
単量体抗体Fc
本発明は、単量体Fcポリペプチド、およびこうしたポリペプチドを作製しそして用いる方法に関する。ポリペプチドは、CH3領域中の1またはそれより多い疎水性接合部分残基の極性アミノ酸での置換を含む。

【発明の詳細な説明】
【背景技術】
【0001】
[0001]抗体は、侵襲する非自己分子に対する防御において、中心的な役割を果たす。抗体が、pH依存方式で、新生Fc受容体(FcRn)と相互作用する能力によって、血清半減期が延長される(GhetieおよびWard 2000)。抗体のこのユニークな特徴のため、Fc融合分子を操作することによって、血清における療法タンパク質またはペプチドの半減期を延長することが可能になる。天然存在IgG抗体および操作Fc融合分子は二価であり、そして単一特異性である。これは、Fcがホモ二量体の性質を持つためである。特定の療法適用のため、抗体または抗体のFc断片によって与えられるすべての正の特質を保持するが、単量体Fcを操作することによって、一価特異性を達成することが望ましいであろう。
【0002】
[0002]抗体は、IgG、IgA、IgE、IgM、およびIgDを含むタンパク質の免疫グロブリンクラスに属する。ヒト血清において最も豊富な免疫グロブリンクラスはIgGであり、その模式的構造を図1に示す(Deisenhofer 1981; Huber 1984; Roux 1999)。IgG構造は、4つの鎖、2つの軽鎖および2つの重鎖を有し;各軽鎖は2つのドメインを有し、そして各重鎖は4つのドメインを有する。抗原結合部位は、可変軽(VL)鎖および可変重(VH)鎖ドメイン、ならびに定常軽(LC)鎖および定常重(CH1)鎖ドメインを含有するFab領域(抗原結合断片)中に位置する。抗体のFc(結晶化可能断片)断片は、重鎖のCH2およびCH3ドメイン領域を含有する。IgG分子は、ヒンジ領域でジスルフィド結合(−S−S−)によって一緒に保持される2つの重鎖、および2つの軽鎖を有する、ヘテロ四量体と見なされることも可能である。ヒンジ・ジスルフィド結合の数は、免疫グロブリンサブクラス間で多様である(PapadeaおよびCheck 1989)。FcRn結合部位は、抗体のFc領域中に位置し(Martin、Westら 2001)、そしてしたがって抗体の血清半減期延長特性は、Fc断片中に保持される。Fc領域は、単独では、CH2およびCH3ドメインを含む重鎖のホモ二量体と考えられうる。
【先行技術文献】
【非特許文献】
【0003】
【非特許文献1】GhetieおよびWard 2000
【非特許文献2】Deisenhofer 1981
【非特許文献3】Huber 1984
【非特許文献4】Roux 1999
【非特許文献5】PapadeaおよびCheck 1989
【非特許文献6】Martin、Westら 2001
【発明の概要】
【発明が解決しようとする課題】
【0004】
[0003]本明細書に提供するのは、ポリペプチドがホモ二量体を形成する能力を有意に減少させる、CH3接合部分ドメイン中の改変を含有するFcポリペプチドである。好ましい態様において、二量体化の減少はほぼ100%である。好ましくは、Fcポリペプチドには、CH3ホモ二量体形成に静電的に好ましくない、1またはそれより多い荷電アミノ酸、および1またはそれより多い疎水性CH3接合部分残基の極性アミノ酸残基、例えばスレオニンでの置換が含まれる。
【0005】
[0004]特定の態様において、CH3ドメインは、392位および/または409位で、負に荷電されたアミノ酸、例えばアスパラギン酸を含む、CH3ホモ二量体形成に静電的に好ましくない改変、ならびにY349、L351、L368、V397、L398、F405、およびY407からなる群より選択される、1またはそれより多い置換された疎水性接合部分残基を有する、ヒトIgG CH3ドメインである。
【0006】
[0005]単量体Fcポリペプチドは、抗体CH1ドメインをさらに含んでもよいし、または抗体重鎖内で構成される。特定の態様において、単量体抗体は、単量体重鎖および軽鎖を含み、本質的に半抗体(half−antibody)を生成する。単量体重鎖は、ジスルフィド結合形成を防止するため、1またはそれより多い突然変異システイン残基を有してもよい。特に有用なシステイン突然変異は、重鎖ヒンジ領域中のものである。
【0007】
[0006]本発明の1つの側面において、ポリペプチドは、野生型CH3ドメインを含むポリペプチドに比較して、ホモ二量体を形成する能力が減少した、抗体CH3ドメインを含む。好ましいポリペプチドは、抗体のCH3ドメインであって、1またはそれより多い荷電アミノ酸が、CH3ホモ二量体形成に静電的に好ましくないアミノ酸で置換され、そして1またはそれより多い疎水性接合部分残基が、極性アミノ酸で置換されるように、野生型CH3ドメインと異なるアミノ酸配列を含む、前記CH3ドメインを含む。
【0008】
[0007]本発明の他の側面は、単量体Fcポリペプチドをコードする核酸、こうした核酸を含む発現ベクター、およびこうした発現ベクターを含有する宿主細胞である。
[0008]本発明の態様には、単量体Fcポリペプチドを調製する方法がさらに含まれる。好ましい態様において、こうした方法は、単量体Fcポリペプチドをコードする核酸を含む宿主細胞を、単量体Fcポリペプチドが発現される条件下で培養し、そして次いで、宿主細胞培養から単量体Fcポリペプチドを回収する工程を含む。
【図面の簡単な説明】
【0009】
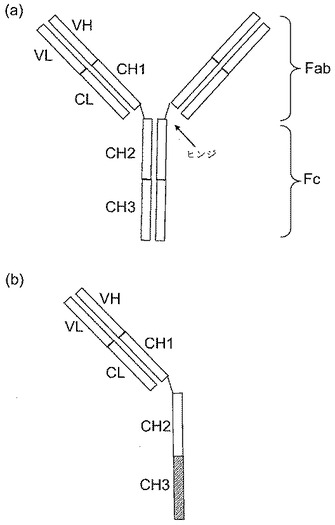
【図1】[0009](a)示すドメインを含むIgG1抗体の模式図。IgG1抗体は、2つの重鎖(より長い長さ)および2つの軽鎖(より短い長さ)を含むY型の四量体である。2つの重鎖は、ヒンジ領域で、ジスルフィド結合(−S−S−)によって一緒に連結される。Fab−抗原結合断片、Fc−結晶化可能断片、VL−可変軽鎖ドメイン、VH−可変重鎖ドメイン、CL−定常(配列変動なし)軽鎖ドメイン、CH1−定常重鎖ドメイン1、CH2−定常重鎖ドメイン2、CH3−定常重鎖ドメイン3。(b)一価抗体の模式図−単量体Fcに融合したFab。この場合、CH3ドメイン接合部分は、アミノ酸突然変異を通じて修飾される。
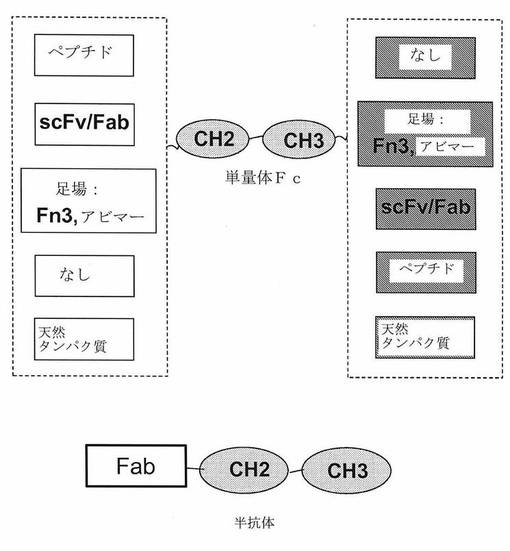
【図2】[0010]図は、単量体(または一価)Fcを含む態様のいくつかを示す。これらには、単量体FcのNおよびC末端両方への融合が含まれる。Fcは、二量体化を伴わなくてさえ、またはFabドメインがなくてさえ、FcRn受容体と相互作用する能力を保持し、単量体Fcと融合しているタンパク質/ドメインの、より長い血清半減期を導く。scFv−一本鎖可変ドメイン断片。
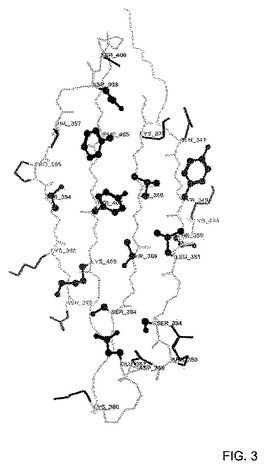
【図3】[0011]ドメイン−ドメイン相互作用に関与する残基を示した、CH3ドメイン接合部分構造。距離カットオフ法を用いて、接合部分残基を同定した。構造的に保存されそして埋め込まれた(buried)(溶媒アクセス可能表面積≦10%)残基を球棒モデルで示す。溶媒に暴露されるかまたは構造的に保存されていない残基を、棒提示で示す。分析は、高解像度(1.65Å)で決定されている、IgG1結晶構造(PDBコード: 1L6X)に基づく(Idusogie, Prestaら 2000)。
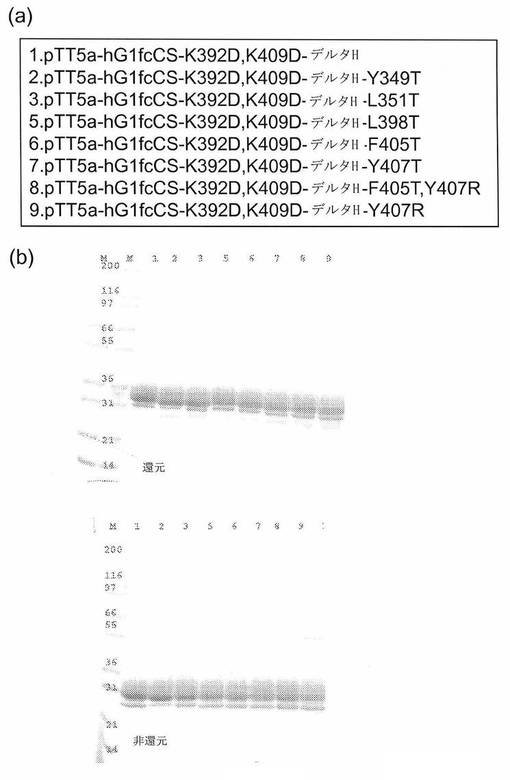
【図4】[0012](a)FcのCH3ドメイン接合部分における突然変異を含めて設計した構築物のリスト、および(b)8つの精製Fc突然変異体タンパク質に関するGELCODETM Blue染色試薬で染色したSDS−PAGEクーマシーゲル。表2の突然変異体4はタンパク質の量が不十分であったため、含めなかった。構築物は、これらのFc構築物がヒンジ・ジスルフィドを欠くという事実のため、還元および非還元ゲル中を同様に移動する。言い換えると、ここでの目的は単量体Fc重鎖を達成することであるため、IgG分子とは異なり、2つの重鎖を連結する重鎖間ジスルフィドはない。
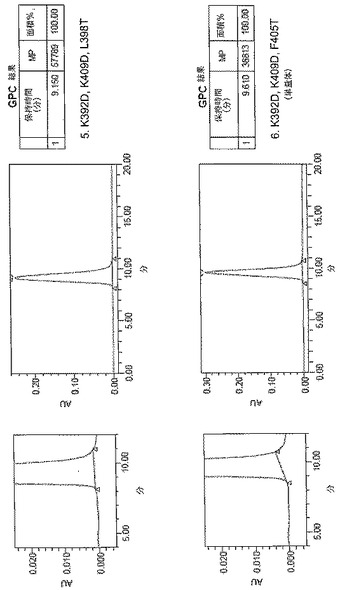
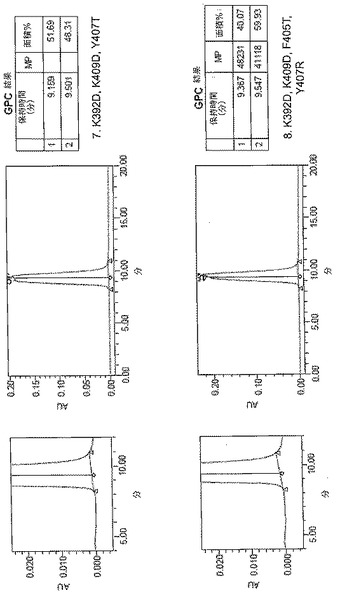
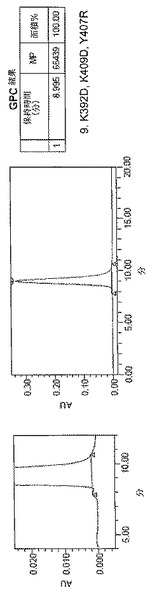
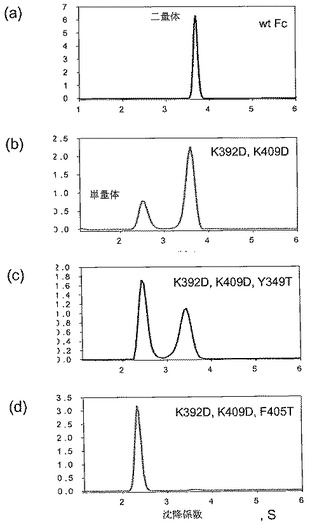
【図5A】[0013]図5A〜5E。表2に列挙する9つの構築物すべてに関するサイズ排除クロマトグラフィー(SEC)プロファイル。例示的な単量体SECプロファイルを示す構築物を「単量体」と標識する。
【図5B】[0013]図5A〜5E。表2に列挙する9つの構築物すべてに関するサイズ排除クロマトグラフィー(SEC)プロファイル。例示的な単量体SECプロファイルを示す構築物を「単量体」と標識する。
【図5C】[0013]図5A〜5E。表2に列挙する9つの構築物すべてに関するサイズ排除クロマトグラフィー(SEC)プロファイル。例示的な単量体SECプロファイルを示す構築物を「単量体」と標識する。
【図5D】[0013]図5A〜5E。表2に列挙する9つの構築物すべてに関するサイズ排除クロマトグラフィー(SEC)プロファイル。例示的な単量体SECプロファイルを示す構築物を「単量体」と標識する。
【図5E】[0013]図5A〜5E。表2に列挙する9つの構築物すべてに関するサイズ排除クロマトグラフィー(SEC)プロファイル。例示的な単量体SECプロファイルを示す構築物を「単量体」と標識する。
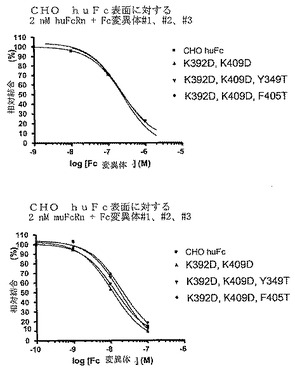
【図6】[0014]野生型および突然変異体Fc構築物に対するヒトおよびマウスFcRn結合のBIACORE分析。
【図7】[0015]分析用超遠心(AUC)を用いたタンパク質サイズ分析。
【図8】[0016]ヒトおよびマウスIgGサブクラスのCH3ドメインの配列比較。同定されたCH3ドメイン接合部分残基(24、表1;「*」によって示す)は非常に保存される。したがって、ヒトIgG1 Fcにおける単量体化を導く突然変異を他のIgGサブクラスに、ならびにヒト以外の種に拡張することも可能である。(a)において、ヒトIgG1、IgG2、IgG3、およびIgG4由来の配列は、それぞれ、配列番号1、配列番号2、配列番号3、および配列番号4に対応する。(b)において、ヒトIgG1、マウスIgG1、マウスIgG2a、マウスIgG2b、およびマウスIgG3由来の配列は、それぞれ、配列番号5、配列番号6、配列番号7、配列番号8、および配列番号9に対応する。
【図9】[0017]実施例2に記載するような、マウスにおける多様なFc融合分子の薬物動体学。
【発明を実施するための形態】
【0010】
[0018]野生型Fcは天然でホモ二量体であり、そしてこの特徴は、2つのCH3ドメイン間に存在する、強い高アフィニティ相互作用によって駆動される。本明細書に記載するのは、単量体Fc分子、およびこうした分子を作製しそして用いる方法である。用語「Fc」は、典型的には、ポリペプチドのホモ二量体として考えられるが、本発明のポリペプチドのユニークな特性のため、本明細書で用いる用語にはまた、重鎖のFc部分に対応するアミノ酸配列を含む、例えばCH2およびCH3ドメインを含有する、単量体ポリペプチドも含まれるであろう。
【0011】
[0019]本明細書に記載する方法は、CH3ドメイン接合部分で残基を置換することによって、CH3/CH3会合を完全に破壊して、なお分子の安定性を維持し、こうして単量体Fcを達成することが可能であることを立証する。改変されたFcの単量体の性質は、例えば、サイズ排除クロマトグラフィー(SEC)および分析用超遠心(AUC)によって評価可能である。置換は2つの事柄を達成する−1つはCH3ドメインのホモ二量体形成を妨害することであり、そしてもう一方は、Fcの単量体型を安定化させることである。
【0012】
[0020]CH3−CH3接合部分を構成するアミノ酸を同定するための方法論は、WO2009089004に開示される。構造に基づく検索アルゴリズム(YeおよびGodzik 2004)を用いて、タンパク質データバンク(PDB)(Bernstein, Koetzleら 1977)より、Fc領域に対応する座標を有する総数48の抗体結晶構造を同定した。同定されたFc結晶構造を調べると、最高解像度で決定された構造が、Z34C(PDBコード:1L6X)と呼ばれる、プロテインA由来のBドメインの最小型に結合したRITUXIMABのFc断片に対応することが明らかになった。寄託されるFc単量体座標および結晶対称性を用いて、1L6Xの生物学的Fcホモ二量体構造を生成した。2つの方法を用いて、CH3−CH3ドメイン相互作用に関与する残基を同定した:(i)距離限界基準によって決定されるような接触および(ii)溶媒アクセス可能表面積分析。
【0013】
[0021]接触に基づく方法にしたがって、その側鎖重原子が、第二の鎖における任意の残基の重原子から、明記する限界より近くに位置する残基として、接合部分残基を定義する。4.5Åの距離限界が好ましいが、接合部分残基を同定するため、より長い距離限界(例えば、5.5Å)もまた使用可能である(BaharおよびJerningan 1997)。
【0014】
[0022]表1は、4.5Åの距離限界を用いた、接触基準法に基づいて同定された24の接合部分残基を列挙する。構造保存に関して、これらの残基をさらに調べた。この目的のため、PDBから同定される48のFc結晶構造を重ね合わせて、そして側鎖重原子に関して、標準偏差を計算することによって分析した。図3は、構造的に保存された位、埋め込まれた位(%ASA≦10)、および暴露された位(%ASA>10)(%ASAは、アミノ酸の標準的ASAに対する、観察されるASAの比を指す(LeeおよびRichards 1971))とともに、CH3ドメイン接合部分を示す。配列比較を通じて、ヒトおよびマウスIgGサブクラス間、ならびに他のIgクラス間での接合部分残基の保存もまた調べた(図8)。
【0015】
[0023]抗体のFc部分に対する多様な置換または突然変異を本明細書に記載する。こうした変異は、KabatのEU番号付けスキームに基づく野生型抗体重鎖におけるその位のアミノ酸の後、その位に置換されるアミノ酸によって示される。例えば、EUの349位のチロシンがスレオニンで置換される際、これは「Y349T」と称される。「野生型配列」によって、動物種、例えばヒト内で天然に存在するアミノ酸の配列を意味する。野生型配列は、集団内の個体間でわずかに変化することも可能であり、例えば多様な免疫グロブリン鎖に関する異なるアレルが当該技術分野に知られる。
【0016】
[0024]ホモ二量体形成を防止するため、CH3−CH3接合部分を構成する1またはそれより多い残基を、相互作用が静電的に好ましくなくなるように、荷電アミノ酸で置換する。好ましい態様において、接合部分中の正荷電アミノ酸、例えばリジン、アルギニン、またはヒスチジンを、負荷電アミノ酸、例えばアスパラギン酸またはグルタミン酸で置換し、そして/または接合部分中の負荷電アミノ酸を正荷電アミノ酸で置換する。例としてヒトIgGを用いると、反対の電荷に変化させてもよい接合部分内の荷電残基には、R355、D356、E357、K370、K392、D399、K409、およびK439が含まれる。特定の好ましい態様において、接合部分内の2またはそれより多い荷電残基を反対の電荷に変化させる。例示的な分子には、K392DおよびK409D突然変異を含むもの、ならびにD399KおよびD356K突然変異を含むものが含まれる。
【0017】
[0025]単量体型において、ポリペプチドの安定性を保持するため、CH3−CH3接合部分を構成する1またはそれより多い疎水性残基を、小さい極性アミノ酸で置換する。例としてヒトIgGを用いると、CH3−CH3接合部分の巨大疎水性残基には、Y349、L351、L368、L398、V397、F405、およびY407が含まれる。小さい極性アミノ酸残基には、アスパラギン、システイン、グルタミン、セリン、およびスレオニンが含まれる。
【0018】
[0026]実施例において、CH3ドメイン接合部分で近くに位置する正荷電Lys残基2つをAspに突然変異させた。次いで、これらの2つのLysからAspへの突然変異の背景において、構造的に保存された巨大疎水性残基に対して、スレオニン・スキャニング突然変異誘発を行った。スレオニンでの多様な置換とともにK392D K409D突然変異を含むFc分子を、単量体形成に関して分析した。例示的な単量体Fc分子には、K392D K409D Y349T置換を有するもの、およびK392D K409D F405T置換を有するものが含まれる。
【0019】
[0027]Fc単量体の半分子の性質により、熱安定性は、高アフィニティCH3−CH3ドメイン相互作用を有するFc二量体のものより低い。熱安定性を増加させるため、CH2およびCH3ドメインにおいて、1またはそれより多いドメイン内ジスルフィド結合を導入してもよい。1またはそれより多い以下のアミノ酸対を突然変異させることによって、ジスルフィド結合を導入してもよい。
【0020】
【化1−1】
【0021】
【化1−2】
【0022】
[0028]抗体がpH依存性様式で新生Fc受容体(FcRn)と相互作用する能力によって、血清半減期が延長される。好ましい態様において、本発明の単量体Fc分子は、野生型Fcポリペプチドより優れていないとしても同様の、FcRnへの結合能を保持する(図6)。実施例2に示すように、本発明の単量体Fc分子は、抗体によって示される延長された血清半減期を保持することも可能であり、そしてしたがって、単量体Fcポリペプチドに共有結合した、例えば融合したポリペプチドの血清半減期を延長させるのに有用である。単量体Fcを操作して、FcRnに対するアフィニティを増加させ、それによってさらに循環中の分子の半減期を増加させる、1またはそれより多いさらなる突然変異を含有するようにすることも可能である。こうしたさらなる突然変異には、限定されるわけではないが、M252Y/S254T/T256E、M428L/N434S、T250Q/M428L、N434H、T307Q/N434A、T307Q/N434S、T307Q/E380A/N434S、およびV308P/N434Sが含まれる。
【0023】
[0029]本発明の組成物および方法は、本明細書に開示する例示的なアレルの変異体には限定されず、本明細書に開示する例示的なアレルに対して、少なくとも70%、少なくとも75%、少なくとも80%、少なくとも81%、少なくとも82%、少なくとも83%、少なくとも84%、少なくとも85%、少なくとも86%、少なくとも87%、少なくとも88%、少なくとも89%、少なくとも90%、少なくとも91%、少なくとも92%、少なくとも93%、少なくとも94%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、および少なくとも99%の同一性を有するものが含まれる。本発明のCH3含有分子の特性を野生型ヒトCH3含有分子のものに比較する目的のため、野生型配列は、図8(a)配列番号1〜4(それぞれ、IgG1、IgG2、IgG3、およびIgG4)に示すものである。
【0024】
[0030]単量体Fc含有分子の生成は、IgG Fcに基づくものに限定されず、IgA、IgE、IgD、およびIgMを含む他の免疫グロブリンサブクラスのFc領域にも適用可能であることが意図される。
【0025】
[0031]Fcドメインを含有する実質的に任意の分子が、本発明の単量体Fcドメインを含んでもよい。こうした分子の例を図2に示す。図2に示すように、多様なペプチドをFcのN末端またはC末端に融合させるかまたはコンジュゲート化してもよい。特定の態様において、Fc含有分子をFabに融合させて、半抗体を生成する。細胞、例えばCHO細胞において、単量体Fcを含む重鎖および軽鎖を組換え的に発現させることによって、こうした半抗体を生成してもよい。重鎖は1またはそれより多いさらなる突然変異を含有してもよい。特定の態様において、重鎖はさらに、ヒンジ領域中に1またはそれより多いシステイン残基の突然変異を含む(Allenら, Biochemistry. 2009 May 5;48(17):3755−66)。
【0026】
[0032]本発明のFcポリペプチドは、野生型Fc分子に比較した際、二量体化減少を示す。したがって、本発明の態様には、抗体またはFc融合分子を含む組成物であって、前記抗体またはFc融合分子が示すFc−Fc二量体化の量が、15%未満、14%未満、13%未満、12%未満、11%未満、10%未満、9%未満、8%未満、7%未満、6%未満、5%未満、4%未満、3%未満、2%未満、または1%未満である、前記組成物が含まれる。当該技術分野に知られるいくつかの技術によって、二量体化を測定してもよい。二量体化を測定するのに好ましい方法には、サイズ排除クロマトグラフィー(SEC)、分析用超遠心(AUC)、動的光散乱(DLS)、および未変性PAGEが含まれる。
【0027】
[0033]本明細書記載のFc単量体分子は、療法タンパク質またはドメインの半減期を延長するのに有用である。Fc単量体療法剤で治療可能な疾患には、炎症、癌、代謝性障害等が含まれることも可能である。潜在的な融合ターゲットには、天然タンパク質結合ドメイン(例えばIL−1Ra、TIMP3、SHKペプチド、EPO、G−CSF)、抗体断片(例えばFab、scFv、ディアボディ、可変ドメイン由来結合剤)、代替足場由来タンパク質結合ドメイン(例えばFn3変異体、アンキリン・リピート変異体、センチリン(centyrin)変異体、アビマー(avimer))および特異的抗原を認識するペプチドが含まれてもよい。Fc単量体融合タンパク質は、サイズが小さく、したがって、組織に浸透する能力がより優れている可能性があるという利点を有する。Fc単量体融合タンパク質は、ターゲット結合の一価性が好ましい場合、特に有用でありうる。多価抗体を用いてターゲティングされた際にアゴニズムに感受性である細胞表面分子をターゲティングする際には、こうした一価性がしばしば好ましい。
【0028】
定義
[0034]本明細書において、別に定義しない限り、本発明と関連して用いられる科学的および技術的用語は、一般の当業者に一般的に理解される意味を有するものとする。さらに、文脈によって別に必要とされない限り、単数形の用語は複数のものを含み、そして複数形の用語は単数形を含むものとする。一般的に、本明細書に記載する、細胞および組織培養、分子生物学、免疫学、微生物学、遺伝学、ならびにタンパク質および核酸化学およびハイブリダイゼーションと関連して用いられる術語、ならびにそれらの技術は、当該技術分野に周知であり、そして一般的に用いられるものである。本発明の方法および技術は、別に示さない限り、一般的に、当該技術分野に周知の慣用法にしたがって、そして本明細書全体で引用され、そして論じられる、多様な一般的な参考文献およびより特異的な参考文献に記載されるように、行われる。例えば、本明細書に援用される、Sambrookら Molecular Cloning: A Laboratory Manual, 第2版, Cold Spring Harbor Laboratory Press, ニューヨーク州コールドスプリングハーバー(1989)、ならびにAusubelら, Current Protocols in Molecular Biology, Greene Publishing Associates(1992)、ならびにHarlowおよびLane Antibodies: A Laboratory Manual Cold Spring Harbor Laboratory Press, ニューヨーク州コールドスプリングハーバー(1990)を参照されたい。酵素反応および精製技術は、当該技術分野において一般的に達成されるように、または本明細書に記載するように、製造者の指定にしたがって行われる。本明細書記載の分析化学、合成有機化学、ならびに医学的および薬学的化学と関連して用いられる専門用語、ならびにこうした化学の実験法および技術は、当該技術分野に周知であり、そして一般的に知られるものである。化学合成、化学分析、薬学的調製、配合、および送達、ならびに患者の治療には、標準的技術を用いてもよい。
【0029】
[0035]以下の用語は、別に示さない限り、以下の意味を有すると理解されるべきである:用語「単離分子」は(分子が、例えばポリペプチド、ポリヌクレオチド、または抗体である場合)、その起源または派生供給源によって、(1)天然状態で該分子に付随する、天然に関連する構成要素と関連していないか、(2)同じ種由来の他の分子を実質的に含まないか、(3)異なる種由来の細胞によって発現されるか、または(4)天然には存在しない分子である。したがって、化学的に合成されるか、または天然に由来する細胞とは異なる細胞系において発現される分子は、天然に関連する構成要素から「単離されている」であろう。分子はまた、当該技術分野に周知の精製技術を用いた単離によって、天然に関連する構成要素を実質的に含まないようにされてもよい。当該技術分野に周知のいくつかの手段によって、分子純度または均一性をアッセイしてもよい。例えば、当該技術分野に周知の技術を用いて、ポリアクリルアミドゲル電気泳動を用い、そしてゲルを染色してポリペプチドを視覚化して、ポリペプチド試料の純度をアッセイしてもよい。特定の目的のため、HPLCまたは当該技術分野に周知の精製のための他の手段を用いることによって、より高い解像度を提供してもよい。
【0030】
[0036]ポリヌクレオチドおよびポリペプチド配列は、標準的な1文字または3文字略記を用いて示される。別に示さない限り、ポリペプチド配列は、アミノ末端を左側に、そしてカルボキシ末端を右側に有し、そして一本鎖核酸配列、および二本鎖核酸配列の上部鎖は、5’端を左に、そして3’端を右に有する。特定のポリペプチドまたはポリヌクレオチド配列はまた、参照配列とどのように異なるかを説明することによって記載されうる。
【0031】
[0037]用語「ペプチド」、「ポリペプチド」および「タンパク質」は、各々、ペプチド結合によって互いに連結された2またはそれより多いアミノ酸残基を含む分子を指す。これらの用語は、例えば天然および人工的タンパク質、タンパク質断片、およびタンパク質配列のポリペプチド類似体(突然変異タンパク質(mutein)、変異体、および融合タンパク質など)、ならびに翻訳後、あるいは別の共有的または非共有的修飾タンパク質を含む。ペプチド、ポリペプチド、またはタンパク質は、単量体性または多量体性であってもよい。
【0032】
[0038]用語「ポリペプチド断片」は、本明細書において、対応する全長タンパク質に比較した際、アミノ末端および/またはカルボキシ末端に欠失を有するポリペプチドを指す。断片は、例えば、少なくとも長さ5、6、7、8、9、10、11、12、13、14、15、20、50、70、80、90、100、150、200、250、300、350、または400アミノ酸であってもよい。断片はまた、例えば、最大で、長さ1,000、750、500、250、200、175、150、125、100、90、80、70、60、50、40、30、20、15、14、13、12、11、または10アミノ酸であってもよい。断片は、さらに、どちらかまたは両方の端に、1またはそれより多いさらなるアミノ酸、例えば異なる天然存在タンパク質または人工的アミノ酸配列由来のアミノ酸配列を含んでもよい。
【0033】
[0039]本発明のポリペプチドには、例えば:(1)タンパク質分解に対する感受性を減少させ、(2)酸化に対する感受性を減少させ、(3)タンパク質複合体を形成するための結合アフィニティを改変し、(4)結合アフィニティを改変し、そして(4)他の物理化学特性または機能特性を与えるかまたはこうした特性を修正するように、いずれかの方式で、そしていずれかの理由のために修飾されているポリペプチドが含まれる。類似体には、ポリペプチドの突然変異タンパク質が含まれる。例えば、単数または複数のアミノ酸置換(例えば、保存的アミノ酸置換)を天然存在配列において(例えば、分子間接触を形成するドメイン(単数または複数)外のポリペプチドの部分において)行うことも可能である。「保存的アミノ酸置換」は、親配列の構造特徴を実質的に変化させないものである(例えば置換アミノ酸は、親配列に存在するらせんを中断させるか、あるいは親配列を特徴付けるかまたはその機能に必要な他のタイプの二次構造を破壊する傾向があってはならない)。当該技術分野に認識されるポリペプチド二次構造および三次構造の例が、Proteins, Structures and Molecular Principles(Creighton監修, W.H. Freeman and Company, ニューヨーク(1984)); Introduction to Protein Structure(C. BrandenおよびJ. Tooze監修, Garland Publishing, ニューヨーク州ニューヨーク(1991));およびThorntonら Nature 354:105(1991)に記載され、これらは各々、本明細書に援用される。
【0034】
[0040]ポリペプチドの「変異体」は、別のポリペプチド配列に比較して、1またはそれより多いアミノ酸残基がアミノ酸配列内で挿入され、欠失され、そして/または置換された、アミノ酸配列を含む。本発明の変異体には、変異体CH2またはCH3ドメインを含むものが含まれる。特定の態様において、変異体は、Fc分子中に存在した際、1またはそれより多いFcγRに対するポリペプチドのアフィニティを増加させる1またはそれより多い突然変異を含む。こうした変異体は、抗体依存性細胞仲介性細胞傷害性の増加を示す。こうしたものを提供する変異体の例は、米国特許第7,317,091号に記載される。
【0035】
[0041]他の変異体には、CH3ドメイン含有ポリペプチドがホモ二量体形成する能力を減少させるものが含まれる。こうしたFc変異体の例が、米国特許第5,731,168号および第7,183,076号に記載される。さらなる例が、08年1月7日出願の共同所有米国仮出願第61/019,569号および08年12月5日出願の第61/120,305号(どちらもその全体が本明細書に援用される)に記載される。
【0036】
[0042]ポリペプチドの「誘導体」は、例えば別の化学部分、例えばポリエチレングリコール、細胞傷害剤、アルブミン(例えばヒト血清アルブミン)などへのコンジュゲート化、リン酸化、およびグリコシル化を介して、化学的に修飾されているポリペプチド(例えば抗体)である。別に示さない限り、用語「抗体」には、2つの全長重鎖および2つの全長軽鎖を含む抗体に加えて、その誘導体、変異体、断片、および突然変異タンパク質が含まれ、それらの例を以下に記載する。
【0037】
[0043]用語「ヒト抗体」には、ヒト免疫グロブリン配列に由来する1またはそれより多い可変領域および定常領域を有する抗体すべてが含まれる。1つの態様において、可変ドメインおよび定常ドメインのすべてがヒト免疫グロブリン配列に由来する(完全ヒト抗体)。これらの抗体は、多様な方法で調製可能であり、その例を以下に記載し、これらには、ヒト重鎖および/または軽鎖をコードする遺伝子に由来する抗体を発現するように遺伝子修飾されたマウスの、関心対象の抗原での免疫を通じたものが含まれる。特定の態様において、ヒト抗体の重鎖は、CH3ドメインにおいて、重鎖が二量体化する能力が減少するように改変される。
【0038】
[0044]ヒト化抗体は、ヒト被験体に投与された際、非ヒト種抗体に比較すると、免疫応答を誘導する可能性がより低く、そして/またはより重度でない免疫応答を誘導するように、1またはそれより多いアミノ酸置換、欠失、および/または付加によって、非ヒト種に由来する抗体の配列と異なる配列を有する。1つの態様において、非ヒト種抗体の重鎖および/または軽鎖のフレームワークおよび定常ドメイン中の特定のアミノ酸を突然変異させて、ヒト化抗体を産生する。別の態様において、ヒト抗体由来の定常ドメイン(単数または複数)を、非ヒト種の可変ドメイン(単数または複数)に融合させる。ヒト化抗体をどのように作製するかの例を、米国特許第6,054,297号、第5,886,152号、および第5,877,293号に見出すことも可能である。
【0039】
[0045]用語「キメラ抗体」は、1つの抗体由来の1またはそれより多い領域、および1またはそれより多い他の抗体由来の1またはそれより多い他の領域を含有する抗体を指す。キメラ抗体の1つの例において、重鎖および/または軽鎖の部分は、特定の種由来であるか、または特定の抗体クラスもしくはサブクラスに属する抗体と同一であるか、該抗体に相同であるか、または該抗体に由来する一方、鎖(単数または複数)の残りは、別の種由来であるか、または別の抗体クラスもしくはサブクラスに属する抗体(単数または複数)と同一であるか、該抗体に相同であるか、または該抗体に由来する。やはり含まれるのは、所望の生物学的活性を示す、こうした抗体の断片である。
【0040】
[0046]抗体の断片または類似体は、本明細書の解説にしたがって、そして当該技術分野に周知の技術を用いて、一般の当業者によって、容易に調製可能である。断片または類似体の好ましいアミノ末端およびカルボキシ末端は、機能ドメインの境界近傍に存在する。公共のまたは私有の(proprietary)配列データベースに、ヌクレオチドおよび/またはアミノ酸配列データを比較することによって、構造ドメインおよび機能ドメインを同定することも可能である。コンピュータ比較法を用いて、既知の構造および/または機能を持つ他のタンパク質に存在する配列モチーフまたは予測されるタンパク質コンホメーションドメインを同定してもよい。既知の三次元構造にフォールディングするタンパク質配列を同定する方法が知られる。例えばBowieら, 1991, Science 253:164を参照されたい。
【0041】
[0047]「CDR移植抗体」は、特定の種またはアイソタイプの抗体由来の1またはそれより多いCDR、および同じまたは異なる種またはアイソタイプの別の抗体のフレームワークを含む抗体である。
【0042】
[0048]2つのポリヌクレオチドまたは2つのポリペプチド配列の「同一性パーセント」は、デフォルト・パラメータを用い、GAPコンピュータ・プログラム(GCGウィスコンシン・パッケージ、バージョン10.3(Accelrys、カリフォルニア州サンディエゴ)の一部)を用いて、配列を比較することによって決定される。
【0043】
[0049]用語「ポリヌクレオチド」、「オリゴヌクレオチド」および「核酸」は、本明細書全体を通じて交換可能に用いられ、そしてDNA分子(例えばcDNAまたはゲノムDNA)、RNA分子(例えばmRNA)、ヌクレオチド類似体(例えばペプチド核酸および非天然存在ヌクレオチド類似体)を用いて生成されるDNAまたはRNAの類似体、およびそれらのハイブリッドを含む。核酸分子は、一本鎖または二本鎖であることも可能である。1つの態様において、本発明の核酸分子は、抗体またはFc融合体、およびその誘導体、突然変異タンパク質、または変異体をコードする、隣接オープンリーディングフレームを含む。
【0044】
[0050]2つの一本鎖ポリヌクレオチドは、ギャップを導入することなく、そしていずれの配列の5’端または3’端にも、対形成しないヌクレオチドを伴わずに、一方のポリヌクレオチド中のすべてのヌクレオチドが、他方のポリヌクレオチド中の相補的ヌクレオチドと反対であるように、逆平行配向で整列可能であるならば、互いに「相補体」である。ポリヌクレオチドは、中程度にストリンジェントな条件下で、2つのポリヌクレオチドが互いにハイブリダイズ可能であるならば、別のポリヌクレオチドに「相補的」である。したがって、ポリヌクレオチドは、別のポリヌクレオチドの相補体であることなく、該ポリヌクレオチドに相補的であることも可能である。
【0045】
[0051]「ベクター」は、連結された別の核酸を、細胞内に導入するために使用可能な核酸である。ベクターの1つのタイプは「プラスミド」であり、その内部にさらなる核酸セグメントを連結可能な、直鎖または環状二本鎖DNA分子を指す。別のタイプのベクターはウイルスベクター(例えば複製不全レトロウイルス、アデノウイルス、およびアデノ随伴ウイルス)であり、ここで、さらなるDNAセグメントをウイルスゲノム内に導入可能である。特定のベクターは、導入された宿主細胞において、自律的に複製可能である(例えば細菌複製起点を含む細菌ベクターおよびエピソーム哺乳動物ベクター)。宿主細胞内への導入に際して、宿主細胞のゲノム内に他のベクター(例えば非エピソーム哺乳動物ベクター)を組み込んで、そしてそれによって宿主ゲノムと一緒に複製させる。「発現ベクター」は、選択したポリヌクレオチドの発現を指示することも可能なベクターのタイプである。
【0046】
[0052]ヌクレオチド配列は、制御配列が該ヌクレオチド配列の発現(例えば発現のレベル、時期、または位置)に影響を及ぼすならば、該制御配列に「機能可能であるように連結されて」いる。「制御配列」は、機能可能であるように連結されている核酸の発現(例えば発現のレベル、時期、または位置)に影響を及ぼす核酸である。制御配列は、例えば、制御される核酸に対して直接、あるいは1またはそれより多い他の分子(例えば制御配列および/または核酸に結合するポリペプチド)の作用を通じて、その効果を発揮しうる。制御配列の例には、プロモーター、エンハンサーおよび他の発現調節要素(例えばポリアデニル化シグナル)が含まれる。制御配列のさらなる例は、例えば、Goeddel, 1990, Gene Expression Technology: Methods in Enzymology 185, Academic Press, カリフォルニア州サンディエゴ、およびBaronら, 1995, Nucleic Acids Res. 23:3605−06に記載される。
【0047】
[0053]「宿主細胞」は、核酸、例えば本発明の核酸を発現するために使用可能な細胞である。宿主細胞は、原核生物、例えば大腸菌(E.coli)であってもよいし、または真核生物、例えば単細胞真核生物(例えば酵母(yeast)または他の真菌)、植物細胞(例えばタバコ(tobacco)またはトマト(tomato)植物細胞)、動物細胞(例えばヒト細胞、サル細胞、ハムスター細胞、ラット細胞、マウス細胞、または昆虫細胞)またはハイブリドーマであってもよい。例示的な宿主細胞には、チャイニーズハムスター卵巣(CHO)細胞株、またはDHFRが欠損しているCHO株DXB−11(Urlaubら, 1980, Proc. Natl. Acad. Sci. USA 77:4216−20を参照されたい)、血清不含培地中で増殖するCHO細胞株(Rasmussenら, 1998, Cytotechnology 28:31を参照されたい)、DXB−11 CHO細胞の誘導体であるCS−9細胞、およびAM−1/D細胞(米国特許第6,210,924号に記載される)を含むその誘導体が含まれる。他のCHO細胞株には、CHO−K1(ATCC#CCL−61)、EM9(ATCC#CRL−1861)、およびUV20(ATCC#CRL−1862)が含まれる。他の宿主細胞の例には、サル腎臓細胞のCOS−7株(ATCC CRL 1651)(Gluzmanら, 1981, Cell 23:175を参照されたい)、L細胞、C127細胞、3T3細胞(ATCC CCL 163)、HeLa細胞、BHK(ATCC CRL 10)細胞株、アフリカミドリザル(African green monkey)腎臓細胞株CV1(ATCC CCL 70)由来のCV1/EBNA細胞株(McMahanら, 1991, EMBO J. 10:2821を参照されたい)、ヒト胚性腎細胞、例えば293、293 EBNAまたはMSR 293、ヒト上皮A431細胞、ヒトColo205細胞、他の形質転換霊長類細胞株、正常二倍体細胞、初代組織のin vitro培養由来の細胞株、初代外植片、HL−60、U937、HaKまたはJurkat細胞が含まれる。典型的には、宿主細胞は、その後、宿主細胞で発現可能なポリペプチドをコードする核酸で形質転換またはトランスフェクションされることが可能な培養細胞である。
【0048】
[0054]句「組換え宿主細胞」を用いて、発現しようとする核酸で形質転換されているかまたはトランスフェクションされている宿主細胞を示すことも可能である。宿主細胞はまた、核酸を含むが、機能可能であるように核酸と連結されるように制御配列が宿主細胞内に導入されない限り、所望のレベルで該核酸を発現しない細胞であってもよい。用語、宿主細胞は、特定の対象の細胞だけでなく、こうした細胞の子孫または潜在的な子孫も指すことが理解される。例えば、突然変異または環境的影響によって、続く世代で特定の修飾が起こりうるため、こうした子孫は、実際、親細胞と同一でない可能性もあるが、なお、本明細書において、この用語の範囲内に含まれる。
【0049】
薬学的組成物
[0055]本発明のポリペプチドは、薬学的組成物内に配合するのに特に有用である。こうした組成物は、生理学的に許容されうるキャリアー、賦形剤または希釈剤などの1またはそれより多いさらなる構成要素を含む。場合によって、組成物は、例えば以下に記載するような、1またはそれより多い生理学的活性剤をさらに含む。多様な特定の態様において、組成物は、1またはそれより多い本発明の単量体抗体および/またはFc融合タンパク質に加えて、1、2、3、4、5、または6の生理学的活性剤を含む。
【0050】
[0056]1つの態様において、薬学的組成物は、本発明の単量体抗体および/またはFc融合タンパク質を、緩衝剤、アスコルビン酸などの酸化防止剤、低分子量ポリペプチド(10アミノ酸未満を有するものなど)、タンパク質、アミノ酸、グルコース、スクロースまたはデキストリンなどの炭水化物、EDTAなどのキレート剤、グルタチオン、安定化剤、ならびに賦形剤からなる群より選択される1またはそれより多い物質とともに、含む。中性緩衝生理食塩水または同種血清アルブミンと混合された生理食塩水が、適切な希釈剤の例である。適切な産業標準にしたがって、ベンジルアルコールなどの保存剤もまた添加してもよい。組成物は、適切な賦形剤溶液(例えばスクロース)を希釈剤として用いた凍結乾燥物として配合されてもよい。適切な構成要素は、使用する投薬量および濃度で、レシピエントに非毒性である。薬学的配合物において使用可能な構成要素のさらなる例が、Remington’s Pharmaceutical Sciences, 第16版(1980)および第20版(2000), Mack Publishing Company, ペンシルバニア州イーストンに提示される。
【0051】
[0057]開業医に使用されるためのキットには、本明細書に論じる任意の状態を治療する際に使用するための、1またはそれより多い本発明の単量体抗体および/またはFc融合タンパク質、ならびにラベルまたは他の使用説明書が含まれる。1つの態様において、キットには、1またはそれより多い単量体抗体および/またはFc融合タンパク質の無菌調製物が含まれ、該調製物は、上に開示するような組成物の形であってもよく、そして1またはそれより多いバイアル中にあってもよい。
【0052】
[0058]投薬量および投与頻度は、投与経路、使用する特定の単量体抗体および/またはFc融合タンパク質、治療しようとする疾患の性質および重症度、状態が急性または慢性であるか、ならびに被験体のサイズおよび全身状態などの要因に応じて、多様でありうる。関連技術において知られる方法によって、例えば用量の段階的増大研究を伴いうる臨床試験において、適切な投薬量を決定してもよい。
【0053】
[0059]本発明の単量体抗体および/またはFc融合タンパク質を、例えば1回または1回より多く、例えばある期間に渡って定期的な間隔で、投与してもよい。特定の態様において、単量体抗体および/またはFc融合タンパク質を、少なくとも1ヶ月に1回またはそれより多く、例えば1ヶ月、2ヶ月、または3ヶ月、あるいはさらに無期限に渡って、投与する。慢性状態を治療するためには、一般的に、長期治療が最も有効である。しかし、急性状態を治療するため、より短い期間、例えば1〜6週間の投与で十分である可能性もある。一般的に、患者が、選択した単数または複数の指標に関して、ベースラインを超えた、医学的に適切な度合いの改善を示すまで、単量体抗体および/またはFc融合タンパク質を投与する。
【0054】
[0060]関連分野で理解されるように、本発明の単量体抗体および/またはFc融合タンパク質を含む薬学的組成物を、適応症に適した方式で、被験体に投与する。限定されるわけではないが、非経口、局所、または吸入によるものを含む、任意の適切な技術によって、薬学的組成物を投与してもよい。注射する場合、薬学的組成物を、例えば、動脈内、静脈内、筋内、病巣内、腹腔内または皮下経路を介して、ボーラス注射によって、あるいは連続注入によって、投与してもよい。局在化投与、例えば疾患部位または傷害部位での投与が意図され、経皮送達および移植物からの持続放出も同様である。吸入による送達には、例えば、鼻または経口吸入、ネブライザーの使用、エアロゾル型での単量体抗体および/またはFc融合タンパク質の吸入等が含まれる。
【実施例】
【0055】
実施例1
[0061]距離限界基準を用いて、CH3−−−CH3ドメイン相互作用に関与する残基を同定した。CH3ドメイン接合部分に位置する24の残基があった(表1)。入手可能な既知のFc抗体結晶構造を用いて、側鎖構造保存に関して、これらの24残基を調べた(図3)。この分析によって、疎水性残基のいくつかに関して、高い構造保存が明らかになった。計算的方法(PokalaおよびHandel 2005)を用いて、24の接合部分位およびL398(接合部分残基K392と接触する残基)を、アラニンで、一度に一残基ずつ突然変異させることによって、つのCH3ドメイン間の会合自由エネルギーもまた計算した。計算および構造保存分析によって、24残基のうち6残基が、CH3ドメイン二量体形成に大きく寄与することが明らかになった。これらの6位を分析して、置換の影響を決定した。
【0056】
表1:第一の鎖(A)中のCH3ドメイン接合部分残基および第二の鎖(B)中の側鎖接触残基のリストa
【0057】
【表1−1】
【0058】
【表1−2】
【0059】
aCH3−CH3ドメイン相互作用に2倍対称性が存在するため、各対の相互作用は、構造中、2回示される(例えば、Ser A 364−Leu B 368’およびLeu A 439−Ser B 364’)。しかし、Leu A 351−Pro B 352’、Leu A 351−Pro B 353’、Lys A 392−Leu B 398’、Val A 397−Thr B 393’、およびTyr A 407−Ser B 408’対は、側鎖−主鎖接触を伴うため、これらは1回しか示されない。
【0060】
[0062]6位各々でのスレオニン置換に対応するFc突然変異タンパク質を生成した(表2)。さらに、F405T Y407RおよびY407R Fc突然変異体もまた生成した。完全単量体Fc形成の可能性を増進させるため、CH3/CH3弱体化Fc変異体の背景で、6つの位で突然変異を生成し、ここで、リジン392およびリジン409をアスパラギン酸に突然変異させる(K392D K409D Fc)。
【0061】
表2
【0062】
【表2−1】
【0063】
【表2−2】
【0064】
ND;未決定。
[0063]Multi−QuikChange部位特異的突然変異誘発(Stratagene)を用いて、ヒトIgG1 Fc K392D K409D中、表2(#2〜#9)に列挙する突然変異を生成した。用いたオリゴプライマーは以下の通りである:
【0065】
【化2】
【0066】
[0064]予期される突然変異をDNA配列決定によって確認した。pTT5一過性哺乳動物発現ベクターを用いて、親(hu IgG1 Fc K409D K392D)および突然変異体Fcタンパク質を293E細胞中で発現させた。標準的プロテインAクロマトグラフィー(5mlカラム、Pierce)を用いて、Fcタンパク質を精製した。TOSO46mm SW3000カラム(TOSO Biosciences LLC、PA)を用いて、Fcタンパク質均一性分析(SEC)を行った。280nmの吸収を測定し、そして1mg/ml=1.74 OD280を用いて計算することによって、タンパク質濃度を決定した。
【0067】
[0065]各試料の場合において、5μgのタンパク質を非還元SDSまたは還元SDS試料緩衝液(Invitroten)で処理し、4〜20% TGゲル(Invitrogen)上で泳動し、そしてGel−code試薬(Pierce)を用いて染色した。
【0068】
[0066]BIAcore3000装置を用いた、Fc突然変異体#1(Fc K392D−K409D、二量体)、#2(Fc K392D−K409D−Y349T、単量体SECプロファイル)および#6(Fc K392D−K409D−Y349T、単量体SECプロファイル)へのヒトおよびマウスFcRn結合のBIAcore分析。CHO細胞で産生したhuFcを、CM5チップ上のフォローセル(follow cell)2(Fc2)上に固定した。Fc1をバックグラウンド対照として用いた。2nMヒトFcRnを1、10、100nMの示したFc変異体と1時間インキュベーションした後、CHO huFc表面に注入した。固定されたCHO huFcへのFcRnの結合減少は、溶液中のFc変異体にFcRnが結合している指標となる。2nMマウスFcRnを0.1、1、10nMのFc変異体と1時間インキュベーションした後、CHO huFc表面に注入した。
【0069】
[0067]突然変異体Fcに関するタンパク質濃度は、それぞれ、0.4および0.6mg/mlであった。Beckmann Coulter ProteomeLab XL−1装置によって、PBS中の試料を分析した。50,000rpmで沈降速度実験を行い、水晶ウインドウを含む二重セクター中心セル・アセンブリー中、280nmの吸光度によって追跡した。間に遅延を含まず、20℃でスキャンを収集した。SEDFIT、バージョン9.4を用いて、AUC−SVデータを分析した。AUC−SV分析において、非線形最小二乗適合中、摩擦比(frictional ration)、時間不変ノイズ(time invariant noise)、およびメニスカス値(meniscus value)が浮遊する(float)ことを可能にした。
【0070】
実施例2
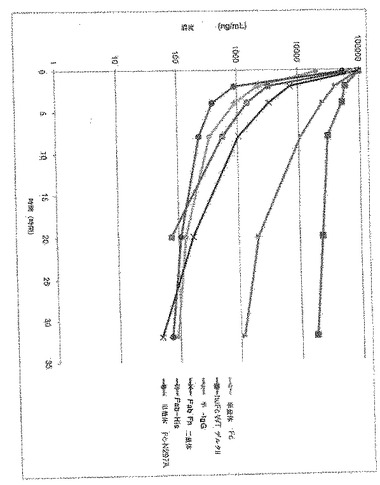
本実施例は、正常マウスにおける多様な分子量の単量体Fc構築物のPKパラメータを示す。単量体Fcは、K392D−K409D−Y349T突然変異を含んだ。単量体Fc N297Aは、グリコシル化部位を除去するため、N297にさらなる突然変異を有する単量体Fcを含んだ。半IgGは、単量体Fcに融合したFabを含んだ。Fab FnFnは、フィブロネクチン二量体に融合したFabを含んだ。Fab Hisは、ヒスチジンタグに融合したFabであった。huFc WTデルタHは、ヒンジ領域を除いたWT Fc二量体を含んだ。54匹のSCIDマウスを以下の処置群に割り当てた。すべてのマウスには、10mg/kg IVを投与し、そして0.25、2、4、8、20および32時間で血清を収集した。
【0071】
表3
【0072】
【表3】
【0073】
分析法のまとめ:
単量体Fc構築物の定量化のため、マイクロタイタープレート(Maxisorp、Nunc)を、1、2および6群に関しては、ヤギ抗ヒトIgG Fcγ特異的(Jackson カタログ#109−005−098)で、または3、4および5群に関しては、ヤギ抗ヒトIgG F(ab’)2特異的(Jacksonカタログ#109−005−097)のいずれかでコーティングした。PBST中の10%NFDM(脱脂粉乳)でブロッキングした後、前処理後、標準、品質管理試料(QC)、および試験試料をNFDM/PBST中、50の希釈係数でインキュベーションした。PBST緩衝液で洗浄することによって、未結合構築物を除去した。次に、1、2および6群において、西洋ワサビペルオキシダーゼ標識ヤギ抗ヒトIgG Fcγ特異的(Jaksonカタログ#109−035−098)を添加して、捕捉されたFc構築物を検出する一方、3、4および5群において、ヤギ抗ヒトIgG F(ab’)2特異的(Jacksonカタログ#109−036−097)を用いて、捕捉されたF(ab’)2構築物を検出した。最終洗浄工程後、TMB基質溶液(1:1テトラメチルベンジジンおよびペルオキシド、Kirkegaard & Perry Laboratories)を添加して、そしてリン酸で反応停止した。光学密度(OD)を450〜650nmの波長で決定した。4パラメータ・ロジスティックモデルにしたがって回帰した、同時に分析される標準曲線に対して、ワトソン・ソフトウェアが仲介する比較を通じて、QCおよび未知の標本に関して、OD値の濃度への変換を達成した。
【0074】
表4−−血清標本に関するデータ
【0075】
【表4】
【0076】
表5
【0077】
【表5】
【0078】
本実施例は、正常マウスPKパラメータ、すなわちFcRn相互作用の度合い、〜60kDaの腎臓クリアランス閾値より高いまたは低い多様な分子量の単量体Fc構築物の度合いを示す。二量体CH2−CH3ドメインからなるhuFc WTデルタH構築物は、クリアランス閾値より低い分子量であるにもかかわらず、すべての他の構築物に比較して、最大の曝露/AUCを示した。Fab Fc単量体からなる半IgG分子は、二量体Fcの曝露の35%を示したが、Fab−フィブロネクチン二量体からなる、サイズが一致したFab Fc二量体に比較して、3.5倍高いAUCを有し、この結果は、半減期、そして最終的には曝露を増加させる際のFcの役割を立証した。Fab単独および単量体Fc構築物、N連結付加部位の除去のため、グリコシル化を欠くWTまたはN297A変異体いずれかは、すべて、迅速なクリアランスおよびhuFC WTデルタH構築物よりも17〜38倍低い最小AUC値を示した。
【0079】
参考文献
【0080】
【化3−1】
【0081】
【化3−2】
【背景技術】
【0001】
[0001]抗体は、侵襲する非自己分子に対する防御において、中心的な役割を果たす。抗体が、pH依存方式で、新生Fc受容体(FcRn)と相互作用する能力によって、血清半減期が延長される(GhetieおよびWard 2000)。抗体のこのユニークな特徴のため、Fc融合分子を操作することによって、血清における療法タンパク質またはペプチドの半減期を延長することが可能になる。天然存在IgG抗体および操作Fc融合分子は二価であり、そして単一特異性である。これは、Fcがホモ二量体の性質を持つためである。特定の療法適用のため、抗体または抗体のFc断片によって与えられるすべての正の特質を保持するが、単量体Fcを操作することによって、一価特異性を達成することが望ましいであろう。
【0002】
[0002]抗体は、IgG、IgA、IgE、IgM、およびIgDを含むタンパク質の免疫グロブリンクラスに属する。ヒト血清において最も豊富な免疫グロブリンクラスはIgGであり、その模式的構造を図1に示す(Deisenhofer 1981; Huber 1984; Roux 1999)。IgG構造は、4つの鎖、2つの軽鎖および2つの重鎖を有し;各軽鎖は2つのドメインを有し、そして各重鎖は4つのドメインを有する。抗原結合部位は、可変軽(VL)鎖および可変重(VH)鎖ドメイン、ならびに定常軽(LC)鎖および定常重(CH1)鎖ドメインを含有するFab領域(抗原結合断片)中に位置する。抗体のFc(結晶化可能断片)断片は、重鎖のCH2およびCH3ドメイン領域を含有する。IgG分子は、ヒンジ領域でジスルフィド結合(−S−S−)によって一緒に保持される2つの重鎖、および2つの軽鎖を有する、ヘテロ四量体と見なされることも可能である。ヒンジ・ジスルフィド結合の数は、免疫グロブリンサブクラス間で多様である(PapadeaおよびCheck 1989)。FcRn結合部位は、抗体のFc領域中に位置し(Martin、Westら 2001)、そしてしたがって抗体の血清半減期延長特性は、Fc断片中に保持される。Fc領域は、単独では、CH2およびCH3ドメインを含む重鎖のホモ二量体と考えられうる。
【先行技術文献】
【非特許文献】
【0003】
【非特許文献1】GhetieおよびWard 2000
【非特許文献2】Deisenhofer 1981
【非特許文献3】Huber 1984
【非特許文献4】Roux 1999
【非特許文献5】PapadeaおよびCheck 1989
【非特許文献6】Martin、Westら 2001
【発明の概要】
【発明が解決しようとする課題】
【0004】
[0003]本明細書に提供するのは、ポリペプチドがホモ二量体を形成する能力を有意に減少させる、CH3接合部分ドメイン中の改変を含有するFcポリペプチドである。好ましい態様において、二量体化の減少はほぼ100%である。好ましくは、Fcポリペプチドには、CH3ホモ二量体形成に静電的に好ましくない、1またはそれより多い荷電アミノ酸、および1またはそれより多い疎水性CH3接合部分残基の極性アミノ酸残基、例えばスレオニンでの置換が含まれる。
【0005】
[0004]特定の態様において、CH3ドメインは、392位および/または409位で、負に荷電されたアミノ酸、例えばアスパラギン酸を含む、CH3ホモ二量体形成に静電的に好ましくない改変、ならびにY349、L351、L368、V397、L398、F405、およびY407からなる群より選択される、1またはそれより多い置換された疎水性接合部分残基を有する、ヒトIgG CH3ドメインである。
【0006】
[0005]単量体Fcポリペプチドは、抗体CH1ドメインをさらに含んでもよいし、または抗体重鎖内で構成される。特定の態様において、単量体抗体は、単量体重鎖および軽鎖を含み、本質的に半抗体(half−antibody)を生成する。単量体重鎖は、ジスルフィド結合形成を防止するため、1またはそれより多い突然変異システイン残基を有してもよい。特に有用なシステイン突然変異は、重鎖ヒンジ領域中のものである。
【0007】
[0006]本発明の1つの側面において、ポリペプチドは、野生型CH3ドメインを含むポリペプチドに比較して、ホモ二量体を形成する能力が減少した、抗体CH3ドメインを含む。好ましいポリペプチドは、抗体のCH3ドメインであって、1またはそれより多い荷電アミノ酸が、CH3ホモ二量体形成に静電的に好ましくないアミノ酸で置換され、そして1またはそれより多い疎水性接合部分残基が、極性アミノ酸で置換されるように、野生型CH3ドメインと異なるアミノ酸配列を含む、前記CH3ドメインを含む。
【0008】
[0007]本発明の他の側面は、単量体Fcポリペプチドをコードする核酸、こうした核酸を含む発現ベクター、およびこうした発現ベクターを含有する宿主細胞である。
[0008]本発明の態様には、単量体Fcポリペプチドを調製する方法がさらに含まれる。好ましい態様において、こうした方法は、単量体Fcポリペプチドをコードする核酸を含む宿主細胞を、単量体Fcポリペプチドが発現される条件下で培養し、そして次いで、宿主細胞培養から単量体Fcポリペプチドを回収する工程を含む。
【図面の簡単な説明】
【0009】
【図1】[0009](a)示すドメインを含むIgG1抗体の模式図。IgG1抗体は、2つの重鎖(より長い長さ)および2つの軽鎖(より短い長さ)を含むY型の四量体である。2つの重鎖は、ヒンジ領域で、ジスルフィド結合(−S−S−)によって一緒に連結される。Fab−抗原結合断片、Fc−結晶化可能断片、VL−可変軽鎖ドメイン、VH−可変重鎖ドメイン、CL−定常(配列変動なし)軽鎖ドメイン、CH1−定常重鎖ドメイン1、CH2−定常重鎖ドメイン2、CH3−定常重鎖ドメイン3。(b)一価抗体の模式図−単量体Fcに融合したFab。この場合、CH3ドメイン接合部分は、アミノ酸突然変異を通じて修飾される。
【図2】[0010]図は、単量体(または一価)Fcを含む態様のいくつかを示す。これらには、単量体FcのNおよびC末端両方への融合が含まれる。Fcは、二量体化を伴わなくてさえ、またはFabドメインがなくてさえ、FcRn受容体と相互作用する能力を保持し、単量体Fcと融合しているタンパク質/ドメインの、より長い血清半減期を導く。scFv−一本鎖可変ドメイン断片。
【図3】[0011]ドメイン−ドメイン相互作用に関与する残基を示した、CH3ドメイン接合部分構造。距離カットオフ法を用いて、接合部分残基を同定した。構造的に保存されそして埋め込まれた(buried)(溶媒アクセス可能表面積≦10%)残基を球棒モデルで示す。溶媒に暴露されるかまたは構造的に保存されていない残基を、棒提示で示す。分析は、高解像度(1.65Å)で決定されている、IgG1結晶構造(PDBコード: 1L6X)に基づく(Idusogie, Prestaら 2000)。
【図4】[0012](a)FcのCH3ドメイン接合部分における突然変異を含めて設計した構築物のリスト、および(b)8つの精製Fc突然変異体タンパク質に関するGELCODETM Blue染色試薬で染色したSDS−PAGEクーマシーゲル。表2の突然変異体4はタンパク質の量が不十分であったため、含めなかった。構築物は、これらのFc構築物がヒンジ・ジスルフィドを欠くという事実のため、還元および非還元ゲル中を同様に移動する。言い換えると、ここでの目的は単量体Fc重鎖を達成することであるため、IgG分子とは異なり、2つの重鎖を連結する重鎖間ジスルフィドはない。
【図5A】[0013]図5A〜5E。表2に列挙する9つの構築物すべてに関するサイズ排除クロマトグラフィー(SEC)プロファイル。例示的な単量体SECプロファイルを示す構築物を「単量体」と標識する。
【図5B】[0013]図5A〜5E。表2に列挙する9つの構築物すべてに関するサイズ排除クロマトグラフィー(SEC)プロファイル。例示的な単量体SECプロファイルを示す構築物を「単量体」と標識する。
【図5C】[0013]図5A〜5E。表2に列挙する9つの構築物すべてに関するサイズ排除クロマトグラフィー(SEC)プロファイル。例示的な単量体SECプロファイルを示す構築物を「単量体」と標識する。
【図5D】[0013]図5A〜5E。表2に列挙する9つの構築物すべてに関するサイズ排除クロマトグラフィー(SEC)プロファイル。例示的な単量体SECプロファイルを示す構築物を「単量体」と標識する。
【図5E】[0013]図5A〜5E。表2に列挙する9つの構築物すべてに関するサイズ排除クロマトグラフィー(SEC)プロファイル。例示的な単量体SECプロファイルを示す構築物を「単量体」と標識する。
【図6】[0014]野生型および突然変異体Fc構築物に対するヒトおよびマウスFcRn結合のBIACORE分析。
【図7】[0015]分析用超遠心(AUC)を用いたタンパク質サイズ分析。
【図8】[0016]ヒトおよびマウスIgGサブクラスのCH3ドメインの配列比較。同定されたCH3ドメイン接合部分残基(24、表1;「*」によって示す)は非常に保存される。したがって、ヒトIgG1 Fcにおける単量体化を導く突然変異を他のIgGサブクラスに、ならびにヒト以外の種に拡張することも可能である。(a)において、ヒトIgG1、IgG2、IgG3、およびIgG4由来の配列は、それぞれ、配列番号1、配列番号2、配列番号3、および配列番号4に対応する。(b)において、ヒトIgG1、マウスIgG1、マウスIgG2a、マウスIgG2b、およびマウスIgG3由来の配列は、それぞれ、配列番号5、配列番号6、配列番号7、配列番号8、および配列番号9に対応する。
【図9】[0017]実施例2に記載するような、マウスにおける多様なFc融合分子の薬物動体学。
【発明を実施するための形態】
【0010】
[0018]野生型Fcは天然でホモ二量体であり、そしてこの特徴は、2つのCH3ドメイン間に存在する、強い高アフィニティ相互作用によって駆動される。本明細書に記載するのは、単量体Fc分子、およびこうした分子を作製しそして用いる方法である。用語「Fc」は、典型的には、ポリペプチドのホモ二量体として考えられるが、本発明のポリペプチドのユニークな特性のため、本明細書で用いる用語にはまた、重鎖のFc部分に対応するアミノ酸配列を含む、例えばCH2およびCH3ドメインを含有する、単量体ポリペプチドも含まれるであろう。
【0011】
[0019]本明細書に記載する方法は、CH3ドメイン接合部分で残基を置換することによって、CH3/CH3会合を完全に破壊して、なお分子の安定性を維持し、こうして単量体Fcを達成することが可能であることを立証する。改変されたFcの単量体の性質は、例えば、サイズ排除クロマトグラフィー(SEC)および分析用超遠心(AUC)によって評価可能である。置換は2つの事柄を達成する−1つはCH3ドメインのホモ二量体形成を妨害することであり、そしてもう一方は、Fcの単量体型を安定化させることである。
【0012】
[0020]CH3−CH3接合部分を構成するアミノ酸を同定するための方法論は、WO2009089004に開示される。構造に基づく検索アルゴリズム(YeおよびGodzik 2004)を用いて、タンパク質データバンク(PDB)(Bernstein, Koetzleら 1977)より、Fc領域に対応する座標を有する総数48の抗体結晶構造を同定した。同定されたFc結晶構造を調べると、最高解像度で決定された構造が、Z34C(PDBコード:1L6X)と呼ばれる、プロテインA由来のBドメインの最小型に結合したRITUXIMABのFc断片に対応することが明らかになった。寄託されるFc単量体座標および結晶対称性を用いて、1L6Xの生物学的Fcホモ二量体構造を生成した。2つの方法を用いて、CH3−CH3ドメイン相互作用に関与する残基を同定した:(i)距離限界基準によって決定されるような接触および(ii)溶媒アクセス可能表面積分析。
【0013】
[0021]接触に基づく方法にしたがって、その側鎖重原子が、第二の鎖における任意の残基の重原子から、明記する限界より近くに位置する残基として、接合部分残基を定義する。4.5Åの距離限界が好ましいが、接合部分残基を同定するため、より長い距離限界(例えば、5.5Å)もまた使用可能である(BaharおよびJerningan 1997)。
【0014】
[0022]表1は、4.5Åの距離限界を用いた、接触基準法に基づいて同定された24の接合部分残基を列挙する。構造保存に関して、これらの残基をさらに調べた。この目的のため、PDBから同定される48のFc結晶構造を重ね合わせて、そして側鎖重原子に関して、標準偏差を計算することによって分析した。図3は、構造的に保存された位、埋め込まれた位(%ASA≦10)、および暴露された位(%ASA>10)(%ASAは、アミノ酸の標準的ASAに対する、観察されるASAの比を指す(LeeおよびRichards 1971))とともに、CH3ドメイン接合部分を示す。配列比較を通じて、ヒトおよびマウスIgGサブクラス間、ならびに他のIgクラス間での接合部分残基の保存もまた調べた(図8)。
【0015】
[0023]抗体のFc部分に対する多様な置換または突然変異を本明細書に記載する。こうした変異は、KabatのEU番号付けスキームに基づく野生型抗体重鎖におけるその位のアミノ酸の後、その位に置換されるアミノ酸によって示される。例えば、EUの349位のチロシンがスレオニンで置換される際、これは「Y349T」と称される。「野生型配列」によって、動物種、例えばヒト内で天然に存在するアミノ酸の配列を意味する。野生型配列は、集団内の個体間でわずかに変化することも可能であり、例えば多様な免疫グロブリン鎖に関する異なるアレルが当該技術分野に知られる。
【0016】
[0024]ホモ二量体形成を防止するため、CH3−CH3接合部分を構成する1またはそれより多い残基を、相互作用が静電的に好ましくなくなるように、荷電アミノ酸で置換する。好ましい態様において、接合部分中の正荷電アミノ酸、例えばリジン、アルギニン、またはヒスチジンを、負荷電アミノ酸、例えばアスパラギン酸またはグルタミン酸で置換し、そして/または接合部分中の負荷電アミノ酸を正荷電アミノ酸で置換する。例としてヒトIgGを用いると、反対の電荷に変化させてもよい接合部分内の荷電残基には、R355、D356、E357、K370、K392、D399、K409、およびK439が含まれる。特定の好ましい態様において、接合部分内の2またはそれより多い荷電残基を反対の電荷に変化させる。例示的な分子には、K392DおよびK409D突然変異を含むもの、ならびにD399KおよびD356K突然変異を含むものが含まれる。
【0017】
[0025]単量体型において、ポリペプチドの安定性を保持するため、CH3−CH3接合部分を構成する1またはそれより多い疎水性残基を、小さい極性アミノ酸で置換する。例としてヒトIgGを用いると、CH3−CH3接合部分の巨大疎水性残基には、Y349、L351、L368、L398、V397、F405、およびY407が含まれる。小さい極性アミノ酸残基には、アスパラギン、システイン、グルタミン、セリン、およびスレオニンが含まれる。
【0018】
[0026]実施例において、CH3ドメイン接合部分で近くに位置する正荷電Lys残基2つをAspに突然変異させた。次いで、これらの2つのLysからAspへの突然変異の背景において、構造的に保存された巨大疎水性残基に対して、スレオニン・スキャニング突然変異誘発を行った。スレオニンでの多様な置換とともにK392D K409D突然変異を含むFc分子を、単量体形成に関して分析した。例示的な単量体Fc分子には、K392D K409D Y349T置換を有するもの、およびK392D K409D F405T置換を有するものが含まれる。
【0019】
[0027]Fc単量体の半分子の性質により、熱安定性は、高アフィニティCH3−CH3ドメイン相互作用を有するFc二量体のものより低い。熱安定性を増加させるため、CH2およびCH3ドメインにおいて、1またはそれより多いドメイン内ジスルフィド結合を導入してもよい。1またはそれより多い以下のアミノ酸対を突然変異させることによって、ジスルフィド結合を導入してもよい。
【0020】
【化1−1】
【0021】
【化1−2】
【0022】
[0028]抗体がpH依存性様式で新生Fc受容体(FcRn)と相互作用する能力によって、血清半減期が延長される。好ましい態様において、本発明の単量体Fc分子は、野生型Fcポリペプチドより優れていないとしても同様の、FcRnへの結合能を保持する(図6)。実施例2に示すように、本発明の単量体Fc分子は、抗体によって示される延長された血清半減期を保持することも可能であり、そしてしたがって、単量体Fcポリペプチドに共有結合した、例えば融合したポリペプチドの血清半減期を延長させるのに有用である。単量体Fcを操作して、FcRnに対するアフィニティを増加させ、それによってさらに循環中の分子の半減期を増加させる、1またはそれより多いさらなる突然変異を含有するようにすることも可能である。こうしたさらなる突然変異には、限定されるわけではないが、M252Y/S254T/T256E、M428L/N434S、T250Q/M428L、N434H、T307Q/N434A、T307Q/N434S、T307Q/E380A/N434S、およびV308P/N434Sが含まれる。
【0023】
[0029]本発明の組成物および方法は、本明細書に開示する例示的なアレルの変異体には限定されず、本明細書に開示する例示的なアレルに対して、少なくとも70%、少なくとも75%、少なくとも80%、少なくとも81%、少なくとも82%、少なくとも83%、少なくとも84%、少なくとも85%、少なくとも86%、少なくとも87%、少なくとも88%、少なくとも89%、少なくとも90%、少なくとも91%、少なくとも92%、少なくとも93%、少なくとも94%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、および少なくとも99%の同一性を有するものが含まれる。本発明のCH3含有分子の特性を野生型ヒトCH3含有分子のものに比較する目的のため、野生型配列は、図8(a)配列番号1〜4(それぞれ、IgG1、IgG2、IgG3、およびIgG4)に示すものである。
【0024】
[0030]単量体Fc含有分子の生成は、IgG Fcに基づくものに限定されず、IgA、IgE、IgD、およびIgMを含む他の免疫グロブリンサブクラスのFc領域にも適用可能であることが意図される。
【0025】
[0031]Fcドメインを含有する実質的に任意の分子が、本発明の単量体Fcドメインを含んでもよい。こうした分子の例を図2に示す。図2に示すように、多様なペプチドをFcのN末端またはC末端に融合させるかまたはコンジュゲート化してもよい。特定の態様において、Fc含有分子をFabに融合させて、半抗体を生成する。細胞、例えばCHO細胞において、単量体Fcを含む重鎖および軽鎖を組換え的に発現させることによって、こうした半抗体を生成してもよい。重鎖は1またはそれより多いさらなる突然変異を含有してもよい。特定の態様において、重鎖はさらに、ヒンジ領域中に1またはそれより多いシステイン残基の突然変異を含む(Allenら, Biochemistry. 2009 May 5;48(17):3755−66)。
【0026】
[0032]本発明のFcポリペプチドは、野生型Fc分子に比較した際、二量体化減少を示す。したがって、本発明の態様には、抗体またはFc融合分子を含む組成物であって、前記抗体またはFc融合分子が示すFc−Fc二量体化の量が、15%未満、14%未満、13%未満、12%未満、11%未満、10%未満、9%未満、8%未満、7%未満、6%未満、5%未満、4%未満、3%未満、2%未満、または1%未満である、前記組成物が含まれる。当該技術分野に知られるいくつかの技術によって、二量体化を測定してもよい。二量体化を測定するのに好ましい方法には、サイズ排除クロマトグラフィー(SEC)、分析用超遠心(AUC)、動的光散乱(DLS)、および未変性PAGEが含まれる。
【0027】
[0033]本明細書記載のFc単量体分子は、療法タンパク質またはドメインの半減期を延長するのに有用である。Fc単量体療法剤で治療可能な疾患には、炎症、癌、代謝性障害等が含まれることも可能である。潜在的な融合ターゲットには、天然タンパク質結合ドメイン(例えばIL−1Ra、TIMP3、SHKペプチド、EPO、G−CSF)、抗体断片(例えばFab、scFv、ディアボディ、可変ドメイン由来結合剤)、代替足場由来タンパク質結合ドメイン(例えばFn3変異体、アンキリン・リピート変異体、センチリン(centyrin)変異体、アビマー(avimer))および特異的抗原を認識するペプチドが含まれてもよい。Fc単量体融合タンパク質は、サイズが小さく、したがって、組織に浸透する能力がより優れている可能性があるという利点を有する。Fc単量体融合タンパク質は、ターゲット結合の一価性が好ましい場合、特に有用でありうる。多価抗体を用いてターゲティングされた際にアゴニズムに感受性である細胞表面分子をターゲティングする際には、こうした一価性がしばしば好ましい。
【0028】
定義
[0034]本明細書において、別に定義しない限り、本発明と関連して用いられる科学的および技術的用語は、一般の当業者に一般的に理解される意味を有するものとする。さらに、文脈によって別に必要とされない限り、単数形の用語は複数のものを含み、そして複数形の用語は単数形を含むものとする。一般的に、本明細書に記載する、細胞および組織培養、分子生物学、免疫学、微生物学、遺伝学、ならびにタンパク質および核酸化学およびハイブリダイゼーションと関連して用いられる術語、ならびにそれらの技術は、当該技術分野に周知であり、そして一般的に用いられるものである。本発明の方法および技術は、別に示さない限り、一般的に、当該技術分野に周知の慣用法にしたがって、そして本明細書全体で引用され、そして論じられる、多様な一般的な参考文献およびより特異的な参考文献に記載されるように、行われる。例えば、本明細書に援用される、Sambrookら Molecular Cloning: A Laboratory Manual, 第2版, Cold Spring Harbor Laboratory Press, ニューヨーク州コールドスプリングハーバー(1989)、ならびにAusubelら, Current Protocols in Molecular Biology, Greene Publishing Associates(1992)、ならびにHarlowおよびLane Antibodies: A Laboratory Manual Cold Spring Harbor Laboratory Press, ニューヨーク州コールドスプリングハーバー(1990)を参照されたい。酵素反応および精製技術は、当該技術分野において一般的に達成されるように、または本明細書に記載するように、製造者の指定にしたがって行われる。本明細書記載の分析化学、合成有機化学、ならびに医学的および薬学的化学と関連して用いられる専門用語、ならびにこうした化学の実験法および技術は、当該技術分野に周知であり、そして一般的に知られるものである。化学合成、化学分析、薬学的調製、配合、および送達、ならびに患者の治療には、標準的技術を用いてもよい。
【0029】
[0035]以下の用語は、別に示さない限り、以下の意味を有すると理解されるべきである:用語「単離分子」は(分子が、例えばポリペプチド、ポリヌクレオチド、または抗体である場合)、その起源または派生供給源によって、(1)天然状態で該分子に付随する、天然に関連する構成要素と関連していないか、(2)同じ種由来の他の分子を実質的に含まないか、(3)異なる種由来の細胞によって発現されるか、または(4)天然には存在しない分子である。したがって、化学的に合成されるか、または天然に由来する細胞とは異なる細胞系において発現される分子は、天然に関連する構成要素から「単離されている」であろう。分子はまた、当該技術分野に周知の精製技術を用いた単離によって、天然に関連する構成要素を実質的に含まないようにされてもよい。当該技術分野に周知のいくつかの手段によって、分子純度または均一性をアッセイしてもよい。例えば、当該技術分野に周知の技術を用いて、ポリアクリルアミドゲル電気泳動を用い、そしてゲルを染色してポリペプチドを視覚化して、ポリペプチド試料の純度をアッセイしてもよい。特定の目的のため、HPLCまたは当該技術分野に周知の精製のための他の手段を用いることによって、より高い解像度を提供してもよい。
【0030】
[0036]ポリヌクレオチドおよびポリペプチド配列は、標準的な1文字または3文字略記を用いて示される。別に示さない限り、ポリペプチド配列は、アミノ末端を左側に、そしてカルボキシ末端を右側に有し、そして一本鎖核酸配列、および二本鎖核酸配列の上部鎖は、5’端を左に、そして3’端を右に有する。特定のポリペプチドまたはポリヌクレオチド配列はまた、参照配列とどのように異なるかを説明することによって記載されうる。
【0031】
[0037]用語「ペプチド」、「ポリペプチド」および「タンパク質」は、各々、ペプチド結合によって互いに連結された2またはそれより多いアミノ酸残基を含む分子を指す。これらの用語は、例えば天然および人工的タンパク質、タンパク質断片、およびタンパク質配列のポリペプチド類似体(突然変異タンパク質(mutein)、変異体、および融合タンパク質など)、ならびに翻訳後、あるいは別の共有的または非共有的修飾タンパク質を含む。ペプチド、ポリペプチド、またはタンパク質は、単量体性または多量体性であってもよい。
【0032】
[0038]用語「ポリペプチド断片」は、本明細書において、対応する全長タンパク質に比較した際、アミノ末端および/またはカルボキシ末端に欠失を有するポリペプチドを指す。断片は、例えば、少なくとも長さ5、6、7、8、9、10、11、12、13、14、15、20、50、70、80、90、100、150、200、250、300、350、または400アミノ酸であってもよい。断片はまた、例えば、最大で、長さ1,000、750、500、250、200、175、150、125、100、90、80、70、60、50、40、30、20、15、14、13、12、11、または10アミノ酸であってもよい。断片は、さらに、どちらかまたは両方の端に、1またはそれより多いさらなるアミノ酸、例えば異なる天然存在タンパク質または人工的アミノ酸配列由来のアミノ酸配列を含んでもよい。
【0033】
[0039]本発明のポリペプチドには、例えば:(1)タンパク質分解に対する感受性を減少させ、(2)酸化に対する感受性を減少させ、(3)タンパク質複合体を形成するための結合アフィニティを改変し、(4)結合アフィニティを改変し、そして(4)他の物理化学特性または機能特性を与えるかまたはこうした特性を修正するように、いずれかの方式で、そしていずれかの理由のために修飾されているポリペプチドが含まれる。類似体には、ポリペプチドの突然変異タンパク質が含まれる。例えば、単数または複数のアミノ酸置換(例えば、保存的アミノ酸置換)を天然存在配列において(例えば、分子間接触を形成するドメイン(単数または複数)外のポリペプチドの部分において)行うことも可能である。「保存的アミノ酸置換」は、親配列の構造特徴を実質的に変化させないものである(例えば置換アミノ酸は、親配列に存在するらせんを中断させるか、あるいは親配列を特徴付けるかまたはその機能に必要な他のタイプの二次構造を破壊する傾向があってはならない)。当該技術分野に認識されるポリペプチド二次構造および三次構造の例が、Proteins, Structures and Molecular Principles(Creighton監修, W.H. Freeman and Company, ニューヨーク(1984)); Introduction to Protein Structure(C. BrandenおよびJ. Tooze監修, Garland Publishing, ニューヨーク州ニューヨーク(1991));およびThorntonら Nature 354:105(1991)に記載され、これらは各々、本明細書に援用される。
【0034】
[0040]ポリペプチドの「変異体」は、別のポリペプチド配列に比較して、1またはそれより多いアミノ酸残基がアミノ酸配列内で挿入され、欠失され、そして/または置換された、アミノ酸配列を含む。本発明の変異体には、変異体CH2またはCH3ドメインを含むものが含まれる。特定の態様において、変異体は、Fc分子中に存在した際、1またはそれより多いFcγRに対するポリペプチドのアフィニティを増加させる1またはそれより多い突然変異を含む。こうした変異体は、抗体依存性細胞仲介性細胞傷害性の増加を示す。こうしたものを提供する変異体の例は、米国特許第7,317,091号に記載される。
【0035】
[0041]他の変異体には、CH3ドメイン含有ポリペプチドがホモ二量体形成する能力を減少させるものが含まれる。こうしたFc変異体の例が、米国特許第5,731,168号および第7,183,076号に記載される。さらなる例が、08年1月7日出願の共同所有米国仮出願第61/019,569号および08年12月5日出願の第61/120,305号(どちらもその全体が本明細書に援用される)に記載される。
【0036】
[0042]ポリペプチドの「誘導体」は、例えば別の化学部分、例えばポリエチレングリコール、細胞傷害剤、アルブミン(例えばヒト血清アルブミン)などへのコンジュゲート化、リン酸化、およびグリコシル化を介して、化学的に修飾されているポリペプチド(例えば抗体)である。別に示さない限り、用語「抗体」には、2つの全長重鎖および2つの全長軽鎖を含む抗体に加えて、その誘導体、変異体、断片、および突然変異タンパク質が含まれ、それらの例を以下に記載する。
【0037】
[0043]用語「ヒト抗体」には、ヒト免疫グロブリン配列に由来する1またはそれより多い可変領域および定常領域を有する抗体すべてが含まれる。1つの態様において、可変ドメインおよび定常ドメインのすべてがヒト免疫グロブリン配列に由来する(完全ヒト抗体)。これらの抗体は、多様な方法で調製可能であり、その例を以下に記載し、これらには、ヒト重鎖および/または軽鎖をコードする遺伝子に由来する抗体を発現するように遺伝子修飾されたマウスの、関心対象の抗原での免疫を通じたものが含まれる。特定の態様において、ヒト抗体の重鎖は、CH3ドメインにおいて、重鎖が二量体化する能力が減少するように改変される。
【0038】
[0044]ヒト化抗体は、ヒト被験体に投与された際、非ヒト種抗体に比較すると、免疫応答を誘導する可能性がより低く、そして/またはより重度でない免疫応答を誘導するように、1またはそれより多いアミノ酸置換、欠失、および/または付加によって、非ヒト種に由来する抗体の配列と異なる配列を有する。1つの態様において、非ヒト種抗体の重鎖および/または軽鎖のフレームワークおよび定常ドメイン中の特定のアミノ酸を突然変異させて、ヒト化抗体を産生する。別の態様において、ヒト抗体由来の定常ドメイン(単数または複数)を、非ヒト種の可変ドメイン(単数または複数)に融合させる。ヒト化抗体をどのように作製するかの例を、米国特許第6,054,297号、第5,886,152号、および第5,877,293号に見出すことも可能である。
【0039】
[0045]用語「キメラ抗体」は、1つの抗体由来の1またはそれより多い領域、および1またはそれより多い他の抗体由来の1またはそれより多い他の領域を含有する抗体を指す。キメラ抗体の1つの例において、重鎖および/または軽鎖の部分は、特定の種由来であるか、または特定の抗体クラスもしくはサブクラスに属する抗体と同一であるか、該抗体に相同であるか、または該抗体に由来する一方、鎖(単数または複数)の残りは、別の種由来であるか、または別の抗体クラスもしくはサブクラスに属する抗体(単数または複数)と同一であるか、該抗体に相同であるか、または該抗体に由来する。やはり含まれるのは、所望の生物学的活性を示す、こうした抗体の断片である。
【0040】
[0046]抗体の断片または類似体は、本明細書の解説にしたがって、そして当該技術分野に周知の技術を用いて、一般の当業者によって、容易に調製可能である。断片または類似体の好ましいアミノ末端およびカルボキシ末端は、機能ドメインの境界近傍に存在する。公共のまたは私有の(proprietary)配列データベースに、ヌクレオチドおよび/またはアミノ酸配列データを比較することによって、構造ドメインおよび機能ドメインを同定することも可能である。コンピュータ比較法を用いて、既知の構造および/または機能を持つ他のタンパク質に存在する配列モチーフまたは予測されるタンパク質コンホメーションドメインを同定してもよい。既知の三次元構造にフォールディングするタンパク質配列を同定する方法が知られる。例えばBowieら, 1991, Science 253:164を参照されたい。
【0041】
[0047]「CDR移植抗体」は、特定の種またはアイソタイプの抗体由来の1またはそれより多いCDR、および同じまたは異なる種またはアイソタイプの別の抗体のフレームワークを含む抗体である。
【0042】
[0048]2つのポリヌクレオチドまたは2つのポリペプチド配列の「同一性パーセント」は、デフォルト・パラメータを用い、GAPコンピュータ・プログラム(GCGウィスコンシン・パッケージ、バージョン10.3(Accelrys、カリフォルニア州サンディエゴ)の一部)を用いて、配列を比較することによって決定される。
【0043】
[0049]用語「ポリヌクレオチド」、「オリゴヌクレオチド」および「核酸」は、本明細書全体を通じて交換可能に用いられ、そしてDNA分子(例えばcDNAまたはゲノムDNA)、RNA分子(例えばmRNA)、ヌクレオチド類似体(例えばペプチド核酸および非天然存在ヌクレオチド類似体)を用いて生成されるDNAまたはRNAの類似体、およびそれらのハイブリッドを含む。核酸分子は、一本鎖または二本鎖であることも可能である。1つの態様において、本発明の核酸分子は、抗体またはFc融合体、およびその誘導体、突然変異タンパク質、または変異体をコードする、隣接オープンリーディングフレームを含む。
【0044】
[0050]2つの一本鎖ポリヌクレオチドは、ギャップを導入することなく、そしていずれの配列の5’端または3’端にも、対形成しないヌクレオチドを伴わずに、一方のポリヌクレオチド中のすべてのヌクレオチドが、他方のポリヌクレオチド中の相補的ヌクレオチドと反対であるように、逆平行配向で整列可能であるならば、互いに「相補体」である。ポリヌクレオチドは、中程度にストリンジェントな条件下で、2つのポリヌクレオチドが互いにハイブリダイズ可能であるならば、別のポリヌクレオチドに「相補的」である。したがって、ポリヌクレオチドは、別のポリヌクレオチドの相補体であることなく、該ポリヌクレオチドに相補的であることも可能である。
【0045】
[0051]「ベクター」は、連結された別の核酸を、細胞内に導入するために使用可能な核酸である。ベクターの1つのタイプは「プラスミド」であり、その内部にさらなる核酸セグメントを連結可能な、直鎖または環状二本鎖DNA分子を指す。別のタイプのベクターはウイルスベクター(例えば複製不全レトロウイルス、アデノウイルス、およびアデノ随伴ウイルス)であり、ここで、さらなるDNAセグメントをウイルスゲノム内に導入可能である。特定のベクターは、導入された宿主細胞において、自律的に複製可能である(例えば細菌複製起点を含む細菌ベクターおよびエピソーム哺乳動物ベクター)。宿主細胞内への導入に際して、宿主細胞のゲノム内に他のベクター(例えば非エピソーム哺乳動物ベクター)を組み込んで、そしてそれによって宿主ゲノムと一緒に複製させる。「発現ベクター」は、選択したポリヌクレオチドの発現を指示することも可能なベクターのタイプである。
【0046】
[0052]ヌクレオチド配列は、制御配列が該ヌクレオチド配列の発現(例えば発現のレベル、時期、または位置)に影響を及ぼすならば、該制御配列に「機能可能であるように連結されて」いる。「制御配列」は、機能可能であるように連結されている核酸の発現(例えば発現のレベル、時期、または位置)に影響を及ぼす核酸である。制御配列は、例えば、制御される核酸に対して直接、あるいは1またはそれより多い他の分子(例えば制御配列および/または核酸に結合するポリペプチド)の作用を通じて、その効果を発揮しうる。制御配列の例には、プロモーター、エンハンサーおよび他の発現調節要素(例えばポリアデニル化シグナル)が含まれる。制御配列のさらなる例は、例えば、Goeddel, 1990, Gene Expression Technology: Methods in Enzymology 185, Academic Press, カリフォルニア州サンディエゴ、およびBaronら, 1995, Nucleic Acids Res. 23:3605−06に記載される。
【0047】
[0053]「宿主細胞」は、核酸、例えば本発明の核酸を発現するために使用可能な細胞である。宿主細胞は、原核生物、例えば大腸菌(E.coli)であってもよいし、または真核生物、例えば単細胞真核生物(例えば酵母(yeast)または他の真菌)、植物細胞(例えばタバコ(tobacco)またはトマト(tomato)植物細胞)、動物細胞(例えばヒト細胞、サル細胞、ハムスター細胞、ラット細胞、マウス細胞、または昆虫細胞)またはハイブリドーマであってもよい。例示的な宿主細胞には、チャイニーズハムスター卵巣(CHO)細胞株、またはDHFRが欠損しているCHO株DXB−11(Urlaubら, 1980, Proc. Natl. Acad. Sci. USA 77:4216−20を参照されたい)、血清不含培地中で増殖するCHO細胞株(Rasmussenら, 1998, Cytotechnology 28:31を参照されたい)、DXB−11 CHO細胞の誘導体であるCS−9細胞、およびAM−1/D細胞(米国特許第6,210,924号に記載される)を含むその誘導体が含まれる。他のCHO細胞株には、CHO−K1(ATCC#CCL−61)、EM9(ATCC#CRL−1861)、およびUV20(ATCC#CRL−1862)が含まれる。他の宿主細胞の例には、サル腎臓細胞のCOS−7株(ATCC CRL 1651)(Gluzmanら, 1981, Cell 23:175を参照されたい)、L細胞、C127細胞、3T3細胞(ATCC CCL 163)、HeLa細胞、BHK(ATCC CRL 10)細胞株、アフリカミドリザル(African green monkey)腎臓細胞株CV1(ATCC CCL 70)由来のCV1/EBNA細胞株(McMahanら, 1991, EMBO J. 10:2821を参照されたい)、ヒト胚性腎細胞、例えば293、293 EBNAまたはMSR 293、ヒト上皮A431細胞、ヒトColo205細胞、他の形質転換霊長類細胞株、正常二倍体細胞、初代組織のin vitro培養由来の細胞株、初代外植片、HL−60、U937、HaKまたはJurkat細胞が含まれる。典型的には、宿主細胞は、その後、宿主細胞で発現可能なポリペプチドをコードする核酸で形質転換またはトランスフェクションされることが可能な培養細胞である。
【0048】
[0054]句「組換え宿主細胞」を用いて、発現しようとする核酸で形質転換されているかまたはトランスフェクションされている宿主細胞を示すことも可能である。宿主細胞はまた、核酸を含むが、機能可能であるように核酸と連結されるように制御配列が宿主細胞内に導入されない限り、所望のレベルで該核酸を発現しない細胞であってもよい。用語、宿主細胞は、特定の対象の細胞だけでなく、こうした細胞の子孫または潜在的な子孫も指すことが理解される。例えば、突然変異または環境的影響によって、続く世代で特定の修飾が起こりうるため、こうした子孫は、実際、親細胞と同一でない可能性もあるが、なお、本明細書において、この用語の範囲内に含まれる。
【0049】
薬学的組成物
[0055]本発明のポリペプチドは、薬学的組成物内に配合するのに特に有用である。こうした組成物は、生理学的に許容されうるキャリアー、賦形剤または希釈剤などの1またはそれより多いさらなる構成要素を含む。場合によって、組成物は、例えば以下に記載するような、1またはそれより多い生理学的活性剤をさらに含む。多様な特定の態様において、組成物は、1またはそれより多い本発明の単量体抗体および/またはFc融合タンパク質に加えて、1、2、3、4、5、または6の生理学的活性剤を含む。
【0050】
[0056]1つの態様において、薬学的組成物は、本発明の単量体抗体および/またはFc融合タンパク質を、緩衝剤、アスコルビン酸などの酸化防止剤、低分子量ポリペプチド(10アミノ酸未満を有するものなど)、タンパク質、アミノ酸、グルコース、スクロースまたはデキストリンなどの炭水化物、EDTAなどのキレート剤、グルタチオン、安定化剤、ならびに賦形剤からなる群より選択される1またはそれより多い物質とともに、含む。中性緩衝生理食塩水または同種血清アルブミンと混合された生理食塩水が、適切な希釈剤の例である。適切な産業標準にしたがって、ベンジルアルコールなどの保存剤もまた添加してもよい。組成物は、適切な賦形剤溶液(例えばスクロース)を希釈剤として用いた凍結乾燥物として配合されてもよい。適切な構成要素は、使用する投薬量および濃度で、レシピエントに非毒性である。薬学的配合物において使用可能な構成要素のさらなる例が、Remington’s Pharmaceutical Sciences, 第16版(1980)および第20版(2000), Mack Publishing Company, ペンシルバニア州イーストンに提示される。
【0051】
[0057]開業医に使用されるためのキットには、本明細書に論じる任意の状態を治療する際に使用するための、1またはそれより多い本発明の単量体抗体および/またはFc融合タンパク質、ならびにラベルまたは他の使用説明書が含まれる。1つの態様において、キットには、1またはそれより多い単量体抗体および/またはFc融合タンパク質の無菌調製物が含まれ、該調製物は、上に開示するような組成物の形であってもよく、そして1またはそれより多いバイアル中にあってもよい。
【0052】
[0058]投薬量および投与頻度は、投与経路、使用する特定の単量体抗体および/またはFc融合タンパク質、治療しようとする疾患の性質および重症度、状態が急性または慢性であるか、ならびに被験体のサイズおよび全身状態などの要因に応じて、多様でありうる。関連技術において知られる方法によって、例えば用量の段階的増大研究を伴いうる臨床試験において、適切な投薬量を決定してもよい。
【0053】
[0059]本発明の単量体抗体および/またはFc融合タンパク質を、例えば1回または1回より多く、例えばある期間に渡って定期的な間隔で、投与してもよい。特定の態様において、単量体抗体および/またはFc融合タンパク質を、少なくとも1ヶ月に1回またはそれより多く、例えば1ヶ月、2ヶ月、または3ヶ月、あるいはさらに無期限に渡って、投与する。慢性状態を治療するためには、一般的に、長期治療が最も有効である。しかし、急性状態を治療するため、より短い期間、例えば1〜6週間の投与で十分である可能性もある。一般的に、患者が、選択した単数または複数の指標に関して、ベースラインを超えた、医学的に適切な度合いの改善を示すまで、単量体抗体および/またはFc融合タンパク質を投与する。
【0054】
[0060]関連分野で理解されるように、本発明の単量体抗体および/またはFc融合タンパク質を含む薬学的組成物を、適応症に適した方式で、被験体に投与する。限定されるわけではないが、非経口、局所、または吸入によるものを含む、任意の適切な技術によって、薬学的組成物を投与してもよい。注射する場合、薬学的組成物を、例えば、動脈内、静脈内、筋内、病巣内、腹腔内または皮下経路を介して、ボーラス注射によって、あるいは連続注入によって、投与してもよい。局在化投与、例えば疾患部位または傷害部位での投与が意図され、経皮送達および移植物からの持続放出も同様である。吸入による送達には、例えば、鼻または経口吸入、ネブライザーの使用、エアロゾル型での単量体抗体および/またはFc融合タンパク質の吸入等が含まれる。
【実施例】
【0055】
実施例1
[0061]距離限界基準を用いて、CH3−−−CH3ドメイン相互作用に関与する残基を同定した。CH3ドメイン接合部分に位置する24の残基があった(表1)。入手可能な既知のFc抗体結晶構造を用いて、側鎖構造保存に関して、これらの24残基を調べた(図3)。この分析によって、疎水性残基のいくつかに関して、高い構造保存が明らかになった。計算的方法(PokalaおよびHandel 2005)を用いて、24の接合部分位およびL398(接合部分残基K392と接触する残基)を、アラニンで、一度に一残基ずつ突然変異させることによって、つのCH3ドメイン間の会合自由エネルギーもまた計算した。計算および構造保存分析によって、24残基のうち6残基が、CH3ドメイン二量体形成に大きく寄与することが明らかになった。これらの6位を分析して、置換の影響を決定した。
【0056】
表1:第一の鎖(A)中のCH3ドメイン接合部分残基および第二の鎖(B)中の側鎖接触残基のリストa
【0057】
【表1−1】
【0058】
【表1−2】
【0059】
aCH3−CH3ドメイン相互作用に2倍対称性が存在するため、各対の相互作用は、構造中、2回示される(例えば、Ser A 364−Leu B 368’およびLeu A 439−Ser B 364’)。しかし、Leu A 351−Pro B 352’、Leu A 351−Pro B 353’、Lys A 392−Leu B 398’、Val A 397−Thr B 393’、およびTyr A 407−Ser B 408’対は、側鎖−主鎖接触を伴うため、これらは1回しか示されない。
【0060】
[0062]6位各々でのスレオニン置換に対応するFc突然変異タンパク質を生成した(表2)。さらに、F405T Y407RおよびY407R Fc突然変異体もまた生成した。完全単量体Fc形成の可能性を増進させるため、CH3/CH3弱体化Fc変異体の背景で、6つの位で突然変異を生成し、ここで、リジン392およびリジン409をアスパラギン酸に突然変異させる(K392D K409D Fc)。
【0061】
表2
【0062】
【表2−1】
【0063】
【表2−2】
【0064】
ND;未決定。
[0063]Multi−QuikChange部位特異的突然変異誘発(Stratagene)を用いて、ヒトIgG1 Fc K392D K409D中、表2(#2〜#9)に列挙する突然変異を生成した。用いたオリゴプライマーは以下の通りである:
【0065】
【化2】
【0066】
[0064]予期される突然変異をDNA配列決定によって確認した。pTT5一過性哺乳動物発現ベクターを用いて、親(hu IgG1 Fc K409D K392D)および突然変異体Fcタンパク質を293E細胞中で発現させた。標準的プロテインAクロマトグラフィー(5mlカラム、Pierce)を用いて、Fcタンパク質を精製した。TOSO46mm SW3000カラム(TOSO Biosciences LLC、PA)を用いて、Fcタンパク質均一性分析(SEC)を行った。280nmの吸収を測定し、そして1mg/ml=1.74 OD280を用いて計算することによって、タンパク質濃度を決定した。
【0067】
[0065]各試料の場合において、5μgのタンパク質を非還元SDSまたは還元SDS試料緩衝液(Invitroten)で処理し、4〜20% TGゲル(Invitrogen)上で泳動し、そしてGel−code試薬(Pierce)を用いて染色した。
【0068】
[0066]BIAcore3000装置を用いた、Fc突然変異体#1(Fc K392D−K409D、二量体)、#2(Fc K392D−K409D−Y349T、単量体SECプロファイル)および#6(Fc K392D−K409D−Y349T、単量体SECプロファイル)へのヒトおよびマウスFcRn結合のBIAcore分析。CHO細胞で産生したhuFcを、CM5チップ上のフォローセル(follow cell)2(Fc2)上に固定した。Fc1をバックグラウンド対照として用いた。2nMヒトFcRnを1、10、100nMの示したFc変異体と1時間インキュベーションした後、CHO huFc表面に注入した。固定されたCHO huFcへのFcRnの結合減少は、溶液中のFc変異体にFcRnが結合している指標となる。2nMマウスFcRnを0.1、1、10nMのFc変異体と1時間インキュベーションした後、CHO huFc表面に注入した。
【0069】
[0067]突然変異体Fcに関するタンパク質濃度は、それぞれ、0.4および0.6mg/mlであった。Beckmann Coulter ProteomeLab XL−1装置によって、PBS中の試料を分析した。50,000rpmで沈降速度実験を行い、水晶ウインドウを含む二重セクター中心セル・アセンブリー中、280nmの吸光度によって追跡した。間に遅延を含まず、20℃でスキャンを収集した。SEDFIT、バージョン9.4を用いて、AUC−SVデータを分析した。AUC−SV分析において、非線形最小二乗適合中、摩擦比(frictional ration)、時間不変ノイズ(time invariant noise)、およびメニスカス値(meniscus value)が浮遊する(float)ことを可能にした。
【0070】
実施例2
本実施例は、正常マウスにおける多様な分子量の単量体Fc構築物のPKパラメータを示す。単量体Fcは、K392D−K409D−Y349T突然変異を含んだ。単量体Fc N297Aは、グリコシル化部位を除去するため、N297にさらなる突然変異を有する単量体Fcを含んだ。半IgGは、単量体Fcに融合したFabを含んだ。Fab FnFnは、フィブロネクチン二量体に融合したFabを含んだ。Fab Hisは、ヒスチジンタグに融合したFabであった。huFc WTデルタHは、ヒンジ領域を除いたWT Fc二量体を含んだ。54匹のSCIDマウスを以下の処置群に割り当てた。すべてのマウスには、10mg/kg IVを投与し、そして0.25、2、4、8、20および32時間で血清を収集した。
【0071】
表3
【0072】
【表3】
【0073】
分析法のまとめ:
単量体Fc構築物の定量化のため、マイクロタイタープレート(Maxisorp、Nunc)を、1、2および6群に関しては、ヤギ抗ヒトIgG Fcγ特異的(Jackson カタログ#109−005−098)で、または3、4および5群に関しては、ヤギ抗ヒトIgG F(ab’)2特異的(Jacksonカタログ#109−005−097)のいずれかでコーティングした。PBST中の10%NFDM(脱脂粉乳)でブロッキングした後、前処理後、標準、品質管理試料(QC)、および試験試料をNFDM/PBST中、50の希釈係数でインキュベーションした。PBST緩衝液で洗浄することによって、未結合構築物を除去した。次に、1、2および6群において、西洋ワサビペルオキシダーゼ標識ヤギ抗ヒトIgG Fcγ特異的(Jaksonカタログ#109−035−098)を添加して、捕捉されたFc構築物を検出する一方、3、4および5群において、ヤギ抗ヒトIgG F(ab’)2特異的(Jacksonカタログ#109−036−097)を用いて、捕捉されたF(ab’)2構築物を検出した。最終洗浄工程後、TMB基質溶液(1:1テトラメチルベンジジンおよびペルオキシド、Kirkegaard & Perry Laboratories)を添加して、そしてリン酸で反応停止した。光学密度(OD)を450〜650nmの波長で決定した。4パラメータ・ロジスティックモデルにしたがって回帰した、同時に分析される標準曲線に対して、ワトソン・ソフトウェアが仲介する比較を通じて、QCおよび未知の標本に関して、OD値の濃度への変換を達成した。
【0074】
表4−−血清標本に関するデータ
【0075】
【表4】
【0076】
表5
【0077】
【表5】
【0078】
本実施例は、正常マウスPKパラメータ、すなわちFcRn相互作用の度合い、〜60kDaの腎臓クリアランス閾値より高いまたは低い多様な分子量の単量体Fc構築物の度合いを示す。二量体CH2−CH3ドメインからなるhuFc WTデルタH構築物は、クリアランス閾値より低い分子量であるにもかかわらず、すべての他の構築物に比較して、最大の曝露/AUCを示した。Fab Fc単量体からなる半IgG分子は、二量体Fcの曝露の35%を示したが、Fab−フィブロネクチン二量体からなる、サイズが一致したFab Fc二量体に比較して、3.5倍高いAUCを有し、この結果は、半減期、そして最終的には曝露を増加させる際のFcの役割を立証した。Fab単独および単量体Fc構築物、N連結付加部位の除去のため、グリコシル化を欠くWTまたはN297A変異体いずれかは、すべて、迅速なクリアランスおよびhuFC WTデルタH構築物よりも17〜38倍低い最小AUC値を示した。
【0079】
参考文献
【0080】
【化3−1】
【0081】
【化3−2】
【特許請求の範囲】
【請求項1】
CH2およびCH3ドメインを含む単量体Fcポリペプチドであって、前記CH3ドメインが:
(a)CH3ホモ二量体形成に静電的に好ましくない2またはそれより多い荷電アミノ酸;および
(b)極性アミノ酸残基での1またはそれより多い疎水性接合部分残基のアミノ酸置換
を含む、前記単量体Fcポリペプチド。
【請求項2】
極性アミノ酸残基がスレオニンである、請求項1の単量体Fcポリペプチド。
【請求項3】
CH3ドメインがIgG CH3ドメインである、請求項1の単量体Fcポリペプチド。
【請求項4】
疎水性接合部分残基が、Y349、L351、L368、V397、L398、F405、およびY407からなる群より選択される、請求項3の単量体Fc。
【請求項5】
疎水性接合部分残基がY349である、請求項4の単量体Fc。
【請求項6】
スレオニンがY349に対して置換される、請求項5の単量体Fc。
【請求項7】
疎水性接合部分残基がF405である、請求項4の単量体Fc。
【請求項8】
スレオニンがF405に対して置換される、請求項7の単量体Fc。
【請求項9】
前記CH3ドメインが、392位で負に荷電されたアミノ酸を含む、請求項3の単量体Fcポリペプチド。
【請求項10】
負に荷電されたアミノ酸がアスパラギン酸である、請求項9の単量体Fcポリペプチド。
【請求項11】
前記CH3ドメインが、409位で負に荷電されたアミノ酸を含む、請求項3の単量体Fcポリペプチド。
【請求項12】
負に荷電されたアミノ酸がアスパラギン酸である、請求項11の単量体Fcポリペプチド。
【請求項13】
前記CH3ドメインが、392位および409位で負に荷電されたアミノ酸を含む、請求項3の単量体Fcポリペプチド。
【請求項14】
負に荷電されたアミノ酸がアスパラギン酸である、請求項13の単量体Fcポリペプチド。
【請求項15】
疎水性接合部分残基に対して置換されたスレオニンをさらに含む、請求項14の単量体Fcポリペプチド。
【請求項16】
疎水性接合部分残基が、Y349、L351、L368、V397、L398、F405、およびY407からなる群より選択される、請求項15の単量体Fcポリペプチド。
【請求項17】
疎水性接合部分残基がY349である、請求項16の単量体Fcポリペプチド。
【請求項18】
抗体可変ドメインを含む、請求項1の単量体Fcポリペプチド。
【請求項19】
CH1ドメインを含む、請求項18の単量体Fcポリペプチド。
【請求項20】
抗体重鎖を含む、請求項19の単量体Fcポリペプチド。
【請求項21】
請求項20の単量体Fcポリペプチドおよび抗体軽鎖を含む、単量体抗体。
【請求項22】
請求項1の単量体Fcポリペプチドをコードする、単離核酸。
【請求項23】
請求項1の単量体Fcポリペプチドをコードする核酸を含む、発現ベクター。
【請求項24】
請求項1の単量体Fcポリペプチドをコードする核酸を含む、宿主細胞。
【請求項25】
単量体Fcポリペプチドを調製する方法であって:
(a)請求項1の単量体Fcポリペプチドをコードする核酸を含む宿主細胞を、前記単量体Fcポリペプチドが発現される条件下で培養し;そして
(b)宿主細胞培養から単量体Fcポリペプチドを回収する
工程を含む、前記方法。
【請求項26】
抗体のCH3ドメインを含むポリペプチドであって、CH3ドメインが:
(a)1またはそれより多い荷電アミノ酸が、CH3ホモ二量体形成に静電的に好ましくないアミノ酸で置換されており;そして
(b)1またはそれより多い疎水性接合部分残基が、極性アミノ酸で置換されている
ここで、野生型CH3ドメインを含むポリペプチドに比較して、ポリペプチドがホモ二量体を形成する能力が減少する
ように、野生型CH3ドメインと異なるポリペプチド配列を含む、前記ポリペプチド。
【請求項1】
CH2およびCH3ドメインを含む単量体Fcポリペプチドであって、前記CH3ドメインが:
(a)CH3ホモ二量体形成に静電的に好ましくない2またはそれより多い荷電アミノ酸;および
(b)極性アミノ酸残基での1またはそれより多い疎水性接合部分残基のアミノ酸置換
を含む、前記単量体Fcポリペプチド。
【請求項2】
極性アミノ酸残基がスレオニンである、請求項1の単量体Fcポリペプチド。
【請求項3】
CH3ドメインがIgG CH3ドメインである、請求項1の単量体Fcポリペプチド。
【請求項4】
疎水性接合部分残基が、Y349、L351、L368、V397、L398、F405、およびY407からなる群より選択される、請求項3の単量体Fc。
【請求項5】
疎水性接合部分残基がY349である、請求項4の単量体Fc。
【請求項6】
スレオニンがY349に対して置換される、請求項5の単量体Fc。
【請求項7】
疎水性接合部分残基がF405である、請求項4の単量体Fc。
【請求項8】
スレオニンがF405に対して置換される、請求項7の単量体Fc。
【請求項9】
前記CH3ドメインが、392位で負に荷電されたアミノ酸を含む、請求項3の単量体Fcポリペプチド。
【請求項10】
負に荷電されたアミノ酸がアスパラギン酸である、請求項9の単量体Fcポリペプチド。
【請求項11】
前記CH3ドメインが、409位で負に荷電されたアミノ酸を含む、請求項3の単量体Fcポリペプチド。
【請求項12】
負に荷電されたアミノ酸がアスパラギン酸である、請求項11の単量体Fcポリペプチド。
【請求項13】
前記CH3ドメインが、392位および409位で負に荷電されたアミノ酸を含む、請求項3の単量体Fcポリペプチド。
【請求項14】
負に荷電されたアミノ酸がアスパラギン酸である、請求項13の単量体Fcポリペプチド。
【請求項15】
疎水性接合部分残基に対して置換されたスレオニンをさらに含む、請求項14の単量体Fcポリペプチド。
【請求項16】
疎水性接合部分残基が、Y349、L351、L368、V397、L398、F405、およびY407からなる群より選択される、請求項15の単量体Fcポリペプチド。
【請求項17】
疎水性接合部分残基がY349である、請求項16の単量体Fcポリペプチド。
【請求項18】
抗体可変ドメインを含む、請求項1の単量体Fcポリペプチド。
【請求項19】
CH1ドメインを含む、請求項18の単量体Fcポリペプチド。
【請求項20】
抗体重鎖を含む、請求項19の単量体Fcポリペプチド。
【請求項21】
請求項20の単量体Fcポリペプチドおよび抗体軽鎖を含む、単量体抗体。
【請求項22】
請求項1の単量体Fcポリペプチドをコードする、単離核酸。
【請求項23】
請求項1の単量体Fcポリペプチドをコードする核酸を含む、発現ベクター。
【請求項24】
請求項1の単量体Fcポリペプチドをコードする核酸を含む、宿主細胞。
【請求項25】
単量体Fcポリペプチドを調製する方法であって:
(a)請求項1の単量体Fcポリペプチドをコードする核酸を含む宿主細胞を、前記単量体Fcポリペプチドが発現される条件下で培養し;そして
(b)宿主細胞培養から単量体Fcポリペプチドを回収する
工程を含む、前記方法。
【請求項26】
抗体のCH3ドメインを含むポリペプチドであって、CH3ドメインが:
(a)1またはそれより多い荷電アミノ酸が、CH3ホモ二量体形成に静電的に好ましくないアミノ酸で置換されており;そして
(b)1またはそれより多い疎水性接合部分残基が、極性アミノ酸で置換されている
ここで、野生型CH3ドメインを含むポリペプチドに比較して、ポリペプチドがホモ二量体を形成する能力が減少する
ように、野生型CH3ドメインと異なるポリペプチド配列を含む、前記ポリペプチド。
【図1】

【図3】
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図6】
【図7】
【図8】

【図9】
【図2】
【図4】
【図3】
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図6】
【図7】
【図8】
【図9】
【図2】
【図4】
【公表番号】特表2013−511281(P2013−511281A)
【公表日】平成25年4月4日(2013.4.4)
【国際特許分類】
【出願番号】特願2012−540134(P2012−540134)
【出願日】平成22年11月22日(2010.11.22)
【国際出願番号】PCT/US2010/057662
【国際公開番号】WO2011/063348
【国際公開日】平成23年5月26日(2011.5.26)
【出願人】(500203709)アムジェン インコーポレイテッド (76)
【Fターム(参考)】
【公表日】平成25年4月4日(2013.4.4)
【国際特許分類】
【出願日】平成22年11月22日(2010.11.22)
【国際出願番号】PCT/US2010/057662
【国際公開番号】WO2011/063348
【国際公開日】平成23年5月26日(2011.5.26)
【出願人】(500203709)アムジェン インコーポレイテッド (76)
【Fターム(参考)】
[ Back to top ]