
抗旋毛虫抗体によって認識されるポリペプチドとその使用法
本発明は、抗旋毛虫抗体によって認識される新規なポリペプチドに関するものである。さらに、本発明は、抗旋毛虫抗体の検出ならびに旋毛虫症の予防を目的とする前記ポリペプチドの利用法にも関するものである。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、旋毛虫症の診断および予防の分野における、寄生虫である「旋毛虫」において同定された新規の抗原の利用法に関するものである。
【背景技術】
【0002】
旋毛虫症は、寄生虫である「旋毛虫」に感染した肉の消費に関連した人獣共通感染症の一つである(MURRELL et al.,2000)。
【0003】
「双器綱」のこの線虫は、旋毛虫科に属し、該旋毛虫科は、系統発生学的に異なる二つのグループと関連する8つの種と3つの遺伝子型を含んでおり、該二つのグループとは、一つは哺乳動物に感染する、被嚢を形成する旋毛虫であり(T.spiralis、T.nativa、T.britovi、T.murrelli、T.nelsoni)、もう一つは、哺乳動物、鳥類および爬虫類に感染する、被嚢を形成しない旋毛虫(T.pseudospiralis、T.papuae、T.zimbabwensis)である(GASSER et al.,2004)。これらすべての種は人に感染する可能性がある。
【0004】
寄生虫の生活環は自動異宿主性である。該生活環は、連続的に終宿主(成長した寄生虫のキャリア)でもあり中間宿主(感染幼虫のキャリア)でもある、同一の宿主で全て展開される(BOIREAU et al.,2002)。感染幼虫の一つの宿主からもう一つ宿主への移行は、新しいサイクルを行うために必要である。この移行は、幼虫に感染した生の肉または十分に焼かれていない肉を摂取することによって起こる。消化の際、該幼虫は解放され、そして腸上皮に侵入し、そこで性別のある成虫(Ad)へと脱皮する。受精した雌は、次に新生L1幼虫(L1NN)を生み出し、該幼虫はリンパおよび血液の流れを介して横紋筋に達する。これらのL1NN幼虫は筋肉細胞に侵入し(L1M:筋肉L1幼虫の感染進行段階)、被嚢を形成する旋毛虫の場合には厚い保護性のコラーゲン嚢で、被嚢を形成しない旋毛虫の場合には非常に薄い保護性のコラーゲン嚢で覆われている、フィーダー細胞への脱分化を引き起こす。
【0005】
旋毛虫症は動物においては無症状であるが、ヒトへの感染は、初期の腸の段階では嘔吐感を伴う下痢、嘔吐および激しい腹痛として現れるのに対し、筋肉への侵入段階に関連する症状は、熱、顔のむくみおよび筋肉痛の合併を特徴としている(CAPO & DESPOMMIER、1996)。目、肺、胃腸、心臓および神経系での発症も、進行が致死的となり得る旋毛虫症のこの診断指標に加えることができる。患者における筋肉痛の持続を代表とする感染の慢性的な特徴は、フィーダー細胞内部で寄生虫が生存していることに関連している。
【0006】
駆虫薬によるヒトの旋毛虫症の特異的な治療は、あらゆる寄生段階に対する対処、特にL1M幼虫の周りにコラーゲンの保護嚢が形成される前の対処が可能になるよう感染の診断が早期に行われると、より効果的である(FOURESTIE et al.,1988)。
【0007】
疫学的なデータによって、ブタに代表される家畜における感染のサイクルも支えている野生動物の数多くの種を伴う伝染様式に関連する、あらゆる地帯における寄生虫の地理的分布が示されている(DUPOUY−CAMET,2000)。
【0008】
ヒト旋毛虫症の流行、すなわち突発的または再発的な人獣共通感染症の流行は、食習慣および必ずしも有効ではない衛生管理の問題のために、世界中の公衆衛生における現実的な問題となっている(MURRELL & POZIO,2000)。これらの流行は、豚およびイノシシの肉ならびに馬肉に主として関与している(BOIREAU et al.,2000)。
【0009】
したがって、ヒトの汚染の予防は、肉の中心まで火を入れることと、飼育条件および/または動物(豚、馬、イノシシおよび旋毛虫に敏感なその他の野生動物種)の旋毛虫症の管理を向上させることを介して行われる(BOIREAU et al.,2002)。
【0010】
旋毛虫症のスクリーニング技術は二つに分けられる。すなわち、1)トリキノスコープ(肉片の顕微鏡観察)による、すなわち筋肉サンプルの人工的な消化の後の、L1M幼虫の直接的な検出と、2)旋毛虫抗原に対する抗体の検出を可能にする、さまざまな免疫学的方法による間接的な検出である。
【0011】
寄生虫の各発達段階、すなわち成虫(Ad)、新生幼虫(L1NN)および筋肉幼虫(L1M)は、特定の抗原プロフィールに対応している。
【0012】
現状で免疫診断に用いられているものは、L1M段階の幼虫に由来する抗原調製物である。実際、二つの先行する段階AdおよびL1NNの抗原成分は精製するのが難しく、これら二つの段階の一方および/または他方に関連する免疫優性の抗原は現在までに特定できていない。
【0013】
主として、幼虫の溶解、溶解物の遠心分離、そして上清の回収によって得られた全可溶性抗原の調製物、あるいは、より頻繁には、排泄/分泌抗原(E/S抗原)が用いられている。
【0014】
排泄−分泌抗原はL1M幼虫を培地で生育させる際に産生される。該抗原はスティコソームと称される特定の器官に由来し、該器官は、およそ50の円盤状の細胞であるスティコサイトで構成されている。スティコサイトは粒状物を含んでおり、その内容物は寄生虫の食道の管腔にある小管によって排出されるものである。この内容物は非常に抗原性が高く、排泄分泌抗原の一部を構成する。これらの抗原はタンパク質の複雑な混合物を形成し、該混合物はとりわけ、特定の糖分子を有する糖タンパク質の群(TSL1抗原と称される)を含んでおり、該糖分子は、旋毛虫のみで知られておりかつこの寄生虫のすべての種に存在しているベータ・チベロースである。
【0015】
排泄−分泌抗原の調製物は、現状では旋毛虫症の免疫診断材料の基準として用いられており、Trichinella spiralisのL1M幼虫の培地から得られる。18〜20時間の培養の後、培地を濾過によって回収し、そして濃縮する(GAMBLE et al.,1983、GAMBLE et al.,1988)。
【0016】
全可溶性抗原の調製物は、主な不都合としては特異性が欠けている。その他の寄生虫症と交差した抗原性反応が頻繁に観察されている。排泄−分泌抗原によってより良い特異性を得ることが可能となる。しかし、いずれの場合にも、抗原の標準化されたロットを大量に生産するのは難しい。
【0017】
ベータ・チベロースを含む糖の構造は、E/S抗原調製物の免疫優性のエピトープに相当し(REASON et al.,1994、米国特許第5541075号明細書および米国特許第5707817号明細書)、化学的に合成されており、旋毛虫の免疫診断のために利用することが提案されている。
【0018】
この試薬は良好な特異性を有しているが、その感度はE/S抗原調製物よりも低いようである。さらに、化学的方法によるこの構造の合成は、実施するのにコストが高く、負担の大きいものになっている。
【0019】
旋毛虫の血清学的診断の領域でぶつかるもう一つの問題は、検出の「ブラインド・ウィンドウ」が存在することであり、該ブラインド・ウィンドウは感染の早い段階に対応し、偽陰性の結果として現れる。さらに、馬では、感染後の25週目には抗体の漸次的な消失が観察されている。
【特許文献1】米国特許第5541075号明細書
【特許文献2】米国特許第5707817号明細書
【発明の開示】
【発明が解決しようとする課題】
【0020】
本発明者らは、旋毛虫による感染の早い段階に関連し、旋毛虫症の血清学的診断に利用可能な免疫優性の抗原を特定することで、ヒトだけでなく動物において、早期の、特異的な、かつ旋毛虫への感染に敏感な検出が得られる手段を提供することに取り組んだ。この目的において、本発明者らは、L1NN段階および/またはAd段階の旋毛虫によって発現される遺伝子産物の中に、所望の抗原特性を有するタンパク質が存在するかどうか研究した。
【0021】
この枠組みにおいて、本発明者らは、旋毛虫のL1NN段階で特異的に発現するタンパク質の一部をなすTrichinella spiralisのタンパク質が免疫優勢性の抗原を構成し、該タンパク質によって、旋毛虫に対する液性応答の早期検出が可能になることと、該タンパク質がさらに、旋毛虫のさまざまな種の間で保持されていることを発見した。
【0022】
このタンパク質を以下ではNBL1と称することにする。このタンパク質をコードしているcDNAの完全な配列ならびにこの配列から推定されるポリペプチド配列は、それぞれAF331160およびAAK16520という番号のもと、Genbankでアクセス可能であり(あるいはSwissprot Q9BJL7)、これらの配列は「セリンプロテアーゼSS2、新生幼虫に固有」という註が付けられている。また、これらの配列は、添付の配列表において、SEQ ID NO:1とSEQ ID NO:2という番号で再掲されている。このタンパク質をコードしているcDNAの部分配列ならびにこの配列から推定されるポリペプチド配列は、AY491941およびAAR36900という番号のもと、それぞれGenbankでアクセス可能である(あるいはSwissprot Q6RUJ3)。
【0023】
さらに、本発明者らは、NBL1に対する液性応答に関連する免疫反応性がこのタンパク質のC末端部分に局在していることを示し、この反応性を担う免疫優性のエピトープを特定した。
【0024】
他方では、本発明者らは、T.spiralisのAdとL1NNを混合した早期段階のcDNAライブラリーから、以下では411と称する新たな遺伝子を特定した。
【0025】
この遺伝子の配列は、添付の配列リストにおいて、SEQ ID NO:3という番号で示されており、該遺伝子配列の翻訳産物の配列はSEQ ID NO:4という番号で示されている。この遺伝子の翻訳産物は、タンパク質Tp21−3と称され、T.pseudospiralisで同定されているE/S抗原(AAF79206、NAGANO et al.,2001)と関連性を有し(78.7%の同一性)、さらに、T.spiralisの推定ORF17.20の翻訳産物(AAB48489)とも関連性を有し、該産物とは86.6%の同一性を有する。
【0026】
また、411遺伝子の翻訳産物によって、早期段階で、旋毛虫のさまざまな種に対する液性応答を検出することが可能となる。
【0027】
さらに、実験を行ったところ、他の抗原によってもE/S抗原によっても少なくとも感染後(pi)15日〜30日の間には検出されなかった旋毛虫に感染した動物を、NBL1抗原および411抗原のそれぞれによって検出することが可能であった。
【0028】
したがって、NBL1抗原(または該抗原の免疫優性のエピトープ)と411抗原を組み合わせることによって、診断の感度、とりわけ感染の初期段階(感染後15日〜20日)に対する感度を向上させることが可能となる。
【課題を解決するための手段】
【0029】
したがって、本発明は生体標本において抗旋毛虫抗体を検出するための試薬としての、抗旋毛虫抗体によって認識される抗原ポリペプチドの利用法を対象としており、該使用法は、前記ポリペプチドが、
a)配列PSSGSRPTYP(SEQ ID NO:5)によって定義されるNBL1抗原の免疫優性のエピトープを含んだポリペプチドと、
b)以下で411抗原とも称されるポリペプチドであり、配列SEQ ID NO:4の25〜175番目のアミノ酸を含むか(411タンパク質の成長した形に相当する)、あるいは、配列SEQ ID NO:4の25〜175番目のアミノ酸配列と少なくとも80%、好ましくはより高く、少なくとも85%、90%、または95%の同一性を有する配列を含むポリペプチド、
から選択されることを特徴としている。
【0030】
より特徴的には、本発明は、生体サンプルにおける抗旋毛虫抗体の存否の検出方法を対象としており、該方法は、
−前記生体サンプルを、前記生体サンプルに場合によっては存在する抗旋毛虫抗体との抗原/抗体複合体の形成を可能にする条件のもとで、上記で定義した一つもしくは複数のポリペプチドa)および/または一つもしくは複数のポリペプチドb)と接触させることと、
−あらゆる適切な手段によって、場合によっては形成される抗原/抗体複合体を検出すること、
を含むことを特徴としている。
【0031】
一般的には、前記生体標本は血清標本である。該血清標本は、旋毛虫に感染しうる種に属する、この寄生虫の存否を検出しようとするあらゆる対象(哺乳動物、鳥類または爬虫類)から得ることができる。有利には、哺乳動物、例えば飼育動物またはヒトの患者から得られる標本である。
【0032】
有利には、上記で定義した一つもしくは複数のポリペプチドa)および/または一つもしくは複数のポリペプチドb)を含む混合物が用いられる。
【0033】
この組み合わせによって、とりわけ、個別に用いた各ポリペプチドに比べて反応性の幅を広げることが可能となる。
【0034】
また、アクセス番号Genbank AAK16520で識別される全NBL1抗原と、アクセス番号Genbank AAR36900で識別されるその断片を除き、上記で定義したポリペプチドa)はそれ自体で本発明の対象の一部をなす。
【0035】
本発明にしたがったこれらのポリペプチドのうち、とりわけ、配列PSSGSRPTYPSSGSR(SEQ ID NO:6)、配列PSSGSRPTYPYTGSR(SEQ ID NO:7)、配列RPTSPSSGSRPTYPS(SEQ ID NO:8)のうち一つまたは複数を含んだポリペプチドが挙げられる。
【0036】
これには、例えばNBL1抗原のC末端領域の断片も含まれ、とりわけ、配列SEQ ID NO:2の326〜459番目のアミノ酸に対応する、
ENSPEGTVKWASKEDSPVDLSTASRPTNPYTGSRPTSPSSGSRPTYPSSGSRPTSPSSGSRPTYPSSGSRPTYPSSGSRPTYPYTGSRPTPQKPVFPSYQKYPPAVQKYIDSLPSGTQGTLEYTVTQNGVTTTT(SEQ ID NO:11)、
を含んだ断片が挙げられ、またこの配列SEQ ID NO:11の小断片、とりわけ、配列SEQ ID NO:2の363〜409番目のアミノ酸に対応する、
PSSGSRPTYPSSGSRPTSPSSGSRPTYPSSGSRPTYPSSGSRPTYP(SEQ ID NO:9)という配列を含んだ小断片も含まれ、より特徴的には、配列SEQ ID NO:2の349〜415番目のアミノ酸に対応する、
SRPTNPYTGSRPTSPSSGSRPTYPSSGSRPTSPSSGSRPTYPSSGSRPTYPSSGSRPTYPYTGSRPT(SEQ ID NO:10)という配列を含んだ小断片が含まれる。
【0037】
上記で定義したポリペプチドb)は、アクセス番号GenBank AAF79206およびAAB48489で識別されるものを除いて、それ自体で本発明の対象の一部をなす。好ましいポリペプチドはとりわけ、配列SEQ ID NO:4のポリペプチド、または、配列SEQ ID NO:4の25〜175番目のアミノ酸に対応するポリペプチド、ならびに、配列SEQ ID NO:4または配列SEQ ID NO:4の25〜175番目のアミノ酸配列との間に少なくとも90%、好ましくは少なくとも95%の同一性を有するポリペプチドである。
【0038】
とりわけ、本発明は、配列PSSGSRPTYP(SEQ ID NO:5)の一つもしくは複数のコピー、またはこの配列を含むNBL1抗原の断片の一つもしくは複数のコピー、および/または、上記で定義したポリペプチドb)の一つもしくは複数のコピーを含む、場合によっては一つもしくは複数の他の異種配列と融合しているキメラポリペプチドを含む。
【0039】
また、本発明は、本発明にしたがったポリペプチドをコードするポリヌクレオチド、ならびに、前記ポリヌクレオチドを含んだ組換えベクター、および前記ベクターによって形質転換した宿主細胞も対象としている。
【0040】
また、本発明は上記で定義したような、一つまたは複数のポリペプチドa)および一つまたは複数のポリペプチドb)を含んだ組成物、ならびに、前記ポリペプチドをコードする一つまたは複数のポリヌクレオチドを含んだ組成物も対象としている。
【0041】
上記で定義したポリペプチドa)およびb)は、それ自体既知である、抗体検出のさまざまな方法の範囲で用いることができる。例えば、とりわけ、ELISAタイプの方法(直接、間接またはサンドイッチ)、ビーズへのミクロ凝集方法、ならびに、免疫標識に組み合わせた電気泳動転写法などが挙げられる。
【0042】
また、本発明は生体サンプルにおける抗旋毛虫抗体の存否を検出するために必要な物のセットも対象としており、これは、上記で定義した一つもしくは複数のポリペプチドa)および/または一つもしくは複数のポリペプチドb)、ならびに場合によっては、抗原/抗体複合体の形成を可能にする反応培地を構築するための適切な緩衝液および試薬、ならびに選択的に、前記抗原/抗体複合体の検出手段を含むことを特徴としている。
【0043】
有利には、前記セットは、固体の担体の上に固定されている、上記で定義したポリペプチドa)および/またはポリペプチドb)を含んでいる。利用可能な固体の担体の非限定的な例としては、マイクロタイタープレート、ビーズ、マイクロビーズまたは微粒子、試験紙などが挙げられる。
【0044】
また、前記セットは、一つまたは複数の陰性血清および一つまたは複数の陽性血清のような、コントロールサンプルも含むことができる。
【0045】
また、本発明は、前記ポリペプチドに対する特異的な抗体を調製するための、上記で定義したポリペプチドa)またはポリペプチドb)の利用法も対象としている。
【0046】
これらのポリペプチドは、それ自体既知である、さまざまな抗体の調製方法の範囲で利用することができる。例えば、該ポリペプチドは(場合によっては適切なアジュバントの添加後に)動物の免疫付与に利用することができる。また、該ポリペプチドは、アフィニティークロマトグラフィーの担体にグラフトすることで、生体液から目的のポリペプチドに対する特異的な抗体を精製できるようにすることができる。生体液とは、例えば目的とするポリペプチドで事前に免疫付与した動物の血清あるいはハイブリドーマの上清とすることができ、また、旋毛虫に感染した動物の血清であり、目的とするポリペプチドに対する特異的な抗体の亜集団を単離するための血清とすることもできる。
【0047】
また、本発明には、上記で定義したポリペプチドa)またはポリペプチドb)に対するあらゆる特異的な抗体が含まれる。それはポリクローナル抗体またはモノクローナル抗体とすることができる。好ましい抗体は、PSSGSRPTYP(SEQ ID NO:5)のエピトープを認識する抗体である。
【0048】
ポリペプチドに対する特異的な抗体は、それ自体既知であるさまざまな技術によって、とりわけ、目的とするポリペプチドで動物を免疫付与すること(場合によっては適切なアジュバントを添加する)と、その血清(ポリクローナル抗体の産生用)またはリンパ球細胞(モノクローナル抗体の産生用)を回収することを含む、従来の方法によって得ることができる。
【0049】
上記で定義したポリペプチドa)およびb)ならびにこれらのポリペプチドをコードするポリヌクレオチドは、免疫原性組成物、とりわけ抗旋毛虫ワクチンを調製するために利用することができる。
【0050】
また、本発明は、上記で定義した一つもしくは複数のポリペプチドa)および/または一つもしくは複数のポリペプチドb)を含んだ免疫原性組成物、あるいは、免疫応答の向上を可能にする一つまたは複数のアジュバントと結びついている、前記ポリペプチドをコードする一つまたは複数のポリヌクレオチドを含んだ免疫原性組成物も対象としている。
【0051】
本発明にしたがった免疫原性組成物の好ましい実施態様によると、それはワクチンである。
【0052】
ペプチドの免疫原性の向上を可能にする非常に多様なアジュバントはそれ自体当業者には知られており、例えば、ミョウバン(水酸化アルミニウム)、完全フロイントアジュバントまたは不完全フロイントアジュバント(IFA)、リポソーム、ならびにウィロソーム(再構成されたウイルスエンベロープ)、ムラミン酸のペプチド誘導体などが挙げられる。ワクチンの場合、当然、薬学的に許容可能なアジュバントが選択されることになり、好ましいアジュバントの例としては、例えばMONTANIDE ISA 70およびMONTANIDE ISA 775という名称でSEPPIC社によって市販されており、かつ欧州特許第480982号明細書、欧州特許第825875号明細書、米国特許第5422109号明細書、米国特許第6251407号明細書、米国特許第6610309号明細書にも記載されているアジュバントのような、「油中水型」エマルジョンタイプのアジュバントが挙げられる。
【0053】
場合によっては、とりわけ短いペプチド(30個以下のアミノ酸)の場合、前記ポリペプチドはキャリアタンパク質と結合させることができる。
【0054】
キャリアタンパク質の例としては、とりわけKLH(スカシ貝ヘモシアニン、Keyhole limpet hemocyanin)、ウシ血清アルブミン(BSA)、オボアルブミン、破傷風トキソイド、ジフテリアトキソイドが挙げられる。また、同一ペプチドの複数のコピーを互いに組み合わせることで、また場合によってはキメラポリペプチドの形でその他のペプチドエピトープと組み合わせることで、あるいは、例えばポリリジンのようなポリマー鎖を介して、マルチエピトープの組成物を形成することもできる。
【0055】
免疫原としてポリヌクレオチドを使うのであれば、免疫原組成物は、投与すべきポリヌクレオチドが挿入された組換えベクターの形をとることができる。例えば、ポックスウイルス、アデノウイルス、レトロウイルス、レンチウイルス、ヘルペスウイルス、AAV(アデノ随伴ウイルス)などのウイルスベクターを用いることができる。また、該組成物は、前記ポリヌクレオチドを含む一つまたは複数の発現ベクターによって形質転換された、非病原性バクテリアの形をとることもできる。また、ポリヌクレオチドを裸のDNAの形で直接投与するか、あるいは、リポソームに包入することもできる。ワクチンの場合、好ましくは非病原性バクテリア(例えば乳酸菌、あるいは大腸菌またはサルモネラ菌の非病原性株)、または、ワクチンウイルス株から派生したベクター、例えば仮性狂犬病(オーエスキー病)ウイルスのワクチン株から派生したベクターが用いられることになる。
【発明を実施するための最良の形態】
【0056】
本発明は、旋毛虫症の早期の免疫診断を目的とした、NBL1抗原および411抗原の利用法を示した実施例に関する、以下の説明の補足によってよりよく理解されるものである。
【0057】
図1:タンパク質THX−NBL1(Cterm)の配列。
NBL1に由来する配列を太字で示し、pET102プラスミドに由来する配列は斜体で示している。
【0058】
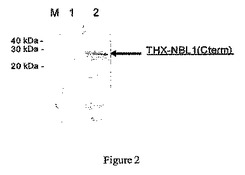
図2:タンパク質THX−NBL1(Cterm)の免疫反応性。
M:分子量マーカー、1:陰性のブタの血清、2:20000匹のT.spiralisのL1Mで実験的に感染させたブタの血清。
【0059】
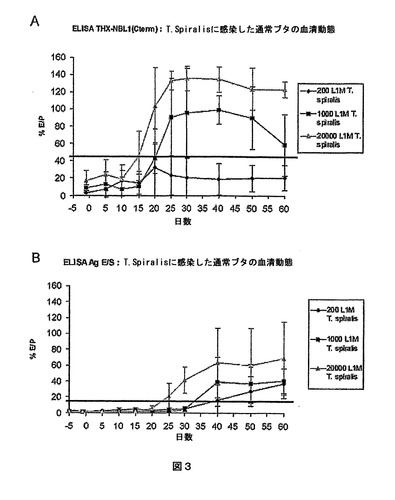
図3:T.spiralisに感染させたブタの血清においてELISA THX−NBL1(Cterm)またはELISA Ag E/Sによって検出された抗旋毛虫抗体の出現動態の比較。
A:ELISA THX−NBL1(Cterm)による検出、B:ELISA Ag E/Sによる検出。
横軸:感染後の日数、縦軸:反応性のパーセント。ELISAによる検出限界は黒い線で示している(ELISA THX−NBL1(Cterm)については44%、ELISA Ag E/Sについては14%)。サンプル/ポジティブコントロール比の平均および標準偏差は感染したブタの各群について示している。
【0060】
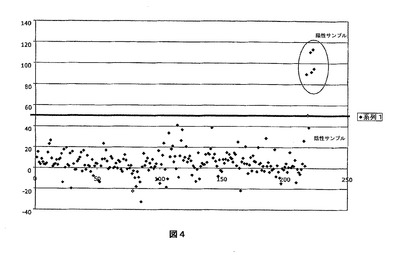
図4:ELISA NBL1(Cterm)の特異性。
横軸:産業用のブタと野外飼育のブタに由来する230の陰性サンプルのコホート。5つの陽性サンプルは実験的に感染させたブタの血清に由来している。限界値は陰性サンプルの平均値の2倍に等しい。縦軸:反応性のパーセント。
【0061】
図5:タンパク質THX−411の配列。
411に由来する配列は太字で示している。プラスミドに由来する配列は斜体で示している。
【0062】
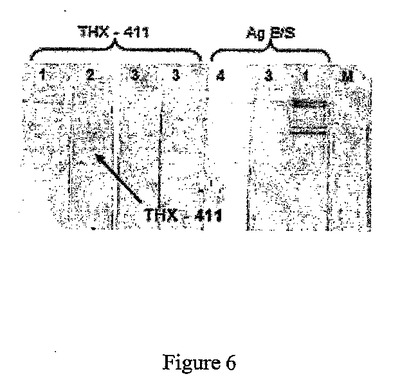
図6:タンパク質THX−411の免疫反応性。
1:感染後50日の陽性血清、2:感染後30日の陽性血清、3:5日前の陰性血清、4:共役型コントロール。M:分子量マーカー。
【0063】
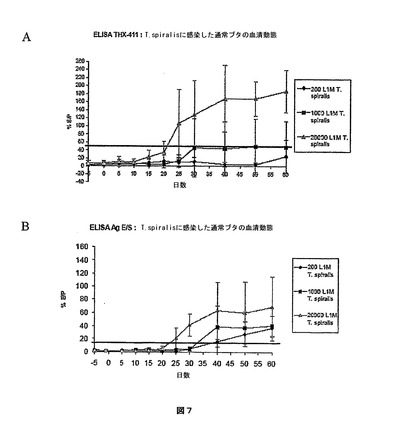
図7:T.spiralisに感染したブタの血清においてELISA THX−411またはELISA Ag E/Sによって検出した抗旋毛虫抗体の出現動態の比較。
A:ELISA THX−411による検出、B:ELISA Ag E/Sによる検出。
横軸:感染後の日数、縦軸:反応性のパーセント。ELISAによる検出限界は黒い線で示している(ELISA THX−411では52%、ELISA Ag E/Sでは14%)。サンプル/ポジティブコントロール比の平均と標準偏差は感染したブタの各群について示している。
【実施例1】
【0064】
実施例1:NBL1のC末端部分を含んだ、組換えタンパク質THX−NBL1(Cterm)の産生。
Trichinella spiralisのL1NNのcDNAライブラリーに対する免疫スクリーニングを、T.spiralisの10000匹のL1Mによって実験的に感染させた後35日目に得られたブタの血清を用いて行った。この血清によって認識されたクローンのシーケンシングによって、それらのうち大半が同一のタンパク質をコードしていることが判定できた。
【0065】
このタンパク質は推定セリンプロテアーゼであり、そのcDNA配列および推定されるアミノ酸配列は、それぞれ、AF331160およびAAK16520という番号のもとGenbankで入手可能であり、また、ここではSEQ ID NO:1およびSEQ ID NO:2の番号で再掲している。該タンパク質はここではNBL1と称することにする。
【0066】
タンパク質のC末端部分を、NBL1CtermF(5’−CACCGAAAATTCTCCTGAAGGA−3’)(SEQ ID NO:12)とNBL1CtermR(5’−TGTTGTTGTAGTAACTCC−3’)(SEQ ID NO:13)オリゴヌクレオチド、そしてDNAポリメラーゼAccuPrime Pfx DNA polymerase(Invitrogen社)を用いて増幅し、製造者(Invitrogen社)の推奨にしたがって発現キット「Champion pET102 Directional TOPO」を用いて、プラスミドpET102D/topoでクローニングした。
【0067】
得られた組換えプラスミドはpET102−NBL1(Cterm)と称するもので、291個のアミノ酸からなり、C末端の位置にポリヒスチジンタグを有している、チオレドキシンNBL1(Cterm)(THX−NBL1(Cterm))融合タンパク質をコードしている。この融合タンパク質の配列は図1に示している。
【0068】
融合タンパク質THX−NBL1(Cterm)は、プラスミドpET102−NBL1(Cterm)によって形質転換した大腸菌BL21Star(DE3)、BL21(DE3)pLys(Invitrogen社)で発現させ、そして、提供者(Qiagen)によって推奨される手順を用いて、Ni−NTAカラム(Ni−NTAスピン・カラム・キット、Ni−NTAビーズ)上で、変性条件下でのアフィニティークロマトグラフィーによって精製した。
【0069】
精製された融合タンパク質THX−NBL1(Cterm)は、変性条件下での電気泳動(SDS−PAGE)の後、31.1kDaの予期されたサイズのバンドの形で現れた。
【0070】
旋毛虫症に感染していないブタの血清、および感染後60日目に採取された、20000匹のT.spiralisのL1M幼虫で感染させたブタの血清に対する、タンパク質THX−NBL1(Cterm)の免疫反応性を、ウェスタンブロッティングによって分析した。
【0071】
変性条件下の電気泳動の後(SDS−PAGE)、提供者(Amersham)の指示にしたがってタンパク質をHybond P膜(PVDF)でエレクトロブロッティングした。膜は、TBS−T(20mM、pH7.5のTris−HCl、150mMのNaCl、0.1%のTween20)と5%の脱脂乳において1時間にわたってプレハイブリダイゼーションした。1分間にわたるTBS−Tによる2回の洗浄の後、そして5分間にわたるTBS−Tでの3回の洗浄の後、膜を、TBSで1/200に希釈したブタの血清を用いて1時間インキュベートした。洗浄後、膜を、1/30000に希釈した、アルカリフォスファターゼ(A1192、Sigma社)で標識した抗ブタIgGウサギ二次抗体を用いて20分間インキュベートし、次に、標識を視認できるようにするために、30分間にわたって純粋なNBT/BCIP基質(E116、Interchim社)を用いてインキュベートした。
【0072】
結果は図2に示している。
【0073】
T.spiralisに感染したブタの血清によるTHX−NBL1(Cterm)の非常に強い免疫反応性が観察される。
【実施例2】
【0074】
実施例2:旋毛虫に対する液性応答を検出するためのTHX−NBL1(Cterm)の利用法。
上記実施例1に記載したように調製したタンパク質THX−NBL1(Cterm)を、T.spiralisの排泄/分泌抗原(E/S)と比較して、旋毛虫に感染したブタに由来する血清に対する間接ELISA法によって評価した。
【0075】
対照のE/S抗原をGAMBLEら(1988)に記載の手順にしたがって調製した。該抗原は、1%のピルビン酸塩、15%のウシ胎児血清(SVF)、1%のL−グルタミン、100U/mlのペニシリン、および100μg/mlのストレプトマイシンを含むRPMI1640において24時間にわたって維持されたL1M幼虫の培養物の上清から得た。
【0076】
ELISA検定のために、E/S抗原については1.25μg/ml、THX−NBL1(Cterm)については2μg/mlの割合で1×PBS緩衝液で希釈した抗原を、96ウェルプレート(MediSorpプレート、NUNC社)において4℃で一晩インキュベートした。3回の洗浄後(1×PBS、0.05%のTween20)、プレートを、洗浄液で2%に希釈した脱脂乳溶液で1時間にわたって室温で飽和させた。
【0077】
各ウェルに、0.05%のTween20を加えた、1×PBS緩衝液で1/20に希釈した100μlのブタの血清を置いた。37℃での30分間のインキュベーションと、続く3回の洗浄の後(1×PBS、0.05%のTween20)、各ウェルに、0.05%のTween20を加えた、1×PBS緩衝液で1/32000に希釈した100μlの共役溶液(Gタンパク質−ペルオキシダーゼ(P−8170、Sigma社))を置いた。37℃での30分間の新たなインキュベーションと、続く3回の洗浄の後(1×PBS、0.05%のTween20)、各ウェルに100μlの基質溶液(3,3’,5,5’テトラメチルベンジジン−過酸化水素:TMB3)を置いた。暗闇において室温で20分にわたるインキュベーションの後、100μl/ウェルで0.5MのH2SO4を添加することで反応を止めた。プレートの読み取りは450nmでの吸光度の測定によって行った。
【0078】
ELISAのプレートの読み取り結果は、
E/P%=(サンプルのOD−ネガティブコントロールのOD/ポジティブコントロールのOD−ネガティブコントロールのOD)×100
という、ポジティブコントロールの血清に対するサンプル血清の反応性のパーセントの形で示される。
【0079】
陽性の限界値(基準となる陰性サンプルの平均値の2倍に等しい)は、ELISA Ag E/Sについては14%、ELISA THX−NBL1(Cterm)については44%である。
【0080】
200、1000または20000匹のT.spiralisのL1M幼虫で実験的に感染させた通常のブタの血清において、ELISA THX−NBL1(Cterm)またはELISA Ag E/Sによって検出された抗旋毛虫抗体の出現動態を比較した。
【0081】
結果は図3に示している。
【0082】
THX−NBL1(Cterm)抗原によって、T.spiralisに対する液性応答の容量依存的な検出が可能となった。また、ウェスタンブロッティングによるリニアエピトープの検出に伴うELISA検定による立体構造エピトープの検出は、旋毛虫のNBL1タンパク質の免疫優性を証明している。
【0083】
ELISA THX−NBL1(Cterm)は感染後25日からセロコンバージョンを検出したのに対し、ELISA Ag E/Sは10日遅れてしかセロコンバージョンを検出しなかった。
【0084】
また、T.spiralisおよび欧州で特定されている旋毛虫の他の三種、T.nativa、T.britoviおよびT.pseudospiralisによって誘発された液性応答のELISA Ag E/Sによる検出とELISA THX−NBL1(Cterm)による検出も比較した。
【0085】
結果は以下の表Iにまとめている。
【0086】
【表1】

【0087】
これらの結果の全体は、ELISA THX−NBL1(Cterm)によって、強く感染した動物については感染後15日目から液性応答の特に早い検出が可能となり、中間的な量である1000匹のL1Mで感染させた動物についてはわずかに遅い検出(感染後25日目)が可能となることを示している。同様の結果は、同じ手順にしたがって感染した微生物を有するブタでも得られた。ELISA Ag E/Sで検出された最も早いセロコンバージョンは感染後25日目だった。2つのELISA検定で得られた結果の比較は、ELISA THX−NBL1(Cterm)の利用によるT.spiralisの診断が早さという点で5〜20日有利であることを証明している(表I)。さらに、ELISA Ag E/SとELISA THX−NBL1(Cterm)で一度に診断したT.spiralisに感染させた動物では、すべてについて、ELISA THX−NBL1(Cterm)での血清の検出ウィンドウが縮小した。ELISA THX−NBL1(Cterm)の感度は、この動物実験のうちの7/9の通常のブタ、すなわち20000匹のT.spiralisのL1Mで感染させた3/3のブタ、1000匹のL1Mで感染させた3/3のブタ、そして200匹のL1Mによってのみ感染した1/3のブタに対する有効なスクリーニングによって証明された。検出は、これらの動物におけるT.spiralisの筋肉寄生虫の量に関連しており、その量は、200匹のL1Mで実験的に感染させたブタについては平均でグラムあたり3匹の幼虫(LpG)、1000匹のL1Mで実験的に感染させたブタについては43LpG、そして20000匹のL1Mで実験的に感染させたブタについては538LpGの変動がある。
【0088】
欧州で特定されている旋毛虫の他の三種、T.nativa、T.britoviおよびT.pseudospiralisによって誘発された液性応答も、ELISA THX−NBL1(Cterm)によって容量依存的に検出されており、このことは、旋毛虫属におけるNBL1の遺伝子的保存および抗原的保存(免疫優性)、ひいては旋毛虫症の広い範囲での診断に対する利点を証明している。診断の感度と早さ(感染後15日目)が確認されている。セロコンバージョンのウィンドウは、ELISA THX−NBL1(Cterm)では5日〜45日で縮小し、T.spiralisの感染と同様に、ELISA Ag E/SとELISA THX−NBL1(Cterm)で一度に診断した動物の全体では、ELISA THX−NBL1(Cterm)での血清の検出ウィンドウが縮小した。さらに、T.nativa種による感染分析では、筋肉寄生虫の量が平均して7×10−4〜0.1LpGでしかなかったにも関わらず6/9の動物(そのうち2頭のブタはELISA Ag E/Sではスクリーニングされていない)を診断したELISA THX−NBL1(Cterm)の強い感度が証明された(対するELISA Ag E/Sでは5/9である)。
【0089】
ELISA THX−NBL1(Cterm)の特異性は、各感染実験の前および感染後10日までに動物の全体から採取した血清によって証明された。野外飼育のブタの200以上の血清を、分子の特異性を示すために用いた。旋毛虫に対して陰性のブタはいずれもELISA THX−NBL1(Cterm)に反応しなかった。
【0090】
これらの結果は図4に示している。
【実施例3】
【0091】
実施例3:NBL1の免疫優性のエピトープの特定。
NBL1のC末端部分から推定されるアミノ酸配列のin silico分析を行い、最も抗原性の強い領域を予測できるようにした。
【0092】
in silico分析の結果によって、NBL1のC末端部分の113アミノ酸(327番アミノ酸から440番アミノ酸)をカバーするオーバーラップした11のペプチドを選択することになった。これらのペプチド配列を以下に示す。
N5EM1:NH2−NSPEGTVKWASKEDS−CONH2(SEQ ID NO:14)
N5EM2:NH2−ASKEDSPVDLSTASR−CONH2(SEQ ID NO:15)
N5EM3:NH2−LSTASRPTNPYTGSR−CONH2(SEQ ID NO:16)
N5EM4:NH2−PYTGSRPTSPSSGSR−CONH2(SEQ ID NO:17)
N5EM5:NH2−PSSGSRPTYPSSGSR−CONH2(SEQ ID NO:6)
N5EM6:NH2−PSSGSRPTSPSSGSR−CONH2(SEQ ID NO:18)
N5EM7:NH2−PSSGSRPTYPYTGSR−CONH2(SEQ ID NO:7)
N5EM8:NH2−PYTGSRPTPQKPVFP−CONH2(SEQ ID NO:19)
N5EM9:NH2−QKPVFPSYQKYPPAV−CONH2(SEQ ID NO:20)
N5EM10:NH2−KYPPAVQKYIDSLPS−CONH2(SEQ ID NO:21)
N5EM11:NH2−RPTSPSSGSRPTYPS−CONH2(SEQ ID NO:8)
【0093】
前もってビオチン化したペプチドを、ストレプトアビジンで事前に処理したプレートでインキュベートすることを除き(1×PBS内で2μg/ml、100μl/ウェル)、実施例2で記載したものと同一の手順を用いて、20000匹のT.spiralisのL1M幼虫で感染させ、感染後60日目に回収したブタの血清に対するN5EMペプチドの抗原性を間接ELISA法によって評価した。
【0094】
三つの免疫反応性ペプチド(N5EM5、7および11)が検出された。免疫反応性ペプチドの一次配列の分析によって、共通した10個のアミノ酸モチーフ(PSSGSRPTYP)の存在が明らかになった(SEQ ID NO:5)。また、このモチーフはNBL1タンパク質の全配列において4回現れる。他方では、6アミノ酸のオーバーラップしたペプチドを介したエピトープマッピングによって、ペプチド内部のリニアエピトープの免疫反応性に関して、単一のアミノ酸、チロシンが最も重要であることを明らかにすることができた。
【0095】
最後に、補助実験によって、感度および早さという点で、N5EM11がN5EM5およびN5EM7と比べて最も反応が良いペプチドであることが証明された。
【0096】
このペプチドを、THX−NBL1(Cterm)について実施例2で記載したように、T.spiralisのE/S抗原と比較した。
【0097】
結果は以下の表IIにまとめている。
【0098】
【表2】
【0099】
これらの結果は、ELISA N5EM11によってT.spiralisへの穏やかな感染を検出することが可能となることを示している。さらに、感染後20日目以降の早期検出、すなわち、ELISA Ag E/Sで可能であるよりも5〜10日早い検出が得られた。
【0100】
また、検出ピーク後に抗N5EM11抗体の割合の減少も観察されており、このことは、この早い性質を裏付けており、また、最近の感染日を判定するために有効である。T.spiralisで感染させ、ELISA N5EM11ペプチドによって診断したブタの全体は、部分的にはELISA Ag E/Sで観察されたウィンドウと類似しているが、ほとんどの場合に縮小したセロコンバージョンのウィンドウを呈している。
【0101】
また、N5EM11は、T.nativa、T.britoviおよびT.pseudospiralisに感染したブタの血清と交差した抗原反応を呈しており、このことは、旋毛虫のL1NN段階におけるこのペプチドの免疫優性を証明している。交差したこれらの抗原反応により、ELISA Ag E/Sより5〜30日早く検出を得ることができる。
【0102】
しかし、T.pseudospiralisによって感染させ二つのテストで同時に検出した動物では検出が遅いという点において、ELISA Ag E/Sと比べて感度の低下が見られた。逆に、ELISA N5EM11によって、T.nativaによって感染し、筋肉寄生虫の量が残っているにも関わらずELISA Ag E/Sではスクリーニングされなかった動物を診断することが可能となっている。
【0103】
ELISA N5EM11の特異性は感染実験の前および感染後10日目までの300頭の動物から採取した血清によって評価した。この特異性は99%を超えている(結果は図示せず)。
【実施例4】
【0104】
411抗原の同定と単離。
411のcDNAクローンをT.spiralisの早期侵入段階AdおよびL1NNのcDNAライブラリーから選別した。
【0105】
このcDNAクローンの核酸配列ならびに推定されたポリペプチド配列を判別し、それぞれ、配列表においてSEQ ID NO:3とSEQ ID NO:4という番号で示した。411のオープンリーディングフレームは20kDaの推定タンパク質をコードしている。シグナルペプチドを除いて、いかなるタンパク質領域も同定されなかった。
【0106】
411の完全なオープンリーディングフレームとGenbankで入手可能な配列との比較は、その411が、T.pseudospiralisで同定されている排泄/分泌タンパク質Tp21−3(AAF79206、NAGANO et al.,2001)とは78.7%の同一性があり、PolvereおよびDespommierによって提示されているT.spiralisの推定ORF17.20(AAB48489)の配列とは86.6%の同一性があることを示している。この配列の比較によって、411は旋毛虫属に共通したこの遺伝子ファミリーの新しいメンバーとして同定される。
【0107】
411の完全なオープンリーディングフレームを、オリゴヌクレオチド411F(5’−CACCCGAGAAAACATGCAT−3’)(SEQ ID NO:22)および411R(5’−TCCATTCAATTTTGCGTCAC−3’)(SEQ ID NO:23)、DNAポリメラーゼAccuPrime Pfx DNA polymerase(Invitrogen社)を用いて増幅し、製造者(Invitrogen社)の推奨にしたがって発現キット「Champion pET102 Directional TOPO」を用い、プラスミドpET102D/topoにおいてクローニングした。
【0108】
得られた組換えプラスミドはpET102−411と称するものであり、予測分子量が36.7kDaの330アミノ酸のチオレドキシン−411融合タンパク質(THX−411)をコードし、C末端の位置にポリヒスチジンタグを有している。この融合タンパク質の配列は図5に示している。
【0109】
融合タンパク質THX−411は、プラスミドpET102−411によって形質転換した大腸菌BL21Star(DE3)、BL21(DE3)pLys(Invitrogen社)の中で発現させ、提供者(Qiagen)によって推奨される手順を用い、Ni−NTAカラム(Ni−NTAスピン・カラム・キット、Ni−NTAビーズ)上で、変性条件下でのアフィニティークロマトグラフィーによって精製した。
【0110】
精製された融合タンパク質THX−411は、変性条件下での電気泳動(SDS−PAGE)の後、予想された36.7kDaのサイズのバンドの形で現れた。
【0111】
旋毛虫症に感染していないブタの血清と、20000匹のT.spiralisのL1M幼虫による感染後30日目および50日目の同一のブタの血清に対するタンパク質THX−411の免疫反応性を、対照のE/S抗原(上記実施例2に記載したように調製)の免疫反応性と比較した。分析はウェスタンブロッティングにより、上記実施例1に記載したものと同一の手順を用いて行った。
【0112】
結果は図6に示している。
【0113】
T.spiralisに感染したブタの血清に対するTHX−411の非常に強い免疫反応性と、抗411抗体の早期検出(感染後30日目)が観察される。軽いノイズも観察されるが、該ノイズは、THX−411の調製物中に残留していた高分子量の細菌タンパク質と、血清中に存在する抗大腸菌抗体との間の交差反応に起因している。
【実施例5】
【0114】
旋毛虫に対する液性応答を検出するためのTHX−411の利用法。
上記実施例4に記載したように調製したタンパク質を、旋毛虫で感染させたブタに由来する血清に対する間接ELISA法により、T.spiralisの排泄/分泌抗原(E/S)との比較で評価した。
【0115】
T.spiralisの200、1000、20000匹のL1M幼虫で実験的に感染させた通常のブタの血清において、ELISA THX−411またはELISA Ag E/Sによって検出された抗旋毛虫抗体の出現動態を比較した。また、ELISA Ag E/SとELISA THX−411による、T.spiralisと、欧州で特定されている他の三種の旋毛虫、T.nativa、T.britoviおよびT.pseudospiralisによって誘発された液性応答の検出も比較した。
【0116】
用いた手順は実施例2で記載したものと同一である。THX−411抗原は2μg/ml、そしてE/S抗原は1.25μg/mlで用いた。
【0117】
陽性の限界はELISA Ag E/Sでは14%、ELISA THX−411では52%である。
【0118】
結果は図7および下の表IIIに示している。
【0119】
【表3】
【0120】
これらの結果の全体は、411抗原によってT.spiralisに対する液性応答の容量依存的な検出が可能となることを示している。また、ウェスタンブロッティングによるリニアエピトープの検出に伴うこのELISA検定による立体構造エピトープの検出は、旋毛虫の411タンパク質の免疫優性を証明している。
【0121】
THX−411組換えタンパク質によって、T.spiralisに対する抗体の特に早い検出が可能となる(20000匹のL1Mで強く感染させた動物については感染後20日目から)。セロコンバージョンは液性応答のプロファイルを伴っており、該液性応答は高い力価を有し、感染後60日まで保持される。中程度のおよび少ない寄生虫の量で感染させた動物のセロコンバージョンはもっと遅く、それぞれ感染後30日目と60日目に検出された。ELISA Ag E/Sによって検出された最も早いセロコンバージョンは感染後25日目である。
【0122】
ELISA Ag E/SとELISA THX−411によって同時に診断された、T.spiralisで感染させた動物は、それらのうち2/5については、ELISA THX−411での血清の検出ウィンドウが縮小し、早さという点で5〜10日有利である。ELISA THX−411の感度は、この動物実験のうちの5/9の通常のブタ、すなわち、20000匹のT.spiralisのL1Mで感染させた3/3のブタ、1000匹のL1Mで感染させた1/3のブタ、そして200匹のL1Mのみで感染させた1/3のブタに対する有効スクリーニングによって証明された。検出はこれらの動物におけるT.spiralisの筋肉寄生虫の量に関連しており、その量は、200匹のL1Mで実験的に感染させたブタについては平均でグラムあたり3匹の幼虫(LpG)、1000匹のL1Mで感染させたブタについては43LpG、そして20000匹のL1Mで感染させたブタについては538LpGの変動がある。
【0123】
T.nativa、T.britoviおよびT.pseudospiralisによって誘発された液性応答もELISA THX−411によって検出されており(20000匹のL1Mで感染させた動物の10/12)、このことは、旋毛虫属における411の遺伝子的保存と抗原的保存、ひいては旋毛虫症の広い範囲での診断に対する利点を証明している。診断の早さは感染後20日目以降で確認されている。セロコンバージョンのウィンドウはELISA 411では5日〜20日で縮小し、T.spiralisによる感染と同様に、ELISA Ag E/SとELISA THX−411で同時に診断した動物の50%〜100%は、ELISA THX−411での血清の検出ウィンドウが縮小した。T.nativa属による感染の分析は、ELISA THX−411の鋭い感度を証明しており、ELISA THX−411は、筋肉寄生虫の量が平均で7×10−4〜0.1LpGしかなかったにも関わらず、6/9の動物を診断している(対するELISA Ag E/Sでは5/9である)。さらに、T.nativa属によって発生した感染力が顕著に弱いにも関わらず、同様の液性応答のプロファイルがT.spiralisとT.nativaによって感染された動物で得られており、このことは、T.nativaの411タンパク質の非常に強い免疫原性と、凍結に対して耐性のあるこの種を検出するための該タンパク質の利用性を示唆している。
【0124】
ELISA THX−411の特異性を、感染実験の前および感染後10日までに150頭の動物から採取した血清を用いて評価した。この特異性は99%を超える(結果は図示せず)。
【0125】
結論。
NBL1抗原および411抗原は免疫優性の抗原を構成し、旋毛虫属内に保持されている。これらの抗原(NBL1のC末端、NBL1のペプチドエピトープN5EM11、411抗原)を用いたELISA検定は、旋毛虫に対する99%を超える特異性を有し、また該検定によって、欧州で特定されている4種の旋毛虫によって引き起こされるブタの旋毛虫症に対する早い診断が可能となる(感染後15〜60日)。
【0126】
精製され、タンパク質のC末端部分に位置するNBL1の免疫優性のエピトープを含有する組換えタンパク質THX−NBL1(Cterm)を用いたELISA検定は、旋毛虫に特異的で感度があり、また該検定によって、ELISA Ag E/Sと比べて5〜45日という鋭い早さでブタの旋毛虫症を診断することが可能となる。さらに、この検定によって、ELISA Ag E/SとELISA THX−NBL1(Cterm)によって同時に診断した動物の100%で、ELISA Ag E/Sに比べて縮小された血清の検出ウィンドウを得ることが可能となった。
【0127】
精製された組換えタンパク質THX−411を用いるELISA検定は、現状では少し低い感度で(診断した動物の17/36)、ELISA Ag E/Sによって検出される液性応答の動態を再現する。しかし、この新しいELISAの感度によって、ELISA Ag E/Sによってスクリーニングされないブタの診断が可能となった。ELISA THX−411によって、ELISA Ag E/Sと比べて5〜20日という鋭い早さでブタの旋毛虫症を診断することを可能にすることができる。さらに、ELISA Ag E/SとELISA THX−411によって同時に診断した動物の50%〜100%で、ELISA THX−411によって血清の検出ウィンドウが縮小した。
【0128】
これらの結果の全体は、NBL1抗体および411抗体をE/S抗原の代わりに、あるいはE/S抗原を補完するものとして、旋毛虫症の早期の血清診断に用いることができることを示している。さらに、旋毛虫のこれら二つの新しい抗原を組み合わせることにより、追加の動物診断によって、ELISAの感度を向上させることが可能となる。
【0129】
N5EM11タイプの免疫優性のペプチドを伴うNBL1(Cterm)と411の組み合わせによって、旋毛虫の診断感度を向上させることが可能となる(検出された動物の24/36)。
【0130】
411を伴うNBL1の追加の効果は、411によって得られる動物検出の利点、ならびに、411によって得られる動物のより速い検出として現れる。これら三種(NBL1、411、N5EM11)は、2つまたは3つの抗原を介した抗体の同時検出によってELISA検定を時間の上で安定させるが、動物の数という点では追加の利点はない。
【0131】
しかし、感染後30日未満という早期診断が二つの抗原NBL1と411によって得られる。
【0132】
参考文献
MURRELL et al., The systematics of the genus Trichinella with a key to species, Vet Parasitol, 93, 293−307, (2000).
GASSER et al., Nonisotopic single−strand conformation polymorphism analysis of sequence variability in ribosomal DNA expansion segments within the genus Trichinella (Nematoda: Adenophorea), Electrophoresis, 25, 3357−3364, (2004).
BOIREAU et al., Risques parasitaires lies aux aliments d’origine animale, Revue francaise des laboratoires, 71−89, (2002).
CAPO & DESPOMMIER, Clinical aspects of infection with Trichinella spp, Clin Microbiol Rev, 9, 47−54, (1996).
FOURESTIE et al., Randomized trial of albendazole versus tiabendazole plus flubendazole during an outbreak of human trichinellosis, Parasitol Res, 75, 36−41, (1988).
DUPOUY−CAMET, Trichinellosis: a worldwide zoonosis, Vet Parasitol, 93, 191−200, (2000).
MURRELL & POZIO, Trichinellosis: the zoonosis that won’t go quietly, Int J Parasitol, 30, 1339−1349, (2000).
BOIREAU et al., Trichinella in horses: a low frequency infection with high human risk, Vet Parasitol, 93, 309−320, (2000).
GAMBLE et al., Diagnosis of swine trichinosis by enzyme−linked immunosorbent assay (ELISA) using an excretory−secretory antigen, Vet Parasitol, 13, 349−361, (1983).
GAMBLE et al., Evaluation of excretory−secretory antigens for the serodiagnosis of swine trichinellosis, Vet Parasitol, 30, 131−137, (1988).
REASON et al., Novel tyvelose−containing tri− and tetra− antennary N−glycans in the immunodominant antigens of the intracellular parasite Trichinella spiralis, Glycobiology, 4, 593−603, (1994).
NAGANO et al., Molecular cloning and characterization of a 21 kDa protein secreted from Trichinella pseudospiralis, J Helminthol, 75, 273−278, (2001).
【図面の簡単な説明】
【0133】
【図1】タンパク質THX−NBL1(Cterm)の配列を示す図である。
【図2】タンパク質THX−NBL1(Cterm)の免疫反応性を示す図である。
【図3】T.spiralisに感染させたブタの血清においてELISA THX−NBL1(Cterm)またはELISA Ag E/Sによって検出した抗旋毛虫抗体の出現動態の比較を示す図である。
【図4】ELISA NBL1(Cterm)の特異性を示す図である。
【図5】タンパク質THX−411の配列を示す図である。
【図6】タンパク質THX−411の免疫反応性を示す図である。
【図7】T.spiralisに感染したブタの血清においてELISA THX−411またはELISA Ag E/Sによって検出した抗旋毛虫抗体の出現動態の比較を示す図である。
【技術分野】
【0001】
本発明は、旋毛虫症の診断および予防の分野における、寄生虫である「旋毛虫」において同定された新規の抗原の利用法に関するものである。
【背景技術】
【0002】
旋毛虫症は、寄生虫である「旋毛虫」に感染した肉の消費に関連した人獣共通感染症の一つである(MURRELL et al.,2000)。
【0003】
「双器綱」のこの線虫は、旋毛虫科に属し、該旋毛虫科は、系統発生学的に異なる二つのグループと関連する8つの種と3つの遺伝子型を含んでおり、該二つのグループとは、一つは哺乳動物に感染する、被嚢を形成する旋毛虫であり(T.spiralis、T.nativa、T.britovi、T.murrelli、T.nelsoni)、もう一つは、哺乳動物、鳥類および爬虫類に感染する、被嚢を形成しない旋毛虫(T.pseudospiralis、T.papuae、T.zimbabwensis)である(GASSER et al.,2004)。これらすべての種は人に感染する可能性がある。
【0004】
寄生虫の生活環は自動異宿主性である。該生活環は、連続的に終宿主(成長した寄生虫のキャリア)でもあり中間宿主(感染幼虫のキャリア)でもある、同一の宿主で全て展開される(BOIREAU et al.,2002)。感染幼虫の一つの宿主からもう一つ宿主への移行は、新しいサイクルを行うために必要である。この移行は、幼虫に感染した生の肉または十分に焼かれていない肉を摂取することによって起こる。消化の際、該幼虫は解放され、そして腸上皮に侵入し、そこで性別のある成虫(Ad)へと脱皮する。受精した雌は、次に新生L1幼虫(L1NN)を生み出し、該幼虫はリンパおよび血液の流れを介して横紋筋に達する。これらのL1NN幼虫は筋肉細胞に侵入し(L1M:筋肉L1幼虫の感染進行段階)、被嚢を形成する旋毛虫の場合には厚い保護性のコラーゲン嚢で、被嚢を形成しない旋毛虫の場合には非常に薄い保護性のコラーゲン嚢で覆われている、フィーダー細胞への脱分化を引き起こす。
【0005】
旋毛虫症は動物においては無症状であるが、ヒトへの感染は、初期の腸の段階では嘔吐感を伴う下痢、嘔吐および激しい腹痛として現れるのに対し、筋肉への侵入段階に関連する症状は、熱、顔のむくみおよび筋肉痛の合併を特徴としている(CAPO & DESPOMMIER、1996)。目、肺、胃腸、心臓および神経系での発症も、進行が致死的となり得る旋毛虫症のこの診断指標に加えることができる。患者における筋肉痛の持続を代表とする感染の慢性的な特徴は、フィーダー細胞内部で寄生虫が生存していることに関連している。
【0006】
駆虫薬によるヒトの旋毛虫症の特異的な治療は、あらゆる寄生段階に対する対処、特にL1M幼虫の周りにコラーゲンの保護嚢が形成される前の対処が可能になるよう感染の診断が早期に行われると、より効果的である(FOURESTIE et al.,1988)。
【0007】
疫学的なデータによって、ブタに代表される家畜における感染のサイクルも支えている野生動物の数多くの種を伴う伝染様式に関連する、あらゆる地帯における寄生虫の地理的分布が示されている(DUPOUY−CAMET,2000)。
【0008】
ヒト旋毛虫症の流行、すなわち突発的または再発的な人獣共通感染症の流行は、食習慣および必ずしも有効ではない衛生管理の問題のために、世界中の公衆衛生における現実的な問題となっている(MURRELL & POZIO,2000)。これらの流行は、豚およびイノシシの肉ならびに馬肉に主として関与している(BOIREAU et al.,2000)。
【0009】
したがって、ヒトの汚染の予防は、肉の中心まで火を入れることと、飼育条件および/または動物(豚、馬、イノシシおよび旋毛虫に敏感なその他の野生動物種)の旋毛虫症の管理を向上させることを介して行われる(BOIREAU et al.,2002)。
【0010】
旋毛虫症のスクリーニング技術は二つに分けられる。すなわち、1)トリキノスコープ(肉片の顕微鏡観察)による、すなわち筋肉サンプルの人工的な消化の後の、L1M幼虫の直接的な検出と、2)旋毛虫抗原に対する抗体の検出を可能にする、さまざまな免疫学的方法による間接的な検出である。
【0011】
寄生虫の各発達段階、すなわち成虫(Ad)、新生幼虫(L1NN)および筋肉幼虫(L1M)は、特定の抗原プロフィールに対応している。
【0012】
現状で免疫診断に用いられているものは、L1M段階の幼虫に由来する抗原調製物である。実際、二つの先行する段階AdおよびL1NNの抗原成分は精製するのが難しく、これら二つの段階の一方および/または他方に関連する免疫優性の抗原は現在までに特定できていない。
【0013】
主として、幼虫の溶解、溶解物の遠心分離、そして上清の回収によって得られた全可溶性抗原の調製物、あるいは、より頻繁には、排泄/分泌抗原(E/S抗原)が用いられている。
【0014】
排泄−分泌抗原はL1M幼虫を培地で生育させる際に産生される。該抗原はスティコソームと称される特定の器官に由来し、該器官は、およそ50の円盤状の細胞であるスティコサイトで構成されている。スティコサイトは粒状物を含んでおり、その内容物は寄生虫の食道の管腔にある小管によって排出されるものである。この内容物は非常に抗原性が高く、排泄分泌抗原の一部を構成する。これらの抗原はタンパク質の複雑な混合物を形成し、該混合物はとりわけ、特定の糖分子を有する糖タンパク質の群(TSL1抗原と称される)を含んでおり、該糖分子は、旋毛虫のみで知られておりかつこの寄生虫のすべての種に存在しているベータ・チベロースである。
【0015】
排泄−分泌抗原の調製物は、現状では旋毛虫症の免疫診断材料の基準として用いられており、Trichinella spiralisのL1M幼虫の培地から得られる。18〜20時間の培養の後、培地を濾過によって回収し、そして濃縮する(GAMBLE et al.,1983、GAMBLE et al.,1988)。
【0016】
全可溶性抗原の調製物は、主な不都合としては特異性が欠けている。その他の寄生虫症と交差した抗原性反応が頻繁に観察されている。排泄−分泌抗原によってより良い特異性を得ることが可能となる。しかし、いずれの場合にも、抗原の標準化されたロットを大量に生産するのは難しい。
【0017】
ベータ・チベロースを含む糖の構造は、E/S抗原調製物の免疫優性のエピトープに相当し(REASON et al.,1994、米国特許第5541075号明細書および米国特許第5707817号明細書)、化学的に合成されており、旋毛虫の免疫診断のために利用することが提案されている。
【0018】
この試薬は良好な特異性を有しているが、その感度はE/S抗原調製物よりも低いようである。さらに、化学的方法によるこの構造の合成は、実施するのにコストが高く、負担の大きいものになっている。
【0019】
旋毛虫の血清学的診断の領域でぶつかるもう一つの問題は、検出の「ブラインド・ウィンドウ」が存在することであり、該ブラインド・ウィンドウは感染の早い段階に対応し、偽陰性の結果として現れる。さらに、馬では、感染後の25週目には抗体の漸次的な消失が観察されている。
【特許文献1】米国特許第5541075号明細書
【特許文献2】米国特許第5707817号明細書
【発明の開示】
【発明が解決しようとする課題】
【0020】
本発明者らは、旋毛虫による感染の早い段階に関連し、旋毛虫症の血清学的診断に利用可能な免疫優性の抗原を特定することで、ヒトだけでなく動物において、早期の、特異的な、かつ旋毛虫への感染に敏感な検出が得られる手段を提供することに取り組んだ。この目的において、本発明者らは、L1NN段階および/またはAd段階の旋毛虫によって発現される遺伝子産物の中に、所望の抗原特性を有するタンパク質が存在するかどうか研究した。
【0021】
この枠組みにおいて、本発明者らは、旋毛虫のL1NN段階で特異的に発現するタンパク質の一部をなすTrichinella spiralisのタンパク質が免疫優勢性の抗原を構成し、該タンパク質によって、旋毛虫に対する液性応答の早期検出が可能になることと、該タンパク質がさらに、旋毛虫のさまざまな種の間で保持されていることを発見した。
【0022】
このタンパク質を以下ではNBL1と称することにする。このタンパク質をコードしているcDNAの完全な配列ならびにこの配列から推定されるポリペプチド配列は、それぞれAF331160およびAAK16520という番号のもと、Genbankでアクセス可能であり(あるいはSwissprot Q9BJL7)、これらの配列は「セリンプロテアーゼSS2、新生幼虫に固有」という註が付けられている。また、これらの配列は、添付の配列表において、SEQ ID NO:1とSEQ ID NO:2という番号で再掲されている。このタンパク質をコードしているcDNAの部分配列ならびにこの配列から推定されるポリペプチド配列は、AY491941およびAAR36900という番号のもと、それぞれGenbankでアクセス可能である(あるいはSwissprot Q6RUJ3)。
【0023】
さらに、本発明者らは、NBL1に対する液性応答に関連する免疫反応性がこのタンパク質のC末端部分に局在していることを示し、この反応性を担う免疫優性のエピトープを特定した。
【0024】
他方では、本発明者らは、T.spiralisのAdとL1NNを混合した早期段階のcDNAライブラリーから、以下では411と称する新たな遺伝子を特定した。
【0025】
この遺伝子の配列は、添付の配列リストにおいて、SEQ ID NO:3という番号で示されており、該遺伝子配列の翻訳産物の配列はSEQ ID NO:4という番号で示されている。この遺伝子の翻訳産物は、タンパク質Tp21−3と称され、T.pseudospiralisで同定されているE/S抗原(AAF79206、NAGANO et al.,2001)と関連性を有し(78.7%の同一性)、さらに、T.spiralisの推定ORF17.20の翻訳産物(AAB48489)とも関連性を有し、該産物とは86.6%の同一性を有する。
【0026】
また、411遺伝子の翻訳産物によって、早期段階で、旋毛虫のさまざまな種に対する液性応答を検出することが可能となる。
【0027】
さらに、実験を行ったところ、他の抗原によってもE/S抗原によっても少なくとも感染後(pi)15日〜30日の間には検出されなかった旋毛虫に感染した動物を、NBL1抗原および411抗原のそれぞれによって検出することが可能であった。
【0028】
したがって、NBL1抗原(または該抗原の免疫優性のエピトープ)と411抗原を組み合わせることによって、診断の感度、とりわけ感染の初期段階(感染後15日〜20日)に対する感度を向上させることが可能となる。
【課題を解決するための手段】
【0029】
したがって、本発明は生体標本において抗旋毛虫抗体を検出するための試薬としての、抗旋毛虫抗体によって認識される抗原ポリペプチドの利用法を対象としており、該使用法は、前記ポリペプチドが、
a)配列PSSGSRPTYP(SEQ ID NO:5)によって定義されるNBL1抗原の免疫優性のエピトープを含んだポリペプチドと、
b)以下で411抗原とも称されるポリペプチドであり、配列SEQ ID NO:4の25〜175番目のアミノ酸を含むか(411タンパク質の成長した形に相当する)、あるいは、配列SEQ ID NO:4の25〜175番目のアミノ酸配列と少なくとも80%、好ましくはより高く、少なくとも85%、90%、または95%の同一性を有する配列を含むポリペプチド、
から選択されることを特徴としている。
【0030】
より特徴的には、本発明は、生体サンプルにおける抗旋毛虫抗体の存否の検出方法を対象としており、該方法は、
−前記生体サンプルを、前記生体サンプルに場合によっては存在する抗旋毛虫抗体との抗原/抗体複合体の形成を可能にする条件のもとで、上記で定義した一つもしくは複数のポリペプチドa)および/または一つもしくは複数のポリペプチドb)と接触させることと、
−あらゆる適切な手段によって、場合によっては形成される抗原/抗体複合体を検出すること、
を含むことを特徴としている。
【0031】
一般的には、前記生体標本は血清標本である。該血清標本は、旋毛虫に感染しうる種に属する、この寄生虫の存否を検出しようとするあらゆる対象(哺乳動物、鳥類または爬虫類)から得ることができる。有利には、哺乳動物、例えば飼育動物またはヒトの患者から得られる標本である。
【0032】
有利には、上記で定義した一つもしくは複数のポリペプチドa)および/または一つもしくは複数のポリペプチドb)を含む混合物が用いられる。
【0033】
この組み合わせによって、とりわけ、個別に用いた各ポリペプチドに比べて反応性の幅を広げることが可能となる。
【0034】
また、アクセス番号Genbank AAK16520で識別される全NBL1抗原と、アクセス番号Genbank AAR36900で識別されるその断片を除き、上記で定義したポリペプチドa)はそれ自体で本発明の対象の一部をなす。
【0035】
本発明にしたがったこれらのポリペプチドのうち、とりわけ、配列PSSGSRPTYPSSGSR(SEQ ID NO:6)、配列PSSGSRPTYPYTGSR(SEQ ID NO:7)、配列RPTSPSSGSRPTYPS(SEQ ID NO:8)のうち一つまたは複数を含んだポリペプチドが挙げられる。
【0036】
これには、例えばNBL1抗原のC末端領域の断片も含まれ、とりわけ、配列SEQ ID NO:2の326〜459番目のアミノ酸に対応する、
ENSPEGTVKWASKEDSPVDLSTASRPTNPYTGSRPTSPSSGSRPTYPSSGSRPTSPSSGSRPTYPSSGSRPTYPSSGSRPTYPYTGSRPTPQKPVFPSYQKYPPAVQKYIDSLPSGTQGTLEYTVTQNGVTTTT(SEQ ID NO:11)、
を含んだ断片が挙げられ、またこの配列SEQ ID NO:11の小断片、とりわけ、配列SEQ ID NO:2の363〜409番目のアミノ酸に対応する、
PSSGSRPTYPSSGSRPTSPSSGSRPTYPSSGSRPTYPSSGSRPTYP(SEQ ID NO:9)という配列を含んだ小断片も含まれ、より特徴的には、配列SEQ ID NO:2の349〜415番目のアミノ酸に対応する、
SRPTNPYTGSRPTSPSSGSRPTYPSSGSRPTSPSSGSRPTYPSSGSRPTYPSSGSRPTYPYTGSRPT(SEQ ID NO:10)という配列を含んだ小断片が含まれる。
【0037】
上記で定義したポリペプチドb)は、アクセス番号GenBank AAF79206およびAAB48489で識別されるものを除いて、それ自体で本発明の対象の一部をなす。好ましいポリペプチドはとりわけ、配列SEQ ID NO:4のポリペプチド、または、配列SEQ ID NO:4の25〜175番目のアミノ酸に対応するポリペプチド、ならびに、配列SEQ ID NO:4または配列SEQ ID NO:4の25〜175番目のアミノ酸配列との間に少なくとも90%、好ましくは少なくとも95%の同一性を有するポリペプチドである。
【0038】
とりわけ、本発明は、配列PSSGSRPTYP(SEQ ID NO:5)の一つもしくは複数のコピー、またはこの配列を含むNBL1抗原の断片の一つもしくは複数のコピー、および/または、上記で定義したポリペプチドb)の一つもしくは複数のコピーを含む、場合によっては一つもしくは複数の他の異種配列と融合しているキメラポリペプチドを含む。
【0039】
また、本発明は、本発明にしたがったポリペプチドをコードするポリヌクレオチド、ならびに、前記ポリヌクレオチドを含んだ組換えベクター、および前記ベクターによって形質転換した宿主細胞も対象としている。
【0040】
また、本発明は上記で定義したような、一つまたは複数のポリペプチドa)および一つまたは複数のポリペプチドb)を含んだ組成物、ならびに、前記ポリペプチドをコードする一つまたは複数のポリヌクレオチドを含んだ組成物も対象としている。
【0041】
上記で定義したポリペプチドa)およびb)は、それ自体既知である、抗体検出のさまざまな方法の範囲で用いることができる。例えば、とりわけ、ELISAタイプの方法(直接、間接またはサンドイッチ)、ビーズへのミクロ凝集方法、ならびに、免疫標識に組み合わせた電気泳動転写法などが挙げられる。
【0042】
また、本発明は生体サンプルにおける抗旋毛虫抗体の存否を検出するために必要な物のセットも対象としており、これは、上記で定義した一つもしくは複数のポリペプチドa)および/または一つもしくは複数のポリペプチドb)、ならびに場合によっては、抗原/抗体複合体の形成を可能にする反応培地を構築するための適切な緩衝液および試薬、ならびに選択的に、前記抗原/抗体複合体の検出手段を含むことを特徴としている。
【0043】
有利には、前記セットは、固体の担体の上に固定されている、上記で定義したポリペプチドa)および/またはポリペプチドb)を含んでいる。利用可能な固体の担体の非限定的な例としては、マイクロタイタープレート、ビーズ、マイクロビーズまたは微粒子、試験紙などが挙げられる。
【0044】
また、前記セットは、一つまたは複数の陰性血清および一つまたは複数の陽性血清のような、コントロールサンプルも含むことができる。
【0045】
また、本発明は、前記ポリペプチドに対する特異的な抗体を調製するための、上記で定義したポリペプチドa)またはポリペプチドb)の利用法も対象としている。
【0046】
これらのポリペプチドは、それ自体既知である、さまざまな抗体の調製方法の範囲で利用することができる。例えば、該ポリペプチドは(場合によっては適切なアジュバントの添加後に)動物の免疫付与に利用することができる。また、該ポリペプチドは、アフィニティークロマトグラフィーの担体にグラフトすることで、生体液から目的のポリペプチドに対する特異的な抗体を精製できるようにすることができる。生体液とは、例えば目的とするポリペプチドで事前に免疫付与した動物の血清あるいはハイブリドーマの上清とすることができ、また、旋毛虫に感染した動物の血清であり、目的とするポリペプチドに対する特異的な抗体の亜集団を単離するための血清とすることもできる。
【0047】
また、本発明には、上記で定義したポリペプチドa)またはポリペプチドb)に対するあらゆる特異的な抗体が含まれる。それはポリクローナル抗体またはモノクローナル抗体とすることができる。好ましい抗体は、PSSGSRPTYP(SEQ ID NO:5)のエピトープを認識する抗体である。
【0048】
ポリペプチドに対する特異的な抗体は、それ自体既知であるさまざまな技術によって、とりわけ、目的とするポリペプチドで動物を免疫付与すること(場合によっては適切なアジュバントを添加する)と、その血清(ポリクローナル抗体の産生用)またはリンパ球細胞(モノクローナル抗体の産生用)を回収することを含む、従来の方法によって得ることができる。
【0049】
上記で定義したポリペプチドa)およびb)ならびにこれらのポリペプチドをコードするポリヌクレオチドは、免疫原性組成物、とりわけ抗旋毛虫ワクチンを調製するために利用することができる。
【0050】
また、本発明は、上記で定義した一つもしくは複数のポリペプチドa)および/または一つもしくは複数のポリペプチドb)を含んだ免疫原性組成物、あるいは、免疫応答の向上を可能にする一つまたは複数のアジュバントと結びついている、前記ポリペプチドをコードする一つまたは複数のポリヌクレオチドを含んだ免疫原性組成物も対象としている。
【0051】
本発明にしたがった免疫原性組成物の好ましい実施態様によると、それはワクチンである。
【0052】
ペプチドの免疫原性の向上を可能にする非常に多様なアジュバントはそれ自体当業者には知られており、例えば、ミョウバン(水酸化アルミニウム)、完全フロイントアジュバントまたは不完全フロイントアジュバント(IFA)、リポソーム、ならびにウィロソーム(再構成されたウイルスエンベロープ)、ムラミン酸のペプチド誘導体などが挙げられる。ワクチンの場合、当然、薬学的に許容可能なアジュバントが選択されることになり、好ましいアジュバントの例としては、例えばMONTANIDE ISA 70およびMONTANIDE ISA 775という名称でSEPPIC社によって市販されており、かつ欧州特許第480982号明細書、欧州特許第825875号明細書、米国特許第5422109号明細書、米国特許第6251407号明細書、米国特許第6610309号明細書にも記載されているアジュバントのような、「油中水型」エマルジョンタイプのアジュバントが挙げられる。
【0053】
場合によっては、とりわけ短いペプチド(30個以下のアミノ酸)の場合、前記ポリペプチドはキャリアタンパク質と結合させることができる。
【0054】
キャリアタンパク質の例としては、とりわけKLH(スカシ貝ヘモシアニン、Keyhole limpet hemocyanin)、ウシ血清アルブミン(BSA)、オボアルブミン、破傷風トキソイド、ジフテリアトキソイドが挙げられる。また、同一ペプチドの複数のコピーを互いに組み合わせることで、また場合によってはキメラポリペプチドの形でその他のペプチドエピトープと組み合わせることで、あるいは、例えばポリリジンのようなポリマー鎖を介して、マルチエピトープの組成物を形成することもできる。
【0055】
免疫原としてポリヌクレオチドを使うのであれば、免疫原組成物は、投与すべきポリヌクレオチドが挿入された組換えベクターの形をとることができる。例えば、ポックスウイルス、アデノウイルス、レトロウイルス、レンチウイルス、ヘルペスウイルス、AAV(アデノ随伴ウイルス)などのウイルスベクターを用いることができる。また、該組成物は、前記ポリヌクレオチドを含む一つまたは複数の発現ベクターによって形質転換された、非病原性バクテリアの形をとることもできる。また、ポリヌクレオチドを裸のDNAの形で直接投与するか、あるいは、リポソームに包入することもできる。ワクチンの場合、好ましくは非病原性バクテリア(例えば乳酸菌、あるいは大腸菌またはサルモネラ菌の非病原性株)、または、ワクチンウイルス株から派生したベクター、例えば仮性狂犬病(オーエスキー病)ウイルスのワクチン株から派生したベクターが用いられることになる。
【発明を実施するための最良の形態】
【0056】
本発明は、旋毛虫症の早期の免疫診断を目的とした、NBL1抗原および411抗原の利用法を示した実施例に関する、以下の説明の補足によってよりよく理解されるものである。
【0057】
図1:タンパク質THX−NBL1(Cterm)の配列。
NBL1に由来する配列を太字で示し、pET102プラスミドに由来する配列は斜体で示している。
【0058】
図2:タンパク質THX−NBL1(Cterm)の免疫反応性。
M:分子量マーカー、1:陰性のブタの血清、2:20000匹のT.spiralisのL1Mで実験的に感染させたブタの血清。
【0059】
図3:T.spiralisに感染させたブタの血清においてELISA THX−NBL1(Cterm)またはELISA Ag E/Sによって検出された抗旋毛虫抗体の出現動態の比較。
A:ELISA THX−NBL1(Cterm)による検出、B:ELISA Ag E/Sによる検出。
横軸:感染後の日数、縦軸:反応性のパーセント。ELISAによる検出限界は黒い線で示している(ELISA THX−NBL1(Cterm)については44%、ELISA Ag E/Sについては14%)。サンプル/ポジティブコントロール比の平均および標準偏差は感染したブタの各群について示している。
【0060】
図4:ELISA NBL1(Cterm)の特異性。
横軸:産業用のブタと野外飼育のブタに由来する230の陰性サンプルのコホート。5つの陽性サンプルは実験的に感染させたブタの血清に由来している。限界値は陰性サンプルの平均値の2倍に等しい。縦軸:反応性のパーセント。
【0061】
図5:タンパク質THX−411の配列。
411に由来する配列は太字で示している。プラスミドに由来する配列は斜体で示している。
【0062】
図6:タンパク質THX−411の免疫反応性。
1:感染後50日の陽性血清、2:感染後30日の陽性血清、3:5日前の陰性血清、4:共役型コントロール。M:分子量マーカー。
【0063】
図7:T.spiralisに感染したブタの血清においてELISA THX−411またはELISA Ag E/Sによって検出した抗旋毛虫抗体の出現動態の比較。
A:ELISA THX−411による検出、B:ELISA Ag E/Sによる検出。
横軸:感染後の日数、縦軸:反応性のパーセント。ELISAによる検出限界は黒い線で示している(ELISA THX−411では52%、ELISA Ag E/Sでは14%)。サンプル/ポジティブコントロール比の平均と標準偏差は感染したブタの各群について示している。
【実施例1】
【0064】
実施例1:NBL1のC末端部分を含んだ、組換えタンパク質THX−NBL1(Cterm)の産生。
Trichinella spiralisのL1NNのcDNAライブラリーに対する免疫スクリーニングを、T.spiralisの10000匹のL1Mによって実験的に感染させた後35日目に得られたブタの血清を用いて行った。この血清によって認識されたクローンのシーケンシングによって、それらのうち大半が同一のタンパク質をコードしていることが判定できた。
【0065】
このタンパク質は推定セリンプロテアーゼであり、そのcDNA配列および推定されるアミノ酸配列は、それぞれ、AF331160およびAAK16520という番号のもとGenbankで入手可能であり、また、ここではSEQ ID NO:1およびSEQ ID NO:2の番号で再掲している。該タンパク質はここではNBL1と称することにする。
【0066】
タンパク質のC末端部分を、NBL1CtermF(5’−CACCGAAAATTCTCCTGAAGGA−3’)(SEQ ID NO:12)とNBL1CtermR(5’−TGTTGTTGTAGTAACTCC−3’)(SEQ ID NO:13)オリゴヌクレオチド、そしてDNAポリメラーゼAccuPrime Pfx DNA polymerase(Invitrogen社)を用いて増幅し、製造者(Invitrogen社)の推奨にしたがって発現キット「Champion pET102 Directional TOPO」を用いて、プラスミドpET102D/topoでクローニングした。
【0067】
得られた組換えプラスミドはpET102−NBL1(Cterm)と称するもので、291個のアミノ酸からなり、C末端の位置にポリヒスチジンタグを有している、チオレドキシンNBL1(Cterm)(THX−NBL1(Cterm))融合タンパク質をコードしている。この融合タンパク質の配列は図1に示している。
【0068】
融合タンパク質THX−NBL1(Cterm)は、プラスミドpET102−NBL1(Cterm)によって形質転換した大腸菌BL21Star(DE3)、BL21(DE3)pLys(Invitrogen社)で発現させ、そして、提供者(Qiagen)によって推奨される手順を用いて、Ni−NTAカラム(Ni−NTAスピン・カラム・キット、Ni−NTAビーズ)上で、変性条件下でのアフィニティークロマトグラフィーによって精製した。
【0069】
精製された融合タンパク質THX−NBL1(Cterm)は、変性条件下での電気泳動(SDS−PAGE)の後、31.1kDaの予期されたサイズのバンドの形で現れた。
【0070】
旋毛虫症に感染していないブタの血清、および感染後60日目に採取された、20000匹のT.spiralisのL1M幼虫で感染させたブタの血清に対する、タンパク質THX−NBL1(Cterm)の免疫反応性を、ウェスタンブロッティングによって分析した。
【0071】
変性条件下の電気泳動の後(SDS−PAGE)、提供者(Amersham)の指示にしたがってタンパク質をHybond P膜(PVDF)でエレクトロブロッティングした。膜は、TBS−T(20mM、pH7.5のTris−HCl、150mMのNaCl、0.1%のTween20)と5%の脱脂乳において1時間にわたってプレハイブリダイゼーションした。1分間にわたるTBS−Tによる2回の洗浄の後、そして5分間にわたるTBS−Tでの3回の洗浄の後、膜を、TBSで1/200に希釈したブタの血清を用いて1時間インキュベートした。洗浄後、膜を、1/30000に希釈した、アルカリフォスファターゼ(A1192、Sigma社)で標識した抗ブタIgGウサギ二次抗体を用いて20分間インキュベートし、次に、標識を視認できるようにするために、30分間にわたって純粋なNBT/BCIP基質(E116、Interchim社)を用いてインキュベートした。
【0072】
結果は図2に示している。
【0073】
T.spiralisに感染したブタの血清によるTHX−NBL1(Cterm)の非常に強い免疫反応性が観察される。
【実施例2】
【0074】
実施例2:旋毛虫に対する液性応答を検出するためのTHX−NBL1(Cterm)の利用法。
上記実施例1に記載したように調製したタンパク質THX−NBL1(Cterm)を、T.spiralisの排泄/分泌抗原(E/S)と比較して、旋毛虫に感染したブタに由来する血清に対する間接ELISA法によって評価した。
【0075】
対照のE/S抗原をGAMBLEら(1988)に記載の手順にしたがって調製した。該抗原は、1%のピルビン酸塩、15%のウシ胎児血清(SVF)、1%のL−グルタミン、100U/mlのペニシリン、および100μg/mlのストレプトマイシンを含むRPMI1640において24時間にわたって維持されたL1M幼虫の培養物の上清から得た。
【0076】
ELISA検定のために、E/S抗原については1.25μg/ml、THX−NBL1(Cterm)については2μg/mlの割合で1×PBS緩衝液で希釈した抗原を、96ウェルプレート(MediSorpプレート、NUNC社)において4℃で一晩インキュベートした。3回の洗浄後(1×PBS、0.05%のTween20)、プレートを、洗浄液で2%に希釈した脱脂乳溶液で1時間にわたって室温で飽和させた。
【0077】
各ウェルに、0.05%のTween20を加えた、1×PBS緩衝液で1/20に希釈した100μlのブタの血清を置いた。37℃での30分間のインキュベーションと、続く3回の洗浄の後(1×PBS、0.05%のTween20)、各ウェルに、0.05%のTween20を加えた、1×PBS緩衝液で1/32000に希釈した100μlの共役溶液(Gタンパク質−ペルオキシダーゼ(P−8170、Sigma社))を置いた。37℃での30分間の新たなインキュベーションと、続く3回の洗浄の後(1×PBS、0.05%のTween20)、各ウェルに100μlの基質溶液(3,3’,5,5’テトラメチルベンジジン−過酸化水素:TMB3)を置いた。暗闇において室温で20分にわたるインキュベーションの後、100μl/ウェルで0.5MのH2SO4を添加することで反応を止めた。プレートの読み取りは450nmでの吸光度の測定によって行った。
【0078】
ELISAのプレートの読み取り結果は、
E/P%=(サンプルのOD−ネガティブコントロールのOD/ポジティブコントロールのOD−ネガティブコントロールのOD)×100
という、ポジティブコントロールの血清に対するサンプル血清の反応性のパーセントの形で示される。
【0079】
陽性の限界値(基準となる陰性サンプルの平均値の2倍に等しい)は、ELISA Ag E/Sについては14%、ELISA THX−NBL1(Cterm)については44%である。
【0080】
200、1000または20000匹のT.spiralisのL1M幼虫で実験的に感染させた通常のブタの血清において、ELISA THX−NBL1(Cterm)またはELISA Ag E/Sによって検出された抗旋毛虫抗体の出現動態を比較した。
【0081】
結果は図3に示している。
【0082】
THX−NBL1(Cterm)抗原によって、T.spiralisに対する液性応答の容量依存的な検出が可能となった。また、ウェスタンブロッティングによるリニアエピトープの検出に伴うELISA検定による立体構造エピトープの検出は、旋毛虫のNBL1タンパク質の免疫優性を証明している。
【0083】
ELISA THX−NBL1(Cterm)は感染後25日からセロコンバージョンを検出したのに対し、ELISA Ag E/Sは10日遅れてしかセロコンバージョンを検出しなかった。
【0084】
また、T.spiralisおよび欧州で特定されている旋毛虫の他の三種、T.nativa、T.britoviおよびT.pseudospiralisによって誘発された液性応答のELISA Ag E/Sによる検出とELISA THX−NBL1(Cterm)による検出も比較した。
【0085】
結果は以下の表Iにまとめている。
【0086】
【表1】
【0087】
これらの結果の全体は、ELISA THX−NBL1(Cterm)によって、強く感染した動物については感染後15日目から液性応答の特に早い検出が可能となり、中間的な量である1000匹のL1Mで感染させた動物についてはわずかに遅い検出(感染後25日目)が可能となることを示している。同様の結果は、同じ手順にしたがって感染した微生物を有するブタでも得られた。ELISA Ag E/Sで検出された最も早いセロコンバージョンは感染後25日目だった。2つのELISA検定で得られた結果の比較は、ELISA THX−NBL1(Cterm)の利用によるT.spiralisの診断が早さという点で5〜20日有利であることを証明している(表I)。さらに、ELISA Ag E/SとELISA THX−NBL1(Cterm)で一度に診断したT.spiralisに感染させた動物では、すべてについて、ELISA THX−NBL1(Cterm)での血清の検出ウィンドウが縮小した。ELISA THX−NBL1(Cterm)の感度は、この動物実験のうちの7/9の通常のブタ、すなわち20000匹のT.spiralisのL1Mで感染させた3/3のブタ、1000匹のL1Mで感染させた3/3のブタ、そして200匹のL1Mによってのみ感染した1/3のブタに対する有効なスクリーニングによって証明された。検出は、これらの動物におけるT.spiralisの筋肉寄生虫の量に関連しており、その量は、200匹のL1Mで実験的に感染させたブタについては平均でグラムあたり3匹の幼虫(LpG)、1000匹のL1Mで実験的に感染させたブタについては43LpG、そして20000匹のL1Mで実験的に感染させたブタについては538LpGの変動がある。
【0088】
欧州で特定されている旋毛虫の他の三種、T.nativa、T.britoviおよびT.pseudospiralisによって誘発された液性応答も、ELISA THX−NBL1(Cterm)によって容量依存的に検出されており、このことは、旋毛虫属におけるNBL1の遺伝子的保存および抗原的保存(免疫優性)、ひいては旋毛虫症の広い範囲での診断に対する利点を証明している。診断の感度と早さ(感染後15日目)が確認されている。セロコンバージョンのウィンドウは、ELISA THX−NBL1(Cterm)では5日〜45日で縮小し、T.spiralisの感染と同様に、ELISA Ag E/SとELISA THX−NBL1(Cterm)で一度に診断した動物の全体では、ELISA THX−NBL1(Cterm)での血清の検出ウィンドウが縮小した。さらに、T.nativa種による感染分析では、筋肉寄生虫の量が平均して7×10−4〜0.1LpGでしかなかったにも関わらず6/9の動物(そのうち2頭のブタはELISA Ag E/Sではスクリーニングされていない)を診断したELISA THX−NBL1(Cterm)の強い感度が証明された(対するELISA Ag E/Sでは5/9である)。
【0089】
ELISA THX−NBL1(Cterm)の特異性は、各感染実験の前および感染後10日までに動物の全体から採取した血清によって証明された。野外飼育のブタの200以上の血清を、分子の特異性を示すために用いた。旋毛虫に対して陰性のブタはいずれもELISA THX−NBL1(Cterm)に反応しなかった。
【0090】
これらの結果は図4に示している。
【実施例3】
【0091】
実施例3:NBL1の免疫優性のエピトープの特定。
NBL1のC末端部分から推定されるアミノ酸配列のin silico分析を行い、最も抗原性の強い領域を予測できるようにした。
【0092】
in silico分析の結果によって、NBL1のC末端部分の113アミノ酸(327番アミノ酸から440番アミノ酸)をカバーするオーバーラップした11のペプチドを選択することになった。これらのペプチド配列を以下に示す。
N5EM1:NH2−NSPEGTVKWASKEDS−CONH2(SEQ ID NO:14)
N5EM2:NH2−ASKEDSPVDLSTASR−CONH2(SEQ ID NO:15)
N5EM3:NH2−LSTASRPTNPYTGSR−CONH2(SEQ ID NO:16)
N5EM4:NH2−PYTGSRPTSPSSGSR−CONH2(SEQ ID NO:17)
N5EM5:NH2−PSSGSRPTYPSSGSR−CONH2(SEQ ID NO:6)
N5EM6:NH2−PSSGSRPTSPSSGSR−CONH2(SEQ ID NO:18)
N5EM7:NH2−PSSGSRPTYPYTGSR−CONH2(SEQ ID NO:7)
N5EM8:NH2−PYTGSRPTPQKPVFP−CONH2(SEQ ID NO:19)
N5EM9:NH2−QKPVFPSYQKYPPAV−CONH2(SEQ ID NO:20)
N5EM10:NH2−KYPPAVQKYIDSLPS−CONH2(SEQ ID NO:21)
N5EM11:NH2−RPTSPSSGSRPTYPS−CONH2(SEQ ID NO:8)
【0093】
前もってビオチン化したペプチドを、ストレプトアビジンで事前に処理したプレートでインキュベートすることを除き(1×PBS内で2μg/ml、100μl/ウェル)、実施例2で記載したものと同一の手順を用いて、20000匹のT.spiralisのL1M幼虫で感染させ、感染後60日目に回収したブタの血清に対するN5EMペプチドの抗原性を間接ELISA法によって評価した。
【0094】
三つの免疫反応性ペプチド(N5EM5、7および11)が検出された。免疫反応性ペプチドの一次配列の分析によって、共通した10個のアミノ酸モチーフ(PSSGSRPTYP)の存在が明らかになった(SEQ ID NO:5)。また、このモチーフはNBL1タンパク質の全配列において4回現れる。他方では、6アミノ酸のオーバーラップしたペプチドを介したエピトープマッピングによって、ペプチド内部のリニアエピトープの免疫反応性に関して、単一のアミノ酸、チロシンが最も重要であることを明らかにすることができた。
【0095】
最後に、補助実験によって、感度および早さという点で、N5EM11がN5EM5およびN5EM7と比べて最も反応が良いペプチドであることが証明された。
【0096】
このペプチドを、THX−NBL1(Cterm)について実施例2で記載したように、T.spiralisのE/S抗原と比較した。
【0097】
結果は以下の表IIにまとめている。
【0098】
【表2】
【0099】
これらの結果は、ELISA N5EM11によってT.spiralisへの穏やかな感染を検出することが可能となることを示している。さらに、感染後20日目以降の早期検出、すなわち、ELISA Ag E/Sで可能であるよりも5〜10日早い検出が得られた。
【0100】
また、検出ピーク後に抗N5EM11抗体の割合の減少も観察されており、このことは、この早い性質を裏付けており、また、最近の感染日を判定するために有効である。T.spiralisで感染させ、ELISA N5EM11ペプチドによって診断したブタの全体は、部分的にはELISA Ag E/Sで観察されたウィンドウと類似しているが、ほとんどの場合に縮小したセロコンバージョンのウィンドウを呈している。
【0101】
また、N5EM11は、T.nativa、T.britoviおよびT.pseudospiralisに感染したブタの血清と交差した抗原反応を呈しており、このことは、旋毛虫のL1NN段階におけるこのペプチドの免疫優性を証明している。交差したこれらの抗原反応により、ELISA Ag E/Sより5〜30日早く検出を得ることができる。
【0102】
しかし、T.pseudospiralisによって感染させ二つのテストで同時に検出した動物では検出が遅いという点において、ELISA Ag E/Sと比べて感度の低下が見られた。逆に、ELISA N5EM11によって、T.nativaによって感染し、筋肉寄生虫の量が残っているにも関わらずELISA Ag E/Sではスクリーニングされなかった動物を診断することが可能となっている。
【0103】
ELISA N5EM11の特異性は感染実験の前および感染後10日目までの300頭の動物から採取した血清によって評価した。この特異性は99%を超えている(結果は図示せず)。
【実施例4】
【0104】
411抗原の同定と単離。
411のcDNAクローンをT.spiralisの早期侵入段階AdおよびL1NNのcDNAライブラリーから選別した。
【0105】
このcDNAクローンの核酸配列ならびに推定されたポリペプチド配列を判別し、それぞれ、配列表においてSEQ ID NO:3とSEQ ID NO:4という番号で示した。411のオープンリーディングフレームは20kDaの推定タンパク質をコードしている。シグナルペプチドを除いて、いかなるタンパク質領域も同定されなかった。
【0106】
411の完全なオープンリーディングフレームとGenbankで入手可能な配列との比較は、その411が、T.pseudospiralisで同定されている排泄/分泌タンパク質Tp21−3(AAF79206、NAGANO et al.,2001)とは78.7%の同一性があり、PolvereおよびDespommierによって提示されているT.spiralisの推定ORF17.20(AAB48489)の配列とは86.6%の同一性があることを示している。この配列の比較によって、411は旋毛虫属に共通したこの遺伝子ファミリーの新しいメンバーとして同定される。
【0107】
411の完全なオープンリーディングフレームを、オリゴヌクレオチド411F(5’−CACCCGAGAAAACATGCAT−3’)(SEQ ID NO:22)および411R(5’−TCCATTCAATTTTGCGTCAC−3’)(SEQ ID NO:23)、DNAポリメラーゼAccuPrime Pfx DNA polymerase(Invitrogen社)を用いて増幅し、製造者(Invitrogen社)の推奨にしたがって発現キット「Champion pET102 Directional TOPO」を用い、プラスミドpET102D/topoにおいてクローニングした。
【0108】
得られた組換えプラスミドはpET102−411と称するものであり、予測分子量が36.7kDaの330アミノ酸のチオレドキシン−411融合タンパク質(THX−411)をコードし、C末端の位置にポリヒスチジンタグを有している。この融合タンパク質の配列は図5に示している。
【0109】
融合タンパク質THX−411は、プラスミドpET102−411によって形質転換した大腸菌BL21Star(DE3)、BL21(DE3)pLys(Invitrogen社)の中で発現させ、提供者(Qiagen)によって推奨される手順を用い、Ni−NTAカラム(Ni−NTAスピン・カラム・キット、Ni−NTAビーズ)上で、変性条件下でのアフィニティークロマトグラフィーによって精製した。
【0110】
精製された融合タンパク質THX−411は、変性条件下での電気泳動(SDS−PAGE)の後、予想された36.7kDaのサイズのバンドの形で現れた。
【0111】
旋毛虫症に感染していないブタの血清と、20000匹のT.spiralisのL1M幼虫による感染後30日目および50日目の同一のブタの血清に対するタンパク質THX−411の免疫反応性を、対照のE/S抗原(上記実施例2に記載したように調製)の免疫反応性と比較した。分析はウェスタンブロッティングにより、上記実施例1に記載したものと同一の手順を用いて行った。
【0112】
結果は図6に示している。
【0113】
T.spiralisに感染したブタの血清に対するTHX−411の非常に強い免疫反応性と、抗411抗体の早期検出(感染後30日目)が観察される。軽いノイズも観察されるが、該ノイズは、THX−411の調製物中に残留していた高分子量の細菌タンパク質と、血清中に存在する抗大腸菌抗体との間の交差反応に起因している。
【実施例5】
【0114】
旋毛虫に対する液性応答を検出するためのTHX−411の利用法。
上記実施例4に記載したように調製したタンパク質を、旋毛虫で感染させたブタに由来する血清に対する間接ELISA法により、T.spiralisの排泄/分泌抗原(E/S)との比較で評価した。
【0115】
T.spiralisの200、1000、20000匹のL1M幼虫で実験的に感染させた通常のブタの血清において、ELISA THX−411またはELISA Ag E/Sによって検出された抗旋毛虫抗体の出現動態を比較した。また、ELISA Ag E/SとELISA THX−411による、T.spiralisと、欧州で特定されている他の三種の旋毛虫、T.nativa、T.britoviおよびT.pseudospiralisによって誘発された液性応答の検出も比較した。
【0116】
用いた手順は実施例2で記載したものと同一である。THX−411抗原は2μg/ml、そしてE/S抗原は1.25μg/mlで用いた。
【0117】
陽性の限界はELISA Ag E/Sでは14%、ELISA THX−411では52%である。
【0118】
結果は図7および下の表IIIに示している。
【0119】
【表3】
【0120】
これらの結果の全体は、411抗原によってT.spiralisに対する液性応答の容量依存的な検出が可能となることを示している。また、ウェスタンブロッティングによるリニアエピトープの検出に伴うこのELISA検定による立体構造エピトープの検出は、旋毛虫の411タンパク質の免疫優性を証明している。
【0121】
THX−411組換えタンパク質によって、T.spiralisに対する抗体の特に早い検出が可能となる(20000匹のL1Mで強く感染させた動物については感染後20日目から)。セロコンバージョンは液性応答のプロファイルを伴っており、該液性応答は高い力価を有し、感染後60日まで保持される。中程度のおよび少ない寄生虫の量で感染させた動物のセロコンバージョンはもっと遅く、それぞれ感染後30日目と60日目に検出された。ELISA Ag E/Sによって検出された最も早いセロコンバージョンは感染後25日目である。
【0122】
ELISA Ag E/SとELISA THX−411によって同時に診断された、T.spiralisで感染させた動物は、それらのうち2/5については、ELISA THX−411での血清の検出ウィンドウが縮小し、早さという点で5〜10日有利である。ELISA THX−411の感度は、この動物実験のうちの5/9の通常のブタ、すなわち、20000匹のT.spiralisのL1Mで感染させた3/3のブタ、1000匹のL1Mで感染させた1/3のブタ、そして200匹のL1Mのみで感染させた1/3のブタに対する有効スクリーニングによって証明された。検出はこれらの動物におけるT.spiralisの筋肉寄生虫の量に関連しており、その量は、200匹のL1Mで実験的に感染させたブタについては平均でグラムあたり3匹の幼虫(LpG)、1000匹のL1Mで感染させたブタについては43LpG、そして20000匹のL1Mで感染させたブタについては538LpGの変動がある。
【0123】
T.nativa、T.britoviおよびT.pseudospiralisによって誘発された液性応答もELISA THX−411によって検出されており(20000匹のL1Mで感染させた動物の10/12)、このことは、旋毛虫属における411の遺伝子的保存と抗原的保存、ひいては旋毛虫症の広い範囲での診断に対する利点を証明している。診断の早さは感染後20日目以降で確認されている。セロコンバージョンのウィンドウはELISA 411では5日〜20日で縮小し、T.spiralisによる感染と同様に、ELISA Ag E/SとELISA THX−411で同時に診断した動物の50%〜100%は、ELISA THX−411での血清の検出ウィンドウが縮小した。T.nativa属による感染の分析は、ELISA THX−411の鋭い感度を証明しており、ELISA THX−411は、筋肉寄生虫の量が平均で7×10−4〜0.1LpGしかなかったにも関わらず、6/9の動物を診断している(対するELISA Ag E/Sでは5/9である)。さらに、T.nativa属によって発生した感染力が顕著に弱いにも関わらず、同様の液性応答のプロファイルがT.spiralisとT.nativaによって感染された動物で得られており、このことは、T.nativaの411タンパク質の非常に強い免疫原性と、凍結に対して耐性のあるこの種を検出するための該タンパク質の利用性を示唆している。
【0124】
ELISA THX−411の特異性を、感染実験の前および感染後10日までに150頭の動物から採取した血清を用いて評価した。この特異性は99%を超える(結果は図示せず)。
【0125】
結論。
NBL1抗原および411抗原は免疫優性の抗原を構成し、旋毛虫属内に保持されている。これらの抗原(NBL1のC末端、NBL1のペプチドエピトープN5EM11、411抗原)を用いたELISA検定は、旋毛虫に対する99%を超える特異性を有し、また該検定によって、欧州で特定されている4種の旋毛虫によって引き起こされるブタの旋毛虫症に対する早い診断が可能となる(感染後15〜60日)。
【0126】
精製され、タンパク質のC末端部分に位置するNBL1の免疫優性のエピトープを含有する組換えタンパク質THX−NBL1(Cterm)を用いたELISA検定は、旋毛虫に特異的で感度があり、また該検定によって、ELISA Ag E/Sと比べて5〜45日という鋭い早さでブタの旋毛虫症を診断することが可能となる。さらに、この検定によって、ELISA Ag E/SとELISA THX−NBL1(Cterm)によって同時に診断した動物の100%で、ELISA Ag E/Sに比べて縮小された血清の検出ウィンドウを得ることが可能となった。
【0127】
精製された組換えタンパク質THX−411を用いるELISA検定は、現状では少し低い感度で(診断した動物の17/36)、ELISA Ag E/Sによって検出される液性応答の動態を再現する。しかし、この新しいELISAの感度によって、ELISA Ag E/Sによってスクリーニングされないブタの診断が可能となった。ELISA THX−411によって、ELISA Ag E/Sと比べて5〜20日という鋭い早さでブタの旋毛虫症を診断することを可能にすることができる。さらに、ELISA Ag E/SとELISA THX−411によって同時に診断した動物の50%〜100%で、ELISA THX−411によって血清の検出ウィンドウが縮小した。
【0128】
これらの結果の全体は、NBL1抗体および411抗体をE/S抗原の代わりに、あるいはE/S抗原を補完するものとして、旋毛虫症の早期の血清診断に用いることができることを示している。さらに、旋毛虫のこれら二つの新しい抗原を組み合わせることにより、追加の動物診断によって、ELISAの感度を向上させることが可能となる。
【0129】
N5EM11タイプの免疫優性のペプチドを伴うNBL1(Cterm)と411の組み合わせによって、旋毛虫の診断感度を向上させることが可能となる(検出された動物の24/36)。
【0130】
411を伴うNBL1の追加の効果は、411によって得られる動物検出の利点、ならびに、411によって得られる動物のより速い検出として現れる。これら三種(NBL1、411、N5EM11)は、2つまたは3つの抗原を介した抗体の同時検出によってELISA検定を時間の上で安定させるが、動物の数という点では追加の利点はない。
【0131】
しかし、感染後30日未満という早期診断が二つの抗原NBL1と411によって得られる。
【0132】
参考文献
MURRELL et al., The systematics of the genus Trichinella with a key to species, Vet Parasitol, 93, 293−307, (2000).
GASSER et al., Nonisotopic single−strand conformation polymorphism analysis of sequence variability in ribosomal DNA expansion segments within the genus Trichinella (Nematoda: Adenophorea), Electrophoresis, 25, 3357−3364, (2004).
BOIREAU et al., Risques parasitaires lies aux aliments d’origine animale, Revue francaise des laboratoires, 71−89, (2002).
CAPO & DESPOMMIER, Clinical aspects of infection with Trichinella spp, Clin Microbiol Rev, 9, 47−54, (1996).
FOURESTIE et al., Randomized trial of albendazole versus tiabendazole plus flubendazole during an outbreak of human trichinellosis, Parasitol Res, 75, 36−41, (1988).
DUPOUY−CAMET, Trichinellosis: a worldwide zoonosis, Vet Parasitol, 93, 191−200, (2000).
MURRELL & POZIO, Trichinellosis: the zoonosis that won’t go quietly, Int J Parasitol, 30, 1339−1349, (2000).
BOIREAU et al., Trichinella in horses: a low frequency infection with high human risk, Vet Parasitol, 93, 309−320, (2000).
GAMBLE et al., Diagnosis of swine trichinosis by enzyme−linked immunosorbent assay (ELISA) using an excretory−secretory antigen, Vet Parasitol, 13, 349−361, (1983).
GAMBLE et al., Evaluation of excretory−secretory antigens for the serodiagnosis of swine trichinellosis, Vet Parasitol, 30, 131−137, (1988).
REASON et al., Novel tyvelose−containing tri− and tetra− antennary N−glycans in the immunodominant antigens of the intracellular parasite Trichinella spiralis, Glycobiology, 4, 593−603, (1994).
NAGANO et al., Molecular cloning and characterization of a 21 kDa protein secreted from Trichinella pseudospiralis, J Helminthol, 75, 273−278, (2001).
【図面の簡単な説明】
【0133】
【図1】タンパク質THX−NBL1(Cterm)の配列を示す図である。
【図2】タンパク質THX−NBL1(Cterm)の免疫反応性を示す図である。
【図3】T.spiralisに感染させたブタの血清においてELISA THX−NBL1(Cterm)またはELISA Ag E/Sによって検出した抗旋毛虫抗体の出現動態の比較を示す図である。
【図4】ELISA NBL1(Cterm)の特異性を示す図である。
【図5】タンパク質THX−411の配列を示す図である。
【図6】タンパク質THX−411の免疫反応性を示す図である。
【図7】T.spiralisに感染したブタの血清においてELISA THX−411またはELISA Ag E/Sによって検出した抗旋毛虫抗体の出現動態の比較を示す図である。
【特許請求の範囲】
【請求項1】
生体標本において抗旋毛虫抗体を検出する試薬としての、抗旋毛虫抗体によって認識される抗原ポリペプチドの利用方法であり、前記ポリペプチドがNBL1抗原の免疫優性のエピトープを含み、該エピトープが配列PSSGSRPTYP(SEQ ID NO:5)によって定義されることを特徴とする利用法。
【請求項2】
配列SEQ ID NO:4の25〜175番目のアミノ酸を含むかまたは配列SEQ ID NO:4の25〜175番目のアミノ酸配列と少なくとも70%の同一性を有する配列を含む一つもしくは複数のポリペプチドと随意に組み合わせた、請求項1で定義した一つまたは複数のポリペプチドを含む混合物を用いることを特徴とする、請求項1に記載の利用方法。
【請求項3】
アクセス番号GenBank AAK16520で特定されるポリペプチドおよびアクセス番号GenBank AAR36900で特定されるポリペプチドを除く、請求項1で定義した抗旋毛虫抗体によって認識される抗原ポリペプチド。
【請求項4】
配列PSSGSRPTYPSSGSR(SEQ ID NO:6)、配列PSSGSRPTYPYTGSR(SEQ ID NO:7)、配列RPTSPSSGSRPTYPS(SEQ ID NO:8)の一つまたは複数の配列を含む、請求項3に記載の抗原ポリペプチド。
【請求項5】
配列SEQ ID NO:2の363〜409番目のアミノ酸を含む、請求項4に記載の抗原ポリペプチド。
【請求項6】
請求項1で定義したポリペプチドの一つまたは複数のコピーを、場合によっては一つまたは複数の異種配列と融合している、請求項2で定義したポリペプチドの一つまたは複数のコピーと随意に組み合わせて含んでいるキメラポリペプチドで構成されることを特徴とする、請求項3に記載の抗原ポリペプチド。
【請求項7】
請求項3〜請求項6のいずれか一つに記載の抗原ポリペプチドをコードするポリヌクレオチド。
【請求項8】
請求項7に記載のポリヌクレオチドを一つまたは複数含む組換えベクター。
【請求項9】
請求項8に記載の組換えベクターによって形質転換された宿主細胞。
【請求項10】
抗体の産生に利用可能な組成物を得るための、請求項1で定義したポリペプチドの利用方法。
【請求項11】
請求項1で定義したポリペプチドを特異的に認識する抗体。
【請求項12】
エピトープPSSGSRPTYP(SEQ ID NO:5)を認識することを特徴とする、請求項11に記載の抗体。
【請求項13】
生体サンプル中で抗旋毛虫抗体の存否を検出する方法であり、
−前記生体サンプルを、前記サンプルの中に場合によっては存在する抗旋毛虫抗体との抗原/抗体複合体の形成を可能にする条件のもとで、請求項2で定義した一つまたは複数のポリペプチドと随意に組み合わせた、請求項1で定義した一つまたは複数のポリペプチドと接触させることと、
−あらゆる適切な手段によって、場合によっては形成される抗原/抗体複合体を検出すること、
を含んでいることを特徴とする検出方法。
【請求項14】
生体サンプル中で抗旋毛虫抗体の存否を検出するために必要な物のセットであり、請求項2で定義した一つまたは複数のポリペプチドと随意に組み合わせた、請求項1で定義した一つまたは複数のポリペプチドを含み、そして、場合によっては、抗原/抗体複合体の形成を可能にする反応培地を構築するための適切な緩衝液および試薬、そして随意に前記抗原/抗体複合体の検出手段を含んでいることを特徴とするセット。
【請求項15】
前記ポリペプチドが固体の担体上に固定されていることを特徴とする、請求項14に記載のセット。
【請求項16】
免疫応答を向上させることを可能にする一つまたは複数のアジュバントと結びついている、
請求項2で定義した一つまたは複数のポリペプチドと随意に組み合わせた、請求項1で定義した一つまたは複数のポリペプチド、
あるいは、請求項2で定義した前記ポリペプチドをコードする一つまたは複数のポリヌクレオチドと随意に組み合わせた、請求項1で定義した前記ポリペプチドをコードする一つまたは複数のポリヌクレオチド
を含む免疫原性組成物。
【請求項17】
ワクチンであることを特徴とする、請求項16に記載の免疫原性組成物。
【請求項1】
生体標本において抗旋毛虫抗体を検出する試薬としての、抗旋毛虫抗体によって認識される抗原ポリペプチドの利用方法であり、前記ポリペプチドがNBL1抗原の免疫優性のエピトープを含み、該エピトープが配列PSSGSRPTYP(SEQ ID NO:5)によって定義されることを特徴とする利用法。
【請求項2】
配列SEQ ID NO:4の25〜175番目のアミノ酸を含むかまたは配列SEQ ID NO:4の25〜175番目のアミノ酸配列と少なくとも70%の同一性を有する配列を含む一つもしくは複数のポリペプチドと随意に組み合わせた、請求項1で定義した一つまたは複数のポリペプチドを含む混合物を用いることを特徴とする、請求項1に記載の利用方法。
【請求項3】
アクセス番号GenBank AAK16520で特定されるポリペプチドおよびアクセス番号GenBank AAR36900で特定されるポリペプチドを除く、請求項1で定義した抗旋毛虫抗体によって認識される抗原ポリペプチド。
【請求項4】
配列PSSGSRPTYPSSGSR(SEQ ID NO:6)、配列PSSGSRPTYPYTGSR(SEQ ID NO:7)、配列RPTSPSSGSRPTYPS(SEQ ID NO:8)の一つまたは複数の配列を含む、請求項3に記載の抗原ポリペプチド。
【請求項5】
配列SEQ ID NO:2の363〜409番目のアミノ酸を含む、請求項4に記載の抗原ポリペプチド。
【請求項6】
請求項1で定義したポリペプチドの一つまたは複数のコピーを、場合によっては一つまたは複数の異種配列と融合している、請求項2で定義したポリペプチドの一つまたは複数のコピーと随意に組み合わせて含んでいるキメラポリペプチドで構成されることを特徴とする、請求項3に記載の抗原ポリペプチド。
【請求項7】
請求項3〜請求項6のいずれか一つに記載の抗原ポリペプチドをコードするポリヌクレオチド。
【請求項8】
請求項7に記載のポリヌクレオチドを一つまたは複数含む組換えベクター。
【請求項9】
請求項8に記載の組換えベクターによって形質転換された宿主細胞。
【請求項10】
抗体の産生に利用可能な組成物を得るための、請求項1で定義したポリペプチドの利用方法。
【請求項11】
請求項1で定義したポリペプチドを特異的に認識する抗体。
【請求項12】
エピトープPSSGSRPTYP(SEQ ID NO:5)を認識することを特徴とする、請求項11に記載の抗体。
【請求項13】
生体サンプル中で抗旋毛虫抗体の存否を検出する方法であり、
−前記生体サンプルを、前記サンプルの中に場合によっては存在する抗旋毛虫抗体との抗原/抗体複合体の形成を可能にする条件のもとで、請求項2で定義した一つまたは複数のポリペプチドと随意に組み合わせた、請求項1で定義した一つまたは複数のポリペプチドと接触させることと、
−あらゆる適切な手段によって、場合によっては形成される抗原/抗体複合体を検出すること、
を含んでいることを特徴とする検出方法。
【請求項14】
生体サンプル中で抗旋毛虫抗体の存否を検出するために必要な物のセットであり、請求項2で定義した一つまたは複数のポリペプチドと随意に組み合わせた、請求項1で定義した一つまたは複数のポリペプチドを含み、そして、場合によっては、抗原/抗体複合体の形成を可能にする反応培地を構築するための適切な緩衝液および試薬、そして随意に前記抗原/抗体複合体の検出手段を含んでいることを特徴とするセット。
【請求項15】
前記ポリペプチドが固体の担体上に固定されていることを特徴とする、請求項14に記載のセット。
【請求項16】
免疫応答を向上させることを可能にする一つまたは複数のアジュバントと結びついている、
請求項2で定義した一つまたは複数のポリペプチドと随意に組み合わせた、請求項1で定義した一つまたは複数のポリペプチド、
あるいは、請求項2で定義した前記ポリペプチドをコードする一つまたは複数のポリヌクレオチドと随意に組み合わせた、請求項1で定義した前記ポリペプチドをコードする一つまたは複数のポリヌクレオチド
を含む免疫原性組成物。
【請求項17】
ワクチンであることを特徴とする、請求項16に記載の免疫原性組成物。
【図1】


【図2】
【図3】
【図4】
【図5】

【図6】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公表番号】特表2009−525740(P2009−525740A)
【公表日】平成21年7月16日(2009.7.16)
【国際特許分類】
【出願番号】特願2008−553794(P2008−553794)
【出願日】平成19年2月7日(2007.2.7)
【国際出願番号】PCT/FR2007/000221
【国際公開番号】WO2007/090960
【国際公開日】平成19年8月16日(2007.8.16)
【出願人】(507417628)アンスティテュ ナシオナル ドゥ ラ ルシェルシュ アグロノミック (4)
【氏名又は名称原語表記】INSTITUT NATIONAL DE LA RECHERCHE AGRONOMIQUE
【出願人】(508238853)アジャンス フランセーズ ドゥ セキュリテ サニテール デ ザリマン (1)
【氏名又は名称原語表記】AGENCE FRANCAISE DE SECURITE SANITAIRE DES ALIMENTS
【出願人】(508238912)ジーリン ユニヴァーシティー (1)
【氏名又は名称原語表記】JILIN UNIVERSITY
【Fターム(参考)】
【公表日】平成21年7月16日(2009.7.16)
【国際特許分類】
【出願日】平成19年2月7日(2007.2.7)
【国際出願番号】PCT/FR2007/000221
【国際公開番号】WO2007/090960
【国際公開日】平成19年8月16日(2007.8.16)
【出願人】(507417628)アンスティテュ ナシオナル ドゥ ラ ルシェルシュ アグロノミック (4)
【氏名又は名称原語表記】INSTITUT NATIONAL DE LA RECHERCHE AGRONOMIQUE
【出願人】(508238853)アジャンス フランセーズ ドゥ セキュリテ サニテール デ ザリマン (1)
【氏名又は名称原語表記】AGENCE FRANCAISE DE SECURITE SANITAIRE DES ALIMENTS
【出願人】(508238912)ジーリン ユニヴァーシティー (1)
【氏名又は名称原語表記】JILIN UNIVERSITY
【Fターム(参考)】
[ Back to top ]