
生産性改善法
【課題】
γ−シクロデキストリン合成酵素、マルトースホスホリラーゼおよびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を大量かつ安定に生産しうる方法を提供することを目的とする。
【解決手段】
γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を菌体外に大量かつ安定に生産しうる宿主−ベクター系により上記課題を解決する。
γ−シクロデキストリン合成酵素、マルトースホスホリラーゼおよびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を大量かつ安定に生産しうる方法を提供することを目的とする。
【解決手段】
γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を菌体外に大量かつ安定に生産しうる宿主−ベクター系により上記課題を解決する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を工業的に生産する方法に関する。さらに詳細には、本発明は、この方法に使用する、転写を調節する塩基配列、シグナルペプチドをコードする遺伝子、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼおよびトレハロースホスホリラーゼからなる群から選択される1種の酵素をコードする遺伝子、およびアミノアシルtRNA合成酵素をコードする遺伝子を含む組換えベクター、該組換えベクターを導入した宿主細胞、ならびにγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法に関する。
【背景技術】
【0002】
γ−シクロデキストリンの合成には、γ−シクロデキストリンを主生成物とするシクロデキストリン・グルカノトランスフェラーゼ(以下、「γ−シクロデキストリン合成酵素」と呼ぶ)を澱粉等に作用させる方法が知られている(たとえば、特許文献1を参照)。特許文献1によれば、外来遺伝子を含む組換えベクターを宿主細胞に導入する宿主−ベクター系を利用してγ−シクロデキストリン合成酵素を取得している。具体的には、γ−シクロデキストリン合成酵素をコードする遺伝子をアンピシリン耐性遺伝子を含むT−ベクターにライゲーションし、大腸菌(Escherichia coli)を形質転換して培養することにより、γ−シクロデキストリン合成酵素を取得している。
【0003】
一方、トレハロースの合成には、マルトースホスホリラーゼおよびトレハロースホスホリラーゼをマルトースに作用させる方法が知られている(たとえば、特許文献2を参照)。特許文献2でも宿主−ベクター系を利用してマルトースホスホリラーゼおよびトレハロースホスホリラーゼを取得している。具体的には、マルトースホスホリラーゼおよびトレハロースホスホリラーゼをコードする遺伝子を適当なベクターに挿入し、大腸菌または枯草菌(Bacillus subtilis)を形質転換してアンピシリンまたはテトラサイクリンなどを含む培地で培養することにより、マルトースホスホリラーゼおよびトレハロースホスホリラーゼを取得している。
【0004】
上記のγ−シクロデキストリン合成酵素の生産方法とマルトースホスホリラーゼおよびトレハロースホスホリラーゼの生産方法は、大腸菌を宿主細胞とする宿主−ベクター系を利用している(たとえば、特許文献1および2を参照)。大腸菌は、世代時間が約20分と短く、様々な糖類を資化して増殖することができる。さらに大腸菌に適した多くのプラスミドベクターも開発されており、大腸菌を宿主細胞とする宿主−ベクター系は、実験室での組換えタンパク質の生産において一応の成果を期待できる。
【0005】
しかしながら、大腸菌の組換えタンパク質を菌体内に蓄積するという性質のために、大腸菌自体の生育が阻害され、さらに菌体内のタンパク質分解酵素の作用により組換えタンパク質の機能が失いやすいという、組換えタンパク質の大量かつ安定に生産する上で不利な点があった。さらに、組換えタンパク質を取得するためには菌体を回収して破砕し、かつ、発熱性物質の一種であるリポポリサッカライドを注意深く除去しなければならない。このために、組換えタンパク質の採取・精製のための工程が頻雑になるという問題も生じていた。
【0006】
そこで、これらの問題を解決する手段として、枯草菌を利用した宿主−ベクター系が知られている(たとえば、特許文献2および非特許文献3を参照)。特許文献2によれば、菌体内よりも菌体外の培養上清中に多くのマルトースホスホリラーゼおよびトレハロースホスホリラーゼを得ている。しかし、取得される酵素量は少なく、大量生産にまでは至っていない。
【0007】
一方、上記のγ−シクロデキストリン合成酵素の生産方法とマルトースホスホリラーゼおよびトレハロースホスホリラーゼの生産方法は、それぞれの酵素遺伝子を抗生物質耐性遺伝子とともに挿入した組換えベクターを利用している。これにより、抗生物質耐性遺伝子に対応する抗生物質を培養培地中に加えることで、抗生物質耐性遺伝子含有組換えベクターを維持する細胞は選択され、該組換えベクターを維持しない細胞は選択されず、したがって排除される。
【0008】
すなわち、抗生物質耐性遺伝子を組換えベクターに挿入する方法は、組換えベクターを宿主細胞内に安定に維持させることができ、実験室で通常行なわれている。しかし、下記の理由により工業的スケールの生産では望ましくない。
(i)抗生物質耐性菌株の使用は、環境に対して危険を呈する可能性がある。
(ii)培養中に必要な抗生物質の量は生産コストを有意に増加させる。
(iii)抗生物質の使用は、ヒト及び動物向けに用いられる物質の生産においては回避すべきである。
【0009】
そこで、抗生物質耐性遺伝子を用いずに組換えベクターを安定化させる方法として、染色体の栄養要求突然変異を補う方法が知られている(非特許文献1および2)。しかしながら、染色体の栄養要求突然変異を補って組換えベクターを安定化させる方法は、宿主細菌が必要とする栄養を培地に加えずに発酵を行う必要があるため、発酵培地の組成を厳しく制限する必要がある。しかも、栄養共生によって、組換えベクターの消失後もなお細胞が増殖しうる可能性がある。
【特許文献1】特開2003−102489号公報
【特許文献2】国際公開第05/003343号パンフレット
【非特許文献1】Genetics. 1989 May;122(1):19-27
【非特許文献2】Curr Genet. 1989 Sep;16(3):159-63
【非特許文献3】Ohta Y, et al., Biosci. Biotechnol. Biochem. 2004 ,68,1073-1081
【発明の開示】
【発明が解決しようとする課題】
【0010】
本発明者らは、枯草菌を宿主細胞として用いた宿主−ベクター系により、γ−シクロデキストリン合成酵素を大量に生産することを試みた。しかし、上記したマルトースホスホリラーゼおよびトレハロースホスホリラーゼの生産と同じく、γ−シクロデキストリン合成酵素についても、枯草菌を宿主細胞として用いたこれまでの宿主−ベクター系では菌体外に大量に分泌することができないことを見出した。
【0011】
そこで、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼまたはトレハロースホスホリラーゼを菌体外に大量に分泌することができる、新たな宿主−ベクター系の提供が強く望まれている。
【0012】
一方、組換えベクターを維持する宿主細胞を選択する方法について、抗生物質耐性遺伝子を組み込む方法および染色体の栄養要求突然変異を補う方法のいずれもが、培地の特殊な処理に依存している。かかる制約は、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼまたはトレハロースホスホリラーゼの生産において、発酵工程の費用の増大、および生産性向上のためにとり得る選択自由度の制限を招く。
【0013】
そこで、抗生物質を用いることなく、かつ培地組成に制約されずに組換えベクターを維持しうる、他の選択方法が強く望まれている。
【0014】
したがって、本発明の目的は、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼまたはトレハロースホスホリラーゼを菌体外に大量に分泌しうる、新たな宿主−ベクター系を提供することにある。
【0015】
さらに、抗生物質を用いることなく、かつ培地組成に制約されずに組換えベクターを維持する新たな手段を提供することにより、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼまたはトレハロースホスホリラーゼを大量かつ安定に生産しうる方法を提供することを目的とする。
【課題を解決するための手段】
【0016】
上記課題を解決するために種々検討を行い、宿主細胞をバチルス属細菌として、組換えベクターに組み込む最適な転写を調節する塩基配列(以下、「遺伝子調節領域」と呼ぶ)とシグナルペプチドをコードする遺伝子(以下、「シグナルペプチド遺伝子」と呼ぶ)の組み合わせを検討し、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を菌体外に大量に分泌することができる宿主−ベクター系を得ることができ、本発明を完成するに至った。
【0017】
さらに、アミノアシルtRNA合成酵素遺伝子を保有する染色体外遺伝子と該アミノアシルtRNA合成酵素活性に欠損を生じさせた染色体変異宿主細胞との相補性に基づくことで、抗生物質を用いることなく、かつ培地組成に制約されずにγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を含む組換えベクターを維持することに成功し、上記した酵素を大量かつ安定に生産しうる方法を見出し、本発明を完成させた。アミノアシルtRNA合成酵素遺伝子を保有する染色体外遺伝子を含有した細胞のみが生存を確保されることに基づく。
【0018】
すなわち、本発明は以下のとおりである。
<1>γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、かつ
アミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターであって、
アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞において用いられる前記組換えベクター。
<2>下記(a)〜(c)の塩基配列または遺伝子を有する組換えベクター:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;および
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子。
<3>下記(a)〜(d)の塩基配列または遺伝子を有し、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞において用いられる組換えベクター:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;および
(d)発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子。
<4>アミノアシルtRNA合成酵素が、トリプトファニルtRNA合成酵素、アラニルtRNA合成酵素、アルギニルtRNA合成酵素、アスパルギニルtRNA合成酵素、アスパルチルtRNA合成酵素、システイニルtRNA合成酵素、グルタミンtRNA合成酵素、グルタメートtRNA合成酵素、グリシンtRNA合成酵素、ヒスチジルtRNA合成酵素、イソロイシルtRNA合成酵素、ロイシルtRNA合成酵素、リシンtRNA合成酵素、メチオニルtRNA合成酵素、フェニルアラニンtRNA合成酵素、プロリルtRNA合成酵素、セリルtRNA合成酵素、トレオニルtRNA合成酵素、チロシルtRNA合成酵素、またはバリルtRNA合成酵素である、<1>または<3>に記載の組換えベクター。
<5>γ−シクロデキストリン合成酵素をコードする遺伝子が、配列番号3に記載のアミノ酸配列をコードする遺伝子である、<1>〜<4>のいずれかに記載の組換えベクター。
<6>マルトースホスホリラーゼをコードする遺伝子が、配列番号4に記載のアミノ酸配列をコードする遺伝子である、<1>〜<4>のいずれかに記載の組換えベクター。
<7>トレハロースホスホリラーゼをコードする遺伝子が、配列番号5に記載のアミノ酸配列をコードする遺伝子である、<1>〜<4>のいずれかに記載の組換えベクター。
<8>組換えベクターは、プラスミド、バクテリオファージ、またはレトロトランスポゾンの形態を有する<1>〜<7>のいずれかに記載の組換えベクター。
<9>アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞であって、
γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、かつ
アミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターで形質転換して用いられる前記宿主細胞。
<10>下記(a)〜(c)の塩基配列または遺伝子を有する組換えベクターで形質転換して用いられる宿主細胞:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;および
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子。
<11>アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞であって、
下記(a)〜(d)の塩基配列または遺伝子を有する組換えベクターで形質転換して用いられる前記宿主細胞:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;および
(d)発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子。
<12>アミノアシルtRNA合成酵素が、トリプトファニルtRNA合成酵素、アラニルtRNA合成酵素、アルギニルtRNA合成酵素、アスパルギニルtRNA合成酵素、アスパルチルtRNA合成酵素、システイニルtRNA合成酵素、グルタミンtRNA合成酵素、グルタメートtRNA合成酵素、グリシンtRNA合成酵素、ヒスチジルtRNA合成酵素、イソロイシルtRNA合成酵素、ロイシルtRNA合成酵素、リシンtRNA合成酵素、メチオニルtRNA合成酵素、フェニルアラニンtRNA合成酵素、プロリルtRNA合成酵素、セリルtRNA合成酵素、トレオニルtRNA合成酵素、チロシルtRNA合成酵素、またはバリルtRNA合成酵素である、<9>または<11>に記載の宿主細胞。
<13>γ−シクロデキストリン合成酵素をコードする遺伝子が、配列番号3に記載のアミノ酸配列をコードする遺伝子である、<9>〜<12>のいずれかに記載の宿主細胞。
<14>マルトースホスホリラーゼをコードする遺伝子が、配列番号4に記載のアミノ酸配列をコードする遺伝子である、<9>〜<12>のいずれかに記載の宿主細胞。
<15>トレハロースホスホリラーゼをコードする遺伝子が、配列番号5に記載のアミノ酸配列をコードする遺伝子である、<9>〜<12>のいずれかに記載の宿主細胞。
<16>宿主細胞が細菌である<9>〜<15>のいずれかに記載の宿主細胞。
<17>細菌がバチルス属細菌である、<16>に記載の宿主細胞。
<18>下記(i)〜(iv)の工程を有する、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法:
(i)γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、さらにアミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターを用意する工程;
(ii)アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞を用意する工程;
(iii)前記宿主細胞を前記組換えベクターで形質転換して形質転換体を得る工程;および
(iv)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
<19>下記(i)〜(iii)の工程を有する、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法:
(i)下記[a]〜[c]の塩基配列または遺伝子を有する組換えベクターを用意する工程、
[a]配列番号1に記載の塩基配列、
[b]前記[a]の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子、および
[c]前記[b]の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;
(ii)前記組換えベクターで宿主細胞を形質転換して形質転換体を得る工程;および
(iii)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
<20>下記(i)〜(iv)の工程を有する、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法:
(i)下記[a]〜[d]の塩基配列または遺伝子を有する組換えベクターを用意する工程、
[a]配列番号1に記載の塩基配列、
[b]前記[a]の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子、
[c]前記[b]の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子、および
[d]発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子;
(ii)アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞を用意する工程;
(iii)前記宿主細胞を前記組換えベクターで形質転換して形質転換体を得る工程;および
(iv)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
<21>アミノアシルtRNA合成酵素が、トリプトファニルtRNA合成酵素、アラニルtRNA合成酵素、アルギニルtRNA合成酵素、アスパルギニルtRNA合成酵素、アスパルチルtRNA合成酵素、システイニルtRNA合成酵素、グルタミンtRNA合成酵素、グルタメートtRNA合成酵素、グリシンtRNA合成酵素、ヒスチジルtRNA合成酵素、イソロイシルtRNA合成酵素、ロイシルtRNA合成酵素、リシンtRNA合成酵素、メチオニルtRNA合成酵素、フェニルアラニンtRNA合成酵素、プロリルtRNA合成酵素、セリルtRNA合成酵素、トレオニルtRNA合成酵素、チロシルtRNA合成酵素、またはバリルtRNA合成酵素である、<18>または<20>に記載の方法。
<22>γ−シクロデキストリン合成酵素をコードする遺伝子が、配列番号3に記載のアミノ酸配列をコードする遺伝子である、<18>〜<21>のいずれかに記載の方法。
<23>マルトースホスホリラーゼをコードする遺伝子が、配列番号4に記載のアミノ酸配列をコードする遺伝子である、<18>〜<21>のいずれかに記載の方法。
<24>トレハロースホスホリラーゼをコードする遺伝子が、配列番号5に記載のアミノ酸配列をコードする遺伝子である、<18>〜<21>のいずれかに記載の方法。
<25>宿主細胞が細菌である<18>〜<24>のいずれかに記載の方法。
<26>細菌がバチルス属細菌である<25>に記載の方法。
<27>γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される1種の酵素の遺伝子が、前記遺伝子と95%以上の相同性を有し、かつ、前記選択される酵素の活性を有するタンパク質をコードする遺伝子である、<5>〜<8>のいずれかに記載の組換えベクター。
<28>γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される1種の酵素の遺伝子が、前記遺伝子と95%以上の相同性を有し、かつ、前記選択される酵素の活性を有するタンパク質をコードする遺伝子である、<13>〜<17>のいずれかに記載の宿主細胞。
<29>γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される1種の酵素の遺伝子が、前記遺伝子と95%以上の相同性を有し、かつ、前記選択される酵素の活性を有するタンパク質をコードする遺伝子である、<22>〜<26>のいずれかに記載の方法。
【発明の効果】
【0019】
本発明によれば、最適な遺伝子調節領域およびシグナルペプチド遺伝子の組み合わせを組換えベクターに含めることで、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を菌体外に大量に分泌することができる。
【0020】
さらに、抗生物質を用いることなく、かつ特定成分を含有しないよう制限を加えた培地を用いることなくγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を含む組換えベクターを維持することができ、その結果、工業的な生産においても、醗酵槽内での培養が終了するまで細胞中の組換えベクターを安定化させることができ、上記した酵素を工業的規模で大量かつ安定に生産することができる。
【発明を実施するための最良の形態】
【0021】
(A)本発明の組換えベクター
本発明の組換えベクターの一つの形態は、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、かつアミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターであって、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞において用いられる前記組換えベクター(以下、「組換えベクターA」と呼ぶ)である。
【0022】
本発明の組換えベクターの一つの形態は、下記(a)〜(c)の塩基配列または遺伝子を有する組換えベクター(以下、「組換えベクターB」と呼ぶ)である:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;および
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子。
【0023】
本発明の組換えベクターの一つの形態は、下記(a)〜(d)の塩基配列または遺伝子を有し、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞において用いられる組換えベクター(以下、「組換えベクターC」と呼ぶ)である:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;および
(d)発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子。
【0024】
本発明の組換えベクターAおよびCは、アミノアシルtRNA合成酵素遺伝子をコードする遺伝子を発現可能な状態で含有する。該組換えベクターAおよびCは、アミノアシルtRNA合成酵素遺伝子をコードする遺伝子を、所望の宿主細胞内で発現可能な状態で含んでおり、当該宿主細胞を形質転換するために使用される。
【0025】
アミノアシルtRNA合成酵素は、20種類のアミノ酸に対応するよう、20数種類(一部は数種類ある)存在し、例えばアラニンに対するアミノアシルtRNA合成酵素はアラニルtRNA合成酵素(alanyl−tRNA synthetaseまたはalanine−tRNA synthetase)と呼ばれている。アミノアシルtRNA合成酵素は、具体的には、例えば、トリプトファニルtRNA合成酵素、アラニルtRNA合成酵素、アルギニルtRNA合成酵素、アスパルギニルtRNA合成酵素、アスパルチルtRNA合成酵素、システイニルtRNA合成酵素、グルタミンtRNA合成酵素、グルタメートtRNA合成酵素、グリシンtRNA合成酵素、ヒスチジルtRNA合成酵素、イソロイシルtRNA合成酵素、ロイシルtRNA合成酵素、リシンtRNA合成酵素、メチオニルtRNA合成酵素、フェニルアラニンtRNA合成酵素、プロリルtRNA合成酵素、セリルtRNA合成酵素、トレオニルtRNA合成酵素、チロシルtRNA合成酵素、またはバリルtRNA合成酵素である。
【0026】
トリプトファンに対応するアミノアシルtRNA合成酵素をトリプトファニルtRNA合成酵素と呼び、トリプトファニルtRNA合成酵素が欠失している細胞株は、トリプトファンを含むタンパクの合成を行うことができないため増殖することができないことになる。
【0027】
アミノアシルtRNA合成酵素遺伝子は原核生物から真核生物に至るあらゆる生物に含まれる。例えば、枯草菌には、alanyl−tRNA synthetase(配列番号6)、arginyl−tRNA synthetase(配列番号7)、asparaginyl−tRNA synthetase(配列番号8)、aspartyl−tRNA synthetase(配列番号9)、cysteinyl−tRNA synthetase(配列番号10)、glutamyl−tRNA synthetase(配列番号11)、glutamyl−tRNA amidotransferase, subunit a(配列番号12)、glycine tRNA synthetase, alpha subunit(配列番号13)、glycine tRNA synthetase, beta subunit(配列番号14)、histidyl tRNA synthetase, hisS(配列番号15)、isoleucyl−tRNA synthetase(配列番号16)、leucyl−tRNA synthetase(配列番号17)、lysine tRNA synthetase, constitutive(配列番号18)、histidyl tRNA synthetase,hisZ(配列番号19)、methionyl−tRNA synthetase(配列番号20)、phenylalanine tRNA synthetase, alpha subunit(配列番号21)、phenylalanine tRNA synthetase, beta subunit(配列番号22)、similar to phenylalanyl−tRNA synthetase, ytpR (配列番号23)、prolyl−tRNA synthetase(配列番号24)、seryl−tRNA synthetase, also charges selenocysteinyl−tRNA with serine(配列番号25)、threonyl−tRNA synthetase, major(配列番号26)、threonyl−tRNA synthetase, minor(配列番号27)、tryptophanyl−tRNA synthetase(配列番号28)、tyrosyl−tRNA synthetase, major(配列番号29)、tyrosyl−tRNA synthetase, minor (配列番号30)、valyl−tRNA synthetase(配列番号31)、glutamyl−tRNA amidotransferase, subunit b(配列番号32)、glutamyl−tRNA amidotransferase, subunit c(配列番号33)などが含まれることが知られている。
【0028】
本発明の組換えベクターAおよびCは、アミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有するものである。したがって、該組換えベクターAおよびCについては、アミノアシルtRNA合成酵素遺伝子以外に、この遺伝子を発現可能な状態とするために、宿主として枯草菌(Bacillus subtilis)などの細菌を使用する場合は、たとえば、プロモーター、オペレーター領域(プロモーター、オペレーターおよびリボゾーム結合領域(SD領域)を含む)、開始コドン、選択マーカー遺伝子、終止コドン、ターミネーター領域、及びプラスミド複製可能単位などを有する。
【0029】
選択マーカー遺伝子としては、特に制限されず、たとえば、ジヒドロ葉酸レダクターゼ(DHFR)遺伝子またはシゾサッカロマイセス・ポンベTPI遺伝子等のようなその補体が宿主細胞に欠けている遺伝子、またはたとえばアンピシリン、カナマイシン、テトラサイクリン、クロラムフェニコール、シクロヘキシミド、テトラマイシン、ネオマイシンもしくはヒグロマイシンのような薬剤耐性遺伝子を挙げることができる。
【0030】
本発明の組換えベクターAおよびCは、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞において用いられる。この宿主細胞については後述する。
【0031】
本発明の組換えベクターは、ベクターの種類は特に限定されず、例えば、宿主細胞内で自立的に複製することが可能なもの(例えばプラスミド等)でもよいし、あるいは、宿主細胞に導入された際に宿主細胞のゲノムに組み込まれ、組み込まれた染色体と共に複製されるものであってもよい。好ましくは、本発明で用いるベクターは、プラスミド、バクテリオファージ、またはレトロトランスポゾンである。
【0032】
宿主細胞内で自立的に複製することが可能なベクターの具体例としては、pRS413、pRS415、pRS416、YCp50、pAUR112またはpAUR123などのYCp型大腸菌−酵母シャトルベクター;pRS403、pRS404、pRS405、pRS406、pAUR101またはpAUR135などのYIp型大腸菌−酵母シャトルベクター;大腸菌由来のプラスミド(たとえば、pBR322、pBR325、pUC18、pUC19、pUC119、pTV118N、pTV119N、pBluescript、pHSG298、pHSG396またはpTrc99AなどのColE系プラスミド;pACYC177またはpACYC184などのp1A系プラスミド;pMW118、pMW119、pMW218またはpMW219などのpSC101系プラスミドなど);枯草菌由来のプラスミド(例えばpUB110、pTP5など);pHY300PLKなどの大腸菌−枯草菌シャトルベクターを挙げることができる。またファージベクターとして、λファージ(たとえば、Charon 4A、Charon21A、EMBL4、λgt100、gt11、zap)、ψX174、M13mp18、M13mp19などを挙げることができる。レトロトランスポゾンとしてはTy因子などを挙げることができる。また、融合タンパク質として発現する発現ベクター、例えばpGEXシリーズ(ファルマシア製)、pMALシリーズ(Biolabs社製)を使用することもできる。
【0033】
本発明の組換えベクターは、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有する。
【0034】
本明細書でいう「遺伝子を発現可能な状態」、または単に「発現可能な状態」とは、遺伝子の情報がmRNAに転写されるためのRNAポリメラーゼ認識部位およびmRNAの情報が該遺伝子の発現産物であるタンパク質等に翻訳されるためのリボゾーム結合領域が該遺伝子の開始コドンの上流に存在する状態を意味する。要するに、該遺伝子の開始コドンから終止コドンまでをmRNAに転写されるために必要な配列が開始コドンより上流に存在し、さらにそのmRNAの情報がタンパク質等に翻訳されるための配列が開始コドンより上流に存在するということになる。ここでいう「開始コドン」とは本来染色体上における該遺伝子の開始コドン配列に限らず、開始コドンとして機能するコドンであればいずれでもよい。またここでいうリボゾーム結合領域とは、例えば原核生物の5´−aaaggagg−3´からなるコンセンサス配列を有する配列に集約されることなく、リボゾームが認識できる配列であれば良く、本来染色体上で該遺伝子のリボゾーム結合領域としての配列に限らず、リボゾームが認識できる配列であれば用いることができる。
【0035】
本明細書でいう「γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子」とは、γ−シクロデキストリン合成酵素をコードする遺伝子、マルトースホスホリラーゼをコードする遺伝子、またはトレハロースホスホリラーゼをコードする遺伝子の単一物だけでなく、各遺伝子を組み合わせた複合物も含む。
【0036】
本発明の組換えベクターの構築は、DNA組換えの一般的な方法、たとえば、Molecular Cloning (1989) (Cold Spring Harbor Lab.)に記載される方法にしたがって行うことができる。
【0037】
本発明の組換えベクターBおよびCは、遺伝子調節領域として配列番号1に記載の塩基配列を有し、シグナルペプチド遺伝子として配列番号2に記載のアミノ酸配列をコードする遺伝子を有する。
【0038】
配列番号1に記載の塩基配列を含むDNA断片は、たとえば、バチルス・エスピー JAMB750(Bacillus sp. JAMB750)株の染色体DNAにランダム変異を導入することにより取得することができる。配列番号1に記載の塩基配列を含むDNA断片を取得するその他の方法としては、たとえば、配列番号1に記載の塩基配列の情報に基づいて、化学合成、遺伝子工学的手法または突然変異誘発などの通常知られる任意の方法で作製することができる。
【0039】
本明細書にいう「シグナルペプチド」とは、シグナルペプチドをコードする塩基配列の下流に存在する遺伝子の遺伝子産物のアミノ末端に付加して、該遺伝子産物を細胞外に分泌しうるペプチドを意味する。また、「遺伝子産物を細胞外に分泌しうる」とは、上記した通り、シグナルペプチドが遺伝子産物である組換えタンパク質のアミノ末端側に付加し、組換えタンパク質の小胞体膜への付着および膜通過等を導き、最終的に細胞外に組換えタンパク質を分泌しうる作用を意味する。
【0040】
本明細書にいう「配列番号1の塩基配列の下流に直接的または間接的に連結された」とは、配列番号1に記載の塩基配列の3´末端側に0〜1000程度の塩基を介して連結されることを意味する。
【0041】
シグナルペプチド遺伝子を含むDNA断片は、たとえば、配列番号2に記載の塩基配列の情報に基づいて、化学合成、遺伝子工学的手法または突然変異誘発などの通常知られる任意の方法で作製することができる。化学合成法としては、たとえば、ホスホアミダイト法を応用することにより自動合成しうる。なお、配列番号2に記載のシグナルペプチドは、バチルス属細菌の分泌するタンパク質群のアミノ末端側のアミノ酸配列情報から統計処理してデザインして得られた数十候補のアミノ酸配列から、組換えタンパク質生産性を評価し選択したものである。本配列は1〜数個のアミノ酸残基を付加、欠失または置換しても利用できる。
【0042】
本発明の組換えベクターBおよびCは、遺伝子調節領域およびその下流に直接的または間接的に連結されたシグナルペプチド遺伝子、ならびにその下流に直接的に連結されたγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を含み、該組換えベクターを宿主細胞として枯草菌に導入し形質転換することにより、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素の発現を促進し、さらに発現する酵素を菌体外に分泌しうる。
【0043】
本明細書にいう「配列番号2に記載のアミノ酸配列をコードする遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子」とは、上記したシグナルペプチド遺伝子の3´末端側に連結されたγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子である。
【0044】
本明細書にいう「γ−シクロデキストリン合成酵素をコードする遺伝子」は、特に限定されるものではないが、好ましくは配列番号3に記載の塩基配列からなる遺伝子である。配列番号3に記載の塩基配列からなる遺伝子は、たとえば、特許文献1の記載に基いてバチルス・クラーキ 7364(Bacillus clarkii 7364)株から取得される。配列番号3に記載の塩基配列からなる遺伝子を取得するその他の方法としては、配列番号3に記載の塩基配列の情報に基づいて、化学合成、遺伝子工学的手法または突然変異誘発などの通常知られる任意の方法で作製することができる。
【0045】
本明細書にいう「γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される1種の酵素の遺伝子が、前記遺伝子と95%以上の相同性を有し、かつ、前記選択される酵素の活性を有するタンパク質をコードする遺伝子」とは、たとえば、γ−シクロデキストリン合成酵素をコードする遺伝子を選択した場合に、γ−シクロデキストリン合成酵素をコードする遺伝子に1〜数個の塩基の欠失、置換、逆位、付加および/または挿入を有し、該遺伝子と95%以上の相同性があり、かつ、発現産物がγ−シクロデキストリン合成酵素活性を有する酵素である遺伝子を意味する。
【0046】
本明細書にいう「γ−シクロデキストリン合成酵素」は、糖類に作用し主生成物としてγ−シクロデキストリンを生成するシクロデキストリン・グルカノトランスフェラーゼ活性を有する酵素であればよく、好ましくは配列番号3に記載の遺伝子が発現して得られる酵素である。
【0047】
本明細書にいう「マルトースホスホリラーゼをコードする遺伝子」は、特に限定されるものではないが、好ましくは配列番号4に記載の塩基配列からなる遺伝子である。配列番号4に記載の塩基配列からなる遺伝子は、たとえば、特許文献2の記載に基づいてパエニバチルス・エスピー SH−55(Paenibacillus sp. SH−55)株から取得される。配列番号4に記載の塩基配列からなる遺伝子を取得するその他の方法としては、配列番号4に記載の塩基配列に記載の情報に基づいて、化学合成、遺伝子工学的手法または突然変異誘発などの通常知られる任意の方法で作製することができる。
【0048】
本明細書にいう「マルトースホスホリラーゼ」は、マルトース中のα−1,4−グルコピラノシド結合を可逆的に加リン酸分解し、グルコースとβ−D−グルコース−1−リン酸を生成することができればよく、好ましくは、配列番号4に記載の遺伝子が発現して得られる酵素である。
【0049】
本明細書にいう「トレハロースホスホリラーゼをコードする遺伝子」は、特に限定されるものではないが、好ましくは配列番号5に記載の塩基配列からなる遺伝子である。配列番号5に記載の塩基配列からなる遺伝子は、たとえば、特許文献2の記載に基いてパエニバチルス・エスピー SH−55株から取得される。配列番号5に記載の塩基配列からなる遺伝子を取得するその他の方法としては、配列番号5に記載の塩基配列に記載の塩基配列の情報に基づいて、化学合成、遺伝子工学的手法または突然変異誘発などの通常知られる任意の方法で作製することができる。
【0050】
本明細書にいう「トレハロースホスホリラーゼ」は、トレハロース中のα−1,1−グルコピラノシド結合を可逆的に加リン酸分解し、グルコースおよびβ−D−グルコース−1−リン酸を生成すればよく、好ましくは、配列番号5に記載の遺伝子が発現して得られる酵素である。
【0051】
本発明の組換えベクターCの具体例を、図1〜3として示す。
【0052】
(B)本発明の宿主細胞
本発明の宿主細胞の一つの形態は、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞であって、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、かつアミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターで形質転換して用いられる前記宿主細胞(以下、「宿主細胞A」と呼ぶ)である。
【0053】
本発明の宿主細胞の一つの形態は、下記(a)〜(c)の塩基配列または遺伝子を有する組換えベクターで形質転換して用いられる宿主細胞(以下、「宿主細胞B」と呼ぶ)である:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;および
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子。
【0054】
本発明の宿主細胞の一つの形態は、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞であって、下記(a)〜(d)の塩基配列または遺伝子を有する組換えベクターで形質転換して用いられる前記宿主細胞(以下、「宿主細胞C」と呼ぶ)である:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;および
(d)発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子。
【0055】
一般に、細胞は、自己の増殖のためにアミノアシルtRNA合成酵素をコードする染色体遺伝子を有している。それに対して、本発明の宿主細胞AおよびCでは、細胞が本来有している、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させるか、あるいはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させる。アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損または該遺伝子発現の低下は、以下の方法で行うことができる。
【0056】
[遺伝子欠損(不活性化)]
不活性化の語は染色体遺伝子のうち1以上の機能発現を妨げる方法を含んでいる。不活性化は核酸遺伝子配列における欠失、置換(例えば変異)、阻害、及び/または挿入などによって行われる。いくつかの実施態様において、変異微生物は好ましくは安定かつ不可逆的な不活性化を生じる1以上の遺伝子の不活性化を含む。
【0057】
(挿入による不活性化)
ここで用いる“挿入配列”の語は微生物染色体内に導入されたDNA配列を言う。いくつかの実施態様において、挿入配列は形質転換される細胞のゲノム中に既に存在する、または存在しない配列であってもよい(すなわち、相同または異種配列である)。さらに別の実施態様において、挿入配列は選択マーカーを含むことがある。さらなる実施態様において、挿入配列は2つの相同ボックスを含むことがある。
【0058】
ここで用いる“相同ボックス”とは微生物染色体の配列と相同な核酸配列をいう。より具体的には、相同ボックスは、本発明に従って不活性化される遺伝子に隣接するコード領域または遺伝子の一部と約80〜100%配列同一性、約90〜100%配列同一性または約95%〜100%配列同一性を有する上流または下流領域をいう。これらの配列は微生物染色体の一部が置換されることを目的とする。本発明を限定するものではないが、相同ボックスは約1塩基対(bp)〜200キロ塩基(kb)を含む。好ましくは、相同ボックスは約1bp〜10.0kb;1bp〜5.0kb;1bp〜2.5kb;1bp〜1.0kb及び0.25kb〜2.5kbを含む。また、相同ボックスは約10.0kb、5.0kb、2.5kb、2.0kb、1.5kb、1.0kb、0.5kb、0.25kb及び0.1kbを含む。いくつかの実施態様において、選択マーカーの5’及び3’末端は相同ボックスに隣接し、ここで相同ボックスは当該遺伝子のコード領域にすぐ隣接した核酸配列を含む。
【0059】
ここで用いる“選択マーカー”の語は宿主細胞において発現できるヌクレオチド配列をいい、選択マーカーの発現により対応する選択因子の存在下、または必須栄養素の不存在下で発現遺伝子を含む細胞は成長できるようになる。ここで用いる“選択可能マーカー”及び“選択マーカー”の語は宿主細胞中で発現できる核酸(例えば遺伝子)を言い、ベクターを含むこれら宿主の選択が簡単にできるようにする。当該選択マーカーの例としては、限定されないが、抗微生物剤などがある。従って、“選択マーカー”とは、宿主細胞が目的の挿入DNAを取り込んだ、またはその他の反応が起こった印を与える遺伝子をいう。通常、選択マーカーは、形質転換時に外来配列を受け取らなかった細胞と外来DNAを含む細胞との区別を可能にするために宿主細胞に抗菌耐性または代謝利点を与える遺伝子をいう。選択マーカーは抗微生物耐性マーカーなどが挙げられる(例えば、ampR;phleoR;specR;kanR;eryR;tetR;cmpR;及びneoR;例えばGuerot−Fleury,Gene,167:335−337[1995];Palmeros et al.、Gene 247:255−264[2000];及びTrieu−Cuot et al.、Gene、23:331−341[1983]を参照)。いくつかの特に好ましい実施態様において、本発明はクロラムフェニコール耐性遺伝子を提供する(例えば、pC194上に存在する遺伝子、及びバチルス・リケニフォルミスゲノム中に存在する耐性遺伝子)。この耐性遺伝子は本発明及び染色体上統合されたカセット及び統合プラスミドの染色体増幅に関する実施態様において特に有用である。
【0060】
例えば、トリプトファニルtRNA合成酵素遺伝子(trpS)が不活性化される遺伝子である場合、DNA構築体は選択マーカーを挿入配列として含みその活性を阻害されるtrpS遺伝子を有する。選択マーカーはtrpSコード配列部分の内側に挿入する。DNA構築体は宿主染色体中のtrpS遺伝子に本質的に同一配列を有し、二重交差の場合、trpS遺伝子は選択マーカーの挿入により不活性化される。
【0061】
(欠失による不活性化)
いくつかの好ましい実施態様において、不活性化は欠失により達成される。いくつかの好ましい実施態様において、遺伝子は相同組換えにより欠失する。例えば、いくつかの実施態様において、欠失する遺伝子がtrpSの場合、相同ボックスの両側に隣接する選択マーカーを有する挿入配列を含むDNA構築体を用いる。相同ボックスは染色体trpS遺伝子の核酸フランキング領域に相同なヌクレオチド配列を含む。DNA構築体は微生物宿主染色体の相同配列と一致し、二重交差の場合、trpS遺伝子は宿主染色体から切除される。
【0062】
ここで用いる遺伝子の“欠失”の語は、コード配列全体の欠失、コード配列の一部の欠失、またはフランキング領域を含むコード配列の欠失をいう。欠失は染色体に残った配列が欠失前の生物活性を発揮しないのであれば部分的であってもよい。コード配列のフランキング領域は約1bp〜約500bpを5´及び3´末端に含むことができる。フランキング領域は500bpよりも大きくてもよいが、好ましくは本発明に従って不活性化または欠失され得るその他の遺伝子を当該領域に含まない。最終的には、欠失遺伝子は事実上、非機能的である。簡単に言えば、“欠失”の語は1以上のヌクレオチドまたはアミノ酸残基がそれぞれ除去された(すなわち、存在しない)、ヌクレオチドまたはアミノ酸配列の変化として定義される。
【0063】
ここで用いる“フランキング配列”とは対象の配列の上流または下流にある配列をいう(例えば、遺伝子A−B−Cは、遺伝子BはA及びC遺伝子配列にフランキング(側面に位置)している)。好ましい実施態様において、挿入配列は相同ボックスの両側の側面に位置する。いくつかの実施態様において、フランキング配列は一方の側(3´または5´)にのみ存在するが、好ましい実施態様において、配列の両側の側面に位置する。各相同ボックスの配列は微生物染色体中の配列に相同である。これらの配列は微生物染色体に新しい構築体が統合されること、及び微生物染色体の一部が挿入配列により置換されることを目的とする。好ましい実施態様において、選択マーカーの5´及び3´末端は不活性化染色体断片の一部を含むポリヌクレオチド配列の側面に位置する。
【0064】
(変異による不活性化)
別の実施態様において、不活性化は遺伝子の変異により生じる。遺伝子を変異する方法はさまざまな方法があり、限定されないが、部位特異的変異、ランダム変異発生、及びgapped−duplex法が挙げられる(例えばMoring et al.、Biotech.2:646[1984];及びKramer et al.、Nucleic Acids Res.,12:9441[1984]を参照)。
【0065】
好ましい実施態様において、変異は少なくとも染色体上の1つのコドンにおいて突然変異誘発を用いて生じる。さらなる実施態様において、変異DNA配列は野生型配列と40%以上、45%以上、50%以上、55%以上、60%以上、65%以上、70%以上、75%以上、80%以上、85%以上、90%以上、95%以上または98%以上の相同性を有する。別の実施態様において、変異DNAは公知の変異誘発手順、例えば、UV照射、ニトロソグアニジン等の化学変異剤を用いてin vivoで生じる。
【0066】
アミノアシルtRNA合成酵素活性に欠損を生じさせる染色体変異の内でも、アミノアシルtRNA合成酵素遺伝子自体の欠損を用いることが望ましい。しかしアミノアシルtRNA合成酵素生合成を調節する他の遺伝子も、対応する遺伝子を保有する染色体外遺伝子を用いる限り使用できる。アミノアシルtRNA合成酵素遺伝子の欠損株の場合、染色体外遺伝子に挿入されるべき遺伝子は該当するアミノアシルtRNA合成酵素遺伝子が相応しい。
【0067】
アミノアシルtRNA合成酵素を有する染色体外遺伝子と染色体の変異されたアミノアシルtRNA合成酵素遺伝子との間の相同性組換えの可能性を回避するためには、目的のアミノアシルtRNA合成酵素遺伝子を染色体から実質的に欠失させることが望ましい。
【0068】
本発明の宿主細胞Aを形質転換する際に用いられる組換えベクターは、上記した本発明の組換えベクターAと同様のものである。一方、本発明の宿主細胞Cを形質転換する際に用いられる組換えベクターは、上記した本発明の組換えベクターCと同様のものである。
【0069】
アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞は、アミノアシルtRNA合成酵素をコードする染色体遺伝子が発現できない、あるいは発現量が不十分であるため、そのままではもはや増殖できない。
【0070】
それに対して、アミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクター(本発明の組換えベクターAまたはC)を含むことで、増殖を維持できる。即ち、アミノアシルtRNA合成酵素の欠失は、染色体外遺伝子上に該アミノアシルtRNA合成酵素遺伝子を導入することにより相補することができる。しかし、もしも細胞が該染色体外遺伝子を失うと、該細胞はもはや増殖できなくなる。
【0071】
したがって、本発明の宿主細胞AまたはCは、上記関係の下に、培地に抗生物質や特定の栄養要求性を満たす栄養分を含めることなく組換えベクターAまたはCを安定して宿主細胞に維持することができる。そのため、本発明の宿主細胞AまたはCは、工業的スケールの生産に使用される任意の培地に適用できるという利点を有する。
【0072】
さらに、本発明の宿主細胞AおよびCは、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現させるために用いられる。本発明の宿主細胞Aは、前記アミノアシルtRNA合成酵素をコードする遺伝子およびγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有する組換えベクターを含むものであり、該組換えベクターを安定して宿主細胞に維持することができる。そして、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現させて、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素の生産に用いることができる。
【0073】
本発明の宿主細胞Bは、本発明の組換えベクターBを導入し形質転換して用いられる宿主細胞であり、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現させるために用いられる。
【0074】
本発明の宿主細胞は細菌であればよく、好ましいのは枯草菌であり、さらに好ましくは枯草菌のISW1214株、BD170株、または168株等である。しかし、本発明の組換えベクターを安定に保持し、複製することができるものであれば、枯草菌以外のバチルス(Bacillus)属細菌などを使用してもよい。
【0075】
本明細書でいう「バチルス属」とは、通常知られるバチルス属に含まれるすべての種類を含み限定されないが、たとえば、バチルス・ズブチリス(Bacillus subtilis)、バチルス・リケニフォルミス(Bacillus licheniformis)、バチルス・レンタス(Bacillus lentus)、バチルス・ステアロサーモフィラス(Bacillus stearothermophilus)、バチルス・アルカロフィラス(Bacillus alkalophilus)、バチルス・アミロリケファシエンス(Bacillus amyloliquefaciens)、バチルス・セレウス(Bacillus cereus)、バチルス・プミルス(Bacillus pumilus)、バチルス・クラウジイ(Bacillus clausii)、バチルス・ハロデュランス(Bacillus halodurans)、バチルス・メガテリウム(Bacillus megaterium)、バチルス・コアグランス(Bacillus coagulans)、バチルス・サーキュランス(Bacillus circulans)、およびバチルス・チューリンゲンシス(Bacillus thuringiensis)などを意味する。なお、バチルス属は分類上再編成を受け続けており、当該属は再分類された種も含むものとする。たとえば、ジオバチルス(Geobacillus)属、アルカリバチルス(Alkalibacillus)属、アンフィバチルス(Amphibacillus)属 、アミロバチルス(Amylobacillus)属、アノキシバチルス(Anoxybacillus)属 、ゴリバチルス(Goribacillus)属、セラシバチルス(Cerasibacillus)属、グラシリバチルス(Gracilibacillus)属、ハロバチルス(Halolactibacillus)属、ハロアルカリバチルス(Halalkalibacillus)属、フィロバチルス(Filobacillus)属、ジョーガリバチルス(Jeotgalibacillus)属、サリバチルス(Salibacillus)属、オーシャノバチルス(Oceanobacillus)属、マリニバチルス(Marinibacillus)属、リシニバチルス(Lysinibacillus)属、レンチバチルス(Lentibacillus)属、ウレーバチルス(Ureibacillus)属、サリニバチルス(Salinibacillus)属、ポンチバチルス(Pontibacillus)属、ピシバチルス(Piscibacillus)属、パラリオバチルス(Paraliobacillus)属、ヴァージバチルス(Virgibacillus)属、サルシューギニバチルス(Salsuginibacillus)属、テニューイバチルス(Tenuibacillus)属、タラソバチルス(Thalassobacillus)属、サームアルカリバチルス(Thermalkalibacillus)属、チューメバチルス(Tumebacillus)属などがあり、これらもまた本明細書にいうバチルス属に含まれるものとする。
【0076】
本発明の組換えベクターを宿主細胞に導入する方法はとくに限定されないが、たとえば、コンピテントセル法、プロトプラスト法、エレクトロポレーション法、カルシウムイオン法、リポフェクション法等を用いることができる。宿主細胞が枯草菌である場合は、コンピテントセル法またはプロトプラスト法が好ましい。
【0077】
(C)γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法。
【0078】
本発明のγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法の一つの態様は、下記(i)〜(iv)の工程を有する方法(以下、「酵素生産方法A」と呼ぶ)である:
(i)γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、さらにアミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターを用意する工程;
(ii)アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異宿主細胞を用意する工程;
(iii)前記変異宿主細胞を前記組換えベクターで形質転換して形質転換体を得る工程;および
(iv)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
【0079】
本発明のγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法の一つの態様は、下記(i)〜(iii)の工程を有する方法(以下、「酵素生産方法B」と呼ぶ)である:
(i)下記[a]〜[c]の塩基配列または遺伝子を有する組換えベクターを用意する工程、
[a]配列番号1に記載の塩基配列、
[b]前記[a]の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子、および
[c]前記[b]の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;
(ii)前記組換えベクターで宿主細胞を形質転換して形質転換体を得る工程;および
(iii)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
【0080】
本発明のγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法の一つの態様は、下記(i)〜(iv)の工程を有する方法(以下、「酵素生産方法C」と呼ぶ)である:
(i)下記[a]〜[d]の塩基配列または遺伝子を有する組換えベクターを用意する工程、
[a]配列番号1に記載の塩基配列、
[b]前記[a]の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子、
[c]前記[b]の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子、および
[d]発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子;
(ii)アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞を用意する工程;
(iii)前記宿主細胞を前記組換えベクターで形質転換して形質転換体を得る工程;および
(iv)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
【0081】
酵素生産方法Aにおける組換えベクターを用意する工程では、アミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で挿入し、かつ、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で挿入した組換えベクターを用意する。さらに、酵素生産方法Cにおける組換えベクターを用意する工程では、アミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で挿入し、かつ、配列番号1に記載の塩基配列、その塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子およびその遺伝子の下流に直接的に連結されたγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を挿入した組換えベクターを用意する。上記組換えベクターは、先に説明した本願発明の組換えベクターAまたはCである。一方、酵素生産方法Bにおける組換えベクターを用意する工程では、配列番号1に記載の塩基配列、その塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子およびその遺伝子の下流に直接的に連結されたγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を挿入した組換えベクターを用意する。
【0082】
遺伝子の挿入は常法によって行うことができ、たとえば、Molecular Cloning (1989) (Cold Spring Harbor Lab.)に記載される方法に従って行うことができる。いずれの遺伝子も発現可能な状態で挿入する。そのため、各々の遺伝子の組換えベクターへの挿入位置は組換えベクターの複製に関与していない場所であれば基本的にはどこでも良く、使用するベクターによって挿入位置は異なる。使用できる組換えベクターは上記に挙げた通りであり、たとえば、枯草菌由来のプラスミド(例えばpUB110、pTP5など);pHY300PLKなどの大腸菌−枯草菌シャトルベクターを挙げることができる。
【0083】
例えば大腸菌−枯草菌のシャトルベクターであるpHY300PLKプラスミドを例に取れば、マルチクローニング部位であるEcoRI−BamHI間に配列番号1のDNA断片と配列番号2のアミノ酸配列をコードするDNA断片を連結させたDNA断片を、EcoRI部位にアミノアシルtRNA合成酵素遺伝子を、同じくマルチクローニング部位BamHI部位にγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を挿入することなどが可能である。以下に、本発明の組換えベクターCを用意する工程を簡単に説明する。
【0084】
配列番号1のDNA断片と配列番号2のアミノ酸配列をコードするDNA断片を連結させたDNA断片をpHY300PLKのEcoRI−BamHI間に挿入したプラスミドを構築する。次に、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を有する生物の染色体DNAを鋳型にPCR増幅を用いて該酵素の活性や安定性に欠かせない領域をコードするDNA断片を増幅し、これを上記した構築済みプラスミドを制限酵素BamHIで消化することによって得られたDNA 断片とライゲーション反応を行い、得られたライゲーション反応混合物を用いて大腸菌を形質転換してpHY300PLKのBamHI部位に目的の通りプロモーター部位から終止コドンまでを含む該遺伝子が挿入されているプラスミドを含む形質転換体を選択し、この形質転換体からプラスミドを調製する。続いて上記の通り調製できたプラスミドを制限酵素EcoRIで消化し、本プラスミドを切断する。枯草菌168(Bacillus subtilis 168)株の染色体DNAを鋳型にPCR増幅を用いてプロモーター部位から終止コドンまでを含むアミノアシルtRNA合成酵素遺伝子を増幅し、EcoRIで切断された構築済みプラスミド由来DNA断片とライゲーション反応を行い、得られたライゲーション反応混合物を用いて大腸菌を形質転換して構築済みプラスミドのEcoRI部位に目的の通りアミノアシルtRNA合成酵素遺伝子が挿入されているプラスミドを含む形質転換体を選択し、この形質転換体から目的プラスミドを調製することができる。
【0085】
酵素生産方法AおよびCにおける宿主細胞を用意する工程では、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞を用意する。この宿主細胞は、前記で説明した本発明の宿主細胞AまたはCである。その調製方法は、以下のとおりである。
【0086】
宿主細胞の調製の手順としては、宿主染色体上に存在する標的遺伝子を計画的に削除又は不活性化する方法のほか、ランダムな遺伝子の削除又は不活性化変異を与え、その後適当な方法により遺伝子解析またはタンパク質生産性の評価を行う方法が挙げられる。
【0087】
標的とする遺伝子を削除又は不活性化するには、例えば相同組換えによる方法を用いればよい。すなわち、標的遺伝子の一部を含むDNA断片を適当なプラスミドベクターにクローニングして得られる環状の組換えプラスミドまたは直鎖上のDNA断片を宿主細胞内に取り込ませ、標的遺伝子の一部領域に於ける相同組換えによって親微生物ゲノム上の標的遺伝子を分断して不活性化することが可能である。あるいは、塩基置換や塩基挿入等による不活性化変異を導入した標的遺伝子、又は標的遺伝子の外側領域を含むが標的遺伝子を含まない直鎖状のDNA断片等をPCR等の方法によって構築し、これを親微生物細胞内に取り込ませて親微生物ゲノムの標的遺伝子内の変異箇所の外側2ヶ所、又は標的遺伝子外側の2ヶ所の領域で2回交差の相同組換えを起こさせることにより、ゲノム上の標的遺伝子を削除或いは不活性化した遺伝子断片と置換することが可能である。
【0088】
本発明の酵素生産方法の形質転換体を得る工程は、上記した組換えベクターを用意する工程で調製した組換えベクターを用いて、宿主細胞または上記した宿主細胞を得る工程で調製した宿主細胞を形質転換する。組換えベクターによる宿主細胞の形質転換も常法により実施できる。
【0089】
組換えベクターの宿主細胞への導入(形質転換)方法は、特に制限されず、トランスフォーメーション法、トランスフェクション法、コンピテント細胞法、エレクトロポレーションなど、導入する宿主細胞の種類や組換えベクターの形態に応じて、適宜選択することができる。
【0090】
ここで用いる、“宿主細胞”とは新しく導入するDNA配列のための宿主または発現媒体として作用する能力を有する細胞をいう。
【0091】
なお、組換えベクターの宿主細胞内での存在様式は、特に制限されず、染色体中に挿入されて、あるいは置換されて組み込まれてもよいし、またプラスミド状態で存在していてもよい。
【0092】
なお、本発明の組換えベクターAまたはCに含まれるアミノアシルtRNA合成酵素をコードする染色体遺伝子は、本発明の宿主細胞AまたはCにおいて、欠損させられたアミノアシルtRNA合成酵素をコードする染色体遺伝子、または宿主細胞が成育できない程度に発現を低下させられたアミノアシルtRNA合成酵素をコードする染色体遺伝子と同種の遺伝子である。例えば、本発明の宿主細胞Aにおいて、欠損させられたアミノアシルtRNA合成酵素をコードする染色体遺伝子、または宿主細胞が成育できない程度に発現を低下させられたアミノアシルtRNA合成酵素をコードする染色体遺伝子が、アラニルtRNA合成酵素遺伝子である場合、本発明の組換えベクターAに含まれるのもアラニルtRNA合成酵素遺伝子である。
【0093】
本発明の酵素生産方法におけるγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程では、上記した形質転換体を得る工程で得られる形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子がコードする酵素を生産する。
【0094】
形質転換体の培養は、宿主細胞の種類に応じて、適切な培地を用い、適切な培養条件の下で実施される。宿主細胞としては、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子がコードする酵素の生産が可能なものであればよく、野生型のものでも変異を施したものでものよい。好ましくは、枯草菌などのバチルス属細菌である。
【0095】
例えば、宿主細胞が枯草菌の場合、培養に用いる培地の種類としては、適当な窒素源、炭素源、ミネラルを含み、本発明の枯草菌宿主が生育し、目的タンパク質を生産することができるものであればよい。例えば、組換え枯草菌168株によって、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子がコードする酵素を生産する場合、炭素源として、ぶどう糖や果糖等の単糖類、しょ糖、麦芽糖等の二糖類又は可溶性澱粉等の多糖類等を配合した培地や、窒素源として、ペプトン類、大豆エキス、酵母エキス、魚肉エキス、コーンスティープリカー( C S L )、金属塩等を配合した培地等を用いることができる。
【0096】
また、培地のpH は、用いる組換え生物体が生育しうる範囲のpHであれば良いが、例えば枯草菌の場合pH 6.0〜8.0に調整するのが好適であり、培養条件は、15〜42℃ 、好ましくは28〜37℃で2〜7日間振盪または、通気撹拌培養すればよい。
【0097】
このようにして得られた培養物から、本発明の組換えベクターに挿入された酵素遺伝子が発現した組換えタンパク質を採取する。該組換えタンパク質は通常形質転換体外に蓄積される。そこで、形質転換体外に蓄積された該組換えタンパク質を培養上清から一般のタンパク質の採取の手段に準じて採取する。たとえば、通常知られる手段によって形質転換体を除いた後に培養上清を組換えタンパク質含有物として用いることができる。
【0098】
ただし、宿主細胞の種類によっては組換えタンパク質を形質転換体内または形質転換体の細胞膜内に蓄積する場合もあり得る。その場合は、特に限定はされないが、たとえば、有機溶剤やリゾチームのような酵素によって形質転換体を溶解する方法、および、超音波破砕法、フレンチプレス法、ガラスビーズ破砕法、ダイノミル破砕法等の細胞破砕法で得られた形質転換体の細胞破砕物および/または培養物を遠心分離法、ろ過法等の操作によって形質転換体と培養上清に分離する。このようにして得られた培養上清を、組換えタンパク質含有物として用いることができる。また、分離した形質転換体をそのまま組換えタンパク質含有物とすることもできる。
【0099】
上記組換えタンパク質含有物は、そのままで使用することもできるが、必要に応じて、たとえば塩析法、沈澱法、透析法、限外濾過法等の通常知られる方法を単独または組み合わせることにより工業用途の濃縮組換えタンパク質含有物を調製できる。
【0100】
さらに前記濃縮組換えタンパク質含有物を、たとえば、イオン交換クロマトグラフィー、等電点クロマトグラフィー、疎水性クロマトグラフィー、ゲル濾過クロマトグラフィー、吸着クロマトグラフィー、アフィニティークロマトグラフィー、逆相クロマトグラフィー、レジンカラム法等の通常知られる単離・精製法の組合せに供すことにより、精製組換えタンパク質を得ることができる。
【実施例】
【0101】
以下、本発明を実施例によりさらに詳細に説明する。
【0102】
[実施例1]枯草菌へのトリプトファニルtRNA合成酵素遺伝子を有するプラスミドの導入
トリプトファニルtRNA合成酵素遺伝子を保有する染色体外遺伝子であるプラスミドpDATS14を以下の方法で構築した。まず、汎用枯草菌−大腸菌のシャトルプラスミドであるpHY300PLK(ヤクルト社製)を鋳型として、プライマーA(配列番号34)とB(配列番号35)を用いてPCRによる増幅を行った。得られた増幅断片を制限酵素XhoIで消化した後、両末端を連結することにより環状プラスミドとした。本プラスミドを用いて大腸菌HB101株を形質転換した。形質転換体からプラスミドを調製しpDA2とした。一方、枯草菌ISW1214(Bacillus subtilis ISW1214)株の染色体を鋳型とし、プライマーC(配列番号36)とD(配列番号37)を用いてトリプトファニルtRNA合成酵素遺伝子とその近傍の領域を含むDNA断片を得た。本DNA断片を制限酵素XhoIで消化し、予め制限酵素XhoIで消化したpDA2と連結した。これを用いて大腸菌HB101株を形質転換した。得られた形質転換体からプラスミドを調製し、pDATS14と命名した。次にpDATS14を用いて、2種のプロテアーゼ(アルカリプロテアーゼE、中性プロテアーゼE)に欠陥を有する枯草菌ISW1214株由来変異株(参考文献 Appl.Microbiol. Biotechnol 65:583-592 (2004)Hatada,Y.ら)を、プロトプラスト形質転換法(参考文献Mol. Gen. Genet. 168:111-115 (1979) Chang, S.とCohen, S. N.)により形質転換し、染色体外にトリプトファニルtRNA合成酵素遺伝子を保有する枯草菌形質転換体を得た。
【0103】
[実施例2]枯草菌の染色体トリプトファニルtRNA合成酵素遺伝子を破壊するためのDNA断片の構築
枯草菌ISW1214株の染色体DNAを鋳型とし、プライマーE(配列番号38)とF(配列番号39)を用いてトリプトファニルtRNA合成酵素遺伝子とその近傍の領域を含む約2.5kbのDNA断片を得た。これを制限酵素EcoRIおよびSalIで消化した。EcoRIおよびSalIで予め消化した汎用性プラスミドpUC18に連結し、プラスミドpTSAFを得た。一方、汎用性プラスミドであるpC194を鋳型として、プライマーG(配列番号40)とH(配列番号41)を用いてスタフィロコッカス由来クロラムフェニコール耐性遺伝子とその近傍領域を含む約1.6kbのDNA断片を得た。さらに、pTSAFを鋳型としてプライマーI(配列番号42)とJ(配列番号43)を用いてPCR反応を行いDNA断片を得た。本DNA断片とクロラムフェニコール耐性遺伝子とその近傍領域を含む約1.6kbのDNA断片を制限酵素BamHIおよびXbaIで消化した後連結し、大腸菌HB101株を形質転換した。形質転換体からプラスミドを調製し、トリプトファニルtRNA合成酵素遺伝子破壊用プラスミドpINTTSとした。pINTTSを鋳型としてプライマーK(配列番号44)とL(配列番号45)を用いてPCR反応を行い直鎖状のトリプトファニルtRNA合成酵素遺伝子破壊用DNA断片を得た。
【0104】
[実施例3]枯草菌の染色体トリプトファニルtRNA合成酵素遺伝子の破壊
上記トリプトファニルtRNA合成酵素遺伝子破壊用DNA断片を用いて、コンピテントセル法を用いて実施例1で得た染色体外にトリプトファニルtRNA合成酵素遺伝子を保有する形質転換体をさらに形質転換し、クロラムフェニコール耐性を獲得した形質転換体を得た。得られた形質転換体から染色体DNAを調製した。本染色体DNAに対して、プライマーM(配列番号46)とN(配列番号47)、あるいはO(配列番号48)とP(配列番号49)を用いてPCR反応を行い、染色体のトリプトファニルtRNA合成酵素遺伝子がコードされていた領域がトリプトファニルtRNA合成酵素遺伝子破壊用DNA断片由来のクロラムフェニコール耐性遺伝子と置換されていることを確認し、本菌株をトリプトファニルtRNA合成酵素遺伝子欠損株であるDTS1451(pDATS14)株とした。
【0105】
[実施例4]DTS1451株へのカナマイシン耐性遺伝子を有するプラスミドの導入
枯草菌ISW1214株の染色体を鋳型とし、プライマーCとDを用いてトリプトファニルtRNA合成酵素遺伝子とその近傍の領域を含むDNA断片を得た。本DNA断片を制限酵素XhoIで消化し、予め制限酵素XhoIで消化したpDA2と連結した。これを用いて、大腸菌HB101株を形質転換した。得られた形質転換体からプラスミドを調製し、pDATS13と命名した。汎用性プラスミドpUB110を鋳型とし、プライマーQ(配列番号50)とR(配列番号51)を用いてカナマイシン耐性遺伝子とその近傍の領域を含むDNA断片を得た。本DNA断片をEcoRIで消化し、pDATS14を鋳型としてプライマーS(配列番号52)とT(配列番号53)を用いてPCRで増幅して、さらにEcoRIで消化することにより得られたDNA断片と連結した。これを用いて、大腸菌HB101株を形質転換した。得られたカナマイシン耐性を有する形質転換体からプラスミドを調製し、プラスミドをpDATSKと命名した。pDATSKを用いてpDATS14を有するDTS1451株をプロトプラスト形質転換法を用いて形質転換し、カナマイシン30μg/mLを含む再生培地(組成:コハク酸ナトリウム 8%、寒天 1%、カザミノ酸 0.5%、酵母エキス 0.5%、リン酸2水素カリウム 0.15%、リン酸水素2カリウム 0.35%、グルコース 0.5%、硫酸マグネシウム 0.4%、牛血清アルブミン 0.01%、メチオニン 0.001%、およびロイシン 0.001%)による選抜を行った。得られた形質転換体をカナマイシン入りLB寒天培地およびテトラサイクリン7.5μg/mLを含む再生培地に植菌し、カナマイシン耐性かつテトラサイクリン感受性となった株をDTS1451(pDATSK)と命名した。
【0106】
[実施例5]遺伝子調節領域DNA断片の探索用プラスミドの構築
遺伝子調節領域DNA断片検索用ベクターpPTCFを以下の方法で構築した。
寄託番号がFERM BP−8320である、マイクロバルバイファー・エスピー JAMB−A7(Microbulbifer sp. JAMB−A7)株由来の染色体DNAを鋳型に用いて、プライマーU(配列番号54)およびプライマーW(配列番号55)を用いてPCRによる増幅を行った。得られたDNA断片を制限酵素、EcoRIで消化した後、予めEcoRIで消化したpHY300PLKと連結することにより環状プラスミドを構築した。本プラスミドを用いて大腸菌HB101株を形質転換した。形質転換体からプラスミドを調製し、寒天分解酵素遺伝子の上流にpHY300PLK由来のポリリンカー部位を有するプラスミドをpPTCFと命名した。
【0107】
[実施例6]遺伝子調節領域DNA断片の取得
Sau3AIを用いて、寄託番号がFERM AP−20227である、バチルス・エスピー JAMB750(Bacillus sp. JAMB750)株の染色体DNAを消化後、これをBamHIで切断したプラスミドpPTCFと混ぜてT4 DNAリガーゼを用いてライゲーション反応を行った。得られたライゲーション液を用いて枯草菌を形質転換した。再生培地にはDM3培地(組成:コハク酸ナトリウム 8%、寒天 1%、カザミノ酸 0.5%、酵母エキス 0.5%、リン酸2水素カリウム 0.15%、リン酸水素2カリウム 0.35%、グルコース 0.5%、硫酸マグネシウム 0.4%、牛血清アルブミン 0.01%、メチオニン 0.001%、ロイシン 0.001%、テトラサイクリン 7.5μg/ml)を用いた。形質転換体の周囲に寒天の窪みを形成した形質転換体を選び、約2,000個の形質転換体を液体培地にて培養(培地組成はポリペプトンS 3%、魚肉エキス 0.5%、酵母エキス 0.05%、リン酸2水素カリウム 0.1%、マルトース 4%、硫酸マグネシウム 0.02%、塩化カルシウム 0.05%、テトラサイクリン 7.5μg/ml、30℃で48時間振とう)した。得られた培養液毎に超音波破砕(Handy sonic model UR−20P、TOMY SEIKO CO.使用)を行い、取得された超音波破砕物の寒天分解酵素活性を測定した。その中で最も活性の高かったものを選択し、この形質転換体の持つプラスミドをpCDAG1と命名した。
【0108】
[実施例7]プラスミドpJEXOPT2の構築
寄託番号がDSM17297である、サラッソモナス・エスピー JAMB−A33(Thalassomonas sp. JAMB−A33)株由来の染色体DNAを鋳型にプライマーX(配列番号56)とプライマーY(配列番号57)を用いてPCR増幅を行い約0.3kbpから成るDNA断片を取得した。このDNA断片を制限酵素HindIIIで切断した。続いて枯草菌−大腸菌のシャトルプラスミドであるpHY300PLK(ヤクルト社製)を制限酵素HindIIIで切断し、上記のDNA断片とリガーゼ反応により連結した。本プラスミドを用いて大腸菌HB101株を形質転換した。形質転換体からプラスミドを調製しpHYTERと命名した。
【0109】
プラスミドpJEXOPT2を以下の方法で構築した。プラスミドpCDAG1を鋳型にしてリン酸化プライマーZ(配列番号58)およびリン酸化プライマーA1(配列番号59)を用いてPCRによる増幅を行った。得られたPCR断片をpUC18のSmaI部位に導入した。構築されたプラスミドを鋳型にして、プライマーZおよびプライマーA1を用いてDiversify PCR Random Mutagenesis Kit( クロンテック社)を用い、キットのプロトコールにしたがって、PCR増幅断片中にランダムに変異を導入した。このランダム変異操作を5回繰り返した。得られたPCR産物を鋳型にプライマーZおよびリン酸化プライマーA1を用いてPCR増幅を行った。その後T4DNAポリメラーゼを用いてPCR産物の末端の平滑化を行った。一方、合成一本鎖DNA断片A(配列番号74)と合成一本鎖DNA断片B(配列番号75)とをアニーリングさせ二本鎖DNAを形成させた。得られたDNA断片をリン酸化した後に、上記のPCR増幅断片とリガーゼ反応により結合した。得られたDNA断片を制限酵素EcoRI、BamHIで消化した後、EcoRIとBamHIで消化したpHYTERと連結することにより環状プラスミド群を構築した。本プラスミド群を用いて大腸菌HB101株を形質転換した。得られた数千あまりの形質転換体からまとめてプラスミド群を調製した。これをプラスミド群Aとした。
【0110】
次にβアガラーゼ生産菌である、寄託番号がFERM BP−8321である、マイクロバルバイファー・エスピー JAMB−A94(Microbulbifer sp. JAMB−A94)株の染色体を鋳型とし、プライマーB1(配列番号60)およびC1(配列番号61)を用いてβアガラーゼ酵素遺伝子DNA断片をPCR増幅した。これを制限酵素BamHIで消化した。プラスミド群Aを制限酵素BamHIで切断し、リガーゼによってβアガラーゼ酵素遺伝子と連結した。これらを大腸菌HB101株を形質転換した。βアガラーゼ活性を示す形質転換体からプラスミドを調製し、これらのプラスミドを用いて枯草菌ISW1214株を形質転換し、テトラサイクリン入り再生培地による選抜を行った。得られた形質転換体を液体培地(ポリペプトンS 3%、魚肉エキス 0.5%、酵母エキス 0.05%、リン酸水素カリウム 0.1%、マルトース 4%、塩化マグネシウム 0.02%、塩化カルシウム 0.05%、テトラサイクリン 15μg/ml)中で30℃、72時間、130rpmにて撹拌培養を行い、得られた培養上清に含まれるβアガラーゼ活性を測定した。その結果、著量のβアガラーゼを生産できるプラスミドpJEXOPT2を取得できた。
【0111】
[実施例8]発現ベクターによるγ−CGTaseの分泌生産
γ−CGTase生産菌であるバチルス・クラーキー 7364(Bacillus clarkii 7364)株由来のγ−CGTase遺伝子(J Biochem(Tokyo). 2003 133:317-324. Takada, M.)をコードする遺伝子配列を含むプラスミドpGFT−01(特許公開番号2003−102489)を鋳型として、プライマーD1(配列番号62)およびE1(配列番号63)を用いてPCR反応を行い、該酵素をコードするDNA断片を得て、これを制限酵素BamHIとSalIで消化した。pJEXOPT2を制限酵素BamHIとSalIで切断し、プラスミドのオリジンを有する方のDNA断片と、リガーゼによってγ−CGTase遺伝子と連結した。これらを大腸菌HB101株に導入して形質転換させた。γ−CGTase活性を示す形質転換体からプラスミドを調製し、pJEXOPT2Gと命名した。次に、枯草菌ISW1214株の染色体を鋳型とし、プライマーF1(配列番号64)とG1(配列番号65)を用いてトリプトファニルtRNA合成酵素遺伝子とその近傍の領域を含むPCR増幅DNA断片を得た。本DNA断片を制限酵素EcoRIで消化し、予め制限酵素EcoRIで消化したpJEXOPT2Gと連結した。これを用いて大腸菌HB101株を形質転換した。得られた形質転換体からプラスミドを調製し、pJEXOPT2GSと命名した。pJEXOPT2GSを用いてDTS1451(pDATSK)を形質転換し、テトラサイクリン 7.5μg/mL再生培地による選抜を行った。得られた形質転換体をカナマイシン入りLB寒天培地およびテトラサイクリン 7.5μg/mLを含む再生培地に植菌し、γ−CGTase活性を示し、さらにカナマイシン感受性かつテトラサイクリン耐性となった株をDTS1451(pJEXOPT2GS)と命名した。DTS1451(pJEXOPT2GS)を液体培地中(ポリペプトンS 5%、魚肉エキス 0.5%、酵母エキス 0.05%、リン酸水素カリウム 0.1%、マルトース 4%、塩化マグネシウム 0.01%、塩化カルシウム 0.1%、抗生物質を含有しない)で30℃、96時間、130rpmにて撹拌培養を行い、得られた培養上清に含まれるγ−CGTase活性を測定した。その結果、培養液1Lあたり約1.4gのγ−CGTase酵素の生産が確認できた。
【0112】
[実施例9]発現ベクターによるマルトースホスホリラーゼ生産
マルトースホスホリラーゼ生産菌である寄託番号FERM−BP8420のパエニバチルス・エスピーSH−55(Paenibacillus sp. SH−55)株由来のマルトースホスホリラーゼ遺伝子を含む組換え体プラスミドpRSMP1(国際公開番号WO2005/00343号公報記載)を組換え大腸菌からGFXPlasmid prep Kit(GEヘルスケアバイオサイエンス株式会社)を用いることにより精製した。得られた組換え体プラスミドを鋳型として、プライマーH1(配列番号66)とプライマーI1(配列番号67)を用い、PCRを行い、マルトースホスホリラーゼ遺伝子を含むDNA断片を取得した。これを制限酵素BamHIとXhoIで消化した。pJEXOPT2を制限酵素BamHIとSalIで切断し、プラスミドのオリジンを有する方のDNA断片とリガーゼによってマルトースホスホリラーゼ遺伝子と連結した。これらを大腸菌HB101株を形質転換した。マルトースホスホリラーゼ活性を示す形質転換体からプラスミドを調製し、pJEXOPT2Mと命名した。枯草菌株ISW1214株の染色体を鋳型とし、プライマーJ1(配列番号68)とK1(配列番号69)を用いてトリプトファニルtRNA合成酵素遺伝子とその近傍の領域を含むPCR増幅DNA断片を得た。本DNA断片を制限酵素EcoRIで消化し、予め制限酵素EcoRIで消化したpJEXOPT2Mと連結した。これを用いて大腸菌HB101株を形質転換した。得られた形質転換体からプラスミドを調製し、pJEXOPT2MSと命名した。pJEXOPT2MSを用いてDTS1451(pDATSK)を形質転換し、テトラサイクリン 7.5μg/mL再生培地による選抜を行った。得られた形質転換体をカナマイシン入りLB寒天培地およびテトラサイクリン 7.5μg/mLを含む再生培地に植菌し、マルトースホスホリラーゼ活性を示し、さらにカナマイシン感受性かつテトラサイクリン耐性となった株をDTS1451(pJEXOPT2MS)と命名した。DTS1451(pJEXOPT2MS)を液体培地中(ポリペプトンS 5%、魚肉エキス 0.5%、酵母エキス 0.05%、リン酸水素カリウム 0.1%、マルトース 4%、塩化マグネシウム 0.01%、塩化カルシウム 0.1%、抗生物質を含有しない)で30℃、96時間、130rpmにて撹拌培養を行い、得られた培養上清に含まれるマルトースホスホリラーゼ活性を測定した。その結果、培養液1Lあたり約0.5gのマルトースホスホリラーゼの生産が確認できた。
【0113】
[実施例10]発現ベクターによるトレハロースホスホリラーゼ生産
トレハロースホスホリラーゼ生産菌であるパエニバチルス・エスピーSH−55株由来のトレハロースホスホリラーゼ遺伝子を含む組換え体プラスミドpRSTP1(国際公開番号WO2005/00343号公報記載)を組換え大腸菌からGFXPlasmid prep Kit(GEヘルスケアバイオサイエンス株式会社)を用いることにより精製した。得られた組換え体プラスミドを鋳型として、プライマーL1(配列番号70)とプライマーM1(配列番号71)を用い、PCRを行い、トレハロースホスホリラーゼ遺伝子を含むDNA断片を取得した。これを制限酵素BamHIとXhoIで消化した。pJEXOPT2を制限酵素BamHIとSalIで切断し、プラスミドのオリジンを有する方のDNA断片とリガーゼによってトレハロースホスホリラーゼ遺伝子と連結した。これらを大腸菌HB101株を形質転換した。トレハロースホスホリラーゼ活性を示す形質転換体からプラスミドを調製し、pJEXOPT2Tと命名した。枯草菌ISW1214株の染色体を鋳型とし、プライマーN1(配列番号72)とO1(配列番号73)を用いてトリプトファニルtRNA合成酵素遺伝子とその近傍の領域を含むPCR増幅DNA断片を得た。本DNA断片を制限酵素EcoRIで消化し、予め制限酵素EcoRIで消化したpJEXOPT2Tと連結した。これを用いて大腸菌HB101株を形質転換した。得られた形質転換体からプラスミドを調製し、pJEXOPT2TSと命名した。pJEXOPT2TSを用いてDTS1451(pDATSK)を形質転換し、テトラサイクリン 7.5μg/mL再生培地による選抜を行った。得られた形質転換体をカナマイシン入りLB寒天培地およびテトラサイクリン 7.5μg/mLを含む再生培地に植菌し、トレハロースホスホリラーゼ活性を示し、さらにカナマイシン感受性かつテトラサイクリン耐性となった株をDTS1451(pJEXOPT2TS)と命名した。DTS1451(pJEXOPT2TS)を液体培地中(ポリペプトンS 5%、魚肉エキス 0.5%、酵母エキス 0.05%、リン酸水素カリウム 0.1%、マルトース 4%、塩化マグネシウム 0.01%、塩化カルシウム 0.1%、抗生物質を含有しない)で30℃、96時間、130rpmにて撹拌培養を行い、得られた培養上清に含まれるトレハロースホスホリラーゼ活性を測定した。その結果、培養液1Lあたり約1.2gのトレハロースホスホリラーゼの生産が確認できた。
【図面の簡単な説明】
【0114】
【図1】γ−シクロデキストリン合成酵素(γ−CGTase)発現ベクターの具体例。
【図2】マルトースホスホリラーゼ発現ベクターの具体例。
【図3】トレハロースホスホリラーゼ発現ベクターの具体例。
【技術分野】
【0001】
本発明は、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を工業的に生産する方法に関する。さらに詳細には、本発明は、この方法に使用する、転写を調節する塩基配列、シグナルペプチドをコードする遺伝子、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼおよびトレハロースホスホリラーゼからなる群から選択される1種の酵素をコードする遺伝子、およびアミノアシルtRNA合成酵素をコードする遺伝子を含む組換えベクター、該組換えベクターを導入した宿主細胞、ならびにγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法に関する。
【背景技術】
【0002】
γ−シクロデキストリンの合成には、γ−シクロデキストリンを主生成物とするシクロデキストリン・グルカノトランスフェラーゼ(以下、「γ−シクロデキストリン合成酵素」と呼ぶ)を澱粉等に作用させる方法が知られている(たとえば、特許文献1を参照)。特許文献1によれば、外来遺伝子を含む組換えベクターを宿主細胞に導入する宿主−ベクター系を利用してγ−シクロデキストリン合成酵素を取得している。具体的には、γ−シクロデキストリン合成酵素をコードする遺伝子をアンピシリン耐性遺伝子を含むT−ベクターにライゲーションし、大腸菌(Escherichia coli)を形質転換して培養することにより、γ−シクロデキストリン合成酵素を取得している。
【0003】
一方、トレハロースの合成には、マルトースホスホリラーゼおよびトレハロースホスホリラーゼをマルトースに作用させる方法が知られている(たとえば、特許文献2を参照)。特許文献2でも宿主−ベクター系を利用してマルトースホスホリラーゼおよびトレハロースホスホリラーゼを取得している。具体的には、マルトースホスホリラーゼおよびトレハロースホスホリラーゼをコードする遺伝子を適当なベクターに挿入し、大腸菌または枯草菌(Bacillus subtilis)を形質転換してアンピシリンまたはテトラサイクリンなどを含む培地で培養することにより、マルトースホスホリラーゼおよびトレハロースホスホリラーゼを取得している。
【0004】
上記のγ−シクロデキストリン合成酵素の生産方法とマルトースホスホリラーゼおよびトレハロースホスホリラーゼの生産方法は、大腸菌を宿主細胞とする宿主−ベクター系を利用している(たとえば、特許文献1および2を参照)。大腸菌は、世代時間が約20分と短く、様々な糖類を資化して増殖することができる。さらに大腸菌に適した多くのプラスミドベクターも開発されており、大腸菌を宿主細胞とする宿主−ベクター系は、実験室での組換えタンパク質の生産において一応の成果を期待できる。
【0005】
しかしながら、大腸菌の組換えタンパク質を菌体内に蓄積するという性質のために、大腸菌自体の生育が阻害され、さらに菌体内のタンパク質分解酵素の作用により組換えタンパク質の機能が失いやすいという、組換えタンパク質の大量かつ安定に生産する上で不利な点があった。さらに、組換えタンパク質を取得するためには菌体を回収して破砕し、かつ、発熱性物質の一種であるリポポリサッカライドを注意深く除去しなければならない。このために、組換えタンパク質の採取・精製のための工程が頻雑になるという問題も生じていた。
【0006】
そこで、これらの問題を解決する手段として、枯草菌を利用した宿主−ベクター系が知られている(たとえば、特許文献2および非特許文献3を参照)。特許文献2によれば、菌体内よりも菌体外の培養上清中に多くのマルトースホスホリラーゼおよびトレハロースホスホリラーゼを得ている。しかし、取得される酵素量は少なく、大量生産にまでは至っていない。
【0007】
一方、上記のγ−シクロデキストリン合成酵素の生産方法とマルトースホスホリラーゼおよびトレハロースホスホリラーゼの生産方法は、それぞれの酵素遺伝子を抗生物質耐性遺伝子とともに挿入した組換えベクターを利用している。これにより、抗生物質耐性遺伝子に対応する抗生物質を培養培地中に加えることで、抗生物質耐性遺伝子含有組換えベクターを維持する細胞は選択され、該組換えベクターを維持しない細胞は選択されず、したがって排除される。
【0008】
すなわち、抗生物質耐性遺伝子を組換えベクターに挿入する方法は、組換えベクターを宿主細胞内に安定に維持させることができ、実験室で通常行なわれている。しかし、下記の理由により工業的スケールの生産では望ましくない。
(i)抗生物質耐性菌株の使用は、環境に対して危険を呈する可能性がある。
(ii)培養中に必要な抗生物質の量は生産コストを有意に増加させる。
(iii)抗生物質の使用は、ヒト及び動物向けに用いられる物質の生産においては回避すべきである。
【0009】
そこで、抗生物質耐性遺伝子を用いずに組換えベクターを安定化させる方法として、染色体の栄養要求突然変異を補う方法が知られている(非特許文献1および2)。しかしながら、染色体の栄養要求突然変異を補って組換えベクターを安定化させる方法は、宿主細菌が必要とする栄養を培地に加えずに発酵を行う必要があるため、発酵培地の組成を厳しく制限する必要がある。しかも、栄養共生によって、組換えベクターの消失後もなお細胞が増殖しうる可能性がある。
【特許文献1】特開2003−102489号公報
【特許文献2】国際公開第05/003343号パンフレット
【非特許文献1】Genetics. 1989 May;122(1):19-27
【非特許文献2】Curr Genet. 1989 Sep;16(3):159-63
【非特許文献3】Ohta Y, et al., Biosci. Biotechnol. Biochem. 2004 ,68,1073-1081
【発明の開示】
【発明が解決しようとする課題】
【0010】
本発明者らは、枯草菌を宿主細胞として用いた宿主−ベクター系により、γ−シクロデキストリン合成酵素を大量に生産することを試みた。しかし、上記したマルトースホスホリラーゼおよびトレハロースホスホリラーゼの生産と同じく、γ−シクロデキストリン合成酵素についても、枯草菌を宿主細胞として用いたこれまでの宿主−ベクター系では菌体外に大量に分泌することができないことを見出した。
【0011】
そこで、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼまたはトレハロースホスホリラーゼを菌体外に大量に分泌することができる、新たな宿主−ベクター系の提供が強く望まれている。
【0012】
一方、組換えベクターを維持する宿主細胞を選択する方法について、抗生物質耐性遺伝子を組み込む方法および染色体の栄養要求突然変異を補う方法のいずれもが、培地の特殊な処理に依存している。かかる制約は、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼまたはトレハロースホスホリラーゼの生産において、発酵工程の費用の増大、および生産性向上のためにとり得る選択自由度の制限を招く。
【0013】
そこで、抗生物質を用いることなく、かつ培地組成に制約されずに組換えベクターを維持しうる、他の選択方法が強く望まれている。
【0014】
したがって、本発明の目的は、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼまたはトレハロースホスホリラーゼを菌体外に大量に分泌しうる、新たな宿主−ベクター系を提供することにある。
【0015】
さらに、抗生物質を用いることなく、かつ培地組成に制約されずに組換えベクターを維持する新たな手段を提供することにより、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼまたはトレハロースホスホリラーゼを大量かつ安定に生産しうる方法を提供することを目的とする。
【課題を解決するための手段】
【0016】
上記課題を解決するために種々検討を行い、宿主細胞をバチルス属細菌として、組換えベクターに組み込む最適な転写を調節する塩基配列(以下、「遺伝子調節領域」と呼ぶ)とシグナルペプチドをコードする遺伝子(以下、「シグナルペプチド遺伝子」と呼ぶ)の組み合わせを検討し、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を菌体外に大量に分泌することができる宿主−ベクター系を得ることができ、本発明を完成するに至った。
【0017】
さらに、アミノアシルtRNA合成酵素遺伝子を保有する染色体外遺伝子と該アミノアシルtRNA合成酵素活性に欠損を生じさせた染色体変異宿主細胞との相補性に基づくことで、抗生物質を用いることなく、かつ培地組成に制約されずにγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を含む組換えベクターを維持することに成功し、上記した酵素を大量かつ安定に生産しうる方法を見出し、本発明を完成させた。アミノアシルtRNA合成酵素遺伝子を保有する染色体外遺伝子を含有した細胞のみが生存を確保されることに基づく。
【0018】
すなわち、本発明は以下のとおりである。
<1>γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、かつ
アミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターであって、
アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞において用いられる前記組換えベクター。
<2>下記(a)〜(c)の塩基配列または遺伝子を有する組換えベクター:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;および
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子。
<3>下記(a)〜(d)の塩基配列または遺伝子を有し、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞において用いられる組換えベクター:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;および
(d)発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子。
<4>アミノアシルtRNA合成酵素が、トリプトファニルtRNA合成酵素、アラニルtRNA合成酵素、アルギニルtRNA合成酵素、アスパルギニルtRNA合成酵素、アスパルチルtRNA合成酵素、システイニルtRNA合成酵素、グルタミンtRNA合成酵素、グルタメートtRNA合成酵素、グリシンtRNA合成酵素、ヒスチジルtRNA合成酵素、イソロイシルtRNA合成酵素、ロイシルtRNA合成酵素、リシンtRNA合成酵素、メチオニルtRNA合成酵素、フェニルアラニンtRNA合成酵素、プロリルtRNA合成酵素、セリルtRNA合成酵素、トレオニルtRNA合成酵素、チロシルtRNA合成酵素、またはバリルtRNA合成酵素である、<1>または<3>に記載の組換えベクター。
<5>γ−シクロデキストリン合成酵素をコードする遺伝子が、配列番号3に記載のアミノ酸配列をコードする遺伝子である、<1>〜<4>のいずれかに記載の組換えベクター。
<6>マルトースホスホリラーゼをコードする遺伝子が、配列番号4に記載のアミノ酸配列をコードする遺伝子である、<1>〜<4>のいずれかに記載の組換えベクター。
<7>トレハロースホスホリラーゼをコードする遺伝子が、配列番号5に記載のアミノ酸配列をコードする遺伝子である、<1>〜<4>のいずれかに記載の組換えベクター。
<8>組換えベクターは、プラスミド、バクテリオファージ、またはレトロトランスポゾンの形態を有する<1>〜<7>のいずれかに記載の組換えベクター。
<9>アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞であって、
γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、かつ
アミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターで形質転換して用いられる前記宿主細胞。
<10>下記(a)〜(c)の塩基配列または遺伝子を有する組換えベクターで形質転換して用いられる宿主細胞:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;および
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子。
<11>アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞であって、
下記(a)〜(d)の塩基配列または遺伝子を有する組換えベクターで形質転換して用いられる前記宿主細胞:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;および
(d)発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子。
<12>アミノアシルtRNA合成酵素が、トリプトファニルtRNA合成酵素、アラニルtRNA合成酵素、アルギニルtRNA合成酵素、アスパルギニルtRNA合成酵素、アスパルチルtRNA合成酵素、システイニルtRNA合成酵素、グルタミンtRNA合成酵素、グルタメートtRNA合成酵素、グリシンtRNA合成酵素、ヒスチジルtRNA合成酵素、イソロイシルtRNA合成酵素、ロイシルtRNA合成酵素、リシンtRNA合成酵素、メチオニルtRNA合成酵素、フェニルアラニンtRNA合成酵素、プロリルtRNA合成酵素、セリルtRNA合成酵素、トレオニルtRNA合成酵素、チロシルtRNA合成酵素、またはバリルtRNA合成酵素である、<9>または<11>に記載の宿主細胞。
<13>γ−シクロデキストリン合成酵素をコードする遺伝子が、配列番号3に記載のアミノ酸配列をコードする遺伝子である、<9>〜<12>のいずれかに記載の宿主細胞。
<14>マルトースホスホリラーゼをコードする遺伝子が、配列番号4に記載のアミノ酸配列をコードする遺伝子である、<9>〜<12>のいずれかに記載の宿主細胞。
<15>トレハロースホスホリラーゼをコードする遺伝子が、配列番号5に記載のアミノ酸配列をコードする遺伝子である、<9>〜<12>のいずれかに記載の宿主細胞。
<16>宿主細胞が細菌である<9>〜<15>のいずれかに記載の宿主細胞。
<17>細菌がバチルス属細菌である、<16>に記載の宿主細胞。
<18>下記(i)〜(iv)の工程を有する、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法:
(i)γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、さらにアミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターを用意する工程;
(ii)アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞を用意する工程;
(iii)前記宿主細胞を前記組換えベクターで形質転換して形質転換体を得る工程;および
(iv)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
<19>下記(i)〜(iii)の工程を有する、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法:
(i)下記[a]〜[c]の塩基配列または遺伝子を有する組換えベクターを用意する工程、
[a]配列番号1に記載の塩基配列、
[b]前記[a]の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子、および
[c]前記[b]の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;
(ii)前記組換えベクターで宿主細胞を形質転換して形質転換体を得る工程;および
(iii)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
<20>下記(i)〜(iv)の工程を有する、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法:
(i)下記[a]〜[d]の塩基配列または遺伝子を有する組換えベクターを用意する工程、
[a]配列番号1に記載の塩基配列、
[b]前記[a]の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子、
[c]前記[b]の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子、および
[d]発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子;
(ii)アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞を用意する工程;
(iii)前記宿主細胞を前記組換えベクターで形質転換して形質転換体を得る工程;および
(iv)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
<21>アミノアシルtRNA合成酵素が、トリプトファニルtRNA合成酵素、アラニルtRNA合成酵素、アルギニルtRNA合成酵素、アスパルギニルtRNA合成酵素、アスパルチルtRNA合成酵素、システイニルtRNA合成酵素、グルタミンtRNA合成酵素、グルタメートtRNA合成酵素、グリシンtRNA合成酵素、ヒスチジルtRNA合成酵素、イソロイシルtRNA合成酵素、ロイシルtRNA合成酵素、リシンtRNA合成酵素、メチオニルtRNA合成酵素、フェニルアラニンtRNA合成酵素、プロリルtRNA合成酵素、セリルtRNA合成酵素、トレオニルtRNA合成酵素、チロシルtRNA合成酵素、またはバリルtRNA合成酵素である、<18>または<20>に記載の方法。
<22>γ−シクロデキストリン合成酵素をコードする遺伝子が、配列番号3に記載のアミノ酸配列をコードする遺伝子である、<18>〜<21>のいずれかに記載の方法。
<23>マルトースホスホリラーゼをコードする遺伝子が、配列番号4に記載のアミノ酸配列をコードする遺伝子である、<18>〜<21>のいずれかに記載の方法。
<24>トレハロースホスホリラーゼをコードする遺伝子が、配列番号5に記載のアミノ酸配列をコードする遺伝子である、<18>〜<21>のいずれかに記載の方法。
<25>宿主細胞が細菌である<18>〜<24>のいずれかに記載の方法。
<26>細菌がバチルス属細菌である<25>に記載の方法。
<27>γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される1種の酵素の遺伝子が、前記遺伝子と95%以上の相同性を有し、かつ、前記選択される酵素の活性を有するタンパク質をコードする遺伝子である、<5>〜<8>のいずれかに記載の組換えベクター。
<28>γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される1種の酵素の遺伝子が、前記遺伝子と95%以上の相同性を有し、かつ、前記選択される酵素の活性を有するタンパク質をコードする遺伝子である、<13>〜<17>のいずれかに記載の宿主細胞。
<29>γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される1種の酵素の遺伝子が、前記遺伝子と95%以上の相同性を有し、かつ、前記選択される酵素の活性を有するタンパク質をコードする遺伝子である、<22>〜<26>のいずれかに記載の方法。
【発明の効果】
【0019】
本発明によれば、最適な遺伝子調節領域およびシグナルペプチド遺伝子の組み合わせを組換えベクターに含めることで、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を菌体外に大量に分泌することができる。
【0020】
さらに、抗生物質を用いることなく、かつ特定成分を含有しないよう制限を加えた培地を用いることなくγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を含む組換えベクターを維持することができ、その結果、工業的な生産においても、醗酵槽内での培養が終了するまで細胞中の組換えベクターを安定化させることができ、上記した酵素を工業的規模で大量かつ安定に生産することができる。
【発明を実施するための最良の形態】
【0021】
(A)本発明の組換えベクター
本発明の組換えベクターの一つの形態は、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、かつアミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターであって、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞において用いられる前記組換えベクター(以下、「組換えベクターA」と呼ぶ)である。
【0022】
本発明の組換えベクターの一つの形態は、下記(a)〜(c)の塩基配列または遺伝子を有する組換えベクター(以下、「組換えベクターB」と呼ぶ)である:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;および
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子。
【0023】
本発明の組換えベクターの一つの形態は、下記(a)〜(d)の塩基配列または遺伝子を有し、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞において用いられる組換えベクター(以下、「組換えベクターC」と呼ぶ)である:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;および
(d)発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子。
【0024】
本発明の組換えベクターAおよびCは、アミノアシルtRNA合成酵素遺伝子をコードする遺伝子を発現可能な状態で含有する。該組換えベクターAおよびCは、アミノアシルtRNA合成酵素遺伝子をコードする遺伝子を、所望の宿主細胞内で発現可能な状態で含んでおり、当該宿主細胞を形質転換するために使用される。
【0025】
アミノアシルtRNA合成酵素は、20種類のアミノ酸に対応するよう、20数種類(一部は数種類ある)存在し、例えばアラニンに対するアミノアシルtRNA合成酵素はアラニルtRNA合成酵素(alanyl−tRNA synthetaseまたはalanine−tRNA synthetase)と呼ばれている。アミノアシルtRNA合成酵素は、具体的には、例えば、トリプトファニルtRNA合成酵素、アラニルtRNA合成酵素、アルギニルtRNA合成酵素、アスパルギニルtRNA合成酵素、アスパルチルtRNA合成酵素、システイニルtRNA合成酵素、グルタミンtRNA合成酵素、グルタメートtRNA合成酵素、グリシンtRNA合成酵素、ヒスチジルtRNA合成酵素、イソロイシルtRNA合成酵素、ロイシルtRNA合成酵素、リシンtRNA合成酵素、メチオニルtRNA合成酵素、フェニルアラニンtRNA合成酵素、プロリルtRNA合成酵素、セリルtRNA合成酵素、トレオニルtRNA合成酵素、チロシルtRNA合成酵素、またはバリルtRNA合成酵素である。
【0026】
トリプトファンに対応するアミノアシルtRNA合成酵素をトリプトファニルtRNA合成酵素と呼び、トリプトファニルtRNA合成酵素が欠失している細胞株は、トリプトファンを含むタンパクの合成を行うことができないため増殖することができないことになる。
【0027】
アミノアシルtRNA合成酵素遺伝子は原核生物から真核生物に至るあらゆる生物に含まれる。例えば、枯草菌には、alanyl−tRNA synthetase(配列番号6)、arginyl−tRNA synthetase(配列番号7)、asparaginyl−tRNA synthetase(配列番号8)、aspartyl−tRNA synthetase(配列番号9)、cysteinyl−tRNA synthetase(配列番号10)、glutamyl−tRNA synthetase(配列番号11)、glutamyl−tRNA amidotransferase, subunit a(配列番号12)、glycine tRNA synthetase, alpha subunit(配列番号13)、glycine tRNA synthetase, beta subunit(配列番号14)、histidyl tRNA synthetase, hisS(配列番号15)、isoleucyl−tRNA synthetase(配列番号16)、leucyl−tRNA synthetase(配列番号17)、lysine tRNA synthetase, constitutive(配列番号18)、histidyl tRNA synthetase,hisZ(配列番号19)、methionyl−tRNA synthetase(配列番号20)、phenylalanine tRNA synthetase, alpha subunit(配列番号21)、phenylalanine tRNA synthetase, beta subunit(配列番号22)、similar to phenylalanyl−tRNA synthetase, ytpR (配列番号23)、prolyl−tRNA synthetase(配列番号24)、seryl−tRNA synthetase, also charges selenocysteinyl−tRNA with serine(配列番号25)、threonyl−tRNA synthetase, major(配列番号26)、threonyl−tRNA synthetase, minor(配列番号27)、tryptophanyl−tRNA synthetase(配列番号28)、tyrosyl−tRNA synthetase, major(配列番号29)、tyrosyl−tRNA synthetase, minor (配列番号30)、valyl−tRNA synthetase(配列番号31)、glutamyl−tRNA amidotransferase, subunit b(配列番号32)、glutamyl−tRNA amidotransferase, subunit c(配列番号33)などが含まれることが知られている。
【0028】
本発明の組換えベクターAおよびCは、アミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有するものである。したがって、該組換えベクターAおよびCについては、アミノアシルtRNA合成酵素遺伝子以外に、この遺伝子を発現可能な状態とするために、宿主として枯草菌(Bacillus subtilis)などの細菌を使用する場合は、たとえば、プロモーター、オペレーター領域(プロモーター、オペレーターおよびリボゾーム結合領域(SD領域)を含む)、開始コドン、選択マーカー遺伝子、終止コドン、ターミネーター領域、及びプラスミド複製可能単位などを有する。
【0029】
選択マーカー遺伝子としては、特に制限されず、たとえば、ジヒドロ葉酸レダクターゼ(DHFR)遺伝子またはシゾサッカロマイセス・ポンベTPI遺伝子等のようなその補体が宿主細胞に欠けている遺伝子、またはたとえばアンピシリン、カナマイシン、テトラサイクリン、クロラムフェニコール、シクロヘキシミド、テトラマイシン、ネオマイシンもしくはヒグロマイシンのような薬剤耐性遺伝子を挙げることができる。
【0030】
本発明の組換えベクターAおよびCは、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞において用いられる。この宿主細胞については後述する。
【0031】
本発明の組換えベクターは、ベクターの種類は特に限定されず、例えば、宿主細胞内で自立的に複製することが可能なもの(例えばプラスミド等)でもよいし、あるいは、宿主細胞に導入された際に宿主細胞のゲノムに組み込まれ、組み込まれた染色体と共に複製されるものであってもよい。好ましくは、本発明で用いるベクターは、プラスミド、バクテリオファージ、またはレトロトランスポゾンである。
【0032】
宿主細胞内で自立的に複製することが可能なベクターの具体例としては、pRS413、pRS415、pRS416、YCp50、pAUR112またはpAUR123などのYCp型大腸菌−酵母シャトルベクター;pRS403、pRS404、pRS405、pRS406、pAUR101またはpAUR135などのYIp型大腸菌−酵母シャトルベクター;大腸菌由来のプラスミド(たとえば、pBR322、pBR325、pUC18、pUC19、pUC119、pTV118N、pTV119N、pBluescript、pHSG298、pHSG396またはpTrc99AなどのColE系プラスミド;pACYC177またはpACYC184などのp1A系プラスミド;pMW118、pMW119、pMW218またはpMW219などのpSC101系プラスミドなど);枯草菌由来のプラスミド(例えばpUB110、pTP5など);pHY300PLKなどの大腸菌−枯草菌シャトルベクターを挙げることができる。またファージベクターとして、λファージ(たとえば、Charon 4A、Charon21A、EMBL4、λgt100、gt11、zap)、ψX174、M13mp18、M13mp19などを挙げることができる。レトロトランスポゾンとしてはTy因子などを挙げることができる。また、融合タンパク質として発現する発現ベクター、例えばpGEXシリーズ(ファルマシア製)、pMALシリーズ(Biolabs社製)を使用することもできる。
【0033】
本発明の組換えベクターは、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有する。
【0034】
本明細書でいう「遺伝子を発現可能な状態」、または単に「発現可能な状態」とは、遺伝子の情報がmRNAに転写されるためのRNAポリメラーゼ認識部位およびmRNAの情報が該遺伝子の発現産物であるタンパク質等に翻訳されるためのリボゾーム結合領域が該遺伝子の開始コドンの上流に存在する状態を意味する。要するに、該遺伝子の開始コドンから終止コドンまでをmRNAに転写されるために必要な配列が開始コドンより上流に存在し、さらにそのmRNAの情報がタンパク質等に翻訳されるための配列が開始コドンより上流に存在するということになる。ここでいう「開始コドン」とは本来染色体上における該遺伝子の開始コドン配列に限らず、開始コドンとして機能するコドンであればいずれでもよい。またここでいうリボゾーム結合領域とは、例えば原核生物の5´−aaaggagg−3´からなるコンセンサス配列を有する配列に集約されることなく、リボゾームが認識できる配列であれば良く、本来染色体上で該遺伝子のリボゾーム結合領域としての配列に限らず、リボゾームが認識できる配列であれば用いることができる。
【0035】
本明細書でいう「γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子」とは、γ−シクロデキストリン合成酵素をコードする遺伝子、マルトースホスホリラーゼをコードする遺伝子、またはトレハロースホスホリラーゼをコードする遺伝子の単一物だけでなく、各遺伝子を組み合わせた複合物も含む。
【0036】
本発明の組換えベクターの構築は、DNA組換えの一般的な方法、たとえば、Molecular Cloning (1989) (Cold Spring Harbor Lab.)に記載される方法にしたがって行うことができる。
【0037】
本発明の組換えベクターBおよびCは、遺伝子調節領域として配列番号1に記載の塩基配列を有し、シグナルペプチド遺伝子として配列番号2に記載のアミノ酸配列をコードする遺伝子を有する。
【0038】
配列番号1に記載の塩基配列を含むDNA断片は、たとえば、バチルス・エスピー JAMB750(Bacillus sp. JAMB750)株の染色体DNAにランダム変異を導入することにより取得することができる。配列番号1に記載の塩基配列を含むDNA断片を取得するその他の方法としては、たとえば、配列番号1に記載の塩基配列の情報に基づいて、化学合成、遺伝子工学的手法または突然変異誘発などの通常知られる任意の方法で作製することができる。
【0039】
本明細書にいう「シグナルペプチド」とは、シグナルペプチドをコードする塩基配列の下流に存在する遺伝子の遺伝子産物のアミノ末端に付加して、該遺伝子産物を細胞外に分泌しうるペプチドを意味する。また、「遺伝子産物を細胞外に分泌しうる」とは、上記した通り、シグナルペプチドが遺伝子産物である組換えタンパク質のアミノ末端側に付加し、組換えタンパク質の小胞体膜への付着および膜通過等を導き、最終的に細胞外に組換えタンパク質を分泌しうる作用を意味する。
【0040】
本明細書にいう「配列番号1の塩基配列の下流に直接的または間接的に連結された」とは、配列番号1に記載の塩基配列の3´末端側に0〜1000程度の塩基を介して連結されることを意味する。
【0041】
シグナルペプチド遺伝子を含むDNA断片は、たとえば、配列番号2に記載の塩基配列の情報に基づいて、化学合成、遺伝子工学的手法または突然変異誘発などの通常知られる任意の方法で作製することができる。化学合成法としては、たとえば、ホスホアミダイト法を応用することにより自動合成しうる。なお、配列番号2に記載のシグナルペプチドは、バチルス属細菌の分泌するタンパク質群のアミノ末端側のアミノ酸配列情報から統計処理してデザインして得られた数十候補のアミノ酸配列から、組換えタンパク質生産性を評価し選択したものである。本配列は1〜数個のアミノ酸残基を付加、欠失または置換しても利用できる。
【0042】
本発明の組換えベクターBおよびCは、遺伝子調節領域およびその下流に直接的または間接的に連結されたシグナルペプチド遺伝子、ならびにその下流に直接的に連結されたγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を含み、該組換えベクターを宿主細胞として枯草菌に導入し形質転換することにより、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素の発現を促進し、さらに発現する酵素を菌体外に分泌しうる。
【0043】
本明細書にいう「配列番号2に記載のアミノ酸配列をコードする遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子」とは、上記したシグナルペプチド遺伝子の3´末端側に連結されたγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子である。
【0044】
本明細書にいう「γ−シクロデキストリン合成酵素をコードする遺伝子」は、特に限定されるものではないが、好ましくは配列番号3に記載の塩基配列からなる遺伝子である。配列番号3に記載の塩基配列からなる遺伝子は、たとえば、特許文献1の記載に基いてバチルス・クラーキ 7364(Bacillus clarkii 7364)株から取得される。配列番号3に記載の塩基配列からなる遺伝子を取得するその他の方法としては、配列番号3に記載の塩基配列の情報に基づいて、化学合成、遺伝子工学的手法または突然変異誘発などの通常知られる任意の方法で作製することができる。
【0045】
本明細書にいう「γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される1種の酵素の遺伝子が、前記遺伝子と95%以上の相同性を有し、かつ、前記選択される酵素の活性を有するタンパク質をコードする遺伝子」とは、たとえば、γ−シクロデキストリン合成酵素をコードする遺伝子を選択した場合に、γ−シクロデキストリン合成酵素をコードする遺伝子に1〜数個の塩基の欠失、置換、逆位、付加および/または挿入を有し、該遺伝子と95%以上の相同性があり、かつ、発現産物がγ−シクロデキストリン合成酵素活性を有する酵素である遺伝子を意味する。
【0046】
本明細書にいう「γ−シクロデキストリン合成酵素」は、糖類に作用し主生成物としてγ−シクロデキストリンを生成するシクロデキストリン・グルカノトランスフェラーゼ活性を有する酵素であればよく、好ましくは配列番号3に記載の遺伝子が発現して得られる酵素である。
【0047】
本明細書にいう「マルトースホスホリラーゼをコードする遺伝子」は、特に限定されるものではないが、好ましくは配列番号4に記載の塩基配列からなる遺伝子である。配列番号4に記載の塩基配列からなる遺伝子は、たとえば、特許文献2の記載に基づいてパエニバチルス・エスピー SH−55(Paenibacillus sp. SH−55)株から取得される。配列番号4に記載の塩基配列からなる遺伝子を取得するその他の方法としては、配列番号4に記載の塩基配列に記載の情報に基づいて、化学合成、遺伝子工学的手法または突然変異誘発などの通常知られる任意の方法で作製することができる。
【0048】
本明細書にいう「マルトースホスホリラーゼ」は、マルトース中のα−1,4−グルコピラノシド結合を可逆的に加リン酸分解し、グルコースとβ−D−グルコース−1−リン酸を生成することができればよく、好ましくは、配列番号4に記載の遺伝子が発現して得られる酵素である。
【0049】
本明細書にいう「トレハロースホスホリラーゼをコードする遺伝子」は、特に限定されるものではないが、好ましくは配列番号5に記載の塩基配列からなる遺伝子である。配列番号5に記載の塩基配列からなる遺伝子は、たとえば、特許文献2の記載に基いてパエニバチルス・エスピー SH−55株から取得される。配列番号5に記載の塩基配列からなる遺伝子を取得するその他の方法としては、配列番号5に記載の塩基配列に記載の塩基配列の情報に基づいて、化学合成、遺伝子工学的手法または突然変異誘発などの通常知られる任意の方法で作製することができる。
【0050】
本明細書にいう「トレハロースホスホリラーゼ」は、トレハロース中のα−1,1−グルコピラノシド結合を可逆的に加リン酸分解し、グルコースおよびβ−D−グルコース−1−リン酸を生成すればよく、好ましくは、配列番号5に記載の遺伝子が発現して得られる酵素である。
【0051】
本発明の組換えベクターCの具体例を、図1〜3として示す。
【0052】
(B)本発明の宿主細胞
本発明の宿主細胞の一つの形態は、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞であって、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、かつアミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターで形質転換して用いられる前記宿主細胞(以下、「宿主細胞A」と呼ぶ)である。
【0053】
本発明の宿主細胞の一つの形態は、下記(a)〜(c)の塩基配列または遺伝子を有する組換えベクターで形質転換して用いられる宿主細胞(以下、「宿主細胞B」と呼ぶ)である:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;および
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子。
【0054】
本発明の宿主細胞の一つの形態は、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞であって、下記(a)〜(d)の塩基配列または遺伝子を有する組換えベクターで形質転換して用いられる前記宿主細胞(以下、「宿主細胞C」と呼ぶ)である:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;および
(d)発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子。
【0055】
一般に、細胞は、自己の増殖のためにアミノアシルtRNA合成酵素をコードする染色体遺伝子を有している。それに対して、本発明の宿主細胞AおよびCでは、細胞が本来有している、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させるか、あるいはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させる。アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損または該遺伝子発現の低下は、以下の方法で行うことができる。
【0056】
[遺伝子欠損(不活性化)]
不活性化の語は染色体遺伝子のうち1以上の機能発現を妨げる方法を含んでいる。不活性化は核酸遺伝子配列における欠失、置換(例えば変異)、阻害、及び/または挿入などによって行われる。いくつかの実施態様において、変異微生物は好ましくは安定かつ不可逆的な不活性化を生じる1以上の遺伝子の不活性化を含む。
【0057】
(挿入による不活性化)
ここで用いる“挿入配列”の語は微生物染色体内に導入されたDNA配列を言う。いくつかの実施態様において、挿入配列は形質転換される細胞のゲノム中に既に存在する、または存在しない配列であってもよい(すなわち、相同または異種配列である)。さらに別の実施態様において、挿入配列は選択マーカーを含むことがある。さらなる実施態様において、挿入配列は2つの相同ボックスを含むことがある。
【0058】
ここで用いる“相同ボックス”とは微生物染色体の配列と相同な核酸配列をいう。より具体的には、相同ボックスは、本発明に従って不活性化される遺伝子に隣接するコード領域または遺伝子の一部と約80〜100%配列同一性、約90〜100%配列同一性または約95%〜100%配列同一性を有する上流または下流領域をいう。これらの配列は微生物染色体の一部が置換されることを目的とする。本発明を限定するものではないが、相同ボックスは約1塩基対(bp)〜200キロ塩基(kb)を含む。好ましくは、相同ボックスは約1bp〜10.0kb;1bp〜5.0kb;1bp〜2.5kb;1bp〜1.0kb及び0.25kb〜2.5kbを含む。また、相同ボックスは約10.0kb、5.0kb、2.5kb、2.0kb、1.5kb、1.0kb、0.5kb、0.25kb及び0.1kbを含む。いくつかの実施態様において、選択マーカーの5’及び3’末端は相同ボックスに隣接し、ここで相同ボックスは当該遺伝子のコード領域にすぐ隣接した核酸配列を含む。
【0059】
ここで用いる“選択マーカー”の語は宿主細胞において発現できるヌクレオチド配列をいい、選択マーカーの発現により対応する選択因子の存在下、または必須栄養素の不存在下で発現遺伝子を含む細胞は成長できるようになる。ここで用いる“選択可能マーカー”及び“選択マーカー”の語は宿主細胞中で発現できる核酸(例えば遺伝子)を言い、ベクターを含むこれら宿主の選択が簡単にできるようにする。当該選択マーカーの例としては、限定されないが、抗微生物剤などがある。従って、“選択マーカー”とは、宿主細胞が目的の挿入DNAを取り込んだ、またはその他の反応が起こった印を与える遺伝子をいう。通常、選択マーカーは、形質転換時に外来配列を受け取らなかった細胞と外来DNAを含む細胞との区別を可能にするために宿主細胞に抗菌耐性または代謝利点を与える遺伝子をいう。選択マーカーは抗微生物耐性マーカーなどが挙げられる(例えば、ampR;phleoR;specR;kanR;eryR;tetR;cmpR;及びneoR;例えばGuerot−Fleury,Gene,167:335−337[1995];Palmeros et al.、Gene 247:255−264[2000];及びTrieu−Cuot et al.、Gene、23:331−341[1983]を参照)。いくつかの特に好ましい実施態様において、本発明はクロラムフェニコール耐性遺伝子を提供する(例えば、pC194上に存在する遺伝子、及びバチルス・リケニフォルミスゲノム中に存在する耐性遺伝子)。この耐性遺伝子は本発明及び染色体上統合されたカセット及び統合プラスミドの染色体増幅に関する実施態様において特に有用である。
【0060】
例えば、トリプトファニルtRNA合成酵素遺伝子(trpS)が不活性化される遺伝子である場合、DNA構築体は選択マーカーを挿入配列として含みその活性を阻害されるtrpS遺伝子を有する。選択マーカーはtrpSコード配列部分の内側に挿入する。DNA構築体は宿主染色体中のtrpS遺伝子に本質的に同一配列を有し、二重交差の場合、trpS遺伝子は選択マーカーの挿入により不活性化される。
【0061】
(欠失による不活性化)
いくつかの好ましい実施態様において、不活性化は欠失により達成される。いくつかの好ましい実施態様において、遺伝子は相同組換えにより欠失する。例えば、いくつかの実施態様において、欠失する遺伝子がtrpSの場合、相同ボックスの両側に隣接する選択マーカーを有する挿入配列を含むDNA構築体を用いる。相同ボックスは染色体trpS遺伝子の核酸フランキング領域に相同なヌクレオチド配列を含む。DNA構築体は微生物宿主染色体の相同配列と一致し、二重交差の場合、trpS遺伝子は宿主染色体から切除される。
【0062】
ここで用いる遺伝子の“欠失”の語は、コード配列全体の欠失、コード配列の一部の欠失、またはフランキング領域を含むコード配列の欠失をいう。欠失は染色体に残った配列が欠失前の生物活性を発揮しないのであれば部分的であってもよい。コード配列のフランキング領域は約1bp〜約500bpを5´及び3´末端に含むことができる。フランキング領域は500bpよりも大きくてもよいが、好ましくは本発明に従って不活性化または欠失され得るその他の遺伝子を当該領域に含まない。最終的には、欠失遺伝子は事実上、非機能的である。簡単に言えば、“欠失”の語は1以上のヌクレオチドまたはアミノ酸残基がそれぞれ除去された(すなわち、存在しない)、ヌクレオチドまたはアミノ酸配列の変化として定義される。
【0063】
ここで用いる“フランキング配列”とは対象の配列の上流または下流にある配列をいう(例えば、遺伝子A−B−Cは、遺伝子BはA及びC遺伝子配列にフランキング(側面に位置)している)。好ましい実施態様において、挿入配列は相同ボックスの両側の側面に位置する。いくつかの実施態様において、フランキング配列は一方の側(3´または5´)にのみ存在するが、好ましい実施態様において、配列の両側の側面に位置する。各相同ボックスの配列は微生物染色体中の配列に相同である。これらの配列は微生物染色体に新しい構築体が統合されること、及び微生物染色体の一部が挿入配列により置換されることを目的とする。好ましい実施態様において、選択マーカーの5´及び3´末端は不活性化染色体断片の一部を含むポリヌクレオチド配列の側面に位置する。
【0064】
(変異による不活性化)
別の実施態様において、不活性化は遺伝子の変異により生じる。遺伝子を変異する方法はさまざまな方法があり、限定されないが、部位特異的変異、ランダム変異発生、及びgapped−duplex法が挙げられる(例えばMoring et al.、Biotech.2:646[1984];及びKramer et al.、Nucleic Acids Res.,12:9441[1984]を参照)。
【0065】
好ましい実施態様において、変異は少なくとも染色体上の1つのコドンにおいて突然変異誘発を用いて生じる。さらなる実施態様において、変異DNA配列は野生型配列と40%以上、45%以上、50%以上、55%以上、60%以上、65%以上、70%以上、75%以上、80%以上、85%以上、90%以上、95%以上または98%以上の相同性を有する。別の実施態様において、変異DNAは公知の変異誘発手順、例えば、UV照射、ニトロソグアニジン等の化学変異剤を用いてin vivoで生じる。
【0066】
アミノアシルtRNA合成酵素活性に欠損を生じさせる染色体変異の内でも、アミノアシルtRNA合成酵素遺伝子自体の欠損を用いることが望ましい。しかしアミノアシルtRNA合成酵素生合成を調節する他の遺伝子も、対応する遺伝子を保有する染色体外遺伝子を用いる限り使用できる。アミノアシルtRNA合成酵素遺伝子の欠損株の場合、染色体外遺伝子に挿入されるべき遺伝子は該当するアミノアシルtRNA合成酵素遺伝子が相応しい。
【0067】
アミノアシルtRNA合成酵素を有する染色体外遺伝子と染色体の変異されたアミノアシルtRNA合成酵素遺伝子との間の相同性組換えの可能性を回避するためには、目的のアミノアシルtRNA合成酵素遺伝子を染色体から実質的に欠失させることが望ましい。
【0068】
本発明の宿主細胞Aを形質転換する際に用いられる組換えベクターは、上記した本発明の組換えベクターAと同様のものである。一方、本発明の宿主細胞Cを形質転換する際に用いられる組換えベクターは、上記した本発明の組換えベクターCと同様のものである。
【0069】
アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞は、アミノアシルtRNA合成酵素をコードする染色体遺伝子が発現できない、あるいは発現量が不十分であるため、そのままではもはや増殖できない。
【0070】
それに対して、アミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクター(本発明の組換えベクターAまたはC)を含むことで、増殖を維持できる。即ち、アミノアシルtRNA合成酵素の欠失は、染色体外遺伝子上に該アミノアシルtRNA合成酵素遺伝子を導入することにより相補することができる。しかし、もしも細胞が該染色体外遺伝子を失うと、該細胞はもはや増殖できなくなる。
【0071】
したがって、本発明の宿主細胞AまたはCは、上記関係の下に、培地に抗生物質や特定の栄養要求性を満たす栄養分を含めることなく組換えベクターAまたはCを安定して宿主細胞に維持することができる。そのため、本発明の宿主細胞AまたはCは、工業的スケールの生産に使用される任意の培地に適用できるという利点を有する。
【0072】
さらに、本発明の宿主細胞AおよびCは、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現させるために用いられる。本発明の宿主細胞Aは、前記アミノアシルtRNA合成酵素をコードする遺伝子およびγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有する組換えベクターを含むものであり、該組換えベクターを安定して宿主細胞に維持することができる。そして、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現させて、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素の生産に用いることができる。
【0073】
本発明の宿主細胞Bは、本発明の組換えベクターBを導入し形質転換して用いられる宿主細胞であり、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現させるために用いられる。
【0074】
本発明の宿主細胞は細菌であればよく、好ましいのは枯草菌であり、さらに好ましくは枯草菌のISW1214株、BD170株、または168株等である。しかし、本発明の組換えベクターを安定に保持し、複製することができるものであれば、枯草菌以外のバチルス(Bacillus)属細菌などを使用してもよい。
【0075】
本明細書でいう「バチルス属」とは、通常知られるバチルス属に含まれるすべての種類を含み限定されないが、たとえば、バチルス・ズブチリス(Bacillus subtilis)、バチルス・リケニフォルミス(Bacillus licheniformis)、バチルス・レンタス(Bacillus lentus)、バチルス・ステアロサーモフィラス(Bacillus stearothermophilus)、バチルス・アルカロフィラス(Bacillus alkalophilus)、バチルス・アミロリケファシエンス(Bacillus amyloliquefaciens)、バチルス・セレウス(Bacillus cereus)、バチルス・プミルス(Bacillus pumilus)、バチルス・クラウジイ(Bacillus clausii)、バチルス・ハロデュランス(Bacillus halodurans)、バチルス・メガテリウム(Bacillus megaterium)、バチルス・コアグランス(Bacillus coagulans)、バチルス・サーキュランス(Bacillus circulans)、およびバチルス・チューリンゲンシス(Bacillus thuringiensis)などを意味する。なお、バチルス属は分類上再編成を受け続けており、当該属は再分類された種も含むものとする。たとえば、ジオバチルス(Geobacillus)属、アルカリバチルス(Alkalibacillus)属、アンフィバチルス(Amphibacillus)属 、アミロバチルス(Amylobacillus)属、アノキシバチルス(Anoxybacillus)属 、ゴリバチルス(Goribacillus)属、セラシバチルス(Cerasibacillus)属、グラシリバチルス(Gracilibacillus)属、ハロバチルス(Halolactibacillus)属、ハロアルカリバチルス(Halalkalibacillus)属、フィロバチルス(Filobacillus)属、ジョーガリバチルス(Jeotgalibacillus)属、サリバチルス(Salibacillus)属、オーシャノバチルス(Oceanobacillus)属、マリニバチルス(Marinibacillus)属、リシニバチルス(Lysinibacillus)属、レンチバチルス(Lentibacillus)属、ウレーバチルス(Ureibacillus)属、サリニバチルス(Salinibacillus)属、ポンチバチルス(Pontibacillus)属、ピシバチルス(Piscibacillus)属、パラリオバチルス(Paraliobacillus)属、ヴァージバチルス(Virgibacillus)属、サルシューギニバチルス(Salsuginibacillus)属、テニューイバチルス(Tenuibacillus)属、タラソバチルス(Thalassobacillus)属、サームアルカリバチルス(Thermalkalibacillus)属、チューメバチルス(Tumebacillus)属などがあり、これらもまた本明細書にいうバチルス属に含まれるものとする。
【0076】
本発明の組換えベクターを宿主細胞に導入する方法はとくに限定されないが、たとえば、コンピテントセル法、プロトプラスト法、エレクトロポレーション法、カルシウムイオン法、リポフェクション法等を用いることができる。宿主細胞が枯草菌である場合は、コンピテントセル法またはプロトプラスト法が好ましい。
【0077】
(C)γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法。
【0078】
本発明のγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法の一つの態様は、下記(i)〜(iv)の工程を有する方法(以下、「酵素生産方法A」と呼ぶ)である:
(i)γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、さらにアミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターを用意する工程;
(ii)アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異宿主細胞を用意する工程;
(iii)前記変異宿主細胞を前記組換えベクターで形質転換して形質転換体を得る工程;および
(iv)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
【0079】
本発明のγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法の一つの態様は、下記(i)〜(iii)の工程を有する方法(以下、「酵素生産方法B」と呼ぶ)である:
(i)下記[a]〜[c]の塩基配列または遺伝子を有する組換えベクターを用意する工程、
[a]配列番号1に記載の塩基配列、
[b]前記[a]の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子、および
[c]前記[b]の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;
(ii)前記組換えベクターで宿主細胞を形質転換して形質転換体を得る工程;および
(iii)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
【0080】
本発明のγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法の一つの態様は、下記(i)〜(iv)の工程を有する方法(以下、「酵素生産方法C」と呼ぶ)である:
(i)下記[a]〜[d]の塩基配列または遺伝子を有する組換えベクターを用意する工程、
[a]配列番号1に記載の塩基配列、
[b]前記[a]の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子、
[c]前記[b]の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子、および
[d]発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子;
(ii)アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞を用意する工程;
(iii)前記宿主細胞を前記組換えベクターで形質転換して形質転換体を得る工程;および
(iv)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
【0081】
酵素生産方法Aにおける組換えベクターを用意する工程では、アミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で挿入し、かつ、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で挿入した組換えベクターを用意する。さらに、酵素生産方法Cにおける組換えベクターを用意する工程では、アミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で挿入し、かつ、配列番号1に記載の塩基配列、その塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子およびその遺伝子の下流に直接的に連結されたγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を挿入した組換えベクターを用意する。上記組換えベクターは、先に説明した本願発明の組換えベクターAまたはCである。一方、酵素生産方法Bにおける組換えベクターを用意する工程では、配列番号1に記載の塩基配列、その塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子およびその遺伝子の下流に直接的に連結されたγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を挿入した組換えベクターを用意する。
【0082】
遺伝子の挿入は常法によって行うことができ、たとえば、Molecular Cloning (1989) (Cold Spring Harbor Lab.)に記載される方法に従って行うことができる。いずれの遺伝子も発現可能な状態で挿入する。そのため、各々の遺伝子の組換えベクターへの挿入位置は組換えベクターの複製に関与していない場所であれば基本的にはどこでも良く、使用するベクターによって挿入位置は異なる。使用できる組換えベクターは上記に挙げた通りであり、たとえば、枯草菌由来のプラスミド(例えばpUB110、pTP5など);pHY300PLKなどの大腸菌−枯草菌シャトルベクターを挙げることができる。
【0083】
例えば大腸菌−枯草菌のシャトルベクターであるpHY300PLKプラスミドを例に取れば、マルチクローニング部位であるEcoRI−BamHI間に配列番号1のDNA断片と配列番号2のアミノ酸配列をコードするDNA断片を連結させたDNA断片を、EcoRI部位にアミノアシルtRNA合成酵素遺伝子を、同じくマルチクローニング部位BamHI部位にγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を挿入することなどが可能である。以下に、本発明の組換えベクターCを用意する工程を簡単に説明する。
【0084】
配列番号1のDNA断片と配列番号2のアミノ酸配列をコードするDNA断片を連結させたDNA断片をpHY300PLKのEcoRI−BamHI間に挿入したプラスミドを構築する。次に、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を有する生物の染色体DNAを鋳型にPCR増幅を用いて該酵素の活性や安定性に欠かせない領域をコードするDNA断片を増幅し、これを上記した構築済みプラスミドを制限酵素BamHIで消化することによって得られたDNA 断片とライゲーション反応を行い、得られたライゲーション反応混合物を用いて大腸菌を形質転換してpHY300PLKのBamHI部位に目的の通りプロモーター部位から終止コドンまでを含む該遺伝子が挿入されているプラスミドを含む形質転換体を選択し、この形質転換体からプラスミドを調製する。続いて上記の通り調製できたプラスミドを制限酵素EcoRIで消化し、本プラスミドを切断する。枯草菌168(Bacillus subtilis 168)株の染色体DNAを鋳型にPCR増幅を用いてプロモーター部位から終止コドンまでを含むアミノアシルtRNA合成酵素遺伝子を増幅し、EcoRIで切断された構築済みプラスミド由来DNA断片とライゲーション反応を行い、得られたライゲーション反応混合物を用いて大腸菌を形質転換して構築済みプラスミドのEcoRI部位に目的の通りアミノアシルtRNA合成酵素遺伝子が挿入されているプラスミドを含む形質転換体を選択し、この形質転換体から目的プラスミドを調製することができる。
【0085】
酵素生産方法AおよびCにおける宿主細胞を用意する工程では、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞を用意する。この宿主細胞は、前記で説明した本発明の宿主細胞AまたはCである。その調製方法は、以下のとおりである。
【0086】
宿主細胞の調製の手順としては、宿主染色体上に存在する標的遺伝子を計画的に削除又は不活性化する方法のほか、ランダムな遺伝子の削除又は不活性化変異を与え、その後適当な方法により遺伝子解析またはタンパク質生産性の評価を行う方法が挙げられる。
【0087】
標的とする遺伝子を削除又は不活性化するには、例えば相同組換えによる方法を用いればよい。すなわち、標的遺伝子の一部を含むDNA断片を適当なプラスミドベクターにクローニングして得られる環状の組換えプラスミドまたは直鎖上のDNA断片を宿主細胞内に取り込ませ、標的遺伝子の一部領域に於ける相同組換えによって親微生物ゲノム上の標的遺伝子を分断して不活性化することが可能である。あるいは、塩基置換や塩基挿入等による不活性化変異を導入した標的遺伝子、又は標的遺伝子の外側領域を含むが標的遺伝子を含まない直鎖状のDNA断片等をPCR等の方法によって構築し、これを親微生物細胞内に取り込ませて親微生物ゲノムの標的遺伝子内の変異箇所の外側2ヶ所、又は標的遺伝子外側の2ヶ所の領域で2回交差の相同組換えを起こさせることにより、ゲノム上の標的遺伝子を削除或いは不活性化した遺伝子断片と置換することが可能である。
【0088】
本発明の酵素生産方法の形質転換体を得る工程は、上記した組換えベクターを用意する工程で調製した組換えベクターを用いて、宿主細胞または上記した宿主細胞を得る工程で調製した宿主細胞を形質転換する。組換えベクターによる宿主細胞の形質転換も常法により実施できる。
【0089】
組換えベクターの宿主細胞への導入(形質転換)方法は、特に制限されず、トランスフォーメーション法、トランスフェクション法、コンピテント細胞法、エレクトロポレーションなど、導入する宿主細胞の種類や組換えベクターの形態に応じて、適宜選択することができる。
【0090】
ここで用いる、“宿主細胞”とは新しく導入するDNA配列のための宿主または発現媒体として作用する能力を有する細胞をいう。
【0091】
なお、組換えベクターの宿主細胞内での存在様式は、特に制限されず、染色体中に挿入されて、あるいは置換されて組み込まれてもよいし、またプラスミド状態で存在していてもよい。
【0092】
なお、本発明の組換えベクターAまたはCに含まれるアミノアシルtRNA合成酵素をコードする染色体遺伝子は、本発明の宿主細胞AまたはCにおいて、欠損させられたアミノアシルtRNA合成酵素をコードする染色体遺伝子、または宿主細胞が成育できない程度に発現を低下させられたアミノアシルtRNA合成酵素をコードする染色体遺伝子と同種の遺伝子である。例えば、本発明の宿主細胞Aにおいて、欠損させられたアミノアシルtRNA合成酵素をコードする染色体遺伝子、または宿主細胞が成育できない程度に発現を低下させられたアミノアシルtRNA合成酵素をコードする染色体遺伝子が、アラニルtRNA合成酵素遺伝子である場合、本発明の組換えベクターAに含まれるのもアラニルtRNA合成酵素遺伝子である。
【0093】
本発明の酵素生産方法におけるγ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程では、上記した形質転換体を得る工程で得られる形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子がコードする酵素を生産する。
【0094】
形質転換体の培養は、宿主細胞の種類に応じて、適切な培地を用い、適切な培養条件の下で実施される。宿主細胞としては、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子がコードする酵素の生産が可能なものであればよく、野生型のものでも変異を施したものでものよい。好ましくは、枯草菌などのバチルス属細菌である。
【0095】
例えば、宿主細胞が枯草菌の場合、培養に用いる培地の種類としては、適当な窒素源、炭素源、ミネラルを含み、本発明の枯草菌宿主が生育し、目的タンパク質を生産することができるものであればよい。例えば、組換え枯草菌168株によって、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子がコードする酵素を生産する場合、炭素源として、ぶどう糖や果糖等の単糖類、しょ糖、麦芽糖等の二糖類又は可溶性澱粉等の多糖類等を配合した培地や、窒素源として、ペプトン類、大豆エキス、酵母エキス、魚肉エキス、コーンスティープリカー( C S L )、金属塩等を配合した培地等を用いることができる。
【0096】
また、培地のpH は、用いる組換え生物体が生育しうる範囲のpHであれば良いが、例えば枯草菌の場合pH 6.0〜8.0に調整するのが好適であり、培養条件は、15〜42℃ 、好ましくは28〜37℃で2〜7日間振盪または、通気撹拌培養すればよい。
【0097】
このようにして得られた培養物から、本発明の組換えベクターに挿入された酵素遺伝子が発現した組換えタンパク質を採取する。該組換えタンパク質は通常形質転換体外に蓄積される。そこで、形質転換体外に蓄積された該組換えタンパク質を培養上清から一般のタンパク質の採取の手段に準じて採取する。たとえば、通常知られる手段によって形質転換体を除いた後に培養上清を組換えタンパク質含有物として用いることができる。
【0098】
ただし、宿主細胞の種類によっては組換えタンパク質を形質転換体内または形質転換体の細胞膜内に蓄積する場合もあり得る。その場合は、特に限定はされないが、たとえば、有機溶剤やリゾチームのような酵素によって形質転換体を溶解する方法、および、超音波破砕法、フレンチプレス法、ガラスビーズ破砕法、ダイノミル破砕法等の細胞破砕法で得られた形質転換体の細胞破砕物および/または培養物を遠心分離法、ろ過法等の操作によって形質転換体と培養上清に分離する。このようにして得られた培養上清を、組換えタンパク質含有物として用いることができる。また、分離した形質転換体をそのまま組換えタンパク質含有物とすることもできる。
【0099】
上記組換えタンパク質含有物は、そのままで使用することもできるが、必要に応じて、たとえば塩析法、沈澱法、透析法、限外濾過法等の通常知られる方法を単独または組み合わせることにより工業用途の濃縮組換えタンパク質含有物を調製できる。
【0100】
さらに前記濃縮組換えタンパク質含有物を、たとえば、イオン交換クロマトグラフィー、等電点クロマトグラフィー、疎水性クロマトグラフィー、ゲル濾過クロマトグラフィー、吸着クロマトグラフィー、アフィニティークロマトグラフィー、逆相クロマトグラフィー、レジンカラム法等の通常知られる単離・精製法の組合せに供すことにより、精製組換えタンパク質を得ることができる。
【実施例】
【0101】
以下、本発明を実施例によりさらに詳細に説明する。
【0102】
[実施例1]枯草菌へのトリプトファニルtRNA合成酵素遺伝子を有するプラスミドの導入
トリプトファニルtRNA合成酵素遺伝子を保有する染色体外遺伝子であるプラスミドpDATS14を以下の方法で構築した。まず、汎用枯草菌−大腸菌のシャトルプラスミドであるpHY300PLK(ヤクルト社製)を鋳型として、プライマーA(配列番号34)とB(配列番号35)を用いてPCRによる増幅を行った。得られた増幅断片を制限酵素XhoIで消化した後、両末端を連結することにより環状プラスミドとした。本プラスミドを用いて大腸菌HB101株を形質転換した。形質転換体からプラスミドを調製しpDA2とした。一方、枯草菌ISW1214(Bacillus subtilis ISW1214)株の染色体を鋳型とし、プライマーC(配列番号36)とD(配列番号37)を用いてトリプトファニルtRNA合成酵素遺伝子とその近傍の領域を含むDNA断片を得た。本DNA断片を制限酵素XhoIで消化し、予め制限酵素XhoIで消化したpDA2と連結した。これを用いて大腸菌HB101株を形質転換した。得られた形質転換体からプラスミドを調製し、pDATS14と命名した。次にpDATS14を用いて、2種のプロテアーゼ(アルカリプロテアーゼE、中性プロテアーゼE)に欠陥を有する枯草菌ISW1214株由来変異株(参考文献 Appl.Microbiol. Biotechnol 65:583-592 (2004)Hatada,Y.ら)を、プロトプラスト形質転換法(参考文献Mol. Gen. Genet. 168:111-115 (1979) Chang, S.とCohen, S. N.)により形質転換し、染色体外にトリプトファニルtRNA合成酵素遺伝子を保有する枯草菌形質転換体を得た。
【0103】
[実施例2]枯草菌の染色体トリプトファニルtRNA合成酵素遺伝子を破壊するためのDNA断片の構築
枯草菌ISW1214株の染色体DNAを鋳型とし、プライマーE(配列番号38)とF(配列番号39)を用いてトリプトファニルtRNA合成酵素遺伝子とその近傍の領域を含む約2.5kbのDNA断片を得た。これを制限酵素EcoRIおよびSalIで消化した。EcoRIおよびSalIで予め消化した汎用性プラスミドpUC18に連結し、プラスミドpTSAFを得た。一方、汎用性プラスミドであるpC194を鋳型として、プライマーG(配列番号40)とH(配列番号41)を用いてスタフィロコッカス由来クロラムフェニコール耐性遺伝子とその近傍領域を含む約1.6kbのDNA断片を得た。さらに、pTSAFを鋳型としてプライマーI(配列番号42)とJ(配列番号43)を用いてPCR反応を行いDNA断片を得た。本DNA断片とクロラムフェニコール耐性遺伝子とその近傍領域を含む約1.6kbのDNA断片を制限酵素BamHIおよびXbaIで消化した後連結し、大腸菌HB101株を形質転換した。形質転換体からプラスミドを調製し、トリプトファニルtRNA合成酵素遺伝子破壊用プラスミドpINTTSとした。pINTTSを鋳型としてプライマーK(配列番号44)とL(配列番号45)を用いてPCR反応を行い直鎖状のトリプトファニルtRNA合成酵素遺伝子破壊用DNA断片を得た。
【0104】
[実施例3]枯草菌の染色体トリプトファニルtRNA合成酵素遺伝子の破壊
上記トリプトファニルtRNA合成酵素遺伝子破壊用DNA断片を用いて、コンピテントセル法を用いて実施例1で得た染色体外にトリプトファニルtRNA合成酵素遺伝子を保有する形質転換体をさらに形質転換し、クロラムフェニコール耐性を獲得した形質転換体を得た。得られた形質転換体から染色体DNAを調製した。本染色体DNAに対して、プライマーM(配列番号46)とN(配列番号47)、あるいはO(配列番号48)とP(配列番号49)を用いてPCR反応を行い、染色体のトリプトファニルtRNA合成酵素遺伝子がコードされていた領域がトリプトファニルtRNA合成酵素遺伝子破壊用DNA断片由来のクロラムフェニコール耐性遺伝子と置換されていることを確認し、本菌株をトリプトファニルtRNA合成酵素遺伝子欠損株であるDTS1451(pDATS14)株とした。
【0105】
[実施例4]DTS1451株へのカナマイシン耐性遺伝子を有するプラスミドの導入
枯草菌ISW1214株の染色体を鋳型とし、プライマーCとDを用いてトリプトファニルtRNA合成酵素遺伝子とその近傍の領域を含むDNA断片を得た。本DNA断片を制限酵素XhoIで消化し、予め制限酵素XhoIで消化したpDA2と連結した。これを用いて、大腸菌HB101株を形質転換した。得られた形質転換体からプラスミドを調製し、pDATS13と命名した。汎用性プラスミドpUB110を鋳型とし、プライマーQ(配列番号50)とR(配列番号51)を用いてカナマイシン耐性遺伝子とその近傍の領域を含むDNA断片を得た。本DNA断片をEcoRIで消化し、pDATS14を鋳型としてプライマーS(配列番号52)とT(配列番号53)を用いてPCRで増幅して、さらにEcoRIで消化することにより得られたDNA断片と連結した。これを用いて、大腸菌HB101株を形質転換した。得られたカナマイシン耐性を有する形質転換体からプラスミドを調製し、プラスミドをpDATSKと命名した。pDATSKを用いてpDATS14を有するDTS1451株をプロトプラスト形質転換法を用いて形質転換し、カナマイシン30μg/mLを含む再生培地(組成:コハク酸ナトリウム 8%、寒天 1%、カザミノ酸 0.5%、酵母エキス 0.5%、リン酸2水素カリウム 0.15%、リン酸水素2カリウム 0.35%、グルコース 0.5%、硫酸マグネシウム 0.4%、牛血清アルブミン 0.01%、メチオニン 0.001%、およびロイシン 0.001%)による選抜を行った。得られた形質転換体をカナマイシン入りLB寒天培地およびテトラサイクリン7.5μg/mLを含む再生培地に植菌し、カナマイシン耐性かつテトラサイクリン感受性となった株をDTS1451(pDATSK)と命名した。
【0106】
[実施例5]遺伝子調節領域DNA断片の探索用プラスミドの構築
遺伝子調節領域DNA断片検索用ベクターpPTCFを以下の方法で構築した。
寄託番号がFERM BP−8320である、マイクロバルバイファー・エスピー JAMB−A7(Microbulbifer sp. JAMB−A7)株由来の染色体DNAを鋳型に用いて、プライマーU(配列番号54)およびプライマーW(配列番号55)を用いてPCRによる増幅を行った。得られたDNA断片を制限酵素、EcoRIで消化した後、予めEcoRIで消化したpHY300PLKと連結することにより環状プラスミドを構築した。本プラスミドを用いて大腸菌HB101株を形質転換した。形質転換体からプラスミドを調製し、寒天分解酵素遺伝子の上流にpHY300PLK由来のポリリンカー部位を有するプラスミドをpPTCFと命名した。
【0107】
[実施例6]遺伝子調節領域DNA断片の取得
Sau3AIを用いて、寄託番号がFERM AP−20227である、バチルス・エスピー JAMB750(Bacillus sp. JAMB750)株の染色体DNAを消化後、これをBamHIで切断したプラスミドpPTCFと混ぜてT4 DNAリガーゼを用いてライゲーション反応を行った。得られたライゲーション液を用いて枯草菌を形質転換した。再生培地にはDM3培地(組成:コハク酸ナトリウム 8%、寒天 1%、カザミノ酸 0.5%、酵母エキス 0.5%、リン酸2水素カリウム 0.15%、リン酸水素2カリウム 0.35%、グルコース 0.5%、硫酸マグネシウム 0.4%、牛血清アルブミン 0.01%、メチオニン 0.001%、ロイシン 0.001%、テトラサイクリン 7.5μg/ml)を用いた。形質転換体の周囲に寒天の窪みを形成した形質転換体を選び、約2,000個の形質転換体を液体培地にて培養(培地組成はポリペプトンS 3%、魚肉エキス 0.5%、酵母エキス 0.05%、リン酸2水素カリウム 0.1%、マルトース 4%、硫酸マグネシウム 0.02%、塩化カルシウム 0.05%、テトラサイクリン 7.5μg/ml、30℃で48時間振とう)した。得られた培養液毎に超音波破砕(Handy sonic model UR−20P、TOMY SEIKO CO.使用)を行い、取得された超音波破砕物の寒天分解酵素活性を測定した。その中で最も活性の高かったものを選択し、この形質転換体の持つプラスミドをpCDAG1と命名した。
【0108】
[実施例7]プラスミドpJEXOPT2の構築
寄託番号がDSM17297である、サラッソモナス・エスピー JAMB−A33(Thalassomonas sp. JAMB−A33)株由来の染色体DNAを鋳型にプライマーX(配列番号56)とプライマーY(配列番号57)を用いてPCR増幅を行い約0.3kbpから成るDNA断片を取得した。このDNA断片を制限酵素HindIIIで切断した。続いて枯草菌−大腸菌のシャトルプラスミドであるpHY300PLK(ヤクルト社製)を制限酵素HindIIIで切断し、上記のDNA断片とリガーゼ反応により連結した。本プラスミドを用いて大腸菌HB101株を形質転換した。形質転換体からプラスミドを調製しpHYTERと命名した。
【0109】
プラスミドpJEXOPT2を以下の方法で構築した。プラスミドpCDAG1を鋳型にしてリン酸化プライマーZ(配列番号58)およびリン酸化プライマーA1(配列番号59)を用いてPCRによる増幅を行った。得られたPCR断片をpUC18のSmaI部位に導入した。構築されたプラスミドを鋳型にして、プライマーZおよびプライマーA1を用いてDiversify PCR Random Mutagenesis Kit( クロンテック社)を用い、キットのプロトコールにしたがって、PCR増幅断片中にランダムに変異を導入した。このランダム変異操作を5回繰り返した。得られたPCR産物を鋳型にプライマーZおよびリン酸化プライマーA1を用いてPCR増幅を行った。その後T4DNAポリメラーゼを用いてPCR産物の末端の平滑化を行った。一方、合成一本鎖DNA断片A(配列番号74)と合成一本鎖DNA断片B(配列番号75)とをアニーリングさせ二本鎖DNAを形成させた。得られたDNA断片をリン酸化した後に、上記のPCR増幅断片とリガーゼ反応により結合した。得られたDNA断片を制限酵素EcoRI、BamHIで消化した後、EcoRIとBamHIで消化したpHYTERと連結することにより環状プラスミド群を構築した。本プラスミド群を用いて大腸菌HB101株を形質転換した。得られた数千あまりの形質転換体からまとめてプラスミド群を調製した。これをプラスミド群Aとした。
【0110】
次にβアガラーゼ生産菌である、寄託番号がFERM BP−8321である、マイクロバルバイファー・エスピー JAMB−A94(Microbulbifer sp. JAMB−A94)株の染色体を鋳型とし、プライマーB1(配列番号60)およびC1(配列番号61)を用いてβアガラーゼ酵素遺伝子DNA断片をPCR増幅した。これを制限酵素BamHIで消化した。プラスミド群Aを制限酵素BamHIで切断し、リガーゼによってβアガラーゼ酵素遺伝子と連結した。これらを大腸菌HB101株を形質転換した。βアガラーゼ活性を示す形質転換体からプラスミドを調製し、これらのプラスミドを用いて枯草菌ISW1214株を形質転換し、テトラサイクリン入り再生培地による選抜を行った。得られた形質転換体を液体培地(ポリペプトンS 3%、魚肉エキス 0.5%、酵母エキス 0.05%、リン酸水素カリウム 0.1%、マルトース 4%、塩化マグネシウム 0.02%、塩化カルシウム 0.05%、テトラサイクリン 15μg/ml)中で30℃、72時間、130rpmにて撹拌培養を行い、得られた培養上清に含まれるβアガラーゼ活性を測定した。その結果、著量のβアガラーゼを生産できるプラスミドpJEXOPT2を取得できた。
【0111】
[実施例8]発現ベクターによるγ−CGTaseの分泌生産
γ−CGTase生産菌であるバチルス・クラーキー 7364(Bacillus clarkii 7364)株由来のγ−CGTase遺伝子(J Biochem(Tokyo). 2003 133:317-324. Takada, M.)をコードする遺伝子配列を含むプラスミドpGFT−01(特許公開番号2003−102489)を鋳型として、プライマーD1(配列番号62)およびE1(配列番号63)を用いてPCR反応を行い、該酵素をコードするDNA断片を得て、これを制限酵素BamHIとSalIで消化した。pJEXOPT2を制限酵素BamHIとSalIで切断し、プラスミドのオリジンを有する方のDNA断片と、リガーゼによってγ−CGTase遺伝子と連結した。これらを大腸菌HB101株に導入して形質転換させた。γ−CGTase活性を示す形質転換体からプラスミドを調製し、pJEXOPT2Gと命名した。次に、枯草菌ISW1214株の染色体を鋳型とし、プライマーF1(配列番号64)とG1(配列番号65)を用いてトリプトファニルtRNA合成酵素遺伝子とその近傍の領域を含むPCR増幅DNA断片を得た。本DNA断片を制限酵素EcoRIで消化し、予め制限酵素EcoRIで消化したpJEXOPT2Gと連結した。これを用いて大腸菌HB101株を形質転換した。得られた形質転換体からプラスミドを調製し、pJEXOPT2GSと命名した。pJEXOPT2GSを用いてDTS1451(pDATSK)を形質転換し、テトラサイクリン 7.5μg/mL再生培地による選抜を行った。得られた形質転換体をカナマイシン入りLB寒天培地およびテトラサイクリン 7.5μg/mLを含む再生培地に植菌し、γ−CGTase活性を示し、さらにカナマイシン感受性かつテトラサイクリン耐性となった株をDTS1451(pJEXOPT2GS)と命名した。DTS1451(pJEXOPT2GS)を液体培地中(ポリペプトンS 5%、魚肉エキス 0.5%、酵母エキス 0.05%、リン酸水素カリウム 0.1%、マルトース 4%、塩化マグネシウム 0.01%、塩化カルシウム 0.1%、抗生物質を含有しない)で30℃、96時間、130rpmにて撹拌培養を行い、得られた培養上清に含まれるγ−CGTase活性を測定した。その結果、培養液1Lあたり約1.4gのγ−CGTase酵素の生産が確認できた。
【0112】
[実施例9]発現ベクターによるマルトースホスホリラーゼ生産
マルトースホスホリラーゼ生産菌である寄託番号FERM−BP8420のパエニバチルス・エスピーSH−55(Paenibacillus sp. SH−55)株由来のマルトースホスホリラーゼ遺伝子を含む組換え体プラスミドpRSMP1(国際公開番号WO2005/00343号公報記載)を組換え大腸菌からGFXPlasmid prep Kit(GEヘルスケアバイオサイエンス株式会社)を用いることにより精製した。得られた組換え体プラスミドを鋳型として、プライマーH1(配列番号66)とプライマーI1(配列番号67)を用い、PCRを行い、マルトースホスホリラーゼ遺伝子を含むDNA断片を取得した。これを制限酵素BamHIとXhoIで消化した。pJEXOPT2を制限酵素BamHIとSalIで切断し、プラスミドのオリジンを有する方のDNA断片とリガーゼによってマルトースホスホリラーゼ遺伝子と連結した。これらを大腸菌HB101株を形質転換した。マルトースホスホリラーゼ活性を示す形質転換体からプラスミドを調製し、pJEXOPT2Mと命名した。枯草菌株ISW1214株の染色体を鋳型とし、プライマーJ1(配列番号68)とK1(配列番号69)を用いてトリプトファニルtRNA合成酵素遺伝子とその近傍の領域を含むPCR増幅DNA断片を得た。本DNA断片を制限酵素EcoRIで消化し、予め制限酵素EcoRIで消化したpJEXOPT2Mと連結した。これを用いて大腸菌HB101株を形質転換した。得られた形質転換体からプラスミドを調製し、pJEXOPT2MSと命名した。pJEXOPT2MSを用いてDTS1451(pDATSK)を形質転換し、テトラサイクリン 7.5μg/mL再生培地による選抜を行った。得られた形質転換体をカナマイシン入りLB寒天培地およびテトラサイクリン 7.5μg/mLを含む再生培地に植菌し、マルトースホスホリラーゼ活性を示し、さらにカナマイシン感受性かつテトラサイクリン耐性となった株をDTS1451(pJEXOPT2MS)と命名した。DTS1451(pJEXOPT2MS)を液体培地中(ポリペプトンS 5%、魚肉エキス 0.5%、酵母エキス 0.05%、リン酸水素カリウム 0.1%、マルトース 4%、塩化マグネシウム 0.01%、塩化カルシウム 0.1%、抗生物質を含有しない)で30℃、96時間、130rpmにて撹拌培養を行い、得られた培養上清に含まれるマルトースホスホリラーゼ活性を測定した。その結果、培養液1Lあたり約0.5gのマルトースホスホリラーゼの生産が確認できた。
【0113】
[実施例10]発現ベクターによるトレハロースホスホリラーゼ生産
トレハロースホスホリラーゼ生産菌であるパエニバチルス・エスピーSH−55株由来のトレハロースホスホリラーゼ遺伝子を含む組換え体プラスミドpRSTP1(国際公開番号WO2005/00343号公報記載)を組換え大腸菌からGFXPlasmid prep Kit(GEヘルスケアバイオサイエンス株式会社)を用いることにより精製した。得られた組換え体プラスミドを鋳型として、プライマーL1(配列番号70)とプライマーM1(配列番号71)を用い、PCRを行い、トレハロースホスホリラーゼ遺伝子を含むDNA断片を取得した。これを制限酵素BamHIとXhoIで消化した。pJEXOPT2を制限酵素BamHIとSalIで切断し、プラスミドのオリジンを有する方のDNA断片とリガーゼによってトレハロースホスホリラーゼ遺伝子と連結した。これらを大腸菌HB101株を形質転換した。トレハロースホスホリラーゼ活性を示す形質転換体からプラスミドを調製し、pJEXOPT2Tと命名した。枯草菌ISW1214株の染色体を鋳型とし、プライマーN1(配列番号72)とO1(配列番号73)を用いてトリプトファニルtRNA合成酵素遺伝子とその近傍の領域を含むPCR増幅DNA断片を得た。本DNA断片を制限酵素EcoRIで消化し、予め制限酵素EcoRIで消化したpJEXOPT2Tと連結した。これを用いて大腸菌HB101株を形質転換した。得られた形質転換体からプラスミドを調製し、pJEXOPT2TSと命名した。pJEXOPT2TSを用いてDTS1451(pDATSK)を形質転換し、テトラサイクリン 7.5μg/mL再生培地による選抜を行った。得られた形質転換体をカナマイシン入りLB寒天培地およびテトラサイクリン 7.5μg/mLを含む再生培地に植菌し、トレハロースホスホリラーゼ活性を示し、さらにカナマイシン感受性かつテトラサイクリン耐性となった株をDTS1451(pJEXOPT2TS)と命名した。DTS1451(pJEXOPT2TS)を液体培地中(ポリペプトンS 5%、魚肉エキス 0.5%、酵母エキス 0.05%、リン酸水素カリウム 0.1%、マルトース 4%、塩化マグネシウム 0.01%、塩化カルシウム 0.1%、抗生物質を含有しない)で30℃、96時間、130rpmにて撹拌培養を行い、得られた培養上清に含まれるトレハロースホスホリラーゼ活性を測定した。その結果、培養液1Lあたり約1.2gのトレハロースホスホリラーゼの生産が確認できた。
【図面の簡単な説明】
【0114】
【図1】γ−シクロデキストリン合成酵素(γ−CGTase)発現ベクターの具体例。
【図2】マルトースホスホリラーゼ発現ベクターの具体例。
【図3】トレハロースホスホリラーゼ発現ベクターの具体例。
【特許請求の範囲】
【請求項1】
γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、かつ
アミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターであって、
アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞において用いられる前記組換えベクター。
【請求項2】
下記(a)〜(c)の塩基配列または遺伝子を有する組換えベクター:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;および
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子。
【請求項3】
下記(a)〜(d)の塩基配列または遺伝子を有し、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞において用いられる組換えベクター:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;および
(d)発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子。
【請求項4】
アミノアシルtRNA合成酵素が、トリプトファニルtRNA合成酵素、アラニルtRNA合成酵素、アルギニルtRNA合成酵素、アスパルギニルtRNA合成酵素、アスパルチルtRNA合成酵素、システイニルtRNA合成酵素、グルタミンtRNA合成酵素、グルタメートtRNA合成酵素、グリシンtRNA合成酵素、ヒスチジルtRNA合成酵素、イソロイシルtRNA合成酵素、ロイシルtRNA合成酵素、リシンtRNA合成酵素、メチオニルtRNA合成酵素、フェニルアラニンtRNA合成酵素、プロリルtRNA合成酵素、セリルtRNA合成酵素、トレオニルtRNA合成酵素、チロシルtRNA合成酵素、またはバリルtRNA合成酵素である、請求項1または3に記載の組換えベクター。
【請求項5】
γ−シクロデキストリン合成酵素をコードする遺伝子が、配列番号3に記載のアミノ酸配列をコードする遺伝子である、請求項1〜4のいずれか1項に記載の組換えベクター。
【請求項6】
マルトースホスホリラーゼをコードする遺伝子が、配列番号4に記載のアミノ酸配列をコードする遺伝子である、請求項1〜4のいずれか1項に記載の組換えベクター。
【請求項7】
トレハロースホスホリラーゼをコードする遺伝子が、配列番号5に記載のアミノ酸配列をコードする遺伝子である、請求項1〜4のいずれか1項に記載の組換えベクター。
【請求項8】
組換えベクターは、プラスミド、バクテリオファージ、またはレトロトランスポゾンの形態を有する請求項1〜7のいずれか1項に記載の組換えベクター。
【請求項9】
アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞であって、
γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、かつ
アミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターで形質転換して用いられる前記宿主細胞。
【請求項10】
下記(a)〜(c)の塩基配列または遺伝子を有する組換えベクターで形質転換して用いられる宿主細胞:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;および
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子。
【請求項11】
アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞であって、
下記(a)〜(d)の塩基配列または遺伝子を有する組換えベクターで形質転換して用いられる前記宿主細胞:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;および
(d)発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子。
【請求項12】
アミノアシルtRNA合成酵素が、トリプトファニルtRNA合成酵素、アラニルtRNA合成酵素、アルギニルtRNA合成酵素、アスパルギニルtRNA合成酵素、アスパルチルtRNA合成酵素、システイニルtRNA合成酵素、グルタミンtRNA合成酵素、グルタメートtRNA合成酵素、グリシンtRNA合成酵素、ヒスチジルtRNA合成酵素、イソロイシルtRNA合成酵素、ロイシルtRNA合成酵素、リシンtRNA合成酵素、メチオニルtRNA合成酵素、フェニルアラニンtRNA合成酵素、プロリルtRNA合成酵素、セリルtRNA合成酵素、トレオニルtRNA合成酵素、チロシルtRNA合成酵素、またはバリルtRNA合成酵素である、請求項9または11に記載の宿主細胞。
【請求項13】
γ−シクロデキストリン合成酵素をコードする遺伝子が、配列番号3に記載のアミノ酸配列をコードする遺伝子である、請求項9〜12のいずれか1項に記載の宿主細胞。
【請求項14】
マルトースホスホリラーゼをコードする遺伝子が、配列番号4に記載のアミノ酸配列をコードする遺伝子である、請求項9〜12のいずれか1項に記載の宿主細胞。
【請求項15】
トレハロースホスホリラーゼをコードする遺伝子が、配列番号5に記載のアミノ酸配列をコードする遺伝子である、請求項9〜12のいずれか1項に記載の宿主細胞。
【請求項16】
宿主細胞が細菌である請求項9〜15のいずれか1項に記載の宿主細胞。
【請求項17】
細菌がバチルス属細菌である、請求項16に記載の宿主細胞。
【請求項18】
下記(i)〜(iv)の工程を有する、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法:
(i)γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、さらにアミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターを用意する工程;
(ii)アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞を用意する工程;
(iii)前記宿主細胞を前記組換えベクターで形質転換して形質転換体を得る工程;および
(iv)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
【請求項19】
下記(i)〜(iii)の工程を有する、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法:
(i)下記[a]〜[c]の塩基配列または遺伝子を有する組換えベクターを用意する工程、
[a]配列番号1に記載の塩基配列、
[b]前記[a]の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子、および
[c]前記[b]の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;
(ii)前記組換えベクターで宿主細胞を形質転換して形質転換体を得る工程;および
(iii)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
【請求項20】
下記(i)〜(iv)の工程を有する、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法:
(i)下記[a]〜[d]の塩基配列または遺伝子を有する組換えベクターを用意する工程、
[a]配列番号1に記載の塩基配列、
[b]前記[a]の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子、
[c]前記[b]の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子、および
[d]発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子;
(ii)アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞を用意する工程;
(iii)前記宿主細胞を前記組換えベクターで形質転換して形質転換体を得る工程;および
(iv)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
【請求項21】
アミノアシルtRNA合成酵素が、トリプトファニルtRNA合成酵素、アラニルtRNA合成酵素、アルギニルtRNA合成酵素、アスパルギニルtRNA合成酵素、アスパルチルtRNA合成酵素、システイニルtRNA合成酵素、グルタミンtRNA合成酵素、グルタメートtRNA合成酵素、グリシンtRNA合成酵素、ヒスチジルtRNA合成酵素、イソロイシルtRNA合成酵素、ロイシルtRNA合成酵素、リシンtRNA合成酵素、メチオニルtRNA合成酵素、フェニルアラニンtRNA合成酵素、プロリルtRNA合成酵素、セリルtRNA合成酵素、トレオニルtRNA合成酵素、チロシルtRNA合成酵素、またはバリルtRNA合成酵素である、請求項18または20に記載の方法。
【請求項22】
γ−シクロデキストリン合成酵素をコードする遺伝子が、配列番号3に記載のアミノ酸配列をコードする遺伝子である、請求項18〜21のいずれか1項に記載の方法。
【請求項23】
マルトースホスホリラーゼをコードする遺伝子が、配列番号4に記載のアミノ酸配列をコードする遺伝子である、請求項18〜21のいずれか1項に記載の方法。
【請求項24】
トレハロースホスホリラーゼをコードする遺伝子が、配列番号5に記載のアミノ酸配列をコードする遺伝子である、請求項18〜21のいずれか1項に記載の方法。
【請求項25】
宿主細胞が細菌である請求項18〜24のいずれか1項に記載の方法。
【請求項26】
細菌がバチルス属細菌である請求項25に記載の方法。
【請求項27】
γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される1種の酵素の遺伝子が、前記遺伝子と95%以上の相同性を有し、かつ、前記選択される酵素の活性を有するタンパク質をコードする遺伝子である、請求項5〜8のいずれか1項に記載の組換えベクター。
【請求項28】
γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される1種の酵素の遺伝子が、前記遺伝子と95%以上の相同性を有し、かつ、前記選択される酵素の活性を有するタンパク質をコードする遺伝子である、請求項13〜17のいずれか1項に記載の宿主細胞。
【請求項29】
γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される1種の酵素の遺伝子が、前記遺伝子と95%以上の相同性を有し、かつ、前記選択される酵素の活性を有するタンパク質をコードする遺伝子である、請求項22〜26のいずれか1項に記載の方法。
【請求項1】
γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、かつ
アミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターであって、
アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞において用いられる前記組換えベクター。
【請求項2】
下記(a)〜(c)の塩基配列または遺伝子を有する組換えベクター:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;および
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子。
【請求項3】
下記(a)〜(d)の塩基配列または遺伝子を有し、アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞において用いられる組換えベクター:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;および
(d)発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子。
【請求項4】
アミノアシルtRNA合成酵素が、トリプトファニルtRNA合成酵素、アラニルtRNA合成酵素、アルギニルtRNA合成酵素、アスパルギニルtRNA合成酵素、アスパルチルtRNA合成酵素、システイニルtRNA合成酵素、グルタミンtRNA合成酵素、グルタメートtRNA合成酵素、グリシンtRNA合成酵素、ヒスチジルtRNA合成酵素、イソロイシルtRNA合成酵素、ロイシルtRNA合成酵素、リシンtRNA合成酵素、メチオニルtRNA合成酵素、フェニルアラニンtRNA合成酵素、プロリルtRNA合成酵素、セリルtRNA合成酵素、トレオニルtRNA合成酵素、チロシルtRNA合成酵素、またはバリルtRNA合成酵素である、請求項1または3に記載の組換えベクター。
【請求項5】
γ−シクロデキストリン合成酵素をコードする遺伝子が、配列番号3に記載のアミノ酸配列をコードする遺伝子である、請求項1〜4のいずれか1項に記載の組換えベクター。
【請求項6】
マルトースホスホリラーゼをコードする遺伝子が、配列番号4に記載のアミノ酸配列をコードする遺伝子である、請求項1〜4のいずれか1項に記載の組換えベクター。
【請求項7】
トレハロースホスホリラーゼをコードする遺伝子が、配列番号5に記載のアミノ酸配列をコードする遺伝子である、請求項1〜4のいずれか1項に記載の組換えベクター。
【請求項8】
組換えベクターは、プラスミド、バクテリオファージ、またはレトロトランスポゾンの形態を有する請求項1〜7のいずれか1項に記載の組換えベクター。
【請求項9】
アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞であって、
γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、かつ
アミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターで形質転換して用いられる前記宿主細胞。
【請求項10】
下記(a)〜(c)の塩基配列または遺伝子を有する組換えベクターで形質転換して用いられる宿主細胞:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;および
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子。
【請求項11】
アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞であって、
下記(a)〜(d)の塩基配列または遺伝子を有する組換えベクターで形質転換して用いられる前記宿主細胞:
(a)配列番号1に記載の塩基配列;
(b)前記(a)の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子;
(c)前記(b)の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;および
(d)発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子。
【請求項12】
アミノアシルtRNA合成酵素が、トリプトファニルtRNA合成酵素、アラニルtRNA合成酵素、アルギニルtRNA合成酵素、アスパルギニルtRNA合成酵素、アスパルチルtRNA合成酵素、システイニルtRNA合成酵素、グルタミンtRNA合成酵素、グルタメートtRNA合成酵素、グリシンtRNA合成酵素、ヒスチジルtRNA合成酵素、イソロイシルtRNA合成酵素、ロイシルtRNA合成酵素、リシンtRNA合成酵素、メチオニルtRNA合成酵素、フェニルアラニンtRNA合成酵素、プロリルtRNA合成酵素、セリルtRNA合成酵素、トレオニルtRNA合成酵素、チロシルtRNA合成酵素、またはバリルtRNA合成酵素である、請求項9または11に記載の宿主細胞。
【請求項13】
γ−シクロデキストリン合成酵素をコードする遺伝子が、配列番号3に記載のアミノ酸配列をコードする遺伝子である、請求項9〜12のいずれか1項に記載の宿主細胞。
【請求項14】
マルトースホスホリラーゼをコードする遺伝子が、配列番号4に記載のアミノ酸配列をコードする遺伝子である、請求項9〜12のいずれか1項に記載の宿主細胞。
【請求項15】
トレハロースホスホリラーゼをコードする遺伝子が、配列番号5に記載のアミノ酸配列をコードする遺伝子である、請求項9〜12のいずれか1項に記載の宿主細胞。
【請求項16】
宿主細胞が細菌である請求項9〜15のいずれか1項に記載の宿主細胞。
【請求項17】
細菌がバチルス属細菌である、請求項16に記載の宿主細胞。
【請求項18】
下記(i)〜(iv)の工程を有する、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法:
(i)γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子を発現可能な状態で有し、さらにアミノアシルtRNA合成酵素をコードする遺伝子を発現可能な状態で有する組換えベクターを用意する工程;
(ii)アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞を用意する工程;
(iii)前記宿主細胞を前記組換えベクターで形質転換して形質転換体を得る工程;および
(iv)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
【請求項19】
下記(i)〜(iii)の工程を有する、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法:
(i)下記[a]〜[c]の塩基配列または遺伝子を有する組換えベクターを用意する工程、
[a]配列番号1に記載の塩基配列、
[b]前記[a]の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子、および
[c]前記[b]の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子;
(ii)前記組換えベクターで宿主細胞を形質転換して形質転換体を得る工程;および
(iii)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
【請求項20】
下記(i)〜(iv)の工程を有する、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する方法:
(i)下記[a]〜[d]の塩基配列または遺伝子を有する組換えベクターを用意する工程、
[a]配列番号1に記載の塩基配列、
[b]前記[a]の塩基配列の下流に直接的または間接的に連結された配列番号2に記載のアミノ酸配列をコードする遺伝子、
[c]前記[b]の遺伝子の下流に直接的に連結された、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素をコードする遺伝子、および
[d]発現可能な状態にあるアミノアシルtRNA合成酵素をコードする遺伝子;
(ii)アミノアシルtRNA合成酵素をコードする染色体遺伝子を欠損させた変異を有する宿主細胞またはアミノアシルtRNA合成酵素をコードする染色体遺伝子の発現を宿主細胞が成育できない程度に低下させた変異を有する宿主細胞を用意する工程;
(iii)前記宿主細胞を前記組換えベクターで形質転換して形質転換体を得る工程;および
(iv)前記形質転換体を培養して、γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される少なくとも1種の酵素を生産する工程。
【請求項21】
アミノアシルtRNA合成酵素が、トリプトファニルtRNA合成酵素、アラニルtRNA合成酵素、アルギニルtRNA合成酵素、アスパルギニルtRNA合成酵素、アスパルチルtRNA合成酵素、システイニルtRNA合成酵素、グルタミンtRNA合成酵素、グルタメートtRNA合成酵素、グリシンtRNA合成酵素、ヒスチジルtRNA合成酵素、イソロイシルtRNA合成酵素、ロイシルtRNA合成酵素、リシンtRNA合成酵素、メチオニルtRNA合成酵素、フェニルアラニンtRNA合成酵素、プロリルtRNA合成酵素、セリルtRNA合成酵素、トレオニルtRNA合成酵素、チロシルtRNA合成酵素、またはバリルtRNA合成酵素である、請求項18または20に記載の方法。
【請求項22】
γ−シクロデキストリン合成酵素をコードする遺伝子が、配列番号3に記載のアミノ酸配列をコードする遺伝子である、請求項18〜21のいずれか1項に記載の方法。
【請求項23】
マルトースホスホリラーゼをコードする遺伝子が、配列番号4に記載のアミノ酸配列をコードする遺伝子である、請求項18〜21のいずれか1項に記載の方法。
【請求項24】
トレハロースホスホリラーゼをコードする遺伝子が、配列番号5に記載のアミノ酸配列をコードする遺伝子である、請求項18〜21のいずれか1項に記載の方法。
【請求項25】
宿主細胞が細菌である請求項18〜24のいずれか1項に記載の方法。
【請求項26】
細菌がバチルス属細菌である請求項25に記載の方法。
【請求項27】
γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される1種の酵素の遺伝子が、前記遺伝子と95%以上の相同性を有し、かつ、前記選択される酵素の活性を有するタンパク質をコードする遺伝子である、請求項5〜8のいずれか1項に記載の組換えベクター。
【請求項28】
γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される1種の酵素の遺伝子が、前記遺伝子と95%以上の相同性を有し、かつ、前記選択される酵素の活性を有するタンパク質をコードする遺伝子である、請求項13〜17のいずれか1項に記載の宿主細胞。
【請求項29】
γ−シクロデキストリン合成酵素、マルトースホスホリラーゼ、およびトレハロースホスホリラーゼからなる群から選択される1種の酵素の遺伝子が、前記遺伝子と95%以上の相同性を有し、かつ、前記選択される酵素の活性を有するタンパク質をコードする遺伝子である、請求項22〜26のいずれか1項に記載の方法。
【図1】


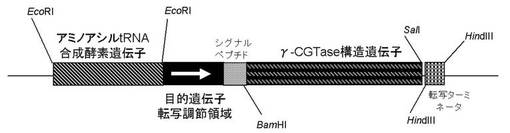
【図2】
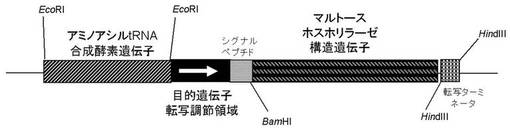
【図3】
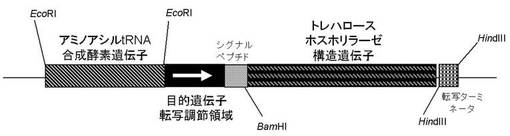
【図2】
【図3】
【公開番号】特開2009−17842(P2009−17842A)
【公開日】平成21年1月29日(2009.1.29)
【国際特許分類】
【出願番号】特願2007−183937(P2007−183937)
【出願日】平成19年7月13日(2007.7.13)
【出願人】(504194878)独立行政法人海洋研究開発機構 (110)
【出願人】(000231453)日本食品化工株式会社 (68)
【Fターム(参考)】
【公開日】平成21年1月29日(2009.1.29)
【国際特許分類】
【出願日】平成19年7月13日(2007.7.13)
【出願人】(504194878)独立行政法人海洋研究開発機構 (110)
【出願人】(000231453)日本食品化工株式会社 (68)
【Fターム(参考)】
[ Back to top ]