
ヘパリン結合タンパク質の組み換え体産生
異種性の宿主細胞において産生される再び折り畳まれたヘパリン結合タンパク質を回収及び精製するための方法には、デキストラン硫酸などのポリ陰イオン類とともに可溶化したタンパク質をインキュベートする工程が含まれる。

【発明の詳細な説明】
【発明の開示】
【0001】
(関連出願)
本出願は、本明細書中にその全体が援用される2005年12月22日に出願の米国特許仮出願第60/753615号および2006年7月14日に出願の米国特許仮出願第60/807432号の優先権を主張する。
【0002】
(発明の分野)
本発明は、細胞培養で生産されるヘパリン-結合タンパク質を得る方法に関する。本発明は、原核生物宿主細胞で産生され、これらの細胞中、典型的には周辺質又は細胞内腔に存在する再び折り畳まれた(refolded)ヘパリン結合タンパク質の回収及び精製に関する方法を包含する。また、原核生物の宿主細胞で産生されるヘパリン結合タンパク質は、可溶タンパク質又は可溶性と不溶性のタンパク質の混合物として提供されてもよい。
【0003】
(発明の背景)
天然に生じ、生物学的に活性な多様なポリペプチドがヘパリンを結合することは公知である。このようなヘパリン-結合ポリペプチドには、サイトカイン、例として、血小板因子4及びIL-8(Barber等, (1972) Biochim. Biophys. Acta, 286:312-329;Handin等, (1976) J. Biol. Chem., 251:4273-422;Loscalzo等, (1985) Arch. Biochem. Biophys. 240:446-455;Zucker等, (1989) Proc. Natl. Acad. Sci. USA, 86:7571-7574;Talpas等, (1991) Biochim. Biophys. Acta, 1078:208-218;Webb等, (1993) Proc. Natl. Acad. Sci. USA, 90:7158-7162)、ヘパリン-結合増殖因子(Burgess and Maciag, (1989) Annu. Rev. Biochem., 58:576-606;Klagsbrun, (1989) Prog. Growth Factor Res., 1:207-235)、例として、上皮細胞増殖因子(EGF);血小板由来増殖因子(PDGF);塩基性線維芽細胞増殖因子(bFGF);酸性線維芽細胞増殖因子(aFGF);血管性内皮増殖因子(VEGF);及び、肝細胞増殖因子(HGF) (Liu等, (1992) Gastrointest. Liver Physiol. 26: G642-G649);そして、セレクチン、例えばL-セレクチン、E-セレクチンおよびP-セレクチン (Norgard-Sumnicht等, (1993) Science, 261:480-483)が含まれる。また、Munoz and Linhardt., (2004) Arterioscler Thromb Vasc Biol., 24:1549-1557を参照のこと。
【0004】
国際公開第95/07097号には、VEGFのようなヘパリン結合増殖因子を含むヘパリン結合タンパク質の製剤とともに、治療的使用のための精製された天然のヘパリン又は他のポリ陰イオン化合物が記載される。オリゴ糖および様々な他のポリ陰イオン化合物由来のヘパリンは、ヘパリン結合増殖因子のための活性な立体構造を安定させることが示されており(Barzu等, (1989) J. Cell. Physiol. 140:538-548;Dabora等, (1991) J. Biol. Chem. 266:23627-23640)、ヘパリン親和性クロマトグラフィは、様々な精製様式で用いられている(一般に、国際公開第96/02562号を参照)。
【0005】
哺乳類起源のヘパリン結合タンパク質の多くは組み換え技術によって産生され、臨床的に関係がある(Munoz and Linhardt, (2004) Arterioscler Thromb Vasc Biol., 24:1549-1557;Favard等 (1996) Diabetes and Metabolism 22(4): 268-73;Matsuda等, (1995) J. Biochem. 118(3): 643-9;Roberts等, (1995) Brain Research 699(1): 51-61)。例えば、VEGFは血管内皮細胞の強力な分裂促進因子である。血管透過性因子(VPF)も知られている。Dvorak等, (1995) Am. J. Pathol. 146:1029-39を参照。VEGFは、胚性脈管構造の発達である脈管形成と、既に存在する血管からの新たな血管の形成プロセスである血管形成の両方に重要な働きがある。例えば、Ferrara, (2004) Endocrine Reviews 25(4): 581-611;Risau等, (1988) Dev. Biol., 125:441-450;Zachary, (1998) Intl. J. Biochem Cell Bio 30:1169-1174;Neufeld等, (1999) FASEB J. 13:9-22;Ferrara (1999) J. Mol. Med. 77: 527-543;及び、Ferrara and Davis-Smyth, (1997) Endocri. Rev. 18: 4-25を参照。VEGFの臨床応用には、例えば、創傷治癒を促進する際(例えば、2006年6月16日出願の「Wound Healing」と題する国際公開第91/02058号(Attorney Docket No. P2358R1)を参照)、肝臓(例えば国際公開第2003/0103581号参照)、骨(例えば国際公開第2003/094617号参照)などの組織成長及び修復の際のように、新規な毛細管床の成長があるものが含まれる。また、Ferrara, (2004) Endocrine Reviews 25(4): 581-611を参照。
【0006】
典型的には、治療的に関係のある組み換えタンパク質は多くの宿主生物において産生される。ほとんどのタンパク質は、CHO細胞などの真核生物宿主においてそれらの天然の形状で発現されうる。一般に、動物細胞培養は、最大細胞密度を達成するために長い成長期間を必要とし、最終的に、原核生物の細胞培養より低細胞密度となる(Cleland, J. (1993) ACS Symposium Series 526, Protein Folding: In Vivo and In Vitro, American Chemical Society)。さらに、動物細胞培養は、所望のタンパク質の回収に干渉しうる成長成分を含有する高価な培地を必要とすることが多い。細菌宿主発現系により、組み換えタンパク質の製造規模産生の費用効果的な代替法が提供される。組み換えタンパク質の一般的な細菌発現については多くの米国特許出願が存在し、その中には、米国特許第4565785号;同第4673641号;同第4795706号;及び同第4710473号などがある。製造方法の主要な利点は、遠心分離法又は精密ろ過法によって生成物を細胞性構成成分から容易に単離することができることである。例えばKipriyanov and Little, (1999) Molecular Biotechnology, 12: 173-201;及びSkerra and Pluckthun, (1988) Science, 240: 1038-1040を参照のこと。
【0007】
酸性線維芽細胞増殖因子、塩基性線維芽細胞増殖因子及び血管内皮細胞増殖因子などの組み換えヘパリン結合増殖因子は、細菌を含む多くの供給源から回収し精製されている(Salter D.H.等, (1996) Labor. Invest. 74(2): 546-556 (VEGF);Siemeister等, (1996) Biochem. Biophys. Res. Commun. 222(2): 249-55 (VEGF);Cao等, (1996) J. Biol. Chem. 261(6): 3154-62 (VEGF);Yang等, (1994) Gaojishu Tongxun, 4:28-31 (VEGF);Anspach等, (1995) J. Chromatogr. A 711(1): 129-139 (aFGF及びbFGF);Gaulandris (1994) J Cell. Physiol. 161(1): 149-59 (bFGF);Estape and Rinas (1996) Biotech. Tech. 10(7): 481-484 (bFGF);McDonald等, (1995) FASEB J. 9(3): A410 (bFGF))。しかしながら、大腸菌などの細菌発現系はタンパク質の適切な再折り畳み(refolding)を促す細胞の機構を欠いており、一般に培養培地にほとんどのタンパク質を分泌しない。細菌性宿主細胞において発現される組み換えタンパク質は、密度の高い、部分的に折り畳まれた(folded)タンパク質と誤って折り畳まれた(misfolded)還元タンパク質からなる封入体(inclusion body)として見られることが多い。この形状において、組み換えタンパク質は、通常不活性である。例えば、VEGFの優れた活性型は、2つの165-アミノ酸ポリペプチド(VEGF-165)のホモ二量体である。この構造において、各サブユニットは、7対の鎖内ジスルフィド結合と2つのサブユニットの共有結合に影響するさらに2対を含む(Ferrara等, (1991) J. Cell. Biochem. 47:211-218)。天然の立体構造は、容易にヘパリンを結合することが示されている非常に基本的なドメインを含む(上掲のFerrara et al (1991))。VEGFの共有結合性二量体化は有効なレセプター結合と生物学的活性に必要とされる(Potgens等, (1994) J. Biol. Chem. 269:32879-32885;Claffey等, (1995) Biochim. et Biophys. Acta 1246:1-9)。細菌性生成物は潜在的にいくつかの誤って折り畳まれ、ジスルフィドが攪乱された中間生成物を含有する。
【0008】
さらに、再折り畳みは、誤って折り畳まれ、ジスルフィド結合した二量体、三量体および多量体を生産することが多い。(Morris等, (1990) Biochem. J., 268:803-806;Toren等, (1988) Anal. Biochem., 169:287-299)。この会合現象は、特により高いタンパク質濃度のタンパク質再折り畳みに非常に共通しており、部分的に折り畳まれた中間生成物の疎水的相互作用による会合に関与していることが多いようである(Cleland and Wang, (1990) Biochemistry, 29:11072-11078)。
【0009】
誤った折り畳みは発酵又は単離手順の間に細胞中で起こる。周辺質又は細胞内腔から回収されるタンパク質は可溶化され、可溶性タンパク質は天然の状態に再び折り畳まれなければならない。正しく、生物学的に活性な立体構造にタンパク質を再び折り畳むためのインビトロの方法は、機能的なタンパク質を得るために必須である。封入体から回収されたタンパク質の典型的な以降のプロセシングには、高濃度の尿素などの変性剤に封入体を溶解し、再折り畳みが生じるように変性剤を希釈することを含む(米国特許第4512922号;同第4511502号;及び同第4511503号を参照)。また、例えばRudolph and Lilie, (1996) FASEB J. 10:49-56;及びFischer等, (1993), Biotechnology and Bioengineering, 41:3-13を参照。このような回収方法は普遍的な適用法であると理解でき、封入体からの生物学的に活性な組み換えタンパク質の回収にわずかな変更も考慮される。これらの方法は、VEGFなどのヘパリン結合タンパク質に応用されている(Siemeister等 (1996) supra)。これらの方法は、その他の安定化作用により組み換えタンパク質を生物学的に活性な立体構造にする前にランダムなジスルフィド結合を取り除くことを目的とし、不適当に折り畳まれた中間生成物を取り除かないか、又は適切に折り畳まれた生成物の均質な集団を提供する。
【0010】
逆ミセル又はイオン交換クロマトグラフィを用いて、ミセル内に単一のタンパク質を封入するか又は単一のタンパク質を樹脂上で単離し、その後変性剤を除去することによって変性したタンパク質の再折り畳みを促している(Hagen等, (1990) Biotechnol. Bioeng. 35:966-975; Creighton (1985) in Protein Structure Folding and Design (Oxender, D.L. Ed.) pp.249-251, New York: Alan R. Liss, Inc.)。これらの方法は、タンパク質凝集を予防し、適当な再折り畳みを促す際に有用であった。再折り畳みの速度又は程度を変えるために、立体構造に特異的な再折り畳みを、タンパク質の天然の構造に対するリガンドおよび抗体によって行った(Biotechnology, (Rehm H.-J., and Reed G. Eds.) pp 528-555, New York, VCH)のCleland and Wang, (1993))。例えば、クレアチンキナーゼは、天然の構造に対する抗体がある場合に再び折り畳まれる(Morris等, (1987) Biochem. J. 248:53-57)。抗体に加えて、リガンドおよび補助因子を用いて再折り畳みを促進している。これらの分子は、おそらく、天然タンパク質の形成後に折り畳まれるタンパク質と相互に作用するようである。したがって、折り畳みの平衡は天然の状態に「制御」されうる。例えば、フェリシトクロムcの再折り畳みの速度は、ヘム鉄の軸位置に対して外部のリガンドによって上げられた(Brems and Stellwagon, (1983) J. Biol. Chem. 258:3655-3661)。また、シャペロンタンパク質は、タンパク質の折り畳みによって促すために用いられている。例としてBaneyx, (1999) Current Opinion in Biotechnology, 10:411-421を参照。
【0011】
例えば、効率が良く経済的な細菌細胞培養物におけるヘパリン結合タンパク質の製造のために、宿主細胞培養物からヘパリン結合タンパク質を折り畳む及び/又は回収する新規のより効率の良い方法が求められている。そのような方法により、生物学的に不活性な中間生成物の除去又は低減や高度に精製された生物学的に活性な適切に再び折り畳まれたタンパク質の回収の改善が可能となり、普遍的にタンパク質産生の製造規模に応用可能である。以下の開示内容を参照すると明らかなように、本発明はこれら及び他の必要性に応えるものである。
【0012】
(発明の概要)
本発明は、再び折り畳まれたヘパリン結合タンパク質の細胞培養物からの回収及び精製のための方法を提供する。特に、本発明は、原核生物の宿主細胞、例えば細菌細胞からヘパリン結合タンパク質を回収する方法を提供する。例えば、方法は、(a) 不溶性ヘパリン結合タンパク質を該細菌細胞の周辺質又は細胞内腔から単離する;(b) カオトロピック剤と還元剤を含む第一緩衝溶液に単離された不溶性ヘパリン結合タンパク質を可溶化する、そして(c) ヘパリン結合タンパク質の再折り畳みが生じる時間と条件で、カオトロピック剤と硫酸化ポリ陰イオン剤を含む第二緩衝溶液中で、可溶化させたヘパリン結合タンパク質をインキュベートする;そして、(d) 該再び折り畳まれたヘパリン結合タンパク質を回収する、工程を含んでなり、このとき硫酸化ポリ陰イオン剤とインキュベートしなかった場合と比べて再び折り畳まれたヘパリン結合タンパク質の回収がおよそ2〜10倍に増加する。一実施態様では、第二緩衝溶液はさらにアルギニンを含む。一実施態様では、第二緩衝溶液はさらにシステイン又は軽度の還元剤を含む。
【0013】
本発明の一実施態様では、例えば、回収された生物学的に活性な再び折り畳まれたタンパク質のタンパク質濃度において2〜8倍の増加、又は回収された生物学的に活性な再び折り畳まれたタンパク質のタンパク質濃度において2〜5倍の増加、又は回収された生物学的に活性な再び折り畳まれたタンパク質のタンパク質濃度において3〜5倍の増加、又は回収された生物学的に活性な再び折り畳まれたタンパク質のタンパク質濃度において2〜3倍の増加がある。本発明の他の実施態様では、生物学的に活性な再び折り畳まれたタンパク質の回収されたタンパク質濃度において、例えば2.0倍、2.5倍、2.8倍、3.0倍、5倍、6倍、7.0倍、8倍、9倍などより大きい増加がある。本発明の一実施態様では、生物学的に活性な再び折り畳まれたVEGFのタンパク質濃度において3〜5倍の増加がある。
【0014】
概して、本発明の方法は、ヘパリン結合タンパク質、特にヘパリン結合増殖因子および、特に血管内皮細胞増殖因子(VEGF)に適用可能である。本発明のある実施態様では、硫酸化ポリ陰イオン剤は、およそ3000から10000ダルトンの間である。一実施態様では、生産プロセスにおいて利用される硫酸化ポリ陰イオン剤は、デキストラン硫酸、硫酸ナトリウム又はヘパリン硫酸塩である。一態様では、デキストラン硫酸は、3000ダルトンから10000ダルトンの間である。
【0015】
本明細書中に記述するように、本発明はさらに、ヘパリン結合タンパク質の精製のみ、又はヘパリン結合タンパク質の回収と関連した精製のための工程及び方法を提供する。特定の実施態様では、精製法は、前記の再び折り畳まれたヘパリン結合タンパク質を、ヒドロキシアパタイトクロマトグラフィ支持体;第一疎水性相互作用クロマトグラフィ支持体、陽イオンクロマトグラフィ支持体および第二疎水性相互作用クロマトグラフィ支持体に接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出させることを含む。他の実施態様では、精製法は、前記の再び折り畳まれたヘパリン結合タンパク質を、陽イオン交換支持体;第一疎水性相互作用クロマトグラフィ支持体、及びイオン交換又は混合媒体クロマトグラフィ支持体に接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出させることを含む。回収工程のための工程は、例えばいずれかの順序で順次、又はクロマトグラフィ支持体の順番を変えて行ってもよい。本発明の特定の実施態様では、製造規模又は工業規模の細胞培養物から再び折り畳まれたヘパリン結合タンパク質を回収する及び精製するための方法を提供する。
【0016】
(詳細な説明)
定義
「ヘパリン」(ヘパリンの酸とも称される)は非常に硫酸化した、直鎖の陰イオン性ムコ多糖の不均一な基であり、グリコサミノグリカンと称される。他も存在しうるが、ヘパリンの主要な糖質は、α-L-イズロン酸2-硫酸塩、2-デオキシ-2-スルファミノ-α-グルコース6-硫酸塩、β-D-グルクロン酸、2-アセタミド-2-デオキシ-α-D-グルコースおよびL-イズロン酸である。これら及び場合によって他の糖質は様々な大きさのポリマーを形成するグリコシド結合によって連結される。その共有結合した硫酸塩とカルボン酸基の存在のために、ヘパリンは非常に酸性である。起源及び測定法によって、ヘパリンの分子量はおよそ3000からおよそ20000ダルトンに変化する。
天然のヘパリンは、いくつかの哺乳類種の様々な組織、特に肝臓および肺や肥満細胞の構成要素である。ヘパリンおよびヘパリン塩類(ヘパリンナトリウム)は市販されており、主に様々な臨床状態において抗凝血物質として用いられる。
【0017】
「デキストラン硫酸」は、主要な構造がD‐グルコースのポリマーであるデキストランの硫酸塩である。グルコース及び場合によって他の糖質は、様々な大きさのポリマーを形成するα-D(1-6)グリコシド結合によって連結される。共有結合した硫酸塩の存在のために、デキストラン硫酸は非常に酸性である。通常、硫黄含量は10%より少なくなく、典型的におよそ15%−20%で、グルコース分子当たり3未満の硫酸基となる。デキストラン硫酸の平均分子量はおよそ1000からおよそ40,000,000ダルトンである。本発明で使用可能なデキストラン硫酸の実施例は、微生物、例としてロイコノストック・メッセンテロイデス(Leuconostoc mesenteroides)およびロイコノストック デキストラニクム(L. dextranicum)から産生されるデキストランの硫酸塩を含む。
【0018】
本発明の範囲内で用いられる「ポリ陰イオン剤」は、他の「ポリ陰イオン剤」、例として、硫酸ナトリウム、硫酸ヘパリン、ヘパラン硫酸、硫酸ペントサン(ポリ)、デキストラン、デキストラン硫酸、ヒアルロン酸、コンドロイチン、コンドロイチン硫酸、デルマタン硫酸およびケラタン硫酸を含む、ヘパリン結合タンパク質に結合可能である市販の精製された天然のヘパリン調製物および化合物を記述すると定める。「硫酸化されたポリ陰イオン剤」、例えば、多糖の硫酸塩誘導体、例としてヘパリン硫酸、デキストラン硫酸、米国特許第5314872号に記載のバシラス‐マセランスなどの微生物によって産生されるシクロデキストリンの硫酸塩、並びにβ-1,3グルカン硫酸塩などの他のグルカンの硫酸塩、アルカリゲネス属又はアグロバクテリウムに属する微生物によって産生されるβ-1,3グルカン、およびコンドロイチン硫酸並びに硫酸化ヘパリン断片は、特に本発明の前後関係の範囲内で有用である。
【0019】
上述した薬剤は一般に利用可能であり、当業者に明らかである。例えば、硫酸化ヘパリン断片は、ゲル浸透クロマトグラフィによって分画化されたヘパリン由来のオリゴ糖のライブラリから得られうる。親和性分画され、ヘパリン由来のオリゴ糖の調製は、Ishihara等, (1993) J. Biol. Chem., 268:4675-4683によって報告された。これらのオリゴ糖は市販のブタヘパリンから調整し、亜硝酸により部分的に脱重合し、水素化ホウ素により還元し、ゲル浸透クロマトグラフィによって分画化した。ジ-、テトラ-、ヘキサ-、オクタ-、及びデカ糖類の結果として生じたプールをセファロースTM4Bに共有結合させたヒト組み換えbFGFの親和性カラムに順にアプライし、さらに、塩化ナトリウムの勾配に応じてこのカラムからの溶出に従ってサブプールに分画した。これにより、Hexa-1からHexa-5と称する5つのプールが生じ、その構造および生物学的活性をさらに評価した。Hexa-5Cの構造とその500-MHzのNMRスペクトルは、Tyrell等, (1993) J. Biol. Chem., 268:4684-4689の図4に示される。このヘキサ糖類は、[IdoA(2-OSO3)α1-4GlcNSO3(6-OSO3)α1-4]2IdoA(2-OSO3)α1-4AManR(6-OSO3)の構造を有する。上記のすべてのヘパリン由来のオリゴ糖、並びに他のヘパリン様オリゴ糖は、本発明に好適であり、本発明に従って用いられうる。本発明のある実施態様では、よりユニットが大きなヘパリンのヘキサ糖類及びポリ糖類(例えば、ヘプタ-、オクタ-、ノナ-、デカ糖類)が用いられる。さらに、例えば高い程度の硫酸化により正味の陰電荷が大きい、ヘパリン由来又はヘパリン様のオリゴ糖が、有利に使われる。
【0020】
本明細書中で用いられる「ヘパリン-結合タンパク質」又は「HPB」なる用語は、ヘパリンを結合することができるポリペプチドを指す(上記に定義されたように)。定義には、成熟、プレ、プレ-プロ、及びプロの形態の天然及び組み換えて産生させたヘパリン-結合タンパク質が含まれる。ヘパリン結合タンパク質の典型的な例は、限定するものではないが、上皮細胞増殖因子(EGF)、血小板由来増殖因子(PDGF)、塩基性線維芽細胞増殖因子(bFGF)、酸性線維芽細胞増殖因子(aFGF)、血管内皮細胞増殖因子(VEGF)、肝増殖因子(HGF)(散乱係数、SFとしても知られる)、及び神経成長因子(NGF)、IL-8などを含む、「ヘパリン結合増殖因子」である。
【0021】
本明細書中で用いられるように「血管内皮細胞増殖因子」又は「VEGF」は、Castor, C.W.,等, (1991) Methods in Enzymol. 198:391-405において開示されるアミノ酸配列を有するウシ下垂体濾胞性細胞から本来得られる哺乳類の増殖因子、並びに限定するものではないがHouck等, (1991) Mol. Endocrin. 5:1806-1814において報告されるヒトVEGFアミノ酸配列を含む、対応する天然のVEGFの定性的生物学的活性を有するこの機能的誘導体を指す。また、Leung等 (1989) Science, 246:1306、及び、Robinson & Stringer, (2001) Journal of Cell Science, 144(5): 853-865、米国特許第5332671号を参照。VEGFの主な形態は、7つの分子内ジスルフィド結合と2つの分子間ジスルフィド結合を形成する16のシステイン残基を有する165アミノ酸ホモ二量体である。オルタナティブスプライシングは、121、145、165、189および206のアミノ酸からなる複数のヒトVEGFポリペプチドの形成に関係しているが、VEGF121変異体は、他の変異体のヘパリン結合ドメインを欠いているため、本明細書に記載のヘパリン結合タンパク質の定義に入らない。VEGFのすべてのアイソフォームは共通のアミノ末端ドメインを共有するが、分子のカルボキシル末端部分の長さが異なる。好ましいVEGFの活性型であるVEGF165は、各単量体のアミノ酸残基Cys26-Cys68;Cys57-Cys104;Cys61-Cys102;Cys117-Cys135;Cys120-Cys137;Cys139-Cys;158;Cys146-Cys160の間にジスルフィド結合を有する。図11を参照。また、例えば、Keck等, (1997) Archives of Biochemistry and Biophysics 344(1): 103-113を参照。VEGF165分子は、アミノ末端レセプター-結合ドメイン(アミノ酸1−110ジスルフィド結合ホモ二量体)とカルボキシル末端ヘパリン-結合ドメイン(残基111−165)の2つのドメインから成る。例えば、Keyt等, (1996) J. Biol. Chem., 271(13): 7788-7795を参照。本発明のある実施態様では、単離して、精製されるVEGF165は、残基75(Asn)でグリコシル化されていない。例えば、Yang等, (1998) Journal of Pharm. & Experimental Therapeutics, 284:103-110を参照。本発明のある実施態様では、単離して、精製されるVEGF165は、残基Asn10で実質的に脱アミド化されていない。本発明のある実施態様では、単離して、精製されるVEGF165は、脱アミド化された(残基Asn10で)タンパク質と脱アミド化されていないタンパク質の混合であり、典型的にはほとんどのタンパク質は脱アミド化されていない。VEGF165はホモ二量体であるので、脱アミド化は一方又は両方のポリペプチド鎖で起こりうる。
【0022】
本明細書中で用いる「適切に折り畳まれた」又は「生物学的に活性な」VEGF又は他のHBPなどは、生物学的に活性な立体構造を有する分子を指す。当業者は、誤って折り畳まれた中間生成物及びジスルフィドスクランブル中間生成物は生物学的活性を有しうると理解するであろう。この場合、適切に折り畳まれた又は生物学的に活性なVEGF又はHBPは、VEGF(上記した)又は他のHBPの天然の折り畳みパターンに対応する。例えば、適切に折り畳まれたVEGFは、二量体分子の2つの分子間ジスルフィド結合に加え、上記のジスルフィド対を有するが、他の中間生成物は細菌細胞培養物によって産生されうる(図1及び3A−3D)。適切に折り畳まれたVEGFでは、各単量体の同じ残基(Cys51とCys60)の間で2つの分子間ジスルフィド結合が生じる。例えば国際公開第98/16551号特許を参照。VEGFの生物学的活性には、限定するものではないが、例えば、血管透過性の促進、血管内皮細胞の増殖促進、VEGFレセプターへの結合、VEGFレセプターによる結合とシグナル伝達(例えば、Keyt等, (1996) Journal of Biological Chemistry, 271(10): 5638-5646を参照)、血管新生の誘導などが含まれる。
【0023】
「精製された」又は「純粋なHBP」なる用語は、組み換え産生及び特に原核生物又は細菌の細胞培養物に見られるような、通常付随する物資を欠く物質を指す。したがって、この用語は、DNA、宿主細胞タンパク質又は元の環境に付随する他の分子の混入がない組み換えHBPを指す。この用語は、少なくともおよそ75%、少なくともおよそ80%、少なくともおよそ85%、少なくともおよそ90%、少なくともおよそ95%又は少なくともおよそ98%又はそれ以上の程度の純度を指す。
【0024】
「封入体」又は「屈折小体」なる用語は、対象の凝集されたポリペプチドの密度の高い細胞内質量を指し、それはすべての細胞構成成分を含む総細胞タンパク質の主な部分を成す。すべてではないが場合によって、ポリペプチドのこれらの凝集塊は、1000倍まで拡大した位相差顕微鏡下で細胞の中に可視の明るいスポットとして認識されうる。
【0025】
本明細書中で用いられる「誤って折り畳まれた」タンパク質なる用語は、屈折小体内に含有される沈殿したないしは凝集したポリペプチドを指す。本明細書中で用いられる「不溶性の」又は「誤って折り畳まれた」VEGF又は他のHBPは、原核生物の宿主細胞の周辺質又は細胞内腔内に含有されるか、またはさもなくば、付随する原核生物の宿主細胞である、沈殿したないしは凝集したVEGFを指し、ミスマッチ又は形成されていないジスルフィド結合を有する生物学的に不活性な立体構造であると思われる。一般に、不溶性HBPは屈折小体に含有される必要はなく、すなわち、位相差顕微鏡下で可視化されることもあるし可視化されないこともある。
【0026】
本明細書中で用いられる「カオトロピック剤」は、水溶液中に適切な濃度であると、ポリペプチドを水溶媒質に可溶化させるために、その表面の変質によってポリペプチドの空間配位又は立体構造を変えることが可能である化合物を指す。変質は、例えば水和作用、溶媒環境又は溶媒-表面相互作用の状態を変えることで生じうる。カオトロピック剤の濃度はその強度および有効性に直接作用する。強力な変性カオトロピック溶液は、溶液中で、タンパク質二次構造を効果的に取り除く溶液中に存在するポリペプチドを効率的にほどく(unfold)高濃度のカオトロピック剤を含む。アンフォールディング(unfolding)は相対的に広範囲であるが、可逆性である。中程度の変性カオトロピック溶液は、溶液中に十分な濃度であると、ポリペプチドは部分的にしか折り畳まれない。内在性又は類似の生理学的条件下でその活性型を操作する場合、それ自体の空間的立体構造に、どんなに立体構造がねじれていようとポリペプチドは溶液中に溶ける中間生成物を経ると考えられているのでカオトロピック剤の例には、塩酸グアニジン、尿素および水酸化物、例えばナトリウム又は水酸化カリウムが含まれる。カオトロピック剤には、これらの試薬の組合せ、例として、尿素又は塩酸グアニジンと水酸化物の混合が含まれる。
【0027】
本明細書中で用いられる「還元剤」は、水溶液中に適切な濃度であると、フリーなスルフヒドリル基を維持して、分子内又は分子間ジスルフィド結合が化学的に崩壊される化合物を指す。適切な還元剤の代表的な例には、ジチオトレイトール(DTT)、ジチオエリトリトール(DTE)、β-メルカプトエタノール(BME)、システイン、システアミン、チオグリコレート、グルタチオン、トリス[2-カルボキシエチル]ホスフィン(TCEP)、及び水素化ホウ素ナトリウムなどがある。
【0028】
本明細書中で用いられる「緩衝溶液」はその酸-塩基コンジュゲート構成成分の作用によってpHの変化に耐える溶液を指す。
【0029】
本明細書中での目的のための「細菌」には真正細菌及び古細菌が含まれる。本発明のある実施態様では、グラム陽性細菌及びグラム陰性細菌を含む真正細菌は本明細書中に記載の方法及びプロセスに用いられる。本発明のある実施態様では、グラム陰性細菌、例えば腸内細菌科が用いられる。腸内細菌科に属する細菌の例は、エシェリキア属、エンテロバクター属、エルウィニア属(Erwinia)、クレブシエラ属、プロテウス属、サルモネラ属、セラシア属、及びシゲラ属を含む。その他の種類の適切な細菌としては、アゾトバクタ、シュードモナス、根粒菌属、ビトレオシラ(Vitreoscilla)、及びパラコッカス(Paracoccus)が挙げられる。本発明のある実施態様では、大腸菌が用いられる。適切な大腸菌宿主には、大腸菌W3110(ATCC 27,325)、大腸菌294(ATCC 31,446)、大腸菌B及び大腸菌X1776(ATCC 31,537)が含まれる。これらの例は、限定的なものではなく例示であり、W3110が一例である。上述の細菌の任意の突然変異細胞も使用できる。当然ながら、バクテリウム属の細胞におけるレプリコンの複製可能性を考慮しながら適切な細菌を選択することが必要である。例えば、pBR322、pBR325,pACYC177、又はpKN410等の周知のプラスミドを使用してレプリコンを供給するとき、大腸菌、セラシア属、又はサルモネラ属を宿主として適切に使用することができる。さらに、好適な細菌宿主細胞の例については以下を参照のこと。
【0030】
本明細書で使用する「細胞」「細胞系」「株」、及び「細胞培養物」という表現は、互いに交換可能に使用され、そのような名称は全て子孫を含んでいる。したがって、「形質転換体」及び「形質転換された細胞」は初代の対象細胞及び、継代回数には関係なく、それに由来する培養物を含んでいる。また、故意又は偶然の変異のために、全ての子孫はDNA含量が正確に一致しないかもしれないことも理解される。最初に形質転換された細胞においてスクリーニングされるのと同じ機能又は生物学的活性を有する突然変異子孫が含まれる。明確な区別が意図される場合には、それは文脈から明らかになるだろう。
【0031】
本明細書中で用いられる場合、一般に、「ポリペプチド」はおよそ10より多いアミノ酸を有する任意の細胞源由来のペプチド及びタンパク質を指す。「異種の」ポリペプチドは、大腸菌によって産生されるヒトタンパク質のような、利用される宿主細胞にとって外来のポリペプチドである。異種ポリペプチドは原核生物のものでも真核生物のものでもよいが、好ましくは真核生物のものであり、より好ましくは哺乳類、最も好ましくはヒトのものである。本発明のある実施態様では、組み換えて産生されるか、又は組み換えポリペプチドである。
【0032】
ヘパリン結合タンパク質
ヘパリン結合タンパク質の単離
不溶性の誤って折り畳まれたヘパリン結合タンパク質(HBP)は、当分野の多くの標準的な技術のいずれかによりタンパク質を発現する原核生物の宿主細胞から単離される。例えば、不溶性のHBPは、適切な単離バッファ中で、細胞を適切なイオン強度のバッファに曝露して殆どの宿主タンパク質を可溶化させるか(しかし対象のタンパク質は実質的に不溶性である)、又は封入体ないしは周辺質又は細胞内腔のタンパク質を放出させ、例えば遠心分離による回収に利用できるようにするために細胞を破壊することにより、単離される。この技術は周知であり、例えば米国特許第4511503号に記載されている。Kleid等は均質化の後に遠心分離をすることによる屈折小体の精製を開示している(Kleid等, (1984) in Developments, Industrial Microbiology, (Society for Industrial Microbiology, Arlington, VA) 25:217-235)。例としてFischer等, (1993) Biotechnology and Bioengineering 41:3-13も参照のこと。
【0033】
米国特許第5410026号は、封入体からのタンパク質の回収のための典型的な方法を記載しており、以下のようにまとめられる。原核生物細胞を好適なバッファに懸濁する。典型的に、バッファはpH5〜9、又はおよそ6〜8に緩衝するために好適な緩衝剤と塩からなる。NaClなどの任意の好適な塩は、緩衝した溶液の十分なイオン強度を維持するために有用である。典型的には、約0.01から2M、又は0.1から0.2Mのイオン強度を用いる。細胞は、この緩衝液中に懸濁され、ついで、例えば機械的方法、例えばホモジナイザー(Manton-Gaulinプレス、マイクロフルイダイザー、又はNiro-Soavi)、フレンチプレス、ビーズミル、又は音波オシレーター、あるいは化学的又は酵素的方法のような通常用いられている技術を使用して破壊又は溶解する。
【0034】
細胞破壊の化学的ないしは酵素的方法の例には、細菌壁を溶解するリゾチームの使用を伴うスフェロプラスティング(Neuら, Biochem.Biophys.Res.Comm.,17:215(1964))、及びポリペプチドの放出のために生細胞を高緊張度の溶液と低緊張度の冷水洗浄で処理することを含む浸透圧ショック(Neuら, J.Biol.Chem.,240:3685-3692(1965))が含まれる。一般的に、発酵培養物の分析スケール容量に含まれる細菌の破壊には超音波処理が用いられる。大きなスケールでは、高圧ホモジナイゼーションが典型的に用いられる。
【0035】
細胞を破壊した後、典型的に、懸濁液を低速、一般的にはおよそ500〜15000×gで遠心し(本発明のある実施態様では、およそ12000×gが用いられる)、標準的な遠心では、不溶性タンパク質のすべてが実質的にすべてペレット状になるのに十分な時間遠心する。この時間は、遠心する容量並びに遠心設定に応じて単に決定されうる。典型的におよそ10分間〜0.5時間が不溶性タンパク質をペレット状にするために十分である。ある実施態様では、懸濁液を12000×gで10分間遠心する。
【0036】
その結果生じたペレットは不溶性タンパク質分画の実質的にすべてを含む。細胞破壊プロセスが完全でないならば、ペレットはインタクトな細胞又は破壊された細胞断片も含有し得る。細胞破壊の完全性は、ペレットを少量の同じバッファ中に懸濁し、懸濁液を位相差顕微鏡で調べることにより検査することができる。破壊細胞断片もしくは全細胞の存在は、断片又は細胞及び随伴する非屈折性(non-refractile)ポリペプチドを除去するために更なる超音波処理又は破壊の他の手段が必要であることを示している。そのような更なる破壊後に、必要ならば、懸濁液を再び遠心分離し、ペレットを回収し、再懸濁し、再分析する。このプロセスを、視覚検査でペレット化材料中に破壊細胞断片がないことが明らかになるか、更なる処理でも得られるペレットのサイズを低減できなくなるまで繰り返す。
【0037】
不溶性タンパク質が細胞内又は周辺質のスペースのいずれにあっても、上記のプロセスは実施できる。本発明のある実施態様では、ヘパリン結合タンパク質を単離するために本明細書中に示した条件は、周辺質スペース又は細胞内スペースに沈殿した封入体と、特にVEGFに関する。しかしながら、この方法及び手順は、以降全体にわたって述べるように一般にわずかな変更があるヘパリン結合タンパク質に適用することができると考えられる。本発明のある実施態様では、この方法及び手順は、HBPの製造又は工業規模の産生、再折り畳み、及び精製に適用することができる。
【0038】
ヘパリン結合タンパク質の再折り畳み
単離した不溶性の誤って折り畳まれたヘパリン結合タンパク質は、ヘパリン結合タンパク質を実質的に可溶化するために十分な量のカオトロピック剤と還元剤を含む第一緩衝溶液中でインキュベートする。このインキュベートは、いくつか又は実質的にすべてのヘパリン結合タンパク質の可溶化とアンフォールド(unfolding)を可能とする濃度、インキュベート時間、及びインキュベート温度の条件下で行う。
【0039】
緩衝溶液中における可溶化の程度の測定は、濁度決定法により、還元されたSDS-PAGEゲル上の遠心分離後に上清とペレットとの間の分画を分析することにより、プロテインアッセイ(例えば、バイオ-ラッドタンパク質アッセイキット(例えば、Pierce, Bio-Radなど))により、又はHPLCにより簡便に実施される。
【0040】
第一緩衝溶液は、バッファのpH範囲を少なくともおよそ7.0に維持するために好適な緩衝剤を含み、典型的な範囲は7.5−10.5である。ある実施態様では、VEGFのpHはpH8.0である。この後の範囲のpHをもたらす適切なバッファの例としては、トリス-HCl(トリス[ヒドロキシメチル]アミノメタン)、HEPPS (N-[2-ヒドロキシエチル]ピペラジン-N'-[3-プロパン-スルホン酸])、HEPES (N-[2-ヒドロキシエチル]ピペラジン-N'-[2-エタンスルホン酸])、CAPSO(3-[シクロヘキシルアミノ]-2-ヒドロキシ-1-プロパンスルホン酸)、AMP(2-アミノ-2-メチル-1-プロパノール)、CAPS(3-[シクロヘキシルアミノ]-1-プロパンスルホン酸)、CHES(2-[N-シクロヘキシルアミノ]エタンスルホン酸)、グリシン、及び酢酸ナトリウムが含まれる。本発明のある実施態様では、本明細書中のバッファはおよそpH8.0のHEPPSである。更なる実施態様では、例えばHEPPSなどのバッファは硫酸化される。
【0041】
本発明の実施に好適なカオトロピック剤には、例えば、尿素及びグアニジン又はチオシアン酸の塩類、例えば尿素、塩酸グアニジン、チオシアン酸ナトリウムなどが含まれる。バッファに存在するために必要なカオトロピック剤の量は、溶液中のHBPをアンフォールドするために十分な量である。本発明のある実施態様では、カオトロピックは、およそ4モルと10モルの間で存在する。本発明の一実施態様では、カオトロピック剤は、およそ5〜8M、又はおよそ7Mの尿素である。他の例では、カオトロピック剤は、およそ6〜8Mの塩酸グアニジンである。
【0042】
適切な還元剤の例には、限定するものではないが、ジチオトレイトール(DTT)、ジチオエリトリトール(DTE)、β-メルカプトエタノール(BME)、システイン、DTEなどがある。バッファに存在すべき還元剤の量は、還元剤とカオトロピック剤の種類、用いるバッファの種類とpH、溶液に取り込まれる又は導入される酸素の量、及びバッファ中のタンパク質の濃度に主に依存するであろう。例えば、4〜8M 尿素を含有するpH7.0〜10.0の緩衝溶液中0.5〜1.5mg/mlのタンパク質の場合、還元剤は、例えば、およそ1〜15mMの濃度のDTT、又はおよそ0.2〜2mMの濃度のBME、又はおよそ2〜10mMの濃度のシステインである。一実施態様では、還元剤は、およそ0.5からおよそ4mM、又は2〜4mMのDTTである。図9はVEGFの抽出に対する尿素とDTTの効果を示す。ピーク3のVEGFは、適切に折り畳まれた生物学的に活性なVEGFを示す。一実施態様では、還元剤はおよそ10mMのDTTである。単一の還元剤又は還元剤の組合せが、本明細書中のバッファで用いられうる。
【0043】
緩衝溶液中のタンパク質の濃度は、タンパク質が光学密度によって測定すると実質的に溶解されている程度でなければならない。用いる正確な量は、例えば、緩衝溶液中の他の成分の濃度と種類、特にタンパク質濃度、還元剤及びバッファのpHに依存するであろう。本発明の一実施態様では、ヘパリン結合タンパク質の濃度は、1ml当たり0.5〜5.5mg又は1.5〜5.0mg/mlの範囲である。典型的に、可溶化は、少なくともおよそ1〜24時間のおよそ0〜45℃、又はおよそ20〜40℃、又はおよそ23〜37℃、又はおよそ25〜37℃、又はおよそ25℃で、少なくともおよそ1〜24時間かけて行う。一実施態様では、可溶化は、室温で少なくともおよそ2時間行う。一般的に、温度は、塩、還元剤およびカオトロピック剤のレベルによって見かけ上影響を受けない。
【0044】
ポリペプチドが可溶化されると、上記のカオトロピック剤と硫酸化ポリ陰イオン剤を、ヘパリン結合タンパク質の再折り畳みが可能なカオトロピック剤の濃度で含有する第二緩衝溶液に置く、又は希釈する。
一般に、可溶性の誤って折り畳まれたタンパク質のこの第二インキュベーションの条件は、タンパク質のいくらか又は実質的又は完全な再折り畳みが起こるようにする。正確な条件は、例えばバッファのpHや、存在するのであれば、硫酸化ポリ陰イオン剤及びカオトロピック剤及び還元剤の種類と濃度に依存するであろう。培養温度は通常、およそ0〜40℃、又は10〜40℃であり、インキュベートは通常、再折り畳みに効果を示すように少なくともおよそ1時間行う。ある実施態様では、反応は、例えばおよそ15〜37℃、又は20〜30℃で、少なくともおよそ6時間、少なくともおよそ10時間、又はおよそ10から48時間の間、又はおよそ15から20時間の間、又は6から20時間の間、又は12から24時間の間で行う。
【0045】
再折り畳みの程度は、HPBの放射線免疫アッセイ(RIA)力価、又は例えばPOROS HE2/Mカラム(PerSeptive BioResearch Products)又は他の好適なヘパリン親和性カラムを用いた高性能液体クロマトグラフィ(HPLC)分析によって、好適に測定される。RIA力価の増加又は適切に折り畳まれたHBPピークサイズは、バッファ中に存在する適切に折り畳まれた、生物学的に活性なHPBの量の増加と直接相関する。RIA又はHPLCによって測定されるように、インキュベートは、回収された誤って折り畳まれたHPBに対する適切に折り畳まれたHPBの比率を最大にするように行う。
【0046】
一実施態様では、適切に折り畳まれたVEGFの質と量は、ヘパリン-結合アッセイを用いて評価される。希釈したヘパリン結合タンパク質を含有する試料は、例えばPOROS HE2/M(4.6×100mm、PerSeptive BioResearch Products, Cambridge, MA)又は他の好適なヘパリン親和性カラムに流す。例えば、ヘパリン親和性カラムは、0.15M 塩化ナトリウムを含有する10mM リン酸ナトリウム、pH7に平衡化した。1ml/分又は2ml/分の流速で、カラムは、0.15〜2M 塩化ナトリウムの直線濃度勾配を含む10mM リン酸ナトリウム、pH7を用いて10分間にわたって溶出する。溶出物を280nmでモニターした。一実施態様では、タンパク質は、生物学的に活性な適切に再び折り畳まれたHBPに対応する単一のピークに回収される。本発明の一実施態様では、適切に再び折り畳まれたHBPを測定するためのアッセイはRPHPLCである。場合によってジスルフィド結合はペプチドマップによって確認することができる。また、円二色性は、2&3D構造/折畳みを決定するために使われてもよい。
【0047】
第二緩衝溶液のためのバッファは、第一緩衝溶液のために上記のもののいずれか、VEGFを再び折り畳むためには例えばおよそ50mMの濃度で、例えばHEPPS pH8.0であってもよい。ポリペプチドは、再折り畳みバッファにて、例えば少なくとも5倍、又は少なくともおよそ10倍、およそ20倍、又はおよそ40倍に希釈されてもよい。あるいは、ポリペプチドは再折り畳みバッファに対して透析されてもよい。
第二緩衝溶液は、HPBの再折り畳みが起こる濃度のカオトロピック剤を含む。通常、カオトロピックはおよそ0.5から2モルの間で存在する。本発明の一実施態様では、本明細書中のカオトロピック剤は、およそ0.5〜2M、0.5〜2M 、又はおよそ1Mの尿素である。一実施態様では、カオトロピック剤はおよそ1.3M 濃度の尿素である。本発明の他の実施態様では、カオトロピック剤はおよそ1Mの塩酸グアニジンである。図10はVEGFの再折り畳みに対する尿素と還元剤DTTの効果を示す。ピーク3のVEGFは適切に折り畳まれた生物学的に活性なVEGFを指す。
【0048】
示すように、場合によって溶液も還元剤を含む。還元剤は、およそ0.5からおよそ10mMのシステイン、0.1〜1.0mMのDTT、及び/又はおよそ0.2mM未満のBMEの濃度範囲での可溶化工程について上記のものから好適に選択される。本発明の一実施態様では、還元剤はおよそ0.5〜2mMのDTTである。本発明の一実施態様では、還元剤はおよそ0.5mMのDTTである。 適切な還元剤の例には、限定するものではないが、例えばジチオトレイトール(DTT)、β-メルカプトエタノール(BME)、システイン、DTEなどが含まれる。DTTおよびBMEは、通常はヘパリン結合タンパク質のために本明細書中で提供される手順と関連して用いられうるのに対して、VEGFの回収のためには本明細書中に記載のようにおよそ0.1からおよそ10mMのシステインと、およそ0.1からおよそ1.0mMのDTTの組み合わせが一例である。
【0049】
再折り畳み工程には、可溶化されたタンパク質の完全な再折り畳みを達成するために十分な濃度で、硫酸化されたポリ陰イオン剤が含まれる。好適なポリ陰イオン剤の例は上記のもの、例えば硫酸化ポリ陰イオン剤、例えばヘパリン硫酸、デキストラン硫酸、ヘパリン硫酸及びコンドロイチン硫酸並びに硫酸化ヘパリン断片を有する上記のポリ糖類の硫酸誘導体である。本発明の関係において用いるヘパリン硫酸では、通常、分子量は、およそ3000から10000ダルトンの間、又はおよそ3000から6000ダルトンの間である。
【0050】
本発明の一実施態様では、デキストラン硫酸は本発明の関係において使用される。本発明で使用されるデキストラン硫酸のような硫酸化されたポリ陰イオン剤又は他の薬剤の分子量は、回収される特定のヘパリン結合タンパク質のサイズに依存する。通常、およそ3000から10000ダルトンの間のデキストラン硫酸が使用される。本発明の一実施態様では、およそ5000ダルトンから10000ダルトンの間のデキストラン硫酸が、例えばVEGFの回収のために使われる。他の実施態様では、およそ5000から8000ダルトンの間のデキストラン硫酸が、HBPの回収のために使われる。図3A−3Dは、ヘパリン親和性クロマトグラフィによって分析した、様々な濃度及び分子量のデキストラン硫酸(図3A−C)およびヘパリン(図3D)によるVEGFの回収を示す。ピーク3は適切に折り畳まれたVEGFに対応する。
【0051】
使用されるポリ陰イオン化合物の濃度は、回収されるタンパク質およびその濃度および折り畳みバッファの温度とpHなどの条件に依存する。代表的な濃度は、およそ50〜500mMの間の硫酸ナトリウム、およそ10〜200μg/mlの6000ダルトンのヘパリンなどの低分子量ヘパリン(Sigma Chemical Co.)、およそ10〜200μg/mlの間のブタヘパリンI-Aなどの高分子量ヘパリン(Sigma Chemical Co.)、及びおよそ10〜400μg/mlの間又はおよそ10〜200μg/mlの間のデキストラン硫酸である。
【0052】
場合によって、再折り畳みバッファは、TRITONTMX-100、NONIDETTMP-40、TWEENTMシリーズ及びBRIJTMシリーズなどの様々な非イオン性界面活性剤のいずれかなどの付加的な薬剤を含みうる。非イオン性界面活性剤はおよそ0.01%〜1.0%の間で存在する。ある例では、非イオン性界面活性剤の濃度はおよそ0.025%から0.05%の間、又はおよそ0.05%である。
場合によって、陽性荷電アミノ酸、例えばアルギニン(例えばL-アルギニン/HCl)、リジンなどは再折り畳みバッファに存在してもよい。本発明のある実施態様では、アルギニンの濃度は、例えばおよそ0〜1000mM、又はおよそ25〜750mM、又はおよそ50〜500mM、又はおよそ50〜250mM、又はおよそ100mM終濃度などである。本発明のある実施態様では、タンパク質は、0.5〜3M 尿素、0〜30mg/L デキストラン硫酸、0〜0.2%トリトンX-100、2〜15mM システイン、0.1〜1mM DTTおよび0〜750mM アルギニン終濃度を含む、pH7.0〜9.0の緩衝溶液中にある。一実施態様では、50mM HEPPSが使われる。一実施態様では、再折り畳みバッファ溶液の終濃度は、1M 尿素、50mM HEPPS、15mg/L デキストラン硫酸、0.05%トリトンX-100、7.5mM システイン、100mM アルギニン、pH8.0である。一実施態様では、再折り畳みバッファ溶液の終濃度は、1.3M 尿素、50mM HEPPS、15mg/L デキストラン硫酸、0.05%トリトンX-100、7.5mM システイン、0.5mM DTT、100mM アルギニン、pH8.0である。
【0053】
ヘパリン結合タンパク質の回収と精製
培養物からのヘパリン結合タンパク質の回収と精製は、例えば、塩と溶媒分別法、コロイド性物質による吸着、ゲル濾過、イオン交換クロマトグラフィ、親和性クロマトグラフィ、免疫親和性クロマトグラフィ、電気泳動法および高性能液体クロマトグラフィ(HPLC)などの様々な方法及び公知の手法をタンパク質の分離のために実施することができるが、前記の再び折り畳まれたヘパリン結合タンパク質を、ヒドロキシアパタイトクロマトグラフィ支持体;第一疎水性相互作用クロマトグラフィ支持体、陽イオンクロマトグラフィ支持体および第二疎水性相互作用クロマトグラフィ支持体に順次接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出させることを含む4工程のクロマトグラフィの手順の例が記載される。あるいは、前記の再び折り畳まれたヘパリン結合タンパク質を陽イオン交換支持体;疎水性相互作用クロマトグラフィ支持体、およびイオン交換クロマトグラフィ支持体に順次接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出することを含む他のクロマトグラフィ手順が記載される。いずれかの手順の工程はいずれの順序でも行うことができると理解される。本発明の一実施態様では、工程は順次行われる。
【0054】
ヘパリン結合タンパク質の更なる回収と精製における適切な第一工程は、典型的に、ヘパリン結合タンパク質の濃縮と試料体積の減少をもたらす。例えば、上記の第二インキュベート工程により、回収されたヘパリン結合タンパク質の容量が大幅に増加し、再折り畳みバッファでのタンパク質が同時に希釈される。適切な第一クロマトグラフィ支持体は、回収されたヘパリン結合タンパク質の容量の減少をもたらし、望ましくない混在タンパク質からタンパク質をいくらか精製する。好適な第一クロマトグラフィ工程には、溶出され、第一疎水性相互作用クロマトグラフィ支持体に直接流し込まれうるクロマトグラフィ支持体が含まれる。例えば、ヘパリン結合タンパク質が疎水性相互作用クロマトグラフィ支持体を流すために適切な高塩濃度に溶出されうるクロマトグラフィ支持体が使われる。
【0055】
例示的な第一クロマトグラフィ支持体には、限定するものではないが、ヒドロキシアパタイトクロマトグラフィ支持体、例えばCHTセラミックI型およびII型(形式的にMacroPrepセラミックとして知られる)、バイオゲルHT、バイオゲルHTP、Biorad, Hercules, CA、など;銅、ニッケルなどの固定した金属イオンの不活性な樹脂からなる金属キレート化クロマトグラフィ支持体;並びに非誘導体化シリカゲルが含まれる。本発明の一実施態様では、VEGFの精製および回収のための第一クロマトグラフィ支持体は、ヒドロキシアパタイトクロマトグラフィ支持体である。本発明の他の一実施態様では、VEGFの精製および回収のための第一クロマトグラフィ支持体は、陽イオン交換支持体であり、例えば更に詳細に後述する。
【0056】
第一クロマトグラフィ支持体からの溶出は当分野の標準的な手法にしたがって達成される。好適な溶出条件とバッファは、下記のような第一疎水性相互作用クロマトグラフィ支持体に直接溶出したHPBの負荷(loading)を促すであろう。
疎水性相互作用クロマトグラフィは当分野で周知であり、「クロマトグラフィ支持体」に付着した疎水性リガンドと相互作用する分子の疎水性部分の相互作用に基づく。基質にカップリングした疎水性リガンドは、HICクロマトグラフィ支持体、HICゲル又はHICカラムなどとさまざまに称される。さらに、タンパク質とHICとの相互作用の強度は、タンパク質上の極性表面に対する非極性表面の割合に依存するだけでなく、同様に非極性表面の関与にも依存する。
【0057】
多くの基質は、HICカラムの調製に使用されてもよい。アガロースが最も広く用いられるが、シリカや有機ポリマーの樹脂も用いられてもよい。有用な疎水性リガンドには、限定するものではないが、ブチル-、プロピル-、又はオクチル-などのおよそ2からおよそ10の炭素原子を有するアルキル基、又はフェニルなどのアリル基が含まれる。ゲルやカラムのための従来のHIC支持体は、製品名ブチル-SEPHAROSETM、フェニル-SEPHAROSETM CL-4B、オクチルSEPHAROSETM FF及びフェニルSEPHAROSETM FFとしてGE Healthcare, Uppsala, Swedenから、製品名TOYOPEARLTM ブチル650M (フラクトゲルTSK ブチル-650)又はTSK-GELフェニル5PWとしてTosoh Corporation, Tokyo, Japanなどの供給元から市販されうる。一実施態様では、VEGFの精製および回収は、ブチル-アガロースである第一HICクロマトグラフィ支持体と、フェニルアガロースである第二疎水性クロマトグラフィ支持体である。他の実施態様では、第一HICクロマトグラフィ支持体はフェニルアガロースである。
【0058】
リガンド密度は、タンパク質の相互作用の強度だけでなく同様にカラムの許容量に影響する重要なパラメーターである。市販のフェニル又はオクチルフェニルゲルのリガンド密度はおよそ5〜40μmol/mlゲル板である。ゲル容量は、対象の特定のタンパク質並びにpH、温度および塩濃度に依存するが、一般に、3〜20mg/mlゲルの範囲内であると思われる。
特定のゲルの選択は当業者により決定されうる。通常、タンパク質およびHICリガンドの相互作用の強度はアルキルリガンドの鎖長によって増加するが、およそ4からおよそ8の炭素原子を有するリガンドがほとんどの分離に適する。フェニル基はペンチル基とほぼ同じ疎水性を有するが、選択性はタンパク質の芳香族基とのπ-π相互作用の可能性のために異なりうる。
【0059】
HICカラムに対するタンパク質の吸着は高塩濃度によるものであるが、有効濃度はタンパク質の性質及び選択された特定のHICリガンドに応じて広範囲で変化しうる。一般におよそ1から4Mの間の塩濃度が有用である。
順に又は勾配の形態のいずれであっても、HIC支持体からの溶出は、例として、a) 塩濃度を変える、b) 溶媒の極性を変える、又はc) 界面活性剤を加えるなどの様々な方法で達成されうる。塩濃度を下げることによって、吸着したタンパク質は疎水性を増すに従って溶出される。極性の変化は、エチレングリコール又はイソプロパノールなどの溶媒の添加とこれによる疎水性相互作用の強度の低減に影響されうる。界面活性剤は、タンパク質のディスプレーサーとして機能し、膜タンパク質の精製と関連して主に使われていた。
【0060】
様々な陰イオン性成分は、クロマトグラフィの陽イオン支持体を形成するために基質に接着されうる。陰イオン性成分には、カルボキシメチル、硫黄エチル基、スルホプロピル基、リン酸およびスルホン酸(S)が含まれる。セルロースのイオン交換樹脂、例として、SE52 SE53、SE92、CM32、CM52、CM92、P11、DE23、DE32、DE52、EXPRESS IONTMSおよびEXPRESS IONTMCは、Whatman LTD, Maidstone Kent U.Kから入手可能である。SEPHADEXTMおよびSEPHAROSETMベースの、相互結合のイオン交換体は、製品名CM SEPHADEXTMC-25、CM SEPHADEXTMC-50、及びSP SEPHADEXTMC-25 SP SEPHADEXTMC-50及びSP-SEPHAROSETM高性能、SP-SEPHAROSETM高速流量、SP-SEPHAROSE XL、CM-SEPHAROSETM高速流量、及びCM-SEPHAROSETMCL-6Bとしても知られており、すべてGE Healthcareから入手できる。本発明を行うためのイオン交換体の例には、限定するものではないが、例えば製品名MACROPREP、例えば、BioRad, Hercules, CAのMACROPREPTMS支持体、MACROPREPTMハイS支持体およびMACROPREPTMCM支持体のイオン交換体が含まれる。
【0061】
陽イオンクロマトグラフィ支持体からの溶出は、通常、塩濃度を増やすことによって達成される。イオンカラムからの溶出物は塩の添加を伴い、上記のように、HICは塩濃度に増強されるので、HIC工程の後にイオン工程又は他の塩工程を取り入れることもある。本発明の一実施態様では、陽イオン交換クロマトグラフィの工程はHIC工程に先行する。
【0062】
VEGFを精製するための方法の例は以下に記載しており、例えば実施例VおよびVIを参照のこと。再折り畳みの後、プール中の不溶性物質はデプス濾過によって除去される。次いで、浄化したプールを、5-mM HEPPS/0.05% TRITONTMX100/pH8で平衡化したセラミックヒドロキシアパタイト(Bio Rad, Hercules, CA)に流す。非結合タンパク質は平衡バッファにて洗浄して除去し、50mM HEPPS/0.05% TRITONTMX100/0.15M リン酸ナトリウム/pH8の均一濃度工程を用いてVEGFを溶出する。VEGFのプールを、50mM HEPPS/0.05% TRITONTMX100/0.15M リン酸ナトリウム/pH8で平衡化したブチルセファロースTM高速流量のカラム(GE Healthcare, Uppsala, Sweden)に流す。カラムは平衡化バッファにて洗浄し、VEGFはカラム溶出物に集まる。ブチルセファロースTMプールを、50mM HEPES/pH8にて平衡化したMacro PrepハイSのカラム(BioRad, Hercules, CA)に流す。溶出物を洗浄した後、280nmの吸光度を規準とし、2カラム容量の50mM HEPES/0.25M 塩化ナトリウム/pH8にて洗浄する。VEGFは、線形の、8カラム容量の0.25〜0.75M 塩化ナトリウムの濃度勾配を含む50mM HEPES/pH8を用いて溶出する。分画を回収し、ヘパリン-結合アッセイによって測定されるように、適切に折り畳まれたVEGFを含むものをプールする。
【0063】
Macro PrepハイSプールは、等量の50mM HEPES/0.8M クエン酸ナトリウム/pH7.5にて調整する。次いで、調整したプールを、50mM HEPES/0.4M クエン酸ナトリウム/pH7.5にて平衡化したフェニル5PW TSKのカラム(Tosohaas, Montgomeryville, PA)に流す。平衡バッファをカラムに通して非結合タンパク質を洗浄した後、10カラム容量の0.4−0M クエン酸ナトリウム勾配を含む50mM HEPES、pH7.5を用いてVEGFを溶出する。SDS‐ポリアクリルアミドゲル電気泳動によって分画をアッセイし、十分な純度のVEGFを含有するものをプールする。
【0064】
宿主細胞におけるヘパリン結合タンパク質の発現
簡潔に言えば、自律複製が可能な発現ベクターと宿主の原核生物細胞ゲノムと関連するタンパク質発現を宿主細胞に導入する。適する発現ベクターの構築は、本明細書中に記載のヘパリン結合タンパク質のヌクレオチド配列を含め、当分野で周知である。例えば、Sambrook等, Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press (Cold Spring Harbor, New York) (2001);Ausubel等, Short Protocols in Molecular Biology, Current Protocols John Wiley and Sons (New Jersey) (2002);及びBaneyx, (1999) Current Opinion in Biotechnology, 10:411-421を参照。細菌を含む適切な原核生物細胞、発現ベクターは、例えばAmerican Type Culture Collection (ATCC), Rockville, Marylandにより市販されている。原核細胞、特に細菌細胞培養物の大規模増殖のための方法は当分野で周知であり、これらの方法が本発明の関係において用いられうる。
【0065】
例えば、原核生物宿主細胞に対象のヘパリン結合タンパク質をコードする発現ないしはクローニングベクターを形質移入し、プロモーターの誘導、形質転換体の選別、又は所望の配列をコードする遺伝子の増幅に応じて変更した従来の栄養培地中で培養する。対象のポリペプチドをコードする核酸は、対象のポリペプチド(一又は複数)をコードする限りにおいて、好ましくは任意の供給源由来のRNA、cDNA又はゲノムDNAである。微生物宿主において異種性ポリペプチド(その変異体を含む)の発現に適する核酸を選別するための方法は周知である。ポリペプチドをコードする核酸分子は当分野で公知の様々な方法によって調整する。例えば、VEGFをコードするDNAは、例えば、VEGFをコードする遺伝子に特異的に結合することができるオリゴヌクレオチドプローブを用いて、単離し、配列決定する。
【0066】
異種の核酸(例えば、cDNA又はゲノムDNA)は、適切なプロモーターの制御の下、微生物における発現のための複製可能なベクターへ適切に挿入される。多くのベクターは、かかる目的のために利用可能であり、適当なベクターの選択は、主としてベクター内に挿入されるべき核酸のサイズ、及びベクターにより形質転換される特定の宿主細胞に依存する。各ベクターは、それが適合する特定の宿主細胞に依存する種々の成分を含む。特定の宿主の種類に依存し、一般にベクター成分には、限定はしないが、以下の一又は複数が含まれる:シグナル配列、複製開始点、一又は複数のマーカー遺伝子、プロモーター、及び転写終結配列。
【0067】
一般には、宿主細胞と適合性のある種に由来するレプリコン及び制御配列を含むプラスミドベクターが、微生物宿主との関連で用いられる。そのベクターは、通常、複製部位、並びに形質転換細胞において表現型の選択を提供可能なマーキング配列を保持する。例えば、大腸菌は、典型的には、E.coli種由来のプラスミドであるpBR322を使って形質転換される(例えば、Bolivar等、Gene, 2: 95 (1977)参照)。pBR322は、アンピシリン及びテトラサイクリン耐性の遺伝子を含んでおり、よって形質転換細胞を同定するための簡単な手段を提供する。そのpBR322プラスミド、もしくは他の微生物プラスミド又はファージもまた、選択マーカー遺伝子の発現のために宿主によって使用され得るプロモーターを含むか、又は含むよう改変される。
【0068】
(i) シグナル配列
本発明のポリペプチドは、直接産生されるだけではなく、典型的にはシグナル配列あるいは成熟ポリペプチドないしはポリペプチドのN末端に特異的切断部位を有する他のポリペプチドである異種性ポリペプチドとの融合ポリペプチドとしても産生される。典型的には選択された異種性シグナル配列は、宿主細胞によって認識されプロセシングされる(すなわち、シグナルペプチダーゼによって切断される)ものである。天然のポリペプチドシグナル配列を認識せずプロセシングしない原核生物宿主細胞に対しては、シグナル配列は、例えばアルカリホスファターゼ、ペニシリナーゼ、lppあるいは熱安定なエンテロトキシンIIリーダーの群から選択される原核生物シグナル配列により置換できる。
【0069】
(ii) 複製開始点成分
発現ベクターは、一又は複数の選択された宿主細胞においてベクターの複製を可能にする核酸配列を含む。そのような配列は様々な細菌についてよく知られている。プラスミドpBR322に由来する複製開始点は大腸菌などの大部分のグラム陰性細菌に好適である。
【0070】
(iii) 選択遺伝子成分
通常、発現ベクターは、選択可能マーカーとも称される選択遺伝子を含む。この遺伝子は、選択培地中で増殖する形質転換された宿主細胞の生存又は増殖に必要なタンパク質をコードする。選択遺伝子を含むベクターで形質転換されない宿主細胞は、培地中で生存できない。この選択可能マーカーは、本発明で利用され、定義されるような遺伝学的マーカーとは異なる。典型的な選択遺伝子は、(a)例えば、アンピシリン、ネオマイシン、メトトレキセートあるいはテトラサイクリンのような抗生物質又はその他の毒素に耐性を付与し、(b)遺伝学的マーカー(一又は複数)の存在によって誘導される欠陥以外の栄養要求性欠陥を補い、又は(c)例えばバチラス菌に対するD-アラニンラセマーゼをコードする遺伝子のような、複合培地から得られない重要な栄養素を供給する、タンパク質をコードする。
【0071】
選択技術の一例においては、宿主細胞の増殖を抑止する薬物が用いられる。この場合、対象の核酸で首尾よく形質転換したこれらの細胞は、抗薬物性を付与し、選択工程を生存するポリペプチドを産生する。このような優性選択の例としては、薬物ネオマイシン(Southern等, J. Molec. Appl. Genet, 1:327 (1982))、ミコフェノール酸(Mulligan等, Science, 209:1422 (1980))又はハイグロマイシン(Sugden等, Mol. Cell. Biol., 5:410-413 (1985))が使用される。上述の3つの例は、各々、適当な薬剤であるG418又はネオマイシン(ジェネティシン)、xgpt(ミコフェノール酸)、又はハイグロマイシンに対する耐性を伝達するために、真核生物での制御下で、細菌性遺伝子を利用する。
【0072】
(iv) プロモーター成分
対象のヘパリン結合タンパク質を産生するための発現ベクターは、宿主微生物によって認識され、対象のポリペプチドをコードする核酸と作用可能に連結される適切なプロモーターを含む。原核生物の宿主での使用に好適なプロモーターはβ-ラクタマーゼ及びラクトースプロモーター系(Chang等, Nature, 275:615 (1978); Goeddel等, Nature, 281:544 (1979))、アラビノースプロモータシステム(Guzman等, J. Bacteriol., 174:7716-7728 (1992))、アルカリホスファターゼ、トリプトファン(trp)プロモーター系(Goeddel, Nucleic Acids Res., 8:4057 (1980);欧州特許第36776号)、及びハイブリッドプロモーター、例えばtacプロモーター(deBoer 等, Proc. Natl. Acad. Sci. USA, 80:21-25 (1983))を含む。しかし、他の既知の細菌性プロモーターも適当である。それらのヌクレオチド配列は公表されており、それにより、任意の必要な制限酵素部位を供給するためのリンカー又はアダプターを用いて、当業者は対象のポリペプチドをコードするDNAにそれらを作用可能に連結する(Siebenlist等, Cell, 20:269 (1980))ことが可能となる。また、上記のSambrook等;及び上記のAusubel等も参照のこと。
【0073】
また、細菌のシステムで使用されるプロモーターも、通常、対象のポリペプチドをコードするDNAと作用可能に結合したシャイン・ダルガルノ(S.D.)配列を有する。該プロモーターは、制限酵素による切断により細菌由来のDNAから取り外すことができ、所望のDNAを含むベクター中へ挿入することができる。
【0074】
(v) ベクターの構築及び分析
一又は複数の上に列挙した成分を含む適切なベクターの構築には標準的なライゲーション技術を用いる。必要とされるプラスミドの生成のために、単離されたプラスミド又はDNA断片を切断させ、整え、そして望ましい型に再ライゲーションする。
構築されたプラスミド中において正しい配列であることを確認する分析のために、ライゲーション混合物を用いて、大腸菌K12菌株294(ATCC31446)又は他の株を形質転換し、適当な場合にはアンピシリン又はテトラサイクリン耐性によって、成功した形質転換細胞を選択する。形質転換細胞からプラスミドを調製し、制限エンドヌクレアーゼ消化により分析し、及び/又はSanger等, Proc. Natl. Acad. Sci. USA, 74:5463-5467 (1977)又はMessing等, Nucleic Acids Res., 9:309 (1981)の方法により、又はMaxam等, Methods in Enzymology, 65:499(1980)の方法により配列決定を行った。また、上記のSambrook等;及び上記のAusubel等も参照のこと。
対象のヘパリン結合タンパク質をコードする核酸は、宿主細胞に挿入される。これは、典型的に、上記の発現ベクターにて宿主細胞を形質転換させ、様々なプロモーターを誘導するために適するように変更した従来の栄養培地中で培養することによって達成される。
【0075】
宿主細胞の培養
対象のヘパリン結合タンパク質を発現するために用いる好適な原核生物細胞は当分野で周知である。典型的に、封入体の形態で、又は周辺質ないしは細胞内腔に大量に組み換えタンパク質を発現する宿主細胞が用いられる。適切な原核生物には、例として、真正細菌、例えばグラム陰性又はグラム陽性生物体、例えば大腸菌、桿菌、例えば枯草菌、シュードモナス属種、例えば、緑膿菌、ネズミチフス菌又はセラチア‐マルセスセンスなどがある。大腸菌宿主の一例は大腸菌294(ATCC 31,446)である。また、他の細胞株、例として、大腸菌B、大腸菌X1776(ATCC 31,537)および大腸菌W3110(ATCC 27,325)も好適である。これらの例は限定するものでなく例示である。株W3110は、組換えDNA産生発酵のための共通の宿主株であるので典型的な宿主である。本発明のある態様では、宿主細胞は最小量のタンパク質分解酵素を分泌する。例えば、株W3110は、タンパク質をコードする遺伝子に遺伝子変異を起こすように修飾してもよく、このような宿主の例には1995年4月25日発行の米国特許第5410026号に記載の大腸菌W3110株1A2、27A7、27B4及び27C7が含まれる。例えば、VEGFの産生のための株は、49B3と称される、遺伝子型tonAΔ ptr3 phoAΔE15 Δ(argF-lac)169 degP41 ilvgを有する大腸菌株W3110である。他の例では、VEGFの産生のための株は、遺伝子型ΔfhuA (ΔtonA) ptr3,lacIq,lacL8,ΔompT Δ(nmpC-fepE) ΔdegP ilvG+を有する大腸菌株(62A7)である。また、例えば国際公開第2004/092393号の頁23〜24にわたる表を参照のこと。
【0076】
対象のヘパリン結合タンパク質を生産するために使用される原核細胞は、当技術分野で既知の培地において成長させたもので、選択された宿主細胞の培養に適しており、一般的にSambrook等, Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press (Cold Spring Harbor, New York) (2001)に記載の培地を含む。細菌に適した培地には、AP5培地、栄養ブロス、Luria-Bertani(LB)ブロス、Neidhardt's最少培地、及びC.R.A.P.最少又は完全培地に必要な栄養を補ったものが含まれるが、これらに限定されるものではない。好ましい実施形態では、培地は発現ベクターの構造に基づいて選択された選択剤も含み、よって発現ベクターを含む原核細胞を選択的に成長させることができる。例えば、アンピシリン耐性遺伝子を発現する細胞を成長させるために、培地にアンピシリンを加える。炭素、窒素、及び無機リン酸塩の供給源に加え、必要な補充物は、単独で、或いは、別の補充物又は複合窒素源等の培地と組み合わせて導入され、適切な濃度で含有される。場合によっては、培養培地は、グルタチオン、システイン、システアミン、チオグリコール酸、ジチオエリスリトール、及びジチオスレイトールからなる群から選択された1又は複数の還元剤を含んでよい。
【0077】
適切な培地の例は、米国特許第5304472号及び同第5342763号に見ることができる。C.R.A.P.リン酸塩制限培地は、3.57gの(NH4)2(SO4)、0.71gのクエン酸Na-2H2O、1.07gのKCl、5,36gの酵母菌抽出物(認証済み)、5.36gのHycaseSFTM−Sheffieldからなり、KOHによりpHを7.3に、脱イオン化H2Oによりqsを872mlに調整したものをオートクレーブしてから55℃に冷却し、110mlの1M MOPS(pH7.3)、11mlの50%グルコース、7mlの1M MgSO4を補ったものである。次いでカルベニシリンを50μg/mlの濃度で導入培地に添加してもよい。
【0078】
原核宿主細胞を適切な温度で培養する。例えば、大腸菌の成長に好ましい温度は約20〜約39℃、又は約25℃〜約37℃、又は約30℃である。
アルカリホスフェートプロモーターが使用される場合、本発明の対象のポリペプチドの生産のために使用される大腸菌細胞は、例えばSambrook等, Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press (Cold Spring Harbor, New York) (2001)に一般的に記載されるようにアルカリホスフェートプロモーターが部分的に又は完全に誘導され得る適切な培地中で培養される。必要とされる培養は、無機リン酸塩の非存在下又はリン酸塩欠乏レベル下では、決して起こらない。第一に、培地はタンパク質合成の誘導レベル以上であり、細菌の増殖に十分な量の無機リン酸塩を含む。細胞が増殖し、リン酸塩を利用する場合、培地中のリン酸塩レベルを低下させ、それによりポリペプチドの合成の誘導を引き起こす。
【0079】
プロモーターが、起こるべき誘導に関し誘導性プロモーターである場合、典型的に細胞はある最適な濃度、例えば、高い細胞濃度過程を用い、点誘導が開始される(例えば、誘導因子の添加、培地成分の除去により)約200のA550が達成されるまで培養され、対象のポリペプチドをコードする遺伝子の発現を誘導する。
必要ないかなる補充物も、単独で、或いは又は別の補充物ないしは複合窒素源等の培地と組み合わせて導入し、当業者に既知の適切な濃度で含めることができる。培地のpHは、主に宿主生物に応じて、およそ5〜9とすることができる。大腸菌については、pHはおよそ6.8〜およそ7.4であり、又はおよそ7.0である。
【0080】
ヘパリン結合タンパク質の製剤
例えば本明細書中に記載の方法を用いて回収したポリペプチドは、薬学的に受容可能な担体に調製化してもよく、様々な診断、治療、又は該分子について公知の他の使用のために用いられる。例として、本明細書中に記載のVEGFは、酵素免疫アッセイなどのイムノアッセイに使われてもよい。本明細書中に記載の方法を用いて得られたヘパリン結合タンパク質についての治療的使用も考慮される。例として、増殖因子又はホルモン、例えばVEGFは所望のように増殖を促すために用いられうる。例として、VEGFは、例えば急性の創傷(例えば熱傷、外科的創傷、通常の創傷など)又は慢性の創傷(例えば糖尿病性潰瘍、褥瘡、褥瘡性潰瘍、静脈潰瘍など)の創傷治癒の促進、発毛促進、組織増殖及び修復(例えば骨、肝臓など)などのために用いられうる。
【0081】
ヘパリン結合タンパク質の治療製剤は、任意の薬学的に許容可能な担体、賦形剤又は安定剤と所望の純度を有する分子、例えばポリペプチドを混合することによって(Remington's Pharmaceutical Sciences, 16版, Osol,A編 [1980])、凍結乾燥製剤又は水溶液の形態で保存用に調製される。許容できる担体、賦形剤又は安定剤は、用いる投与量及び濃度では細胞に対して無毒性であり、リン酸、クエン酸及び他の有機酸等の緩衝液;アスコルビン酸及びメチオニンを含む抗酸化剤;防腐剤(例えば、オクタデシルジメチルベンジルアンモニウムクロリド;塩化ヘキサメトニウム;塩化ベンザルコニウム、塩化ベンゼトニウム;フェノール、ブチル又はベンジルアルコール;アルキルパラベン類、例えばメチル又はプロピルパラベン;カテコール;レゾルシノール;シクロヘキサノール;3-ペンタノール;及びm-クレゾール);低分子量(残基数10個未満)ポリペプチド;血清アルブミン、ゼラチン又は免疫グロブリン等のタンパク質;ポリビニルピロリドン等の親水性重合体;グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン、又はリシン等のアミノ酸;グルコース、マンノース又はデキストリン等の単糖類、二糖類及び他の炭水化物;EDTA等のキレート剤;スクロース、マンニトール、トレハロース又はソルビトール等の糖類;ナトリウム等の塩形成対イオン;金属錯体(例えばZn-タンパク質錯体);及び/又はTWEENTM、PLURONICSTM又はポリエチレングリコール(PEG)等の非イオン性界面活性剤を含む。
【0082】
ある実施態様では、インビボ投与のために使用される製剤は無菌である。これは滅菌濾過膜による濾過によって達成される。HBPは、凍結乾燥された形態、又は、水溶液又はゲル形態で保存されうる。より高い又はより低いpH値でも好適な場合があるが、HBP調製物のpHはおよそ5〜8まででありうる。賦形剤、担体、又は安定剤の使用によりHBPの塩類が形成されうることは理解されるであろう。
典型的に創傷治癒のために、HBPは部位特異的運搬用に調製される。局所に適応される場合、HBPは担体及び/又はアジュバントなどの他の成分と適切に組み合わされる。前記の他の成分の性質に制限はないが、ただし意図する投与のために効果的かつ薬学的に受容可能でなければならず、組成物の活性成分の活性を有意に分解することができないものである。精製されたコラーゲンの有無にかかわらず、適切な溶媒の例には、軟膏、クリーム、ゲル、噴霧又は懸濁液が含まれる。また、組成物は、場合によって液体又は半流動体形態で、滅菌包帯、経皮パッチ、絆創膏及び包帯にしみ込ませてもよい。
【0083】
ゲル製剤を得るため、液体組成物に調製されるHBPは、有効量の水溶性多糖類又は合成ポリマー、例としてポリエチレングリコールと混合して、適切な粘性のゲルを形成して、局所に適応してもよい。使われうる多糖類には、例えば、エーテル化したセルロース誘導体などのセルロース誘導体、例えば、アルキルセルロース、ヒドロキシアルキルセルロース、及びアルキルヒドロキシアルキルセルロース、例えばメチルセルロース、ヒドロキシエチルセルロース、カルボキシメチルセルロース、ヒドロキシプロピルメチルセルロース及びヒドロキシプロピルセルロース;澱粉及び分画された澱粉;寒天;アルギン酸及びアルギナート;アラビアゴム;プルラン;アガロース;カラギーナン;デキストラン;デキストリン;フルクタン;イヌリン;マンナン;キシラン;アラビナン;キトサン;グリコーゲン;グルカン;及び、合成バイオポリマー;並びに、キサンタンガムなどのガム;ガーゴム;ローカストビーンガム;アラビアゴム;トラガカントゴム;及び、インドゴム;及び、その誘導体及び混合物が含まれる。本発明のある実施態様では、本明細書中のゲル化剤は、例えば、生物系に不活性、非中毒性、調製がしやすく、及び/又はあまり流れやすくないか粘性がなくて、その中に含まれるHBPを不安定にしないものである。
【0084】
ある実施態様では、多糖は、エーテル化されたセルロース誘導体であって、他の実施態様では、十分に限定され、精製され、USPに列挙されるもの、例えばメチルセルロース及びヒドロキシアルキルセルロース誘導体、例として、ヒドロキシプロピルセルロース、ヒドロキシエチルセルロース及びヒドロキシプロピルメチルセルロースである。一実施態様では、メチルセルロースが多糖である。メチルセルロースがゲルに使用される場合、典型的には、ゲルのおよそ2〜5%、又はおよそ3%、又はおよそ4%、又はおよそ5%を含んでなり、HBPはゲルの1ml当たりおよそ300〜1000mgの量で存在する。
ゲル化のために有用なポリエチレングリコールは、一般的に、適当な粘性を得るために低分子量と高分子量のポリエチレングリコールの混合物である。この目的のために、例えば、分子量400〜600のポリエチレングリコールと分子量1500のポリエチレングリコールとの混合物は、ペーストを得るために適切な比率で混合される場合に有効である。
【0085】
多糖類及びポリエチレングリコールに用いる「水溶性」なる用語はコロイド溶液及び分散液を含むことを意味する。通常、セルロース誘導体の溶解性はエーテル基の置換の程度で決定され、本明細書において有用な安定化誘導体は誘導体を水溶性にするためにセルロース鎖のアンヒドログルコース単位当たりの該エーテル基を十分な量有しなければならない。一般的に、アンヒドログルコース単位当たり少なくとも0.35のエーテル基のエーテル置換の程度が十分である。さらに、セルロース誘導体は、アルカリ金属塩、例えばLi、Na、K又はCs塩類の形態であってもよい。
【0086】
例えば、Remington's Pharmaceutical Sciences 18th edition, Gennaro, A. Ed. (1995)を参照。またJohnson等, Nat. Med., 2:795-799 (1996);Yasuda, Biomed. Ther., 27:1221-1223 (1993);Hora等, Bio/Technology, 8:755-758 (1990);Vaccine Design: The Subunit and Adjuvant Approach, Powell and Newman, eds, (Plenum Press: New York, 1995)のCleland, "Design and Production of Single Immunization Vaccines Using Polylactide Polyglycolide Microsphere Systems," pp. 439-462;国際公開第97/03692号、国際公開第96/40072号、国際公開第96/07399号;米国特許第5654010号;DE第3218121号;Epstein等, (1985) Proc. Natl. Acad. Sci. USA, 82:3688-3692;Hwang等, (1980) Proc. Natl. Acad. Sci. USA, 77:40304034;欧州特許第52322号;欧州特許第36676号;欧州特許第88046号;欧州特許第143949号;欧州特許第142641号;日本特許出願83-118008;米国特許第4485045号及び同第4544545号;及び欧州特許第102324号も参照のこと。
以下の実施例は、限定するものではなく例示するためのものである。
【実施例】
【0087】
実施例I:大腸菌において発現した組み換えヒトVEGF
組み換えヒトVEGFを大腸菌において発現させた。合成の間、タンパク質は細胞膜周辺腔に分泌され、屈折小体として蓄積された。したがって、タンパク質の抽出および再折り畳みを達成するために研究を行った。これらの研究により、ポリ陰イオン剤を付加することなく標準的な回収技術を用いて少なくとも3種のVEGF(図1)が単離されたことが明らかとなった。天然のVEGFを用いた研究により、ヘパリン添加がカオトロプ-及びチオール-誘導性の変性剤に対する耐性を増すことが示された(図2)。さらに、ヘパリンは、小規模再折り畳み実験において適切に再折り畳みされたVEGFの量を有意に増やした。この結果を大規模な方法に応用するために、ヘパリンに構造上類似する分子であるデキストラン硫酸の存在下においてVEGFの再折り畳みを可能にした条件を探した。デキストラン硫酸の添加により、適切に折り畳まれた生物学的に活性なVEGFの収率がコントロールと比べて3〜5倍に増加した。
【0088】
方法
VEGF165発現のためのプラスミド−プラスミドpVEGF171は大腸菌周辺質におけるヒトVEGF165の発現のために設定した(例えば、Leung等, (1989) Science, 246:1306-1309を参照)。VEGFコード配列の転写は、アルカリホスファターゼ(AP)プロモーターの厳格なコントロール下にあのに対して(例えば、Kikuchi等, (1981) Nucleic Acids Research, 9:5671-8を参照)、翻訳開始に必要な配列はシャイン-ダルガーノ領域にある(例えば、Yanofsky等, (1981) Nucleic Acids Research, 9:6647-68を参照)。VEGFコード配列は、その後の大腸菌周辺質への分泌のために細菌性耐熱性毒素II(STII)シグナル配列の下流に融合した(例えば、Lee等, (1983) Infect. Immun. 42:264-8;及びPicken等, (1983) Infect. Immun. 42:269-75を参照)。STIIシグナル配列のコドン修飾により翻訳レベルが調節され、その結果、周辺質におけるVEGFが最適レベルに集積した(例えば、Simmons and Yansura, (1996) Nature Biotechnoloy, 14:629-34を参照)。転写終結区に対するラムダ(例えば、Scholtissek and Grosse, (1987) Nucleic Acids Research 15:3185を参照)はVEGF翻訳終止コドンの下流に位置した。複製起点と、アンピシリン及びテトラサイクリン耐性遺伝子は、プラスミドpBR322から得た。例えば、Bolivar等, (1977) Gene 2:95-113を参照。
【0089】
細胞均質化と屈折小体の調製−回収した大腸菌細胞を凍結し、−70℃に保存した。細胞は、BTUX (centrifuge, Alfa laval)遠心にて回収し、BEPEX(大きいサイズのフリーザ)に凍結保存した。細胞は、5容量の50mM HEPES/150mM NaCl/5mM EDTA pH7.5(5L/kgペレット)に懸濁し、モデル15M実験室ホモジナイザーGaulin 15M(小型)又はM3(大型) (Gaulin Corporation, Everett, MA) にて均質化した。次いで、細胞懸濁液を等量の同じバッファにて希釈し、BTPX205の(Alfa Laval Separation AB(Tumba, Sweden)連続速度遠心にて遠心分離して屈折小体を回収した。中間生成物スケール遠心機はSA1を使用した。あるいは、細胞を均質にし、BEPEXに凍結して再水和することなく直接ペレットを回収することもできる。
【0090】
実施例II:大腸菌-I(Escherichia coli-I)において発現させた組み換えヒトVEGFの抽出及び再折り畳み
方法
抽出と再折り畳み−屈折性ペレットは、5Lのバッファ/kgペレットで、7M 尿素/50mM HEPPS/pH8(終濃度)を含有する抽出バッファに懸濁した。次いで、固形ジチオトレイトールを、4mM終濃度となるように3.7g/kgペレットで加えた。例として、VEGFの抽出に対する尿素とDTTの効果についての図9を参照のこと。懸濁液は20℃で1〜2時間かけて完全に混合した。pHは、50%水酸化ナトリウム(w/w)にてpH8.0に調整してもよい。抽出バッファの容量当たり、19容量の再折り畳みバッファを加えて再折り畳みを生じさせた。再折り畳みバッファには、50mM HEPPS/1 M−2M 尿素/2−5mM システイン/0.05%−0.2%TRITONTMX100/pH8、終濃度を含めた。例えば、再折り畳みの間に存在する尿素およびDTT濃度の効果についての図10を参照のこと。示すように、デキストラン硫酸、ヘパリン又は硫酸ナトリウムを加えた。再折り畳みインキュベーションは室温で4〜24時間行った。場合によって、インキュベートは室温で最大およそ48時間行うことができる。折畳みは、SDS-PAGEおよび/またはヘパリンHPLCにてモニターした。生成物は、Cuno90SPフィルターによるデプス濾過(depth filtration)の後に0.45μm濾過を行って浄化した。
【0091】
ヘパリン-結合HPLCアッセイ−適切に再び折り畳まれたVEGFの質および量は、固定されたヘパリンを含有するカラムを使用して決定した。カラムPOROS HE2/M(4.6×100mm、HE2/M、PerSeptive BioResearch Products, Cambridge, MA)は0.15M 塩化ナトリウムを含有する10mM リン酸ナトリウム、pH7にて平衡化した。1ml/分又は2ml/分の流速で、カラムは、10分かけて、0.15〜2M 塩化ナトリウムを含む平衡バッファから直線濃度勾配を使用して溶出した。あるアッセイでは、16分で溶出した。溶出物を280nmでモニターした。一般的に、ほとんどのタンパク質は空隙容量で溶出し、3種のVEGFが同定されうる。最も高い親和性で、最も溶出が遅い種を適切に折り畳まれたVEGFとして同定し、その後「ピーク3のVEGF」と同定した。
【0092】
結果
システインが媒介する変性からVEGFを保護するヘパリン−天然のVEGFに10mM システインを加えると、適切に折り畳まれた分子が大きく減少した(図2)。この変性は、20mMほどのわずかな濃度の2つの異なる形態のヘパリンを加えると阻止された。
【0093】
表Ia
VEGF再折り畳みに対するヘパリンとデキストラン硫酸の効果
表の値は、屈折性ペレット(g)当たりのピーク3の形成されたVEGF(mg)の量である。各々の添加の濃度は示すとおりである。*平均コントロール(5.6+5.0=5.3)
【0094】
表Ib
VEGF再折り畳みに対する硫酸ナトリウムの効果
表の値は、屈折性ペレット(g)当たりのピーク3の形成されたVEGF(mg)の量である。硫酸ナトリウムの濃度は示すとおりである。
【0095】
表II
VEGF再折り畳みに対するヘパリンとデキストラン硫酸の効果
表の値は、屈折性ペレット(g)当たりのピーク3の形成されたVEGF(mg)の量である。
【0096】
まとめ
再折り畳み収率を増加させるヘパリンおよびデキストラン硫酸−上記の変性に対する保護的特性のため、VEGFの再折り畳みに対する様々な異なる形態の硫酸化ポリマーの効果を調べた。表Ia(及び図5)に示すように、低分子量及び高分子量の形態のヘパリンは再び折り畳まれたVEGFの収率をおよそ3倍に増加した。表Ib(及び図6)に示すように、硫酸ナトリウムは再び折り畳まれたVEGFの収率をおよそ2倍に増加した。また、10Kd形態のデキストラン硫酸は再折り畳み収率の増加に有効であったが、調査した濃度の高い範囲では基質阻害が生じた。更なる研究により、5Kd、8Kd及び10Kdの形態のデキストラン硫酸はすべて、再折り畳みの際のVEGFの収率を有意に増加したことが示された。図7を参照。また図8も参照。
【0097】
実施例III:VEGFの回収に対する異なるバッファとTRITONTMX-100の効果
結果
【0098】
まとめ
実施例I、II及びIIIのデータをまとめると、ヘパリン-結合増殖因子であるVEGFが再び折り畳まれる際にヘパリン硫酸塩又はデキストラン硫酸のいずれかを含めると収率が有意に(2〜5倍)改善することと、回収条件が示唆される。この方法は工業規模で成功裏に実施された。この方法はヘパリンを結合する他の基本的な増殖因子および他のタンパク質の再折り畳みに応用することができると期待される。
【0099】
実施例IV:大腸菌-II(Escherichia coli-II)において発現される組み換えヒトVEGFの
方法
抽出と再折り畳み−屈折性ペレットを抽出バッファに懸濁した。この抽出バッファの終濃度は5Lのバッファ/kgペレットで7M 尿素、2−30mM DTT(例えば10mM DTT)、50mM HEPPS/pH7−9(例えばpH8)とした。懸濁液は室温で1〜2時間かけて完全に混合した。抽出バッファの容量当たり、19容量の再折り畳みバッファを加えて再折り畳みを生じさせた。再折り畳みバッファは、終濃度として1M又は1.3M尿素、2−15mM システイン(例えば7.5mM システイン)、0.5mM DTT、0−0.75M アルギニン(例えば100mM アルギニン)、15mg/Lのデキストラン硫酸、50mM HEPPS、0.05%TRITONTMX100/pH8を含有した。荷電アミノ酸の存在下における再折り畳みに対する効果について、例えば図12を参照のこと。ここでは、ヒスチジンの添加によりアミノ酸の付加がない場合と同じ効果が生じた。再折り畳みインキュベーションは室温で12〜24時間行った。場合によって、インキュベートは室温で最大およそ48時間行うことができる。場合によって、再折り畳みの間に空気又は酸素を添加してもよい(0.3〜1cc/分/L)。折畳みは、SDS-PAGEおよび/またはヘパリンHPLCにてモニターした。生成物は、Cuno90SPフィルターによるデプス濾過(depth filtration)の後に0.45μm濾過を行って浄化した。
【0100】
抽出および再折り畳みの工程の全体の希釈物は1:100とした。抽出及び再折り畳みの工程の全体の希釈の、例えば1:100へ、1:200への増加により活性なVEGFの総量が増加したが、濃度は低い。図13を参照。
再折り畳みの効率は、二量体/単量体の量を測定することによって決定することができ、このとき単量体は、C18逆相HPLCカラムで測定することができ、二量体形成は、ヘパリンカラムクロマトグラフィ又はSP-5PW陽イオン交換クロマトグラフィアッセイで測定することができる。
【0101】
実施例V:大規模な再折り畳み
最適化の再折り畳み条件の拡張性を試験するため、小規模(0.1L)、中規模(1L)及び試験的設備(250L〜400L)のスケールでの再折り畳みの動態を調べるために実験を行った。図4に示すように、大規模スケールの再折り畳みの動態はより小さいスケールと違いがなく、再び折り畳まれたVEGFの最終力価はわずかに増加した。これらのデータから、デキストラン硫酸による再折り畳みの拡張性が示された。さらに、生成物は、Cuno90SPフィルターによるデプス濾過(depth filtration)の後に0.45μm濾過を行って浄化した。
【0102】
実施例VI:再折り畳み後のrhVEGFの精製I
MacroPrepセラミックヒドロキシアパタイトクロマトグラフィ−再折り畳みの後、プール中の不溶性物質をデプス濾過によって除去した。次いで、浄化したプールを、50mM HEPPS/0.05% TRITONTMX100/pH8で平衡化したセラミックヒドロキシアパタイトカラム(35D×31H=30L) (Bio Rad, Hercules, CA)に流した。非結合タンパク質は平衡バッファにて洗浄して除去し、50mM HEPPS/0.05% TRITONTMX100/0.15M リン酸ナトリウム/pH8の均一濃度工程を用いてVEGFを溶出した。流速は120cm/時とした。プールする分画を分画のヘパリンHPLC分析にて測定した。
【0103】
ブチルセファロースTM高速流量クロマトグラフィ−VEGFのプールを、50mM HEPPS/0.05% TRITONTMX100/0.15M リン酸ナトリウム/pH8で平衡化したブチルセファロースTM高速流量のカラム(35D×26H=25L)(GE Healthcare, Uppsala, Sweden)に流した。流速は100cm/時とした。カラムは平衡化バッファにて洗浄し、VEGFはカラム溶出物に集まった。分画を集め、タンパク質含有分画をプールし、A280nmを測定した。
【0104】
Macro PrepハイSクロマトグラフィ−ブチルセファロースTMプールを、50mM HEPES/pH8にて平衡化したMacro PrepハイSのカラム(30D×39H=27 L)(BioRad, Hercules, CA)に流した。溶出物を洗浄した後、280nmの吸光度を規準とし、2カラム容量の50mM HEPES/0.25M 塩化ナトリウム/pH8にて洗浄した。VEGFは、線形の、8カラム容量の0.25〜0.75M 塩化ナトリウムの濃度勾配を含む50mM HEPES/pH8を用いて溶出した。流速を75〜200cm/時とした。分画を回収し、ヘパリン-結合アッセイ、例えばヘパリンHPLCによって測定したように、適切に折り畳まれたVEGFを含むものをプールした。
【0105】
フェニル5PW TSKクロマトグラフィ−Macro PrepハイSプールは、等量の50mM HEPES/0.8M クエン酸ナトリウム/pH7.5にて調整した。次いで、調整したプールを、50mM HEPES/0.4 M クエン酸ナトリウム/pH7.5にて平衡化したフェニル5PW TSKのカラム(18D×43H=11L)(Tosohaas, Montgomeryville, PA)に流した。平衡バッファをカラムに通して非結合タンパク質を洗浄した後、10カラム容量の0.4−0M クエン酸ナトリウム勾配を含む50mM HEPES、pH7.5を用いてVEGFを溶出した。SDS‐ポリアクリルアミドゲル電気泳動によって分画をアッセイし、十分な純度のVEGFを含有するものをプールした。
【0106】
限外濾過/ダイアフィルトレーション−プールしたVEGFを5kD再生セルロースメンブレン(G30619);Unit Pellicon;Feed Rate 17.1L/分に限外濾過した。ポリソルベート20にてメンブレンを調整した。プールしたVEGFは、6g/L(UF1)の濃度で限外濾過した。試料は、5mM コハク酸ナトリウム/275mM トレハロース/pH5.0と7−14DV(Diavolume)にてダイアフィルトレートした。最終製剤は、5mg/mlの濃度の、5mM コハク酸ナトリウム/275mM トレハロース/0.01%ポリソルベート20/pH5.0であった。
【0107】
実施例VII:再折り畳み後のrhVEGFの精製II
陽イオン交換液体クロマトグラフィ−再折り畳みの後、プール中の不溶性物質をデプス濾過によって除去することができる。再び折り畳まれたプールは、pH5.0−7.5、及び2〜6.5mS/cmに調整する。一実施態様では、プールはpH6.5および5mS/cmに調整している。次いで、再び折り畳まれたプールは、スルホプロピル高度負荷カラム(sulfopropyl extreme load column、SPXL)に流し、漸増塩濃度勾配により溶出した。プールした分画を、分画のヘパリンHPLC分析で測定することができる。
【0108】
疎水性相互作用カラム(HIC):VEGFのSPXL溶出プールを、フェニルTSKクロマトグラフィカラム(Tosohaas, Montgomeryville, PA)に流すために、50mS/cmに調整してもよい。分画を集め、タンパク質含有分画をプールする。
【0109】
IEX又は混合モード:フェニルTSKプールを、イオン交換クロマトグラフィ(IEX)又は混合モードクロマトグラフィのカラムに流してもよい。分画を集め、本明細書に記載のアッセイによって決定した適切に折り畳まれたVEGFを含むものをプールした。
【0110】
限外濾過/ダイアフィルトレーション−プールしたVEGFを5kD再生セルロースメンブレン(G30619);Unit Pellicon;Feed Rate 17.1L/分に限外濾過してもよい。例えば、ポリソルベート20にてメンブレンを調整する。プールしたVEGFは、6g/L(UF1)の濃度で限外濾過する。試料は、5mM コハク酸ナトリウム/275mM トレハロース/pH5.0と7−14DV(Diavolume)にてダイアフィルトレートする。
【0111】
本明細書に記載の方法及びプロセスでは、最終純度及び/又は活性は、ペプチドマッピング、ジスルフィドマッピング、SDS-PAGE(還元及び非還元の両方)、円二色性、リムルスアメーバ様細胞可溶化液(LAL)、ヘパリンクロマトグラフィ、ヘパリンHPLC(例えば、ヘパリンHPLCを用いてVEGF二量体濃度を決定することができる)、逆相(rp)HPLCクロマトグラフィ(例えば、rpHPLCを用いてVEGF単量体の濃度を決定することができる)、ヘパリン結合、レセプター結合(例えば、VEGF、例えばKDRレセプター結合-Bioanalytic R&D、及び/又はFlt1レセプター結合に対して)、SEC分析、細胞アッセイ、HUVEC作用強度アッセイ、VEGF抗体を用いたELISA、質量分析などによって評価することができる。
【0112】
本明細書に記載の寄託、実施例及び実施態様は例示的な目的のみであり、この見地の様々な修飾及び変更は当業者に示唆されており、本出願の精神及び権限と添付の特許請求の範囲内に包含されることは理解されるものである。本明細書において引用されるすべての刊行物、参考文献、特許および特許出願は、すべての目的のために出典明記によりその全体がここに援用される。
【図面の簡単な説明】
【0113】
【図1】POROS HE2/Mカラム(4.6×100mm、PerSeptive BioResearch Products, Cambridge, MA)に流した細菌株W3110により産生されたVEGFのクロマトグラフを示す。例えば、POROS HE/2Mカラムは、0.15M 塩化ナトリウムを含有する10mM リン酸ナトリウム、pH7にて平衡化している。カラムは、10分かけて、0.15〜2M 塩化ナトリウムを含む10mM リン酸ナトリウム、pH7から直線濃度勾配を使用して溶出する。溶出物を280nmでモニターする。それぞれのピークで回収されたタンパク質はVEGFに対応するが、ピーク3のみは生物学的に活性な適切に再折り畳みされたVEGFに対応する。
【図2】ヘパリンによる天然の適切に折り畳まれたVEGFの安定化を表すグラフを示す。VEGFは、5mM EDTA、0.2M NaClおよび10mM システインを含有する50mM HEPES、pH8に懸濁する。
【図3A−3D】細菌株W3110によって産生され、それぞれ12μg/mlのデキストラン硫酸5000ダルトン(図3A)、12μg/mlのデキストラン硫酸8000ダルトン(図3B)、又は12μg/mlのデキストラン硫酸10000ダルトン(図3C)、又は25μg/mlのヘパリン(図3D)3000ダルトンとともにインキュベートし、POROS HE2/Mカラム(4.6×100 mm、PerSeptive BioResearch Products, Cambridge, MA)に流したVEGFのクロマトグラフを示す。例えば、カラムは、0.15M 塩化ナトリウムを含有する10mM リン酸ナトリウム、pH7にて平衡化している。カラムは、10分かけて、0.15〜2M 塩化ナトリウムを含む10mM リン酸ナトリウム、pH7から直線濃度勾配を使用して溶出する。溶出物を280nmでモニターする。それぞれのピークで回収されたタンパク質はVEGFに対応するが、ピーク3のみは生物学的に活性な適切に再折り畳みされたVEGFに対応する。
【図4】VEGFの再折り畳みに対する大きさの影響を示す。
【図5】VEGFの再折り畳みに対する低分子量(MW)及び高MW、及びデキストラン硫酸、10000ダルトンの効果を示す。ピーク3は、生物学的に活性な適切に再び折り畳まれたVEGFに対応する。
【図6】VEGFの再折り畳みに対する硫酸ナトリウムの影響を示す。ピーク3は、生物学的に活性な適切に再び折り畳まれたVEGFに対応する。
【図7】VEGFの再折り畳みに対する低分子量(MW)及び高MW、及びデキストラン硫酸、5000ダルトン、8000ダルトン、及び10000ダルトンの効果を示す。ピーク3は、生物学的に活性な適切に再び折り畳まれたVEGFに対応する。
【図8】VEGFの再折り畳みに対するヘパリンとデキストラン硫酸の効果を示す。ピーク3は、生物学的に活性な適切に再び折り畳まれたVEGFに対応する。
【図9】細菌封入体からのVEGFの抽出に対する尿素とDTTの効果を示す。
【図10】VEGFの再折り畳みに対する尿素とDTT濃度の影響を示す。
【図11】VEGF165のアミノ酸配列(配列番号:1)とともにジスルフィド結合を示す。
【図12】荷電アミノ酸の存在の効果を示す。第二緩衝溶液中0.75M濃度のアルギニンおよびリジンは有益であるのに対して、ヒスチジンはヒスチジンがない緩衝溶液と比較して、ほとんど相加的効果を有さない。さらに、アルギニンは0.1〜1M濃度で類似の効果を有することが示された。
【図13】%再折り畳み効率における希釈の効果を示すが、希釈が増すにつれて総VEGF濃度は低くなり、%再折り畳み効率は希釈が増すと高くなる。
【発明の開示】
【0001】
(関連出願)
本出願は、本明細書中にその全体が援用される2005年12月22日に出願の米国特許仮出願第60/753615号および2006年7月14日に出願の米国特許仮出願第60/807432号の優先権を主張する。
【0002】
(発明の分野)
本発明は、細胞培養で生産されるヘパリン-結合タンパク質を得る方法に関する。本発明は、原核生物宿主細胞で産生され、これらの細胞中、典型的には周辺質又は細胞内腔に存在する再び折り畳まれた(refolded)ヘパリン結合タンパク質の回収及び精製に関する方法を包含する。また、原核生物の宿主細胞で産生されるヘパリン結合タンパク質は、可溶タンパク質又は可溶性と不溶性のタンパク質の混合物として提供されてもよい。
【0003】
(発明の背景)
天然に生じ、生物学的に活性な多様なポリペプチドがヘパリンを結合することは公知である。このようなヘパリン-結合ポリペプチドには、サイトカイン、例として、血小板因子4及びIL-8(Barber等, (1972) Biochim. Biophys. Acta, 286:312-329;Handin等, (1976) J. Biol. Chem., 251:4273-422;Loscalzo等, (1985) Arch. Biochem. Biophys. 240:446-455;Zucker等, (1989) Proc. Natl. Acad. Sci. USA, 86:7571-7574;Talpas等, (1991) Biochim. Biophys. Acta, 1078:208-218;Webb等, (1993) Proc. Natl. Acad. Sci. USA, 90:7158-7162)、ヘパリン-結合増殖因子(Burgess and Maciag, (1989) Annu. Rev. Biochem., 58:576-606;Klagsbrun, (1989) Prog. Growth Factor Res., 1:207-235)、例として、上皮細胞増殖因子(EGF);血小板由来増殖因子(PDGF);塩基性線維芽細胞増殖因子(bFGF);酸性線維芽細胞増殖因子(aFGF);血管性内皮増殖因子(VEGF);及び、肝細胞増殖因子(HGF) (Liu等, (1992) Gastrointest. Liver Physiol. 26: G642-G649);そして、セレクチン、例えばL-セレクチン、E-セレクチンおよびP-セレクチン (Norgard-Sumnicht等, (1993) Science, 261:480-483)が含まれる。また、Munoz and Linhardt., (2004) Arterioscler Thromb Vasc Biol., 24:1549-1557を参照のこと。
【0004】
国際公開第95/07097号には、VEGFのようなヘパリン結合増殖因子を含むヘパリン結合タンパク質の製剤とともに、治療的使用のための精製された天然のヘパリン又は他のポリ陰イオン化合物が記載される。オリゴ糖および様々な他のポリ陰イオン化合物由来のヘパリンは、ヘパリン結合増殖因子のための活性な立体構造を安定させることが示されており(Barzu等, (1989) J. Cell. Physiol. 140:538-548;Dabora等, (1991) J. Biol. Chem. 266:23627-23640)、ヘパリン親和性クロマトグラフィは、様々な精製様式で用いられている(一般に、国際公開第96/02562号を参照)。
【0005】
哺乳類起源のヘパリン結合タンパク質の多くは組み換え技術によって産生され、臨床的に関係がある(Munoz and Linhardt, (2004) Arterioscler Thromb Vasc Biol., 24:1549-1557;Favard等 (1996) Diabetes and Metabolism 22(4): 268-73;Matsuda等, (1995) J. Biochem. 118(3): 643-9;Roberts等, (1995) Brain Research 699(1): 51-61)。例えば、VEGFは血管内皮細胞の強力な分裂促進因子である。血管透過性因子(VPF)も知られている。Dvorak等, (1995) Am. J. Pathol. 146:1029-39を参照。VEGFは、胚性脈管構造の発達である脈管形成と、既に存在する血管からの新たな血管の形成プロセスである血管形成の両方に重要な働きがある。例えば、Ferrara, (2004) Endocrine Reviews 25(4): 581-611;Risau等, (1988) Dev. Biol., 125:441-450;Zachary, (1998) Intl. J. Biochem Cell Bio 30:1169-1174;Neufeld等, (1999) FASEB J. 13:9-22;Ferrara (1999) J. Mol. Med. 77: 527-543;及び、Ferrara and Davis-Smyth, (1997) Endocri. Rev. 18: 4-25を参照。VEGFの臨床応用には、例えば、創傷治癒を促進する際(例えば、2006年6月16日出願の「Wound Healing」と題する国際公開第91/02058号(Attorney Docket No. P2358R1)を参照)、肝臓(例えば国際公開第2003/0103581号参照)、骨(例えば国際公開第2003/094617号参照)などの組織成長及び修復の際のように、新規な毛細管床の成長があるものが含まれる。また、Ferrara, (2004) Endocrine Reviews 25(4): 581-611を参照。
【0006】
典型的には、治療的に関係のある組み換えタンパク質は多くの宿主生物において産生される。ほとんどのタンパク質は、CHO細胞などの真核生物宿主においてそれらの天然の形状で発現されうる。一般に、動物細胞培養は、最大細胞密度を達成するために長い成長期間を必要とし、最終的に、原核生物の細胞培養より低細胞密度となる(Cleland, J. (1993) ACS Symposium Series 526, Protein Folding: In Vivo and In Vitro, American Chemical Society)。さらに、動物細胞培養は、所望のタンパク質の回収に干渉しうる成長成分を含有する高価な培地を必要とすることが多い。細菌宿主発現系により、組み換えタンパク質の製造規模産生の費用効果的な代替法が提供される。組み換えタンパク質の一般的な細菌発現については多くの米国特許出願が存在し、その中には、米国特許第4565785号;同第4673641号;同第4795706号;及び同第4710473号などがある。製造方法の主要な利点は、遠心分離法又は精密ろ過法によって生成物を細胞性構成成分から容易に単離することができることである。例えばKipriyanov and Little, (1999) Molecular Biotechnology, 12: 173-201;及びSkerra and Pluckthun, (1988) Science, 240: 1038-1040を参照のこと。
【0007】
酸性線維芽細胞増殖因子、塩基性線維芽細胞増殖因子及び血管内皮細胞増殖因子などの組み換えヘパリン結合増殖因子は、細菌を含む多くの供給源から回収し精製されている(Salter D.H.等, (1996) Labor. Invest. 74(2): 546-556 (VEGF);Siemeister等, (1996) Biochem. Biophys. Res. Commun. 222(2): 249-55 (VEGF);Cao等, (1996) J. Biol. Chem. 261(6): 3154-62 (VEGF);Yang等, (1994) Gaojishu Tongxun, 4:28-31 (VEGF);Anspach等, (1995) J. Chromatogr. A 711(1): 129-139 (aFGF及びbFGF);Gaulandris (1994) J Cell. Physiol. 161(1): 149-59 (bFGF);Estape and Rinas (1996) Biotech. Tech. 10(7): 481-484 (bFGF);McDonald等, (1995) FASEB J. 9(3): A410 (bFGF))。しかしながら、大腸菌などの細菌発現系はタンパク質の適切な再折り畳み(refolding)を促す細胞の機構を欠いており、一般に培養培地にほとんどのタンパク質を分泌しない。細菌性宿主細胞において発現される組み換えタンパク質は、密度の高い、部分的に折り畳まれた(folded)タンパク質と誤って折り畳まれた(misfolded)還元タンパク質からなる封入体(inclusion body)として見られることが多い。この形状において、組み換えタンパク質は、通常不活性である。例えば、VEGFの優れた活性型は、2つの165-アミノ酸ポリペプチド(VEGF-165)のホモ二量体である。この構造において、各サブユニットは、7対の鎖内ジスルフィド結合と2つのサブユニットの共有結合に影響するさらに2対を含む(Ferrara等, (1991) J. Cell. Biochem. 47:211-218)。天然の立体構造は、容易にヘパリンを結合することが示されている非常に基本的なドメインを含む(上掲のFerrara et al (1991))。VEGFの共有結合性二量体化は有効なレセプター結合と生物学的活性に必要とされる(Potgens等, (1994) J. Biol. Chem. 269:32879-32885;Claffey等, (1995) Biochim. et Biophys. Acta 1246:1-9)。細菌性生成物は潜在的にいくつかの誤って折り畳まれ、ジスルフィドが攪乱された中間生成物を含有する。
【0008】
さらに、再折り畳みは、誤って折り畳まれ、ジスルフィド結合した二量体、三量体および多量体を生産することが多い。(Morris等, (1990) Biochem. J., 268:803-806;Toren等, (1988) Anal. Biochem., 169:287-299)。この会合現象は、特により高いタンパク質濃度のタンパク質再折り畳みに非常に共通しており、部分的に折り畳まれた中間生成物の疎水的相互作用による会合に関与していることが多いようである(Cleland and Wang, (1990) Biochemistry, 29:11072-11078)。
【0009】
誤った折り畳みは発酵又は単離手順の間に細胞中で起こる。周辺質又は細胞内腔から回収されるタンパク質は可溶化され、可溶性タンパク質は天然の状態に再び折り畳まれなければならない。正しく、生物学的に活性な立体構造にタンパク質を再び折り畳むためのインビトロの方法は、機能的なタンパク質を得るために必須である。封入体から回収されたタンパク質の典型的な以降のプロセシングには、高濃度の尿素などの変性剤に封入体を溶解し、再折り畳みが生じるように変性剤を希釈することを含む(米国特許第4512922号;同第4511502号;及び同第4511503号を参照)。また、例えばRudolph and Lilie, (1996) FASEB J. 10:49-56;及びFischer等, (1993), Biotechnology and Bioengineering, 41:3-13を参照。このような回収方法は普遍的な適用法であると理解でき、封入体からの生物学的に活性な組み換えタンパク質の回収にわずかな変更も考慮される。これらの方法は、VEGFなどのヘパリン結合タンパク質に応用されている(Siemeister等 (1996) supra)。これらの方法は、その他の安定化作用により組み換えタンパク質を生物学的に活性な立体構造にする前にランダムなジスルフィド結合を取り除くことを目的とし、不適当に折り畳まれた中間生成物を取り除かないか、又は適切に折り畳まれた生成物の均質な集団を提供する。
【0010】
逆ミセル又はイオン交換クロマトグラフィを用いて、ミセル内に単一のタンパク質を封入するか又は単一のタンパク質を樹脂上で単離し、その後変性剤を除去することによって変性したタンパク質の再折り畳みを促している(Hagen等, (1990) Biotechnol. Bioeng. 35:966-975; Creighton (1985) in Protein Structure Folding and Design (Oxender, D.L. Ed.) pp.249-251, New York: Alan R. Liss, Inc.)。これらの方法は、タンパク質凝集を予防し、適当な再折り畳みを促す際に有用であった。再折り畳みの速度又は程度を変えるために、立体構造に特異的な再折り畳みを、タンパク質の天然の構造に対するリガンドおよび抗体によって行った(Biotechnology, (Rehm H.-J., and Reed G. Eds.) pp 528-555, New York, VCH)のCleland and Wang, (1993))。例えば、クレアチンキナーゼは、天然の構造に対する抗体がある場合に再び折り畳まれる(Morris等, (1987) Biochem. J. 248:53-57)。抗体に加えて、リガンドおよび補助因子を用いて再折り畳みを促進している。これらの分子は、おそらく、天然タンパク質の形成後に折り畳まれるタンパク質と相互に作用するようである。したがって、折り畳みの平衡は天然の状態に「制御」されうる。例えば、フェリシトクロムcの再折り畳みの速度は、ヘム鉄の軸位置に対して外部のリガンドによって上げられた(Brems and Stellwagon, (1983) J. Biol. Chem. 258:3655-3661)。また、シャペロンタンパク質は、タンパク質の折り畳みによって促すために用いられている。例としてBaneyx, (1999) Current Opinion in Biotechnology, 10:411-421を参照。
【0011】
例えば、効率が良く経済的な細菌細胞培養物におけるヘパリン結合タンパク質の製造のために、宿主細胞培養物からヘパリン結合タンパク質を折り畳む及び/又は回収する新規のより効率の良い方法が求められている。そのような方法により、生物学的に不活性な中間生成物の除去又は低減や高度に精製された生物学的に活性な適切に再び折り畳まれたタンパク質の回収の改善が可能となり、普遍的にタンパク質産生の製造規模に応用可能である。以下の開示内容を参照すると明らかなように、本発明はこれら及び他の必要性に応えるものである。
【0012】
(発明の概要)
本発明は、再び折り畳まれたヘパリン結合タンパク質の細胞培養物からの回収及び精製のための方法を提供する。特に、本発明は、原核生物の宿主細胞、例えば細菌細胞からヘパリン結合タンパク質を回収する方法を提供する。例えば、方法は、(a) 不溶性ヘパリン結合タンパク質を該細菌細胞の周辺質又は細胞内腔から単離する;(b) カオトロピック剤と還元剤を含む第一緩衝溶液に単離された不溶性ヘパリン結合タンパク質を可溶化する、そして(c) ヘパリン結合タンパク質の再折り畳みが生じる時間と条件で、カオトロピック剤と硫酸化ポリ陰イオン剤を含む第二緩衝溶液中で、可溶化させたヘパリン結合タンパク質をインキュベートする;そして、(d) 該再び折り畳まれたヘパリン結合タンパク質を回収する、工程を含んでなり、このとき硫酸化ポリ陰イオン剤とインキュベートしなかった場合と比べて再び折り畳まれたヘパリン結合タンパク質の回収がおよそ2〜10倍に増加する。一実施態様では、第二緩衝溶液はさらにアルギニンを含む。一実施態様では、第二緩衝溶液はさらにシステイン又は軽度の還元剤を含む。
【0013】
本発明の一実施態様では、例えば、回収された生物学的に活性な再び折り畳まれたタンパク質のタンパク質濃度において2〜8倍の増加、又は回収された生物学的に活性な再び折り畳まれたタンパク質のタンパク質濃度において2〜5倍の増加、又は回収された生物学的に活性な再び折り畳まれたタンパク質のタンパク質濃度において3〜5倍の増加、又は回収された生物学的に活性な再び折り畳まれたタンパク質のタンパク質濃度において2〜3倍の増加がある。本発明の他の実施態様では、生物学的に活性な再び折り畳まれたタンパク質の回収されたタンパク質濃度において、例えば2.0倍、2.5倍、2.8倍、3.0倍、5倍、6倍、7.0倍、8倍、9倍などより大きい増加がある。本発明の一実施態様では、生物学的に活性な再び折り畳まれたVEGFのタンパク質濃度において3〜5倍の増加がある。
【0014】
概して、本発明の方法は、ヘパリン結合タンパク質、特にヘパリン結合増殖因子および、特に血管内皮細胞増殖因子(VEGF)に適用可能である。本発明のある実施態様では、硫酸化ポリ陰イオン剤は、およそ3000から10000ダルトンの間である。一実施態様では、生産プロセスにおいて利用される硫酸化ポリ陰イオン剤は、デキストラン硫酸、硫酸ナトリウム又はヘパリン硫酸塩である。一態様では、デキストラン硫酸は、3000ダルトンから10000ダルトンの間である。
【0015】
本明細書中に記述するように、本発明はさらに、ヘパリン結合タンパク質の精製のみ、又はヘパリン結合タンパク質の回収と関連した精製のための工程及び方法を提供する。特定の実施態様では、精製法は、前記の再び折り畳まれたヘパリン結合タンパク質を、ヒドロキシアパタイトクロマトグラフィ支持体;第一疎水性相互作用クロマトグラフィ支持体、陽イオンクロマトグラフィ支持体および第二疎水性相互作用クロマトグラフィ支持体に接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出させることを含む。他の実施態様では、精製法は、前記の再び折り畳まれたヘパリン結合タンパク質を、陽イオン交換支持体;第一疎水性相互作用クロマトグラフィ支持体、及びイオン交換又は混合媒体クロマトグラフィ支持体に接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出させることを含む。回収工程のための工程は、例えばいずれかの順序で順次、又はクロマトグラフィ支持体の順番を変えて行ってもよい。本発明の特定の実施態様では、製造規模又は工業規模の細胞培養物から再び折り畳まれたヘパリン結合タンパク質を回収する及び精製するための方法を提供する。
【0016】
(詳細な説明)
定義
「ヘパリン」(ヘパリンの酸とも称される)は非常に硫酸化した、直鎖の陰イオン性ムコ多糖の不均一な基であり、グリコサミノグリカンと称される。他も存在しうるが、ヘパリンの主要な糖質は、α-L-イズロン酸2-硫酸塩、2-デオキシ-2-スルファミノ-α-グルコース6-硫酸塩、β-D-グルクロン酸、2-アセタミド-2-デオキシ-α-D-グルコースおよびL-イズロン酸である。これら及び場合によって他の糖質は様々な大きさのポリマーを形成するグリコシド結合によって連結される。その共有結合した硫酸塩とカルボン酸基の存在のために、ヘパリンは非常に酸性である。起源及び測定法によって、ヘパリンの分子量はおよそ3000からおよそ20000ダルトンに変化する。
天然のヘパリンは、いくつかの哺乳類種の様々な組織、特に肝臓および肺や肥満細胞の構成要素である。ヘパリンおよびヘパリン塩類(ヘパリンナトリウム)は市販されており、主に様々な臨床状態において抗凝血物質として用いられる。
【0017】
「デキストラン硫酸」は、主要な構造がD‐グルコースのポリマーであるデキストランの硫酸塩である。グルコース及び場合によって他の糖質は、様々な大きさのポリマーを形成するα-D(1-6)グリコシド結合によって連結される。共有結合した硫酸塩の存在のために、デキストラン硫酸は非常に酸性である。通常、硫黄含量は10%より少なくなく、典型的におよそ15%−20%で、グルコース分子当たり3未満の硫酸基となる。デキストラン硫酸の平均分子量はおよそ1000からおよそ40,000,000ダルトンである。本発明で使用可能なデキストラン硫酸の実施例は、微生物、例としてロイコノストック・メッセンテロイデス(Leuconostoc mesenteroides)およびロイコノストック デキストラニクム(L. dextranicum)から産生されるデキストランの硫酸塩を含む。
【0018】
本発明の範囲内で用いられる「ポリ陰イオン剤」は、他の「ポリ陰イオン剤」、例として、硫酸ナトリウム、硫酸ヘパリン、ヘパラン硫酸、硫酸ペントサン(ポリ)、デキストラン、デキストラン硫酸、ヒアルロン酸、コンドロイチン、コンドロイチン硫酸、デルマタン硫酸およびケラタン硫酸を含む、ヘパリン結合タンパク質に結合可能である市販の精製された天然のヘパリン調製物および化合物を記述すると定める。「硫酸化されたポリ陰イオン剤」、例えば、多糖の硫酸塩誘導体、例としてヘパリン硫酸、デキストラン硫酸、米国特許第5314872号に記載のバシラス‐マセランスなどの微生物によって産生されるシクロデキストリンの硫酸塩、並びにβ-1,3グルカン硫酸塩などの他のグルカンの硫酸塩、アルカリゲネス属又はアグロバクテリウムに属する微生物によって産生されるβ-1,3グルカン、およびコンドロイチン硫酸並びに硫酸化ヘパリン断片は、特に本発明の前後関係の範囲内で有用である。
【0019】
上述した薬剤は一般に利用可能であり、当業者に明らかである。例えば、硫酸化ヘパリン断片は、ゲル浸透クロマトグラフィによって分画化されたヘパリン由来のオリゴ糖のライブラリから得られうる。親和性分画され、ヘパリン由来のオリゴ糖の調製は、Ishihara等, (1993) J. Biol. Chem., 268:4675-4683によって報告された。これらのオリゴ糖は市販のブタヘパリンから調整し、亜硝酸により部分的に脱重合し、水素化ホウ素により還元し、ゲル浸透クロマトグラフィによって分画化した。ジ-、テトラ-、ヘキサ-、オクタ-、及びデカ糖類の結果として生じたプールをセファロースTM4Bに共有結合させたヒト組み換えbFGFの親和性カラムに順にアプライし、さらに、塩化ナトリウムの勾配に応じてこのカラムからの溶出に従ってサブプールに分画した。これにより、Hexa-1からHexa-5と称する5つのプールが生じ、その構造および生物学的活性をさらに評価した。Hexa-5Cの構造とその500-MHzのNMRスペクトルは、Tyrell等, (1993) J. Biol. Chem., 268:4684-4689の図4に示される。このヘキサ糖類は、[IdoA(2-OSO3)α1-4GlcNSO3(6-OSO3)α1-4]2IdoA(2-OSO3)α1-4AManR(6-OSO3)の構造を有する。上記のすべてのヘパリン由来のオリゴ糖、並びに他のヘパリン様オリゴ糖は、本発明に好適であり、本発明に従って用いられうる。本発明のある実施態様では、よりユニットが大きなヘパリンのヘキサ糖類及びポリ糖類(例えば、ヘプタ-、オクタ-、ノナ-、デカ糖類)が用いられる。さらに、例えば高い程度の硫酸化により正味の陰電荷が大きい、ヘパリン由来又はヘパリン様のオリゴ糖が、有利に使われる。
【0020】
本明細書中で用いられる「ヘパリン-結合タンパク質」又は「HPB」なる用語は、ヘパリンを結合することができるポリペプチドを指す(上記に定義されたように)。定義には、成熟、プレ、プレ-プロ、及びプロの形態の天然及び組み換えて産生させたヘパリン-結合タンパク質が含まれる。ヘパリン結合タンパク質の典型的な例は、限定するものではないが、上皮細胞増殖因子(EGF)、血小板由来増殖因子(PDGF)、塩基性線維芽細胞増殖因子(bFGF)、酸性線維芽細胞増殖因子(aFGF)、血管内皮細胞増殖因子(VEGF)、肝増殖因子(HGF)(散乱係数、SFとしても知られる)、及び神経成長因子(NGF)、IL-8などを含む、「ヘパリン結合増殖因子」である。
【0021】
本明細書中で用いられるように「血管内皮細胞増殖因子」又は「VEGF」は、Castor, C.W.,等, (1991) Methods in Enzymol. 198:391-405において開示されるアミノ酸配列を有するウシ下垂体濾胞性細胞から本来得られる哺乳類の増殖因子、並びに限定するものではないがHouck等, (1991) Mol. Endocrin. 5:1806-1814において報告されるヒトVEGFアミノ酸配列を含む、対応する天然のVEGFの定性的生物学的活性を有するこの機能的誘導体を指す。また、Leung等 (1989) Science, 246:1306、及び、Robinson & Stringer, (2001) Journal of Cell Science, 144(5): 853-865、米国特許第5332671号を参照。VEGFの主な形態は、7つの分子内ジスルフィド結合と2つの分子間ジスルフィド結合を形成する16のシステイン残基を有する165アミノ酸ホモ二量体である。オルタナティブスプライシングは、121、145、165、189および206のアミノ酸からなる複数のヒトVEGFポリペプチドの形成に関係しているが、VEGF121変異体は、他の変異体のヘパリン結合ドメインを欠いているため、本明細書に記載のヘパリン結合タンパク質の定義に入らない。VEGFのすべてのアイソフォームは共通のアミノ末端ドメインを共有するが、分子のカルボキシル末端部分の長さが異なる。好ましいVEGFの活性型であるVEGF165は、各単量体のアミノ酸残基Cys26-Cys68;Cys57-Cys104;Cys61-Cys102;Cys117-Cys135;Cys120-Cys137;Cys139-Cys;158;Cys146-Cys160の間にジスルフィド結合を有する。図11を参照。また、例えば、Keck等, (1997) Archives of Biochemistry and Biophysics 344(1): 103-113を参照。VEGF165分子は、アミノ末端レセプター-結合ドメイン(アミノ酸1−110ジスルフィド結合ホモ二量体)とカルボキシル末端ヘパリン-結合ドメイン(残基111−165)の2つのドメインから成る。例えば、Keyt等, (1996) J. Biol. Chem., 271(13): 7788-7795を参照。本発明のある実施態様では、単離して、精製されるVEGF165は、残基75(Asn)でグリコシル化されていない。例えば、Yang等, (1998) Journal of Pharm. & Experimental Therapeutics, 284:103-110を参照。本発明のある実施態様では、単離して、精製されるVEGF165は、残基Asn10で実質的に脱アミド化されていない。本発明のある実施態様では、単離して、精製されるVEGF165は、脱アミド化された(残基Asn10で)タンパク質と脱アミド化されていないタンパク質の混合であり、典型的にはほとんどのタンパク質は脱アミド化されていない。VEGF165はホモ二量体であるので、脱アミド化は一方又は両方のポリペプチド鎖で起こりうる。
【0022】
本明細書中で用いる「適切に折り畳まれた」又は「生物学的に活性な」VEGF又は他のHBPなどは、生物学的に活性な立体構造を有する分子を指す。当業者は、誤って折り畳まれた中間生成物及びジスルフィドスクランブル中間生成物は生物学的活性を有しうると理解するであろう。この場合、適切に折り畳まれた又は生物学的に活性なVEGF又はHBPは、VEGF(上記した)又は他のHBPの天然の折り畳みパターンに対応する。例えば、適切に折り畳まれたVEGFは、二量体分子の2つの分子間ジスルフィド結合に加え、上記のジスルフィド対を有するが、他の中間生成物は細菌細胞培養物によって産生されうる(図1及び3A−3D)。適切に折り畳まれたVEGFでは、各単量体の同じ残基(Cys51とCys60)の間で2つの分子間ジスルフィド結合が生じる。例えば国際公開第98/16551号特許を参照。VEGFの生物学的活性には、限定するものではないが、例えば、血管透過性の促進、血管内皮細胞の増殖促進、VEGFレセプターへの結合、VEGFレセプターによる結合とシグナル伝達(例えば、Keyt等, (1996) Journal of Biological Chemistry, 271(10): 5638-5646を参照)、血管新生の誘導などが含まれる。
【0023】
「精製された」又は「純粋なHBP」なる用語は、組み換え産生及び特に原核生物又は細菌の細胞培養物に見られるような、通常付随する物資を欠く物質を指す。したがって、この用語は、DNA、宿主細胞タンパク質又は元の環境に付随する他の分子の混入がない組み換えHBPを指す。この用語は、少なくともおよそ75%、少なくともおよそ80%、少なくともおよそ85%、少なくともおよそ90%、少なくともおよそ95%又は少なくともおよそ98%又はそれ以上の程度の純度を指す。
【0024】
「封入体」又は「屈折小体」なる用語は、対象の凝集されたポリペプチドの密度の高い細胞内質量を指し、それはすべての細胞構成成分を含む総細胞タンパク質の主な部分を成す。すべてではないが場合によって、ポリペプチドのこれらの凝集塊は、1000倍まで拡大した位相差顕微鏡下で細胞の中に可視の明るいスポットとして認識されうる。
【0025】
本明細書中で用いられる「誤って折り畳まれた」タンパク質なる用語は、屈折小体内に含有される沈殿したないしは凝集したポリペプチドを指す。本明細書中で用いられる「不溶性の」又は「誤って折り畳まれた」VEGF又は他のHBPは、原核生物の宿主細胞の周辺質又は細胞内腔内に含有されるか、またはさもなくば、付随する原核生物の宿主細胞である、沈殿したないしは凝集したVEGFを指し、ミスマッチ又は形成されていないジスルフィド結合を有する生物学的に不活性な立体構造であると思われる。一般に、不溶性HBPは屈折小体に含有される必要はなく、すなわち、位相差顕微鏡下で可視化されることもあるし可視化されないこともある。
【0026】
本明細書中で用いられる「カオトロピック剤」は、水溶液中に適切な濃度であると、ポリペプチドを水溶媒質に可溶化させるために、その表面の変質によってポリペプチドの空間配位又は立体構造を変えることが可能である化合物を指す。変質は、例えば水和作用、溶媒環境又は溶媒-表面相互作用の状態を変えることで生じうる。カオトロピック剤の濃度はその強度および有効性に直接作用する。強力な変性カオトロピック溶液は、溶液中で、タンパク質二次構造を効果的に取り除く溶液中に存在するポリペプチドを効率的にほどく(unfold)高濃度のカオトロピック剤を含む。アンフォールディング(unfolding)は相対的に広範囲であるが、可逆性である。中程度の変性カオトロピック溶液は、溶液中に十分な濃度であると、ポリペプチドは部分的にしか折り畳まれない。内在性又は類似の生理学的条件下でその活性型を操作する場合、それ自体の空間的立体構造に、どんなに立体構造がねじれていようとポリペプチドは溶液中に溶ける中間生成物を経ると考えられているのでカオトロピック剤の例には、塩酸グアニジン、尿素および水酸化物、例えばナトリウム又は水酸化カリウムが含まれる。カオトロピック剤には、これらの試薬の組合せ、例として、尿素又は塩酸グアニジンと水酸化物の混合が含まれる。
【0027】
本明細書中で用いられる「還元剤」は、水溶液中に適切な濃度であると、フリーなスルフヒドリル基を維持して、分子内又は分子間ジスルフィド結合が化学的に崩壊される化合物を指す。適切な還元剤の代表的な例には、ジチオトレイトール(DTT)、ジチオエリトリトール(DTE)、β-メルカプトエタノール(BME)、システイン、システアミン、チオグリコレート、グルタチオン、トリス[2-カルボキシエチル]ホスフィン(TCEP)、及び水素化ホウ素ナトリウムなどがある。
【0028】
本明細書中で用いられる「緩衝溶液」はその酸-塩基コンジュゲート構成成分の作用によってpHの変化に耐える溶液を指す。
【0029】
本明細書中での目的のための「細菌」には真正細菌及び古細菌が含まれる。本発明のある実施態様では、グラム陽性細菌及びグラム陰性細菌を含む真正細菌は本明細書中に記載の方法及びプロセスに用いられる。本発明のある実施態様では、グラム陰性細菌、例えば腸内細菌科が用いられる。腸内細菌科に属する細菌の例は、エシェリキア属、エンテロバクター属、エルウィニア属(Erwinia)、クレブシエラ属、プロテウス属、サルモネラ属、セラシア属、及びシゲラ属を含む。その他の種類の適切な細菌としては、アゾトバクタ、シュードモナス、根粒菌属、ビトレオシラ(Vitreoscilla)、及びパラコッカス(Paracoccus)が挙げられる。本発明のある実施態様では、大腸菌が用いられる。適切な大腸菌宿主には、大腸菌W3110(ATCC 27,325)、大腸菌294(ATCC 31,446)、大腸菌B及び大腸菌X1776(ATCC 31,537)が含まれる。これらの例は、限定的なものではなく例示であり、W3110が一例である。上述の細菌の任意の突然変異細胞も使用できる。当然ながら、バクテリウム属の細胞におけるレプリコンの複製可能性を考慮しながら適切な細菌を選択することが必要である。例えば、pBR322、pBR325,pACYC177、又はpKN410等の周知のプラスミドを使用してレプリコンを供給するとき、大腸菌、セラシア属、又はサルモネラ属を宿主として適切に使用することができる。さらに、好適な細菌宿主細胞の例については以下を参照のこと。
【0030】
本明細書で使用する「細胞」「細胞系」「株」、及び「細胞培養物」という表現は、互いに交換可能に使用され、そのような名称は全て子孫を含んでいる。したがって、「形質転換体」及び「形質転換された細胞」は初代の対象細胞及び、継代回数には関係なく、それに由来する培養物を含んでいる。また、故意又は偶然の変異のために、全ての子孫はDNA含量が正確に一致しないかもしれないことも理解される。最初に形質転換された細胞においてスクリーニングされるのと同じ機能又は生物学的活性を有する突然変異子孫が含まれる。明確な区別が意図される場合には、それは文脈から明らかになるだろう。
【0031】
本明細書中で用いられる場合、一般に、「ポリペプチド」はおよそ10より多いアミノ酸を有する任意の細胞源由来のペプチド及びタンパク質を指す。「異種の」ポリペプチドは、大腸菌によって産生されるヒトタンパク質のような、利用される宿主細胞にとって外来のポリペプチドである。異種ポリペプチドは原核生物のものでも真核生物のものでもよいが、好ましくは真核生物のものであり、より好ましくは哺乳類、最も好ましくはヒトのものである。本発明のある実施態様では、組み換えて産生されるか、又は組み換えポリペプチドである。
【0032】
ヘパリン結合タンパク質
ヘパリン結合タンパク質の単離
不溶性の誤って折り畳まれたヘパリン結合タンパク質(HBP)は、当分野の多くの標準的な技術のいずれかによりタンパク質を発現する原核生物の宿主細胞から単離される。例えば、不溶性のHBPは、適切な単離バッファ中で、細胞を適切なイオン強度のバッファに曝露して殆どの宿主タンパク質を可溶化させるか(しかし対象のタンパク質は実質的に不溶性である)、又は封入体ないしは周辺質又は細胞内腔のタンパク質を放出させ、例えば遠心分離による回収に利用できるようにするために細胞を破壊することにより、単離される。この技術は周知であり、例えば米国特許第4511503号に記載されている。Kleid等は均質化の後に遠心分離をすることによる屈折小体の精製を開示している(Kleid等, (1984) in Developments, Industrial Microbiology, (Society for Industrial Microbiology, Arlington, VA) 25:217-235)。例としてFischer等, (1993) Biotechnology and Bioengineering 41:3-13も参照のこと。
【0033】
米国特許第5410026号は、封入体からのタンパク質の回収のための典型的な方法を記載しており、以下のようにまとめられる。原核生物細胞を好適なバッファに懸濁する。典型的に、バッファはpH5〜9、又はおよそ6〜8に緩衝するために好適な緩衝剤と塩からなる。NaClなどの任意の好適な塩は、緩衝した溶液の十分なイオン強度を維持するために有用である。典型的には、約0.01から2M、又は0.1から0.2Mのイオン強度を用いる。細胞は、この緩衝液中に懸濁され、ついで、例えば機械的方法、例えばホモジナイザー(Manton-Gaulinプレス、マイクロフルイダイザー、又はNiro-Soavi)、フレンチプレス、ビーズミル、又は音波オシレーター、あるいは化学的又は酵素的方法のような通常用いられている技術を使用して破壊又は溶解する。
【0034】
細胞破壊の化学的ないしは酵素的方法の例には、細菌壁を溶解するリゾチームの使用を伴うスフェロプラスティング(Neuら, Biochem.Biophys.Res.Comm.,17:215(1964))、及びポリペプチドの放出のために生細胞を高緊張度の溶液と低緊張度の冷水洗浄で処理することを含む浸透圧ショック(Neuら, J.Biol.Chem.,240:3685-3692(1965))が含まれる。一般的に、発酵培養物の分析スケール容量に含まれる細菌の破壊には超音波処理が用いられる。大きなスケールでは、高圧ホモジナイゼーションが典型的に用いられる。
【0035】
細胞を破壊した後、典型的に、懸濁液を低速、一般的にはおよそ500〜15000×gで遠心し(本発明のある実施態様では、およそ12000×gが用いられる)、標準的な遠心では、不溶性タンパク質のすべてが実質的にすべてペレット状になるのに十分な時間遠心する。この時間は、遠心する容量並びに遠心設定に応じて単に決定されうる。典型的におよそ10分間〜0.5時間が不溶性タンパク質をペレット状にするために十分である。ある実施態様では、懸濁液を12000×gで10分間遠心する。
【0036】
その結果生じたペレットは不溶性タンパク質分画の実質的にすべてを含む。細胞破壊プロセスが完全でないならば、ペレットはインタクトな細胞又は破壊された細胞断片も含有し得る。細胞破壊の完全性は、ペレットを少量の同じバッファ中に懸濁し、懸濁液を位相差顕微鏡で調べることにより検査することができる。破壊細胞断片もしくは全細胞の存在は、断片又は細胞及び随伴する非屈折性(non-refractile)ポリペプチドを除去するために更なる超音波処理又は破壊の他の手段が必要であることを示している。そのような更なる破壊後に、必要ならば、懸濁液を再び遠心分離し、ペレットを回収し、再懸濁し、再分析する。このプロセスを、視覚検査でペレット化材料中に破壊細胞断片がないことが明らかになるか、更なる処理でも得られるペレットのサイズを低減できなくなるまで繰り返す。
【0037】
不溶性タンパク質が細胞内又は周辺質のスペースのいずれにあっても、上記のプロセスは実施できる。本発明のある実施態様では、ヘパリン結合タンパク質を単離するために本明細書中に示した条件は、周辺質スペース又は細胞内スペースに沈殿した封入体と、特にVEGFに関する。しかしながら、この方法及び手順は、以降全体にわたって述べるように一般にわずかな変更があるヘパリン結合タンパク質に適用することができると考えられる。本発明のある実施態様では、この方法及び手順は、HBPの製造又は工業規模の産生、再折り畳み、及び精製に適用することができる。
【0038】
ヘパリン結合タンパク質の再折り畳み
単離した不溶性の誤って折り畳まれたヘパリン結合タンパク質は、ヘパリン結合タンパク質を実質的に可溶化するために十分な量のカオトロピック剤と還元剤を含む第一緩衝溶液中でインキュベートする。このインキュベートは、いくつか又は実質的にすべてのヘパリン結合タンパク質の可溶化とアンフォールド(unfolding)を可能とする濃度、インキュベート時間、及びインキュベート温度の条件下で行う。
【0039】
緩衝溶液中における可溶化の程度の測定は、濁度決定法により、還元されたSDS-PAGEゲル上の遠心分離後に上清とペレットとの間の分画を分析することにより、プロテインアッセイ(例えば、バイオ-ラッドタンパク質アッセイキット(例えば、Pierce, Bio-Radなど))により、又はHPLCにより簡便に実施される。
【0040】
第一緩衝溶液は、バッファのpH範囲を少なくともおよそ7.0に維持するために好適な緩衝剤を含み、典型的な範囲は7.5−10.5である。ある実施態様では、VEGFのpHはpH8.0である。この後の範囲のpHをもたらす適切なバッファの例としては、トリス-HCl(トリス[ヒドロキシメチル]アミノメタン)、HEPPS (N-[2-ヒドロキシエチル]ピペラジン-N'-[3-プロパン-スルホン酸])、HEPES (N-[2-ヒドロキシエチル]ピペラジン-N'-[2-エタンスルホン酸])、CAPSO(3-[シクロヘキシルアミノ]-2-ヒドロキシ-1-プロパンスルホン酸)、AMP(2-アミノ-2-メチル-1-プロパノール)、CAPS(3-[シクロヘキシルアミノ]-1-プロパンスルホン酸)、CHES(2-[N-シクロヘキシルアミノ]エタンスルホン酸)、グリシン、及び酢酸ナトリウムが含まれる。本発明のある実施態様では、本明細書中のバッファはおよそpH8.0のHEPPSである。更なる実施態様では、例えばHEPPSなどのバッファは硫酸化される。
【0041】
本発明の実施に好適なカオトロピック剤には、例えば、尿素及びグアニジン又はチオシアン酸の塩類、例えば尿素、塩酸グアニジン、チオシアン酸ナトリウムなどが含まれる。バッファに存在するために必要なカオトロピック剤の量は、溶液中のHBPをアンフォールドするために十分な量である。本発明のある実施態様では、カオトロピックは、およそ4モルと10モルの間で存在する。本発明の一実施態様では、カオトロピック剤は、およそ5〜8M、又はおよそ7Mの尿素である。他の例では、カオトロピック剤は、およそ6〜8Mの塩酸グアニジンである。
【0042】
適切な還元剤の例には、限定するものではないが、ジチオトレイトール(DTT)、ジチオエリトリトール(DTE)、β-メルカプトエタノール(BME)、システイン、DTEなどがある。バッファに存在すべき還元剤の量は、還元剤とカオトロピック剤の種類、用いるバッファの種類とpH、溶液に取り込まれる又は導入される酸素の量、及びバッファ中のタンパク質の濃度に主に依存するであろう。例えば、4〜8M 尿素を含有するpH7.0〜10.0の緩衝溶液中0.5〜1.5mg/mlのタンパク質の場合、還元剤は、例えば、およそ1〜15mMの濃度のDTT、又はおよそ0.2〜2mMの濃度のBME、又はおよそ2〜10mMの濃度のシステインである。一実施態様では、還元剤は、およそ0.5からおよそ4mM、又は2〜4mMのDTTである。図9はVEGFの抽出に対する尿素とDTTの効果を示す。ピーク3のVEGFは、適切に折り畳まれた生物学的に活性なVEGFを示す。一実施態様では、還元剤はおよそ10mMのDTTである。単一の還元剤又は還元剤の組合せが、本明細書中のバッファで用いられうる。
【0043】
緩衝溶液中のタンパク質の濃度は、タンパク質が光学密度によって測定すると実質的に溶解されている程度でなければならない。用いる正確な量は、例えば、緩衝溶液中の他の成分の濃度と種類、特にタンパク質濃度、還元剤及びバッファのpHに依存するであろう。本発明の一実施態様では、ヘパリン結合タンパク質の濃度は、1ml当たり0.5〜5.5mg又は1.5〜5.0mg/mlの範囲である。典型的に、可溶化は、少なくともおよそ1〜24時間のおよそ0〜45℃、又はおよそ20〜40℃、又はおよそ23〜37℃、又はおよそ25〜37℃、又はおよそ25℃で、少なくともおよそ1〜24時間かけて行う。一実施態様では、可溶化は、室温で少なくともおよそ2時間行う。一般的に、温度は、塩、還元剤およびカオトロピック剤のレベルによって見かけ上影響を受けない。
【0044】
ポリペプチドが可溶化されると、上記のカオトロピック剤と硫酸化ポリ陰イオン剤を、ヘパリン結合タンパク質の再折り畳みが可能なカオトロピック剤の濃度で含有する第二緩衝溶液に置く、又は希釈する。
一般に、可溶性の誤って折り畳まれたタンパク質のこの第二インキュベーションの条件は、タンパク質のいくらか又は実質的又は完全な再折り畳みが起こるようにする。正確な条件は、例えばバッファのpHや、存在するのであれば、硫酸化ポリ陰イオン剤及びカオトロピック剤及び還元剤の種類と濃度に依存するであろう。培養温度は通常、およそ0〜40℃、又は10〜40℃であり、インキュベートは通常、再折り畳みに効果を示すように少なくともおよそ1時間行う。ある実施態様では、反応は、例えばおよそ15〜37℃、又は20〜30℃で、少なくともおよそ6時間、少なくともおよそ10時間、又はおよそ10から48時間の間、又はおよそ15から20時間の間、又は6から20時間の間、又は12から24時間の間で行う。
【0045】
再折り畳みの程度は、HPBの放射線免疫アッセイ(RIA)力価、又は例えばPOROS HE2/Mカラム(PerSeptive BioResearch Products)又は他の好適なヘパリン親和性カラムを用いた高性能液体クロマトグラフィ(HPLC)分析によって、好適に測定される。RIA力価の増加又は適切に折り畳まれたHBPピークサイズは、バッファ中に存在する適切に折り畳まれた、生物学的に活性なHPBの量の増加と直接相関する。RIA又はHPLCによって測定されるように、インキュベートは、回収された誤って折り畳まれたHPBに対する適切に折り畳まれたHPBの比率を最大にするように行う。
【0046】
一実施態様では、適切に折り畳まれたVEGFの質と量は、ヘパリン-結合アッセイを用いて評価される。希釈したヘパリン結合タンパク質を含有する試料は、例えばPOROS HE2/M(4.6×100mm、PerSeptive BioResearch Products, Cambridge, MA)又は他の好適なヘパリン親和性カラムに流す。例えば、ヘパリン親和性カラムは、0.15M 塩化ナトリウムを含有する10mM リン酸ナトリウム、pH7に平衡化した。1ml/分又は2ml/分の流速で、カラムは、0.15〜2M 塩化ナトリウムの直線濃度勾配を含む10mM リン酸ナトリウム、pH7を用いて10分間にわたって溶出する。溶出物を280nmでモニターした。一実施態様では、タンパク質は、生物学的に活性な適切に再び折り畳まれたHBPに対応する単一のピークに回収される。本発明の一実施態様では、適切に再び折り畳まれたHBPを測定するためのアッセイはRPHPLCである。場合によってジスルフィド結合はペプチドマップによって確認することができる。また、円二色性は、2&3D構造/折畳みを決定するために使われてもよい。
【0047】
第二緩衝溶液のためのバッファは、第一緩衝溶液のために上記のもののいずれか、VEGFを再び折り畳むためには例えばおよそ50mMの濃度で、例えばHEPPS pH8.0であってもよい。ポリペプチドは、再折り畳みバッファにて、例えば少なくとも5倍、又は少なくともおよそ10倍、およそ20倍、又はおよそ40倍に希釈されてもよい。あるいは、ポリペプチドは再折り畳みバッファに対して透析されてもよい。
第二緩衝溶液は、HPBの再折り畳みが起こる濃度のカオトロピック剤を含む。通常、カオトロピックはおよそ0.5から2モルの間で存在する。本発明の一実施態様では、本明細書中のカオトロピック剤は、およそ0.5〜2M、0.5〜2M 、又はおよそ1Mの尿素である。一実施態様では、カオトロピック剤はおよそ1.3M 濃度の尿素である。本発明の他の実施態様では、カオトロピック剤はおよそ1Mの塩酸グアニジンである。図10はVEGFの再折り畳みに対する尿素と還元剤DTTの効果を示す。ピーク3のVEGFは適切に折り畳まれた生物学的に活性なVEGFを指す。
【0048】
示すように、場合によって溶液も還元剤を含む。還元剤は、およそ0.5からおよそ10mMのシステイン、0.1〜1.0mMのDTT、及び/又はおよそ0.2mM未満のBMEの濃度範囲での可溶化工程について上記のものから好適に選択される。本発明の一実施態様では、還元剤はおよそ0.5〜2mMのDTTである。本発明の一実施態様では、還元剤はおよそ0.5mMのDTTである。 適切な還元剤の例には、限定するものではないが、例えばジチオトレイトール(DTT)、β-メルカプトエタノール(BME)、システイン、DTEなどが含まれる。DTTおよびBMEは、通常はヘパリン結合タンパク質のために本明細書中で提供される手順と関連して用いられうるのに対して、VEGFの回収のためには本明細書中に記載のようにおよそ0.1からおよそ10mMのシステインと、およそ0.1からおよそ1.0mMのDTTの組み合わせが一例である。
【0049】
再折り畳み工程には、可溶化されたタンパク質の完全な再折り畳みを達成するために十分な濃度で、硫酸化されたポリ陰イオン剤が含まれる。好適なポリ陰イオン剤の例は上記のもの、例えば硫酸化ポリ陰イオン剤、例えばヘパリン硫酸、デキストラン硫酸、ヘパリン硫酸及びコンドロイチン硫酸並びに硫酸化ヘパリン断片を有する上記のポリ糖類の硫酸誘導体である。本発明の関係において用いるヘパリン硫酸では、通常、分子量は、およそ3000から10000ダルトンの間、又はおよそ3000から6000ダルトンの間である。
【0050】
本発明の一実施態様では、デキストラン硫酸は本発明の関係において使用される。本発明で使用されるデキストラン硫酸のような硫酸化されたポリ陰イオン剤又は他の薬剤の分子量は、回収される特定のヘパリン結合タンパク質のサイズに依存する。通常、およそ3000から10000ダルトンの間のデキストラン硫酸が使用される。本発明の一実施態様では、およそ5000ダルトンから10000ダルトンの間のデキストラン硫酸が、例えばVEGFの回収のために使われる。他の実施態様では、およそ5000から8000ダルトンの間のデキストラン硫酸が、HBPの回収のために使われる。図3A−3Dは、ヘパリン親和性クロマトグラフィによって分析した、様々な濃度及び分子量のデキストラン硫酸(図3A−C)およびヘパリン(図3D)によるVEGFの回収を示す。ピーク3は適切に折り畳まれたVEGFに対応する。
【0051】
使用されるポリ陰イオン化合物の濃度は、回収されるタンパク質およびその濃度および折り畳みバッファの温度とpHなどの条件に依存する。代表的な濃度は、およそ50〜500mMの間の硫酸ナトリウム、およそ10〜200μg/mlの6000ダルトンのヘパリンなどの低分子量ヘパリン(Sigma Chemical Co.)、およそ10〜200μg/mlの間のブタヘパリンI-Aなどの高分子量ヘパリン(Sigma Chemical Co.)、及びおよそ10〜400μg/mlの間又はおよそ10〜200μg/mlの間のデキストラン硫酸である。
【0052】
場合によって、再折り畳みバッファは、TRITONTMX-100、NONIDETTMP-40、TWEENTMシリーズ及びBRIJTMシリーズなどの様々な非イオン性界面活性剤のいずれかなどの付加的な薬剤を含みうる。非イオン性界面活性剤はおよそ0.01%〜1.0%の間で存在する。ある例では、非イオン性界面活性剤の濃度はおよそ0.025%から0.05%の間、又はおよそ0.05%である。
場合によって、陽性荷電アミノ酸、例えばアルギニン(例えばL-アルギニン/HCl)、リジンなどは再折り畳みバッファに存在してもよい。本発明のある実施態様では、アルギニンの濃度は、例えばおよそ0〜1000mM、又はおよそ25〜750mM、又はおよそ50〜500mM、又はおよそ50〜250mM、又はおよそ100mM終濃度などである。本発明のある実施態様では、タンパク質は、0.5〜3M 尿素、0〜30mg/L デキストラン硫酸、0〜0.2%トリトンX-100、2〜15mM システイン、0.1〜1mM DTTおよび0〜750mM アルギニン終濃度を含む、pH7.0〜9.0の緩衝溶液中にある。一実施態様では、50mM HEPPSが使われる。一実施態様では、再折り畳みバッファ溶液の終濃度は、1M 尿素、50mM HEPPS、15mg/L デキストラン硫酸、0.05%トリトンX-100、7.5mM システイン、100mM アルギニン、pH8.0である。一実施態様では、再折り畳みバッファ溶液の終濃度は、1.3M 尿素、50mM HEPPS、15mg/L デキストラン硫酸、0.05%トリトンX-100、7.5mM システイン、0.5mM DTT、100mM アルギニン、pH8.0である。
【0053】
ヘパリン結合タンパク質の回収と精製
培養物からのヘパリン結合タンパク質の回収と精製は、例えば、塩と溶媒分別法、コロイド性物質による吸着、ゲル濾過、イオン交換クロマトグラフィ、親和性クロマトグラフィ、免疫親和性クロマトグラフィ、電気泳動法および高性能液体クロマトグラフィ(HPLC)などの様々な方法及び公知の手法をタンパク質の分離のために実施することができるが、前記の再び折り畳まれたヘパリン結合タンパク質を、ヒドロキシアパタイトクロマトグラフィ支持体;第一疎水性相互作用クロマトグラフィ支持体、陽イオンクロマトグラフィ支持体および第二疎水性相互作用クロマトグラフィ支持体に順次接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出させることを含む4工程のクロマトグラフィの手順の例が記載される。あるいは、前記の再び折り畳まれたヘパリン結合タンパク質を陽イオン交換支持体;疎水性相互作用クロマトグラフィ支持体、およびイオン交換クロマトグラフィ支持体に順次接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出することを含む他のクロマトグラフィ手順が記載される。いずれかの手順の工程はいずれの順序でも行うことができると理解される。本発明の一実施態様では、工程は順次行われる。
【0054】
ヘパリン結合タンパク質の更なる回収と精製における適切な第一工程は、典型的に、ヘパリン結合タンパク質の濃縮と試料体積の減少をもたらす。例えば、上記の第二インキュベート工程により、回収されたヘパリン結合タンパク質の容量が大幅に増加し、再折り畳みバッファでのタンパク質が同時に希釈される。適切な第一クロマトグラフィ支持体は、回収されたヘパリン結合タンパク質の容量の減少をもたらし、望ましくない混在タンパク質からタンパク質をいくらか精製する。好適な第一クロマトグラフィ工程には、溶出され、第一疎水性相互作用クロマトグラフィ支持体に直接流し込まれうるクロマトグラフィ支持体が含まれる。例えば、ヘパリン結合タンパク質が疎水性相互作用クロマトグラフィ支持体を流すために適切な高塩濃度に溶出されうるクロマトグラフィ支持体が使われる。
【0055】
例示的な第一クロマトグラフィ支持体には、限定するものではないが、ヒドロキシアパタイトクロマトグラフィ支持体、例えばCHTセラミックI型およびII型(形式的にMacroPrepセラミックとして知られる)、バイオゲルHT、バイオゲルHTP、Biorad, Hercules, CA、など;銅、ニッケルなどの固定した金属イオンの不活性な樹脂からなる金属キレート化クロマトグラフィ支持体;並びに非誘導体化シリカゲルが含まれる。本発明の一実施態様では、VEGFの精製および回収のための第一クロマトグラフィ支持体は、ヒドロキシアパタイトクロマトグラフィ支持体である。本発明の他の一実施態様では、VEGFの精製および回収のための第一クロマトグラフィ支持体は、陽イオン交換支持体であり、例えば更に詳細に後述する。
【0056】
第一クロマトグラフィ支持体からの溶出は当分野の標準的な手法にしたがって達成される。好適な溶出条件とバッファは、下記のような第一疎水性相互作用クロマトグラフィ支持体に直接溶出したHPBの負荷(loading)を促すであろう。
疎水性相互作用クロマトグラフィは当分野で周知であり、「クロマトグラフィ支持体」に付着した疎水性リガンドと相互作用する分子の疎水性部分の相互作用に基づく。基質にカップリングした疎水性リガンドは、HICクロマトグラフィ支持体、HICゲル又はHICカラムなどとさまざまに称される。さらに、タンパク質とHICとの相互作用の強度は、タンパク質上の極性表面に対する非極性表面の割合に依存するだけでなく、同様に非極性表面の関与にも依存する。
【0057】
多くの基質は、HICカラムの調製に使用されてもよい。アガロースが最も広く用いられるが、シリカや有機ポリマーの樹脂も用いられてもよい。有用な疎水性リガンドには、限定するものではないが、ブチル-、プロピル-、又はオクチル-などのおよそ2からおよそ10の炭素原子を有するアルキル基、又はフェニルなどのアリル基が含まれる。ゲルやカラムのための従来のHIC支持体は、製品名ブチル-SEPHAROSETM、フェニル-SEPHAROSETM CL-4B、オクチルSEPHAROSETM FF及びフェニルSEPHAROSETM FFとしてGE Healthcare, Uppsala, Swedenから、製品名TOYOPEARLTM ブチル650M (フラクトゲルTSK ブチル-650)又はTSK-GELフェニル5PWとしてTosoh Corporation, Tokyo, Japanなどの供給元から市販されうる。一実施態様では、VEGFの精製および回収は、ブチル-アガロースである第一HICクロマトグラフィ支持体と、フェニルアガロースである第二疎水性クロマトグラフィ支持体である。他の実施態様では、第一HICクロマトグラフィ支持体はフェニルアガロースである。
【0058】
リガンド密度は、タンパク質の相互作用の強度だけでなく同様にカラムの許容量に影響する重要なパラメーターである。市販のフェニル又はオクチルフェニルゲルのリガンド密度はおよそ5〜40μmol/mlゲル板である。ゲル容量は、対象の特定のタンパク質並びにpH、温度および塩濃度に依存するが、一般に、3〜20mg/mlゲルの範囲内であると思われる。
特定のゲルの選択は当業者により決定されうる。通常、タンパク質およびHICリガンドの相互作用の強度はアルキルリガンドの鎖長によって増加するが、およそ4からおよそ8の炭素原子を有するリガンドがほとんどの分離に適する。フェニル基はペンチル基とほぼ同じ疎水性を有するが、選択性はタンパク質の芳香族基とのπ-π相互作用の可能性のために異なりうる。
【0059】
HICカラムに対するタンパク質の吸着は高塩濃度によるものであるが、有効濃度はタンパク質の性質及び選択された特定のHICリガンドに応じて広範囲で変化しうる。一般におよそ1から4Mの間の塩濃度が有用である。
順に又は勾配の形態のいずれであっても、HIC支持体からの溶出は、例として、a) 塩濃度を変える、b) 溶媒の極性を変える、又はc) 界面活性剤を加えるなどの様々な方法で達成されうる。塩濃度を下げることによって、吸着したタンパク質は疎水性を増すに従って溶出される。極性の変化は、エチレングリコール又はイソプロパノールなどの溶媒の添加とこれによる疎水性相互作用の強度の低減に影響されうる。界面活性剤は、タンパク質のディスプレーサーとして機能し、膜タンパク質の精製と関連して主に使われていた。
【0060】
様々な陰イオン性成分は、クロマトグラフィの陽イオン支持体を形成するために基質に接着されうる。陰イオン性成分には、カルボキシメチル、硫黄エチル基、スルホプロピル基、リン酸およびスルホン酸(S)が含まれる。セルロースのイオン交換樹脂、例として、SE52 SE53、SE92、CM32、CM52、CM92、P11、DE23、DE32、DE52、EXPRESS IONTMSおよびEXPRESS IONTMCは、Whatman LTD, Maidstone Kent U.Kから入手可能である。SEPHADEXTMおよびSEPHAROSETMベースの、相互結合のイオン交換体は、製品名CM SEPHADEXTMC-25、CM SEPHADEXTMC-50、及びSP SEPHADEXTMC-25 SP SEPHADEXTMC-50及びSP-SEPHAROSETM高性能、SP-SEPHAROSETM高速流量、SP-SEPHAROSE XL、CM-SEPHAROSETM高速流量、及びCM-SEPHAROSETMCL-6Bとしても知られており、すべてGE Healthcareから入手できる。本発明を行うためのイオン交換体の例には、限定するものではないが、例えば製品名MACROPREP、例えば、BioRad, Hercules, CAのMACROPREPTMS支持体、MACROPREPTMハイS支持体およびMACROPREPTMCM支持体のイオン交換体が含まれる。
【0061】
陽イオンクロマトグラフィ支持体からの溶出は、通常、塩濃度を増やすことによって達成される。イオンカラムからの溶出物は塩の添加を伴い、上記のように、HICは塩濃度に増強されるので、HIC工程の後にイオン工程又は他の塩工程を取り入れることもある。本発明の一実施態様では、陽イオン交換クロマトグラフィの工程はHIC工程に先行する。
【0062】
VEGFを精製するための方法の例は以下に記載しており、例えば実施例VおよびVIを参照のこと。再折り畳みの後、プール中の不溶性物質はデプス濾過によって除去される。次いで、浄化したプールを、5-mM HEPPS/0.05% TRITONTMX100/pH8で平衡化したセラミックヒドロキシアパタイト(Bio Rad, Hercules, CA)に流す。非結合タンパク質は平衡バッファにて洗浄して除去し、50mM HEPPS/0.05% TRITONTMX100/0.15M リン酸ナトリウム/pH8の均一濃度工程を用いてVEGFを溶出する。VEGFのプールを、50mM HEPPS/0.05% TRITONTMX100/0.15M リン酸ナトリウム/pH8で平衡化したブチルセファロースTM高速流量のカラム(GE Healthcare, Uppsala, Sweden)に流す。カラムは平衡化バッファにて洗浄し、VEGFはカラム溶出物に集まる。ブチルセファロースTMプールを、50mM HEPES/pH8にて平衡化したMacro PrepハイSのカラム(BioRad, Hercules, CA)に流す。溶出物を洗浄した後、280nmの吸光度を規準とし、2カラム容量の50mM HEPES/0.25M 塩化ナトリウム/pH8にて洗浄する。VEGFは、線形の、8カラム容量の0.25〜0.75M 塩化ナトリウムの濃度勾配を含む50mM HEPES/pH8を用いて溶出する。分画を回収し、ヘパリン-結合アッセイによって測定されるように、適切に折り畳まれたVEGFを含むものをプールする。
【0063】
Macro PrepハイSプールは、等量の50mM HEPES/0.8M クエン酸ナトリウム/pH7.5にて調整する。次いで、調整したプールを、50mM HEPES/0.4M クエン酸ナトリウム/pH7.5にて平衡化したフェニル5PW TSKのカラム(Tosohaas, Montgomeryville, PA)に流す。平衡バッファをカラムに通して非結合タンパク質を洗浄した後、10カラム容量の0.4−0M クエン酸ナトリウム勾配を含む50mM HEPES、pH7.5を用いてVEGFを溶出する。SDS‐ポリアクリルアミドゲル電気泳動によって分画をアッセイし、十分な純度のVEGFを含有するものをプールする。
【0064】
宿主細胞におけるヘパリン結合タンパク質の発現
簡潔に言えば、自律複製が可能な発現ベクターと宿主の原核生物細胞ゲノムと関連するタンパク質発現を宿主細胞に導入する。適する発現ベクターの構築は、本明細書中に記載のヘパリン結合タンパク質のヌクレオチド配列を含め、当分野で周知である。例えば、Sambrook等, Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press (Cold Spring Harbor, New York) (2001);Ausubel等, Short Protocols in Molecular Biology, Current Protocols John Wiley and Sons (New Jersey) (2002);及びBaneyx, (1999) Current Opinion in Biotechnology, 10:411-421を参照。細菌を含む適切な原核生物細胞、発現ベクターは、例えばAmerican Type Culture Collection (ATCC), Rockville, Marylandにより市販されている。原核細胞、特に細菌細胞培養物の大規模増殖のための方法は当分野で周知であり、これらの方法が本発明の関係において用いられうる。
【0065】
例えば、原核生物宿主細胞に対象のヘパリン結合タンパク質をコードする発現ないしはクローニングベクターを形質移入し、プロモーターの誘導、形質転換体の選別、又は所望の配列をコードする遺伝子の増幅に応じて変更した従来の栄養培地中で培養する。対象のポリペプチドをコードする核酸は、対象のポリペプチド(一又は複数)をコードする限りにおいて、好ましくは任意の供給源由来のRNA、cDNA又はゲノムDNAである。微生物宿主において異種性ポリペプチド(その変異体を含む)の発現に適する核酸を選別するための方法は周知である。ポリペプチドをコードする核酸分子は当分野で公知の様々な方法によって調整する。例えば、VEGFをコードするDNAは、例えば、VEGFをコードする遺伝子に特異的に結合することができるオリゴヌクレオチドプローブを用いて、単離し、配列決定する。
【0066】
異種の核酸(例えば、cDNA又はゲノムDNA)は、適切なプロモーターの制御の下、微生物における発現のための複製可能なベクターへ適切に挿入される。多くのベクターは、かかる目的のために利用可能であり、適当なベクターの選択は、主としてベクター内に挿入されるべき核酸のサイズ、及びベクターにより形質転換される特定の宿主細胞に依存する。各ベクターは、それが適合する特定の宿主細胞に依存する種々の成分を含む。特定の宿主の種類に依存し、一般にベクター成分には、限定はしないが、以下の一又は複数が含まれる:シグナル配列、複製開始点、一又は複数のマーカー遺伝子、プロモーター、及び転写終結配列。
【0067】
一般には、宿主細胞と適合性のある種に由来するレプリコン及び制御配列を含むプラスミドベクターが、微生物宿主との関連で用いられる。そのベクターは、通常、複製部位、並びに形質転換細胞において表現型の選択を提供可能なマーキング配列を保持する。例えば、大腸菌は、典型的には、E.coli種由来のプラスミドであるpBR322を使って形質転換される(例えば、Bolivar等、Gene, 2: 95 (1977)参照)。pBR322は、アンピシリン及びテトラサイクリン耐性の遺伝子を含んでおり、よって形質転換細胞を同定するための簡単な手段を提供する。そのpBR322プラスミド、もしくは他の微生物プラスミド又はファージもまた、選択マーカー遺伝子の発現のために宿主によって使用され得るプロモーターを含むか、又は含むよう改変される。
【0068】
(i) シグナル配列
本発明のポリペプチドは、直接産生されるだけではなく、典型的にはシグナル配列あるいは成熟ポリペプチドないしはポリペプチドのN末端に特異的切断部位を有する他のポリペプチドである異種性ポリペプチドとの融合ポリペプチドとしても産生される。典型的には選択された異種性シグナル配列は、宿主細胞によって認識されプロセシングされる(すなわち、シグナルペプチダーゼによって切断される)ものである。天然のポリペプチドシグナル配列を認識せずプロセシングしない原核生物宿主細胞に対しては、シグナル配列は、例えばアルカリホスファターゼ、ペニシリナーゼ、lppあるいは熱安定なエンテロトキシンIIリーダーの群から選択される原核生物シグナル配列により置換できる。
【0069】
(ii) 複製開始点成分
発現ベクターは、一又は複数の選択された宿主細胞においてベクターの複製を可能にする核酸配列を含む。そのような配列は様々な細菌についてよく知られている。プラスミドpBR322に由来する複製開始点は大腸菌などの大部分のグラム陰性細菌に好適である。
【0070】
(iii) 選択遺伝子成分
通常、発現ベクターは、選択可能マーカーとも称される選択遺伝子を含む。この遺伝子は、選択培地中で増殖する形質転換された宿主細胞の生存又は増殖に必要なタンパク質をコードする。選択遺伝子を含むベクターで形質転換されない宿主細胞は、培地中で生存できない。この選択可能マーカーは、本発明で利用され、定義されるような遺伝学的マーカーとは異なる。典型的な選択遺伝子は、(a)例えば、アンピシリン、ネオマイシン、メトトレキセートあるいはテトラサイクリンのような抗生物質又はその他の毒素に耐性を付与し、(b)遺伝学的マーカー(一又は複数)の存在によって誘導される欠陥以外の栄養要求性欠陥を補い、又は(c)例えばバチラス菌に対するD-アラニンラセマーゼをコードする遺伝子のような、複合培地から得られない重要な栄養素を供給する、タンパク質をコードする。
【0071】
選択技術の一例においては、宿主細胞の増殖を抑止する薬物が用いられる。この場合、対象の核酸で首尾よく形質転換したこれらの細胞は、抗薬物性を付与し、選択工程を生存するポリペプチドを産生する。このような優性選択の例としては、薬物ネオマイシン(Southern等, J. Molec. Appl. Genet, 1:327 (1982))、ミコフェノール酸(Mulligan等, Science, 209:1422 (1980))又はハイグロマイシン(Sugden等, Mol. Cell. Biol., 5:410-413 (1985))が使用される。上述の3つの例は、各々、適当な薬剤であるG418又はネオマイシン(ジェネティシン)、xgpt(ミコフェノール酸)、又はハイグロマイシンに対する耐性を伝達するために、真核生物での制御下で、細菌性遺伝子を利用する。
【0072】
(iv) プロモーター成分
対象のヘパリン結合タンパク質を産生するための発現ベクターは、宿主微生物によって認識され、対象のポリペプチドをコードする核酸と作用可能に連結される適切なプロモーターを含む。原核生物の宿主での使用に好適なプロモーターはβ-ラクタマーゼ及びラクトースプロモーター系(Chang等, Nature, 275:615 (1978); Goeddel等, Nature, 281:544 (1979))、アラビノースプロモータシステム(Guzman等, J. Bacteriol., 174:7716-7728 (1992))、アルカリホスファターゼ、トリプトファン(trp)プロモーター系(Goeddel, Nucleic Acids Res., 8:4057 (1980);欧州特許第36776号)、及びハイブリッドプロモーター、例えばtacプロモーター(deBoer 等, Proc. Natl. Acad. Sci. USA, 80:21-25 (1983))を含む。しかし、他の既知の細菌性プロモーターも適当である。それらのヌクレオチド配列は公表されており、それにより、任意の必要な制限酵素部位を供給するためのリンカー又はアダプターを用いて、当業者は対象のポリペプチドをコードするDNAにそれらを作用可能に連結する(Siebenlist等, Cell, 20:269 (1980))ことが可能となる。また、上記のSambrook等;及び上記のAusubel等も参照のこと。
【0073】
また、細菌のシステムで使用されるプロモーターも、通常、対象のポリペプチドをコードするDNAと作用可能に結合したシャイン・ダルガルノ(S.D.)配列を有する。該プロモーターは、制限酵素による切断により細菌由来のDNAから取り外すことができ、所望のDNAを含むベクター中へ挿入することができる。
【0074】
(v) ベクターの構築及び分析
一又は複数の上に列挙した成分を含む適切なベクターの構築には標準的なライゲーション技術を用いる。必要とされるプラスミドの生成のために、単離されたプラスミド又はDNA断片を切断させ、整え、そして望ましい型に再ライゲーションする。
構築されたプラスミド中において正しい配列であることを確認する分析のために、ライゲーション混合物を用いて、大腸菌K12菌株294(ATCC31446)又は他の株を形質転換し、適当な場合にはアンピシリン又はテトラサイクリン耐性によって、成功した形質転換細胞を選択する。形質転換細胞からプラスミドを調製し、制限エンドヌクレアーゼ消化により分析し、及び/又はSanger等, Proc. Natl. Acad. Sci. USA, 74:5463-5467 (1977)又はMessing等, Nucleic Acids Res., 9:309 (1981)の方法により、又はMaxam等, Methods in Enzymology, 65:499(1980)の方法により配列決定を行った。また、上記のSambrook等;及び上記のAusubel等も参照のこと。
対象のヘパリン結合タンパク質をコードする核酸は、宿主細胞に挿入される。これは、典型的に、上記の発現ベクターにて宿主細胞を形質転換させ、様々なプロモーターを誘導するために適するように変更した従来の栄養培地中で培養することによって達成される。
【0075】
宿主細胞の培養
対象のヘパリン結合タンパク質を発現するために用いる好適な原核生物細胞は当分野で周知である。典型的に、封入体の形態で、又は周辺質ないしは細胞内腔に大量に組み換えタンパク質を発現する宿主細胞が用いられる。適切な原核生物には、例として、真正細菌、例えばグラム陰性又はグラム陽性生物体、例えば大腸菌、桿菌、例えば枯草菌、シュードモナス属種、例えば、緑膿菌、ネズミチフス菌又はセラチア‐マルセスセンスなどがある。大腸菌宿主の一例は大腸菌294(ATCC 31,446)である。また、他の細胞株、例として、大腸菌B、大腸菌X1776(ATCC 31,537)および大腸菌W3110(ATCC 27,325)も好適である。これらの例は限定するものでなく例示である。株W3110は、組換えDNA産生発酵のための共通の宿主株であるので典型的な宿主である。本発明のある態様では、宿主細胞は最小量のタンパク質分解酵素を分泌する。例えば、株W3110は、タンパク質をコードする遺伝子に遺伝子変異を起こすように修飾してもよく、このような宿主の例には1995年4月25日発行の米国特許第5410026号に記載の大腸菌W3110株1A2、27A7、27B4及び27C7が含まれる。例えば、VEGFの産生のための株は、49B3と称される、遺伝子型tonAΔ ptr3 phoAΔE15 Δ(argF-lac)169 degP41 ilvgを有する大腸菌株W3110である。他の例では、VEGFの産生のための株は、遺伝子型ΔfhuA (ΔtonA) ptr3,lacIq,lacL8,ΔompT Δ(nmpC-fepE) ΔdegP ilvG+を有する大腸菌株(62A7)である。また、例えば国際公開第2004/092393号の頁23〜24にわたる表を参照のこと。
【0076】
対象のヘパリン結合タンパク質を生産するために使用される原核細胞は、当技術分野で既知の培地において成長させたもので、選択された宿主細胞の培養に適しており、一般的にSambrook等, Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press (Cold Spring Harbor, New York) (2001)に記載の培地を含む。細菌に適した培地には、AP5培地、栄養ブロス、Luria-Bertani(LB)ブロス、Neidhardt's最少培地、及びC.R.A.P.最少又は完全培地に必要な栄養を補ったものが含まれるが、これらに限定されるものではない。好ましい実施形態では、培地は発現ベクターの構造に基づいて選択された選択剤も含み、よって発現ベクターを含む原核細胞を選択的に成長させることができる。例えば、アンピシリン耐性遺伝子を発現する細胞を成長させるために、培地にアンピシリンを加える。炭素、窒素、及び無機リン酸塩の供給源に加え、必要な補充物は、単独で、或いは、別の補充物又は複合窒素源等の培地と組み合わせて導入され、適切な濃度で含有される。場合によっては、培養培地は、グルタチオン、システイン、システアミン、チオグリコール酸、ジチオエリスリトール、及びジチオスレイトールからなる群から選択された1又は複数の還元剤を含んでよい。
【0077】
適切な培地の例は、米国特許第5304472号及び同第5342763号に見ることができる。C.R.A.P.リン酸塩制限培地は、3.57gの(NH4)2(SO4)、0.71gのクエン酸Na-2H2O、1.07gのKCl、5,36gの酵母菌抽出物(認証済み)、5.36gのHycaseSFTM−Sheffieldからなり、KOHによりpHを7.3に、脱イオン化H2Oによりqsを872mlに調整したものをオートクレーブしてから55℃に冷却し、110mlの1M MOPS(pH7.3)、11mlの50%グルコース、7mlの1M MgSO4を補ったものである。次いでカルベニシリンを50μg/mlの濃度で導入培地に添加してもよい。
【0078】
原核宿主細胞を適切な温度で培養する。例えば、大腸菌の成長に好ましい温度は約20〜約39℃、又は約25℃〜約37℃、又は約30℃である。
アルカリホスフェートプロモーターが使用される場合、本発明の対象のポリペプチドの生産のために使用される大腸菌細胞は、例えばSambrook等, Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Laboratory Press (Cold Spring Harbor, New York) (2001)に一般的に記載されるようにアルカリホスフェートプロモーターが部分的に又は完全に誘導され得る適切な培地中で培養される。必要とされる培養は、無機リン酸塩の非存在下又はリン酸塩欠乏レベル下では、決して起こらない。第一に、培地はタンパク質合成の誘導レベル以上であり、細菌の増殖に十分な量の無機リン酸塩を含む。細胞が増殖し、リン酸塩を利用する場合、培地中のリン酸塩レベルを低下させ、それによりポリペプチドの合成の誘導を引き起こす。
【0079】
プロモーターが、起こるべき誘導に関し誘導性プロモーターである場合、典型的に細胞はある最適な濃度、例えば、高い細胞濃度過程を用い、点誘導が開始される(例えば、誘導因子の添加、培地成分の除去により)約200のA550が達成されるまで培養され、対象のポリペプチドをコードする遺伝子の発現を誘導する。
必要ないかなる補充物も、単独で、或いは又は別の補充物ないしは複合窒素源等の培地と組み合わせて導入し、当業者に既知の適切な濃度で含めることができる。培地のpHは、主に宿主生物に応じて、およそ5〜9とすることができる。大腸菌については、pHはおよそ6.8〜およそ7.4であり、又はおよそ7.0である。
【0080】
ヘパリン結合タンパク質の製剤
例えば本明細書中に記載の方法を用いて回収したポリペプチドは、薬学的に受容可能な担体に調製化してもよく、様々な診断、治療、又は該分子について公知の他の使用のために用いられる。例として、本明細書中に記載のVEGFは、酵素免疫アッセイなどのイムノアッセイに使われてもよい。本明細書中に記載の方法を用いて得られたヘパリン結合タンパク質についての治療的使用も考慮される。例として、増殖因子又はホルモン、例えばVEGFは所望のように増殖を促すために用いられうる。例として、VEGFは、例えば急性の創傷(例えば熱傷、外科的創傷、通常の創傷など)又は慢性の創傷(例えば糖尿病性潰瘍、褥瘡、褥瘡性潰瘍、静脈潰瘍など)の創傷治癒の促進、発毛促進、組織増殖及び修復(例えば骨、肝臓など)などのために用いられうる。
【0081】
ヘパリン結合タンパク質の治療製剤は、任意の薬学的に許容可能な担体、賦形剤又は安定剤と所望の純度を有する分子、例えばポリペプチドを混合することによって(Remington's Pharmaceutical Sciences, 16版, Osol,A編 [1980])、凍結乾燥製剤又は水溶液の形態で保存用に調製される。許容できる担体、賦形剤又は安定剤は、用いる投与量及び濃度では細胞に対して無毒性であり、リン酸、クエン酸及び他の有機酸等の緩衝液;アスコルビン酸及びメチオニンを含む抗酸化剤;防腐剤(例えば、オクタデシルジメチルベンジルアンモニウムクロリド;塩化ヘキサメトニウム;塩化ベンザルコニウム、塩化ベンゼトニウム;フェノール、ブチル又はベンジルアルコール;アルキルパラベン類、例えばメチル又はプロピルパラベン;カテコール;レゾルシノール;シクロヘキサノール;3-ペンタノール;及びm-クレゾール);低分子量(残基数10個未満)ポリペプチド;血清アルブミン、ゼラチン又は免疫グロブリン等のタンパク質;ポリビニルピロリドン等の親水性重合体;グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン、又はリシン等のアミノ酸;グルコース、マンノース又はデキストリン等の単糖類、二糖類及び他の炭水化物;EDTA等のキレート剤;スクロース、マンニトール、トレハロース又はソルビトール等の糖類;ナトリウム等の塩形成対イオン;金属錯体(例えばZn-タンパク質錯体);及び/又はTWEENTM、PLURONICSTM又はポリエチレングリコール(PEG)等の非イオン性界面活性剤を含む。
【0082】
ある実施態様では、インビボ投与のために使用される製剤は無菌である。これは滅菌濾過膜による濾過によって達成される。HBPは、凍結乾燥された形態、又は、水溶液又はゲル形態で保存されうる。より高い又はより低いpH値でも好適な場合があるが、HBP調製物のpHはおよそ5〜8まででありうる。賦形剤、担体、又は安定剤の使用によりHBPの塩類が形成されうることは理解されるであろう。
典型的に創傷治癒のために、HBPは部位特異的運搬用に調製される。局所に適応される場合、HBPは担体及び/又はアジュバントなどの他の成分と適切に組み合わされる。前記の他の成分の性質に制限はないが、ただし意図する投与のために効果的かつ薬学的に受容可能でなければならず、組成物の活性成分の活性を有意に分解することができないものである。精製されたコラーゲンの有無にかかわらず、適切な溶媒の例には、軟膏、クリーム、ゲル、噴霧又は懸濁液が含まれる。また、組成物は、場合によって液体又は半流動体形態で、滅菌包帯、経皮パッチ、絆創膏及び包帯にしみ込ませてもよい。
【0083】
ゲル製剤を得るため、液体組成物に調製されるHBPは、有効量の水溶性多糖類又は合成ポリマー、例としてポリエチレングリコールと混合して、適切な粘性のゲルを形成して、局所に適応してもよい。使われうる多糖類には、例えば、エーテル化したセルロース誘導体などのセルロース誘導体、例えば、アルキルセルロース、ヒドロキシアルキルセルロース、及びアルキルヒドロキシアルキルセルロース、例えばメチルセルロース、ヒドロキシエチルセルロース、カルボキシメチルセルロース、ヒドロキシプロピルメチルセルロース及びヒドロキシプロピルセルロース;澱粉及び分画された澱粉;寒天;アルギン酸及びアルギナート;アラビアゴム;プルラン;アガロース;カラギーナン;デキストラン;デキストリン;フルクタン;イヌリン;マンナン;キシラン;アラビナン;キトサン;グリコーゲン;グルカン;及び、合成バイオポリマー;並びに、キサンタンガムなどのガム;ガーゴム;ローカストビーンガム;アラビアゴム;トラガカントゴム;及び、インドゴム;及び、その誘導体及び混合物が含まれる。本発明のある実施態様では、本明細書中のゲル化剤は、例えば、生物系に不活性、非中毒性、調製がしやすく、及び/又はあまり流れやすくないか粘性がなくて、その中に含まれるHBPを不安定にしないものである。
【0084】
ある実施態様では、多糖は、エーテル化されたセルロース誘導体であって、他の実施態様では、十分に限定され、精製され、USPに列挙されるもの、例えばメチルセルロース及びヒドロキシアルキルセルロース誘導体、例として、ヒドロキシプロピルセルロース、ヒドロキシエチルセルロース及びヒドロキシプロピルメチルセルロースである。一実施態様では、メチルセルロースが多糖である。メチルセルロースがゲルに使用される場合、典型的には、ゲルのおよそ2〜5%、又はおよそ3%、又はおよそ4%、又はおよそ5%を含んでなり、HBPはゲルの1ml当たりおよそ300〜1000mgの量で存在する。
ゲル化のために有用なポリエチレングリコールは、一般的に、適当な粘性を得るために低分子量と高分子量のポリエチレングリコールの混合物である。この目的のために、例えば、分子量400〜600のポリエチレングリコールと分子量1500のポリエチレングリコールとの混合物は、ペーストを得るために適切な比率で混合される場合に有効である。
【0085】
多糖類及びポリエチレングリコールに用いる「水溶性」なる用語はコロイド溶液及び分散液を含むことを意味する。通常、セルロース誘導体の溶解性はエーテル基の置換の程度で決定され、本明細書において有用な安定化誘導体は誘導体を水溶性にするためにセルロース鎖のアンヒドログルコース単位当たりの該エーテル基を十分な量有しなければならない。一般的に、アンヒドログルコース単位当たり少なくとも0.35のエーテル基のエーテル置換の程度が十分である。さらに、セルロース誘導体は、アルカリ金属塩、例えばLi、Na、K又はCs塩類の形態であってもよい。
【0086】
例えば、Remington's Pharmaceutical Sciences 18th edition, Gennaro, A. Ed. (1995)を参照。またJohnson等, Nat. Med., 2:795-799 (1996);Yasuda, Biomed. Ther., 27:1221-1223 (1993);Hora等, Bio/Technology, 8:755-758 (1990);Vaccine Design: The Subunit and Adjuvant Approach, Powell and Newman, eds, (Plenum Press: New York, 1995)のCleland, "Design and Production of Single Immunization Vaccines Using Polylactide Polyglycolide Microsphere Systems," pp. 439-462;国際公開第97/03692号、国際公開第96/40072号、国際公開第96/07399号;米国特許第5654010号;DE第3218121号;Epstein等, (1985) Proc. Natl. Acad. Sci. USA, 82:3688-3692;Hwang等, (1980) Proc. Natl. Acad. Sci. USA, 77:40304034;欧州特許第52322号;欧州特許第36676号;欧州特許第88046号;欧州特許第143949号;欧州特許第142641号;日本特許出願83-118008;米国特許第4485045号及び同第4544545号;及び欧州特許第102324号も参照のこと。
以下の実施例は、限定するものではなく例示するためのものである。
【実施例】
【0087】
実施例I:大腸菌において発現した組み換えヒトVEGF
組み換えヒトVEGFを大腸菌において発現させた。合成の間、タンパク質は細胞膜周辺腔に分泌され、屈折小体として蓄積された。したがって、タンパク質の抽出および再折り畳みを達成するために研究を行った。これらの研究により、ポリ陰イオン剤を付加することなく標準的な回収技術を用いて少なくとも3種のVEGF(図1)が単離されたことが明らかとなった。天然のVEGFを用いた研究により、ヘパリン添加がカオトロプ-及びチオール-誘導性の変性剤に対する耐性を増すことが示された(図2)。さらに、ヘパリンは、小規模再折り畳み実験において適切に再折り畳みされたVEGFの量を有意に増やした。この結果を大規模な方法に応用するために、ヘパリンに構造上類似する分子であるデキストラン硫酸の存在下においてVEGFの再折り畳みを可能にした条件を探した。デキストラン硫酸の添加により、適切に折り畳まれた生物学的に活性なVEGFの収率がコントロールと比べて3〜5倍に増加した。
【0088】
方法
VEGF165発現のためのプラスミド−プラスミドpVEGF171は大腸菌周辺質におけるヒトVEGF165の発現のために設定した(例えば、Leung等, (1989) Science, 246:1306-1309を参照)。VEGFコード配列の転写は、アルカリホスファターゼ(AP)プロモーターの厳格なコントロール下にあのに対して(例えば、Kikuchi等, (1981) Nucleic Acids Research, 9:5671-8を参照)、翻訳開始に必要な配列はシャイン-ダルガーノ領域にある(例えば、Yanofsky等, (1981) Nucleic Acids Research, 9:6647-68を参照)。VEGFコード配列は、その後の大腸菌周辺質への分泌のために細菌性耐熱性毒素II(STII)シグナル配列の下流に融合した(例えば、Lee等, (1983) Infect. Immun. 42:264-8;及びPicken等, (1983) Infect. Immun. 42:269-75を参照)。STIIシグナル配列のコドン修飾により翻訳レベルが調節され、その結果、周辺質におけるVEGFが最適レベルに集積した(例えば、Simmons and Yansura, (1996) Nature Biotechnoloy, 14:629-34を参照)。転写終結区に対するラムダ(例えば、Scholtissek and Grosse, (1987) Nucleic Acids Research 15:3185を参照)はVEGF翻訳終止コドンの下流に位置した。複製起点と、アンピシリン及びテトラサイクリン耐性遺伝子は、プラスミドpBR322から得た。例えば、Bolivar等, (1977) Gene 2:95-113を参照。
【0089】
細胞均質化と屈折小体の調製−回収した大腸菌細胞を凍結し、−70℃に保存した。細胞は、BTUX (centrifuge, Alfa laval)遠心にて回収し、BEPEX(大きいサイズのフリーザ)に凍結保存した。細胞は、5容量の50mM HEPES/150mM NaCl/5mM EDTA pH7.5(5L/kgペレット)に懸濁し、モデル15M実験室ホモジナイザーGaulin 15M(小型)又はM3(大型) (Gaulin Corporation, Everett, MA) にて均質化した。次いで、細胞懸濁液を等量の同じバッファにて希釈し、BTPX205の(Alfa Laval Separation AB(Tumba, Sweden)連続速度遠心にて遠心分離して屈折小体を回収した。中間生成物スケール遠心機はSA1を使用した。あるいは、細胞を均質にし、BEPEXに凍結して再水和することなく直接ペレットを回収することもできる。
【0090】
実施例II:大腸菌-I(Escherichia coli-I)において発現させた組み換えヒトVEGFの抽出及び再折り畳み
方法
抽出と再折り畳み−屈折性ペレットは、5Lのバッファ/kgペレットで、7M 尿素/50mM HEPPS/pH8(終濃度)を含有する抽出バッファに懸濁した。次いで、固形ジチオトレイトールを、4mM終濃度となるように3.7g/kgペレットで加えた。例として、VEGFの抽出に対する尿素とDTTの効果についての図9を参照のこと。懸濁液は20℃で1〜2時間かけて完全に混合した。pHは、50%水酸化ナトリウム(w/w)にてpH8.0に調整してもよい。抽出バッファの容量当たり、19容量の再折り畳みバッファを加えて再折り畳みを生じさせた。再折り畳みバッファには、50mM HEPPS/1 M−2M 尿素/2−5mM システイン/0.05%−0.2%TRITONTMX100/pH8、終濃度を含めた。例えば、再折り畳みの間に存在する尿素およびDTT濃度の効果についての図10を参照のこと。示すように、デキストラン硫酸、ヘパリン又は硫酸ナトリウムを加えた。再折り畳みインキュベーションは室温で4〜24時間行った。場合によって、インキュベートは室温で最大およそ48時間行うことができる。折畳みは、SDS-PAGEおよび/またはヘパリンHPLCにてモニターした。生成物は、Cuno90SPフィルターによるデプス濾過(depth filtration)の後に0.45μm濾過を行って浄化した。
【0091】
ヘパリン-結合HPLCアッセイ−適切に再び折り畳まれたVEGFの質および量は、固定されたヘパリンを含有するカラムを使用して決定した。カラムPOROS HE2/M(4.6×100mm、HE2/M、PerSeptive BioResearch Products, Cambridge, MA)は0.15M 塩化ナトリウムを含有する10mM リン酸ナトリウム、pH7にて平衡化した。1ml/分又は2ml/分の流速で、カラムは、10分かけて、0.15〜2M 塩化ナトリウムを含む平衡バッファから直線濃度勾配を使用して溶出した。あるアッセイでは、16分で溶出した。溶出物を280nmでモニターした。一般的に、ほとんどのタンパク質は空隙容量で溶出し、3種のVEGFが同定されうる。最も高い親和性で、最も溶出が遅い種を適切に折り畳まれたVEGFとして同定し、その後「ピーク3のVEGF」と同定した。
【0092】
結果
システインが媒介する変性からVEGFを保護するヘパリン−天然のVEGFに10mM システインを加えると、適切に折り畳まれた分子が大きく減少した(図2)。この変性は、20mMほどのわずかな濃度の2つの異なる形態のヘパリンを加えると阻止された。
【0093】
表Ia
VEGF再折り畳みに対するヘパリンとデキストラン硫酸の効果
表の値は、屈折性ペレット(g)当たりのピーク3の形成されたVEGF(mg)の量である。各々の添加の濃度は示すとおりである。*平均コントロール(5.6+5.0=5.3)
【0094】
表Ib
VEGF再折り畳みに対する硫酸ナトリウムの効果
表の値は、屈折性ペレット(g)当たりのピーク3の形成されたVEGF(mg)の量である。硫酸ナトリウムの濃度は示すとおりである。
【0095】
表II
VEGF再折り畳みに対するヘパリンとデキストラン硫酸の効果
表の値は、屈折性ペレット(g)当たりのピーク3の形成されたVEGF(mg)の量である。
【0096】
まとめ
再折り畳み収率を増加させるヘパリンおよびデキストラン硫酸−上記の変性に対する保護的特性のため、VEGFの再折り畳みに対する様々な異なる形態の硫酸化ポリマーの効果を調べた。表Ia(及び図5)に示すように、低分子量及び高分子量の形態のヘパリンは再び折り畳まれたVEGFの収率をおよそ3倍に増加した。表Ib(及び図6)に示すように、硫酸ナトリウムは再び折り畳まれたVEGFの収率をおよそ2倍に増加した。また、10Kd形態のデキストラン硫酸は再折り畳み収率の増加に有効であったが、調査した濃度の高い範囲では基質阻害が生じた。更なる研究により、5Kd、8Kd及び10Kdの形態のデキストラン硫酸はすべて、再折り畳みの際のVEGFの収率を有意に増加したことが示された。図7を参照。また図8も参照。
【0097】
実施例III:VEGFの回収に対する異なるバッファとTRITONTMX-100の効果
結果
【0098】
まとめ
実施例I、II及びIIIのデータをまとめると、ヘパリン-結合増殖因子であるVEGFが再び折り畳まれる際にヘパリン硫酸塩又はデキストラン硫酸のいずれかを含めると収率が有意に(2〜5倍)改善することと、回収条件が示唆される。この方法は工業規模で成功裏に実施された。この方法はヘパリンを結合する他の基本的な増殖因子および他のタンパク質の再折り畳みに応用することができると期待される。
【0099】
実施例IV:大腸菌-II(Escherichia coli-II)において発現される組み換えヒトVEGFの
方法
抽出と再折り畳み−屈折性ペレットを抽出バッファに懸濁した。この抽出バッファの終濃度は5Lのバッファ/kgペレットで7M 尿素、2−30mM DTT(例えば10mM DTT)、50mM HEPPS/pH7−9(例えばpH8)とした。懸濁液は室温で1〜2時間かけて完全に混合した。抽出バッファの容量当たり、19容量の再折り畳みバッファを加えて再折り畳みを生じさせた。再折り畳みバッファは、終濃度として1M又は1.3M尿素、2−15mM システイン(例えば7.5mM システイン)、0.5mM DTT、0−0.75M アルギニン(例えば100mM アルギニン)、15mg/Lのデキストラン硫酸、50mM HEPPS、0.05%TRITONTMX100/pH8を含有した。荷電アミノ酸の存在下における再折り畳みに対する効果について、例えば図12を参照のこと。ここでは、ヒスチジンの添加によりアミノ酸の付加がない場合と同じ効果が生じた。再折り畳みインキュベーションは室温で12〜24時間行った。場合によって、インキュベートは室温で最大およそ48時間行うことができる。場合によって、再折り畳みの間に空気又は酸素を添加してもよい(0.3〜1cc/分/L)。折畳みは、SDS-PAGEおよび/またはヘパリンHPLCにてモニターした。生成物は、Cuno90SPフィルターによるデプス濾過(depth filtration)の後に0.45μm濾過を行って浄化した。
【0100】
抽出および再折り畳みの工程の全体の希釈物は1:100とした。抽出及び再折り畳みの工程の全体の希釈の、例えば1:100へ、1:200への増加により活性なVEGFの総量が増加したが、濃度は低い。図13を参照。
再折り畳みの効率は、二量体/単量体の量を測定することによって決定することができ、このとき単量体は、C18逆相HPLCカラムで測定することができ、二量体形成は、ヘパリンカラムクロマトグラフィ又はSP-5PW陽イオン交換クロマトグラフィアッセイで測定することができる。
【0101】
実施例V:大規模な再折り畳み
最適化の再折り畳み条件の拡張性を試験するため、小規模(0.1L)、中規模(1L)及び試験的設備(250L〜400L)のスケールでの再折り畳みの動態を調べるために実験を行った。図4に示すように、大規模スケールの再折り畳みの動態はより小さいスケールと違いがなく、再び折り畳まれたVEGFの最終力価はわずかに増加した。これらのデータから、デキストラン硫酸による再折り畳みの拡張性が示された。さらに、生成物は、Cuno90SPフィルターによるデプス濾過(depth filtration)の後に0.45μm濾過を行って浄化した。
【0102】
実施例VI:再折り畳み後のrhVEGFの精製I
MacroPrepセラミックヒドロキシアパタイトクロマトグラフィ−再折り畳みの後、プール中の不溶性物質をデプス濾過によって除去した。次いで、浄化したプールを、50mM HEPPS/0.05% TRITONTMX100/pH8で平衡化したセラミックヒドロキシアパタイトカラム(35D×31H=30L) (Bio Rad, Hercules, CA)に流した。非結合タンパク質は平衡バッファにて洗浄して除去し、50mM HEPPS/0.05% TRITONTMX100/0.15M リン酸ナトリウム/pH8の均一濃度工程を用いてVEGFを溶出した。流速は120cm/時とした。プールする分画を分画のヘパリンHPLC分析にて測定した。
【0103】
ブチルセファロースTM高速流量クロマトグラフィ−VEGFのプールを、50mM HEPPS/0.05% TRITONTMX100/0.15M リン酸ナトリウム/pH8で平衡化したブチルセファロースTM高速流量のカラム(35D×26H=25L)(GE Healthcare, Uppsala, Sweden)に流した。流速は100cm/時とした。カラムは平衡化バッファにて洗浄し、VEGFはカラム溶出物に集まった。分画を集め、タンパク質含有分画をプールし、A280nmを測定した。
【0104】
Macro PrepハイSクロマトグラフィ−ブチルセファロースTMプールを、50mM HEPES/pH8にて平衡化したMacro PrepハイSのカラム(30D×39H=27 L)(BioRad, Hercules, CA)に流した。溶出物を洗浄した後、280nmの吸光度を規準とし、2カラム容量の50mM HEPES/0.25M 塩化ナトリウム/pH8にて洗浄した。VEGFは、線形の、8カラム容量の0.25〜0.75M 塩化ナトリウムの濃度勾配を含む50mM HEPES/pH8を用いて溶出した。流速を75〜200cm/時とした。分画を回収し、ヘパリン-結合アッセイ、例えばヘパリンHPLCによって測定したように、適切に折り畳まれたVEGFを含むものをプールした。
【0105】
フェニル5PW TSKクロマトグラフィ−Macro PrepハイSプールは、等量の50mM HEPES/0.8M クエン酸ナトリウム/pH7.5にて調整した。次いで、調整したプールを、50mM HEPES/0.4 M クエン酸ナトリウム/pH7.5にて平衡化したフェニル5PW TSKのカラム(18D×43H=11L)(Tosohaas, Montgomeryville, PA)に流した。平衡バッファをカラムに通して非結合タンパク質を洗浄した後、10カラム容量の0.4−0M クエン酸ナトリウム勾配を含む50mM HEPES、pH7.5を用いてVEGFを溶出した。SDS‐ポリアクリルアミドゲル電気泳動によって分画をアッセイし、十分な純度のVEGFを含有するものをプールした。
【0106】
限外濾過/ダイアフィルトレーション−プールしたVEGFを5kD再生セルロースメンブレン(G30619);Unit Pellicon;Feed Rate 17.1L/分に限外濾過した。ポリソルベート20にてメンブレンを調整した。プールしたVEGFは、6g/L(UF1)の濃度で限外濾過した。試料は、5mM コハク酸ナトリウム/275mM トレハロース/pH5.0と7−14DV(Diavolume)にてダイアフィルトレートした。最終製剤は、5mg/mlの濃度の、5mM コハク酸ナトリウム/275mM トレハロース/0.01%ポリソルベート20/pH5.0であった。
【0107】
実施例VII:再折り畳み後のrhVEGFの精製II
陽イオン交換液体クロマトグラフィ−再折り畳みの後、プール中の不溶性物質をデプス濾過によって除去することができる。再び折り畳まれたプールは、pH5.0−7.5、及び2〜6.5mS/cmに調整する。一実施態様では、プールはpH6.5および5mS/cmに調整している。次いで、再び折り畳まれたプールは、スルホプロピル高度負荷カラム(sulfopropyl extreme load column、SPXL)に流し、漸増塩濃度勾配により溶出した。プールした分画を、分画のヘパリンHPLC分析で測定することができる。
【0108】
疎水性相互作用カラム(HIC):VEGFのSPXL溶出プールを、フェニルTSKクロマトグラフィカラム(Tosohaas, Montgomeryville, PA)に流すために、50mS/cmに調整してもよい。分画を集め、タンパク質含有分画をプールする。
【0109】
IEX又は混合モード:フェニルTSKプールを、イオン交換クロマトグラフィ(IEX)又は混合モードクロマトグラフィのカラムに流してもよい。分画を集め、本明細書に記載のアッセイによって決定した適切に折り畳まれたVEGFを含むものをプールした。
【0110】
限外濾過/ダイアフィルトレーション−プールしたVEGFを5kD再生セルロースメンブレン(G30619);Unit Pellicon;Feed Rate 17.1L/分に限外濾過してもよい。例えば、ポリソルベート20にてメンブレンを調整する。プールしたVEGFは、6g/L(UF1)の濃度で限外濾過する。試料は、5mM コハク酸ナトリウム/275mM トレハロース/pH5.0と7−14DV(Diavolume)にてダイアフィルトレートする。
【0111】
本明細書に記載の方法及びプロセスでは、最終純度及び/又は活性は、ペプチドマッピング、ジスルフィドマッピング、SDS-PAGE(還元及び非還元の両方)、円二色性、リムルスアメーバ様細胞可溶化液(LAL)、ヘパリンクロマトグラフィ、ヘパリンHPLC(例えば、ヘパリンHPLCを用いてVEGF二量体濃度を決定することができる)、逆相(rp)HPLCクロマトグラフィ(例えば、rpHPLCを用いてVEGF単量体の濃度を決定することができる)、ヘパリン結合、レセプター結合(例えば、VEGF、例えばKDRレセプター結合-Bioanalytic R&D、及び/又はFlt1レセプター結合に対して)、SEC分析、細胞アッセイ、HUVEC作用強度アッセイ、VEGF抗体を用いたELISA、質量分析などによって評価することができる。
【0112】
本明細書に記載の寄託、実施例及び実施態様は例示的な目的のみであり、この見地の様々な修飾及び変更は当業者に示唆されており、本出願の精神及び権限と添付の特許請求の範囲内に包含されることは理解されるものである。本明細書において引用されるすべての刊行物、参考文献、特許および特許出願は、すべての目的のために出典明記によりその全体がここに援用される。
【図面の簡単な説明】
【0113】
【図1】POROS HE2/Mカラム(4.6×100mm、PerSeptive BioResearch Products, Cambridge, MA)に流した細菌株W3110により産生されたVEGFのクロマトグラフを示す。例えば、POROS HE/2Mカラムは、0.15M 塩化ナトリウムを含有する10mM リン酸ナトリウム、pH7にて平衡化している。カラムは、10分かけて、0.15〜2M 塩化ナトリウムを含む10mM リン酸ナトリウム、pH7から直線濃度勾配を使用して溶出する。溶出物を280nmでモニターする。それぞれのピークで回収されたタンパク質はVEGFに対応するが、ピーク3のみは生物学的に活性な適切に再折り畳みされたVEGFに対応する。
【図2】ヘパリンによる天然の適切に折り畳まれたVEGFの安定化を表すグラフを示す。VEGFは、5mM EDTA、0.2M NaClおよび10mM システインを含有する50mM HEPES、pH8に懸濁する。
【図3A−3D】細菌株W3110によって産生され、それぞれ12μg/mlのデキストラン硫酸5000ダルトン(図3A)、12μg/mlのデキストラン硫酸8000ダルトン(図3B)、又は12μg/mlのデキストラン硫酸10000ダルトン(図3C)、又は25μg/mlのヘパリン(図3D)3000ダルトンとともにインキュベートし、POROS HE2/Mカラム(4.6×100 mm、PerSeptive BioResearch Products, Cambridge, MA)に流したVEGFのクロマトグラフを示す。例えば、カラムは、0.15M 塩化ナトリウムを含有する10mM リン酸ナトリウム、pH7にて平衡化している。カラムは、10分かけて、0.15〜2M 塩化ナトリウムを含む10mM リン酸ナトリウム、pH7から直線濃度勾配を使用して溶出する。溶出物を280nmでモニターする。それぞれのピークで回収されたタンパク質はVEGFに対応するが、ピーク3のみは生物学的に活性な適切に再折り畳みされたVEGFに対応する。
【図4】VEGFの再折り畳みに対する大きさの影響を示す。
【図5】VEGFの再折り畳みに対する低分子量(MW)及び高MW、及びデキストラン硫酸、10000ダルトンの効果を示す。ピーク3は、生物学的に活性な適切に再び折り畳まれたVEGFに対応する。
【図6】VEGFの再折り畳みに対する硫酸ナトリウムの影響を示す。ピーク3は、生物学的に活性な適切に再び折り畳まれたVEGFに対応する。
【図7】VEGFの再折り畳みに対する低分子量(MW)及び高MW、及びデキストラン硫酸、5000ダルトン、8000ダルトン、及び10000ダルトンの効果を示す。ピーク3は、生物学的に活性な適切に再び折り畳まれたVEGFに対応する。
【図8】VEGFの再折り畳みに対するヘパリンとデキストラン硫酸の効果を示す。ピーク3は、生物学的に活性な適切に再び折り畳まれたVEGFに対応する。
【図9】細菌封入体からのVEGFの抽出に対する尿素とDTTの効果を示す。
【図10】VEGFの再折り畳みに対する尿素とDTT濃度の影響を示す。
【図11】VEGF165のアミノ酸配列(配列番号:1)とともにジスルフィド結合を示す。
【図12】荷電アミノ酸の存在の効果を示す。第二緩衝溶液中0.75M濃度のアルギニンおよびリジンは有益であるのに対して、ヒスチジンはヒスチジンがない緩衝溶液と比較して、ほとんど相加的効果を有さない。さらに、アルギニンは0.1〜1M濃度で類似の効果を有することが示された。
【図13】%再折り畳み効率における希釈の効果を示すが、希釈が増すにつれて総VEGF濃度は低くなり、%再折り畳み効率は希釈が増すと高くなる。
【特許請求の範囲】
【請求項1】
原核生物の細胞培養物からヘパリン結合タンパク質を回収するための方法であって、
(a) 該原核生物の細胞培養物の周辺質から該ヘパリン結合タンパク質を単離する;
(b) カオトロピック剤と還元剤を含む第一緩衝溶液中で単離されたヘパリン結合タンパク質を変性させる;
(c) ヘパリン結合タンパク質の再折り畳みが生じる時間と条件で、カオトロピック剤と硫酸化ポリ陰イオン剤を含む第二緩衝溶液中で、変性したヘパリン結合タンパク質をインキュベートする;そして、
(d) 該再び折り畳まれたヘパリン結合タンパク質を回収する
工程を含んでなり、このとき硫酸化ポリ陰イオン剤とインキュベートしなかった場合と比べて再び折り畳まれたヘパリン結合タンパク質の回収がおよそ2〜5倍に増加する方法。
【請求項2】
前記ヘパリン結合タンパク質がヘパリン結合増殖因子である、請求項1に記載の方法。
【請求項3】
前記ヘパリン結合増殖因子が血管内皮細胞増殖因子(VEGF)である、請求項2に記載の方法。
【請求項4】
前記VEGFがVEGF165である、請求項3に記載の方法。
【請求項5】
前記硫酸化ポリ陰イオン剤がおよそ3000ダルトンから10000ダルトンの間である、請求項1に記載の方法。
【請求項6】
前記第二緩衝溶液がデキストラン硫酸を含む、請求項3に記載の方法。
【請求項7】
前記第二緩衝溶液が硫酸ナトリウムを含む、請求項3に記載の方法。
【請求項8】
前記第二緩衝溶液がヘパリンを含む、請求項3に記載の方法。
【請求項9】
前記デキストラン硫酸がおよそ8000から10000ダルトンの間である、請求項6に記載の方法。
【請求項10】
前記第一および第二の緩衝溶液がHEPPS pH8.0を含む、請求項3に記載の方法。
【請求項11】
さらに、前記第二緩衝溶液が還元剤を含む、請求項1に記載の方法。
【請求項12】
前記第二緩衝溶液の還元剤がシステインとDTTの組合せを含む、請求項11に記載の方法。
【請求項13】
さらに、前記第二緩衝溶液が非イオン性界面活性剤を含む、請求項1に記載の方法。
【請求項14】
さらに、前記第二緩衝溶液がアルギニンおよび/またはリジンを含む、請求項1に記載の方法。
【請求項15】
前記の回収工程(d)が、前記の再び折り畳まれたヘパリン結合タンパク質を、ヒドロキシアパタイトクロマトグラフィ支持体、第一疎水性相互作用クロマトグラフィ支持体;陽イオンクロマトグラフィ支持体、および第二疎水性相互作用クロマトグラフィ支持体に順次接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出させることを含む、請求項1に記載の方法。
【請求項16】
前記第一及び第二の疎水性相互作用クロマトグラフィ支持体がブチル-、プロピル-、オクチル-及びアリル-アガロース樹脂からなる群から選択される、請求項15に記載の方法。
【請求項17】
前記第一疎水性相互作用クロマトグラフィ支持体がブチル-アガロース支持体であり、前記第二疎水性相互作用クロマトグラフィ支持体がフェニル-アガロース支持体樹脂である、請求項15に記載の方法。
【請求項18】
前記回収工程(d)が、前記の再び折り畳まれたヘパリン結合タンパク質を、陽イオン交換支持体;疎水性相互作用クロマトグラフィ支持体、およびイオン交換クロマトグラフィ支持体に順次接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出することを含む、請求項1に記載の方法。
【請求項19】
原核生物の細胞培養物からヘパリン結合タンパク質を回収するための方法であって、
(a) 該ヘパリン結合タンパク質を該原核生物の細胞培養物の周辺質から単離する;
(b) カオトロピック剤と還元剤を含む第一緩衝溶液中で単離した該ヘパリン結合タンパク質を変性させる;
(c) ヘパリン結合タンパク質の再折り畳みが生じる時間と条件で、カオトロピック剤と硫酸化ポリ陰イオン剤を含む第二緩衝溶液中で、変性したヘパリン結合タンパク質をインキュベートし、このとき硫酸化ポリ陰イオン剤とインキュベートしなかった場合と比べて再び折り畳まれたヘパリン結合タンパク質の回収がおよそ2〜5倍に増加するものであり;そして、
(d) 再び折り畳まれた該ヘパリン結合タンパク質を、ヒドロキシアパタイトクロマトグラフィ支持体、第一疎水性相互作用クロマトグラフィ支持体、陽イオンクロマトグラフィ支持体、および第二疎水性相互作用クロマトグラフィ支持体に順次接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出させる
という工程を含んでなる方法。
【請求項20】
再び折り畳まれたヘパリン結合タンパク質を、ヒドロキシアパタイトクロマトグラフィ支持体、第一疎水性相互作用クロマトグラフィ支持体、陽イオンクロマトグラフィ支持体、および第二疎水性相互作用クロマトグラフィ支持体に順次接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出させる工程を含んでなる、ヘパリン結合タンパク質を精製するための方法。
【請求項21】
原核生物の細胞培養物からヘパリン結合タンパク質を回収するための方法であって、
(a) 該ヘパリン結合タンパク質を該原核生物の細胞培養物の周辺質から単離する;
(b) カオトロピック剤と還元剤を含む第一緩衝溶液中で単離した該ヘパリン結合タンパク質を変性させる;
(c) ヘパリン結合タンパク質の再折り畳みが生じる時間と条件で、カオトロピック剤と硫酸化ポリ陰イオン剤を含む第二緩衝溶液中で、変性したヘパリン結合タンパク質をインキュベートし、このとき硫酸化ポリ陰イオン剤とインキュベートしなかった場合と比べて再び折り畳まれたヘパリン結合タンパク質の回収がおよそ2〜5倍に増加するものであり;そして、
(d) 再び折り畳まれた該ヘパリン結合タンパク質を、陽イオン交換支持体;疎水性相互作用クロマトグラフィ支持体、およびイオン交換クロマトグラフィ支持体に順次接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出させる
という工程を含んでなる方法。
【請求項22】
前記ポリ陰イオン剤が、およそ3000ダルトンから10000ダルトンの間である、請求項19又は21に記載の方法。
【請求項23】
再び折り畳まれたヘパリン結合タンパク質を、陽イオン交換支持体;疎水性相互作用クロマトグラフィ支持体、およびイオン交換クロマトグラフィ支持体に順次接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出させる工程を含んでなる、ヘパリン結合タンパク質を精製するための方法。
【請求項1】
原核生物の細胞培養物からヘパリン結合タンパク質を回収するための方法であって、
(a) 該原核生物の細胞培養物の周辺質から該ヘパリン結合タンパク質を単離する;
(b) カオトロピック剤と還元剤を含む第一緩衝溶液中で単離されたヘパリン結合タンパク質を変性させる;
(c) ヘパリン結合タンパク質の再折り畳みが生じる時間と条件で、カオトロピック剤と硫酸化ポリ陰イオン剤を含む第二緩衝溶液中で、変性したヘパリン結合タンパク質をインキュベートする;そして、
(d) 該再び折り畳まれたヘパリン結合タンパク質を回収する
工程を含んでなり、このとき硫酸化ポリ陰イオン剤とインキュベートしなかった場合と比べて再び折り畳まれたヘパリン結合タンパク質の回収がおよそ2〜5倍に増加する方法。
【請求項2】
前記ヘパリン結合タンパク質がヘパリン結合増殖因子である、請求項1に記載の方法。
【請求項3】
前記ヘパリン結合増殖因子が血管内皮細胞増殖因子(VEGF)である、請求項2に記載の方法。
【請求項4】
前記VEGFがVEGF165である、請求項3に記載の方法。
【請求項5】
前記硫酸化ポリ陰イオン剤がおよそ3000ダルトンから10000ダルトンの間である、請求項1に記載の方法。
【請求項6】
前記第二緩衝溶液がデキストラン硫酸を含む、請求項3に記載の方法。
【請求項7】
前記第二緩衝溶液が硫酸ナトリウムを含む、請求項3に記載の方法。
【請求項8】
前記第二緩衝溶液がヘパリンを含む、請求項3に記載の方法。
【請求項9】
前記デキストラン硫酸がおよそ8000から10000ダルトンの間である、請求項6に記載の方法。
【請求項10】
前記第一および第二の緩衝溶液がHEPPS pH8.0を含む、請求項3に記載の方法。
【請求項11】
さらに、前記第二緩衝溶液が還元剤を含む、請求項1に記載の方法。
【請求項12】
前記第二緩衝溶液の還元剤がシステインとDTTの組合せを含む、請求項11に記載の方法。
【請求項13】
さらに、前記第二緩衝溶液が非イオン性界面活性剤を含む、請求項1に記載の方法。
【請求項14】
さらに、前記第二緩衝溶液がアルギニンおよび/またはリジンを含む、請求項1に記載の方法。
【請求項15】
前記の回収工程(d)が、前記の再び折り畳まれたヘパリン結合タンパク質を、ヒドロキシアパタイトクロマトグラフィ支持体、第一疎水性相互作用クロマトグラフィ支持体;陽イオンクロマトグラフィ支持体、および第二疎水性相互作用クロマトグラフィ支持体に順次接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出させることを含む、請求項1に記載の方法。
【請求項16】
前記第一及び第二の疎水性相互作用クロマトグラフィ支持体がブチル-、プロピル-、オクチル-及びアリル-アガロース樹脂からなる群から選択される、請求項15に記載の方法。
【請求項17】
前記第一疎水性相互作用クロマトグラフィ支持体がブチル-アガロース支持体であり、前記第二疎水性相互作用クロマトグラフィ支持体がフェニル-アガロース支持体樹脂である、請求項15に記載の方法。
【請求項18】
前記回収工程(d)が、前記の再び折り畳まれたヘパリン結合タンパク質を、陽イオン交換支持体;疎水性相互作用クロマトグラフィ支持体、およびイオン交換クロマトグラフィ支持体に順次接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出することを含む、請求項1に記載の方法。
【請求項19】
原核生物の細胞培養物からヘパリン結合タンパク質を回収するための方法であって、
(a) 該ヘパリン結合タンパク質を該原核生物の細胞培養物の周辺質から単離する;
(b) カオトロピック剤と還元剤を含む第一緩衝溶液中で単離した該ヘパリン結合タンパク質を変性させる;
(c) ヘパリン結合タンパク質の再折り畳みが生じる時間と条件で、カオトロピック剤と硫酸化ポリ陰イオン剤を含む第二緩衝溶液中で、変性したヘパリン結合タンパク質をインキュベートし、このとき硫酸化ポリ陰イオン剤とインキュベートしなかった場合と比べて再び折り畳まれたヘパリン結合タンパク質の回収がおよそ2〜5倍に増加するものであり;そして、
(d) 再び折り畳まれた該ヘパリン結合タンパク質を、ヒドロキシアパタイトクロマトグラフィ支持体、第一疎水性相互作用クロマトグラフィ支持体、陽イオンクロマトグラフィ支持体、および第二疎水性相互作用クロマトグラフィ支持体に順次接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出させる
という工程を含んでなる方法。
【請求項20】
再び折り畳まれたヘパリン結合タンパク質を、ヒドロキシアパタイトクロマトグラフィ支持体、第一疎水性相互作用クロマトグラフィ支持体、陽イオンクロマトグラフィ支持体、および第二疎水性相互作用クロマトグラフィ支持体に順次接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出させる工程を含んでなる、ヘパリン結合タンパク質を精製するための方法。
【請求項21】
原核生物の細胞培養物からヘパリン結合タンパク質を回収するための方法であって、
(a) 該ヘパリン結合タンパク質を該原核生物の細胞培養物の周辺質から単離する;
(b) カオトロピック剤と還元剤を含む第一緩衝溶液中で単離した該ヘパリン結合タンパク質を変性させる;
(c) ヘパリン結合タンパク質の再折り畳みが生じる時間と条件で、カオトロピック剤と硫酸化ポリ陰イオン剤を含む第二緩衝溶液中で、変性したヘパリン結合タンパク質をインキュベートし、このとき硫酸化ポリ陰イオン剤とインキュベートしなかった場合と比べて再び折り畳まれたヘパリン結合タンパク質の回収がおよそ2〜5倍に増加するものであり;そして、
(d) 再び折り畳まれた該ヘパリン結合タンパク質を、陽イオン交換支持体;疎水性相互作用クロマトグラフィ支持体、およびイオン交換クロマトグラフィ支持体に順次接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出させる
という工程を含んでなる方法。
【請求項22】
前記ポリ陰イオン剤が、およそ3000ダルトンから10000ダルトンの間である、請求項19又は21に記載の方法。
【請求項23】
再び折り畳まれたヘパリン結合タンパク質を、陽イオン交換支持体;疎水性相互作用クロマトグラフィ支持体、およびイオン交換クロマトグラフィ支持体に順次接触させ、選択的に各支持体から該ヘパリン結合タンパク質を溶出させる工程を含んでなる、ヘパリン結合タンパク質を精製するための方法。
【図1】

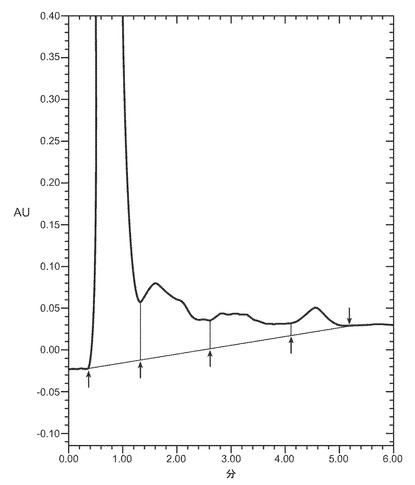
【図2】
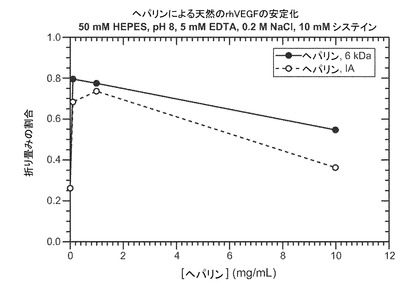
【図3A】
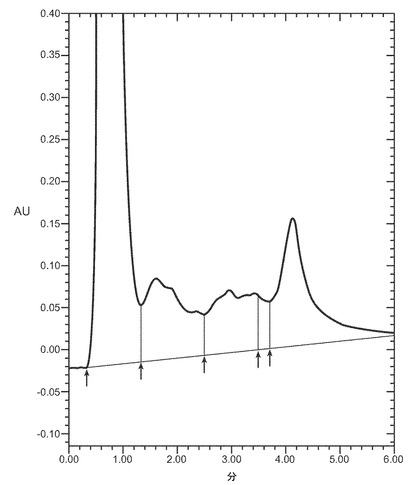
【図3B】
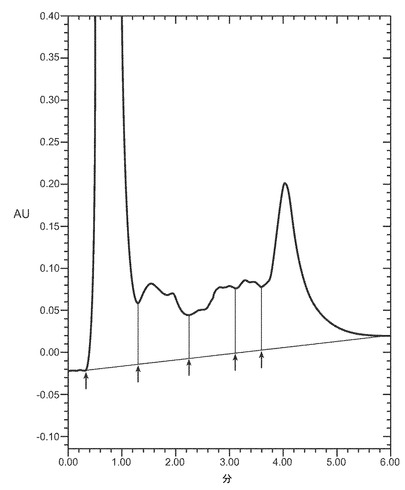
【図3C】
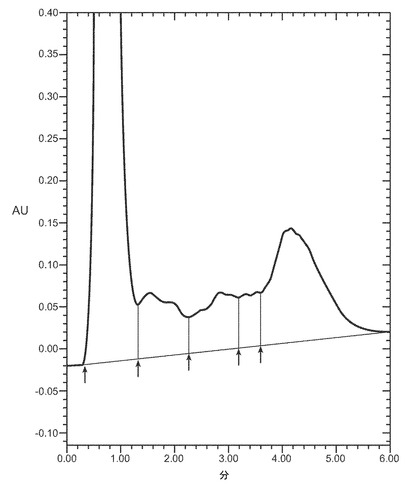
【図3D】
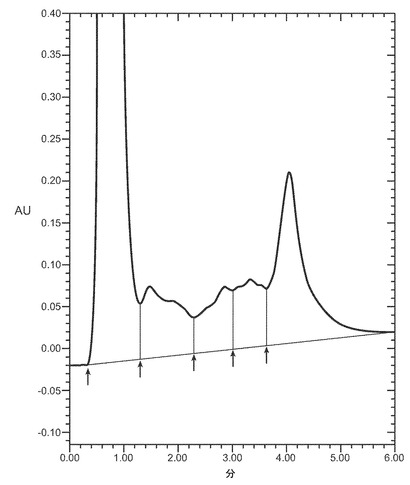
【図4】
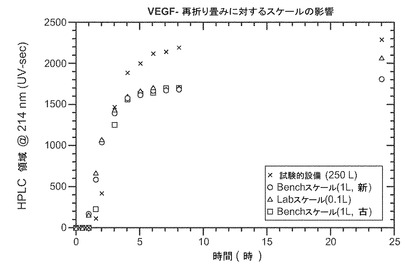
【図5】
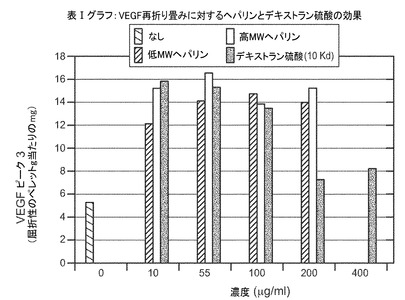
【図6】
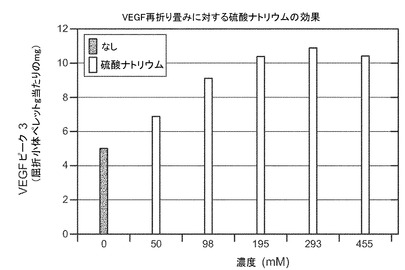
【図7】
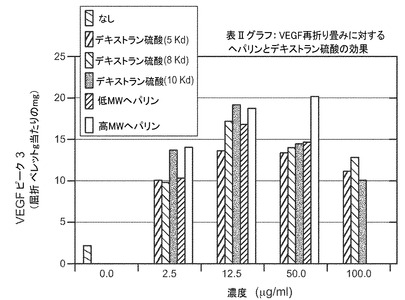
【図8】
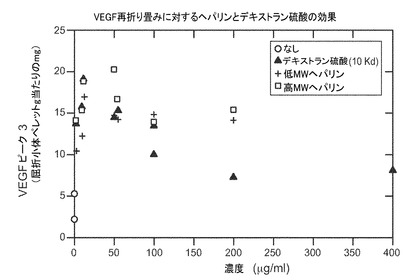
【図9】

【図10】
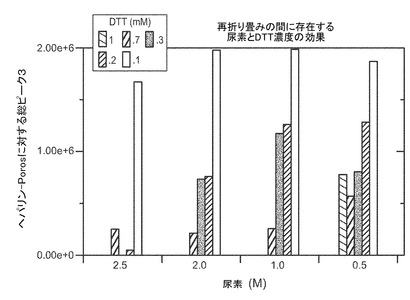
【図11】
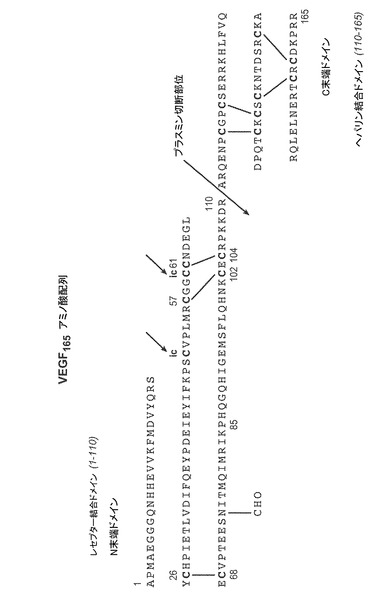
【図12】
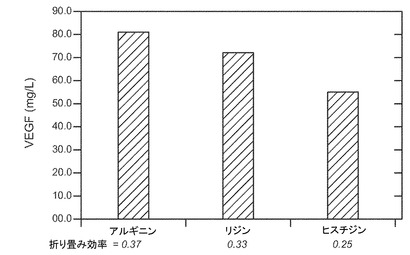
【図13】
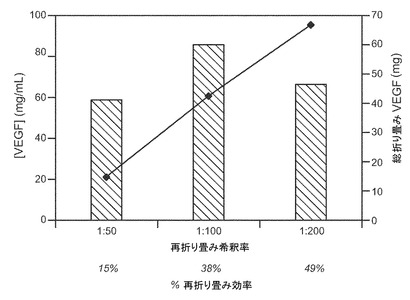
【図2】
【図3A】
【図3B】
【図3C】
【図3D】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公表番号】特表2009−521232(P2009−521232A)
【公表日】平成21年6月4日(2009.6.4)
【国際特許分類】
【出願番号】特願2008−547733(P2008−547733)
【出願日】平成18年12月19日(2006.12.19)
【国際出願番号】PCT/US2006/062320
【国際公開番号】WO2007/130154
【国際公開日】平成19年11月15日(2007.11.15)
【出願人】(596168317)ジェネンテック・インコーポレーテッド (372)
【氏名又は名称原語表記】GENENTECH,INC.
【Fターム(参考)】
【公表日】平成21年6月4日(2009.6.4)
【国際特許分類】
【出願日】平成18年12月19日(2006.12.19)
【国際出願番号】PCT/US2006/062320
【国際公開番号】WO2007/130154
【国際公開日】平成19年11月15日(2007.11.15)
【出願人】(596168317)ジェネンテック・インコーポレーテッド (372)
【氏名又は名称原語表記】GENENTECH,INC.
【Fターム(参考)】
[ Back to top ]