
変異体のスクリーニング方法
【課題】高分子基質を結合する高分子基質結合部位を有する酵素について高活性変異体を効率的に取得できるスクリーニング方法を提供する。
【解決手段】高分子基質に結合するトンネル状又は溝状の基質結合部位を有する酵素につき、基質相互作用を有する可能性のある部位のアミノ酸をアラニン置換した2種類以上の一次変異体を調製する工程と、前記一次変異体につき、変異による酵素活性の上昇の有無をスクリーニングして、変異前より酵素活性が上昇した前記一次変異体のアラニン置換部位を変異導入候補部位として選択する工程と、を備える、スクリーニング方法とする。
【解決手段】高分子基質に結合するトンネル状又は溝状の基質結合部位を有する酵素につき、基質相互作用を有する可能性のある部位のアミノ酸をアラニン置換した2種類以上の一次変異体を調製する工程と、前記一次変異体につき、変異による酵素活性の上昇の有無をスクリーニングして、変異前より酵素活性が上昇した前記一次変異体のアラニン置換部位を変異導入候補部位として選択する工程と、を備える、スクリーニング方法とする。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、変異体のスクリーニング方法に関し、特に、高分子基質に作用するプロセッシブ酵素について高活性な変異体を効率的に取得するスクリーニング方法に関する。
【背景技術】
【0002】
多くの酵素の基質結合部位は鍵穴状であり、酵素のごく一部に限定されていることが多い。一方、セルロースなどの高分子基質に作用する酵素の中には、高分子基質の鎖がはまり込むトンネル状又は溝状の相対的に長い基質結合部位を有する酵素があることが知られている。さらに、当該基質結合部位に高分子基質を保持しつつ触媒反応を行う酵素(プロセッシブ酵)があることも知られている。こうした酵素における広範囲にわたる基質結合部位においては、その一部が活性部位として機能していると考えられる。
【0003】
鍵穴状の基質結合部位を有する酵素について高活性な変異体を得る方法としては、たとえば、ある程度ランダムに複数の部位に部位特異的に各種のアミノ酸置換変異を導入する方法や、立体構造から活性に影響を及ぼす変異導入候補部位を選抜し部位特異的にアミノ酸変異を導入する方法等が一般に採用される。そして、こうした方法によりある程度高活性な変異体を取得できることが期待される。
【0004】
また、酵素活性部位を特定するのにアラニンスキャニングという手法がある(非特許文献1)。この手法は、活性部位周辺のアミノ酸をアラニンに置換した変異体を作製して、当該変異体のタンパク質の特異的活性が低下するかどうか検出し、活性が低下する場合には、当該部位は、酵素活性に強く関与する部位であると判定する手法である。
【0005】
【非特許文献1】Cunningham,B. C. and J. A. Wells, Science 244(4908):1081-1085,1989
【発明の開示】
【発明が解決しようとする課題】
【0006】
しかしながら、トンネル状等の基質結合部位を有する酵素につき、上記のような手法を試みても高活性変異体を取得するのは困難であった。すなわち、各種のアミノ酸置換変異の導入を試みる場合、広い範囲の基質結合部位にわたって多数の部位を置換対象としなければならず、かかる作業は膨大な時間と労力を有するものであった。また、基質結合部位が広い範囲にわたるため、置換導入部位を決定することも困難であった。
【0007】
以上のように、高分子基質を結合するトンネル状又は溝状の基質結合部位を有する酵素において高活性変異体を得るのはきわめて困難であった。
【0008】
そこで、本発明は、トンネル状又は溝状の基質結合部位を有する酵素について高活性変異体を効率的に取得できるスクリーニング方法を提供することを一つの目的とする。
【課題を解決するための手段】
【0009】
本発明者らは、上記課題について種々検討したところ、まず、大まかに予め選択した基質と相互作用する可能性のある部位に対してアラニンスキャニングを実施し、当該アラニンスキャニングにより活性上昇傾向のある部位を変異導入候補部位として選抜し、当該変異導入候補部位に各種アミノ酸置換を導入することにより、高活性な変異体を極めて効率的に取得できることを見出し、本発明を完成した。すなわち、本発明によれば以下の手段が提供される。
【0010】
本発明によれば、高分子基質を結合するトンネル状又は溝状の基質結合部位を有する酵素につき、基質相互作用を有する可能性のある部位のアミノ酸をアラニン置換した2種類以上の一次変異体を調製する工程と、前記一次変異体につき、変異による酵素活性の上昇の有無をスクリーニングして、変異前より酵素活性が上昇した前記一次変異体のアラニン置換部位を変異導入候補部位として選択する工程と、を備える、変異体のスクリーニング方法が提供される。
【0011】
本発明によれば、前記基質相互作用を有する部位は、前記酵素の前記基質結合部位の入口又は出口もしくはこれらの近傍に位置される部位を含む、前記スクリーニング方法も提供される。さらに、前記変異導入候補部位のそれぞれにつき、当該部位の本来のアミノ酸残基を2種類以上の他のアミノ酸に置換して得られる2種類以上の二次変異体を調製する工程と、前記二次変異体につき、変異による酵素活性の上昇の有無をスクリーニングして、変異前より酵素活性が上昇した前記二次変異体の変異導入部位のアミノ酸及び部位を置換アミノ酸候補及び置換候補部位として選択する工程と、を備える、前記スクリーニング方法も提供される。
【0012】
前記高分子基質は、セルロースであってもよく、前記酵素は、セロビオヒドロラーゼであってもよく、Phanerocaete chrysosporium由来のセロビオヒドロラーゼであってもよい。また、セロビオヒドロラーゼは、配列番号2で表されるアミノ酸配列又はこれに相当するアミノ酸配列を有し、前記変異導入候補部位は、配列番号2で表されるアミノ酸配列におけるW38位、W40位及びY47位又はこれらに相当する部位から選択されるものであってもよい。前記酵素は、エンドグルカナーゼであってもよく、Phanerocaete chrysosporium由来のエンドグルカナーゼであってもよい。前記エンドグルカナーゼは配列番号4で表されるアミノ酸配列又はこれに相当するアミノ酸配列を有し、前記変異導入候補部位は、配列番号4で表されるアミノ酸配列におけるY7位及びP134位又はこれらに相当する部位から選択されるものであってもよい。
【0013】
本発明によれば、配列番号2で表されるアミノ酸配列又はこれに相当するアミノ酸配列において、W38F、W40N、W40D、W40E、W40I、W40G、W40R、W40Q、W40S、W40Y、W40T、W40V、W40H、W40F、W40P、W40M、W40K、W40L、W40A、Y47N、Y47D、Y47E、Y47I、Y47R、Y47Q、Y47S、Y47W、Y47T、Y47H、Y47F、Y47P、Y47M、Y47K、Y47L及びY47Aからなる群から選択されるいずれかの変異又はこれに相当する変異を有するアミノ酸配列を有し、セロビオヒドロラーゼ活性を有するタンパク質が提供される。前記変異は、W40N、W40D、W40E、W40I、W40S、W40T、W40V、W40M、W40K及びW40A並びにこれらに相当する変異からなる群から選択されるいずれであってもよい。
【0014】
本発明によれば、配列番号2で表されるアミノ酸配列又はこれに相当するアミノ酸配列において、W38F、W40N、W40D、W40E、W40I、W40G、W40R、W40Q、W40S、W40Y、W40T、W40V、W40H、W40F、W40P、W40M、W40K、W40L、W40A、Y47N、Y47D、Y47E、Y47I、Y47R、Y47Q、Y47S、Y47W、Y47T、Y47H、Y47F、Y47P、Y47M、Y47K、Y47L及びY47Aからなる群から選択されるいずれかの変異又はこれに相当する変異を有するアミノ酸配列を有し、セロビオヒドロラーゼ活性を有するタンパク質をコードするポリヌクレオチドが提供される。
【発明を実施するための最良の形態】
【0015】
本発明は、高分子基質を結合する基質結合部位を有する酵素の高活性変異体を取得するスクリーニング方法及び変異体に関する。本発明のスクリーニング方法によれば、高分子基質を結合するトンネル状又は溝状の基質結合部位を有する酵素であっても、アラニン置換により酵素活性が上昇傾向にある部位を変異導入候補部位とし、当該変異導入候補部位にアミノ酸置換を導入した変異体を取得することで、ランダムな変異導入や多数に渡る部位特異的変異導入を回避でき、効率的に高活性な変異体を得ることができる。
【0016】
部位特異的にアラニン置換するアラニンスキャニングは、活性部位の特定に一般的に用いられる。すなわち、アラニン置換により活性が低下する部位は活性に寄与することがわかる。一方、アラニン置換により活性が向上する部位の位置づけは明らかではない。本発明者らは、高分子基質に結合するトンネル状や溝状等の基質結合部位を有する酵素において、基質相互作用を有する可能性の部位をアラニン置換し、アラニン置換により酵素活性が上昇する部位を、トンネル状の基質結合部位に対する基質の通過性に関与する変異導入候補部位として特定し、さらに当該部位に他のアミノ酸残基を導入することにより、高活性な変異体が得られることがわかった。以下、本発明の各種実施形態について詳細に説明する。
【0017】
(高分子基質結合部位を有する酵素の変異体のスクリーニング方法)
本発明のスクリーニング方法は、高分子基質結合部位を有する酵素につき、基質相互作用を有する可能性のある部位をアラニン置換した2種類以上の一次変異体を取得する工程を備えている。
【0018】
高分子基質結合部位を有する酵素は、トンネル状又は溝状の基質結合部位を有している。こうした基質結合部位は、高分子鎖がはまり込む構造となっているため、比較的細長い構造を有している。こうした酵素としては、例えば、セルロースを基質とするグリコシドヒドロラーゼファミリー(GHF)6、7、48に属するセルローゼであるセロビオヒドロラーゼが挙げられる。これらの酵素は、セルロースあるいはセロオリゴ糖などの高分子基質を結合するトンネル状の基質結合部位を有するプロセッシブ酵素であるとされている。また、GHF5,9に属するセロビオヒドロラーゼも溝状の基質結合部位を有しているとされている。さらにまた、GHF5、8、9、10、12、26、44、45、45、51、61及び74に属するエンドグルカナーゼも挙げられる。さらに、GHFに分類されるほかの高分子基質を分解する酵素としては、ヘミセルラーゼ及びアミラーゼが挙げられる。さらにまた、GHFに属さないが高分子基質を結合するトンネル状又は溝状の高分子基質結合部位を有する酵素としては、プロテアーゼ、エステラーゼ、リアーゼ及びトランスフェラーゼが挙げられる。
【0019】
本発明のスクリーニング方法は、高分子基質結合部位を有する酵素のなかでもプロセッシブ酵素のスクリーニングに適している。プロセッシブ酵素においては、高分子基質と強く相互作用が酵素活性に影響が大きいと考えられるからである。なお、プロセッシブ酵素とは、より詳細には、酵素反応を通じて酵素が基質をと分離することを要しない酵素である。例えば、プロセッシブ酵素としては、セルロースを基質とするグリコシドヒドロラーゼファミリー6、7、48に属するセルローゼであるセロビオヒドロラーゼが挙げられる。
【0020】
こうした酵素としては、例えば、各種菌に由来するセルラーゼが挙げられる。なかでも、GHF6に属する、Cellulomonas fimiなどのCellulomonas属、Humicola insolens、Humicola insolensなどのHumicola属、Trichoderma virideなどのTrichoderma属;GHF7に属するAspergillus aculeatus、Aspergillus nigar、Aspergillus oryzaeなどのAspergillus属、Phanerochaete chrysosporiumなどのPhanerocaete属、Acremonium thermophilumなどのAcremonium属、 Chaetomium thermophilumなどのChaetomium属、Trichoderma属;GHF48に属するClostridium acetobutylicum、Clostridium cellulolyticum、Clostridium cellulovorans、Clostridium josui、Clostridium thermocellumなどのClostridium属の菌由来のセロビオヒドロラーゼを含むセルラーゼが挙げられる。
【0021】
(一次変異体の調製工程)
まず、高分子基質結合部位を有する酵素において、基質相互作用を有する可能性のある部位をアラニン置換して一次変異体を調製する。基質相互作用を有する可能性のある部位(アラニン置換部位)は、立体構造が解明されている適当な他の参照用の高分子基質結合部位を有する酵素を選択し、その立体構造等から高分子基質と相互作用してその安定化に寄与していると考えられるアミノ酸残基を選択し、さらに、改変対象の高分子基質結合部位を有する酵素のアミノ酸配列から当該アミノ酸残基に相当するアミノ酸残基を抽出することにより決定できる。
【0022】
参照用の酵素は、アミノ酸配列において一定以上の同一性を有し同種活性を有する酵素とすることが好ましい。参照用酵素としては、例えば、改変対象がセルラーゼであれば、同一のGHFに属するセルラーゼが挙げられる。なお、参照用酵素としては、以下に記載するように、立体構造及び活性部位等が研究されている酵素等を用いることができる。
【0023】
参照用酵素の立体構造においてトンネル状又は溝状等の高分子基質結合部位において基質との相互作用に関与するアミノ酸残基(アラニン置換部位)を特定するには、立体構造モデルを構築する方法と既知の立体構造モデルを用いる方法とが挙げられる。立体構造モデルを構築する方法は、改変対象の酵素のアミノ酸配列をPSI−BLAST等のプログラムに入力し、類似性の高いアミノ酸配列の酵素を検索する。最も類似の酵素の構造を参照し、InsightII、Discovery studio等のソフトにより、ホモロジーモデリングを行うことにより、構造を予測する。予測した構造モデルを見ながら基質結合部位周辺のアミノ酸を抽出し、これらのアミノ酸を、アラニン置換部位とすることができる。
【0024】
また、立体構造モデルを構築しないで基質との相互作用に関与するアミノ酸残基を特定するには、例えば、同じGHFの酵素のうち、PDB等の立体構造データベースに構造がある酵素を検索する。選んだ構造既知の酵素のアミノ酸配列と、ターゲット酵素のアミノ酸配列をCLUSTAL W、GENETYX等のソフトに入力しアライメントする。構造既知の酵素の構造を見ながら基質結合部位周辺のアミノ酸を抽出し、そのアミノ酸とアライメントされた改変対象酵素のアミノ酸を、アラニン置換部位とすることができる。
【0025】
アラニン置換部位の個数は、特に限定しないが、必要に応じた数とすればよい。典型的には、数個以上、好ましくは10個以上から30個程度以下とすることができる。
【0026】
アラニン置換部位は、トンネル状又は溝状の基質結合部位の中央寄りの部分よりも入り口又は出口もしくはこれらの近傍から選択することが好ましい。本発明者らによる知見によれば、高分子基質結合部位を有する酵素におけるかかる部位のアミノ酸残基をアラニン置換することで、置換前に比べて基質結合部位への基質のアクセシビリティが大きく変化する可能性が高いと考えられる。そして、アラニン置換により酵素活性が向上するとき、当該部位の本来のアミノ酸残基は触媒作用を抑制している可能性があると考えられ、当該部位の他のアミノ酸への置換により活性が向上する可能性が高いと考えられる。
【0027】
決定したアラニン置換部位から選択される1種又は2種以上の部位をアラニン置換して一次変異体を調製する。それぞれの一次変異体は、複数のアラニン置換がなされていてもよいが、好ましくは、それぞれの一次変異体につき、一つのアミノ残基についてのみアラニン置換がなされている。部位特異的にアラニン置換された一次変異体を得るには、公知の部位特異的変異導入法を用いることができる。例えば、部位特異的にアラニン置換を導入するための変異型遺伝子は、Kunkel法やGapped duplex法などの公知の手法又はこれに準ずる方法により、例えば変異 導入用キット(例えばMutant-K(TAKARA社製)やMutant-G(TAKARA社製)など)を用いて、あるいは、TAKARA社のLA PCR in vitro Mutagenesisシリーズキットを用いて作製することができる。また、遺伝子の塩基配列を参照すれば、Molecular Cloning [Sambrookら編集, 15, Site-directed Mutagenesis of Cloned DNA, 15.3〜15.113, Cold Spring Harbor Lab. Press, New York (1989)]等の文献の記載に従って、当業者であれば格別の困難性なしに選択し実施することができる。また、当業者であれば、遺伝子の塩基配列を基にして、当該塩基配列から1以上(1又は数個以上)の塩基の置換、欠失、挿入又は付加を人為的に行う技術(部位特異的突然変異誘発)については、実施例に示すように、オーバーラップPCRを用いるほか、Proc. Natl. Acad. Sci. USA, 81, 5662-5666 (1984); WO85/00817; Nature, 316, 601-605 (1985); Gene, 34, 315-323 (1985); Nucleic Acids Res., 13, 4431-4442 (1985); Proc. Natl. Acad. Sci. USA, 79, 6409-6413 (1982); Science, 224, 1431-1433 (1984) 等に記載の技術に従って実施することができる。
【0028】
また、部位特異的変異導入法により鋳型DNAを取得後は、大腸菌等を用いた各種タンパク質の合成方法によりDNA上の情報に基づくタンパク質を取得できるが、好ましくは公知の無細胞タンパク質合成法を用いる。無細胞タンパク質合成系を用いることで、著しく作業時間と労働強度を低下させることができる。無細胞タンパク質合成系としては、公知の各種キットを用いて実施できるほか、特開2003−116590号公報、特開2006−61080号公報等に記載の方法を利用できる。無細胞タンパク質合成系を用いることにより、例えば、同時にスキャンするアラニン置換部位が10個〜100個程度のとき、従来なら5日から2週間程度要していたのを、0.5日〜2日程度で終了させることができる。さらに、同時スキャン部位が400個程度の場合、大腸菌による菌体生産による場合には60日間程度要するところを、無細胞タンパク質合成系によれば4日程度で達成することができる。
【0029】
(一次変異体のスクリーニング工程)
次に、一次変異体についてのスクリーニング工程(一次スクリーニング工程)を実施する。一次スクリーニング工程では、得られた一次変異体につき、変異による酵素活性の上昇の有無をスクリーニングして、変異前よりも活性が上昇した前記一次変異体におけるアラニン置換部位を変異導入候補部位として決定することができる。アラニン置換により酵素活性が向上したとき、当該部位は置換前のアミノ酸残基が酵素活性に抑制的に作用している可能性があり、当該アラニン置換部位をさらに他のアミノ酸残基に置換することにより、酵素活性の向上が期待できる。なお、酵素活性の測定方法は、改変対象酵素の種類によって適宜決定される。また、活性の上昇程度は、特に限定しないが、好ましくは30%以上、より好ましくは60%以上、さらに好ましくは100%以上、いっそう好ましくは200%以上である。
【0030】
以上の工程により、高分子基質結合部位を有する酵素における変異導入候補部位が迅速に決定される。従来、一次的な変異導入候補部位の決定に、多大な時間と労力を費やしており、基質結合部位が広範囲にわたるプロセッシブ酵素の場合、多数個のアミノ酸残基につき部位特異的変異導入を試みる必要があった。しかしながら、アラニンスキャニングをこうした酵素の基質結合部位の変異導入候補部位の決定に利用することで従来の作業を著しく簡素化して導入候補部位を決定できるようになった。
【0031】
本発明においては、一次スクリーニング工程を実施した上、抽出した置換変異導入候補部位に各種のアミノ酸を導入した二次変異体を調製し、これら二次変異体につき二次スクリーニング工程を実施する。このため、高活性な変異体のスクリーニングのための二次変異体を調製する作業及びスクリーニングサ行を著しく省力化し短時間化することができる。例えば、高分子基質結合部位が数十個のアミノ酸残基を含むとき、これらの全てのアミノ酸残基につき飽和アミノ酸変異を導入すると数百〜千数百程度の変異遺伝子及び変異体タンパク質を調製し、これらにつき酵素活性を測定することになる。これに対し、本発明の一次変異体のスクリーニング工程で数十個のアミノ酸残基から例えば数分の1から数十分の1程度に置換変異導入候補部位を絞ることができる。そして、これらの候補部位に飽和アミノ酸変異を導入することにより、変異遺伝子及び変異体のライブラリーサイズも数分の1〜数十分の1程度に縮小される。このようなライブラリ縮小効果は、広範囲にわたる高分子基質結合部位を有する酵素において極めて有効である。このようなライブラリーサイズの縮小効果に加えて、一次変異体及び二次変異体の調製に無細胞タンパク質合成系を用いることで、より一層スクリーニングを省力化することができる。
【0032】
(二次変異体の調製工程)
一次スクリーニング工程実施後には、さらに、二次変異体の調製工程と二次変異体についてのスクリーニング工程(二次スクリーニング工程)を備えることで、より具体的に置換アミノ酸候補及び置換候補部位を決定できる。二次変異体の調製工程では、一次スクリーニング工程において決定した変異導入候補部位に、ランダムにあるいは一定の規則に従い、本来のアミノ酸残基以外のアミノ酸残基を導入した二次変異体を調製することができる。決定された各変異導入候補部位につき、置換するアミノ酸は、本来のアミノ酸残基以外の全てのアミノ酸残基(全19種類)の全部又は一部であるが、少なくとも2種類以上の他のアミノ酸に置換することが好ましい。一部のアミノ酸に置換するとき、例えば、保存的アミノ酸置換の手法を採用できる。
【0033】
それぞれの二次変異体は、複数の変異導入候補部位において他のアミノ酸に置換されていてもよいが、各二次変異体は、それぞれ一つの変異導入候補部位においてのみ置換がなされていることが好ましい。二次変異体は、一つの置換候補部位につき複数種類のアミノ酸置換がなされた複数種類とすることが好ましい。さらに、こうした置換候補部位毎に複数種類の二次変異体を調製することが好ましい。
【0034】
二次変異体を調製するには、一次変異体の調製時と同様、公知の部位特異的変異導入法から適宜選択した手法を用いればよく、タンパク質合成にあたっては無細胞タンパク質合成系を用いることが好ましい。
【0035】
(二次変異体のスクリーニング工程)
次に、二次スクリーニング工程を実施する。二次スクリーニング工程では、得られた二次変異体につき、変異による酵素活性の上昇の有無をスクリーニングして、変異前よりも活性が上昇した前記二次変異体における置換アミノ酸及び置換部位を、置換アミノ酸候補及び置換候補部位として決定できる。酵素活性の測定方法は、改変対象酵素の種類によって適宜決定される。
【0036】
こうして得られた置換アミノ酸候補及び置換候補部位を、2種類以上組み合わせるなどの進化分子工学的手法を用いることで、より高活性な高分子基質結合部位を有する酵素を得ることができる。
【0037】
例えば、改変対象酵素が、Phanerocaete chrysosporium由来のセロビオヒドロラーゼなどのセロビオヒドロラーゼの場合、配列番号2で表されるアミノ酸配列又はこれに相当するアミノ酸配列(相当アミノ酸配列)を有し、配列番号2で表されるアミノ酸配列におけるW38位、W40位及びY47位から選択される1種又は2種以上又は当該相相当するアミノ酸配列これらに相当する部位から選択される1種又は2種以上を置換候補部位として二次変異体を調製し、二次変異体のスクリーニングを実施することができる。
【0038】
また、改変対象酵素は、Phanerocaete chrysosporium由来のエンドグルカナーゼなどのエンドグルカナーゼである場合、配列番号4で表されるアミノ酸配列又はこれに相当するアミノ酸配列(相当アミノ酸配列)を有し、配列番号4で表されるアミノ酸配列におけるY7位及びP134位から選択される1種又は2種以上又は当該相相当するアミノ酸配列におけるこれらに相当する部位から選択される1種又は2種以上を置換候補部位として二次変異体を調製し、二次変異体のスクリーニングを実施することもできる。
【0039】
改変対象酵素は上記のとおり、必ずしも配列番号2又は4で表されるアミノ酸配列と完全に一致するアミノ酸配列を有していなくともよく、これらに相当するアミノ酸配列(相当アミノ酸配列)を有していればよい。相当アミノ酸配列とは、BLASTなどの配列相同性検索において配列番号2又は4で表されるアミノ酸配列と同一性が40%以上、より好ましくは50%以上、さらに好ましくは60%以上、一層好ましくは70%以上、より一層好ましくは80%以上、さらに一層好ましくは90%以上、最も好ましくは95%以上のアミノ酸配列である。改変対象酵素が、こうしたアミノ酸配列を有する場合、ペアワイズアラインメントなどの手法により配列番号2又は配列番号4で表されるアミノ酸配列とアラインメントしたとき、上記置換候補部位に対応するものとして検出される部位を置換候補部位として二次変異体を調製することができる。
【0040】
本明細書において同一性とは、当該技術で知られているとおり、配列を比較することにより決定される、2以上のタンパク質あるいは2以上のポリヌクレオチドの間の関係である。当該技術で“同一性 ”とは、タンパク質またはポリヌクレオチド配列の間の適合によって、あるいは場合によっては、一続きのそのような配列間の適合によって決定されるような、タンパク質またはポリヌクレオチド配列の間の配列相関性の程度を意味する。“同一性 ”および“類似性”は、既知の方法により容易に決定できる。同一性 を決定する好ましい方法は、試験する配列間で最も長く適合するように設計される。同一性 および類似性を決定するための方法は、公に利用可能なプログラムにコードされている。相同性決定には、AltschulらによるBLAST (Basic Local Alignment Search Tool) プログラム(たとえば、Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ., J. Mol. Biol., 215: p403-410 (1990), Altschyl SF, Madden TL, Schaffer AA, Zhang J, Miller W, Lipman DJ., Nucleic Acids Res. 25: p3389-3402 (1997))を利用し決定することができる。BLASTのようなソフトウェアを用いる場合、デフォルト値を用いるのが好ましい。BLAST検索に一般的に用いられる主な初期条件は、以下の通りであるが、これに限定されない。
【0041】
(変異体タンパク質)
本発明のタンパク質は、配列番号2で表されるアミノ酸配列において、W38F、W40N、W40D、W40E、W40I、W40G、W40R、W40Q、W40S、W40Y、W40T、W40V、W40H、W40F、W40P、W40M、W40K、W40L、W40A、Y47N、Y47D、Y47E、Y47I、Y47R、Y47Q、Y47S、Y47W、Y47T、Y47H、Y47F、Y47P、Y47M、Y47K、Y47L及びY47Aからなる群から選択されるいずれかの変異を有するアミノ酸配列を有し、セロビオヒドロラーゼ活性を有するタンパク質である。これらのタンパク質は、親タンパク質である配列番号2で表されるアミノ酸配列からなるタンパク質、すなわち、Phanerocaete chrysosporium由来のCBH Iよりもよりもセルロースを分解する活性が高い。したがって、セルロース系バイオマスなどのセルロース含有材料の有効利用に有利に用いることができる。これらの変異体のなかでも、特に、W40N、W40D、W40E、W40I、W40S、W40T、W40V、W40M、W40K及びW40Aのいずれかがより高い活性を有している。これらの変異体は、親タンパク質よりも還元糖量に基づく酵素活性測定系において約2倍以上の活性を有している。
【0042】
変異体タンパク質は、配列番号2に相当するアミノ酸配列(相当アミノ酸配列)において、配列番号2で表されるアミノ酸配列において上記各変異に相当する変異のいずれかを有し、セロビオヒドロラーゼ活性を有するタンパク質であってもよい。すなわち、変異体タンパク質は、必ずしも配列番号2で表されるアミノ酸配列と完全に一致するアミノ酸配列において上記変異のいずれかを有するものでなくてもよい。すなわち、変異体タンパク質は、BLASTなどの配列相同性検索において、配列番号2で表されるアミノ酸配列と同一性が40%以上、より好ましくは50%以上、さらに好ましくは60%以上、一層好ましくは70%以上、より一層好ましくは80%以上、さらに一層好ましくは90%以上、最も好ましくは95%以上のアミノ酸配列を、上記の相当アミノ酸配列とし、この配列をペアワイズアラインメントなどの手法により配列番号2で表されるアミノ酸配列とアラインメントしたとき、上記変異部位に対応するものとして検出される部位のいずれかにおいて対応するアミノ酸置換を有するものであってもよい。
【0043】
こうした変異体タンパク質は、公知の方法によって取得できる。例えば、大腸菌、酵母等を用いた菌体によるタンパク質発現系、無細胞タンパク質合成系、形質転換した動植物を用いたタンパク質発現系などを用いて取得することができる。酵母等の細胞表層に提示させる形態で発現させることもできる。タンパク質の細胞表層提示技術は、当業者であれば、Cell surface engineering of yeast: construction of arming yeast with biocatalyst. J. Biosci. Bioeng., 90, 125-136 (2000)等に基づき実施することができる。
【0044】
(本発明のポリヌクレオチド)
本発明のポリヌクレオチドは、本発明の変異体タンパク質をコードしている。すなわち、配列番号2で表されるアミノ酸配列又はこれに相当するアミノ酸配列において、W38F、W40N、W40D、W40E、W40I、W40G、W40R、W40Q、W40S、W40Y、W40T、W40V、W40H、W40F、W40P、W40M、W40K、W40L、W40A、Y47N、Y47D、Y47E、Y47I、Y47R、Y47Q、Y47S、Y47W、Y47T、Y47H、Y47F、Y47P、Y47M、Y47K、Y47L及びY47Aからなる群から選択されるいずれかの変異又はこれに相当する変異を有するアミノ酸配列を有し、セロビオヒドロラーゼ活性を有するタンパク質をコードしている。
【0045】
本発明のポリヌクレオチドは、DNAの形態(たとえば、cDNAおよびクローニングによって得られるか、あるいは合成的に生成されるゲノムDNAを含む)であってもよく、RNA(たとえばmRNA)の形態であってもよい。該ポリヌクレオチドは、二本鎖であっても、一本鎖であってもよい。二本鎖の場合は、二本鎖DNA、二本鎖RNAまたはDNA:RNAのハイブリッドであってもよい。一本鎖の場合は、センス鎖(コード鎖としても知られる)であっても、アンチセンス鎖(非コード鎖としても知られる)であってもよい。
【0046】
本発明のポリヌクレオチドは、すでに説明したように、配列番号2で表されるアミノ酸配列をコードする配列番号1で表される塩基配列のDNA等の特定箇所に公知の各種方法によって変異を導入すること等によって得ることができる。また、配列番号1で表される塩基配列に対して、配列番号1とは異なるコドン用法を用いて得られる同一のアミノ酸配列をコードするDNAに対して変異を導入してもよい。
【0047】
以下、本発明を、実施例を挙げて具体的に説明するが、本発明はこれらの実施例に限定されるものではない。
【実施例1】
【0048】
(アラニン置換導入部位の決定)
白色腐朽菌Phanerocaete chrysosporium由来のセロビオヒドロラーゼIIであるCBH I(PcCBH7C)とエンドグルカナーゼであるEG(PcEGIII)に種々の変異を導入して、そのセルロース分解活性を向上させるため、まず、変異導入に適した箇所を探索した。すなわち、PcCBH7Cのアミノ酸配列(配列番号2 )とPcEGIIIのアミノ酸配列(配列番号4)それぞれに類似構造を持つ別のセルラーゼの立体構造から推測された、酵素活性を示す際にセルロース鎖と相互作用しその安定化に寄与していると考えられるアミノ酸残基を選択した。そして、選択した部位のアミノ酸残基をまずアラニンに置換し、それらの中でも特に変異導入による活性変化への影響が大きいと思われるアミノ酸残基の位置を特定することとした。
【0049】
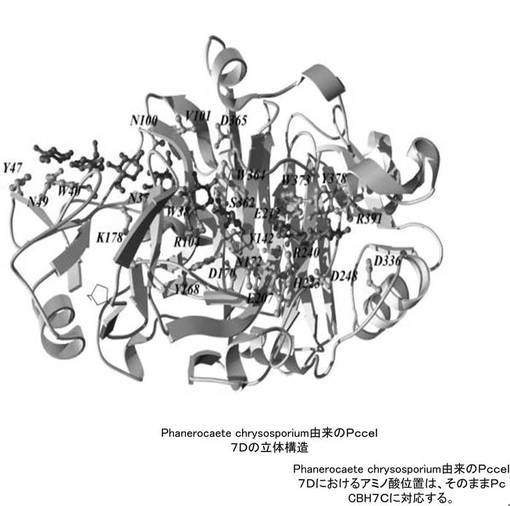
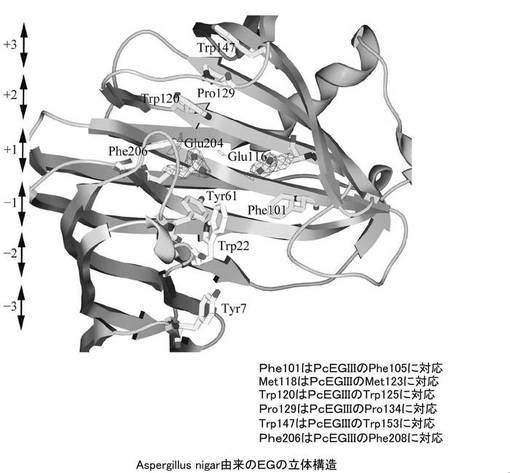
PcCBH7Cの参照構造として、同じPhanerocaete chrysosporium由来のセロビオヒドロラーゼIであるCBH I(図1A参照)とした。また、PcEG IIIについては、Aspergillus nigar由来のEGIII(図1B参照)を参照構造とした。これらについてはいずれもPDB(http://www.rcsb.org/pdb/home/home.do)において立体構造を取得できる。参照の結果、これより、PcCBH7Cについては、以下の表(表1)に示す24個のアミノ酸残基をアラニン置換することとした。また、PcEGIllについては、以下の表(表2)に示す10個のアミノ酸残基をアラニンに置換することとした。
【0050】
【表1】

【表2】
【実施例2】
【0051】
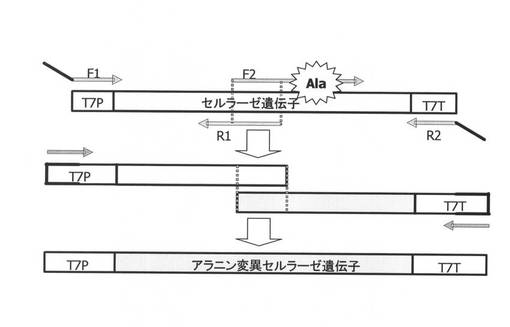
(オーバーラッピングPCRによるセルラーゼ遺伝子へのアラニン変異の導入)
(PcCBH7C遺伝子)
オーバーラッピングPCRによるセルラーゼ遺伝子へのアラニン変異の導入のスキームを図2に示す。プラスミドpRPccel7C(Pccel7C遺伝子、配列番号1)の両端をNdelおよびEcoRlで切断し、pRSETBにクローニングして作成)を鋳型とし、表3に示すF3−K4プライマーと各変異導入部位につき個別のリバースプライマーを用いてPCRを行うことによりT7プロモーターとRBSを含んだ断片を増幅した。また、表3に示すR3−K4(と各変異導入部位につき個別のフォワードプライマーを用いてPCRを行うことにより各変異導入部位へのアラニン変異とT7ターミネーターを含む断片を増幅した。なお、PCRの反応液組成は、以下の通りとした。反応条件は、94℃で3分後、94℃で10秒、60℃で10秒及び72℃で30秒を1サイクルとして25サイクル実施し、72℃で7分とした。
【0052】
【表3A】
【表3B】
【0053】
(PCR反応液)
10×PfuUItra(登録商標)IIバッファー 2.0μ1
2.5mMdNTPs 2.0μ1
鋳型DNA(1ng/μ1) 1.0μ1
10μMフォワードプライマー 1.0μ1
10μMリバースプライマー 1.0μl
5U/μlPfuUltra(登録商標)II Fusion HS DNAポリメラーゼ(Strategene製) 0.2μl
S.W. 12.8μl
合計 20μl
【0054】
(PcEGIII遺伝子)
プラスミドpET−PcEG(ppET」PcEG(pET23b(+)vetQrのNdel、XhoサイトにPcEGIII(配列番号3)の全長遺伝子を挿入したもの)を鋳型とし、PcCBH7Cの場合と同様にF3−K4プライマーと表4に示した各変異導入部位につき個別のリバースプラィマーを用いてPCRを行うことによりT7プロモーターとRBSを含んだ断片を増幅し、R3−K4と表4に示した各変異導入部位につき個別のフォワードプライマーを用いてPCRを行うことにより各変異導入部位へのアラニン変異とT7ターミネーターを含む断片を増幅した。PCR条件は、PcCBH7C遺伝子の場合と同様とした。
【0055】
【表4】
【0056】
得られた各PCR産物につき、アガロースゲル電気泳動を行い、MinEluteGelExtraction Kitを用いてアガロースゲルから精製した。アガロースゲルから精製した2種類の断片を、図2に示すオーバーラッピングPCRを行うことにより連結させた。PCR反応液の組成は、以下のとおりとした。反応条件は、94℃で3分後、94℃で15秒、55℃で30秒、及び72℃で90秒を1サイクルとして10サイクル実施後に、プライマーK4を添加し、引き続き、94℃で15秒、55℃で30秒、72℃で90秒を1サイクルとして10サイクル実施後、72℃で7分の条件でPCRを行った。この際、オーバーラップさせる各断片の濃度はNanoDrop(商標)ND−1000分光光度計によって測定し、各々が10ngずつPCR反応系に含まれるよう調整した。こうして作製したアラニン変異導入セルラーゼ遺伝子につき、表5に示すプライマーを用いて塩基配列の確認を行った。
【0057】
(オーバーラッピングPCR反応液の組成)
10xLA Taq(登録商標)Buffer 2.0μl
2.5mMdNTPs 3.2μl
25mM MgCl2 2.0μl
フォワード断片(10ng/μl) 1.0μl
リバース断片(10ng/μl) 1.0μl
20μMK4プライマー 1.0μl
5U/μl LA Taq DNAポリメラーゼ(タカラバイオ製)
0.2μl
S.W. 9.6μl
合計 20μl
【0058】
【表5】
【0059】
また、PcCBH7C及びPcEGIIlの各野生型遺伝子は、それぞれプラスミドpRPccel7C及びpE下PcEGを鋳型とし、プライマーにF3−K4とR3−K4を用いてPCRを行うことによって作成した。PCR反応液の組成は、以下のとおりとした。
反応条件は、94℃で3分後、94℃で15秒、55℃で15秒、72℃で30秒を1サイクルとして25サイクル実施後、72℃で7分とした。作製した各変異遺伝子と野生型遺伝子は、Qubit(商標)fluorometerを用いてそのDNA濃度を測定した。
【0060】
(PCR反応液)
10×PfuUltra(商標)IIバッファー2.0μ1
2.5mM dNTPs 2.0μ1
鋳型(1ng/μ1) 1.0μ1
10μM F3K4プライマー 1.0μ1
10μM R3K4プライマー 1.0μ1
5U/μl PfuUltra(商標)II FusionHSDNAポリメラーゼ
0.2μ1
S.W. 12.8μ1
合計 20μl
【実施例3】
【0061】
(無細胞タンパク質合成系によるセルラーゼの発現)
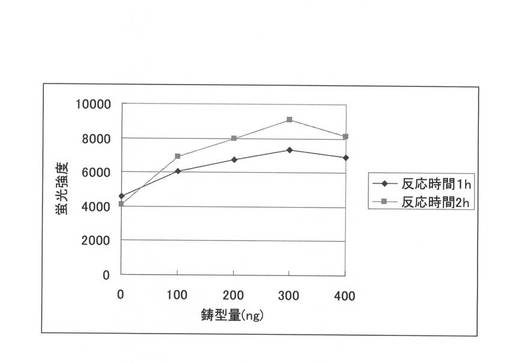
実施例2で作製したPcCBH7Cの野生型のPCR産物を、大腸菌由来無細胞蛋白質合成の鋳型とし、その鋳型量を0ng,100ng、200ng、300ng、及び400ngと変化させた。そして、それぞれの鋳型量で合成した蛋白質の酵素活性を測定し、無細胞蛋白質合成系に必要な鋳型量を決定した。鋳型量は、約200〜300ngが最適であった(図3)。さらに、PD1(Humicola insolens由来)及び還元型グルタチオン(GSH)、酸化型グルタチオン(GSSG)を、以下の組成の転写・翻訳共役反応液に加えて混合し、26℃で、6時間無細胞蛋白質合成を行った。
【0062】
(転写・翻訳共役反応液)
PDI 0.15μ1
10mM GSH 0.3μ1
0.4M 酢酸マグネシウム 0.6μ1
5M 酢酸カリウム 0.6μl
0.1mg/mlリファンピシリン 3.0μ1
LM(20a.a.) 7.5μ1
10mg/ml CK 0.45μ1
2.3mg/ml T7RNAポリメラーゼ 1.0μ1
(pAR1219;Davanloo et al.,1984で形質転換したE.coli BL21から精製)
100mMGSSG 0.3μ1
S30抽出物(特開2006−61080) 8.5μ1
鋳型+S.W. 7.6μ1
合計 30μ1
【0063】
なお、無細胞合成反応試薬の終濃度は、LM;LowMolecular weight mixture(56.4mM トリス−酢酸(pH7.4)、1.22mM ATP(pH7.0)、各0.85mM GUC ミックス(GTP、UTP、CTP)、50mM クレアチンリン酸2ナトリウム、各0.32mM 20アミノ酸ミックス、4%(w/v)ポリエチレングリコール6000、34.6μg/ml 葉酸、35.9mM 酢酸アンモニウム、0.17mg/ml E.coli tRNA、28.3%(v/v)S30抽出物)、0.15mg/mlクレアチンキナーゼ、7.7μg/ml T7RNAポリメラーゼ、10μg/m1リファンピシリン、10mM 酢酸マグネシウム、100mM 酢酸カリウムであった。
【0064】
また、オートラジオグラフィによる蛋白質発現量の確認を行った。無細胞蛋白質合成系の組成液の1つであるLMに含まれる20種類のアミノ酸を、ロイシン以外の19アミノ酸に14C−ロイシンを加えたものに変更し、26℃で6時間無細胞蛋白質合成を行った。合成反応液の組成は、以下のとおりとした。
【0065】
(無細胞タンパク質合成反応液の組成)
PDI 0.15μl
10mM GSH 0.3μl
0.4M 酢酸マグネシウム 0.6μl
0.5M 酢酸カリウム 0.6μl
0.1mg/m1リファンピシリン 3.0μl
LM(19アミノ酸) 7.5μl
10mg/mlCK 0.45μl
2.3mg/mlT7RNAポリメラーゼ 1.0μl
100mM GSSG 0.3μ1
14C−ロイシン 2.0μ1
(Amersham−PhamaciaBiotech製)
S30抽出物 8.5μl
鋳型DNA+S.W 5.6μl
合計 30μl
【0066】
無細胞蛋白質合成反応終了液に対し15,000rpmで5分間遠心を行い、反応終了液を上清画分と沈澱画分に分けた。この上清画分と沈殿画分の各々に60%アセトンを9倍量加え、氷上で5分間静置した後、4℃、15,000rpmで5分間遠心を行い、上清を除いた上で7.5μ1のS.W.に再懸濁した。等量の2xSDS−PAGEサンプルバッファーと混合し、煮沸したものをサンプルとしてSDS−PAGEを行った。SDS−PAGE終了後のゲルを、ゲルドライヤーを用いて80℃、1時間乾燥させた後、イメージングプレートに挟み12時間以上置いてRIを転写させた。転写後のイメージングプレートをBAS IP MAGAZINEにセットし、BAS2000で14C−ロイシンの取り込み量を解析した。
【0067】
PcCBH7C及びPcEGIIIのアラニン変異導入遺伝子と野生型遺伝子の無細胞蛋白質合成系による発現と、オートラジオグラフィによる発現量の確認を行った。結果を図4、図5及び図6に示す。PcCBH7Cではいずれの変異体及び野生型についてもその大部分が上清画分に発現されており、発現量の差もほとんど見られなかった。上清画分には可溶性の蛋白質が見出されるので、発現されたPcCBH7Cの大部分はフォールディングの不具合等は起こらずに、活性を持った正常な構造で発現されているものと考えられる。一方PcEGlllについては、各変異体間でやや大きな差が見られ、上清画分と沈殿画分への発現量の比率が各変異体によって異なっていた。発現量には差がないことから、発現後の蛋白質のフォールディング過程が原因ではないかと考えられた。
【実施例4】
【0068】
(PcCBH7C及びPcEGHIIIの酵素活性の測定)
実施例3で得られた反応終了液につき、酵素活性を測定した。PcCBH7Cの酵素活性の基質として、不溶性の高分子セルロースであるPASC(リン酸膨潤アビセル)と、可溶性のセロビオースに4−methylumbelliferylが付加された蛍光基質であるMUC(4−methylumbelliferyl−β−D−cellobioside)を用いた。PASCは高分子セルロースであるため、セルロース鎖は基質結合ポケットの溝の全域に入り込む形で結合し反応する。よって、アラニン置換によるポケット全域への影響を見ることができる。MUCは低分子であるため、G1〜G3結合部位周辺への影響のみを見ることができる。また、PcEGIIIの酵素活性の基質として、可溶性基質のCMC(力ルボキシメチル化セルロース;SIGMA−ALDRICH製)とResorufinceIIobioside(MarkerGeneTMFIuorescentCellulaseAssayKit;MGTInc.社製)を用いた。詳細を以下に示す。
【0069】
PASCによる酵素反応系は、1%PASC 50mM酢酸バッファー(pH5.0)250μl,50mM酢酸バッファー(pH5.0)225μl、無細胞蛋白質合成反応終了液25μl,計500μlの組成で、45℃、24時間酵素反応を行った。PASCとの反応性を上げるためアシストチューブを横にし、振邊しながら反応させた。その後、反応液を用いて生成した還元糖の定量(ソモギーネルソン法又はTZ assay法)を行った。ソモギー・ネルソン法は、反応液サンプル25μ1にソモギー銅液50μ1を加えよく混合させた。サンプルとは別に0〜1mg/mlのグルコース水溶液を調製し同様に以下の操作を行った。サーマルサイクラーを用いて、10g℃で30分間反応させた後、ネルソン液50μ1を加えよく混合した。3,000rpm、3分間遠心した後、上清50μ1を150μ1のS.W.を入れた96穴プレートに加え、プレートリーダーFusionで540nmの吸光度を測定した。測定結果からグルコース検量線を作成し、生成した還元糖量をグルコース濃度に換算し求めた。また、TZassay法は(Chong, K. et al., Journal of Biochemical and Biophysical Methods, 111 - 109(1985))は、以下のとおりとした。すなわち、TZassayバッファ195μlを分注し、反応液サンプル5μlを加えてゆっくり混合した。これを100℃で3分間ヒート処理した後に氷冷し、660nmの吸光度を測定した。グルコースの検量線を作製し、グルコース換算の還元糖量を得た。
【0070】
MUCによる酵素反応系は、2mMMUC/50mM酢酸バッファー(pH5.0)50μ1、50mM酢酸バッファー(pH5.0)50μl、無細胞蛋白質合成反応終了液20μl、計100μlの組成で、96穴プレートにおいて40℃、1時間酵素反応を行った。その後、プレートリーダーFusionでEx/Em=330/515nmの蛍光強度を測定した。
【0071】
CMCによる酵素反応系は、エッペンドルフチューブを用いて、1%CMC/50mM酢酸バッファー(pH5.0)50μ1,50mM酢酸バッファー(pH5.O)45μ1,無細胞蛋白質合成反応終了液5μ1、計100μ1の組成で、45℃で1時間酵素反応を行った。CMCとの反応性を上げるためエッペンドルフチューブを横にし、振とうしながら反応させた。その後、反応液を用いて生成した還元糖の定量を、前述のソモギー・ネルソン法によって行った。
【0072】
Resorufin cellobiosideによる酵素反応系は、0.5mM Resorufin cellobioside/100mM酢酸バッファー(pH6.0)50μ1、100mM酢酸バッファー(pH6.0)25μl、無細胞蛋白質合成反応終了液25μ1、計100μlの組成で、96穴プレートにおいて40℃で1時間酵素反応を行った。その後、プレートリーダーFusionでEx/Em=540/590nmの蛍光強度を測定した。
【0073】
(PcCBH7C及びそのアラニン置換変異体の酵素活性)
無細胞タンパク質合成系によって発現させたPcCBH7Cの野生型と各アラニン変異体24サンプルの酵素活性をPASCとMUCを基質として用いて行った。その活性測定結果を図7に示す。図7に示すように、大部分のアラニン変異体ではその酵素活性が野生型よりも低下した。ここで、PASCに対する活性測定結果とMUCに対する活性測定結果の傾向が若干異なっていることがわかるが、これはおそらくこの二つの基質の構造の違いに起因するものと考えられる。PASCが非晶性のβ−1,4−グリコシド結合で連結したグルコースの鎖状の構造を持つのに対し、MUCは2分子のグルコースがβ−1,4−グリコシド結合でつながった2糖のセロビオシドに分子量176の大きな蛍光基であるメチルウンベリフェロンが結合した構造を持つ。PcCBH7Cは2枚のβシートからなるトンネル構造の中央に活性中心を持っている。このトンネルにセルロース鎖が入り、トンネルの内側に面するアミノ酸残基との相互作用により安定化されつつ活性中心に移動し、グリコシド結合を切断されるのであるが、MUCはその蛍光基の存在の為にこのトンネルヘの侵入及びトンネル内でのアミノ酸残基との相互作用に基づく安定化に通常とは異なる寄与が生まれ、PASCに対する酵素活性測定結果との間に相違が生じたのではないかと考えている。
【0074】
さらに、38位のTrp、40位のTrp、47位のTyrをアラニンに置換した変異体では、その酵素活性が野生型よりも向上した。これらの3個のアミノ酸残基は、立体構造においては上述したセルロース鎖と相互作用するトンネル構造の入りロ付近に存在している。TrpやPhe、Tyrのような芳香族の側鎖を持ったアミノ酸は、このトンネル構造の内部によく見られ、疎水性相互作用や水素結合によってセルロース鎖と相互作用する。今回これらの酵素活性に重要であると思われるアミノ酸残基をアラニンに置換したのであるが、Trp38、Trp40、Tyr47の3つの変異体の酵素活性が野生型よりも向上したのは、アラニンに置換することで側鎖による影響が著しく減少し、その結果としてトンネル構造の入りロを広めることとなり、セルロース鎖がトンネル構造に侵入しやすくなったためではないかと考えられる。しかし、これらの芳香族側鎖をもつアミノ酸残基がトンネル構造内での相互作用に重要な働きをしていることは間違いないようであり、この3つ以外の、よりトンネル構造の内部に存在するTyr142やTrp364のようなアミノ酸残基をアラニンに置換した変異体では、その酵素活性は野生型に比べて著しく低下していた。また、これ以外にトンネル構造の出ロ付近にも芳香族側鎖を持つアミノ酸残基が存在しており、これらはセルロース鎖の分解によって生じたセロビオースの酵素外への迅速な排出に関与し、このセロビオースによる酵素活性の阻害を防ぐ働きをしていると考えられている。今回の結果からもこの説にあてはまる傾向が見られ、Trp373やTyr378をアラニンに置換した変異体では酵素活性が著しく低下していた。
【実施例5】
【0075】
(PcEGIII及びそのアラニン置換変異体の酵素活性)
PcEGIIIは、図6の結果のように各変異体間で蛋白質の発現量に差が見られたので、まずBAS2000に付属の画像解析ソフトImage Gougeでその放射線強度を計測し、その値を元に各変異体の比活性を算出した。その結果が図8である。Tyr7を除くほぼ全ての変異体で、その酵素活性がいずれの基質に対しても激減した。CBHの結果とは異なる、このような大幅な酵素活性の減少の原因について考察を加えた。EGの立体構造は2枚のβシートからなるものの、上述のCBHのトンネル構造とは異なりU字型の構造をとっている。このU字型構造により、EGはCBHには無いセルロース鎖へのエンド型の分解作用を持つことが可能となっている。U字型であるため、EGにはトンネル構造のような密閉感は無く、セルロース鎖との相互作用に働くアミノ酸残基たちもある程度外部にさらされた状況にある。つまりこれらのアミノ酸残基はその多くが協同的に、外部からの酵素内へのセルロース鎖の取り込みに関与しているのではないかと考えることができ、この理由により各部位における1残基のアミノ酸変異でも大幅な酵素活性の低下が生じたのではないかと考えている。この仮説に基づくと、このU字型の雨どい構造の両端部分へのアラニン変異導入ならば、活性中心からはかなり離れた位置に存在し、CBHのように活性中心へとセルロース鎖を送り届ける必要もないため、それほど酵素活性に影響を与えないのではないかと推測されるが、実際に活性測定結果を見るとこのU字構造の両末端に位置するTyr7とPro134ヘアラニン変異を導入したものでは、その酵素活性の低下が他の8部位ほど大幅でないことがわかった。
【実施例6】
【0076】
(Phanerocaete chrysosporium由来のセロビオヒドロラーゼの38位、40位及47位に対する飽和変異)
セロビオヒドロラーゼのアラニンスキャニングの結果より、pc由来のCBHの38位、40位及び47位の変異体で酵素活性が向上したことから、これら3部位に対して、実施例2に準じてオーバーラッピングPCRによって飽和変異を導入し、実施例3に準じて各変異体を作製した。変異導入のために使用したプライマーを表6に示した。変異導入部位は大文字で表記し、変異アミノ酸は1文字表記で示した。表6のプライマーを用いた各変異導入断片作製のためのPCR反応液の組成を表7を示す。PCR反応条件は、実施例2の断片作製時と同様に条件とした。オーバーラッピングPCR反応液の組成及び反応条件は、実施例のオーバーラッピングPCRと同様の条件とした。また、pcCBH7c野生型の遺伝子は、実施例2と同様にして作製した。
【0077】
【表6】
[表7]
(PCR反応液)
10×Pyrobest(登録商標)バッファーII 2.0μ1
2.5mMdNTPs 2.0μ1
鋳型DNA(1ng/μ1) 1.0μ1
10μM フォワードプライマー 1.0μ1
10μM リバースプライマー 1.0μl
5U/μlPyrobest(登録商標)DNAポリメラーゼ
0.4μl
S.W. 12.6μl
合計 20μl
【0078】
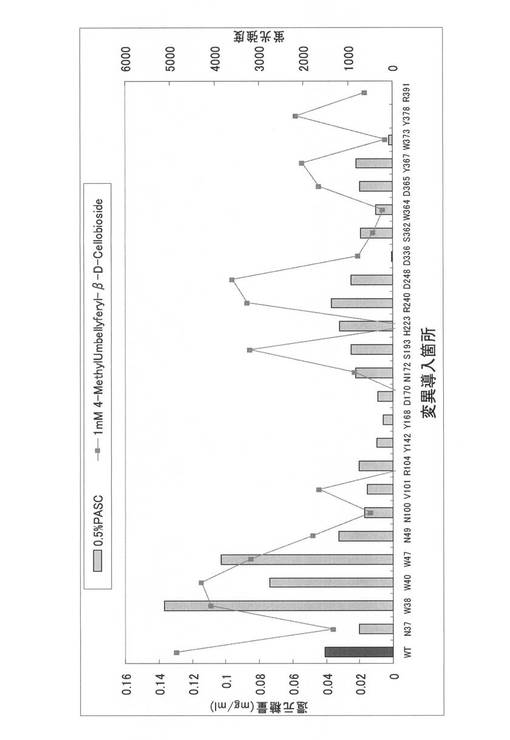
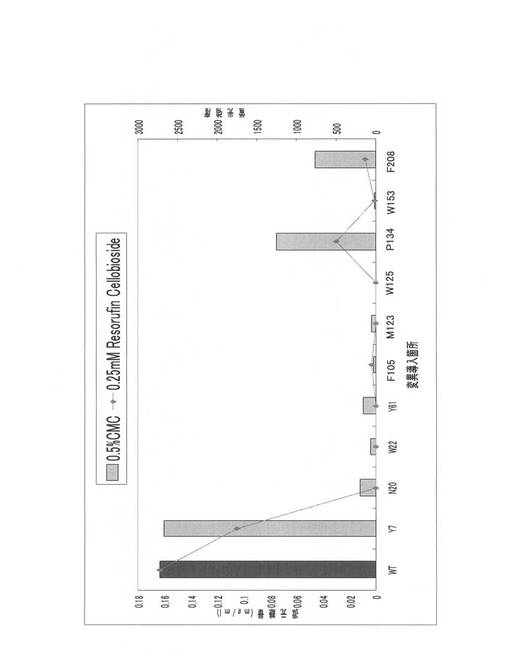
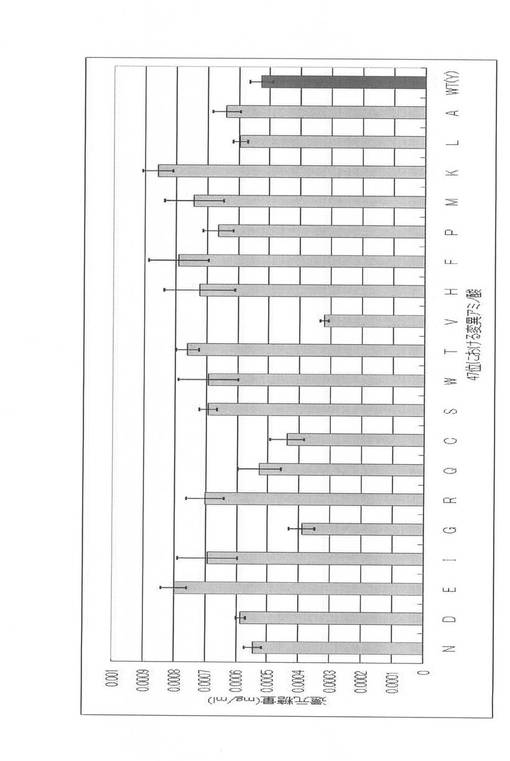
得られたPCR産物の各1μlのアガロースゲル電気泳動結果を図9に示す。W30、W40、Y47変異体のオーバーラップPCR産物は、鋳型量が200〜300ngになるように調製し、実施例3に記載の大腸菌由来無細胞タンパク合成系で発現させた。発現させた変異体を、実施例5と同様にして基質(0.5%PASC)と反応させ、ソモギー・ネルソン法によって活性測定を行い、野性型(WT)との酵素活性を比較した。各変異体の活性測定結果をそれぞれ図10〜図12に示した。
【0079】
図10〜図12に示すように、各位の飽和変異体中、野生型よりも高活性を示す変異体を高頻度で見出すことができた。すなわち、W38F、W40N、W40D、W40E、W40I、W40G、W40R、W40Q、W40S、W40Y、W40T、W40V、W40H、W40F、W40P、W40M、W40K、W40L、W40A、Y47N、Y47D、Y47E、Y47I、Y47R、Y47Q、Y47S、Y47W、Y47T、Y47H、Y47F、Y47P、Y47M、Y47K、Y47L及びY47Aであった。W38位については、アラニン変異型であるW38Aが野生型よりも低い酵素活性を呈していたが、W40位及びY47位においては、多くの変異体について野生型よりも高活性であった。特に、W40N、W40D、W40E、W40I、W40S、W40T、W40V、W40M、W40K及びW40Aにおいて野生型の約2倍以上の活性を認めた。
【0080】
以上のことから、セルロースなどの高分子を基質とする酵素、特に、プロセッセッシブ酵素に分類される酵素等、予想される基質結合部位が多数にわたるときには、効率的に変異体を得るには、あらかじめアラニンスキャニングにより、アラニン変異によって前記高分子基質に対する酵素活性が少なくとも低下しない箇所を変異候補部位としてスクリーニングし、当該変異候補部位を標的として複数のアミノ酸変異を導入した複数種類の変異体を作製することにより、きわめて効率的に高活性な変異体が得られることがわかった。
【図面の簡単な説明】
【0081】
【図1A】PcCBH7Cの参照構造としたPhanerocaete chrysosporium由来のセロビオヒドロラーゼIであるCBH Iの立体構造を示す図である。
【図1B】PcEG IIIの参照構造としたAspergillus nigar由来のEGIIIの立体構造を示す図である。
【図2】オーバーラッピングPCRによるセルラーゼ遺伝子へのアラニン変異の導入のスキームを示す図である。
【図3】無細胞タンパク質合成系における鋳型量の検討結果を示す図である。
【図4】PcCBH7Cの上清画分のオートラジオグラフィによる発現量の測定結果を示す図である。
【図5】PcCBH7Cの沈殿画分のオートラジオグラフィによる発現量の測定結果を示す図である。
【図6】PcEGIIIの上清画分と沈殿画分のオートラジオグラフィによる発現量の測定結果を示す図である。上段に上清画分の結果を示し、下段に沈殿画分の結果を示す。
【図7】PcCBH7Cのアラニン置換変異体の酵素活性を比較する図である。
【図8】PcEGIIIのアラニン置換変異体の酵素活性を比較する図である。
【図9】PCR産物のアガロースゲル電気泳動結果を示す図である。
【図10】W38位における各種アミノ酸置換変異体の活性を比較する図である。
【図11】W40位における各種アミノ酸置換変異体の活性を比較する図である。
【図12】Y47位における各種アミノ酸置換変異体の活性を比較する図である。
【配列表フリーテキスト】
【0082】
配列番号5〜143:プライマー
【技術分野】
【0001】
本発明は、変異体のスクリーニング方法に関し、特に、高分子基質に作用するプロセッシブ酵素について高活性な変異体を効率的に取得するスクリーニング方法に関する。
【背景技術】
【0002】
多くの酵素の基質結合部位は鍵穴状であり、酵素のごく一部に限定されていることが多い。一方、セルロースなどの高分子基質に作用する酵素の中には、高分子基質の鎖がはまり込むトンネル状又は溝状の相対的に長い基質結合部位を有する酵素があることが知られている。さらに、当該基質結合部位に高分子基質を保持しつつ触媒反応を行う酵素(プロセッシブ酵)があることも知られている。こうした酵素における広範囲にわたる基質結合部位においては、その一部が活性部位として機能していると考えられる。
【0003】
鍵穴状の基質結合部位を有する酵素について高活性な変異体を得る方法としては、たとえば、ある程度ランダムに複数の部位に部位特異的に各種のアミノ酸置換変異を導入する方法や、立体構造から活性に影響を及ぼす変異導入候補部位を選抜し部位特異的にアミノ酸変異を導入する方法等が一般に採用される。そして、こうした方法によりある程度高活性な変異体を取得できることが期待される。
【0004】
また、酵素活性部位を特定するのにアラニンスキャニングという手法がある(非特許文献1)。この手法は、活性部位周辺のアミノ酸をアラニンに置換した変異体を作製して、当該変異体のタンパク質の特異的活性が低下するかどうか検出し、活性が低下する場合には、当該部位は、酵素活性に強く関与する部位であると判定する手法である。
【0005】
【非特許文献1】Cunningham,B. C. and J. A. Wells, Science 244(4908):1081-1085,1989
【発明の開示】
【発明が解決しようとする課題】
【0006】
しかしながら、トンネル状等の基質結合部位を有する酵素につき、上記のような手法を試みても高活性変異体を取得するのは困難であった。すなわち、各種のアミノ酸置換変異の導入を試みる場合、広い範囲の基質結合部位にわたって多数の部位を置換対象としなければならず、かかる作業は膨大な時間と労力を有するものであった。また、基質結合部位が広い範囲にわたるため、置換導入部位を決定することも困難であった。
【0007】
以上のように、高分子基質を結合するトンネル状又は溝状の基質結合部位を有する酵素において高活性変異体を得るのはきわめて困難であった。
【0008】
そこで、本発明は、トンネル状又は溝状の基質結合部位を有する酵素について高活性変異体を効率的に取得できるスクリーニング方法を提供することを一つの目的とする。
【課題を解決するための手段】
【0009】
本発明者らは、上記課題について種々検討したところ、まず、大まかに予め選択した基質と相互作用する可能性のある部位に対してアラニンスキャニングを実施し、当該アラニンスキャニングにより活性上昇傾向のある部位を変異導入候補部位として選抜し、当該変異導入候補部位に各種アミノ酸置換を導入することにより、高活性な変異体を極めて効率的に取得できることを見出し、本発明を完成した。すなわち、本発明によれば以下の手段が提供される。
【0010】
本発明によれば、高分子基質を結合するトンネル状又は溝状の基質結合部位を有する酵素につき、基質相互作用を有する可能性のある部位のアミノ酸をアラニン置換した2種類以上の一次変異体を調製する工程と、前記一次変異体につき、変異による酵素活性の上昇の有無をスクリーニングして、変異前より酵素活性が上昇した前記一次変異体のアラニン置換部位を変異導入候補部位として選択する工程と、を備える、変異体のスクリーニング方法が提供される。
【0011】
本発明によれば、前記基質相互作用を有する部位は、前記酵素の前記基質結合部位の入口又は出口もしくはこれらの近傍に位置される部位を含む、前記スクリーニング方法も提供される。さらに、前記変異導入候補部位のそれぞれにつき、当該部位の本来のアミノ酸残基を2種類以上の他のアミノ酸に置換して得られる2種類以上の二次変異体を調製する工程と、前記二次変異体につき、変異による酵素活性の上昇の有無をスクリーニングして、変異前より酵素活性が上昇した前記二次変異体の変異導入部位のアミノ酸及び部位を置換アミノ酸候補及び置換候補部位として選択する工程と、を備える、前記スクリーニング方法も提供される。
【0012】
前記高分子基質は、セルロースであってもよく、前記酵素は、セロビオヒドロラーゼであってもよく、Phanerocaete chrysosporium由来のセロビオヒドロラーゼであってもよい。また、セロビオヒドロラーゼは、配列番号2で表されるアミノ酸配列又はこれに相当するアミノ酸配列を有し、前記変異導入候補部位は、配列番号2で表されるアミノ酸配列におけるW38位、W40位及びY47位又はこれらに相当する部位から選択されるものであってもよい。前記酵素は、エンドグルカナーゼであってもよく、Phanerocaete chrysosporium由来のエンドグルカナーゼであってもよい。前記エンドグルカナーゼは配列番号4で表されるアミノ酸配列又はこれに相当するアミノ酸配列を有し、前記変異導入候補部位は、配列番号4で表されるアミノ酸配列におけるY7位及びP134位又はこれらに相当する部位から選択されるものであってもよい。
【0013】
本発明によれば、配列番号2で表されるアミノ酸配列又はこれに相当するアミノ酸配列において、W38F、W40N、W40D、W40E、W40I、W40G、W40R、W40Q、W40S、W40Y、W40T、W40V、W40H、W40F、W40P、W40M、W40K、W40L、W40A、Y47N、Y47D、Y47E、Y47I、Y47R、Y47Q、Y47S、Y47W、Y47T、Y47H、Y47F、Y47P、Y47M、Y47K、Y47L及びY47Aからなる群から選択されるいずれかの変異又はこれに相当する変異を有するアミノ酸配列を有し、セロビオヒドロラーゼ活性を有するタンパク質が提供される。前記変異は、W40N、W40D、W40E、W40I、W40S、W40T、W40V、W40M、W40K及びW40A並びにこれらに相当する変異からなる群から選択されるいずれであってもよい。
【0014】
本発明によれば、配列番号2で表されるアミノ酸配列又はこれに相当するアミノ酸配列において、W38F、W40N、W40D、W40E、W40I、W40G、W40R、W40Q、W40S、W40Y、W40T、W40V、W40H、W40F、W40P、W40M、W40K、W40L、W40A、Y47N、Y47D、Y47E、Y47I、Y47R、Y47Q、Y47S、Y47W、Y47T、Y47H、Y47F、Y47P、Y47M、Y47K、Y47L及びY47Aからなる群から選択されるいずれかの変異又はこれに相当する変異を有するアミノ酸配列を有し、セロビオヒドロラーゼ活性を有するタンパク質をコードするポリヌクレオチドが提供される。
【発明を実施するための最良の形態】
【0015】
本発明は、高分子基質を結合する基質結合部位を有する酵素の高活性変異体を取得するスクリーニング方法及び変異体に関する。本発明のスクリーニング方法によれば、高分子基質を結合するトンネル状又は溝状の基質結合部位を有する酵素であっても、アラニン置換により酵素活性が上昇傾向にある部位を変異導入候補部位とし、当該変異導入候補部位にアミノ酸置換を導入した変異体を取得することで、ランダムな変異導入や多数に渡る部位特異的変異導入を回避でき、効率的に高活性な変異体を得ることができる。
【0016】
部位特異的にアラニン置換するアラニンスキャニングは、活性部位の特定に一般的に用いられる。すなわち、アラニン置換により活性が低下する部位は活性に寄与することがわかる。一方、アラニン置換により活性が向上する部位の位置づけは明らかではない。本発明者らは、高分子基質に結合するトンネル状や溝状等の基質結合部位を有する酵素において、基質相互作用を有する可能性の部位をアラニン置換し、アラニン置換により酵素活性が上昇する部位を、トンネル状の基質結合部位に対する基質の通過性に関与する変異導入候補部位として特定し、さらに当該部位に他のアミノ酸残基を導入することにより、高活性な変異体が得られることがわかった。以下、本発明の各種実施形態について詳細に説明する。
【0017】
(高分子基質結合部位を有する酵素の変異体のスクリーニング方法)
本発明のスクリーニング方法は、高分子基質結合部位を有する酵素につき、基質相互作用を有する可能性のある部位をアラニン置換した2種類以上の一次変異体を取得する工程を備えている。
【0018】
高分子基質結合部位を有する酵素は、トンネル状又は溝状の基質結合部位を有している。こうした基質結合部位は、高分子鎖がはまり込む構造となっているため、比較的細長い構造を有している。こうした酵素としては、例えば、セルロースを基質とするグリコシドヒドロラーゼファミリー(GHF)6、7、48に属するセルローゼであるセロビオヒドロラーゼが挙げられる。これらの酵素は、セルロースあるいはセロオリゴ糖などの高分子基質を結合するトンネル状の基質結合部位を有するプロセッシブ酵素であるとされている。また、GHF5,9に属するセロビオヒドロラーゼも溝状の基質結合部位を有しているとされている。さらにまた、GHF5、8、9、10、12、26、44、45、45、51、61及び74に属するエンドグルカナーゼも挙げられる。さらに、GHFに分類されるほかの高分子基質を分解する酵素としては、ヘミセルラーゼ及びアミラーゼが挙げられる。さらにまた、GHFに属さないが高分子基質を結合するトンネル状又は溝状の高分子基質結合部位を有する酵素としては、プロテアーゼ、エステラーゼ、リアーゼ及びトランスフェラーゼが挙げられる。
【0019】
本発明のスクリーニング方法は、高分子基質結合部位を有する酵素のなかでもプロセッシブ酵素のスクリーニングに適している。プロセッシブ酵素においては、高分子基質と強く相互作用が酵素活性に影響が大きいと考えられるからである。なお、プロセッシブ酵素とは、より詳細には、酵素反応を通じて酵素が基質をと分離することを要しない酵素である。例えば、プロセッシブ酵素としては、セルロースを基質とするグリコシドヒドロラーゼファミリー6、7、48に属するセルローゼであるセロビオヒドロラーゼが挙げられる。
【0020】
こうした酵素としては、例えば、各種菌に由来するセルラーゼが挙げられる。なかでも、GHF6に属する、Cellulomonas fimiなどのCellulomonas属、Humicola insolens、Humicola insolensなどのHumicola属、Trichoderma virideなどのTrichoderma属;GHF7に属するAspergillus aculeatus、Aspergillus nigar、Aspergillus oryzaeなどのAspergillus属、Phanerochaete chrysosporiumなどのPhanerocaete属、Acremonium thermophilumなどのAcremonium属、 Chaetomium thermophilumなどのChaetomium属、Trichoderma属;GHF48に属するClostridium acetobutylicum、Clostridium cellulolyticum、Clostridium cellulovorans、Clostridium josui、Clostridium thermocellumなどのClostridium属の菌由来のセロビオヒドロラーゼを含むセルラーゼが挙げられる。
【0021】
(一次変異体の調製工程)
まず、高分子基質結合部位を有する酵素において、基質相互作用を有する可能性のある部位をアラニン置換して一次変異体を調製する。基質相互作用を有する可能性のある部位(アラニン置換部位)は、立体構造が解明されている適当な他の参照用の高分子基質結合部位を有する酵素を選択し、その立体構造等から高分子基質と相互作用してその安定化に寄与していると考えられるアミノ酸残基を選択し、さらに、改変対象の高分子基質結合部位を有する酵素のアミノ酸配列から当該アミノ酸残基に相当するアミノ酸残基を抽出することにより決定できる。
【0022】
参照用の酵素は、アミノ酸配列において一定以上の同一性を有し同種活性を有する酵素とすることが好ましい。参照用酵素としては、例えば、改変対象がセルラーゼであれば、同一のGHFに属するセルラーゼが挙げられる。なお、参照用酵素としては、以下に記載するように、立体構造及び活性部位等が研究されている酵素等を用いることができる。
【0023】
参照用酵素の立体構造においてトンネル状又は溝状等の高分子基質結合部位において基質との相互作用に関与するアミノ酸残基(アラニン置換部位)を特定するには、立体構造モデルを構築する方法と既知の立体構造モデルを用いる方法とが挙げられる。立体構造モデルを構築する方法は、改変対象の酵素のアミノ酸配列をPSI−BLAST等のプログラムに入力し、類似性の高いアミノ酸配列の酵素を検索する。最も類似の酵素の構造を参照し、InsightII、Discovery studio等のソフトにより、ホモロジーモデリングを行うことにより、構造を予測する。予測した構造モデルを見ながら基質結合部位周辺のアミノ酸を抽出し、これらのアミノ酸を、アラニン置換部位とすることができる。
【0024】
また、立体構造モデルを構築しないで基質との相互作用に関与するアミノ酸残基を特定するには、例えば、同じGHFの酵素のうち、PDB等の立体構造データベースに構造がある酵素を検索する。選んだ構造既知の酵素のアミノ酸配列と、ターゲット酵素のアミノ酸配列をCLUSTAL W、GENETYX等のソフトに入力しアライメントする。構造既知の酵素の構造を見ながら基質結合部位周辺のアミノ酸を抽出し、そのアミノ酸とアライメントされた改変対象酵素のアミノ酸を、アラニン置換部位とすることができる。
【0025】
アラニン置換部位の個数は、特に限定しないが、必要に応じた数とすればよい。典型的には、数個以上、好ましくは10個以上から30個程度以下とすることができる。
【0026】
アラニン置換部位は、トンネル状又は溝状の基質結合部位の中央寄りの部分よりも入り口又は出口もしくはこれらの近傍から選択することが好ましい。本発明者らによる知見によれば、高分子基質結合部位を有する酵素におけるかかる部位のアミノ酸残基をアラニン置換することで、置換前に比べて基質結合部位への基質のアクセシビリティが大きく変化する可能性が高いと考えられる。そして、アラニン置換により酵素活性が向上するとき、当該部位の本来のアミノ酸残基は触媒作用を抑制している可能性があると考えられ、当該部位の他のアミノ酸への置換により活性が向上する可能性が高いと考えられる。
【0027】
決定したアラニン置換部位から選択される1種又は2種以上の部位をアラニン置換して一次変異体を調製する。それぞれの一次変異体は、複数のアラニン置換がなされていてもよいが、好ましくは、それぞれの一次変異体につき、一つのアミノ残基についてのみアラニン置換がなされている。部位特異的にアラニン置換された一次変異体を得るには、公知の部位特異的変異導入法を用いることができる。例えば、部位特異的にアラニン置換を導入するための変異型遺伝子は、Kunkel法やGapped duplex法などの公知の手法又はこれに準ずる方法により、例えば変異 導入用キット(例えばMutant-K(TAKARA社製)やMutant-G(TAKARA社製)など)を用いて、あるいは、TAKARA社のLA PCR in vitro Mutagenesisシリーズキットを用いて作製することができる。また、遺伝子の塩基配列を参照すれば、Molecular Cloning [Sambrookら編集, 15, Site-directed Mutagenesis of Cloned DNA, 15.3〜15.113, Cold Spring Harbor Lab. Press, New York (1989)]等の文献の記載に従って、当業者であれば格別の困難性なしに選択し実施することができる。また、当業者であれば、遺伝子の塩基配列を基にして、当該塩基配列から1以上(1又は数個以上)の塩基の置換、欠失、挿入又は付加を人為的に行う技術(部位特異的突然変異誘発)については、実施例に示すように、オーバーラップPCRを用いるほか、Proc. Natl. Acad. Sci. USA, 81, 5662-5666 (1984); WO85/00817; Nature, 316, 601-605 (1985); Gene, 34, 315-323 (1985); Nucleic Acids Res., 13, 4431-4442 (1985); Proc. Natl. Acad. Sci. USA, 79, 6409-6413 (1982); Science, 224, 1431-1433 (1984) 等に記載の技術に従って実施することができる。
【0028】
また、部位特異的変異導入法により鋳型DNAを取得後は、大腸菌等を用いた各種タンパク質の合成方法によりDNA上の情報に基づくタンパク質を取得できるが、好ましくは公知の無細胞タンパク質合成法を用いる。無細胞タンパク質合成系を用いることで、著しく作業時間と労働強度を低下させることができる。無細胞タンパク質合成系としては、公知の各種キットを用いて実施できるほか、特開2003−116590号公報、特開2006−61080号公報等に記載の方法を利用できる。無細胞タンパク質合成系を用いることにより、例えば、同時にスキャンするアラニン置換部位が10個〜100個程度のとき、従来なら5日から2週間程度要していたのを、0.5日〜2日程度で終了させることができる。さらに、同時スキャン部位が400個程度の場合、大腸菌による菌体生産による場合には60日間程度要するところを、無細胞タンパク質合成系によれば4日程度で達成することができる。
【0029】
(一次変異体のスクリーニング工程)
次に、一次変異体についてのスクリーニング工程(一次スクリーニング工程)を実施する。一次スクリーニング工程では、得られた一次変異体につき、変異による酵素活性の上昇の有無をスクリーニングして、変異前よりも活性が上昇した前記一次変異体におけるアラニン置換部位を変異導入候補部位として決定することができる。アラニン置換により酵素活性が向上したとき、当該部位は置換前のアミノ酸残基が酵素活性に抑制的に作用している可能性があり、当該アラニン置換部位をさらに他のアミノ酸残基に置換することにより、酵素活性の向上が期待できる。なお、酵素活性の測定方法は、改変対象酵素の種類によって適宜決定される。また、活性の上昇程度は、特に限定しないが、好ましくは30%以上、より好ましくは60%以上、さらに好ましくは100%以上、いっそう好ましくは200%以上である。
【0030】
以上の工程により、高分子基質結合部位を有する酵素における変異導入候補部位が迅速に決定される。従来、一次的な変異導入候補部位の決定に、多大な時間と労力を費やしており、基質結合部位が広範囲にわたるプロセッシブ酵素の場合、多数個のアミノ酸残基につき部位特異的変異導入を試みる必要があった。しかしながら、アラニンスキャニングをこうした酵素の基質結合部位の変異導入候補部位の決定に利用することで従来の作業を著しく簡素化して導入候補部位を決定できるようになった。
【0031】
本発明においては、一次スクリーニング工程を実施した上、抽出した置換変異導入候補部位に各種のアミノ酸を導入した二次変異体を調製し、これら二次変異体につき二次スクリーニング工程を実施する。このため、高活性な変異体のスクリーニングのための二次変異体を調製する作業及びスクリーニングサ行を著しく省力化し短時間化することができる。例えば、高分子基質結合部位が数十個のアミノ酸残基を含むとき、これらの全てのアミノ酸残基につき飽和アミノ酸変異を導入すると数百〜千数百程度の変異遺伝子及び変異体タンパク質を調製し、これらにつき酵素活性を測定することになる。これに対し、本発明の一次変異体のスクリーニング工程で数十個のアミノ酸残基から例えば数分の1から数十分の1程度に置換変異導入候補部位を絞ることができる。そして、これらの候補部位に飽和アミノ酸変異を導入することにより、変異遺伝子及び変異体のライブラリーサイズも数分の1〜数十分の1程度に縮小される。このようなライブラリ縮小効果は、広範囲にわたる高分子基質結合部位を有する酵素において極めて有効である。このようなライブラリーサイズの縮小効果に加えて、一次変異体及び二次変異体の調製に無細胞タンパク質合成系を用いることで、より一層スクリーニングを省力化することができる。
【0032】
(二次変異体の調製工程)
一次スクリーニング工程実施後には、さらに、二次変異体の調製工程と二次変異体についてのスクリーニング工程(二次スクリーニング工程)を備えることで、より具体的に置換アミノ酸候補及び置換候補部位を決定できる。二次変異体の調製工程では、一次スクリーニング工程において決定した変異導入候補部位に、ランダムにあるいは一定の規則に従い、本来のアミノ酸残基以外のアミノ酸残基を導入した二次変異体を調製することができる。決定された各変異導入候補部位につき、置換するアミノ酸は、本来のアミノ酸残基以外の全てのアミノ酸残基(全19種類)の全部又は一部であるが、少なくとも2種類以上の他のアミノ酸に置換することが好ましい。一部のアミノ酸に置換するとき、例えば、保存的アミノ酸置換の手法を採用できる。
【0033】
それぞれの二次変異体は、複数の変異導入候補部位において他のアミノ酸に置換されていてもよいが、各二次変異体は、それぞれ一つの変異導入候補部位においてのみ置換がなされていることが好ましい。二次変異体は、一つの置換候補部位につき複数種類のアミノ酸置換がなされた複数種類とすることが好ましい。さらに、こうした置換候補部位毎に複数種類の二次変異体を調製することが好ましい。
【0034】
二次変異体を調製するには、一次変異体の調製時と同様、公知の部位特異的変異導入法から適宜選択した手法を用いればよく、タンパク質合成にあたっては無細胞タンパク質合成系を用いることが好ましい。
【0035】
(二次変異体のスクリーニング工程)
次に、二次スクリーニング工程を実施する。二次スクリーニング工程では、得られた二次変異体につき、変異による酵素活性の上昇の有無をスクリーニングして、変異前よりも活性が上昇した前記二次変異体における置換アミノ酸及び置換部位を、置換アミノ酸候補及び置換候補部位として決定できる。酵素活性の測定方法は、改変対象酵素の種類によって適宜決定される。
【0036】
こうして得られた置換アミノ酸候補及び置換候補部位を、2種類以上組み合わせるなどの進化分子工学的手法を用いることで、より高活性な高分子基質結合部位を有する酵素を得ることができる。
【0037】
例えば、改変対象酵素が、Phanerocaete chrysosporium由来のセロビオヒドロラーゼなどのセロビオヒドロラーゼの場合、配列番号2で表されるアミノ酸配列又はこれに相当するアミノ酸配列(相当アミノ酸配列)を有し、配列番号2で表されるアミノ酸配列におけるW38位、W40位及びY47位から選択される1種又は2種以上又は当該相相当するアミノ酸配列これらに相当する部位から選択される1種又は2種以上を置換候補部位として二次変異体を調製し、二次変異体のスクリーニングを実施することができる。
【0038】
また、改変対象酵素は、Phanerocaete chrysosporium由来のエンドグルカナーゼなどのエンドグルカナーゼである場合、配列番号4で表されるアミノ酸配列又はこれに相当するアミノ酸配列(相当アミノ酸配列)を有し、配列番号4で表されるアミノ酸配列におけるY7位及びP134位から選択される1種又は2種以上又は当該相相当するアミノ酸配列におけるこれらに相当する部位から選択される1種又は2種以上を置換候補部位として二次変異体を調製し、二次変異体のスクリーニングを実施することもできる。
【0039】
改変対象酵素は上記のとおり、必ずしも配列番号2又は4で表されるアミノ酸配列と完全に一致するアミノ酸配列を有していなくともよく、これらに相当するアミノ酸配列(相当アミノ酸配列)を有していればよい。相当アミノ酸配列とは、BLASTなどの配列相同性検索において配列番号2又は4で表されるアミノ酸配列と同一性が40%以上、より好ましくは50%以上、さらに好ましくは60%以上、一層好ましくは70%以上、より一層好ましくは80%以上、さらに一層好ましくは90%以上、最も好ましくは95%以上のアミノ酸配列である。改変対象酵素が、こうしたアミノ酸配列を有する場合、ペアワイズアラインメントなどの手法により配列番号2又は配列番号4で表されるアミノ酸配列とアラインメントしたとき、上記置換候補部位に対応するものとして検出される部位を置換候補部位として二次変異体を調製することができる。
【0040】
本明細書において同一性とは、当該技術で知られているとおり、配列を比較することにより決定される、2以上のタンパク質あるいは2以上のポリヌクレオチドの間の関係である。当該技術で“同一性 ”とは、タンパク質またはポリヌクレオチド配列の間の適合によって、あるいは場合によっては、一続きのそのような配列間の適合によって決定されるような、タンパク質またはポリヌクレオチド配列の間の配列相関性の程度を意味する。“同一性 ”および“類似性”は、既知の方法により容易に決定できる。同一性 を決定する好ましい方法は、試験する配列間で最も長く適合するように設計される。同一性 および類似性を決定するための方法は、公に利用可能なプログラムにコードされている。相同性決定には、AltschulらによるBLAST (Basic Local Alignment Search Tool) プログラム(たとえば、Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ., J. Mol. Biol., 215: p403-410 (1990), Altschyl SF, Madden TL, Schaffer AA, Zhang J, Miller W, Lipman DJ., Nucleic Acids Res. 25: p3389-3402 (1997))を利用し決定することができる。BLASTのようなソフトウェアを用いる場合、デフォルト値を用いるのが好ましい。BLAST検索に一般的に用いられる主な初期条件は、以下の通りであるが、これに限定されない。
【0041】
(変異体タンパク質)
本発明のタンパク質は、配列番号2で表されるアミノ酸配列において、W38F、W40N、W40D、W40E、W40I、W40G、W40R、W40Q、W40S、W40Y、W40T、W40V、W40H、W40F、W40P、W40M、W40K、W40L、W40A、Y47N、Y47D、Y47E、Y47I、Y47R、Y47Q、Y47S、Y47W、Y47T、Y47H、Y47F、Y47P、Y47M、Y47K、Y47L及びY47Aからなる群から選択されるいずれかの変異を有するアミノ酸配列を有し、セロビオヒドロラーゼ活性を有するタンパク質である。これらのタンパク質は、親タンパク質である配列番号2で表されるアミノ酸配列からなるタンパク質、すなわち、Phanerocaete chrysosporium由来のCBH Iよりもよりもセルロースを分解する活性が高い。したがって、セルロース系バイオマスなどのセルロース含有材料の有効利用に有利に用いることができる。これらの変異体のなかでも、特に、W40N、W40D、W40E、W40I、W40S、W40T、W40V、W40M、W40K及びW40Aのいずれかがより高い活性を有している。これらの変異体は、親タンパク質よりも還元糖量に基づく酵素活性測定系において約2倍以上の活性を有している。
【0042】
変異体タンパク質は、配列番号2に相当するアミノ酸配列(相当アミノ酸配列)において、配列番号2で表されるアミノ酸配列において上記各変異に相当する変異のいずれかを有し、セロビオヒドロラーゼ活性を有するタンパク質であってもよい。すなわち、変異体タンパク質は、必ずしも配列番号2で表されるアミノ酸配列と完全に一致するアミノ酸配列において上記変異のいずれかを有するものでなくてもよい。すなわち、変異体タンパク質は、BLASTなどの配列相同性検索において、配列番号2で表されるアミノ酸配列と同一性が40%以上、より好ましくは50%以上、さらに好ましくは60%以上、一層好ましくは70%以上、より一層好ましくは80%以上、さらに一層好ましくは90%以上、最も好ましくは95%以上のアミノ酸配列を、上記の相当アミノ酸配列とし、この配列をペアワイズアラインメントなどの手法により配列番号2で表されるアミノ酸配列とアラインメントしたとき、上記変異部位に対応するものとして検出される部位のいずれかにおいて対応するアミノ酸置換を有するものであってもよい。
【0043】
こうした変異体タンパク質は、公知の方法によって取得できる。例えば、大腸菌、酵母等を用いた菌体によるタンパク質発現系、無細胞タンパク質合成系、形質転換した動植物を用いたタンパク質発現系などを用いて取得することができる。酵母等の細胞表層に提示させる形態で発現させることもできる。タンパク質の細胞表層提示技術は、当業者であれば、Cell surface engineering of yeast: construction of arming yeast with biocatalyst. J. Biosci. Bioeng., 90, 125-136 (2000)等に基づき実施することができる。
【0044】
(本発明のポリヌクレオチド)
本発明のポリヌクレオチドは、本発明の変異体タンパク質をコードしている。すなわち、配列番号2で表されるアミノ酸配列又はこれに相当するアミノ酸配列において、W38F、W40N、W40D、W40E、W40I、W40G、W40R、W40Q、W40S、W40Y、W40T、W40V、W40H、W40F、W40P、W40M、W40K、W40L、W40A、Y47N、Y47D、Y47E、Y47I、Y47R、Y47Q、Y47S、Y47W、Y47T、Y47H、Y47F、Y47P、Y47M、Y47K、Y47L及びY47Aからなる群から選択されるいずれかの変異又はこれに相当する変異を有するアミノ酸配列を有し、セロビオヒドロラーゼ活性を有するタンパク質をコードしている。
【0045】
本発明のポリヌクレオチドは、DNAの形態(たとえば、cDNAおよびクローニングによって得られるか、あるいは合成的に生成されるゲノムDNAを含む)であってもよく、RNA(たとえばmRNA)の形態であってもよい。該ポリヌクレオチドは、二本鎖であっても、一本鎖であってもよい。二本鎖の場合は、二本鎖DNA、二本鎖RNAまたはDNA:RNAのハイブリッドであってもよい。一本鎖の場合は、センス鎖(コード鎖としても知られる)であっても、アンチセンス鎖(非コード鎖としても知られる)であってもよい。
【0046】
本発明のポリヌクレオチドは、すでに説明したように、配列番号2で表されるアミノ酸配列をコードする配列番号1で表される塩基配列のDNA等の特定箇所に公知の各種方法によって変異を導入すること等によって得ることができる。また、配列番号1で表される塩基配列に対して、配列番号1とは異なるコドン用法を用いて得られる同一のアミノ酸配列をコードするDNAに対して変異を導入してもよい。
【0047】
以下、本発明を、実施例を挙げて具体的に説明するが、本発明はこれらの実施例に限定されるものではない。
【実施例1】
【0048】
(アラニン置換導入部位の決定)
白色腐朽菌Phanerocaete chrysosporium由来のセロビオヒドロラーゼIIであるCBH I(PcCBH7C)とエンドグルカナーゼであるEG(PcEGIII)に種々の変異を導入して、そのセルロース分解活性を向上させるため、まず、変異導入に適した箇所を探索した。すなわち、PcCBH7Cのアミノ酸配列(配列番号2 )とPcEGIIIのアミノ酸配列(配列番号4)それぞれに類似構造を持つ別のセルラーゼの立体構造から推測された、酵素活性を示す際にセルロース鎖と相互作用しその安定化に寄与していると考えられるアミノ酸残基を選択した。そして、選択した部位のアミノ酸残基をまずアラニンに置換し、それらの中でも特に変異導入による活性変化への影響が大きいと思われるアミノ酸残基の位置を特定することとした。
【0049】
PcCBH7Cの参照構造として、同じPhanerocaete chrysosporium由来のセロビオヒドロラーゼIであるCBH I(図1A参照)とした。また、PcEG IIIについては、Aspergillus nigar由来のEGIII(図1B参照)を参照構造とした。これらについてはいずれもPDB(http://www.rcsb.org/pdb/home/home.do)において立体構造を取得できる。参照の結果、これより、PcCBH7Cについては、以下の表(表1)に示す24個のアミノ酸残基をアラニン置換することとした。また、PcEGIllについては、以下の表(表2)に示す10個のアミノ酸残基をアラニンに置換することとした。
【0050】
【表1】
【表2】
【実施例2】
【0051】
(オーバーラッピングPCRによるセルラーゼ遺伝子へのアラニン変異の導入)
(PcCBH7C遺伝子)
オーバーラッピングPCRによるセルラーゼ遺伝子へのアラニン変異の導入のスキームを図2に示す。プラスミドpRPccel7C(Pccel7C遺伝子、配列番号1)の両端をNdelおよびEcoRlで切断し、pRSETBにクローニングして作成)を鋳型とし、表3に示すF3−K4プライマーと各変異導入部位につき個別のリバースプライマーを用いてPCRを行うことによりT7プロモーターとRBSを含んだ断片を増幅した。また、表3に示すR3−K4(と各変異導入部位につき個別のフォワードプライマーを用いてPCRを行うことにより各変異導入部位へのアラニン変異とT7ターミネーターを含む断片を増幅した。なお、PCRの反応液組成は、以下の通りとした。反応条件は、94℃で3分後、94℃で10秒、60℃で10秒及び72℃で30秒を1サイクルとして25サイクル実施し、72℃で7分とした。
【0052】
【表3A】
【表3B】
【0053】
(PCR反応液)
10×PfuUItra(登録商標)IIバッファー 2.0μ1
2.5mMdNTPs 2.0μ1
鋳型DNA(1ng/μ1) 1.0μ1
10μMフォワードプライマー 1.0μ1
10μMリバースプライマー 1.0μl
5U/μlPfuUltra(登録商標)II Fusion HS DNAポリメラーゼ(Strategene製) 0.2μl
S.W. 12.8μl
合計 20μl
【0054】
(PcEGIII遺伝子)
プラスミドpET−PcEG(ppET」PcEG(pET23b(+)vetQrのNdel、XhoサイトにPcEGIII(配列番号3)の全長遺伝子を挿入したもの)を鋳型とし、PcCBH7Cの場合と同様にF3−K4プライマーと表4に示した各変異導入部位につき個別のリバースプラィマーを用いてPCRを行うことによりT7プロモーターとRBSを含んだ断片を増幅し、R3−K4と表4に示した各変異導入部位につき個別のフォワードプライマーを用いてPCRを行うことにより各変異導入部位へのアラニン変異とT7ターミネーターを含む断片を増幅した。PCR条件は、PcCBH7C遺伝子の場合と同様とした。
【0055】
【表4】
【0056】
得られた各PCR産物につき、アガロースゲル電気泳動を行い、MinEluteGelExtraction Kitを用いてアガロースゲルから精製した。アガロースゲルから精製した2種類の断片を、図2に示すオーバーラッピングPCRを行うことにより連結させた。PCR反応液の組成は、以下のとおりとした。反応条件は、94℃で3分後、94℃で15秒、55℃で30秒、及び72℃で90秒を1サイクルとして10サイクル実施後に、プライマーK4を添加し、引き続き、94℃で15秒、55℃で30秒、72℃で90秒を1サイクルとして10サイクル実施後、72℃で7分の条件でPCRを行った。この際、オーバーラップさせる各断片の濃度はNanoDrop(商標)ND−1000分光光度計によって測定し、各々が10ngずつPCR反応系に含まれるよう調整した。こうして作製したアラニン変異導入セルラーゼ遺伝子につき、表5に示すプライマーを用いて塩基配列の確認を行った。
【0057】
(オーバーラッピングPCR反応液の組成)
10xLA Taq(登録商標)Buffer 2.0μl
2.5mMdNTPs 3.2μl
25mM MgCl2 2.0μl
フォワード断片(10ng/μl) 1.0μl
リバース断片(10ng/μl) 1.0μl
20μMK4プライマー 1.0μl
5U/μl LA Taq DNAポリメラーゼ(タカラバイオ製)
0.2μl
S.W. 9.6μl
合計 20μl
【0058】
【表5】
【0059】
また、PcCBH7C及びPcEGIIlの各野生型遺伝子は、それぞれプラスミドpRPccel7C及びpE下PcEGを鋳型とし、プライマーにF3−K4とR3−K4を用いてPCRを行うことによって作成した。PCR反応液の組成は、以下のとおりとした。
反応条件は、94℃で3分後、94℃で15秒、55℃で15秒、72℃で30秒を1サイクルとして25サイクル実施後、72℃で7分とした。作製した各変異遺伝子と野生型遺伝子は、Qubit(商標)fluorometerを用いてそのDNA濃度を測定した。
【0060】
(PCR反応液)
10×PfuUltra(商標)IIバッファー2.0μ1
2.5mM dNTPs 2.0μ1
鋳型(1ng/μ1) 1.0μ1
10μM F3K4プライマー 1.0μ1
10μM R3K4プライマー 1.0μ1
5U/μl PfuUltra(商標)II FusionHSDNAポリメラーゼ
0.2μ1
S.W. 12.8μ1
合計 20μl
【実施例3】
【0061】
(無細胞タンパク質合成系によるセルラーゼの発現)
実施例2で作製したPcCBH7Cの野生型のPCR産物を、大腸菌由来無細胞蛋白質合成の鋳型とし、その鋳型量を0ng,100ng、200ng、300ng、及び400ngと変化させた。そして、それぞれの鋳型量で合成した蛋白質の酵素活性を測定し、無細胞蛋白質合成系に必要な鋳型量を決定した。鋳型量は、約200〜300ngが最適であった(図3)。さらに、PD1(Humicola insolens由来)及び還元型グルタチオン(GSH)、酸化型グルタチオン(GSSG)を、以下の組成の転写・翻訳共役反応液に加えて混合し、26℃で、6時間無細胞蛋白質合成を行った。
【0062】
(転写・翻訳共役反応液)
PDI 0.15μ1
10mM GSH 0.3μ1
0.4M 酢酸マグネシウム 0.6μ1
5M 酢酸カリウム 0.6μl
0.1mg/mlリファンピシリン 3.0μ1
LM(20a.a.) 7.5μ1
10mg/ml CK 0.45μ1
2.3mg/ml T7RNAポリメラーゼ 1.0μ1
(pAR1219;Davanloo et al.,1984で形質転換したE.coli BL21から精製)
100mMGSSG 0.3μ1
S30抽出物(特開2006−61080) 8.5μ1
鋳型+S.W. 7.6μ1
合計 30μ1
【0063】
なお、無細胞合成反応試薬の終濃度は、LM;LowMolecular weight mixture(56.4mM トリス−酢酸(pH7.4)、1.22mM ATP(pH7.0)、各0.85mM GUC ミックス(GTP、UTP、CTP)、50mM クレアチンリン酸2ナトリウム、各0.32mM 20アミノ酸ミックス、4%(w/v)ポリエチレングリコール6000、34.6μg/ml 葉酸、35.9mM 酢酸アンモニウム、0.17mg/ml E.coli tRNA、28.3%(v/v)S30抽出物)、0.15mg/mlクレアチンキナーゼ、7.7μg/ml T7RNAポリメラーゼ、10μg/m1リファンピシリン、10mM 酢酸マグネシウム、100mM 酢酸カリウムであった。
【0064】
また、オートラジオグラフィによる蛋白質発現量の確認を行った。無細胞蛋白質合成系の組成液の1つであるLMに含まれる20種類のアミノ酸を、ロイシン以外の19アミノ酸に14C−ロイシンを加えたものに変更し、26℃で6時間無細胞蛋白質合成を行った。合成反応液の組成は、以下のとおりとした。
【0065】
(無細胞タンパク質合成反応液の組成)
PDI 0.15μl
10mM GSH 0.3μl
0.4M 酢酸マグネシウム 0.6μl
0.5M 酢酸カリウム 0.6μl
0.1mg/m1リファンピシリン 3.0μl
LM(19アミノ酸) 7.5μl
10mg/mlCK 0.45μl
2.3mg/mlT7RNAポリメラーゼ 1.0μl
100mM GSSG 0.3μ1
14C−ロイシン 2.0μ1
(Amersham−PhamaciaBiotech製)
S30抽出物 8.5μl
鋳型DNA+S.W 5.6μl
合計 30μl
【0066】
無細胞蛋白質合成反応終了液に対し15,000rpmで5分間遠心を行い、反応終了液を上清画分と沈澱画分に分けた。この上清画分と沈殿画分の各々に60%アセトンを9倍量加え、氷上で5分間静置した後、4℃、15,000rpmで5分間遠心を行い、上清を除いた上で7.5μ1のS.W.に再懸濁した。等量の2xSDS−PAGEサンプルバッファーと混合し、煮沸したものをサンプルとしてSDS−PAGEを行った。SDS−PAGE終了後のゲルを、ゲルドライヤーを用いて80℃、1時間乾燥させた後、イメージングプレートに挟み12時間以上置いてRIを転写させた。転写後のイメージングプレートをBAS IP MAGAZINEにセットし、BAS2000で14C−ロイシンの取り込み量を解析した。
【0067】
PcCBH7C及びPcEGIIIのアラニン変異導入遺伝子と野生型遺伝子の無細胞蛋白質合成系による発現と、オートラジオグラフィによる発現量の確認を行った。結果を図4、図5及び図6に示す。PcCBH7Cではいずれの変異体及び野生型についてもその大部分が上清画分に発現されており、発現量の差もほとんど見られなかった。上清画分には可溶性の蛋白質が見出されるので、発現されたPcCBH7Cの大部分はフォールディングの不具合等は起こらずに、活性を持った正常な構造で発現されているものと考えられる。一方PcEGlllについては、各変異体間でやや大きな差が見られ、上清画分と沈殿画分への発現量の比率が各変異体によって異なっていた。発現量には差がないことから、発現後の蛋白質のフォールディング過程が原因ではないかと考えられた。
【実施例4】
【0068】
(PcCBH7C及びPcEGHIIIの酵素活性の測定)
実施例3で得られた反応終了液につき、酵素活性を測定した。PcCBH7Cの酵素活性の基質として、不溶性の高分子セルロースであるPASC(リン酸膨潤アビセル)と、可溶性のセロビオースに4−methylumbelliferylが付加された蛍光基質であるMUC(4−methylumbelliferyl−β−D−cellobioside)を用いた。PASCは高分子セルロースであるため、セルロース鎖は基質結合ポケットの溝の全域に入り込む形で結合し反応する。よって、アラニン置換によるポケット全域への影響を見ることができる。MUCは低分子であるため、G1〜G3結合部位周辺への影響のみを見ることができる。また、PcEGIIIの酵素活性の基質として、可溶性基質のCMC(力ルボキシメチル化セルロース;SIGMA−ALDRICH製)とResorufinceIIobioside(MarkerGeneTMFIuorescentCellulaseAssayKit;MGTInc.社製)を用いた。詳細を以下に示す。
【0069】
PASCによる酵素反応系は、1%PASC 50mM酢酸バッファー(pH5.0)250μl,50mM酢酸バッファー(pH5.0)225μl、無細胞蛋白質合成反応終了液25μl,計500μlの組成で、45℃、24時間酵素反応を行った。PASCとの反応性を上げるためアシストチューブを横にし、振邊しながら反応させた。その後、反応液を用いて生成した還元糖の定量(ソモギーネルソン法又はTZ assay法)を行った。ソモギー・ネルソン法は、反応液サンプル25μ1にソモギー銅液50μ1を加えよく混合させた。サンプルとは別に0〜1mg/mlのグルコース水溶液を調製し同様に以下の操作を行った。サーマルサイクラーを用いて、10g℃で30分間反応させた後、ネルソン液50μ1を加えよく混合した。3,000rpm、3分間遠心した後、上清50μ1を150μ1のS.W.を入れた96穴プレートに加え、プレートリーダーFusionで540nmの吸光度を測定した。測定結果からグルコース検量線を作成し、生成した還元糖量をグルコース濃度に換算し求めた。また、TZassay法は(Chong, K. et al., Journal of Biochemical and Biophysical Methods, 111 - 109(1985))は、以下のとおりとした。すなわち、TZassayバッファ195μlを分注し、反応液サンプル5μlを加えてゆっくり混合した。これを100℃で3分間ヒート処理した後に氷冷し、660nmの吸光度を測定した。グルコースの検量線を作製し、グルコース換算の還元糖量を得た。
【0070】
MUCによる酵素反応系は、2mMMUC/50mM酢酸バッファー(pH5.0)50μ1、50mM酢酸バッファー(pH5.0)50μl、無細胞蛋白質合成反応終了液20μl、計100μlの組成で、96穴プレートにおいて40℃、1時間酵素反応を行った。その後、プレートリーダーFusionでEx/Em=330/515nmの蛍光強度を測定した。
【0071】
CMCによる酵素反応系は、エッペンドルフチューブを用いて、1%CMC/50mM酢酸バッファー(pH5.0)50μ1,50mM酢酸バッファー(pH5.O)45μ1,無細胞蛋白質合成反応終了液5μ1、計100μ1の組成で、45℃で1時間酵素反応を行った。CMCとの反応性を上げるためエッペンドルフチューブを横にし、振とうしながら反応させた。その後、反応液を用いて生成した還元糖の定量を、前述のソモギー・ネルソン法によって行った。
【0072】
Resorufin cellobiosideによる酵素反応系は、0.5mM Resorufin cellobioside/100mM酢酸バッファー(pH6.0)50μ1、100mM酢酸バッファー(pH6.0)25μl、無細胞蛋白質合成反応終了液25μ1、計100μlの組成で、96穴プレートにおいて40℃で1時間酵素反応を行った。その後、プレートリーダーFusionでEx/Em=540/590nmの蛍光強度を測定した。
【0073】
(PcCBH7C及びそのアラニン置換変異体の酵素活性)
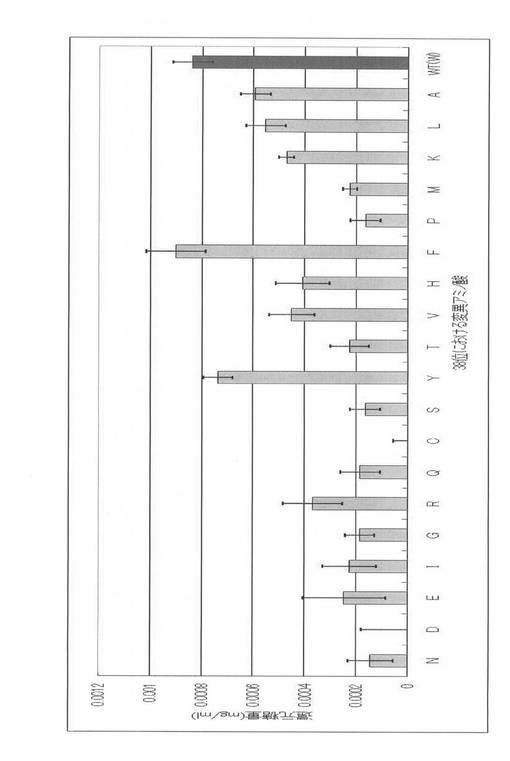
無細胞タンパク質合成系によって発現させたPcCBH7Cの野生型と各アラニン変異体24サンプルの酵素活性をPASCとMUCを基質として用いて行った。その活性測定結果を図7に示す。図7に示すように、大部分のアラニン変異体ではその酵素活性が野生型よりも低下した。ここで、PASCに対する活性測定結果とMUCに対する活性測定結果の傾向が若干異なっていることがわかるが、これはおそらくこの二つの基質の構造の違いに起因するものと考えられる。PASCが非晶性のβ−1,4−グリコシド結合で連結したグルコースの鎖状の構造を持つのに対し、MUCは2分子のグルコースがβ−1,4−グリコシド結合でつながった2糖のセロビオシドに分子量176の大きな蛍光基であるメチルウンベリフェロンが結合した構造を持つ。PcCBH7Cは2枚のβシートからなるトンネル構造の中央に活性中心を持っている。このトンネルにセルロース鎖が入り、トンネルの内側に面するアミノ酸残基との相互作用により安定化されつつ活性中心に移動し、グリコシド結合を切断されるのであるが、MUCはその蛍光基の存在の為にこのトンネルヘの侵入及びトンネル内でのアミノ酸残基との相互作用に基づく安定化に通常とは異なる寄与が生まれ、PASCに対する酵素活性測定結果との間に相違が生じたのではないかと考えている。
【0074】
さらに、38位のTrp、40位のTrp、47位のTyrをアラニンに置換した変異体では、その酵素活性が野生型よりも向上した。これらの3個のアミノ酸残基は、立体構造においては上述したセルロース鎖と相互作用するトンネル構造の入りロ付近に存在している。TrpやPhe、Tyrのような芳香族の側鎖を持ったアミノ酸は、このトンネル構造の内部によく見られ、疎水性相互作用や水素結合によってセルロース鎖と相互作用する。今回これらの酵素活性に重要であると思われるアミノ酸残基をアラニンに置換したのであるが、Trp38、Trp40、Tyr47の3つの変異体の酵素活性が野生型よりも向上したのは、アラニンに置換することで側鎖による影響が著しく減少し、その結果としてトンネル構造の入りロを広めることとなり、セルロース鎖がトンネル構造に侵入しやすくなったためではないかと考えられる。しかし、これらの芳香族側鎖をもつアミノ酸残基がトンネル構造内での相互作用に重要な働きをしていることは間違いないようであり、この3つ以外の、よりトンネル構造の内部に存在するTyr142やTrp364のようなアミノ酸残基をアラニンに置換した変異体では、その酵素活性は野生型に比べて著しく低下していた。また、これ以外にトンネル構造の出ロ付近にも芳香族側鎖を持つアミノ酸残基が存在しており、これらはセルロース鎖の分解によって生じたセロビオースの酵素外への迅速な排出に関与し、このセロビオースによる酵素活性の阻害を防ぐ働きをしていると考えられている。今回の結果からもこの説にあてはまる傾向が見られ、Trp373やTyr378をアラニンに置換した変異体では酵素活性が著しく低下していた。
【実施例5】
【0075】
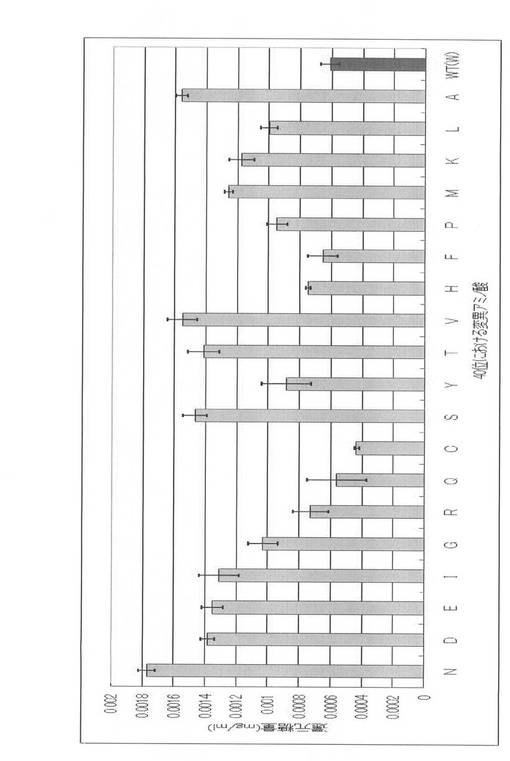
(PcEGIII及びそのアラニン置換変異体の酵素活性)
PcEGIIIは、図6の結果のように各変異体間で蛋白質の発現量に差が見られたので、まずBAS2000に付属の画像解析ソフトImage Gougeでその放射線強度を計測し、その値を元に各変異体の比活性を算出した。その結果が図8である。Tyr7を除くほぼ全ての変異体で、その酵素活性がいずれの基質に対しても激減した。CBHの結果とは異なる、このような大幅な酵素活性の減少の原因について考察を加えた。EGの立体構造は2枚のβシートからなるものの、上述のCBHのトンネル構造とは異なりU字型の構造をとっている。このU字型構造により、EGはCBHには無いセルロース鎖へのエンド型の分解作用を持つことが可能となっている。U字型であるため、EGにはトンネル構造のような密閉感は無く、セルロース鎖との相互作用に働くアミノ酸残基たちもある程度外部にさらされた状況にある。つまりこれらのアミノ酸残基はその多くが協同的に、外部からの酵素内へのセルロース鎖の取り込みに関与しているのではないかと考えることができ、この理由により各部位における1残基のアミノ酸変異でも大幅な酵素活性の低下が生じたのではないかと考えている。この仮説に基づくと、このU字型の雨どい構造の両端部分へのアラニン変異導入ならば、活性中心からはかなり離れた位置に存在し、CBHのように活性中心へとセルロース鎖を送り届ける必要もないため、それほど酵素活性に影響を与えないのではないかと推測されるが、実際に活性測定結果を見るとこのU字構造の両末端に位置するTyr7とPro134ヘアラニン変異を導入したものでは、その酵素活性の低下が他の8部位ほど大幅でないことがわかった。
【実施例6】
【0076】
(Phanerocaete chrysosporium由来のセロビオヒドロラーゼの38位、40位及47位に対する飽和変異)
セロビオヒドロラーゼのアラニンスキャニングの結果より、pc由来のCBHの38位、40位及び47位の変異体で酵素活性が向上したことから、これら3部位に対して、実施例2に準じてオーバーラッピングPCRによって飽和変異を導入し、実施例3に準じて各変異体を作製した。変異導入のために使用したプライマーを表6に示した。変異導入部位は大文字で表記し、変異アミノ酸は1文字表記で示した。表6のプライマーを用いた各変異導入断片作製のためのPCR反応液の組成を表7を示す。PCR反応条件は、実施例2の断片作製時と同様に条件とした。オーバーラッピングPCR反応液の組成及び反応条件は、実施例のオーバーラッピングPCRと同様の条件とした。また、pcCBH7c野生型の遺伝子は、実施例2と同様にして作製した。
【0077】
【表6】
[表7]
(PCR反応液)
10×Pyrobest(登録商標)バッファーII 2.0μ1
2.5mMdNTPs 2.0μ1
鋳型DNA(1ng/μ1) 1.0μ1
10μM フォワードプライマー 1.0μ1
10μM リバースプライマー 1.0μl
5U/μlPyrobest(登録商標)DNAポリメラーゼ
0.4μl
S.W. 12.6μl
合計 20μl
【0078】
得られたPCR産物の各1μlのアガロースゲル電気泳動結果を図9に示す。W30、W40、Y47変異体のオーバーラップPCR産物は、鋳型量が200〜300ngになるように調製し、実施例3に記載の大腸菌由来無細胞タンパク合成系で発現させた。発現させた変異体を、実施例5と同様にして基質(0.5%PASC)と反応させ、ソモギー・ネルソン法によって活性測定を行い、野性型(WT)との酵素活性を比較した。各変異体の活性測定結果をそれぞれ図10〜図12に示した。
【0079】
図10〜図12に示すように、各位の飽和変異体中、野生型よりも高活性を示す変異体を高頻度で見出すことができた。すなわち、W38F、W40N、W40D、W40E、W40I、W40G、W40R、W40Q、W40S、W40Y、W40T、W40V、W40H、W40F、W40P、W40M、W40K、W40L、W40A、Y47N、Y47D、Y47E、Y47I、Y47R、Y47Q、Y47S、Y47W、Y47T、Y47H、Y47F、Y47P、Y47M、Y47K、Y47L及びY47Aであった。W38位については、アラニン変異型であるW38Aが野生型よりも低い酵素活性を呈していたが、W40位及びY47位においては、多くの変異体について野生型よりも高活性であった。特に、W40N、W40D、W40E、W40I、W40S、W40T、W40V、W40M、W40K及びW40Aにおいて野生型の約2倍以上の活性を認めた。
【0080】
以上のことから、セルロースなどの高分子を基質とする酵素、特に、プロセッセッシブ酵素に分類される酵素等、予想される基質結合部位が多数にわたるときには、効率的に変異体を得るには、あらかじめアラニンスキャニングにより、アラニン変異によって前記高分子基質に対する酵素活性が少なくとも低下しない箇所を変異候補部位としてスクリーニングし、当該変異候補部位を標的として複数のアミノ酸変異を導入した複数種類の変異体を作製することにより、きわめて効率的に高活性な変異体が得られることがわかった。
【図面の簡単な説明】
【0081】
【図1A】PcCBH7Cの参照構造としたPhanerocaete chrysosporium由来のセロビオヒドロラーゼIであるCBH Iの立体構造を示す図である。
【図1B】PcEG IIIの参照構造としたAspergillus nigar由来のEGIIIの立体構造を示す図である。
【図2】オーバーラッピングPCRによるセルラーゼ遺伝子へのアラニン変異の導入のスキームを示す図である。
【図3】無細胞タンパク質合成系における鋳型量の検討結果を示す図である。
【図4】PcCBH7Cの上清画分のオートラジオグラフィによる発現量の測定結果を示す図である。
【図5】PcCBH7Cの沈殿画分のオートラジオグラフィによる発現量の測定結果を示す図である。
【図6】PcEGIIIの上清画分と沈殿画分のオートラジオグラフィによる発現量の測定結果を示す図である。上段に上清画分の結果を示し、下段に沈殿画分の結果を示す。
【図7】PcCBH7Cのアラニン置換変異体の酵素活性を比較する図である。
【図8】PcEGIIIのアラニン置換変異体の酵素活性を比較する図である。
【図9】PCR産物のアガロースゲル電気泳動結果を示す図である。
【図10】W38位における各種アミノ酸置換変異体の活性を比較する図である。
【図11】W40位における各種アミノ酸置換変異体の活性を比較する図である。
【図12】Y47位における各種アミノ酸置換変異体の活性を比較する図である。
【配列表フリーテキスト】
【0082】
配列番号5〜143:プライマー
【特許請求の範囲】
【請求項1】
高分子基質を結合するトンネル状又は溝状の基質結合部位を有する酵素につき、基質相互作用を有する可能性のある部位のアミノ酸をアラニン置換した2種類以上の一次変異体を調製する工程と、
前記一次変異体につき、変異による酵素活性の上昇の有無をスクリーニングして、変異前より酵素活性が上昇した前記一次変異体のアラニン置換部位を変異導入候補部位として選択する工程と、
を備える、変異体のスクリーニング方法。
【請求項2】
前記基質相互作用を有する部位は、前記プロセッシブ酵素の前記基質結合部位の入口又は出口もしくはこれらの近傍に位置される部位を含む、請求項1に記載の方法。
【請求項3】
さらに、
前記変異導入候補部位のそれぞれにつき、当該部位の本来のアミノ酸残基を2種類以上の他のアミノ酸に置換して得られる2種類以上の二次変異体を調製する工程と、
前記二次変異体につき、変異による酵素活性の上昇の有無をスクリーニングして、変異前より酵素活性が上昇した前記二次変異体の変異導入部位のアミノ酸及び部位を置換アミノ酸候補及び置換候補部位として選択する工程と、
を備える、請求項1又は2に記載のスクリーニング方法。
【請求項4】
前記高分子基質は、セルロースである、請求項1〜3のいずれかに記載のスクリーニング方法。
【請求項5】
前記酵素は、セロビオヒドロラーゼである、請求項4に記載のスクリーニング方法。
【請求項6】
前記Phanerocaete chrysosporium由来のセロビオヒドロラーゼである、請求項5に記載のスクリーニング方法。
【請求項7】
前記セロビオヒドロラーゼは、配列番号2で表されるアミノ酸配列又はこれに相当するアミノ酸配列を有し、前記変異導入候補部位は、配列番号2で表されるアミノ酸配列におけるW38位、W40位及びY47位又はこれに相当する部位から選択される、請求項5又は6に記載のスクリーニング方法。
【請求項8】
前記酵素は、エンドグルカナーゼである、請求項4に記載のスクリーニング方法。
【請求項9】
前記エンドグルカナーゼは、Phanerocaete chrysosporium由来のエンドグルカナーゼである、請求項8に記載のスクリーニング方法。
【請求項10】
前記エンドグルカナーゼは配列番号4で表されるアミノ酸配列又はこれに相当するアミノ酸配列を有し、前記前記変異導入候補部位は、配列番号4で表されるアミノ酸配列におけるY7位及びP134位又はこれらに相当する部位から選択される、請求項8又は9に記載のスクリーニング方法。
【請求項11】
配列番号2で表されるアミノ酸配列又はこれに相当するアミノ酸配列において、W38F、W40N、W40D、W40E、W40I、W40G、W40R、W40Q、W40S、W40Y、W40T、W40V、W40H、W40F、W40P、W40M、W40K、W40L、W40A、Y47N、Y47D、Y47E、Y47I、Y47R、Y47Q、Y47S、Y47W、Y47T、Y47H、Y47F、Y47P、Y47M、Y47K、Y47L及びY47Aからなる群から選択されるいずれかの変異又はこれに相当する変異を有するアミノ酸配列を有し、セロビオヒドロラーゼ活性を有するタンパク質。
【請求項12】
前記変異は、W40N、W40D、W40E、W40I、W40S、W40T、W40V、W40M、W40K及びW40A並びにこれらに相当する変異からなる群から選択されるいずれである、請求項11に記載のタンパク質。
【請求項13】
配列番号2で表されるアミノ酸配列又はこれに相当するアミノ酸配列において、W38F、W40N、W40D、W40E、W40I、W40G、W40R、W40Q、W40S、W40Y、W40T、W40V、W40H、W40F、W40P、W40M、W40K、W40L、W40A、Y47N、Y47D、Y47E、Y47I、Y47R、Y47Q、Y47S、Y47W、Y47T、Y47H、Y47F、Y47P、Y47M、Y47K、Y47L及びY47Aからなる群から選択されるいずれかの変異又はこれに相当する変異を有するアミノ酸配列を有し、セロビオヒドロラーゼ活性を有するタンパク質をコードするポリヌクレオチド。
【請求項1】
高分子基質を結合するトンネル状又は溝状の基質結合部位を有する酵素につき、基質相互作用を有する可能性のある部位のアミノ酸をアラニン置換した2種類以上の一次変異体を調製する工程と、
前記一次変異体につき、変異による酵素活性の上昇の有無をスクリーニングして、変異前より酵素活性が上昇した前記一次変異体のアラニン置換部位を変異導入候補部位として選択する工程と、
を備える、変異体のスクリーニング方法。
【請求項2】
前記基質相互作用を有する部位は、前記プロセッシブ酵素の前記基質結合部位の入口又は出口もしくはこれらの近傍に位置される部位を含む、請求項1に記載の方法。
【請求項3】
さらに、
前記変異導入候補部位のそれぞれにつき、当該部位の本来のアミノ酸残基を2種類以上の他のアミノ酸に置換して得られる2種類以上の二次変異体を調製する工程と、
前記二次変異体につき、変異による酵素活性の上昇の有無をスクリーニングして、変異前より酵素活性が上昇した前記二次変異体の変異導入部位のアミノ酸及び部位を置換アミノ酸候補及び置換候補部位として選択する工程と、
を備える、請求項1又は2に記載のスクリーニング方法。
【請求項4】
前記高分子基質は、セルロースである、請求項1〜3のいずれかに記載のスクリーニング方法。
【請求項5】
前記酵素は、セロビオヒドロラーゼである、請求項4に記載のスクリーニング方法。
【請求項6】
前記Phanerocaete chrysosporium由来のセロビオヒドロラーゼである、請求項5に記載のスクリーニング方法。
【請求項7】
前記セロビオヒドロラーゼは、配列番号2で表されるアミノ酸配列又はこれに相当するアミノ酸配列を有し、前記変異導入候補部位は、配列番号2で表されるアミノ酸配列におけるW38位、W40位及びY47位又はこれに相当する部位から選択される、請求項5又は6に記載のスクリーニング方法。
【請求項8】
前記酵素は、エンドグルカナーゼである、請求項4に記載のスクリーニング方法。
【請求項9】
前記エンドグルカナーゼは、Phanerocaete chrysosporium由来のエンドグルカナーゼである、請求項8に記載のスクリーニング方法。
【請求項10】
前記エンドグルカナーゼは配列番号4で表されるアミノ酸配列又はこれに相当するアミノ酸配列を有し、前記前記変異導入候補部位は、配列番号4で表されるアミノ酸配列におけるY7位及びP134位又はこれらに相当する部位から選択される、請求項8又は9に記載のスクリーニング方法。
【請求項11】
配列番号2で表されるアミノ酸配列又はこれに相当するアミノ酸配列において、W38F、W40N、W40D、W40E、W40I、W40G、W40R、W40Q、W40S、W40Y、W40T、W40V、W40H、W40F、W40P、W40M、W40K、W40L、W40A、Y47N、Y47D、Y47E、Y47I、Y47R、Y47Q、Y47S、Y47W、Y47T、Y47H、Y47F、Y47P、Y47M、Y47K、Y47L及びY47Aからなる群から選択されるいずれかの変異又はこれに相当する変異を有するアミノ酸配列を有し、セロビオヒドロラーゼ活性を有するタンパク質。
【請求項12】
前記変異は、W40N、W40D、W40E、W40I、W40S、W40T、W40V、W40M、W40K及びW40A並びにこれらに相当する変異からなる群から選択されるいずれである、請求項11に記載のタンパク質。
【請求項13】
配列番号2で表されるアミノ酸配列又はこれに相当するアミノ酸配列において、W38F、W40N、W40D、W40E、W40I、W40G、W40R、W40Q、W40S、W40Y、W40T、W40V、W40H、W40F、W40P、W40M、W40K、W40L、W40A、Y47N、Y47D、Y47E、Y47I、Y47R、Y47Q、Y47S、Y47W、Y47T、Y47H、Y47F、Y47P、Y47M、Y47K、Y47L及びY47Aからなる群から選択されるいずれかの変異又はこれに相当する変異を有するアミノ酸配列を有し、セロビオヒドロラーゼ活性を有するタンパク質をコードするポリヌクレオチド。
【図1A】

【図1B】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
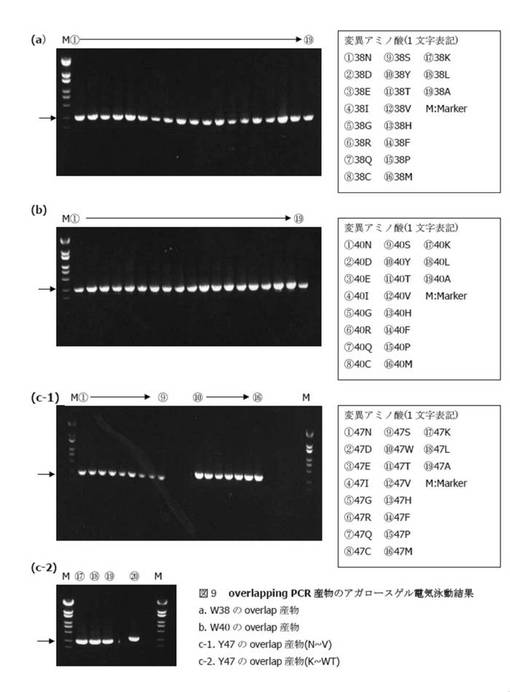
【図10】
【図11】
【図12】
【図1B】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公開番号】特開2010−46034(P2010−46034A)
【公開日】平成22年3月4日(2010.3.4)
【国際特許分類】
【出願番号】特願2008−214714(P2008−214714)
【出願日】平成20年8月22日(2008.8.22)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成19年度、独立行政法人新エネルギー・産業技術総合開発機構 微生物機能を活用した環境調和型製造基盤技術開発の委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(000003609)株式会社豊田中央研究所 (4,200)
【出願人】(504139662)国立大学法人名古屋大学 (996)
【Fターム(参考)】
【公開日】平成22年3月4日(2010.3.4)
【国際特許分類】
【出願日】平成20年8月22日(2008.8.22)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成19年度、独立行政法人新エネルギー・産業技術総合開発機構 微生物機能を活用した環境調和型製造基盤技術開発の委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(000003609)株式会社豊田中央研究所 (4,200)
【出願人】(504139662)国立大学法人名古屋大学 (996)
【Fターム(参考)】
[ Back to top ]