
ヒト胚性幹細胞の分化
本発明は、多能性幹細胞からの、インスリン産生細胞への分化を促進させるための方法を提供する。詳細には、本発明は、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞を産生する方法を提供する。

【発明の詳細な説明】
【技術分野】
【0001】
本出願は、米国特許仮出願第61/289,671号(2009年12月23日出願)の利益を主張し、その全体が参照により本明細書に組み込まれる。
本発明は、多能性幹細胞からのインスリン産生細胞への分化を促進させるための方法を提供する。詳細には、本発明は、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞を産生する方法を提供する。
【背景技術】
【0002】
I型糖尿病の細胞置換療法の進歩及び移植可能なランゲルハンス島の不足により、生着に適したインスリン産生細胞、すなわちβ細胞の供給源の開発に注目が集まっている。1つの手法は、例えば、胚性幹細胞のような多能性幹細胞から機能性のβ細胞を生成することである。
【0003】
脊椎動物の胚発生において、多能性細胞は、原腸形成として公知のプロセスにより3つの胚葉(外胚葉、中胚葉、及び内胚葉)を含む細胞のグループを生じる。例えば、甲状腺、胸腺、膵臓、腸、及び肝臓などの組織は、内胚葉から中間ステージを経て発達する。このプロセスにおける中間ステージは、胚体内胚葉(definitive endoderm)の形成である。胚体内胚葉細胞は、HNF3β、GATA4、MIXL1、CXCR4及びSOX17などの多数のマーカーを発現する。
【0004】
膵臓の形成は、胚体内胚葉が膵臓内胚葉へと分化することにより生じる。膵臓内胚葉の細胞は膵臓−十二指腸ホメオボックス遺伝子、PDX1を発現する。PDX1が存在しない場合、膵臓は、腹側芽及び背側芽の形成より先に発達が進行しない。したがって、PDX1の発現は、膵臓器官形成において重要な工程として特徴付けられる。成熟した膵臓は、他の細胞型間で外分泌組織及び内分泌組織を含有する。外分泌組織及び内分泌組織は、膵臓内胚葉の分化によって生じる。
【0005】
島細胞の特徴を保持する細胞が、マウスの胚細胞から誘導されたことが報告されている。例えば、Lumelskyら(Science 292:1389,2001)は、膵島と同様のインスリン分泌構造へのマウスの胚性幹細胞の分化を報告している。Soriaら(Diabetes 49:157,2000)は、ストレプトゾトシン誘発糖尿病のマウスにおいて、マウスの胚性幹細胞から誘導されたインスリン分泌細胞が血糖を正常化することを報告している。
【0006】
一例において、Horiら(PNAS 99:16105,2002)は、ホスホイノシチド3−キナーゼ(LY294002)の阻害剤でマウス胚性幹細胞を処理することにより、β細胞に類似した細胞が生じたことを開示している。
【0007】
別の例において、Blyszczukら(PNAS 100:998,2003)は、Pax4を構成的に発現するマウス胚性幹細胞からのインスリン産生細胞の生成を報告している。
【0008】
Micallefらは、レチノイン酸が、胚性幹細胞のPDX1陽性膵臓内胚葉の形成に対する関与を制御できることを報告している。レチノイン酸は、胚における原腸形成の終了時に対応する期間中の、胚性幹細胞分化の4日目に培養液に添加すると、Pdx1発現の誘発に最も効果的である(Diabetes 54:301、2005)。
【0009】
Miyazakiらは、Pdx1を過剰発現するマウス胚性幹細胞株を報告している。この結果は、外因性のPdx1発現が、得られた分化細胞内でインスリン、ソマトスタチン、グルコキナーゼ、ニューロゲニン3、p48、Pax6、及びHnf6遺伝子の発現を明らかに増加させたことを示している(Diabetes 53:1030,2004)。
【0010】
Skoudyらは、マウス胚性幹細胞内で、アクチビンA(TGFβスーパーファミリーのメンバー)が、膵臓外分泌遺伝子(p48及びアミラーゼ)、並びに内分泌遺伝子(Pdx1、インスリン及びグルカゴン)の発現を上方制御することを報告している。最大の効果は、1nMアクチビンAを使用した場合に認められた。また、インスリン及びPdx1のmRNA発現レベルはレチノイン酸に影響されないが、3nM FGF7処理により、Pdx1の転写物量が増加する結果となることが認められた(Biochem.J.379:749,2004)。
【0011】
Shirakiらは、PDX1陽性細胞への胚性幹細胞の分化を特異的に増加させる増殖因子の効果を研究した。Shirakiらは、TGFβ2によってPDX1陽性細胞がより高い比率で再現性良く得られたことを観察した(Genes Cells.2005 Jun;10(6):503〜16)。
【0012】
Gordonらは、血清の非存在下、かつアクチビンとWntシグナル伝達阻害剤の存在下での、マウス胚性幹細胞からの短尾奇形[陽性]/HNF−3β[陽性]内胚葉細胞への誘導を示した(米国特許第2006/0003446A1号)。
【0013】
Gordonら(PNAS,103巻、16806ページ、2006)は、「Wnt及びTGF−β/nodal/アクチビンの同時シグナル伝達が前原始線条の形成には必要であった」と述べている。
【0014】
しかしながら、胚性幹細胞発達のマウスモデルは、例えばヒトなどのより高等な哺乳動物における発達プログラムを正確には模倣しない恐れがある。
【0015】
Thomsonらは、ヒト胚盤胞から胚性幹細胞を単離した(Science 282:114,1998)。同時に、Gearhart及び共同研究者は、胎児生殖腺組織から、ヒト胚生殖(hEG)細胞株を誘導した(Shamblottら、Proc.Natl.Acad.Sci.USA 95:13726、1998)。白血病阻害因子(LIF)と共に培養するのみで分化を阻止し得るマウス胚性幹細胞とは異なり、ヒト胚性幹細胞は、非常に特殊な条件下で維持する必要がある(米国特許第6,200,806号、国際公開第99/20741号;同第01/51616号)。
【0016】
D’Amourらは、高濃度のアクチビン及び低濃度の血清の存在下で、ヒト胚性幹細胞由来の胚体内胚葉の濃縮化された培養物が調製されたことを述べている(Nature Biotechnology 2005)。これらの細胞を、マウスの腎臓被膜下に移植することにより、内胚葉性器官の特徴の一部を有するより成熟した細胞への分化が得られた。ヒト胚性幹細胞由来の胚体内胚葉細胞は、FGF−10の添加後、PDX1陽性細胞に更に分化することができる(米国特許出願公開第2005/0266554A1号)。
【0017】
D’Amourら(Nature Biotechnology−24,1392〜1401(2006))は、「我々はヒト胚性幹(hES)細胞を、膵臓ホルモンインスリン、グルカゴン、ソマトスタチン、膵臓ポリペプチド及びグレリンを合成できる内分泌細胞へと変換する分化プロセスを開発した。このプロセスは、胚体内胚葉、腸管内胚葉、膵臓内胚葉及び内分泌前駆体が、内分泌ホルモンを発現する細胞へと向かう段階に類似した段階を介して細胞を指向させることにより、in vivoでの膵臓器官形成を模倣する」と述べている。
【0018】
別の例において、Fiskらは、ヒト胚性幹細胞から膵島細胞を産生するシステムを報告している(米国特許出願公開第2006/0040387A1号)。この場合、分化経路は3つのステージに分割された。先ず、ヒト胚性幹細胞を、酪酸ナトリウムとアクチビンAとの組み合わせを用いて内胚葉に分化させた。次に、細胞をノギンなどのTGFβアンタゴニストとEGF又はベータセルリンとの組み合わせと培養して、PDX1陽性細胞を生成する。最終分化は、ニコチンアミドにより誘導した。
【0019】
一例において、Benvenistryらは、「我々は、PDX1の過剰発現が、膵臓に多く見られる遺伝子の発現を上昇させたことを結論付ける。インスリン発現の誘導には、in vivoでのみ存在する更なるシグナルを必要とする可能性がある」と述べている(Benvenistryら、Stem Cells 2006;24:1923〜1930)。
【0020】
別の例において、Grapin−Bottonらは次のように述べている。「Ngn3を初期活性化させると、ほぼ例外なくグルカゴン+細胞を誘導し、膵臓前駆細胞プールを消費した。E11.5からPDX−1前駆細胞はコンピテントになり、インスリン[陽性]でPP[陽性]の細胞へと分化した」(Johansson KAら、Developmental Cell 12,457〜465,March 2007)。
【0021】
例えば、Diezらは、次のように述べている。「9及び10週において、ほとんどのグルカゴン陽性細胞はインスリンを共発現したが、これらのステージにおいて明確にインスリンのみの細胞がはっきりと検出可能であった。インスリン及びグルカゴンを共発現する細胞は、全試験期間中(9〜21週)に観察されたが、全インスリン及びグルカゴン発現細胞のごく一部にすぎなかった」(J Histochem Cytochem.2009 Sep;57(9):811〜24.2009 Apr 13)。
【0022】
一例として、Chenらは、「(−)−インドラクタムV[(ILV)]は、プロテインキナーゼCシグナル伝達を活性化し、内胚葉系に既に確定したhESCの膵臓特殊化に向かわせる...ILV及びレチノイン酸は、関連メカニズムを介して機能する...ILVは、レチノイン酸よりも、高いPDX−1発現細胞誘導(PDX−1を発現する細胞の比率)を示す」(Nature Chemical Biology 5,195〜196(April 2009)doi:10.1038/nchembio0409−195)と述べている。
【0023】
Lyttleらは、「NKX6−1がインスリン細胞のみと共局在したことは、NKX6−1がヒトβ細胞発生に独占的に関与することを示す」(Diabetologia 2008 Jul:51(7):1169〜80,2008)と述べている。
【発明の概要】
【発明が解決しようとする課題】
【0024】
したがって、β細胞に更によく類似する機能性インスリン発現細胞を生成するための、in vitro法開発の必要性が著しく今も残っている。本発明は、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞集団を生成することにより、ヒト胚性幹細胞からインスリン発現細胞への分化効率を改善させるという代替アプローチを用いる。
【課題を解決するための手段】
【0025】
一実施形態では、本発明は、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞集団を提供する。
【0026】
一実施形態では、本発明は、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞集団へと、多能性幹細胞集団を分化させる方法を提供し、この方法は以下の工程を包含する。
a.多能性幹細胞を培養する工程、
b.多能性幹細胞を、胚体内胚葉系(definitive endoderm lineage)に特徴的なマーカーを発現する細胞へと分化させる工程、
c.胚体内胚葉系に特徴的なマーカーを発現する細胞を、膵臓内胚葉系に特徴的なマーカーを発現する細胞へと分化させる工程、
d.膵臓内胚葉系に特徴的なマーカーを発現する細胞を、プロテインキナーゼC活性化因子を追加した培地で処理することにより、膵臓内胚葉系に特徴的なマーカーを発現する細胞を、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞へと分化させる工程。
【図面の簡単な説明】
【0027】
【図1】本発明の細胞におけるインスリン及びグルカゴンの発現に対するTPB処理の影響を示す。パネルa及びbは、TPBで処理した細胞のそれぞれインスリン及びグルカゴンの発現を示す。対照細胞集団をパネルc及びdに示す。
【図2】本発明の方法に従って処理された細胞における、インスリン及びグルカゴンの発現に対する様々な濃度のTPBの影響を示す。パネルa〜dは、示される濃度のTPBで処理した細胞集団のインスリン及びグルカゴンの発現を示す。
【図3】本発明の方法に従って処理された細胞における、インスリン及びグルカゴンの発現に対するプロテインキナーゼC阻害剤の影響を示す。パネルaは、TPBで処理した細胞のインスリン及びグルカゴンの発現を表し、パネルcは対応するDAPI染色を表す。パネルbは、TPB及びGO 6976で処理した細胞のインスリン及びグルカゴンの発現を表し、パネルdは対応するDAPI染色を表す。
【図4】本発明の方法に従って処理された細胞における、インスリンの発現に対する様々なプロテインキナーゼC阻害剤の影響を示す。パネルaは、TPBで処理した細胞のインスリンの発現を示す。パネルbは、ILVで処理した細胞のインスリンの発現を示す。パネルcは、PMAで処理した細胞のインスリンの発現を示す。
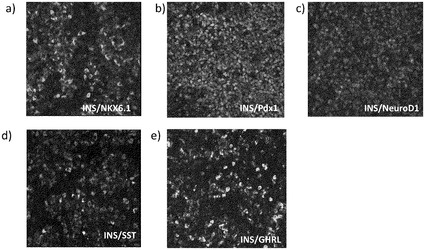
【図5】本発明の方法に従って処理された細胞における、膵臓内分泌系に特徴的なマーカーの発現を示す。これらのパネルは、インスリン及びNKX6.1(パネルa)、インスリン及びPDX1(パネルb)、インスリン及びNEUROD1(パネルc)、インスリン及びソマトスタチン(パネルd)、並びにインスリン及びグレリン(パネルe)の発現を表す。
【図6】本発明の方法に従って処理された細胞における、インスリン及びグルカゴンの発現を示す。パネルa〜cは、1%のB27、50ng/mLのFGF7、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、100ng/mLのノギン、20ng/mLのアクチビンA、及びp38キナーゼ阻害剤(米国特許第6,214,830号に開示されるもの、2.5μM)を添加したDMEM(高グルコース)で4日間にわたって処理した細胞(ステージ3、処理8、実施例2)における、インスリン発現(パネルa)、グルカゴン発現(パネルb)、及びDAPI染色(パネルc)を示す。パネルd〜fは、1%のB27、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、及び100ng/mLのノギンを添加したDMEM(高グルコース)で4日間にわたって処理した細胞(ステージ3、処理9、実施例2)における、インスリン発現(パネルd)、グルカゴン発現(パネルe)、及びDAPI染色(パネルf)を示す。
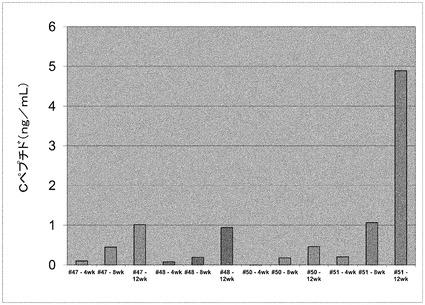
【図7】本発明の細胞の投与4、8、及び12週間後の(SCID)−beige(Bg)マウスにおいて、グルコース負荷に続いてヒトCペプチドが検出されたことを示す。
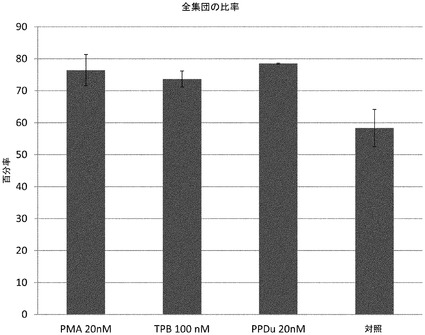
【図8】記載される濃度の様々なプロテインキナーゼC阻害剤処理後に得られた、PDX1及びNKX6.1を共発現する細胞の比率を示す。
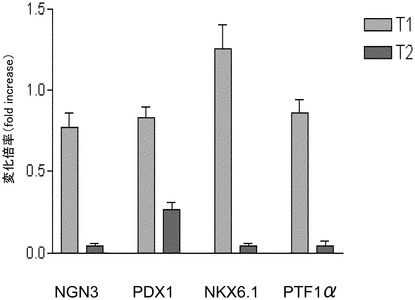
【図9】実施例6に記載の方法に従って処理した細胞の、NGN3、PDX1、NKX6.1、及びPTF1α発現を示す。
【発明を実施するための形態】
【0028】
開示を明確にするため、及び限定ではなく、本発明の「発明を実施するための形態」を、本発明の特定の特徴、実施形態又は応用を説明若しくは図示した以下の小項目に分ける。
【0029】
定義
幹細胞は、単一の細胞レベルにおいて自己再生及び分化して、自己再生前駆細胞、非再生前駆細胞、及び最終分化細胞を含む、後代細胞を産生する、能力で定義される未分化細胞である。幹細胞は、複数の胚葉(内胚葉、中胚葉、及び外胚葉)から様々な細胞系の機能的細胞へとin vitroで分化する能力、並びに移植後に複数の胚葉の組織を生じ、及び胚盤胞への注入後、全部ではないとしても大部分の組織に実質的に寄与する能力によっても特徴付けられる。
【0030】
幹細胞は、発生能によって、(1)全胚及び胚体外細胞型を生じる能力を意味する全能性、(2)全胚細胞型を生じる能力を意味する多能性、(3)細胞系の小集合を生じるが、すべて特定の組織、器官、又は生理学的システム内で生じる能力を意味する複能性(例えば、造血幹細胞(HSC)は、HSC(自己再生)、血液細胞に限定された寡能性前駆細胞、並びに血液の通常の構成要素である全細胞型及び要素(例えば、血小板)を含む後代を産生できる)、(4)複能性幹細胞と比較して、より限定された細胞系統の小集合を生じる能力を意味する寡能性、並びに(5)1つの細胞系(例えば、精子形成幹細胞)を生じる能力を意味する単能性に分類される。
【0031】
分化は、非特殊化(「未確定」)又は低特殊化細胞が、例えば神経細胞又は筋細胞などの特殊化細胞の特徴を得るプロセスである。分化細胞又は分化を誘導された細胞は、細胞系内でより特殊化した(「確定した」)状況を呈している細胞である。分化プロセスに適用された際の用語「確定した」は、通常の環境下で特定の細胞型又は細胞型の小集合への分化を続け、かつ通常の環境下で異なる細胞型に分化したり、又は低分化細胞型に戻ったりすることができない地点まで、分化経路において進行した細胞を指す。脱分化は、細胞が細胞系内で低特殊化(又は確定)した状況に戻るプロセスを指す。本明細書で使用するとき、細胞系は、細胞の遺伝、すなわちその細胞がどの細胞から来たか、またどの細胞を生じ得るかを規定する。細胞系は、細胞を発生及び分化の遺伝的スキーム内に配置する。系特異的なマーカーは、対象とする系の細胞の表現型に特異的に関連した特徴を指し、未確定の細胞の対象とする系への分化を評価する際に使用可能である。
【0032】
本明細書で使用するとき、「胚体内胚葉系に特徴的なマーカーを発現する細胞」又は「ステージ1細胞」又は「ステージ1」とは、以下のマーカー、すなわち、SOX17、GATA4、HNF3β、GSC、CER1、Nodal、FGF8、短尾奇形、Mix−様ホメオボックスタンパク質、FGF4、CD48、eomesodermin(EOMES)、DKK4、FGF17、GATA6、CXCR4、C−Kit、CD99、又はOTX2のうちの少なくとも1つを発現する細胞を指す。胚体内胚葉系に特徴的なマーカーを発現する細胞には、原始線条前駆体細胞、原始線条細胞、中内胚葉細胞、及び胚体内胚葉細胞が挙げられる。
【0033】
本明細書で使用するとき、「膵臓内胚葉系に特徴的なマーカーを発現する細胞」とは、以下のマーカー、すなわち、PDX1、NKX6.1、HNF1β、PTF1α、HNF6、HNF4α、SOX9、HB9、又はPROX1のうちの少なくとも1つを発現する細胞を指す。膵臓内胚葉系に特徴的なマーカーを発現する細胞には、膵臓内胚葉細胞、原腸管細胞、及び後部前腸細胞が挙げられる。
【0034】
本明細書で使用するとき、「胚体内胚葉」は、原腸形成中、胚盤葉上層から生じ、胃腸管及びその誘導体を形成する細胞の特徴を保持する細胞を指す。胚体内胚葉細胞は、以下のマーカー、すなわち、HNF3β、GATA4、SOX17、Cerberus、OTX2、goosecoid、C−Kit、CD99、及びMIXL1を発現する。
【0035】
本明細書で使用するとき、「マーカー」は、対象とする細胞で差異的に発現される核酸又はポリペプチド分子である。この文脈において、差次的な発現は、陽性マーカーのレベルの上昇、及び陰性マーカーのレベルの減少を意味する。マーカー核酸又はポリペプチドの検出可能なレベルは、他の細胞と比較して対象とする細胞内で充分高いか又は低く、そのため当該技術分野において既知の多様な方法のいずれかを使用して、対象とする細胞を他の細胞から識別及び区別することができる。
【0036】
本明細書で使用するとき、「膵臓内分泌細胞」又は「膵臓ホルモン発現細胞」又は「膵臓内分泌系に特徴的なマーカーを発現する細胞」とは、以下のホルモン、すなわち、インスリン、グルカゴン、ソマトスタチン、及び膵臓ポリペプチドのうちの少なくとも1つを発現することが可能な細胞を指す。
【0037】
多能性幹細胞の単離、増殖及び培養
多能性幹細胞の特徴付け
多能性幹細胞は、ステージ特異的胚抗原(SSEA)3及び4、並びにTra−1−60及びTra−1−81と呼ばれる抗体を使用して検出可能なマーカーのうちの1つ以上を発現する(Thomsonら,Science 282:1145,1998)。多能性幹細胞をin vitroで分化させると、SSEA−4、Tra−1−60、及びTra−1−81(存在する場合)の発現が減少し、SSEA−1の発現が上昇する。未分化の多能性幹細胞は通常アルカリホスファターゼ活性を有し、細胞を4%パラホルムアルデヒドで固定した後、製造業者(Vector Laboratories,Burlingame Calif.)の説明書に従ってVector Redを基質として発生させることによって検出可能である。未分化の多能性幹細胞は、OCT4及びTERTも通常発現し、RT−PCRにより検出される。
【0038】
増殖した多能性幹細胞の別の望ましい表現型は、3つの胚葉のすべて、すなわち、内胚葉、中胚葉、及び外胚葉組織の細胞に分化する能力である。多能性幹細胞の多能性は、例えば、細胞を重症複合免疫不全症(SCID)マウスに注入し、形成される奇形腫を4%パラホルムアルデヒドで固定し、次いでこれを3つの胚葉由来の細胞型の根拠について組織学的に調べることによって確認することができる。あるいは、胚様体を形成させ、この胚様体を3つの胚葉に関連したマーカーの存在に関して評価することにより、多能性を決定してもよい。
【0039】
増殖した多能性幹細胞株を、標準的なGバンド法を使用して核型決定し、確立された対応する霊長類種の核型と比較してもよい。細胞が正倍数体であり、全ヒト染色体が存在し、かつ著しく変更されてはいないことを意味する、「正常な核型」を有する細胞を得ることが望ましい。
【0040】
多能性幹細胞源
使用が可能な多能性幹細胞の種類としては、妊娠期間中の任意の時期(必ずしもではないが、通常は妊娠約10〜12週よりも前)に採取した前胚性組織(例えば、胚盤胞など)、胚性組織又は胎児組織などの、妊娠後に形成される組織に由来する多能性細胞の樹立株が含まれる。非限定的な例は、例えばヒト胚性幹細胞株H1、H7、及びH9(WiCell)などのヒト胚性幹細胞又はヒト胚生殖細胞の確立株である。それらの細胞の最初の樹立又は安定化中に本開示の組成物を使用することも想定され、その場合、供給源となる細胞は、供給源となる組織から直接採取した一次多能性細胞であろう。フィーダー細胞の不在下で既に培養された多能性幹細胞集団から採取した細胞も好適である。例えば、BG01v(BresaGen(Athens,GA))などの変異ヒト胚性幹細胞株も好適である。
【0041】
一実施形態では、ヒト胚性幹細胞は、Thomsonら(米国特許第5,843,780号、Science 282:1145,1998;Curr.Top.Dev.Biol.38:133 ff.,1998;Proc.Natl.Acad.Sci.U.S.A.92:7844,1995)により記載されるように調製される。
【0042】
多能性幹細胞の培養
一実施形態では、多能性幹細胞は、典型的にはフィーダー細胞の層上で培養され、このフィーダー細胞は、多能性幹細胞を様々な方法で支持する。あるいは、多能性幹細胞を、フィーダー細胞を本質的に含まないにも関わらず、細胞を実質的に分化させることなく多能性幹細胞の増殖を支持するような培養システム中で培養する。フィーダー細胞不含培養における多能性幹細胞の、分化を伴わない増殖は、あらかじめ別の細胞型を培養することにより馴化培地を使用して支持される。あるいは、フィーダー細胞不含培養における多能性幹細胞の、分化を伴わない増殖は、既知組成培地を使用して支持される。
【0043】
例えば、Reubinoffら(Nature Biotechnology 18:399〜404(2000))及びThompsonら(Science 6 November 1998:282巻、5391号、1145〜1147ページ)は、マウス胚性線維芽細胞のフィーダー細胞層を用いたヒト胚盤胞からの多能性幹細胞株の培養について開示している。
【0044】
Richardsら(Stem Cells 21:546〜556,2003)は、11種類の異なるヒトの成人、胎児、及び新生児フィーダー細胞層のパネルを、それらのヒト多能性幹細胞の培養を支持する能力に関して評価した。Richardsらは、「成人の皮膚線維芽フィーダー細胞上で培養したヒト胚性幹細胞系は、ヒト胚性幹細胞の形態を有し、多能性を維持する」と述べている。
【0045】
米国特許出願公開第20020072117号は、フィーダーを含まない培養において霊長類の多能性幹細胞の増殖を支持する培地を生成する細胞系を開示している。使用される細胞系は、胚性組織から得られるか又は胚性幹細胞から分化された間葉系及び線維芽細胞様の細胞系である。米国特許出願公開第20020072117号は、また、この細胞系の、1次フィーダー細胞層としての使用も開示している。
【0046】
別の例において、Wangら(Stem Cells 23:1221〜1227,2005)は、ヒト胚性幹細胞由来のフィーダー細胞層上でのヒト多能性幹細胞の長期間増殖のための方法を開示している。
【0047】
別の例において、Stojkovicら(Stem Cells 2005 23:306〜314,2005)は、ヒト胚性幹細胞の自発的分化に由来するフィーダー細胞システムを開示している。
【0048】
更なる例において、Miyamotoら(Stem Cells 22:433〜440,2004)は、ヒト胎盤から得たフィーダー細胞源を開示している。
【0049】
Amitら(Biol.Reprod 68:2150〜2156,2003)は、ヒト包皮に由来するフィーダー細胞層を開示している。
【0050】
別の例において、Inzunzaら(Stem Cells 23:544〜549,2005)は、ヒト出生後包皮線維芽細胞からのフィーダー細胞層を開示している。
【0051】
米国特許第6642048号は、フィーダーを含まない培養における霊長類の多能性幹(pPS)細胞の増殖を支持する培地、及びこうした培地の製造に有用な細胞系を開示している。米国特許第6642048号は、「本発明は、胚性組織から得られるか、あるいは胚性幹細胞から分化した間葉系及び線維芽細胞様の細胞系を含む。本開示では、こうした細胞系を誘導し、培地を調整し、この馴化培地を用いて幹細胞を増殖させるための方法を説明及び図示する」と述べている。
【0052】
別の例において、国際公開第2005014799号は、哺乳動物細胞の維持、増殖及び分化のための馴化培地を開示している。国際公開第2005014799号は、「本発明に基づいて調整される培地は、マウス細胞、特にMMH(Metマウス肝細胞)と呼ばれる分化かつ不死化したトランスジェニック肝細胞の細胞分泌活性によって馴化される」と述べている。
【0053】
別の例において、Xuら(Stem Cells 22:972〜980,2004)は、ヒトテロメラーゼ逆転写酵素を過剰発現するように遺伝子改変されたヒト胚性幹細胞由来細胞誘導体から得られた馴化培地を開示している。
【0054】
別の例において、米国特許出願公開第20070010011号は、多能性幹細胞を維持するための既知組成培地を開示している。
【0055】
代替的な培養システムは、胚性幹細胞の増殖を促進することが可能な増殖因子を追加した無血清培地を使用している。例えば、Cheonら(BioReprod DOI:10.1095/biolreprod.105.046870,October 19,2005)は、胚性幹細胞の自己再生の誘発が可能な異なる増殖因子を追加した非馴化血清補充(SR)培地中に胚性幹細胞が維持されている、フィーダーを含まずかつ血清を含まない培養システムを開示している。
【0056】
別の例において、Levensteinら(Stem Cells 24:568〜574,2006)は、bFGFを追加した培地を使用して、線維芽細胞又は馴化培地の非存在において、ヒト胚性幹細胞を長期間培養する方法を開示している。
【0057】
別の例において、米国特許出願公開第20050148070号は、血清及び繊維芽細胞フィーダー細胞を含まない合成培地中でのヒト胚性幹細胞の培養方法を開示しており、この方法は、アルブミン、アミノ酸、ビタミン、ミネラル、少なくとも1つのトランスフェリン又はトランスフェリン代用物質、少なくとも1つのインスリン又はインスリン代用物質を含有する培地中で幹細胞を培養する工程を含み、この培地は、哺乳動物胎児血清を本質的に含有せず、線維芽細胞増殖因子シグナル伝達受容体を活性化することが可能な少なくとも約100ng/mLの線維芽細胞増殖因子を含有し、増殖因子は線維芽細胞フィーダー層以外の供給源からも供給され、培地はフィーダー細胞又は条件培地なしで未分化状態の幹細胞の増殖を支持するものである。
【0058】
別の例において、米国特許出願公開第20050233446号は、未分化の霊長類始原幹細胞などの幹細胞の培養に有用な合成培地を開示している。溶液では、培地は培養されている幹細胞と比較して実質的に等張である。所定の培養において、特定の培地は、基本培地及び実質的に未分化の始原幹細胞の増殖の支持に必要な量のbFGF、インスリン、及びアスコルビン酸を含む。
【0059】
別の例として、米国特許第6800480号は、「一実施形態では、実質的に未分化状態の霊長類由来の始原幹細胞を増殖させるための細胞培地であって、霊長類由来の始原幹細胞の増殖を支持する上で効果的な低浸透圧、低エンドトキシンの基本培地を含む、細胞培地を提供する。この基本培地は、霊長類由来の始原幹細胞の増殖を支持する上で効果的な栄養素血清、並びにフィーダー細胞及びフィーダー細胞から誘導される細胞外マトリックス成分からなる群から選択される基質と組み合わされる。培地は更に、非必須アミノ酸、抗酸化剤、並びにヌクレオシド及びピルビン酸塩からなる群から選択される第1の増殖因子を含む」と述べている。
【0060】
別の例において、米国特許出願公開第20050244962号は、「一態様において本発明は、霊長類の胚性幹細胞を培養する方法を提供する。1つの方法は、哺乳動物の胎児血清を本質的に含まない(好ましくはあらゆる動物の血清をも本質的に含まない)培地中で、線維芽細胞フィーダー層以外の供給源から供給される線維芽細胞増殖因子の存在下で、幹細胞を培養する。好ましい形態では、十分な量の線維芽細胞増殖因子を添加することによって、幹細胞の培養を維持するために従来必要とされていた線維芽細胞フィーダー層の必要性がなくなる」と述べている。
【0061】
更なる例において、国際公開第2005065354号は、フィーダー及び血清を本質的に含まない合成等張培地を開示しており、この培地は、a.基本培地と、b.実質的に未分化の哺乳類幹細胞の増殖を支持するのに十分な量のbFGFと、c.実質的に未分化の哺乳類幹細胞の増殖を支持するのに十分な量のインスリンと、d.実質的に未分化の哺乳類幹細胞の増殖を支持するのに十分な量のアスコルビン酸と、を含む。
【0062】
別の例において、国際公開第2005086845号は、細胞を未分化な状態に維持するのに充分な量の、トランスフォーミング増殖因子β(TGF−β)ファミリータンパク質の構成員、線維芽細胞増殖因子(FGF)ファミリータンパク質の構成員、又はニコチンアミド(NIC)に、所望の結果を得るのに充分な時間幹細胞を曝露することを含む、未分化の幹細胞を維持するための方法を開示している。
【0063】
多能性幹細胞は、好適な培養基材上に播くことができる。一実施形態では、好適な培養基質は、例えば基底膜から誘導されたもの、又は接着分子受容体−リガンド結合の一部を形成し得るものなどの細胞外マトリックス成分である。一実施形態において、好適な培養基質は、MATRIGEL(登録商標)(Becton Dickenson)である。MATRIGEL(登録商標)は、Engelbreth−Holm Swarm腫瘍細胞由来の可溶性製剤であって、室温でゲル化して再構成基底膜を形成する。
【0064】
他の細胞外マトリックス成分及び成分混合物は代替物として好適である。これには、増殖させる細胞型に応じて、ラミニン、フィブロネクチン、プロテオグリカン、エンタクチン、ヘパラン硫塩、及び同様物を単独で又は様々な組み合わせで含み得る。
【0065】
多能性幹細胞は、細胞の生存、増殖、及び所望の特徴の維持を促進する培地の存在下で、基質上に好適に分布させることで播いてもよい。これらすべての特徴は、播種分布に細心の注意を払うことから利益が得られ、かつ当業者により容易に決定可能である。
【0066】
好適な培地は、以下の成分、すなわち例えば、ダルベッコ改変イーグル培地(DMEM)、Gibco # 11965−092;ノックアウトダルベッコ改変イーグル培地(KO DMEM)、Gibco #10829−018;ハムF12/50% DMEM基本培地;200mM L−グルタミン、Gibco#15039−027;非必須アミノ酸溶液、Gibco # 11140−050;β−メルカプトエタノール、Sigma # M7522;ヒト組み換え塩基性線維芽細胞増殖因子(bFGF)、Gibco # 13256−029等から調製することもできる。
【0067】
多能性幹細胞からの、膵臓内分泌系に特徴的なマーカーを発現する細胞の形成
一実施形態では、本発明は、多能性幹細胞から、膵臓内胚葉系に特徴的なマーカーを発現する細胞を産生するための方法を提供し、この方法は以下の工程a、b、c及びdを包含する:
a.多能性幹細胞を培養する工程、
b.多能性幹細胞を、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させる工程、
c.胚体内胚葉系に特徴的なマーカーを発現する細胞を、膵臓内胚葉系に特徴的なマーカーを発現する細胞へと分化させる工程、
d.膵臓内胚葉系に特徴的なマーカーを発現する細胞を、膵内分泌系に特徴的なマーカーを発現する細胞へと分化させる工程。
【0068】
本発明の一態様では、膵臓内分泌系に特徴的なマーカーを発現する細胞は、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する。
【0069】
胚体内胚葉系に特徴的なマーカーを発現する細胞への多能性幹細胞の分化
胚体内胚葉系に特徴的なマーカーを発現する細胞の形成は、以下の特定のプロトコルの前後に、マーカーの存在に関して試験することにより決定され得る。多能性幹細胞は、典型的にはそのようなマーカーを最小量発現する。したがって、多能性細胞の分化は、細胞がそれらの発現を開始した際に検出される。
【0070】
多能性幹細胞は、当該技術分野のいかなる方法、又は本発明で提案されるいかなる方法によって胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させてもよい。
【0071】
例えば、多能性幹細胞は、D’Amourら、Nature Biotechnology 23,1534〜1541(2005)に開示される方法に従って、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0072】
例えば、多能性幹細胞は、Shinozakiら、Development 131,1651〜1662(2004)により開示される方法に従って、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0073】
例えば、多能性幹細胞は、McLeanら、Stem Cells 25,29〜38(2007)により開示される方法に従って、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0074】
例えば、多能性幹細胞は、D’Amourら、Nature Biotechnology 24,1392〜1401(2006)に開示される方法に従って、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0075】
例えば、多能性幹細胞は、アクチビンAを含む培地中、血清の非存在下で多能性幹細胞を培養し、次いで細胞をアクチビンA及び血清と培養し、次いで細胞をアクチビンA及び異なる濃度の血清と培養することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。この方法の一例は、Nature Biotechnology 23,1534〜1541(2005)に開示されている。
【0076】
例えば、多能性幹細胞は、アクチビンAを含む培地中、血清の非存在下で多能性幹細胞を培養し、次いで細胞をアクチビンAと、別の濃度の血清と共に培養することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。この方法の例は、D’Amourら、Nature Biotechnology(2005)に開示されている。
【0077】
例えば、多能性幹細胞は、アクチビンA及びWntリガンドを含む培地中、血清の非存在下で多能性幹細胞を培養し、次いでWntリガンドを除去し、細胞をアクチビンAと、血清と共に培養することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。この方法の例は、Nature Biotechnology 24,1392〜1401(2006)に開示されている。
【0078】
例えば、多能性幹細胞は、LifeScan,Inc.に譲渡された米国特許出願第11/736,908号に開示される方法に従って多能性幹細胞を処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0079】
例えば、多能性幹細胞は、LifeScan,Inc.に譲渡された米国特許出願第11/779,311号に開示される方法に従って多能性幹細胞を処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0080】
例えば、多能性幹細胞は、米国特許出願第60/990,529号に開示される方法に従って多能性幹細胞を処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0081】
例えば、多能性幹細胞は、米国特許出願第61/076,889号に開示される方法に従って多能性幹細胞を処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0082】
例えば、多能性幹細胞は、米国特許出願第61/076,900号に開示される方法に従って多能性幹細胞を処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0083】
例えば、多能性幹細胞は、米国特許出願第61/076,908号に開示される方法に従って多能性幹細胞を処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0084】
例えば、多能性幹細胞は、米国特許出願第61/076,915号に開示される方法に従って多能性幹細胞を処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0085】
膵臓内胚葉系に特徴的なマーカーを発現する細胞への、胚体内胚葉系に特徴的なマーカーを発現する細胞の分化
胚体内胚葉系に特徴的なマーカーを発現する細胞は、当該技術分野の任意の方法、又は本発明で提案する任意の方法により、膵臓内胚葉系に特徴的なマーカーを発現する細胞へと分化され得る。
【0086】
例えば、胚体内胚葉系に特徴的なマーカーを発現する細胞は、D’Amourら、Nature Biotechnol.24:1392〜1401,2006に開示されている方法に従って、膵臓内胚葉系に特徴的なマーカーを発現する細胞へと分化され得る。
【0087】
一実施形態では、膵臓内胚葉系に特徴的なマーカーを発現する細胞は、PDX1、NKX6.1を共発現するが、最小量のCDX2及びNGN3を発現しない。
【0088】
一実施形態では、胚体内胚葉系に特徴的なマーカーを発現する細胞は、胚体内胚葉系に特徴的なマーカーを発現する細胞を、FGF7を追加した第1培地で培養し、次いで細胞を、FGF7、BMP阻害能を有する因子、TGFβ受容体アゴニスト、レチノイン酸、及びヘッジホッグシグナル経路阻害剤を追加した第2培地で培養することにより、PDX1、NKX6.1を共発現するが、最小量のCDX2及びNGN3を発現しない、膵臓内胚葉系に特徴的なマーカーを発現する細胞へと分化させる。
【0089】
一実施形態では、FGF7は、約50pg/mL〜約50μg/mLの濃度で使用することができる。一実施形態では、FGF7は、50ng/mLの濃度で使用することができる。
【0090】
一実施形態では、BMP阻害能を有する因子はノギンである。ノギンは、約500ng/mL〜約500μg/mLの濃度で使用することができる。一実施形態では、ノギンは100ng/mLの濃度で使用される。
【0091】
一実施形態では、TGFβ受容体アゴニストは、アクチビンA、アクチビンB、TGFβ−I、TGFβ−II、GDF−8、及びGDF−11からなる群から選択される。
【0092】
アクチビンAは、約2ng/mL〜100ng/mLの濃度で使用することができる。一実施形態では、アクチビンAは20ng/mLの濃度で使用される。別の実施形態では、アクチビンAは50ng/mLの濃度で使用される。
【0093】
アクチビンBは、約2ng/mL〜100ng/mLの濃度で使用することができる。一実施形態では、アクチビンBは20ng/mLの濃度で使用される。別の実施形態では、アクチビンBは50ng/mLの濃度で使用される。
【0094】
TGFβ−Iは、約2ng/mL〜100ng/mLの濃度で使用することができる。一実施形態では、TGFβ−Iは20ng/mLの濃度で使用される。別の実施形態では、TGFβ−Iは50ng/mLの濃度で使用される。
【0095】
TGFβ−IIは、約2ng/mL〜100ng/nLの濃度で使用することができる。一実施形態では、TGFβ−IIは20ng/mLの濃度で使用される。別の実施形態では、TGFβ−IIは50ng/mLの濃度で使用される。
【0096】
GDF−8は、約2ng/mL〜100ng/mLの濃度で使用することができる。一実施形態では、GDF−8は20ng/mLの濃度で使用される。別の実施形態では、GDF−8は50ng/mLの濃度で使用される。
【0097】
GDF−11は、約2ng/mL〜100ng/mLの濃度で使用することができる。一実施形態では、GDF−11は20ng/mLの濃度で使用される。別の実施形態では、GDF−11は50ng/mLの濃度で使用される。
【0098】
レチノイン酸は、約1nM〜約1mMの濃度で使用することができる。一実施形態では、レチノイン酸は1μMの濃度で使用される。
【0099】
一実施形態では、ヘッジホッグシグナル経路阻害剤はシクロパミン−KAADである。シクロパミン−KAADは、約0.025μM〜約2.5μMの濃度で使用することができる。一実施形態では、シクロパミン−KAADは0.25μMの濃度で使用される。
【0100】
分化効率は、処理した細胞集団を、胚体内胚葉系に特徴的なマーカーを発現する細胞により発現されたタンパク質マーカーを特異的に認識する薬剤(抗体など)に曝露することにより測定することができる。
【0101】
培養又は単離された細胞中のタンパク質及び核酸マーカーの発現を評価する方法は、当該技術分野において標準技術である。これらには、定量的逆転写酵素ポリメラーゼ連鎖反応(RT−PCR)、ノーザンブロット、in situハイブリダイゼーション(例えば、Current Protocols in Molecular Biology(Ausubelら編集、2001 supplement)参照)、並びにイムノアッセイ、例えば切片材料の免疫組織化学的分析、ウェスタンブロッティング、及び完全細胞で利用できるマーカーについてのフローサイトメトリー分析(FACS)(例えば、Harlow及びLane,Using Antibodies:A Laboratory Manual,New York:Cold Spring Harbor Laboratory Press(1998)参照)が挙げられる。
【0102】
多能性幹細胞の特徴は当業者に周知であり、多能性幹細胞の更なる特徴は、継続して同定されている。多能性幹細胞のマーカーとして、例えば、以下のもの、すなわち、ABCG2、cripto、FOXD3、CONNEXIN43、CONNEXIN45、OCT4、SOX2、Nanog、hTERT、UTF1、ZFP42、SSEA−3、SSEA−4、Tra 1−60、Tra 1−81の1つ以上の発現が挙げられる。
【0103】
多能性幹細胞を本発明の方法で処理した後、処理した細胞集団を、胚体内胚葉系に特徴的なマーカーを発現する細胞により発現される、例えばCXCR4などのタンパク質マーカーを特異的に認識する薬剤(抗体など)に曝露することにより、分化した細胞を精製することができる。
【0104】
本発明で用いるのに好適な多能性幹細胞としては、例えば、ヒト胚性幹細胞株H9(NIHコード:WA09)、ヒト胚性幹細胞株H1(NIHコード:WA01)、ヒト胚性幹細胞株H7(NIHコード:WA07)、及びヒト胚性幹細胞株SA002(Cellartis,Sweden)が挙げられる。多能性細胞に特徴的な以下のマーカー、すなわち、ABCG2、cripto、CD9、FOXD3、CONNEXIN43、CONNEXIN45、OCT4、SOX2、Nanog、hTERT、UTF1、ZFP42、SSEA−3、SSEA−4、Tra 1−60、及びTra 1−81のうちのすくなくとも1つを発現する細胞も本発明で用いるのに好適である。
【0105】
胚体内胚葉系に特徴的なマーカーは、SOX17、GATA4、HNF3β、GSC、CER1、Nodal、FGF8、短尾奇形、Mix様ホメオボックスタンパク質、FGF4、CD48、エオメソダーミン(EOMES)、DKK4、FGF17、GATA6、CXCR4、C−Kit、CD99、及びOTX2からなる群から選択される。本発明での使用に好適なのは、胚体内胚葉系に特徴的なマーカーのうちの少なくとも1つを発現する細胞である。本発明の一態様では、胚体内胚葉系に特徴的なマーカーを発現する細胞は、原始線条前駆体細胞である。別の態様では、胚体内胚葉系に特徴的なマーカーを発現する細胞は、中内胚葉細胞である。別の態様では、胚体内胚葉系に特徴的なマーカーを発現する細胞は、胚体内胚葉細胞である。
【0106】
膵臓内胚葉系に特徴的なマーカーは、PDX1、NKX6.1、HNF1β、PTF1α、HNF6、HNF4α、SOX9、HB9、及びPROX1からなる群から選択される。本発明での使用に好適なのは、膵臓内胚葉系に特徴的な少なくとも1つのマーカーを発現する細胞である。本発明の一態様において、膵臓内胚葉系に特徴的なマーカーを発現する細胞は、膵臓内胚葉細胞である。
【0107】
膵臓内分泌系のマーカーを発現する細胞への、膵臓内胚葉系に特徴的なマーカーを発現する細胞の分化
一実施形態では、膵臓内胚葉系に特徴的なマーカーを発現する細胞は、膵臓内分泌系に特徴的なマーカーを発現する細胞に更に分化する。
【0108】
一実施形態では、膵臓内胚葉系に特徴的なマーカーを発現する細胞は、PDX1、NKX6.1を共発現するが、最小量のCDX2及びNGN3を発現しない。
【0109】
一実施形態では、膵臓内分泌系に特徴的なマーカーを発現する細胞は、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する。
【0110】
一実施形態では、膵臓内胚葉系に特徴的なマーカーを発現する細胞を、BMP阻害能を有する因子、TGFβ受容体シグナル伝達阻害剤、及びプロテインキナーゼC活性化因子を追加した培地で培養することにより、膵臓内胚葉系に特徴的なマーカーを発現する細胞を、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞へと分化させる。
【0111】
一実施形態では、BMP阻害能を有する因子はノギンである。ノギンは、約500ng/mL〜約500μg/mLの濃度で使用することができる。一実施形態では、ノギンは100ng/mLの濃度で使用される。
【0112】
一実施形態では、TGFβ受容体シグナル伝達阻害剤は、ALK5の阻害剤である。一実施形態では、ALK5の阻害剤は、ALK5阻害剤IIである。ALK5阻害剤IIは、約0.1μM〜約10μMの濃度で使用することができる。一実施形態では、ALK5阻害剤IIは1μMの濃度で使用される。
【0113】
一実施形態では、プロテインキナーゼC活性化因子は、(2S,5S)−(E,E)−8−(5−(4−(トリフルオロメチル)フェニル)−2,4−ペンタジエノイルアミノ(pentadiemoylamino))ベンゾラクタム、インドラクタムV、及びホルボール−12−ミリステート−13−アセテートからなる群から選択される。一実施形態では、プロテインキナーゼC活性化因子は、(2S,5S)−(E,E)−8−(5−(4−(トリフルオロメチル)フェニル)−2,4−ペンタジエノイルアミノ(pentadiemoylamino))ベンゾラクタムである。(2S,5S)−(E,E)−8−(5−(4−(トリフルオロメチル)フェニル)−2,4−ペンタジエノイルアミノ(pentadiemoylamino))ベンゾラクタムは、約20nM〜約500nMの濃度で使用することができる。(2S,5S)−(E,E)−8−(5−(4−(トリフルオロメチル)フェニル)−2,4−ペンタジエノイルアミノ(pentadiemoylamino))ベンゾラクタム、インドラクタムV、及びホルボール−12−ミリステート−13−アセテートは、本明細書で「TPB」と称される。
【0114】
膵臓内分泌系に特徴的なマーカーは、NEUROD、ISL1、PDX1、NKX6.1、NKX2.2、PAX4、及びPAX6からなる群から選択される。一実施形態では、膵臓内分泌系に特徴的なマーカーを発現する細胞は、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する。
【0115】
治療
一態様では、本発明は、I型糖尿病に罹患しているか、あるいはI型糖尿病を発症するリスクを有する患者を治療する方法を提供する。一実施形態では、この方法は、多能性幹細胞を培養する工程と、多能性幹細胞を、膵臓内分泌系に特徴的なマーカーを発現する細胞へとin vitroで分化させる工程と、膵臓内分泌系に特徴的なマーカーを発現する細胞を患者に移植する工程と、を含む。
【0116】
更に別の態様では、本発明は、II型糖尿病に罹患しているか、あるいはII型糖尿病を発症するリスクを有する患者を治療する方法を提供する。一実施形態では、この方法は、多能性幹細胞を培養する工程と、多能性幹細胞を、膵臓内分泌系に特徴的なマーカーを発現する細胞へとin vitroで分化させる工程と、膵臓内分泌系に特徴的なマーカーを発現する細胞を患者に移植する工程と、を含む。
【0117】
適切であるならば、移植した細胞の生存及び機能を亢進する医薬品又は生理活性物質で患者を更に処置してもよい。それらの薬剤には、例えば、特に、インスリン、TGF−β1、2、及び3を含むTGF−βファミリーのメンバー、骨形成タンパク質(BMP−2、−3、−4、−5、−6、−7、−11、−12、及び−13)、線維芽細胞増殖因子−1及び−2、血小板由来増殖因子−AA及び−BB、多血小板血漿、インスリン増殖因子(IGF−I、II)、増殖分化因子(GDF−5、−6、−7、−8、−10、−15)、血管内皮細胞由来増殖因子(VEGF)、プレイオトロフィン、エンドセリンを挙げてもよい。他の医薬化合物には、例えば、ニコチンアミド、グルカゴン様ペプチド−I(GLP−1)及びII、GLP−1及び2模倣体(mimetibody)、エキセンディン−4、レチノイン酸、副甲状腺ホルモン、例えば米国特許出願公開第2004/0209901号及び同第2004/0132729号に開示される化合物のようなMAPK阻害剤などが挙げられる。
【0118】
多能性幹細胞は、レシピエントに移植する前にインスリン産生細胞へと分化させてもよい。具体的な実施形態では、多能性幹細胞は、レシピエントに移植する前にβ細胞へと完全に分化させる。あるいは多能性幹細胞を、未分化又は一部が分化した状態でレシピエントに移植してもよい。更なる分化をレシピエント内で行ってもよい。
【0119】
胚体内胚葉細胞、又は代替的には膵臓内胚葉細胞、又は代替的にはβ細胞を、分散した細胞として移植してもよく、あるいは肝門静脈内に注入され得るクラスターに形成してもよい。あるいは、細胞を、生体適合性の分解性ポリマー支持体、多孔性の非分解性デバイス中で提供してもよく、又は宿主免疫応答から保護されるよう封入してもよい。細胞を、レシピエントの適切な部位内に移植してもよい。移植部位には、例えば肝臓、天然の膵臓、腎被膜下空間、網、腹膜、漿膜下空間、腸、胃、又は皮下ポケットが挙げられる。
【0120】
移植された細胞の更なる分化、生存又は活性を向上するために、増殖因子、抗酸化剤又は抗炎症剤などの追加の因子を、細胞の投与前、投与と同時、又は投与後に投与してもよい。特定の実施形態では、増殖因子は、投与された細胞をin vivoで分化させるように使用される。これらの因子は、内在性細胞により分泌され、投与された細胞にin situで曝露されてもよい。移植された細胞を誘導して、当該技術分野で既知の内因性の及び外因性の投与された増殖因子の任意の組み合わせにより分化させることができる。
【0121】
移植に使用する細胞の量は、患者の状態及び治療に対する応答を含む、多数の様々な要因に依存し、当業者により決定され得る。
【0122】
一態様では、本発明は、糖尿病に罹患しているか、あるいは糖尿病を発症するリスクを有する患者を治療する方法を提供する。本方法は、多能性幹細胞を培養し、培養した細胞をβ細胞系にin vitroで分化させ、この細胞を3次元支持体に埋め込むことを含む。細胞は、患者に移植する前に、この支持体上にin vitroで維持してもよい。あるいは、細胞を含む支持体を、in vitroで更に培養することなく直接患者に移植してもよい。所望により、支持体を、埋め込まれた細胞の生存及び機能を促進する少なくとも1つの医薬品と共に組み込んでもよい。
【0123】
本発明の目的のために使用するのに好適な支持体材料には、組織修復に有用な組織鋳型、導管、バリア、及びリザーバが挙げられる。より詳細には、発泡体、スポンジ、ゲル、ヒドロゲル、織物、及び不織構造の形態を有する合成及び天然材料であって、in vitro及びin vivoで使用されて、生物組織を再構築又は再生し、また走化性薬剤を送達して組織増殖を誘発する材料が、本発明の方法の実施における使用に好適である。例えば、米国特許第5,770,417号、同第6,022,743号、同第5,567,612号、同第5,759,830号、同第6,626,950号、同第6,534,084号、同第6,306,424号、同第6,365,149号、同第6,599,323号、同第6,656,488号、米国特許出願公開第2004/0062753 A1号、米国特許第4,557,264号及び同第6,333,029号に開示されている材料を参照のこと。
【0124】
医薬品が組み込まれた支持体を形成するために、薬剤を、支持体を形成するのに先立ち、ポリマー溶液と混合することもできる。あるいは加工された支持体上に、医薬品を好ましくは医薬担体の存在下で被覆してもよい。医薬品は、液体、超微粒子状固体、又は任意の他の適切な物理的形態として存在し得る。あるいは医薬品の放出速度を変更するために、支持体に賦形剤を加えてもよい。別の実施形態では、抗炎症性化合物である少なくとも1種の医薬化合物、例えば米国特許第6,509,369号に開示されている化合物を支持体に組み込む。
【0125】
支持体には、抗アポトーシス化合物である少なくとも1種の医薬化合物、例えば米国特許第6,793,945号に開示されている化合物を組み込んでもよい。
【0126】
支持体には、線維症阻害剤である少なくとも1種の医薬化合物、例えば米国特許第6,331,298号に開示されている化合物も組み込まれ得る。
【0127】
支持体には、血管新生を促進させることができる少なくとも1種の医薬化合物、例えば米国特許出願公開第2004/0220393号及び同第2004/0209901号に開示されている化合物も組み込まれ得る。
【0128】
支持体には、免疫抑制化合物である少なくとも1種の医薬化合物、例えば米国特許出願公開第2004/0171623号に開示されている化合物も組み込まれ得る。
【0129】
支持体には、例えば、特に、TGF−β1、2、及び3を含むTGF−βファミリーのメンバー、骨形成タンパク質(BMP−2、−3、−4、−5、−6、−7、−11、−12、及び−13)、線維芽細胞増殖因子−1及び−2、血小板由来増殖因子−AA及び−BB、多血小板血漿、インスリン増殖因子(IGF−I、II)、増殖分化因子(GDF−5、−6、−8、−10、−15)、血管内皮細胞由来増殖因子(VEGF)、プレイオトロフィン、エンドセリンなどの増殖因子である、少なくとも1種の医薬化合物も組み込まれ得る。他の医薬化合物には、例えば、ニコチンアミド、低酸素誘導因子1−α、グルカゴン様ペプチド−I(GLP−1)、GLP−1及びGLP−2疑似体、並びにII、エキセンディン4、ノダル、ノギン、NGF、レチノイン酸、副甲状腺ホルモン、テネイシン−C、トロポエラスチン、トロンビン由来ペプチド、カテリシジン、デフェンシン、ラミニン、フィブロネクチン及びビトロネクチンなどの接着性細胞外マトリックスタンパク質の細胞−及びヘパリン−結合ドメインを含む生物学的ペプチド、例えば米国特許出願公開第2004/0209901号及び同第2004/0132729号に開示されている化合物などのMAPK阻害剤を挙げることができる。
【0130】
骨格内への本発明の細胞の組み込みは、細胞を骨格上に単に沈着させることにより達成できる。細胞は、単に拡散により骨格内に入ることができる(J.Pediatr.Surg.23(1 Pt 2):3〜9(1988))。細胞播種の効率を向上させるために、いくつかの他の手法が開発されている。例えば、軟骨細胞をポリグリコール酸骨格上に播種する際に、スピナーフラスコが使用されている(Biotechnol.Prog.14(2):193〜202(1998))。細胞播種のための他の手法は遠心法の使用であり、これは播種する細胞に与えるストレスを最小にし、かつ播種効率を高める。例えば、Yangらは、遠心分離細胞固定化法(CCI)と呼ばれる細胞播種法を開発した(J.Biomed.Mater.Res.55(3):379〜86(2001))。
【0131】
本発明を以下の実施例によって更に説明するが、本発明はこれらの実施例により限定されるものではない。
【実施例】
【0132】
(実施例1)
インスリン及びNKX6.1、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞集団の形成
ヒト胚性幹細胞株H1細胞を、MATRIGEL(登録商標)(1:30希釈)(BD Biosciences;カタログ# 356231)をコーティングしたディッシュと0.2%のFBS、100ng/mLのアクチビンA(PeproTech;カタログ#120−14)、及び20ng/mLのWNT−3a(R&D Systems;カタログ# 1324−WN/CF)を添加したRPMI培地(Invitrogen;カタログ#:22400)により1日培養した後で、0.5%のFBS及び100ng/mLのアクチビンAを添加したRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで、
a.2%のFBS及び50ng/mLのFGF7(PeproTech;カタログ#100−19)を添加したDMEM/F12(Invitrogen;カタログ# 11330−032)で3日間にわたって処理し(ステージ2)、次いで、
b.1%のB27、50ng/mLのFGF7、0.25μMのシクロパミン−KAAD(Calbiochem;カタログ# 239804)、及び100ng/mLのノギン(R&D Systems;カタログ# 3344−NG)を添加したDMEM(高グルコース)(Invitrogen;カタログ# 10569)で4日間にわたって培養し(ステージ3)、次いで
c.1%のB27(Invitrogen;カタログ# 0791)、100ng/mLのノギン、1μMのALK5阻害剤II(Axxora;カタログ# ALX−270−445)、及び500nMのTBP((2S,5S)−(E,E)−8−(5−(4−(トリフルオロメチル)フェニル)−2,4−ペンタジエノイルアミノ(pentadiemoylamino))ベンゾラクタム)(Calbiochem;カタログ# 565740)を添加したDMEM(高グルコース)で6日間にわたって培養する(ステージ4)。
【0133】
対照として、別の細胞集団を、1%のB27、100ng/mLのノギン、及び1μMのALK5阻害剤IIを添加したDMEM(高グルコース)で6日間にわたって処理した(ステージ4、対照群)。
【0134】
図1に示すように、ステージ4におけるTBP処理により、インスリン発現細胞が増加する結果となった(図1、パネルa)。なお、これらインスリン発現細胞の約60%が単独の内分泌ホルモン発現細胞であり、これらの細胞はインスリンを発現し、グルカゴン、ソマトスタチン及びグレリン(図1、パネルa及びb、図5、パネルd及びe)を発現しなかった。TBP処理を受けた培養においては、グルカゴン発現細胞も確認された。ほとんどのグルカゴン発現細胞は、インスリンも共発現した(図1、パネルa及びb)。対照群について、細胞の大部分はインスリン及びグルカゴンを共発現した(図1、パネルc及びd)。
【0135】
異なる実験において、ヒト胚性幹細胞株H1細胞を、MATRIGEL(登録商標)(1:30希釈)をコーティングしたディッシュと、0.2%のFBS、100ng/mLのアクチビンA、及び20ng/mLのWNT−3aを添加したRPMI培地により1日培養し、次いで0.5%のFBS及び100ng/mLのアクチビンAを添加したRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
a.2%のFBS及び50ng/mLのFGF7を添加したDMEM/F12で3日間にわたって処理し(ステージ2)、次いで
b.1%のB27、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、及び100ng/mLのノギンを添加したDMEM(高グルコース)で4日間にわたって処理し(ステージ3)、次いで
c.処理1:1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び500nMのTBPを添加したDMEM(高グルコース)で6日間(ステージ4、処理1)、又は
d.処理2:1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び100nMのTBPを添加したDMEM(高グルコース)で6日間(ステージ4、処理2)、又は
e.処理3:1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び20nMのTBPを添加したDMEM(高グルコース)で6日間(ステージ4、処理3)、又は
f.処理4:1%のB27、100ng/mLのノギン、及び1μMのALK5阻害剤IIを添加したDMEM(高グルコース)で6日間(ステージ4、処理4)にわたって処理した。
【0136】
免疫細胞化学的分析を用い、本発明の細胞の形成に対する様々な濃度のTPBの影響を評価した。インスリン単独発現細胞数の著しい増加が、500nM及び100nMのTPB処理群の両方(図2、パネルa及びb)において観察された。FACS分析では、両方の処理によりin vitroにおいて12%のインスリン単独発現細胞をもたらし、この集団の15%がNKX6.1も発現していたことが確認された(表1−NKX6.1/INS発現細胞は総集団の2.4%であった)。対照群と同様に、20nMのTPBにおいては、ほとんどの細胞がインスリン及びグルカゴンを共発現した(図2、パネルc及びd)。
【0137】
【表1】
【0138】
内分泌ホルモン発現細胞形成への作用が、プロテインキナーゼCの活性化を介してなされたことを更に確認するため、ヒト胚性幹細胞株H1の別の細胞集団を、MATRIGEL(登録商標)(1:30希釈)をコーティングしたディッシュと0.2%のFBS、100ng/mLのアクチビンA、及び20ng/mL WNT−3aを添加したRPMI培地により1日培養した後で、0.5%のFBS及び100ng/mLのアクチビンAを添加したRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
a.2%のFBS及び50ng/mLのFGF7を添加したDMEM/F12で3日間にわたって処理し(ステージ2)、次いで
b.1%のB27、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、及び100ng/mLのノギンを添加したDMEM(高グルコース)で4日間にわたって処理し(ステージ3)、次いで
c.処理5:1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び500nMのTBPを添加したDMEM(高グルコース)で6日間(ステージ4、処理5)、又は
d.処理6:1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、500nMのTPB、及び5μMのGO 6976を添加したDMEM(高グルコース)で6日間(ステージ4、処理6)、又は
e.処理7:1%のB27、100ng/mLのノギン、及び1μMのALK5阻害剤IIを添加したDMEM(高グルコース)で処理し(ステージ4、処理7)、次いで
f.1%のB27を添加したDMEM(高グルコース)で4日間にわたって処理した(ステージ5)。
【0139】
GO 6976は、Ca2+依存性プロテインキナーゼCのイソ型を選択的に阻害するといわれている。TPBのみ(図3、パネルa)、並びにTPB及びGO 6976(図3、パネルb)を与えられた培養液において、膵臓内分泌系に特徴的なマーカーを発現する細胞の数が著しく減少する。FACS分析では、TPB処理(処理6)が、30.6%のシナプトフィジン発現細胞、12%のインスリン単独発現細胞、及び4.6%のグルカゴン発現細胞をもたらしたことが確認された。一方、TBP及びGO 6976処理(処理7)は、10.6%のシナプトフィジン発現細胞と、検出できないレベルのインスリン単独発現細胞をもたらした(表2)。処理6と処理7との間で、観察された総細胞数に違いはなかった(図3、パネルc及びd参照、処理6及び処理7の総細胞数を表すDAPI染色を示す)。これらの結果は、プロテインキナーゼCのシグナル伝達が、膵臓内分泌系に特徴的なマーカーを発現する細胞の形成に重要であり得ることを示唆する。
【0140】
その他のプロテインキナーゼC活性化因子も調べた。これらはインドラクタムV(ILV)(Axxora;カタログ# ALX−420−011−C300)及びホルボール−12−ミリステート−13−アセテート(PMA)(Calbiochem;カタログ#524400)とした。しかし、TPBのみがインスリン単独発現細胞の形成を示した(図4、パネルa)。ILV(図4、パネルb)及びPMA(図4、パネルc)の両方が500nMにおいて、6日後にインスリン及びグルカゴンを共発現する細胞をもたらした。FACS分析では、TPB処理が、12%のインスリン単独発現細胞、及び4.6%のグルカゴン発現細胞、並びに7.1%のインスリン及びグルカゴンを共発現する細胞をもたらしたことが確認された。一方、ILV処理は、3%のインスリン単独発現細胞、及び12%のグルカゴン細胞、並びに12%のインスリン及びグルカゴンを共発現する細胞をもたらした(表2)。免疫細胞化学的分析により、TPBで処理した培養液において、インスリン発現細胞の20%がNKX6.1(図5、パネルa)及びPDX1(図5、パネルb)を共発現したことが示された。インスリン発現細胞の大部分は、内分泌腺マーカー(maker)であるNEURODを共発現した(図5、パネルc)。ソマトスタチン又はグレリン(GHRL)を共発現したインスリン発現細胞はほとんどなかった(図5、パネルd及びe)。
【0141】
【表2】
【0142】
(実施例2)
インスリン及びNKX6.1、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞集団を形成する別法
異なる実験において、ヒト胚性幹細胞株H1細胞を、MATRIGEL(登録商標)(1:30希釈)をコーティングしたディッシュと、0.2%のFBS、100ng/mLのアクチビンA、及び20ng/mL WNT−3aを添加したRPMI培地により1日培養し、次いで0.5%のFBS及び100ng/mLのアクチビンAを添加したRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
a.2%のFBS及び50ng/mLのFGF7を添加したDMEM/F12で3日間にわたって処理し(ステージ2)、次いで
b.処理8:1%のB27、50ng/mLのFGF7、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、100ng/mLのノギン、20ng/mLのアクチビンA、及びp38キナーゼ阻害剤(米国特許第6,214,830号に開示されるもの、2.5μM)を添加したDMEM(高グルコース)で4日間(ステージ3、処理8)、又は
c.処理9:1%のB27、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、及び100ng/mLのノギンを添加したDMEM(高グルコース)で4日間(ステージ3、処理9)、次いで
d.1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び500nMのTPBを添加したDMEM(高グルコース)で6日間にわたって処理した(ステージ4)。
【0143】
ステージ3、処理8により、PDX1及びNKX6.1を共発現したが、CDX2及びNGN3を共発現しなかった膵臓内胚葉系に特徴的なマーカーを発現する細胞集団の形成をもたらした。一方、ステージ3、処理9では、PDX1、NKX6.1、及びNGN3を共発現した膵臓内胚葉系に特徴的なマーカーを発現する細胞集団の形成をもたらした。これらの細胞集団に対するプロテインキナーゼC活性化因子処理の影響を調べた(上記ステージ4)。
【0144】
FACS分析を行い、インスリン単独陽性細胞、グルカゴン単独陽性細胞、インスリン/グルカゴン二重陽性細胞、NKX6.1陽性細胞を発現する細胞、インスリン/NKX6.1陽性細胞、及びシナプトフィジン陽性細胞(膵臓内分泌腺マーカー)の比率を明らかにした。
【0145】
表3に示すように、処理8で形成された細胞集団は、シナプトフィジン発現で表される、より大部分の内分泌細胞を生じ、総細胞集団の49.7%はシナプトフィジンを発現した。総集団の27.8%はインスリン単独陽性細胞であった。
【0146】
一方、処理9で形成された細胞集団が生じたシナプトフィジン発現細胞は、たった25.7%であった。総集団の7.6%はインスリン単独発現細胞であった。両方の処理で観察されたグルカゴン単独発現細胞に有意差はなく、グルカゴン発現細胞の比率はインスリン発現細胞より有意に低かった。
【0147】
かなりの量のインスリン発現細胞はまた、NKX6.1も共発現した。処理8を受けた細胞集団では、総集団の11%がインスリン及びNKX6.1を発現した。処理9を受けた細胞集団では、総集団の2%がインスリン及びNKX6.1を発現した。
【0148】
免疫蛍光分析によりこれを確認した(図6)。処理8は、処理9に比べてインスリン発現細胞を増加させた(図6、パネルa及びd)。ほとんどのグルカゴン発現細胞は多ホルモン細胞であった(図6、パネルa、b、d及びe)。これらの結果は、処理8で生じた細胞集団(PDX1及びNKX6.1を共発現したが、CDX2及びNGN3は共発現しなかった膵臓内胚葉系に特徴的なマーカー(maker)を発現する細胞)は、本発明の方法により、より効率的に誘導され成熟した機能性インスリン発現細胞になれることを示唆する。
【0149】
【表3】
【0150】
(実施例3)
インスリン及びNKX6.1、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞集団を形成する別法
別の実験において、ヒト胚性幹細胞株H1細胞を、MATRIGEL(登録商標)(1:30希釈)をコーティングしたディッシュと、0.2%のFBS、100ng/mLのアクチビンA、及び20ng/mLのWNT−3aを添加したRPMI培地により1日培養し、次いで0.5%のFBS及び100ng/mLのアクチビンAを添加したRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
a.2%のFBS及び50ng/mLのFGF7を添加したDMEM/F12で3日間にわたって処理し(ステージ2)、次いで
b.1%のB27、50ng/mLのFGF7、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、100ng/mLのノギン、20ng/mLのアクチビンA、及びp38キナーゼ阻害剤(JNJ3026582、2.5μM)を添加したDMEM(高グルコース)で4日間にわたって処理し(ステージ3)、次いで
c.処理10:1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び500nMのTPBを添加したDMEM(高グルコース)で6日間(ステージ4、処理10)、又は
d.処理11:1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び500nMのTPBを添加したDMEM(高グルコース)で9日間(ステージ4、処理11)、又は
e.処理12:1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び500nMのTPBを添加したDMEM(高グルコース)で12日間にわたって処理した(ステージ4、処理12)。
【0151】
表4に示すように、プロテインキナーゼC活性化因子による処理期間を、9日間(処理11)又は12日間(処理12)のいずれかに延長したとき、更なる効果はみられなかった。インスリン単独発現細胞は、処理10で6日間処理後、総集団の27.8%であった。反対に、インスリン発現細胞は、9日間処理(処理11)後10%に、12日間処理(処理12)後更に4%に減少した。同時に、インスリン及びNKX6.1を共発現する細胞集団の総比率も、処理の延長後に著しく下がった。これらの結果は、ノギン、Alk5阻害剤II、及びプロテインキナーゼC活性化因子による6日間処理は、本発明の細胞の形成に十分であったことを示唆した。
【0152】
【表4】
【0153】
(実施例4)
重症複合免疫不全(SCID)−beige(Bg)マウスへの、本発明の細胞の移植
ヒト胚性幹細胞株H1細胞を、MATRIGEL(登録商標)(1:30希釈)をコーティングしたディッシュと、0.2%のFBS、100ng/mLのアクチビンA、及び20ng/mLのWNT−3aを添加したRPMI培地により1日培養し、次いで0.5%のFBS及び100ng/mLのアクチビンAを添加したRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
a.2%のFBS及び50ng/mLのFGF7を添加したDMEM/F12で3日間にわたって処理し(ステージ2)、次いで
b.1%のB27、50ng/mLのFGF7、0.25μMのシクロパミン−KAAD、及び100ng/mLのノギンを添加したDMEM(高グルコース)で4日間にわたって処理し(ステージ3)、次いで
c.1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び500nMのTBPを添加したDMEM(高グルコース)で6日間にわたって処理した(ステージ4)。
【0154】
ステージ4の終了時に、1mLのガラス製ピペットを用いて細胞を機械的に回収し、続いて非接着性プレートに移し一晩培養した。得られた細胞集合体を回収し、5百万個の細胞を含有する集合体を免疫不全マウス(SCID/Bg、動物番号47、48、49、50、及び51)の腎臓被膜に移植した。図7を参照されたい。
【0155】
4週間後、これらの移植物中のインスリン産生細胞の機能性を、動物にグルコースを投与しインスリン分泌を誘導することにより試験した。動物を約15〜20時間にわたって絶食させ、その後、後眼窩から血液サンプルを採取した(処理前グルコース)。次いで各動物に、約3g/kgのグルコース/30%のデキストロース溶液を腹腔内注射により投与し、グルコース注入の約60分後に血液を採取した。非常に感受性の高い、ヒト特異的なCペプチドELISAプレート(カタログ番号80−CPTHU−E01、Alpco Diagnostics,NH)を使用して、マウス血清を用いて血中ヒトCペプチドを検出した。ヒトCペプチドの検出は、インスリン分泌が移植細胞に由来していることを示す。
【0156】
移植後4週目程の早期に、動物血清中のヒトCペプチドを検出した。ヒトCペプチドは経時的に増加した。移植データを図7にまとめる。1ヶ月の終了時点で、試験群の60%の動物において、グルコース投与に応答するヒトCペプチド(0.2ng/mL未満)が検出できた。4週間後、4例中3例のマウスで、ヒトCペプチドのグルコース刺激化血清濃度が5〜10倍上昇した。移植12週間後、移植マウスにおけるヒトcペプチドの平均グルコース刺激化血清濃度は、1mg/mLより高かった(n=4)。
【0157】
(実施例5)
PDX1及びNKX6.1を共発現する、膵臓内胚葉系に特徴的なマーカーを発現する細胞集団を形成する別法
簡潔に述べると、ヒト胚性幹細胞株H1細胞を、MATRIGEL(登録商標)(1:30希釈)をコーティングしたディッシュと、0.2%のFBS、100ng/mLのアクチビンA、及び20ng/mLのWNT−3aを追加したRPMI培地により1日培養した後で、0.5%のFBS及び100ng/mLのアクチビンAを追加したRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
a.2%のFBS及び50ng/mLのFGF7を添加したDMEM/F12で3日間にわたって処理し(ステージ2)、次いで
b.1%のB27、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、及び100ng/mLのノギンを添加したDMEM(高グルコース)で4日間にわたって処理し(ステージ3)、次いで
c.1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、20nM PMA、又は100nMのTPB、又は20nMのホルボール−12,13−ジブチラート(PDBu)(Calbioche、カタログ#524390)を添加したDMEM(高グルコース)で6日間にわたって処理した(ステージ4)。
【0158】
対照として、別の細胞集団を、1%のB27、100ng/mLのノギン、及び1μMのALK5阻害剤IIを追加したDMEM(高グルコース)で6日間にわたって処理した(ステージ4)。
【0159】
ステージ4の6日目に培養液を2回サンプリングし、IN Cell Analyzer 1000(GE Healthcare)を用いて撮像した。ウェルあたり100視野の撮像を得て、バイオアッセイ及び続く染色手順の間の任意の細胞ロスを補正した。IN Cell Developer Toolbox 1.7(GE Healthcare)ソフトウェアを用いて、各ウェルから総細胞数、総PDX1発現細胞、及び総NKX6.1発現細胞の測定値を得た。各複製データセットについて平均及び標準偏差を算出した。総PDX1及びNKX6.1タンパク質発現細胞は、総細胞集団の比率として報告した。図8に示すように、プロテインキナーゼC活性化因子処理群では、対照処理で得られたサンプルと比較して低い有効濃度(約20nM)において、NKX6.1/PDX1発現細胞集団が劇的に増加した。ステージ4の6日目までに、プロテインキナーゼC活性化因子又は対照処理のいずれかを受けた細胞集団では、集団の92%±4%がPDX1を発現した。プロテインキナーゼC活性化因子処理群では、75%±5%のPDX1発現細胞がNKX6.1を発現した。しかし、ノギン及びTGFβ受容体阻害剤のみで処理された集団(対照)では、PDX1発現細胞の58%±5%のみがNKX6.1を発現した。プロテインキナーゼC活性化因子の存在下では、20%のNKX6.1発現細胞は、増殖マーカー、EdU(Click−iT(登録商標)EdU Imaging Kit、Invitrogen、カタログ#C10337)に対して共に陽性であった。
【0160】
この実験では、プロテインキナーゼC活性化因子を比較的低い有効濃度(〜20nM)でノギン及びTGFβ受容体阻害剤と組み合わせて用いると、Nkx6.1発現の上方制御、並びにPDX1及びNKX6.1を発現する細胞比率の増加を促進できることを示す。
【0161】
(実施例6)
膵臓内胚葉系に特徴的なマーカーを発現する細胞の、プロテインキナーゼC活性化因子による処理
簡潔に述べると、ヒト胚性幹細胞株H1細胞を、MATRIGEL(登録商標)(1:30希釈)をコーティングしたディッシュと、0.2%のFBS、100ng/mLのアクチビンA、及び20ng/mLのWNT−3aを追加したRPMI培地により1日培養した後で、0.5%のFBS、及び100ng/mLのアクチビンAを追加したRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
a.2%のFBS及び50ng/mLのFGF7を添加したDMEM/F12で3日間にわたって処理し(ステージ2)、次いで
b.T1:1%のB27、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、100ng/mLのノギン、及び50ng/mLのFGF10を添加したDMEM(高グルコース)で4日間(ステージ3、T1)、又は
T2:1%のB27、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、100ng/mLのノギン、50ng/mLのFGF10、及び100nMのTPBを添加したDMEM(高グルコース)で4日間にわたって処理し(ステージ3、T2)、次いで
c.1%のB27、100ng/mLのノギン、及び1μMのALK5阻害剤IIを添加したDMEM(高グルコース)で6日間にわたって処理した(ステージ4)。
【0162】
図9に示すように、対照群(T1)と比較してTPBで処理した細胞(T2)において、膵臓内胚葉マーカーであるPDX1、NKX6.1、及びPTF1αの顕著な下方制御が観察された。NKX6.1は免疫組織化学的に検出できなかった。これらのデータは、ステージ3におけるプロテインキナーゼ活性化因子処理により、PDX1/NKX6.1共発現細胞の生成が促進されなかったことを示唆する。
【0163】
本明細書の全体を通じて引用した刊行物は、その全容を本明細書に援用するものである。以上、本発明の様々な態様を実施例及び好ましい実施形態を参照して説明したが、本発明の範囲は、上記の説明文によってではなく、特許法の原則の下で適宜解釈される以下の「特許請求の範囲」によって定義されるものである点は認識されるであろう。
【技術分野】
【0001】
本出願は、米国特許仮出願第61/289,671号(2009年12月23日出願)の利益を主張し、その全体が参照により本明細書に組み込まれる。
本発明は、多能性幹細胞からのインスリン産生細胞への分化を促進させるための方法を提供する。詳細には、本発明は、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞を産生する方法を提供する。
【背景技術】
【0002】
I型糖尿病の細胞置換療法の進歩及び移植可能なランゲルハンス島の不足により、生着に適したインスリン産生細胞、すなわちβ細胞の供給源の開発に注目が集まっている。1つの手法は、例えば、胚性幹細胞のような多能性幹細胞から機能性のβ細胞を生成することである。
【0003】
脊椎動物の胚発生において、多能性細胞は、原腸形成として公知のプロセスにより3つの胚葉(外胚葉、中胚葉、及び内胚葉)を含む細胞のグループを生じる。例えば、甲状腺、胸腺、膵臓、腸、及び肝臓などの組織は、内胚葉から中間ステージを経て発達する。このプロセスにおける中間ステージは、胚体内胚葉(definitive endoderm)の形成である。胚体内胚葉細胞は、HNF3β、GATA4、MIXL1、CXCR4及びSOX17などの多数のマーカーを発現する。
【0004】
膵臓の形成は、胚体内胚葉が膵臓内胚葉へと分化することにより生じる。膵臓内胚葉の細胞は膵臓−十二指腸ホメオボックス遺伝子、PDX1を発現する。PDX1が存在しない場合、膵臓は、腹側芽及び背側芽の形成より先に発達が進行しない。したがって、PDX1の発現は、膵臓器官形成において重要な工程として特徴付けられる。成熟した膵臓は、他の細胞型間で外分泌組織及び内分泌組織を含有する。外分泌組織及び内分泌組織は、膵臓内胚葉の分化によって生じる。
【0005】
島細胞の特徴を保持する細胞が、マウスの胚細胞から誘導されたことが報告されている。例えば、Lumelskyら(Science 292:1389,2001)は、膵島と同様のインスリン分泌構造へのマウスの胚性幹細胞の分化を報告している。Soriaら(Diabetes 49:157,2000)は、ストレプトゾトシン誘発糖尿病のマウスにおいて、マウスの胚性幹細胞から誘導されたインスリン分泌細胞が血糖を正常化することを報告している。
【0006】
一例において、Horiら(PNAS 99:16105,2002)は、ホスホイノシチド3−キナーゼ(LY294002)の阻害剤でマウス胚性幹細胞を処理することにより、β細胞に類似した細胞が生じたことを開示している。
【0007】
別の例において、Blyszczukら(PNAS 100:998,2003)は、Pax4を構成的に発現するマウス胚性幹細胞からのインスリン産生細胞の生成を報告している。
【0008】
Micallefらは、レチノイン酸が、胚性幹細胞のPDX1陽性膵臓内胚葉の形成に対する関与を制御できることを報告している。レチノイン酸は、胚における原腸形成の終了時に対応する期間中の、胚性幹細胞分化の4日目に培養液に添加すると、Pdx1発現の誘発に最も効果的である(Diabetes 54:301、2005)。
【0009】
Miyazakiらは、Pdx1を過剰発現するマウス胚性幹細胞株を報告している。この結果は、外因性のPdx1発現が、得られた分化細胞内でインスリン、ソマトスタチン、グルコキナーゼ、ニューロゲニン3、p48、Pax6、及びHnf6遺伝子の発現を明らかに増加させたことを示している(Diabetes 53:1030,2004)。
【0010】
Skoudyらは、マウス胚性幹細胞内で、アクチビンA(TGFβスーパーファミリーのメンバー)が、膵臓外分泌遺伝子(p48及びアミラーゼ)、並びに内分泌遺伝子(Pdx1、インスリン及びグルカゴン)の発現を上方制御することを報告している。最大の効果は、1nMアクチビンAを使用した場合に認められた。また、インスリン及びPdx1のmRNA発現レベルはレチノイン酸に影響されないが、3nM FGF7処理により、Pdx1の転写物量が増加する結果となることが認められた(Biochem.J.379:749,2004)。
【0011】
Shirakiらは、PDX1陽性細胞への胚性幹細胞の分化を特異的に増加させる増殖因子の効果を研究した。Shirakiらは、TGFβ2によってPDX1陽性細胞がより高い比率で再現性良く得られたことを観察した(Genes Cells.2005 Jun;10(6):503〜16)。
【0012】
Gordonらは、血清の非存在下、かつアクチビンとWntシグナル伝達阻害剤の存在下での、マウス胚性幹細胞からの短尾奇形[陽性]/HNF−3β[陽性]内胚葉細胞への誘導を示した(米国特許第2006/0003446A1号)。
【0013】
Gordonら(PNAS,103巻、16806ページ、2006)は、「Wnt及びTGF−β/nodal/アクチビンの同時シグナル伝達が前原始線条の形成には必要であった」と述べている。
【0014】
しかしながら、胚性幹細胞発達のマウスモデルは、例えばヒトなどのより高等な哺乳動物における発達プログラムを正確には模倣しない恐れがある。
【0015】
Thomsonらは、ヒト胚盤胞から胚性幹細胞を単離した(Science 282:114,1998)。同時に、Gearhart及び共同研究者は、胎児生殖腺組織から、ヒト胚生殖(hEG)細胞株を誘導した(Shamblottら、Proc.Natl.Acad.Sci.USA 95:13726、1998)。白血病阻害因子(LIF)と共に培養するのみで分化を阻止し得るマウス胚性幹細胞とは異なり、ヒト胚性幹細胞は、非常に特殊な条件下で維持する必要がある(米国特許第6,200,806号、国際公開第99/20741号;同第01/51616号)。
【0016】
D’Amourらは、高濃度のアクチビン及び低濃度の血清の存在下で、ヒト胚性幹細胞由来の胚体内胚葉の濃縮化された培養物が調製されたことを述べている(Nature Biotechnology 2005)。これらの細胞を、マウスの腎臓被膜下に移植することにより、内胚葉性器官の特徴の一部を有するより成熟した細胞への分化が得られた。ヒト胚性幹細胞由来の胚体内胚葉細胞は、FGF−10の添加後、PDX1陽性細胞に更に分化することができる(米国特許出願公開第2005/0266554A1号)。
【0017】
D’Amourら(Nature Biotechnology−24,1392〜1401(2006))は、「我々はヒト胚性幹(hES)細胞を、膵臓ホルモンインスリン、グルカゴン、ソマトスタチン、膵臓ポリペプチド及びグレリンを合成できる内分泌細胞へと変換する分化プロセスを開発した。このプロセスは、胚体内胚葉、腸管内胚葉、膵臓内胚葉及び内分泌前駆体が、内分泌ホルモンを発現する細胞へと向かう段階に類似した段階を介して細胞を指向させることにより、in vivoでの膵臓器官形成を模倣する」と述べている。
【0018】
別の例において、Fiskらは、ヒト胚性幹細胞から膵島細胞を産生するシステムを報告している(米国特許出願公開第2006/0040387A1号)。この場合、分化経路は3つのステージに分割された。先ず、ヒト胚性幹細胞を、酪酸ナトリウムとアクチビンAとの組み合わせを用いて内胚葉に分化させた。次に、細胞をノギンなどのTGFβアンタゴニストとEGF又はベータセルリンとの組み合わせと培養して、PDX1陽性細胞を生成する。最終分化は、ニコチンアミドにより誘導した。
【0019】
一例において、Benvenistryらは、「我々は、PDX1の過剰発現が、膵臓に多く見られる遺伝子の発現を上昇させたことを結論付ける。インスリン発現の誘導には、in vivoでのみ存在する更なるシグナルを必要とする可能性がある」と述べている(Benvenistryら、Stem Cells 2006;24:1923〜1930)。
【0020】
別の例において、Grapin−Bottonらは次のように述べている。「Ngn3を初期活性化させると、ほぼ例外なくグルカゴン+細胞を誘導し、膵臓前駆細胞プールを消費した。E11.5からPDX−1前駆細胞はコンピテントになり、インスリン[陽性]でPP[陽性]の細胞へと分化した」(Johansson KAら、Developmental Cell 12,457〜465,March 2007)。
【0021】
例えば、Diezらは、次のように述べている。「9及び10週において、ほとんどのグルカゴン陽性細胞はインスリンを共発現したが、これらのステージにおいて明確にインスリンのみの細胞がはっきりと検出可能であった。インスリン及びグルカゴンを共発現する細胞は、全試験期間中(9〜21週)に観察されたが、全インスリン及びグルカゴン発現細胞のごく一部にすぎなかった」(J Histochem Cytochem.2009 Sep;57(9):811〜24.2009 Apr 13)。
【0022】
一例として、Chenらは、「(−)−インドラクタムV[(ILV)]は、プロテインキナーゼCシグナル伝達を活性化し、内胚葉系に既に確定したhESCの膵臓特殊化に向かわせる...ILV及びレチノイン酸は、関連メカニズムを介して機能する...ILVは、レチノイン酸よりも、高いPDX−1発現細胞誘導(PDX−1を発現する細胞の比率)を示す」(Nature Chemical Biology 5,195〜196(April 2009)doi:10.1038/nchembio0409−195)と述べている。
【0023】
Lyttleらは、「NKX6−1がインスリン細胞のみと共局在したことは、NKX6−1がヒトβ細胞発生に独占的に関与することを示す」(Diabetologia 2008 Jul:51(7):1169〜80,2008)と述べている。
【発明の概要】
【発明が解決しようとする課題】
【0024】
したがって、β細胞に更によく類似する機能性インスリン発現細胞を生成するための、in vitro法開発の必要性が著しく今も残っている。本発明は、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞集団を生成することにより、ヒト胚性幹細胞からインスリン発現細胞への分化効率を改善させるという代替アプローチを用いる。
【課題を解決するための手段】
【0025】
一実施形態では、本発明は、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞集団を提供する。
【0026】
一実施形態では、本発明は、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞集団へと、多能性幹細胞集団を分化させる方法を提供し、この方法は以下の工程を包含する。
a.多能性幹細胞を培養する工程、
b.多能性幹細胞を、胚体内胚葉系(definitive endoderm lineage)に特徴的なマーカーを発現する細胞へと分化させる工程、
c.胚体内胚葉系に特徴的なマーカーを発現する細胞を、膵臓内胚葉系に特徴的なマーカーを発現する細胞へと分化させる工程、
d.膵臓内胚葉系に特徴的なマーカーを発現する細胞を、プロテインキナーゼC活性化因子を追加した培地で処理することにより、膵臓内胚葉系に特徴的なマーカーを発現する細胞を、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞へと分化させる工程。
【図面の簡単な説明】
【0027】
【図1】本発明の細胞におけるインスリン及びグルカゴンの発現に対するTPB処理の影響を示す。パネルa及びbは、TPBで処理した細胞のそれぞれインスリン及びグルカゴンの発現を示す。対照細胞集団をパネルc及びdに示す。
【図2】本発明の方法に従って処理された細胞における、インスリン及びグルカゴンの発現に対する様々な濃度のTPBの影響を示す。パネルa〜dは、示される濃度のTPBで処理した細胞集団のインスリン及びグルカゴンの発現を示す。
【図3】本発明の方法に従って処理された細胞における、インスリン及びグルカゴンの発現に対するプロテインキナーゼC阻害剤の影響を示す。パネルaは、TPBで処理した細胞のインスリン及びグルカゴンの発現を表し、パネルcは対応するDAPI染色を表す。パネルbは、TPB及びGO 6976で処理した細胞のインスリン及びグルカゴンの発現を表し、パネルdは対応するDAPI染色を表す。
【図4】本発明の方法に従って処理された細胞における、インスリンの発現に対する様々なプロテインキナーゼC阻害剤の影響を示す。パネルaは、TPBで処理した細胞のインスリンの発現を示す。パネルbは、ILVで処理した細胞のインスリンの発現を示す。パネルcは、PMAで処理した細胞のインスリンの発現を示す。
【図5】本発明の方法に従って処理された細胞における、膵臓内分泌系に特徴的なマーカーの発現を示す。これらのパネルは、インスリン及びNKX6.1(パネルa)、インスリン及びPDX1(パネルb)、インスリン及びNEUROD1(パネルc)、インスリン及びソマトスタチン(パネルd)、並びにインスリン及びグレリン(パネルe)の発現を表す。
【図6】本発明の方法に従って処理された細胞における、インスリン及びグルカゴンの発現を示す。パネルa〜cは、1%のB27、50ng/mLのFGF7、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、100ng/mLのノギン、20ng/mLのアクチビンA、及びp38キナーゼ阻害剤(米国特許第6,214,830号に開示されるもの、2.5μM)を添加したDMEM(高グルコース)で4日間にわたって処理した細胞(ステージ3、処理8、実施例2)における、インスリン発現(パネルa)、グルカゴン発現(パネルb)、及びDAPI染色(パネルc)を示す。パネルd〜fは、1%のB27、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、及び100ng/mLのノギンを添加したDMEM(高グルコース)で4日間にわたって処理した細胞(ステージ3、処理9、実施例2)における、インスリン発現(パネルd)、グルカゴン発現(パネルe)、及びDAPI染色(パネルf)を示す。
【図7】本発明の細胞の投与4、8、及び12週間後の(SCID)−beige(Bg)マウスにおいて、グルコース負荷に続いてヒトCペプチドが検出されたことを示す。
【図8】記載される濃度の様々なプロテインキナーゼC阻害剤処理後に得られた、PDX1及びNKX6.1を共発現する細胞の比率を示す。
【図9】実施例6に記載の方法に従って処理した細胞の、NGN3、PDX1、NKX6.1、及びPTF1α発現を示す。
【発明を実施するための形態】
【0028】
開示を明確にするため、及び限定ではなく、本発明の「発明を実施するための形態」を、本発明の特定の特徴、実施形態又は応用を説明若しくは図示した以下の小項目に分ける。
【0029】
定義
幹細胞は、単一の細胞レベルにおいて自己再生及び分化して、自己再生前駆細胞、非再生前駆細胞、及び最終分化細胞を含む、後代細胞を産生する、能力で定義される未分化細胞である。幹細胞は、複数の胚葉(内胚葉、中胚葉、及び外胚葉)から様々な細胞系の機能的細胞へとin vitroで分化する能力、並びに移植後に複数の胚葉の組織を生じ、及び胚盤胞への注入後、全部ではないとしても大部分の組織に実質的に寄与する能力によっても特徴付けられる。
【0030】
幹細胞は、発生能によって、(1)全胚及び胚体外細胞型を生じる能力を意味する全能性、(2)全胚細胞型を生じる能力を意味する多能性、(3)細胞系の小集合を生じるが、すべて特定の組織、器官、又は生理学的システム内で生じる能力を意味する複能性(例えば、造血幹細胞(HSC)は、HSC(自己再生)、血液細胞に限定された寡能性前駆細胞、並びに血液の通常の構成要素である全細胞型及び要素(例えば、血小板)を含む後代を産生できる)、(4)複能性幹細胞と比較して、より限定された細胞系統の小集合を生じる能力を意味する寡能性、並びに(5)1つの細胞系(例えば、精子形成幹細胞)を生じる能力を意味する単能性に分類される。
【0031】
分化は、非特殊化(「未確定」)又は低特殊化細胞が、例えば神経細胞又は筋細胞などの特殊化細胞の特徴を得るプロセスである。分化細胞又は分化を誘導された細胞は、細胞系内でより特殊化した(「確定した」)状況を呈している細胞である。分化プロセスに適用された際の用語「確定した」は、通常の環境下で特定の細胞型又は細胞型の小集合への分化を続け、かつ通常の環境下で異なる細胞型に分化したり、又は低分化細胞型に戻ったりすることができない地点まで、分化経路において進行した細胞を指す。脱分化は、細胞が細胞系内で低特殊化(又は確定)した状況に戻るプロセスを指す。本明細書で使用するとき、細胞系は、細胞の遺伝、すなわちその細胞がどの細胞から来たか、またどの細胞を生じ得るかを規定する。細胞系は、細胞を発生及び分化の遺伝的スキーム内に配置する。系特異的なマーカーは、対象とする系の細胞の表現型に特異的に関連した特徴を指し、未確定の細胞の対象とする系への分化を評価する際に使用可能である。
【0032】
本明細書で使用するとき、「胚体内胚葉系に特徴的なマーカーを発現する細胞」又は「ステージ1細胞」又は「ステージ1」とは、以下のマーカー、すなわち、SOX17、GATA4、HNF3β、GSC、CER1、Nodal、FGF8、短尾奇形、Mix−様ホメオボックスタンパク質、FGF4、CD48、eomesodermin(EOMES)、DKK4、FGF17、GATA6、CXCR4、C−Kit、CD99、又はOTX2のうちの少なくとも1つを発現する細胞を指す。胚体内胚葉系に特徴的なマーカーを発現する細胞には、原始線条前駆体細胞、原始線条細胞、中内胚葉細胞、及び胚体内胚葉細胞が挙げられる。
【0033】
本明細書で使用するとき、「膵臓内胚葉系に特徴的なマーカーを発現する細胞」とは、以下のマーカー、すなわち、PDX1、NKX6.1、HNF1β、PTF1α、HNF6、HNF4α、SOX9、HB9、又はPROX1のうちの少なくとも1つを発現する細胞を指す。膵臓内胚葉系に特徴的なマーカーを発現する細胞には、膵臓内胚葉細胞、原腸管細胞、及び後部前腸細胞が挙げられる。
【0034】
本明細書で使用するとき、「胚体内胚葉」は、原腸形成中、胚盤葉上層から生じ、胃腸管及びその誘導体を形成する細胞の特徴を保持する細胞を指す。胚体内胚葉細胞は、以下のマーカー、すなわち、HNF3β、GATA4、SOX17、Cerberus、OTX2、goosecoid、C−Kit、CD99、及びMIXL1を発現する。
【0035】
本明細書で使用するとき、「マーカー」は、対象とする細胞で差異的に発現される核酸又はポリペプチド分子である。この文脈において、差次的な発現は、陽性マーカーのレベルの上昇、及び陰性マーカーのレベルの減少を意味する。マーカー核酸又はポリペプチドの検出可能なレベルは、他の細胞と比較して対象とする細胞内で充分高いか又は低く、そのため当該技術分野において既知の多様な方法のいずれかを使用して、対象とする細胞を他の細胞から識別及び区別することができる。
【0036】
本明細書で使用するとき、「膵臓内分泌細胞」又は「膵臓ホルモン発現細胞」又は「膵臓内分泌系に特徴的なマーカーを発現する細胞」とは、以下のホルモン、すなわち、インスリン、グルカゴン、ソマトスタチン、及び膵臓ポリペプチドのうちの少なくとも1つを発現することが可能な細胞を指す。
【0037】
多能性幹細胞の単離、増殖及び培養
多能性幹細胞の特徴付け
多能性幹細胞は、ステージ特異的胚抗原(SSEA)3及び4、並びにTra−1−60及びTra−1−81と呼ばれる抗体を使用して検出可能なマーカーのうちの1つ以上を発現する(Thomsonら,Science 282:1145,1998)。多能性幹細胞をin vitroで分化させると、SSEA−4、Tra−1−60、及びTra−1−81(存在する場合)の発現が減少し、SSEA−1の発現が上昇する。未分化の多能性幹細胞は通常アルカリホスファターゼ活性を有し、細胞を4%パラホルムアルデヒドで固定した後、製造業者(Vector Laboratories,Burlingame Calif.)の説明書に従ってVector Redを基質として発生させることによって検出可能である。未分化の多能性幹細胞は、OCT4及びTERTも通常発現し、RT−PCRにより検出される。
【0038】
増殖した多能性幹細胞の別の望ましい表現型は、3つの胚葉のすべて、すなわち、内胚葉、中胚葉、及び外胚葉組織の細胞に分化する能力である。多能性幹細胞の多能性は、例えば、細胞を重症複合免疫不全症(SCID)マウスに注入し、形成される奇形腫を4%パラホルムアルデヒドで固定し、次いでこれを3つの胚葉由来の細胞型の根拠について組織学的に調べることによって確認することができる。あるいは、胚様体を形成させ、この胚様体を3つの胚葉に関連したマーカーの存在に関して評価することにより、多能性を決定してもよい。
【0039】
増殖した多能性幹細胞株を、標準的なGバンド法を使用して核型決定し、確立された対応する霊長類種の核型と比較してもよい。細胞が正倍数体であり、全ヒト染色体が存在し、かつ著しく変更されてはいないことを意味する、「正常な核型」を有する細胞を得ることが望ましい。
【0040】
多能性幹細胞源
使用が可能な多能性幹細胞の種類としては、妊娠期間中の任意の時期(必ずしもではないが、通常は妊娠約10〜12週よりも前)に採取した前胚性組織(例えば、胚盤胞など)、胚性組織又は胎児組織などの、妊娠後に形成される組織に由来する多能性細胞の樹立株が含まれる。非限定的な例は、例えばヒト胚性幹細胞株H1、H7、及びH9(WiCell)などのヒト胚性幹細胞又はヒト胚生殖細胞の確立株である。それらの細胞の最初の樹立又は安定化中に本開示の組成物を使用することも想定され、その場合、供給源となる細胞は、供給源となる組織から直接採取した一次多能性細胞であろう。フィーダー細胞の不在下で既に培養された多能性幹細胞集団から採取した細胞も好適である。例えば、BG01v(BresaGen(Athens,GA))などの変異ヒト胚性幹細胞株も好適である。
【0041】
一実施形態では、ヒト胚性幹細胞は、Thomsonら(米国特許第5,843,780号、Science 282:1145,1998;Curr.Top.Dev.Biol.38:133 ff.,1998;Proc.Natl.Acad.Sci.U.S.A.92:7844,1995)により記載されるように調製される。
【0042】
多能性幹細胞の培養
一実施形態では、多能性幹細胞は、典型的にはフィーダー細胞の層上で培養され、このフィーダー細胞は、多能性幹細胞を様々な方法で支持する。あるいは、多能性幹細胞を、フィーダー細胞を本質的に含まないにも関わらず、細胞を実質的に分化させることなく多能性幹細胞の増殖を支持するような培養システム中で培養する。フィーダー細胞不含培養における多能性幹細胞の、分化を伴わない増殖は、あらかじめ別の細胞型を培養することにより馴化培地を使用して支持される。あるいは、フィーダー細胞不含培養における多能性幹細胞の、分化を伴わない増殖は、既知組成培地を使用して支持される。
【0043】
例えば、Reubinoffら(Nature Biotechnology 18:399〜404(2000))及びThompsonら(Science 6 November 1998:282巻、5391号、1145〜1147ページ)は、マウス胚性線維芽細胞のフィーダー細胞層を用いたヒト胚盤胞からの多能性幹細胞株の培養について開示している。
【0044】
Richardsら(Stem Cells 21:546〜556,2003)は、11種類の異なるヒトの成人、胎児、及び新生児フィーダー細胞層のパネルを、それらのヒト多能性幹細胞の培養を支持する能力に関して評価した。Richardsらは、「成人の皮膚線維芽フィーダー細胞上で培養したヒト胚性幹細胞系は、ヒト胚性幹細胞の形態を有し、多能性を維持する」と述べている。
【0045】
米国特許出願公開第20020072117号は、フィーダーを含まない培養において霊長類の多能性幹細胞の増殖を支持する培地を生成する細胞系を開示している。使用される細胞系は、胚性組織から得られるか又は胚性幹細胞から分化された間葉系及び線維芽細胞様の細胞系である。米国特許出願公開第20020072117号は、また、この細胞系の、1次フィーダー細胞層としての使用も開示している。
【0046】
別の例において、Wangら(Stem Cells 23:1221〜1227,2005)は、ヒト胚性幹細胞由来のフィーダー細胞層上でのヒト多能性幹細胞の長期間増殖のための方法を開示している。
【0047】
別の例において、Stojkovicら(Stem Cells 2005 23:306〜314,2005)は、ヒト胚性幹細胞の自発的分化に由来するフィーダー細胞システムを開示している。
【0048】
更なる例において、Miyamotoら(Stem Cells 22:433〜440,2004)は、ヒト胎盤から得たフィーダー細胞源を開示している。
【0049】
Amitら(Biol.Reprod 68:2150〜2156,2003)は、ヒト包皮に由来するフィーダー細胞層を開示している。
【0050】
別の例において、Inzunzaら(Stem Cells 23:544〜549,2005)は、ヒト出生後包皮線維芽細胞からのフィーダー細胞層を開示している。
【0051】
米国特許第6642048号は、フィーダーを含まない培養における霊長類の多能性幹(pPS)細胞の増殖を支持する培地、及びこうした培地の製造に有用な細胞系を開示している。米国特許第6642048号は、「本発明は、胚性組織から得られるか、あるいは胚性幹細胞から分化した間葉系及び線維芽細胞様の細胞系を含む。本開示では、こうした細胞系を誘導し、培地を調整し、この馴化培地を用いて幹細胞を増殖させるための方法を説明及び図示する」と述べている。
【0052】
別の例において、国際公開第2005014799号は、哺乳動物細胞の維持、増殖及び分化のための馴化培地を開示している。国際公開第2005014799号は、「本発明に基づいて調整される培地は、マウス細胞、特にMMH(Metマウス肝細胞)と呼ばれる分化かつ不死化したトランスジェニック肝細胞の細胞分泌活性によって馴化される」と述べている。
【0053】
別の例において、Xuら(Stem Cells 22:972〜980,2004)は、ヒトテロメラーゼ逆転写酵素を過剰発現するように遺伝子改変されたヒト胚性幹細胞由来細胞誘導体から得られた馴化培地を開示している。
【0054】
別の例において、米国特許出願公開第20070010011号は、多能性幹細胞を維持するための既知組成培地を開示している。
【0055】
代替的な培養システムは、胚性幹細胞の増殖を促進することが可能な増殖因子を追加した無血清培地を使用している。例えば、Cheonら(BioReprod DOI:10.1095/biolreprod.105.046870,October 19,2005)は、胚性幹細胞の自己再生の誘発が可能な異なる増殖因子を追加した非馴化血清補充(SR)培地中に胚性幹細胞が維持されている、フィーダーを含まずかつ血清を含まない培養システムを開示している。
【0056】
別の例において、Levensteinら(Stem Cells 24:568〜574,2006)は、bFGFを追加した培地を使用して、線維芽細胞又は馴化培地の非存在において、ヒト胚性幹細胞を長期間培養する方法を開示している。
【0057】
別の例において、米国特許出願公開第20050148070号は、血清及び繊維芽細胞フィーダー細胞を含まない合成培地中でのヒト胚性幹細胞の培養方法を開示しており、この方法は、アルブミン、アミノ酸、ビタミン、ミネラル、少なくとも1つのトランスフェリン又はトランスフェリン代用物質、少なくとも1つのインスリン又はインスリン代用物質を含有する培地中で幹細胞を培養する工程を含み、この培地は、哺乳動物胎児血清を本質的に含有せず、線維芽細胞増殖因子シグナル伝達受容体を活性化することが可能な少なくとも約100ng/mLの線維芽細胞増殖因子を含有し、増殖因子は線維芽細胞フィーダー層以外の供給源からも供給され、培地はフィーダー細胞又は条件培地なしで未分化状態の幹細胞の増殖を支持するものである。
【0058】
別の例において、米国特許出願公開第20050233446号は、未分化の霊長類始原幹細胞などの幹細胞の培養に有用な合成培地を開示している。溶液では、培地は培養されている幹細胞と比較して実質的に等張である。所定の培養において、特定の培地は、基本培地及び実質的に未分化の始原幹細胞の増殖の支持に必要な量のbFGF、インスリン、及びアスコルビン酸を含む。
【0059】
別の例として、米国特許第6800480号は、「一実施形態では、実質的に未分化状態の霊長類由来の始原幹細胞を増殖させるための細胞培地であって、霊長類由来の始原幹細胞の増殖を支持する上で効果的な低浸透圧、低エンドトキシンの基本培地を含む、細胞培地を提供する。この基本培地は、霊長類由来の始原幹細胞の増殖を支持する上で効果的な栄養素血清、並びにフィーダー細胞及びフィーダー細胞から誘導される細胞外マトリックス成分からなる群から選択される基質と組み合わされる。培地は更に、非必須アミノ酸、抗酸化剤、並びにヌクレオシド及びピルビン酸塩からなる群から選択される第1の増殖因子を含む」と述べている。
【0060】
別の例において、米国特許出願公開第20050244962号は、「一態様において本発明は、霊長類の胚性幹細胞を培養する方法を提供する。1つの方法は、哺乳動物の胎児血清を本質的に含まない(好ましくはあらゆる動物の血清をも本質的に含まない)培地中で、線維芽細胞フィーダー層以外の供給源から供給される線維芽細胞増殖因子の存在下で、幹細胞を培養する。好ましい形態では、十分な量の線維芽細胞増殖因子を添加することによって、幹細胞の培養を維持するために従来必要とされていた線維芽細胞フィーダー層の必要性がなくなる」と述べている。
【0061】
更なる例において、国際公開第2005065354号は、フィーダー及び血清を本質的に含まない合成等張培地を開示しており、この培地は、a.基本培地と、b.実質的に未分化の哺乳類幹細胞の増殖を支持するのに十分な量のbFGFと、c.実質的に未分化の哺乳類幹細胞の増殖を支持するのに十分な量のインスリンと、d.実質的に未分化の哺乳類幹細胞の増殖を支持するのに十分な量のアスコルビン酸と、を含む。
【0062】
別の例において、国際公開第2005086845号は、細胞を未分化な状態に維持するのに充分な量の、トランスフォーミング増殖因子β(TGF−β)ファミリータンパク質の構成員、線維芽細胞増殖因子(FGF)ファミリータンパク質の構成員、又はニコチンアミド(NIC)に、所望の結果を得るのに充分な時間幹細胞を曝露することを含む、未分化の幹細胞を維持するための方法を開示している。
【0063】
多能性幹細胞は、好適な培養基材上に播くことができる。一実施形態では、好適な培養基質は、例えば基底膜から誘導されたもの、又は接着分子受容体−リガンド結合の一部を形成し得るものなどの細胞外マトリックス成分である。一実施形態において、好適な培養基質は、MATRIGEL(登録商標)(Becton Dickenson)である。MATRIGEL(登録商標)は、Engelbreth−Holm Swarm腫瘍細胞由来の可溶性製剤であって、室温でゲル化して再構成基底膜を形成する。
【0064】
他の細胞外マトリックス成分及び成分混合物は代替物として好適である。これには、増殖させる細胞型に応じて、ラミニン、フィブロネクチン、プロテオグリカン、エンタクチン、ヘパラン硫塩、及び同様物を単独で又は様々な組み合わせで含み得る。
【0065】
多能性幹細胞は、細胞の生存、増殖、及び所望の特徴の維持を促進する培地の存在下で、基質上に好適に分布させることで播いてもよい。これらすべての特徴は、播種分布に細心の注意を払うことから利益が得られ、かつ当業者により容易に決定可能である。
【0066】
好適な培地は、以下の成分、すなわち例えば、ダルベッコ改変イーグル培地(DMEM)、Gibco # 11965−092;ノックアウトダルベッコ改変イーグル培地(KO DMEM)、Gibco #10829−018;ハムF12/50% DMEM基本培地;200mM L−グルタミン、Gibco#15039−027;非必須アミノ酸溶液、Gibco # 11140−050;β−メルカプトエタノール、Sigma # M7522;ヒト組み換え塩基性線維芽細胞増殖因子(bFGF)、Gibco # 13256−029等から調製することもできる。
【0067】
多能性幹細胞からの、膵臓内分泌系に特徴的なマーカーを発現する細胞の形成
一実施形態では、本発明は、多能性幹細胞から、膵臓内胚葉系に特徴的なマーカーを発現する細胞を産生するための方法を提供し、この方法は以下の工程a、b、c及びdを包含する:
a.多能性幹細胞を培養する工程、
b.多能性幹細胞を、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させる工程、
c.胚体内胚葉系に特徴的なマーカーを発現する細胞を、膵臓内胚葉系に特徴的なマーカーを発現する細胞へと分化させる工程、
d.膵臓内胚葉系に特徴的なマーカーを発現する細胞を、膵内分泌系に特徴的なマーカーを発現する細胞へと分化させる工程。
【0068】
本発明の一態様では、膵臓内分泌系に特徴的なマーカーを発現する細胞は、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する。
【0069】
胚体内胚葉系に特徴的なマーカーを発現する細胞への多能性幹細胞の分化
胚体内胚葉系に特徴的なマーカーを発現する細胞の形成は、以下の特定のプロトコルの前後に、マーカーの存在に関して試験することにより決定され得る。多能性幹細胞は、典型的にはそのようなマーカーを最小量発現する。したがって、多能性細胞の分化は、細胞がそれらの発現を開始した際に検出される。
【0070】
多能性幹細胞は、当該技術分野のいかなる方法、又は本発明で提案されるいかなる方法によって胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させてもよい。
【0071】
例えば、多能性幹細胞は、D’Amourら、Nature Biotechnology 23,1534〜1541(2005)に開示される方法に従って、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0072】
例えば、多能性幹細胞は、Shinozakiら、Development 131,1651〜1662(2004)により開示される方法に従って、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0073】
例えば、多能性幹細胞は、McLeanら、Stem Cells 25,29〜38(2007)により開示される方法に従って、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0074】
例えば、多能性幹細胞は、D’Amourら、Nature Biotechnology 24,1392〜1401(2006)に開示される方法に従って、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0075】
例えば、多能性幹細胞は、アクチビンAを含む培地中、血清の非存在下で多能性幹細胞を培養し、次いで細胞をアクチビンA及び血清と培養し、次いで細胞をアクチビンA及び異なる濃度の血清と培養することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。この方法の一例は、Nature Biotechnology 23,1534〜1541(2005)に開示されている。
【0076】
例えば、多能性幹細胞は、アクチビンAを含む培地中、血清の非存在下で多能性幹細胞を培養し、次いで細胞をアクチビンAと、別の濃度の血清と共に培養することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。この方法の例は、D’Amourら、Nature Biotechnology(2005)に開示されている。
【0077】
例えば、多能性幹細胞は、アクチビンA及びWntリガンドを含む培地中、血清の非存在下で多能性幹細胞を培養し、次いでWntリガンドを除去し、細胞をアクチビンAと、血清と共に培養することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。この方法の例は、Nature Biotechnology 24,1392〜1401(2006)に開示されている。
【0078】
例えば、多能性幹細胞は、LifeScan,Inc.に譲渡された米国特許出願第11/736,908号に開示される方法に従って多能性幹細胞を処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0079】
例えば、多能性幹細胞は、LifeScan,Inc.に譲渡された米国特許出願第11/779,311号に開示される方法に従って多能性幹細胞を処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0080】
例えば、多能性幹細胞は、米国特許出願第60/990,529号に開示される方法に従って多能性幹細胞を処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0081】
例えば、多能性幹細胞は、米国特許出願第61/076,889号に開示される方法に従って多能性幹細胞を処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0082】
例えば、多能性幹細胞は、米国特許出願第61/076,900号に開示される方法に従って多能性幹細胞を処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0083】
例えば、多能性幹細胞は、米国特許出願第61/076,908号に開示される方法に従って多能性幹細胞を処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0084】
例えば、多能性幹細胞は、米国特許出願第61/076,915号に開示される方法に従って多能性幹細胞を処理することによって、胚体内胚葉系に特徴的なマーカーを発現する細胞へと分化させることができる。
【0085】
膵臓内胚葉系に特徴的なマーカーを発現する細胞への、胚体内胚葉系に特徴的なマーカーを発現する細胞の分化
胚体内胚葉系に特徴的なマーカーを発現する細胞は、当該技術分野の任意の方法、又は本発明で提案する任意の方法により、膵臓内胚葉系に特徴的なマーカーを発現する細胞へと分化され得る。
【0086】
例えば、胚体内胚葉系に特徴的なマーカーを発現する細胞は、D’Amourら、Nature Biotechnol.24:1392〜1401,2006に開示されている方法に従って、膵臓内胚葉系に特徴的なマーカーを発現する細胞へと分化され得る。
【0087】
一実施形態では、膵臓内胚葉系に特徴的なマーカーを発現する細胞は、PDX1、NKX6.1を共発現するが、最小量のCDX2及びNGN3を発現しない。
【0088】
一実施形態では、胚体内胚葉系に特徴的なマーカーを発現する細胞は、胚体内胚葉系に特徴的なマーカーを発現する細胞を、FGF7を追加した第1培地で培養し、次いで細胞を、FGF7、BMP阻害能を有する因子、TGFβ受容体アゴニスト、レチノイン酸、及びヘッジホッグシグナル経路阻害剤を追加した第2培地で培養することにより、PDX1、NKX6.1を共発現するが、最小量のCDX2及びNGN3を発現しない、膵臓内胚葉系に特徴的なマーカーを発現する細胞へと分化させる。
【0089】
一実施形態では、FGF7は、約50pg/mL〜約50μg/mLの濃度で使用することができる。一実施形態では、FGF7は、50ng/mLの濃度で使用することができる。
【0090】
一実施形態では、BMP阻害能を有する因子はノギンである。ノギンは、約500ng/mL〜約500μg/mLの濃度で使用することができる。一実施形態では、ノギンは100ng/mLの濃度で使用される。
【0091】
一実施形態では、TGFβ受容体アゴニストは、アクチビンA、アクチビンB、TGFβ−I、TGFβ−II、GDF−8、及びGDF−11からなる群から選択される。
【0092】
アクチビンAは、約2ng/mL〜100ng/mLの濃度で使用することができる。一実施形態では、アクチビンAは20ng/mLの濃度で使用される。別の実施形態では、アクチビンAは50ng/mLの濃度で使用される。
【0093】
アクチビンBは、約2ng/mL〜100ng/mLの濃度で使用することができる。一実施形態では、アクチビンBは20ng/mLの濃度で使用される。別の実施形態では、アクチビンBは50ng/mLの濃度で使用される。
【0094】
TGFβ−Iは、約2ng/mL〜100ng/mLの濃度で使用することができる。一実施形態では、TGFβ−Iは20ng/mLの濃度で使用される。別の実施形態では、TGFβ−Iは50ng/mLの濃度で使用される。
【0095】
TGFβ−IIは、約2ng/mL〜100ng/nLの濃度で使用することができる。一実施形態では、TGFβ−IIは20ng/mLの濃度で使用される。別の実施形態では、TGFβ−IIは50ng/mLの濃度で使用される。
【0096】
GDF−8は、約2ng/mL〜100ng/mLの濃度で使用することができる。一実施形態では、GDF−8は20ng/mLの濃度で使用される。別の実施形態では、GDF−8は50ng/mLの濃度で使用される。
【0097】
GDF−11は、約2ng/mL〜100ng/mLの濃度で使用することができる。一実施形態では、GDF−11は20ng/mLの濃度で使用される。別の実施形態では、GDF−11は50ng/mLの濃度で使用される。
【0098】
レチノイン酸は、約1nM〜約1mMの濃度で使用することができる。一実施形態では、レチノイン酸は1μMの濃度で使用される。
【0099】
一実施形態では、ヘッジホッグシグナル経路阻害剤はシクロパミン−KAADである。シクロパミン−KAADは、約0.025μM〜約2.5μMの濃度で使用することができる。一実施形態では、シクロパミン−KAADは0.25μMの濃度で使用される。
【0100】
分化効率は、処理した細胞集団を、胚体内胚葉系に特徴的なマーカーを発現する細胞により発現されたタンパク質マーカーを特異的に認識する薬剤(抗体など)に曝露することにより測定することができる。
【0101】
培養又は単離された細胞中のタンパク質及び核酸マーカーの発現を評価する方法は、当該技術分野において標準技術である。これらには、定量的逆転写酵素ポリメラーゼ連鎖反応(RT−PCR)、ノーザンブロット、in situハイブリダイゼーション(例えば、Current Protocols in Molecular Biology(Ausubelら編集、2001 supplement)参照)、並びにイムノアッセイ、例えば切片材料の免疫組織化学的分析、ウェスタンブロッティング、及び完全細胞で利用できるマーカーについてのフローサイトメトリー分析(FACS)(例えば、Harlow及びLane,Using Antibodies:A Laboratory Manual,New York:Cold Spring Harbor Laboratory Press(1998)参照)が挙げられる。
【0102】
多能性幹細胞の特徴は当業者に周知であり、多能性幹細胞の更なる特徴は、継続して同定されている。多能性幹細胞のマーカーとして、例えば、以下のもの、すなわち、ABCG2、cripto、FOXD3、CONNEXIN43、CONNEXIN45、OCT4、SOX2、Nanog、hTERT、UTF1、ZFP42、SSEA−3、SSEA−4、Tra 1−60、Tra 1−81の1つ以上の発現が挙げられる。
【0103】
多能性幹細胞を本発明の方法で処理した後、処理した細胞集団を、胚体内胚葉系に特徴的なマーカーを発現する細胞により発現される、例えばCXCR4などのタンパク質マーカーを特異的に認識する薬剤(抗体など)に曝露することにより、分化した細胞を精製することができる。
【0104】
本発明で用いるのに好適な多能性幹細胞としては、例えば、ヒト胚性幹細胞株H9(NIHコード:WA09)、ヒト胚性幹細胞株H1(NIHコード:WA01)、ヒト胚性幹細胞株H7(NIHコード:WA07)、及びヒト胚性幹細胞株SA002(Cellartis,Sweden)が挙げられる。多能性細胞に特徴的な以下のマーカー、すなわち、ABCG2、cripto、CD9、FOXD3、CONNEXIN43、CONNEXIN45、OCT4、SOX2、Nanog、hTERT、UTF1、ZFP42、SSEA−3、SSEA−4、Tra 1−60、及びTra 1−81のうちのすくなくとも1つを発現する細胞も本発明で用いるのに好適である。
【0105】
胚体内胚葉系に特徴的なマーカーは、SOX17、GATA4、HNF3β、GSC、CER1、Nodal、FGF8、短尾奇形、Mix様ホメオボックスタンパク質、FGF4、CD48、エオメソダーミン(EOMES)、DKK4、FGF17、GATA6、CXCR4、C−Kit、CD99、及びOTX2からなる群から選択される。本発明での使用に好適なのは、胚体内胚葉系に特徴的なマーカーのうちの少なくとも1つを発現する細胞である。本発明の一態様では、胚体内胚葉系に特徴的なマーカーを発現する細胞は、原始線条前駆体細胞である。別の態様では、胚体内胚葉系に特徴的なマーカーを発現する細胞は、中内胚葉細胞である。別の態様では、胚体内胚葉系に特徴的なマーカーを発現する細胞は、胚体内胚葉細胞である。
【0106】
膵臓内胚葉系に特徴的なマーカーは、PDX1、NKX6.1、HNF1β、PTF1α、HNF6、HNF4α、SOX9、HB9、及びPROX1からなる群から選択される。本発明での使用に好適なのは、膵臓内胚葉系に特徴的な少なくとも1つのマーカーを発現する細胞である。本発明の一態様において、膵臓内胚葉系に特徴的なマーカーを発現する細胞は、膵臓内胚葉細胞である。
【0107】
膵臓内分泌系のマーカーを発現する細胞への、膵臓内胚葉系に特徴的なマーカーを発現する細胞の分化
一実施形態では、膵臓内胚葉系に特徴的なマーカーを発現する細胞は、膵臓内分泌系に特徴的なマーカーを発現する細胞に更に分化する。
【0108】
一実施形態では、膵臓内胚葉系に特徴的なマーカーを発現する細胞は、PDX1、NKX6.1を共発現するが、最小量のCDX2及びNGN3を発現しない。
【0109】
一実施形態では、膵臓内分泌系に特徴的なマーカーを発現する細胞は、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する。
【0110】
一実施形態では、膵臓内胚葉系に特徴的なマーカーを発現する細胞を、BMP阻害能を有する因子、TGFβ受容体シグナル伝達阻害剤、及びプロテインキナーゼC活性化因子を追加した培地で培養することにより、膵臓内胚葉系に特徴的なマーカーを発現する細胞を、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞へと分化させる。
【0111】
一実施形態では、BMP阻害能を有する因子はノギンである。ノギンは、約500ng/mL〜約500μg/mLの濃度で使用することができる。一実施形態では、ノギンは100ng/mLの濃度で使用される。
【0112】
一実施形態では、TGFβ受容体シグナル伝達阻害剤は、ALK5の阻害剤である。一実施形態では、ALK5の阻害剤は、ALK5阻害剤IIである。ALK5阻害剤IIは、約0.1μM〜約10μMの濃度で使用することができる。一実施形態では、ALK5阻害剤IIは1μMの濃度で使用される。
【0113】
一実施形態では、プロテインキナーゼC活性化因子は、(2S,5S)−(E,E)−8−(5−(4−(トリフルオロメチル)フェニル)−2,4−ペンタジエノイルアミノ(pentadiemoylamino))ベンゾラクタム、インドラクタムV、及びホルボール−12−ミリステート−13−アセテートからなる群から選択される。一実施形態では、プロテインキナーゼC活性化因子は、(2S,5S)−(E,E)−8−(5−(4−(トリフルオロメチル)フェニル)−2,4−ペンタジエノイルアミノ(pentadiemoylamino))ベンゾラクタムである。(2S,5S)−(E,E)−8−(5−(4−(トリフルオロメチル)フェニル)−2,4−ペンタジエノイルアミノ(pentadiemoylamino))ベンゾラクタムは、約20nM〜約500nMの濃度で使用することができる。(2S,5S)−(E,E)−8−(5−(4−(トリフルオロメチル)フェニル)−2,4−ペンタジエノイルアミノ(pentadiemoylamino))ベンゾラクタム、インドラクタムV、及びホルボール−12−ミリステート−13−アセテートは、本明細書で「TPB」と称される。
【0114】
膵臓内分泌系に特徴的なマーカーは、NEUROD、ISL1、PDX1、NKX6.1、NKX2.2、PAX4、及びPAX6からなる群から選択される。一実施形態では、膵臓内分泌系に特徴的なマーカーを発現する細胞は、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する。
【0115】
治療
一態様では、本発明は、I型糖尿病に罹患しているか、あるいはI型糖尿病を発症するリスクを有する患者を治療する方法を提供する。一実施形態では、この方法は、多能性幹細胞を培養する工程と、多能性幹細胞を、膵臓内分泌系に特徴的なマーカーを発現する細胞へとin vitroで分化させる工程と、膵臓内分泌系に特徴的なマーカーを発現する細胞を患者に移植する工程と、を含む。
【0116】
更に別の態様では、本発明は、II型糖尿病に罹患しているか、あるいはII型糖尿病を発症するリスクを有する患者を治療する方法を提供する。一実施形態では、この方法は、多能性幹細胞を培養する工程と、多能性幹細胞を、膵臓内分泌系に特徴的なマーカーを発現する細胞へとin vitroで分化させる工程と、膵臓内分泌系に特徴的なマーカーを発現する細胞を患者に移植する工程と、を含む。
【0117】
適切であるならば、移植した細胞の生存及び機能を亢進する医薬品又は生理活性物質で患者を更に処置してもよい。それらの薬剤には、例えば、特に、インスリン、TGF−β1、2、及び3を含むTGF−βファミリーのメンバー、骨形成タンパク質(BMP−2、−3、−4、−5、−6、−7、−11、−12、及び−13)、線維芽細胞増殖因子−1及び−2、血小板由来増殖因子−AA及び−BB、多血小板血漿、インスリン増殖因子(IGF−I、II)、増殖分化因子(GDF−5、−6、−7、−8、−10、−15)、血管内皮細胞由来増殖因子(VEGF)、プレイオトロフィン、エンドセリンを挙げてもよい。他の医薬化合物には、例えば、ニコチンアミド、グルカゴン様ペプチド−I(GLP−1)及びII、GLP−1及び2模倣体(mimetibody)、エキセンディン−4、レチノイン酸、副甲状腺ホルモン、例えば米国特許出願公開第2004/0209901号及び同第2004/0132729号に開示される化合物のようなMAPK阻害剤などが挙げられる。
【0118】
多能性幹細胞は、レシピエントに移植する前にインスリン産生細胞へと分化させてもよい。具体的な実施形態では、多能性幹細胞は、レシピエントに移植する前にβ細胞へと完全に分化させる。あるいは多能性幹細胞を、未分化又は一部が分化した状態でレシピエントに移植してもよい。更なる分化をレシピエント内で行ってもよい。
【0119】
胚体内胚葉細胞、又は代替的には膵臓内胚葉細胞、又は代替的にはβ細胞を、分散した細胞として移植してもよく、あるいは肝門静脈内に注入され得るクラスターに形成してもよい。あるいは、細胞を、生体適合性の分解性ポリマー支持体、多孔性の非分解性デバイス中で提供してもよく、又は宿主免疫応答から保護されるよう封入してもよい。細胞を、レシピエントの適切な部位内に移植してもよい。移植部位には、例えば肝臓、天然の膵臓、腎被膜下空間、網、腹膜、漿膜下空間、腸、胃、又は皮下ポケットが挙げられる。
【0120】
移植された細胞の更なる分化、生存又は活性を向上するために、増殖因子、抗酸化剤又は抗炎症剤などの追加の因子を、細胞の投与前、投与と同時、又は投与後に投与してもよい。特定の実施形態では、増殖因子は、投与された細胞をin vivoで分化させるように使用される。これらの因子は、内在性細胞により分泌され、投与された細胞にin situで曝露されてもよい。移植された細胞を誘導して、当該技術分野で既知の内因性の及び外因性の投与された増殖因子の任意の組み合わせにより分化させることができる。
【0121】
移植に使用する細胞の量は、患者の状態及び治療に対する応答を含む、多数の様々な要因に依存し、当業者により決定され得る。
【0122】
一態様では、本発明は、糖尿病に罹患しているか、あるいは糖尿病を発症するリスクを有する患者を治療する方法を提供する。本方法は、多能性幹細胞を培養し、培養した細胞をβ細胞系にin vitroで分化させ、この細胞を3次元支持体に埋め込むことを含む。細胞は、患者に移植する前に、この支持体上にin vitroで維持してもよい。あるいは、細胞を含む支持体を、in vitroで更に培養することなく直接患者に移植してもよい。所望により、支持体を、埋め込まれた細胞の生存及び機能を促進する少なくとも1つの医薬品と共に組み込んでもよい。
【0123】
本発明の目的のために使用するのに好適な支持体材料には、組織修復に有用な組織鋳型、導管、バリア、及びリザーバが挙げられる。より詳細には、発泡体、スポンジ、ゲル、ヒドロゲル、織物、及び不織構造の形態を有する合成及び天然材料であって、in vitro及びin vivoで使用されて、生物組織を再構築又は再生し、また走化性薬剤を送達して組織増殖を誘発する材料が、本発明の方法の実施における使用に好適である。例えば、米国特許第5,770,417号、同第6,022,743号、同第5,567,612号、同第5,759,830号、同第6,626,950号、同第6,534,084号、同第6,306,424号、同第6,365,149号、同第6,599,323号、同第6,656,488号、米国特許出願公開第2004/0062753 A1号、米国特許第4,557,264号及び同第6,333,029号に開示されている材料を参照のこと。
【0124】
医薬品が組み込まれた支持体を形成するために、薬剤を、支持体を形成するのに先立ち、ポリマー溶液と混合することもできる。あるいは加工された支持体上に、医薬品を好ましくは医薬担体の存在下で被覆してもよい。医薬品は、液体、超微粒子状固体、又は任意の他の適切な物理的形態として存在し得る。あるいは医薬品の放出速度を変更するために、支持体に賦形剤を加えてもよい。別の実施形態では、抗炎症性化合物である少なくとも1種の医薬化合物、例えば米国特許第6,509,369号に開示されている化合物を支持体に組み込む。
【0125】
支持体には、抗アポトーシス化合物である少なくとも1種の医薬化合物、例えば米国特許第6,793,945号に開示されている化合物を組み込んでもよい。
【0126】
支持体には、線維症阻害剤である少なくとも1種の医薬化合物、例えば米国特許第6,331,298号に開示されている化合物も組み込まれ得る。
【0127】
支持体には、血管新生を促進させることができる少なくとも1種の医薬化合物、例えば米国特許出願公開第2004/0220393号及び同第2004/0209901号に開示されている化合物も組み込まれ得る。
【0128】
支持体には、免疫抑制化合物である少なくとも1種の医薬化合物、例えば米国特許出願公開第2004/0171623号に開示されている化合物も組み込まれ得る。
【0129】
支持体には、例えば、特に、TGF−β1、2、及び3を含むTGF−βファミリーのメンバー、骨形成タンパク質(BMP−2、−3、−4、−5、−6、−7、−11、−12、及び−13)、線維芽細胞増殖因子−1及び−2、血小板由来増殖因子−AA及び−BB、多血小板血漿、インスリン増殖因子(IGF−I、II)、増殖分化因子(GDF−5、−6、−8、−10、−15)、血管内皮細胞由来増殖因子(VEGF)、プレイオトロフィン、エンドセリンなどの増殖因子である、少なくとも1種の医薬化合物も組み込まれ得る。他の医薬化合物には、例えば、ニコチンアミド、低酸素誘導因子1−α、グルカゴン様ペプチド−I(GLP−1)、GLP−1及びGLP−2疑似体、並びにII、エキセンディン4、ノダル、ノギン、NGF、レチノイン酸、副甲状腺ホルモン、テネイシン−C、トロポエラスチン、トロンビン由来ペプチド、カテリシジン、デフェンシン、ラミニン、フィブロネクチン及びビトロネクチンなどの接着性細胞外マトリックスタンパク質の細胞−及びヘパリン−結合ドメインを含む生物学的ペプチド、例えば米国特許出願公開第2004/0209901号及び同第2004/0132729号に開示されている化合物などのMAPK阻害剤を挙げることができる。
【0130】
骨格内への本発明の細胞の組み込みは、細胞を骨格上に単に沈着させることにより達成できる。細胞は、単に拡散により骨格内に入ることができる(J.Pediatr.Surg.23(1 Pt 2):3〜9(1988))。細胞播種の効率を向上させるために、いくつかの他の手法が開発されている。例えば、軟骨細胞をポリグリコール酸骨格上に播種する際に、スピナーフラスコが使用されている(Biotechnol.Prog.14(2):193〜202(1998))。細胞播種のための他の手法は遠心法の使用であり、これは播種する細胞に与えるストレスを最小にし、かつ播種効率を高める。例えば、Yangらは、遠心分離細胞固定化法(CCI)と呼ばれる細胞播種法を開発した(J.Biomed.Mater.Res.55(3):379〜86(2001))。
【0131】
本発明を以下の実施例によって更に説明するが、本発明はこれらの実施例により限定されるものではない。
【実施例】
【0132】
(実施例1)
インスリン及びNKX6.1、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞集団の形成
ヒト胚性幹細胞株H1細胞を、MATRIGEL(登録商標)(1:30希釈)(BD Biosciences;カタログ# 356231)をコーティングしたディッシュと0.2%のFBS、100ng/mLのアクチビンA(PeproTech;カタログ#120−14)、及び20ng/mLのWNT−3a(R&D Systems;カタログ# 1324−WN/CF)を添加したRPMI培地(Invitrogen;カタログ#:22400)により1日培養した後で、0.5%のFBS及び100ng/mLのアクチビンAを添加したRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで、
a.2%のFBS及び50ng/mLのFGF7(PeproTech;カタログ#100−19)を添加したDMEM/F12(Invitrogen;カタログ# 11330−032)で3日間にわたって処理し(ステージ2)、次いで、
b.1%のB27、50ng/mLのFGF7、0.25μMのシクロパミン−KAAD(Calbiochem;カタログ# 239804)、及び100ng/mLのノギン(R&D Systems;カタログ# 3344−NG)を添加したDMEM(高グルコース)(Invitrogen;カタログ# 10569)で4日間にわたって培養し(ステージ3)、次いで
c.1%のB27(Invitrogen;カタログ# 0791)、100ng/mLのノギン、1μMのALK5阻害剤II(Axxora;カタログ# ALX−270−445)、及び500nMのTBP((2S,5S)−(E,E)−8−(5−(4−(トリフルオロメチル)フェニル)−2,4−ペンタジエノイルアミノ(pentadiemoylamino))ベンゾラクタム)(Calbiochem;カタログ# 565740)を添加したDMEM(高グルコース)で6日間にわたって培養する(ステージ4)。
【0133】
対照として、別の細胞集団を、1%のB27、100ng/mLのノギン、及び1μMのALK5阻害剤IIを添加したDMEM(高グルコース)で6日間にわたって処理した(ステージ4、対照群)。
【0134】
図1に示すように、ステージ4におけるTBP処理により、インスリン発現細胞が増加する結果となった(図1、パネルa)。なお、これらインスリン発現細胞の約60%が単独の内分泌ホルモン発現細胞であり、これらの細胞はインスリンを発現し、グルカゴン、ソマトスタチン及びグレリン(図1、パネルa及びb、図5、パネルd及びe)を発現しなかった。TBP処理を受けた培養においては、グルカゴン発現細胞も確認された。ほとんどのグルカゴン発現細胞は、インスリンも共発現した(図1、パネルa及びb)。対照群について、細胞の大部分はインスリン及びグルカゴンを共発現した(図1、パネルc及びd)。
【0135】
異なる実験において、ヒト胚性幹細胞株H1細胞を、MATRIGEL(登録商標)(1:30希釈)をコーティングしたディッシュと、0.2%のFBS、100ng/mLのアクチビンA、及び20ng/mLのWNT−3aを添加したRPMI培地により1日培養し、次いで0.5%のFBS及び100ng/mLのアクチビンAを添加したRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
a.2%のFBS及び50ng/mLのFGF7を添加したDMEM/F12で3日間にわたって処理し(ステージ2)、次いで
b.1%のB27、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、及び100ng/mLのノギンを添加したDMEM(高グルコース)で4日間にわたって処理し(ステージ3)、次いで
c.処理1:1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び500nMのTBPを添加したDMEM(高グルコース)で6日間(ステージ4、処理1)、又は
d.処理2:1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び100nMのTBPを添加したDMEM(高グルコース)で6日間(ステージ4、処理2)、又は
e.処理3:1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び20nMのTBPを添加したDMEM(高グルコース)で6日間(ステージ4、処理3)、又は
f.処理4:1%のB27、100ng/mLのノギン、及び1μMのALK5阻害剤IIを添加したDMEM(高グルコース)で6日間(ステージ4、処理4)にわたって処理した。
【0136】
免疫細胞化学的分析を用い、本発明の細胞の形成に対する様々な濃度のTPBの影響を評価した。インスリン単独発現細胞数の著しい増加が、500nM及び100nMのTPB処理群の両方(図2、パネルa及びb)において観察された。FACS分析では、両方の処理によりin vitroにおいて12%のインスリン単独発現細胞をもたらし、この集団の15%がNKX6.1も発現していたことが確認された(表1−NKX6.1/INS発現細胞は総集団の2.4%であった)。対照群と同様に、20nMのTPBにおいては、ほとんどの細胞がインスリン及びグルカゴンを共発現した(図2、パネルc及びd)。
【0137】
【表1】
【0138】
内分泌ホルモン発現細胞形成への作用が、プロテインキナーゼCの活性化を介してなされたことを更に確認するため、ヒト胚性幹細胞株H1の別の細胞集団を、MATRIGEL(登録商標)(1:30希釈)をコーティングしたディッシュと0.2%のFBS、100ng/mLのアクチビンA、及び20ng/mL WNT−3aを添加したRPMI培地により1日培養した後で、0.5%のFBS及び100ng/mLのアクチビンAを添加したRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
a.2%のFBS及び50ng/mLのFGF7を添加したDMEM/F12で3日間にわたって処理し(ステージ2)、次いで
b.1%のB27、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、及び100ng/mLのノギンを添加したDMEM(高グルコース)で4日間にわたって処理し(ステージ3)、次いで
c.処理5:1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び500nMのTBPを添加したDMEM(高グルコース)で6日間(ステージ4、処理5)、又は
d.処理6:1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、500nMのTPB、及び5μMのGO 6976を添加したDMEM(高グルコース)で6日間(ステージ4、処理6)、又は
e.処理7:1%のB27、100ng/mLのノギン、及び1μMのALK5阻害剤IIを添加したDMEM(高グルコース)で処理し(ステージ4、処理7)、次いで
f.1%のB27を添加したDMEM(高グルコース)で4日間にわたって処理した(ステージ5)。
【0139】
GO 6976は、Ca2+依存性プロテインキナーゼCのイソ型を選択的に阻害するといわれている。TPBのみ(図3、パネルa)、並びにTPB及びGO 6976(図3、パネルb)を与えられた培養液において、膵臓内分泌系に特徴的なマーカーを発現する細胞の数が著しく減少する。FACS分析では、TPB処理(処理6)が、30.6%のシナプトフィジン発現細胞、12%のインスリン単独発現細胞、及び4.6%のグルカゴン発現細胞をもたらしたことが確認された。一方、TBP及びGO 6976処理(処理7)は、10.6%のシナプトフィジン発現細胞と、検出できないレベルのインスリン単独発現細胞をもたらした(表2)。処理6と処理7との間で、観察された総細胞数に違いはなかった(図3、パネルc及びd参照、処理6及び処理7の総細胞数を表すDAPI染色を示す)。これらの結果は、プロテインキナーゼCのシグナル伝達が、膵臓内分泌系に特徴的なマーカーを発現する細胞の形成に重要であり得ることを示唆する。
【0140】
その他のプロテインキナーゼC活性化因子も調べた。これらはインドラクタムV(ILV)(Axxora;カタログ# ALX−420−011−C300)及びホルボール−12−ミリステート−13−アセテート(PMA)(Calbiochem;カタログ#524400)とした。しかし、TPBのみがインスリン単独発現細胞の形成を示した(図4、パネルa)。ILV(図4、パネルb)及びPMA(図4、パネルc)の両方が500nMにおいて、6日後にインスリン及びグルカゴンを共発現する細胞をもたらした。FACS分析では、TPB処理が、12%のインスリン単独発現細胞、及び4.6%のグルカゴン発現細胞、並びに7.1%のインスリン及びグルカゴンを共発現する細胞をもたらしたことが確認された。一方、ILV処理は、3%のインスリン単独発現細胞、及び12%のグルカゴン細胞、並びに12%のインスリン及びグルカゴンを共発現する細胞をもたらした(表2)。免疫細胞化学的分析により、TPBで処理した培養液において、インスリン発現細胞の20%がNKX6.1(図5、パネルa)及びPDX1(図5、パネルb)を共発現したことが示された。インスリン発現細胞の大部分は、内分泌腺マーカー(maker)であるNEURODを共発現した(図5、パネルc)。ソマトスタチン又はグレリン(GHRL)を共発現したインスリン発現細胞はほとんどなかった(図5、パネルd及びe)。
【0141】
【表2】
【0142】
(実施例2)
インスリン及びNKX6.1、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞集団を形成する別法
異なる実験において、ヒト胚性幹細胞株H1細胞を、MATRIGEL(登録商標)(1:30希釈)をコーティングしたディッシュと、0.2%のFBS、100ng/mLのアクチビンA、及び20ng/mL WNT−3aを添加したRPMI培地により1日培養し、次いで0.5%のFBS及び100ng/mLのアクチビンAを添加したRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
a.2%のFBS及び50ng/mLのFGF7を添加したDMEM/F12で3日間にわたって処理し(ステージ2)、次いで
b.処理8:1%のB27、50ng/mLのFGF7、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、100ng/mLのノギン、20ng/mLのアクチビンA、及びp38キナーゼ阻害剤(米国特許第6,214,830号に開示されるもの、2.5μM)を添加したDMEM(高グルコース)で4日間(ステージ3、処理8)、又は
c.処理9:1%のB27、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、及び100ng/mLのノギンを添加したDMEM(高グルコース)で4日間(ステージ3、処理9)、次いで
d.1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び500nMのTPBを添加したDMEM(高グルコース)で6日間にわたって処理した(ステージ4)。
【0143】
ステージ3、処理8により、PDX1及びNKX6.1を共発現したが、CDX2及びNGN3を共発現しなかった膵臓内胚葉系に特徴的なマーカーを発現する細胞集団の形成をもたらした。一方、ステージ3、処理9では、PDX1、NKX6.1、及びNGN3を共発現した膵臓内胚葉系に特徴的なマーカーを発現する細胞集団の形成をもたらした。これらの細胞集団に対するプロテインキナーゼC活性化因子処理の影響を調べた(上記ステージ4)。
【0144】
FACS分析を行い、インスリン単独陽性細胞、グルカゴン単独陽性細胞、インスリン/グルカゴン二重陽性細胞、NKX6.1陽性細胞を発現する細胞、インスリン/NKX6.1陽性細胞、及びシナプトフィジン陽性細胞(膵臓内分泌腺マーカー)の比率を明らかにした。
【0145】
表3に示すように、処理8で形成された細胞集団は、シナプトフィジン発現で表される、より大部分の内分泌細胞を生じ、総細胞集団の49.7%はシナプトフィジンを発現した。総集団の27.8%はインスリン単独陽性細胞であった。
【0146】
一方、処理9で形成された細胞集団が生じたシナプトフィジン発現細胞は、たった25.7%であった。総集団の7.6%はインスリン単独発現細胞であった。両方の処理で観察されたグルカゴン単独発現細胞に有意差はなく、グルカゴン発現細胞の比率はインスリン発現細胞より有意に低かった。
【0147】
かなりの量のインスリン発現細胞はまた、NKX6.1も共発現した。処理8を受けた細胞集団では、総集団の11%がインスリン及びNKX6.1を発現した。処理9を受けた細胞集団では、総集団の2%がインスリン及びNKX6.1を発現した。
【0148】
免疫蛍光分析によりこれを確認した(図6)。処理8は、処理9に比べてインスリン発現細胞を増加させた(図6、パネルa及びd)。ほとんどのグルカゴン発現細胞は多ホルモン細胞であった(図6、パネルa、b、d及びe)。これらの結果は、処理8で生じた細胞集団(PDX1及びNKX6.1を共発現したが、CDX2及びNGN3は共発現しなかった膵臓内胚葉系に特徴的なマーカー(maker)を発現する細胞)は、本発明の方法により、より効率的に誘導され成熟した機能性インスリン発現細胞になれることを示唆する。
【0149】
【表3】
【0150】
(実施例3)
インスリン及びNKX6.1、並びに最小量のグルカゴンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞集団を形成する別法
別の実験において、ヒト胚性幹細胞株H1細胞を、MATRIGEL(登録商標)(1:30希釈)をコーティングしたディッシュと、0.2%のFBS、100ng/mLのアクチビンA、及び20ng/mLのWNT−3aを添加したRPMI培地により1日培養し、次いで0.5%のFBS及び100ng/mLのアクチビンAを添加したRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
a.2%のFBS及び50ng/mLのFGF7を添加したDMEM/F12で3日間にわたって処理し(ステージ2)、次いで
b.1%のB27、50ng/mLのFGF7、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、100ng/mLのノギン、20ng/mLのアクチビンA、及びp38キナーゼ阻害剤(JNJ3026582、2.5μM)を添加したDMEM(高グルコース)で4日間にわたって処理し(ステージ3)、次いで
c.処理10:1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び500nMのTPBを添加したDMEM(高グルコース)で6日間(ステージ4、処理10)、又は
d.処理11:1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び500nMのTPBを添加したDMEM(高グルコース)で9日間(ステージ4、処理11)、又は
e.処理12:1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び500nMのTPBを添加したDMEM(高グルコース)で12日間にわたって処理した(ステージ4、処理12)。
【0151】
表4に示すように、プロテインキナーゼC活性化因子による処理期間を、9日間(処理11)又は12日間(処理12)のいずれかに延長したとき、更なる効果はみられなかった。インスリン単独発現細胞は、処理10で6日間処理後、総集団の27.8%であった。反対に、インスリン発現細胞は、9日間処理(処理11)後10%に、12日間処理(処理12)後更に4%に減少した。同時に、インスリン及びNKX6.1を共発現する細胞集団の総比率も、処理の延長後に著しく下がった。これらの結果は、ノギン、Alk5阻害剤II、及びプロテインキナーゼC活性化因子による6日間処理は、本発明の細胞の形成に十分であったことを示唆した。
【0152】
【表4】
【0153】
(実施例4)
重症複合免疫不全(SCID)−beige(Bg)マウスへの、本発明の細胞の移植
ヒト胚性幹細胞株H1細胞を、MATRIGEL(登録商標)(1:30希釈)をコーティングしたディッシュと、0.2%のFBS、100ng/mLのアクチビンA、及び20ng/mLのWNT−3aを添加したRPMI培地により1日培養し、次いで0.5%のFBS及び100ng/mLのアクチビンAを添加したRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
a.2%のFBS及び50ng/mLのFGF7を添加したDMEM/F12で3日間にわたって処理し(ステージ2)、次いで
b.1%のB27、50ng/mLのFGF7、0.25μMのシクロパミン−KAAD、及び100ng/mLのノギンを添加したDMEM(高グルコース)で4日間にわたって処理し(ステージ3)、次いで
c.1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、及び500nMのTBPを添加したDMEM(高グルコース)で6日間にわたって処理した(ステージ4)。
【0154】
ステージ4の終了時に、1mLのガラス製ピペットを用いて細胞を機械的に回収し、続いて非接着性プレートに移し一晩培養した。得られた細胞集合体を回収し、5百万個の細胞を含有する集合体を免疫不全マウス(SCID/Bg、動物番号47、48、49、50、及び51)の腎臓被膜に移植した。図7を参照されたい。
【0155】
4週間後、これらの移植物中のインスリン産生細胞の機能性を、動物にグルコースを投与しインスリン分泌を誘導することにより試験した。動物を約15〜20時間にわたって絶食させ、その後、後眼窩から血液サンプルを採取した(処理前グルコース)。次いで各動物に、約3g/kgのグルコース/30%のデキストロース溶液を腹腔内注射により投与し、グルコース注入の約60分後に血液を採取した。非常に感受性の高い、ヒト特異的なCペプチドELISAプレート(カタログ番号80−CPTHU−E01、Alpco Diagnostics,NH)を使用して、マウス血清を用いて血中ヒトCペプチドを検出した。ヒトCペプチドの検出は、インスリン分泌が移植細胞に由来していることを示す。
【0156】
移植後4週目程の早期に、動物血清中のヒトCペプチドを検出した。ヒトCペプチドは経時的に増加した。移植データを図7にまとめる。1ヶ月の終了時点で、試験群の60%の動物において、グルコース投与に応答するヒトCペプチド(0.2ng/mL未満)が検出できた。4週間後、4例中3例のマウスで、ヒトCペプチドのグルコース刺激化血清濃度が5〜10倍上昇した。移植12週間後、移植マウスにおけるヒトcペプチドの平均グルコース刺激化血清濃度は、1mg/mLより高かった(n=4)。
【0157】
(実施例5)
PDX1及びNKX6.1を共発現する、膵臓内胚葉系に特徴的なマーカーを発現する細胞集団を形成する別法
簡潔に述べると、ヒト胚性幹細胞株H1細胞を、MATRIGEL(登録商標)(1:30希釈)をコーティングしたディッシュと、0.2%のFBS、100ng/mLのアクチビンA、及び20ng/mLのWNT−3aを追加したRPMI培地により1日培養した後で、0.5%のFBS及び100ng/mLのアクチビンAを追加したRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
a.2%のFBS及び50ng/mLのFGF7を添加したDMEM/F12で3日間にわたって処理し(ステージ2)、次いで
b.1%のB27、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、及び100ng/mLのノギンを添加したDMEM(高グルコース)で4日間にわたって処理し(ステージ3)、次いで
c.1%のB27、100ng/mLのノギン、1μMのALK5阻害剤II、20nM PMA、又は100nMのTPB、又は20nMのホルボール−12,13−ジブチラート(PDBu)(Calbioche、カタログ#524390)を添加したDMEM(高グルコース)で6日間にわたって処理した(ステージ4)。
【0158】
対照として、別の細胞集団を、1%のB27、100ng/mLのノギン、及び1μMのALK5阻害剤IIを追加したDMEM(高グルコース)で6日間にわたって処理した(ステージ4)。
【0159】
ステージ4の6日目に培養液を2回サンプリングし、IN Cell Analyzer 1000(GE Healthcare)を用いて撮像した。ウェルあたり100視野の撮像を得て、バイオアッセイ及び続く染色手順の間の任意の細胞ロスを補正した。IN Cell Developer Toolbox 1.7(GE Healthcare)ソフトウェアを用いて、各ウェルから総細胞数、総PDX1発現細胞、及び総NKX6.1発現細胞の測定値を得た。各複製データセットについて平均及び標準偏差を算出した。総PDX1及びNKX6.1タンパク質発現細胞は、総細胞集団の比率として報告した。図8に示すように、プロテインキナーゼC活性化因子処理群では、対照処理で得られたサンプルと比較して低い有効濃度(約20nM)において、NKX6.1/PDX1発現細胞集団が劇的に増加した。ステージ4の6日目までに、プロテインキナーゼC活性化因子又は対照処理のいずれかを受けた細胞集団では、集団の92%±4%がPDX1を発現した。プロテインキナーゼC活性化因子処理群では、75%±5%のPDX1発現細胞がNKX6.1を発現した。しかし、ノギン及びTGFβ受容体阻害剤のみで処理された集団(対照)では、PDX1発現細胞の58%±5%のみがNKX6.1を発現した。プロテインキナーゼC活性化因子の存在下では、20%のNKX6.1発現細胞は、増殖マーカー、EdU(Click−iT(登録商標)EdU Imaging Kit、Invitrogen、カタログ#C10337)に対して共に陽性であった。
【0160】
この実験では、プロテインキナーゼC活性化因子を比較的低い有効濃度(〜20nM)でノギン及びTGFβ受容体阻害剤と組み合わせて用いると、Nkx6.1発現の上方制御、並びにPDX1及びNKX6.1を発現する細胞比率の増加を促進できることを示す。
【0161】
(実施例6)
膵臓内胚葉系に特徴的なマーカーを発現する細胞の、プロテインキナーゼC活性化因子による処理
簡潔に述べると、ヒト胚性幹細胞株H1細胞を、MATRIGEL(登録商標)(1:30希釈)をコーティングしたディッシュと、0.2%のFBS、100ng/mLのアクチビンA、及び20ng/mLのWNT−3aを追加したRPMI培地により1日培養した後で、0.5%のFBS、及び100ng/mLのアクチビンAを追加したRPMI培地で更に2日間にわたって処理し(ステージ1)、次いで
a.2%のFBS及び50ng/mLのFGF7を添加したDMEM/F12で3日間にわたって処理し(ステージ2)、次いで
b.T1:1%のB27、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、100ng/mLのノギン、及び50ng/mLのFGF10を添加したDMEM(高グルコース)で4日間(ステージ3、T1)、又は
T2:1%のB27、0.25μMのシクロパミン−KAAD、2μMのレチノイン酸(RA)、100ng/mLのノギン、50ng/mLのFGF10、及び100nMのTPBを添加したDMEM(高グルコース)で4日間にわたって処理し(ステージ3、T2)、次いで
c.1%のB27、100ng/mLのノギン、及び1μMのALK5阻害剤IIを添加したDMEM(高グルコース)で6日間にわたって処理した(ステージ4)。
【0162】
図9に示すように、対照群(T1)と比較してTPBで処理した細胞(T2)において、膵臓内胚葉マーカーであるPDX1、NKX6.1、及びPTF1αの顕著な下方制御が観察された。NKX6.1は免疫組織化学的に検出できなかった。これらのデータは、ステージ3におけるプロテインキナーゼ活性化因子処理により、PDX1/NKX6.1共発現細胞の生成が促進されなかったことを示唆する。
【0163】
本明細書の全体を通じて引用した刊行物は、その全容を本明細書に援用するものである。以上、本発明の様々な態様を実施例及び好ましい実施形態を参照して説明したが、本発明の範囲は、上記の説明文によってではなく、特許法の原則の下で適宜解釈される以下の「特許請求の範囲」によって定義されるものである点は認識されるであろう。
【特許請求の範囲】
【請求項1】
NKX6.1及びインスリンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞集団であって、前記集団の10%未満の細胞がグルカゴンを発現する、細胞集団。
【請求項2】
前記細胞の少なくとも30%がNKX6.1を発現する、請求項1に記載の細胞集団。
【請求項3】
前記細胞の少なくとも40%がNKX.6.1を発現する、請求項1に記載の細胞集団。
【請求項4】
前記細胞の少なくとも50%がNKX6.1を発現する、請求項1に記載の細胞集団。
【請求項5】
前記細胞の少なくとも60%がNKX6.1を発現する、請求項1に記載の細胞集団。
【請求項6】
前記細胞の少なくとも5%がインスリンを発現する、請求項1に記載の細胞集団。
【請求項7】
NKX6.1及びインスリンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞集団を生成する方法であって、前記集団の10%未満の細胞がグルカゴンを発現し、前記方法が、
a.多能性幹細胞を培養する工程と、
b.前記多能性幹細胞を、胚体内胚葉系(definitive endoderm lineage)に特徴的なマーカーを発現する細胞へと分化させる工程と、
c.前記胚体内胚葉系に特徴的なマーカーを発現する細胞を、膵臓内胚葉系に特徴的なマーカーを発現する細胞へと分化させる工程と、
d.前記膵臓内胚葉系に特徴的なマーカーを発現する細胞を、BMP阻害能を有する因子、TGFβ受容体シグナル伝達阻害剤、及びプロテインキナーゼC活性化因子を追加した培地で培養することにより、前記膵臓内胚葉系に特徴的なマーカーを発現する細胞を、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する、前記膵臓内分泌系に特徴的なマーカーを発現する細胞へと分化させる工程と、を含む、方法。
【請求項1】
NKX6.1及びインスリンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞集団であって、前記集団の10%未満の細胞がグルカゴンを発現する、細胞集団。
【請求項2】
前記細胞の少なくとも30%がNKX6.1を発現する、請求項1に記載の細胞集団。
【請求項3】
前記細胞の少なくとも40%がNKX.6.1を発現する、請求項1に記載の細胞集団。
【請求項4】
前記細胞の少なくとも50%がNKX6.1を発現する、請求項1に記載の細胞集団。
【請求項5】
前記細胞の少なくとも60%がNKX6.1を発現する、請求項1に記載の細胞集団。
【請求項6】
前記細胞の少なくとも5%がインスリンを発現する、請求項1に記載の細胞集団。
【請求項7】
NKX6.1及びインスリンを共発現する、膵臓内分泌系に特徴的なマーカーを発現する細胞集団を生成する方法であって、前記集団の10%未満の細胞がグルカゴンを発現し、前記方法が、
a.多能性幹細胞を培養する工程と、
b.前記多能性幹細胞を、胚体内胚葉系(definitive endoderm lineage)に特徴的なマーカーを発現する細胞へと分化させる工程と、
c.前記胚体内胚葉系に特徴的なマーカーを発現する細胞を、膵臓内胚葉系に特徴的なマーカーを発現する細胞へと分化させる工程と、
d.前記膵臓内胚葉系に特徴的なマーカーを発現する細胞を、BMP阻害能を有する因子、TGFβ受容体シグナル伝達阻害剤、及びプロテインキナーゼC活性化因子を追加した培地で培養することにより、前記膵臓内胚葉系に特徴的なマーカーを発現する細胞を、NKX6.1及びインスリン、並びに最小量のグルカゴンを共発現する、前記膵臓内分泌系に特徴的なマーカーを発現する細胞へと分化させる工程と、を含む、方法。
【図1】


【図2】

【図3】

【図4】

【図5】
【図6】

【図7】
【図8】
【図9】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2013−515480(P2013−515480A)
【公表日】平成25年5月9日(2013.5.9)
【国際特許分類】
【出願番号】特願2012−546058(P2012−546058)
【出願日】平成22年12月16日(2010.12.16)
【国際出願番号】PCT/US2010/060756
【国際公開番号】WO2011/079017
【国際公開日】平成23年6月30日(2011.6.30)
【出願人】(509087759)ヤンセン バイオテツク,インコーポレーテツド (77)
【Fターム(参考)】
【公表日】平成25年5月9日(2013.5.9)
【国際特許分類】
【出願日】平成22年12月16日(2010.12.16)
【国際出願番号】PCT/US2010/060756
【国際公開番号】WO2011/079017
【国際公開日】平成23年6月30日(2011.6.30)
【出願人】(509087759)ヤンセン バイオテツク,インコーポレーテツド (77)
【Fターム(参考)】
[ Back to top ]