/またはJセグメント、ならびに/あるいはC領域セグメントの動物のゲノムへの組み込みが含まれる。同様に、ヒト化Ig軽鎖遺伝子座の作成には、1つ以上のVおよび/またはJセグメント、ならびに/あるいはC領域セグメントの動物のゲノムへの組み込みが含まれる。
【0081】
使用されるアプローチに応じて、ヒトIg遺伝子セグメント(単数または複数)を、動物の内因性のIg遺伝子座が通常存在する染色体位置に、あるいは、動物の異なる遺伝子座に組み込むことができる。染色体の位置にはかかわらず、本発明のヒト化Ig遺伝子座は、非ヒト動物中で、遺伝子再構成、および遺伝子変換、および超変異を受ける能力を有しており、それによってヒト化Ig分子の多様化されたレパートリーを生じる。遺伝子再構成および遺伝子変換を受ける能力を有しているIg遺伝子座はまた、「機能的な」Ig遺伝子座とも呼ばれ、機能的なIg遺伝子座によって作成される多様性を有している抗体はまた、「機能的な」抗体または抗体分子の「機能的な」レパートリーとも呼ばれる。
【0082】
さらなる局面においては、本発明により、非霊長類動物のIg重鎖または軽鎖遺伝子中の複数のコード領域の間隔を隔てるスペーサー配列に由来するか、あるいは、2つ以上のそのようなスペーサー配列のコンセンサス配列に由来する、少なくとも約20個の連続しているヌクレオチドを含むヌクレオチド配列が隣接している、ヒトIg遺伝子セグメントを含む核酸分子が提供される。隣接配列中のスペーサー配列に由来するセクションとしての隣接配列は、同一であっても、異なっていてもよい。スペーサー配列に由来するか、あるいは2つ以上のスペーサー配列のコンセンサス配列に由来する、連続しているヌクレオチドは、ヒトIg遺伝子セグメントに対して直接融合させることができる。あるいは、ヒトIg遺伝子セグメントと、少なくとも1つのスペーサーを起源とするヌクレオチド配列との間の介在配列である場合もある。したがって、例えば、ヒトV遺伝子セグメントの5’末端の隣接配列にはプロモーター領域が含まれ、これは、ヒトV遺伝子セグメントに対して直接連結させられ、少なくとも20ヌクレオチドのスペーサー配列に由来するヌクレオチドのストレッチからのその間隔が隔てられる。
【0083】
さらに別の局面においては、本発明は、ヒト重鎖V、D、および/またはJ遺伝子セグメント、ならびに/あるいはC領域セグメントが、もとの非ヒト動物の免疫グロブリン遺伝子中と同じ立体配置で存在し、非霊長類動物のIg重鎖または軽鎖遺伝子中のコード領域の間隔を隔てるイントロン配列に由来する少なくとも約20個の連続しているヌクレオチドを含む配列によって間隔を隔てられている、ヒト化Ig重鎖遺伝子座に関する。別の実施形態においては、本発明により、ヒト軽鎖C領域セグメント、および/またはJ遺伝子セグメント、および/またはV領域セグメントが、もとの非ヒト動物の免疫グロブリン遺伝子と同じ立体配置で非ヒト動物(例えば、霊長類以外)のイントロン配列によって間隔を隔てられている、ヒト化軽鎖遺伝子座が提供される。特定の実施形態においては、スペーサー配列は、ヒト以外(霊長類以外)の動物の非コード配列(例えば、イントロン配列)に基づいて設計される。1つの実施形態においては、スペーサーは、ヒト以外(霊長類以外)の動物に由来する適切な非コード配列を保持し得る。あるいは、構築を簡単にするために、高度に相同である非コード(イントロン)配列に基づいて設計されるコンセンサス配列が設計され、複数のヒト重鎖または軽鎖の遺伝子セグメントの調製のための同じ型のスペーサー配列として使用される。
【0084】
本発明により、具体的には、ヒト化免疫グロブリン遺伝子座の構築に有用な、単離された核酸配列およびベクターが提供される。
【0085】
1つの実施形態においては、ヒト化されるIg遺伝子座を含むDNAフラグメントは、遺伝子変換により抗体多様性を生じる動物(例えば、ウサギおよびニワトリ)から単離される。このような大きなDNAフラグメントは、ヒト以外、例えば、非霊長類動物のゲノムDNAから調製することができる、プラスミド、コスミド、酵母人工染色体(YAC)、または細菌の人工染色体(BAC)などのライブラリーをスクリーニングすることによって単離することができる。動物のC領域の全体を、1つのプラスミドまたはコスミドクローンに含めることができ、これは続いて、ヒト化される。YACクローンには、2メガ塩基までのDNAフラグメントを持たせることができ、したがって、動物の重鎖遺伝子座全体、またはその大きな部分を1つのYACクローン中に単離することができ、また、1つのYACクローン中に含ませるように再構築することもできる。BACクローンは、より小さい大きさ(約150〜450kb)のDNAフラグメントを持つことができる。しかし、Ig遺伝子座の重複するフラグメントを含む複数のBACクローンを別々にヒト化して、その後、動物のレシピエント細胞に一緒に注入することができる。ここで、重複しているフラグメントは、レシピエント動物の細胞中で組み換わって連続しているIg遺伝子座を生じる。
【0086】
ヒトIg遺伝子セグメントは、DNAフラグメントの連結、または相同組み換えによるDNAフラグメントの挿入を含む種々の方法によって、ベクター(例えば、BACクローン)上のIg遺伝子座に組み込むことができる。ヒトIg遺伝子セグメントの組み込みは、ヒトIg遺伝子セグメントがトランス遺伝子中で宿主動物配列に動作可能であるように連結されて、機能的なヒト化Ig遺伝子座、すなわち、遺伝子再構成、および遺伝子変換、および超変異が可能であるIg遺伝子座を生じ、これによってヒト化抗体の多様化されたレパートリーの生産を導くように、行われる。
【0087】
1つの実施形態においては、ヒトIg遺伝子セグメントは、相同組み換えによってIg遺伝子座に組み込むことができる。相同組み換えは、細菌、酵母、および他の細胞中で、高頻度の相同組み換え事象で行うことができる。例えば、酵母細胞は、動物のIg遺伝子座またはその大きな部分を含むYACで形質転換される。続いて、そのような酵母細胞は、さらに、本明細書中で上記に記載されるような組み換えベクターで形質転換される。組み換えベクターは、5’隣接配列と3’隣接配列に連結されたヒトIg遺伝子セグメントを有している。組み換えベクター中の5’隣接配列と3’隣接配列は、YAC上の動物のIg遺伝子セグメントの隣接配列と相同である。相同組み換えの結果、YAC上の動物のIg遺伝子セグメントは、ヒトIg遺伝子セグメントで置き換えられる。あるいは、大腸菌(E.coli)のような細菌細胞は、動物のIg遺伝子座またはその大きな部分を含むBACで形質転換される。そのような細菌細胞は、さらに、5’隣接配列と3’隣接配列に連結されているヒトIg遺伝子セグメントを有している組み換えベクターで形質転換される。組み換えベクター中の5’隣接配列と3’隣接配列は、相同組み換えを媒介し、組み換えベクター上のヒトIg遺伝子セグメントと、BAC上の動物のIg遺伝子セグメントとを交換する。ヒト化YACとBACは、細胞から容易に単離することができ、トランスジェニック動物の作成に使用することができる。
【0088】
本発明のさらなる局面においては、ヒト化免疫グロブリンを生産することができるトランスジェニック動物を作成する方法が提供される。
【0089】
本発明にしたがって、ヒト化免疫グロブリンを作成することができるトランスジェニック動物が、レシピエント細胞または動物の細胞中に、ヒト化Ig遺伝子座を有している本明細書中の上記に記載されている1つ以上のトランスジェニックベクターを導入し、遺伝子修飾されたレシピエント細胞(単数または複数)から動物へと導くことによって、作成することができる。
【0090】
レシピエント細胞は、例えば、非ヒト動物に由来するものであり得る。非ヒト動物は、遺伝子変換、および/または超変異によって抗体多様性を生じる動物、例えば、鳥類(例えば、ニワトリ)、ウサギ、ウシなどである。このような動物においては、3’近接V遺伝子セグメントは、免疫グロブリンの生産のために優先的に使用される。動物の3’近接V遺伝子セグメントの置き換えによるか、または3’近接V遺伝子セグメントの極近くに置くことによるかのいずれかによる、トランスジェニックベクター上のIg遺伝子座へのヒトV遺伝子セグメントの組み込みにより、免疫グロブリンの大部分において、ヒトV領域ポリペプチド配列の発現が生じる。あるいは、再構成されたヒトV(D)Jセグメントを、トランスジェニックベクター上の免疫グロブリン遺伝子座のJ遺伝子座中に挿入することもできる。
【0091】
ヒト化Ig遺伝子座を含むトランスジェニックベクターは、レシピエント細胞(単数または複数)に導入され、その後、ランダムな組み込みによって、または標的化された組み込みによって、レシピエント細胞(単数または複数)のゲノム中に組み込まれる。
【0092】
ランダムな組み込みのために、ヒト化Ig遺伝子座を含むトランスジェニックベクターを、標準的なトランスジェニック技術によって動物のレシピエント細胞中に導入することができる。例えば、トランスジェニックベクターを、受精卵の前核に直接注入することができる。トランスジェニックベクターは、また、卵母細胞の受精の前に、精子をトランスジェニックベクターとともにインキュベーションすることによって導入することもできる。トランスジェニック動物は、受精卵から発生させることができる。トランスジェニックベクターを導入することに代わる別の方法は、胚性幹細胞をトランスフェクトし、その後、遺伝子修飾された胚性幹細胞を胚発生させることによる。あるいは、トランスジェニックベクター(裸のもの、または促進試薬と組み合わせたもの)を、発生しつつある胚に直接注入することができる。最終的には、キメラであるトランスジェニック動物が、トランスジェニック動物の少なくともいくつかの体細胞のゲノム中に組み込まれたヒト化Igトランス遺伝子を含む胚から作成される。
【0093】
特定の実施形態においては、ヒト化Ig遺伝子座を含むトランス遺伝子が、内因性の免疫グロブリン遺伝子の発現が欠損している動物株に由来するレシピエント細胞(例えば、受精卵、または発生しつつある胚)のゲノム中にランダムに組み込まれる。このような動物株の使用により、ヒト化トランスジェニックIg遺伝子座からの免疫グロブリン分子の優先的な発現が可能になる。このような動物の例としては、AliciaおよびBasileaウサギ株、さらには、ガンマグロブリン血症(Agammaglobinemic)ニワトリ株、さらには、免疫グロブリンノックアウトマウスが挙げられる。あるいは、ヒト化免疫グロブリントランス遺伝子または遺伝子座を有しているトランスジェニック動物を、内因性免疫グロブリンの発現が欠損している動物株と交配させることができる。内因性Ig遺伝子座の欠損と、ヒト化トランスジェニックIg遺伝子座についてホモ接合型である子孫を得ることができる。
【0094】
標的化された組み換えのためには、トランスジェニックベクターを、胚性幹細胞、またはすでに分化した体細胞のような、適切な動物のレシピエント細胞中に導入することができる。以後、トランス遺伝子が動物のゲノムに組み込まれており、相同組み換えによって対応する内因性のIg遺伝子座が置き換えられている細胞を、標準的な方法によって選択することができる。例えば、Kuroiwa et al,Nature Genetics 2004年6月6日を参照のこと。選択された細胞は、その後、徐核された核導入ユニット細胞、例えば、卵母細胞または胚性幹細胞、全能性のきちんとした機能を有している新生児を形成することができる細胞と融合させられる。融合は、十分に確立されている従来技術にしたがって行われる。卵母細胞の徐核と、核の導入は、また、注入用ピペットを使用する顕微手術によって行うこともできる。(例えば、Wakayama et al.,Nature(1998)394:369を参照のこと)。得られた卵細胞は、その後、適切な培地中で培養されて、トランスジェニック動物を作成するために同調させたレシピエントに導入される。あるいは、選択された遺伝子修飾された細胞を、発生しつつある胚に注入して、その後、キメラ動物中へと発生させることもできる。
【0095】
さらに、本発明にしたがって、ヒト化免疫グロブリンを生産することができるトランスジェニック動物を、また、レシピエント細胞(単数または複数)中に、内因性のIg遺伝子セグメントの隣接配列に相同である5’隣接配列と3’隣接配列に連結されたヒトIg遺伝子セグメントを有している、本明細書中で上記に記載されている1つ以上の組み換えベクターを導入し、内因性のIg遺伝子セグメントが相同組み換えによってヒトIg遺伝子セグメントによって置き換えられている細胞を選択し、そして選択された遺伝子修飾されたレシピエント細胞(単数または複数)から動物へと誘導することによって、作成することもできる。
【0096】
トランスジェニックベクターの標的挿入と同様に、このアプローチにおいてレシピエントとしての使用に適切な細胞としては、胚性幹細胞、またはすでに分化した体細胞が挙げられる。ヒトIg遺伝子セグメントを有している組み換えベクターを、例えば、トランスフェクションのような、任意の適切な手段によってそのようなレシピエント細胞中に導入することができる。以後、ヒトIg遺伝子セグメントが相同組み換えによって対応する内因性Ig遺伝子セグメントと置き換わっている細胞を、標準的な方法によって選択することができる。これらの遺伝子修飾された細胞を、トランスジェニック動物のクローニングのための核導入手順において、核ドナー細胞とすることができる。あるいは、選択された遺伝子修飾された胚性幹細胞を、発生しつつある胚に注入することができる。これは続いて、キメラ動物へと発達させることができる。
【0097】
上記の方法のいずれかによって作成されたトランスジェニック動物は、本発明の別の実施形態を形成する。トランスジェニック動物は、少なくとも1つ、すなわち、1つ以上の、ヒト化Ig遺伝子座をゲノム中に有しており、この遺伝子座から、ヒト化抗体の機能的なレパートリーが生産される。
【0098】
特異的な実施形態においては、本発明により、ゲノム中に1つ以上のヒト化Ig遺伝子座を有しているトランスジェニックウサギが提供される。本発明のトランスジェニックウサギは、ヒト化Ig遺伝子座を再構成および遺伝子変換することができ、ヒト化された交代の機能的なレパートリーを発現することができる。
【0099】
別の特異的な実施形態においては、本発明により、1つ以上のヒト化Ig遺伝子座をゲノム中に有しているトランスジェニックニワトリが提供される。本発明のトランスジェニックニワトリは、ヒト化Ig遺伝子座を再構成および遺伝子変換することができ、ヒト化抗体の機能的なレパートリーを発現することができる。別の特異的な実施形態においては、本発明により、1つ以上のヒト化V領域をゲノム中に有しているトランスジェニックマウスが提供される。ヒト化V領域には、非ヒトスペーサー配列が隣接している少なくとも2つのヒトV遺伝子セグメントが含まれる。トランスジェニックマウスは、ヒトVエレメントを再構成することができ、抗体の機能的なレパートリーを発現することができる。
【0100】
一旦、多様化させられたヒト化免疫グロブリン分子を生産することができる非ヒトトランスジェニック動物が作成されると、抗原に対するヒト化免疫グロブリンおよびヒト化抗体調製物を、抗原で動物を免疫化することによって容易に得ることができる。種々の抗原を使用して、トランスジェニックである宿主動物を免疫化することができる。このような抗原としては、微生物、例えば、ウイルス、および単細胞微生物(例えば、細菌および真菌)、生存している微生物、弱毒化された微生物、または死滅させた微生物、微生物のフラグメント、あるいは、微生物から単離された抗原性分子が挙げられるが、これらに限定されない。
【0101】
動物の免疫化に使用される例示的な細菌抗原としては、莢膜多糖類であるタイプ5および8のような、黄色ブドウ球菌(Staphylococcus aureus)から精製された抗原、α毒素、付着因子結合タンパク質、コラーゲン結合タンパク質、およびフィブロネクチン結合タンパク質のような感染性因子の組み換えバージョンが挙げられる。例示的な細菌抗原としては、また、黄色ブドウ球菌(S.aureus)、シュードモナス・アエルギノーサ(Pseudomonas aeruginosa)、腸球菌(enterococcus)、腸内細菌(enterobacter)、および肺炎桿菌(Klebsiella pneumoniae)の弱毒化されたバージョン、あるいは、これらの細菌細胞に由来する培養上清が挙げられる。免疫化に使用することができる他の細菌抗原としては、精製されたリポ多糖(LPS)、莢膜抗原、莢膜多糖類、および/または外膜タンパク質の組み換えバージョン、フィブロネクチン結合タンパク質、ならびに、シュードモナス・アエルギノーサ(Psuedomonas aeruginosa)、超球菌(enterococcus)、腸内細菌(enterobacter)、および肺炎桿菌(Klebsiella pneumoniae)由来の内毒素および外毒素が挙げられる。
【0102】
真菌に対する抗体の作成のための例示的な抗原としては、真菌の弱毒化されたバージョン、またはその外膜タンパク質が挙げられる。真菌としては、カンジダ・アルビカンス(Candida albicans)、カンジダ・パラシロシ(Candida parapsilosis)、カンジタ・トロピカリス(Candida tropicalis)、およびクリプトコッカス・ネオホルマンス(Cryptococcus neoformans)が挙げられるが、これらに限定されない。
【0103】
ウイルスに対する抗体を作成するための免疫化に使用される例示的な抗原としては、ウイルスの外膜タンパク質および弱毒化させられたバージョンが挙げられる。ウイルスとして、呼吸器合胞体ウイルス(respiratory syncytial virus)(RSV)(特に、F−タンパク質)、C型肝炎ウイルス(HCV)、B型肝炎ウイルス(HBV)、サイトメガロウイルス(CMV)、EBV、およびHSVが含まれるが、これらに限定されない。
【0104】
治療用抗体を、単離された腫瘍細胞または腫瘍細胞株;腫瘍関連抗原(これには、Her−2−neu抗原(それに対する抗体が乳ガンの処置に有用である);CD19、CD20、CD22、およびCD53抗原(それに対する抗体がB細胞リンパ腫の処置に有用である);(3)前立腺特異的膜抗原(PMSA)(それに対する抗体が前立腺ガンの処置に有用である)、および17−1A分子(それに対する抗体が結腸ガンの処置に有用である)が含まれるが、これらに限定されない)でトランスジェニック動物を免疫化することによって、ガンの処置のために作成することができる。
【0105】
抗原は、任意の従来の様式でトランスジェニック宿主動物に、アジュバントとともに、またはアジュバントなしで投与することができ、予め決定されたスケジュールに従って投与することができる。
【0106】
免疫化後、免疫化されたトランスジェニック動物に由来する血清または乳汁を、抗原に特異的な薬学的等級のポリクローナル抗体を精製するために分画することができる。トランスジェニックである鳥類の場合には、抗体は、また、卵黄を分画することによっても得ることができる。濃縮され、精製された免疫グロブリン画分は、クロマトグラフィー(アフィニティー、イオン交換、ゲル濾過など)、硫酸アンモニウムのような塩、エタノールのような有機溶媒、またはポリエチレングリコールのような高分子を用いる選択的な沈降によって、得ることができる。
【0107】
モノクローナル抗体を作成するために、脾細胞が、免疫化されたトランスジェニック動物から単離され、これは、ハイブリドーマの生産のための形質転換細胞株との細胞融合において使用されるか、または、抗体をコードするcDNAが、標準的な分子生物学的技術によってクローニングされ、トランスフェクトされた細胞中で発現させられるかのいずれかが行われる。モノクローナル抗体を作成するための手順は、当該分野で十分に確立されている。例えば、欧州特許第0 583 980 A1号(“Method For Generating Monoclonal Antibodies From Rabbits”)、米国特許第4,977,081号(“Stable Rabbit−Mouse Hybridomas And Secretion Products Thereof”)、WO97/16537(“Stable Chicken B−cell Line And Method of Use Thereof”)、およびEP
0 491 057 B1(“Hybridoma Which Produces Avian Specific Immunoglobulin G”)を参照のこと。これらの開示は、引用により本明細書中に組み入れられる。クローニングされたcDNA分子からのモノクローナル抗体の生体外での生産は、Andris−Widhopf et al.,“Methods for the generation of chicken monoclonal antibody fragments by phage display”、J Immunol Methods 242:159(2000)によって、そして、Burton,D.R.,“Phage display”Immunotechnology 1:87(1995)によって記載されている。それらの開示は、引用により本明細書中に組み入れられる。
【0108】
B細胞のような、本発明のトランスジェニック動物に由来する細胞、または抗原に対して免疫化されたトランスジェニック動物から確立された細胞株もまた、本発明の一部である。
【0109】
本発明のさらなる局面においては、疾患の処置に望ましい、精製されたヒト化抗体組成物、好ましくは、ヒト化ポリクローナル抗体組成物を投与することにより、霊長類、具体的には、ヒト患者の疾患を処置するための方法が提供される。
【0110】
本発明の別の局面においては、精製されたモノクローナル抗体またはポリクローナル抗体が、霊長類、特に、ヒトへの投与に適している適切な薬学的担体と混合させられて、薬学的組成物が提供される。本発明の薬学的組成物において使用することができる薬学的に許容される担体は、任意の全ての溶媒、分散媒体、等張化剤などであり得る。任意の従来の媒体、試薬、希釈剤、または担体は、レシピエントに対して、またはその中に含まれる抗体の治療有効性に対して不利益をもたらさない限りは、本発明の薬学的組成物中でのその使用は適切である。担体は、液体、半固体(例えば、ペースト状)、または固体の担体であり得る。担体の例としては、油、水、生理食塩溶液、アルコール、糖、ゲル、脂質、リポソーム、樹脂、多孔性基質、結合剤、増量剤、コーティング剤、保存料など、あるいはそれらの組み合わせが挙げられる。
【0111】
投与に使用されるヒト化ポリクローナル抗体組成物は、通常、0.1から100mg/mlまでの免疫グロブリン濃度、より一般的には、1から10mg/mlまでの濃度のポリクローナル抗体の集団を含むことを特徴とする。抗体組成物には、種々のイソ型の免疫グロブリンが含まれる場合がある。あるいは、抗体組成物には、1つにイソ型の抗体のみが含まれる場合も、また、複数の選択されたイソ型の抗体が含まれる場合もある。
【0112】
ほとんどの場合においては、抗体組成物は、修飾されていない免疫グロブリン、すなわち、さらに例えば、化学的もしくは酵素的に、修飾されることなく動物から、調製されたヒト化抗体から構成される。あるいは、免疫グロブリン画分に、酵素消化(例えば、ペプシン、パパイン、プラスミン、グリコシダーゼ、ヌクレアーゼなどで)、加熱などのような処理が行われる場合もあり、そして/また、さらに分画される場合もある。
【0113】
抗体組成物は、通常は、血管系に投与され、適切な静脈に入れられたカテーテルを通じて、注射または注入によって静脈内に投与することが好都合である。抗体組成物は、適切な速度で、通常は、約10分から約24時間の範囲で、さらに一般的には、約30分から約6時間の範囲で、患者が順応できる液体の速度によって投与される。有効投与量の投与は、1回の注入の中に存在させることができ、また、一連の注入の中に存在させることもできる。反復注入は、抗体調製物の半減期および臨床的な適応症に応じて、1日に1回、1週間に1回、1ヶ月に1回、または3ヶ月ごとに1回投与することができる。上皮表面への塗布については、抗体組成物は、意図される最終的な結果をもたらすために十分な量で処置が必要な表面に塗布され、そして必要な場合には繰り返すことができる。さらに、抗体は、例えば、筋肉内のボーラス注射として投与することができ、これには、その後、例えば、注入による持続的な投与が続く場合があるが、これは必ずしも必要ではない。
【0114】
抗体組成物は、疾患の原因となるか、あるいは、望ましくないかまたは異常な免疫反応を誘発する、ヒトの体組織中の抗原性物質に結合し、それを中和するために使用することができる。「抗原性物質」は、本明細書中では、タンパク質を含む任意の可溶性または細胞表面結合分子、さらには、細胞、または、疾患を引き起こす微生物、あるいは、少なくとも抗体に結合することができ、好ましくは、免疫反応を刺激することもできる因子を含むように定義される。
【0115】
単剤療法として、または化学療法と組み合わせた、感染性因子に対する抗体組成物の投与により、感染性粒子の排除がもたらされる。抗体の単独での投与により、感染性粒子の数は、10倍から100倍、より一般的には、1000倍以上減少する。同様に、単剤療法として、または化学療法との組み合わせにおいて使用された、悪性疾患を有している患者における抗体療法により、悪性細胞の数は、通常、10倍から100倍、あるいは、さらに1000倍減少する。治療は、感染性粒子、悪性細胞などの完全な排除が確実となるまでの期間、繰り返すことができる。いくつかの症例においては、抗体調製物での治療が、検出できる量の感染性粒子または望ましくない細胞が存在しなくなるまでの期間、続けられる。同様に、免疫反応の調節のための抗体療法の使用は、1回の治療用抗体の投与から構成される場合も、また、複数回の投与から構成される場合もある。治療は、任意の疾患の兆候がなくなるまでの期間、続けられ得る。
【0116】
この処置は、感染性疾患または悪性腫瘍を阻害するために十分な投与量で、化学療法と組み合わせて使用することができる。自己免疫疾患の患者または移植のレシピエントにおいては、抗体療法を、免疫反応を阻害するために十分な投与量の免疫抑制療法と組み合わせて使用することもできる。本発明はさらに、以下の実施例によって説明されるが、実施例は決して限定ではない。
【実施例】
【0117】
(実施例1)
(ウサギ免疫グロブリン遺伝子座を含むBACクローンの単離および配列決定)
高分子量のDNAをa2b5雄ウサギから単離した。ウサギを安楽死させ、脾臓と腎臓を取り出し、氷冷したPBS中でリンスした。脂肪と結合組織を除去し、別々に処理した。臓器をフラグメントになるように切り、予め冷却しておいたDounceホモジナイザーにおいてホモジナイズした。上清を冷却した50mlのファルコンチューブに移し、冷却したPBSと混合し、大きな組織の破片を2分間の間、底に沈ませた。上清中の細胞を、200gで4℃で10分間かけてペレット状にし、PBSで1回洗浄し、1mlのPBS中に再度懸濁させ、数えた。5×106、5×107、および5×108個の細胞のセットを、CHEF哺乳動物ゲノムDNAプラグキット(CHEF Mammalian Genomic DNA Plug Kit)(BIORAD)を使用してアガロースプラグに包埋した。HindIIIでの部分的消化の条件を最適化するために、プラグを5個の同じフラグメントに切断し、1から10単位のHindIIIで種々の時間の間、種々の温度で消化した。最良の結果は、4℃で3時間、または37℃で25分間の、2単位のHindIIIを用いた場合に得られた。消化したDNAを、以下のパラメーターを使用してパルスフィールドゲル電気泳動(Pulse Field Gel Electrophoresis)(PFGE)装置上でダブルサイズ分画した:6時間の逆行、15秒間のスイッチ時間;6時間前方へ進める、15秒間のスイッチ時間;20時間前方へ進める、90秒間のスイッチ時間;200V、14℃。所望される大きさの部分的に消化されたDNAを有しているゲルの領域を切り取り、DNAをゲレース(gelase)を使用して単離した。11ngの挿入フラグメントを、1ngのHindIIIで消化したpBELOBAC11と連結し、DH10B細胞中にエレクトロポレーションした。得られたコロニーの1%についてNotIを使用して大きさを測定し、平均の挿入フラグメントの大きさが124kbであることを明らかにした。1×105個のクローンをナイロンフィルター(Nylon filter)上にスポットし、特異的プローブとのハイブリダイゼーションによってスクリーニングした。
【0118】
スクリーニングのためのプローブを、ウサギ由来のゲノムDNAを使用してPCRによって増幅し、pBlueScriptにクローニング氏、配列決定によって確認した。プライマー対(配列番号193〜208、表2)を、公開されている配列にしたがって設計した。ウサギ重鎖および軽鎖免疫グロブリン遺伝子座を提示するいくつかのBACを単離し、マップした(図1および5)。BACs 219D23 219D23(GenBank Acc.No.AY386695)、225P18(GenBank Acc.No.AY386697)、27N5(GenBank Acc.No.AY386696)、38A2(GenBank Acc.No.AY386694)、179L1(GenBank Acc.No.AY495827)、215M22(GenBank Acc.No.AY495826)、19(GenBank Acc.No.AY495828)、およびフォスミドFos15B(GenBank Acc.No.AY3866968)を配列決定した。配列決定のためのショットガンライブラリーを、pCR−Blunt中に、1.5〜2kbの大きさの挿入フラグメントを含めて構築した。配列分析には、STADENパッケージ(Roger Staden,Cambridge,UK)を使用した。ソフトウェアによってプレギャップを調節し、gap4を組み立てとギャップを閉じるために使用した。配列の特質クリッピング(quality clipping)のために、PHRED(Washington University)とSTADENパッケージを組み合わせた。
【0119】
(実施例2)
(ヒト化ウサギ免疫グロブリン重鎖遺伝子座の構築)
ウサギの免疫グロブリン重鎖遺伝子座の配列を含むBACおよびフォスミドクローンを、定常領域、可変領域、および結合遺伝子セグメント、あるいは、3’エンハンサー領域に特異的なプローブを使用してゲノムDNAライブラリーから単離した。単離したBAC(図1)27N5(GenBank Acc.No.AY386696)、219D23(GenBank Acc.No.AY386695)、225P18(GenBank
Acc.No.AY386697)、38A2(GenBank Acc.No.AY386694)、フォスミドFos15B(GenBank Acc.No.AY3866968)を配列決定した(Ros et al.,Gene 330,49−59)。
【0120】
選択した免疫グロブリンコード配列を、ETクローニングによる大腸菌(E.coli)中での相同組み換えにより、対応するヒトの部分と交換した(E−Chiang Lee et al.,Genomics 73,56−65(2001);Daiguan
Yu et al.,PNAS 97,5978−5983(2000);Muyrers et al.,Nucleic Acids Research 27,1555−1557(1999);Zhang et al.,Nature Biotechnology 18,1314−1317(2000))。
【0121】
あるいは、DNAフラグメントを、生体外での連結によって組み換え、その後、大腸菌(E.coli)を形質転換した。BACおよび/またはFos15B、あるいはその一部を、生体外での連結および形質転換、ETクローニングによって、あるいはCreリコンビナーゼによって媒介される組み込みによって結合させた。
【0122】
ETクローニングのために、標的配列を含むベクターを、誘導性のλファージ組み換え酵素Redα、Redβおよびγを含むストレプトマイシン耐性大腸菌(E.coli)株に形質転換した。これらの組み換えタンパク質を、同時トランスフェクトさせたプラスミドから(プラスミドpSC101を有しているDH10B大腸菌(E.coli)細胞)、またはゲノムに組み込まれたλプロファージから(DY380大腸菌(E.coli)株)のいずれかから発現させた。ETクローニング手順には、2回の相同組み換え工程を含めた。
【0123】
最初の工程では、標的遺伝子座が、選択−対抗選択(selection−counter selection)カセット(例えば、neo−rpsL、これは、ネオマイシンに対する耐性(neo)と、ストレプトマイシンに対する感受性(rpsL)を与える)によって置き換えた。neo−耐性コロニーの単離後、相同組み換えによる選択カセットの挿入を、制限酵素分析と部分的な配列決定によって確認した。
【0124】
第2の工程では、rpsL−neo選択カセットを、新しい配列と交換した。ストレプトマイシン耐性クローンを制限分析と配列決定によって分析した。ETクローニング手順に使用したフラグメントには、20から50bpの長さの隣接配列が含まれており、これは、標的配列と同じであった。連結に使用した配列は、その3’末端と5’末端に適切な制限酵素部位を有していた。これらの部位は、もともと備わっている部位であるか、またはそれらは、適切な部位を含むプライマーを使用してPCRによって導入されたものであるかのいずれかであった。
【0125】
あるいは、配列を合成によって作成した。
【0126】
ヒト化重鎖を、BAC 219D23中のウサギJH、Cμ、およびBAC 27N5中のCγの、対応するヒト部分での、ETクローニングにより置き換えによって構築した。ETクローニング手順に使用したヒト配列は、ヒトゲノムDNAからPCRによって増幅した。
【0127】
ヒトCμ、Cγ、およびJH遺伝子セグメントを、ウサギの標的配列に相同である50bpのプライマー(配列番号209〜214、表2)を使用して増幅した。
【0128】
BACクローン225P18のクローン219D23との連結、およびBAC 27N5のフォスミド15Bとの連結の後、連結させた構築物を大腸菌(E.coli)に形質転換し、Creリコンビナーゼによって媒介される挿入によって繋いだ。これによって、18個のウサギの可変遺伝子、ウサギD領域、ヒトJ領域、ヒトCμ、ヒトCγ、ウサギCε、ウサギCα4、および3’エンハンサーエレメントから構成される機能的な遺伝子座が得られた。
【0129】
トランスジェニック動物の作成のために、ヒト化BACクローンを、3個の重複しているBAC(225P18と219D23とBAC27N5)、または2つの重複している結合させたBAC(225P18−229D23と、BAC27N5−フォスミド15B)を同時にか、または別々のいずれかで、あるいは、1つのBACとして(225P18−219D23−27N5−フォスミド15B)注入した。トランス遺伝子を有している創始動物をPCRによって同定した。
【0130】
(実施例3)
(合成のフラグメントを用いたヒト化免疫グロブリン重鎖遺伝子座の構築)
ヒトV配列とウサギのスペーサーを有している、ユニット1、ユニット2、ユニット3、およびユニット4と示される4個のフラグメント(図13、配列番号189〜192)を化学的に合成した。それぞれのフラグメントには、5’に、AscI制限エンドヌクレアーゼ認識部位が、3’にはlox71 Creリコンビナーゼ認識配列と、その後ろにFseIとMluI制限酵素認識配列が隣接している。ユニット1は、ウサギのスペーサーI29−30、I3−4、I2−3、およびIl−2の3’側半分(I1−2B)によって間隔を隔てられているヒトVH3−49、VH3−11、VH3−7、およびVH3−15可変遺伝子から構成されている。ユニット2は、ウサギのスペーサーI1−2A、(I1−2の5’側半分)、I7−8、I6−7、およびI4−5の3’側半分(I4−5B)によって間隔を隔てられているヒトVH3−48、VH3−43、およびVH3−64から構成されている。ユニット3は、ウサギのスペーサー配列I4−5B、I26−27、I11−12、およびI17−18によって間隔を隔てられているヒトVH3−74、VH3−30、およびVH3−9から構成されている。
【0131】
ユニット3は、上記に加えて、上流のFlpリコンビナーゼ認識標的(FRT)配列の隣接と、その後ろに、上記のAscI部位の前にSglfI制限エンドヌクレアーゼ認識配列を有している。
【0132】
ユニット4は、ウサギのスペーサーI1−2が5’側に隣接しているヒトVH3−23遺伝子、lox66 Creリコンビナーゼ標的配列と、AscIエンドヌクレアーゼ認識配列を有しており、3’には、IV−C(5’側半分)ウサギスペーサー配列が隣接しており、その後ろにMluIエンドヌクレアーゼ認識配列が続く。
【0133】
ゲンタマイシン選択カセットを、AscIとFseI部位を含むプライマー配列番号215および216(表2)を使用してPCR増幅し、AscI、FseI、およびMluIエンドヌクレアーゼ認識部位を含む修飾されたクローニング部位を有しているpGEMベクター(配列番号217および218(表2)を使用してPCRによって修飾したpGEM.Genta)に連結した。
【0134】
ユニット1、2、および3を、pGEM.Genta(Promega)ベクターにクローニングした。
【0135】
ユニット4を、HindIIIで直鎖状にしたカスタマイズしたpBELOBAC11(NEB)ベクターにサブクローニングし、PCR増幅した。正方向プライマー(配列番号219、表2)は、HindIII、PacI、およびAatIIについての制限部位を有しており、逆方向プライマー(配列番号220)は、BamHI、MluI、およびAscIについての制限部位を有している。プライマーは、pBELOBAC11クロラムフェニコール選択カセットを欠失させるように設計した。さらに、ネオマイシン選択カセットを、BamHIとHindIII制限部位を有しているプライマー配列番号221および222(表2)を用いてPCR増幅し、修飾したpBELOBAC11ベクター(pBB11.1)に連結した。
【0136】
ユニット1〜4を、記載されているようにcreによって媒介される組み換えによって組み立てた(Mejia et al.,Genomics 70(2)165−70(2000))。最初に、ユニット1をカスタマイズしたpGEM.Gentaベクターにクローニングし、FseIで消化し、その後、連結によって再度環化させた。このベクターではない(vectorless)構築物で、pBB11.1.Unit4とp706−Creプラスミドを有している大腸菌(E.coli)を形質転換した。ユニット1とPBB11.1.unit4との組み換えの後、ポジティブクローン(ユニット4/1)を、カナマイシンとゲンタマイシンを含む培地上で選択した。クローンを、種々の酵素を用いる制限分析によって特徴付けた。
【0137】
ユニット2の組み換えのために、ユニット4/1挿入フラグメントを、AscIとPacIでの二重消化によって切り出し、pBELOBAC11中に修飾したリンカーとともにクローニングした(pBB11.2:プライマー配列番号223および224(表2)を使用してPCRによって増幅した)。
【0138】
pBELOBAC11を、HindIIIで直鎖状にし、PacIおよびAatIIエンドヌクレアーゼ認識部位をコードする正方向プライマーと、MluIおよびNotIエンドヌクレアーゼ認識部位、ならびに、lox66 Creリコンビナーゼ標的部位をコードする逆方向プライマーを用いて、PCR増幅した。ユニット1/4の連結のために、pBB11.2ベクターをMluIとPacIで開環させた。
【0139】
pGEM.Genta.Unit2を、pGEM.Genta.Unit1について記載したように、環状のベクターではない構築物へと変換し、pBB11.2.Unit4/1と、生体内でのCreによって媒介される組み換えによって繋いだ。続いて、得られた構築物pBB11.2.Unit4/1/2を、ETクローニングによって野生型のloxp部位をlox66標的部位で置き換えることによる、ユニット3とのCreによって媒介される組み換えのために調製した(Muyrers et al.,Nucleic Acids Research 27,1555−1557(1999);Muyers et al.,Trends Biochem.Sci.26(5):325−31(2001))。クロラムフェニコール選択カセットを、BAC標的配列に相同である50bpの配列を含むプライマー(配列番号225および226、表2)を用いてPCRによって増幅した。逆方向プライマーには、lox66部位を含めた。ゲルから精製したPCR産物を、標的BACと、さらに相同組み換えに必要なpSC101プラスミドを有している細胞に形質転換した。ポジティブクローンを、クロラムフェニコールで選択し、制限分析と配列決定によって確認した。pGEM.Genta.Unit3を、ユニット1および2について上記に記載したように生体内での組み換えのために調製し、受容体BAC、さらには、p706−Creプラスミドを有している細胞に形質転換した。ポジティブクローンpBB11.2.Unit4/1/2/3を、ゲンタマイシンを用いて選択し、制限分析によって確認した。pBB11.2.Unit4/1/2/3を、さらにETクローニングによって、lox71標的部位を生じるように修飾した。その後、pBB11.2.Unit4/1/2/3を、BAC 219D23、27N5、およびFos15Bに由来するフラグメントに繋いだ。
【0140】
(実施例4)
(PCR増幅したフラグメントを用いるヒト化免疫グロブリン重鎖遺伝子座の構築)
ヒトVHエレメントを、ゲノムDNA(ClonTech)とプライマー配列番号227〜248(表2)を使用して増幅した。PCR産物を、ゲル電気泳動によって分析し、GENECLEANキット(Q−Biogen)を使用してゲル精製した。その後、増幅産物を、Zero−Blunt TOPO(登録商標)(Invitrogen)に、製造業者の説明書にしたがってサブクローニングした。全ての増幅したVエレメントの配列を確認した。ヒト化V領域の構築のために、Vエレメントを、鋳型としてのプラスミドDNAと、プライマー配列番号249〜270(表2)を使用して増幅した。正方向プライマーには、AscI部位、その後ろに、ウサギスプライシング部位を含めた。逆方向プライマーには、ウサギの組み換えシグナル配列(RSS)と、MluI制限部位を含めた。PCR産物を、GENECLEANキットを使用してゲル精製した。
【0141】
ヒトVHs、V3−33、V3−74、V3−49、V3−21、V3−48、V3−73、V3−7、およびV3−Dは、PCRによっては単離できず、これらは、化学合成した(BlueHeron,Bothel,WA)。制限部位とウサギの調節配列を、合成の間に付加した。
【0142】
ウサギのスペーサー配列を、鋳型としてのBAC 38A2および225P18と、プライマー配列番号271〜288(表2)を使用して増幅した。BAC 225P18をNheIで二重消化し、41kbのフラグメントをゲル精製した。このフラグメントを、スペーサーV1−2、V2−3、V3−4、V4−5、およびV5−6の増幅のための鋳型とした。
【0143】
BAC 225P18を、BstBIで消化し、スペーサーV6−7とV7−8の鋳型をゲル精製した。BAC 38A2のPacIとRsrIIでの二重消化により、スペーサーV21−22とV22−23の鋳型のゲル精製ができた。
【0144】
増幅したスペーサー配列をゲル精製し、XL−PCR−TOPO(登録商標)(Invitrogen)に、製造業者の説明書にしたがってサブクローニングした。
【0145】
VHエレメントとウサギのスペーサー配列を、修飾したpGEM(Promega)とpBS(Strategene)ベクターにサブクローニングした。pGEMベクターをNotIとHindIIIで切断し、FseI、AscI、およびMluI部位を含む化学合成したオリゴヌクレオチド配列(オリゴ1:配列番号289;オリゴ2:配列番号290;表2)に連結させた。ベクターpBSをSacIとKpnIで切断し、制限部位FseI、AscI、およびMluIを含む化学合成したオリゴヌクレオチド配列(オリゴ1:配列番号291;オリゴ2:配列番号292;表2)と連結させた。
【0146】
ゲンタマイシンとネオマイシン選択カセットを、FseIまたはAscI部位を有するプライマー(配列番号293〜296、表2)を使用して増幅し、修飾したpGEMとpBSベクターに連結した。
【0147】
最終的な構築物を、修飾したpBeloBACIIベクター中で組み立てた。pBeloBACIIベクターを、BamHIとHindIIIで開環し、クローニング部位を、FseI、AscI、MluI部位を含むように、化学合成したオリゴヌクレオチド配列(オリゴ1:配列番号297;オリゴ2:配列番号298;表2)を使用して修飾した。
【0148】
BAC 219D23を、ETクローニングを使用して制限部位を導入することによって修飾した(Muyrers et al.,Nucleic Acids Research 27,1555−1557(1999);Muyers et al.,Trends Biochem.Sci.26(5):325−31(2001))。ネオマイシン選択カセットを、プライマー配列番号299および配列番号300(表2)を用いて増幅した。正方向プライマーにはFseI部位を含め、逆方向プライマーにはAscI部位を含めた。
【0149】
精製したPCR産物で、BAC 219D23と、相同組み換えに必要なプラスミドpSC101を有している大腸菌(E.coli)を形質転換した。カセットとBAC中の標的部位の相同組み換えの後、導入した制限部位を制限分析によって確認した。その後、修飾したBAC 219D23をFseIとMluIで消化し、得られた17kbのフラグメント(FseI−Neo−AscIカセット)を、PFGEによって分離し、電気的溶出によって精製した。この精製したフラグメントを、FseIとMluIで開環させた修飾したpBeloBAC IIベクターと連結させた。
【0150】
ヒトVHエレメントをコードする精製したDNAフラグメントを、AscIとMluIで開環させた修飾したpGEM.neoベクターと連結させた。同様に、スペーサー配列を、修飾したpGEM.gentaベクターにサブクローニングした。その後、スペーサー配列を有しているpGEM.gentaベクターをFseIとMluIで切断し、挿入フラグメントを、FseIとAscIで開環させたpGEM.neo.VHベクターと連結させた。この工程を数回繰り返して、いくつかのスペーサーとVHセグメントから構成されるフラグメントを組み立てた。このようなフラグメントをFseIとMluIで切り出し、FseIとAscIで直鎖状にした修飾したpBeloBACIIベクターと連結させた。これらのプロセスを、大きな免疫グロブリンV領域を組み立てるために繰り返した(図6)。ヒト化重鎖遺伝子座を、トランスジェニック動物の作成に使用した。
【0151】
(実施例5)
(ヒト化Cκとヒト化再構成VJを含む、ヒト化ウサギ軽鎖遺伝子座の構築)
ウサギのゲノムBACライブラリーのスクリーニングにより、ウサギの軽鎖κ1遺伝子セグメントを含む3個のBAC(215M22、179L1、および196O2;それぞれ、Gene Bank Acc.No.AY495826、AY495827、およびAY495828)が得られた。ウサギのCκ1を、記載されている(E−Chiang
Lee et al.,Genomics 73,56−65(2001);Daiguan Yu et al.,PNAS 97,5978−5983(2000);Muyrers et al.,Nucleic Acids Research 27,1555−1557(1999);Zhang et al.,Nature Biotechnology 18,1314−1317(2000))ようなETクローニングによって、ヒトCκアロタイプKm3と交換した。ヒトCκ(アロタイプKm3)を、標的配列に相同である50bpの配列を含むプライマー(配列番号301および302、表2)を用いてPCRによって増幅した。相同アームを、ウサギの生殖細胞系列κ(b5;GenBank Accession No.K01363)の公開されている配列に基づいて設計し、Cκのイントロン−エキソン境界と適合させた。BAC 179L1中でのウサギのCκのヒトCκへの交換を、配列決定によって確認した。
【0152】
BAC 179L1−huCκを2回のETクローニングによって修飾した。ネオマイシン選択カセットを、BAC 179L1に相同である50bpの配列を含むプライマー(配列番号303および304、表2)を用いて増幅した。正方向プライマーには、さらに、i−CeuIメガヌクレアーゼ部位を含めた。PCR産物をETクローニングに使用した。ポジティブクローンをネオマイシンで選択し、制限酵素消化と配列決定によって正確であることをチェックした。ゼオシン選択カセットを、BAC 179L1に相同である50bpの配列を含むプライマー(配列番号305および306、表2)を用いて増幅した。正方向プライマーには、さらに、i−SceIメガヌクレアーゼ部位を含めた。PCR産物をETクローニングに使用した。ポジティブクローンをゼオシンで選択し、制限酵素消化と配列決定によって正確であることをチェックした。
【0153】
BAC 215M22を1回のETクローニングによって修飾した。ゲンタマイシン耐性遺伝子を、BAC 215M22に相同である50bpの配列を含むプライマー(配列番号307および308、表2)を用いて増幅した。正方向プライマーには、さらに、i−CeuIメガヌクレアーゼ部位を含め、逆方向プライマーには、i−SceIメガヌクレアーゼ部位を含めた。PCR産物をETクローニングに使用した。得られたクローンをゲンタマイシンで選択し、制限酵素消化と配列決定によって分析した。
【0154】
修飾したBAC 179L1および225M22を、i−CeuIとi−SceIで切断した。98kbのフラグメントと、132kbのフラグメントを精製し、連結させた。得られたクローンを、カナマイシンとクロラムフェニコールで選択し、制限酵素消化、i−SceIとi−CeuI制限部位を含む領域のPCR、および配列決定によって正確であることをチェックした。得られたBACを、179−215−huCκと呼んだ。
【0155】
BAC 179−215−huCκのウサギのJκ1とJκ2を、ETクローニングによって、合成のヒトの再構成したVκJκ遺伝子と置き換えた。ウサギのプロモーター、ウサギのリーダー、ウサギのイントロンと、ヒトVκJκ遺伝子を有しているDNAフラグメントを化学合成した。合成のヒトVJのコドン使用は、ウサギのVκ遺伝子に最も高く相同であるDNA配列となるように最適化した。
【0156】
合成のヒトVJを、BAC 179L1に相同である50bpの配列を含む正方向プライマー(配列番号309、表2)と、ゲンタマイシン耐性遺伝子に相同である配列およびFRT部位を含む逆方向プライマー(配列番号310、表2)を用いてPCR増幅した。ゲンタマイシン耐性遺伝子を、FRT部位を含む正方向プライマー(配列番号311、表2)と、BAC 179L1に相同である50bpおよびFRT部位を有している逆方向プライマー(配列312、表2)を用いて増幅した。合成のヒトVJとゲンタマイシン耐性遺伝子とを、合成のヒトVJ遺伝子についての正方向プライマーと、ゲンタマイシン耐性遺伝子についての逆方向プライマーを使用する重複伸張(overlap extension)PCRによって結合させた。得られたフラグメントをETクローニングに使用した。ポジティブクローンをゲンタマイシンで選択し、制限酵素消化と配列決定によって正確であることを確認した。
【0157】
ゲンタマイシン耐性遺伝子を、Flpリコンビナーゼの発現を通じて、部位特異的組み換えによって除去した。組み換え後、1つのFRTが残った。このFRT部位をETクローニングによって欠失させた。合成のヒトVJに由来する232bpのフラグメントを、(プライマー配列番号313および314(表2)を使用して)PCRによって増幅し、ETクローニングに使用した。得られたコロニーを、FRT部位が存在しないことについて(プライマー配列番号315および316(表2)を使用して)PCRによってスクリーニングし、配列決定によって確認した。
【0158】
BAC 179−215−huCκのネオマイシン耐性遺伝子をETクローニングによって置き換えた。ゲンタマイシン耐性(pRep−Genta;Genebridges)遺伝子を、BAC 179−215−huCκに相同である50bpの配列を含むプライマー(配列番号317および318、表2)を用いてPCRによって増幅した。正方向プライマーには、さらに、loxP部位、attB部位、およびPvuI制限部位を持たせておいた。得られたクローンをゲンタマイシンで選択し、制限酵素消化と配列決定によって正確であることをチェックした。
【0159】
得られたBAC(rLC3−B、図8)を、トランスジェニック動物の作成に使用した。
【0160】
(実施例6)
(複数のヒトVκエレメント、ニワトリのスペーサーエレメント、および再構成されたヒトVJを含む、ヒト化ウサギ軽鎖遺伝子座の構築)
ウサギのゲノムBACライブラリーのスクリーニングにより、ウサギの軽鎖κ1遺伝子セグメントを含む3個のBAC(215M22、179L1、および196O2;それぞれ、Gene Bank Acc.No.AY495826、AY495827、およびAY495828)が得られた。ウサギのCκ1を、記載されている(E−Chiang
Lee et al.,Genomics 73,56−65(2001);Daiguan Yu et al.,PNAS 97,5978−5983(2000);Muyrers et al.,Nucleic Acids Research 27,1555−1557(1999);Zhang et al.,Nature Biotechnology 18,1314−1317(2000))ようなETクローニングによって、ヒトCκアロタイプKm3と交換した。ヒトCκ(アロタイプKm3)を、標的配列に相同である50bpの配列を含むプライマー(配列番号301および302、表2)を用いてPCRによって増幅した。相同アームを、ウサギの生殖細胞系列κ(b5;GenBank Accession No.K01363)の公開されている配列に基づいて設計し、Cκのイントロン−エキソン境界と適合させた。BAC 179L1中でのウサギのCκのヒトCκへの交換を、配列決定によって確認した。
【0161】
2つのDNAフラグメント、17個のヒトのV偽遺伝子と18個のニワトリのスペーサー配列を含むユニット1(12,235bp、図12a、配列番号187)と、リーダーを伴う機能的な再構成されたヒトκVJ遺伝子フラグメント、11個のヒトV偽遺伝子、12個のニワトリのスペーサー配列、イントロン1、およびイントロン2の一部を含むユニット2(13,283bp、図12b、配列番号188)を化学合成し、ベクターpBR322にクローニングした。
【0162】
ユニット1およびユニット2を、制限酵素NgoMIVとAsiSI、またはNgoMIVとAscIでそれぞれ消化し、3個のフラグメントの連結により修飾されたリンカーを有しているpBELOBAC11に連結させた。修飾されたリンカーには、BsiWI制限部位、FRT5部位、rpsL−Neoカセット、AscI部位、およびAsiSI部位が含まれている。リンカーフラグメントを、高品質のポリメラーゼ(Roche)、プライマーCE_1_001_012904(配列番号319、表2)、および鋳型としてのプラスミドpRpsL−Neo(Genebridges)を用いて増幅した。その後、増幅した産物を、pBELOBAC11のBamHIとHindIII部位に連結した。ユニット1とユニット2の連結のために、修飾されたpBELOBAC11をAsiSIとAscIを用いて開環させた。ポジティブクローン(pBELOBAC11 Unit1/2)を制限酵素消化によってチェックした。
【0163】
BAC 179L1(GENBANK Acc.No.AY495827)を、ETクローニングによる2つの修飾された選択カセットの挿入によって修飾した。カセット1は、BAC 179L1に相同である50bpの配列とAscI部位を逆方向プライマー中に含むプライマー(配列番号321および322、表2)を用いて増幅したゲンタマイシン耐性遺伝子である。カセット2は、BAC 179L1に相同である50bpの配列、attB部位、FRT5部位、およびBsiWI部位を正方向プライマー中に含むプライマー(配列番号323および324、表2)を用いて増幅したrps1−Neo選択カセットである。
【0164】
精製したPCR産物で、BACと、相同組み換えに必要なプラスミドpSC101を有している大腸菌(E.coli)を形質転換した。相同組み換えの後、BACの修飾がうまくできたことを、制限消化分析、サザンブロット、および配列決定によって確認した。
【0165】
修飾したBAC 179L1を、制限酵素AscIとBsiWIで切り出した。ヒトCκを含むフラグメントを精製し、同じ制限酵素で開環させたpBELOBAC11 Unit1/2と連結させた。ポジティブクローンを、制限酵素消化によってチェックした。最終的な構築物(図12c)を、トランスジェニック動物の作成のために使用した。
【0166】
(実施例7)
(複数のヒトVκエレメント、ニワトリのスペーサー、および再構成されていないヒトJκ遺伝子座を含む、ヒト化ウサギ軽鎖遺伝子座の構築)
実施例6に記載した構築物を、以下のようなETクローニングによって修飾した。再構成された機能的なVJ配列を、機能的な組み換えシグナル配列(RSS)が隣接している機能的なV1と交換した。RSSを、pBELOBAC11 Unit1/2のV1に相同である50bpの配列を含む正方向プライマー(配列番号325、表2)、および、AscI制限酵素部位を含み、ゲンタマイシン耐性遺伝子に対する相同性を有している逆方向プライマー(配列番号326、表2)を用いて、BAC 179L1からPCR増幅した。ゲンタマイシン耐性遺伝子を、RSSの増幅に使用した逆方向プライマーに相同である配列を含む正方向プライマー(配列番号327、表2)、およびpBELOBAC11
Unit1/2に相同である50bpの配列と、BsiWI制限酵素部位を含む逆方向プライマー(配列番号328、表2)を用いて増幅した。
【0167】
RSSとゲンタマイシン耐性遺伝子を、RSSの増幅のための正方向プライマーと、ゲンタマイシン耐性遺伝子の増幅のための逆方向プライマーを使用する、重複伸張PCRによって結合させた。得られたフラグメントを、ETクローニングによりpBELOBAC11 Unit1/2を修飾するために使用した。ポジティブクローンをゲンタマイシンで選択し、制限酵素消化と配列決定によって分析した。
【0168】
ヒトのCκを有しているBAC 179L1を、ETクローニングによってさらに修飾した。カナマイシン選択カセットを、BAC 179L1に相同である50bpの配列を含むプライマー(配列番号329および330、表2)を用いて増幅した。逆方向プライマーにはまた、AscI制限酵素部位とFRT部位を含めておいた。PCR産物をETクローニングに使用した。アンピシリン選択カセットを、BAC 179L1に相同である50bpの配列を含むプライマー(配列番号331および332、表2)を用いて増幅した。正方向プライマーには、また、attB部位、AsiSI制限酵素部位、およびFRT5部位も含めておいた。逆方向プライマーには、BsiWI制限酵素部位とFRT部位を含めておいた。PCR産物をETクローニングに使用した。ヒトJ領域を、BAC 179L1に相同である50bpの配列を含むプライマー(配列番号333および334、表2)を用いて、ヒトのゲノムDNAから増幅した。PCR産物をETクローニングに使用した。得られたクローンを、制限酵素消化と配列決定によって分析した。
【0169】
ポジティブクローンをAscIとBsiWIで切断した。得られたフラグメントを精製し、同じ制限酵素で切断した修飾したpBELOBAC11 Unit1/2に連結した。ポジティブクローンをアンピシリンで選択し、制限酵素消化と配列決定によって分析した。正しいクローンを、トランスジェニック動物を作成するために使用した。
【0170】
(実施例8)
(複数のヒトVκエレメントを含むヒト化ウサギ軽鎖遺伝子座の構築)
ウサギのゲノムBACライブラリーのスクリーニングにより、ウサギの軽鎖κ1遺伝子セグメントを含む3個のBAC(215M22、179L1、および196O2;それぞれ、Gene Bank Acc.No.AY495826、AY495827、およびAY495828)が得られた。ウサギのCκ1を、記載されている(E−Chiang
Lee et al.,Genomics 73,56−65(2001);Daiguan Yu et al.,PNAS 97,5978−5983(2000);Muyrers et al.,Nucleic Acids Research 27,1555−1557(1999);Zhang et al.,Nature Biotechnology 18,1314−1317(2000))ようなETクローニングによって、ヒトCκアロタイプKm3と交換した。ヒトCκ(アロタイプKm3)を、標的配列に相同である50bpの配列を含むプライマー(配列番号301および302、表2)を用いてPCRによって増幅した。相同アームを、ウサギの生殖細胞系列κ(b5;GenBank Accession No.K01363)の公開されている配列に基づいて設計し、Cκのイントロン−エキソン境界と適合させた。BAC 179L1中でのウサギのCκのヒトCκへの交換を、配列決定によって確認した。
【0171】
Vκ1ファミリー(O2、L8、L4、A30、L11、L1、L5、L15、O8、L19、L12、A20、O4、L14、L23、L9、A4、L24、O6、L22、A9、A25、A15、O9)のヒトVκエレメントを、プライマー(配列番号335〜382、表2)、および鋳型としてのヒトゲノムDNAを使用してPCRによって増幅した。
【0172】
増幅産物をゲル電気泳動によって分析し、GENECLEAN(Q−Biogen)を使用してゲル精製し、Zero−Blunt TOPO(登録商標)ベクター(Invitrogen)にサブクローニングし、配列決定した。再構成されたヒトVκ(O2)Jκ(J4)エレメントが、PCR増幅によって生じ、これをサブクローニングし、配列決定した。ヒトVκエレメントをウサギのスペーサーと結合させるために、ヒトVκエレメントを、プラスミドDNAを鋳型として使用して、プライマー(配列番号383〜430、表2)を用いて、PCRによって増幅した。プライマーには、AscI部位またはMluI部位を含めた。
【0173】
ウサギのスペーサー配列を、プライマー配列番号431〜450(表2)を使用してPCRによって増幅した。BAC 179L1および215M22を、SpeI、NheI、AclI、SfoI、MluI、およびSalI/XhoIで消化した。フラグメントをゲル精製し、増幅鋳型として使用した。
【0174】
5’末端に存在するスペーサー配列を、FRTとattB部位を含む上流オリゴヌクレオチドによって増幅した。PCR産物を、GENECLEANキットを使用してゲル精製し、XL−PCR−TOPO(登録商標)(Invitrogen)に、製造業者の説明書にしたがってサブクローニングした。
【0175】
ヒトVκエレメントとウサギのスペーサー配列を、実施例4に記載したように修飾したpGEM(Promega)にクローニングした。
【0176】
ヒトVκと修飾したpGEM.gentaベクターを、AscIとMluIで消化し、これらを連結させた。同様に、ウサギのスペーサー配列をpGEM.neoにクローニングした。その後、pGEM.neoVκをFseIとAscIで切断し、FseIとMluIで切り出したpGem.genta.spacerの精製した挿入フラグメントと連結させた。AscIとMluIの相補末端の連結により、それらの制限酵素部位は破壊され、いくつかのVκとスペーサーエレメントを含むVκ遺伝子座の構築のために、AscIとMluIを繰り返し使用することができるようになる。ヒト化BAC 179L1および215M22のフラグメントと、ヒト化Vκ領域から構成される最終的な構築物を、pBeloBACの中で組み立てた。BAC 179L1および215M22を修飾し、結合させた。その後、BAC 179L1−215M22−huCκを、ETクローニングによってさらに修飾した。制限酵素部位、選択マーカー、およびさらなる機能的部位を含む2つのカセットを、図7に示すようにETクローニングによってベクター中に挿入した。カセットの増幅のために使用したプライマー(配列番号451〜454)を、表2に列挙する。
【0177】
最終的な構築物を組み立てるために、ヒトVエレメント、ウサギのスペーサーエレメント、および耐性マーカーから構成されるユニットを、pGEMからFseIとMluIで切り出し、FseIとAscIで消化したBAC 179L1−215M22と連結させた。その後、耐性マーカーを、ヒトVエレメント、ウサギのスペーサーエレメント、および別の耐性マーカーから構成される新しい挿入フラグメントと置き換えた。数回繰り返した後、最終的な構築物は、ウサギのスペーサー配列によって隔てられた多くのVκセグメント(L8、L4、A30、L11、L1、L5、L15、O8、L19、L12、A20、O4、L14、L23、L9、A4、L24、O6、L22、A9、A25、A15、O9)から構成される。ヒト化軽鎖遺伝子座を、トランスジェニック動物の作成のために使用する。
【0178】
(実施例9)
(ニワトリ重鎖遺伝子座スペーサー配列を有するヒト化重鎖遺伝子座の構築)
ニワトリのスペーサー配列によって隔てられている、ウサギD領域、ヒトJ領域、ヒトCμ、ヒトCγ、ウサギCα4、ウサギ3’αエンハンサー、およびヒトVHエレメント(プロモーター、および7マー/9マーの配列を含む)を含む、合成のヒト化重鎖遺伝子座を構築する。
【0179】
修飾したウサギBAC 27N5(「実施例2」を参照のこと)を、ETクローニングによってさらに修飾した。この構築物には、ヒト化CμおよびCγと、2つの1つずつしか存在しないBsiWIおよびAsiSI制限部位が、α−4膜エキソンの下流に含まれている。DNAを、オリゴヌクレオチド配列番号455および456(表2)を用いて増幅した。
【0180】
フォスミドFos15BをNheIで消化し、3’αエンハンサーを含む得られた13kbのフラグメントを、挿入フラグメントがBsiWIおよびAsiSI部位と隣接するように、クローニングベクター中にサブクローニングした。その後、挿入フラグメントを、BsiWIおよびAsiSIで切断し、修飾したBAC 27N5と連結させてBAC
27N5Fosを形成させた。
【0181】
BAC 219D23中のウサギJ領域を、ETクローニングによって対応するヒトJ領域と交換した。ヒトJ領域を、プライマー配列番号457および458(表2)を使用してPCRによって増幅した。
【0182】
1つずつしか存在しない制限酵素部位を、BAC 219D23のD領域の上流(A)とCμの上流(B)に挿入した。BAC 27N5Fos中では、制限部位Aを、リンカー領域の上流に挿入し、Bを、BAC 219D23に相同である配列の中に挿入した。酵素AおよびBでの消化後、ヒトJ領域とウサギD領域を含むフラグメントを単離し、BAC 27N5Fosと連結させて、BAC 219D23/27N5Fosを作成した。
【0183】
ニワトリの重鎖スペーサー配列を、ニワトリ重鎖V偽遺伝子(Mansikka et
al.,J Immunol 145(11),3601−3609(1990)、Reynaud et al.,Cell 59(1),171−183(1989))に特異的なプライマー(配列番号459および460、表2)を使用して、PCRによってニワトリのゲノムDNAから増幅した。あるいは、スペーサー配列を化学合成した。
【0184】
PCR産物をゲル精製し、pTOPO(Invitrogen)にクローニングし、配列決定した。
【0185】
ヒト重鎖可変エレメントを、GENBANKに公開されている配列(例えば、Acc.No.NG_001019)にしたがって設計したプライマーを用いてPCRによって増幅したか、あるいは化学合成した。ヒトVエレメントには、ヒトプロモーター領域、ヒトリーダー配列、リーダーとVをコードする領域との間のヒトのイントロン、ヒトVをコードする領域、およびヒトの組み換えシグナル配列が含まれている。増幅したか、または合成したフラグメントには、特異的な制限エンドヌクレアーゼ認識部位が隣接している。ニワトリのスペーサー配列とヒトのVエレメントを、ヒト化免疫グロブリン遺伝子座を含む1つまたはいくつかの大きなDNAフラグメントとなるように結合させた。構築物を、トランスジェニック動物を作成するために使用した。
【0186】
(実施例10)
(ヒトVエレメントおよび非ヒトスペーサー配列を含む(プロモーター領域とRSSは含まない)ヒト化免疫グロブリン遺伝子座の構築)
非ヒトゲノムDNAを用いて作成したBACライブラリーを、免疫グロブリンに特異的なプローブを用いてスクリーニングし、重鎖および軽鎖免疫グロブリンC、J、およびD領域を含むBACクローンを同定した。BACクローンを修飾して、制限酵素部位を含ませた。ヒト重鎖および軽鎖可変エレメントを、GenBank(例えば、Acc.No.NG_001019)に公開されている配列にしたがって設計したプライマーを使用してPCRによって増幅した。配列は、ゲノムDNAから増幅したか、または化学合成した。ヒトVエレメントには、ヒトのプロモーター領域、ヒトのリーダー配列、リーダーとVをコードする領域との間のヒトのイントロン、ヒトVをコードする領域、およびヒトの組み換えシグナル配列(RSS)が含まれている。増幅したか、または合成したフラグメントは、末端に特異的な制限エンドヌクレアーゼ認識部位を有している。非ヒトスペーサー配列は、PCR増幅したか、または化学合成した。非ヒトスペーサー配列とヒトのVエレメントを、ヒト化免疫グロブリン遺伝子座を含む1つまたはいくつかの大きなDNAフラグメントとなるように結合させた。構築物を、トランスジェニック動物を作成するために使用した。ニワトリのスペーサー配列を用いたヒト化V領域の構築の例を、図10に示す。
【0187】
(実施例11)
(ヒトVエレメントおよび非ヒトスペーサー配列を含むヒト化免疫グロブリン遺伝子座の構築)
非ヒトゲノムDNAを用いて作成したBACライブラリーを、免疫グロブリンに特異的なプローブを用いてスクリーニングし、重鎖および軽鎖免疫グロブリンC、J、およびD領域を含むBACクローンを同定した。BACクローンを修飾して、制限酵素部位を含ませた。ヒト重鎖および軽鎖可変エレメントを、GenBank(例えば、Acc.No.NG_001019)に公開されている配列にしたがって設計したプライマーを使用してPCRによって増幅した。配列は、ゲノムDNAから増幅したか、または化学合成した。ヒトVエレメントには、ヒトVをコードする領域が含まれている。非ヒトスペーサー配列は、PCRによって増幅したか、または化学合成した。これには、組み換えシグナル配列、スペーサー配列、プロモーター領域、リーダー配列、およびリーダーとVをコードする領域との間のイントロンが含まれている。このような非ヒトスペーサー配列を、ヒトのVエレメントと、1つまたはいくつかの大きなDNAフラグメントとなるように結合させ、トランスジェニック動物を作成するために使用した。マウスまたはウサギのスペーサー配列を用いたヒト化V領域の構築の例を、図11に示す。
【0188】
(実施例12)
(ヒト化免疫グロブリンを発現するトランスジェニックウサギ)
トランスジェニックウサギを、Fan et al.(Pathol.Int.49:583−594,1999)に記載されているように作成した。簡単に説明すると、雌のウサギを標準的な方法を使用して過剰排卵させ、雄のウサギと交配させた。前核段階の受精卵を卵管から回収し、20%のウシ胎児血清を補充したダルベッコのリン酸緩衝化生理食塩水のような適切な培養液に入れた。ヒト化免疫グロブリン遺伝子座を含むBACを、1対のマニピュレーターの力を借りて雄の前核にマイクロインジェクションした。形態学的に生存している受精卵を、偽妊娠させたウサギの卵管に移した。偽妊娠は、ヒトの絨毛膜性腺刺激ホルモン(hCG)の注射によって誘導した。ヒト化軽鎖構築物を受精卵の4645個の前核に注入した後、4043個の卵母細胞を132匹のレシピエントに移した。全部で253匹の生存性の子孫が産まれ、そのうちの11匹がトランスジェニックであった。ヒトκ軽鎖の発現を、ヒトκ軽鎖特異的試薬(例えば、マウスの抗ヒトκ、Southern Biotech,9220−01;ヤギの抗ヒトκ、Southern Biotech 2063−08)を使用してELISAによって検出した。
【0189】
ヒト化重鎖構築物を、4083個の受精卵の前核に注入した。3485個の卵母細胞を119匹のレシピエントに移し、433匹の子孫が生まれた。PCRおよびFISHによる分析により、これらの動物のうちの20匹がトランスジェニックであることが明らかになった。創始動物の血液中のヒト化重鎖を、ヒトIgM/IgGに特異的な抗体(例えば、ウサギの抗ヒトIgM、Rockland 609−4131;ウサギの抗ヒトIgM、Rockland 609−4631;ウサギの抗ヒトIgG、Pierce 31142、ウサギの抗ヒトIgG、Southern Biotech 6145−08;ウサギの抗ヒトIgG、Pierce 31784)を使用してELISAによって検出した。
【0190】
ヒト化κ、μ、およびγ鎖を検出するサンドイッチ型ELISAを、標準的な手順を使用して行った。簡単に説明すると、マイクロタイタープレートを捕捉抗体でコーティングし、希釈した血清試料とともにインキュベートした。結合したヒト免疫グロブリンを、二次標識抗体と、ペルオキシダーゼ−ストレプトアビジン結合体(Sigma、S2438)を使用して検出した。
【0191】
ヒト化重鎖免疫グロブリン鎖とヒト化軽鎖免疫グロブリン鎖の両方を発現する二重トランスジェニックである動物を、創始動物を交配させることによって作成した。
【0192】
(実施例13)
(ヒト化免疫グロブリンを発現するトランスジェニックマウス)
トランスジェニックマウスを、Nagy et al.,(Manipulating
the Mouse Embryo:A Laboratory Manual;Cold Spring Harbor Laboratory Press,New York,2003)に記載されているように作成した。簡単に説明すると、雌のマウスを標準的な方法を使用して過剰排卵させ、雄のマウスと交配させた。前核段階の受精卵を卵管から回収し、M2培地のような適切な培養液に入れた。ヒト化免疫グロブリン遺伝子座を含むBACを、1対のマニピュレーターの力を借りて雄の前核にマイクロインジェクションした。形態学的に生存している受精卵を、偽妊娠させた雌のマウスの卵管に移した。偽妊娠は無精子症の雄との交配によって誘導した。ヒト化軽鎖構築物を受精卵の1325個の受精卵の前核への注入の後、787個の卵母細胞を29匹のレシピエントに移した。全部で55匹の生存性の子孫が産まれ、そのうちの11匹がトランスジェニックであった。
【0193】
ヒト化重鎖構築物を、1050個の受精卵の前核に注入した。650個の卵母細胞を25匹のレシピエントに移し、64匹の生存している子孫が生まれた。PCRによる分析から、これらの動物のうちの19匹がトランスジェニックであることが明らかになった。
【0194】
ヒト化重鎖免疫グロブリン鎖とヒト化軽鎖免疫グロブリン鎖の両方を発現する二重トランスジェニックである動物を、創始動物を交配させることによって作成した。ヒト化κ、μ、およびγ鎖の発現を、標準的な手順を使用してELISAによって検出した。簡単に説明すると、マイクロタイタープレートを捕捉抗体でコーティングし、希釈した血清試料とともにインキュベートした。結合したヒト免疫グロブリンを、二次標識抗体と、ペルオキシダーゼ−ストレプトアビジン結合体(Sigma、S2438)を使用して検出した。
【0195】
明細書を通じて引用された全ての参考文献は、引用により本明細書中に明確に組み入れられる。本発明は、特定の実施例を参照することによって説明されるが、それらに限定されない。当業者であれば、種々の変更はバリエーションが、本発明の本質から逸脱することなく可能であることを認識するであろう。全てのこのような変更とバリエーションが、本明細書の範囲に具体的に含まれる。
【0196】
【表1−1】
【0197】
【表1−2】
【0198】
【表1−3】
【0199】
【表1−4】
【0200】
【表1−5】
【0201】
【表2−1】
【0202】
【表2−2】
【0203】
【表2−3】
【0204】
【表2−4】
【0205】
【表2−5】
【0206】
【表2−6】
【0207】
【表2−7】
【0208】
【表2−8】
【0209】
【表2−9】
【特許請求の範囲】
【請求項1】
ヌクレオチド配列が隣接しているヒト免疫グロブリン遺伝子セグメントを含む単離された核酸分子であって、ここで、該隣接配列は同じであっても異なっていてもよく、該隣接配列は、主に遺伝子変換および/もしくは超変異によって抗体多様性を生じる動物の免疫グロブリン重鎖遺伝子あるいは免疫グロブリン軽鎖遺伝子に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列、または2つ以上の該スペーサー配列のコンセンサス配列に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列を含む、単離された核酸分子。
【請求項2】
前記隣接配列が、主に遺伝子変換および/もしくは超変異によって抗体多様性を生じる動物の免疫グロブリン重鎖遺伝子あるいは免疫グロブリン軽鎖遺伝子に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列、または2つ以上の該スペーサー配列のコンセンサス配列に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列から本質的に構成されている、請求項1に記載の単離された核酸分子。
【請求項3】
前記隣接配列が、少なくとも約50個の連続しているヌクレオチドを含む、請求項1に記載の単離された核酸分子。
【請求項4】
前記隣接配列が同じである、請求項1に記載の単離された核酸分子。
【請求項5】
前記隣接配列が異なっている、請求項1に記載の単離された核酸分子。
【請求項6】
前記ヒト免疫グロブリン遺伝子セグメントが、重鎖V遺伝子セグメント、重鎖D遺伝子セグメントまたは重鎖J遺伝子セグメントである、請求項1に記載の単離された核酸分子。
【請求項7】
前記ヒト免疫グロブリン遺伝子セグメントが、重鎖Vセグメントである、請求項6に記載の単離された核酸分子。
【請求項8】
前記ヒト免疫グロブリン遺伝子セグメントが、VH3ファミリー、VH1ファミリー、VH5ファミリー、またはVH4ファミリーのメンバーである、請求項7に記載の単離された核酸分子。
【請求項9】
前記ヒト免疫グロブリン遺伝子セグメントが、軽鎖Vセグメントまたは軽鎖Jセグメントである、請求項1に記載の単離された核酸分子。
【請求項10】
前記ヒト免疫グロブリン遺伝子セグメントが、軽鎖Vセグメントである、請求項9に記載の単離された核酸分子。
【請求項11】
前記ヒト免疫グロブリン遺伝子セグメントが、κ軽鎖遺伝子セグメントである、請求項10に記載の単離された核酸分子。
【請求項12】
前記ヒト免疫グロブリン遺伝子セグメントが、Vκ1ファミリー、Vκ3ファミリーまたはVκ4ファミリーのメンバーである、請求項11に記載の単離された核酸分子。
【請求項13】
前記ヒト免疫グロブリン遺伝子セグメントが、λ軽鎖遺伝子セグメントである、請求項10に記載の単離された核酸分子。
【請求項14】
前記ヒト免疫グロブリン遺伝子セグメントが、Vλ1ファミリー、Vλ2ファミリー、またはVλ3ファミリーのメンバーである、請求項13に記載の単離された核酸分子。
【請求項15】
前記ヒト免疫グロブリン遺伝子セグメントが、重鎖Cセグメントである、請求項1に記載の単離された核酸分子。
【請求項16】
前記ヒト免疫グロブリン遺伝子セグメントが、軽鎖Cセグメントである、請求項1に記載の単離された核酸分子。
【請求項17】
主に遺伝子変換および/または超変異によって抗体多様性を生じる前記動物が、ウサギ、ニワトリ、ブタ、ヒツジ、ヤギおよびウシからなる群より選択される、請求項1に記載の単離された核酸分子。
【請求項18】
前記隣接配列が、配列番号1〜185からなる群より選択される配列に由来するか、または2つ以上の該配列のコンセンサス配列に由来する、少なくとも20個の連続しているヌクレオチドを含むヌクレオチド配列からなる郡より選択される、請求項1に記載の単離された核酸分子。
【請求項19】
前記ヒト免疫グロブリン遺伝子セグメントが、重鎖Vセグメント、重鎖Dセグメントまたは重鎖Jセグメントである、請求項1に記載の単離された核酸分子。
【請求項20】
前記隣接配列が、配列番号1〜56からなる群より選択される配列に由来するか、または2つ以上の該配列のコンセンサス配列に由来する、少なくとも20個の連続しているヌクレオチドを含む、請求項19に記載の単離された核酸分子。
【請求項21】
前記ヒト免疫グロブリン遺伝子セグメントが、軽鎖Vセグメントである、請求項1に記載の単離された核酸分子。
【請求項22】
前記隣接配列が、配列番号107〜184から選択される配列に由来するか、または2つ以上の該配列のコンセンサス配列に由来する、少なくとも20個の連続しているヌクレオチドを含む、請求項21に記載の単離された核酸分子。
【請求項23】
前記ヒト免疫グロブリン遺伝子セグメントが、重鎖Cセグメントである、請求項1に記載の単離された核酸分子。
【請求項24】
前記隣接配列が、配列番号57〜104から選択される配列に由来するか、または2つ以上の該配列のコンセンサス配列に由来する、少なくとも20個の連続しているヌクレオチドが含まれる、請求項23に記載の単離された核酸分子。
【請求項25】
前記ヒト免疫グロブリン遺伝子セグメントが、軽鎖Cセグメントである、請求項1に記載の単離された核酸分子。
【請求項26】
前記隣接配列が、配列番号105、106、184、185から選択される配列に由来するか、または2つ以上の該配列のコンセンサス配列に由来する、少なくとも20個の連続しているヌクレオチドを含む、請求項25に記載の単離された核酸分子。
【請求項27】
請求項1または請求項26に記載の核酸分子を含む組み換えベクター。
【請求項28】
ヒト化IgG遺伝子座を含むトランスジェニックベクターであって、該ヒト化Ig遺伝子座が、主に遺伝子変換および/もしくは超変異によって抗体多様性を生じる動物のIg遺伝子座またはIg遺伝子座の一部に由来し、該ヒト化Ig遺伝子座は、複数のIg遺伝子セグメントを含み、ここで:
(a)少なくとも1つの該遺伝子セグメントが、主に遺伝子変換および/もしくは超変異によって抗体多様性を生じる該動物の免疫グロブリン重鎖遺伝子あるいは免疫グロブリン軽鎖遺伝子に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列、または2つ以上の該スペーサー配列のコンセンサス配列に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列を含むヌクレオチド配列が隣接しているヒトIg遺伝子セグメントであり;
(b)該遺伝子セグメントが、再構成されていない立体配置、部分的に再構成された立体配置、または完全に再構成された立体配置において並列しており;そして
(c)該ヒト化Ig遺伝子座が、遺伝子再構成を受けることができ、必要に応じて、遺伝子変換および/または超変異を受けることができ、そして該非霊長類動物において、ヒト化免疫グロブリンのレパートリーを生じることができる、
トランスジェニックベクター。
【請求項29】
前記スペーサー配列が、配列番号1から185からなる群、または2つ以上の該配列のコンセンサス配列から選択される、請求項28に記載のトランスジェニックベクター。
【請求項30】
主に遺伝子変換によって抗体多様性を生じる前記動物が、ウサギ、ブタ、ニワトリ、ヒツジ、ヤギおよびウシからなる群より選択される、請求項29に記載のトランスジェニックベクター。
【請求項31】
前記ヒト化Ig遺伝子座が重鎖遺伝子座であり、該重鎖遺伝子座は、少なくとも1つのV遺伝子セグメント、少なくとも1つのD遺伝子セグメント、少なくとも1つのJ遺伝子セグメント、および少なくとも1つの定常領域遺伝子セグメントを含む、請求項30に記載のトランスジェニックベクター。
【請求項32】
前記定常領域遺伝子セグメントが、ヒト重鎖定常領域遺伝子セグメントである、請求項31に記載のトランスジェニックベクター。
【請求項33】
前記ヒト重鎖定常領域セグメントが、Cγおよび/またはCμセグメントである、請求項32に記載のトランスジェニックベクター。
【請求項34】
約5〜100個のV遺伝子セグメント、および少なくとも1つのヒトV遺伝子セグメントを含む、請求項31に記載のトランスジェニックベクター。
【請求項35】
前記ヒト化Ig遺伝子座が軽鎖遺伝子座であり、該軽鎖遺伝子座は、少なくとも1つのV遺伝子セグメント、少なくとも1つのJ遺伝子セグメント、および少なくとも1つの定常領域遺伝子セグメントを含む、請求項30に記載のトランスジェニックベクター。
【請求項36】
前記定常領域遺伝子セグメントがヒト軽鎖定常領域遺伝子セグメントである、請求項35に記載のトランスジェニックベクター。
【請求項37】
前記ヒト軽鎖定常領域遺伝子セグメントがCλまたはCκである、請求項36に記載のトランスジェニックベクター。
【請求項38】
約5〜100個のV遺伝子セグメント、および少なくとも1つのヒトV遺伝子セグメントを含む、請求項36に記載のトランスジェニックベクター。
【請求項39】
前記ヒトV遺伝子セグメントが、再構成された立体配置においてJ遺伝子セグメントのすぐ5’にある、請求項36に記載のトランスジェニックベクター。
【請求項40】
5’から3’の方向で、5’ヌクレオチド配列、ヒト免疫グロブリン遺伝子セグメント、および3’ヌクレオチド配列からなる2つ以上のユニットを含む核酸分子であって、ここで、該5’ヌクレオチド配列および該3’ヌクレオチド配列は同じであっても異なっていてもよく、主に遺伝子変換および/もしくは超変異によって抗体多様性を生じる動物の免疫グロブリン重鎖遺伝子あるいは免疫グロブリン軽鎖遺伝子に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列、または2つ以上の該スペーサー配列のコンセンサス配列に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列を含む、核酸分子。
【請求項41】
前記スペーサー配列が、配列番号1〜185からなる群、または2つ以上の該スペーサー配列のコンセンサス配列から選択される、請求項40に記載の核酸分子。
【請求項42】
前記ユニットが同じである、請求項41に記載の核酸分子。
【請求項43】
前記ユニットが異なる、請求項41に記載の核酸分子。
【請求項44】
主に遺伝子変換および/または超変異によって抗体多様性を生じる動物中でヒト化抗体の機能的なレパートリーを生じることができるヒト化免疫グロブリン遺伝子座を含むトランスジェニックベクターを作製するための方法であって、該方法は、以下:
(a)少なくとも1つのV遺伝子セグメント、少なくとも1つのJ遺伝子セグメント、および少なくとも1つの定常領域遺伝子セグメントを含む該動物に由来するIg遺伝子座またはその一部を含むDNAフラグメントを得る工程;ならびに
(b)ヒト化Ig遺伝子座を生じるように、工程(a)のDNAフラグメント中に少なくとも1つのヒトIg遺伝子セグメントを組み込む工程であって、ここで、ヒトIg遺伝子セグメントは、主に遺伝子変換および/もしくは超変異によって抗体多様性を生じる動物の免疫グロブリン重鎖遺伝子あるいは免疫グロブリン軽鎖遺伝子に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列、または2つ以上のそのようなスペーサー配列のコンセンサス配列に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列を含むヌクレオチド配列が隣接しており;ここで、(i)該遺伝子セグメントが、再構成されていない立体配置、部分的に再構成された立体配置、または完全に再構成された立体配置において並列しており;そして(ii)ヒト化Ig遺伝子座が遺伝子再構成を受けることができ、必要に応じて、遺伝子変換および/または超変異を受けることができ、そして該動物中でヒト化免疫グロブリンのレパートリーを生じることができる、工程、
を包含する、方法。
【請求項45】
請求項28に記載のトランスジェニックベクター中に存在するヒト化免疫グロブリン遺伝子座を含む、主に遺伝子変換および/または超変異によって抗体多様性を生じるトランスジェニック動物。
【請求項46】
本質的に完全な内因性の調節機構および抗体産生機構を保存している、請求項45に記載のトランスジェニック動物。
【請求項47】
トランスジェニックのウサギ、ニワトリ、ブタ、ヒツジ、ヤギ、およびウシからなる群より選択される、請求項46に記載のトランスジェニック動物。
【請求項48】
請求項45に記載のトランスジェニック動物に由来するB細胞。
【請求項49】
請求項45に記載のトランスジェニック動物を使用して産生される、ヒト化免疫グロブリン。
【請求項50】
抗体である、請求項49に記載のヒト化免疫グロブリン。
【請求項51】
ポリクローナル抗体である、請求項50に記載のヒト化免疫グロブリン。
【請求項52】
モノクローナル抗体である、請求項50に記載のヒト化免疫グロブリン。
【請求項53】
請求項49に記載のヒト化免疫グロブリンを含む、抗体調製物。
【請求項54】
請求項49に記載のヒト化免疫グロブリンを含む、薬学的組成物。
【請求項55】
ヌクレオチド配列が隣接している2つ以上のヒト免疫グロブリン重鎖または軽鎖の可変(V)領域遺伝子セグメントを含む単離された核酸分子であって、ここで、該隣接配列は同じであっても異なっていてもよく、該隣接配列は、非霊長類動物の免疫グロブリン重鎖遺伝子もしくは免疫グロブリン軽鎖遺伝子に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列、または2つ以上の該スペーサー配列のコンセンサス配列に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列を含む、単離された核酸分子。
【請求項56】
前記スペーサー配列が、配列番号1〜185からなる群より選択される、請求項55に記載の単離された核酸分子。
【請求項57】
前記スペーサー配列が、齧歯類の免疫グロブリンスペーサー配列から選択される、請求項55に記載の単離された核酸分子。
【請求項58】
前記齧歯類が、マウスまたはラットである、請求項57に記載の単離された核酸、請求項55または請求項56に記載の単離された核酸を含むトランスジェニックベクター。
【請求項1】
ヌクレオチド配列が隣接しているヒト免疫グロブリン遺伝子セグメントを含む単離された核酸分子であって、ここで、該隣接配列は同じであっても異なっていてもよく、該隣接配列は、主に遺伝子変換および/もしくは超変異によって抗体多様性を生じる動物の免疫グロブリン重鎖遺伝子あるいは免疫グロブリン軽鎖遺伝子に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列、または2つ以上の該スペーサー配列のコンセンサス配列に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列を含む、単離された核酸分子。
【請求項2】
前記隣接配列が、主に遺伝子変換および/もしくは超変異によって抗体多様性を生じる動物の免疫グロブリン重鎖遺伝子あるいは免疫グロブリン軽鎖遺伝子に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列、または2つ以上の該スペーサー配列のコンセンサス配列に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列から本質的に構成されている、請求項1に記載の単離された核酸分子。
【請求項3】
前記隣接配列が、少なくとも約50個の連続しているヌクレオチドを含む、請求項1に記載の単離された核酸分子。
【請求項4】
前記隣接配列が同じである、請求項1に記載の単離された核酸分子。
【請求項5】
前記隣接配列が異なっている、請求項1に記載の単離された核酸分子。
【請求項6】
前記ヒト免疫グロブリン遺伝子セグメントが、重鎖V遺伝子セグメント、重鎖D遺伝子セグメントまたは重鎖J遺伝子セグメントである、請求項1に記載の単離された核酸分子。
【請求項7】
前記ヒト免疫グロブリン遺伝子セグメントが、重鎖Vセグメントである、請求項6に記載の単離された核酸分子。
【請求項8】
前記ヒト免疫グロブリン遺伝子セグメントが、VH3ファミリー、VH1ファミリー、VH5ファミリー、またはVH4ファミリーのメンバーである、請求項7に記載の単離された核酸分子。
【請求項9】
前記ヒト免疫グロブリン遺伝子セグメントが、軽鎖Vセグメントまたは軽鎖Jセグメントである、請求項1に記載の単離された核酸分子。
【請求項10】
前記ヒト免疫グロブリン遺伝子セグメントが、軽鎖Vセグメントである、請求項9に記載の単離された核酸分子。
【請求項11】
前記ヒト免疫グロブリン遺伝子セグメントが、κ軽鎖遺伝子セグメントである、請求項10に記載の単離された核酸分子。
【請求項12】
前記ヒト免疫グロブリン遺伝子セグメントが、Vκ1ファミリー、Vκ3ファミリーまたはVκ4ファミリーのメンバーである、請求項11に記載の単離された核酸分子。
【請求項13】
前記ヒト免疫グロブリン遺伝子セグメントが、λ軽鎖遺伝子セグメントである、請求項10に記載の単離された核酸分子。
【請求項14】
前記ヒト免疫グロブリン遺伝子セグメントが、Vλ1ファミリー、Vλ2ファミリー、またはVλ3ファミリーのメンバーである、請求項13に記載の単離された核酸分子。
【請求項15】
前記ヒト免疫グロブリン遺伝子セグメントが、重鎖Cセグメントである、請求項1に記載の単離された核酸分子。
【請求項16】
前記ヒト免疫グロブリン遺伝子セグメントが、軽鎖Cセグメントである、請求項1に記載の単離された核酸分子。
【請求項17】
主に遺伝子変換および/または超変異によって抗体多様性を生じる前記動物が、ウサギ、ニワトリ、ブタ、ヒツジ、ヤギおよびウシからなる群より選択される、請求項1に記載の単離された核酸分子。
【請求項18】
前記隣接配列が、配列番号1〜185からなる群より選択される配列に由来するか、または2つ以上の該配列のコンセンサス配列に由来する、少なくとも20個の連続しているヌクレオチドを含むヌクレオチド配列からなる郡より選択される、請求項1に記載の単離された核酸分子。
【請求項19】
前記ヒト免疫グロブリン遺伝子セグメントが、重鎖Vセグメント、重鎖Dセグメントまたは重鎖Jセグメントである、請求項1に記載の単離された核酸分子。
【請求項20】
前記隣接配列が、配列番号1〜56からなる群より選択される配列に由来するか、または2つ以上の該配列のコンセンサス配列に由来する、少なくとも20個の連続しているヌクレオチドを含む、請求項19に記載の単離された核酸分子。
【請求項21】
前記ヒト免疫グロブリン遺伝子セグメントが、軽鎖Vセグメントである、請求項1に記載の単離された核酸分子。
【請求項22】
前記隣接配列が、配列番号107〜184から選択される配列に由来するか、または2つ以上の該配列のコンセンサス配列に由来する、少なくとも20個の連続しているヌクレオチドを含む、請求項21に記載の単離された核酸分子。
【請求項23】
前記ヒト免疫グロブリン遺伝子セグメントが、重鎖Cセグメントである、請求項1に記載の単離された核酸分子。
【請求項24】
前記隣接配列が、配列番号57〜104から選択される配列に由来するか、または2つ以上の該配列のコンセンサス配列に由来する、少なくとも20個の連続しているヌクレオチドが含まれる、請求項23に記載の単離された核酸分子。
【請求項25】
前記ヒト免疫グロブリン遺伝子セグメントが、軽鎖Cセグメントである、請求項1に記載の単離された核酸分子。
【請求項26】
前記隣接配列が、配列番号105、106、184、185から選択される配列に由来するか、または2つ以上の該配列のコンセンサス配列に由来する、少なくとも20個の連続しているヌクレオチドを含む、請求項25に記載の単離された核酸分子。
【請求項27】
請求項1または請求項26に記載の核酸分子を含む組み換えベクター。
【請求項28】
ヒト化IgG遺伝子座を含むトランスジェニックベクターであって、該ヒト化Ig遺伝子座が、主に遺伝子変換および/もしくは超変異によって抗体多様性を生じる動物のIg遺伝子座またはIg遺伝子座の一部に由来し、該ヒト化Ig遺伝子座は、複数のIg遺伝子セグメントを含み、ここで:
(a)少なくとも1つの該遺伝子セグメントが、主に遺伝子変換および/もしくは超変異によって抗体多様性を生じる該動物の免疫グロブリン重鎖遺伝子あるいは免疫グロブリン軽鎖遺伝子に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列、または2つ以上の該スペーサー配列のコンセンサス配列に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列を含むヌクレオチド配列が隣接しているヒトIg遺伝子セグメントであり;
(b)該遺伝子セグメントが、再構成されていない立体配置、部分的に再構成された立体配置、または完全に再構成された立体配置において並列しており;そして
(c)該ヒト化Ig遺伝子座が、遺伝子再構成を受けることができ、必要に応じて、遺伝子変換および/または超変異を受けることができ、そして該非霊長類動物において、ヒト化免疫グロブリンのレパートリーを生じることができる、
トランスジェニックベクター。
【請求項29】
前記スペーサー配列が、配列番号1から185からなる群、または2つ以上の該配列のコンセンサス配列から選択される、請求項28に記載のトランスジェニックベクター。
【請求項30】
主に遺伝子変換によって抗体多様性を生じる前記動物が、ウサギ、ブタ、ニワトリ、ヒツジ、ヤギおよびウシからなる群より選択される、請求項29に記載のトランスジェニックベクター。
【請求項31】
前記ヒト化Ig遺伝子座が重鎖遺伝子座であり、該重鎖遺伝子座は、少なくとも1つのV遺伝子セグメント、少なくとも1つのD遺伝子セグメント、少なくとも1つのJ遺伝子セグメント、および少なくとも1つの定常領域遺伝子セグメントを含む、請求項30に記載のトランスジェニックベクター。
【請求項32】
前記定常領域遺伝子セグメントが、ヒト重鎖定常領域遺伝子セグメントである、請求項31に記載のトランスジェニックベクター。
【請求項33】
前記ヒト重鎖定常領域セグメントが、Cγおよび/またはCμセグメントである、請求項32に記載のトランスジェニックベクター。
【請求項34】
約5〜100個のV遺伝子セグメント、および少なくとも1つのヒトV遺伝子セグメントを含む、請求項31に記載のトランスジェニックベクター。
【請求項35】
前記ヒト化Ig遺伝子座が軽鎖遺伝子座であり、該軽鎖遺伝子座は、少なくとも1つのV遺伝子セグメント、少なくとも1つのJ遺伝子セグメント、および少なくとも1つの定常領域遺伝子セグメントを含む、請求項30に記載のトランスジェニックベクター。
【請求項36】
前記定常領域遺伝子セグメントがヒト軽鎖定常領域遺伝子セグメントである、請求項35に記載のトランスジェニックベクター。
【請求項37】
前記ヒト軽鎖定常領域遺伝子セグメントがCλまたはCκである、請求項36に記載のトランスジェニックベクター。
【請求項38】
約5〜100個のV遺伝子セグメント、および少なくとも1つのヒトV遺伝子セグメントを含む、請求項36に記載のトランスジェニックベクター。
【請求項39】
前記ヒトV遺伝子セグメントが、再構成された立体配置においてJ遺伝子セグメントのすぐ5’にある、請求項36に記載のトランスジェニックベクター。
【請求項40】
5’から3’の方向で、5’ヌクレオチド配列、ヒト免疫グロブリン遺伝子セグメント、および3’ヌクレオチド配列からなる2つ以上のユニットを含む核酸分子であって、ここで、該5’ヌクレオチド配列および該3’ヌクレオチド配列は同じであっても異なっていてもよく、主に遺伝子変換および/もしくは超変異によって抗体多様性を生じる動物の免疫グロブリン重鎖遺伝子あるいは免疫グロブリン軽鎖遺伝子に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列、または2つ以上の該スペーサー配列のコンセンサス配列に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列を含む、核酸分子。
【請求項41】
前記スペーサー配列が、配列番号1〜185からなる群、または2つ以上の該スペーサー配列のコンセンサス配列から選択される、請求項40に記載の核酸分子。
【請求項42】
前記ユニットが同じである、請求項41に記載の核酸分子。
【請求項43】
前記ユニットが異なる、請求項41に記載の核酸分子。
【請求項44】
主に遺伝子変換および/または超変異によって抗体多様性を生じる動物中でヒト化抗体の機能的なレパートリーを生じることができるヒト化免疫グロブリン遺伝子座を含むトランスジェニックベクターを作製するための方法であって、該方法は、以下:
(a)少なくとも1つのV遺伝子セグメント、少なくとも1つのJ遺伝子セグメント、および少なくとも1つの定常領域遺伝子セグメントを含む該動物に由来するIg遺伝子座またはその一部を含むDNAフラグメントを得る工程;ならびに
(b)ヒト化Ig遺伝子座を生じるように、工程(a)のDNAフラグメント中に少なくとも1つのヒトIg遺伝子セグメントを組み込む工程であって、ここで、ヒトIg遺伝子セグメントは、主に遺伝子変換および/もしくは超変異によって抗体多様性を生じる動物の免疫グロブリン重鎖遺伝子あるいは免疫グロブリン軽鎖遺伝子に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列、または2つ以上のそのようなスペーサー配列のコンセンサス配列に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列を含むヌクレオチド配列が隣接しており;ここで、(i)該遺伝子セグメントが、再構成されていない立体配置、部分的に再構成された立体配置、または完全に再構成された立体配置において並列しており;そして(ii)ヒト化Ig遺伝子座が遺伝子再構成を受けることができ、必要に応じて、遺伝子変換および/または超変異を受けることができ、そして該動物中でヒト化免疫グロブリンのレパートリーを生じることができる、工程、
を包含する、方法。
【請求項45】
請求項28に記載のトランスジェニックベクター中に存在するヒト化免疫グロブリン遺伝子座を含む、主に遺伝子変換および/または超変異によって抗体多様性を生じるトランスジェニック動物。
【請求項46】
本質的に完全な内因性の調節機構および抗体産生機構を保存している、請求項45に記載のトランスジェニック動物。
【請求項47】
トランスジェニックのウサギ、ニワトリ、ブタ、ヒツジ、ヤギ、およびウシからなる群より選択される、請求項46に記載のトランスジェニック動物。
【請求項48】
請求項45に記載のトランスジェニック動物に由来するB細胞。
【請求項49】
請求項45に記載のトランスジェニック動物を使用して産生される、ヒト化免疫グロブリン。
【請求項50】
抗体である、請求項49に記載のヒト化免疫グロブリン。
【請求項51】
ポリクローナル抗体である、請求項50に記載のヒト化免疫グロブリン。
【請求項52】
モノクローナル抗体である、請求項50に記載のヒト化免疫グロブリン。
【請求項53】
請求項49に記載のヒト化免疫グロブリンを含む、抗体調製物。
【請求項54】
請求項49に記載のヒト化免疫グロブリンを含む、薬学的組成物。
【請求項55】
ヌクレオチド配列が隣接している2つ以上のヒト免疫グロブリン重鎖または軽鎖の可変(V)領域遺伝子セグメントを含む単離された核酸分子であって、ここで、該隣接配列は同じであっても異なっていてもよく、該隣接配列は、非霊長類動物の免疫グロブリン重鎖遺伝子もしくは免疫グロブリン軽鎖遺伝子に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列、または2つ以上の該スペーサー配列のコンセンサス配列に由来する少なくとも約20個の連続しているヌクレオチドのスペーサー配列を含む、単離された核酸分子。
【請求項56】
前記スペーサー配列が、配列番号1〜185からなる群より選択される、請求項55に記載の単離された核酸分子。
【請求項57】
前記スペーサー配列が、齧歯類の免疫グロブリンスペーサー配列から選択される、請求項55に記載の単離された核酸分子。
【請求項58】
前記齧歯類が、マウスまたはラットである、請求項57に記載の単離された核酸、請求項55または請求項56に記載の単離された核酸を含むトランスジェニックベクター。
【図9b−1】


【図9b−2】

【図9b−3】

【図9b−4】

【図9b−5】

【図9b−6】

【図9b−7】

【図9b−8】

【図9b−9】

【図9b−10】

【図12a−1】

【図12a−2】

【図12a−3】

【図12a−4】

【図12b−1】

【図12b−2】

【図12b−3】

【図12b−4】

【図13a−1】

【図13a−2】

【図13b−1】

【図13b−2】

【図13b−3】

【図13b−4】

【図13b−5】

【図13c−1】

【図13c−2】

【図13c−3】

【図13c−4】

【図13d−1】

【図13d−2】

【図13d−3】

【図13d−4】

【図13d−5】

【図1】
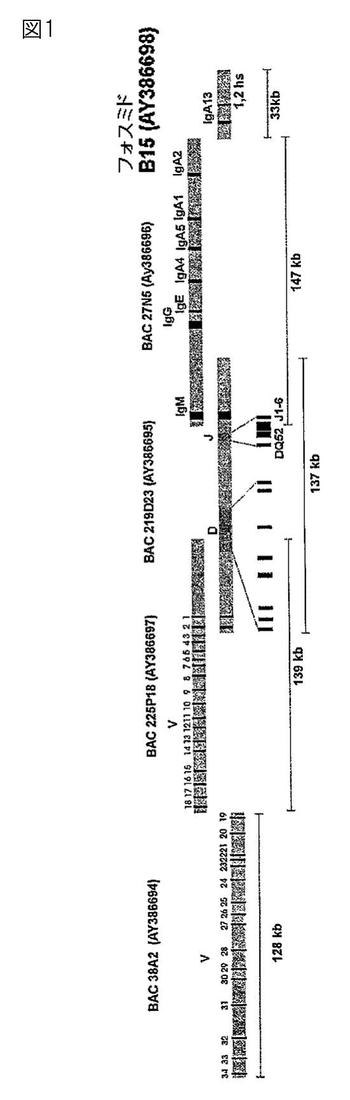
【図2】
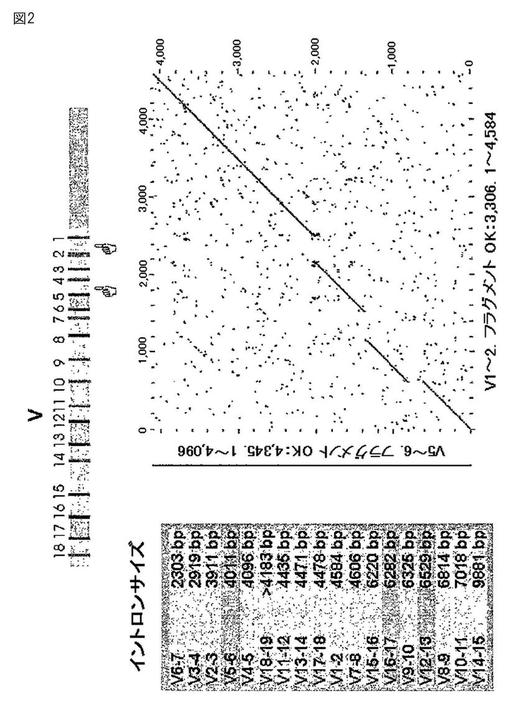
【図3】
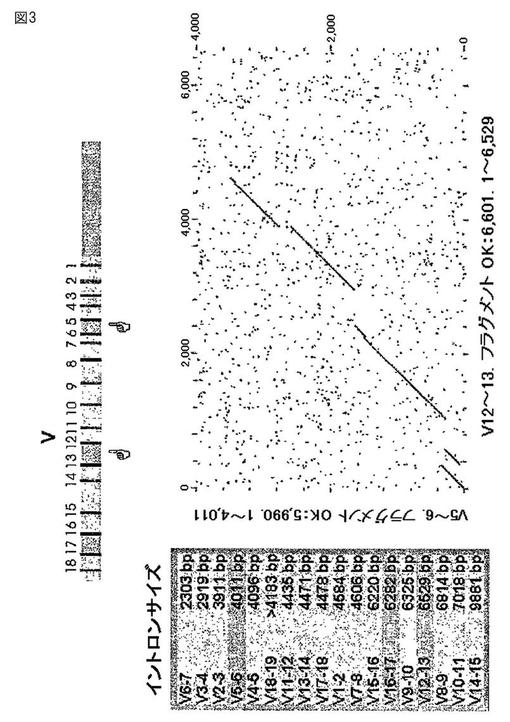
【図4】
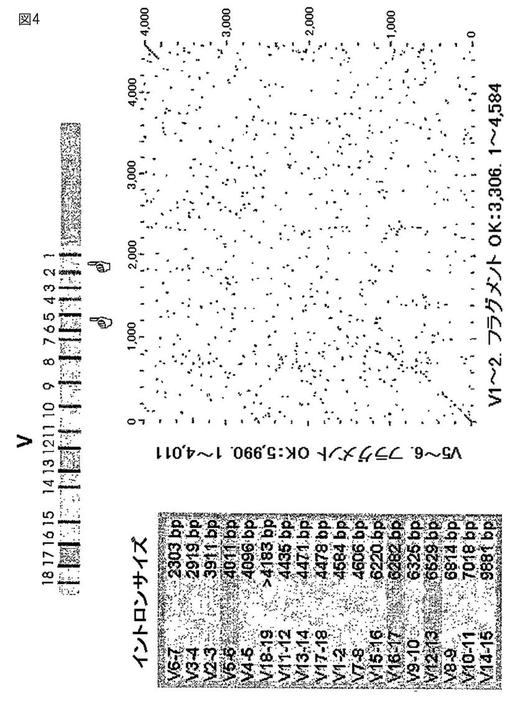
【図5】
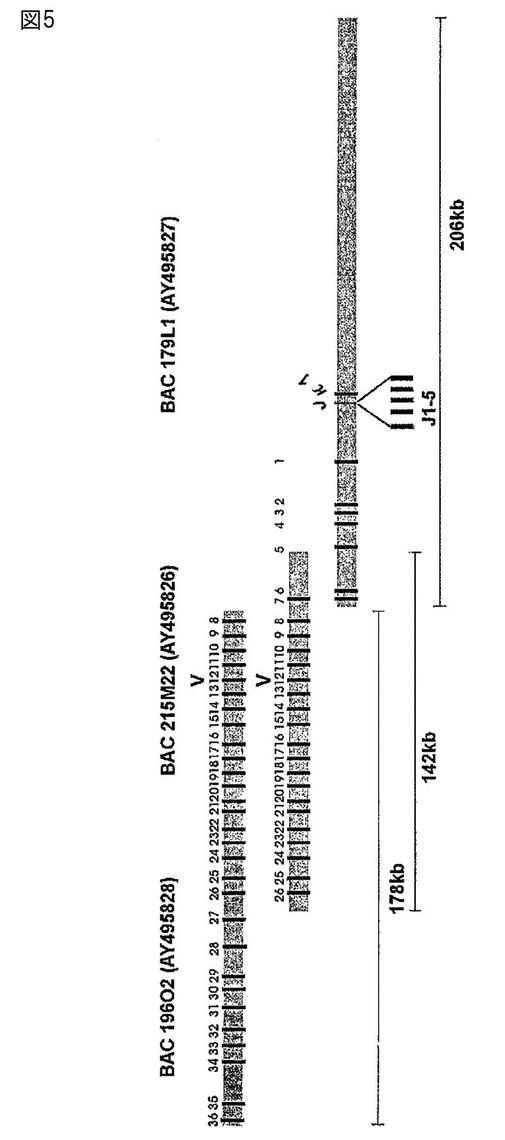
【図6】
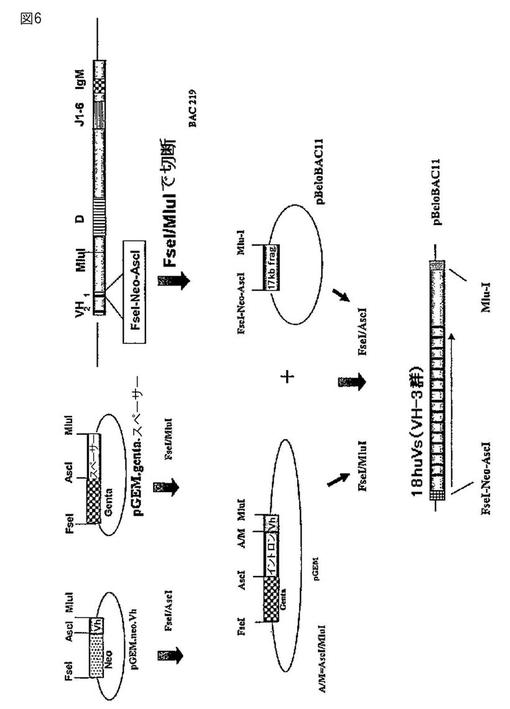
【図7】
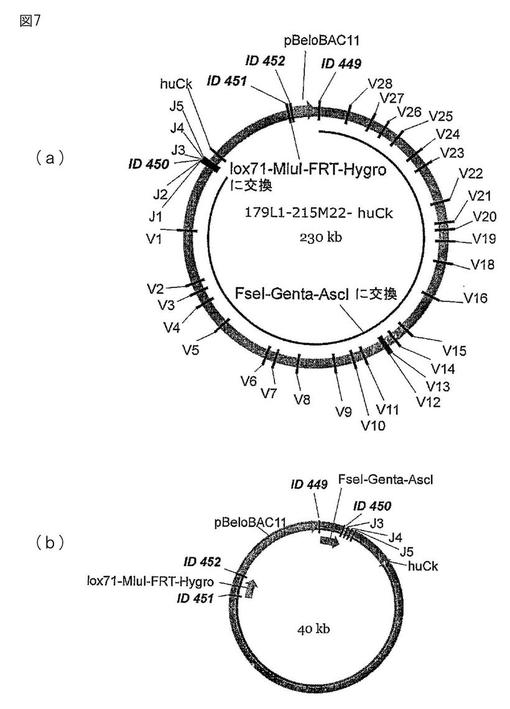
【図8】
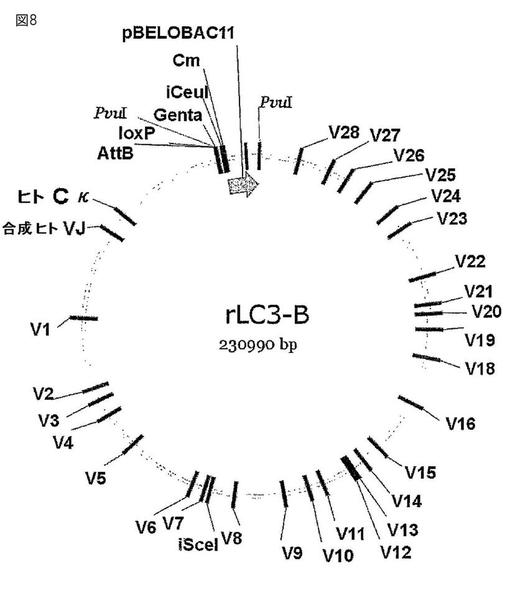
【図9a】
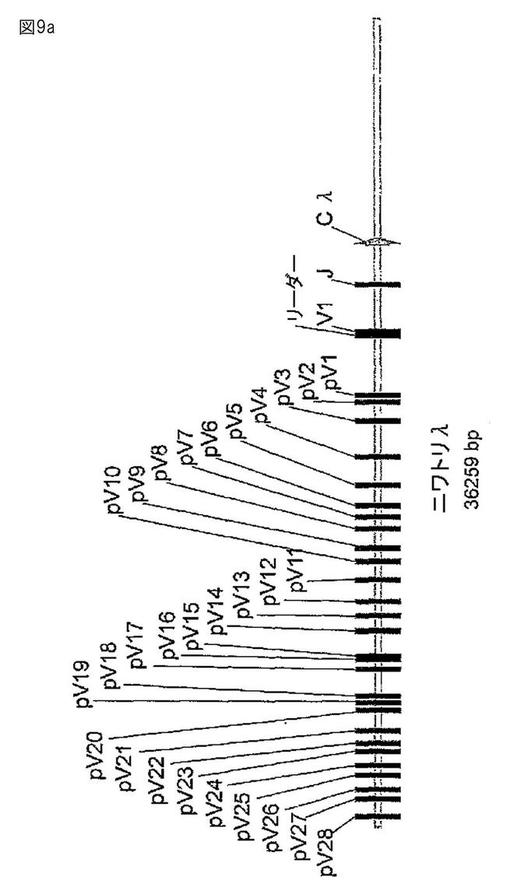
【図10】
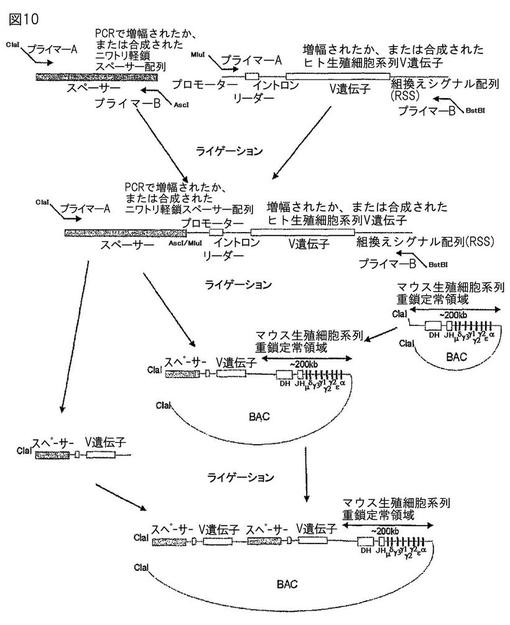
【図11】
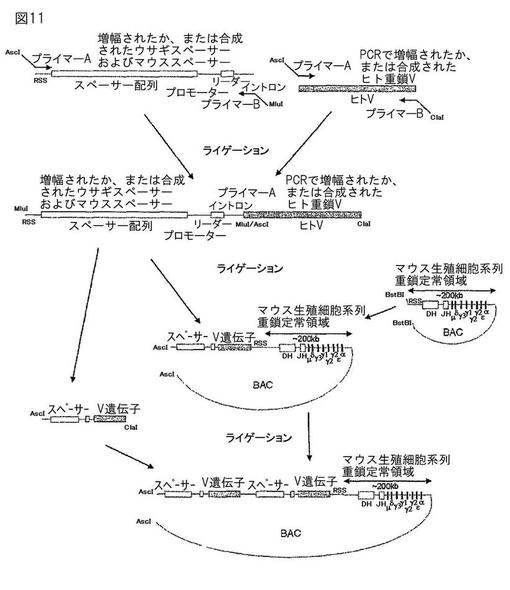
【図12c】
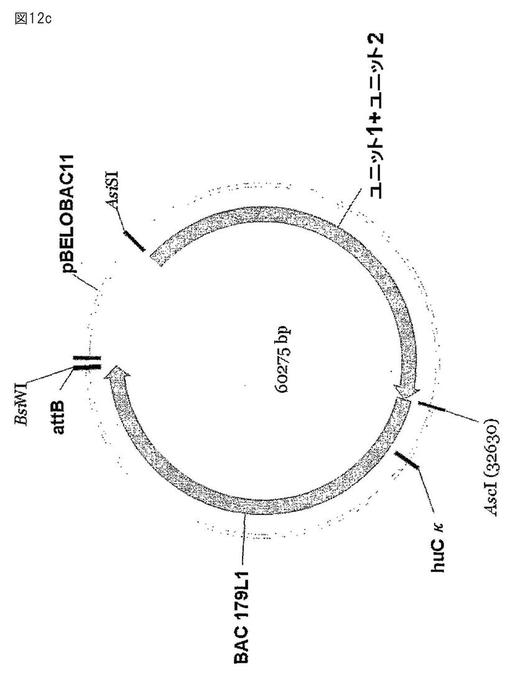
【図13e】
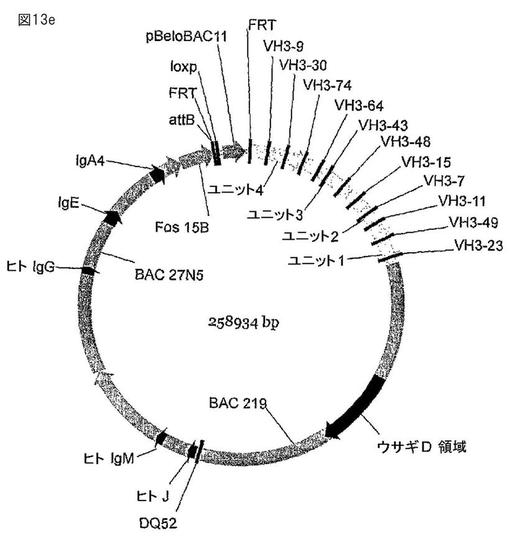
【図9b−2】
【図9b−3】
【図9b−4】
【図9b−5】
【図9b−6】
【図9b−7】
【図9b−8】
【図9b−9】
【図9b−10】
【図12a−1】
【図12a−2】
【図12a−3】
【図12a−4】
【図12b−1】
【図12b−2】
【図12b−3】
【図12b−4】
【図13a−1】
【図13a−2】
【図13b−1】
【図13b−2】
【図13b−3】
【図13b−4】
【図13b−5】
【図13c−1】
【図13c−2】
【図13c−3】
【図13c−4】
【図13d−1】
【図13d−2】
【図13d−3】
【図13d−4】
【図13d−5】
【図1】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9a】
【図10】
【図11】
【図12c】
【図13e】
【公開番号】特開2011−182801(P2011−182801A)
【公開日】平成23年9月22日(2011.9.22)
【国際特許分類】
【外国語出願】
【出願番号】特願2011−131898(P2011−131898)
【出願日】平成23年6月14日(2011.6.14)
【分割の表示】特願2006−520411(P2006−520411)の分割
【原出願日】平成16年7月15日(2004.7.15)
【出願人】(506011504)セラピューティック ヒューマン ポリクローナルズ, インコーポレイテッド (6)
【Fターム(参考)】
【公開日】平成23年9月22日(2011.9.22)
【国際特許分類】
【出願番号】特願2011−131898(P2011−131898)
【出願日】平成23年6月14日(2011.6.14)
【分割の表示】特願2006−520411(P2006−520411)の分割
【原出願日】平成16年7月15日(2004.7.15)
【出願人】(506011504)セラピューティック ヒューマン ポリクローナルズ, インコーポレイテッド (6)
【Fターム(参考)】
[ Back to top ]