
免疫原性グリコペプチド、スクリーニング、製造及び使用
【課題】病原性微生物により引き起こされる感染の予防接種及び診断に有用な病原性微生物由来免疫原性グリコペプチド、その選択方法及び調製方法を提供する。
【解決手段】少なくとも1つの中性アミノ酸は二糖又は三糖に結合し、少なくとも15%のアミノ酸はプロリンで、その1つは中性アミノ酸の位置に対して−1〜−4位に位置し、クラスIIMHC分子で示され、由来する天然型グリコペプチドでの免疫化で誘導されるCD4+Tリンパ球により特異的に同定されるが、同一配列を有する非グリコシル化ペプチドでの免疫化で誘導されるCD4+Tリンパ球により同定されず、同定されるCD4+Tリンパ球の増殖と該リンパ球によるサイトカインの分泌を誘導できる、14〜25アミノ酸からなるグリコシル化Tエピトープからなる免疫原性グリコペプチド。
【解決手段】少なくとも1つの中性アミノ酸は二糖又は三糖に結合し、少なくとも15%のアミノ酸はプロリンで、その1つは中性アミノ酸の位置に対して−1〜−4位に位置し、クラスIIMHC分子で示され、由来する天然型グリコペプチドでの免疫化で誘導されるCD4+Tリンパ球により特異的に同定されるが、同一配列を有する非グリコシル化ペプチドでの免疫化で誘導されるCD4+Tリンパ球により同定されず、同定されるCD4+Tリンパ球の増殖と該リンパ球によるサイトカインの分泌を誘導できる、14〜25アミノ酸からなるグリコシル化Tエピトープからなる免疫原性グリコペプチド。
【発明の詳細な説明】
【技術分野】
【0001】
この発明は、病原性微生物のようなもの(細菌又は真菌)による免疫化及び感染診断に使用できる、病原性微生物由来の免疫原性グリコペプチド及びその選択ならびに製造方法に関する。
【背景技術】
【0002】
これらの感染を予防し、治療するために実行されている手段は、まず、感染をモニターし、治療できるスクリーニングと、次いで免疫化からなる。
ヒト医学で最も深刻な感染の一例としてエム.ツベルクローシスでの感染を挙げて、これらの手段を以降に例示する。具体的には、正常な免疫反応を有するエム.ツベルクローシスに感染した個体の5〜10%が、深刻な疾患(結核)に進行する;この頻度は、免疫反応を欠いている個体(HIV感染、免疫抑制剤での治療など)ではさらに高い。
【0003】
診 断
現在利用可能な種々の技術には、以下が挙げられる:
− 結核を正確に診断するのに最も厳密な手段であるエム.ツベルクローシスの純粋培養物の生産。それは、肺結核症例の2/3を診断できる穏やかな高感度技術である。結果は、最低3〜4週間後にのみ、時には2ヶ月培養した後にのみ利用できる。標識した前駆体を用いる培養技術の使用はこれらの遅れを短縮できるが、にもかかわらず依然として相当な期間である。培養によるエム.ツベルクローシスのこの検出は、肺結核のためにすら時には入手しにくく、症例の約1/3が生物学的追認を受けない細菌含有試料を要する。時折り、この検査は、肺外形態の疾患に対する特殊な医学的介入(脳脊髄液の腰椎穿刺又はリンパ節バイオプシー)を要する。
【0004】
− 分子遺伝学に基づく微生物学的技術(PCR)は、細菌含有試料を得るという同一の要件に直面している。さらに、試料中にPCR反応阻害剤が存在するために、その由来を調節できず、時にこれらの技術を使用できない。技術は、一般的な実施には有効ではない。
− 現在、診断用途に適合した感度と特異性を有する血清学的診断法はない。
【0005】
− ツベルクリンに対する反応は、個体が感作され、エム.ツベルクローシスに感染しているか、又はBCGで免疫化されていることを示す。ツベルクリンは、実際エム.ツベルクローシス抗原の混合物であり、したがって、エム.ツベルクローシスでの感染とBCGでの免疫化とを識別することができない。というのは、ワクチン抗原とエム.ツベルクローシスとで交差反応が極めて多いからである。さらに、ツベルクリンに対するこの反応は、進行中の疾患である結核をエム.ツベルクローシスでの感染と識別することができない。
【0006】
ワクチン
BCGでの免疫化は、一次感染(エム.ツベルクローシスの初期増殖)、特にこれらの細菌の二次転移を制御することができる。それは、おそらく現在利用できる有効な治療がない潜伏期の感染の発症を減じるのに寄与している。BCGは、特に副作用なく、結核に対して30億人以上の個体を免疫化するために使用されている。BCGでの免疫化中、毒性が弱毒化されたこれらの細菌の局所増殖がある。細胞性免疫が、誘発される。それは、マイコバクテリアのタンパク質又は抗原に対する遅延型過敏症(HSR)(ツベルクリンに対する反応)を引き起こし、エム.ツベルクローシスでの感染に対する耐性を増した。これらの2つの免疫反応(HSR-型感作及び耐性増加)は、マイコバクテリア抗原と反応するTリンパ球によって支持される。
【0007】
BCGは、急性型感染(例えば子供の結核性脳膜炎)に対しても保護する。その有効性は、大人では多岐にわたる。BCGとツベルクローシス複合体に属さない他のマイコバクテリアとの交差反応性の存在、及びマイコバクテリウム・ツベルクローシスのある免疫原性抗原のBCGゲノムでの不在、又は感染中のこれらの抗原に対する異なる発現プロフィルは、多岐にわたるBCGの有効性を説明し得る。
さらに、BCGは、毒性が弱毒化された生株である。したがって、それは、免疫抑制された個体、特にヒト免疫不全ウイルス(HIV)に感染したことが確認された個体での使用を妨げる残留病原力を有している。
【0008】
これらの感染により効果的に対抗するために、これらの感染の原因である病原性微生物に対して保護する抗原に基づいて、診断上の手段とワクチン、特に危険性のない「サブユニット」ワクチンを有することが賢明であろう。
強力な保護免疫反応を誘導できるこれらの病原性微生物の分子を見出すために、この意味において幾つかの研究が行われている。つまり、J. Hessら(C.R. Acad. Sci. Paris, 1999, 322: 953-958)は、結核に対するワクチンとして使用できる抗原が有すべき性質を概説している。その概説では、単一抗原よりむしろ予備選択した抗原の組み合わせを用いることが重要視されている。特に、種々の株に高度に保存される領域の存在、毒性株と弱毒性株の遺伝子発現プロフィルの相違、免疫反応のエフェクター細胞(B、CD4+ T、CD8+ Tリンパ球)に関する反応性又は主要組織適合性複合体(MHC)の大多数のHLA分子へのこれらの抗原の結合能力などの基準に基づくこれらの抗原の選択が推奨されている。
【0009】
これらの抗原の幾つかは、シー.アルビカンス(C. albicans)のマンノタンパク質のような表面抗原形態(Buurmanら、PNAS, 1998, 95, 7670-7675)、又はエム.ツベルクローシスでの分泌抗原形態: MPT59 (30 kDa)、85A (32 kDa)、MPT64 (23 kDa)、hsp71 (71 kfla)、MPT51 (24 kDa)、MPT63 (16 kDa)及びESAT-6 (6 kDa) (Andersen, Infect. Immun., 1994, 62, 2536-2544;Horwitzら、PNAS, 1995, 92, 1530-1534)のいずれかで存在する。これらのエム.ツベルクローシス抗原は、CD4+ Tリンパ球によって優先的に認識されるので、免疫化組成物の潜在的な候補物としてすでに提案されている(Andersenら、上記;Horwitzら、上記)。
【0010】
また、MHCクラスII分子によって提示され、かつ特異的CD4+ Tリンパ球によって認識されるエピトープを含むペプチドをエム.ツベルクローシス抗原から単離することが提案されている;このようなエピトープは、特に2つのタンパク質について報告されている: ESAT-6 (Olsenら、Eur. J. Immunol., 2000, 30, 1724-1732)及びMPT-39 (Mustafaら、Inf. Immunol., 2000, 68, 3933-3940)。
【0011】
幾つかの観察が、発明者によって以前になされている(Romainら、Inf. Immun., 1993, 61, 742-750;Romainら、Proc. Natl. Acad. Sci. USA 1993, 90: 5322-5326):
− 生細菌のみが保護免疫を誘導でき、死菌細菌も保護なしに免疫反応を誘導する;
− 培養培地では、細菌の成長中に細菌によって放出され、生細菌で免疫化された動物の免疫系によって認識されるタンパク質は、これらが死菌細菌での免疫化後にあまり認識されないか、又は全く認識されないので、存在する。
【0012】
選択についてこの二重の基準を用いて、2つの新たなタンパク質を精製した。Apa、又はMPT-32 あるいは45/47 kDa 抗原複合体と命名されるエム.ツベルクローシスによって分泌されるタンパク質は、Rv 1860遺伝子産物である(Laqueyrerieら、Infect. Immun. 1995, 63: 4003-4010)。第二分子は、Rv 1796遺伝子によってエンコードされる推定上のセリンプロテアーゼの内部ペプチドである。
抗原として天然のApaタンパク質を用い、発明者らは、培養物にツベルクローシス群の細菌(エム.ツベルクローシス、エム.ボビス及びBCG)によって分泌されるタンパク質のわずか2%を示すこのタンパク質が、エム.ツベルクローシスに感染しているか、又はBCGで免疫化された動物を由来とする特異的な CD4+ Tリンパ球によって極めて有効に認識される免疫優勢抗原であることを以前に示している(Romainら、 Inf. Immun., 1999, 67, 5567-5572;Horn ら、J. Biol. Chem., 1999, 274, 32023-32030)。
【0013】
これらの同一の研究において、発明者らは、Apaのマンノシル化が、このタンパク質の抗原活性に必須であることも示した:
− α-マンノシダーゼ又はトリフルオロメタンスルホン酸(TFMS)で天然のApaを処理することによって、又はグリコシル化できない細菌(イー.コリ)でApaを発現することによって得られるApaの脱マンノシル化は、100倍の抗原性損失を伴う。
− エム.ツベルクローシスにって産生されるApaとわずかに異なるマンノース組成を全体に有するマイコバクテリウム・スメグマティス(Mycobacterium smegmatis)によって産生されるグリコシル化Apaは、約10/1に低下する抗原活性を有する。
【0014】
さらに、このエム.ツベルクローシスのApa分子は、ジマンノース(T10及びT18)、マンノース(T27)、 マンノース、ジマンノース又はトリマンノース(T277)で、α-(1,2)型のグリコシド結合を介して4個のトレオニン残基(T10、T18、T27 及びT277)に結合した6〜9個のマンノース残基を含むことが報告されている(Dobosら、J. Bacteriol., 1996, 178, 2498-2506)。モノ-、ジ-又はトリ-マンノースを含むこの糖類の構造は、酵母、特にカンジダ・アルビカンス(Candida albicans)由来のマンノタンパク質の構造に似ており、長いオリゴマンノース鎖を有するエフ.メニゴセプティカム(F.meningosepticum)由来タンパク質の構造と異なることに留意すべきである。
【0015】
脱マンノシル化後にみられるApa抗原性の損失は、この抗原の食作用とプロセシング、あるいは CD4+ Tリンパ球による後者の認識の低下に起因している可能性がある。具体的には、ヘキソースに特異的に結合するマクロファージ及び樹状細胞、特にシー.アルビカンス由来マンノタンパク質及びマイコバクテリア由来のリポアラビノマンナンのようなマンノリピドのマンノースレセプターは、ペプチド/クラスII MHC分子複合体形態のこれらの細胞表面に存在する抗原の食作用とプロセシングで役割を果たしている(Stahlら、Current Opinion in Immunology, 1998, 10, 50-55)。また、(N-末端位のリジン残基についてマンノシル化された)マンノシル化ペプチドは、同一の配列を持つ非グリコシル化ペプチドよりも有効に樹状細胞によって食菌され、処理されることも分かっている(Tanら、Eur. J. Immunol., 1997, 27, 2426-2435)。
【0016】
ニワトリのリソチームモデルでは、この抗原のTエピトープを構成するペプチドのグリコシル化類似体であるペプチドが、このグリコシル化エピトープを特異的に認識するCD4+ Tリンパ球を誘導できることが示されている(Deckら、J. Immunol., 1995, 155, 1074-1078)。しかし、CD4+ Tリンパ球によって特異的に認識されるグリコシル化Tエピトープは病原性微生物(細菌/真菌)由来の天然抗原で同定されないので、CD4+ Tリンパ球によるこれらの病原性微生物由来抗原の認識におけるグリコシル化の重要性は依然として立証されべきものである。
さらに、これは、抗原のグリコシル化に関するエム.ツベルクローシスApaと一般的な知見に関するデータであるにもかかわらず、免疫原性組成物又は免疫化組成物及び/又は診断試験において有効に使用できる、これらの病原性微生物のO-グリコシル化タンパク質由来抗原を製造する可能性は、現在のところない。
【0017】
本質的に:
− これらの微生物によって産生されるタンパク質のごく少量の%を示す活性タンパク質は、これらの病原性剤を大量に扱うため危険な方法を用いて、極めて低い収率で精製される、
− 異種発現系(グリコシル化できない真核細胞又は細菌)で産生されるタンパク質は、低い抗原活性を有する、
− エム.スメグマティスのような同種の発現系で産生されるタンパク質は、許容される抗原活性を有するが、それらは複雑な方法を用いては不十分な量で産生される。
【先行技術文献】
【非特許文献】
【0018】
【非特許文献1】J. Hessら(C.R. Acad. Sci. Paris, 1999, 322: 953-958)
【非特許文献2】(Buurmanら、PNAS, 1998, 95, 7670-7675)
【非特許文献3】Andersen, Infect. Immun., 1994, 62, 2536-2544
【非特許文献4】Horwitzら、PNAS, 1995, 92, 1530-1534
【非特許文献5】Olsenら、Eur. J. Immunol., 2000, 30, 1724-1732
【非特許文献6】Mustafaら、Inf. Immunol., 2000, 68, 3933-3940
【非特許文献7】Romainら、Inf. Immun., 1993, 61, 742-750
【非特許文献8】Romainら、Proc. Natl. Acad. Sci. USA 1993, 90: 5322-5326
【非特許文献9】Laqueyrerieら、Infect. Immun. 1995, 63: 4003-4010
【非特許文献10】Romainら、Inf. Immun., 1999, 67, 5567-5572
【非特許文献11】Horn ら、J. Biol. Chem., 1999, 274, 32023-32030
【非特許文献12】Dobosら、J. Bacteriol., 1996, 178, 2498-2506
【非特許文献13】Stahlら、Current Opinion in Immunology, 1998, 10, 50-55
【非特許文献14】Tanら、Eur. J. Immunol., 1997, 27, 2426-2435
【非特許文献15】Deckら、J. Immunol., 1995, 155, 1074-1078
【発明の概要】
【発明が解決しようとする課題】
【0019】
この結果、発明者らは、単独で又は他の剤と組み合わせて投与する際に、一方で免疫抑制された個体(生ワクチンの使用に関連した危険性の消失)を含む全ての個体に使用できるワクチンを構成でき、他方で診断目的で使用できる、保護体液性及び/又は細胞性免疫反応を誘導できる免疫優勢抗原を製造することを目的とした。
脱グリコシル化された天然タンパク質又はイー.コリで産生される組換えタンパク質に少なくとも等しく、さもなければ、より大きい抗原活性を示すグリコタンパク質を合成する病原性微生物(特にマイコバクテリア)由来のあるグリコペプチドが見出された。
また、これらのグリコペプチドを大量に製造するための実施しやすい手段を開発することが、本発明の目的である。
【課題を解決するための手段】
【0020】
本発明の対象は、
a1)少なくとも1つの中性アミノ酸は二糖又は三糖に結合し(グリコシド結合)、アミノ酸の少なくとも15%はプロリンで、プロリンの1つは中性アミノ酸の位置に対して-1〜-4位に位置する、病原性微生物由来で、14〜25アミノ酸からなるグリコシル化Tエピトープから本質的になり、
− クラスII MHC分子で提示され、
− 由来する天然のグリコタンパク質での免疫化で誘導される CD4+ Tリンパ球によって特異的に認識されるが、同一の配列を有する非グリコシル化ペプチドでの免疫化で誘導されるCD4+ Tリンパ球によっては認識されず、かつ
− それらを認識するCD4+ Tリンパ球の増殖と該リンパ球によるサイトカインの分泌を誘導できる
グリコペプチド、及び
b1) Dobosら(J. Bacteriol., 1996, 178, 2498-2506)によって記載されるApa由来の、配列SEQ ID NO:11のグリコペプチドを除く、a1)に定義されるグリコペプチドの配列を含む15〜39アミノ酸の配列を有するグリコペプチド
からなる群から選択される免疫原性グリコペプチドである。
【0021】
グリコシル化Tエピトープから本質的になるこれらのグリコペプチドは、このグリコシル化Tエピトープを介してCD4+ Tリンパ球によって認識される。具体的には、ツベルクローシス群の生細菌での免疫化後、同一配列を有する非グリコシル化ペプチドに特異的なTリンパ球よりもこれらのグリコペプチドに特異的なTリンパ球がかなり多い。
有利には、グリコペプチドは、同一配列を有する対照のペプチドよりも少なくとも10倍、好ましくは少なくとも30倍大きい抗原活性を有する。
【0022】
このグリペプチドは、以下の利点を有する:
− 保護細胞型免疫反応、あるいは体液性反応の誘導、及び免疫抑制個体における抗原としての潜在的な使用、
− 病原性微生物に特異的な多数のTリンパ球によって認識されるための、従来の抗原(弱毒化生微生物の培養物、該培養物から調製される抗原混合物又は非グリコシル化ペプチド)に少なくとも等しく、さもなければより大きい抗原活性、
− 他の微生物、特に他の異型性マイコバクテリアとの交差反応性の問題を除き、免疫化と病原性微生物の診断の有効性を増すことができる、極めて狭い特異性;特に、より具体的には、病原性微生物に排他的に存在するそのオリゴ糖残基は、CD4+ Tリンパ球によって認識されるTエピトープの定義に必須な方法で寄与する;つまり、それらは、そのタンパク質の幾つかをO-グリコシル化できる病原性微生物(ツベルクローシス複合体細菌、フラボバクテリウム・メニンゴセプティカム(Flavobacterium meningosepticum)、カンジダ・アルビカンスなど)での免疫化及び感染を診断するのに特異的な抗原を構成する、
【0023】
− 全体に非病原性(apathogenic)であるための免疫抑制個体での使用、
− 大量での潜在的な生産、
− 進行中の用量とワクチンの有効性についてのより良い標準化、
− 貯蔵と使用の容易化。
グリコペプチドの有利な具体例によれば、中性アミノ酸は、セリンとトレオニンからなる群から選択される。
グリコペプチドのこの具体例の有利なアレンジによれば、それらは、二糖又は三糖に結合した1〜7個のトレオニン残基を含む。
グリコペプチドの別の有利な具体例によれば、二糖又は三糖は、ヘキソース、好ましくはマンノースの二量体又は三量体である。
【0024】
グリコペプチドのさらに別の有利な具体例によれば、グリコシド結合はα-(1,2)結合である。
さらに別の有利な具体例によれば、グリコペプチドは、タンパク質をO-グリコシル化できる病原性微生物、好ましくはマイコバクテリウム・ツベルクローシス又はカンジダ・アルビカンス由来である。
この発明によれば、グリコペプチドは、好ましくは
− エム.ツベルクローシス(Genbank 番号X80268)のApaタンパク質、又は
− エム.ツベルクローシス株H37Rv (Sanger bank)のゲノム配列の注釈に関し、Rv 1796遺伝子によってエンコードされるRv1796タンパク質
に由来する。
【0025】
好ましくは、グリコペプチドは、
− 配列(SEQ ID NO:1)が、Apaタンパク質の配列の1〜39位から伸長する配列であり、SEQ ID NO:1の10、18及び27位のトレオニン残基の少なくとも1つがグリコシド結合を介して二糖又は三糖に結合する、39アミノ酸のグリコペプチド、
− 配列(SEQ ID NO:2)が、Apaタンパク質の配列(C-末端配列)の261〜286位から伸長する配列であり、SEQ ID NO:2の17位のトレオニン残基がグリコシド結合を介して二糖又は三糖に結合する26アミノ酸のグリコペプチド、及び
− 配列(SEQ ID NO:3)がRv 1796 タンパク質の配列の169〜203位から伸長する配列であり、SEQ ID NO:3の4、5、7、13、15、23及び25位のトレオニン残基の少なくとも1つがグリコシド結合を介して二糖又は三糖に結合する35アミノ酸のグリコペプチド
からなる群から選択される。
【0026】
本発明の対象は、
− グリコシド結合を介して二糖又は三糖に結合したグリコシル化中性アミノ酸を溶液中に調製し、
− グリコペプチドのペプチド配列を産生するのに必要なアミノ酸及び上記で得たグリコシル化中性アミノ酸を用いて、固体支持体上でグリコペプチドを合成し、かつ
− 固体支持体からグリコペプチドを切断する
工程からなることを特徴とする、上記のグリコペプチドの合成方法でもある。
【0027】
この方法の有利な具体例によれば、中性アミノ酸は、セリン及びトレオニンからなる群から選択される。
この具体例の有利なアレンジによれば、グリコペプチドが、配列(Tは、2又は3個のグリコシド残基で官能化されたO-グリコシル化トレオニンを示し、かつAcはアセテート官能基を示す):
SEQ ID NO:1:
H2N-DPEPAPPVPTTAASPPSTAAAPPAPATPVAPPPPAAANT-CONH2
SEQ ID NO:2:
AcNH-PAPAPAPAGEVAPTPTTPTPQRTLPA-COOH
SEQ ID NO:3:
AcNH-TIPTTETPPPPQTVTLSPVPPQTVTVIPAPPPEEG-CONH2
を有する際、
【0028】
その方法は、
i)2又は3個のグリコシド残基で官能化されたO-グリコシル化トレオニンを溶液中に調製し、
ii)これらの配列を製造するために必要なアミノ酸及び工程i)で得たO-グリコシル化トレオニンを用いて、固体支持体上で、上記の配列SEQ ID NO:1、SEQ ID NO:2 及びSEQ ID NO:3に相当するペプチドを合成し、
iii)固体支持体からペプチドを切断し、かつ
iv)化学合成により、ペプチドSEQ ID NO:1及びSEQ ID NO:3のC-末端にアミド官能性を、またペプチドSEQ ID NO:2及びSEQ ID NO:3のN-末端にアセテート官能性を導入する
工程からなる。
【0029】
したがって、ペプチドSEQ ID NO:1、SEQ ID NO:2及びSEQ ID NO:3の合成は、そのあいだにグリコシル化アミノ酸が導入される従来の固相ペプチド合成に相当する。固相ペプチド合成の分野で知られているように、使用されるアミノ酸は適当に保護され、必要であれば、ペプチド配列に他のものが導入される前に活性化される。同様に、トレオニンによって担持されるグリコシド残基に存在するヒドロキシルは、ペプチド合成のあいだに適切に保護される必要がある。
いったんペプチド合成が行われると、ペプチドは、固体支持体から分離され、脱保護される。それらは、逆相高速液体クロマトグラフィーによって精製することができる。
【0030】
この具体例の有利なアレンジによれば、工程i)で製造されるO-グリコシル化トレオニンによって担持されるグリコシド結合はヘキソース、好ましくはマンノースであり、マンノース残基は、有利にはα-(1,2)結合を介して互いに結合される。
【0031】
このアレンジの有利な態様によれば、マンノース残基で官能化されるトレオニンは、以下のように製造される:
a2)式(I)及び(II)
【0032】
【化1】

[式中、P1及びP2は、同一又は異なっていてもよく、ヒドロキシル官能性を保護する基を示し、かつXは、臭素原子のような活性化官能性を示す]のマンノース誘導体の製造:
b2)2つのマンノース残基からなり、式(III):
【0033】
【化2】
[式中、P1、P2及びXは、式(I)及び(II)に関して定義されるとおり]
に相当する活性化誘導体の生産をもたらす、式(I)の誘導体と式(II)の誘導体との反応、次いで得られた化合物の活性化、
【0034】
c2)任意に、3つのマンノース残基からなり、式(IV):
【0035】
【化3】
[式中、P1、P2及びXは、式(I)及び(II)に関して定義されるとおり]
に相当する活性化誘導体の産生をもたらす、a2)に定義される式(I)のマンノース誘導体での式(III)の化合物の反応、次いで得られた化合物の活性化、及び
d2)それぞれ式(VI)又は(VII):
【0036】
【化4】
[式中、P1、P2、P3及びP4は上記のとおり]
のグリコシル化トレオニンの生産をもたらす、式(III)の化合物又は式(IV)の化合物の、式(V):
【0037】
【化5】
[式中、P3は、第一級アミン官能性を保護する基を示し、かつP4はヒドロキシル官能性を保護する基を示す]
の適当に保護されたトレオニンとの縮合。
【0038】
保護基P1、P2、P3及びP4は、研究Protective Groups in Organic Synthesis (T.W. GREENE 及びP.G.M. WUTS、第2版1991年、J. WILEY and Sons)に記載される基から選択してもよい。例として、また非制限に、P1とP2はアセチル又はベンゾイル基を示してもよく、P3はFmoc (9-フルオレニルメトキシカルボニル)基を示してもよく、かつP4はペンタフルオロフェニル基を示してもよい。
【0039】
この発明の対象は、上記のように、本発明によるグリコペプチドを合成する方法に付随して有利に行ってもよい、病原性微生物のタンパク質のペプチド配列を用いて免疫原性グリコペプチドを選択し、スクリーニングするための方法であって、方法は、少なくとも
a3)二糖又は三糖に結合した少なくとも1つの中性アミノ酸及びプロリンの1つが中性アミノ酸の位置に対して-1〜-4位に位置する少なくとも15%のプロリンを含む少なくとも1つの14〜25アミノ酸配列を、タンパク質のペプチド配列において、またそれから、検索し、選択し、
b3)上記の合成法にしたがって、工程a3)で選択されるグリコペプチドを調製し、かつ
c3)抗原活性が、同一配列を有する対照のペプチドよりも少なくとも10倍、好ましくは少なくとも30倍大きいグリコペプチドを選択する
工程からなることを特徴とする。
【0040】
このスクリーニング方法の有利な具体例によれば、工程a3)に先立って、少なくとも1つの抗原性グリコタンパク質を予備選択する工程が含まれる。
このスクリーニング方法の別の有利な具体例によれば、工程c3)では、グリコペプチドの抗原活性は、弱毒化病原性微生物又は病原性微生物の抗原画分で免疫化された動物のCD4+ Tリンパ球の活性を測定して評価される。
【0041】
Tリンパ球の活性化は、Current protocols in Immunology (John E. Coligan, 2000, Wiley and son Inc, Library of Congress, USA)に記載されるような従来の免疫技術を用いて立証することができる。例として、リンパ球増殖アッセイ、活性化CD4+ Tリンパ球によって合成されるサイトカイン(タンパク質又はmRNA)のためのアッセイ(イムノアッセイ(ELISA)又はRT-PCR型の重合鎖反応)、又はエム.ツベルクローシスの場合には、遅延型過敏症アッセイが挙げられる。
また、この発明は、上記のような選択及びスクリーニング方法を用いて得られるグリコペプチドを包含する。
【0042】
また、この発明の対象は、免疫原性組成物もしくは免疫化組成物又は診断試薬の製造のための、本発明による少なくとも1つのグリコペプチド又は配列SEQ ID NO:11のグリコペプチドの使用である。
病原性微生物、特にエム.ツベルクローシスでの感染によって誘導される細胞性及び/又は体液性免疫を極めて特異的に検出する本発明によるグリコペプチドは、当業者にそれ自体知られており、細胞性免疫を検出できるいずれかの技術によって結核の診断に有利に用いることができる。例として、T-リンパ球増殖アッセイ及びCD4+ Tリンパ球、特にγ-IFNに特異的なサイトカインのための免疫酵素アッセイが挙げられる。
【0043】
また、本発明の対象は、少なくとも1つの医薬的に許容される賦形剤と組み合わさった、上記の少なくとも1つのグリコペプチドからなることを特徴とする、体液性及び/又は細胞性免疫を誘導できる免疫原性組成物である。
体液性又は細胞性免疫反応の設定でのCD4+ Tリンパ球とCD8+ Tリンパ球つまりBリンパ球との協同作用のために、本発明のグリコペプチドは、抗原に対する免疫化の有効性を増すために、いずれかの他の抗原用の輸送タンパク質(担体)として有利に用いることができる。この抗原/担体の組み合わせは、免疫化抗原に特異的なB及びTリンパ球の選択と増幅を有利に容易にすることができる。
【0044】
本発明の対象は、少なくとも1つの医薬的に許容される賦形剤及び任意に少なくとも1つのアジュバントと組み合わさった、上記の少なくとも1つのグリコペプチドからなることを特徴とする、体液性及び/又は細胞性免疫を誘発し得る免疫化組成物である。
免疫原性組成物又は免疫化組成物の有利な具体例によれば、グリコペプチドは、少なくとも1つのBエピトープ、CD4+型の1つのTエピトープ又はCD8+型の1つのTエピトープからなるタンパク質又はタンパク質フラグメントと組み合わさっている。
【0045】
本発明の目的のため、タンパク質の配列に対して、用語「Bエピトープ」、「CD4+型Tエピトープ」及び「CD8+型Tエピトープ」は、それぞれ抗体、CD4+リンパ球Tレセプター及びCD8+リンパ球Tレセプターに結合し得るこの配列のフラグメントを意味することを意図する。
本発明の目的のため、表現「タンパク質とグリコペプチドの組み合わせ」は、いずれかの物理的手段又は化学的手段、例えばグリコペプチドの配列とタンパク質又はタンパク質フラグメントの配列との融合物の発現による混合と結合の双方を意味することを意図する。
用いられるアジュバントは、従来使用されるアジュバントである;有利には、それらは水酸化アルミニウムとスクアレンからなる群から選択される。
【0046】
グリコペプチドは、ペプチドの免疫原性を増すことのできる、当業者にはそれ自体公知のいずれかの他の手段と任意に組み合わせてもよい。例として、Wilkinsonら、1999, Eur. J. Immunol., 29, 2788-2796により記載されるような分枝状多量体ペプチドを生産できる担体ペプチドへの結合が挙げられる。
本発明の対象は、本発明によるグリコペプチドの1以上に対することを特徴とする抗体でもある。
抗体の有利な具体例によれば、それらは、モノクローナル抗体及びポリクローナル抗体から選択される。
【0047】
本発明の対象は、本発明によるグリコペプチドと抗体からなる群から選択されることを特徴とする診断試薬でもある。
また、本発明の対象は、病原性微生物に感染している恐れのある患者由来の生物学的試料を上記の診断試薬(場合によって抗体又はグリコペプチド)と接触させ、生物学的試料複合体中に存在する抗体/微生物又は試料複合体に存在するグリコペプチド/抗体の形成を検出することからなることを特徴とする、病原性微生物での感染の検出方法である。
【0048】
上記のアレンジに加えて、この発明は、この発明の具体例ならびに添付の図面に言及する以下の記載から明らかになるであろう多くのアレンジをさらに含む:
【図面の簡単な説明】
【0049】
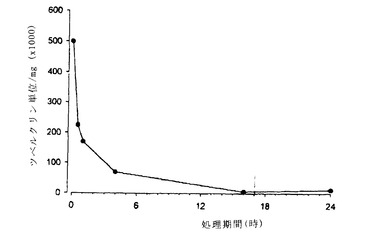
【図1】図1は、遅延型過敏症アッセイを用いる、エム.ツベルクローシスから精製した天然のApaの抗原活性測定を、α-マンノシダーゼによるApaタンパク質の消化動態の関数として示す。結果は、時間の関数としてタンパク質mg当たりのツベルクリン単位で示す。
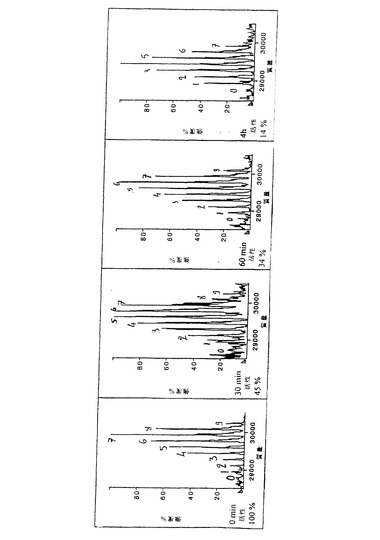
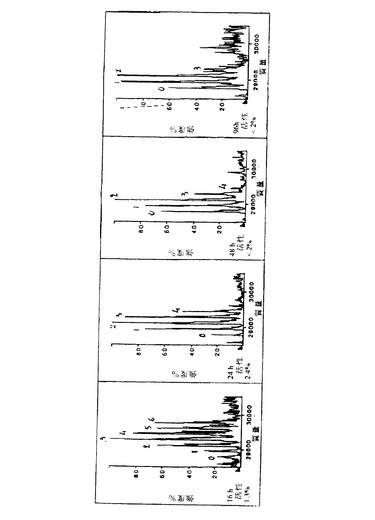
【図2a】図2は、α-マンノシダーゼでのApaタンパク質の消化動態の関数として、Apa分子のマンノース組成の質量分析を示す。Apaタンパク質の各ピークに相当するマンノース残基数を示し、Apa消化産物の全体的な抗原活性を研究した種々の時間で示す。
【図2b】図2は、α-マンノシダーゼでのApaタンパク質の消化動態の関数として、Apa分子のマンノース組成の質量分析を示す。Apaタンパク質の各ピークに相当するマンノース残基数を示し、Apa消化産物の全体的な抗原活性を研究した種々の時間で示す。
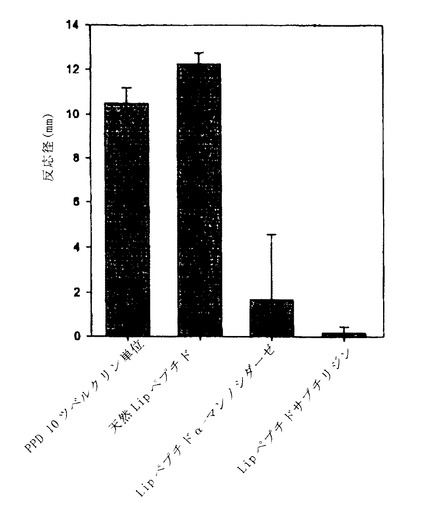
【図3】図3は、遅延型過敏症アッセイを用いる、Rv 1796タンパク質由来のLipと命名されるグリコペプチド(SEQ ID NO:3)の抗原活性測定を示す。エム.ツベルクローシス(PPD)由来の標準的な精製タンパク質を0.1ml中0.25μg用量で正の対照として用いる。Lipペプチドを0.1ml中0.02μg用量で用いる。α-マンノシダーゼ又はサブチリシンで処理したLipペプチドは、同一用量ではネガティブである。結果を紅斑反応直径により示す。
【図4】図4は、インビトロのリンパ球増殖アッセイを用いる Lipペプチドの抗原活性を示す。Tリンパ球によるグリコシル化Lipペプチド(天然Lip)の認識を、脱グリコシル化ペプチド(Lip + α-マンノシダーゼ)又は抗-T-リンパ球CD4+レセプター抗体と組み合わさったLipペプチド(Lip + 抗Cd4)の認識と比較する。
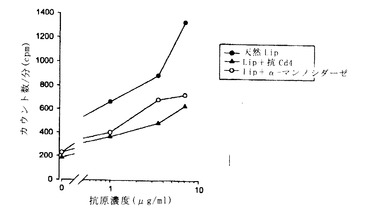
【図5】図5は、遅延型過敏症アッセイを用いる、エム.ツベルクローシスから精製した天然のApa(天然Apa)又はイー.コリで産生される脱グリコシル化組換えApa(イー.コリrApa)の抗原活性測定を、モルモットの免疫化の関数として示す。後者を、皮内注射される生BCG又はサイトメガロウイルス初期プロモーターのコントロール下に位置するApaのコード配列を含むプラスミドpAG831もしくはpAG832で予め免疫化した。プラスミドでのモルモットの免疫化は、遅延型過敏症反応によって発現され得る感作を生ずる。この反応を生ずるには2つの型の抗原は等価であるが、BCGでの免疫化後には、グリコシル化された天然のApaのみが抗原性である。
【図6】図6は、α-(1,2)結合を介して結合する2又は3個のマンノース残基からなる単位の調製を示す。
【図7a】図7(7a及び7b)は、2又は3個のマンノース単位で官能化されたトレオニンの調製を示す。
【図7b】図7(7a及び7b)は、2又は3個のマンノース単位で官能化されたトレオニンの調製を示す。
【実施例】
【0050】
実施例1:Apaタンパク質の抗原性におけるオリゴサッカライド残基数の重要性
1.材料と方法
a)α-マンノシダーゼでの消化によるApaの制限された脱グリコシル化
上記のHornらによって記載されるプロトコルにしたがって、エム.ツベルクローシスの培養上清から精製した450μgのApaタンパク質を、450μl量の緩衝液A (100 mM CH3COO-Na+、2mM ZnCl2)に希釈する。
最初の時点で、75μlのApaタンパク質溶液を除き、25μlの緩衝液Aに希釈し、対照として凍結する。1mg/ml (3IU/ml, Oxford Glycosciences)のα-マンノシダーゼ125μlを375μlのApa溶液に加え、500μlの反応容量を37℃でインキュベートする。30分、1時間、4時間、16時間及び24時間後、100μlの反応物を除き、-20℃で凍結する。
【0051】
b)消化産物の精製
100μlの試料を90℃で2分加熱し、次いで 急冷し、真空下で乾燥させ、水中0.1%のトリフルオロ酢酸300μl(溶液B)に再懸濁する。
Apa消化産物を、90分で溶液B中の0〜90%アセトニトリル勾配を用いて逆相クロマトグラフィーカラム(Ressource RPC, Pharmacia)でα-マンノシダーゼから分離する。Apaを、51.5%±0.5%のアセトニトリルに相当する時間t=68分でカラムから溶出する。Apaに相当する画分を回収し、凍結乾燥し、水中5%のブタノール溶液(溶液C)に再懸濁し、次いで真空下で乾燥させる。精製した試料を次いで溶液C 100μlに再懸濁する。
【0052】
c)Apa消化産物の生化学分析
各試料のオリゴサッカライド組成を、上記のHornらに記載される条件下で質量分析により分析する。
各試料中に存在するタンパク質の相対量を評価するために、210 nmでの吸光度を測定する。
次いで、試料を乾燥し、その濃度を滴定緩衝液(緩衝液D:PBS、0.9% NaCl、0.05% Tween 80)中で1 mg/mlに調整する。
【0053】
d)遅延型過敏症アッセイにおける、α-マンノシダーゼでのApaの制限された消化産物の抗原活性の生物学的滴定
2回の注射点で生BCG 2mgを皮内注射することによりあらかじめ3ヶ月免疫化したモルモットでの遅延型過敏症アッセイを用いて、抗原活性を測定する。
各試料を緩衝液Dで2μg/mlの濃度に希釈し、この希釈液100μl (0.2μg)を2頭のあらかじめ免疫化したモルモットのバッチに皮内注射する。
【0054】
動物の種々のバッチは以下のとおりである:
・バッチ1:緩衝液D 100μlを投与した負の対照
・バッチ2:Apa t=0
・バッチ3:Apa t=30分
・バッチ4:Apa t=1時間
・バッチ5:Apa t=4時間
・バッチ6:Apa t=16時間
・バッチ7:Apa t=24時間
・バッチ8:正の対照(10ツベルクリン単位(TU)に相当するマイコバクテリウム・ツベルクローシス(PPD)由来の標準的な精製タンパク質0.25μg)
注射から24時間後、紅斑反応直径の平均を、種々の動物バッチについて測定し、試料のツベルクリン力価をPPD標準に対して測定する。
【0055】
2.結果
結果を図1と2に示す。
α-マンノシダーゼでの消化動態の関数としてApaの抗原活性分析(図1)は、Apaの抗原活性が、α-マンノシダーゼでの消化中に徐々に失われることを示している:1時間で66%、4時間で86%及び長い消化には97〜99%。
種々の消化時間で得た生成物のマンノース組成の分析(図2)は、
・ 天然のApa分子が、6〜8マンノース残基を有し、かつ
・ 3〜6マンノース残基が依然としてあるApa分子は、その抗原活性の86%を失うことを示している。
【0056】
Apaのオリゴマンノース組成は、ジマンノース(T10及びT18)、マンノース(T27)、マンノース、ジマノース又はトリマンノース (T277)、上記のDobosらのとおりであることが分かっている。さらに、α-マンノシダーゼは、エキソマンノシダーゼである。
その結果、結果は、
・ Apaの4オリゴマンノース鎖の1又は2個の末端マンノースの損失は、抗原活性の劇的な損失をもたらし、かつ
・ Apaの抗原性は、グリコシル化トレオニン残基の1以上に対するジマンノース又はトリマンノースの存在に関連している
ことを示している。
【0057】
実施例2:エム.ツベルクローシスのLipグリコペプチドの立証
材料と方法
a)グリコペプチドの精製
a1)粗物質の調製
マイコバクテリウム・ツベルクローシス(H37Rv)株の細菌を、Sauton合成培地(culture medium, H. Cassagne, 1961, Institut Pasteur編集、2巻、242頁)で20日間培養する。10,000 Daより大きい分子量分子のみを保持するように、培地に分泌された分子を限外ろ過膜(PM10, AMICON)で濃縮し、次いで凍結乾燥する。約10 gの凍結乾燥物が、培養培地60Lについて得られる。
【0058】
a2)分子ろ過(工程1)
調製用カラムを、Sup75 プレップグレードマトリクス(Pharmacia)で充填する。この50×750 mmのカラムを、1ml/minの流速でリン酸緩衝液(50 mM Na2/K PO4, pH 7.1;100 mM NaCl;4%ブタノール)で平衡化する。上記の粗物質を、100ml当たり10gの最終濃度で平衡化緩衝液に溶解し、4時間43,000 gで遠心分離し、次いで0.22μmのろ紙で濾過して清澄にする。13 mlを注入し、280 nmでの吸光度を介して検出される種々の画分をPM10膜上で濃縮し、次いで凍結乾燥する。
溶出画分700〜800 mlは極めて抗原性である:遅延型過敏症は、生BCGで免疫化したモルモットで認められる;この画分は、他方、熱不活化BCGで免疫化したモルモットでは比較的不活性である。
【0059】
a3)イオン交換(工程2)
24×250 mmのPharmacia Source 15Q調製用カラム(15μm)を、8barの最大圧を用いて流速5ml/minで20 mM tris/HCl、pH8.4% ブタノール緩衝液で平衡化する。当初の緩衝液10mlに溶解した上記の画分500mgを注入した後、同一緩衝液で0〜150mMの直線状のNaCl勾配を適用する。溶出した画分を280 nmの吸光度で検出し、PM10膜で濃縮し、次いで凍結乾燥する。
40〜75 mM NaClで溶出された画分は、極めて抗原性である;遅延型過敏症が生BCGで免疫化されたモルモットで認められる;この画分は、他方、熱不活化BCGで免疫化したモルモットで比較的不活性である。
【0060】
a4)C8カラムでの逆相(工程3)
4.6×100 mmのPharmacia RPCカラム(逆相カラム) Resource 15RPCを、流速1 ml/minで20 mM CH3COO-NH4+ 緩衝液、pH6.5を用いて平衡化する。0〜90%の非直線状アセトニトリル勾配を、緩衝液2ml中の上記画分10mgをカラムに注入した後に適用する。溶出した画分を280 nmで検出し、次いで凍結乾燥前に40℃で真空下に濃縮する。
アセトニトリル濃度18〜22%の溶出画分は、生BCGワクチンで免疫化したモルモットでの遅延型過敏症を顕在化する点で極めて抗原性であり、熱不活化BCGで免疫化したモルモットでは比較的不活性である。
【0061】
a5)C18カラムでの逆相(工程4)
C18逆相マイクロボア(microbore)カラム(Browlec lab. 1×250 mm)を、20 mM CH3COO-NH4+ 緩衝液、pH6.5を用いて1ml/minの流速で平衡化する。0〜90%の非直線アセトニトリル勾配を、カラムに上記の画分を注入した後、適用する。
220 nmでのみ検出された画分を約11%濃度のアセトニトリルで溶出する。この画分(3mg)は、生細菌で免疫化したモルモットでの遅延型過敏症反応の顕在化に極めて活性で、熱不活化BCGで免疫化したモルモットでは比較的不活性である。
【0062】
b)精製グリコペプチドの生化学的分析
最終精製工程で得た画分を、製造者の指示に従って改変エドマン技術(Applied Biosystems 473A)を用いてシークエンスした。
各試料の組成を、上記のHornらによって記載される条件下で質量分析(MALDI-TOF 分光計)により分析する。
c)α-マンノシダーゼでのグリコペプチドの消化
上記の精製グリコペプチド9μgを65μgの100 mM CH3COO-Na+緩衝液、pH5に溶解し、次いで、3μlの1mg/ml α-マンノシダーゼ溶液(Oxford Glyco System)、つまり90 mUのα-マンノシダーゼを加える。全てを消化するように反応を24時間37℃でインキュベートし、次いで得られた生成物を真空下で乾燥する。
【0063】
d)ペプチドのサブチリジンでの消化
上記の精製グリコペプチド690 ngを、5μの100 mM炭酸アンモニウム緩衝液pH8に溶解し、次いで1μlの100μg/ml サブチリジン溶液、つまり約100 ngを加える。反応を24時間37℃でインキュベートし、次いで反応生成物を真空下で乾燥させ、滴定緩衝液(緩衝液D)に溶解する。
e)遅延型過敏症アッセイを用いるグリコペプチドの抗原活性の生物学的滴定
未消化又はα-マンノシダーゼもしくはサブチリジンで消化した上記の精製グリコペプチド0.02μgを、実施例1に記載のプロトコルにしたがってあらかじめ免疫化したモルモットのバッチに注射する。結果を、紅斑反応直径の値で示す。対照は、10TUに相当する0.25μgのPPDからなる。
f)インビトロのリンパ球増殖アッセイを用いるグリコペプチドの抗原活性測定
アッセイの条件は、上記のHornらの記載する条件である。
【0064】
2.結果
a)Lip グリコペプチドの精製及び生化学的分析
精製グリコペプチドについて行った質量測定は、測定が162質量単位値異なると、マンノースでおそらくグリコシル化される複雑な分子の存在を示す。α-マンノシダーゼで処理したペプチドの質量に相当する6951 Daの質量は、これらの分子の最少質量としてみなされる。
精製グリコペプチドのN-末端配列は、主要配列 TIPTT... と少数配列IPTTE...の存在を示している。
これらの結果は、配列(SEQ ID NO:3)がエム.ツベルクローシスの Rv1796遺伝子によってエンコードされるタンパク質由来ペプチドのN-末端フラグメントの配列であり、Sanger bankのエム.ツベルクローシス株H37Rvのゲノム配列の注釈に関し、該タンパク質の169〜239位に伸長している、Lipと称されるマンノシル化グリコペプチドに適合している。
【0065】
b)遅延型過敏症アッセイを用いるLipグリコペプチドの抗原活性測定
グリコペプチドは、生細菌で免疫化したモルモットの遅延型過敏症反応の顕在化に極めて活性であり、他方、熱不活化BCGで免疫化したモルモットでは比較的不活性である。
グリコペプチドの抗原活性は、精製工程中に増す:
− 工程1:得られた画分は、生BCGで免疫化したモルモットで180,000 TU/mg及び熱不活化BCGで免疫化したモルモットで10,000 TU/mgの活性を有する。
− 工程2:得られた画分は、生BCGで免疫化したモルモットでは900,000 TU/mg及び熱不活化BCGで免疫化したモルモットでは30,000 TU/mgの活性を有する。
− 工程3:精製画分は、生BCGで免疫化したモルモットでは1,000,000 TU/mgより大きく、熱不活化BCGで免疫化したモルモットでは30,000 TU/mg未満の活性を有する。
【0066】
図3に示す結果は、
− 37℃で24時間のα-マンノシダーゼの作用は、95%より高い抗原活性の損失を引き起こす:脱グリコシル化後、画分は、1,000,000 TU/mgの活性から30,000 TU/mg未満の活性に低下した、
− サブチリジンの作用は、抗原活性を廃する、及び
− 等量のタンパク質で、Lipグリコペプチドは、マイコバクテリウム・ツベルクローシス(PPD)由来の標準的な精製タンパク質より少なくとも10倍活性が高い、ことを示している。
【0067】
c)インビトロのリンパ球増殖アッセイを用いるLipグリコペプチドの抗原活性測定
図4に示す結果は、T-リンパ球増殖がペプチド濃度に依存していることを示している。この増殖は、Tリンパ球がCD4分子に対する抗体で処理される際、又はグリコペプチドがα-マンノースで処理される際、重要ではない。
【0068】
実施例3:Apaタンパク質をエンコードする裸のDNAでの免疫化による、Tエピトープの定義におけるApaのオリゴサッカライド残基の役割の立証
1.材料と方法
a)Apaタンパク質をエンコードする配列を含むプラスミドの構築
サイトメガロウイルス初期プロモーター配列を含むプラスミド pS65T (Clontech)を、制限酵素NheIとBspEIで切断し、クレノウ酵素で修復し、次いで連結してプラスミドpAG800を得る。
プラスミドpAG800を酵素ApaIで切断して、それ自体にハイブリダイズしたオリゴヌクレオチド12M48 (5' CAACGTTGGGCC 3';SEQ ID NO:4)と連結し、プラスミドpAG802を得る。
【0069】
シグナル配列を欠くApaのコード配列を含む875塩基対のフラグメントを、オリゴヌクレオチド22M42 (5' TCCCAAGCTTTTGGTAGCCG 3';SEQ ID NO:5)と33M44 (5' CTAGGATCCACCATGCCGGAGCCAGCGCCCCCG 3';SEQ ID NO:6)を用いて、プラスミドpLA34-2 (Laqueyrerie, 1995, Infect. Immun., 63, 4003-4010)からポリメラーゼ連鎖反応(PCR)により増幅する。
オリゴヌクレオチド33M44を、コザック型のコンセンサス翻訳開始部位を含むように合成した(Nucl. Acids Res., 1987, 15, 8125-8148)。PCRで得たフラグメントをBamHIとEcoRVで切断し、BgIIIとSmaIで切断したプラスミドpAG802に挿入し、プラスミドpAG803を得る。これらの操作中、オリゴヌクレオチド配列5' CAACGTTGGGCC 3'が失われる;Psp1046 ISSと称されるこの配列は、配列IL-12p40 ISS と同様に、免疫反応を増す免疫刺激配列であると考えられる(Lipford GBら、1997, Eur. J. Immunol., 27, 3420-3426)。
【0070】
Psp1046 ISS配列を、オリゴヌクレオチド25M46 (5' GATCCCCCCCCAACGTTCCCCCCCG 3';SEQ ID NO:8)とハイブリダイズしたオリゴヌクレオチド25M45 (5' GATCCGGGGGGGAACGTTGGGGGGG 3';SEQ ID NO:7)をクローニングすることによってプラスミドpAG803のBamHI部位に挿入し、プラスミドpAG831を得る。
IL-12p40 ISS配列を、オリゴヌクレオチド24M64 (5' GGGCCCTTGGAACGTCATAGCGCT 3';SEQ ID NO:10)とハイブリダイズしたオリゴヌクレオチド24M63 (5' AGCGCTATGACGTTCCAAGGGCCC 3';SEQ ID NO:9)をクローニングすることによって、プラスミドpAG803のBamHI部位に挿入し、プラスミドpAG832を得る。
【0071】
エシェリキア・コリ株XL1 Blueを形質転換後、上記のプラスミドを、カナマイシン25μg/mlを含むLB培養培地(Sambrookら、Molecular cloning: A laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989)で増幅する。細菌溶解物をTriton X-114 (1%)で処理することによりエンドトキシンを除く先の工程の後、プラスミドDNAを製造者の指示に従ってMaxiPrep QIA フィルターカラム(QIAGEN)で精製する。
【0072】
b)プラスミドpAG831とpAG832でのモルモットの免疫化
わき腹に2回皮内注射することにより、上記のように調製し、精製したプラスミドpAG831又は pAG832のDNA 50μgで体重300〜400gのモルモット(Hartley)を免疫化する。
対照は、実施例1又は実施例2に記載される条件下で生BCGで免疫化したモルモットの群からなる。
【0073】
c)遅延型過敏症アッセイを用いる、裸のDNAで免疫化したモルモットにおける真核細胞により産生されるApaタンパク質の抗原活性測定
免疫化から1〜2ヶ月後に、遅延型過敏症反応を、天然のApaタンパク質又はエシェリキア・コリの形質転換株で産生される組換えApaタンパク質に対して測定し、タンパク質を上記のHornらに記載されるプロトコルにしたがって精製する。
天然のApa及び組換えApaを、滴定緩衝液(緩衝液D) 100μl中0.2μg用量で皮内注射する。抗原活性を、実施例2に記載されるように測定する。
【0074】
2.結果
図5に示す結果は以下のとおりである:
・ 真核プロモーターの制御下でApaのコード配列を含むプラスミドpAG831又はpAG832で免疫化したモルモットは、天然のApaタンパク質に対する免疫反応(抗原とまとめられる際には、遅延型過敏症アッセイ又はインビトロT-リンパ球増殖アッセイを用いて測定できる抗体及びCD4+型のT-反応)を多くの場合に生ずる。天然のApa抗原に相当する動物では、酵素経路を介して脱グリコシル化される抗原又はイー.コリ由来の非グリコシル化組換え抗原に対するCD4+ Tリンパ球の反応は、グリコシル化天然抗原で見られる反応と同一の強度である。
・ 他方、生BCGで免疫化したモルモットは、天然のApaに対してのみ遅延型過敏症反応を示す。これらの動物は、上記のようにイー.コリで産生される非グリコシル化組換えApaに対し、反応を生じないか、又はかなり低下した反応を生ずる。
【0075】
これらの結果は、以下のことを示している:
1)Apaをエンコードする裸のDNAで免疫化した動物で認められる結果は、マクロファージ又は樹状細胞によるApaタンパク質の食菌及び提示能力が、天然又は組換えの(非グリコシル化)Apaタンパク質と同一であることを示している、
2)生BCGで免疫化した動物で認められる結果と上記の結果の組み合わせは、脱グリコシル化Apaタンパク質に対する反応の不在は、マクロファージ又は樹状細胞によるその提示能力の低下ではなく、CD4+ Tリンパ球による認識不在に起因していることを示している。その結果、エム.ツベルクローシス又は生BCGによって産生されるような、天然型のApa又はLipタンパク質の側鎖のオリゴマンノース残基は、CD4+ Tリンパ球によって認識されるTエピトープの構成で役割を果たしている。
【0076】
実施例4:グリコシル化ペプチドSEQ ID NO:1、SEQ ID NO:2 及びSEQ ID NO:3の調製
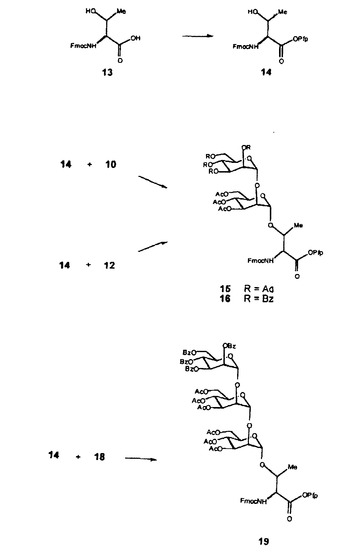
1)グリコシル化シントン15、16及び19の調製
ペプチド合成の前に、グリコシル化シントン、つまり2又は3個のマンノース残基で官能化したトレオニンを調製する。
・ 化合物5及び8の調製(図6)
化合物5及び8の調製は、それぞれH. FRANZYKら(J. Chem. Soc. Perkin Trans. 1, 1995, 2883-2898)及びR.K. NESSら(J. Am. Chem. Soc. Perkin, 1950, 72, 2200-2205)に記載されている。
市販の過アセチル化マンノース1(つまり、1,2,3,4,5-ペンタ-O-アセチル-α-D-マンノピラノース)を、A. LEVENEら(J. Biol. Chem., 1931, 90, 89-98)によって記載されるように、酢酸中の臭化水素の作用によりアノマー位で臭素化する。活性化中間体2を、2,6-ジメチルピリジン/メタノール混合物でオルソエステル3に環化する。10%水性トリフルオロ酢酸/アセトニトリル混合物中で0℃での酸加水分解によるオルソエステルの位置選択的な開環は、1,3,4,6-テトラ-O-アセチル-α-D-マンノピラノース(5)を生じる。位置異性体4も単離される。
【0077】
市販のマンノース6は、ピリジン中の塩化ベンゾイルの作用により7に過ベンゾイル化される。後者は、酢酸中の臭化水素の作用により8に活性化される。しかし、このプロトコルと以下の方法では、臭化水素の作用による以外の活性化方法、例えば当業者に公知の方法を用いてもよい。
・ 二糖10及び12の調製(図7a)
化合物10及び12の調製は、それぞれA. JANSSONら(J. Chem. Soc. Perkin Trans. 1, 1992, 1699-1707)及びH. FRANZYKら(同節)に記載されている。化合物2及び5は、ジクロロメタン中でトリフルオロメタンスルホネート銀(又はいずれかの他の縮合反応プロモーター)の存在下で縮合され、過アセチル化二糖9を生じ、次いで酢酸中の臭化水素の作用により臭素化前駆体に活性化する。同一のプロトコルによれば、化合物5及び8は、縮合されて化合物11を生じ、それ自体12に活性化される。
【0078】
・ 三糖18の調製(図7a)
活性化二糖12を、ジクロロメタン中のトリフルオロメタンスルホネート銀の存在下で単糖アクセプター5に縮合し、過アセチル化三糖17を生じ、次いでこれを酢酸中の臭化水素の作用により臭素化前駆体18に活性化する。
・ 2つのマンノース単位を有するシントン15ならびに16、及び3つのマンノース単位を有するシントン19の調製(図7b)
I. SCHONら(Synthesis, 1986, 303-305)に記載されるように、第一級アミン官能基がFmoc基で保護されている市販のトレオニン13の酸官能性を、ジシクロヘキシルカルボジイミド(DCCI)の存在下でペンタフルオロフェノール(pfp)の作用によりエステルの形態で阻害し、アクセプター前駆体14を生じる。
【0079】
シントン15及び16の調製は、それぞれA. JANSSONら(同節)及びH. FRANZYKら(同節)により記載されている。ジクロロメタン中のトリフルオロメタンスルホネート銀の存在下で行なわれる活性化二糖10及び12との化合物14の縮合は、それぞれシントン15及び16を生じる。同じプロトコルによれば、化合物14と活性三糖18との縮合は、シントン19を生じる。
【0080】
2)グリコシル化ペプチドSEQ ID NO:1、SEQ ID NO:2及びSEQ ID NO:3の調製
ペプチドは、Fmoc化学を用いて固相で合成される。ペプチド合成は、所望の配列を生産する一方、ペンタフルオロフェノールの活性化エステルの形態のグリコシル化シントン(シントン15、16及び19)を挿入するために必要なアミノ酸を用いて自動合成機行われる。
使用されるシントンに応じて、2つのマンノース残基で官能化(ペプチド合成中のシントン15及び/又は16の挿入)されるトレオニンからなるペプチド又は3つのマンノース残基で官能化(ペプチド合成中のシントン19の挿入)されるトレオニンからなるペプチド又は2つのマンノースで官能化されるトレオニンと33のマンノース残基で官能化(ペプチド合成中のシントン19及び15及び/又は16の挿入)されるトレオニンの双方を含むペプチドのいずれかが得られる。
【0081】
合成の最後に、トリフルオロ酢酸を用いて固体支持体からペプチドを切断し、種々のアミノ酸及びマンノースのヒドロキシル官能性を脱保護した後、ペプチドを逆相高速液体クロマトグラフィー(HPLC)により精製する。その構造を、当業者に公知の技術、例えば質量分析及びアミノ酸分析を用いて制御する。
次に、当業者に公知の有機化学技術を用いる化学合成により、アミド官能性(ペプチドSEQ ID NO:1及びSEQ ID NO:3のC-末端位)ならびにアセテート官能性(ペプチドSEQ ID NO:2及びSEQ ID NO:3のN-末端位)を導入する。
【0082】
実施例5:イー.コリで産生されるApaペプチドでの免疫化による、Tエピトープの定義におけるApaのオリゴサッカライド残基の役割の立証
1)材料と方法
当業者に周知のイー.コリにおける組換えタンパク質のクローニング、発現及び精製の従来技術(例えば、Current Protocols in Molecular Biology (Frederick M. AUSUBEL, 2000, Wiley and Son Inc, Library of Congress, USA)に記載されるプロトコル参照)にしたがって、Apaの250〜280位に相当するペプチドをボルデテラ・ペルツシス(Bordetella pertussis)のサイクラーゼのフラグメントとの融合体の形態でイー.コリで産生した。
体重300〜400gの5頭のHartleyモルモットの3群を、アジュバント溶液0.1ml中のこの精製Apaペプチド20μgを用いて、1ヶ月空けて2回皮内注射し、免疫化した。
【0083】
実施例1に記載される条件下で生BCGで予め4ヶ月間免疫化した4頭のモルモット3群を、対照として用いる。
免疫化から1〜2ヶ月後、遅延過敏症反応を、天然のApaタンパク質、イー.コリで産生される組換えApaタンパク質及び実施例1に記載されるようにして調製された脱グリコシル化Apaタンパク質に関して、実施例3に定義する条件下で測定した。
【0084】
2)結果
Apa融合ペプチド又は生BCGで免疫化したモルモットの遅延過敏症反応を、天然のApaタンパク質、イー.コリで産生される組換えApaタンパク質及び脱グリコシル化Apaタンパク質に関して測定した。紅斑反応の直径(mm)で表された結果を、以下の表1に示す:
【0085】
【表1】
【0086】
上記の表1に示すように、生BCGで免疫化したモルモットで見られる遅延過敏症反応は、天然のApa分子の注射後に著しい。反応は、化学的に脱グリコシル化した分子又はイー.コリで産生される分子の注射後に極めて弱いか、あるいはない。他方、ボルデテラ・ペルツシスのサイクラーゼフラグメントとApa分子の内部フラグメントとの融合に相当する組換え分子で免疫化したモルモットについては、感作は、天然分子又は脱グリコシル化分子に対して同一である。
【0087】
これらの結果は、Apa分子のグリコシル化Tエピトープが、生細菌で免疫化されたモルモットによって選択的に認識されることを示している。また、結果は、モルモットによる脱グリコシル化分子の認識の不在又は低下は、これらの分子の低下した固有の抗原性に伴わないことを示している。
上記から明らかであるように、この発明は、より具体的に記載したにすぎないその実施方法、調製及び適用にいかなる方法でも制限されない;逆に、本発明は、本発明の概要又は範囲を逸脱しない限り当業者に起こりうる全ての変形を包含する。
【技術分野】
【0001】
この発明は、病原性微生物のようなもの(細菌又は真菌)による免疫化及び感染診断に使用できる、病原性微生物由来の免疫原性グリコペプチド及びその選択ならびに製造方法に関する。
【背景技術】
【0002】
これらの感染を予防し、治療するために実行されている手段は、まず、感染をモニターし、治療できるスクリーニングと、次いで免疫化からなる。
ヒト医学で最も深刻な感染の一例としてエム.ツベルクローシスでの感染を挙げて、これらの手段を以降に例示する。具体的には、正常な免疫反応を有するエム.ツベルクローシスに感染した個体の5〜10%が、深刻な疾患(結核)に進行する;この頻度は、免疫反応を欠いている個体(HIV感染、免疫抑制剤での治療など)ではさらに高い。
【0003】
診 断
現在利用可能な種々の技術には、以下が挙げられる:
− 結核を正確に診断するのに最も厳密な手段であるエム.ツベルクローシスの純粋培養物の生産。それは、肺結核症例の2/3を診断できる穏やかな高感度技術である。結果は、最低3〜4週間後にのみ、時には2ヶ月培養した後にのみ利用できる。標識した前駆体を用いる培養技術の使用はこれらの遅れを短縮できるが、にもかかわらず依然として相当な期間である。培養によるエム.ツベルクローシスのこの検出は、肺結核のためにすら時には入手しにくく、症例の約1/3が生物学的追認を受けない細菌含有試料を要する。時折り、この検査は、肺外形態の疾患に対する特殊な医学的介入(脳脊髄液の腰椎穿刺又はリンパ節バイオプシー)を要する。
【0004】
− 分子遺伝学に基づく微生物学的技術(PCR)は、細菌含有試料を得るという同一の要件に直面している。さらに、試料中にPCR反応阻害剤が存在するために、その由来を調節できず、時にこれらの技術を使用できない。技術は、一般的な実施には有効ではない。
− 現在、診断用途に適合した感度と特異性を有する血清学的診断法はない。
【0005】
− ツベルクリンに対する反応は、個体が感作され、エム.ツベルクローシスに感染しているか、又はBCGで免疫化されていることを示す。ツベルクリンは、実際エム.ツベルクローシス抗原の混合物であり、したがって、エム.ツベルクローシスでの感染とBCGでの免疫化とを識別することができない。というのは、ワクチン抗原とエム.ツベルクローシスとで交差反応が極めて多いからである。さらに、ツベルクリンに対するこの反応は、進行中の疾患である結核をエム.ツベルクローシスでの感染と識別することができない。
【0006】
ワクチン
BCGでの免疫化は、一次感染(エム.ツベルクローシスの初期増殖)、特にこれらの細菌の二次転移を制御することができる。それは、おそらく現在利用できる有効な治療がない潜伏期の感染の発症を減じるのに寄与している。BCGは、特に副作用なく、結核に対して30億人以上の個体を免疫化するために使用されている。BCGでの免疫化中、毒性が弱毒化されたこれらの細菌の局所増殖がある。細胞性免疫が、誘発される。それは、マイコバクテリアのタンパク質又は抗原に対する遅延型過敏症(HSR)(ツベルクリンに対する反応)を引き起こし、エム.ツベルクローシスでの感染に対する耐性を増した。これらの2つの免疫反応(HSR-型感作及び耐性増加)は、マイコバクテリア抗原と反応するTリンパ球によって支持される。
【0007】
BCGは、急性型感染(例えば子供の結核性脳膜炎)に対しても保護する。その有効性は、大人では多岐にわたる。BCGとツベルクローシス複合体に属さない他のマイコバクテリアとの交差反応性の存在、及びマイコバクテリウム・ツベルクローシスのある免疫原性抗原のBCGゲノムでの不在、又は感染中のこれらの抗原に対する異なる発現プロフィルは、多岐にわたるBCGの有効性を説明し得る。
さらに、BCGは、毒性が弱毒化された生株である。したがって、それは、免疫抑制された個体、特にヒト免疫不全ウイルス(HIV)に感染したことが確認された個体での使用を妨げる残留病原力を有している。
【0008】
これらの感染により効果的に対抗するために、これらの感染の原因である病原性微生物に対して保護する抗原に基づいて、診断上の手段とワクチン、特に危険性のない「サブユニット」ワクチンを有することが賢明であろう。
強力な保護免疫反応を誘導できるこれらの病原性微生物の分子を見出すために、この意味において幾つかの研究が行われている。つまり、J. Hessら(C.R. Acad. Sci. Paris, 1999, 322: 953-958)は、結核に対するワクチンとして使用できる抗原が有すべき性質を概説している。その概説では、単一抗原よりむしろ予備選択した抗原の組み合わせを用いることが重要視されている。特に、種々の株に高度に保存される領域の存在、毒性株と弱毒性株の遺伝子発現プロフィルの相違、免疫反応のエフェクター細胞(B、CD4+ T、CD8+ Tリンパ球)に関する反応性又は主要組織適合性複合体(MHC)の大多数のHLA分子へのこれらの抗原の結合能力などの基準に基づくこれらの抗原の選択が推奨されている。
【0009】
これらの抗原の幾つかは、シー.アルビカンス(C. albicans)のマンノタンパク質のような表面抗原形態(Buurmanら、PNAS, 1998, 95, 7670-7675)、又はエム.ツベルクローシスでの分泌抗原形態: MPT59 (30 kDa)、85A (32 kDa)、MPT64 (23 kDa)、hsp71 (71 kfla)、MPT51 (24 kDa)、MPT63 (16 kDa)及びESAT-6 (6 kDa) (Andersen, Infect. Immun., 1994, 62, 2536-2544;Horwitzら、PNAS, 1995, 92, 1530-1534)のいずれかで存在する。これらのエム.ツベルクローシス抗原は、CD4+ Tリンパ球によって優先的に認識されるので、免疫化組成物の潜在的な候補物としてすでに提案されている(Andersenら、上記;Horwitzら、上記)。
【0010】
また、MHCクラスII分子によって提示され、かつ特異的CD4+ Tリンパ球によって認識されるエピトープを含むペプチドをエム.ツベルクローシス抗原から単離することが提案されている;このようなエピトープは、特に2つのタンパク質について報告されている: ESAT-6 (Olsenら、Eur. J. Immunol., 2000, 30, 1724-1732)及びMPT-39 (Mustafaら、Inf. Immunol., 2000, 68, 3933-3940)。
【0011】
幾つかの観察が、発明者によって以前になされている(Romainら、Inf. Immun., 1993, 61, 742-750;Romainら、Proc. Natl. Acad. Sci. USA 1993, 90: 5322-5326):
− 生細菌のみが保護免疫を誘導でき、死菌細菌も保護なしに免疫反応を誘導する;
− 培養培地では、細菌の成長中に細菌によって放出され、生細菌で免疫化された動物の免疫系によって認識されるタンパク質は、これらが死菌細菌での免疫化後にあまり認識されないか、又は全く認識されないので、存在する。
【0012】
選択についてこの二重の基準を用いて、2つの新たなタンパク質を精製した。Apa、又はMPT-32 あるいは45/47 kDa 抗原複合体と命名されるエム.ツベルクローシスによって分泌されるタンパク質は、Rv 1860遺伝子産物である(Laqueyrerieら、Infect. Immun. 1995, 63: 4003-4010)。第二分子は、Rv 1796遺伝子によってエンコードされる推定上のセリンプロテアーゼの内部ペプチドである。
抗原として天然のApaタンパク質を用い、発明者らは、培養物にツベルクローシス群の細菌(エム.ツベルクローシス、エム.ボビス及びBCG)によって分泌されるタンパク質のわずか2%を示すこのタンパク質が、エム.ツベルクローシスに感染しているか、又はBCGで免疫化された動物を由来とする特異的な CD4+ Tリンパ球によって極めて有効に認識される免疫優勢抗原であることを以前に示している(Romainら、 Inf. Immun., 1999, 67, 5567-5572;Horn ら、J. Biol. Chem., 1999, 274, 32023-32030)。
【0013】
これらの同一の研究において、発明者らは、Apaのマンノシル化が、このタンパク質の抗原活性に必須であることも示した:
− α-マンノシダーゼ又はトリフルオロメタンスルホン酸(TFMS)で天然のApaを処理することによって、又はグリコシル化できない細菌(イー.コリ)でApaを発現することによって得られるApaの脱マンノシル化は、100倍の抗原性損失を伴う。
− エム.ツベルクローシスにって産生されるApaとわずかに異なるマンノース組成を全体に有するマイコバクテリウム・スメグマティス(Mycobacterium smegmatis)によって産生されるグリコシル化Apaは、約10/1に低下する抗原活性を有する。
【0014】
さらに、このエム.ツベルクローシスのApa分子は、ジマンノース(T10及びT18)、マンノース(T27)、 マンノース、ジマンノース又はトリマンノース(T277)で、α-(1,2)型のグリコシド結合を介して4個のトレオニン残基(T10、T18、T27 及びT277)に結合した6〜9個のマンノース残基を含むことが報告されている(Dobosら、J. Bacteriol., 1996, 178, 2498-2506)。モノ-、ジ-又はトリ-マンノースを含むこの糖類の構造は、酵母、特にカンジダ・アルビカンス(Candida albicans)由来のマンノタンパク質の構造に似ており、長いオリゴマンノース鎖を有するエフ.メニゴセプティカム(F.meningosepticum)由来タンパク質の構造と異なることに留意すべきである。
【0015】
脱マンノシル化後にみられるApa抗原性の損失は、この抗原の食作用とプロセシング、あるいは CD4+ Tリンパ球による後者の認識の低下に起因している可能性がある。具体的には、ヘキソースに特異的に結合するマクロファージ及び樹状細胞、特にシー.アルビカンス由来マンノタンパク質及びマイコバクテリア由来のリポアラビノマンナンのようなマンノリピドのマンノースレセプターは、ペプチド/クラスII MHC分子複合体形態のこれらの細胞表面に存在する抗原の食作用とプロセシングで役割を果たしている(Stahlら、Current Opinion in Immunology, 1998, 10, 50-55)。また、(N-末端位のリジン残基についてマンノシル化された)マンノシル化ペプチドは、同一の配列を持つ非グリコシル化ペプチドよりも有効に樹状細胞によって食菌され、処理されることも分かっている(Tanら、Eur. J. Immunol., 1997, 27, 2426-2435)。
【0016】
ニワトリのリソチームモデルでは、この抗原のTエピトープを構成するペプチドのグリコシル化類似体であるペプチドが、このグリコシル化エピトープを特異的に認識するCD4+ Tリンパ球を誘導できることが示されている(Deckら、J. Immunol., 1995, 155, 1074-1078)。しかし、CD4+ Tリンパ球によって特異的に認識されるグリコシル化Tエピトープは病原性微生物(細菌/真菌)由来の天然抗原で同定されないので、CD4+ Tリンパ球によるこれらの病原性微生物由来抗原の認識におけるグリコシル化の重要性は依然として立証されべきものである。
さらに、これは、抗原のグリコシル化に関するエム.ツベルクローシスApaと一般的な知見に関するデータであるにもかかわらず、免疫原性組成物又は免疫化組成物及び/又は診断試験において有効に使用できる、これらの病原性微生物のO-グリコシル化タンパク質由来抗原を製造する可能性は、現在のところない。
【0017】
本質的に:
− これらの微生物によって産生されるタンパク質のごく少量の%を示す活性タンパク質は、これらの病原性剤を大量に扱うため危険な方法を用いて、極めて低い収率で精製される、
− 異種発現系(グリコシル化できない真核細胞又は細菌)で産生されるタンパク質は、低い抗原活性を有する、
− エム.スメグマティスのような同種の発現系で産生されるタンパク質は、許容される抗原活性を有するが、それらは複雑な方法を用いては不十分な量で産生される。
【先行技術文献】
【非特許文献】
【0018】
【非特許文献1】J. Hessら(C.R. Acad. Sci. Paris, 1999, 322: 953-958)
【非特許文献2】(Buurmanら、PNAS, 1998, 95, 7670-7675)
【非特許文献3】Andersen, Infect. Immun., 1994, 62, 2536-2544
【非特許文献4】Horwitzら、PNAS, 1995, 92, 1530-1534
【非特許文献5】Olsenら、Eur. J. Immunol., 2000, 30, 1724-1732
【非特許文献6】Mustafaら、Inf. Immunol., 2000, 68, 3933-3940
【非特許文献7】Romainら、Inf. Immun., 1993, 61, 742-750
【非特許文献8】Romainら、Proc. Natl. Acad. Sci. USA 1993, 90: 5322-5326
【非特許文献9】Laqueyrerieら、Infect. Immun. 1995, 63: 4003-4010
【非特許文献10】Romainら、Inf. Immun., 1999, 67, 5567-5572
【非特許文献11】Horn ら、J. Biol. Chem., 1999, 274, 32023-32030
【非特許文献12】Dobosら、J. Bacteriol., 1996, 178, 2498-2506
【非特許文献13】Stahlら、Current Opinion in Immunology, 1998, 10, 50-55
【非特許文献14】Tanら、Eur. J. Immunol., 1997, 27, 2426-2435
【非特許文献15】Deckら、J. Immunol., 1995, 155, 1074-1078
【発明の概要】
【発明が解決しようとする課題】
【0019】
この結果、発明者らは、単独で又は他の剤と組み合わせて投与する際に、一方で免疫抑制された個体(生ワクチンの使用に関連した危険性の消失)を含む全ての個体に使用できるワクチンを構成でき、他方で診断目的で使用できる、保護体液性及び/又は細胞性免疫反応を誘導できる免疫優勢抗原を製造することを目的とした。
脱グリコシル化された天然タンパク質又はイー.コリで産生される組換えタンパク質に少なくとも等しく、さもなければ、より大きい抗原活性を示すグリコタンパク質を合成する病原性微生物(特にマイコバクテリア)由来のあるグリコペプチドが見出された。
また、これらのグリコペプチドを大量に製造するための実施しやすい手段を開発することが、本発明の目的である。
【課題を解決するための手段】
【0020】
本発明の対象は、
a1)少なくとも1つの中性アミノ酸は二糖又は三糖に結合し(グリコシド結合)、アミノ酸の少なくとも15%はプロリンで、プロリンの1つは中性アミノ酸の位置に対して-1〜-4位に位置する、病原性微生物由来で、14〜25アミノ酸からなるグリコシル化Tエピトープから本質的になり、
− クラスII MHC分子で提示され、
− 由来する天然のグリコタンパク質での免疫化で誘導される CD4+ Tリンパ球によって特異的に認識されるが、同一の配列を有する非グリコシル化ペプチドでの免疫化で誘導されるCD4+ Tリンパ球によっては認識されず、かつ
− それらを認識するCD4+ Tリンパ球の増殖と該リンパ球によるサイトカインの分泌を誘導できる
グリコペプチド、及び
b1) Dobosら(J. Bacteriol., 1996, 178, 2498-2506)によって記載されるApa由来の、配列SEQ ID NO:11のグリコペプチドを除く、a1)に定義されるグリコペプチドの配列を含む15〜39アミノ酸の配列を有するグリコペプチド
からなる群から選択される免疫原性グリコペプチドである。
【0021】
グリコシル化Tエピトープから本質的になるこれらのグリコペプチドは、このグリコシル化Tエピトープを介してCD4+ Tリンパ球によって認識される。具体的には、ツベルクローシス群の生細菌での免疫化後、同一配列を有する非グリコシル化ペプチドに特異的なTリンパ球よりもこれらのグリコペプチドに特異的なTリンパ球がかなり多い。
有利には、グリコペプチドは、同一配列を有する対照のペプチドよりも少なくとも10倍、好ましくは少なくとも30倍大きい抗原活性を有する。
【0022】
このグリペプチドは、以下の利点を有する:
− 保護細胞型免疫反応、あるいは体液性反応の誘導、及び免疫抑制個体における抗原としての潜在的な使用、
− 病原性微生物に特異的な多数のTリンパ球によって認識されるための、従来の抗原(弱毒化生微生物の培養物、該培養物から調製される抗原混合物又は非グリコシル化ペプチド)に少なくとも等しく、さもなければより大きい抗原活性、
− 他の微生物、特に他の異型性マイコバクテリアとの交差反応性の問題を除き、免疫化と病原性微生物の診断の有効性を増すことができる、極めて狭い特異性;特に、より具体的には、病原性微生物に排他的に存在するそのオリゴ糖残基は、CD4+ Tリンパ球によって認識されるTエピトープの定義に必須な方法で寄与する;つまり、それらは、そのタンパク質の幾つかをO-グリコシル化できる病原性微生物(ツベルクローシス複合体細菌、フラボバクテリウム・メニンゴセプティカム(Flavobacterium meningosepticum)、カンジダ・アルビカンスなど)での免疫化及び感染を診断するのに特異的な抗原を構成する、
【0023】
− 全体に非病原性(apathogenic)であるための免疫抑制個体での使用、
− 大量での潜在的な生産、
− 進行中の用量とワクチンの有効性についてのより良い標準化、
− 貯蔵と使用の容易化。
グリコペプチドの有利な具体例によれば、中性アミノ酸は、セリンとトレオニンからなる群から選択される。
グリコペプチドのこの具体例の有利なアレンジによれば、それらは、二糖又は三糖に結合した1〜7個のトレオニン残基を含む。
グリコペプチドの別の有利な具体例によれば、二糖又は三糖は、ヘキソース、好ましくはマンノースの二量体又は三量体である。
【0024】
グリコペプチドのさらに別の有利な具体例によれば、グリコシド結合はα-(1,2)結合である。
さらに別の有利な具体例によれば、グリコペプチドは、タンパク質をO-グリコシル化できる病原性微生物、好ましくはマイコバクテリウム・ツベルクローシス又はカンジダ・アルビカンス由来である。
この発明によれば、グリコペプチドは、好ましくは
− エム.ツベルクローシス(Genbank 番号X80268)のApaタンパク質、又は
− エム.ツベルクローシス株H37Rv (Sanger bank)のゲノム配列の注釈に関し、Rv 1796遺伝子によってエンコードされるRv1796タンパク質
に由来する。
【0025】
好ましくは、グリコペプチドは、
− 配列(SEQ ID NO:1)が、Apaタンパク質の配列の1〜39位から伸長する配列であり、SEQ ID NO:1の10、18及び27位のトレオニン残基の少なくとも1つがグリコシド結合を介して二糖又は三糖に結合する、39アミノ酸のグリコペプチド、
− 配列(SEQ ID NO:2)が、Apaタンパク質の配列(C-末端配列)の261〜286位から伸長する配列であり、SEQ ID NO:2の17位のトレオニン残基がグリコシド結合を介して二糖又は三糖に結合する26アミノ酸のグリコペプチド、及び
− 配列(SEQ ID NO:3)がRv 1796 タンパク質の配列の169〜203位から伸長する配列であり、SEQ ID NO:3の4、5、7、13、15、23及び25位のトレオニン残基の少なくとも1つがグリコシド結合を介して二糖又は三糖に結合する35アミノ酸のグリコペプチド
からなる群から選択される。
【0026】
本発明の対象は、
− グリコシド結合を介して二糖又は三糖に結合したグリコシル化中性アミノ酸を溶液中に調製し、
− グリコペプチドのペプチド配列を産生するのに必要なアミノ酸及び上記で得たグリコシル化中性アミノ酸を用いて、固体支持体上でグリコペプチドを合成し、かつ
− 固体支持体からグリコペプチドを切断する
工程からなることを特徴とする、上記のグリコペプチドの合成方法でもある。
【0027】
この方法の有利な具体例によれば、中性アミノ酸は、セリン及びトレオニンからなる群から選択される。
この具体例の有利なアレンジによれば、グリコペプチドが、配列(Tは、2又は3個のグリコシド残基で官能化されたO-グリコシル化トレオニンを示し、かつAcはアセテート官能基を示す):
SEQ ID NO:1:
H2N-DPEPAPPVPTTAASPPSTAAAPPAPATPVAPPPPAAANT-CONH2
SEQ ID NO:2:
AcNH-PAPAPAPAGEVAPTPTTPTPQRTLPA-COOH
SEQ ID NO:3:
AcNH-TIPTTETPPPPQTVTLSPVPPQTVTVIPAPPPEEG-CONH2
を有する際、
【0028】
その方法は、
i)2又は3個のグリコシド残基で官能化されたO-グリコシル化トレオニンを溶液中に調製し、
ii)これらの配列を製造するために必要なアミノ酸及び工程i)で得たO-グリコシル化トレオニンを用いて、固体支持体上で、上記の配列SEQ ID NO:1、SEQ ID NO:2 及びSEQ ID NO:3に相当するペプチドを合成し、
iii)固体支持体からペプチドを切断し、かつ
iv)化学合成により、ペプチドSEQ ID NO:1及びSEQ ID NO:3のC-末端にアミド官能性を、またペプチドSEQ ID NO:2及びSEQ ID NO:3のN-末端にアセテート官能性を導入する
工程からなる。
【0029】
したがって、ペプチドSEQ ID NO:1、SEQ ID NO:2及びSEQ ID NO:3の合成は、そのあいだにグリコシル化アミノ酸が導入される従来の固相ペプチド合成に相当する。固相ペプチド合成の分野で知られているように、使用されるアミノ酸は適当に保護され、必要であれば、ペプチド配列に他のものが導入される前に活性化される。同様に、トレオニンによって担持されるグリコシド残基に存在するヒドロキシルは、ペプチド合成のあいだに適切に保護される必要がある。
いったんペプチド合成が行われると、ペプチドは、固体支持体から分離され、脱保護される。それらは、逆相高速液体クロマトグラフィーによって精製することができる。
【0030】
この具体例の有利なアレンジによれば、工程i)で製造されるO-グリコシル化トレオニンによって担持されるグリコシド結合はヘキソース、好ましくはマンノースであり、マンノース残基は、有利にはα-(1,2)結合を介して互いに結合される。
【0031】
このアレンジの有利な態様によれば、マンノース残基で官能化されるトレオニンは、以下のように製造される:
a2)式(I)及び(II)
【0032】
【化1】
[式中、P1及びP2は、同一又は異なっていてもよく、ヒドロキシル官能性を保護する基を示し、かつXは、臭素原子のような活性化官能性を示す]のマンノース誘導体の製造:
b2)2つのマンノース残基からなり、式(III):
【0033】
【化2】
[式中、P1、P2及びXは、式(I)及び(II)に関して定義されるとおり]
に相当する活性化誘導体の生産をもたらす、式(I)の誘導体と式(II)の誘導体との反応、次いで得られた化合物の活性化、
【0034】
c2)任意に、3つのマンノース残基からなり、式(IV):
【0035】
【化3】
[式中、P1、P2及びXは、式(I)及び(II)に関して定義されるとおり]
に相当する活性化誘導体の産生をもたらす、a2)に定義される式(I)のマンノース誘導体での式(III)の化合物の反応、次いで得られた化合物の活性化、及び
d2)それぞれ式(VI)又は(VII):
【0036】
【化4】
[式中、P1、P2、P3及びP4は上記のとおり]
のグリコシル化トレオニンの生産をもたらす、式(III)の化合物又は式(IV)の化合物の、式(V):
【0037】
【化5】
[式中、P3は、第一級アミン官能性を保護する基を示し、かつP4はヒドロキシル官能性を保護する基を示す]
の適当に保護されたトレオニンとの縮合。
【0038】
保護基P1、P2、P3及びP4は、研究Protective Groups in Organic Synthesis (T.W. GREENE 及びP.G.M. WUTS、第2版1991年、J. WILEY and Sons)に記載される基から選択してもよい。例として、また非制限に、P1とP2はアセチル又はベンゾイル基を示してもよく、P3はFmoc (9-フルオレニルメトキシカルボニル)基を示してもよく、かつP4はペンタフルオロフェニル基を示してもよい。
【0039】
この発明の対象は、上記のように、本発明によるグリコペプチドを合成する方法に付随して有利に行ってもよい、病原性微生物のタンパク質のペプチド配列を用いて免疫原性グリコペプチドを選択し、スクリーニングするための方法であって、方法は、少なくとも
a3)二糖又は三糖に結合した少なくとも1つの中性アミノ酸及びプロリンの1つが中性アミノ酸の位置に対して-1〜-4位に位置する少なくとも15%のプロリンを含む少なくとも1つの14〜25アミノ酸配列を、タンパク質のペプチド配列において、またそれから、検索し、選択し、
b3)上記の合成法にしたがって、工程a3)で選択されるグリコペプチドを調製し、かつ
c3)抗原活性が、同一配列を有する対照のペプチドよりも少なくとも10倍、好ましくは少なくとも30倍大きいグリコペプチドを選択する
工程からなることを特徴とする。
【0040】
このスクリーニング方法の有利な具体例によれば、工程a3)に先立って、少なくとも1つの抗原性グリコタンパク質を予備選択する工程が含まれる。
このスクリーニング方法の別の有利な具体例によれば、工程c3)では、グリコペプチドの抗原活性は、弱毒化病原性微生物又は病原性微生物の抗原画分で免疫化された動物のCD4+ Tリンパ球の活性を測定して評価される。
【0041】
Tリンパ球の活性化は、Current protocols in Immunology (John E. Coligan, 2000, Wiley and son Inc, Library of Congress, USA)に記載されるような従来の免疫技術を用いて立証することができる。例として、リンパ球増殖アッセイ、活性化CD4+ Tリンパ球によって合成されるサイトカイン(タンパク質又はmRNA)のためのアッセイ(イムノアッセイ(ELISA)又はRT-PCR型の重合鎖反応)、又はエム.ツベルクローシスの場合には、遅延型過敏症アッセイが挙げられる。
また、この発明は、上記のような選択及びスクリーニング方法を用いて得られるグリコペプチドを包含する。
【0042】
また、この発明の対象は、免疫原性組成物もしくは免疫化組成物又は診断試薬の製造のための、本発明による少なくとも1つのグリコペプチド又は配列SEQ ID NO:11のグリコペプチドの使用である。
病原性微生物、特にエム.ツベルクローシスでの感染によって誘導される細胞性及び/又は体液性免疫を極めて特異的に検出する本発明によるグリコペプチドは、当業者にそれ自体知られており、細胞性免疫を検出できるいずれかの技術によって結核の診断に有利に用いることができる。例として、T-リンパ球増殖アッセイ及びCD4+ Tリンパ球、特にγ-IFNに特異的なサイトカインのための免疫酵素アッセイが挙げられる。
【0043】
また、本発明の対象は、少なくとも1つの医薬的に許容される賦形剤と組み合わさった、上記の少なくとも1つのグリコペプチドからなることを特徴とする、体液性及び/又は細胞性免疫を誘導できる免疫原性組成物である。
体液性又は細胞性免疫反応の設定でのCD4+ Tリンパ球とCD8+ Tリンパ球つまりBリンパ球との協同作用のために、本発明のグリコペプチドは、抗原に対する免疫化の有効性を増すために、いずれかの他の抗原用の輸送タンパク質(担体)として有利に用いることができる。この抗原/担体の組み合わせは、免疫化抗原に特異的なB及びTリンパ球の選択と増幅を有利に容易にすることができる。
【0044】
本発明の対象は、少なくとも1つの医薬的に許容される賦形剤及び任意に少なくとも1つのアジュバントと組み合わさった、上記の少なくとも1つのグリコペプチドからなることを特徴とする、体液性及び/又は細胞性免疫を誘発し得る免疫化組成物である。
免疫原性組成物又は免疫化組成物の有利な具体例によれば、グリコペプチドは、少なくとも1つのBエピトープ、CD4+型の1つのTエピトープ又はCD8+型の1つのTエピトープからなるタンパク質又はタンパク質フラグメントと組み合わさっている。
【0045】
本発明の目的のため、タンパク質の配列に対して、用語「Bエピトープ」、「CD4+型Tエピトープ」及び「CD8+型Tエピトープ」は、それぞれ抗体、CD4+リンパ球Tレセプター及びCD8+リンパ球Tレセプターに結合し得るこの配列のフラグメントを意味することを意図する。
本発明の目的のため、表現「タンパク質とグリコペプチドの組み合わせ」は、いずれかの物理的手段又は化学的手段、例えばグリコペプチドの配列とタンパク質又はタンパク質フラグメントの配列との融合物の発現による混合と結合の双方を意味することを意図する。
用いられるアジュバントは、従来使用されるアジュバントである;有利には、それらは水酸化アルミニウムとスクアレンからなる群から選択される。
【0046】
グリコペプチドは、ペプチドの免疫原性を増すことのできる、当業者にはそれ自体公知のいずれかの他の手段と任意に組み合わせてもよい。例として、Wilkinsonら、1999, Eur. J. Immunol., 29, 2788-2796により記載されるような分枝状多量体ペプチドを生産できる担体ペプチドへの結合が挙げられる。
本発明の対象は、本発明によるグリコペプチドの1以上に対することを特徴とする抗体でもある。
抗体の有利な具体例によれば、それらは、モノクローナル抗体及びポリクローナル抗体から選択される。
【0047】
本発明の対象は、本発明によるグリコペプチドと抗体からなる群から選択されることを特徴とする診断試薬でもある。
また、本発明の対象は、病原性微生物に感染している恐れのある患者由来の生物学的試料を上記の診断試薬(場合によって抗体又はグリコペプチド)と接触させ、生物学的試料複合体中に存在する抗体/微生物又は試料複合体に存在するグリコペプチド/抗体の形成を検出することからなることを特徴とする、病原性微生物での感染の検出方法である。
【0048】
上記のアレンジに加えて、この発明は、この発明の具体例ならびに添付の図面に言及する以下の記載から明らかになるであろう多くのアレンジをさらに含む:
【図面の簡単な説明】
【0049】
【図1】図1は、遅延型過敏症アッセイを用いる、エム.ツベルクローシスから精製した天然のApaの抗原活性測定を、α-マンノシダーゼによるApaタンパク質の消化動態の関数として示す。結果は、時間の関数としてタンパク質mg当たりのツベルクリン単位で示す。
【図2a】図2は、α-マンノシダーゼでのApaタンパク質の消化動態の関数として、Apa分子のマンノース組成の質量分析を示す。Apaタンパク質の各ピークに相当するマンノース残基数を示し、Apa消化産物の全体的な抗原活性を研究した種々の時間で示す。
【図2b】図2は、α-マンノシダーゼでのApaタンパク質の消化動態の関数として、Apa分子のマンノース組成の質量分析を示す。Apaタンパク質の各ピークに相当するマンノース残基数を示し、Apa消化産物の全体的な抗原活性を研究した種々の時間で示す。
【図3】図3は、遅延型過敏症アッセイを用いる、Rv 1796タンパク質由来のLipと命名されるグリコペプチド(SEQ ID NO:3)の抗原活性測定を示す。エム.ツベルクローシス(PPD)由来の標準的な精製タンパク質を0.1ml中0.25μg用量で正の対照として用いる。Lipペプチドを0.1ml中0.02μg用量で用いる。α-マンノシダーゼ又はサブチリシンで処理したLipペプチドは、同一用量ではネガティブである。結果を紅斑反応直径により示す。
【図4】図4は、インビトロのリンパ球増殖アッセイを用いる Lipペプチドの抗原活性を示す。Tリンパ球によるグリコシル化Lipペプチド(天然Lip)の認識を、脱グリコシル化ペプチド(Lip + α-マンノシダーゼ)又は抗-T-リンパ球CD4+レセプター抗体と組み合わさったLipペプチド(Lip + 抗Cd4)の認識と比較する。
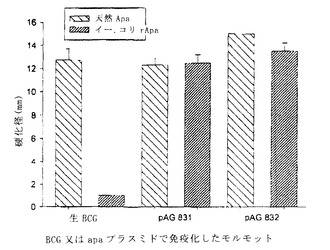
【図5】図5は、遅延型過敏症アッセイを用いる、エム.ツベルクローシスから精製した天然のApa(天然Apa)又はイー.コリで産生される脱グリコシル化組換えApa(イー.コリrApa)の抗原活性測定を、モルモットの免疫化の関数として示す。後者を、皮内注射される生BCG又はサイトメガロウイルス初期プロモーターのコントロール下に位置するApaのコード配列を含むプラスミドpAG831もしくはpAG832で予め免疫化した。プラスミドでのモルモットの免疫化は、遅延型過敏症反応によって発現され得る感作を生ずる。この反応を生ずるには2つの型の抗原は等価であるが、BCGでの免疫化後には、グリコシル化された天然のApaのみが抗原性である。
【図6】図6は、α-(1,2)結合を介して結合する2又は3個のマンノース残基からなる単位の調製を示す。
【図7a】図7(7a及び7b)は、2又は3個のマンノース単位で官能化されたトレオニンの調製を示す。
【図7b】図7(7a及び7b)は、2又は3個のマンノース単位で官能化されたトレオニンの調製を示す。
【実施例】
【0050】
実施例1:Apaタンパク質の抗原性におけるオリゴサッカライド残基数の重要性
1.材料と方法
a)α-マンノシダーゼでの消化によるApaの制限された脱グリコシル化
上記のHornらによって記載されるプロトコルにしたがって、エム.ツベルクローシスの培養上清から精製した450μgのApaタンパク質を、450μl量の緩衝液A (100 mM CH3COO-Na+、2mM ZnCl2)に希釈する。
最初の時点で、75μlのApaタンパク質溶液を除き、25μlの緩衝液Aに希釈し、対照として凍結する。1mg/ml (3IU/ml, Oxford Glycosciences)のα-マンノシダーゼ125μlを375μlのApa溶液に加え、500μlの反応容量を37℃でインキュベートする。30分、1時間、4時間、16時間及び24時間後、100μlの反応物を除き、-20℃で凍結する。
【0051】
b)消化産物の精製
100μlの試料を90℃で2分加熱し、次いで 急冷し、真空下で乾燥させ、水中0.1%のトリフルオロ酢酸300μl(溶液B)に再懸濁する。
Apa消化産物を、90分で溶液B中の0〜90%アセトニトリル勾配を用いて逆相クロマトグラフィーカラム(Ressource RPC, Pharmacia)でα-マンノシダーゼから分離する。Apaを、51.5%±0.5%のアセトニトリルに相当する時間t=68分でカラムから溶出する。Apaに相当する画分を回収し、凍結乾燥し、水中5%のブタノール溶液(溶液C)に再懸濁し、次いで真空下で乾燥させる。精製した試料を次いで溶液C 100μlに再懸濁する。
【0052】
c)Apa消化産物の生化学分析
各試料のオリゴサッカライド組成を、上記のHornらに記載される条件下で質量分析により分析する。
各試料中に存在するタンパク質の相対量を評価するために、210 nmでの吸光度を測定する。
次いで、試料を乾燥し、その濃度を滴定緩衝液(緩衝液D:PBS、0.9% NaCl、0.05% Tween 80)中で1 mg/mlに調整する。
【0053】
d)遅延型過敏症アッセイにおける、α-マンノシダーゼでのApaの制限された消化産物の抗原活性の生物学的滴定
2回の注射点で生BCG 2mgを皮内注射することによりあらかじめ3ヶ月免疫化したモルモットでの遅延型過敏症アッセイを用いて、抗原活性を測定する。
各試料を緩衝液Dで2μg/mlの濃度に希釈し、この希釈液100μl (0.2μg)を2頭のあらかじめ免疫化したモルモットのバッチに皮内注射する。
【0054】
動物の種々のバッチは以下のとおりである:
・バッチ1:緩衝液D 100μlを投与した負の対照
・バッチ2:Apa t=0
・バッチ3:Apa t=30分
・バッチ4:Apa t=1時間
・バッチ5:Apa t=4時間
・バッチ6:Apa t=16時間
・バッチ7:Apa t=24時間
・バッチ8:正の対照(10ツベルクリン単位(TU)に相当するマイコバクテリウム・ツベルクローシス(PPD)由来の標準的な精製タンパク質0.25μg)
注射から24時間後、紅斑反応直径の平均を、種々の動物バッチについて測定し、試料のツベルクリン力価をPPD標準に対して測定する。
【0055】
2.結果
結果を図1と2に示す。
α-マンノシダーゼでの消化動態の関数としてApaの抗原活性分析(図1)は、Apaの抗原活性が、α-マンノシダーゼでの消化中に徐々に失われることを示している:1時間で66%、4時間で86%及び長い消化には97〜99%。
種々の消化時間で得た生成物のマンノース組成の分析(図2)は、
・ 天然のApa分子が、6〜8マンノース残基を有し、かつ
・ 3〜6マンノース残基が依然としてあるApa分子は、その抗原活性の86%を失うことを示している。
【0056】
Apaのオリゴマンノース組成は、ジマンノース(T10及びT18)、マンノース(T27)、マンノース、ジマノース又はトリマンノース (T277)、上記のDobosらのとおりであることが分かっている。さらに、α-マンノシダーゼは、エキソマンノシダーゼである。
その結果、結果は、
・ Apaの4オリゴマンノース鎖の1又は2個の末端マンノースの損失は、抗原活性の劇的な損失をもたらし、かつ
・ Apaの抗原性は、グリコシル化トレオニン残基の1以上に対するジマンノース又はトリマンノースの存在に関連している
ことを示している。
【0057】
実施例2:エム.ツベルクローシスのLipグリコペプチドの立証
材料と方法
a)グリコペプチドの精製
a1)粗物質の調製
マイコバクテリウム・ツベルクローシス(H37Rv)株の細菌を、Sauton合成培地(culture medium, H. Cassagne, 1961, Institut Pasteur編集、2巻、242頁)で20日間培養する。10,000 Daより大きい分子量分子のみを保持するように、培地に分泌された分子を限外ろ過膜(PM10, AMICON)で濃縮し、次いで凍結乾燥する。約10 gの凍結乾燥物が、培養培地60Lについて得られる。
【0058】
a2)分子ろ過(工程1)
調製用カラムを、Sup75 プレップグレードマトリクス(Pharmacia)で充填する。この50×750 mmのカラムを、1ml/minの流速でリン酸緩衝液(50 mM Na2/K PO4, pH 7.1;100 mM NaCl;4%ブタノール)で平衡化する。上記の粗物質を、100ml当たり10gの最終濃度で平衡化緩衝液に溶解し、4時間43,000 gで遠心分離し、次いで0.22μmのろ紙で濾過して清澄にする。13 mlを注入し、280 nmでの吸光度を介して検出される種々の画分をPM10膜上で濃縮し、次いで凍結乾燥する。
溶出画分700〜800 mlは極めて抗原性である:遅延型過敏症は、生BCGで免疫化したモルモットで認められる;この画分は、他方、熱不活化BCGで免疫化したモルモットでは比較的不活性である。
【0059】
a3)イオン交換(工程2)
24×250 mmのPharmacia Source 15Q調製用カラム(15μm)を、8barの最大圧を用いて流速5ml/minで20 mM tris/HCl、pH8.4% ブタノール緩衝液で平衡化する。当初の緩衝液10mlに溶解した上記の画分500mgを注入した後、同一緩衝液で0〜150mMの直線状のNaCl勾配を適用する。溶出した画分を280 nmの吸光度で検出し、PM10膜で濃縮し、次いで凍結乾燥する。
40〜75 mM NaClで溶出された画分は、極めて抗原性である;遅延型過敏症が生BCGで免疫化されたモルモットで認められる;この画分は、他方、熱不活化BCGで免疫化したモルモットで比較的不活性である。
【0060】
a4)C8カラムでの逆相(工程3)
4.6×100 mmのPharmacia RPCカラム(逆相カラム) Resource 15RPCを、流速1 ml/minで20 mM CH3COO-NH4+ 緩衝液、pH6.5を用いて平衡化する。0〜90%の非直線状アセトニトリル勾配を、緩衝液2ml中の上記画分10mgをカラムに注入した後に適用する。溶出した画分を280 nmで検出し、次いで凍結乾燥前に40℃で真空下に濃縮する。
アセトニトリル濃度18〜22%の溶出画分は、生BCGワクチンで免疫化したモルモットでの遅延型過敏症を顕在化する点で極めて抗原性であり、熱不活化BCGで免疫化したモルモットでは比較的不活性である。
【0061】
a5)C18カラムでの逆相(工程4)
C18逆相マイクロボア(microbore)カラム(Browlec lab. 1×250 mm)を、20 mM CH3COO-NH4+ 緩衝液、pH6.5を用いて1ml/minの流速で平衡化する。0〜90%の非直線アセトニトリル勾配を、カラムに上記の画分を注入した後、適用する。
220 nmでのみ検出された画分を約11%濃度のアセトニトリルで溶出する。この画分(3mg)は、生細菌で免疫化したモルモットでの遅延型過敏症反応の顕在化に極めて活性で、熱不活化BCGで免疫化したモルモットでは比較的不活性である。
【0062】
b)精製グリコペプチドの生化学的分析
最終精製工程で得た画分を、製造者の指示に従って改変エドマン技術(Applied Biosystems 473A)を用いてシークエンスした。
各試料の組成を、上記のHornらによって記載される条件下で質量分析(MALDI-TOF 分光計)により分析する。
c)α-マンノシダーゼでのグリコペプチドの消化
上記の精製グリコペプチド9μgを65μgの100 mM CH3COO-Na+緩衝液、pH5に溶解し、次いで、3μlの1mg/ml α-マンノシダーゼ溶液(Oxford Glyco System)、つまり90 mUのα-マンノシダーゼを加える。全てを消化するように反応を24時間37℃でインキュベートし、次いで得られた生成物を真空下で乾燥する。
【0063】
d)ペプチドのサブチリジンでの消化
上記の精製グリコペプチド690 ngを、5μの100 mM炭酸アンモニウム緩衝液pH8に溶解し、次いで1μlの100μg/ml サブチリジン溶液、つまり約100 ngを加える。反応を24時間37℃でインキュベートし、次いで反応生成物を真空下で乾燥させ、滴定緩衝液(緩衝液D)に溶解する。
e)遅延型過敏症アッセイを用いるグリコペプチドの抗原活性の生物学的滴定
未消化又はα-マンノシダーゼもしくはサブチリジンで消化した上記の精製グリコペプチド0.02μgを、実施例1に記載のプロトコルにしたがってあらかじめ免疫化したモルモットのバッチに注射する。結果を、紅斑反応直径の値で示す。対照は、10TUに相当する0.25μgのPPDからなる。
f)インビトロのリンパ球増殖アッセイを用いるグリコペプチドの抗原活性測定
アッセイの条件は、上記のHornらの記載する条件である。
【0064】
2.結果
a)Lip グリコペプチドの精製及び生化学的分析
精製グリコペプチドについて行った質量測定は、測定が162質量単位値異なると、マンノースでおそらくグリコシル化される複雑な分子の存在を示す。α-マンノシダーゼで処理したペプチドの質量に相当する6951 Daの質量は、これらの分子の最少質量としてみなされる。
精製グリコペプチドのN-末端配列は、主要配列 TIPTT... と少数配列IPTTE...の存在を示している。
これらの結果は、配列(SEQ ID NO:3)がエム.ツベルクローシスの Rv1796遺伝子によってエンコードされるタンパク質由来ペプチドのN-末端フラグメントの配列であり、Sanger bankのエム.ツベルクローシス株H37Rvのゲノム配列の注釈に関し、該タンパク質の169〜239位に伸長している、Lipと称されるマンノシル化グリコペプチドに適合している。
【0065】
b)遅延型過敏症アッセイを用いるLipグリコペプチドの抗原活性測定
グリコペプチドは、生細菌で免疫化したモルモットの遅延型過敏症反応の顕在化に極めて活性であり、他方、熱不活化BCGで免疫化したモルモットでは比較的不活性である。
グリコペプチドの抗原活性は、精製工程中に増す:
− 工程1:得られた画分は、生BCGで免疫化したモルモットで180,000 TU/mg及び熱不活化BCGで免疫化したモルモットで10,000 TU/mgの活性を有する。
− 工程2:得られた画分は、生BCGで免疫化したモルモットでは900,000 TU/mg及び熱不活化BCGで免疫化したモルモットでは30,000 TU/mgの活性を有する。
− 工程3:精製画分は、生BCGで免疫化したモルモットでは1,000,000 TU/mgより大きく、熱不活化BCGで免疫化したモルモットでは30,000 TU/mg未満の活性を有する。
【0066】
図3に示す結果は、
− 37℃で24時間のα-マンノシダーゼの作用は、95%より高い抗原活性の損失を引き起こす:脱グリコシル化後、画分は、1,000,000 TU/mgの活性から30,000 TU/mg未満の活性に低下した、
− サブチリジンの作用は、抗原活性を廃する、及び
− 等量のタンパク質で、Lipグリコペプチドは、マイコバクテリウム・ツベルクローシス(PPD)由来の標準的な精製タンパク質より少なくとも10倍活性が高い、ことを示している。
【0067】
c)インビトロのリンパ球増殖アッセイを用いるLipグリコペプチドの抗原活性測定
図4に示す結果は、T-リンパ球増殖がペプチド濃度に依存していることを示している。この増殖は、Tリンパ球がCD4分子に対する抗体で処理される際、又はグリコペプチドがα-マンノースで処理される際、重要ではない。
【0068】
実施例3:Apaタンパク質をエンコードする裸のDNAでの免疫化による、Tエピトープの定義におけるApaのオリゴサッカライド残基の役割の立証
1.材料と方法
a)Apaタンパク質をエンコードする配列を含むプラスミドの構築
サイトメガロウイルス初期プロモーター配列を含むプラスミド pS65T (Clontech)を、制限酵素NheIとBspEIで切断し、クレノウ酵素で修復し、次いで連結してプラスミドpAG800を得る。
プラスミドpAG800を酵素ApaIで切断して、それ自体にハイブリダイズしたオリゴヌクレオチド12M48 (5' CAACGTTGGGCC 3';SEQ ID NO:4)と連結し、プラスミドpAG802を得る。
【0069】
シグナル配列を欠くApaのコード配列を含む875塩基対のフラグメントを、オリゴヌクレオチド22M42 (5' TCCCAAGCTTTTGGTAGCCG 3';SEQ ID NO:5)と33M44 (5' CTAGGATCCACCATGCCGGAGCCAGCGCCCCCG 3';SEQ ID NO:6)を用いて、プラスミドpLA34-2 (Laqueyrerie, 1995, Infect. Immun., 63, 4003-4010)からポリメラーゼ連鎖反応(PCR)により増幅する。
オリゴヌクレオチド33M44を、コザック型のコンセンサス翻訳開始部位を含むように合成した(Nucl. Acids Res., 1987, 15, 8125-8148)。PCRで得たフラグメントをBamHIとEcoRVで切断し、BgIIIとSmaIで切断したプラスミドpAG802に挿入し、プラスミドpAG803を得る。これらの操作中、オリゴヌクレオチド配列5' CAACGTTGGGCC 3'が失われる;Psp1046 ISSと称されるこの配列は、配列IL-12p40 ISS と同様に、免疫反応を増す免疫刺激配列であると考えられる(Lipford GBら、1997, Eur. J. Immunol., 27, 3420-3426)。
【0070】
Psp1046 ISS配列を、オリゴヌクレオチド25M46 (5' GATCCCCCCCCAACGTTCCCCCCCG 3';SEQ ID NO:8)とハイブリダイズしたオリゴヌクレオチド25M45 (5' GATCCGGGGGGGAACGTTGGGGGGG 3';SEQ ID NO:7)をクローニングすることによってプラスミドpAG803のBamHI部位に挿入し、プラスミドpAG831を得る。
IL-12p40 ISS配列を、オリゴヌクレオチド24M64 (5' GGGCCCTTGGAACGTCATAGCGCT 3';SEQ ID NO:10)とハイブリダイズしたオリゴヌクレオチド24M63 (5' AGCGCTATGACGTTCCAAGGGCCC 3';SEQ ID NO:9)をクローニングすることによって、プラスミドpAG803のBamHI部位に挿入し、プラスミドpAG832を得る。
【0071】
エシェリキア・コリ株XL1 Blueを形質転換後、上記のプラスミドを、カナマイシン25μg/mlを含むLB培養培地(Sambrookら、Molecular cloning: A laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989)で増幅する。細菌溶解物をTriton X-114 (1%)で処理することによりエンドトキシンを除く先の工程の後、プラスミドDNAを製造者の指示に従ってMaxiPrep QIA フィルターカラム(QIAGEN)で精製する。
【0072】
b)プラスミドpAG831とpAG832でのモルモットの免疫化
わき腹に2回皮内注射することにより、上記のように調製し、精製したプラスミドpAG831又は pAG832のDNA 50μgで体重300〜400gのモルモット(Hartley)を免疫化する。
対照は、実施例1又は実施例2に記載される条件下で生BCGで免疫化したモルモットの群からなる。
【0073】
c)遅延型過敏症アッセイを用いる、裸のDNAで免疫化したモルモットにおける真核細胞により産生されるApaタンパク質の抗原活性測定
免疫化から1〜2ヶ月後に、遅延型過敏症反応を、天然のApaタンパク質又はエシェリキア・コリの形質転換株で産生される組換えApaタンパク質に対して測定し、タンパク質を上記のHornらに記載されるプロトコルにしたがって精製する。
天然のApa及び組換えApaを、滴定緩衝液(緩衝液D) 100μl中0.2μg用量で皮内注射する。抗原活性を、実施例2に記載されるように測定する。
【0074】
2.結果
図5に示す結果は以下のとおりである:
・ 真核プロモーターの制御下でApaのコード配列を含むプラスミドpAG831又はpAG832で免疫化したモルモットは、天然のApaタンパク質に対する免疫反応(抗原とまとめられる際には、遅延型過敏症アッセイ又はインビトロT-リンパ球増殖アッセイを用いて測定できる抗体及びCD4+型のT-反応)を多くの場合に生ずる。天然のApa抗原に相当する動物では、酵素経路を介して脱グリコシル化される抗原又はイー.コリ由来の非グリコシル化組換え抗原に対するCD4+ Tリンパ球の反応は、グリコシル化天然抗原で見られる反応と同一の強度である。
・ 他方、生BCGで免疫化したモルモットは、天然のApaに対してのみ遅延型過敏症反応を示す。これらの動物は、上記のようにイー.コリで産生される非グリコシル化組換えApaに対し、反応を生じないか、又はかなり低下した反応を生ずる。
【0075】
これらの結果は、以下のことを示している:
1)Apaをエンコードする裸のDNAで免疫化した動物で認められる結果は、マクロファージ又は樹状細胞によるApaタンパク質の食菌及び提示能力が、天然又は組換えの(非グリコシル化)Apaタンパク質と同一であることを示している、
2)生BCGで免疫化した動物で認められる結果と上記の結果の組み合わせは、脱グリコシル化Apaタンパク質に対する反応の不在は、マクロファージ又は樹状細胞によるその提示能力の低下ではなく、CD4+ Tリンパ球による認識不在に起因していることを示している。その結果、エム.ツベルクローシス又は生BCGによって産生されるような、天然型のApa又はLipタンパク質の側鎖のオリゴマンノース残基は、CD4+ Tリンパ球によって認識されるTエピトープの構成で役割を果たしている。
【0076】
実施例4:グリコシル化ペプチドSEQ ID NO:1、SEQ ID NO:2 及びSEQ ID NO:3の調製
1)グリコシル化シントン15、16及び19の調製
ペプチド合成の前に、グリコシル化シントン、つまり2又は3個のマンノース残基で官能化したトレオニンを調製する。
・ 化合物5及び8の調製(図6)
化合物5及び8の調製は、それぞれH. FRANZYKら(J. Chem. Soc. Perkin Trans. 1, 1995, 2883-2898)及びR.K. NESSら(J. Am. Chem. Soc. Perkin, 1950, 72, 2200-2205)に記載されている。
市販の過アセチル化マンノース1(つまり、1,2,3,4,5-ペンタ-O-アセチル-α-D-マンノピラノース)を、A. LEVENEら(J. Biol. Chem., 1931, 90, 89-98)によって記載されるように、酢酸中の臭化水素の作用によりアノマー位で臭素化する。活性化中間体2を、2,6-ジメチルピリジン/メタノール混合物でオルソエステル3に環化する。10%水性トリフルオロ酢酸/アセトニトリル混合物中で0℃での酸加水分解によるオルソエステルの位置選択的な開環は、1,3,4,6-テトラ-O-アセチル-α-D-マンノピラノース(5)を生じる。位置異性体4も単離される。
【0077】
市販のマンノース6は、ピリジン中の塩化ベンゾイルの作用により7に過ベンゾイル化される。後者は、酢酸中の臭化水素の作用により8に活性化される。しかし、このプロトコルと以下の方法では、臭化水素の作用による以外の活性化方法、例えば当業者に公知の方法を用いてもよい。
・ 二糖10及び12の調製(図7a)
化合物10及び12の調製は、それぞれA. JANSSONら(J. Chem. Soc. Perkin Trans. 1, 1992, 1699-1707)及びH. FRANZYKら(同節)に記載されている。化合物2及び5は、ジクロロメタン中でトリフルオロメタンスルホネート銀(又はいずれかの他の縮合反応プロモーター)の存在下で縮合され、過アセチル化二糖9を生じ、次いで酢酸中の臭化水素の作用により臭素化前駆体に活性化する。同一のプロトコルによれば、化合物5及び8は、縮合されて化合物11を生じ、それ自体12に活性化される。
【0078】
・ 三糖18の調製(図7a)
活性化二糖12を、ジクロロメタン中のトリフルオロメタンスルホネート銀の存在下で単糖アクセプター5に縮合し、過アセチル化三糖17を生じ、次いでこれを酢酸中の臭化水素の作用により臭素化前駆体18に活性化する。
・ 2つのマンノース単位を有するシントン15ならびに16、及び3つのマンノース単位を有するシントン19の調製(図7b)
I. SCHONら(Synthesis, 1986, 303-305)に記載されるように、第一級アミン官能基がFmoc基で保護されている市販のトレオニン13の酸官能性を、ジシクロヘキシルカルボジイミド(DCCI)の存在下でペンタフルオロフェノール(pfp)の作用によりエステルの形態で阻害し、アクセプター前駆体14を生じる。
【0079】
シントン15及び16の調製は、それぞれA. JANSSONら(同節)及びH. FRANZYKら(同節)により記載されている。ジクロロメタン中のトリフルオロメタンスルホネート銀の存在下で行なわれる活性化二糖10及び12との化合物14の縮合は、それぞれシントン15及び16を生じる。同じプロトコルによれば、化合物14と活性三糖18との縮合は、シントン19を生じる。
【0080】
2)グリコシル化ペプチドSEQ ID NO:1、SEQ ID NO:2及びSEQ ID NO:3の調製
ペプチドは、Fmoc化学を用いて固相で合成される。ペプチド合成は、所望の配列を生産する一方、ペンタフルオロフェノールの活性化エステルの形態のグリコシル化シントン(シントン15、16及び19)を挿入するために必要なアミノ酸を用いて自動合成機行われる。
使用されるシントンに応じて、2つのマンノース残基で官能化(ペプチド合成中のシントン15及び/又は16の挿入)されるトレオニンからなるペプチド又は3つのマンノース残基で官能化(ペプチド合成中のシントン19の挿入)されるトレオニンからなるペプチド又は2つのマンノースで官能化されるトレオニンと33のマンノース残基で官能化(ペプチド合成中のシントン19及び15及び/又は16の挿入)されるトレオニンの双方を含むペプチドのいずれかが得られる。
【0081】
合成の最後に、トリフルオロ酢酸を用いて固体支持体からペプチドを切断し、種々のアミノ酸及びマンノースのヒドロキシル官能性を脱保護した後、ペプチドを逆相高速液体クロマトグラフィー(HPLC)により精製する。その構造を、当業者に公知の技術、例えば質量分析及びアミノ酸分析を用いて制御する。
次に、当業者に公知の有機化学技術を用いる化学合成により、アミド官能性(ペプチドSEQ ID NO:1及びSEQ ID NO:3のC-末端位)ならびにアセテート官能性(ペプチドSEQ ID NO:2及びSEQ ID NO:3のN-末端位)を導入する。
【0082】
実施例5:イー.コリで産生されるApaペプチドでの免疫化による、Tエピトープの定義におけるApaのオリゴサッカライド残基の役割の立証
1)材料と方法
当業者に周知のイー.コリにおける組換えタンパク質のクローニング、発現及び精製の従来技術(例えば、Current Protocols in Molecular Biology (Frederick M. AUSUBEL, 2000, Wiley and Son Inc, Library of Congress, USA)に記載されるプロトコル参照)にしたがって、Apaの250〜280位に相当するペプチドをボルデテラ・ペルツシス(Bordetella pertussis)のサイクラーゼのフラグメントとの融合体の形態でイー.コリで産生した。
体重300〜400gの5頭のHartleyモルモットの3群を、アジュバント溶液0.1ml中のこの精製Apaペプチド20μgを用いて、1ヶ月空けて2回皮内注射し、免疫化した。
【0083】
実施例1に記載される条件下で生BCGで予め4ヶ月間免疫化した4頭のモルモット3群を、対照として用いる。
免疫化から1〜2ヶ月後、遅延過敏症反応を、天然のApaタンパク質、イー.コリで産生される組換えApaタンパク質及び実施例1に記載されるようにして調製された脱グリコシル化Apaタンパク質に関して、実施例3に定義する条件下で測定した。
【0084】
2)結果
Apa融合ペプチド又は生BCGで免疫化したモルモットの遅延過敏症反応を、天然のApaタンパク質、イー.コリで産生される組換えApaタンパク質及び脱グリコシル化Apaタンパク質に関して測定した。紅斑反応の直径(mm)で表された結果を、以下の表1に示す:
【0085】
【表1】
【0086】
上記の表1に示すように、生BCGで免疫化したモルモットで見られる遅延過敏症反応は、天然のApa分子の注射後に著しい。反応は、化学的に脱グリコシル化した分子又はイー.コリで産生される分子の注射後に極めて弱いか、あるいはない。他方、ボルデテラ・ペルツシスのサイクラーゼフラグメントとApa分子の内部フラグメントとの融合に相当する組換え分子で免疫化したモルモットについては、感作は、天然分子又は脱グリコシル化分子に対して同一である。
【0087】
これらの結果は、Apa分子のグリコシル化Tエピトープが、生細菌で免疫化されたモルモットによって選択的に認識されることを示している。また、結果は、モルモットによる脱グリコシル化分子の認識の不在又は低下は、これらの分子の低下した固有の抗原性に伴わないことを示している。
上記から明らかであるように、この発明は、より具体的に記載したにすぎないその実施方法、調製及び適用にいかなる方法でも制限されない;逆に、本発明は、本発明の概要又は範囲を逸脱しない限り当業者に起こりうる全ての変形を包含する。
【特許請求の範囲】
【請求項1】
a1)少なくとも1つの中性アミノ酸は二糖又は三糖に結合し(グリコシド結合)、アミノ酸の少なくとも15%はプロリンで、プロリンの1つは中性アミノ酸の位置に対して−1〜−4位に位置する、病原性微生物由来で、14〜25アミノ酸からなるグリコシル化Tエピトープから本質的になり、
− クラスII MHC分子で提示され、
− 由来する天然のグリコタンパク質での免疫化で誘導される CD4+ Tリンパ球によって特異的に認識されるが、同一の配列を有する非グリコシル化ペプチドでの免疫化で誘導されるCD4+ Tリンパ球によっては認識されず、かつ
− それらを認識するCD4+ Tリンパ球の増殖と該リンパ球によるサイトカインの分泌を誘導できる
グリコペプチド、及び
b1)配列SEQ ID NO:11のグリコペプチドを除く、a1)に定義されるグリコペプチドの配列を含む15〜39アミノ酸の配列を有するグリコペプチドからなる群から選択されるが、
ただし、
− 配列(SEQ ID NO:1)が、Apaタンパク質の配列の1〜39位から伸長する配列であり、SEQ ID NO:1の10、18及び27位のトレオニン残基の少なくとも1つがグリコシド結合を介して二糖又は三糖に結合する、39アミノ酸のグリコペプチド、
− 配列(SEQ ID NO:2)が、Apaタンパク質の配列(C-末端配列)の261〜286位から伸長する配列であり、SEQ ID NO:2の17位のトレオニン残基がグリコシド結合を介して二糖又は三糖に結合する26アミノ酸のグリコペプチド、及び
− 配列(SEQ ID NO:3)がRv 1796 タンパク質の配列の169〜203位から伸長する配列であり、SEQ ID NO:3の4、5、7、13、15、23及び25位のトレオニン残基の少なくとも1つがグリコシド結合を介して二糖又は三糖に結合する35アミノ酸のグリコペプチド
を除く、免疫原性グリコペプチド。
【請求項2】
中性アミノ酸が、セリンとトレオニンからなる群から選択されることを特徴とする請求項1に記載の免疫原性グリコペプチド。
【請求項3】
二糖又は三糖に結合した1〜7個のトレオニン残基を含むことを特徴とする請求項1又は請求項2に記載のグリコペプチド。
【請求項4】
二糖又は三糖が、ヘキソースの二量体又は三量体であることを特徴とする請求項1〜3のいずれか1つに記載のグリコペプチド。
【請求項5】
ヘキソースがマンノースであることを特徴とする請求項4に記載のグリコペプチド。
【請求項6】
グリコシド結合が、α-(1,2)-結合であることを特徴とする請求項1〜5のいずれか1つに記載のグリコペプチド。
【請求項7】
病原性微生物が、タンパク質をO-グリコシル化できることを特徴とする請求項1〜6のいずれか1つに記載のグリコペプチド。
【請求項8】
病原性微生物が、マイコバクテリウム・ツベルクローシス又はカンジダ・アルビカンスであることを特徴とする請求項7に記載のグリコペプチド。
【請求項9】
エム.ツベルクローシス(Genbank 番号X80268)のApaタンパク質由来、又はエム.ツベルクローシス株H37Rvのゲノム配列の注釈に関し、Rv 1796遺伝子によってエンコードされるRv1796タンパク質由来であることを特徴とする、請求項1〜8のいずれか1つに記載のグリコペプチド。
【請求項10】
− グリコシド結合を介して二糖又は三糖に結合したグリコシル化中性アミノ酸を溶液中に調製し、
− グリコペプチドのペプチド配列を産生するのに必要なアミノ酸及び上記で得た中性アミノ酸を用いて、固体支持体上でグリコペプチドを合成し、かつ
− 固体支持体からグリコペプチドを切断する
工程からなることを特徴とする、請求項1〜9のいずれか1つに記載のグリコペプチドの合成方法。
【請求項11】
中性アミノ酸が、セリン及びトレオニンからなる群から選択されることを特徴とする請求項10に記載の方法。
【請求項12】
トレオニンによって担持されるグリコシド残基が、ヘキソース、好ましくはマンノースであることを特徴とする請求項11に記載の方法。
【請求項13】
マンノース残基で官能化されるトレオニンが、
a2)式(I)及び(II)
【化1】
[式中、P1及びP2は、同一又は異なっていてもよく、ヒドロキシル官能性を保護する基を示し、かつXは、臭素原子のような活性化官能性を示す]のマンノース誘導体の製造:
b2)2つのマンノース残基からなり、式(III):
【化2】
[式中、P1、P2及びXは、式(I)及び(II)に関して定義されるとおり]
に相当する活性化誘導体の生産をもたらす、式(I)の誘導体と式(II)の誘導体との反応、次いで得られた化合物の活性化、
c2)任意に、3つのマンノース残基からなり、式(IV):
【化3】
[式中、P1、P2及びXは、式(I)及び(II)に関して定義されるとおり]
に相当する活性化誘導体の産生をもたらす、a2)に定義される式(I)のマンノース誘導体での式(III)の化合物の反応、次いで得られた化合物の活性化、及び
d2)それぞれ式(VI)又は(VII):
【化4】
[式中、P1、P2、P3及びP4は上記のとおり]
のグリコシル化トレオニンの生産をもたらす、式(III)の化合物又は式(IV)の化合物の、式(V):
【化5】
[式中、P3は、第一級アミン官能性を保護する基を示し、かつP4はヒドロキシル官能性を保護する基を示す]
の適当に保護されたトレオニンとの縮合
工程を用いて調製されることを特徴とする請求項12に記載の方法。
【請求項14】
少なくとも
a3)二糖又は三糖に結合した少なくとも1つの中性アミノ酸及びプロリンの1つが中性アミノ酸の位置に対して−1〜−4位に位置する少なくとも15%のプロリンを含む少なくとも1つの14〜25アミノ酸配列を、タンパク質のペプチド配列において、またそれから、検索し、選択し、
b3)請求項10〜13に記載の方法にしたがって、工程a3)で選択されるグリコペプチドを調製し、かつ
c3)抗原活性が、同一配列を有する対照のペプチドよりも少なくとも10倍、好ましくは少なくとも30倍大きいグリコペプチドを選択する
工程からなることを特徴とする、病原性微生物のタンパク質のペプチド配列を用いて免疫原性グリコペプチドを選択し、スクリーニングするための方法。
【請求項15】
工程a3)に先立って、少なくとも1つの抗原性グリコタンパク質を予備選択する工程を含むことを特徴とする請求項14に記載の方法。
【請求項16】
中性アミノ酸が、セリンとトレオニンからなる群から選択されることを特徴とする請求項14に記載の方法。
【請求項17】
工程c3)で、グリコペプチドの抗原活性が、弱毒化病原性微生物又は病原性微生物の抗原画分で免疫化された動物のCD4+ Tリンパ球の活性を測定して評価されることを特徴とする請求項14に記載の方法。
【請求項18】
請求項10〜17のいずれか1つに記載の方法を用いて得られることを特徴とするグリコペプチド。
【請求項19】
免疫原性組成物もしくは免疫化組成物又は診断試薬の製造のための、請求項1〜9もしくは18のいずれか1つに記載のグリコペプチド又は配列SEQ ID NO:11のグリコペプチドからなる群から選択される少なくとも1つのグリコペプチドの使用。
【請求項20】
少なくとも1つの医薬的に許容される賦形剤と組み合わさった、請求項1〜9もしくは18のいずれか1つに記載の少なくとも1つのグリコペプチドを含むことを特徴とする、体液性及び/又は細胞性免疫を誘導できる免疫原性組成物。
【請求項21】
少なくとも1つの医薬的に許容される賦形剤及び任意に少なくとも1つのアジュバントと組み合わさった、請求項1〜9又は18のいずれか1つに記載の少なくとも1つのグリコペプチドからなることを特徴とする、体液性及び/又は細胞性免疫を誘発し得る免疫化組成物。
【請求項22】
グリコペプチドが、少なくとも1つのBエピトープ、CD4+型の1つのTエピトープ又はCD8+型の1つのTエピトープからなるタンパク質又はタンパク質フラグメントと組み合わさっていることを特徴とする、請求項20又は請求項21に記載の免疫原性組成物又は免疫化組成物。
【請求項23】
請求項1〜9又は18のいずれか1つに記載の1以上のグリコペプチドに対することを特徴とする抗体。
【請求項24】
請求項1〜9もしくは18のいずれか1つに記載するグリコペプチド又は請求項23に記載の抗体からなることを特徴とする診断試薬。
【請求項25】
病原性微生物に感染している恐れのある患者由来の生物学的試料を請求項24に記載の診断試薬と接触させ、生物学的試料複合体中に存在する抗体/微生物又は試料複合体に存在するグリコペプチド/抗体の形成を検出することからなることを特徴とする、病原性微生物での感染の検出方法。
【請求項1】
a1)少なくとも1つの中性アミノ酸は二糖又は三糖に結合し(グリコシド結合)、アミノ酸の少なくとも15%はプロリンで、プロリンの1つは中性アミノ酸の位置に対して−1〜−4位に位置する、病原性微生物由来で、14〜25アミノ酸からなるグリコシル化Tエピトープから本質的になり、
− クラスII MHC分子で提示され、
− 由来する天然のグリコタンパク質での免疫化で誘導される CD4+ Tリンパ球によって特異的に認識されるが、同一の配列を有する非グリコシル化ペプチドでの免疫化で誘導されるCD4+ Tリンパ球によっては認識されず、かつ
− それらを認識するCD4+ Tリンパ球の増殖と該リンパ球によるサイトカインの分泌を誘導できる
グリコペプチド、及び
b1)配列SEQ ID NO:11のグリコペプチドを除く、a1)に定義されるグリコペプチドの配列を含む15〜39アミノ酸の配列を有するグリコペプチドからなる群から選択されるが、
ただし、
− 配列(SEQ ID NO:1)が、Apaタンパク質の配列の1〜39位から伸長する配列であり、SEQ ID NO:1の10、18及び27位のトレオニン残基の少なくとも1つがグリコシド結合を介して二糖又は三糖に結合する、39アミノ酸のグリコペプチド、
− 配列(SEQ ID NO:2)が、Apaタンパク質の配列(C-末端配列)の261〜286位から伸長する配列であり、SEQ ID NO:2の17位のトレオニン残基がグリコシド結合を介して二糖又は三糖に結合する26アミノ酸のグリコペプチド、及び
− 配列(SEQ ID NO:3)がRv 1796 タンパク質の配列の169〜203位から伸長する配列であり、SEQ ID NO:3の4、5、7、13、15、23及び25位のトレオニン残基の少なくとも1つがグリコシド結合を介して二糖又は三糖に結合する35アミノ酸のグリコペプチド
を除く、免疫原性グリコペプチド。
【請求項2】
中性アミノ酸が、セリンとトレオニンからなる群から選択されることを特徴とする請求項1に記載の免疫原性グリコペプチド。
【請求項3】
二糖又は三糖に結合した1〜7個のトレオニン残基を含むことを特徴とする請求項1又は請求項2に記載のグリコペプチド。
【請求項4】
二糖又は三糖が、ヘキソースの二量体又は三量体であることを特徴とする請求項1〜3のいずれか1つに記載のグリコペプチド。
【請求項5】
ヘキソースがマンノースであることを特徴とする請求項4に記載のグリコペプチド。
【請求項6】
グリコシド結合が、α-(1,2)-結合であることを特徴とする請求項1〜5のいずれか1つに記載のグリコペプチド。
【請求項7】
病原性微生物が、タンパク質をO-グリコシル化できることを特徴とする請求項1〜6のいずれか1つに記載のグリコペプチド。
【請求項8】
病原性微生物が、マイコバクテリウム・ツベルクローシス又はカンジダ・アルビカンスであることを特徴とする請求項7に記載のグリコペプチド。
【請求項9】
エム.ツベルクローシス(Genbank 番号X80268)のApaタンパク質由来、又はエム.ツベルクローシス株H37Rvのゲノム配列の注釈に関し、Rv 1796遺伝子によってエンコードされるRv1796タンパク質由来であることを特徴とする、請求項1〜8のいずれか1つに記載のグリコペプチド。
【請求項10】
− グリコシド結合を介して二糖又は三糖に結合したグリコシル化中性アミノ酸を溶液中に調製し、
− グリコペプチドのペプチド配列を産生するのに必要なアミノ酸及び上記で得た中性アミノ酸を用いて、固体支持体上でグリコペプチドを合成し、かつ
− 固体支持体からグリコペプチドを切断する
工程からなることを特徴とする、請求項1〜9のいずれか1つに記載のグリコペプチドの合成方法。
【請求項11】
中性アミノ酸が、セリン及びトレオニンからなる群から選択されることを特徴とする請求項10に記載の方法。
【請求項12】
トレオニンによって担持されるグリコシド残基が、ヘキソース、好ましくはマンノースであることを特徴とする請求項11に記載の方法。
【請求項13】
マンノース残基で官能化されるトレオニンが、
a2)式(I)及び(II)
【化1】
[式中、P1及びP2は、同一又は異なっていてもよく、ヒドロキシル官能性を保護する基を示し、かつXは、臭素原子のような活性化官能性を示す]のマンノース誘導体の製造:
b2)2つのマンノース残基からなり、式(III):
【化2】
[式中、P1、P2及びXは、式(I)及び(II)に関して定義されるとおり]
に相当する活性化誘導体の生産をもたらす、式(I)の誘導体と式(II)の誘導体との反応、次いで得られた化合物の活性化、
c2)任意に、3つのマンノース残基からなり、式(IV):
【化3】
[式中、P1、P2及びXは、式(I)及び(II)に関して定義されるとおり]
に相当する活性化誘導体の産生をもたらす、a2)に定義される式(I)のマンノース誘導体での式(III)の化合物の反応、次いで得られた化合物の活性化、及び
d2)それぞれ式(VI)又は(VII):
【化4】
[式中、P1、P2、P3及びP4は上記のとおり]
のグリコシル化トレオニンの生産をもたらす、式(III)の化合物又は式(IV)の化合物の、式(V):
【化5】
[式中、P3は、第一級アミン官能性を保護する基を示し、かつP4はヒドロキシル官能性を保護する基を示す]
の適当に保護されたトレオニンとの縮合
工程を用いて調製されることを特徴とする請求項12に記載の方法。
【請求項14】
少なくとも
a3)二糖又は三糖に結合した少なくとも1つの中性アミノ酸及びプロリンの1つが中性アミノ酸の位置に対して−1〜−4位に位置する少なくとも15%のプロリンを含む少なくとも1つの14〜25アミノ酸配列を、タンパク質のペプチド配列において、またそれから、検索し、選択し、
b3)請求項10〜13に記載の方法にしたがって、工程a3)で選択されるグリコペプチドを調製し、かつ
c3)抗原活性が、同一配列を有する対照のペプチドよりも少なくとも10倍、好ましくは少なくとも30倍大きいグリコペプチドを選択する
工程からなることを特徴とする、病原性微生物のタンパク質のペプチド配列を用いて免疫原性グリコペプチドを選択し、スクリーニングするための方法。
【請求項15】
工程a3)に先立って、少なくとも1つの抗原性グリコタンパク質を予備選択する工程を含むことを特徴とする請求項14に記載の方法。
【請求項16】
中性アミノ酸が、セリンとトレオニンからなる群から選択されることを特徴とする請求項14に記載の方法。
【請求項17】
工程c3)で、グリコペプチドの抗原活性が、弱毒化病原性微生物又は病原性微生物の抗原画分で免疫化された動物のCD4+ Tリンパ球の活性を測定して評価されることを特徴とする請求項14に記載の方法。
【請求項18】
請求項10〜17のいずれか1つに記載の方法を用いて得られることを特徴とするグリコペプチド。
【請求項19】
免疫原性組成物もしくは免疫化組成物又は診断試薬の製造のための、請求項1〜9もしくは18のいずれか1つに記載のグリコペプチド又は配列SEQ ID NO:11のグリコペプチドからなる群から選択される少なくとも1つのグリコペプチドの使用。
【請求項20】
少なくとも1つの医薬的に許容される賦形剤と組み合わさった、請求項1〜9もしくは18のいずれか1つに記載の少なくとも1つのグリコペプチドを含むことを特徴とする、体液性及び/又は細胞性免疫を誘導できる免疫原性組成物。
【請求項21】
少なくとも1つの医薬的に許容される賦形剤及び任意に少なくとも1つのアジュバントと組み合わさった、請求項1〜9又は18のいずれか1つに記載の少なくとも1つのグリコペプチドからなることを特徴とする、体液性及び/又は細胞性免疫を誘発し得る免疫化組成物。
【請求項22】
グリコペプチドが、少なくとも1つのBエピトープ、CD4+型の1つのTエピトープ又はCD8+型の1つのTエピトープからなるタンパク質又はタンパク質フラグメントと組み合わさっていることを特徴とする、請求項20又は請求項21に記載の免疫原性組成物又は免疫化組成物。
【請求項23】
請求項1〜9又は18のいずれか1つに記載の1以上のグリコペプチドに対することを特徴とする抗体。
【請求項24】
請求項1〜9もしくは18のいずれか1つに記載するグリコペプチド又は請求項23に記載の抗体からなることを特徴とする診断試薬。
【請求項25】
病原性微生物に感染している恐れのある患者由来の生物学的試料を請求項24に記載の診断試薬と接触させ、生物学的試料複合体中に存在する抗体/微生物又は試料複合体に存在するグリコペプチド/抗体の形成を検出することからなることを特徴とする、病原性微生物での感染の検出方法。
【図1】

【図2a】
【図2b】
【図3】
【図4】
【図5】
【図6】
【図7a】
【図7b】
【図2a】
【図2b】
【図3】
【図4】
【図5】
【図6】
【図7a】
【図7b】
【公開番号】特開2010−235606(P2010−235606A)
【公開日】平成22年10月21日(2010.10.21)
【国際特許分類】
【出願番号】特願2010−88027(P2010−88027)
【出願日】平成22年4月6日(2010.4.6)
【分割の表示】特願2002−552001(P2002−552001)の分割
【原出願日】平成13年12月20日(2001.12.20)
【出願人】(501474748)インスティティ・パスツール (27)
【氏名又は名称原語表記】INSTITUT PASTEUR
【住所又は居所原語表記】28,rue du Docteur Roux,F−75724 Paris Cedex 15 FRANCE
【Fターム(参考)】
【公開日】平成22年10月21日(2010.10.21)
【国際特許分類】
【出願日】平成22年4月6日(2010.4.6)
【分割の表示】特願2002−552001(P2002−552001)の分割
【原出願日】平成13年12月20日(2001.12.20)
【出願人】(501474748)インスティティ・パスツール (27)
【氏名又は名称原語表記】INSTITUT PASTEUR
【住所又は居所原語表記】28,rue du Docteur Roux,F−75724 Paris Cedex 15 FRANCE
【Fターム(参考)】
[ Back to top ]