
前癌と癌細胞用のマーカー、及び前記細胞の増殖阻害方法
【課題】前癌と癌細胞用のマーカー、及び前記細胞の増殖阻害方法の提供。
【解決手段】正常、前癌又は癌細胞の中で異なった形で発現するキメラRNAと称されるヒトミトコンドリアRNAの新規なファミリー。前記キメラRNAを標的にするオリゴヌクレオチド。研究用、癌診断及び癌治療用における、前記オリゴヌクレオチド又はその類似物の使用。前記オリゴヌクレオチドの、センス又はアンチセンスミトコンドリアキメラRNAでハイブリダイズすることによって、前記オリゴヌクレオチドをし、正常に増殖する細胞、前癌細胞及び癌細胞を識別する方法。
【解決手段】正常、前癌又は癌細胞の中で異なった形で発現するキメラRNAと称されるヒトミトコンドリアRNAの新規なファミリー。前記キメラRNAを標的にするオリゴヌクレオチド。研究用、癌診断及び癌治療用における、前記オリゴヌクレオチド又はその類似物の使用。前記オリゴヌクレオチドの、センス又はアンチセンスミトコンドリアキメラRNAでハイブリダイズすることによって、前記オリゴヌクレオチドをし、正常に増殖する細胞、前癌細胞及び癌細胞を識別する方法。
【発明の詳細な説明】
【技術分野】
【0001】
1.本発明の技術分野
本発明は、2003年5月21日付けで出願された米国仮特許出願第60/47210
6の利益を主張する。
本発明は、ヒトのミトコンドリアキメラRNAと称される、ヒトミトコンドリアRNA
の新規なファミリーに関連する癌治療法、癌診断及び研究試薬に関する。
特に、本発明は、ヒトミトコンドリアRNAを標的とするオリゴヌクレオチドに関する
。本発明のオリゴヌクレオチドは、癌細胞死を誘発するキメラRNAにハイブリタイズす
る。本発明により提供される組成物と方法は、新規な癌治療として有効である。更に、こ
のオリゴヌクレオチドは、休止期及び増殖期の正常細胞、前癌および癌細胞においてミト
コンドリアキメラRNAの発現が異なることに基づき癌と前癌細胞の診断に用いることが
できる。
【背景技術】
【0002】
2.本発明の背景技術
2.1 ミトコンドリア
ミトコンドリアは、酸化性りん酸化反応により必須分子であるアデノシン三りん酸(A
TP)を製造する細胞内小器官である。1665塩基対のヒトミトコンドリア(mtDN
A)は、2つのリボゾームRNA、22転移RNA(tRNAs)、及び同じ数のポリペ
プチドをエンコードしている13オープンリーデングフレーム(ORF)をエンコードし
ている(Clayton,Hum Reprod. Suppl 2:11-17, 2000; Taanman, Biochim. Biophys. Act
a,1410:103-123, 1999)。
G+T塩基組成物の成分を基に、mtDNAの2つの鎖は、ボイヤント密度(浮遊密度)
が異なり、セシウムクロライド勾配(gradient)を変性して(denaturating)分離される
。重鎖(又はH鎖)は、2つのリボゾームRNAs(12Sと16S)、14tRNAs、並びに、
ND1、ND2、ND3、ND4L、ND4、ND5、COXI、COX II、COX III、ATP6、ATP8及びCyt bに対応
する12ポリペプチドをエンコードしている。
軽鎖(又はL鎖)は、8tRNAs及びコンプレックスNADハイドロゼナーゼNS6
をコードしている(Clayton,Hum Reprod. Suppl 2:11-17, 2000; Taanman, Biochim. Bio
phys. Acta,1410:103-123, 1999)。
【0003】
多くのmtDNAは、置換ループ又はDループと呼ばれる短い3つの鎖構造を含んでいる
。ヒトにおいて1,006の塩基対である、この領域は、フェニルアラニンのtRNA(tRN
APhe)及びプロリンのtRNA(tRNAPro)の遺伝子に接しており、L鎖に相補してH鎖を置
換する短核酸鎖を含んでいる(Clayton, Hum Reprod. Suppl 2:11-17, 2000; Taanman, Bi
ochim. Biophys.Acta, 1410:103-123, 1999)。
この領域は、mtDNA発現のための主なコントロール・サイトとして発展しており、リ
ーデング鎖又は複製のH鎖起点、及びH鎖(HSP)とL鎖(LSP)の転写のための主なプロモー
ターを含んでいる。
HSPとLSP(約150bp)は隣接しているにもかかわらず、これらの調節要素は、ミト
コンドリア疾病を有する患者のモデルを利用するインビボ(体内)(Chinnery and Turnbu
ll, Mol. Med.Today, 6:425432, 2000)と同様に、インビトロ(体外)でも機能的に独立
である(Shueyand Attardi, J Biol Chem. 260:1952-1958, 1985; Taanman, Biochim. Bi
ophys. Acta,1410:103-123, 1999)。
【0004】
双方の鎖は、ポリシトロンRNAsとして転写され、これらはその後個々のmRNA、t
RNA及びrRNAをリリースするためにプロセスされる(Taanman, Biochim. Biophys.
Acta,1410:103-123, 1999)。
ヒトでは、ミトコンドリアのRNAポリメラーゼは、イースト・ミトコンドリアRNA
ポリメラーゼの配列、及びいくつかのバクテリオファージのRNAポリメラーゼと著しい
相同性を有する1,230のアミノ酸の蛋白質である(Tirantiら,Hum Mol Genet. 6:615-
625, 1997)。
更に、転写要因のファミリーは、哺乳類のmtDNA転写として本質的で、DNA結合蛋
白の高移動性グループ(HMG)-boxファミリーのメンバーである、ミトコンドリア転
写因子A又はTFMAなどで特徴づけらている(Parisi and Clayton, Science. 252:965-
969, 1991)。
最近、2つの独立した報告書で、ヒトおよびマウスにおける新規な転写因子TFB1M
とTFB2Mの特徴について記述されている(McCulloch et al., Mol. Cell Biol. 22:11
16-1125, 2002;Falkenberg et al., Nat Genet. 31:289-294, 2002; Rantanen et al.,
Mamm Genome.14:1-6, 2003)。
mtDNA転写に包含されるcis-とtrans-活動因子についてなされた相当の進歩にもかか
わらず、機能の詳細については、十分には理解されていない。
【0005】
2.2 ミトコンドリアおよびアポトーシス
ミトコンドリアは、細胞がよくコントロールされた又はプログラムされた方法で細胞が
死滅する基本的な生物学のプロセスである、アポトーシス(細胞死)における中心的な役
割を果たす。この細胞自殺プログラムは、進化の間、そして後生動物の成熟したホメオス
タシスには本質的である。アポトーシスは、不必要な、損傷した、突然変異した、そして
老化した細胞を撲滅するために活性化される(Meier et al., Nature 407:796-801, 2000)
。アポトーシスの不規則化(disregulation)は、いくつかの病理学の発表で示されてい
る。
このように、アポトーシスの異常な抑制は、腫瘍形成の特徴で、この場合、広範なアポ
トーシスが発作、敗血性ショック及び神経性変性不調(neurodegerative disorders)の
ような急性の疾病と関係づけられている。
現在、アポトーシスのプロセスは、外因性と内因性経路として知られている2つの主な
経路について述べられている(Zornig et al., Biochim.Biophys. Acta, 1551:F1-F37, 20
01)。
外因性経路は、異なるリガンドが死のレセプターに結合することにより、細胞膜で開始
されるプロセスである(Krammer, Nature 407:789-795, 2000; Zornigら, Biochim. Bioph
ys. Acta,1551:F1-f37, 2001)。
【0006】
カスパーゼは、アポトーシスにおいて蛋白分解性カスケードに関与する。カスパーゼは
、蛋白分解性成熟又はアポトーシス誘導プロセッシングを受ける不活性プレカーサーたん
ぱく質として合成される(Zornig et al., Biochim. Biophys. Acta, 1551:F1-F37, 2001)
。
しかしながら、より最近、いくつかの実験証拠は、リソソームプロテアーゼがアポトー
シス性損傷後に蛋白質分解の代替となる経路を構成することを示している(Guicciardi et
al.,Oncogene, 23:2881-2890, 2004)。
他方、ヒトのオンコプロテイン(癌タンパク)Bcl-2に同種のアンチ−アポトーシ
ス性蛋白が記述されている。この蛋白は、アンチ−アポトーシス(Bcl-2、Bcl-XL、Bcl-w
)あるいはプロ−アポトーシス(Bax、Bak、Bim、Bid等)であるたんぱく質のファミリー
に属する(Zorninget al., Biochim. Biophys. Acta, 1551:F1-F37, 2001)。
【0007】
ミトコンドリアは、特にアポトーシス性プロセスの間の初期に影響を受ける、そして現
在、これらは、細胞死の中心的なコーディネーターとして認識されている(Boya et al.,
Biochem.Biophys. Res. Commun. 304:575-581, 2003; Ferri and Kroemer, Nature Cell
Biol.3:E255-E263, 2001; Zornig et al., Biochim. Biophys. Acta, 1551:F1-F37, 20
01)。
いくつかのプロ−アポトーシスシグナルと損傷経路は、Bcl-2蛋白の制御下にある
ミトコンドリア膜浸透、現象を誘発するミトコンドリアに集中する(Boya et al., Bioche
m. Biophys.Res. Commun. 304:575-581, 2003; Zornig et al., Biochim. Biophys. Act
a, 1551:F1-F37,2001)。
細胞がアポトーシス傷害を受けた後に、トランス−膜ポテンシャル(Dym )は、消失して
、外部のミトコンドリア膜の完全な浸透が起こり、そしてその結果、有害なミトコンドリ
ア内部膜蛋白の漏出が起こる。
ミトコンドリア蛋白質の漏出の最初の例は、シトクロムcの放出であった(Liu et al.,
Apoptosis,6:453-462, 2001)。
シトクロムcがサイトゾル中に存在すると、それは、アポプトゾームと名付けられた、
カスパーゼを活性化する錯体の組み立てを促進する。
シトクロムcは、Apaf-1(アポトーシスプロテアーゼ活性因子-1)に結合して、d
ATO/ATPの錯体への結合と、Apaf-1のオリゴマー化を促進する(Adrain et al
., 1999;Benedict et al., 2000)。
Apaf-1のオリゴマー化は、前駆体の蛋白質分解活性化を触媒するプロ-カスパーゼ
-9の補充と活性プロ-カスパーゼ-9の生成を可能にする(Adrainet al., J. Biol. Chem
.274:20855-20860, 1999; Benedict et al., J. Biol. Chem., 275:8461-8468, 2000)。
【0008】
アポトーシス蛋白質の細胞基質抑制剤のファミリーは、記述されており、XIAP、c-
IAP1及びc-IAP2として知られている。これらの蛋白質は、処理されたカスパーゼ
-3及びカスパーゼ-9に結合して抑制し、そしてその結果、分解のカスケードを停止する
。しかし、細胞は、更にこのアンチ−アポトーシス経路をバイパスするための対抗メカニ
ズムを含んでいる。
アポトーシスを被る細胞において、カスパーゼは、Smac(カスパーゼの第2のミト
コンドリアの活性剤)あるいはDIABLO(蛋白質をLowplに結び付けるDirect IAP)と
して知られている蛋白質のIAPに結合することにより、この抑制の影響から解放される
(Verhagen etal., Apoptosis, 7:163-166, 2002)。
IAPに結合することによって、Smac/DIABLOは、IAPから活性カスパー
ゼを置換し、その結果細胞死を促進する。
HtrA2として知られている他の蛋白質は、アポトーシス損傷後に、ミトコンドリア
から細胞基質に開放される、この場合、たんぱく質は、Smac/DIABLOと同じ方
法でIAPに結合する、そしてそれによりカスパーゼ活性を促進する(Verhagen et al.,
Apoptosis,7:163-166, 2002; Martins et al., 2001; Suzuki et al., Mol. Cell, 8:61
3-621, 2001;Hedge et al., Apoptosis, 7:123-132, 2002)。
【0009】
アポトーシス誘発因子(AIF)は、アポトーシスカスケードの他の成分である。
アポトーシスの誘発後、AIFは、細胞基質と核に移動する。核では、AIFが周辺の
クロマチン凝縮およびDNA断片化(fragmentation)を引き起こす。AIFは、末梢の
クロマチル縮合とDNA断片化を誘発する。AIFは、更にDym消失とホスファチジルセ
リン露出のようなアポトーシスのいくつかの特徴を引き起こす(Zornig et al., Biochim.
Biophys. Acta,1551:F1-F37, 2001)。
AIFのアポトーシス活性を規制するように思われる因子は、熱ショック蛋白質70であ
る(Ravagnanet al., Nature Cell Biol. 3:839-843, 2001)。
AIFのように、ミトコンドリアを出て、核に移動する他のミトコンドリア因子は、エ
ンドヌクレアーゼG(endonuclease G)かエンドG(Endo G)である。
核では、エンドGがカスパーゼ抑制剤の存在下でもDNA断片化を発生する(Li et al.,
Nature,412:95-99, 2001)。
エンドGは、CAD(カスパーゼに活性化されたDNAse)のように、その活性化がカス
パーゼに極めて依存しているヌクレアーゼと同様の様式で作用する(Samejima et al., J.
Biol.Chem., 276:45427-45432, 2001)。
【0010】
2.3 癌および前癌
癌は、細胞の悪性疾患で、その独自の特徴、すなわち細胞サイクルの通常の制御を失っ
て不規則に成長し、分化が欠如し、及び他の細胞組織に進入して転移する。発癌は、正常
細胞が悪性細胞に変化することによるプロセスである。
発癌は、開始が遺伝的事象で始まり、その後に初期腺腫の形成が促進される期間に改変
された細胞が選択的に増加する多段階プロセスである。
連続的な発癌補助作用がないと、腺腫は消退して消滅する。
第二の遺伝的事象で、少量の発癌補助作用を受けた腺腫は、後発の腺腫形成が進行して
、その後悪性の変化をたどる(McKinnell et al., “The Biology Basis of Cancer”, Ch
. 3, 1998)。
【0011】
癌の病因は複雑で、細胞サイクル規則の改変、染色体異常、及び染色体破損を含んでい
る。発癌性ウィルス、化学品、放射線(紫外線あるいは電離放射線)、及び免疫障害のよ
うなタイプの感染性因子は、発癌の主な原因であると考えられている(McKinnell et al.,
“TheBiological Basis of Cancer, Ch.3, 1998)。
【0012】
癌もミトコンドリアの機能障害と関係のあることは長い間提案されてきた。
これらの理論のうちの1つは、ミトコンドリアの突然変異が細胞形質転換と癌の主要な
原因であるとする提案である(Warburg, 1956; Carew and Huang, Mol. Cancer, 1:1-12,
2002)。
ミトコンドリアDNA(mtDNA)の変質は、血液作用による腫瘍(Clayton and Vinogra
d, Nature,216:652-657, 1967)、及び乳癌(Tan et al., 2002; Parrella et al., 2001)
において報告されている。
いくつかの部位のmtDNAの突然変異と欠失(deletions)は、また患者において、直
腸癌、前立腺癌、卵巣癌、胃癌、膵臓癌、肝細胞性癌、食道癌、腎臓癌、甲状腺癌および
脳腫瘍で明らかにされてきている(reviewd by Carew and Huang, Mol. Cancer, 1:1-12,
2002)。
一般的に、腫瘍タイプと関係のない癌において、mtDNA変質の2つの主な特徴がある
と推定される。大部分の突然変異は、TからCとGからAへの塩基の転移である。第二に
、突然変異が生ずる特別の遺伝子における多様性はあるが、D−ループは、ほとんどの腺
腫タイプにおいてmtDNAの最も頻繁な体細胞の突然変異領域になると思われる。
【0013】
前癌とは、悪性細胞に進化または分化する形質転換細胞として定義される。いくつかの
例は、DNAまたはRNAのオンコウィルスによって変化した細胞である。
本発明は、ミトコンドリアRNAの新規なファミリー、及び診断法と癌治療を目標とし
てこれらのRNAの使用に関する。
本発明は、組成物と方法を提供する、そしてこのことは、腫瘍細胞又は前悪性細胞もし
くは腫瘍形成ウィルスで変化した細胞と正常細胞とを区別するのに有用である。
特に、以下に詳述するように、本発明は、前癌及び癌細胞と正常細胞とを区別する診断
検査のための組成物と方法を提供する。
本発明の他の態様では、多くの又は選択的な癌細胞死を誘発するための組成物と方法が
提供される。それ故、本発明は、研究のみならず、癌および前癌の診断と治療に用いるこ
とが可能である、組成物及び方法を提供する。
【発明の概要】
【課題を解決するための手段】
【0014】
3.発明の概要
本発明は、正常な休止及び増殖する細胞、前癌細胞及び癌細胞において特異的に発現さ
れる、ミトコンドリアキメラRNAと称される、新規なヒトミトコンドリアRNAのファ
ミリーを検出するのに有用な組成物及び方法を対象とする。
【0015】
3.1 センスミトコンドリアキメラRNA
本発明の1の態様において、16SミトコンドリアリボゾームRNA(SEQ ID NO 1)の
5’末端に共有結合している815ヌクレオチドの逆方向反復配列(inverted repeat)
を含むミトコンドリアキメラRNAを検出するための組成物及び方法が提供される。
この逆方向反復配列は、mtDNAの16S遺伝子のL−鎖から転写されるRNAの81
5ヌクレオチドのフラグメントに相当する。
このように、この新規なRNAの合成は、mtDNAの16SのL―鎖とH−鎖の転写を
必要とする。mtDNAの両鎖の転写は、異なるプロモーターに規制されるので、我々は、
ミトコンドリアキメラRNA(SEQ ID NO 1)として、本発明におけるこの新規なRNAに
ついて言及する。更に、815ヌクレオチドの逆方向反復配列は“センス”16SのRN
A(H−鎖から転写された)に結合されるので、我々は、“センスミトコンドリアキメラ
RNA”として、この新規なRNAについて言及する。
【0016】
本発明は、培養細胞、細胞サンプル及び組織切片(tissue sections)におけるセンス
ミトコンドリアキメラRNAの発現を検出する組成物と方法を提供する。
この検出は、insituハイブリダイゼーション、対応するcDNAの合成とPCRによる
増殖、転写媒介増殖(transcription mediated amplification)(TMA)(Comanor et
al., J. ClinVirol., 28:14-26, 2003)もしくはノーザンブロット、又は当技術分野の技
術者に自明である他の方法により行われる。
【0017】
本発明の1の側面によると、in situ ハイブリダイゼーションアッセイは、センスミト
コンドリアキメラRNAが正常の増殖期の細胞、異なる腫瘍型の人体生検に存在する腫瘍
細胞におけると同様に培養中の腫瘍細胞に発現することを示す。センスミトコンドリアキ
メラRNAは、正常の休止期の細胞には発現しない。
本発明の他の態様によると、パピローマ・ウイルス(乳頭腫ウィルス)16又は18に
より形質転換(transform)した細胞中の第二の新規なセンスミトコンドリアキメラRN
Aを検出する方法と構成が提供される(Hausen, Biochim. Biophys. Acta, 1288:F55-F78,
1996)。
これらの形質転換した細胞において、16SミトコンドリアRNAの5’末端に共有結
合している754ヌクレオチドの逆方向反復配列からなる、新規なセンスミトコンドリア
キメラRNAが発現する(SEQ ID NO 2)。
このRNAは、正常な増殖細胞中又は腫瘍細胞中には存在しない。本方法と組成物は、
また16SミトコンドリアRNA(SEQ ID NO 3)の5’末端に共有結合している694
ヌクレオチドの逆方向反復配列からなる第三のセンスミトコンドリアキメラRNAがHT
LV−1で形質転換した細胞中に存在することを実証する。
【0018】
3.2 アンチセンスのミトコンドリアキメラRNA
本発明は、更に正常な増殖細胞がSEQ ID NO 4、SEQ ID NO 5、SEQ ID NO 6に対応する
アンチセンスミトコンドリアキメラRNAを発現することを示す方法と組成物を提供する
。
これらの転写産物は、16SミトコンドリアリボゾームのRNA(L−鎖から転写され
た)の5’末端に結合している、それ故にアンチセンスのミトコンドリアRNA名である
、可変の長さ(H−鎖から転写された)の逆方向反復配列を含む。
アンチセンスミトコンドリアキメラRNAの発現は、形質転換した又は前癌細胞中のみ
ならず、異なるタイプの腫瘍のヒト生検中に存在する腫瘍細胞中と同様に、増殖中の腫瘍
細胞系中で下方制御(down regulate)される。
従って、本発明は、通常の増殖細胞を癌細胞と前癌細胞から区別するセンスとアンチセ
ンスのミトコンドリアキメラRNAの発現を検出する方法と組成物を提供し、それ故に悪
性細胞と癌のための新規なマーカーを提供する。
【0019】
3.3 癌療法
本発明の他の態様では、方法と組成物は、センスとアンチセンスミトコンドリアキメラ
RNAを阻害(interfere)するために提供される。
1つの好ましい態様は、低コピー数のこの転写産物を含む腫瘍細胞中のアンチセンスの
ミトコンドリアキメラRNAを阻害することである。この阻害は、その配列がアンチセン
スのミトコンドリアキメラRNA(SEQ ID NO 4及び/又はSEQ ID NO 5 及び/又は SEQ I
D NO 6)の配列に相補している、オリゴヌクレオチド又はオリゴヌクレオチド類似体を用
いて行われる。1又はそれ以上のこれらの相補的なオリゴヌクオチドを用いた異なるタイ
プの腫瘍細胞の処理は、細胞死又はアポトーシスを誘発する。
【0020】
これらのオリゴヌクレオチドは、少なくとも15核酸塩基がSEQ ID NO 4及び/又はSEQ
ID NO 5及び/又はSEQID NO 6に相補する15ないし50のヌクレオチドの合成物であ
る。
これらの相補的なオリゴヌクレオチドの例は、SEQ ID NOS 9〜98の中で示される。
ヒトリンパ球(正常な休止細胞)又はフィトヘムアグルチニン(植物性血球凝集素)で
活性化されたヒトリンパ球(正常な増殖細胞)の処理は、アンチセンスミトコンドリアキメ
ラRNAの配列に相補するオリゴヌクレオチドを用いて同条件で処理した後にはアポトー
シスを受けないので、アポトーシスの誘発は、選択的である。
もし、腫瘍細胞がセンスミトコンドリアキメラRNA(SEQ ID NO 1及び/又はSEQ ID
NO 2及び/又はSEQ IDNO 3)を標的又は相補するオリゴヌクレオチドで処理すると、細
胞死又はアポトーシスの誘発が減じられる。
例えば、本願発明は以下の項目を提供する。
(項目1)
アンチセンスミトコンドリアキメラDNAの分離された核酸分子であって、かつSEQ ID
NO 4、SEQID NO 5又はSEQ ID NO 6のヌクレオチド配列を含む核酸分子。
(項目2)
項目1に記載のSEQID NO 4、SEQ ID NO 5又はSEQ ID NO 6のアンチセンスのミトコン
ドリアキメラDNAに相補する、長さで10ないし50核酸塩基の合成物又はオリゴヌク
レオチド。
(項目3)
前記合成物又はオリゴヌクレオチドが、長さ10から50核酸塩基で、SEQ ID NOが9
から98で構成されるグループから選択される配列の少なくとも10核酸塩基部分を含む
、項目2に記載の合成物。
(項目4)
前記合成物又はオリゴヌクレオチドが、長さ10から50までの核酸塩基で、前癌およ
び癌細胞死を誘発する、SEQ ID NOが9から98で構成されるグループから選択される配
列の少なくとも10核酸塩基部を含む、項目2に記載の合成物。
(項目5)
前記オリゴヌクレオチドがSEQ ID NO 4、SEQ ID NO 5又はSEQ ID NO 6のアンチセンス
ミトコンドリアのキメラRNAでハイブリダイズされて、前癌及び癌細胞死を誘発する、
項目2に記載の合成物。
(項目6)
前記オリゴヌクレオチドが少なくとも1つの修飾されたインターヌクレオシド連鎖を含
む、項目2に記載の合成物。
(項目7)
前記修飾されたインターヌクレオシド連鎖がホスフォロチオエート、メチルホスフォネ
ート、ホスフォロジチオエート、ホスフォロトリエステル、ペプチド性核酸、及びロック
された核酸連鎖から構成されるグループから選択された、項目6に記載の合成物。
(項目8)
前記合成物が少なくとも1つの修飾された糖部分を含む、項目2に記載の合成物。
(項目9)
前記修飾された糖部分が2’-Oメトキシエチル、2’-Oメチル及びモルフォリノ基から
構成されるグループから選択される、項目8に記載の合成物。
(項目10)
前記オリゴヌクレオチドが少なくとも1の修飾された核酸塩基を含む、項目2に記載
の合成物。
(項目11)
修飾された核酸塩基が5-メチルシトシンである、項目10に記載の合成物。
(項目12)
前記オリゴヌクレオチドが、ホスフォロチオエート連鎖で結合されたDNA残基を含む
キメラ核酸であり、該DNA残基がホスフォロチオエート連鎖で共に結合された2’-Oメ
トキシエチル又は2’-OメチルRNA残基の少なくとも3つの残基により5’及び3’サ
イドで側面に位置されている、項目2に記載の合成物。
(項目13)
前記合成物がリボザイムである、項目2に記載の合成物。
(項目14)
前記合成物が低分子干渉RNAすなわちsiRNAである、項目2に記載の合成物。
(項目15)
センスミトコンドリアキメラDNAの分離された核酸分子であって、かつSEQ ID NO 1
、SEQID NO 2又はSEQ ID NO 3のヌクレオチド配列を含む、核酸分子。
(項目16)
項目15のSEQID NO 1、SEQ ID NQ.2又はSEQ ID NO 3のセンスミトコンドリアキメ
ラDNAを標的とする、長さ10から50核酸塩基の合成物又はオリゴヌクレオチド。
(項目17)
前記合成物又はオリゴヌクレオチドがSEQ ID NO 99〜197から構成されるグループから
選択された配列の少なくとも10核酸塩基部を含み、長さで50核酸塩基以下である、請
求項16に記載の合成物。
(項目18)
前記合成物又はオリゴヌクレオチドが前癌又は癌細胞死を誘発するSEQ ID NO 99〜197
から構成されるグループから選択された配列の少なくとも10核酸塩基部を含み、長さで
50核酸塩基以下である、項目16に記載の合成物。
(項目19)
前記オリゴヌクレオチドがSEQ ID NO 1、SEQ ID NO 2又はSEQ ID NO 3のセンスミトコ
ンドリアキメラDNAでハイブリダイズされていて、前癌又は癌細胞死を誘発する、請求
項16に記載の合成物。
(項目20)
前記オリゴヌクレオチドが少なくとも1の修飾されたインターヌクレオシド連鎖を含む
、項目16に記載の合成物。
(項目21)
前記修飾インターヌクレオシド連鎖がホスフォロチオエート、メチルホスフォネート、
ホスフォロジチオエート、ホスフォロトリエステル、ペプチド性核酸、及びロックされた
核酸連鎖から構成されるグループから選択された、項目20に記載の合成物。
(項目22)
前記合成物が少なくとも1つの修飾された糖部分を含む、項目16に記載の合成物。
(項目23)
前記修飾された糖部分が2’-Oメトキシエチル、2’-Oメチル及びモルフォリノ基から
構成されるグループから選択されている、項目23に記載の合成物。
(項目24)
前記オリゴヌクレオチドが少なくとも1の修飾された核酸塩基を含む、項目16に記
載の合成物。
(項目25)
前記修飾された核酸塩基が5-メチルシトシンである、項目24に記載の合成物。
(項目26)
前記オリゴヌクレオチドが、ホスフォロチオエート連鎖で結合されたDNA残基を含む
キメラ核酸であり、該DNA残基がホスフォロチオエート連鎖で結合された2’-Oメトキ
シエチル又は2’-OメチルRNA残基の少なくとも3つの残基により5’及び3’サイド
で側面に位置されている、項目16に記載の合成物。
(項目27)
前記合成物がリボザイムである、項目16に記載の合成物。
(項目28)
前記合成物が低分子干渉RNA又はsiRNAである、項目16に記載の合成物。
(項目29)
SEQID NO 4、SEQ ID NO 5もしくはSEQ ID NO 6のアンチセンスミトコンドリアキメラ
RNA及び/又はSEQ ID NO 1、SEQ ID NO 2もしくはSEQ ID NO 3のセンスミトコンドリ
アキメラRNAに相補する、長さ10から50核酸塩基の1以上のオリゴヌクレオチド、
並びに薬剤の受け入れ可能なキャリアー又は希釈剤を含む、薬剤組成物。
(項目30)
前記キャリアー又は希釈剤が乳剤、リポソーム、浸透促進剤、界面活性剤、キレート試
薬、キャリアーを含む、項目29に記載の薬剤組成物。
(項目31)
経口、口腔、非経口もしくは直腸投与、又は吸入もしくは通気による肺投与用の、請求
項29に記載の薬剤組成物。
(項目32)
SEQID NO 4、SEQ ID NO 5もしくはSEQ ID NO 6のアンチセンスミトコンドリアキメラ
RNA及び/又はSEQ ID NO 1、SEQ ID NO 2もしくはSEQ ID NO 3のセンスミトコンドリ
アキメラRNAに相補する、長さ10から50核酸塩基の合成物で、癌もしくは前癌細胞
又は組織と接触させることを含む、細胞又は組織中における前癌又は癌細胞死の誘発方法
。
(項目33)
SEQID NO 4、SEQ ID NO 5もしくはSEQ ID NO 6のアンチセンスミトコンドリアキメラ
RNA及び/又はSEQ ID NO 1、SEQ ID NO 2もしくはSEQ ID NO 3のセンスミトコンドリ
アキメラRNAに相補する、長さ10から50核酸塩基の2以上の合成物で、細胞又は組
織と接触させることを含む、細胞又は組織中における前癌又は癌細胞死の誘発方法。
(項目34)
前癌又は癌細胞死を誘発することが可能である、SEQ ID NO 4、SEQ ID NO 5もしくはSE
Q ID NO 6のアンチセンスミトコンドリアキメラRNA及び/又はSEQID NO 1、SEQ ID N
O 2もしくはSEQID NO 3のセンスミトコンドリアキメラRNAに相補する、長さ10から
50核酸塩基の合成物の治療に有効な量をヒト又は動物に投与することを含む、前癌又は
癌を有する疑いのあるヒト又は動物の治療方法。
(項目35)
SEQID NO 4、SEQ ID NO 5もしくはSEQ ID NO 6のアンチセンスミトコンドリアキメラ
RNA及び/又はSEQ ID NO 1、SEQ ID NO 2もしくはSEQ ID NO 3のセンスミトコンドリ
アキメラRNAに相補する、長さ10から50核酸塩基の2以上の合成物の治療に有効な
量をヒト又は動物に投与することを含む、前癌又は癌細胞死の誘発することが可能である
、前癌又は癌を有する疑いのあるヒト又は動物の治療方法。
(項目36)
前記合成物又は複数の合成物が癌の治療に対して化学療法薬と併用して投与される、請
求項34に記載の方法
(項目37)
癌の化学療法薬治療の効能改善方法であって、前癌又は癌細胞死を誘発する、SEQ ID N
O 4、SEQ IDNO 5もしくはSEQ ID NO 6のアンチセンスミトコンドリアキメラRNA及び
/又はSEQID NO 1、SEQ ID NO 2もしくはSEQ ID NO 3のセンスミトコンドリアキメラR
NAに相補する、長さ10から50核酸塩基の合成物又は複数の合成物を化学療法薬治療
と併用して投与することを含む、改善方法。
(項目38)
前記合成物又は複数の合成物が少なくとも1つの修飾されたインターヌクレオシド連鎖
を含む、オリゴヌクレオチドである項目32に記載の方法。
(項目39)
前記修飾されたインターヌクレオシド連鎖がホスフォロチオエート、メチルホスフォネ
ート、ホスフォロジチオエート、ホスフォロトリエステル、ペプチド性核酸、及びロック
された核酸連鎖から構成されるグループから選択された、項目38に記載の方法。
(項目40)
前記合成物又は複数の合成物が少なくとも1つの修飾された糖部分を含む、項目32
に記載の方法。
(項目41)
前記修飾された糖部分が2’-Oメトキシエチル、2’-Oメチル及びモルフォリノ基から
構成されるグループから選択される、項目40に記載の方法。
(項目42)
哺乳類の細胞において、SEQ ID NO 4、SEQ ID NO 5、SEQ ID NO 6のアンチセンスミト
コンドリアキメラRNA及びSEQ ID NO 1、SEQ ID NO 2、SEQ ID NO 3のセンスミトコン
ドリアキメラRNAの存在又は不存在を決定することによる正常、前癌又は癌細胞を識別
するハイブリダイゼーション法であって、該方法は、下記a)ないしc)を含むハイブリ
ダイゼーション法。
a)サンプルを、少なくとも1つの検出可能なプローブが標的ミトコンドリアキメラRN
A配列にハイブリダイズするプローブとして、SEQ ID NO 4、SEQ ID NO 5もしくはSEQ ID
NO 6のアンチセンスミトコンドリアキメラRNA及び/又はSEQID NO 1、SEQ ID NO 2
もしくはSEQID NO 3のセンスミトコンドリアキメラRNAに相補する長さ10から50
核酸塩基の2又はそれ以上の独立して検出可能な又は標識された合成物と接触させる
b)検出可能な合成物の標的配列へのハイブリダイゼーションの検出、同定、又は定量
c)正常、前癌又は癌細胞における、センス及びアンチセンスミトコンドリアキメラRN
Aの異なる発現のパターンで得られた結果の相互の関連付けと比較
(項目43)
個々に独立して検出可能なプローブがオリゴヌクレオチドである、項目42に記載の
方法。
(項目44)
前記プローブがDNA及び/又はRNAの10から50残基のオリゴヌクレオチドを含
む、項目42に記載の方法。
(項目45)
前記プローブがSEQ ID NO 1、SEQ ID NO 2、SEQ ID NO 3、SEQ ID NO 4、SEQ ID NO 5
あるいはSEQID NO 6のRNAの領域で少なくとも95%がホモロジーである10から5
0残基のオリゴヌクレオチドを含む、項目44に記載の方法。
(項目46)
前記オリゴヌクレオチドが少なくとも1つの修飾されたインターヌクレオシド連鎖を含
む項目45に記載の方法。
(項目47)
前記修飾されたインターヌクレオシド連鎖がホスフォロチオエート、メチルホスフォネ
ート、ホスフォロジチオエート、ホスフォロトリエステル、ペプチド性核酸、及びロック
された核酸連鎖から構成されるグループから選択されている、項目46に記載の方法。
(項目48)
前記オリゴヌクレオチドが少なくとも1つの修飾された糖部分を含む、項目48に記
載の方法。
(項目49)
前記修飾された糖部分が2’-Oメトキシエチル、2’-Oメチル及びモルフォリノ基から
構成されるグループから選択されている、項目48に記載の方法。
(項目50)
前記オリゴヌクレオチドが放射能、又はジゴキシゲニン、ビオチン、蛍光色素のグルー
プから選択されたハプテンで標識されている、項目45に記載の方法。
(項目51)
前記検出方法がアンチビオチン抗体、アンチジゴキシゲニン抗体及びアンチフルオレセ
イン抗体を含む、項目42に記載の方法。
(項目52)
前記抗体が抗体とアルカリホスファターゼ又はペルオキシダーゼと接合している、請求
項51に記載の方法。
(項目53)
前記検出方法がストレプトアビジン-アルカリホスファターゼ接合と増殖試薬を含む、
項目42に記載の方法。
(項目54)
センスミトコンドリアRNAを標的にする前記オリゴヌクレオチドが蛍光色素で標識さ
れており、またアンチセンスミトコンドリアRNAを標的にする前記アンチセンスミトコ
ンドリアRNAが異なる発光の蛍光色素で標識されている、項目42に記載の方法。
(項目55)
前記検出方法が異なる蛍光色素の発光の検出を含み、蛍光顕微鏡もしくは共焦点顕微鏡
、レーザースキャナ装置、又は流量血球計算器で検出される、項目54に記載の方法。
(項目56)
前記サンプルが正常な休止又は増殖細胞、培養下の前癌及び癌細胞のサンプルを含む、
項目42に記載の方法。
(項目57)
前記サンプルが癌を有する疑いのある患者からの細胞を含む、項目42に記載の方法
。
(項目58)
前記サンプルが固定された又は冷凍のヒトの生体からの組織切片である、項目42に
記載の方法。
(項目59)
前記サンプルが尿、肺洗浄、唾液、コロノサイト、血液塗抹標本、骨髄の塗抹標本、腹
水、血液、又はリンパ液循環転移細胞中に存在する細胞を含む、項目42に記載の方法
。
(項目60)
項目42の方法による、正常なヒト増殖細胞及び形質転換又は悪性ヒト細胞を識別す
るのに有用なinsituハイブリダイゼーション用キットであって、下記A)ないしd)を
含むキット。
a)センス及び/又はアンチセンスミトコンドリアキメラRNAを標的にする1又はそれ
以上の検出可能な、標識されたオリゴヌクレオチド
b)標識検出のために接合された抗体試薬
c)ハイブリダイゼーションと洗浄緩衝液
d)固定されたヒト正常増殖細胞の1又はそれ以上のスライド、及び対照として、固定さ
れ形質転換された又は悪性ヒト細胞の1又はそれ以上のスライド
(項目61)
前記オリゴヌクレオチドの標識がジゴキシゲニン、バイオチン、フルオレセインのグル
ープから選択されたハプテンを含む、項目60のキット。
(項目62)
前記検出方法がアンチビオチン抗体、アンチジゴキシゲニン抗体及びアンチフルオレセ
イン抗体を含む項目60に記載のキット。
(項目63)
前記抗体がアルカリホスファターゼ又はペルオキシダーゼと接合した抗体である、請求
項60に記載のキット。
(項目64)
前記検出方法が抗ビオチン抗体、抗ジゴキシゲニン抗体又は抗フルオレセイン抗体に対
する抗体と接合したアルカリホスファターゼ又はペルオキシダーゼ、及び増殖試薬を含む
、項目60に記載のキット。
(項目65)
前記検出方法がストレプトアビジン−アルカリホスファターゼ接合及び増殖試薬を含む
、項目60に記載のキット。
(項目66)
前記増殖試薬がアルカリホスファターゼニトロブルーテトラゾリウム用基質又はペルオ
キシダーゼ用基質である、項目60に記載のキット。
(項目67)
サンプルの中にある細胞質、細胞核、及び細胞の核小体中のアンチセンス(SEQ ID NO
4、5及び6)又はセンス(SEQID NO 1、2及び3)ミトコンドリアキメラRNAの細胞局在
化を決定するinsituハイブリダイゼーション法であって、下記a)とb)を含むin situ
ハイブリダイゼーション法。
a)サンプルを、少なくとも1つの検出可能なプローブが標識ミトコンドリアキメラRN
A配列にハイブリタイズするプローブとして、SEQ ID NO 4、SEQ ID NO 5もしくはSEQ ID
NO 6のアンチセンスミトコンドリアキメラRNA及び/又はSEQID NO 1、SEQ ID NO 2
もしくはSEQID NO 3のセンスミトコンドリアキメラRNAに相補する長さ10から50
核酸塩基の1又はそれ以上の独立して検出可能な合成物又は標識された合成物と接触させ
る
b)検出可能な合成物の標的配列へのハイブリダイゼーションの検出、同定、又は定量
(項目68)
項目67の方法によるサンプルの中に存在する細胞質、細胞核、及び細胞の核小体中
のアンチセンス(SEQ ID NO 4,5及び6)又はセンス(SEQ ID NO 1、2及び3)ミトコンド
リアキメラRNAの細胞局在化を決定するin situハイブリダイゼーション用キットであ
って、下記a)ないしd)を含むキット。
a)センス又はアンチセンスミトコンドリアキメラRNAを標的にする、標識された1又
はそれ以上の検出可能なオリゴヌクレオチド
b)標識検出のために接合された抗体試薬
c)ハイブリダイゼーションと洗浄緩衝液
d)固定されたヒト正常増殖細胞の1又はそれ以上のスライド、及び対照として固定され
形質転換された又は悪性のヒト細胞の1又はそれ以上のスライド
(項目69)
前記センス及びアンチセンスミトコンドリアキメラRNAの検出、同定及び定量が以下
の手順のいずれかにより行われる、項目42に記載の方法。
insituハイブリダイゼーション又は蛍光in situハイブリダイゼーション、ドットブロ
ット、ノーザンブロット及びPCRによるcDNA増殖又はTMAによるRNA増殖
(項目70)
前記合成物又は複数の合成物が少なくとも1の修飾されたインターヌクレオシド連鎖を
含むオリゴヌクレオチドである、項目33に記載の方法。
(項目71)
前記合成物又は複数の合成物が少なくとも1の修飾されたインターヌクレオシド連鎖を
含むオリゴヌクレオチドである、項目34に記載の方法。
(項目72)
前記合成物又は複数の合成物が少なくとも1の修飾されたインターヌクレオシド連鎖を
含むオリゴヌクレオチドである、項目35に記載の方法。
(項目73)
前記合成物又は複数の合成物が少なくとも1つの修飾された糖部分を含む、項目33
に記載の方法。
(項目74)
前記合成物又は複数の合成物が少なくとも1つの修飾された糖部分を含む、項目34
に記載の方法。
(項目75)
前記合成物又は複数の合成物が少なくとも1つの修飾された糖部分を含む、項目35
に記載の方法。
(項目76)
前記合成物又は複数の合成物が少なくとも1つの修飾された糖部分を含む、項目36
に記載の方法。
【図面の簡単な説明】
【0021】
【図1】図1A、B及びCは、SEQ ID NO 1, SEQ ID NO 2及びSEQ ID NO 3に相当するセンスミトコンドリアキメラRNAの構造を示す線画である。 矢印は、ピースによりRNAを増殖するのに用いるプライマーの相対的位置を示す。 線の下部の矢印は、リバースプライマーで、線の先頭の矢は、フォワードプライマーである。プライマー1は、16Sミトコンドリアの5’末端に隣接して位置付けされる。黒線は、センス16Sミトコンドリアに相当する、一方、点線は、アンチセンス16Sミトコンドリアに相当する。
【0022】
【図2】図2は、図1Aに示したプライマー1とプライマー3の間で得られた増殖産物を示すアガロースゲル電気泳動である。 増殖は、鋳型としてSiHa(癌細胞)、HPV16(HFK698)で形質転換されたヒト角化細胞、あるいはHTLV-1で形質転換されたBリンパ細胞を用いて、PT−CRにより実行された。SiHa細胞からのRNAで、210bpの1つの単一アンプリコンだけが得られ、SEQID NO 1のセグメントに相当する。HPV16で形質転換されたヒト角化細胞の全RNAにおいて、210bpのアンプリコンに加えて、150bpの2番目のアンプリコンが得られた、SEQ ID NO 2に相当する。 HTLV-1で形質転換された細胞からのRNAで、210bp及び150bpのアンプリコンに加えて、3番目のアンプリコンが得られ、SEQ ID NO 3に相当する。
【0023】
【図3】図3は、SEQ ID NO 4、SEQ ID NO 5及びSEQ ID NO 6に相当するアンチセンスミトコンドリアキメラRNAの構造を示す線画である。矢印は、増殖に用いられたプライマーを示す。プライマー1は、アンチセンスの16SミトコンドリアRNAの5’末端に隣接して位置付けされる。 これらの転写産物の配列を得るための方策は、図1に記載されたのと同様である。
【0024】
【図4A】図4A及び図4Bは、培養液中でいくつかの腫瘍細胞を用いて行われたin situハイブリダイゼーション分析を示す。 細胞は、センスミトコンドリアキメラRNAに相補するオリゴヌクレオチドプローブでハイブリダイズされ、そしてジゴキシゲニンでラベル表示された(左パネル)。 更に、細胞は、ジゴキシゲニンでラベル表示されたアンチセンスミトコンドリアキメラRNAに相補するオリゴヌクレオチドプローブで並行してハイブリダイズされた(右パネル)。細胞系は左側で識別される。
【図4B】図4A及び図4Bは、培養液中でいくつかの腫瘍細胞を用いて行われたin situハイブリダイゼーション分析を示す。 細胞は、センスミトコンドリアキメラRNAに相補するオリゴヌクレオチドプローブでハイブリダイズされ、そしてジゴキシゲニンでラベル表示された(左パネル)。 更に、細胞は、ジゴキシゲニンでラベル表示されたアンチセンスミトコンドリアキメラRNAに相補するオリゴヌクレオチドプローブで並行してハイブリダイズされた(右パネル)。細胞系は左側で識別される。
【0025】
【図5A】図5Aは、異なる腫瘍細胞タイプに相当するヒトの生検のいくつかのセクションのin situハイブリダイゼーションである。 腫瘍セクションは、センスミトコンドリアキメラRNAに相補するオリゴヌクレオチドプローブでハイブリダイズされ、ジゴキシゲニンでラベル表示された(左パネル)。 更に、並列の腫瘍セクションは、ジゴキシゲニンでラベル表示されたアンチセンスセンスミトコンドリアキメラRNAに相補するオリゴヌクレオチドプローブでハイブリダイズされた(右パネル)。
【図5B】図5Bは、ジゴキシゲニンでラベル表示されたセンスミトコンドリアキメラRNAに相補するオリゴヌクレオチドプローブを用いて行われた、異なるヒト腫瘍細胞のin situハイブリダイゼーションである。
【0026】
【図6】図6は、正常増殖細胞のin situハイブリダイゼーションである。 サンプルは、センス又はアンチセンスミトコンドリアキメラRNAを標的にしたプローブでハイブリダイズし、ジゴキシゲニンでラベル表示された。 強いハイブリダイゼーションシグナルは、双方のプローブで得られた、1つはセンスミトコンドリアキメラRNAに相補し(左パネル)、同様に他はアンチセンスミトコンドリアキメラRNAに相補する(右パネル)。 組織あるいは細胞は、左側で識別される。
【0027】
【図7】図7は、ミトゲンPHAで増殖するために刺激されたヒトリンパ細胞における発現の変化を示すための免疫細胞化学及びin situハイブリダイゼーションである。 PHAで刺激48時間後、リンパ細胞は、抗原Ki-67及びPCNAを表現する。 これらの抗原は、コントロール中又は休止リンパ細胞中では発現されない(左パネル)。前記in situハイブリダイゼーションは、センス及びアンチセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオチドで行ない、ジゴキシゲニンでラベル表示された。刺激を受けたリンパ細胞は、アンチセンスミトコンドリアキメラRNAだけでなくセンスミトコンドリアキメラRNAを過剰発現した(右パネル)。
【0028】
【図8】図8は、核小体中でセンスミトコンドリアキメラRNAの局在化を示す腫瘍細胞のinsituハイブリダイゼーションである。 細胞あるいは腫瘍セクションは、左側で示される。
【0029】
【図9】図9は、アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオチドプローブで処理された腫瘍HL-60細胞に生じる変化示す蛍光顕微鏡検査である。A、B、C及びDは、活性化されたカスパーゼに高い親和性で結合する化合物(VAD-fmok)で染色されることを示す。この化合物は、フルオレスセインで標識される。アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオチドプローブは、(B,Cと対比して)同じ方法でも薬剤スタウロスポリンよりも高いカスパーゼ活性を誘発する。 活性化されたカスパーゼは、対照として、コントロール未処理細胞(A)又は12SミトコンドリアRNAを標的にするオリゴヌクレオチドプローブで処理された細胞(D)中では検出されなかった。EとFは、DAPIでHL-60細胞の染色を示す。 コントロール細胞(未処理)は、核の均質な染色を示す(E)、一方、アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオチドプローブで処理された細胞は、広範な核の断片化を示す(F)。
【0030】
【図10】図10は、休止及び増殖リンパ細胞の異なる処理条件後のアポトーシスを起こした細胞の割合を示す。 アポトーシスは、DAPI染色により休止中の細胞又はPHAに刺激されたリンパ細胞で測定された。バー1及び2は、未処理細胞に相当する。コントロール(1)又はPHAに刺激されたリンパ細胞(2)の低い自然のアポトーシスが観察された。同様の低いレベルのアポトーシスは、休止中のリンパ細胞又はアンチセンスミトコンドリアキメラRNAを標的にする15uMのオリゴヌクレオチドプローブで15時間処理されたPHA刺激リンパ細胞で観察された、アポトーシスは正常細胞中では誘発されないことを示した。対照として、休止中の細胞とPHAに刺激されたリンパ細胞は、スタウロスポリンで処理された。これらの条件下では、休止中のリンパ細胞(5)あるいはPHAに刺激されたリンパ細胞(6)の約90%がアポトーシスをこうむる。
【発明を実施するための形態】
【0031】
4.発明の詳細な説明
4.1 ヒトミトコンドリアキメラRNAファミリー
本発明は、ヒトの細胞がヒトのミトコンドリアキメラRNAと称される、新規なミトコ
ンドリアRNAのファミリーを発現するという驚くべき発見に基づくものである。
【0032】
これらの転写産物に1つは、センスミトコンドリアキメラRNAと名付けられた、ミト
コンドリアの16SリボゾームRNAの5’末端に共有結合している815ヌクレオチド
の長い逆方向反復配列が含まれる。この長い逆方向反復配列は、長い二重鎖幹と50ヌク
レオチドを形成する、位置51から866の16SリボゾームRNAに十分に相補してい
る。
図1Aに示すように、815塩基対のステムは、逆方向転写酵素(reverse transcript
ase)が相当するcDNAを合成するのに重要な問題を示す。従って、新しい戦略は、図
1Aに例示するRT−PCRによりこのRNAを増殖するのに用いられる。
各重なり合うフラグメントの配列を得た後に、それらは、SEQ ID No1(図1A)に示すセ
ンスミトコンドリアキメラRNAの完全な配列を得るためにコンティグ(contigs)とし
て組み立てられる。
【0033】
この発明の他の側面は、癌遺伝子のヒトの乳頭腫ウィルス(パピローマ・ウイルス)1
6又は18で形質転換した細胞中に発現する他の新規なセンスミトコンドリアキメラRN
Aの発見である。
人間の包皮角化細胞(Human foreskin keratinocytes)(HFK)は、HPV16又は18
により感染した(Hausen,Biochim. Biophys. Acta, 1288:F55-F78, 1996)。
この感染は、HFKの形質転換又は不死化を引き起こす。
しかしながら、これらの細胞は、関連するSiHa細胞(HPV16に感染した)又は
HeLa細胞(HPV18に感染した)のような発癌性ではない。
これらの細胞は、SiHaとHeLa細胞に類似するセンスミトコンドリアキメラRN
A(SEQ ID NO1)を発現する。
しかしながら、形質転換した細胞は、更に16SリボゾームRNA(図1B)(SEQ ID No
2)に結合する754ヌクレオチドの逆方向反復配列を含む、他の第二のセンスミトコン
ドリアキメラRNAを発現する。この新しいセンスミトコンドリアキメラRNAは、正常
なヒトの細胞(HFK)中もしくは発癌性細胞(SiHa又はHeLa細胞)中では下方制御される
か、又は発現しない。
【0034】
この発明の別の態様で、我々は、HTLV-1(Kobayashi et al., EMBO J., 3:1339-13
43, 1984)で形質転換した細胞中で第三のセンスミトコンドリアキメラRNAの発現を決
定した。
HTLV-1に感染したMT-2細胞は、センスミトコンドリアキメラRNA(SEQID NO
1)とHPV16あるいは18(SEQID NO 2)で形質転換された細胞中に発現するセンスミ
トコンドリアキメラRNAを発現する。
これらの転写産物に加えて、HTLV-1で感染された細胞は、16SリボゾームRN
Aの5’末端に結合する694ヌクレオチドの逆方向反復配列を含む、第三のセンスミト
コンドリアキメラRNAを発現する。
この新規なRNA(図1C)(SEQID NO 3)は、正常な増殖する細胞の中で、癌細胞の中
で、あるいはHPV16あるいは18で形質転換されたHFKの中では発現されない。
【0035】
前節に記載したヒトの包皮角化細胞(forskin keratinocytes)(HFK)のような正常の
増殖する細胞は、センスミトコンドリアキメラRNA(図6) (SEQ ID NO 1)を過剰発現(
over express)する。
フィトヘムアグルチニン(phytohaemagglutinin)(PHA)のようなミトゲンで活性化され
たヒトのリンパ細胞は、細胞周期のSフェーズに入って、DNAの合成を始める(Yu et a
l., J. Biol.Chem., 266:7588-7595, 1991)。
増殖する細胞として、リンパ細胞は、Ki-67のような増殖に関連して、細胞核抗原
またはPCNAを増殖する抗原を発現する(Bantis et al., Cytopathology, 15:25-31, 2
004)。
脾臓の胚の中心にあるリンパ細胞、精原細胞、及び胚の細胞のような他の増殖する細胞
は、またセンスミトコンドリアキメラRNA(SEQ ID NO 1)(図4)を過剰発現する。対照的
に、活性化されないリンパ細胞、又は筋細胞のような非増殖細胞は、センスミトコンドリ
アキメラRNA(図7)を発現しない。
【0036】
本発明の他の態様で、腫瘍細胞と正常に増殖する細胞とを区別する方法が提供される。
以前に記述したように、腫瘍細胞と正常に増殖する細胞は、SEQ ID NO 1に記述したセン
スミトコンドリアキメラRNAを過剰発現する。
更に、HPVおよびHTLV-1で感染された特別の状況において、付加的なキメラR
NAが見出される(SEQ ID NO 2及びSEQ ID NO 3)。
しかしながら、本発明は、アンチセンスミトコンドリアキメラRNAを過剰発現すると
いう驚くべき発見に基づくものである。
アンチセンスミトコンドリアキメラRNAの発現は、PHA(図7)で活性化されたヒト
のリンパ細胞中で、正常なHFKで及び他の正常な増殖する細胞(図6)中で確認された。
この発明の他の驚くべき発見は、正常に増殖する細胞と異なり、腫瘍細胞がアンチセン
スミトコンドリアキメラRNAを発現しないか、又はその生産を下方制御するということ
である(図6及び図7と図4を比較する)。
【0037】
RT-PCRによってオーバーラップしているフラグメントに基づくキメラRNAを増
殖するために同様の方策を用いて、アンチセンスミトコンドリアキメラRNAの構造が決
定された(図3)。
コンティグにおけるその配列と組み立ては、アンチセンスの16Sミトコンドリアリボ
ゾームRNA(図3A、B及びC)(SEQID NO 4、SEQ ID NO 5、SEQ ID NO 6)の5’末端
に結合している異なる長さの逆方向反復配列を含むアンチセンスミトコンドリアキメラR
NAの複雑なファミリーを明らかにする。
その配列は、またこれらのRNAにおける二重鎖構造又はステムの生成、及び7、96
及び451ヌクレオチド(図3A、B及びC、SEQ ID NO 4、SEQ ID NO 5、SEQ ID NO 6)
をそれぞれ有するループの生成を明らかにする。
【0038】
本発明の他の態様では、腫瘍ウィルスによって細胞の発癌性形質転換を生じさせる方法
と組成物が提供される。HeLa細胞(HPV 18に感染した)又はSiHa細胞(HPV 16に感
染した)は、センスミトコンドリアキメラRNAを過剰発現する、しかしアンチセンスミ
トコンドリアキメラRNAの発現を下方制御する。他方、正常な増殖細胞であるHFKは
、アンチセンスミトコンドリアキメラRNAと同様にセンスミトコンドリアキメラRNA
の双方を過剰発現する。
HPV16あるいはHPV18でHFKの形質転換の後に、細胞は、腫瘍表現型を得、
これらはセンスミトコンドリアキメラRNAを過剰発現し、そしてアンチセンスミトコン
ドリアキメラRNAの発現を下方制御する。センスミトコンドリアキメラRNAの過剰発
現及びアンチセンスミトコンドリアキメラRNAの発現の下方制御は、in situハイブリ
ダイゼーション、RT-PCRによるRNAの増殖、又は当該分野の技術者によってよく
知られた方法によるRNAを決定する他の方法を用いることにより決定される。これらの
方法と組成物は、他の腫瘍ウィルスで又は形質転換又は発癌性を誘発する複合物により、
キメラRNA細胞ファミリーの発現における変化を決定するのに用いられる(McKinnell e
t al., “Thebiological basis of Cancer, Cambridge University Press1998)。
【0039】
4.2 癌および前癌診断
本発明により、方法と組成物は、生体サンプル中にセンスミトコンドリアキメラRNA
とアンチセンスミトコンドリアキメラRNAの存在を検出するために提供される。
1つの好ましい態様では、検出は、insituハイブリダイゼーションにより行われる。
生体サンプルの細胞中のセンスミトコンドリアキメラRNAとアンチセンスミトコンド
リアキメラRNAの検出は、細胞が正常な増殖中の細胞であることを示す。
他の態様において、腫瘍細胞によるin situハイブリダイゼーションの結果は、センス
ミトコンドリアキメラRNAの発現と、アンチセンスミトコンドリアキメラRNAの下方
制御又は不存在を示す。
もし、生体サンプルが非増殖の正常細胞を含む場合、in situハイブリダイゼーション
は、センスミトコンドリアキメラRNAとアンチセンスミトコンドリアキメラRNAのい
ずれも発現しないことを示す。
【0040】
生体サンプルは、培養中の又は血液スミア(smears)もしくは骨髄スミア中の正常細胞
(休止又は増殖する細胞)、培養中の腫瘍細胞、及び腫瘍ウィルスで形質転換された正常
細胞として理解される。
更に、生体細胞は、膀胱もしくは腎臓癌を有する疑いのある患者からの尿もしくは膀胱
洗浄から得られた細胞、又は頭と首の癌を有する疑いのある患者の唾液からの細胞、肺癌
を有する疑いのある患者の管支肺胞からの細胞を含む。また、生体サンプルは、白血病を
有する疑いのある患者の血液からの細胞スミア、又は転移を有する疑いのある患者の血液
、リンパ液、リンパ節からの細胞スミアを含む。
【0041】
本発明による生体サンプルは、当分野の技術者に良く知られた技術である、組織病理学
の分析のため急速冷凍した組織あるいは細胞サンプルの使用を含む。
あるいは、生体サンプルは、当分野でよく知られた多種多様の固定プロトコル(Frederi
ck et al,Current Protocols In Molecular Biology, Volume 2, Unit 14, Frederick M
. Ausubul etal. edS., 1995; Celis, Cell Biology, A Laboratoty Handbook, Julio E
. Celis, ed.,1994)により行うことができる化学処理の使用により固定される薄片の生
体検査になりうる。
生体サンプルは、また化学的修飾、ホルマリン処理又は当該分野でよく知られた他の固
定がなされていない非固定の生物由来物質であってもよい。
【0042】
あるいは、insituハイブリダイゼーションは、パラフィン又は他の包埋ができるポリ
マーのような物質に包埋された生体サンプルの使用により行うことができる。
包埋により得られたブロックは、ミクロトームで約4ないし10μmの厚みの切片にす
る。この切片は、ポリリシン又はムッセル(mussel)接着性蛋白質のような技術分野で知
られた接着性物質で被覆されてガラス又はポラスチックスライドに乗せられる(Burzio et
al.,Curr. Opin. Biotechnol., 8:309-312, 1997)。
【0043】
in situハイブリダイゼーションは、この分野の技術者によく知られた方法で行われる
。例えば、細胞中でセンスミトコンドリアキメラRNA(SEQ ID NO 1, SEQ ID NO 2, SE
Q ID NO 3)、又はアンチセンスミトコンドリアキメラRNA(SEQID NO 4, SEQ ID NO 5
, SEQ ID NO 6)の1又は2以上の配列を標的にした1又は2以上の標識されたプローブか
らなるハイブリダイゼーション溶液は、ハイブリダイゼーション条件下で細胞と接触され
る。そのハイブリダイゼーションシグナルは、それから正常の又は制御された癌及び前癌
細胞から予め決定されたハイブリダイゼーションパターンと比較される。
【0044】
ここに使用されているように、in situハイブリダイゼーションを行うために標識され
たプローブは、RNA、DNA、又は当該分野で知られている任意の方法により製造され
うる合成核酸である。
合成核酸は、インビトロで転写されたリボプローブ又はPCRフラグメントを含む。
この発明の好ましい態様において、合成された相補的なオリゴヌクレオチドが用いられ
る。相補的なオリゴヌクレオチドプローブは、少なくとも長さ約10のヌクレオチド、よ
り好ましくは少なくとも約14、特に好ましくは少なくとも長さ18のヌクレオチドであ
る。当業者は、長さが10ヌクレオチドから、センスミトコンドリアキメラRNA又はア
ンチセンスミトコンドリアキメラRNAにハイブリダイゼーションが可能である長さにま
で拡張できることを理解する。ここでの好ましい態様において、長さは、約30ヌクレオ
チド、より好ましくは約25ヌクレオチド、そしと最も好ましくは10から50ヌクレオ
チドである。
より長いプロービング核酸もまた使用できる。プローブの配列は、少なくとも95%が
SEQ ID NO 1、SEQID NO 2、SEQ ID NO 3、SEQ ID NO 4、SEQ ID NO 5及びSEQ ID NO 6に
リストされた配列に同種である。
【0045】
本発明の相補的なオリゴヌクレオチドプローブは、一般にリン酸ジエステル(phosphod
iester)結合を含む、しかし、ある場合には、限定されるものではないが、ホスフォロチ
オエート(phosphorothioate)(Mag et al., Nucleic Acids Res. 19:1437-1441, 1991;
及び米国特許番号5,644,048),ペプチド性核酸(peptide nucleic acid)又はPNA(Eg
holm, Nature,365:566-568, 1993; 及び米国特許番号6,656,687), phosphoramide (Beau
cage, MethodsMol. Biol. 20:33-61, 1993), ホスフォアミド(phosphoramide) (Beauc
age, MethodsMol. Biol. 20:33-61, 1993), ホスフォロジチオエート(phosphorodithio
ate)(Capaldi etal., Nucleic Acids Res., 28:E40, 2000)からなるインターヌクレオ
シド連鎖を交互に含むオリゴヌクレオチドプローブ類似体も含まれる。
他の相補的なオリゴヌクレオチド類似体は、限定されるものではないが、モルフォリノ
(morpholino)(Summerton, Biochim. Biophys. Acta, 1489:141-158, 1999)、ロックさ
れたオリゴヌクレオチド(locked oligonucleotides) (Wahlestedt wt al., Proc. Natl
. Acad. Sci.US, 97:5633-5638, 2000)、ペプチド性核酸(peptidic nucleic acids)又
はPNA(Nielsen et al., 1993; Hyrup and Nielsen, 1996)又は2-O-(2-メトキシ)エチ
ル変性5’及び3’末端オリゴヌクレオチド(2-o-(2-methoxy) ethyl modified 5’ and
3’ endoligonucleotides)(McKay et al., J. Biol. Chem., 274:1715-1722, 1999)な
どを含む。
これらの引例のすべては、参照することにより明らかに本明細書に組み込まれる。
核酸は、デオキシリボ−とリボ−ヌクレオチドのいかなる結合、及びウラシル、アデニ
ン、チミン、シトシン、グアニン、イノシン、キササナイン(xathanine)、ハイポキサ
ナイン(hypoxathanine)、イソシトシン、イソグアニン(isoguanine)等を含むいかな
る塩基の結合を含めることができる。
【0046】
本発明の他の態様では、核酸またはオリゴヌクレオチドプローブは、センスミトコンド
リアキメラRNA又はアンチセンスミトコンドリアキメラRNAによるハイブリダイゼー
ションを検出するために標識される。プローブは、その技術分野で知られている任意の方
法により検出可能なマーカーにより標識される。
プローブを標識する方法は、ランダムのプライミング、エンドラベリング、PCR、及
びニックトランスレーションを含む。
酵素の標識付けは、核酸ポリメラーゼ、3つの未標識ヌクレオチド、及び標識が直接付
けられるか、標識するためにリンカーアーム(linker arm)を含むか、又はハプテンもし
くは標識が付けられた結合分子が結合するかもしれない他の分子に付けられる4番めのヌ
クレオチドの存在下で行なわれる。
適切な直接の標識は、.sup.32 P、.sup.3 Hおよび.sup.35 Sのような放射性のラベル、
並びに蛍光性のマーカーのような非放射性の標識を含む。
好ましい蛍光色素(fluorophores)は、5(6)カルボキシフルオレセン[5(6)carboxyfluo
rescein]、6-((7-アミノの-4-メチルコウマリン-3-アセチル)アミノ)ヘキサノイック
酸[6-((7-amino-4-methylcoumarin-3-acetyl)amino)hexanoic acid]、5(及び6)-カ
ルボキシ-X-ローダミン[5(and6)-carboxy-X-rhodamine]、シアニン2(Cy2)染料[Cyanin
e 2 (Cy2) Dye]、シアニン3(Cy3)染料[Cyanine3 (Cy3) Dye]、シアニン3.5(Cy3.5
)染料[Cyanine 3.5(Cy3.5) Dye]、シアニン5(Cy5)染料[Cyanine 5 (Cy5) Dye]、シア
ニン5.5(Cy5.5)染料[Cyanine 5.5 (Cy5.5) Dye]、シアニン7(Cy7)染料[Cyanine 7
(Cy7) Dye]、シアニン9(Cy9)染料[Cyanine9 (Cy9) Dye]、(シアニン染料2、3、3.5
、5および5.5は、Amersham,Arlington Heights, III からのNHSエステルとして入手可能
である。)、又はアレクサ染料[Alexa dyes](Alexa 488、Alexa 532、Alexa 556、Alexa
590等を含む。)(MolecularProbes, Eugene, Oreg.)を含む。
【0047】
プローブは、ハプテン又は他の分子に共有結合しているヌクレオチドに結合することに
より直接的に標識される。好ましいハプテンは、限定されるものではないが、5(6)-カ
ルボキシフルオレセイン[5(6)-carboxyfluorescein]、2,4-ジニトロフェニルジオキシ
ゼニン[2,4-dinitrophenyl、digoxigenin]、及びビオチンを含み、そしてそのハプテン又
は他の分子に対する標識された抗体でプローブの検出を行う。
ビオチンの場合、検出は、検出可能なラベルに接合しているアビディン(avidin)ある
いはストレプトアビジン(streptavidin)により行うことができる。
抗体、ストレプトアビジンおよびアビディンは、蛍光性のマーカーで、あるいはそれら
を検出可能にするアルカリホスファターゼかホースラディッシュ・ペルオキシダーゼのよ
うな酵素のマーカーに接合される。
接合されたストレプトアビジン(streptavidin)、アビディン及び抗体アンチジゴキシゲ
ニン(digoxigenin)は、ベクター研究所(バーリンゲーム,カリフォルニア)およびボエリン
ゲルマンハイム(BoehringerMannheim)(インディアナポリス、インディア)のような会社
から商業的に入手可能である。
他の具体例では、抗体又はストレプトアビジンは、多くの安定した蛍光放射量子ドット
に接合される(Wuet al., Nature Biotechnol. 21:41-46, 2003)。
【0048】
抗体とストレプトアビジンに結合している酵素は、酵素に対する基質(substrate)を
提供することにより、熱量反応を通して検出することができる。
種々の基質の存在において、異なる色は、反応により生成され、そしてこれらの色は、
多数のプローブを別々に検知するために視覚化することができる。
当該分野で公知のいかなる基質も使用できる。
アルカリ性ホスファターゼに対する好ましい基板は、5-ブロモ-4-クロロ-3-インド
リルホスフェーテ(5-bromo-4-chloro-3-indolylphosphate)(BCIP)及びニトロブルーテト
ラゾリウム(nitroblue tetrazolium)(NBT)を含む。
ホースラディシュ・ペルオキシダーゼ(horseradish peroxidase)用の好ましい基質は
、ジアミノベンゾエート(diaminobenzoate)(DAB)である。
その分野の当業者は、更にその他の酵素活性を用いることが可能であることを理解する
。
【0049】
本発明の他の具体例では、正確で複製可能な結果を達成するin situハイブリダイゼー
ションを行うための条件が記述される。
核酸ハイブリダイゼーションの分野の技術者は、ハイブリダイゼーションの厳格性をコ
ントロールするために一般に使用される因子がホルムアミド濃度あるいは他の化学的変性
試薬、塩濃度又は可変のイオン強度、ハイブリダイゼーション温度、界面活性剤濃度、p
Hとカオトロピック剤(chaotropic agents)の存在又は不存在、を含むことを認識する
であろう。
これらの厳格因子は、それによりキメラRNAに対するオリゴヌクレオチドプローブの
ハイブリダイゼーションの厳格さをコントロールするために調整することができる。分析
用の最適の厳格さは、所望の厳格さの程度が達成されるまで、各厳格因子の実験によって
実験的に決定される。
【0050】
最良のinsituハイブリダイゼーションを制御するための他の条件は、例えば、標的の
キメラRNAよりも生体サンプル中に存在する、成分に対するプローブの非特定結合をブ
ロックする化学薬品の使用である。
ブロッキング試薬(遮断薬)は、限定されるものではないが、RNA、DNAあるいは
標識されていないオリゴヌクレオチドである。
ハイブリダイゼーション溶液中で結合しているブロッキング試薬は、標識されたプロー
ブの非特定結合を制御する、そしてそれ故分析のシグナル対ノイス比を増加する。
本発明の他の側面において、プローブは、センス又はアンチセンスのミトコンドリアR
NAの配列に相補する配列を有する(SEQ ID NO 1, SEQ ID NO 2, SEQ ID NO 3, SEQ ID N
O 4, SEQ ID NO5 and SEQ ID N 6 参照)
【0051】
生化学的配列の固定は、また実験的に決定されるin situハイブリダイゼーションの重
要な側面である。グルタルアルデヒドのような高度に架橋された固定剤は、標的のセンス
ミトコンドリアキメラRNA又はアンチセンスミトコンドリアキメラRNAへのプローブ
のアクセスがブロックされるので、推奨されない。
この発明の好ましい方法は、冷凍のサンプルも好ましいが、ホルマリンで固定された生
体サンプルである。センスミトコンドリアキメラRNA又はアンチセンスミトコンドリア
キメラRNAを標識されたプローブに触れさせるために、付加的な操作を用いることがで
きる。
例えば、生体サンプルは、プローブが標的キメラRNAにアクセスするのをブロックす
る蛋白質を除去するためにプロテイナーゼKで消化することができる。
in situハイブリダイゼーションに先立って、生体サンプルをプロテイナーゼK又は他
のプロテアーゼで処理することは、当該分野の技術者によく知られている。
【0052】
前述したように、キメラRNAとアンチセンスキメラRNA中の長い逆方向反復配列は
、高度に安定な二重鎖の生成を誘発する。キメラRNAの単一鎖領域の第二の構造を有す
るこれらの構造は、プローブが標的キメラRNAにアクセスするのにバリヤーを構成する
。
従って、本発明の他の側面では、生体サンプルは、キメラRNAを変性するために、室
温で10分間0.2MのHClで処理される。
該サンプルは、ここに記述するin situハイブリダイゼーションプロトコルを適用する
前に、pH7.4の緩衝溶液で数回洗浄することにより急速に中和される。
ここに記述する、通常のルーチン実験およびその開示の下にこの分野の技術者は、ここ
に記述する方法と組成物を利用して分析を行うための適切なハイブリゼイション条件を容
易に決めることができる。適切なin situハイブリダイゼーション条件は、in situハイブ
リダイゼーション操作を行うのに適切なこれらの条件である。
従って、適切なinsitu ハイブリダイゼーション条件は、ここにおける開示と引例(場
合によっては更なるルーチン実験を用いて)を用いて明らかになる。
【0053】
本発明の他の具体例において、in situハイブリダイゼーションにより決められたよう
にセンスミトコンドリアキメラRNAの局在化(localization)は、癌を持つ患者の予後
と管理に重要な情報である。腫瘍細胞において、センスミトコンドリアキメラRNAは、
遅発エンドソーム/リソソームに付随する細胞質中に最も見出される。しかし、核小体中
の局在化は、また、ある細胞中にも見出される。
ヒトの生体検査における腫瘍細胞の存在において、そのハイブリダイゼーションのシグ
ナルは、センスミトコンドリアキメラRNAが細胞質中にのみ、又は細胞質と核小体中も
しくは細胞質と細胞核中に存在することを示す。
従って、異なる局在化は、重要な予後の重要性を持つこともよそうされる。
好ましい具体例において、例えば乳房、大腸及び前立腺腫瘍からのヒトの生検のパネル
は、キメラRNAを検出するためのin situハイブリダイゼーションにより研究されうる
。
肯定的なハイブリダイゼーションシグナルと共に(プローブがどのように標識されたか
には左右されず)、細胞内局在化(唯一細胞質、細胞質及び核小体、又は細胞質及び細胞
核)は、各腫瘍中で確立され、そしてその結果は各患者の生存と比較された。
【0054】
この発明の他の側面において、正常な及び/又は腫瘍細胞を含む個別細胞の混合物は、
蛍光色素(fluorochrome)で標識され、センスミトコンドリアキメラRNAとアンチセン
スミトコンドリアキメラRNAに相補するオリゴヌクレオチドプローブを有する検査液中
でハイブリダイゼーションされる。
例えば、センスミトコンドリアキメラRNAを標的にしているプローブは、ローダミン
で標識表示され、そしてセンスミトコンドリアキメラRNAを標的にしているプローブは
、アレクサ(Alexa)488で標識表示されうる。
以前に記述した条件の下にハイブリダイゼーションと洗浄後に、細胞は、細胞内の標識
表示するフローサイトメトリー(flow cytometry)により分析することができる。
【0055】
本発明の好ましい態様は、腫瘍細胞中のキメラRNAの特定の局在化について得られた
情報が予後診断の重要な更なる情報を提供するので、in situハイブリダイゼーションに
用いることである。
本発明の更に他の態様において、代替分子法は、正常、前癌及び癌細胞中でのキメラR
NAの発現及びセンスとアンチセンスキメラRNAの差次的発現を検出するのに用いられ
る。これらの別法は、限定されるものではないがノーザンブロット、ドットブロット、キ
メラRNAとアンチセンスキメラRNAに対するオリゴヌクレオチド配列、RT−PCR
によるRNAの増殖、インビトロ転移介在性増殖又はTMAによるRNAの増殖、S1又
はリボヌクレアーゼ(RNA分解酵素)アッセイ、等を含む。
【0056】
本発明の1つの態様で、センスミトコンドリアキメラRNAは、16SリボソームRN
Aの5’末端部を含む領域、又は逆方向反復配列の部分的もしくは全領域の増殖により得
られたプローブを用いて診断目的で検出することができる。
図1に示すように、リバースプライマーは例えばプライマー1(SEQ ID NO 139)にす
ることができ、そしてフォワードプライマーは、プライマー3,4,5,6又は7(SEQ ID
NOS129,11,106,102,63)にすることができる。他の部分に位置するプライマーもまた使
用することができ、そしてこれらは該分野の技術者により容易にデザインできる。この発
明の他の側面で、逆転写活性を有する酵素とヘキサマー又はより長鎖のようなランダムプ
ライマーにより合成されるcDNAは、当該分野の技術者によく知られている。
【0057】
得られた210、350、500もしくは850bp、又は他の部分に位置するプライ
マーを用いて得られた他のサイズのアンプリコン(amplicons、単位複製配列)は、アガ
ロースゲル又はポリアクリルアミドゲル中の電気泳動(Sambrook et al., 1989)及び臭化
エチジウム又は他の挿入染料を用いて染色することにより検出できる。
このアンプリコンは、メーカーの指示書により精製される。
【0058】
ミトコンドリアキメラRNAの検出は、ノーザンブロットにより行うことができる(Sam
brook et al.,1989)。
アガロースゲル中のRNAの分離後に、フラグメントは、当該分野の技術者によく知ら
れた操作により膜(ニトロセルロースまたはナイロン)に転送される(Sambrook et al.,
1989)。膜を調べるために、センスミトコンドリアキメラRNAの1000から125の
位置に相当する250bpのフラグメントは、増殖される。
アンプリコンは、製造指示書により精製され(Wizard, Promega)、そして10ナノグラ
ムが第2の増殖のために鋳型として用いられる。
この増殖は、PCR(Invitrogen)と32P-a-dCTP (Amerscham)の5マイクロキュリーの
鎖混合物で行う。放射性の増殖フラグメントは、95℃で10分間インキュベートにより
変性し、変性されたプローブは、ハイブリダイゼーション混合物に添加される。膜は、6
5℃で16時間、ハイブリダイズされて、その後2倍のSSC緩衝液で2度、60℃で0
.5倍のSSCで2度、そして45℃で0.2倍のSSCで洗浄される(Sambrook et al.
, 1989)。洗浄された膜は、−70℃で一晩X線フィルムに曝される(Sambrooket al., 1
989)。
膜上のハイブリダイゼーションシグナルは、16SリボゾームRNA(1559のヌクレオ
チド)と815ヌクレオチドの逆方向反復配列に相当するサイズである2,400のヌクレ
オチドの主な成分に相当する。
【0059】
本発明の他の態様で、センスミトコンドリアキメラRNAの一部は、細胞又は細胞の組
織から抽出された全RNAのリボヌクレアーゼ(RNA分解酵素)の消化の後に検出され
る。
二重鎖構造又はセンスミトコンドリアキメラRNAのステムは、リボヌクレアーゼによ
る消化に耐性がある。
細胞又は細胞の組織からトリゾール(TriZol) (Invitrogen)で抽出された全RNAは、
2倍のSSCの小容量中に溶解される。溶液は、50μg/mlの最終濃度でRNA分解
酵素A(Sigma)を用いてインキュベートされる。
25℃で30分後に、ヌクレアーゼに対するRNA耐性は、トリゾールで抽出され、−
20℃で一晩イソプロパノールを用いて沈殿させる。ヌクレアーゼに対するRNA耐性は
、蒸留されたDEPC処理水に溶解し、そしてRT-PCR増殖のために鋳型として用い
られる。
16SリボゾームRNAの位置55および790を標的とするプライマーを用いて行っ
た増殖は、センスミトコンドリアキメラRNA(SEQ ID NO 1)のステムと同じ配列で10
0%同一性を示す配列と約730塩基対のフラグメントを生ずる。
その一方、キメラRNAで3’ハーフに相補する単一鎖、又は12Sミトコンドリアリ
ボゾームRNA、又は18SリボゾームRNAもしくはGAPDHに対するmRNAは、
リボヌクレアーゼAで処理することにより全体的に消化され、そしてそれ故、これらのR
NAを標的にするプライマーが用いられた場合に、増殖生成物は、得られない。
【0060】
本発明の他の態様では、全RNAがリボヌクレアーゼAで処理された後、得られたセン
スミトコンドリアキメラRNAのステムは、ノーザンブロットにより検出できる。ヌクレ
アーゼに耐性を示し、トリゾールによる抽出とイソプロピルアルコールを用いた沈殿によ
り回収されるRNAは、アガロースゲル中の電気泳動によって分離される。転移の後に、
膜は、前述したプローブでブロットされ、そしてセンスミトコンドリアキメラRNAをノ
ーザンブロット(Sambrooket al., 1989)するのに用いられる。
【0061】
他の態様では、この発明は、センスミトコンドリアキメラRNA又はアンチセンスミト
コンドリアキメラRNAを生体サンプル中でアッセイを検出するのに適したキッドを教示
する。キメラRNAの検出に適した、一般的で好ましい態様、組成物及び方法が提供され
、in situハイブリダイゼーションによるアンチセンスキメラRNAは、既に本明細書で
記述された。
好ましいオリゴヌクレオチドプローブ配列は、限定されるものではないがリストに記載
されている。更に、オリゴヌクレオチドプローブを使用するのに適した方法あるいはサン
プル中のキメラRNAもしくはアンチセンスキメラRNAを検出するキッドのオリゴヌク
レオチドプローブは、既に本明細書で記述された。
【0062】
本発明のキットは、1つ以上のオリゴヌクレオチドプローブと、サンプル中のセンスミ
トコンドリアキメラRNA又はアンチセンスミトコンドリアキメラRNAを検出するため
に用いるinsituハイブリダイゼーションを行うために選択された試薬又は組成物からな
る。2以上のオリゴヌクレオチドの各セットは独立して検知可能な構成部分で好ましくは
ラベル化され、そして生体サンプルの個々の細胞において、センスミトコンドリアキメラ
RNA又はセンスミトコンドリアキメラRNAが検出される。
好ましい態様において、センスミトコンドリアキメラRNA又はアンチセンスミトコン
ドリアキメラRNAを検出するのに用いるキットのオリゴヌクレオチドプローブは、異な
るハプテンで各セットごとに標識される。このハプテンは、ビオチン、ジゴキシゲニン、
あるいは異なる酵素(例えばアルカリホスファターゼあるいはペルオキシダーゼ)で標識し
た抗体あるいはストレプトアビジンでin situハイブリダイゼーション方法で認識できる
フルオレセイン(蛍光色素)とすることができる。
他の方法として、プローブの各セットの各オリゴヌクレオチドプローブは、独立して検
出可能な蛍光性のグループで標識できる。
例えば、センスミトコンドリアキメラRNAを検出するためのオリゴヌクレオチドプロ
ーブのセットは、ローダミンで標識できる、一方、アンチセンスミトコンドリアキメラR
NAを検出するためのオリゴヌクレオチドプローブのセットは、アレクサ488(Alexa 4
88)で標識できる。
更に、生体サンプルの細胞中のキメラRNA又はアンチセンスキメラRNAの局在化を
決定する方法が提供される。
更に、本発明の組成物と方法は、共焦点顕微鏡分析を用いて異なる細胞小器官の特定の
マーカーでキメラRNA又はアンチセンスキメラRNAの共−局在化(co-localization
)を決定するのに使用することができる。
【0063】
ここに提供される組成物および方法は、前癌および癌の検知と診断に特に有効であると
考えられる。
ここに提供されるような用語「癌」は、以下の確認されている特異な条件のいずれか1
により悩まされる細胞を含んでいる。
これらは、急性慢性である骨髄性白血病、急性慢性であるリンパ芽球の白血病、多発性
骨髄腫、ホジキン病、非ホジキンのリンパ腫あるいは悪性リンパ腫;胃癌、食道癌か腺癌
、膵臓送管の腺癌、インスリノーマ(膵島細胞腫)、グルカゴン産生腫瘍(glucagonoma
)、ガストリノーマ(gastrinoma)、小腸腺癌(small bowel adenocarcinoma)、結腸直
腸癌;肝細胞性の癌、肝細胞性の腺腫;カルチノイド、腎臓腺癌のような尿生殖路、ウイル
ム腫瘍(Wilm'stumor)、膀胱および尿道癌および前立腺癌、精上皮腫のような精巣癌、
奇形腫、奇形癌、間質細胞癌腫; 子宮内膜の癌、子宮頚癌、卵巣癌、外陰癌と膣癌、セル
トリ-L-アイディグ(Sertoli-L-eydig)細胞癌、黒色腫および卵管癌;肺、歯槽と細気管
支の癌;脳腫瘍; 皮膚悪性黒色腫、基底細胞癌、扁平上皮細胞癌及びカルポシ(Karposi)肉
腫を含む。
繊維肉腫、心臓の血管肉腫と横紋筋肉腫、及び他の悪性癌は、当該分野の技術者によく
知られている。
【0064】
4.3 癌および前癌療法
化学療法薬は、癌細胞増殖および存続に影響を与える一連の細胞応答を含む。
これらの細胞応答の研究された最良のものは、アポトーシスであり、抗癌剤が共通のア
ポトーシス経路を活性化することにより腫瘍細胞を殺すことができることは現在明らかで
ある。。
不運にも、これらの薬は、さらに急速に分裂する骨髄の正常細胞、正常な造血と腸の細
胞および髪の毛マトリックスのケラチン生成細胞にも影響を与える(McKinnell et al., T
he biological Basisof cancer, 1998; Komarov et al., Science 285:1733-1737, 1999
; Johnstone etal., Cell 108:153-164, 2002)。
【0065】
他方、多くの腫瘍細胞が細胞死開始因子、調整因子及び細胞死の遂行因子を変化させる
、これらは、異なる癌タイプの腫瘍細胞がなぜ種々の化学療法薬と放射線に耐性を示すこ
とになるかを説明している。
この変化は、固有の経路の因子、ポストミトコンドリアル事象及び細胞死の固有の経路
について報告されている(Rampino et al., Science 275:967-969, 1997; Vogelstein et
al., Nature408: 307-310, 2000; Teitz et al., Nature Med. 6:529-535, 2000; Reed,
J. Clin.Oncol., 17:2941-2953, 1999; Johnston et al., Cell 108:153-164, 2002)。
従って、癌治療法のパラダイムは、腫瘍細胞の細胞死の選択的誘引となる操作、正常な
増殖細胞を変化させない操作、及び異なる細胞死経路を変化又は突然変異させる要因を回
避する操作である。
【0066】
本発明の構成および方法は、正常な増殖細胞と同様のレベルで、腫瘍及び前腫瘍細胞が
センスミトコンドリアキメラRNAを過剰発現するという発見に基づく。
これらの転写産物の構造は、図1、図2、及びSEQ ID NO 1、SEQ ID NO 2、SEQ ID NO
3、SEQ ID NO 4、SEQID NO 5およびSEQ ID NO 6における対応する配列である。
【0067】
その一方、他の驚きの発見は、前腫瘍及び腫瘍細胞がセンスミトコンドリアキメラRN
Aを過剰発現し、そしてアンチセンスミトコンドリアキメラRNAを下方制御する、とい
うことである。前腫瘍及び腫瘍細胞におけるアンチセンスミトコンドリアキメラRNAの
合成の抑制と阻害は、癌細胞と正常増殖細胞間の表現型の新たな差異を構成する、これは
、本発明の主な態様に1つとして考えられる。
さらに、異なる癌タイプのヒトの生検における癌はまた、培養中の癌と同じ表現型を示
す(図5Aおよび5B)。
【0068】
センスミトコンドリアキメラRNAとアンチセンスミトコンドリアキメラRNAの機能
は明確ではないが、密接な相関関係はこれらのRNAの発現と細胞増殖間に存在する。例
えば、肝臓、腎臓および脾臓のような細胞組織中の、Ki-67、PCNAあるいはリン酸
化されたヒストンH3(histone H3)のような抗原を意味するようなもので定義される正常
な増殖する細胞は、アンチセンスミトコンドリアキメラRNAだけでなくセンスミトコン
ドリアキメラRNAを過剰発現する。
Ki-67又はPCNAを発現しない同様の細胞組織の非増殖細胞において、センスミト
コンドリアキメラRNAとアンチセンスミトコンドリアキメラRNAは発現しない。更に
、図7に示すように、ミトゲンPHAで活性化されたヒトのリンパ球は、DNAを合成し
、増殖する抗原Ki-67とPCNAを発現する。
この活性化されたリンパ球は、またアンチセンスミトコンドリアキメラRNAだけでな
くセンスミトコンドリアキメラRNAを過剰発現する(図7)。
対照的に、休止しているリンパ球又は非活性リンパ球は、センスミトコンドリアキメラ
RNAもアンチセンスミトコンドリアキメラRNAも発現しない。
【0069】
本発明の1つの基本部分である、前述の発見は、正常の増殖細胞がセンスとアンチセン
スミトコンドリアキメラRNAを発現する期間、腫瘍細胞はセンスミトコンドリアキメラ
RNAを発現し、そしてアンチセンスミトコンドリアキメラRNAの発現を下方制御する
、ことを示す。
細胞増殖におけるこれらのRNAの機能を理解するために、培養中の癌細胞は、センス
ミトコンドリアキメラRNA(SEQ ID NO 1、SEQ ID NO 2、SEQ ID NO 3)又はアンチセン
スミトコンドリアキメラRNA(SEQ ID NO 4、SEQ ID NO 5、SEQ ID NO 6)を標的にする
アンチセンスオリゴヌクレオチドで処理された。
他の驚くべき発見である結果は、これらの条件下で細胞は、予定された細胞死又はアポ
トーシスをこうむることを示した。
センス又はアンチセンスミトコンドリアキメラRNAに相補するオリゴヌクレオチドで
6ないし15時間処理され処理後、細胞の75〜96%は、アポトーシス(表2参照)を被
る。
処理された細胞で観察された変化は、クロマチン凝縮、核断片化、DNA断片化、カス
パーゼ活性及び細胞膜の変化プロセスであった。
4またはそれ以上のミスマッチ又はスクランブルオリゴヌクレオチドを有する制御オリ
ゴヌクレオチドは、アポトーシスを誘発しなかった。
【0070】
また、センス又はアンチセンス12SミトコンドリアRNAを標的とする、又はmRN
AもしくはミトコンドリアND1サブユニットのアンチセンス転写産物を標的とするオリ
ゴヌクレオチドで処理しても細胞は影響を受けなかった。
一般に、アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオチドは
、同じ濃度ではセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオチドより
もより効果的であった。
このことは、予期された結果であった、なぜなら腫瘍細胞は、センスミトコンドリアキ
メラRNAを過剰発現し、そしてそれ故細胞内でこの転写産物のすべてのコピーを阻害す
る細胞内のオリゴヌクレオチド濃度に到達ことが困難だからである。
他方、腫瘍細胞は、アンチセンスミトコンドリアキメラRNAを下方制御するので、細
胞あたりのコピー数が低くこのRNAで阻害することが容易になる。
【0071】
アポトーシスの誘発は、腫瘍細胞に対してまた選択的である。休止しているヒトリンパ
球又はPHAで48時間活性化したヒトのリンパ球は、アンチセンスミトコンドリアキメ
ラRNAに相補する、又はセンスミトコンドリアキメラRNAを標識にするオリゴヌクレ
オチドで、一晩処理をしても、また相補的オリゴヌクレオチド(15uM)の高用量で処
理しても影響されない。
【0072】
アンチセンスミトコンドリアキメラRNA(SEQ ID NO 4、SEQ ID NO 5、SEQ ID NO 6)
を標的にする相補的オリゴヌクレオチドで処理することによるアポトーシスの誘発は、限
定されるものではないが前骨髄球性白血病細胞HL-60、急性リンパ芽球の白血病MOLT-
4、T−リンパ球白血病細胞、Jurkat、T細胞白血病、Devernelle、又はBリンパ腫、N
SO/2又は骨髄腫、HeLa細胞、DU145、PC-3、Caco-2、Hep-2及
びHepG2で達成される。
治療の効かない(化学療法又は放射線治療)腫瘍細胞のパラダイムと見なすことができ
る、MCF/7(乳癌)とメラノーマの2つの細胞は、アンチセンスミトコンドリアキメ
ラRNA(SEQID NO 4、SEQ ID NO 5、SEQ ID NO 6)を標的とする相補的オリゴヌクレオ
チドで15時間処理するとき、80%以上アポトーシスを受ける。
より低いアポーシス結果は、センスキメラRNA(SEQ ID NO 1)に相補するオリゴヌク
レオチドで得られた。
すでに記載したように、4つのミスマッチ又はスクランブルオリゴヌクレオチドを有す
るオリゴヌクレオチドは、細胞死を誘発しない。
【0073】
以下の記載は、治療標的として、センスキメラRNAとアンチセンスキメラRNAを用
いて癌を処理するための方法と組成物である。
好ましい態様は、限定されるものでないが、アンチセンスキメラRNAに相補するオリ
ゴヌクレオチドを用いて癌を処理するための方法と組成物である。
この治療の結果は、処理された対象で少なくとも健康に良い利益を生ずる、この利益は
、癌の場合に癌の沈静化に限定されないが、癌の症状の緩和及び癌の転移拡散の制御を含
む。このようなすべての方法は、腫瘍細胞において、及び正常細胞においてはマイナーな
効果でアポトーシスの誘発を伴う。
特定のRNAを標的する相補的なオリゴヌクレオチドは、多くのさまざまなmRNAの
発現又は対応するたんぱく質の合成を少なくするかなくす為に用いられてきた(e.g. Vick
ers et al., J.Biol. Chem., 278:7108-7118, 2003)。
現在、異なる化学的性質を有する約42のアンチセンスオリゴヌクレオチドは、可能性
のある薬剤としてスクリーニングされている(Stephens and Rivers, Curr. Opin. Mol. T
herapeut.,5:118-122, 2003) (参照例:U.S. Pat. Nos. 5,801,154; 6,576,759; 6,720,
413; 6,573,050and 6,673,917)。
これらの引例のすべては、参照することにより明らかに本明細書に組み込まれる
【0074】
この発明の1つの側面において、アンチセンスキメラRNAを標的にする1以上のオリゴ
ヌクレオチドが用いられる。
2つ以上の相補的なオリゴヌクレオチドの使用は、より有効で、ある程度の相乗作用を
示す。
【0075】
本発明のオリゴヌクレオチドは、アンチセンスミトコンドリアキメラRNA又はセンスミ
トコンドリアキメラRNAに相補することができる。
相補的なオリゴヌクレオチドは、アンチセンスミトコンドリアキメラRNA又はセンス
ミトコンドリアキメラRNAに結合し、それらの機能を妨げる。
絶対的な相補性は、好ましいが、要求はされない。
ここで記述するように、一部のRNAに相補するオリゴヌクレオチド配列は、RNAで
ハイブリダイズして安定な二重鎖を形成するに十分な相補性を有する配列を意味する。ハ
イブリダイズする能力は、両者の相補の程度とオリゴヌクレオチドの長さによる。一般に
、ハイブリダイズする核酸が長いほど、RNAでのミスマッチの増加した塩基を多く含む
が、それでも安定な二重鎖を形成する。
当該分野の技術者は、ハイブリダイズ化された錯体の融点を決定する標準操作の使用に
よりミスマッチの許容程度を確認することができる。
【0076】
一般に、異なる蛋白質をmRNAでハイブリダイズするための相補的オリゴヌクレオチ
ドは、AUGスタートコドンの相補物を含むmRNAの5’未翻訳領域、又はより効果的
な3’未翻訳領域を標的にする。
mRNAコード領域に相補するオリゴヌクレオチドは、翻訳の効果が低い抑制剤である
(前の引例を参照)。
センスミトコンドリアキメラRNAとアンチセンスミトコンドリアキメラRNAは、非
コードRNAであり、それ故オリゴヌクレオチドの標的領域は、これらの転写産物のいか
なる領域とも相補することができる。最も効果的な領域は、各30ヌクレオチドでデザイ
ンされた相補的オリゴヌクレオチドを用いてアンチセンス又はセンスミトコンドリアキメ
ラRNAの配列をスキャニングすることにより決定されたアンチセンスミトコンドリアキ
メラRNAの単一鎖セグメントの周りに局在化される。
この分野の技術者は、SEQ ID NO 2、SEQ ID NO 3、SEQ ID NO 4、SEQ ID NO 5、及びSE
Q ID NO 6の転写産物の完全な配列内の他の配列が代替の相補的なオリゴヌクレオチドを
デザインするための標的であることを理解する。。
【0077】
アンチセンスミトコンドリアキメラRNA又はセンスミトコンドリアキメラRNAを標
的とし、本発明によると腫瘍細胞死を誘発することになるこの相補的なオリゴヌクレオチ
ドは、一般にナチュラルなリン酸ジエステル結合と異なるバックボーンを含む。このオリ
ゴヌクレオチドは、限定されないが、ホスフォロチオエーテ(Mag et al., Nucleic Acids
Res.19:1437-1441, 1991; and U.S. Pat. No. 5,644,048)、ペプチド核酸又はPNA(E
gholm, Nature,365:566-568, 1993; and U.S. Pat. No. 6,656,687)、ホスフォアミド(B
eaucage,Methods Mol. Biol. 20:33-61, 1993)、ホスフォロチジオエーテ(Capaldi et a
l., NucleicAcids Res., 28:E40, 2000)を含む、交互に並ぶインターヌクレオシド結合
を有することができる。
他のオリゴヌクレオチド誘導体は、限定されないがモルホリノ(Summerton, Biochim. B
iophys. Acta,1489:141-158, 1999)、固定されたオリゴヌクレオチド(Wahlestedt wt al
., Proc. Natl.Acad. Sci. US, 97:5633-5638, 2000)、ペプチド核酸又はPNA(Nielse
n et al., 1993;Hyrup and Nielsen, 1996)又は2-o-(2-メトキシ)エチル修飾5’と3
’末端オリゴヌクレオチド(McKay et al., J. Biol. Chem., 274:1715-1722, 1999)のよ
うなものを含む。
これらの引例のすべては、参照することにより明らかに本明細書に組み込まれる。
核酸は、デオキシリボ−とリボ核酸のいかなる結合、及びウラシル、アデニン、チミン
、シトシン、グアニン、イノシン、キササニン(xathanine)、ハイポキササニン(hypoxat
hanine)イソシトシン、イソグアニンを含む、塩基のいかなる結合も含むことができる。
【0078】
本発明による相補的オリゴヌクレオチドは、限定されないが
5-フルオロウラシル、5-ブロモウラシル、5-クロロウラシル、5-ヨードウラシル、
ヒポキサンチン、キサンチン、4-アセチルシトシン、
5-(カルボキシヒドロキシメチル)ウラシル、
5-カルボキシメチルアミノメチル-2-チオウリジン、
5-カルボキシメチルアミノメチルウラシル、ジヒドロウラシル、
ベータD-ガラクトシルキオシン(beta-D-galactosylqueosine)、イノシン、
N6-イソペンテニルアデニン、1-メチルグアニン、1-メチルイノシン、
2、2-ジメチルグアニン、2-メチルアデニン、2-メチルグアニン、3-メチルシトシン
、5-メチルシトシン、N6アデニン、7-メチルグアニン、
5-メチルアミノメチルウラシル、5-メトキシアミノメチル-2-チオウラシル、
ベータD−マノシルキオシン(beta-D-mannosylqueosine)、
5'-メトキシカルボキシメチルウラシル、5-メトシキウラシル、
2-メチルチオ-N6-イソペンテニルアデニン、ウラシル-5-オキシアセテック酸(V)
、ワイブトキソシン(wybutoxosine)、プソイドウラシル、キオシン(queosine)、
2-チオシトシン(thiocytosine)、5-メチル-2-チオウラシル、2-チオウラシル、
4-チオウラシル、5-メチルウラシル、ウラシル-5-オキシアセテックアシッド メチル
エステル、ウラシル-5-オキシアセテック酸(V)、5-メチル-2-チオウラシル、3-(
3-アミノ-3-N-2-カルボキシプロピル)ウラシル、
(acp3)w及び2,6-ジアミノプリン
を含むグループから選択された少なくとも1つの修飾塩基部分を含む。
【0079】
相補的なオリゴヌクレオチドは、更に限定されないがアラビノース、2-フルオロアラ
ビノーズ、キシルロース及びヘキソースを含むグループから選択された少なくとも1つの
修飾糖部分を含むことができる。
【0080】
本発明の他の態様では、相補的なオリゴヌクレオチドが、アンチセンスミトコンドリア
キメラRNAのいかなる領域とも、又はセンスミトコンドリアキメラRNAのいかなる領
域ともハイブリダイズするようにデザインされる。
相補的なオリゴヌクレオチドは、長さで少なくとも10のヌクレオチドでなければなら
ず、長さで10から50のヌクレオチド範囲の相補的オリゴヌクレオチドが好ましい。
特定の側面では、相補的なオリゴヌクレオチドが少なくとも12のヌクレオチド、少な
くとも18のヌクレオチド、少なくとも22のヌクレオチド、少なくとも30のヌクレオ
チド、少なくとも50のヌクレオチドである。
【0081】
In vivoだけでなくinvitro実験においても、アンチセンス又は相補的オリゴヌクレオ
チドの非特定の生物学的効果で、アンチセンスミトコンドリアキメラRNA又はセンスミ
トコンドリアキメラRNAの機能を用いてアンチセンス阻害を識別する制御を利用する実
験を考慮することは重要である。
従って、オリゴヌクレオチドのデザインは、参照することにより明らかに本発明に組み
込まれる、米国特許番号 6,673,917で報告されているように動物細胞に有毒効果があるCp
Gトラック、5' GGGGトラックおよび他の配列の存在は、避けなければならない。
更に、配列5'CGTTAの存在は、報告されているアンチセンスでない効果を避けられる(T
idd et al.,Nucleic Acids Res. 28:2242-2250, 2000)。
【0082】
本発明の他の態様では、癌を有する動物又は病人の治療薬として、センスミトコンドリ
アキメラRNAを標的とする又はアンチセンスミトコンドリアキメラRNAを標的とする
相補的オリゴヌクレオチドは、抗癌治療剤と放射線療法に対する活性を誘発することがで
きる。
増感又は過敏性として知られている誘発される感度は、抗癌治療又は放射線療法に対す
る耐性を示す腫瘍細胞中で測定される。
抗癌剤は、これらの当該分野で使用中であり既に知られている、又は未発見の薬剤から
なる。従来の化学療法薬の中には、アルキル化剤、代謝拮抗物質、抗生物質および抗微小
管薬がある。
これらの薬のいくつかの例はシスプラチン、メトトレキサート、ドキソルビシン,
ダクチノマイシン、マイトマイシン、シクロホスファミドなどである。
この発明の他の側面で、癌を有する動物又は病人をアンチセンスミトコンドリアキメラ
RNA及び/又はセンスミトコンドリアキメラRNAを標的にした相補的オリゴヌクレオ
チドでの処理と共に又は処理後に、患者は、放射線療法で処理される、この場合、前記放
射線療法は、紫外線、ガンマ線、アルファ粒子、ベータ粒子、X線および電子線を含んで
いる。
【0083】
この発明の他の側面で、アンチセンスミトコンドリアキメラRNA又はセンスミトコン
ドリアキメラRNAの機能で腫瘍細胞死の阻害は、RNA阻害又はRNAサイレンシング
によりもたらされる。
過去6年にわたって、RNA阻害(RNAi)は、哺乳類細胞中の遺伝子サイレンシングに対
する新規で有望なアプローチとして浮上してきている(Elbashir et al., Nature 411:494
-498, 2001;McManus et al., Nature Rev. Genet. 3:737-747, 2002)。
長さで約19から21のヌクレオチドの合成的に合成された二重鎖のRNA分子は、m
RNAの分解とその後の蛋白質ノックダウンをもたらす、特にそれらの相補的標的mRN
Aにハイブリダイズする。
いくつかの異なる遺伝子は、小さなRNA又はsiRNA阻害により成功裡にサイレン化
される(Lu etal., Curr. Opin. Mol. Ther. 5:225-234, 2003.; Wacheck et al., Oligo
nucleotides13:393-400, 2003)。
アンチセンスミトコンドリアキメラRNA又はセンスミトコンドリアキメラRNAを標
的とする19から21のヌクレオチドの合成された二重鎖のRNAは、これらの転写産物
を分解して、腫瘍細胞死を誘発する。
当該分野の技術者は、siRNAの配列がアンチセンスミトコンドリアキメラRNA又は
センスミトコンドリアキメラRNA(SEQ ID NO 1, SEQ ID NO 2, SEQ ID NO 3, SEQ ID N
O 4, SEQ ID NO5, and SEQ ID NO 6)のいかなる領域にも相補する必要があることは理解
するだろう。
【0084】
この発明の他の態様で、リボザイムは、腫瘍細胞死を誘発するためにアンチセンスミト
コンドリアキメラRNA又はセンスミトコンドリアキメラRNAを阻害するのに用いるこ
とができる。
リボザイムの配列は、腫瘍細胞死を引き起こすのにより効果的な転写産物の特定の領域
を開裂するために、アンチセンスミトコンドリアキメラRNA(SEQ ID NO 4, SEQ ID NO
5, SEQ ID NO 6)又はセンスミトコンドリアキメラRNA(SEQID NO 1, SEQ ID NO 2, SE
Q ID NO 3)の配列によりデザインされなければならない。
リボザイムは、RNAの特定の開裂を触媒することが可能な酵素RNA分子である(Ros
si, Curr.Biology 4:469-471, 1994)。
【0085】
リボザイム作用のメカニズムは、標的RNAに相補するリボザイム分子の配列の特定の
ハイブリダイゼーションを含み、その後エンドヌクレアーゼ的に切断する。
リボザイム分子の構成は、標的遺伝子mRNAに相補する1つ以上の配列を含んでいる
と予想され、mRNA開裂の原因であることがよく知られ、また米国特許番号5,093,246
(これは参照によってその全部がここに組み入れられる)に記述された触媒配列を含んで
いると思われる。
本発明の範囲内において、ハンマーヘッドリボザイム分子は、アンチセンスミトコンド
リアキメラRNA又はセンスミトコンドリアキメラRNAのエンドヌクレアーゼ的切断を
特異的及び効果的に触媒するように作り出されている。
ハンマーヘッドリボザイムの構造と生産は、当該分野でよく知られており、また記述さ
れている(Haseloffet al., Gene, 82:43-52, 1989)。
本発明のリボザイムは、更にRNAエンドリボヌクレアーゼを含んでいる(Zaug et al.
, Science,224:574-578, 1984)。
【0086】
遺伝子治療は、癌にかかっている患者か、癌の予防あるいは禁止が望ましい患者への核
酸の投与によって行なわれる癌の治療あるいは予防に言及する。
本発明のこの態様では、細胞内で生産された治療用核酸は、腫瘍細胞死を誘発するこれ
らのミトコンドリア転移産物の機能を阻害又は禁止することによる治療の影響を調節する
、アンチセンスミトコンドリアキメラRNA又はセンスミトコンドリアキメラRNAを標
的にする相補的RNAである。
従って、1つの好ましいアプローチは、アンチセンスRNAの転移産物がRNAポリメ
ラーゼII又はIIIの強いプロモーターの制御下で置換された、組み換えられたDNAの利
用である。
相補的RNAをコード化する配列は、ほ乳類、好ましくはヒト細胞中で作用するその技
術分野で知られているいかなるプロモーターでも発現する。
【0087】
このようなプロモーターは、限定されるものでないが、SV4初期プロモーター領域(B
enoist andChambon, Nature 290:304-310, 1981)、ヘルペスチミジンキナーゼプロモー
ター(Wagneret al., Proc. Natl. Acad. Sci. U.S.A. 78:1441-1445, 1981)、メタロチ
オネイン遺伝子の調節塩基配列(Brinster et al., Nature 296:39-42, 1982)、ラウス肉
腫ウイルス(Yamamotoet al., Cell 22:787-797, 1980)、等を含む。
相補的RNAを生産する組み換えられたDNA構造は、限定されるものでないが、アデ
ノウィルス・ベクター、アデノ関連性ウィルス・ベクター、単純ヘルペスウィルス・ベク
ター、ワクシニアウィルス・ベクター、及びレトロウィルス・ベクターを含むウィルス・
ベクターにすることができる。
当該分野の技術者によく知られている方法を使用して、このベクターは、製薬組成物と
して標的腫瘍細胞中に導入される。
【0088】
薬学的に受理可能なキャリヤーにおける相補的核酸(相補的オリゴヌクレオチド、siR
NA、リボザイム、又はウィルス・ベクター)の効果的量からなる、本発明の薬剤組成物
は、アンチセンスミトコンドリアキメラRNA又はセンスミトコンドリアキメラRNAの
作用を阻害するために癌患者に投与され、腫瘍細胞のアポトーシスを誘発する。
相補的な核酸は、薬剤組成物中に配合される、この薬剤組成物は、キャリヤー、希釈剤
、バッファー、予防薬、界面活性剤、ポリエチレンイミド(PEI)、リポソームあるい
はこの分野で周知の他の脂質製剤を含めることができる。
前記薬剤組成物は、局所適用、経口、非経口又は直腸投与によって投与される。
非経口投与は、静脈、皮下、腹腔内、又は筋肉注射、又は吸入又は通気による肺内投与
を含んでいる。
【0089】
本発明の組成物は、治療学、診断学、予防処置、そして研究試薬およびキットとして利
用することができる。
ここに提供される組成物及び方法は、癌の治療に特に役立つと考えられる。
ここに提供される用語「癌」は、次の識別された特異な条件のうちのいずれか1つで悩
まされる細胞を含んでいる。
これらは、急性慢性である脊髄性白血病、急性慢性であるリンパ芽球白血病、多発性骨
髄腫、ホジキン病、非ホジキンのリンパ腫あるいは悪性リンパ腫;胃癌、食道癌又は腺癌
、膵臓送管の腺癌、インシュリノーマ、グルカゴン産生腫瘍、多発性胃潰瘍、
小さな腸腺癌、結腸直腸癌;肝細胞性の癌、肝細胞性の腺腫;類癌腫、腎臓腺癌のような
尿生殖器官、ウイルム腫瘍(Wilm's tumor)、膀胱および尿道癌及び前立腺腺癌、精上皮
腫のような精巣癌、奇形腫、奇形癌、間質細胞癌腫;子宮内膜の癌、子宮頚癌、卵巣癌、
外陰癌と膣癌、セルトリ-L-アイディグ(Sertoli-L-eydig)細胞癌、黒色腫および卵管
癌;肺、歯槽と細気管支の癌;脳腫瘍;皮膚悪性黒色腫、基底細胞癌、扁平上皮細胞癌及
びカルポシ(Karposi)肉腫を含む。
繊維肉腫、心臓の血管肉腫と横紋筋肉腫、及び他の悪性癌は、当該分野の熟練技術者に
よく知られている。
ここに提供される用語「癌細胞」は、上記の識別された特異な条件のうちのいずれか1
つで悩まされる細胞を含んでいる。
次の例は、本発明の様々な側面の実行のために最良の方法を述べるだけでなく、上記の
記述された発明を使用する方法について記述する役割を果たす。
これらの例は、本発明の範囲を少しも制限することはなく、むしろこれらは実例となる目
的のために示されることが理解される。
【実施例】
【0090】
[実施例1]
センスミトコンドリアヒトキメラRNAの分離と配列
(図1A、SEQ ID NO1)
最初の実験は、推定のヒトセンスミトコンドリアキメラRNAがより複雑で安定な、マ
ウスのキメラRNAである第二の構造を含むことを示した(Villegas et al., DNA & Cell
Biol.19:579-588, 2000; Villegas et al., Nucleic Acids Res. 30:1895-1901, 2002)
。
それ故、またマウスミトコンドリアキメラRNAの第二の構造に基づき、理論的なヒト
のセンスミトコンドリアキメラRNAの第二の構造が、推定された(図1A)。
理論的なヒトの転写産物は、未知の長さ(図1A)のループを形成するアンチセンス16
SミトコンドリアRNAのフラグメントに5’末端で結合しているセンス16Sミトコン
ドリアRNAの完全な配列を含んでいた。
アンチセンス16SミトコンドリアRNAのセグメントは、センス16Sミトコンドリ
アRNAに十分に相補した、またそれゆえセンス16S転写産物の5’末端に結合する逆
方向反復配列に相当する。この構造に基づき、プライマーは、RT−PCRによるこの推
定の転写産物を増殖するようにデザインされた。
1つのリバースプライマーは、ヒトセンス16SミトコンドリアRNAの5’末端から
11から31に位置、又は理論的ループの始点に位置していた(プライマー1、図1A)
(SEQ IDNO 1)。
【0091】
用いたフォワードプライマーの配列は、センス16SミトコンドリアRNAの位置21
3−234の配列で、図1Aの中のプライマー3に相当する。
HeLa細胞、HL−60、Du145、MCF/7及びヒトのリンパ細胞を含むヒト
の組織と細胞からRNAの増殖は、プライマー1と3(図1A)を用いてRT−PCRに
よりPHA(実施例7参照)で刺激され、約210bpの固有のアンプリコン(図2)を産
出する。
RT−PCRは、前記した方法で実施した(Villegas et al., DNA & Cell Biol. 19:57
9-588, 2000;Villegas et al., Nucleic Acids Res. 30:1895-1901, 2002)。
個々のヒトの組織あるいは細胞からのアンプリコンは、クローン化された、また両鎖は
配列された。
すべての場合に、216bpの同一鎖が得られ、センス16SミトコンドリアRNAの
5’末端の最初の31ヌクレチドに結合している184ヌクレチドの逆方向反復配列を含
んでいた。
その後、我々は、逆方向反復配列が184ヌクレチドより長いか、またアンチセンス1
6SミトコンドリアRNA(図1A)の5’末端に向かって更に拡張しているかを決定し
た。
前記したHeLa細胞又は他の細胞からのcDNAは、前記したようにループに位置す
るリバースプライマー1と、推定のより長い逆方向反復配列の5’末端に向かってウォー
クするプライマー4から7との間で増殖された(図1A)。
このアプローチを用いて、プライマー1が、プライマー4、5及び6とそれぞれ結合す
るのに使用されたとき、およそ500、700及び800bpの増殖フラグメントが得ら
れた。
【0092】
他方、cDNAがプライマー1とプライマー7の間で増殖されたとき、増殖産物は得ら
れなかった、これは、逆方向反復配列の5’末端がプライマー7とプライマー8の間であ
ったためと推定される(以下を参照)。
前記800bpのアンプリコンの完全な配列は、センス16SミトコンドリアRNA(S
EQ ID NO 1)(図1A)の最初の31ヌクレオチドに結合する769ヌクレオチドの逆方向
反復配列を示す。
センス16SミトコンドリアRNAに結合する逆方向反復配列の3’末端で配列は、2
16bpのアンプリコンの同じ領域に見出されるものと同一である。このことは、重要で
ある、なぜなら両者の場合に我々は、同じRNAを増殖していたからである。
更に、配列は、アンチセンス16SミトコンドリアRNAの3’末端のその50ヌクレ
オチドがセンス16SミトコンドリアRNAの逆方向反復配列中には見当たらない、こと
を示した。
すべて、これらの結果は、二重鎖構造が逆方向反復配列と後者の51の位置で始まるセ
ンス16SミトコンドリアRNAとの間で形成され、また50ヌクレオチドの推定ループ
を形成することを示唆する。
【0093】
ループのサイズを確認するために、ヒトのcDNAは、逆方向反復配列の3’末端に位
置するフォワードプライマー2と、センス16SミトコンドリアRNA(図1A)の位置
213−234でまたリバースであるプライマー3との間でPCRにより増殖された。お
よそ240bpのアンプリコンが得られた、またこの配列は、センス16Sミトコンドリ
アRNAの最初の234ヌクレオチドが逆方向反復配列の3’末端の最後の25ヌクレオ
チドに結合していたことを示した。逆方向反復配列の25ヌクレオチドの配列は、センス
16SミトコンドリアRNAの位置51から75(図1A)に十分に相補していた。
【0094】
プライマー1と6で得られたアンプリコンの配列、及びプライマー2と3で得られたア
ンプリコンの配列がコンティグのように組い立てられる場合には、ヒトセンスミトコンド
リアキメラRNAの新たな構造は、50ヌクレオチドのループと少なくとも769bpの
二重鎖構造であることを確認した(図1A)(SEQ ID NO 1参照)。
【0095】
二重鎖RNAは、RNaseAによって消化されないので、ヒトセンスミトコンドリアキメ
ラRNAのステムは、この酵素に耐性がなければならない。一方、ループ又は二重鎖を越
えて拡張したセンス16SミトコンドリアRNA鎖の3’の領域は、酵素により消化され
なければならない。
HeLa細胞又は他の細胞からのRNAは、RNase A(50μg/ml)によって消化され
、続いてフェノール抽出された、そしてヌクレアーゼ耐性物質はエタノール沈殿により回
収された。
消化されたRNAからのcDNAは、図1Aに示すプライマーを用いてPCRにより増
殖された。
プライマー1と6で得られた約800bpのアンプリコンは、ループが酵素により消化
されたことを示すRNaseA消化後には増殖されなかった。図1Aに示したようにプライマ
ー10と11で得られた360bpのアンプリコンと同じであった。
すべて、これらの結果は、ステムを超えて拡張したセンスミトコンドリアキメラRNA
の3’領域だけでなくループも酵素により消化されたことを示した。
センスミトコンドリアキメラRNAの二重鎖の構造に一致しまたプライマー8と6で得
られた750bpアンプリコンの増殖は、RNase A消化により影響されなかった。
リボ核酸消化に耐性を示す二重鎖フラグメントの配列は、予期されたステムの配列と同
一であった。
【0096】
センスミトコンドリアキメラRNAの逆方向反復配列の5’末端を決めるために、RNas
e A消化後に得られた転写産物のステムは、5’RACE分析に用いられた。
逆方向反復配列の5’末端決定は、メーカーのインストラクション(Invitrogen)に従
って行われた。
この結果は、逆方向反復配列がプライマー1と6を用いたセンスミトコンドリアキメラ
RNAの増殖後に得られたアンプリコンの5’末端から46の付加的ヌクレオチドに伸び
ていることを示した。
要約すると、815ヌクレオチドの逆方向反復配列は、16SミトコンドリアRNAの
最初の865ヌクレオチドの5’末端に結合している。この転写産物の配列は、ヒト16
Sミトコンドリア遺伝子(HとL鎖)(SEQ ID NO 1)と99.8%の同一性を示した。二
重鎖ステムの両端の5’末端は、5’RACEにより確認された。
【0097】
上記の結果は、センスミトコンドリアキメラRNAが815塩基対のステムあるいは二
重鎖構造と50ヌクレオチドを含んでいたことを示した。
しかしながら、これらの結果は、逆方向反復配列が完全な16SミトコンドリアRNA
に結合していることを立証するものではない。3’末端から完全なcDNAの合成のよう
な従来のアプローチの使用は役に立たない、なぜなら転写産物の二重鎖構造は、Tthを
含む逆転写酵素に対し克服できない問題を示すからである(Myers and Gelfand, Biochemi
stry30:7661-7666, 1991)。
もし815ヌクレオチドの逆方向反復配列が16SミトコンドリアRNAの1559ヌ
クレオチドに結合したら、2.3Kbの転写産物が予期できる。
HeLa細胞、HL−60及びMCF/7細胞から全RNAのノーザンブロット分析は
、32Pで標識されたプローブを用いて行われ、センスミトコンドリアキメラRNAの二
重鎖構造を標的にした。その結果は、1.6Kbのバンドを除いて、センスミトコンドリ
アキメラRNAとセンス16SミトコンドリアRNAにそれぞれ対応する約2.4Kbの
バンドを示した。
もし前記RNAがノーザンブロットより先にRNase Aで消化されたら、およそ0.8K
bのシングルハイブリダイゼーションバンドが得られた、これはセンスミトコンドリアキ
メラRNAのステムのサイズに相当する。
すべて、これらの結果は、前記センスミトコンドリアキメラRNAが完全なセンス16
SミトコンドリアRNAの5’末端に結合した815ヌクレオチドの逆方向反復配列を含
んでいて、SEQID NO 1に相当していた、ことを強く実証した。
【0098】
逆方向反復配列とセンス16SミトコンドリアRNA間の結合領域を、ヌクレオチドプ
ローブを用いて具体的に検出することが可能である。
プローブは、逆方向反復配列の3’末端とセンス16SミトコンドリアRNA間の結合
点の各サイドで7から10のヌクレオチドを含まなければならない。
このオリゴヌクレオチドは、in situハイブリダイゼーション、又はRT−PCRもし
くはこの新規なRNAを検出するための分野の技術者によく知られた任意の方法による増
殖に用いることができる。
【0099】
センスミトコンドリアキメラRNAは、正常な増殖細胞(ヒトの包皮角化細胞、脾臓、
PHAで刺激されたリンパ球、マウス胚)中、前癌細胞(HPV16又は18で形質転換
された角化細胞、HTLV−1で形質転換されたMT−2細胞)中に、そして癌細胞中に
存在する。正常な休止細胞中には存在しない。これらの結果の要約は、表1(実施例4)
に示されている。
【0100】
[実施例2]
乳頭腫ウィルスで形質転換されたヒトの角化細胞は、新規なセンスミトコンドリアキメ
ラRNAを合成する(図1B、SEQID NO 2)
ヒトの包皮角化細胞(HFK)は、ヒトの乳頭腫ウィルス16(HPV16)に既に感染
した細胞溶解物を用いた培養により形質転換された。
この細胞は、3部のK−SFM、1部のDMEM培地(Invitrogen)、5ng/mlの
下垂体抽出物、及び10%の子牛胎児血清で培養された。
培養条件は、37℃で5%CO2であった。24時間の感染後、形質転換されたHFK
は、が新しいフラスコに転送され、同じ条件下で培養された。
この後に、細胞(HFK698)は、1:4から1:3の分離比率を使用して、3日ごとに新し
い培養フラスコに連続的に転送された。
19継代後に、細胞(HPV16で形質転換されたHFK698)は、記載したように集菌され(Haus
en,Biochim.Biophys. Acta, 1288:F55-F78, 1996)、10分間の300×gでの遠心分離
で集められ、そして塩性のリン酸塩バッファー(PBS)で2度洗浄された。
全RNAは、トリゾール(Invitrogene)で洗浄された細胞から抽出された。
約200ナノグラムのRNAは、実施例1に記載したようにランダムのヘキサマー(he
xamers)でcDNAを合成するために使用された。
【0101】
cDNAは、図1Aに記述されているように、リバースプライマー1とフォワードプラ
イマー3を用いてPCRにより増殖された。
この増殖プロトコルは、予期した210bpのアンプリコンを生じた、ここでセンス1
6SミトコンドリアRNAの最初の31ヌクレオチドは、実施例1に記述したように18
4ヌクレオチドの逆方向反復配列に結合していた。
この増殖製品の電気泳動分析は、センスミトコンドリアキメラRNAに相当する210
塩基対のアンプリコンと、図2に示すように約150塩基対の他の増殖フラグメントの存
在を示した。この新しいフラグメント(SEQ ID NO 2)の完全な存在は、センス16Sミト
コンドリアRNAの5’ 末端から最初の31のヌクレオチドが121ヌクレオチドの逆
方向反復配列に結合されており、これはSEQ ID NO 1のセンスミトコンドリアキメラRN
Aの逆方向反復配列と比較すると、63ヌクレオチドより短い。
【0102】
このより短い逆方向反復配列は、ミトコンドリアキメラRNAの構造においてRNA9
6ヌクレオチド(図1B)のより長いループを生成する。
配列の残部は、SEQID NO 1と同一である。
この新規なミトコンドリアキメラRNAは、SiHa細胞(図4A)には存在しない、こ
れは、PHAで刺激されたヒトリンパ球のような増殖型細胞中にも存在しない、HPVで
形質転換された発癌性細胞である(実施例参照)。
同様の結果は、HPV16又はHPV18で形質転換された又は不滅化された(しかし
発癌性ではない)細胞は、前−悪性細胞と考えられ、それ故新規なセンスミトコンドリア
キメラRNAは、前−悪性細胞の新規なポテンシャルマーカーである。
【0103】
16SミトコンドリアRNAに結合しているSEQ ID NO 2の逆方向反復配列の3’末端
の配列がSEQID NO 1の同じ領域とは異なるので、オリゴヌクレオチドプローブは、この
転写産物の特別の検出に用いることができる。
このプローブは、逆方向反復配列の3’末端とSEQ ID NO 7のオリゴヌクレオチドのよ
うなセンス16SミトコンドリアRNAの始めとの間の結合点の各端で7ないし10のヌ
クレオチドを含まなければならない。
このオリゴヌクレオチドは、in situハイブリダイゼーションもしくはRT−PCRに
よる増殖、又は前癌細胞のこの新規な特別のマーカーを検出する、この分野の技術者によ
く知られた任意の方法に用いることができる。
【0104】
[実施例3]
HTLV-1で形質転換されたヒトMT-2細胞は、第3の新規なセンスミトコンドリアキメラR
NAの発現を誘発する(図1C、SEQ ID NO 3)。
HTLV-1で形質転換されたヒトMT-2細胞は、記述された方法で培養された(Kobayashiet
al., EMBO J.,3:1339-1343, 1984)。
細胞は集菌され、300×gで10分間遠心分離され、PBSで2度洗浄された。最後
の細胞ペレットは、実施例1で記述したようにトリゾールで抽出された。
cDNAは、RNAを鋳型として用いてランダムヘキサマーで合成された、またcDN
Aは、リバースプライマー1および図1Aに記述されるようなフォワードプライマー3を
使用して、PCRにより増殖された。
以前に記述したように、この増殖プロトコルは、実施例1に記述したセンスミトコンド
リアキメラRNAに相当する、184ヌクレオチドの逆方向反復配列に結合したセンス1
6SミトコンドリアRNAの最初の31ヌクレオチドを含む210塩基対のアンプリコン
を生ずる。
増殖産物の電気泳動分析は、すでに記述された210塩基対のアンプリコンの存在に加
えて、約150塩基対のバンドを示した(図2を参照)。
【0105】
この150塩基対のアンプリコンの配列は、HPV16あるいはHPV18(SEQ ID NO
2)で形質転換された細胞中で発現する第2のセンスミトコンドリアキメラRNAに相当
する、実施例2で記述したアンプリコンの配列に同じである。
更に、新しい増殖産物は、約100bpであることが見出された(図2)。
この第3のアンプリコンの配列は、センス16SミトコンドリアRNAの5’末端に結
合して、167ヌクレオチドのループを発生する、61ヌクレオチドの逆方向反復配列を
示した(図1C;SEQID NO 3)。
この新規なアンプリコンは、正常細胞中に、腫瘍細胞中にまたEPV16あるいは18
で形質転換された細胞中には存在しなかった。
従って、この新規なセンスミトコンドリアキメラRNAは、発癌性レトロウイルスで形
質転換された細胞のポテンシャルマーカーである。
【0106】
16SミトコンドリアRNAに結合しているSEQ ID NO 3の逆方向反復配列の3’末端
の配列は、SEQID NO 1及びSEQ ID NO 2の同じ領域とは異なるので、オリゴヌクレオチド
プローブは、この転写産物の特定の検出に用いることができる。
このプローブは、逆方向反復配列の3’末端とSEQ ID NO 8のオリゴヌクレオチドのよ
うなセンス16SミトコンドリアRNAの始まりとの間の結合点の各端で7から10のヌ
クレオチドに及ばなければならない。
このオリゴヌクレオチドは、in situハイブリダイゼーションもしくはRT−PCRに
よる増殖、又はレトロウィルス腫瘍ウィルスで形質転換された細胞のこの特別なマーカー
を検出する、この分野の技術者によく知られた任意の方法に用いることができる。
【0107】
[実施例4]
ヒトアンチセンスミトコンドリアキメラRNAの構造
我々の最初の実験は、アンチセンスミトコンドリアキメラRNAに相当するキメラRN
Aの第二のファミリーは、いくつかの細胞研究に存在することを示した。
ヒトアンチセンスミトコンドリアキメラRNAの構造を決定するために、センスミトコ
ンドリアキメラRNAに用いられる方法が採用された(図1)。
理論的なアンチセンスミトコンドリアキメラRNAは、アンチセンス16Sミトコンド
リアRNAの5’末端に結合している逆方向反復配列としてのセンス16Sミトコンドリ
アRNAのフラグメントを含んでいた。
後者のRNAは、ミトコンドリアDNAのL−鎖から転写され、そして16Sミトコン
ドリア遺伝子に相当する(図3)。このRNAを増殖するために、リバースプライマーは
アンチセンス16SミトコンドリアRNAの5’末端に接近してハイブリダイズされた、
またフォワードプライマーは、逆方向反復配列(図3)の推定のフラグメントの異なる位
置でハイブリダイズされた。
PHAで48時間刺激されたヒトリンパ球からの全RNAは、鋳型として用いられた。
cDNAは、実施例1に記載したようにランダムヘキサマーで合成された。
【0108】
PCRによるcDNAの増殖は、アンチセンス16SミトコンドリアRNAの5’末端
の始めに接近した位置でのリバースプライマー(プライマー1、図3)及び逆方向反復配
列(図3)上に位置する異なるフォワードプライマーで行われた。
3つの主なアンプリコンだけが得られた、これは逆方向反復配列のサイズとループのサ
イズで異なっていた。
これらのアンプリコンは、精製され、配列が決定された。これらのアンチセンスミトコ
ンドリアキメラRNAの1つは、365ヌクレオチドの逆方向反復配列と17のヌクレオ
チド(SEQ IDNO 4)のループを含んでいる。
他のRNAは、96ヌクレオチドのループと189ヌクレオチド(SEQ ID NO 5)の逆方
向反復配列を含んでいる。
更に、アンチセンスミトコンドリアキメラRNAの他の種は、296ヌクレオチドの逆
方向反復配列と451ヌクレオチド(SEQ ID NO 6)のループを含んでいる。
3つのアンチセンスミトコンドリアキメラRNAのすべての配列は、ミトコンドリアD
NA遺伝子(HとL鎖)の配列と99.8%同一であった。
【0109】
以下の例において示される結果は、前腫瘍及び腫瘍細胞と、アンチセンスミトコンドリ
アキメラRNAの発現にかかわる正常な増殖細胞との間に大きな違いがあることを示す。
増殖する細胞はすべてセンスミトコンドリアキメラRNAを過剰発現する。
しかしながら、一方正常な増殖細胞もまたセンスミトコンドリアキメラRNAを発現す
るが、これらの転写産物は、腫瘍細胞中では下方制御される。非増殖細胞又は休止細胞は
共にミトコンドリアキメラRNAを発現しない。
従って、これらの異なる発現は、発癌の新規で有効なマーカーであることを示し、これ
はin situハイブリダイゼーション、ノーザンブロット分析、RT−PCRもしくはTM
A又はこの分野の技術者に知られている他の技術により検出できる。
センスとアンチセンスミトコンドリアキメラRNAの異なる発現の要約は、表1に示さ
れている。
【0110】
【表1】

【0111】
[実施例5]
腫瘍細胞系は、センスミトコンドリアキメラRNA(SEQ ID NO 1)を過剰発現し、またア
ンチセンスミトコンドリアキメラRNA(SEQ ID NO 4,5及び6)の発現を下方制御する。
in situハイブリダイゼーションは、培養中の腫瘍細胞系でセンスミトコンドリアキメ
ラRNAの発現を決定するのに用いられた。in situハイブリダイゼーションに対し、粘
着性腫瘍細胞は、8−ウエルのチャンバースライド(Lab-TekO, NUNC)中で、アメリカン組
織培養収集(AmericanTissue Culture Collection)(ATCC)により推奨された適切な培地
と条件を用いて37℃で24ないし48時間培養された。
非粘着性細胞(e.g.HL-60, Jurkat and Ramos)に対しては、小さなフラスコ中で37℃
で48時間培養された。細胞は、300×gで10分間遠心分離により回収され、PBS
の小さな容量中に再懸濁され、そして10から20ulのアリコートは、あらかじめポリ
リシン又はマッスル(mussel)から精製された接着タンパク質でコートされたガラススラ
イド上に塗布された(Burzioet al, Curr. Opin. Biotechnol., 8:309-312, 1997)。細胞
は室温で30分間乾燥された。
【0112】
前記細胞は、PBSで3回洗浄され、4%のパラホルムアルデヒドで室温下10分間固
定された。スライドはその後PBSで5分間3度洗浄され、0.2N HClで室温下10
分間インキュベートされた。
該細胞は、更に3度、最初は室温下にPBSで、その後2×SSCで10分間(2×SSC
:0.3MのNaCl、30mMクエン酸ナトリウム、pH7.0)洗浄された(Sambrook et al., 1989)。
プレハイブリダイゼーションは、4×SSC、10%硫酸デキストラン、150μg/ml
酵母tRNA及びニシン精液DNA、50%のホルムアミド及び1×Denhardt溶液(0.2mg
/mlのFicollタイプ400、0、2mg/mlのポリビニルピロリジン、0.2mg/mlのBSA)を含む溶液
中で37℃で30分間行われた。ハイブリダイゼーションは、センスとアンチセンスミト
コンドリアキメラRNAを標的にする3.5pmolのプローブを含む同じプレハイブリ
ダイゼーション混合物中37℃で15時間行った。
前記プローブは、センスとアンチセンスミトコンドリアキメラRNAの配列の異なる領
域をターゲットにする20以上のデオキシヌクレオチドを含んでいた(SEQ ID NO 99〜197
及びSEQ IDNO 9〜98をそれぞれ参照)。プローブは、既に記述したように予めジゴキシゲ
ニン-11-dUTP(Roche)とターミナル・トランスフェラーゼ(Promega)を用いて3’末端でラ
ベル表示された(Villegaset al., DNA & Cell Biol., 19:579-588, 2000)。
過剰のプローブを除去するために、スライドは、室温で、最初2×SSCで10分間及び1
×SSCで10分間洗浄された。その後、サンプルは室温で0.2×SSCで45℃30分間、及
び最後に0.2×SSCで10分間洗浄された。
【0113】
ハイブリダイゼーション後、細胞はブロッキング緩衝液中(1%BSA、0.3%トリトンX-100
in PBS)中で30分間インキュベートされ、その後予めブロッキング緩衝液中で1:50
0に希釈され、アルカリ性ホスファターゼに結合しているアンチ−ジゴキシゲニン単クロ
ーン抗体を用いて室温で2時間インキュベートされた。
最後に、スライドは、PBSで2度洗われた、そして呈色反応は以前に記載したようにBC
IP/NBT基質混合物(DAKO)を用いて実行された(Villegas et al., DNA & Cell Biol.
, 19:579-588,2000)。同様の操作は、フルオレスセインまたはローダミンに接合される
アンチ−ジゴキシゲニン抗体を使用して、FISHに対して行われた。
【0114】
図4Aと4Bに示されるように、SEQ ID NO 63に対応するジゴキシゲニンでラベル表示
したプローブを用いたin situハイブリダイゼーションは、ヒト腫瘍細胞がセンスミトコ
ンドリアキメラRNAを過剰発現することを示す。
ジゴキシゲニンでラベル表示した、SEQ ID NO 64に対応するセンスプローブを用いたin
situハイブリダイゼーションは、アンチセンスミトコンドリアキメラRNAの発現を下
方制御を示して、ネガティブであった(図4A及びB)。同じ結果は、センスあるいはアン
チセンスミトコンドリアキメラRNAの他の領域を標的にするオリゴヌクレオチドプロー
ブで得られた。
【0115】
[実施例6]
ヒト生体中の腫瘍細胞はセンスミトコンドリアキメラRNA(SEQ ID NO 1)を過剰発現
し、アンチセンスミトコンドリアキメラRNA(SEQ ID NOS 4、5及び6)を下方制御する。
ヒトの生体は、病理学者あるいはDAKOからの組織アレーから入手した
分析されたほとんどのサンプルはパラフィンに埋め込まれていたか、またはホルマリン
で固定されていた。他の組織サンプルは、Boiun固定剤で固定されていた、また他のサン
プルは、新鮮な凍結組織切片であった。
約4ないし8μmの組織切片は、予めポリリシン又はムラサキイガイ(Aulacomya ater)
から精製された接着性ポリフェノール蛋白質でコートされたガラススライド上に固定され
た(Burzioet al., Curr. Opin. Biotechnol., 8:309-312, 1997)。
パラフィンに包埋された組織切片は、60℃で1時間インキュベートされた、そしてパ
ラフィンは、キシロールで各15分間3度洗浄により除去された。
切片は、空気乾燥され、PBSで4回洗浄された。その後、切片は、0.2N HCl
を用いて、室温で10分間インキュベートされた、そしてPBSで十分に洗浄された。後
で、サンプルは、実施例4に記述されたプロトコルに従って、ジゴキシゲニンでラベル表
示されたアンチセンスプローブを用いてin situハイブリダイゼーションに供された。パ
ラレルセクションはセンスミトコンドリアキメラRNAの同じ領域に対応するセンスプロ
ーブでハイブリダイズされた。
【0116】
図5Aに示すように、胸、子宮頚部、膀胱の腫瘍及び肺の癌は、転移産物の強い存在を
示して、センスミトコンドリアキメラRNAを標的にするアンチセンスプローブで強く染
色されることを示した。他方、アンチセンスミトコンドリアキメラRNAを標的にするプ
ローブを用いたinsituハイブリダイゼーションは、この転写産物(図5A)の下方制御
を示してネガティブであった。
他の腫瘍細胞もまたセンスミトコンドリアキメラRNAを過剰発現し、そしてアンチセ
ンスミトコンドリアキメラRNAの発現を下方制御する(図5B)。
【0117】
[実施例7]
正常増殖細胞はセンス及びアンチセンストコンドリアキメラRNAを過剰発現する。
実施例5及び6に記述されたin situハイブリダイゼーションで同じプロトコルを使用
して、センスミトコンドリアキメラRNAの発現は、増殖細胞で決定した。
図6に示すように、HFK細胞、精原細胞、脾臓細胞、及び増殖型マウス胚は、センス
ミトコンドリアキメラRNAの過剰発現を示唆する、強いハイブリダイゼーションシグナ
ルを示した。
対照的に、脳、筋肉及び肝臓の細胞のような非増殖細胞は、センスミトコンドリアキメ
ラRNAが発現しないか又はこれらの細胞で下方制御されることを示唆して、シグナルを
示さなかった。
【0118】
しかしながら、驚くべき結果は、in situハイブリダイゼーションがアンチセンスミト
コンドリアキメラRNAを標的にするプローブで行われたとき、強いシグナルも観察され
た(図6)。
並行して分析されたいくつかの制御は、これらのプローブのハイブリダイゼーションシ
グナルが技術上の原因によらないことを示した。
もしinsituハイブリダイゼーションがジゴキシゲニンで非ラベル表示されたものを除
いて、過剰(50ないし100倍)の同じプローブと共にラベル表示されたプローブで行
われたら、ハイブリダイゼーションシグナルは消失した。
ハイブリダイゼーションに先立ちサンプルがリボヌクレアーゼAで一晩インキュベート
された場合、ハイブリダイゼーションシグナルは消失した。
また、ハイブリダイゼーションが4つのミスマッチを有するアンチセンスミトコンドリ
アキメラRNAを標的にするラベル表示されたプローブで行われた場合、ハイブリダイゼ
ーションシグナルは観察されなかった。
【0119】
[実施例8]
フィトヘムアグルチニン(PHA)で刺激された正常なヒトのリンパ細胞は、センス及びア
ンチセンスミトコンドリアキメラRNAを過剰発現する。健康なドナーから5mlの血液
がEDTAで集められた。この血液は、0.9%NaClの1容積で希釈され、該混合物
は、遠心分離機チューブ中の5mlのHistopaque-1077(Sigma)に適用された。このチュー
ブは室温下で20分間800×gで遠心分離された。
分裂期の白血球が集められ、0.9%のNaClの2容積で薄められ、室温で10分間、
250×gで遠心分離された。集められた細胞は、けん濁され、200mMグルタミン、
10mM非必須アミノ酸、ペニシリン、子牛の胎児血清を欠いているストレプトマイシン
が追加されたRPMI1640倍地で2度洗浄された。
最後の沈殿物は、10%子牛の胎児血清を含む同じ培地で再けん濁され、1ml当たり
のヒトリンパ球の数は、ノイバウエルチャンバー中の顕微鏡で数えて決定された。
【0120】
ヒトのリンパ球は、上記の追加されたRPMI1640倍地に10%の子牛の胎児血清
を添加した96ウエルのマイクロリッタープレート中で37℃、5%CO2の条件下で培
養された。1ウエル当たり約30,000のリンパ球は、10ug/mlの(細胞増殖を誘発す
る)ミトゲンPHA存在、非存在下で培養された(Yu et al., J. Biol. Chem., 266:7588
-7595, 1991)。
PHAで処理後48ないし72時間後、細胞は、H3−チミジン又はBrdUの取り込
みにより立証されたように活発にDNA合成にたずさわった(Yu et al., J. Biol. Chem.
,266:7588-7595, 1991)。また、PHAで刺激された48時間後に、リンパ球は、増殖細胞
核抗原又はPCNA及びKi−67のように、細胞増殖の他のマーカーを過剰発現した(B
antis et al.,Cytopathology, 15:25-31, 2004) (図7)。
休止中又はコントロール・リンパ細胞は、これらの抗原を発現しなかった(図7)。
【0121】
刺激されたリンパ球がセンスミトコンドリアキメラRNAを発現する否かを決定するた
めに、細胞は、ジゴキシゲニンでラベル表示され、センスミトコンドリアキメラRNAを
標的にするオリゴヌクレオチドプローブでin situハイブリダイズされた。
使用されたinsituハイブリダイゼーションプロトコルは、実施例5に記載された。
この転写産物の過剰発現を示唆する、強いハイブリダイゼーションシグナルが得られた
(図7)。ハイブリダイゼーションシグナルは、腫瘍細胞又は他の正常増殖細胞で観察され
たシグナルと同程度であった(図7を図4AとB、図5AとBと比較)。
ハイブリダイゼーションシグナルは、PHAなしでインキュベートされたコントロール
リンパ球には観察されなかった(図7)。
【0122】
このinsituハイブリダイゼーションがジゴキシゲニンでラベル表示され、アンチセン
スミトコンドリアキメラRNAを標的にするセンスオリゴヌクレオチドを用いて行なわれ
た場合、同様に強いハイブリダイゼーションシグナルが得られた(図7)。
いくつかのコントロールは、ハイブリダイゼーションシグナルが技術上の原因よる可能
性を否定するために行なわれた。
ハイブリダイゼーションシグナルは、もしin situハイブリダイゼーションが過剰(5
0ないし100倍)の(ジゴキシゲニンでラベル表示されていない)同一センスプローブ
と共にセンスラベル表示プローブで行なわれるときには、消失する。
ハイブリダイゼーションに先立ちサンプルをリボヌクレアーゼAで一晩インキュベート
すると、ハイブリダイゼーションシグナルは、消失する。
更に、ハイブリダイゼーションが4つのミスマッチを有するセンスプローブで行なうと
、ハイブリダイゼーションシグナルは、観察されない。対照的に、刺激を受けていないリ
ンパ球のinsituハイブリダイゼーションは、ハイブリダイゼーションシグナルを示さな
い(図7)。
結論として、増殖するために刺激された正常なヒトリンパ細胞は、センスミトコンドリ
アキメラRNA及びアンチセンスミトコンドリアキメラRNAの双方を過剰発現する。こ
れらの転写産物は、休止細胞中では発現しない。
【0123】
[実施例9]
センスミトコンドリアキメラRNAは、正常な及び腫瘍細胞中で異なる局在化を示す。
実施例5と6に記述したin situハイブリダイゼーションは、ヒトの生検の腫瘍細胞中
だけでなくいくつかの腫瘍細胞系においても、センスミトコンドリアキメラRNAは、細
胞質中で選択的に局在化される。しかしながら、ある腫瘍生検において、核中の転写産物
の明らかな局在化が見出された(図4A、B)。
【0124】
驚くべき発見は、核中のセンスミトコンドリアキメラRNAの局在化であった。
実施例5に記載したように実施されたin situハイブリダイゼーションは、HeLaと
SiHa細胞の核中にポジティブなハイブリダイゼーションシグナルを示した(図8)。
ハイブリダイゼーションシグナルは、HPV16で形質転換されたHFKの核中で強くな
った(図8)。核の局在化は、また胸腫瘍及び横紋筋肉腫からの腫瘍細胞中にも見出され
た(図8)。
【0125】
共−局在化(co-localization)の研究は、細胞質で局在化したセンスミトコンドリア
キメラRNAがミトコンドリアの外側で後期エンドゾーム/リゾソームと結合することを
示した。もし共−局在化の研究が分子プローブ(Mitotrack)又は抗体アンチ−シトクロム
c(Promega)もしくはアンチ−エンドヌクレアーゼG(Chemicon)のようなミトコンドリア
のマーカーで行なわれる場合、in situハイブリダイゼーションは、弱い共−局在化を示
した。
しかしながら、完全な共−局在化は、リソトラック(Lysotrack)(Molecular Probes)又
はアンチ-ランプ-2(anti-Lamp-2)(BD Pharmigen)もしくはアンチカプサイシンD(anti-
cathepsin D)(Zymed)のような後期エンドゾーム/リゾソームの免疫細胞学でハイブリダ
イゼーションシグナル中に見出された。
【0126】
HeLa細胞は、実施例5に記述したようなジゴキシゲニンでラベル表示されたオリゴ
ヌクレオチドプローブでin situハイブリダイズされた。
ポスト−ハイブリダイゼーションと洗浄操作後、細胞は、ローダミン(Roche)でラベル
表示されたアンチ−ジゴキシゲニン及びフルオレスセイン(BD Pharmingen)でラベル表示
されたanti-Lamp-2antibodyでインキュベートされた。暗所で室温下に3時間インキュベ
ートした後、スライドは洗浄され、マウントされてツァィス共焦点顕微鏡で分析された。
リソトラック(Lysotrack)又はアンチ−カテプシンD抗体がリソソーム分画として用
いられたとき、ランプ-2(Lamp-2)の局在化によるハイブリダイゼーションシグナルの
明らかな共−局在化が得られた。同様のハイブリダイゼーションシグナルの共−局在化結
果が得られた。
我々が知っている限り、これは、RNA(特にミトコンドリア転写産物)が細胞のリソ
ソームに結合されたことを示す最初の報告書である。
腫瘍細胞中のセンスミトコンドリアキメラRNAの局在化の決定は、癌を有する患者に
対する重要な予後の値となる。一般に、正常な増殖細胞中でセンス及びアンチセンスミト
コンドリアキメラRNAは、主に核中に局在化される。
【0127】
[実施例10]
アンチセンスミトコンドリアキメラRNAを標的にするアンチセンスオリゴヌクレオチ
ドを用いて腫瘍細胞をin vitro処理すると細胞死を誘発する。
HL-60細胞は、ATCCによって推奨された最適条件の下で培養される。
約30,000個の細胞が96―ウエルのマイクロタイタープレートで培養された。
センス又はアンチセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオチド
(2uM)が加えられた。細胞の浸透性を増すために、オリゴヌクレオチドは、リプフェ
クタミン(lipofectamin)もしくはオリゴフェクタミン(oligofectamin)(Invitrogen)
又はポリエチレンイミド(PEI)を有する混合物中に添加された( Exgen TM500, Fermenta
s)。細胞に特に非毒性であるPEIがより好ましい、
細胞は、オリゴヌクレオチドで6時間インキュベートされる、そして細胞残存の割合は
、トリパンブルーへの浸透性で決定される。
オリゴヌクレオチドで6時間インキュベートした後、細胞の主要な割合が死滅した。
しかしながら、アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオ
チドは、細胞死を誘発するのに更に有効であった(細胞死の15%に対し約90%)。
【0128】
他方、細胞がセンスもしくはアンチセンス12SミトコンドリアRNA又はND1サブ
ニットのmRNAを標的にするオリゴヌクレオチド、又はスクランブルオリゴヌクレオチ
ドもしくは4つのミスマッチを有するオリゴヌクレオチド(これらのすべてはコントロー
ルとして使用された)で処理されたとき、細胞死は生じなかった。
これらの研究に用いられたオリゴヌクレオチドは、5’末端で最初の5ヌクレオチドま
た3’末端で最初の5ヌクレオチドにホスフォロチオエートリンケージを含んでいる。平
均して、10の中央のヌクレオチドは、ホスフォジエステル結合を含んでいる。これらの
オリゴヌクレオチドを備えた細胞の処理がDNA断片化を引き起こすかどうか確証するた
めに、HL-60細胞は、オリゴヌクレオチドで6時間、以前に記述されたと同じ条件下でイ
ンキュベートされた。
約30,000個のHL-60細胞が、96ウエルのマイクロチッタープレート上で、200ul
のIDMEMプラス10%の子牛胎児血清中で、センスミトコンドリアキメラRNAを標
的とする又はアンチセンスミトコンドリアキメラRNAを標的とする1uMのオリゴヌク
レオチドと共に培養された。
【0129】
PEIを含む混合物中に添加されたオリゴヌクレオチドの化学は、前節に記載したのと
同様である。オリゴヌクレオチドで6時間定温放置した後、細胞は、TUNEL分析(DeadEnd
の比色定量のTUNELシステム、Promega)を用いてDNA断片化のために分析された。
表2に示されるように、アンチセンスミトコンドリアキメラRNAを標的とするオリゴ
ヌクレオチドで処理後、約96%の細胞がDNA断片化を示した。同様のDNA断片化率
は薬物スタウロスポリンで得られた。スクランブルオリゴヌクレオチド又はミスマッチを
有するオリゴヌクレオチドは効力を示さなかった。
対照的に、センスミトコンドリアキメラRNAを標的とするオリゴヌクレオチドで処理
されると、約20%の腫瘍細胞はアンチセンスミトコンドリアキメラRNAの発現を下方
制御し、そしてその結果これらの細胞は、この転写産物の少ないコピー数をもたらす(表
2)。
従って、細胞死は、アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌク
レオチドでより効率的に誘発される。
これらの結果は、腫瘍細胞中のアンチセンスミトコンドリアキメラRNAの少ないコピ
ー数は、治療目的になる、ことを強く示唆する。
【0130】
【表2】
【0131】
別の研究で、我々は、アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌ
クレオチドで細胞を処理するとカスパーゼ活性を誘発する、ことを決定した。
カスパーゼは、プログラムされた細胞死あるいはアポトーシスに活発に伴う、蛋白質分
解を生ずる酵素である。HL-60は、アンチセンスミトコンドリアキメラRNAを標的
にするオリゴヌクレオチド、又はスタウロスポリンを用いて、以前に記述された培養条件
下で6時間、インキュベートされた。
その後、フルオレセインで結合したVAD-fmk(CaspaCe FITC-VAD-FMK、Promega)を
培養に加え、37℃で30分間インキュベートされた。 VAD-fmokは、カスパーゼの強い
抑制剤で、非常に高い親和性でプロテアーゼに結合する(Gracia-Calvo et al., J. Biol.
Chem.,273:32608-32613, 1998)。
細胞は、遠心分離によって洗われ、標本化されて蛍光顕微鏡で観察された。
ズ9に示すように、アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌク
レオチドで処理したHL-60細胞は、スタウロスポリンで達成された活性化と同様のレ
ベルで、カスパーゼ活性を誘発した。12SミトコンドリアRNAを標的にしたアンチセ
ンスのオリゴヌクレオチドでは、カスパーゼ活性は得られなかった。
【0132】
アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオチドで処理した
細胞は、またアポトーシスに一致する他の変化を示す。電子顕微鏡分析は、核断片化およ
びクロマチン凝縮を示した。核断片化もDAPIで核を染色することにより実証された。
アンチセンスミトコンドリアキメラRNAを標的にするこれらのオリゴヌクレオチドで
処理後、細胞は、DAPI染色で示されたように(図9E及び9F)、核断片化を受ける。
【0133】
[実施例11]
アンチセンストコンドリアキメラRNAに相補するオリゴヌクレオチドで処理されると、
他の癌細胞も細胞死を被る。
他の癌細胞は、実施例10に記載したプロトコルに従い、アンチセンストコンドリアキ
メラRNAに相補するオリゴヌクレオチドで処理された。この細胞は、ATCCのリコメ
ンドに従い、それらの最適条件下でインキュベートし、そして2uMオリゴヌクレオチド
がPEIと共に実験の初期段階で添加された。
6時間後に、オリゴヌクレオチドの2回目の添加を同様の濃度で実行し、結果は、実験
の開始の後15時間で決定された。細胞死は、DAPI染色と断片化された核の細胞数を
数えることにより決定された。表3に示すように、オリゴヌクレオチドで処理された細胞
の70%以上がアポトーシスを被った。
薬剤処理に全くの耐性を示すことが知られている、黒色腫細胞、リンパ腫細胞および胸
癌細胞MCF/7は、非常に高い割合でアポトーシスを被ることに気づくことは重要であ
る(表3)。
【0134】
【表3】
【0135】
アポトーシスを誘発する際にオリゴヌクレオチドに対するより有効な標的となる転写産
物の領域があるかを決定するために、次の実験を行った。アポトーシスの誘発は、アンチ
センストコンドリアキメラRNAの5’末端から始まっておよそ各30ヌクレオチドを標
的とした、約20ヌクレオチドのアンチセンスオリゴヌクレオチドを有するHeLa、H
E-60及びMCF/7細胞において研究された。
時間0では、1uMオリゴヌクレオチドがPEIと共に加えられた、そしてこの処理は
6時間後に繰り返された。処理開始後15時間でアポトーシスを被る細胞の割合は、DA
PI染色と断片化された核を有する細胞を数えることにより決定された。
殆どのオリゴヌクレオチドは、変わりうる程度のアポトーシスを誘発したが、アンチセ
ンスミトコンドリアキメラRNAの単一鎖領域は、細胞死を誘発するよい標的であった。
推定二重鎖又はアンチセンスミトコンドリアキメラRNAのループ構造を標的にするオリ
ゴヌクレオチドは、それほど有効ではない
【0136】
アポトーシスは、またトリパンブルー染色、ヨウ化プロピジウム染色、アネキシン(an
exine)免疫化学により決定することができる。 これらの技術において、細胞は、蛍光
顕微鏡又はフロー・サイトメトリーにより分析することができる。DNA断片化は、TUNE
L、あるいはDNAラダーを示す電気泳動によって測定されうる。
ウエスタンブロット分析もまた、カスパーゼ、poly(ADP Rib)合成等のような蛋白質の
プロセッシングを決定するために使用することができる。
【0137】
[実施例12]
アンチセンスミトコンドリアキメラRNAに相補するオリゴヌクレオチドで正常増殖また
は休止細胞の処理は、アポトーシスに効果がない。以前に記載したように、正常増殖細胞
は、アンチセンスミトコンドリアキメラRNAと同様にセンスミトコンドリアキメラRN
Aを過剰発現する。
他方、休止細胞は、これらの転写産物を発言しない。
それ故、アンチセンスミトコンドリアキメラRNAに相補するオリゴヌクレオチドが正
常細胞において細胞死を誘発するかを決定することは重要である。ヒトリンパ細胞は、実
施例8に記述したように10ug/mlのPHAで48時間刺激された。
並行して、コントロールリンパ細胞がPHAなしで48時間インキュベートされた。
48時間の培養では、PEI(実施例10参照)と混合した15uMのオリゴヌクレオチ
ドは、刺激されそしてコントロールされたリンパ細胞に添加された、そして更に15時間
インキュベートされた。オリゴヌクレオチドの濃度は、以前の実験(1−2uM)に用い
た濃度よりも10フォールド高かった。
刺激されそしてコントロールされたリンパ細胞の他の例は、同じ期間の間、0.4uM
スタウロスポリンで処理された。実験の終わりに、細胞死は、トリパンブルー染色又はD
API染色のいずれかにより測定された。
図10に示すように、コントロールリンパ細胞またはオリゴヌクレオチドを含まない1
5時間のインキュベートでPHA刺激されたリンパ細胞は、異なる実験で7から10%間
の変動で自然におこるアポトーシスと同様のレベルを示した。
【0138】
同様の結果は、オリゴヌクレオチドのより低い(1-2 uM)濃度で得られた。
更に、15uMアンチセンスオリゴヌクレオチドで15時間インキュベートしたコント
ロール及び刺激されたリンパ細胞は、同じ低レベルのアポトーシス(およそ10%)を示
した(図10)。
対照的に、コントロールリンパ細胞又はPHA刺激され、スタウロスポリンで15時間
インキュベートされたリンパ細胞は、80%以上の細胞がアポトーシスを被ることを示し
た(図10)。これは非常に重要な結果である、なぜならヒトリンパ細胞のような正常休
止細胞又は正常増殖細胞がアンチセンスミトコンドリアキメラRNAに相補するオリゴヌ
クレオチドによりアポトーシスを誘発するのに抵抗性であるからである。言いかえれば、
アンチセンスミトコンドリアキメラRNAで阻害することにより、腫瘍細胞中のアポトー
シスの誘発が癌に対する選択的治療アプローチとなるからである。
【0139】
引用文献
1.Adrain et al.,“Regulation of apoptotic protease activating factor-1 oligomer
ization andapoptosis by the WD-40 repeat region”, J Biol Chem. 274:20855-20860
, 1999.
2.Bantis etal., “Expression of p129, Ki-67 and PCNA as proliferation markers i
n imprintsmears of prostate carcinoma and their prognostic value”, Cytopatholo
gy, 15:25-31,2004.
3.Beaucage, “Oligodeoxyribonucleotidessynthesis. Phosphoramidite approach”,
Methods Mol.Biol. 20:33-61, 1993.
4.Benedict etal., “Expression and functional analysis of Apaf-1 isoforms. Extr
a Wd-40 repeatis required for cytochrome c binding and regulated activation of
procaspase-9”,J Biol Chem. 275:8461-8468, 2000.
5.Benoist andChambon, “In vivo requirement of the SV40 early promoter region”
, Nature290:304-310, 1981.
6.Boya et al., “Mitochondrion-targetedapoptosis regulators of viral origin”,
Biochem.Biophys. Res. Commun. 304:575-581, 2003.
7.Brinster etal., “Regulation of metallothionein-thymidine kinase fusion plasm
ids injectedinto mouse eggs”, Nature 296:39-42, 1982.
8.Burzio etal., “Enviromental bioadhesion: themes and applications”, Curr. Op
in.Biotechnol., 8:309-312, 1997.
9.Capaldi etal., “Highly efficient solid phase synthesis of oligonucleotide an
alogs containingphosphorodithioate linkages”, Nucleic Acids Res. 28:E40, 2000.
.
10.Carew andHuang, “Mitochondrial defects in cancer”, Mol. Cancer, 1:1-12, 20
02.
11.Celis, “CellBiology, A Laboratoty Handbook”, Julio E. Celis, ed., 1994
12.Chinnery andTurnbull, “Mitochondrial DNA mutations in the pathogenesis of h
uman disease”,Mol. Med. Today, 6:425-432, 2000.
13.Clayton andVinograd, “Circular dimer and catenate forms of mitochondrial DN
A in humanluekaemic leucocytes”, Nature, 216:652-657, 1967.
14.Clayton andVinograd, “Complex mitochondrialDNA in leukemic and normal human
myeloidcells”, Proc. Natl. Acad. Sci. US, 62:1077-1084, 1969.
15.Clayton, “Transcriptionand replication of mitochondrial DNA”, Hum Reprod.
Suppl2:11-17, 2000.
16.Comanor etal., “Successful HCV genotyping of previously failed and low vira
l loadspecimens using an HCV RNA qualitative assay based on transcription-media
tedamplification in conjunction with the line probe assay.”, J. Clin Virol., 2
8:14-26,2003.
17.Egholm etal., “PNA hybridizes to complementary oligonucleotides obeying the
Watson-Crickhydrogen-bonding rules”, Nature, 365:566-568, 1993.
18.Elbashir etal., “Duplexes of 21-nucleotides RNAs mediate RNA interference i
n cultured mammaliancells,” Nature 411:494-498, 2001.
19.Falkenberget al., “Mitochondrial transcription factors B1 and B2 activate t
ranscription ofhuman mtDNA”, Nat Genet. 31:289-294, 2002.
20.Ferri andKroemer, “Organelle-specific initiation of cell death pathways”,
Nature CellBiol. 3:E255-263, 2001.
21.Frederick etal., in “Current Protocols In Molecular Biology”, Volume 2, Un
it 14,Frederick M. Ausubul et al. eds., 1995.
22.Gracia-Calvoet al., “Inhibition of human caspases by peptide-based and macr
omolecularinhibitors”, J. Biol. Chem., 273:32608-32613, 1998.
23.Guicciardiet al., “Lysosomesin cell death”, Oncogene, 23:2881-2890, 2004.
24.Haseloff etal., “Sequence required for self-catalysed cleavage of the satel
lite RNA oftabacco ringspot virus”, Gene, 82:43-52, 1989.
25.Hausen, “Papillomavirus infection- a major cause of human cancer,” Biochim
.Biophys. Acta,1288:F55-F78, 1996.
26.Hedge etal., “Commitment to apoptosis induced by tumour necrosis factor-alp
ha is dependenton caspase activity”, Apoptosis,7:123-132, 2002.
27.Hyrup andNielsen, “Peptide nucleic acids (PNA): synthesis,properties and po
tentialapplications”, Bioorg. Med. Chem., 4:5-23, 1996.
28.Johnstone etal., “Apoptosis: a link between cancer genetics and chemotherap
y”, Cell108:153-164, 2002.
29.Kobayashi etal., “Genomic structure of HTLV: detection of defective genome
and itsamplification in MT2 cells”, EMBO J., 3:1339-1343, 1984
30.Komarov etal., “A chemical inhibitor of p53 that protect mice from the side
effectsof cancer therapy”, Science 285:1733-1737, 1999.
31.Krammer, “CD95’sdeadly mission in immune system”, Nature 407:789-795, 200
0.
32.Li et al., “EndonucleaseG is an apoptotic DNase when released from mitochon
dria”, Nature,412:95-99, 2001.
33.Liu et al., “Syntheticpeptides and non-peptidic molecules as probes of stru
cture andfunction of Bcl-2 family proteins and modulators of apoptosis”, Apopt
osis,6:453-462, 2001..
34.Lu et al., “siRNA-mediatetedantitumorogenesis for drug target validation an
d therapeutics,”Curr. Opin. Mol. Ther. 5:225-234, 2003.
35.Mag et al., “Synthesisand selective cleavage of an oligodeoxynucleotide con
taining abridged internucleotide 5'-phosphorothioate linkage”,
36.Martins etal., “The serine protease Omi/HtrA2 regulates apoptosis by bindin
g XIAP througha reaper-like motif”, J. Biol. Chem. 277:439-444, 2002.
37.McCulloch etal., “A human mitochondrial transcription factor is related to
RNA adenine methyltransferasesand binds S-adenosylmethionine”, Mol. Cell Biol.
22:1116-1125,2002.
38.McKay etal., “Characterization of a potent and specific class of antisense
oligonucleotideinhibitor of human protein kinase C-a expression”, J. Biol. Ch
em.,274:1715-1722, 1999.
39.McKinnell etal., “The Biological Basis of Cancer, (R:G:McKinnell, R:E: Parc
hment, A:O:Perantoni, G:B: Pierce, eds.), Ch.3, Cambridge University Press, UK,
1998
40.McManus etal., “Gene silincing in mammals by small interfering RNAs,” Natu
re Rev.Genet.3: 737-747, 2002.
41.Meier etal., “Apoptosis in development”, Nature 407:796-801, 2000.
42.Myers etal., “Reverse transcription and DNA amplification by a Thermus ther
mophilus DNApolymerase”, Biochemistry, 30:7661-7666, 1991.
43.Nielsen etal., “Peptide nucleic acids (PNA): oligonucleotide analogs with a
polyamidebackbone”, in S. Crooke, B. Lebleu (eds.) Antisense Research and App
lications, Ch9, CRC Press, Boca Raton, Florida, pp. 363-373, 1993.
44.Parisi and Clayton,“Similarity of human mitochondrial transcription factor
1 to highmobility group proteins”, Science. 252:965-969, 1991.
45.Parrella etal., “Detection of mitochondrial DNA mutations in primary breast
cancerand fine-needle aspirates”, Cancer Res., 61:7623-7626, 2001.
46.Rampino etal., “Somatic frameshift mutations in the BAX gene in colon cance
rs of themicrosatellite mutator phenotype”, Science, 275:967-969, 1997.
47.Rantanen et al.,“Characterization of the mouse genes for mitochondrial tran
scriptionfactors B1 and B2”, Mamm Genome. 14:1-6, 2003.
48.Ravagnan etal., “Heat-shock protein 70 antagonizes apoptosis-inducing facto
r”,
Nat Cell Biol.3:839-843, 2001.
49.Reed, “Dysregulationof apoptosis in cancer”, J. Clin. Oncol., 17:2941-2953
, 1999.
50.Rossi, “Practicalribozymes. Making ribozymes work in cells”, Curr Biol. 4:
469-471, 1994.
51.ambrook etal., Molecular Cloning. A Laboratory Manual, (Sambrook, J., Frits
ch, E.T. andManiatis, T. Eds.,) 2nd Edn, Cold Spring Harbor Laboratory, Cold S
pring Harbor,NY. 1989.
52.Samejima etal., “CAD/DFF40 nuclease is dispensable for high molecular weigh
t DNA cleavageand stage I chromatin condensation in apoptosis”, J Biol Chem. 2
76:45427-45432,2001.
53.Shuey andAttardi, “Characterization of an RNA polymerase activity from HeLa
cellmitochondria, which initiates transcription at the heavy strand rRNA promo
ter and thelight strand promoter in human mitochondrial DNA”, J Biol Chem. 260
:1952-1958,1985.
54.Stephens andRivers, “Antisense oligonucleotide therapy in cancer”, Curr. O
pin. Mol.Therapeut., 5:118-122, 2003
55.Summerton, “Morpholinoantisense oligomers: a case for an Rnase H-independen
t structural type”,Biochim. Biophys. Acta, 1489:141-158, 1999.
56.Suzuki etal., “A serine protease, Htra2, is<released from the mitochondria
and interactswith xiap, inducing cell death”, Mol. Cell, 8:613-621, 2001.
57.Taanman, “The mitochondrial genome: structure, transcription, translation a
nd replication”, Biochim. Biophys. Acta, 1410:103-123, 1999.
58.Tan et al., “Comprehensivescanning of somatic mitochondrial DNA mutations i
n breast cancer”,Cancer Res., 62:972-076, 2002.
59.Teitz etal., “Caspase 8 is deleted or silenced preferentially in childhood
neuroblastomaswith amplification of MYCN”, Nature Med. 6:529-535, 2000.
60.Tidd et al.,“Oligodeoxynucleotide 5mers containing a 5’-CpG induce apoptos
is through a mitochondrialmechanism in T lymphocytic leukemia cells,” Nucleic
Acids Res. ,28:2242-2250 (2000).
61.Tiranti etal., “Identification of the gene encoding the human mitochondrial
RNApolymerase (h-mtRPOL) by cyberscreening of the Expressed Sequence Tags data
base”, Hum MolGenet. 6:615-625, 1997.
62.Verhagen etal., “Cell death regulation by the mammalian IAP antagonist Diab
lo/Smac”,Apoptosis, 7:163-166, 2002.
63.Vickers etal., “Efficient reduction of target RNAs by small interfering RNA
and RnaseH-dependent antisense agents”, J. Biol. Chem., 278:7108-7118, 2003.
64.Villegas etal., “A novel chimeric mitochondrial RNA localized in the nucleu
s of mousesperm”, DNA & Cell Biol. 19:579-588, 2000.
65.Villegas etal., “A putative RNA editing from U to C in a mouse mitochondria
l transcript”,Nucleic Acids Res. 30:1895-1901, 2002.
66.Vogelsteinet al., “Surfing the p53 network”, Nature 408: 307-310, 2000.
67.Wacheck etal., “ Small interfering RNA targeting Bcl-2 sensitizes malignant
melanoma,”Oligonucleotides 13:393-400, 2003.
68.Wagner etal., “Nucleotide sequence of the thymidine kinase gene of herpes s
implex virustype 1”, Proc. Natl. Acad. Sci. U.S.A. 78:1441-1445, 1981.
69.Wahlestedtet al., “Potent and nontoxic antisense oligonucleotides containin
g lockednucleic acids”, Proc. Natl. Acad. Sci. US, 97:5633-5638, 2000.
70.Warburg, “Onthe origin of cancer cells”, Science, 123:309-314, 1956.
71.Wu et al., “Immunofluorescentlabelingof cancer marker Her2 and other cellul
ar targets withsemiconductor quantum dots,” Nature Biotechnol. 21:41-46, 2003.
72.Yamamoto etal., “Identification of a functional promoter in the long termin
al repeat ofRous sarcoma virus”, Cell 22:787-797, 1980
73.Yu et al., “VitaminD receptor expression in human lymphocytes. Signal requi
rements andcharacterization by western blots and DNA sequencing”, J. Biol. Che
m.,266:7588-7595, 1991.
74.Zaug et al.,“A labile phosphodiester bond at the ligationjunctionin a circu
lar interveningsequence RNA”, Science, 224:574-578, 1984.
75.Zornig etal., “Apoptosis regulators and their role in tumorogenesis”, Bioc
him.Biophys.Acta, 1551:F1-F37, 2001.
【化1】
【化2】
【化3】
【化4】
【化5】
【化6】
【化7】
【化8】
【化9】
【化10】
【化11】
【化12】
【化13】
【化14】
【化15】
【化16】
【化17】
【化18】
【化19】
【化20】
【化21】
【化22】
【化23】
【化24】
【化25】
【化26】
【化27】
【化28】
【化29】
【化30】
【化31】
【化32】
【化33】
【化34】
【化35】
【技術分野】
【0001】
1.本発明の技術分野
本発明は、2003年5月21日付けで出願された米国仮特許出願第60/47210
6の利益を主張する。
本発明は、ヒトのミトコンドリアキメラRNAと称される、ヒトミトコンドリアRNA
の新規なファミリーに関連する癌治療法、癌診断及び研究試薬に関する。
特に、本発明は、ヒトミトコンドリアRNAを標的とするオリゴヌクレオチドに関する
。本発明のオリゴヌクレオチドは、癌細胞死を誘発するキメラRNAにハイブリタイズす
る。本発明により提供される組成物と方法は、新規な癌治療として有効である。更に、こ
のオリゴヌクレオチドは、休止期及び増殖期の正常細胞、前癌および癌細胞においてミト
コンドリアキメラRNAの発現が異なることに基づき癌と前癌細胞の診断に用いることが
できる。
【背景技術】
【0002】
2.本発明の背景技術
2.1 ミトコンドリア
ミトコンドリアは、酸化性りん酸化反応により必須分子であるアデノシン三りん酸(A
TP)を製造する細胞内小器官である。1665塩基対のヒトミトコンドリア(mtDN
A)は、2つのリボゾームRNA、22転移RNA(tRNAs)、及び同じ数のポリペ
プチドをエンコードしている13オープンリーデングフレーム(ORF)をエンコードし
ている(Clayton,Hum Reprod. Suppl 2:11-17, 2000; Taanman, Biochim. Biophys. Act
a,1410:103-123, 1999)。
G+T塩基組成物の成分を基に、mtDNAの2つの鎖は、ボイヤント密度(浮遊密度)
が異なり、セシウムクロライド勾配(gradient)を変性して(denaturating)分離される
。重鎖(又はH鎖)は、2つのリボゾームRNAs(12Sと16S)、14tRNAs、並びに、
ND1、ND2、ND3、ND4L、ND4、ND5、COXI、COX II、COX III、ATP6、ATP8及びCyt bに対応
する12ポリペプチドをエンコードしている。
軽鎖(又はL鎖)は、8tRNAs及びコンプレックスNADハイドロゼナーゼNS6
をコードしている(Clayton,Hum Reprod. Suppl 2:11-17, 2000; Taanman, Biochim. Bio
phys. Acta,1410:103-123, 1999)。
【0003】
多くのmtDNAは、置換ループ又はDループと呼ばれる短い3つの鎖構造を含んでいる
。ヒトにおいて1,006の塩基対である、この領域は、フェニルアラニンのtRNA(tRN
APhe)及びプロリンのtRNA(tRNAPro)の遺伝子に接しており、L鎖に相補してH鎖を置
換する短核酸鎖を含んでいる(Clayton, Hum Reprod. Suppl 2:11-17, 2000; Taanman, Bi
ochim. Biophys.Acta, 1410:103-123, 1999)。
この領域は、mtDNA発現のための主なコントロール・サイトとして発展しており、リ
ーデング鎖又は複製のH鎖起点、及びH鎖(HSP)とL鎖(LSP)の転写のための主なプロモー
ターを含んでいる。
HSPとLSP(約150bp)は隣接しているにもかかわらず、これらの調節要素は、ミト
コンドリア疾病を有する患者のモデルを利用するインビボ(体内)(Chinnery and Turnbu
ll, Mol. Med.Today, 6:425432, 2000)と同様に、インビトロ(体外)でも機能的に独立
である(Shueyand Attardi, J Biol Chem. 260:1952-1958, 1985; Taanman, Biochim. Bi
ophys. Acta,1410:103-123, 1999)。
【0004】
双方の鎖は、ポリシトロンRNAsとして転写され、これらはその後個々のmRNA、t
RNA及びrRNAをリリースするためにプロセスされる(Taanman, Biochim. Biophys.
Acta,1410:103-123, 1999)。
ヒトでは、ミトコンドリアのRNAポリメラーゼは、イースト・ミトコンドリアRNA
ポリメラーゼの配列、及びいくつかのバクテリオファージのRNAポリメラーゼと著しい
相同性を有する1,230のアミノ酸の蛋白質である(Tirantiら,Hum Mol Genet. 6:615-
625, 1997)。
更に、転写要因のファミリーは、哺乳類のmtDNA転写として本質的で、DNA結合蛋
白の高移動性グループ(HMG)-boxファミリーのメンバーである、ミトコンドリア転
写因子A又はTFMAなどで特徴づけらている(Parisi and Clayton, Science. 252:965-
969, 1991)。
最近、2つの独立した報告書で、ヒトおよびマウスにおける新規な転写因子TFB1M
とTFB2Mの特徴について記述されている(McCulloch et al., Mol. Cell Biol. 22:11
16-1125, 2002;Falkenberg et al., Nat Genet. 31:289-294, 2002; Rantanen et al.,
Mamm Genome.14:1-6, 2003)。
mtDNA転写に包含されるcis-とtrans-活動因子についてなされた相当の進歩にもかか
わらず、機能の詳細については、十分には理解されていない。
【0005】
2.2 ミトコンドリアおよびアポトーシス
ミトコンドリアは、細胞がよくコントロールされた又はプログラムされた方法で細胞が
死滅する基本的な生物学のプロセスである、アポトーシス(細胞死)における中心的な役
割を果たす。この細胞自殺プログラムは、進化の間、そして後生動物の成熟したホメオス
タシスには本質的である。アポトーシスは、不必要な、損傷した、突然変異した、そして
老化した細胞を撲滅するために活性化される(Meier et al., Nature 407:796-801, 2000)
。アポトーシスの不規則化(disregulation)は、いくつかの病理学の発表で示されてい
る。
このように、アポトーシスの異常な抑制は、腫瘍形成の特徴で、この場合、広範なアポ
トーシスが発作、敗血性ショック及び神経性変性不調(neurodegerative disorders)の
ような急性の疾病と関係づけられている。
現在、アポトーシスのプロセスは、外因性と内因性経路として知られている2つの主な
経路について述べられている(Zornig et al., Biochim.Biophys. Acta, 1551:F1-F37, 20
01)。
外因性経路は、異なるリガンドが死のレセプターに結合することにより、細胞膜で開始
されるプロセスである(Krammer, Nature 407:789-795, 2000; Zornigら, Biochim. Bioph
ys. Acta,1551:F1-f37, 2001)。
【0006】
カスパーゼは、アポトーシスにおいて蛋白分解性カスケードに関与する。カスパーゼは
、蛋白分解性成熟又はアポトーシス誘導プロセッシングを受ける不活性プレカーサーたん
ぱく質として合成される(Zornig et al., Biochim. Biophys. Acta, 1551:F1-F37, 2001)
。
しかしながら、より最近、いくつかの実験証拠は、リソソームプロテアーゼがアポトー
シス性損傷後に蛋白質分解の代替となる経路を構成することを示している(Guicciardi et
al.,Oncogene, 23:2881-2890, 2004)。
他方、ヒトのオンコプロテイン(癌タンパク)Bcl-2に同種のアンチ−アポトーシ
ス性蛋白が記述されている。この蛋白は、アンチ−アポトーシス(Bcl-2、Bcl-XL、Bcl-w
)あるいはプロ−アポトーシス(Bax、Bak、Bim、Bid等)であるたんぱく質のファミリー
に属する(Zorninget al., Biochim. Biophys. Acta, 1551:F1-F37, 2001)。
【0007】
ミトコンドリアは、特にアポトーシス性プロセスの間の初期に影響を受ける、そして現
在、これらは、細胞死の中心的なコーディネーターとして認識されている(Boya et al.,
Biochem.Biophys. Res. Commun. 304:575-581, 2003; Ferri and Kroemer, Nature Cell
Biol.3:E255-E263, 2001; Zornig et al., Biochim. Biophys. Acta, 1551:F1-F37, 20
01)。
いくつかのプロ−アポトーシスシグナルと損傷経路は、Bcl-2蛋白の制御下にある
ミトコンドリア膜浸透、現象を誘発するミトコンドリアに集中する(Boya et al., Bioche
m. Biophys.Res. Commun. 304:575-581, 2003; Zornig et al., Biochim. Biophys. Act
a, 1551:F1-F37,2001)。
細胞がアポトーシス傷害を受けた後に、トランス−膜ポテンシャル(Dym )は、消失して
、外部のミトコンドリア膜の完全な浸透が起こり、そしてその結果、有害なミトコンドリ
ア内部膜蛋白の漏出が起こる。
ミトコンドリア蛋白質の漏出の最初の例は、シトクロムcの放出であった(Liu et al.,
Apoptosis,6:453-462, 2001)。
シトクロムcがサイトゾル中に存在すると、それは、アポプトゾームと名付けられた、
カスパーゼを活性化する錯体の組み立てを促進する。
シトクロムcは、Apaf-1(アポトーシスプロテアーゼ活性因子-1)に結合して、d
ATO/ATPの錯体への結合と、Apaf-1のオリゴマー化を促進する(Adrain et al
., 1999;Benedict et al., 2000)。
Apaf-1のオリゴマー化は、前駆体の蛋白質分解活性化を触媒するプロ-カスパーゼ
-9の補充と活性プロ-カスパーゼ-9の生成を可能にする(Adrainet al., J. Biol. Chem
.274:20855-20860, 1999; Benedict et al., J. Biol. Chem., 275:8461-8468, 2000)。
【0008】
アポトーシス蛋白質の細胞基質抑制剤のファミリーは、記述されており、XIAP、c-
IAP1及びc-IAP2として知られている。これらの蛋白質は、処理されたカスパーゼ
-3及びカスパーゼ-9に結合して抑制し、そしてその結果、分解のカスケードを停止する
。しかし、細胞は、更にこのアンチ−アポトーシス経路をバイパスするための対抗メカニ
ズムを含んでいる。
アポトーシスを被る細胞において、カスパーゼは、Smac(カスパーゼの第2のミト
コンドリアの活性剤)あるいはDIABLO(蛋白質をLowplに結び付けるDirect IAP)と
して知られている蛋白質のIAPに結合することにより、この抑制の影響から解放される
(Verhagen etal., Apoptosis, 7:163-166, 2002)。
IAPに結合することによって、Smac/DIABLOは、IAPから活性カスパー
ゼを置換し、その結果細胞死を促進する。
HtrA2として知られている他の蛋白質は、アポトーシス損傷後に、ミトコンドリア
から細胞基質に開放される、この場合、たんぱく質は、Smac/DIABLOと同じ方
法でIAPに結合する、そしてそれによりカスパーゼ活性を促進する(Verhagen et al.,
Apoptosis,7:163-166, 2002; Martins et al., 2001; Suzuki et al., Mol. Cell, 8:61
3-621, 2001;Hedge et al., Apoptosis, 7:123-132, 2002)。
【0009】
アポトーシス誘発因子(AIF)は、アポトーシスカスケードの他の成分である。
アポトーシスの誘発後、AIFは、細胞基質と核に移動する。核では、AIFが周辺の
クロマチン凝縮およびDNA断片化(fragmentation)を引き起こす。AIFは、末梢の
クロマチル縮合とDNA断片化を誘発する。AIFは、更にDym消失とホスファチジルセ
リン露出のようなアポトーシスのいくつかの特徴を引き起こす(Zornig et al., Biochim.
Biophys. Acta,1551:F1-F37, 2001)。
AIFのアポトーシス活性を規制するように思われる因子は、熱ショック蛋白質70であ
る(Ravagnanet al., Nature Cell Biol. 3:839-843, 2001)。
AIFのように、ミトコンドリアを出て、核に移動する他のミトコンドリア因子は、エ
ンドヌクレアーゼG(endonuclease G)かエンドG(Endo G)である。
核では、エンドGがカスパーゼ抑制剤の存在下でもDNA断片化を発生する(Li et al.,
Nature,412:95-99, 2001)。
エンドGは、CAD(カスパーゼに活性化されたDNAse)のように、その活性化がカス
パーゼに極めて依存しているヌクレアーゼと同様の様式で作用する(Samejima et al., J.
Biol.Chem., 276:45427-45432, 2001)。
【0010】
2.3 癌および前癌
癌は、細胞の悪性疾患で、その独自の特徴、すなわち細胞サイクルの通常の制御を失っ
て不規則に成長し、分化が欠如し、及び他の細胞組織に進入して転移する。発癌は、正常
細胞が悪性細胞に変化することによるプロセスである。
発癌は、開始が遺伝的事象で始まり、その後に初期腺腫の形成が促進される期間に改変
された細胞が選択的に増加する多段階プロセスである。
連続的な発癌補助作用がないと、腺腫は消退して消滅する。
第二の遺伝的事象で、少量の発癌補助作用を受けた腺腫は、後発の腺腫形成が進行して
、その後悪性の変化をたどる(McKinnell et al., “The Biology Basis of Cancer”, Ch
. 3, 1998)。
【0011】
癌の病因は複雑で、細胞サイクル規則の改変、染色体異常、及び染色体破損を含んでい
る。発癌性ウィルス、化学品、放射線(紫外線あるいは電離放射線)、及び免疫障害のよ
うなタイプの感染性因子は、発癌の主な原因であると考えられている(McKinnell et al.,
“TheBiological Basis of Cancer, Ch.3, 1998)。
【0012】
癌もミトコンドリアの機能障害と関係のあることは長い間提案されてきた。
これらの理論のうちの1つは、ミトコンドリアの突然変異が細胞形質転換と癌の主要な
原因であるとする提案である(Warburg, 1956; Carew and Huang, Mol. Cancer, 1:1-12,
2002)。
ミトコンドリアDNA(mtDNA)の変質は、血液作用による腫瘍(Clayton and Vinogra
d, Nature,216:652-657, 1967)、及び乳癌(Tan et al., 2002; Parrella et al., 2001)
において報告されている。
いくつかの部位のmtDNAの突然変異と欠失(deletions)は、また患者において、直
腸癌、前立腺癌、卵巣癌、胃癌、膵臓癌、肝細胞性癌、食道癌、腎臓癌、甲状腺癌および
脳腫瘍で明らかにされてきている(reviewd by Carew and Huang, Mol. Cancer, 1:1-12,
2002)。
一般的に、腫瘍タイプと関係のない癌において、mtDNA変質の2つの主な特徴がある
と推定される。大部分の突然変異は、TからCとGからAへの塩基の転移である。第二に
、突然変異が生ずる特別の遺伝子における多様性はあるが、D−ループは、ほとんどの腺
腫タイプにおいてmtDNAの最も頻繁な体細胞の突然変異領域になると思われる。
【0013】
前癌とは、悪性細胞に進化または分化する形質転換細胞として定義される。いくつかの
例は、DNAまたはRNAのオンコウィルスによって変化した細胞である。
本発明は、ミトコンドリアRNAの新規なファミリー、及び診断法と癌治療を目標とし
てこれらのRNAの使用に関する。
本発明は、組成物と方法を提供する、そしてこのことは、腫瘍細胞又は前悪性細胞もし
くは腫瘍形成ウィルスで変化した細胞と正常細胞とを区別するのに有用である。
特に、以下に詳述するように、本発明は、前癌及び癌細胞と正常細胞とを区別する診断
検査のための組成物と方法を提供する。
本発明の他の態様では、多くの又は選択的な癌細胞死を誘発するための組成物と方法が
提供される。それ故、本発明は、研究のみならず、癌および前癌の診断と治療に用いるこ
とが可能である、組成物及び方法を提供する。
【発明の概要】
【課題を解決するための手段】
【0014】
3.発明の概要
本発明は、正常な休止及び増殖する細胞、前癌細胞及び癌細胞において特異的に発現さ
れる、ミトコンドリアキメラRNAと称される、新規なヒトミトコンドリアRNAのファ
ミリーを検出するのに有用な組成物及び方法を対象とする。
【0015】
3.1 センスミトコンドリアキメラRNA
本発明の1の態様において、16SミトコンドリアリボゾームRNA(SEQ ID NO 1)の
5’末端に共有結合している815ヌクレオチドの逆方向反復配列(inverted repeat)
を含むミトコンドリアキメラRNAを検出するための組成物及び方法が提供される。
この逆方向反復配列は、mtDNAの16S遺伝子のL−鎖から転写されるRNAの81
5ヌクレオチドのフラグメントに相当する。
このように、この新規なRNAの合成は、mtDNAの16SのL―鎖とH−鎖の転写を
必要とする。mtDNAの両鎖の転写は、異なるプロモーターに規制されるので、我々は、
ミトコンドリアキメラRNA(SEQ ID NO 1)として、本発明におけるこの新規なRNAに
ついて言及する。更に、815ヌクレオチドの逆方向反復配列は“センス”16SのRN
A(H−鎖から転写された)に結合されるので、我々は、“センスミトコンドリアキメラ
RNA”として、この新規なRNAについて言及する。
【0016】
本発明は、培養細胞、細胞サンプル及び組織切片(tissue sections)におけるセンス
ミトコンドリアキメラRNAの発現を検出する組成物と方法を提供する。
この検出は、insituハイブリダイゼーション、対応するcDNAの合成とPCRによる
増殖、転写媒介増殖(transcription mediated amplification)(TMA)(Comanor et
al., J. ClinVirol., 28:14-26, 2003)もしくはノーザンブロット、又は当技術分野の技
術者に自明である他の方法により行われる。
【0017】
本発明の1の側面によると、in situ ハイブリダイゼーションアッセイは、センスミト
コンドリアキメラRNAが正常の増殖期の細胞、異なる腫瘍型の人体生検に存在する腫瘍
細胞におけると同様に培養中の腫瘍細胞に発現することを示す。センスミトコンドリアキ
メラRNAは、正常の休止期の細胞には発現しない。
本発明の他の態様によると、パピローマ・ウイルス(乳頭腫ウィルス)16又は18に
より形質転換(transform)した細胞中の第二の新規なセンスミトコンドリアキメラRN
Aを検出する方法と構成が提供される(Hausen, Biochim. Biophys. Acta, 1288:F55-F78,
1996)。
これらの形質転換した細胞において、16SミトコンドリアRNAの5’末端に共有結
合している754ヌクレオチドの逆方向反復配列からなる、新規なセンスミトコンドリア
キメラRNAが発現する(SEQ ID NO 2)。
このRNAは、正常な増殖細胞中又は腫瘍細胞中には存在しない。本方法と組成物は、
また16SミトコンドリアRNA(SEQ ID NO 3)の5’末端に共有結合している694
ヌクレオチドの逆方向反復配列からなる第三のセンスミトコンドリアキメラRNAがHT
LV−1で形質転換した細胞中に存在することを実証する。
【0018】
3.2 アンチセンスのミトコンドリアキメラRNA
本発明は、更に正常な増殖細胞がSEQ ID NO 4、SEQ ID NO 5、SEQ ID NO 6に対応する
アンチセンスミトコンドリアキメラRNAを発現することを示す方法と組成物を提供する
。
これらの転写産物は、16SミトコンドリアリボゾームのRNA(L−鎖から転写され
た)の5’末端に結合している、それ故にアンチセンスのミトコンドリアRNA名である
、可変の長さ(H−鎖から転写された)の逆方向反復配列を含む。
アンチセンスミトコンドリアキメラRNAの発現は、形質転換した又は前癌細胞中のみ
ならず、異なるタイプの腫瘍のヒト生検中に存在する腫瘍細胞中と同様に、増殖中の腫瘍
細胞系中で下方制御(down regulate)される。
従って、本発明は、通常の増殖細胞を癌細胞と前癌細胞から区別するセンスとアンチセ
ンスのミトコンドリアキメラRNAの発現を検出する方法と組成物を提供し、それ故に悪
性細胞と癌のための新規なマーカーを提供する。
【0019】
3.3 癌療法
本発明の他の態様では、方法と組成物は、センスとアンチセンスミトコンドリアキメラ
RNAを阻害(interfere)するために提供される。
1つの好ましい態様は、低コピー数のこの転写産物を含む腫瘍細胞中のアンチセンスの
ミトコンドリアキメラRNAを阻害することである。この阻害は、その配列がアンチセン
スのミトコンドリアキメラRNA(SEQ ID NO 4及び/又はSEQ ID NO 5 及び/又は SEQ I
D NO 6)の配列に相補している、オリゴヌクレオチド又はオリゴヌクレオチド類似体を用
いて行われる。1又はそれ以上のこれらの相補的なオリゴヌクオチドを用いた異なるタイ
プの腫瘍細胞の処理は、細胞死又はアポトーシスを誘発する。
【0020】
これらのオリゴヌクレオチドは、少なくとも15核酸塩基がSEQ ID NO 4及び/又はSEQ
ID NO 5及び/又はSEQID NO 6に相補する15ないし50のヌクレオチドの合成物であ
る。
これらの相補的なオリゴヌクレオチドの例は、SEQ ID NOS 9〜98の中で示される。
ヒトリンパ球(正常な休止細胞)又はフィトヘムアグルチニン(植物性血球凝集素)で
活性化されたヒトリンパ球(正常な増殖細胞)の処理は、アンチセンスミトコンドリアキメ
ラRNAの配列に相補するオリゴヌクレオチドを用いて同条件で処理した後にはアポトー
シスを受けないので、アポトーシスの誘発は、選択的である。
もし、腫瘍細胞がセンスミトコンドリアキメラRNA(SEQ ID NO 1及び/又はSEQ ID
NO 2及び/又はSEQ IDNO 3)を標的又は相補するオリゴヌクレオチドで処理すると、細
胞死又はアポトーシスの誘発が減じられる。
例えば、本願発明は以下の項目を提供する。
(項目1)
アンチセンスミトコンドリアキメラDNAの分離された核酸分子であって、かつSEQ ID
NO 4、SEQID NO 5又はSEQ ID NO 6のヌクレオチド配列を含む核酸分子。
(項目2)
項目1に記載のSEQID NO 4、SEQ ID NO 5又はSEQ ID NO 6のアンチセンスのミトコン
ドリアキメラDNAに相補する、長さで10ないし50核酸塩基の合成物又はオリゴヌク
レオチド。
(項目3)
前記合成物又はオリゴヌクレオチドが、長さ10から50核酸塩基で、SEQ ID NOが9
から98で構成されるグループから選択される配列の少なくとも10核酸塩基部分を含む
、項目2に記載の合成物。
(項目4)
前記合成物又はオリゴヌクレオチドが、長さ10から50までの核酸塩基で、前癌およ
び癌細胞死を誘発する、SEQ ID NOが9から98で構成されるグループから選択される配
列の少なくとも10核酸塩基部を含む、項目2に記載の合成物。
(項目5)
前記オリゴヌクレオチドがSEQ ID NO 4、SEQ ID NO 5又はSEQ ID NO 6のアンチセンス
ミトコンドリアのキメラRNAでハイブリダイズされて、前癌及び癌細胞死を誘発する、
項目2に記載の合成物。
(項目6)
前記オリゴヌクレオチドが少なくとも1つの修飾されたインターヌクレオシド連鎖を含
む、項目2に記載の合成物。
(項目7)
前記修飾されたインターヌクレオシド連鎖がホスフォロチオエート、メチルホスフォネ
ート、ホスフォロジチオエート、ホスフォロトリエステル、ペプチド性核酸、及びロック
された核酸連鎖から構成されるグループから選択された、項目6に記載の合成物。
(項目8)
前記合成物が少なくとも1つの修飾された糖部分を含む、項目2に記載の合成物。
(項目9)
前記修飾された糖部分が2’-Oメトキシエチル、2’-Oメチル及びモルフォリノ基から
構成されるグループから選択される、項目8に記載の合成物。
(項目10)
前記オリゴヌクレオチドが少なくとも1の修飾された核酸塩基を含む、項目2に記載
の合成物。
(項目11)
修飾された核酸塩基が5-メチルシトシンである、項目10に記載の合成物。
(項目12)
前記オリゴヌクレオチドが、ホスフォロチオエート連鎖で結合されたDNA残基を含む
キメラ核酸であり、該DNA残基がホスフォロチオエート連鎖で共に結合された2’-Oメ
トキシエチル又は2’-OメチルRNA残基の少なくとも3つの残基により5’及び3’サ
イドで側面に位置されている、項目2に記載の合成物。
(項目13)
前記合成物がリボザイムである、項目2に記載の合成物。
(項目14)
前記合成物が低分子干渉RNAすなわちsiRNAである、項目2に記載の合成物。
(項目15)
センスミトコンドリアキメラDNAの分離された核酸分子であって、かつSEQ ID NO 1
、SEQID NO 2又はSEQ ID NO 3のヌクレオチド配列を含む、核酸分子。
(項目16)
項目15のSEQID NO 1、SEQ ID NQ.2又はSEQ ID NO 3のセンスミトコンドリアキメ
ラDNAを標的とする、長さ10から50核酸塩基の合成物又はオリゴヌクレオチド。
(項目17)
前記合成物又はオリゴヌクレオチドがSEQ ID NO 99〜197から構成されるグループから
選択された配列の少なくとも10核酸塩基部を含み、長さで50核酸塩基以下である、請
求項16に記載の合成物。
(項目18)
前記合成物又はオリゴヌクレオチドが前癌又は癌細胞死を誘発するSEQ ID NO 99〜197
から構成されるグループから選択された配列の少なくとも10核酸塩基部を含み、長さで
50核酸塩基以下である、項目16に記載の合成物。
(項目19)
前記オリゴヌクレオチドがSEQ ID NO 1、SEQ ID NO 2又はSEQ ID NO 3のセンスミトコ
ンドリアキメラDNAでハイブリダイズされていて、前癌又は癌細胞死を誘発する、請求
項16に記載の合成物。
(項目20)
前記オリゴヌクレオチドが少なくとも1の修飾されたインターヌクレオシド連鎖を含む
、項目16に記載の合成物。
(項目21)
前記修飾インターヌクレオシド連鎖がホスフォロチオエート、メチルホスフォネート、
ホスフォロジチオエート、ホスフォロトリエステル、ペプチド性核酸、及びロックされた
核酸連鎖から構成されるグループから選択された、項目20に記載の合成物。
(項目22)
前記合成物が少なくとも1つの修飾された糖部分を含む、項目16に記載の合成物。
(項目23)
前記修飾された糖部分が2’-Oメトキシエチル、2’-Oメチル及びモルフォリノ基から
構成されるグループから選択されている、項目23に記載の合成物。
(項目24)
前記オリゴヌクレオチドが少なくとも1の修飾された核酸塩基を含む、項目16に記
載の合成物。
(項目25)
前記修飾された核酸塩基が5-メチルシトシンである、項目24に記載の合成物。
(項目26)
前記オリゴヌクレオチドが、ホスフォロチオエート連鎖で結合されたDNA残基を含む
キメラ核酸であり、該DNA残基がホスフォロチオエート連鎖で結合された2’-Oメトキ
シエチル又は2’-OメチルRNA残基の少なくとも3つの残基により5’及び3’サイド
で側面に位置されている、項目16に記載の合成物。
(項目27)
前記合成物がリボザイムである、項目16に記載の合成物。
(項目28)
前記合成物が低分子干渉RNA又はsiRNAである、項目16に記載の合成物。
(項目29)
SEQID NO 4、SEQ ID NO 5もしくはSEQ ID NO 6のアンチセンスミトコンドリアキメラ
RNA及び/又はSEQ ID NO 1、SEQ ID NO 2もしくはSEQ ID NO 3のセンスミトコンドリ
アキメラRNAに相補する、長さ10から50核酸塩基の1以上のオリゴヌクレオチド、
並びに薬剤の受け入れ可能なキャリアー又は希釈剤を含む、薬剤組成物。
(項目30)
前記キャリアー又は希釈剤が乳剤、リポソーム、浸透促進剤、界面活性剤、キレート試
薬、キャリアーを含む、項目29に記載の薬剤組成物。
(項目31)
経口、口腔、非経口もしくは直腸投与、又は吸入もしくは通気による肺投与用の、請求
項29に記載の薬剤組成物。
(項目32)
SEQID NO 4、SEQ ID NO 5もしくはSEQ ID NO 6のアンチセンスミトコンドリアキメラ
RNA及び/又はSEQ ID NO 1、SEQ ID NO 2もしくはSEQ ID NO 3のセンスミトコンドリ
アキメラRNAに相補する、長さ10から50核酸塩基の合成物で、癌もしくは前癌細胞
又は組織と接触させることを含む、細胞又は組織中における前癌又は癌細胞死の誘発方法
。
(項目33)
SEQID NO 4、SEQ ID NO 5もしくはSEQ ID NO 6のアンチセンスミトコンドリアキメラ
RNA及び/又はSEQ ID NO 1、SEQ ID NO 2もしくはSEQ ID NO 3のセンスミトコンドリ
アキメラRNAに相補する、長さ10から50核酸塩基の2以上の合成物で、細胞又は組
織と接触させることを含む、細胞又は組織中における前癌又は癌細胞死の誘発方法。
(項目34)
前癌又は癌細胞死を誘発することが可能である、SEQ ID NO 4、SEQ ID NO 5もしくはSE
Q ID NO 6のアンチセンスミトコンドリアキメラRNA及び/又はSEQID NO 1、SEQ ID N
O 2もしくはSEQID NO 3のセンスミトコンドリアキメラRNAに相補する、長さ10から
50核酸塩基の合成物の治療に有効な量をヒト又は動物に投与することを含む、前癌又は
癌を有する疑いのあるヒト又は動物の治療方法。
(項目35)
SEQID NO 4、SEQ ID NO 5もしくはSEQ ID NO 6のアンチセンスミトコンドリアキメラ
RNA及び/又はSEQ ID NO 1、SEQ ID NO 2もしくはSEQ ID NO 3のセンスミトコンドリ
アキメラRNAに相補する、長さ10から50核酸塩基の2以上の合成物の治療に有効な
量をヒト又は動物に投与することを含む、前癌又は癌細胞死の誘発することが可能である
、前癌又は癌を有する疑いのあるヒト又は動物の治療方法。
(項目36)
前記合成物又は複数の合成物が癌の治療に対して化学療法薬と併用して投与される、請
求項34に記載の方法
(項目37)
癌の化学療法薬治療の効能改善方法であって、前癌又は癌細胞死を誘発する、SEQ ID N
O 4、SEQ IDNO 5もしくはSEQ ID NO 6のアンチセンスミトコンドリアキメラRNA及び
/又はSEQID NO 1、SEQ ID NO 2もしくはSEQ ID NO 3のセンスミトコンドリアキメラR
NAに相補する、長さ10から50核酸塩基の合成物又は複数の合成物を化学療法薬治療
と併用して投与することを含む、改善方法。
(項目38)
前記合成物又は複数の合成物が少なくとも1つの修飾されたインターヌクレオシド連鎖
を含む、オリゴヌクレオチドである項目32に記載の方法。
(項目39)
前記修飾されたインターヌクレオシド連鎖がホスフォロチオエート、メチルホスフォネ
ート、ホスフォロジチオエート、ホスフォロトリエステル、ペプチド性核酸、及びロック
された核酸連鎖から構成されるグループから選択された、項目38に記載の方法。
(項目40)
前記合成物又は複数の合成物が少なくとも1つの修飾された糖部分を含む、項目32
に記載の方法。
(項目41)
前記修飾された糖部分が2’-Oメトキシエチル、2’-Oメチル及びモルフォリノ基から
構成されるグループから選択される、項目40に記載の方法。
(項目42)
哺乳類の細胞において、SEQ ID NO 4、SEQ ID NO 5、SEQ ID NO 6のアンチセンスミト
コンドリアキメラRNA及びSEQ ID NO 1、SEQ ID NO 2、SEQ ID NO 3のセンスミトコン
ドリアキメラRNAの存在又は不存在を決定することによる正常、前癌又は癌細胞を識別
するハイブリダイゼーション法であって、該方法は、下記a)ないしc)を含むハイブリ
ダイゼーション法。
a)サンプルを、少なくとも1つの検出可能なプローブが標的ミトコンドリアキメラRN
A配列にハイブリダイズするプローブとして、SEQ ID NO 4、SEQ ID NO 5もしくはSEQ ID
NO 6のアンチセンスミトコンドリアキメラRNA及び/又はSEQID NO 1、SEQ ID NO 2
もしくはSEQID NO 3のセンスミトコンドリアキメラRNAに相補する長さ10から50
核酸塩基の2又はそれ以上の独立して検出可能な又は標識された合成物と接触させる
b)検出可能な合成物の標的配列へのハイブリダイゼーションの検出、同定、又は定量
c)正常、前癌又は癌細胞における、センス及びアンチセンスミトコンドリアキメラRN
Aの異なる発現のパターンで得られた結果の相互の関連付けと比較
(項目43)
個々に独立して検出可能なプローブがオリゴヌクレオチドである、項目42に記載の
方法。
(項目44)
前記プローブがDNA及び/又はRNAの10から50残基のオリゴヌクレオチドを含
む、項目42に記載の方法。
(項目45)
前記プローブがSEQ ID NO 1、SEQ ID NO 2、SEQ ID NO 3、SEQ ID NO 4、SEQ ID NO 5
あるいはSEQID NO 6のRNAの領域で少なくとも95%がホモロジーである10から5
0残基のオリゴヌクレオチドを含む、項目44に記載の方法。
(項目46)
前記オリゴヌクレオチドが少なくとも1つの修飾されたインターヌクレオシド連鎖を含
む項目45に記載の方法。
(項目47)
前記修飾されたインターヌクレオシド連鎖がホスフォロチオエート、メチルホスフォネ
ート、ホスフォロジチオエート、ホスフォロトリエステル、ペプチド性核酸、及びロック
された核酸連鎖から構成されるグループから選択されている、項目46に記載の方法。
(項目48)
前記オリゴヌクレオチドが少なくとも1つの修飾された糖部分を含む、項目48に記
載の方法。
(項目49)
前記修飾された糖部分が2’-Oメトキシエチル、2’-Oメチル及びモルフォリノ基から
構成されるグループから選択されている、項目48に記載の方法。
(項目50)
前記オリゴヌクレオチドが放射能、又はジゴキシゲニン、ビオチン、蛍光色素のグルー
プから選択されたハプテンで標識されている、項目45に記載の方法。
(項目51)
前記検出方法がアンチビオチン抗体、アンチジゴキシゲニン抗体及びアンチフルオレセ
イン抗体を含む、項目42に記載の方法。
(項目52)
前記抗体が抗体とアルカリホスファターゼ又はペルオキシダーゼと接合している、請求
項51に記載の方法。
(項目53)
前記検出方法がストレプトアビジン-アルカリホスファターゼ接合と増殖試薬を含む、
項目42に記載の方法。
(項目54)
センスミトコンドリアRNAを標的にする前記オリゴヌクレオチドが蛍光色素で標識さ
れており、またアンチセンスミトコンドリアRNAを標的にする前記アンチセンスミトコ
ンドリアRNAが異なる発光の蛍光色素で標識されている、項目42に記載の方法。
(項目55)
前記検出方法が異なる蛍光色素の発光の検出を含み、蛍光顕微鏡もしくは共焦点顕微鏡
、レーザースキャナ装置、又は流量血球計算器で検出される、項目54に記載の方法。
(項目56)
前記サンプルが正常な休止又は増殖細胞、培養下の前癌及び癌細胞のサンプルを含む、
項目42に記載の方法。
(項目57)
前記サンプルが癌を有する疑いのある患者からの細胞を含む、項目42に記載の方法
。
(項目58)
前記サンプルが固定された又は冷凍のヒトの生体からの組織切片である、項目42に
記載の方法。
(項目59)
前記サンプルが尿、肺洗浄、唾液、コロノサイト、血液塗抹標本、骨髄の塗抹標本、腹
水、血液、又はリンパ液循環転移細胞中に存在する細胞を含む、項目42に記載の方法
。
(項目60)
項目42の方法による、正常なヒト増殖細胞及び形質転換又は悪性ヒト細胞を識別す
るのに有用なinsituハイブリダイゼーション用キットであって、下記A)ないしd)を
含むキット。
a)センス及び/又はアンチセンスミトコンドリアキメラRNAを標的にする1又はそれ
以上の検出可能な、標識されたオリゴヌクレオチド
b)標識検出のために接合された抗体試薬
c)ハイブリダイゼーションと洗浄緩衝液
d)固定されたヒト正常増殖細胞の1又はそれ以上のスライド、及び対照として、固定さ
れ形質転換された又は悪性ヒト細胞の1又はそれ以上のスライド
(項目61)
前記オリゴヌクレオチドの標識がジゴキシゲニン、バイオチン、フルオレセインのグル
ープから選択されたハプテンを含む、項目60のキット。
(項目62)
前記検出方法がアンチビオチン抗体、アンチジゴキシゲニン抗体及びアンチフルオレセ
イン抗体を含む項目60に記載のキット。
(項目63)
前記抗体がアルカリホスファターゼ又はペルオキシダーゼと接合した抗体である、請求
項60に記載のキット。
(項目64)
前記検出方法が抗ビオチン抗体、抗ジゴキシゲニン抗体又は抗フルオレセイン抗体に対
する抗体と接合したアルカリホスファターゼ又はペルオキシダーゼ、及び増殖試薬を含む
、項目60に記載のキット。
(項目65)
前記検出方法がストレプトアビジン−アルカリホスファターゼ接合及び増殖試薬を含む
、項目60に記載のキット。
(項目66)
前記増殖試薬がアルカリホスファターゼニトロブルーテトラゾリウム用基質又はペルオ
キシダーゼ用基質である、項目60に記載のキット。
(項目67)
サンプルの中にある細胞質、細胞核、及び細胞の核小体中のアンチセンス(SEQ ID NO
4、5及び6)又はセンス(SEQID NO 1、2及び3)ミトコンドリアキメラRNAの細胞局在
化を決定するinsituハイブリダイゼーション法であって、下記a)とb)を含むin situ
ハイブリダイゼーション法。
a)サンプルを、少なくとも1つの検出可能なプローブが標識ミトコンドリアキメラRN
A配列にハイブリタイズするプローブとして、SEQ ID NO 4、SEQ ID NO 5もしくはSEQ ID
NO 6のアンチセンスミトコンドリアキメラRNA及び/又はSEQID NO 1、SEQ ID NO 2
もしくはSEQID NO 3のセンスミトコンドリアキメラRNAに相補する長さ10から50
核酸塩基の1又はそれ以上の独立して検出可能な合成物又は標識された合成物と接触させ
る
b)検出可能な合成物の標的配列へのハイブリダイゼーションの検出、同定、又は定量
(項目68)
項目67の方法によるサンプルの中に存在する細胞質、細胞核、及び細胞の核小体中
のアンチセンス(SEQ ID NO 4,5及び6)又はセンス(SEQ ID NO 1、2及び3)ミトコンド
リアキメラRNAの細胞局在化を決定するin situハイブリダイゼーション用キットであ
って、下記a)ないしd)を含むキット。
a)センス又はアンチセンスミトコンドリアキメラRNAを標的にする、標識された1又
はそれ以上の検出可能なオリゴヌクレオチド
b)標識検出のために接合された抗体試薬
c)ハイブリダイゼーションと洗浄緩衝液
d)固定されたヒト正常増殖細胞の1又はそれ以上のスライド、及び対照として固定され
形質転換された又は悪性のヒト細胞の1又はそれ以上のスライド
(項目69)
前記センス及びアンチセンスミトコンドリアキメラRNAの検出、同定及び定量が以下
の手順のいずれかにより行われる、項目42に記載の方法。
insituハイブリダイゼーション又は蛍光in situハイブリダイゼーション、ドットブロ
ット、ノーザンブロット及びPCRによるcDNA増殖又はTMAによるRNA増殖
(項目70)
前記合成物又は複数の合成物が少なくとも1の修飾されたインターヌクレオシド連鎖を
含むオリゴヌクレオチドである、項目33に記載の方法。
(項目71)
前記合成物又は複数の合成物が少なくとも1の修飾されたインターヌクレオシド連鎖を
含むオリゴヌクレオチドである、項目34に記載の方法。
(項目72)
前記合成物又は複数の合成物が少なくとも1の修飾されたインターヌクレオシド連鎖を
含むオリゴヌクレオチドである、項目35に記載の方法。
(項目73)
前記合成物又は複数の合成物が少なくとも1つの修飾された糖部分を含む、項目33
に記載の方法。
(項目74)
前記合成物又は複数の合成物が少なくとも1つの修飾された糖部分を含む、項目34
に記載の方法。
(項目75)
前記合成物又は複数の合成物が少なくとも1つの修飾された糖部分を含む、項目35
に記載の方法。
(項目76)
前記合成物又は複数の合成物が少なくとも1つの修飾された糖部分を含む、項目36
に記載の方法。
【図面の簡単な説明】
【0021】
【図1】図1A、B及びCは、SEQ ID NO 1, SEQ ID NO 2及びSEQ ID NO 3に相当するセンスミトコンドリアキメラRNAの構造を示す線画である。 矢印は、ピースによりRNAを増殖するのに用いるプライマーの相対的位置を示す。 線の下部の矢印は、リバースプライマーで、線の先頭の矢は、フォワードプライマーである。プライマー1は、16Sミトコンドリアの5’末端に隣接して位置付けされる。黒線は、センス16Sミトコンドリアに相当する、一方、点線は、アンチセンス16Sミトコンドリアに相当する。
【0022】
【図2】図2は、図1Aに示したプライマー1とプライマー3の間で得られた増殖産物を示すアガロースゲル電気泳動である。 増殖は、鋳型としてSiHa(癌細胞)、HPV16(HFK698)で形質転換されたヒト角化細胞、あるいはHTLV-1で形質転換されたBリンパ細胞を用いて、PT−CRにより実行された。SiHa細胞からのRNAで、210bpの1つの単一アンプリコンだけが得られ、SEQID NO 1のセグメントに相当する。HPV16で形質転換されたヒト角化細胞の全RNAにおいて、210bpのアンプリコンに加えて、150bpの2番目のアンプリコンが得られた、SEQ ID NO 2に相当する。 HTLV-1で形質転換された細胞からのRNAで、210bp及び150bpのアンプリコンに加えて、3番目のアンプリコンが得られ、SEQ ID NO 3に相当する。
【0023】
【図3】図3は、SEQ ID NO 4、SEQ ID NO 5及びSEQ ID NO 6に相当するアンチセンスミトコンドリアキメラRNAの構造を示す線画である。矢印は、増殖に用いられたプライマーを示す。プライマー1は、アンチセンスの16SミトコンドリアRNAの5’末端に隣接して位置付けされる。 これらの転写産物の配列を得るための方策は、図1に記載されたのと同様である。
【0024】
【図4A】図4A及び図4Bは、培養液中でいくつかの腫瘍細胞を用いて行われたin situハイブリダイゼーション分析を示す。 細胞は、センスミトコンドリアキメラRNAに相補するオリゴヌクレオチドプローブでハイブリダイズされ、そしてジゴキシゲニンでラベル表示された(左パネル)。 更に、細胞は、ジゴキシゲニンでラベル表示されたアンチセンスミトコンドリアキメラRNAに相補するオリゴヌクレオチドプローブで並行してハイブリダイズされた(右パネル)。細胞系は左側で識別される。
【図4B】図4A及び図4Bは、培養液中でいくつかの腫瘍細胞を用いて行われたin situハイブリダイゼーション分析を示す。 細胞は、センスミトコンドリアキメラRNAに相補するオリゴヌクレオチドプローブでハイブリダイズされ、そしてジゴキシゲニンでラベル表示された(左パネル)。 更に、細胞は、ジゴキシゲニンでラベル表示されたアンチセンスミトコンドリアキメラRNAに相補するオリゴヌクレオチドプローブで並行してハイブリダイズされた(右パネル)。細胞系は左側で識別される。
【0025】
【図5A】図5Aは、異なる腫瘍細胞タイプに相当するヒトの生検のいくつかのセクションのin situハイブリダイゼーションである。 腫瘍セクションは、センスミトコンドリアキメラRNAに相補するオリゴヌクレオチドプローブでハイブリダイズされ、ジゴキシゲニンでラベル表示された(左パネル)。 更に、並列の腫瘍セクションは、ジゴキシゲニンでラベル表示されたアンチセンスセンスミトコンドリアキメラRNAに相補するオリゴヌクレオチドプローブでハイブリダイズされた(右パネル)。
【図5B】図5Bは、ジゴキシゲニンでラベル表示されたセンスミトコンドリアキメラRNAに相補するオリゴヌクレオチドプローブを用いて行われた、異なるヒト腫瘍細胞のin situハイブリダイゼーションである。
【0026】
【図6】図6は、正常増殖細胞のin situハイブリダイゼーションである。 サンプルは、センス又はアンチセンスミトコンドリアキメラRNAを標的にしたプローブでハイブリダイズし、ジゴキシゲニンでラベル表示された。 強いハイブリダイゼーションシグナルは、双方のプローブで得られた、1つはセンスミトコンドリアキメラRNAに相補し(左パネル)、同様に他はアンチセンスミトコンドリアキメラRNAに相補する(右パネル)。 組織あるいは細胞は、左側で識別される。
【0027】
【図7】図7は、ミトゲンPHAで増殖するために刺激されたヒトリンパ細胞における発現の変化を示すための免疫細胞化学及びin situハイブリダイゼーションである。 PHAで刺激48時間後、リンパ細胞は、抗原Ki-67及びPCNAを表現する。 これらの抗原は、コントロール中又は休止リンパ細胞中では発現されない(左パネル)。前記in situハイブリダイゼーションは、センス及びアンチセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオチドで行ない、ジゴキシゲニンでラベル表示された。刺激を受けたリンパ細胞は、アンチセンスミトコンドリアキメラRNAだけでなくセンスミトコンドリアキメラRNAを過剰発現した(右パネル)。
【0028】
【図8】図8は、核小体中でセンスミトコンドリアキメラRNAの局在化を示す腫瘍細胞のinsituハイブリダイゼーションである。 細胞あるいは腫瘍セクションは、左側で示される。
【0029】
【図9】図9は、アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオチドプローブで処理された腫瘍HL-60細胞に生じる変化示す蛍光顕微鏡検査である。A、B、C及びDは、活性化されたカスパーゼに高い親和性で結合する化合物(VAD-fmok)で染色されることを示す。この化合物は、フルオレスセインで標識される。アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオチドプローブは、(B,Cと対比して)同じ方法でも薬剤スタウロスポリンよりも高いカスパーゼ活性を誘発する。 活性化されたカスパーゼは、対照として、コントロール未処理細胞(A)又は12SミトコンドリアRNAを標的にするオリゴヌクレオチドプローブで処理された細胞(D)中では検出されなかった。EとFは、DAPIでHL-60細胞の染色を示す。 コントロール細胞(未処理)は、核の均質な染色を示す(E)、一方、アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオチドプローブで処理された細胞は、広範な核の断片化を示す(F)。
【0030】
【図10】図10は、休止及び増殖リンパ細胞の異なる処理条件後のアポトーシスを起こした細胞の割合を示す。 アポトーシスは、DAPI染色により休止中の細胞又はPHAに刺激されたリンパ細胞で測定された。バー1及び2は、未処理細胞に相当する。コントロール(1)又はPHAに刺激されたリンパ細胞(2)の低い自然のアポトーシスが観察された。同様の低いレベルのアポトーシスは、休止中のリンパ細胞又はアンチセンスミトコンドリアキメラRNAを標的にする15uMのオリゴヌクレオチドプローブで15時間処理されたPHA刺激リンパ細胞で観察された、アポトーシスは正常細胞中では誘発されないことを示した。対照として、休止中の細胞とPHAに刺激されたリンパ細胞は、スタウロスポリンで処理された。これらの条件下では、休止中のリンパ細胞(5)あるいはPHAに刺激されたリンパ細胞(6)の約90%がアポトーシスをこうむる。
【発明を実施するための形態】
【0031】
4.発明の詳細な説明
4.1 ヒトミトコンドリアキメラRNAファミリー
本発明は、ヒトの細胞がヒトのミトコンドリアキメラRNAと称される、新規なミトコ
ンドリアRNAのファミリーを発現するという驚くべき発見に基づくものである。
【0032】
これらの転写産物に1つは、センスミトコンドリアキメラRNAと名付けられた、ミト
コンドリアの16SリボゾームRNAの5’末端に共有結合している815ヌクレオチド
の長い逆方向反復配列が含まれる。この長い逆方向反復配列は、長い二重鎖幹と50ヌク
レオチドを形成する、位置51から866の16SリボゾームRNAに十分に相補してい
る。
図1Aに示すように、815塩基対のステムは、逆方向転写酵素(reverse transcript
ase)が相当するcDNAを合成するのに重要な問題を示す。従って、新しい戦略は、図
1Aに例示するRT−PCRによりこのRNAを増殖するのに用いられる。
各重なり合うフラグメントの配列を得た後に、それらは、SEQ ID No1(図1A)に示すセ
ンスミトコンドリアキメラRNAの完全な配列を得るためにコンティグ(contigs)とし
て組み立てられる。
【0033】
この発明の他の側面は、癌遺伝子のヒトの乳頭腫ウィルス(パピローマ・ウイルス)1
6又は18で形質転換した細胞中に発現する他の新規なセンスミトコンドリアキメラRN
Aの発見である。
人間の包皮角化細胞(Human foreskin keratinocytes)(HFK)は、HPV16又は18
により感染した(Hausen,Biochim. Biophys. Acta, 1288:F55-F78, 1996)。
この感染は、HFKの形質転換又は不死化を引き起こす。
しかしながら、これらの細胞は、関連するSiHa細胞(HPV16に感染した)又は
HeLa細胞(HPV18に感染した)のような発癌性ではない。
これらの細胞は、SiHaとHeLa細胞に類似するセンスミトコンドリアキメラRN
A(SEQ ID NO1)を発現する。
しかしながら、形質転換した細胞は、更に16SリボゾームRNA(図1B)(SEQ ID No
2)に結合する754ヌクレオチドの逆方向反復配列を含む、他の第二のセンスミトコン
ドリアキメラRNAを発現する。この新しいセンスミトコンドリアキメラRNAは、正常
なヒトの細胞(HFK)中もしくは発癌性細胞(SiHa又はHeLa細胞)中では下方制御される
か、又は発現しない。
【0034】
この発明の別の態様で、我々は、HTLV-1(Kobayashi et al., EMBO J., 3:1339-13
43, 1984)で形質転換した細胞中で第三のセンスミトコンドリアキメラRNAの発現を決
定した。
HTLV-1に感染したMT-2細胞は、センスミトコンドリアキメラRNA(SEQID NO
1)とHPV16あるいは18(SEQID NO 2)で形質転換された細胞中に発現するセンスミ
トコンドリアキメラRNAを発現する。
これらの転写産物に加えて、HTLV-1で感染された細胞は、16SリボゾームRN
Aの5’末端に結合する694ヌクレオチドの逆方向反復配列を含む、第三のセンスミト
コンドリアキメラRNAを発現する。
この新規なRNA(図1C)(SEQID NO 3)は、正常な増殖する細胞の中で、癌細胞の中
で、あるいはHPV16あるいは18で形質転換されたHFKの中では発現されない。
【0035】
前節に記載したヒトの包皮角化細胞(forskin keratinocytes)(HFK)のような正常の
増殖する細胞は、センスミトコンドリアキメラRNA(図6) (SEQ ID NO 1)を過剰発現(
over express)する。
フィトヘムアグルチニン(phytohaemagglutinin)(PHA)のようなミトゲンで活性化され
たヒトのリンパ細胞は、細胞周期のSフェーズに入って、DNAの合成を始める(Yu et a
l., J. Biol.Chem., 266:7588-7595, 1991)。
増殖する細胞として、リンパ細胞は、Ki-67のような増殖に関連して、細胞核抗原
またはPCNAを増殖する抗原を発現する(Bantis et al., Cytopathology, 15:25-31, 2
004)。
脾臓の胚の中心にあるリンパ細胞、精原細胞、及び胚の細胞のような他の増殖する細胞
は、またセンスミトコンドリアキメラRNA(SEQ ID NO 1)(図4)を過剰発現する。対照的
に、活性化されないリンパ細胞、又は筋細胞のような非増殖細胞は、センスミトコンドリ
アキメラRNA(図7)を発現しない。
【0036】
本発明の他の態様で、腫瘍細胞と正常に増殖する細胞とを区別する方法が提供される。
以前に記述したように、腫瘍細胞と正常に増殖する細胞は、SEQ ID NO 1に記述したセン
スミトコンドリアキメラRNAを過剰発現する。
更に、HPVおよびHTLV-1で感染された特別の状況において、付加的なキメラR
NAが見出される(SEQ ID NO 2及びSEQ ID NO 3)。
しかしながら、本発明は、アンチセンスミトコンドリアキメラRNAを過剰発現すると
いう驚くべき発見に基づくものである。
アンチセンスミトコンドリアキメラRNAの発現は、PHA(図7)で活性化されたヒト
のリンパ細胞中で、正常なHFKで及び他の正常な増殖する細胞(図6)中で確認された。
この発明の他の驚くべき発見は、正常に増殖する細胞と異なり、腫瘍細胞がアンチセン
スミトコンドリアキメラRNAを発現しないか、又はその生産を下方制御するということ
である(図6及び図7と図4を比較する)。
【0037】
RT-PCRによってオーバーラップしているフラグメントに基づくキメラRNAを増
殖するために同様の方策を用いて、アンチセンスミトコンドリアキメラRNAの構造が決
定された(図3)。
コンティグにおけるその配列と組み立ては、アンチセンスの16Sミトコンドリアリボ
ゾームRNA(図3A、B及びC)(SEQID NO 4、SEQ ID NO 5、SEQ ID NO 6)の5’末端
に結合している異なる長さの逆方向反復配列を含むアンチセンスミトコンドリアキメラR
NAの複雑なファミリーを明らかにする。
その配列は、またこれらのRNAにおける二重鎖構造又はステムの生成、及び7、96
及び451ヌクレオチド(図3A、B及びC、SEQ ID NO 4、SEQ ID NO 5、SEQ ID NO 6)
をそれぞれ有するループの生成を明らかにする。
【0038】
本発明の他の態様では、腫瘍ウィルスによって細胞の発癌性形質転換を生じさせる方法
と組成物が提供される。HeLa細胞(HPV 18に感染した)又はSiHa細胞(HPV 16に感
染した)は、センスミトコンドリアキメラRNAを過剰発現する、しかしアンチセンスミ
トコンドリアキメラRNAの発現を下方制御する。他方、正常な増殖細胞であるHFKは
、アンチセンスミトコンドリアキメラRNAと同様にセンスミトコンドリアキメラRNA
の双方を過剰発現する。
HPV16あるいはHPV18でHFKの形質転換の後に、細胞は、腫瘍表現型を得、
これらはセンスミトコンドリアキメラRNAを過剰発現し、そしてアンチセンスミトコン
ドリアキメラRNAの発現を下方制御する。センスミトコンドリアキメラRNAの過剰発
現及びアンチセンスミトコンドリアキメラRNAの発現の下方制御は、in situハイブリ
ダイゼーション、RT-PCRによるRNAの増殖、又は当該分野の技術者によってよく
知られた方法によるRNAを決定する他の方法を用いることにより決定される。これらの
方法と組成物は、他の腫瘍ウィルスで又は形質転換又は発癌性を誘発する複合物により、
キメラRNA細胞ファミリーの発現における変化を決定するのに用いられる(McKinnell e
t al., “Thebiological basis of Cancer, Cambridge University Press1998)。
【0039】
4.2 癌および前癌診断
本発明により、方法と組成物は、生体サンプル中にセンスミトコンドリアキメラRNA
とアンチセンスミトコンドリアキメラRNAの存在を検出するために提供される。
1つの好ましい態様では、検出は、insituハイブリダイゼーションにより行われる。
生体サンプルの細胞中のセンスミトコンドリアキメラRNAとアンチセンスミトコンド
リアキメラRNAの検出は、細胞が正常な増殖中の細胞であることを示す。
他の態様において、腫瘍細胞によるin situハイブリダイゼーションの結果は、センス
ミトコンドリアキメラRNAの発現と、アンチセンスミトコンドリアキメラRNAの下方
制御又は不存在を示す。
もし、生体サンプルが非増殖の正常細胞を含む場合、in situハイブリダイゼーション
は、センスミトコンドリアキメラRNAとアンチセンスミトコンドリアキメラRNAのい
ずれも発現しないことを示す。
【0040】
生体サンプルは、培養中の又は血液スミア(smears)もしくは骨髄スミア中の正常細胞
(休止又は増殖する細胞)、培養中の腫瘍細胞、及び腫瘍ウィルスで形質転換された正常
細胞として理解される。
更に、生体細胞は、膀胱もしくは腎臓癌を有する疑いのある患者からの尿もしくは膀胱
洗浄から得られた細胞、又は頭と首の癌を有する疑いのある患者の唾液からの細胞、肺癌
を有する疑いのある患者の管支肺胞からの細胞を含む。また、生体サンプルは、白血病を
有する疑いのある患者の血液からの細胞スミア、又は転移を有する疑いのある患者の血液
、リンパ液、リンパ節からの細胞スミアを含む。
【0041】
本発明による生体サンプルは、当分野の技術者に良く知られた技術である、組織病理学
の分析のため急速冷凍した組織あるいは細胞サンプルの使用を含む。
あるいは、生体サンプルは、当分野でよく知られた多種多様の固定プロトコル(Frederi
ck et al,Current Protocols In Molecular Biology, Volume 2, Unit 14, Frederick M
. Ausubul etal. edS., 1995; Celis, Cell Biology, A Laboratoty Handbook, Julio E
. Celis, ed.,1994)により行うことができる化学処理の使用により固定される薄片の生
体検査になりうる。
生体サンプルは、また化学的修飾、ホルマリン処理又は当該分野でよく知られた他の固
定がなされていない非固定の生物由来物質であってもよい。
【0042】
あるいは、insituハイブリダイゼーションは、パラフィン又は他の包埋ができるポリ
マーのような物質に包埋された生体サンプルの使用により行うことができる。
包埋により得られたブロックは、ミクロトームで約4ないし10μmの厚みの切片にす
る。この切片は、ポリリシン又はムッセル(mussel)接着性蛋白質のような技術分野で知
られた接着性物質で被覆されてガラス又はポラスチックスライドに乗せられる(Burzio et
al.,Curr. Opin. Biotechnol., 8:309-312, 1997)。
【0043】
in situハイブリダイゼーションは、この分野の技術者によく知られた方法で行われる
。例えば、細胞中でセンスミトコンドリアキメラRNA(SEQ ID NO 1, SEQ ID NO 2, SE
Q ID NO 3)、又はアンチセンスミトコンドリアキメラRNA(SEQID NO 4, SEQ ID NO 5
, SEQ ID NO 6)の1又は2以上の配列を標的にした1又は2以上の標識されたプローブか
らなるハイブリダイゼーション溶液は、ハイブリダイゼーション条件下で細胞と接触され
る。そのハイブリダイゼーションシグナルは、それから正常の又は制御された癌及び前癌
細胞から予め決定されたハイブリダイゼーションパターンと比較される。
【0044】
ここに使用されているように、in situハイブリダイゼーションを行うために標識され
たプローブは、RNA、DNA、又は当該分野で知られている任意の方法により製造され
うる合成核酸である。
合成核酸は、インビトロで転写されたリボプローブ又はPCRフラグメントを含む。
この発明の好ましい態様において、合成された相補的なオリゴヌクレオチドが用いられ
る。相補的なオリゴヌクレオチドプローブは、少なくとも長さ約10のヌクレオチド、よ
り好ましくは少なくとも約14、特に好ましくは少なくとも長さ18のヌクレオチドであ
る。当業者は、長さが10ヌクレオチドから、センスミトコンドリアキメラRNA又はア
ンチセンスミトコンドリアキメラRNAにハイブリダイゼーションが可能である長さにま
で拡張できることを理解する。ここでの好ましい態様において、長さは、約30ヌクレオ
チド、より好ましくは約25ヌクレオチド、そしと最も好ましくは10から50ヌクレオ
チドである。
より長いプロービング核酸もまた使用できる。プローブの配列は、少なくとも95%が
SEQ ID NO 1、SEQID NO 2、SEQ ID NO 3、SEQ ID NO 4、SEQ ID NO 5及びSEQ ID NO 6に
リストされた配列に同種である。
【0045】
本発明の相補的なオリゴヌクレオチドプローブは、一般にリン酸ジエステル(phosphod
iester)結合を含む、しかし、ある場合には、限定されるものではないが、ホスフォロチ
オエート(phosphorothioate)(Mag et al., Nucleic Acids Res. 19:1437-1441, 1991;
及び米国特許番号5,644,048),ペプチド性核酸(peptide nucleic acid)又はPNA(Eg
holm, Nature,365:566-568, 1993; 及び米国特許番号6,656,687), phosphoramide (Beau
cage, MethodsMol. Biol. 20:33-61, 1993), ホスフォアミド(phosphoramide) (Beauc
age, MethodsMol. Biol. 20:33-61, 1993), ホスフォロジチオエート(phosphorodithio
ate)(Capaldi etal., Nucleic Acids Res., 28:E40, 2000)からなるインターヌクレオ
シド連鎖を交互に含むオリゴヌクレオチドプローブ類似体も含まれる。
他の相補的なオリゴヌクレオチド類似体は、限定されるものではないが、モルフォリノ
(morpholino)(Summerton, Biochim. Biophys. Acta, 1489:141-158, 1999)、ロックさ
れたオリゴヌクレオチド(locked oligonucleotides) (Wahlestedt wt al., Proc. Natl
. Acad. Sci.US, 97:5633-5638, 2000)、ペプチド性核酸(peptidic nucleic acids)又
はPNA(Nielsen et al., 1993; Hyrup and Nielsen, 1996)又は2-O-(2-メトキシ)エチ
ル変性5’及び3’末端オリゴヌクレオチド(2-o-(2-methoxy) ethyl modified 5’ and
3’ endoligonucleotides)(McKay et al., J. Biol. Chem., 274:1715-1722, 1999)な
どを含む。
これらの引例のすべては、参照することにより明らかに本明細書に組み込まれる。
核酸は、デオキシリボ−とリボ−ヌクレオチドのいかなる結合、及びウラシル、アデニ
ン、チミン、シトシン、グアニン、イノシン、キササナイン(xathanine)、ハイポキサ
ナイン(hypoxathanine)、イソシトシン、イソグアニン(isoguanine)等を含むいかな
る塩基の結合を含めることができる。
【0046】
本発明の他の態様では、核酸またはオリゴヌクレオチドプローブは、センスミトコンド
リアキメラRNA又はアンチセンスミトコンドリアキメラRNAによるハイブリダイゼー
ションを検出するために標識される。プローブは、その技術分野で知られている任意の方
法により検出可能なマーカーにより標識される。
プローブを標識する方法は、ランダムのプライミング、エンドラベリング、PCR、及
びニックトランスレーションを含む。
酵素の標識付けは、核酸ポリメラーゼ、3つの未標識ヌクレオチド、及び標識が直接付
けられるか、標識するためにリンカーアーム(linker arm)を含むか、又はハプテンもし
くは標識が付けられた結合分子が結合するかもしれない他の分子に付けられる4番めのヌ
クレオチドの存在下で行なわれる。
適切な直接の標識は、.sup.32 P、.sup.3 Hおよび.sup.35 Sのような放射性のラベル、
並びに蛍光性のマーカーのような非放射性の標識を含む。
好ましい蛍光色素(fluorophores)は、5(6)カルボキシフルオレセン[5(6)carboxyfluo
rescein]、6-((7-アミノの-4-メチルコウマリン-3-アセチル)アミノ)ヘキサノイック
酸[6-((7-amino-4-methylcoumarin-3-acetyl)amino)hexanoic acid]、5(及び6)-カ
ルボキシ-X-ローダミン[5(and6)-carboxy-X-rhodamine]、シアニン2(Cy2)染料[Cyanin
e 2 (Cy2) Dye]、シアニン3(Cy3)染料[Cyanine3 (Cy3) Dye]、シアニン3.5(Cy3.5
)染料[Cyanine 3.5(Cy3.5) Dye]、シアニン5(Cy5)染料[Cyanine 5 (Cy5) Dye]、シア
ニン5.5(Cy5.5)染料[Cyanine 5.5 (Cy5.5) Dye]、シアニン7(Cy7)染料[Cyanine 7
(Cy7) Dye]、シアニン9(Cy9)染料[Cyanine9 (Cy9) Dye]、(シアニン染料2、3、3.5
、5および5.5は、Amersham,Arlington Heights, III からのNHSエステルとして入手可能
である。)、又はアレクサ染料[Alexa dyes](Alexa 488、Alexa 532、Alexa 556、Alexa
590等を含む。)(MolecularProbes, Eugene, Oreg.)を含む。
【0047】
プローブは、ハプテン又は他の分子に共有結合しているヌクレオチドに結合することに
より直接的に標識される。好ましいハプテンは、限定されるものではないが、5(6)-カ
ルボキシフルオレセイン[5(6)-carboxyfluorescein]、2,4-ジニトロフェニルジオキシ
ゼニン[2,4-dinitrophenyl、digoxigenin]、及びビオチンを含み、そしてそのハプテン又
は他の分子に対する標識された抗体でプローブの検出を行う。
ビオチンの場合、検出は、検出可能なラベルに接合しているアビディン(avidin)ある
いはストレプトアビジン(streptavidin)により行うことができる。
抗体、ストレプトアビジンおよびアビディンは、蛍光性のマーカーで、あるいはそれら
を検出可能にするアルカリホスファターゼかホースラディッシュ・ペルオキシダーゼのよ
うな酵素のマーカーに接合される。
接合されたストレプトアビジン(streptavidin)、アビディン及び抗体アンチジゴキシゲ
ニン(digoxigenin)は、ベクター研究所(バーリンゲーム,カリフォルニア)およびボエリン
ゲルマンハイム(BoehringerMannheim)(インディアナポリス、インディア)のような会社
から商業的に入手可能である。
他の具体例では、抗体又はストレプトアビジンは、多くの安定した蛍光放射量子ドット
に接合される(Wuet al., Nature Biotechnol. 21:41-46, 2003)。
【0048】
抗体とストレプトアビジンに結合している酵素は、酵素に対する基質(substrate)を
提供することにより、熱量反応を通して検出することができる。
種々の基質の存在において、異なる色は、反応により生成され、そしてこれらの色は、
多数のプローブを別々に検知するために視覚化することができる。
当該分野で公知のいかなる基質も使用できる。
アルカリ性ホスファターゼに対する好ましい基板は、5-ブロモ-4-クロロ-3-インド
リルホスフェーテ(5-bromo-4-chloro-3-indolylphosphate)(BCIP)及びニトロブルーテト
ラゾリウム(nitroblue tetrazolium)(NBT)を含む。
ホースラディシュ・ペルオキシダーゼ(horseradish peroxidase)用の好ましい基質は
、ジアミノベンゾエート(diaminobenzoate)(DAB)である。
その分野の当業者は、更にその他の酵素活性を用いることが可能であることを理解する
。
【0049】
本発明の他の具体例では、正確で複製可能な結果を達成するin situハイブリダイゼー
ションを行うための条件が記述される。
核酸ハイブリダイゼーションの分野の技術者は、ハイブリダイゼーションの厳格性をコ
ントロールするために一般に使用される因子がホルムアミド濃度あるいは他の化学的変性
試薬、塩濃度又は可変のイオン強度、ハイブリダイゼーション温度、界面活性剤濃度、p
Hとカオトロピック剤(chaotropic agents)の存在又は不存在、を含むことを認識する
であろう。
これらの厳格因子は、それによりキメラRNAに対するオリゴヌクレオチドプローブの
ハイブリダイゼーションの厳格さをコントロールするために調整することができる。分析
用の最適の厳格さは、所望の厳格さの程度が達成されるまで、各厳格因子の実験によって
実験的に決定される。
【0050】
最良のinsituハイブリダイゼーションを制御するための他の条件は、例えば、標的の
キメラRNAよりも生体サンプル中に存在する、成分に対するプローブの非特定結合をブ
ロックする化学薬品の使用である。
ブロッキング試薬(遮断薬)は、限定されるものではないが、RNA、DNAあるいは
標識されていないオリゴヌクレオチドである。
ハイブリダイゼーション溶液中で結合しているブロッキング試薬は、標識されたプロー
ブの非特定結合を制御する、そしてそれ故分析のシグナル対ノイス比を増加する。
本発明の他の側面において、プローブは、センス又はアンチセンスのミトコンドリアR
NAの配列に相補する配列を有する(SEQ ID NO 1, SEQ ID NO 2, SEQ ID NO 3, SEQ ID N
O 4, SEQ ID NO5 and SEQ ID N 6 参照)
【0051】
生化学的配列の固定は、また実験的に決定されるin situハイブリダイゼーションの重
要な側面である。グルタルアルデヒドのような高度に架橋された固定剤は、標的のセンス
ミトコンドリアキメラRNA又はアンチセンスミトコンドリアキメラRNAへのプローブ
のアクセスがブロックされるので、推奨されない。
この発明の好ましい方法は、冷凍のサンプルも好ましいが、ホルマリンで固定された生
体サンプルである。センスミトコンドリアキメラRNA又はアンチセンスミトコンドリア
キメラRNAを標識されたプローブに触れさせるために、付加的な操作を用いることがで
きる。
例えば、生体サンプルは、プローブが標的キメラRNAにアクセスするのをブロックす
る蛋白質を除去するためにプロテイナーゼKで消化することができる。
in situハイブリダイゼーションに先立って、生体サンプルをプロテイナーゼK又は他
のプロテアーゼで処理することは、当該分野の技術者によく知られている。
【0052】
前述したように、キメラRNAとアンチセンスキメラRNA中の長い逆方向反復配列は
、高度に安定な二重鎖の生成を誘発する。キメラRNAの単一鎖領域の第二の構造を有す
るこれらの構造は、プローブが標的キメラRNAにアクセスするのにバリヤーを構成する
。
従って、本発明の他の側面では、生体サンプルは、キメラRNAを変性するために、室
温で10分間0.2MのHClで処理される。
該サンプルは、ここに記述するin situハイブリダイゼーションプロトコルを適用する
前に、pH7.4の緩衝溶液で数回洗浄することにより急速に中和される。
ここに記述する、通常のルーチン実験およびその開示の下にこの分野の技術者は、ここ
に記述する方法と組成物を利用して分析を行うための適切なハイブリゼイション条件を容
易に決めることができる。適切なin situハイブリダイゼーション条件は、in situハイブ
リダイゼーション操作を行うのに適切なこれらの条件である。
従って、適切なinsitu ハイブリダイゼーション条件は、ここにおける開示と引例(場
合によっては更なるルーチン実験を用いて)を用いて明らかになる。
【0053】
本発明の他の具体例において、in situハイブリダイゼーションにより決められたよう
にセンスミトコンドリアキメラRNAの局在化(localization)は、癌を持つ患者の予後
と管理に重要な情報である。腫瘍細胞において、センスミトコンドリアキメラRNAは、
遅発エンドソーム/リソソームに付随する細胞質中に最も見出される。しかし、核小体中
の局在化は、また、ある細胞中にも見出される。
ヒトの生体検査における腫瘍細胞の存在において、そのハイブリダイゼーションのシグ
ナルは、センスミトコンドリアキメラRNAが細胞質中にのみ、又は細胞質と核小体中も
しくは細胞質と細胞核中に存在することを示す。
従って、異なる局在化は、重要な予後の重要性を持つこともよそうされる。
好ましい具体例において、例えば乳房、大腸及び前立腺腫瘍からのヒトの生検のパネル
は、キメラRNAを検出するためのin situハイブリダイゼーションにより研究されうる
。
肯定的なハイブリダイゼーションシグナルと共に(プローブがどのように標識されたか
には左右されず)、細胞内局在化(唯一細胞質、細胞質及び核小体、又は細胞質及び細胞
核)は、各腫瘍中で確立され、そしてその結果は各患者の生存と比較された。
【0054】
この発明の他の側面において、正常な及び/又は腫瘍細胞を含む個別細胞の混合物は、
蛍光色素(fluorochrome)で標識され、センスミトコンドリアキメラRNAとアンチセン
スミトコンドリアキメラRNAに相補するオリゴヌクレオチドプローブを有する検査液中
でハイブリダイゼーションされる。
例えば、センスミトコンドリアキメラRNAを標的にしているプローブは、ローダミン
で標識表示され、そしてセンスミトコンドリアキメラRNAを標的にしているプローブは
、アレクサ(Alexa)488で標識表示されうる。
以前に記述した条件の下にハイブリダイゼーションと洗浄後に、細胞は、細胞内の標識
表示するフローサイトメトリー(flow cytometry)により分析することができる。
【0055】
本発明の好ましい態様は、腫瘍細胞中のキメラRNAの特定の局在化について得られた
情報が予後診断の重要な更なる情報を提供するので、in situハイブリダイゼーションに
用いることである。
本発明の更に他の態様において、代替分子法は、正常、前癌及び癌細胞中でのキメラR
NAの発現及びセンスとアンチセンスキメラRNAの差次的発現を検出するのに用いられ
る。これらの別法は、限定されるものではないがノーザンブロット、ドットブロット、キ
メラRNAとアンチセンスキメラRNAに対するオリゴヌクレオチド配列、RT−PCR
によるRNAの増殖、インビトロ転移介在性増殖又はTMAによるRNAの増殖、S1又
はリボヌクレアーゼ(RNA分解酵素)アッセイ、等を含む。
【0056】
本発明の1つの態様で、センスミトコンドリアキメラRNAは、16SリボソームRN
Aの5’末端部を含む領域、又は逆方向反復配列の部分的もしくは全領域の増殖により得
られたプローブを用いて診断目的で検出することができる。
図1に示すように、リバースプライマーは例えばプライマー1(SEQ ID NO 139)にす
ることができ、そしてフォワードプライマーは、プライマー3,4,5,6又は7(SEQ ID
NOS129,11,106,102,63)にすることができる。他の部分に位置するプライマーもまた使
用することができ、そしてこれらは該分野の技術者により容易にデザインできる。この発
明の他の側面で、逆転写活性を有する酵素とヘキサマー又はより長鎖のようなランダムプ
ライマーにより合成されるcDNAは、当該分野の技術者によく知られている。
【0057】
得られた210、350、500もしくは850bp、又は他の部分に位置するプライ
マーを用いて得られた他のサイズのアンプリコン(amplicons、単位複製配列)は、アガ
ロースゲル又はポリアクリルアミドゲル中の電気泳動(Sambrook et al., 1989)及び臭化
エチジウム又は他の挿入染料を用いて染色することにより検出できる。
このアンプリコンは、メーカーの指示書により精製される。
【0058】
ミトコンドリアキメラRNAの検出は、ノーザンブロットにより行うことができる(Sam
brook et al.,1989)。
アガロースゲル中のRNAの分離後に、フラグメントは、当該分野の技術者によく知ら
れた操作により膜(ニトロセルロースまたはナイロン)に転送される(Sambrook et al.,
1989)。膜を調べるために、センスミトコンドリアキメラRNAの1000から125の
位置に相当する250bpのフラグメントは、増殖される。
アンプリコンは、製造指示書により精製され(Wizard, Promega)、そして10ナノグラ
ムが第2の増殖のために鋳型として用いられる。
この増殖は、PCR(Invitrogen)と32P-a-dCTP (Amerscham)の5マイクロキュリーの
鎖混合物で行う。放射性の増殖フラグメントは、95℃で10分間インキュベートにより
変性し、変性されたプローブは、ハイブリダイゼーション混合物に添加される。膜は、6
5℃で16時間、ハイブリダイズされて、その後2倍のSSC緩衝液で2度、60℃で0
.5倍のSSCで2度、そして45℃で0.2倍のSSCで洗浄される(Sambrook et al.
, 1989)。洗浄された膜は、−70℃で一晩X線フィルムに曝される(Sambrooket al., 1
989)。
膜上のハイブリダイゼーションシグナルは、16SリボゾームRNA(1559のヌクレオ
チド)と815ヌクレオチドの逆方向反復配列に相当するサイズである2,400のヌクレ
オチドの主な成分に相当する。
【0059】
本発明の他の態様で、センスミトコンドリアキメラRNAの一部は、細胞又は細胞の組
織から抽出された全RNAのリボヌクレアーゼ(RNA分解酵素)の消化の後に検出され
る。
二重鎖構造又はセンスミトコンドリアキメラRNAのステムは、リボヌクレアーゼによ
る消化に耐性がある。
細胞又は細胞の組織からトリゾール(TriZol) (Invitrogen)で抽出された全RNAは、
2倍のSSCの小容量中に溶解される。溶液は、50μg/mlの最終濃度でRNA分解
酵素A(Sigma)を用いてインキュベートされる。
25℃で30分後に、ヌクレアーゼに対するRNA耐性は、トリゾールで抽出され、−
20℃で一晩イソプロパノールを用いて沈殿させる。ヌクレアーゼに対するRNA耐性は
、蒸留されたDEPC処理水に溶解し、そしてRT-PCR増殖のために鋳型として用い
られる。
16SリボゾームRNAの位置55および790を標的とするプライマーを用いて行っ
た増殖は、センスミトコンドリアキメラRNA(SEQ ID NO 1)のステムと同じ配列で10
0%同一性を示す配列と約730塩基対のフラグメントを生ずる。
その一方、キメラRNAで3’ハーフに相補する単一鎖、又は12Sミトコンドリアリ
ボゾームRNA、又は18SリボゾームRNAもしくはGAPDHに対するmRNAは、
リボヌクレアーゼAで処理することにより全体的に消化され、そしてそれ故、これらのR
NAを標的にするプライマーが用いられた場合に、増殖生成物は、得られない。
【0060】
本発明の他の態様では、全RNAがリボヌクレアーゼAで処理された後、得られたセン
スミトコンドリアキメラRNAのステムは、ノーザンブロットにより検出できる。ヌクレ
アーゼに耐性を示し、トリゾールによる抽出とイソプロピルアルコールを用いた沈殿によ
り回収されるRNAは、アガロースゲル中の電気泳動によって分離される。転移の後に、
膜は、前述したプローブでブロットされ、そしてセンスミトコンドリアキメラRNAをノ
ーザンブロット(Sambrooket al., 1989)するのに用いられる。
【0061】
他の態様では、この発明は、センスミトコンドリアキメラRNA又はアンチセンスミト
コンドリアキメラRNAを生体サンプル中でアッセイを検出するのに適したキッドを教示
する。キメラRNAの検出に適した、一般的で好ましい態様、組成物及び方法が提供され
、in situハイブリダイゼーションによるアンチセンスキメラRNAは、既に本明細書で
記述された。
好ましいオリゴヌクレオチドプローブ配列は、限定されるものではないがリストに記載
されている。更に、オリゴヌクレオチドプローブを使用するのに適した方法あるいはサン
プル中のキメラRNAもしくはアンチセンスキメラRNAを検出するキッドのオリゴヌク
レオチドプローブは、既に本明細書で記述された。
【0062】
本発明のキットは、1つ以上のオリゴヌクレオチドプローブと、サンプル中のセンスミ
トコンドリアキメラRNA又はアンチセンスミトコンドリアキメラRNAを検出するため
に用いるinsituハイブリダイゼーションを行うために選択された試薬又は組成物からな
る。2以上のオリゴヌクレオチドの各セットは独立して検知可能な構成部分で好ましくは
ラベル化され、そして生体サンプルの個々の細胞において、センスミトコンドリアキメラ
RNA又はセンスミトコンドリアキメラRNAが検出される。
好ましい態様において、センスミトコンドリアキメラRNA又はアンチセンスミトコン
ドリアキメラRNAを検出するのに用いるキットのオリゴヌクレオチドプローブは、異な
るハプテンで各セットごとに標識される。このハプテンは、ビオチン、ジゴキシゲニン、
あるいは異なる酵素(例えばアルカリホスファターゼあるいはペルオキシダーゼ)で標識し
た抗体あるいはストレプトアビジンでin situハイブリダイゼーション方法で認識できる
フルオレセイン(蛍光色素)とすることができる。
他の方法として、プローブの各セットの各オリゴヌクレオチドプローブは、独立して検
出可能な蛍光性のグループで標識できる。
例えば、センスミトコンドリアキメラRNAを検出するためのオリゴヌクレオチドプロ
ーブのセットは、ローダミンで標識できる、一方、アンチセンスミトコンドリアキメラR
NAを検出するためのオリゴヌクレオチドプローブのセットは、アレクサ488(Alexa 4
88)で標識できる。
更に、生体サンプルの細胞中のキメラRNA又はアンチセンスキメラRNAの局在化を
決定する方法が提供される。
更に、本発明の組成物と方法は、共焦点顕微鏡分析を用いて異なる細胞小器官の特定の
マーカーでキメラRNA又はアンチセンスキメラRNAの共−局在化(co-localization
)を決定するのに使用することができる。
【0063】
ここに提供される組成物および方法は、前癌および癌の検知と診断に特に有効であると
考えられる。
ここに提供されるような用語「癌」は、以下の確認されている特異な条件のいずれか1
により悩まされる細胞を含んでいる。
これらは、急性慢性である骨髄性白血病、急性慢性であるリンパ芽球の白血病、多発性
骨髄腫、ホジキン病、非ホジキンのリンパ腫あるいは悪性リンパ腫;胃癌、食道癌か腺癌
、膵臓送管の腺癌、インスリノーマ(膵島細胞腫)、グルカゴン産生腫瘍(glucagonoma
)、ガストリノーマ(gastrinoma)、小腸腺癌(small bowel adenocarcinoma)、結腸直
腸癌;肝細胞性の癌、肝細胞性の腺腫;カルチノイド、腎臓腺癌のような尿生殖路、ウイル
ム腫瘍(Wilm'stumor)、膀胱および尿道癌および前立腺癌、精上皮腫のような精巣癌、
奇形腫、奇形癌、間質細胞癌腫; 子宮内膜の癌、子宮頚癌、卵巣癌、外陰癌と膣癌、セル
トリ-L-アイディグ(Sertoli-L-eydig)細胞癌、黒色腫および卵管癌;肺、歯槽と細気管
支の癌;脳腫瘍; 皮膚悪性黒色腫、基底細胞癌、扁平上皮細胞癌及びカルポシ(Karposi)肉
腫を含む。
繊維肉腫、心臓の血管肉腫と横紋筋肉腫、及び他の悪性癌は、当該分野の技術者によく
知られている。
【0064】
4.3 癌および前癌療法
化学療法薬は、癌細胞増殖および存続に影響を与える一連の細胞応答を含む。
これらの細胞応答の研究された最良のものは、アポトーシスであり、抗癌剤が共通のア
ポトーシス経路を活性化することにより腫瘍細胞を殺すことができることは現在明らかで
ある。。
不運にも、これらの薬は、さらに急速に分裂する骨髄の正常細胞、正常な造血と腸の細
胞および髪の毛マトリックスのケラチン生成細胞にも影響を与える(McKinnell et al., T
he biological Basisof cancer, 1998; Komarov et al., Science 285:1733-1737, 1999
; Johnstone etal., Cell 108:153-164, 2002)。
【0065】
他方、多くの腫瘍細胞が細胞死開始因子、調整因子及び細胞死の遂行因子を変化させる
、これらは、異なる癌タイプの腫瘍細胞がなぜ種々の化学療法薬と放射線に耐性を示すこ
とになるかを説明している。
この変化は、固有の経路の因子、ポストミトコンドリアル事象及び細胞死の固有の経路
について報告されている(Rampino et al., Science 275:967-969, 1997; Vogelstein et
al., Nature408: 307-310, 2000; Teitz et al., Nature Med. 6:529-535, 2000; Reed,
J. Clin.Oncol., 17:2941-2953, 1999; Johnston et al., Cell 108:153-164, 2002)。
従って、癌治療法のパラダイムは、腫瘍細胞の細胞死の選択的誘引となる操作、正常な
増殖細胞を変化させない操作、及び異なる細胞死経路を変化又は突然変異させる要因を回
避する操作である。
【0066】
本発明の構成および方法は、正常な増殖細胞と同様のレベルで、腫瘍及び前腫瘍細胞が
センスミトコンドリアキメラRNAを過剰発現するという発見に基づく。
これらの転写産物の構造は、図1、図2、及びSEQ ID NO 1、SEQ ID NO 2、SEQ ID NO
3、SEQ ID NO 4、SEQID NO 5およびSEQ ID NO 6における対応する配列である。
【0067】
その一方、他の驚きの発見は、前腫瘍及び腫瘍細胞がセンスミトコンドリアキメラRN
Aを過剰発現し、そしてアンチセンスミトコンドリアキメラRNAを下方制御する、とい
うことである。前腫瘍及び腫瘍細胞におけるアンチセンスミトコンドリアキメラRNAの
合成の抑制と阻害は、癌細胞と正常増殖細胞間の表現型の新たな差異を構成する、これは
、本発明の主な態様に1つとして考えられる。
さらに、異なる癌タイプのヒトの生検における癌はまた、培養中の癌と同じ表現型を示
す(図5Aおよび5B)。
【0068】
センスミトコンドリアキメラRNAとアンチセンスミトコンドリアキメラRNAの機能
は明確ではないが、密接な相関関係はこれらのRNAの発現と細胞増殖間に存在する。例
えば、肝臓、腎臓および脾臓のような細胞組織中の、Ki-67、PCNAあるいはリン酸
化されたヒストンH3(histone H3)のような抗原を意味するようなもので定義される正常
な増殖する細胞は、アンチセンスミトコンドリアキメラRNAだけでなくセンスミトコン
ドリアキメラRNAを過剰発現する。
Ki-67又はPCNAを発現しない同様の細胞組織の非増殖細胞において、センスミト
コンドリアキメラRNAとアンチセンスミトコンドリアキメラRNAは発現しない。更に
、図7に示すように、ミトゲンPHAで活性化されたヒトのリンパ球は、DNAを合成し
、増殖する抗原Ki-67とPCNAを発現する。
この活性化されたリンパ球は、またアンチセンスミトコンドリアキメラRNAだけでな
くセンスミトコンドリアキメラRNAを過剰発現する(図7)。
対照的に、休止しているリンパ球又は非活性リンパ球は、センスミトコンドリアキメラ
RNAもアンチセンスミトコンドリアキメラRNAも発現しない。
【0069】
本発明の1つの基本部分である、前述の発見は、正常の増殖細胞がセンスとアンチセン
スミトコンドリアキメラRNAを発現する期間、腫瘍細胞はセンスミトコンドリアキメラ
RNAを発現し、そしてアンチセンスミトコンドリアキメラRNAの発現を下方制御する
、ことを示す。
細胞増殖におけるこれらのRNAの機能を理解するために、培養中の癌細胞は、センス
ミトコンドリアキメラRNA(SEQ ID NO 1、SEQ ID NO 2、SEQ ID NO 3)又はアンチセン
スミトコンドリアキメラRNA(SEQ ID NO 4、SEQ ID NO 5、SEQ ID NO 6)を標的にする
アンチセンスオリゴヌクレオチドで処理された。
他の驚くべき発見である結果は、これらの条件下で細胞は、予定された細胞死又はアポ
トーシスをこうむることを示した。
センス又はアンチセンスミトコンドリアキメラRNAに相補するオリゴヌクレオチドで
6ないし15時間処理され処理後、細胞の75〜96%は、アポトーシス(表2参照)を被
る。
処理された細胞で観察された変化は、クロマチン凝縮、核断片化、DNA断片化、カス
パーゼ活性及び細胞膜の変化プロセスであった。
4またはそれ以上のミスマッチ又はスクランブルオリゴヌクレオチドを有する制御オリ
ゴヌクレオチドは、アポトーシスを誘発しなかった。
【0070】
また、センス又はアンチセンス12SミトコンドリアRNAを標的とする、又はmRN
AもしくはミトコンドリアND1サブユニットのアンチセンス転写産物を標的とするオリ
ゴヌクレオチドで処理しても細胞は影響を受けなかった。
一般に、アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオチドは
、同じ濃度ではセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオチドより
もより効果的であった。
このことは、予期された結果であった、なぜなら腫瘍細胞は、センスミトコンドリアキ
メラRNAを過剰発現し、そしてそれ故細胞内でこの転写産物のすべてのコピーを阻害す
る細胞内のオリゴヌクレオチド濃度に到達ことが困難だからである。
他方、腫瘍細胞は、アンチセンスミトコンドリアキメラRNAを下方制御するので、細
胞あたりのコピー数が低くこのRNAで阻害することが容易になる。
【0071】
アポトーシスの誘発は、腫瘍細胞に対してまた選択的である。休止しているヒトリンパ
球又はPHAで48時間活性化したヒトのリンパ球は、アンチセンスミトコンドリアキメ
ラRNAに相補する、又はセンスミトコンドリアキメラRNAを標識にするオリゴヌクレ
オチドで、一晩処理をしても、また相補的オリゴヌクレオチド(15uM)の高用量で処
理しても影響されない。
【0072】
アンチセンスミトコンドリアキメラRNA(SEQ ID NO 4、SEQ ID NO 5、SEQ ID NO 6)
を標的にする相補的オリゴヌクレオチドで処理することによるアポトーシスの誘発は、限
定されるものではないが前骨髄球性白血病細胞HL-60、急性リンパ芽球の白血病MOLT-
4、T−リンパ球白血病細胞、Jurkat、T細胞白血病、Devernelle、又はBリンパ腫、N
SO/2又は骨髄腫、HeLa細胞、DU145、PC-3、Caco-2、Hep-2及
びHepG2で達成される。
治療の効かない(化学療法又は放射線治療)腫瘍細胞のパラダイムと見なすことができ
る、MCF/7(乳癌)とメラノーマの2つの細胞は、アンチセンスミトコンドリアキメ
ラRNA(SEQID NO 4、SEQ ID NO 5、SEQ ID NO 6)を標的とする相補的オリゴヌクレオ
チドで15時間処理するとき、80%以上アポトーシスを受ける。
より低いアポーシス結果は、センスキメラRNA(SEQ ID NO 1)に相補するオリゴヌク
レオチドで得られた。
すでに記載したように、4つのミスマッチ又はスクランブルオリゴヌクレオチドを有す
るオリゴヌクレオチドは、細胞死を誘発しない。
【0073】
以下の記載は、治療標的として、センスキメラRNAとアンチセンスキメラRNAを用
いて癌を処理するための方法と組成物である。
好ましい態様は、限定されるものでないが、アンチセンスキメラRNAに相補するオリ
ゴヌクレオチドを用いて癌を処理するための方法と組成物である。
この治療の結果は、処理された対象で少なくとも健康に良い利益を生ずる、この利益は
、癌の場合に癌の沈静化に限定されないが、癌の症状の緩和及び癌の転移拡散の制御を含
む。このようなすべての方法は、腫瘍細胞において、及び正常細胞においてはマイナーな
効果でアポトーシスの誘発を伴う。
特定のRNAを標的する相補的なオリゴヌクレオチドは、多くのさまざまなmRNAの
発現又は対応するたんぱく質の合成を少なくするかなくす為に用いられてきた(e.g. Vick
ers et al., J.Biol. Chem., 278:7108-7118, 2003)。
現在、異なる化学的性質を有する約42のアンチセンスオリゴヌクレオチドは、可能性
のある薬剤としてスクリーニングされている(Stephens and Rivers, Curr. Opin. Mol. T
herapeut.,5:118-122, 2003) (参照例:U.S. Pat. Nos. 5,801,154; 6,576,759; 6,720,
413; 6,573,050and 6,673,917)。
これらの引例のすべては、参照することにより明らかに本明細書に組み込まれる
【0074】
この発明の1つの側面において、アンチセンスキメラRNAを標的にする1以上のオリゴ
ヌクレオチドが用いられる。
2つ以上の相補的なオリゴヌクレオチドの使用は、より有効で、ある程度の相乗作用を
示す。
【0075】
本発明のオリゴヌクレオチドは、アンチセンスミトコンドリアキメラRNA又はセンスミ
トコンドリアキメラRNAに相補することができる。
相補的なオリゴヌクレオチドは、アンチセンスミトコンドリアキメラRNA又はセンス
ミトコンドリアキメラRNAに結合し、それらの機能を妨げる。
絶対的な相補性は、好ましいが、要求はされない。
ここで記述するように、一部のRNAに相補するオリゴヌクレオチド配列は、RNAで
ハイブリダイズして安定な二重鎖を形成するに十分な相補性を有する配列を意味する。ハ
イブリダイズする能力は、両者の相補の程度とオリゴヌクレオチドの長さによる。一般に
、ハイブリダイズする核酸が長いほど、RNAでのミスマッチの増加した塩基を多く含む
が、それでも安定な二重鎖を形成する。
当該分野の技術者は、ハイブリダイズ化された錯体の融点を決定する標準操作の使用に
よりミスマッチの許容程度を確認することができる。
【0076】
一般に、異なる蛋白質をmRNAでハイブリダイズするための相補的オリゴヌクレオチ
ドは、AUGスタートコドンの相補物を含むmRNAの5’未翻訳領域、又はより効果的
な3’未翻訳領域を標的にする。
mRNAコード領域に相補するオリゴヌクレオチドは、翻訳の効果が低い抑制剤である
(前の引例を参照)。
センスミトコンドリアキメラRNAとアンチセンスミトコンドリアキメラRNAは、非
コードRNAであり、それ故オリゴヌクレオチドの標的領域は、これらの転写産物のいか
なる領域とも相補することができる。最も効果的な領域は、各30ヌクレオチドでデザイ
ンされた相補的オリゴヌクレオチドを用いてアンチセンス又はセンスミトコンドリアキメ
ラRNAの配列をスキャニングすることにより決定されたアンチセンスミトコンドリアキ
メラRNAの単一鎖セグメントの周りに局在化される。
この分野の技術者は、SEQ ID NO 2、SEQ ID NO 3、SEQ ID NO 4、SEQ ID NO 5、及びSE
Q ID NO 6の転写産物の完全な配列内の他の配列が代替の相補的なオリゴヌクレオチドを
デザインするための標的であることを理解する。。
【0077】
アンチセンスミトコンドリアキメラRNA又はセンスミトコンドリアキメラRNAを標
的とし、本発明によると腫瘍細胞死を誘発することになるこの相補的なオリゴヌクレオチ
ドは、一般にナチュラルなリン酸ジエステル結合と異なるバックボーンを含む。このオリ
ゴヌクレオチドは、限定されないが、ホスフォロチオエーテ(Mag et al., Nucleic Acids
Res.19:1437-1441, 1991; and U.S. Pat. No. 5,644,048)、ペプチド核酸又はPNA(E
gholm, Nature,365:566-568, 1993; and U.S. Pat. No. 6,656,687)、ホスフォアミド(B
eaucage,Methods Mol. Biol. 20:33-61, 1993)、ホスフォロチジオエーテ(Capaldi et a
l., NucleicAcids Res., 28:E40, 2000)を含む、交互に並ぶインターヌクレオシド結合
を有することができる。
他のオリゴヌクレオチド誘導体は、限定されないがモルホリノ(Summerton, Biochim. B
iophys. Acta,1489:141-158, 1999)、固定されたオリゴヌクレオチド(Wahlestedt wt al
., Proc. Natl.Acad. Sci. US, 97:5633-5638, 2000)、ペプチド核酸又はPNA(Nielse
n et al., 1993;Hyrup and Nielsen, 1996)又は2-o-(2-メトキシ)エチル修飾5’と3
’末端オリゴヌクレオチド(McKay et al., J. Biol. Chem., 274:1715-1722, 1999)のよ
うなものを含む。
これらの引例のすべては、参照することにより明らかに本明細書に組み込まれる。
核酸は、デオキシリボ−とリボ核酸のいかなる結合、及びウラシル、アデニン、チミン
、シトシン、グアニン、イノシン、キササニン(xathanine)、ハイポキササニン(hypoxat
hanine)イソシトシン、イソグアニンを含む、塩基のいかなる結合も含むことができる。
【0078】
本発明による相補的オリゴヌクレオチドは、限定されないが
5-フルオロウラシル、5-ブロモウラシル、5-クロロウラシル、5-ヨードウラシル、
ヒポキサンチン、キサンチン、4-アセチルシトシン、
5-(カルボキシヒドロキシメチル)ウラシル、
5-カルボキシメチルアミノメチル-2-チオウリジン、
5-カルボキシメチルアミノメチルウラシル、ジヒドロウラシル、
ベータD-ガラクトシルキオシン(beta-D-galactosylqueosine)、イノシン、
N6-イソペンテニルアデニン、1-メチルグアニン、1-メチルイノシン、
2、2-ジメチルグアニン、2-メチルアデニン、2-メチルグアニン、3-メチルシトシン
、5-メチルシトシン、N6アデニン、7-メチルグアニン、
5-メチルアミノメチルウラシル、5-メトキシアミノメチル-2-チオウラシル、
ベータD−マノシルキオシン(beta-D-mannosylqueosine)、
5'-メトキシカルボキシメチルウラシル、5-メトシキウラシル、
2-メチルチオ-N6-イソペンテニルアデニン、ウラシル-5-オキシアセテック酸(V)
、ワイブトキソシン(wybutoxosine)、プソイドウラシル、キオシン(queosine)、
2-チオシトシン(thiocytosine)、5-メチル-2-チオウラシル、2-チオウラシル、
4-チオウラシル、5-メチルウラシル、ウラシル-5-オキシアセテックアシッド メチル
エステル、ウラシル-5-オキシアセテック酸(V)、5-メチル-2-チオウラシル、3-(
3-アミノ-3-N-2-カルボキシプロピル)ウラシル、
(acp3)w及び2,6-ジアミノプリン
を含むグループから選択された少なくとも1つの修飾塩基部分を含む。
【0079】
相補的なオリゴヌクレオチドは、更に限定されないがアラビノース、2-フルオロアラ
ビノーズ、キシルロース及びヘキソースを含むグループから選択された少なくとも1つの
修飾糖部分を含むことができる。
【0080】
本発明の他の態様では、相補的なオリゴヌクレオチドが、アンチセンスミトコンドリア
キメラRNAのいかなる領域とも、又はセンスミトコンドリアキメラRNAのいかなる領
域ともハイブリダイズするようにデザインされる。
相補的なオリゴヌクレオチドは、長さで少なくとも10のヌクレオチドでなければなら
ず、長さで10から50のヌクレオチド範囲の相補的オリゴヌクレオチドが好ましい。
特定の側面では、相補的なオリゴヌクレオチドが少なくとも12のヌクレオチド、少な
くとも18のヌクレオチド、少なくとも22のヌクレオチド、少なくとも30のヌクレオ
チド、少なくとも50のヌクレオチドである。
【0081】
In vivoだけでなくinvitro実験においても、アンチセンス又は相補的オリゴヌクレオ
チドの非特定の生物学的効果で、アンチセンスミトコンドリアキメラRNA又はセンスミ
トコンドリアキメラRNAの機能を用いてアンチセンス阻害を識別する制御を利用する実
験を考慮することは重要である。
従って、オリゴヌクレオチドのデザインは、参照することにより明らかに本発明に組み
込まれる、米国特許番号 6,673,917で報告されているように動物細胞に有毒効果があるCp
Gトラック、5' GGGGトラックおよび他の配列の存在は、避けなければならない。
更に、配列5'CGTTAの存在は、報告されているアンチセンスでない効果を避けられる(T
idd et al.,Nucleic Acids Res. 28:2242-2250, 2000)。
【0082】
本発明の他の態様では、癌を有する動物又は病人の治療薬として、センスミトコンドリ
アキメラRNAを標的とする又はアンチセンスミトコンドリアキメラRNAを標的とする
相補的オリゴヌクレオチドは、抗癌治療剤と放射線療法に対する活性を誘発することがで
きる。
増感又は過敏性として知られている誘発される感度は、抗癌治療又は放射線療法に対す
る耐性を示す腫瘍細胞中で測定される。
抗癌剤は、これらの当該分野で使用中であり既に知られている、又は未発見の薬剤から
なる。従来の化学療法薬の中には、アルキル化剤、代謝拮抗物質、抗生物質および抗微小
管薬がある。
これらの薬のいくつかの例はシスプラチン、メトトレキサート、ドキソルビシン,
ダクチノマイシン、マイトマイシン、シクロホスファミドなどである。
この発明の他の側面で、癌を有する動物又は病人をアンチセンスミトコンドリアキメラ
RNA及び/又はセンスミトコンドリアキメラRNAを標的にした相補的オリゴヌクレオ
チドでの処理と共に又は処理後に、患者は、放射線療法で処理される、この場合、前記放
射線療法は、紫外線、ガンマ線、アルファ粒子、ベータ粒子、X線および電子線を含んで
いる。
【0083】
この発明の他の側面で、アンチセンスミトコンドリアキメラRNA又はセンスミトコン
ドリアキメラRNAの機能で腫瘍細胞死の阻害は、RNA阻害又はRNAサイレンシング
によりもたらされる。
過去6年にわたって、RNA阻害(RNAi)は、哺乳類細胞中の遺伝子サイレンシングに対
する新規で有望なアプローチとして浮上してきている(Elbashir et al., Nature 411:494
-498, 2001;McManus et al., Nature Rev. Genet. 3:737-747, 2002)。
長さで約19から21のヌクレオチドの合成的に合成された二重鎖のRNA分子は、m
RNAの分解とその後の蛋白質ノックダウンをもたらす、特にそれらの相補的標的mRN
Aにハイブリダイズする。
いくつかの異なる遺伝子は、小さなRNA又はsiRNA阻害により成功裡にサイレン化
される(Lu etal., Curr. Opin. Mol. Ther. 5:225-234, 2003.; Wacheck et al., Oligo
nucleotides13:393-400, 2003)。
アンチセンスミトコンドリアキメラRNA又はセンスミトコンドリアキメラRNAを標
的とする19から21のヌクレオチドの合成された二重鎖のRNAは、これらの転写産物
を分解して、腫瘍細胞死を誘発する。
当該分野の技術者は、siRNAの配列がアンチセンスミトコンドリアキメラRNA又は
センスミトコンドリアキメラRNA(SEQ ID NO 1, SEQ ID NO 2, SEQ ID NO 3, SEQ ID N
O 4, SEQ ID NO5, and SEQ ID NO 6)のいかなる領域にも相補する必要があることは理解
するだろう。
【0084】
この発明の他の態様で、リボザイムは、腫瘍細胞死を誘発するためにアンチセンスミト
コンドリアキメラRNA又はセンスミトコンドリアキメラRNAを阻害するのに用いるこ
とができる。
リボザイムの配列は、腫瘍細胞死を引き起こすのにより効果的な転写産物の特定の領域
を開裂するために、アンチセンスミトコンドリアキメラRNA(SEQ ID NO 4, SEQ ID NO
5, SEQ ID NO 6)又はセンスミトコンドリアキメラRNA(SEQID NO 1, SEQ ID NO 2, SE
Q ID NO 3)の配列によりデザインされなければならない。
リボザイムは、RNAの特定の開裂を触媒することが可能な酵素RNA分子である(Ros
si, Curr.Biology 4:469-471, 1994)。
【0085】
リボザイム作用のメカニズムは、標的RNAに相補するリボザイム分子の配列の特定の
ハイブリダイゼーションを含み、その後エンドヌクレアーゼ的に切断する。
リボザイム分子の構成は、標的遺伝子mRNAに相補する1つ以上の配列を含んでいる
と予想され、mRNA開裂の原因であることがよく知られ、また米国特許番号5,093,246
(これは参照によってその全部がここに組み入れられる)に記述された触媒配列を含んで
いると思われる。
本発明の範囲内において、ハンマーヘッドリボザイム分子は、アンチセンスミトコンド
リアキメラRNA又はセンスミトコンドリアキメラRNAのエンドヌクレアーゼ的切断を
特異的及び効果的に触媒するように作り出されている。
ハンマーヘッドリボザイムの構造と生産は、当該分野でよく知られており、また記述さ
れている(Haseloffet al., Gene, 82:43-52, 1989)。
本発明のリボザイムは、更にRNAエンドリボヌクレアーゼを含んでいる(Zaug et al.
, Science,224:574-578, 1984)。
【0086】
遺伝子治療は、癌にかかっている患者か、癌の予防あるいは禁止が望ましい患者への核
酸の投与によって行なわれる癌の治療あるいは予防に言及する。
本発明のこの態様では、細胞内で生産された治療用核酸は、腫瘍細胞死を誘発するこれ
らのミトコンドリア転移産物の機能を阻害又は禁止することによる治療の影響を調節する
、アンチセンスミトコンドリアキメラRNA又はセンスミトコンドリアキメラRNAを標
的にする相補的RNAである。
従って、1つの好ましいアプローチは、アンチセンスRNAの転移産物がRNAポリメ
ラーゼII又はIIIの強いプロモーターの制御下で置換された、組み換えられたDNAの利
用である。
相補的RNAをコード化する配列は、ほ乳類、好ましくはヒト細胞中で作用するその技
術分野で知られているいかなるプロモーターでも発現する。
【0087】
このようなプロモーターは、限定されるものでないが、SV4初期プロモーター領域(B
enoist andChambon, Nature 290:304-310, 1981)、ヘルペスチミジンキナーゼプロモー
ター(Wagneret al., Proc. Natl. Acad. Sci. U.S.A. 78:1441-1445, 1981)、メタロチ
オネイン遺伝子の調節塩基配列(Brinster et al., Nature 296:39-42, 1982)、ラウス肉
腫ウイルス(Yamamotoet al., Cell 22:787-797, 1980)、等を含む。
相補的RNAを生産する組み換えられたDNA構造は、限定されるものでないが、アデ
ノウィルス・ベクター、アデノ関連性ウィルス・ベクター、単純ヘルペスウィルス・ベク
ター、ワクシニアウィルス・ベクター、及びレトロウィルス・ベクターを含むウィルス・
ベクターにすることができる。
当該分野の技術者によく知られている方法を使用して、このベクターは、製薬組成物と
して標的腫瘍細胞中に導入される。
【0088】
薬学的に受理可能なキャリヤーにおける相補的核酸(相補的オリゴヌクレオチド、siR
NA、リボザイム、又はウィルス・ベクター)の効果的量からなる、本発明の薬剤組成物
は、アンチセンスミトコンドリアキメラRNA又はセンスミトコンドリアキメラRNAの
作用を阻害するために癌患者に投与され、腫瘍細胞のアポトーシスを誘発する。
相補的な核酸は、薬剤組成物中に配合される、この薬剤組成物は、キャリヤー、希釈剤
、バッファー、予防薬、界面活性剤、ポリエチレンイミド(PEI)、リポソームあるい
はこの分野で周知の他の脂質製剤を含めることができる。
前記薬剤組成物は、局所適用、経口、非経口又は直腸投与によって投与される。
非経口投与は、静脈、皮下、腹腔内、又は筋肉注射、又は吸入又は通気による肺内投与
を含んでいる。
【0089】
本発明の組成物は、治療学、診断学、予防処置、そして研究試薬およびキットとして利
用することができる。
ここに提供される組成物及び方法は、癌の治療に特に役立つと考えられる。
ここに提供される用語「癌」は、次の識別された特異な条件のうちのいずれか1つで悩
まされる細胞を含んでいる。
これらは、急性慢性である脊髄性白血病、急性慢性であるリンパ芽球白血病、多発性骨
髄腫、ホジキン病、非ホジキンのリンパ腫あるいは悪性リンパ腫;胃癌、食道癌又は腺癌
、膵臓送管の腺癌、インシュリノーマ、グルカゴン産生腫瘍、多発性胃潰瘍、
小さな腸腺癌、結腸直腸癌;肝細胞性の癌、肝細胞性の腺腫;類癌腫、腎臓腺癌のような
尿生殖器官、ウイルム腫瘍(Wilm's tumor)、膀胱および尿道癌及び前立腺腺癌、精上皮
腫のような精巣癌、奇形腫、奇形癌、間質細胞癌腫;子宮内膜の癌、子宮頚癌、卵巣癌、
外陰癌と膣癌、セルトリ-L-アイディグ(Sertoli-L-eydig)細胞癌、黒色腫および卵管
癌;肺、歯槽と細気管支の癌;脳腫瘍;皮膚悪性黒色腫、基底細胞癌、扁平上皮細胞癌及
びカルポシ(Karposi)肉腫を含む。
繊維肉腫、心臓の血管肉腫と横紋筋肉腫、及び他の悪性癌は、当該分野の熟練技術者に
よく知られている。
ここに提供される用語「癌細胞」は、上記の識別された特異な条件のうちのいずれか1
つで悩まされる細胞を含んでいる。
次の例は、本発明の様々な側面の実行のために最良の方法を述べるだけでなく、上記の
記述された発明を使用する方法について記述する役割を果たす。
これらの例は、本発明の範囲を少しも制限することはなく、むしろこれらは実例となる目
的のために示されることが理解される。
【実施例】
【0090】
[実施例1]
センスミトコンドリアヒトキメラRNAの分離と配列
(図1A、SEQ ID NO1)
最初の実験は、推定のヒトセンスミトコンドリアキメラRNAがより複雑で安定な、マ
ウスのキメラRNAである第二の構造を含むことを示した(Villegas et al., DNA & Cell
Biol.19:579-588, 2000; Villegas et al., Nucleic Acids Res. 30:1895-1901, 2002)
。
それ故、またマウスミトコンドリアキメラRNAの第二の構造に基づき、理論的なヒト
のセンスミトコンドリアキメラRNAの第二の構造が、推定された(図1A)。
理論的なヒトの転写産物は、未知の長さ(図1A)のループを形成するアンチセンス16
SミトコンドリアRNAのフラグメントに5’末端で結合しているセンス16Sミトコン
ドリアRNAの完全な配列を含んでいた。
アンチセンス16SミトコンドリアRNAのセグメントは、センス16Sミトコンドリ
アRNAに十分に相補した、またそれゆえセンス16S転写産物の5’末端に結合する逆
方向反復配列に相当する。この構造に基づき、プライマーは、RT−PCRによるこの推
定の転写産物を増殖するようにデザインされた。
1つのリバースプライマーは、ヒトセンス16SミトコンドリアRNAの5’末端から
11から31に位置、又は理論的ループの始点に位置していた(プライマー1、図1A)
(SEQ IDNO 1)。
【0091】
用いたフォワードプライマーの配列は、センス16SミトコンドリアRNAの位置21
3−234の配列で、図1Aの中のプライマー3に相当する。
HeLa細胞、HL−60、Du145、MCF/7及びヒトのリンパ細胞を含むヒト
の組織と細胞からRNAの増殖は、プライマー1と3(図1A)を用いてRT−PCRに
よりPHA(実施例7参照)で刺激され、約210bpの固有のアンプリコン(図2)を産
出する。
RT−PCRは、前記した方法で実施した(Villegas et al., DNA & Cell Biol. 19:57
9-588, 2000;Villegas et al., Nucleic Acids Res. 30:1895-1901, 2002)。
個々のヒトの組織あるいは細胞からのアンプリコンは、クローン化された、また両鎖は
配列された。
すべての場合に、216bpの同一鎖が得られ、センス16SミトコンドリアRNAの
5’末端の最初の31ヌクレチドに結合している184ヌクレチドの逆方向反復配列を含
んでいた。
その後、我々は、逆方向反復配列が184ヌクレチドより長いか、またアンチセンス1
6SミトコンドリアRNA(図1A)の5’末端に向かって更に拡張しているかを決定し
た。
前記したHeLa細胞又は他の細胞からのcDNAは、前記したようにループに位置す
るリバースプライマー1と、推定のより長い逆方向反復配列の5’末端に向かってウォー
クするプライマー4から7との間で増殖された(図1A)。
このアプローチを用いて、プライマー1が、プライマー4、5及び6とそれぞれ結合す
るのに使用されたとき、およそ500、700及び800bpの増殖フラグメントが得ら
れた。
【0092】
他方、cDNAがプライマー1とプライマー7の間で増殖されたとき、増殖産物は得ら
れなかった、これは、逆方向反復配列の5’末端がプライマー7とプライマー8の間であ
ったためと推定される(以下を参照)。
前記800bpのアンプリコンの完全な配列は、センス16SミトコンドリアRNA(S
EQ ID NO 1)(図1A)の最初の31ヌクレオチドに結合する769ヌクレオチドの逆方向
反復配列を示す。
センス16SミトコンドリアRNAに結合する逆方向反復配列の3’末端で配列は、2
16bpのアンプリコンの同じ領域に見出されるものと同一である。このことは、重要で
ある、なぜなら両者の場合に我々は、同じRNAを増殖していたからである。
更に、配列は、アンチセンス16SミトコンドリアRNAの3’末端のその50ヌクレ
オチドがセンス16SミトコンドリアRNAの逆方向反復配列中には見当たらない、こと
を示した。
すべて、これらの結果は、二重鎖構造が逆方向反復配列と後者の51の位置で始まるセ
ンス16SミトコンドリアRNAとの間で形成され、また50ヌクレオチドの推定ループ
を形成することを示唆する。
【0093】
ループのサイズを確認するために、ヒトのcDNAは、逆方向反復配列の3’末端に位
置するフォワードプライマー2と、センス16SミトコンドリアRNA(図1A)の位置
213−234でまたリバースであるプライマー3との間でPCRにより増殖された。お
よそ240bpのアンプリコンが得られた、またこの配列は、センス16Sミトコンドリ
アRNAの最初の234ヌクレオチドが逆方向反復配列の3’末端の最後の25ヌクレオ
チドに結合していたことを示した。逆方向反復配列の25ヌクレオチドの配列は、センス
16SミトコンドリアRNAの位置51から75(図1A)に十分に相補していた。
【0094】
プライマー1と6で得られたアンプリコンの配列、及びプライマー2と3で得られたア
ンプリコンの配列がコンティグのように組い立てられる場合には、ヒトセンスミトコンド
リアキメラRNAの新たな構造は、50ヌクレオチドのループと少なくとも769bpの
二重鎖構造であることを確認した(図1A)(SEQ ID NO 1参照)。
【0095】
二重鎖RNAは、RNaseAによって消化されないので、ヒトセンスミトコンドリアキメ
ラRNAのステムは、この酵素に耐性がなければならない。一方、ループ又は二重鎖を越
えて拡張したセンス16SミトコンドリアRNA鎖の3’の領域は、酵素により消化され
なければならない。
HeLa細胞又は他の細胞からのRNAは、RNase A(50μg/ml)によって消化され
、続いてフェノール抽出された、そしてヌクレアーゼ耐性物質はエタノール沈殿により回
収された。
消化されたRNAからのcDNAは、図1Aに示すプライマーを用いてPCRにより増
殖された。
プライマー1と6で得られた約800bpのアンプリコンは、ループが酵素により消化
されたことを示すRNaseA消化後には増殖されなかった。図1Aに示したようにプライマ
ー10と11で得られた360bpのアンプリコンと同じであった。
すべて、これらの結果は、ステムを超えて拡張したセンスミトコンドリアキメラRNA
の3’領域だけでなくループも酵素により消化されたことを示した。
センスミトコンドリアキメラRNAの二重鎖の構造に一致しまたプライマー8と6で得
られた750bpアンプリコンの増殖は、RNase A消化により影響されなかった。
リボ核酸消化に耐性を示す二重鎖フラグメントの配列は、予期されたステムの配列と同
一であった。
【0096】
センスミトコンドリアキメラRNAの逆方向反復配列の5’末端を決めるために、RNas
e A消化後に得られた転写産物のステムは、5’RACE分析に用いられた。
逆方向反復配列の5’末端決定は、メーカーのインストラクション(Invitrogen)に従
って行われた。
この結果は、逆方向反復配列がプライマー1と6を用いたセンスミトコンドリアキメラ
RNAの増殖後に得られたアンプリコンの5’末端から46の付加的ヌクレオチドに伸び
ていることを示した。
要約すると、815ヌクレオチドの逆方向反復配列は、16SミトコンドリアRNAの
最初の865ヌクレオチドの5’末端に結合している。この転写産物の配列は、ヒト16
Sミトコンドリア遺伝子(HとL鎖)(SEQ ID NO 1)と99.8%の同一性を示した。二
重鎖ステムの両端の5’末端は、5’RACEにより確認された。
【0097】
上記の結果は、センスミトコンドリアキメラRNAが815塩基対のステムあるいは二
重鎖構造と50ヌクレオチドを含んでいたことを示した。
しかしながら、これらの結果は、逆方向反復配列が完全な16SミトコンドリアRNA
に結合していることを立証するものではない。3’末端から完全なcDNAの合成のよう
な従来のアプローチの使用は役に立たない、なぜなら転写産物の二重鎖構造は、Tthを
含む逆転写酵素に対し克服できない問題を示すからである(Myers and Gelfand, Biochemi
stry30:7661-7666, 1991)。
もし815ヌクレオチドの逆方向反復配列が16SミトコンドリアRNAの1559ヌ
クレオチドに結合したら、2.3Kbの転写産物が予期できる。
HeLa細胞、HL−60及びMCF/7細胞から全RNAのノーザンブロット分析は
、32Pで標識されたプローブを用いて行われ、センスミトコンドリアキメラRNAの二
重鎖構造を標的にした。その結果は、1.6Kbのバンドを除いて、センスミトコンドリ
アキメラRNAとセンス16SミトコンドリアRNAにそれぞれ対応する約2.4Kbの
バンドを示した。
もし前記RNAがノーザンブロットより先にRNase Aで消化されたら、およそ0.8K
bのシングルハイブリダイゼーションバンドが得られた、これはセンスミトコンドリアキ
メラRNAのステムのサイズに相当する。
すべて、これらの結果は、前記センスミトコンドリアキメラRNAが完全なセンス16
SミトコンドリアRNAの5’末端に結合した815ヌクレオチドの逆方向反復配列を含
んでいて、SEQID NO 1に相当していた、ことを強く実証した。
【0098】
逆方向反復配列とセンス16SミトコンドリアRNA間の結合領域を、ヌクレオチドプ
ローブを用いて具体的に検出することが可能である。
プローブは、逆方向反復配列の3’末端とセンス16SミトコンドリアRNA間の結合
点の各サイドで7から10のヌクレオチドを含まなければならない。
このオリゴヌクレオチドは、in situハイブリダイゼーション、又はRT−PCRもし
くはこの新規なRNAを検出するための分野の技術者によく知られた任意の方法による増
殖に用いることができる。
【0099】
センスミトコンドリアキメラRNAは、正常な増殖細胞(ヒトの包皮角化細胞、脾臓、
PHAで刺激されたリンパ球、マウス胚)中、前癌細胞(HPV16又は18で形質転換
された角化細胞、HTLV−1で形質転換されたMT−2細胞)中に、そして癌細胞中に
存在する。正常な休止細胞中には存在しない。これらの結果の要約は、表1(実施例4)
に示されている。
【0100】
[実施例2]
乳頭腫ウィルスで形質転換されたヒトの角化細胞は、新規なセンスミトコンドリアキメ
ラRNAを合成する(図1B、SEQID NO 2)
ヒトの包皮角化細胞(HFK)は、ヒトの乳頭腫ウィルス16(HPV16)に既に感染
した細胞溶解物を用いた培養により形質転換された。
この細胞は、3部のK−SFM、1部のDMEM培地(Invitrogen)、5ng/mlの
下垂体抽出物、及び10%の子牛胎児血清で培養された。
培養条件は、37℃で5%CO2であった。24時間の感染後、形質転換されたHFK
は、が新しいフラスコに転送され、同じ条件下で培養された。
この後に、細胞(HFK698)は、1:4から1:3の分離比率を使用して、3日ごとに新し
い培養フラスコに連続的に転送された。
19継代後に、細胞(HPV16で形質転換されたHFK698)は、記載したように集菌され(Haus
en,Biochim.Biophys. Acta, 1288:F55-F78, 1996)、10分間の300×gでの遠心分離
で集められ、そして塩性のリン酸塩バッファー(PBS)で2度洗浄された。
全RNAは、トリゾール(Invitrogene)で洗浄された細胞から抽出された。
約200ナノグラムのRNAは、実施例1に記載したようにランダムのヘキサマー(he
xamers)でcDNAを合成するために使用された。
【0101】
cDNAは、図1Aに記述されているように、リバースプライマー1とフォワードプラ
イマー3を用いてPCRにより増殖された。
この増殖プロトコルは、予期した210bpのアンプリコンを生じた、ここでセンス1
6SミトコンドリアRNAの最初の31ヌクレオチドは、実施例1に記述したように18
4ヌクレオチドの逆方向反復配列に結合していた。
この増殖製品の電気泳動分析は、センスミトコンドリアキメラRNAに相当する210
塩基対のアンプリコンと、図2に示すように約150塩基対の他の増殖フラグメントの存
在を示した。この新しいフラグメント(SEQ ID NO 2)の完全な存在は、センス16Sミト
コンドリアRNAの5’ 末端から最初の31のヌクレオチドが121ヌクレオチドの逆
方向反復配列に結合されており、これはSEQ ID NO 1のセンスミトコンドリアキメラRN
Aの逆方向反復配列と比較すると、63ヌクレオチドより短い。
【0102】
このより短い逆方向反復配列は、ミトコンドリアキメラRNAの構造においてRNA9
6ヌクレオチド(図1B)のより長いループを生成する。
配列の残部は、SEQID NO 1と同一である。
この新規なミトコンドリアキメラRNAは、SiHa細胞(図4A)には存在しない、こ
れは、PHAで刺激されたヒトリンパ球のような増殖型細胞中にも存在しない、HPVで
形質転換された発癌性細胞である(実施例参照)。
同様の結果は、HPV16又はHPV18で形質転換された又は不滅化された(しかし
発癌性ではない)細胞は、前−悪性細胞と考えられ、それ故新規なセンスミトコンドリア
キメラRNAは、前−悪性細胞の新規なポテンシャルマーカーである。
【0103】
16SミトコンドリアRNAに結合しているSEQ ID NO 2の逆方向反復配列の3’末端
の配列がSEQID NO 1の同じ領域とは異なるので、オリゴヌクレオチドプローブは、この
転写産物の特別の検出に用いることができる。
このプローブは、逆方向反復配列の3’末端とSEQ ID NO 7のオリゴヌクレオチドのよ
うなセンス16SミトコンドリアRNAの始めとの間の結合点の各端で7ないし10のヌ
クレオチドを含まなければならない。
このオリゴヌクレオチドは、in situハイブリダイゼーションもしくはRT−PCRに
よる増殖、又は前癌細胞のこの新規な特別のマーカーを検出する、この分野の技術者によ
く知られた任意の方法に用いることができる。
【0104】
[実施例3]
HTLV-1で形質転換されたヒトMT-2細胞は、第3の新規なセンスミトコンドリアキメラR
NAの発現を誘発する(図1C、SEQ ID NO 3)。
HTLV-1で形質転換されたヒトMT-2細胞は、記述された方法で培養された(Kobayashiet
al., EMBO J.,3:1339-1343, 1984)。
細胞は集菌され、300×gで10分間遠心分離され、PBSで2度洗浄された。最後
の細胞ペレットは、実施例1で記述したようにトリゾールで抽出された。
cDNAは、RNAを鋳型として用いてランダムヘキサマーで合成された、またcDN
Aは、リバースプライマー1および図1Aに記述されるようなフォワードプライマー3を
使用して、PCRにより増殖された。
以前に記述したように、この増殖プロトコルは、実施例1に記述したセンスミトコンド
リアキメラRNAに相当する、184ヌクレオチドの逆方向反復配列に結合したセンス1
6SミトコンドリアRNAの最初の31ヌクレオチドを含む210塩基対のアンプリコン
を生ずる。
増殖産物の電気泳動分析は、すでに記述された210塩基対のアンプリコンの存在に加
えて、約150塩基対のバンドを示した(図2を参照)。
【0105】
この150塩基対のアンプリコンの配列は、HPV16あるいはHPV18(SEQ ID NO
2)で形質転換された細胞中で発現する第2のセンスミトコンドリアキメラRNAに相当
する、実施例2で記述したアンプリコンの配列に同じである。
更に、新しい増殖産物は、約100bpであることが見出された(図2)。
この第3のアンプリコンの配列は、センス16SミトコンドリアRNAの5’末端に結
合して、167ヌクレオチドのループを発生する、61ヌクレオチドの逆方向反復配列を
示した(図1C;SEQID NO 3)。
この新規なアンプリコンは、正常細胞中に、腫瘍細胞中にまたEPV16あるいは18
で形質転換された細胞中には存在しなかった。
従って、この新規なセンスミトコンドリアキメラRNAは、発癌性レトロウイルスで形
質転換された細胞のポテンシャルマーカーである。
【0106】
16SミトコンドリアRNAに結合しているSEQ ID NO 3の逆方向反復配列の3’末端
の配列は、SEQID NO 1及びSEQ ID NO 2の同じ領域とは異なるので、オリゴヌクレオチド
プローブは、この転写産物の特定の検出に用いることができる。
このプローブは、逆方向反復配列の3’末端とSEQ ID NO 8のオリゴヌクレオチドのよ
うなセンス16SミトコンドリアRNAの始まりとの間の結合点の各端で7から10のヌ
クレオチドに及ばなければならない。
このオリゴヌクレオチドは、in situハイブリダイゼーションもしくはRT−PCRに
よる増殖、又はレトロウィルス腫瘍ウィルスで形質転換された細胞のこの特別なマーカー
を検出する、この分野の技術者によく知られた任意の方法に用いることができる。
【0107】
[実施例4]
ヒトアンチセンスミトコンドリアキメラRNAの構造
我々の最初の実験は、アンチセンスミトコンドリアキメラRNAに相当するキメラRN
Aの第二のファミリーは、いくつかの細胞研究に存在することを示した。
ヒトアンチセンスミトコンドリアキメラRNAの構造を決定するために、センスミトコ
ンドリアキメラRNAに用いられる方法が採用された(図1)。
理論的なアンチセンスミトコンドリアキメラRNAは、アンチセンス16Sミトコンド
リアRNAの5’末端に結合している逆方向反復配列としてのセンス16Sミトコンドリ
アRNAのフラグメントを含んでいた。
後者のRNAは、ミトコンドリアDNAのL−鎖から転写され、そして16Sミトコン
ドリア遺伝子に相当する(図3)。このRNAを増殖するために、リバースプライマーは
アンチセンス16SミトコンドリアRNAの5’末端に接近してハイブリダイズされた、
またフォワードプライマーは、逆方向反復配列(図3)の推定のフラグメントの異なる位
置でハイブリダイズされた。
PHAで48時間刺激されたヒトリンパ球からの全RNAは、鋳型として用いられた。
cDNAは、実施例1に記載したようにランダムヘキサマーで合成された。
【0108】
PCRによるcDNAの増殖は、アンチセンス16SミトコンドリアRNAの5’末端
の始めに接近した位置でのリバースプライマー(プライマー1、図3)及び逆方向反復配
列(図3)上に位置する異なるフォワードプライマーで行われた。
3つの主なアンプリコンだけが得られた、これは逆方向反復配列のサイズとループのサ
イズで異なっていた。
これらのアンプリコンは、精製され、配列が決定された。これらのアンチセンスミトコ
ンドリアキメラRNAの1つは、365ヌクレオチドの逆方向反復配列と17のヌクレオ
チド(SEQ IDNO 4)のループを含んでいる。
他のRNAは、96ヌクレオチドのループと189ヌクレオチド(SEQ ID NO 5)の逆方
向反復配列を含んでいる。
更に、アンチセンスミトコンドリアキメラRNAの他の種は、296ヌクレオチドの逆
方向反復配列と451ヌクレオチド(SEQ ID NO 6)のループを含んでいる。
3つのアンチセンスミトコンドリアキメラRNAのすべての配列は、ミトコンドリアD
NA遺伝子(HとL鎖)の配列と99.8%同一であった。
【0109】
以下の例において示される結果は、前腫瘍及び腫瘍細胞と、アンチセンスミトコンドリ
アキメラRNAの発現にかかわる正常な増殖細胞との間に大きな違いがあることを示す。
増殖する細胞はすべてセンスミトコンドリアキメラRNAを過剰発現する。
しかしながら、一方正常な増殖細胞もまたセンスミトコンドリアキメラRNAを発現す
るが、これらの転写産物は、腫瘍細胞中では下方制御される。非増殖細胞又は休止細胞は
共にミトコンドリアキメラRNAを発現しない。
従って、これらの異なる発現は、発癌の新規で有効なマーカーであることを示し、これ
はin situハイブリダイゼーション、ノーザンブロット分析、RT−PCRもしくはTM
A又はこの分野の技術者に知られている他の技術により検出できる。
センスとアンチセンスミトコンドリアキメラRNAの異なる発現の要約は、表1に示さ
れている。
【0110】
【表1】
【0111】
[実施例5]
腫瘍細胞系は、センスミトコンドリアキメラRNA(SEQ ID NO 1)を過剰発現し、またア
ンチセンスミトコンドリアキメラRNA(SEQ ID NO 4,5及び6)の発現を下方制御する。
in situハイブリダイゼーションは、培養中の腫瘍細胞系でセンスミトコンドリアキメ
ラRNAの発現を決定するのに用いられた。in situハイブリダイゼーションに対し、粘
着性腫瘍細胞は、8−ウエルのチャンバースライド(Lab-TekO, NUNC)中で、アメリカン組
織培養収集(AmericanTissue Culture Collection)(ATCC)により推奨された適切な培地
と条件を用いて37℃で24ないし48時間培養された。
非粘着性細胞(e.g.HL-60, Jurkat and Ramos)に対しては、小さなフラスコ中で37℃
で48時間培養された。細胞は、300×gで10分間遠心分離により回収され、PBS
の小さな容量中に再懸濁され、そして10から20ulのアリコートは、あらかじめポリ
リシン又はマッスル(mussel)から精製された接着タンパク質でコートされたガラススラ
イド上に塗布された(Burzioet al, Curr. Opin. Biotechnol., 8:309-312, 1997)。細胞
は室温で30分間乾燥された。
【0112】
前記細胞は、PBSで3回洗浄され、4%のパラホルムアルデヒドで室温下10分間固
定された。スライドはその後PBSで5分間3度洗浄され、0.2N HClで室温下10
分間インキュベートされた。
該細胞は、更に3度、最初は室温下にPBSで、その後2×SSCで10分間(2×SSC
:0.3MのNaCl、30mMクエン酸ナトリウム、pH7.0)洗浄された(Sambrook et al., 1989)。
プレハイブリダイゼーションは、4×SSC、10%硫酸デキストラン、150μg/ml
酵母tRNA及びニシン精液DNA、50%のホルムアミド及び1×Denhardt溶液(0.2mg
/mlのFicollタイプ400、0、2mg/mlのポリビニルピロリジン、0.2mg/mlのBSA)を含む溶液
中で37℃で30分間行われた。ハイブリダイゼーションは、センスとアンチセンスミト
コンドリアキメラRNAを標的にする3.5pmolのプローブを含む同じプレハイブリ
ダイゼーション混合物中37℃で15時間行った。
前記プローブは、センスとアンチセンスミトコンドリアキメラRNAの配列の異なる領
域をターゲットにする20以上のデオキシヌクレオチドを含んでいた(SEQ ID NO 99〜197
及びSEQ IDNO 9〜98をそれぞれ参照)。プローブは、既に記述したように予めジゴキシゲ
ニン-11-dUTP(Roche)とターミナル・トランスフェラーゼ(Promega)を用いて3’末端でラ
ベル表示された(Villegaset al., DNA & Cell Biol., 19:579-588, 2000)。
過剰のプローブを除去するために、スライドは、室温で、最初2×SSCで10分間及び1
×SSCで10分間洗浄された。その後、サンプルは室温で0.2×SSCで45℃30分間、及
び最後に0.2×SSCで10分間洗浄された。
【0113】
ハイブリダイゼーション後、細胞はブロッキング緩衝液中(1%BSA、0.3%トリトンX-100
in PBS)中で30分間インキュベートされ、その後予めブロッキング緩衝液中で1:50
0に希釈され、アルカリ性ホスファターゼに結合しているアンチ−ジゴキシゲニン単クロ
ーン抗体を用いて室温で2時間インキュベートされた。
最後に、スライドは、PBSで2度洗われた、そして呈色反応は以前に記載したようにBC
IP/NBT基質混合物(DAKO)を用いて実行された(Villegas et al., DNA & Cell Biol.
, 19:579-588,2000)。同様の操作は、フルオレスセインまたはローダミンに接合される
アンチ−ジゴキシゲニン抗体を使用して、FISHに対して行われた。
【0114】
図4Aと4Bに示されるように、SEQ ID NO 63に対応するジゴキシゲニンでラベル表示
したプローブを用いたin situハイブリダイゼーションは、ヒト腫瘍細胞がセンスミトコ
ンドリアキメラRNAを過剰発現することを示す。
ジゴキシゲニンでラベル表示した、SEQ ID NO 64に対応するセンスプローブを用いたin
situハイブリダイゼーションは、アンチセンスミトコンドリアキメラRNAの発現を下
方制御を示して、ネガティブであった(図4A及びB)。同じ結果は、センスあるいはアン
チセンスミトコンドリアキメラRNAの他の領域を標的にするオリゴヌクレオチドプロー
ブで得られた。
【0115】
[実施例6]
ヒト生体中の腫瘍細胞はセンスミトコンドリアキメラRNA(SEQ ID NO 1)を過剰発現
し、アンチセンスミトコンドリアキメラRNA(SEQ ID NOS 4、5及び6)を下方制御する。
ヒトの生体は、病理学者あるいはDAKOからの組織アレーから入手した
分析されたほとんどのサンプルはパラフィンに埋め込まれていたか、またはホルマリン
で固定されていた。他の組織サンプルは、Boiun固定剤で固定されていた、また他のサン
プルは、新鮮な凍結組織切片であった。
約4ないし8μmの組織切片は、予めポリリシン又はムラサキイガイ(Aulacomya ater)
から精製された接着性ポリフェノール蛋白質でコートされたガラススライド上に固定され
た(Burzioet al., Curr. Opin. Biotechnol., 8:309-312, 1997)。
パラフィンに包埋された組織切片は、60℃で1時間インキュベートされた、そしてパ
ラフィンは、キシロールで各15分間3度洗浄により除去された。
切片は、空気乾燥され、PBSで4回洗浄された。その後、切片は、0.2N HCl
を用いて、室温で10分間インキュベートされた、そしてPBSで十分に洗浄された。後
で、サンプルは、実施例4に記述されたプロトコルに従って、ジゴキシゲニンでラベル表
示されたアンチセンスプローブを用いてin situハイブリダイゼーションに供された。パ
ラレルセクションはセンスミトコンドリアキメラRNAの同じ領域に対応するセンスプロ
ーブでハイブリダイズされた。
【0116】
図5Aに示すように、胸、子宮頚部、膀胱の腫瘍及び肺の癌は、転移産物の強い存在を
示して、センスミトコンドリアキメラRNAを標的にするアンチセンスプローブで強く染
色されることを示した。他方、アンチセンスミトコンドリアキメラRNAを標的にするプ
ローブを用いたinsituハイブリダイゼーションは、この転写産物(図5A)の下方制御
を示してネガティブであった。
他の腫瘍細胞もまたセンスミトコンドリアキメラRNAを過剰発現し、そしてアンチセ
ンスミトコンドリアキメラRNAの発現を下方制御する(図5B)。
【0117】
[実施例7]
正常増殖細胞はセンス及びアンチセンストコンドリアキメラRNAを過剰発現する。
実施例5及び6に記述されたin situハイブリダイゼーションで同じプロトコルを使用
して、センスミトコンドリアキメラRNAの発現は、増殖細胞で決定した。
図6に示すように、HFK細胞、精原細胞、脾臓細胞、及び増殖型マウス胚は、センス
ミトコンドリアキメラRNAの過剰発現を示唆する、強いハイブリダイゼーションシグナ
ルを示した。
対照的に、脳、筋肉及び肝臓の細胞のような非増殖細胞は、センスミトコンドリアキメ
ラRNAが発現しないか又はこれらの細胞で下方制御されることを示唆して、シグナルを
示さなかった。
【0118】
しかしながら、驚くべき結果は、in situハイブリダイゼーションがアンチセンスミト
コンドリアキメラRNAを標的にするプローブで行われたとき、強いシグナルも観察され
た(図6)。
並行して分析されたいくつかの制御は、これらのプローブのハイブリダイゼーションシ
グナルが技術上の原因によらないことを示した。
もしinsituハイブリダイゼーションがジゴキシゲニンで非ラベル表示されたものを除
いて、過剰(50ないし100倍)の同じプローブと共にラベル表示されたプローブで行
われたら、ハイブリダイゼーションシグナルは消失した。
ハイブリダイゼーションに先立ちサンプルがリボヌクレアーゼAで一晩インキュベート
された場合、ハイブリダイゼーションシグナルは消失した。
また、ハイブリダイゼーションが4つのミスマッチを有するアンチセンスミトコンドリ
アキメラRNAを標的にするラベル表示されたプローブで行われた場合、ハイブリダイゼ
ーションシグナルは観察されなかった。
【0119】
[実施例8]
フィトヘムアグルチニン(PHA)で刺激された正常なヒトのリンパ細胞は、センス及びア
ンチセンスミトコンドリアキメラRNAを過剰発現する。健康なドナーから5mlの血液
がEDTAで集められた。この血液は、0.9%NaClの1容積で希釈され、該混合物
は、遠心分離機チューブ中の5mlのHistopaque-1077(Sigma)に適用された。このチュー
ブは室温下で20分間800×gで遠心分離された。
分裂期の白血球が集められ、0.9%のNaClの2容積で薄められ、室温で10分間、
250×gで遠心分離された。集められた細胞は、けん濁され、200mMグルタミン、
10mM非必須アミノ酸、ペニシリン、子牛の胎児血清を欠いているストレプトマイシン
が追加されたRPMI1640倍地で2度洗浄された。
最後の沈殿物は、10%子牛の胎児血清を含む同じ培地で再けん濁され、1ml当たり
のヒトリンパ球の数は、ノイバウエルチャンバー中の顕微鏡で数えて決定された。
【0120】
ヒトのリンパ球は、上記の追加されたRPMI1640倍地に10%の子牛の胎児血清
を添加した96ウエルのマイクロリッタープレート中で37℃、5%CO2の条件下で培
養された。1ウエル当たり約30,000のリンパ球は、10ug/mlの(細胞増殖を誘発す
る)ミトゲンPHA存在、非存在下で培養された(Yu et al., J. Biol. Chem., 266:7588
-7595, 1991)。
PHAで処理後48ないし72時間後、細胞は、H3−チミジン又はBrdUの取り込
みにより立証されたように活発にDNA合成にたずさわった(Yu et al., J. Biol. Chem.
,266:7588-7595, 1991)。また、PHAで刺激された48時間後に、リンパ球は、増殖細胞
核抗原又はPCNA及びKi−67のように、細胞増殖の他のマーカーを過剰発現した(B
antis et al.,Cytopathology, 15:25-31, 2004) (図7)。
休止中又はコントロール・リンパ細胞は、これらの抗原を発現しなかった(図7)。
【0121】
刺激されたリンパ球がセンスミトコンドリアキメラRNAを発現する否かを決定するた
めに、細胞は、ジゴキシゲニンでラベル表示され、センスミトコンドリアキメラRNAを
標的にするオリゴヌクレオチドプローブでin situハイブリダイズされた。
使用されたinsituハイブリダイゼーションプロトコルは、実施例5に記載された。
この転写産物の過剰発現を示唆する、強いハイブリダイゼーションシグナルが得られた
(図7)。ハイブリダイゼーションシグナルは、腫瘍細胞又は他の正常増殖細胞で観察され
たシグナルと同程度であった(図7を図4AとB、図5AとBと比較)。
ハイブリダイゼーションシグナルは、PHAなしでインキュベートされたコントロール
リンパ球には観察されなかった(図7)。
【0122】
このinsituハイブリダイゼーションがジゴキシゲニンでラベル表示され、アンチセン
スミトコンドリアキメラRNAを標的にするセンスオリゴヌクレオチドを用いて行なわれ
た場合、同様に強いハイブリダイゼーションシグナルが得られた(図7)。
いくつかのコントロールは、ハイブリダイゼーションシグナルが技術上の原因よる可能
性を否定するために行なわれた。
ハイブリダイゼーションシグナルは、もしin situハイブリダイゼーションが過剰(5
0ないし100倍)の(ジゴキシゲニンでラベル表示されていない)同一センスプローブ
と共にセンスラベル表示プローブで行なわれるときには、消失する。
ハイブリダイゼーションに先立ちサンプルをリボヌクレアーゼAで一晩インキュベート
すると、ハイブリダイゼーションシグナルは、消失する。
更に、ハイブリダイゼーションが4つのミスマッチを有するセンスプローブで行なうと
、ハイブリダイゼーションシグナルは、観察されない。対照的に、刺激を受けていないリ
ンパ球のinsituハイブリダイゼーションは、ハイブリダイゼーションシグナルを示さな
い(図7)。
結論として、増殖するために刺激された正常なヒトリンパ細胞は、センスミトコンドリ
アキメラRNA及びアンチセンスミトコンドリアキメラRNAの双方を過剰発現する。こ
れらの転写産物は、休止細胞中では発現しない。
【0123】
[実施例9]
センスミトコンドリアキメラRNAは、正常な及び腫瘍細胞中で異なる局在化を示す。
実施例5と6に記述したin situハイブリダイゼーションは、ヒトの生検の腫瘍細胞中
だけでなくいくつかの腫瘍細胞系においても、センスミトコンドリアキメラRNAは、細
胞質中で選択的に局在化される。しかしながら、ある腫瘍生検において、核中の転写産物
の明らかな局在化が見出された(図4A、B)。
【0124】
驚くべき発見は、核中のセンスミトコンドリアキメラRNAの局在化であった。
実施例5に記載したように実施されたin situハイブリダイゼーションは、HeLaと
SiHa細胞の核中にポジティブなハイブリダイゼーションシグナルを示した(図8)。
ハイブリダイゼーションシグナルは、HPV16で形質転換されたHFKの核中で強くな
った(図8)。核の局在化は、また胸腫瘍及び横紋筋肉腫からの腫瘍細胞中にも見出され
た(図8)。
【0125】
共−局在化(co-localization)の研究は、細胞質で局在化したセンスミトコンドリア
キメラRNAがミトコンドリアの外側で後期エンドゾーム/リゾソームと結合することを
示した。もし共−局在化の研究が分子プローブ(Mitotrack)又は抗体アンチ−シトクロム
c(Promega)もしくはアンチ−エンドヌクレアーゼG(Chemicon)のようなミトコンドリア
のマーカーで行なわれる場合、in situハイブリダイゼーションは、弱い共−局在化を示
した。
しかしながら、完全な共−局在化は、リソトラック(Lysotrack)(Molecular Probes)又
はアンチ-ランプ-2(anti-Lamp-2)(BD Pharmigen)もしくはアンチカプサイシンD(anti-
cathepsin D)(Zymed)のような後期エンドゾーム/リゾソームの免疫細胞学でハイブリダ
イゼーションシグナル中に見出された。
【0126】
HeLa細胞は、実施例5に記述したようなジゴキシゲニンでラベル表示されたオリゴ
ヌクレオチドプローブでin situハイブリダイズされた。
ポスト−ハイブリダイゼーションと洗浄操作後、細胞は、ローダミン(Roche)でラベル
表示されたアンチ−ジゴキシゲニン及びフルオレスセイン(BD Pharmingen)でラベル表示
されたanti-Lamp-2antibodyでインキュベートされた。暗所で室温下に3時間インキュベ
ートした後、スライドは洗浄され、マウントされてツァィス共焦点顕微鏡で分析された。
リソトラック(Lysotrack)又はアンチ−カテプシンD抗体がリソソーム分画として用
いられたとき、ランプ-2(Lamp-2)の局在化によるハイブリダイゼーションシグナルの
明らかな共−局在化が得られた。同様のハイブリダイゼーションシグナルの共−局在化結
果が得られた。
我々が知っている限り、これは、RNA(特にミトコンドリア転写産物)が細胞のリソ
ソームに結合されたことを示す最初の報告書である。
腫瘍細胞中のセンスミトコンドリアキメラRNAの局在化の決定は、癌を有する患者に
対する重要な予後の値となる。一般に、正常な増殖細胞中でセンス及びアンチセンスミト
コンドリアキメラRNAは、主に核中に局在化される。
【0127】
[実施例10]
アンチセンスミトコンドリアキメラRNAを標的にするアンチセンスオリゴヌクレオチ
ドを用いて腫瘍細胞をin vitro処理すると細胞死を誘発する。
HL-60細胞は、ATCCによって推奨された最適条件の下で培養される。
約30,000個の細胞が96―ウエルのマイクロタイタープレートで培養された。
センス又はアンチセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオチド
(2uM)が加えられた。細胞の浸透性を増すために、オリゴヌクレオチドは、リプフェ
クタミン(lipofectamin)もしくはオリゴフェクタミン(oligofectamin)(Invitrogen)
又はポリエチレンイミド(PEI)を有する混合物中に添加された( Exgen TM500, Fermenta
s)。細胞に特に非毒性であるPEIがより好ましい、
細胞は、オリゴヌクレオチドで6時間インキュベートされる、そして細胞残存の割合は
、トリパンブルーへの浸透性で決定される。
オリゴヌクレオチドで6時間インキュベートした後、細胞の主要な割合が死滅した。
しかしながら、アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオ
チドは、細胞死を誘発するのに更に有効であった(細胞死の15%に対し約90%)。
【0128】
他方、細胞がセンスもしくはアンチセンス12SミトコンドリアRNA又はND1サブ
ニットのmRNAを標的にするオリゴヌクレオチド、又はスクランブルオリゴヌクレオチ
ドもしくは4つのミスマッチを有するオリゴヌクレオチド(これらのすべてはコントロー
ルとして使用された)で処理されたとき、細胞死は生じなかった。
これらの研究に用いられたオリゴヌクレオチドは、5’末端で最初の5ヌクレオチドま
た3’末端で最初の5ヌクレオチドにホスフォロチオエートリンケージを含んでいる。平
均して、10の中央のヌクレオチドは、ホスフォジエステル結合を含んでいる。これらの
オリゴヌクレオチドを備えた細胞の処理がDNA断片化を引き起こすかどうか確証するた
めに、HL-60細胞は、オリゴヌクレオチドで6時間、以前に記述されたと同じ条件下でイ
ンキュベートされた。
約30,000個のHL-60細胞が、96ウエルのマイクロチッタープレート上で、200ul
のIDMEMプラス10%の子牛胎児血清中で、センスミトコンドリアキメラRNAを標
的とする又はアンチセンスミトコンドリアキメラRNAを標的とする1uMのオリゴヌク
レオチドと共に培養された。
【0129】
PEIを含む混合物中に添加されたオリゴヌクレオチドの化学は、前節に記載したのと
同様である。オリゴヌクレオチドで6時間定温放置した後、細胞は、TUNEL分析(DeadEnd
の比色定量のTUNELシステム、Promega)を用いてDNA断片化のために分析された。
表2に示されるように、アンチセンスミトコンドリアキメラRNAを標的とするオリゴ
ヌクレオチドで処理後、約96%の細胞がDNA断片化を示した。同様のDNA断片化率
は薬物スタウロスポリンで得られた。スクランブルオリゴヌクレオチド又はミスマッチを
有するオリゴヌクレオチドは効力を示さなかった。
対照的に、センスミトコンドリアキメラRNAを標的とするオリゴヌクレオチドで処理
されると、約20%の腫瘍細胞はアンチセンスミトコンドリアキメラRNAの発現を下方
制御し、そしてその結果これらの細胞は、この転写産物の少ないコピー数をもたらす(表
2)。
従って、細胞死は、アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌク
レオチドでより効率的に誘発される。
これらの結果は、腫瘍細胞中のアンチセンスミトコンドリアキメラRNAの少ないコピ
ー数は、治療目的になる、ことを強く示唆する。
【0130】
【表2】
【0131】
別の研究で、我々は、アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌ
クレオチドで細胞を処理するとカスパーゼ活性を誘発する、ことを決定した。
カスパーゼは、プログラムされた細胞死あるいはアポトーシスに活発に伴う、蛋白質分
解を生ずる酵素である。HL-60は、アンチセンスミトコンドリアキメラRNAを標的
にするオリゴヌクレオチド、又はスタウロスポリンを用いて、以前に記述された培養条件
下で6時間、インキュベートされた。
その後、フルオレセインで結合したVAD-fmk(CaspaCe FITC-VAD-FMK、Promega)を
培養に加え、37℃で30分間インキュベートされた。 VAD-fmokは、カスパーゼの強い
抑制剤で、非常に高い親和性でプロテアーゼに結合する(Gracia-Calvo et al., J. Biol.
Chem.,273:32608-32613, 1998)。
細胞は、遠心分離によって洗われ、標本化されて蛍光顕微鏡で観察された。
ズ9に示すように、アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌク
レオチドで処理したHL-60細胞は、スタウロスポリンで達成された活性化と同様のレ
ベルで、カスパーゼ活性を誘発した。12SミトコンドリアRNAを標的にしたアンチセ
ンスのオリゴヌクレオチドでは、カスパーゼ活性は得られなかった。
【0132】
アンチセンスミトコンドリアキメラRNAを標的にするオリゴヌクレオチドで処理した
細胞は、またアポトーシスに一致する他の変化を示す。電子顕微鏡分析は、核断片化およ
びクロマチン凝縮を示した。核断片化もDAPIで核を染色することにより実証された。
アンチセンスミトコンドリアキメラRNAを標的にするこれらのオリゴヌクレオチドで
処理後、細胞は、DAPI染色で示されたように(図9E及び9F)、核断片化を受ける。
【0133】
[実施例11]
アンチセンストコンドリアキメラRNAに相補するオリゴヌクレオチドで処理されると、
他の癌細胞も細胞死を被る。
他の癌細胞は、実施例10に記載したプロトコルに従い、アンチセンストコンドリアキ
メラRNAに相補するオリゴヌクレオチドで処理された。この細胞は、ATCCのリコメ
ンドに従い、それらの最適条件下でインキュベートし、そして2uMオリゴヌクレオチド
がPEIと共に実験の初期段階で添加された。
6時間後に、オリゴヌクレオチドの2回目の添加を同様の濃度で実行し、結果は、実験
の開始の後15時間で決定された。細胞死は、DAPI染色と断片化された核の細胞数を
数えることにより決定された。表3に示すように、オリゴヌクレオチドで処理された細胞
の70%以上がアポトーシスを被った。
薬剤処理に全くの耐性を示すことが知られている、黒色腫細胞、リンパ腫細胞および胸
癌細胞MCF/7は、非常に高い割合でアポトーシスを被ることに気づくことは重要であ
る(表3)。
【0134】
【表3】
【0135】
アポトーシスを誘発する際にオリゴヌクレオチドに対するより有効な標的となる転写産
物の領域があるかを決定するために、次の実験を行った。アポトーシスの誘発は、アンチ
センストコンドリアキメラRNAの5’末端から始まっておよそ各30ヌクレオチドを標
的とした、約20ヌクレオチドのアンチセンスオリゴヌクレオチドを有するHeLa、H
E-60及びMCF/7細胞において研究された。
時間0では、1uMオリゴヌクレオチドがPEIと共に加えられた、そしてこの処理は
6時間後に繰り返された。処理開始後15時間でアポトーシスを被る細胞の割合は、DA
PI染色と断片化された核を有する細胞を数えることにより決定された。
殆どのオリゴヌクレオチドは、変わりうる程度のアポトーシスを誘発したが、アンチセ
ンスミトコンドリアキメラRNAの単一鎖領域は、細胞死を誘発するよい標的であった。
推定二重鎖又はアンチセンスミトコンドリアキメラRNAのループ構造を標的にするオリ
ゴヌクレオチドは、それほど有効ではない
【0136】
アポトーシスは、またトリパンブルー染色、ヨウ化プロピジウム染色、アネキシン(an
exine)免疫化学により決定することができる。 これらの技術において、細胞は、蛍光
顕微鏡又はフロー・サイトメトリーにより分析することができる。DNA断片化は、TUNE
L、あるいはDNAラダーを示す電気泳動によって測定されうる。
ウエスタンブロット分析もまた、カスパーゼ、poly(ADP Rib)合成等のような蛋白質の
プロセッシングを決定するために使用することができる。
【0137】
[実施例12]
アンチセンスミトコンドリアキメラRNAに相補するオリゴヌクレオチドで正常増殖また
は休止細胞の処理は、アポトーシスに効果がない。以前に記載したように、正常増殖細胞
は、アンチセンスミトコンドリアキメラRNAと同様にセンスミトコンドリアキメラRN
Aを過剰発現する。
他方、休止細胞は、これらの転写産物を発言しない。
それ故、アンチセンスミトコンドリアキメラRNAに相補するオリゴヌクレオチドが正
常細胞において細胞死を誘発するかを決定することは重要である。ヒトリンパ細胞は、実
施例8に記述したように10ug/mlのPHAで48時間刺激された。
並行して、コントロールリンパ細胞がPHAなしで48時間インキュベートされた。
48時間の培養では、PEI(実施例10参照)と混合した15uMのオリゴヌクレオチ
ドは、刺激されそしてコントロールされたリンパ細胞に添加された、そして更に15時間
インキュベートされた。オリゴヌクレオチドの濃度は、以前の実験(1−2uM)に用い
た濃度よりも10フォールド高かった。
刺激されそしてコントロールされたリンパ細胞の他の例は、同じ期間の間、0.4uM
スタウロスポリンで処理された。実験の終わりに、細胞死は、トリパンブルー染色又はD
API染色のいずれかにより測定された。
図10に示すように、コントロールリンパ細胞またはオリゴヌクレオチドを含まない1
5時間のインキュベートでPHA刺激されたリンパ細胞は、異なる実験で7から10%間
の変動で自然におこるアポトーシスと同様のレベルを示した。
【0138】
同様の結果は、オリゴヌクレオチドのより低い(1-2 uM)濃度で得られた。
更に、15uMアンチセンスオリゴヌクレオチドで15時間インキュベートしたコント
ロール及び刺激されたリンパ細胞は、同じ低レベルのアポトーシス(およそ10%)を示
した(図10)。
対照的に、コントロールリンパ細胞又はPHA刺激され、スタウロスポリンで15時間
インキュベートされたリンパ細胞は、80%以上の細胞がアポトーシスを被ることを示し
た(図10)。これは非常に重要な結果である、なぜならヒトリンパ細胞のような正常休
止細胞又は正常増殖細胞がアンチセンスミトコンドリアキメラRNAに相補するオリゴヌ
クレオチドによりアポトーシスを誘発するのに抵抗性であるからである。言いかえれば、
アンチセンスミトコンドリアキメラRNAで阻害することにより、腫瘍細胞中のアポトー
シスの誘発が癌に対する選択的治療アプローチとなるからである。
【0139】
引用文献
1.Adrain et al.,“Regulation of apoptotic protease activating factor-1 oligomer
ization andapoptosis by the WD-40 repeat region”, J Biol Chem. 274:20855-20860
, 1999.
2.Bantis etal., “Expression of p129, Ki-67 and PCNA as proliferation markers i
n imprintsmears of prostate carcinoma and their prognostic value”, Cytopatholo
gy, 15:25-31,2004.
3.Beaucage, “Oligodeoxyribonucleotidessynthesis. Phosphoramidite approach”,
Methods Mol.Biol. 20:33-61, 1993.
4.Benedict etal., “Expression and functional analysis of Apaf-1 isoforms. Extr
a Wd-40 repeatis required for cytochrome c binding and regulated activation of
procaspase-9”,J Biol Chem. 275:8461-8468, 2000.
5.Benoist andChambon, “In vivo requirement of the SV40 early promoter region”
, Nature290:304-310, 1981.
6.Boya et al., “Mitochondrion-targetedapoptosis regulators of viral origin”,
Biochem.Biophys. Res. Commun. 304:575-581, 2003.
7.Brinster etal., “Regulation of metallothionein-thymidine kinase fusion plasm
ids injectedinto mouse eggs”, Nature 296:39-42, 1982.
8.Burzio etal., “Enviromental bioadhesion: themes and applications”, Curr. Op
in.Biotechnol., 8:309-312, 1997.
9.Capaldi etal., “Highly efficient solid phase synthesis of oligonucleotide an
alogs containingphosphorodithioate linkages”, Nucleic Acids Res. 28:E40, 2000.
.
10.Carew andHuang, “Mitochondrial defects in cancer”, Mol. Cancer, 1:1-12, 20
02.
11.Celis, “CellBiology, A Laboratoty Handbook”, Julio E. Celis, ed., 1994
12.Chinnery andTurnbull, “Mitochondrial DNA mutations in the pathogenesis of h
uman disease”,Mol. Med. Today, 6:425-432, 2000.
13.Clayton andVinograd, “Circular dimer and catenate forms of mitochondrial DN
A in humanluekaemic leucocytes”, Nature, 216:652-657, 1967.
14.Clayton andVinograd, “Complex mitochondrialDNA in leukemic and normal human
myeloidcells”, Proc. Natl. Acad. Sci. US, 62:1077-1084, 1969.
15.Clayton, “Transcriptionand replication of mitochondrial DNA”, Hum Reprod.
Suppl2:11-17, 2000.
16.Comanor etal., “Successful HCV genotyping of previously failed and low vira
l loadspecimens using an HCV RNA qualitative assay based on transcription-media
tedamplification in conjunction with the line probe assay.”, J. Clin Virol., 2
8:14-26,2003.
17.Egholm etal., “PNA hybridizes to complementary oligonucleotides obeying the
Watson-Crickhydrogen-bonding rules”, Nature, 365:566-568, 1993.
18.Elbashir etal., “Duplexes of 21-nucleotides RNAs mediate RNA interference i
n cultured mammaliancells,” Nature 411:494-498, 2001.
19.Falkenberget al., “Mitochondrial transcription factors B1 and B2 activate t
ranscription ofhuman mtDNA”, Nat Genet. 31:289-294, 2002.
20.Ferri andKroemer, “Organelle-specific initiation of cell death pathways”,
Nature CellBiol. 3:E255-263, 2001.
21.Frederick etal., in “Current Protocols In Molecular Biology”, Volume 2, Un
it 14,Frederick M. Ausubul et al. eds., 1995.
22.Gracia-Calvoet al., “Inhibition of human caspases by peptide-based and macr
omolecularinhibitors”, J. Biol. Chem., 273:32608-32613, 1998.
23.Guicciardiet al., “Lysosomesin cell death”, Oncogene, 23:2881-2890, 2004.
24.Haseloff etal., “Sequence required for self-catalysed cleavage of the satel
lite RNA oftabacco ringspot virus”, Gene, 82:43-52, 1989.
25.Hausen, “Papillomavirus infection- a major cause of human cancer,” Biochim
.Biophys. Acta,1288:F55-F78, 1996.
26.Hedge etal., “Commitment to apoptosis induced by tumour necrosis factor-alp
ha is dependenton caspase activity”, Apoptosis,7:123-132, 2002.
27.Hyrup andNielsen, “Peptide nucleic acids (PNA): synthesis,properties and po
tentialapplications”, Bioorg. Med. Chem., 4:5-23, 1996.
28.Johnstone etal., “Apoptosis: a link between cancer genetics and chemotherap
y”, Cell108:153-164, 2002.
29.Kobayashi etal., “Genomic structure of HTLV: detection of defective genome
and itsamplification in MT2 cells”, EMBO J., 3:1339-1343, 1984
30.Komarov etal., “A chemical inhibitor of p53 that protect mice from the side
effectsof cancer therapy”, Science 285:1733-1737, 1999.
31.Krammer, “CD95’sdeadly mission in immune system”, Nature 407:789-795, 200
0.
32.Li et al., “EndonucleaseG is an apoptotic DNase when released from mitochon
dria”, Nature,412:95-99, 2001.
33.Liu et al., “Syntheticpeptides and non-peptidic molecules as probes of stru
cture andfunction of Bcl-2 family proteins and modulators of apoptosis”, Apopt
osis,6:453-462, 2001..
34.Lu et al., “siRNA-mediatetedantitumorogenesis for drug target validation an
d therapeutics,”Curr. Opin. Mol. Ther. 5:225-234, 2003.
35.Mag et al., “Synthesisand selective cleavage of an oligodeoxynucleotide con
taining abridged internucleotide 5'-phosphorothioate linkage”,
36.Martins etal., “The serine protease Omi/HtrA2 regulates apoptosis by bindin
g XIAP througha reaper-like motif”, J. Biol. Chem. 277:439-444, 2002.
37.McCulloch etal., “A human mitochondrial transcription factor is related to
RNA adenine methyltransferasesand binds S-adenosylmethionine”, Mol. Cell Biol.
22:1116-1125,2002.
38.McKay etal., “Characterization of a potent and specific class of antisense
oligonucleotideinhibitor of human protein kinase C-a expression”, J. Biol. Ch
em.,274:1715-1722, 1999.
39.McKinnell etal., “The Biological Basis of Cancer, (R:G:McKinnell, R:E: Parc
hment, A:O:Perantoni, G:B: Pierce, eds.), Ch.3, Cambridge University Press, UK,
1998
40.McManus etal., “Gene silincing in mammals by small interfering RNAs,” Natu
re Rev.Genet.3: 737-747, 2002.
41.Meier etal., “Apoptosis in development”, Nature 407:796-801, 2000.
42.Myers etal., “Reverse transcription and DNA amplification by a Thermus ther
mophilus DNApolymerase”, Biochemistry, 30:7661-7666, 1991.
43.Nielsen etal., “Peptide nucleic acids (PNA): oligonucleotide analogs with a
polyamidebackbone”, in S. Crooke, B. Lebleu (eds.) Antisense Research and App
lications, Ch9, CRC Press, Boca Raton, Florida, pp. 363-373, 1993.
44.Parisi and Clayton,“Similarity of human mitochondrial transcription factor
1 to highmobility group proteins”, Science. 252:965-969, 1991.
45.Parrella etal., “Detection of mitochondrial DNA mutations in primary breast
cancerand fine-needle aspirates”, Cancer Res., 61:7623-7626, 2001.
46.Rampino etal., “Somatic frameshift mutations in the BAX gene in colon cance
rs of themicrosatellite mutator phenotype”, Science, 275:967-969, 1997.
47.Rantanen et al.,“Characterization of the mouse genes for mitochondrial tran
scriptionfactors B1 and B2”, Mamm Genome. 14:1-6, 2003.
48.Ravagnan etal., “Heat-shock protein 70 antagonizes apoptosis-inducing facto
r”,
Nat Cell Biol.3:839-843, 2001.
49.Reed, “Dysregulationof apoptosis in cancer”, J. Clin. Oncol., 17:2941-2953
, 1999.
50.Rossi, “Practicalribozymes. Making ribozymes work in cells”, Curr Biol. 4:
469-471, 1994.
51.ambrook etal., Molecular Cloning. A Laboratory Manual, (Sambrook, J., Frits
ch, E.T. andManiatis, T. Eds.,) 2nd Edn, Cold Spring Harbor Laboratory, Cold S
pring Harbor,NY. 1989.
52.Samejima etal., “CAD/DFF40 nuclease is dispensable for high molecular weigh
t DNA cleavageand stage I chromatin condensation in apoptosis”, J Biol Chem. 2
76:45427-45432,2001.
53.Shuey andAttardi, “Characterization of an RNA polymerase activity from HeLa
cellmitochondria, which initiates transcription at the heavy strand rRNA promo
ter and thelight strand promoter in human mitochondrial DNA”, J Biol Chem. 260
:1952-1958,1985.
54.Stephens andRivers, “Antisense oligonucleotide therapy in cancer”, Curr. O
pin. Mol.Therapeut., 5:118-122, 2003
55.Summerton, “Morpholinoantisense oligomers: a case for an Rnase H-independen
t structural type”,Biochim. Biophys. Acta, 1489:141-158, 1999.
56.Suzuki etal., “A serine protease, Htra2, is<released from the mitochondria
and interactswith xiap, inducing cell death”, Mol. Cell, 8:613-621, 2001.
57.Taanman, “The mitochondrial genome: structure, transcription, translation a
nd replication”, Biochim. Biophys. Acta, 1410:103-123, 1999.
58.Tan et al., “Comprehensivescanning of somatic mitochondrial DNA mutations i
n breast cancer”,Cancer Res., 62:972-076, 2002.
59.Teitz etal., “Caspase 8 is deleted or silenced preferentially in childhood
neuroblastomaswith amplification of MYCN”, Nature Med. 6:529-535, 2000.
60.Tidd et al.,“Oligodeoxynucleotide 5mers containing a 5’-CpG induce apoptos
is through a mitochondrialmechanism in T lymphocytic leukemia cells,” Nucleic
Acids Res. ,28:2242-2250 (2000).
61.Tiranti etal., “Identification of the gene encoding the human mitochondrial
RNApolymerase (h-mtRPOL) by cyberscreening of the Expressed Sequence Tags data
base”, Hum MolGenet. 6:615-625, 1997.
62.Verhagen etal., “Cell death regulation by the mammalian IAP antagonist Diab
lo/Smac”,Apoptosis, 7:163-166, 2002.
63.Vickers etal., “Efficient reduction of target RNAs by small interfering RNA
and RnaseH-dependent antisense agents”, J. Biol. Chem., 278:7108-7118, 2003.
64.Villegas etal., “A novel chimeric mitochondrial RNA localized in the nucleu
s of mousesperm”, DNA & Cell Biol. 19:579-588, 2000.
65.Villegas etal., “A putative RNA editing from U to C in a mouse mitochondria
l transcript”,Nucleic Acids Res. 30:1895-1901, 2002.
66.Vogelsteinet al., “Surfing the p53 network”, Nature 408: 307-310, 2000.
67.Wacheck etal., “ Small interfering RNA targeting Bcl-2 sensitizes malignant
melanoma,”Oligonucleotides 13:393-400, 2003.
68.Wagner etal., “Nucleotide sequence of the thymidine kinase gene of herpes s
implex virustype 1”, Proc. Natl. Acad. Sci. U.S.A. 78:1441-1445, 1981.
69.Wahlestedtet al., “Potent and nontoxic antisense oligonucleotides containin
g lockednucleic acids”, Proc. Natl. Acad. Sci. US, 97:5633-5638, 2000.
70.Warburg, “Onthe origin of cancer cells”, Science, 123:309-314, 1956.
71.Wu et al., “Immunofluorescentlabelingof cancer marker Her2 and other cellul
ar targets withsemiconductor quantum dots,” Nature Biotechnol. 21:41-46, 2003.
72.Yamamoto etal., “Identification of a functional promoter in the long termin
al repeat ofRous sarcoma virus”, Cell 22:787-797, 1980
73.Yu et al., “VitaminD receptor expression in human lymphocytes. Signal requi
rements andcharacterization by western blots and DNA sequencing”, J. Biol. Che
m.,266:7588-7595, 1991.
74.Zaug et al.,“A labile phosphodiester bond at the ligationjunctionin a circu
lar interveningsequence RNA”, Science, 224:574-578, 1984.
75.Zornig etal., “Apoptosis regulators and their role in tumorogenesis”, Bioc
him.Biophys.Acta, 1551:F1-F37, 2001.
【化1】
【化2】
【化3】
【化4】
【化5】
【化6】
【化7】
【化8】
【化9】
【化10】
【化11】
【化12】
【化13】
【化14】
【化15】
【化16】
【化17】
【化18】
【化19】
【化20】
【化21】
【化22】
【化23】
【化24】
【化25】
【化26】
【化27】
【化28】
【化29】
【化30】
【化31】
【化32】
【化33】
【化34】
【化35】
【特許請求の範囲】
【請求項1】
本願明細書に記載された発明。
【請求項1】
本願明細書に記載された発明。
【図1】


【図2】
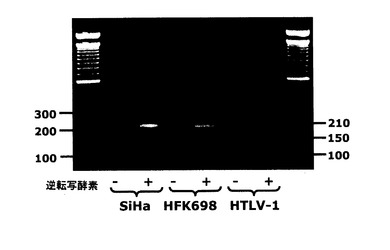
【図3】
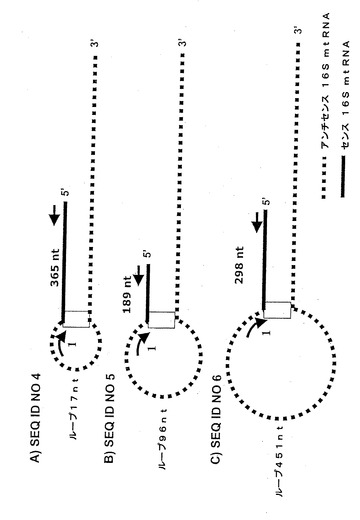
【図4A】

【図4B】
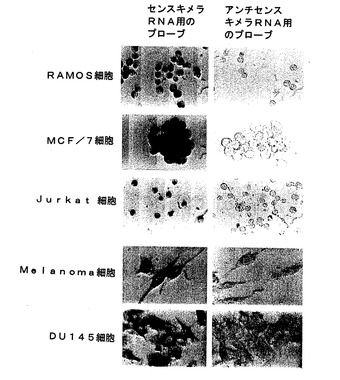
【図5A】

【図5B】
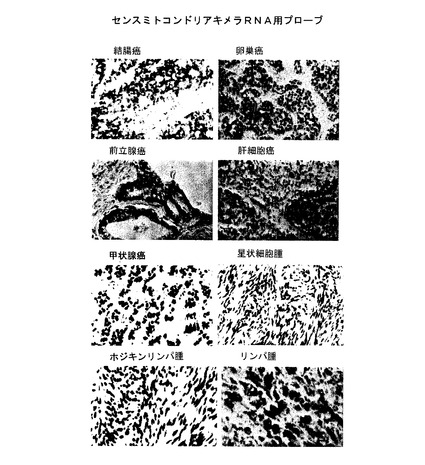
【図6】
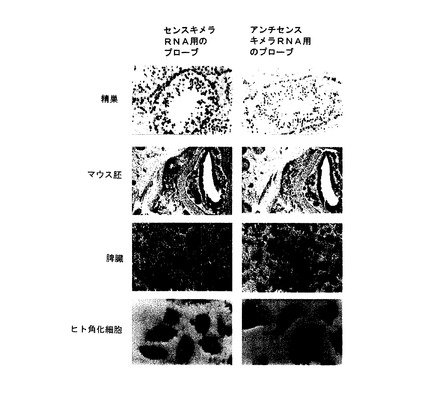
【図7】

【図8】
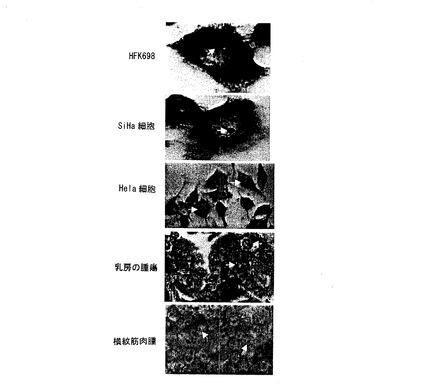
【図9】

【図10】

【図2】
【図3】
【図4A】
【図4B】
【図5A】
【図5B】
【図6】
【図7】
【図8】
【図9】
【図10】
【公開番号】特開2013−66478(P2013−66478A)
【公開日】平成25年4月18日(2013.4.18)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−269587(P2012−269587)
【出願日】平成24年12月10日(2012.12.10)
【分割の表示】特願2006−533267(P2006−533267)の分割
【原出願日】平成16年5月21日(2004.5.21)
【出願人】(310008941)アンデス バイオテクノロジーズ ソシエダード アノニマ (1)
【氏名又は名称原語表記】ANDES BIOTECHNOLOGIES S.A.
【住所又は居所原語表記】Avda. Zanartu 1482,Nunoa,Santiago,Chile
【Fターム(参考)】
【公開日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願番号】特願2012−269587(P2012−269587)
【出願日】平成24年12月10日(2012.12.10)
【分割の表示】特願2006−533267(P2006−533267)の分割
【原出願日】平成16年5月21日(2004.5.21)
【出願人】(310008941)アンデス バイオテクノロジーズ ソシエダード アノニマ (1)
【氏名又は名称原語表記】ANDES BIOTECHNOLOGIES S.A.
【住所又は居所原語表記】Avda. Zanartu 1482,Nunoa,Santiago,Chile
【Fターム(参考)】
[ Back to top ]