
多種サブタイプFIVワクチン
【課題】多種サブタイプFIVワクチンを用いて、広範囲のFIV株による感染からネコを守るための新規の方法および組成物に関する。
【解決手段】無細胞ウイルス全体またはウイルス感染細胞株のいずれかを含む多種サブタイプFIVワクチンについて記述する。本ワクチン組成物によるネコのワクチン接種法についても記述する。本発明の方法および組成物に従ってワクチン接種したネコは、同種または異種FIV株でチャレンジした場合、FIVに対して防御的液性および細胞性免疫反応を示す。本発明はまた、FIV易感染性である新規のネコ科細胞株およびその利用法にも関する。
【解決手段】無細胞ウイルス全体またはウイルス感染細胞株のいずれかを含む多種サブタイプFIVワクチンについて記述する。本ワクチン組成物によるネコのワクチン接種法についても記述する。本発明の方法および組成物に従ってワクチン接種したネコは、同種または異種FIV株でチャレンジした場合、FIVに対して防御的液性および細胞性免疫反応を示す。本発明はまた、FIV易感染性である新規のネコ科細胞株およびその利用法にも関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は米国国立衛生研究所助成金番号NIH AI30904による助成を受けた研究プロジェクトの下で政府の支援を受けて行われた。米国政府は本発明に一定の権利を保有する。
【背景技術】
【0002】
発明の背景
飼いネコは、ネコ白血病ウイルス(FeLV)、ネコ肉腫ウイルス(FeSV)、内因型C型オンコロナウイルス(RD-114)、およびネコ合胞体形成ウイルス(FeSFV)を含むいくつかのレトロウイルスによる感染症にかかりやすい。これらの中で、FeLVは最も重要な病原体で、リンパ網内系および骨髄新生物、貧血、免疫媒介疾患、ならびにヒト後天性免疫不全症候群(AIDS)と同様の免疫不全症候群を含む多様な症状を引き起こす。最近では、FeLV-AIDSと呼ばれる特定の複製欠損FeLV変異株が免疫抑制特性により一層関連している。
【0003】
ネコTリンパ球親和性レンチウイルス(現在では、ネコ免疫不全ウイルス、FIVと呼ばれている)の発見は、ペダーセンら(非特許文献1)によって最初に報告された。FIVの特徴は、ヤマモトら(非特許文献2);ヤマモトら(非特許文献3);およびアックリーら(非特許文献4)において報告されている。血清疫学的データにより、FIV感染が世界中の飼いネコおよび野生のネコに固有であることが示された。流産、脱毛、貧血、結膜炎、慢性鼻炎、腸炎、歯肉炎、血便排泄、神経異常、歯周炎、および脂漏性皮膚炎を含む広く多様な症状は、FIVによる感染症に関連する。FIVに感染した飼いネコの顕著な免疫学的特徴は、ネコのCD4+末梢血リンパ球の慢性的および進行性枯渇、CD4:CD8細胞比の低下、およびいくつかの症例ではCD8保有リンパ球の増加である。分子、生化学、および免疫病理学的特徴に基づき、ネコのFIV感染症は今や、FeLV-FAIDSより良いネコのAIDSモデルであると思われる。
【0004】
FIVのクローニングおよび配列分析は、オルムステッドら(非特許文献5);オルムステッドら(非特許文献6);およびタルボットら(非特許文献7)において報告されている。ホージー&ジャレット(非特許文献8)は、FIVに感染したネコの血清学的反応について記述している。FIVウイルスサブタイプは、それぞれの株によって誘発された交叉中和抗体の濃度に基づく免疫型に従って分類することができる(マーフィー&キングスバリー(非特許文献9))。近年、ヌクレオチド配列の相同性に基づく遺伝子型に従って、ウイルスがサブタイプに分類された。HIVおよびFIVサブタイプ分類は遺伝子型に基づくが(ソドラら(非特許文献10);リグビーら(非特許文献11);およびルワジーら(非特許文献12))、サブタイプの遺伝子型と免疫型の間の相関に関してはほとんどわかっていない。FIVウイルス単離体は現在、4つのFIVサブタイプに分類されている:A、B、CおよびD(カキヌマら(非特許文献13))。感染性単離体および感染性分子クローンは、サブタイプCを除く全てのFIVサブタイプについて記述されている(ソドラら(非特許文献10))。サブタイプCのFIVは、カナダのネコの細胞DNAから唯一同定されている(ソドラら(非特許文献10);リグビーら(非特許文献11);カキヌマら(非特許文献13))。
【0005】
FIVワクチンを開発する上で主に難しい点は、異なるサブタイプまたはクレードからのフィールド単離体を含む広範囲のFIV株に対して有効なワクチンアプローチを特定することであった。FIVに対するワクチン予防は、単一株のワクチンを用いた場合、同種およびわずかに異種の株に対しては予防できたが、中程度から大きく異なる種の株によるチャレンジでは予防できない(ジョンソンら(非特許文献14);ヤマモトら(非特許文献15))。したがって、依然として多数のFIVサブタイプから防御することのできるワクチンが必要とされている。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Pedersen, N.C., E.W. Ho, M.L. Brown, J.K. Yamaoto (1987) “Isolation of a T-lymphotropic virus from domestic cats with an immunodeficiency-like syndrome,” Science 235:790-793
【非特許文献2】Yamamoto, J.K., N.C. Pedersen, E.W. Ho, T. Okuda, G.H. Theilen (1988a) “Feline immunodeficiency syndrome - a comparison between feline T- lymphotropic lentivirus and feline leukemia virus,” Leukemia, December Supplement 2:204S-215S.
【非特許文献3】Yamamoto, J.K., E. Sparger, E.W. Ho, P.H. Andersen, T.P. O'Connor, C.P. Mandell, L. Lowenstine, N.C. Pedersen (1988) “Pathogenesis of experimentally induced feline immunodeficiency virus infection in cats,” Am. J. Vet. Res. 49:1246-1258.
【非特許文献4】Ackley, C.D., J.K. Yamamoto, N.B. Levy, N.C. Pedersen, M.D. Cooper (1990) “Immunologic abnormalities in pathogen-free cats experimentally infected with feline immunodeficiency virus,” J. Virol. 64:5652-5655.
【非特許文献5】Olmsted, R.A., A.K. Barnes, J.K. Yamamoto, V.M. Hirsch, R.H. Purcell, P.R. Johnson (1989) “Molecular cloning of feline immunodeficiency virus,” Proc. Nat. Acad. Sci. 86:2448-2452
【非特許文献6】Olmsted, R.A., V.M. Hirsch, R.H. Purcell, P.R. Johnson (1989) “Nucleotide sequence analysis of feline immunodeficiency virus: Genome organization and relationship to other lentivirus,” Proc. Natl. Acad. Sci. USA 86:8088-8092.
【非特許文献7】Talbott, R.L., E.E. Sparger, K.M. Lovelace, W.M. Fitch, N.C. Pedersen, P.A. Luciw, J.H. Elder (1989) “Nucleotide sequence and genomic organization of feline immunodeficiency virus,” Proc. Natl. Acad. Sci. USA 86:5743-5747.
【非特許文献8】Hosie, M.J., O. Jarrett (1990) “Serological responses of cats to feline immunodeficiency virus,” AIDS 4:215-220.
【非特許文献9】Murphy, F., D.W. Kingsbury (1990) “Virus Taxonomy,” In Fields Virology, 2nd Ed., B.N. Fields, D.M. Knipe et al., eds, Raven Press, New York, Chapter 2, pp. 9-36.
【非特許文献10】Sodora, D.L., E.G. Shpaer, B.E. Kitchell, S.W. Dow, E.A. Hoover, J.I. Mullins (1994) “Identification of three feline immunodeficiency virus (FIV) env gene subtype and comparison of the FIV and human immunodeficiency virus type 1 evolutionary patterns,” J Virol. 68:2230-2238.
【非特許文献11】Rigby, M.A, E.C. Holmes, M. Pistello, A. Mackay, A.J. Leigh-Brown, J.C. Neil (1993) “Evolution of structural proteins of feline immunodeficiency virus: molecular epidemiology and evidence of selection for change,” J. Gen. Virol. 74:425-436.
【非特許文献12】Louwagie, J., F.E. McCutchan, M. Peeters, T.P. Brennan, E. Sanders-Buell, G.A. Eddy, G. van den Grosen, K. Fransen, G.M. Gershy-Damet, R. Deleys, D.S. Burke (1993) “Phylogenetic analysis of gag genes from 70 international HIV-1 isolates provides evidence for multiple genotypes,” AIDS 7:769-780.
【非特許文献13】Kakinuma, S., K. Motokawa, T. Hohdatsu, J.K. Yamamoto, H. Koyama, H. Hashimoto (1995) “Nucleotide Sequence of Feline Immunodeficiency Virus: Classification of Japanese Isolates into Two Subtypes Which Are Distinct from Non-Japanese Subtypes,” Journal of Virology 69(6):3639-3646.
【非特許文献14】Johnson, C.M., B.A. Torres, H. Koyama, J.K. Yamamoto (1994) “FIV as a model for AIDS vaccination,” AIDS Res. Hum. Retroviruses 10:225-228.
【非特許文献15】Yamamoto, J.K., T. Hohdatsu, R.A. Olmsted, R. Pu, H. Louie, H. Zochlinski, V. Acevedo, H.M. Johnson, G.A. Soulds, M.B. Gardner (1993) “Experimental vaccine protection against homologous and heterologous strains of feline immunodeficiency virus,” J. Virol. 67:601-605.
【発明の概要】
【0007】
発明の簡単な概要
本発明は、宿主動物におけるFIV感染症に対して広範囲の防御的免疫を誘発するワクチンに関する。特に、本発明は異なるFIVサブタイプから単離された無細胞ウイルス単離株、またはそれぞれが異なるサブタイプの異なるプロトタイプFIVウイルスに感染した細胞株の組み合わせ、を用いて調製される多種サブタイプFIVワクチンに関する。本発明のFIVワクチンを接種したネコは、同種および異種FIV株に対して液性および細胞性の免疫反応を引き起こす。
【0008】
本発明はまた、多種のFIVサブタイプに易感染性である新規のネコ科細胞株にも関する。本発明の細胞株は、本発明の方法によるFIVワクチンにおける使用と共に、多種のFIVサブタイプの増殖および産生に有用である。さらに、本細胞株はまた、ネコ抗血清のFIVウイルス中和アッセイにおいてネコ末梢血単核球(PBMC)の代わりに用いることができる。
【0009】
配列の簡単な説明
配列番号:1は、SV-V3-2と呼ばれるFIV表面エンベロープペプチドのアミノ酸配列である。
配列番号:2は、TM-C1と呼ばれるFIV膜通過ペプチドのアミノ酸配列である。
配列番号:3は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:4は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:5は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:6は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:7は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:8は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:9は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:10は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:11は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:12は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:13は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:14は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:15は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:16は、FIV PCRプライマーのヌクレオチド配列である。
【図面の簡単な説明】
【0010】
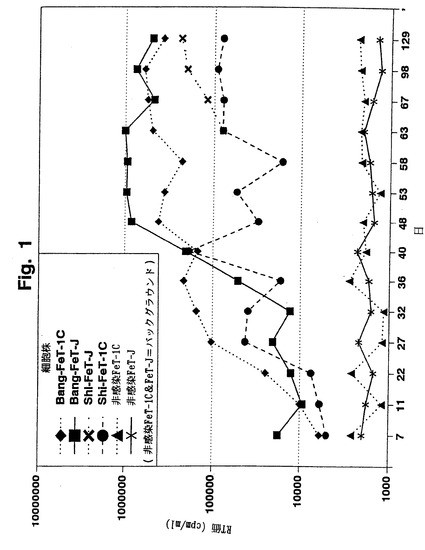
【図1】図1は、これらのFIV株でFeT-1CおよびFeT-J細胞株を感染させた後に産生されたFIVBangおよびFIVShiの逆転写酵素(RT)濃度を示す。
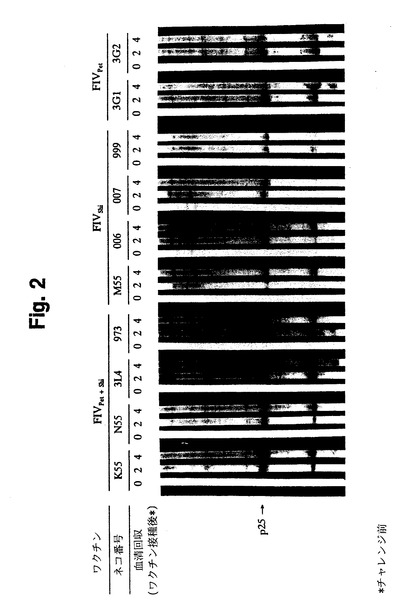
【図2】図2は、イムノブロットによって検出された、2種サブタイプワクチン接種ネコの抗FIV抗体とFIV蛋白質との免疫反応を示す。各ブロット上の数値は、血清を検査した時点での動物に行ったワクチン接種回数を表す。
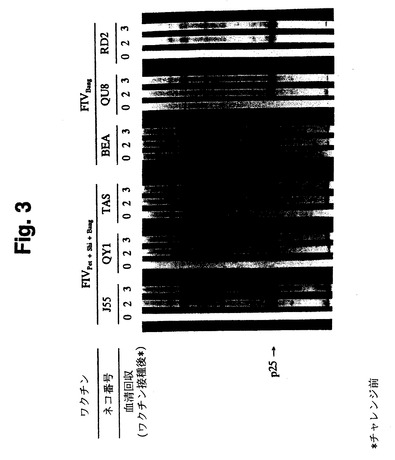
【図3】図3は、イムノブロットによって検出された、3種サブタイプワクチン接種ネコの抗FIV抗体とFIV蛋白質との免疫反応を示す。各ブロット上の数値は、血清を検査した時点での動物に行ったワクチン接種回数を表す。
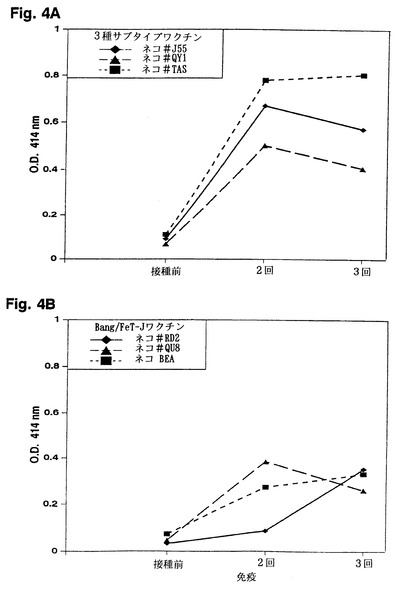
【図4】図4は、ELISAによって検出した、3種サブタイプワクチン接種ネコから得た抗FIV抗体とFIV SU-V3-2ペプチドとの免疫反応性を示す。
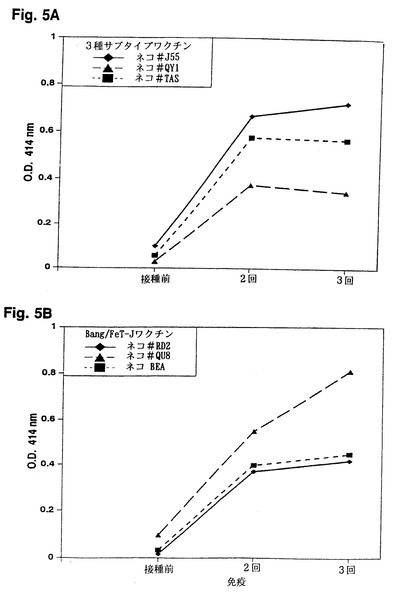
【図5】図5は、ELISAによって検出した、3種サブタイプワクチン接種ネコから得た抗FIV抗体とFIV TM-C1ペプチドとの免疫反応性を示す。
【図6】図6は、FIVPet(AP)、FIVDix(AD)、FIVUK8(AU)、FIVBang(BB)、FIVAom1(BA)、およびFIVShi(DS)のいずれかに感染したネコから得た血清の交叉中和抗体価を示す。感染前(カラム1)、感染後6ヶ月(カラム2)、および感染後12ヶ月(カラム3)での血清を、FeT-1C-細胞株においてサブタイプA FIVPet、サブタイプB FIVBang、およびサブタイプD FIVShiに関して試験した。1株当たり少なくとも3匹のネコを試験し、結果は各株の代表的なネコから得たVN力価を示す。VNアッセイに関しては、初代培養PBMCを用いても同様の結果が得られた。
【発明を実施するための形態】
【0011】
発明の詳細な開示
本発明は、感受性宿主動物におけるFIV感染症に対する防御的免疫の誘導に有用な新規の方法およびワクチン組成物に関する。本明細書に記載のワクチン組成物は、宿主動物に投与すると、FIVの同種および異種株による感染症に対して防御的液性および細胞性免疫反応を誘導する。ワクチン組成物は、無細胞FIVウイルス単離株を含んでいてもFIV-感染細胞株を含んでいてもよい。好ましい態様において、本発明のワクチン組成物は、異なる2つのFIVサブタイプからのFIV株を含む。好ましくは、ワクチン組成物は、各株が異なるFIVサブタイプである3つのFIV株を含む。より好ましくは、FIVサブタイプA、サブタイプBおよびサブタイプDのそれぞれから少なくとも1つのFIV株がワクチン組成物に含まれる。
【0012】
特定の態様において、ワクチン組成物はFIVPet-およびFIVShi-感染細胞株を含む。もう一つの態様において、ワクチン組成物はFIVPet-、FIVBang-、およびFIVShi-感染細胞株を含む。FIVサブタイプの全てまたは一部を表すその他のFIV株の使用は、本発明において特に企図されるところである。例えば、FIVサブタイプAプロトタイプウイルスを提供する目的で、FIVDixまたはFIVUK8をFIVPetに加えて、またはFIVPetの代わりに、ワクチン組成物に含めることができる。FIVサブタイプBおよびDプロトタイプウイルスに関しても、その他のFIV株による同様の追加または置換を行うことができる。
【0013】
本明細書に記載のように、本明細書のワクチン組成物は、FIV感染細胞株、または無細胞ウイルスおよび感染細胞株の組み合わせと共に、無細胞ウイルス全体、またはウイルス、FIV蛋白質およびポリペプチドの一部を含んでいてもよい。FIV感染細胞株からなるワクチン組成物は、それぞれが異なるFIVサブタイプに感染した多数の細胞株を含んでいてもよい。本発明のワクチン組成物はまた、例えばFIV env、gag/pro、またはenv-gag/proを含む組換えウイルスベクターに基づくFIV構築物を含む。組換え型ベクター/FIV構築物を調製するために用いることができるいかなる適当なウイルスベクターも、本発明での使用が企図される。例えば、アデノウイルス、鳥類ポックスウイルス、ネコヘルペスウイルス、ワクシニア、カナリアポックス、昆虫ポックス、ブタポックス、および当技術分野で既知のその他のウイルスに由来するウイルスベクターを、本発明の組成物および方法と共に用いることができる。FIV成分をコードし発現する組換えポリヌクレオチドベクターは、当技術分野で既知の標準的な遺伝子操作技術を用いて構築することができる。さらに、本明細書に記載の様々なワクチン組成物は、個別に用いることも、互いに組み合わせて用いることもできる。例えば、動物の一次免疫には、単一または複数のサブタイプ成分を有する、組換えベクターに基づくFIV構築物を用い、二次免疫には不活性化FIV-感染細胞株からなるワクチン組成物を用いてもよい。本発明のワクチン組成物によるその他の免疫プロトコルは、当業者には明らかで、本発明の範囲内であると企図される。
【0014】
本明細書に特に記載された多種サブタイプFIVワクチンのネコにおける免疫原性および有効性を調べた。本ワクチン組成物でワクチン接種した特定病原体非保有(SPF)ネコの、同種および異種FIV株によるチャレンジ前後の液性および細胞性免疫をモニターした。液性免疫は、ウイルス中和(VN)抗体活性の測定によってモニターし、細胞性免疫は細胞障害性Tリンパ球(CTL)活性の測定によってモニターした。ワクチン接種ネコの血清および免疫細胞の同種および異種FIV株に対するVNおよびCTL活性をそれぞれインビトロで試験し、本ワクチンがFIV感染に対する広範囲の防御を誘発できることを証明した。本発明の指示に従い、異なるFIVサブタイプからのプロトタイプウイルス単離株を組み合わせることによって、または異なるサブタイプのプロトタイプウイルスに感染させた個々の細胞を組み合わせることによって、有効な多種サブタイプFIVワクチンを産生することができる。
【0015】
本明細書に特に例示した株に加えて全てのFIV株は、本発明での使用が企図される。多くのFIV単離株が文献に記述されており、当業者に既知である。FIVPetは米国特許第5,037,753号に記述されている。記述されているその他のFIV単離体は、当業者によって定法を用いて感染したネコから容易に単離することができる。FIVの単離および培養法は、参照として本明細書に組み入れられる米国特許第5,037,753号および第5,118,602号に記述されている。
【0016】
本明細書に例示されている新規の細胞株は、本発明のワクチン接種法および組成物において用いることができる。末梢血単核細胞を含むFIV株易感染性のその他の細胞または細胞株もまた、本発明での使用が企図される。
【0017】
FIVウイルス蛋白質の天然ポリペプチド、組換えポリペプチドまたは合成ポリペプチド、およびそのペプチド断片もまた、本方法によるワクチン組成物として用いることができる。好ましい態様において、多種のFIVサブタイプに由来するFIVポリペプチドをワクチン組成物に結合させ、これを用いて宿主動物にワクチン接種する。例えば、異なるサブタイプからの少なくとも2つのプロトタイプFIV株からのFIVエンベロープ糖蛋白質に基づくポリペプチドをワクチンに組み合わせることができる。ポリペプチドは一つの株に対して同種であってもよく、そのアミノ酸配列が少なくとも2つの明確なFIVサブタイプからの接合または結合ポリペプチドに由来する「ハイブリッド」または「キメラ」ポリペプチドを含むものであってもよい。FIVポリペプチドの調製手順は当業者に周知である。例えば、FIVポリペプチドは固相合成法を用いて合成することができる(メリフィールド(Merrifield)、1963)。FIVポリペプチドはまた、FIV蛋白質またはペプチドをコードするポリペプチド分子が、細菌、酵母、または哺乳類細胞株などの宿主細胞で発現されるよう、DNA組換え技術を用いて産生することができ、当技術分野の標準的な技術を用いて発現した蛋白質を精製することができる。
【0018】
本発明はまた、FIVに対して易感染性である新規のネコ科T細胞株にも関する。インターロイキン-2(IL-2)依存的細胞および非依存的細胞の両者を特に例示する。FeT-1CおよびFeT-Jと呼ばれる細胞株を本明細書に記述する。FeT-1C細胞株はIL-2依存的であり、FeT-J細胞株はIL-2非依存的である。本発明の細胞株は、インビトロでのFIVウイルス株の増殖および産生と共に、ネコのFIV免疫の媒体を提供するのに有用である。IL-2依存的FeT-1CおよびIL-2非依存的FeT-J非感染細胞株をいずれも培養液中の逆転写酵素(RT)活性についておよびPCRによるFIVプロウイルス配列について20回以上試験し、FIV陰性であることを確認した。FeT-J細胞株は、FIVShi、FIVDix、FIVUK8、FIVPetおよびFIVBangを含む調べた全てのFIV株に高度に感染可能であったが、FIVShiに直接感染させることはよ比較的困難であった。
【0019】
本発明はさらに、本発明の細胞株によって産生された細胞性産物にも関する。細胞性産物は当業者に既知の方法を用いて単離および検出することができる。細胞株に対する抗体は、既知の方法を用いて産生させることができ、本発明に企図される。
【0020】
FeT-1C(ATCC寄託番号CRL 11968)、およびFeT-J(ATCC寄託番号CTL 11967)と呼ばれるFIV非感染細胞株はいずれも、1995年8月24日に、米国メリーランド州ロックビルのアメリカンタイプカルチャーコレクション(ATCC)に寄託された。FIVBang-(ATCC寄託番号11975)およびFIVShi-(ATCC寄託番号11976)感染細胞株は、1995年8月25日にアメリカンタイプカルチャーコレクションに寄託された。
【0021】
本培養物は、本特許出願の係属中、特許庁長官により37 CFR 1.14および35 U.S.C.122の下で資格を与えられると決定された者は、寄託物を利用することができるよう保証される条件下で寄託されている。寄託物は、本出願の写しまたはその所産が提出されている国の外国特許法の要請に応じて利用することができるだろう。しかし、寄託物の利用は、政府の決定によって付与された特許権の適用の制限において本発明を実施する認可とはならないことを理解すべきである。
【0022】
さらに、本培養寄託物は、微生物の寄託に関するブダペスト条約(Budapest Treaty for the Deposit of Microorgamisms)の要件に従って保存され、公的に利用可能となる、すなわち、寄託物は、寄託試料の最も最近の分譲請求から少なくとも5年間、そしていずれにしても、寄託日から少なくとも30年間、または培養物の開示が発行されるあらゆる特許の有効期間にわたって、細心の注意を払って保存され、汚染を免れる。寄託者は、寄託物の状態のために、寄託所が請求時に試料を分譲できない場合には、寄託物を交換する義務を理解している。本培養寄託物の公衆への利用可能性を制限するものは全て、これを開示する特許が付与された際に、変更不可能に取り除かれるであろう。
【0023】
本発明の方法によれば、本明細書に記載のFIVワクチン組成物は、易感染性宿主、典型的には飼いネコに有効な量および方法で投与され、その後のチャレンジまたはFIVによる宿主の感染に対する防御的免疫を誘導する。ワクチンは、例えば皮下、腹腔内、または筋肉内注射によって非経口的に投与されるのが一般的である。その他の適当な投与法は経口または鼻腔内投与である。通常、ワクチンは宿主に少なくとも2回、各投与間に1週間以上の間隔をあけて投与する。しかし、ワクチンの初回および追加投与に関するその他のレジメは企図されており、実施者の判断および処置する特定の宿主動物に依る。
【0024】
本発明のワクチン組成物は、当業者に周知の方法によって調製することができる。例えば、ワクチンは注射可能の形態、例えば溶液または懸濁液として調製することが一般的である。ワクチンは投与剤形と融和するように、受容者に治療的有効量かつ免疫誘発量で投与する。特定のワクチン製剤の至適投与量および投与パターンは、当業者によって容易に決定することができる。
【0025】
ワクチン製剤におけるウイルスおよび細胞は、当技術分野で既知の方法を用いて不活化または弱毒化してもよい。例えば、ウイルス全体および感染細胞をパラホルムアルデヒド、ホルマリン、フェノール、UV光線、温度上昇等に暴露することによって不活化または弱毒化することができる。ワクチン用量における無細胞FIV全体の量は通常、約0.1 mg〜約5 mgの範囲内で、より通常は約0.2 mg〜約2 mgの範囲内である。FIV感染細胞株からなるワクチン製剤の投与量は通常、約106〜約108細胞/用量であり、より一般的には5×106〜約7.5×107細胞/用量である。ウイルスまたは細胞は、投与直前にアジュバントと混合するのが一般的である。ワクチン製剤において用いられるアジュバントは、トレオニルムラミルジペプチド(MDP)(バイアーズら(Byars)、1987)またはフロイトの完全および不完全アジュバントの混合液のいずれかであることが一般的である。ミョウバンなどの、本発明の方法およびワクチンでの使用に適当なその他の多様なアジュバントは、当技術分野で周知であり、本発明での使用が企図される。
【0026】
本発明はさらに、本発明の非感染細胞株を用いて、試料中のウイルス中和(VN)抗体を解析する新規の方法にも関する。一定数の継代後に使用できなくなり、FeT-1CまたはFeT-J細胞ほど容易に増殖しないPBMCとは異なり、FeT-1CおよびFeT-J細胞は、確立された細胞株で、今後の使用のために容易に凍結保存することができる。異なるSPFネコから得られたPBMCは細胞増殖速度およびFIV易感染性が個々に異なるため、FeT-1C細胞を用いたVNアッセイから得られた結果は、PBMCを用いたVNアッセイより再現性が高い。さらに、VNアッセイ用のPBMCは、PBMCを用いたインビトロVNアッセイに影響を及ぼす可能性がある、起こりうるインビボ感染を排除するために、無菌室収容および維持を必要とするSPFネコから得なければならない。このように、異なるサブタイプのFIVに容易に感染させることが可能なFeT-1Cのようなネコ科細胞株は、VNアッセイにおいてPBMCの都合の良い代用とすることができる。
【0027】
本明細書では、FIV株について以下の略語を用いる:
株(サブタイプ) 略語
Petaluma(A) FIVPet
Dixon(A) FIVDix
UK8(A) FIVUK8
Bangston(B) FIVBang
Aomori-1(B) FIVAom1
Aomori-2(B) FIVAom2
Shizuoka(D) FIVShi
【0028】
材料および方法
細胞培養。全ての懸濁細胞株は10%熱不活化仔ウシ胎児血清(FCS)を含むPRMI 1640培地で培養した。10 mM HEPES(N-2-ヒドロキシエチルピペラジン-n'-2-エタンスルホン酸)、2 mM L-グルタミン、50 μg/mlゲンタマイシンおよび5×10−5 M 2-メルカプトエタノール。IL-2依存的細胞にはヒト組換えIL-2(Cetus Corporation、エメリービル、カリフォルニア)100 U/mlを補足した。懸濁細胞は、0.5〜4×106細胞/mlの細胞濃度で継代し、週2回新鮮な培地で再度培養した。単層細胞は全て、細胞初濃度2×106細胞/mlで週2回継代した。FIV感染細胞からの組織培養液(TCF)を週2回回収し、3000 rpmで1時間遠心して、残留細胞を除去し、試験時に直ちに使用する予定のTCFに関しては、-20℃または-70℃で保存した。FIV-易感染性細胞(1×106細胞/ml)は約30,000 cpm/mlの逆転写酵素(RT)活性を有するFIVに感染させた。
【0029】
FIVの精製。FIV感染細胞株の組織培養液は、個々に2000〜3000 rpmで1時間遠心して細胞を除去した。TCF中のウイルスは16,000 rpmで2時間超遠心してペレットにし、まず10/50%(w/v)不連続蔗糖勾配で、次に10/50%連続蔗糖勾配で超遠心して精製した(ペダーセンら(Pedersen)、1987;ヤマモトら(Yamamoto)、1988)。ウイルス単離株のそれぞれを18時間1.25%滅菌パラホルムアルデヒド(0.22 μm濾過滅菌)で不活化し、その後滅菌PBSで十分に透析した。不活化ウイルスは、滅菌PBSで500 μg/mlの濃度に希釈し、各株の250 μg/0.5 mlを滅菌マイクロフュージチューブに移して-70℃で保存した。不活化FIV株は室温で解凍し、不活化ウイルス250 μgの0.5 ml滅菌PBS溶液を免疫直前にアジュバント0.5 mlと混合した。FIV-感染細胞株は、個別に18時間1.25%滅菌パラホルムアルデヒドで不活化し、滅菌PBSで3回洗浄し、滅菌チューブ中で約5.0×107細胞/mlの濃度に再懸濁して4℃で保存した。典型的には、不活化感染細胞約2.5×107細胞/mlの0.5 ml PBS溶液を免疫直前にアジュバント0.5 mlと混合した。トレオニルムラミルジペプチド(MDP MF75.2アジュバント;Chiron Corporation、エメリービル、CA)250 μg/0.5 mlをアジュバントとして用いた。
【0030】
CTLアッセイ。末梢血単核細胞(PBMC)を、10日間のFIV感染前にコンカナバリンA(Con A)で3日間刺激した(ソングら(Song, 1992))。これらの細胞をCTLアッセイの標的細胞として用いた。CTL活性は、Con A刺激PBMCをUV-および放射線不活化FIV-感染自家PBMCと共に5日間培養することによって生じた。これらの細胞を刺激エフェクター細胞として用いた。解析当日に標的細胞をNa51CrO4 50 μCiで1〜3時間標識して、3回洗浄し、次に固定数の標識標的細胞(5×104細胞/ウェル)をマイクロタイタープレートに加えた。エフェクター細胞を様々なエフェクター/標的細胞比(例えば、100:1、50:1、および10:1)で3連ずつ加えた。プレートを400 rpmで1分遠心し、37℃で4時間インキュベートした。対照51Cr-標識標的細胞を洗浄剤で溶解し、最大放出値を得た。被験試料ウェルから上清を収集し、ガンマカウンターを用いて放射能を定量した。エフェクター細胞の非存在下で51Cr-標識標的細胞をインキュベートすることによって、自然発生放出を定量した。特異的細胞障害百分率は以下のように計算した:
【数1】

【0031】
イムノブロットおよび酵素結合イムノソルベントアッセイ(ELISA) ヤマモトら(Yamamoto, 1993)の記述のように、蔗糖勾配精製ウイルスをイムノブロットアッセイの基質として用いた。感染細胞の組織培養液からのFIVPetは低速遠心(2000 rpmで45分)で不純物を除去し、超遠心(16,000 rpmで2時間)によって濃縮し、10/50%(w/v)連続蔗糖勾配上での超遠心によって精製した。この技法で精製したウイルスをイムノブロットアッセイの基質として用いた。
【0032】
これまでに記述されたイムノブロット技法(ヤマモトら(Yamamoto)、1991a)の改変を用いた。ウイルスブロット片は、ウイルスを0.1%SDSで可溶化し、その後10%SDSポリアクリルアミドゲル上で電気泳動し、ニトロセルロース膜上に電気泳動的に移動させることによって調製した。ワクチン接種ネコからの血清試料を緩衝液3(0.15 M塩化ナトリウム、0.001 Mエジチ酸、0.05 Mトリス塩基、0.05%ツイーン20、および0.1%ウシ血清アルブミン)で1:50に希釈し、イムノブロットプレートの個々のウェル中でウイルスブロット片と共に37℃で18時間インキュベートした。ブロット片は洗浄液(0.15 M NaClおよび0.05%ツイーン20脱イオン水溶液)で個々に洗浄し、ビオチン化抗ネコIgG(Vector Laboratories、バーリンゲイム、CA)と共に37℃で1時間インキュベートし、洗浄液で3回洗浄した。次にブロット片を個々に、ホースラディッシュペルオキシダーゼ結合ストレプトアビジン(ベクター・ラボラトリーズ)と共に30分インキュベートした。十分な洗浄後、各片を新鮮な基質溶液(0.05%ジアミノベンジジン、400 μg/ml NiCl2、および0.01%H2O2の0.1 Mトリス緩衝溶液、pH 7.4)と共に室温でインキュベートした。目に見えるバンドが確立した後、過剰量の蒸留水を加えて反応を停止させ、ブロット片を乾燥させた。次に、イムノブロット上のバンドの分子量は、先にアミドブラックで染色した片上の分子量標準物質の移動距離とそれらの移動距離を比較して決定した。各イムノブロット分析には診断的評価の内部対照として陽性および陰性対照血清を含めた。
【0033】
ウイルス抗原特異的ELISAは既に記述されている(ヤマモトら(Yamamoto)、1991a;ヤマモトら(Yamamoto)、1993)。蔗糖勾配精製FIVPetおよび表面エンベロープ(SU)ならびにFIVPetの保存(C)および可変(V)領域双方の膜通過(TM)ペプチドは、96ウェルイムノロンプレート(Dynatic Laboratories, Inc.、シャンティリー、VA)上に重炭酸緩衝液(pH 9.6)と共に250 ng/ウェルで12〜18時間、37℃でコーティングし、ELISAの基質として用いた。SU-V3-2ペプチドのアミノ酸配列は:Gly Ser Trp Phe Arg Ala Ile Ser Ser Trp Lys Gln Arg Asn Arg Trp Glu Trp Arg Pro Asp Phe(配列番号:1);およびTM-C1ペプチドのアミノ酸配列は:Gln Glu Leu Gly Cys Asn Gln Asn Gln Phe Phe Cys Lys Ile(配列番号:2)である。合成ペプチドは、FMOCペプチド合成化学(マガジンら(Magazine)、1988)を用いて、バイオサーチ9500ペプチドシンセサイザー(Biosearch、サンラファエル、CA)で合成した。合成ペプチドの純度は、逆相高速液体クロマトグラフィー上での単一のピークの存在によって決定し、ピーク試料について実施したアミノ酸配列分析によって確認した。
【0034】
ペプチドコーティングプレートは、使用直前に緩衝液3で1回洗浄した。血清試料を緩衝液3で1:200に希釈し、FIV抗原コーティングウェル中で37℃で1時間インキュベートし、6回洗浄した。ウェルを洗浄液で洗浄し、ビオチン化抗ネコIgG(Vector Laboratories、バーリンゲイム、CA)と共に37℃で1時間インキュベートし、6回洗浄し、ホースラディッシュペルオキシダーゼ結合ストレプトアビジン(ベクター・ラボラトリーズ)と共に37℃で1時間インキュベートした。次にウェルを洗浄液で6回洗浄して、ELISA基質溶液(0.005%テトラメチルベンジジンおよび0.015%H2O2の0.96%クエン酸溶液)と共に室温でインキュベートした。既知のFIV陽性ネコ血清からなる連続希釈標準物質に可視反応色が確立すれば、0.1 Mフッ化水素酸で反応を停止させた。バイオラッドELISAリーダー(バイオラッド・ラボラトリーズ、ハーキュルズ、CA)で光学濃度414 nmでの吸光度を測定した。
【0035】
ポリメラーゼ連鎖反応(PCR)。感染細胞のプロウイルスDNA濃度は、同じまたは異なるサブタイプからの多種FIV株を識別するために最近開発された分別PCRによってモニターした(オカダら(Okada)、1994)。PCRの感度を増加させる手段として、表1に示すネステッドPCRプライマーセットを用いた。PCRは2段階反応で行い、最初はオカダら(Okada, 1994)が記述した条件下で1対の外側のプライマー(全てのFIV株に共通)で行った。PCRの第二段階では、内側のプライマー(各FIV株に特異的)を用いて第一段階産物の1/25を増幅した。ネステッドPCRを用いて、FIVPet、FIVUK8、FIVBang、FIVAom1、FIVAom2、およびFIVShiに感染した細胞を互いに区別することができる。
【表1】
【0036】
プロウイルスDNAの細胞当たりのおおよその量は、既知の細胞数から抽出したDNAの希釈を変化させて半定量的PCRによって定量した。例えば、105個の細胞をDNA抽出に用いた場合、DNA調製物を10−5希釈すれば、細胞1個当たりに存在するDNAにほぼ相当することになる。これらの変化させたDNA希釈液についてPCRを実施し、陽性PCR結果を示す最終希釈をエンドポイント希釈と見なす。エンドポイント希釈に相当する細胞数を用いて、以下の式に従って所定の細胞調製物中のウイルス感染細胞の百分率を決定した:
【数2】
式中、Zはエンドポイント希釈に相当する細胞数である。
【0037】
逆転写酵素(RT)アッセイ。RNA-依存的DNAポリメラーゼ(RT)の有無は、本質的にレイら(Rey)の記載に従い細胞培養上清において解析した。FIVを検出するRTアッセイは、外因性鋳型プライマーとしてポリ(rA)-オリゴ(dT12−18)、異なる4つのデオキシリボヌクレオチド3リン酸、2価陽イオンとしてMg++と共に20 mM KCl、および1試料当たり5μCi[3H]-標識チミジン3リン酸を用いた。5μCi[3H]TTPは、ベックマンLS250シンチレーションカウンター(Beckman Instruments, Inc.、パロアルト、CA)でシンチレーション液混合物(キシレン1対研究産物国際生物分解計数シンチラント9の割合)を用いて、平均総カウント1,200,000 cpmを生じた。その結果、調べる試料のRT値は1,200,000 cpm/ml未満となると考えられる。
【0038】
ウイルス中和アッセイ。株およびサブタイプ特異的VNアッセイを開発する計略は記述されている(オカダら(Okada)、1994)。ネコ末梢血単核細胞(PBMC)(4×105細胞/ml)またはFIV易感染性FeT-1C細胞(2×105細胞/ml)を加える前に、熱不活化血清の連続希釈物を24ウェルプレート中で37℃で45分、各FIV株の100 TCID50と共にインキュベートした。培養3日後に、細胞をHank's緩衝塩溶液で1回洗浄し、培養から残留ウイルスを除去し、次に細胞を新鮮な培養培地(10%熱不活化仔ウシ胎児血清、10 mM HEPES緩衝液、50μg/mlゲンタマイシン、5×10−5 M 2-メルカプトエタノール、および100 単位/mlヒト組換えIL-2を含むRPMI-1640)に再懸濁した。細胞のウイルス感染は、培養9、12、15、および18日目に回収した培養液のMg++依存的RTアッセイによってモニターした。RT活性が、SPF血清からなる感染対照培養の25%以下である場合、血清はVN抗体陽性と見なした。
【0039】
以下は、本発明の実施に関する最良の形態を含む手順を図示する実施例である。これらの実施例は、制限的なものと解釈してはならない。百分率は全て重量で表し、溶媒混合比は、他に明記していなければ全て容積である。
【実施例1】
【0040】
実施例1−FIV感染細胞株。
IL-2依存的FeT-1Mクローンの母株であるFeT-1Cと呼ばれる新規インターロイキン-2(IL-2)依存的ネコ科T細胞株を用いて、FIVPet、FIVDix、FIVUK8、FIVBang、FIVAom2、またはFIVShiのいずれかに慢性的に感染させた個々の細胞株を確立した。FeT-1Mクローン(FIV-Fet1Mとも呼ばれる)は、本明細書に参照として組み入れられる米国特許第5,275,813号に記述されており、これを用いて、FIVPetを慢性的に産生するIL-2非依存的細胞株FL-4(これも米国特許第5,275,813号に記述されている)を産生した。FeT-1C細胞株は、FIVサブタイプA、B、およびDの異なる単離体に高度に感染可能である。FeT-1C細胞株を長期間継代すると、その感染性、特にFIVサブタイプDに対する感染性は低下する;したがって、至適FIV感染率を得るため、またはVNアッセイで使用するためには、継代回数は約35回未満とすること。半定量的PCRおよびウイルスコア抗原分析により、FIVに暴露された細胞株は全て、個々のFIV株に有意に感染することが示された。
【0041】
FIV易感染性のIL-2非依存的ネコ科細胞株も同様に、FeT-1C細胞から作製した。FeT-Jと呼ばれるこの細胞株は、FIV感染培地または細胞を用いて共培養によってFIVに感染させることができる。例えば、FIVBang感染FeT-1C細胞株を、IL-2の非存在下で非感染FeT-J細胞と共培養して、IL-2非依存的FIVBang感染FeT-J細胞株(Bang/FeT-Jと呼ぶ)を確立した。共培養による感染法では、Bang/FeT-1C細胞を非感染FeT-J細胞と約2:1〜約10:1(感染:非感染)の比で混合した。細胞混合物はIL-2の非存在下で培地中で数日間培養し、FeT-1C細胞を死亡させた。残った細胞は、FIVBang感染FeT-J細胞から成った。このように、FIV感染FeT-1C細胞を用いてFeT-J細胞を感染させ、異なるFIVサブタイプに感染したIL-2非依存的FeT-J細胞株を確立することができる。FIV感染FeT-1C細胞との共培養法の結果、異なるFIVサブタイプを中等度から高レベル産生するIL-2非依存的FeT-J細胞株が得られた。
【0042】
FeT-1C細胞株をまたFIVShiに感染させ、十分に継代すると、Shi/FeT-1Cと呼ばれるIL-2依存的細胞株が得られた。Shi/FeT-1C細胞株を後に、IL-2の非存在下でFeT-Jと共培養し、得られたIL-2非依存的FIVShi-感染細胞株をShi/FeT-Jと呼んだ。IL-2非依存的Shi/FeT-J細胞株は、IL-2依存的Shi/FeT-1C細胞株より高レベルのFIVShiを産生する(図1)。
【0043】
FIVBangに感染させたFeT-J細胞株の作製はまた、FeT-1C細胞株を用いずに実施した。FeT-J細胞株を無細胞FIVBang接種体に直接感染させ、IL-2を用いずに十分に継代した。得られたIL-2非依存的FIVBang産生細胞株をBang/FeT-Jと命名した。Bang/FeT-J細胞株は、FeT-1C細胞株をFIVBangに感染させて作製したIL-2依存的Bang/FeT-1C細胞株より高レベルのFIVBangを産生した。
【実施例2】
【0044】
実施例2−多種サブタイプFIVワクチン。
FIV感染細胞は遠心によって上清から除去し、不活化し、そしてワクチンとして用いた。同様に、FIVウイルス全体は、超遠心によって感染した無細胞上清から沈殿させ、不活化した。感染細胞およびウイルスはいずれも、5℃で1.25%パラホルムアルデヒドで24時間処置して不活化し、その後それぞれ、十分に洗浄、またはPBSで透析した。この方法は、免疫原性を失うことなくFIVを効率よく不活化する。本方法によって産生されたFIV免疫原は、防御免疫の誘導に非常に有効である(ヤマモトら(Yamamoto)、1993;ヤマモトら(Yamamoto)、1991a;ヤマモトら(Yamamoto)、1991b)。弱毒化したウイルス単離体もまた、本発明のワクチン組成物に用いることができると企図される。
【0045】
FIVShi感染FeT-1C細胞株をFIVPet株に重複感染させると、多種サブタイプFIV(すなわち多種サブタイプA/D FeT-1C細胞株)に感染した単一の細胞株が得られるが、共感染の2ヶ月以内に、FIVShiプロウイルス濃度は50%から5%未満に減少し、一方FIVPetプロウイルス濃度は同時に約50%に増加した。このように、FIVワクチンとして用いるための、多種サブタイプFIVに感染させた単一細胞株の維持は、本発明の好ましい態様ではない。
【0046】
その結果として、本発明の一つの態様において、ワクチン組成物は、各株が異なるFIVサブタイプに感染した2つの個々の細胞株から作製した。特定の態様において、2種サブタイプのFIVワクチン組成物は、FIVサブタイプA感染細胞株(Pet/FL-4)とFIVサブタイプD感染細胞株(Shi/FeT-1C)との組み合わせから成った。AサブタイプおよびDサブタイプ感染細胞株は記述のように不活化し、等しい細胞数(MDP 250 μg中に各2.5×107細胞)を混合し、これを用いてネコを免疫した。SPFネコ3匹を不活化Pet/FL-4細胞でワクチン接種し、別のネコ4匹を不活化Shi/FeT-1C細胞でワクチン接種した(2.5×107細胞/接種量)。一連の4回のワクチン接種後、2種サブタイプ(Pet/FL-4およびShi/FeT-1C)ワクチンは、調べた両FIV株に対して有意なVN抗体価を含む抗FIV抗体を誘導した(図2および表2、試験I)。2種サブタイプ(Pet/FL-4およびShi/FeT-1C)ワクチン接種ネコ4匹に、FIVBang(50 CID50)をチャレンジした。Pet/FL-4ワクチン接種ネコ3匹全てとShi/FeT-1Cワクチン接種ネコ2匹にFIVBangの50 CID50量をチャレンジした。残る2匹のShi/FeT-1Cワクチン接種ネコには、FIVShiの50 CID50量をチャレンジした。
【0047】
2種サブタイプワクチン接種ネコは全て、感染後(pi)6週目でのウイルス単離およびPBMCのPCRにより、FIVBang陰性であったが、偽免疫ネコは全て、感染後6週目でのウイルス単離およびPCRにより、FIVBangまたはFIVShiのいずれかに対して陽性であった(表2、試験I)。対照的に、Pet/FL-4ワクチン接種およびShi/FeT-1Cワクチン接種群のネコそれぞれ1匹ずつにFIVBangをチャレンジすると、FIVBang陽性であった。予想されたように、FIVShiワクチンを接種し、その後FIVShiをチャレンジしたネコは全て、感染後6週目でFIVShiに対して陰性であった。このように、特に例示した2種サブタイプワクチンは、異種FIVBangチャレンジに対してと同様、同種FIVShiに対する感染も防御または遅らせた。
【0048】
2種サブタイプワクチンを接種したネコ(Pet/FL-4細胞およびShi/FeT-1C細胞)は、2回目の免疫後、ウイルスコア蛋白質p25(FIV p28とも呼ばれる)に対して特異的なFIV抗体を産生した(図2)。他のウイルス抗原に対するより高い抗体価は、3〜4回の免疫後に示された。FIVPetに対するVN抗体は、2回の免疫後に産生されたが、FIVShiに対するVN抗体は4回の免疫後に産生された(表4)。FIVPetおよびFIVShiに対するCTL反応は、調べた全てのネコにおいて3回の免疫で検出され(表3)、両株に対するより強いCTL反応は4回の免疫後に得られた。さらに、調べたネコ3匹のうち2匹が、4回の免疫後FIVBangに対するCTL反応を示した。結果は、4回のワクチン接種後、2種サブタイプワクチンは、FIVPetおよびFIVShiに対して強いCTL反応を誘導し(表3)、両FIV株に対してVN抗体価を含む高いFIV抗体を誘導した。
【0049】
不活化Shi/FeT-1C細胞で免疫したネコは、2回の免疫後ウイルスコア蛋白質p25に対して特異的なFIV抗体を産生し、3回の免疫後その他のウイルス抗原に対する抗体を産生した(図2)。これらのネコでは、FIVShiに対するVN抗体は4回の免疫後に産生されたが、FIVPetに対するVN抗体は免疫の過程において検出されなかった。Shi/FeT-1Cワクチン接種ネコはいずれも、4回の免疫後に限ってFIVShiに対するCTL反応を起こしたが、FIVPetに対するCTL反応は4回の免疫後でも起こさなかった(表3)。
【0050】
不活化Pet/FL-4細胞で免疫したネコは、2回の免疫後にp25に対する抗体を産生し(図2)、2〜3回の免疫後FIVPetに対するVN抗体を含む他のウイルス抗原に対する抗体を産生した(表4)。Pet/FL-4細胞で免疫したネコに検出される唯一のCTL反応は、FIVPetに対するものであった。全体的に、2種サブタイプワクチンは、1種サブタイプワクチンより、速やかにかつ高いVN抗体価、および両FIV株に対するCTL反応を誘導した。偽免疫SPFネコは、ウイルス抗体、VN抗体、または抗FIV CTL反応を示さなかった。
【表2】
【表3】
【表4】
【0051】
好ましい態様において、本発明のワクチン組成物は、各細胞株が異なるFIVサブタイプ(AまたはBもしくはD)からのウイルス株に感染している3つの細胞株から調製された3種サブタイプFIVワクチンからなる。特定病原体不在ネコ3匹を3種サブタイプ(FIVPet+FIVBang+FIVShi)ワクチンで免疫した。その他のネコは1種サブタイプFIVBangワクチンで免疫して、ワクチン成分としてマクロファージ親和性FIVBangの免疫原性を評価した。VN抗体価の結果は、3種サブタイプ(FIVPet+FIVBang+FIVShi)と1種類サブタイプFIVBangワクチンはいずれも、2回の免疫後においても高い抗ウイルス抗体価を誘発した(表2、試験IIおよび表5)。このように、リンパ球親和性およびマクロファージ親和性FIVは、本発明のワクチン組成物の成分として用いることができる。
【0052】
不活化Pet/FL-4、不活化Bang/FeT-J、および不活化Shi/FeT-1C細胞の組み合わせ(全体でMDP 250 μg中に各2.5×107細胞)で免疫したSPFネコ3匹は、ウイルスコア蛋白質p25およびFIV SUおよびTMエンベロープ蛋白質を含むその他のウイルス抗原に特異的なFIV抗体を2回の免疫後に産生した(図3、4、5)。FIVPet、FIVBang、およびFIVShiに対するVN抗体は、2回の免疫後まもなく大多数のネコに産生され、3回目の免疫により全てのネコに産生された(表5)。さらに、ネコ1匹が3回の免疫後にFIVUK8に交叉反応するVN抗体を産生した。不活化Bang/FeT-J細胞のみで免疫したSPFネコ4匹は、ウイルスコア蛋白質p25およびその他のウイルス抗原に特異的な抗体を産生した(図3)。これらのネコではFIVBangに対するVN抗体は2回の免疫後に産生された(表5)が、FIVPetおよびFIVUK8に対するVN抗体は免疫過程において検出されなかった。3種サブタイプFIVワクチン(Pet/FL-4、Bang/FeT-JおよびShi/FeT-1C細胞)で3回免疫したネコのFIV A、B、およびDサブタイプ標的細胞に対するCTL反応を表6に示す。調べたこれら3つのFIVサブタイプ全てに対してCTL反応が検出された。このように、3種サブタイプワクチンは、1種サブタイプワクチンより幅広いCTL反応ならびに、より速やかかつより高いVN抗体価およびSU-エンベロープ抗体価を誘導した。非感染FeT-Jまたは偽免疫SPFネコも、ウイルス抗体またはVN抗体を産生しなかった。
【表5】
【表6】
【実施例3】
【0053】
実施例3−FIVサブタイプに対するVN抗体。
FIVに対するVN抗体のアッセイはまた、本発明のFeT-1C細胞を用いて開発された。FIVPet感染ネコおよび不活化Pet/FL-4細胞または不活化FIVPetウイルスでワクチン接種したSPFネコの血清を、本明細書に記述のVNアッセイ法に従って、FeT-1C細胞またはPBMCのいずれかを用いてVN抗体価を調べた。ワクチンを接種せず、FIVに感染していないSPFネコ2匹の血清を対照血清として用いた。ワクチン接種およびFIV感染ネコの血清は1000以上の高いVN抗体価を示したが、非ワクチン接種SPFネコの血清は検出可能なVN抗体価を示さなかった。FeT-1Cに基づくVNアッセイは、ネコの初代培養PBMCを用いて得られた結果と同等のVN抗体価結果を示す(表6)。この知見は、FeT-1C細胞を用いたVNアッセイでのVN抗体価が、PBMCを用いたVNアッセイに関して得られた結果と相関することを証明している。したがって、FeT-1C細胞は、全てのFIVサブタイプに感染させることが可能で、組織培養において容易に増殖させることが可能なため、FIVに関する標準的なVNアッセイにおいてPBMCの代わりに都合良く用いることができる。
【表7】
【実施例4】
【0054】
実施例4−FIV株の免疫タイピング
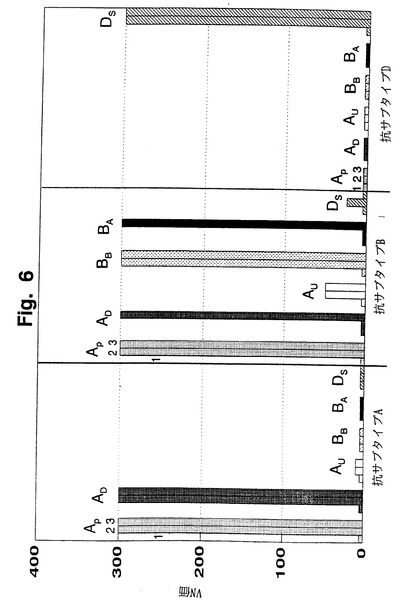
FIVサブタイプがFIV免疫型を反映するか否かを評価するために、FeT-1C細胞を用いてインビトロ試験を実施した。免疫タイピングはワクチン防御におけるVN抗体の役割を理解する上で重要である。FIVサブタイプA株(FIVPet、FIVDix、FIVUK8)、サブタイプB(FIVBang、FIVAom1)、およびサブタイプD(FIVShi)に感染させたネコの抗血清を、VNアッセイにおいてFeT-1C細胞を用いてインビトロでこれらの株を中和する能力に関して調べた(図6)。試験抗血清は全て、対応する同種FIV株に対する中和活性を示した。サブタイプA株であるFIVPetは、FIVDixに感染させたネコの抗血清によって有意に交叉中和された。FIVPetは、表面エンベロープ糖蛋白質(Env)領域がFIVDixと約9%異なる。FIVサブタイプAに感染させたネコの抗血清はサブタイプBのFIVBangと交叉中和したが、サブタイプD FIVShiを中和しなかった。サブタイプBおよびD株に感染させたネコの抗血清のみが、同種サブタイプ内の他のFIV株を交叉中和した。さらに、FIVUK8に感染させたネコの抗血清はFIVBangを中和したが、サブタイプA内のFIV株を中和しなかった。FIVUK8はサブタイプAに分類されているが(ソドラら(Sodora)、1994;リグビーら(Rigby)、1993;カキヌマら(Kakinuma)、1995)が、これらの結果は、FIVUK8に対する抗血清がサブタイプB株を認識するがサブタイプA株を認識しないことを示唆しており、不活化FIVPetワクチンがFIVUK8およびFIVShiに対して無効である理由を説明する可能性がある(ジョンソンら(Johnson)、1994)。このように、遺伝子型と免疫型との間には緩やかな相関が存在する。遺伝子型分析によりFIV株分類が可能となったが、交叉中和抗体試験はFIV株の免疫原性を反映し、これは、ワクチンによって誘発される広範囲の液性防御における重要なパラメーターである。
【実施例5】
【0055】
実施例5−FIV細胞親和性
感染FeT-1Cおよび感染FeT-J細胞株から得たFIV株の細胞指向性を、初代培養PBMCから得たFIV株の細胞指向性と比較した(表8)。FIV単離体2個、FIVUK8およびFIVBangはいずれも等しくリンパ球親和性およびマクロファージ親和性であるが、FIVShiは非常にリンパ球親和性である。FIVPetは、マクロファージ親和性よりリンパ球親和性で、その細胞親和性はその細胞源によって有意に影響を受けなかった。FIVBangのマクロファージ親和性は、ウイルスの細胞源によって影響を受けなかった。感染FeT-1C細胞株からのFIV株の細胞親和性は、初代培養PBMCから得られた細胞親和性と同等であるため、FeT-1C細胞内で増殖するウイルスは、VNアッセイの接種体として用いることができ、治療および予防的アプローチを評価するための試験では、インビボ接種体として用いることもできる。
【表8】
【0056】
本明細書に記載の実施例および態様は例示目的のためのみであり、その様々な改変または変更は当業者に示唆され、本出願および付属の請求の範囲の範囲内に含まれるものと理解すべきである。
【技術分野】
【0001】
本発明は米国国立衛生研究所助成金番号NIH AI30904による助成を受けた研究プロジェクトの下で政府の支援を受けて行われた。米国政府は本発明に一定の権利を保有する。
【背景技術】
【0002】
発明の背景
飼いネコは、ネコ白血病ウイルス(FeLV)、ネコ肉腫ウイルス(FeSV)、内因型C型オンコロナウイルス(RD-114)、およびネコ合胞体形成ウイルス(FeSFV)を含むいくつかのレトロウイルスによる感染症にかかりやすい。これらの中で、FeLVは最も重要な病原体で、リンパ網内系および骨髄新生物、貧血、免疫媒介疾患、ならびにヒト後天性免疫不全症候群(AIDS)と同様の免疫不全症候群を含む多様な症状を引き起こす。最近では、FeLV-AIDSと呼ばれる特定の複製欠損FeLV変異株が免疫抑制特性により一層関連している。
【0003】
ネコTリンパ球親和性レンチウイルス(現在では、ネコ免疫不全ウイルス、FIVと呼ばれている)の発見は、ペダーセンら(非特許文献1)によって最初に報告された。FIVの特徴は、ヤマモトら(非特許文献2);ヤマモトら(非特許文献3);およびアックリーら(非特許文献4)において報告されている。血清疫学的データにより、FIV感染が世界中の飼いネコおよび野生のネコに固有であることが示された。流産、脱毛、貧血、結膜炎、慢性鼻炎、腸炎、歯肉炎、血便排泄、神経異常、歯周炎、および脂漏性皮膚炎を含む広く多様な症状は、FIVによる感染症に関連する。FIVに感染した飼いネコの顕著な免疫学的特徴は、ネコのCD4+末梢血リンパ球の慢性的および進行性枯渇、CD4:CD8細胞比の低下、およびいくつかの症例ではCD8保有リンパ球の増加である。分子、生化学、および免疫病理学的特徴に基づき、ネコのFIV感染症は今や、FeLV-FAIDSより良いネコのAIDSモデルであると思われる。
【0004】
FIVのクローニングおよび配列分析は、オルムステッドら(非特許文献5);オルムステッドら(非特許文献6);およびタルボットら(非特許文献7)において報告されている。ホージー&ジャレット(非特許文献8)は、FIVに感染したネコの血清学的反応について記述している。FIVウイルスサブタイプは、それぞれの株によって誘発された交叉中和抗体の濃度に基づく免疫型に従って分類することができる(マーフィー&キングスバリー(非特許文献9))。近年、ヌクレオチド配列の相同性に基づく遺伝子型に従って、ウイルスがサブタイプに分類された。HIVおよびFIVサブタイプ分類は遺伝子型に基づくが(ソドラら(非特許文献10);リグビーら(非特許文献11);およびルワジーら(非特許文献12))、サブタイプの遺伝子型と免疫型の間の相関に関してはほとんどわかっていない。FIVウイルス単離体は現在、4つのFIVサブタイプに分類されている:A、B、CおよびD(カキヌマら(非特許文献13))。感染性単離体および感染性分子クローンは、サブタイプCを除く全てのFIVサブタイプについて記述されている(ソドラら(非特許文献10))。サブタイプCのFIVは、カナダのネコの細胞DNAから唯一同定されている(ソドラら(非特許文献10);リグビーら(非特許文献11);カキヌマら(非特許文献13))。
【0005】
FIVワクチンを開発する上で主に難しい点は、異なるサブタイプまたはクレードからのフィールド単離体を含む広範囲のFIV株に対して有効なワクチンアプローチを特定することであった。FIVに対するワクチン予防は、単一株のワクチンを用いた場合、同種およびわずかに異種の株に対しては予防できたが、中程度から大きく異なる種の株によるチャレンジでは予防できない(ジョンソンら(非特許文献14);ヤマモトら(非特許文献15))。したがって、依然として多数のFIVサブタイプから防御することのできるワクチンが必要とされている。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Pedersen, N.C., E.W. Ho, M.L. Brown, J.K. Yamaoto (1987) “Isolation of a T-lymphotropic virus from domestic cats with an immunodeficiency-like syndrome,” Science 235:790-793
【非特許文献2】Yamamoto, J.K., N.C. Pedersen, E.W. Ho, T. Okuda, G.H. Theilen (1988a) “Feline immunodeficiency syndrome - a comparison between feline T- lymphotropic lentivirus and feline leukemia virus,” Leukemia, December Supplement 2:204S-215S.
【非特許文献3】Yamamoto, J.K., E. Sparger, E.W. Ho, P.H. Andersen, T.P. O'Connor, C.P. Mandell, L. Lowenstine, N.C. Pedersen (1988) “Pathogenesis of experimentally induced feline immunodeficiency virus infection in cats,” Am. J. Vet. Res. 49:1246-1258.
【非特許文献4】Ackley, C.D., J.K. Yamamoto, N.B. Levy, N.C. Pedersen, M.D. Cooper (1990) “Immunologic abnormalities in pathogen-free cats experimentally infected with feline immunodeficiency virus,” J. Virol. 64:5652-5655.
【非特許文献5】Olmsted, R.A., A.K. Barnes, J.K. Yamamoto, V.M. Hirsch, R.H. Purcell, P.R. Johnson (1989) “Molecular cloning of feline immunodeficiency virus,” Proc. Nat. Acad. Sci. 86:2448-2452
【非特許文献6】Olmsted, R.A., V.M. Hirsch, R.H. Purcell, P.R. Johnson (1989) “Nucleotide sequence analysis of feline immunodeficiency virus: Genome organization and relationship to other lentivirus,” Proc. Natl. Acad. Sci. USA 86:8088-8092.
【非特許文献7】Talbott, R.L., E.E. Sparger, K.M. Lovelace, W.M. Fitch, N.C. Pedersen, P.A. Luciw, J.H. Elder (1989) “Nucleotide sequence and genomic organization of feline immunodeficiency virus,” Proc. Natl. Acad. Sci. USA 86:5743-5747.
【非特許文献8】Hosie, M.J., O. Jarrett (1990) “Serological responses of cats to feline immunodeficiency virus,” AIDS 4:215-220.
【非特許文献9】Murphy, F., D.W. Kingsbury (1990) “Virus Taxonomy,” In Fields Virology, 2nd Ed., B.N. Fields, D.M. Knipe et al., eds, Raven Press, New York, Chapter 2, pp. 9-36.
【非特許文献10】Sodora, D.L., E.G. Shpaer, B.E. Kitchell, S.W. Dow, E.A. Hoover, J.I. Mullins (1994) “Identification of three feline immunodeficiency virus (FIV) env gene subtype and comparison of the FIV and human immunodeficiency virus type 1 evolutionary patterns,” J Virol. 68:2230-2238.
【非特許文献11】Rigby, M.A, E.C. Holmes, M. Pistello, A. Mackay, A.J. Leigh-Brown, J.C. Neil (1993) “Evolution of structural proteins of feline immunodeficiency virus: molecular epidemiology and evidence of selection for change,” J. Gen. Virol. 74:425-436.
【非特許文献12】Louwagie, J., F.E. McCutchan, M. Peeters, T.P. Brennan, E. Sanders-Buell, G.A. Eddy, G. van den Grosen, K. Fransen, G.M. Gershy-Damet, R. Deleys, D.S. Burke (1993) “Phylogenetic analysis of gag genes from 70 international HIV-1 isolates provides evidence for multiple genotypes,” AIDS 7:769-780.
【非特許文献13】Kakinuma, S., K. Motokawa, T. Hohdatsu, J.K. Yamamoto, H. Koyama, H. Hashimoto (1995) “Nucleotide Sequence of Feline Immunodeficiency Virus: Classification of Japanese Isolates into Two Subtypes Which Are Distinct from Non-Japanese Subtypes,” Journal of Virology 69(6):3639-3646.
【非特許文献14】Johnson, C.M., B.A. Torres, H. Koyama, J.K. Yamamoto (1994) “FIV as a model for AIDS vaccination,” AIDS Res. Hum. Retroviruses 10:225-228.
【非特許文献15】Yamamoto, J.K., T. Hohdatsu, R.A. Olmsted, R. Pu, H. Louie, H. Zochlinski, V. Acevedo, H.M. Johnson, G.A. Soulds, M.B. Gardner (1993) “Experimental vaccine protection against homologous and heterologous strains of feline immunodeficiency virus,” J. Virol. 67:601-605.
【発明の概要】
【0007】
発明の簡単な概要
本発明は、宿主動物におけるFIV感染症に対して広範囲の防御的免疫を誘発するワクチンに関する。特に、本発明は異なるFIVサブタイプから単離された無細胞ウイルス単離株、またはそれぞれが異なるサブタイプの異なるプロトタイプFIVウイルスに感染した細胞株の組み合わせ、を用いて調製される多種サブタイプFIVワクチンに関する。本発明のFIVワクチンを接種したネコは、同種および異種FIV株に対して液性および細胞性の免疫反応を引き起こす。
【0008】
本発明はまた、多種のFIVサブタイプに易感染性である新規のネコ科細胞株にも関する。本発明の細胞株は、本発明の方法によるFIVワクチンにおける使用と共に、多種のFIVサブタイプの増殖および産生に有用である。さらに、本細胞株はまた、ネコ抗血清のFIVウイルス中和アッセイにおいてネコ末梢血単核球(PBMC)の代わりに用いることができる。
【0009】
配列の簡単な説明
配列番号:1は、SV-V3-2と呼ばれるFIV表面エンベロープペプチドのアミノ酸配列である。
配列番号:2は、TM-C1と呼ばれるFIV膜通過ペプチドのアミノ酸配列である。
配列番号:3は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:4は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:5は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:6は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:7は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:8は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:9は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:10は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:11は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:12は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:13は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:14は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:15は、FIV PCRプライマーのヌクレオチド配列である。
配列番号:16は、FIV PCRプライマーのヌクレオチド配列である。
【図面の簡単な説明】
【0010】
【図1】図1は、これらのFIV株でFeT-1CおよびFeT-J細胞株を感染させた後に産生されたFIVBangおよびFIVShiの逆転写酵素(RT)濃度を示す。
【図2】図2は、イムノブロットによって検出された、2種サブタイプワクチン接種ネコの抗FIV抗体とFIV蛋白質との免疫反応を示す。各ブロット上の数値は、血清を検査した時点での動物に行ったワクチン接種回数を表す。
【図3】図3は、イムノブロットによって検出された、3種サブタイプワクチン接種ネコの抗FIV抗体とFIV蛋白質との免疫反応を示す。各ブロット上の数値は、血清を検査した時点での動物に行ったワクチン接種回数を表す。
【図4】図4は、ELISAによって検出した、3種サブタイプワクチン接種ネコから得た抗FIV抗体とFIV SU-V3-2ペプチドとの免疫反応性を示す。
【図5】図5は、ELISAによって検出した、3種サブタイプワクチン接種ネコから得た抗FIV抗体とFIV TM-C1ペプチドとの免疫反応性を示す。
【図6】図6は、FIVPet(AP)、FIVDix(AD)、FIVUK8(AU)、FIVBang(BB)、FIVAom1(BA)、およびFIVShi(DS)のいずれかに感染したネコから得た血清の交叉中和抗体価を示す。感染前(カラム1)、感染後6ヶ月(カラム2)、および感染後12ヶ月(カラム3)での血清を、FeT-1C-細胞株においてサブタイプA FIVPet、サブタイプB FIVBang、およびサブタイプD FIVShiに関して試験した。1株当たり少なくとも3匹のネコを試験し、結果は各株の代表的なネコから得たVN力価を示す。VNアッセイに関しては、初代培養PBMCを用いても同様の結果が得られた。
【発明を実施するための形態】
【0011】
発明の詳細な開示
本発明は、感受性宿主動物におけるFIV感染症に対する防御的免疫の誘導に有用な新規の方法およびワクチン組成物に関する。本明細書に記載のワクチン組成物は、宿主動物に投与すると、FIVの同種および異種株による感染症に対して防御的液性および細胞性免疫反応を誘導する。ワクチン組成物は、無細胞FIVウイルス単離株を含んでいてもFIV-感染細胞株を含んでいてもよい。好ましい態様において、本発明のワクチン組成物は、異なる2つのFIVサブタイプからのFIV株を含む。好ましくは、ワクチン組成物は、各株が異なるFIVサブタイプである3つのFIV株を含む。より好ましくは、FIVサブタイプA、サブタイプBおよびサブタイプDのそれぞれから少なくとも1つのFIV株がワクチン組成物に含まれる。
【0012】
特定の態様において、ワクチン組成物はFIVPet-およびFIVShi-感染細胞株を含む。もう一つの態様において、ワクチン組成物はFIVPet-、FIVBang-、およびFIVShi-感染細胞株を含む。FIVサブタイプの全てまたは一部を表すその他のFIV株の使用は、本発明において特に企図されるところである。例えば、FIVサブタイプAプロトタイプウイルスを提供する目的で、FIVDixまたはFIVUK8をFIVPetに加えて、またはFIVPetの代わりに、ワクチン組成物に含めることができる。FIVサブタイプBおよびDプロトタイプウイルスに関しても、その他のFIV株による同様の追加または置換を行うことができる。
【0013】
本明細書に記載のように、本明細書のワクチン組成物は、FIV感染細胞株、または無細胞ウイルスおよび感染細胞株の組み合わせと共に、無細胞ウイルス全体、またはウイルス、FIV蛋白質およびポリペプチドの一部を含んでいてもよい。FIV感染細胞株からなるワクチン組成物は、それぞれが異なるFIVサブタイプに感染した多数の細胞株を含んでいてもよい。本発明のワクチン組成物はまた、例えばFIV env、gag/pro、またはenv-gag/proを含む組換えウイルスベクターに基づくFIV構築物を含む。組換え型ベクター/FIV構築物を調製するために用いることができるいかなる適当なウイルスベクターも、本発明での使用が企図される。例えば、アデノウイルス、鳥類ポックスウイルス、ネコヘルペスウイルス、ワクシニア、カナリアポックス、昆虫ポックス、ブタポックス、および当技術分野で既知のその他のウイルスに由来するウイルスベクターを、本発明の組成物および方法と共に用いることができる。FIV成分をコードし発現する組換えポリヌクレオチドベクターは、当技術分野で既知の標準的な遺伝子操作技術を用いて構築することができる。さらに、本明細書に記載の様々なワクチン組成物は、個別に用いることも、互いに組み合わせて用いることもできる。例えば、動物の一次免疫には、単一または複数のサブタイプ成分を有する、組換えベクターに基づくFIV構築物を用い、二次免疫には不活性化FIV-感染細胞株からなるワクチン組成物を用いてもよい。本発明のワクチン組成物によるその他の免疫プロトコルは、当業者には明らかで、本発明の範囲内であると企図される。
【0014】
本明細書に特に記載された多種サブタイプFIVワクチンのネコにおける免疫原性および有効性を調べた。本ワクチン組成物でワクチン接種した特定病原体非保有(SPF)ネコの、同種および異種FIV株によるチャレンジ前後の液性および細胞性免疫をモニターした。液性免疫は、ウイルス中和(VN)抗体活性の測定によってモニターし、細胞性免疫は細胞障害性Tリンパ球(CTL)活性の測定によってモニターした。ワクチン接種ネコの血清および免疫細胞の同種および異種FIV株に対するVNおよびCTL活性をそれぞれインビトロで試験し、本ワクチンがFIV感染に対する広範囲の防御を誘発できることを証明した。本発明の指示に従い、異なるFIVサブタイプからのプロトタイプウイルス単離株を組み合わせることによって、または異なるサブタイプのプロトタイプウイルスに感染させた個々の細胞を組み合わせることによって、有効な多種サブタイプFIVワクチンを産生することができる。
【0015】
本明細書に特に例示した株に加えて全てのFIV株は、本発明での使用が企図される。多くのFIV単離株が文献に記述されており、当業者に既知である。FIVPetは米国特許第5,037,753号に記述されている。記述されているその他のFIV単離体は、当業者によって定法を用いて感染したネコから容易に単離することができる。FIVの単離および培養法は、参照として本明細書に組み入れられる米国特許第5,037,753号および第5,118,602号に記述されている。
【0016】
本明細書に例示されている新規の細胞株は、本発明のワクチン接種法および組成物において用いることができる。末梢血単核細胞を含むFIV株易感染性のその他の細胞または細胞株もまた、本発明での使用が企図される。
【0017】
FIVウイルス蛋白質の天然ポリペプチド、組換えポリペプチドまたは合成ポリペプチド、およびそのペプチド断片もまた、本方法によるワクチン組成物として用いることができる。好ましい態様において、多種のFIVサブタイプに由来するFIVポリペプチドをワクチン組成物に結合させ、これを用いて宿主動物にワクチン接種する。例えば、異なるサブタイプからの少なくとも2つのプロトタイプFIV株からのFIVエンベロープ糖蛋白質に基づくポリペプチドをワクチンに組み合わせることができる。ポリペプチドは一つの株に対して同種であってもよく、そのアミノ酸配列が少なくとも2つの明確なFIVサブタイプからの接合または結合ポリペプチドに由来する「ハイブリッド」または「キメラ」ポリペプチドを含むものであってもよい。FIVポリペプチドの調製手順は当業者に周知である。例えば、FIVポリペプチドは固相合成法を用いて合成することができる(メリフィールド(Merrifield)、1963)。FIVポリペプチドはまた、FIV蛋白質またはペプチドをコードするポリペプチド分子が、細菌、酵母、または哺乳類細胞株などの宿主細胞で発現されるよう、DNA組換え技術を用いて産生することができ、当技術分野の標準的な技術を用いて発現した蛋白質を精製することができる。
【0018】
本発明はまた、FIVに対して易感染性である新規のネコ科T細胞株にも関する。インターロイキン-2(IL-2)依存的細胞および非依存的細胞の両者を特に例示する。FeT-1CおよびFeT-Jと呼ばれる細胞株を本明細書に記述する。FeT-1C細胞株はIL-2依存的であり、FeT-J細胞株はIL-2非依存的である。本発明の細胞株は、インビトロでのFIVウイルス株の増殖および産生と共に、ネコのFIV免疫の媒体を提供するのに有用である。IL-2依存的FeT-1CおよびIL-2非依存的FeT-J非感染細胞株をいずれも培養液中の逆転写酵素(RT)活性についておよびPCRによるFIVプロウイルス配列について20回以上試験し、FIV陰性であることを確認した。FeT-J細胞株は、FIVShi、FIVDix、FIVUK8、FIVPetおよびFIVBangを含む調べた全てのFIV株に高度に感染可能であったが、FIVShiに直接感染させることはよ比較的困難であった。
【0019】
本発明はさらに、本発明の細胞株によって産生された細胞性産物にも関する。細胞性産物は当業者に既知の方法を用いて単離および検出することができる。細胞株に対する抗体は、既知の方法を用いて産生させることができ、本発明に企図される。
【0020】
FeT-1C(ATCC寄託番号CRL 11968)、およびFeT-J(ATCC寄託番号CTL 11967)と呼ばれるFIV非感染細胞株はいずれも、1995年8月24日に、米国メリーランド州ロックビルのアメリカンタイプカルチャーコレクション(ATCC)に寄託された。FIVBang-(ATCC寄託番号11975)およびFIVShi-(ATCC寄託番号11976)感染細胞株は、1995年8月25日にアメリカンタイプカルチャーコレクションに寄託された。
【0021】
本培養物は、本特許出願の係属中、特許庁長官により37 CFR 1.14および35 U.S.C.122の下で資格を与えられると決定された者は、寄託物を利用することができるよう保証される条件下で寄託されている。寄託物は、本出願の写しまたはその所産が提出されている国の外国特許法の要請に応じて利用することができるだろう。しかし、寄託物の利用は、政府の決定によって付与された特許権の適用の制限において本発明を実施する認可とはならないことを理解すべきである。
【0022】
さらに、本培養寄託物は、微生物の寄託に関するブダペスト条約(Budapest Treaty for the Deposit of Microorgamisms)の要件に従って保存され、公的に利用可能となる、すなわち、寄託物は、寄託試料の最も最近の分譲請求から少なくとも5年間、そしていずれにしても、寄託日から少なくとも30年間、または培養物の開示が発行されるあらゆる特許の有効期間にわたって、細心の注意を払って保存され、汚染を免れる。寄託者は、寄託物の状態のために、寄託所が請求時に試料を分譲できない場合には、寄託物を交換する義務を理解している。本培養寄託物の公衆への利用可能性を制限するものは全て、これを開示する特許が付与された際に、変更不可能に取り除かれるであろう。
【0023】
本発明の方法によれば、本明細書に記載のFIVワクチン組成物は、易感染性宿主、典型的には飼いネコに有効な量および方法で投与され、その後のチャレンジまたはFIVによる宿主の感染に対する防御的免疫を誘導する。ワクチンは、例えば皮下、腹腔内、または筋肉内注射によって非経口的に投与されるのが一般的である。その他の適当な投与法は経口または鼻腔内投与である。通常、ワクチンは宿主に少なくとも2回、各投与間に1週間以上の間隔をあけて投与する。しかし、ワクチンの初回および追加投与に関するその他のレジメは企図されており、実施者の判断および処置する特定の宿主動物に依る。
【0024】
本発明のワクチン組成物は、当業者に周知の方法によって調製することができる。例えば、ワクチンは注射可能の形態、例えば溶液または懸濁液として調製することが一般的である。ワクチンは投与剤形と融和するように、受容者に治療的有効量かつ免疫誘発量で投与する。特定のワクチン製剤の至適投与量および投与パターンは、当業者によって容易に決定することができる。
【0025】
ワクチン製剤におけるウイルスおよび細胞は、当技術分野で既知の方法を用いて不活化または弱毒化してもよい。例えば、ウイルス全体および感染細胞をパラホルムアルデヒド、ホルマリン、フェノール、UV光線、温度上昇等に暴露することによって不活化または弱毒化することができる。ワクチン用量における無細胞FIV全体の量は通常、約0.1 mg〜約5 mgの範囲内で、より通常は約0.2 mg〜約2 mgの範囲内である。FIV感染細胞株からなるワクチン製剤の投与量は通常、約106〜約108細胞/用量であり、より一般的には5×106〜約7.5×107細胞/用量である。ウイルスまたは細胞は、投与直前にアジュバントと混合するのが一般的である。ワクチン製剤において用いられるアジュバントは、トレオニルムラミルジペプチド(MDP)(バイアーズら(Byars)、1987)またはフロイトの完全および不完全アジュバントの混合液のいずれかであることが一般的である。ミョウバンなどの、本発明の方法およびワクチンでの使用に適当なその他の多様なアジュバントは、当技術分野で周知であり、本発明での使用が企図される。
【0026】
本発明はさらに、本発明の非感染細胞株を用いて、試料中のウイルス中和(VN)抗体を解析する新規の方法にも関する。一定数の継代後に使用できなくなり、FeT-1CまたはFeT-J細胞ほど容易に増殖しないPBMCとは異なり、FeT-1CおよびFeT-J細胞は、確立された細胞株で、今後の使用のために容易に凍結保存することができる。異なるSPFネコから得られたPBMCは細胞増殖速度およびFIV易感染性が個々に異なるため、FeT-1C細胞を用いたVNアッセイから得られた結果は、PBMCを用いたVNアッセイより再現性が高い。さらに、VNアッセイ用のPBMCは、PBMCを用いたインビトロVNアッセイに影響を及ぼす可能性がある、起こりうるインビボ感染を排除するために、無菌室収容および維持を必要とするSPFネコから得なければならない。このように、異なるサブタイプのFIVに容易に感染させることが可能なFeT-1Cのようなネコ科細胞株は、VNアッセイにおいてPBMCの都合の良い代用とすることができる。
【0027】
本明細書では、FIV株について以下の略語を用いる:
株(サブタイプ) 略語
Petaluma(A) FIVPet
Dixon(A) FIVDix
UK8(A) FIVUK8
Bangston(B) FIVBang
Aomori-1(B) FIVAom1
Aomori-2(B) FIVAom2
Shizuoka(D) FIVShi
【0028】
材料および方法
細胞培養。全ての懸濁細胞株は10%熱不活化仔ウシ胎児血清(FCS)を含むPRMI 1640培地で培養した。10 mM HEPES(N-2-ヒドロキシエチルピペラジン-n'-2-エタンスルホン酸)、2 mM L-グルタミン、50 μg/mlゲンタマイシンおよび5×10−5 M 2-メルカプトエタノール。IL-2依存的細胞にはヒト組換えIL-2(Cetus Corporation、エメリービル、カリフォルニア)100 U/mlを補足した。懸濁細胞は、0.5〜4×106細胞/mlの細胞濃度で継代し、週2回新鮮な培地で再度培養した。単層細胞は全て、細胞初濃度2×106細胞/mlで週2回継代した。FIV感染細胞からの組織培養液(TCF)を週2回回収し、3000 rpmで1時間遠心して、残留細胞を除去し、試験時に直ちに使用する予定のTCFに関しては、-20℃または-70℃で保存した。FIV-易感染性細胞(1×106細胞/ml)は約30,000 cpm/mlの逆転写酵素(RT)活性を有するFIVに感染させた。
【0029】
FIVの精製。FIV感染細胞株の組織培養液は、個々に2000〜3000 rpmで1時間遠心して細胞を除去した。TCF中のウイルスは16,000 rpmで2時間超遠心してペレットにし、まず10/50%(w/v)不連続蔗糖勾配で、次に10/50%連続蔗糖勾配で超遠心して精製した(ペダーセンら(Pedersen)、1987;ヤマモトら(Yamamoto)、1988)。ウイルス単離株のそれぞれを18時間1.25%滅菌パラホルムアルデヒド(0.22 μm濾過滅菌)で不活化し、その後滅菌PBSで十分に透析した。不活化ウイルスは、滅菌PBSで500 μg/mlの濃度に希釈し、各株の250 μg/0.5 mlを滅菌マイクロフュージチューブに移して-70℃で保存した。不活化FIV株は室温で解凍し、不活化ウイルス250 μgの0.5 ml滅菌PBS溶液を免疫直前にアジュバント0.5 mlと混合した。FIV-感染細胞株は、個別に18時間1.25%滅菌パラホルムアルデヒドで不活化し、滅菌PBSで3回洗浄し、滅菌チューブ中で約5.0×107細胞/mlの濃度に再懸濁して4℃で保存した。典型的には、不活化感染細胞約2.5×107細胞/mlの0.5 ml PBS溶液を免疫直前にアジュバント0.5 mlと混合した。トレオニルムラミルジペプチド(MDP MF75.2アジュバント;Chiron Corporation、エメリービル、CA)250 μg/0.5 mlをアジュバントとして用いた。
【0030】
CTLアッセイ。末梢血単核細胞(PBMC)を、10日間のFIV感染前にコンカナバリンA(Con A)で3日間刺激した(ソングら(Song, 1992))。これらの細胞をCTLアッセイの標的細胞として用いた。CTL活性は、Con A刺激PBMCをUV-および放射線不活化FIV-感染自家PBMCと共に5日間培養することによって生じた。これらの細胞を刺激エフェクター細胞として用いた。解析当日に標的細胞をNa51CrO4 50 μCiで1〜3時間標識して、3回洗浄し、次に固定数の標識標的細胞(5×104細胞/ウェル)をマイクロタイタープレートに加えた。エフェクター細胞を様々なエフェクター/標的細胞比(例えば、100:1、50:1、および10:1)で3連ずつ加えた。プレートを400 rpmで1分遠心し、37℃で4時間インキュベートした。対照51Cr-標識標的細胞を洗浄剤で溶解し、最大放出値を得た。被験試料ウェルから上清を収集し、ガンマカウンターを用いて放射能を定量した。エフェクター細胞の非存在下で51Cr-標識標的細胞をインキュベートすることによって、自然発生放出を定量した。特異的細胞障害百分率は以下のように計算した:
【数1】
【0031】
イムノブロットおよび酵素結合イムノソルベントアッセイ(ELISA) ヤマモトら(Yamamoto, 1993)の記述のように、蔗糖勾配精製ウイルスをイムノブロットアッセイの基質として用いた。感染細胞の組織培養液からのFIVPetは低速遠心(2000 rpmで45分)で不純物を除去し、超遠心(16,000 rpmで2時間)によって濃縮し、10/50%(w/v)連続蔗糖勾配上での超遠心によって精製した。この技法で精製したウイルスをイムノブロットアッセイの基質として用いた。
【0032】
これまでに記述されたイムノブロット技法(ヤマモトら(Yamamoto)、1991a)の改変を用いた。ウイルスブロット片は、ウイルスを0.1%SDSで可溶化し、その後10%SDSポリアクリルアミドゲル上で電気泳動し、ニトロセルロース膜上に電気泳動的に移動させることによって調製した。ワクチン接種ネコからの血清試料を緩衝液3(0.15 M塩化ナトリウム、0.001 Mエジチ酸、0.05 Mトリス塩基、0.05%ツイーン20、および0.1%ウシ血清アルブミン)で1:50に希釈し、イムノブロットプレートの個々のウェル中でウイルスブロット片と共に37℃で18時間インキュベートした。ブロット片は洗浄液(0.15 M NaClおよび0.05%ツイーン20脱イオン水溶液)で個々に洗浄し、ビオチン化抗ネコIgG(Vector Laboratories、バーリンゲイム、CA)と共に37℃で1時間インキュベートし、洗浄液で3回洗浄した。次にブロット片を個々に、ホースラディッシュペルオキシダーゼ結合ストレプトアビジン(ベクター・ラボラトリーズ)と共に30分インキュベートした。十分な洗浄後、各片を新鮮な基質溶液(0.05%ジアミノベンジジン、400 μg/ml NiCl2、および0.01%H2O2の0.1 Mトリス緩衝溶液、pH 7.4)と共に室温でインキュベートした。目に見えるバンドが確立した後、過剰量の蒸留水を加えて反応を停止させ、ブロット片を乾燥させた。次に、イムノブロット上のバンドの分子量は、先にアミドブラックで染色した片上の分子量標準物質の移動距離とそれらの移動距離を比較して決定した。各イムノブロット分析には診断的評価の内部対照として陽性および陰性対照血清を含めた。
【0033】
ウイルス抗原特異的ELISAは既に記述されている(ヤマモトら(Yamamoto)、1991a;ヤマモトら(Yamamoto)、1993)。蔗糖勾配精製FIVPetおよび表面エンベロープ(SU)ならびにFIVPetの保存(C)および可変(V)領域双方の膜通過(TM)ペプチドは、96ウェルイムノロンプレート(Dynatic Laboratories, Inc.、シャンティリー、VA)上に重炭酸緩衝液(pH 9.6)と共に250 ng/ウェルで12〜18時間、37℃でコーティングし、ELISAの基質として用いた。SU-V3-2ペプチドのアミノ酸配列は:Gly Ser Trp Phe Arg Ala Ile Ser Ser Trp Lys Gln Arg Asn Arg Trp Glu Trp Arg Pro Asp Phe(配列番号:1);およびTM-C1ペプチドのアミノ酸配列は:Gln Glu Leu Gly Cys Asn Gln Asn Gln Phe Phe Cys Lys Ile(配列番号:2)である。合成ペプチドは、FMOCペプチド合成化学(マガジンら(Magazine)、1988)を用いて、バイオサーチ9500ペプチドシンセサイザー(Biosearch、サンラファエル、CA)で合成した。合成ペプチドの純度は、逆相高速液体クロマトグラフィー上での単一のピークの存在によって決定し、ピーク試料について実施したアミノ酸配列分析によって確認した。
【0034】
ペプチドコーティングプレートは、使用直前に緩衝液3で1回洗浄した。血清試料を緩衝液3で1:200に希釈し、FIV抗原コーティングウェル中で37℃で1時間インキュベートし、6回洗浄した。ウェルを洗浄液で洗浄し、ビオチン化抗ネコIgG(Vector Laboratories、バーリンゲイム、CA)と共に37℃で1時間インキュベートし、6回洗浄し、ホースラディッシュペルオキシダーゼ結合ストレプトアビジン(ベクター・ラボラトリーズ)と共に37℃で1時間インキュベートした。次にウェルを洗浄液で6回洗浄して、ELISA基質溶液(0.005%テトラメチルベンジジンおよび0.015%H2O2の0.96%クエン酸溶液)と共に室温でインキュベートした。既知のFIV陽性ネコ血清からなる連続希釈標準物質に可視反応色が確立すれば、0.1 Mフッ化水素酸で反応を停止させた。バイオラッドELISAリーダー(バイオラッド・ラボラトリーズ、ハーキュルズ、CA)で光学濃度414 nmでの吸光度を測定した。
【0035】
ポリメラーゼ連鎖反応(PCR)。感染細胞のプロウイルスDNA濃度は、同じまたは異なるサブタイプからの多種FIV株を識別するために最近開発された分別PCRによってモニターした(オカダら(Okada)、1994)。PCRの感度を増加させる手段として、表1に示すネステッドPCRプライマーセットを用いた。PCRは2段階反応で行い、最初はオカダら(Okada, 1994)が記述した条件下で1対の外側のプライマー(全てのFIV株に共通)で行った。PCRの第二段階では、内側のプライマー(各FIV株に特異的)を用いて第一段階産物の1/25を増幅した。ネステッドPCRを用いて、FIVPet、FIVUK8、FIVBang、FIVAom1、FIVAom2、およびFIVShiに感染した細胞を互いに区別することができる。
【表1】
【0036】
プロウイルスDNAの細胞当たりのおおよその量は、既知の細胞数から抽出したDNAの希釈を変化させて半定量的PCRによって定量した。例えば、105個の細胞をDNA抽出に用いた場合、DNA調製物を10−5希釈すれば、細胞1個当たりに存在するDNAにほぼ相当することになる。これらの変化させたDNA希釈液についてPCRを実施し、陽性PCR結果を示す最終希釈をエンドポイント希釈と見なす。エンドポイント希釈に相当する細胞数を用いて、以下の式に従って所定の細胞調製物中のウイルス感染細胞の百分率を決定した:
【数2】
式中、Zはエンドポイント希釈に相当する細胞数である。
【0037】
逆転写酵素(RT)アッセイ。RNA-依存的DNAポリメラーゼ(RT)の有無は、本質的にレイら(Rey)の記載に従い細胞培養上清において解析した。FIVを検出するRTアッセイは、外因性鋳型プライマーとしてポリ(rA)-オリゴ(dT12−18)、異なる4つのデオキシリボヌクレオチド3リン酸、2価陽イオンとしてMg++と共に20 mM KCl、および1試料当たり5μCi[3H]-標識チミジン3リン酸を用いた。5μCi[3H]TTPは、ベックマンLS250シンチレーションカウンター(Beckman Instruments, Inc.、パロアルト、CA)でシンチレーション液混合物(キシレン1対研究産物国際生物分解計数シンチラント9の割合)を用いて、平均総カウント1,200,000 cpmを生じた。その結果、調べる試料のRT値は1,200,000 cpm/ml未満となると考えられる。
【0038】
ウイルス中和アッセイ。株およびサブタイプ特異的VNアッセイを開発する計略は記述されている(オカダら(Okada)、1994)。ネコ末梢血単核細胞(PBMC)(4×105細胞/ml)またはFIV易感染性FeT-1C細胞(2×105細胞/ml)を加える前に、熱不活化血清の連続希釈物を24ウェルプレート中で37℃で45分、各FIV株の100 TCID50と共にインキュベートした。培養3日後に、細胞をHank's緩衝塩溶液で1回洗浄し、培養から残留ウイルスを除去し、次に細胞を新鮮な培養培地(10%熱不活化仔ウシ胎児血清、10 mM HEPES緩衝液、50μg/mlゲンタマイシン、5×10−5 M 2-メルカプトエタノール、および100 単位/mlヒト組換えIL-2を含むRPMI-1640)に再懸濁した。細胞のウイルス感染は、培養9、12、15、および18日目に回収した培養液のMg++依存的RTアッセイによってモニターした。RT活性が、SPF血清からなる感染対照培養の25%以下である場合、血清はVN抗体陽性と見なした。
【0039】
以下は、本発明の実施に関する最良の形態を含む手順を図示する実施例である。これらの実施例は、制限的なものと解釈してはならない。百分率は全て重量で表し、溶媒混合比は、他に明記していなければ全て容積である。
【実施例1】
【0040】
実施例1−FIV感染細胞株。
IL-2依存的FeT-1Mクローンの母株であるFeT-1Cと呼ばれる新規インターロイキン-2(IL-2)依存的ネコ科T細胞株を用いて、FIVPet、FIVDix、FIVUK8、FIVBang、FIVAom2、またはFIVShiのいずれかに慢性的に感染させた個々の細胞株を確立した。FeT-1Mクローン(FIV-Fet1Mとも呼ばれる)は、本明細書に参照として組み入れられる米国特許第5,275,813号に記述されており、これを用いて、FIVPetを慢性的に産生するIL-2非依存的細胞株FL-4(これも米国特許第5,275,813号に記述されている)を産生した。FeT-1C細胞株は、FIVサブタイプA、B、およびDの異なる単離体に高度に感染可能である。FeT-1C細胞株を長期間継代すると、その感染性、特にFIVサブタイプDに対する感染性は低下する;したがって、至適FIV感染率を得るため、またはVNアッセイで使用するためには、継代回数は約35回未満とすること。半定量的PCRおよびウイルスコア抗原分析により、FIVに暴露された細胞株は全て、個々のFIV株に有意に感染することが示された。
【0041】
FIV易感染性のIL-2非依存的ネコ科細胞株も同様に、FeT-1C細胞から作製した。FeT-Jと呼ばれるこの細胞株は、FIV感染培地または細胞を用いて共培養によってFIVに感染させることができる。例えば、FIVBang感染FeT-1C細胞株を、IL-2の非存在下で非感染FeT-J細胞と共培養して、IL-2非依存的FIVBang感染FeT-J細胞株(Bang/FeT-Jと呼ぶ)を確立した。共培養による感染法では、Bang/FeT-1C細胞を非感染FeT-J細胞と約2:1〜約10:1(感染:非感染)の比で混合した。細胞混合物はIL-2の非存在下で培地中で数日間培養し、FeT-1C細胞を死亡させた。残った細胞は、FIVBang感染FeT-J細胞から成った。このように、FIV感染FeT-1C細胞を用いてFeT-J細胞を感染させ、異なるFIVサブタイプに感染したIL-2非依存的FeT-J細胞株を確立することができる。FIV感染FeT-1C細胞との共培養法の結果、異なるFIVサブタイプを中等度から高レベル産生するIL-2非依存的FeT-J細胞株が得られた。
【0042】
FeT-1C細胞株をまたFIVShiに感染させ、十分に継代すると、Shi/FeT-1Cと呼ばれるIL-2依存的細胞株が得られた。Shi/FeT-1C細胞株を後に、IL-2の非存在下でFeT-Jと共培養し、得られたIL-2非依存的FIVShi-感染細胞株をShi/FeT-Jと呼んだ。IL-2非依存的Shi/FeT-J細胞株は、IL-2依存的Shi/FeT-1C細胞株より高レベルのFIVShiを産生する(図1)。
【0043】
FIVBangに感染させたFeT-J細胞株の作製はまた、FeT-1C細胞株を用いずに実施した。FeT-J細胞株を無細胞FIVBang接種体に直接感染させ、IL-2を用いずに十分に継代した。得られたIL-2非依存的FIVBang産生細胞株をBang/FeT-Jと命名した。Bang/FeT-J細胞株は、FeT-1C細胞株をFIVBangに感染させて作製したIL-2依存的Bang/FeT-1C細胞株より高レベルのFIVBangを産生した。
【実施例2】
【0044】
実施例2−多種サブタイプFIVワクチン。
FIV感染細胞は遠心によって上清から除去し、不活化し、そしてワクチンとして用いた。同様に、FIVウイルス全体は、超遠心によって感染した無細胞上清から沈殿させ、不活化した。感染細胞およびウイルスはいずれも、5℃で1.25%パラホルムアルデヒドで24時間処置して不活化し、その後それぞれ、十分に洗浄、またはPBSで透析した。この方法は、免疫原性を失うことなくFIVを効率よく不活化する。本方法によって産生されたFIV免疫原は、防御免疫の誘導に非常に有効である(ヤマモトら(Yamamoto)、1993;ヤマモトら(Yamamoto)、1991a;ヤマモトら(Yamamoto)、1991b)。弱毒化したウイルス単離体もまた、本発明のワクチン組成物に用いることができると企図される。
【0045】
FIVShi感染FeT-1C細胞株をFIVPet株に重複感染させると、多種サブタイプFIV(すなわち多種サブタイプA/D FeT-1C細胞株)に感染した単一の細胞株が得られるが、共感染の2ヶ月以内に、FIVShiプロウイルス濃度は50%から5%未満に減少し、一方FIVPetプロウイルス濃度は同時に約50%に増加した。このように、FIVワクチンとして用いるための、多種サブタイプFIVに感染させた単一細胞株の維持は、本発明の好ましい態様ではない。
【0046】
その結果として、本発明の一つの態様において、ワクチン組成物は、各株が異なるFIVサブタイプに感染した2つの個々の細胞株から作製した。特定の態様において、2種サブタイプのFIVワクチン組成物は、FIVサブタイプA感染細胞株(Pet/FL-4)とFIVサブタイプD感染細胞株(Shi/FeT-1C)との組み合わせから成った。AサブタイプおよびDサブタイプ感染細胞株は記述のように不活化し、等しい細胞数(MDP 250 μg中に各2.5×107細胞)を混合し、これを用いてネコを免疫した。SPFネコ3匹を不活化Pet/FL-4細胞でワクチン接種し、別のネコ4匹を不活化Shi/FeT-1C細胞でワクチン接種した(2.5×107細胞/接種量)。一連の4回のワクチン接種後、2種サブタイプ(Pet/FL-4およびShi/FeT-1C)ワクチンは、調べた両FIV株に対して有意なVN抗体価を含む抗FIV抗体を誘導した(図2および表2、試験I)。2種サブタイプ(Pet/FL-4およびShi/FeT-1C)ワクチン接種ネコ4匹に、FIVBang(50 CID50)をチャレンジした。Pet/FL-4ワクチン接種ネコ3匹全てとShi/FeT-1Cワクチン接種ネコ2匹にFIVBangの50 CID50量をチャレンジした。残る2匹のShi/FeT-1Cワクチン接種ネコには、FIVShiの50 CID50量をチャレンジした。
【0047】
2種サブタイプワクチン接種ネコは全て、感染後(pi)6週目でのウイルス単離およびPBMCのPCRにより、FIVBang陰性であったが、偽免疫ネコは全て、感染後6週目でのウイルス単離およびPCRにより、FIVBangまたはFIVShiのいずれかに対して陽性であった(表2、試験I)。対照的に、Pet/FL-4ワクチン接種およびShi/FeT-1Cワクチン接種群のネコそれぞれ1匹ずつにFIVBangをチャレンジすると、FIVBang陽性であった。予想されたように、FIVShiワクチンを接種し、その後FIVShiをチャレンジしたネコは全て、感染後6週目でFIVShiに対して陰性であった。このように、特に例示した2種サブタイプワクチンは、異種FIVBangチャレンジに対してと同様、同種FIVShiに対する感染も防御または遅らせた。
【0048】
2種サブタイプワクチンを接種したネコ(Pet/FL-4細胞およびShi/FeT-1C細胞)は、2回目の免疫後、ウイルスコア蛋白質p25(FIV p28とも呼ばれる)に対して特異的なFIV抗体を産生した(図2)。他のウイルス抗原に対するより高い抗体価は、3〜4回の免疫後に示された。FIVPetに対するVN抗体は、2回の免疫後に産生されたが、FIVShiに対するVN抗体は4回の免疫後に産生された(表4)。FIVPetおよびFIVShiに対するCTL反応は、調べた全てのネコにおいて3回の免疫で検出され(表3)、両株に対するより強いCTL反応は4回の免疫後に得られた。さらに、調べたネコ3匹のうち2匹が、4回の免疫後FIVBangに対するCTL反応を示した。結果は、4回のワクチン接種後、2種サブタイプワクチンは、FIVPetおよびFIVShiに対して強いCTL反応を誘導し(表3)、両FIV株に対してVN抗体価を含む高いFIV抗体を誘導した。
【0049】
不活化Shi/FeT-1C細胞で免疫したネコは、2回の免疫後ウイルスコア蛋白質p25に対して特異的なFIV抗体を産生し、3回の免疫後その他のウイルス抗原に対する抗体を産生した(図2)。これらのネコでは、FIVShiに対するVN抗体は4回の免疫後に産生されたが、FIVPetに対するVN抗体は免疫の過程において検出されなかった。Shi/FeT-1Cワクチン接種ネコはいずれも、4回の免疫後に限ってFIVShiに対するCTL反応を起こしたが、FIVPetに対するCTL反応は4回の免疫後でも起こさなかった(表3)。
【0050】
不活化Pet/FL-4細胞で免疫したネコは、2回の免疫後にp25に対する抗体を産生し(図2)、2〜3回の免疫後FIVPetに対するVN抗体を含む他のウイルス抗原に対する抗体を産生した(表4)。Pet/FL-4細胞で免疫したネコに検出される唯一のCTL反応は、FIVPetに対するものであった。全体的に、2種サブタイプワクチンは、1種サブタイプワクチンより、速やかにかつ高いVN抗体価、および両FIV株に対するCTL反応を誘導した。偽免疫SPFネコは、ウイルス抗体、VN抗体、または抗FIV CTL反応を示さなかった。
【表2】
【表3】
【表4】
【0051】
好ましい態様において、本発明のワクチン組成物は、各細胞株が異なるFIVサブタイプ(AまたはBもしくはD)からのウイルス株に感染している3つの細胞株から調製された3種サブタイプFIVワクチンからなる。特定病原体不在ネコ3匹を3種サブタイプ(FIVPet+FIVBang+FIVShi)ワクチンで免疫した。その他のネコは1種サブタイプFIVBangワクチンで免疫して、ワクチン成分としてマクロファージ親和性FIVBangの免疫原性を評価した。VN抗体価の結果は、3種サブタイプ(FIVPet+FIVBang+FIVShi)と1種類サブタイプFIVBangワクチンはいずれも、2回の免疫後においても高い抗ウイルス抗体価を誘発した(表2、試験IIおよび表5)。このように、リンパ球親和性およびマクロファージ親和性FIVは、本発明のワクチン組成物の成分として用いることができる。
【0052】
不活化Pet/FL-4、不活化Bang/FeT-J、および不活化Shi/FeT-1C細胞の組み合わせ(全体でMDP 250 μg中に各2.5×107細胞)で免疫したSPFネコ3匹は、ウイルスコア蛋白質p25およびFIV SUおよびTMエンベロープ蛋白質を含むその他のウイルス抗原に特異的なFIV抗体を2回の免疫後に産生した(図3、4、5)。FIVPet、FIVBang、およびFIVShiに対するVN抗体は、2回の免疫後まもなく大多数のネコに産生され、3回目の免疫により全てのネコに産生された(表5)。さらに、ネコ1匹が3回の免疫後にFIVUK8に交叉反応するVN抗体を産生した。不活化Bang/FeT-J細胞のみで免疫したSPFネコ4匹は、ウイルスコア蛋白質p25およびその他のウイルス抗原に特異的な抗体を産生した(図3)。これらのネコではFIVBangに対するVN抗体は2回の免疫後に産生された(表5)が、FIVPetおよびFIVUK8に対するVN抗体は免疫過程において検出されなかった。3種サブタイプFIVワクチン(Pet/FL-4、Bang/FeT-JおよびShi/FeT-1C細胞)で3回免疫したネコのFIV A、B、およびDサブタイプ標的細胞に対するCTL反応を表6に示す。調べたこれら3つのFIVサブタイプ全てに対してCTL反応が検出された。このように、3種サブタイプワクチンは、1種サブタイプワクチンより幅広いCTL反応ならびに、より速やかかつより高いVN抗体価およびSU-エンベロープ抗体価を誘導した。非感染FeT-Jまたは偽免疫SPFネコも、ウイルス抗体またはVN抗体を産生しなかった。
【表5】
【表6】
【実施例3】
【0053】
実施例3−FIVサブタイプに対するVN抗体。
FIVに対するVN抗体のアッセイはまた、本発明のFeT-1C細胞を用いて開発された。FIVPet感染ネコおよび不活化Pet/FL-4細胞または不活化FIVPetウイルスでワクチン接種したSPFネコの血清を、本明細書に記述のVNアッセイ法に従って、FeT-1C細胞またはPBMCのいずれかを用いてVN抗体価を調べた。ワクチンを接種せず、FIVに感染していないSPFネコ2匹の血清を対照血清として用いた。ワクチン接種およびFIV感染ネコの血清は1000以上の高いVN抗体価を示したが、非ワクチン接種SPFネコの血清は検出可能なVN抗体価を示さなかった。FeT-1Cに基づくVNアッセイは、ネコの初代培養PBMCを用いて得られた結果と同等のVN抗体価結果を示す(表6)。この知見は、FeT-1C細胞を用いたVNアッセイでのVN抗体価が、PBMCを用いたVNアッセイに関して得られた結果と相関することを証明している。したがって、FeT-1C細胞は、全てのFIVサブタイプに感染させることが可能で、組織培養において容易に増殖させることが可能なため、FIVに関する標準的なVNアッセイにおいてPBMCの代わりに都合良く用いることができる。
【表7】
【実施例4】
【0054】
実施例4−FIV株の免疫タイピング
FIVサブタイプがFIV免疫型を反映するか否かを評価するために、FeT-1C細胞を用いてインビトロ試験を実施した。免疫タイピングはワクチン防御におけるVN抗体の役割を理解する上で重要である。FIVサブタイプA株(FIVPet、FIVDix、FIVUK8)、サブタイプB(FIVBang、FIVAom1)、およびサブタイプD(FIVShi)に感染させたネコの抗血清を、VNアッセイにおいてFeT-1C細胞を用いてインビトロでこれらの株を中和する能力に関して調べた(図6)。試験抗血清は全て、対応する同種FIV株に対する中和活性を示した。サブタイプA株であるFIVPetは、FIVDixに感染させたネコの抗血清によって有意に交叉中和された。FIVPetは、表面エンベロープ糖蛋白質(Env)領域がFIVDixと約9%異なる。FIVサブタイプAに感染させたネコの抗血清はサブタイプBのFIVBangと交叉中和したが、サブタイプD FIVShiを中和しなかった。サブタイプBおよびD株に感染させたネコの抗血清のみが、同種サブタイプ内の他のFIV株を交叉中和した。さらに、FIVUK8に感染させたネコの抗血清はFIVBangを中和したが、サブタイプA内のFIV株を中和しなかった。FIVUK8はサブタイプAに分類されているが(ソドラら(Sodora)、1994;リグビーら(Rigby)、1993;カキヌマら(Kakinuma)、1995)が、これらの結果は、FIVUK8に対する抗血清がサブタイプB株を認識するがサブタイプA株を認識しないことを示唆しており、不活化FIVPetワクチンがFIVUK8およびFIVShiに対して無効である理由を説明する可能性がある(ジョンソンら(Johnson)、1994)。このように、遺伝子型と免疫型との間には緩やかな相関が存在する。遺伝子型分析によりFIV株分類が可能となったが、交叉中和抗体試験はFIV株の免疫原性を反映し、これは、ワクチンによって誘発される広範囲の液性防御における重要なパラメーターである。
【実施例5】
【0055】
実施例5−FIV細胞親和性
感染FeT-1Cおよび感染FeT-J細胞株から得たFIV株の細胞指向性を、初代培養PBMCから得たFIV株の細胞指向性と比較した(表8)。FIV単離体2個、FIVUK8およびFIVBangはいずれも等しくリンパ球親和性およびマクロファージ親和性であるが、FIVShiは非常にリンパ球親和性である。FIVPetは、マクロファージ親和性よりリンパ球親和性で、その細胞親和性はその細胞源によって有意に影響を受けなかった。FIVBangのマクロファージ親和性は、ウイルスの細胞源によって影響を受けなかった。感染FeT-1C細胞株からのFIV株の細胞親和性は、初代培養PBMCから得られた細胞親和性と同等であるため、FeT-1C細胞内で増殖するウイルスは、VNアッセイの接種体として用いることができ、治療および予防的アプローチを評価するための試験では、インビボ接種体として用いることもできる。
【表8】
【0056】
本明細書に記載の実施例および態様は例示目的のためのみであり、その様々な改変または変更は当業者に示唆され、本出願および付属の請求の範囲の範囲内に含まれるものと理解すべきである。
【特許請求の範囲】
【請求項1】
FIV感受性動物において複数のFIVサブタイプに対して免疫反応を誘発することが可能なFIV免疫源を含むワクチン組成物。
【請求項2】
組換えウイルスベクターFIV構築物、多種FIVサブタイプに由来するFIVポリペプチド、多種の無細胞FIVウイルス全体、およびそれぞれが異なるFIVサブタイプ由来のFIV株に感染している多種の細胞株からなる群より選択される、請求項1記載のワクチン組成物。
【請求項3】
ワクチンを宿主動物に投与する前に、FIVウイルスまたはFIV感染細胞株を、該ウイルスまたは該細胞株が不活化されるよう処置する、請求項2記載のワクチン組成物。
【請求項4】
ワクチンを宿主動物に投与する前に、FIVウイルスまたはFIV感染細胞株を、該ウイルスまたは該細胞株が弱毒化されるよう処置する、請求項2記載のワクチン組成物。
【請求項5】
複数のFIVサブタイプに対して免疫反応を誘発することが可能なワクチン組成物の有効量を宿主に投与することを含む、感受性宿主動物においてFIV感染に対する防御的免疫反応を誘導する方法。
【請求項6】
ワクチン組成物が、組換えウイルスベクターFIV構築物、多種FIVサブタイプに由来するFIVポリペプチド、多種の無細胞FIVウイルス全体、およびそれぞれが異なるFIVサブタイプのFIV株に感染している多種の細胞株からなる群より選択される、請求項5記載の方法。
【請求項7】
FIVウイルスまたはFIV感染細胞株が、ワクチンを宿主動物へ投与する前に、不活化されるよう処置される、請求項6記載の方法。
【請求項8】
FIVウイルスまたはFIV感染細胞株が、ワクチンを宿主動物へ投与する前に、弱毒化されるよう処置される、請求項6記載の方法。
【請求項9】
FIVサブタイプが、サブタイプA、B、CおよびDからなる群より選択される、請求項5記載の方法。
【請求項10】
少なくとも一次免疫が組換えウイルスベクターFIV構築物を投与することを含み、その後、組換えウイルスベクターFIV構築物、FIVポリペプチド、無細胞FIVウイルス全体、およびFIV感染細胞株からなる群より選択されるワクチン組成物で追加免疫が行われる、請求項5記載の方法。
【請求項11】
細胞株が、少なくとも1つのFIVサブタイプによる注射に感受性であり、該FIVサブタイプが、サブタイプA、B、CおよびDからなる群より選択される、ネコ科由来T細胞株。
【請求項12】
FeT-1Cと称される、請求項11記載の細胞株。
【請求項13】
細胞株が、FIVDix、FIVUK8、FIVBang、FIVAom1、FIVAom2、FIVPet、およびFIVShiからなる群より選択されるFIVウイルス株の少なくとも1つに感染する、請求項11記載の細胞株。
【請求項14】
IL-2非依存的である、請求項11記載の細胞株。
【請求項15】
FIVDix、FIVUK8、FIVBang、FIVAom1、FIVAom2、FIVPet、およびFIVShiからなる群より選択されるFIVウイルス株の少なくとも1つに感染する、請求項14記載の細胞株。
【請求項16】
FeT-Jと称される、請求項14記載の細胞株。
【請求項17】
試料をFIVに接触させ、次に請求項10記載の細胞株を該試料中で有効な時間培養し、該細胞を新鮮な培養培地で培養し、次に該培養培地中の逆転写酵素活性量を定量することを含む、試料中のFIVウイルス中和抗体の検出または定量法。
【請求項18】
細胞株が、FeT-1CおよびFeT-Jと称される細胞株からなる群より選択される、請求項17記載の方法。
【請求項1】
FIV感受性動物において複数のFIVサブタイプに対して免疫反応を誘発することが可能なFIV免疫源を含むワクチン組成物。
【請求項2】
組換えウイルスベクターFIV構築物、多種FIVサブタイプに由来するFIVポリペプチド、多種の無細胞FIVウイルス全体、およびそれぞれが異なるFIVサブタイプ由来のFIV株に感染している多種の細胞株からなる群より選択される、請求項1記載のワクチン組成物。
【請求項3】
ワクチンを宿主動物に投与する前に、FIVウイルスまたはFIV感染細胞株を、該ウイルスまたは該細胞株が不活化されるよう処置する、請求項2記載のワクチン組成物。
【請求項4】
ワクチンを宿主動物に投与する前に、FIVウイルスまたはFIV感染細胞株を、該ウイルスまたは該細胞株が弱毒化されるよう処置する、請求項2記載のワクチン組成物。
【請求項5】
複数のFIVサブタイプに対して免疫反応を誘発することが可能なワクチン組成物の有効量を宿主に投与することを含む、感受性宿主動物においてFIV感染に対する防御的免疫反応を誘導する方法。
【請求項6】
ワクチン組成物が、組換えウイルスベクターFIV構築物、多種FIVサブタイプに由来するFIVポリペプチド、多種の無細胞FIVウイルス全体、およびそれぞれが異なるFIVサブタイプのFIV株に感染している多種の細胞株からなる群より選択される、請求項5記載の方法。
【請求項7】
FIVウイルスまたはFIV感染細胞株が、ワクチンを宿主動物へ投与する前に、不活化されるよう処置される、請求項6記載の方法。
【請求項8】
FIVウイルスまたはFIV感染細胞株が、ワクチンを宿主動物へ投与する前に、弱毒化されるよう処置される、請求項6記載の方法。
【請求項9】
FIVサブタイプが、サブタイプA、B、CおよびDからなる群より選択される、請求項5記載の方法。
【請求項10】
少なくとも一次免疫が組換えウイルスベクターFIV構築物を投与することを含み、その後、組換えウイルスベクターFIV構築物、FIVポリペプチド、無細胞FIVウイルス全体、およびFIV感染細胞株からなる群より選択されるワクチン組成物で追加免疫が行われる、請求項5記載の方法。
【請求項11】
細胞株が、少なくとも1つのFIVサブタイプによる注射に感受性であり、該FIVサブタイプが、サブタイプA、B、CおよびDからなる群より選択される、ネコ科由来T細胞株。
【請求項12】
FeT-1Cと称される、請求項11記載の細胞株。
【請求項13】
細胞株が、FIVDix、FIVUK8、FIVBang、FIVAom1、FIVAom2、FIVPet、およびFIVShiからなる群より選択されるFIVウイルス株の少なくとも1つに感染する、請求項11記載の細胞株。
【請求項14】
IL-2非依存的である、請求項11記載の細胞株。
【請求項15】
FIVDix、FIVUK8、FIVBang、FIVAom1、FIVAom2、FIVPet、およびFIVShiからなる群より選択されるFIVウイルス株の少なくとも1つに感染する、請求項14記載の細胞株。
【請求項16】
FeT-Jと称される、請求項14記載の細胞株。
【請求項17】
試料をFIVに接触させ、次に請求項10記載の細胞株を該試料中で有効な時間培養し、該細胞を新鮮な培養培地で培養し、次に該培養培地中の逆転写酵素活性量を定量することを含む、試料中のFIVウイルス中和抗体の検出または定量法。
【請求項18】
細胞株が、FeT-1CおよびFeT-Jと称される細胞株からなる群より選択される、請求項17記載の方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2013−79286(P2013−79286A)
【公開日】平成25年5月2日(2013.5.2)
【国際特許分類】
【出願番号】特願2013−12043(P2013−12043)
【出願日】平成25年1月25日(2013.1.25)
【分割の表示】特願2007−121532(P2007−121532)の分割
【原出願日】平成8年8月23日(1996.8.23)
【出願人】(507371168)ユニバーシティ オブ フロリダ リサーチ ファンデーション インコーポレーティッド (38)
【出願人】(506115514)ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア (87)
【Fターム(参考)】
【公開日】平成25年5月2日(2013.5.2)
【国際特許分類】
【出願日】平成25年1月25日(2013.1.25)
【分割の表示】特願2007−121532(P2007−121532)の分割
【原出願日】平成8年8月23日(1996.8.23)
【出願人】(507371168)ユニバーシティ オブ フロリダ リサーチ ファンデーション インコーポレーティッド (38)
【出願人】(506115514)ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア (87)
【Fターム(参考)】
[ Back to top ]