
ファゴピリトールの調製とその使用
【課題】ファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3の単利、およびファゴピリトールを用いる方法の提供。
【解決手段】ファゴピリトールA1、ファゴピリトールA2、ファゴピリトールA3、さらに、ファゴピリトールB1、ファゴピリトールB2、ファゴピリトールB3、およびD-カイロ-イノシトールの二つまたはそれ以上を含み、その少なくとも一つが単離されたファゴピリトールA1、単離されたファゴピリトールA3、または単離されたファゴピリトールB3である組成物についても、その使用法を提示する。原料とするソバから実質的に純粋なファゴピリトールA1、ファゴピリトールA3、ファゴピリトールB3、またはその混合物を調製する方法についても明らかにする。ファゴピリトールはその投与が糖尿病の治療に用いられうる薬学的組成物を調製するために用いることができる。
【解決手段】ファゴピリトールA1、ファゴピリトールA2、ファゴピリトールA3、さらに、ファゴピリトールB1、ファゴピリトールB2、ファゴピリトールB3、およびD-カイロ-イノシトールの二つまたはそれ以上を含み、その少なくとも一つが単離されたファゴピリトールA1、単離されたファゴピリトールA3、または単離されたファゴピリトールB3である組成物についても、その使用法を提示する。原料とするソバから実質的に純粋なファゴピリトールA1、ファゴピリトールA3、ファゴピリトールB3、またはその混合物を調製する方法についても明らかにする。ファゴピリトールはその投与が糖尿病の治療に用いられうる薬学的組成物を調製するために用いることができる。
【発明の詳細な説明】
【技術分野】
【0001】
本出願は、参照として本明細書に組み入れられる、1997年5月7日に提出された米国特許仮出願第60/045,927号の恩典を主張する、1998年5月6日に提出された米国特許出願第09/073,467号の一部継続出願である。
【0002】
発明の分野
本発明は、ファゴピリトールおよびファゴピリトールを用いる方法に関する。
【背景技術】
【0003】
発明の背景
真性糖尿病は、世界保健機構によって流行の割合に達していると認識されている主な世界規模の問題である。これは現在、先進諸国における死因の第4位であり、工業化が進む諸国では急速に増加しつつある疾患である。全世界の糖尿病発生率の推計は、1985年の3000万人から1994年での1億人を超えるまでに増加した。真性糖尿病は、糖質代謝の欠損によって引き起こされ、血液および尿に異常に大量のグルコースが存在することを特徴とする疾患である。真性糖尿病は、最終的に眼、腎臓、心臓、および脚の障害に至ることがあり、妊娠を危険に曝しうる。
【0004】
真性糖尿病は通常二つの種類に分類される。I型またはインスリン依存型真性糖尿病(「IDDM」)は、主に小児および若い成人に発症することからこれまで若年性糖尿病と呼ばれており、自己免疫疾患の一つに関係している。発症と進行が急激であり、これは全症例の約10〜15%を占める。II型またはインスリン非依存型真性糖尿病(「NIDDM」)は、これまで成人型糖尿病と呼ばれており、通常年齢40歳以上の人に認められ、進行は遅い。これはその初期段階で臨床的に病的な状態を伴わないことがしばしばあり、その代わりに血中グルコースレベルまたは尿中グルコースレベルによって検出される。
【0005】
糖尿病は、単一の障害というより多数の原因を有する障害群であると見なされる。ヒトの膵臓は、グルコースの体組織への流入およびその利用を促進して、それによって生体活動のエネルギーを供給するインスリンと呼ばれるホルモンを分泌する。しかし、糖尿病の人では、グルコースの流入が損なわれ、その結果産生されるインスリン量の欠乏または標的細胞の変化のいずれかが起こる。そのため、糖が血液中に蓄積して尿中に排泄される。I型糖尿病では、問題はほぼ常にインスリン産生の重度または全体的な減少である。II型糖尿病では、膵臓はしばしばかなりの量のインスリンを産生するが、このホルモンは組織によるグルコースの利用を促進することができない。
【0006】
適切な処置を行えば、ほとんどの糖尿病患者は血糖値を正常またはほぼ正常範囲に維持する。これによって、患者は通常の生活を送ることができ、疾患のいくつかの長期的な結末を予防することができる。インスリンをほとんどまたは全く産生しないI型糖尿病の場合、治療はインスリン注射を必要とする。II型糖尿病の場合、患者のほとんどが少なくとも中等度に体重過剰であるため、治療は食事制限、体重減少、および運動に基づく。体重の減少は、組織におけるインスリン耐性状態を部分的に逆転させるように思われる。II型患者の血糖値がそれでも高ければ、医師は、治療レジメにインスリン注射を加えてもよい。多くの場合、インスリン注射を必要とするのは、インスリンの欠乏によるためではなく、インスリンメディエータであるガラクトサミン-D-カイロ-イノシトールの欠乏のために患者ではインスリンの効率よい利用能が減少しているためである。
【0007】
注射による投与に関連した不快感の他に、インスリンの用量を制御する問題も存在する。インスリンの過量投与によって生じる低血糖症は振せん、冷や汗、立毛、低体温症、および頭痛を引き起こすことがあり、混乱、幻覚、奇異行動を伴い、最終的に痙攣および昏睡を伴うことがある。したがって、インスリン注射にたよることなく糖尿病患者の血糖値を制御できれば都合がよいと考えられる。本発明は、そのような制御を提供することに向けられる。
【発明の概要】
【0008】
発明の概要
本発明は、単離されたファゴピリトールA1、単離されたファゴピリトールA3、および単離されたファゴピリトールB3に関する。
【0009】
本発明はまた、ファゴピリトールA1、ファゴピリトールA2、ファゴピリトールA3、ファゴピリトールB1、ファゴピリトールB2、ファゴピリトールB3、およびD-カイロ-イノシトールの二つまたはそれ以上を含む組成物にも向けられる。組成物は、単離されたファゴピリトールA1、単離されたファゴピリトールA3、または単離されたファゴピリトールB3の少なくとも一つを含む。
【0010】
本発明は、ファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3からなる群より選択される実質的に純粋なファゴピリトールにも関する。
【0011】
もう一つの局面において、本発明は、ファゴピリトールA1、ファゴピリトールA3、ファゴピリトールB3、およびその混合物からなる群より選択される材料を調製する方法に向けられる。ソバを、粗抽出物を産生するために有効な条件で溶媒に接触させる。粗抽出物は、非ファゴピリトール材料ならびにファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3からなる群より選択される一つまたは複数のファゴピリトールを含む。次に、非ファゴピリトール材料を一つまたは複数のファゴピリトールから分離する。
【0012】
本発明の単離されたファゴピリトールは、同様に薬学的担体を含む薬学的組成物において用いることができる。この薬学的組成物または本発明の実質的に純粋なファゴピリトールまたは本発明の単離されたファゴピリトールは、糖尿病を治療するために患者に投与することができる。
【図面の簡単な説明】
【0013】
【図1】ソバ種子の成分を示す略図である。Aは成熟ソバ痩果の垂直方向の断面である。Bは成熟ソバ痩果の水平方向の断面である。Cはひき割り成熟ソバ(殻を除いたソバ痩果)の水平方向断面である。Dはその粉砕断片および破断平面を示すひき割り成熟ソバの水平断面である。
【図2】エタノール:水(1:1、v:v)によって成熟ソバ種子の胚組織から抽出したトリメチルシリル化可溶性成分のガスクロマトグラムである。ピークは以下のように表示される:(a)D-カイロ-イノシトール、(b)ミオ-イノシトール、(c)フェニルαD-グルコシド(内部標準)、(d)ショ糖、(e)ファゴピリトールA1、(f)ファゴピリトールB1、(g)ガラクチノール、(h)ラフィノース、(i)不明物質、(j)不明物質、(k)ファゴピリトールA2、(l)ファゴピリトールB2、(m)不明物質(おそらくジガラクトシルミオ-イノシトール)、(n)スタキオース、および(o)ファゴピリトールA3。
【図3】エタノール:水(1:1、容積:容積)によって20 DAPソバ種子の胚組織から抽出したトリメチルシリル化可溶性成分のガスクロマトグラムである。Aは加水分解前のガスクロマトグラムである;Bはαガラクトシダーゼ酵素による23時間の加水分解後のガスクロマトグラムである;およびCは2Nトリフルオロ酢酸(「TFA」)による70℃で3時間の酸加水分解後のガスクロマトグラムである。ピークは以下のように同定される:(a)D-カイロ-イノシトール、(b)ミオ-イノシトール、(c)フェニルα-D-グルコシド(内部対照)、(d)ショ糖、(e)ファゴピリトールA1、(f)ファゴピリトールB1、(g)ガラクチノール、(k)ファゴピリトールA2、(l)ファゴピリトールB2、(1)ガラクトース、(2)グルコース、および(3)果糖。グルコース、果糖、およびガラクトースのトリメチルシリル(「TMS」)誘導体は、アノマー型の糖を明確なTMS産物として捕獲する。
【図4】ソバ種子からのファゴピリトールの画分を示すグラフである。ひき割り成熟ソバを50%エタノールによって抽出して、80℃で45分間加熱し、加水分解酵素を不活化して、分子量カットオフ10,000のフィルターを通過させ、可溶性画分を蒸発乾固させた。残査をアセトニトリル:水(75:25、容積:容積)5mlに溶解して、3-アミノプロピルを9%官能化させたシリカゲルを充填した9×290 mmのカラム上でクロマトグラフィーを行った。成分をアセトニトリル:水(75:25、容積:容積)によって0.4 ml/分の流速で溶出し、3-アミノプロピルを9%官能化したシリカゲルを充填した9×290 mmのカラム上でクロマトグラフィーを行った。成分は、アセトニトリル:水(画分1〜70に関して75:25、容積:容積;および画分71〜90に関して60:40のアセトニトリル:水)によって0.4 ml/分で溶出した。各5ml画分の1mlを蒸発乾固させて、トリメチルシリルイミダゾール(「TMSI」)によって誘導体化して、ガスクロマトグラフィーによって分析した。選択した画分1ml試料を、酵素的および/または酸加水分解のために蒸発乾固させた。各画分における成分は以下のように同定される:(a)カイロ-イノシトール+ミオ-イノシトール+ショ糖、(b)ファゴピリトールA1、(c)不明物質、(d)ファゴピリトールB1、(e)ファゴピリトールA2、および(f)ファゴピリトールB2。
【図5】一連のガラクチノールの構造式を示す。酵素ガラクチノールシンターゼは、UDP-ガラクトースからのα-ガラクトシルを、D-ミオ-イノシトールの3位に転移して、ガラクチノール(O-α-D-ガラクトピラノシル-(1→3)-D-ミオイノシトール)を形成する。ガラクチノールは、ガラクトース供与体として作用して、多くの種子においてラフィノースシリーズのオリゴ糖を形成する。ガラクチノールはまた、アクセプターとして作用して、カラスノエンドウ(Vicia sativa L.)および他の豆果の種子において認められるジガラクトシルミオ-イノシトール(O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→3)-D-ミオイノシトール)を形成することができる(参照として本明細書に組み入れられる、ペテク(Petek)ら、「カラスノエンドウの種子由来のミオ-イノシトールの二つのガラクトシドの単離(Isolation of Two Galactosides of myo-Inositol from Vetch Seeds)」、C.R. Acad. Sci.(パリ)Serie D:Sci. Nat. 263:195〜197(1966)(ペテクI)、およびペテク(Petek)ら、「カラスノエンドウ種子の発芽における、αガラクトシダーゼの精製および特性(Purification and Properties of α-Galactosidase in Germinating Vicia sativa seeds.)」、Eur. J. Biochem. 8:395〜402(1969)(ペテクII))。少量の高次オリゴマー(トリガラクトシルミオ-イノシトールおよびテトラガラクトシルミオ-イノシトール)が、豆果種子に存在することがある(参照として本明細書に組み入れられる、カートワ(Courtois)ら、「単糖、オリゴ糖、およびポリオールの分布(Distribution of Monosaccharides, Oligosaccharides and Polyols)」、ハーボーン(Harborne)ら編、「マメ科植物の化学分類(Chemotaxonomy of the Leguminosae)」、ニューヨーク、アカデミック出版社、207〜229頁(1971)(「カートワ」))。
【図6】受粉後(「DAP」)6日〜32日に収穫した殻を除去した「マンカン」ソバ種子の発芽と乾燥耐性とを示すグラフであり、そのまま(○)または飽和LiCl溶液上で相対湿度(「RH」)12%で急速に乾燥後(●)発芽させた。発芽は湿らせたペーパータオル上で25℃で10日目に記録した。
【図7】1日目93%RH、2日目87%RH、3日目75%RH、4日目51%RH、5日目45%RH、および6日目33%RHでインキュベートした場合の、「マンカン」ソバの未成熟(10 DAP)なひき割り種子の緩速乾燥誘導耐性を示すグラフである。新鮮な種子と、連続的に低いRH環境で1〜6日までインキュベートした(様々な塩の飽和溶液上で)後の種子を、12%RHおよび22℃で1週間急速乾燥させた。乾燥後、種子を25℃でペーパータオル上で発芽させた。7日目に発芽した種子は、正常な幼根の成長を示し、乾燥に耐性であると考えられた。Aは、緩速乾燥時の日数の関数としての乾燥の耐性(●)および50%発芽するまでの日数(○)を示す。Bは緩速乾燥時の日数の関数としての胚あたりのシクリトールであるD-カイロ-イノシトール(●)およびミオ-イノシトール(■)の量を示す。Cは、緩速乾燥時の日数の関数としての胚あたりのガラクトシルシクリトールであるファゴピリトールB1(●)、ファゴピリトールA1(黒三角)、およびガラクチノール(■)の量を示す。Dは、緩速乾燥時の日数の関数としての胚あたりのジガラクトシルシクリトールであるファゴピリトールB2(●)、ファゴピリトールA2(黒三角)、およびジガラクトシル-ミオ-イノシトール(□)の量を示す。Eは緩速乾燥時の日数の関数としてのショ糖(●)の量を示す。
【図8】新鮮な10 DAPの未成熟な殻を除去した「マンカン」ソバ種子を高い相対湿度(98%)でインキュベートした日数の関数としての様々な特性を示すグラフである。Aは、高い相対湿度での日数の関数としての乾燥耐性(●)および50%発芽するまでの日数(○)を示す。Bは、高い相対湿度での日数の関数としての胚あたりのシクリトールであるD-カイロ-イノシトール(●)およびミオ-イノシトール(■)の量を示す。Cは、高い相対湿度での日数の関数としての胚あたりのガラクトシルシクリトールであるファゴピリトールB1(●)、ファゴピリトールA1(黒三角)、およびガラクチノール(■)の量を示す。Dは、高い相対湿度での日数の関数としての胚あたりのジガラクトシルおよびトリガラクトシルシクリトールであるファゴピリトールA2(黒三角)、ファゴピリトールB2(●)、ファゴピリトールA3(○)およびジガラクトシルミオ-イノシトール(□)の量を示す。Eは、高い相対湿度での日数の関数としての胚あたりのショ糖(●)の量を示す。
【図9A】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての乾燥耐性(●)および発芽開点始(○)を示す。
【図9B】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての軸の長さ(●)を示す。
【図9C】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての軸あたりのシクリトールであるD-カイロ-イノシトール(●)およびミオ-イノシトール(○)の量を示す。
【図9D】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての子葉あたりのシクリトールであるD-カイロ-イノシトール(●)およびミオ-イノシトール(○)の量を示す。
【図9E】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての軸あたりのファゴピリトールB1(●)、ファゴピリトールA1(○)、ファゴピリトールB2(■)、およびファゴピリトールA2(□)の量を示す。
【図9F】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての子葉あたりのファゴピリトールB1(●)、ファゴピリトールA1(○)、ファゴピリトールB2(■)、およびファゴピリトールA2(□)の量を示す。
【図9G】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての軸あたりのショ糖(●)の量を示す。
【図9H】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての子葉あたりのショ糖の量を示す。
【図9I】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての軸あたりのグルコース(●)、果糖(○)、および不明物質(■)の量を示す。
【図9J】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての子葉あたりのマルトース(●)、マルトトリオース(○)、およびマリトテトラオース(■)の量を示す。
【図10】3-アミノプロピル基官能性シリカゲル上で分離した様々な画分に存在する様々なファゴピリトールの量のグラフである。カラムにローディングした材料は、P2カラムから回収した濃縮ファゴピリトールA3画分であった。
【図11】3-アミノプロピル基官能性シリカゲル上で分離した様々な画分に存在する様々なファゴピリトールの量のグラフである。カラムにローディングした材料は、P2カラムから選択されたP2画分であった。
【図12】3-アミノプロピル基官能性シリカゲル上で分離した様々な画分に存在する様々なファゴピリトールの量のグラフである。カラムにローディングした材料は、全ひき割り粉(バーケットミルズ社)50 g由来の抽出物であった。
【図13】3-アミノプロピル基官能性シリカゲル上で分離した様々な画分に存在する様々なファゴピリトールの量のグラフである。カラムにローディングした材料は、全ひき割り粉(バーケットミルズ社)50 gからの抽出物であった。
【図14】3-アミノプロピル基官能性シリカゲル上で分離した様々な画分に存在する様々なファゴピリトールの量のグラフである。カラムにローディングした材料は、全ひき割り粉(バーケットミルズ社)50 gからの抽出物であった。
【図15】3-アミノプロピル基官能性シリカゲル上で分離した様々な画分に存在する様々なファゴピリトールの量のグラフである。カラムにローディングした材料は、軽ひき割り粉(ミンダック社)50 gからの抽出物であった。
【図16】エタノール:水(v/v)によって段階的に溶出したファゴピリトールを含む予備植物抽出物(20 g総量の糖)の炭素-セライトカラム(cc38)クロマトグラフィーを示す。
【図17】ジガラクトシルミオ-イノシトール(DGMI)、ファゴピリトールA2(A2)、ファゴピリトールB3(B3)、トリガラクトシルミオ-イノシトール(T、TGMI)、およびファゴピリトールA3(A3)の溶出を示す図16からの縮尺を拡大したプロットを示す。
【図18】関係するピークを図示するために縮尺を拡大してプロットしたファゴピリトールA3(A3、38.23分)、ファゴピリトールB3(B3、39.00分)、およびトリガラクトシルミオ-イノシトール(TGMI、39.46分)を示すガスクロマトグラムである。
【図19】それぞれファゴピリトールA3(A3;37.83 分)、ファゴピリトールB3(B3;38.50分)、およびトリガラクトシルミオ-イノシトール(TGMI;38.97分)を濃縮した画分のピークの分離を示すGCクロマトグラムを三つ重ね合わせた軌跡を示す。
【発明を実施するための形態】
【0014】
発明の詳細な説明
本発明は、ファゴピリトールに関する。ファゴピリトールは本明細書において用いられる一般的な用語であり、特定されないα-ガラクトシルD-カイロ-イノシトールまたはその塩もしくは誘導体を意味する。より詳しく述べると、本発明は単離されたファゴピリトールA1、特に以下の式Iを有する複数のファゴピリトールA1およびその塩に関する:
【化4】

【0015】
本発明はまた、単離されたファゴピリトールA3、特に以下の式IIを有する複数のファゴピリトールA3に関する:
【化5】
【0016】
本発明はさらに、単離されたファゴピリトールB3、特に以下の式IIIを有する複数のファゴピリトールB3ならびにその塩および誘導体に関する:
【化6】
【0017】
本発明の複数のファゴピリトールA1、A3およびB3の塩は、水酸化金属または金属アルコキシド、水酸化アンモニウムまたはアミン(例えば、トリエチルアミンのような3級アミン)のような、ファゴピリトールのヒドロキシル基の一つまたは複数のpKa以上のpKa(すなわち、-log Ka)を有する塩基との反応産物となりうる。例としての塩は、リチウム塩、ナトリウム塩、およびカリウム塩のようなアルカリ金属塩、カルシウム塩およびバリウム塩のようなアルカリ土類金属塩、アンモニウム塩、スルホニウム塩、ならびにホスホニウム塩である。
【0018】
本発明の複数のファゴピリトールA1、A3、およびB3の誘導体には、例えば、ハロゲン化アルカリのような陽性荷電を有する炭素を有する化合物とファゴピリトールとの反応産物が含まれ、この場合誘導体はファゴピリトールのエーテルとなり、またはハロゲン化カルボン酸(例えば塩化アセチル)もしくは無水物(例えば、無水酢酸)との反応産物が含まれ、この場合誘導体はファゴピリトールのエステル(例えばアセテート)となる。
【0019】
本明細書において用いられるように、単離されたファゴピリトールは、それが自然界に存在する他のソバ成分を実質的に含まないファゴピリトールである。本発明の単離されたファゴピリトールは、例えば、ソバまたはその他の天然資源からの抽出と共に化学合成を含む様々な方法によって調製することができると理解すべきである。
【0020】
本発明はまた、好ましくは式I、IIおよびIIIをそれぞれ有する実質的に純粋なファゴピリトールA1、実質的に純粋なファゴピリトールA3、および実質的に純粋なファゴピリトールB3、その塩および誘導体にも関する。本発明はまた、実質的に純粋なファゴピリトールA2およびその塩にも関し、特に複数のファゴピリトールA2は以下の式IVを有する:
【化7】
【0021】
本発明はさらに、実質的に純粋なファゴピリトールB1、特に以下の式Vを有するファゴピリトールならびにその塩および誘導体にも関する:
【化8】
本発明はまた、実質的に純粋なファゴピリトールB2、特に以下の式VIを有するファゴピリトールと共にその塩および誘導体にも関する:
【化9】
本明細書において言及されるように、実質的に純粋とは、ガラクチノール、ミオ-イノシトール、ジガラクトシルミオ-イノシトール、フィチン、芳香性物質(例えば、ポリフェノールおよび色素、ならびに他の着色芳香性物質)、細胞壁粒子、タンパク質、ならびに酸(例えば、有機酸、核酸、およびアミノ酸)およびその塩のような他の成分または材料を実質的に含まないことを意味する。典型的に、実質的に純粋なファゴピリトールは、約98%を超える純度または約95%〜約99%の純度のような、95%を超える純度を有するファゴピリトールである。
【0022】
上記のように、本発明のファゴピリトールは、その調製方法によって制限されない。特に好ましい一つの方法は、ソバからの本発明のファゴピリトールの精製を含む。簡単に説明すると、非ファゴピリトール材料と、ファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3からなる群より選択される一つまたは複数のファゴピリトールとを含む粗抽出物を産生するために有効な条件で、ソバを溶媒に接触させる。次に、一つまたは複数のファゴピリトールを非ファゴピリトール材料から分離する。この過程は下記により詳細に説明する。
【0023】
ソバ植物(ファゴピルム・エスクレンツム(Fagopyrum esculentum))の任意の部分を、本発明のファゴピリトールを調製するために用いることができる。
【0024】
開発中のソバ種子の構造は、参照として本明細書に組み入れられる、オーベンドルフ(Obendorf)ら、「開発中のソバ種子の構造および化学的組成(Structure and Chemical Composition of Developing Buckwheat Seed)」、244〜251頁、ヤニク(Janick)ら編、「新たな穀物(New Crops)」、ニューヨーク、ジョンウィリー・アンド・サンズ(1993)(「オーベンドルフ」)中、によって論評され説明されている。成熟したソバ種子の写真は、参照として本明細書に組み入れられる、マーシャル(Marshall)ら、「ソバ:説明、育種、生産、および利用(Buckwheat:Description, Breeding, Production, and Utilization)」、Advances in Cereal Science and Technology 5:157〜210(1982)(「マーシャル」))およびポメランズ(Pomeranz)、「ソバ:構造、組成、および利用(Buckwheat:Structure, Composition, and Utilization)」、CRC Critical Reviews in Food Science and Nutrition 19:213〜258(1983))による論評に示されている。後者の論評における構造情報のほとんどは、参照として本明細書に組み入れられる、ポメランズ(Pomeranz)、「ソバ穀粒の走査型電子顕微鏡写真(Scanning Electron Microscopy of the Buckwheat Kernel)」、Cereal Chemistry 49:23〜25(1972))によるソバ穀粒の走査型電子顕微鏡研究から得ている。胚は、図1Aに略図で示すように、ソバ痩果(成熟したソバ果の名称)の先端の胚の幼根(根)に存在する。参照として本明細書に組み入れられる、オーベンドルフによって説明されるように、子葉は、受粉後8日目までに軟らかい(ほとんど液体)内乳から伸張して内乳の底に達する。痩果は、小柄によって親植物の総状花序(花序または花集合体の名称)に結合する。軸および子葉を含む胚は、内乳の中を生育し続け、内胚葉の端部で巻いて、受粉後16日目で最高乾燥重量に達する。内乳は、受粉後24〜28日までに乾燥物質(ほとんどがデンプン)を蓄積し続ける。さらに数日後、ソバ穀粒(痩果または果実)が成熟して乾燥する。
【0025】
成熟ソバ痩果の水平断面図(図1Aにおいて「断面図」と記した線)を図1Bに略図で示す。この「階層(top-douwn)」図は痩果の外層として外殻(果皮または果実の外皮)を示す(図1Bおよび1Cにおける斜線入りの黒い部分)。外殻は通常黒または暗茶色の硬い繊維様構造である。外殻は粉砕の際に除去される最初の層であり、それによって殻が除去されたひき割り果(果皮のない真の「種子」)を生じる(図1C)。殻を除去したひき割り果の外側の部分は、「皮膜」または種皮(種皮とも呼ばれる)である。これは、細胞壁が肥厚した種皮細胞と核の残遺物(外乳;厚さ細胞1〜3個分)で構成される繊維層である。内乳における細胞の最も外側の層は糊粉層と呼ばれ、通常、成熟穀粒において種皮と核組織に接着する。糊粉は、中心内乳細胞より厚みのある細胞壁を有する小さい細胞の単層であり、デンプンを含まず、種子を乾燥後もなおも「生存」している。内乳細胞の残遺物は、より薄い細胞壁を有し、参照として本明細書に組み入れられる、マーシャル(Marshall)において説明されるようにデンプン顆粒が充填されている。
【0026】
ソバ胚は軸(中心部の円)と、内乳の中を辺縁部まで巻いて、曲がって種皮近傍の内胚葉の外側部分に沿って続く二つの薄い葉状の子葉とを有する(図1Bおよび1C;胚の構造は黒色、内乳構造は白色、および種皮または「皮膜」は灰色)。成熟穀粒では、子葉は内乳をほぼ取り巻く。胚(軸プラス子葉)は、成熟乾燥穀粒においても生存している。発芽後、胚はソバの実生を形成する。成熟乾燥穀粒では、胚と糊粉層の細胞は、ソバ穀粒における全てのファゴピリトールとD-カイロ-イノシトールとを含む。胚(糊粉層と共に)は、ほとんどのタンパク質と、油、フィチン、無機質、ビタミン、およびルチンのほぼ全てを含むが、デンプンは微量に過ぎない。ほぼ全てのデンプンは内乳に存在するが、内乳の最も外側の層である糊粉層にはデンプンは存在しない。
【0027】
全ひき割り粉は、画分を分離しないですりつぶした全ひき割りソバ(図1C)を含む。外皮が繊維様性質であるために、ひき割りソバの粉砕時により大きな断片が通常形成される。破断平面は通常厚みのある細胞壁を有する層の内側である。小麦、オート麦、およびオオムギの乾燥粉砕物と同様に、ふすま画分は果皮(種皮を含む)、核残遺物、糊粉、および糊粉下層を含む。破断平面は、これらの穀粒において、壁がより厚みのある糊粉下層と壁がより薄い中心内乳のあいだである。同様に、ソバの糊粉層は通常、粉砕時に「ふすま」または「皮膜」画分に接着する。ふすま画分では、「種皮」(種皮としても知られる)または「皮膜」は、二つの子葉の外側に接着して子葉のその部分が引き裂かれて、ふすま画分と共に分離される(図1Dにおける理論的破断平面の略図を参照のこと)。必ずしも全ての皮膜が、接着した子葉組織を有するわけではない。ほとんどの内乳は、粉砕時に粉を形成する軟らかい薄い壁の細胞を有する。いくつかの種子において、内乳中心部の子葉は、全くまばらで、「純粋な」子葉組織のより大きな部分に剥がれ落ちる可能性がある。子葉と軸のこれらのより大きな断片は、ふすま画分にも分離される可能性があるが、軟らかい胚組織の多くは微粉にされてデンプンに富む明るい色の粉画分と共に分離する。これらの理由から、ふすま画分はファゴピリトールとD-カイロ-イノシトールに富む原料となりうる。同様に、明るい色の粉画分は、内乳に埋もれた胚画分からのファゴピリトールを含むが、その濃度は大量のデンプンによって希釈されるために低い。内乳の前に胚が成熟するために、胚の大きさは成熟穀粒において内乳の大きさより小さく変動する。より小さい種子(より少ない内乳)では、子葉は種皮により多く接触しているのに対し、より大きな種子(より多い内乳)では、子葉は種皮全体にあまり接触していない。画分において外殻を有しない「精製されたふすま」は、ファゴピリトール、無機質、タンパク質、および油に富む。
【0028】
この説明にとって重要であるのは、胚細胞(子葉と軸)および糊粉層の細胞がショ糖、ファゴピリトール、ルチン、フィチン、ほとんどの無機質、ほとんどのビタミン、ほぼ全ての油、および高濃度のタンパク質を含む細胞である点である。胚および糊粉層の細胞は、乾燥種子における「生存」細胞であるが、糊粉下、中心内乳、および種皮(種皮または皮膜)の細胞は死亡する。外殻(果皮)の細胞も同様に乾燥種子では死んでいる。このように、胚と糊粉層細胞に富む任意の画分をこれらの成分に富むと考えられる。中心内乳細胞は主にデンプンであり、したがって、明るい色の粉画分はデンプンに富む。しかし、胚が中心内乳を横切るため、胚の多くは小さい断片に破壊されて、明るい色の粉画分に入るが、暗い色の粉画分(外皮も含む)または全ひき割り粉にも存在すると考えられる。
【0029】
本発明のファゴピリトールを調製するために用いられるソバ植物の好ましい部分は、ソバ胚、特に胚軸、または「ふすま」もしくは「精製ふすま」粉砕画分を含む、胚、子葉、軸、または糊粉層細胞に富む粉砕画分である。容易に利用できることから、市販の特選ソバ粉(好ましくは水分約12.3%を含む)のようなソバ粉、または市販の全白ひき割り粉から調製した全ひき割り粉は、ファゴピリトールを単離するための優れたソバ原料であることが判明している。または、粉はソバ種子、好ましくは殻を除去したソバ種子から産生することができる。ソバの起源にかかわりなく、ソバ粉のようなソバを微粉に粉砕すれば、好ましくは100目のふるいを通過するような粉末に粉砕すれば、抽出は改善される。ソバ粉は生または脱脂粉となりえて、脱脂はアルカン溶媒、好ましくはヘキサンのような脂肪酸のための溶媒と生ソバとを混合することによって行う。脱脂は、初回抽出段階の効率を増加させるが、一般的に脱脂粉を用いた場合にはその後の濾過およびクロマトグラフィーがより難しくなる。
【0030】
次に、粉を適した溶媒によって抽出する。抽出過程にとって適した溶媒には、水、アルコール、および水/アルコール混合物が含まれる。水抽出物は、おそらくタンパク質およびデンプンの水中の溶解度が高いために、典型的により懸濁材料を含む。アルコールおよびアルコール/水抽出物は一般的に懸濁材料をあまり含まず、抽出の際の微生物および内因性酵素活性を最小限にする。アルコールおよびアルコール/水抽出において用いられる適したアルコールには、メタノール、エタノール、イソプロパノール、およびn-プロパノールが含まれ、エタノールが好ましい。水/エタノール混合物は好ましい抽出溶媒である。これらの中で、好ましい溶媒混合物は、エタノール対水の容量比が約1:2〜約2:1、より好ましくは約1:1である混合物である。
【0031】
抽出は、ソバ粉またはその他のソバ原料を適量の抽出溶媒と共に、好ましくは攪拌しながら混合することによって行われる。攪拌は、ブレンダー、好ましくは「高剪断力」のポリトロンヘッドを有するブレンダー、パドルミキサー、または剪断ミキサーのような任意の適した手段によって行うことができる。攪拌装置としてブレンダーを用いる場合、ブレンダーのスイッチを10秒間入れて10秒間切るサイクルを1分または2分間繰り返すと良好な結果が得られる。粉の単位重量あたり用いられる抽出溶媒の容積は、本発明の実践にあたって特に重要ではない。典型的に、溶媒/粉比(ml/g)約1:1〜約200:1が適しており、溶媒/粉比約10:1〜約50:1が好ましく、溶媒/粉比約20:1が最も好ましい。
【0032】
抽出を行う温度は、本発明の実践にとって特に重要ではない。しかし、熱い抽出後では、混合物は濃厚になりすぎて(ソバに存在するデンプンの膨張のため)濾過または遠心を効率よく行うことができず、抽出されるファゴピリトールの容量が少なくなる。約4℃〜ほぼ室温(約25℃)までの温度での低温抽出、好ましくは約10℃〜ほぼ室温までの温度、最も好ましくはほぼ室温での低温抽出が一般的に最も高い収率を生じる。
【0033】
抽出過程の時間は、攪拌の種類および激しさ、抽出を行う容器の形状、溶媒対粉比、抽出過程を行う温度、抽出される材料の容積、抽出される材料の粉砕の細かさ(粒子径)等を含む多くの要因に依存する。好ましくは、抽出は均一な材料が得られるように十分な時間行う。典型的に、約15秒〜約1時間、好ましくは約1分〜約5分、より好ましくは約1分〜約2分が有効である。
【0034】
抽出されたファゴピリトールの収率はしばしば、抽出過程を1回、2回、またはそれ以上繰り返すことによって増加させることができる。典型的に、最初の抽出では存在するファゴピリトールの85%〜95%が抽出される。2回目の抽出を行うと、さらに8%〜10%収率が増加して、3回目の抽出によって一般的にさらに1%収率が上昇する。
【0035】
第一のおよび選択的にさらなる抽出段階を行った後、ホモジネートを沈殿させて、溶媒を除去する。溶媒の除去は任意の適した手段によって行ってよく、濾過、特に限外濾過または遠心が最もよい結果を生じる。次に、粗抽出物(すなわち、遠心後の上清または濾過後の濾液)を好ましくは、ソバに存在してファゴピリトールを加水分解する可能性がある任意のα-ガラクトシダーゼおよび/またはその他の酵素を不活化するために十分な温度に加熱する。一般的に、不活化は、粗抽出物を約70℃〜約90℃、好ましくは約80℃の温度に加熱することによって行うことができる。冷却後、粗抽出物を好ましくは濾過によって透明にし、蒸発によって濃縮した後、限外濾過によってさらに透明にする。粗抽出物の組成は、抽出過程およびその後の濾過、遠心、不活化、蒸発、または透明化の際に、ガス液体クロマトグラフィー、高速液体クロマトグラフィー(「HPLC」)、分光光度分析、色素結合タンパク質アッセイ、および原子吸収分析を含む標準的な方法によってモニターすることができる。必要であれば、ファゴピリトールの濃縮画分をエタノール沈殿によって画分して、高次オリゴマー型の望ましくない可溶性糖質(例えば、五糖および高次糖)を除去することができる。
【0036】
ファゴピリトールの回収はさらに、細胞壁粒子および大きなタンパク質を、例えば粗抽出物を、約5000〜約20,000、好ましくは約10,000の分子量カットオフ濾紙に通過させて除去することによって、さらに増強することができる。
【0037】
さらにまたは代わりは、ファゴピリトールは例えば、サイバーン樹脂(アフテックインク、ロチェスター、ニューヨーク州から入手可能)、ポリビニルポリピロリドン(「PVPP」)(例えば、ポリクラー-VT、ISPテクノロジーズインク、ウェイン、ニュージャージー州から販売)、ベントナイト(例えば、KWKベントナイト凝集粉、プレスク島ワインセラーズ、ノースイースト、ペンシルバニア州から販売)、および珪藻土粉末(例えば、セライトコーポレーション、ラモック、カリフォルニア州から販売)のような食品等級のイオン交換樹脂によって非糖質の混入物質を除去することによってさらに精製することができる。説明すると、荷電材料は、粗抽出物をアンバーライトERA-94イオン交換樹脂およびドウェックス50Wx4イオン交換樹脂(いずれもシグマケミカル社、セントルイスから市販されている)のようなイオン交換材料に通過させることによって、粗抽出物から除去することができる。ファゴピリトールのさらなる精製は、粗抽出物を、ポリビニルピロリドン(「PVP」)または活性炭のような芳香性物質を吸収する材料を含むカラムに通過させることによって、ポリフェノール材料、芳香性色素、およびその他の茶色の混入物質のような芳香性物質を除去することによって行うことができる。次に、濃縮粗抽出物を冷やすか、もしくはエタノールのようなアルコールを濃縮抽出物に加えること、またはその双方によってファゴピリトールを沈殿させることができる。
【0038】
次に、得られたファゴピリトールと他の糖との混合物は、例えば、クロマトグラフィーによってその成分に分離することができる。例えば、クロマトグラフィーによる分離は、アミン官能性シリカゲル上で、もしくは炭素セライトカラムを用いることによって、またはその両者を用いて行うことができる。一般的に、水溶性の低分子量着色混入物を除去するためには炭素セライトカラムのほうがよい。
【0039】
好ましいアミン官能性シリカゲルである3-アミノプロピル官能性シリカゲルは、シグマ-アルドリッチ社(ミルウォーキー、ウィスコンシン州)から販売されている。シリカゲルの官能化(functionalization)の程度は本発明の実践にとって重要ではない。好ましくは、このシリカゲルは約5%〜約15%官能化しており、より好ましくは約9%官能化している。粗抽出物をローディングして様々なファゴピリトールを3-アミノプロピル官能性シリカゲルから溶出するために適した溶媒は、当業者によって容易に確認することができる。好ましくは、分離はアセトニトリル/水の溶離剤を用いて行う。より好ましくは、粗抽出物はアセトニトリル:水(70:30、v:v)溶液としてローディングして、70:30(v:v)アセトニトリル:水、60:40(v:v)アセトニトリル:水、および次に50:50(v:v)アセトニトリル:水によって段階的に溶出する。溶出の後、3-アミノプロピル官能性シリカゲルは60:40(v:v)アセトニトリル:水によって再生することができる。一般的に、3-アミノプロピル官能性シリカゲルを用いる場合の溶出の順序は以下の通りである:単糖類、D-カイロ-イノシトール、ミオ-イノシトール、ショ糖、トレハロース、ファゴピリトールA1、ファゴピリトールB1、ガラクチノール、ファゴピリトールA2、ファゴピリトールB2、ジガラクトシルミオ-イノシトール、ファゴピリトールA3、ファゴピリトールB3、およびトリガラクトシルミオ-イノシトール。
【0040】
セライト炭素カラムを用いてクロマトグラフィーによる分離を行う場合、適したカラムには、炭素(例えば、ダルコG60、J.T.ベイカー、フィリップスバーグ、ニュージャージー州)とセライト(例えば、セライト545-AW、スペルコ、ベルフォンテ、ペンシルバニア州)との混合物を含むカラムが含まれる。一般的に、カラム材料は炭素とセライトとを蒸留水でスラリーにすることによって調製する。次に、適した大きさのカラムにスラリーを充填して、充填したカラムを蒸留水で洗浄する。粗抽出物は、任意の適した溶媒を用いてローディングすることができ、水が好ましい。炭素セライトカラムは、例えば、水、アルコール(例えば、エタノール)、またはその組み合わせによって溶出することができる。説明すると、溶出は50%エタノール(すなわち、エタノールと水の50:50(v/v)混合物)を用いて行うことができる。または、溶出は蒸留水でまず溶出した後、エタノール含有量を増加させた一連のエタノールと水との混合物によって溶出することによって段階的に行うことができる。例えば、カラムをまず蒸留水によって溶出して次に5%エタノール、次に10%エタノール、次に20%エタノール、次に30%エタノール、次に40%エタノール、そして次に50%エタノールによって溶出することができる。場合によっては、一つまたは複数の溶出段階を省略することができる。例えば溶出は、30%および40%エタノール溶出を行わずに、20%エタノール溶液から50%溶出に直接進行することができる。または、溶出は水/エタノール勾配溶媒系を用いて行うことができる。一般的に炭素-セライトを用いる場合の溶出順序は以下の通りである:D-カイロ-イノシトール、ミオ-イノシトール、単糖類、ファゴピリトールB1、ガラクチノール、ファゴピリトールA1、トレハロース、ファゴピリトールB2、ショ糖、ジガラクトシル-ミオ-イノシトール、ファゴピリトールA2、ファゴピリトールB3、トリガラクトシルミオ-イノシトールおよびファゴピリトールA3。
【0041】
様々な画分の組成は、画分からの少量を蒸発乾固させ、トリメチルシリルイミダゾール(「TMSI」)のピリジン溶液(1:1)によって誘導体化してトリメチルシリル(「TMS」)誘導体化糖質を作製して、例えば高分解能ガスクロマトグラフィーによってTMS誘導体化糖質を分析することができる。一つの特定のファゴピリトールを含む画分をプールして蒸発乾固させることによって、実質的に純粋なファゴピリトールが得られる。ファゴピリトールの混合物を含む画分を再度クロマトグラフィーを行ってその分離を行うことができる。
【0042】
本発明はまた、二つまたはそれ以上のファゴピリトールA1、ファゴピリトールA2、ファゴピリトールA3、ファゴピリトールB1、ファゴピリトールB2、ファゴピリトールB3、およびD-カイロ-イノシトールを含む組成物にも関する。好ましくは、この組成物には、少なくとも一つの単離されたファゴピリトールA1、ファゴピリトールA3、またはファゴピリトールB3が含まれる。例としての組成物は、単離されたファゴピリトールA1と単離されたファゴピリトールA2、単離されたファゴピリトールB2と単離されたファゴピリトールB3、D-カイロ-イノシトールと単離されたファゴピリトールB3等を含む組成物である。好ましくは、組成物は、一つまたは複数のガラクチノール、ミオ-イノシトール、ジガラクトシルミオ-イノシトール、フィチン、芳香性物質(例えば、ポリフェノール、色素、およびその他の着色芳香性物質)、細胞壁粒子、タンパク質、ならびに酸(例えば、有機酸、核酸、およびアミノ酸)およびその塩を実質的に含まない。ファゴピリトールの混合物は、研究室でのインビトロ条件でヒト便の細菌の存在下で6時間以内に分解された。したがって、ファゴピリトールは消化管における細菌によって消化されて、遊離のD-カイロ-イノシトールが放出されて取り込まれると考えられており、単量体または二量体の場合、消化管の細胞によって取り込まれる可能性がある。
【0043】
ファゴピリトールおよび組成物は、上記の単離されたファゴピリトール、実質的に純粋なファゴピリトール、または組成物をイヌ、ネコ、ラット、マウス、およびヒトを含む哺乳類のような患者に投与することによって、そのような患者における糖尿病を治療するために有用である。上記のファゴピリトールおよび組成物はまた、多嚢胞性卵巣症候群を治療するために有用となる可能性がある(参照として本明細書に組み入れられる、ネスラー(Nestler)ら、New England J. of Med. 340:1314〜1320(1999))。例えば、本発明の実質的に純粋なファゴピリトール、組成物、または一つもしくはそれ以上の単離されたファゴピリトールは、単独で投与することができ、または本発明の単離されたファゴピリトールは適した薬学的担体または希釈剤と共に投与することができる。希釈剤または担体成分は、それらが本発明のファゴピリトールまたは組成物の治療効果を減少させないように選択すべきである。適した薬学的組成物には、薬学的担体と、例えば単離されたファゴピリトールA1、単離されたファゴピリトールA3、または単離されたファゴピリトールB3とを含む組成物が含まれる。薬学的組成物はさらに、ファゴピリトールA2、ファゴピリトールB1、ファゴピリトールB2、または三つ全てを含みうる。
【0044】
本明細書に記載のファゴピリトールおよび組成物は、所望の用途に適当な適した任意の形状で構成することが可能である:例えば、経口、非経口、または局所投与。非経口投与の例には、脳室内、脳内、筋肉内、静脈内、腹腔内、直腸内、および皮下投与である。好ましい投与経路は経口である。ファゴピリトールが局所または非経口投与される場合には、ファゴピリトールは予め加水分解することが好ましい。
【0045】
経口で用いるために適した投与剤形には、錠剤、分散粉末、顆粒、カプセル剤、懸濁剤、シロップ剤、およびエリキシル剤が含まれる。錠剤の不活性希釈剤および担体には、例えば炭酸カルシウム、炭酸ナトリウム、乳糖およびタルクが含まれる。錠剤はまた、デンプンおよびアルギン酸のような造粒および崩壊物質;デンプン、ゼラチンおよびアカシアのような結合剤;およびステアリン酸マグネシウム、ステアリン酸およびタルクのような潤滑剤が含まれる。錠剤は、非コーティングであってもよく、または崩壊および吸収を遅延させるために既知の技術によってコーティングしてもよい。カプセル剤において用いてもよい不活性希釈剤および担体には、例えば、炭酸カルシウム、リン酸カルシウム、およびカオリンが含まれる。懸濁剤、シロップ剤、およびエリキシル剤はメチルセルロース、トラガカント、アルギン酸ナトリウムのような従来の賦形剤;レシチンおよびステアリン酸ポリオキシエチレンのような湿潤剤;ならびにエチル-p-ヒドロキシベンゾエートのような保存剤を含んでもよい。非経口投与に適した投与剤形には、溶液、懸濁液、分散剤、乳剤等が含まれる。それらはまた、使用直前に滅菌注射用媒体に溶解または懸濁することができる滅菌固形組成物の形で製造してもよい。それらは、当技術分野で既知の懸濁剤または分散剤を含んでもよい。
【0046】
経口投与の場合、固形または液体単位投与剤形を調製することができる。錠剤のような固形組成物を調製する場合、上記に開示したように適したファゴピリトールまたは組成物を、タルク、ステアリン酸マグネシウム、リン酸二カルシウム、ケイ酸アルミニウムマグネシウム、硫酸カルシウム、デンプン、乳糖、アカシア、メチルセルロース、および薬学的希釈剤または単体と機能的に類似の材料のような従来の成分と混合する。カプセル剤は、開示のファゴピリトールまたは組成物を不活性な薬学的希釈剤と混合して、混合物を適当な大きさの硬いゼラチンカプセルに充填することによって調製される。軟ゼラチンカプセルは、ファゴピリトールまたは組成物と、許容される植物油、軽白色鉱油、または他の不活性油とのスラリーを機械的に封入することによって調製される。
【0047】
シロップ剤、エリキシル剤および懸濁剤のような経口投与のための液体単位投与剤形を調製することができる。水溶性型は、糖、芳香性着香料および保存剤と共に水性媒体に溶解してシロップ剤を形成することができる。エリキシル剤は、芳香性着香料と共に、糖およびサッカリンのような適した甘味料と共に水性アルコール(エタノール)溶媒を用いることによって調製する。懸濁剤は、アカシア、トラガカント、メチルセルロース等のような懸濁剤の助けを借りてシロップ溶媒と共に調製することができる。
【0048】
ファゴピリトールまたは組成物を経口投与する場合、適した1日用量は、参照として本明細書に組み入れられる、ラーナー(Larner)らの米国特許第5,124,360号に記載のような、遊離のD-カイロ-イノシトールの適した用量に基づくことができる。抽出されたファゴピリトールの約半分がD-カイロ-イノシトールであり、ほとんどが結合型のD-カイロ-イノシトールであり、遊離のD-カイロ-イノシトールは少量であると考えられる。したがって、ファゴピリトールの適した用量は、D-カイロ-イノシトールの適した用量の約2倍である。典型的に、経口投与の場合、適した1日用量は、被験者の体重1kgあたり約5 mg〜約200 mgのファゴピリトールまたは組成物である。
【0049】
または、本発明のファゴピリトールは、食品に入れて経口的に摂取することができる。例えば、ファゴピリトールは、パン、ロールパン、およびその他の食品において、精製型で、またはソバのふすまの形で組み入れて、ファゴピリトールを摂取するための食品を作製することができる。抽出したファゴピリトールによってパン、ロールパン、およびその他の食品を栄養強化することは、日常的な食事に大量のファゴピリトールを取り込む方法を提供しうる。パンを調製するための適した技法は、例えば、参照として本明細書に組み入れられる、ブラウン(Brown、「タサヤラのパンの本(The Tassajara Bread Book)」、ボストン:シャンバラ出版(1986))に見出される。
【0050】
非経口投与の場合、液体単位用量剤形は上記のファゴピリトールまたは組成物と、水が好ましい滅菌溶媒とを利用して調製される。用いる溶媒および濃度に応じて、ファゴピリトールまたは組成物は、溶媒に懸濁または溶解することができる。溶液を調製する場合、ファゴピリトールまたは組成物は、注射するために水に溶解して、濾過滅菌してから適したバイアルまたはアンプルに充填して密封することができる。都合のよいことに、局所麻酔剤、保存剤、および緩衝剤のようなアジュバントは溶媒に溶解することができる。安定性を増強するために、液体単位用量剤形は、バイアルに充填して、水分を真空下で除去した後凍結することができる。次に、凍結乾燥粉末をバイアルに密封して、使用前に溶液にするために注射用水のバイアルを添付して供給する。非経口懸濁剤は、ファゴピリトールまたは組成物を溶解する代わりに溶媒に懸濁して、実質的に同じように調製するが、この場合、濾過滅菌を行うことができない。ファゴピリトールまたは組成物は滅菌溶媒に懸濁する前にエチレンオキサイドに曝露することによって滅菌することができる。ファゴピリトールまたは組成物の均一な分布を促進するために、界面活性剤または湿潤剤を非経口懸濁液に含めることが都合がよい。非経口投与量は、1日あたり被験者の体重1kgあたり約5 mg〜約200 mgの範囲のファゴピリトールまたは組成物となりうる。好ましくは、非経口投与の1日用量は、経口投与では、ファゴピリトールからのガラクトースが消化管の微生物によって消費されるが、非経口投与ではガラクトースは血糖値に関与することを考慮して、被験者の体重1kgあたりの用量よりかなり低いと考えられる。
【0051】
または、ファゴピリトールもしくは組成物は、従来の方法を用いて徐放性製剤に組み入れて、外科的に埋め込むことができる。適した徐放性マトリクスには、エチレン酢酸ビニルと他の生体適合性ポリマーからなるマトリクスが含まれる。
【0052】
局所投与の場合、上記のファゴピリトールを含むリン脂質小胞のような担体は、皮膚を通してファゴピリトールの取り込みを促進する可能性がある。
【0053】
上記のように、ファゴピリトールは、消化管において細菌によって消化されて遊離のD-カイロ-イノシトールを放出し、これが取り込まれると考えられる。D-カイロ-イノシトールは抗酸化剤であり、より詳しくはヒドロキシルラジカルスキャベンジャーであることが知られている。したがって、本発明のファゴピリトールおよび組成物はまた、本発明のファゴピリトールおよび組成物は、好ましくは本発明のファゴピリトールおよび組成物を被験者に経口投与することによって、抗酸化剤であるD-カイロ-イノシトールの原料として用いることができる。
【0054】
本発明は、以下の実施例によってさらに説明する。
【実施例】
【0055】
実施例
実施例1-材料および方法
植物材料
ソバ普通種(ファゴピルム・エスクレンツムモエンク栽培品種「マンカン」(Fagopyrum esculentum Moench cv. 'Mancan')、タデ科)の胚を、種子の発達中および成熟種子における可溶性糖質およびシクリトールに関して分析した。ソバ普通種の栽培品種である「マノー(Manor)」および「タータリー(Tartary)」ソバ(ファゴピルム・タータリクム(L)ガートン(Fagopyrum tartaricum(L.)Gaertn))の成熟種子も同様に分析した。ソバ種子は温室で、既に記載されているように昼間(14時間)24℃、夜(10時間)18℃で生育させた(参照として本明細書に組み入れられる、ホーボビッツ(Horbowicz)ら、「種子発達の際のソバの内乳と胚のステロールと脂肪酸の変化(Changes in Sterols and Fatty Acids of Buckwheat Endosperm and Embryo During Seed Development)」、J. Agric. Food Chem. 40:745〜750(1992))。1000ワットのシルバニアハロゲン化金属ランプからの白熱光約740 μmol m-2 s-1によって、自然の太陽光を毎日14時間供給した。胚は6〜32 DAPおよび成熟時に収穫した種子から採取した。温度の効果を評価するために、4週齢の植物を、種子が成熟するまで温室から18℃と25℃の一定温度の生育チャンバーに移した。植物は毎日約300 μmol m-2 s-1の蛍光を16時間与えた。ピン型の花を有する植物と房状の花を有する植物を、生育チャンバーまたは温室内で同じ温度条件で正統法の他家受粉によって手で受粉させてピン×房、および房×ピンを作製した。新鮮および乾燥重量、水、および水分濃度は、温室で生育させた植物について果皮、殻を除去したひき割り果(種子)、胚、および内乳に関して処置毎に種子6個〜10個を調べる実験3回の平均値±SEとして表記する。軸および子葉組織における可溶性成分を分析する場合、それぞれ胚10個の同じもの3個を軸および子葉画分に関して分けた。内乳および胚画分を分析する場合、3つの成熟、乾燥、および手で胚および内乳画分に分離されるひき割り(果皮を除去した)種子の3回の実験を行った。内乳画分は種皮を含むが果皮は含まない。栽培品種と種を比較するために、それぞれひき割り果3個の実験3回を分析した。
【0056】
糖質およびシクリトール標準物質
グルコース、果糖、ガラクトース、ソルビトール、マルトース、マルトトリオース、マルトテトラオース、ミオ-イノシトール、ショ糖、ラフィノース、スタキオース、フェノールα-D-グルコシド(内部標準)、ケルセチン、およびルチンは、シグマケミカル社(セントルイス、ミズーリ州)から購入した。ガラクチノールはクオ(T.M. Kuo)氏(ピオリア、イリノイ州)から贈与された;D-オノニトール、セクオイトール、およびL-(+)-ボメシトールはローウス(F.A. Loewus)氏(プルマン、ワシントン州)から贈与された;D-カイロ-イノシトールとD-ピニトールは、アンギアル(S.J. Angyal)氏(ケニントン、ニューサウスウェールズ州、オーストラリア)から贈与された;O-α-D-ガラクトピラノシル-(1→2)-4-O-メチル-D-カイロ-イノシトール(ガラクトピニトールA)、O-α-D-ガラクトピラノシル-(1→3)-3-O-メチル-D-カイロ-イノシトール(ガラクトピニトールB)、D-ピニトール、およびD-カイロ-イノシトールは、ストリーター(J.G. Streeter)氏(ウースター、オハイオ州)から贈与された;ベルバスコース、D-カイロ-イノシトール、L-カイロ-イノシトール、およびD-ピニトールは、ヴィールシュ(P. Wiirsch)氏(ローザンヌ、スイス)から贈与された;ベルバスコースおよびD-カイロ-イノシトールは、アダムスおよびジェンセン(P. AdamsおよびR.G. Jensen)両氏(ツーソン、アリゾナ州)から贈与された;ならびにD-カイロ-イノシトールおよびL-カイロ-イノシトールはリヒター(A. Richter)氏から贈与された(ウィーン、オーストリア)。
【0057】
可溶性糖質の抽出と分析
20 DAPの胚7個、または発達しつつある(6〜32 DAP)の胚5個または10個、および成熟乾燥種子の同じもの3個からの軸および子葉組織を分離して、重量を測定し、内部標準としてフェニルα-D-グルコシド100 μgを含むエタノール:水(1:1、v:v)0.6 mlによって乳鉢で十分にホモジナイズした。ホモジネートおよびすすぎ液を1.5 ml微量遠心管に合わせて、80℃で45分間加熱して、内因性の酵素を不活化して、ベックマンマイクロフュージEにおいて最高速で20分間遠心した。残査を再度抽出して。合わせた上清を分子量10,000のカットオフフィルターに通過させた。少量の濾液をシリル化バイアルに移して、窒素ガス蒸気中で蒸発乾固させた。残査を、乾燥器において五酸化リン上で一晩保存して、微量の水を除去した。乾燥残査を、堅くキャップをしたシリル化バイアル(スペルコ、ベルフォンテ、カリフォルニア州)においてアルミニウムブロックヒーターにおいて70℃で30分シリル化混合物(TMSI:ピリジン、1:1、v:v)200 μlによって誘導体化した後、22℃に冷却した。TMSIによって誘導体化した可溶性糖質1μlを、水素炎イオン化検出器とヒューレットパッカード3396A積分器とを備えたヒューレット・パッカード5890シリーズIIガスクロマトグラフのスプリットモード注入器に注入した。可溶性糖質は、DB-1毛細管カラム(長さ15 、内径0.25 mm、被膜の厚さ0.25 μm;J&Wサイエンティフィック、フォルソム、カリフォルニア州)において分析して、このカラムはプログラムされた初回温度150℃、3℃/分で200℃となるように調節し、7℃/分で325℃となるように調節し、そして325℃で20分間維持した。注入器部分は、335℃で操作して、検出器は350℃で操作した。担体ガスはヘリウムで、3.0 ml/分、スプリット1:50であった。検出器のガスは水素30 ml/分、および空気300 ml/分であった。
【0058】
可溶性糖質の量は、標準曲線からの外挿によって決定し、内部標準であるフェニルα-D-グルコシドのピーク面積に対するそれぞれの既知の糖またはシクリトールのピーク面積の比を各成分の既知量に対してプロットした。標準物質は全て、アノマー型をTMS誘導体として捕獲するために分析前にエタノール:水(1:1、v:v)に溶解した。不明物質の糖質の量は最も近い既知の標準物質による計算によって推定した。量は、各実験3回の平均値±平均値のSEとして表記する。
【0059】
ソバ種子のファゴピリトールの画分
エタノール:水(1:1、v:v)で抽出した可溶性成分を蒸発乾固させた。残査をアセトニトリル:水(75:25、v:v)5mlに溶解して、9%官能化した3-アミノプロピル官能性シリカゲル(アルドリッチケミカル社、ミルウォーキー、ウィスコンシン州)を充填した9×290 mmカラム上でクロマトグラフィーを行った。成分は、アセトニトリル:水(画分<70に関しては75:25、v:v;そして画分>70に関しては60:40、v:v)によって流速0.4 ml/分で溶出した。各5ml画分の1mlを蒸発乾固させて、上記のようにガスクロマトグラフィーによって分析した。選択した画分の1ml試料を酵素および/または酸加水分解のために蒸発乾固した。
【0060】
酵素的消化
緑色コーヒー豆のα-ガラクトシダーゼ(EC 3.2.1.22)(ベーリンガーマンハイム社、インジアナポリス、インジアナ州)の3.2 M硫酸アンモニウム溶液1mlを蒸留水1000 mlに対して一晩透析して、シリル化反応を妨害して多様な結果を生じる硫酸アンモニウムを除去した。透析した酵素の0.1 ml水溶液を糖質またはシクリトール水溶液0.1 mlに加えて、37℃でインキュベートした。24時間後、無水エタノール0.2 mlおよび内部標準100 μgを加えた。混合物を90℃で5分間加熱して酵素を不活化し、分子量10,000のカットオフフィルターを通して遠心した。濾液を窒素ガス蒸気下で蒸発乾固させて、シリル化して、上記のようにガスクロマトグラフィーによって分析した。
【0061】
酸加水分解
選択した化合物の乾燥試料を2N TFAによって70℃で3時間加水分解した。化合物および加水分解産物は、ガスクロマトグラフィー、またはガスクロマトグラフィー/質量分析(「GC-MS」)および/またはプロトン核磁気共鳴(「1H-NMR」)によって、真正標準物質と比較することによって同定した。
【0062】
乾燥耐性
未成熟な種子を6〜32 DAPで植物から採取して、殻を除去して(果皮を除去する)、そのまま発芽させるか、または乾燥耐性に関して調べた(ブラックマン(Blackman)ら、「発達途中の大豆種子の乾燥耐性における、成熟タンパク質と糖(Maturation Proteins and Sugars in Dessiccation Tolerance of Developing Soybean Seeds)」、Plant Physiol. 100:225〜230(1992)(「ブラックマンI」)、参照として本明細書に組み入れられる)。殻を除去したひき割り果を12%相対湿度(「RH」)でLiClの飽和濃度に対して25℃で急速乾燥させた。6日後、種子を水の上での高湿度で一晩湿らせて、湿らせたペーパータオル上で25℃で発芽させた。7日目での発芽率および50%発芽するまでの日数を報告する。
【0063】
未成熟な種子は10 DAPで収穫して、殻を除去して、種子を連続的に、密封したデシケータ内でKNO3、Na2CO3、NaCl、Mg(NO3)2、K2CO3、およびMgCl2の飽和塩濃度上でそれぞれ、より低い相対湿度(92%、87%、75%、51%、45%、および32%RH)に毎日移すことによって6日間ゆっくり乾燥させた(ブラックマン(Blackman)ら、「大豆の乾燥耐性に関連した成熟タンパク質(Maturation Proteins associated with Dessiccation Tolerance in Soybean)」、Plant Physiol. 96:868〜874(1991)(「ブラックマンII」)、参照として本明細書に組み入れられる)。殻を除去したひき割り果(種子)を12%RHに7日間移して、その後乾燥させた種子(湿らせた後)を湿らせたペーパータオル上で25℃で7日間発芽させることによって、乾燥耐性に関して試験した。高い相対湿度制御処置の場合、未成熟な種子は6日間水を加えたデシケータに入れた(ブラックマンII、参照として本明細書に組み入れられる)後、乾燥耐性に関して調べた。緩速乾燥過程の各段階後または高相対湿度では毎日、胚を採取して可溶性の糖質およびシクリトールに関してアッセイした。
【0064】
「マンカン」商業用保存種子の成熟種子をペーパータオル上で0時間、6時間、12時間、18時間、24時間、36時間、および48時間発芽させた後、上記のようにLiClの飽和溶液上で12%RHで急速に乾燥させた。湿らせたペーパータオル上で25℃で7日間再水和させた後、正常な幼根の生育を有する実生を乾燥耐性であると分類した。軸または子葉は、可溶性糖質およびシクリトールに関してアッセイした。
【0065】
実施例2-成熟種子の胚におけるオリゴ糖とシクリトール
「マンカン」の市販の登録保存種子からの成熟乾燥ソバ種子の胚から抽出した可溶性糖とシクリトールの典型的なガスクロマトグラムを図2に示す。ソバ胚はそれらがラフィノースシリーズのオリゴ糖ではなくてガラクトシルシクリトール(糖アルコール)を蓄積することから、独特であると考えられている。種の名称ファゴピルム(Fagopyrum)にちなんでファゴピリトールB1(図2、ピークfを参照のこと)と命名された主な可溶性成分は、ソバ胚において全可溶性ショ糖およびシクリトールを40%まで蓄積した。ショ糖(図2、ピークd)は、全可溶性糖およびシクリトールの別の42%を占めた。
【0066】
ファゴピリトールB1は、シクリトールD-カイロ-イノシトールのα-ガラクトシル誘導体であり、大豆種子における少ない成分であるO-α-D-ガラクトピラノシル-(1→2)-D-カイロ-イノシトールと同じ保持時間を有する(シュバイツァー(Schweizer)ら、「大豆の三つのα-D-ガラクトピラノシルシクリトールの、精製および構造決定(Purification and Structure Determination of Three α-D-galactopyranosylcyclitols from Soya Beans)」、Carbohydr. Res. 95:61〜71(1981)およびトレイトラー(Traitler)ら、「毛細管カラム上でのオンカラムインジェクションによる、ガスクロマトグラフィーによる糖の分離(Gas Chromatographic Separation of Sugars by On-Column Injection on Glass Capillary Column)」、J. High Resolut. Chromatogr. Chromatogr. Commun. 7:558〜562(1984)、参照として本明細書に組み入れられる)。二つの異なるガラクトシルカイロ-イノシトール異性体であるファゴピリトールA1とファゴピリトールB1(図2、ピークeおよびf)、および二つの異なるジガラクトシルカイロ-イノシトール異性体であるファゴピリトールA2とファゴピリトールB2(図2、ピークkおよびl)とがソバ胚に存在することが証拠によって示唆された。さらに、トリガラクトシルカイロ-イノシトールである少量のファゴピリトールA3(図2、ピークo)が市販の保存種子に検出された。5個のうち少なくと3個がこれまで報告されていなかった。誘導体化されていない二つの遊離のシクリトールであるD-カイロ-イノシトールとミオ-イノシトール(図2、ピークaおよびb)がソバに認められた。ピニトールと、ガラクトピニトールA、ガラクトピニトールBおよびシセリトールを含むピニトールのいくつかのガラクトシル誘導体とを蓄積する(ホーボビッツ(Horbowicz)、「種子の乾燥耐性と保存性;膨満によって産生されるオリゴ糖とシクリトール-論評と調査(Seed Desiccation Tolerance and Storability:Dependence on Flatulence-Producing Oligosaccharides and Cyclitols-Review and Survey)」、Seed Sci. Res. 4:385〜405(1994)(「ホーボビッツ」)、参照として本明細書に組み入れられる)多くのマメ科の種子とは異なり、ソバ種子は、ピニトールまたはピニトール誘導体を含まなかった。ソバは、ガラクチノール(O-α-D-ガラクトピラノシル-(1→3)-D-カイロ-イノシトール)(図2、ピークg)、またはジガラクトシルミオ-イノシトール(O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→3)-D-ミオ-イノシトール)(図2、ピークm)(ペテクIおよびカートワ、参照として本明細書に組み入れられる)のごく少量を蓄積したに過ぎなかった。トウモロコシまたは大豆およびルピナスのような他の豆果とは異なり(ホーボビッツ(Horbowicz)、参照として本明細書に組み入れられる)、ソバ胚によるラフィノース(O-α-D-ガラクトピラノシル-(1→6)-α-D-ガラクトピラノシル-(1→2)-β-D-フルクトフラノシド)(図2、ピークh)の蓄積は非常に少量で、スタキオース(O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→6)-α-D-グルコピラノシル-(1→2)-β-D-フルクトフラノシド)(全可溶性成分の1%未満)(図2、ピークn)の蓄積はごく少量であった。還元糖(グルコース、果糖、ガラクトース)は、種子発達および成熟のあいだソバ胚に蓄積されなかった。
【0067】
ファゴピリトールピークの同一性の証拠を図3A〜Cに示す。加水分解前、ファゴピリトールは容易に明らかである(図3A、ピークe、f、k、l)。α位のガラクトシル結合を加水分解するα-ガラクトシダーゼによって加水分解すると、ピークe〜lは消失したのに対し(図3B)、D-カイロ-イノシトール(図3B、ピークa)は大きく増加し、ミオ-イノシトール(ピークb)はわずかに増加するが、他の遊離のシクリトールは認められない。ショ糖(図3B、ピークd)は、有意に変化しないが、ガラクトースのアノマー型(図3B、「1」と記されたピーク)が出現する。ラフィノースおよびスタキオースは微量存在したに過ぎないための、これらの糖の加水分解によるショ糖およびガラクトースはほとんど認められない。したがって、ほぼ全てのガラクトースがガラクトシルシクリトールから放出された。ミオ-イノシトールはガラクチノール(図3A、ピークg)および微量のジガラクトシルミオ-イノシトール(図2、ピークm)の加水分解によってわずかに増加する(図3B、ピークb)。ファゴピリトールはβ-ガラクトシダーゼ(EC 3.2.1.23)に対して37℃で24時間耐性であり、ガラクトースは放出されなかった。このパターンは、図3Aにおけるピークe、f、kおよびlがα-ガラクトシル結合のみを含むことを確認する。ピークl(ファゴピリトールB2)は、成熟テンサイ(ベータ・ブルガリス(Beta Vulgaris L.))種子の胚から抽出した、O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→2)-D-カイロ-イノシトールと同一の保持時間を示した(シオミ(Shiomi)ら、「テンサイの球状種子からの新しいジガラクトシルシクリトール(A New Digalactosyl Cyclitol from Seed Balls of Sugar Beet)」、Agric. Biol. Chem. 52:1587〜1588(1988)、参照として本明細書に組み入れられる)。強い酸(2N TFA)による加水分解後、ショ糖も同様に加水分解されてグルコース(図3C、「2」と記された二つのピーク)および果糖(図3C、「3」と記された三つのピーク)のアノマー型が得られた。加水分解後に内部標準(ピークc)を加えた。ガラクチノールに関して修正して、ファゴピリトールA2およびファゴピリトールB2がジガラクトシルカイロ-イノシトールであると仮定すると、組織抽出物の酸加水分解から生じた産物は、ファゴピリトールA1プラスファゴピリトールB1(ピークeおよびf)に関してD-カイロ-イノシトール1.00に対してガラクトース残基0.93の計算比を有し、双方がガラクトシルカイロ-イノシトールである証拠を提供する。
【0068】
さらに確認するために、ファゴピリトールを液体カラムクロマトグラフィーによって分離した(図4)。ファゴピリトールB1(図4、ピークd)のαガラクトシダーゼ加水分解産物は、ガラクトースの回収に関して修正後、D-カイロ-イノシトール1.00に対してガラクトース1.08の比を示し、ファゴピリトールB1がαガラクトシルD-カイロ-イノシトールであることを確認した。図4のピークfのTMS-産物のガスクロマトグラフィー分析によって、ファゴピリトールB2に対応する一つのピークを認めた。2N TFAによって部分的に加水分解すると、ファゴピリトールB2、ファゴピリトールB1、ガラクトシルガラクトース、D-カイロ-イノシトール、およびガラクトースに対応するピークを認めた。ガラクトースの回収に関して修正すると、D-カイロ-イノシトール1.00に対してガラクトース残基1.93の比を認め、ファゴピリトールB2がジガラクトシルD-カイロ-イノシトールであることを確認する。ファゴピリトールにおけるガラクトースのα結合はまた、1H-NMRによっても確認した。ガラクトースに対するD-カイロ-イノシトールの比およびピークの位置は、ピークeおよびfがおそらくガラクトシルD-カイロ-イノシトールの異なる位置異性体であること、そしてピークkおよびlが対応するジガラクトシルD-カイロ-イノシトールであることを示唆している。GC-MS分析は、ファゴピリトールA1およびファゴピリトールB1が密接に関連しているが異なる断片化パターンを有することを証明している。ガラクチノールシリーズの構造は図5に示す。
【0069】
ファゴピリトールB1およびファゴピリトールA1におけるD-カイロ-イノシトールの存在は、高分解能ガスクロマトグラフィーおよびキラル毛細管カラムを用いて行った。真正のD-カイロ-イノシトールおよびL-カイロ-イノシトール標準物質、ならびに純粋なファゴピリトールB1および純粋なファゴピリトールA1の酸加水分解産物(2N TFA、70℃で3時間)を、無水ペンタフルオロプロピオン酸(「PFPN」):アセトニトリル(1:1、v/v)によって誘導体化した。誘導体産物を真正のD-カイロ-イノシトールおよびL-カイロ-イノシトール標準物質と共に、カイラル毛細管カラムを備えた高分解能ガスクロマトグラフィー(キラシル-バル、アルテック社、ディアフィールド、イリノイ州;長さ25 m、内径0.25 mm、被膜の厚さ0.16 gin)を用いて100℃で等温的に溶出させて同時にクロマトグラフィーを行った。注入口の温度は130℃で水素炎イオン化検出器(「FID」)の温度は130℃であった。ファゴピリトールB1およびファゴピリトールA1におけるD-カイロ-イノシトールは、真正のD-カイロ-イノシトール(保持時間4.6分)と共に同時クロマトグラフされたが、真正のL-カイロ-イノシトールは(保持時間、4.9分)同時にクロマトグラフされなかった。ファゴピリトールB1およびファゴピリトールA1は、保持時間4.6分でD-カイロ-イノシトールのみを有することが確認された。
【0070】
さらに、ガラクトシル環、D-カイロ-イノシトール環の正確な構造、およびD-カイロ-イノシトール環上の2位の炭素に対するガラクトースのアノマー炭素のα結合を、純粋なファゴピリトールB1を用いたNMR実験によって確認した。スゼンシンスキ(Szczencinski)ら、「ソバ種子由来のファゴピリトールB1の構造に関するNMR研究(NMR Investigation of the Structure of Fagopyritol B1 from Buckwheat Seeds)」、Bulletin of the Polish Academy of Sciences Chemistry 40:9〜13(1998)、参照として本明細書に組み入れられる)を参照のこと。
【0071】
実施例3-種子発達および成熟の際の糖およびシクリトールの蓄積
ソバ胚は、12 DAPで最大水分含有量を蓄積するが、最大新鮮重量は16 DAPで起こり、16 DAP以降は胚の乾燥物質にこれ以上の有意な増加を認めない(表1)。種子成熟の後期段階で認められた胚乾燥物質のわずかな減少は、内乳全体に絡み合って(参照として本明細書に組み入れられる、マーシャル(Marshall)ら、「ソバ:説明、育種、生産、および利用(Buckwheat:Description, Breeding, Production, and Utilization)」、Adv. Cereal Sci. Tech. 5:157〜210(1982)に示されるように)乾燥種子において種皮に接着している全ての子葉残遺物を物理的に除去することが難しいことを反映している。
【0072】
(表1)
【0073】
表2は、種子発達および成熟の様々な段階で、ソバ胚に存在する様々な糖質およびシクリトールの量を示す(μg/胚の単位で表す)。果糖およびグルコースは6および8DAPでの非常に若い胚にのみ存在し、胚発達および成熟の後期には存在しなかった(表2)。D-カイロ-イノシトールおよびミオ-イノシトールは、胚発達および成熟を通して最も初期の段階から存在した(表2)のに対し、ピニトールまたは他のO-メチルシクリトールは胚発達および成熟の任意の段階において検出されなかった。ショ糖は8〜10 DAPのあいだに劇的に増加し(表2)、胚の非常に急速な生育の始まりと一致して(表1)、16 DAPまではこれより遅い速度で増加し続けた(表2)。ファゴピリトールB1およびその異性体型であるファゴピリトールA1は、12 DAPで最初に検出され、12〜16 DAPのあいだに非常に急速に蓄積された(表2)。ファゴピリトールA2およびファゴピリトールB2は、16 DAPで最初に検出されて、20 DAPまで急速に蓄積した。ファゴピリトールA3は、胚乾燥の初期段階(50〜35%水分濃度;表1)のあいだに20 DAP(表2)で蓄積した。全可溶性糖およびシクリトールは2相性で蓄積された:最初に胚の急速な発達およびショ糖の蓄積に関連して8〜10 DAPで、第二にファゴピリトールB1の急速な蓄積に関連して12〜16 DAPで(表2)蓄積された。
【0074】
(表2)
a ガラクチノール標準物質に基づいて計算。
b ラフィノース標準物質に基づいて計算。
c スタキオース標準物質に基づいて計算。
d ショ糖対ファゴピリトールB1の比およびショ糖対非ショ糖の比は、平均値に基づいて計算した。
【0075】
実施例4-ソバ種子における糖とシクリトールの分布
軸は、20 DAPにおいて胚の新鮮重量の8%であることが判明したが、子葉は、胚の新鮮重量の92%であった(表3)。全ての成分の濃度は、子葉より軸において高かった(表3)。ショ糖、シクリトール、およびその誘導体は、ソバ胚の軸では乾燥重量の15%に蓄積したのに対し子葉の乾燥重量の8%であった(表3)。軸組織におけるファゴピリトールB1濃度は、子葉での濃度の倍であり、ショ糖対ファゴピリトールB1の比は、子葉における1.64と比較して軸において1.07(w:w)であった。膨満によって産生されるオリゴ糖である、ラフィノースおよびスタキオースは軸組織に存在したが、子葉ではごく微量に過ぎなかった(表3)。ガラクチノールはショ糖(DP-2)にガラクトースを供与して、ラフィノース(ガラクトシルショ糖、DP-3)を形成し、ラフィノースにガラクトースを供与してスタキオース(ガラクトシルショ糖、DP-4)を形成する。
【0076】
(表3)
a ガラクチノール標準物質に基づいて計算。
b ラフィノース標準物質に基づいて計算。不明物質は、保持時間の比がフェニルα-D-グルコシドに対して2.44である。
c スタキオース標準物質に基づいて計算。
d ショ糖対ファゴピリトールB1の比およびショ糖対非ショ糖の比は、平均値に基づいて計算した。
【0077】
糊粉層および親の種皮-核(スパーモダーム(spermoderm))組織のふすま画分を含む内乳画分は、ひき割り果の74%であり、胚はひき割り果の26%であった(表4)。胚画分は全可溶性糖質およびシクリトールの71.4%を含み、内乳画分は28.6%を含んだ。胚は、内乳画分(76 μg)の5倍ものファゴピリトールB1(403 μg)を含んだ(表4)。内乳画分に存在する糖、シクリトール、およびシクリトール誘導体のほとんどはおそらく糊粉層に存在した。いくつかは、成熟乾燥ひき割り果の内乳画分から完全に分離できなかった子葉の断片であった可能性がある。内乳画分における可溶性成分の濃度は、デンプンが主な貯蔵成分であるために低かった。
【0078】
(表4)
a ガラクチノール標準物質に基づいて計算。
b ラフィノース標準物質に基づいて計算。
c スタキオース標準物質に基づいて計算。
【0079】
実施例5-シクリトールおよび可溶性糖質における栽培品種と種の差
成熟ひき割り果における全可溶性糖質およびシクリトールの濃度は、「マンカン」および「マノー」のソバ普通種(表5)および「タータリー」ソバにおいてほぼ同じであった。ショ糖濃度はひき割り果における三つ全ての遺伝子型において同じであった。ファゴピリトールB1は「マンカン」において濃度が最高であり、「マノー」では低く、「タータリー」では「マンカン」の20%に過ぎなかった。「タータリー」には、「マンカン」または「マノー」において認められない不明化合物2個が含まれたが、「マノー」では、より高い濃度のファゴピリトールA2、ファゴピリトールB2、およびファゴピリトールA3を示した(表5)。三つの遺伝子型は同じ環境条件で生育しなかったために、差の一部は生産環境に関連する可能性がある。しかし、「タータリー」ソバのひき割り果において不明物質1の濃度が高かったこととファゴピリトールB1の濃度が低かったことは、おそらく遺伝的な差の反映であった。「マノー」ソバのひき割り果において、より高次のガラクトシルオリゴマーがより高濃度へシフトしたことは環境の影響を反映した可能性がある。
【0080】
(表5)
a ガラクチノール標準物質に基づいて計算。
b ラフィノース標準物質に基づいて計算。
c スタキオース標準物質に基づいて計算。
【0081】
フラボノール、ケルセチン、およびルチンに対する感受性は本明細書において用いたアッセイ方法では全く低かった。ソバの葉に一般的に存在するルチンは、「マンカン」または「マノー」栽培品種のいずれかのソバ普通種の成熟種子のひき割り果においてごく微量検出されたに過ぎなかった。対照的に、「タータリー」ソバの成熟種子のひき割り果は、ルチン標準物質によって計算し、そして試料を加水分解後ケルセチンおよびラムノース標準物質によって計算すると、乾燥重量1gあたりルチン約7.14 mgを含んだ。ルチンの酸加水分解産物は、ケルセチン、ラムノース、およびグルコースであった。ケルセチンおよびラムノースは、「タータリー」ソバのひき割り果からの抽出物を酸加水分解(2N TFAで70℃で3時間)した後に検出されたが、ソバ普通種の胚からの抽出物の酸加水分解産物にはいずれも認めなかった。グルコースもまたショ糖の酸加水分解産物であり、したがって、組織抽出物におけるルチンのアッセイのために用いることができなかった。胚は、成熟乾燥「タータリー」ソバ種子の内乳から分離することができなかったため、胚組織5個を分析できなかった。
【0082】
実施例6-ソバひき割り果、痩果、および粉砕画分における総ファゴピリトール、遊離のD-カイロ-イノシトール、および総D-カイロ-イノシトール
粉砕画分において、「精製ふすま」画分は種皮に接着する胚画分を高濃度に有した。殻が存在しない場合、この画分は、粉砕画分の中でもファゴピリトールとD-カイロ-イノシトールの最高濃度を示した(表6)。「精製ふすま」における総ファゴピリトールおよび総D-カイロ-イノシトール濃度は、次に最も高い粉砕画分(ふすま#1)の濃度の2倍であった。総ファゴピリトールおよび総D-カイロ-イノシトールの濃度は、粉砕画分において、その粉砕画分における胚組織の濃度の減少の程度に応じて減少した(表6)。この減少順序は「精製ふすま」>「ふすま#1」(殻付き)>「ふすま#2」(殻はより少ない)>全ひき割り粉>「特選粉」(デンプンが多い)>「ひび割れ」(胚断片を小数含む)であった。「ひび割れ」画分は粉砕および抽出することが非常に難しかった。「ひび割れ」画分のデンプン質の内乳は、乾燥していた部分的に発芽した種子に類似していた。乾燥種子を湿らせると、αアミラーゼが、子葉からデンプンの多い内乳に放出され、デンプンを消化し始めた。この粘着性のデンプンは、乾燥時に非常に堅くなった。「ひび割れ」のいくつかは、収穫前に植物において再度湿った種子に由来すると仮定された。ファゴピリトールは再湿潤の12〜18時間以内に消失し、デンプンの分解はその時点で既に始まっていた。類推すると、ひび割れの画分は、少数の胚画分を有し、存在するそれらの画分は、種子を収穫前に再度湿らせるとファゴピリトールが減少している可能性がある。
【0083】
(表6)
【0084】
実施例7-糖とシクリトールの蓄積に及ぼす温度の影響
胚(子葉および軸を合わせて)の糖組成は、18℃および25℃で生育させた植物から収穫した成熟種子から採取した胚に関して報告されている。ソバ胚における主な可溶性糖質は、ショ糖とファゴピリトールB1であり、これらは18℃では全体の85%を占めた(表7)。胚の乾燥重量は二つの温度で類似であったため、内乳は、18℃では25℃より種子あたり平均で15 mg多い乾燥重量を蓄積した。胚の可溶性糖質に及ぼす温度の影響は全く興味深かった。18℃で増殖させた種子の胚は、25℃で生育させた場合と比較して、ショ糖が減少し、ファゴピリトールB1が2.5倍多く、スタキオースは3倍多かった。ファゴピリトールA2およびファゴピリトールB2は25℃ではより高いレベルに蓄積した(表7)。温度がしばしば25℃を超える温室で生育させると、ファゴピリトールA3は20 DAPで検出可能であった(表2)。このパターンは、ファゴピリトールB1の蓄積には低温が都合がよいのに対し、ファゴピリトールのより高次ガラクトシルオリゴマーの蓄積には高温が都合がよいことを示唆している。スタキオースに関しては逆のパターンを認めた。スタキオースの蓄積は、低温で都合がよく(表7)、温室でのより暖かい温度で成熟させた種子からの胚では検出されなかった(表2)。
【0085】
(表7)
a 成熟乾燥種子は、胚切除前に473 mg(18℃)および32±3 mg(25℃)であった;3回の実験。
b ガラクチノール標準物質に基づいて計算。
c ラフィノース標準物質に基づいて計算。不明物質は、フェニルα-D-グルコシドと比較して保持時間の比2.44を示す。
【0086】
実施例8-胚発達の際の乾燥耐性とファゴピリトールB1との関連
ソバ種子における発達途中の胚は、12 DAPで乾燥耐性となった(図6)。新たに収穫した種子は、高度の休眠を示した(サミミー(Samimy)ら、「ソバ普通種における種子休眠(Seed Dormancy in Common Buckwheat(Fagopyruni esculentum Moench)」、Plant Varieties and Seeds 7:17〜22(1994)、参照として本明細書に組み入れられる)。25℃で湿らせたペーパータオル上で16日後も発芽しなかった種子は休眠していると見なされ、20および24 DAPでの植物種子(表2)の場合の76%〜87%ほどのファゴピリトールB1(表8)を含み、休眠がファゴピリトールB1の動員を遅らせることを示唆している。12 DAP(図6)での乾燥耐性の発達は12〜16 DAPでファゴピリトールB1の蓄積に関連して起こった(表2)。
【0087】
(表8)
a 新鮮な種子を20または24 DARで収穫した。25℃で湿らせたペーパータオル上で16日後も発芽しなかった種子は休眠していると見なされる。
b ガラクチノール標準物質に基づいて計算。
c ラフィノース標準物質に基づいて計算。不明物質は、フェニルα-D-グルコシドに対する保持時間の比が2.49および2.51であった。
【0088】
未成熟なソバ種子を10 DAPで収穫すると、乾燥耐性は、制御された相対湿度下(図7A)での緩速乾燥によって誘導された(図7A)。未成熟な種子の緩速乾燥時の乾燥耐性の獲得(早期成熟または誘導成熟)は、緩速乾燥の初日におけるファゴピリトールB1の蓄積に密接に関連した(図7C)。D-カイロ-イノシトールは、緩速乾燥の初日のあいだに急速に減少した(図7B)。ミオ-イノシトールは一定のままであったが(図7B)、ガラクチノールは2日目に増加した(図7C)。少量のファゴピリトールA2、ファゴピリトールB2、およびジガラクトシルミオ-イノシトールが5日目に蓄積した(図7D)。ショ糖は、初日のあいだに減少し、その後一定であった(図7E)。
【0089】
同様に、高い相対湿度で維持すると、未成熟な10 DAPの殻を除去したソバ種子は、ファゴピリトールB1(図8C)およびD-カイロ-イノシトール(図8B)の蓄積に関連して1日後に乾燥耐性となった(図8A)。ガラクチノールは、高い相対湿度では2日目に増加した(図8C)。少量のファゴピリトールA2およびファゴピリトールB2が2〜5日に蓄積し、ジガラクトシルミオ-イノシトールは2〜6日に蓄積した(図8D)。ファゴピリトールA3は、6日後に限って検出された。ショ糖は初日以降徐々に増加した(図8E)。このパターンは、大豆種子の軸とは対照的であり、大豆種子は、高い相対湿度でインキュベートしても乾燥耐性とならず、高い相対湿度でインキュベートしてもスタキオースを蓄積しない(ブラックマンI、参照として本明細書に組み入れられる)。ファゴピリトールB1は、非常にマイルドな乾燥条件で蓄積することができるがラフィノースシリーズのオリゴ糖の蓄積はより多くの乾燥を必要とするように思われる。
【0090】
実施例9-発芽の際の乾燥耐性の喪失に対するファゴピリトールB1の関係
乾燥耐性の喪失に対するファゴピリトールB1の喪失の関係(12%相対湿度および22℃での急速乾燥)は、殻を除去した成熟ソバ種子(「マンカン」)の発芽の際の軸および子葉組織において測定した。吸水開始後6時間以内にファゴピリトールB1は、軸組織から消失し始めた。12時間までに低いレベルのみが残り、発芽18時間までに、ファゴピリトールは全て微量の高次オリゴマーを除いて完全に消失した(図9E)。ファゴピリトールB1の消失は、再水和時の胚軸-幼根軸(一次根の生育)の「正常な」生育によって測定した急速な発芽の開始および乾燥耐性の喪失に密接に関連した(図9A)。ショ糖レベルは減少した後48時間後に増加して、胚軸から糖が動員されていることを反映した(図9C)。ショ糖:ファゴピリトールB1比は、乾燥耐性の喪失および発芽の開始に関連して12時間後非常に急速に増加した。ショ糖とは対照的に、D-カイロ-イノシトールは12時間後に増加して、ファゴピリトールの加水分解を反映する(図9C)。ガラクトースはファゴピリトールの動員の際に検出されなかったが、明らかに他の産物に急速に変換された。果糖およびグルコースは、軸組織が急速に生育しつつある(図9I)18時間後に軸組織に存在した(図9B)。果糖およびグルコースは軸組織に存在する唯一の還元糖であった(図9I)。
【0091】
発芽のあいだ、マルトース、マルトトリオース、およびマルトテトラオース(図9J)は、増加したショ糖レベルと共に胚軸組織に存在し(図9H)、デンプンの分解を反映した。D-カイロ-イノシトールは、発芽の6〜18時間のあいだに5倍増加し(図9B)、胚軸におけるファゴピリトールの分解を反映した(図9B)。胚軸におけるD-カイロ-イノシトールの総量(遊離および結合)は発芽の際に減少し、軸組織への輸送の発生または他の産物への代謝を示唆した。発芽の18〜48時間で胚軸におけるショ糖の量が多いことは、内乳におけるデンプンの動員および胚軸による加水分解産物の吸収を反映した(図9H〜J)。胚軸組織は内乳に埋もれ、発芽の際にデンプンの多い内乳に取り巻かれた。0時間では胚軸におけるファゴピリトールB1および全可溶性糖質(図9F)のレベルが低いことは、乾燥種子の外種皮(種皮)に接する外側の子葉の一部が、成熟乾燥種子からの胚組織の剥離の際に失われたことを示した。
【0092】
実施例10-画分実験の材料
殻を除去した種子100個の試料(約2.5 g)を粉砕して、エタノール:水(50:50)によって抽出した。抽出物を繰り返し遠心して、容積数mlに濃縮して、分子量10,000のカットオフフィルターを通過させた。エタノールを濃縮試料に加えて、冷凍庫に入れて、粗抽出物からファゴピリトールA3を濃縮することを目的としてファゴピリトールの沈殿を促進した。ほとんどの場合、調製物からの試料ではファゴピリトールA3が失われるか、または検出レベル以下であった。
【0093】
ショ糖の約97%がインベルターゼによって除去された。グルコース、果糖、およびガラクトースは増加して、ラフィノースおよびスタキオースは、おそらくインベルターゼ酵素調製物が十分に純粋でなかったためになおも存在した。グルコースおよび果糖を異なる方法によって破壊する試みは、部分的に成功したに過ぎなかった。
【0094】
実施例11-P2ゲルの画分
いくつかの組の抽出物をプールして濃縮し、遠心および限外濾過によって透明にした。種子1081個を合わせた抽出物を3mlに濃縮した。可溶性糖質の濃縮調製物をバイオゲルP2ゲルカラム(3m×40 cm)にローディングして、脱気した水によって65℃で溶出した。初期画分は茶色であり、その後の画分は黄色であった。糖およびファゴピリトールのほとんどが5ml画分3本に溶出され、なおも茶色の材料が混入していた。これらの画分をプールして濃縮し、第二のバイオゲルP2ゲルカラムに20℃でローディングした。ファゴピリトールA3に富む画分をプールした。ファゴピリトールA1およびB1に富む画分をプールした;そしてファゴピリトールA1、B1、A2、およびB2ならびにショ糖に富む画分をプールした。カラムの流速は、茶色の材料(おそらくポリフェノール材料)の量が増加するにつれて減少した。
【0095】
種子1300個からの抽出物(約35 g)を遠心によって透明にして、濃縮し、分子量10,000のカットオフフィルターを通過させた。1.5 mlに濃縮した後、試料を限外濾過して、濃縮した抽出物をP2カラム上でクロマトグラフィーを行った。それぞれの抽出物からのファゴピリトールA3に富む画分および他のファゴピリトールに富む画分を合わせた。遠心前に沸騰すると、デンプンの膨張が起こり、抽出可能な容積が減少して、抽出物の着色が増加する。いずれのカラムも異なるファゴピリトールを効率よく分離できなかった。
【0096】
実施例12-3-アミノプロピル官能性シリカゲルカラムでの画分
0.9 cm×30 cmの3-アミノプロピル結合(9%官能化)シリカゲル(シグマ-アルドリッチ社)をその後の実験におけるファゴピリトールの画分に用いた。P2ゲルカラムからの濃縮画分を最初の5個のアミノカラムにローディングした。
【0097】
カラム3に関して、P2ゲルカラム上で部分精製した後のファゴピリトールA3に富む試料を蒸発乾固させて、水2mlに溶解して、PVPカラムを通過させた。溶離剤を蒸発乾固させて、アセトニトリル:水(70:30)7mlに溶解して、0.9 cm×30 cmの3-アミノプロピル官能性シリカゲルカラムにローディングした。アセトニトリル:水(70:30)350 ml、アセトニトリル:水(60:40)150 ml、およびアセトニトリル:水(50:50)50 mlによって段階的な溶出を行った。カラムは、アセトニトリル:水(60:40)30 mlおよびアセトニトリル:水(75:25)30 mlによって再生した。画分(5ml)を回収して、回収した画分からの試料0.5 mlを蒸発乾固させて、TMSI:ピリジン(1:1)によって誘導体化した。TMS-糖質は高分解能のガスクロマトグラフィーによって分析した。ファゴピリトールA3は他のファゴピリトールから明らかに分離された(図10)。「純粋な」ファゴピリトールA3を含むプールした画分を蒸発乾固させて、微量の混入物を除去するために再度クロマトグラフィーを行うために保存した。「純粋な」ファゴピリトールA2または「純粋な」ファゴピリトールB2の画分をプールした。ファゴピリトールA2およびB2の混合物の画分も同様にリサイクルするためにプールした。
【0098】
カラム4に関して、P2ゲルクロマトグラフィー後のファゴピリトールの混合物を含む試料をアミノプロピル官能性シリカゲル上でさらに分離した(図11)。カラム4に類似の量のファゴピリトールA3をローディングしたが、他のファゴピリトールのためにカラムにローディングする全量が多くなった。ファゴピリトールA2およびB2は混合したままであったが、いくつかの「純粋な」ファゴピリトールA1、B1、およびA3画分は、さらに精製するためにプールした。
【0099】
実施例13-炭素セライトカラム上での画分
粗抽出物の混合物を、ソバ精製ふすまの25 g試料3個から調製したところ、可溶性糖質を全量で1.2 g含み、そのうち540 mgがファゴピリトールであった。精製ふすまを50%エタノールによって抽出し、エタノールをロータリーエバポレーションによって除去して、抽出物をポリビニルポリピロリドン(「PVPP」)およびアンバーライトIRN-150イオン交換樹脂を含むカラムの中に通過させた。次に、抽出物を酵母(2 mg酵母/精製ふすま1g)によって消化して、ショ糖の97%を除去した。この処置の副産物は二糖類であるトレハロース(O-α-D-グルコピラノシル-(1→1)-O-α-D-グルコシド)であった。酵母処置抽出物を分子量10,000のカットオフフィルター(アミコン社)によって濾過して、凍結乾燥した。次に、合わせた抽出物を蒸留水9mlに再度溶解して炭素セライトカラムの上部に適用した。カラムは炭素(「ダルコ-G60」、J.T.ベイカー、フィリップスバーグ、ニュージャージー州)とセライト545-AW(スペルコ、ベルフォンテ、ペンシルバニア州)とを重量で等しい比率で含む混合物を含んだ。炭素セライト混合物は、蒸留水において3.3 cmのEDカラム、高さ17 cmに充填したスラリーで、充填したカラムを蒸留水1Lによって洗浄した。カラムを4℃で蒸留水800 ml、5%エタノール1000 ml、10%エタノール1000 ml、20%エタノール250 ml、および50%エタノール200 mlによって段階的に溶出した。溶離剤をフラスコに集めて、これを1時間毎に手で交換した。平均すると、フラスコは容積37 mlを含んだ。画分に含まれるファゴピリトール濃度は、高分解能のガスクロマトグラフィーによって分析した。単糖類、グルコース、および果糖(酵母処理により大量に存在する)、ならびにミオ-イノシトールおよびD-カイロ-イノシトールは、水画分に溶出した。ファゴピリトールB1は5%エタノールによってカラムから溶出したがガラクチノールが混入していた。ファゴピリトールA1およびB2は10%エタノールによって溶出されたがそれぞれ、トレハロースとショ糖が混入した(酵母によって除去されなかった少量)。ファゴピリトールA2は20%エタノール溶離剤に含まれ、ファゴピリトールA3は50%エタノール溶出の段階までカラムに残っていた。着色混合物を除去する場合、炭素-セライトカラムは、3-アミノプロピル官能性シリカゲルカラムより有効であった。
【0100】
実施例14-粉の高温対低温抽出
全白ひき割り粉(バーケットミルズ社)からの粉(100目のふるいを通したもの、C&Nラボラトリーミル)を、粉に対する様々な溶媒比で抽出して、「高剪断」ポリトロンヘッドを備えたワーリングブレンダーを用いて高温抽出または低温抽出を行った。ブレンダーは10秒間のオンと10秒間のオフを1分または2分間行った。高温抽出の後、混合物は濃厚すぎて(デンプンの膨張による)濾過または遠心を効率よくできず、抽出されたファゴピリトールの容量は少なくなった。低温抽出(寒冷または室温)では、最高の収率が得られた。最適な抽出は、溶媒/粉の比率が20:1(例えば、粉25 gをエタノール:水(50:50)500 mlに)であった。ファゴピリトールの相対的な収率は、同じ25 g試料の第一、第二、および第三の抽出に関して約90%、9%、および1%であった。低温抽出物の遠心後の上清または濾過後の濾液を80℃に加熱して、粉に存在して抽出の際にファゴピリトールを加水分解する可能性があるαガラクトシダーゼおよび他の酵素を不活化した。これらの酵素は、エタノール:水(50:50)中では不活性であったが、エタノールが濃縮中に蒸発した後の水相では活性化されるようになる可能性がある。
【0101】
実施例15-脱脂粉を用いた効果
いくつかの試料は抽出前にヘキサンによって脱脂した。これらの実験において、PVPカラムおよびイオン交換カラム段階は限外濾過の後に行った。初期段階は容易に進行するように思われたが、限外濾過はなおも遅く、全技法を行うには非常に時間を浪費して面倒であった。技法を単純にして有効性を改善するために、非脱脂粉についてさらなる実験を行った。
【0102】
実施例16-水対エタノール:水(50:50)による抽出
ミンダック社製の粉(各200 mg)を、遠心管において蒸留水2mlまたはエタノール:水(50:50)によって抽出した。ファゴピリトールの収率は水またはエタノール:水(50:50)による抽出後同じであった(表9)。水抽出物は曇っているように思われ、より懸濁した材料を含んだ。この知見は、おそらくタンパク質およびデンプンの水に対する溶解度が高いことに関連した。エタノール抽出物はあまり懸濁していない材料を含み、抽出の際の微生物および内因性の酵素活性も最小限であった。エタノール:水(50:50)をその後の抽出において用いた。
【0103】
(表9)
【0104】
実施例17-バーケットミルズ社製とミンダック社製の粉の比較
ショ糖およびファゴピリトールB1は双方の原料において主要な成分であった(表10)。バーケットミルズ社製は、全ひき割り粉に由来し、ミンダック社製のより精製粉より、総可溶性糖質の収率が約2倍高かった。ショ糖は、双方の原料において総可溶性糖質の約60%であった。バーケットミルズ社の抽出物は、赤みがかった茶色であったのに対し、ミンダック社の抽出物は曇った白色であった。ファゴピリトールA2、B2、およびA3は、ミンダック社製の粉のこの試料において検出されなかった。
【0105】
(表10)
【0106】
実施例18-ショ糖およびファゴピリトールの酵母消化
ミンダック社製の粉の試料25 mgをエタノール:水(50:50)500 mlによって抽出して、遠心し、加熱してエタノールを蒸発させて濾過した。ケーキ酵母(8500 mg)を水抽出物に加えて、22℃で0〜48時間インキュベートした。ショ糖の約97%が最初の6時間以内に消化されたが、いくつかのファゴピリトールが失われ、D-カイロ-イノシトールは増加した(表11)。ミオ-イノシトールが大きく増加したことは意外であり、おそらく粉におけるフィチンの分解を反映した。酵母による消化前にイオン交換樹脂によってフィチンを除去すれば、より良好な結果が得られると思われる。ショ糖と比較すると、ファゴピリトールは酵母によってかなりゆっくり消化された。
【0107】
(表11)
【0108】
もう一つの酵母消化実験において、抽出物をイオン交換カラムであるPVPカラムに通過させて、アミコン限外濾過装置を通過させてから酵母を加えた。この実験は乾燥顆粒酵母(200 mg)を用いて行った。結果は、ショ糖が最初の6時間以内に消化され、そのあいだファゴピリトールが比較的安定であるように思われることを示した(表12)。当初不明であった材料は、最初の6時間に増加してその後ゆっくり分解した。この不明物質は、真正トレハロース(誘導体化後)との同時クロマトグラフィーおよびガスクロマトグラフィー/質量分析によってトレハロースであると同定された。DB1およびDB5カラム上での保持時間および不明物質の質量分析は、真正のトレハロースと同一であった。α,α'-トレハロースは、グルコース(O-α-D-グルコピラノシル-(1-1')-O-α-D-グルコシド)の二量体である。
【0109】
(表12)
【0110】
バーケットミルズ社製の粉を用いて第三の実験においてより少ない量の酵母を調べた。ショ糖約97%が最初の2時間以内に酵母50 mgによって消化され、グルコースの倍加を伴った(表13)。ファゴピリトールは酵母とのインキュベーションのあいだ比較的安定であるように思われた。産生されたトレハロースも同様に低いレベルであることからより低い量の酵母が好ましい。ファゴピリトールから高分子量混入物質より単糖類を除去するほうがおそらくより容易である。
【0111】
(表13)
【0112】
実施例19-精製段階
表14は、バーケットミルズ社製の全ひき割りソバ粉の2回抽出した25 g試料2個から、単離の際の異なる段階(抽出、加熱、および遠心、ならびに蒸発)でのファゴピリトール分析の結果を示す。連続的な精製段階のあいだ、高分子量ファゴピリトールは容易に分解するように思われた。これは、部分的に試料がますます純粋になったことと、部分的にファゴピリトールA3がより容易に分解されるより濃縮された試料になったことによる考えられる。結果は、抽出技法の短縮も望ましいかも知れないことを示した。低温抽出では、より少量を用いて、なおも良好な抽出物の収率を得ることができる。
【0113】
(表14)
#1および#2は、抽出直後に650 ml抽出物2個から得た試料0.5 mlによる。
#3および#4は、加熱および遠心後の350 ml抽出物2個から得た試料0.5 mlによる。
#5は、蒸発および遠心後の抽出物250 mlから得た試料0.5 mlによる。
【0114】
第一および第二の抽出によって抽出されたファゴピリトールの量を表15に示す。全ひき割りソバ粉(25 g、バーケットミルズ社)を、第一の抽出ではエタノール:水(50:50)320 mlによって、そして第二の抽出ではエタノール:水(50:50)200 mlによって抽出した。表15に示されるように、第一の抽出は、可溶性糖質の約90%を除去した。第二の抽出はより多くのファゴピリトールA1を除去したが、より大きなファゴピリトールは検出可能なレベルで除去されなかった。これらの結果は、抽出をより低い容量での一段階に短縮すれば、抽出時間が短縮されるが、ファゴピリトールA1を十分に回収しない可能性があることを示している。
【0115】
(表15)
【0116】
ファゴピリトール粗抽出物は、3-アミノプロピル官能性シリカゲルカラム上で二量体、三量体、および四量体に分離することができた(図12)。カラムは、ファゴピリトールA3を回収するためにオーバーローディングした。オーバーローディングにもかかわらず、かなりの(しかし全てではない)精製が得られた。ファゴピリトールAおよびファゴピリトールBは、カラムをオーバーローディングすると通常重なり合うことが認められた。しかし、注意して行えば、カラムをオーバーローディングした場合でも比較的純粋な画分を得ることができる(図13)。
【0117】
限外濾過装置の前にPVPカラムを置くと、アミコンの分子量10,000カットオフフィルターを通しての抽出物の通過時間が短縮された。図14および図15によって示されるように、これはまた、二量体、三量体、および四量体ファゴピリトールの分解を改善した。
【0118】
実施例20-ヒト便菌叢によるファゴピリトール濃縮ソバ抽出物のインビトロ発酵
発酵試験は、乾燥顆粒パン酵母によって予め処置したファゴピリトール濃縮バーケットミルズ社製ソバ抽出物によって行った。酵母細胞を遠心(ソルバル冷蔵遠心機を用いて19K rpmで10分間)および限外濾過(アミコン濾過装置、分子量10,000カットオフ)によって除去した。ファゴピリトール水抽出物は、滅菌して他の全てのガスを除去するために二酸化炭素下で沸騰させた。
【0119】
便接種物および全ての試料を調製するあいだ、二酸化炭素ガスの流れの下で作業することによって嫌気的条件を維持した。発酵ボトルはゴム製の栓と金属製のキャップによって密封した。
【0120】
ファゴピリトール基質を最初に調製した。次に、ブランクを除き、全てのボトルに1mlを入れた(1試料あたり2個にさらに対照1個とブランク1個とを含む4個を、0時間、6時間、12時間、および24時間に行った)。次に、栄養培地を調製した。対照ボトルには、接種物の代わりに培地1mlを入れた。残りの培地を用いて、新鮮な便を1:12(w/v)に希釈して、参照として本明細書に組み入れられる、バリー(Barry)ら、「ルミナールおよびヒト便菌叢による、中性単糖類のインビトロ発酵(In Vitro Fermentation of Neutral Monosaccharides by Ruminal and Human Fecal Microflora)」、Acta Vet Scand 86(補則):93〜95(1989)およびギロン(Guillon)ら、「ヒト便細菌によるエンドウおよびリンゴ繊維の発酵の、残留繊維の特徴付け(Characterization of Residual Fibers from Fermentation of Pea and Apple Fibers by Human Fecal Bacteria)」、J. Sci. Food Agric. 68:521〜529(1995)に記載のように調製した。全ての試料およびブランクに、ゴム製の栓を通してシリンジによって便接種物1mlを接種した。
【0121】
発酵のために40℃の水浴を用いて、明記した時間にボトルを開けて空気を入れることによって発酵を中止し、直ちに凍結した。
【0122】
試料は、融解後1回遠心した後(微量遠心機、8000 rpm)、分子量10,000カットオフの微量遠心管において再度遠心することによって調製した。栄養培地を含むそれらの試料に存在する塩は、それらがTMS誘導体化を妨害することから、各試料をアンバーライト混合床樹脂の中に通過させることによって除去した。当初の抽出物の対照試料2個も同様に、ファゴピリトールが処置によって除去されないことを保証するために樹脂の中を通過させた。
【0123】
各(750 μl)少量を、シリル化バイアルにおいて内部標準(フェニルα-D-グルコピラノシド)100 μgと共に窒素ガスの流れの下で乾燥させた;バイアルをP205上で一晩デシケータに入れた。
【0124】
トリメチルシリル誘導体は、1:1ピリジン:TMS溶液100 μlを加えて、70℃で30分加熱することによって調製した。
【0125】
最後に誘導体化試料をGC(HP 5890シリーズII:DB-1カラム;内径0.25 mm、長さ15 m、被膜の厚さ0.25 μm)において分析した。
【0126】
便の細菌は、試料から全てのファゴピリトールを除去した。イノシトール化合物およびトレハロースも同様に細菌によって代謝された。ピーク2個(保持時間約3.1および3.4)は、全ての試料および対照クロマトグラムについて繰り返し存在し、このことは便細菌によって代謝されない酵母処置の副産物がソバ抽出物に存在することを示している;これらの同じピークは当初の抽出物の分析においても示された。
【0127】
内部標準物質は全ての分析において示され、誘導体化が成功したことを示した。表16は、対照および試料におけるイノシトールおよびファゴピリトールの濃度(mg/750 μlで報告)を示す。
【0128】
(表16)
【0129】
実施例21-ファゴピリトールのパイロットプラント産生技法
粉砕
ソバふすま試料を、アルパイングラインダー(アウグスバーグ、ドイツ)を低速で速いフィードに設定して粉砕した。ふすま50ポンド(22.6 kg)の試料10個をすりつぶして、各試料は完了して粒子径の変化に至るまでに約1.5時間を要した。
【0130】
抽出
すりつぶしたふすま試料を、備え付けの風圧駆動攪拌機(リーメタルプロダクツ、フィリップスバーグ、ペンシルバニア州)を有する100ガロンの蒸気被覆バットに入れた。ふすま試料は、50%エタノール(95%エタノール35ガロン、および蒸留水31ガロン)66ガロンによって抽出して、室温で最高速度で1時間攪拌した。混合物を1時間沈降させ、沈降した後、上清をプラスチックホースを通してサイフォンによって30ガロンの車輪付きステンレススチールドラム缶に移した。
【0131】
操作1および2において、さらに50%エタノール15ガロン(95%エタノール8ガロンと蒸留水7ガロン)を加えて残っている固体をすすいだ。これを沈降させて、上清をサイフォンで除去した。操作3〜10では、最初の抽出によって上清を採取する技術が改善されたためにすすぎ段階を省略した。これは、透明な上清を珪藻土プレートフィルター(「DEPF」)を通してまず濾過する段階、そして次にバットの底の泥を、プレートフィルターが目詰まりするまで可能な限り多く濾過する段階を含む。
【0132】
蒸発
操作1で合わせた上清を、容量20ガロンのドラム型真空蒸気ケトル(ゴレン、シカゴ、イリノイ州)において60℃、20 psi真空で蒸発させた。しかし、抽出物の実質的な比率は、真空排管を通しての泡立ちのために失われた。
【0133】
操作2では、エタノールを100ガロンの蒸気被覆バットにおいて加熱することによって蒸発させた。この方法は時間がかかるものの(丸1日かかる)有効であり、エタノールヒュームがパイロットプラントに放出された。
【0134】
操作3からの抽出物7ガロンについて、蒸発させるために容量20ガロンのドラム型真空蒸気ケトルを用いて試行を行ったが、あらゆる抽出物の喪失を目に見えるようにして、したがって温度調節によって減少できるようにプレキシガラス片を備えた排管を用いた。この過程は遅く、試料の過度の泡立ちを抑えるために温度を50℃に低下させなければならず、抽出物は17%エタノールに蒸発されたに過ぎなかった。
【0135】
操作3の残り、ならびに操作4、5、および6はそれぞれ、ユニペクチン濃縮器(ユニペクチンAG、チューリッヒ、スイス)を用いて濃縮した。この真空補助過程によって温度は35〜40℃に維持され、これを一連の長くて細いパイプを通して流すことによって抽出物の表面積を増加することによって操作することが可能であった。ユニペクチンは、用いるために適度に速かったものの、低温を維持したために、泥状の茶色の物質の形成を減少させず、各抽出物を濃縮器に4回通過させても、エタノール濃度は5.5%に減少したに過ぎなかった。抽出物のエタノール含有量は、蒸気被覆25ガロンバットを沸騰させることによって1〜2%に減少した(リーメタルプロダクツ、フィリップスバーグ、ペンシルバニア州)。
【0136】
操作7〜10の蒸発に関して、操作2に記載したように100ガロンの蒸気被覆バットを用いて、エタノール蒸気をパイロットプラントに放出した。エタノール濃度は、モデル360沸点測定装置(ジュジャージン-サラーソン、パリ、フランス)を用いて測定した。
【0137】
抽出物の保存
ファゴピリトールをエタノール中に含む抽出物を+2℃で保存して、水性抽出物(蒸発段階後)を-20℃で保存した。抽出物はプラスチック4または蓋付の5ガロン容器に保存した。
【0138】
珪藻土プレートフィルター
珪藻土プレートフィルター(「DEPF」)(T.シュライバー社、ハリソン、ニュージャージー州)は、プラスチックホルダーと濾紙片(フィルトレーションアンリミテッド、アクロン、ニューヨーク州)をいずれかの側に備えた金属プレート6枚(7インチ×7インチ)からなり、全て、きつく互いに圧縮されている。DEPFは使用するために、水と混合したプレコート120 g(プレートあたり20 g)の中に送ることによって調製した。DEPFを通過した水が透明であれば、プレコートをその場に置いた。ボディフィードをファゴピリトール抽出物の中に攪拌した。次に、試料をボディフィードと共に、ボディフィードが懸濁液を維持するように絶えず攪拌しながらDEPFの中を通過させた。プレコートまたはボディフィードとして用いられる珪藻土粉末は、セライト503、セライト535、セライト577、およびハイフロースーパーセル(「HFSC」)(セライトコーポレーション、ロンポック、カリフォルニア州)であった。特に明記していなければ、用いた組み合わせは、プレコート(プレートあたり20 g)としてセライト577、そしてボディフィードとしてHFSC(抽出物の重量に対して乾燥粉末の0.25%重量)であった。
【0139】
PVPP処置
パイロットプラント抽出において用いたポリビニルポリピロリドン(PVPP)は、ポリクラー-VT(ISPテクノロジーズ社、ウェイン、ニュージャージー州)であった。抽出物を蒸発によって濃縮した後にPVPP処置を行う場合、PVPP 0.75 kg(特に明記していない限り)を25ガロンの蒸気被覆バットにおいて抽出物に加えて、風圧駆動攪拌器によって30分間攪拌した(ガスト社、ベントンハーバー、ミシガン州)。これによっていくつかの着色が除去されたが、PVPPをカラムにローディングして抽出物をその中に通過させた実験室の規模の場合ほど有効ではなかった。ワイン産業において用いられるように処置時間を3時間(操作2)に増加しても、得られた抽出物の色は認識できるほど改善しなかった。
【0140】
操作7〜10に関して、PVPP 1.5 kgを、蒸発前に100ガロンバットにおいてファゴピリトールのエタノール抽出物に加えた。混合物を30分攪拌して、さらに30分間沈降させて、上清を除去した。蒸発後、抽出物は泥状の茶色(酸化および縮合フェノール類)というよりむしろ濃い黄色(可溶性プロアントシアニジン)となった。上記のように0.75 kg PVPP処置はまた、抽出物をさらにきれいにするために蒸発後に用いた。操作7〜10は、その結果着色混入物質に関して他の抽出物よりかなりきれいであった。
【0141】
最初にPVPPを沈降させて、DEPFのために上清を除去し、PVPPおよび底にある関連する液体を捨てた。次に、PVPPを含む全抽出物をDEPFに送ると、PVPPは、目詰まりを起こすことなくDEPFによって保持された。
【0142】
ベントナイト処置
ベントナイトはワイン産業において、ワインにおいてプロアントシアニジンに反応する不安定なタンパク質によって引き起こされるくもりを透明にするために用いられる。ベントナイト(プレスク島ワインセラーズ、ノースイースト、ペンシルバニア州)は、少量の蒸留水と共に混合する(特に明記していない限り試料1ガロンあたり3g)ことによって濃いスラリーを形成することによって調製した。ブレンダーを用いて塊を破壊した。2〜3時間後、少量の抽出物をベントナイトスラリーと共に混合した後、これを抽出物の残りに加えて、数分攪拌した。処置は約24時間低温で行って、フロキュレーションを起こさせて、タンパク質を沈殿させた。上清を注意深く注いだ。PVPPとは対照的に、ベントナイトを除去するためにDEPFを用いることは、DEPFが容易に目詰まりするために適切ではなかった。
【0143】
イオン交換処置
サイバーン樹脂(アフテック社、ロチェスター、ニューヨーク州)を加えて(1.8 kg)30分間攪拌した。100目のふるいを通過させることによって試料から樹脂を除去した。
【0144】
酵母処理
ショ糖を除去するために酵母による抽出物の処置を操作2で行った。抽出物を32℃に加温して、乾燥顆粒パン酵母40 gを加えて、時折攪拌しながら1時間発酵させた。77℃で15分加熱して酵母を殺した。この方法は、酵母はショ糖の除去には有効であったが、これはトレハロースを産生し、トレハロースはファゴピリトールのその後の精製時にファゴピリトールA1から分離することが難しいため中断した。
【0145】
限外濾過
分子量10,000のカットオフ中空線維フィルターカートリッジ2個を限外濾過ユニット(「UF」)(ロミコン社、ウィルミントン、マサチューセッツ州)に適合させた。
【0146】
操作1は抽出過程において初期にUFを通過させ、大量のエタノール抽出物を速やかに濾過した。約5ガロンのボイド容量が存在するため、UFについて大量の液体を用いなければならないことはこの操作から明白であった。同様に、UFはほとんどのエタノールが抽出物から除去された後に用いなければならず、濾紙は50%エタノールの存在下でわずかにカールするが、なおも再利用可能である。濾過と使用後の濾紙の洗浄はいずれも時間と労力がかかり、そのため労力を省くために、それぞれを個々に行うより他の抽出物を合わせて最終的なUF処置を行った。
【0147】
したがって、操作2〜10からの抽出物を合わせて限外濾過を行った。温度は、濾紙が総抽出物65ガロンを濾過するために必要な1.5時間のあいだ27℃を維持した。10,000ダルトンより小さい粒子は濾紙を通過するが、より大きな粒子は、カートリッジの中をまっすぐ通過して、再循環した。大量の微粒子を含む老廃物約5ガロンが蓄積した。100ガロンのバットに集められた濾過された抽出物は、透明で薄い黄色であった。色はおそらく、特に操作2〜6で限外濾過によって濾過されずに逃れてPVPP、ベントナイト、またはイオン交換樹脂に吸収された可溶性の低分子量プロアントシアニジンの存在を反映する。
【0148】
実施例22-ファゴピリトールA2、ファゴピリトールA3、およびファゴピリトールB2の精製とNMRによる分子構造
ソバからのファゴピリトールの抽出
ソバ普通種(ファゴピルム・エスクレンツムモエンク栽培品種「マノー」(Fagopyrum esculentum Moench cv. 'Manor')種子はミンダックグロワーズ社(グランドフォークス、ノースダコタ州)から提供された。ソバふすまは、EtOH:H2O 1:1(v/v)によってパイロットプラントの規模で抽出し(実施例21を参照)、抽出物は珪藻土フィルターを通して濾過して、蒸発させて容量を減少させ、EtOHを除去して、ポリビニルポリピロリドン(PVPP)、ベントナイト、およびイオン交換樹脂によって処置して、非糖質成分を除去した。可溶性の糖質抽出物は、最終的に分子量10,000のカットオフ中空線維フィルターカートリッジを通して濾過して、蒸発させて容積を減少させた。
【0149】
ファゴピリトールの精製
精製は、炭素(ダルコG60;J.T.ベイカー社、フィリップスバーグ、ニュージャージー州)とセライト545-AW(スペルコ、ベルフォンテ、ペンシルバニア州)の1:1(w/w)静止相での調整的クロマトグラフィーによって実施した(ウィスラー(Whisler)ら、J. Amer. Chem. Soc. 72:677〜679(1950)、参照として本明細書に組み入れられる)。カラムに新しくスラリーを水と共に充填して、凍結乾燥した試料を最小量の水に溶解してローディングし、可溶性糖質をEtOH:H2O(v/v)を段階的に増加させて4℃で溶出し、20 ml画分に回収した。カラム溶離剤試料を定期的に採取して、フェニルαD-グルコシドを内部標準として加えて、乾燥試料をN-トリメチルシリルイミダゾール:ピリジンの1:1(v/v)混合物によって誘導体化して、水素炎イオン化検出器、スプリットモードインジェクタ(1:50)、およびHP-1MS毛細管カラム(15 m×0.25 mm内径、被膜の厚み0.25 μm)を備えたヒューレットパッカード6890 GCによって分析した(ホーボビッツ(Horbowicz)、参照として本明細書に組み入れられる)。GCはプログラムされた初回温度150℃で操作して、3℃/分で200℃に、7℃/分で325℃になるように調節し、325℃で20分間維持した。注入口は335℃で操作して、検出器は350℃で操作した。担体ガスは2.5 ml/分の窒素であった。
【0150】
ファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3の精製に関しては、ソバふすま抽出物を吸着床25×900 mmのクロマトグラフィーを行った。水2.2 L、4%EtOH 2.5 L、5%EtOH 3 Lによって連続的に溶出した後、ファゴピリトールB2は7%EtOH 2Lによって、ファゴピリトールA2は12%EtOH 1Lによって、そしてファゴピリトールA3は20%EtOH 1Lによって溶出された。ファゴピリトールB2またはファゴピリトールA2、またはファゴピリトールA3を含む画分を凍結乾燥して再度クロマトグラフィーを行った。
【0151】
ファゴピリトールB2の画分を、カンジダ・ユチリス(Candida utilis)由来のインベルターゼ(EC 3.2.1.26)によって22℃で1時間処置してショ糖を加水分解した後、吸着床15×450 mmのクロマトグラフィーを行った。5%EtOH1Lによって混入物質を溶出した後、ファゴピリトールB2が6%EtOHによって溶出された。
【0152】
ファゴピリトールA2は、吸着床15×500 mm上でクロマトグラフィーを行った。少量の混入物質がH2O 0.5 L、6%EtOH 1L、および8%EtOH 0.5 Lによって連続的に溶出され、ファゴピリトールA2は9%EtOHによって溶出された。
【0153】
ソバふすま抽出物の異なる4つのカラムからファゴピリトールA3を含む画分を合わせて、再度クロマトグラフィーを行った(19×350 mm)。ファゴピリトールA3は14%EtOHと共に溶出して、その後12%EtOHによる溶出によって混入物を除去した。
【0154】
酸および酵素的加水分解
ファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3の試料を、1M CF3CO2Hによって80℃で16時間加水分解して蒸発乾固させた。D-ガラクトースとD-カイロ-イノシトールとのモル比は、トリメチルシリル産物のGC分析後に計算した。ファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3の試料(100 μg)を、1.25単位の脱塩緑色コーヒー豆α-D-ガラクトシダーゼ(EC 3.2.1.22)の水溶液200 μlと共に22℃でインキュベートするか、または0.5単位ウシ肝β-D-ガラクトシダーゼ(EC 3.2.1.23)の水溶液200 μlと共に37℃で24時間インキュベートした。酵素タンパク質を濾過(分子量10,000のカットオフフィルター)によって除去し、試料を乾燥させ、産物をトリメチルシリル誘導体のGC分析によってアッセイした。ラフィノースおよび乳糖(100 μg)はそれぞれ、αまたはβ-D-ガラクトシダーゼのみによって加水分解して、双方の酵素が活性であることを確認した。
【0155】
カイロ-イノシトールのカイラリティの分析
真正D-カイロ-イノシトールおよびL-カイロ-イノシトール標準物質(アルドリッチ社、ミルウォーキー、ウィスコンシン州)およびファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3の酸加水分解産物(2M CF3CO2H、70℃で3時間)を、無水ペンタフルオロプロピオン酸:アセトニトリル(1:1、v/v)によって誘導体化した(オストルンド(Ostlund)、Proc. Natl. Acad. Sci. USA 90:9988〜9992(1993))。誘導体化産物は、ヒューレットパッカード5890シリーズ11 GCによって、100℃で等温的にキラシル-バルカイラル毛細管カラム(25×0.25 mm内径、被膜の厚さ0.16 μm)を用いるGCによって、誘導体化標準物質と共に同時クロマトグラフされた(リービット(Leavitt)ら、Carbohydr. Res. 103:203〜212(1982))。注入口の温度は130℃であり、FID温度は130℃であった。担体ガスはヘリウム3ml/分、スプリット1:50であった。
【0156】
NMR分析
精製試料(ファゴピリトールB2 41 mg、ファゴピリトールA2 13.5 mg、ファゴピリトールA3 34.5 mg)を、分析するために99.96%D2O(ケンブリッジアイソトープ研究所、アンドバー、マサチューセッツ州)700 μlに溶解した。バリアン(パロアルト、カリフォルニア州)VXRS-400を、1D 13C NMRおよびDEPT(分極転移のひずみのない増強)に関して、100.5 MHzで、25℃で外部炭素標準物質として1,4-ジオキサン(δc 67.4)を用いて行った。他の全てのNMR実験は、残留HOD(δH 4.63)に対して25℃で標準化した、バリアンユニティ500上で499.9 MHzで行った。これらは1D 1H NMRであり、二倍量フィルター相関分光法(double-quantum filtered correlated spectroscopy、DQCOSY)、勾配増強異核種多量相関(gradient-enhanced heteronuclear multiple quantum correlation、GHMQC)、同核種2D J-分光法(homonuclear 2D J-spectroscopy、HOM2DJ)、核オーバーハウザー交換分光法(Nuclear Overhauser and Exchange Spectroscopy、NOESY)、および勾配増強異核種多重結合相関(gradient-enhanced heteronuclear multiple bond correlation、GHMBC)であった。データを分析してバリアンVNMRソフトウェア、バージョン6.1Aを用いてシミュレーションを行った。
【0157】
結果
ソバふすまを50%EtOHによって抽出し、ファゴピリトールを炭素-セライトの1:1(w/w)混合物によってカラムクロマトグラフィーを用いて精製した。ファゴピリトールB2は7%EtOHによって溶出された画分から精製され、ファゴピリトールA2は、9%EtOHによって溶出され、ファゴピリトールA3は、14%EtOHによって溶出された。ファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3の酸加水分解からのカイロ-イノシトールのペル(ペンタフルオロプロピオネート)は、カイラルカラムにおいてD-カイロ-イノシトールと共に同時クロマトグラフされたが、L-カイロ-イノシトールとは同時クロマトグラフされなかった。ファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3の酸加水分解によって、D-ガラクトースとD-カイロ-イノシトールを、ファゴピリトールB2に関して2.02:1.00の比で、ファゴピリトールA2に関して2.03:1.00の比で、そしてファゴピリトールA3に関して2.98:1.00の比で生じた。ファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3の部分的加水分解産物のGC分析によって、ファゴピリトールB2からファゴピリトールB1、ファゴピリトールA2からファゴピリトールA1、そしてファゴピリトールA3からファゴピリトールA1、D-カイロ-イノシトール、ならびにファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3からガラクトースが同定された。ファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3は、α-D-ガラクトシダーゼによって加水分解されたが、β-D-ガラクトシダーゼによっては加水分解されず、全てのD-ガラクトピラノシル残基に関してαアノマー結合であることを証明している。
【0158】
ファゴピリトールB2の13C NMRスペクトルによって、炭素共鳴18個が同定され、1H NMRスペクトルによって水素共鳴20個が同定された。炭素18個の化学シフトはそれぞれ、GHMQCを用いて水素20個の化学シフトと結合していた。炭素の化学シフト4個を除く全てが69 ppm〜76 ppmのあいだであった。炭素共鳴2個は、99.02および95.74 ppmで低磁場への化学シフトを示し、これは置換アノマー炭素の特徴であった。炭素2個は62.03および67.72 ppmで高磁場に存在して、これらはDEPTによってCH2炭素であると同定された。水素原子3個を除く全てが3.5〜3.9 ppmでの化学シフトを有した。最も低磁場の水素2個(δ4.81およびδ4.99)はアノマー水素に特徴的な化学シフトを有し、GHMQCによってアノマー炭素(それぞれ、δ99.02およびδ95.74)に結合していることが示された。これらは、DQCOSYによるガラクトーススピン系内での連結度を追跡するための開始点として作用した。高磁場のCH2炭素に水素共鳴を割り当てると、二つのガラクトース残基の相違が明らかになった;CH2炭素の一つ(δ62.03)は偶発的に等価の共鳴(δ3.59)を有する水素2個を有し、他のCH2炭素(δ67.72)は等価でない共鳴(δ3.72およびδ3.54)を有した。CH2水素のあいだに等価でない共鳴が存在することは、この炭素にさらに置換が存在することを示唆しており;このように、等価でない水素を含むスピン系(δ3.54、H-6'a;δ3.72、H-6'b)を、カイロ-イノシトールに近位の(最も近い)ガラクトースに割り当てて、等価のプロトン(δ3.59、H-6"a、H-6"b)を含むスピン系を遠位のガラクトース残基に割り当てた(表17)。
【0159】
(表17)ファゴピリトールB2(1)とファゴピリトールA2(2)の、1Hおよび13C NMR(25℃、D2O)化学シフトおよびプロトン-プロトンカップリング定数
a、b、d、e 割り当ては逆転させてもよい
c 予測値;実験データのシミュレーションによって確認
【0160】
この割り当てのさらなる証拠はC-6'(δ67.72)に由来し、これはC-6"(δ62.03)と比較すると5.5 ppmによってデシールドされており、隣接するC-5'(δ69.98)は、C-5"(δ71.71)と比較して少量が高磁場側にシフトしていることから、C-6'での置換を示している。1D 1Hスペクトル(表17)から計算したカップリング定数(J1',2'、3.9 Hz;J1",2"、3.9 Hz)が小さいことから、双方のガラクトシル環に関してαアノマー結合が確認された。
【0161】
ファゴピリトールB2に関して、全てが3.47〜4.08 ppmのあいだの残っている6個の1H共鳴を、Dカイロ-イノシトールに割り当てて、連結度をDQCOSY実験から表した。それぞれの水素が結合している炭素をGHMQCから推定した。シクリトール環におけるこれらの炭素(δ75.58)の最も低磁場は、ガラクトースとの結合点であった。置換炭素(δ75.58)に結合した水素(δ3.73、H-2)について認められたカップリング定数(J1,2 3.0 Hz;J2,3 10.0 Hz)、ならびにH-1(J1,2 3.0 Hz;J1,6 4.1 Hz)およびH-3(J2,3 10.0 Hz;J3,4 9,6 Hz)と割り当てられた隣接する水素について認められた定数から、隣接する軸結合および赤道結合を有する軸水素(すなわち、H-2またはH-5)に3.73 ppmが割り当てられることを確認した。カイロ-イノシトール環の対称性により、C-2またはC-5での置換の結果、等価な構造が得られ、より小さい数が割り当てられる。したがって、ファゴピリトールB2は、O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→2)-D-カイロ-イノシトールである。
【0162】
ファゴピリトールA2に関する13C NMRスペクトルから、炭素共鳴18個が同定され、1H NMRスペクトルによって水素共鳴20個が同定された。ファゴピリトールA2の二つのガラクトシル残基は、ファゴピリトールB2(上記)に関する記述と同じように解明し、化学シフトとカップリング定数とはファゴピリトールB2と同等であった(表17)。ファゴピリトールA2に関して、D-カイロ-イノシトール環における置換炭素(δ81.78)上での水素(δ3.54)は、1H NMRおよび同核種2D J分光法(「HOM2DJ」)実験の双方から、唯一の大きなカップリング定数(J2,3 9.8 Hz;J3,4 9.8 Hz)を有することが認められ、トランス-アキシャル近接水素(すなわち、H-3またはH-4)であることを示している。カイロ-イノシトール環が対称であるために、C-3またはC-4での置換は同等の構造を生じ、より小さい数を割り当てる。D-カイロ-イノシトール残基の結合点は、ファゴピリトールA2においてファゴピリトールA1と同様にC-3であった。C-1'(δ100.33)に関する唯一の見かけのGHMBC相互作用はH-3(δ3.54)であり、D-カイロ-イノシトール環のH-2およびH-4に軸結合水素が存在することを確認した。同様に、C-3とH-1'との相互作用は(1'→3)結合を確認したが、H-2'は環に対して軸結合であるためC-3とH-2'では確認されなかった。ファゴピリトールA2は、O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→3)-D-カイロ-イノシトールである。
【0163】
ファゴピリトールAシリーズとファゴピリトールBシリーズとのあいだには、置換炭素の化学シフトに明確な差を認めることができ、これは非アノマー炭素の最も低磁場に存在する。Bシリーズでは、これは76 ppmであるのに対し、Aシリーズでは置換炭素は6ppmさらに低磁場で82 ppmであった。ファゴピリトールA3が82.04 ppm(C-3)で炭素共鳴を含むことから、このパターンによって、トリガラクトシルD-カイロ-イノシトールであるファゴピリトールA3がファゴピリトールAシリーズのメンバーであることが定義された(表18)。
【0164】
(表18)ファゴピリトールA3の、1Hおよび13C NMR(25℃、D2O)化学シフトならびに1Hカップリング定数
a、b、d、e 割り当ては逆転させてもよい
c 予測値;実験データのシミュレーションによって確認
【0165】
さらに、置換炭素(δ82.04)に結合した水素は、1H NMRおよびHOM2DJから計算すると、大きなカップリング定数(J2,3 10.1 Hz、J3,4 10.1 Hz)を有する化学シフト3.59 ppm(H-3)を有することが認められた。この化合物は、これまでファゴピリトールBシリーズのトリガラクトシルD-カイロ-イノシトールであると予想されていた(ホーボビッツおよびホーボビッツ(HorbowiczおよびHorbowicz)ら、Planta、205:1〜11(1998)、参照として本明細書に組み入れられる);その理由はソバ種子は、ファゴピリトールA1より多くのファゴピリトールB1、そしてファゴピリトールA2より多くのファゴピリトールB2を有するためであり、同じパターンがトリガラクトシルD-カイロ-イノシトールについても予測された。しかし、このトリガラクトシルD-カイロ-イノシトールはO-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→3)-D-カイロ-イノシトールであると決定され、慣用名ファゴピリトールA3を割付された。
【0166】
四つの環を重ね合わせると、ファゴピリトールA3のスペクトルは複雑であった。水素27個のほとんどが1D 1HおよびHOM2DJスペクトルから同定されたが、DQCOSYを用いて、水素がガラクトースまたはD-カイロ-イノシトールに属すると割り当てた。三つのガラクトース環の2位、3位、および4位の水素は、環のあいだの共鳴が非常に類似であるか、または同一であったため、水素を個々の環に直ちに割り当てることができなかった。水素の三つの対(δ3.50およびδ3.78;δ3.57およびδ3.72;δ3.60)は、DEPTによってCH2であると同定された炭素(δ67.61;δ67.26;δ61.95)とのGHMQC実験におけるその相互作用によって、ガラクトース残基の6位であると同定された。一対の水素は偶発的に等価であり(δ3.60)、最も高磁場の炭素(δ61.95)に結合し、H-6'"aおよびH-6'"bと割り当てられた。これらの水素(δ3.60)は、DQCOSYによって3.87 ppmで(H-5'")水素と相互作用した。H-5'"との相互作用を通して、98.85 ppmでのアノマー炭素をC-1'"と割り当てた。この炭素(δ98.85)も同様に、CH2水素と相互作用して化学シフトが3.72および3.57 ppmであり、H-6'"aおよびH-6'"bと割り当てられた。同様に、99.05 ppm(C-1")での炭素は4.02 ppm(H-5")での水素、ならびに3.78 ppmおよび3.50 ppm(H-6'aおよびH-6'b)での水素と相互作用した。C-1'(δ100.36)とD-カイロ-イノシトールのH-3(δ3.59)のみとのGHMBC相互作用によって、H-3が置換点(C-3)で軸結合のH-2およびH-4と共に炭素に結合していることを確認した。これらの割り当てを行った後、残りの割り当ては四つのスピン系のそれぞれについて行うことができる。最後に、GHMQCを用いて炭素を割り当てた。69.95 ppm〜70.23 ppmのあいだに起こる炭素共鳴24個中8個では、絶対的な割り当ては不可能であり、おそらく変更可能な多くの割り当てを与えた(表18)。
【0167】
ファゴピリトールBシリーズのD-カイロ-イノシトール環における水素間のカップリング定数をその1D 1H NMRスペクトルから計算することは可能であった。しかし、ファゴピリトールAシリーズにおいて、H-4とH-5の化学シフトは一致し、H-4とH-5との強いカップリングによって複雑な多重線を生じ、この中から6.5 Hz以上のカップリング定数を発見することは不可能である。ファゴピリトールBシリーズに関して測定した定数に基づいてD-カイロ-イノシトールに関して予想されるカップリング定数によって、ファゴピリトールA2またはファゴピリトールA3におけるD-カイロ-イノシトールについて認められる1H化学シフトのシミュレーションを行うと、ファゴピリトールAシリーズにおいて認められたものと一致するH-4およびH-5の多重線が得られた。これによって、ファゴピリトールA2に関してシミュレーションされたカップリング定数(J4,5 9.8 Hz)(表17)およびファゴピリトールA3に関してシミュレーションされた定数(J4,5 10.1 Hz)(表18)が確認された。
【0168】
ファゴピリトールBシリーズのメンバーは、これまで他の種において同定されている。ソバ(ホーボビッツ(Horbowicz)ら、Planta, 205:1〜11(1998);ズクゼシンスキ(Szczecinski)ら、Bull. Pol. Acad. Sci. Chem. 46:9〜13(1998)、参照として本明細書に組み入れられる)の他に、ファゴピリトールB1は大豆の種子(シュバイツァー(Schweizer)ら、Carbohydr. Res. 95:61〜71(1981)、参照として本明細書に組み入れられる)、ルピナス、レンズマメ、およびヒヨコマメ(ケメナー(Quemener)ら、Phytochem. 22:1745〜1751(1983)、参照として本明細書に組み入れられる)、およびホホバマメ(オガワ(Ogawa)ら、Carbohydr. Res. 302:219〜221(1997)、参照として本明細書に組み入れられる)に存在する。ファゴピリトールB1の化学合成が報告されている(コルニエンコ(Kornienko)ら、Carbohydr. res. 310:141〜144(1998)、参照として本明細書に組み入れられる)。ファゴピリトールB1は、テンサイの球状種子において化学的方法によって同定された(シオミ(Shiomi)ら、Agric. Biol. Chem. 52:1587〜1588(1988)、参照として本明細書に組み入れられる)が、その絶対的な構造は本発明において初めて示されている。ファゴピリトールA2およびファゴピリトールA3は、ファゴピリトールA1と共に、ソバの種子におけるガラクトシルシクリトールの新規シリーズのメンバーである。
【0169】
実施例23-ファゴピリトールA1およびファゴピリトールB1の精製とNMRによる分子構造
ソバからのファゴピリトールの抽出。ソバ普通種(ファゴピルム・エスクレンツムモエンク栽培品種「マノー」(Fagopyrum esculentum Moench cv. 'Manor')種子のふすま粉砕画分は、ミンダックグロワーズ社(グランドフォークス、ノースダコタ州)から提供された。ソバふすまは、EtOH:H2O 1:1(v/v)によってパイロットプラントの規模で抽出し(実施例21を参照)、抽出物を珪藻土フィルターに通して濾過して、蒸発させて容量を減少させ、EtOHを除去して、ポリビニルポリピロリドン(PVPP)、ベントナイト、およびイオン交換樹脂(強陰イオン樹脂と陽イオン樹脂とを混合)によって処置して、非糖質成分を除去した。可溶性の糖質抽出物は、最終的に分子量10,000のカットオフ中空線維フィルターカートリッジを通して濾過して蒸発させて容積を減少させた。
【0170】
ファゴピリトールの精製
精製は、炭素(ダルコG60;J.T.ベイカー社、フィリップスバーグ、ニュージャージー州)とセライト545-AW(スペルコ、ベルフォンテ、ペンシルバニア州)との1:1(w/w)静止相での調整的クロマトグラフィーによって実施した(ウィスラー(Whisler)ら、J. Amer. Chem. Soc. 72:677〜679(1950)、参照として本明細書に組み入れられる)。カラムに新しくスラリーを水と共に充填して、凍結乾燥した試料を最小量の水に溶解してローディングし、可溶性糖質をEtOH:H2O(v/v)を段階的に増加させて4℃で溶出し、20 ml画分に回収した。カラム溶離剤試料を定期的に採取して、フェニルαD-グルコシドを内部標準として加えて、乾燥試料をN-トリメチルシリルイミダゾール:ピリジンの1:1(v/v)混合物によって誘導体化して、水素炎イオン化検出器、スプリットモードインジェクタ(1:50)、およびHP-1MS毛細管カラム(15 m×0.25 mm内径、被膜の厚み0.25 μm)を備えたヒューレットパッカード6890 GCによって分析した。GCはプログラムされた初回温度150℃で操作して、3℃/分で200℃に、7℃/分で325℃になるように調節し、325℃で20分間維持した。注入口は335℃で操作して、検出器は350℃で操作した。担体ガスは2.5 ml/分の窒素であった。
【0171】
ファゴピリトールA1を精製するため、可溶性糖質(5%ファゴピリトールA1)約1.8 gを含むソバふすま抽出物を、炭素-セライトの吸着床25×900 mmでクロマトグラフィーを行った。水2.2 Lによって溶出させて単糖類を除去して、4%EtOH 2.5 Lによって溶出させてガラクチノールとファゴピリトールB1とを除去した後、ファゴピリトールA1は5%EtOHによって溶出され、これを凍結乾燥して、再度クロマトグラフィー(吸着床25×550 mm)を行った。最初にファゴピリトールB1を4%EtOHによって除去した後にファゴピリトールA1を5%EtOHによって溶出することによってファゴピリトールA1(95%純粋)を得た。ファゴピリトールA1を含む画分に、再クロマトグラフィーを3回行い(吸着床10×370 mm;水によって溶出した後5%EtOHによって溶出)、NMR分析のために純粋なファゴピリトールA1を得た。
【0172】
ファゴピリトールB1を精製するために、ソバふすま抽出物をパン酵母と共にインキュベートしてショ糖を除去した。可溶性糖質(25%ファゴピリトールB1)3.15 gを含む試料を吸着床43×360 mmの炭素-セライト上でクロマトグラフィーを行った。水1.5 Lによって溶出して単糖類を除去した後、ファゴピリトールB1およびガラクチノールが4%EtOHによって溶出された。ファゴピリトールB1を含む画分を凍結乾燥して25×900 mmのカラム上で再度クロマトグラフィーを行った。ガラクチノールはファゴピリトールB1の初回大量と共に溶出して、ファゴピリトールB1溶離剤の最後部からの画分を合わせると、NMR分析にとって純粋なファゴピリトールB1が得られた。
【0173】
D-カイロ-イノシトールは、部分的に精製したファゴピリトールB1(ファゴピリトールB1 106 mg;ファゴピリトールA1 14 mg;ガラクチノール2.5 mg)を4M HCl中で95℃で1時間加水分解して、NaOHによって中和し、得られた混合物を炭素-セライト(25×900 mm)上でクロマトグラフィーを行い、水によって溶出することによって得られた。
【0174】
酸および酵素的加水分解
ファゴピリトールA1およびファゴピリトールB1の試料を1M CF3CO2Hによって80℃で16時間加水分解して蒸発乾固した。D-ガラクトースとD-カイロ-イノシトールのモル比は、トリメチルシリル産物のGC分析後に計算した。ファゴピリトールA1とファゴピリトールB1の試料(200 μg)を1.25単位の脱塩した緑色コーヒー豆α-D-ガラクトシダーゼ(EC 3.2.1.22)の水溶液200 μlと共に22℃で、または0.5単位ウシ肝β-D-ガラクトシダーゼ(EC 3.2.1.23)の水溶液200 μlと共に37℃で24時間インキュベートした。酵素タンパク質は、濾過(分子量10,000のカットオフフィルター)によって除去し、試料を乾燥させて、産物をトリメチルシリル誘導体のGC分析によってアッセイした。ラフィノースおよび乳糖(100 μg)をそれぞれ、αまたはβ-D-ガラクトシダーゼによって加水分解して、双方の酵素が活性であることを確認した。
【0175】
カイラルカラムGCによるD-カイロ-イノシトールとL-カイロ-イノシトールの分離
真正のD-カイロ-イノシトールおよびL-カイロ-イノシトール標準物質(ウィーン大学のリヒター(A. Richter)氏からの寄贈)および純粋なファゴピリトールA1または純粋なファゴピリトールB1の酸加水分解産物(2M CF3CO2H、70℃で3時間)を無水ペンタフルオロプロピオン酸:アセトニトリル(1:1、v/v)によって誘導体化した(オストルンド(Ostlund)、Proc. Natl. Acad. Sci. USA 90:9988〜9992(1993))。誘導体化産物は、ヒューレットパッカード5890シリーズ11 GCによって100℃で等温的にキラシル-バルカイラル毛細管カラム(25×0.25 mm内径、被膜の厚さ0.16 μm)を用いるGCによって、誘導体化標準物質と共に同時クロマトグラフされた(リービット(Leavitt)ら、Carbohydr. Res. 103:203〜212(1982))。注入口の温度は130℃であり、FID温度は130℃であった。担体ガスはヘリウム3ml/分、スプリット1:50であった。
【0176】
NMR分析
精製試料(ファゴピリトールA1 14.4 mg、ファゴピリトールB1 21 mg;D-カイロ-イノシトール9 mg)を、分析するために99.96%D2O 700 μlに溶解した。バリアンVXRS-400を、1D 13C NMRに関して100.5 MHzで、25℃で外部炭素標準物質として1,4-ジオキサン(δC 67.4)を用いて行った。他の全てのNMR実験は、残留HOD(δH 4.63)に対して25℃で標準化したバリアンユニティ500(パロアルト、カリフォルニア州)において499.9 MHzで行った。これらは1D 1H NMR、二倍量フィルターCOSY、勾配増強HMQC、同核種2D J-分光法(HOM2DJ)、NOESY、および勾配増強HMBCであった。バリアンVNMRソフトウェア、バージョン6.1Aを用いてデータを分析してシミュレーションを行った。
【0177】
GC-MS分析
ファゴピリトールA1およびファゴピリトールB1を含むソバ種子からのトリメチルシリル誘導体化糖質を、HP 5970質量分析器にカップリングさせたヒューレットパッカード5890ガスクロマトグラフにおいてDB-5毛細管カラム(30 m×0.25 mm内径、被膜の厚さ0.25 μm)上でクロマトグラフィーを行った。温度プログラムは140℃で4分であり、270℃まで3℃/分で増加させ、270℃で20分間維持した。担体ガスはヘリウム1ml/分であった。誘導体化ファゴピリトールA1およびファゴピリトールB1に対応するGCピークの質量スペクトルを、真正D-カイロ-イノシトールおよびD-ガラクトースのトリメチルシリル誘導体の質量スペクトルと比較した。
【0178】
結果
ソバふすまを50%EtOHによって抽出して、得られた抽出物を炭素-セライト1:1(w/w)上でクロマトグラフィーを行い、ファゴピリトールA1を精製した。比較のために、ファゴピリトールB1も同様に同じ試料から精製した。ファゴピリトールA1およびファゴピリトールB1の酸加水分解からのカイロ-イノシトールのペル(ペンタフルオロプロピオネート)は、真正のD-カイロ-イノシトールと共にカイラルカラム上で同時クロマトグラフされたが、真正のL-カイロ-イノシトールとは同時クロマトグラフされなかった。ファゴピリトールA1およびファゴピリトールB1の加水分解は、D-ガラクトースとD-カイロ-イノシトールとを、ファゴピリトールA1に関してモル比1.01:1.00で、そしてファゴピリトールB1に関して1.03:1.00で生じた。遊離のD-カイロ-イノシトールは、ファゴピリトールA1とファゴピリトールB1の混合物の酸加水分解の後に炭素-セライトカラム上で精製することによって得た。ファゴピリトールA1およびファゴピリトールB1は、α-D-ガラクトシダーゼによって加水分解され、β-D-ガラクトシダーゼによって加水分解されず、α-D-ガラクトシル結合が存在することを証明している。
【0179】
ファゴピリトールA1の13C NMRスペクトルから、炭素共鳴12個が同定され、1H NMRスペクトルから1H共鳴13個が同定された。ファゴピリトールA1における炭素12個の化学シフトは、GHMQCを用いると水素13個の化学シフトに関連した。D-ガラクトピラノシル残基の特徴的な低磁場のアノマー炭素(δ100.26)およびアノマー1H(δ5.16)を、スピン系における連結度に関するDQCOSYの分析の開始点として利用した。ガラクトシル残基上の二つの一致するH-6'水素も同様に有用なマーカーであり、1Hスペクトルにおいてδ3.55で二重強度を有する二重線によって明らかに同定可能であり、13Cスペクトルにおいて最も高磁場の炭素(δ61.80)に結合した。ガラクトシル環の残りの共鳴の割り当ては簡単であった。ガラクトース残基の1Hシグナルを割り当てれば(表19)、残りの6個の共鳴はD-カイロ-イノシトール環に割り当てることができると考えられる。
【0180】
(表19)D2O中で25℃でのファゴピリトールA1、ファゴピリトールB1、および遊離のD-カイロ-イノシトールの、1Hおよび13C NMR化学シフト
a 割り当ては逆転させてもよい。
【0181】
3.54 ppmでの水素は、遊離のD-カイロ-イノシトールに関する共鳴と比較して8〜10 ppm低磁場にシフトした共鳴を有する低磁場の炭素(δ81.77)に結合し(表19)、それによってD-カイロ-イノシトール環上でのガラクトース置換点が同定された。さらに、GHMBAおよびNOESY実験から、3.54での水素と、ガラクトースのアノマー水素(δ5.16;H-1')およびアノマー炭素(δ100.26:C-1')との明らかな相互作用が示され、結合点を確認した。
【0182】
隣接する水素間のカップリング定数は1D 1Hシグナルから計算した。HOM2DJは、重なり合うシグナルから形成された領域3.5〜3.9 ppmにおける多重線を分離するために役立った。得られたカップリング定数(表20)から、ファゴピリトールA1のガラクトース環に関して行った割り当てを確認した。
【0183】
(表20)ファゴピリトールA1、ファゴピリトールB1、および遊離のD-カイロ-イノシトールの、1H-NMRスペクトルデータに由来する推定カップリング定数
a 予測値;実験データのシミュレーションによって確認した。
【0184】
特に、小さいカップリング定数J1',2' 3.9 Hzは、α-アノマー結合を確認した。D-カイロ-イノシトール環の結合点でのδ3.54での水素はカップリング定数9.7 Hzを示し、双方の隣接する水素とトランスジアキシャル関係にあることを示した。したがって、ファゴピリトールA1に構造O-α-D-ガラクトピラノシル-(1→3)-D-カイロ-イノシトールを割り当てた。ファゴピリトールA1は、推定のインスリンメディエータ(ラーナー(Larner)ら、Biochem. Biophys. Res. Comm. 151:1416〜1426(1988);ベルリン(Berlin)ら、Tetrahedron Lett. 31:1109〜1112(1990)、参照として本明細書に組み入れられる)に関連する、O-(2-アミノ-2-デオキシ)-α-D-ガラクトピラノシル-(1→3)-D-カイロ-イノシトール(ベルリン(Berlin)ら、Tetrahedron Lett. 31:1109〜1112(1990)、参照として本明細書に組み入れられる)と等配電子である。
【0185】
D-カイロ-イノシトール環の隣接する水素間のカップリング定数は環の番号をつける際に重要であった。しかし、ファゴピリトールA1のH-4およびH-5のカップリング定数は容易に区別できなかった。これらの同一の化学シフト(δ3.61;H-4およびH-5)を有する二つの強くカップリングした水素の強い相互作用は、見かけの最大スプリットが6.8 Hzである有する複雑な多重線を生じた。D-カイロ-イノシトールにおける近接するトランスジアキシャル水素の3つの対(H-2、H-3、H-4、H-5)は、O-α-D-ガラクトピラノシル-(1→2)-D-カイロ-イノシトール(ファゴピリトールB1、表20)におけるD-カイロ-イノシトールについて認められたように、9〜10 Hzのカップリング定数を示すはずである(ズクゼシンスキ(Szczecinski)ら、Bull. Pol. Acad. Sci. Chem. 46:9〜13(1998)、参照として本明細書に組み入れられる)。この矛盾は、化学シフト(表19)および実験スペクトルラインデータから計算したカップリング定数(表20)を用いて、しかしJ4,5に関しては予測値9.7 Hzを用いて、C-置換D-カイロ-イノシトールに関する1Hスペクトルのシミュレーションによって解決した。シミュレーションしたスペクトルと実験1Hスペクトルとは同一であり、H-4およびH-5のあいだの強い相互作用の影響によって、見かけの最大スプリット6.8 Hzの多重線が3.61 ppmで得られた。このように、J4,5は、シミュレーションに基づくと9.7 Hz(表20)として与えられる。D-カイロ-イノシトール環の水素の割り当ては、ファゴピリトールA1のH-3でのガラクトシル置換が、ファゴピリトールA1の隣接するH-4(δ3.61)を、それがH-5(δ3.61)と一致するようにファゴピリトールB1(δ3.44)およびD-カイロ-イノシトール(δ3.41)より低磁場方向にシフトさせることから、ファゴピリトールB1に関してはファゴピリトールA1より容易であった。ファゴピリトールB1では、H-4(δ3.44)は、H-5(δ3.60)の高磁場であり、これによってトランスジアキシャル相互作用の大きなカップリング定数(J4,5 9.7 Hz)をファゴピリトールB1に関して認めることができる(表20)。シミュレーションはまた、遊離のD-カイロ-イノシトールにおける水素間のカップリングの解明においても有用であった。D-カイロ-イノシトールの1H NMRスペクトルは、それぞれの多重線が同一の水素2個を表す三つの多重線パターンを示し、その2組は、対内部でカップリングを有した。計算した化学シフトおよびカップリング定数のシミュレーションによって、D-カイロ-イノシトールに関して認められたスペクトルと一致したスペクトルが得られ、表20に示すカップリング定数の計算値を確認した。
【0186】
ファゴピリトールA1およびファゴピリトールB1は、2つの異なるシリーズのファゴピリトールオリゴマーを表す位置異性体である。ファゴピリトールB1であるO-α-D-ガラクトピラノシル-(1→2)-D-カイロ-イノシトールの13Cおよび/または1H共鳴は、ソバ(ズクゼシンスキ(Szczecinski)ら、Bull. Pol. Acad. Sci. Chem. 46:9〜13(1998)、参照として本明細書に組み入れられる)、大豆(シュバイツァー(Schweizer)ら、Carbohydr. Res. 95:61〜71(1981)、参照として本明細書に組み入れられる)、およびホホバマメ(オガワ(Ogawa)ら、Carbohydr. Res. 302:219〜221(1997))におけるこの構造に関して報告された値と厳密に一致した。ガラクトピラノシル環の共鳴も同様に、ガラクトシルオノニトール(リヒター(Richter)ら、J. Nat. Prod. 60:749〜751(1997)、参照として本明細書に組み入れられる)およびアノマー炭素でα置換されたガラクトースを含む他のオリゴ糖(ボック(Bock)ら、Adv. Carbohydr. Chem. Biochem. 42:193〜225(1984)、参照として本明細書に組み入れられる)の共鳴と厳密に一致した。
【0187】
ファゴピリトールA1とファゴピリトールB1のトリメチルシリル産物の質量スペクトルはm/z 73、103、129、147、191、204、217、305、318、319、および361でイオン断片ピークと類似であった。m/z 305/318の断片の量の比は、ファゴピリトールA1に関して0.9、およびファゴピリトールB1に関して2.6であったが、m/z 318/319の断片の量の比は、ファゴピリトールA1に関して2.2、およびファゴピリトールB1に関して0.9であった。これらの量の比は、GC-MSによるファゴピリトールA1とファゴピリトールB1のトリメチルシリル誘導体を区別する可能性がある。
【0188】
実施例24-ファゴピリトールB3の単離と特徴付け
ソバからのファゴピリトールの抽出
ソバ普通種(ファゴピルム・エスクレンツムモエンク栽培品種「マノー」(Fagopyrum esculentum Moench cv. 'Manor')種子のふすま粉砕画分は、ミンダックグロワーズ社(グランドフォークス、ノースダコタ州)から提供された。ソバのふすまを、EtOH:H2O1:1(v/v)によってパイロットプラントの規模で抽出し(実施例21を参照)、抽出物を珪藻土フィルターに通して濾過して、蒸発させて容量を減少させ、EtOHを除去して、ポリビニルポリピロリドン(PVPP)、ベントナイト、およびイオン交換樹脂によって処置して、非糖質成分を除去した。可溶性の糖質抽出物は、最後に分子量10,000のカットオフ中空線維フィルターカートリッジを通して濾過して蒸発させて容積を減少させた。
【0189】
ファゴピリトールの精製
精製は、炭素(ダルコG60;J.T.ベイカー社、フィリップスバーグ、ニュージャージー州)とセライト545-AW(スペルコ、ベルフォンテ、ペンシルバニア州)との1:1(w/w)静止相での調整的クロマトグラフィーによって実施した(ウィスラーおよびデュルソ(WhislerおよびDurso)、1950)。炭素320 gとセライト320 gとの混合物を水と共に混合して、薄いペースト状にして100 mm×300 mmカラムに吸着床の厚さが200 mm(吸着床の容積1570 ml)となるように入れた。カラムには糖全体の約20 gを含むパイロットプラント抽出物220をローディングした。可溶性糖質は、ガラス蒸留水4L、2%エタノール4L、4%エタノール4L、5%エタノール4L、8%エタノール4L、10%エタノール4L、14%エタノール4L、20%エタノール2L、および50%エタノール2Lによって4℃で段階的に溶出して、500 mL画分に手動で回収した。流速は水での500 ml/時間から50%エタノールでの約175 ml/時間であった。容量(速度)は画分の収集容器にとっては多すぎるために、画分を手動で収集する必要があった。各画分の試料(200 μl)を採取してフェニル-α-D-グルコシド(25 μg)を内部標準物質として加え、乾燥した試料をN-トリメチルシリルイミダゾール:ピリジン、1:1(v/v)によって誘導体化して、水素炎イオン化検出器、スプリットモードインジェクタ(1:50)、およびHP-1MS毛細管カラム(15 m×0.25 mm内径、被膜の厚み0.25 μm)を備えたヒューレットパッカード6890 GCによって分析した(ホーボビッツ(Horbowicz)、参照として本明細書に組み入れられる)。GCはプログラムされた初回温度150℃で操作して、3℃/分で200℃に、7℃/分で325℃になるように調節し、325℃で20分間維持した。注入口は335℃で操作して、検出器は350℃で操作した。担体ガスは2.5 ml/分の窒素であった。
【0190】
結果
カラム2つを流した(cc37およびcc38(上記のように精製溶出プロトコールによって))。カラムcc38に関する溶出配列プロフィールを図16に示す。D-カイロ-イノシトール(カイロ)およびミオ-イノシトール(ミオ)は水と共に溶出され、果糖(Fru)およびグルコース(Glc)は2%エタノールと共に溶出され、ファゴピリトールB1は4%エタノール、ファゴピリトールA1は5%エタノール、過剰量のショ糖は8%エタノール、ファゴピリトールB2は10%エタノールによって溶出されてその後にDGMIが溶出され、ファゴピリトールA2は14%エタノール、ファゴピリトールB3およびTGMIは20%エタノールおよび50%エタノールによって溶出されてその後にファゴピリトールA3が溶出された。50%エタノール溶液は、ファゴピリトールA3画分を濃縮するために役立った。ファゴピリトールB3として暫定的に同定されたピークは、ファゴピリトールA3の前に溶出した(図16)。
【0191】
ファゴピリトールB3は、予想通りファゴピリトールA3の前に溶出した。暫定的にトリガラクトシルミオ-イノシトールであると同定されたもう一つの化合物(TGMI:図17において「T」と記す)は、ファゴピリトールB3の尾部で溶出し、同様に、ジガラクトシル-ミオ-イノシトール(DGMI)はファゴピリトールB2の尾部に溶出した。
【0192】
ファゴピリトールを含む画分(図16から)を凍結乾燥した。類似の混入物質を含む画分をプールして、さらなる炭素-セライトカラムクロマトグラフィーによってさらなる精製のために濃縮した。カラムcc37(示していない)から、ファゴピリトールB3およびTGMIと同定されたピークを含む2つの画分(60および61)を凍結乾燥して、プールし、濃縮して、高分解能ガスクロマトグラフィーによって分析した。この濃縮画分(60+61)のガスクロマトグラムからのデータを、関係するピークを強調するために拡大尺度で図18において再度プロットした。ファゴピリトールA3は保持時間38.27分(A3)、ファゴピリトールB3は保持時間39.01分(B3)、およびトリガラクトシルミオ-イノシトールは保持時間39.46分(TGMI)であった(図18;表21)。
【0193】
(表21)典型的なGCクロマトグラム*上でのTMSI-誘導体化糖の保持時間
*乾燥試料をN-トリメチルシリルイミダゾール:ピリジン1:1(v/v)によって誘導体化して、3℃/分、7℃/分で325℃まで調節し、325℃で20分維持したHe上で分析した。注入口は335℃で操作して、検出器は350℃で操作した。担体ガスは2.5 ml/分の窒素であった。
【0194】
このクロマトグラムはGCの「後ろの」カラムで実施した。「前の」カラムで実施すると、保持時間は約0.3分長かった。
【0195】
ファゴピリトールの混合物でのファゴピリトールB3は、α-ガラクトシダーゼによって完全に加水分解された。ファゴピリトールB3に富むが、ファゴピリトールA3およびトリガラクトシルミオ-イノシトール(TGMI)も含む画分は、トリフルオロ酢酸によって完全に加水分解されてガラクトースとシクリトールとなった。ガラクトース対シクリトールのモル比は3:1であった。部分的加水分解後の産物の分析は、ファゴピリトールB2(O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→2)-D-カイロ-イノシトール)およびファゴピリトールB1(O-α-D-ガラクトピラノシル-(1→2)-D-カイロ-イノシトール)の存在を同定し(ファゴピリトールB2およびファゴピリトールB1は、非加水分解試料には存在しなかった)、それによって、ファゴピリトールB3がO-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→2)-D-カイロ-イノシトールと同一であることを確認した。
【0196】
実施例25-ファゴピリトールAおよびBシリーズの単離と特徴付け
ソバのファゴピリトールの二つの完全なシリーズは、同定して特徴を調べられている。ファゴピリトールAシリーズはファゴピリトールA1、ファゴピリトールA2、およびファゴピリトールA3からなる。ファゴピリトールBシリーズはファゴピリトールB1、ファゴピリトールB2、およびファゴピリトールB3からなる。全てはαガラクトシダーゼによって加水分解されるが、βガラクトシダーゼによっては加水分解されず、全てのファゴピリトールがα-ガラクトシドであることを確認する(表22)。
【0197】
(表22)ファゴピリトールAシリーズおよびファゴピリトールBシリーズオリゴガラクトシドの加水分解産物
【0198】
全てはトリフルオロ酢酸によってガラクトースとD-カイロ-イノシトールに完全に加水分解される。高分解能ガスクロマトグラフィーによるカイラル毛細管カラム上の酸加水分解産物の分析から、ファゴピリトールにD-カイロ-イノシトールが存在することが確認された(表22)。ガラクトース対D-カイロ-イノシトールのモル比は、ファゴピリトールA1およびファゴピリトールB1に関して1:1、ファゴピリトールA2およびファゴピリトールB2に関して2:1、ならびにファゴピリトールA3およびファゴピリトールB3に関して3:1であった(表22)。ファゴピリトールA1、ファゴピリトールB1、およびD-カイロ-イノシトールの結合および絶対配置はNMRによって決定し、ファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3も同様に決定した。ファゴピリトールA1は推定のインスリンメディエータと等配電子である。実施例24はファゴピリトールB3の単離および同定を図示する。ファゴピリトールB3は、ソバ種子における少量成分である。ファゴピリトールA3:ファゴピリトールB3の定量的な比は約10:1である。
【0199】
実施例26-オリゴガラクトシドのガラクチノールシリーズにおけるDGMIおよびTGMIの同定に関する証拠
ガラクチノールは、ラフィノースシリーズのオリゴ糖(ラフィノース、スタキオース、バーバスコース)、ガラクトピニトール、およびおそらくファゴピリトールの生合成に対する重要なガラクトシルドナーであるO-α-D-ガラクトピラノシル(1→1)-L-ミオ-イノシトールである。ガラクチノールシリーズのメンバーは、ガラクチノール、ジガラクトシルミオ-イノシトール、トリガラクトシルミオ-イノシトール、およびテトラガラクトシルミオ-イノシトール(ホーボビッツ&ペテクI、参照として本明細書に組み入れられる)であると提唱されている。ガラクチノールの絶対配置は、NMRによって決定されている。DGMIは、NMRによって分析されていない。ガラクチノールシリーズのトリおよびテトラガラクトシルメンバーが提唱されているが、これまで単離または特徴付けがなされていなかった。
【0200】
ソバ種子は、ガラクチノール、ジガラクトシルミオ-イノシトール(DGMI)、およびトリガラクトシルミオ-イノシトール(TGMI)を含む。DGMIは、炭素-セライトクロマトグラフィーの際に、ファゴピリトールB2とファゴピリトールA2のあいだに10%エタノールによって溶出される(実施例24を参照のこと)。HP-1 MS毛細管カラム上でのガスクロマトグラフィーでは、DGMIのピークは、ファゴピリトールB2に関するピークより約0.5 分長い保持時間を有した(表21を参照のこと)。DGMIは、α-ガラクトシダーゼによって完全に加水分解されるが、β-ガラクトシダーゼでは加水分解されず、DGMIがαガラクトシドであることを確認する(表23)。
【0201】
(表23)ガラクチノールシリーズのオリゴガラクトシドの加水分解産物
【0202】
DGMIは完全に加水分解されてガラクトースとミオ-イノシトールをモル比で2.08:1.00を生じた(表23)。部分的加水分解の後、同定された産物は、ガラクチノール、ガラクトビオース(gal-gal)、D-ガラクトース、およびミオ-イノシトール出合った。これらの結果から、DGMIの構造はO-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→1)-L-ミオ-イノシトール(O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→3)-D-ミオ-イノシトールとしても知られる)であることが確認された。
【0203】
トリガラクトシルミオ-イノシトール(TGMI)は、炭素-セライトクロマトグラフィーの際にファゴピリトールB3とファゴピリトールA3のあいだに20%エタノールによって溶出される(実施例24を参照のこと)。HP-1MS毛細管カラム上でのガスクロマトグラフィーでは、TGMIのピークはファゴピリトールB3のピークより約0.45分長い保持時間を有する(表21;図19)。TGMIはα-ガラクトシダーゼおよびトリフルオロ酢酸によって完全に加水分解された。ガラクトース対シクリトールのモル比は1:3であった。TGMIに富み、同様にファゴピリトールB3およびファゴピリトールA3(しかし、ガラクチノール、DGMI、またはミオ-イノシトールではない)を含む画分の部分的加水分解後の産物の分析を行うと、ガラクチノールとミオ-イノシトールが共に検出された(表23)。これらの結果は、トリガラクトシルミオ-イノシトールがO-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→1)-L-ミオイノシトール(同様に、O-α-D-ガラクトピラノシル-(1→6)O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→3)-D-ミオ-イノシトールとしても知られる)であると同定する。
【0204】
本発明は、説明する目的で詳細に説明してきたが、そのような詳細はその目的のために提供するのであって、当業者は変更を行うことができ、それらも以下の請求の範囲によって定義される本発明の趣旨および範囲に含まれると理解すべきである。
【技術分野】
【0001】
本出願は、参照として本明細書に組み入れられる、1997年5月7日に提出された米国特許仮出願第60/045,927号の恩典を主張する、1998年5月6日に提出された米国特許出願第09/073,467号の一部継続出願である。
【0002】
発明の分野
本発明は、ファゴピリトールおよびファゴピリトールを用いる方法に関する。
【背景技術】
【0003】
発明の背景
真性糖尿病は、世界保健機構によって流行の割合に達していると認識されている主な世界規模の問題である。これは現在、先進諸国における死因の第4位であり、工業化が進む諸国では急速に増加しつつある疾患である。全世界の糖尿病発生率の推計は、1985年の3000万人から1994年での1億人を超えるまでに増加した。真性糖尿病は、糖質代謝の欠損によって引き起こされ、血液および尿に異常に大量のグルコースが存在することを特徴とする疾患である。真性糖尿病は、最終的に眼、腎臓、心臓、および脚の障害に至ることがあり、妊娠を危険に曝しうる。
【0004】
真性糖尿病は通常二つの種類に分類される。I型またはインスリン依存型真性糖尿病(「IDDM」)は、主に小児および若い成人に発症することからこれまで若年性糖尿病と呼ばれており、自己免疫疾患の一つに関係している。発症と進行が急激であり、これは全症例の約10〜15%を占める。II型またはインスリン非依存型真性糖尿病(「NIDDM」)は、これまで成人型糖尿病と呼ばれており、通常年齢40歳以上の人に認められ、進行は遅い。これはその初期段階で臨床的に病的な状態を伴わないことがしばしばあり、その代わりに血中グルコースレベルまたは尿中グルコースレベルによって検出される。
【0005】
糖尿病は、単一の障害というより多数の原因を有する障害群であると見なされる。ヒトの膵臓は、グルコースの体組織への流入およびその利用を促進して、それによって生体活動のエネルギーを供給するインスリンと呼ばれるホルモンを分泌する。しかし、糖尿病の人では、グルコースの流入が損なわれ、その結果産生されるインスリン量の欠乏または標的細胞の変化のいずれかが起こる。そのため、糖が血液中に蓄積して尿中に排泄される。I型糖尿病では、問題はほぼ常にインスリン産生の重度または全体的な減少である。II型糖尿病では、膵臓はしばしばかなりの量のインスリンを産生するが、このホルモンは組織によるグルコースの利用を促進することができない。
【0006】
適切な処置を行えば、ほとんどの糖尿病患者は血糖値を正常またはほぼ正常範囲に維持する。これによって、患者は通常の生活を送ることができ、疾患のいくつかの長期的な結末を予防することができる。インスリンをほとんどまたは全く産生しないI型糖尿病の場合、治療はインスリン注射を必要とする。II型糖尿病の場合、患者のほとんどが少なくとも中等度に体重過剰であるため、治療は食事制限、体重減少、および運動に基づく。体重の減少は、組織におけるインスリン耐性状態を部分的に逆転させるように思われる。II型患者の血糖値がそれでも高ければ、医師は、治療レジメにインスリン注射を加えてもよい。多くの場合、インスリン注射を必要とするのは、インスリンの欠乏によるためではなく、インスリンメディエータであるガラクトサミン-D-カイロ-イノシトールの欠乏のために患者ではインスリンの効率よい利用能が減少しているためである。
【0007】
注射による投与に関連した不快感の他に、インスリンの用量を制御する問題も存在する。インスリンの過量投与によって生じる低血糖症は振せん、冷や汗、立毛、低体温症、および頭痛を引き起こすことがあり、混乱、幻覚、奇異行動を伴い、最終的に痙攣および昏睡を伴うことがある。したがって、インスリン注射にたよることなく糖尿病患者の血糖値を制御できれば都合がよいと考えられる。本発明は、そのような制御を提供することに向けられる。
【発明の概要】
【0008】
発明の概要
本発明は、単離されたファゴピリトールA1、単離されたファゴピリトールA3、および単離されたファゴピリトールB3に関する。
【0009】
本発明はまた、ファゴピリトールA1、ファゴピリトールA2、ファゴピリトールA3、ファゴピリトールB1、ファゴピリトールB2、ファゴピリトールB3、およびD-カイロ-イノシトールの二つまたはそれ以上を含む組成物にも向けられる。組成物は、単離されたファゴピリトールA1、単離されたファゴピリトールA3、または単離されたファゴピリトールB3の少なくとも一つを含む。
【0010】
本発明は、ファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3からなる群より選択される実質的に純粋なファゴピリトールにも関する。
【0011】
もう一つの局面において、本発明は、ファゴピリトールA1、ファゴピリトールA3、ファゴピリトールB3、およびその混合物からなる群より選択される材料を調製する方法に向けられる。ソバを、粗抽出物を産生するために有効な条件で溶媒に接触させる。粗抽出物は、非ファゴピリトール材料ならびにファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3からなる群より選択される一つまたは複数のファゴピリトールを含む。次に、非ファゴピリトール材料を一つまたは複数のファゴピリトールから分離する。
【0012】
本発明の単離されたファゴピリトールは、同様に薬学的担体を含む薬学的組成物において用いることができる。この薬学的組成物または本発明の実質的に純粋なファゴピリトールまたは本発明の単離されたファゴピリトールは、糖尿病を治療するために患者に投与することができる。
【図面の簡単な説明】
【0013】
【図1】ソバ種子の成分を示す略図である。Aは成熟ソバ痩果の垂直方向の断面である。Bは成熟ソバ痩果の水平方向の断面である。Cはひき割り成熟ソバ(殻を除いたソバ痩果)の水平方向断面である。Dはその粉砕断片および破断平面を示すひき割り成熟ソバの水平断面である。
【図2】エタノール:水(1:1、v:v)によって成熟ソバ種子の胚組織から抽出したトリメチルシリル化可溶性成分のガスクロマトグラムである。ピークは以下のように表示される:(a)D-カイロ-イノシトール、(b)ミオ-イノシトール、(c)フェニルαD-グルコシド(内部標準)、(d)ショ糖、(e)ファゴピリトールA1、(f)ファゴピリトールB1、(g)ガラクチノール、(h)ラフィノース、(i)不明物質、(j)不明物質、(k)ファゴピリトールA2、(l)ファゴピリトールB2、(m)不明物質(おそらくジガラクトシルミオ-イノシトール)、(n)スタキオース、および(o)ファゴピリトールA3。
【図3】エタノール:水(1:1、容積:容積)によって20 DAPソバ種子の胚組織から抽出したトリメチルシリル化可溶性成分のガスクロマトグラムである。Aは加水分解前のガスクロマトグラムである;Bはαガラクトシダーゼ酵素による23時間の加水分解後のガスクロマトグラムである;およびCは2Nトリフルオロ酢酸(「TFA」)による70℃で3時間の酸加水分解後のガスクロマトグラムである。ピークは以下のように同定される:(a)D-カイロ-イノシトール、(b)ミオ-イノシトール、(c)フェニルα-D-グルコシド(内部対照)、(d)ショ糖、(e)ファゴピリトールA1、(f)ファゴピリトールB1、(g)ガラクチノール、(k)ファゴピリトールA2、(l)ファゴピリトールB2、(1)ガラクトース、(2)グルコース、および(3)果糖。グルコース、果糖、およびガラクトースのトリメチルシリル(「TMS」)誘導体は、アノマー型の糖を明確なTMS産物として捕獲する。
【図4】ソバ種子からのファゴピリトールの画分を示すグラフである。ひき割り成熟ソバを50%エタノールによって抽出して、80℃で45分間加熱し、加水分解酵素を不活化して、分子量カットオフ10,000のフィルターを通過させ、可溶性画分を蒸発乾固させた。残査をアセトニトリル:水(75:25、容積:容積)5mlに溶解して、3-アミノプロピルを9%官能化させたシリカゲルを充填した9×290 mmのカラム上でクロマトグラフィーを行った。成分をアセトニトリル:水(75:25、容積:容積)によって0.4 ml/分の流速で溶出し、3-アミノプロピルを9%官能化したシリカゲルを充填した9×290 mmのカラム上でクロマトグラフィーを行った。成分は、アセトニトリル:水(画分1〜70に関して75:25、容積:容積;および画分71〜90に関して60:40のアセトニトリル:水)によって0.4 ml/分で溶出した。各5ml画分の1mlを蒸発乾固させて、トリメチルシリルイミダゾール(「TMSI」)によって誘導体化して、ガスクロマトグラフィーによって分析した。選択した画分1ml試料を、酵素的および/または酸加水分解のために蒸発乾固させた。各画分における成分は以下のように同定される:(a)カイロ-イノシトール+ミオ-イノシトール+ショ糖、(b)ファゴピリトールA1、(c)不明物質、(d)ファゴピリトールB1、(e)ファゴピリトールA2、および(f)ファゴピリトールB2。
【図5】一連のガラクチノールの構造式を示す。酵素ガラクチノールシンターゼは、UDP-ガラクトースからのα-ガラクトシルを、D-ミオ-イノシトールの3位に転移して、ガラクチノール(O-α-D-ガラクトピラノシル-(1→3)-D-ミオイノシトール)を形成する。ガラクチノールは、ガラクトース供与体として作用して、多くの種子においてラフィノースシリーズのオリゴ糖を形成する。ガラクチノールはまた、アクセプターとして作用して、カラスノエンドウ(Vicia sativa L.)および他の豆果の種子において認められるジガラクトシルミオ-イノシトール(O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→3)-D-ミオイノシトール)を形成することができる(参照として本明細書に組み入れられる、ペテク(Petek)ら、「カラスノエンドウの種子由来のミオ-イノシトールの二つのガラクトシドの単離(Isolation of Two Galactosides of myo-Inositol from Vetch Seeds)」、C.R. Acad. Sci.(パリ)Serie D:Sci. Nat. 263:195〜197(1966)(ペテクI)、およびペテク(Petek)ら、「カラスノエンドウ種子の発芽における、αガラクトシダーゼの精製および特性(Purification and Properties of α-Galactosidase in Germinating Vicia sativa seeds.)」、Eur. J. Biochem. 8:395〜402(1969)(ペテクII))。少量の高次オリゴマー(トリガラクトシルミオ-イノシトールおよびテトラガラクトシルミオ-イノシトール)が、豆果種子に存在することがある(参照として本明細書に組み入れられる、カートワ(Courtois)ら、「単糖、オリゴ糖、およびポリオールの分布(Distribution of Monosaccharides, Oligosaccharides and Polyols)」、ハーボーン(Harborne)ら編、「マメ科植物の化学分類(Chemotaxonomy of the Leguminosae)」、ニューヨーク、アカデミック出版社、207〜229頁(1971)(「カートワ」))。
【図6】受粉後(「DAP」)6日〜32日に収穫した殻を除去した「マンカン」ソバ種子の発芽と乾燥耐性とを示すグラフであり、そのまま(○)または飽和LiCl溶液上で相対湿度(「RH」)12%で急速に乾燥後(●)発芽させた。発芽は湿らせたペーパータオル上で25℃で10日目に記録した。
【図7】1日目93%RH、2日目87%RH、3日目75%RH、4日目51%RH、5日目45%RH、および6日目33%RHでインキュベートした場合の、「マンカン」ソバの未成熟(10 DAP)なひき割り種子の緩速乾燥誘導耐性を示すグラフである。新鮮な種子と、連続的に低いRH環境で1〜6日までインキュベートした(様々な塩の飽和溶液上で)後の種子を、12%RHおよび22℃で1週間急速乾燥させた。乾燥後、種子を25℃でペーパータオル上で発芽させた。7日目に発芽した種子は、正常な幼根の成長を示し、乾燥に耐性であると考えられた。Aは、緩速乾燥時の日数の関数としての乾燥の耐性(●)および50%発芽するまでの日数(○)を示す。Bは緩速乾燥時の日数の関数としての胚あたりのシクリトールであるD-カイロ-イノシトール(●)およびミオ-イノシトール(■)の量を示す。Cは、緩速乾燥時の日数の関数としての胚あたりのガラクトシルシクリトールであるファゴピリトールB1(●)、ファゴピリトールA1(黒三角)、およびガラクチノール(■)の量を示す。Dは、緩速乾燥時の日数の関数としての胚あたりのジガラクトシルシクリトールであるファゴピリトールB2(●)、ファゴピリトールA2(黒三角)、およびジガラクトシル-ミオ-イノシトール(□)の量を示す。Eは緩速乾燥時の日数の関数としてのショ糖(●)の量を示す。
【図8】新鮮な10 DAPの未成熟な殻を除去した「マンカン」ソバ種子を高い相対湿度(98%)でインキュベートした日数の関数としての様々な特性を示すグラフである。Aは、高い相対湿度での日数の関数としての乾燥耐性(●)および50%発芽するまでの日数(○)を示す。Bは、高い相対湿度での日数の関数としての胚あたりのシクリトールであるD-カイロ-イノシトール(●)およびミオ-イノシトール(■)の量を示す。Cは、高い相対湿度での日数の関数としての胚あたりのガラクトシルシクリトールであるファゴピリトールB1(●)、ファゴピリトールA1(黒三角)、およびガラクチノール(■)の量を示す。Dは、高い相対湿度での日数の関数としての胚あたりのジガラクトシルおよびトリガラクトシルシクリトールであるファゴピリトールA2(黒三角)、ファゴピリトールB2(●)、ファゴピリトールA3(○)およびジガラクトシルミオ-イノシトール(□)の量を示す。Eは、高い相対湿度での日数の関数としての胚あたりのショ糖(●)の量を示す。
【図9A】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての乾燥耐性(●)および発芽開点始(○)を示す。
【図9B】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての軸の長さ(●)を示す。
【図9C】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての軸あたりのシクリトールであるD-カイロ-イノシトール(●)およびミオ-イノシトール(○)の量を示す。
【図9D】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての子葉あたりのシクリトールであるD-カイロ-イノシトール(●)およびミオ-イノシトール(○)の量を示す。
【図9E】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての軸あたりのファゴピリトールB1(●)、ファゴピリトールA1(○)、ファゴピリトールB2(■)、およびファゴピリトールA2(□)の量を示す。
【図9F】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての子葉あたりのファゴピリトールB1(●)、ファゴピリトールA1(○)、ファゴピリトールB2(■)、およびファゴピリトールA2(□)の量を示す。
【図9G】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての軸あたりのショ糖(●)の量を示す。
【図9H】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての子葉あたりのショ糖の量を示す。
【図9I】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての軸あたりのグルコース(●)、果糖(○)、および不明物質(■)の量を示す。
【図9J】殻を除去した成熟「マンカン」ソバ種子の湿らせたペーパータオル上で25℃での発芽0〜48時間における乾燥耐性の喪失および糖類およびシクリトールの変化を示すグラフである。25℃で湿らせたペーパータオル上での発芽の時間の関数としての子葉あたりのマルトース(●)、マルトトリオース(○)、およびマリトテトラオース(■)の量を示す。
【図10】3-アミノプロピル基官能性シリカゲル上で分離した様々な画分に存在する様々なファゴピリトールの量のグラフである。カラムにローディングした材料は、P2カラムから回収した濃縮ファゴピリトールA3画分であった。
【図11】3-アミノプロピル基官能性シリカゲル上で分離した様々な画分に存在する様々なファゴピリトールの量のグラフである。カラムにローディングした材料は、P2カラムから選択されたP2画分であった。
【図12】3-アミノプロピル基官能性シリカゲル上で分離した様々な画分に存在する様々なファゴピリトールの量のグラフである。カラムにローディングした材料は、全ひき割り粉(バーケットミルズ社)50 g由来の抽出物であった。
【図13】3-アミノプロピル基官能性シリカゲル上で分離した様々な画分に存在する様々なファゴピリトールの量のグラフである。カラムにローディングした材料は、全ひき割り粉(バーケットミルズ社)50 gからの抽出物であった。
【図14】3-アミノプロピル基官能性シリカゲル上で分離した様々な画分に存在する様々なファゴピリトールの量のグラフである。カラムにローディングした材料は、全ひき割り粉(バーケットミルズ社)50 gからの抽出物であった。
【図15】3-アミノプロピル基官能性シリカゲル上で分離した様々な画分に存在する様々なファゴピリトールの量のグラフである。カラムにローディングした材料は、軽ひき割り粉(ミンダック社)50 gからの抽出物であった。
【図16】エタノール:水(v/v)によって段階的に溶出したファゴピリトールを含む予備植物抽出物(20 g総量の糖)の炭素-セライトカラム(cc38)クロマトグラフィーを示す。
【図17】ジガラクトシルミオ-イノシトール(DGMI)、ファゴピリトールA2(A2)、ファゴピリトールB3(B3)、トリガラクトシルミオ-イノシトール(T、TGMI)、およびファゴピリトールA3(A3)の溶出を示す図16からの縮尺を拡大したプロットを示す。
【図18】関係するピークを図示するために縮尺を拡大してプロットしたファゴピリトールA3(A3、38.23分)、ファゴピリトールB3(B3、39.00分)、およびトリガラクトシルミオ-イノシトール(TGMI、39.46分)を示すガスクロマトグラムである。
【図19】それぞれファゴピリトールA3(A3;37.83 分)、ファゴピリトールB3(B3;38.50分)、およびトリガラクトシルミオ-イノシトール(TGMI;38.97分)を濃縮した画分のピークの分離を示すGCクロマトグラムを三つ重ね合わせた軌跡を示す。
【発明を実施するための形態】
【0014】
発明の詳細な説明
本発明は、ファゴピリトールに関する。ファゴピリトールは本明細書において用いられる一般的な用語であり、特定されないα-ガラクトシルD-カイロ-イノシトールまたはその塩もしくは誘導体を意味する。より詳しく述べると、本発明は単離されたファゴピリトールA1、特に以下の式Iを有する複数のファゴピリトールA1およびその塩に関する:
【化4】
【0015】
本発明はまた、単離されたファゴピリトールA3、特に以下の式IIを有する複数のファゴピリトールA3に関する:
【化5】
【0016】
本発明はさらに、単離されたファゴピリトールB3、特に以下の式IIIを有する複数のファゴピリトールB3ならびにその塩および誘導体に関する:
【化6】
【0017】
本発明の複数のファゴピリトールA1、A3およびB3の塩は、水酸化金属または金属アルコキシド、水酸化アンモニウムまたはアミン(例えば、トリエチルアミンのような3級アミン)のような、ファゴピリトールのヒドロキシル基の一つまたは複数のpKa以上のpKa(すなわち、-log Ka)を有する塩基との反応産物となりうる。例としての塩は、リチウム塩、ナトリウム塩、およびカリウム塩のようなアルカリ金属塩、カルシウム塩およびバリウム塩のようなアルカリ土類金属塩、アンモニウム塩、スルホニウム塩、ならびにホスホニウム塩である。
【0018】
本発明の複数のファゴピリトールA1、A3、およびB3の誘導体には、例えば、ハロゲン化アルカリのような陽性荷電を有する炭素を有する化合物とファゴピリトールとの反応産物が含まれ、この場合誘導体はファゴピリトールのエーテルとなり、またはハロゲン化カルボン酸(例えば塩化アセチル)もしくは無水物(例えば、無水酢酸)との反応産物が含まれ、この場合誘導体はファゴピリトールのエステル(例えばアセテート)となる。
【0019】
本明細書において用いられるように、単離されたファゴピリトールは、それが自然界に存在する他のソバ成分を実質的に含まないファゴピリトールである。本発明の単離されたファゴピリトールは、例えば、ソバまたはその他の天然資源からの抽出と共に化学合成を含む様々な方法によって調製することができると理解すべきである。
【0020】
本発明はまた、好ましくは式I、IIおよびIIIをそれぞれ有する実質的に純粋なファゴピリトールA1、実質的に純粋なファゴピリトールA3、および実質的に純粋なファゴピリトールB3、その塩および誘導体にも関する。本発明はまた、実質的に純粋なファゴピリトールA2およびその塩にも関し、特に複数のファゴピリトールA2は以下の式IVを有する:
【化7】
【0021】
本発明はさらに、実質的に純粋なファゴピリトールB1、特に以下の式Vを有するファゴピリトールならびにその塩および誘導体にも関する:
【化8】
本発明はまた、実質的に純粋なファゴピリトールB2、特に以下の式VIを有するファゴピリトールと共にその塩および誘導体にも関する:
【化9】
本明細書において言及されるように、実質的に純粋とは、ガラクチノール、ミオ-イノシトール、ジガラクトシルミオ-イノシトール、フィチン、芳香性物質(例えば、ポリフェノールおよび色素、ならびに他の着色芳香性物質)、細胞壁粒子、タンパク質、ならびに酸(例えば、有機酸、核酸、およびアミノ酸)およびその塩のような他の成分または材料を実質的に含まないことを意味する。典型的に、実質的に純粋なファゴピリトールは、約98%を超える純度または約95%〜約99%の純度のような、95%を超える純度を有するファゴピリトールである。
【0022】
上記のように、本発明のファゴピリトールは、その調製方法によって制限されない。特に好ましい一つの方法は、ソバからの本発明のファゴピリトールの精製を含む。簡単に説明すると、非ファゴピリトール材料と、ファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3からなる群より選択される一つまたは複数のファゴピリトールとを含む粗抽出物を産生するために有効な条件で、ソバを溶媒に接触させる。次に、一つまたは複数のファゴピリトールを非ファゴピリトール材料から分離する。この過程は下記により詳細に説明する。
【0023】
ソバ植物(ファゴピルム・エスクレンツム(Fagopyrum esculentum))の任意の部分を、本発明のファゴピリトールを調製するために用いることができる。
【0024】
開発中のソバ種子の構造は、参照として本明細書に組み入れられる、オーベンドルフ(Obendorf)ら、「開発中のソバ種子の構造および化学的組成(Structure and Chemical Composition of Developing Buckwheat Seed)」、244〜251頁、ヤニク(Janick)ら編、「新たな穀物(New Crops)」、ニューヨーク、ジョンウィリー・アンド・サンズ(1993)(「オーベンドルフ」)中、によって論評され説明されている。成熟したソバ種子の写真は、参照として本明細書に組み入れられる、マーシャル(Marshall)ら、「ソバ:説明、育種、生産、および利用(Buckwheat:Description, Breeding, Production, and Utilization)」、Advances in Cereal Science and Technology 5:157〜210(1982)(「マーシャル」))およびポメランズ(Pomeranz)、「ソバ:構造、組成、および利用(Buckwheat:Structure, Composition, and Utilization)」、CRC Critical Reviews in Food Science and Nutrition 19:213〜258(1983))による論評に示されている。後者の論評における構造情報のほとんどは、参照として本明細書に組み入れられる、ポメランズ(Pomeranz)、「ソバ穀粒の走査型電子顕微鏡写真(Scanning Electron Microscopy of the Buckwheat Kernel)」、Cereal Chemistry 49:23〜25(1972))によるソバ穀粒の走査型電子顕微鏡研究から得ている。胚は、図1Aに略図で示すように、ソバ痩果(成熟したソバ果の名称)の先端の胚の幼根(根)に存在する。参照として本明細書に組み入れられる、オーベンドルフによって説明されるように、子葉は、受粉後8日目までに軟らかい(ほとんど液体)内乳から伸張して内乳の底に達する。痩果は、小柄によって親植物の総状花序(花序または花集合体の名称)に結合する。軸および子葉を含む胚は、内乳の中を生育し続け、内胚葉の端部で巻いて、受粉後16日目で最高乾燥重量に達する。内乳は、受粉後24〜28日までに乾燥物質(ほとんどがデンプン)を蓄積し続ける。さらに数日後、ソバ穀粒(痩果または果実)が成熟して乾燥する。
【0025】
成熟ソバ痩果の水平断面図(図1Aにおいて「断面図」と記した線)を図1Bに略図で示す。この「階層(top-douwn)」図は痩果の外層として外殻(果皮または果実の外皮)を示す(図1Bおよび1Cにおける斜線入りの黒い部分)。外殻は通常黒または暗茶色の硬い繊維様構造である。外殻は粉砕の際に除去される最初の層であり、それによって殻が除去されたひき割り果(果皮のない真の「種子」)を生じる(図1C)。殻を除去したひき割り果の外側の部分は、「皮膜」または種皮(種皮とも呼ばれる)である。これは、細胞壁が肥厚した種皮細胞と核の残遺物(外乳;厚さ細胞1〜3個分)で構成される繊維層である。内乳における細胞の最も外側の層は糊粉層と呼ばれ、通常、成熟穀粒において種皮と核組織に接着する。糊粉は、中心内乳細胞より厚みのある細胞壁を有する小さい細胞の単層であり、デンプンを含まず、種子を乾燥後もなおも「生存」している。内乳細胞の残遺物は、より薄い細胞壁を有し、参照として本明細書に組み入れられる、マーシャル(Marshall)において説明されるようにデンプン顆粒が充填されている。
【0026】
ソバ胚は軸(中心部の円)と、内乳の中を辺縁部まで巻いて、曲がって種皮近傍の内胚葉の外側部分に沿って続く二つの薄い葉状の子葉とを有する(図1Bおよび1C;胚の構造は黒色、内乳構造は白色、および種皮または「皮膜」は灰色)。成熟穀粒では、子葉は内乳をほぼ取り巻く。胚(軸プラス子葉)は、成熟乾燥穀粒においても生存している。発芽後、胚はソバの実生を形成する。成熟乾燥穀粒では、胚と糊粉層の細胞は、ソバ穀粒における全てのファゴピリトールとD-カイロ-イノシトールとを含む。胚(糊粉層と共に)は、ほとんどのタンパク質と、油、フィチン、無機質、ビタミン、およびルチンのほぼ全てを含むが、デンプンは微量に過ぎない。ほぼ全てのデンプンは内乳に存在するが、内乳の最も外側の層である糊粉層にはデンプンは存在しない。
【0027】
全ひき割り粉は、画分を分離しないですりつぶした全ひき割りソバ(図1C)を含む。外皮が繊維様性質であるために、ひき割りソバの粉砕時により大きな断片が通常形成される。破断平面は通常厚みのある細胞壁を有する層の内側である。小麦、オート麦、およびオオムギの乾燥粉砕物と同様に、ふすま画分は果皮(種皮を含む)、核残遺物、糊粉、および糊粉下層を含む。破断平面は、これらの穀粒において、壁がより厚みのある糊粉下層と壁がより薄い中心内乳のあいだである。同様に、ソバの糊粉層は通常、粉砕時に「ふすま」または「皮膜」画分に接着する。ふすま画分では、「種皮」(種皮としても知られる)または「皮膜」は、二つの子葉の外側に接着して子葉のその部分が引き裂かれて、ふすま画分と共に分離される(図1Dにおける理論的破断平面の略図を参照のこと)。必ずしも全ての皮膜が、接着した子葉組織を有するわけではない。ほとんどの内乳は、粉砕時に粉を形成する軟らかい薄い壁の細胞を有する。いくつかの種子において、内乳中心部の子葉は、全くまばらで、「純粋な」子葉組織のより大きな部分に剥がれ落ちる可能性がある。子葉と軸のこれらのより大きな断片は、ふすま画分にも分離される可能性があるが、軟らかい胚組織の多くは微粉にされてデンプンに富む明るい色の粉画分と共に分離する。これらの理由から、ふすま画分はファゴピリトールとD-カイロ-イノシトールに富む原料となりうる。同様に、明るい色の粉画分は、内乳に埋もれた胚画分からのファゴピリトールを含むが、その濃度は大量のデンプンによって希釈されるために低い。内乳の前に胚が成熟するために、胚の大きさは成熟穀粒において内乳の大きさより小さく変動する。より小さい種子(より少ない内乳)では、子葉は種皮により多く接触しているのに対し、より大きな種子(より多い内乳)では、子葉は種皮全体にあまり接触していない。画分において外殻を有しない「精製されたふすま」は、ファゴピリトール、無機質、タンパク質、および油に富む。
【0028】
この説明にとって重要であるのは、胚細胞(子葉と軸)および糊粉層の細胞がショ糖、ファゴピリトール、ルチン、フィチン、ほとんどの無機質、ほとんどのビタミン、ほぼ全ての油、および高濃度のタンパク質を含む細胞である点である。胚および糊粉層の細胞は、乾燥種子における「生存」細胞であるが、糊粉下、中心内乳、および種皮(種皮または皮膜)の細胞は死亡する。外殻(果皮)の細胞も同様に乾燥種子では死んでいる。このように、胚と糊粉層細胞に富む任意の画分をこれらの成分に富むと考えられる。中心内乳細胞は主にデンプンであり、したがって、明るい色の粉画分はデンプンに富む。しかし、胚が中心内乳を横切るため、胚の多くは小さい断片に破壊されて、明るい色の粉画分に入るが、暗い色の粉画分(外皮も含む)または全ひき割り粉にも存在すると考えられる。
【0029】
本発明のファゴピリトールを調製するために用いられるソバ植物の好ましい部分は、ソバ胚、特に胚軸、または「ふすま」もしくは「精製ふすま」粉砕画分を含む、胚、子葉、軸、または糊粉層細胞に富む粉砕画分である。容易に利用できることから、市販の特選ソバ粉(好ましくは水分約12.3%を含む)のようなソバ粉、または市販の全白ひき割り粉から調製した全ひき割り粉は、ファゴピリトールを単離するための優れたソバ原料であることが判明している。または、粉はソバ種子、好ましくは殻を除去したソバ種子から産生することができる。ソバの起源にかかわりなく、ソバ粉のようなソバを微粉に粉砕すれば、好ましくは100目のふるいを通過するような粉末に粉砕すれば、抽出は改善される。ソバ粉は生または脱脂粉となりえて、脱脂はアルカン溶媒、好ましくはヘキサンのような脂肪酸のための溶媒と生ソバとを混合することによって行う。脱脂は、初回抽出段階の効率を増加させるが、一般的に脱脂粉を用いた場合にはその後の濾過およびクロマトグラフィーがより難しくなる。
【0030】
次に、粉を適した溶媒によって抽出する。抽出過程にとって適した溶媒には、水、アルコール、および水/アルコール混合物が含まれる。水抽出物は、おそらくタンパク質およびデンプンの水中の溶解度が高いために、典型的により懸濁材料を含む。アルコールおよびアルコール/水抽出物は一般的に懸濁材料をあまり含まず、抽出の際の微生物および内因性酵素活性を最小限にする。アルコールおよびアルコール/水抽出において用いられる適したアルコールには、メタノール、エタノール、イソプロパノール、およびn-プロパノールが含まれ、エタノールが好ましい。水/エタノール混合物は好ましい抽出溶媒である。これらの中で、好ましい溶媒混合物は、エタノール対水の容量比が約1:2〜約2:1、より好ましくは約1:1である混合物である。
【0031】
抽出は、ソバ粉またはその他のソバ原料を適量の抽出溶媒と共に、好ましくは攪拌しながら混合することによって行われる。攪拌は、ブレンダー、好ましくは「高剪断力」のポリトロンヘッドを有するブレンダー、パドルミキサー、または剪断ミキサーのような任意の適した手段によって行うことができる。攪拌装置としてブレンダーを用いる場合、ブレンダーのスイッチを10秒間入れて10秒間切るサイクルを1分または2分間繰り返すと良好な結果が得られる。粉の単位重量あたり用いられる抽出溶媒の容積は、本発明の実践にあたって特に重要ではない。典型的に、溶媒/粉比(ml/g)約1:1〜約200:1が適しており、溶媒/粉比約10:1〜約50:1が好ましく、溶媒/粉比約20:1が最も好ましい。
【0032】
抽出を行う温度は、本発明の実践にとって特に重要ではない。しかし、熱い抽出後では、混合物は濃厚になりすぎて(ソバに存在するデンプンの膨張のため)濾過または遠心を効率よく行うことができず、抽出されるファゴピリトールの容量が少なくなる。約4℃〜ほぼ室温(約25℃)までの温度での低温抽出、好ましくは約10℃〜ほぼ室温までの温度、最も好ましくはほぼ室温での低温抽出が一般的に最も高い収率を生じる。
【0033】
抽出過程の時間は、攪拌の種類および激しさ、抽出を行う容器の形状、溶媒対粉比、抽出過程を行う温度、抽出される材料の容積、抽出される材料の粉砕の細かさ(粒子径)等を含む多くの要因に依存する。好ましくは、抽出は均一な材料が得られるように十分な時間行う。典型的に、約15秒〜約1時間、好ましくは約1分〜約5分、より好ましくは約1分〜約2分が有効である。
【0034】
抽出されたファゴピリトールの収率はしばしば、抽出過程を1回、2回、またはそれ以上繰り返すことによって増加させることができる。典型的に、最初の抽出では存在するファゴピリトールの85%〜95%が抽出される。2回目の抽出を行うと、さらに8%〜10%収率が増加して、3回目の抽出によって一般的にさらに1%収率が上昇する。
【0035】
第一のおよび選択的にさらなる抽出段階を行った後、ホモジネートを沈殿させて、溶媒を除去する。溶媒の除去は任意の適した手段によって行ってよく、濾過、特に限外濾過または遠心が最もよい結果を生じる。次に、粗抽出物(すなわち、遠心後の上清または濾過後の濾液)を好ましくは、ソバに存在してファゴピリトールを加水分解する可能性がある任意のα-ガラクトシダーゼおよび/またはその他の酵素を不活化するために十分な温度に加熱する。一般的に、不活化は、粗抽出物を約70℃〜約90℃、好ましくは約80℃の温度に加熱することによって行うことができる。冷却後、粗抽出物を好ましくは濾過によって透明にし、蒸発によって濃縮した後、限外濾過によってさらに透明にする。粗抽出物の組成は、抽出過程およびその後の濾過、遠心、不活化、蒸発、または透明化の際に、ガス液体クロマトグラフィー、高速液体クロマトグラフィー(「HPLC」)、分光光度分析、色素結合タンパク質アッセイ、および原子吸収分析を含む標準的な方法によってモニターすることができる。必要であれば、ファゴピリトールの濃縮画分をエタノール沈殿によって画分して、高次オリゴマー型の望ましくない可溶性糖質(例えば、五糖および高次糖)を除去することができる。
【0036】
ファゴピリトールの回収はさらに、細胞壁粒子および大きなタンパク質を、例えば粗抽出物を、約5000〜約20,000、好ましくは約10,000の分子量カットオフ濾紙に通過させて除去することによって、さらに増強することができる。
【0037】
さらにまたは代わりは、ファゴピリトールは例えば、サイバーン樹脂(アフテックインク、ロチェスター、ニューヨーク州から入手可能)、ポリビニルポリピロリドン(「PVPP」)(例えば、ポリクラー-VT、ISPテクノロジーズインク、ウェイン、ニュージャージー州から販売)、ベントナイト(例えば、KWKベントナイト凝集粉、プレスク島ワインセラーズ、ノースイースト、ペンシルバニア州から販売)、および珪藻土粉末(例えば、セライトコーポレーション、ラモック、カリフォルニア州から販売)のような食品等級のイオン交換樹脂によって非糖質の混入物質を除去することによってさらに精製することができる。説明すると、荷電材料は、粗抽出物をアンバーライトERA-94イオン交換樹脂およびドウェックス50Wx4イオン交換樹脂(いずれもシグマケミカル社、セントルイスから市販されている)のようなイオン交換材料に通過させることによって、粗抽出物から除去することができる。ファゴピリトールのさらなる精製は、粗抽出物を、ポリビニルピロリドン(「PVP」)または活性炭のような芳香性物質を吸収する材料を含むカラムに通過させることによって、ポリフェノール材料、芳香性色素、およびその他の茶色の混入物質のような芳香性物質を除去することによって行うことができる。次に、濃縮粗抽出物を冷やすか、もしくはエタノールのようなアルコールを濃縮抽出物に加えること、またはその双方によってファゴピリトールを沈殿させることができる。
【0038】
次に、得られたファゴピリトールと他の糖との混合物は、例えば、クロマトグラフィーによってその成分に分離することができる。例えば、クロマトグラフィーによる分離は、アミン官能性シリカゲル上で、もしくは炭素セライトカラムを用いることによって、またはその両者を用いて行うことができる。一般的に、水溶性の低分子量着色混入物を除去するためには炭素セライトカラムのほうがよい。
【0039】
好ましいアミン官能性シリカゲルである3-アミノプロピル官能性シリカゲルは、シグマ-アルドリッチ社(ミルウォーキー、ウィスコンシン州)から販売されている。シリカゲルの官能化(functionalization)の程度は本発明の実践にとって重要ではない。好ましくは、このシリカゲルは約5%〜約15%官能化しており、より好ましくは約9%官能化している。粗抽出物をローディングして様々なファゴピリトールを3-アミノプロピル官能性シリカゲルから溶出するために適した溶媒は、当業者によって容易に確認することができる。好ましくは、分離はアセトニトリル/水の溶離剤を用いて行う。より好ましくは、粗抽出物はアセトニトリル:水(70:30、v:v)溶液としてローディングして、70:30(v:v)アセトニトリル:水、60:40(v:v)アセトニトリル:水、および次に50:50(v:v)アセトニトリル:水によって段階的に溶出する。溶出の後、3-アミノプロピル官能性シリカゲルは60:40(v:v)アセトニトリル:水によって再生することができる。一般的に、3-アミノプロピル官能性シリカゲルを用いる場合の溶出の順序は以下の通りである:単糖類、D-カイロ-イノシトール、ミオ-イノシトール、ショ糖、トレハロース、ファゴピリトールA1、ファゴピリトールB1、ガラクチノール、ファゴピリトールA2、ファゴピリトールB2、ジガラクトシルミオ-イノシトール、ファゴピリトールA3、ファゴピリトールB3、およびトリガラクトシルミオ-イノシトール。
【0040】
セライト炭素カラムを用いてクロマトグラフィーによる分離を行う場合、適したカラムには、炭素(例えば、ダルコG60、J.T.ベイカー、フィリップスバーグ、ニュージャージー州)とセライト(例えば、セライト545-AW、スペルコ、ベルフォンテ、ペンシルバニア州)との混合物を含むカラムが含まれる。一般的に、カラム材料は炭素とセライトとを蒸留水でスラリーにすることによって調製する。次に、適した大きさのカラムにスラリーを充填して、充填したカラムを蒸留水で洗浄する。粗抽出物は、任意の適した溶媒を用いてローディングすることができ、水が好ましい。炭素セライトカラムは、例えば、水、アルコール(例えば、エタノール)、またはその組み合わせによって溶出することができる。説明すると、溶出は50%エタノール(すなわち、エタノールと水の50:50(v/v)混合物)を用いて行うことができる。または、溶出は蒸留水でまず溶出した後、エタノール含有量を増加させた一連のエタノールと水との混合物によって溶出することによって段階的に行うことができる。例えば、カラムをまず蒸留水によって溶出して次に5%エタノール、次に10%エタノール、次に20%エタノール、次に30%エタノール、次に40%エタノール、そして次に50%エタノールによって溶出することができる。場合によっては、一つまたは複数の溶出段階を省略することができる。例えば溶出は、30%および40%エタノール溶出を行わずに、20%エタノール溶液から50%溶出に直接進行することができる。または、溶出は水/エタノール勾配溶媒系を用いて行うことができる。一般的に炭素-セライトを用いる場合の溶出順序は以下の通りである:D-カイロ-イノシトール、ミオ-イノシトール、単糖類、ファゴピリトールB1、ガラクチノール、ファゴピリトールA1、トレハロース、ファゴピリトールB2、ショ糖、ジガラクトシル-ミオ-イノシトール、ファゴピリトールA2、ファゴピリトールB3、トリガラクトシルミオ-イノシトールおよびファゴピリトールA3。
【0041】
様々な画分の組成は、画分からの少量を蒸発乾固させ、トリメチルシリルイミダゾール(「TMSI」)のピリジン溶液(1:1)によって誘導体化してトリメチルシリル(「TMS」)誘導体化糖質を作製して、例えば高分解能ガスクロマトグラフィーによってTMS誘導体化糖質を分析することができる。一つの特定のファゴピリトールを含む画分をプールして蒸発乾固させることによって、実質的に純粋なファゴピリトールが得られる。ファゴピリトールの混合物を含む画分を再度クロマトグラフィーを行ってその分離を行うことができる。
【0042】
本発明はまた、二つまたはそれ以上のファゴピリトールA1、ファゴピリトールA2、ファゴピリトールA3、ファゴピリトールB1、ファゴピリトールB2、ファゴピリトールB3、およびD-カイロ-イノシトールを含む組成物にも関する。好ましくは、この組成物には、少なくとも一つの単離されたファゴピリトールA1、ファゴピリトールA3、またはファゴピリトールB3が含まれる。例としての組成物は、単離されたファゴピリトールA1と単離されたファゴピリトールA2、単離されたファゴピリトールB2と単離されたファゴピリトールB3、D-カイロ-イノシトールと単離されたファゴピリトールB3等を含む組成物である。好ましくは、組成物は、一つまたは複数のガラクチノール、ミオ-イノシトール、ジガラクトシルミオ-イノシトール、フィチン、芳香性物質(例えば、ポリフェノール、色素、およびその他の着色芳香性物質)、細胞壁粒子、タンパク質、ならびに酸(例えば、有機酸、核酸、およびアミノ酸)およびその塩を実質的に含まない。ファゴピリトールの混合物は、研究室でのインビトロ条件でヒト便の細菌の存在下で6時間以内に分解された。したがって、ファゴピリトールは消化管における細菌によって消化されて、遊離のD-カイロ-イノシトールが放出されて取り込まれると考えられており、単量体または二量体の場合、消化管の細胞によって取り込まれる可能性がある。
【0043】
ファゴピリトールおよび組成物は、上記の単離されたファゴピリトール、実質的に純粋なファゴピリトール、または組成物をイヌ、ネコ、ラット、マウス、およびヒトを含む哺乳類のような患者に投与することによって、そのような患者における糖尿病を治療するために有用である。上記のファゴピリトールおよび組成物はまた、多嚢胞性卵巣症候群を治療するために有用となる可能性がある(参照として本明細書に組み入れられる、ネスラー(Nestler)ら、New England J. of Med. 340:1314〜1320(1999))。例えば、本発明の実質的に純粋なファゴピリトール、組成物、または一つもしくはそれ以上の単離されたファゴピリトールは、単独で投与することができ、または本発明の単離されたファゴピリトールは適した薬学的担体または希釈剤と共に投与することができる。希釈剤または担体成分は、それらが本発明のファゴピリトールまたは組成物の治療効果を減少させないように選択すべきである。適した薬学的組成物には、薬学的担体と、例えば単離されたファゴピリトールA1、単離されたファゴピリトールA3、または単離されたファゴピリトールB3とを含む組成物が含まれる。薬学的組成物はさらに、ファゴピリトールA2、ファゴピリトールB1、ファゴピリトールB2、または三つ全てを含みうる。
【0044】
本明細書に記載のファゴピリトールおよび組成物は、所望の用途に適当な適した任意の形状で構成することが可能である:例えば、経口、非経口、または局所投与。非経口投与の例には、脳室内、脳内、筋肉内、静脈内、腹腔内、直腸内、および皮下投与である。好ましい投与経路は経口である。ファゴピリトールが局所または非経口投与される場合には、ファゴピリトールは予め加水分解することが好ましい。
【0045】
経口で用いるために適した投与剤形には、錠剤、分散粉末、顆粒、カプセル剤、懸濁剤、シロップ剤、およびエリキシル剤が含まれる。錠剤の不活性希釈剤および担体には、例えば炭酸カルシウム、炭酸ナトリウム、乳糖およびタルクが含まれる。錠剤はまた、デンプンおよびアルギン酸のような造粒および崩壊物質;デンプン、ゼラチンおよびアカシアのような結合剤;およびステアリン酸マグネシウム、ステアリン酸およびタルクのような潤滑剤が含まれる。錠剤は、非コーティングであってもよく、または崩壊および吸収を遅延させるために既知の技術によってコーティングしてもよい。カプセル剤において用いてもよい不活性希釈剤および担体には、例えば、炭酸カルシウム、リン酸カルシウム、およびカオリンが含まれる。懸濁剤、シロップ剤、およびエリキシル剤はメチルセルロース、トラガカント、アルギン酸ナトリウムのような従来の賦形剤;レシチンおよびステアリン酸ポリオキシエチレンのような湿潤剤;ならびにエチル-p-ヒドロキシベンゾエートのような保存剤を含んでもよい。非経口投与に適した投与剤形には、溶液、懸濁液、分散剤、乳剤等が含まれる。それらはまた、使用直前に滅菌注射用媒体に溶解または懸濁することができる滅菌固形組成物の形で製造してもよい。それらは、当技術分野で既知の懸濁剤または分散剤を含んでもよい。
【0046】
経口投与の場合、固形または液体単位投与剤形を調製することができる。錠剤のような固形組成物を調製する場合、上記に開示したように適したファゴピリトールまたは組成物を、タルク、ステアリン酸マグネシウム、リン酸二カルシウム、ケイ酸アルミニウムマグネシウム、硫酸カルシウム、デンプン、乳糖、アカシア、メチルセルロース、および薬学的希釈剤または単体と機能的に類似の材料のような従来の成分と混合する。カプセル剤は、開示のファゴピリトールまたは組成物を不活性な薬学的希釈剤と混合して、混合物を適当な大きさの硬いゼラチンカプセルに充填することによって調製される。軟ゼラチンカプセルは、ファゴピリトールまたは組成物と、許容される植物油、軽白色鉱油、または他の不活性油とのスラリーを機械的に封入することによって調製される。
【0047】
シロップ剤、エリキシル剤および懸濁剤のような経口投与のための液体単位投与剤形を調製することができる。水溶性型は、糖、芳香性着香料および保存剤と共に水性媒体に溶解してシロップ剤を形成することができる。エリキシル剤は、芳香性着香料と共に、糖およびサッカリンのような適した甘味料と共に水性アルコール(エタノール)溶媒を用いることによって調製する。懸濁剤は、アカシア、トラガカント、メチルセルロース等のような懸濁剤の助けを借りてシロップ溶媒と共に調製することができる。
【0048】
ファゴピリトールまたは組成物を経口投与する場合、適した1日用量は、参照として本明細書に組み入れられる、ラーナー(Larner)らの米国特許第5,124,360号に記載のような、遊離のD-カイロ-イノシトールの適した用量に基づくことができる。抽出されたファゴピリトールの約半分がD-カイロ-イノシトールであり、ほとんどが結合型のD-カイロ-イノシトールであり、遊離のD-カイロ-イノシトールは少量であると考えられる。したがって、ファゴピリトールの適した用量は、D-カイロ-イノシトールの適した用量の約2倍である。典型的に、経口投与の場合、適した1日用量は、被験者の体重1kgあたり約5 mg〜約200 mgのファゴピリトールまたは組成物である。
【0049】
または、本発明のファゴピリトールは、食品に入れて経口的に摂取することができる。例えば、ファゴピリトールは、パン、ロールパン、およびその他の食品において、精製型で、またはソバのふすまの形で組み入れて、ファゴピリトールを摂取するための食品を作製することができる。抽出したファゴピリトールによってパン、ロールパン、およびその他の食品を栄養強化することは、日常的な食事に大量のファゴピリトールを取り込む方法を提供しうる。パンを調製するための適した技法は、例えば、参照として本明細書に組み入れられる、ブラウン(Brown、「タサヤラのパンの本(The Tassajara Bread Book)」、ボストン:シャンバラ出版(1986))に見出される。
【0050】
非経口投与の場合、液体単位用量剤形は上記のファゴピリトールまたは組成物と、水が好ましい滅菌溶媒とを利用して調製される。用いる溶媒および濃度に応じて、ファゴピリトールまたは組成物は、溶媒に懸濁または溶解することができる。溶液を調製する場合、ファゴピリトールまたは組成物は、注射するために水に溶解して、濾過滅菌してから適したバイアルまたはアンプルに充填して密封することができる。都合のよいことに、局所麻酔剤、保存剤、および緩衝剤のようなアジュバントは溶媒に溶解することができる。安定性を増強するために、液体単位用量剤形は、バイアルに充填して、水分を真空下で除去した後凍結することができる。次に、凍結乾燥粉末をバイアルに密封して、使用前に溶液にするために注射用水のバイアルを添付して供給する。非経口懸濁剤は、ファゴピリトールまたは組成物を溶解する代わりに溶媒に懸濁して、実質的に同じように調製するが、この場合、濾過滅菌を行うことができない。ファゴピリトールまたは組成物は滅菌溶媒に懸濁する前にエチレンオキサイドに曝露することによって滅菌することができる。ファゴピリトールまたは組成物の均一な分布を促進するために、界面活性剤または湿潤剤を非経口懸濁液に含めることが都合がよい。非経口投与量は、1日あたり被験者の体重1kgあたり約5 mg〜約200 mgの範囲のファゴピリトールまたは組成物となりうる。好ましくは、非経口投与の1日用量は、経口投与では、ファゴピリトールからのガラクトースが消化管の微生物によって消費されるが、非経口投与ではガラクトースは血糖値に関与することを考慮して、被験者の体重1kgあたりの用量よりかなり低いと考えられる。
【0051】
または、ファゴピリトールもしくは組成物は、従来の方法を用いて徐放性製剤に組み入れて、外科的に埋め込むことができる。適した徐放性マトリクスには、エチレン酢酸ビニルと他の生体適合性ポリマーからなるマトリクスが含まれる。
【0052】
局所投与の場合、上記のファゴピリトールを含むリン脂質小胞のような担体は、皮膚を通してファゴピリトールの取り込みを促進する可能性がある。
【0053】
上記のように、ファゴピリトールは、消化管において細菌によって消化されて遊離のD-カイロ-イノシトールを放出し、これが取り込まれると考えられる。D-カイロ-イノシトールは抗酸化剤であり、より詳しくはヒドロキシルラジカルスキャベンジャーであることが知られている。したがって、本発明のファゴピリトールおよび組成物はまた、本発明のファゴピリトールおよび組成物は、好ましくは本発明のファゴピリトールおよび組成物を被験者に経口投与することによって、抗酸化剤であるD-カイロ-イノシトールの原料として用いることができる。
【0054】
本発明は、以下の実施例によってさらに説明する。
【実施例】
【0055】
実施例
実施例1-材料および方法
植物材料
ソバ普通種(ファゴピルム・エスクレンツムモエンク栽培品種「マンカン」(Fagopyrum esculentum Moench cv. 'Mancan')、タデ科)の胚を、種子の発達中および成熟種子における可溶性糖質およびシクリトールに関して分析した。ソバ普通種の栽培品種である「マノー(Manor)」および「タータリー(Tartary)」ソバ(ファゴピルム・タータリクム(L)ガートン(Fagopyrum tartaricum(L.)Gaertn))の成熟種子も同様に分析した。ソバ種子は温室で、既に記載されているように昼間(14時間)24℃、夜(10時間)18℃で生育させた(参照として本明細書に組み入れられる、ホーボビッツ(Horbowicz)ら、「種子発達の際のソバの内乳と胚のステロールと脂肪酸の変化(Changes in Sterols and Fatty Acids of Buckwheat Endosperm and Embryo During Seed Development)」、J. Agric. Food Chem. 40:745〜750(1992))。1000ワットのシルバニアハロゲン化金属ランプからの白熱光約740 μmol m-2 s-1によって、自然の太陽光を毎日14時間供給した。胚は6〜32 DAPおよび成熟時に収穫した種子から採取した。温度の効果を評価するために、4週齢の植物を、種子が成熟するまで温室から18℃と25℃の一定温度の生育チャンバーに移した。植物は毎日約300 μmol m-2 s-1の蛍光を16時間与えた。ピン型の花を有する植物と房状の花を有する植物を、生育チャンバーまたは温室内で同じ温度条件で正統法の他家受粉によって手で受粉させてピン×房、および房×ピンを作製した。新鮮および乾燥重量、水、および水分濃度は、温室で生育させた植物について果皮、殻を除去したひき割り果(種子)、胚、および内乳に関して処置毎に種子6個〜10個を調べる実験3回の平均値±SEとして表記する。軸および子葉組織における可溶性成分を分析する場合、それぞれ胚10個の同じもの3個を軸および子葉画分に関して分けた。内乳および胚画分を分析する場合、3つの成熟、乾燥、および手で胚および内乳画分に分離されるひき割り(果皮を除去した)種子の3回の実験を行った。内乳画分は種皮を含むが果皮は含まない。栽培品種と種を比較するために、それぞれひき割り果3個の実験3回を分析した。
【0056】
糖質およびシクリトール標準物質
グルコース、果糖、ガラクトース、ソルビトール、マルトース、マルトトリオース、マルトテトラオース、ミオ-イノシトール、ショ糖、ラフィノース、スタキオース、フェノールα-D-グルコシド(内部標準)、ケルセチン、およびルチンは、シグマケミカル社(セントルイス、ミズーリ州)から購入した。ガラクチノールはクオ(T.M. Kuo)氏(ピオリア、イリノイ州)から贈与された;D-オノニトール、セクオイトール、およびL-(+)-ボメシトールはローウス(F.A. Loewus)氏(プルマン、ワシントン州)から贈与された;D-カイロ-イノシトールとD-ピニトールは、アンギアル(S.J. Angyal)氏(ケニントン、ニューサウスウェールズ州、オーストラリア)から贈与された;O-α-D-ガラクトピラノシル-(1→2)-4-O-メチル-D-カイロ-イノシトール(ガラクトピニトールA)、O-α-D-ガラクトピラノシル-(1→3)-3-O-メチル-D-カイロ-イノシトール(ガラクトピニトールB)、D-ピニトール、およびD-カイロ-イノシトールは、ストリーター(J.G. Streeter)氏(ウースター、オハイオ州)から贈与された;ベルバスコース、D-カイロ-イノシトール、L-カイロ-イノシトール、およびD-ピニトールは、ヴィールシュ(P. Wiirsch)氏(ローザンヌ、スイス)から贈与された;ベルバスコースおよびD-カイロ-イノシトールは、アダムスおよびジェンセン(P. AdamsおよびR.G. Jensen)両氏(ツーソン、アリゾナ州)から贈与された;ならびにD-カイロ-イノシトールおよびL-カイロ-イノシトールはリヒター(A. Richter)氏から贈与された(ウィーン、オーストリア)。
【0057】
可溶性糖質の抽出と分析
20 DAPの胚7個、または発達しつつある(6〜32 DAP)の胚5個または10個、および成熟乾燥種子の同じもの3個からの軸および子葉組織を分離して、重量を測定し、内部標準としてフェニルα-D-グルコシド100 μgを含むエタノール:水(1:1、v:v)0.6 mlによって乳鉢で十分にホモジナイズした。ホモジネートおよびすすぎ液を1.5 ml微量遠心管に合わせて、80℃で45分間加熱して、内因性の酵素を不活化して、ベックマンマイクロフュージEにおいて最高速で20分間遠心した。残査を再度抽出して。合わせた上清を分子量10,000のカットオフフィルターに通過させた。少量の濾液をシリル化バイアルに移して、窒素ガス蒸気中で蒸発乾固させた。残査を、乾燥器において五酸化リン上で一晩保存して、微量の水を除去した。乾燥残査を、堅くキャップをしたシリル化バイアル(スペルコ、ベルフォンテ、カリフォルニア州)においてアルミニウムブロックヒーターにおいて70℃で30分シリル化混合物(TMSI:ピリジン、1:1、v:v)200 μlによって誘導体化した後、22℃に冷却した。TMSIによって誘導体化した可溶性糖質1μlを、水素炎イオン化検出器とヒューレットパッカード3396A積分器とを備えたヒューレット・パッカード5890シリーズIIガスクロマトグラフのスプリットモード注入器に注入した。可溶性糖質は、DB-1毛細管カラム(長さ15 、内径0.25 mm、被膜の厚さ0.25 μm;J&Wサイエンティフィック、フォルソム、カリフォルニア州)において分析して、このカラムはプログラムされた初回温度150℃、3℃/分で200℃となるように調節し、7℃/分で325℃となるように調節し、そして325℃で20分間維持した。注入器部分は、335℃で操作して、検出器は350℃で操作した。担体ガスはヘリウムで、3.0 ml/分、スプリット1:50であった。検出器のガスは水素30 ml/分、および空気300 ml/分であった。
【0058】
可溶性糖質の量は、標準曲線からの外挿によって決定し、内部標準であるフェニルα-D-グルコシドのピーク面積に対するそれぞれの既知の糖またはシクリトールのピーク面積の比を各成分の既知量に対してプロットした。標準物質は全て、アノマー型をTMS誘導体として捕獲するために分析前にエタノール:水(1:1、v:v)に溶解した。不明物質の糖質の量は最も近い既知の標準物質による計算によって推定した。量は、各実験3回の平均値±平均値のSEとして表記する。
【0059】
ソバ種子のファゴピリトールの画分
エタノール:水(1:1、v:v)で抽出した可溶性成分を蒸発乾固させた。残査をアセトニトリル:水(75:25、v:v)5mlに溶解して、9%官能化した3-アミノプロピル官能性シリカゲル(アルドリッチケミカル社、ミルウォーキー、ウィスコンシン州)を充填した9×290 mmカラム上でクロマトグラフィーを行った。成分は、アセトニトリル:水(画分<70に関しては75:25、v:v;そして画分>70に関しては60:40、v:v)によって流速0.4 ml/分で溶出した。各5ml画分の1mlを蒸発乾固させて、上記のようにガスクロマトグラフィーによって分析した。選択した画分の1ml試料を酵素および/または酸加水分解のために蒸発乾固した。
【0060】
酵素的消化
緑色コーヒー豆のα-ガラクトシダーゼ(EC 3.2.1.22)(ベーリンガーマンハイム社、インジアナポリス、インジアナ州)の3.2 M硫酸アンモニウム溶液1mlを蒸留水1000 mlに対して一晩透析して、シリル化反応を妨害して多様な結果を生じる硫酸アンモニウムを除去した。透析した酵素の0.1 ml水溶液を糖質またはシクリトール水溶液0.1 mlに加えて、37℃でインキュベートした。24時間後、無水エタノール0.2 mlおよび内部標準100 μgを加えた。混合物を90℃で5分間加熱して酵素を不活化し、分子量10,000のカットオフフィルターを通して遠心した。濾液を窒素ガス蒸気下で蒸発乾固させて、シリル化して、上記のようにガスクロマトグラフィーによって分析した。
【0061】
酸加水分解
選択した化合物の乾燥試料を2N TFAによって70℃で3時間加水分解した。化合物および加水分解産物は、ガスクロマトグラフィー、またはガスクロマトグラフィー/質量分析(「GC-MS」)および/またはプロトン核磁気共鳴(「1H-NMR」)によって、真正標準物質と比較することによって同定した。
【0062】
乾燥耐性
未成熟な種子を6〜32 DAPで植物から採取して、殻を除去して(果皮を除去する)、そのまま発芽させるか、または乾燥耐性に関して調べた(ブラックマン(Blackman)ら、「発達途中の大豆種子の乾燥耐性における、成熟タンパク質と糖(Maturation Proteins and Sugars in Dessiccation Tolerance of Developing Soybean Seeds)」、Plant Physiol. 100:225〜230(1992)(「ブラックマンI」)、参照として本明細書に組み入れられる)。殻を除去したひき割り果を12%相対湿度(「RH」)でLiClの飽和濃度に対して25℃で急速乾燥させた。6日後、種子を水の上での高湿度で一晩湿らせて、湿らせたペーパータオル上で25℃で発芽させた。7日目での発芽率および50%発芽するまでの日数を報告する。
【0063】
未成熟な種子は10 DAPで収穫して、殻を除去して、種子を連続的に、密封したデシケータ内でKNO3、Na2CO3、NaCl、Mg(NO3)2、K2CO3、およびMgCl2の飽和塩濃度上でそれぞれ、より低い相対湿度(92%、87%、75%、51%、45%、および32%RH)に毎日移すことによって6日間ゆっくり乾燥させた(ブラックマン(Blackman)ら、「大豆の乾燥耐性に関連した成熟タンパク質(Maturation Proteins associated with Dessiccation Tolerance in Soybean)」、Plant Physiol. 96:868〜874(1991)(「ブラックマンII」)、参照として本明細書に組み入れられる)。殻を除去したひき割り果(種子)を12%RHに7日間移して、その後乾燥させた種子(湿らせた後)を湿らせたペーパータオル上で25℃で7日間発芽させることによって、乾燥耐性に関して試験した。高い相対湿度制御処置の場合、未成熟な種子は6日間水を加えたデシケータに入れた(ブラックマンII、参照として本明細書に組み入れられる)後、乾燥耐性に関して調べた。緩速乾燥過程の各段階後または高相対湿度では毎日、胚を採取して可溶性の糖質およびシクリトールに関してアッセイした。
【0064】
「マンカン」商業用保存種子の成熟種子をペーパータオル上で0時間、6時間、12時間、18時間、24時間、36時間、および48時間発芽させた後、上記のようにLiClの飽和溶液上で12%RHで急速に乾燥させた。湿らせたペーパータオル上で25℃で7日間再水和させた後、正常な幼根の生育を有する実生を乾燥耐性であると分類した。軸または子葉は、可溶性糖質およびシクリトールに関してアッセイした。
【0065】
実施例2-成熟種子の胚におけるオリゴ糖とシクリトール
「マンカン」の市販の登録保存種子からの成熟乾燥ソバ種子の胚から抽出した可溶性糖とシクリトールの典型的なガスクロマトグラムを図2に示す。ソバ胚はそれらがラフィノースシリーズのオリゴ糖ではなくてガラクトシルシクリトール(糖アルコール)を蓄積することから、独特であると考えられている。種の名称ファゴピルム(Fagopyrum)にちなんでファゴピリトールB1(図2、ピークfを参照のこと)と命名された主な可溶性成分は、ソバ胚において全可溶性ショ糖およびシクリトールを40%まで蓄積した。ショ糖(図2、ピークd)は、全可溶性糖およびシクリトールの別の42%を占めた。
【0066】
ファゴピリトールB1は、シクリトールD-カイロ-イノシトールのα-ガラクトシル誘導体であり、大豆種子における少ない成分であるO-α-D-ガラクトピラノシル-(1→2)-D-カイロ-イノシトールと同じ保持時間を有する(シュバイツァー(Schweizer)ら、「大豆の三つのα-D-ガラクトピラノシルシクリトールの、精製および構造決定(Purification and Structure Determination of Three α-D-galactopyranosylcyclitols from Soya Beans)」、Carbohydr. Res. 95:61〜71(1981)およびトレイトラー(Traitler)ら、「毛細管カラム上でのオンカラムインジェクションによる、ガスクロマトグラフィーによる糖の分離(Gas Chromatographic Separation of Sugars by On-Column Injection on Glass Capillary Column)」、J. High Resolut. Chromatogr. Chromatogr. Commun. 7:558〜562(1984)、参照として本明細書に組み入れられる)。二つの異なるガラクトシルカイロ-イノシトール異性体であるファゴピリトールA1とファゴピリトールB1(図2、ピークeおよびf)、および二つの異なるジガラクトシルカイロ-イノシトール異性体であるファゴピリトールA2とファゴピリトールB2(図2、ピークkおよびl)とがソバ胚に存在することが証拠によって示唆された。さらに、トリガラクトシルカイロ-イノシトールである少量のファゴピリトールA3(図2、ピークo)が市販の保存種子に検出された。5個のうち少なくと3個がこれまで報告されていなかった。誘導体化されていない二つの遊離のシクリトールであるD-カイロ-イノシトールとミオ-イノシトール(図2、ピークaおよびb)がソバに認められた。ピニトールと、ガラクトピニトールA、ガラクトピニトールBおよびシセリトールを含むピニトールのいくつかのガラクトシル誘導体とを蓄積する(ホーボビッツ(Horbowicz)、「種子の乾燥耐性と保存性;膨満によって産生されるオリゴ糖とシクリトール-論評と調査(Seed Desiccation Tolerance and Storability:Dependence on Flatulence-Producing Oligosaccharides and Cyclitols-Review and Survey)」、Seed Sci. Res. 4:385〜405(1994)(「ホーボビッツ」)、参照として本明細書に組み入れられる)多くのマメ科の種子とは異なり、ソバ種子は、ピニトールまたはピニトール誘導体を含まなかった。ソバは、ガラクチノール(O-α-D-ガラクトピラノシル-(1→3)-D-カイロ-イノシトール)(図2、ピークg)、またはジガラクトシルミオ-イノシトール(O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→3)-D-ミオ-イノシトール)(図2、ピークm)(ペテクIおよびカートワ、参照として本明細書に組み入れられる)のごく少量を蓄積したに過ぎなかった。トウモロコシまたは大豆およびルピナスのような他の豆果とは異なり(ホーボビッツ(Horbowicz)、参照として本明細書に組み入れられる)、ソバ胚によるラフィノース(O-α-D-ガラクトピラノシル-(1→6)-α-D-ガラクトピラノシル-(1→2)-β-D-フルクトフラノシド)(図2、ピークh)の蓄積は非常に少量で、スタキオース(O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→6)-α-D-グルコピラノシル-(1→2)-β-D-フルクトフラノシド)(全可溶性成分の1%未満)(図2、ピークn)の蓄積はごく少量であった。還元糖(グルコース、果糖、ガラクトース)は、種子発達および成熟のあいだソバ胚に蓄積されなかった。
【0067】
ファゴピリトールピークの同一性の証拠を図3A〜Cに示す。加水分解前、ファゴピリトールは容易に明らかである(図3A、ピークe、f、k、l)。α位のガラクトシル結合を加水分解するα-ガラクトシダーゼによって加水分解すると、ピークe〜lは消失したのに対し(図3B)、D-カイロ-イノシトール(図3B、ピークa)は大きく増加し、ミオ-イノシトール(ピークb)はわずかに増加するが、他の遊離のシクリトールは認められない。ショ糖(図3B、ピークd)は、有意に変化しないが、ガラクトースのアノマー型(図3B、「1」と記されたピーク)が出現する。ラフィノースおよびスタキオースは微量存在したに過ぎないための、これらの糖の加水分解によるショ糖およびガラクトースはほとんど認められない。したがって、ほぼ全てのガラクトースがガラクトシルシクリトールから放出された。ミオ-イノシトールはガラクチノール(図3A、ピークg)および微量のジガラクトシルミオ-イノシトール(図2、ピークm)の加水分解によってわずかに増加する(図3B、ピークb)。ファゴピリトールはβ-ガラクトシダーゼ(EC 3.2.1.23)に対して37℃で24時間耐性であり、ガラクトースは放出されなかった。このパターンは、図3Aにおけるピークe、f、kおよびlがα-ガラクトシル結合のみを含むことを確認する。ピークl(ファゴピリトールB2)は、成熟テンサイ(ベータ・ブルガリス(Beta Vulgaris L.))種子の胚から抽出した、O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→2)-D-カイロ-イノシトールと同一の保持時間を示した(シオミ(Shiomi)ら、「テンサイの球状種子からの新しいジガラクトシルシクリトール(A New Digalactosyl Cyclitol from Seed Balls of Sugar Beet)」、Agric. Biol. Chem. 52:1587〜1588(1988)、参照として本明細書に組み入れられる)。強い酸(2N TFA)による加水分解後、ショ糖も同様に加水分解されてグルコース(図3C、「2」と記された二つのピーク)および果糖(図3C、「3」と記された三つのピーク)のアノマー型が得られた。加水分解後に内部標準(ピークc)を加えた。ガラクチノールに関して修正して、ファゴピリトールA2およびファゴピリトールB2がジガラクトシルカイロ-イノシトールであると仮定すると、組織抽出物の酸加水分解から生じた産物は、ファゴピリトールA1プラスファゴピリトールB1(ピークeおよびf)に関してD-カイロ-イノシトール1.00に対してガラクトース残基0.93の計算比を有し、双方がガラクトシルカイロ-イノシトールである証拠を提供する。
【0068】
さらに確認するために、ファゴピリトールを液体カラムクロマトグラフィーによって分離した(図4)。ファゴピリトールB1(図4、ピークd)のαガラクトシダーゼ加水分解産物は、ガラクトースの回収に関して修正後、D-カイロ-イノシトール1.00に対してガラクトース1.08の比を示し、ファゴピリトールB1がαガラクトシルD-カイロ-イノシトールであることを確認した。図4のピークfのTMS-産物のガスクロマトグラフィー分析によって、ファゴピリトールB2に対応する一つのピークを認めた。2N TFAによって部分的に加水分解すると、ファゴピリトールB2、ファゴピリトールB1、ガラクトシルガラクトース、D-カイロ-イノシトール、およびガラクトースに対応するピークを認めた。ガラクトースの回収に関して修正すると、D-カイロ-イノシトール1.00に対してガラクトース残基1.93の比を認め、ファゴピリトールB2がジガラクトシルD-カイロ-イノシトールであることを確認する。ファゴピリトールにおけるガラクトースのα結合はまた、1H-NMRによっても確認した。ガラクトースに対するD-カイロ-イノシトールの比およびピークの位置は、ピークeおよびfがおそらくガラクトシルD-カイロ-イノシトールの異なる位置異性体であること、そしてピークkおよびlが対応するジガラクトシルD-カイロ-イノシトールであることを示唆している。GC-MS分析は、ファゴピリトールA1およびファゴピリトールB1が密接に関連しているが異なる断片化パターンを有することを証明している。ガラクチノールシリーズの構造は図5に示す。
【0069】
ファゴピリトールB1およびファゴピリトールA1におけるD-カイロ-イノシトールの存在は、高分解能ガスクロマトグラフィーおよびキラル毛細管カラムを用いて行った。真正のD-カイロ-イノシトールおよびL-カイロ-イノシトール標準物質、ならびに純粋なファゴピリトールB1および純粋なファゴピリトールA1の酸加水分解産物(2N TFA、70℃で3時間)を、無水ペンタフルオロプロピオン酸(「PFPN」):アセトニトリル(1:1、v/v)によって誘導体化した。誘導体産物を真正のD-カイロ-イノシトールおよびL-カイロ-イノシトール標準物質と共に、カイラル毛細管カラムを備えた高分解能ガスクロマトグラフィー(キラシル-バル、アルテック社、ディアフィールド、イリノイ州;長さ25 m、内径0.25 mm、被膜の厚さ0.16 gin)を用いて100℃で等温的に溶出させて同時にクロマトグラフィーを行った。注入口の温度は130℃で水素炎イオン化検出器(「FID」)の温度は130℃であった。ファゴピリトールB1およびファゴピリトールA1におけるD-カイロ-イノシトールは、真正のD-カイロ-イノシトール(保持時間4.6分)と共に同時クロマトグラフされたが、真正のL-カイロ-イノシトールは(保持時間、4.9分)同時にクロマトグラフされなかった。ファゴピリトールB1およびファゴピリトールA1は、保持時間4.6分でD-カイロ-イノシトールのみを有することが確認された。
【0070】
さらに、ガラクトシル環、D-カイロ-イノシトール環の正確な構造、およびD-カイロ-イノシトール環上の2位の炭素に対するガラクトースのアノマー炭素のα結合を、純粋なファゴピリトールB1を用いたNMR実験によって確認した。スゼンシンスキ(Szczencinski)ら、「ソバ種子由来のファゴピリトールB1の構造に関するNMR研究(NMR Investigation of the Structure of Fagopyritol B1 from Buckwheat Seeds)」、Bulletin of the Polish Academy of Sciences Chemistry 40:9〜13(1998)、参照として本明細書に組み入れられる)を参照のこと。
【0071】
実施例3-種子発達および成熟の際の糖およびシクリトールの蓄積
ソバ胚は、12 DAPで最大水分含有量を蓄積するが、最大新鮮重量は16 DAPで起こり、16 DAP以降は胚の乾燥物質にこれ以上の有意な増加を認めない(表1)。種子成熟の後期段階で認められた胚乾燥物質のわずかな減少は、内乳全体に絡み合って(参照として本明細書に組み入れられる、マーシャル(Marshall)ら、「ソバ:説明、育種、生産、および利用(Buckwheat:Description, Breeding, Production, and Utilization)」、Adv. Cereal Sci. Tech. 5:157〜210(1982)に示されるように)乾燥種子において種皮に接着している全ての子葉残遺物を物理的に除去することが難しいことを反映している。
【0072】
(表1)
【0073】
表2は、種子発達および成熟の様々な段階で、ソバ胚に存在する様々な糖質およびシクリトールの量を示す(μg/胚の単位で表す)。果糖およびグルコースは6および8DAPでの非常に若い胚にのみ存在し、胚発達および成熟の後期には存在しなかった(表2)。D-カイロ-イノシトールおよびミオ-イノシトールは、胚発達および成熟を通して最も初期の段階から存在した(表2)のに対し、ピニトールまたは他のO-メチルシクリトールは胚発達および成熟の任意の段階において検出されなかった。ショ糖は8〜10 DAPのあいだに劇的に増加し(表2)、胚の非常に急速な生育の始まりと一致して(表1)、16 DAPまではこれより遅い速度で増加し続けた(表2)。ファゴピリトールB1およびその異性体型であるファゴピリトールA1は、12 DAPで最初に検出され、12〜16 DAPのあいだに非常に急速に蓄積された(表2)。ファゴピリトールA2およびファゴピリトールB2は、16 DAPで最初に検出されて、20 DAPまで急速に蓄積した。ファゴピリトールA3は、胚乾燥の初期段階(50〜35%水分濃度;表1)のあいだに20 DAP(表2)で蓄積した。全可溶性糖およびシクリトールは2相性で蓄積された:最初に胚の急速な発達およびショ糖の蓄積に関連して8〜10 DAPで、第二にファゴピリトールB1の急速な蓄積に関連して12〜16 DAPで(表2)蓄積された。
【0074】
(表2)
a ガラクチノール標準物質に基づいて計算。
b ラフィノース標準物質に基づいて計算。
c スタキオース標準物質に基づいて計算。
d ショ糖対ファゴピリトールB1の比およびショ糖対非ショ糖の比は、平均値に基づいて計算した。
【0075】
実施例4-ソバ種子における糖とシクリトールの分布
軸は、20 DAPにおいて胚の新鮮重量の8%であることが判明したが、子葉は、胚の新鮮重量の92%であった(表3)。全ての成分の濃度は、子葉より軸において高かった(表3)。ショ糖、シクリトール、およびその誘導体は、ソバ胚の軸では乾燥重量の15%に蓄積したのに対し子葉の乾燥重量の8%であった(表3)。軸組織におけるファゴピリトールB1濃度は、子葉での濃度の倍であり、ショ糖対ファゴピリトールB1の比は、子葉における1.64と比較して軸において1.07(w:w)であった。膨満によって産生されるオリゴ糖である、ラフィノースおよびスタキオースは軸組織に存在したが、子葉ではごく微量に過ぎなかった(表3)。ガラクチノールはショ糖(DP-2)にガラクトースを供与して、ラフィノース(ガラクトシルショ糖、DP-3)を形成し、ラフィノースにガラクトースを供与してスタキオース(ガラクトシルショ糖、DP-4)を形成する。
【0076】
(表3)
a ガラクチノール標準物質に基づいて計算。
b ラフィノース標準物質に基づいて計算。不明物質は、保持時間の比がフェニルα-D-グルコシドに対して2.44である。
c スタキオース標準物質に基づいて計算。
d ショ糖対ファゴピリトールB1の比およびショ糖対非ショ糖の比は、平均値に基づいて計算した。
【0077】
糊粉層および親の種皮-核(スパーモダーム(spermoderm))組織のふすま画分を含む内乳画分は、ひき割り果の74%であり、胚はひき割り果の26%であった(表4)。胚画分は全可溶性糖質およびシクリトールの71.4%を含み、内乳画分は28.6%を含んだ。胚は、内乳画分(76 μg)の5倍ものファゴピリトールB1(403 μg)を含んだ(表4)。内乳画分に存在する糖、シクリトール、およびシクリトール誘導体のほとんどはおそらく糊粉層に存在した。いくつかは、成熟乾燥ひき割り果の内乳画分から完全に分離できなかった子葉の断片であった可能性がある。内乳画分における可溶性成分の濃度は、デンプンが主な貯蔵成分であるために低かった。
【0078】
(表4)
a ガラクチノール標準物質に基づいて計算。
b ラフィノース標準物質に基づいて計算。
c スタキオース標準物質に基づいて計算。
【0079】
実施例5-シクリトールおよび可溶性糖質における栽培品種と種の差
成熟ひき割り果における全可溶性糖質およびシクリトールの濃度は、「マンカン」および「マノー」のソバ普通種(表5)および「タータリー」ソバにおいてほぼ同じであった。ショ糖濃度はひき割り果における三つ全ての遺伝子型において同じであった。ファゴピリトールB1は「マンカン」において濃度が最高であり、「マノー」では低く、「タータリー」では「マンカン」の20%に過ぎなかった。「タータリー」には、「マンカン」または「マノー」において認められない不明化合物2個が含まれたが、「マノー」では、より高い濃度のファゴピリトールA2、ファゴピリトールB2、およびファゴピリトールA3を示した(表5)。三つの遺伝子型は同じ環境条件で生育しなかったために、差の一部は生産環境に関連する可能性がある。しかし、「タータリー」ソバのひき割り果において不明物質1の濃度が高かったこととファゴピリトールB1の濃度が低かったことは、おそらく遺伝的な差の反映であった。「マノー」ソバのひき割り果において、より高次のガラクトシルオリゴマーがより高濃度へシフトしたことは環境の影響を反映した可能性がある。
【0080】
(表5)
a ガラクチノール標準物質に基づいて計算。
b ラフィノース標準物質に基づいて計算。
c スタキオース標準物質に基づいて計算。
【0081】
フラボノール、ケルセチン、およびルチンに対する感受性は本明細書において用いたアッセイ方法では全く低かった。ソバの葉に一般的に存在するルチンは、「マンカン」または「マノー」栽培品種のいずれかのソバ普通種の成熟種子のひき割り果においてごく微量検出されたに過ぎなかった。対照的に、「タータリー」ソバの成熟種子のひき割り果は、ルチン標準物質によって計算し、そして試料を加水分解後ケルセチンおよびラムノース標準物質によって計算すると、乾燥重量1gあたりルチン約7.14 mgを含んだ。ルチンの酸加水分解産物は、ケルセチン、ラムノース、およびグルコースであった。ケルセチンおよびラムノースは、「タータリー」ソバのひき割り果からの抽出物を酸加水分解(2N TFAで70℃で3時間)した後に検出されたが、ソバ普通種の胚からの抽出物の酸加水分解産物にはいずれも認めなかった。グルコースもまたショ糖の酸加水分解産物であり、したがって、組織抽出物におけるルチンのアッセイのために用いることができなかった。胚は、成熟乾燥「タータリー」ソバ種子の内乳から分離することができなかったため、胚組織5個を分析できなかった。
【0082】
実施例6-ソバひき割り果、痩果、および粉砕画分における総ファゴピリトール、遊離のD-カイロ-イノシトール、および総D-カイロ-イノシトール
粉砕画分において、「精製ふすま」画分は種皮に接着する胚画分を高濃度に有した。殻が存在しない場合、この画分は、粉砕画分の中でもファゴピリトールとD-カイロ-イノシトールの最高濃度を示した(表6)。「精製ふすま」における総ファゴピリトールおよび総D-カイロ-イノシトール濃度は、次に最も高い粉砕画分(ふすま#1)の濃度の2倍であった。総ファゴピリトールおよび総D-カイロ-イノシトールの濃度は、粉砕画分において、その粉砕画分における胚組織の濃度の減少の程度に応じて減少した(表6)。この減少順序は「精製ふすま」>「ふすま#1」(殻付き)>「ふすま#2」(殻はより少ない)>全ひき割り粉>「特選粉」(デンプンが多い)>「ひび割れ」(胚断片を小数含む)であった。「ひび割れ」画分は粉砕および抽出することが非常に難しかった。「ひび割れ」画分のデンプン質の内乳は、乾燥していた部分的に発芽した種子に類似していた。乾燥種子を湿らせると、αアミラーゼが、子葉からデンプンの多い内乳に放出され、デンプンを消化し始めた。この粘着性のデンプンは、乾燥時に非常に堅くなった。「ひび割れ」のいくつかは、収穫前に植物において再度湿った種子に由来すると仮定された。ファゴピリトールは再湿潤の12〜18時間以内に消失し、デンプンの分解はその時点で既に始まっていた。類推すると、ひび割れの画分は、少数の胚画分を有し、存在するそれらの画分は、種子を収穫前に再度湿らせるとファゴピリトールが減少している可能性がある。
【0083】
(表6)
【0084】
実施例7-糖とシクリトールの蓄積に及ぼす温度の影響
胚(子葉および軸を合わせて)の糖組成は、18℃および25℃で生育させた植物から収穫した成熟種子から採取した胚に関して報告されている。ソバ胚における主な可溶性糖質は、ショ糖とファゴピリトールB1であり、これらは18℃では全体の85%を占めた(表7)。胚の乾燥重量は二つの温度で類似であったため、内乳は、18℃では25℃より種子あたり平均で15 mg多い乾燥重量を蓄積した。胚の可溶性糖質に及ぼす温度の影響は全く興味深かった。18℃で増殖させた種子の胚は、25℃で生育させた場合と比較して、ショ糖が減少し、ファゴピリトールB1が2.5倍多く、スタキオースは3倍多かった。ファゴピリトールA2およびファゴピリトールB2は25℃ではより高いレベルに蓄積した(表7)。温度がしばしば25℃を超える温室で生育させると、ファゴピリトールA3は20 DAPで検出可能であった(表2)。このパターンは、ファゴピリトールB1の蓄積には低温が都合がよいのに対し、ファゴピリトールのより高次ガラクトシルオリゴマーの蓄積には高温が都合がよいことを示唆している。スタキオースに関しては逆のパターンを認めた。スタキオースの蓄積は、低温で都合がよく(表7)、温室でのより暖かい温度で成熟させた種子からの胚では検出されなかった(表2)。
【0085】
(表7)
a 成熟乾燥種子は、胚切除前に473 mg(18℃)および32±3 mg(25℃)であった;3回の実験。
b ガラクチノール標準物質に基づいて計算。
c ラフィノース標準物質に基づいて計算。不明物質は、フェニルα-D-グルコシドと比較して保持時間の比2.44を示す。
【0086】
実施例8-胚発達の際の乾燥耐性とファゴピリトールB1との関連
ソバ種子における発達途中の胚は、12 DAPで乾燥耐性となった(図6)。新たに収穫した種子は、高度の休眠を示した(サミミー(Samimy)ら、「ソバ普通種における種子休眠(Seed Dormancy in Common Buckwheat(Fagopyruni esculentum Moench)」、Plant Varieties and Seeds 7:17〜22(1994)、参照として本明細書に組み入れられる)。25℃で湿らせたペーパータオル上で16日後も発芽しなかった種子は休眠していると見なされ、20および24 DAPでの植物種子(表2)の場合の76%〜87%ほどのファゴピリトールB1(表8)を含み、休眠がファゴピリトールB1の動員を遅らせることを示唆している。12 DAP(図6)での乾燥耐性の発達は12〜16 DAPでファゴピリトールB1の蓄積に関連して起こった(表2)。
【0087】
(表8)
a 新鮮な種子を20または24 DARで収穫した。25℃で湿らせたペーパータオル上で16日後も発芽しなかった種子は休眠していると見なされる。
b ガラクチノール標準物質に基づいて計算。
c ラフィノース標準物質に基づいて計算。不明物質は、フェニルα-D-グルコシドに対する保持時間の比が2.49および2.51であった。
【0088】
未成熟なソバ種子を10 DAPで収穫すると、乾燥耐性は、制御された相対湿度下(図7A)での緩速乾燥によって誘導された(図7A)。未成熟な種子の緩速乾燥時の乾燥耐性の獲得(早期成熟または誘導成熟)は、緩速乾燥の初日におけるファゴピリトールB1の蓄積に密接に関連した(図7C)。D-カイロ-イノシトールは、緩速乾燥の初日のあいだに急速に減少した(図7B)。ミオ-イノシトールは一定のままであったが(図7B)、ガラクチノールは2日目に増加した(図7C)。少量のファゴピリトールA2、ファゴピリトールB2、およびジガラクトシルミオ-イノシトールが5日目に蓄積した(図7D)。ショ糖は、初日のあいだに減少し、その後一定であった(図7E)。
【0089】
同様に、高い相対湿度で維持すると、未成熟な10 DAPの殻を除去したソバ種子は、ファゴピリトールB1(図8C)およびD-カイロ-イノシトール(図8B)の蓄積に関連して1日後に乾燥耐性となった(図8A)。ガラクチノールは、高い相対湿度では2日目に増加した(図8C)。少量のファゴピリトールA2およびファゴピリトールB2が2〜5日に蓄積し、ジガラクトシルミオ-イノシトールは2〜6日に蓄積した(図8D)。ファゴピリトールA3は、6日後に限って検出された。ショ糖は初日以降徐々に増加した(図8E)。このパターンは、大豆種子の軸とは対照的であり、大豆種子は、高い相対湿度でインキュベートしても乾燥耐性とならず、高い相対湿度でインキュベートしてもスタキオースを蓄積しない(ブラックマンI、参照として本明細書に組み入れられる)。ファゴピリトールB1は、非常にマイルドな乾燥条件で蓄積することができるがラフィノースシリーズのオリゴ糖の蓄積はより多くの乾燥を必要とするように思われる。
【0090】
実施例9-発芽の際の乾燥耐性の喪失に対するファゴピリトールB1の関係
乾燥耐性の喪失に対するファゴピリトールB1の喪失の関係(12%相対湿度および22℃での急速乾燥)は、殻を除去した成熟ソバ種子(「マンカン」)の発芽の際の軸および子葉組織において測定した。吸水開始後6時間以内にファゴピリトールB1は、軸組織から消失し始めた。12時間までに低いレベルのみが残り、発芽18時間までに、ファゴピリトールは全て微量の高次オリゴマーを除いて完全に消失した(図9E)。ファゴピリトールB1の消失は、再水和時の胚軸-幼根軸(一次根の生育)の「正常な」生育によって測定した急速な発芽の開始および乾燥耐性の喪失に密接に関連した(図9A)。ショ糖レベルは減少した後48時間後に増加して、胚軸から糖が動員されていることを反映した(図9C)。ショ糖:ファゴピリトールB1比は、乾燥耐性の喪失および発芽の開始に関連して12時間後非常に急速に増加した。ショ糖とは対照的に、D-カイロ-イノシトールは12時間後に増加して、ファゴピリトールの加水分解を反映する(図9C)。ガラクトースはファゴピリトールの動員の際に検出されなかったが、明らかに他の産物に急速に変換された。果糖およびグルコースは、軸組織が急速に生育しつつある(図9I)18時間後に軸組織に存在した(図9B)。果糖およびグルコースは軸組織に存在する唯一の還元糖であった(図9I)。
【0091】
発芽のあいだ、マルトース、マルトトリオース、およびマルトテトラオース(図9J)は、増加したショ糖レベルと共に胚軸組織に存在し(図9H)、デンプンの分解を反映した。D-カイロ-イノシトールは、発芽の6〜18時間のあいだに5倍増加し(図9B)、胚軸におけるファゴピリトールの分解を反映した(図9B)。胚軸におけるD-カイロ-イノシトールの総量(遊離および結合)は発芽の際に減少し、軸組織への輸送の発生または他の産物への代謝を示唆した。発芽の18〜48時間で胚軸におけるショ糖の量が多いことは、内乳におけるデンプンの動員および胚軸による加水分解産物の吸収を反映した(図9H〜J)。胚軸組織は内乳に埋もれ、発芽の際にデンプンの多い内乳に取り巻かれた。0時間では胚軸におけるファゴピリトールB1および全可溶性糖質(図9F)のレベルが低いことは、乾燥種子の外種皮(種皮)に接する外側の子葉の一部が、成熟乾燥種子からの胚組織の剥離の際に失われたことを示した。
【0092】
実施例10-画分実験の材料
殻を除去した種子100個の試料(約2.5 g)を粉砕して、エタノール:水(50:50)によって抽出した。抽出物を繰り返し遠心して、容積数mlに濃縮して、分子量10,000のカットオフフィルターを通過させた。エタノールを濃縮試料に加えて、冷凍庫に入れて、粗抽出物からファゴピリトールA3を濃縮することを目的としてファゴピリトールの沈殿を促進した。ほとんどの場合、調製物からの試料ではファゴピリトールA3が失われるか、または検出レベル以下であった。
【0093】
ショ糖の約97%がインベルターゼによって除去された。グルコース、果糖、およびガラクトースは増加して、ラフィノースおよびスタキオースは、おそらくインベルターゼ酵素調製物が十分に純粋でなかったためになおも存在した。グルコースおよび果糖を異なる方法によって破壊する試みは、部分的に成功したに過ぎなかった。
【0094】
実施例11-P2ゲルの画分
いくつかの組の抽出物をプールして濃縮し、遠心および限外濾過によって透明にした。種子1081個を合わせた抽出物を3mlに濃縮した。可溶性糖質の濃縮調製物をバイオゲルP2ゲルカラム(3m×40 cm)にローディングして、脱気した水によって65℃で溶出した。初期画分は茶色であり、その後の画分は黄色であった。糖およびファゴピリトールのほとんどが5ml画分3本に溶出され、なおも茶色の材料が混入していた。これらの画分をプールして濃縮し、第二のバイオゲルP2ゲルカラムに20℃でローディングした。ファゴピリトールA3に富む画分をプールした。ファゴピリトールA1およびB1に富む画分をプールした;そしてファゴピリトールA1、B1、A2、およびB2ならびにショ糖に富む画分をプールした。カラムの流速は、茶色の材料(おそらくポリフェノール材料)の量が増加するにつれて減少した。
【0095】
種子1300個からの抽出物(約35 g)を遠心によって透明にして、濃縮し、分子量10,000のカットオフフィルターを通過させた。1.5 mlに濃縮した後、試料を限外濾過して、濃縮した抽出物をP2カラム上でクロマトグラフィーを行った。それぞれの抽出物からのファゴピリトールA3に富む画分および他のファゴピリトールに富む画分を合わせた。遠心前に沸騰すると、デンプンの膨張が起こり、抽出可能な容積が減少して、抽出物の着色が増加する。いずれのカラムも異なるファゴピリトールを効率よく分離できなかった。
【0096】
実施例12-3-アミノプロピル官能性シリカゲルカラムでの画分
0.9 cm×30 cmの3-アミノプロピル結合(9%官能化)シリカゲル(シグマ-アルドリッチ社)をその後の実験におけるファゴピリトールの画分に用いた。P2ゲルカラムからの濃縮画分を最初の5個のアミノカラムにローディングした。
【0097】
カラム3に関して、P2ゲルカラム上で部分精製した後のファゴピリトールA3に富む試料を蒸発乾固させて、水2mlに溶解して、PVPカラムを通過させた。溶離剤を蒸発乾固させて、アセトニトリル:水(70:30)7mlに溶解して、0.9 cm×30 cmの3-アミノプロピル官能性シリカゲルカラムにローディングした。アセトニトリル:水(70:30)350 ml、アセトニトリル:水(60:40)150 ml、およびアセトニトリル:水(50:50)50 mlによって段階的な溶出を行った。カラムは、アセトニトリル:水(60:40)30 mlおよびアセトニトリル:水(75:25)30 mlによって再生した。画分(5ml)を回収して、回収した画分からの試料0.5 mlを蒸発乾固させて、TMSI:ピリジン(1:1)によって誘導体化した。TMS-糖質は高分解能のガスクロマトグラフィーによって分析した。ファゴピリトールA3は他のファゴピリトールから明らかに分離された(図10)。「純粋な」ファゴピリトールA3を含むプールした画分を蒸発乾固させて、微量の混入物を除去するために再度クロマトグラフィーを行うために保存した。「純粋な」ファゴピリトールA2または「純粋な」ファゴピリトールB2の画分をプールした。ファゴピリトールA2およびB2の混合物の画分も同様にリサイクルするためにプールした。
【0098】
カラム4に関して、P2ゲルクロマトグラフィー後のファゴピリトールの混合物を含む試料をアミノプロピル官能性シリカゲル上でさらに分離した(図11)。カラム4に類似の量のファゴピリトールA3をローディングしたが、他のファゴピリトールのためにカラムにローディングする全量が多くなった。ファゴピリトールA2およびB2は混合したままであったが、いくつかの「純粋な」ファゴピリトールA1、B1、およびA3画分は、さらに精製するためにプールした。
【0099】
実施例13-炭素セライトカラム上での画分
粗抽出物の混合物を、ソバ精製ふすまの25 g試料3個から調製したところ、可溶性糖質を全量で1.2 g含み、そのうち540 mgがファゴピリトールであった。精製ふすまを50%エタノールによって抽出し、エタノールをロータリーエバポレーションによって除去して、抽出物をポリビニルポリピロリドン(「PVPP」)およびアンバーライトIRN-150イオン交換樹脂を含むカラムの中に通過させた。次に、抽出物を酵母(2 mg酵母/精製ふすま1g)によって消化して、ショ糖の97%を除去した。この処置の副産物は二糖類であるトレハロース(O-α-D-グルコピラノシル-(1→1)-O-α-D-グルコシド)であった。酵母処置抽出物を分子量10,000のカットオフフィルター(アミコン社)によって濾過して、凍結乾燥した。次に、合わせた抽出物を蒸留水9mlに再度溶解して炭素セライトカラムの上部に適用した。カラムは炭素(「ダルコ-G60」、J.T.ベイカー、フィリップスバーグ、ニュージャージー州)とセライト545-AW(スペルコ、ベルフォンテ、ペンシルバニア州)とを重量で等しい比率で含む混合物を含んだ。炭素セライト混合物は、蒸留水において3.3 cmのEDカラム、高さ17 cmに充填したスラリーで、充填したカラムを蒸留水1Lによって洗浄した。カラムを4℃で蒸留水800 ml、5%エタノール1000 ml、10%エタノール1000 ml、20%エタノール250 ml、および50%エタノール200 mlによって段階的に溶出した。溶離剤をフラスコに集めて、これを1時間毎に手で交換した。平均すると、フラスコは容積37 mlを含んだ。画分に含まれるファゴピリトール濃度は、高分解能のガスクロマトグラフィーによって分析した。単糖類、グルコース、および果糖(酵母処理により大量に存在する)、ならびにミオ-イノシトールおよびD-カイロ-イノシトールは、水画分に溶出した。ファゴピリトールB1は5%エタノールによってカラムから溶出したがガラクチノールが混入していた。ファゴピリトールA1およびB2は10%エタノールによって溶出されたがそれぞれ、トレハロースとショ糖が混入した(酵母によって除去されなかった少量)。ファゴピリトールA2は20%エタノール溶離剤に含まれ、ファゴピリトールA3は50%エタノール溶出の段階までカラムに残っていた。着色混合物を除去する場合、炭素-セライトカラムは、3-アミノプロピル官能性シリカゲルカラムより有効であった。
【0100】
実施例14-粉の高温対低温抽出
全白ひき割り粉(バーケットミルズ社)からの粉(100目のふるいを通したもの、C&Nラボラトリーミル)を、粉に対する様々な溶媒比で抽出して、「高剪断」ポリトロンヘッドを備えたワーリングブレンダーを用いて高温抽出または低温抽出を行った。ブレンダーは10秒間のオンと10秒間のオフを1分または2分間行った。高温抽出の後、混合物は濃厚すぎて(デンプンの膨張による)濾過または遠心を効率よくできず、抽出されたファゴピリトールの容量は少なくなった。低温抽出(寒冷または室温)では、最高の収率が得られた。最適な抽出は、溶媒/粉の比率が20:1(例えば、粉25 gをエタノール:水(50:50)500 mlに)であった。ファゴピリトールの相対的な収率は、同じ25 g試料の第一、第二、および第三の抽出に関して約90%、9%、および1%であった。低温抽出物の遠心後の上清または濾過後の濾液を80℃に加熱して、粉に存在して抽出の際にファゴピリトールを加水分解する可能性があるαガラクトシダーゼおよび他の酵素を不活化した。これらの酵素は、エタノール:水(50:50)中では不活性であったが、エタノールが濃縮中に蒸発した後の水相では活性化されるようになる可能性がある。
【0101】
実施例15-脱脂粉を用いた効果
いくつかの試料は抽出前にヘキサンによって脱脂した。これらの実験において、PVPカラムおよびイオン交換カラム段階は限外濾過の後に行った。初期段階は容易に進行するように思われたが、限外濾過はなおも遅く、全技法を行うには非常に時間を浪費して面倒であった。技法を単純にして有効性を改善するために、非脱脂粉についてさらなる実験を行った。
【0102】
実施例16-水対エタノール:水(50:50)による抽出
ミンダック社製の粉(各200 mg)を、遠心管において蒸留水2mlまたはエタノール:水(50:50)によって抽出した。ファゴピリトールの収率は水またはエタノール:水(50:50)による抽出後同じであった(表9)。水抽出物は曇っているように思われ、より懸濁した材料を含んだ。この知見は、おそらくタンパク質およびデンプンの水に対する溶解度が高いことに関連した。エタノール抽出物はあまり懸濁していない材料を含み、抽出の際の微生物および内因性の酵素活性も最小限であった。エタノール:水(50:50)をその後の抽出において用いた。
【0103】
(表9)
【0104】
実施例17-バーケットミルズ社製とミンダック社製の粉の比較
ショ糖およびファゴピリトールB1は双方の原料において主要な成分であった(表10)。バーケットミルズ社製は、全ひき割り粉に由来し、ミンダック社製のより精製粉より、総可溶性糖質の収率が約2倍高かった。ショ糖は、双方の原料において総可溶性糖質の約60%であった。バーケットミルズ社の抽出物は、赤みがかった茶色であったのに対し、ミンダック社の抽出物は曇った白色であった。ファゴピリトールA2、B2、およびA3は、ミンダック社製の粉のこの試料において検出されなかった。
【0105】
(表10)
【0106】
実施例18-ショ糖およびファゴピリトールの酵母消化
ミンダック社製の粉の試料25 mgをエタノール:水(50:50)500 mlによって抽出して、遠心し、加熱してエタノールを蒸発させて濾過した。ケーキ酵母(8500 mg)を水抽出物に加えて、22℃で0〜48時間インキュベートした。ショ糖の約97%が最初の6時間以内に消化されたが、いくつかのファゴピリトールが失われ、D-カイロ-イノシトールは増加した(表11)。ミオ-イノシトールが大きく増加したことは意外であり、おそらく粉におけるフィチンの分解を反映した。酵母による消化前にイオン交換樹脂によってフィチンを除去すれば、より良好な結果が得られると思われる。ショ糖と比較すると、ファゴピリトールは酵母によってかなりゆっくり消化された。
【0107】
(表11)
【0108】
もう一つの酵母消化実験において、抽出物をイオン交換カラムであるPVPカラムに通過させて、アミコン限外濾過装置を通過させてから酵母を加えた。この実験は乾燥顆粒酵母(200 mg)を用いて行った。結果は、ショ糖が最初の6時間以内に消化され、そのあいだファゴピリトールが比較的安定であるように思われることを示した(表12)。当初不明であった材料は、最初の6時間に増加してその後ゆっくり分解した。この不明物質は、真正トレハロース(誘導体化後)との同時クロマトグラフィーおよびガスクロマトグラフィー/質量分析によってトレハロースであると同定された。DB1およびDB5カラム上での保持時間および不明物質の質量分析は、真正のトレハロースと同一であった。α,α'-トレハロースは、グルコース(O-α-D-グルコピラノシル-(1-1')-O-α-D-グルコシド)の二量体である。
【0109】
(表12)
【0110】
バーケットミルズ社製の粉を用いて第三の実験においてより少ない量の酵母を調べた。ショ糖約97%が最初の2時間以内に酵母50 mgによって消化され、グルコースの倍加を伴った(表13)。ファゴピリトールは酵母とのインキュベーションのあいだ比較的安定であるように思われた。産生されたトレハロースも同様に低いレベルであることからより低い量の酵母が好ましい。ファゴピリトールから高分子量混入物質より単糖類を除去するほうがおそらくより容易である。
【0111】
(表13)
【0112】
実施例19-精製段階
表14は、バーケットミルズ社製の全ひき割りソバ粉の2回抽出した25 g試料2個から、単離の際の異なる段階(抽出、加熱、および遠心、ならびに蒸発)でのファゴピリトール分析の結果を示す。連続的な精製段階のあいだ、高分子量ファゴピリトールは容易に分解するように思われた。これは、部分的に試料がますます純粋になったことと、部分的にファゴピリトールA3がより容易に分解されるより濃縮された試料になったことによる考えられる。結果は、抽出技法の短縮も望ましいかも知れないことを示した。低温抽出では、より少量を用いて、なおも良好な抽出物の収率を得ることができる。
【0113】
(表14)
#1および#2は、抽出直後に650 ml抽出物2個から得た試料0.5 mlによる。
#3および#4は、加熱および遠心後の350 ml抽出物2個から得た試料0.5 mlによる。
#5は、蒸発および遠心後の抽出物250 mlから得た試料0.5 mlによる。
【0114】
第一および第二の抽出によって抽出されたファゴピリトールの量を表15に示す。全ひき割りソバ粉(25 g、バーケットミルズ社)を、第一の抽出ではエタノール:水(50:50)320 mlによって、そして第二の抽出ではエタノール:水(50:50)200 mlによって抽出した。表15に示されるように、第一の抽出は、可溶性糖質の約90%を除去した。第二の抽出はより多くのファゴピリトールA1を除去したが、より大きなファゴピリトールは検出可能なレベルで除去されなかった。これらの結果は、抽出をより低い容量での一段階に短縮すれば、抽出時間が短縮されるが、ファゴピリトールA1を十分に回収しない可能性があることを示している。
【0115】
(表15)
【0116】
ファゴピリトール粗抽出物は、3-アミノプロピル官能性シリカゲルカラム上で二量体、三量体、および四量体に分離することができた(図12)。カラムは、ファゴピリトールA3を回収するためにオーバーローディングした。オーバーローディングにもかかわらず、かなりの(しかし全てではない)精製が得られた。ファゴピリトールAおよびファゴピリトールBは、カラムをオーバーローディングすると通常重なり合うことが認められた。しかし、注意して行えば、カラムをオーバーローディングした場合でも比較的純粋な画分を得ることができる(図13)。
【0117】
限外濾過装置の前にPVPカラムを置くと、アミコンの分子量10,000カットオフフィルターを通しての抽出物の通過時間が短縮された。図14および図15によって示されるように、これはまた、二量体、三量体、および四量体ファゴピリトールの分解を改善した。
【0118】
実施例20-ヒト便菌叢によるファゴピリトール濃縮ソバ抽出物のインビトロ発酵
発酵試験は、乾燥顆粒パン酵母によって予め処置したファゴピリトール濃縮バーケットミルズ社製ソバ抽出物によって行った。酵母細胞を遠心(ソルバル冷蔵遠心機を用いて19K rpmで10分間)および限外濾過(アミコン濾過装置、分子量10,000カットオフ)によって除去した。ファゴピリトール水抽出物は、滅菌して他の全てのガスを除去するために二酸化炭素下で沸騰させた。
【0119】
便接種物および全ての試料を調製するあいだ、二酸化炭素ガスの流れの下で作業することによって嫌気的条件を維持した。発酵ボトルはゴム製の栓と金属製のキャップによって密封した。
【0120】
ファゴピリトール基質を最初に調製した。次に、ブランクを除き、全てのボトルに1mlを入れた(1試料あたり2個にさらに対照1個とブランク1個とを含む4個を、0時間、6時間、12時間、および24時間に行った)。次に、栄養培地を調製した。対照ボトルには、接種物の代わりに培地1mlを入れた。残りの培地を用いて、新鮮な便を1:12(w/v)に希釈して、参照として本明細書に組み入れられる、バリー(Barry)ら、「ルミナールおよびヒト便菌叢による、中性単糖類のインビトロ発酵(In Vitro Fermentation of Neutral Monosaccharides by Ruminal and Human Fecal Microflora)」、Acta Vet Scand 86(補則):93〜95(1989)およびギロン(Guillon)ら、「ヒト便細菌によるエンドウおよびリンゴ繊維の発酵の、残留繊維の特徴付け(Characterization of Residual Fibers from Fermentation of Pea and Apple Fibers by Human Fecal Bacteria)」、J. Sci. Food Agric. 68:521〜529(1995)に記載のように調製した。全ての試料およびブランクに、ゴム製の栓を通してシリンジによって便接種物1mlを接種した。
【0121】
発酵のために40℃の水浴を用いて、明記した時間にボトルを開けて空気を入れることによって発酵を中止し、直ちに凍結した。
【0122】
試料は、融解後1回遠心した後(微量遠心機、8000 rpm)、分子量10,000カットオフの微量遠心管において再度遠心することによって調製した。栄養培地を含むそれらの試料に存在する塩は、それらがTMS誘導体化を妨害することから、各試料をアンバーライト混合床樹脂の中に通過させることによって除去した。当初の抽出物の対照試料2個も同様に、ファゴピリトールが処置によって除去されないことを保証するために樹脂の中を通過させた。
【0123】
各(750 μl)少量を、シリル化バイアルにおいて内部標準(フェニルα-D-グルコピラノシド)100 μgと共に窒素ガスの流れの下で乾燥させた;バイアルをP205上で一晩デシケータに入れた。
【0124】
トリメチルシリル誘導体は、1:1ピリジン:TMS溶液100 μlを加えて、70℃で30分加熱することによって調製した。
【0125】
最後に誘導体化試料をGC(HP 5890シリーズII:DB-1カラム;内径0.25 mm、長さ15 m、被膜の厚さ0.25 μm)において分析した。
【0126】
便の細菌は、試料から全てのファゴピリトールを除去した。イノシトール化合物およびトレハロースも同様に細菌によって代謝された。ピーク2個(保持時間約3.1および3.4)は、全ての試料および対照クロマトグラムについて繰り返し存在し、このことは便細菌によって代謝されない酵母処置の副産物がソバ抽出物に存在することを示している;これらの同じピークは当初の抽出物の分析においても示された。
【0127】
内部標準物質は全ての分析において示され、誘導体化が成功したことを示した。表16は、対照および試料におけるイノシトールおよびファゴピリトールの濃度(mg/750 μlで報告)を示す。
【0128】
(表16)
【0129】
実施例21-ファゴピリトールのパイロットプラント産生技法
粉砕
ソバふすま試料を、アルパイングラインダー(アウグスバーグ、ドイツ)を低速で速いフィードに設定して粉砕した。ふすま50ポンド(22.6 kg)の試料10個をすりつぶして、各試料は完了して粒子径の変化に至るまでに約1.5時間を要した。
【0130】
抽出
すりつぶしたふすま試料を、備え付けの風圧駆動攪拌機(リーメタルプロダクツ、フィリップスバーグ、ペンシルバニア州)を有する100ガロンの蒸気被覆バットに入れた。ふすま試料は、50%エタノール(95%エタノール35ガロン、および蒸留水31ガロン)66ガロンによって抽出して、室温で最高速度で1時間攪拌した。混合物を1時間沈降させ、沈降した後、上清をプラスチックホースを通してサイフォンによって30ガロンの車輪付きステンレススチールドラム缶に移した。
【0131】
操作1および2において、さらに50%エタノール15ガロン(95%エタノール8ガロンと蒸留水7ガロン)を加えて残っている固体をすすいだ。これを沈降させて、上清をサイフォンで除去した。操作3〜10では、最初の抽出によって上清を採取する技術が改善されたためにすすぎ段階を省略した。これは、透明な上清を珪藻土プレートフィルター(「DEPF」)を通してまず濾過する段階、そして次にバットの底の泥を、プレートフィルターが目詰まりするまで可能な限り多く濾過する段階を含む。
【0132】
蒸発
操作1で合わせた上清を、容量20ガロンのドラム型真空蒸気ケトル(ゴレン、シカゴ、イリノイ州)において60℃、20 psi真空で蒸発させた。しかし、抽出物の実質的な比率は、真空排管を通しての泡立ちのために失われた。
【0133】
操作2では、エタノールを100ガロンの蒸気被覆バットにおいて加熱することによって蒸発させた。この方法は時間がかかるものの(丸1日かかる)有効であり、エタノールヒュームがパイロットプラントに放出された。
【0134】
操作3からの抽出物7ガロンについて、蒸発させるために容量20ガロンのドラム型真空蒸気ケトルを用いて試行を行ったが、あらゆる抽出物の喪失を目に見えるようにして、したがって温度調節によって減少できるようにプレキシガラス片を備えた排管を用いた。この過程は遅く、試料の過度の泡立ちを抑えるために温度を50℃に低下させなければならず、抽出物は17%エタノールに蒸発されたに過ぎなかった。
【0135】
操作3の残り、ならびに操作4、5、および6はそれぞれ、ユニペクチン濃縮器(ユニペクチンAG、チューリッヒ、スイス)を用いて濃縮した。この真空補助過程によって温度は35〜40℃に維持され、これを一連の長くて細いパイプを通して流すことによって抽出物の表面積を増加することによって操作することが可能であった。ユニペクチンは、用いるために適度に速かったものの、低温を維持したために、泥状の茶色の物質の形成を減少させず、各抽出物を濃縮器に4回通過させても、エタノール濃度は5.5%に減少したに過ぎなかった。抽出物のエタノール含有量は、蒸気被覆25ガロンバットを沸騰させることによって1〜2%に減少した(リーメタルプロダクツ、フィリップスバーグ、ペンシルバニア州)。
【0136】
操作7〜10の蒸発に関して、操作2に記載したように100ガロンの蒸気被覆バットを用いて、エタノール蒸気をパイロットプラントに放出した。エタノール濃度は、モデル360沸点測定装置(ジュジャージン-サラーソン、パリ、フランス)を用いて測定した。
【0137】
抽出物の保存
ファゴピリトールをエタノール中に含む抽出物を+2℃で保存して、水性抽出物(蒸発段階後)を-20℃で保存した。抽出物はプラスチック4または蓋付の5ガロン容器に保存した。
【0138】
珪藻土プレートフィルター
珪藻土プレートフィルター(「DEPF」)(T.シュライバー社、ハリソン、ニュージャージー州)は、プラスチックホルダーと濾紙片(フィルトレーションアンリミテッド、アクロン、ニューヨーク州)をいずれかの側に備えた金属プレート6枚(7インチ×7インチ)からなり、全て、きつく互いに圧縮されている。DEPFは使用するために、水と混合したプレコート120 g(プレートあたり20 g)の中に送ることによって調製した。DEPFを通過した水が透明であれば、プレコートをその場に置いた。ボディフィードをファゴピリトール抽出物の中に攪拌した。次に、試料をボディフィードと共に、ボディフィードが懸濁液を維持するように絶えず攪拌しながらDEPFの中を通過させた。プレコートまたはボディフィードとして用いられる珪藻土粉末は、セライト503、セライト535、セライト577、およびハイフロースーパーセル(「HFSC」)(セライトコーポレーション、ロンポック、カリフォルニア州)であった。特に明記していなければ、用いた組み合わせは、プレコート(プレートあたり20 g)としてセライト577、そしてボディフィードとしてHFSC(抽出物の重量に対して乾燥粉末の0.25%重量)であった。
【0139】
PVPP処置
パイロットプラント抽出において用いたポリビニルポリピロリドン(PVPP)は、ポリクラー-VT(ISPテクノロジーズ社、ウェイン、ニュージャージー州)であった。抽出物を蒸発によって濃縮した後にPVPP処置を行う場合、PVPP 0.75 kg(特に明記していない限り)を25ガロンの蒸気被覆バットにおいて抽出物に加えて、風圧駆動攪拌器によって30分間攪拌した(ガスト社、ベントンハーバー、ミシガン州)。これによっていくつかの着色が除去されたが、PVPPをカラムにローディングして抽出物をその中に通過させた実験室の規模の場合ほど有効ではなかった。ワイン産業において用いられるように処置時間を3時間(操作2)に増加しても、得られた抽出物の色は認識できるほど改善しなかった。
【0140】
操作7〜10に関して、PVPP 1.5 kgを、蒸発前に100ガロンバットにおいてファゴピリトールのエタノール抽出物に加えた。混合物を30分攪拌して、さらに30分間沈降させて、上清を除去した。蒸発後、抽出物は泥状の茶色(酸化および縮合フェノール類)というよりむしろ濃い黄色(可溶性プロアントシアニジン)となった。上記のように0.75 kg PVPP処置はまた、抽出物をさらにきれいにするために蒸発後に用いた。操作7〜10は、その結果着色混入物質に関して他の抽出物よりかなりきれいであった。
【0141】
最初にPVPPを沈降させて、DEPFのために上清を除去し、PVPPおよび底にある関連する液体を捨てた。次に、PVPPを含む全抽出物をDEPFに送ると、PVPPは、目詰まりを起こすことなくDEPFによって保持された。
【0142】
ベントナイト処置
ベントナイトはワイン産業において、ワインにおいてプロアントシアニジンに反応する不安定なタンパク質によって引き起こされるくもりを透明にするために用いられる。ベントナイト(プレスク島ワインセラーズ、ノースイースト、ペンシルバニア州)は、少量の蒸留水と共に混合する(特に明記していない限り試料1ガロンあたり3g)ことによって濃いスラリーを形成することによって調製した。ブレンダーを用いて塊を破壊した。2〜3時間後、少量の抽出物をベントナイトスラリーと共に混合した後、これを抽出物の残りに加えて、数分攪拌した。処置は約24時間低温で行って、フロキュレーションを起こさせて、タンパク質を沈殿させた。上清を注意深く注いだ。PVPPとは対照的に、ベントナイトを除去するためにDEPFを用いることは、DEPFが容易に目詰まりするために適切ではなかった。
【0143】
イオン交換処置
サイバーン樹脂(アフテック社、ロチェスター、ニューヨーク州)を加えて(1.8 kg)30分間攪拌した。100目のふるいを通過させることによって試料から樹脂を除去した。
【0144】
酵母処理
ショ糖を除去するために酵母による抽出物の処置を操作2で行った。抽出物を32℃に加温して、乾燥顆粒パン酵母40 gを加えて、時折攪拌しながら1時間発酵させた。77℃で15分加熱して酵母を殺した。この方法は、酵母はショ糖の除去には有効であったが、これはトレハロースを産生し、トレハロースはファゴピリトールのその後の精製時にファゴピリトールA1から分離することが難しいため中断した。
【0145】
限外濾過
分子量10,000のカットオフ中空線維フィルターカートリッジ2個を限外濾過ユニット(「UF」)(ロミコン社、ウィルミントン、マサチューセッツ州)に適合させた。
【0146】
操作1は抽出過程において初期にUFを通過させ、大量のエタノール抽出物を速やかに濾過した。約5ガロンのボイド容量が存在するため、UFについて大量の液体を用いなければならないことはこの操作から明白であった。同様に、UFはほとんどのエタノールが抽出物から除去された後に用いなければならず、濾紙は50%エタノールの存在下でわずかにカールするが、なおも再利用可能である。濾過と使用後の濾紙の洗浄はいずれも時間と労力がかかり、そのため労力を省くために、それぞれを個々に行うより他の抽出物を合わせて最終的なUF処置を行った。
【0147】
したがって、操作2〜10からの抽出物を合わせて限外濾過を行った。温度は、濾紙が総抽出物65ガロンを濾過するために必要な1.5時間のあいだ27℃を維持した。10,000ダルトンより小さい粒子は濾紙を通過するが、より大きな粒子は、カートリッジの中をまっすぐ通過して、再循環した。大量の微粒子を含む老廃物約5ガロンが蓄積した。100ガロンのバットに集められた濾過された抽出物は、透明で薄い黄色であった。色はおそらく、特に操作2〜6で限外濾過によって濾過されずに逃れてPVPP、ベントナイト、またはイオン交換樹脂に吸収された可溶性の低分子量プロアントシアニジンの存在を反映する。
【0148】
実施例22-ファゴピリトールA2、ファゴピリトールA3、およびファゴピリトールB2の精製とNMRによる分子構造
ソバからのファゴピリトールの抽出
ソバ普通種(ファゴピルム・エスクレンツムモエンク栽培品種「マノー」(Fagopyrum esculentum Moench cv. 'Manor')種子はミンダックグロワーズ社(グランドフォークス、ノースダコタ州)から提供された。ソバふすまは、EtOH:H2O 1:1(v/v)によってパイロットプラントの規模で抽出し(実施例21を参照)、抽出物は珪藻土フィルターを通して濾過して、蒸発させて容量を減少させ、EtOHを除去して、ポリビニルポリピロリドン(PVPP)、ベントナイト、およびイオン交換樹脂によって処置して、非糖質成分を除去した。可溶性の糖質抽出物は、最終的に分子量10,000のカットオフ中空線維フィルターカートリッジを通して濾過して、蒸発させて容積を減少させた。
【0149】
ファゴピリトールの精製
精製は、炭素(ダルコG60;J.T.ベイカー社、フィリップスバーグ、ニュージャージー州)とセライト545-AW(スペルコ、ベルフォンテ、ペンシルバニア州)の1:1(w/w)静止相での調整的クロマトグラフィーによって実施した(ウィスラー(Whisler)ら、J. Amer. Chem. Soc. 72:677〜679(1950)、参照として本明細書に組み入れられる)。カラムに新しくスラリーを水と共に充填して、凍結乾燥した試料を最小量の水に溶解してローディングし、可溶性糖質をEtOH:H2O(v/v)を段階的に増加させて4℃で溶出し、20 ml画分に回収した。カラム溶離剤試料を定期的に採取して、フェニルαD-グルコシドを内部標準として加えて、乾燥試料をN-トリメチルシリルイミダゾール:ピリジンの1:1(v/v)混合物によって誘導体化して、水素炎イオン化検出器、スプリットモードインジェクタ(1:50)、およびHP-1MS毛細管カラム(15 m×0.25 mm内径、被膜の厚み0.25 μm)を備えたヒューレットパッカード6890 GCによって分析した(ホーボビッツ(Horbowicz)、参照として本明細書に組み入れられる)。GCはプログラムされた初回温度150℃で操作して、3℃/分で200℃に、7℃/分で325℃になるように調節し、325℃で20分間維持した。注入口は335℃で操作して、検出器は350℃で操作した。担体ガスは2.5 ml/分の窒素であった。
【0150】
ファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3の精製に関しては、ソバふすま抽出物を吸着床25×900 mmのクロマトグラフィーを行った。水2.2 L、4%EtOH 2.5 L、5%EtOH 3 Lによって連続的に溶出した後、ファゴピリトールB2は7%EtOH 2Lによって、ファゴピリトールA2は12%EtOH 1Lによって、そしてファゴピリトールA3は20%EtOH 1Lによって溶出された。ファゴピリトールB2またはファゴピリトールA2、またはファゴピリトールA3を含む画分を凍結乾燥して再度クロマトグラフィーを行った。
【0151】
ファゴピリトールB2の画分を、カンジダ・ユチリス(Candida utilis)由来のインベルターゼ(EC 3.2.1.26)によって22℃で1時間処置してショ糖を加水分解した後、吸着床15×450 mmのクロマトグラフィーを行った。5%EtOH1Lによって混入物質を溶出した後、ファゴピリトールB2が6%EtOHによって溶出された。
【0152】
ファゴピリトールA2は、吸着床15×500 mm上でクロマトグラフィーを行った。少量の混入物質がH2O 0.5 L、6%EtOH 1L、および8%EtOH 0.5 Lによって連続的に溶出され、ファゴピリトールA2は9%EtOHによって溶出された。
【0153】
ソバふすま抽出物の異なる4つのカラムからファゴピリトールA3を含む画分を合わせて、再度クロマトグラフィーを行った(19×350 mm)。ファゴピリトールA3は14%EtOHと共に溶出して、その後12%EtOHによる溶出によって混入物を除去した。
【0154】
酸および酵素的加水分解
ファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3の試料を、1M CF3CO2Hによって80℃で16時間加水分解して蒸発乾固させた。D-ガラクトースとD-カイロ-イノシトールとのモル比は、トリメチルシリル産物のGC分析後に計算した。ファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3の試料(100 μg)を、1.25単位の脱塩緑色コーヒー豆α-D-ガラクトシダーゼ(EC 3.2.1.22)の水溶液200 μlと共に22℃でインキュベートするか、または0.5単位ウシ肝β-D-ガラクトシダーゼ(EC 3.2.1.23)の水溶液200 μlと共に37℃で24時間インキュベートした。酵素タンパク質を濾過(分子量10,000のカットオフフィルター)によって除去し、試料を乾燥させ、産物をトリメチルシリル誘導体のGC分析によってアッセイした。ラフィノースおよび乳糖(100 μg)はそれぞれ、αまたはβ-D-ガラクトシダーゼのみによって加水分解して、双方の酵素が活性であることを確認した。
【0155】
カイロ-イノシトールのカイラリティの分析
真正D-カイロ-イノシトールおよびL-カイロ-イノシトール標準物質(アルドリッチ社、ミルウォーキー、ウィスコンシン州)およびファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3の酸加水分解産物(2M CF3CO2H、70℃で3時間)を、無水ペンタフルオロプロピオン酸:アセトニトリル(1:1、v/v)によって誘導体化した(オストルンド(Ostlund)、Proc. Natl. Acad. Sci. USA 90:9988〜9992(1993))。誘導体化産物は、ヒューレットパッカード5890シリーズ11 GCによって、100℃で等温的にキラシル-バルカイラル毛細管カラム(25×0.25 mm内径、被膜の厚さ0.16 μm)を用いるGCによって、誘導体化標準物質と共に同時クロマトグラフされた(リービット(Leavitt)ら、Carbohydr. Res. 103:203〜212(1982))。注入口の温度は130℃であり、FID温度は130℃であった。担体ガスはヘリウム3ml/分、スプリット1:50であった。
【0156】
NMR分析
精製試料(ファゴピリトールB2 41 mg、ファゴピリトールA2 13.5 mg、ファゴピリトールA3 34.5 mg)を、分析するために99.96%D2O(ケンブリッジアイソトープ研究所、アンドバー、マサチューセッツ州)700 μlに溶解した。バリアン(パロアルト、カリフォルニア州)VXRS-400を、1D 13C NMRおよびDEPT(分極転移のひずみのない増強)に関して、100.5 MHzで、25℃で外部炭素標準物質として1,4-ジオキサン(δc 67.4)を用いて行った。他の全てのNMR実験は、残留HOD(δH 4.63)に対して25℃で標準化した、バリアンユニティ500上で499.9 MHzで行った。これらは1D 1H NMRであり、二倍量フィルター相関分光法(double-quantum filtered correlated spectroscopy、DQCOSY)、勾配増強異核種多量相関(gradient-enhanced heteronuclear multiple quantum correlation、GHMQC)、同核種2D J-分光法(homonuclear 2D J-spectroscopy、HOM2DJ)、核オーバーハウザー交換分光法(Nuclear Overhauser and Exchange Spectroscopy、NOESY)、および勾配増強異核種多重結合相関(gradient-enhanced heteronuclear multiple bond correlation、GHMBC)であった。データを分析してバリアンVNMRソフトウェア、バージョン6.1Aを用いてシミュレーションを行った。
【0157】
結果
ソバふすまを50%EtOHによって抽出し、ファゴピリトールを炭素-セライトの1:1(w/w)混合物によってカラムクロマトグラフィーを用いて精製した。ファゴピリトールB2は7%EtOHによって溶出された画分から精製され、ファゴピリトールA2は、9%EtOHによって溶出され、ファゴピリトールA3は、14%EtOHによって溶出された。ファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3の酸加水分解からのカイロ-イノシトールのペル(ペンタフルオロプロピオネート)は、カイラルカラムにおいてD-カイロ-イノシトールと共に同時クロマトグラフされたが、L-カイロ-イノシトールとは同時クロマトグラフされなかった。ファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3の酸加水分解によって、D-ガラクトースとD-カイロ-イノシトールを、ファゴピリトールB2に関して2.02:1.00の比で、ファゴピリトールA2に関して2.03:1.00の比で、そしてファゴピリトールA3に関して2.98:1.00の比で生じた。ファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3の部分的加水分解産物のGC分析によって、ファゴピリトールB2からファゴピリトールB1、ファゴピリトールA2からファゴピリトールA1、そしてファゴピリトールA3からファゴピリトールA1、D-カイロ-イノシトール、ならびにファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3からガラクトースが同定された。ファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3は、α-D-ガラクトシダーゼによって加水分解されたが、β-D-ガラクトシダーゼによっては加水分解されず、全てのD-ガラクトピラノシル残基に関してαアノマー結合であることを証明している。
【0158】
ファゴピリトールB2の13C NMRスペクトルによって、炭素共鳴18個が同定され、1H NMRスペクトルによって水素共鳴20個が同定された。炭素18個の化学シフトはそれぞれ、GHMQCを用いて水素20個の化学シフトと結合していた。炭素の化学シフト4個を除く全てが69 ppm〜76 ppmのあいだであった。炭素共鳴2個は、99.02および95.74 ppmで低磁場への化学シフトを示し、これは置換アノマー炭素の特徴であった。炭素2個は62.03および67.72 ppmで高磁場に存在して、これらはDEPTによってCH2炭素であると同定された。水素原子3個を除く全てが3.5〜3.9 ppmでの化学シフトを有した。最も低磁場の水素2個(δ4.81およびδ4.99)はアノマー水素に特徴的な化学シフトを有し、GHMQCによってアノマー炭素(それぞれ、δ99.02およびδ95.74)に結合していることが示された。これらは、DQCOSYによるガラクトーススピン系内での連結度を追跡するための開始点として作用した。高磁場のCH2炭素に水素共鳴を割り当てると、二つのガラクトース残基の相違が明らかになった;CH2炭素の一つ(δ62.03)は偶発的に等価の共鳴(δ3.59)を有する水素2個を有し、他のCH2炭素(δ67.72)は等価でない共鳴(δ3.72およびδ3.54)を有した。CH2水素のあいだに等価でない共鳴が存在することは、この炭素にさらに置換が存在することを示唆しており;このように、等価でない水素を含むスピン系(δ3.54、H-6'a;δ3.72、H-6'b)を、カイロ-イノシトールに近位の(最も近い)ガラクトースに割り当てて、等価のプロトン(δ3.59、H-6"a、H-6"b)を含むスピン系を遠位のガラクトース残基に割り当てた(表17)。
【0159】
(表17)ファゴピリトールB2(1)とファゴピリトールA2(2)の、1Hおよび13C NMR(25℃、D2O)化学シフトおよびプロトン-プロトンカップリング定数
a、b、d、e 割り当ては逆転させてもよい
c 予測値;実験データのシミュレーションによって確認
【0160】
この割り当てのさらなる証拠はC-6'(δ67.72)に由来し、これはC-6"(δ62.03)と比較すると5.5 ppmによってデシールドされており、隣接するC-5'(δ69.98)は、C-5"(δ71.71)と比較して少量が高磁場側にシフトしていることから、C-6'での置換を示している。1D 1Hスペクトル(表17)から計算したカップリング定数(J1',2'、3.9 Hz;J1",2"、3.9 Hz)が小さいことから、双方のガラクトシル環に関してαアノマー結合が確認された。
【0161】
ファゴピリトールB2に関して、全てが3.47〜4.08 ppmのあいだの残っている6個の1H共鳴を、Dカイロ-イノシトールに割り当てて、連結度をDQCOSY実験から表した。それぞれの水素が結合している炭素をGHMQCから推定した。シクリトール環におけるこれらの炭素(δ75.58)の最も低磁場は、ガラクトースとの結合点であった。置換炭素(δ75.58)に結合した水素(δ3.73、H-2)について認められたカップリング定数(J1,2 3.0 Hz;J2,3 10.0 Hz)、ならびにH-1(J1,2 3.0 Hz;J1,6 4.1 Hz)およびH-3(J2,3 10.0 Hz;J3,4 9,6 Hz)と割り当てられた隣接する水素について認められた定数から、隣接する軸結合および赤道結合を有する軸水素(すなわち、H-2またはH-5)に3.73 ppmが割り当てられることを確認した。カイロ-イノシトール環の対称性により、C-2またはC-5での置換の結果、等価な構造が得られ、より小さい数が割り当てられる。したがって、ファゴピリトールB2は、O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→2)-D-カイロ-イノシトールである。
【0162】
ファゴピリトールA2に関する13C NMRスペクトルから、炭素共鳴18個が同定され、1H NMRスペクトルによって水素共鳴20個が同定された。ファゴピリトールA2の二つのガラクトシル残基は、ファゴピリトールB2(上記)に関する記述と同じように解明し、化学シフトとカップリング定数とはファゴピリトールB2と同等であった(表17)。ファゴピリトールA2に関して、D-カイロ-イノシトール環における置換炭素(δ81.78)上での水素(δ3.54)は、1H NMRおよび同核種2D J分光法(「HOM2DJ」)実験の双方から、唯一の大きなカップリング定数(J2,3 9.8 Hz;J3,4 9.8 Hz)を有することが認められ、トランス-アキシャル近接水素(すなわち、H-3またはH-4)であることを示している。カイロ-イノシトール環が対称であるために、C-3またはC-4での置換は同等の構造を生じ、より小さい数を割り当てる。D-カイロ-イノシトール残基の結合点は、ファゴピリトールA2においてファゴピリトールA1と同様にC-3であった。C-1'(δ100.33)に関する唯一の見かけのGHMBC相互作用はH-3(δ3.54)であり、D-カイロ-イノシトール環のH-2およびH-4に軸結合水素が存在することを確認した。同様に、C-3とH-1'との相互作用は(1'→3)結合を確認したが、H-2'は環に対して軸結合であるためC-3とH-2'では確認されなかった。ファゴピリトールA2は、O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→3)-D-カイロ-イノシトールである。
【0163】
ファゴピリトールAシリーズとファゴピリトールBシリーズとのあいだには、置換炭素の化学シフトに明確な差を認めることができ、これは非アノマー炭素の最も低磁場に存在する。Bシリーズでは、これは76 ppmであるのに対し、Aシリーズでは置換炭素は6ppmさらに低磁場で82 ppmであった。ファゴピリトールA3が82.04 ppm(C-3)で炭素共鳴を含むことから、このパターンによって、トリガラクトシルD-カイロ-イノシトールであるファゴピリトールA3がファゴピリトールAシリーズのメンバーであることが定義された(表18)。
【0164】
(表18)ファゴピリトールA3の、1Hおよび13C NMR(25℃、D2O)化学シフトならびに1Hカップリング定数
a、b、d、e 割り当ては逆転させてもよい
c 予測値;実験データのシミュレーションによって確認
【0165】
さらに、置換炭素(δ82.04)に結合した水素は、1H NMRおよびHOM2DJから計算すると、大きなカップリング定数(J2,3 10.1 Hz、J3,4 10.1 Hz)を有する化学シフト3.59 ppm(H-3)を有することが認められた。この化合物は、これまでファゴピリトールBシリーズのトリガラクトシルD-カイロ-イノシトールであると予想されていた(ホーボビッツおよびホーボビッツ(HorbowiczおよびHorbowicz)ら、Planta、205:1〜11(1998)、参照として本明細書に組み入れられる);その理由はソバ種子は、ファゴピリトールA1より多くのファゴピリトールB1、そしてファゴピリトールA2より多くのファゴピリトールB2を有するためであり、同じパターンがトリガラクトシルD-カイロ-イノシトールについても予測された。しかし、このトリガラクトシルD-カイロ-イノシトールはO-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→3)-D-カイロ-イノシトールであると決定され、慣用名ファゴピリトールA3を割付された。
【0166】
四つの環を重ね合わせると、ファゴピリトールA3のスペクトルは複雑であった。水素27個のほとんどが1D 1HおよびHOM2DJスペクトルから同定されたが、DQCOSYを用いて、水素がガラクトースまたはD-カイロ-イノシトールに属すると割り当てた。三つのガラクトース環の2位、3位、および4位の水素は、環のあいだの共鳴が非常に類似であるか、または同一であったため、水素を個々の環に直ちに割り当てることができなかった。水素の三つの対(δ3.50およびδ3.78;δ3.57およびδ3.72;δ3.60)は、DEPTによってCH2であると同定された炭素(δ67.61;δ67.26;δ61.95)とのGHMQC実験におけるその相互作用によって、ガラクトース残基の6位であると同定された。一対の水素は偶発的に等価であり(δ3.60)、最も高磁場の炭素(δ61.95)に結合し、H-6'"aおよびH-6'"bと割り当てられた。これらの水素(δ3.60)は、DQCOSYによって3.87 ppmで(H-5'")水素と相互作用した。H-5'"との相互作用を通して、98.85 ppmでのアノマー炭素をC-1'"と割り当てた。この炭素(δ98.85)も同様に、CH2水素と相互作用して化学シフトが3.72および3.57 ppmであり、H-6'"aおよびH-6'"bと割り当てられた。同様に、99.05 ppm(C-1")での炭素は4.02 ppm(H-5")での水素、ならびに3.78 ppmおよび3.50 ppm(H-6'aおよびH-6'b)での水素と相互作用した。C-1'(δ100.36)とD-カイロ-イノシトールのH-3(δ3.59)のみとのGHMBC相互作用によって、H-3が置換点(C-3)で軸結合のH-2およびH-4と共に炭素に結合していることを確認した。これらの割り当てを行った後、残りの割り当ては四つのスピン系のそれぞれについて行うことができる。最後に、GHMQCを用いて炭素を割り当てた。69.95 ppm〜70.23 ppmのあいだに起こる炭素共鳴24個中8個では、絶対的な割り当ては不可能であり、おそらく変更可能な多くの割り当てを与えた(表18)。
【0167】
ファゴピリトールBシリーズのD-カイロ-イノシトール環における水素間のカップリング定数をその1D 1H NMRスペクトルから計算することは可能であった。しかし、ファゴピリトールAシリーズにおいて、H-4とH-5の化学シフトは一致し、H-4とH-5との強いカップリングによって複雑な多重線を生じ、この中から6.5 Hz以上のカップリング定数を発見することは不可能である。ファゴピリトールBシリーズに関して測定した定数に基づいてD-カイロ-イノシトールに関して予想されるカップリング定数によって、ファゴピリトールA2またはファゴピリトールA3におけるD-カイロ-イノシトールについて認められる1H化学シフトのシミュレーションを行うと、ファゴピリトールAシリーズにおいて認められたものと一致するH-4およびH-5の多重線が得られた。これによって、ファゴピリトールA2に関してシミュレーションされたカップリング定数(J4,5 9.8 Hz)(表17)およびファゴピリトールA3に関してシミュレーションされた定数(J4,5 10.1 Hz)(表18)が確認された。
【0168】
ファゴピリトールBシリーズのメンバーは、これまで他の種において同定されている。ソバ(ホーボビッツ(Horbowicz)ら、Planta, 205:1〜11(1998);ズクゼシンスキ(Szczecinski)ら、Bull. Pol. Acad. Sci. Chem. 46:9〜13(1998)、参照として本明細書に組み入れられる)の他に、ファゴピリトールB1は大豆の種子(シュバイツァー(Schweizer)ら、Carbohydr. Res. 95:61〜71(1981)、参照として本明細書に組み入れられる)、ルピナス、レンズマメ、およびヒヨコマメ(ケメナー(Quemener)ら、Phytochem. 22:1745〜1751(1983)、参照として本明細書に組み入れられる)、およびホホバマメ(オガワ(Ogawa)ら、Carbohydr. Res. 302:219〜221(1997)、参照として本明細書に組み入れられる)に存在する。ファゴピリトールB1の化学合成が報告されている(コルニエンコ(Kornienko)ら、Carbohydr. res. 310:141〜144(1998)、参照として本明細書に組み入れられる)。ファゴピリトールB1は、テンサイの球状種子において化学的方法によって同定された(シオミ(Shiomi)ら、Agric. Biol. Chem. 52:1587〜1588(1988)、参照として本明細書に組み入れられる)が、その絶対的な構造は本発明において初めて示されている。ファゴピリトールA2およびファゴピリトールA3は、ファゴピリトールA1と共に、ソバの種子におけるガラクトシルシクリトールの新規シリーズのメンバーである。
【0169】
実施例23-ファゴピリトールA1およびファゴピリトールB1の精製とNMRによる分子構造
ソバからのファゴピリトールの抽出。ソバ普通種(ファゴピルム・エスクレンツムモエンク栽培品種「マノー」(Fagopyrum esculentum Moench cv. 'Manor')種子のふすま粉砕画分は、ミンダックグロワーズ社(グランドフォークス、ノースダコタ州)から提供された。ソバふすまは、EtOH:H2O 1:1(v/v)によってパイロットプラントの規模で抽出し(実施例21を参照)、抽出物を珪藻土フィルターに通して濾過して、蒸発させて容量を減少させ、EtOHを除去して、ポリビニルポリピロリドン(PVPP)、ベントナイト、およびイオン交換樹脂(強陰イオン樹脂と陽イオン樹脂とを混合)によって処置して、非糖質成分を除去した。可溶性の糖質抽出物は、最終的に分子量10,000のカットオフ中空線維フィルターカートリッジを通して濾過して蒸発させて容積を減少させた。
【0170】
ファゴピリトールの精製
精製は、炭素(ダルコG60;J.T.ベイカー社、フィリップスバーグ、ニュージャージー州)とセライト545-AW(スペルコ、ベルフォンテ、ペンシルバニア州)との1:1(w/w)静止相での調整的クロマトグラフィーによって実施した(ウィスラー(Whisler)ら、J. Amer. Chem. Soc. 72:677〜679(1950)、参照として本明細書に組み入れられる)。カラムに新しくスラリーを水と共に充填して、凍結乾燥した試料を最小量の水に溶解してローディングし、可溶性糖質をEtOH:H2O(v/v)を段階的に増加させて4℃で溶出し、20 ml画分に回収した。カラム溶離剤試料を定期的に採取して、フェニルαD-グルコシドを内部標準として加えて、乾燥試料をN-トリメチルシリルイミダゾール:ピリジンの1:1(v/v)混合物によって誘導体化して、水素炎イオン化検出器、スプリットモードインジェクタ(1:50)、およびHP-1MS毛細管カラム(15 m×0.25 mm内径、被膜の厚み0.25 μm)を備えたヒューレットパッカード6890 GCによって分析した。GCはプログラムされた初回温度150℃で操作して、3℃/分で200℃に、7℃/分で325℃になるように調節し、325℃で20分間維持した。注入口は335℃で操作して、検出器は350℃で操作した。担体ガスは2.5 ml/分の窒素であった。
【0171】
ファゴピリトールA1を精製するため、可溶性糖質(5%ファゴピリトールA1)約1.8 gを含むソバふすま抽出物を、炭素-セライトの吸着床25×900 mmでクロマトグラフィーを行った。水2.2 Lによって溶出させて単糖類を除去して、4%EtOH 2.5 Lによって溶出させてガラクチノールとファゴピリトールB1とを除去した後、ファゴピリトールA1は5%EtOHによって溶出され、これを凍結乾燥して、再度クロマトグラフィー(吸着床25×550 mm)を行った。最初にファゴピリトールB1を4%EtOHによって除去した後にファゴピリトールA1を5%EtOHによって溶出することによってファゴピリトールA1(95%純粋)を得た。ファゴピリトールA1を含む画分に、再クロマトグラフィーを3回行い(吸着床10×370 mm;水によって溶出した後5%EtOHによって溶出)、NMR分析のために純粋なファゴピリトールA1を得た。
【0172】
ファゴピリトールB1を精製するために、ソバふすま抽出物をパン酵母と共にインキュベートしてショ糖を除去した。可溶性糖質(25%ファゴピリトールB1)3.15 gを含む試料を吸着床43×360 mmの炭素-セライト上でクロマトグラフィーを行った。水1.5 Lによって溶出して単糖類を除去した後、ファゴピリトールB1およびガラクチノールが4%EtOHによって溶出された。ファゴピリトールB1を含む画分を凍結乾燥して25×900 mmのカラム上で再度クロマトグラフィーを行った。ガラクチノールはファゴピリトールB1の初回大量と共に溶出して、ファゴピリトールB1溶離剤の最後部からの画分を合わせると、NMR分析にとって純粋なファゴピリトールB1が得られた。
【0173】
D-カイロ-イノシトールは、部分的に精製したファゴピリトールB1(ファゴピリトールB1 106 mg;ファゴピリトールA1 14 mg;ガラクチノール2.5 mg)を4M HCl中で95℃で1時間加水分解して、NaOHによって中和し、得られた混合物を炭素-セライト(25×900 mm)上でクロマトグラフィーを行い、水によって溶出することによって得られた。
【0174】
酸および酵素的加水分解
ファゴピリトールA1およびファゴピリトールB1の試料を1M CF3CO2Hによって80℃で16時間加水分解して蒸発乾固した。D-ガラクトースとD-カイロ-イノシトールのモル比は、トリメチルシリル産物のGC分析後に計算した。ファゴピリトールA1とファゴピリトールB1の試料(200 μg)を1.25単位の脱塩した緑色コーヒー豆α-D-ガラクトシダーゼ(EC 3.2.1.22)の水溶液200 μlと共に22℃で、または0.5単位ウシ肝β-D-ガラクトシダーゼ(EC 3.2.1.23)の水溶液200 μlと共に37℃で24時間インキュベートした。酵素タンパク質は、濾過(分子量10,000のカットオフフィルター)によって除去し、試料を乾燥させて、産物をトリメチルシリル誘導体のGC分析によってアッセイした。ラフィノースおよび乳糖(100 μg)をそれぞれ、αまたはβ-D-ガラクトシダーゼによって加水分解して、双方の酵素が活性であることを確認した。
【0175】
カイラルカラムGCによるD-カイロ-イノシトールとL-カイロ-イノシトールの分離
真正のD-カイロ-イノシトールおよびL-カイロ-イノシトール標準物質(ウィーン大学のリヒター(A. Richter)氏からの寄贈)および純粋なファゴピリトールA1または純粋なファゴピリトールB1の酸加水分解産物(2M CF3CO2H、70℃で3時間)を無水ペンタフルオロプロピオン酸:アセトニトリル(1:1、v/v)によって誘導体化した(オストルンド(Ostlund)、Proc. Natl. Acad. Sci. USA 90:9988〜9992(1993))。誘導体化産物は、ヒューレットパッカード5890シリーズ11 GCによって100℃で等温的にキラシル-バルカイラル毛細管カラム(25×0.25 mm内径、被膜の厚さ0.16 μm)を用いるGCによって、誘導体化標準物質と共に同時クロマトグラフされた(リービット(Leavitt)ら、Carbohydr. Res. 103:203〜212(1982))。注入口の温度は130℃であり、FID温度は130℃であった。担体ガスはヘリウム3ml/分、スプリット1:50であった。
【0176】
NMR分析
精製試料(ファゴピリトールA1 14.4 mg、ファゴピリトールB1 21 mg;D-カイロ-イノシトール9 mg)を、分析するために99.96%D2O 700 μlに溶解した。バリアンVXRS-400を、1D 13C NMRに関して100.5 MHzで、25℃で外部炭素標準物質として1,4-ジオキサン(δC 67.4)を用いて行った。他の全てのNMR実験は、残留HOD(δH 4.63)に対して25℃で標準化したバリアンユニティ500(パロアルト、カリフォルニア州)において499.9 MHzで行った。これらは1D 1H NMR、二倍量フィルターCOSY、勾配増強HMQC、同核種2D J-分光法(HOM2DJ)、NOESY、および勾配増強HMBCであった。バリアンVNMRソフトウェア、バージョン6.1Aを用いてデータを分析してシミュレーションを行った。
【0177】
GC-MS分析
ファゴピリトールA1およびファゴピリトールB1を含むソバ種子からのトリメチルシリル誘導体化糖質を、HP 5970質量分析器にカップリングさせたヒューレットパッカード5890ガスクロマトグラフにおいてDB-5毛細管カラム(30 m×0.25 mm内径、被膜の厚さ0.25 μm)上でクロマトグラフィーを行った。温度プログラムは140℃で4分であり、270℃まで3℃/分で増加させ、270℃で20分間維持した。担体ガスはヘリウム1ml/分であった。誘導体化ファゴピリトールA1およびファゴピリトールB1に対応するGCピークの質量スペクトルを、真正D-カイロ-イノシトールおよびD-ガラクトースのトリメチルシリル誘導体の質量スペクトルと比較した。
【0178】
結果
ソバふすまを50%EtOHによって抽出して、得られた抽出物を炭素-セライト1:1(w/w)上でクロマトグラフィーを行い、ファゴピリトールA1を精製した。比較のために、ファゴピリトールB1も同様に同じ試料から精製した。ファゴピリトールA1およびファゴピリトールB1の酸加水分解からのカイロ-イノシトールのペル(ペンタフルオロプロピオネート)は、真正のD-カイロ-イノシトールと共にカイラルカラム上で同時クロマトグラフされたが、真正のL-カイロ-イノシトールとは同時クロマトグラフされなかった。ファゴピリトールA1およびファゴピリトールB1の加水分解は、D-ガラクトースとD-カイロ-イノシトールとを、ファゴピリトールA1に関してモル比1.01:1.00で、そしてファゴピリトールB1に関して1.03:1.00で生じた。遊離のD-カイロ-イノシトールは、ファゴピリトールA1とファゴピリトールB1の混合物の酸加水分解の後に炭素-セライトカラム上で精製することによって得た。ファゴピリトールA1およびファゴピリトールB1は、α-D-ガラクトシダーゼによって加水分解され、β-D-ガラクトシダーゼによって加水分解されず、α-D-ガラクトシル結合が存在することを証明している。
【0179】
ファゴピリトールA1の13C NMRスペクトルから、炭素共鳴12個が同定され、1H NMRスペクトルから1H共鳴13個が同定された。ファゴピリトールA1における炭素12個の化学シフトは、GHMQCを用いると水素13個の化学シフトに関連した。D-ガラクトピラノシル残基の特徴的な低磁場のアノマー炭素(δ100.26)およびアノマー1H(δ5.16)を、スピン系における連結度に関するDQCOSYの分析の開始点として利用した。ガラクトシル残基上の二つの一致するH-6'水素も同様に有用なマーカーであり、1Hスペクトルにおいてδ3.55で二重強度を有する二重線によって明らかに同定可能であり、13Cスペクトルにおいて最も高磁場の炭素(δ61.80)に結合した。ガラクトシル環の残りの共鳴の割り当ては簡単であった。ガラクトース残基の1Hシグナルを割り当てれば(表19)、残りの6個の共鳴はD-カイロ-イノシトール環に割り当てることができると考えられる。
【0180】
(表19)D2O中で25℃でのファゴピリトールA1、ファゴピリトールB1、および遊離のD-カイロ-イノシトールの、1Hおよび13C NMR化学シフト
a 割り当ては逆転させてもよい。
【0181】
3.54 ppmでの水素は、遊離のD-カイロ-イノシトールに関する共鳴と比較して8〜10 ppm低磁場にシフトした共鳴を有する低磁場の炭素(δ81.77)に結合し(表19)、それによってD-カイロ-イノシトール環上でのガラクトース置換点が同定された。さらに、GHMBAおよびNOESY実験から、3.54での水素と、ガラクトースのアノマー水素(δ5.16;H-1')およびアノマー炭素(δ100.26:C-1')との明らかな相互作用が示され、結合点を確認した。
【0182】
隣接する水素間のカップリング定数は1D 1Hシグナルから計算した。HOM2DJは、重なり合うシグナルから形成された領域3.5〜3.9 ppmにおける多重線を分離するために役立った。得られたカップリング定数(表20)から、ファゴピリトールA1のガラクトース環に関して行った割り当てを確認した。
【0183】
(表20)ファゴピリトールA1、ファゴピリトールB1、および遊離のD-カイロ-イノシトールの、1H-NMRスペクトルデータに由来する推定カップリング定数
a 予測値;実験データのシミュレーションによって確認した。
【0184】
特に、小さいカップリング定数J1',2' 3.9 Hzは、α-アノマー結合を確認した。D-カイロ-イノシトール環の結合点でのδ3.54での水素はカップリング定数9.7 Hzを示し、双方の隣接する水素とトランスジアキシャル関係にあることを示した。したがって、ファゴピリトールA1に構造O-α-D-ガラクトピラノシル-(1→3)-D-カイロ-イノシトールを割り当てた。ファゴピリトールA1は、推定のインスリンメディエータ(ラーナー(Larner)ら、Biochem. Biophys. Res. Comm. 151:1416〜1426(1988);ベルリン(Berlin)ら、Tetrahedron Lett. 31:1109〜1112(1990)、参照として本明細書に組み入れられる)に関連する、O-(2-アミノ-2-デオキシ)-α-D-ガラクトピラノシル-(1→3)-D-カイロ-イノシトール(ベルリン(Berlin)ら、Tetrahedron Lett. 31:1109〜1112(1990)、参照として本明細書に組み入れられる)と等配電子である。
【0185】
D-カイロ-イノシトール環の隣接する水素間のカップリング定数は環の番号をつける際に重要であった。しかし、ファゴピリトールA1のH-4およびH-5のカップリング定数は容易に区別できなかった。これらの同一の化学シフト(δ3.61;H-4およびH-5)を有する二つの強くカップリングした水素の強い相互作用は、見かけの最大スプリットが6.8 Hzである有する複雑な多重線を生じた。D-カイロ-イノシトールにおける近接するトランスジアキシャル水素の3つの対(H-2、H-3、H-4、H-5)は、O-α-D-ガラクトピラノシル-(1→2)-D-カイロ-イノシトール(ファゴピリトールB1、表20)におけるD-カイロ-イノシトールについて認められたように、9〜10 Hzのカップリング定数を示すはずである(ズクゼシンスキ(Szczecinski)ら、Bull. Pol. Acad. Sci. Chem. 46:9〜13(1998)、参照として本明細書に組み入れられる)。この矛盾は、化学シフト(表19)および実験スペクトルラインデータから計算したカップリング定数(表20)を用いて、しかしJ4,5に関しては予測値9.7 Hzを用いて、C-置換D-カイロ-イノシトールに関する1Hスペクトルのシミュレーションによって解決した。シミュレーションしたスペクトルと実験1Hスペクトルとは同一であり、H-4およびH-5のあいだの強い相互作用の影響によって、見かけの最大スプリット6.8 Hzの多重線が3.61 ppmで得られた。このように、J4,5は、シミュレーションに基づくと9.7 Hz(表20)として与えられる。D-カイロ-イノシトール環の水素の割り当ては、ファゴピリトールA1のH-3でのガラクトシル置換が、ファゴピリトールA1の隣接するH-4(δ3.61)を、それがH-5(δ3.61)と一致するようにファゴピリトールB1(δ3.44)およびD-カイロ-イノシトール(δ3.41)より低磁場方向にシフトさせることから、ファゴピリトールB1に関してはファゴピリトールA1より容易であった。ファゴピリトールB1では、H-4(δ3.44)は、H-5(δ3.60)の高磁場であり、これによってトランスジアキシャル相互作用の大きなカップリング定数(J4,5 9.7 Hz)をファゴピリトールB1に関して認めることができる(表20)。シミュレーションはまた、遊離のD-カイロ-イノシトールにおける水素間のカップリングの解明においても有用であった。D-カイロ-イノシトールの1H NMRスペクトルは、それぞれの多重線が同一の水素2個を表す三つの多重線パターンを示し、その2組は、対内部でカップリングを有した。計算した化学シフトおよびカップリング定数のシミュレーションによって、D-カイロ-イノシトールに関して認められたスペクトルと一致したスペクトルが得られ、表20に示すカップリング定数の計算値を確認した。
【0186】
ファゴピリトールA1およびファゴピリトールB1は、2つの異なるシリーズのファゴピリトールオリゴマーを表す位置異性体である。ファゴピリトールB1であるO-α-D-ガラクトピラノシル-(1→2)-D-カイロ-イノシトールの13Cおよび/または1H共鳴は、ソバ(ズクゼシンスキ(Szczecinski)ら、Bull. Pol. Acad. Sci. Chem. 46:9〜13(1998)、参照として本明細書に組み入れられる)、大豆(シュバイツァー(Schweizer)ら、Carbohydr. Res. 95:61〜71(1981)、参照として本明細書に組み入れられる)、およびホホバマメ(オガワ(Ogawa)ら、Carbohydr. Res. 302:219〜221(1997))におけるこの構造に関して報告された値と厳密に一致した。ガラクトピラノシル環の共鳴も同様に、ガラクトシルオノニトール(リヒター(Richter)ら、J. Nat. Prod. 60:749〜751(1997)、参照として本明細書に組み入れられる)およびアノマー炭素でα置換されたガラクトースを含む他のオリゴ糖(ボック(Bock)ら、Adv. Carbohydr. Chem. Biochem. 42:193〜225(1984)、参照として本明細書に組み入れられる)の共鳴と厳密に一致した。
【0187】
ファゴピリトールA1とファゴピリトールB1のトリメチルシリル産物の質量スペクトルはm/z 73、103、129、147、191、204、217、305、318、319、および361でイオン断片ピークと類似であった。m/z 305/318の断片の量の比は、ファゴピリトールA1に関して0.9、およびファゴピリトールB1に関して2.6であったが、m/z 318/319の断片の量の比は、ファゴピリトールA1に関して2.2、およびファゴピリトールB1に関して0.9であった。これらの量の比は、GC-MSによるファゴピリトールA1とファゴピリトールB1のトリメチルシリル誘導体を区別する可能性がある。
【0188】
実施例24-ファゴピリトールB3の単離と特徴付け
ソバからのファゴピリトールの抽出
ソバ普通種(ファゴピルム・エスクレンツムモエンク栽培品種「マノー」(Fagopyrum esculentum Moench cv. 'Manor')種子のふすま粉砕画分は、ミンダックグロワーズ社(グランドフォークス、ノースダコタ州)から提供された。ソバのふすまを、EtOH:H2O1:1(v/v)によってパイロットプラントの規模で抽出し(実施例21を参照)、抽出物を珪藻土フィルターに通して濾過して、蒸発させて容量を減少させ、EtOHを除去して、ポリビニルポリピロリドン(PVPP)、ベントナイト、およびイオン交換樹脂によって処置して、非糖質成分を除去した。可溶性の糖質抽出物は、最後に分子量10,000のカットオフ中空線維フィルターカートリッジを通して濾過して蒸発させて容積を減少させた。
【0189】
ファゴピリトールの精製
精製は、炭素(ダルコG60;J.T.ベイカー社、フィリップスバーグ、ニュージャージー州)とセライト545-AW(スペルコ、ベルフォンテ、ペンシルバニア州)との1:1(w/w)静止相での調整的クロマトグラフィーによって実施した(ウィスラーおよびデュルソ(WhislerおよびDurso)、1950)。炭素320 gとセライト320 gとの混合物を水と共に混合して、薄いペースト状にして100 mm×300 mmカラムに吸着床の厚さが200 mm(吸着床の容積1570 ml)となるように入れた。カラムには糖全体の約20 gを含むパイロットプラント抽出物220をローディングした。可溶性糖質は、ガラス蒸留水4L、2%エタノール4L、4%エタノール4L、5%エタノール4L、8%エタノール4L、10%エタノール4L、14%エタノール4L、20%エタノール2L、および50%エタノール2Lによって4℃で段階的に溶出して、500 mL画分に手動で回収した。流速は水での500 ml/時間から50%エタノールでの約175 ml/時間であった。容量(速度)は画分の収集容器にとっては多すぎるために、画分を手動で収集する必要があった。各画分の試料(200 μl)を採取してフェニル-α-D-グルコシド(25 μg)を内部標準物質として加え、乾燥した試料をN-トリメチルシリルイミダゾール:ピリジン、1:1(v/v)によって誘導体化して、水素炎イオン化検出器、スプリットモードインジェクタ(1:50)、およびHP-1MS毛細管カラム(15 m×0.25 mm内径、被膜の厚み0.25 μm)を備えたヒューレットパッカード6890 GCによって分析した(ホーボビッツ(Horbowicz)、参照として本明細書に組み入れられる)。GCはプログラムされた初回温度150℃で操作して、3℃/分で200℃に、7℃/分で325℃になるように調節し、325℃で20分間維持した。注入口は335℃で操作して、検出器は350℃で操作した。担体ガスは2.5 ml/分の窒素であった。
【0190】
結果
カラム2つを流した(cc37およびcc38(上記のように精製溶出プロトコールによって))。カラムcc38に関する溶出配列プロフィールを図16に示す。D-カイロ-イノシトール(カイロ)およびミオ-イノシトール(ミオ)は水と共に溶出され、果糖(Fru)およびグルコース(Glc)は2%エタノールと共に溶出され、ファゴピリトールB1は4%エタノール、ファゴピリトールA1は5%エタノール、過剰量のショ糖は8%エタノール、ファゴピリトールB2は10%エタノールによって溶出されてその後にDGMIが溶出され、ファゴピリトールA2は14%エタノール、ファゴピリトールB3およびTGMIは20%エタノールおよび50%エタノールによって溶出されてその後にファゴピリトールA3が溶出された。50%エタノール溶液は、ファゴピリトールA3画分を濃縮するために役立った。ファゴピリトールB3として暫定的に同定されたピークは、ファゴピリトールA3の前に溶出した(図16)。
【0191】
ファゴピリトールB3は、予想通りファゴピリトールA3の前に溶出した。暫定的にトリガラクトシルミオ-イノシトールであると同定されたもう一つの化合物(TGMI:図17において「T」と記す)は、ファゴピリトールB3の尾部で溶出し、同様に、ジガラクトシル-ミオ-イノシトール(DGMI)はファゴピリトールB2の尾部に溶出した。
【0192】
ファゴピリトールを含む画分(図16から)を凍結乾燥した。類似の混入物質を含む画分をプールして、さらなる炭素-セライトカラムクロマトグラフィーによってさらなる精製のために濃縮した。カラムcc37(示していない)から、ファゴピリトールB3およびTGMIと同定されたピークを含む2つの画分(60および61)を凍結乾燥して、プールし、濃縮して、高分解能ガスクロマトグラフィーによって分析した。この濃縮画分(60+61)のガスクロマトグラムからのデータを、関係するピークを強調するために拡大尺度で図18において再度プロットした。ファゴピリトールA3は保持時間38.27分(A3)、ファゴピリトールB3は保持時間39.01分(B3)、およびトリガラクトシルミオ-イノシトールは保持時間39.46分(TGMI)であった(図18;表21)。
【0193】
(表21)典型的なGCクロマトグラム*上でのTMSI-誘導体化糖の保持時間
*乾燥試料をN-トリメチルシリルイミダゾール:ピリジン1:1(v/v)によって誘導体化して、3℃/分、7℃/分で325℃まで調節し、325℃で20分維持したHe上で分析した。注入口は335℃で操作して、検出器は350℃で操作した。担体ガスは2.5 ml/分の窒素であった。
【0194】
このクロマトグラムはGCの「後ろの」カラムで実施した。「前の」カラムで実施すると、保持時間は約0.3分長かった。
【0195】
ファゴピリトールの混合物でのファゴピリトールB3は、α-ガラクトシダーゼによって完全に加水分解された。ファゴピリトールB3に富むが、ファゴピリトールA3およびトリガラクトシルミオ-イノシトール(TGMI)も含む画分は、トリフルオロ酢酸によって完全に加水分解されてガラクトースとシクリトールとなった。ガラクトース対シクリトールのモル比は3:1であった。部分的加水分解後の産物の分析は、ファゴピリトールB2(O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→2)-D-カイロ-イノシトール)およびファゴピリトールB1(O-α-D-ガラクトピラノシル-(1→2)-D-カイロ-イノシトール)の存在を同定し(ファゴピリトールB2およびファゴピリトールB1は、非加水分解試料には存在しなかった)、それによって、ファゴピリトールB3がO-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→2)-D-カイロ-イノシトールと同一であることを確認した。
【0196】
実施例25-ファゴピリトールAおよびBシリーズの単離と特徴付け
ソバのファゴピリトールの二つの完全なシリーズは、同定して特徴を調べられている。ファゴピリトールAシリーズはファゴピリトールA1、ファゴピリトールA2、およびファゴピリトールA3からなる。ファゴピリトールBシリーズはファゴピリトールB1、ファゴピリトールB2、およびファゴピリトールB3からなる。全てはαガラクトシダーゼによって加水分解されるが、βガラクトシダーゼによっては加水分解されず、全てのファゴピリトールがα-ガラクトシドであることを確認する(表22)。
【0197】
(表22)ファゴピリトールAシリーズおよびファゴピリトールBシリーズオリゴガラクトシドの加水分解産物
【0198】
全てはトリフルオロ酢酸によってガラクトースとD-カイロ-イノシトールに完全に加水分解される。高分解能ガスクロマトグラフィーによるカイラル毛細管カラム上の酸加水分解産物の分析から、ファゴピリトールにD-カイロ-イノシトールが存在することが確認された(表22)。ガラクトース対D-カイロ-イノシトールのモル比は、ファゴピリトールA1およびファゴピリトールB1に関して1:1、ファゴピリトールA2およびファゴピリトールB2に関して2:1、ならびにファゴピリトールA3およびファゴピリトールB3に関して3:1であった(表22)。ファゴピリトールA1、ファゴピリトールB1、およびD-カイロ-イノシトールの結合および絶対配置はNMRによって決定し、ファゴピリトールB2、ファゴピリトールA2、およびファゴピリトールA3も同様に決定した。ファゴピリトールA1は推定のインスリンメディエータと等配電子である。実施例24はファゴピリトールB3の単離および同定を図示する。ファゴピリトールB3は、ソバ種子における少量成分である。ファゴピリトールA3:ファゴピリトールB3の定量的な比は約10:1である。
【0199】
実施例26-オリゴガラクトシドのガラクチノールシリーズにおけるDGMIおよびTGMIの同定に関する証拠
ガラクチノールは、ラフィノースシリーズのオリゴ糖(ラフィノース、スタキオース、バーバスコース)、ガラクトピニトール、およびおそらくファゴピリトールの生合成に対する重要なガラクトシルドナーであるO-α-D-ガラクトピラノシル(1→1)-L-ミオ-イノシトールである。ガラクチノールシリーズのメンバーは、ガラクチノール、ジガラクトシルミオ-イノシトール、トリガラクトシルミオ-イノシトール、およびテトラガラクトシルミオ-イノシトール(ホーボビッツ&ペテクI、参照として本明細書に組み入れられる)であると提唱されている。ガラクチノールの絶対配置は、NMRによって決定されている。DGMIは、NMRによって分析されていない。ガラクチノールシリーズのトリおよびテトラガラクトシルメンバーが提唱されているが、これまで単離または特徴付けがなされていなかった。
【0200】
ソバ種子は、ガラクチノール、ジガラクトシルミオ-イノシトール(DGMI)、およびトリガラクトシルミオ-イノシトール(TGMI)を含む。DGMIは、炭素-セライトクロマトグラフィーの際に、ファゴピリトールB2とファゴピリトールA2のあいだに10%エタノールによって溶出される(実施例24を参照のこと)。HP-1 MS毛細管カラム上でのガスクロマトグラフィーでは、DGMIのピークは、ファゴピリトールB2に関するピークより約0.5 分長い保持時間を有した(表21を参照のこと)。DGMIは、α-ガラクトシダーゼによって完全に加水分解されるが、β-ガラクトシダーゼでは加水分解されず、DGMIがαガラクトシドであることを確認する(表23)。
【0201】
(表23)ガラクチノールシリーズのオリゴガラクトシドの加水分解産物
【0202】
DGMIは完全に加水分解されてガラクトースとミオ-イノシトールをモル比で2.08:1.00を生じた(表23)。部分的加水分解の後、同定された産物は、ガラクチノール、ガラクトビオース(gal-gal)、D-ガラクトース、およびミオ-イノシトール出合った。これらの結果から、DGMIの構造はO-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→1)-L-ミオ-イノシトール(O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→3)-D-ミオ-イノシトールとしても知られる)であることが確認された。
【0203】
トリガラクトシルミオ-イノシトール(TGMI)は、炭素-セライトクロマトグラフィーの際にファゴピリトールB3とファゴピリトールA3のあいだに20%エタノールによって溶出される(実施例24を参照のこと)。HP-1MS毛細管カラム上でのガスクロマトグラフィーでは、TGMIのピークはファゴピリトールB3のピークより約0.45分長い保持時間を有する(表21;図19)。TGMIはα-ガラクトシダーゼおよびトリフルオロ酢酸によって完全に加水分解された。ガラクトース対シクリトールのモル比は1:3であった。TGMIに富み、同様にファゴピリトールB3およびファゴピリトールA3(しかし、ガラクチノール、DGMI、またはミオ-イノシトールではない)を含む画分の部分的加水分解後の産物の分析を行うと、ガラクチノールとミオ-イノシトールが共に検出された(表23)。これらの結果は、トリガラクトシルミオ-イノシトールがO-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→1)-L-ミオイノシトール(同様に、O-α-D-ガラクトピラノシル-(1→6)O-α-D-ガラクトピラノシル-(1→6)-O-α-D-ガラクトピラノシル-(1→3)-D-ミオ-イノシトールとしても知られる)であると同定する。
【0204】
本発明は、説明する目的で詳細に説明してきたが、そのような詳細はその目的のために提供するのであって、当業者は変更を行うことができ、それらも以下の請求の範囲によって定義される本発明の趣旨および範囲に含まれると理解すべきである。
【特許請求の範囲】
【請求項1】
ファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3からなる群より選択される、単離されたファゴピリトール。
【請求項2】
以下の式を有するファゴピリトールA1である、請求項1記載の単離されたファゴピリトール:
【化1】
【請求項3】
以下の式を有するファゴピリトールA3である、請求項1記載の単離されたファゴピリトール:
【化2】
【請求項4】
以下の式を有するファゴピリトールB3である、請求項1記載の単離されたファゴピリトール:
【化3】
【請求項5】
一つまたは複数のガラクチノール、ミオ-イノシトール、ジガラクトシルミオ-イノシトール、フィチン、芳香性物質、細胞壁粒子、タンパク質、有機酸、アミノ酸、核酸、およびその塩を実質的に含まない、請求項1記載の単離されたファゴピリトール。
【請求項6】
ファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3からなる群より選択される、実質的に純粋なファゴピリトール。
【請求項7】
約95%から約99%純粋である、請求項6記載の実質的に純粋なファゴピリトール。
【請求項8】
ファゴピリトールA1である、請求項6記載の実質的に純粋なファゴピリトール。
【請求項9】
ファゴピリトールA3である、請求項6記載の実質的に純粋なファゴピリトール。
【請求項10】
ファゴピリトールB3である、請求項6記載の実質的に純粋なファゴピリトール。
【請求項11】
二つまたはそれ以上のファゴピリトールA1、ファゴピリトールA2、ファゴピリトールA3、ファゴピリトールB1、ファゴピリトールB2、ファゴピリトールB3、およびD-カイロ(chiro)-イノシトールを含み、請求項1記載の単離されたファゴピリトールの少なくとも一つを含む組成物。
【請求項12】
一つまたは複数のガラクチノール、ミオ-イノシトール、ジガラクトシルミオ-イノシトール、フィチン、芳香性物質、細胞壁粒子、タンパク質、ならびに有機酸、アミノ酸、核酸、およびその塩を実質的に含まない、請求項11記載の組成物。
【請求項13】
以下を含む、II型糖尿病の治療に用いられる薬学的組成物:
薬学的担体、および
請求項1記載の一つまたは複数の単離されたファゴピリトール。
【請求項14】
ファゴピリトールA2、ファゴピリトールB1、ファゴピリトールB2、およびその組み合わせからなる群より選択される、ファゴピリトールをさらに含む、請求項13記載の薬学的組成物。
【請求項15】
本質的には薬学的担体および請求項1記載の一つまたは複数の単離されたファゴピリトールからなる、請求項13記載の薬学的組成物。
【請求項16】
以下を含む、患者におけるII型糖尿病を治療する方法:
請求項1記載の一つまたは複数の単離されたファゴピリトールの有効量を、患者に投与する段階。
【請求項17】
以下を含む、患者におけるII型糖尿病を治療する方法:
請求項11記載の組成物の有効量を、患者に投与する段階。
【請求項18】
以下を含む、患者におけるII型糖尿病を治療する方法:
請求項6記載の実質的に純粋なファゴピリトールの有効量を、患者に投与する段階。
【請求項19】
以下を含む、患者におけるII型糖尿病を治療する方法:
請求項13記載の薬学的組成物の有効量を、患者に投与する段階。
【請求項20】
以下の段階を含む、ファゴピリトールA1、ファゴピリトールA3、ファゴピリトールB3、またはその混合物を調製する方法:
非ファゴピリトール材料、ならびにファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3からなる群より選択される、一つまたは複数のファゴピリトールを含む粗抽出物を産生するために有効な条件下で、ソバを溶媒に接触させる段階;ならびに
非ファゴピリトール材料を一つまたは複数のファゴピリトールから分離する段階。
【請求項21】
ソバがソバ種子胚を含む、請求項20記載の方法。
【請求項22】
ソバがソバ粉である、請求項20記載の方法。
【請求項23】
非ファゴピリトール材料が、一つまたは複数の細胞壁粒子、大きなタンパク質、荷電材料、および芳香性物質を含む、請求項20記載の方法。
【請求項24】
分離する段階が以下の段階を含む、請求項20記載の方法:
粗抽出物を濃縮するために有効な条件下で、溶媒を蒸発させる段階。
【請求項25】
分離する段階が以下の段階を含む、請求項20記載の方法:
細胞壁粒子および大きなタンパク質を粗抽出物から除去するために有効な条件下で、粗抽出物を分子量カットオフフィルターに通過させる段階。
【請求項26】
分離する段階が以下の段階を含む、請求項20記載の方法:
荷電材料を粗抽出物から除去するために有効な条件下で、粗抽出物をイオン交換材料に通過させる段階。
【請求項27】
分離する段階が以下の段階を含む、請求項20記載の方法:
芳香性物質を粗抽出物から除去するために有効な条件下で、粗抽出物をポリビニルピロリドンカラムまたは活性炭カラムに通過させる段階。
【請求項28】
一つまたは複数のファゴピリトールが少なくとも二つのファゴピリトールを含み、該方法がさらに以下の段階を含む、請求項20記載の方法:
少なくとも二つのファゴピリトールからファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3の一つを分離する段階。
【請求項29】
少なくとも二つのファゴピリトールからファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3の一つを分離する段階が、クロマトグラフィーによって行われる、請求項28記載の方法。
【請求項30】
少なくとも二つのファゴピリトールからファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3の一つを分離する段階が、アミン官能性(amine-functionalized)シリカゲル上で行われる、請求項29記載の方法。
【請求項31】
少なくとも二つのファゴピリトールからファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3の一つを分離する段階が、アセトニトリル:水の溶離剤を用いて行われる、請求項29記載の方法。
【請求項1】
ファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3からなる群より選択される、単離されたファゴピリトール。
【請求項2】
以下の式を有するファゴピリトールA1である、請求項1記載の単離されたファゴピリトール:
【化1】
【請求項3】
以下の式を有するファゴピリトールA3である、請求項1記載の単離されたファゴピリトール:
【化2】
【請求項4】
以下の式を有するファゴピリトールB3である、請求項1記載の単離されたファゴピリトール:
【化3】
【請求項5】
一つまたは複数のガラクチノール、ミオ-イノシトール、ジガラクトシルミオ-イノシトール、フィチン、芳香性物質、細胞壁粒子、タンパク質、有機酸、アミノ酸、核酸、およびその塩を実質的に含まない、請求項1記載の単離されたファゴピリトール。
【請求項6】
ファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3からなる群より選択される、実質的に純粋なファゴピリトール。
【請求項7】
約95%から約99%純粋である、請求項6記載の実質的に純粋なファゴピリトール。
【請求項8】
ファゴピリトールA1である、請求項6記載の実質的に純粋なファゴピリトール。
【請求項9】
ファゴピリトールA3である、請求項6記載の実質的に純粋なファゴピリトール。
【請求項10】
ファゴピリトールB3である、請求項6記載の実質的に純粋なファゴピリトール。
【請求項11】
二つまたはそれ以上のファゴピリトールA1、ファゴピリトールA2、ファゴピリトールA3、ファゴピリトールB1、ファゴピリトールB2、ファゴピリトールB3、およびD-カイロ(chiro)-イノシトールを含み、請求項1記載の単離されたファゴピリトールの少なくとも一つを含む組成物。
【請求項12】
一つまたは複数のガラクチノール、ミオ-イノシトール、ジガラクトシルミオ-イノシトール、フィチン、芳香性物質、細胞壁粒子、タンパク質、ならびに有機酸、アミノ酸、核酸、およびその塩を実質的に含まない、請求項11記載の組成物。
【請求項13】
以下を含む、II型糖尿病の治療に用いられる薬学的組成物:
薬学的担体、および
請求項1記載の一つまたは複数の単離されたファゴピリトール。
【請求項14】
ファゴピリトールA2、ファゴピリトールB1、ファゴピリトールB2、およびその組み合わせからなる群より選択される、ファゴピリトールをさらに含む、請求項13記載の薬学的組成物。
【請求項15】
本質的には薬学的担体および請求項1記載の一つまたは複数の単離されたファゴピリトールからなる、請求項13記載の薬学的組成物。
【請求項16】
以下を含む、患者におけるII型糖尿病を治療する方法:
請求項1記載の一つまたは複数の単離されたファゴピリトールの有効量を、患者に投与する段階。
【請求項17】
以下を含む、患者におけるII型糖尿病を治療する方法:
請求項11記載の組成物の有効量を、患者に投与する段階。
【請求項18】
以下を含む、患者におけるII型糖尿病を治療する方法:
請求項6記載の実質的に純粋なファゴピリトールの有効量を、患者に投与する段階。
【請求項19】
以下を含む、患者におけるII型糖尿病を治療する方法:
請求項13記載の薬学的組成物の有効量を、患者に投与する段階。
【請求項20】
以下の段階を含む、ファゴピリトールA1、ファゴピリトールA3、ファゴピリトールB3、またはその混合物を調製する方法:
非ファゴピリトール材料、ならびにファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3からなる群より選択される、一つまたは複数のファゴピリトールを含む粗抽出物を産生するために有効な条件下で、ソバを溶媒に接触させる段階;ならびに
非ファゴピリトール材料を一つまたは複数のファゴピリトールから分離する段階。
【請求項21】
ソバがソバ種子胚を含む、請求項20記載の方法。
【請求項22】
ソバがソバ粉である、請求項20記載の方法。
【請求項23】
非ファゴピリトール材料が、一つまたは複数の細胞壁粒子、大きなタンパク質、荷電材料、および芳香性物質を含む、請求項20記載の方法。
【請求項24】
分離する段階が以下の段階を含む、請求項20記載の方法:
粗抽出物を濃縮するために有効な条件下で、溶媒を蒸発させる段階。
【請求項25】
分離する段階が以下の段階を含む、請求項20記載の方法:
細胞壁粒子および大きなタンパク質を粗抽出物から除去するために有効な条件下で、粗抽出物を分子量カットオフフィルターに通過させる段階。
【請求項26】
分離する段階が以下の段階を含む、請求項20記載の方法:
荷電材料を粗抽出物から除去するために有効な条件下で、粗抽出物をイオン交換材料に通過させる段階。
【請求項27】
分離する段階が以下の段階を含む、請求項20記載の方法:
芳香性物質を粗抽出物から除去するために有効な条件下で、粗抽出物をポリビニルピロリドンカラムまたは活性炭カラムに通過させる段階。
【請求項28】
一つまたは複数のファゴピリトールが少なくとも二つのファゴピリトールを含み、該方法がさらに以下の段階を含む、請求項20記載の方法:
少なくとも二つのファゴピリトールからファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3の一つを分離する段階。
【請求項29】
少なくとも二つのファゴピリトールからファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3の一つを分離する段階が、クロマトグラフィーによって行われる、請求項28記載の方法。
【請求項30】
少なくとも二つのファゴピリトールからファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3の一つを分離する段階が、アミン官能性(amine-functionalized)シリカゲル上で行われる、請求項29記載の方法。
【請求項31】
少なくとも二つのファゴピリトールからファゴピリトールA1、ファゴピリトールA3、およびファゴピリトールB3の一つを分離する段階が、アセトニトリル:水の溶離剤を用いて行われる、請求項29記載の方法。
【図1】

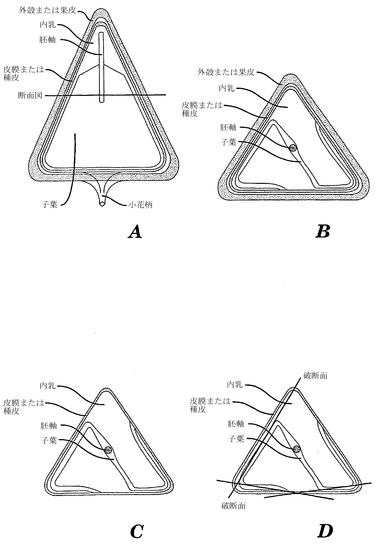
【図2】
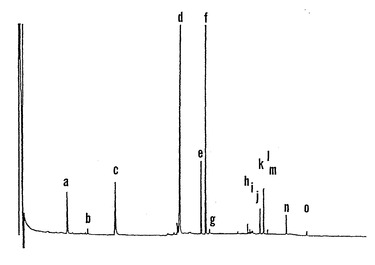
【図3】
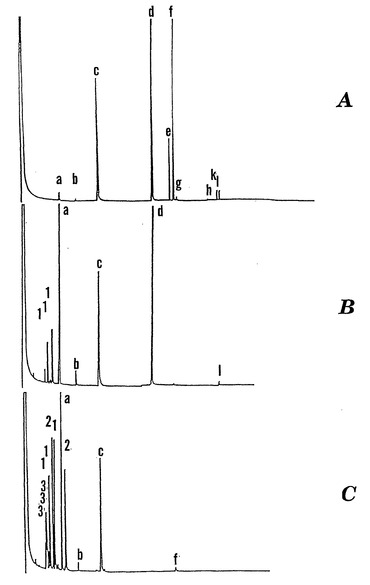
【図4】
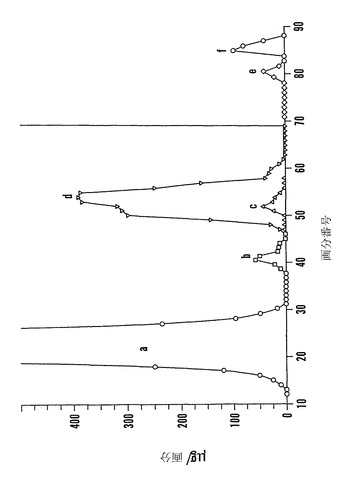
【図5】
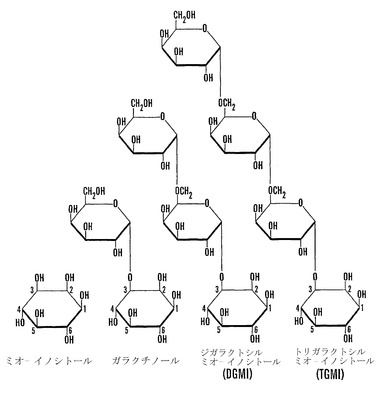
【図6】
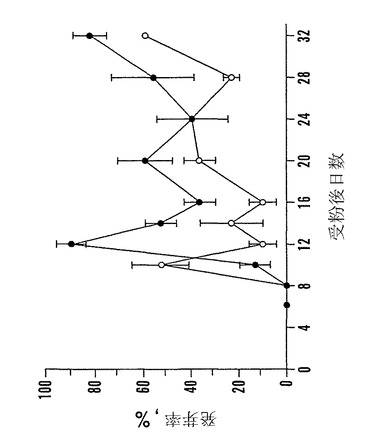
【図7】
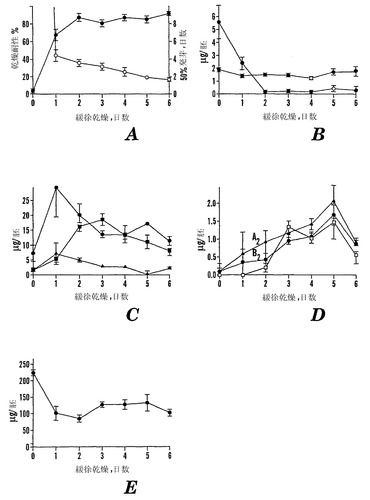
【図8】
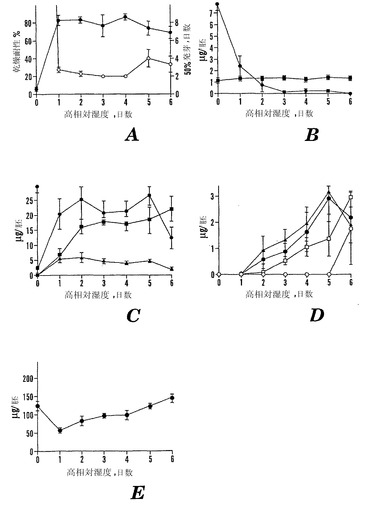
【図9A】
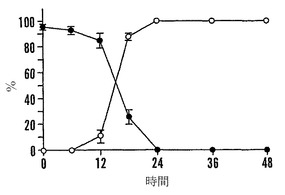
【図9B】
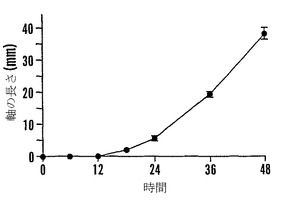
【図9C】
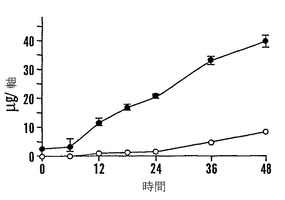
【図9D】
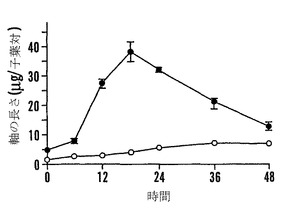
【図9E】
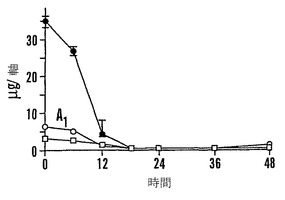
【図9F】
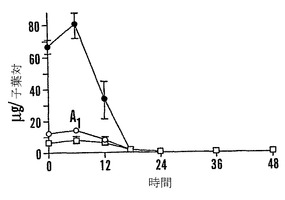
【図9G】
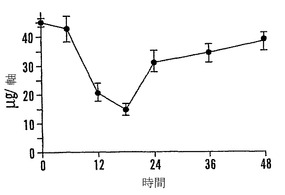
【図9H】
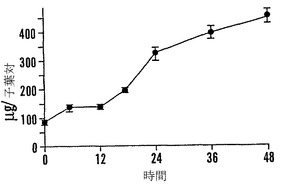
【図9I】
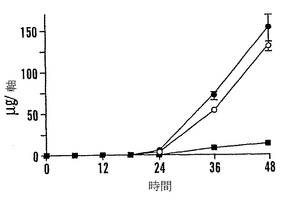
【図9J】
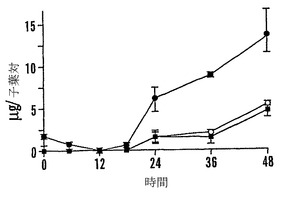
【図10】
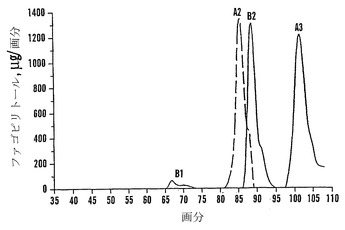
【図11】
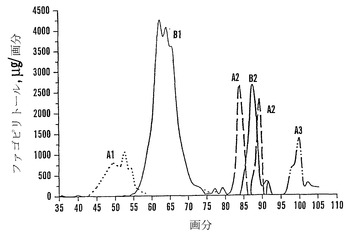
【図12】
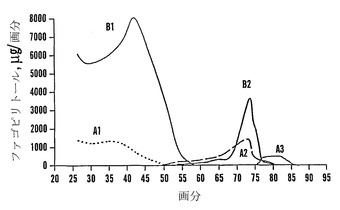
【図13】
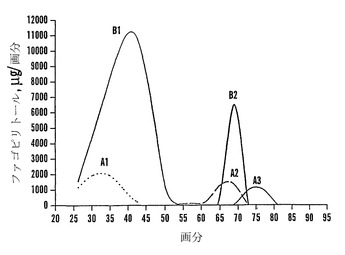
【図14】
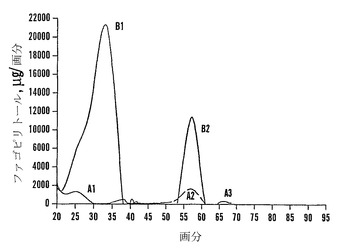
【図15】
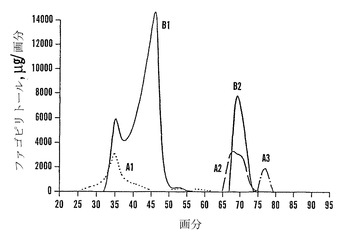
【図16】
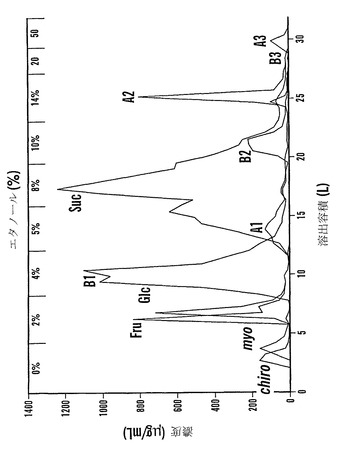
【図17】
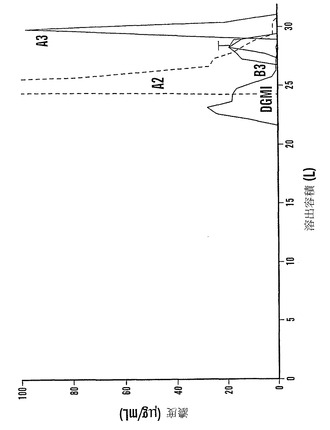
【図18】
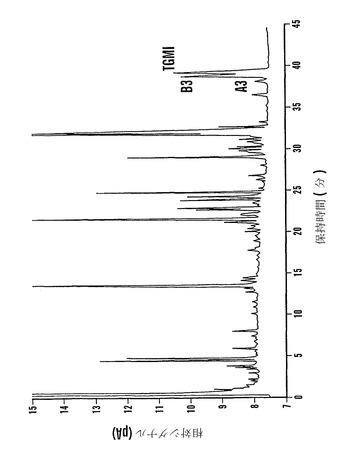
【図19】
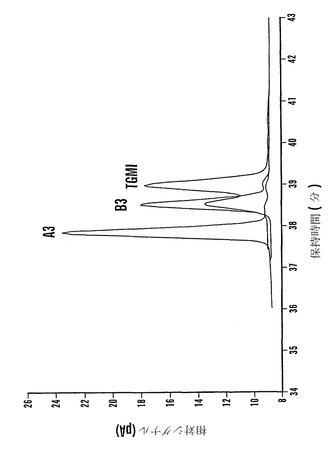
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9A】
【図9B】
【図9C】
【図9D】
【図9E】
【図9F】
【図9G】
【図9H】
【図9I】
【図9J】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【公開番号】特開2013−10782(P2013−10782A)
【公開日】平成25年1月17日(2013.1.17)
【国際特許分類】
【出願番号】特願2012−196803(P2012−196803)
【出願日】平成24年9月7日(2012.9.7)
【分割の表示】特願2001−585776(P2001−585776)の分割
【原出願日】平成13年5月22日(2001.5.22)
【出願人】(500056220)コーネル リサーチ ファンデーション インク. (7)
【Fターム(参考)】
【公開日】平成25年1月17日(2013.1.17)
【国際特許分類】
【出願日】平成24年9月7日(2012.9.7)
【分割の表示】特願2001−585776(P2001−585776)の分割
【原出願日】平成13年5月22日(2001.5.22)
【出願人】(500056220)コーネル リサーチ ファンデーション インク. (7)
【Fターム(参考)】
[ Back to top ]