
微生物のDNA情報に基づいたメタン発酵の運転管理
【課題】有機性廃棄物のメタン発酵系におけるメタン発酵効率の低下を予測し、安定なメタン発酵を可能にするための運転管理法を提供することを目的とする。
【解決手段】本発明は、メタン発酵効率の低下に先駆けて増殖するか又は該低下と同時に増殖する特定の真正細菌をそのDNA情報に基づいて検出及び定量することによるモニタリング方法、及び該方法に使用されるプライマーセットに関する。本発明によれば、特定の真正細菌の関与に基づいて、メタン発酵の失敗を客観的に予測し、安定的なメタン発酵を達成させるための運転管理の方法を提供することができる。
【解決手段】本発明は、メタン発酵効率の低下に先駆けて増殖するか又は該低下と同時に増殖する特定の真正細菌をそのDNA情報に基づいて検出及び定量することによるモニタリング方法、及び該方法に使用されるプライマーセットに関する。本発明によれば、特定の真正細菌の関与に基づいて、メタン発酵の失敗を客観的に予測し、安定的なメタン発酵を達成させるための運転管理の方法を提供することができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、有機性廃棄物のメタン発酵系におけるメタン発酵効率をモニタリングする方法、及び該方法に使用されるプライマーセットに関する。より具体的には、本発明は、有機性廃棄物のメタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下と同時に増殖する特定の真正細菌をそのDNA情報に基づいて検出及び定量することを特徴とするメタン発酵の運転管理に関する。
【背景技術】
【0002】
メタン発酵は、有機性廃水を処理する方法の1つで、ランニングコストが安く、さらにバイオガスとしてエネルギー回収ができ、余剰汚泥発生量も少ないという特徴を有する優れた方法として知られている(非特許文献1参照)。メタン発酵法の歴史は古く、嫌気性微生物による高濃度有機性廃水や汚泥消化等の処理技術として開発されてきた。近年では、高濃度有機性廃水等に使われた第一世代に代わって、UASB法(上向流嫌気性汚泥床法)の開発に伴う第二世代のメタン発酵法が実績を増やしている。UASB法とは、0.5〜2mm程度に自己造粒した高密度グラニュール菌体(真正細菌及び古細菌を含む)を利用して、高濃度基質の高負荷高速運転を可能としたリアクターを利用する方法である(非特許文献1及び2参照)。グラニュール菌体は沈降性が良いことから、それまでの懸濁型メタン発酵に見られるような気液固分離の欠点を解決できる手法である。
【0003】
メタン発酵は、多くの微生物の共生により有機物を分解し、その過程で生成する有機酸などをメタンに還元する方法であって、主として3段階で進行すると考えられている。複雑な有機物は、第1段階の酸生成過程(液化過程)で酸生成菌類の作用により、単糖類、アミノ酸などの低分子量の物質を経て、酢酸、プロピオン酸、ギ酸、酪酸などの低級脂肪酸、乳酸及びエタノールなどになる。第2段階においては、酢酸以外の低級脂肪酸、乳酸及びエタノールは、水素生成細菌により水素及び酢酸に変換され、最終の第3段階において基質特異性の強いメタン生成古細菌群により、メタン及び二酸化炭素などに分解される。ここで、酢酸及びプロピオン酸などの低級脂肪酸はメタン発酵プロセスにおける主要な中間生成物であり、酸生成菌類及び水素生成細菌などの真正細菌がその役割を担っている(非特許文献3参照)。
【0004】
メタン発酵においては、その原料となる有機性廃棄物の供給量(有機物負荷)が大きくなると、供給量に応じてメタンガスの発生量も増加するが、有機物負荷が過剰になると、メタン生成に関与する真正細菌及び古細菌の活性が著しく低下し、メタンガスが全く生成されなくなる(メタン発酵が失敗する)。有機物負荷の過剰によりメタン生成が行われなくなると、メタン発酵を復帰させることは非常に困難であるという問題点がある。この限界の負荷量をリアクター内への流速や有機物濃度などから決定することは困難であり、これまで限界の負荷量は熟練のオペレータの判断に任されていた。これまで、メタン発酵が失敗した後に有用な微生物が死滅又は減少するという報告がなされている(特許文献1参照)。しかしながら、特許文献1に開示された方法は、メタン発酵の失敗を後付け的に説明することはできても、メタン発酵の失敗を予測し、未然に予防することができない。また、メタン発酵が失敗するとき、又は失敗したときには、リアクター内のpHが低下している(pH低下はメタン発酵失敗の原因であり、結果である、すなわち負のフィードバックと考えられる)ということは経験的によく知られている。しかしながら、メタン発酵が失敗しなくても一時的にpH低下が起こることも周知な現象である。よって、pH低下に基づいて、メタン発酵失敗の予兆とすることはできない。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特表2006−325581
【非特許文献】
【0006】
【非特許文献1】松本豊、中崎清彦共著、『化学工学の進歩第36集 環境調和型エネルギーシステム』(社団法人化学工学会編集)、「4.3 メタン発酵技術」(140〜156頁)、2002年、槇書店
【非特許文献2】松尾幸彦他、『用水と廃水』、「UASB法を用いたPEG排水の高速処理」、Vol.43、No.12、1060−1066(2001)
【非特許文献3】重松亨ら、『生物工学会誌』、第87巻、第12号、570〜596頁、2009年
【発明の概要】
【発明が解決しようとする課題】
【0007】
これまで、メタン発酵の失敗を客観的に予測し、メタン発酵の運転管理を行う手法は開発されていない。したがって、本発明の目的は、特定微生物の関与に基づいて、メタン発酵の失敗を客観的に予測し、安定的なメタン発酵を達成させるための運転管理の方法を提供することにある。
【課題を解決するための手段】
【0008】
上記実情に鑑み、本発明者らは、メタン発酵が失敗するときに、いわゆる悪玉菌ともいうべき、かかる失敗を誘発する特定の微生物を同定することに成功し、該微生物の増殖をメタン発酵中にモニタリングすることにより、メタン発酵の運転管理を首尾よく行うことができることを見出し、本発明を完成するに至った。
【0009】
すなわち、本発明は、以下の通りである。
[1]有機性廃棄物のメタン発酵系におけるメタン発酵効率をモニタリングする方法であって、下記のプライマーセット:
(a)配列番号1に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド;及び
(b)配列番号2に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド
を用いてポリメラーゼ連鎖反応(PCR)を行い、該メタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下と同時に増殖する真正細菌の16S rDNA量又は16S rRNA量を測定することを含むモニタリング方法。
[2]前記真正細菌が、配列番号3に示される塩基配列を含む16S rDNAを有する、上記[1]に記載のモニタリング方法。
[3]有機性廃棄物が、食品廃棄物、糞尿、動植物性残さ、古紙、及び汚泥からなる群から選択される、上記[1]又は[2]に記載のモニタリング方法。
[4]前記食品廃棄物がシロップ廃液である、上記[3]に記載のモニタリング方法。
[5]有機性廃棄物のメタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下の開始とともに増殖する真正細菌を検出又は定量するためのプライマーセットであって、ここで、該プライマーセットは、
(a)配列番号1に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド;及び
(b)配列番号2に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド
であるプライマーセット。
【発明の効果】
【0010】
本発明は、有機性廃棄物の有効利用を目的としたメタン発酵において、メタン発酵が失敗に至る状態の変化を特定の微生物のDNA情報に基づいて客観的に予測することにより、メタン発酵の運転管理が可能となる。
【図面の簡単な説明】
【0011】
【図1】バンドB6全長の塩基配列を示す。
【図2】Anaerobic syntrophic bacterium NE23−3の16S rRNAの部分塩基配列を示す。
【図3】バンドB6を有する真正細菌を含む系統分布図を示す。
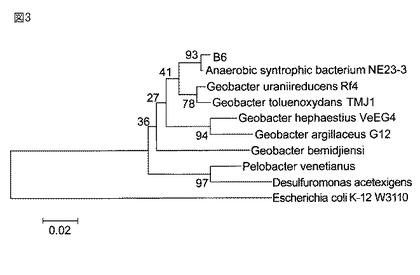
【図4】有機性廃棄物の供給量(TOC濃度)を経時的に変化させたパターンを示す。
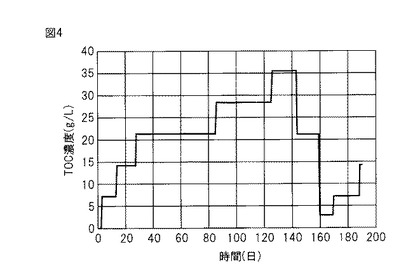
【図5】リアルタイムPCR法に使用したプライマーを用いた検量線を示す。
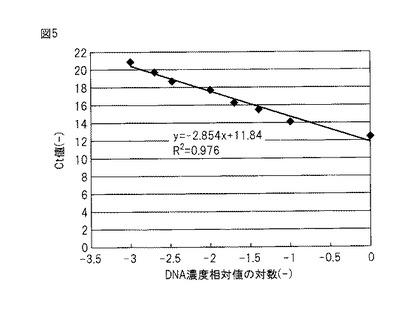
【図6】(A)TOC濃度を経時的に変化させた場合に、メタン発酵中に現れる微生物に固有の16S rRNAを示す。(B)TOC濃度の変化に対応させたメタンガスの発生量を示す。(C)バンドB6の経時的なDNA濃度の相対的変化を示す。
【発明を実施するための形態】
【0012】
以下、本発明の説明のために、好ましい実施形態に関して詳述する。
前述の通り、本発明は、有機性廃棄物のメタン発酵系におけるメタン発酵効率をモニタリングする方法であって、下記のプライマーセット:
(a)配列番号1に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド;及び
(b)配列番号2に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド
を用いてポリメラーゼ連鎖反応(PCR)を行い、該メタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下と同時に増殖する真正細菌の16S rDNA量又は16S rRNA量を測定することを含むモニタリング方法に関する。
【0013】
(1)メタン発酵効率の低下に関与する真正細菌の遺伝子のクローニング
本発明のモニタリング方法において標的とする真正細菌は、Anaerobic syntrophic bacterium NE23−3に近縁の真正細菌であって、固有の16S rRNA(又は16S rDNA)を有する。本発明者らは、メタン発酵中に有機物負荷を徐々に増大させていくと、ある量(「限界負荷量」という。)を超えた時点からメタン発酵効率の低下が見られ、この限界負荷量の前後からある種の真正細菌が増加することを見出し、メタン発酵中のこの真正細菌の16S rRNA(又は16S rDNA)(本明細書において、その部分塩基配列を「バンドB6」と呼ぶ。)量をリアルタイムPCRで測定することにより、メタン発酵効率のモニタリングに成功し、本発明を完成させた(後述する実施例4参照、図6)。なお、バンドB6の塩基配列を図1に示す(配列番号3)。
【0014】
バンドB6は、近縁のAnaerobic syntrophic bacterium NE23−3(GenBank Accession No.AB231802)の16S rRNA(又は16S rDNA)の部分塩基配列(配列番号4)の情報を利用することにより同定された(実施例3参照)。バンドB6を有する真正細菌、及び上記NE23−3株はいずれも未同定であり、基準株(type strain)が存在しない。本発明のモニタリング方法においては、モニタリングの標的となる真正細菌の固有の16S rRNA(又は16S rDNA)の一部であるバンドB6の部分配列(配列番号3のヌクレオチド118−255)を検出の対象とする。
【0015】
本発明のモニタリング方法において使用されるバンドB6の同定は、例えば、以下のようにして行うことができる。簡単には、真正細菌から核酸を抽出後、PCRで増幅して、アガロースゲル電気泳動等の分子量により核酸断片を篩い分ける方法で分離し、特定のバンドを切り出す方法等の常法に従って核酸を単離することができる。メタン発酵に関与する真正細菌は複数種含まれるため、最初に特定の真正細菌における固有の16S rRNA遺伝子を同定することが必要である。まず、グラニュール菌体に含まれる微生物群(「微生物叢」)の16S rRNAをPCR−DGGE法によって分離及び検出する。具体的には、メタン発酵中のリアクター内溶液をサンプリングし、グラニュール状メタン菌を遠心分離によって回収した後、溶菌し、メタン菌に含まれるタンパク質及び脂質をクロロホルムの添加によって抽出・除去する。続いて、エタノールの添加により核酸のみを回収する。その後、真正細菌由来のDNAを検出するために汎用されている以下のプライマーセット(以下、「プライマーセットI」と称する場合がある。):
357f(フォワード):5’−CCTACGGGAGGCAGCAG−3’(配列番号5)(配列番号4のヌクレオチド324−340に対応)
517r(リバース):5’−ATTACCGCGGCTGCTGG−3’(配列番号6)(配列番号4のヌクレオチド518−502に対応)
を用いて、該核酸をポリメラーゼ連鎖反応(PCR)法により増幅する。上記プライマーセットIは、真正細菌を含む大部分の原核生物の16S rRNA遺伝子に結合するとされるユニバーサルプライマーであって、大腸菌の16S rRNA遺伝子の塩基配列を基準にすると、プライマー357f及び517rは、それぞれヌクレオチド341−357及び517−534に対応する。なお、上記プライマー357fには、GCクランプ(5’−CGCCCGCCGCGCGCGGCGGGCGGGGCGGGGGCACGGGGGG−3’)(配列番号7)が付加されている。また、核酸を増幅するための方法としては、PCR法(Saiki R.K.ら,Science,230,1350−1354(1985))に限定されず、当業者であれば、ライゲース連鎖反応(LCR)(Wu D.Y.ら,Genomics,4,560−569(1989))、及び転写に基づく増幅(Kwoh D.Y.ら,Proc.Natl.Acad.Sci.USA,86,1173−1177(1989))等の温度循環を必要とする反応、並びに鎖置換反応(SDA)(Walker G.T.ら,Proc.Natl.Acad.Sci.USA,89,392−396(1992); Walker G.T.ら,Nuc.Acids Res.,20,1691−1696(1992))等を用いることができることは容易に理解される。なお、本発明のモニタリング方法では、メタン発酵効率をリアルタイムで測定する必要性からPCR法を利用することが好ましい。PCR反応条件は、例えば、2本鎖DNAの熱変性を90〜98℃で5秒〜10分間行い、アニーリングを50〜70℃で10秒〜2分間行い、伸長反応を60〜75℃で10秒〜10分間行う。任意に、最終伸長反応を伸長反応と同じ温度で10秒〜15分間行う。熱変性−アニーリング−伸長反応のサイクルを5〜50回行う。
【0016】
次に、上記で核酸を増幅させたPCR増幅産物をアガロースゲル又はポリアクリルアミドゲルで電気泳動することによって分離する。電気泳動によるPCR増幅産物の分離は常法に従って容易に行うことができる。電気泳動で使用するゲルは、核酸を変性させるための変性剤を含み、核酸の分離に適した所定の濃度勾配を有するポリアクリルアミドゲルであることが好ましい。また、電気泳動に使用される泳動溶液及び電圧等の泳動条件は適宜調整することができる。上記のPCR−DGGE法を使用することにより、各真正細菌に該当する核酸のバンドの濃淡から、可視的に真正細菌の経時的な増減をモニタリングすることができる。ここで、ゲル上のバンドの検出は、限定されないが、エチジウムブロマイド含有溶液を用いてゲルを染色することにより行う。
【0017】
さらに、本発明のモニタリング方法で対象とされる真正細菌は、限界負荷量の前後において出現する細菌であり、上記の電気泳動上の経時的なバンドの濃淡の変化において限界負荷量の前後に出現したバンド(固有の16S rRNA遺伝子に対応するDNA断片)を切り出し、塩基配列を解析することにより、メタン発酵効率の低下に関与する真正細菌の遺伝子をクローニングすることができる。ゲルから切り出されたDNA断片には、目的以外のDNA断片が混在している可能性があるため、所望のDNA断片を得るためにさらに分離及び精製することが好ましい。一般に、PCR産物をクローニングするため、ベクター(例えば、pCR4、pMW、pBluescript、pBR322、pUC、pDEL系などのプラスミドベクター、M13mpなどのファージベクター、Charomid9系などのコスミドベクター)に、PCR産物をライゲーションさせ、コンピテントセルに形質転換させる。ベクターにはさらにプロモーター、エンハンサー、ターミネーターなどを適宜含めることができる。形質転換したコンピテントセルを培養してコロニーを形成させ、塩基配列の解読のためのサンプルとする。その後、所望のDNA断片の塩基配列を決定するが、塩基配列の解読は、従来法を用いればよく、例えば、Sanger法、サイクルシークエンス法による自動シークエンサーを利用することができる。自動シークエンサーのシークエンス反応には、プライマーを蛍光標識したDye primer法、ddNTPを蛍光標識したDye terminator法、さらに基質のdNTPに標識したInternal−label法があるが、いずれの方法でもよい。
【0018】
本発明のバンドB6のクローニングにおいては、所望のDNA断片を得るために、PCRによるDNA合成の際に付加されるデオキシアデノシン(dA)を利用したTAクローニング法を用いることができる。TAクローニング法は、3’→5’エキソヌクレア−ゼ活性を有するDNAポリメラーゼを利用したクローニング法であり、まず、このDNAポリメラーゼを用いて得られたPCR産物の平滑末端をTaqDNAポリメラーゼ等のpolI型酵素によって処理することによって、3’末端にアデニン又はチミンを付加させる。一方、ベクターにdTTP又はtaqDNAポリメラーゼを反応させることによってTAベクターを作製する。次に、上記のように処理した核酸断片とベクターをライゲーションさせて宿主細胞に形質転換し、所望のDNAをクローニングする方法である。より具体的には、上記でゲルから切り出されたDNA断片をまず上記のプライマーセットIを使用し、PCRにより増幅する。次に、得られたPCR産物をTAクローニング用のプラスミドに組込み、このプラスミドを大腸菌に形質転換する。次に、PCR産物が組み込まれたプラスミドによって大腸菌に形質転換されたことを確認するために、アンピシリン等の抗生物質による選択を行う。その後、プラスミドDNAを抽出し、プライマーセットIを用いて所望のPCR産物を増幅させ、さらに、上記と同様にDGGEを行い、バンド位置を確認した後、バンドB6の部分塩基配列を決定することができる。本明細書においては、プライマーセットIを用いて決定されたバンドB6の部分塩基配列(配列番号3のヌクレオチド18−178)から、DDBJ(http://www.ddbj.nig.ac.jp/Welcome−j.html)を用いた相同性検索を行い、最も近縁な微生物として、Anaerobic syntrophic bacterium NE23−3(GenBank Accession No.AB231802)を特定した(実施例3参照)。上述の通り、NE23−3株は、未同定であり、基準株が存在しない。得られたバンドB6の部分塩基配列(配列番号3のヌクレオチド18−178)とNE23−3株の部分塩基配列の一部(配列番号4のヌクレオチド341−501)とを比較すると、両者は一致していた。なお、プライマーセットIは、引用文献(Pedro,S.M.ら,J.Biosci.Bioeng.,92,159−164(2001))に記載され、特に変更せずに使用した。また、後述するように、バンドB6の塩基配列としてヌクレオチド1−255を決定した(配列番号3、図1参照)。
【0019】
(2)モニタリング方法
本発明によれば、有機性廃棄物のメタン発酵系におけるメタン発酵効率をモニタリングする方法が提供され、より具体的には、下記のプライマーセット:
(a)配列番号1に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド;及び
(b)配列番号2に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド
を用いてポリメラーゼ連鎖反応(PCR)を行い、該メタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下と同時に増殖する真正細菌の16S rDNA量又は16S rRNA量を測定することを含むモニタリング方法である。
【0020】
本明細書で使用するとき、用語「有機性廃棄物」とは、一般に、生物学的分解によって環境中に直接還元することができる廃棄物を指し、限定されないが、生ごみなどの食品廃棄物、糞尿、動植物性残さ、古紙、及び汚泥などが挙げられる。食品廃棄物には、一般廃棄物(事業系及び家庭系を問わない)及び産業廃棄物が含まれる。具体的には、食品廃棄物の例は、限定されないが、生ごみ、シロップ廃液、おから、排乳、コーヒーかす、茶かすなどが挙げられる。
【0021】
本発明によれば、メタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下と同時に増殖する真正細菌のバンドB6(16S rRNA)量を測定するために使用されるバンドB6に特異的なプライマーセットを提供することができる。本発明においては、下記のプライマーセットII:
GMP478F(フォワード):5’−CGGGTGCTAATATCATCTGCGC−3’(配列番号1)(配列番号3のヌクレオチド118−139に対応)
GMP575R(リバース):5’−ATCAAACCGCCTACACGCGC−3’(配列番号2)(配列番号3のヌクレオチド255−236に対応)
を設計し、メタン発酵効率のモニタリングに使用した。大腸菌の16S rRNA遺伝子の塩基配列を基準にすると、プライマーGMP478F及びGMP575Rは、それぞれヌクレオチド457−478及び575−594に対応する。本発明のモニタリング方法では、上記プライマーセットII以外に、各プライマーに実質的に同一な塩基配列からなるオリゴヌクレオチドを使用することができる。ここで、本明細書で使用するとき、語句「実質的に同一な塩基配列」とは、対応する上記プライマーの塩基配列と90%以上、95%以上、98%以上、若しくは99%以上の配列同一性を有する塩基配列、又は上記プライマーの塩基配列の塩基の複数個、好ましくは5個、より好ましくは4個、さらに好ましくは3個、さらにより好ましくは2個、最も好ましくは1個が欠失、付加及び/又は置換されている塩基配列をいう。
【0022】
プライマーの設計は以下の通りである。上述した通り、バンドB6は、プライマーセットI(プライマー357Fと517R)に含まれる塩基配列の情報からAnaerobic syntrophic bacterium NE23−3(Accession No.AB231802)に最も近縁であることが判明した(図3参照)。そこで、NE23−3株の塩基配列情報(配列番号4、図2)に基づいて、BioEdit v7.0.9 General Informationを用いて、NE23−3株とその他の微生物群における塩基配列の相同性を確認し、塩基配列中で保存性の低い領域を検索して、特異性の高いプライマーをFastPCR(URL:http://www.primerdigital.com/fastpcr.html)の推奨結果を参考に設計した。なお、プライマーの設計時は、一般的にいわれている以下の条件を満たすように留意した。増幅サイズは80〜150塩基(増幅効率を100%にするため)、プライマー長さは17〜25塩基(長過ぎるとアニーリング効率が低下する)GC含量は40〜60%(45〜55%が望ましい)とし、偏りがないように、部分的にGC、ATリッチができないように注意する、T/C、A/Gが連続しないように注意する、プライマー内部での3塩基以上の相補的配列を避ける、プライマー間での3塩基以上の相補的配列を避ける、プライマー3’末端が2塩基相補する配列を避ける。プライマーの標的塩基配列への特異性を確認するため、所定のPCR反応条件下で定量的PCR法(例えば、SYBRグリーンを利用したリアルタイムPCR法、蛍光プローブ法、競合PCR法)を行い、増幅曲線分析及び融解曲線分析を行ってバンドB6に相当する微生物濃度を定量することができる。
【0023】
上記で設計したプライマーセットIIの使用により、メタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下と同時に増殖する真正細菌の16S rRNA(又は16S rDNA)(一部にバンドB6遺伝子を含む)を検出し、例えば、リアルタイムPCR法によってバンドB6を経時的に定量することができる。なお、バンドB6を有する真正細菌(菌体)は、バンドB6の塩基配列を含む約1500塩基の16S領域を有するが、現時点では完全に配列決定されていない。また、このバンドB6は、メタン発酵効率の低下と関連付けて見出されたが、バンドB6を有する真正細菌は、メタン発酵系においてメタン発酵効率の低下の開始前、開始と同時、又は開始後に増殖してメタン発酵系に出現する細菌である。本発明のモニタリング方法においては、メタン発酵効率の低下を予測することを目的としているため、メタン発酵効率の低下に先駆けて(低下の開始前に)増殖するか又は該低下と同時に増殖するバンドB6を有する真正細菌を検出されることが望ましい。ここで、本明細書で使用するとき、「先駆けて」とは、メタン発酵効率の低下開始の少なくとも2〜3週間前、1週間前、数日前、1日前、12時間前、6時間前、3時間前、1時間前、又は30分前に、該真正細菌が増殖を開始するか又はバンドB6が検出される場合をいう。検出されるバンドB6は、遺伝子(DNA若しくはmRNA)の形態であるか又はタンパク質の形態であってもよい。
【0024】
本発明のモニタリング方法によれば、プライマーセットIIを用いたリアルタイムPCRを行った後、バンドB6に対応する菌体を含んでいるメタン発酵過程で採取したグラニュールサンプルを連続希釈し、この希釈率に応じた菌体数に対するバンドB6の検量線を作成することができる。これにより、目的とする真正細菌の試料中での有無、その数量を精度よく知ることができる。また、本明細書で使用するとき、メタン発酵効率の「低下」とは、有機性廃棄物の限界負荷量を超えて添加された場合に、メタン発酵系に有機性廃棄物を添加することによって発生していたメタンガスの量が減少している、又は有機性廃棄物を添加したとしてもメタンガスが発生しないことを意味する。限界定負荷量を超えたことによるメタン発酵効率の低下は、メタンガスの発生量が5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、又は100%減少した場合を指す。本発明のモニタリング方法を適用することにより、メタン発酵効率の低下に先駆けて又は低下と同時に増加するバンドB6量をリアルタイムで定量することにより、メタン発酵が失敗に至る状態の変化を予測することができるため、添加する有機性廃棄物の量を調節することによって、安定なメタン発酵が可能になる。また、本発明によれば、本発明のモニタリング方法を応用したバイオセンサーを提供することができる。より具体的には、前述のプライマーセットIIが電極に固定化されたバイオセンサーを用いることにより、リアクター内の発酵液中のバンドB6量を検出及び定量することにより、リアクターに供給する有機性廃棄物の量を調節することができる。
以下、実施例により本発明をさらに詳細に説明するが、本発明はこれらの実施例に限定されない。
【実施例】
【0025】
実施例1:UASBメタン発酵
メタン発酵にはUASB法を使用し、原料の有機性廃棄物としては、共同出願人である山梨缶詰株式会社の工場において缶詰製造過程で排出される平均2.2トン/日のシロップ廃液を用いた。使用したシロップ廃液は、主成分がスクロースであり、糖度が15.2ブリックス、全有機炭素(TOC)濃度が約71g/Lであった。メタン発酵には、外寸法φ2500×H5000mmのUASB発酵槽を用いた。発酵液容積は20m3であった。実験開始時にビール工場のUASB発酵槽で形成されたグラニュール汚泥を15m3充填した。また、実験開始から2日間は馴致期間とし、シロップ廃液を供給せずに温度を40℃に維持した。実験開始から2日以降にシロップ廃液の供給を始めるが、シロップ廃液をそのままUASB発酵槽へ投入すると負荷が高すぎるため、調整槽において水で適宣希釈し、pHが7.5付近になるように苛性ソーダを用いて調整した。その後、温調槽で39℃まで昇温した後にUASB発酵槽に導いた。水で希釈したシロップ廃液の濃度を段階的に変化させたが、およそ4〜5ヶ月後にはシロップ廃液を等量の水で希釈し、TOC濃度が35.5g/Lとなるように負荷を増加させた(図4参照)。なお、145日目以降は再び負荷量を低下させた。また、運転開始49日以降にはメタン発酵を担う微生物の活性を高める目的で金属塩CoCl2、MgCl2、MnCl2、NiCl2をそれぞれ20gずつ(供給するシロップ廃液の1ppm程度に相当)を添加し、さらに、64日以降にはC/N比が15になるように調整槽にて尿素を添加した。ガス発生量は発酵槽上部に取り付けたガス出口の流量計(山武、気体用マスフローメーター、CMS0500)で計測し、pHは槽内のpHメータ(YOKOGAWA、pH100)で計測した。なお、発生するガスは、1日1回、テドラーバッグに捕集し、その中に含まれるメタンガス濃度をガスクロマトグラフ分析装置(ヤナコ、G−1011、カラム MS13X+MS5A)を用いて測定した。メタン発酵の設定温度を40℃とし、グラニュールを含む発酵液サンプルを発酵槽下部から1700mm、壁面から750mmのサンプリング口から所定の時間間隔で、約200mLを採取して、微生物叢の解析に用いた。
【0026】
実施例2:グラニュール粒子からのDNA抽出及びPCR増幅
採取した発酵液サンプル中のグラニュールの約1gを滅菌したスパーテルですくい取り、30mLの滅菌水で洗浄してグラニュール表面に付着している微生物を除去した。その後、その0.3gを使用して、Isoil for Beads Beating kit(Nippon Gene Co.,Ltd.)によりDNAを抽出した。次に、グラニュールから抽出したDNAには、真正細菌及び古細菌に由来するDNAが含まれるが、真正細菌に由来するDNAを増幅させるために、前述のプライマーセットI(プライマー357fと517r)を用いた。増幅条件を以下の表1に示す。
【0027】
【表1】

【0028】
使用したPCR増幅装置はPTC−100 thermal cycler(TaKaRa Shuzo CO.,Ltd.,Kyoto,Japan)であった。また、PCR反応液の組成を表2に示す。
【0029】
【表2】
【0030】
実施例3:バンドB6の分離及び同定
真正細菌からバンドB6を検出するためにDGGE(変性剤濃度勾配ゲル電気泳動)法を用いた。DGGEにはThe Dcode Universal Mutation Detection system(Bio−Rad、Hercules、Calif.)を使用した。まず、グラニュールから抽出した真正細菌由来のDNAを増幅したPCR産物を変性剤の濃度勾配が30%から60%(変性剤100%では7Mの尿素と40%[wt/vol]のホルムアミドを含む)の厚さ0.8mmのポリアクリルアミドゲル(10%[wt/vol]アクリルアミドとビスアクリルアミドの割合は37.5:1)を用いて、1.0×Tris−acetate−EDTA(TAE)溶液中で60℃、200V、3.5時間の条件で電気泳動を行った。なお、電気泳動の後、200mLの1.0×TAE溶液と20μLのエチジウムブロマイド溶液を混合した溶液にゲルを浸し、30分間染色した後、紫外線下で可視化した。
【0031】
DGGE法で検出されたポリアクリルアミドゲル中のバンドからDNA断片を抽出・回収した。抽出したDNA断片から所望のDNA断片を分離及び精製するために、TAクローニング法を用いた。まず、抽出されたDNA断片は、前述のプライマーセットIを用いて、上記と同一の増幅条件を用いて増幅させた。得られたPCR増幅産物は、TOPO TA Cloning Kit(Invitrogen Copration,USA)を用いてプラスミドにライゲーション後、大腸菌に形質転換した。この形質転換した大腸菌を、アンピシリンを添加したLB寒天培地に塗沫し、カラーセレクションを行った。カラーセレクションによりPCR産物をライゲーションしたプラスミドを有することが確認された大腸菌については、まず、下記に示すプライマーセットIII:
M13 F(5’−GTAAAACGACGGCCAG−3’)(配列番号8)
M13 R(5’−CAGGAAACAGCTATGAC−3’)(配列番号9)
を用いてDNAを増幅し、その増幅産物について、さらにプライマーセットIを用いてPCRを行い、再度DGGEを行ってバンドの出現位置を確認した。なお、プライマーセットIIIを用いたPCR条件を以下の表3に示す。
【0032】
【表3】
【0033】
その後、確認された所望のDNA断片はWizard(登録商標)SV Gel and PCR Clean−UP system(Promega Copration,USA)を用いて精製した。精製したPCR増幅物はBigDye terminator kit(Perkin Elmer Japan,Applied Biosystems Division)を用いてシーケンス反応を行った。塩基配列はABI Prism 310 Genetic Analyzer(Applied Biosystems)で解析した。決定した塩基配列については、DDBJ(http://www.ddbj.nig.ac.jp/Welcome−j.html)に登録されている配列に対して相同性検索を行い最も近縁な微生物名を決定した。
【0034】
実施例4:リアルタイムPCR法を用いたDNA濃度の定量
一般に嫌気性菌は増殖が遅く、生成するコロニーも小さいので寒天平板上でのコロニーアピアランスによる識別が困難であることから、メタン発酵のような複数の微生物が共存する複合微生物の系で特定の微生物の濃度を選択的に定量するためには、その微生物に特徴的な遺伝子の配列を検出するリアルタイムPCR法を有効に使用することができる。そこで、バンドB6を特異的に検出するプライマーセットを設計し、これを用いてメタン発酵槽のグラニュール中の真正細菌に含まれるバンドB6を定量した。ここで使用するリアルタイムPCR法は、PCRによるDNAの増幅量をリアルタイムでモニタリングし、測定される蛍光強度の閾値と増幅曲線が交わるサイクル数(Ct値(Threshold Cycle))と、初期鋳型量、すなわち初期微生物濃度の間に直線関係があることを利用して、微生物濃度を定量する方法である。
【0035】
(1)プライマーの設計
バンドB6は、前述のプライマー357Fと517Rに含まれる塩基配列の情報からAnaerobic syntrophic bacterium NE23−3(Accession No.AB231802)に最も近縁であることが判明した。NE23−3株の塩基配列情報(図2)をもとにBioEdit v7.0.9 General Informationを用いて、NE23−3株とその他の微生物群における塩基配列の相同性を確認し、塩基配列の中で保存性の低い領域を検索して、特異性の高いプライマーをFastPCR(URL:http://www.primerdigital.com/fastpcr.html)の推奨結果を参考に設計した。なお、プライマーの設計時は、一般的にいわれている以下の条件を満たすように留意した。増幅サイズは80〜150塩基(増幅効率を100%にするため)、プライマー長さは17〜25塩基(長過ぎるとアニーリング効率が低下する)GC含量は40〜60%(45〜55%が望ましい)とし、偏りがないように、部分的にGC、ATリッチができないように注意する、T/C、A/Gが連続しないように注意する、プライマー内部での3 塩基以上の相補的配列を避ける、プライマー間での3塩基以上の相補的配列を避ける、プライマー3’末端が2塩基相補する配列を避ける。設計したプライマー(プライマーセットII)の塩基配列は前述の通りである。
【0036】
(2)設計したプライマーの特異性の検討
設計したプライマーセットIIが目的菌に選択的であることをBioEdit v7.0.9 General Informationを用いて、プライマーの塩基配列と目的菌以外の微生物群の塩基配列とを比較することで確認するとともに、実際のメタン発酵グラニュールサンプルに設計したプライマー(プライマーセットII)を作用させて、増幅される複数のDNA断片の塩基配列がすべて同一であることを確かめた(データ示さず)。増幅されるDNA断片の塩基配列が同一であれば、設計したプライマーが類似の塩基配列を有する複数種類の微生物を検出する可能性は極めて小さくなる。まず、メタン発酵グラニュールサンプルにプライマーセットIIを加えてPCRによりDNA断片を増幅させた。なお、PCR条件を以下の表4に示す。
【0037】
【表4】
【0038】
次に、増幅したDNA断片をTAクローニング法により大腸菌に形質転換した。続いてPCR産物がライゲーションされたプラスミドを持つことが確認された大腸菌にプライマーセットIII(前述)を加えて、PCR増幅し、精製した。その後、BigDye terminator kit(Perkin Elmer Japan,Applied Biosystems Division)を用いてシーケンス反応を行った。塩基配列はABI Prism 310 Genetic Analyzer(Applied Biosystems)で解析した。目的としたPCR産物がライゲーションされたプラスミドを持つことが確認された大腸菌コロニーが12個得られた。これらのすべてについてPCR産物の塩基配列を解析したところ、塩基配列は完全に同一であり、設計したプライマーが類似の塩基配列を有する複数種類の微生物を検出している確率は極めて小さいことが判明した。また、このプライマーセットIIで増幅したDNA領域は、バンドB6の同定に用いたDNA領域と同じ遺伝子上に存在することを確かめた。
【0039】
(3)設計したプライマーを用いた検量線の作成
メタン発酵グラニュールサンプルを適宜希釈して、設計したプライマーの定量性を確認した。メタン発酵グラニュールサンプルは発酵開始125日目のものを用いた。リアルタイムPCR装置はSmart Cycler II(タカラバイオ株式会社)を用い、PCR反応液は、プライマーセットIIとSYBR Premix Ex Taq(タカラバイオ株式会社)を用いて、下記の表5のように調製した。
【0040】
【表5】
【0041】
増幅条件は最初に95℃、10秒の変性をおこなった後、95℃、5秒(変性)、60℃、20秒(アニーリング/伸長)を40サイクルとした(表6参照)。作製した検量線を図5に示す。検量線のR2値は0.976であり、定量に用いるのに十分な精度であった。
【0042】
【表6】
【0043】
(4)リアルタイムPCR法によるバンドB6に対応する微生物の定量
メタン発酵グラニュールサンプルからのDNA抽出物にリアルタイムPCR法を適用してバンドB6に対応する菌体濃度を測定した。用いたプライマーセットとPCR条件は、検量線を作成したときと同様とした。なお、菌体濃度はメタン発酵125日後の菌体濃度が最大になるときを1とした相対濃度で示した(図6参照)。
【技術分野】
【0001】
本発明は、有機性廃棄物のメタン発酵系におけるメタン発酵効率をモニタリングする方法、及び該方法に使用されるプライマーセットに関する。より具体的には、本発明は、有機性廃棄物のメタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下と同時に増殖する特定の真正細菌をそのDNA情報に基づいて検出及び定量することを特徴とするメタン発酵の運転管理に関する。
【背景技術】
【0002】
メタン発酵は、有機性廃水を処理する方法の1つで、ランニングコストが安く、さらにバイオガスとしてエネルギー回収ができ、余剰汚泥発生量も少ないという特徴を有する優れた方法として知られている(非特許文献1参照)。メタン発酵法の歴史は古く、嫌気性微生物による高濃度有機性廃水や汚泥消化等の処理技術として開発されてきた。近年では、高濃度有機性廃水等に使われた第一世代に代わって、UASB法(上向流嫌気性汚泥床法)の開発に伴う第二世代のメタン発酵法が実績を増やしている。UASB法とは、0.5〜2mm程度に自己造粒した高密度グラニュール菌体(真正細菌及び古細菌を含む)を利用して、高濃度基質の高負荷高速運転を可能としたリアクターを利用する方法である(非特許文献1及び2参照)。グラニュール菌体は沈降性が良いことから、それまでの懸濁型メタン発酵に見られるような気液固分離の欠点を解決できる手法である。
【0003】
メタン発酵は、多くの微生物の共生により有機物を分解し、その過程で生成する有機酸などをメタンに還元する方法であって、主として3段階で進行すると考えられている。複雑な有機物は、第1段階の酸生成過程(液化過程)で酸生成菌類の作用により、単糖類、アミノ酸などの低分子量の物質を経て、酢酸、プロピオン酸、ギ酸、酪酸などの低級脂肪酸、乳酸及びエタノールなどになる。第2段階においては、酢酸以外の低級脂肪酸、乳酸及びエタノールは、水素生成細菌により水素及び酢酸に変換され、最終の第3段階において基質特異性の強いメタン生成古細菌群により、メタン及び二酸化炭素などに分解される。ここで、酢酸及びプロピオン酸などの低級脂肪酸はメタン発酵プロセスにおける主要な中間生成物であり、酸生成菌類及び水素生成細菌などの真正細菌がその役割を担っている(非特許文献3参照)。
【0004】
メタン発酵においては、その原料となる有機性廃棄物の供給量(有機物負荷)が大きくなると、供給量に応じてメタンガスの発生量も増加するが、有機物負荷が過剰になると、メタン生成に関与する真正細菌及び古細菌の活性が著しく低下し、メタンガスが全く生成されなくなる(メタン発酵が失敗する)。有機物負荷の過剰によりメタン生成が行われなくなると、メタン発酵を復帰させることは非常に困難であるという問題点がある。この限界の負荷量をリアクター内への流速や有機物濃度などから決定することは困難であり、これまで限界の負荷量は熟練のオペレータの判断に任されていた。これまで、メタン発酵が失敗した後に有用な微生物が死滅又は減少するという報告がなされている(特許文献1参照)。しかしながら、特許文献1に開示された方法は、メタン発酵の失敗を後付け的に説明することはできても、メタン発酵の失敗を予測し、未然に予防することができない。また、メタン発酵が失敗するとき、又は失敗したときには、リアクター内のpHが低下している(pH低下はメタン発酵失敗の原因であり、結果である、すなわち負のフィードバックと考えられる)ということは経験的によく知られている。しかしながら、メタン発酵が失敗しなくても一時的にpH低下が起こることも周知な現象である。よって、pH低下に基づいて、メタン発酵失敗の予兆とすることはできない。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特表2006−325581
【非特許文献】
【0006】
【非特許文献1】松本豊、中崎清彦共著、『化学工学の進歩第36集 環境調和型エネルギーシステム』(社団法人化学工学会編集)、「4.3 メタン発酵技術」(140〜156頁)、2002年、槇書店
【非特許文献2】松尾幸彦他、『用水と廃水』、「UASB法を用いたPEG排水の高速処理」、Vol.43、No.12、1060−1066(2001)
【非特許文献3】重松亨ら、『生物工学会誌』、第87巻、第12号、570〜596頁、2009年
【発明の概要】
【発明が解決しようとする課題】
【0007】
これまで、メタン発酵の失敗を客観的に予測し、メタン発酵の運転管理を行う手法は開発されていない。したがって、本発明の目的は、特定微生物の関与に基づいて、メタン発酵の失敗を客観的に予測し、安定的なメタン発酵を達成させるための運転管理の方法を提供することにある。
【課題を解決するための手段】
【0008】
上記実情に鑑み、本発明者らは、メタン発酵が失敗するときに、いわゆる悪玉菌ともいうべき、かかる失敗を誘発する特定の微生物を同定することに成功し、該微生物の増殖をメタン発酵中にモニタリングすることにより、メタン発酵の運転管理を首尾よく行うことができることを見出し、本発明を完成するに至った。
【0009】
すなわち、本発明は、以下の通りである。
[1]有機性廃棄物のメタン発酵系におけるメタン発酵効率をモニタリングする方法であって、下記のプライマーセット:
(a)配列番号1に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド;及び
(b)配列番号2に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド
を用いてポリメラーゼ連鎖反応(PCR)を行い、該メタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下と同時に増殖する真正細菌の16S rDNA量又は16S rRNA量を測定することを含むモニタリング方法。
[2]前記真正細菌が、配列番号3に示される塩基配列を含む16S rDNAを有する、上記[1]に記載のモニタリング方法。
[3]有機性廃棄物が、食品廃棄物、糞尿、動植物性残さ、古紙、及び汚泥からなる群から選択される、上記[1]又は[2]に記載のモニタリング方法。
[4]前記食品廃棄物がシロップ廃液である、上記[3]に記載のモニタリング方法。
[5]有機性廃棄物のメタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下の開始とともに増殖する真正細菌を検出又は定量するためのプライマーセットであって、ここで、該プライマーセットは、
(a)配列番号1に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド;及び
(b)配列番号2に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド
であるプライマーセット。
【発明の効果】
【0010】
本発明は、有機性廃棄物の有効利用を目的としたメタン発酵において、メタン発酵が失敗に至る状態の変化を特定の微生物のDNA情報に基づいて客観的に予測することにより、メタン発酵の運転管理が可能となる。
【図面の簡単な説明】
【0011】
【図1】バンドB6全長の塩基配列を示す。
【図2】Anaerobic syntrophic bacterium NE23−3の16S rRNAの部分塩基配列を示す。
【図3】バンドB6を有する真正細菌を含む系統分布図を示す。
【図4】有機性廃棄物の供給量(TOC濃度)を経時的に変化させたパターンを示す。
【図5】リアルタイムPCR法に使用したプライマーを用いた検量線を示す。
【図6】(A)TOC濃度を経時的に変化させた場合に、メタン発酵中に現れる微生物に固有の16S rRNAを示す。(B)TOC濃度の変化に対応させたメタンガスの発生量を示す。(C)バンドB6の経時的なDNA濃度の相対的変化を示す。
【発明を実施するための形態】
【0012】
以下、本発明の説明のために、好ましい実施形態に関して詳述する。
前述の通り、本発明は、有機性廃棄物のメタン発酵系におけるメタン発酵効率をモニタリングする方法であって、下記のプライマーセット:
(a)配列番号1に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド;及び
(b)配列番号2に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド
を用いてポリメラーゼ連鎖反応(PCR)を行い、該メタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下と同時に増殖する真正細菌の16S rDNA量又は16S rRNA量を測定することを含むモニタリング方法に関する。
【0013】
(1)メタン発酵効率の低下に関与する真正細菌の遺伝子のクローニング
本発明のモニタリング方法において標的とする真正細菌は、Anaerobic syntrophic bacterium NE23−3に近縁の真正細菌であって、固有の16S rRNA(又は16S rDNA)を有する。本発明者らは、メタン発酵中に有機物負荷を徐々に増大させていくと、ある量(「限界負荷量」という。)を超えた時点からメタン発酵効率の低下が見られ、この限界負荷量の前後からある種の真正細菌が増加することを見出し、メタン発酵中のこの真正細菌の16S rRNA(又は16S rDNA)(本明細書において、その部分塩基配列を「バンドB6」と呼ぶ。)量をリアルタイムPCRで測定することにより、メタン発酵効率のモニタリングに成功し、本発明を完成させた(後述する実施例4参照、図6)。なお、バンドB6の塩基配列を図1に示す(配列番号3)。
【0014】
バンドB6は、近縁のAnaerobic syntrophic bacterium NE23−3(GenBank Accession No.AB231802)の16S rRNA(又は16S rDNA)の部分塩基配列(配列番号4)の情報を利用することにより同定された(実施例3参照)。バンドB6を有する真正細菌、及び上記NE23−3株はいずれも未同定であり、基準株(type strain)が存在しない。本発明のモニタリング方法においては、モニタリングの標的となる真正細菌の固有の16S rRNA(又は16S rDNA)の一部であるバンドB6の部分配列(配列番号3のヌクレオチド118−255)を検出の対象とする。
【0015】
本発明のモニタリング方法において使用されるバンドB6の同定は、例えば、以下のようにして行うことができる。簡単には、真正細菌から核酸を抽出後、PCRで増幅して、アガロースゲル電気泳動等の分子量により核酸断片を篩い分ける方法で分離し、特定のバンドを切り出す方法等の常法に従って核酸を単離することができる。メタン発酵に関与する真正細菌は複数種含まれるため、最初に特定の真正細菌における固有の16S rRNA遺伝子を同定することが必要である。まず、グラニュール菌体に含まれる微生物群(「微生物叢」)の16S rRNAをPCR−DGGE法によって分離及び検出する。具体的には、メタン発酵中のリアクター内溶液をサンプリングし、グラニュール状メタン菌を遠心分離によって回収した後、溶菌し、メタン菌に含まれるタンパク質及び脂質をクロロホルムの添加によって抽出・除去する。続いて、エタノールの添加により核酸のみを回収する。その後、真正細菌由来のDNAを検出するために汎用されている以下のプライマーセット(以下、「プライマーセットI」と称する場合がある。):
357f(フォワード):5’−CCTACGGGAGGCAGCAG−3’(配列番号5)(配列番号4のヌクレオチド324−340に対応)
517r(リバース):5’−ATTACCGCGGCTGCTGG−3’(配列番号6)(配列番号4のヌクレオチド518−502に対応)
を用いて、該核酸をポリメラーゼ連鎖反応(PCR)法により増幅する。上記プライマーセットIは、真正細菌を含む大部分の原核生物の16S rRNA遺伝子に結合するとされるユニバーサルプライマーであって、大腸菌の16S rRNA遺伝子の塩基配列を基準にすると、プライマー357f及び517rは、それぞれヌクレオチド341−357及び517−534に対応する。なお、上記プライマー357fには、GCクランプ(5’−CGCCCGCCGCGCGCGGCGGGCGGGGCGGGGGCACGGGGGG−3’)(配列番号7)が付加されている。また、核酸を増幅するための方法としては、PCR法(Saiki R.K.ら,Science,230,1350−1354(1985))に限定されず、当業者であれば、ライゲース連鎖反応(LCR)(Wu D.Y.ら,Genomics,4,560−569(1989))、及び転写に基づく増幅(Kwoh D.Y.ら,Proc.Natl.Acad.Sci.USA,86,1173−1177(1989))等の温度循環を必要とする反応、並びに鎖置換反応(SDA)(Walker G.T.ら,Proc.Natl.Acad.Sci.USA,89,392−396(1992); Walker G.T.ら,Nuc.Acids Res.,20,1691−1696(1992))等を用いることができることは容易に理解される。なお、本発明のモニタリング方法では、メタン発酵効率をリアルタイムで測定する必要性からPCR法を利用することが好ましい。PCR反応条件は、例えば、2本鎖DNAの熱変性を90〜98℃で5秒〜10分間行い、アニーリングを50〜70℃で10秒〜2分間行い、伸長反応を60〜75℃で10秒〜10分間行う。任意に、最終伸長反応を伸長反応と同じ温度で10秒〜15分間行う。熱変性−アニーリング−伸長反応のサイクルを5〜50回行う。
【0016】
次に、上記で核酸を増幅させたPCR増幅産物をアガロースゲル又はポリアクリルアミドゲルで電気泳動することによって分離する。電気泳動によるPCR増幅産物の分離は常法に従って容易に行うことができる。電気泳動で使用するゲルは、核酸を変性させるための変性剤を含み、核酸の分離に適した所定の濃度勾配を有するポリアクリルアミドゲルであることが好ましい。また、電気泳動に使用される泳動溶液及び電圧等の泳動条件は適宜調整することができる。上記のPCR−DGGE法を使用することにより、各真正細菌に該当する核酸のバンドの濃淡から、可視的に真正細菌の経時的な増減をモニタリングすることができる。ここで、ゲル上のバンドの検出は、限定されないが、エチジウムブロマイド含有溶液を用いてゲルを染色することにより行う。
【0017】
さらに、本発明のモニタリング方法で対象とされる真正細菌は、限界負荷量の前後において出現する細菌であり、上記の電気泳動上の経時的なバンドの濃淡の変化において限界負荷量の前後に出現したバンド(固有の16S rRNA遺伝子に対応するDNA断片)を切り出し、塩基配列を解析することにより、メタン発酵効率の低下に関与する真正細菌の遺伝子をクローニングすることができる。ゲルから切り出されたDNA断片には、目的以外のDNA断片が混在している可能性があるため、所望のDNA断片を得るためにさらに分離及び精製することが好ましい。一般に、PCR産物をクローニングするため、ベクター(例えば、pCR4、pMW、pBluescript、pBR322、pUC、pDEL系などのプラスミドベクター、M13mpなどのファージベクター、Charomid9系などのコスミドベクター)に、PCR産物をライゲーションさせ、コンピテントセルに形質転換させる。ベクターにはさらにプロモーター、エンハンサー、ターミネーターなどを適宜含めることができる。形質転換したコンピテントセルを培養してコロニーを形成させ、塩基配列の解読のためのサンプルとする。その後、所望のDNA断片の塩基配列を決定するが、塩基配列の解読は、従来法を用いればよく、例えば、Sanger法、サイクルシークエンス法による自動シークエンサーを利用することができる。自動シークエンサーのシークエンス反応には、プライマーを蛍光標識したDye primer法、ddNTPを蛍光標識したDye terminator法、さらに基質のdNTPに標識したInternal−label法があるが、いずれの方法でもよい。
【0018】
本発明のバンドB6のクローニングにおいては、所望のDNA断片を得るために、PCRによるDNA合成の際に付加されるデオキシアデノシン(dA)を利用したTAクローニング法を用いることができる。TAクローニング法は、3’→5’エキソヌクレア−ゼ活性を有するDNAポリメラーゼを利用したクローニング法であり、まず、このDNAポリメラーゼを用いて得られたPCR産物の平滑末端をTaqDNAポリメラーゼ等のpolI型酵素によって処理することによって、3’末端にアデニン又はチミンを付加させる。一方、ベクターにdTTP又はtaqDNAポリメラーゼを反応させることによってTAベクターを作製する。次に、上記のように処理した核酸断片とベクターをライゲーションさせて宿主細胞に形質転換し、所望のDNAをクローニングする方法である。より具体的には、上記でゲルから切り出されたDNA断片をまず上記のプライマーセットIを使用し、PCRにより増幅する。次に、得られたPCR産物をTAクローニング用のプラスミドに組込み、このプラスミドを大腸菌に形質転換する。次に、PCR産物が組み込まれたプラスミドによって大腸菌に形質転換されたことを確認するために、アンピシリン等の抗生物質による選択を行う。その後、プラスミドDNAを抽出し、プライマーセットIを用いて所望のPCR産物を増幅させ、さらに、上記と同様にDGGEを行い、バンド位置を確認した後、バンドB6の部分塩基配列を決定することができる。本明細書においては、プライマーセットIを用いて決定されたバンドB6の部分塩基配列(配列番号3のヌクレオチド18−178)から、DDBJ(http://www.ddbj.nig.ac.jp/Welcome−j.html)を用いた相同性検索を行い、最も近縁な微生物として、Anaerobic syntrophic bacterium NE23−3(GenBank Accession No.AB231802)を特定した(実施例3参照)。上述の通り、NE23−3株は、未同定であり、基準株が存在しない。得られたバンドB6の部分塩基配列(配列番号3のヌクレオチド18−178)とNE23−3株の部分塩基配列の一部(配列番号4のヌクレオチド341−501)とを比較すると、両者は一致していた。なお、プライマーセットIは、引用文献(Pedro,S.M.ら,J.Biosci.Bioeng.,92,159−164(2001))に記載され、特に変更せずに使用した。また、後述するように、バンドB6の塩基配列としてヌクレオチド1−255を決定した(配列番号3、図1参照)。
【0019】
(2)モニタリング方法
本発明によれば、有機性廃棄物のメタン発酵系におけるメタン発酵効率をモニタリングする方法が提供され、より具体的には、下記のプライマーセット:
(a)配列番号1に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド;及び
(b)配列番号2に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド
を用いてポリメラーゼ連鎖反応(PCR)を行い、該メタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下と同時に増殖する真正細菌の16S rDNA量又は16S rRNA量を測定することを含むモニタリング方法である。
【0020】
本明細書で使用するとき、用語「有機性廃棄物」とは、一般に、生物学的分解によって環境中に直接還元することができる廃棄物を指し、限定されないが、生ごみなどの食品廃棄物、糞尿、動植物性残さ、古紙、及び汚泥などが挙げられる。食品廃棄物には、一般廃棄物(事業系及び家庭系を問わない)及び産業廃棄物が含まれる。具体的には、食品廃棄物の例は、限定されないが、生ごみ、シロップ廃液、おから、排乳、コーヒーかす、茶かすなどが挙げられる。
【0021】
本発明によれば、メタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下と同時に増殖する真正細菌のバンドB6(16S rRNA)量を測定するために使用されるバンドB6に特異的なプライマーセットを提供することができる。本発明においては、下記のプライマーセットII:
GMP478F(フォワード):5’−CGGGTGCTAATATCATCTGCGC−3’(配列番号1)(配列番号3のヌクレオチド118−139に対応)
GMP575R(リバース):5’−ATCAAACCGCCTACACGCGC−3’(配列番号2)(配列番号3のヌクレオチド255−236に対応)
を設計し、メタン発酵効率のモニタリングに使用した。大腸菌の16S rRNA遺伝子の塩基配列を基準にすると、プライマーGMP478F及びGMP575Rは、それぞれヌクレオチド457−478及び575−594に対応する。本発明のモニタリング方法では、上記プライマーセットII以外に、各プライマーに実質的に同一な塩基配列からなるオリゴヌクレオチドを使用することができる。ここで、本明細書で使用するとき、語句「実質的に同一な塩基配列」とは、対応する上記プライマーの塩基配列と90%以上、95%以上、98%以上、若しくは99%以上の配列同一性を有する塩基配列、又は上記プライマーの塩基配列の塩基の複数個、好ましくは5個、より好ましくは4個、さらに好ましくは3個、さらにより好ましくは2個、最も好ましくは1個が欠失、付加及び/又は置換されている塩基配列をいう。
【0022】
プライマーの設計は以下の通りである。上述した通り、バンドB6は、プライマーセットI(プライマー357Fと517R)に含まれる塩基配列の情報からAnaerobic syntrophic bacterium NE23−3(Accession No.AB231802)に最も近縁であることが判明した(図3参照)。そこで、NE23−3株の塩基配列情報(配列番号4、図2)に基づいて、BioEdit v7.0.9 General Informationを用いて、NE23−3株とその他の微生物群における塩基配列の相同性を確認し、塩基配列中で保存性の低い領域を検索して、特異性の高いプライマーをFastPCR(URL:http://www.primerdigital.com/fastpcr.html)の推奨結果を参考に設計した。なお、プライマーの設計時は、一般的にいわれている以下の条件を満たすように留意した。増幅サイズは80〜150塩基(増幅効率を100%にするため)、プライマー長さは17〜25塩基(長過ぎるとアニーリング効率が低下する)GC含量は40〜60%(45〜55%が望ましい)とし、偏りがないように、部分的にGC、ATリッチができないように注意する、T/C、A/Gが連続しないように注意する、プライマー内部での3塩基以上の相補的配列を避ける、プライマー間での3塩基以上の相補的配列を避ける、プライマー3’末端が2塩基相補する配列を避ける。プライマーの標的塩基配列への特異性を確認するため、所定のPCR反応条件下で定量的PCR法(例えば、SYBRグリーンを利用したリアルタイムPCR法、蛍光プローブ法、競合PCR法)を行い、増幅曲線分析及び融解曲線分析を行ってバンドB6に相当する微生物濃度を定量することができる。
【0023】
上記で設計したプライマーセットIIの使用により、メタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下と同時に増殖する真正細菌の16S rRNA(又は16S rDNA)(一部にバンドB6遺伝子を含む)を検出し、例えば、リアルタイムPCR法によってバンドB6を経時的に定量することができる。なお、バンドB6を有する真正細菌(菌体)は、バンドB6の塩基配列を含む約1500塩基の16S領域を有するが、現時点では完全に配列決定されていない。また、このバンドB6は、メタン発酵効率の低下と関連付けて見出されたが、バンドB6を有する真正細菌は、メタン発酵系においてメタン発酵効率の低下の開始前、開始と同時、又は開始後に増殖してメタン発酵系に出現する細菌である。本発明のモニタリング方法においては、メタン発酵効率の低下を予測することを目的としているため、メタン発酵効率の低下に先駆けて(低下の開始前に)増殖するか又は該低下と同時に増殖するバンドB6を有する真正細菌を検出されることが望ましい。ここで、本明細書で使用するとき、「先駆けて」とは、メタン発酵効率の低下開始の少なくとも2〜3週間前、1週間前、数日前、1日前、12時間前、6時間前、3時間前、1時間前、又は30分前に、該真正細菌が増殖を開始するか又はバンドB6が検出される場合をいう。検出されるバンドB6は、遺伝子(DNA若しくはmRNA)の形態であるか又はタンパク質の形態であってもよい。
【0024】
本発明のモニタリング方法によれば、プライマーセットIIを用いたリアルタイムPCRを行った後、バンドB6に対応する菌体を含んでいるメタン発酵過程で採取したグラニュールサンプルを連続希釈し、この希釈率に応じた菌体数に対するバンドB6の検量線を作成することができる。これにより、目的とする真正細菌の試料中での有無、その数量を精度よく知ることができる。また、本明細書で使用するとき、メタン発酵効率の「低下」とは、有機性廃棄物の限界負荷量を超えて添加された場合に、メタン発酵系に有機性廃棄物を添加することによって発生していたメタンガスの量が減少している、又は有機性廃棄物を添加したとしてもメタンガスが発生しないことを意味する。限界定負荷量を超えたことによるメタン発酵効率の低下は、メタンガスの発生量が5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、又は100%減少した場合を指す。本発明のモニタリング方法を適用することにより、メタン発酵効率の低下に先駆けて又は低下と同時に増加するバンドB6量をリアルタイムで定量することにより、メタン発酵が失敗に至る状態の変化を予測することができるため、添加する有機性廃棄物の量を調節することによって、安定なメタン発酵が可能になる。また、本発明によれば、本発明のモニタリング方法を応用したバイオセンサーを提供することができる。より具体的には、前述のプライマーセットIIが電極に固定化されたバイオセンサーを用いることにより、リアクター内の発酵液中のバンドB6量を検出及び定量することにより、リアクターに供給する有機性廃棄物の量を調節することができる。
以下、実施例により本発明をさらに詳細に説明するが、本発明はこれらの実施例に限定されない。
【実施例】
【0025】
実施例1:UASBメタン発酵
メタン発酵にはUASB法を使用し、原料の有機性廃棄物としては、共同出願人である山梨缶詰株式会社の工場において缶詰製造過程で排出される平均2.2トン/日のシロップ廃液を用いた。使用したシロップ廃液は、主成分がスクロースであり、糖度が15.2ブリックス、全有機炭素(TOC)濃度が約71g/Lであった。メタン発酵には、外寸法φ2500×H5000mmのUASB発酵槽を用いた。発酵液容積は20m3であった。実験開始時にビール工場のUASB発酵槽で形成されたグラニュール汚泥を15m3充填した。また、実験開始から2日間は馴致期間とし、シロップ廃液を供給せずに温度を40℃に維持した。実験開始から2日以降にシロップ廃液の供給を始めるが、シロップ廃液をそのままUASB発酵槽へ投入すると負荷が高すぎるため、調整槽において水で適宣希釈し、pHが7.5付近になるように苛性ソーダを用いて調整した。その後、温調槽で39℃まで昇温した後にUASB発酵槽に導いた。水で希釈したシロップ廃液の濃度を段階的に変化させたが、およそ4〜5ヶ月後にはシロップ廃液を等量の水で希釈し、TOC濃度が35.5g/Lとなるように負荷を増加させた(図4参照)。なお、145日目以降は再び負荷量を低下させた。また、運転開始49日以降にはメタン発酵を担う微生物の活性を高める目的で金属塩CoCl2、MgCl2、MnCl2、NiCl2をそれぞれ20gずつ(供給するシロップ廃液の1ppm程度に相当)を添加し、さらに、64日以降にはC/N比が15になるように調整槽にて尿素を添加した。ガス発生量は発酵槽上部に取り付けたガス出口の流量計(山武、気体用マスフローメーター、CMS0500)で計測し、pHは槽内のpHメータ(YOKOGAWA、pH100)で計測した。なお、発生するガスは、1日1回、テドラーバッグに捕集し、その中に含まれるメタンガス濃度をガスクロマトグラフ分析装置(ヤナコ、G−1011、カラム MS13X+MS5A)を用いて測定した。メタン発酵の設定温度を40℃とし、グラニュールを含む発酵液サンプルを発酵槽下部から1700mm、壁面から750mmのサンプリング口から所定の時間間隔で、約200mLを採取して、微生物叢の解析に用いた。
【0026】
実施例2:グラニュール粒子からのDNA抽出及びPCR増幅
採取した発酵液サンプル中のグラニュールの約1gを滅菌したスパーテルですくい取り、30mLの滅菌水で洗浄してグラニュール表面に付着している微生物を除去した。その後、その0.3gを使用して、Isoil for Beads Beating kit(Nippon Gene Co.,Ltd.)によりDNAを抽出した。次に、グラニュールから抽出したDNAには、真正細菌及び古細菌に由来するDNAが含まれるが、真正細菌に由来するDNAを増幅させるために、前述のプライマーセットI(プライマー357fと517r)を用いた。増幅条件を以下の表1に示す。
【0027】
【表1】
【0028】
使用したPCR増幅装置はPTC−100 thermal cycler(TaKaRa Shuzo CO.,Ltd.,Kyoto,Japan)であった。また、PCR反応液の組成を表2に示す。
【0029】
【表2】
【0030】
実施例3:バンドB6の分離及び同定
真正細菌からバンドB6を検出するためにDGGE(変性剤濃度勾配ゲル電気泳動)法を用いた。DGGEにはThe Dcode Universal Mutation Detection system(Bio−Rad、Hercules、Calif.)を使用した。まず、グラニュールから抽出した真正細菌由来のDNAを増幅したPCR産物を変性剤の濃度勾配が30%から60%(変性剤100%では7Mの尿素と40%[wt/vol]のホルムアミドを含む)の厚さ0.8mmのポリアクリルアミドゲル(10%[wt/vol]アクリルアミドとビスアクリルアミドの割合は37.5:1)を用いて、1.0×Tris−acetate−EDTA(TAE)溶液中で60℃、200V、3.5時間の条件で電気泳動を行った。なお、電気泳動の後、200mLの1.0×TAE溶液と20μLのエチジウムブロマイド溶液を混合した溶液にゲルを浸し、30分間染色した後、紫外線下で可視化した。
【0031】
DGGE法で検出されたポリアクリルアミドゲル中のバンドからDNA断片を抽出・回収した。抽出したDNA断片から所望のDNA断片を分離及び精製するために、TAクローニング法を用いた。まず、抽出されたDNA断片は、前述のプライマーセットIを用いて、上記と同一の増幅条件を用いて増幅させた。得られたPCR増幅産物は、TOPO TA Cloning Kit(Invitrogen Copration,USA)を用いてプラスミドにライゲーション後、大腸菌に形質転換した。この形質転換した大腸菌を、アンピシリンを添加したLB寒天培地に塗沫し、カラーセレクションを行った。カラーセレクションによりPCR産物をライゲーションしたプラスミドを有することが確認された大腸菌については、まず、下記に示すプライマーセットIII:
M13 F(5’−GTAAAACGACGGCCAG−3’)(配列番号8)
M13 R(5’−CAGGAAACAGCTATGAC−3’)(配列番号9)
を用いてDNAを増幅し、その増幅産物について、さらにプライマーセットIを用いてPCRを行い、再度DGGEを行ってバンドの出現位置を確認した。なお、プライマーセットIIIを用いたPCR条件を以下の表3に示す。
【0032】
【表3】
【0033】
その後、確認された所望のDNA断片はWizard(登録商標)SV Gel and PCR Clean−UP system(Promega Copration,USA)を用いて精製した。精製したPCR増幅物はBigDye terminator kit(Perkin Elmer Japan,Applied Biosystems Division)を用いてシーケンス反応を行った。塩基配列はABI Prism 310 Genetic Analyzer(Applied Biosystems)で解析した。決定した塩基配列については、DDBJ(http://www.ddbj.nig.ac.jp/Welcome−j.html)に登録されている配列に対して相同性検索を行い最も近縁な微生物名を決定した。
【0034】
実施例4:リアルタイムPCR法を用いたDNA濃度の定量
一般に嫌気性菌は増殖が遅く、生成するコロニーも小さいので寒天平板上でのコロニーアピアランスによる識別が困難であることから、メタン発酵のような複数の微生物が共存する複合微生物の系で特定の微生物の濃度を選択的に定量するためには、その微生物に特徴的な遺伝子の配列を検出するリアルタイムPCR法を有効に使用することができる。そこで、バンドB6を特異的に検出するプライマーセットを設計し、これを用いてメタン発酵槽のグラニュール中の真正細菌に含まれるバンドB6を定量した。ここで使用するリアルタイムPCR法は、PCRによるDNAの増幅量をリアルタイムでモニタリングし、測定される蛍光強度の閾値と増幅曲線が交わるサイクル数(Ct値(Threshold Cycle))と、初期鋳型量、すなわち初期微生物濃度の間に直線関係があることを利用して、微生物濃度を定量する方法である。
【0035】
(1)プライマーの設計
バンドB6は、前述のプライマー357Fと517Rに含まれる塩基配列の情報からAnaerobic syntrophic bacterium NE23−3(Accession No.AB231802)に最も近縁であることが判明した。NE23−3株の塩基配列情報(図2)をもとにBioEdit v7.0.9 General Informationを用いて、NE23−3株とその他の微生物群における塩基配列の相同性を確認し、塩基配列の中で保存性の低い領域を検索して、特異性の高いプライマーをFastPCR(URL:http://www.primerdigital.com/fastpcr.html)の推奨結果を参考に設計した。なお、プライマーの設計時は、一般的にいわれている以下の条件を満たすように留意した。増幅サイズは80〜150塩基(増幅効率を100%にするため)、プライマー長さは17〜25塩基(長過ぎるとアニーリング効率が低下する)GC含量は40〜60%(45〜55%が望ましい)とし、偏りがないように、部分的にGC、ATリッチができないように注意する、T/C、A/Gが連続しないように注意する、プライマー内部での3 塩基以上の相補的配列を避ける、プライマー間での3塩基以上の相補的配列を避ける、プライマー3’末端が2塩基相補する配列を避ける。設計したプライマー(プライマーセットII)の塩基配列は前述の通りである。
【0036】
(2)設計したプライマーの特異性の検討
設計したプライマーセットIIが目的菌に選択的であることをBioEdit v7.0.9 General Informationを用いて、プライマーの塩基配列と目的菌以外の微生物群の塩基配列とを比較することで確認するとともに、実際のメタン発酵グラニュールサンプルに設計したプライマー(プライマーセットII)を作用させて、増幅される複数のDNA断片の塩基配列がすべて同一であることを確かめた(データ示さず)。増幅されるDNA断片の塩基配列が同一であれば、設計したプライマーが類似の塩基配列を有する複数種類の微生物を検出する可能性は極めて小さくなる。まず、メタン発酵グラニュールサンプルにプライマーセットIIを加えてPCRによりDNA断片を増幅させた。なお、PCR条件を以下の表4に示す。
【0037】
【表4】
【0038】
次に、増幅したDNA断片をTAクローニング法により大腸菌に形質転換した。続いてPCR産物がライゲーションされたプラスミドを持つことが確認された大腸菌にプライマーセットIII(前述)を加えて、PCR増幅し、精製した。その後、BigDye terminator kit(Perkin Elmer Japan,Applied Biosystems Division)を用いてシーケンス反応を行った。塩基配列はABI Prism 310 Genetic Analyzer(Applied Biosystems)で解析した。目的としたPCR産物がライゲーションされたプラスミドを持つことが確認された大腸菌コロニーが12個得られた。これらのすべてについてPCR産物の塩基配列を解析したところ、塩基配列は完全に同一であり、設計したプライマーが類似の塩基配列を有する複数種類の微生物を検出している確率は極めて小さいことが判明した。また、このプライマーセットIIで増幅したDNA領域は、バンドB6の同定に用いたDNA領域と同じ遺伝子上に存在することを確かめた。
【0039】
(3)設計したプライマーを用いた検量線の作成
メタン発酵グラニュールサンプルを適宜希釈して、設計したプライマーの定量性を確認した。メタン発酵グラニュールサンプルは発酵開始125日目のものを用いた。リアルタイムPCR装置はSmart Cycler II(タカラバイオ株式会社)を用い、PCR反応液は、プライマーセットIIとSYBR Premix Ex Taq(タカラバイオ株式会社)を用いて、下記の表5のように調製した。
【0040】
【表5】
【0041】
増幅条件は最初に95℃、10秒の変性をおこなった後、95℃、5秒(変性)、60℃、20秒(アニーリング/伸長)を40サイクルとした(表6参照)。作製した検量線を図5に示す。検量線のR2値は0.976であり、定量に用いるのに十分な精度であった。
【0042】
【表6】
【0043】
(4)リアルタイムPCR法によるバンドB6に対応する微生物の定量
メタン発酵グラニュールサンプルからのDNA抽出物にリアルタイムPCR法を適用してバンドB6に対応する菌体濃度を測定した。用いたプライマーセットとPCR条件は、検量線を作成したときと同様とした。なお、菌体濃度はメタン発酵125日後の菌体濃度が最大になるときを1とした相対濃度で示した(図6参照)。
【特許請求の範囲】
【請求項1】
有機性廃棄物のメタン発酵系におけるメタン発酵効率をモニタリングする方法であって、下記のプライマーセット:
(a)配列番号1に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド;及び
(b)配列番号2に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド
を用いてポリメラーゼ連鎖反応(PCR)を行い、該メタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下と同時に増殖する真正細菌の16S rDNA量又は16S rRNA量を測定することを含むモニタリング方法。
【請求項2】
前記真正細菌が、配列番号3に示される塩基配列を含む16S rDNAを有する、請求項1に記載のモニタリング方法。
【請求項3】
有機性廃棄物が、食品廃棄物、糞尿、動植物残さ、古紙、及び汚泥からなる群から選択される、請求項1又は2に記載のモニタリング方法。
【請求項4】
前記食品廃棄物がシロップ廃液である、請求項3に記載のモニタリング方法。
【請求項5】
有機性廃棄物のメタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下の開始とともに増殖する真正細菌を検出又は定量するためのプライマーセットであって、ここで、該プライマーセットは、
(a)配列番号1に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド;及び
(b)配列番号2に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド
であるプライマーセット。
【請求項1】
有機性廃棄物のメタン発酵系におけるメタン発酵効率をモニタリングする方法であって、下記のプライマーセット:
(a)配列番号1に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド;及び
(b)配列番号2に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド
を用いてポリメラーゼ連鎖反応(PCR)を行い、該メタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下と同時に増殖する真正細菌の16S rDNA量又は16S rRNA量を測定することを含むモニタリング方法。
【請求項2】
前記真正細菌が、配列番号3に示される塩基配列を含む16S rDNAを有する、請求項1に記載のモニタリング方法。
【請求項3】
有機性廃棄物が、食品廃棄物、糞尿、動植物残さ、古紙、及び汚泥からなる群から選択される、請求項1又は2に記載のモニタリング方法。
【請求項4】
前記食品廃棄物がシロップ廃液である、請求項3に記載のモニタリング方法。
【請求項5】
有機性廃棄物のメタン発酵系においてメタン発酵効率の低下に先駆けて増殖するか又は該低下の開始とともに増殖する真正細菌を検出又は定量するためのプライマーセットであって、ここで、該プライマーセットは、
(a)配列番号1に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド;及び
(b)配列番号2に示される塩基配列又はこれと実質的に同一の塩基配列からなるオリゴヌクレオチド
であるプライマーセット。
【図1】


【図2】

【図3】
【図4】
【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2012−249558(P2012−249558A)
【公開日】平成24年12月20日(2012.12.20)
【国際特許分類】
【出願番号】特願2011−123529(P2011−123529)
【出願日】平成23年6月1日(2011.6.1)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 化学工学会第76年会研究発表講演要旨集(発行所:公益社団法人 化学工学会 発行日:平成23年2月22日)
【出願人】(304021417)国立大学法人東京工業大学 (1,821)
【出願人】(511133679)山梨罐詰株式会社 (1)
【Fターム(参考)】
【公開日】平成24年12月20日(2012.12.20)
【国際特許分類】
【出願日】平成23年6月1日(2011.6.1)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 化学工学会第76年会研究発表講演要旨集(発行所:公益社団法人 化学工学会 発行日:平成23年2月22日)
【出願人】(304021417)国立大学法人東京工業大学 (1,821)
【出願人】(511133679)山梨罐詰株式会社 (1)
【Fターム(参考)】
[ Back to top ]