
ビタミンD類水酸化酵素の改良
【課題】Vdh及びVdhと構造的に関連したVdh-K1などの水酸化酵素を改変することによりビタミンD類の水酸化反応における副反応を抑制し、さらに水酸化酵素の比活性を改善して水酸化ビタミンD誘導体を効率よく生産する方法を提供する。
【解決手段】ビタミンD類の水酸化において活性が増強され、望ましくない副反応が低減したポリペプチド(酵素)をコードするDNA、そのDNAによりコードされる改変型ビタミンD類水酸化酵素、そのDNAを含む自律複製性または組み込み複製性の組換えベクター、そのDNAを含む形質転換体、及びその形質転換体を培地で培養し、その培養液から水酸化ビタミンD誘導体を採取する水酸化ビタミンD誘導体の製造方法。
【解決手段】ビタミンD類の水酸化において活性が増強され、望ましくない副反応が低減したポリペプチド(酵素)をコードするDNA、そのDNAによりコードされる改変型ビタミンD類水酸化酵素、そのDNAを含む自律複製性または組み込み複製性の組換えベクター、そのDNAを含む形質転換体、及びその形質転換体を培地で培養し、その培養液から水酸化ビタミンD誘導体を採取する水酸化ビタミンD誘導体の製造方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ビタミンD類の水酸化に関与するポリペプチド(ビタミンD類水酸化酵素)をコードする塩基配列を改変することにより得られる、ビタミンD類の水酸化活性が増強され、あるいは副反応が抑制されたビタミンD類水酸化酵素、その酵素をコードするDNA、そのDNAを含むベクター、そのベクターを含む形質転換体、及びその形質転換体を用いた水酸化ビタミンD誘導体の製造方法に関する。
【背景技術】
【0002】
ビタミンDは、主に高等生物において必須とされる脂溶性ビタミン群でコレステロールから生合成される。その生理作用はカルシウムの吸収促進や代謝促進、細胞分化誘導、免疫調節作用など多岐にわたり、生体において重要な役割を果たしている。
【0003】
ヒトにおけるビタミンDの生合成の主要な経路は、まずコレステロールから7−デヒドロコレステロール(プロビタミンD3)が合成された後、皮膚において紫外線と熱反応からビタミンD3がつくられる。ビタミンD3は肝臓のミトコンドリアに局在するシトクロムP450(例えば、CYP27A1)によって25位の水酸化を受けて25−ヒドロキシビタミンD3となる。その後、腎臓の近位尿細管において別のシトクロムP450(例えば、CYP27B1)によって、さらに1α位の水酸化を受けて1α,25−ジヒドロキシビタミンD3(活性化型ビタミンD3)が作られる。この物質が細胞内の受容体(レセプター)に結合することによって、生理作用発現に関わる核内の特定の遺伝子の発現調節を行うことが分かっている。
【0004】
従って、肝臓または腎臓が機能不全に陥ると、正常なビタミンD代謝が阻害されることがある。そのような患者では血中の25-ヒドロキシビタミンD3や1α,25−ジヒドロキシビタミンD3が極度に低下することがあり、治療のためには25-ヒドロキシビタミンD3や1α,25−ジヒドロキシビタミンD3を投与して補う必要がある。
【0005】
一方、先天的に、ビタミンD3の水酸化に関与するシトクロムP450遺伝子に変異があることに起因する水酸化機能不全により1α,25−ジヒドロキシビタミンD3が不足して起こるくる病が知られている。のみならず、後天的なくる病や骨粗鬆症を含めて、1α,25−ジヒドロキシビタミンD3が不足することで起こる疾患の治療薬として、1α,25−ジヒドロキシビタミンD3及び同様な生理作用をもつ類縁化合物の重要性は非常に高い。
【0006】
さらに、ビタミンD類はその多様な生理活性から様々な誘導体が薬物の候補として研究され、抗腫瘍薬、乾癬治療薬、免疫賦活薬などの開発にも期待がもたれている。このような観点からビタミンD類を誘導体化する一手法としても水酸化は重要な反応である。この場合、水酸化の位置は1α位及び25位に限定されず、他の位置の水酸化修飾にも薬効を発現させるための高い必要性が生じることがある。このように医薬品開発の課題として水酸化ビタミンD誘導体の製造法を改良し、より効率的で安価に水酸化ビタミンD誘導体を製造して供給することが医薬産業から強く望まれてきた。
【0007】
ビタミンD類の中で特に生理活性が強く、価値の高い1α,25−ジヒドロキシビタミンD3について言及すると、その製造手法としては、有機合成法、ヒトのシトクロムP450を利用する方法、微生物を利用する方法等が挙げられる。このうち有機合成法ではコレステロールから約20工程を経て合成する製法が知られている(非特許文献1及び2参照)。またヒトのシトクロムP450を利用する方法では、関与する遺伝子、発現する技術、シトクロムP450酵素の性質に関する生物学的・生化学的知見の蓄積がある(非特許文献3参照)。しかし、これら2つのいずれの方法も高い製造コストや低い生産性の問題から、実用化には極めて不向きである。
【0008】
微生物を利用する方法としては放線菌シュードノカルディア・オートトロフィカ(Pseudonocardia autotrophica)を変換菌として用いたビタミンD3から1α,25−ジヒドロキシビタミンD3の製造方法が知られており、この方法は産業的に実用化されている(非特許文献4、非特許文献5、特許文献1及び特許文献2参照)。この方法は、シュードノカルディア(Pseudonocardia)属放線菌を発酵槽において培養した後、ビタミンD3を反応基質として培養液に添加し、菌体のもつ水酸化酵素の作用で反応を進行させて、その結果蓄積する1α,25−ジヒドロキシビタミンD3を培養液から回収・精製して行うものである。
【0009】
この方法においては、変換微生物として生育時間の長い放線菌を用いるため製造期間が長期になり、さらにこの変換菌がもつビタミンD3変換作用の中には目的の水酸化反応のみならず、望ましくない副反応も存在する。その副反応として例えば、26位あるいは24位の水酸化などが挙げられる。また基質であるビタミンD3や目的生成物である1α,25−ジヒドロキシビタミンD3に対する分解活性も存在する。特に1α,25−ジヒドロキシビタミンD3の分解は、目的物の蓄積量に直接的に関わる極めて重要な問題である。これに加えて、副産物の存在も変換反応後の培養液からの1α,25−ジヒドロキシビタミンD3の回収・精製工程を多くし、結果的に目的物の収量を低下させる。
【0010】
このような問題点を解決するため、変換菌である放線菌シュードノカルディア・オートトロフィカからビタミンD類の水酸化酵素(「Vdh」または人為的変異が導入されていないことを強調するため「野生型Vdh」とも言う。)をコードする遺伝子(「vdh」または「野生型vdh」と言う。)を取得し、これを生育の速い大腸菌に組込んで得られる形質転換体を用いてビタミンD類を水酸化し、水酸化ビタミンD誘導体を効率的に生産する方法が提案されている(特許文献3)。さらにその方法において、vdh遺伝子を改変して水酸化活性を高めた改変酵素をコードする遺伝子を利用する、より効果的な方法も提案されている(非特許文献6)。非特許文献6によると、その遺伝子(vdh-K1)がコードする改変酵素Vdh-K1は野生型Vdh酵素にアミノ酸配列レベルで以下の4種の変異を導入した酵素である。
【0011】
(1) 70番目のスレオニンがアルギニンに置き換わる変異、
(2) 156番目のバリンがロイシンに置き換わる変異、
(3) 216番目のグルタミン酸がメチオニンに置き換わる変異、
(4) 384番目のグルタミン酸がアルギニンに置き換わる変異。
【0012】
Vdh-K1はVD3に対する25位水酸化活性が野生型酵素の約20倍比活性が上昇したことが知られている。vdh-K1遺伝子を大腸菌に形質転換して得られた株を変換菌として用いることで比較的短期間で効率よくビタミンD類を水酸化することができるが、産業利用の観点から、反応特異性が不十分で副生物である26位水酸化体の産生を抑制できない問題点は残っており、さらに水酸化活性においてもより高い水準が求められている。したがって、副生物である26位水酸化体の産生を抑制させるとともにより一層の活性上昇が実現することでVdhまたはVdh-K1の高機能化を実現し、水酸化ビタミンD誘導体の効率的な製造方法を確立することが強く望まれていた。
【先行技術文献】
【特許文献】
【0013】
【特許文献1】特開平2-469号公報
【特許文献2】特開平2-231089号公報
【特許文献3】国際公開第2007/138894号パンフレット
【非特許文献】
【0014】
【非特許文献1】Kametani T., Furuyama H., Synthesis Med. Res. Rev. 7, 147-171 (1987)
【非特許文献2】Zunk G-D., Okamura W. H. Norman A. W., Chem. Rev. 95, 1877-1952 (1995)
【非特許文献3】T. Sakakiらの総説, Frontiers in Bioscience, 10, 119-134, 2005
【非特許文献4】J. Sasaki et. al, Applied Microbiology and Biotechnology, 38, 152-157, 1992
【非特許文献5】K. Takeda et. al, J. Ferment. Bioeng., 78, 380-382, 1994
【非特許文献6】Y. Fujii et. al, Biochem. Biophys. Res. Commun., 385, 170-175, 2009)
【発明の概要】
【発明が解決しようとする課題】
【0015】
本発明は、活性や副反応抑制においてビタミンD類の水酸化機能が向上したビタミンD類水酸化酵素、当該酵素をコードするDNA、該DNA含むベクター、及び該ベクターを含む形質転換体を提供することを課題とする。
さらに本発明は、Vdh及びVdhと構造的に関連したVdh-K1などの水酸化酵素を改変することでビタミンD類の副反応を抑制し、望ましくはさらに水酸化酵素の比活性を改善して水酸化ビタミンD誘導体を効率よく生産する方法を提供することにある。
【課題を解決するための手段】
【0016】
本発明者らは、前述の課題を解決するために、これまでに知見が得られていない副反応低減や活性上昇に関与する新規な変異酵素を取得することを目的として鋭意研究を実施した。まず、非特許文献6に開示されたシュードノカルディア・オートトロフィカNBRC12743株由来のvdh遺伝子を改変して得られたvdh-K1遺伝子に対して、その塩基配列中のアミノ酸コドンにランダムに変異を加え、DNA変異体のライブラリーを調製した。次いでそれらの変異体がコードする変異酵素タンパク質を大腸菌において発現させ、それらのクローンのビタミンD類の水酸化活性を測定することで、25位水酸化活性に対する26位水酸化活性の比率の低下した改変遺伝子を選択し、変異が加えられた部位(改変部位)を特定した。次にこれらの改変部位を組み合わせて、さらに機能が高められた改変遺伝子を調製した。その後、これらの改変遺伝子を含むベクターを大腸菌に組み込んで26位水酸化活性が抑制された水酸化酵素または25位水酸化活性が増強された水酸化酵素を発現させて酵素の機能を評価した結果、Vdh-K1に比較して水酸化ビタミンD誘導体を効率よく生産できる改変型ビタミンD類水酸化酵素を見出し、本発明を完成した。
【0017】
すなわち、本発明は以下の[1]〜[12]に関する。
[1] 配列番号2または4に記載のアミノ酸配列において、下記の(1)〜(4)からなる群より選択される少なくとも1つの改変部位を有する改変アミノ酸配列からなる改変型ビタミンD類水酸化酵素:
(1)88位のイソロイシンがバリンに置換された改変部位、
(2)171位のロイシンがバリンに置換された改変部位、
(3)177位のアラニンがアルギニンに置換された改変部位、
(4)240位のトレオニンがセリンに置換された改変部位。
[2] 前記[1]に記載の改変アミノ酸配列において、(1)〜(4)からなる群より選択される少なくとも1つの改変部位のほかに、さらに1または数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有する改変型ビタミンD類水酸化酵素。
[3] 前記[1]に記載の改変アミノ酸配列と90%以上の相同性を有し、(1)〜(4)からなる群より選択される少なくとも1つの改変部位を含むアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有する改変型ビタミンD類水酸化酵素。
[4] 配列番号2または4に記載のアミノ酸配列において、88位のイソロイシンがバリンに、及び171位のロイシンがバリンにそれぞれ置換された改変アミノ酸配列からなる前記[1]に記載の改変型ビタミンD類水酸化酵素。
[5] 前記[4]に記載の改変アミノ酸配列において、88位及び171位のアミノ酸置換のほかに、さらに1または数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有する改変型ビタミンD類水酸化酵素。
[6] 前記[4]に記載の改変アミノ酸配列と90%以上の相同性を有し、88位及び171位のアミノ酸置換を含むアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有する改変型ビタミンD類水酸化酵素。
[7] 前記[1]〜[6]のいずれか1項に記載の改変型ビタミンD類水酸化酵素をコードするDNA。
[8] 配列番号1または3に記載の塩基配列において、下記の(1)〜(4)からなる群より選択される少なくとも1つの改変部位を有する改変塩基配列からなる前記[1]に記載の改変型ビタミンD類水酸化酵素をコードするDNA:
(1)262〜264番目の塩基配列ATCが、バリンをコードするコドンに置換された改変部位、
(2)511〜513番目の塩基配列CTGが、バリンをコードするコドンに置換された改変部位、
(3)529〜531番目の塩基配列GCCが、アルギニンをコードするコドンに置換された改変部位、
(4)718〜720番目の塩基配列ACGが、セリンをコードするコドンに置換された改変部位。
[9] 配列番号1または3に記載の塩基配列において、262〜264番目の塩基配列ATCがバリンをコードするコドンに、及び511〜513番目の塩基配列CTGがバリンをコードするコドンにそれぞれ置換された改変塩基配列からなる前記[8]に記載のDNA。
[10] 前記[7]〜[9]のいずれか1項に記載のDNAを含む組換えベクター。
[11] 前記[10]に記載のベクターを含む形質転換体。
[12] 前記[11]に記載の形質転換体を培養し、培養中または培養後の形質転換体とビタミンD類とを接触させ、産出された水酸化ビタミンD誘導体を採取することを特徴とする水酸化ビタミンD誘導体の製造方法。
【発明の効果】
【0018】
本発明は、活性や副反応抑制においてビタミンD類の水酸化機能が向上したビタミンD類水酸化酵素及びそれをコードするDNA、そのDNAを形質転換した細胞、その細胞を用いた高効率な水酸化ビタミンD類の製造方法に関する。本発明によって、ビタミンD3の25位水酸化能が向上した酵素、ビタミンD3の26位水酸化能が抑制した酵素等、産業上有用な特徴的性質をもつビタミンD類の水酸化酵素及び該酵素をコードするDNAを取得できる。さらに、それらのDNAを担持するベクター、そのベクターで大腸菌やシュードノカルディア・オートトロフィカ等を形質転換した形質転換体を作製し、その形質転換体を用いて水酸化ビタミンD誘導体を効率よく製造することができる。従って、本発明により、産業上極めて有用で、かつ効率のよい水酸化ビタミンD誘導体の製造方法が提供される。
【図面の簡単な説明】
【0019】
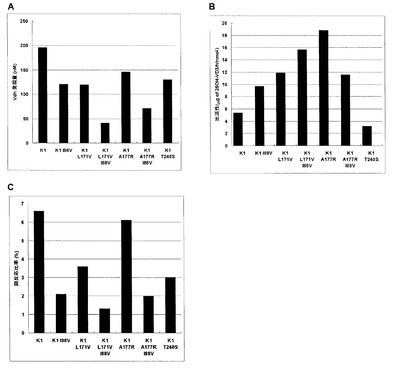
【図1】Aは大腸菌におけるVdh-K1各改変体の発現量を示したグラフである。Bは大腸菌において発現させた改変体の比活性を示したグラフである。Cは改変体の25位水酸化ビタミンD3量に対する26位水酸化ビタミンD3量の比率を示したグラフである。
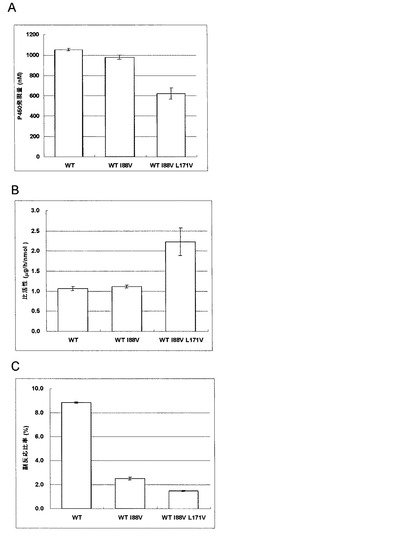
【図2】Aは野生型Vdh改変体の発現量を示したグラフである。Bは野生型Vdh改変体の比活性を示したグラフである。Cは野生型Vdh改変体の25位水酸化ビタミンD3量に対する26位水酸化ビタミンD3量の比率を示したグラフである。
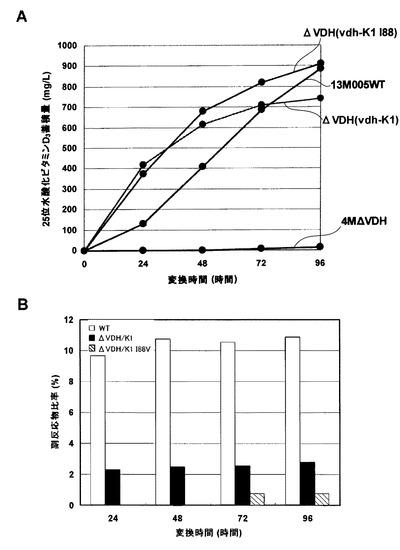
【図3】Aはシュードノカルディア・オートトロフィカの培地中における25位水酸化ビタミンD3の蓄積量を示したグラフである。Bは培地中の25位水酸化ビタミンD3量に対する26位水酸化ビタミンD3量の比率を示したグラフである。
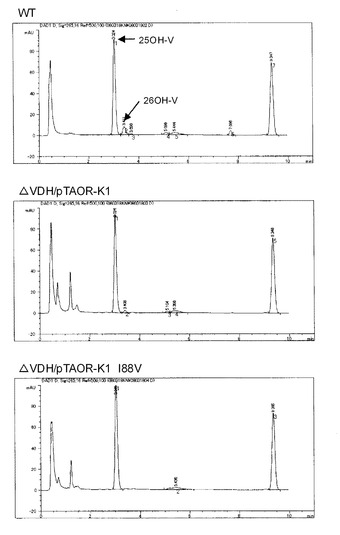
【図4】シュードノカルディア・オートトロフィカ親株及び形質転換株によるビタミンD3変換試験を行った時のHPLCのクロマトグラムを示した図である。
【発明を実施するための形態】
【0020】
以下に本願明細書において記載する用語、記号等の意義を説明し、本発明を詳細に説明する。
本願明細書において用いる「ビタミンD類」とは、本発明の水酸化酵素により基質として認識され、1α位、25位、その他の炭素に一原子酸素添加を行うことが可能なものを意味し、以下の化合物を包含する。
【0021】
(1)ビタミンD3、ビタミンD2などのビタミンD類、
(2)ビタミンD類の前駆体、代謝物、その他の誘導体(この中には、25−ヒドロキシビタミンD3、1α−ヒドロキシビタミンD3などが含まれる。)、
(3)コレスタン系のステロイド類またはその修飾化合物(この中には、コレステロール、7−デヒドロコレステロールなどが含まれる。)。
【0022】
本明細書において用いる「水酸化ビタミンD誘導体」とは、本発明の水酸化酵素によって、ビタミンD類の1α位、25位、その他の位置の炭素原子に1酸素原子が添加された化合物を意味する。
【0023】
また、本発明において「ビタミンD類の水酸化酵素活性」とは、ビタミンD類を水酸化し、水酸化ビタミンD誘導体へ変換する酵素活性を意味する。「ビタミンD類の水酸化酵素」とはVdhまたはVdh-K1などのVdhと構造的かつ機能的に相同性を有する酵素を意味する。Vdh及びVdh-K1はそれぞれ配列番号4及び2で表されるアミノ酸配列からなるポリペプチドである。vdh遺伝子及びvdh-K1遺伝子はそれぞれ配列番号3及び1で表されるポリヌクレオチドからなるDNAである。
【0024】
さらに、本発明において「改変型酵素」、「改変アミノ酸配列」、「DNA改変体」、「改変塩基配列」とは、構成アミノ酸または塩基の置換、削除、付加、挿入などにより修飾された酵素、アミノ酸配列、DNA、塩基配列を意味する。
【0025】
以下、本発明の実施の形態について詳細に説明する。
[本発明に係る酵素及びそれをコードするDNA]
本発明に係る改変型ビタミンD類水酸化酵素は、野生型のビタミンD類水酸化酵素Vdh及び改変酵素Vdh-K1の特定のアミノ酸配列を改変することによって得られたものであり、当該酵素を水酸化ビタミンD誘導体の製造に用いた場合、改変前の酵素と比較して、その副反応を有意に抑制し、所望の水酸化ビタミンD誘導体の製造効率を向上させることを可能とするものである。
本発明に係る酵素及びそれをコードするDNAは、以下の方法で取得された。
本発明者らは、非特許文献6に記載された情報に従って適宜設計されたプライマーを用いてPCRを行い、ビタミンD類の水酸化酵素活性を有するポリペプチドをコードするvdh-K1遺伝子(配列番号1)を含むDNA断片を取得した。次いで、フェレドキシンをコードするDNA、フェレドキシン還元酵素をコードするDNA及び大腸菌のリボゾーム結合部位として有効とされるDNA(SD配列)をすべて含むプラスミドにこの断片を連結させ、大腸菌においてビタミンD類の水酸化酵素活性を発現しうるプラスミドを作製した。得られたプラスミドを用いて、PCRにより配列番号1で示されるDNAにランダムな変異が入ったDNA断片を増幅し、ビタミンD水酸化酵素発現用プラスミドに連結させ、DNA改変体を含むプラスミドのライブラリーを作製した。次いでこれらライブラリーのプラスミドで形質転換した大腸菌がもつビタミンD類の水酸化酵素活性をスクリーニングすることにより、特徴的な水酸化活性をもつ複数のポリペプチド(改変型ビタミンD類水酸化酵素)及びそれらをコードするDNAを見出したものである。Vdh-K1遺伝子は403残基のポリペプチドからなるVdh-K1をコードしているが、開始コドン(メチオニン)を除く402残基のうちいずれか1つの残基が置換された改変体を作るには、各残基ごとに元のアミノ酸を除く19種類のアミノ酸に置換する。これをその残基の「アミノ酸総置換」と言う。この結果、調製可能な改変体は7638種に及ぶ。現在の技術レベルではこれらのすべてを調製し試験するには多大な労力と技術的な熟練が必要であり、さらに網羅的に試験した結果、必ず所望の改変体が得られる確証はない。本発明者らは高機能化したビタミンD類の水酸化酵素改変体を獲得するため、このような困難なスクリーニングを遂行し、本発明の改変型酵素構築の基本情報となる有効変異についての情報を得るに至った。
【0026】
こうして得られた本発明のビタミンD類水酸化酵素及び当該酵素をコードするDNAは、下記(a)のアミノ酸配列からなる改変型ビタミンD類水酸化酵素、及び下記(a)のアミノ酸配列からなる改変型ビタミンD類水酸化酵素をコードするDNAである。
(a)配列番号2または4に記載のアミノ酸配列において、以下の(1)〜(4)からなる群より選択される少なくとも1つの改変部位を有する改変アミノ酸配列:
(1)88位のイソロイシンがバリンに置換された改変部位、
(2)171位のロイシンがバリンに置換された改変部位、
(3)177位のアラニンがアルギニンに置換された改変部位、
(4)240位のトレオニンがセリンに置換された改変部位。
【0027】
前記(a)のアミノ酸配列からなる改変型ビタミンD類水酸化酵素をコードするDNAの中でも下記のDNAが好ましい。
配列番号1または3に記載の塩基配列において、以下の(1)〜(4)からなる群より選択される少なくとも1つの改変部位を有する改変塩基配列からなるDNA:
(1)262〜264番目の塩基配列ATCが、バリンをコードするコドンに置換された改変部位、
(2)511〜513番目の塩基配列CTGが、バリンをコードするコドンに置換された改変部位、
(3)529〜531番目の塩基配列GCCが、アルギニンをコードするコドンに置換された改変部位、
(4)718〜720番目の塩基配列ACGが、セリンをコードするコドンに置換された改変部位。
【0028】
ここで、バリンをコードするコドンは、具体的にはGTT、GTG、GTA、GTGであり、アルギニンをコードするコドンは、具体的にはCGT、CGC、CGA、CGG、AGA、AGGであり、セリンをコードするコドンは、具体的にはTCT、TCC、TCA、TCG、AGT、AGCである。
【0029】
これらのアミノ酸改変による優れた効果は、以下の(ア)及び(イ)にまとめることができる。
(ア)改変型ビタミンD類水酸化酵素自体のビタミンD類水酸化能が増強されている(水酸化能の比活性が高くなっている)。
(イ)改変型ビタミンD類水酸化酵素自体のビタミンD類26位水酸化能が抑制されている(25位に対する26位水酸化活性の比率が低くなっている)。
【0030】
なお、上記の各DNA改変体がもつ個々の特長は以下のとおりである。
1)「88位のイソロイシンがバリンに置換された改変部位を有する改変アミノ酸配列をコードするDNA」または「塩基262から塩基264にかけての配列ATCを、バリンをコードするコドンに改変した改変部位を有する改変塩基配列」を組み込んだ形質転換体は、ビタミンD3の変換反応において、26−ヒドロキシビタミンD3の生成量が減少する。
【0031】
2)「171位のロイシンがバリンに置換された改変部位を有する改変アミノ酸配列をコードするDNA」または「塩基511から塩基513にかけての配列CTGを、バリンをコードするコドンに改変した改変部位を有する改変塩基配列」を組み込んだ形質転換体は、ビタミンD3の変換反応において、26−ヒドロキシビタミンD3の生成量が減少する。そして、そのDNA改変体がコードするポリペプチドは、ビタミンD3の25位水酸化能の比活性が高い。
【0032】
3)「177位のアラニンがアルギニンに置換された改変部位を有する改変アミノ酸配列をコードするDNA」または「塩基529から塩基531にかけての配列GCCを、アルギニンをコードするコドンに改変した改変部位を有する改変塩基配列」を組み込んだ形質転換体は、ビタミンD3の変換反応において、対応する25-ヒドロキシ体の生成量が増大する。そして、そのDNA改変体がコードするポリペプチドは、ビタミンD3の25位水酸化能の比活性が高い。
【0033】
4)「240位のトレオニンがセリンに置換された改変部位を有する改変アミノ酸配列をコードするDNA」または「塩基718から塩基720にかけての配列ACGを、セリンをコードするコドンに改変した改変部位を有する改変塩基配列」を組み込んだ形質転換体は、ビタミンD3の変換反応において、26−ヒドロキシビタミンD3の生成量が減少する。
【0034】
上述のように、本発明において「ビタミンD類水酸化酵素」とはVdh、またはVdh-K1等のVdhと構造的かつ機能的に相同性を有する酵素を包含するものである。したがって、本発明に係る改変型ビタミンD類水酸化酵素を得るために用いられるビタミンD類水酸化酵素としては、vdh-K1遺伝子(配列番号1)がコードするビタミンD類水酸化酵素Vdh-K1(配列番号2)、及び野生型vdh遺伝子(配列番号3)がコードするビタミンD類水酸化酵素Vdh(配列番号4)のほか、vdh-K1遺伝子または野生型vdh遺伝子と高い相同性を示し、かつ同じ機能をもつ遺伝子がコードする酵素を用いることも可能である。
【0035】
本発明において、「vdh-K1遺伝子または野生型vdh遺伝子と高い相同性を示し、かつ同じ機能をもつ遺伝子がコードするビタミンD類水酸化酵素」とは、下記(b)または(c)のアミノ酸配列からなるビタミンD類水酸化酵素を意味する。
(b)配列番号2または4に記載のアミノ酸配列において、1または数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有するアミノ酸配列、
(c)配列番号2または4に記載のアミノ酸配列と90%以上の相同性を有するアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有するアミノ酸配列。
【0036】
すなわち、本発明の酵素には、上記(b)または(c)のアミノ酸配列において、前記(a)と同様の改変部位、すなわち(1)88位のイソロイシンがバリンに置換された改変部位、(2)171位のロイシンがバリンに置換された改変部位、(3)177位のアラニンがアルギニンに置換された改変部位、及び(4)240位のトレオニンがセリンに置換された改変部位からなる群より選択される少なくとも1つの改変部位を有するアミノ酸配列からなるビタミンD類水酸化酵素が包含される。
【0037】
本発明において、「1または数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列」における「1または数個」の範囲は特に限定されないが、好ましくは、1〜40個であり、より好ましくは1〜20個であり、特に好ましくは1〜10個である。
また、アミノ酸配列の相同性については、例えば、Genetyx(Genetyx corporation)のHomology Search機能により求めることができる。本発明において、アミノ酸配列の相同性の範囲は、90%以上であることが好ましく、95%以上であることがより好ましく、98%以上であることが特に好ましい。
【0038】
「野生型vdh遺伝子と高い相同性を示し、かつ同じ機能をもつ遺伝子」として、具体的には、特許文献3の表1に記載されているシュードノカルディア・オートトロフィカやシュードノカルディア・サターネア(Pseudonocardia saturnea)の様々な株から由来するvdhに相同な遺伝子が挙げられる。
【0039】
さらに上記のDNA改変体の改変部位を組み合わせて、さらに個々の機能が強化された改変型ビタミンD類水酸化酵素及び当該酵素をコードするDNAを取得することもできる。そのような酵素及びDNAの具体例として下記(1)〜(3)のアミノ酸配列からなる改変型ビタミンD類水酸化酵素、及び下記(1)〜(3)のアミノ酸配列からなる改変型ビタミンD類水酸化酵素をコードするDNAが挙げられる。
【0040】
(1)配列番号2または4に記載のアミノ酸配列において、88位のイソロイシンがバリンに、及び171位のロイシンがバリンにそれぞれ置換された改変アミノ酸配列、
(2)(1)の改変アミノ酸配列において、前記88位及び171位のアミノ酸置換のほかに、さらに1または数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有するアミノ酸配列、
(3)(1)の改変アミノ酸配列と90%以上の相同性を有し前記88位及び171位のアミノ酸置換を含むアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有するアミノ酸配列。
【0041】
上記のDNA改変体は、大腸菌において発現したポリペプチドが示すビタミンD3の25位水酸化能の比活性が野生型Vdhに対して高い(約2倍)のに加え、ビタミンD3の25位水酸化能に対する26位水酸化能の比率が低い(約1/5)という特長がある。
【0042】
前記(1)のアミノ酸配列からなる改変型ビタミンD類水酸化酵素をコードするDNAの中でも、配列番号1または3に記載の塩基配列において、262〜264番目の塩基配列ATCがバリンをコードするコドンに、及び511〜513番目の塩基配列CTGがバリンをコードするコドンにそれぞれ置換された改変塩基配列からなるDNAが好ましい。
【0043】
本発明の酵素をコードするDNAの取得方法は特に限定されない。前述したとおり、シュードノカルディア・オートトロフィカNBRC12743株のゲノムDNAを鋳型とし、PCRにより配列番号1の配列を含むDNAを取得し、これを用いてエラープローン(error-prone)PCRにより複数の変異を加えたDNA改変体を作製して、それらで形質転換された大腸菌の水酸化活性をスクリーニングして取得することができる。
【0044】
例えば、上記したvdh遺伝子断片を鋳型として使用し、配列番号3に示したvdh遺伝子のオープンリーディングフレームの全長を増幅できるように設計した1対のプライマーを用いてPCRを行う。反応液中のマンガンイオン濃度やデオキシグアノシン5'−三リン酸(dGTP)の濃度を変化させることにより変異の誘発を操作できる。PCRの反応条件は適宜設定することができ、例えば、94℃で30秒間(変性)、55℃で30秒〜1分間(アニーリング)、72℃で2分間(伸長)からなる反応工程を1サイクルとして、例えば25サイクル行った後、72℃で7分間反応させる条件などを挙げることができる。増幅されたDNA断片にはランダムに変異が導入されており、適当な宿主中で増幅可能なベクター中にクローニングすることができる。
【0045】
また、改変部位を指定して特定の塩基部分を置換することも可能である。例えば、置換したいアミノ酸をコードするコドンを中央に含むプライマー(変異−F)とそれに対する相補配列を含むプライマー(変異−R)を作製する。vdh遺伝子の5'末端から下流のDNA断片を増幅するプライマー(VDH-F)とプライマー(変異−R)のペアで完全長vdh遺伝子断片を鋳型としてPCRを行いvdh遺伝子の5'末端から置換挿入部位までを含むDNA断片を増幅する。また、vdh遺伝子の3'末端から上流のDNA断片を増幅するプライマー(VDH-R)とプライマー(変異−F)のペアで完全長vdh遺伝子断片を鋳型としてPCRを行い置換挿入部位からvdh遺伝子の3'末端までを含むDNA断片を増幅する。次にVDH-FとVDH-Rのプライマーペアを用いて上記で調製したvdh遺伝子の5'末端から置換挿入部位までを含むDNA断片と置換挿入部位からvdh遺伝子の3'末端までを含むDNA断片を鋳型としてPCRを行い、置換が導入された完全長vdh遺伝子断片を増幅する。その他、本明細書において開示した配列情報に基づいて全合成することも可能である。
【0046】
上記したプライマーの調製、DNAライブラリーの構築、DNAライブラリーのスクリーニング、並びに目的遺伝子のクローニングなどの操作は当業者に既知であり、例えば、モレキュラークローニング第2版、Current Protocols in Molecular Biology, Supplement 1〜38, John Wiley & Sons (1987-1997)等に記載の方法に準じて行うことができる。
【0047】
本発明のポリペプチド(酵素)の取得方法は特に制限されず、化学合成により合成したポリペプチドでもよいし、遺伝子組み換え技術により作製したものでもよい。遺伝子組み換え技術により作製する場合には、先ず、本明細書に上記したとおり、当該酵素をコードするDNA改変体を取得する。このDNA改変体を適当な発現系に導入することにより、本発明の酵素を産生することができる。発現系での酵素の発現については本明細書中において後記する。
【0048】
[本発明の組み換えベクター]
本発明に係るDNAは適当なベクター中に挿入して使用することができる。本発明で用いるベクターの種類は特に限定されず、例えば、自律的に複製するベクター(例えば、プラスミド等)でもよいし、あるいは、宿主細胞に導入された際に宿主細胞のゲノムに組み込まれ、組み込まれた染色体と共に複製されるものであってもよい。発現ベクターにおいて本発明のDNAは、転写に必要な要素(例えば、プロモーター等)が機能的に連結されている。プロモーターは宿主細胞において転写活性を示すDNA配列であり、宿主の種類に応じて適宜選択することができる。具体的には、宿主として大腸菌を用いる場合、例えば、ベクターとしてpET vector、pUC19等を用い、プロモーターとしてT7 promoter、lac promoter等を用いることができる。
【0049】
[本発明の形質転換体及びそれを用いた組み換えポリペプチドの製造]
本発明に係るDNAまたは組み換えベクターを適当な宿主に導入することによって形質転換体を作製することができる。本発明のDNAまたは組み換えベクターが導入される宿主細胞は、本発明の遺伝子を発現できれば任意の細胞でよく、細菌、酵母、真菌及び高等真核細胞等が挙げられる。細菌細胞の例としては、バチルス(Bacillus)またはストレプトマイセス(Streptomyces)等のグラム陽性菌または大腸菌等のグラム陰性菌が挙げられる。これら細菌の形質転換は、塩化カルシウム法、プロトプラスト法、エレクトロポレーション法または公知の方法で調製が可能なコンピテント細胞を用いることにより行えばよい。例えば、エレクトロポレーション法は以下のように行うことができる。外来遺伝子が挿入されたプラスミドをコンピテント細胞の懸濁液に加え、この懸濁液をエレクトロポレーション法専用のキュベットに入れ、そのキュベットに高電圧の電気パルスをかける。その後選択培地で培養し、平板寒天培地上で形質転換体を単離する。
【0050】
酵母細胞の例としては、サッカロマイセス(Saccharomyces)またはシゾサッカロマイセス(Schizosaccharomyces)に属する細胞が挙げられ、例えば、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)またはサッカロマイセス・クルイベリ(Saccharomyces kluyveri)等が挙げられる。酵母宿主への組み換えベクターの導入方法としては、例えば、エレクトロポレーション法、スフェロプラスト法、酢酸リチウム法等を挙げられる。真菌細胞の例としては、糸状菌、例えばアスペルギルス、ニューロスポラ、フザリウム、またはトリコデルマに属する細胞が挙げられる。宿主細胞として糸状菌を用いる場合、DNA構築物を宿主染色体に組み込んで組換え宿主細胞を得ることにより形質転換を行うことができる。DNA構築物の宿主染色体への組み込みは、公知の方法に従い、例えば相同組換えまたは異種組換えにより行うことができる。特に、製造効率の点から、本発明においては形質転換の宿主として、シュードノカルディア(Pseudonocardia)属細菌、大腸菌、ロードコッカス(Rhodococcus)属細菌を用いることが好ましい。
【0051】
上記の形質転換体は、導入された遺伝子の発現を可能にする条件下で適切な栄養培地中で培養する。形質転換体の培養物から、本発明のポリペプチドを単離精製するには、通常のポリペプチドの単離、精製法を用いればよい。
【0052】
例えば、本発明のポリペプチドが、細胞内に溶解状態で発現した場合には、培養終了後、細胞を遠心分離により回収し水系緩衝液に懸濁後、超音波破砕機等により細胞を破砕し、無細胞抽出液を得る。該無細胞抽出液を遠心分離することにより得られた上清から、通常のポリペプチドの単離精製法、即ち、溶媒抽出法、硫安等による塩析法、脱塩法、有機溶媒による沈殿法、ジエチルアミノエチル(DEAE)セファロース等のレジンを用いた陰イオン交換クロマトグラフィー法、SP-Sepharose FF(アマシャムバイオサイエンス社製)等のレジンを用いた陽イオン交換クロマトグラフィー法、ブチルセファロース、フェニルセファロース等のレジンを用いた疎水性クロマトグラフィー法、分子ふるいを用いたゲルろ過法、アフィニティークロマトグラフィ一法、クロマトフォーカシング法、等電点電気泳動等の電気泳動法等の手法を単独あるいは組み合わせて用い、精製標品を得ることができる。
【0053】
[水酸化ビタミンD誘導体の製造方法]
本発明は、ビタミンD類の水酸化酵素機能が向上した改変型ビタミンD類水酸化酵素をコードするDNAを導入した形質転換体の存在下でビタミンD類を水酸化させることを含む、水酸化ビタミンD誘導体の製造方法を包含する。具体的には、本発明に係る製造方法は、改変型ビタミンD類水酸化酵素をコードするDNAを含む組換えベクターによって形質転換された形質転換体を培養し、培養中または培養後において、当該形質転換体とビタミンD類とを接触させてビタミンD類を水酸化させ、産出された水酸化ビタミンD誘導体を採取することを特徴とする水酸化ビタミンD誘導体の製造方法である。
【0054】
このような形質転換体の存在下でビタミンD類を水酸化させるのに好適な条件は、以下のとおりである。
まず、形質転換体中のビタミンD水酸化酵素関連DNA(ビタミンD類の水酸化酵素をコードするDNA、フェレドキシンをコードするDNA及びフェレドキシン還元酵素をコードするDNA)を必要に応じて誘導物質を添加して菌体内で発現させる。これらのDNAが発現した菌体を添加したビタミンD類と接触させ、変換反応をさせる。変換反応の温度は、形質転換体の至適温度を考慮して、適宜決定できる。また、反応時間も水酸化ビタミンD誘導体への変換率(反応の進行度合い)等を考慮して、適宜決定することができる。例えば、20〜31℃で、1〜5日が好適である。さらに、反応様式は、バッチ式でも連続式でも、いずれの形式でも実施することができる。
【0055】
生成した水酸化ビタミンD誘導体の単離及び精製は、一般に微生物代謝産物をその培養液から単離するために用いられる分離、精製の方法が利用できる。例えば、メタノール、エタノール、アセトン、ブタノール、酢酸エチル、酢酸ブチル、クロロホルム、トルエン等を用いた有機溶媒抽出、ダイヤイオンHP-20などの疎水性吸着樹脂を用いた吸脱着処理、セファデックスLH-20等を用いたゲルろ過クロマトグラフィー、活性炭、シリカゲル等による吸着クロマトグラフィー、もしくは薄層クロマトグラフィーによる吸脱着処理、あるいは逆相カラム等を用いた高速液体クロマトグラフィー等の公知のあらゆる方法がこれにあたる。また、ここに示した方法に特に限定されるものではない。これらの方法を単独あるいは任意の順序に組み合わせ、また反復して用いることにより、目的の水酸化ビタミンD誘導体を単離精製することができる。
【実施例】
【0056】
以下、本発明について具体例を挙げてより詳細に説明するが、本発明をこれらの例に限定することを意図するものではない。なお、下記の例中のパーセント(%)は、培地の説明においては質量パーセント、HPLCの移動相の説明においては、容量パーセントを示す。
【0057】
実施例1:高機能化したVdh-K1の改変体の取得
(1)88番目の残基のアミノ酸総置換ライブラリーの作製
88、171、177または240番目のアミノ酸残基のうち、いずれか1箇所のアミノ酸が総置換されたvdh-K1遺伝子(配列番号1)を増幅するためプライマーを設計した。88番目の残基のアミノ酸総置換にはプライマーI88sat-F(配列番号11)とI88sat-R(配列番号12)を設計した。2組のプライマー、(1):Vdh-1F(配列番号5)とI88sat-R及び(2):Vdh-1R(配列番号6)とI88sat-Fを用いて、pET-aciBC-SD-VdhK1(非特許文献6)を鋳型とし、PCR反応を行って、それぞれ約0.25 kb及び約0.95 kbの2つのDNA断片を得た。これらのDNA断片を反応液からWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収精製した。続いて、完全長の遺伝子を増幅するためにプライマーVdh-1FとVdh-1Rを用い、上記で調製した2つのDNA断片の等モル混合物をテンプレートとしてPCR反応を行い、1.2 kbpのDNA断片を得た。このDNA断片を、反応液からWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収精製した後、制限酵素NdeIとBglIIにより37℃で15時間消化し、Wizard SV Gel and PCR Clean-Up System(プロメガ社)で精製した後、DNA Ligation kit ver 2.1(タカラバイオ社)を用いてプラスミドpETaciBC(特許文献3)のNde I-Bam HI部位に連結し、大腸菌DH5α株(タカラバイオ社)を形質転換した。カルベニシリン(50μg/ml)を含むLB寒天培地に出現した大腸菌形質転換株約1000コロニーに2 mlのLB培地を加えて、寒天培地上でスプレッダーを用いて菌体を懸濁した。この菌体懸濁液からWizard Plus SV Minipreps DNA Purification system(プロメガ社)を用いてプラスミドを抽出した。このプラスミドには88番目の残基が任意のアミノ酸に置換されたvdh-K1遺伝子が挿入されていると考えられ、Vdh-K1の88番目のアミノ酸残基の総置換ライブラリー(pETaciBC-SD-Vdh-88sat)として用いた。
【0058】
(2)171、177または240番目の残基のアミノ酸総置換ライブラリーの作製
前述(1)に記載した方法と同様にして、171及び177番目の残基のアミノ酸総置換ライブラリーを調製した。この時、171番目の残基のアミノ酸総置換にはプライマーL171sat-F(配列番号13)とL171sat-R(配列番号14)、177番目の残基のアミノ酸総置換にはプライマーA177sat-F(配列番号15)とA177sat-R(配列番号16)、240番目の残基のアミノ酸総置換にはプライマーT240sat-F(配列番号17)とT240sat-R(配列番号18)を用いた。この結果、作製したライブラリーをそれぞれVdh-K1の171番目のアミノ酸残基の総置換ライブラリー(pETaciBC-SD-Vdh-171sat)、Vdh-K1の177番目のアミノ酸残基の総置換ライブラリー(pETaciBC-SD-Vdh-177sat)、Vdh-K1の240番目のアミノ酸残基の総置換ライブラリー(pETaciBC-SD-Vdh-240sat)と名付けた。
【0059】
(3)88番目のアミノ酸残基の総置換ライブラリーを用いた副反応抑制クローンのスクリーニング
作製したVdh-K1の88番目のアミノ酸残基総置換ライブラリー(pETaciBC-SD-Vdh-88sat)で大腸菌BL21(DE3)を形質転換して大腸菌BL21(DE3)/pETaciBC-SD-Vdh-88sat株のコロニーを多数得た。96穴ディープウエルプレート(IWAKI社)2枚の各ウエルにカルベニシリン(50μg/ml)を含む1 mlのTB培地を入れ、大腸菌BL21(DE3)/pETaciBC-SD-Vdh-88satを1穴あたり1コロニー植菌した。コントロールとして、各プレートにpETaciBC-SD-VdhK1を8株植菌した。プレート用シェーカー(SCF-98-8:いわしや社)にプレートをセットし、30℃、800 rpmで16時間培養した。レプリケーターを用いて、96穴ディープウエルプレート(IWAKI社)2枚の各ウエル中にカルベニシリン(50μg/ml)とOvernight autoinduction system 1(メルク社)を含む1 mlのTB培地へ植菌した。前培養の96穴ディープウエルプレートはそのまま−80℃で凍結保存した。プレート用シェーカー(SCF-98-8:いわしや社)にプレートをセットし25℃、800 rpmで24時間培養した。3500 rpm、10分間の遠心により菌体を沈殿させ、上清を取り除いた。500μlのCV2反応液(1.0% (w/v) partially methylated β-cyclodextrin, 500 mg/L ビタミンD3, 5.0% (v/v) DMSO)を加え、プレート用シェーカー(SCF-98-8: いわしや社)にプレートをセットし28℃、800 rpmで24時間変換反応をした。各500μlのメタノールを添加し、反応を止め、800 rpmで5分間撹拌した。4000 rpm、10分間の遠心により菌体を沈殿させ、上澄みをHPLC分析用サンプルとして保存した。他のアミノ酸総置換ライブラリーも同様にHPLC分析用サンプルを調製した。HPLCの分析条件を以下に示す。
【0060】
分析装置:Agilent 100 series、
カラム:J' sphere ODS-H80(YMC, Inc.), 75 mm×4.6 mmI.D.、
移動相:A;イオン交換水、
B;アセトニトリル、
グラジエント時間設定:0分 移動相A/B =20:80、
4.3分 移動相A/B =20:80、
4.5分 移動相A/B =0:100、
9.5分 移動相A/B =0:100、
9.7分 移動相A/B =20:80、
10.0分 移動相A/B =20:80、
流速:1.5 ml/分、
検出:UV 265 nm、
インジェクション容量:10 μl、
カラム温度:40℃、
分析時間:10分、
保持時間:25−ヒドロキシビタミンD3 3.2分、
26−ヒドロキシビタミンD3 3.6分、
ビタミンD3 9.2分。
【0061】
上記の条件で変換サンプルを分析し、25−ヒドロキシビタミンD3が生成していることを確認し、副反応物比率が減少しているクローンを選抜した。副反応物比率は以下の式で求めた。
【数1】

【0062】
上記に示したHPLC分析条件により26−ヒドロキシビタミンD3生成比がvdh-K1発現株より減少した4クローンについてアンピシリン(50μg/ml)を含む2 ml LB液体培地で培養し、増殖した形質転換大腸菌からWizard Plus SV Minipreps DNA Purification system(プロメガ社)を用いてプラスミドDNAを精製した。得られたプラスミドを鋳型として改変遺伝子の塩基配列を決定した。塩基配列の解析には、DNA塩基配列解析装置(アプライドバイオシシテム;3130)を用い、配列決定用プライマー(配列番号7,8,9,10)を使用してダイターミネーター・サイクル・シークエンス法で解析した。
【0063】
アミノ酸総置換ライブラリーをスクリーニングした結果、Vdh-K1の88番目の残基のイソロイシンがバリンに置換された改変体を発現する大腸菌形質転換株が副反応を抑制すると示唆された。さらにVdh-K1の171、177及び240番目の残基のアミノ酸総置換ライブラリーのスクリーニングをそれぞれ行った結果、171番目の残基のロイシンがバリンに、177番目のアラニンがアルギニンにまたは240番目のトレオニンがセリンに置換された改変体を発現する大腸菌形質転換体が副反応を抑制することが示唆された。
【0064】
【表1】
【0065】
実施例2:高機能化したVdh-K1改変体の比活性測定
変異の導入により大腸菌での発現量が変化することが考えられたため、単位酵素あたりのビタミンD3水酸化活性を測定した。上記の基質認識に関わるアミノ酸のアミノ酸総置換ライブラリーをスクリーニングした結果、88番目の残基のイソロイシンがバリンに置換したアミノ酸置換体が副反応抑制に寄与した。これらの改変Vdh-K1を発現させるプラスミド(pETaciBC-SD-Vdh-K1 I88V)を導入し、大腸菌BL21(DE3)を形質転換した。得られた大腸菌BL21(DE3)形質転換株を50μg/mlのカルベニシリンを含む2 mlのTB培地に植菌した。30℃、220 rpmで14時間培養後、カルベニシリンとOvernight autoinduction system 1(メルク社)を含む25 mlのTB培地へ250μlの前培養液を接種した。25℃、220 rpmで24時間培養後、2 mlの培養液を試験管にとり、3500 rpm、10分間の遠心により菌体を沈殿させて上澄みを捨て、1 mlのCV2反応液を加えた。28℃、220 rpmで6時間変換反応を行い1.5 mlの酢酸エチルを添加して反応を止めた。30秒間ボルテックスで撹拌し、3500 rpm、10分間の遠心により水相と酢酸エチル相に分離させた。酢酸エチル相をとり、残った水相に0.75 mlの酢酸エチルを加えた。30秒間ボルテックスで撹拌し、3500 rpm、10分間の遠心により水相と酢酸エチル相に分離させた。酢酸エチル相をとり、先程得た酢酸エチル相と合わせて、減圧濃縮機により乾固した。乾固物に200μlのメタノールを添加し、溶解後、HPLC分析用バイアルへ移して、分析まで遮光4℃で保存した。
【0066】
変換試験に使用しなかった培養液から4300 rpm、10分間の遠心により菌体を沈殿として得た。上澄みを捨て、培養液に対して5分の1量のCV緩衝液(50 mMリン酸カリウム緩衝液pH 7.4, 10% (v/v)グリセロール)を加え、5倍濃縮細胞懸濁液を調製した。2 mlの5倍濃縮細胞懸濁液に68μl 10X Bug Buster(ノバジェン社)、2μl Benzonase(ノバジェン社)、200μl 40 mg/ml Lysozymeを加え、30℃で20分間緩やかに振とうした。7000 rpm、10分間の遠心により上清に無細胞抽出液を得た。この無細胞抽出液をキャップ付き試験管2本に750μlずつ分け、一方の無細胞抽出液に一酸化炭素を通した。次に、両方の無細胞抽出液にハイドロサルファイトナトリウムを少量(1-2 mg)添加した。一酸化炭素を通していないサンプルの400 nmから500 nmの吸収スペクトルをベースラインとして、スペクトルフォトメーター(HITACHI社、U-3310 SpectrophotoMeter)で一酸化炭素を通したサンプルの400 nmから500 nmの吸収をスキャンした。その結果、シトクロムP450に特徴的な450 nmのピークを観察し、その吸収から一酸化炭素結合型で還元状態Vdhの分子吸光係数を1 mMあたり91として培養液中のVdh改変体の発現量を計算した。
【0067】
前記変換試験サンプルのHPLC解析による25−ヒドロキシビタミンD3の定量結果と一酸化炭素結合スペクトル解析より得られたVdhまたはその改変体の発現量から、1 nmolのVdhまたはその改変体が1時間あたりに産生する25−ヒドロキシビタミンD3量を算出した。この値を比較することによりVdh-K1の88番目の残基のイソロイシンがバリンに置換したアミノ酸置換体(Vdh-K1I88V)における比活性の評価を行った。表2と図1にVdh-K1I88Vの比活性測定の結果を示した。Vdh発現量」はVdhまたはその改変体を発現させた大腸菌形質転換株の培養液中の濃度(nM)で示し、25-ヒドロキシビタミンD3生成量は休止菌体反応を6時間行った1 mlの反応液中の25-ヒドロキシビタミンD3濃度(mg/L)で示した。比活性として、1 nmolのVdhまたはその改変体が1時間で生成する25−ヒドロキシビタミンD3量(μg/hr/nmol of Vdh)で示し、コントロール(Vdh-K1)に対する比活性の相対活性を示した。Vdh-K1I88Vの比活性は、Vdh-K1の比活性の1.8倍を示し、単位酵素あたりのVD3水酸化活性は増加した。さらに、副反応比率は2.1%まで顕著に減少した。大腸菌において、Vdh-K1I88Vは、ビタミンD3変換時の副反応抑制に寄与した。同様にVdh-K1の171番目の残基のロイシンがバリンに置換されたアミノ酸置換体(Vdh-K1L171V)、177番目の残基のアラニンがアルギニンに置換されたアミノ酸置換体(Vdh-K1A177R)及び240番目の残基トレオニンがセリンに置換されたアミノ酸置換体(Vdh-K1T240S)における比活性の評価を行った。Vdh-K1L171VはVdh-K1よりも比活性が高いのに加え、副反応比率は3.6%に低減した。Vdh-K1A177RはVdh-K1の約3.5倍の比活性を示した。Vdh-K1A177Rの副反応比率はVdh-K1と同等であった。Vdh-K1T240SはVdh-K1よりも副反応比率は低減したが、比活性がVdh-K1の約60%に低下した。
【0068】
実施例3:有効変異の二重化によるVdh-K1改変体の調製と比活性測定
上記に見出した有効変異の重複効果を調べるため、Vdh-K1I88Vに171番目の残基のロイシンがバリンに置換した変異を導入するためのプライマーL171V-F(配列番号19)とL171V-R(配列番号20)を設計した。pET-aciBC-SD-VdhK1 I88Vを鋳型とし、2組のプライマー、(i):Vdh-1FとL171V-R及び(ii):Vdh-1RとL171V-Fを用いてPCR反応を行って、それぞれ約0.51kb及び約0.69 kbの2つのDNA断片を得た。これらのDNA断片をWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収した。続いて、完全長の遺伝子を増幅するためにプライマーVdh-1FとVdh-1Rを用い、上記で調製した2つのDNA断片の等モル混合物をテンプレートとしてPCR反応を行い、1.2 kbpのDNA断片を得た。このDNA断片を、反応液からWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収精製した後、制限酵素NdeIとBglIIにより37℃で15時間消化し、Wizard SV Gel and PCR Clean-Up System(プロメガ社)で精製した後、DNA Ligation kit ver 2.1(タカラバイオ社)を用いてプラスミドpETaciBCのNde I-Bam HI部位に連結し、大腸菌DH5α株を形質転換した。その後、カルベニシリン(50μg/ml)を含むLB寒天培地を用いて、形質転換された大腸菌を選択した。形質転換株からコロニーを分離し、カルベニシリン(50μg/ml)を含む2 mlのLB培地で増殖させた。増殖菌体からWizard Plus SV Minipreps DNA Purification system(プロメガ社)を用いてプラスミドpET-aciBC-SD-VdhK1 I88V L171Vを調製した。
【0069】
Vdh-K1I88Vに177番目の残基のアラニンがアルギニンに置換した変異を導入するためA177R-F(配列番号21)とA177R-R(配列番号22)を設計した。pET-aciBC-SD-VdhK1 I88Vを鋳型とし、2組のプライマー、(i):Vdh-1FとA177R-R及び(ii): Vdh-1RとA177R-Fを用いてPCR反応を行ってそれぞれ約0.53 kb及び約0.67 kbの2つのDNA断片を得た。これらのDNA断片をWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収した。続いて、完全長の遺伝子を増幅するためにプライマーVdh-1FとVdh-1Rを用い、上記で調製した2つのDNA断片の等モル混合物をテンプレートとしてPCR反応を行い、1.2 kbpのDNA断片を得た。このDNA断片を、反応液からWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収精製した後、制限酵素NdeIとBglIIにより37℃で15時間消化し、Wizard SV Gel and PCR Clean-Up System(プロメガ社)で精製した後、DNA Ligation kit ver 2.1(タカラバイオ社)を用いてプラスミドpETaciBCのNde I-Bam HI部位に連結し、大腸菌DH5α株を形質転換した。その後、カルベニシリン(50μg/ml)を含むLB寒天培地を用いて、形質転換された大腸菌を選択した。形質転換株からコロニーを分離し、カルベニシリン(50μg/ml)を含む2 mlのLB培地で増殖させた。増殖菌体からWizard Plus SV Minipreps DNA Purification system(プロメガ社)を用いてプラスミドpET-aciBC-SD-vdhK1 I88V A177Rを調製した。
【0070】
前述の方法で調製したプラスミド(pET-aciBC-SD-VdhK1 I88V L171V及びpET-aciBC-SD-VdhK1 I88V A177R)で大腸菌BL21(DE3)を形質転換してそれぞれBL21(DE3)/ pETaciBC-SD-VdhK1 I88V L171V及びBL21(DE3)/ pETaciBC-SD-VdhK1 I88V A177R株を得た。これらアミノ酸置換体の比活性及び副反応比率を親株と比較した結果、BL21(DE3)/ pETaciBC-SD-VdhK1 I88V A177R株は重複効果を示さなかったが、pETaciBC-SD-VdhK1 I88V L171Vは変異を重複することにより、単独変異の副反応比率よりもさらに低減することができた(表2及び図1)。このように大腸菌において、Vdh-K1の88番目の残基のイソロイシンがバリンに置換され、かつ171番目の残基のロイシンがバリンに置換されたアミノ酸置換体はVdh-K1の26位水酸化活性を低減するのに有効な変異であることが明らかとなった。
【0071】
【表2】
【0072】
実施例4:Vdh改変体の調製と比活性測定
(1)88番目の残基をバリンへ置換したVdhをコードする遺伝子の調製
上記に見出した副反応を抑制する有効変異の野生型Vdhに対する効果を調べるため、プライマーI88V-F(配列番号24)とI88V-R(配列番号25)を合成した。pET-aciBC-SD-vdh(非特許文献6)を鋳型として、2組のプライマー、(i):Vdh-1FとI88V-R 及び(ii):Vdh-1RとI88V-Fを用いてPCR反応を行ってそれぞれ約0.25 kb及び約0.95 kbの2つのDNA断片を得た。これらのDNA断片をPCR反応液からWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収精製した。続いて、完全長の遺伝子を増幅するためにプライマーVdh-1FとVdh-1Rを用い、上記で調製した2つのDNA断片の等モル混合物をテンプレートとしてPCR反応を行い、1.2 kbpのDNA断片を得た。このDNA断片を、反応液からWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収精製した後、制限酵素NdeIとBglIIにより37℃で15時間消化し、Wizard SV Gel and PCR Clean-Up System(プロメガ社)で精製した後、DNA Ligation kit ver 2.1(タカラバイオ社)を用いてプラスミドpETaciBC(非特許文献6)のNde I-Bam HI部位に連結し、大腸菌DH5α株を形質転換した。その後、カルベニシリン(50μg/ml)を含むLB寒天培地を用いて、形質転換された大腸菌を選択した。形質転換株からコロニーを分離し、カルベニシリン(50μg/ml)を含む2 mlのLB培地で増殖させた。増殖菌体からWizard Plus SV Minipreps DNA Purification system(プロメガ社)を用いてプラスミドpET-aciBC-SD-vdhI88Vを調製した。
【0073】
(2)88番目及び171番目の残基をバリンへ置換したVdhをコードする遺伝子の調製
前記(1)で調製した遺伝子にさらに171残基目のアミノ酸置換の変異を導入するため、pET-aciBC-SD-vdh I88Vを鋳型とし、2組のプライマー、(i):Vdh-1FとL171V-R 及び(ii):Vdh-1RとL171V-Fを用いてPCR反応を行ってそれぞれ約0.51 kb及び約0.69 kbの2つのDNA断片を得た。これらのDNA断片をWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収した。続いて、完全長の遺伝子を増幅するためにプライマーVdh-1FとVdh-1Rを用い、上記で調製した2つのDNA断片の等モル混合物をテンプレートとしてPCR反応を行い、1.2 kbpのDNA断片を得た。このDNA断片を、反応液からWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収精製した後、制限酵素NdeIとBglIIにより37℃で15時間消化し、Wizard SV Gel and PCR Clean-Up System(プロメガ社)で精製した後、DNA Ligation kit ver 2.1(タカラバイオ社)を用いてプラスミドpETaciBCのNde I-Bam HI部位に連結し、大腸菌DH5α株を形質転換した。その後、カルベニシリン(50μg/ml)を含むLB寒天培地を用いて、形質転換された大腸菌を選択した。形質転換株からコロニーを分離し、カルベニシリン(50μg/ml)を含む2 mlのLB培地で増殖させた。増殖菌体からWizard Plus SV Minipreps DNA Purification system(プロメガ社)を用いてプラスミドpET-aciBC-SD-vdhI88V L171Vプラスミドを調製した。
【0074】
(3)Vdh改変体の比活性測定
pET-aciBC-SD-vdh(非特許文献6)、pET-aciBC-SD-vdhI88V及びpET-aciBC-SD-vdh I88V L171Vプラスミドでそれぞれ大腸菌BL21(DE3)を形質転換して形質転換株を得た。アミノ酸置換体の比活性及び副反応比率を親株と比較した結果、野生型Vdhの88残基目のイソロイシンがバリンに置換されたアミノ酸置換体は野生型Vdhの25位水酸化活性を維持しながら、25位水酸化活性に対する26位水酸化活性比を2.5%に低減した。さらに、野生型Vdhの88残基目のイソロイシンがバリンに置換され、かつ171残基目のロイシンがバリンに置換されたアミノ酸置換体はVdh I88Vよりも酵素あたりの25位水酸化活性が高いのに加えて、25位水酸化活性に対する26位水酸化活性比は1.5%まで低減した(図2)。
【0075】
実施例5:シュードノカルディア・オートトロフィカにおけるvdh遺伝子の破壊
(1)vdh遺伝子破壊用プラスミドpTYM18-2の作製
プラスミドpTYM18(尾仲らJ. Anibiot. 56, 950-956, 2003)を制限酵素Eco RI及びBgl IIで消化して得られる約3.3 kbのDNA断片を精製し、その末端をDNA Blunting Kit(タカラバイオ)で平滑化した後、DNA Ligation Kit(タカラバイオ)で自己環状化した。この反応液で大腸菌Escherichia coli DH5αを形質転換して得られたクローンからプラスミドDNAを調製してpTYM18-2を得た。
【0076】
(2)シュードノカルディア・オートトロフィカ vdh遺伝子破壊株の分離
特許文献3記載のvdh遺伝子を担持するプラスミドpVdh-camABを鋳型にして以下2種のプライマー、
(i) Vdh236Spe-F 配列番号26:5'-GCACTAGTGCCACACGCTGCCCGAGGACCAG-3'
(ii) Vdh1039Bam-R 配列番号27:5'-GCGGATCCGTGCCCGAAGAACACCCCGCC-3'
により約820bからなる野生型vdh遺伝子の一部断片を増幅した。このDNAを制限酵素Spe I及びBam HIで消化し、精製した後、pTYM18-2のXba I-Bam HI制限酵素部位へクローニングしてプラスミドpTYM-VDHを構築した。このプラスミドを接合用大腸菌Escherichia coli S17-1株に形質転換し、カナマイシン25μg/mlを含む2 mlのLB培地に植菌して30℃で20時間培養した。この培養液200μlをLB培地10 mlに植菌して30℃で3時間培養した。この培養液をLB培地で2回洗浄した後、LB培地3 mlに懸濁した。一方で、TSB培地にて28℃で3日間増殖させたシュードノカルディア・オートトロフィカの培養液500μlをTSB培地10 mlに植菌して28℃で20時間培養した。この増殖菌体を滅菌水で2回洗浄した後、滅菌水500μlに懸濁した。この懸濁液30μlに対し前述の大腸菌S17-1株(Escherichia coli ATCC47055)の懸濁液500μlを混合し、ISP4平板培地(寒天濃度1.5%)へ塗布した。この培地を30℃で20時間培養した。リボスタマイシン200μg/ml及びナリジクス酸25μg/mlを含む2 mlのSNA(ソフトニュートリエントアガー)を平板上に重層した。この培地を30℃で2週間培養し生育してきた菌株をvdh遺伝子の破壊株として分離した。
【0077】
実施例6:Vdh-K1 I88Vアミノ酸置換体遺伝子のシュードノカルディア・オートトロフィカでの発現
Vdh-K1 I88Vアミノ酸置換体遺伝子を増幅するため、Vdh-2R(配列番号23)を合成した。プライマーVdh-1FとVdh-2Rを用い、pETaciBC-SD-Vdh-K1 I88Vを鋳型にしてPCR反応を行った。その結果増幅した約1.2 kbpのDNA断片を反応液からWizard SV Gel and PCR Clean-Up System プロメガ社)によって回収精製した後、制限酵素Nde IとNhe Iにより37℃で10時間消化しWizard SV Gel and PCR Clean-Up System(プロメガ社)で精製した。予め制限酵素NdeI及びNheIで消化して、vdh遺伝子断片を切り離したpTAOR3-vdh(シュードノカルディア・オートトロフィカ発現ベクター、特願2008-259317特許、平成20年10月6日出願)のベクターDNA断片に、このDNA断片を、DNA Ligation kit ver 2.1(タカラバイオ社)を用いて連結し、大腸菌DH5α株(タカラバイオ社)を形質転換した。その後、カルベニシリン(50μg/ml)を含むLB培地を用いて、形質転換された大腸菌を増殖させた。増殖菌体からWizard Plus SV Minipreps DNA Purification system(プロメガ社)を用いてプラスミドを抽出した。このプラスミドをpTAOR-K1I88Vと名付けた。Vdh-K1 アミノ酸置換体についても上記と同様に構築し、pTAOR-K1と名付けた。得られたプラスミドを鋳型としてvdh-K1の改変体の塩基配列の決定をDNA塩基配列解析装置(アプライドバイオシシテム;3130)で用いて行った。配列決定用プライマーを使用してダイターミネーター・サイクル・シークエンス法で解析し、正しいDNA配列が含まれていることを確認した。
【0078】
pTAOR-K1I88VまたはpTAOR-K1プラスミドで大腸菌S17-1株(Escherichia coli ATCC47055)を形質転換し、アプラマイシン(60μg/ml)を含むLB寒天培地で選抜し、pTAOR-K1 I88V/S17-1またはpTAOR-K1/S17-1株を得た。pTAOR-K1 I88VまたはpTAOR-K1プラスミドをカナマイシン遺伝子で前記vdh遺伝子破壊シュードノカルディア・オートトロフィカ株に接合伝達により導入した。導入方法を以下に示す。
【0079】
シュードノカルディア・オートトロフィカの凍結菌体液をカナマイシン(200μg/ml)を含むLB培地を用いて、30℃、3日間培養した。培養液500μlを遠心分離(7000 rpm, 30秒間)し、培養菌体を集菌した。菌体を500μlのLB培地に懸濁し、この培養液をシュードノカルディア・オートトロフィカ細胞懸濁液とした。さらに、大腸菌pTAOR-K1 I88V/S17-1株をアプラマイシン(60μg/ml)を含むLB培地を用いて、30℃、24 h培養した。培養液200μlを遠心分離(7000 rpm, 30秒間)し、培養菌体を集菌した。菌体を200μlのLB培地に懸濁し、この培養液を大腸菌細胞懸濁液とした。シュードノカルディア・オートトロフィカ及び大腸菌細胞懸濁液を混和後、遠心分離(7000 rpm, 30秒間)した。上清を500μl除き、残り200μlのLB培地で菌体を懸濁後、薬剤を含まないLB寒天培地に塗布し、30℃、24 h培養した。寒天プレートに2 mlのLB液体培地を加え、寒天培地に生育した菌体を懸濁後、マイクロチューブに懸濁液を移した。懸濁液をカナマイシン(200μg/ml)、アプラマイシン(24μg/ml)及びナリジクス酸(50μg/ml)を含むLB寒天培地にスプレッドし、30℃、一週間培養した。生育したコロニーを再度カナマイシン(200μg/ml)、アプラマイシン(24μg/ml)及びナリジクス酸(50μg/ml)を含むLB寒天培地に画線し耐性を確認した。得られた菌株をpTAOR-K1 I88VまたはpTAOR-K1株とした。
【0080】
得られたシュードノカルディア・オートトロフィカ形質転換株、シュードノカルディア・オートトロフィカ vdh破壊株及びvdh遺伝子が破壊されていないシュードノカルディア・オートトロフィカ親株をビタミンD3変換試験に供した。培地組成は以下の通りである。
(前培養培地組成)
グルコース 1.5%
酵母エキス 0.3%
NaCl 0.4%
CaCO3 0.2%
ポリペプトン 1.5%
pH7.4
(本培養培地組成)
グルコース 2.0%
酵母エキス 0.5%
NaCl 0.04%
CaCO3 0.3%
ポリペプトン 1.0%
大豆ペプトン 1.0%
K2HPO4 0.04%
pH7.4
【0081】
試験株を100 mlの前培養液に植菌した。シュードノカルディア・オートトロフィカ形質転換株及びシュードノカルディア・オートトロフィカ vdh破壊株用培地にはカナマイシン(200μg/ml)を添加した。28℃、220 rpm、2日間培養後、100 mlの本培養培地へ1 mlの前培養液を接種した。28℃、220 rpm、24 h培養後、培養液に1 mlのアセトンを添加した。さらに24時間培養し、ビタミンD3溶液(終濃度2.0% (w/v) partially methylated β-cyclodextrin,1000μg/L ビタミンD3, 2.0% (v/v)エタノール)を添加した。以後、24 時間ごとに 1 mlサンプリングを行い、サンプリングと同時にビタミンD3溶液(終濃度2.0% (w/v) partially methylated β-cyclodextrin, 1000μg/L ビタミンD3, 2.0% (v/v)エタノール)を添加した。サンプリングした培養液はメタノールで10倍希釈し、遠心上清をHPLC分析用サンプルとした。HPLCの分析条件を以下に示す。
【0082】
分析装置:Agilent 100 series、
カラム:J' sphere ODS-H80 (YMC, Inc.), 75 mm×4.6 mmI.D.、
移動相:A; イオン交換水、
B; アセトニトリル、
グラジエント時間設定:0分 移動相A/B =20:80、
4.3分 移動相A/B =20:80、
4.5分 移動相A/B =0:100、
9.5分 移動相A/B =0:100、
9.7分 移動相A/B =20:80、
10.0分 移動相A/B =20:80、
流速:1.5 ml/分、
検出:UV 265 nm、
インジェクション容量:10 μl、
カラム温度:40℃、
分析時間:10分、
保持時間:25−ヒドロキシビタミンD3 3.2分、
26−ヒドロキシビタミンD3 3.6分、
ビタミンD3 9.2分。
【0083】
まず、上記の条件でサンプルを分析し、形質転換株培養液中に25−ヒドロキシビタミンD3及び26−ヒドロキシビタミンD3が蓄積していることを確認した。次に培養時間経過による25−ヒドロキシビタミンD3蓄積濃度及び副反応物比率(%)を求めた。結果を表3及び図3に示す。親株とアミノ酸置換体の形質転換株は25−ヒドロキシビタミンD3の最終蓄積量に大きな差が認められなかった。副反応物比率は親株が約10%程度であるのに対し、pTAOR-K1形質転換株は2.5%程度、pTAOR-K1 I88V形質転換株は1%以下に副反応物比率が低減した。HPLCクロマトチャート上でも10倍希釈で測定すると、26−ヒドロキシビタミンD3のピークは検出限界以下になった。結果を図4に示す。以上より、I88Vアミノ酸置換体はシュードノカルディア・オートトロフィカにおいてもVdhの副反応抑制に顕著な効果があった。
【0084】
【表3】
【技術分野】
【0001】
本発明は、ビタミンD類の水酸化に関与するポリペプチド(ビタミンD類水酸化酵素)をコードする塩基配列を改変することにより得られる、ビタミンD類の水酸化活性が増強され、あるいは副反応が抑制されたビタミンD類水酸化酵素、その酵素をコードするDNA、そのDNAを含むベクター、そのベクターを含む形質転換体、及びその形質転換体を用いた水酸化ビタミンD誘導体の製造方法に関する。
【背景技術】
【0002】
ビタミンDは、主に高等生物において必須とされる脂溶性ビタミン群でコレステロールから生合成される。その生理作用はカルシウムの吸収促進や代謝促進、細胞分化誘導、免疫調節作用など多岐にわたり、生体において重要な役割を果たしている。
【0003】
ヒトにおけるビタミンDの生合成の主要な経路は、まずコレステロールから7−デヒドロコレステロール(プロビタミンD3)が合成された後、皮膚において紫外線と熱反応からビタミンD3がつくられる。ビタミンD3は肝臓のミトコンドリアに局在するシトクロムP450(例えば、CYP27A1)によって25位の水酸化を受けて25−ヒドロキシビタミンD3となる。その後、腎臓の近位尿細管において別のシトクロムP450(例えば、CYP27B1)によって、さらに1α位の水酸化を受けて1α,25−ジヒドロキシビタミンD3(活性化型ビタミンD3)が作られる。この物質が細胞内の受容体(レセプター)に結合することによって、生理作用発現に関わる核内の特定の遺伝子の発現調節を行うことが分かっている。
【0004】
従って、肝臓または腎臓が機能不全に陥ると、正常なビタミンD代謝が阻害されることがある。そのような患者では血中の25-ヒドロキシビタミンD3や1α,25−ジヒドロキシビタミンD3が極度に低下することがあり、治療のためには25-ヒドロキシビタミンD3や1α,25−ジヒドロキシビタミンD3を投与して補う必要がある。
【0005】
一方、先天的に、ビタミンD3の水酸化に関与するシトクロムP450遺伝子に変異があることに起因する水酸化機能不全により1α,25−ジヒドロキシビタミンD3が不足して起こるくる病が知られている。のみならず、後天的なくる病や骨粗鬆症を含めて、1α,25−ジヒドロキシビタミンD3が不足することで起こる疾患の治療薬として、1α,25−ジヒドロキシビタミンD3及び同様な生理作用をもつ類縁化合物の重要性は非常に高い。
【0006】
さらに、ビタミンD類はその多様な生理活性から様々な誘導体が薬物の候補として研究され、抗腫瘍薬、乾癬治療薬、免疫賦活薬などの開発にも期待がもたれている。このような観点からビタミンD類を誘導体化する一手法としても水酸化は重要な反応である。この場合、水酸化の位置は1α位及び25位に限定されず、他の位置の水酸化修飾にも薬効を発現させるための高い必要性が生じることがある。このように医薬品開発の課題として水酸化ビタミンD誘導体の製造法を改良し、より効率的で安価に水酸化ビタミンD誘導体を製造して供給することが医薬産業から強く望まれてきた。
【0007】
ビタミンD類の中で特に生理活性が強く、価値の高い1α,25−ジヒドロキシビタミンD3について言及すると、その製造手法としては、有機合成法、ヒトのシトクロムP450を利用する方法、微生物を利用する方法等が挙げられる。このうち有機合成法ではコレステロールから約20工程を経て合成する製法が知られている(非特許文献1及び2参照)。またヒトのシトクロムP450を利用する方法では、関与する遺伝子、発現する技術、シトクロムP450酵素の性質に関する生物学的・生化学的知見の蓄積がある(非特許文献3参照)。しかし、これら2つのいずれの方法も高い製造コストや低い生産性の問題から、実用化には極めて不向きである。
【0008】
微生物を利用する方法としては放線菌シュードノカルディア・オートトロフィカ(Pseudonocardia autotrophica)を変換菌として用いたビタミンD3から1α,25−ジヒドロキシビタミンD3の製造方法が知られており、この方法は産業的に実用化されている(非特許文献4、非特許文献5、特許文献1及び特許文献2参照)。この方法は、シュードノカルディア(Pseudonocardia)属放線菌を発酵槽において培養した後、ビタミンD3を反応基質として培養液に添加し、菌体のもつ水酸化酵素の作用で反応を進行させて、その結果蓄積する1α,25−ジヒドロキシビタミンD3を培養液から回収・精製して行うものである。
【0009】
この方法においては、変換微生物として生育時間の長い放線菌を用いるため製造期間が長期になり、さらにこの変換菌がもつビタミンD3変換作用の中には目的の水酸化反応のみならず、望ましくない副反応も存在する。その副反応として例えば、26位あるいは24位の水酸化などが挙げられる。また基質であるビタミンD3や目的生成物である1α,25−ジヒドロキシビタミンD3に対する分解活性も存在する。特に1α,25−ジヒドロキシビタミンD3の分解は、目的物の蓄積量に直接的に関わる極めて重要な問題である。これに加えて、副産物の存在も変換反応後の培養液からの1α,25−ジヒドロキシビタミンD3の回収・精製工程を多くし、結果的に目的物の収量を低下させる。
【0010】
このような問題点を解決するため、変換菌である放線菌シュードノカルディア・オートトロフィカからビタミンD類の水酸化酵素(「Vdh」または人為的変異が導入されていないことを強調するため「野生型Vdh」とも言う。)をコードする遺伝子(「vdh」または「野生型vdh」と言う。)を取得し、これを生育の速い大腸菌に組込んで得られる形質転換体を用いてビタミンD類を水酸化し、水酸化ビタミンD誘導体を効率的に生産する方法が提案されている(特許文献3)。さらにその方法において、vdh遺伝子を改変して水酸化活性を高めた改変酵素をコードする遺伝子を利用する、より効果的な方法も提案されている(非特許文献6)。非特許文献6によると、その遺伝子(vdh-K1)がコードする改変酵素Vdh-K1は野生型Vdh酵素にアミノ酸配列レベルで以下の4種の変異を導入した酵素である。
【0011】
(1) 70番目のスレオニンがアルギニンに置き換わる変異、
(2) 156番目のバリンがロイシンに置き換わる変異、
(3) 216番目のグルタミン酸がメチオニンに置き換わる変異、
(4) 384番目のグルタミン酸がアルギニンに置き換わる変異。
【0012】
Vdh-K1はVD3に対する25位水酸化活性が野生型酵素の約20倍比活性が上昇したことが知られている。vdh-K1遺伝子を大腸菌に形質転換して得られた株を変換菌として用いることで比較的短期間で効率よくビタミンD類を水酸化することができるが、産業利用の観点から、反応特異性が不十分で副生物である26位水酸化体の産生を抑制できない問題点は残っており、さらに水酸化活性においてもより高い水準が求められている。したがって、副生物である26位水酸化体の産生を抑制させるとともにより一層の活性上昇が実現することでVdhまたはVdh-K1の高機能化を実現し、水酸化ビタミンD誘導体の効率的な製造方法を確立することが強く望まれていた。
【先行技術文献】
【特許文献】
【0013】
【特許文献1】特開平2-469号公報
【特許文献2】特開平2-231089号公報
【特許文献3】国際公開第2007/138894号パンフレット
【非特許文献】
【0014】
【非特許文献1】Kametani T., Furuyama H., Synthesis Med. Res. Rev. 7, 147-171 (1987)
【非特許文献2】Zunk G-D., Okamura W. H. Norman A. W., Chem. Rev. 95, 1877-1952 (1995)
【非特許文献3】T. Sakakiらの総説, Frontiers in Bioscience, 10, 119-134, 2005
【非特許文献4】J. Sasaki et. al, Applied Microbiology and Biotechnology, 38, 152-157, 1992
【非特許文献5】K. Takeda et. al, J. Ferment. Bioeng., 78, 380-382, 1994
【非特許文献6】Y. Fujii et. al, Biochem. Biophys. Res. Commun., 385, 170-175, 2009)
【発明の概要】
【発明が解決しようとする課題】
【0015】
本発明は、活性や副反応抑制においてビタミンD類の水酸化機能が向上したビタミンD類水酸化酵素、当該酵素をコードするDNA、該DNA含むベクター、及び該ベクターを含む形質転換体を提供することを課題とする。
さらに本発明は、Vdh及びVdhと構造的に関連したVdh-K1などの水酸化酵素を改変することでビタミンD類の副反応を抑制し、望ましくはさらに水酸化酵素の比活性を改善して水酸化ビタミンD誘導体を効率よく生産する方法を提供することにある。
【課題を解決するための手段】
【0016】
本発明者らは、前述の課題を解決するために、これまでに知見が得られていない副反応低減や活性上昇に関与する新規な変異酵素を取得することを目的として鋭意研究を実施した。まず、非特許文献6に開示されたシュードノカルディア・オートトロフィカNBRC12743株由来のvdh遺伝子を改変して得られたvdh-K1遺伝子に対して、その塩基配列中のアミノ酸コドンにランダムに変異を加え、DNA変異体のライブラリーを調製した。次いでそれらの変異体がコードする変異酵素タンパク質を大腸菌において発現させ、それらのクローンのビタミンD類の水酸化活性を測定することで、25位水酸化活性に対する26位水酸化活性の比率の低下した改変遺伝子を選択し、変異が加えられた部位(改変部位)を特定した。次にこれらの改変部位を組み合わせて、さらに機能が高められた改変遺伝子を調製した。その後、これらの改変遺伝子を含むベクターを大腸菌に組み込んで26位水酸化活性が抑制された水酸化酵素または25位水酸化活性が増強された水酸化酵素を発現させて酵素の機能を評価した結果、Vdh-K1に比較して水酸化ビタミンD誘導体を効率よく生産できる改変型ビタミンD類水酸化酵素を見出し、本発明を完成した。
【0017】
すなわち、本発明は以下の[1]〜[12]に関する。
[1] 配列番号2または4に記載のアミノ酸配列において、下記の(1)〜(4)からなる群より選択される少なくとも1つの改変部位を有する改変アミノ酸配列からなる改変型ビタミンD類水酸化酵素:
(1)88位のイソロイシンがバリンに置換された改変部位、
(2)171位のロイシンがバリンに置換された改変部位、
(3)177位のアラニンがアルギニンに置換された改変部位、
(4)240位のトレオニンがセリンに置換された改変部位。
[2] 前記[1]に記載の改変アミノ酸配列において、(1)〜(4)からなる群より選択される少なくとも1つの改変部位のほかに、さらに1または数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有する改変型ビタミンD類水酸化酵素。
[3] 前記[1]に記載の改変アミノ酸配列と90%以上の相同性を有し、(1)〜(4)からなる群より選択される少なくとも1つの改変部位を含むアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有する改変型ビタミンD類水酸化酵素。
[4] 配列番号2または4に記載のアミノ酸配列において、88位のイソロイシンがバリンに、及び171位のロイシンがバリンにそれぞれ置換された改変アミノ酸配列からなる前記[1]に記載の改変型ビタミンD類水酸化酵素。
[5] 前記[4]に記載の改変アミノ酸配列において、88位及び171位のアミノ酸置換のほかに、さらに1または数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有する改変型ビタミンD類水酸化酵素。
[6] 前記[4]に記載の改変アミノ酸配列と90%以上の相同性を有し、88位及び171位のアミノ酸置換を含むアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有する改変型ビタミンD類水酸化酵素。
[7] 前記[1]〜[6]のいずれか1項に記載の改変型ビタミンD類水酸化酵素をコードするDNA。
[8] 配列番号1または3に記載の塩基配列において、下記の(1)〜(4)からなる群より選択される少なくとも1つの改変部位を有する改変塩基配列からなる前記[1]に記載の改変型ビタミンD類水酸化酵素をコードするDNA:
(1)262〜264番目の塩基配列ATCが、バリンをコードするコドンに置換された改変部位、
(2)511〜513番目の塩基配列CTGが、バリンをコードするコドンに置換された改変部位、
(3)529〜531番目の塩基配列GCCが、アルギニンをコードするコドンに置換された改変部位、
(4)718〜720番目の塩基配列ACGが、セリンをコードするコドンに置換された改変部位。
[9] 配列番号1または3に記載の塩基配列において、262〜264番目の塩基配列ATCがバリンをコードするコドンに、及び511〜513番目の塩基配列CTGがバリンをコードするコドンにそれぞれ置換された改変塩基配列からなる前記[8]に記載のDNA。
[10] 前記[7]〜[9]のいずれか1項に記載のDNAを含む組換えベクター。
[11] 前記[10]に記載のベクターを含む形質転換体。
[12] 前記[11]に記載の形質転換体を培養し、培養中または培養後の形質転換体とビタミンD類とを接触させ、産出された水酸化ビタミンD誘導体を採取することを特徴とする水酸化ビタミンD誘導体の製造方法。
【発明の効果】
【0018】
本発明は、活性や副反応抑制においてビタミンD類の水酸化機能が向上したビタミンD類水酸化酵素及びそれをコードするDNA、そのDNAを形質転換した細胞、その細胞を用いた高効率な水酸化ビタミンD類の製造方法に関する。本発明によって、ビタミンD3の25位水酸化能が向上した酵素、ビタミンD3の26位水酸化能が抑制した酵素等、産業上有用な特徴的性質をもつビタミンD類の水酸化酵素及び該酵素をコードするDNAを取得できる。さらに、それらのDNAを担持するベクター、そのベクターで大腸菌やシュードノカルディア・オートトロフィカ等を形質転換した形質転換体を作製し、その形質転換体を用いて水酸化ビタミンD誘導体を効率よく製造することができる。従って、本発明により、産業上極めて有用で、かつ効率のよい水酸化ビタミンD誘導体の製造方法が提供される。
【図面の簡単な説明】
【0019】
【図1】Aは大腸菌におけるVdh-K1各改変体の発現量を示したグラフである。Bは大腸菌において発現させた改変体の比活性を示したグラフである。Cは改変体の25位水酸化ビタミンD3量に対する26位水酸化ビタミンD3量の比率を示したグラフである。
【図2】Aは野生型Vdh改変体の発現量を示したグラフである。Bは野生型Vdh改変体の比活性を示したグラフである。Cは野生型Vdh改変体の25位水酸化ビタミンD3量に対する26位水酸化ビタミンD3量の比率を示したグラフである。
【図3】Aはシュードノカルディア・オートトロフィカの培地中における25位水酸化ビタミンD3の蓄積量を示したグラフである。Bは培地中の25位水酸化ビタミンD3量に対する26位水酸化ビタミンD3量の比率を示したグラフである。
【図4】シュードノカルディア・オートトロフィカ親株及び形質転換株によるビタミンD3変換試験を行った時のHPLCのクロマトグラムを示した図である。
【発明を実施するための形態】
【0020】
以下に本願明細書において記載する用語、記号等の意義を説明し、本発明を詳細に説明する。
本願明細書において用いる「ビタミンD類」とは、本発明の水酸化酵素により基質として認識され、1α位、25位、その他の炭素に一原子酸素添加を行うことが可能なものを意味し、以下の化合物を包含する。
【0021】
(1)ビタミンD3、ビタミンD2などのビタミンD類、
(2)ビタミンD類の前駆体、代謝物、その他の誘導体(この中には、25−ヒドロキシビタミンD3、1α−ヒドロキシビタミンD3などが含まれる。)、
(3)コレスタン系のステロイド類またはその修飾化合物(この中には、コレステロール、7−デヒドロコレステロールなどが含まれる。)。
【0022】
本明細書において用いる「水酸化ビタミンD誘導体」とは、本発明の水酸化酵素によって、ビタミンD類の1α位、25位、その他の位置の炭素原子に1酸素原子が添加された化合物を意味する。
【0023】
また、本発明において「ビタミンD類の水酸化酵素活性」とは、ビタミンD類を水酸化し、水酸化ビタミンD誘導体へ変換する酵素活性を意味する。「ビタミンD類の水酸化酵素」とはVdhまたはVdh-K1などのVdhと構造的かつ機能的に相同性を有する酵素を意味する。Vdh及びVdh-K1はそれぞれ配列番号4及び2で表されるアミノ酸配列からなるポリペプチドである。vdh遺伝子及びvdh-K1遺伝子はそれぞれ配列番号3及び1で表されるポリヌクレオチドからなるDNAである。
【0024】
さらに、本発明において「改変型酵素」、「改変アミノ酸配列」、「DNA改変体」、「改変塩基配列」とは、構成アミノ酸または塩基の置換、削除、付加、挿入などにより修飾された酵素、アミノ酸配列、DNA、塩基配列を意味する。
【0025】
以下、本発明の実施の形態について詳細に説明する。
[本発明に係る酵素及びそれをコードするDNA]
本発明に係る改変型ビタミンD類水酸化酵素は、野生型のビタミンD類水酸化酵素Vdh及び改変酵素Vdh-K1の特定のアミノ酸配列を改変することによって得られたものであり、当該酵素を水酸化ビタミンD誘導体の製造に用いた場合、改変前の酵素と比較して、その副反応を有意に抑制し、所望の水酸化ビタミンD誘導体の製造効率を向上させることを可能とするものである。
本発明に係る酵素及びそれをコードするDNAは、以下の方法で取得された。
本発明者らは、非特許文献6に記載された情報に従って適宜設計されたプライマーを用いてPCRを行い、ビタミンD類の水酸化酵素活性を有するポリペプチドをコードするvdh-K1遺伝子(配列番号1)を含むDNA断片を取得した。次いで、フェレドキシンをコードするDNA、フェレドキシン還元酵素をコードするDNA及び大腸菌のリボゾーム結合部位として有効とされるDNA(SD配列)をすべて含むプラスミドにこの断片を連結させ、大腸菌においてビタミンD類の水酸化酵素活性を発現しうるプラスミドを作製した。得られたプラスミドを用いて、PCRにより配列番号1で示されるDNAにランダムな変異が入ったDNA断片を増幅し、ビタミンD水酸化酵素発現用プラスミドに連結させ、DNA改変体を含むプラスミドのライブラリーを作製した。次いでこれらライブラリーのプラスミドで形質転換した大腸菌がもつビタミンD類の水酸化酵素活性をスクリーニングすることにより、特徴的な水酸化活性をもつ複数のポリペプチド(改変型ビタミンD類水酸化酵素)及びそれらをコードするDNAを見出したものである。Vdh-K1遺伝子は403残基のポリペプチドからなるVdh-K1をコードしているが、開始コドン(メチオニン)を除く402残基のうちいずれか1つの残基が置換された改変体を作るには、各残基ごとに元のアミノ酸を除く19種類のアミノ酸に置換する。これをその残基の「アミノ酸総置換」と言う。この結果、調製可能な改変体は7638種に及ぶ。現在の技術レベルではこれらのすべてを調製し試験するには多大な労力と技術的な熟練が必要であり、さらに網羅的に試験した結果、必ず所望の改変体が得られる確証はない。本発明者らは高機能化したビタミンD類の水酸化酵素改変体を獲得するため、このような困難なスクリーニングを遂行し、本発明の改変型酵素構築の基本情報となる有効変異についての情報を得るに至った。
【0026】
こうして得られた本発明のビタミンD類水酸化酵素及び当該酵素をコードするDNAは、下記(a)のアミノ酸配列からなる改変型ビタミンD類水酸化酵素、及び下記(a)のアミノ酸配列からなる改変型ビタミンD類水酸化酵素をコードするDNAである。
(a)配列番号2または4に記載のアミノ酸配列において、以下の(1)〜(4)からなる群より選択される少なくとも1つの改変部位を有する改変アミノ酸配列:
(1)88位のイソロイシンがバリンに置換された改変部位、
(2)171位のロイシンがバリンに置換された改変部位、
(3)177位のアラニンがアルギニンに置換された改変部位、
(4)240位のトレオニンがセリンに置換された改変部位。
【0027】
前記(a)のアミノ酸配列からなる改変型ビタミンD類水酸化酵素をコードするDNAの中でも下記のDNAが好ましい。
配列番号1または3に記載の塩基配列において、以下の(1)〜(4)からなる群より選択される少なくとも1つの改変部位を有する改変塩基配列からなるDNA:
(1)262〜264番目の塩基配列ATCが、バリンをコードするコドンに置換された改変部位、
(2)511〜513番目の塩基配列CTGが、バリンをコードするコドンに置換された改変部位、
(3)529〜531番目の塩基配列GCCが、アルギニンをコードするコドンに置換された改変部位、
(4)718〜720番目の塩基配列ACGが、セリンをコードするコドンに置換された改変部位。
【0028】
ここで、バリンをコードするコドンは、具体的にはGTT、GTG、GTA、GTGであり、アルギニンをコードするコドンは、具体的にはCGT、CGC、CGA、CGG、AGA、AGGであり、セリンをコードするコドンは、具体的にはTCT、TCC、TCA、TCG、AGT、AGCである。
【0029】
これらのアミノ酸改変による優れた効果は、以下の(ア)及び(イ)にまとめることができる。
(ア)改変型ビタミンD類水酸化酵素自体のビタミンD類水酸化能が増強されている(水酸化能の比活性が高くなっている)。
(イ)改変型ビタミンD類水酸化酵素自体のビタミンD類26位水酸化能が抑制されている(25位に対する26位水酸化活性の比率が低くなっている)。
【0030】
なお、上記の各DNA改変体がもつ個々の特長は以下のとおりである。
1)「88位のイソロイシンがバリンに置換された改変部位を有する改変アミノ酸配列をコードするDNA」または「塩基262から塩基264にかけての配列ATCを、バリンをコードするコドンに改変した改変部位を有する改変塩基配列」を組み込んだ形質転換体は、ビタミンD3の変換反応において、26−ヒドロキシビタミンD3の生成量が減少する。
【0031】
2)「171位のロイシンがバリンに置換された改変部位を有する改変アミノ酸配列をコードするDNA」または「塩基511から塩基513にかけての配列CTGを、バリンをコードするコドンに改変した改変部位を有する改変塩基配列」を組み込んだ形質転換体は、ビタミンD3の変換反応において、26−ヒドロキシビタミンD3の生成量が減少する。そして、そのDNA改変体がコードするポリペプチドは、ビタミンD3の25位水酸化能の比活性が高い。
【0032】
3)「177位のアラニンがアルギニンに置換された改変部位を有する改変アミノ酸配列をコードするDNA」または「塩基529から塩基531にかけての配列GCCを、アルギニンをコードするコドンに改変した改変部位を有する改変塩基配列」を組み込んだ形質転換体は、ビタミンD3の変換反応において、対応する25-ヒドロキシ体の生成量が増大する。そして、そのDNA改変体がコードするポリペプチドは、ビタミンD3の25位水酸化能の比活性が高い。
【0033】
4)「240位のトレオニンがセリンに置換された改変部位を有する改変アミノ酸配列をコードするDNA」または「塩基718から塩基720にかけての配列ACGを、セリンをコードするコドンに改変した改変部位を有する改変塩基配列」を組み込んだ形質転換体は、ビタミンD3の変換反応において、26−ヒドロキシビタミンD3の生成量が減少する。
【0034】
上述のように、本発明において「ビタミンD類水酸化酵素」とはVdh、またはVdh-K1等のVdhと構造的かつ機能的に相同性を有する酵素を包含するものである。したがって、本発明に係る改変型ビタミンD類水酸化酵素を得るために用いられるビタミンD類水酸化酵素としては、vdh-K1遺伝子(配列番号1)がコードするビタミンD類水酸化酵素Vdh-K1(配列番号2)、及び野生型vdh遺伝子(配列番号3)がコードするビタミンD類水酸化酵素Vdh(配列番号4)のほか、vdh-K1遺伝子または野生型vdh遺伝子と高い相同性を示し、かつ同じ機能をもつ遺伝子がコードする酵素を用いることも可能である。
【0035】
本発明において、「vdh-K1遺伝子または野生型vdh遺伝子と高い相同性を示し、かつ同じ機能をもつ遺伝子がコードするビタミンD類水酸化酵素」とは、下記(b)または(c)のアミノ酸配列からなるビタミンD類水酸化酵素を意味する。
(b)配列番号2または4に記載のアミノ酸配列において、1または数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有するアミノ酸配列、
(c)配列番号2または4に記載のアミノ酸配列と90%以上の相同性を有するアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有するアミノ酸配列。
【0036】
すなわち、本発明の酵素には、上記(b)または(c)のアミノ酸配列において、前記(a)と同様の改変部位、すなわち(1)88位のイソロイシンがバリンに置換された改変部位、(2)171位のロイシンがバリンに置換された改変部位、(3)177位のアラニンがアルギニンに置換された改変部位、及び(4)240位のトレオニンがセリンに置換された改変部位からなる群より選択される少なくとも1つの改変部位を有するアミノ酸配列からなるビタミンD類水酸化酵素が包含される。
【0037】
本発明において、「1または数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列」における「1または数個」の範囲は特に限定されないが、好ましくは、1〜40個であり、より好ましくは1〜20個であり、特に好ましくは1〜10個である。
また、アミノ酸配列の相同性については、例えば、Genetyx(Genetyx corporation)のHomology Search機能により求めることができる。本発明において、アミノ酸配列の相同性の範囲は、90%以上であることが好ましく、95%以上であることがより好ましく、98%以上であることが特に好ましい。
【0038】
「野生型vdh遺伝子と高い相同性を示し、かつ同じ機能をもつ遺伝子」として、具体的には、特許文献3の表1に記載されているシュードノカルディア・オートトロフィカやシュードノカルディア・サターネア(Pseudonocardia saturnea)の様々な株から由来するvdhに相同な遺伝子が挙げられる。
【0039】
さらに上記のDNA改変体の改変部位を組み合わせて、さらに個々の機能が強化された改変型ビタミンD類水酸化酵素及び当該酵素をコードするDNAを取得することもできる。そのような酵素及びDNAの具体例として下記(1)〜(3)のアミノ酸配列からなる改変型ビタミンD類水酸化酵素、及び下記(1)〜(3)のアミノ酸配列からなる改変型ビタミンD類水酸化酵素をコードするDNAが挙げられる。
【0040】
(1)配列番号2または4に記載のアミノ酸配列において、88位のイソロイシンがバリンに、及び171位のロイシンがバリンにそれぞれ置換された改変アミノ酸配列、
(2)(1)の改変アミノ酸配列において、前記88位及び171位のアミノ酸置換のほかに、さらに1または数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有するアミノ酸配列、
(3)(1)の改変アミノ酸配列と90%以上の相同性を有し前記88位及び171位のアミノ酸置換を含むアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有するアミノ酸配列。
【0041】
上記のDNA改変体は、大腸菌において発現したポリペプチドが示すビタミンD3の25位水酸化能の比活性が野生型Vdhに対して高い(約2倍)のに加え、ビタミンD3の25位水酸化能に対する26位水酸化能の比率が低い(約1/5)という特長がある。
【0042】
前記(1)のアミノ酸配列からなる改変型ビタミンD類水酸化酵素をコードするDNAの中でも、配列番号1または3に記載の塩基配列において、262〜264番目の塩基配列ATCがバリンをコードするコドンに、及び511〜513番目の塩基配列CTGがバリンをコードするコドンにそれぞれ置換された改変塩基配列からなるDNAが好ましい。
【0043】
本発明の酵素をコードするDNAの取得方法は特に限定されない。前述したとおり、シュードノカルディア・オートトロフィカNBRC12743株のゲノムDNAを鋳型とし、PCRにより配列番号1の配列を含むDNAを取得し、これを用いてエラープローン(error-prone)PCRにより複数の変異を加えたDNA改変体を作製して、それらで形質転換された大腸菌の水酸化活性をスクリーニングして取得することができる。
【0044】
例えば、上記したvdh遺伝子断片を鋳型として使用し、配列番号3に示したvdh遺伝子のオープンリーディングフレームの全長を増幅できるように設計した1対のプライマーを用いてPCRを行う。反応液中のマンガンイオン濃度やデオキシグアノシン5'−三リン酸(dGTP)の濃度を変化させることにより変異の誘発を操作できる。PCRの反応条件は適宜設定することができ、例えば、94℃で30秒間(変性)、55℃で30秒〜1分間(アニーリング)、72℃で2分間(伸長)からなる反応工程を1サイクルとして、例えば25サイクル行った後、72℃で7分間反応させる条件などを挙げることができる。増幅されたDNA断片にはランダムに変異が導入されており、適当な宿主中で増幅可能なベクター中にクローニングすることができる。
【0045】
また、改変部位を指定して特定の塩基部分を置換することも可能である。例えば、置換したいアミノ酸をコードするコドンを中央に含むプライマー(変異−F)とそれに対する相補配列を含むプライマー(変異−R)を作製する。vdh遺伝子の5'末端から下流のDNA断片を増幅するプライマー(VDH-F)とプライマー(変異−R)のペアで完全長vdh遺伝子断片を鋳型としてPCRを行いvdh遺伝子の5'末端から置換挿入部位までを含むDNA断片を増幅する。また、vdh遺伝子の3'末端から上流のDNA断片を増幅するプライマー(VDH-R)とプライマー(変異−F)のペアで完全長vdh遺伝子断片を鋳型としてPCRを行い置換挿入部位からvdh遺伝子の3'末端までを含むDNA断片を増幅する。次にVDH-FとVDH-Rのプライマーペアを用いて上記で調製したvdh遺伝子の5'末端から置換挿入部位までを含むDNA断片と置換挿入部位からvdh遺伝子の3'末端までを含むDNA断片を鋳型としてPCRを行い、置換が導入された完全長vdh遺伝子断片を増幅する。その他、本明細書において開示した配列情報に基づいて全合成することも可能である。
【0046】
上記したプライマーの調製、DNAライブラリーの構築、DNAライブラリーのスクリーニング、並びに目的遺伝子のクローニングなどの操作は当業者に既知であり、例えば、モレキュラークローニング第2版、Current Protocols in Molecular Biology, Supplement 1〜38, John Wiley & Sons (1987-1997)等に記載の方法に準じて行うことができる。
【0047】
本発明のポリペプチド(酵素)の取得方法は特に制限されず、化学合成により合成したポリペプチドでもよいし、遺伝子組み換え技術により作製したものでもよい。遺伝子組み換え技術により作製する場合には、先ず、本明細書に上記したとおり、当該酵素をコードするDNA改変体を取得する。このDNA改変体を適当な発現系に導入することにより、本発明の酵素を産生することができる。発現系での酵素の発現については本明細書中において後記する。
【0048】
[本発明の組み換えベクター]
本発明に係るDNAは適当なベクター中に挿入して使用することができる。本発明で用いるベクターの種類は特に限定されず、例えば、自律的に複製するベクター(例えば、プラスミド等)でもよいし、あるいは、宿主細胞に導入された際に宿主細胞のゲノムに組み込まれ、組み込まれた染色体と共に複製されるものであってもよい。発現ベクターにおいて本発明のDNAは、転写に必要な要素(例えば、プロモーター等)が機能的に連結されている。プロモーターは宿主細胞において転写活性を示すDNA配列であり、宿主の種類に応じて適宜選択することができる。具体的には、宿主として大腸菌を用いる場合、例えば、ベクターとしてpET vector、pUC19等を用い、プロモーターとしてT7 promoter、lac promoter等を用いることができる。
【0049】
[本発明の形質転換体及びそれを用いた組み換えポリペプチドの製造]
本発明に係るDNAまたは組み換えベクターを適当な宿主に導入することによって形質転換体を作製することができる。本発明のDNAまたは組み換えベクターが導入される宿主細胞は、本発明の遺伝子を発現できれば任意の細胞でよく、細菌、酵母、真菌及び高等真核細胞等が挙げられる。細菌細胞の例としては、バチルス(Bacillus)またはストレプトマイセス(Streptomyces)等のグラム陽性菌または大腸菌等のグラム陰性菌が挙げられる。これら細菌の形質転換は、塩化カルシウム法、プロトプラスト法、エレクトロポレーション法または公知の方法で調製が可能なコンピテント細胞を用いることにより行えばよい。例えば、エレクトロポレーション法は以下のように行うことができる。外来遺伝子が挿入されたプラスミドをコンピテント細胞の懸濁液に加え、この懸濁液をエレクトロポレーション法専用のキュベットに入れ、そのキュベットに高電圧の電気パルスをかける。その後選択培地で培養し、平板寒天培地上で形質転換体を単離する。
【0050】
酵母細胞の例としては、サッカロマイセス(Saccharomyces)またはシゾサッカロマイセス(Schizosaccharomyces)に属する細胞が挙げられ、例えば、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)またはサッカロマイセス・クルイベリ(Saccharomyces kluyveri)等が挙げられる。酵母宿主への組み換えベクターの導入方法としては、例えば、エレクトロポレーション法、スフェロプラスト法、酢酸リチウム法等を挙げられる。真菌細胞の例としては、糸状菌、例えばアスペルギルス、ニューロスポラ、フザリウム、またはトリコデルマに属する細胞が挙げられる。宿主細胞として糸状菌を用いる場合、DNA構築物を宿主染色体に組み込んで組換え宿主細胞を得ることにより形質転換を行うことができる。DNA構築物の宿主染色体への組み込みは、公知の方法に従い、例えば相同組換えまたは異種組換えにより行うことができる。特に、製造効率の点から、本発明においては形質転換の宿主として、シュードノカルディア(Pseudonocardia)属細菌、大腸菌、ロードコッカス(Rhodococcus)属細菌を用いることが好ましい。
【0051】
上記の形質転換体は、導入された遺伝子の発現を可能にする条件下で適切な栄養培地中で培養する。形質転換体の培養物から、本発明のポリペプチドを単離精製するには、通常のポリペプチドの単離、精製法を用いればよい。
【0052】
例えば、本発明のポリペプチドが、細胞内に溶解状態で発現した場合には、培養終了後、細胞を遠心分離により回収し水系緩衝液に懸濁後、超音波破砕機等により細胞を破砕し、無細胞抽出液を得る。該無細胞抽出液を遠心分離することにより得られた上清から、通常のポリペプチドの単離精製法、即ち、溶媒抽出法、硫安等による塩析法、脱塩法、有機溶媒による沈殿法、ジエチルアミノエチル(DEAE)セファロース等のレジンを用いた陰イオン交換クロマトグラフィー法、SP-Sepharose FF(アマシャムバイオサイエンス社製)等のレジンを用いた陽イオン交換クロマトグラフィー法、ブチルセファロース、フェニルセファロース等のレジンを用いた疎水性クロマトグラフィー法、分子ふるいを用いたゲルろ過法、アフィニティークロマトグラフィ一法、クロマトフォーカシング法、等電点電気泳動等の電気泳動法等の手法を単独あるいは組み合わせて用い、精製標品を得ることができる。
【0053】
[水酸化ビタミンD誘導体の製造方法]
本発明は、ビタミンD類の水酸化酵素機能が向上した改変型ビタミンD類水酸化酵素をコードするDNAを導入した形質転換体の存在下でビタミンD類を水酸化させることを含む、水酸化ビタミンD誘導体の製造方法を包含する。具体的には、本発明に係る製造方法は、改変型ビタミンD類水酸化酵素をコードするDNAを含む組換えベクターによって形質転換された形質転換体を培養し、培養中または培養後において、当該形質転換体とビタミンD類とを接触させてビタミンD類を水酸化させ、産出された水酸化ビタミンD誘導体を採取することを特徴とする水酸化ビタミンD誘導体の製造方法である。
【0054】
このような形質転換体の存在下でビタミンD類を水酸化させるのに好適な条件は、以下のとおりである。
まず、形質転換体中のビタミンD水酸化酵素関連DNA(ビタミンD類の水酸化酵素をコードするDNA、フェレドキシンをコードするDNA及びフェレドキシン還元酵素をコードするDNA)を必要に応じて誘導物質を添加して菌体内で発現させる。これらのDNAが発現した菌体を添加したビタミンD類と接触させ、変換反応をさせる。変換反応の温度は、形質転換体の至適温度を考慮して、適宜決定できる。また、反応時間も水酸化ビタミンD誘導体への変換率(反応の進行度合い)等を考慮して、適宜決定することができる。例えば、20〜31℃で、1〜5日が好適である。さらに、反応様式は、バッチ式でも連続式でも、いずれの形式でも実施することができる。
【0055】
生成した水酸化ビタミンD誘導体の単離及び精製は、一般に微生物代謝産物をその培養液から単離するために用いられる分離、精製の方法が利用できる。例えば、メタノール、エタノール、アセトン、ブタノール、酢酸エチル、酢酸ブチル、クロロホルム、トルエン等を用いた有機溶媒抽出、ダイヤイオンHP-20などの疎水性吸着樹脂を用いた吸脱着処理、セファデックスLH-20等を用いたゲルろ過クロマトグラフィー、活性炭、シリカゲル等による吸着クロマトグラフィー、もしくは薄層クロマトグラフィーによる吸脱着処理、あるいは逆相カラム等を用いた高速液体クロマトグラフィー等の公知のあらゆる方法がこれにあたる。また、ここに示した方法に特に限定されるものではない。これらの方法を単独あるいは任意の順序に組み合わせ、また反復して用いることにより、目的の水酸化ビタミンD誘導体を単離精製することができる。
【実施例】
【0056】
以下、本発明について具体例を挙げてより詳細に説明するが、本発明をこれらの例に限定することを意図するものではない。なお、下記の例中のパーセント(%)は、培地の説明においては質量パーセント、HPLCの移動相の説明においては、容量パーセントを示す。
【0057】
実施例1:高機能化したVdh-K1の改変体の取得
(1)88番目の残基のアミノ酸総置換ライブラリーの作製
88、171、177または240番目のアミノ酸残基のうち、いずれか1箇所のアミノ酸が総置換されたvdh-K1遺伝子(配列番号1)を増幅するためプライマーを設計した。88番目の残基のアミノ酸総置換にはプライマーI88sat-F(配列番号11)とI88sat-R(配列番号12)を設計した。2組のプライマー、(1):Vdh-1F(配列番号5)とI88sat-R及び(2):Vdh-1R(配列番号6)とI88sat-Fを用いて、pET-aciBC-SD-VdhK1(非特許文献6)を鋳型とし、PCR反応を行って、それぞれ約0.25 kb及び約0.95 kbの2つのDNA断片を得た。これらのDNA断片を反応液からWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収精製した。続いて、完全長の遺伝子を増幅するためにプライマーVdh-1FとVdh-1Rを用い、上記で調製した2つのDNA断片の等モル混合物をテンプレートとしてPCR反応を行い、1.2 kbpのDNA断片を得た。このDNA断片を、反応液からWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収精製した後、制限酵素NdeIとBglIIにより37℃で15時間消化し、Wizard SV Gel and PCR Clean-Up System(プロメガ社)で精製した後、DNA Ligation kit ver 2.1(タカラバイオ社)を用いてプラスミドpETaciBC(特許文献3)のNde I-Bam HI部位に連結し、大腸菌DH5α株(タカラバイオ社)を形質転換した。カルベニシリン(50μg/ml)を含むLB寒天培地に出現した大腸菌形質転換株約1000コロニーに2 mlのLB培地を加えて、寒天培地上でスプレッダーを用いて菌体を懸濁した。この菌体懸濁液からWizard Plus SV Minipreps DNA Purification system(プロメガ社)を用いてプラスミドを抽出した。このプラスミドには88番目の残基が任意のアミノ酸に置換されたvdh-K1遺伝子が挿入されていると考えられ、Vdh-K1の88番目のアミノ酸残基の総置換ライブラリー(pETaciBC-SD-Vdh-88sat)として用いた。
【0058】
(2)171、177または240番目の残基のアミノ酸総置換ライブラリーの作製
前述(1)に記載した方法と同様にして、171及び177番目の残基のアミノ酸総置換ライブラリーを調製した。この時、171番目の残基のアミノ酸総置換にはプライマーL171sat-F(配列番号13)とL171sat-R(配列番号14)、177番目の残基のアミノ酸総置換にはプライマーA177sat-F(配列番号15)とA177sat-R(配列番号16)、240番目の残基のアミノ酸総置換にはプライマーT240sat-F(配列番号17)とT240sat-R(配列番号18)を用いた。この結果、作製したライブラリーをそれぞれVdh-K1の171番目のアミノ酸残基の総置換ライブラリー(pETaciBC-SD-Vdh-171sat)、Vdh-K1の177番目のアミノ酸残基の総置換ライブラリー(pETaciBC-SD-Vdh-177sat)、Vdh-K1の240番目のアミノ酸残基の総置換ライブラリー(pETaciBC-SD-Vdh-240sat)と名付けた。
【0059】
(3)88番目のアミノ酸残基の総置換ライブラリーを用いた副反応抑制クローンのスクリーニング
作製したVdh-K1の88番目のアミノ酸残基総置換ライブラリー(pETaciBC-SD-Vdh-88sat)で大腸菌BL21(DE3)を形質転換して大腸菌BL21(DE3)/pETaciBC-SD-Vdh-88sat株のコロニーを多数得た。96穴ディープウエルプレート(IWAKI社)2枚の各ウエルにカルベニシリン(50μg/ml)を含む1 mlのTB培地を入れ、大腸菌BL21(DE3)/pETaciBC-SD-Vdh-88satを1穴あたり1コロニー植菌した。コントロールとして、各プレートにpETaciBC-SD-VdhK1を8株植菌した。プレート用シェーカー(SCF-98-8:いわしや社)にプレートをセットし、30℃、800 rpmで16時間培養した。レプリケーターを用いて、96穴ディープウエルプレート(IWAKI社)2枚の各ウエル中にカルベニシリン(50μg/ml)とOvernight autoinduction system 1(メルク社)を含む1 mlのTB培地へ植菌した。前培養の96穴ディープウエルプレートはそのまま−80℃で凍結保存した。プレート用シェーカー(SCF-98-8:いわしや社)にプレートをセットし25℃、800 rpmで24時間培養した。3500 rpm、10分間の遠心により菌体を沈殿させ、上清を取り除いた。500μlのCV2反応液(1.0% (w/v) partially methylated β-cyclodextrin, 500 mg/L ビタミンD3, 5.0% (v/v) DMSO)を加え、プレート用シェーカー(SCF-98-8: いわしや社)にプレートをセットし28℃、800 rpmで24時間変換反応をした。各500μlのメタノールを添加し、反応を止め、800 rpmで5分間撹拌した。4000 rpm、10分間の遠心により菌体を沈殿させ、上澄みをHPLC分析用サンプルとして保存した。他のアミノ酸総置換ライブラリーも同様にHPLC分析用サンプルを調製した。HPLCの分析条件を以下に示す。
【0060】
分析装置:Agilent 100 series、
カラム:J' sphere ODS-H80(YMC, Inc.), 75 mm×4.6 mmI.D.、
移動相:A;イオン交換水、
B;アセトニトリル、
グラジエント時間設定:0分 移動相A/B =20:80、
4.3分 移動相A/B =20:80、
4.5分 移動相A/B =0:100、
9.5分 移動相A/B =0:100、
9.7分 移動相A/B =20:80、
10.0分 移動相A/B =20:80、
流速:1.5 ml/分、
検出:UV 265 nm、
インジェクション容量:10 μl、
カラム温度:40℃、
分析時間:10分、
保持時間:25−ヒドロキシビタミンD3 3.2分、
26−ヒドロキシビタミンD3 3.6分、
ビタミンD3 9.2分。
【0061】
上記の条件で変換サンプルを分析し、25−ヒドロキシビタミンD3が生成していることを確認し、副反応物比率が減少しているクローンを選抜した。副反応物比率は以下の式で求めた。
【数1】
【0062】
上記に示したHPLC分析条件により26−ヒドロキシビタミンD3生成比がvdh-K1発現株より減少した4クローンについてアンピシリン(50μg/ml)を含む2 ml LB液体培地で培養し、増殖した形質転換大腸菌からWizard Plus SV Minipreps DNA Purification system(プロメガ社)を用いてプラスミドDNAを精製した。得られたプラスミドを鋳型として改変遺伝子の塩基配列を決定した。塩基配列の解析には、DNA塩基配列解析装置(アプライドバイオシシテム;3130)を用い、配列決定用プライマー(配列番号7,8,9,10)を使用してダイターミネーター・サイクル・シークエンス法で解析した。
【0063】
アミノ酸総置換ライブラリーをスクリーニングした結果、Vdh-K1の88番目の残基のイソロイシンがバリンに置換された改変体を発現する大腸菌形質転換株が副反応を抑制すると示唆された。さらにVdh-K1の171、177及び240番目の残基のアミノ酸総置換ライブラリーのスクリーニングをそれぞれ行った結果、171番目の残基のロイシンがバリンに、177番目のアラニンがアルギニンにまたは240番目のトレオニンがセリンに置換された改変体を発現する大腸菌形質転換体が副反応を抑制することが示唆された。
【0064】
【表1】
【0065】
実施例2:高機能化したVdh-K1改変体の比活性測定
変異の導入により大腸菌での発現量が変化することが考えられたため、単位酵素あたりのビタミンD3水酸化活性を測定した。上記の基質認識に関わるアミノ酸のアミノ酸総置換ライブラリーをスクリーニングした結果、88番目の残基のイソロイシンがバリンに置換したアミノ酸置換体が副反応抑制に寄与した。これらの改変Vdh-K1を発現させるプラスミド(pETaciBC-SD-Vdh-K1 I88V)を導入し、大腸菌BL21(DE3)を形質転換した。得られた大腸菌BL21(DE3)形質転換株を50μg/mlのカルベニシリンを含む2 mlのTB培地に植菌した。30℃、220 rpmで14時間培養後、カルベニシリンとOvernight autoinduction system 1(メルク社)を含む25 mlのTB培地へ250μlの前培養液を接種した。25℃、220 rpmで24時間培養後、2 mlの培養液を試験管にとり、3500 rpm、10分間の遠心により菌体を沈殿させて上澄みを捨て、1 mlのCV2反応液を加えた。28℃、220 rpmで6時間変換反応を行い1.5 mlの酢酸エチルを添加して反応を止めた。30秒間ボルテックスで撹拌し、3500 rpm、10分間の遠心により水相と酢酸エチル相に分離させた。酢酸エチル相をとり、残った水相に0.75 mlの酢酸エチルを加えた。30秒間ボルテックスで撹拌し、3500 rpm、10分間の遠心により水相と酢酸エチル相に分離させた。酢酸エチル相をとり、先程得た酢酸エチル相と合わせて、減圧濃縮機により乾固した。乾固物に200μlのメタノールを添加し、溶解後、HPLC分析用バイアルへ移して、分析まで遮光4℃で保存した。
【0066】
変換試験に使用しなかった培養液から4300 rpm、10分間の遠心により菌体を沈殿として得た。上澄みを捨て、培養液に対して5分の1量のCV緩衝液(50 mMリン酸カリウム緩衝液pH 7.4, 10% (v/v)グリセロール)を加え、5倍濃縮細胞懸濁液を調製した。2 mlの5倍濃縮細胞懸濁液に68μl 10X Bug Buster(ノバジェン社)、2μl Benzonase(ノバジェン社)、200μl 40 mg/ml Lysozymeを加え、30℃で20分間緩やかに振とうした。7000 rpm、10分間の遠心により上清に無細胞抽出液を得た。この無細胞抽出液をキャップ付き試験管2本に750μlずつ分け、一方の無細胞抽出液に一酸化炭素を通した。次に、両方の無細胞抽出液にハイドロサルファイトナトリウムを少量(1-2 mg)添加した。一酸化炭素を通していないサンプルの400 nmから500 nmの吸収スペクトルをベースラインとして、スペクトルフォトメーター(HITACHI社、U-3310 SpectrophotoMeter)で一酸化炭素を通したサンプルの400 nmから500 nmの吸収をスキャンした。その結果、シトクロムP450に特徴的な450 nmのピークを観察し、その吸収から一酸化炭素結合型で還元状態Vdhの分子吸光係数を1 mMあたり91として培養液中のVdh改変体の発現量を計算した。
【0067】
前記変換試験サンプルのHPLC解析による25−ヒドロキシビタミンD3の定量結果と一酸化炭素結合スペクトル解析より得られたVdhまたはその改変体の発現量から、1 nmolのVdhまたはその改変体が1時間あたりに産生する25−ヒドロキシビタミンD3量を算出した。この値を比較することによりVdh-K1の88番目の残基のイソロイシンがバリンに置換したアミノ酸置換体(Vdh-K1I88V)における比活性の評価を行った。表2と図1にVdh-K1I88Vの比活性測定の結果を示した。Vdh発現量」はVdhまたはその改変体を発現させた大腸菌形質転換株の培養液中の濃度(nM)で示し、25-ヒドロキシビタミンD3生成量は休止菌体反応を6時間行った1 mlの反応液中の25-ヒドロキシビタミンD3濃度(mg/L)で示した。比活性として、1 nmolのVdhまたはその改変体が1時間で生成する25−ヒドロキシビタミンD3量(μg/hr/nmol of Vdh)で示し、コントロール(Vdh-K1)に対する比活性の相対活性を示した。Vdh-K1I88Vの比活性は、Vdh-K1の比活性の1.8倍を示し、単位酵素あたりのVD3水酸化活性は増加した。さらに、副反応比率は2.1%まで顕著に減少した。大腸菌において、Vdh-K1I88Vは、ビタミンD3変換時の副反応抑制に寄与した。同様にVdh-K1の171番目の残基のロイシンがバリンに置換されたアミノ酸置換体(Vdh-K1L171V)、177番目の残基のアラニンがアルギニンに置換されたアミノ酸置換体(Vdh-K1A177R)及び240番目の残基トレオニンがセリンに置換されたアミノ酸置換体(Vdh-K1T240S)における比活性の評価を行った。Vdh-K1L171VはVdh-K1よりも比活性が高いのに加え、副反応比率は3.6%に低減した。Vdh-K1A177RはVdh-K1の約3.5倍の比活性を示した。Vdh-K1A177Rの副反応比率はVdh-K1と同等であった。Vdh-K1T240SはVdh-K1よりも副反応比率は低減したが、比活性がVdh-K1の約60%に低下した。
【0068】
実施例3:有効変異の二重化によるVdh-K1改変体の調製と比活性測定
上記に見出した有効変異の重複効果を調べるため、Vdh-K1I88Vに171番目の残基のロイシンがバリンに置換した変異を導入するためのプライマーL171V-F(配列番号19)とL171V-R(配列番号20)を設計した。pET-aciBC-SD-VdhK1 I88Vを鋳型とし、2組のプライマー、(i):Vdh-1FとL171V-R及び(ii):Vdh-1RとL171V-Fを用いてPCR反応を行って、それぞれ約0.51kb及び約0.69 kbの2つのDNA断片を得た。これらのDNA断片をWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収した。続いて、完全長の遺伝子を増幅するためにプライマーVdh-1FとVdh-1Rを用い、上記で調製した2つのDNA断片の等モル混合物をテンプレートとしてPCR反応を行い、1.2 kbpのDNA断片を得た。このDNA断片を、反応液からWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収精製した後、制限酵素NdeIとBglIIにより37℃で15時間消化し、Wizard SV Gel and PCR Clean-Up System(プロメガ社)で精製した後、DNA Ligation kit ver 2.1(タカラバイオ社)を用いてプラスミドpETaciBCのNde I-Bam HI部位に連結し、大腸菌DH5α株を形質転換した。その後、カルベニシリン(50μg/ml)を含むLB寒天培地を用いて、形質転換された大腸菌を選択した。形質転換株からコロニーを分離し、カルベニシリン(50μg/ml)を含む2 mlのLB培地で増殖させた。増殖菌体からWizard Plus SV Minipreps DNA Purification system(プロメガ社)を用いてプラスミドpET-aciBC-SD-VdhK1 I88V L171Vを調製した。
【0069】
Vdh-K1I88Vに177番目の残基のアラニンがアルギニンに置換した変異を導入するためA177R-F(配列番号21)とA177R-R(配列番号22)を設計した。pET-aciBC-SD-VdhK1 I88Vを鋳型とし、2組のプライマー、(i):Vdh-1FとA177R-R及び(ii): Vdh-1RとA177R-Fを用いてPCR反応を行ってそれぞれ約0.53 kb及び約0.67 kbの2つのDNA断片を得た。これらのDNA断片をWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収した。続いて、完全長の遺伝子を増幅するためにプライマーVdh-1FとVdh-1Rを用い、上記で調製した2つのDNA断片の等モル混合物をテンプレートとしてPCR反応を行い、1.2 kbpのDNA断片を得た。このDNA断片を、反応液からWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収精製した後、制限酵素NdeIとBglIIにより37℃で15時間消化し、Wizard SV Gel and PCR Clean-Up System(プロメガ社)で精製した後、DNA Ligation kit ver 2.1(タカラバイオ社)を用いてプラスミドpETaciBCのNde I-Bam HI部位に連結し、大腸菌DH5α株を形質転換した。その後、カルベニシリン(50μg/ml)を含むLB寒天培地を用いて、形質転換された大腸菌を選択した。形質転換株からコロニーを分離し、カルベニシリン(50μg/ml)を含む2 mlのLB培地で増殖させた。増殖菌体からWizard Plus SV Minipreps DNA Purification system(プロメガ社)を用いてプラスミドpET-aciBC-SD-vdhK1 I88V A177Rを調製した。
【0070】
前述の方法で調製したプラスミド(pET-aciBC-SD-VdhK1 I88V L171V及びpET-aciBC-SD-VdhK1 I88V A177R)で大腸菌BL21(DE3)を形質転換してそれぞれBL21(DE3)/ pETaciBC-SD-VdhK1 I88V L171V及びBL21(DE3)/ pETaciBC-SD-VdhK1 I88V A177R株を得た。これらアミノ酸置換体の比活性及び副反応比率を親株と比較した結果、BL21(DE3)/ pETaciBC-SD-VdhK1 I88V A177R株は重複効果を示さなかったが、pETaciBC-SD-VdhK1 I88V L171Vは変異を重複することにより、単独変異の副反応比率よりもさらに低減することができた(表2及び図1)。このように大腸菌において、Vdh-K1の88番目の残基のイソロイシンがバリンに置換され、かつ171番目の残基のロイシンがバリンに置換されたアミノ酸置換体はVdh-K1の26位水酸化活性を低減するのに有効な変異であることが明らかとなった。
【0071】
【表2】
【0072】
実施例4:Vdh改変体の調製と比活性測定
(1)88番目の残基をバリンへ置換したVdhをコードする遺伝子の調製
上記に見出した副反応を抑制する有効変異の野生型Vdhに対する効果を調べるため、プライマーI88V-F(配列番号24)とI88V-R(配列番号25)を合成した。pET-aciBC-SD-vdh(非特許文献6)を鋳型として、2組のプライマー、(i):Vdh-1FとI88V-R 及び(ii):Vdh-1RとI88V-Fを用いてPCR反応を行ってそれぞれ約0.25 kb及び約0.95 kbの2つのDNA断片を得た。これらのDNA断片をPCR反応液からWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収精製した。続いて、完全長の遺伝子を増幅するためにプライマーVdh-1FとVdh-1Rを用い、上記で調製した2つのDNA断片の等モル混合物をテンプレートとしてPCR反応を行い、1.2 kbpのDNA断片を得た。このDNA断片を、反応液からWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収精製した後、制限酵素NdeIとBglIIにより37℃で15時間消化し、Wizard SV Gel and PCR Clean-Up System(プロメガ社)で精製した後、DNA Ligation kit ver 2.1(タカラバイオ社)を用いてプラスミドpETaciBC(非特許文献6)のNde I-Bam HI部位に連結し、大腸菌DH5α株を形質転換した。その後、カルベニシリン(50μg/ml)を含むLB寒天培地を用いて、形質転換された大腸菌を選択した。形質転換株からコロニーを分離し、カルベニシリン(50μg/ml)を含む2 mlのLB培地で増殖させた。増殖菌体からWizard Plus SV Minipreps DNA Purification system(プロメガ社)を用いてプラスミドpET-aciBC-SD-vdhI88Vを調製した。
【0073】
(2)88番目及び171番目の残基をバリンへ置換したVdhをコードする遺伝子の調製
前記(1)で調製した遺伝子にさらに171残基目のアミノ酸置換の変異を導入するため、pET-aciBC-SD-vdh I88Vを鋳型とし、2組のプライマー、(i):Vdh-1FとL171V-R 及び(ii):Vdh-1RとL171V-Fを用いてPCR反応を行ってそれぞれ約0.51 kb及び約0.69 kbの2つのDNA断片を得た。これらのDNA断片をWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収した。続いて、完全長の遺伝子を増幅するためにプライマーVdh-1FとVdh-1Rを用い、上記で調製した2つのDNA断片の等モル混合物をテンプレートとしてPCR反応を行い、1.2 kbpのDNA断片を得た。このDNA断片を、反応液からWizard SV Gel and PCR Clean-Up System(プロメガ社)によって回収精製した後、制限酵素NdeIとBglIIにより37℃で15時間消化し、Wizard SV Gel and PCR Clean-Up System(プロメガ社)で精製した後、DNA Ligation kit ver 2.1(タカラバイオ社)を用いてプラスミドpETaciBCのNde I-Bam HI部位に連結し、大腸菌DH5α株を形質転換した。その後、カルベニシリン(50μg/ml)を含むLB寒天培地を用いて、形質転換された大腸菌を選択した。形質転換株からコロニーを分離し、カルベニシリン(50μg/ml)を含む2 mlのLB培地で増殖させた。増殖菌体からWizard Plus SV Minipreps DNA Purification system(プロメガ社)を用いてプラスミドpET-aciBC-SD-vdhI88V L171Vプラスミドを調製した。
【0074】
(3)Vdh改変体の比活性測定
pET-aciBC-SD-vdh(非特許文献6)、pET-aciBC-SD-vdhI88V及びpET-aciBC-SD-vdh I88V L171Vプラスミドでそれぞれ大腸菌BL21(DE3)を形質転換して形質転換株を得た。アミノ酸置換体の比活性及び副反応比率を親株と比較した結果、野生型Vdhの88残基目のイソロイシンがバリンに置換されたアミノ酸置換体は野生型Vdhの25位水酸化活性を維持しながら、25位水酸化活性に対する26位水酸化活性比を2.5%に低減した。さらに、野生型Vdhの88残基目のイソロイシンがバリンに置換され、かつ171残基目のロイシンがバリンに置換されたアミノ酸置換体はVdh I88Vよりも酵素あたりの25位水酸化活性が高いのに加えて、25位水酸化活性に対する26位水酸化活性比は1.5%まで低減した(図2)。
【0075】
実施例5:シュードノカルディア・オートトロフィカにおけるvdh遺伝子の破壊
(1)vdh遺伝子破壊用プラスミドpTYM18-2の作製
プラスミドpTYM18(尾仲らJ. Anibiot. 56, 950-956, 2003)を制限酵素Eco RI及びBgl IIで消化して得られる約3.3 kbのDNA断片を精製し、その末端をDNA Blunting Kit(タカラバイオ)で平滑化した後、DNA Ligation Kit(タカラバイオ)で自己環状化した。この反応液で大腸菌Escherichia coli DH5αを形質転換して得られたクローンからプラスミドDNAを調製してpTYM18-2を得た。
【0076】
(2)シュードノカルディア・オートトロフィカ vdh遺伝子破壊株の分離
特許文献3記載のvdh遺伝子を担持するプラスミドpVdh-camABを鋳型にして以下2種のプライマー、
(i) Vdh236Spe-F 配列番号26:5'-GCACTAGTGCCACACGCTGCCCGAGGACCAG-3'
(ii) Vdh1039Bam-R 配列番号27:5'-GCGGATCCGTGCCCGAAGAACACCCCGCC-3'
により約820bからなる野生型vdh遺伝子の一部断片を増幅した。このDNAを制限酵素Spe I及びBam HIで消化し、精製した後、pTYM18-2のXba I-Bam HI制限酵素部位へクローニングしてプラスミドpTYM-VDHを構築した。このプラスミドを接合用大腸菌Escherichia coli S17-1株に形質転換し、カナマイシン25μg/mlを含む2 mlのLB培地に植菌して30℃で20時間培養した。この培養液200μlをLB培地10 mlに植菌して30℃で3時間培養した。この培養液をLB培地で2回洗浄した後、LB培地3 mlに懸濁した。一方で、TSB培地にて28℃で3日間増殖させたシュードノカルディア・オートトロフィカの培養液500μlをTSB培地10 mlに植菌して28℃で20時間培養した。この増殖菌体を滅菌水で2回洗浄した後、滅菌水500μlに懸濁した。この懸濁液30μlに対し前述の大腸菌S17-1株(Escherichia coli ATCC47055)の懸濁液500μlを混合し、ISP4平板培地(寒天濃度1.5%)へ塗布した。この培地を30℃で20時間培養した。リボスタマイシン200μg/ml及びナリジクス酸25μg/mlを含む2 mlのSNA(ソフトニュートリエントアガー)を平板上に重層した。この培地を30℃で2週間培養し生育してきた菌株をvdh遺伝子の破壊株として分離した。
【0077】
実施例6:Vdh-K1 I88Vアミノ酸置換体遺伝子のシュードノカルディア・オートトロフィカでの発現
Vdh-K1 I88Vアミノ酸置換体遺伝子を増幅するため、Vdh-2R(配列番号23)を合成した。プライマーVdh-1FとVdh-2Rを用い、pETaciBC-SD-Vdh-K1 I88Vを鋳型にしてPCR反応を行った。その結果増幅した約1.2 kbpのDNA断片を反応液からWizard SV Gel and PCR Clean-Up System プロメガ社)によって回収精製した後、制限酵素Nde IとNhe Iにより37℃で10時間消化しWizard SV Gel and PCR Clean-Up System(プロメガ社)で精製した。予め制限酵素NdeI及びNheIで消化して、vdh遺伝子断片を切り離したpTAOR3-vdh(シュードノカルディア・オートトロフィカ発現ベクター、特願2008-259317特許、平成20年10月6日出願)のベクターDNA断片に、このDNA断片を、DNA Ligation kit ver 2.1(タカラバイオ社)を用いて連結し、大腸菌DH5α株(タカラバイオ社)を形質転換した。その後、カルベニシリン(50μg/ml)を含むLB培地を用いて、形質転換された大腸菌を増殖させた。増殖菌体からWizard Plus SV Minipreps DNA Purification system(プロメガ社)を用いてプラスミドを抽出した。このプラスミドをpTAOR-K1I88Vと名付けた。Vdh-K1 アミノ酸置換体についても上記と同様に構築し、pTAOR-K1と名付けた。得られたプラスミドを鋳型としてvdh-K1の改変体の塩基配列の決定をDNA塩基配列解析装置(アプライドバイオシシテム;3130)で用いて行った。配列決定用プライマーを使用してダイターミネーター・サイクル・シークエンス法で解析し、正しいDNA配列が含まれていることを確認した。
【0078】
pTAOR-K1I88VまたはpTAOR-K1プラスミドで大腸菌S17-1株(Escherichia coli ATCC47055)を形質転換し、アプラマイシン(60μg/ml)を含むLB寒天培地で選抜し、pTAOR-K1 I88V/S17-1またはpTAOR-K1/S17-1株を得た。pTAOR-K1 I88VまたはpTAOR-K1プラスミドをカナマイシン遺伝子で前記vdh遺伝子破壊シュードノカルディア・オートトロフィカ株に接合伝達により導入した。導入方法を以下に示す。
【0079】
シュードノカルディア・オートトロフィカの凍結菌体液をカナマイシン(200μg/ml)を含むLB培地を用いて、30℃、3日間培養した。培養液500μlを遠心分離(7000 rpm, 30秒間)し、培養菌体を集菌した。菌体を500μlのLB培地に懸濁し、この培養液をシュードノカルディア・オートトロフィカ細胞懸濁液とした。さらに、大腸菌pTAOR-K1 I88V/S17-1株をアプラマイシン(60μg/ml)を含むLB培地を用いて、30℃、24 h培養した。培養液200μlを遠心分離(7000 rpm, 30秒間)し、培養菌体を集菌した。菌体を200μlのLB培地に懸濁し、この培養液を大腸菌細胞懸濁液とした。シュードノカルディア・オートトロフィカ及び大腸菌細胞懸濁液を混和後、遠心分離(7000 rpm, 30秒間)した。上清を500μl除き、残り200μlのLB培地で菌体を懸濁後、薬剤を含まないLB寒天培地に塗布し、30℃、24 h培養した。寒天プレートに2 mlのLB液体培地を加え、寒天培地に生育した菌体を懸濁後、マイクロチューブに懸濁液を移した。懸濁液をカナマイシン(200μg/ml)、アプラマイシン(24μg/ml)及びナリジクス酸(50μg/ml)を含むLB寒天培地にスプレッドし、30℃、一週間培養した。生育したコロニーを再度カナマイシン(200μg/ml)、アプラマイシン(24μg/ml)及びナリジクス酸(50μg/ml)を含むLB寒天培地に画線し耐性を確認した。得られた菌株をpTAOR-K1 I88VまたはpTAOR-K1株とした。
【0080】
得られたシュードノカルディア・オートトロフィカ形質転換株、シュードノカルディア・オートトロフィカ vdh破壊株及びvdh遺伝子が破壊されていないシュードノカルディア・オートトロフィカ親株をビタミンD3変換試験に供した。培地組成は以下の通りである。
(前培養培地組成)
グルコース 1.5%
酵母エキス 0.3%
NaCl 0.4%
CaCO3 0.2%
ポリペプトン 1.5%
pH7.4
(本培養培地組成)
グルコース 2.0%
酵母エキス 0.5%
NaCl 0.04%
CaCO3 0.3%
ポリペプトン 1.0%
大豆ペプトン 1.0%
K2HPO4 0.04%
pH7.4
【0081】
試験株を100 mlの前培養液に植菌した。シュードノカルディア・オートトロフィカ形質転換株及びシュードノカルディア・オートトロフィカ vdh破壊株用培地にはカナマイシン(200μg/ml)を添加した。28℃、220 rpm、2日間培養後、100 mlの本培養培地へ1 mlの前培養液を接種した。28℃、220 rpm、24 h培養後、培養液に1 mlのアセトンを添加した。さらに24時間培養し、ビタミンD3溶液(終濃度2.0% (w/v) partially methylated β-cyclodextrin,1000μg/L ビタミンD3, 2.0% (v/v)エタノール)を添加した。以後、24 時間ごとに 1 mlサンプリングを行い、サンプリングと同時にビタミンD3溶液(終濃度2.0% (w/v) partially methylated β-cyclodextrin, 1000μg/L ビタミンD3, 2.0% (v/v)エタノール)を添加した。サンプリングした培養液はメタノールで10倍希釈し、遠心上清をHPLC分析用サンプルとした。HPLCの分析条件を以下に示す。
【0082】
分析装置:Agilent 100 series、
カラム:J' sphere ODS-H80 (YMC, Inc.), 75 mm×4.6 mmI.D.、
移動相:A; イオン交換水、
B; アセトニトリル、
グラジエント時間設定:0分 移動相A/B =20:80、
4.3分 移動相A/B =20:80、
4.5分 移動相A/B =0:100、
9.5分 移動相A/B =0:100、
9.7分 移動相A/B =20:80、
10.0分 移動相A/B =20:80、
流速:1.5 ml/分、
検出:UV 265 nm、
インジェクション容量:10 μl、
カラム温度:40℃、
分析時間:10分、
保持時間:25−ヒドロキシビタミンD3 3.2分、
26−ヒドロキシビタミンD3 3.6分、
ビタミンD3 9.2分。
【0083】
まず、上記の条件でサンプルを分析し、形質転換株培養液中に25−ヒドロキシビタミンD3及び26−ヒドロキシビタミンD3が蓄積していることを確認した。次に培養時間経過による25−ヒドロキシビタミンD3蓄積濃度及び副反応物比率(%)を求めた。結果を表3及び図3に示す。親株とアミノ酸置換体の形質転換株は25−ヒドロキシビタミンD3の最終蓄積量に大きな差が認められなかった。副反応物比率は親株が約10%程度であるのに対し、pTAOR-K1形質転換株は2.5%程度、pTAOR-K1 I88V形質転換株は1%以下に副反応物比率が低減した。HPLCクロマトチャート上でも10倍希釈で測定すると、26−ヒドロキシビタミンD3のピークは検出限界以下になった。結果を図4に示す。以上より、I88Vアミノ酸置換体はシュードノカルディア・オートトロフィカにおいてもVdhの副反応抑制に顕著な効果があった。
【0084】
【表3】
【特許請求の範囲】
【請求項1】
配列番号2または4に記載のアミノ酸配列において、下記の(1)〜(4)からなる群より選択される少なくとも1つの改変部位を有する改変アミノ酸配列からなる改変型ビタミンD類水酸化酵素:
(1)88位のイソロイシンがバリンに置換された改変部位、
(2)171位のロイシンがバリンに置換された改変部位、
(3)177位のアラニンがアルギニンに置換された改変部位、
(4)240位のトレオニンがセリンに置換された改変部位。
【請求項2】
請求項1に記載の改変アミノ酸配列において、(1)〜(4)からなる群より選択される少なくとも1つの改変部位のほかに、さらに1または数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有する改変型ビタミンD類水酸化酵素。
【請求項3】
請求項1に記載の改変アミノ酸配列と90%以上の相同性を有し、(1)〜(4)からなる群より選択される少なくとも1つの改変部位を含むアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有する改変型ビタミンD類水酸化酵素。
【請求項4】
配列番号2または4に記載のアミノ酸配列において、88位のイソロイシンがバリンに、及び171位のロイシンがバリンにそれぞれ置換された改変アミノ酸配列からなる請求項1に記載の改変型ビタミンD類水酸化酵素。
【請求項5】
請求項4に記載の改変アミノ酸配列において、88位及び171位のアミノ酸置換のほかに、さらに1または数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有する改変型ビタミンD類水酸化酵素。
【請求項6】
請求項4に記載の改変アミノ酸配列と90%以上の相同性を有し、88位及び171位のアミノ酸置換を含むアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有する改変型ビタミンD類水酸化酵素。
【請求項7】
請求項1〜6のいずれか1項に記載の改変型ビタミンD類水酸化酵素をコードするDNA。
【請求項8】
配列番号1または3に記載の塩基配列において、下記の(1)〜(4)からなる群より選択される少なくとも1つの改変部位を有する改変塩基配列からなる請求項1に記載の改変型ビタミンD類水酸化酵素をコードするDNA:
(1)262〜264番目の塩基配列ATCが、バリンをコードするコドンに置換された改変部位、
(2)511〜513番目の塩基配列CTGが、バリンをコードするコドンに置換された改変部位、
(3)529〜531番目の塩基配列GCCが、アルギニンをコードするコドンに置換された改変部位、
(4)718〜720番目の塩基配列ACGが、セリンをコードするコドンに置換された改変部位。
【請求項9】
配列番号1または3に記載の塩基配列において、262〜264番目の塩基配列ATCがバリンをコードするコドンに、及び511〜513番目の塩基配列CTGがバリンをコードするコドンにそれぞれ置換された改変塩基配列からなる請求項8に記載のDNA。
【請求項10】
請求項7〜9のいずれか1項に記載のDNAを含む組換えベクター。
【請求項11】
請求項10に記載のベクターを含む形質転換体。
【請求項12】
請求項11に記載の形質転換体を培養し、培養中または培養後の形質転換体とビタミンD類とを接触させ、産出された水酸化ビタミンD誘導体を採取することを特徴とする水酸化ビタミンD誘導体の製造方法。
【請求項1】
配列番号2または4に記載のアミノ酸配列において、下記の(1)〜(4)からなる群より選択される少なくとも1つの改変部位を有する改変アミノ酸配列からなる改変型ビタミンD類水酸化酵素:
(1)88位のイソロイシンがバリンに置換された改変部位、
(2)171位のロイシンがバリンに置換された改変部位、
(3)177位のアラニンがアルギニンに置換された改変部位、
(4)240位のトレオニンがセリンに置換された改変部位。
【請求項2】
請求項1に記載の改変アミノ酸配列において、(1)〜(4)からなる群より選択される少なくとも1つの改変部位のほかに、さらに1または数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有する改変型ビタミンD類水酸化酵素。
【請求項3】
請求項1に記載の改変アミノ酸配列と90%以上の相同性を有し、(1)〜(4)からなる群より選択される少なくとも1つの改変部位を含むアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有する改変型ビタミンD類水酸化酵素。
【請求項4】
配列番号2または4に記載のアミノ酸配列において、88位のイソロイシンがバリンに、及び171位のロイシンがバリンにそれぞれ置換された改変アミノ酸配列からなる請求項1に記載の改変型ビタミンD類水酸化酵素。
【請求項5】
請求項4に記載の改変アミノ酸配列において、88位及び171位のアミノ酸置換のほかに、さらに1または数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有する改変型ビタミンD類水酸化酵素。
【請求項6】
請求項4に記載の改変アミノ酸配列と90%以上の相同性を有し、88位及び171位のアミノ酸置換を含むアミノ酸配列からなり、かつビタミンD類水酸化酵素活性を有する改変型ビタミンD類水酸化酵素。
【請求項7】
請求項1〜6のいずれか1項に記載の改変型ビタミンD類水酸化酵素をコードするDNA。
【請求項8】
配列番号1または3に記載の塩基配列において、下記の(1)〜(4)からなる群より選択される少なくとも1つの改変部位を有する改変塩基配列からなる請求項1に記載の改変型ビタミンD類水酸化酵素をコードするDNA:
(1)262〜264番目の塩基配列ATCが、バリンをコードするコドンに置換された改変部位、
(2)511〜513番目の塩基配列CTGが、バリンをコードするコドンに置換された改変部位、
(3)529〜531番目の塩基配列GCCが、アルギニンをコードするコドンに置換された改変部位、
(4)718〜720番目の塩基配列ACGが、セリンをコードするコドンに置換された改変部位。
【請求項9】
配列番号1または3に記載の塩基配列において、262〜264番目の塩基配列ATCがバリンをコードするコドンに、及び511〜513番目の塩基配列CTGがバリンをコードするコドンにそれぞれ置換された改変塩基配列からなる請求項8に記載のDNA。
【請求項10】
請求項7〜9のいずれか1項に記載のDNAを含む組換えベクター。
【請求項11】
請求項10に記載のベクターを含む形質転換体。
【請求項12】
請求項11に記載の形質転換体を培養し、培養中または培養後の形質転換体とビタミンD類とを接触させ、産出された水酸化ビタミンD誘導体を採取することを特徴とする水酸化ビタミンD誘導体の製造方法。
【図1】

【図2】
【図3】
【図4】
【図2】
【図3】
【図4】
【公開番号】特開2011−115078(P2011−115078A)
【公開日】平成23年6月16日(2011.6.16)
【国際特許分類】
【出願番号】特願2009−274179(P2009−274179)
【出願日】平成21年12月2日(2009.12.2)
【出願人】(000001915)メルシャン株式会社 (48)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成23年6月16日(2011.6.16)
【国際特許分類】
【出願日】平成21年12月2日(2009.12.2)
【出願人】(000001915)メルシャン株式会社 (48)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]