
桂皮酸の製造方法
【課題】桂皮酸を効率よく製造する方法を提供すること。
【解決手段】発明は、桂皮酸の製造方法を提供し、この方法は、フェニルアラニンアンモニアリアーゼ高発現放線菌を液体培地で培養する工程を含む。
【解決手段】発明は、桂皮酸の製造方法を提供し、この方法は、フェニルアラニンアンモニアリアーゼ高発現放線菌を液体培地で培養する工程を含む。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、桂皮酸の製造方法に関する。
【背景技術】
【0002】
桂皮酸は、芳香族不飽和カルボン酸に分類される有機化合物であり、IUPAC系統名は3-フェニル-2-プロペン酸である。β-フェニルアクリル酸とも表される。桂皮酸は、植物界に広く存在している。
【0003】
桂皮酸は、医薬品や化学品の原料、食品用の香料などとして有用であり、その需要は大きい。桂皮酸は、従来、化学法によって製造されてきた。例えば、桂皮酸は、シンナムアルデヒドの酸化によって生成することができる。工業的には、ベンズアルデヒドおよび無水酢酸に酢酸カリウムを作用させるパーキン反応によって製造され得る。
【0004】
天然に存在する桂皮酸は、フェニルアラニンが酵素フェニルアラニンアンモニアリアーゼ(以下、「PAL」ともいう)によって脱アミノ化されることによって生成する。フェニルアラニンアンモニアリアーゼは、主として植物に見出されていたが、ポリケチド生産性放線菌であるストレプトマイセス・マリチマス(Streptomyces maritimus)にも存在することが見出された(非特許文献1)。非特許文献1では、ストレプトマイセス・マリチマスのベンゾイル−CoAを中間体とするII型ポリケチド合成系(エンテロシン生合成)の遺伝子クラスター由来のencPが、フェニルアラニンアンモニアリアーゼをコードする遺伝子であることが報告されている。
【0005】
放線菌、特にストレプトマイセス属(Streptomyces)放線菌は、抗生物質および免疫抑制剤に代表される医薬品および医薬品中間体をはじめ、種々の有用物質の生産菌として世界中で広く利用されており、遺伝子組換え技術を用いた有用物質の生産においても、宿主として利用が期待されている。例えば、ストレプトベルティシリウム・シナモネウム(Streptoverticillium cinnamoneum)由来のホスホリパーゼD(PLD)のプロモーターを含む領域を放線菌−大腸菌シャトルベクターpUC702に組み込み、ストレプトマイセス属放線菌のストレプトマイセス・リビダンス(Streptomyces lividans)でタンパク質を発現させると、タンパク質の収量が多くなることが記載されている(非特許文献2)。
【0006】
放線菌の代謝経路に関して、ストレプトマイセス・セリカラー(Streptomyces coelicolor)がグリセロールを資化できることが知られている(非特許文献3)。ストレプトマイセス・リビダンスは、アミラーゼおよびキシラナーゼなどの糖化酵素を保持しているとの報告がある(アミラーゼ:非特許文献4、キシラナーゼ:非特許文献5)。
【0007】
微生物による桂皮酸の生産について、非特許文献6には、赤色酵母のロドスポリジウム・トルロイデス(Rhodosporidium toruloides)由来のフェニルアラニンアンモニアリアーゼをコードする遺伝子を導入したシュードモナス・プチダ(Pseudomonas putida)S12株による桂皮酸の生産が報告されている。非特許文献6では、培養に用いる好適な株として、上記遺伝子を導入した後ランダム変異誘発に供し、m−フルオロフェニルアラニン含有培地で選抜した株が用いられ、さらに炭素源としてのグルコースまたはグリセロールの使用および窒素制限下のバッチ培養が検討されている。非特許文献1には、encP遺伝子をストレプトマイセス・セリカラーに導入して得られた形質転換株をR2YE寒天培地で増殖させた際に桂皮酸の合成が見られたことが報告されている。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Xiangら, J. Biol. Chem., 2002年, 277(36)巻, 32505-32509頁
【非特許文献2】Oginoら, Appl. Microbiol. Biotechnol., 2004年, 64(6)巻, 823-828頁
【非特許文献3】Smithら, Mol. Gen. Genet., 1988年, 211巻, 129-137頁
【非特許文献4】Yinら、Gene.,1997年,197(1-2)巻,37-45頁
【非特許文献5】Bielyら、Biochim.Biophys.Acta.,1993年、1162(3)巻、246-254頁
【非特許文献6】Nijkampら, Appl. Microbiol. Biotechnol., 2005年, 69巻, 170-177頁
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明の目的は、桂皮酸を効率よく製造する方法を提供することである。
【課題を解決するための手段】
【0010】
本発明は、桂皮酸の製造方法を提供し、この方法は、フェニルアラニンアンモニアリアーゼ高発現放線菌を液体培地で培養する工程を含む。
【0011】
1つの実施態様では、上記フェニルアラニンアンモニアリアーゼ高発現放線菌は、フェニルアラニンアンモニアリアーゼをコードする遺伝子を導入して得られる形質転換放線菌である。上記遺伝子は高発現型プロモーターの制御下で発現される。
【0012】
さらなる実施態様では、上記高発現型プロモーターは、ホスホリパーゼDプロモーターである。
【0013】
別の実施態様では、上記フェニルアラニンアンモニアリアーゼ高発現放線菌は、ストレプトマイセス属に属する放線菌である。
【0014】
さらに別の実施態様では、上記液体培地は、フェニルアラニンを含む。
【0015】
本発明はまた、ストレプトマイセス・マリチマス由来encP遺伝子を導入して得られる形質転換ストレプトマイセス・リビダンスを提供し、該形質転換ストレプトマイセス・リビダンスでは、該encP遺伝子がホスホリパーゼDプロモーターの制御下で発現されている。
【発明の効果】
【0016】
本発明によれば、桂皮酸を効率よく製造する方法が提供される。
【図面の簡単な説明】
【0017】
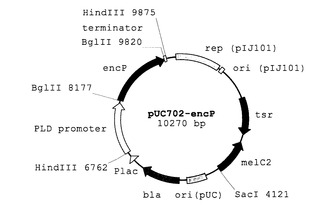
【図1】フェニルアラニンアンモニアリアーゼ高発現系プラスミドベクターpUC702-encPを示す模式図である。
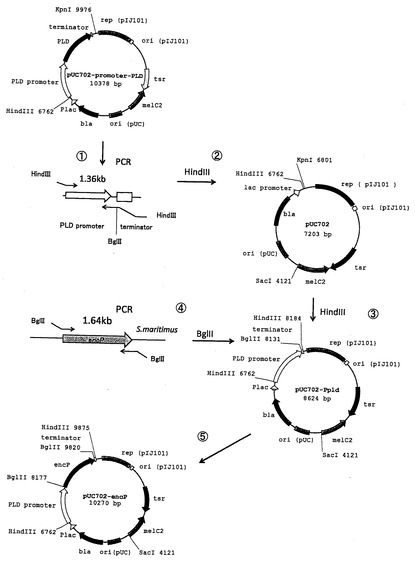
【図2】フェニルアラニンアンモニアリアーゼ高発現系プラスミドベクターpUC702-encPの構築スキームの一例を示す模式図である。
【図3】pUC702-encP形質転換ストレプトマイセス・リビダンスの培養における乾燥菌体重量およびグルコース濃度の経時変化を示すグラフである。
【図4】pUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸濃度およびL-フェニルアラニン濃度の経時変化を示すグラフである。
【図5】種々の量の窒素源を用いたpUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸生産量の経時変化を示すグラフである。
【図6】炭素源としてグリセロールまたは生デンプンを含む培地を用いたpUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸生産量の経時変化を示すグラフである。
【図7】炭素源として生デンプン、可溶デンプンまたはキシランを含む培地を用いたpUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸生産量の経時変化を示すグラフである。
【図8】炭素源としてキシロースまたはアラビノースを含む、あるいは炭素源未添加の培地を用いたpUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸生産量の経時変化を示すグラフである。
【発明を実施するための形態】
【0018】
本発明は、桂皮酸の製造方法を提供し、この方法は、フェニルアラニンアンモニアリアーゼ高発現放線菌を液体培地で培養する工程を含む。
【0019】
本明細書で「フェニルアラニンアンモニアリアーゼ高発現放線菌」とは、酵素フェニルアラニンアンモニアリアーゼ(PAL)の発現量もしくは活性が高い放線菌をいう。PALの発現量もしくは活性は、以下に説明する液体培養で桂皮酸を生産し得る程度が好ましい。PAL高発現放線菌は、以下に説明する液体培養で桂皮酸を生産できればよい。
【0020】
フェニルアラニンアンモニアリアーゼ高発現放線菌には、PALをコードする遺伝子を導入して得られた形質転換放線菌が含まれる。好ましくは、PALをコードする遺伝子は、高発現型プロモーターの制御下に発現し得る。
【0021】
PALは、フェニルアラニン(特にL-フェニルアラニン)からの桂皮酸の生成を触媒する酵素である。PALをコードする遺伝子は、その発現によりPALの活性があればよく、他の活性(例えば、チロシンアンモニアリアーゼ(TAL)活性)を併せ持つものであってもよい。TALをコードする遺伝子として知られるものであっても、その発現によりPAL活性を有する遺伝子は用いられ得、本明細書中では、まとめて「PALをコードする遺伝子」という。PALをコードする遺伝子は、細菌、酵母、植物などのいずれに由来する遺伝子であってもよい。放線菌由来の遺伝子が特に好ましく、例えば、ストレプトマイセス・マリチマス由来encP遺伝子が挙げられる。ストレプトマイセス・マリチマス由来encP遺伝子の塩基配列は、GenBankにエンテロシン生合成遺伝子クラスターとして登録されている(Accession Number AF254925)。PALをコードする遺伝子としてはさらに、例えば、光合成細菌のロドバクター(例えば、Rhodobacter capsulatus(Kyndtら、FEBS Lett.,2002年、512巻、240-244頁)、Rhodobacter sphaeroides(Xueら, J. Ind. Microbiol. Biotechnol., 2007年, 34巻, 599-604頁))、赤色酵母のロドトルラ(例えば、Rhodotorula glutinis(Vannelliaら, Metabolic Engineering, 2007年, 9巻, 142-151頁)、Rhodotorula mucilaginosa(NCBI番号X13094)、およびロドスポリジウム・トルロイデス(Rhodosporidium toruloides)(非特許文献6))、植物(例えば、シロイヌナズナ(Arabidopsis thaliana(Olsenら, Journal of Plant Physiology, 2008年, 165巻, 1491-1499頁))、サルビア(Salvia miltiorrhiza(Songら, Mol. Biol. Rep., 2009年, 36巻, 939-952頁))、タケ(Bambusa oldhamii(Hsiehら, Protein Expression and Purification, 2010年, 71巻, 224-230頁))、リコリス(Lycoris radiata)(NCBI番号FJ603650)、パパイヤ(Carica papaya)(NCBI番号GQ365163)、ダイズ(Glycine max)(NCBI番号 Gene ID:100302735)、シソ(Perilla frutescens)(NCBI番号HQ388347)、パセリ(Petroselmum crispum(Schulzら, FEBS Letters, 1989年, 258巻, 335-338頁)))、さらにBrassica juncea(NCBI番号GQ505065)、Trifolium subterraneum(NCBI番号M91192)、Urochloa maxima(NCBI番号EU741934)、Liriodendron tulipifera(NCBI番号EU190449)、Stylosanthes guianensis(NCBI番号EU404160)、Fagopyrum dibotrys(NCBI番号HM628904)などに由来するPALをコードする遺伝子が挙げられる。これらの遺伝子は、PAL活性を有する限り、天然または人工的に変異を受けたものであってもよい。
【0022】
PALをコードする遺伝子は、公知の塩基配列情報に基づいて調製したプライマーまたはプローブを用いてPCRまたはハイブリダイゼーションによって調製され得る。ストレプトマイセス・マリチマス由来encP遺伝子は、例えば、ストレプトマイセス・マリチマス(Streptomyces maritimus)DSM 41777株(ドイツ細胞バンク:Deutsche Sammlung Von Mikroorganismen Und Zellkulturen(DSMZ))のゲノムDNAを鋳型として、encP遺伝子の塩基配列情報に基づいて設計され得るプライマー対を用いるPCRによって調製され得る。
【0023】
PALをコードする遺伝子を用いて遺伝子発現カセットを構築し得る。遺伝子発現カセットは、その遺伝子の発現を調節するプロモーター、ターミネーター、オペレーター、エンハンサーなどのいわゆる調節因子を含み得る。調節因子は、発現させる遺伝子自身のものであっても、他の遺伝子由来のものであってもよく、当業者によって適宜選択され得る。例えば、プロモーターとしては高発現型プロモーターが好ましく、例えば、ホスホリパーゼD(PLD)プロモーター(特に、ストレプトベルティシリウム・シナモネウム(Streptoverticillium cinnamoneum)由来のPLDプロモーター:非特許文献2)が挙げられる。発現カセットは、遺伝子発現の目的に応じて、必要な機能配列(例えば、シャイン・ダルガルノ配列(SD配列))をさらに含むこともできる。発現カセットは、必要に応じてリンカーも含み得る。
【0024】
各種塩基配列を含むDNAの合成および連結は、当業者が通常用い得る手法で行われ得る。
【0025】
PALをコードする遺伝子または該遺伝子を含む発現カセットは、プラスミドの形態のベクターに挿入され得る。DNAの取得の簡易化の点から、放線菌と大腸菌とのシャトルベクターであることが好ましい。必要に応じて、ベクターは、上述したような調節配列を含み得る。ベクターは、例えば、放線菌および大腸菌の複製開始点および選択マーカー(例えば、放線菌についてチオストレプトン耐性遺伝子、および大腸菌についてアンピシリン耐性遺伝子(例えばβ-ラクタマーゼ遺伝子))などを有し得る。プラスミドベクターの例としては、pUC702(非特許文献2)などが挙げられる。
【0026】
本発明においては、PAL高発現放線菌の作製のために、例えば、encP遺伝子をPLDプロモーターの制御下に配置する放線菌と大腸菌とのシャトルベクターが用いられ得る。このようなベクターとして調製された例として、pUC702-encPを挙げる。図1は、フェニルアラニンアンモニアリアーゼ高発現系プラスミドベクターpUC702-encPを示す模式図である。図1の記号は以下の通りである:「PLD promoter」ストレプトベティシリウム・シナモネウム由来ホスホリパーゼD遺伝子プロモーター領域;「encP」ストレプトマイセス・マリチマス由来フェニルアラニンアンモニアリアーゼ遺伝子;「terminator」ストレプトベティシリウム・シナモネウム由来ホスホリパーゼD遺伝子ターミネーター領域;「rep(pIJ101)」放線菌プラスミドpIJ101の複製領域;「ori(pIJ101)」放線菌プラスミドpIJ101の複製起点;「tsr」チオストレプトン耐性遺伝子;「melC2」チロシナーゼ遺伝子;「ori(pUC)」大腸菌プラスミドpUC19複製起点;「bla」アンピシリン耐性遺伝子(β-ラクタマーゼ遺伝子);「Plac」lacプロモーター領域。図2は、フェニルアラニンアンモニアリアーゼ高発現系プラスミドベクターpUC702-encPの構築スキームの一例を示す模式図である。図2に示されるように、大腸菌−放線菌シャトルベクターであるpUC702ベクター(約7.2kb)のHindIII部位に、pUC702-promoter-PLD(非特許文献2)よりPCRで増幅したPLDのプロモーターおよびターミネーターを含む約1.4kbの領域(好ましくは、PLDのリボゾーム結合部位(Ribosome Binding Site (RBS))(シャイン・ダルガルノ配列(SD配列))をさらに含む)をHindIIIで消化後組み込むことにより、pUC702-Ppld(約8.6kb)を得る。続いて、PCRにより取得した約1.6kbのencP遺伝子断片(好ましくは、encP遺伝子およびそのSD配列を含む)およびpUC702-PpldをBglIIで消化後連結し、pUC702-encP(約10.2kb)を得る。pUC702-encPでは、encP遺伝子断片が、PLDプロモーターの下流に位置するPLDのRBS配列(SD配列)の直後かつターミネーターの上流に配置されている。
【0027】
本明細書で遺伝子の「導入」とは、細胞の中に遺伝子またはDNAを導入するだけでなく、発現させることも意味する。「形質転換」は、細胞の中に遺伝子またはDNAを導入して発現させることにより宿主の遺伝的形質を変えること、またはその操作をいう。遺伝子またはDNAの導入、または形質転換のために、当業者が通常用いる手法が用いられ得、放線菌に関しては、例えば、プロトプラスト法が用いられ得る。導入されるDNAは、プラスミドの形態で存在してもよく、あるいは宿主の染色体に取り込まれてもよく、または宿主の遺伝子と相同組換えを起こして染色体に組み込まれてもよい。
【0028】
放線菌としては、好ましくは、ストレプトマイセス属(Streptomyces)の微生物、例えば、ストレプトマイセス・リビダンス(Streptomyces lividans)、ストレプトマイセス・バイオラセオルバー(Streptomyces violaceoruber)、ストレプトマイセス・セリカラー(Streptomyces coelicolor)、ストレプトマイセス・アベルミチリス(Streptomyces avermitilis)、ストレプトマイセス・グリセウス(Streptomyces griseus)などが挙げられる。放線菌は、例えば、微生物寄託分譲機関から分譲を受けることによって入手され得る。例えば、ストレプトマイセス・リビダンス1326株が、独立行政法人 製品評価技術基盤機構 バイオテクノロジー本部 生物遺伝資源部門(NBRC)から分譲を受けられ得る(NBRC番号15675)。
【0029】
放線菌にPALをコードする遺伝子が導入され、形質転換されたかどうかは、選択マーカーによる選択に加え、例えばL-フェニルアラニンからのまたはグルコースからの桂皮酸の生産を調べることによって確認することができる。
【0030】
PAL高発現放線菌を液体培養して桂皮酸を製造し得る。本明細書中では、このための培養を便宜上「本培養」という。PAL高発現放線菌は、好ましくは、上記液体培養の前に、予め培養して細胞数を増大させ得る(本明細書中では、この培養を便宜上「前培養」という)。
【0031】
前培養の培養条件は、放線菌(例えば、ストレプトマイセス属)が良好に増殖し得る限り限定されない。前培養では、例えば、放線菌の増殖用培地として通常用いられる培地(例えば、トリプケースソイブイヨン(TSB培地:これは、カゼインの膵液消化物(通常、17g/L)、大豆ミールのパパイン消化物(通常、3g/L)、グルコース(通常、2.5g/L)、塩化ナトリウム(通常、5g/L)およびリン酸二カリウム(通常、2.5g/L)を含む;例えば、Becton, Dickinson and Co.,より市販され得る)が用いられ得る。培地には、形質転換放線菌の選択のため、チオストレプトン(Ts)のような抗生物質が添加され得る。培養温度(例えば28℃)、培養時間(例えば24〜48時間)、培地のpH(例えば6〜7)などは、適宜設定され得る。
【0032】
桂皮酸製造のための本培養では、PAL高発現放線菌の培養培地として、放線菌の培養のために通常用いられ得る液体培地が用いられる。このような液体培地は、放線菌の培養に通常用いられる培地の成分を含有し得、炭素(C)源および窒素(N)源を含み、そして必要に応じてミネラル、ビタミン、バッファーなどをさらに含み得る。炭素源としては、放線菌が資化し得る糖類または糖アルコールが用いられ得る。例えば、グルコース、グリセロール、キシロース、アラビノース、デンプン(生デンプン、可溶デンプン)、キシランなどが、炭素源として用いられ得る。生デンプン、グリセロール、およびキシロースが好ましい。窒素源としては、各種の無機および有機アンモニウム塩類(例えば、アンモニア、塩化アンモニウム、硫酸アンモニウム、炭酸アンモニウム、酢酸アンモニウム、尿素、および他の窒素含有物質)、複数のアミノ酸を混合物として含有する天然物およびその抽出物、ならびにその加水分解物(例えば、トリプトン、ペプトン、NZ−アミン、肉エキス、酵母エキス、カザミノ酸、ホエー、廃糖蜜、コーンスチープリカー、カゼイン加水分解物、大豆粉末、フィッシュミール、およびこれらの消化物)が用いられ得る。炭素源および窒素源の量、または炭素源と窒素源との比は特に限定されないが、窒素源の量は、放線菌の培養に通常用いられる場合に比べて多いものであってもよい。
【0033】
本培養のために、例えば、上述のトリプケースソイブイヨン(TSB培地)が用いられ得る。トリプケースソイブイヨン(TSB培地)に、炭素源の糖または糖アルコール(例えば、上記)を例えば0〜15g/Lまで、または0〜30g/Lで、および窒素源(例えば、上記)を例えば0〜50g/L(トリプトン換算量)で添加され得る。
【0034】
本培養用の液体培地は、フェニルアラニン(特にL-フェニルアラニン)を含有し得る。本培養用の液体培地中のフェニルアラニン(特にL-フェニルアラニン)は、放線菌培養培地およびその添加成分(例えば、トリプケースソイブイヨン(TSB培地)、トリプトンなど)に由来するものであっても、あるいはこれらとは別に該液体培地に添加されるものであってもよい。
【0035】
本培養用の培地にも、導入した遺伝子の脱落を防ぐ、培地の汚染を防止するなどのため、チオストレプトン(Ts)のような抗生物質が添加され得る。
【0036】
本培養の培養条件は、放線菌(例えば、ストレプトマイセス属)が良好に増殖し、桂皮酸が生産される限り限定されない。本培養の培養温度は、例えば26〜28℃、好ましくは28℃である。本培養の培養期間は、3日以上であり得、好ましくは、3〜8日間、より好ましくは6〜8日間である。培地のpHは、例えば5〜7.5、好ましくは6〜7である。PAL高発現放線菌は、好気条件下で好適に培養され得る。
【0037】
培養液からの桂皮酸の回収、分離および精製は常法に従って行われる。例えば、培養液から遠心分離によって菌体を除去して得られた培養上清をそのまままたは濃縮して桂皮酸として使用することもできるが、各種クロマトグラフィーなどの公知の精製手段に供することによってさらに純度の高い桂皮酸を得ることができる。
【0038】
以下、実施例を挙げて本発明を説明するが、本発明はこの実施例によって限定されるものではない。
【実施例】
【0039】
大腸菌からのプラスミド調製を含め、遺伝子組換え操作技術全般はMolecular Cloning A Laboratory Manual 2nd Edition (1989)に従った。PCR全般にはKOD DNA polymerase (東洋紡績株式会社製)を使用し、目的DNAのゲルからの抽出にはWizard SV Gel Extraction Kit(Promega社製)を使用した。入手先を特に記載していない材料は、当業者が通常入手可能なものを使用し得る。
【0040】
(実施例1:桂皮酸製造用ベクターの構築)
PCRの鋳型にするため、ストレプトマイセス・マリチマス(Streptomyces maritimus)DSM 41777株(ドイツ細胞バンク:Deutsche Sammlung Von Mikroorganismen Und Zellkulturen(DSMZ))のゲノムDNAを、PowerSoil DNA Isolation Kit(Mo Bio Laboratories社製)を用いて調製した。
【0041】
ホスホリパーゼD(PLD)のプロモーターおよびターミネーターを含むプラスミドベクターを、以下のように調製した。ストレプトベティシリウム・シナモネウム由来のPLDの発現カセットが挿入されている大腸菌放線菌シャトルベクタープラスミドpUC702-promoter-PLD(非特許文献2)を鋳型として、フォワードプライマー(配列番号1)およびリバースプライマー(配列番号2)を使用し、PCRを行った。フォワードプライマーは、5’から3’の方向に、付加配列−HindIII制限部位−PLDプロモーターの5’末端側領域を含む断片、リバースプライマーは、5’から3’の方向に、HindIII制限部位−PLDターミネーター−BglII制限部位−PLDのリボゾーム結合部位(RBS)−PLDプロモーターの3’末端側領域を含む(オープンリーディングフレーム(ORF)に対して順方向に表す場合、PLDプロモーターの3’末端側領域-PLDのリボゾーム結合部位(RBS)-BglII制限部位-PLDターミネーター-HindIIIに相当する)断片となるように設計した。このPCR増幅により得られた断片をHindIIIで切断し、PLDのプロモーターおよびターミネーターを含む1.36kbのDNA断片を得た。この1.36kbのDNA断片は、PLDプロモーターの下流にPLDのRBS(SD配列)をさらに含み、PLDのRBS(SD配列)のすぐ下流にBglII制限部位を含む。次に、同様にHindIIIで消化した大腸菌放線菌シャトルベクタープラスミドpUC702(非特許文献2)に、このDNA断片を組み込んだ。得られたプラスミドをpUC702-Ppldと命名した(図2)。
【0042】
PCRにより、ストレプトマイセス・マリチマスDSM 41777株ゲノムDNAを鋳型として、フォワードプライマー(配列番号3)およびリバースプライマー(配列番号4)を使用し、フェニルアラニンアンモニアリアーゼ(PAL)をコードする遺伝子encPの増幅を行った。プライマー対は、開始コドンの約40bp上流から下流に増幅するように設計した。これは、開始コドンの10bp程度上流に存在する推定シャイン・ダルガルノ配列(SD配列)を含むようにするためである。また、フォワードプライマーおよびリバースプライマーとも、BglII制限部位を含むようにもした。上記PCRにより、encP遺伝子およびその上流配列(40bp)を含む1.64kbのDNA断片を得た(図2)。
【0043】
上記PCRにより得られたencP遺伝子およびその上流配列を含むDNA断片をBglIIで消化し、pUC702-PpldのBglII部位間に挿入し、PLDプロモーターの下流に位置するPLDのSD配列の直後かつターミネーターの上流に配置した(図2)。得られたプラスミドで大腸菌Novablue(Novagen, メルク株式会社製)を形質転換し、大腸菌内で増幅した目的プラスミドを抽出した。大腸菌Novablueの形質転換体の選択は、アンピシリンを含むLB固形培地上に大腸菌を塗布して行い、そしてプラスミドの抽出は、Labo PassTM Mini Plasmid DNA Purification kit(COSMO Genentech社製)を用いて行った。構築したプラスミドの塩基配列は、ABI PRISM BigDye Terminator v3.1 Cycle Sequencing KitによるPCRおよびXTerminator TM Solutionによるプライマー除去を行った後に、3130 Pop-7TMポリマーを用いてABI PRISM 3130xl Genetic Analyzer(Applied Biosystems)により解読して確認した。これにより得られたプラスミドをpUC702-encPと命名した(図2)。
【0044】
(実施例2:放線菌の形質転換)
実施例1で調製した桂皮酸製造用ベクタープラスミドpUC702-encPを用いて、放線菌ストレプトマイセス・リビダンス(Streptomyces lividans)1326株(独立行政法人 製品評価技術基盤機構 バイオテクノロジー本部 生物遺伝資源部門(NBRC):NBRC番号15675)を形質転換した。放線菌の形質転換方法は、Plactical Streptomyces Genetics(The John Innes Foundation, 2000)に従い、プロトプラスト-PEG法により行った。放線菌の形質転換体の選択は、チオストレプトン(Ts, 50μg/mL)軟寒天を重層したR2YE培地(Genetic Manipulation of Streptomyces-A Laboratory Manual, 著者:D.A.Hopwoodら,the John Innes Foundation発行 1985年)上で30℃にて5日間放線菌を培養して行った。
【0045】
(実施例3:形質転換放線菌による桂皮酸の生成におけるフェニルアラニン添加の影響)
<前培養>
実施例2において得られた形質転換体コロニーの1つ(pUC702-encP形質転換ストレプトマイセス・リビダンス)を、チオストレプトン(Ts)5μg/mLを含むトリプケースソイブイヨン(TSB:Becton, Dickinson and Co.,製)培地5mLを入れた試験管中で、培養温度28℃にて振とう速度180rpmの条件で3日間、培養した。
【0046】
<本培養>
本培養のための培地として、以下のいずれかを用いた:(1)グルコース15g/Lおよびトリプトン15g/Lを添加した、Ts(5μg/mL)を含むTSB培地(以下、「培地1」);(2)培地1にフェニルアラニン10mM添加した培地(以下、「培地2」);および(3)培地1にフェニルアラニン100mM添加した培地(以下、「培地3」)。各培地100mLを含む500mL容量の坂口フラスコ中で、培養温度28℃にて振とう速度200rpmの条件でBioShaker G-BR-200(TAITEC)を用いて8日間培養した。
【0047】
<生産物の分析>
培養開始から24時間毎に培養液をサンプリングし、菌体濃度(OD600)を測定した。サンプリングした培養液を遠心して培養上清を回収した。
【0048】
培養上清を高速液体クロマトグラフィー(HPLC)にて分析し、桂皮酸、L-フェニルアラニン、およびグルコース量を測定した。分析サンプルとして、pUC702-encP形質転換ストレプトマイセス・リビダンスの培養上清を、リン酸ナトリウム緩衝液で10倍希釈して用いた。桂皮酸については、Cosmosil 5C18-MS II Packed column(4.6mm×250mm、ナカライテスク製)を用いた高速液体クロマトグラフィーシステムLC Prominence(島津製作所製)により、リン酸ナトリウムバッファー(pH2.5):アセトニトリル=7:3(v/v)の組成の溶離液を用いて、カラム温度40℃および流速1.2mL/分で分離し、紫外部吸光検出系SPD-20A(島津製作所製)によって254nmにて検出した。フェニルアラニンについては、Cosmosil Chorester(4.6mm×250mm、ナカライテスク)を用いて、40%(v/v)アセトニトリル−0.01%(v/v)酢酸を溶媒とし、カラム温度40℃および流速0.6mL/分で分離し、254nmにて検出した。グルコースについては、Shim-pack SPR-Pb(7.8mm×250mm、Shimadzu)を用いて、水を溶媒とし、カラム温度80℃および流速0.6mL/分で分離し、示差屈折計RID-10A(島津製作所)を用いて検出した。
【0049】
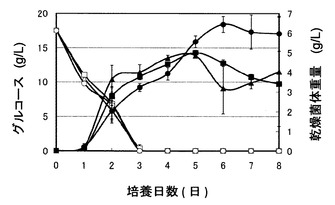
図3は、pUC702-encP形質転換ストレプトマイセス・リビダンスの培養における乾燥菌体重量およびグルコース濃度の経時変化を示すグラフである。図3のグラフの右の縦軸は乾燥菌体重量(g/L)を表し、左の縦軸はグルコース濃度(g/L)を表し、横軸は培養時間(日)を表す。図3の記号は以下の通りである:黒三角、培地1で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスの乾燥菌体重量;黒四角、培地2で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスの乾燥菌体重量;黒丸、培地3で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスの乾燥菌体重量;白三角、培地1で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスのグルコース濃度;白四角、培地2で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスのグルコース濃度;白丸、培地3で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスのグルコース濃度。フェニルアラニンの添加の有無にかかわらず糖(グルコース)はほぼ同速度にて3日間で消費された。フェニルアラニンの添加の有無にかかわらず菌体の増殖は良好であった。
【0050】
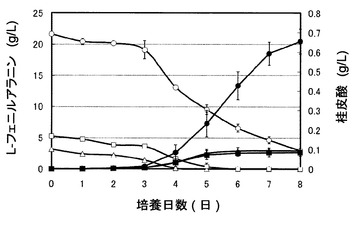
図4は、pUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸濃度およびL-フェニルアラニン濃度の経時変化を示すグラフである。図4のグラフの右の縦軸は桂皮酸濃度(g/L)を表し、左の縦軸はL-フェニルアラニン濃度(g/L)を表し、横軸は培養日数(日)を表す。図4の記号は以下の通りである:黒三角、培地1で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスの桂皮酸濃度;黒四角、培地2で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスの桂皮酸濃度;黒丸、培地3で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスの桂皮酸濃度;白三角、培地1で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスのL-フェニルアラニン濃度;白四角、培地2で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスのL-フェニルアラニン濃度;白丸、培地3で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスのL-フェニルアラニン濃度。培地1(グルコース含有培地、フェニルアラニン無添加)、培地2(フェニルアラニン10mM添加)、および培地3(フェニルアラニン100mM添加)のいずれの場合も桂皮酸が生成した。フェニルアラニンは培養日数の経過と共に減少した。図4において、培地1での培養にて培養0時間で検出されたフェニルアラニンは、TSB培地およびトリプトンに由来する遊離アミノ酸によるものである。桂皮酸の生成は、図3に見られる糖の消費開始時点から始まっていた。桂皮酸生産量は、フェニルアラニン100mM添加の場合が最も大きく、8日間で約0.65g/Lであった。
【0051】
(実施例4:種々の量の窒素源を用いた形質転換放線菌による桂皮酸の生成)
本培養のための培地として、以下のいずれかを用いたこと以外は、実施例3と同様に、pUC702-encP形質転換ストレプトマイセス・リビダンスの前培養および本培養を行った:Ts(5mg/L)を含むTSB培地に、(1)グルコース15g/Lおよびトリプトン15g/Lを添加;(2)グルコース15g/Lおよびトリプトン30g/Lを添加;または(3)グルコース15g/Lおよびトリプトン50g/Lを添加。コントロールとして、PALをコードする遺伝子を含まないpUC702を用いて実施例2に記載のように形質転換したストレプトマイセス・リビダンスもまた培養に用いた。
【0052】
培養後の培養液から21,880×gにて20分間遠心分離することにより培養上清を分離し、これを高速液体クロマトグラフィー(HPLC)にて分析し、桂皮酸生産量を測定した。桂皮酸について、Choresterカラム(4.6mm×250mm、ナカライテスク製)を用いた高速液体クロマトグラフィーシステム(島津製作所製)により、リン酸ナトリウムバッファー(pH2.5):アセトニトリル=7:3(v/v)の組成の溶離液を用いて、カラム温度30℃および流速1.2mL/分で分離し、紫外部吸光検出系SPD-20A(島津製作所製)によって254nmにて検出した。
【0053】
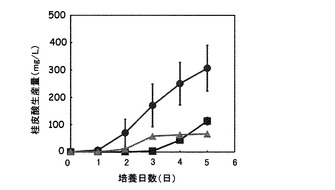
図5は、種々の量の窒素源を含む培地を用いたpUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸生産量の経時変化を示すグラフである。図5の縦軸は桂皮酸生産量(g/L)を表し、横軸は培養日数(日)を表す。図5の記号は以下の通りである:黒三角、(1)の培地で培養したコントロールのpUC702形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒菱形、(1)の培地で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒四角、(2)の培地で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒丸、(3)の培地で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量。
【0054】
図5に示されるように、pUC702-encP形質転換ストレプトマイセス・リビダンスは、培養開始時の窒素源のトリプトンの量を増大させるにつれて、桂皮酸生産量を増大した。本実施例では、培養開始時のトリプトン量を50g/Lより増大しても、桂皮酸生産量は増大しなかった(データは示さず)。
【0055】
他方、コントロールのpUC702形質転換ストレプトマイセス・リビダンスは、培養の間に桂皮酸を全く生産しなかった。本実施例および実施例3の結果から、ストレプトマイセス・リビダンスのPALの発現が、桂皮酸の生産に関与することが確認された。
【0056】
(実施例5:種々の炭素源を用いた形質転換放線菌による桂皮酸の生成)
本培養のための培地として、以下のいずれかを用いたこと以外は、実施例3と同様に、pUC702-encP形質転換ストレプトマイセス・リビダンスの前培養および本培養を行った:Ts(5mg/L)を含むTSB培地に、(1)グリセロール15g/Lおよびトリプトン15g/Lを添加;(2)グリセロール30g/Lおよびトリプトン50g/Lを添加;(3)生デンプン(和光純薬工業株式会社製)15g/Lおよびトリプトン50g/Lを添加;(4)可溶デンプン(和光純薬工業株式会社製)15g/Lおよびトリプトン50g/Lを添加;(5)キシラン(SIGMA-ALDRICH Inc.製)15g/Lおよびトリプトン50g/Lを添加;(6)キシロース15g/Lおよびトリプトン50g/Lを添加;(7)アラビノース15g/Lおよびトリプトン50g/Lを添加;または(8)トリプトン50g/Lのみを添加(「炭素源未添加」)。桂皮酸生産量の測定は、実施例4と同様に行った。
【0057】
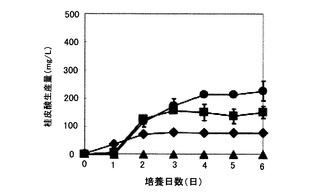
図6は、炭素源としてグリセロールまたは生デンプンを含む培地を用いたpUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸生産量の経時変化を示すグラフである。図6の縦軸は桂皮酸生産量(g/L)を表し、横軸は培養日数(日)を表す。図6の記号は以下の通りである:黒菱形、(1)の培地(炭素源グリセロール)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒丸、(2)の培地(炭素源グリセロール)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒四角、(3)の培地(炭素源生デンプン)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量。
【0058】
図6に示されるように、pUC702-encP形質転換ストレプトマイセス・リビダンスは、炭素源としてグルコースの代わりにグリセロールを用いた場合でも桂皮酸を生産した。同量のトリプトン添加の場合(15g/L)で比較すると、グリセロール(15g/L添加、図6)は、グルコース(15g/L添加、図5)よりも1.5倍高い桂皮酸生産量を示した。炭素源としてグルコースの代わりに生デンプンを用いた場合も、pUC702-encP形質転換ストレプトマイセス・リビダンスは桂皮酸を生産し、同量のトリプトン添加の場合(50g/L)で比較すると、生デンプン(15g/L添加、図6)では、グルコース(15g/L添加、図5)よりも顕著に高い桂皮酸生産量が得られた。
【0059】
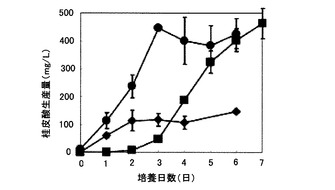
図7は、炭素源として生デンプン、可溶デンプンまたはキシランを含む培地を用いたpUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸生産量の経時変化を示すグラフである。図7の縦軸は桂皮酸生産量(g/L)を表し、横軸は培養日数(日)を表す。図7の記号は以下の通りである:黒丸、(3)の培地(炭素源生デンプン)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒三角、(4)の培地(炭素源可溶デンプン)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒菱形、(5)の培地(炭素源キシラン)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量。
【0060】
図7に示されるように、pUC702-encP形質転換ストレプトマイセス・リビダンスは、炭素源としてグルコースの代わりにデンプン(生デンプン、可溶デンプン)またはキシランを用いた場合でも桂皮酸を生産し、特に生デンプンを用いた場合で桂皮酸の生産量は高く、生産量は培養5日後で300mg/Lを越え、培養7日後には460mg/Lの生産量に達した。
【0061】
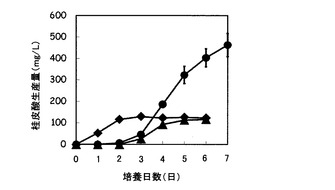
図8は、炭素源としてキシロースまたはアラビノースを含む、あるいは炭素源未添加の培地を用いたpUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸生産量の経時変化を示すグラフである。図8の縦軸は桂皮酸生産量(g/L)を表し、横軸は培養日数(日)を表す。図8の記号は以下の通りである:黒丸、(6)の培地(炭素源キシロース)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒四角、(7)の培地(炭素源アラビノース)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒三角、(8)の培地(炭素源添加せず)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量。
【0062】
図8に示されるように、pUC702-encP形質転換ストレプトマイセス・リビダンスは、炭素源としてグルコースの代わりにキシロースまたはアラビノースを用いた場合でも桂皮酸を生産した。特にキシロースを用いた場合で桂皮酸の生産量は高く、生産量は培養5日後で300mg/Lに達した。炭素源未添加の(8)の培地でも桂皮酸の生産はみられたが、その生産量は100mg/Lにも届かず、それ以上増大することなく停滞した。
【産業上の利用可能性】
【0063】
本発明によって桂皮酸を効率的に製造でき、製造された桂皮酸は、医薬品あるいは化成品原料として、食品用の香料として好適に利用できる。また、本発明によれば、生物による安全な方法で桂皮酸を生産することができる。また、原料物質(初期物質)としてグルコース以外にも種々の炭素源、特に、生デンプンなどの植物バイオマスに由来する炭素源を使用できるので、製造コストを抑えることができる。
【技術分野】
【0001】
本発明は、桂皮酸の製造方法に関する。
【背景技術】
【0002】
桂皮酸は、芳香族不飽和カルボン酸に分類される有機化合物であり、IUPAC系統名は3-フェニル-2-プロペン酸である。β-フェニルアクリル酸とも表される。桂皮酸は、植物界に広く存在している。
【0003】
桂皮酸は、医薬品や化学品の原料、食品用の香料などとして有用であり、その需要は大きい。桂皮酸は、従来、化学法によって製造されてきた。例えば、桂皮酸は、シンナムアルデヒドの酸化によって生成することができる。工業的には、ベンズアルデヒドおよび無水酢酸に酢酸カリウムを作用させるパーキン反応によって製造され得る。
【0004】
天然に存在する桂皮酸は、フェニルアラニンが酵素フェニルアラニンアンモニアリアーゼ(以下、「PAL」ともいう)によって脱アミノ化されることによって生成する。フェニルアラニンアンモニアリアーゼは、主として植物に見出されていたが、ポリケチド生産性放線菌であるストレプトマイセス・マリチマス(Streptomyces maritimus)にも存在することが見出された(非特許文献1)。非特許文献1では、ストレプトマイセス・マリチマスのベンゾイル−CoAを中間体とするII型ポリケチド合成系(エンテロシン生合成)の遺伝子クラスター由来のencPが、フェニルアラニンアンモニアリアーゼをコードする遺伝子であることが報告されている。
【0005】
放線菌、特にストレプトマイセス属(Streptomyces)放線菌は、抗生物質および免疫抑制剤に代表される医薬品および医薬品中間体をはじめ、種々の有用物質の生産菌として世界中で広く利用されており、遺伝子組換え技術を用いた有用物質の生産においても、宿主として利用が期待されている。例えば、ストレプトベルティシリウム・シナモネウム(Streptoverticillium cinnamoneum)由来のホスホリパーゼD(PLD)のプロモーターを含む領域を放線菌−大腸菌シャトルベクターpUC702に組み込み、ストレプトマイセス属放線菌のストレプトマイセス・リビダンス(Streptomyces lividans)でタンパク質を発現させると、タンパク質の収量が多くなることが記載されている(非特許文献2)。
【0006】
放線菌の代謝経路に関して、ストレプトマイセス・セリカラー(Streptomyces coelicolor)がグリセロールを資化できることが知られている(非特許文献3)。ストレプトマイセス・リビダンスは、アミラーゼおよびキシラナーゼなどの糖化酵素を保持しているとの報告がある(アミラーゼ:非特許文献4、キシラナーゼ:非特許文献5)。
【0007】
微生物による桂皮酸の生産について、非特許文献6には、赤色酵母のロドスポリジウム・トルロイデス(Rhodosporidium toruloides)由来のフェニルアラニンアンモニアリアーゼをコードする遺伝子を導入したシュードモナス・プチダ(Pseudomonas putida)S12株による桂皮酸の生産が報告されている。非特許文献6では、培養に用いる好適な株として、上記遺伝子を導入した後ランダム変異誘発に供し、m−フルオロフェニルアラニン含有培地で選抜した株が用いられ、さらに炭素源としてのグルコースまたはグリセロールの使用および窒素制限下のバッチ培養が検討されている。非特許文献1には、encP遺伝子をストレプトマイセス・セリカラーに導入して得られた形質転換株をR2YE寒天培地で増殖させた際に桂皮酸の合成が見られたことが報告されている。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Xiangら, J. Biol. Chem., 2002年, 277(36)巻, 32505-32509頁
【非特許文献2】Oginoら, Appl. Microbiol. Biotechnol., 2004年, 64(6)巻, 823-828頁
【非特許文献3】Smithら, Mol. Gen. Genet., 1988年, 211巻, 129-137頁
【非特許文献4】Yinら、Gene.,1997年,197(1-2)巻,37-45頁
【非特許文献5】Bielyら、Biochim.Biophys.Acta.,1993年、1162(3)巻、246-254頁
【非特許文献6】Nijkampら, Appl. Microbiol. Biotechnol., 2005年, 69巻, 170-177頁
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明の目的は、桂皮酸を効率よく製造する方法を提供することである。
【課題を解決するための手段】
【0010】
本発明は、桂皮酸の製造方法を提供し、この方法は、フェニルアラニンアンモニアリアーゼ高発現放線菌を液体培地で培養する工程を含む。
【0011】
1つの実施態様では、上記フェニルアラニンアンモニアリアーゼ高発現放線菌は、フェニルアラニンアンモニアリアーゼをコードする遺伝子を導入して得られる形質転換放線菌である。上記遺伝子は高発現型プロモーターの制御下で発現される。
【0012】
さらなる実施態様では、上記高発現型プロモーターは、ホスホリパーゼDプロモーターである。
【0013】
別の実施態様では、上記フェニルアラニンアンモニアリアーゼ高発現放線菌は、ストレプトマイセス属に属する放線菌である。
【0014】
さらに別の実施態様では、上記液体培地は、フェニルアラニンを含む。
【0015】
本発明はまた、ストレプトマイセス・マリチマス由来encP遺伝子を導入して得られる形質転換ストレプトマイセス・リビダンスを提供し、該形質転換ストレプトマイセス・リビダンスでは、該encP遺伝子がホスホリパーゼDプロモーターの制御下で発現されている。
【発明の効果】
【0016】
本発明によれば、桂皮酸を効率よく製造する方法が提供される。
【図面の簡単な説明】
【0017】
【図1】フェニルアラニンアンモニアリアーゼ高発現系プラスミドベクターpUC702-encPを示す模式図である。
【図2】フェニルアラニンアンモニアリアーゼ高発現系プラスミドベクターpUC702-encPの構築スキームの一例を示す模式図である。
【図3】pUC702-encP形質転換ストレプトマイセス・リビダンスの培養における乾燥菌体重量およびグルコース濃度の経時変化を示すグラフである。
【図4】pUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸濃度およびL-フェニルアラニン濃度の経時変化を示すグラフである。
【図5】種々の量の窒素源を用いたpUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸生産量の経時変化を示すグラフである。
【図6】炭素源としてグリセロールまたは生デンプンを含む培地を用いたpUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸生産量の経時変化を示すグラフである。
【図7】炭素源として生デンプン、可溶デンプンまたはキシランを含む培地を用いたpUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸生産量の経時変化を示すグラフである。
【図8】炭素源としてキシロースまたはアラビノースを含む、あるいは炭素源未添加の培地を用いたpUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸生産量の経時変化を示すグラフである。
【発明を実施するための形態】
【0018】
本発明は、桂皮酸の製造方法を提供し、この方法は、フェニルアラニンアンモニアリアーゼ高発現放線菌を液体培地で培養する工程を含む。
【0019】
本明細書で「フェニルアラニンアンモニアリアーゼ高発現放線菌」とは、酵素フェニルアラニンアンモニアリアーゼ(PAL)の発現量もしくは活性が高い放線菌をいう。PALの発現量もしくは活性は、以下に説明する液体培養で桂皮酸を生産し得る程度が好ましい。PAL高発現放線菌は、以下に説明する液体培養で桂皮酸を生産できればよい。
【0020】
フェニルアラニンアンモニアリアーゼ高発現放線菌には、PALをコードする遺伝子を導入して得られた形質転換放線菌が含まれる。好ましくは、PALをコードする遺伝子は、高発現型プロモーターの制御下に発現し得る。
【0021】
PALは、フェニルアラニン(特にL-フェニルアラニン)からの桂皮酸の生成を触媒する酵素である。PALをコードする遺伝子は、その発現によりPALの活性があればよく、他の活性(例えば、チロシンアンモニアリアーゼ(TAL)活性)を併せ持つものであってもよい。TALをコードする遺伝子として知られるものであっても、その発現によりPAL活性を有する遺伝子は用いられ得、本明細書中では、まとめて「PALをコードする遺伝子」という。PALをコードする遺伝子は、細菌、酵母、植物などのいずれに由来する遺伝子であってもよい。放線菌由来の遺伝子が特に好ましく、例えば、ストレプトマイセス・マリチマス由来encP遺伝子が挙げられる。ストレプトマイセス・マリチマス由来encP遺伝子の塩基配列は、GenBankにエンテロシン生合成遺伝子クラスターとして登録されている(Accession Number AF254925)。PALをコードする遺伝子としてはさらに、例えば、光合成細菌のロドバクター(例えば、Rhodobacter capsulatus(Kyndtら、FEBS Lett.,2002年、512巻、240-244頁)、Rhodobacter sphaeroides(Xueら, J. Ind. Microbiol. Biotechnol., 2007年, 34巻, 599-604頁))、赤色酵母のロドトルラ(例えば、Rhodotorula glutinis(Vannelliaら, Metabolic Engineering, 2007年, 9巻, 142-151頁)、Rhodotorula mucilaginosa(NCBI番号X13094)、およびロドスポリジウム・トルロイデス(Rhodosporidium toruloides)(非特許文献6))、植物(例えば、シロイヌナズナ(Arabidopsis thaliana(Olsenら, Journal of Plant Physiology, 2008年, 165巻, 1491-1499頁))、サルビア(Salvia miltiorrhiza(Songら, Mol. Biol. Rep., 2009年, 36巻, 939-952頁))、タケ(Bambusa oldhamii(Hsiehら, Protein Expression and Purification, 2010年, 71巻, 224-230頁))、リコリス(Lycoris radiata)(NCBI番号FJ603650)、パパイヤ(Carica papaya)(NCBI番号GQ365163)、ダイズ(Glycine max)(NCBI番号 Gene ID:100302735)、シソ(Perilla frutescens)(NCBI番号HQ388347)、パセリ(Petroselmum crispum(Schulzら, FEBS Letters, 1989年, 258巻, 335-338頁)))、さらにBrassica juncea(NCBI番号GQ505065)、Trifolium subterraneum(NCBI番号M91192)、Urochloa maxima(NCBI番号EU741934)、Liriodendron tulipifera(NCBI番号EU190449)、Stylosanthes guianensis(NCBI番号EU404160)、Fagopyrum dibotrys(NCBI番号HM628904)などに由来するPALをコードする遺伝子が挙げられる。これらの遺伝子は、PAL活性を有する限り、天然または人工的に変異を受けたものであってもよい。
【0022】
PALをコードする遺伝子は、公知の塩基配列情報に基づいて調製したプライマーまたはプローブを用いてPCRまたはハイブリダイゼーションによって調製され得る。ストレプトマイセス・マリチマス由来encP遺伝子は、例えば、ストレプトマイセス・マリチマス(Streptomyces maritimus)DSM 41777株(ドイツ細胞バンク:Deutsche Sammlung Von Mikroorganismen Und Zellkulturen(DSMZ))のゲノムDNAを鋳型として、encP遺伝子の塩基配列情報に基づいて設計され得るプライマー対を用いるPCRによって調製され得る。
【0023】
PALをコードする遺伝子を用いて遺伝子発現カセットを構築し得る。遺伝子発現カセットは、その遺伝子の発現を調節するプロモーター、ターミネーター、オペレーター、エンハンサーなどのいわゆる調節因子を含み得る。調節因子は、発現させる遺伝子自身のものであっても、他の遺伝子由来のものであってもよく、当業者によって適宜選択され得る。例えば、プロモーターとしては高発現型プロモーターが好ましく、例えば、ホスホリパーゼD(PLD)プロモーター(特に、ストレプトベルティシリウム・シナモネウム(Streptoverticillium cinnamoneum)由来のPLDプロモーター:非特許文献2)が挙げられる。発現カセットは、遺伝子発現の目的に応じて、必要な機能配列(例えば、シャイン・ダルガルノ配列(SD配列))をさらに含むこともできる。発現カセットは、必要に応じてリンカーも含み得る。
【0024】
各種塩基配列を含むDNAの合成および連結は、当業者が通常用い得る手法で行われ得る。
【0025】
PALをコードする遺伝子または該遺伝子を含む発現カセットは、プラスミドの形態のベクターに挿入され得る。DNAの取得の簡易化の点から、放線菌と大腸菌とのシャトルベクターであることが好ましい。必要に応じて、ベクターは、上述したような調節配列を含み得る。ベクターは、例えば、放線菌および大腸菌の複製開始点および選択マーカー(例えば、放線菌についてチオストレプトン耐性遺伝子、および大腸菌についてアンピシリン耐性遺伝子(例えばβ-ラクタマーゼ遺伝子))などを有し得る。プラスミドベクターの例としては、pUC702(非特許文献2)などが挙げられる。
【0026】
本発明においては、PAL高発現放線菌の作製のために、例えば、encP遺伝子をPLDプロモーターの制御下に配置する放線菌と大腸菌とのシャトルベクターが用いられ得る。このようなベクターとして調製された例として、pUC702-encPを挙げる。図1は、フェニルアラニンアンモニアリアーゼ高発現系プラスミドベクターpUC702-encPを示す模式図である。図1の記号は以下の通りである:「PLD promoter」ストレプトベティシリウム・シナモネウム由来ホスホリパーゼD遺伝子プロモーター領域;「encP」ストレプトマイセス・マリチマス由来フェニルアラニンアンモニアリアーゼ遺伝子;「terminator」ストレプトベティシリウム・シナモネウム由来ホスホリパーゼD遺伝子ターミネーター領域;「rep(pIJ101)」放線菌プラスミドpIJ101の複製領域;「ori(pIJ101)」放線菌プラスミドpIJ101の複製起点;「tsr」チオストレプトン耐性遺伝子;「melC2」チロシナーゼ遺伝子;「ori(pUC)」大腸菌プラスミドpUC19複製起点;「bla」アンピシリン耐性遺伝子(β-ラクタマーゼ遺伝子);「Plac」lacプロモーター領域。図2は、フェニルアラニンアンモニアリアーゼ高発現系プラスミドベクターpUC702-encPの構築スキームの一例を示す模式図である。図2に示されるように、大腸菌−放線菌シャトルベクターであるpUC702ベクター(約7.2kb)のHindIII部位に、pUC702-promoter-PLD(非特許文献2)よりPCRで増幅したPLDのプロモーターおよびターミネーターを含む約1.4kbの領域(好ましくは、PLDのリボゾーム結合部位(Ribosome Binding Site (RBS))(シャイン・ダルガルノ配列(SD配列))をさらに含む)をHindIIIで消化後組み込むことにより、pUC702-Ppld(約8.6kb)を得る。続いて、PCRにより取得した約1.6kbのencP遺伝子断片(好ましくは、encP遺伝子およびそのSD配列を含む)およびpUC702-PpldをBglIIで消化後連結し、pUC702-encP(約10.2kb)を得る。pUC702-encPでは、encP遺伝子断片が、PLDプロモーターの下流に位置するPLDのRBS配列(SD配列)の直後かつターミネーターの上流に配置されている。
【0027】
本明細書で遺伝子の「導入」とは、細胞の中に遺伝子またはDNAを導入するだけでなく、発現させることも意味する。「形質転換」は、細胞の中に遺伝子またはDNAを導入して発現させることにより宿主の遺伝的形質を変えること、またはその操作をいう。遺伝子またはDNAの導入、または形質転換のために、当業者が通常用いる手法が用いられ得、放線菌に関しては、例えば、プロトプラスト法が用いられ得る。導入されるDNAは、プラスミドの形態で存在してもよく、あるいは宿主の染色体に取り込まれてもよく、または宿主の遺伝子と相同組換えを起こして染色体に組み込まれてもよい。
【0028】
放線菌としては、好ましくは、ストレプトマイセス属(Streptomyces)の微生物、例えば、ストレプトマイセス・リビダンス(Streptomyces lividans)、ストレプトマイセス・バイオラセオルバー(Streptomyces violaceoruber)、ストレプトマイセス・セリカラー(Streptomyces coelicolor)、ストレプトマイセス・アベルミチリス(Streptomyces avermitilis)、ストレプトマイセス・グリセウス(Streptomyces griseus)などが挙げられる。放線菌は、例えば、微生物寄託分譲機関から分譲を受けることによって入手され得る。例えば、ストレプトマイセス・リビダンス1326株が、独立行政法人 製品評価技術基盤機構 バイオテクノロジー本部 生物遺伝資源部門(NBRC)から分譲を受けられ得る(NBRC番号15675)。
【0029】
放線菌にPALをコードする遺伝子が導入され、形質転換されたかどうかは、選択マーカーによる選択に加え、例えばL-フェニルアラニンからのまたはグルコースからの桂皮酸の生産を調べることによって確認することができる。
【0030】
PAL高発現放線菌を液体培養して桂皮酸を製造し得る。本明細書中では、このための培養を便宜上「本培養」という。PAL高発現放線菌は、好ましくは、上記液体培養の前に、予め培養して細胞数を増大させ得る(本明細書中では、この培養を便宜上「前培養」という)。
【0031】
前培養の培養条件は、放線菌(例えば、ストレプトマイセス属)が良好に増殖し得る限り限定されない。前培養では、例えば、放線菌の増殖用培地として通常用いられる培地(例えば、トリプケースソイブイヨン(TSB培地:これは、カゼインの膵液消化物(通常、17g/L)、大豆ミールのパパイン消化物(通常、3g/L)、グルコース(通常、2.5g/L)、塩化ナトリウム(通常、5g/L)およびリン酸二カリウム(通常、2.5g/L)を含む;例えば、Becton, Dickinson and Co.,より市販され得る)が用いられ得る。培地には、形質転換放線菌の選択のため、チオストレプトン(Ts)のような抗生物質が添加され得る。培養温度(例えば28℃)、培養時間(例えば24〜48時間)、培地のpH(例えば6〜7)などは、適宜設定され得る。
【0032】
桂皮酸製造のための本培養では、PAL高発現放線菌の培養培地として、放線菌の培養のために通常用いられ得る液体培地が用いられる。このような液体培地は、放線菌の培養に通常用いられる培地の成分を含有し得、炭素(C)源および窒素(N)源を含み、そして必要に応じてミネラル、ビタミン、バッファーなどをさらに含み得る。炭素源としては、放線菌が資化し得る糖類または糖アルコールが用いられ得る。例えば、グルコース、グリセロール、キシロース、アラビノース、デンプン(生デンプン、可溶デンプン)、キシランなどが、炭素源として用いられ得る。生デンプン、グリセロール、およびキシロースが好ましい。窒素源としては、各種の無機および有機アンモニウム塩類(例えば、アンモニア、塩化アンモニウム、硫酸アンモニウム、炭酸アンモニウム、酢酸アンモニウム、尿素、および他の窒素含有物質)、複数のアミノ酸を混合物として含有する天然物およびその抽出物、ならびにその加水分解物(例えば、トリプトン、ペプトン、NZ−アミン、肉エキス、酵母エキス、カザミノ酸、ホエー、廃糖蜜、コーンスチープリカー、カゼイン加水分解物、大豆粉末、フィッシュミール、およびこれらの消化物)が用いられ得る。炭素源および窒素源の量、または炭素源と窒素源との比は特に限定されないが、窒素源の量は、放線菌の培養に通常用いられる場合に比べて多いものであってもよい。
【0033】
本培養のために、例えば、上述のトリプケースソイブイヨン(TSB培地)が用いられ得る。トリプケースソイブイヨン(TSB培地)に、炭素源の糖または糖アルコール(例えば、上記)を例えば0〜15g/Lまで、または0〜30g/Lで、および窒素源(例えば、上記)を例えば0〜50g/L(トリプトン換算量)で添加され得る。
【0034】
本培養用の液体培地は、フェニルアラニン(特にL-フェニルアラニン)を含有し得る。本培養用の液体培地中のフェニルアラニン(特にL-フェニルアラニン)は、放線菌培養培地およびその添加成分(例えば、トリプケースソイブイヨン(TSB培地)、トリプトンなど)に由来するものであっても、あるいはこれらとは別に該液体培地に添加されるものであってもよい。
【0035】
本培養用の培地にも、導入した遺伝子の脱落を防ぐ、培地の汚染を防止するなどのため、チオストレプトン(Ts)のような抗生物質が添加され得る。
【0036】
本培養の培養条件は、放線菌(例えば、ストレプトマイセス属)が良好に増殖し、桂皮酸が生産される限り限定されない。本培養の培養温度は、例えば26〜28℃、好ましくは28℃である。本培養の培養期間は、3日以上であり得、好ましくは、3〜8日間、より好ましくは6〜8日間である。培地のpHは、例えば5〜7.5、好ましくは6〜7である。PAL高発現放線菌は、好気条件下で好適に培養され得る。
【0037】
培養液からの桂皮酸の回収、分離および精製は常法に従って行われる。例えば、培養液から遠心分離によって菌体を除去して得られた培養上清をそのまままたは濃縮して桂皮酸として使用することもできるが、各種クロマトグラフィーなどの公知の精製手段に供することによってさらに純度の高い桂皮酸を得ることができる。
【0038】
以下、実施例を挙げて本発明を説明するが、本発明はこの実施例によって限定されるものではない。
【実施例】
【0039】
大腸菌からのプラスミド調製を含め、遺伝子組換え操作技術全般はMolecular Cloning A Laboratory Manual 2nd Edition (1989)に従った。PCR全般にはKOD DNA polymerase (東洋紡績株式会社製)を使用し、目的DNAのゲルからの抽出にはWizard SV Gel Extraction Kit(Promega社製)を使用した。入手先を特に記載していない材料は、当業者が通常入手可能なものを使用し得る。
【0040】
(実施例1:桂皮酸製造用ベクターの構築)
PCRの鋳型にするため、ストレプトマイセス・マリチマス(Streptomyces maritimus)DSM 41777株(ドイツ細胞バンク:Deutsche Sammlung Von Mikroorganismen Und Zellkulturen(DSMZ))のゲノムDNAを、PowerSoil DNA Isolation Kit(Mo Bio Laboratories社製)を用いて調製した。
【0041】
ホスホリパーゼD(PLD)のプロモーターおよびターミネーターを含むプラスミドベクターを、以下のように調製した。ストレプトベティシリウム・シナモネウム由来のPLDの発現カセットが挿入されている大腸菌放線菌シャトルベクタープラスミドpUC702-promoter-PLD(非特許文献2)を鋳型として、フォワードプライマー(配列番号1)およびリバースプライマー(配列番号2)を使用し、PCRを行った。フォワードプライマーは、5’から3’の方向に、付加配列−HindIII制限部位−PLDプロモーターの5’末端側領域を含む断片、リバースプライマーは、5’から3’の方向に、HindIII制限部位−PLDターミネーター−BglII制限部位−PLDのリボゾーム結合部位(RBS)−PLDプロモーターの3’末端側領域を含む(オープンリーディングフレーム(ORF)に対して順方向に表す場合、PLDプロモーターの3’末端側領域-PLDのリボゾーム結合部位(RBS)-BglII制限部位-PLDターミネーター-HindIIIに相当する)断片となるように設計した。このPCR増幅により得られた断片をHindIIIで切断し、PLDのプロモーターおよびターミネーターを含む1.36kbのDNA断片を得た。この1.36kbのDNA断片は、PLDプロモーターの下流にPLDのRBS(SD配列)をさらに含み、PLDのRBS(SD配列)のすぐ下流にBglII制限部位を含む。次に、同様にHindIIIで消化した大腸菌放線菌シャトルベクタープラスミドpUC702(非特許文献2)に、このDNA断片を組み込んだ。得られたプラスミドをpUC702-Ppldと命名した(図2)。
【0042】
PCRにより、ストレプトマイセス・マリチマスDSM 41777株ゲノムDNAを鋳型として、フォワードプライマー(配列番号3)およびリバースプライマー(配列番号4)を使用し、フェニルアラニンアンモニアリアーゼ(PAL)をコードする遺伝子encPの増幅を行った。プライマー対は、開始コドンの約40bp上流から下流に増幅するように設計した。これは、開始コドンの10bp程度上流に存在する推定シャイン・ダルガルノ配列(SD配列)を含むようにするためである。また、フォワードプライマーおよびリバースプライマーとも、BglII制限部位を含むようにもした。上記PCRにより、encP遺伝子およびその上流配列(40bp)を含む1.64kbのDNA断片を得た(図2)。
【0043】
上記PCRにより得られたencP遺伝子およびその上流配列を含むDNA断片をBglIIで消化し、pUC702-PpldのBglII部位間に挿入し、PLDプロモーターの下流に位置するPLDのSD配列の直後かつターミネーターの上流に配置した(図2)。得られたプラスミドで大腸菌Novablue(Novagen, メルク株式会社製)を形質転換し、大腸菌内で増幅した目的プラスミドを抽出した。大腸菌Novablueの形質転換体の選択は、アンピシリンを含むLB固形培地上に大腸菌を塗布して行い、そしてプラスミドの抽出は、Labo PassTM Mini Plasmid DNA Purification kit(COSMO Genentech社製)を用いて行った。構築したプラスミドの塩基配列は、ABI PRISM BigDye Terminator v3.1 Cycle Sequencing KitによるPCRおよびXTerminator TM Solutionによるプライマー除去を行った後に、3130 Pop-7TMポリマーを用いてABI PRISM 3130xl Genetic Analyzer(Applied Biosystems)により解読して確認した。これにより得られたプラスミドをpUC702-encPと命名した(図2)。
【0044】
(実施例2:放線菌の形質転換)
実施例1で調製した桂皮酸製造用ベクタープラスミドpUC702-encPを用いて、放線菌ストレプトマイセス・リビダンス(Streptomyces lividans)1326株(独立行政法人 製品評価技術基盤機構 バイオテクノロジー本部 生物遺伝資源部門(NBRC):NBRC番号15675)を形質転換した。放線菌の形質転換方法は、Plactical Streptomyces Genetics(The John Innes Foundation, 2000)に従い、プロトプラスト-PEG法により行った。放線菌の形質転換体の選択は、チオストレプトン(Ts, 50μg/mL)軟寒天を重層したR2YE培地(Genetic Manipulation of Streptomyces-A Laboratory Manual, 著者:D.A.Hopwoodら,the John Innes Foundation発行 1985年)上で30℃にて5日間放線菌を培養して行った。
【0045】
(実施例3:形質転換放線菌による桂皮酸の生成におけるフェニルアラニン添加の影響)
<前培養>
実施例2において得られた形質転換体コロニーの1つ(pUC702-encP形質転換ストレプトマイセス・リビダンス)を、チオストレプトン(Ts)5μg/mLを含むトリプケースソイブイヨン(TSB:Becton, Dickinson and Co.,製)培地5mLを入れた試験管中で、培養温度28℃にて振とう速度180rpmの条件で3日間、培養した。
【0046】
<本培養>
本培養のための培地として、以下のいずれかを用いた:(1)グルコース15g/Lおよびトリプトン15g/Lを添加した、Ts(5μg/mL)を含むTSB培地(以下、「培地1」);(2)培地1にフェニルアラニン10mM添加した培地(以下、「培地2」);および(3)培地1にフェニルアラニン100mM添加した培地(以下、「培地3」)。各培地100mLを含む500mL容量の坂口フラスコ中で、培養温度28℃にて振とう速度200rpmの条件でBioShaker G-BR-200(TAITEC)を用いて8日間培養した。
【0047】
<生産物の分析>
培養開始から24時間毎に培養液をサンプリングし、菌体濃度(OD600)を測定した。サンプリングした培養液を遠心して培養上清を回収した。
【0048】
培養上清を高速液体クロマトグラフィー(HPLC)にて分析し、桂皮酸、L-フェニルアラニン、およびグルコース量を測定した。分析サンプルとして、pUC702-encP形質転換ストレプトマイセス・リビダンスの培養上清を、リン酸ナトリウム緩衝液で10倍希釈して用いた。桂皮酸については、Cosmosil 5C18-MS II Packed column(4.6mm×250mm、ナカライテスク製)を用いた高速液体クロマトグラフィーシステムLC Prominence(島津製作所製)により、リン酸ナトリウムバッファー(pH2.5):アセトニトリル=7:3(v/v)の組成の溶離液を用いて、カラム温度40℃および流速1.2mL/分で分離し、紫外部吸光検出系SPD-20A(島津製作所製)によって254nmにて検出した。フェニルアラニンについては、Cosmosil Chorester(4.6mm×250mm、ナカライテスク)を用いて、40%(v/v)アセトニトリル−0.01%(v/v)酢酸を溶媒とし、カラム温度40℃および流速0.6mL/分で分離し、254nmにて検出した。グルコースについては、Shim-pack SPR-Pb(7.8mm×250mm、Shimadzu)を用いて、水を溶媒とし、カラム温度80℃および流速0.6mL/分で分離し、示差屈折計RID-10A(島津製作所)を用いて検出した。
【0049】
図3は、pUC702-encP形質転換ストレプトマイセス・リビダンスの培養における乾燥菌体重量およびグルコース濃度の経時変化を示すグラフである。図3のグラフの右の縦軸は乾燥菌体重量(g/L)を表し、左の縦軸はグルコース濃度(g/L)を表し、横軸は培養時間(日)を表す。図3の記号は以下の通りである:黒三角、培地1で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスの乾燥菌体重量;黒四角、培地2で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスの乾燥菌体重量;黒丸、培地3で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスの乾燥菌体重量;白三角、培地1で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスのグルコース濃度;白四角、培地2で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスのグルコース濃度;白丸、培地3で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスのグルコース濃度。フェニルアラニンの添加の有無にかかわらず糖(グルコース)はほぼ同速度にて3日間で消費された。フェニルアラニンの添加の有無にかかわらず菌体の増殖は良好であった。
【0050】
図4は、pUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸濃度およびL-フェニルアラニン濃度の経時変化を示すグラフである。図4のグラフの右の縦軸は桂皮酸濃度(g/L)を表し、左の縦軸はL-フェニルアラニン濃度(g/L)を表し、横軸は培養日数(日)を表す。図4の記号は以下の通りである:黒三角、培地1で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスの桂皮酸濃度;黒四角、培地2で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスの桂皮酸濃度;黒丸、培地3で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスの桂皮酸濃度;白三角、培地1で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスのL-フェニルアラニン濃度;白四角、培地2で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスのL-フェニルアラニン濃度;白丸、培地3で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスのL-フェニルアラニン濃度。培地1(グルコース含有培地、フェニルアラニン無添加)、培地2(フェニルアラニン10mM添加)、および培地3(フェニルアラニン100mM添加)のいずれの場合も桂皮酸が生成した。フェニルアラニンは培養日数の経過と共に減少した。図4において、培地1での培養にて培養0時間で検出されたフェニルアラニンは、TSB培地およびトリプトンに由来する遊離アミノ酸によるものである。桂皮酸の生成は、図3に見られる糖の消費開始時点から始まっていた。桂皮酸生産量は、フェニルアラニン100mM添加の場合が最も大きく、8日間で約0.65g/Lであった。
【0051】
(実施例4:種々の量の窒素源を用いた形質転換放線菌による桂皮酸の生成)
本培養のための培地として、以下のいずれかを用いたこと以外は、実施例3と同様に、pUC702-encP形質転換ストレプトマイセス・リビダンスの前培養および本培養を行った:Ts(5mg/L)を含むTSB培地に、(1)グルコース15g/Lおよびトリプトン15g/Lを添加;(2)グルコース15g/Lおよびトリプトン30g/Lを添加;または(3)グルコース15g/Lおよびトリプトン50g/Lを添加。コントロールとして、PALをコードする遺伝子を含まないpUC702を用いて実施例2に記載のように形質転換したストレプトマイセス・リビダンスもまた培養に用いた。
【0052】
培養後の培養液から21,880×gにて20分間遠心分離することにより培養上清を分離し、これを高速液体クロマトグラフィー(HPLC)にて分析し、桂皮酸生産量を測定した。桂皮酸について、Choresterカラム(4.6mm×250mm、ナカライテスク製)を用いた高速液体クロマトグラフィーシステム(島津製作所製)により、リン酸ナトリウムバッファー(pH2.5):アセトニトリル=7:3(v/v)の組成の溶離液を用いて、カラム温度30℃および流速1.2mL/分で分離し、紫外部吸光検出系SPD-20A(島津製作所製)によって254nmにて検出した。
【0053】
図5は、種々の量の窒素源を含む培地を用いたpUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸生産量の経時変化を示すグラフである。図5の縦軸は桂皮酸生産量(g/L)を表し、横軸は培養日数(日)を表す。図5の記号は以下の通りである:黒三角、(1)の培地で培養したコントロールのpUC702形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒菱形、(1)の培地で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒四角、(2)の培地で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒丸、(3)の培地で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量。
【0054】
図5に示されるように、pUC702-encP形質転換ストレプトマイセス・リビダンスは、培養開始時の窒素源のトリプトンの量を増大させるにつれて、桂皮酸生産量を増大した。本実施例では、培養開始時のトリプトン量を50g/Lより増大しても、桂皮酸生産量は増大しなかった(データは示さず)。
【0055】
他方、コントロールのpUC702形質転換ストレプトマイセス・リビダンスは、培養の間に桂皮酸を全く生産しなかった。本実施例および実施例3の結果から、ストレプトマイセス・リビダンスのPALの発現が、桂皮酸の生産に関与することが確認された。
【0056】
(実施例5:種々の炭素源を用いた形質転換放線菌による桂皮酸の生成)
本培養のための培地として、以下のいずれかを用いたこと以外は、実施例3と同様に、pUC702-encP形質転換ストレプトマイセス・リビダンスの前培養および本培養を行った:Ts(5mg/L)を含むTSB培地に、(1)グリセロール15g/Lおよびトリプトン15g/Lを添加;(2)グリセロール30g/Lおよびトリプトン50g/Lを添加;(3)生デンプン(和光純薬工業株式会社製)15g/Lおよびトリプトン50g/Lを添加;(4)可溶デンプン(和光純薬工業株式会社製)15g/Lおよびトリプトン50g/Lを添加;(5)キシラン(SIGMA-ALDRICH Inc.製)15g/Lおよびトリプトン50g/Lを添加;(6)キシロース15g/Lおよびトリプトン50g/Lを添加;(7)アラビノース15g/Lおよびトリプトン50g/Lを添加;または(8)トリプトン50g/Lのみを添加(「炭素源未添加」)。桂皮酸生産量の測定は、実施例4と同様に行った。
【0057】
図6は、炭素源としてグリセロールまたは生デンプンを含む培地を用いたpUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸生産量の経時変化を示すグラフである。図6の縦軸は桂皮酸生産量(g/L)を表し、横軸は培養日数(日)を表す。図6の記号は以下の通りである:黒菱形、(1)の培地(炭素源グリセロール)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒丸、(2)の培地(炭素源グリセロール)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒四角、(3)の培地(炭素源生デンプン)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量。
【0058】
図6に示されるように、pUC702-encP形質転換ストレプトマイセス・リビダンスは、炭素源としてグルコースの代わりにグリセロールを用いた場合でも桂皮酸を生産した。同量のトリプトン添加の場合(15g/L)で比較すると、グリセロール(15g/L添加、図6)は、グルコース(15g/L添加、図5)よりも1.5倍高い桂皮酸生産量を示した。炭素源としてグルコースの代わりに生デンプンを用いた場合も、pUC702-encP形質転換ストレプトマイセス・リビダンスは桂皮酸を生産し、同量のトリプトン添加の場合(50g/L)で比較すると、生デンプン(15g/L添加、図6)では、グルコース(15g/L添加、図5)よりも顕著に高い桂皮酸生産量が得られた。
【0059】
図7は、炭素源として生デンプン、可溶デンプンまたはキシランを含む培地を用いたpUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸生産量の経時変化を示すグラフである。図7の縦軸は桂皮酸生産量(g/L)を表し、横軸は培養日数(日)を表す。図7の記号は以下の通りである:黒丸、(3)の培地(炭素源生デンプン)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒三角、(4)の培地(炭素源可溶デンプン)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒菱形、(5)の培地(炭素源キシラン)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量。
【0060】
図7に示されるように、pUC702-encP形質転換ストレプトマイセス・リビダンスは、炭素源としてグルコースの代わりにデンプン(生デンプン、可溶デンプン)またはキシランを用いた場合でも桂皮酸を生産し、特に生デンプンを用いた場合で桂皮酸の生産量は高く、生産量は培養5日後で300mg/Lを越え、培養7日後には460mg/Lの生産量に達した。
【0061】
図8は、炭素源としてキシロースまたはアラビノースを含む、あるいは炭素源未添加の培地を用いたpUC702-encP形質転換ストレプトマイセス・リビダンスの培養における桂皮酸生産量の経時変化を示すグラフである。図8の縦軸は桂皮酸生産量(g/L)を表し、横軸は培養日数(日)を表す。図8の記号は以下の通りである:黒丸、(6)の培地(炭素源キシロース)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒四角、(7)の培地(炭素源アラビノース)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量;黒三角、(8)の培地(炭素源添加せず)で培養したpUC702-encP形質転換ストレプトマイセス・リビダンスによる桂皮酸生産量。
【0062】
図8に示されるように、pUC702-encP形質転換ストレプトマイセス・リビダンスは、炭素源としてグルコースの代わりにキシロースまたはアラビノースを用いた場合でも桂皮酸を生産した。特にキシロースを用いた場合で桂皮酸の生産量は高く、生産量は培養5日後で300mg/Lに達した。炭素源未添加の(8)の培地でも桂皮酸の生産はみられたが、その生産量は100mg/Lにも届かず、それ以上増大することなく停滞した。
【産業上の利用可能性】
【0063】
本発明によって桂皮酸を効率的に製造でき、製造された桂皮酸は、医薬品あるいは化成品原料として、食品用の香料として好適に利用できる。また、本発明によれば、生物による安全な方法で桂皮酸を生産することができる。また、原料物質(初期物質)としてグルコース以外にも種々の炭素源、特に、生デンプンなどの植物バイオマスに由来する炭素源を使用できるので、製造コストを抑えることができる。
【特許請求の範囲】
【請求項1】
桂皮酸の製造方法であって、
フェニルアラニンアンモニアリアーゼ高発現放線菌を液体培地で培養する工程を含む、方法。
【請求項2】
前記フェニルアラニンアンモニアリアーゼ高発現放線菌が、フェニルアラニンアンモニアリアーゼをコードする遺伝子を導入して得られる形質転換放線菌であり、該遺伝子が高発現型プロモーターの制御下で発現される、請求項1に記載の方法。
【請求項3】
前記高発現型プロモーターが、ホスホリパーゼDプロモーターである、請求項2に記載の方法。
【請求項4】
前記フェニルアラニンアンモニアリアーゼ高発現放線菌が、ストレプトマイセス属に属する放線菌である、請求項1から3のいずれかに記載の方法。
【請求項5】
前記液体培地が、フェニルアラニンを含む、請求項1から4のいずれかに記載の方法。
【請求項6】
ストレプトマイセス・マリチマス由来encP遺伝子を導入して得られる形質転換ストレプトマイセス・リビダンスであって、該encP遺伝子がホスホリパーゼDプロモーターの制御下で発現されている、形質転換ストレプトマイセス・リビダンス。
【請求項1】
桂皮酸の製造方法であって、
フェニルアラニンアンモニアリアーゼ高発現放線菌を液体培地で培養する工程を含む、方法。
【請求項2】
前記フェニルアラニンアンモニアリアーゼ高発現放線菌が、フェニルアラニンアンモニアリアーゼをコードする遺伝子を導入して得られる形質転換放線菌であり、該遺伝子が高発現型プロモーターの制御下で発現される、請求項1に記載の方法。
【請求項3】
前記高発現型プロモーターが、ホスホリパーゼDプロモーターである、請求項2に記載の方法。
【請求項4】
前記フェニルアラニンアンモニアリアーゼ高発現放線菌が、ストレプトマイセス属に属する放線菌である、請求項1から3のいずれかに記載の方法。
【請求項5】
前記液体培地が、フェニルアラニンを含む、請求項1から4のいずれかに記載の方法。
【請求項6】
ストレプトマイセス・マリチマス由来encP遺伝子を導入して得られる形質転換ストレプトマイセス・リビダンスであって、該encP遺伝子がホスホリパーゼDプロモーターの制御下で発現されている、形質転換ストレプトマイセス・リビダンス。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2011−223981(P2011−223981A)
【公開日】平成23年11月10日(2011.11.10)
【国際特許分類】
【出願番号】特願2010−287418(P2010−287418)
【出願日】平成22年12月24日(2010.12.24)
【出願人】(504150450)国立大学法人神戸大学 (421)
【出願人】(000003001)帝人株式会社 (1,209)
【Fターム(参考)】
【公開日】平成23年11月10日(2011.11.10)
【国際特許分類】
【出願日】平成22年12月24日(2010.12.24)
【出願人】(504150450)国立大学法人神戸大学 (421)
【出願人】(000003001)帝人株式会社 (1,209)
【Fターム(参考)】
[ Back to top ]