
上皮細胞増殖因子受容体に対するヒトモノクローナル抗体
【課題】ヒト上皮細胞増殖因子受容体(EGF-r)に対する抗体によって固形癌を抗体療法により治療するための、ヒトモノクローナル抗体の提供。
【解決手段】ヒト上皮細胞増殖因子受容体(EGF-r)に対する完全なヒトモノクローナル抗体が提供される。重鎖および軽鎖免疫グロブリン分子をコードするヌクレオチド配列および該分子を含むアミノ酸配列、特にCDR1からCDR3までの近接する重鎖および軽鎖配列に相当する配列が提供される。そのような免疫グロブリン分子およびモノクローナル抗体を発現するハイブリドーマも提供される。
【解決手段】ヒト上皮細胞増殖因子受容体(EGF-r)に対する完全なヒトモノクローナル抗体が提供される。重鎖および軽鎖免疫グロブリン分子をコードするヌクレオチド配列および該分子を含むアミノ酸配列、特にCDR1からCDR3までの近接する重鎖および軽鎖配列に相当する配列が提供される。そのような免疫グロブリン分子およびモノクローナル抗体を発現するハイブリドーマも提供される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明により、ヒト上皮細胞増殖因子受容体(EGF-r)に対する相補性決定領域に及ぶ完全なヒト近接(contiguous)重鎖および軽鎖配列モノクローナル抗体が提供される。重鎖および軽鎖免疫グロブリン分子をコードするヌクレオチド配列およびこれらの分子を含むアミノ酸配列、特に(CDR')、中でもCDR1からCDR3までに相当する配列が提供される。そのような免疫グロブリン分子およびモノクローナル抗体を発現するハイブリドーマも提供する。
【背景技術】
【0002】
EGF-rは、多くのタイプのヒト固形癌上に過剰発現されることが示されている。メンデルソーン(Mendelsohn)、Cancer Cells 7:359(1989)(非特許文献1)、メンデルソーン(Mendelsohn)、Cancer Biology 1:339〜344(1990)(非特許文献2)、モジュタエジ&ディーン(Modjtahedi and Dean)、Int'l J.Oncology 4:277〜296(1994)(非特許文献3)。例えば、EGF-rの過剰発現は特定の肺、乳房、結腸、胃、脳、膀胱、頭頸部、卵巣、および前立腺の癌腫に認められている[モジュタエジ&ディーン(Modjtahedi and Dean)、Int'l J.Oncology 4:277〜296(1994)(非特許文献3)]。上皮細胞増殖因子(EGF)およびトランスフォーミング増殖因子-α(TGF-α)はいずれもEGF-rに結合し、細胞増殖を引き起こし、腫瘍を増殖させることが示されている。
【0003】
このように、EGF、TGF-α、およびEGF-rに対する抗体はEGF-rを発現するまたは過剰発現する腫瘍の治療において有用となる可能性があることが一部のグループにより提唱された[メンデルソーン(Mendelsohn)、Cancer Cells 7:359(1989)(非特許文献1)、メンデルソーン(Mendelsohn)、Cancer Biology 1:339〜344(1990)(非特許文献2)、モジュタエジ&ディーン(Modjtahedi and Dean)、Int'l J.Oncology 4:277〜296(1994)(非特許文献3)、トシら(Tosi)、Int'l J.Cancer 62:643〜650(1995)(非特許文献4)]。実際に、抗EGF-r抗体はEGFおよびTGF-αの受容体への結合を遮断しながら、腫瘍細胞の増殖を阻害するように思われることが証明された。しかし、同時に抗EGF-r抗体はEGFおよびTGF-α非依存的細胞増殖についてはこれを阻害しないように思われた[モジュタエジ&ディーン(Modjtahedi and Dean)、Int'l J.Oncology 4:277〜296(1994)(非特許文献3)]。
【0004】
これらの知見を考慮して、EGF-rに対する多くのマウスおよびラットモノクローナル抗体を作製し、インビトロおよびインビボでそれらが腫瘍細胞の増殖を阻害するか否かを調べた[モジュタエジ&ディーン(Modjtahedi and Dean)、Int'l J.Oncology 4:277〜296(1994)(非特許文献3)]。臨床においておそらく開発が最も進行している抗体はC225と名付けられたキメラ抗体で、これはマウスの可変領域とヒトIgG1定常領域とを有する[モジュタエジ&ディーン(Modjtahedi and Dean)、Int'l J.Oncology 4:277〜296(1994)(非特許文献3)]。C225抗体に基づく225と呼ばれるマウス抗体は、カリフォルニア大学およびローラー社(Rorer)によって開発された。その開示が参照として本明細書に組み入れられる、米国特許第4,943,533号(特許文献1)および欧州特許第359,282号(特許文献2)を参照のこと。C225抗体は、インビトロでEGF-媒介腫瘍細胞の増殖を阻害し、インビボでヌードマウスにおいてヒト腫瘍形成を阻害することが示された。その上、該抗体は特定の化学療法剤と相乗的に作用して、インビボで異種移植マウスモデルにおいてヒト腫瘍を根治させるように作用するように思われた[モジュタエジ&ディーン(Modjtahedi and Dean)、Int'l J.Oncology 4:277〜296(1994)(非特許文献3)]。
【0005】
イムクローン社(ImClone)はC225と名付けられた抗EGF-r抗体を用いてヒト臨床試験を実施している。C225に関して、頭頸部、前立腺および肺癌患者におけるフェーズIおよびフェーズI/II臨床試験は、実施されたかまたは現在進行中のようである。フェーズI臨床試験では、免疫無防備状態(immunocompromised)の患者を含む症例においても、多数回の注射によって、およびおそらく400mg/m2までの用量で毒性が認められなかった。該試験は約5〜約200 mg/m2までの5用量を含む用量増加試験として実施され、化学療法(すなわち、ドキソルビシン、アドリアマイシン、タキソール、およびシスプラチン)と併用して行われた。これらの試験において得られた明らかな安全性データと共に、研究からの予備的な結果によって、前立腺癌患者の80%において腫瘍の退縮に関する何らかの証拠が示されるように思われる。
【0006】
しかし、これらの上記抗体のそれぞれはマウスまたはラットの可変および/または定常領域を有する。そのようなマウスまたはラットに由来する蛋白質が存在すれば、抗体の急速な一掃が起こり得る、または患者が抗体に対する免疫応答を起こしうる。マウスまたはラット由来の抗体の使用を避けるために、齧歯類が完全なヒト抗体を産生するように、ヒトの抗体機能を齧歯類に導入できるのではないかと考えられた。
【0007】
メガベースサイズのヒト遺伝子座をYAC中にクローニングおよび再構築し、それらをマウス生殖細胞系に導入しうることにより、極めて大きく、または大まかにマッピングされた遺伝子座の機能的構成要素を解明するための、ならびにヒト疾患の有用なモデルを作製するための、強力な手法が提供される。さらに、このような技術を利用してマウス遺伝子座をそれらのヒト等価物に置換することにより、発生過程におけるヒト遺伝子産物の発現および調節、それらの他の系との連絡、ならびに疾患の誘導および進行におけるそれらの関与に関して比類のない洞察が得られると考えられる。
【0008】
このような方法の1つの重要な実用的応用は、マウス体液性免疫系の「ヒト化」である。内因性Ig遺伝子が不活性化されたマウスにヒト免疫グロブリン(Ig)遺伝子座を導入することにより、抗体のプログラム化された発現および構築の根底にある機序に加えて、B細胞の発生におけるそれらの役割をも検討する機会が提供される。さらに、このような戦略により、ヒト疾患の抗体療法という可能性の実現へ向けての重要な里程標となる、完全ヒトモノクローナル抗体(Mab)を生産するための理想的な供給源が提供されると考えられる。完全ヒト抗体は、マウスまたはマウス由来のMabに固有の免疫原性およびアレルギー性反応を最小限に抑え、それにより、投与される抗体の有効性および安全性を高めると考えられる。完全ヒト抗体の使用は、抗体の反復投与を必要とする、炎症、自己免疫および癌などの慢性的および再発性のヒト疾患の治療に、実質的な利益を提供すると考えられる。
【0009】
この目標に向けての1つの手法は、マウス抗体の産生に欠陥があってヒトIg遺伝子座の大きな断片を有するマウス系統を、組換え操作によって作製することであり、これは、このようなマウスが、マウス抗体を生じずに多様なレパトア(repertoire)のヒト抗体を産生するであろうとの予測に立つものであった。大きなヒトIg断片は、可変遺伝子の広範な多様性に加えて、抗体の産生および発現の適切な調節も保持すると考えられる。抗体の多様化および選択のため、ならびにヒトタンパク質に対する免疫寛容を失わせるためのマウス機構を開発すれば、これらのマウス系統で再現されたヒト抗体レパトアはヒト抗原を含む関心対象の任意の抗原に対する高親和性抗体を生じるはずである。ハイブリドーマ技術を用いることにより、望ましい特異性をもつ抗原特異的ヒトMabは容易に製造および選択しうると考えられる。
【0010】
この一般的な方法は、本発明者らが1994年に発表した最初のXenoMouse(登録商標)系統の作出との関連で示された。グリーン(Green)ら、Nature Genetics 7:13〜21(1994)(非特許文献5)参照。XenoMouse(登録商標)系統は、可変および定常領域のコア配列を含む、ヒト重鎖およびκ軽鎖遺伝子座のそれぞれ245kbおよび190kbのサイズの生殖細胞系コンフィギュレーション断片を有する酵母人工染色体(YAC)を用いて操作された。同上。ヒトIgを含むYACは、抗体の再配列および発現の双方に関してマウス系への適合性があることが実証されており、不活性化されたマウスIg遺伝子の置換も可能であった。これは、それらがB細胞発生の誘導および完全ヒト抗体の成人様ヒトレパトアの生成、ならびに抗原特異的ヒトMabの産生の能力をもつことによって示された。また、これらの結果は、比較的多数のV遺伝子、付加的な調節要素、およびヒトIg定常領域を含むヒトIg遺伝子座のかなりの部分を導入することにより、感染および免疫化に対するヒト体液性応答の特徴である実質的に完全なレパトアが再現される可能性があることも示唆した。グリーンら(Green)は最近研究を拡大して、メガベースの大きさの生殖細胞系列構造のヒト重鎖座およびκ軽鎖座のYAC断片をそれぞれ導入することによって、ヒト抗体のレパトア(repertoire)の約80%以上を導入した。メンデスら(Mendez)、Nature Genetics 15:146〜156(1997)(非特許文献6)および本明細書に参照として組み入れられる、1996年12月3日に提出された米国特許出願第08/759,620号(特許文献3)を参照のこと。
【0011】
このような手法は、1990年1月12日に提出された米国特許出願第07/466,008号(特許文献4)、1990年11月8日に提出された第07/610,515号(特許文献5)、1992年7月24日に提出された第07/919,297号(特許文献6)、1992年7月30日に提出された第07/922,649号(特許文献7)、1993年3月15日に提出された第08/031,801号(特許文献8)、1993年8月27日に提出された第08/112,848号(特許文献9)、1994年4月28日に提出された第08/234,145号(特許文献10)、1995年1月20日に提出された第08/376,279号(特許文献11)、1995年4月27日に提出された第08/430,938号(特許文献12)、1995年6月5日に提出された第08/464,584号(特許文献13)、1995年6月5日に提出された第08/464,582号(特許文献14)、1995年6月5日に提出された第08/463,191号(特許文献15)、1995年6月5日に提出された第08/462,837号(特許文献16)、1995年6月5日に提出された第08/486,853号(特許文献17)、1995年6月5日に提出された第08/486,857号(特許文献18)、1995年6月5日に提出された第08/486,859号(特許文献19)、1995年6月5日に提出された第08/462,513号(特許文献20)、1996年10月2日に提出された第08/724,752号(特許文献21)、および1996年12月3日に提出された第08/759,620号(特許文献3)という米国特許出願においてさらに考察および描写されている。メンデズ(Mendez)ら、Nature Genetics 15:146-156(1997)(非特許文献6)も参照のこと。また、1996年6月12日に認可が発行されたEP 0 463 151 B1(特許文献22)、1994年2月3日に発行された国際特許出願WO 94/02602号(特許文献23)、1996年10月31日に発行された国際特許出願WO 96/34096号(特許文献24)、および1996年4月29日に提出されたPCT/US96/05928(特許文献25)も参照されたい。上記に引用したそれぞれの特許、特許出願、および参考文献の開示は、その全体が参照として本明細書に組み入れられる。
【0012】
1つの代替的なアプローチにおいて、ジェンファームインターナショナル社(GenPharm International,Inc.)を含む他者は、「ミニ遺伝子座(minilocus)」法を用いている。ミニ遺伝子座法では、Ig遺伝子座からの複数の小片(個々の遺伝子)を含めることにより、外来性Ig遺伝子座を模倣する。このため、1つまたは複数のVH遺伝子、1つまたは複数のDH遺伝子、1つまたは複数のJH遺伝子、μ定常領域、および第2の定常領域(好ましくはγ定常領域)が、動物に挿入するための構築物(construct)中に形成される。この手法はスラニ(Surani)らに対する米国特許第5,545,807号(特許文献26)、いずれもロンバーグ(Lonberg)およびケイ(Kay)に対する米国特許第5,545,806号(特許文献27)および第5,625,825号(特許文献28)、ならびにいずれもジェンファームインターナショナル社(GenPharm International,Inc.)による米国特許出願である1990年8月29日に提出された第07/574,748号(特許文献29)、1990年8月31日に提出された第07/575,962号(特許文献30)、1991年12月17日に提出された第07/810,279号(特許文献31)、1992年3月18日に提出された第07/853,408号(特許文献32)、1992年6月23日に提出された第07/904,068号(特許文献33)、1992年12月16日に提出された第07/990,860号(特許文献34)、1993年4月26日に提出された第08/053,131号(特許文献35)、1993年7月22日に提出された第08/096,762号(特許文献36)、1993年11月18日に提出された第08/155,301号(特許文献37)、1993年12月3日に提出された第08/161,739号(特許文献38)、1993年12月10日に提出された第08/165,699号(特許文献39)、1994年3月9日に提出された第08/209,741号(特許文献40)に記載されており、それらの開示は参照として本明細書に組み入れられる。また、その開示の全体が参照として本明細書に組み入れられる1994年11月10日に発行された国際公開公報第94/25585号(特許文献41)、1993年7月24日に発行された国際公開公報第93/12227号(特許文献42)、1992年12月23日に発行された国際公開公報第92/22645号(特許文献43)、1992年3月19日に発行された国際公開公報第92/03918号(特許文献44)も参照されたい。さらに、その開示の全体が参照として本明細書に組み入れられるテイラー(Taylor)ら、1992(非特許文献7)、チェン(Chen)ら、1993(非特許文献8)、トゥアイロン(Tuaillon)ら、1993(非特許文献9)、チョイ(Choi)ら、1993(非特許文献10)、ロンバーグ(Lonberg)ら(1994)(非特許文献11)、テイラーら(1994)(非特許文献12)、およびトゥアイロンら(1995)(非特許文献13)も参照されたい。
【0013】
上記に引用し、英国医学研究会議(Medical Research Council)(「MRC」)に譲渡されたスラニ(Surani)らの発明者らは、ミニ遺伝子座法の使用により、Ig遺伝子座を有するトランスジェニックマウスを作製した。上記に引用したジェンファームインターナショナル社の業績の発明者らであるロンバーグおよびケイは、本発明者の先例にならって、内因性マウスIg遺伝子座の不活性化とスラニらの業績の実質的な重複物との組み合わせを提案した。
【0014】
ミニ遺伝子座法の1つの利点は、Ig遺伝子座の一部を含む構築物の作製および動物への導入を迅速に行いうることである。しかしこれを相殺する程度に、ミニ遺伝子座法には理論的にみて、少数のV、DおよびJ遺伝子を含めることによって導入される多様性が不十分であるとの重大な欠点がある。事実、発表された研究はこの懸念を裏づけるように思われる。ミニ遺伝子座法を用いて作製された動物のB細胞発生および抗体産生は阻害されていると考えられる。したがって、本発明を取り囲む研究は一貫して、より広範な多様性の実現を目的として、および動物の免疫レパトアを再構築するための努力において、Ig遺伝子座の大部分を導入することを目的としている。
【0015】
ヒト抗マウス抗体(HAMA)反応のために、業界はキメラまたはそれ以外のヒト化抗体を作製するようになった。C225抗体はヒト定常領域とマウス可変領域とを有するキメラ抗体であるが、特に抗体の長期的または多用量使用時には、特定のヒト抗キメラ抗体(HACA)反応が認められるであろうと予想される。
このように、HAMAまたはHACA反応に関する懸念および/または作用を無用のものとするために、C225と比較して同程度または増強した活性を有する、EGF-rに対する完全なヒト抗体を提供することが望ましいと考えられる。
【先行技術文献】
【特許文献】
【0016】
【特許文献1】米国特許第4,943,533号
【特許文献2】欧州特許第359,282号
【特許文献3】米国特許出願第08/759,620号
【特許文献4】米国特許出願第07/466,008号
【特許文献5】米国特許出願第07/610,515号
【特許文献6】米国特許出願第07/919,297号
【特許文献7】米国特許出願第07/922,649号
【特許文献8】米国特許出願第08/031,801号
【特許文献9】米国特許出願第08/112,848号
【特許文献10】米国特許出願第08/234,145号
【特許文献11】米国特許出願第08/376,279号
【特許文献12】米国特許出願第08/430,938号
【特許文献13】米国特許出願第08/464,584号
【特許文献14】米国特許出願第08/464,582号
【特許文献15】米国特許出願第08/463,191号
【特許文献16】米国特許出願第08/462,837号
【特許文献17】米国特許出願第08/486,853号
【特許文献18】米国特許出願第08/486,857号
【特許文献19】米国特許出願第08/486,859号
【特許文献20】米国特許出願第08/462,513号
【特許文献21】米国特許出願第08/724,752号
【特許文献22】EP 0 463 151 B1
【特許文献23】国際特許出願WO 94/02602号
【特許文献24】国際特許出願WO 96/34096号
【特許文献25】PCT/US96/05928
【特許文献26】米国特許第5,545,807号
【特許文献27】米国特許第5,545,806号
【特許文献28】米国特許第5,625,825号
【特許文献29】米国特許出願第07/574,748号
【特許文献30】米国特許出願第07/575,962号
【特許文献31】米国特許出願第07/810,279号
【特許文献32】米国特許出願第07/853,408号
【特許文献33】米国特許出願第07/904,068号
【特許文献34】米国特許出願第07/990,860号
【特許文献35】米国特許出願第08/053,131号
【特許文献36】米国特許出願第08/096,762号
【特許文献37】米国特許出願第08/155,301号
【特許文献38】米国特許出願第08/161,739号
【特許文献39】米国特許出願第08/165,699号
【特許文献40】米国特許出願第08/209,741号
【特許文献41】国際公開公報第94/25585号
【特許文献42】国際公開公報第93/12227号
【特許文献43】国際公開公報第92/22645号
【特許文献44】国際公開公報第92/03918号
【非特許文献】
【0017】
【非特許文献1】メンデルソーン(Mendelsohn)、Cancer Cells 7:359(1989)
【非特許文献2】メンデルソーン(Mendelsohn)、Cancer Biology 1:339〜344(1990)
【非特許文献3】モジュタエジ&ディーン(Modjtahedi and Dean)、Int'l J.Oncology 4:277〜296(1994)
【非特許文献4】トシら(Tosi)、Int'l J.Cancer 62:643〜650(1995)
【非特許文献5】グリーン(Green)ら、Nature Genetics 7:13〜21(1994)
【非特許文献6】メンデスら(Mendez)、Nature Genetics 15:146〜156(1997)
【非特許文献7】Taylorら、"A transgenic mouse that expresses a diversity of human sequence heavy and light chain immunoglobulins." Nucleic Acids Research 20:6287-6295(1992)
【非特許文献8】Chenら、"Immunoglobulin gene rearrangement in B-cell deficient mice generated by targeted deletion of the JH locus," International Immunology 5:647-656(1993)
【非特許文献9】Tuaillonら、"Human immunoglobulin heavy-chain minilocus recombination in transgenic mice:gene-segment use in m and g transcripts." Proc.Natl.Acad.Sci.USA 90:3720-3724(1993)
【非特許文献10】Choiら、"Transgenic mice containing a human heavy chain immunoglobulin gene fragment cloned in a yeast artificial cnromosome" Nature Genetics 4:117-123(1993)
【非特許文献11】Lonbergら、"Antigen-specific human antibodies from mice comprising four distinct genetic modifications." Nature 368:856-859(1994)
【非特許文献12】Taylorら、"Human immunoglobulin transgenes undergo rearrangement,somatic mutation and class switching in mice that lack endogenous IgM." International Immunology 6:579-591(1994)
【非特許文献13】Tuaillonら、"Analysis of direct and inverted DJH rearrangements in a human Ig heavy chain transgenic minilocus" J.Immunol.154:6453-6465(1995)
【発明の概要】
【0018】
本発明の第一の局面に従って、配列の一部がヒトVH 4ファミリー遺伝子ならびに図2、6、10、14、18、22、26および30に示すヌクレオチド配列によって表されるその変異体のいずれかによってコードされる、重鎖可変領域アミノ酸配列を含む、上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、重鎖可変領域アミノ酸配列は、10位の残基においてアスパラギン酸アミノ酸置換を含む。
【0019】
本発明の第2の局面に従って、その配列の一部がヒトVH 4〜31遺伝子ならびに図2、6、10、14および18に示すヌクレオチド配列によって表されるその変異体のいずれかによってコードされる、重鎖可変領域アミノ酸配列を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、重鎖可変領域は配列番号:23に表されるようにCDR1からCDR3までの近接配列を含む。好ましい態様において、抗体は、配列番号:24によって表される配列を含む軽鎖可変領域をさらに含む。好ましい態様において、重鎖可変領域は配列番号:25に表されるように、CDR1からCDR3までの近接配列を含む。好ましい態様において、抗体は配列番号:26によって表される配列を含む軽鎖可変領域をさらに含む。好ましい態様において、重鎖可変領域は配列番号:27に表されるようにCDR1からCDR3までの近接配列を含む。好ましい態様において、抗体は配列番号:28によって表される配列を含む軽鎖可変領域をさらに含む。好ましい態様において、重鎖可変領域は配列番号:29に表されるようにCDR1からCDR3までの近接配列を含む。好ましい態様において、抗体は配列番号:30によって表される配列を含む軽鎖可変領域をさらに含む。好ましい態様において、重鎖可変領域は配列番号:31に表されるようにCDR1からCDR3までの近接配列を含む。好ましい態様において、抗体は配列番号:32によって表される配列を含む軽鎖可変領域をさらに含む。
【0020】
本発明の第3の局面に従って、その配列の一部がヒトVH 4〜61遺伝子ならびに図22、26、および30に示すヌクレオチド配列によって表されるその変異体のいずれかによってコードされる重鎖可変領域アミノ酸配列を含む、上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、重鎖可変領域は配列番号:33に表されるようにCDR1からCDR3までの近接配列を含む。好ましい態様において抗体は、配列番号:34によって表される配列を含む軽鎖可変領域をさらに含む。好ましい態様において、重鎖可変領域は配列番号:35に表されるようにCDR1からCDR3までの近接配列を含む。好ましい態様において、抗体は配列番号:36によって表される配列を含む軽鎖可変領域をさらに含む。好ましい態様において、重鎖可変領域は配列番号:37に表されるようにCDR1からCDR3までの近接配列を含む、好ましい態様において抗体は、配列番号:38によって表される配列を含む軽鎖可変領域をさらに含む。
【0021】
本発明の第4の局面において、その配列の一部がヒトVK Iファミリー遺伝子ならびに図4、8、12、16、20、24、28、および32に示すヌクレオチド配列によって表されるその変異体のいずれかによってコードされる軽鎖可変領域アミノ酸配列を含む、上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、軽鎖可変領域は配列番号:24によって表される配列を含む。好ましい態様において、軽鎖可変領域は配列番号:26によって表される配列を含む。好ましい態様において、軽鎖可変領域は配列番号:28によって表される配列を含む。好ましい態様において、軽鎖可変領域は配列番号:30によって表される配列を含む。好ましい態様において、軽鎖可変領域は配列番号:32によって表される配列を含む。好ましい態様において、軽鎖可変領域は配列番号:34によって表される配列を含む。好ましい態様において、軽鎖可変領域は配列番号:36によって表される配列を含む。好ましい態様において、軽鎖可変領域は配列番号:38によって表される配列を含む。
【0022】
本発明の第5の局面に従って、配列番号:23に表されるようにCDR1からCDR3までの近接配列を含む重鎖可変領域を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、抗体は配列番号:24によって表される配列を含む軽鎖可変領域をさらに含む。
【0023】
本発明の第6の局面において、配列番号:25に表されるようにCDR1からCDR3までの近接配列を含む重鎖可変領域を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、抗体は配列番号:26によって表される配列を含む軽鎖可変領域をさらに含む。
【0024】
本発明の第7の局面において、配列番号:27に表されるようにCDR1からCDR3までの近接配列を含む重鎖可変領域を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、抗体は配列番号:28によって表される配列を含む軽鎖可変領域をさらに含む。
【0025】
本発明の第8の局面において、配列番号:29に表されるようにCDR1からCDR3までの近接配列を含む重鎖可変領域を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、抗体は配列番号:30によって表される配列を含む軽鎖可変領域をさらに含む。
【0026】
本発明の第9の局面において、配列番号:31に表されるようにCDR1からCDR3までの近接配列を含む重鎖可変領域を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、抗体は配列番号:32によって表される配列を含む軽鎖可変領域をさらに含む。
【0027】
本発明の第10の局面において、配列番号:33に表されるようにCDR1からCDR3までの近接配列を含む重鎖可変領域を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、抗体は配列番号:34によって表される配列を含む軽鎖可変領域をさらに含む。
【0028】
本発明の第11の局面において、配列番号:35に表されるようにCDR1からCDR3までの近接配列を含む重鎖可変領域を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、抗体は配列番号:36によって表される配列を含む軽鎖可変領域をさらに含む。
【0029】
本発明の第12の局面において、配列番号:37に表されるようにCDR1からCDR3までの近接配列を含む重鎖可変領域を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、抗体は配列番号:38によって表される配列を含む軽鎖可変領域をさらに含む。
【0030】
本発明の第13の局面に従って、上皮細胞増殖因子受容体に対する抗体によって固形腫瘍を治療する方法において、固形腫瘍を有する患者に本発明の前述の抗体の一つを投与することを含む改善が提供される。
本発明は例えば、以下の項目を提供する:
(項目1)
配列の一部がヒトVH 4ファミリー遺伝子および図2、6、10、14、18、22、26および30に示すヌクレオチド配列によって表されるその変異体のいずれかによってコードされる、重鎖可変領域アミノ酸配列を含む上皮細胞増殖因子受容体に対する抗体。
(項目2)
重鎖可変領域アミノ酸配列が残基10においてアスパラギン酸アミノ酸置換を含む、項目1記載の抗体。
(項目3)
配列の一部がヒトVH 4〜31遺伝子および図2、6、10、14、および18に示すヌクレオチド配列によって表されるその変異体のいずれかによってコードされる、重鎖可変領域アミノ酸配列を含む上皮細胞増殖因子受容体に対する抗体。
(項目4)
重鎖可変領域が配列番号:23に示すCDR1からCDR3までの近接配列を含む、項目3記載の抗体。
(項目5)
配列番号:24によって表される配列を含む軽鎖可変領域をさらに含む、項目4記載の抗体。
(項目6)
重鎖可変領域が配列番号:25に示すCDR1からCDR3までの近接配列を含む、項目3記載の抗体。
(項目7)
配列番号:26によって表される配列を含む軽鎖可変領域をさらに含む、項目6記載の抗体。
(項目8)
重鎖可変領域が配列番号:27に示すCDR1からCDR3までの近接配列を含む、項目3記載の抗体。
(項目9)
配列番号:28によって表される配列を含む軽鎖可変領域をさらに含む、項目8記載の抗体。
(項目10)
重鎖可変領域が配列番号:29に示すCDR1からCDR3までの近接配列を含む、項目3記載の抗体。
(項目11)
配列番号:30によって表される配列を含む軽鎖可変領域をさらに含む、項目10記載の抗体。
(項目12)
重鎖可変領域が配列番号:31に示すCDR1からCDR3までの近接配列を含む、項目3記載の抗体。
(項目13)
配列番号:32によって表される配列を含む軽鎖可変領域をさらに含む、項目12記載の抗体。
(項目14)
配列の一部がヒトVH 4〜61遺伝子および図22、26、および30に示すヌクレオチド配列によって表されるその変異体のいずれかによってコードされる、重鎖可変領域アミノ酸配列を含む上皮細胞増殖因子受容体に対する抗体。
(項目15)
重鎖可変領域が配列番号:33に示すCDR1からCDR3までの近接配列を含む、項目14記載の抗体。
(項目16)
配列番号:34によって表される配列を含む軽鎖可変領域をさらに含む、項目15記載の抗体。
(項目17)
重鎖可変領域が配列番号:35に示すCDR1からCDR3までの近接配列を含む、項目14記載の抗体。
(項目18)
配列番号:36によって表される配列を含む軽鎖可変領域をさらに含む、項目17記載の抗体。
(項目19)
重鎖可変領域が配列番号:37に示すCDR1からCDR3までの近接配列を含む、項目14記載の抗体。
(項目20)
配列番号:38によって表される配列を含む軽鎖可変領域をさらに含む、項目19記載の抗体。
(項目21)
配列の一部がヒトVK Iファミリー遺伝子および図4、8、12、16、20、24、28、および32に示されるヌクレオチド配列によって表されるその変異体のいずれかによってコードされる、軽鎖可変領域アミノ酸配列を含む、上皮細胞増殖因子受容体に対する抗体。
(項目22)
軽鎖可変領域が配列番号:24によって表される配列を含む、項目21記載の抗体。
(項目23)
軽鎖可変領域が配列番号:26によって表される配列を含む、項目21記載の抗体。
(項目24)
軽鎖可変領域が配列番号:28によって表される配列を含む、項目21記載の抗体。
(項目25)
軽鎖可変領域が配列番号:30によって表される配列を含む、項目21記載の抗体。
(項目26)
軽鎖可変領域が配列番号:32によって表される配列を含む、項目21記載の抗体。
(項目27)
軽鎖可変領域が配列番号:34によって表される配列を含む、項目21記載の抗体。
(項目28)
軽鎖可変領域が配列番号:36によって表される配列を含む、項目21記載の抗体。
(項目29)
軽鎖可変領域が配列番号:38によって表される配列を含む、項目21記載の抗体。
(項目30)
配列番号:23に示すCDR1からCDR3までの近接配列を含む重鎖可変領域を含む、上皮細胞増殖因子受容体に対する抗体。
(項目31)
配列番号:24によって表される配列を含む軽鎖可変領域をさらに含む、項目30記載の抗体。
(項目32)
配列番号:25に示すCDR1からCDR3までの近接配列を含む重鎖可変領域を含む、上皮細胞増殖因子受容体に対する抗体。
(項目33)
配列番号:26によって表される配列を含む軽鎖可変領域をさらに含む、項目32記載の抗体。
(項目34)
配列番号:27に示すCDR1からCDR3までの近接配列を含む重鎖可変領域を含む、上皮細胞増殖因子受容体に対する抗体。
(項目35)
配列番号:28によって表される配列を含む軽鎖可変領域をさらに含む、項目34記載の抗体。
(項目36)
配列番号:29に示すCDR1からCDR3までの近接配列を含む重鎖可変領域を含む、上皮細胞増殖因子受容体に対する抗体。
(項目37)
配列番号:30によって表される配列を含む軽鎖可変領域をさらに含む、項目36記載の抗体。
(項目38)
配列番号:31に示すCDR1からCDR3までの近接配列を含む重鎖可変領域を含む、上皮細胞増殖因子受容体に対する抗体。
(項目39)
配列番号:32によって表される配列を含む軽鎖可変領域をさらに含む、項目38記載の抗体。
(項目40)
配列番号:33に示すCDR1からCDR3までの近接配列を含む重鎖可変領域を含む、上皮細胞増殖因子受容体に対する抗体。
(項目41)
配列番号:34によって表される配列を含む軽鎖可変領域をさらに含む、項目40記載の抗体。
(項目42)
配列番号:35に示すCDR1からCDR3までの近接配列を含む重鎖可変領域を含む、上皮細胞増殖因子受容体に対する抗体。
(項目43)
配列番号:36によって表される配列を含む軽鎖可変領域をさらに含む、項目42記載の抗体。
(項目44)
配列番号:37に示すCDR1からCDR3までの近接配列を含む重鎖可変領域を含む、上皮細胞増殖因子受容体に対する抗体。
(項目45)
配列番号:38によって表される配列を含む軽鎖可変領域をさらに含む、項目44記載の抗体。
(項目46)
上皮細胞増殖因子受容体に対する抗体によって固形癌を治療する方法において、固形癌を有する患者に項目1〜45のいずれか一項記載の抗体を投与することを含む改善。
【図面の簡単な説明】
【0031】
【図1】ハイブリドーマE1.1によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE1.1分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列(contiguous sequence)を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図2】ハイブリドーマE1.1からクローニングした図1の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図3】ハイブリドーマE1.1によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE1.1分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図4】ハイブリドーマE1.1からクローニングした図3のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図5】ハイブリドーマE2.4によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE2.4分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図6】ハイブリドーマE2.4からクローニングした図5の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図7】ハイブリドーマE2.4によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE2.4分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図8】ハイブリドーマE2.4からクローニングした図7のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図9】ハイブリドーマE2.5によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE2.5分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図10】ハイブリドーマE2.5からクローニングした図9の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図11】ハイブリドーマE2.5によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE2.5分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図12】ハイブリドーマE2.5からクローニングした図11のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図13】ハイブリドーマE6.2によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE6.2分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図14】ハイブリドーマE6.2からクローニングした図13の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図15】ハイブリドーマE6.2によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE6.2分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図16】ハイブリドーマE6.2からクローニングした図15のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図17】ハイブリドーマE6.4によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE6.4分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図18】ハイブリドーマE6.2からクローニングした図17の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図19】ハイブリドーマE6.4によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE6.4分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図20】ハイブリドーマE6.4からクローニングした図19のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図21】ハイブリドーマE2.11によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜61によってコードされる配列とE2.11分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図22】ハイブリドーマE2.11からクローニングした図21の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図23】ハイブリドーマE2.11によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE2.11分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図24】ハイブリドーマE2.11からクローニングした図23のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図25】ハイブリドーマE6.3によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜61によってコードされる配列とE6.3分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図26】ハイブリドーマE6.3からクローニングした図25の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図27】ハイブリドーマE6.3によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE6.3分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図28】ハイブリドーマE6.3からクローニングした図27のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図29】ハイブリドーマE7.6.3によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜61によってコードされる配列とE7.6.3分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図30】ハイブリドーマE7.6.3からクローニングした図29の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図31】ハイブリドーマE7.6.3によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE7.6.3分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図32】ハイブリドーマE7.6.3からクローニングした図31のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図33】特異的抗EGF-r抗体重鎖アミノ酸配列と、特定の抗体の重鎖をコードする特定のVH遺伝子のアミノ酸配列との比較を示す。
【図34】特異的抗EGF-r抗体軽鎖アミノ酸配列と、特定の抗体の軽鎖をコードする特定のVK遺伝子のアミノ酸配列との比較を示す。
【図35】インビトロでヒト抗EGF-r抗体によるヒト類上皮癌A431細胞に対するEGF結合の遮断を示し、(□)は本発明の抗EGF-r抗体によって得られた結果を示し、(●)は、マウスモノクローナル抗体225によって得られた結果を示し、および(黒上向き三角)は、対照の非特異的ヒトIgG2抗体によって得られた結果を示す。
【図36】インビトロでヒト抗EGF-r抗体によるヒト類上皮癌A431細胞に対するEGF結合の阻害を示し、(□)は、マウスモノクローナル抗体225によって得られた結果を示し、(○)はマウスモノクローナル抗体528によって得られた結果を示し、(黒下向き三角)は本発明のE1.1抗体を用いて得られた結果を示し、(黒上向き三角)は本発明のE2.4抗体を用いて得られた結果を示し、(黒右向き三角)は、本発明のE2.5抗体を用いて得られた結果を示し、(黒左向き三角)は本発明のE2.6抗体を用いて得られた結果を示し、(◆)は本発明のE2.11抗体を用いて得られた結果を示し、および(四角で囲まれた×印)は、対照の、非特異的ヒトIgG2抗体を用いて得られた結果を示す。
【図37】インビトロでヒト抗EGF-r抗体によるヒト類上皮癌A431細胞に対するTGF-α結合の阻害を示し、(□)は、マウスモノクローナル抗体225によって得られた結果を示し、(◆)は本発明のE6.2抗体を用いて得られた結果を示し、(●)は本発明のE6.3抗体を用いて得られた結果を示し、(黒上向き三角)は本発明のE7.2抗体を用いて得られた結果を示し、(黒四角)は本発明のE7.10抗体を用いて得られた結果を示し、(黒下向き三角)はE7.6.3を用いて得られた結果を示し、および(○で囲まれた十字)は対照の非特異的ヒトIgG2抗体を用いて得られた結果を示す。
【図38】インビトロでヒト抗EGF-r抗体によるヒト結腸癌SW948細胞に対するEGF結合の阻害を示し、(●)は、本発明の抗EGF-r抗体によって得られた結果を示し、(□)はマウスモノクローナル抗体225によって得られた結果を示し、および(黒上向き三角)は対照の非特異的ヒトIgG2抗体によって得られた結果を示す。
【図39】ゼノマウスII株に由来するヒト抗EGF-r抗体がインビトロでSW948細胞の増殖を阻害することを示し、(●)は、本発明の抗EGF-r抗体によって得られた結果を示し、(□)は、マウスモノクローナル抗体225によって得られた結果を示し、および(黒上向き三角)は対照の非特異的ヒトIgG2抗体によって得られた結果を示す。
【図40】インビボで本発明のヒト抗EGF-r抗体を用いることによる、ヌードマウスにおけるヒト類上皮癌A431細胞増殖の阻害を示す。図において、(黒上向き三角)は、本発明のヒト抗EGF-r抗体1mgの投与によって得られた結果を示し、(黒下向き三角)は本発明のヒト抗EGF-r抗体0.2 mgの投与によって得られた結果を示し、(□)は、対照の非特異的ヒトIgG2抗体によって得られた結果を示し、および(○)は、対照として燐酸緩衝生理食塩液を用いた場合に得られた結果を示す。
【図41】インビボで本発明のヒト抗EGF-r抗体を用いることによる、ヌードマウスにおける類上皮癌形成の阻害に関連したデータを示し、19日目での腫瘍の発生率を示す。
【図42】インビボで本発明のヒト抗EGF-r抗体を用いることによる、ヌードマウスにおける類上皮癌形成の阻害に関連したデータを示し、120日目での腫瘍の発生率を示す。
【図43】インビボにおいて本発明のヒト抗EGF-r抗体を用いることによって、ヌードマウスにおいて確立されたヒト類上皮腫瘍の根治に関連したデータを示す。図において、(△)は本発明のヒト抗EGF-r抗体(E7.6.3)各1mgを多数回投与した場合に得られた結果を示し、(×)は、ドキソルビシン各125μgを2回投与した場合に得られた結果を示し、(XX)は本発明のヒト抗EGF-r抗体(E7.6.3)各1 mgの多数回投与をドキソルビシン各125 μgの2回投与と併用した場合に得られた結果を示し、(黒四角)は対照の非特異的ヒトIgG2抗体によって得られた結果を示し、および(◆)は対照として燐酸緩衝生理食塩液を用いた場合に得られた結果を示す。
【図44】インビボで本発明のヒト抗EGF-r抗体を用いることによってヌードマウスにおいて確立されたヒト類上皮腫瘍の根治に関連したデータを示す。図において、(◆)は本発明のヒト抗EGF-r抗体(E2.5)の各0.5mgを多数回投与した場合に得られた結果を示し、(黒四角)はドキソルビシンの各125 μgを2回投与することによって得られた結果を示し、(△)は本発明のヒト抗EGF-r抗体(E2.5)の各0.5mgの多数回投与をドキソルビシン各125 μgの2回投与と併用した場合に得られた結果を示し、(×)は、対照として燐酸緩衝生理食塩液を用いた場合に得られた結果を示し、および(XX)は、対照の非特異的ヒトIgG2抗体を1mgの用量で用いた場合に得られた結果を示す。
【発明を実施するための形態】
【0032】
好ましい態様の詳細な説明
本発明に従って、ヒト上皮細胞増殖因子受容体(EGF-r)に対する完全なヒトモノクローナル抗体を提供する。重鎖および軽鎖免疫グロブリン分子をコードするヌクレオチド配列、ならびにこれらの分子を含むアミノ酸配列、特にCDR1からCDR3までの近接する重鎖および軽鎖配列に相当する配列が提供される。そのような免疫グロブリン分子およびモノクローナル抗体を発現するハイブリドーマもまた提供する。
【0033】
定義
特に明記していない限り、本発明に関連して用いられる科学技術用語は、当業者によって一般的に理解される意味を有する。さらに、本文において特に要求していなければ、単数の用語には複数の用語が含まれ、複数の用語には単数の用語が含まれる。一般に、本明細書に記述の細胞および組織培養、分子生物学、蛋白質およびオリゴまたはポリヌクレオチド化学、およびハイブリダイゼーションに関連して用いられる用語体系は、当技術分野において周知であり一般的に用いられる。組換え型DNA、オリゴヌクレオチド合成、ならびに組織培養および形質転換(例えば電気穿孔、リポフェクション)に関しては標準的な技法が用いられる。酵素反応および精製技法は、製造元の説明書に従って、もしくは当技術分野で一般的に実施されるように、または本明細書に記述されるように実施する。前述の技術および技法は、概して当技術分野で周知の従来の方法に従って、ならびに本明細書を通じて引用され考察される、様々な一般的およびより詳細な引用文献において記述されているように実施する。例えば、本明細書に参照として組み入れられる、サムブルックら(Sambrook)、「分子クローニング:実験マニュアル(Molecular Cloning:A Laboratory Manual)」(第二版、Cold Spring Harbor Laboratory Press,Cold Spring Harbor、ニューヨーク(1989))を参照のこと。本明細書に記述の分析化学、合成有機化学、ならびに医化学および製薬化学に関連して用いられる用語体系、ならびにそれらの実験方法および技術は、当技術分野において周知であり、一般的に用いられる。化学合成、化学分析、薬学的調製、製剤および輸送ならびに患者の治療に関しては標準的な技法を用いる。
【0034】
本開示に従って用いられるように、以下の用語は、特に明記していない限り、以下の意味を有すると理解される:
本明細書において用いられる「単離されたポリヌクレオチド」という用語は、その起源のために「単離されたポリヌクレオチド」が(1)「単離されたポリヌクレオチド」が本来認められるポリヌクレオチドの全体もしくは一部と会合していない、(2)本来結合していないポリヌクレオチドに機能的に結合している、または(3)より大きい配列の一部として天然では起こらない、ゲノム、cDNAもしくは合成起源またはその何らかの組合せのポリヌクレオチドを意味する。
【0035】
本明細書において用いられる「単離された蛋白質」という用語は、その起源もしくは由来源のために、「単離された蛋白質」が(1)天然で認められる蛋白質とは会合していない、(2)同じ起源からのその他の蛋白質を含まない、例えば、マウス蛋白質を含まない、(3)異なる種の細胞によって発現される、または(4)天然に起こらない、cDNA、組換え型RNAもしくは合成起源またはその何らかの組合せの蛋白質を意味する。
【0036】
「ポリペプチド」という用語は、本明細書においてポリペプチド配列の本来の蛋白質、断片または類似体を指す一般的用語として用いられる。したがって、本来の蛋白質、断片および類似体はポリペプチド属に含まれる種である。本発明に係る好ましいポリペプチドは、図1、5、9、13、17、21、25、および29によって表されるヒト重鎖免疫グロブリン分子ならびに図3、7、11、15、19、23、27、および31によって表されるヒトカッパ軽鎖免疫グロブリン分子と共に、重鎖免疫グロブリン分子と、カッパ軽鎖免疫グロブリン分子のような軽鎖免疫グロブリン分子とを含む組合せによって形成された抗体分子、ならびにその逆と共に、その断片および類似体を含む。
【0037】
本明細書においてある物質に適用して用いられる「天然」という用語はある物質が天然に認められるという事実を指す。例えば、天然の起源から単離することができる有機体(ウイルスを含む)に存在し、且つ実験室もしくはそれ以外の場所でヒトによって意図的に改変されていない、ポリペプチドまたはポリヌクレオチド配列は、天然である。
【0038】
本明細書において用いられる「機能的に結合された」という用語は、その意図する方法においてそれらが機能する関係にある、と記述された成分の位置を意味する。コード配列に「機能的に結合された」調節配列は、コード配列の発現が調節配列と適合する条件下で得られるようにライゲーションされる。
【0039】
本明細書において用いられる「調節配列」という用語は、それに対してライゲーションされるコード配列の発現およびプロセシングを行うために必要なポリヌクレオチド配列を意味する。そのような調節配列の性質は宿主生物に依って異なる;原核生物では、該調節配列は一般に、プロモーター、リボソーム結合部位、および転写終結配列を含む;真核生物では、一般に該調節配列はプロモーターおよび転写終結配列を含む。「調節配列」という用語は、その存在が発現およびプロセシングにとって必須である成分を最少でも全て含むものと解釈され、その存在が有利となるさらなる成分、例えばリーダー配列および融合パートナー配列を含むこともできる。
【0040】
本明細書において言及される「ポリヌクレオチド」という用語は、長さが少なくとも10塩基のヌクレオチドのポリマー型、リボヌクレオチドもしくはデオキシヌクレオチドのいずれか、またはいずれかのタイプのヌクレオチドの改変型を意味する。この用語にはDNAの一本鎖および二本鎖型が含まれる。
【0041】
本明細書において言及する「オリゴヌクレオチド」という用語には、天然のヌクレオチド、ならびに天然および非天然のオリゴヌクレオチド結合によって結合された改変型ヌクレオチドが含まれる。オリゴヌクレオチドは一般に長さ200塩基以下を含むポリヌクレオチドサブセットである。好ましくは、オリゴヌクレオチドは長さが10〜60塩基であり、最も好ましくは長さが12、13、14、15、16、17、18、19、または20〜40塩基である。オリゴヌクレオチドは通常、例えばプローブに用いる場合には一本鎖である;しかし、例えば遺伝子変異体の構築に用いる場合にはオリゴヌクレオチドは二本鎖であってもよい。本発明のオリゴヌクレオチドは、センスオリゴヌクレオチドまたはアンチセンスオリゴヌクレオチドのいずれであってもよい。
【0042】
本明細書において用いられる「天然ヌクレオチド」という用語には、デオキシリボヌクレオチドおよびリボヌクレオチドが含まれる。本明細書における「改変型ヌクレオチド」という用語には、改変されたまたは置換された糖基等を有するヌクレオチドが含まれる。本明細書において用いられる「オリゴヌクレオチド結合」という用語には、ホスホロチオ酸エステル、ホスホロジチオ酸エステル、ホスホロセレン酸エステル、ホスホロジセセレン酸エステル、ホスホロアニロチオ酸エステル、ホスホロアニラデート、ホスホロアミデート等のオリゴヌクレオチド結合が含まれる。例えば、その開示が本明細書に参照として組み入れられる、ラプランシェら(Laplanche)、Nucl.Acids Res.14:9081(1986);ステックら(Stec)、J.Am.Chem.Soc.106:6077(1984);スタインら(Stein)、Nucl.Acads Res.16:3209(1988);ツォンら(Zon)、AntiCancer Drug Design 6:539(1991);ツォンら(Zon)、「オリゴヌクレオチドと類似体:実践的アプローチ(Oligonucleotides and Analogues:A Practical Approach)」、87〜108頁(エックスタイン(F.Eckstein)編、Oxford University Press,Oxford,England(1991));ステックら(Stec)、米国特許第5,151,510号;ウールマン&ペイマン(Uhlman and Peyman)、Reviews 90:543(1990)を参照のこと。オリゴヌクレオチドは必要に応じて、検出のための標識を含むことができる。
【0043】
本明細書において用いられる「選択的にハイブリダイズする」という用語は、検出可能且つ特異的に結合することを意味する。本発明のポリヌクレオチド、オリゴヌクレオチド、およびその断片は、非特異的核酸に対する検出可能な結合の認識量を最小限にするようなハイブリダイゼーションおよび洗浄条件下で、核酸鎖と選択的にハイブリダイズする。高ストリンジェンシー条件は、当技術分野で既知であり、本明細書において考察するように、選択的なハイブリダイゼーション条件を得るために用いることができる。一般に、本発明のポリヌクレオチド、オリゴヌクレオチドおよび断片と、対象核酸配列との間の核酸配列の相同性は少なくとも80%、より典型的には少なくとも85%、90%、95%、99%、および100%のような相同性の増加が好ましい。それらの配列間に部分的または完全な同一性が認められれば、2つのアミノ酸配列は相同である。例えば、85%相同性とは、2つの配列を最大に合致するように配置した場合にアミノ酸の85%が同一であることを意味する。ギャップ(対合させる2つの配列のいずれかにおける)によって、対合が最大となるようにする;ギャップの長さは5以下であることが好ましく、2以下であることがより好ましい。または好ましくは、2つの蛋白質の配列(または長さが少なくともアミノ酸30個である、それらに由来するポリペプチド配列)は、本明細書においてこの用語を用いるように、変異データマトリクスおよびギャップペナルティ6以上を用いたプログラムALIGNを用いた場合に配置スコアが(標準偏差の単位で)5以上であれば、相同である。デイホフ(Dayhoff,M.O.)、「蛋白質の配列と構造の図解(Atlas of Protein Sequence and Structure)」、101〜110頁(第5巻、National Biomedical Reseach Foundation(1972))およびこの巻の補足2、1〜10頁を参照のこと。2つの配列またはその一部はより好ましくは、ALIGNプログラムを用いて最適に配置した場合に、そのアミノ酸が50%以上同一であれば、または50%同一に等しければ相同である。「に相当する」という用語は、本明細書において、あるポリヌクレオチド配列が、参照ポリヌクレオチド配列の全てもしくは一部と相同である(すなわち、同一であるが厳密に進化的に関連していない)、またはポリペプチド配列が参照ポリペプチド配列と同一であることを意味するように用いられる。対照区別して、「と相補的である」という用語は、本明細書において、相補的配列が参照ポリヌクレオチド配列の全てまたは一部と相同であることを意味するように用いられる。説明すると、ヌクレオチド配列「TATAC」は、参照配列「TATAC」に対応し、参照配列「GTATA」と相補的である。
【0044】
以下の用語は、2つ以上のポリヌクレオチドまたはアミノ酸配列の間の配列関係を記述するために用いられる:「参照配列」、「比較ウィンドウ」、「配列同一性」、「配列同一性の割合」、および「実質的な同一性」。「参照配列」とは、配列比較の根拠として用いられる明確に定められた配列である;参照配列はより大きい配列のサブセット、例えば全長cDNAのセグメントもしくは配列表に示されている遺伝子配列であってもよく、または完全なcDNAもしくは遺伝子配列を含んでもよい。一般に、参照配列は長さが少なくともヌクレオチド18個またはアミノ酸6個、しばしば長さが少なくともヌクレオチド24個またはアミノ酸8個、および多くの場合長さが少なくともヌクレオチド48個またはアミノ酸16個である。2つのポリヌクレオチドまたはアミノ酸配列はそれぞれ、(1)2つの分子の間で類似である配列(すなわち完全なポリヌクレオチドまたはアミノ酸配列の一部)を含んでもよく、および(2)2つのポリヌクレオチドまたはアミノ酸配列間で異なる配列をさらに含んでもよいため、2つ(以上)の分子間の配列比較は典型的に、配列類似性の局所領域を特定および比較するために、「比較ウィンドウ」の範囲で2つの分子の配列を比較することによって行われる。本明細書において用いられる「比較ウィンドウ」とは、ポリヌクレオチド配列またはアミノ酸配列を、近接ヌクレオチド少なくとも18個またはアミノ酸6個の配列の参照配列と比較する場合、および比較ウィンドウにおけるポリヌクレオチド配列の一部が、2つの配列を最適に配置した場合に参照配列(付加または欠失を含まない)と比較して20%未満の付加、欠失、置換等(すなわちギャップ)を含む場合の、近接ヌクレオチド少なくとも18個の位置またはアミノ酸6個という概念的部分を指す。比較ウィンドウを配置させるための配列の最適な配置は、スミス&ウォーターマン(Smith and Waterman)、Adv.Appl.Math.2:482(1981)の局所相同性アルゴリズム、ニードルマン&ビュンシュ(Needleman and Wunsch)、J.Mol.Biol.48:443(1970)の相同性配置アルゴリズム、ペアソン&リップマン(Pearson and Lipman)、Proc.Natl.Acad.Sci.(U.S.A.) 85:2444(1988)の類似性方法に関する研究、これらのアルゴリズムのコンピューターによる実行(ウィスコンシン・ジェネティックス・ソフトウェアパッケージ7.0(ジェネティクス・コンピューターグループ、575 Science Dr.Madison,Wis.)、ジーンワークス、またはマックベクターソフトウェアパッケージにおけるGAP、BESTFIT、FASTAおよびTFASTA)、または肉眼での検分によって行ってもよく、様々な方法によって得られた最適な配置(すなわち比較ウィンドウの範囲で最高の相同性百分率が得られる)を選択する。
【0045】
「配列同一性」という用語は2つのポリヌクレオチドまたはアミノ酸配列が、比較ウィンドウの範囲で同一である(すなわち、ヌクレオチド・ヌクレオチドまたは残基・残基毎に)ことを意味する。「配列同一性の割合」という用語は、比較ウィンドウの範囲で2つの最適に配置された配列を比較し、同一核酸塩基(例えば、A、T、C、G、UもしくはI)または残基が双方の配列に存在する位置の数を決定して対合した位置の数を得て、対合した位置の数を比較ウィンドウにおける位置の総数で除して、および結果を100倍して配列同一性の割合を算出することによって計算される。本明細書において用いられる「実質的に同一」という用語は、ポリヌクレオチドまたはアミノ酸が、参照配列と比較して、少なくともヌクレオチド18(アミノ酸6個)位の比較ウィンドウ、しばしば少なくともヌクレオチド24〜48(アミノ酸8〜16個)位のウィンドウの範囲で少なくとも85%の配列同一性を有する、好ましくは少なくとも90〜95%の配列同一性を有する、より一般的には少なくとも99%の配列同一性を有する配列を含む場合であって、参照配列を、比較ウィンドウの範囲において参照配列の全体の20%未満である欠失または付加を含んでもよい配列と比較することによって配列同一性の割合が計算される場合の、ポリヌクレオチドまたはアミノ酸配列の特徴を指す。参照配列はより大きい配列のサブセットであってもよい。
【0046】
本明細書において用いられるように、通常のアミノ酸20個およびその略語は、従来の用法に従う。本明細書に参照として組み入れられる、「免疫学−合成(Immunology- A Synthesis)」(第二版、ゴルブ&グレン(E.S.Golub and D.R.Gren)編、Sinauer Associates、サンダーランド、マサチューセッツ州(1991))を参照のこと。通常のアミノ酸20個の立体異性体(例えば、D-アミノ酸)、α、α-2置換アミノ酸のような非天然型アミノ酸、N-アルキルアミノ酸、乳酸、およびその他の異形のアミノ酸もまた、本発明のポリペプチドの適した成分となってもよい。異形アミノ酸の例には:4-ヒドロキシプロリン、Y-カルボキシグルタミン酸塩、ε-N,N,N-トリメチルリジン、ε-N-アセチルリジン、O-ホスホセリン、N-アセチルセリン、N-ホルミルメチオニン、3-メチルヒスチジン、5-ヒドロキシリジン、σ-N-メチルアルギニン、ならびにその他の類似のアミノ酸およびイミノ酸(例えば、4-ヒドロキシプロリン)が含まれる。本明細書において用いられるポリペプチドの表記において、標準的な用法および慣例に従って、左側の方向がアミノ末端の方向であり、右側の方向がカルボキシ末端の方向である。
【0047】
同様に、特に明記していなければ、一本鎖ポリヌクレオチド配列の左側は5'末端である;二本鎖ポリヌクレオチド配列の左側の方向は5'方向と呼ばれる。新生RNA転写物の5'から3'への付加方向を転写方向と呼ぶ;RNAと同じ配列を有するDNA鎖上の配列領域であってRNA転写物の5'末端に対して5'方向にある領域を、「上流配列」と呼び;RNAと同じ配列を有するDNA鎖上の配列領域であって、RNA転写物の3'末端に対して3'方向にある領域を「下流配列」と呼ぶ。
【0048】
ポリペプチドに適用されるように、「実質的に同一」という用語は、2つのポリペプチド配列を、例えば、デフォルトギャップ加重を用いたプログラムGAPまたはBESTFITによって最適に配置した場合に、それらが少なくとも80%の配列同一性を有する、好ましくは少なくとも90%の配列同一性を有する、より好ましくは少なくとも95%の配列同一性を有する、および最も好ましくは少なくとも99%の配列同一性を有することを意味する。同一でない残基の位置は、保存的アミノ酸置換によって異なることが好ましい。保存的アミノ酸置換は類似の側鎖を有する残基の相互交換を指す。例えば、脂肪族側鎖を有するアミノ酸のグループは、グリシン、アラニン、バリン、ロイシン、およびイソロイシンである;脂肪族ヒドロキシル側鎖を有するアミノ酸のグループはセリンおよびトレオニンである;アミド含有側鎖を有するアミノ酸のグループは、アスパラギンおよびグルタミンである;芳香族側鎖を有するアミノ酸のグループは、フェニルアラニン、チロシン、およびトリプトファンである;塩基性側鎖を有するアミノ酸のグループはリジン、アルギニン、およびヒスチジンである;ならびに硫黄含有側鎖を有するアミノ酸のグループはシステインおよびメチオニンである。好ましい保存的アミノ酸置換グループは:バリン・ロイシン・イソロイシン、フェニルアラニン・チロシン、リジン・アルギニン、アラニン・バリン、グルタミン酸・アスパラギン酸、およびアスパラギン・グルタミンである。
【0049】
本明細書において考察するように、抗体または免疫グロブリン分子のアミノ酸配列における小さい変化は、アミノ酸配列の変化が少なくとも75%、より好ましくは少なくとも80%、90%、95%、および最も好ましくは99%を維持している限り、本発明に含まれるものとする。特に、保存的アミノ酸置換について考察する。保存的置換はその側鎖において関連するアミノ酸のファミリー内に起こる置換である。遺伝的にコードされるアミノ酸は一般に以下のファミリーに分類される:(1)酸性=アスパラギン酸、グルタミン酸;(2)塩基性=リジン、アルギニン、ヒスチジン;(3)非極性=アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン;および(4)非荷電極性=グリシン、アスパラギン、グルタミン、システイン、セリン、トレオニン、チロシン。より好ましいファミリーは:セリンおよびトレオニンは脂肪族ヒドロキシファミリーである;アスパラギンおよびグルタミンはアミド含有ファミリーである;アラニン、バリン、ロイシン、およびイソロイシンは脂肪族ファミリーである;ならびにフェニルアラニン、トリプトファン、およびチロシンは芳香族ファミリーである。例えば、ロイシンのイソロイシンまたはバリンへの1箇所置換、アスパラギン酸のグルタミン酸への置換、トレオニンのセリンへの置換、または一つのアミノ酸の、構造的に関連するアミノ酸への同様な置換は、特に置換がフレームワーク部位内のアミノ酸を含まなければ、得られた分子の結合もしくは特性に主要な影響を及ぼさないであろうと十分に予想される。アミノ酸が変化した結果機能的ペプチドが得られるか否かは、ポリペプチド誘導体の特異的活性をアッセイすることによって容易に決定することができる。アッセイは本明細書において詳細に記述する。抗体または免疫グロブリン分子の断片または類似体は、当業者によって容易に調製することができる。断片または類似体の好ましいアミノ末端およびカルボキシ末端は、機能的ドメインの境界付近で起こる。構造的および機能的ドメインは、ヌクレオチドおよび/またはアミノ酸配列データを、公共または私有の配列データベースと比較することによって同定することができる。好ましくは、コンピューターによる比較法を用いて、既知構造および/または機能のその他の蛋白質において起こる配列モチーフまたは推定蛋白質構造ドメインを同定する。既知の3次元構造の中に折り畳まれる蛋白質配列を同定する方法は公知である(ボウイら(Bowie)、Science 253:164(1991))。このように、前述の例は、当業者が本発明に従って構造および機能的ドメインを定義するために用いてもよい配列モチーフおよび構造の配座を認識することができることを示している。
【0050】
好ましいアミノ酸置換は:(1)蛋白質溶解に対する感受性を低下させる、(2)酸化に対する感受性を低下させる、(3)蛋白質複合体を形成する結合親和性を変化させる、(4)結合親和性を変化させる、および(5)そのような類似体の他の物理化学的または機能的特性を付与または改変する、置換である。類似体は天然に存在するペプチド配列以外の配列の様々な変異体を含むことができる。例えば、単一または多数のアミノ酸置換(好ましくは保存的アミノ酸置換)は、天然に存在する配列において起こってもよい(好ましくは、分子間接触を形成するドメインの外側のポリペプチドの一部において)。保存的アミノ酸置換は、親配列の構造的特徴を実質的に変化させてはならない(例えば、置換アミノ酸は親配列において起こるヘリックスを切断する傾向があってはならない、または親配列の特徴となる他のタイプの二次構造を乱す傾向があってはならない)。当技術分野において認識されるポリペプチドの二次および三次構造の例は、それぞれ本明細書に参照として組み入れられる、「蛋白質、構造および分子の基本(Proteins,Structures and Molecular Principles)」(クレイトン(Creighton)編、W.H.Freeman and Company,ニューヨーク(1984));「蛋白質構造入門(Introduction to Protein Structure)」(ブランデン&トーズ(C.Branden and J.Tooze)編、Garland Publishing,ニューヨーク、ニューヨーク州(1991));およびソーントンら(Thornton)、Nature 354:105(1991)を参照のこと。
【0051】
本明細書において用いられる「ポリペプチド断片」という用語は、アミノ末端および/またはカルボキシ末端の欠失を有するが、残りのアミノ酸配列は、例えば全長のcDNA配列から導出した天然に存在する配列における対応する位置において同一であるポリペプチドを指す。断片は典型的に、長さが少なくともアミノ酸5、6、8または10個、好ましくは少なくとも14アミノ酸長、より好ましくは少なくとも20アミノ酸長、通常少なくとも50アミノ酸長、およびさらにより好ましくは少なくとも70アミノ酸長である。本明細書において用いられる「類似体」という用語は、推定アミノ酸配列の一部と実質的な同一性を有し、以下の特性の少なくとも1つ有する、少なくとも25個のアミノ酸のセグメントを含むポリペプチドを意味する:(1)適当な結合条件下でのEGF-rに対する特異的結合、(2)その受容体に対するEGFの結合能、または(3)インビトロもしくはインビボでEGF-rを発現する細胞増殖の阻害能。典型的に、ポリペプチド類似体は、天然に存在する配列に関して保存的アミノ酸置換(または付加もしくは欠失)を含む。類似体は典型的に、長さが少なくともアミノ酸20個、好ましくは長さが少なくともアミノ酸50個以上であり、しばしば全長の天然に存在するポリペプチドと同じ長さとなることができる。
【0052】
ペプチド類似体は一般的に、製薬産業において鋳型ペプチドと類似の特性を有する非ペプチド薬として用いられる。これらのタイプの非ペプチド化合物は「ペプチド模倣体」または「ペプチド擬似体(peptidomimetics)」と呼ばれる。フォーシェル(Fauchere)、J Adv.Drug Res.15:29(1986);ベベール&フライディンガー(Veber and Freidinger)、TINS 392(1985);およびエバンスら(Evans)、J.Med.Chem.30:1229(1987)が、参照として本明細書に組み入れられる。そのような化合物はしばしば、コンピューターによる分子モデリングを用いて開発される。同等の治療または予防効果を生じるために、治療的に有用なペプチドと構造的に類似のペプチド模倣体を用いてもよい。一般に、ペプチド擬似体は、ヒト抗体のような模範ポリペプチド(すなわち生化学特性または薬学活性を有するポリペプチド)と構造的に類似であるが、当技術分野で周知の方法によって、1つ以上のペプチド結合が選択的に以下からなる群より選択される結合に置換されている:−−CH2NH−−、−−CH2S−−、−−CH2−CH2−−、−−CH=CH−−(シスおよびトランス)、−−COCH2−−、−−CH(OH)CH2−−、および−CH2SO−−。コンセンサス配列のアミノ酸1個以上を同じタイプのD-アミノ酸に系統的に置換(例えば、L-リジンの代わりにD-リジン)することによって、より安定なペプチドを作製してもよい。さらに、当技術分野で既知の方法によって(本明細書に参照として組み入れられる、リゾ&ギエラッシュ(Rizo and Gierasch)、Ann.Rev.Biochem.61:387(1992));例えば、ペプチドを環状にする分子内ジスルフィド架橋を形成することができる内部システイン残基を加えることによって、コンセンサス配列または実質的に同一なコンセンサス配列変異を含む不自然なペプチドを産生させてもよい。
【0053】
「抗体」または「抗体ペプチド」とは、無傷の抗体、または特異的結合のために無傷の抗体と競合するその結合断片を指す。結合断片は、DNA組換え技法によって、または無傷抗体の酵素的もしくは化学的開裂によって産生される。結合断片には、Fab、Fab'、F(ab')2、Fv、および一本鎖抗体が含まれる。「二重特異性」、または「双機能性」抗体以外の抗体は、その結合部位のそれぞれが同一であると理解される。抗体は、抗体の過剰量が対受容体に結合した受容体の量を少なくとも約20%、40%、60%または80%、およびより通常約85%以上減少させる場合に(インビトロ競合結合アッセイにおいて測定する)、対受容体に対する受容体の接着を実質的に阻害する。
【0054】
「エピトープ」という用語は、免疫グロブリンまたはT-細胞受容体に特異的に結合することができるいかなる蛋白質決定因子も含む。エピトープ決定因子は通常、アミノ酸または糖側鎖のような分子の化学的に活性な表面での集合体を含み、通常、特異的電荷特性と共に、特異的な三次元構造特性を有する。抗体は、解離定数が≦1μM、好ましくは≦100nM、および最も好ましくは≦10 nMであれば、抗原に特異的に結合すると言える。
「物質」という用語は本明細書において、化学化合物、化学化合物の混合物、生物学的巨大分子、または生物材料からの抽出物を指す。
【0055】
本明細書において用いるように、「標識」または「標識された」という用語は、例えば放射性標識アミノ酸を組み入れることによって、または印をつけたアビジンによって検出することができるビオチン化部分をポリペプチドに接着させることによって(例えば、蛍光マーカーまたは光学もしくは比色法によって検出することができる酵素活性を含むストレプトアビジン)、検出可能なマーカーを組み入れることを意味する。特定の状況では、標識またはマーカーもまた治療的となりうる。ポリペプチドおよび糖蛋白質を標識する様々な方法は、当技術分野において既知であり、用いてもよい。ポリペプチドの標識の例は、以下を含むがこれらに限定されない:放射性同位元素または放射性核種(例えば、3H、14C、15N、35S、90Y、99Tc、111In、125I、131I)、蛍光標識(例えば、FITC、ローダミン、ランタニド燐)、酵素標識(例えば、ホースラディッシュ・ペルオキシダーゼ、β-ガラクトシダーゼ、ルシフェラーゼ、アルカリフォスファターゼ)、化学発光物質、ビオチニル基、二次リポーター(例えば、ロイシンジッパー対配列、二次抗体の結合部位、金属結合ドメイン、エピトープタグ)によって認識される既定のポリペプチドエピトープ。いくつかの態様において、標識は起こりうる立体妨害を減少させるために、様々な長さのスペーサーアームによって結合する。
【0056】
本明細書で用いられる「薬学的物質または薬物」という用語は、患者に適切に投与した場合に望ましい治療効果を誘導することができる化学化合物または組成物を指す。本明細書におけるその他の化学的用語は、本明細書において参照として組み入れられる、マグローヒル化学用語辞典(パーカー(Parker,S.)編、McGraw-Hill、サンフランシスコ(1985))に説明されるように、当技術分野における従来の用途に従って用いられる。
【0057】
「抗癌剤」という用語は、本明細書において、ヒトの新生物、特に癌腫、肉腫、リンパ腫、または白血病のような悪性(癌様)病変の発生または進行を阻害する機能的特性を有する物質を指すために用いられる。転移の阻害はしばしば抗新生物薬の特性である。
【0058】
本明細書において用いるように、「実質的に純粋」とは、目的とする種が最も多く存在する種であることを意味する(すなわち、モルベースで組成物における他のいかなる個々の種より豊富である)、および好ましくは、実質的に精製された分画は、目的とする種が存在する全ての巨大分子種の少なくとも約50%(モルベースで)を含む組成物である。一般に、実質的に純粋な組成物は、組成物中に存在する全ての巨大分子種の約80%以上、より好ましくは約85%、90%、95%、および99%を含む。最も好ましくは、組成物が本質的に単一の巨大分子種からなるように、目的とする種を本質的に均一になるまで精製する(組成物中の汚染種は、従来の検出法によって検出できない)。
患者という用語にはヒトおよび獣医学での被験動物が含まれる。
【0059】
抗体の構造
基本抗体構造単位は4量体からなることが知られている。各4量体はポリペプチド鎖の2つの同一の対からなり、それぞれの対は1つの「軽」鎖(約25 kDa)および1つの「重」鎖(約50〜70kDa)を有する。各鎖のアミノ末端部分は、主に抗原認識を担うアミノ酸約100〜110個以上の可変領域を含む。各鎖のカルボキシ末端部分は、イフェクター機能を主に担う定常領域を定義する。ヒト軽鎖はカッパおよびラムダ軽鎖として分類される。重鎖は、ミュー、デルタ、ガンマ、アルファまたはイプシロンとして分類され、抗体のアイソタイプはそれぞれIgM、IgD、IgAおよびIgEと定義される。軽鎖および重鎖の内部で、可変領域および定常領域はアミノ酸約12個以上の「J」領域によって結合し、重鎖はまたアミノ酸約10個以上の「D」領域を含む。一般的に、「基礎免疫学(Fundamental Immunology)」(ポール(Paul,W.)編、第二版、Raven Press、ニューヨーク(1989))(参照として本明細書に組み入れられる)を参照のこと。各軽鎖/重鎖対の可変領域は、抗体結合部位を形成する。
このように、無傷の抗体は二つの結合部位を有する。双機能性または二重特異性抗体の場合を除いて、2つの抗体結合部位は同一である。
【0060】
鎖は全て、相補性決定領域またはCDRとも呼ばれる3つの超可変領域によって結合した比較的保存されたフレームワーク領域(FR)の同じ一般構造を示す。各対の2つの鎖からのCDRはフレームワーク領域によって整列し、特異的エピトープとの結合が可能となる。N末端からC末端に向けて、軽鎖および重鎖はいずれもドメインFR1、CDR1、FR2、CDR2、FR3、CDR3およびFR4を含む。各ドメインに対するアミノ酸の割付は、「カバットの免疫学的に重要な蛋白質の配列(Kabat Sequences of Proteins of Immunological Interest)」(国立衛生研究所、ベセスダ、メリーランド州(1987および1991))、またはコシア&レスク(Chothia and Lesk)、J.Mol.Biol.196:901〜917(1987);コシアら(Chothia)、Nature 342:878〜883(1989)の定義に従っている。
【0061】
二重特異性または双機能抗体は、異なる2つの重鎖/軽鎖対および異なる2つの結合部位を有する人工ハイブリッド抗体である。二重特異性抗体は、ハイブリドーマの融合またはFab'断片の結合を含む多様な方法によって産生することができる。例えば、ソングシビライ&ラックマン(Songsivilai and Lachman)、Clin.Exp.Immunol.79:315〜321(1990)、コステルニーら(Kostelny)、J.Immunol.148:1547〜1553(1992)を参照のこと。二重特異性抗体の産生は、従来の抗体の産生と比較して比較的大きな労働力を要するプロセスであり、一般に二重特異性抗体に関して収率および純度の程度は低い。二重特異性抗体は1つの結合部位を有する断片の形では存在しない(例えば、Fab、Fab'、およびFv)。
【0062】
抗体の調製
本発明に係る抗体は、好ましくはヒト抗体産生ゲノムの実質的な一部が挿入されているが、内因性のマウス抗体の産生は欠損しているトランスジェニックマウスを使用することによって調製する。次に、そのようなマウスはヒト免疫グロブリン分子および抗体を産生することができるが、マウス免疫グロブリン分子および抗体の産生を欠損している。これらを得るために用いられる技術は、特許、出願、および本明細書の背景に開示されている引用文献に開示されている。しかし特に、マウスのトランスジェニック産生およびその抗体の好ましい態様は、その開示が参照として本明細書に組み入れられる、1996年12月3日に提出された米国特許出願第08/759,620号に開示されている。同様に、その開示が本明細書に参照として組み入れられる、メンデス(Mendez)、Nature Genetics 15:146〜156(1997)を参照のこと。
【0063】
そのような技術を用いることによって、本発明者らは多様な抗原に対する完全なヒトモノクローナル抗体を産生した。本質的に、本発明者らは、目的の抗原でゼノマウス(XenoMouse(登録商標))系マウスを免疫し、抗体を発現するマウスからリンパ球(B細胞など)を回収し、そのような回収された細胞を骨髄系の細胞株と融合させて、不死化ハイブリドーマ細胞株を調製し、該ハイブリドーマ細胞株をスクリーニングして選択し、目的の抗原に対して特異的な抗体を産生するハイブリドーマ細胞株を特定する。本発明者らは、本発明に従ってこれらの技法をEGF-rに特異的な抗体を調製するために使用した。本明細書において、本発明者らは、EGF-rに対して特異的な抗体を産生するハイブリドーマ細胞株8個の産生について記述する。さらに、そのような抗体の重鎖および軽鎖のヌクレオチドおよびアミノ酸配列解析を含む、そのような細胞株によって産生された抗体の特徴分析を提供する。
【0064】
本明細書に記述のハイブリドーマ細胞株はE1.1、E2.4、E2.5、E6.2、E6.4、E2.11、E6.3、およびE7.6.3と命名される。前述の細胞株によって産生されたそれぞれの抗体は、ヒトカッパ軽鎖を有する完全なヒトIgG2重鎖である。一般に、本発明の抗体は非常に高い親和性を有し、固相および液相のいずれかによって測定すると、典型的に約10-9〜約10-11MのKdを有する。
【0065】
認識されるように、本発明の抗体はハイブリドーマ細胞株以外の細胞株において発現させることができる。特定の抗体をコードする配列は、適した哺乳類宿主細胞の形質転換に用いることができる。形質転換は、例えば、ウイルス(またはウイルスベクター)にポリヌクレオチドをパッケージングし、および宿主細胞を該ウイルス(またはベクター)によって形質導入することを含む、宿主細胞にポリヌクレオチドを導入するためのいかなる既知の方法によっても、または米国特許第4,399,216号、第4,912,040号、第4,740,461号、および第4,959,455号(これらの特許は参照として本明細書に組み入れられる)によって示されるように、当技術分野で既知のトランスフェクション技法によって行うことができる。用いる形質転換技法は、形質転換すべき宿主に依存する。異種ポリヌクレオチドを哺乳類細胞に導入する方法は、当技術分野で周知であり、デキストラン媒介トランスフェクション、リン酸カルシウム沈降法、ポリブレン媒介トランスフェクション、プロトプラスト融合、電気穿孔、リポソームへのポリヌクレオチドの封入、および核へのDNAの直接マイクロインジェクションを含む。
【0066】
発現のための宿主として利用できる哺乳類細胞株は当技術分野で周知であり、チャイニーズハムスター卵巣(CHO)細胞、Hela細胞、ベビーハムスター腎(BHK)細胞、サル腎(COS)細胞、ヒト肝細胞癌細胞(例えば、Hep G2)および多くのその他の細胞株を含むがこれらに限定されない、アメリカンタイプカルチャーコレクション(ATCC)から入手できる多くの不死化細胞株を含む。特に好ましい細胞株は、どの細胞株が高発現レベルを有するか、および構成的なEGF-r結合特性を有する抗体を産生するか否かを決定することによって選択される。
【0067】
本発明の抗体は、EGFおよびTGF-αの、その受容体であるEGF-rに対する結合の強力な阻害剤である。それらの結果を実施例5および6において考察し、図35〜38に示す。それらの結果と一致して、図39に示し、実施例7において考察するように、本発明の抗体はインビトロで特定のヒト癌細胞株の増殖をも阻害する。本発明の抗体はまた、インビボで特定のヒト癌の増殖を予防する。それらの結果を図40〜42に示し、実施例8において考察する。実施例9において、本発明者らは、本発明の抗体を少なくとも抗癌剤と併用すると、動物において既に存在する腫瘍が根治することを示した。その上、単剤療法(すなわち抗癌剤と併用しないで)としての抗体療法は、先行技術、例えば抗体225を使用する場合では不可能であるように思われたが、本発明の抗体を用いれば可能であるように思われる。それらの結果を実施例9において考察し、図43〜44に示す。
【0068】
本発明に従って示された結果から、本発明の抗体は、EGF-rに対する現在の治療的抗体、例えば225と比べて、本発明をより有効なものとする特定の特性を有することが示される。イムクローン社(ImClon)によって臨床開発された225抗体は親和性が2×10-10MのキメラIgG1抗体であり、これは抗癌剤との併用療法では有効であるように思われるが、単剤療法ではあまり有効でないように思われる。対照的に、本発明の抗体(特に本発明のE2.5およびE7.6.3抗体)は有意に高い親和性(E2.5:1.6×10-11M;E7.6.3:5.7×10-11 M)を有し、抗癌剤との併用療法のみならず、単剤療法でもC225抗体より低い用量で有効であるように思われる。
【実施例】
【0069】
実施した実験および得られた結果を含む以下の実施例は、説明目的のためにのみ提供されるのであって、本発明を制限するものと解釈してはならない。
【0070】
実施例1
抗EGF-r抗体産生ハイブリドーマの作製
本発明の抗体は、本実施例に従って調製、選択および解析した。
免疫およびハイブリドーマの作製 ゼノマウス(XenoMice)(8〜10週齢)に、リン酸緩衝塩類溶液(PBS)中に再懸濁した2×107個のA431(ATCC CRL-7907)細胞の腹腔内投与による免疫化を施した。この量の投与を3回繰り返した。融合4日前に、マウスにPBS中にて抗原または細胞の最終注射を行った。免疫化マウスからの脾臓およびリンパ節のリンパ球を、非分泌型骨髄腫NSO-bcl2系(RayおよびDiamond、1994)と融合させ、以前に記載されている通り(GalfreおよびMilstein、1981)にHAT選択にかけた。EGF-r特異的ヒトIgG2κ(下記に検出されるように)抗体を全てが分泌するハイブリドーマの大きい集団を回収した。実施例2に示すように、パネルから選択した特定の抗体を225抗体との競合能に関して選択した。
【0071】
ELISAアッセイ: マウス血清中およびハイブリドーマ上清中の抗原特異的抗体を測定するためのELISAは、抗体を捕捉するためにA431細胞からアフィニティー精製されたEGF-r(Sigma、E-3641)を用いて、記載されている通り(Coliganら、1994)に実施した。ヒトおよびマウスの免疫グロブリンの濃度は以下の捕捉抗体を用いて測定した:ヒトガンマ、カッパ、ミューIgについてはそれぞれウサギ抗ヒトIgG(Southern Biotechnology、6145-01)、ヤギ抗ヒトIgκ(Vector Laboratories、AI-3060)、マウス抗ヒトIgM(CGI/ATCC、HB-57)、ならびにマウスガンマ、カッパ、ミューおよびラムダIgの捕捉のためにはそれぞれヤギ抗マウスIgG(Caltag、M 30100)、ヤギ抗マウスIgκ(Southern Biotechnology、1050-01)、ヤギ抗マウスIgM(Southern Biotechnology、1020-01)およびヤギ抗マウスλ(Southern Biotechnology、1060-01)。ELISA実験に用いた検出用抗体は、ヤギ抗マウスIgG-HRP(Caltag、M-30107)、ヤギ抗マウスIgκ-HRT(Caltag、M 33007)、マウス抗ヒトIgG2-HRP(Southern Biotechnology、9070-05)、マウス抗ヒトIgM-HRP(Southern Biotechnology、9020-05)およびヤギ抗ヒトκ-ビオチン(Vector、BA-3060)である。ヒトおよびマウスのIgの定量化のために用いた標準は以下の通りである:ヒトIgG2κ(Calbiochem、400122)、ヒトIgMκ(Cappel、13000)、マウスIgGκ(Cappel 55939)、マウスIgMκ(Sigma、M-3795)およびマウスIgG3λ(Sigma、M-9019)。
【0072】
BIAcoreによる完全ヒトMabの親和定数の決定: 製造者によって概説された標準的な手順によりBIAcore 2000装置を用いて、精製ヒトモノクローナル抗体、Fab断片またはハイブリドーマ上清の親和性の測定をプラズモン共鳴によって実施した。
抗体の速度論的分析は、センサー表面上に低密度に固定された抗原を用いて行った。A431細胞膜から精製された可溶性EGF-r(Sigma、E-3641)を228RUの表面密度で概して用いた。解離速度(kd)および会合速度(ka)は、製造者によって提供されたソフトウエア(BIAevaluation 2.1)を用いて決定した。
【0073】
ELISAによる溶液中の親和定数の決定:ELISAによって溶液中での抗体結合親和性を決定するために、EGF-rに対するモノクローナル抗体の様々な濃度を平行に達するまで一定濃度のEGF-rと共にインキュベートした。その後、反応溶液中の遊離のEGF-rの濃度を間接ELISAによって決定した。よって、3.0×10-11M〜2.7×10-7 Mの濃度のモノクローナル抗体を、0.5%BSAを含むPBS 200μlにおいて4×10-10 Mの濃度のEGF-rと共に室温で15時間インキュベートした。インキュベート後、同じモノクローナル抗体を予めコーティングした(100μl/ウェル、コーティング緩衝液2μg/mlで)96穴マイクロタイタープレートのウェルに各混合液70μlを移し、室温で15分間インキュベートした。洗浄緩衝液で洗浄した後、EGF-r蛋白質の炭水化物に結合するマウス抗EGF-r-HRPによって、プレート上に保持されたEGF-rを検出した。EGF-rの濃度をその標準物質から計算し、これを用いて最初の抗原抗体反応溶液中での結合および遊離の抗体を計算した。各モノクローナル抗体のEGF-rに対する結合親和性はスキャッチャード分析を用いて計算した。
【0074】
受容体結合アッセイ法: EGF受容体結合アッセイ法は、PBS結合緩衝液中、様々な濃度の抗体と共に30分間4℃でインキュベートされたA431細胞またはSW948細胞を用いて実施した(0.4×106細胞/ウェル)。0.1nM[125I]EGF(Amersham,IM-196)または[125I]TGF-α(Amersham)を各ウェルに加え、そのプレートを90分間4℃でインキュベートした。該プレートを5回洗浄し、風乾し、シンチレーションカウンターで計測した。抗EGF-rマウス抗体225および528(Calbiochem)を対照として用いた。
【0075】
実施例2
m225抗体による抗EGF-r抗体の共選択
上記のように、抗体225はEGFおよびTGF-αのEGF-rに対する結合に対して高い親和性を有し、これを有効に阻害することが示された。したがって、本発明者らは、競合アッセイにおいて抗体225を用いて、本発明に従って調製したEGF-rに対するヒト抗体を選択すれば、それに対して225抗体が結合する同一または類似のエピトープに対する抗体が選択されるであろうと予想した。
【0076】
したがって、本発明者らは、A431細胞膜(シグマ社、E-3641)から精製した可溶性EGF-rを抗体225によって前処置し、その後本発明の抗体で処置するバイアコア(BIAcore)アッセイを行った。本発明の抗体が結合しなければ、本発明のそのような抗体を上記のように結合親和性に関してスクリーニングした。
以下の表において、このようにして選択した特定の抗体の親和性測定を提供する:
【0077】
【表1】

表から認められるように、このようにして選択された抗体は、例外的に高い親和性および結合定数を有する。
【0078】
実施例3
本発明に従って調製した抗EGF-r抗体の構造
以下の考察において、本発明に従って調製した抗体に関連した構造的情報を提供する。
本発明に従って産生された抗体の構造を分析するため、本発明者らは特定のハイブリドーマから重鎖および軽鎖断片をコードする遺伝子をクローニングした。遺伝子のクローニングおよびシークエンシングは以下のように実施した:
【0079】
ファストトラック(Fast-Track)・キット(Invitrogen)を用いて、免疫したゼノマウスに由来するハイブリドーマ細胞約2×105個からポリ(A)+ mRNAを単離した。ランダムプライミングしたcDNAを産生した後にPCRを行った。ヒトVHまたはヒトVKファミリー特異的可変領域プライマー(マークスら(Marks)、1991)または万能ヒトVHプライマー、MG-30(CAGGTGCAGCTGGAGCAGTCIGG)(配列番号:1)を、ヒトCγ2定常領域に対して特異的なプライマー(MG-40d;5'-GCTGAGGGAGTAGAGTCCTGAGGA-3')(配列番号:2)、またはCK定常領域(hkP2;既にグリーンら(Green)、1994が記述したように)と共に用いた。ハイブリドーマからのヒトMab-由来重鎖およびカッパ鎖転写物の配列は、上記のプライマーを用いてポリ(A)+ RNAから産生したPCR産物の直接シークエンシングによって得た。PCR産物はまた、TAクローニングキット(インビトロゲン社)を用いてpCRIIの中にクローニングし、プリスム色素ターミネーター・シークエンシングキットおよびABI 377シークエンシング装置を用いて双方の鎖をシークエンシングした。配列は全て、マックベクター(MacVector)およびジーンワークス(GeneWorks)ソフトウェアプログラムを用いて、「V BASE配列ディレクトリ」(トムリンソンら(Tomlinson)、MRC 蛋白質操作センター、ケンブリッジ、イギリス)に配置することによって分析した。
【0080】
ハイブリドーマE1.1
ハイブリドーマE1.1によって分泌された抗体はヒトカッパ軽鎖を有するヒトIgG2抗体を含む。抗体をそのアミノ酸配列と共に、その重鎖および軽鎖の遺伝子使用度に関連した構造情報に関して分析した。このように、重鎖VH、DおよびJHならびに軽鎖VKおよびJK遺伝子使用度を分析して、コードされる産物と特定の遺伝子使用度との相違点についても分析した。よって、ハイブリドーマE1.1によって分泌された抗体は以下の遺伝子使用度を有することが証明された:
VH −4〜31
D −2
JH −5
VK −018
JK −4
【0081】
V BASE配列ディレクトリにおいて報告されるように、VH 4〜31遺伝子によってコードされるアミノ酸配列を以下のように決定した:
VSGGSISSGGYYWSWIRQHPGKGLEWIGYIYYSGSTYYNPSLKSRVTISVDTSKNQFSLKLSSVTAADTAVYYCAR(配列番号:19)
V BASE配列ディレクトリにおいて報告されるように、VK(018)遺伝子によってコードされるアミノ酸配列を以下のように決定した:
TITCQASQDISNYLNWYQQKPGKAPKLLIYDASNLETGVPSRFSGSGSGTDFTFTISSLQPEDIATYYCQQYDNLP(配列番号:20)
【0082】
重鎖および軽鎖に関するアミノ酸およびヌクレオチド配列情報は図1〜4と共に下記に示す。図1は、ハイブリドーマE1.1によって分泌された重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE1.1分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【0083】
図2は、ハイブリドーマE1.1からクローニングした図1の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
図3は、ハイブリドーマE1.1によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE1.1分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
図4は、ハイブリドーマE1.1からクローニングした図3のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【0084】
ハイブリドーマE2.4
ハイブリドーマE2.4によって分泌された抗体は、ヒトカッパ軽鎖を有するヒトIgG2抗体を含む。抗体をそのアミノ酸配列と共にその重鎖および軽鎖遺伝子使用度に関連した構造情報に関して分析した。このように、重鎖VH、DおよびJHならびに軽鎖VKおよびJK遺伝子使用度を分析して、コードされる産物と特定の遺伝子使用度との相違点についても分析した。よって、ハイブリドーマE2.4によって分泌された抗体は以下の遺伝子使用度を有することが証明された:
VH −4〜31
D −A1/A4
JH −3
VK −018
JK −4
【0085】
重鎖および軽鎖に関するアミノ酸およびヌクレオチド配列情報は図5〜8と共に下記に提供する。図5は、ハイブリドーマE2.4によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE2.4分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【0086】
図6は、ハイブリドーマE2.4からクローニングした図5の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
図7は、ハイブリドーマE2.4によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE2.4分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
図8は、ハイブリドーマE2.4からクローニングした図7のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【0087】
ハイブリドーマE2.5
ハイブリドーマE2.5によって分泌された抗体は、ヒトカッパ軽鎖を有するヒトIgG2抗体を含む。抗体をそのアミノ酸配列と共に、その重鎖および軽鎖の遺伝子使用度に関連した構造情報に関して分析した。このように、重鎖VH、DおよびJHならびに軽鎖VKおよびJK遺伝子の利用を分析して、コードされる産物と特定の遺伝子使用度との相違点についても分析した。よって、ハイブリドーマE2.5によって分泌された抗体は以下の遺伝子使用度を有することが証明された:
VH −4〜31
D −XP1/21〜10
JH −4
VK −018
JK −2
【0088】
重鎖および軽鎖に関するアミノ酸およびヌクレオチド配列情報は図9〜12と共に下記に示す。図9は、ハイブリドーマE2.5によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE2.5分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【0089】
図10は、ハイブリドーマE2.5からクローニングした図9の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
図11は、ハイブリドーマE2.5によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE2.5分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
図12は、ハイブリドーマE2.5からクローニングした図11のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【0090】
ハイブリドーマE6.2
ハイブリドーマE6.2によって分泌された抗体はヒトカッパ軽鎖を有するヒトIgG2抗体を含む。抗体をそのアミノ酸配列と共に、その重鎖および軽鎖の遺伝子使用度に関連した構造情報に関して分析した。このように、重鎖VH、DおよびJHならびに軽鎖VKおよびJK遺伝子使用度を分析して、コードされる産物と特定の遺伝子使用度との相違点についても分析した。よって、ハイブリドーマE6.2によって分泌された抗体は以下の遺伝子使用度を有することが証明された:
VH −4〜31
D −?(CNTCCCTT)
JH −6
VK −018
JK −1
【0091】
重鎖および軽鎖に関するアミノ酸およびヌクレオチド配列情報は図13〜16と共に下記に示す。図13は、ハイブリドーマE6.2によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE6.2分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【0092】
図14は、ハイブリドーマE6.2からクローニングした図13の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
図15は、ハイブリドーマE6.2によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE6.2分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
図16は、ハイブリドーマE6.2からクローニングした図15のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【0093】
ハイブリドーマE6.4
ハイブリドーマE6.4によって分泌された抗体はヒトカッパ軽鎖を有するヒトIgG2抗体を含む。抗体をそのアミノ酸配列と共に、その重鎖および軽鎖の遺伝子使用度に関連した構造情報に関して分析した。このように、重鎖VH、DおよびJHならびに軽鎖VKおよびJK遺伝子使用度を分析して、コードされる産物と特定の遺伝子使用度との相違点についても分析した。よって、ハイブリドーマE6.4によって分泌された抗体は以下の遺伝子使用度を有することが証明された:
VH −4〜31
D −A1/A4
JH −4
VK −012
JK −2
【0094】
V BASE配列ディレクトリにおいて報告されるように、VK012遺伝子によってコードされるアミノ酸配列を以下のように決定した:
TITCRASQSISSYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSYSTP(配列番号:21)
【0095】
重鎖および軽鎖に関するアミノ酸およびヌクレオチド配列情報は図17〜20と共に下記に示す。図17は、ハイブリドーマE6.4によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE6.4分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【0096】
図18は、ハイブリドーマE6.4からクローニングした図17の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
図19は、ハイブリドーマE6.4によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子012によってコードされる配列とE6.4分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
図20は、ハイブリドーマE6.4からクローニングした図19のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【0097】
ハイブリドーマE2.11
ハイブリドーマE2.11によって分泌された抗体はヒトカッパ軽鎖を有するヒトIgG2抗体を含む。抗体をそのアミノ酸配列と共に、その重鎖および軽鎖の遺伝子使用度に関連した構造情報に関して分析した。このように、重鎖VH、DおよびJHならびに軽鎖VKおよびJK遺伝子使用度を分析して、コードされる産物と特定の遺伝子使用度との相違点についても分析した。よって、ハイブリドーマE2.11によって分泌された抗体は以下の遺伝子使用度を有することが証明された:
VH −4〜61
D −XP1/21〜10
JH −4
VK −018
JK −4
【0098】
V BASE配列ディレクトリにおいて報告されるように、VH4〜61遺伝子によってコードされるアミノ酸配列を以下のように決定した:
VSGGSVSSGSYYWSWIRQPPGKGLEWIGYIYYSGSTNYNPSLKSRVTISVDTSKNQFSLKLSSVTAADTAVYYCAR(配列番号:22)
【0099】
重鎖および軽鎖に関するアミノ酸およびヌクレオチド配列情報は図21〜24と共に下記に示す。図21は、ハイブリドーマE2.11によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜61によってコードされる配列とE2.11分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【0100】
図22は、ハイブリドーマE2.11からクローニングした図21の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
図23は、ハイブリドーマE2.11によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE2.11分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
図24は、ハイブリドーマE2.11からクローニングした図23のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【0101】
ハイブリドーマE6.3
ハイブリドーマE6.3によって分泌された抗体はヒトカッパ軽鎖を有するヒトIgG2抗体を含む。抗体をそのアミノ酸配列と共に、その重鎖および軽鎖の遺伝子使用度に関連した構造情報に関して分析した。このように、重鎖VH、DおよびJHならびに軽鎖VKおよびJK遺伝子の利用を分析して、コードされる産物と特定の遺伝子使用度との相違点についても分析した。よって、ハイブリドーマE6.3によって分泌された抗体は以下の遺伝子使用度を有することが証明された:
VH −4〜61
D −1〜2rc
JH −4
VK −018
JK −4
【0102】
重鎖および軽鎖に関するアミノ酸およびヌクレオチド配列情報は図25〜28と共に下記に示す。図25は、ハイブリドーマE6.3によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜61によってコードされる配列とE6.3分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【0103】
図26は、ハイブリドーマE6.3からクローニングした図25の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
図27は、ハイブリドーマE6.3によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE6.3分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
図28は、ハイブリドーマE6.3からクローニングした図27のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【0104】
ハイブリドーマE7.6.3
ハイブリドーマE7.6.3によって分泌された抗体はヒトカッパ軽鎖を有するヒトIgG2抗体を含む。抗体をそのアミノ酸配列と共に、その重鎖および軽鎖の遺伝子使用度に関連した構造情報に関して分析した。このように、重鎖VH、DおよびJHならびに軽鎖VKおよびJK遺伝子使用度を分析して、コードされる産物と特定の遺伝子使用度との相違点についても分析した。よって、ハイブリドーマE7.6.3によって分泌された抗体は以下の遺伝子使用度を有することが証明された:
VH −4〜61
D −XP4rc〜XP1
JH −3
VK −018
JK −4
【0105】
重鎖および軽鎖に関するアミノ酸およびヌクレオチド配列情報は図29〜32と共に下記に示す。図29は、ハイブリドーマE7.6.3によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜61によってコードされる配列とE7.6.3分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【0106】
図30は、ハイブリドーマE7.6.3からクローニングした図29の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
図31は、ハイブリドーマE7.6.3によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE7.6.3分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
図32は、ハイブリドーマE7.6.3からクローニングした図31のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【0107】
実施例4
重鎖および軽鎖アミノ酸置換の分析
図33は、特異的抗EGF-r抗体重鎖アミノ酸配列と、特定の抗体の重鎖をコードする特定のVH遺伝子のアミノ酸配列との比較を示す。図34は、特異的抗EGF-r抗体軽鎖アミノ酸配列と、特定の抗体の軽鎖をコードする特定のVK遺伝子のアミノ酸配列との類似比較を示す。認められるように、重鎖および軽鎖配列には極めて保存されたアミノ酸置換がいくつか存在する。特に、抗体の重鎖では、重鎖分子の全てがVH 4ファミリー遺伝子によってコードされ、VH 4〜31によってコードされる抗体では10位にグリシンを有し、VH 4〜61によってコードされる抗体では10位にセリンを有するが、これらはそれぞれアスパラギン酸に置換されている。VH 4〜31重鎖の場合と同様に、1つを除いて全ての抗体が7位のセリンのアスパラギンへの置換を含む。35位ではそれほど優勢な置換ではないが同様の置換が認められ、VH4〜31によってコードされる抗体の2つおよびVH4〜61によってコードされる抗体の2つにおいてセリンがアスパラギンに置換されている。同様に、VH4〜31によってコードされる抗体の2つおよびVH4〜61によってコードされる抗体の2つでは、28位に置換が存在し、それぞれの場合についてチロシンがセリン(E2.4)またはヒスチジン(E6.4、E2.11、およびE7.6.3)に置換されている。抗体の5つ、すなわちVH4〜31によってコードされる抗体の3つ、およびVH4〜61によってコードされる抗体の2つは、50位でバリンの代わりにロイシン(E2.4およびE2.11)またはイソロイシン(E2.5、E6.2、およびE7.6.3)を有する。
【0108】
カッパ軽鎖アミノ酸配列では、配列は全てVK Iファミリー遺伝子によってコードされ、分子の7個は018遺伝子によってコードされ、1個(E6.4)は012遺伝子によってコードされている。図34において、E6.4分子を他の分子と共に018遺伝子産物と比較するとわかるように、012遺伝子産物と018遺伝子産物との間には高度の相同性が存在する。E6.4分子は図19に示すように、012遺伝子産物と比較して2箇所の置換を有するに過ぎず、018遺伝子産物と比較して13箇所の置換を有するに過ぎない。抗体は全て、CDR3の74位で置換を有し、アスパラギンがセリン(E1.1、E2.5、E2.11、およびE6.3)、ヒスチジン(E2.4、E6.2、およびE7.6.3)、またはアルギニン(E6.4)に置換されている。残りの置換はあまり高度に保存されてはいない。しかし、多くの抗体はCDR内部に置換を有するように思われる。しかし、非常に親和性の高い抗体であるE7.6.3がCDR3にまさに隣接するまでの軽鎖アミノ酸配列、およびCDR3内にアミノ酸置換を有しないことに注目することは興味深い。
【0109】
上記のように確認されたアミノ酸置換はそれぞれCDRに非常に近位でまたはCDRの内部に存在することが認識されると思われる。そのような置換はEGF受容体分子に対する抗体の結合に基づいて何らかの作用を有すると思われる。さらに、そのような置換は抗体の親和性に有意な作用を及ぼしうる。
既に考察したように、抗EGF-r抗体は、特定の抗腫瘍活性を有することが示されている。以下の実験は、本発明の抗体がそのような抗腫瘍活性を有するか否かを決定するために実施した。
【0110】
実施例5
インビトロでヒト抗EGF-r抗体によるヒト類上皮癌A431細胞に対するEGFおよびTGF-αの結合の遮断
インビトロアッセイを実施して、本発明の抗体がヒト癌細胞株に対するEGFの結合を遮断することができるか否かを明らかにした。上記のように既に抗癌活性が証明されたマウスモノクローナル抗体225と、本発明の抗体の結合とを比較する実験を実施した。
【0111】
この実施例ではヒト類上皮癌A431細胞株を使用した。A431細胞株はそのEGF-rの発現レベルが高いこと(約2×106個EGF-r分子/細胞)が知られている。したがって、全ての結合部位を飽和するためにはより高濃度の抗EGF-r抗体を必要とする。本実施例から得られた結果を図35に示す。図において、インビトロでヒト抗EGF-r抗体によるヒト類上皮癌A431細胞に対するI125標識EGF結合の遮断が示される。図において、(□)は本発明の抗EGF-r抗体(E7.6.3)によって得られた結果を示し、(●)は、マウスモノクローナル抗体225によって得られた結果を示し、および(黒上向き三角)は、対照の非特異的ヒトIgG2抗体によって得られた結果を示す。
【0112】
図36は、インビトロで本発明のヒト抗EGF-r抗体集団によるヒト類上皮癌A431細胞に対するEGF結合の阻害を225、528および非特異的ヒトIgG2対照と比較して示す。図において、(□)は、マウスモノクローナル抗体225によって得られた結果を示し、(○)はマウスモノクローナル抗体528によって得られた結果を示し、(黒下向き三角)は本発明のE1.1抗体を用いて得られた結果を示し、(黒上向き三角)は本発明のE2.4抗体を用いて得られた結果を示し、(黒右向き三角)は、本発明のE2.5抗体を用いて得た結果を示し、(黒左向き三角)は本発明のE2.6抗体を用いて得られた結果を示し、(◆)は本発明のE2.11抗体を用いて得られた結果を示し、且つ(四角で囲んだ×印)は、対照の、非特異的ヒトIgG2抗体を用いて得られた結果を示す。
【0113】
結果は、本発明の抗体が、A431細胞上の表面に発現されたEGF-rに対するEGF結合を225および528抗体より良好に遮断する可能性があることを示している。本発明の抗体は8nM濃度で結合を阻害し始めるように思われるのに対し、225抗体では10 nM濃度であった。
【0114】
TGF-α結合の阻害に関しては、本発明の抗体を用いることによって225抗体と比較して類似の有効性が認められた。図37は、インビトロでヒト抗EGF-r抗体によるヒト類上皮癌A431細胞に対するTGF-α結合の阻害を示し、(□)は、マウスモノクローナル抗体225によって得られた結果を示し、(◆)は本発明のE6.2抗体を用いて得られた結果を示し、(●)は本発明のE6.3抗体を用いて得られた結果を示し、(黒上向き三角)は本発明のE7.2抗体を用いて得られた結果を示し、(黒四角)は本発明のE7.10抗体を用いて得られた結果を示し、(黒下向き三角)はE7.6.3を用いて得られた結果を示し、且つ(○で囲んだ十字)は対照の非特異的ヒトIgG2抗体を用いて得られた結果を示す。
【0115】
結果は本発明の抗体が、A431細胞の表面に発現されたEGF-rに対するTGF-α結合を225抗体より良好に遮断する可能性があることを示している。本発明の抗体は0.1nM濃度で結合を阻害し始めるように思われるのに対し、225抗体では1nM濃度であった。
【0116】
実施例6
インビトロでヒト抗EGF-r抗体によるヒト結腸腺癌SW948細胞に対するEGF結合の遮断
本発明の抗体がさらにもう一つのヒト癌細胞株に対するEGF結合を遮断できるか否かを明らかにするために、もう一つのインビトロアッセイを実施した。上記のように既に抗癌活性を有することが証明されているマウスモノクローナル抗体225と、本発明の抗体の結合を比較する実験を実施した。
【0117】
本実施例ではヒト結腸腺癌SW948細胞株を用いた。A431細胞株とは対照的に、SW948細胞株はその表面上のEGF-rの発現数が比較的少ない(約40,000分子/細胞)。したがって細胞上の受容体の全ての結合部位を飽和するために必要な抗EGF-r抗体の量は比較的少ない。本実施例の結果を図38に示す。図において、インビトロでヒト抗EGF-r抗体によるヒト結腸腺癌SW948細胞に対するI125標識EGF結合の阻害を示す。図において、(●)は、本発明の抗EGF-r抗体(E7.6.3)によって得られた結果を示し、(□)はマウスモノクローナル抗体225によって得られた結果を示し、且つ(黒上向き三角)は対照の非特異的ヒトIgG2抗体によって得られた結果を示す。
【0118】
結果は本発明の抗体がSW948細胞に対するEGF結合を少なくとも225抗体と同程度に遮断することを示している。実際に、曲線は本発明の抗体ではわずかに改善しており、225抗体と比較すると低濃度で阻害が起こることを示している。
【0119】
実施例7
インビトロでのヒト抗EGF-r抗体によるヒト結腸腺癌SW948細胞増殖の阻害
本発明者らはまた、本発明の抗体が癌細胞増殖を阻害するか否か、および225抗体と比較してどの程度まで阻害するかを明らかにするためにインビトロアッセイを実施した。上記のように抗癌活性を有することが既に証明されているマウスモノクローナル抗体225による阻害と、本発明の抗体による阻害を比較する実験を実施した。
【0120】
本実施例ではヒト結腸腺癌SW948細胞株を用いた。本発明者らが行うと、SW948細胞のみがEGF-依存的細胞増殖を示した。対照的にA431細胞株はインビトロでEGFの存在下で増殖阻害を示した。結果を図39に示し、本発明のヒト抗EGF-r抗体がインビトロでSW948細胞の増殖を阻害することを示す。図において(●)は、本発明の抗EGF-r抗体(E7.6.3)によって得られた結果を示し、(□)は、マウスモノクローナル抗体225によって得られた結果を示し、且つ(黒上向き三角)は対照の非特異的ヒトIgG2抗体によって得られた結果を示す。
【0121】
結果は、本発明の抗体がSW948細胞の増殖を少なくとも225抗体と同程度に阻害することを示す。実際に、本発明の抗体に関して曲線はわずかに改善し、約100 μg/mlで細胞増殖を明らかに100%阻害を示したが、225抗体は同じ用量範囲では80〜90%の阻害レベルで平衡に達するように思われる。
【0122】
実施例8
インビボでのヒト抗EGF-r抗体によるヌードマウスにおけるヒト類上皮癌細胞増殖の阻害
本実験において、本発明者らは本発明の抗体がインビボで腫瘍の細胞増殖を阻害することができるか否かを明らかにしようと試みた。実験において、8週齢のヌードマウスにヒト類上皮癌A431細胞株を皮下接種した。マウスにA431細胞5×106個を注射した。本発明の抗体の2つの用量のうち1つ、または2つの対照のうち1つを、A431細胞接種と同じ日に腹腔内注射した。いずれかの抗体または対照の3つの投与を行った後に、マウスの皮下腫瘍形成および大きさを調べた。用いた抗体の用量は1.0mgまたは0.2 mgのいずれかであった。対照は、燐酸緩衝生理食塩液または非特異的ヒトIgG2抗体のいずれかであった。
【0123】
本実験の結果を図40に示す。図において、インビボで本発明のヒト抗EGF-r抗体を用いることによる、ヌードマウスにおけるヒト類上皮癌細胞増殖の阻害は明らかである。図において、(黒上向き三角)は、本発明のヒト抗EGF-r抗体(E7.6.3)1.0mgの投与によって得られた結果を示し(n=5)、(黒下向き三角)はE7.6.3抗体0.2 mgの投与によって得られた結果を示し(n=4)、(□)は、対照の非特異的ヒトIgG2抗体によって得られた結果を示し(n=6)、且つ(○)は、対照として燐酸緩衝生理食塩液を用いた場合に得られた結果を示す(n=6)。
E7.6.3抗体で処置した動物では腫瘍の増殖は認められなかったが、対照動物は腫瘍細胞を接種した25日以内に有意な腫瘍の増殖が認められた。
【0124】
同じ実験において、本発明の3つの抗体を比較した。結果を図41に示す。本発明の抗体のそれぞれ、すなわちE7.6.3の1 mgをマウス5匹に、0.2 mgを4匹に、E2.5の1mgをマウス3匹に、および0.2 mgをマウス3匹に、ならびにE1.1の1mgをマウス3匹に投与することにより、対照と比較してマウスにヒト類上皮癌形成の阻害が示された。対照動物(PBS処置動物6匹およびヒトIgG2-処置動物6匹を含む)は全て接種の19日以内に有意な腫瘍を発生したが、本発明のヒト抗EGF-r抗体で処置した動物はいずれも接種の19日以内に腫瘍を発生しなかった。
【0125】
図42は、上記の同じ実験の動物を、ヒト類上皮癌の接種後130日間追跡した結果を示す。この実験の結果を図42に示す。図において、対照マウスは全て腫瘍細胞接種の20日以内に腫瘍を発生したことが認められる。対照的に、本発明の抗体で処置した最初のマウスが腫瘍を発症したのは70日目であった。130日までに、腫瘍を発生したのは実験動物15匹中4匹に過ぎなかった。興味深いことに、E2.5抗体の0.2mg用量で処置した実験動物のいずれも試験期間内に腫瘍を発生しなかった。
【0126】
この実施例8に関連した上記実験は、本発明の抗体が、腫瘍細胞株の接種と同時に投与すれば、腫瘍細胞の増殖の開始および腫瘍の開始をほぼ完全に予防するように思われることを証明する。その上、腫瘍細胞増殖に及ぼす作用は長期間持続するように思われることが認められた。
【0127】
実施例9
インビボでヒト抗EGF-r抗体によるヌードマウスにおけるヒト類上皮癌増殖の根治
腫瘍細胞の増殖および/または腫瘍の確立を予防することは、先に述べた実施例に関連して上記で考察したように、治療的な観点から見て肯定的な知見であるが、確立した腫瘍の根治も同様に非常に望ましい。したがって、以下の実験において、本発明の抗体が哺乳類において確立した腫瘍を根治することができるか否かを調べた。225抗体に関して得られたこれまでのデータから、確立された腫瘍を225抗体を用いることによって有効に根治するためには、抗癌剤による治療を補うことが必要であることが示された。このように本発明者らの実験では、単独および抗癌剤治療と併用した場合の双方の抗体治療を調べた。
【0128】
実験において、0日目にヌードマウスにA431ヒト類上皮癌細胞5×106個を皮下接種した。腫瘍が十分に確立してから(大きさ≧0.4 cm3)、マウスを抗体、化学療法剤、および/または対照のいずれかで治療した。治療は5、8、10、14、16、および21日目に開始して継続し、化学療法剤は5および6日のみに投与した。治療には、本発明の抗体(E7.6.3)、抗癌剤ドキソルビシン、および抗体とドキソルビシンの併用が含まれた。対照は燐酸緩衝生理食塩液または非特異的ヒトIgG2抗体であった。各治療群は動物5匹を含んだ。実験から得られたデータを図43に示し、(△)は本発明のヒト抗EGF-r抗体(E7.6.3)1mgを投与した場合に得られた結果を示し、(×)は、ドキソルビシン125μgを投与した場合に得られた結果を示し、(XX)は本発明のヒト抗EGF-r抗体(E7.6.3)1 mgの投与をドキソルビシン125 μgの投与と併用した場合に得られた結果を示し、(黒四角)は対照の非特異的ヒトIgG2抗体によって得られた結果を示し、および(◆)は対照として燐酸緩衝生理食塩液を用いた場合に得られた結果を示す。
【0129】
図から認められるように、E7.6.3抗体をドキソルビシンと併用して投与すると、腫瘍増殖の完全な根治が得られた。さらに、腫瘍の増殖はE7.6.3抗体のみを投与した場合にも完全に停止した。
【0130】
図44に結果を示す同様の実験において、腫瘍の接種後にマウス5匹にE2.5抗体の0.5 mgを5、8、10、14、16、および21日に投与することによって治療し、およびマウス5匹にE2.5抗体の5、8、10、14、16、および21日目の投与と、ドキソルビシン5および6日目の投与の併用によって治療した。図において、(◆)は本発明のヒト抗EGF-r抗体(E2.5)0.5mgを投与した場合に得られた結果を示し、(黒四角)はドキソルビシンの125 μgを投与することによって得られた結果を示し、(△)は本発明のヒト抗EGF-r抗体(E2.5)0.5mgの投与をドキソルビシン125 μgの投与と併用した場合に得られた結果を示し、(×)は、対照として燐酸緩衝生理食塩液を用いた場合に得られた結果を示し、および(XX)は、対照の非特異的ヒトIgG2抗体を用いた場合に得られた結果を示す。
図から認められるように、E2.5抗体を単独で、またはドキソルビシンと併用して投与すると、マウスにおいて腫瘍のほぼ完全な根治が得られた。
【0131】
実施例10
インビボでヒト抗EGF-r抗体を用いることによるヒト癌の治療および診断に関するヒト臨床試験
緒言
本発明の抗体は特定の固形癌の治療に適応される。特にEGF-r発現レベルを含む多くの要因に基づき、以下の腫瘍のタイプが好ましい適応となるように思われる:乳癌、卵巣癌、結腸癌、前立腺癌、膀胱および非小細胞肺癌。これらの適応のそれぞれに関して、3つの臨床経路が明確に臨床的に成功する可能性があるように思われる。
【0132】
補助療法:補助療法において、患者は、本発明の抗体と化学療法剤または抗癌剤および/または放射線療法との併用によって治療する。上記の主な標的は、標準的な第一選択および第二選択療法に発明の抗体を加えることによってプロトコルに基づいて治療されるであろう。プロトコルは、標準的な化学療法の通常の用量を減少させることと共に腫瘍塊の減少によって評価する有効性が得られるようにデザインされる。これらの用量の減少により、化学療法剤の用量関連毒性を減少させることによってさらなるおよび/または長期間の療法が可能となるであろう。先行技術の抗EGF-r抗体は、いくつかの補助的臨床試験において、化学療法もしくは抗癌剤であるアドリアマイシン(C225:進行前立腺癌)、シスプラチン(C225:進行頭頸部癌および肺癌)、タキソール(C225:乳癌)、ならびにドキソルビシン(C225:前臨床)と併用して用いられた、または用いられている。
【0133】
単剤療法:腫瘍の単剤療法に本発明の抗体を利用する場合には、化学療法剤または抗癌剤を用いないで抗体を患者に投与する。本発明の抗体の使用によって得られ、本明細書において考察する前臨床試験の結果から、補助療法および/または単剤療法の双方に関して同程度の結果が示された。その上、単剤療法は明らかに臨床的に広範囲の転移疾患を有する末期段階の癌患者において実施された。患者は疾患に何らかの安定化を示すように思われた。同上。臨床試験は(癌)腫瘍を有する難治性の患者において作用が示されるようにデザインされる。
【0134】
造影剤:放射性核種(例えば、イットリウム(90Y))を本発明の抗体に結合することによって、本発明の放射標識抗体は、診断的造影剤として利用することができると予想される。そのような役割において、本発明の抗体は固形癌のみならずEGF受容体を発現する細胞の転移病変の双方に存在するであろう。本発明の抗体を造影剤として用いる場合には、どの腫瘍が残存するかおよび/または再発するかを決定するために術後の追跡調査と共に術前スクリーニングとして、固形癌の外科治療を補助するために抗体を用いることができる。111In-C225抗体は、切除不能の肺扁平細胞癌を有する患者におけるフェーズIヒト臨床試験において造影剤として用いられている。ディブジら(Divgi)、J.Natl.Cancer Inst.83:97〜104(1991)。患者は標準的な前後方γカメラによって追跡した。前臨床データから、全ての原発病変および大きい転移病変は特定されるが、小さい転移病変(1cm以下)は半数が検出されたに過ぎなかった。
【0135】
用量および投与経路
本発明の抗体の特定の投与法についてはまだ決定していないが、臨床において用いられる同様の産物(イムクローンC255)と比較することによって、特定の投与に関する検討を行うことが可能である。C225抗体は典型的に5〜400mg/m2の範囲の用量を投与し、これより低い用量は安全性試験に関連した場合に限って用いられる。本発明の抗体の親和性はC225抗体より1対数高い。さらに、本発明の抗体は、C225抗体のキメラ特性と比較して完全なヒト抗体であり、このように抗体のクリアランスはより遅くなると予想される。したがって、本発明の抗体を患者に投与する場合には、用量はこれより低く、おそらく50〜300mg/m2の範囲となり、それでもなお有効であろうと予想される。従来のmg/kgで表す用量単位に対して、mg/m2での投与は表面積に基づいた単位であり、幼児から成人までの全ての体格の患者を含むようにデザインされる都合のよい投与単位である。
【0136】
本発明の抗体の輸送には3つの明確な輸送アプローチが有用であると予想される。従来の静脈内輸送はおそらく、ほとんどの腫瘍にとって標準的な輸送技法であろう。しかし、卵巣、胆管、その他の管等の腫瘍のような腹腔における腫瘍に関しては、腫瘍での抗体量を高め、および抗体のクリアランスを最小限にするために、腹腔内投与が都合がよい可能性がある。同様にして、固形癌の中には局所還流に適した血管を有するものがある。局所還流によって腫瘍部位での抗体の高用量を得ることが可能となり、抗体の短期間でのクリアランスが最小限となる。
【0137】
臨床開発計画(CDP)
総説:CDPにより、本発明の抗EGF-r抗体の補助療法、単剤療法に関連した治療、および造影剤としての処置が追求および開発される。まず臨床試験を用いて安全性を証明し、次に投与を繰り返した際の有効性を得るために臨床試験を行う。臨床試験は、標準的な化学療法と標準的な療法プラス本発明の抗体とを比較するオープンラベルとなるであろう。認識されるように、患者の登録に関して利用することができる1つの基準は、生検によって決定した患者の腫瘍のEGF-r発現レベルであってもよい。
【0138】
蛋白質または抗体注入に基づく治療に関して、安全性に関する懸念は主に(i)サイトカイン放出症候群、すなわち血圧低下、発熱、震え、悪寒、(ii)材料に対する免疫原反応の発症(すなわち、ヒト抗体治療に対する患者によるヒト抗体の産生またはHAHA反応)、および(iii)EGF受容体を発現する正常細胞、例えばEGF-rを発現する肝細胞に対する毒性、に関する。標準的な試験および追跡調査を利用して、これらの安全性に関する懸念をそれぞれモニターする。特に、あるとすれば肝臓の障害を評価するために臨床試験の間、肝機能を頻繁にモニターする。
【0139】
ヒト臨床試験:抗EGF-r抗体および化学療法剤との補助療法
フェーズIヒト臨床試験は固形癌、例えば乳癌の治療において、本発明のヒト抗EGF-r抗体の6用量を静脈内投与した場合の安全性を評価するために開始されるであろう。本試験において、シスプラチン、トポテカン、ドキソルビシン、アドリアマイシン、タキソール等のような抗癌剤または化学療法剤に対する補助療法として用いた場合の、本発明の抗体の単回投与の安全性を評価する。臨床試験のデザインには、以下のスケジュールに従って、治療の過程において本発明の抗体の用量を約25mg/m2〜約275 mg/m2に増加させて、本発明の抗体の単回投与を6回行うことが含まれる。
【0140】
【表2】
患者は抗体および化学療法のそれぞれの投与後1週間厳密に追跡調査する。特に、上記の安全性に関する懸念について患者を評価する:(i)サイトカイン放出症候群、(ii)材料に対する免疫原反応の発症(すなわち、患者によるヒト抗体治療に対するヒト抗体の産生、またはHAHA反応)、および(iii)EGF受容体を発現する正常細胞、例えばEGF-rを発現する肝細胞に対する毒性。標準的な試験および追跡調査を利用してこれらの安全性に関する懸念のそれぞれをモニターする。特に、あるとすれば肝臓に対する障害を評価するために、臨床試験において肝機能を頻繁にモニターする。
【0141】
患者はまた、臨床転帰に関して、特にMRIまたはその他の画像によって示される腫瘍塊の減少に関しても評価する。
安全性が証明されることおよび有効性が示されることを仮定して、有効性をさらに探索し、最適な投与を決定するために、フェーズII臨床試験がおそらく開始されるであろう。
【0142】
ヒト臨床試験:ヒト抗EGF-r抗体による単剤療法
本発明の抗体が上記補助臨床試験に関連して安全であるように思われると仮定して、単剤療法に関する有効性および最適な投与を評価するためのヒト臨床試験。そのような臨床試験を行って、患者に対して本発明の抗体と同時に化学療法を投与しないことを除き、上記補助臨床試験の場合と同じ安全性および転帰分析を必要とする。
【0143】
ヒト臨床試験:抗EGF-r抗体による診断的造影
再度、上記の補助療法が上記の安全性基準の中で安全であるように思われると仮定して、本発明の抗体を診断的造影剤として用いることに関するヒト臨床試験を実施することができる。プロトコルはディブジら(Divgi)、J.Nat.Cancer Inst.83:97〜104(1991)に記述されたものと実質的に同様にしてデザインされると予想される。
【0144】
参照としての組み入れ
特許、特許出願、論文、教科書等を含む、本明細書における全ての引用文献、およびそれらに引用される引用文献は、それらが先ではないという程度に、本明細書に参照としてその全文が組み入れられる。さらに、そのような引用文献において引用される引用文献を含む、以下の引用文献もまた、本明細書においてその全文が参照として組み入れられる。
Abertsenら、"Construction and characterization of a yeast artificial chromosome library containing seven haploid human genome equivalents." Proc.Natl.Acad.Sci.87:4256(1990)
Anandら、"Construction of yeast artificial chromosome libraries with large inserts using fractionation by pulsed-field gel electrophoresis." Nucl.Acids Res.17:3425-3433(1989)
Bermanら、"Content and organization of the human Ig VH locus:definition of three new VH families and linkage to the Ig CH locus."(1988) EMB0 J.7:727-738
Brezinschekら、"Analysis of the heavy chain repertoire of human peripheral B-cells using single-cell polymerase chain reaction." J.Immunol.155:190-202(1995)
Brownsteinら、"Isolation of singie-copy human genes from a library of yeast artificial chromosome clones." Science 244:1348-1351(1989)
Bruggemanら、PNAS USA 86:6709-6713 (1989)
Bruggemann ら、"Human antibody production in transgenic mice:expression from 100 kb of the human IgH locus." Eur.J.Immunol.21:1323-1326(1991)
Bruggeman,M.およびNeuberger,M.S.in Methods:A companion to Methods in Enzymology 2:159-165(Lernerら編、Academic Press (1991))
Bruggemann,M.およびNeuberger,M.S."Strategies for expressing human antibody repertoires in transgenic mice." Immunology Today 17:391-397(1996)
Chenら、"Immunoglobulin gene rearrangement in B-cell deficient mice generated by targeted deletion of the JH locus," International Immunology 5:647-656(1993)
Choiら、"Transgenic mice containing a human heavy chain immunoglobulin gene fragment cloned in a yeast artificial cnromosome" Nature Genetics 4:117-123(1993)
Coliganら、Unit 2.1,"Enzyme-linked Immunosorbent assays," in Current protocols in Immunology(1994)
Cook,G.P.およびTomlinson,I.M.,"The human immunoglobulin VH repertoire." Immunology Today 16:237-242(1995)
Coxら、"A directory of human germ-line VX segments reveals a strong bias in their usage." Eur.J.Immunol.24:827-836(1994)
Dariavachら、"The mouse IgH 3'-enhancer." Eur.J.linmunol.21:1499-1504(1901)
Den Dunnenら、"Reconstruction of the 2.4 Mb human DMD-gene by homologous YAC recombination." Human Molecular Genetics 1:19-28(1992)
Feeney,A.J."Lack of N regions in fetal and neonatal mouse immunoglobulin V-D-J junctional sequences." J.Exp.Med.172:137-1390(1990)
Fishwildら、"High-avidity human IgGκ monoclonal antibodies from a novel strain of minilocus transgenic mice." Nature Biotech.14:845-851(1996)
Flanagan,J.G.およびRabbitts,T.H.,"Arrangement of human immunoglobulin heavy chain constan region genes implies evolutionary duplication of a segment containing g,e,and a genes." Nature 300:709-713(1982)
Galfre,G.およびMilstein,C.,"Preparation of monoclonal antibodies:strategies and procedures." Methods Enzymol.73:3-46(1981)
Gemmillら、"Protocols for pulsed field gel electrophoresis:Separation and detection of large DNA molecules." Advances in Genome Biology 1:217-251(1991)
Gillら、"Monoclonal anti-epidermal growth factor receptor antibodies which are inhibitors of epidermal growth factor binding and antagonists of epidermal growth factorstimulated tyrosine protein kinase activity." J.Biol.Chem.259:7755(1984)
Greenら、"Antigen-specific human monoclonal antibodies from mice engineered with human Ig heavy and light chain YACs." Nature Genetics 7:13-21(1994)
Hermansonら、"Rescue of end fragments of yeast artificial chromosomes by homologous recombination in yeast." Nucleic Acids Res.19:4943-4948(1991)
Huberら、"The human immunoglobulin κ locus.Characterization of the partially duplicated L regions." Eur.J.Immunol.23:2860-2967(1993)
Jakobovits,A.,"Humanizing the mouse genome." Current Biology 4:761-763(1994)
Jakobovits,A.,"Production of fully human antibodies by transgenic mice." Current opinion in Biotechnology 6:561-566(1995)
Jakobovitsら、"Germ-line transmission and expression of a human-derived yeast artificial-chromosome." Nature 362:255-258(1993)
Jakobovits,A.ら、"Analysis of homozygous mutant chimeric mice:Deletion of the immunoglobulin heavy-chain joining region blocks B-cell development and antibody production." Proc.Natl.Acad.Sci.USA 90:2551-2555(1993)
Kawamotoら、"Growth stimulation of A431 cells by epidermal growth factor:Identification of high affinity receptors for EGF by an anti-receptor monoclonal antibody." Proc.Nat.Acad.Sci.,USA 80:1337-1341(1983)
Lonbergら、"Antigen-specific human antibodies from mice comprising four distinct genetic modifications." Nature 368:856-859(1994)
Lusti-Marasimhanら、"Mutation of Leu25 and Val27 introduces CC chemokine activity into interleukin-8." J.Biol.Chem.270:2716-2721(1995)
Marksら、"Oligonucleotide primers for pclymerase chain reaction amplification of human immunoglobulin variable genes and design of family-specific oligonucleotide probes." Eur.J.Immunol.21:985-991(1991)
Matsudaら、"Structure and physical map of 64 variable segments in the 3' 0.8-megabase region of the human immunoglobulin heavy-chain locus." Nature Genetics 3:88-94(1993)
Max,E.Molecular genetics of immunoglobulins.Fundamental Immunology.315-382(Paul,WE編、New York:Raven Press (1993))
Mendezら、"A set of YAC targeting vectors for the interconversion of centric and acentric arms." Cold Spring Harbor Laboratory Press,Genome Mapping and Sequencing meeting,163(1993)
Mendezら、"Analysis of the structural integrity of YACs comprising human immunoglobulin genes in yeast and in embryonic stem cells." Genomics 26:294-307(1995)
Ray,S.およびDiamond,B.,"Generation of a fusion partner to sample the repertoire of Splenic B-cells destined for apoptosis." Proc.Natl.Acad.Sci.USA 91:5548-5551(1994)
Satoら、"Biological effects in vitro of monoclonal antibodies to human epidermal growth factor receptors" Mol.Bioi.Med.1:511-529(1983)
Schiestl,R.H.およびGietz,R.D.,"High efficiency transformation of intact yeast cells using stranded nucleic acids as a carrier." Curr.Genet.16:339-346(1989)
Shermanら、"Laboratory Course Manual for Methods in Yeast Genetics."(Cold Spring Harbor Laboratory Press,Cold Spring Harbor,NY (1986))
Silvermanら、"Meiotic recombination between yeast artificial chromosomes yields a single clone containing the entire BCL2 protooncogene." Proc.Nati.Acad.Sci.USA 87:9913-9917(19 )
Srivastava,A.およびSchiessinger,D.,"Vectors for inserting selectable markers in vector arms and human DNA inserts of yeast artificial chromosomes(YACs)." Gene 103:53-59 (1991)
Taylorら、"A transgenic mouse that expresses a diversity of human sequence heavy and light chain immunoglobulins." Nucleic Acids Research 20:6287-6295(1992)
Taylorら、"Human immunoglobulin transgenes undergo rearrangement,somatic mutation and class switching in mice that lack endogenous IgM." International Immunology 6:579-591(1994)
Tuaillonら、"Human immunoglobulin heavy-chain minilocus recombination in transgenic mice:gene-segment use in m and g transcripts." Proc.Natl.Acad.Sci.USA 90:3720-3724(1993)
Tuaillonら、"Analysis of direct and inverted DJH rearrangements in a human Ig heavy chain transgenic minilocus" J.Immunol.154:6453-6465(1995)
Vaughanら、"Human antibodies with subnanomolar affinities isolated from a large non-immunized phage display library." Nature Biotech.14:309-314(1996)
Wagnerら、"The diversity of antigen-specific monoclonal antibodies from transgenic mice bearing human immunoglobulin gene miniloci." Eur.J.Immunol.24:2672-2681(1994)
Weichholdら、"The human immunoglobulin κ locus consists of two copies that are organized in opposite polarity." Genomics 16:503-511(1993)
Yamada,M.ら、"Preferential utilization of specific immunoglobulin heavy chain diversity and joining segments in adult human peripheral blood B lymphocytes." J.Exp.Med.173:395-407(1991)
【0145】
同等性
前述の記述および実施例は、本発明の特定の好ましい態様について詳述し、発明者が意図する最善の様式を記述する。しかし、前述がどれほど詳細に本文において現れようとも、本発明は多くの方法において実践されること、および本発明は添付の請求の範囲およびその同等物に従って解釈されるべきであると認識される。
【技術分野】
【0001】
本発明により、ヒト上皮細胞増殖因子受容体(EGF-r)に対する相補性決定領域に及ぶ完全なヒト近接(contiguous)重鎖および軽鎖配列モノクローナル抗体が提供される。重鎖および軽鎖免疫グロブリン分子をコードするヌクレオチド配列およびこれらの分子を含むアミノ酸配列、特に(CDR')、中でもCDR1からCDR3までに相当する配列が提供される。そのような免疫グロブリン分子およびモノクローナル抗体を発現するハイブリドーマも提供する。
【背景技術】
【0002】
EGF-rは、多くのタイプのヒト固形癌上に過剰発現されることが示されている。メンデルソーン(Mendelsohn)、Cancer Cells 7:359(1989)(非特許文献1)、メンデルソーン(Mendelsohn)、Cancer Biology 1:339〜344(1990)(非特許文献2)、モジュタエジ&ディーン(Modjtahedi and Dean)、Int'l J.Oncology 4:277〜296(1994)(非特許文献3)。例えば、EGF-rの過剰発現は特定の肺、乳房、結腸、胃、脳、膀胱、頭頸部、卵巣、および前立腺の癌腫に認められている[モジュタエジ&ディーン(Modjtahedi and Dean)、Int'l J.Oncology 4:277〜296(1994)(非特許文献3)]。上皮細胞増殖因子(EGF)およびトランスフォーミング増殖因子-α(TGF-α)はいずれもEGF-rに結合し、細胞増殖を引き起こし、腫瘍を増殖させることが示されている。
【0003】
このように、EGF、TGF-α、およびEGF-rに対する抗体はEGF-rを発現するまたは過剰発現する腫瘍の治療において有用となる可能性があることが一部のグループにより提唱された[メンデルソーン(Mendelsohn)、Cancer Cells 7:359(1989)(非特許文献1)、メンデルソーン(Mendelsohn)、Cancer Biology 1:339〜344(1990)(非特許文献2)、モジュタエジ&ディーン(Modjtahedi and Dean)、Int'l J.Oncology 4:277〜296(1994)(非特許文献3)、トシら(Tosi)、Int'l J.Cancer 62:643〜650(1995)(非特許文献4)]。実際に、抗EGF-r抗体はEGFおよびTGF-αの受容体への結合を遮断しながら、腫瘍細胞の増殖を阻害するように思われることが証明された。しかし、同時に抗EGF-r抗体はEGFおよびTGF-α非依存的細胞増殖についてはこれを阻害しないように思われた[モジュタエジ&ディーン(Modjtahedi and Dean)、Int'l J.Oncology 4:277〜296(1994)(非特許文献3)]。
【0004】
これらの知見を考慮して、EGF-rに対する多くのマウスおよびラットモノクローナル抗体を作製し、インビトロおよびインビボでそれらが腫瘍細胞の増殖を阻害するか否かを調べた[モジュタエジ&ディーン(Modjtahedi and Dean)、Int'l J.Oncology 4:277〜296(1994)(非特許文献3)]。臨床においておそらく開発が最も進行している抗体はC225と名付けられたキメラ抗体で、これはマウスの可変領域とヒトIgG1定常領域とを有する[モジュタエジ&ディーン(Modjtahedi and Dean)、Int'l J.Oncology 4:277〜296(1994)(非特許文献3)]。C225抗体に基づく225と呼ばれるマウス抗体は、カリフォルニア大学およびローラー社(Rorer)によって開発された。その開示が参照として本明細書に組み入れられる、米国特許第4,943,533号(特許文献1)および欧州特許第359,282号(特許文献2)を参照のこと。C225抗体は、インビトロでEGF-媒介腫瘍細胞の増殖を阻害し、インビボでヌードマウスにおいてヒト腫瘍形成を阻害することが示された。その上、該抗体は特定の化学療法剤と相乗的に作用して、インビボで異種移植マウスモデルにおいてヒト腫瘍を根治させるように作用するように思われた[モジュタエジ&ディーン(Modjtahedi and Dean)、Int'l J.Oncology 4:277〜296(1994)(非特許文献3)]。
【0005】
イムクローン社(ImClone)はC225と名付けられた抗EGF-r抗体を用いてヒト臨床試験を実施している。C225に関して、頭頸部、前立腺および肺癌患者におけるフェーズIおよびフェーズI/II臨床試験は、実施されたかまたは現在進行中のようである。フェーズI臨床試験では、免疫無防備状態(immunocompromised)の患者を含む症例においても、多数回の注射によって、およびおそらく400mg/m2までの用量で毒性が認められなかった。該試験は約5〜約200 mg/m2までの5用量を含む用量増加試験として実施され、化学療法(すなわち、ドキソルビシン、アドリアマイシン、タキソール、およびシスプラチン)と併用して行われた。これらの試験において得られた明らかな安全性データと共に、研究からの予備的な結果によって、前立腺癌患者の80%において腫瘍の退縮に関する何らかの証拠が示されるように思われる。
【0006】
しかし、これらの上記抗体のそれぞれはマウスまたはラットの可変および/または定常領域を有する。そのようなマウスまたはラットに由来する蛋白質が存在すれば、抗体の急速な一掃が起こり得る、または患者が抗体に対する免疫応答を起こしうる。マウスまたはラット由来の抗体の使用を避けるために、齧歯類が完全なヒト抗体を産生するように、ヒトの抗体機能を齧歯類に導入できるのではないかと考えられた。
【0007】
メガベースサイズのヒト遺伝子座をYAC中にクローニングおよび再構築し、それらをマウス生殖細胞系に導入しうることにより、極めて大きく、または大まかにマッピングされた遺伝子座の機能的構成要素を解明するための、ならびにヒト疾患の有用なモデルを作製するための、強力な手法が提供される。さらに、このような技術を利用してマウス遺伝子座をそれらのヒト等価物に置換することにより、発生過程におけるヒト遺伝子産物の発現および調節、それらの他の系との連絡、ならびに疾患の誘導および進行におけるそれらの関与に関して比類のない洞察が得られると考えられる。
【0008】
このような方法の1つの重要な実用的応用は、マウス体液性免疫系の「ヒト化」である。内因性Ig遺伝子が不活性化されたマウスにヒト免疫グロブリン(Ig)遺伝子座を導入することにより、抗体のプログラム化された発現および構築の根底にある機序に加えて、B細胞の発生におけるそれらの役割をも検討する機会が提供される。さらに、このような戦略により、ヒト疾患の抗体療法という可能性の実現へ向けての重要な里程標となる、完全ヒトモノクローナル抗体(Mab)を生産するための理想的な供給源が提供されると考えられる。完全ヒト抗体は、マウスまたはマウス由来のMabに固有の免疫原性およびアレルギー性反応を最小限に抑え、それにより、投与される抗体の有効性および安全性を高めると考えられる。完全ヒト抗体の使用は、抗体の反復投与を必要とする、炎症、自己免疫および癌などの慢性的および再発性のヒト疾患の治療に、実質的な利益を提供すると考えられる。
【0009】
この目標に向けての1つの手法は、マウス抗体の産生に欠陥があってヒトIg遺伝子座の大きな断片を有するマウス系統を、組換え操作によって作製することであり、これは、このようなマウスが、マウス抗体を生じずに多様なレパトア(repertoire)のヒト抗体を産生するであろうとの予測に立つものであった。大きなヒトIg断片は、可変遺伝子の広範な多様性に加えて、抗体の産生および発現の適切な調節も保持すると考えられる。抗体の多様化および選択のため、ならびにヒトタンパク質に対する免疫寛容を失わせるためのマウス機構を開発すれば、これらのマウス系統で再現されたヒト抗体レパトアはヒト抗原を含む関心対象の任意の抗原に対する高親和性抗体を生じるはずである。ハイブリドーマ技術を用いることにより、望ましい特異性をもつ抗原特異的ヒトMabは容易に製造および選択しうると考えられる。
【0010】
この一般的な方法は、本発明者らが1994年に発表した最初のXenoMouse(登録商標)系統の作出との関連で示された。グリーン(Green)ら、Nature Genetics 7:13〜21(1994)(非特許文献5)参照。XenoMouse(登録商標)系統は、可変および定常領域のコア配列を含む、ヒト重鎖およびκ軽鎖遺伝子座のそれぞれ245kbおよび190kbのサイズの生殖細胞系コンフィギュレーション断片を有する酵母人工染色体(YAC)を用いて操作された。同上。ヒトIgを含むYACは、抗体の再配列および発現の双方に関してマウス系への適合性があることが実証されており、不活性化されたマウスIg遺伝子の置換も可能であった。これは、それらがB細胞発生の誘導および完全ヒト抗体の成人様ヒトレパトアの生成、ならびに抗原特異的ヒトMabの産生の能力をもつことによって示された。また、これらの結果は、比較的多数のV遺伝子、付加的な調節要素、およびヒトIg定常領域を含むヒトIg遺伝子座のかなりの部分を導入することにより、感染および免疫化に対するヒト体液性応答の特徴である実質的に完全なレパトアが再現される可能性があることも示唆した。グリーンら(Green)は最近研究を拡大して、メガベースの大きさの生殖細胞系列構造のヒト重鎖座およびκ軽鎖座のYAC断片をそれぞれ導入することによって、ヒト抗体のレパトア(repertoire)の約80%以上を導入した。メンデスら(Mendez)、Nature Genetics 15:146〜156(1997)(非特許文献6)および本明細書に参照として組み入れられる、1996年12月3日に提出された米国特許出願第08/759,620号(特許文献3)を参照のこと。
【0011】
このような手法は、1990年1月12日に提出された米国特許出願第07/466,008号(特許文献4)、1990年11月8日に提出された第07/610,515号(特許文献5)、1992年7月24日に提出された第07/919,297号(特許文献6)、1992年7月30日に提出された第07/922,649号(特許文献7)、1993年3月15日に提出された第08/031,801号(特許文献8)、1993年8月27日に提出された第08/112,848号(特許文献9)、1994年4月28日に提出された第08/234,145号(特許文献10)、1995年1月20日に提出された第08/376,279号(特許文献11)、1995年4月27日に提出された第08/430,938号(特許文献12)、1995年6月5日に提出された第08/464,584号(特許文献13)、1995年6月5日に提出された第08/464,582号(特許文献14)、1995年6月5日に提出された第08/463,191号(特許文献15)、1995年6月5日に提出された第08/462,837号(特許文献16)、1995年6月5日に提出された第08/486,853号(特許文献17)、1995年6月5日に提出された第08/486,857号(特許文献18)、1995年6月5日に提出された第08/486,859号(特許文献19)、1995年6月5日に提出された第08/462,513号(特許文献20)、1996年10月2日に提出された第08/724,752号(特許文献21)、および1996年12月3日に提出された第08/759,620号(特許文献3)という米国特許出願においてさらに考察および描写されている。メンデズ(Mendez)ら、Nature Genetics 15:146-156(1997)(非特許文献6)も参照のこと。また、1996年6月12日に認可が発行されたEP 0 463 151 B1(特許文献22)、1994年2月3日に発行された国際特許出願WO 94/02602号(特許文献23)、1996年10月31日に発行された国際特許出願WO 96/34096号(特許文献24)、および1996年4月29日に提出されたPCT/US96/05928(特許文献25)も参照されたい。上記に引用したそれぞれの特許、特許出願、および参考文献の開示は、その全体が参照として本明細書に組み入れられる。
【0012】
1つの代替的なアプローチにおいて、ジェンファームインターナショナル社(GenPharm International,Inc.)を含む他者は、「ミニ遺伝子座(minilocus)」法を用いている。ミニ遺伝子座法では、Ig遺伝子座からの複数の小片(個々の遺伝子)を含めることにより、外来性Ig遺伝子座を模倣する。このため、1つまたは複数のVH遺伝子、1つまたは複数のDH遺伝子、1つまたは複数のJH遺伝子、μ定常領域、および第2の定常領域(好ましくはγ定常領域)が、動物に挿入するための構築物(construct)中に形成される。この手法はスラニ(Surani)らに対する米国特許第5,545,807号(特許文献26)、いずれもロンバーグ(Lonberg)およびケイ(Kay)に対する米国特許第5,545,806号(特許文献27)および第5,625,825号(特許文献28)、ならびにいずれもジェンファームインターナショナル社(GenPharm International,Inc.)による米国特許出願である1990年8月29日に提出された第07/574,748号(特許文献29)、1990年8月31日に提出された第07/575,962号(特許文献30)、1991年12月17日に提出された第07/810,279号(特許文献31)、1992年3月18日に提出された第07/853,408号(特許文献32)、1992年6月23日に提出された第07/904,068号(特許文献33)、1992年12月16日に提出された第07/990,860号(特許文献34)、1993年4月26日に提出された第08/053,131号(特許文献35)、1993年7月22日に提出された第08/096,762号(特許文献36)、1993年11月18日に提出された第08/155,301号(特許文献37)、1993年12月3日に提出された第08/161,739号(特許文献38)、1993年12月10日に提出された第08/165,699号(特許文献39)、1994年3月9日に提出された第08/209,741号(特許文献40)に記載されており、それらの開示は参照として本明細書に組み入れられる。また、その開示の全体が参照として本明細書に組み入れられる1994年11月10日に発行された国際公開公報第94/25585号(特許文献41)、1993年7月24日に発行された国際公開公報第93/12227号(特許文献42)、1992年12月23日に発行された国際公開公報第92/22645号(特許文献43)、1992年3月19日に発行された国際公開公報第92/03918号(特許文献44)も参照されたい。さらに、その開示の全体が参照として本明細書に組み入れられるテイラー(Taylor)ら、1992(非特許文献7)、チェン(Chen)ら、1993(非特許文献8)、トゥアイロン(Tuaillon)ら、1993(非特許文献9)、チョイ(Choi)ら、1993(非特許文献10)、ロンバーグ(Lonberg)ら(1994)(非特許文献11)、テイラーら(1994)(非特許文献12)、およびトゥアイロンら(1995)(非特許文献13)も参照されたい。
【0013】
上記に引用し、英国医学研究会議(Medical Research Council)(「MRC」)に譲渡されたスラニ(Surani)らの発明者らは、ミニ遺伝子座法の使用により、Ig遺伝子座を有するトランスジェニックマウスを作製した。上記に引用したジェンファームインターナショナル社の業績の発明者らであるロンバーグおよびケイは、本発明者の先例にならって、内因性マウスIg遺伝子座の不活性化とスラニらの業績の実質的な重複物との組み合わせを提案した。
【0014】
ミニ遺伝子座法の1つの利点は、Ig遺伝子座の一部を含む構築物の作製および動物への導入を迅速に行いうることである。しかしこれを相殺する程度に、ミニ遺伝子座法には理論的にみて、少数のV、DおよびJ遺伝子を含めることによって導入される多様性が不十分であるとの重大な欠点がある。事実、発表された研究はこの懸念を裏づけるように思われる。ミニ遺伝子座法を用いて作製された動物のB細胞発生および抗体産生は阻害されていると考えられる。したがって、本発明を取り囲む研究は一貫して、より広範な多様性の実現を目的として、および動物の免疫レパトアを再構築するための努力において、Ig遺伝子座の大部分を導入することを目的としている。
【0015】
ヒト抗マウス抗体(HAMA)反応のために、業界はキメラまたはそれ以外のヒト化抗体を作製するようになった。C225抗体はヒト定常領域とマウス可変領域とを有するキメラ抗体であるが、特に抗体の長期的または多用量使用時には、特定のヒト抗キメラ抗体(HACA)反応が認められるであろうと予想される。
このように、HAMAまたはHACA反応に関する懸念および/または作用を無用のものとするために、C225と比較して同程度または増強した活性を有する、EGF-rに対する完全なヒト抗体を提供することが望ましいと考えられる。
【先行技術文献】
【特許文献】
【0016】
【特許文献1】米国特許第4,943,533号
【特許文献2】欧州特許第359,282号
【特許文献3】米国特許出願第08/759,620号
【特許文献4】米国特許出願第07/466,008号
【特許文献5】米国特許出願第07/610,515号
【特許文献6】米国特許出願第07/919,297号
【特許文献7】米国特許出願第07/922,649号
【特許文献8】米国特許出願第08/031,801号
【特許文献9】米国特許出願第08/112,848号
【特許文献10】米国特許出願第08/234,145号
【特許文献11】米国特許出願第08/376,279号
【特許文献12】米国特許出願第08/430,938号
【特許文献13】米国特許出願第08/464,584号
【特許文献14】米国特許出願第08/464,582号
【特許文献15】米国特許出願第08/463,191号
【特許文献16】米国特許出願第08/462,837号
【特許文献17】米国特許出願第08/486,853号
【特許文献18】米国特許出願第08/486,857号
【特許文献19】米国特許出願第08/486,859号
【特許文献20】米国特許出願第08/462,513号
【特許文献21】米国特許出願第08/724,752号
【特許文献22】EP 0 463 151 B1
【特許文献23】国際特許出願WO 94/02602号
【特許文献24】国際特許出願WO 96/34096号
【特許文献25】PCT/US96/05928
【特許文献26】米国特許第5,545,807号
【特許文献27】米国特許第5,545,806号
【特許文献28】米国特許第5,625,825号
【特許文献29】米国特許出願第07/574,748号
【特許文献30】米国特許出願第07/575,962号
【特許文献31】米国特許出願第07/810,279号
【特許文献32】米国特許出願第07/853,408号
【特許文献33】米国特許出願第07/904,068号
【特許文献34】米国特許出願第07/990,860号
【特許文献35】米国特許出願第08/053,131号
【特許文献36】米国特許出願第08/096,762号
【特許文献37】米国特許出願第08/155,301号
【特許文献38】米国特許出願第08/161,739号
【特許文献39】米国特許出願第08/165,699号
【特許文献40】米国特許出願第08/209,741号
【特許文献41】国際公開公報第94/25585号
【特許文献42】国際公開公報第93/12227号
【特許文献43】国際公開公報第92/22645号
【特許文献44】国際公開公報第92/03918号
【非特許文献】
【0017】
【非特許文献1】メンデルソーン(Mendelsohn)、Cancer Cells 7:359(1989)
【非特許文献2】メンデルソーン(Mendelsohn)、Cancer Biology 1:339〜344(1990)
【非特許文献3】モジュタエジ&ディーン(Modjtahedi and Dean)、Int'l J.Oncology 4:277〜296(1994)
【非特許文献4】トシら(Tosi)、Int'l J.Cancer 62:643〜650(1995)
【非特許文献5】グリーン(Green)ら、Nature Genetics 7:13〜21(1994)
【非特許文献6】メンデスら(Mendez)、Nature Genetics 15:146〜156(1997)
【非特許文献7】Taylorら、"A transgenic mouse that expresses a diversity of human sequence heavy and light chain immunoglobulins." Nucleic Acids Research 20:6287-6295(1992)
【非特許文献8】Chenら、"Immunoglobulin gene rearrangement in B-cell deficient mice generated by targeted deletion of the JH locus," International Immunology 5:647-656(1993)
【非特許文献9】Tuaillonら、"Human immunoglobulin heavy-chain minilocus recombination in transgenic mice:gene-segment use in m and g transcripts." Proc.Natl.Acad.Sci.USA 90:3720-3724(1993)
【非特許文献10】Choiら、"Transgenic mice containing a human heavy chain immunoglobulin gene fragment cloned in a yeast artificial cnromosome" Nature Genetics 4:117-123(1993)
【非特許文献11】Lonbergら、"Antigen-specific human antibodies from mice comprising four distinct genetic modifications." Nature 368:856-859(1994)
【非特許文献12】Taylorら、"Human immunoglobulin transgenes undergo rearrangement,somatic mutation and class switching in mice that lack endogenous IgM." International Immunology 6:579-591(1994)
【非特許文献13】Tuaillonら、"Analysis of direct and inverted DJH rearrangements in a human Ig heavy chain transgenic minilocus" J.Immunol.154:6453-6465(1995)
【発明の概要】
【0018】
本発明の第一の局面に従って、配列の一部がヒトVH 4ファミリー遺伝子ならびに図2、6、10、14、18、22、26および30に示すヌクレオチド配列によって表されるその変異体のいずれかによってコードされる、重鎖可変領域アミノ酸配列を含む、上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、重鎖可変領域アミノ酸配列は、10位の残基においてアスパラギン酸アミノ酸置換を含む。
【0019】
本発明の第2の局面に従って、その配列の一部がヒトVH 4〜31遺伝子ならびに図2、6、10、14および18に示すヌクレオチド配列によって表されるその変異体のいずれかによってコードされる、重鎖可変領域アミノ酸配列を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、重鎖可変領域は配列番号:23に表されるようにCDR1からCDR3までの近接配列を含む。好ましい態様において、抗体は、配列番号:24によって表される配列を含む軽鎖可変領域をさらに含む。好ましい態様において、重鎖可変領域は配列番号:25に表されるように、CDR1からCDR3までの近接配列を含む。好ましい態様において、抗体は配列番号:26によって表される配列を含む軽鎖可変領域をさらに含む。好ましい態様において、重鎖可変領域は配列番号:27に表されるようにCDR1からCDR3までの近接配列を含む。好ましい態様において、抗体は配列番号:28によって表される配列を含む軽鎖可変領域をさらに含む。好ましい態様において、重鎖可変領域は配列番号:29に表されるようにCDR1からCDR3までの近接配列を含む。好ましい態様において、抗体は配列番号:30によって表される配列を含む軽鎖可変領域をさらに含む。好ましい態様において、重鎖可変領域は配列番号:31に表されるようにCDR1からCDR3までの近接配列を含む。好ましい態様において、抗体は配列番号:32によって表される配列を含む軽鎖可変領域をさらに含む。
【0020】
本発明の第3の局面に従って、その配列の一部がヒトVH 4〜61遺伝子ならびに図22、26、および30に示すヌクレオチド配列によって表されるその変異体のいずれかによってコードされる重鎖可変領域アミノ酸配列を含む、上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、重鎖可変領域は配列番号:33に表されるようにCDR1からCDR3までの近接配列を含む。好ましい態様において抗体は、配列番号:34によって表される配列を含む軽鎖可変領域をさらに含む。好ましい態様において、重鎖可変領域は配列番号:35に表されるようにCDR1からCDR3までの近接配列を含む。好ましい態様において、抗体は配列番号:36によって表される配列を含む軽鎖可変領域をさらに含む。好ましい態様において、重鎖可変領域は配列番号:37に表されるようにCDR1からCDR3までの近接配列を含む、好ましい態様において抗体は、配列番号:38によって表される配列を含む軽鎖可変領域をさらに含む。
【0021】
本発明の第4の局面において、その配列の一部がヒトVK Iファミリー遺伝子ならびに図4、8、12、16、20、24、28、および32に示すヌクレオチド配列によって表されるその変異体のいずれかによってコードされる軽鎖可変領域アミノ酸配列を含む、上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、軽鎖可変領域は配列番号:24によって表される配列を含む。好ましい態様において、軽鎖可変領域は配列番号:26によって表される配列を含む。好ましい態様において、軽鎖可変領域は配列番号:28によって表される配列を含む。好ましい態様において、軽鎖可変領域は配列番号:30によって表される配列を含む。好ましい態様において、軽鎖可変領域は配列番号:32によって表される配列を含む。好ましい態様において、軽鎖可変領域は配列番号:34によって表される配列を含む。好ましい態様において、軽鎖可変領域は配列番号:36によって表される配列を含む。好ましい態様において、軽鎖可変領域は配列番号:38によって表される配列を含む。
【0022】
本発明の第5の局面に従って、配列番号:23に表されるようにCDR1からCDR3までの近接配列を含む重鎖可変領域を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、抗体は配列番号:24によって表される配列を含む軽鎖可変領域をさらに含む。
【0023】
本発明の第6の局面において、配列番号:25に表されるようにCDR1からCDR3までの近接配列を含む重鎖可変領域を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、抗体は配列番号:26によって表される配列を含む軽鎖可変領域をさらに含む。
【0024】
本発明の第7の局面において、配列番号:27に表されるようにCDR1からCDR3までの近接配列を含む重鎖可変領域を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、抗体は配列番号:28によって表される配列を含む軽鎖可変領域をさらに含む。
【0025】
本発明の第8の局面において、配列番号:29に表されるようにCDR1からCDR3までの近接配列を含む重鎖可変領域を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、抗体は配列番号:30によって表される配列を含む軽鎖可変領域をさらに含む。
【0026】
本発明の第9の局面において、配列番号:31に表されるようにCDR1からCDR3までの近接配列を含む重鎖可変領域を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、抗体は配列番号:32によって表される配列を含む軽鎖可変領域をさらに含む。
【0027】
本発明の第10の局面において、配列番号:33に表されるようにCDR1からCDR3までの近接配列を含む重鎖可変領域を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、抗体は配列番号:34によって表される配列を含む軽鎖可変領域をさらに含む。
【0028】
本発明の第11の局面において、配列番号:35に表されるようにCDR1からCDR3までの近接配列を含む重鎖可変領域を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、抗体は配列番号:36によって表される配列を含む軽鎖可変領域をさらに含む。
【0029】
本発明の第12の局面において、配列番号:37に表されるようにCDR1からCDR3までの近接配列を含む重鎖可変領域を含む上皮細胞増殖因子受容体に対する抗体が提供される。好ましい態様において、抗体は配列番号:38によって表される配列を含む軽鎖可変領域をさらに含む。
【0030】
本発明の第13の局面に従って、上皮細胞増殖因子受容体に対する抗体によって固形腫瘍を治療する方法において、固形腫瘍を有する患者に本発明の前述の抗体の一つを投与することを含む改善が提供される。
本発明は例えば、以下の項目を提供する:
(項目1)
配列の一部がヒトVH 4ファミリー遺伝子および図2、6、10、14、18、22、26および30に示すヌクレオチド配列によって表されるその変異体のいずれかによってコードされる、重鎖可変領域アミノ酸配列を含む上皮細胞増殖因子受容体に対する抗体。
(項目2)
重鎖可変領域アミノ酸配列が残基10においてアスパラギン酸アミノ酸置換を含む、項目1記載の抗体。
(項目3)
配列の一部がヒトVH 4〜31遺伝子および図2、6、10、14、および18に示すヌクレオチド配列によって表されるその変異体のいずれかによってコードされる、重鎖可変領域アミノ酸配列を含む上皮細胞増殖因子受容体に対する抗体。
(項目4)
重鎖可変領域が配列番号:23に示すCDR1からCDR3までの近接配列を含む、項目3記載の抗体。
(項目5)
配列番号:24によって表される配列を含む軽鎖可変領域をさらに含む、項目4記載の抗体。
(項目6)
重鎖可変領域が配列番号:25に示すCDR1からCDR3までの近接配列を含む、項目3記載の抗体。
(項目7)
配列番号:26によって表される配列を含む軽鎖可変領域をさらに含む、項目6記載の抗体。
(項目8)
重鎖可変領域が配列番号:27に示すCDR1からCDR3までの近接配列を含む、項目3記載の抗体。
(項目9)
配列番号:28によって表される配列を含む軽鎖可変領域をさらに含む、項目8記載の抗体。
(項目10)
重鎖可変領域が配列番号:29に示すCDR1からCDR3までの近接配列を含む、項目3記載の抗体。
(項目11)
配列番号:30によって表される配列を含む軽鎖可変領域をさらに含む、項目10記載の抗体。
(項目12)
重鎖可変領域が配列番号:31に示すCDR1からCDR3までの近接配列を含む、項目3記載の抗体。
(項目13)
配列番号:32によって表される配列を含む軽鎖可変領域をさらに含む、項目12記載の抗体。
(項目14)
配列の一部がヒトVH 4〜61遺伝子および図22、26、および30に示すヌクレオチド配列によって表されるその変異体のいずれかによってコードされる、重鎖可変領域アミノ酸配列を含む上皮細胞増殖因子受容体に対する抗体。
(項目15)
重鎖可変領域が配列番号:33に示すCDR1からCDR3までの近接配列を含む、項目14記載の抗体。
(項目16)
配列番号:34によって表される配列を含む軽鎖可変領域をさらに含む、項目15記載の抗体。
(項目17)
重鎖可変領域が配列番号:35に示すCDR1からCDR3までの近接配列を含む、項目14記載の抗体。
(項目18)
配列番号:36によって表される配列を含む軽鎖可変領域をさらに含む、項目17記載の抗体。
(項目19)
重鎖可変領域が配列番号:37に示すCDR1からCDR3までの近接配列を含む、項目14記載の抗体。
(項目20)
配列番号:38によって表される配列を含む軽鎖可変領域をさらに含む、項目19記載の抗体。
(項目21)
配列の一部がヒトVK Iファミリー遺伝子および図4、8、12、16、20、24、28、および32に示されるヌクレオチド配列によって表されるその変異体のいずれかによってコードされる、軽鎖可変領域アミノ酸配列を含む、上皮細胞増殖因子受容体に対する抗体。
(項目22)
軽鎖可変領域が配列番号:24によって表される配列を含む、項目21記載の抗体。
(項目23)
軽鎖可変領域が配列番号:26によって表される配列を含む、項目21記載の抗体。
(項目24)
軽鎖可変領域が配列番号:28によって表される配列を含む、項目21記載の抗体。
(項目25)
軽鎖可変領域が配列番号:30によって表される配列を含む、項目21記載の抗体。
(項目26)
軽鎖可変領域が配列番号:32によって表される配列を含む、項目21記載の抗体。
(項目27)
軽鎖可変領域が配列番号:34によって表される配列を含む、項目21記載の抗体。
(項目28)
軽鎖可変領域が配列番号:36によって表される配列を含む、項目21記載の抗体。
(項目29)
軽鎖可変領域が配列番号:38によって表される配列を含む、項目21記載の抗体。
(項目30)
配列番号:23に示すCDR1からCDR3までの近接配列を含む重鎖可変領域を含む、上皮細胞増殖因子受容体に対する抗体。
(項目31)
配列番号:24によって表される配列を含む軽鎖可変領域をさらに含む、項目30記載の抗体。
(項目32)
配列番号:25に示すCDR1からCDR3までの近接配列を含む重鎖可変領域を含む、上皮細胞増殖因子受容体に対する抗体。
(項目33)
配列番号:26によって表される配列を含む軽鎖可変領域をさらに含む、項目32記載の抗体。
(項目34)
配列番号:27に示すCDR1からCDR3までの近接配列を含む重鎖可変領域を含む、上皮細胞増殖因子受容体に対する抗体。
(項目35)
配列番号:28によって表される配列を含む軽鎖可変領域をさらに含む、項目34記載の抗体。
(項目36)
配列番号:29に示すCDR1からCDR3までの近接配列を含む重鎖可変領域を含む、上皮細胞増殖因子受容体に対する抗体。
(項目37)
配列番号:30によって表される配列を含む軽鎖可変領域をさらに含む、項目36記載の抗体。
(項目38)
配列番号:31に示すCDR1からCDR3までの近接配列を含む重鎖可変領域を含む、上皮細胞増殖因子受容体に対する抗体。
(項目39)
配列番号:32によって表される配列を含む軽鎖可変領域をさらに含む、項目38記載の抗体。
(項目40)
配列番号:33に示すCDR1からCDR3までの近接配列を含む重鎖可変領域を含む、上皮細胞増殖因子受容体に対する抗体。
(項目41)
配列番号:34によって表される配列を含む軽鎖可変領域をさらに含む、項目40記載の抗体。
(項目42)
配列番号:35に示すCDR1からCDR3までの近接配列を含む重鎖可変領域を含む、上皮細胞増殖因子受容体に対する抗体。
(項目43)
配列番号:36によって表される配列を含む軽鎖可変領域をさらに含む、項目42記載の抗体。
(項目44)
配列番号:37に示すCDR1からCDR3までの近接配列を含む重鎖可変領域を含む、上皮細胞増殖因子受容体に対する抗体。
(項目45)
配列番号:38によって表される配列を含む軽鎖可変領域をさらに含む、項目44記載の抗体。
(項目46)
上皮細胞増殖因子受容体に対する抗体によって固形癌を治療する方法において、固形癌を有する患者に項目1〜45のいずれか一項記載の抗体を投与することを含む改善。
【図面の簡単な説明】
【0031】
【図1】ハイブリドーマE1.1によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE1.1分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列(contiguous sequence)を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図2】ハイブリドーマE1.1からクローニングした図1の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図3】ハイブリドーマE1.1によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE1.1分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図4】ハイブリドーマE1.1からクローニングした図3のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図5】ハイブリドーマE2.4によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE2.4分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図6】ハイブリドーマE2.4からクローニングした図5の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図7】ハイブリドーマE2.4によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE2.4分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図8】ハイブリドーマE2.4からクローニングした図7のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図9】ハイブリドーマE2.5によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE2.5分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図10】ハイブリドーマE2.5からクローニングした図9の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図11】ハイブリドーマE2.5によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE2.5分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図12】ハイブリドーマE2.5からクローニングした図11のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図13】ハイブリドーマE6.2によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE6.2分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図14】ハイブリドーマE6.2からクローニングした図13の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図15】ハイブリドーマE6.2によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE6.2分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図16】ハイブリドーマE6.2からクローニングした図15のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図17】ハイブリドーマE6.4によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE6.4分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図18】ハイブリドーマE6.2からクローニングした図17の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図19】ハイブリドーマE6.4によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE6.4分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図20】ハイブリドーマE6.4からクローニングした図19のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図21】ハイブリドーマE2.11によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜61によってコードされる配列とE2.11分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図22】ハイブリドーマE2.11からクローニングした図21の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図23】ハイブリドーマE2.11によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE2.11分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図24】ハイブリドーマE2.11からクローニングした図23のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図25】ハイブリドーマE6.3によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜61によってコードされる配列とE6.3分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図26】ハイブリドーマE6.3からクローニングした図25の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図27】ハイブリドーマE6.3によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE6.3分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図28】ハイブリドーマE6.3からクローニングした図27のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図29】ハイブリドーマE7.6.3によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜61によってコードされる配列とE7.6.3分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図30】ハイブリドーマE7.6.3からクローニングした図29の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図31】ハイブリドーマE7.6.3によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE7.6.3分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【図32】ハイブリドーマE7.6.3からクローニングした図31のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【図33】特異的抗EGF-r抗体重鎖アミノ酸配列と、特定の抗体の重鎖をコードする特定のVH遺伝子のアミノ酸配列との比較を示す。
【図34】特異的抗EGF-r抗体軽鎖アミノ酸配列と、特定の抗体の軽鎖をコードする特定のVK遺伝子のアミノ酸配列との比較を示す。
【図35】インビトロでヒト抗EGF-r抗体によるヒト類上皮癌A431細胞に対するEGF結合の遮断を示し、(□)は本発明の抗EGF-r抗体によって得られた結果を示し、(●)は、マウスモノクローナル抗体225によって得られた結果を示し、および(黒上向き三角)は、対照の非特異的ヒトIgG2抗体によって得られた結果を示す。
【図36】インビトロでヒト抗EGF-r抗体によるヒト類上皮癌A431細胞に対するEGF結合の阻害を示し、(□)は、マウスモノクローナル抗体225によって得られた結果を示し、(○)はマウスモノクローナル抗体528によって得られた結果を示し、(黒下向き三角)は本発明のE1.1抗体を用いて得られた結果を示し、(黒上向き三角)は本発明のE2.4抗体を用いて得られた結果を示し、(黒右向き三角)は、本発明のE2.5抗体を用いて得られた結果を示し、(黒左向き三角)は本発明のE2.6抗体を用いて得られた結果を示し、(◆)は本発明のE2.11抗体を用いて得られた結果を示し、および(四角で囲まれた×印)は、対照の、非特異的ヒトIgG2抗体を用いて得られた結果を示す。
【図37】インビトロでヒト抗EGF-r抗体によるヒト類上皮癌A431細胞に対するTGF-α結合の阻害を示し、(□)は、マウスモノクローナル抗体225によって得られた結果を示し、(◆)は本発明のE6.2抗体を用いて得られた結果を示し、(●)は本発明のE6.3抗体を用いて得られた結果を示し、(黒上向き三角)は本発明のE7.2抗体を用いて得られた結果を示し、(黒四角)は本発明のE7.10抗体を用いて得られた結果を示し、(黒下向き三角)はE7.6.3を用いて得られた結果を示し、および(○で囲まれた十字)は対照の非特異的ヒトIgG2抗体を用いて得られた結果を示す。
【図38】インビトロでヒト抗EGF-r抗体によるヒト結腸癌SW948細胞に対するEGF結合の阻害を示し、(●)は、本発明の抗EGF-r抗体によって得られた結果を示し、(□)はマウスモノクローナル抗体225によって得られた結果を示し、および(黒上向き三角)は対照の非特異的ヒトIgG2抗体によって得られた結果を示す。
【図39】ゼノマウスII株に由来するヒト抗EGF-r抗体がインビトロでSW948細胞の増殖を阻害することを示し、(●)は、本発明の抗EGF-r抗体によって得られた結果を示し、(□)は、マウスモノクローナル抗体225によって得られた結果を示し、および(黒上向き三角)は対照の非特異的ヒトIgG2抗体によって得られた結果を示す。
【図40】インビボで本発明のヒト抗EGF-r抗体を用いることによる、ヌードマウスにおけるヒト類上皮癌A431細胞増殖の阻害を示す。図において、(黒上向き三角)は、本発明のヒト抗EGF-r抗体1mgの投与によって得られた結果を示し、(黒下向き三角)は本発明のヒト抗EGF-r抗体0.2 mgの投与によって得られた結果を示し、(□)は、対照の非特異的ヒトIgG2抗体によって得られた結果を示し、および(○)は、対照として燐酸緩衝生理食塩液を用いた場合に得られた結果を示す。
【図41】インビボで本発明のヒト抗EGF-r抗体を用いることによる、ヌードマウスにおける類上皮癌形成の阻害に関連したデータを示し、19日目での腫瘍の発生率を示す。
【図42】インビボで本発明のヒト抗EGF-r抗体を用いることによる、ヌードマウスにおける類上皮癌形成の阻害に関連したデータを示し、120日目での腫瘍の発生率を示す。
【図43】インビボにおいて本発明のヒト抗EGF-r抗体を用いることによって、ヌードマウスにおいて確立されたヒト類上皮腫瘍の根治に関連したデータを示す。図において、(△)は本発明のヒト抗EGF-r抗体(E7.6.3)各1mgを多数回投与した場合に得られた結果を示し、(×)は、ドキソルビシン各125μgを2回投与した場合に得られた結果を示し、(XX)は本発明のヒト抗EGF-r抗体(E7.6.3)各1 mgの多数回投与をドキソルビシン各125 μgの2回投与と併用した場合に得られた結果を示し、(黒四角)は対照の非特異的ヒトIgG2抗体によって得られた結果を示し、および(◆)は対照として燐酸緩衝生理食塩液を用いた場合に得られた結果を示す。
【図44】インビボで本発明のヒト抗EGF-r抗体を用いることによってヌードマウスにおいて確立されたヒト類上皮腫瘍の根治に関連したデータを示す。図において、(◆)は本発明のヒト抗EGF-r抗体(E2.5)の各0.5mgを多数回投与した場合に得られた結果を示し、(黒四角)はドキソルビシンの各125 μgを2回投与することによって得られた結果を示し、(△)は本発明のヒト抗EGF-r抗体(E2.5)の各0.5mgの多数回投与をドキソルビシン各125 μgの2回投与と併用した場合に得られた結果を示し、(×)は、対照として燐酸緩衝生理食塩液を用いた場合に得られた結果を示し、および(XX)は、対照の非特異的ヒトIgG2抗体を1mgの用量で用いた場合に得られた結果を示す。
【発明を実施するための形態】
【0032】
好ましい態様の詳細な説明
本発明に従って、ヒト上皮細胞増殖因子受容体(EGF-r)に対する完全なヒトモノクローナル抗体を提供する。重鎖および軽鎖免疫グロブリン分子をコードするヌクレオチド配列、ならびにこれらの分子を含むアミノ酸配列、特にCDR1からCDR3までの近接する重鎖および軽鎖配列に相当する配列が提供される。そのような免疫グロブリン分子およびモノクローナル抗体を発現するハイブリドーマもまた提供する。
【0033】
定義
特に明記していない限り、本発明に関連して用いられる科学技術用語は、当業者によって一般的に理解される意味を有する。さらに、本文において特に要求していなければ、単数の用語には複数の用語が含まれ、複数の用語には単数の用語が含まれる。一般に、本明細書に記述の細胞および組織培養、分子生物学、蛋白質およびオリゴまたはポリヌクレオチド化学、およびハイブリダイゼーションに関連して用いられる用語体系は、当技術分野において周知であり一般的に用いられる。組換え型DNA、オリゴヌクレオチド合成、ならびに組織培養および形質転換(例えば電気穿孔、リポフェクション)に関しては標準的な技法が用いられる。酵素反応および精製技法は、製造元の説明書に従って、もしくは当技術分野で一般的に実施されるように、または本明細書に記述されるように実施する。前述の技術および技法は、概して当技術分野で周知の従来の方法に従って、ならびに本明細書を通じて引用され考察される、様々な一般的およびより詳細な引用文献において記述されているように実施する。例えば、本明細書に参照として組み入れられる、サムブルックら(Sambrook)、「分子クローニング:実験マニュアル(Molecular Cloning:A Laboratory Manual)」(第二版、Cold Spring Harbor Laboratory Press,Cold Spring Harbor、ニューヨーク(1989))を参照のこと。本明細書に記述の分析化学、合成有機化学、ならびに医化学および製薬化学に関連して用いられる用語体系、ならびにそれらの実験方法および技術は、当技術分野において周知であり、一般的に用いられる。化学合成、化学分析、薬学的調製、製剤および輸送ならびに患者の治療に関しては標準的な技法を用いる。
【0034】
本開示に従って用いられるように、以下の用語は、特に明記していない限り、以下の意味を有すると理解される:
本明細書において用いられる「単離されたポリヌクレオチド」という用語は、その起源のために「単離されたポリヌクレオチド」が(1)「単離されたポリヌクレオチド」が本来認められるポリヌクレオチドの全体もしくは一部と会合していない、(2)本来結合していないポリヌクレオチドに機能的に結合している、または(3)より大きい配列の一部として天然では起こらない、ゲノム、cDNAもしくは合成起源またはその何らかの組合せのポリヌクレオチドを意味する。
【0035】
本明細書において用いられる「単離された蛋白質」という用語は、その起源もしくは由来源のために、「単離された蛋白質」が(1)天然で認められる蛋白質とは会合していない、(2)同じ起源からのその他の蛋白質を含まない、例えば、マウス蛋白質を含まない、(3)異なる種の細胞によって発現される、または(4)天然に起こらない、cDNA、組換え型RNAもしくは合成起源またはその何らかの組合せの蛋白質を意味する。
【0036】
「ポリペプチド」という用語は、本明細書においてポリペプチド配列の本来の蛋白質、断片または類似体を指す一般的用語として用いられる。したがって、本来の蛋白質、断片および類似体はポリペプチド属に含まれる種である。本発明に係る好ましいポリペプチドは、図1、5、9、13、17、21、25、および29によって表されるヒト重鎖免疫グロブリン分子ならびに図3、7、11、15、19、23、27、および31によって表されるヒトカッパ軽鎖免疫グロブリン分子と共に、重鎖免疫グロブリン分子と、カッパ軽鎖免疫グロブリン分子のような軽鎖免疫グロブリン分子とを含む組合せによって形成された抗体分子、ならびにその逆と共に、その断片および類似体を含む。
【0037】
本明細書においてある物質に適用して用いられる「天然」という用語はある物質が天然に認められるという事実を指す。例えば、天然の起源から単離することができる有機体(ウイルスを含む)に存在し、且つ実験室もしくはそれ以外の場所でヒトによって意図的に改変されていない、ポリペプチドまたはポリヌクレオチド配列は、天然である。
【0038】
本明細書において用いられる「機能的に結合された」という用語は、その意図する方法においてそれらが機能する関係にある、と記述された成分の位置を意味する。コード配列に「機能的に結合された」調節配列は、コード配列の発現が調節配列と適合する条件下で得られるようにライゲーションされる。
【0039】
本明細書において用いられる「調節配列」という用語は、それに対してライゲーションされるコード配列の発現およびプロセシングを行うために必要なポリヌクレオチド配列を意味する。そのような調節配列の性質は宿主生物に依って異なる;原核生物では、該調節配列は一般に、プロモーター、リボソーム結合部位、および転写終結配列を含む;真核生物では、一般に該調節配列はプロモーターおよび転写終結配列を含む。「調節配列」という用語は、その存在が発現およびプロセシングにとって必須である成分を最少でも全て含むものと解釈され、その存在が有利となるさらなる成分、例えばリーダー配列および融合パートナー配列を含むこともできる。
【0040】
本明細書において言及される「ポリヌクレオチド」という用語は、長さが少なくとも10塩基のヌクレオチドのポリマー型、リボヌクレオチドもしくはデオキシヌクレオチドのいずれか、またはいずれかのタイプのヌクレオチドの改変型を意味する。この用語にはDNAの一本鎖および二本鎖型が含まれる。
【0041】
本明細書において言及する「オリゴヌクレオチド」という用語には、天然のヌクレオチド、ならびに天然および非天然のオリゴヌクレオチド結合によって結合された改変型ヌクレオチドが含まれる。オリゴヌクレオチドは一般に長さ200塩基以下を含むポリヌクレオチドサブセットである。好ましくは、オリゴヌクレオチドは長さが10〜60塩基であり、最も好ましくは長さが12、13、14、15、16、17、18、19、または20〜40塩基である。オリゴヌクレオチドは通常、例えばプローブに用いる場合には一本鎖である;しかし、例えば遺伝子変異体の構築に用いる場合にはオリゴヌクレオチドは二本鎖であってもよい。本発明のオリゴヌクレオチドは、センスオリゴヌクレオチドまたはアンチセンスオリゴヌクレオチドのいずれであってもよい。
【0042】
本明細書において用いられる「天然ヌクレオチド」という用語には、デオキシリボヌクレオチドおよびリボヌクレオチドが含まれる。本明細書における「改変型ヌクレオチド」という用語には、改変されたまたは置換された糖基等を有するヌクレオチドが含まれる。本明細書において用いられる「オリゴヌクレオチド結合」という用語には、ホスホロチオ酸エステル、ホスホロジチオ酸エステル、ホスホロセレン酸エステル、ホスホロジセセレン酸エステル、ホスホロアニロチオ酸エステル、ホスホロアニラデート、ホスホロアミデート等のオリゴヌクレオチド結合が含まれる。例えば、その開示が本明細書に参照として組み入れられる、ラプランシェら(Laplanche)、Nucl.Acids Res.14:9081(1986);ステックら(Stec)、J.Am.Chem.Soc.106:6077(1984);スタインら(Stein)、Nucl.Acads Res.16:3209(1988);ツォンら(Zon)、AntiCancer Drug Design 6:539(1991);ツォンら(Zon)、「オリゴヌクレオチドと類似体:実践的アプローチ(Oligonucleotides and Analogues:A Practical Approach)」、87〜108頁(エックスタイン(F.Eckstein)編、Oxford University Press,Oxford,England(1991));ステックら(Stec)、米国特許第5,151,510号;ウールマン&ペイマン(Uhlman and Peyman)、Reviews 90:543(1990)を参照のこと。オリゴヌクレオチドは必要に応じて、検出のための標識を含むことができる。
【0043】
本明細書において用いられる「選択的にハイブリダイズする」という用語は、検出可能且つ特異的に結合することを意味する。本発明のポリヌクレオチド、オリゴヌクレオチド、およびその断片は、非特異的核酸に対する検出可能な結合の認識量を最小限にするようなハイブリダイゼーションおよび洗浄条件下で、核酸鎖と選択的にハイブリダイズする。高ストリンジェンシー条件は、当技術分野で既知であり、本明細書において考察するように、選択的なハイブリダイゼーション条件を得るために用いることができる。一般に、本発明のポリヌクレオチド、オリゴヌクレオチドおよび断片と、対象核酸配列との間の核酸配列の相同性は少なくとも80%、より典型的には少なくとも85%、90%、95%、99%、および100%のような相同性の増加が好ましい。それらの配列間に部分的または完全な同一性が認められれば、2つのアミノ酸配列は相同である。例えば、85%相同性とは、2つの配列を最大に合致するように配置した場合にアミノ酸の85%が同一であることを意味する。ギャップ(対合させる2つの配列のいずれかにおける)によって、対合が最大となるようにする;ギャップの長さは5以下であることが好ましく、2以下であることがより好ましい。または好ましくは、2つの蛋白質の配列(または長さが少なくともアミノ酸30個である、それらに由来するポリペプチド配列)は、本明細書においてこの用語を用いるように、変異データマトリクスおよびギャップペナルティ6以上を用いたプログラムALIGNを用いた場合に配置スコアが(標準偏差の単位で)5以上であれば、相同である。デイホフ(Dayhoff,M.O.)、「蛋白質の配列と構造の図解(Atlas of Protein Sequence and Structure)」、101〜110頁(第5巻、National Biomedical Reseach Foundation(1972))およびこの巻の補足2、1〜10頁を参照のこと。2つの配列またはその一部はより好ましくは、ALIGNプログラムを用いて最適に配置した場合に、そのアミノ酸が50%以上同一であれば、または50%同一に等しければ相同である。「に相当する」という用語は、本明細書において、あるポリヌクレオチド配列が、参照ポリヌクレオチド配列の全てもしくは一部と相同である(すなわち、同一であるが厳密に進化的に関連していない)、またはポリペプチド配列が参照ポリペプチド配列と同一であることを意味するように用いられる。対照区別して、「と相補的である」という用語は、本明細書において、相補的配列が参照ポリヌクレオチド配列の全てまたは一部と相同であることを意味するように用いられる。説明すると、ヌクレオチド配列「TATAC」は、参照配列「TATAC」に対応し、参照配列「GTATA」と相補的である。
【0044】
以下の用語は、2つ以上のポリヌクレオチドまたはアミノ酸配列の間の配列関係を記述するために用いられる:「参照配列」、「比較ウィンドウ」、「配列同一性」、「配列同一性の割合」、および「実質的な同一性」。「参照配列」とは、配列比較の根拠として用いられる明確に定められた配列である;参照配列はより大きい配列のサブセット、例えば全長cDNAのセグメントもしくは配列表に示されている遺伝子配列であってもよく、または完全なcDNAもしくは遺伝子配列を含んでもよい。一般に、参照配列は長さが少なくともヌクレオチド18個またはアミノ酸6個、しばしば長さが少なくともヌクレオチド24個またはアミノ酸8個、および多くの場合長さが少なくともヌクレオチド48個またはアミノ酸16個である。2つのポリヌクレオチドまたはアミノ酸配列はそれぞれ、(1)2つの分子の間で類似である配列(すなわち完全なポリヌクレオチドまたはアミノ酸配列の一部)を含んでもよく、および(2)2つのポリヌクレオチドまたはアミノ酸配列間で異なる配列をさらに含んでもよいため、2つ(以上)の分子間の配列比較は典型的に、配列類似性の局所領域を特定および比較するために、「比較ウィンドウ」の範囲で2つの分子の配列を比較することによって行われる。本明細書において用いられる「比較ウィンドウ」とは、ポリヌクレオチド配列またはアミノ酸配列を、近接ヌクレオチド少なくとも18個またはアミノ酸6個の配列の参照配列と比較する場合、および比較ウィンドウにおけるポリヌクレオチド配列の一部が、2つの配列を最適に配置した場合に参照配列(付加または欠失を含まない)と比較して20%未満の付加、欠失、置換等(すなわちギャップ)を含む場合の、近接ヌクレオチド少なくとも18個の位置またはアミノ酸6個という概念的部分を指す。比較ウィンドウを配置させるための配列の最適な配置は、スミス&ウォーターマン(Smith and Waterman)、Adv.Appl.Math.2:482(1981)の局所相同性アルゴリズム、ニードルマン&ビュンシュ(Needleman and Wunsch)、J.Mol.Biol.48:443(1970)の相同性配置アルゴリズム、ペアソン&リップマン(Pearson and Lipman)、Proc.Natl.Acad.Sci.(U.S.A.) 85:2444(1988)の類似性方法に関する研究、これらのアルゴリズムのコンピューターによる実行(ウィスコンシン・ジェネティックス・ソフトウェアパッケージ7.0(ジェネティクス・コンピューターグループ、575 Science Dr.Madison,Wis.)、ジーンワークス、またはマックベクターソフトウェアパッケージにおけるGAP、BESTFIT、FASTAおよびTFASTA)、または肉眼での検分によって行ってもよく、様々な方法によって得られた最適な配置(すなわち比較ウィンドウの範囲で最高の相同性百分率が得られる)を選択する。
【0045】
「配列同一性」という用語は2つのポリヌクレオチドまたはアミノ酸配列が、比較ウィンドウの範囲で同一である(すなわち、ヌクレオチド・ヌクレオチドまたは残基・残基毎に)ことを意味する。「配列同一性の割合」という用語は、比較ウィンドウの範囲で2つの最適に配置された配列を比較し、同一核酸塩基(例えば、A、T、C、G、UもしくはI)または残基が双方の配列に存在する位置の数を決定して対合した位置の数を得て、対合した位置の数を比較ウィンドウにおける位置の総数で除して、および結果を100倍して配列同一性の割合を算出することによって計算される。本明細書において用いられる「実質的に同一」という用語は、ポリヌクレオチドまたはアミノ酸が、参照配列と比較して、少なくともヌクレオチド18(アミノ酸6個)位の比較ウィンドウ、しばしば少なくともヌクレオチド24〜48(アミノ酸8〜16個)位のウィンドウの範囲で少なくとも85%の配列同一性を有する、好ましくは少なくとも90〜95%の配列同一性を有する、より一般的には少なくとも99%の配列同一性を有する配列を含む場合であって、参照配列を、比較ウィンドウの範囲において参照配列の全体の20%未満である欠失または付加を含んでもよい配列と比較することによって配列同一性の割合が計算される場合の、ポリヌクレオチドまたはアミノ酸配列の特徴を指す。参照配列はより大きい配列のサブセットであってもよい。
【0046】
本明細書において用いられるように、通常のアミノ酸20個およびその略語は、従来の用法に従う。本明細書に参照として組み入れられる、「免疫学−合成(Immunology- A Synthesis)」(第二版、ゴルブ&グレン(E.S.Golub and D.R.Gren)編、Sinauer Associates、サンダーランド、マサチューセッツ州(1991))を参照のこと。通常のアミノ酸20個の立体異性体(例えば、D-アミノ酸)、α、α-2置換アミノ酸のような非天然型アミノ酸、N-アルキルアミノ酸、乳酸、およびその他の異形のアミノ酸もまた、本発明のポリペプチドの適した成分となってもよい。異形アミノ酸の例には:4-ヒドロキシプロリン、Y-カルボキシグルタミン酸塩、ε-N,N,N-トリメチルリジン、ε-N-アセチルリジン、O-ホスホセリン、N-アセチルセリン、N-ホルミルメチオニン、3-メチルヒスチジン、5-ヒドロキシリジン、σ-N-メチルアルギニン、ならびにその他の類似のアミノ酸およびイミノ酸(例えば、4-ヒドロキシプロリン)が含まれる。本明細書において用いられるポリペプチドの表記において、標準的な用法および慣例に従って、左側の方向がアミノ末端の方向であり、右側の方向がカルボキシ末端の方向である。
【0047】
同様に、特に明記していなければ、一本鎖ポリヌクレオチド配列の左側は5'末端である;二本鎖ポリヌクレオチド配列の左側の方向は5'方向と呼ばれる。新生RNA転写物の5'から3'への付加方向を転写方向と呼ぶ;RNAと同じ配列を有するDNA鎖上の配列領域であってRNA転写物の5'末端に対して5'方向にある領域を、「上流配列」と呼び;RNAと同じ配列を有するDNA鎖上の配列領域であって、RNA転写物の3'末端に対して3'方向にある領域を「下流配列」と呼ぶ。
【0048】
ポリペプチドに適用されるように、「実質的に同一」という用語は、2つのポリペプチド配列を、例えば、デフォルトギャップ加重を用いたプログラムGAPまたはBESTFITによって最適に配置した場合に、それらが少なくとも80%の配列同一性を有する、好ましくは少なくとも90%の配列同一性を有する、より好ましくは少なくとも95%の配列同一性を有する、および最も好ましくは少なくとも99%の配列同一性を有することを意味する。同一でない残基の位置は、保存的アミノ酸置換によって異なることが好ましい。保存的アミノ酸置換は類似の側鎖を有する残基の相互交換を指す。例えば、脂肪族側鎖を有するアミノ酸のグループは、グリシン、アラニン、バリン、ロイシン、およびイソロイシンである;脂肪族ヒドロキシル側鎖を有するアミノ酸のグループはセリンおよびトレオニンである;アミド含有側鎖を有するアミノ酸のグループは、アスパラギンおよびグルタミンである;芳香族側鎖を有するアミノ酸のグループは、フェニルアラニン、チロシン、およびトリプトファンである;塩基性側鎖を有するアミノ酸のグループはリジン、アルギニン、およびヒスチジンである;ならびに硫黄含有側鎖を有するアミノ酸のグループはシステインおよびメチオニンである。好ましい保存的アミノ酸置換グループは:バリン・ロイシン・イソロイシン、フェニルアラニン・チロシン、リジン・アルギニン、アラニン・バリン、グルタミン酸・アスパラギン酸、およびアスパラギン・グルタミンである。
【0049】
本明細書において考察するように、抗体または免疫グロブリン分子のアミノ酸配列における小さい変化は、アミノ酸配列の変化が少なくとも75%、より好ましくは少なくとも80%、90%、95%、および最も好ましくは99%を維持している限り、本発明に含まれるものとする。特に、保存的アミノ酸置換について考察する。保存的置換はその側鎖において関連するアミノ酸のファミリー内に起こる置換である。遺伝的にコードされるアミノ酸は一般に以下のファミリーに分類される:(1)酸性=アスパラギン酸、グルタミン酸;(2)塩基性=リジン、アルギニン、ヒスチジン;(3)非極性=アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン;および(4)非荷電極性=グリシン、アスパラギン、グルタミン、システイン、セリン、トレオニン、チロシン。より好ましいファミリーは:セリンおよびトレオニンは脂肪族ヒドロキシファミリーである;アスパラギンおよびグルタミンはアミド含有ファミリーである;アラニン、バリン、ロイシン、およびイソロイシンは脂肪族ファミリーである;ならびにフェニルアラニン、トリプトファン、およびチロシンは芳香族ファミリーである。例えば、ロイシンのイソロイシンまたはバリンへの1箇所置換、アスパラギン酸のグルタミン酸への置換、トレオニンのセリンへの置換、または一つのアミノ酸の、構造的に関連するアミノ酸への同様な置換は、特に置換がフレームワーク部位内のアミノ酸を含まなければ、得られた分子の結合もしくは特性に主要な影響を及ぼさないであろうと十分に予想される。アミノ酸が変化した結果機能的ペプチドが得られるか否かは、ポリペプチド誘導体の特異的活性をアッセイすることによって容易に決定することができる。アッセイは本明細書において詳細に記述する。抗体または免疫グロブリン分子の断片または類似体は、当業者によって容易に調製することができる。断片または類似体の好ましいアミノ末端およびカルボキシ末端は、機能的ドメインの境界付近で起こる。構造的および機能的ドメインは、ヌクレオチドおよび/またはアミノ酸配列データを、公共または私有の配列データベースと比較することによって同定することができる。好ましくは、コンピューターによる比較法を用いて、既知構造および/または機能のその他の蛋白質において起こる配列モチーフまたは推定蛋白質構造ドメインを同定する。既知の3次元構造の中に折り畳まれる蛋白質配列を同定する方法は公知である(ボウイら(Bowie)、Science 253:164(1991))。このように、前述の例は、当業者が本発明に従って構造および機能的ドメインを定義するために用いてもよい配列モチーフおよび構造の配座を認識することができることを示している。
【0050】
好ましいアミノ酸置換は:(1)蛋白質溶解に対する感受性を低下させる、(2)酸化に対する感受性を低下させる、(3)蛋白質複合体を形成する結合親和性を変化させる、(4)結合親和性を変化させる、および(5)そのような類似体の他の物理化学的または機能的特性を付与または改変する、置換である。類似体は天然に存在するペプチド配列以外の配列の様々な変異体を含むことができる。例えば、単一または多数のアミノ酸置換(好ましくは保存的アミノ酸置換)は、天然に存在する配列において起こってもよい(好ましくは、分子間接触を形成するドメインの外側のポリペプチドの一部において)。保存的アミノ酸置換は、親配列の構造的特徴を実質的に変化させてはならない(例えば、置換アミノ酸は親配列において起こるヘリックスを切断する傾向があってはならない、または親配列の特徴となる他のタイプの二次構造を乱す傾向があってはならない)。当技術分野において認識されるポリペプチドの二次および三次構造の例は、それぞれ本明細書に参照として組み入れられる、「蛋白質、構造および分子の基本(Proteins,Structures and Molecular Principles)」(クレイトン(Creighton)編、W.H.Freeman and Company,ニューヨーク(1984));「蛋白質構造入門(Introduction to Protein Structure)」(ブランデン&トーズ(C.Branden and J.Tooze)編、Garland Publishing,ニューヨーク、ニューヨーク州(1991));およびソーントンら(Thornton)、Nature 354:105(1991)を参照のこと。
【0051】
本明細書において用いられる「ポリペプチド断片」という用語は、アミノ末端および/またはカルボキシ末端の欠失を有するが、残りのアミノ酸配列は、例えば全長のcDNA配列から導出した天然に存在する配列における対応する位置において同一であるポリペプチドを指す。断片は典型的に、長さが少なくともアミノ酸5、6、8または10個、好ましくは少なくとも14アミノ酸長、より好ましくは少なくとも20アミノ酸長、通常少なくとも50アミノ酸長、およびさらにより好ましくは少なくとも70アミノ酸長である。本明細書において用いられる「類似体」という用語は、推定アミノ酸配列の一部と実質的な同一性を有し、以下の特性の少なくとも1つ有する、少なくとも25個のアミノ酸のセグメントを含むポリペプチドを意味する:(1)適当な結合条件下でのEGF-rに対する特異的結合、(2)その受容体に対するEGFの結合能、または(3)インビトロもしくはインビボでEGF-rを発現する細胞増殖の阻害能。典型的に、ポリペプチド類似体は、天然に存在する配列に関して保存的アミノ酸置換(または付加もしくは欠失)を含む。類似体は典型的に、長さが少なくともアミノ酸20個、好ましくは長さが少なくともアミノ酸50個以上であり、しばしば全長の天然に存在するポリペプチドと同じ長さとなることができる。
【0052】
ペプチド類似体は一般的に、製薬産業において鋳型ペプチドと類似の特性を有する非ペプチド薬として用いられる。これらのタイプの非ペプチド化合物は「ペプチド模倣体」または「ペプチド擬似体(peptidomimetics)」と呼ばれる。フォーシェル(Fauchere)、J Adv.Drug Res.15:29(1986);ベベール&フライディンガー(Veber and Freidinger)、TINS 392(1985);およびエバンスら(Evans)、J.Med.Chem.30:1229(1987)が、参照として本明細書に組み入れられる。そのような化合物はしばしば、コンピューターによる分子モデリングを用いて開発される。同等の治療または予防効果を生じるために、治療的に有用なペプチドと構造的に類似のペプチド模倣体を用いてもよい。一般に、ペプチド擬似体は、ヒト抗体のような模範ポリペプチド(すなわち生化学特性または薬学活性を有するポリペプチド)と構造的に類似であるが、当技術分野で周知の方法によって、1つ以上のペプチド結合が選択的に以下からなる群より選択される結合に置換されている:−−CH2NH−−、−−CH2S−−、−−CH2−CH2−−、−−CH=CH−−(シスおよびトランス)、−−COCH2−−、−−CH(OH)CH2−−、および−CH2SO−−。コンセンサス配列のアミノ酸1個以上を同じタイプのD-アミノ酸に系統的に置換(例えば、L-リジンの代わりにD-リジン)することによって、より安定なペプチドを作製してもよい。さらに、当技術分野で既知の方法によって(本明細書に参照として組み入れられる、リゾ&ギエラッシュ(Rizo and Gierasch)、Ann.Rev.Biochem.61:387(1992));例えば、ペプチドを環状にする分子内ジスルフィド架橋を形成することができる内部システイン残基を加えることによって、コンセンサス配列または実質的に同一なコンセンサス配列変異を含む不自然なペプチドを産生させてもよい。
【0053】
「抗体」または「抗体ペプチド」とは、無傷の抗体、または特異的結合のために無傷の抗体と競合するその結合断片を指す。結合断片は、DNA組換え技法によって、または無傷抗体の酵素的もしくは化学的開裂によって産生される。結合断片には、Fab、Fab'、F(ab')2、Fv、および一本鎖抗体が含まれる。「二重特異性」、または「双機能性」抗体以外の抗体は、その結合部位のそれぞれが同一であると理解される。抗体は、抗体の過剰量が対受容体に結合した受容体の量を少なくとも約20%、40%、60%または80%、およびより通常約85%以上減少させる場合に(インビトロ競合結合アッセイにおいて測定する)、対受容体に対する受容体の接着を実質的に阻害する。
【0054】
「エピトープ」という用語は、免疫グロブリンまたはT-細胞受容体に特異的に結合することができるいかなる蛋白質決定因子も含む。エピトープ決定因子は通常、アミノ酸または糖側鎖のような分子の化学的に活性な表面での集合体を含み、通常、特異的電荷特性と共に、特異的な三次元構造特性を有する。抗体は、解離定数が≦1μM、好ましくは≦100nM、および最も好ましくは≦10 nMであれば、抗原に特異的に結合すると言える。
「物質」という用語は本明細書において、化学化合物、化学化合物の混合物、生物学的巨大分子、または生物材料からの抽出物を指す。
【0055】
本明細書において用いるように、「標識」または「標識された」という用語は、例えば放射性標識アミノ酸を組み入れることによって、または印をつけたアビジンによって検出することができるビオチン化部分をポリペプチドに接着させることによって(例えば、蛍光マーカーまたは光学もしくは比色法によって検出することができる酵素活性を含むストレプトアビジン)、検出可能なマーカーを組み入れることを意味する。特定の状況では、標識またはマーカーもまた治療的となりうる。ポリペプチドおよび糖蛋白質を標識する様々な方法は、当技術分野において既知であり、用いてもよい。ポリペプチドの標識の例は、以下を含むがこれらに限定されない:放射性同位元素または放射性核種(例えば、3H、14C、15N、35S、90Y、99Tc、111In、125I、131I)、蛍光標識(例えば、FITC、ローダミン、ランタニド燐)、酵素標識(例えば、ホースラディッシュ・ペルオキシダーゼ、β-ガラクトシダーゼ、ルシフェラーゼ、アルカリフォスファターゼ)、化学発光物質、ビオチニル基、二次リポーター(例えば、ロイシンジッパー対配列、二次抗体の結合部位、金属結合ドメイン、エピトープタグ)によって認識される既定のポリペプチドエピトープ。いくつかの態様において、標識は起こりうる立体妨害を減少させるために、様々な長さのスペーサーアームによって結合する。
【0056】
本明細書で用いられる「薬学的物質または薬物」という用語は、患者に適切に投与した場合に望ましい治療効果を誘導することができる化学化合物または組成物を指す。本明細書におけるその他の化学的用語は、本明細書において参照として組み入れられる、マグローヒル化学用語辞典(パーカー(Parker,S.)編、McGraw-Hill、サンフランシスコ(1985))に説明されるように、当技術分野における従来の用途に従って用いられる。
【0057】
「抗癌剤」という用語は、本明細書において、ヒトの新生物、特に癌腫、肉腫、リンパ腫、または白血病のような悪性(癌様)病変の発生または進行を阻害する機能的特性を有する物質を指すために用いられる。転移の阻害はしばしば抗新生物薬の特性である。
【0058】
本明細書において用いるように、「実質的に純粋」とは、目的とする種が最も多く存在する種であることを意味する(すなわち、モルベースで組成物における他のいかなる個々の種より豊富である)、および好ましくは、実質的に精製された分画は、目的とする種が存在する全ての巨大分子種の少なくとも約50%(モルベースで)を含む組成物である。一般に、実質的に純粋な組成物は、組成物中に存在する全ての巨大分子種の約80%以上、より好ましくは約85%、90%、95%、および99%を含む。最も好ましくは、組成物が本質的に単一の巨大分子種からなるように、目的とする種を本質的に均一になるまで精製する(組成物中の汚染種は、従来の検出法によって検出できない)。
患者という用語にはヒトおよび獣医学での被験動物が含まれる。
【0059】
抗体の構造
基本抗体構造単位は4量体からなることが知られている。各4量体はポリペプチド鎖の2つの同一の対からなり、それぞれの対は1つの「軽」鎖(約25 kDa)および1つの「重」鎖(約50〜70kDa)を有する。各鎖のアミノ末端部分は、主に抗原認識を担うアミノ酸約100〜110個以上の可変領域を含む。各鎖のカルボキシ末端部分は、イフェクター機能を主に担う定常領域を定義する。ヒト軽鎖はカッパおよびラムダ軽鎖として分類される。重鎖は、ミュー、デルタ、ガンマ、アルファまたはイプシロンとして分類され、抗体のアイソタイプはそれぞれIgM、IgD、IgAおよびIgEと定義される。軽鎖および重鎖の内部で、可変領域および定常領域はアミノ酸約12個以上の「J」領域によって結合し、重鎖はまたアミノ酸約10個以上の「D」領域を含む。一般的に、「基礎免疫学(Fundamental Immunology)」(ポール(Paul,W.)編、第二版、Raven Press、ニューヨーク(1989))(参照として本明細書に組み入れられる)を参照のこと。各軽鎖/重鎖対の可変領域は、抗体結合部位を形成する。
このように、無傷の抗体は二つの結合部位を有する。双機能性または二重特異性抗体の場合を除いて、2つの抗体結合部位は同一である。
【0060】
鎖は全て、相補性決定領域またはCDRとも呼ばれる3つの超可変領域によって結合した比較的保存されたフレームワーク領域(FR)の同じ一般構造を示す。各対の2つの鎖からのCDRはフレームワーク領域によって整列し、特異的エピトープとの結合が可能となる。N末端からC末端に向けて、軽鎖および重鎖はいずれもドメインFR1、CDR1、FR2、CDR2、FR3、CDR3およびFR4を含む。各ドメインに対するアミノ酸の割付は、「カバットの免疫学的に重要な蛋白質の配列(Kabat Sequences of Proteins of Immunological Interest)」(国立衛生研究所、ベセスダ、メリーランド州(1987および1991))、またはコシア&レスク(Chothia and Lesk)、J.Mol.Biol.196:901〜917(1987);コシアら(Chothia)、Nature 342:878〜883(1989)の定義に従っている。
【0061】
二重特異性または双機能抗体は、異なる2つの重鎖/軽鎖対および異なる2つの結合部位を有する人工ハイブリッド抗体である。二重特異性抗体は、ハイブリドーマの融合またはFab'断片の結合を含む多様な方法によって産生することができる。例えば、ソングシビライ&ラックマン(Songsivilai and Lachman)、Clin.Exp.Immunol.79:315〜321(1990)、コステルニーら(Kostelny)、J.Immunol.148:1547〜1553(1992)を参照のこと。二重特異性抗体の産生は、従来の抗体の産生と比較して比較的大きな労働力を要するプロセスであり、一般に二重特異性抗体に関して収率および純度の程度は低い。二重特異性抗体は1つの結合部位を有する断片の形では存在しない(例えば、Fab、Fab'、およびFv)。
【0062】
抗体の調製
本発明に係る抗体は、好ましくはヒト抗体産生ゲノムの実質的な一部が挿入されているが、内因性のマウス抗体の産生は欠損しているトランスジェニックマウスを使用することによって調製する。次に、そのようなマウスはヒト免疫グロブリン分子および抗体を産生することができるが、マウス免疫グロブリン分子および抗体の産生を欠損している。これらを得るために用いられる技術は、特許、出願、および本明細書の背景に開示されている引用文献に開示されている。しかし特に、マウスのトランスジェニック産生およびその抗体の好ましい態様は、その開示が参照として本明細書に組み入れられる、1996年12月3日に提出された米国特許出願第08/759,620号に開示されている。同様に、その開示が本明細書に参照として組み入れられる、メンデス(Mendez)、Nature Genetics 15:146〜156(1997)を参照のこと。
【0063】
そのような技術を用いることによって、本発明者らは多様な抗原に対する完全なヒトモノクローナル抗体を産生した。本質的に、本発明者らは、目的の抗原でゼノマウス(XenoMouse(登録商標))系マウスを免疫し、抗体を発現するマウスからリンパ球(B細胞など)を回収し、そのような回収された細胞を骨髄系の細胞株と融合させて、不死化ハイブリドーマ細胞株を調製し、該ハイブリドーマ細胞株をスクリーニングして選択し、目的の抗原に対して特異的な抗体を産生するハイブリドーマ細胞株を特定する。本発明者らは、本発明に従ってこれらの技法をEGF-rに特異的な抗体を調製するために使用した。本明細書において、本発明者らは、EGF-rに対して特異的な抗体を産生するハイブリドーマ細胞株8個の産生について記述する。さらに、そのような抗体の重鎖および軽鎖のヌクレオチドおよびアミノ酸配列解析を含む、そのような細胞株によって産生された抗体の特徴分析を提供する。
【0064】
本明細書に記述のハイブリドーマ細胞株はE1.1、E2.4、E2.5、E6.2、E6.4、E2.11、E6.3、およびE7.6.3と命名される。前述の細胞株によって産生されたそれぞれの抗体は、ヒトカッパ軽鎖を有する完全なヒトIgG2重鎖である。一般に、本発明の抗体は非常に高い親和性を有し、固相および液相のいずれかによって測定すると、典型的に約10-9〜約10-11MのKdを有する。
【0065】
認識されるように、本発明の抗体はハイブリドーマ細胞株以外の細胞株において発現させることができる。特定の抗体をコードする配列は、適した哺乳類宿主細胞の形質転換に用いることができる。形質転換は、例えば、ウイルス(またはウイルスベクター)にポリヌクレオチドをパッケージングし、および宿主細胞を該ウイルス(またはベクター)によって形質導入することを含む、宿主細胞にポリヌクレオチドを導入するためのいかなる既知の方法によっても、または米国特許第4,399,216号、第4,912,040号、第4,740,461号、および第4,959,455号(これらの特許は参照として本明細書に組み入れられる)によって示されるように、当技術分野で既知のトランスフェクション技法によって行うことができる。用いる形質転換技法は、形質転換すべき宿主に依存する。異種ポリヌクレオチドを哺乳類細胞に導入する方法は、当技術分野で周知であり、デキストラン媒介トランスフェクション、リン酸カルシウム沈降法、ポリブレン媒介トランスフェクション、プロトプラスト融合、電気穿孔、リポソームへのポリヌクレオチドの封入、および核へのDNAの直接マイクロインジェクションを含む。
【0066】
発現のための宿主として利用できる哺乳類細胞株は当技術分野で周知であり、チャイニーズハムスター卵巣(CHO)細胞、Hela細胞、ベビーハムスター腎(BHK)細胞、サル腎(COS)細胞、ヒト肝細胞癌細胞(例えば、Hep G2)および多くのその他の細胞株を含むがこれらに限定されない、アメリカンタイプカルチャーコレクション(ATCC)から入手できる多くの不死化細胞株を含む。特に好ましい細胞株は、どの細胞株が高発現レベルを有するか、および構成的なEGF-r結合特性を有する抗体を産生するか否かを決定することによって選択される。
【0067】
本発明の抗体は、EGFおよびTGF-αの、その受容体であるEGF-rに対する結合の強力な阻害剤である。それらの結果を実施例5および6において考察し、図35〜38に示す。それらの結果と一致して、図39に示し、実施例7において考察するように、本発明の抗体はインビトロで特定のヒト癌細胞株の増殖をも阻害する。本発明の抗体はまた、インビボで特定のヒト癌の増殖を予防する。それらの結果を図40〜42に示し、実施例8において考察する。実施例9において、本発明者らは、本発明の抗体を少なくとも抗癌剤と併用すると、動物において既に存在する腫瘍が根治することを示した。その上、単剤療法(すなわち抗癌剤と併用しないで)としての抗体療法は、先行技術、例えば抗体225を使用する場合では不可能であるように思われたが、本発明の抗体を用いれば可能であるように思われる。それらの結果を実施例9において考察し、図43〜44に示す。
【0068】
本発明に従って示された結果から、本発明の抗体は、EGF-rに対する現在の治療的抗体、例えば225と比べて、本発明をより有効なものとする特定の特性を有することが示される。イムクローン社(ImClon)によって臨床開発された225抗体は親和性が2×10-10MのキメラIgG1抗体であり、これは抗癌剤との併用療法では有効であるように思われるが、単剤療法ではあまり有効でないように思われる。対照的に、本発明の抗体(特に本発明のE2.5およびE7.6.3抗体)は有意に高い親和性(E2.5:1.6×10-11M;E7.6.3:5.7×10-11 M)を有し、抗癌剤との併用療法のみならず、単剤療法でもC225抗体より低い用量で有効であるように思われる。
【実施例】
【0069】
実施した実験および得られた結果を含む以下の実施例は、説明目的のためにのみ提供されるのであって、本発明を制限するものと解釈してはならない。
【0070】
実施例1
抗EGF-r抗体産生ハイブリドーマの作製
本発明の抗体は、本実施例に従って調製、選択および解析した。
免疫およびハイブリドーマの作製 ゼノマウス(XenoMice)(8〜10週齢)に、リン酸緩衝塩類溶液(PBS)中に再懸濁した2×107個のA431(ATCC CRL-7907)細胞の腹腔内投与による免疫化を施した。この量の投与を3回繰り返した。融合4日前に、マウスにPBS中にて抗原または細胞の最終注射を行った。免疫化マウスからの脾臓およびリンパ節のリンパ球を、非分泌型骨髄腫NSO-bcl2系(RayおよびDiamond、1994)と融合させ、以前に記載されている通り(GalfreおよびMilstein、1981)にHAT選択にかけた。EGF-r特異的ヒトIgG2κ(下記に検出されるように)抗体を全てが分泌するハイブリドーマの大きい集団を回収した。実施例2に示すように、パネルから選択した特定の抗体を225抗体との競合能に関して選択した。
【0071】
ELISAアッセイ: マウス血清中およびハイブリドーマ上清中の抗原特異的抗体を測定するためのELISAは、抗体を捕捉するためにA431細胞からアフィニティー精製されたEGF-r(Sigma、E-3641)を用いて、記載されている通り(Coliganら、1994)に実施した。ヒトおよびマウスの免疫グロブリンの濃度は以下の捕捉抗体を用いて測定した:ヒトガンマ、カッパ、ミューIgについてはそれぞれウサギ抗ヒトIgG(Southern Biotechnology、6145-01)、ヤギ抗ヒトIgκ(Vector Laboratories、AI-3060)、マウス抗ヒトIgM(CGI/ATCC、HB-57)、ならびにマウスガンマ、カッパ、ミューおよびラムダIgの捕捉のためにはそれぞれヤギ抗マウスIgG(Caltag、M 30100)、ヤギ抗マウスIgκ(Southern Biotechnology、1050-01)、ヤギ抗マウスIgM(Southern Biotechnology、1020-01)およびヤギ抗マウスλ(Southern Biotechnology、1060-01)。ELISA実験に用いた検出用抗体は、ヤギ抗マウスIgG-HRP(Caltag、M-30107)、ヤギ抗マウスIgκ-HRT(Caltag、M 33007)、マウス抗ヒトIgG2-HRP(Southern Biotechnology、9070-05)、マウス抗ヒトIgM-HRP(Southern Biotechnology、9020-05)およびヤギ抗ヒトκ-ビオチン(Vector、BA-3060)である。ヒトおよびマウスのIgの定量化のために用いた標準は以下の通りである:ヒトIgG2κ(Calbiochem、400122)、ヒトIgMκ(Cappel、13000)、マウスIgGκ(Cappel 55939)、マウスIgMκ(Sigma、M-3795)およびマウスIgG3λ(Sigma、M-9019)。
【0072】
BIAcoreによる完全ヒトMabの親和定数の決定: 製造者によって概説された標準的な手順によりBIAcore 2000装置を用いて、精製ヒトモノクローナル抗体、Fab断片またはハイブリドーマ上清の親和性の測定をプラズモン共鳴によって実施した。
抗体の速度論的分析は、センサー表面上に低密度に固定された抗原を用いて行った。A431細胞膜から精製された可溶性EGF-r(Sigma、E-3641)を228RUの表面密度で概して用いた。解離速度(kd)および会合速度(ka)は、製造者によって提供されたソフトウエア(BIAevaluation 2.1)を用いて決定した。
【0073】
ELISAによる溶液中の親和定数の決定:ELISAによって溶液中での抗体結合親和性を決定するために、EGF-rに対するモノクローナル抗体の様々な濃度を平行に達するまで一定濃度のEGF-rと共にインキュベートした。その後、反応溶液中の遊離のEGF-rの濃度を間接ELISAによって決定した。よって、3.0×10-11M〜2.7×10-7 Mの濃度のモノクローナル抗体を、0.5%BSAを含むPBS 200μlにおいて4×10-10 Mの濃度のEGF-rと共に室温で15時間インキュベートした。インキュベート後、同じモノクローナル抗体を予めコーティングした(100μl/ウェル、コーティング緩衝液2μg/mlで)96穴マイクロタイタープレートのウェルに各混合液70μlを移し、室温で15分間インキュベートした。洗浄緩衝液で洗浄した後、EGF-r蛋白質の炭水化物に結合するマウス抗EGF-r-HRPによって、プレート上に保持されたEGF-rを検出した。EGF-rの濃度をその標準物質から計算し、これを用いて最初の抗原抗体反応溶液中での結合および遊離の抗体を計算した。各モノクローナル抗体のEGF-rに対する結合親和性はスキャッチャード分析を用いて計算した。
【0074】
受容体結合アッセイ法: EGF受容体結合アッセイ法は、PBS結合緩衝液中、様々な濃度の抗体と共に30分間4℃でインキュベートされたA431細胞またはSW948細胞を用いて実施した(0.4×106細胞/ウェル)。0.1nM[125I]EGF(Amersham,IM-196)または[125I]TGF-α(Amersham)を各ウェルに加え、そのプレートを90分間4℃でインキュベートした。該プレートを5回洗浄し、風乾し、シンチレーションカウンターで計測した。抗EGF-rマウス抗体225および528(Calbiochem)を対照として用いた。
【0075】
実施例2
m225抗体による抗EGF-r抗体の共選択
上記のように、抗体225はEGFおよびTGF-αのEGF-rに対する結合に対して高い親和性を有し、これを有効に阻害することが示された。したがって、本発明者らは、競合アッセイにおいて抗体225を用いて、本発明に従って調製したEGF-rに対するヒト抗体を選択すれば、それに対して225抗体が結合する同一または類似のエピトープに対する抗体が選択されるであろうと予想した。
【0076】
したがって、本発明者らは、A431細胞膜(シグマ社、E-3641)から精製した可溶性EGF-rを抗体225によって前処置し、その後本発明の抗体で処置するバイアコア(BIAcore)アッセイを行った。本発明の抗体が結合しなければ、本発明のそのような抗体を上記のように結合親和性に関してスクリーニングした。
以下の表において、このようにして選択した特定の抗体の親和性測定を提供する:
【0077】
【表1】
表から認められるように、このようにして選択された抗体は、例外的に高い親和性および結合定数を有する。
【0078】
実施例3
本発明に従って調製した抗EGF-r抗体の構造
以下の考察において、本発明に従って調製した抗体に関連した構造的情報を提供する。
本発明に従って産生された抗体の構造を分析するため、本発明者らは特定のハイブリドーマから重鎖および軽鎖断片をコードする遺伝子をクローニングした。遺伝子のクローニングおよびシークエンシングは以下のように実施した:
【0079】
ファストトラック(Fast-Track)・キット(Invitrogen)を用いて、免疫したゼノマウスに由来するハイブリドーマ細胞約2×105個からポリ(A)+ mRNAを単離した。ランダムプライミングしたcDNAを産生した後にPCRを行った。ヒトVHまたはヒトVKファミリー特異的可変領域プライマー(マークスら(Marks)、1991)または万能ヒトVHプライマー、MG-30(CAGGTGCAGCTGGAGCAGTCIGG)(配列番号:1)を、ヒトCγ2定常領域に対して特異的なプライマー(MG-40d;5'-GCTGAGGGAGTAGAGTCCTGAGGA-3')(配列番号:2)、またはCK定常領域(hkP2;既にグリーンら(Green)、1994が記述したように)と共に用いた。ハイブリドーマからのヒトMab-由来重鎖およびカッパ鎖転写物の配列は、上記のプライマーを用いてポリ(A)+ RNAから産生したPCR産物の直接シークエンシングによって得た。PCR産物はまた、TAクローニングキット(インビトロゲン社)を用いてpCRIIの中にクローニングし、プリスム色素ターミネーター・シークエンシングキットおよびABI 377シークエンシング装置を用いて双方の鎖をシークエンシングした。配列は全て、マックベクター(MacVector)およびジーンワークス(GeneWorks)ソフトウェアプログラムを用いて、「V BASE配列ディレクトリ」(トムリンソンら(Tomlinson)、MRC 蛋白質操作センター、ケンブリッジ、イギリス)に配置することによって分析した。
【0080】
ハイブリドーマE1.1
ハイブリドーマE1.1によって分泌された抗体はヒトカッパ軽鎖を有するヒトIgG2抗体を含む。抗体をそのアミノ酸配列と共に、その重鎖および軽鎖の遺伝子使用度に関連した構造情報に関して分析した。このように、重鎖VH、DおよびJHならびに軽鎖VKおよびJK遺伝子使用度を分析して、コードされる産物と特定の遺伝子使用度との相違点についても分析した。よって、ハイブリドーマE1.1によって分泌された抗体は以下の遺伝子使用度を有することが証明された:
VH −4〜31
D −2
JH −5
VK −018
JK −4
【0081】
V BASE配列ディレクトリにおいて報告されるように、VH 4〜31遺伝子によってコードされるアミノ酸配列を以下のように決定した:
VSGGSISSGGYYWSWIRQHPGKGLEWIGYIYYSGSTYYNPSLKSRVTISVDTSKNQFSLKLSSVTAADTAVYYCAR(配列番号:19)
V BASE配列ディレクトリにおいて報告されるように、VK(018)遺伝子によってコードされるアミノ酸配列を以下のように決定した:
TITCQASQDISNYLNWYQQKPGKAPKLLIYDASNLETGVPSRFSGSGSGTDFTFTISSLQPEDIATYYCQQYDNLP(配列番号:20)
【0082】
重鎖および軽鎖に関するアミノ酸およびヌクレオチド配列情報は図1〜4と共に下記に示す。図1は、ハイブリドーマE1.1によって分泌された重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE1.1分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【0083】
図2は、ハイブリドーマE1.1からクローニングした図1の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
図3は、ハイブリドーマE1.1によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE1.1分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
図4は、ハイブリドーマE1.1からクローニングした図3のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【0084】
ハイブリドーマE2.4
ハイブリドーマE2.4によって分泌された抗体は、ヒトカッパ軽鎖を有するヒトIgG2抗体を含む。抗体をそのアミノ酸配列と共にその重鎖および軽鎖遺伝子使用度に関連した構造情報に関して分析した。このように、重鎖VH、DおよびJHならびに軽鎖VKおよびJK遺伝子使用度を分析して、コードされる産物と特定の遺伝子使用度との相違点についても分析した。よって、ハイブリドーマE2.4によって分泌された抗体は以下の遺伝子使用度を有することが証明された:
VH −4〜31
D −A1/A4
JH −3
VK −018
JK −4
【0085】
重鎖および軽鎖に関するアミノ酸およびヌクレオチド配列情報は図5〜8と共に下記に提供する。図5は、ハイブリドーマE2.4によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE2.4分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【0086】
図6は、ハイブリドーマE2.4からクローニングした図5の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
図7は、ハイブリドーマE2.4によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE2.4分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
図8は、ハイブリドーマE2.4からクローニングした図7のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【0087】
ハイブリドーマE2.5
ハイブリドーマE2.5によって分泌された抗体は、ヒトカッパ軽鎖を有するヒトIgG2抗体を含む。抗体をそのアミノ酸配列と共に、その重鎖および軽鎖の遺伝子使用度に関連した構造情報に関して分析した。このように、重鎖VH、DおよびJHならびに軽鎖VKおよびJK遺伝子の利用を分析して、コードされる産物と特定の遺伝子使用度との相違点についても分析した。よって、ハイブリドーマE2.5によって分泌された抗体は以下の遺伝子使用度を有することが証明された:
VH −4〜31
D −XP1/21〜10
JH −4
VK −018
JK −2
【0088】
重鎖および軽鎖に関するアミノ酸およびヌクレオチド配列情報は図9〜12と共に下記に示す。図9は、ハイブリドーマE2.5によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE2.5分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【0089】
図10は、ハイブリドーマE2.5からクローニングした図9の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
図11は、ハイブリドーマE2.5によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE2.5分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
図12は、ハイブリドーマE2.5からクローニングした図11のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【0090】
ハイブリドーマE6.2
ハイブリドーマE6.2によって分泌された抗体はヒトカッパ軽鎖を有するヒトIgG2抗体を含む。抗体をそのアミノ酸配列と共に、その重鎖および軽鎖の遺伝子使用度に関連した構造情報に関して分析した。このように、重鎖VH、DおよびJHならびに軽鎖VKおよびJK遺伝子使用度を分析して、コードされる産物と特定の遺伝子使用度との相違点についても分析した。よって、ハイブリドーマE6.2によって分泌された抗体は以下の遺伝子使用度を有することが証明された:
VH −4〜31
D −?(CNTCCCTT)
JH −6
VK −018
JK −1
【0091】
重鎖および軽鎖に関するアミノ酸およびヌクレオチド配列情報は図13〜16と共に下記に示す。図13は、ハイブリドーマE6.2によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE6.2分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【0092】
図14は、ハイブリドーマE6.2からクローニングした図13の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
図15は、ハイブリドーマE6.2によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE6.2分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
図16は、ハイブリドーマE6.2からクローニングした図15のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【0093】
ハイブリドーマE6.4
ハイブリドーマE6.4によって分泌された抗体はヒトカッパ軽鎖を有するヒトIgG2抗体を含む。抗体をそのアミノ酸配列と共に、その重鎖および軽鎖の遺伝子使用度に関連した構造情報に関して分析した。このように、重鎖VH、DおよびJHならびに軽鎖VKおよびJK遺伝子使用度を分析して、コードされる産物と特定の遺伝子使用度との相違点についても分析した。よって、ハイブリドーマE6.4によって分泌された抗体は以下の遺伝子使用度を有することが証明された:
VH −4〜31
D −A1/A4
JH −4
VK −012
JK −2
【0094】
V BASE配列ディレクトリにおいて報告されるように、VK012遺伝子によってコードされるアミノ酸配列を以下のように決定した:
TITCRASQSISSYLNWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSYSTP(配列番号:21)
【0095】
重鎖および軽鎖に関するアミノ酸およびヌクレオチド配列情報は図17〜20と共に下記に示す。図17は、ハイブリドーマE6.4によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜31によってコードされる配列とE6.4分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【0096】
図18は、ハイブリドーマE6.4からクローニングした図17の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
図19は、ハイブリドーマE6.4によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子012によってコードされる配列とE6.4分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
図20は、ハイブリドーマE6.4からクローニングした図19のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【0097】
ハイブリドーマE2.11
ハイブリドーマE2.11によって分泌された抗体はヒトカッパ軽鎖を有するヒトIgG2抗体を含む。抗体をそのアミノ酸配列と共に、その重鎖および軽鎖の遺伝子使用度に関連した構造情報に関して分析した。このように、重鎖VH、DおよびJHならびに軽鎖VKおよびJK遺伝子使用度を分析して、コードされる産物と特定の遺伝子使用度との相違点についても分析した。よって、ハイブリドーマE2.11によって分泌された抗体は以下の遺伝子使用度を有することが証明された:
VH −4〜61
D −XP1/21〜10
JH −4
VK −018
JK −4
【0098】
V BASE配列ディレクトリにおいて報告されるように、VH4〜61遺伝子によってコードされるアミノ酸配列を以下のように決定した:
VSGGSVSSGSYYWSWIRQPPGKGLEWIGYIYYSGSTNYNPSLKSRVTISVDTSKNQFSLKLSSVTAADTAVYYCAR(配列番号:22)
【0099】
重鎖および軽鎖に関するアミノ酸およびヌクレオチド配列情報は図21〜24と共に下記に示す。図21は、ハイブリドーマE2.11によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜61によってコードされる配列とE2.11分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【0100】
図22は、ハイブリドーマE2.11からクローニングした図21の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
図23は、ハイブリドーマE2.11によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE2.11分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
図24は、ハイブリドーマE2.11からクローニングした図23のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【0101】
ハイブリドーマE6.3
ハイブリドーマE6.3によって分泌された抗体はヒトカッパ軽鎖を有するヒトIgG2抗体を含む。抗体をそのアミノ酸配列と共に、その重鎖および軽鎖の遺伝子使用度に関連した構造情報に関して分析した。このように、重鎖VH、DおよびJHならびに軽鎖VKおよびJK遺伝子の利用を分析して、コードされる産物と特定の遺伝子使用度との相違点についても分析した。よって、ハイブリドーマE6.3によって分泌された抗体は以下の遺伝子使用度を有することが証明された:
VH −4〜61
D −1〜2rc
JH −4
VK −018
JK −4
【0102】
重鎖および軽鎖に関するアミノ酸およびヌクレオチド配列情報は図25〜28と共に下記に示す。図25は、ハイブリドーマE6.3によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜61によってコードされる配列とE6.3分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【0103】
図26は、ハイブリドーマE6.3からクローニングした図25の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
図27は、ハイブリドーマE6.3によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE6.3分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
図28は、ハイブリドーマE6.3からクローニングした図27のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【0104】
ハイブリドーマE7.6.3
ハイブリドーマE7.6.3によって分泌された抗体はヒトカッパ軽鎖を有するヒトIgG2抗体を含む。抗体をそのアミノ酸配列と共に、その重鎖および軽鎖の遺伝子使用度に関連した構造情報に関して分析した。このように、重鎖VH、DおよびJHならびに軽鎖VKおよびJK遺伝子使用度を分析して、コードされる産物と特定の遺伝子使用度との相違点についても分析した。よって、ハイブリドーマE7.6.3によって分泌された抗体は以下の遺伝子使用度を有することが証明された:
VH −4〜61
D −XP4rc〜XP1
JH −3
VK −018
JK −4
【0105】
重鎖および軽鎖に関するアミノ酸およびヌクレオチド配列情報は図29〜32と共に下記に示す。図29は、ハイブリドーマE7.6.3によって分泌される重鎖免疫グロブリン分子のアミノ酸配列である。重鎖可変領域遺伝子4〜61によってコードされる配列とE7.6.3分泌重鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
【0106】
図30は、ハイブリドーマE7.6.3からクローニングした図29の重鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
図31は、ハイブリドーマE7.6.3によって分泌されるカッパ軽鎖免疫グロブリン分子のアミノ酸配列である。軽鎖可変領域遺伝子018によってコードされる配列とE7.6.3分泌軽鎖の配列との相違点を太字および拡大フォントで示す。CDR1からCDR3までの近接配列を下線で示し、CDR1、CDR2およびCDR3配列はそれぞれ二重下線で示す。
図32は、ハイブリドーマE7.6.3からクローニングした図31のカッパ軽鎖免疫グロブリン分子をコードするcDNAのヌクレオチド配列である。
【0107】
実施例4
重鎖および軽鎖アミノ酸置換の分析
図33は、特異的抗EGF-r抗体重鎖アミノ酸配列と、特定の抗体の重鎖をコードする特定のVH遺伝子のアミノ酸配列との比較を示す。図34は、特異的抗EGF-r抗体軽鎖アミノ酸配列と、特定の抗体の軽鎖をコードする特定のVK遺伝子のアミノ酸配列との類似比較を示す。認められるように、重鎖および軽鎖配列には極めて保存されたアミノ酸置換がいくつか存在する。特に、抗体の重鎖では、重鎖分子の全てがVH 4ファミリー遺伝子によってコードされ、VH 4〜31によってコードされる抗体では10位にグリシンを有し、VH 4〜61によってコードされる抗体では10位にセリンを有するが、これらはそれぞれアスパラギン酸に置換されている。VH 4〜31重鎖の場合と同様に、1つを除いて全ての抗体が7位のセリンのアスパラギンへの置換を含む。35位ではそれほど優勢な置換ではないが同様の置換が認められ、VH4〜31によってコードされる抗体の2つおよびVH4〜61によってコードされる抗体の2つにおいてセリンがアスパラギンに置換されている。同様に、VH4〜31によってコードされる抗体の2つおよびVH4〜61によってコードされる抗体の2つでは、28位に置換が存在し、それぞれの場合についてチロシンがセリン(E2.4)またはヒスチジン(E6.4、E2.11、およびE7.6.3)に置換されている。抗体の5つ、すなわちVH4〜31によってコードされる抗体の3つ、およびVH4〜61によってコードされる抗体の2つは、50位でバリンの代わりにロイシン(E2.4およびE2.11)またはイソロイシン(E2.5、E6.2、およびE7.6.3)を有する。
【0108】
カッパ軽鎖アミノ酸配列では、配列は全てVK Iファミリー遺伝子によってコードされ、分子の7個は018遺伝子によってコードされ、1個(E6.4)は012遺伝子によってコードされている。図34において、E6.4分子を他の分子と共に018遺伝子産物と比較するとわかるように、012遺伝子産物と018遺伝子産物との間には高度の相同性が存在する。E6.4分子は図19に示すように、012遺伝子産物と比較して2箇所の置換を有するに過ぎず、018遺伝子産物と比較して13箇所の置換を有するに過ぎない。抗体は全て、CDR3の74位で置換を有し、アスパラギンがセリン(E1.1、E2.5、E2.11、およびE6.3)、ヒスチジン(E2.4、E6.2、およびE7.6.3)、またはアルギニン(E6.4)に置換されている。残りの置換はあまり高度に保存されてはいない。しかし、多くの抗体はCDR内部に置換を有するように思われる。しかし、非常に親和性の高い抗体であるE7.6.3がCDR3にまさに隣接するまでの軽鎖アミノ酸配列、およびCDR3内にアミノ酸置換を有しないことに注目することは興味深い。
【0109】
上記のように確認されたアミノ酸置換はそれぞれCDRに非常に近位でまたはCDRの内部に存在することが認識されると思われる。そのような置換はEGF受容体分子に対する抗体の結合に基づいて何らかの作用を有すると思われる。さらに、そのような置換は抗体の親和性に有意な作用を及ぼしうる。
既に考察したように、抗EGF-r抗体は、特定の抗腫瘍活性を有することが示されている。以下の実験は、本発明の抗体がそのような抗腫瘍活性を有するか否かを決定するために実施した。
【0110】
実施例5
インビトロでヒト抗EGF-r抗体によるヒト類上皮癌A431細胞に対するEGFおよびTGF-αの結合の遮断
インビトロアッセイを実施して、本発明の抗体がヒト癌細胞株に対するEGFの結合を遮断することができるか否かを明らかにした。上記のように既に抗癌活性が証明されたマウスモノクローナル抗体225と、本発明の抗体の結合とを比較する実験を実施した。
【0111】
この実施例ではヒト類上皮癌A431細胞株を使用した。A431細胞株はそのEGF-rの発現レベルが高いこと(約2×106個EGF-r分子/細胞)が知られている。したがって、全ての結合部位を飽和するためにはより高濃度の抗EGF-r抗体を必要とする。本実施例から得られた結果を図35に示す。図において、インビトロでヒト抗EGF-r抗体によるヒト類上皮癌A431細胞に対するI125標識EGF結合の遮断が示される。図において、(□)は本発明の抗EGF-r抗体(E7.6.3)によって得られた結果を示し、(●)は、マウスモノクローナル抗体225によって得られた結果を示し、および(黒上向き三角)は、対照の非特異的ヒトIgG2抗体によって得られた結果を示す。
【0112】
図36は、インビトロで本発明のヒト抗EGF-r抗体集団によるヒト類上皮癌A431細胞に対するEGF結合の阻害を225、528および非特異的ヒトIgG2対照と比較して示す。図において、(□)は、マウスモノクローナル抗体225によって得られた結果を示し、(○)はマウスモノクローナル抗体528によって得られた結果を示し、(黒下向き三角)は本発明のE1.1抗体を用いて得られた結果を示し、(黒上向き三角)は本発明のE2.4抗体を用いて得られた結果を示し、(黒右向き三角)は、本発明のE2.5抗体を用いて得た結果を示し、(黒左向き三角)は本発明のE2.6抗体を用いて得られた結果を示し、(◆)は本発明のE2.11抗体を用いて得られた結果を示し、且つ(四角で囲んだ×印)は、対照の、非特異的ヒトIgG2抗体を用いて得られた結果を示す。
【0113】
結果は、本発明の抗体が、A431細胞上の表面に発現されたEGF-rに対するEGF結合を225および528抗体より良好に遮断する可能性があることを示している。本発明の抗体は8nM濃度で結合を阻害し始めるように思われるのに対し、225抗体では10 nM濃度であった。
【0114】
TGF-α結合の阻害に関しては、本発明の抗体を用いることによって225抗体と比較して類似の有効性が認められた。図37は、インビトロでヒト抗EGF-r抗体によるヒト類上皮癌A431細胞に対するTGF-α結合の阻害を示し、(□)は、マウスモノクローナル抗体225によって得られた結果を示し、(◆)は本発明のE6.2抗体を用いて得られた結果を示し、(●)は本発明のE6.3抗体を用いて得られた結果を示し、(黒上向き三角)は本発明のE7.2抗体を用いて得られた結果を示し、(黒四角)は本発明のE7.10抗体を用いて得られた結果を示し、(黒下向き三角)はE7.6.3を用いて得られた結果を示し、且つ(○で囲んだ十字)は対照の非特異的ヒトIgG2抗体を用いて得られた結果を示す。
【0115】
結果は本発明の抗体が、A431細胞の表面に発現されたEGF-rに対するTGF-α結合を225抗体より良好に遮断する可能性があることを示している。本発明の抗体は0.1nM濃度で結合を阻害し始めるように思われるのに対し、225抗体では1nM濃度であった。
【0116】
実施例6
インビトロでヒト抗EGF-r抗体によるヒト結腸腺癌SW948細胞に対するEGF結合の遮断
本発明の抗体がさらにもう一つのヒト癌細胞株に対するEGF結合を遮断できるか否かを明らかにするために、もう一つのインビトロアッセイを実施した。上記のように既に抗癌活性を有することが証明されているマウスモノクローナル抗体225と、本発明の抗体の結合を比較する実験を実施した。
【0117】
本実施例ではヒト結腸腺癌SW948細胞株を用いた。A431細胞株とは対照的に、SW948細胞株はその表面上のEGF-rの発現数が比較的少ない(約40,000分子/細胞)。したがって細胞上の受容体の全ての結合部位を飽和するために必要な抗EGF-r抗体の量は比較的少ない。本実施例の結果を図38に示す。図において、インビトロでヒト抗EGF-r抗体によるヒト結腸腺癌SW948細胞に対するI125標識EGF結合の阻害を示す。図において、(●)は、本発明の抗EGF-r抗体(E7.6.3)によって得られた結果を示し、(□)はマウスモノクローナル抗体225によって得られた結果を示し、且つ(黒上向き三角)は対照の非特異的ヒトIgG2抗体によって得られた結果を示す。
【0118】
結果は本発明の抗体がSW948細胞に対するEGF結合を少なくとも225抗体と同程度に遮断することを示している。実際に、曲線は本発明の抗体ではわずかに改善しており、225抗体と比較すると低濃度で阻害が起こることを示している。
【0119】
実施例7
インビトロでのヒト抗EGF-r抗体によるヒト結腸腺癌SW948細胞増殖の阻害
本発明者らはまた、本発明の抗体が癌細胞増殖を阻害するか否か、および225抗体と比較してどの程度まで阻害するかを明らかにするためにインビトロアッセイを実施した。上記のように抗癌活性を有することが既に証明されているマウスモノクローナル抗体225による阻害と、本発明の抗体による阻害を比較する実験を実施した。
【0120】
本実施例ではヒト結腸腺癌SW948細胞株を用いた。本発明者らが行うと、SW948細胞のみがEGF-依存的細胞増殖を示した。対照的にA431細胞株はインビトロでEGFの存在下で増殖阻害を示した。結果を図39に示し、本発明のヒト抗EGF-r抗体がインビトロでSW948細胞の増殖を阻害することを示す。図において(●)は、本発明の抗EGF-r抗体(E7.6.3)によって得られた結果を示し、(□)は、マウスモノクローナル抗体225によって得られた結果を示し、且つ(黒上向き三角)は対照の非特異的ヒトIgG2抗体によって得られた結果を示す。
【0121】
結果は、本発明の抗体がSW948細胞の増殖を少なくとも225抗体と同程度に阻害することを示す。実際に、本発明の抗体に関して曲線はわずかに改善し、約100 μg/mlで細胞増殖を明らかに100%阻害を示したが、225抗体は同じ用量範囲では80〜90%の阻害レベルで平衡に達するように思われる。
【0122】
実施例8
インビボでのヒト抗EGF-r抗体によるヌードマウスにおけるヒト類上皮癌細胞増殖の阻害
本実験において、本発明者らは本発明の抗体がインビボで腫瘍の細胞増殖を阻害することができるか否かを明らかにしようと試みた。実験において、8週齢のヌードマウスにヒト類上皮癌A431細胞株を皮下接種した。マウスにA431細胞5×106個を注射した。本発明の抗体の2つの用量のうち1つ、または2つの対照のうち1つを、A431細胞接種と同じ日に腹腔内注射した。いずれかの抗体または対照の3つの投与を行った後に、マウスの皮下腫瘍形成および大きさを調べた。用いた抗体の用量は1.0mgまたは0.2 mgのいずれかであった。対照は、燐酸緩衝生理食塩液または非特異的ヒトIgG2抗体のいずれかであった。
【0123】
本実験の結果を図40に示す。図において、インビボで本発明のヒト抗EGF-r抗体を用いることによる、ヌードマウスにおけるヒト類上皮癌細胞増殖の阻害は明らかである。図において、(黒上向き三角)は、本発明のヒト抗EGF-r抗体(E7.6.3)1.0mgの投与によって得られた結果を示し(n=5)、(黒下向き三角)はE7.6.3抗体0.2 mgの投与によって得られた結果を示し(n=4)、(□)は、対照の非特異的ヒトIgG2抗体によって得られた結果を示し(n=6)、且つ(○)は、対照として燐酸緩衝生理食塩液を用いた場合に得られた結果を示す(n=6)。
E7.6.3抗体で処置した動物では腫瘍の増殖は認められなかったが、対照動物は腫瘍細胞を接種した25日以内に有意な腫瘍の増殖が認められた。
【0124】
同じ実験において、本発明の3つの抗体を比較した。結果を図41に示す。本発明の抗体のそれぞれ、すなわちE7.6.3の1 mgをマウス5匹に、0.2 mgを4匹に、E2.5の1mgをマウス3匹に、および0.2 mgをマウス3匹に、ならびにE1.1の1mgをマウス3匹に投与することにより、対照と比較してマウスにヒト類上皮癌形成の阻害が示された。対照動物(PBS処置動物6匹およびヒトIgG2-処置動物6匹を含む)は全て接種の19日以内に有意な腫瘍を発生したが、本発明のヒト抗EGF-r抗体で処置した動物はいずれも接種の19日以内に腫瘍を発生しなかった。
【0125】
図42は、上記の同じ実験の動物を、ヒト類上皮癌の接種後130日間追跡した結果を示す。この実験の結果を図42に示す。図において、対照マウスは全て腫瘍細胞接種の20日以内に腫瘍を発生したことが認められる。対照的に、本発明の抗体で処置した最初のマウスが腫瘍を発症したのは70日目であった。130日までに、腫瘍を発生したのは実験動物15匹中4匹に過ぎなかった。興味深いことに、E2.5抗体の0.2mg用量で処置した実験動物のいずれも試験期間内に腫瘍を発生しなかった。
【0126】
この実施例8に関連した上記実験は、本発明の抗体が、腫瘍細胞株の接種と同時に投与すれば、腫瘍細胞の増殖の開始および腫瘍の開始をほぼ完全に予防するように思われることを証明する。その上、腫瘍細胞増殖に及ぼす作用は長期間持続するように思われることが認められた。
【0127】
実施例9
インビボでヒト抗EGF-r抗体によるヌードマウスにおけるヒト類上皮癌増殖の根治
腫瘍細胞の増殖および/または腫瘍の確立を予防することは、先に述べた実施例に関連して上記で考察したように、治療的な観点から見て肯定的な知見であるが、確立した腫瘍の根治も同様に非常に望ましい。したがって、以下の実験において、本発明の抗体が哺乳類において確立した腫瘍を根治することができるか否かを調べた。225抗体に関して得られたこれまでのデータから、確立された腫瘍を225抗体を用いることによって有効に根治するためには、抗癌剤による治療を補うことが必要であることが示された。このように本発明者らの実験では、単独および抗癌剤治療と併用した場合の双方の抗体治療を調べた。
【0128】
実験において、0日目にヌードマウスにA431ヒト類上皮癌細胞5×106個を皮下接種した。腫瘍が十分に確立してから(大きさ≧0.4 cm3)、マウスを抗体、化学療法剤、および/または対照のいずれかで治療した。治療は5、8、10、14、16、および21日目に開始して継続し、化学療法剤は5および6日のみに投与した。治療には、本発明の抗体(E7.6.3)、抗癌剤ドキソルビシン、および抗体とドキソルビシンの併用が含まれた。対照は燐酸緩衝生理食塩液または非特異的ヒトIgG2抗体であった。各治療群は動物5匹を含んだ。実験から得られたデータを図43に示し、(△)は本発明のヒト抗EGF-r抗体(E7.6.3)1mgを投与した場合に得られた結果を示し、(×)は、ドキソルビシン125μgを投与した場合に得られた結果を示し、(XX)は本発明のヒト抗EGF-r抗体(E7.6.3)1 mgの投与をドキソルビシン125 μgの投与と併用した場合に得られた結果を示し、(黒四角)は対照の非特異的ヒトIgG2抗体によって得られた結果を示し、および(◆)は対照として燐酸緩衝生理食塩液を用いた場合に得られた結果を示す。
【0129】
図から認められるように、E7.6.3抗体をドキソルビシンと併用して投与すると、腫瘍増殖の完全な根治が得られた。さらに、腫瘍の増殖はE7.6.3抗体のみを投与した場合にも完全に停止した。
【0130】
図44に結果を示す同様の実験において、腫瘍の接種後にマウス5匹にE2.5抗体の0.5 mgを5、8、10、14、16、および21日に投与することによって治療し、およびマウス5匹にE2.5抗体の5、8、10、14、16、および21日目の投与と、ドキソルビシン5および6日目の投与の併用によって治療した。図において、(◆)は本発明のヒト抗EGF-r抗体(E2.5)0.5mgを投与した場合に得られた結果を示し、(黒四角)はドキソルビシンの125 μgを投与することによって得られた結果を示し、(△)は本発明のヒト抗EGF-r抗体(E2.5)0.5mgの投与をドキソルビシン125 μgの投与と併用した場合に得られた結果を示し、(×)は、対照として燐酸緩衝生理食塩液を用いた場合に得られた結果を示し、および(XX)は、対照の非特異的ヒトIgG2抗体を用いた場合に得られた結果を示す。
図から認められるように、E2.5抗体を単独で、またはドキソルビシンと併用して投与すると、マウスにおいて腫瘍のほぼ完全な根治が得られた。
【0131】
実施例10
インビボでヒト抗EGF-r抗体を用いることによるヒト癌の治療および診断に関するヒト臨床試験
緒言
本発明の抗体は特定の固形癌の治療に適応される。特にEGF-r発現レベルを含む多くの要因に基づき、以下の腫瘍のタイプが好ましい適応となるように思われる:乳癌、卵巣癌、結腸癌、前立腺癌、膀胱および非小細胞肺癌。これらの適応のそれぞれに関して、3つの臨床経路が明確に臨床的に成功する可能性があるように思われる。
【0132】
補助療法:補助療法において、患者は、本発明の抗体と化学療法剤または抗癌剤および/または放射線療法との併用によって治療する。上記の主な標的は、標準的な第一選択および第二選択療法に発明の抗体を加えることによってプロトコルに基づいて治療されるであろう。プロトコルは、標準的な化学療法の通常の用量を減少させることと共に腫瘍塊の減少によって評価する有効性が得られるようにデザインされる。これらの用量の減少により、化学療法剤の用量関連毒性を減少させることによってさらなるおよび/または長期間の療法が可能となるであろう。先行技術の抗EGF-r抗体は、いくつかの補助的臨床試験において、化学療法もしくは抗癌剤であるアドリアマイシン(C225:進行前立腺癌)、シスプラチン(C225:進行頭頸部癌および肺癌)、タキソール(C225:乳癌)、ならびにドキソルビシン(C225:前臨床)と併用して用いられた、または用いられている。
【0133】
単剤療法:腫瘍の単剤療法に本発明の抗体を利用する場合には、化学療法剤または抗癌剤を用いないで抗体を患者に投与する。本発明の抗体の使用によって得られ、本明細書において考察する前臨床試験の結果から、補助療法および/または単剤療法の双方に関して同程度の結果が示された。その上、単剤療法は明らかに臨床的に広範囲の転移疾患を有する末期段階の癌患者において実施された。患者は疾患に何らかの安定化を示すように思われた。同上。臨床試験は(癌)腫瘍を有する難治性の患者において作用が示されるようにデザインされる。
【0134】
造影剤:放射性核種(例えば、イットリウム(90Y))を本発明の抗体に結合することによって、本発明の放射標識抗体は、診断的造影剤として利用することができると予想される。そのような役割において、本発明の抗体は固形癌のみならずEGF受容体を発現する細胞の転移病変の双方に存在するであろう。本発明の抗体を造影剤として用いる場合には、どの腫瘍が残存するかおよび/または再発するかを決定するために術後の追跡調査と共に術前スクリーニングとして、固形癌の外科治療を補助するために抗体を用いることができる。111In-C225抗体は、切除不能の肺扁平細胞癌を有する患者におけるフェーズIヒト臨床試験において造影剤として用いられている。ディブジら(Divgi)、J.Natl.Cancer Inst.83:97〜104(1991)。患者は標準的な前後方γカメラによって追跡した。前臨床データから、全ての原発病変および大きい転移病変は特定されるが、小さい転移病変(1cm以下)は半数が検出されたに過ぎなかった。
【0135】
用量および投与経路
本発明の抗体の特定の投与法についてはまだ決定していないが、臨床において用いられる同様の産物(イムクローンC255)と比較することによって、特定の投与に関する検討を行うことが可能である。C225抗体は典型的に5〜400mg/m2の範囲の用量を投与し、これより低い用量は安全性試験に関連した場合に限って用いられる。本発明の抗体の親和性はC225抗体より1対数高い。さらに、本発明の抗体は、C225抗体のキメラ特性と比較して完全なヒト抗体であり、このように抗体のクリアランスはより遅くなると予想される。したがって、本発明の抗体を患者に投与する場合には、用量はこれより低く、おそらく50〜300mg/m2の範囲となり、それでもなお有効であろうと予想される。従来のmg/kgで表す用量単位に対して、mg/m2での投与は表面積に基づいた単位であり、幼児から成人までの全ての体格の患者を含むようにデザインされる都合のよい投与単位である。
【0136】
本発明の抗体の輸送には3つの明確な輸送アプローチが有用であると予想される。従来の静脈内輸送はおそらく、ほとんどの腫瘍にとって標準的な輸送技法であろう。しかし、卵巣、胆管、その他の管等の腫瘍のような腹腔における腫瘍に関しては、腫瘍での抗体量を高め、および抗体のクリアランスを最小限にするために、腹腔内投与が都合がよい可能性がある。同様にして、固形癌の中には局所還流に適した血管を有するものがある。局所還流によって腫瘍部位での抗体の高用量を得ることが可能となり、抗体の短期間でのクリアランスが最小限となる。
【0137】
臨床開発計画(CDP)
総説:CDPにより、本発明の抗EGF-r抗体の補助療法、単剤療法に関連した治療、および造影剤としての処置が追求および開発される。まず臨床試験を用いて安全性を証明し、次に投与を繰り返した際の有効性を得るために臨床試験を行う。臨床試験は、標準的な化学療法と標準的な療法プラス本発明の抗体とを比較するオープンラベルとなるであろう。認識されるように、患者の登録に関して利用することができる1つの基準は、生検によって決定した患者の腫瘍のEGF-r発現レベルであってもよい。
【0138】
蛋白質または抗体注入に基づく治療に関して、安全性に関する懸念は主に(i)サイトカイン放出症候群、すなわち血圧低下、発熱、震え、悪寒、(ii)材料に対する免疫原反応の発症(すなわち、ヒト抗体治療に対する患者によるヒト抗体の産生またはHAHA反応)、および(iii)EGF受容体を発現する正常細胞、例えばEGF-rを発現する肝細胞に対する毒性、に関する。標準的な試験および追跡調査を利用して、これらの安全性に関する懸念をそれぞれモニターする。特に、あるとすれば肝臓の障害を評価するために臨床試験の間、肝機能を頻繁にモニターする。
【0139】
ヒト臨床試験:抗EGF-r抗体および化学療法剤との補助療法
フェーズIヒト臨床試験は固形癌、例えば乳癌の治療において、本発明のヒト抗EGF-r抗体の6用量を静脈内投与した場合の安全性を評価するために開始されるであろう。本試験において、シスプラチン、トポテカン、ドキソルビシン、アドリアマイシン、タキソール等のような抗癌剤または化学療法剤に対する補助療法として用いた場合の、本発明の抗体の単回投与の安全性を評価する。臨床試験のデザインには、以下のスケジュールに従って、治療の過程において本発明の抗体の用量を約25mg/m2〜約275 mg/m2に増加させて、本発明の抗体の単回投与を6回行うことが含まれる。
【0140】
【表2】
患者は抗体および化学療法のそれぞれの投与後1週間厳密に追跡調査する。特に、上記の安全性に関する懸念について患者を評価する:(i)サイトカイン放出症候群、(ii)材料に対する免疫原反応の発症(すなわち、患者によるヒト抗体治療に対するヒト抗体の産生、またはHAHA反応)、および(iii)EGF受容体を発現する正常細胞、例えばEGF-rを発現する肝細胞に対する毒性。標準的な試験および追跡調査を利用してこれらの安全性に関する懸念のそれぞれをモニターする。特に、あるとすれば肝臓に対する障害を評価するために、臨床試験において肝機能を頻繁にモニターする。
【0141】
患者はまた、臨床転帰に関して、特にMRIまたはその他の画像によって示される腫瘍塊の減少に関しても評価する。
安全性が証明されることおよび有効性が示されることを仮定して、有効性をさらに探索し、最適な投与を決定するために、フェーズII臨床試験がおそらく開始されるであろう。
【0142】
ヒト臨床試験:ヒト抗EGF-r抗体による単剤療法
本発明の抗体が上記補助臨床試験に関連して安全であるように思われると仮定して、単剤療法に関する有効性および最適な投与を評価するためのヒト臨床試験。そのような臨床試験を行って、患者に対して本発明の抗体と同時に化学療法を投与しないことを除き、上記補助臨床試験の場合と同じ安全性および転帰分析を必要とする。
【0143】
ヒト臨床試験:抗EGF-r抗体による診断的造影
再度、上記の補助療法が上記の安全性基準の中で安全であるように思われると仮定して、本発明の抗体を診断的造影剤として用いることに関するヒト臨床試験を実施することができる。プロトコルはディブジら(Divgi)、J.Nat.Cancer Inst.83:97〜104(1991)に記述されたものと実質的に同様にしてデザインされると予想される。
【0144】
参照としての組み入れ
特許、特許出願、論文、教科書等を含む、本明細書における全ての引用文献、およびそれらに引用される引用文献は、それらが先ではないという程度に、本明細書に参照としてその全文が組み入れられる。さらに、そのような引用文献において引用される引用文献を含む、以下の引用文献もまた、本明細書においてその全文が参照として組み入れられる。
Abertsenら、"Construction and characterization of a yeast artificial chromosome library containing seven haploid human genome equivalents." Proc.Natl.Acad.Sci.87:4256(1990)
Anandら、"Construction of yeast artificial chromosome libraries with large inserts using fractionation by pulsed-field gel electrophoresis." Nucl.Acids Res.17:3425-3433(1989)
Bermanら、"Content and organization of the human Ig VH locus:definition of three new VH families and linkage to the Ig CH locus."(1988) EMB0 J.7:727-738
Brezinschekら、"Analysis of the heavy chain repertoire of human peripheral B-cells using single-cell polymerase chain reaction." J.Immunol.155:190-202(1995)
Brownsteinら、"Isolation of singie-copy human genes from a library of yeast artificial chromosome clones." Science 244:1348-1351(1989)
Bruggemanら、PNAS USA 86:6709-6713 (1989)
Bruggemann ら、"Human antibody production in transgenic mice:expression from 100 kb of the human IgH locus." Eur.J.Immunol.21:1323-1326(1991)
Bruggeman,M.およびNeuberger,M.S.in Methods:A companion to Methods in Enzymology 2:159-165(Lernerら編、Academic Press (1991))
Bruggemann,M.およびNeuberger,M.S."Strategies for expressing human antibody repertoires in transgenic mice." Immunology Today 17:391-397(1996)
Chenら、"Immunoglobulin gene rearrangement in B-cell deficient mice generated by targeted deletion of the JH locus," International Immunology 5:647-656(1993)
Choiら、"Transgenic mice containing a human heavy chain immunoglobulin gene fragment cloned in a yeast artificial cnromosome" Nature Genetics 4:117-123(1993)
Coliganら、Unit 2.1,"Enzyme-linked Immunosorbent assays," in Current protocols in Immunology(1994)
Cook,G.P.およびTomlinson,I.M.,"The human immunoglobulin VH repertoire." Immunology Today 16:237-242(1995)
Coxら、"A directory of human germ-line VX segments reveals a strong bias in their usage." Eur.J.Immunol.24:827-836(1994)
Dariavachら、"The mouse IgH 3'-enhancer." Eur.J.linmunol.21:1499-1504(1901)
Den Dunnenら、"Reconstruction of the 2.4 Mb human DMD-gene by homologous YAC recombination." Human Molecular Genetics 1:19-28(1992)
Feeney,A.J."Lack of N regions in fetal and neonatal mouse immunoglobulin V-D-J junctional sequences." J.Exp.Med.172:137-1390(1990)
Fishwildら、"High-avidity human IgGκ monoclonal antibodies from a novel strain of minilocus transgenic mice." Nature Biotech.14:845-851(1996)
Flanagan,J.G.およびRabbitts,T.H.,"Arrangement of human immunoglobulin heavy chain constan region genes implies evolutionary duplication of a segment containing g,e,and a genes." Nature 300:709-713(1982)
Galfre,G.およびMilstein,C.,"Preparation of monoclonal antibodies:strategies and procedures." Methods Enzymol.73:3-46(1981)
Gemmillら、"Protocols for pulsed field gel electrophoresis:Separation and detection of large DNA molecules." Advances in Genome Biology 1:217-251(1991)
Gillら、"Monoclonal anti-epidermal growth factor receptor antibodies which are inhibitors of epidermal growth factor binding and antagonists of epidermal growth factorstimulated tyrosine protein kinase activity." J.Biol.Chem.259:7755(1984)
Greenら、"Antigen-specific human monoclonal antibodies from mice engineered with human Ig heavy and light chain YACs." Nature Genetics 7:13-21(1994)
Hermansonら、"Rescue of end fragments of yeast artificial chromosomes by homologous recombination in yeast." Nucleic Acids Res.19:4943-4948(1991)
Huberら、"The human immunoglobulin κ locus.Characterization of the partially duplicated L regions." Eur.J.Immunol.23:2860-2967(1993)
Jakobovits,A.,"Humanizing the mouse genome." Current Biology 4:761-763(1994)
Jakobovits,A.,"Production of fully human antibodies by transgenic mice." Current opinion in Biotechnology 6:561-566(1995)
Jakobovitsら、"Germ-line transmission and expression of a human-derived yeast artificial-chromosome." Nature 362:255-258(1993)
Jakobovits,A.ら、"Analysis of homozygous mutant chimeric mice:Deletion of the immunoglobulin heavy-chain joining region blocks B-cell development and antibody production." Proc.Natl.Acad.Sci.USA 90:2551-2555(1993)
Kawamotoら、"Growth stimulation of A431 cells by epidermal growth factor:Identification of high affinity receptors for EGF by an anti-receptor monoclonal antibody." Proc.Nat.Acad.Sci.,USA 80:1337-1341(1983)
Lonbergら、"Antigen-specific human antibodies from mice comprising four distinct genetic modifications." Nature 368:856-859(1994)
Lusti-Marasimhanら、"Mutation of Leu25 and Val27 introduces CC chemokine activity into interleukin-8." J.Biol.Chem.270:2716-2721(1995)
Marksら、"Oligonucleotide primers for pclymerase chain reaction amplification of human immunoglobulin variable genes and design of family-specific oligonucleotide probes." Eur.J.Immunol.21:985-991(1991)
Matsudaら、"Structure and physical map of 64 variable segments in the 3' 0.8-megabase region of the human immunoglobulin heavy-chain locus." Nature Genetics 3:88-94(1993)
Max,E.Molecular genetics of immunoglobulins.Fundamental Immunology.315-382(Paul,WE編、New York:Raven Press (1993))
Mendezら、"A set of YAC targeting vectors for the interconversion of centric and acentric arms." Cold Spring Harbor Laboratory Press,Genome Mapping and Sequencing meeting,163(1993)
Mendezら、"Analysis of the structural integrity of YACs comprising human immunoglobulin genes in yeast and in embryonic stem cells." Genomics 26:294-307(1995)
Ray,S.およびDiamond,B.,"Generation of a fusion partner to sample the repertoire of Splenic B-cells destined for apoptosis." Proc.Natl.Acad.Sci.USA 91:5548-5551(1994)
Satoら、"Biological effects in vitro of monoclonal antibodies to human epidermal growth factor receptors" Mol.Bioi.Med.1:511-529(1983)
Schiestl,R.H.およびGietz,R.D.,"High efficiency transformation of intact yeast cells using stranded nucleic acids as a carrier." Curr.Genet.16:339-346(1989)
Shermanら、"Laboratory Course Manual for Methods in Yeast Genetics."(Cold Spring Harbor Laboratory Press,Cold Spring Harbor,NY (1986))
Silvermanら、"Meiotic recombination between yeast artificial chromosomes yields a single clone containing the entire BCL2 protooncogene." Proc.Nati.Acad.Sci.USA 87:9913-9917(19 )
Srivastava,A.およびSchiessinger,D.,"Vectors for inserting selectable markers in vector arms and human DNA inserts of yeast artificial chromosomes(YACs)." Gene 103:53-59 (1991)
Taylorら、"A transgenic mouse that expresses a diversity of human sequence heavy and light chain immunoglobulins." Nucleic Acids Research 20:6287-6295(1992)
Taylorら、"Human immunoglobulin transgenes undergo rearrangement,somatic mutation and class switching in mice that lack endogenous IgM." International Immunology 6:579-591(1994)
Tuaillonら、"Human immunoglobulin heavy-chain minilocus recombination in transgenic mice:gene-segment use in m and g transcripts." Proc.Natl.Acad.Sci.USA 90:3720-3724(1993)
Tuaillonら、"Analysis of direct and inverted DJH rearrangements in a human Ig heavy chain transgenic minilocus" J.Immunol.154:6453-6465(1995)
Vaughanら、"Human antibodies with subnanomolar affinities isolated from a large non-immunized phage display library." Nature Biotech.14:309-314(1996)
Wagnerら、"The diversity of antigen-specific monoclonal antibodies from transgenic mice bearing human immunoglobulin gene miniloci." Eur.J.Immunol.24:2672-2681(1994)
Weichholdら、"The human immunoglobulin κ locus consists of two copies that are organized in opposite polarity." Genomics 16:503-511(1993)
Yamada,M.ら、"Preferential utilization of specific immunoglobulin heavy chain diversity and joining segments in adult human peripheral blood B lymphocytes." J.Exp.Med.173:395-407(1991)
【0145】
同等性
前述の記述および実施例は、本発明の特定の好ましい態様について詳述し、発明者が意図する最善の様式を記述する。しかし、前述がどれほど詳細に本文において現れようとも、本発明は多くの方法において実践されること、および本発明は添付の請求の範囲およびその同等物に従って解釈されるべきであると認識される。
【特許請求の範囲】
【請求項1】
明細書中に記載の発明。
【請求項1】
明細書中に記載の発明。
【図1】


【図2】
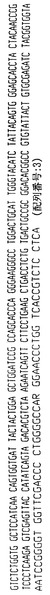
【図3】

【図4】
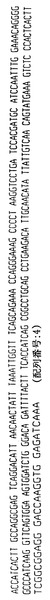
【図5】

【図6】
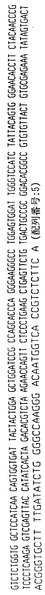
【図7】

【図8】
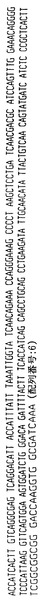
【図9】

【図10】
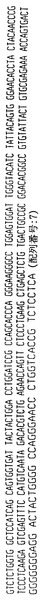
【図11】

【図12】
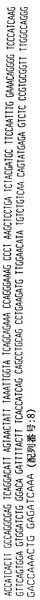
【図13】

【図14】
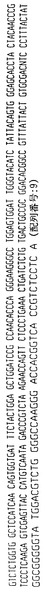
【図15】
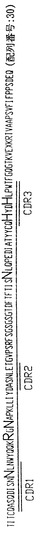
【図16】
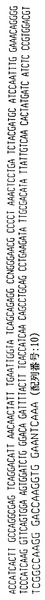
【図17】

【図18】
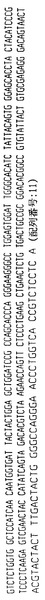
【図19】

【図20】
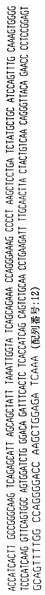
【図21】

【図22】
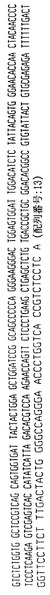
【図23】

【図24】

【図25】
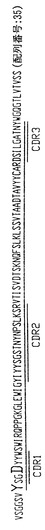
【図26】
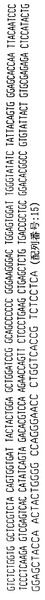
【図27】

【図28】
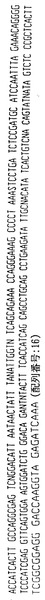
【図29】

【図30】
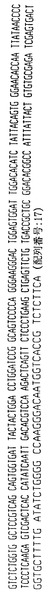
【図31】

【図32】
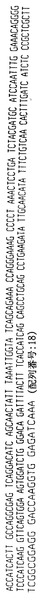
【図33】
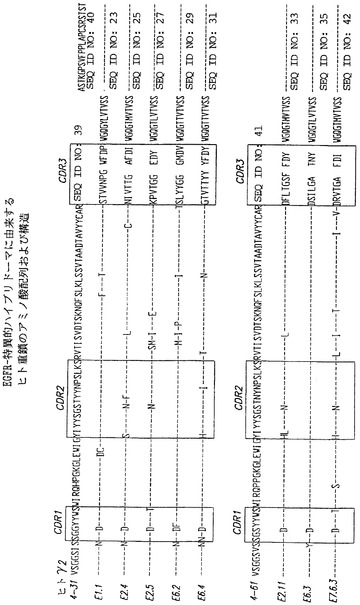
【図34】

【図35】
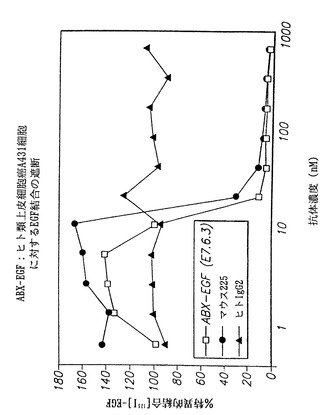
【図36】
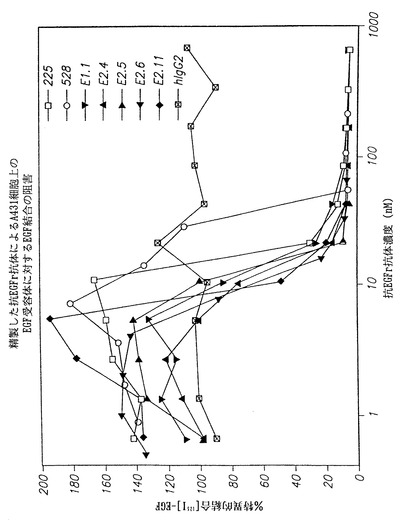
【図37】
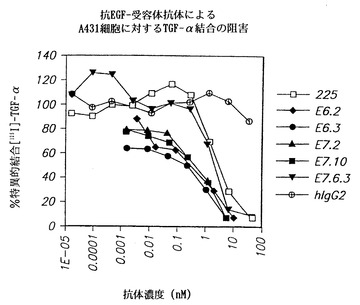
【図38】
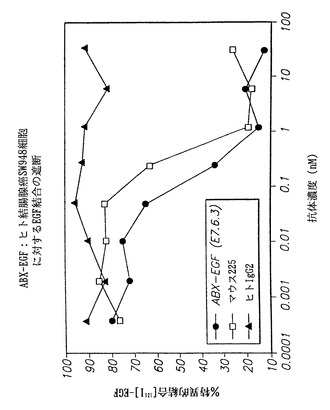
【図39】
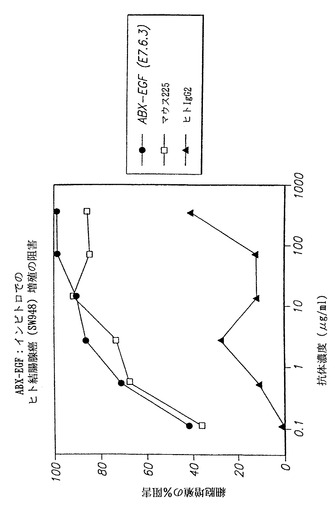
【図40】
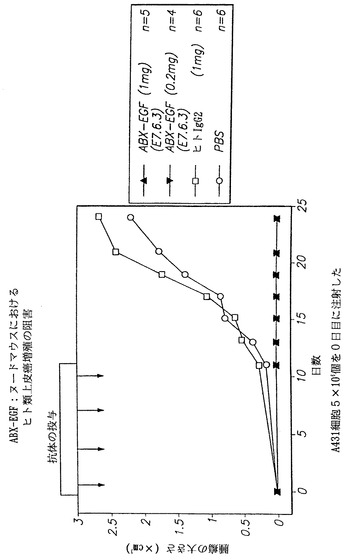
【図41】
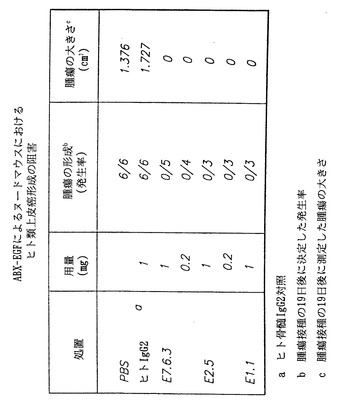
【図42】
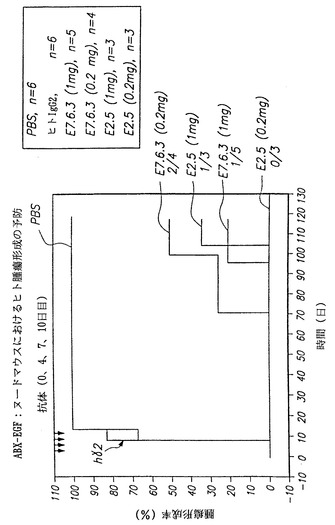
【図43】
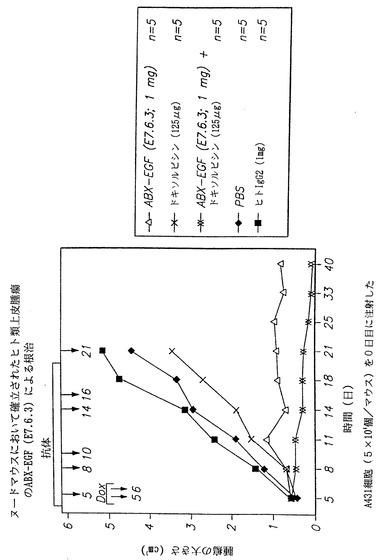
【図44】
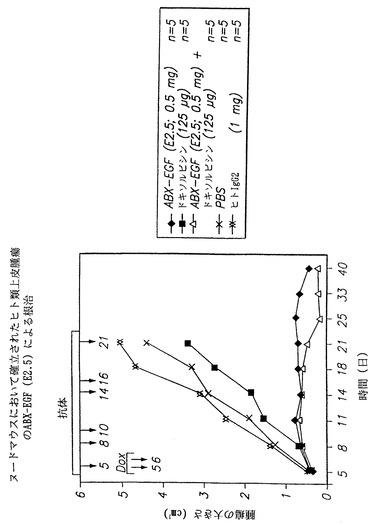
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【図34】
【図35】
【図36】
【図37】
【図38】
【図39】
【図40】
【図41】
【図42】
【図43】
【図44】
【公開番号】特開2012−158601(P2012−158601A)
【公開日】平成24年8月23日(2012.8.23)
【国際特許分類】
【出願番号】特願2012−104486(P2012−104486)
【出願日】平成24年5月1日(2012.5.1)
【分割の表示】特願2010−52859(P2010−52859)の分割
【原出願日】平成10年5月5日(1998.5.5)
【出願人】(398005777)アムジェン フレモント インク. (40)
【Fターム(参考)】
【公開日】平成24年8月23日(2012.8.23)
【国際特許分類】
【出願日】平成24年5月1日(2012.5.1)
【分割の表示】特願2010−52859(P2010−52859)の分割
【原出願日】平成10年5月5日(1998.5.5)
【出願人】(398005777)アムジェン フレモント インク. (40)
【Fターム(参考)】
[ Back to top ]