
光増感性蛍光タンパク質
【課題】単量体型の光増感性蛍光タンパク質、光増感活性が高い光増感性蛍光タンパク質、又はKillerRedとは異なる、励起波長及び蛍光波長を有する光増感性蛍光タンパク質を提供する。
【解決手段】特定のアミノ酸配列からなり、発色団補助光不活性化法に用いることのできる、光増感性蛍光タンパク質又はその機能的等価改変体、および該タンパク質をコードするDNA。また、該タンパク質を用いる光増感性の活性酸素産生方法、さらに標的分子の機能解析方法。
【解決手段】特定のアミノ酸配列からなり、発色団補助光不活性化法に用いることのできる、光増感性蛍光タンパク質又はその機能的等価改変体、および該タンパク質をコードするDNA。また、該タンパク質を用いる光増感性の活性酸素産生方法、さらに標的分子の機能解析方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、光増感性蛍光タンパク質に関する。本発明によれば、発色団補助光不活性化法において、標的タンパク質を効率よく破壊することができる。
【背景技術】
【0002】
生体試料内の任意のタンパク質の機能を調べる方法として、標的タンパク質をコードする遺伝子を破壊する、いわゆる遺伝子ノックアウト法、標的タンパク質をコードするmRNAに結合して分解するRNA干渉法、又は標的タンパク質に対する抗体を作用させることで機能不活性化を行う方法などが知られている。これらの方法はいずれも標的タンパク質が発現している細胞内の全ての領域の標的タンパク質に効果が及ぶことを特徴とする。従って、細胞内の任意の位置に存在する標的タンパク質だけを選択的に不活性化する事は、原理上不可能であった。
【0003】
一方、標的タンパク質の機能部位を光照射依存的に産生された活性酸素による酸化によって時空間的に不活性化し、そのタンパク質の機能解析を行う方法として、発色団補助光不活性化法(chromophore-assisted light inactivation;以下、CALIと称することがある)が知られている。具体的には、発色団補助光不活性化法はマラカイトグリーン、ローダミン誘導体、又はフルオレセイン誘導体などを光増感剤として用い、標的分子(例えば、標的タンパク質)に結合させ、標的分子の機能を抑制することによって、標的分子の機能を解析する方法である。例えば、フルオレセイン誘導体を用いた場合、フルオレセイン誘導体から、発生する一重項酸素(活性酸素)が標的タンパク質の機能を破壊することが知られている。
しかしながら、発色団補助光不活性化法は、通常フルオレセイン標識した標的分子に対する抗体又はフルオレセイン標識した標的分子(例えば、標的タンパク質)をマイクロインジェクション等で細胞に導入する必要があり、操作が煩雑であるという問題点があった。
【0004】
最近、Lukyanovらは、発色団補助光不活性化法に用いることのできる光増感性蛍光タンパク質(CALI色素)である、「KillerRed」を報告した(特許文献1及び非特許文献1)。KillerRedはタンパク質であるため、KillerRedをコードするDNAを含むプラスミドなどを用いることによって、形質転換又はトランスフェクションなどの手法により、光増感性蛍光タンパク質(CALI色素)を細胞内において発現させることが可能になった。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】国際公開第2006/117694号公報
【非特許文献】
【0006】
【非特許文献1】「ネイチャー・バイオテクノロジー(Nature Biotechnology)」(米国)2006年、第24巻、p.95〜99
【非特許文献2】「フェブス・レター(FEBS letter)」(オランダ)2009年、第583巻、p.2839−2842
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、前記KillerRedは、細胞内において二量体を形成するため、多くのタンパク質はKillerRedと融合しただけで、立体障害などによる機能の不活性化が起こり、光照射依存的な標的タンパク質の機能不活性化を行うことができないという問題を有していた。
また、KillerRedは、光増感活性が低いため、活性酸素の産生量がフルオレセインよりも少ないという問題を有していた。
更に、例えば、生体試料内の2つ以上の標的タンパク質を、個別に光照射によって不活性化する場合には、吸光特性の異なる光増感性蛍光タンパク質が必要である。しかしながら、KillerRed以外の吸光スペクトルを有する光増感性蛍光タンパク質は報告されておらず、別の励起波長及び蛍光波長を有する光増感性蛍光タンパク質の開発が望まれていた。
従って、本発明の目的は、単量体型の光増感性蛍光タンパク質を提供することである。また、本発明の別の目的は、光増感活性が高い光増感性蛍光タンパク質を提供することである。更に、本発明の別の目的は、KillerRedとは異なる、励起波長及び蛍光波長を有する光増感性蛍光タンパク質を提供することである。
【0008】
本発明者は、生物個体内、組織内、細胞内、細胞内小器官内、又は水溶液中において、光刺激により活性酸素を効率良く産生することのできる光増感活性の高い光増感性蛍光タンパク質について、鋭意研究した結果、驚くべきことに、KillerRedのアミノ酸配列において、5番目のグリシン(G)をバリン(V)に、147番目のアスパラギン(N)をセリン(S)に、162番目のロイシン(L)をトレオニン(T)に、164番目のフェニルアラニン(F)をトレオニン(T)に、174番目のロイシン(L)をリシン(K)に、そして206番目のメチオニン(M)をトレオニン(T)に置換した変異体であるSupernova−Redは、単量体として細胞内などに存在するようになることを見出した。更に、前記Supernova−Redの68番目のチロシン(Y)をトリプトファン(W)に置換した変異体であるSupernova−Orangeは、一重項酸素の産生量が増加し、励起極大波長及び蛍光極大波長が、それぞれ455nm及び544nmにシフトすることを見出した。
更に、Supernova−Orangeの46番のバリン(V)をアラニン(A)に置換した変異体であるSupernova−Greenは、一重項酸素の産生量がKillerRedと比較して増加し、励起極大波長及び蛍光極大波長が、それぞれ439nm及び496nmにシフトすることを見出した。
本発明は、こうした知見に基づくものである。
【課題を解決するための手段】
【0009】
従って、本発明は、配列表の配列番号2で表されるアミノ酸配列、配列番号4で表されるアミノ酸配列、配列番号6で表されるアミノ酸配列、配列番号9で表されるアミノ酸配列、及び配列番号10で表されるアミノ酸配列、からなる群から選択される少なくとも1つのアミノ酸配列を含む光増感性蛍光タンパク質又はその機能的等価改変体に関する。
本発明の光増感性蛍光タンパク質又はその機能的等価改変体の好ましい態様においては、前記光増感性蛍光タンパク質又はその機能的等価改変体が、他のタンパク質と結合した融合タンパク質である。
また、本発明は、前記光増感性蛍光タンパク質又はその機能的等価改変体をコードするDNAに関する。
本発明のDNAの好ましい態様においては、配列表の配列番号1で表される塩基配列、配列番号3で表される塩基配列、及び配列番号5で表される塩基配列からなる群から選択される少なくとも1つの塩基配列からなるDNAである。
更に、本発明は、前記DNAを含むプラスミドに関する。
更に、本発明は、前記光増感性蛍光タンパク質又はその機能的等価改変体を用いた、光増感性の活性酸素産生方法であって、前記光増感性蛍光タンパク質又はその機能的等価改変体に、励起光を照射することを特徴とする、前記方法に関する。
本発明の光増感性の活性酸素産生方法の好ましい態様においては、前記励起光の波長が、490〜580nm、400〜500nm、及び390〜440nmからなる群から選択される少なくとも1つの波長である。
更に、本発明は、光増感性蛍光タンパク質又はその機能的等価改変体を用いた、標的分子の機能解析方法であって、前記光増感性蛍光タンパク質又はその機能的等価改変体に、励起光を照射することを特徴とする、前記方法に関する。
本発明の標的分子の機能解析方法の好ましい態様においては、前記励起光の波長が、490〜580nm、400〜500nm、及び390〜440nmからなる群から選択される少なくとも1つの波長である。
【発明の効果】
【0010】
本発明の光増感性蛍光タンパク質又はその機能的等価改変体によれば、単量体として細胞内などに存在することが可能であり、多くのタンパク質に対してCALIにおける光増感性蛍光タンパク質として用いることが可能である。
本発明の光増感性蛍光タンパク質及びその機能的等価改変体、特にはSupernova−Orangeによれば、Supernova−Redの68番目のチロシンをトリプトファンに置換(Y68W)することで、励起極大/蛍光極大を455nm/544nm(目視ではオレンジ色)にシフトし、更に、一重項酸素の産生量がKillerRedと比較して増加する。従って、CALIにおいて、増強型光増感性蛍光タンパク質として用いることが可能であり、更にKillerRed、Supernova−Red及びSupernova−Greenなどと同時に用いることにより、2種以上の標的タンパク質の機能相関などを解析することが可能である。
本発明の光増感性蛍光タンパク質及びその機能的等価改変体、特にはSupernova−Greenによれば、Supernova−Orangeの46番のバリンをアラニンに置換することで、励起極大/蛍光極大を439nm/496nm(目視では緑色)にシフトし、一重項酸素の産生量がKillerRedと比較して増加する。従って、CALIにおいて、増強型光増感性蛍光タンパク質として用いることが可能であり、更にKillerRed、Supernova−Red、及びSupernova−Orangeなどと同時に用いることにより、2種以上の標的タンパク質の機能相関などを解析することが可能である。
なお、KillerRedのX線結晶構造解析を行った論文(非特許文献2)によれば、KillerRedの46番のバリンをアラニンに置換(V46A)することは、発色団の光増感作用により発生した活性酸素をKillerRedタンパク質の外部に効率良く放出するようになる可能性が記載されている。
しかしながら、後述の実施例に示すように、Supernova−RedにV46A変異を導入すると、活性酸素の放出量は減少した。一方、Y68W変異が導入されているSupernova−OrangeにV46A変異を導入したSupernova−Greenでは、活性酸素の放出量が2倍以上に増大し、更に蛍光波長がグリーンに変化した。従って、光増感性蛍光タンパク質において、変異の導入により、活性酸素の放出量が増加したり、又は減少したりすることを予想すること、また蛍光波長が変化することは当業者であっても決して容易ではなく、試行錯誤を要するものである。
【図面の簡単な説明】
【0011】
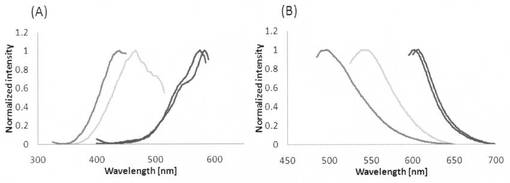
【図1】KillerRed、Supernova−Red、Supernova−Orange、及びSupernova−Greenの励起スペクトル(A)及び蛍光スペクトル(B)を示したグラフである。
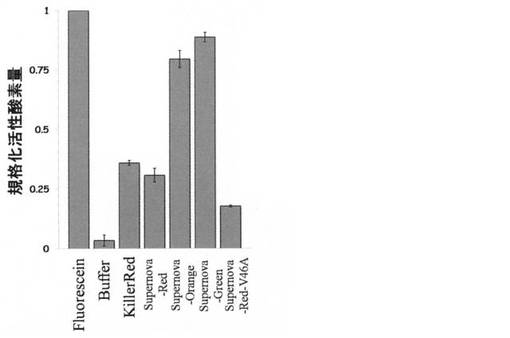
【図2】KillerRed、Supernova−Red、Supernova−Orange、Supernova−Green、及びSupernova−Red−V46Aの活性酸素の産生量を示したグラフである。FITC(フルオレセインイソチアシアネート)は陽性コントロールを示し、Buffer(1xPBS)は陰性コントロールを示している。
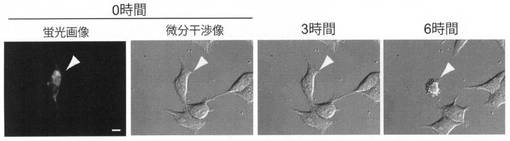
【図3】Supernova−RedをHela細胞に導入し、光照射を行い、0時間、3時間、及び6時間後の細胞の微分干渉画像を示した写真である。Supernova−Redがミトコンドリアに発現している細胞は、6時間後にアポトーシスにより、死滅している。
【発明を実施するための形態】
【0012】
本発明の光増感性蛍光タンパク質は、配列番号2で表されるアミノ酸配列、配列番号4で表されるアミノ酸配列、配列番号6で表されるアミノ酸配列、配列番号9で表されるアミノ酸配列、及び配列番号10で表されるアミノ酸配列からなる群から選択される少なくとも1つのアミノ酸配列を含むタンパク質である。
【0013】
本発明の機能的等価改変体は、前記光増感性蛍光タンパク質の機能的等価改変体である。本明細書において、「機能的等価改変体」とは、そのアミノ酸配列が、元となるタンパク質のアミノ酸配列において1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列であって、しかも、元となるタンパク質と実質的に同じ活性を示すタンパク質を意味する。前記アミノ酸の欠失、置換、又は付加は、例えば10個であり、好ましくは1〜10個、より好ましくは1〜5個、更に好ましくは1〜2個である。
【0014】
本発明の光増感性蛍光タンパク質の第1の実施態様は、配列番号2で表されるアミノ酸配列を含むタンパク質(以下、第1光増感性蛍光タンパク質と称する)又はその機能的等価改変体(以下、第1機能的等価改変体と称する;以下、第1光増感性蛍光タンパク質及び第1機能的等価改変体を、まとめて第1光増感性蛍光タンパク質等と称することがある)であり、配列番号2で表されるアミノ酸配列からなるタンパク質又はその機能的等価改変体であることが好ましい。
第1光増感性蛍光タンパク質は、配列番号8で表されるアミノ酸配列からなる光増感性蛍光タンパク質(すなわち、KillerRed)のアミノ酸配列において、5番目のグリシン(G)をバリン(V)に置換(以下、G5Vと称することがある)、147番目のアスパラギン(N)をセリン(S)に置換(以下、N147Sと称することがある)、162番目のロイシン(L)をトレオニン(T)に置換(以下、L162Tと称することがある)、164番目のフェニルアラニン(F)をトレオニン(T)に置換(以下、F164Tと称することがある)、174番目のロイシン(L)をリシン(K)に置換(以下、L174Kと称することがある)、そして206番目のメチオニン(M)をトレオニン(T)に置換(以下、M206Tと称することがある)したアミノ酸配列を含む光増感性蛍光タンパク質である。代表的な第1光増感性蛍光タンパク質は、配列番号2で表されるアミノ酸配列からなるタンパク質であるSupernova−Redである。
KillerRedは、二量体を形成することが知られているが、これに対してSupernova−Redは、試料中(例えば、生物個体内、組織内、細胞内、細胞内小器官内、又は水溶液中)において、単量体で存在する。また、Supernova−Redの励起極大波長及び蛍光極大波長は、それぞれ579nm及び610nmである。
【0015】
本発明の第1光増感性蛍光タンパク質等において、前記L162T及びF164Tの置換(以下、L162T、及びF164Tの置換を有するKillerRedの変異体を、KillerRed−L162T/F164Tと称する)により、二量体であったKillerRedが、単量体となる。従って、前記KillerRed−L162T/F164Tは、標的タンパク質との融合タンパク質として発現しても、標的タンパク質の機能を阻害しない。一方、L162T及びF164Tの置換により、KillerRed−L162T/F164Tは、KillerRedと比較して585nmにおける吸光及び610nmにおける蛍光が減少した。
しかしながら、前記KillerRed−L162T/F164Tに、更にG5V、N147S、L174K、及びM206Tの置換を導入することにより、KillerRedと同等の蛍光強度を有するSupernova−Redを得ることができた。
【0016】
本発明の第1機能的等価改変体は、実質的にSupernova−Redと同じ活性を有するものである。「Supernova−Redと同じ活性」とは、光照射により活性酸素を産生すること、光照射により蛍光を発生すること、試料中において単量体で存在すること、そして励起極大波長及び蛍光極大波長がそれぞれ約579nm及び610nmであることを意味する。なお、励起極大波長及び蛍光極大波長がそれぞれ約579nm及び610nmであるとは、好ましくは569〜589nm及び600〜620nmであり、より好ましくは574〜584nm及び605〜615nmである。第1機能的等価改変体としては、例えば、前記配列番号2で表されるアミノ酸配列において、第2番目のグリシン及び第3番目のセリンを欠如したタンパク質を挙げることができる。
【0017】
本発明の光増感性蛍光タンパク質の第2の実施態様は、配列番号4で表されるアミノ酸配列を含むタンパク質(以下、第2光増感性蛍光タンパク質と称する)又はその機能的等価改変体(以下、第2機能的等価改変体と称する;以下、第2光増感性蛍光タンパク質及び第2機能的等価改変体を、まとめて第2光増感性蛍光タンパク質等と称することがある)であり、配列番号4で表されるアミノ酸配列からなるタンパク質又はその機能的等価改変体であることが好ましい。第2光増感性蛍光タンパク質は、前記Supernova−Redのアミノ酸配列において、68番目のチロシン(Y)をトリプトファン(W)に置換(以下、Y68Wと称する)したアミノ酸配列を含む光増感性蛍光タンパク質である。代表的な第2光増感性蛍光タンパク質は、配列番号4で表されるアミノ酸配列からなるタンパク質であるSupernova−Orangeである。
Supernova−Orangeの励起極大波長及び蛍光極大波長は、それぞれ455nm及び544nmであり、KillerRedと比較して、一重項酸素の産生量が増加している。
【0018】
本発明の第2光増感性蛍光タンパク質等において、前記L162T及びF164Tの置換により、二量体であったKillerRedが、単量体となり、Y68Wの置換により、励起極大波長及び蛍光極大波長が、それぞれおよそ455nm及び544nmとなる。
本発明の第2機能的等価改変体は、実質的にSupernova−Orangeと同じ活性を有するものである。「Supernova−Orangeと同じ活性」とは、光照射により活性酸素を産生すること、光照射により蛍光を発生すること、試料中において単量体で存在すること、そして励起極大波長及び蛍光極大波長がそれぞれ約455nm及び544nmであることを意味する。なお、励起極大波長及び蛍光極大波長がそれぞれ約455nm及び544nmであるとは、好ましくは445〜465nm及び534〜554nmであり、より好ましくは450〜460nm及び539〜549nmである。第2機能的等価改変体としては、例えば、前記配列番号8で表されるアミノ酸配列において、L162T、F164T、及びY68Wの置換が導入されたタンパク質、及び前記タンパク質において、第2番目のグリシン及び第3番目のセリンを欠如したタンパク質、並びに前記配列番号4で表されるアミノ酸配列において、第2番目のグリシン及び第3番目のセリンを欠如したタンパク質を挙げることができる。
【0019】
本発明の光増感性蛍光タンパク質の第3の実施態様は、配列番号6で表されるアミノ酸配列を含むタンパク質(以下、第3光増感性蛍光タンパク質と称する)又はその機能的等価改変体(以下、第3機能的等価改変体と称する;以下、第3光増感性蛍光タンパク質及び第3機能的等価改変体を、まとめて第3光増感性蛍光タンパク質等と称することがある)であり、配列番号6で表されるアミノ酸配列からなるタンパク質又はその機能的等価改変体であることが好ましい。第3光増感性蛍光タンパク質は、前記Supernova−Orangeのアミノ酸配列において、46番のバリン(V)をアラニン(A)に置換(以下、V46Aと称する)したアミノ酸配列を含む光増感性蛍光タンパク質である。代表的な第3光増感性蛍光タンパク質は、配列番号6で表されるアミノ酸配列からなるタンパク質である、Supernova−Greenである。
Supernova−Greenの励起極大波長及び蛍光極大波長は、それぞれ439nm及び496nmであり、KillerRedと比較して、一重項酸素の産生量が増加している。
【0020】
本発明の第3光増感性蛍光タンパク質等において、前記L162T及びF164Tの置換により二量体であったKillerRedが単量体となり、Y68W及びV46Aの置換により、励起極大波長及び蛍光極大波長が、それぞれ439nm及び496nmとなる。
本発明の第3機能的等価改変体は、実質的にSupernova−Greenと同じ活性を有するものである。「Supernova−Greenと同じ活性」とは、光照射により活性酸素を産生すること、光照射により蛍光を発生すること、試料中において単量体で存在すること、そして励起極大波長及び蛍光極大波長がそれぞれ約439nm及び496nmであることを意味する。なお励起極大波長及び蛍光極大波長がそれぞれ約439nm及び496nmであるとは、好ましくは429〜449nm及び486〜506nmであり、より好ましくは434〜444nm及び591〜501nmである。第3機能的等価改変体としては、例えば、前記配列番号8で表されるアミノ酸配列において、L162T、F164T、Y68W及びV46Aの置換が導入されたタンパク質、及び前記タンパク質において、第2番目のグリシン及び第3番目のセリンを欠如したタンパク質、並びに前記配列番号6で表されるアミノ酸配列において、第2番目のグリシン及び第3番目のセリンを欠如したタンパク質を挙げることができる。
【0021】
本発明の光増感性蛍光タンパク質の第4の実施態様は、配列番号9で表されるアミノ酸配列を含むタンパク質(以下、第4タンパク質と称する)又はその機能的等価改変体(以下、第4機能的等価改変体と称する;以下、第4光増感性蛍光タンパク質及び第4機能的等価改変体を、まとめて第4光増感性蛍光タンパク質等と称することがある)であり、配列番号9で表されるアミノ酸配列からなるタンパク質又はその機能的等価改変体であることが好ましい。
第4光増感性蛍光タンパク質は、配列番号8で表されるアミノ酸配列からなる光増感性蛍光タンパク質(すなわち、KillerRed)のアミノ酸配列において、L162T、F164T、及びN147Sの置換を有するアミノ酸配列を含む光増感性蛍光タンパク質である。代表的な第4光増感性蛍光タンパク質は、配列番号9で表されるアミノ酸配列からなるタンパク質であるKillerRed_L162T_F164T_N147Sである。
KillerRed_L162T_F164T_N147Sの励起極大波長は、512nmであり、蛍光は黄緑色(目視)を示す。
【0022】
本発明の第4光増感性蛍光タンパク質等において、前記L162T及びF164Tの置換により二量体であったKillerRedが単量体となり、N147Sの置換により、励起極大波長が512nmで、蛍光が黄緑色となる。
【0023】
本発明の第4機能的等価改変体は、実質的にKillerRed_L162T_F164T_N147Sと同じ活性を有するものである。「KillerRed_L162T_F164T_N147Sと同じ活性」とは、光照射により活性酸素を産生すること、光照射により蛍光を発生すること、試料中において単量体で存在すること、そして励起極大波長が約512nm及び蛍光が黄緑色であることを意味する。なお、励起極大波長が約512nmであるとは、好ましくは502〜522nmであり、より好ましくは507〜517nmである。第4機能的等価改変体としては、例えば、前記配列番号9で表されるアミノ酸配列において、第2番目のグリシン及び第3番目のセリンを欠如したタンパク質を挙げることができる。
【0024】
本発明の光増感性蛍光タンパク質の第5の実施態様は、配列番号10で表されるアミノ酸配列を含むタンパク質(以下、第5タンパク質と称する)又はその機能的等価改変体(以下、第5機能的等価改変体と称する;以下、第5光増感性蛍光タンパク質及び第5機能的等価改変体を、まとめて第5光増感性蛍光タンパク質等と称することがある)であり、配列番号10で表されるアミノ酸配列からなるタンパク質又はその機能的等価改変体であることが好ましい。
第5光増感性蛍光タンパク質は、配列番号8で表されるアミノ酸配列からなる光増感性蛍光タンパク質(すなわち、KillerRed)のアミノ酸配列において、L162TF164T、G5V、N147S、L174K、M206T、及びV46Aの置換を有するアミノ酸配列を含む光増感性蛍光タンパク質である。代表的な第5光増感性蛍光タンパク質は、配列番号10で表されるアミノ酸配列からなるタンパク質であるSupernova−Red_V46Aである。
Supernova−Red_V46Aの励起極大波長及び蛍光極大波長は、それぞれ509nm及び594nmである。
【0025】
本発明の第5光増感性蛍光タンパク質等において、前記L162T及びF164Tの置換により二量体であったKillerRedが単量体となり、G5V、N147S、L174K、M206T、及びV46Aの置換により、励起極大波長及び蛍光極大波長が、それぞれ509nm及び594nmとなる。
【0026】
本発明の第5機能的等価改変体は、実質的にSupernova−Red_V46Aと同じ活性を有するものである。「Supernova−Red_V46Aと同じ活性」とは、光照射により活性酸素を産生すること、光照射により蛍光を発生すること、試料中において単量体で存在すること、そして励起極大波長及び蛍光極大波長がそれぞれ約509nm及び594nmであることを意味する。なお、励起極大波長及び蛍光極大波長がそれぞれ約509nm及び594nmであるとは、好ましくは499〜519nm及び584〜604nmであり、より好ましくは504〜514nm及び589〜599nmである。第5機能的等価改変体としては、例えば、前記配列番号10で表されるアミノ酸配列において、第2番目のグリシン及び第3番目のセリンを欠如したタンパク質を挙げることができる。
【0027】
本発明の光増感性蛍光タンパク質及び機能的等価改変体(以下、まとめて本発明の光増感性蛍光タンパク質等と称することがある)は、光の照射によって、活性酸素(例えば、一重項酸素)を産生することが可能である。従って、本発明のタンパク質等を細胞内に導入し、適当な励起波長の光を照射することによって、一重項酸素を発生させ、細胞に障害を与え、更には細胞を死滅させることもできる。
また、本発明のタンパク質等は、発色団補助光不活性化法に用いることもできる。具体的には本発明の光増感性蛍光タンパク質等を標的タンパク質に結合させ、光の照射によって活性酸素(例えば、一重項酸素)を産生することによって、その標的タンパク質の機能を抑制、又はその標的タンパク質を破壊することによって、標的タンパク質の機能を解析することができる。
【0028】
本発明による光増感性蛍光タンパク質及び機能的等価改変体は、種々の公知の方法によって得ることができ、例えば、光増感性蛍光タンパク質であるKillerRedをコードするDNAを含むプラスミドベクターを用いて公知の遺伝子工学的手法により調製することができる。より具体的には、部位特異的突然変異誘発法(site−directed mutagenesis)により調製することが可能である。
更に得られたプラスミドを用いて、大腸菌などの宿主細胞を形質転換し、形質転換体を得ることができる。得られた形質転換体(すなわち、本発明による光増感性蛍光タンパク質又は機能的等価改変体をコードするDNAを含むプラスミドを有する形質転換体)を、前記光増感性蛍光タンパク質等の発現が可能な条件下で培養し、タンパク質の分離及び精製に一般的に用いられる方法により、その培養物から光増感性蛍光タンパク質等を分離及び精製することにより調製することができる。前記の分離及び精製方法としては、例えば、光増感性蛍光タンパク質にHisタグを付けニッケルNTAカラムで生成する方法、硫安塩析、イオン交換セルロースを用いるイオン交換カラムクロマトグラフィー、分子篩ゲルを用いる分子篩カラムクロマトグラフィー、プロテインA結合多糖類を用いる親和性カラムクロマトグラフィー、透析、又は凍結乾燥等を挙げることができる。
【0029】
本発明の光増感性蛍光タンパク質等は、他のタンパク質と結合した融合タンパク質とすることができる。融合される他のタンパク質は、光増感性蛍光タンパク質等の活性を完全に阻害しない限り、特に制限されるものではなく、例えば、発色団補助光不活性化法における標的タンパク質、又は標的分子に結合させる結合タンパク質を挙げることができる。
【0030】
前記標的タンパク質は、発色団補助光不活性化法において、機能を解析されるタンパク質である。標的タンパク質を本発明の光増感性蛍光タンパク質等との融合タンパク質とした場合、光の照射により光増感性蛍光タンパク質等から産生された活性酸素が、容易に標的タンパク質を不活化することができる。これは、産生された活性酸素により、半径約10〜50Åに存在する標的分子が特異的に不活化されるためである。本発明の光増感性蛍光タンパク質等は、単量体であるため、標的タンパク質の細胞内における本来の局在性や活性を妨害することが少ない。後述の実施例で示すように、Supernova−Redと、標的タンパク質(例えば、チューブリン、ケラチン、又はCx43)とを、融合タンパク質として発現させた場合、標的タンパク質の局在化に影響を与えなかった。
【0031】
前記結合タンパク質は、発色団補助光不活性化法において、標的分子に光増感性蛍光タンパク質を結合させるタンパク質である。結合タンパク質は、標的分子に結合することのできるタンパク質であれば、特に限定されるものではないが、例えば、抗体及びその抗原結合性フラグメント(例えば、scFv、Fab、又は抗体可変領域全長の遺伝子産物)、レセプター、又はリガンドを挙げることができる。
【0032】
前記融合タンパク質は、公知の遺伝子組み換え技術によって、調整することが可能である。具体的には、光増感性蛍光タンパク質等をコードするDNAと、他のタンパク質をコードするDNAとを、定法に従って、発現用プラスミドに導入する。得られたプラスミドを、形質転換などにより、宿主細胞に導入することによって、融合タンパク質を発現させることが可能である。
【0033】
本発明によるDNAは、本発明の光増感性蛍光タンパク質等をコードする限り、特に限定されるものではなく、例えば、配列表の配列番号1、3、又は5で表される塩基配列からなるDNAを挙げることができる。配列表の配列番号1で表される塩基配列からなる前記DNAは、配列表の配列番号2で表されるアミノ酸配列からなるSupernova−Redをコードする。また、配列表の配列番号3で表される塩基配列からなる前記DNAは、配列表の配列番号4で表されるアミノ酸配列からなるSupernova−Orangeをコードする。更に、配列表の配列番号5で表される塩基配列からなる前記DNAは、配列表の配列番号6で表されるアミノ酸配列からなるSupernova−Greenをコードする。
【0034】
本発明によるプラスミドは、本発明による前記DNAを含む限り、特に限定されるものではなく、例えば、用いる宿主細胞に応じて適宜選択した公知の発現ベクターに、本発明による前記DNAを挿入することにより得られるプラスミドを挙げることができる。
得られたプラスミドを所望の宿主細胞に形質転換することにより、形質転換体を得ることができる。前記形質転換体も、本発明による前記プラスミドを含む限り、特に限定されるものではない。
【0035】
前記宿主細胞としては、例えば、通常使用される公知の微生物、例えば、大腸菌又は酵母(Saccharomyces cerevisiae)、あるいは、公知の培養細胞、例えば、動物細胞(例えば、CHO細胞、HEK−293細胞、又はCOS細胞)又は昆虫細胞(例えば、BmN4細胞)を挙げることができる。
【0036】
また、公知の前記発現ベクターとしては、例えば、大腸菌に対しては、pUC、pTV、pGEX、pKK、又はpTrcHisを;酵母に対しては、pEMBLY又はpYES2を;CHO細胞に対してはpcDNA3又はpMAMneoを;HEK−293細胞に対してはpcDNA3を;COS細胞に対してはpcDNA3を;BmN4細胞に対しては、カイコ核多角体ウイルス(BmNPV)のポリヘドリンプロモーターを有するベクター(例えば、pBK283)を挙げることができる。
【0037】
本発明の光増感性の活性酸素産生方法は、本発明の光増感性蛍光タンパク質等を用いる。前記光増感性蛍光タンパク質等に、励起光を照射することにより、光増感性蛍光タンパク質等から活性酸素が産生される。
本発明の光増感性の活性酸素産生方法によれば、例えば、細胞に前記光増感性蛍光タンパク質等を導入し、そして励起光を照射することにより、光増感性蛍光タンパク質等から活性酸素が産生され、細胞を死滅させることができる。
前記励起光の波長は、本発明の光増感性蛍光タンパク質等から活性酸素を産生させることのできる波長である。本発明の第1光増感性蛍光タンパク質等では、前記励起波長は、490〜620nmが好ましく、540〜580nmがより好ましく、560〜580nmが最も好ましい。本発明の第2光増感性蛍光タンパク質等では、前記励起波長は400〜500nmが好ましく、420〜480nmがより好ましく、440〜460nmが最も好ましい。本発明の第3光増感性蛍光タンパク質等では、前記励起波長は390〜480nmが好ましく、390〜460nmがより好ましく、430〜450nmが最も好ましい。
また前記励起光としては、前記励起波長の光のほか、多光子励起現象にもとづき、前記励起波長のおよそ整数倍の波長を有する光を用いることもできる。
【0038】
本発明の光増感性の活性酸素産生方法によれば、細胞のアポトーシスを誘導することが可能である。アポトーシスを誘導することにより、本発明の光増感性の活性酸素産生方法は、光線力学療法(PDT)に用いることができる。光線力学療法とは、光増感性の物質を腫瘍細胞に取り込ませ、光を照射することにより、細胞を死滅させる腫瘍の治療方法である。より具体的には、腫瘍細胞又は腫瘍組織内の新生血管の内皮細胞内に、本発明の光増感性蛍光タンパク質等を導入し、適当な光を照射することにより、活性酸素を発生させる。この活性酸素により、腫瘍細胞又は腫瘍組織が傷害され、腫瘍が消失するものである。
【0039】
本発明の標的分子の機能解析方法は、本発明の光増感性蛍光タンパク質等を用いる。前記光増感性蛍光タンパク質等に、励起光を照射することにより、光増感性蛍光タンパク質等から活性酸素が産生される。産生された活性酸素は近傍の標的分子を破壊し、それにより破壊された標的分子の機能を解析することが可能である。
前記励起光の波長は、本発明の光増感性蛍光タンパク質等から活性酸素を産生させることのできる波長である。本発明の第1光増感性蛍光タンパク質等では、前記励起波長は、490〜620nmが好ましく、540〜580nmがより好ましく、560〜580nmが最も好ましい。本発明の第2光増感性蛍光タンパク質等では、前記励起波長は400〜500nmが好ましく、420〜480nmがより好ましく、440〜460nmが最も好ましい。本発明の第3光増感性蛍光タンパク質等では、前記励起波長は390〜480nmが好ましく、390〜460nmがより好ましく、430〜450nmが最も好ましい。
また前記励起光としては、前記励起波長の光のほか、多光子励起現象にもとづき、前記励起波長のおよそ整数倍の波長を有する光を用いることもできる。
より具体的には本発明の標的分子の機能解析方法は、発色団補助光不活性化法として、用いることができる。
【0040】
本発明の方法において、標的分子とは、その生理的機能を解明する対象となる生体分子をいい、特にこれらに限定されないが、例えば、タンパク質、ペプチド、炭水化物、脂質、DNA、RNA、糖、シグナル伝達物質などが挙げられる。特には、本発明の方法において好ましい標的分子は、タンパク質であり、酵素、受容体タンパク質、リガンドタンパク質、シグナル伝達タンパク質、転写制御タンパク質、骨格タンパク質、細胞接着タンパク質、スキャホールドタンパク質等を挙げることができる。
【0041】
前記光増感性蛍光タンパク質等が産生する活性酸素は、一重項酸素である。前記一重項酸素は、励起状態の光増感性蛍光タンパク質発色団が項間交差によって励起3重項状態に遷移し、発色団近傍に存在する基底状態の3重項酸素にエネルギーを転移することにより発生する。発生した一重項酸素は、約10〜50Åの半径で光増感性蛍光タンパク質等に結合された標的分子(例えば、標的タンパク質)あるいはその機能性部位を選択的に不活化することができる。このようにして、標的分子自体あるいは標的分子の特定部位を不活性化することによって、その標的物質の生理的機能を解析することができる。例えば、このような不活性化によって、タンパク質の機能性部位の同定、その機能性部位の機能の確認、リガンドの機能の確認、機能性部位のタンパク質寿命に及ぼす影響の確認、機能性部位のタンパク質動態に及ぼす影響の確認、機能性部位のタンパク質フォールディングに及ぼす影響の確認などを行うことができる。
なお、本発明の標的分子の機能解析方法は、in vitro及びin vivoアッセイに、また細胞内外の標的分子に利用可能である。
【0042】
本発明の標的分子の機能解析方法において、光増感性蛍光タンパク質等を標的分子(例えば、タンパク質、炭水化物、脂質、DNA、RNA、糖、シグナル伝達物質)に結合させる方法は、特に限定されず、例えば化学結合によることが好ましい。光増感性蛍光タンパク質等を標的分子に共有結合により結合させることができる。また、光増感性蛍光タンパク質等に標的分子に対する抗体を共有結合により結合させ、その抗体により光増感性蛍光タンパク質等を標的分子に結合させることができる。更に、光増感性蛍光タンパク質等と標的分子に対する抗体とを融合タンパク質として発現させ、その抗体により光増感性蛍光タンパク質等を標的分子に結合させることができる。更には、標的分子がタンパク質の場合は、光増感性蛍光タンパク質等と標的タンパク質とを融合タンパク質として発現させることができる。
【実施例】
【0043】
以下、実施例によって本発明を具体的に説明するが、これらは本発明の範囲を限定するものではない。
【0044】
《実施例1》
本実施例では、KillerRedのアミノ酸配列に変異を導入することにより、単量体として存在することのできる光増感蛍光タンパク質の作製を試みた。
具体的には、KillerRedのアミノ酸配列における5番目のグリシン(G)、147番目のアスパラギン(N)、162番目のロイシン(L)、164番目のフェニルアラニン(F)、174番目のロイシン(L)、206番目のメチオニン(M)を、それぞれ、バリン(V)、セリン(S)、トレオニン(T)、トレオニン(T)、リシン(K)、トレオニン(T)に変異させるために、Sawanoらの方法(Sawano A and Miyawaki A. Nucleic Acid Research 28,E78,2000)に従って、部位特異的突然変異誘発法により、順次変異を導入した。
【0045】
(1)162番目のロイシン(L)のトレオニン(T)への置換
162番目のロイシン(L)をトレオニン(T)に置換するため、以下の変異導入用のプライマーを合成した。
Primer−L162T:5’-GTGCGCCAGACCGCCACCATC-3’(配列番号11)
Primer−L162Tの末端は、T4ポリヌクレオチドキナーゼによってリン酸化した。変異を導入する鋳型プラスミドDNAとしては、KillerRed遺伝子を含むプラスミドベクターを鋳型に用いたポリメラーゼ連鎖反応により、KillerRed遺伝子の上流と下流にそれぞれBamHI及びEcoRIを付加してKillerRed遺伝子を増幅した後、制限酵素BamHI及びEcoRIで切り出し、プラプラスミドpRSETB(インビロロジェン社)に挿入して得られたKillerRed/pRSETBを用いた。
【0046】
突然変異鎖の合成は、以下の通り行った。以下の組成の反応溶液40μLを準備し、PCR法によりin vitroで合成した。
KillerRed/pRSETB 50ng、
Primer−L162T 10pmol、
dNTP 3.75nmol、
PfuDNAポリメラーゼ 1.25U、
PfuDNAリガーゼ 20U
なお、PfuDNAポリメラーゼはSTRATAGENE社製であり、10×Pfuポリメラーゼバッファー中に溶解したものである。また、PfuDNAリガーゼは、STRATAGENE社製のものであり、10×PfuDNAリガーゼバッファー中に溶解したものである。
サーマルサイクラーは以下の通りプログラムした。すなわち、まず、65℃、5分のプレインキュベーションを行ってPfuDNAリガーゼによる鋳型DNAのニックを修復し、その後94℃、1分の最初の変性を行った。続いて、94℃、10秒のDNA変性、55℃、30秒のアニーリング反応及び65℃、10分の伸長・連結反応を1サイクルとして35サイクル行った。最後に75℃、10分のポストインキュベーションを行った。
【0047】
得られたPCR産物を、以下の反応条件に従い、DpnIによって切断し、変異の導入されていないメチル化又はヘミメチル化した鋳型プラスミドDNAを選択的に消化した。具体的には、前記PCR後の反応溶液40μLに1μL(20U)のDpnI(New England BioLab社製)を加え、37℃で2時間インキュベートした。なお、DpnIは〔5’−Gm6ATC−3’〕を認識して2本鎖DNAを切断するエンドヌクレアーゼである。更に、DpnIで処理した反応溶液20.4μLに対して、フェノールクロロホルム抽出を行ってサンプルを精製し、上清をエタノール沈殿して乾燥させた。
【0048】
次に、得られたDNAを用いて、形質転換及び発現を行った。前記1本鎖突然変異導入環状DNA鎖を用いて、ヒートショック法により大腸菌(E.coli)コンピテントセル(JM109(DE3))を形質転換した。そして形質転換した大腸菌細胞を、100μg/mLのアンピシリンを含むLB固体培地で37℃にて培養し、生育した単一コロニーをピックアップして100μg/mLのアンピシリンを含むLB培養液2mL中へ移し、37℃で一晩培養して増殖させた。
162番目のロイシン(L)のトレオニン(T)への置換は、定法に従い、塩基配列のシークエンスを行い、変異の導入されたプラスミドを、KillerRed_L162T/pRSETBと命名した。
【0049】
(2)164番目のフェニルアラニン(F)のトレオニン(T)への置換
KillerRed_L162T/pRSETBの164番目のフェニルアラニン(F)をトレオニン(T)に置換するため、以下の変異導入用のプライマーを合成した。
Primer−F164T:5’-CAGACCGCCACCATCGGCTTC-3’(配列番号12)
164番目のフェニルアラニン(F)をトレオニン(T)への置換は、鋳型プラスミドDNAとして、KillerRed_L162T/pRSETBを用いること、及びプライマーとしてPrimer−F164Tを用いることを除いては、前記(1)の操作を繰り返し、164番目のフェニルアラニン(F)をトレオニン(T)に置換されたタンパク質をコードするDNAを含むプラスミドKillerRed_L162T_F164T/pRSETBを得た。
【0050】
(3)KillerRed_L162T_F164Tの発現
得られたプラスミドKillerRed_L162T_F164T/pRSETBタンパク質は、T7発現系(pRSETB/JM109(DE3))を用いて発現させた。すなわち、大腸菌(E.coli)コンピテントセル(JM109(DE3))をKillerRed_L162T_F164T/pRSETBで形質転換し、100μg/mLのアンピシリンを含むLB固体培地で37℃にて培養した。その後、生育した単一コロニーをピックアップして100μg/mLのアンピシリンを含むLB培養液300mL中へ植菌し、23℃で4日間培養した。得られた細胞をフレンチプレスにより破砕し、得られた上清液から、ニッケルキレートカラム(QIAGEN社製)を用いてポリヒスチジン標識化タンパク質を精製した。更に、この溶出液(100mMイミダゾール、50mM Tris−Cl、pH7.4、300mM NaCl)をPD10脱塩・バッファー交換カラム(GE healthcare Bio−Sciences社製)により、1×PBS(pH7.4)を用いて精製し、タンパク質KillerRed_L162T_F164Tを得た。
【0051】
(4)KillerRed_L162T_F164Tの発色及び単量体化の確認
得られたKillerRed_L162T_F164Tを、pseudo native PAGEにより解析した。KillerRed_L162T_F164Tのバンドは、二量体のKillerRedのバンドと比較すると、低分子量にシフトし、単量体として存在した。
しかしながら、KillerRed_L162T_F164Tは、KillerRedと比較すると、610nmにおける蛍光が減衰した。結果を、表1に示す。
【0052】
(5)G5V、N147S、L174K、及びM206Tの置換の導入
G5V、N147S、L174K、及びM206Tの置換を、KillerRed_L162T/F164Tに導入するため、以下のプライマーを合成した。
Primer−G5V:5’-GGTTCAGAGGTCGGCCCCGCC-3’(配列番号13)
Primer−N147S:5’-CAGACCGCCACCATCGGCTTC-3’(配列番号14)
Primer−L174K:5’-GACGGCGGCAAGATGATGGGC-3’(配列番号15)
Primer−M206T:5’-ACCAAGCAGACGAGGGACACT-3’(配列番号16)
【0053】
5番目のグリシン(G)のバリン(V)への置換は、鋳型プラスミドDNAとして、KillerRed_L162T_F164T/pRSETBを用いること、及びプライマーとしてPrimer−G5Vを用いることを除いては、前記(1)の操作を繰り返し、5番目のグリシン(G)のバリン(V)に置換されたタンパク質をコードするDNAを含むプラスミドKillerRed_L162T_F164T_G5V/pRSETBを得た。
以下、同様の操作により、順次変異を導入し、KillerRedのアミノ酸配列において、147番目のアスパラギン(N)をセリン(S)に、174番目のロイシン(L)をリシン(K)に、そして206番目のメチオニン(M)をトレオニン(T)に置換したタンパク質をコードするDNAを含むプラスミドSupernova−Red/pRSETBを得た。
【0054】
(6)Supernova−Redの発現
Supernova−Redタンパク質は、プラスミドとしてSupernova−Red/pRSETBを用いたことを除いては、前記(3)の操作を繰り返し、精製タンパク質Supernova−Redを得た。
【0055】
《比較例1〜21》
比較例1〜21では、二量体のKillerRedを、単量体にするための変異の導入を試みた。具体的には、各比較例では以下のアミノ酸の置換を行った。
比較例1:126番目のアスパラギンをアルギニン(N126R)に置換した。
比較例2:128番目のアスパラギン酸をトレオニン(D128T)に置換した。
比較例3:126番目のアスパラギンをアルギニンに置換(N126R)し、128番目のアスパラギン酸をトレオニン(N126R/D128T)に置換した。
比較例4:154番目のヒスチジンをグルタミン酸(H154E)に置換した。
比較例5:165番目のイソロイシンをアルギニン(I165R)に置換した。
比較例6:163番目のアラニンをリシン(A163K/I165R)に置換し、165番目のイソロイシンをアルギニン(N126R/D128T)に置換した。
比較例7:176番目のメチオニンをアスパラギン酸(M176D)に置換した。
比較例8:196番目のプロリンをアラニン(P196A)に置換した。
比較例9:198番目のフェニルアラニンをリシン(F198K)に置換した。
比較例10:140番目のグルタミンをアラニン(Q140A)に置換した。
比較例11:26番目のリシンをグルタミン酸(K26E)に置換した。
比較例12:174番目のロイシンをヒスチジン(L174H)に置換した。
比較例13:174番目のロイシンをアルギニン(L174R)に置換した。
比較例14:227番目のバリンをリシン(V227K)に置換した。
比較例15:230番目のイソロイシンをリシン(I230K)に置換した。
比較例16:234番目のイソロイシンをリシン(I234K)に置換した。
比較例17:152番目のフェニルアラニンをトレオニン(F152T)に置換した。
比較例18:162番目のロイシンをリシン(L162K)に置換した。
比較例19:164番目のフェニルアラニンをリシン(F164K)に置換した。
比較例20:167番目のフェニルアラニンをリシン(F167K)に置換した。
比較例21:167番目のフェニルアラニンをトレオニン(F167T)に置換した。
前記各比較例において、それぞれの置換に対応するプライマーを合成し、それぞれのプライマーを用いたことを除いては、前記実施例1の工程(1)の操作を繰り返すことにより、それぞれの変異を有するタンパク質をコードするDNAを含むプラスミドを得た。
得られたプラスミドを用いることを除いては、前記実施例の工程(3)及び(4)の操作を繰り返すことにより、それぞれのタンパク質の発色及び単量体化を確認した。比較例1〜21では、得られたタンパク質は、二量体のままで、単量体化しなかった。結果を纏めて、表1に示す。
【0056】
【表1】

【0057】
《実施例2》
本実施例では、前記タンパク質Supernova−Redの68番目のチロシン(Y)をトリプトファン(W)に置換した。
68番目のチロシン(Y)をトリプトファン(W)に置換するため、以下の変異導入用のプライマーを合成した。
Primer−Y68W:5’-GCCACCTGATCCAGTGGGGCGAGCCC-3’(配列番号17)
鋳型プラスミドDNAとして、Supernova−Red/pRSETBを用いること、及びプライマーとしてPrimer−Y68Wを用いることを除いては、前記実施例1の工程(1)の操作を繰り返し、プラスミドSupernova−Orange/pRSETBを得た。
更に、プラスミドSupernova−Orange/pRSETBを用いることを除いては、前記実施例1の工程(3)の操作を繰り返し、タンパク質Supernova−Orangeを得た。
【0058】
《実施例3》
本実施例では、前記タンパク質Supernova−Orangeの46番目のバリン(V)をアラニン(A)に置換した。
46番目のバリン(V)をアラニン(A)に置換するため、以下の変異導入用のプライマーを合成した。
Primer−V46A:5’-GGCGACTTCAACGCCCACGCCGTG-3’(配列番号18)
鋳型プラスミドDNAとして、Supernova−Orange/pRSETBを用いること、及びプライマーとしてPrimer−V46Aを用いることを除いては、前記実施例1の工程(1)の操作を繰り返し、プラスミドSupernova−Green/pRSETBを得た。
更に、プラスミドSupernova−Green/pRSETBを用いることを除いては、前記実施例1の工程(3)の操作を繰り返し、タンパク質Supernova−Greenを得た。
【0059】
《実施例4》
本実施例では、タンパク質KillerRed_L162T_F164Tに、更にN147Sの置換を導入し、KillerRed_L162T_F164T_N147Sの作製を試みた。
鋳型プラスミドDNAとして、KillerRed_L162T_F164T/pRSETBを用いること、及びプライマーとしてPrimer−V46Aを用いることを除いては、前記実施例1の工程(1)の操作を繰り返し、プラスミドKillerRed_L162T_F164T_V46A/pRSETBを得た。
更に、プラスミドKillerRed_L162T_F164T_V46A/pRSETBを用いることを除いては、前記実施例1の工程(3)の操作を繰り返し、タンパク質KillerRed_L162T_F164T_N147S(配列番号9)を得た。
【0060】
《実施例5》
本実施例では、前記タンパク質Supernova−Redの46番目のバリン(V)をアラニン(A)に置換した。
鋳型プラスミドDNAとして、Supernova−Red/pRSETBを用いること、及びプライマーとしてPrimer−V46Aを用いることを除いては、前記実施例1の工程(1)の操作を繰り返し、プラスミドSupernova−Red_V46A/pRSETBを得た。更に、プラスミドSupernova−Red_V46A/pRSETBを用いることを除いては、前記実施例1の工程(3)の操作を繰り返し、タンパク質Supernova−Red_V46A(配列番号10)を得た。
【0061】
《比較例22〜27》
比較例22〜27では、Supernova−Redの発色団の発色を変化させるための変異の導入を試みた。具体的には、各比較例では、L162T/F164Tの置換に、更に以下のアミノ酸の置換を行った。
比較例1:150番目のヒスチジンをトレオニン(H150T)に置換した。
比較例2:148番目のグルタミン酸をトレオニン(E148T)に置換した。
比較例3:148番目のグルタミン酸をリシン(E148K)に置換した。
比較例4:148番目のグルタミン酸をバリン(E148V)に置換した。
比較例5:226番目のセリンをイソロイシン(S226I)に置換した。
比較例6:148番目のグルタミン酸をバリン(E148V)に置換し、226番目のセリンをイソロイシン(S226I)に置換した。
前記各比較例において、それぞれの置換に対応するプライマーを合成し、それぞれのプライマーを用いたことを除いては、前記実施例1の工程(5)の操作を繰り返すことにより、それぞれの変異を有するタンパク質をコードするDNAを含むプラスミドを得た。
得られたプラスミドを用いることを除いては、前記実施例の工程(3)及び(4)の操作を繰り返すことにより、それぞれのタンパク質の発色及び単量体化を確認した。比較例22〜27で得られたタンパク質は、赤色からの発色のシフトは見られず、また、発色の増強も見られなかった。結果を纏めて表2に示す。
【0062】
【表2】
【0063】
《実施例6》
得られたタンパク質が所望の突然変異を導入したものであることを、該タンパク質の励起波長及び発光波長を測定するとともに既存のタンパク質(KillerRed等)の励起波長及び発光波長と比較することにより確認した。
得られたタンパク質の励起波長を測定し、規格化したものを図1(A)中に曲線で示す。また、得られたタンパク質の蛍光波長を測定し、正規化したものを図1(B)中曲線で示す。なお、図1(A)及び(B)には、比較のためにKillerRed、Supernova−Redについて励起波長及び蛍光波長を併せて示す。図1(A)及び(B)に示すように、上述した方法で変異を導入したタンパク質Supernova−Orangeは励起波長のピークが455nm、蛍光波長のピークが544nmであり、Supernova−Greenは励起波長のピークが439nm、蛍光波長のピークが469nmであることがわかる。これに対してKillerRedの励起波長のピークは585nmであり、蛍光波長のピークは610nm、またSupernova−Redの励起波長のピークは579nmであり、蛍光波長のピークは610nmである。
なお、この表3において、λabsはナノメーター単位での吸光度スペクトルのピークを表している。(ε)は103M−1cm−1単位でのモル吸光係数である。λemはナノメーター単位での蛍光スペクトルのピークを表している。
【0064】
更に、実施例4で得られたKillerRed_L162T_F164T_N147S及び、実施例5で得られたSupernova−Red_V46Aについても、励起波長及び、蛍光波長を測定した。KillerRed_L162T_F164T_N147Sは、励起波長のピークが512nm、そして蛍光が黄緑色であり、Supernova−Red_V46Aは励起波長のピークが509nm、蛍光波長のピークが594nmであった。
【0065】
《実施例7》
また、KillerRed、Supernova−Red、Supernova−Orange及びSupernova−Greenについて活性酸素量の測定を行った。ここではまず、10μM anthracene−9,10−dipropionic acid(以下ADPAと記す)を含む50μSupernova−Red、Supernova−Red、Supernova−Orange及びSupernova−Greenタンパク質溶液をそれぞれ100μLずつ調製した。ADPAは励起極大が380nm、蛍光極大が430nmの一重項酸素検出用試薬である。一重項酸素とは活性酸素の一種で、大変強力な酸化能力を有しており、ADPAが一重項酸素と反応することでADPAは過酸化物へと変化し、430nmにある蛍光ピークが失われる。
得られたサンプル溶液を以下の測定条件に従って、活性酸素測定を行った。すなわち、15μLをテラサキディッシュに添加し、YAGレーザー(LOTIS社、LS−2137U−YAG、周波数10Hz、パルス幅15ns)の第三高調波(355nm)をオプティカルパラメトリック発振器(LOTIS社、LT−2214−OPO)を用いて波長変換し、20mJ/cm2/パルスのエネルギー密度で5分間照射した後、これを1mLの1×PBSで希釈し、分光蛍光光度計F−2500(日立)を用いてADPAの退色率を測定した。レーザーの波長はそれぞれKillerRedとSupernova−Redが580nm、Supernova−Orangeが450nm、Supernova−Greenが440nmである。上述した突然変異を導入した蛍光タンパク質を含む4つの蛍光タンパク質の活性酸素量の比較を図2に示す。図2においてKR各種光増感性蛍光タンパク質によるADPA褪色率、すなわち活性酸素放出量を示している。なお、図2では比較のためフルオレセインイソチオシアネート(FITC)の活性酸素量で規格化した。
【0066】
【表3】
【0067】
《実施例8》
本実施例では、実施例1において得られたSupernova−Redを、チューブリン、ケラチン、又はCx43との融合タンパク質として発現させた。前記Supernova−Red/pRSETBに、PCRで得られたチューブリン、ケラチン、又はCx43をコードするDNAを挿入した。得られたプラスミドを、それぞれtublin−Supernova−Red/pRSETB、keratin−Supernova−Red/pRSETB、及びCx43S−upernova−Red/pRSETBと称する。
得られたプラスミドtublin−Supernova−Red/pRSETB、keratin−Supernova−Red/pRSETB、及びCx43−Supernova−Red/pRSETBを、HeLa細胞にトランスフェクションし、細胞内局在を蛍光顕微鏡で調べた。
チューブリン、ケラチン、又はCx43、それぞれ、細胞の微小管、中間系フィラメント、及びギャップジャンクションに局在しており、それぞれのタンパク質が存在している位置で発現しており、タンパク質の局在化に影響を与えなかった。
【0068】
《実施例9》
本実施例では、前記Supernova−Redにミトコンドリアマトリクス移行シグナルをつなげることで細胞内のミトコンドリアに導入し、光照射によってミトコンドリア機能を不活性化することによりアポトーシスを誘導した。
1μgのプラスミド_Supernova−Red/pcDNA3、を、トランスフェクション試薬(Superfect QIAGEN社)を用いて、35mmガラスボトム皿上で培養しているHela細胞に導入した。2日間培養後、580nmの光を8W/cm2のパワー密度で90秒間照射した後、0時間、3時間、及び6時間の細胞を微分干渉顕微鏡を用いて観察した。Supernova−Redが、ミトコンドリアに発現している細胞は、6時間後には退縮し、死滅した。
【産業上の利用可能性】
【0069】
本発明の光増感性蛍光タンパク質又はその機能的等価改変体は、発色団補助光不活性化法に用いることが可能であり、標的分子の機能解析に用いることができる。
【技術分野】
【0001】
本発明は、光増感性蛍光タンパク質に関する。本発明によれば、発色団補助光不活性化法において、標的タンパク質を効率よく破壊することができる。
【背景技術】
【0002】
生体試料内の任意のタンパク質の機能を調べる方法として、標的タンパク質をコードする遺伝子を破壊する、いわゆる遺伝子ノックアウト法、標的タンパク質をコードするmRNAに結合して分解するRNA干渉法、又は標的タンパク質に対する抗体を作用させることで機能不活性化を行う方法などが知られている。これらの方法はいずれも標的タンパク質が発現している細胞内の全ての領域の標的タンパク質に効果が及ぶことを特徴とする。従って、細胞内の任意の位置に存在する標的タンパク質だけを選択的に不活性化する事は、原理上不可能であった。
【0003】
一方、標的タンパク質の機能部位を光照射依存的に産生された活性酸素による酸化によって時空間的に不活性化し、そのタンパク質の機能解析を行う方法として、発色団補助光不活性化法(chromophore-assisted light inactivation;以下、CALIと称することがある)が知られている。具体的には、発色団補助光不活性化法はマラカイトグリーン、ローダミン誘導体、又はフルオレセイン誘導体などを光増感剤として用い、標的分子(例えば、標的タンパク質)に結合させ、標的分子の機能を抑制することによって、標的分子の機能を解析する方法である。例えば、フルオレセイン誘導体を用いた場合、フルオレセイン誘導体から、発生する一重項酸素(活性酸素)が標的タンパク質の機能を破壊することが知られている。
しかしながら、発色団補助光不活性化法は、通常フルオレセイン標識した標的分子に対する抗体又はフルオレセイン標識した標的分子(例えば、標的タンパク質)をマイクロインジェクション等で細胞に導入する必要があり、操作が煩雑であるという問題点があった。
【0004】
最近、Lukyanovらは、発色団補助光不活性化法に用いることのできる光増感性蛍光タンパク質(CALI色素)である、「KillerRed」を報告した(特許文献1及び非特許文献1)。KillerRedはタンパク質であるため、KillerRedをコードするDNAを含むプラスミドなどを用いることによって、形質転換又はトランスフェクションなどの手法により、光増感性蛍光タンパク質(CALI色素)を細胞内において発現させることが可能になった。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】国際公開第2006/117694号公報
【非特許文献】
【0006】
【非特許文献1】「ネイチャー・バイオテクノロジー(Nature Biotechnology)」(米国)2006年、第24巻、p.95〜99
【非特許文献2】「フェブス・レター(FEBS letter)」(オランダ)2009年、第583巻、p.2839−2842
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、前記KillerRedは、細胞内において二量体を形成するため、多くのタンパク質はKillerRedと融合しただけで、立体障害などによる機能の不活性化が起こり、光照射依存的な標的タンパク質の機能不活性化を行うことができないという問題を有していた。
また、KillerRedは、光増感活性が低いため、活性酸素の産生量がフルオレセインよりも少ないという問題を有していた。
更に、例えば、生体試料内の2つ以上の標的タンパク質を、個別に光照射によって不活性化する場合には、吸光特性の異なる光増感性蛍光タンパク質が必要である。しかしながら、KillerRed以外の吸光スペクトルを有する光増感性蛍光タンパク質は報告されておらず、別の励起波長及び蛍光波長を有する光増感性蛍光タンパク質の開発が望まれていた。
従って、本発明の目的は、単量体型の光増感性蛍光タンパク質を提供することである。また、本発明の別の目的は、光増感活性が高い光増感性蛍光タンパク質を提供することである。更に、本発明の別の目的は、KillerRedとは異なる、励起波長及び蛍光波長を有する光増感性蛍光タンパク質を提供することである。
【0008】
本発明者は、生物個体内、組織内、細胞内、細胞内小器官内、又は水溶液中において、光刺激により活性酸素を効率良く産生することのできる光増感活性の高い光増感性蛍光タンパク質について、鋭意研究した結果、驚くべきことに、KillerRedのアミノ酸配列において、5番目のグリシン(G)をバリン(V)に、147番目のアスパラギン(N)をセリン(S)に、162番目のロイシン(L)をトレオニン(T)に、164番目のフェニルアラニン(F)をトレオニン(T)に、174番目のロイシン(L)をリシン(K)に、そして206番目のメチオニン(M)をトレオニン(T)に置換した変異体であるSupernova−Redは、単量体として細胞内などに存在するようになることを見出した。更に、前記Supernova−Redの68番目のチロシン(Y)をトリプトファン(W)に置換した変異体であるSupernova−Orangeは、一重項酸素の産生量が増加し、励起極大波長及び蛍光極大波長が、それぞれ455nm及び544nmにシフトすることを見出した。
更に、Supernova−Orangeの46番のバリン(V)をアラニン(A)に置換した変異体であるSupernova−Greenは、一重項酸素の産生量がKillerRedと比較して増加し、励起極大波長及び蛍光極大波長が、それぞれ439nm及び496nmにシフトすることを見出した。
本発明は、こうした知見に基づくものである。
【課題を解決するための手段】
【0009】
従って、本発明は、配列表の配列番号2で表されるアミノ酸配列、配列番号4で表されるアミノ酸配列、配列番号6で表されるアミノ酸配列、配列番号9で表されるアミノ酸配列、及び配列番号10で表されるアミノ酸配列、からなる群から選択される少なくとも1つのアミノ酸配列を含む光増感性蛍光タンパク質又はその機能的等価改変体に関する。
本発明の光増感性蛍光タンパク質又はその機能的等価改変体の好ましい態様においては、前記光増感性蛍光タンパク質又はその機能的等価改変体が、他のタンパク質と結合した融合タンパク質である。
また、本発明は、前記光増感性蛍光タンパク質又はその機能的等価改変体をコードするDNAに関する。
本発明のDNAの好ましい態様においては、配列表の配列番号1で表される塩基配列、配列番号3で表される塩基配列、及び配列番号5で表される塩基配列からなる群から選択される少なくとも1つの塩基配列からなるDNAである。
更に、本発明は、前記DNAを含むプラスミドに関する。
更に、本発明は、前記光増感性蛍光タンパク質又はその機能的等価改変体を用いた、光増感性の活性酸素産生方法であって、前記光増感性蛍光タンパク質又はその機能的等価改変体に、励起光を照射することを特徴とする、前記方法に関する。
本発明の光増感性の活性酸素産生方法の好ましい態様においては、前記励起光の波長が、490〜580nm、400〜500nm、及び390〜440nmからなる群から選択される少なくとも1つの波長である。
更に、本発明は、光増感性蛍光タンパク質又はその機能的等価改変体を用いた、標的分子の機能解析方法であって、前記光増感性蛍光タンパク質又はその機能的等価改変体に、励起光を照射することを特徴とする、前記方法に関する。
本発明の標的分子の機能解析方法の好ましい態様においては、前記励起光の波長が、490〜580nm、400〜500nm、及び390〜440nmからなる群から選択される少なくとも1つの波長である。
【発明の効果】
【0010】
本発明の光増感性蛍光タンパク質又はその機能的等価改変体によれば、単量体として細胞内などに存在することが可能であり、多くのタンパク質に対してCALIにおける光増感性蛍光タンパク質として用いることが可能である。
本発明の光増感性蛍光タンパク質及びその機能的等価改変体、特にはSupernova−Orangeによれば、Supernova−Redの68番目のチロシンをトリプトファンに置換(Y68W)することで、励起極大/蛍光極大を455nm/544nm(目視ではオレンジ色)にシフトし、更に、一重項酸素の産生量がKillerRedと比較して増加する。従って、CALIにおいて、増強型光増感性蛍光タンパク質として用いることが可能であり、更にKillerRed、Supernova−Red及びSupernova−Greenなどと同時に用いることにより、2種以上の標的タンパク質の機能相関などを解析することが可能である。
本発明の光増感性蛍光タンパク質及びその機能的等価改変体、特にはSupernova−Greenによれば、Supernova−Orangeの46番のバリンをアラニンに置換することで、励起極大/蛍光極大を439nm/496nm(目視では緑色)にシフトし、一重項酸素の産生量がKillerRedと比較して増加する。従って、CALIにおいて、増強型光増感性蛍光タンパク質として用いることが可能であり、更にKillerRed、Supernova−Red、及びSupernova−Orangeなどと同時に用いることにより、2種以上の標的タンパク質の機能相関などを解析することが可能である。
なお、KillerRedのX線結晶構造解析を行った論文(非特許文献2)によれば、KillerRedの46番のバリンをアラニンに置換(V46A)することは、発色団の光増感作用により発生した活性酸素をKillerRedタンパク質の外部に効率良く放出するようになる可能性が記載されている。
しかしながら、後述の実施例に示すように、Supernova−RedにV46A変異を導入すると、活性酸素の放出量は減少した。一方、Y68W変異が導入されているSupernova−OrangeにV46A変異を導入したSupernova−Greenでは、活性酸素の放出量が2倍以上に増大し、更に蛍光波長がグリーンに変化した。従って、光増感性蛍光タンパク質において、変異の導入により、活性酸素の放出量が増加したり、又は減少したりすることを予想すること、また蛍光波長が変化することは当業者であっても決して容易ではなく、試行錯誤を要するものである。
【図面の簡単な説明】
【0011】
【図1】KillerRed、Supernova−Red、Supernova−Orange、及びSupernova−Greenの励起スペクトル(A)及び蛍光スペクトル(B)を示したグラフである。
【図2】KillerRed、Supernova−Red、Supernova−Orange、Supernova−Green、及びSupernova−Red−V46Aの活性酸素の産生量を示したグラフである。FITC(フルオレセインイソチアシアネート)は陽性コントロールを示し、Buffer(1xPBS)は陰性コントロールを示している。
【図3】Supernova−RedをHela細胞に導入し、光照射を行い、0時間、3時間、及び6時間後の細胞の微分干渉画像を示した写真である。Supernova−Redがミトコンドリアに発現している細胞は、6時間後にアポトーシスにより、死滅している。
【発明を実施するための形態】
【0012】
本発明の光増感性蛍光タンパク質は、配列番号2で表されるアミノ酸配列、配列番号4で表されるアミノ酸配列、配列番号6で表されるアミノ酸配列、配列番号9で表されるアミノ酸配列、及び配列番号10で表されるアミノ酸配列からなる群から選択される少なくとも1つのアミノ酸配列を含むタンパク質である。
【0013】
本発明の機能的等価改変体は、前記光増感性蛍光タンパク質の機能的等価改変体である。本明細書において、「機能的等価改変体」とは、そのアミノ酸配列が、元となるタンパク質のアミノ酸配列において1以上(特には1又は数個)のアミノ酸が欠失、置換、又は付加されたアミノ酸配列であって、しかも、元となるタンパク質と実質的に同じ活性を示すタンパク質を意味する。前記アミノ酸の欠失、置換、又は付加は、例えば10個であり、好ましくは1〜10個、より好ましくは1〜5個、更に好ましくは1〜2個である。
【0014】
本発明の光増感性蛍光タンパク質の第1の実施態様は、配列番号2で表されるアミノ酸配列を含むタンパク質(以下、第1光増感性蛍光タンパク質と称する)又はその機能的等価改変体(以下、第1機能的等価改変体と称する;以下、第1光増感性蛍光タンパク質及び第1機能的等価改変体を、まとめて第1光増感性蛍光タンパク質等と称することがある)であり、配列番号2で表されるアミノ酸配列からなるタンパク質又はその機能的等価改変体であることが好ましい。
第1光増感性蛍光タンパク質は、配列番号8で表されるアミノ酸配列からなる光増感性蛍光タンパク質(すなわち、KillerRed)のアミノ酸配列において、5番目のグリシン(G)をバリン(V)に置換(以下、G5Vと称することがある)、147番目のアスパラギン(N)をセリン(S)に置換(以下、N147Sと称することがある)、162番目のロイシン(L)をトレオニン(T)に置換(以下、L162Tと称することがある)、164番目のフェニルアラニン(F)をトレオニン(T)に置換(以下、F164Tと称することがある)、174番目のロイシン(L)をリシン(K)に置換(以下、L174Kと称することがある)、そして206番目のメチオニン(M)をトレオニン(T)に置換(以下、M206Tと称することがある)したアミノ酸配列を含む光増感性蛍光タンパク質である。代表的な第1光増感性蛍光タンパク質は、配列番号2で表されるアミノ酸配列からなるタンパク質であるSupernova−Redである。
KillerRedは、二量体を形成することが知られているが、これに対してSupernova−Redは、試料中(例えば、生物個体内、組織内、細胞内、細胞内小器官内、又は水溶液中)において、単量体で存在する。また、Supernova−Redの励起極大波長及び蛍光極大波長は、それぞれ579nm及び610nmである。
【0015】
本発明の第1光増感性蛍光タンパク質等において、前記L162T及びF164Tの置換(以下、L162T、及びF164Tの置換を有するKillerRedの変異体を、KillerRed−L162T/F164Tと称する)により、二量体であったKillerRedが、単量体となる。従って、前記KillerRed−L162T/F164Tは、標的タンパク質との融合タンパク質として発現しても、標的タンパク質の機能を阻害しない。一方、L162T及びF164Tの置換により、KillerRed−L162T/F164Tは、KillerRedと比較して585nmにおける吸光及び610nmにおける蛍光が減少した。
しかしながら、前記KillerRed−L162T/F164Tに、更にG5V、N147S、L174K、及びM206Tの置換を導入することにより、KillerRedと同等の蛍光強度を有するSupernova−Redを得ることができた。
【0016】
本発明の第1機能的等価改変体は、実質的にSupernova−Redと同じ活性を有するものである。「Supernova−Redと同じ活性」とは、光照射により活性酸素を産生すること、光照射により蛍光を発生すること、試料中において単量体で存在すること、そして励起極大波長及び蛍光極大波長がそれぞれ約579nm及び610nmであることを意味する。なお、励起極大波長及び蛍光極大波長がそれぞれ約579nm及び610nmであるとは、好ましくは569〜589nm及び600〜620nmであり、より好ましくは574〜584nm及び605〜615nmである。第1機能的等価改変体としては、例えば、前記配列番号2で表されるアミノ酸配列において、第2番目のグリシン及び第3番目のセリンを欠如したタンパク質を挙げることができる。
【0017】
本発明の光増感性蛍光タンパク質の第2の実施態様は、配列番号4で表されるアミノ酸配列を含むタンパク質(以下、第2光増感性蛍光タンパク質と称する)又はその機能的等価改変体(以下、第2機能的等価改変体と称する;以下、第2光増感性蛍光タンパク質及び第2機能的等価改変体を、まとめて第2光増感性蛍光タンパク質等と称することがある)であり、配列番号4で表されるアミノ酸配列からなるタンパク質又はその機能的等価改変体であることが好ましい。第2光増感性蛍光タンパク質は、前記Supernova−Redのアミノ酸配列において、68番目のチロシン(Y)をトリプトファン(W)に置換(以下、Y68Wと称する)したアミノ酸配列を含む光増感性蛍光タンパク質である。代表的な第2光増感性蛍光タンパク質は、配列番号4で表されるアミノ酸配列からなるタンパク質であるSupernova−Orangeである。
Supernova−Orangeの励起極大波長及び蛍光極大波長は、それぞれ455nm及び544nmであり、KillerRedと比較して、一重項酸素の産生量が増加している。
【0018】
本発明の第2光増感性蛍光タンパク質等において、前記L162T及びF164Tの置換により、二量体であったKillerRedが、単量体となり、Y68Wの置換により、励起極大波長及び蛍光極大波長が、それぞれおよそ455nm及び544nmとなる。
本発明の第2機能的等価改変体は、実質的にSupernova−Orangeと同じ活性を有するものである。「Supernova−Orangeと同じ活性」とは、光照射により活性酸素を産生すること、光照射により蛍光を発生すること、試料中において単量体で存在すること、そして励起極大波長及び蛍光極大波長がそれぞれ約455nm及び544nmであることを意味する。なお、励起極大波長及び蛍光極大波長がそれぞれ約455nm及び544nmであるとは、好ましくは445〜465nm及び534〜554nmであり、より好ましくは450〜460nm及び539〜549nmである。第2機能的等価改変体としては、例えば、前記配列番号8で表されるアミノ酸配列において、L162T、F164T、及びY68Wの置換が導入されたタンパク質、及び前記タンパク質において、第2番目のグリシン及び第3番目のセリンを欠如したタンパク質、並びに前記配列番号4で表されるアミノ酸配列において、第2番目のグリシン及び第3番目のセリンを欠如したタンパク質を挙げることができる。
【0019】
本発明の光増感性蛍光タンパク質の第3の実施態様は、配列番号6で表されるアミノ酸配列を含むタンパク質(以下、第3光増感性蛍光タンパク質と称する)又はその機能的等価改変体(以下、第3機能的等価改変体と称する;以下、第3光増感性蛍光タンパク質及び第3機能的等価改変体を、まとめて第3光増感性蛍光タンパク質等と称することがある)であり、配列番号6で表されるアミノ酸配列からなるタンパク質又はその機能的等価改変体であることが好ましい。第3光増感性蛍光タンパク質は、前記Supernova−Orangeのアミノ酸配列において、46番のバリン(V)をアラニン(A)に置換(以下、V46Aと称する)したアミノ酸配列を含む光増感性蛍光タンパク質である。代表的な第3光増感性蛍光タンパク質は、配列番号6で表されるアミノ酸配列からなるタンパク質である、Supernova−Greenである。
Supernova−Greenの励起極大波長及び蛍光極大波長は、それぞれ439nm及び496nmであり、KillerRedと比較して、一重項酸素の産生量が増加している。
【0020】
本発明の第3光増感性蛍光タンパク質等において、前記L162T及びF164Tの置換により二量体であったKillerRedが単量体となり、Y68W及びV46Aの置換により、励起極大波長及び蛍光極大波長が、それぞれ439nm及び496nmとなる。
本発明の第3機能的等価改変体は、実質的にSupernova−Greenと同じ活性を有するものである。「Supernova−Greenと同じ活性」とは、光照射により活性酸素を産生すること、光照射により蛍光を発生すること、試料中において単量体で存在すること、そして励起極大波長及び蛍光極大波長がそれぞれ約439nm及び496nmであることを意味する。なお励起極大波長及び蛍光極大波長がそれぞれ約439nm及び496nmであるとは、好ましくは429〜449nm及び486〜506nmであり、より好ましくは434〜444nm及び591〜501nmである。第3機能的等価改変体としては、例えば、前記配列番号8で表されるアミノ酸配列において、L162T、F164T、Y68W及びV46Aの置換が導入されたタンパク質、及び前記タンパク質において、第2番目のグリシン及び第3番目のセリンを欠如したタンパク質、並びに前記配列番号6で表されるアミノ酸配列において、第2番目のグリシン及び第3番目のセリンを欠如したタンパク質を挙げることができる。
【0021】
本発明の光増感性蛍光タンパク質の第4の実施態様は、配列番号9で表されるアミノ酸配列を含むタンパク質(以下、第4タンパク質と称する)又はその機能的等価改変体(以下、第4機能的等価改変体と称する;以下、第4光増感性蛍光タンパク質及び第4機能的等価改変体を、まとめて第4光増感性蛍光タンパク質等と称することがある)であり、配列番号9で表されるアミノ酸配列からなるタンパク質又はその機能的等価改変体であることが好ましい。
第4光増感性蛍光タンパク質は、配列番号8で表されるアミノ酸配列からなる光増感性蛍光タンパク質(すなわち、KillerRed)のアミノ酸配列において、L162T、F164T、及びN147Sの置換を有するアミノ酸配列を含む光増感性蛍光タンパク質である。代表的な第4光増感性蛍光タンパク質は、配列番号9で表されるアミノ酸配列からなるタンパク質であるKillerRed_L162T_F164T_N147Sである。
KillerRed_L162T_F164T_N147Sの励起極大波長は、512nmであり、蛍光は黄緑色(目視)を示す。
【0022】
本発明の第4光増感性蛍光タンパク質等において、前記L162T及びF164Tの置換により二量体であったKillerRedが単量体となり、N147Sの置換により、励起極大波長が512nmで、蛍光が黄緑色となる。
【0023】
本発明の第4機能的等価改変体は、実質的にKillerRed_L162T_F164T_N147Sと同じ活性を有するものである。「KillerRed_L162T_F164T_N147Sと同じ活性」とは、光照射により活性酸素を産生すること、光照射により蛍光を発生すること、試料中において単量体で存在すること、そして励起極大波長が約512nm及び蛍光が黄緑色であることを意味する。なお、励起極大波長が約512nmであるとは、好ましくは502〜522nmであり、より好ましくは507〜517nmである。第4機能的等価改変体としては、例えば、前記配列番号9で表されるアミノ酸配列において、第2番目のグリシン及び第3番目のセリンを欠如したタンパク質を挙げることができる。
【0024】
本発明の光増感性蛍光タンパク質の第5の実施態様は、配列番号10で表されるアミノ酸配列を含むタンパク質(以下、第5タンパク質と称する)又はその機能的等価改変体(以下、第5機能的等価改変体と称する;以下、第5光増感性蛍光タンパク質及び第5機能的等価改変体を、まとめて第5光増感性蛍光タンパク質等と称することがある)であり、配列番号10で表されるアミノ酸配列からなるタンパク質又はその機能的等価改変体であることが好ましい。
第5光増感性蛍光タンパク質は、配列番号8で表されるアミノ酸配列からなる光増感性蛍光タンパク質(すなわち、KillerRed)のアミノ酸配列において、L162TF164T、G5V、N147S、L174K、M206T、及びV46Aの置換を有するアミノ酸配列を含む光増感性蛍光タンパク質である。代表的な第5光増感性蛍光タンパク質は、配列番号10で表されるアミノ酸配列からなるタンパク質であるSupernova−Red_V46Aである。
Supernova−Red_V46Aの励起極大波長及び蛍光極大波長は、それぞれ509nm及び594nmである。
【0025】
本発明の第5光増感性蛍光タンパク質等において、前記L162T及びF164Tの置換により二量体であったKillerRedが単量体となり、G5V、N147S、L174K、M206T、及びV46Aの置換により、励起極大波長及び蛍光極大波長が、それぞれ509nm及び594nmとなる。
【0026】
本発明の第5機能的等価改変体は、実質的にSupernova−Red_V46Aと同じ活性を有するものである。「Supernova−Red_V46Aと同じ活性」とは、光照射により活性酸素を産生すること、光照射により蛍光を発生すること、試料中において単量体で存在すること、そして励起極大波長及び蛍光極大波長がそれぞれ約509nm及び594nmであることを意味する。なお、励起極大波長及び蛍光極大波長がそれぞれ約509nm及び594nmであるとは、好ましくは499〜519nm及び584〜604nmであり、より好ましくは504〜514nm及び589〜599nmである。第5機能的等価改変体としては、例えば、前記配列番号10で表されるアミノ酸配列において、第2番目のグリシン及び第3番目のセリンを欠如したタンパク質を挙げることができる。
【0027】
本発明の光増感性蛍光タンパク質及び機能的等価改変体(以下、まとめて本発明の光増感性蛍光タンパク質等と称することがある)は、光の照射によって、活性酸素(例えば、一重項酸素)を産生することが可能である。従って、本発明のタンパク質等を細胞内に導入し、適当な励起波長の光を照射することによって、一重項酸素を発生させ、細胞に障害を与え、更には細胞を死滅させることもできる。
また、本発明のタンパク質等は、発色団補助光不活性化法に用いることもできる。具体的には本発明の光増感性蛍光タンパク質等を標的タンパク質に結合させ、光の照射によって活性酸素(例えば、一重項酸素)を産生することによって、その標的タンパク質の機能を抑制、又はその標的タンパク質を破壊することによって、標的タンパク質の機能を解析することができる。
【0028】
本発明による光増感性蛍光タンパク質及び機能的等価改変体は、種々の公知の方法によって得ることができ、例えば、光増感性蛍光タンパク質であるKillerRedをコードするDNAを含むプラスミドベクターを用いて公知の遺伝子工学的手法により調製することができる。より具体的には、部位特異的突然変異誘発法(site−directed mutagenesis)により調製することが可能である。
更に得られたプラスミドを用いて、大腸菌などの宿主細胞を形質転換し、形質転換体を得ることができる。得られた形質転換体(すなわち、本発明による光増感性蛍光タンパク質又は機能的等価改変体をコードするDNAを含むプラスミドを有する形質転換体)を、前記光増感性蛍光タンパク質等の発現が可能な条件下で培養し、タンパク質の分離及び精製に一般的に用いられる方法により、その培養物から光増感性蛍光タンパク質等を分離及び精製することにより調製することができる。前記の分離及び精製方法としては、例えば、光増感性蛍光タンパク質にHisタグを付けニッケルNTAカラムで生成する方法、硫安塩析、イオン交換セルロースを用いるイオン交換カラムクロマトグラフィー、分子篩ゲルを用いる分子篩カラムクロマトグラフィー、プロテインA結合多糖類を用いる親和性カラムクロマトグラフィー、透析、又は凍結乾燥等を挙げることができる。
【0029】
本発明の光増感性蛍光タンパク質等は、他のタンパク質と結合した融合タンパク質とすることができる。融合される他のタンパク質は、光増感性蛍光タンパク質等の活性を完全に阻害しない限り、特に制限されるものではなく、例えば、発色団補助光不活性化法における標的タンパク質、又は標的分子に結合させる結合タンパク質を挙げることができる。
【0030】
前記標的タンパク質は、発色団補助光不活性化法において、機能を解析されるタンパク質である。標的タンパク質を本発明の光増感性蛍光タンパク質等との融合タンパク質とした場合、光の照射により光増感性蛍光タンパク質等から産生された活性酸素が、容易に標的タンパク質を不活化することができる。これは、産生された活性酸素により、半径約10〜50Åに存在する標的分子が特異的に不活化されるためである。本発明の光増感性蛍光タンパク質等は、単量体であるため、標的タンパク質の細胞内における本来の局在性や活性を妨害することが少ない。後述の実施例で示すように、Supernova−Redと、標的タンパク質(例えば、チューブリン、ケラチン、又はCx43)とを、融合タンパク質として発現させた場合、標的タンパク質の局在化に影響を与えなかった。
【0031】
前記結合タンパク質は、発色団補助光不活性化法において、標的分子に光増感性蛍光タンパク質を結合させるタンパク質である。結合タンパク質は、標的分子に結合することのできるタンパク質であれば、特に限定されるものではないが、例えば、抗体及びその抗原結合性フラグメント(例えば、scFv、Fab、又は抗体可変領域全長の遺伝子産物)、レセプター、又はリガンドを挙げることができる。
【0032】
前記融合タンパク質は、公知の遺伝子組み換え技術によって、調整することが可能である。具体的には、光増感性蛍光タンパク質等をコードするDNAと、他のタンパク質をコードするDNAとを、定法に従って、発現用プラスミドに導入する。得られたプラスミドを、形質転換などにより、宿主細胞に導入することによって、融合タンパク質を発現させることが可能である。
【0033】
本発明によるDNAは、本発明の光増感性蛍光タンパク質等をコードする限り、特に限定されるものではなく、例えば、配列表の配列番号1、3、又は5で表される塩基配列からなるDNAを挙げることができる。配列表の配列番号1で表される塩基配列からなる前記DNAは、配列表の配列番号2で表されるアミノ酸配列からなるSupernova−Redをコードする。また、配列表の配列番号3で表される塩基配列からなる前記DNAは、配列表の配列番号4で表されるアミノ酸配列からなるSupernova−Orangeをコードする。更に、配列表の配列番号5で表される塩基配列からなる前記DNAは、配列表の配列番号6で表されるアミノ酸配列からなるSupernova−Greenをコードする。
【0034】
本発明によるプラスミドは、本発明による前記DNAを含む限り、特に限定されるものではなく、例えば、用いる宿主細胞に応じて適宜選択した公知の発現ベクターに、本発明による前記DNAを挿入することにより得られるプラスミドを挙げることができる。
得られたプラスミドを所望の宿主細胞に形質転換することにより、形質転換体を得ることができる。前記形質転換体も、本発明による前記プラスミドを含む限り、特に限定されるものではない。
【0035】
前記宿主細胞としては、例えば、通常使用される公知の微生物、例えば、大腸菌又は酵母(Saccharomyces cerevisiae)、あるいは、公知の培養細胞、例えば、動物細胞(例えば、CHO細胞、HEK−293細胞、又はCOS細胞)又は昆虫細胞(例えば、BmN4細胞)を挙げることができる。
【0036】
また、公知の前記発現ベクターとしては、例えば、大腸菌に対しては、pUC、pTV、pGEX、pKK、又はpTrcHisを;酵母に対しては、pEMBLY又はpYES2を;CHO細胞に対してはpcDNA3又はpMAMneoを;HEK−293細胞に対してはpcDNA3を;COS細胞に対してはpcDNA3を;BmN4細胞に対しては、カイコ核多角体ウイルス(BmNPV)のポリヘドリンプロモーターを有するベクター(例えば、pBK283)を挙げることができる。
【0037】
本発明の光増感性の活性酸素産生方法は、本発明の光増感性蛍光タンパク質等を用いる。前記光増感性蛍光タンパク質等に、励起光を照射することにより、光増感性蛍光タンパク質等から活性酸素が産生される。
本発明の光増感性の活性酸素産生方法によれば、例えば、細胞に前記光増感性蛍光タンパク質等を導入し、そして励起光を照射することにより、光増感性蛍光タンパク質等から活性酸素が産生され、細胞を死滅させることができる。
前記励起光の波長は、本発明の光増感性蛍光タンパク質等から活性酸素を産生させることのできる波長である。本発明の第1光増感性蛍光タンパク質等では、前記励起波長は、490〜620nmが好ましく、540〜580nmがより好ましく、560〜580nmが最も好ましい。本発明の第2光増感性蛍光タンパク質等では、前記励起波長は400〜500nmが好ましく、420〜480nmがより好ましく、440〜460nmが最も好ましい。本発明の第3光増感性蛍光タンパク質等では、前記励起波長は390〜480nmが好ましく、390〜460nmがより好ましく、430〜450nmが最も好ましい。
また前記励起光としては、前記励起波長の光のほか、多光子励起現象にもとづき、前記励起波長のおよそ整数倍の波長を有する光を用いることもできる。
【0038】
本発明の光増感性の活性酸素産生方法によれば、細胞のアポトーシスを誘導することが可能である。アポトーシスを誘導することにより、本発明の光増感性の活性酸素産生方法は、光線力学療法(PDT)に用いることができる。光線力学療法とは、光増感性の物質を腫瘍細胞に取り込ませ、光を照射することにより、細胞を死滅させる腫瘍の治療方法である。より具体的には、腫瘍細胞又は腫瘍組織内の新生血管の内皮細胞内に、本発明の光増感性蛍光タンパク質等を導入し、適当な光を照射することにより、活性酸素を発生させる。この活性酸素により、腫瘍細胞又は腫瘍組織が傷害され、腫瘍が消失するものである。
【0039】
本発明の標的分子の機能解析方法は、本発明の光増感性蛍光タンパク質等を用いる。前記光増感性蛍光タンパク質等に、励起光を照射することにより、光増感性蛍光タンパク質等から活性酸素が産生される。産生された活性酸素は近傍の標的分子を破壊し、それにより破壊された標的分子の機能を解析することが可能である。
前記励起光の波長は、本発明の光増感性蛍光タンパク質等から活性酸素を産生させることのできる波長である。本発明の第1光増感性蛍光タンパク質等では、前記励起波長は、490〜620nmが好ましく、540〜580nmがより好ましく、560〜580nmが最も好ましい。本発明の第2光増感性蛍光タンパク質等では、前記励起波長は400〜500nmが好ましく、420〜480nmがより好ましく、440〜460nmが最も好ましい。本発明の第3光増感性蛍光タンパク質等では、前記励起波長は390〜480nmが好ましく、390〜460nmがより好ましく、430〜450nmが最も好ましい。
また前記励起光としては、前記励起波長の光のほか、多光子励起現象にもとづき、前記励起波長のおよそ整数倍の波長を有する光を用いることもできる。
より具体的には本発明の標的分子の機能解析方法は、発色団補助光不活性化法として、用いることができる。
【0040】
本発明の方法において、標的分子とは、その生理的機能を解明する対象となる生体分子をいい、特にこれらに限定されないが、例えば、タンパク質、ペプチド、炭水化物、脂質、DNA、RNA、糖、シグナル伝達物質などが挙げられる。特には、本発明の方法において好ましい標的分子は、タンパク質であり、酵素、受容体タンパク質、リガンドタンパク質、シグナル伝達タンパク質、転写制御タンパク質、骨格タンパク質、細胞接着タンパク質、スキャホールドタンパク質等を挙げることができる。
【0041】
前記光増感性蛍光タンパク質等が産生する活性酸素は、一重項酸素である。前記一重項酸素は、励起状態の光増感性蛍光タンパク質発色団が項間交差によって励起3重項状態に遷移し、発色団近傍に存在する基底状態の3重項酸素にエネルギーを転移することにより発生する。発生した一重項酸素は、約10〜50Åの半径で光増感性蛍光タンパク質等に結合された標的分子(例えば、標的タンパク質)あるいはその機能性部位を選択的に不活化することができる。このようにして、標的分子自体あるいは標的分子の特定部位を不活性化することによって、その標的物質の生理的機能を解析することができる。例えば、このような不活性化によって、タンパク質の機能性部位の同定、その機能性部位の機能の確認、リガンドの機能の確認、機能性部位のタンパク質寿命に及ぼす影響の確認、機能性部位のタンパク質動態に及ぼす影響の確認、機能性部位のタンパク質フォールディングに及ぼす影響の確認などを行うことができる。
なお、本発明の標的分子の機能解析方法は、in vitro及びin vivoアッセイに、また細胞内外の標的分子に利用可能である。
【0042】
本発明の標的分子の機能解析方法において、光増感性蛍光タンパク質等を標的分子(例えば、タンパク質、炭水化物、脂質、DNA、RNA、糖、シグナル伝達物質)に結合させる方法は、特に限定されず、例えば化学結合によることが好ましい。光増感性蛍光タンパク質等を標的分子に共有結合により結合させることができる。また、光増感性蛍光タンパク質等に標的分子に対する抗体を共有結合により結合させ、その抗体により光増感性蛍光タンパク質等を標的分子に結合させることができる。更に、光増感性蛍光タンパク質等と標的分子に対する抗体とを融合タンパク質として発現させ、その抗体により光増感性蛍光タンパク質等を標的分子に結合させることができる。更には、標的分子がタンパク質の場合は、光増感性蛍光タンパク質等と標的タンパク質とを融合タンパク質として発現させることができる。
【実施例】
【0043】
以下、実施例によって本発明を具体的に説明するが、これらは本発明の範囲を限定するものではない。
【0044】
《実施例1》
本実施例では、KillerRedのアミノ酸配列に変異を導入することにより、単量体として存在することのできる光増感蛍光タンパク質の作製を試みた。
具体的には、KillerRedのアミノ酸配列における5番目のグリシン(G)、147番目のアスパラギン(N)、162番目のロイシン(L)、164番目のフェニルアラニン(F)、174番目のロイシン(L)、206番目のメチオニン(M)を、それぞれ、バリン(V)、セリン(S)、トレオニン(T)、トレオニン(T)、リシン(K)、トレオニン(T)に変異させるために、Sawanoらの方法(Sawano A and Miyawaki A. Nucleic Acid Research 28,E78,2000)に従って、部位特異的突然変異誘発法により、順次変異を導入した。
【0045】
(1)162番目のロイシン(L)のトレオニン(T)への置換
162番目のロイシン(L)をトレオニン(T)に置換するため、以下の変異導入用のプライマーを合成した。
Primer−L162T:5’-GTGCGCCAGACCGCCACCATC-3’(配列番号11)
Primer−L162Tの末端は、T4ポリヌクレオチドキナーゼによってリン酸化した。変異を導入する鋳型プラスミドDNAとしては、KillerRed遺伝子を含むプラスミドベクターを鋳型に用いたポリメラーゼ連鎖反応により、KillerRed遺伝子の上流と下流にそれぞれBamHI及びEcoRIを付加してKillerRed遺伝子を増幅した後、制限酵素BamHI及びEcoRIで切り出し、プラプラスミドpRSETB(インビロロジェン社)に挿入して得られたKillerRed/pRSETBを用いた。
【0046】
突然変異鎖の合成は、以下の通り行った。以下の組成の反応溶液40μLを準備し、PCR法によりin vitroで合成した。
KillerRed/pRSETB 50ng、
Primer−L162T 10pmol、
dNTP 3.75nmol、
PfuDNAポリメラーゼ 1.25U、
PfuDNAリガーゼ 20U
なお、PfuDNAポリメラーゼはSTRATAGENE社製であり、10×Pfuポリメラーゼバッファー中に溶解したものである。また、PfuDNAリガーゼは、STRATAGENE社製のものであり、10×PfuDNAリガーゼバッファー中に溶解したものである。
サーマルサイクラーは以下の通りプログラムした。すなわち、まず、65℃、5分のプレインキュベーションを行ってPfuDNAリガーゼによる鋳型DNAのニックを修復し、その後94℃、1分の最初の変性を行った。続いて、94℃、10秒のDNA変性、55℃、30秒のアニーリング反応及び65℃、10分の伸長・連結反応を1サイクルとして35サイクル行った。最後に75℃、10分のポストインキュベーションを行った。
【0047】
得られたPCR産物を、以下の反応条件に従い、DpnIによって切断し、変異の導入されていないメチル化又はヘミメチル化した鋳型プラスミドDNAを選択的に消化した。具体的には、前記PCR後の反応溶液40μLに1μL(20U)のDpnI(New England BioLab社製)を加え、37℃で2時間インキュベートした。なお、DpnIは〔5’−Gm6ATC−3’〕を認識して2本鎖DNAを切断するエンドヌクレアーゼである。更に、DpnIで処理した反応溶液20.4μLに対して、フェノールクロロホルム抽出を行ってサンプルを精製し、上清をエタノール沈殿して乾燥させた。
【0048】
次に、得られたDNAを用いて、形質転換及び発現を行った。前記1本鎖突然変異導入環状DNA鎖を用いて、ヒートショック法により大腸菌(E.coli)コンピテントセル(JM109(DE3))を形質転換した。そして形質転換した大腸菌細胞を、100μg/mLのアンピシリンを含むLB固体培地で37℃にて培養し、生育した単一コロニーをピックアップして100μg/mLのアンピシリンを含むLB培養液2mL中へ移し、37℃で一晩培養して増殖させた。
162番目のロイシン(L)のトレオニン(T)への置換は、定法に従い、塩基配列のシークエンスを行い、変異の導入されたプラスミドを、KillerRed_L162T/pRSETBと命名した。
【0049】
(2)164番目のフェニルアラニン(F)のトレオニン(T)への置換
KillerRed_L162T/pRSETBの164番目のフェニルアラニン(F)をトレオニン(T)に置換するため、以下の変異導入用のプライマーを合成した。
Primer−F164T:5’-CAGACCGCCACCATCGGCTTC-3’(配列番号12)
164番目のフェニルアラニン(F)をトレオニン(T)への置換は、鋳型プラスミドDNAとして、KillerRed_L162T/pRSETBを用いること、及びプライマーとしてPrimer−F164Tを用いることを除いては、前記(1)の操作を繰り返し、164番目のフェニルアラニン(F)をトレオニン(T)に置換されたタンパク質をコードするDNAを含むプラスミドKillerRed_L162T_F164T/pRSETBを得た。
【0050】
(3)KillerRed_L162T_F164Tの発現
得られたプラスミドKillerRed_L162T_F164T/pRSETBタンパク質は、T7発現系(pRSETB/JM109(DE3))を用いて発現させた。すなわち、大腸菌(E.coli)コンピテントセル(JM109(DE3))をKillerRed_L162T_F164T/pRSETBで形質転換し、100μg/mLのアンピシリンを含むLB固体培地で37℃にて培養した。その後、生育した単一コロニーをピックアップして100μg/mLのアンピシリンを含むLB培養液300mL中へ植菌し、23℃で4日間培養した。得られた細胞をフレンチプレスにより破砕し、得られた上清液から、ニッケルキレートカラム(QIAGEN社製)を用いてポリヒスチジン標識化タンパク質を精製した。更に、この溶出液(100mMイミダゾール、50mM Tris−Cl、pH7.4、300mM NaCl)をPD10脱塩・バッファー交換カラム(GE healthcare Bio−Sciences社製)により、1×PBS(pH7.4)を用いて精製し、タンパク質KillerRed_L162T_F164Tを得た。
【0051】
(4)KillerRed_L162T_F164Tの発色及び単量体化の確認
得られたKillerRed_L162T_F164Tを、pseudo native PAGEにより解析した。KillerRed_L162T_F164Tのバンドは、二量体のKillerRedのバンドと比較すると、低分子量にシフトし、単量体として存在した。
しかしながら、KillerRed_L162T_F164Tは、KillerRedと比較すると、610nmにおける蛍光が減衰した。結果を、表1に示す。
【0052】
(5)G5V、N147S、L174K、及びM206Tの置換の導入
G5V、N147S、L174K、及びM206Tの置換を、KillerRed_L162T/F164Tに導入するため、以下のプライマーを合成した。
Primer−G5V:5’-GGTTCAGAGGTCGGCCCCGCC-3’(配列番号13)
Primer−N147S:5’-CAGACCGCCACCATCGGCTTC-3’(配列番号14)
Primer−L174K:5’-GACGGCGGCAAGATGATGGGC-3’(配列番号15)
Primer−M206T:5’-ACCAAGCAGACGAGGGACACT-3’(配列番号16)
【0053】
5番目のグリシン(G)のバリン(V)への置換は、鋳型プラスミドDNAとして、KillerRed_L162T_F164T/pRSETBを用いること、及びプライマーとしてPrimer−G5Vを用いることを除いては、前記(1)の操作を繰り返し、5番目のグリシン(G)のバリン(V)に置換されたタンパク質をコードするDNAを含むプラスミドKillerRed_L162T_F164T_G5V/pRSETBを得た。
以下、同様の操作により、順次変異を導入し、KillerRedのアミノ酸配列において、147番目のアスパラギン(N)をセリン(S)に、174番目のロイシン(L)をリシン(K)に、そして206番目のメチオニン(M)をトレオニン(T)に置換したタンパク質をコードするDNAを含むプラスミドSupernova−Red/pRSETBを得た。
【0054】
(6)Supernova−Redの発現
Supernova−Redタンパク質は、プラスミドとしてSupernova−Red/pRSETBを用いたことを除いては、前記(3)の操作を繰り返し、精製タンパク質Supernova−Redを得た。
【0055】
《比較例1〜21》
比較例1〜21では、二量体のKillerRedを、単量体にするための変異の導入を試みた。具体的には、各比較例では以下のアミノ酸の置換を行った。
比較例1:126番目のアスパラギンをアルギニン(N126R)に置換した。
比較例2:128番目のアスパラギン酸をトレオニン(D128T)に置換した。
比較例3:126番目のアスパラギンをアルギニンに置換(N126R)し、128番目のアスパラギン酸をトレオニン(N126R/D128T)に置換した。
比較例4:154番目のヒスチジンをグルタミン酸(H154E)に置換した。
比較例5:165番目のイソロイシンをアルギニン(I165R)に置換した。
比較例6:163番目のアラニンをリシン(A163K/I165R)に置換し、165番目のイソロイシンをアルギニン(N126R/D128T)に置換した。
比較例7:176番目のメチオニンをアスパラギン酸(M176D)に置換した。
比較例8:196番目のプロリンをアラニン(P196A)に置換した。
比較例9:198番目のフェニルアラニンをリシン(F198K)に置換した。
比較例10:140番目のグルタミンをアラニン(Q140A)に置換した。
比較例11:26番目のリシンをグルタミン酸(K26E)に置換した。
比較例12:174番目のロイシンをヒスチジン(L174H)に置換した。
比較例13:174番目のロイシンをアルギニン(L174R)に置換した。
比較例14:227番目のバリンをリシン(V227K)に置換した。
比較例15:230番目のイソロイシンをリシン(I230K)に置換した。
比較例16:234番目のイソロイシンをリシン(I234K)に置換した。
比較例17:152番目のフェニルアラニンをトレオニン(F152T)に置換した。
比較例18:162番目のロイシンをリシン(L162K)に置換した。
比較例19:164番目のフェニルアラニンをリシン(F164K)に置換した。
比較例20:167番目のフェニルアラニンをリシン(F167K)に置換した。
比較例21:167番目のフェニルアラニンをトレオニン(F167T)に置換した。
前記各比較例において、それぞれの置換に対応するプライマーを合成し、それぞれのプライマーを用いたことを除いては、前記実施例1の工程(1)の操作を繰り返すことにより、それぞれの変異を有するタンパク質をコードするDNAを含むプラスミドを得た。
得られたプラスミドを用いることを除いては、前記実施例の工程(3)及び(4)の操作を繰り返すことにより、それぞれのタンパク質の発色及び単量体化を確認した。比較例1〜21では、得られたタンパク質は、二量体のままで、単量体化しなかった。結果を纏めて、表1に示す。
【0056】
【表1】
【0057】
《実施例2》
本実施例では、前記タンパク質Supernova−Redの68番目のチロシン(Y)をトリプトファン(W)に置換した。
68番目のチロシン(Y)をトリプトファン(W)に置換するため、以下の変異導入用のプライマーを合成した。
Primer−Y68W:5’-GCCACCTGATCCAGTGGGGCGAGCCC-3’(配列番号17)
鋳型プラスミドDNAとして、Supernova−Red/pRSETBを用いること、及びプライマーとしてPrimer−Y68Wを用いることを除いては、前記実施例1の工程(1)の操作を繰り返し、プラスミドSupernova−Orange/pRSETBを得た。
更に、プラスミドSupernova−Orange/pRSETBを用いることを除いては、前記実施例1の工程(3)の操作を繰り返し、タンパク質Supernova−Orangeを得た。
【0058】
《実施例3》
本実施例では、前記タンパク質Supernova−Orangeの46番目のバリン(V)をアラニン(A)に置換した。
46番目のバリン(V)をアラニン(A)に置換するため、以下の変異導入用のプライマーを合成した。
Primer−V46A:5’-GGCGACTTCAACGCCCACGCCGTG-3’(配列番号18)
鋳型プラスミドDNAとして、Supernova−Orange/pRSETBを用いること、及びプライマーとしてPrimer−V46Aを用いることを除いては、前記実施例1の工程(1)の操作を繰り返し、プラスミドSupernova−Green/pRSETBを得た。
更に、プラスミドSupernova−Green/pRSETBを用いることを除いては、前記実施例1の工程(3)の操作を繰り返し、タンパク質Supernova−Greenを得た。
【0059】
《実施例4》
本実施例では、タンパク質KillerRed_L162T_F164Tに、更にN147Sの置換を導入し、KillerRed_L162T_F164T_N147Sの作製を試みた。
鋳型プラスミドDNAとして、KillerRed_L162T_F164T/pRSETBを用いること、及びプライマーとしてPrimer−V46Aを用いることを除いては、前記実施例1の工程(1)の操作を繰り返し、プラスミドKillerRed_L162T_F164T_V46A/pRSETBを得た。
更に、プラスミドKillerRed_L162T_F164T_V46A/pRSETBを用いることを除いては、前記実施例1の工程(3)の操作を繰り返し、タンパク質KillerRed_L162T_F164T_N147S(配列番号9)を得た。
【0060】
《実施例5》
本実施例では、前記タンパク質Supernova−Redの46番目のバリン(V)をアラニン(A)に置換した。
鋳型プラスミドDNAとして、Supernova−Red/pRSETBを用いること、及びプライマーとしてPrimer−V46Aを用いることを除いては、前記実施例1の工程(1)の操作を繰り返し、プラスミドSupernova−Red_V46A/pRSETBを得た。更に、プラスミドSupernova−Red_V46A/pRSETBを用いることを除いては、前記実施例1の工程(3)の操作を繰り返し、タンパク質Supernova−Red_V46A(配列番号10)を得た。
【0061】
《比較例22〜27》
比較例22〜27では、Supernova−Redの発色団の発色を変化させるための変異の導入を試みた。具体的には、各比較例では、L162T/F164Tの置換に、更に以下のアミノ酸の置換を行った。
比較例1:150番目のヒスチジンをトレオニン(H150T)に置換した。
比較例2:148番目のグルタミン酸をトレオニン(E148T)に置換した。
比較例3:148番目のグルタミン酸をリシン(E148K)に置換した。
比較例4:148番目のグルタミン酸をバリン(E148V)に置換した。
比較例5:226番目のセリンをイソロイシン(S226I)に置換した。
比較例6:148番目のグルタミン酸をバリン(E148V)に置換し、226番目のセリンをイソロイシン(S226I)に置換した。
前記各比較例において、それぞれの置換に対応するプライマーを合成し、それぞれのプライマーを用いたことを除いては、前記実施例1の工程(5)の操作を繰り返すことにより、それぞれの変異を有するタンパク質をコードするDNAを含むプラスミドを得た。
得られたプラスミドを用いることを除いては、前記実施例の工程(3)及び(4)の操作を繰り返すことにより、それぞれのタンパク質の発色及び単量体化を確認した。比較例22〜27で得られたタンパク質は、赤色からの発色のシフトは見られず、また、発色の増強も見られなかった。結果を纏めて表2に示す。
【0062】
【表2】
【0063】
《実施例6》
得られたタンパク質が所望の突然変異を導入したものであることを、該タンパク質の励起波長及び発光波長を測定するとともに既存のタンパク質(KillerRed等)の励起波長及び発光波長と比較することにより確認した。
得られたタンパク質の励起波長を測定し、規格化したものを図1(A)中に曲線で示す。また、得られたタンパク質の蛍光波長を測定し、正規化したものを図1(B)中曲線で示す。なお、図1(A)及び(B)には、比較のためにKillerRed、Supernova−Redについて励起波長及び蛍光波長を併せて示す。図1(A)及び(B)に示すように、上述した方法で変異を導入したタンパク質Supernova−Orangeは励起波長のピークが455nm、蛍光波長のピークが544nmであり、Supernova−Greenは励起波長のピークが439nm、蛍光波長のピークが469nmであることがわかる。これに対してKillerRedの励起波長のピークは585nmであり、蛍光波長のピークは610nm、またSupernova−Redの励起波長のピークは579nmであり、蛍光波長のピークは610nmである。
なお、この表3において、λabsはナノメーター単位での吸光度スペクトルのピークを表している。(ε)は103M−1cm−1単位でのモル吸光係数である。λemはナノメーター単位での蛍光スペクトルのピークを表している。
【0064】
更に、実施例4で得られたKillerRed_L162T_F164T_N147S及び、実施例5で得られたSupernova−Red_V46Aについても、励起波長及び、蛍光波長を測定した。KillerRed_L162T_F164T_N147Sは、励起波長のピークが512nm、そして蛍光が黄緑色であり、Supernova−Red_V46Aは励起波長のピークが509nm、蛍光波長のピークが594nmであった。
【0065】
《実施例7》
また、KillerRed、Supernova−Red、Supernova−Orange及びSupernova−Greenについて活性酸素量の測定を行った。ここではまず、10μM anthracene−9,10−dipropionic acid(以下ADPAと記す)を含む50μSupernova−Red、Supernova−Red、Supernova−Orange及びSupernova−Greenタンパク質溶液をそれぞれ100μLずつ調製した。ADPAは励起極大が380nm、蛍光極大が430nmの一重項酸素検出用試薬である。一重項酸素とは活性酸素の一種で、大変強力な酸化能力を有しており、ADPAが一重項酸素と反応することでADPAは過酸化物へと変化し、430nmにある蛍光ピークが失われる。
得られたサンプル溶液を以下の測定条件に従って、活性酸素測定を行った。すなわち、15μLをテラサキディッシュに添加し、YAGレーザー(LOTIS社、LS−2137U−YAG、周波数10Hz、パルス幅15ns)の第三高調波(355nm)をオプティカルパラメトリック発振器(LOTIS社、LT−2214−OPO)を用いて波長変換し、20mJ/cm2/パルスのエネルギー密度で5分間照射した後、これを1mLの1×PBSで希釈し、分光蛍光光度計F−2500(日立)を用いてADPAの退色率を測定した。レーザーの波長はそれぞれKillerRedとSupernova−Redが580nm、Supernova−Orangeが450nm、Supernova−Greenが440nmである。上述した突然変異を導入した蛍光タンパク質を含む4つの蛍光タンパク質の活性酸素量の比較を図2に示す。図2においてKR各種光増感性蛍光タンパク質によるADPA褪色率、すなわち活性酸素放出量を示している。なお、図2では比較のためフルオレセインイソチオシアネート(FITC)の活性酸素量で規格化した。
【0066】
【表3】
【0067】
《実施例8》
本実施例では、実施例1において得られたSupernova−Redを、チューブリン、ケラチン、又はCx43との融合タンパク質として発現させた。前記Supernova−Red/pRSETBに、PCRで得られたチューブリン、ケラチン、又はCx43をコードするDNAを挿入した。得られたプラスミドを、それぞれtublin−Supernova−Red/pRSETB、keratin−Supernova−Red/pRSETB、及びCx43S−upernova−Red/pRSETBと称する。
得られたプラスミドtublin−Supernova−Red/pRSETB、keratin−Supernova−Red/pRSETB、及びCx43−Supernova−Red/pRSETBを、HeLa細胞にトランスフェクションし、細胞内局在を蛍光顕微鏡で調べた。
チューブリン、ケラチン、又はCx43、それぞれ、細胞の微小管、中間系フィラメント、及びギャップジャンクションに局在しており、それぞれのタンパク質が存在している位置で発現しており、タンパク質の局在化に影響を与えなかった。
【0068】
《実施例9》
本実施例では、前記Supernova−Redにミトコンドリアマトリクス移行シグナルをつなげることで細胞内のミトコンドリアに導入し、光照射によってミトコンドリア機能を不活性化することによりアポトーシスを誘導した。
1μgのプラスミド_Supernova−Red/pcDNA3、を、トランスフェクション試薬(Superfect QIAGEN社)を用いて、35mmガラスボトム皿上で培養しているHela細胞に導入した。2日間培養後、580nmの光を8W/cm2のパワー密度で90秒間照射した後、0時間、3時間、及び6時間の細胞を微分干渉顕微鏡を用いて観察した。Supernova−Redが、ミトコンドリアに発現している細胞は、6時間後には退縮し、死滅した。
【産業上の利用可能性】
【0069】
本発明の光増感性蛍光タンパク質又はその機能的等価改変体は、発色団補助光不活性化法に用いることが可能であり、標的分子の機能解析に用いることができる。
【特許請求の範囲】
【請求項1】
配列表の配列番号2で表されるアミノ酸配列、配列番号4で表されるアミノ酸配列、配列番号6で表されるアミノ酸配列、配列番号9で表されるアミノ酸配列、及び配列番号10で表されるアミノ酸配列、からなる群から選択される少なくとも1つのアミノ酸配列を含む光増感性蛍光タンパク質又はその機能的等価改変体。
【請求項2】
前記光増感性蛍光タンパク質又はその機能的等価改変体が、他のタンパク質と結合した融合タンパク質である、請求項1に記載のタンパク質又はその機能的等価改変体。
【請求項3】
請求項1又は2に記載の光増感性蛍光タンパク質又はその機能的等価改変体をコードするDNA。
【請求項4】
配列表の配列番号1で表される塩基配列、配列番号3で表される塩基配列、及び配列番号5で表される塩基配列からなる群から選択される少なくとも1つの塩基配列からなる、請求項3に記載のDNA。
【請求項5】
請求項3又は4に記載のDNAを含むプラスミド。
【請求項6】
請求項1又は2に記載の光増感性蛍光タンパク質又はその機能的等価改変体を用いた、光増感性の活性酸素産生方法であって、前記光増感性蛍光タンパク質又はその機能的等価改変体に、励起光を照射することを特徴とする、前記方法。
【請求項7】
前記励起光の波長が、490〜580nm、400〜500nm、及び390〜440nmからなる群から選択される少なくとも1つの波長である、請求項6に記載の光増感性の活性酸素産生方法。
【請求項8】
請求項1又は2に記載の光増感性蛍光タンパク質又はその機能的等価改変体を用いた、標的分子の機能解析方法であって、前記光増感性蛍光タンパク質又はその機能的等価改変体に、励起光を照射することを特徴とする、前記方法。
【請求項9】
前記励起光の波長が、490〜580nm、400〜500nm、及び390〜440nmからなる群から選択される少なくとも1つの波長である、請求項8に記載の標的分子の機能解析方法。
【請求項10】
前記標的分子が、タンパク質、DNA、RNA、糖質、及び脂質からなる群から選択される少なくとも1つの標的分子である、請求項9に記載の標的分子の機能解析方法。
【請求項1】
配列表の配列番号2で表されるアミノ酸配列、配列番号4で表されるアミノ酸配列、配列番号6で表されるアミノ酸配列、配列番号9で表されるアミノ酸配列、及び配列番号10で表されるアミノ酸配列、からなる群から選択される少なくとも1つのアミノ酸配列を含む光増感性蛍光タンパク質又はその機能的等価改変体。
【請求項2】
前記光増感性蛍光タンパク質又はその機能的等価改変体が、他のタンパク質と結合した融合タンパク質である、請求項1に記載のタンパク質又はその機能的等価改変体。
【請求項3】
請求項1又は2に記載の光増感性蛍光タンパク質又はその機能的等価改変体をコードするDNA。
【請求項4】
配列表の配列番号1で表される塩基配列、配列番号3で表される塩基配列、及び配列番号5で表される塩基配列からなる群から選択される少なくとも1つの塩基配列からなる、請求項3に記載のDNA。
【請求項5】
請求項3又は4に記載のDNAを含むプラスミド。
【請求項6】
請求項1又は2に記載の光増感性蛍光タンパク質又はその機能的等価改変体を用いた、光増感性の活性酸素産生方法であって、前記光増感性蛍光タンパク質又はその機能的等価改変体に、励起光を照射することを特徴とする、前記方法。
【請求項7】
前記励起光の波長が、490〜580nm、400〜500nm、及び390〜440nmからなる群から選択される少なくとも1つの波長である、請求項6に記載の光増感性の活性酸素産生方法。
【請求項8】
請求項1又は2に記載の光増感性蛍光タンパク質又はその機能的等価改変体を用いた、標的分子の機能解析方法であって、前記光増感性蛍光タンパク質又はその機能的等価改変体に、励起光を照射することを特徴とする、前記方法。
【請求項9】
前記励起光の波長が、490〜580nm、400〜500nm、及び390〜440nmからなる群から選択される少なくとも1つの波長である、請求項8に記載の標的分子の機能解析方法。
【請求項10】
前記標的分子が、タンパク質、DNA、RNA、糖質、及び脂質からなる群から選択される少なくとも1つの標的分子である、請求項9に記載の標的分子の機能解析方法。
【図1】

【図2】
【図3】
【図2】
【図3】
【公開番号】特開2013−78264(P2013−78264A)
【公開日】平成25年5月2日(2013.5.2)
【国際特許分類】
【出願番号】特願2010−22603(P2010−22603)
【出願日】平成22年2月3日(2010.2.3)
【出願人】(504173471)国立大学法人北海道大学 (971)
【Fターム(参考)】
【公開日】平成25年5月2日(2013.5.2)
【国際特許分類】
【出願日】平成22年2月3日(2010.2.3)
【出願人】(504173471)国立大学法人北海道大学 (971)
【Fターム(参考)】
[ Back to top ]