
眼乾燥症候群治療におけるガレクチン組成物及びその使用方法
哺乳類における眼乾燥の薬剤治療方法であり、この方法は、上皮創傷に苦しむ哺乳類に対し治療に有効量のガレクチン‐1タンパク質、ガレクチン‐3、ガレクチン‐7、及び/又はガレクチン‐8タンパク質を提供する過程を備える。薬剤及び眼科系組成物並びにキットを提供し、該キットは、薬学的有効成分としてガレクチン‐1タンパク質、ガレクチン‐3、ガレクチン‐7、及び/又はガレクチン‐8タンパク質を備え、適当な担体又は希釈剤を備える。

【発明の詳細な説明】
【技術分野】
【0001】
眼乾燥症候群治療のためのガレクチンタンパク質の組成物、方法及びキットを提供する。
【背景技術】
【0002】
眼乾燥症候群(DES)はよくある症状で、30から45歳人口の10%もが症状を示し、45歳以上では増加し、人口の20%もが症状を示す。(Schein et al., 1997, Am J Ophthamol 124, 723‐72; Brewitt and Sistani, 2001, Surv of Ophthalmol 45:S119‐S202)。DESは眼球刺激、かすみ目及び視野振動を引き起こし、角膜感染や潰瘍にかかる視力にかかわる危険性を増加する。DESの組織学的な影響としては、異常増殖や分化が挙げられ、眼球表面上皮に、結膜杯細胞密度の減少及び眼球表面上皮による粘液生産の減少及び異常が見られる(Murillo‐Lopez and Pflugfelder, 1996, Dry Eye. In: Krachmer J, Mannis M, Holland E, eds. The Cornea. Mosby, St. Louis, MO. 663‐686; Dursun et al., 2002, Invest Ophthalmol & Vis Sci 43:632‐638)。眼乾燥症は慢性疾患で、症状や兆候は環境要因により大きく影響され、湿度や空気移動などが挙げられるが、読書やコンピューターの使用といった看視作業の度合いにも大きく影響される。
【0003】
DESの典型的な症状としては、灼熱痛、かゆみ、異物感、刺痛、乾燥、光線恐怖症、眼球疲労、及び発赤が挙げられる。眼乾燥症は慢性疾患で、症状や兆候は環境要因により大きく影響され、湿度や空気移動などが挙げられるが、読書やコンピューターの使用といった看視作業の度合いにも大きく影響される(Rheinstrom, 1999, Dry eye. In Yanoff, ed. Ophthalmology. 1st Ed. Editor. Mosley International Ltd, St Louis, MO; Foulks, 2003, The Ocular Surface, 1: 20‐30)。
【0004】
多種多様の人工涙液製品が販売されているが、どれもが症状を一時的に緩和するだけである。今のところ、症状を改善するような治療薬はない。従って、本技術分野では、眼乾燥症候群を治療する、更なる医薬品及び組成物が必要とされている。特に、薬品、組成物及び治療方法が必要とされている。
【0005】
【非特許文献1】Schein et al., 1997, Am J Ophthamol 124, 723‐72
【非特許文献2】Brewitt and Sistani, 2001, Surv of Ophthalmol 45:S119‐S202
【非特許文献3】Murillo‐Lopez and Pflugfelder, 1996, Dry Eye. In: Krachmer J, Mannis M, Holland E, eds. The Cornea. Mosby, St. Lois, MO. 663‐686
【非特許文献4】Dursun et al., 2002, Invest Ophthalmol & Vis Sci 43:632‐638
【非特許文献5】Rheinstrom, 1999, Dry eye. In Yanoff, ed. Ophthalmology. 1st Ed. Editor. Mosley International Ltd, St Louis, MO
【非特許文献6】Foulks, 2003, The Ocular Surface, 1: 20‐30
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明は、治療を必要としてする哺乳類の眼乾燥を治療する方法であって、治療に有効な量のガレクチンタンパク質を哺乳類に投与することを特徴とする。ある実施例において、哺乳類における眼乾燥の予防方法が挙げられ、眼乾燥予防を必要とする哺乳類の同定、及び同定した哺乳類に対し治療に有効なガレクチンタンパク質を投与する工程からなる。従って、該方法は治療又は予防を必要としている哺乳類に対して用いられ、少なくとも、眼上皮創傷である哺乳類、抗ヒスタミン剤の使用前である哺乳類、抗炎症剤の使用前である哺乳類、エクシマーレーザー治療を施す前の哺乳類のいずれかに当てはまる哺乳類の治療又は予防に対し用いられる。例えば、哺乳類にはヒトが挙げられる。更に、ガレクチンタンパク質はガレクチン‐8、ガレクチン‐7もしくはガレクチン‐3から選択される。ある実施例において眼乾燥が持続性症候群である。例えば、眼乾燥は上皮びらんを引き起こし、上皮びらんは更に角膜創傷へとつながる。
【0007】
一般的には、上述の方法で用いられるガレクチン-8タンパク質は、配列番号4又は5のアミノ酸配列を備える。例えば、ガレクチン‐8タンパク質は、配列番号4又は5のアミノ酸配列と実質的に同一のアミノ酸配列を含む。本明細書中で用いられている「実質的に同一」とは、ガレクチン‐8は、配列番号4又は5のアミノ酸配列と、少なくとも60%の同一性、70%の同一性、80%の同一性、又は90%の同一性を持つことを意味する。
【0008】
一般的には、上述の方法で用いられるガレクチン‐7タンパク質は、配列番号2のアミノ酸配列を備える。例えば、ガレクチン‐7タンパク質は、配列番号2のアミノ酸配列と実質的に同一のアミノ酸配列を含む。ガレクチン‐7は、配列番号2のアミノ酸配列と、少なくとも60%の同一性、70%の同一性、80%の同一性、又は90%の同一性を持つ。ある実施例ではガレクチンン‐7タンパク質はガレクチン‐7ガラクシド結合領域を含む。
【0009】
一般的には、上述の方法で用いられるガレクチン‐3タンパク質は、配列番号1のアミノ酸配列を備える。例えば、ガレクチン‐3タンパク質は、配列番号1のアミノ酸配列と実質的に同一のアミノ酸配列を含む。ガレクチン‐3は、配列番号1のアミノ酸配列と、少なくとも60%の同一性、70%の同一性、80%の同一性、又は90%の同一性を持つ。ある実施例ではガレクチンン‐3タンパク質はガレクチン‐3ガラクシド結合領域を含む。
【0010】
本発明の他の実施例としては、眼乾燥の治療促進効果を持つ医薬品組成物が挙げられ、該医薬品組成物は、薬学的に適当な担体もしくは希釈剤、及結膜上皮及び/又は角膜上皮の健全性を促進する十分量のガレクチン‐8タンパク質を備える。本発明の他の実施例としては、眼乾燥の治療に促進効果のある医薬品組成物が挙げられ、該医薬品組成物は、薬学的に適当な担体もしくは希釈剤、並びに結膜上皮及び/又は角膜上皮の健全性を促進する十分量のガレクチン‐7タンパク質を備える。本発明の他の実施例としては、眼乾燥の治療に促進効果のある医薬品組成物が挙げられ、該医薬品組成物は、薬学的に適当な担体もしくは希釈剤、及び結膜上皮及び/又は角膜上皮の健全性を促進する十分量のガレクチン‐3タンパク質を備える。本発明の他の実施例としては、眼乾燥の治療に促進効果のある医薬品組成物が挙げられ、該医薬品組成物は、薬学的に適当な担体もしくは希釈剤、及び結膜上皮及び/又は角膜上皮の健全性を促進する十分量のガレクチン‐8タンパク質を備える。これらの医薬品組成物のいずれかにおいて、眼乾燥が再発性上皮びらんを伴う眼乾燥症である。例えば、眼乾燥症は創傷を引き起こし、該創傷は角膜創傷である。更に眼乾燥は、エクシマーレーザー角膜切除術により引き起こされる。
【課題を解決するための手段】
【0011】
様々な医薬品組成物の実施例において、ガレクチン‐8タンパク質は、配列番号4又は5のアミノ酸配列を含む。又はガレクチン8タンパク質はアミノ酸番号4又は5のアミノ酸配列と実質的に同一なアミノ酸配列を含む。或いは、ガレクチン‐8タンパク質は、ガレクチン‐8N末端領域と,ガレクチン‐8プロリン、グリシン、及びチロシンが豊富な領域とを含む。もしくは、ガレクチン‐8タンパク質は、ガレクチン‐8プロリン、グリシン、及びチロシンが豊富な領域と、ガレクチン‐8ガラクトシド結合領域とを含む。例えば、ガレクチン‐8タンパク質はガレクチン‐8ガラクトシド結合領域を含む。或いは、ガレクチン‐7タンパク質は配列番号2のアミノ酸配列を含む。例えば、ガレクチン‐7タンパク質は配列番号2のアミノ酸配列と実質的に同一なアミノ酸配列を含む。又は、ガレクチン‐7タンパク質はガレクチン‐7ガラクトシド結合領域を含む。或いは、ガレクチン‐3タンパク質は配列番号1のアミノ酸配列を含む。例えば、ガレクチン‐3タンパク質はアミノ酸番号1のアミノ酸配列と実質的に同一なアミノ酸配列を含む。例えば、ガレクチン‐3タンパク質はガレクチン‐3ガラクトシド結合領域を含む。
【0012】
本発明は、更に治療を必要としている哺乳類の眼乾燥症を治療する方法であることを特徴とし、治療に有効量の物質を投与する工程を備え、該物質は角膜表面又は結膜表面上での涙液層の拡散に影響する。また、本発明は、治療を必要としている哺乳類における眼乾燥症の治療方法であることを特徴とし、眼乾燥症に苦しむ哺乳類において治療に有効量の物質を投与する工程からなり、該物質はガレクチン‐8タンパク質の発現に影響する。例えば、該物質とは、配列番号4又は5のアミノ酸配列を持つガレクチン‐8タンパク質を備える。例えば、該物質とは、配列番号4又は5のアミノ酸配列と実質的に同一のアミノ酸配列を持つガレクチン‐8タンパク質を備える。
【発明を実施するための最良の形態】
【0013】
本発明は、様々な特許、科学論文、その他の出版物に言及している。それぞれの事項の内容は参照することにより本明細書に含まれるものとする。またここで言及する全てのウェブサイトの内容(本出願時のもの)についても、それらを参照することにより本明細書に含まれるものとする。
【0014】
本発明の実施例は、炭水化物結合タンパク質であるガレクチンが、角膜及び結膜表皮において涙液層の拡散を促進するという概念に基づき示されている。ガレクチンはガラクト−ス結合タンパク質で、ムチンのO結合したオリゴ糖に見られるGalβ1‐4GlcNAC二糖に高い親和性で結合する。ガレクチンの多くは二価もしくは多価であり、ガレクチンが分泌性ムチンのオリゴ糖鎖と膜貫通型ムチン(その他の糖タンパク質)を結合することにより、涙液層の拡散を促進する。前記膜貫通型ムチン(その他の糖タンパク質)は角膜及び結膜上皮の最端面に存在するものである。
【0015】
本発明は、ガレクチン‐8、ガレクチン‐3及び/又はガレクチン‐7を備える医薬品組成物を提供し、損傷した哺乳類組織の傷の再上皮化を促進する際に有用である。本発明は、更に、上皮損傷した哺乳類組織における治療上の処置方法を提供し、該方法は上皮損傷を被った哺乳類に、治療に有効量のガレクチン‐1、ガレクチン‐3、ガレクチン‐7、ガレクチン‐8、又はガレクチン‐1、‐3、‐7、‐8のうち少なくとも2種類の混合物を投与する工程を備える。ガレクチン‐1、‐3、‐7、‐8の混合物が投与される場合、前述ガレクチンのいずれもが、他のガレクチンの投与前、併用して、もしくは投与後に投与可能である。
【0016】
本発明は、ガレクチン‐3が、損傷に伴って角膜上皮細胞の角膜への移動が上方制御されているという発見を包含する(例1)。また、本発明は、野生型マウスに比べてガレクチン‐3欠損型マウスでは、角膜経上皮エクシマーレーザー創傷及び角膜のアルカリによる火傷の再上皮化が際立って遅くなるという発見も含む(例2)。本発明は更に、ガレクチン‐3欠損型マウスでは、多くの損傷関連遺伝子(例えば、トロイド様タンパク質及びガレクチン‐7)の発現が異常であるという発見も提供している(例3)。更に、本発明は、外因性ガレクチン‐3及びガレクチン‐7が角膜傷の再上皮化を促進することも示している(それぞれ、例4、5)。
【0017】
<ガレクチン>
レクチンはタンパク質で、特に炭水化物に対する結合能、細胞凝集能により明確にされている(例えば、Sharon, Trends Biochem. Sci. 18:221, 1993参照)。レクチンは多種多様の細胞機能を担っていることが示されており、該機能は細胞‐細胞及び細胞‐マトリックス間相互作用を含む。レクチンは植物、無脊椎動物及び哺乳類の広範囲に見られる。動物のレクチンは4つの異なった群に区別されており、1)C型レクチン、2)P型レクチン、3)ガレクチン(以前はS型レクチンと称されていた)、及び4)ペントラキシンが挙げられる(例えば、Barondes et al., J. Biol. Chem. 269:20807, 1994)。
【0018】
全ての哺乳類のガレクチンは、β‐ラクトース及びβ‐ガラクトシド関連因子に対する認識について詳細に解析されている。全ての哺乳類のガレクチンは小さいβガラクトシドに対し同様な親和性を示すのに対し、より複雑な複合多糖に対しては、結合特異性に著しい違いが見られる(Henrick et al., Glycobiology 8;45, 1998; Sato et al., J. Biol. Chem. 267:6983, 1992; Seetharaman et al., J. Bioil. Chem. 273:13047, 1998)。β‐ガラクトシド糖類への結合に加えて、ガレクチンは赤血球凝集活性を持つ。ラミニンは自然界に存在する糖タンパク質で、多数のポリラクトサミン鎖を多く持ち、特定のガレクチンに対する天然リガンドであることが示されている。ラミニンは基底層及び細胞外マトリックスの構成要素であり、細胞外マトリックスは全ての上皮の下にあり、個々の筋肉、脂肪、及びシュワン細胞を取り囲んでいる。細胞と基底層の相互作用は、哺乳類の成長過程において、種々の細胞種の輸送及び/又は分化に影響を与えることが知られている。ガレクチンは膜移動を特定する在来の配列は持たないが、細胞内で分泌され細胞内に存在する。βガラクトシド糖に対する親和性だけでなく、ガレクチンのファミリーメンバーは炭水化物認識領域(CRD、又はタンパク質結合領域とも呼ばれる)における著しい配列類似性を共有する。炭水化物認識領域に関連するアミノ酸残基は、X線結晶学により決定されている(Lobsanov et al, J. Biol. Chem. 267:27034, 1993 and Seethnaraman et al., supra)。ガレクチンは、多岐にわたる生物機能に関与しているとされ、該機能は細胞接着(Copper et al., J. Cell Biol. 115:1437, 1991)、成長制御(Wells et al., Cell 64:91, 1991)、細胞移動(Hughes, Curr. Opin. Struct. Biol. 2:687, 1992)、悪性形質転換(Raz et al., Int. J. Cancer 46:871, 1990)及び免疫反応(Offner et al, J. Neuroimmunol. 28:177, 1990)が挙げられる。現在、12のガレクチンファミリーの真核メンバーが特徴付けられている。
【0019】
<ガレクチン‐3>
ガレクチン‐3ファミリーメンバーのタンパク質(以前はCBP‐35、Mac‐2、L‐34、εBP及びRL‐29として知られていた)は、一般的には約240から270のアミノ酸を含み、分子量は約25から29kDaの範囲である。ガレクチン‐3タンパク質は、一般的には、短いN末端領域と、ガラクトシド結合領域を含むC末端領域と、介在するプロリン、グリシン及びチロシンが豊富な領域とからなる。該介在するプロリン、グリシン及びチロシンが豊富な領域は、7から10の保存アミノ酸配列の繰り返しを含む(Liu et al., Biochemistry 35:6073, 1996及びCherayil et al., Proc. Natl. Acad. Sci. USA, 87:7324, 1990)。縦列反復はコラ−ゲン遺伝子ス−パ−ファミリ−に見られる反復と類似性がある。反復数はガレクチン‐3タンパク質により様々で、異なる種によるガレクチン‐3タンパク質の大きさの違いを説明している。ガレクチン‐3N末端領域は、複合糖質のリガンドを含む面に結合して、該タンパク質を多量体化させる。
【0020】
ガレクチン‐3は様々な炎症細胞(活性型マクロファージ、好塩基球及び肥満細胞)及び多種の組織の上皮及び繊維芽細胞で発現している(Perillo et al., J. Mol. Med. 76:402, 1998)。ガレクチン‐3は細胞表面、細胞外マトリックス(ECM)、細胞質及び細胞核に見られる。ガレクチン‐3は、細胞表面上又はECMで、細胞‐細胞及び細胞‐マトリックス間相互作用を媒介しているとされ、該相互作用は、複合糖質に相補的に結合することでなされる。該複合糖質はECM及び細胞表面分子に多く見られるポリラクトサミン鎖を含む。ガレクチン‐3は、ラミンに結合することで細胞‐マトリックス間接着を抑制しているとされる。細胞核では、ガレクチン‐3は公知の細胞接着分子(例えば、α6β1及びα4β7インテグリン、Warlfield et al., Invasion Metastasis 17:101, 1997及び、Matarrese et al., Int. J. Cancer 85:545, 2000)及びサイトカイン(例えば、IL‐1、Jeng et al., Immunol. Lett. 42: 113, 1994)の発現を制御することで、細胞‐マトリックス間相互作用に非間接的に関与することもできる。腎臓などの特定の器官では、ガレクチン‐3の発現は発達的に制御されており、肺胞上皮細胞及び肝細胞におけるガレクチン‐3の発現レベルでは損傷に伴い上方制御されている。増殖過程では、ガレクチン‐3は特定の細胞種の核内で凝縮することが示されている。ガレクチン‐3の発現は、特定の腫瘍では増加していることから、ガレクチン‐3は転移に関与していると示唆される。実際に、弱転移性細胞系でのガレクチン‐3の過剰発現は、転移能を顕著に増加させる(Raz et al., supra)。
【0021】
ヒトのガレクチン‐3は250アミノ酸長からなり、約26.1kDaの分子量を持つ(配列番号1、図1)。図1、3、5及び7に示してあるように、ヒトガレクチン‐3は次の様な領域、特徴的配列又はその他の構造上の特徴を持つ(PS及びPFの接頭認識番号に関する一般情報は、Sonnhammer et al., Protein 28:405, 1997参照)。配列番号1のアミノ酸残基1から14の周囲にあるN末端領域、配列番号1のアミノ酸残基15から116の周囲にあるプロリン、グリシン及びチロシンが豊富な領域、配列番号1のアミノ酸残基117から247の周囲にあるガラクトシド結合領域、配列番号1のアミノ酸残基181から200の周囲にあるガラプチン特徴的配列(PROSITE番号、PS00309)、配列番号1のアミノ酸残基4から7の周囲にある一つの潜在的なN‐糖鎖形成部位(PROSITE番号、PS00001)、配列番号1のアミノ酸残基137から179及び194から196の周囲にある二つの潜在的なタンパク質キナーゼCリン酸化部位(PROSITE番号、PS00005)、配列番号1のアミノ酸残基6から9及び175から178の周囲にある二つの潜在的なカゼインキナーゼIIリン酸化部位(PROSITE番号、PS00006)、及び、配列番号1のアミノ酸残基24から29、27から32、34から39、43から48、52から57、61から66、65から70及び68から73の周囲にある八つの潜在的なミリストイル化部位(PROSITE番号、PS00008)が挙げられる。
【0022】
本明細書に定義するように、ガレクチン‐3タンパク質は、「ガレクチン‐3N末端領域」、ガレクチン‐3「プロリン、グリシン及びチロシンが豊富な領域」及び/又はガレクチン‐3「ガラクトシド結合領域」を含む。これらの領域は、更に以下のように定義される。
【0023】
本明細書に用いられているように、ガラクチン‐3「N末端領域」には約10から20のアミノ酸、好適には約14のアミノ酸のアミノ酸配列が含まれ、配列番号1の1から14のアミノ酸と少なくとも60%、70%、80%、90%、95%、99%又は100%の同一性を有するものとする。N末端領域は、N‐糖鎖形成部位(PROSITE番号、PS00001)及び/又はカゼインキナーゼIIリン酸化部位(PROSITE番号、PS00006)を含むことが可能である。PROSITEのN‐糖鎖形成部位は、共通配列として、N‐{P}‐[ST]‐{P}を持ち、カゼインキナーゼIIリン酸化部位は、共通配列として、[ST]‐X(2)‐[DE]を持つ。上述の共通配列及び本明細書に記載の他のモチ−フ又は特徴的配列では、IUPAC規格であるアミノ酸の一文字記号が用いられている。様式の各要素はダッシュ(‐)で区切られており、角括弧([])は各位置で許容されている特定の残基を示し、Xは各位置で許容されている任意の残基を示し、丸括弧(())内の番号は付随アミノ酸によって示されている残基数を示す。ある実施例によっては、N末端領域は配列番号1のアミノ酸L7及びL11を含む。図3に示す様に、これらのアミノ酸はガレクチン‐3の様々な哺乳類の種を超えて保存されており、該アミノ酸は触媒及び/又は構造的な役割を果たす。
【0024】
本明細書にあるように、ガレクチン‐3の「プロリン、グリシン及びチロシンが豊富な領域」は約60から140のアミノ酸のアミノ酸配列を含み、より好適には約80から120のアミノ酸、又は約90から110のアミノ酸のアミノ酸配列を含み、配列番号1の15から116アミノ酸と約70%、80%、90%、95%、99%又は100%の同一性を持つ。プロリン、グリシン及びチロシンが豊富な領域は、1,2,3,4,5,6,7又は8のN‐ミリストイル化部位(PROSITE番号、PS00008)を含むことも可能であり、該N‐ミリストイル化部位は共通配列として、G‐{EDRKHPFYW}‐X(2)‐[STAGCN]‐{P}を持つ。ある実施例では、プロリン、グリシン及びチロシンが豊富な領域は配列番号1の以下のアミノ酸及び領域を含む。配列及び領域には、G21、P23、G27、N28、P30、G32、G34、P37、Y41‐P46、G53、Y55‐G57、P61、G62、G66、P72、G73、G77、Y79‐G81、P83、G87、Y89、P90、G99、Y101、P102、P106、Y107、A109、L114及びV116が挙げられる。これらのアミノ酸及び領域は、ガレクチン‐3の様々な哺乳類の種を超えて保存されており、該アミノ及び領域は触媒作用及び/又は構造上の役割を担う。(図3において「*」で示されたアミノ酸を参照)
【0025】
本明細書にあるように、ガレクチン‐3の「ガラクトシド結合領域」は約80から180のアミノ酸のアミノ酸配列を含み、該アミノ酸配列はPFAM(番号3)に登録されている共通配列PF00337との配列比較で、少なくとも150のビットスコアを持つ。好適には、ガレクチン‐3のガラクトシド結合領域は少なくとも約100から160のアミノ酸を含み、より好適には110から150のアミノ酸又は約120から140のアミノ酸を含み、PFAM(番号3)に登録されている共通配列PF00337との配列比較で、少なくとも150、より好適には少なくとも175、更に好適には200以上のビットスコアを持つ。
【0026】
特定の配列とPFAM(番号3)に登録されている共通配列PF00337との配列比較におけるビットスコアを計算するには、対象配列をHMMのPFAMデ−タベ−ス(例えばPFAMデータベース、リリース2.1)で検索し、初期変数には、www.sanger.ac.uk/Sogtware/Pfam.で得られる数値を用いる。PFAMデ−タベ−スの説明は、Sonnhammer et al., supraに記載され、HMMの詳細は例えば、文献(Griboskov et al., Meth. Enzymol. 183:146,1990及びStulz et al., Protein Sci. 2:305, 1993)に記載されている。
【0027】
ガレクチン‐3のガラクトシド結合領域は更に1つの、好ましくは2つのタンパク質キナーゼCリン酸化部位(PROSITE番号、PS00005)、カゼインキナーゼIIリン酸化部位(PROSITE番号、PS00006)及び/又はガラプチン特徴的配列(PROSITE番号、PS00309)を含むことができる。タンパク質キナーゼCリン酸化部位は、次の共通配列、[ST]‐X‐[RK]を持つ。ガラプチン特徴的配列は次の共通配列、W‐[GEK]‐X‐[EQ]‐X‐[KRE]‐X(3、6)‐[PCTF]‐[LIVMF]‐[NQEGSKV]‐X‐[GH]‐X(3)‐[DENKHS]‐[LIVMFC]を持つ。ある実施では、ガレクチン‐3のガラクトシド結合領域は配列番号1の次のアミノ酸及び領域、P117、Y118、L120‐L122、G125、P128、R129、L131‐I134、G136‐V138、N141、N143、R144、L147、F149、R151、G152、D154、A156‐F163、E165、R169‐N174、N179‐G182、E184‐R186、F190‐E193、G195、P197‐K199、Q201‐L203、E205、D207‐Q220、N222、R224、L228、I231、I236、G238‐I240及びL242‐S244を持つ。これらのアミノ酸及び領域は、ガレクチン‐3の様々な哺乳類の種を超えて保存されており、該アミノ酸及び領域は触媒作用及び/又は構造上の役割を担う。(図3に「*」で表記されたアミノ酸を参照)
【0028】
本発明のあるガレクチン‐3タンパク質は、配列番号1に示されるヒトのガレクチン‐3のアミノ酸配列を含む。本発明の他のガレクチン‐3タンパク質は、配列番号1のアミノ酸配列と実質的に同一のアミノ酸配列を持つ。「実質的に同一」とは、本明細書では、十分又は最低限の数のアミノ酸残基を含む第1のアミノ酸を言及し、該アミノ酸残基は、第2のアミノ酸配列の整列アミノ酸配列残基と同一であり、これにより第1及び第2のアミノ酸配列は共通の構造領域及び/又は共通の機能活性を持つことが可能であるとする。例えば、配列番号1と少なくとも60%又は65%の同一性、好ましくは、少なくとも75%の同一性、より好ましくは少なくとも85%、90%、91%、92%、93%、94%、95%、96%、97%、98%又は99%の同一性を持つ共通の構造領域を含むアミノ酸配列は、配列番号1のアミノ酸配列と実質的に同一とされる。特に、配列番号1の特定アミノ酸残基における欠失、付加、置換もしくは変形などの偶発的又は意図的に誘導された変異を含むタンパク質は、本明細書記載の、ガレクチン‐3の定義に含まれる。ここに定義するように、ガレクチン‐3タンパク質は、他の哺乳類種のガレクチン‐3のアミノ酸配列によって示される領域を含むことが可能であることも理解されるであろう。他の哺乳類種とは、ウシ、イヌ、ネコ、ヤギ、ヒツジ、ブタ、ネズミ及びウマを含むが、これらに限定されない。
【0029】
配列間の配列同一性の計算は、以下の様にして行われる。2つのアミノ酸配列の同一性の割合の決定には、最適な比較をするために、まず、配列を整列する(例えば最適な比較では、1つのアミノ酸配列、又は1番目及び2番目両方のアミノ酸配列に相違が取り入れられる)。そして、対応するアミノ酸部位又はヌクレオチド部位のアミノ酸残基を比較する。第一配列の部位が第二配列の対応する部位と同一のアミノ酸残基又はヌクレオチドである場合は、タンパク質はその部位で同一である。2つの配列間の同一性の割合は、配列間で同一な部位数により決まり、相違の数及び各相違の長さを考慮に入れる。該相違の数及び各相違の長さは、2つの配列の最適な配列比較に必要とされる。
【0030】
2つの配列間の配列比較及び同一性の割合の決定は、数学的アルゴリズムを用いてなされる。好適な実施例においては、2つのアミノ酸配列間の同一性の割合は、初期値変数を用い配列比較ソフトウェアプログラムにより決定される。適当なプログラムには、例えば、CLUSTAL W(Thompson et al., Nuc. Acids Research 22:4673, 1994(www.ebi.ac.uk/clustalw))、BL2SEQ(Tatusova and Madden, FEMS Microbiol. Lett. 174:247, 1999(www.ncbi.nlm.nih.gov/blast/bl2seq/bl2.html))、SAGA(Notredame and Higgins, Nuc. Acids Research 24:1515, 1996(igs-server.cnrs-mrs.fr/~cnotred))及びDIALIGN(Morgenstern et al., Bioinformatics 14:290, 1998(bibiserv.techfak.uni-bielefeld.de/dialign))が挙げられる。
【0031】
(www.ncbi.
<ガレクチン‐7>
ガレクチン‐7ファミリーメンバーのタンパク質は、通常は単量体として存在し、130から140のアミノ酸を含み、分子量は約15から16kDaの範囲である(例えば、Magnaldo et al., Develop. Biol. 168:259, 1995及びMadsen et al., J. Biol. Chem. 270:5823, 1995参照)。ガレクチン‐7の発現は、上皮層形成の開始と関連している(Timmons et al., Int. J. Dev. Biol. 43:229, 1999)。ガレクチン‐7は、細胞‐マトリックス間及び細胞‐細胞間相互作用を担っていると考えられている。ガレクチン‐7は細胞と細胞が接触している領域(例えば、ヒト表皮の上層)にみられ、ガレクチン‐7の発現は足場非依存性ケラチン生成細胞では非常に下方制御されており、悪性のケラチン生成細胞系では見られない。ガレクチン‐7は、正常なケラチン生成細胞の維持に必要とされる(Madsen et al., supra参照)。
【0032】
ヒトガレクチン‐7は136アミノ酸を含み、約15.1kDaの分子量を持つ(配列番号2、図2)。図2、4、6及び8に示すように、ヒトガレクチン‐7は次のような領域、特徴的配列、もしくは他の構造的特徴を含む。配列番号2のアミノ酸残基5から135の周囲に位置するガラクトシド結合領域、配列番号2のアミノ酸残基70から89の周囲に位置するガラプチン特徴的配列(PROSITE番号、PS00309)、配列番号2のアミノ酸残基29から32の周囲に位置する1つのN‐糖鎖形成部位(PROSITE番号、PS00001)、配列番号2のアミノ酸残基132から134の周囲に位置する1つのタンパク質キナーゼCリン酸化部位(PROSITE番号、PS00005)、配列番号2のアミノ酸残基9から12の周囲に位置する1つのカゼインキナーゼIIリン酸化部位(PROSITE番号、PS00006)、及び配列番号2のアミノ酸残基13から18及び44から49の周囲に位置する2つのミリストイル化部位(PROSITE番号、PS00008)である。
【0033】
本明細書に定義されている通り、ガレクチン‐7を含む「ガレクチン‐7タンパク質」はガラクトシド結合領域を含む。この領域は、更に次の様に定義されている。
【0034】
本明細書で用いられているように、ガレクチン‐7のガラクトシド結合領域は、約80から180のアミノ酸のアミノ酸配列からなり、PFAM(配列番号3)の共通配列PF00337との配列比較で、少なくとも80のビットスコアを持つ。好適には、ガレクチン‐7のガラクトシド結合領域は少なくとも約100から160のアミノ酸からなり、より好適には約110から150のアミノ酸、又は約120から140のアミノ酸からなり、PFAM(配列番号3)の共通配列、PF00337との配列比較で、少なくとも80のビットスコア、より好適には少なくとも100のビットスコア、更に好適には120以上のビットスコアを持つ。ガレクチン‐7のガラクトシド結合領域は、1つのN‐糖鎖形成部位(PROSITE番号PS00001)、1つのタンパク質キナーゼCリン酸化部位(PROSITE番号PS00005)、1つのカゼインキナーゼIIリン酸化部位(PROSITE番号、PS00006)、1つもしくは2つのミリストイル化部位(PROSITE番号、PS00008)、及び/又はガレプチン特徴的配列(PROSITE番号、PS00309)を含むことが可能である。ある実施例によっては、ガレクチン‐7のガラクトシド結合領域は、配列番号2の次のアミノ酸配列又は領域を含む。M1、S2、H6、K7、L10、P11、G13、R15、G17‐V19、R21‐G24、V26、P27、A30、R32‐Q43、D46‐N63、K65、Q67、G68、W70‐G76、G78、P80‐L90、I92、G97‐K99、V101、G103、D104、Y107、H109、F110、H112、R113、P115、V119、R120、V122‐L130、S132、I135及びF136である。これらのアミノ酸及び領域は、幾つかの哺乳類のガレクチン‐7で保存されており、触媒作用及び/又は構造上の役割を担う(図4に「*」で表記されたアミノ酸を参照)。
【0035】
本発明のあるガレクチン‐7タンパク質は、配列番号2に記載のヒトのガレクチン‐7アミノ酸配列を含む。本発明の他のガレクチン‐7タンパク質は、配列番号2のアミノ酸配列と実質的に同一のアミノ酸配列を含む。特に、偶発的若しくは意図的に誘導された変異、例えば、配列番号2の特定アミノ酸残基で、欠損、付加、置換若しくは修飾などを含むタンパク質は、本発明のガレクチン‐7の定義に当てはまる。ここに定義するように、ガレクチン‐3タンパク質は、他の哺乳類種のガレクチン‐3のアミノ酸配列によって示される領域を含むことが可能であることも理解されるであろう。他の哺乳類種とは、ウシ、イヌ、ネコ、ヤギ、ヒツジ、ブタ、ネズミ及びウマを含むが、これらに限定されない。
【0036】
<ガレクチン‐8>
ガレクチン−8は広範囲に発現しているタンパク質で、例えば、肝臓、心臓、筋肉、腎臓、脾臓、後肢及び脳で見られ、ヒト及びラットのガレクチン‐8遺伝子及びタンパク質の配列が入手可能である(例えば、Hadari, et al., Trends in Glycosci and Glycotechnol. 9: 103-112, 1997参照)。高親水性の特徴、及びムチンのO結合型オリゴ糖に見られるGalβ1‐4GlcNAC二糖との結合能により、該タンパク質は理想的な眼症候群の治療薬とされる。
【0037】
ヒトのガレクチン‐8のアミノ酸配列には2つの型が知られており、316アミノ酸型(受入番号O00214、1997年11月1日作成)及び359アミノ酸型(受入番号Q8TEV1、2002年6月1日作成)がある。これらの配列は、かなりの長さにおいて類似又は同一である一方、全体で見ればいくらかの異なる部分がある。316型アミノ酸配列は、1文字アミノ酸記号を用いて以下に示す(配列番号4)。
【0038】
【表1】
【0039】
長型アミノ酸配列を以下に示す(配列番号5)。
【0040】
【表2】
【0041】
本明細書に定義されている通り、「ガレクチン‐8タンパク質」は、ガレクチン‐8の「N末端領域」、ガレクチン‐8の「プロリン、グリセリン及びチロシンが豊富な領域」、及び/又はガレクチン‐8の「ガラクトシド結合領域」を含むことが可能である。
【0042】
本明細書にあるように、ガレクチン‐8の「N末端領域」は約10から20アミノ酸のアミノ酸配列を含み、好適には約14のアミノ酸で、配列番号もしくは5のアミノ酸と60%、70%、80%、90%、95%、99%、又は100%の同一性を持つ。N末端領域には、N‐糖鎖形成部位(PROSITE番号、PS00001)及び/又はカゼインキナーゼIIリン酸化部位(PROSITE番号、PS00006)が含まれる。PROSITEのN‐糖鎖形成部位は共通配列としてN‐{P}‐[ST]‐{P}を持ち、PROSITEによるカゼインキナーゼIIリン酸化部位は共通配列として[ST]‐X(2)‐[DE]を持つ。上記共通配列には、他のモチーフ又は特徴的配列も含まれる。
【0043】
本明細書では、ガレクチン‐8の「プロリン、グリシン及びチロシンが豊富な領域」は、約60から140のアミノ酸のアミノ酸配列からなり、より好適には、80から120のアミノ酸、もしくは、90から110のアミノ酸で、配列番号4及びは5の各アミノ酸の15から116のアミノ酸と60%、70%、80%、90%、95%、99%、又は100%の同一性を持つ。プロリン、グリシン及びチロシンが豊富な領域は、1,2,3,4,5,6,7又は8のN‐ミリストイル化部位(PROSITE番号、PS0008)を持つことも可能で、ミリストイル化部位は共通配列として、G‐{EDRKHPFYW}‐X(2)‐[STAGCN]‐{P}を持つ。ある実施例では、プロリン、グリシン及びチロシンが豊富な領域は、配列番号4の以下のアミノ酸及び領域を含む。アミノ酸及び領域は、G20、P23、P28、G29、G36、P39及び当業者にとって明確なその他の残基である。これらのアミノ酸及び領域は、幾つかの哺乳類のガレクチン‐8で保存されていて、触媒作用及び構造的役割を担っている。ある実施例では、プロリン、グリシン及びチロシンが豊富な領域は、配列番号5の以下のアミノ酸及び領域を含む。アミノ酸及び領域は、G21、P24、P29、G30、G37、P40及び当業者にとって明確なその他の残基である。
【0044】
本明細書では、ガレクチン‐4の「ガラクトシド結合領域」は約80から180のアミノ酸のアミノ酸配列からなり、PFAM(配列番号3)の共通配列であるPF00337との配列比較で、少なくとも150のビットスコアを持つ。好適には、ガレクチン‐3ガラクトシド結合領域は、少なくとも約100から160のアミノ酸を含み、より好適には、110から150アミノ酸、又は120から140アミノ酸を含み、PFAM(配列番号3)の共通配列であるPF00337との配列比較により、少なくとも150、より好適には少なくとも175、更に好適には200以上のビットスコアを持つ。
【0045】
特定の配列とPFAM(配列番号3)の共通配列であるPF00337との配列比較からビットスコアを計算するには、対象配列をPFAMのデータベースであるHMM(PFAM データベース、リリース 2.1)で検索する。初期値変数は、www.sanger.ac.uk/Software/Pfamで取得可能である。PFAMデータベースの説明は、参考文献(Sonnhammer et al., supra)にあり、HMMの詳細な説明は、例えば、参考文献(Gribskov et al., Meth., Enzymol. 183:146, 1990及びStultz et al., Protein Sci. 2:305, 1993)にある。
【0046】
ガレクチン‐8のガラクトシド結合領域は更に、1の、好適には2のタンパク質キナーゼCリン酸化部位(PROSITE番号、PS00005)、カゼインキナーゼIIリン酸化部位(PROSITE番号PS00006)及び/又はガラプチンの特徴的配列(PROSITE番号、PS00309)を含む。タンパク質キナーゼCリン酸化部位は次の共通配列、[ST]‐X‐[RK]を持つ。ガラプチン特徴的配列は、以下の共通配列、W‐[GEK]‐X‐[EQ]‐X‐[KRE]‐X(3,6)‐[PCTF]‐[LIVMF]‐[NQEGSKV]‐X‐[GH]‐X(3)‐[DENKHS]‐[LIVMFC]を持つ。ある実施例では、ガレクチン‐8のガラクトシド結合領域は配列番号4の次のアミノ酸及び領域、L123‐L124、G126、P131、R128、L140‐I146及び前述した部位と類似したその他の部位を含む。これらのアミノ酸及び領域は、幾つかの哺乳類のガレクチン‐8で保存されており、触媒作用及び構造的役割を担っている(図3に「*」で表記されたアミノ酸参照)。
【0047】
本発明のあるガレクチン‐8タンパク質は、配列番号4及び5記載のヒトガレクチン‐8アミノ酸配列を含む。他の本発明のガレクチン‐8は、配列番号4又は5記載のアミノ酸配列と実質的に同一のアミノ酸配列を含む。実質的に同一」とは、本明細書では、十分又は最低限の数のアミノ酸残基を含む第1のアミノ酸を言及し、該アミノ酸残基は、第2のアミノ酸配列の整列アミノ酸配列残基と同一であり、これにより第1及び第2のアミノ酸配列は共通の構造領域及び/又は共通の機能活性を持つことが可能であるとする。例えば、配列番号4又は5と少なくとも60%又は65%の同一性、好ましくは、少なくとも75%の同一性、より好ましくは少なくとも85%、90%、91%、92%、93%、94%、95%、96%、97%、98%又は99%の同一性を持つ共通の構造領域を含むアミノ酸配列は、配列番号4又は5のアミノ酸配列と実質的に同一とされる。特に、配列番号4又は5の特定アミノ酸残基における欠失、付加、置換もしくは修飾などの偶発的又は意図的に誘導された変異を含むタンパク質は、本明細書記載の、ガレクチン‐8タンパク質の定義に含まれる。ここに定義するように、ガレクチン‐8タンパク質は、他の哺乳類種のガレクチン‐8のアミノ酸配列によって示される領域を含むことが可能であることも理解されるであろう。他の哺乳類種とは、ウシ、イヌ、ネコ、ヤギ、ヒツジ、ブタ、ネズミ及びウマを含むが、これらに限定されない。
【0048】
眼乾燥症候群は、眼科では「風邪」のようなものと考えられていて、眼科医を訪れる患者にみられる2番目に多い病状である。1500万人近くのアメリカ人が眼乾燥に苦しんでいる。該疾患は、涙液層における構造又は機能の欠損により起こる。涙液層は動的構造を持ち、涙液層の生産と代謝回転は健全な角膜表面維持に不可欠である。涙液層は3層の、ムチン(最内側)、水溶性(中間)及び脂質(表面)層から構成される。これらの層いずれかの機能不全により、目乾燥症は引き起こされる。涙液層は最内側にあるムチン層を介して上皮細胞と結合する。ムチン層は、涙液層が角膜全体に、均一に拡散することを促進し、表面を適切に湿らせるために必要である。ムチン分子が眼に接着しないと、通常の水溶性涙液生産でも上皮損傷が起こる。涙液層拡散障害は、眼表面又は涙液に支障をきたし、眼乾燥症を引き起こす可能性がある。最内側ムチン層は、結膜杯細胞及び結膜又は角膜上皮細胞により生産される。眼ムチンは、膜貫通型(例えば、MUCI及びMUCU)又はゲル形成/分泌性のいずれかである(例えば、MUC5AC)。
【0049】
ムチンは非常に巨大な糖タンパク質で、少なくとも質量の半分にO結合型炭水化物を持つ。O結合型炭水化物の豊富な側鎖により、ムチンは非常に親水性であるという特徴を持つ。この親水性の特性により、水溶性層が眼上に均一に拡散することを可能にしていると考えられている。
【0050】
本発明は眼乾燥症候群(DES)の治療におけるガレクチンの治療的使用に基づく。特定の機構には限定されず、該使用はガレクチンが炭水化物結合するという特性に基づく。点眼液の局所的投与により類液の拡散が促進され、これは分泌性ムチンが、結膜及び角膜上の上皮表面において、膜貫通型ムチン及び他の糖タンパク質と結合することで促進される。
【0051】
本明細書の例では、三つの異なった眼乾燥マウスモデルを用いて、ガレクチン液の局所的投与の効果を特定している。一番目のモデルでは、涙液不足は、正常マウスの尾部にスコポラミンパッチを投与する、Pflugfelder‘s Laboratoryの報告にある方法(Dursun et al., 2002, Invest Ophthamol & Vis Sci. 43:632-638及びPflugfelder et al., 2003, The Ocular Surface, 1:31-36)により誘導されている。二番目のモデルでは、MRL/lpr自己免疫性マウスを利用し、これはシェーグレン症候群(Jabs et al., 1991, Invest Ophthamol Vis Sci. 32:371-380及びJabs et al., 1997, Curr Eye Res. 16:909--916)に提唱されているモデルで、涙腺炎症性病変を起こしている。三番目のモデルでは、IL‐1αの注入を用い、動物モデルを作成する。該モデルは、シェーグレン症候群(Zoukhri, D. et al., Invest Ophthalmol Vis Sci 42(5): 925-932参照)モデルである。
【0052】
測定には、生体内解析による涙液生成、涙液クリアランス及び角膜免疫染色が挙げられ、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7およびガレクチン‐8の各種濃度による効果を、これらパラメーターにおいて単独又は併用し、溶媒処理された対照動物と比較して特定する。生体内解析に引き続き、単離された涙腺を用いた生化学的解析、角膜表面の免疫組織学的解析が行われる。更に、スコポロミン処理前に行われるガレクチン液による前処理の効果並びに、ガレクチン及びガレクチン阻害剤であるβ乳糖を併用した治療により推定される効果についても検討される。
【0053】
本明細書の例から得られたデータによると、該モデルによる眼乾燥症において、ガレクチンは、測定可能なパラメーターにおいて候群統計学的に顕著な効果を示した。ガレクチン処理された動物の眼角膜では、偽薬処理された動物と比べて、角膜免疫染色の減少が見られたことから、ガレクチンは、ヒトDESに対しすばらしい治療可能性があるという根拠が得られた。二つの動物モデルは、異なったDESの型を示すので、どちらかのモデルで得られた肯定的な結果はDES治療におけるガレクチンの役割を特定するのに役立ち、臨床治験において焦点を絞る際の指針となる。臨床治験には追加動物実験が含まれ、有益な効果を最大限に得るのに必要なガレクチン濃度、効果の持続期間、並びにガレクチンと角膜及び内在性ムチンとの結合特性を決定する。更に毒性検査が行われ、安全な角膜用量範囲を特定し、臨床試験の承認を受ける前に、米国食品衛生局の必要条件を満たす。
【0054】
涙液の最内側は、結膜杯状細胞、並びに角膜及び結膜上皮細胞によって生産されたムチンにより構成される。ムチンは放出され分泌性ムチンとして存在したり、膜貫通型ムチンとして上皮に付属した状態を保ったりする。これらの分子は糖タンパク質であり、豊富なO結合した炭水化物側鎖を持つ。側鎖によってムチンは非常に親水性な特性を持ち、この特性により、水溶性層が眼上を均一に拡散することが可能となっている。該水溶液層は涙腺細胞により生産され、涙液層の約90%を構成する。水溶液層は主に水から成り、塩、ブドウ糖、リゾチーム、涙特異的プレアルブミン、ラクトフェリン、分泌性免疫グロブリンA、及び他のタンパク質が溶解している。外膜は蒸発を遅らせ、マイボーム腺によって生産された皮脂腺性物質からなる。瞬きにより、涙液層上に該層が拡散される(Rheinstrom SD, 1999, Dry eye. In Yanoff, ed, Ophthalmology. 1st Ed. Editor. Mosley International Ltd, St Louis, MO)。
【0055】
国立眼研究所は、眼乾燥の症状を二つのカテゴリー(水溶性層欠損及び蒸発欠損)に分類している。しかし、臨床症状は、多くの場合二つの発病経路が混合している(涙液生産の減少は、多くの場合欠陥油層の拡散につながり、結果過剰蒸発が起こる。また、マイボーム腺症な非常に多くの場合、低感受性眼乾燥と関連している)。水溶性涙液欠損類は、シューグレン症候群眼乾燥と非シューグレン症候群眼乾燥のサブカテゴリーに分類され、症状の重さの違いにより分類され、シューグレン症候群の炎症の発現が増強されている(Foulks, 2003, The Ocular Surface 1:20-30)。
【0056】
水溶性層欠損は眼乾燥の最も一般的な要因で、通常は涙腺からの涙液分泌の減少によって起こるが、涙液蒸発の増加によっても起こる。分泌減少の原因には、シューグレン症候群、老人性分泌過減、涙腺切除、ビタミンA欠乏、類肉腫及びリンパ腫における免疫涙腺損傷、感覚消失又は運動反射喪失、結膜の瘢痕状態、及びコンタクトレンズによる磨耗が挙げられる(Rolando and Zierhut, Surv Ophthlmol. 45:S203-S210, 2001)。水溶性層における組成の変化も眼表面損傷に関連している。組成の変化には、電解質濃度の増加、成長因子の損失又は炎症性サイトカイニンの存在等が挙げられ、涙液の低代謝回転と共に、眼表面損傷に関連している。
【0057】
杯細胞の欠乏は、従ってムチン量の減少につながり、眼乾燥の多くの様態を伴う。特に、疾患によっては杯細胞損失を引き起こす。これらは、ビタミンA欠損及び瘢痕を残す結膜症を含む。ビタミンAは、眼表面において、杯細胞やムチンの維持に必須とされる。該結膜症には、スティーブンス・ジョンソン症候群、トラコーマ、類天疱瘡及び化学熱傷等がある。局所的薬剤投与や保存料もまた、眼表面及び杯細胞を損傷しうる(Rheinstrom SD, 1999 Dry eye. In Yanoff, ed Ophthalmology. 1st Ed. Editor. Mosley International Ltd, St Louis MO; Abelson et al., 2003, Rev Ophthalmol 10:1)。
【0058】
患者の不快感の評価だけでなく、診断技術は、DESの重症度及び原因の特定に役立つことが証明された。涙液層の非安定性は、非侵襲性な涙液破砕時間測定(TBUT)により評価される。該測定では、蛍光溶液が投与され、涙液層が最初に破壊するまでの時間が、生体顕微鏡によって観測される。涙液生産は、典型的又は改良スキマー試験によって推定される。フルオレセイン、又は類似色素を用いた眼表面染色の度合いは、日常的に用いられ、眼表面交代の重症度が診断されている。これらの色素は、ムチンタンパク質の生産が欠乏した上皮表面、又は上皮細胞膜が露出した上皮表面を染色する。標準評点方式が開発され、損傷の重症度が定量化されている。該試験は、臨床試験において日常的に行われ、侵襲的な眼乾燥治療の治療効果が評価されている(Foulks 2003, The Ocular Surface, 1:20-30)。
【0059】
人口涙液は現在の眼乾燥治療の中心である。様々な商品が入手可能だが、どれもが一過的に症状を軽減するだけである。現在のところ、症状を戻すような治療薬はない。セルロースエステル(メチルセルロース、ヒドロキシプロピルメチルセルロース)又はポリビニルアルコールなどの高分子量重合体を生理食塩水に添加し、人口涙液を開発可能である。他の人口涙液処方は涙液中のムチン組成を模倣して調整されている。研究によると、人口涙液に使われている保存剤にある種の毒性があることが示され、保存料が入っていない、一回分ずつに分かれて小瓶に入った涙液が入手可能となっている(Rheinstrom SD, 1999 Dry eye. In Yanoff, ed Ophthalmology. 1st Ed. Editor. Mosley International Ltd, St Louis MO; Abelson et al., 2003, Rev Ophthalmol 10:1)。
【0060】
外疾患の病理生理学の理解向上に伴い、多くの異なった様式の治療方法が導入されてきた。これらには、涙液脂質膜を標的とした疾患、及び根底にある免疫性又はホルモン性病因が含まれる(Brewitt et al., 2001, Surv of Ophthalmol 45:S119-S202)。治験的治療には、15(S)‐HETE(水酸化エイコサテトラエン酸)(Gamache et al., 2000, Cornea 19:6:S88)等の局所的分泌促進物質、及び合成のP2Y2受容体アゴニスト(Jumblatt et al., 1998, Exp Eye Res 67:341-346)が含まれ、シクロスポリンA及び副腎皮質ステロイドなどの抗炎症物質ももちろん含まれる(Pflugfelder, 2003, The Ocular Surface, 1:31-36)。
【0061】
ムチンは、健全な眼球涙液層の生理機能の維持を担っていることが知られている。ムチンは高分子量糖タンパク質で、タンパク質骨格及び高炭水化物含有量を持っている。ムチン層に関与しているだけでなく、ムチン自体も糖衣、即ち足場構造を形成し、該足場様構造が細胞接着を促進する。ムチンは、眼表面損傷に対し防御の役割も担っている。また、ムチンは涙液層を親水性にする役割を担っている。これにより、涙液層が安定化され、角膜張力が減少し、水溶性層が眼表面上を均一に拡散することが可能となる。このムチン層なしでは、涙液は表面にとどまらず、眼表面は傷つきやすくなる(Abelson et al., 2003, Rev Ophthalmol 10:1)。
【0062】
体内では、膜貫通型及び分泌性の、二種類の原発型ムチンが生産さている。膜貫通型ムチン(MUC1、MUC2及びMUC4)は細胞の脂質二重膜に包埋されている。膜貫通型ムチンは角膜及び結膜の非杯細胞により発現され、分泌性ムチン(MUC5AC及びMUC7)の拡散を促進していると考えられている。該分泌性ムチンは、眼表皮を横断する杯細胞により分泌される。二種類のムチンは共に生涙液層を形成している(Danjo et al., 1998, Invest Ophthalmol Vis Sci 39: 2602-2609及びWatanabe 2002, Cornea 21:S17-S22)。
【0063】
結膜ムチンの欠乏は特定種の眼乾燥疾患の一因となっている。考えられる原因には、杯細胞密度の減少、ムチン分布又は特性の変化、並びにムチンmRNA発現の現象が挙げられる(Gipson et al., 2000, Prog Retinal Eye Res, 16:1:81-98及びGilbard, 2000, 「Dry-eye disorders」, In: Albert M et al., eds. Principals and Practice of Ophthalmology: 2nd Edition, Philadelphia: W.B. Saunders Company: 982-1001)。ビタミンA欠乏、局所用薬剤で、保存料を含む液滴の過剰投与、及び、瘢痕を残す結膜障害は全て杯細胞及び眼表面の損傷になり得る(Danjo et al., 1998, Invest Ophthalmol Vis Sci 39: 2602-2609及びAbelson et al., 2003, Rev Ophthalmol 10:1)。
【0064】
また、眼表面に見られる膜貫通型ムチンは、眼乾燥症の患者では変質している。研究者によると、膜貫通型ムチンであるMUC1等は、眼乾燥症患者のものは糖化されていないが、正常な患者のムチンは糖化されている。構造の変化は、眼表面上のムチン機能を変える可能性があり、眼乾燥症を更に悪化させる可能性がある(Danjo et al., 1998, Invest Ophthalmol Vis Sci 39:2602-2609)。
【0065】
<涙液層機能不全におけるガレクチンの役割>
ガレクチンは炭水化物結合タンパク質で、角膜及び結膜表面上の涙液層拡散を促進させる、理想的な候補となる可能性を持っている。これらのタンパク質は、高い親和性を持ってムチンと結合し、特に、ムチンのO結合炭水化物側鎖と結合する。特定の理論、又は作用機序に基づき結合するのではないが、ガレクチンは、分泌性ムチンが膜貫通型ムチン(又は他の糖タンパク質)と結合することによって、涙液層による角膜及び結膜表面上の拡散を促進させる。ムチン欠損眼では、外来性ガレクチンがムチン結合を延長又は促進させる。更に、ガレクチン自体が角膜表面の炭水化物結合タンパク質と結合することにより、ムチン欠損状態でも、涙液層の均一的な拡散をよりいっそう促進させる。
【0066】
ムチンとガレクチンの相互作用が、眼以外の組織で証明されている。ガレクチンは、ムチンの発現及び生産を、様々なムチンに対する強い結合親和性により調整していることが示されている。ガレクチン‐3はヒト結腸癌細胞由来のムチンと結合し、該ムチンの発現を濃度依存的に調整する(Bresalier et al., 1996, Cancer Res 56: 4354-4357及びDudas et al., 2002, Gastroenterology 118: 1553)。卵巣腫瘍細胞では、ムチン様糖タンパク質であるCA125はガレクチン‐1と結合する(Seelenmeyer et al., 2002, J Cell Sci, 116:1305-1318)。
【0067】
ガレクチンは、広く分布している炭水化物結合タンパク質からなるファミリーを構成し、多くの細胞表面及びECM糖タンパク質に見られるβ‐ガラクトシド含有グリカンとの親和性により特徴付けられている。哺乳類では、現在のところ14メンバーのガレクチンファミリーがあり(ガレクチン‐1から‐14)、炭水化物認識領域(CRD)における構造的類似性により定義されている。ガレクチンは可溶性タンパク質である。細胞内では、ガレクチンは主に細胞質に存在し、ガレクチン‐1及び‐3は増殖細胞の核でも確認されている。細胞外では、ガレクチンは細胞表面及びECMに見られる。ある種の成長因子(例えば、bFGF)及びサイトカイニン(例えば、IL−1)と同様に、ガレクチンは典型的なシグナル配列又は膜貫通領域を持たず、まだ良く理解されていないが、細胞から、非典型的な経路を経て分泌される。ガレクチン‐1、‐3、‐8及び‐9等のガレクチンは、幅広い組織で見られるが、一方、ガレクチン‐4、‐5及び‐6等のガレクチンは組織特異性を示す。ガレクチン‐1及び‐3は最も研究されているガレクチンである。細胞外では、両レクチンは細胞‐細胞間、及び細胞‐マトリックス間接着に関与していると考えられている。該接着は、フィブロネクチン、ラミニン、ビトロネクチン及びインテグリンの独特なイソ型上のオリゴ糖との結合によるものとされている。ガレクチンのほとんどは2つのCRDを持つか二量体として存在し、機能的に二価である。二価の特性により、一方では、そのレクチンの糖タンパク質である分泌性ムチンのオリゴ糖鎖と相互作用が可能であり、他方では、角膜の先端面における膜貫通型ムチン又は糖タンパク質のオリゴ糖鎖と相互作用が可能であり、これにより、眼表面上において涙液層の拡散を促進させている。幾つかの研究により、ガレクチン‐3はマウス及びヒトの角膜表皮で 発現していることが示されている。外来性ガレクチン‐3及び‐7は、マウス動物モデルにおいて角膜傷の再上皮化を促進させ、角膜上皮細胞の輸送に関与している可能性がある(Cao et al, 2002, J Biol Chem. 277:42299-4230及び2003, Arch Ophthalmol, 121: 82-86)。
【0068】
本明細書の例にある予備的な研究では、2つの異なったモデルの角膜傷治癒が用いられ、創傷の再上皮化は、以前に、ガレクチン‐3欠損(gal3-/-)マウスでは野生型(gal3+/+)マウスに比べて著しく遅いことが発見されている。一方、角膜上皮創傷閉塞率は、ガレクチン‐1欠損マウスと野生型と比べて差は見られなかった。外来性ガレクチン‐3及びガレクチン‐7は角膜アルカリ熱傷創傷の再上皮化を、濃度依存的に促進させた。該作用は、競合する糖、即ちβ‐ラクトースにより抑制されたが、関係のない二糖であるショ糖には抑制されなかった(Cao et al, 2002, J Biol Chem. 277:42299-4230及び2003, Arch Ophthalmol, 121: 82-86)。
【実施例】
【0069】
<材料と方法>
以下の材料と方法は、本明細書全般の例で使用される。
【0070】
<治療濃度のガレクチン‐1、‐3、‐7及び‐8を含む人工涙液溶液>
標準的な人工涙液溶液は動物研究に基づくガレクチン濃度で処方されている。該溶液は緩衝液で処理され、pHは中性、等張でCMC及びHMPC等の粘度を増すための増粘剤を含む。理想的な対照溶液は、ガレクチン以外全ての成分を含み、作成される。保存料が上皮損傷を悪化させる為、該溶液は保存用ではない。従って、新しい溶液はサンプルごとに調整され、投与時までに冷凍される。種々の処方には、対照人工涙液を含む。この対照人工涙液は、10μg/mlの各ガレクチン‐1、‐3、‐7若しくは‐8、又は対の組み合わせを含む人工涙液、及び、20μg/mlのガレクチン‐1、‐3、‐7若しくは‐8、又は対の組み合わせを含む人工涙液を含み、さらに、これら人工涙液に0.1Mのβ‐ラクトース及び二糖を含む人工涙液を含む。尚、二糖はガレクチン阻害剤として働く。
【0071】
<DES動物モデル:正常マウスにおいてスコポラミンより誘発される涙液不足>
スコポラミンは抗コリン作用薬で、ムスカリン性コリン受容体を標的とする。経皮スコポラミン(scop)パッチは、ヒトにおいて抗嘔気目的で及び動揺病治療で一般的に使われている。公表された研究(Dursun et al., 2002, Invest Ophthalmol & Vis Sci. 43:632-638)によると、この方法を用いたscop治療により、対照群の涙液生産の20%まで減少させるという、顕著で統計学的に有意な結果が得られた。涙液中のフルオレセインのクリアランス、及び角膜におけるカルボキシフルオレセイン取り込みは、どちらも約3倍となった。更に該動物を送風フード内に置くことにより、これらのパラメーターは一層劇的に増大した。更に、scop+送風フード処理したマウスでは、結膜杯細胞濃度が対照と比べて90%以上も減少した。径皮パッチ送達系により制御され継続的な薬剤送達が可能となるため、涙液生産の減少は24時間以上も続いた。一方、局所的アトロピン投与では、涙液の減少は数時間しか持続しなかった。新しいスコポラミンパッチの再投与により、効果は更に持続する。
【0072】
該例では、C57BL/6系マウス(Charles River Laboratories, Wilmington, MA)が用いられる。該マウスは6から8週齢で性混合である。無麻酔のマウスを手で捕まえ、1インチ部分の尾部中部を電気かみそりで剃る。経皮scopパッチ(Novartis, Summit, NJ)を4つに切り、4分の1を脱毛した尾部中部に投与する。パッチは48時間ごとに張り替えられ、安定した薬剤送達が維持される。動物を1時間、1日3回、実験の間毎日送風フード内に置く。
【0073】
<MRL/lpr自己免疫性マウス>
本実験では、MRL/Mp‐lpr/lpr(MRL/lpr)系のマウス(Jackson Laboratory, Bar Harbor, ME)が用いる。該マウスは類遺伝子亜系で、自己免疫性疾患を発病し、16週齢以内に涙腺炎症性病変を引き起こす。病理学及び組織学的評価に基づき、該マウスはヒトシューグレン症候群(Jabs et al., 1991, Invest Ophthamol Vis Sci. 32:371-380及び2001, Curr Eye Res. 16:909-916)のモデルとして提唱された。本明細書中例に用いられるマウスは、少なくとも16週齢以上で性混合のものである。
【0074】
<動物における涙液生産、涙液クリアランス及び角膜染色の測定>
水溶性類液生産は、フェノールレッドを含侵させた綿糸(Zone-quick, Oasis, Glendora, CA)を用いて測定した。該糸をピンセットで持ち、外眼角内の眼表面に1分間当てる。綿糸上の目盛りを使って、何ミリ綿糸が濡れたかで水性の涙液生産を測定する。
【0075】
涙液破砕時間(TBUT)は、1%ナトリウムフルオレセイン2μLを滴剤として用いて、これを眼表面に投与して測定した。涙液層を隙灯生体顕微鏡で観察し、涙液層が拡散するまでの時間を記録する。
【0076】
フルオレセイン染色はTBUT測定直後に行い、1%ナトリウムフルオレセイン2μLを滴剤として用い、これを眼表面に投与する。投与5分後、隙灯生体顕微鏡を用い、角膜におけるフルオレセイン染色を評価する。フルオレセイン染色は、臨床では角膜表面損傷を判定する為に一般的に用いられている。眼乾燥症の動物の角膜表面はフルオレセイン染色するが、正常の角膜表面は染色しないとされている。フルオレセイン染色の程度は、眼乾燥臨床治験でも使われた尺度と同様の標準F1評価尺度を用いて記述する。
【0077】
<同等の治療>
治療及び測定計画は、短期的効果(ガレクエン治療後2時間)及び長期的効果(ガレクエン治療後14時間まで)が測定できるようにされている。ガレクチンは、1日4回の治療計画に長期的効果があると期待される。治療計画、測定計画又は投与量の変更は、角膜に有益な効果をもたらすために必要なものであれば、本明細書に例と同等のものとする。例えば、20μL/mLのガレクチン治療で反応が見られ、10μL/mLで反応が見られない場合は、追加例はより高い投与量(50μg/mL)を備える。
【0078】
<ガレクチン‐1、‐3、ガレクチン‐7及びガレクチン‐8の製剤>
本発明のガレクチンは、当業者が入手可能である。入手可能なガレクチンには、単離されたタンパク質だけでなく、遺伝子組み換えや合成により生産されたタンパク質が含まれ、固相方法によるタンパク質が例に挙げられる。本発明によると、ガレクチン‐3、ガレクチン‐7又はガレクチン‐8をコードするポリヌクレオチド配列は、組み換えDNA分子として用いられることが可能で、該組換え分子は適当な宿主細胞内で本発明のガレクチン発現を指示する。参考文献(Cherayil et al., supra、Madsen et al., supra 及びHadri et al., supra)には、ヒトガレクチン‐1、‐3、‐7及び‐8夫々のクローニング方法が詳細に記載されている。生物学的に活性を持つガレクチン‐1、ガレクチン‐3、ガレクチン‐7、ガレクチン‐8を発現するために、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7、ガレクチン‐8をエンコードするヌクレオチド配列又は機能的に同等な配列が適当な発現ベクターに挿入される。発現ベクターには、挿入したコードされた配列の転写及び翻訳に必要な要素を含むものとする。当業者に公知の方法を用いて、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7若しくはガレクチン‐8をコードする配列、並びに適当な転写又は翻訳制御を含む発現ベクターを構築してもよい。該方法には、生体外遺伝子組み換え技術、合成技術、並びに生体内組み換え若しくは遺伝子組み換えが含まれる。欠失、付加、又は置換は、当業者に公知の任意の技術を用いて導入することができる。例えば、PCRによる突然変異誘導が挙げられる。これらの技術は、参考文献(Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Press, Plainview, NY, 1989及びAusubel et al, Current Protocols in Molecular Biology, John Wiley & Sons, New York, NY, 1989))に記載されている。様々な発現ベクター/宿主系に導入され、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7又はガレクチン‐8を発現させることが可能である。これらの発現ベクター/宿主系には、組換えバクテリオファージ、プラスミド又はコスミドDNA発現ベクターにより形質転換した細菌等の微生物、酵母発現ベクターにより形質転換した酵母、ウイルス発現ベクター(例えば、バキュロウイルス)に感染した昆虫細胞系、多種の発現ベクター(例えば、カリフラワーモザイクウイルス(CaMV)、タバコモザイクウイルス(TMV))若しくは細菌発現ベクター(例えば、Ti、pBR322又はpET25bプラスミド)により形質転換した植物細胞系、又は動物細胞系が含まれるが、限定されるものではない。或いは、本発明のガレクチンは、化学的手法を用いてガレクチン‐1、ガレクチン‐3、ガレクチン‐7又はガレクチン‐8アミノ酸配列を全体又は一部を合成してもよい。例えば、ペプチド合成は、多様な固相法(Roberge et al., Science 269:202, 1995)を用いて行うことが可能であり、自動合成も実施可能であり、例えば、431Aペプチド合成機(Applied Biosystems of Foster City, CAから入手可能)を用い、機器添付の取扱説明書に従いペプチド合成することができる。
【0079】
<医薬組成物>
本発明の特徴の一つは医薬組成物が提供されていることである。該組成物は、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7及び/又はガレクチン‐8を備え、薬学的に許容可能な担体を任意に備える。実施例によっては、これらの組成は、一又はそれ以上の追加治療剤を任意に備える。実施例によっては、治療剤又は薬剤は以下の群から選択される。該薬剤は増殖因子、抗炎症剤、血管収縮剤、コラゲナーゼ阻害剤、局所ステロイド、マトリックスメタロプロテナーゼ阻害剤、アスコルビン酸、アンジオテンシンII、アンジオテンシンIII、カルレティキュリン、テトラサイクリン、フィブロネクチン、コラーゲン、トロンボスポンジン、形質転換増殖因子(TGF)、ケラチノサイト増殖因子(KGF)、線維眼細胞増殖因子(FGF)、インシュリン様増殖因子(IGF)、上皮増殖因子(EGF)、血小板由来増殖因子(PDGF)、ニューレギュリン(NDF)、肝細胞増殖因子(HGF)ビオチン等のビタミンB及びニアルロン酸からなる。
【0080】
本明細書で用いられている「薬学的に許容可能な担体」は、所望の投与形態により、ありとあらゆる溶媒、希釈剤又は他の液体媒体、分散若しくは懸濁酸、表面活性剤、等張剤、増粘若しくは乳化剤、防腐剤、固体結合剤並びに潤滑剤等からなる。参考文献(Remington's Pharmaceutical Sciences Ed. by Gennaro, Mack Publishing, Easton, PA, 1995)には、医薬品組成物の処方に用いられる多種多様の担体が開示されており、公知の調剤方法が記載されている。薬学的に許容可能な担体の物質例としては、ブドウ糖及びショ糖等の糖類、コーンスターチ及びジャガイモ澱粉等の澱粉、カルボキシメチルセルロースナトリウム、エチルセルロース及び酢酸セルロース等のセルロース並びにその派生物、粉末トラガカント、麦芽、ゼラチン、ココアバター及び座薬ワックス等の賦形剤、ピーナッツ油、綿実油、紅花油、ゴマ油、オリーブ油、コーン油及び大豆油等の油、プロピレングリコール等のグリコール、オレイン酸エチル及びウラリル酸エチル等のエステル、寒天、水酸化マグネシウム及び水酸化アルミニウム等の緩衝剤、アルギン酸、pyrogen-free water、等張食塩水、リンガー溶液、エチルアルコール並びにリン酸緩衝液が挙げられ、また、ラウリルシュウ酸ナトリウム及びステアリン酸マグネシウムであるような非毒性相溶性の滑剤、さらに、着色剤、解除剤、コーティング剤、甘味、香料添加及び香料剤、並びに保存料及び抗酸化剤も、策定者の判断により組成物に含有可能である。
【0081】
<治療に有効な投与量>
他の特徴としては、本発明の治療法によると、眼と医薬品組成物を接触させることによる眼乾燥の治療があげられ、以下に記載する。従って、本発明は眼乾燥の治療方法を提供し、治療に有効量の医薬品組成物を、治療を必要とする対象に投与する過程を備える。該組成は、ガレクチン‐3、ガレクチン‐7及び/又はガレクチン‐8を含む有効成分を備え、投与量及び投与回数は、所望の結果に応じて決められる。当然のことながら、これには本発明の医薬品を治療的処置として投与する過程を含み、この過程により角膜及び結膜表面における涙液層の拡散が促進される。或いは、眼乾燥に伴う合併症を最小限に抑える為の予防手段(例えば、手術中及び/又は手術後、或いは抗ヒスタミン剤と併用した炎症症状の治療中及び/又は治療後の傷洗浄液として)として投与する過程を含む。本発明の実施例によっては、「治療に有効な量」の医薬品組成物とは、眼乾燥治療に有効な量である。本発明の方法によると、該組成物は、眼乾燥治療を促進させるために有効な任意の量を任意の投与経路により投与可能である。従って、本明細書にある「眼乾燥治療を促進させるために有効な量」との表現は、涙液層を促進させる為に十分な量の組成物に言及する。的確な投与量は、個々の医師により、治療する患者を考慮し選択される。投与量と投与は、十分量の有効成分を提供する為に、又は所望の効果を維持するために調節される。考慮すべき更なる要因としては症状の重症度が挙げられ、例えば眼乾燥の程度、症歴、患者の体重、年齢、性別、食習慣、投与時間及び回数、併用する薬剤、反応特異性、並びに治療に対する耐性/反応が挙げられる。長時間作用性医薬品組成物は、1日に数回、毎日、3から4日間、毎週又は2週間に一度、組成物の半減期及びクリアランス時間により投与可能である。
【0082】
本発明の有効成分は、好適には、投与のし易さ及び投与の均一性を考慮して投与単位形態として処方されるものとする。「投与単位形態」との表現は、本明細書では、物理的に独立した単位に言及し、治療対象の患者に適当な有効成分の単位である。しかしながら、本発明組成の1日当たりの使用量は、主治医により医学的に妥当な判断内で決定されるものとする。有効成分の治療に有効な投与量は、先ずは細胞培養試験又は動物モデルを用いて概算され、一般的にはマウス、ウサギ、イヌ又は豚が用いられる。動物モデルは所望の濃度範囲及び投与経路を特定する際にも用いられる。直接投与が投与経路と考えられるが、得られた情報は、ヒトへの投与に有用な投与量及び追加投与経路を特定する為に利用される。治療に有効な投与量とは、症状や状態を回復させる有効成分量のに言及する。有効成分の治療的有効性及び毒性は、培養細胞又は実験動物を用いた標準的な薬学手法により特定可能であり、例えば、ED50(投与量は集団の50%に治療的有効)及びLD50(投与量は集団の50%で致死)が挙げられる。毒性用量と治療有効性用量の比が治療指数とされ、例えばLD50/ED50の比率で表される。大きい治療指数を示す医薬品組成物が好適である。細胞培養解析及び動物実験により得られたデータはヒトに用いられる一連の処方に利用される。
【0083】
<医薬品組成物の投与>
薬学的に許容な適当な担体と共に所望の投与量に処方後、本発明の医薬品組成物は、ヒト及び他の哺乳類に、眼等(粉末、軟膏又は点滴薬により)局所的に、例えば眼に直接投与可能である。代替及び付加経路としては、経口、直腸、非経口、大槽内、膣内、腹腔内、口腔内又は経鼻的経路等が挙げられ、治療対象の症状の重症度により想定される。
【0084】
眼投与のための液状剤形は、緩衝剤及び可溶化剤、水等所望の希釈剤、チモゾール等の保存剤、並びに溶液調製の為の一以上の生重合体又は重合体を含む。生重合体又は重合体には、ポリエチレングリコール、メチルセルロース水酸化プロプル、ヒアルロン酸ナトリウム、ポリアクリル酸ナトリウム又はタマリンドゴム等がある。
【0085】
経口投与のための液状剤形には、限定されるものではないが、薬学的に許容可能な乳剤、微小乳剤、液剤、懸濁剤、シロップ及びエリキシル剤が含まれる。これらの有効成分に加えて、液体剤形は不活性希釈剤を含み、該技術分野で一般的に用いられているものとしては、例えば、水若しくは他の溶媒、可溶化剤又は乳化剤{例えば、エチルアルコール、イソプロピルアルコール、炭酸エステル、酢酸エチル、ベンジルアルコール、安息香酸ベンジル、プロピレングリコール、1,3−ブチレングリコール、ジメチルホルムアミド、油(特に綿実油、落花生油、コーン油、オリーブ油、ひまし油及びゴマ油)、グリセロール及びテトラヒドロフリフリルアルコール等}、ポリエチレングリコール、ソルビタン脂肪酸エステル、並びにこれらの混合物が挙げられる。不活性希釈剤の他には、経口組成物は湿潤剤等の補助剤、乳化剤及び懸濁剤、甘味剤、香料添加剤、並びに香料剤を含むことも可能である。
【0086】
発明の医薬品組成物の局所投与又は経皮投与剤形は、軟膏、ペースト、クリーム、ローション、ゲル、粉末、水溶液、スプレー、吸入剤又はパッチを含む。該有効成分は、無菌状態で混合され、薬学的に許容可能な担体、並びに必要に応じて保存料若しくは緩衝剤が加えられる。例えば、眼又は皮膚感染は水溶性点滴剤、ミスト、乳剤又はクリームにより治療される。薬剤投与は、治療にも予防にもなりうる。予防的処方は潜在性創傷又は創傷の原因に適用され、コンタクトレンズ、コンタクトレンズ洗浄及び水洗液、コンタクトレンズ保存又は運搬容器、コンタクトレンズ取り扱い器具、目薬、手術用洗浄液、点耳薬、眼帯及び目周辺用化粧品(クリーム、ローション、マスカラ、アイライナー及びアイシャドウ等を含む)に用いられる。本発明は、眼科機器、手術用機器、聴覚機器又は開示の組成物を含む製品(ガーゼ、包帯又はストリップ)、並びにこれらの機器若しくは製品を作るための若しくは使うための方法を含む。これらの機器は開示の組成物により被膜し、組成物を浸透させ、結合させ、又は他の方法で処理しても良い。
【0087】
該軟膏、ペースト、クリーム及びゲルは本発明の有効成分だけでなく、動物性若しくは植物性脂肪等の賦形剤、油、ワックス、パラフィン、スターチ、トラガカント、セルロース派生物、ポリエチレングリコール、シリコン、ベントナイト、ケイ酸、タルク並びに酸化亜鉛又はこれらの混合物を含むことが可能である。
【0088】
粉末及びスプレーは、本発明の薬剤に加えて、タルク、ケイ酸、水酸化アルミニウム、ケイ酸カルシウム、ポリアミン粉末又は該薬物の混合物等の賦形剤を含むことが可能である。スプレーは更にクロロフルオロ炭化水素等の噴霧剤を含む。
【0089】
経皮パッチは、体への有効成分供給を制御できるという点で更に優れている。そのような剤形は、化合物を適当な溶剤に溶解又は分注し作られる。吸収促進剤を用いて、化合物が皮膚上に良く行渡らせることも可能である。該速度は、速度調整膜を用いることで、又は、化合物をポリマーマトリックス若しくはゲルに分散させることで制御可能である。
【0090】
注射製剤は、例えば無菌の水溶性又は油性の懸濁液が既知の技術に基づき、分散剤又は湿潤剤及び懸濁剤を用いて処方することができる。該無菌注射製剤は、非毒性で非経口的に許容可能な希釈又は溶媒(例えば1,3−ブタンジオール水溶液等)含有の無菌の注射剤、懸濁剤又は乳剤でもよい。許容可能な溶媒又は溶媒として使用されるものには、水、リンガー水溶液、U.S.P.及び等張塩化ナトリウム溶液がある。更に、通常は無菌の、不揮発性油が溶剤又は懸濁剤として用いられている。この用途には、合成モノ‐又はジグリセリドを含む、どの製品の不揮発性油も用いることが可能である。また、オレイン酸等の脂肪酸が注射製剤に用いられている。注射製剤は滅菌可能で、例えば、細菌保持フィルターを用いた濾過により、又は、事前に滅菌水若しくは他の滅菌注射溶剤に溶解または拡散させるなどして混入した無菌固体組成形状の無菌剤により、滅菌することができる。有効成分の効果を持続させるためには、皮下又は筋肉内注射による有効成分の吸収を遅らせることが好適である。非経口投与による有効成分の吸収を遅らせるために、有効成分を油溶媒に溶解又は懸濁することも可能である。デポー注射形態は、ポリラクチド-ポリグリコライド(polylactide-polyglycolide)であるような生分解性重合体に有効成分を含んだマイクロカプセル剤を形成することにより製造される。有効成分と重合体の割合、及び用いられる重合体の性質により、放出される有効成分の割合は制御可能である。生分解性重合体には、poly(orthoesters)及びpoly(anhydrides)が例として挙げられる。デポー注射製剤は有効成分をリポソーム又はマイクロエマルジョンに封入し調整することも可能で、これらは体組織と適合性がある。
【0091】
直腸又は膣内投与用組成物は好適には座薬で、本発明の有効成分を適当な非刺激性の賦形剤又は担体と混合し調整することが可能である。該賦形剤又は担体には、ココアバター、ポリエチレングリコール又は座薬ワックス等が挙げられ、これらは室温では固体であるが体温で液体になるため、直腸又は膣腔で溶け該有効成分を放出する。
【0092】
経口投与用固体剤形はカプセル剤、錠剤、丸剤、散剤及び顆粒剤を含む。該固体剤形には、有効成分は、少なくとも一の非活性で薬学的許容可能な賦形剤又は担体(クエン酸ナトリウム又は第二リン酸カルシウム等)並びに/又は、
a)注入剤若しくは増量剤(スターチ、ショ糖、ブドウ糖、マニトール及びケイ酸等)、
b)結合剤(例えば、カルボキシメチルセルロース、アルギン酸塩、ゼラチン、ポリビニルピロリジノン、ショ糖及びアカシア等)、
c)保湿剤(グリセロール等)、d)崩壊剤(寒天、炭酸カルシウム、ジャガイモ若しくはタピオカ澱粉、アルギン酸、特定のケイ酸並びに炭酸ナトリウム等)、
e)溶液緩染剤(パラフィン等)、
f)吸収促進剤(第四アンモニア化合物等)、
g)湿潤剤(例えば、セチルアルコール及びモノステアレートグリセロール等)、
h)吸収剤(カオリン及びベントナイト粘土等)、
i)潤滑剤(タルク、ステアリン酸カルシウム、ステアリン酸マグネシウム、固体ポリエチレングリコール、ラウリル硫酸ナトリウム)
及びこれらの混合物と混合されている
【0093】
同類の固体組成物を軟又は硬ゼラチンカプセル状の注入剤として使用することも可能で、高分子ポリエチレングリコール等は言うまでもなく乳糖等の賦形剤が用いられる。固体剤形は、錠剤、糖衣錠、カプセル剤、丸剤及び顆粒剤が、コーティング剤又はシェル剤(腸溶コーティング、放出制御コーティング及び他の薬剤処方技術として公知のコーティング)を用いて調剤される。前記固形剤形状では、有効成分はショ糖又はスターチ等の少なくとも一の非活性希釈剤と混合される。前記剤形は、通常行われている様に、非活性希釈剤以外の追加物質を備えることも可能で、例えば、錠剤潤滑剤並びにステアリン酸マグネシウム及びマイクロクリスタリンセルロース等の他の錠剤補助剤が挙げられる。カプセル剤、錠剤及び丸剤の場合は、剤形は緩衝剤を備えることも可能である。任意に乳白剤を含むことも可能である。乳白剤は腸管の特定部位だけで、又は腸管の特定部位選択的に遅発的に有効成分を放出する。使用可能な包埋組成の例には、ポリマー物質及びワックスが挙げられる。
【0094】
<医薬品組成物の使用>
上述の記載及び例により更に詳細に記載の如く、ガレクチン‐3、ガレクチン‐7及びガレクチン‐8は、涙液層の拡散促進に有用であり、これは該ガレクチンが分泌性ムチンのオリゴ糖鎖を膜貫通型ムチン(又は他の糖タンパク質)と結合させることでなされる。一般的には、ガレクチンは臨床的に眼乾燥の治癒を促進するのに有効と考えられている。
【0095】
一般的には、ガレクチンは、皮膚上皮に限定されずあらゆる皮膚上皮組織、角膜上皮、消化器管の内張り、肺上皮、並びに腎管、血管上皮、子宮及び膣上皮、尿道上皮、又は呼吸器上皮の内表皮に関連した治癒の促進に有用であることが本明細書に示されている。本発明は多種多様の実施例を網羅し、種々の上皮創傷を治癒する。上皮創傷には、外科創傷、切除損傷、疱疹、潰瘍、病変部、剥離、糜爛、裂傷、おでき、傷口、痛み、並びに熱又は化学物質暴露による熱傷が挙げられるが、限定されるものではない。該創傷は健常者のものである場合もあるが、異常な損傷治癒を誘発する症状(糖尿病、角膜ジストロフィー、尿道症、栄養失調、ビタミン欠乏、肥満、感染症、免疫抑制、並びに、ステロイド、放射線治療、非ステロイド系抗炎症薬(NSAID)、抗新生物治療薬及び代謝拮抗物による全身治療に伴う合併症)を持つ患者の創傷でもよい。
【0096】
ガレクチン‐1、‐3及び‐7及び/又は‐8は、本明細書では、例えば、皮膚の損失の後で起こる皮膚の回復の促進に役立つ。或いは、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7及び/又はガレクチン‐8について、本明細書では、創傷を起こしている層に対する植皮の密着性を増強すること、及び創傷を起こしている層で再度上皮形成を促すことが示された。適切な植皮は、自己移植片、人工皮膚、同種移植片、自己植皮片、自己上皮植皮片、無血管性植皮片、ブレア‐ブラウン植皮(Blair‐Brown graft)、骨移植、胚胎組織移植片、真皮移植片、遅延移植片、皮膚移植片、表皮移植片、筋膜移植片、全層植皮、異種移植片、同種移植片、増植性移植片、表層移植片、網状移植片、粘膜移植片、オレア‐ティールシュ植皮片(Ollier-Thiersch graft)、大網移植片、切り張り移植片、茎状移植片、穿通性植皮片、中間層皮膚移植片、分層植皮片を含むが、これらに限定されない。
【0097】
ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、本明細書では、疱疹状皮膚炎を治療するのに役立つ。この疱疹状皮膚炎は、真皮と上皮の接合部で疱疹を形成する。ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、本明細書では、表皮水疱症を治療することに、また、上皮とその下の真皮との密着性の欠損を治療することに役立つ。尚、また、この密着性の欠損は頻発することとなる。ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、開いて痛みを伴う水膨れを治療することに役立つ。これらの治療は、ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8が、これら損傷部の上皮再形成を促進ことによりなしえる。ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、さらに上皮内の細胞間の接着の損失を伴う天疱瘡を治療すること、或いは、真皮と上皮の接合部の細胞間の接着の損失を伴う接合部類天疱瘡を治療することに役立つ。ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、様々な潰瘍を治療することに使用される。前記潰瘍には、糖尿病(性)潰瘍、皮膚潰瘍、褥瘡性潰瘍、動脈潰瘍、及び様々なうっ血性潰瘍が含まれるが、これらに限定されない。
【0098】
本発明は、角膜組織修復を促進するための方法も含むものとする。前記方法は、角膜潰瘍、熱、放射線、泡性角膜炎、角膜潰瘍又は断裂、近視矯正のための光屈折率手術、異物性及び無菌性角膜浸潤、酸又はアルカリ(例えば、フッ化水素酸、ギ酸、無水アンモニア、セメント、フェノール類、或いは他の化学薬剤(黄燐、元素金属、硝酸塩炭化水素、タール))の暴露による化学熱傷、神経栄養角膜症、糖尿病性の角膜症、Thygeson's点状表層角膜炎等の角膜症、ウイルス性角膜炎(メタヘルペス性(metaherpetic)角膜炎、ヘルペス性角膜炎)及び細菌性角膜炎等の角膜炎、格子状ジストロフィー等の角膜ジストロフィー、上皮基底膜ジストロフィー(EBMD)及びフックス内皮ジストロフィーにより引き起こされる角膜上皮欠損を治療することを含む。
【0099】
本明細書における方法により、ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、胃腸の潰瘍を治療することに有用であり、粘膜内層の治癒を助け、また腺性粘膜及び十二指腸粘膜内層をより早く再生させることができる。クローン病及び潰瘍性大腸炎等の炎症性腸疾患は、小腸及び大腸それぞれの粘膜表面の破壊を引き起こす。従って、ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、その粘膜表面を再形成することを促進することにより使用されることができので、より早い治癒を助け、炎症性腸疾患の進行を遅らせることができる。ガレクチン‐1、‐3、‐7及びガレクチン‐8は、ムチンを固定し、ムチンと上皮の先端面を結合させることにより、有害な物質(例えば摂取される有害物質または手術に伴い生じる有害物質)から胃腸管を保護することができる。ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、腸毒性の副作用を減らすために使用される。前記腸毒性は、細菌感染、ウイルス感染、放射線治療、化学療法、或いは他の治療により引き起こされる。ガレクチン‐8、‐3及び/又はガレクチン‐7は、例えば、粘膜炎、食道炎或いは胃炎を、予防的に或いは治療的に予防或いは緩和する。(例えば、経口、食道、腸管、結腸、直腸、肛門潰瘍に関連する損傷が治癒される。)
【0100】
ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は尿路上皮の治癒を促進するのに有益である。尿路上皮細胞を含む組織層は、多くの手段により損傷している。該手段とは、カテーテル法、外科手術、或いはバクテリア感染(例えば、淋病といった、性感染症を引き起こす媒介物による感染)を含む。本発明はまた女性生殖系の組織の治癒の促進方法も包含する。該方法は有効量のガレクチン‐8、‐3及び又はガレクチン‐7を投与することを含む。女性生殖系の組織の損傷は、カンジダ菌、トリコモナス、ガードネレラ、淋病、クラミジア、マイコプラズマ感染症及びその他の性感染症といった感染を含む広範囲の病気により引き起こされる。
【0101】
ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は腎上皮細胞の治療を促進する方法により有益とされる。そしてそれ故に、急性又は慢性腎不全や末期の腎臓病などの腎疾患や病状の緩和や治療に有益でありうる。ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は乳房組織の治療を促進する方法により有益とされる。そしてそれ故に、外科手術、外傷又は癌によって受けた乳房組織の治癒の促進に有益でありうる。ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、脳組織の損傷の治癒や緩和を促進する本明細書に示された方法により有益とされる。該脳組織の損傷とは、外傷、外科手術又は化学薬品によるものである。
【0102】
ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は予防的に投与されて、様々な病理状態により引き起こされる肺への損傷を軽減又は回避することが可能である。例えば、ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は肺胞や細気管支上皮の治療を促進し、急性又は慢性肺損傷を回避、減弱又は治療することが可能である。肺胞の進行性消失をもたらす肺気腫や、気道障害、すなわち煙の吸入や火傷に起因し、細気管支上皮や肺胞の壊死の原因となるものは、ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8を用いて効果的に治療することができる。該ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、化学療法、放射線治療、肺癌、ぜんそく、黒色肺及びその他の肺損傷状況に起因する損傷を治療することができる。
【0103】
当然のことながら、本発明に包含される治療方法は、ヒトの創傷の治療に限定されるものではなく、任意の哺乳類の創傷の治療に用いられてもよい。該哺乳類は、ウシ、イヌ、ネコ、ヤギ、ヒツジ、ブタ、ネズミ及びウマの種を含むが、これらに限定されない。ある種の創傷を治療する場合、使用されたガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、種に自然に発生するガレクチン‐1、‐3、‐7及び/又はガレクチン‐8のアミノ酸配列と実質的に同一であるアミノ酸配列を持つことが好まれるが、これに限定されない。
【0104】
これらの例の中に述べられてきた全ての動物は、眼科研究での動物の使用の、眼科と視覚に関する研究会議の決議、またNIHの実験動物の管理と使用に関する指針による勧告に準拠している。
【0105】
<実施例1:損傷に伴う角膜上皮の移動におけるガレクチン−3の上方制御>
ガレクチン−3の発現レベルが、損傷に伴った治癒角膜の上皮において変わるかどうかを決定するために、生体内において部分的に治癒することができる程度の2mmのエクシマレーザー切除と切除創傷をもつマウスの角膜を、ラットの抗ヒトガレクチン3-mAb M3/38(アメリカ培養コレクション、Rockville, MD)により、免疫染色した。角膜上皮は、層鱗状上皮の原型である。マウスにおいて、角膜上皮は全体の角膜の厚さの20〜25%を構成し、5〜6層の細胞からなる。上皮基底膜の下面には角膜実質を備え、マウスの場合、該角膜実質は全体の角膜の厚さの70〜80%である。切除創傷は、角膜実質に傷をつけずに上皮を除去する。反対に、エクシマレーザー処置は一般的に近視の矯正に用いられ、上皮だけでなくその下面に位置する角膜実質も除去する。
【0106】
スイス・ウェブスターマウス(Taconic Laboratory Animal Service, Germantown, NY)を、1.25%のアバーティン(0.2ml/10kg体重)を用いて筋肉注射によって麻酔した。該アバーティンは、2,2,2トリブロモエタノール2.5gと、2−メチル−2ブタノール(Aldrich, Milwaukee, WI)5mlと、蒸留水195mlを混合して調製した。プロパラカイン点眼薬(Alcon Labs Fort Worth, TXより入手可能なALCAINE(登録商標))を角膜の局所麻酔として用いた。第1群マウスの右目に、APEX PLUS(登録商標)エクシマレーザー(Summit Technology of Waltham、MA)を用いて、経上皮エクシマレーザー切除術を行った(2mmの光学部、42から44μmの切除の深さ、PTKモード)。第2群のマウスの右目に、アルジャー・ブラッシュ(Alger brush (Alger Equipment Company of Lago Vista,TX))を用いて、2mmの切除創傷を作製した。
【0107】
手術に伴い、全ての動物に鎮痛薬として、ブプレノルフィン(0.3mg/mlを0.2ml、Reckitt&Colman Pharmaceuticals, Richmond, VAより入手可能なBUPRENEX(登録商標))を用いて筋肉注射を行った。抗生物質軟膏(Pharmaderm, Melville, NY から入手可能なVETROPOLYCIN(登録商標))を塗ると、角膜は、生体内で16〜18時間後に部分的に治癒された。治癒期間の終了時、上記のように動物に麻酔をかけた後、頸部脱臼させて解剖を行った。解剖後、その眼をホルマリンで2時間固定した後、パラフィンろうに包埋した。組織断片(厚さ5μm)は眼球軸に対して平行な位置で切断した。その断片はキシレンによる処置と、段階的なエタノール溶液(100%、70%、30%)を用いて再水和することによって脱パラフィン化した。免疫染色については、組織断片は、連続して3%のH2O2(37℃、10分)、2.5%の標準的なヤギ血清で培養した。それぞれ、H2O2は内因性のペルオキシダーゼ活性を阻害し、ヤギ血清は非特異的結合を阻害する。その断片は、連続してmAb M3/38(不希釈のハイブリドーマ流動体、1時間)で培養し、抗ラット免疫グロブリン(IgG)を1時間ビオチン化し(1:200、Vector Labs、Burlingame、CA)、アビジンDとビオチンペルオキシダーゼ(Vector Lab)とジアミノベンジジン(DAB)―H2O2試薬(Kirkegaard&Perry Labs, Gaithersburg, MD)との複合体を20時間で新たに調製した。負対照として、関連のないmAbまたは培地のみを用いて断片を処置した。
【0108】
図9に、免疫組織化学的染色したパラフィン断片における標準的な角膜(図9AとB)と治癒角膜(図9CとD)を示す。この図9には、角膜創傷治癒モデルのどちらにおいても、移動性の治癒角膜上皮が、標準的な表皮よりもmAb M3/38によって濃く染色され、詳細には基底細胞層と中間細胞層が濃く染色されていることが示されている。治癒角膜上皮と標準的な角膜上皮のどちらも、免疫染色は細胞−マトリックスの接着部位においてさらに濃く染色されている。標準的な角膜の間質細胞は、mAb M3/38と反応しない一方、治癒角膜の下の前部角膜実質に位置する細胞は、特に移動性上皮の下の領域において、確実にガレクチン−3を発現する。
【0109】
角膜上皮におけるガレクチン−3の免疫活性は、血清を含まないイーグルの最小必須培地において、器官培養を16〜18時間することにより角膜が治癒された場合と類似している。該イーグルの最小必須培地には、非必須アミノ酸、L−グルタミン、抗生物質、0.4%のウシ血清アルブミン(BSA)が含まれる。しかしながら、前部角膜実質は、ガレクチン−3を発現する細胞なしに生体内で治癒され、これにより、生体内で治癒を行う可能性がある角膜実質で見られるガレクチン−3陽性の細胞は、ケラトサイトではなく白血球である可能性が高いことが示唆された。
【0110】
ガレクチン−3の糖質認識領域が、損傷に伴う角膜上皮層の移動に関与するかどうかを決定するために、2mmのエクシマレーザー切除と切除創傷をもつ角膜を、二糖のβ−ラクトースとスクロースの存在下または非存在下で、器官培養を行うことで治癒させた。β−ラクトースがガラクトースを含み、ガレクチンと結合する一方で、スクロースはガラクトースをもたず、ガレクチンと結合しなかった。これらの実験によると、角膜創傷の再上皮形成率は、β−ラクトースの存在下では著しく遅く、一方スクロースは何も影響しなかった。図10で示すとおり、異なる群間の治癒率(少なくとも2つの実験による平均±標準誤差)は、mm2/hで以下のように示される。培地のみは0.088±0.003(N=29)、培地とβ−ラクトースは0.063±0.003(N=19)、培地とスクロースは0.084±0.004(N=10)であった。
【0111】
<実施例2:野生型マウスとガレクチン−3欠損マウスにおける角膜上皮創傷閉塞>
角膜創傷の再上皮形成が、ガレクチン−3欠損マウスで低下するかどうかを決定するために、角膜創傷治癒の4つの異なるモデルを用いた。ガレクチン−3欠損マウス(gal-3-/-)は、Hsu et al., Am J. Pathol 156:1073,2000に記載されたガレクチン−3遺伝子の遮断を対象として生産された。特に、CRDをコードする領域は、ネオマイシン抵抗性遺伝子によって遮断された。これは、0.5kbイントロン4−エクソンの5断片を抗生物質抵抗性遺伝子(neo)に置換することを含む。不活性化されたガレクチン−3遺伝子が、サザンブロット法及びウエスタンブロット法によって確認された。
【0112】
簡潔に説明すると、エクシマレーザー切除術を行った角膜(実施例1で記載したように)又はアルカリによる火傷は、生体内又は生体外で(実施例1で記載したように)部分的に治癒された。アルカリ損傷については、2mmのフィルターディスク(Whatman 50, Whatman International, Maidstone, UK)を、トレフィンを用いることにより作製し、0.5Nの水酸化ナトリウムに浸し、それを第2群のマウスの右目の角膜表面に30秒おいた。その後、該眼を十分なPBSで洗浄した。治癒期間の終了時、創傷範囲をメチレンブルーで染色することにより視覚化した。その後、染色された創傷を標準距離で撮影し、染色された創傷の映像から創傷範囲の輪郭を紙に写しとった。SIGMASCAN(登録商標)のソフトウェア(SPSS Science of Chicago, IL)を用いて、これらの輪郭をデジタル化して定量化した。創傷治癒の異なるモデルのgal-3+/+マウスの創傷閉塞率の分析では、gal-3+/+のmm2/hで示される創傷閉塞率は、エクシマレーザーによって損傷した角膜の方がアルカリによる火傷によって損傷した角膜に比べて、低いことが明らかになった。また、損傷方法に関らず、創傷閉塞率は、生体内で治癒する角膜の方が器官培養で治癒する角膜に比べて高い。図11に示すように、gal-3+/+群間の創傷閉塞率は、エクシマレーザー/生体内群は0.076±0.003mm2/h、エクシマレーザー/生体外群は0.050±0.003mm2/h、アルカリによる火傷/生体内群は0.182±0.003mm2/h、アルカリによる火傷/生体外群は0.106±0.005mm2/hである。それぞれの群の値は、少なくとも2つの実験(各群N=9以上)の平均±標準誤差で示されている。gal-3+/+群とgal-3-/-群の創傷閉塞率を比較すると、角膜がエクシマレーザーによる損傷を受けたかアルカリ処置による損傷を受けたかに関らず、また角膜が生体内又は生体外で治癒されたかに関らず、mm2/hで表される角膜上皮創傷閉塞率は、gal-3-/-群のマウスがgal-3+/+群のマウスと比較して著しく低いことが明らかになった。異なるgal-3-/-群における創傷閉塞率は、エクシマレーザー/生体内群は0.060±0.004mm2/h、エクシマレーザー/生体外群は0.036±0.005mm2/h、アルカリによる火傷/生体内群は0.150±0.008mm2、アルカリによる火傷/生体外群は0.081±0.004mm2/hである。この場合も、すべての値が少なくとも2つの実験(各群N=8以上)の平均±標準誤差で示されている。
【0113】
<実施例3:損傷に伴うガレクチン−3欠損マウスの角膜上皮の移動における遺伝子の発現パターン>
角膜上皮創傷の再上皮形成が、なぜgal-3-/-マウスで乱れるのかを理解する試みとして、gal-3+/+とgal-3-/-の角膜の治癒における遺伝子発現パターンを、cDNAのマイクロアレイを用いて比較し、結果をさらに半定量的RT−PCRによって確かめた。経上皮エクシマレーザー切除(半径2mm)は、gal-3+/+とgal-3-/-のマウスの右目において作製した(実施例1のように)。角膜は、部分的に20〜24時間で生体内で治癒された。治癒期間の終了時、動物を解剖し、角膜を摘出してすぐに液体窒素に入れ、SMART(登録商標)のcDNA技術を用いた遺伝子発現分析のため、Clontech Labolatories, Palo Alto, CAに発送した。簡潔に説明すると、ATLAS(登録商標)のピュアトータルRNAラベリングシステムで定める試薬を用いて全RNAを分離した。30のgal-3+/+とgal-3-/-の角膜から生産されるRNAは、それぞれ3.5μgと2.6μgであった。gal-3+/+とgal-3-/-のマウスの角膜のRNA作製のA260:A280比率は、それぞれ1.48と1.37であった。リボソームRNAの28S:18S比率は、どちらの作製においても1.8であった。これにより、RNA作製量は十分であるということが確かとなった。プローブ作製については、第1のらせん構造cDNAは、RNAを175ng、修飾オリゴ(dT)プライマー(CDSプライマー)、POWERSCRIPT(登録商標)逆転写酵素と、SMART(登録商標)IIオリゴヌクレオチドを用いて合成した。対照群は、逆転写酵素なしにサンプルを培養した。長距離(LD)−PCRによってcDNAを増幅させた。増幅サイクルの最適数を決定するために、試薬製品の一定分量を15、18、21、24サイクル時に集め、アガロースゲル電気泳動によって分析した。最適数の増幅サイクル(例えば23サイクル)を用いて増幅した2重らせんcDNAの生産量は1〜1.6μgであった。増幅したcDNA(500ng)は、クレノウ酵素と33P−αATPを、ATLAS(登録商標)のマイクロアレイ(Clontech)に用いるSMART(登録商標)cDNAプローブ合成用の取扱説明書を用いて、放射性同位体でラベル付けした。ラベルしたプローブをNUCLEOSPIN(登録商標)フィルターによる濾過によって精製し、マウスの1.2k-I ATLAS (登録商標)ナイロンのcDNAマイクロアレイ(Clonetech)とハイブリッド形成した。これは、1200までのマウス遺伝子からなる広域スペクトルアレイである。ハイブリダイゼーションに伴い、その膜を放射線画像解析装置の画面にさらし、結果をATLAS IMAGE (登録商標) 2.0 ソフトウェア (Clonetech)で分析した。データは半定量的RT−PCRによって確認した。
【0114】
RT−PCRとして、全RNAと第1のらせん構造cDNAは、前述した手順によって、gal-3+/+治癒角膜とgal-3-/-の治癒角膜により調製した。PCR増幅は、cDNAを14ng、Clonetechから購入した遺伝子特異性の特別プライマー、及びADVANTAGE (登録商標) 2 PCRキット(Clonetech)によるその他の試薬を用いて50μl量中で行った。アニーリングの温度は68℃とし、試薬はPCR増幅のサイクル数の変化に従った。ハウスキーピング遺伝子の分析として、増幅生成物の一定分量5μlを5サイクルごと及び18サイクル開始時に集める一方、異なる発現遺伝子反応の増幅生成物を1つおきのサイクルごと及び28サイクル開始時に集めた。様々なサイクルで集められた増幅生成物は、1.5%アガロース/エチジウムブロマイドゲルで電気泳動することにより分析した(図12)。これらの実験から、gal-3+/+マウスの治癒角膜と比較して、gal-3-/-マウスの治癒角膜は、ガレクチンー7のmRNA転写産物、その他のガラクトース結合タンパク質及びトロイド様タンパク質(TLL)、メタロプロテイナーゼが顕著に少ない量であることが明らかになった。全体的には、gal-3+/+の治癒角膜と比較して、gal-3-/-の治癒角膜は、ガレクチン−7を約1/12、TLL遺伝子転写産物を約1/14(データ示さず)含んでいた(図12)。様々なハウスキーピング遺伝子のmRNA転写産物の発現レベルは、マイクロアレイ技術(図12)と半定量的RT−PCR(図12、GAPDHはD−グリセルアルデヒド−3−リン酸脱水素酵素、RPS29はリボソームタンパク質S29、ODCはオルニチンデカルボキシラーゼである)によって検出した結果、gal-3+/+治癒とgal-3-/-治癒の両方が類似していた。
【0115】
ガレクチン−7タンパク質の発現レベルがgal-3-/-マウスの治癒角膜でも減少するかどうかを確認するために、gal-3+/+治癒角膜とgal-3-/-の治癒角膜の界面活性剤抽出物を用いたウエスタンブロット分析(図13A)と、gal-3+/+とgal-3-/-マウスの角膜由来のパラフィン断片を用いた抗ガレクチン−7のポリクローナル抗体の免疫組織化学的研究(図13B)を行った。この免疫活性は、激しい(+++)、中程度(++)、弱い(+)、又は負(−)で級付けをした。gal-3-/-治癒角膜上皮の移動が、gal-3+/+治癒角膜上皮の移動と比較して、著しくガレクチン−7の免疫活性が少ないことを発見した(gal-3+/+:+++36/42、++5/42、+1/42以下、gal-3-/-:+++3/42、++26/42、+13/42以下)。また、細胞培養で成長したgal-3-/-マウスの胎児線維芽細胞(MEF)は、gal-3+/+のMEF培養と比較して減少量を示した(図13C)。
【0116】
<実施例4:外因性のガレクチン−3が、野生型マウスとガレクチン−3欠損マウス内で角膜創の上皮再形成を促進する>
角膜上皮創閉塞速度がガレクチン−3欠損マウス(gal-3-/-マウス)内で変動したが(実施例2)、器官培養において、角膜を治療する上皮再形成を外因性ガレクチンが促進させるか否かという点は興味深いところである。本研究において、アルカリによる火傷を負ったgal-3+/+マウスとgal-3-/-マウスの角膜が、無血清培地内で培養された。尚、この培養は、様々な量の組み換え型ガレクチン−3の存在下及び不存在下で行なわれている。
【0117】
上記の如く(Yang et al., Biochemistry 37:4086,1998)、大腸菌内で、組み換え型全長ヒトガレクチン−3が作り出されるとともに精製された。実施例2において説明されたように、アルカリが含浸されたフィルターディスクを用いて、麻酔されたマウスの両目にアルカリ火傷創(2mm直径)が作られた。この創傷の後、マウスは殺傷され、眼球が摘出され、外因性ガレクチン−3の存在下或いは不存在下で、眼球の培養が18時間から20時間行なわれた。
マウスの左目は対象として用いられるものであるが、無血清培地でのみ培養された。右目は、様々なテスト試薬を含む無血清培地内で培養された。このテスト試薬には、以下のものが含まれる。
(i)ガレクチン−3(5μg/mLから20μg/mL)
(ii)ガレクチン−3(10μg/mL)と0.1Mのβ−ラクトース
(iii)ガレクチン−3(10μg/mL)と0.1Mのスクロース
(iv)0.1Mのβ−ラクトース
(v)0.1Mのスクロース
治癒期間の終わりに、実施例2で説明した如く、SIGMASCAN(登録商標)ソフトウェア(イリノイ州シカゴのSPSS Science社製)を用いて、残存する創傷領域が染色され、撮像され、定量化された。各グループは、最低3つの眼球を含み、全ての実験は、少なくとも2回行なわれた。
【0118】
外因性ガレクチン−3は、gal-3-/-マウスの角膜創傷の上皮再形成速度に何ら盈虚を与えなかった(図14A参照)。しかし、外因性ガレクチンは、gal-3+/+マウスの創傷閉塞速度を促進した。この創傷閉塞速度は、10μg/mLから20μg/mLの濃度に依存する(0及び5μg/mLの濃度では、0.090±0.010mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は7以上)であり、10μg/mLでは、0.129±0.010 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は7以上)であり、20μg/mLでは、0.154±0.004 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は7以上)である。
図15に示す如く、gal-3+/+マウスの角膜上皮創傷閉塞に対するガレクチン−3の促進効果は、β−ラクトースにより特に抑制されたがスクロースによっては抑制されなかった(10μg/mLのガレクチン−3に対して、0.127±0.010 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は7以上)であり、ガレクチン−3(10μg/mL)と0.1Mのβ−ラクトースに対して、0.103±0.014 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は7以上)であり、ガレクチン−3(10μg/mL)と0.1Mのスクロースに対して、0.130±0.003 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は7以上)である)。
【0119】
<実施例5:外因性ガレクチン−7が、野生型マウスとガレクチン−3欠損マウスの角膜創傷の上皮再形成を促す>
別個の研究において、cDNAマイクロアレイ(例えば、実施例3に示すような)を用いて、gal-3+/+マウスの正常角膜と治癒角膜の遺伝子発現パターンの比較が行なわれ、治癒角膜においては、ガレクチン−7の発現が著しく向上することが明らかとなった。実施例3で説明した研究と関連して、これらの発見は、gal-3-/-マウスの治癒角膜において、ガレクチン−7の発現が低下することを示している。これに起因して、これらの発見は、外因性ガレクチン−7が器官培養において治癒角膜の上皮再形成を促すか否かを判定する実験設計に繋がっている。
この研究において、アルカリ火傷創を負ったgal-3-/-マウスの角膜が、無血清培地で培養された。この培養は、様々な量の組み換え型ガレクチン−7の存在下及び不存在下で行なわれている。
【0120】
cDNA(ヴァージニア州のマナサスにあるAmerican Type Culture Collection社からESTクローンとして入手可能である)をpET25bプラスミド(ウィスコンシン州マジソンにあるNovagen社から入手可能である)内にクローン化することにより、大腸菌内に組み換え型全長ヒトガレクチン−7が作り出された。
実施例2において説明されたように、アルカリが含浸されたフィルターディスクを用いて、麻酔されたマウスの両目にアルカリ火傷創(2mm直径)が作られた。この創傷の後、マウスは殺傷され、眼球が摘出され、外因性ガレクチン−7の存在下或いは不存在下で、眼球の培養が18時間から20時間行なわれた。
マウスの左目は対象として用いられるものであるが、無血清培地でのみ培養された。右目は、様々なテスト試薬を含む無血清培地内で培養された。このテスト試薬には、以下のものが含まれる。
(i)ガレクチン−7(20μg/mL)
(ii)ガレクチン−7(20μg/mL)と0.1Mのβ−ラクトース
(iii)ガレクチン−7(20μg/mL)と0.1Mのスクロース
治癒期間の終わりに、実施例2で説明した如く、SIGMASCAN(登録商標)ソフトウェア(イリノイ州シカゴのSPSS Science社製)を用いて、残存する創傷領域が染色され、撮像され、定量化された。各グループは、最低6つの眼球を含み、全ての実験は、少なくとも2回行なわれた。
【0121】
図16に示す如く、外因性ガレクチン−7は、創傷閉塞速度を促進させた(培地単独の場合は、0.036±0.006 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は10以上)であり、ガレクチン−7(20μg/mL)の場合は、0.072±0.004 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は10以上)である。
図16に示す如く、角膜上皮創傷閉塞に対するガレクチン−7の促進効果は、β−ラクトースにより特に抑制されたが、スクロースによっては抑制されなかった(20μg/mLのガレクチン−7に対して、0.072±0.004 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は9以上)であり、ガレクチン−7(20μg/mL)と0.1Mのβ−ラクトースに対して、0.050±0.004 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は9以上)であり、ガレクチン−7(20μg/mL)と0.1Mのスクロースに対して、0.079±0.007 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は9以上)である)。
図16に示す如く、外因性ガレクチン−7が、gal-3-/-マウスに代えて、gal-3+/+マウスの損傷した角膜に添加されると、損傷閉塞速度が更に高まった(0.094±0.003 gal-3+/+ mm2/h)。
【0122】
<実施例6:野生型マウスとガレクチン−3欠損マウス内での皮膚上皮創閉塞>
gal-3+/+マウスとgal-3-/-マウスは、1.25%のトリブロモエタノール(Avertin)(0.2mL/10g体重)の腹腔内投与により、麻酔された。レーザ処置の前に、剃刀刃を用いて、背部領域の体毛が剃られた。上皮を透過する6mmの背部の皮膚の創傷が、エキシマレーザ(マサチューセッツ州のワルシャムにあるSummit Technology社製)を用いて作られた。外科的処置の後、抗菌性軟膏が、創傷表面に塗布され、ブプノルフィン(2 mg/kg体重)が、皮下に投与され、術後の痛感を最小限化させている。当該創傷は、部分的に生体内治癒が可能であり、術後、24、48及び72時間で試験が行なわれている。
治癒期間の終わりに、マウスは、1.25%のトリブロモエタノール(Avertin)(0.2mL/10g体重)の腹腔内投与により、再度麻酔され、Sigma Scanソフトウェアを用いて撮像され、その後定量化される。
実験動物の2つのグループ(即ち、gal-3+/+マウスとgal-3-/-マウス)の間で創傷閉塞速度が比較される。実験動物は、二酸化炭素吸入或いはペントバービタルの過投与により、その後殺傷される。
【0123】
<実施例7:皮膚創傷の上皮再形成に対する外因性ガレクチン−3の効果>
実験動物(マウス:57BL/6及び129混合遺伝背景;齢:6週から8週;性別:混合)が、1.25%のトリブロモエタノール(Avertin)(0.2mL/10g体重)の腹腔内投与により麻酔された。
レーザ処置の前に、剃刀刃を用いて、背部領域の体毛が剃られた。上皮を透過する6mmの背部の2つの皮膚上の創傷(各側1つの創傷)が、エキシマレーザー(マサチューセッツ州のワルシャムにあるSummit Technology社製)を用いて作られた。
外科的処置の後、抗菌性軟膏が、創傷表面に塗布され、ブプノルフィン(2 mg/kg体重)が、皮下に投与され、術後の痛感を最小限化させている。当該創傷は、部分的に生体内治癒が可能であり、術後、4時間から6時間おきにガレクチン−3を含有する軟膏が右側の創傷に塗布され、キャリアのみが左側の創傷に塗布された。尚、左側の創傷は対象としての役割を担う。
治癒期間の終わり(24時間から48時間後)に、実験動物は、1.25%のトリブロモエタノール(Avertin)(0.2mL/10g体重)の腹腔内投与により、麻酔され、Sigma Scanソフトウェアを用いて撮像され、その後定量化される。
実験動物の2つのグループ(即ち、ガレクチン−3を用いて治療されたものと対象)の間で創傷閉塞速度が比較される。実験動物は、二酸化炭素吸入或いはペントバービタルの過投与により、その後殺傷される。
【0124】
<実施例8:皮膚創傷の上皮再形成に対する外因性ガレクチン−7の効果>
実験動物(マウス:57BL/6及び129混合遺伝背景;齢:6週から8週;性別:混合)が、1.25%のトリブロモエタノール(Avertin)(0.2mL/10g体重)の腹腔内投与により麻酔された。
レーザ処置の前に、剃刀刃を用いて、背部領域の体毛が剃られた。上皮を透過する6mmの背部の2つの皮膚上の創傷(各側1つの創傷)が、エキシマレーザー(マサチューセッツ州のワルシャムにあるSummit Technology社製)を用いて作られた。
外科的処置の後、抗菌性軟膏が、創傷表面に塗布され、ブプノルフィン(2 mg/kg体重)が、皮下に投与され、術後の痛感を最小限化させている。当該創傷は、部分的に生体内治癒が可能であり、術後、4時間から6時間おきにガレクチン−7を含有する軟膏が右側の創傷に塗布され、キャリアのみが左側の創傷に塗布された。尚、左側の創傷は対象としての役割を担う。
治癒期間の終わり(24時間から48時間後)に、実験動物は、1.25%のトリブロモエタノール(Avertin)(0.2mL/10g体重)の腹腔内投与により、麻酔され、Sigma Scanソフトウェアを用いて撮像され、その後定量化される。
実験動物の2つのグループ(即ち、ガレクチン−7を用いて治療されたものと対象)の間で創傷閉塞速度が比較される。実験動物は、二酸化炭素吸入或いはペントバービタルの過投与により、その後殺傷される。
【0125】
<実施例9:アルビノウサギのドライアイ症状に対する外因性ガレクチン−8或いはガレクチン−1の影響>
実験動物(ウサギ:アルビノ;齢:6週から8週;性別:混合)が、1.25%のトリブロモエタノール(Avertin)(0.2mL/10g体重)の腹腔内投与により麻酔された。1.0%の硫酸アトロピンの日常的な繰り返し滴下(Burgalassi, S., et al., Ophthalm Res 31:229-235, 1997)により、各ウサギにドライアイ症状を引き起こす。Schirmer I test及び蛍光染色後の角膜の試験により、ドライアイ症候群の評価が各実験動物に対してなされる。
【0126】
4時間から6時間おきに、ガレクチン−8の溶液を含有する点眼薬が、各実験動物の右眼に投与される。また、キャリアのみが左目に投与される。尚、左目は対象としての役割を担う。
治癒期間の終わり(24時間から48時間後)に、実験動物は、1.25%のトリブロモエタノール(Avertin)(0.2mL/10g体重)の腹腔内投与により、麻酔され、眼領域がSigma Scanソフトウェアを用いて撮像され、その後定量化される。
眼表面及びドライアイの症状が2つの眼のグループ(即ち、ガレクチン−8により治療された右目と対象となる左目)の間で評価され、これらの表面が比較される。
実験動物は、二酸化炭素吸入或いはペントバービタルの過投与により、その後殺傷され、標準的な毒学的基準により解析がなされた。
【0127】
ヒトと他の脊椎動物(図20参照)の多種のガレクチン−8タンパク質(図18及び配列識別番号:4及び5)のアミノ酸配列は略同一であり、このことは、他の種の哺乳類に当てはまる。同様に、ヒトと他の哺乳類の多種のガレクチン−1タンパク質(図19及び配列識別番号:6)のアミノ酸配列は略同一である。保存部位及び保存変化が少ない部位は、残基が変化し他の機能性ガレクチン−8及び/又は機能性ガレクチン−1タンパク質を得られ、これらタンパク質が、ドライアイ症状及び他の眼症状を治療する能力を備える。
【0128】
<実施例10:スコポラミンモデル:DESに対するガレクチン治療並びにスコポラミン治療前のガレクチン溶液を用いた前治療の効果>
溶液1mL当たり0, 10又は20μgのいずれかの量のガレクチン−1、ガレクチン−3、ガレクチン−7及びガレクチン−8のうち1つ或いは2つを含有する涙液溶液が、以下に示す治療スケジュールに則って投与される。
10μLの滴下量を以って、各実験動物の一方の眼に対する治療が各時点で行なわれる。スコポラミンパッチモデルのグループに対して、10μg/mLのガレクチン溶液が、1日当たり4回、一方の目に投与される。この投与は、スコポラミンパッチの使用直後に開始されるものである。
マウスで構成される治療グループが使用される。治療グループ及び眼は、ランダム化並びにコード化され、無作為に測定が行われる。各実験動物の一方の眼が治療溶液を投与され、他方の眼は対象滴を投与される。測定は、各時点で相互に行われる。
【0129】
<実施例11:スコポラミンモデル:βラクトース、ガレクチン阻害剤とともに投与されるガレクチンの混合治療>
実施例1に示すものと同一の手順が実行された。尚、0.1 Mのβ−ラクトースとともに10μg/mLのガレクチン溶液が用いられ、ガレクチンの効果が、競合するに糖類により抑制されるか否かを判断する。
【0130】
【表3】
【0131】
<実施例12:DESにおけるガレクチン−1、ガレクチン−3、ガレクチン−7及びガレクチン−8の役割:マウスの自己免疫MRL/lpr染色>
治療グループ及び眼は、ランダム化並びにコード化され、無作為に測定が行われる。各実験動物の一方の眼が治療溶液を投与され、他方の眼は対象滴を投与される。測定は、各時点で相互に行われる。
【0132】
【表4】
【0133】
<実施例13:ドライアイ症状を有する被験体に対するガレクチン−3による治療効果>
ドライアイ症状に対するガレクチンの効果を判断するために、インターロイキン−1を用いた動物モデルシステムが用いられ、4つのマウスグループが使用された。各グループは、5つの実験動物からなり、以下のように処置を施された。
3つのグループに対して、第0日目にインターロイキン−1(IL-1)が被検体に注入された。この注入は、シェーグレン症候群のマウスモデル用に創られた標準的手順にしたがって行われた(Zoukhri, D. et al., Invest Ophthalmol Vis Sci 42 (5): 925-932)。対象被検体は当該注入を受けておらず、点眼薬を用いた更なる治療を施されていない。IL-1の注入を受けた被検体は、その後、点眼薬の投与による治療を受けた。この点眼薬には、ガレクチン−3が含まれ、点眼量は、75μg/mL又は150μg/mLであり、1日当たり4回の点眼が行なわれる。この点眼は、第1日目から行なわれる。1つの被検体グループは、同様にバッファのみ(ガレクチンなし)を1日当たり4回投与された。ドライアイの程度は、上述の如く(Zoukhri et al.)、フルオレセインのクリアランスによって、各被検体ごとに評価された。
【0134】
第1日目の治療初期において、涙液分泌液が、フルオレセインのクリアランスにより測定され、75μg/mL或いは150μg/mLの投与量のガレクチン−3それぞれにより治療された被検体において、対象の被検体と比して、改善が見られた。第2日目までに、ガレクチン治療を施された被検体の症状は完全に改善された。実際に、第2日目において、幾分良好な涙液クリアランスが、低投与量75μg/mLの被検体で観察されたことから、これより少ない投与量が最適な投与量である可能性が示されている。図21において、棒グラフの棒(第2日目の治療された被検体に対する)上のアスタリスクは、ドライアイ症状を有する対象の被検体グループであって、バッファ賦形剤のみを投与されたものからのデータと比してこれらのデータが統計学的に有意であることを示している(p<0.05)。結論として、ガレクチン−3は、ドライアイを治療するとともに、1日から2日の治療期間内でこの症状を改善する能力を備えるということができる。哺乳類間の高程度の同一性並びにガレクチンファミリーの異なる型間でのガレクチン機能の同一性及び特異的アミノ酸コンセンサス配列の同一性の存在により、ガレクチン−1、ガレクチン−3、ガレクチン−7及びガレクチン−8のいずれか或いはこれらの組み合わせが、同様にドライアイ及びこれに関連する症状の治療に有効であることが予測できる。
【0135】
<結論>
本明細書では、ガレクチン−3及びガレクチン−7が角膜創傷の再上皮化の役割を担うことが実証された。実施例1では、免疫組織化学の研究により、損傷後にガレクチン−3が角膜上皮の細胞間基質接着部位で高密度存在することが明らかになった。そこは細胞基質の相互作用に影響を与え、細胞移動にも理想的な部位である。実施例2では、角膜創傷の再上皮化の進度がガレクチン−3欠損マウスにおいては、野生型マウスよりも著しく遅れることが示された。実施例3では、損傷後にガレクチン−7の発現がガレクチン−3欠損マウスにおいては、野生型マウスよりも著しく減少することが確認された。実施例4、5では、外因性の組み換え型ガレクチン−3とガレクチン−7が、gal3+/+マウスにおいて角膜創傷の再上皮化を刺激することが示された。実施例6−8では、創傷の再上皮化に際するガレクチンの効果の測定手段を明示した。実施例9−12においては、眼乾燥症候群におけるガレクチンの効果の測定手段を明らかにした。実施例13では、ガレクチン−3の眼乾燥への即効性治療が示された。さらに、実施例1においては、角膜上皮創閉塞速度におけるガレクチン−3の刺激性効果が、ガラクトシド化学構造の競合二糖(β−ラクトース)によって抑制されることが実証された。同時に、科学的に関連性のない二糖(スクロース)の影響を受けないことも示された。本結果として、糖鎖認識部位は角膜に対する外因性ガレクチンの有用な効果に直接関与していることが示された。
【0136】
ガレクチン1、3、7及びガレクチン8が眼乾燥症候群の治療に影響するというメカニズムに関して、いかなる特定の理論にも結びつけるという意図ではないが、以下を提案する。
【0137】
上記の通り、ガレクチン−3は糖鎖ポリラクトサミンを包含する補完的複合等質と結合して、細胞同士の、又は細胞基質の相互作用の仲介をすると考えられている。糖鎖ポリラクトサミンは多数の細胞外基質(ECM)において、又はイソ型のフィブロネクチン、ラミニン及び、インテグリンのような細胞表面分子において見られる。(Liu, Clin. Immunol. 97:79, 2000 及び Perillo, supra) しかしながら、外因性のガレクチン−3がgal3-/-マウスにおいては創傷再上皮化を促進しないという本明細書の所見(例4参照)は、細胞内のガレクチン−3が創傷治癒の過程で有意に貢献しているということを示唆している。細胞内のガレクチン−3は、特定の細胞表面、又は/あるいは、細胞外基質(ECM)受容体の発現において最も影響を与えており、結果として、細胞基質の相互作用や細胞移動にも影響を与えている。この考えはすでに発表された研究と一致しており、その研究ではガレクチン−3は乳癌細胞株内で安定的に過剰発現した。親細胞株はガレクチンをほとんど、あるいはまったく発現しなかった。結果として、α4β7、α6β1インテグリンが増加し、ラミニン、フィブロネクチン、及びビトロネクチンを含む様々な細胞外基質(ECM)分子との接着が強固になった。(Warfield, supra及びMattarese, supra) 別の研究においては(Dudas et al, Gastroenterology 118:1553, 2000)発現ガレクチン−3を形質移入された結腸癌細胞株が、レクチン本体の主要な結合基である特定ムチン、MUC2を増加させた。(Bresalier et al, Cancer Research 56:4354,1996) gal3+/+マウスにおいて創傷再上皮化速度に対する外因性ガレクチン−3の刺激性効果がラクロース阻害であるという事実は、以下の興味深い可能性を示唆している。細胞内ガレクチン−3は、実際にレクチン本体の細胞外基質(ECM)受容体や細胞表面として機能するタンパク質糖鎖付加を制御しているのかもしれない。細胞内ガレクチン−3が核基質に働きかけ、複雑な生物学的過程に影響を与える可能性を有しているということはまた、以下の所見を示唆している。特定の状況下では、レクチンは核内でリボ核タンパク質複合体と結合したり、RNA前駆体のスプライシング因子として作用したりすることが可能である。(Dagher et al., Proc. Natl. Acad. ScL USA 92:1213, 1995). 同様に、ワング(Wang)などによって、ガレクチン−3は前立腺癌細胞において核基質と結合し、又、一本鎖DNAとRNAの双方と結びつくことが実証された。(Wang et al., Biochem. Biophys. Res. Commun. 217:292, 1995).
【0138】
マウスの相補DNAマイクロアレイを用いたgal3+/+及びgal3-/-マウスの角膜治癒の遺伝子発現パターンの分析により、発現したgal3-/-マウスの角膜治癒は野生タイプのマウスよりもガレクチン−7を著しく減少させることが明らかになった。(例3、5参照)ガレクチン−7は1994年に初めて報告され(Barondes, supra)、ガレクチン−3ほど十分に特徴づけられていない。ガレクチン−3とは異なり、ガレクチン−7には顕著な組織特異性がある。動物の成体において、ガレクチン−7の発現は上皮に限られており、上皮は重層化しているか、重層化することになる。(Timmons et al., supra). タンパク質は細胞基質や細胞間の相互作用、及びアポトーシスに関与していると考えられている。(Leonidas, Biochemistry 37:13930, 1998 及び Bernerd et al., Proc. Natl. Acad. Sci. USA 96:11329, 1999). 一般的に、ガレクチン−7の発現とケラチノサイト増殖との間には逆相関が存在しており、ガレクチン−7の発現はSV40ウィルスに形質転換したケラチノサイトで抑制され、表皮性腫瘍に由来する株細胞でも同様に抑制される。外因性ガレクチン−3はgal3-/-角膜では創傷再上皮化を刺激せず、gal3-/-角膜の治癒はガレクチン−7の量を減少させる。この発見により、ガレクチン−3がガレクチン−7を調節することで創傷再上皮化に少なくとも部分的には影響を与えるということが示唆されている。実際に、ガレクチン−3と異なり、ガレクチン−7はラクトースを抑制してgal3-/-角膜で創傷の再上皮化を促進することが発見されている。また、gal3-/-マウスの胚繊維芽細胞は発現してガレクチン−7の量を減少させた。
【0139】
関与するメカニズムにかかわらず、ガレクチン−3とガレクチン−7の双方が角膜創傷の再上皮化を刺激するという所見は、上皮創傷の治療やとりわけ上皮創傷の非治癒に広範な意味を持つ。今日では、角膜の持続性上皮欠損の治療が主要な臨床問題となっている。さらに、移植後の創傷、高齢者の慢性創傷、褥瘡、及び皮膚の静脈うっ血性潰瘍への効果的な治療の必要性が高まっている。細胞増殖を刺激するとして公知の多数の成長因子(例えば、上皮成長細胞因子(EGF)、トランスフォーミング増殖因子(TGF)、繊維芽細胞増殖因子(FGF)、KGF、肝細胞増殖因子(HGF))が角膜での有用性を検査されており、皮膚の上皮創傷治癒においては、全体的に失望させる結果に終わっている。(Eaglstein, Surg. Clin. North Am. 77:689, 1997; Singer 及び Clark, N. Engl. J. Med. 341:738, 1999; Zieske 及び Gipson, pp. 364-372 in "Principle 及び Practice of Ophthalmology" Ed. by D. M. Albert 及び F. A. Jakobiec, W.B. Saunders Company, Philadelphia, PA, 2000; Schultz et al., Eye 8:184, 1994; Kandarakiset al., Am. J. Ophthalmol. 98:411, 1984; and Singh 及び Foster, Am. J. Ophthalmol. 103:802, 1987). 成長因子を用いる上記研究の大部分で、創傷の再上皮化の加速度範囲は今日の研究でガレクチンによって確認される範囲より著しく小さかった。また、上皮細胞成長因子(EGF)のような成長因子として処遇される角膜上皮は過形成のものであり、(Singh 及び Foster, Cornea 8:45, 1989) 明らかに望ましくない状態である。この観点においては、上皮細胞でレクチンの細胞有糸分裂の誘発性は観察されなかったため、臨床的な可能性はガレクチン−3とガレクチン−7の方が成長因子よりも魅力的である。ここ10年以上に渡って、近視の矯正のために角膜特性を修正するエキシマレーザー角膜切除術の可能性が現実味を帯びてきている。上記のような施術が毎週数千も行われ、近視の人々はメガネやコンタクトレンズから解放されている。世界の成人人口の25%−30%が近視であるという事実に着目すれば、年間約50万もの施術がアメリカだけで行われると想定されるだろう。エキシマレーザー手術のあとに、上皮治癒で遅延が生じる場合もある。上記のような遅延は可能な限り避けなければならず、そうでないと角膜が術後濁度や感染性角膜炎、及び角膜腫瘍に罹ってしまう危険性がある。やはり、ガレクチンに基づく治療はそのような症例で創傷の上皮化を促進する。
【0140】
<他の実施例>
本発明の他の具体例については、本明細書で開示された本発明の実施や規格を考慮すれば、同業者には明白である。規格や諸例については例示しただけに過ぎず、本発明の正確な範囲は以下の請求項で示されている。
【図面の簡単な説明】
【0141】
【図1】アミノ酸配列とヒトのガレクチン‐3を示す。(ジェンバンク受入番号BAA22164、配列番号1)
【図2】アミノ酸配列とヒトのガレクチン‐7を示す。(ジェンバンク受入番号I55469、配列番号2)
【図3】ヒトのガレクチン3(配列番号1)と、ウサギのガレクチン‐3のアミノ酸配列(ジェンバンク受入番号JC4300)、ニワトリのガレクチン‐3のアミノ酸配列(ジェンバンク受入番号AAB02856)及びハムスターのガレクチン‐3のアミノ酸配列(ジェンバンク受入番号CAA55479)をクラスタルWを用いて縦に配列したものを示す。図中の最初(上部)の配列はヒトのガレクチン‐3(配列番号1)のアミノ酸1から250、図中の2番目の配列はハムスターのガレクチン‐3のアミノ酸1から245、図中の3番目の配列はウサギのガレクチン‐3のアミノ酸1から242、図中の4番目(下部)の配列はニワトリのガレクチン‐3のアミノ酸1から262である。
【図4】ヒトのガレクチン‐7(配列番号2)とラットのガレクチン‐7(ジェンバンク受入番号P97590)及びマウスのガレクチン‐7(ジェンバンク受入番号O54974)をクラスタルWを用いて縦に配列したものを示す。図中の最初(上部)の配列はラットのガレクチン‐7のアミノ酸1から136、図中の2番目の配列はマウスのガレクチン‐7のアミノ酸1から136、図中3番目(下部)の配列はヒトのガレクチン‐7(配列番号2)のアミノ酸1から136である。
【図5】ヒトのガレクチン‐3(配列番号1)をPROSITEでスキャンした結果の一覧である。
【図6】ヒトのガレクチン‐7(配列番号2)をPROSITEでスキャンした結果の一覧である。
【図7】ヒトのガレクチン‐3のガラクトシド結合領域と、PFAM中の隠れマルコフモデル(HMM)由来の共通アミノ酸配列(PF00337)を縦に配列したものを示す。上部の配列は共通アミノ酸配列(PF00337、配列番号3)であり、また下部のアミノ酸配列は配列番号1のアミノ酸117から247に対応する。
【図8】ヒトのガレクチン‐7のガラクトシド結合領域と、PFAM中の隠れマルコフモデル(HMM)由来の共通アミノ酸配列(PF00337)を縦に配列したものを示す。上部の配列は共通アミノ酸配列(PF00337、配列番号3)であり、また下部のアミノ酸配列は配列番号2のアミノ酸5から135に対応する。
【図9】生体内で治癒可能な程度で角膜を2ミリ剥離し、或いは角膜にエクシマーレーザー創傷を与え、それからパラフィン切片中でガレクチン‐3の免疫反応性を分析した一連の写真を含む。(A)は、メマトキシリンとエオシンで色づけした(i)正常な角膜、(ii)剥離直後の角膜、(iii)エクシマーレーザー創傷を与えた直後の角膜である。(B)は、免疫組織学上色づけした(i)通常のgal-3+/+角膜、(ii)エクシマーレーザー創傷後の治癒gal-3+/+角膜である。免疫組織学上色づけした(iii)通常のgal-3-/-角膜と(iv)エクシマーレーザー創傷後の治癒gal-3-/-角膜である。濃い色は陽性免疫染色を示す。WE、(傷の端(wound edge))、LE(移動した上皮の先端(leading edge of migrating epithelium))、すなわち矢印(上皮)、矢頭(白血球/間質細胞)である。
【図10】創傷を受けた角膜上皮の治癒率に及ぼされるβ‐ラクトース(Lac)とサッカロース(Suc)の影響を図示するグラフである。
【図11】野生型(gal-3+/+)マウス及びガレクチン‐3欠損(gal-3-/-)マウスに、エクシマーレーザー或いはアルカリ処理を用いて、体内又は体外で治癒可能な程度の創傷を与えた角膜上皮の治癒率を図示している。
【図12】ガレクチン‐7の遺伝子発現とハウスキーピング遺伝子の選択における野生型(gal-3+/+)とガレクチン‐3欠損(gal-3-/-)マウス間での差を図示している。(GAPDHはD‐グリセルアルデヒド3リン酸デヒドロゲナーゼ、RPS29はリボソームたんぱく質S29、ODCはオルニチンデカルボキシラーゼである。)該差は、cDNAマイクロアレイ及び半定量PCRにより決定される。
【図13】ガレクチン‐7の発現における野生型(gal-3+/+)マウスとガレクチン‐3欠損(gal-3-/-)マウス間での差を図示している。該差は、(A)ウェスタンブロット分析、(B)免疫組織化学分析、(C)マウスの胚前脳部分を用いることにより決定される。
【図14】(図14Aの)ガレクチン‐3欠損(gal-3-/-)マウス及び(図14Bの)野生型(gal-3+/+)マウスの創傷を受けた角膜上皮の治癒率に及ぼされる外因性ガレクチン‐3の影響を図示するグラフである。
【図15】外因性ガレクチン‐3の存在下で、野生型(gal-3+/+)マウスの創傷を受けた角膜上皮の治癒率に及ぼされるβ‐ラクトース(Lac)とサッカロース(Suc)の影響を図示するグラフである。
【図16】(A)は外因性ガレクチン‐7が単独で、もしくはβ‐ラクトース(Lac)と共に、或いはサッカロース(Suc)と共に用いられた場合の、野生型(gal-3+/+)マウスの創傷を受けた角膜上皮の治癒率に及ぼされる外因性ガレクチン‐7の影響を図示したグラフである。また(B)は、野生型(gal-3+/+)マウス及びガレクチン‐3欠損(gal-3-/-)マウスの創傷を受けた角膜上皮の治癒率に及ぼされる外因性ガレクチン‐7の影響を比較するグラフである。
【図17】ヒトのガレクチン‐1(配列番号6)のアミノ酸配列を図示する。
【図18】ヒトのガレクチン‐8(配列番号4)の短形のアミノ酸配列を図示する。
【図19】ヒトのガレクチン‐1(配列番号6)、ラットのガレクチン‐1、マウスのガレクチン‐1及びハムスターのガレクチン‐1それぞれのアミノ酸配列を示す。しかもこれらは非常に高率な同一性を共有しているので、ガレクチン‐1配列は哺乳類種の間で強力に保存されている。黒い部分は残基の位置を表し、該位置は全ての哺乳類の配列において同一ではない。ラットとヒトは90.4%が同一で、マウスとヒトは88.2%同一で、またハムスターとヒトは91.2%同一である。また、図は非同一の位置にある残基の大部分が保存的な変化をすることを示す。例えば、アルギニンはポジション19でリシンに置換され、ロイシンはポジション24でバリンに、セリンはポジション73でトレオニンに、そしてアスパラギン酸はポジション136でグルタミン酸に置換される。
【図20】ヒトのガレクチン‐8(配列番号4)の短形(316アミノ酸)のそれぞれのアミノ酸配列と、マウスのガレクチン‐8、ラットのガレクチン‐8、ニワトリのガレクチン‐8及びカエルのガレクチン‐8の短形の対応するアミノ酸配列を示す。該アミノ酸配列は、強調部分と共に配列され、ヒトとラットガレクチン‐8の配列に匹敵する非ヒトアミノ酸配列が相違していることを表す。3つの哺乳類のガレクチン‐8アミノ酸配列は、非常に高率の同一性を共有し、また78.2%同一であって、非同一部分(21.8%)の大部分においてアミノ酸変化が保存されている。該アミノ酸変化は、マウス及びラットの類似のポジションにあるチロシンに匹敵するヒトのポジション20にあるフェニルアラニン、ヒトのポジション22にあるグリシンとラットのセリン、ヒトのポジション26にあるグリシンとげっ歯類配列中のアスパラギン酸塩である。さらに、脊椎動物種は43.0%のかなりの同一性を共有しているので、ガレクチン-8配列は哺乳類種の間で高い同一性を持ち、脊椎動物種間両方で保存されている。
【図21】マウスモデル中の眼乾燥症候群にガレクチンを投与した影響を示す棒グラフである。該症候群は時間の関数としての0日目にインターロイキン-1α(図では「IL-1」と示される)を注入して引き起こされたものである。ガレクチン-3は1日目と2日目に、一日4回、目薬を用いて1グループにつき5つの被験物それぞれに投与した。該目薬は75μg/mlまたは150μg/mlの濃度である。対照群の被験物にはIL-1を注入せず、目薬を投与しなかった。「緩衝液」グループ中の被験物には一日4回キャリア媒体のみの治療薬を投与した。Zoukhri,D.et.al.,Invest Ophthalmol Vis Sci 42(5):925−932に述べられているように、フルオレセインクリアランスにより涙の動態が評価される。低い値はより良く涙が流れることを指し示す。2日目のp<0.05の統計学上の有意差を持って対照群のデータと治療を施したグループのデータを比較すると、ガレクチン治療により、ガレクチンを投与されていない制御動物と比較して、涙の動態を正常なグループの涙の動態に回復したことがデータに示している。
【技術分野】
【0001】
眼乾燥症候群治療のためのガレクチンタンパク質の組成物、方法及びキットを提供する。
【背景技術】
【0002】
眼乾燥症候群(DES)はよくある症状で、30から45歳人口の10%もが症状を示し、45歳以上では増加し、人口の20%もが症状を示す。(Schein et al., 1997, Am J Ophthamol 124, 723‐72; Brewitt and Sistani, 2001, Surv of Ophthalmol 45:S119‐S202)。DESは眼球刺激、かすみ目及び視野振動を引き起こし、角膜感染や潰瘍にかかる視力にかかわる危険性を増加する。DESの組織学的な影響としては、異常増殖や分化が挙げられ、眼球表面上皮に、結膜杯細胞密度の減少及び眼球表面上皮による粘液生産の減少及び異常が見られる(Murillo‐Lopez and Pflugfelder, 1996, Dry Eye. In: Krachmer J, Mannis M, Holland E, eds. The Cornea. Mosby, St. Louis, MO. 663‐686; Dursun et al., 2002, Invest Ophthalmol & Vis Sci 43:632‐638)。眼乾燥症は慢性疾患で、症状や兆候は環境要因により大きく影響され、湿度や空気移動などが挙げられるが、読書やコンピューターの使用といった看視作業の度合いにも大きく影響される。
【0003】
DESの典型的な症状としては、灼熱痛、かゆみ、異物感、刺痛、乾燥、光線恐怖症、眼球疲労、及び発赤が挙げられる。眼乾燥症は慢性疾患で、症状や兆候は環境要因により大きく影響され、湿度や空気移動などが挙げられるが、読書やコンピューターの使用といった看視作業の度合いにも大きく影響される(Rheinstrom, 1999, Dry eye. In Yanoff, ed. Ophthalmology. 1st Ed. Editor. Mosley International Ltd, St Louis, MO; Foulks, 2003, The Ocular Surface, 1: 20‐30)。
【0004】
多種多様の人工涙液製品が販売されているが、どれもが症状を一時的に緩和するだけである。今のところ、症状を改善するような治療薬はない。従って、本技術分野では、眼乾燥症候群を治療する、更なる医薬品及び組成物が必要とされている。特に、薬品、組成物及び治療方法が必要とされている。
【0005】
【非特許文献1】Schein et al., 1997, Am J Ophthamol 124, 723‐72
【非特許文献2】Brewitt and Sistani, 2001, Surv of Ophthalmol 45:S119‐S202
【非特許文献3】Murillo‐Lopez and Pflugfelder, 1996, Dry Eye. In: Krachmer J, Mannis M, Holland E, eds. The Cornea. Mosby, St. Lois, MO. 663‐686
【非特許文献4】Dursun et al., 2002, Invest Ophthalmol & Vis Sci 43:632‐638
【非特許文献5】Rheinstrom, 1999, Dry eye. In Yanoff, ed. Ophthalmology. 1st Ed. Editor. Mosley International Ltd, St Louis, MO
【非特許文献6】Foulks, 2003, The Ocular Surface, 1: 20‐30
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明は、治療を必要としてする哺乳類の眼乾燥を治療する方法であって、治療に有効な量のガレクチンタンパク質を哺乳類に投与することを特徴とする。ある実施例において、哺乳類における眼乾燥の予防方法が挙げられ、眼乾燥予防を必要とする哺乳類の同定、及び同定した哺乳類に対し治療に有効なガレクチンタンパク質を投与する工程からなる。従って、該方法は治療又は予防を必要としている哺乳類に対して用いられ、少なくとも、眼上皮創傷である哺乳類、抗ヒスタミン剤の使用前である哺乳類、抗炎症剤の使用前である哺乳類、エクシマーレーザー治療を施す前の哺乳類のいずれかに当てはまる哺乳類の治療又は予防に対し用いられる。例えば、哺乳類にはヒトが挙げられる。更に、ガレクチンタンパク質はガレクチン‐8、ガレクチン‐7もしくはガレクチン‐3から選択される。ある実施例において眼乾燥が持続性症候群である。例えば、眼乾燥は上皮びらんを引き起こし、上皮びらんは更に角膜創傷へとつながる。
【0007】
一般的には、上述の方法で用いられるガレクチン-8タンパク質は、配列番号4又は5のアミノ酸配列を備える。例えば、ガレクチン‐8タンパク質は、配列番号4又は5のアミノ酸配列と実質的に同一のアミノ酸配列を含む。本明細書中で用いられている「実質的に同一」とは、ガレクチン‐8は、配列番号4又は5のアミノ酸配列と、少なくとも60%の同一性、70%の同一性、80%の同一性、又は90%の同一性を持つことを意味する。
【0008】
一般的には、上述の方法で用いられるガレクチン‐7タンパク質は、配列番号2のアミノ酸配列を備える。例えば、ガレクチン‐7タンパク質は、配列番号2のアミノ酸配列と実質的に同一のアミノ酸配列を含む。ガレクチン‐7は、配列番号2のアミノ酸配列と、少なくとも60%の同一性、70%の同一性、80%の同一性、又は90%の同一性を持つ。ある実施例ではガレクチンン‐7タンパク質はガレクチン‐7ガラクシド結合領域を含む。
【0009】
一般的には、上述の方法で用いられるガレクチン‐3タンパク質は、配列番号1のアミノ酸配列を備える。例えば、ガレクチン‐3タンパク質は、配列番号1のアミノ酸配列と実質的に同一のアミノ酸配列を含む。ガレクチン‐3は、配列番号1のアミノ酸配列と、少なくとも60%の同一性、70%の同一性、80%の同一性、又は90%の同一性を持つ。ある実施例ではガレクチンン‐3タンパク質はガレクチン‐3ガラクシド結合領域を含む。
【0010】
本発明の他の実施例としては、眼乾燥の治療促進効果を持つ医薬品組成物が挙げられ、該医薬品組成物は、薬学的に適当な担体もしくは希釈剤、及結膜上皮及び/又は角膜上皮の健全性を促進する十分量のガレクチン‐8タンパク質を備える。本発明の他の実施例としては、眼乾燥の治療に促進効果のある医薬品組成物が挙げられ、該医薬品組成物は、薬学的に適当な担体もしくは希釈剤、並びに結膜上皮及び/又は角膜上皮の健全性を促進する十分量のガレクチン‐7タンパク質を備える。本発明の他の実施例としては、眼乾燥の治療に促進効果のある医薬品組成物が挙げられ、該医薬品組成物は、薬学的に適当な担体もしくは希釈剤、及び結膜上皮及び/又は角膜上皮の健全性を促進する十分量のガレクチン‐3タンパク質を備える。本発明の他の実施例としては、眼乾燥の治療に促進効果のある医薬品組成物が挙げられ、該医薬品組成物は、薬学的に適当な担体もしくは希釈剤、及び結膜上皮及び/又は角膜上皮の健全性を促進する十分量のガレクチン‐8タンパク質を備える。これらの医薬品組成物のいずれかにおいて、眼乾燥が再発性上皮びらんを伴う眼乾燥症である。例えば、眼乾燥症は創傷を引き起こし、該創傷は角膜創傷である。更に眼乾燥は、エクシマーレーザー角膜切除術により引き起こされる。
【課題を解決するための手段】
【0011】
様々な医薬品組成物の実施例において、ガレクチン‐8タンパク質は、配列番号4又は5のアミノ酸配列を含む。又はガレクチン8タンパク質はアミノ酸番号4又は5のアミノ酸配列と実質的に同一なアミノ酸配列を含む。或いは、ガレクチン‐8タンパク質は、ガレクチン‐8N末端領域と,ガレクチン‐8プロリン、グリシン、及びチロシンが豊富な領域とを含む。もしくは、ガレクチン‐8タンパク質は、ガレクチン‐8プロリン、グリシン、及びチロシンが豊富な領域と、ガレクチン‐8ガラクトシド結合領域とを含む。例えば、ガレクチン‐8タンパク質はガレクチン‐8ガラクトシド結合領域を含む。或いは、ガレクチン‐7タンパク質は配列番号2のアミノ酸配列を含む。例えば、ガレクチン‐7タンパク質は配列番号2のアミノ酸配列と実質的に同一なアミノ酸配列を含む。又は、ガレクチン‐7タンパク質はガレクチン‐7ガラクトシド結合領域を含む。或いは、ガレクチン‐3タンパク質は配列番号1のアミノ酸配列を含む。例えば、ガレクチン‐3タンパク質はアミノ酸番号1のアミノ酸配列と実質的に同一なアミノ酸配列を含む。例えば、ガレクチン‐3タンパク質はガレクチン‐3ガラクトシド結合領域を含む。
【0012】
本発明は、更に治療を必要としている哺乳類の眼乾燥症を治療する方法であることを特徴とし、治療に有効量の物質を投与する工程を備え、該物質は角膜表面又は結膜表面上での涙液層の拡散に影響する。また、本発明は、治療を必要としている哺乳類における眼乾燥症の治療方法であることを特徴とし、眼乾燥症に苦しむ哺乳類において治療に有効量の物質を投与する工程からなり、該物質はガレクチン‐8タンパク質の発現に影響する。例えば、該物質とは、配列番号4又は5のアミノ酸配列を持つガレクチン‐8タンパク質を備える。例えば、該物質とは、配列番号4又は5のアミノ酸配列と実質的に同一のアミノ酸配列を持つガレクチン‐8タンパク質を備える。
【発明を実施するための最良の形態】
【0013】
本発明は、様々な特許、科学論文、その他の出版物に言及している。それぞれの事項の内容は参照することにより本明細書に含まれるものとする。またここで言及する全てのウェブサイトの内容(本出願時のもの)についても、それらを参照することにより本明細書に含まれるものとする。
【0014】
本発明の実施例は、炭水化物結合タンパク質であるガレクチンが、角膜及び結膜表皮において涙液層の拡散を促進するという概念に基づき示されている。ガレクチンはガラクト−ス結合タンパク質で、ムチンのO結合したオリゴ糖に見られるGalβ1‐4GlcNAC二糖に高い親和性で結合する。ガレクチンの多くは二価もしくは多価であり、ガレクチンが分泌性ムチンのオリゴ糖鎖と膜貫通型ムチン(その他の糖タンパク質)を結合することにより、涙液層の拡散を促進する。前記膜貫通型ムチン(その他の糖タンパク質)は角膜及び結膜上皮の最端面に存在するものである。
【0015】
本発明は、ガレクチン‐8、ガレクチン‐3及び/又はガレクチン‐7を備える医薬品組成物を提供し、損傷した哺乳類組織の傷の再上皮化を促進する際に有用である。本発明は、更に、上皮損傷した哺乳類組織における治療上の処置方法を提供し、該方法は上皮損傷を被った哺乳類に、治療に有効量のガレクチン‐1、ガレクチン‐3、ガレクチン‐7、ガレクチン‐8、又はガレクチン‐1、‐3、‐7、‐8のうち少なくとも2種類の混合物を投与する工程を備える。ガレクチン‐1、‐3、‐7、‐8の混合物が投与される場合、前述ガレクチンのいずれもが、他のガレクチンの投与前、併用して、もしくは投与後に投与可能である。
【0016】
本発明は、ガレクチン‐3が、損傷に伴って角膜上皮細胞の角膜への移動が上方制御されているという発見を包含する(例1)。また、本発明は、野生型マウスに比べてガレクチン‐3欠損型マウスでは、角膜経上皮エクシマーレーザー創傷及び角膜のアルカリによる火傷の再上皮化が際立って遅くなるという発見も含む(例2)。本発明は更に、ガレクチン‐3欠損型マウスでは、多くの損傷関連遺伝子(例えば、トロイド様タンパク質及びガレクチン‐7)の発現が異常であるという発見も提供している(例3)。更に、本発明は、外因性ガレクチン‐3及びガレクチン‐7が角膜傷の再上皮化を促進することも示している(それぞれ、例4、5)。
【0017】
<ガレクチン>
レクチンはタンパク質で、特に炭水化物に対する結合能、細胞凝集能により明確にされている(例えば、Sharon, Trends Biochem. Sci. 18:221, 1993参照)。レクチンは多種多様の細胞機能を担っていることが示されており、該機能は細胞‐細胞及び細胞‐マトリックス間相互作用を含む。レクチンは植物、無脊椎動物及び哺乳類の広範囲に見られる。動物のレクチンは4つの異なった群に区別されており、1)C型レクチン、2)P型レクチン、3)ガレクチン(以前はS型レクチンと称されていた)、及び4)ペントラキシンが挙げられる(例えば、Barondes et al., J. Biol. Chem. 269:20807, 1994)。
【0018】
全ての哺乳類のガレクチンは、β‐ラクトース及びβ‐ガラクトシド関連因子に対する認識について詳細に解析されている。全ての哺乳類のガレクチンは小さいβガラクトシドに対し同様な親和性を示すのに対し、より複雑な複合多糖に対しては、結合特異性に著しい違いが見られる(Henrick et al., Glycobiology 8;45, 1998; Sato et al., J. Biol. Chem. 267:6983, 1992; Seetharaman et al., J. Bioil. Chem. 273:13047, 1998)。β‐ガラクトシド糖類への結合に加えて、ガレクチンは赤血球凝集活性を持つ。ラミニンは自然界に存在する糖タンパク質で、多数のポリラクトサミン鎖を多く持ち、特定のガレクチンに対する天然リガンドであることが示されている。ラミニンは基底層及び細胞外マトリックスの構成要素であり、細胞外マトリックスは全ての上皮の下にあり、個々の筋肉、脂肪、及びシュワン細胞を取り囲んでいる。細胞と基底層の相互作用は、哺乳類の成長過程において、種々の細胞種の輸送及び/又は分化に影響を与えることが知られている。ガレクチンは膜移動を特定する在来の配列は持たないが、細胞内で分泌され細胞内に存在する。βガラクトシド糖に対する親和性だけでなく、ガレクチンのファミリーメンバーは炭水化物認識領域(CRD、又はタンパク質結合領域とも呼ばれる)における著しい配列類似性を共有する。炭水化物認識領域に関連するアミノ酸残基は、X線結晶学により決定されている(Lobsanov et al, J. Biol. Chem. 267:27034, 1993 and Seethnaraman et al., supra)。ガレクチンは、多岐にわたる生物機能に関与しているとされ、該機能は細胞接着(Copper et al., J. Cell Biol. 115:1437, 1991)、成長制御(Wells et al., Cell 64:91, 1991)、細胞移動(Hughes, Curr. Opin. Struct. Biol. 2:687, 1992)、悪性形質転換(Raz et al., Int. J. Cancer 46:871, 1990)及び免疫反応(Offner et al, J. Neuroimmunol. 28:177, 1990)が挙げられる。現在、12のガレクチンファミリーの真核メンバーが特徴付けられている。
【0019】
<ガレクチン‐3>
ガレクチン‐3ファミリーメンバーのタンパク質(以前はCBP‐35、Mac‐2、L‐34、εBP及びRL‐29として知られていた)は、一般的には約240から270のアミノ酸を含み、分子量は約25から29kDaの範囲である。ガレクチン‐3タンパク質は、一般的には、短いN末端領域と、ガラクトシド結合領域を含むC末端領域と、介在するプロリン、グリシン及びチロシンが豊富な領域とからなる。該介在するプロリン、グリシン及びチロシンが豊富な領域は、7から10の保存アミノ酸配列の繰り返しを含む(Liu et al., Biochemistry 35:6073, 1996及びCherayil et al., Proc. Natl. Acad. Sci. USA, 87:7324, 1990)。縦列反復はコラ−ゲン遺伝子ス−パ−ファミリ−に見られる反復と類似性がある。反復数はガレクチン‐3タンパク質により様々で、異なる種によるガレクチン‐3タンパク質の大きさの違いを説明している。ガレクチン‐3N末端領域は、複合糖質のリガンドを含む面に結合して、該タンパク質を多量体化させる。
【0020】
ガレクチン‐3は様々な炎症細胞(活性型マクロファージ、好塩基球及び肥満細胞)及び多種の組織の上皮及び繊維芽細胞で発現している(Perillo et al., J. Mol. Med. 76:402, 1998)。ガレクチン‐3は細胞表面、細胞外マトリックス(ECM)、細胞質及び細胞核に見られる。ガレクチン‐3は、細胞表面上又はECMで、細胞‐細胞及び細胞‐マトリックス間相互作用を媒介しているとされ、該相互作用は、複合糖質に相補的に結合することでなされる。該複合糖質はECM及び細胞表面分子に多く見られるポリラクトサミン鎖を含む。ガレクチン‐3は、ラミンに結合することで細胞‐マトリックス間接着を抑制しているとされる。細胞核では、ガレクチン‐3は公知の細胞接着分子(例えば、α6β1及びα4β7インテグリン、Warlfield et al., Invasion Metastasis 17:101, 1997及び、Matarrese et al., Int. J. Cancer 85:545, 2000)及びサイトカイン(例えば、IL‐1、Jeng et al., Immunol. Lett. 42: 113, 1994)の発現を制御することで、細胞‐マトリックス間相互作用に非間接的に関与することもできる。腎臓などの特定の器官では、ガレクチン‐3の発現は発達的に制御されており、肺胞上皮細胞及び肝細胞におけるガレクチン‐3の発現レベルでは損傷に伴い上方制御されている。増殖過程では、ガレクチン‐3は特定の細胞種の核内で凝縮することが示されている。ガレクチン‐3の発現は、特定の腫瘍では増加していることから、ガレクチン‐3は転移に関与していると示唆される。実際に、弱転移性細胞系でのガレクチン‐3の過剰発現は、転移能を顕著に増加させる(Raz et al., supra)。
【0021】
ヒトのガレクチン‐3は250アミノ酸長からなり、約26.1kDaの分子量を持つ(配列番号1、図1)。図1、3、5及び7に示してあるように、ヒトガレクチン‐3は次の様な領域、特徴的配列又はその他の構造上の特徴を持つ(PS及びPFの接頭認識番号に関する一般情報は、Sonnhammer et al., Protein 28:405, 1997参照)。配列番号1のアミノ酸残基1から14の周囲にあるN末端領域、配列番号1のアミノ酸残基15から116の周囲にあるプロリン、グリシン及びチロシンが豊富な領域、配列番号1のアミノ酸残基117から247の周囲にあるガラクトシド結合領域、配列番号1のアミノ酸残基181から200の周囲にあるガラプチン特徴的配列(PROSITE番号、PS00309)、配列番号1のアミノ酸残基4から7の周囲にある一つの潜在的なN‐糖鎖形成部位(PROSITE番号、PS00001)、配列番号1のアミノ酸残基137から179及び194から196の周囲にある二つの潜在的なタンパク質キナーゼCリン酸化部位(PROSITE番号、PS00005)、配列番号1のアミノ酸残基6から9及び175から178の周囲にある二つの潜在的なカゼインキナーゼIIリン酸化部位(PROSITE番号、PS00006)、及び、配列番号1のアミノ酸残基24から29、27から32、34から39、43から48、52から57、61から66、65から70及び68から73の周囲にある八つの潜在的なミリストイル化部位(PROSITE番号、PS00008)が挙げられる。
【0022】
本明細書に定義するように、ガレクチン‐3タンパク質は、「ガレクチン‐3N末端領域」、ガレクチン‐3「プロリン、グリシン及びチロシンが豊富な領域」及び/又はガレクチン‐3「ガラクトシド結合領域」を含む。これらの領域は、更に以下のように定義される。
【0023】
本明細書に用いられているように、ガラクチン‐3「N末端領域」には約10から20のアミノ酸、好適には約14のアミノ酸のアミノ酸配列が含まれ、配列番号1の1から14のアミノ酸と少なくとも60%、70%、80%、90%、95%、99%又は100%の同一性を有するものとする。N末端領域は、N‐糖鎖形成部位(PROSITE番号、PS00001)及び/又はカゼインキナーゼIIリン酸化部位(PROSITE番号、PS00006)を含むことが可能である。PROSITEのN‐糖鎖形成部位は、共通配列として、N‐{P}‐[ST]‐{P}を持ち、カゼインキナーゼIIリン酸化部位は、共通配列として、[ST]‐X(2)‐[DE]を持つ。上述の共通配列及び本明細書に記載の他のモチ−フ又は特徴的配列では、IUPAC規格であるアミノ酸の一文字記号が用いられている。様式の各要素はダッシュ(‐)で区切られており、角括弧([])は各位置で許容されている特定の残基を示し、Xは各位置で許容されている任意の残基を示し、丸括弧(())内の番号は付随アミノ酸によって示されている残基数を示す。ある実施例によっては、N末端領域は配列番号1のアミノ酸L7及びL11を含む。図3に示す様に、これらのアミノ酸はガレクチン‐3の様々な哺乳類の種を超えて保存されており、該アミノ酸は触媒及び/又は構造的な役割を果たす。
【0024】
本明細書にあるように、ガレクチン‐3の「プロリン、グリシン及びチロシンが豊富な領域」は約60から140のアミノ酸のアミノ酸配列を含み、より好適には約80から120のアミノ酸、又は約90から110のアミノ酸のアミノ酸配列を含み、配列番号1の15から116アミノ酸と約70%、80%、90%、95%、99%又は100%の同一性を持つ。プロリン、グリシン及びチロシンが豊富な領域は、1,2,3,4,5,6,7又は8のN‐ミリストイル化部位(PROSITE番号、PS00008)を含むことも可能であり、該N‐ミリストイル化部位は共通配列として、G‐{EDRKHPFYW}‐X(2)‐[STAGCN]‐{P}を持つ。ある実施例では、プロリン、グリシン及びチロシンが豊富な領域は配列番号1の以下のアミノ酸及び領域を含む。配列及び領域には、G21、P23、G27、N28、P30、G32、G34、P37、Y41‐P46、G53、Y55‐G57、P61、G62、G66、P72、G73、G77、Y79‐G81、P83、G87、Y89、P90、G99、Y101、P102、P106、Y107、A109、L114及びV116が挙げられる。これらのアミノ酸及び領域は、ガレクチン‐3の様々な哺乳類の種を超えて保存されており、該アミノ及び領域は触媒作用及び/又は構造上の役割を担う。(図3において「*」で示されたアミノ酸を参照)
【0025】
本明細書にあるように、ガレクチン‐3の「ガラクトシド結合領域」は約80から180のアミノ酸のアミノ酸配列を含み、該アミノ酸配列はPFAM(番号3)に登録されている共通配列PF00337との配列比較で、少なくとも150のビットスコアを持つ。好適には、ガレクチン‐3のガラクトシド結合領域は少なくとも約100から160のアミノ酸を含み、より好適には110から150のアミノ酸又は約120から140のアミノ酸を含み、PFAM(番号3)に登録されている共通配列PF00337との配列比較で、少なくとも150、より好適には少なくとも175、更に好適には200以上のビットスコアを持つ。
【0026】
特定の配列とPFAM(番号3)に登録されている共通配列PF00337との配列比較におけるビットスコアを計算するには、対象配列をHMMのPFAMデ−タベ−ス(例えばPFAMデータベース、リリース2.1)で検索し、初期変数には、www.sanger.ac.uk/Sogtware/Pfam.で得られる数値を用いる。PFAMデ−タベ−スの説明は、Sonnhammer et al., supraに記載され、HMMの詳細は例えば、文献(Griboskov et al., Meth. Enzymol. 183:146,1990及びStulz et al., Protein Sci. 2:305, 1993)に記載されている。
【0027】
ガレクチン‐3のガラクトシド結合領域は更に1つの、好ましくは2つのタンパク質キナーゼCリン酸化部位(PROSITE番号、PS00005)、カゼインキナーゼIIリン酸化部位(PROSITE番号、PS00006)及び/又はガラプチン特徴的配列(PROSITE番号、PS00309)を含むことができる。タンパク質キナーゼCリン酸化部位は、次の共通配列、[ST]‐X‐[RK]を持つ。ガラプチン特徴的配列は次の共通配列、W‐[GEK]‐X‐[EQ]‐X‐[KRE]‐X(3、6)‐[PCTF]‐[LIVMF]‐[NQEGSKV]‐X‐[GH]‐X(3)‐[DENKHS]‐[LIVMFC]を持つ。ある実施では、ガレクチン‐3のガラクトシド結合領域は配列番号1の次のアミノ酸及び領域、P117、Y118、L120‐L122、G125、P128、R129、L131‐I134、G136‐V138、N141、N143、R144、L147、F149、R151、G152、D154、A156‐F163、E165、R169‐N174、N179‐G182、E184‐R186、F190‐E193、G195、P197‐K199、Q201‐L203、E205、D207‐Q220、N222、R224、L228、I231、I236、G238‐I240及びL242‐S244を持つ。これらのアミノ酸及び領域は、ガレクチン‐3の様々な哺乳類の種を超えて保存されており、該アミノ酸及び領域は触媒作用及び/又は構造上の役割を担う。(図3に「*」で表記されたアミノ酸を参照)
【0028】
本発明のあるガレクチン‐3タンパク質は、配列番号1に示されるヒトのガレクチン‐3のアミノ酸配列を含む。本発明の他のガレクチン‐3タンパク質は、配列番号1のアミノ酸配列と実質的に同一のアミノ酸配列を持つ。「実質的に同一」とは、本明細書では、十分又は最低限の数のアミノ酸残基を含む第1のアミノ酸を言及し、該アミノ酸残基は、第2のアミノ酸配列の整列アミノ酸配列残基と同一であり、これにより第1及び第2のアミノ酸配列は共通の構造領域及び/又は共通の機能活性を持つことが可能であるとする。例えば、配列番号1と少なくとも60%又は65%の同一性、好ましくは、少なくとも75%の同一性、より好ましくは少なくとも85%、90%、91%、92%、93%、94%、95%、96%、97%、98%又は99%の同一性を持つ共通の構造領域を含むアミノ酸配列は、配列番号1のアミノ酸配列と実質的に同一とされる。特に、配列番号1の特定アミノ酸残基における欠失、付加、置換もしくは変形などの偶発的又は意図的に誘導された変異を含むタンパク質は、本明細書記載の、ガレクチン‐3の定義に含まれる。ここに定義するように、ガレクチン‐3タンパク質は、他の哺乳類種のガレクチン‐3のアミノ酸配列によって示される領域を含むことが可能であることも理解されるであろう。他の哺乳類種とは、ウシ、イヌ、ネコ、ヤギ、ヒツジ、ブタ、ネズミ及びウマを含むが、これらに限定されない。
【0029】
配列間の配列同一性の計算は、以下の様にして行われる。2つのアミノ酸配列の同一性の割合の決定には、最適な比較をするために、まず、配列を整列する(例えば最適な比較では、1つのアミノ酸配列、又は1番目及び2番目両方のアミノ酸配列に相違が取り入れられる)。そして、対応するアミノ酸部位又はヌクレオチド部位のアミノ酸残基を比較する。第一配列の部位が第二配列の対応する部位と同一のアミノ酸残基又はヌクレオチドである場合は、タンパク質はその部位で同一である。2つの配列間の同一性の割合は、配列間で同一な部位数により決まり、相違の数及び各相違の長さを考慮に入れる。該相違の数及び各相違の長さは、2つの配列の最適な配列比較に必要とされる。
【0030】
2つの配列間の配列比較及び同一性の割合の決定は、数学的アルゴリズムを用いてなされる。好適な実施例においては、2つのアミノ酸配列間の同一性の割合は、初期値変数を用い配列比較ソフトウェアプログラムにより決定される。適当なプログラムには、例えば、CLUSTAL W(Thompson et al., Nuc. Acids Research 22:4673, 1994(www.ebi.ac.uk/clustalw))、BL2SEQ(Tatusova and Madden, FEMS Microbiol. Lett. 174:247, 1999(www.ncbi.nlm.nih.gov/blast/bl2seq/bl2.html))、SAGA(Notredame and Higgins, Nuc. Acids Research 24:1515, 1996(igs-server.cnrs-mrs.fr/~cnotred))及びDIALIGN(Morgenstern et al., Bioinformatics 14:290, 1998(bibiserv.techfak.uni-bielefeld.de/dialign))が挙げられる。
【0031】
(www.ncbi.
<ガレクチン‐7>
ガレクチン‐7ファミリーメンバーのタンパク質は、通常は単量体として存在し、130から140のアミノ酸を含み、分子量は約15から16kDaの範囲である(例えば、Magnaldo et al., Develop. Biol. 168:259, 1995及びMadsen et al., J. Biol. Chem. 270:5823, 1995参照)。ガレクチン‐7の発現は、上皮層形成の開始と関連している(Timmons et al., Int. J. Dev. Biol. 43:229, 1999)。ガレクチン‐7は、細胞‐マトリックス間及び細胞‐細胞間相互作用を担っていると考えられている。ガレクチン‐7は細胞と細胞が接触している領域(例えば、ヒト表皮の上層)にみられ、ガレクチン‐7の発現は足場非依存性ケラチン生成細胞では非常に下方制御されており、悪性のケラチン生成細胞系では見られない。ガレクチン‐7は、正常なケラチン生成細胞の維持に必要とされる(Madsen et al., supra参照)。
【0032】
ヒトガレクチン‐7は136アミノ酸を含み、約15.1kDaの分子量を持つ(配列番号2、図2)。図2、4、6及び8に示すように、ヒトガレクチン‐7は次のような領域、特徴的配列、もしくは他の構造的特徴を含む。配列番号2のアミノ酸残基5から135の周囲に位置するガラクトシド結合領域、配列番号2のアミノ酸残基70から89の周囲に位置するガラプチン特徴的配列(PROSITE番号、PS00309)、配列番号2のアミノ酸残基29から32の周囲に位置する1つのN‐糖鎖形成部位(PROSITE番号、PS00001)、配列番号2のアミノ酸残基132から134の周囲に位置する1つのタンパク質キナーゼCリン酸化部位(PROSITE番号、PS00005)、配列番号2のアミノ酸残基9から12の周囲に位置する1つのカゼインキナーゼIIリン酸化部位(PROSITE番号、PS00006)、及び配列番号2のアミノ酸残基13から18及び44から49の周囲に位置する2つのミリストイル化部位(PROSITE番号、PS00008)である。
【0033】
本明細書に定義されている通り、ガレクチン‐7を含む「ガレクチン‐7タンパク質」はガラクトシド結合領域を含む。この領域は、更に次の様に定義されている。
【0034】
本明細書で用いられているように、ガレクチン‐7のガラクトシド結合領域は、約80から180のアミノ酸のアミノ酸配列からなり、PFAM(配列番号3)の共通配列PF00337との配列比較で、少なくとも80のビットスコアを持つ。好適には、ガレクチン‐7のガラクトシド結合領域は少なくとも約100から160のアミノ酸からなり、より好適には約110から150のアミノ酸、又は約120から140のアミノ酸からなり、PFAM(配列番号3)の共通配列、PF00337との配列比較で、少なくとも80のビットスコア、より好適には少なくとも100のビットスコア、更に好適には120以上のビットスコアを持つ。ガレクチン‐7のガラクトシド結合領域は、1つのN‐糖鎖形成部位(PROSITE番号PS00001)、1つのタンパク質キナーゼCリン酸化部位(PROSITE番号PS00005)、1つのカゼインキナーゼIIリン酸化部位(PROSITE番号、PS00006)、1つもしくは2つのミリストイル化部位(PROSITE番号、PS00008)、及び/又はガレプチン特徴的配列(PROSITE番号、PS00309)を含むことが可能である。ある実施例によっては、ガレクチン‐7のガラクトシド結合領域は、配列番号2の次のアミノ酸配列又は領域を含む。M1、S2、H6、K7、L10、P11、G13、R15、G17‐V19、R21‐G24、V26、P27、A30、R32‐Q43、D46‐N63、K65、Q67、G68、W70‐G76、G78、P80‐L90、I92、G97‐K99、V101、G103、D104、Y107、H109、F110、H112、R113、P115、V119、R120、V122‐L130、S132、I135及びF136である。これらのアミノ酸及び領域は、幾つかの哺乳類のガレクチン‐7で保存されており、触媒作用及び/又は構造上の役割を担う(図4に「*」で表記されたアミノ酸を参照)。
【0035】
本発明のあるガレクチン‐7タンパク質は、配列番号2に記載のヒトのガレクチン‐7アミノ酸配列を含む。本発明の他のガレクチン‐7タンパク質は、配列番号2のアミノ酸配列と実質的に同一のアミノ酸配列を含む。特に、偶発的若しくは意図的に誘導された変異、例えば、配列番号2の特定アミノ酸残基で、欠損、付加、置換若しくは修飾などを含むタンパク質は、本発明のガレクチン‐7の定義に当てはまる。ここに定義するように、ガレクチン‐3タンパク質は、他の哺乳類種のガレクチン‐3のアミノ酸配列によって示される領域を含むことが可能であることも理解されるであろう。他の哺乳類種とは、ウシ、イヌ、ネコ、ヤギ、ヒツジ、ブタ、ネズミ及びウマを含むが、これらに限定されない。
【0036】
<ガレクチン‐8>
ガレクチン−8は広範囲に発現しているタンパク質で、例えば、肝臓、心臓、筋肉、腎臓、脾臓、後肢及び脳で見られ、ヒト及びラットのガレクチン‐8遺伝子及びタンパク質の配列が入手可能である(例えば、Hadari, et al., Trends in Glycosci and Glycotechnol. 9: 103-112, 1997参照)。高親水性の特徴、及びムチンのO結合型オリゴ糖に見られるGalβ1‐4GlcNAC二糖との結合能により、該タンパク質は理想的な眼症候群の治療薬とされる。
【0037】
ヒトのガレクチン‐8のアミノ酸配列には2つの型が知られており、316アミノ酸型(受入番号O00214、1997年11月1日作成)及び359アミノ酸型(受入番号Q8TEV1、2002年6月1日作成)がある。これらの配列は、かなりの長さにおいて類似又は同一である一方、全体で見ればいくらかの異なる部分がある。316型アミノ酸配列は、1文字アミノ酸記号を用いて以下に示す(配列番号4)。
【0038】
【表1】
【0039】
長型アミノ酸配列を以下に示す(配列番号5)。
【0040】
【表2】
【0041】
本明細書に定義されている通り、「ガレクチン‐8タンパク質」は、ガレクチン‐8の「N末端領域」、ガレクチン‐8の「プロリン、グリセリン及びチロシンが豊富な領域」、及び/又はガレクチン‐8の「ガラクトシド結合領域」を含むことが可能である。
【0042】
本明細書にあるように、ガレクチン‐8の「N末端領域」は約10から20アミノ酸のアミノ酸配列を含み、好適には約14のアミノ酸で、配列番号もしくは5のアミノ酸と60%、70%、80%、90%、95%、99%、又は100%の同一性を持つ。N末端領域には、N‐糖鎖形成部位(PROSITE番号、PS00001)及び/又はカゼインキナーゼIIリン酸化部位(PROSITE番号、PS00006)が含まれる。PROSITEのN‐糖鎖形成部位は共通配列としてN‐{P}‐[ST]‐{P}を持ち、PROSITEによるカゼインキナーゼIIリン酸化部位は共通配列として[ST]‐X(2)‐[DE]を持つ。上記共通配列には、他のモチーフ又は特徴的配列も含まれる。
【0043】
本明細書では、ガレクチン‐8の「プロリン、グリシン及びチロシンが豊富な領域」は、約60から140のアミノ酸のアミノ酸配列からなり、より好適には、80から120のアミノ酸、もしくは、90から110のアミノ酸で、配列番号4及びは5の各アミノ酸の15から116のアミノ酸と60%、70%、80%、90%、95%、99%、又は100%の同一性を持つ。プロリン、グリシン及びチロシンが豊富な領域は、1,2,3,4,5,6,7又は8のN‐ミリストイル化部位(PROSITE番号、PS0008)を持つことも可能で、ミリストイル化部位は共通配列として、G‐{EDRKHPFYW}‐X(2)‐[STAGCN]‐{P}を持つ。ある実施例では、プロリン、グリシン及びチロシンが豊富な領域は、配列番号4の以下のアミノ酸及び領域を含む。アミノ酸及び領域は、G20、P23、P28、G29、G36、P39及び当業者にとって明確なその他の残基である。これらのアミノ酸及び領域は、幾つかの哺乳類のガレクチン‐8で保存されていて、触媒作用及び構造的役割を担っている。ある実施例では、プロリン、グリシン及びチロシンが豊富な領域は、配列番号5の以下のアミノ酸及び領域を含む。アミノ酸及び領域は、G21、P24、P29、G30、G37、P40及び当業者にとって明確なその他の残基である。
【0044】
本明細書では、ガレクチン‐4の「ガラクトシド結合領域」は約80から180のアミノ酸のアミノ酸配列からなり、PFAM(配列番号3)の共通配列であるPF00337との配列比較で、少なくとも150のビットスコアを持つ。好適には、ガレクチン‐3ガラクトシド結合領域は、少なくとも約100から160のアミノ酸を含み、より好適には、110から150アミノ酸、又は120から140アミノ酸を含み、PFAM(配列番号3)の共通配列であるPF00337との配列比較により、少なくとも150、より好適には少なくとも175、更に好適には200以上のビットスコアを持つ。
【0045】
特定の配列とPFAM(配列番号3)の共通配列であるPF00337との配列比較からビットスコアを計算するには、対象配列をPFAMのデータベースであるHMM(PFAM データベース、リリース 2.1)で検索する。初期値変数は、www.sanger.ac.uk/Software/Pfamで取得可能である。PFAMデータベースの説明は、参考文献(Sonnhammer et al., supra)にあり、HMMの詳細な説明は、例えば、参考文献(Gribskov et al., Meth., Enzymol. 183:146, 1990及びStultz et al., Protein Sci. 2:305, 1993)にある。
【0046】
ガレクチン‐8のガラクトシド結合領域は更に、1の、好適には2のタンパク質キナーゼCリン酸化部位(PROSITE番号、PS00005)、カゼインキナーゼIIリン酸化部位(PROSITE番号PS00006)及び/又はガラプチンの特徴的配列(PROSITE番号、PS00309)を含む。タンパク質キナーゼCリン酸化部位は次の共通配列、[ST]‐X‐[RK]を持つ。ガラプチン特徴的配列は、以下の共通配列、W‐[GEK]‐X‐[EQ]‐X‐[KRE]‐X(3,6)‐[PCTF]‐[LIVMF]‐[NQEGSKV]‐X‐[GH]‐X(3)‐[DENKHS]‐[LIVMFC]を持つ。ある実施例では、ガレクチン‐8のガラクトシド結合領域は配列番号4の次のアミノ酸及び領域、L123‐L124、G126、P131、R128、L140‐I146及び前述した部位と類似したその他の部位を含む。これらのアミノ酸及び領域は、幾つかの哺乳類のガレクチン‐8で保存されており、触媒作用及び構造的役割を担っている(図3に「*」で表記されたアミノ酸参照)。
【0047】
本発明のあるガレクチン‐8タンパク質は、配列番号4及び5記載のヒトガレクチン‐8アミノ酸配列を含む。他の本発明のガレクチン‐8は、配列番号4又は5記載のアミノ酸配列と実質的に同一のアミノ酸配列を含む。実質的に同一」とは、本明細書では、十分又は最低限の数のアミノ酸残基を含む第1のアミノ酸を言及し、該アミノ酸残基は、第2のアミノ酸配列の整列アミノ酸配列残基と同一であり、これにより第1及び第2のアミノ酸配列は共通の構造領域及び/又は共通の機能活性を持つことが可能であるとする。例えば、配列番号4又は5と少なくとも60%又は65%の同一性、好ましくは、少なくとも75%の同一性、より好ましくは少なくとも85%、90%、91%、92%、93%、94%、95%、96%、97%、98%又は99%の同一性を持つ共通の構造領域を含むアミノ酸配列は、配列番号4又は5のアミノ酸配列と実質的に同一とされる。特に、配列番号4又は5の特定アミノ酸残基における欠失、付加、置換もしくは修飾などの偶発的又は意図的に誘導された変異を含むタンパク質は、本明細書記載の、ガレクチン‐8タンパク質の定義に含まれる。ここに定義するように、ガレクチン‐8タンパク質は、他の哺乳類種のガレクチン‐8のアミノ酸配列によって示される領域を含むことが可能であることも理解されるであろう。他の哺乳類種とは、ウシ、イヌ、ネコ、ヤギ、ヒツジ、ブタ、ネズミ及びウマを含むが、これらに限定されない。
【0048】
眼乾燥症候群は、眼科では「風邪」のようなものと考えられていて、眼科医を訪れる患者にみられる2番目に多い病状である。1500万人近くのアメリカ人が眼乾燥に苦しんでいる。該疾患は、涙液層における構造又は機能の欠損により起こる。涙液層は動的構造を持ち、涙液層の生産と代謝回転は健全な角膜表面維持に不可欠である。涙液層は3層の、ムチン(最内側)、水溶性(中間)及び脂質(表面)層から構成される。これらの層いずれかの機能不全により、目乾燥症は引き起こされる。涙液層は最内側にあるムチン層を介して上皮細胞と結合する。ムチン層は、涙液層が角膜全体に、均一に拡散することを促進し、表面を適切に湿らせるために必要である。ムチン分子が眼に接着しないと、通常の水溶性涙液生産でも上皮損傷が起こる。涙液層拡散障害は、眼表面又は涙液に支障をきたし、眼乾燥症を引き起こす可能性がある。最内側ムチン層は、結膜杯細胞及び結膜又は角膜上皮細胞により生産される。眼ムチンは、膜貫通型(例えば、MUCI及びMUCU)又はゲル形成/分泌性のいずれかである(例えば、MUC5AC)。
【0049】
ムチンは非常に巨大な糖タンパク質で、少なくとも質量の半分にO結合型炭水化物を持つ。O結合型炭水化物の豊富な側鎖により、ムチンは非常に親水性であるという特徴を持つ。この親水性の特性により、水溶性層が眼上に均一に拡散することを可能にしていると考えられている。
【0050】
本発明は眼乾燥症候群(DES)の治療におけるガレクチンの治療的使用に基づく。特定の機構には限定されず、該使用はガレクチンが炭水化物結合するという特性に基づく。点眼液の局所的投与により類液の拡散が促進され、これは分泌性ムチンが、結膜及び角膜上の上皮表面において、膜貫通型ムチン及び他の糖タンパク質と結合することで促進される。
【0051】
本明細書の例では、三つの異なった眼乾燥マウスモデルを用いて、ガレクチン液の局所的投与の効果を特定している。一番目のモデルでは、涙液不足は、正常マウスの尾部にスコポラミンパッチを投与する、Pflugfelder‘s Laboratoryの報告にある方法(Dursun et al., 2002, Invest Ophthamol & Vis Sci. 43:632-638及びPflugfelder et al., 2003, The Ocular Surface, 1:31-36)により誘導されている。二番目のモデルでは、MRL/lpr自己免疫性マウスを利用し、これはシェーグレン症候群(Jabs et al., 1991, Invest Ophthamol Vis Sci. 32:371-380及びJabs et al., 1997, Curr Eye Res. 16:909--916)に提唱されているモデルで、涙腺炎症性病変を起こしている。三番目のモデルでは、IL‐1αの注入を用い、動物モデルを作成する。該モデルは、シェーグレン症候群(Zoukhri, D. et al., Invest Ophthalmol Vis Sci 42(5): 925-932参照)モデルである。
【0052】
測定には、生体内解析による涙液生成、涙液クリアランス及び角膜免疫染色が挙げられ、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7およびガレクチン‐8の各種濃度による効果を、これらパラメーターにおいて単独又は併用し、溶媒処理された対照動物と比較して特定する。生体内解析に引き続き、単離された涙腺を用いた生化学的解析、角膜表面の免疫組織学的解析が行われる。更に、スコポロミン処理前に行われるガレクチン液による前処理の効果並びに、ガレクチン及びガレクチン阻害剤であるβ乳糖を併用した治療により推定される効果についても検討される。
【0053】
本明細書の例から得られたデータによると、該モデルによる眼乾燥症において、ガレクチンは、測定可能なパラメーターにおいて候群統計学的に顕著な効果を示した。ガレクチン処理された動物の眼角膜では、偽薬処理された動物と比べて、角膜免疫染色の減少が見られたことから、ガレクチンは、ヒトDESに対しすばらしい治療可能性があるという根拠が得られた。二つの動物モデルは、異なったDESの型を示すので、どちらかのモデルで得られた肯定的な結果はDES治療におけるガレクチンの役割を特定するのに役立ち、臨床治験において焦点を絞る際の指針となる。臨床治験には追加動物実験が含まれ、有益な効果を最大限に得るのに必要なガレクチン濃度、効果の持続期間、並びにガレクチンと角膜及び内在性ムチンとの結合特性を決定する。更に毒性検査が行われ、安全な角膜用量範囲を特定し、臨床試験の承認を受ける前に、米国食品衛生局の必要条件を満たす。
【0054】
涙液の最内側は、結膜杯状細胞、並びに角膜及び結膜上皮細胞によって生産されたムチンにより構成される。ムチンは放出され分泌性ムチンとして存在したり、膜貫通型ムチンとして上皮に付属した状態を保ったりする。これらの分子は糖タンパク質であり、豊富なO結合した炭水化物側鎖を持つ。側鎖によってムチンは非常に親水性な特性を持ち、この特性により、水溶性層が眼上を均一に拡散することが可能となっている。該水溶液層は涙腺細胞により生産され、涙液層の約90%を構成する。水溶液層は主に水から成り、塩、ブドウ糖、リゾチーム、涙特異的プレアルブミン、ラクトフェリン、分泌性免疫グロブリンA、及び他のタンパク質が溶解している。外膜は蒸発を遅らせ、マイボーム腺によって生産された皮脂腺性物質からなる。瞬きにより、涙液層上に該層が拡散される(Rheinstrom SD, 1999, Dry eye. In Yanoff, ed, Ophthalmology. 1st Ed. Editor. Mosley International Ltd, St Louis, MO)。
【0055】
国立眼研究所は、眼乾燥の症状を二つのカテゴリー(水溶性層欠損及び蒸発欠損)に分類している。しかし、臨床症状は、多くの場合二つの発病経路が混合している(涙液生産の減少は、多くの場合欠陥油層の拡散につながり、結果過剰蒸発が起こる。また、マイボーム腺症な非常に多くの場合、低感受性眼乾燥と関連している)。水溶性涙液欠損類は、シューグレン症候群眼乾燥と非シューグレン症候群眼乾燥のサブカテゴリーに分類され、症状の重さの違いにより分類され、シューグレン症候群の炎症の発現が増強されている(Foulks, 2003, The Ocular Surface 1:20-30)。
【0056】
水溶性層欠損は眼乾燥の最も一般的な要因で、通常は涙腺からの涙液分泌の減少によって起こるが、涙液蒸発の増加によっても起こる。分泌減少の原因には、シューグレン症候群、老人性分泌過減、涙腺切除、ビタミンA欠乏、類肉腫及びリンパ腫における免疫涙腺損傷、感覚消失又は運動反射喪失、結膜の瘢痕状態、及びコンタクトレンズによる磨耗が挙げられる(Rolando and Zierhut, Surv Ophthlmol. 45:S203-S210, 2001)。水溶性層における組成の変化も眼表面損傷に関連している。組成の変化には、電解質濃度の増加、成長因子の損失又は炎症性サイトカイニンの存在等が挙げられ、涙液の低代謝回転と共に、眼表面損傷に関連している。
【0057】
杯細胞の欠乏は、従ってムチン量の減少につながり、眼乾燥の多くの様態を伴う。特に、疾患によっては杯細胞損失を引き起こす。これらは、ビタミンA欠損及び瘢痕を残す結膜症を含む。ビタミンAは、眼表面において、杯細胞やムチンの維持に必須とされる。該結膜症には、スティーブンス・ジョンソン症候群、トラコーマ、類天疱瘡及び化学熱傷等がある。局所的薬剤投与や保存料もまた、眼表面及び杯細胞を損傷しうる(Rheinstrom SD, 1999 Dry eye. In Yanoff, ed Ophthalmology. 1st Ed. Editor. Mosley International Ltd, St Louis MO; Abelson et al., 2003, Rev Ophthalmol 10:1)。
【0058】
患者の不快感の評価だけでなく、診断技術は、DESの重症度及び原因の特定に役立つことが証明された。涙液層の非安定性は、非侵襲性な涙液破砕時間測定(TBUT)により評価される。該測定では、蛍光溶液が投与され、涙液層が最初に破壊するまでの時間が、生体顕微鏡によって観測される。涙液生産は、典型的又は改良スキマー試験によって推定される。フルオレセイン、又は類似色素を用いた眼表面染色の度合いは、日常的に用いられ、眼表面交代の重症度が診断されている。これらの色素は、ムチンタンパク質の生産が欠乏した上皮表面、又は上皮細胞膜が露出した上皮表面を染色する。標準評点方式が開発され、損傷の重症度が定量化されている。該試験は、臨床試験において日常的に行われ、侵襲的な眼乾燥治療の治療効果が評価されている(Foulks 2003, The Ocular Surface, 1:20-30)。
【0059】
人口涙液は現在の眼乾燥治療の中心である。様々な商品が入手可能だが、どれもが一過的に症状を軽減するだけである。現在のところ、症状を戻すような治療薬はない。セルロースエステル(メチルセルロース、ヒドロキシプロピルメチルセルロース)又はポリビニルアルコールなどの高分子量重合体を生理食塩水に添加し、人口涙液を開発可能である。他の人口涙液処方は涙液中のムチン組成を模倣して調整されている。研究によると、人口涙液に使われている保存剤にある種の毒性があることが示され、保存料が入っていない、一回分ずつに分かれて小瓶に入った涙液が入手可能となっている(Rheinstrom SD, 1999 Dry eye. In Yanoff, ed Ophthalmology. 1st Ed. Editor. Mosley International Ltd, St Louis MO; Abelson et al., 2003, Rev Ophthalmol 10:1)。
【0060】
外疾患の病理生理学の理解向上に伴い、多くの異なった様式の治療方法が導入されてきた。これらには、涙液脂質膜を標的とした疾患、及び根底にある免疫性又はホルモン性病因が含まれる(Brewitt et al., 2001, Surv of Ophthalmol 45:S119-S202)。治験的治療には、15(S)‐HETE(水酸化エイコサテトラエン酸)(Gamache et al., 2000, Cornea 19:6:S88)等の局所的分泌促進物質、及び合成のP2Y2受容体アゴニスト(Jumblatt et al., 1998, Exp Eye Res 67:341-346)が含まれ、シクロスポリンA及び副腎皮質ステロイドなどの抗炎症物質ももちろん含まれる(Pflugfelder, 2003, The Ocular Surface, 1:31-36)。
【0061】
ムチンは、健全な眼球涙液層の生理機能の維持を担っていることが知られている。ムチンは高分子量糖タンパク質で、タンパク質骨格及び高炭水化物含有量を持っている。ムチン層に関与しているだけでなく、ムチン自体も糖衣、即ち足場構造を形成し、該足場様構造が細胞接着を促進する。ムチンは、眼表面損傷に対し防御の役割も担っている。また、ムチンは涙液層を親水性にする役割を担っている。これにより、涙液層が安定化され、角膜張力が減少し、水溶性層が眼表面上を均一に拡散することが可能となる。このムチン層なしでは、涙液は表面にとどまらず、眼表面は傷つきやすくなる(Abelson et al., 2003, Rev Ophthalmol 10:1)。
【0062】
体内では、膜貫通型及び分泌性の、二種類の原発型ムチンが生産さている。膜貫通型ムチン(MUC1、MUC2及びMUC4)は細胞の脂質二重膜に包埋されている。膜貫通型ムチンは角膜及び結膜の非杯細胞により発現され、分泌性ムチン(MUC5AC及びMUC7)の拡散を促進していると考えられている。該分泌性ムチンは、眼表皮を横断する杯細胞により分泌される。二種類のムチンは共に生涙液層を形成している(Danjo et al., 1998, Invest Ophthalmol Vis Sci 39: 2602-2609及びWatanabe 2002, Cornea 21:S17-S22)。
【0063】
結膜ムチンの欠乏は特定種の眼乾燥疾患の一因となっている。考えられる原因には、杯細胞密度の減少、ムチン分布又は特性の変化、並びにムチンmRNA発現の現象が挙げられる(Gipson et al., 2000, Prog Retinal Eye Res, 16:1:81-98及びGilbard, 2000, 「Dry-eye disorders」, In: Albert M et al., eds. Principals and Practice of Ophthalmology: 2nd Edition, Philadelphia: W.B. Saunders Company: 982-1001)。ビタミンA欠乏、局所用薬剤で、保存料を含む液滴の過剰投与、及び、瘢痕を残す結膜障害は全て杯細胞及び眼表面の損傷になり得る(Danjo et al., 1998, Invest Ophthalmol Vis Sci 39: 2602-2609及びAbelson et al., 2003, Rev Ophthalmol 10:1)。
【0064】
また、眼表面に見られる膜貫通型ムチンは、眼乾燥症の患者では変質している。研究者によると、膜貫通型ムチンであるMUC1等は、眼乾燥症患者のものは糖化されていないが、正常な患者のムチンは糖化されている。構造の変化は、眼表面上のムチン機能を変える可能性があり、眼乾燥症を更に悪化させる可能性がある(Danjo et al., 1998, Invest Ophthalmol Vis Sci 39:2602-2609)。
【0065】
<涙液層機能不全におけるガレクチンの役割>
ガレクチンは炭水化物結合タンパク質で、角膜及び結膜表面上の涙液層拡散を促進させる、理想的な候補となる可能性を持っている。これらのタンパク質は、高い親和性を持ってムチンと結合し、特に、ムチンのO結合炭水化物側鎖と結合する。特定の理論、又は作用機序に基づき結合するのではないが、ガレクチンは、分泌性ムチンが膜貫通型ムチン(又は他の糖タンパク質)と結合することによって、涙液層による角膜及び結膜表面上の拡散を促進させる。ムチン欠損眼では、外来性ガレクチンがムチン結合を延長又は促進させる。更に、ガレクチン自体が角膜表面の炭水化物結合タンパク質と結合することにより、ムチン欠損状態でも、涙液層の均一的な拡散をよりいっそう促進させる。
【0066】
ムチンとガレクチンの相互作用が、眼以外の組織で証明されている。ガレクチンは、ムチンの発現及び生産を、様々なムチンに対する強い結合親和性により調整していることが示されている。ガレクチン‐3はヒト結腸癌細胞由来のムチンと結合し、該ムチンの発現を濃度依存的に調整する(Bresalier et al., 1996, Cancer Res 56: 4354-4357及びDudas et al., 2002, Gastroenterology 118: 1553)。卵巣腫瘍細胞では、ムチン様糖タンパク質であるCA125はガレクチン‐1と結合する(Seelenmeyer et al., 2002, J Cell Sci, 116:1305-1318)。
【0067】
ガレクチンは、広く分布している炭水化物結合タンパク質からなるファミリーを構成し、多くの細胞表面及びECM糖タンパク質に見られるβ‐ガラクトシド含有グリカンとの親和性により特徴付けられている。哺乳類では、現在のところ14メンバーのガレクチンファミリーがあり(ガレクチン‐1から‐14)、炭水化物認識領域(CRD)における構造的類似性により定義されている。ガレクチンは可溶性タンパク質である。細胞内では、ガレクチンは主に細胞質に存在し、ガレクチン‐1及び‐3は増殖細胞の核でも確認されている。細胞外では、ガレクチンは細胞表面及びECMに見られる。ある種の成長因子(例えば、bFGF)及びサイトカイニン(例えば、IL−1)と同様に、ガレクチンは典型的なシグナル配列又は膜貫通領域を持たず、まだ良く理解されていないが、細胞から、非典型的な経路を経て分泌される。ガレクチン‐1、‐3、‐8及び‐9等のガレクチンは、幅広い組織で見られるが、一方、ガレクチン‐4、‐5及び‐6等のガレクチンは組織特異性を示す。ガレクチン‐1及び‐3は最も研究されているガレクチンである。細胞外では、両レクチンは細胞‐細胞間、及び細胞‐マトリックス間接着に関与していると考えられている。該接着は、フィブロネクチン、ラミニン、ビトロネクチン及びインテグリンの独特なイソ型上のオリゴ糖との結合によるものとされている。ガレクチンのほとんどは2つのCRDを持つか二量体として存在し、機能的に二価である。二価の特性により、一方では、そのレクチンの糖タンパク質である分泌性ムチンのオリゴ糖鎖と相互作用が可能であり、他方では、角膜の先端面における膜貫通型ムチン又は糖タンパク質のオリゴ糖鎖と相互作用が可能であり、これにより、眼表面上において涙液層の拡散を促進させている。幾つかの研究により、ガレクチン‐3はマウス及びヒトの角膜表皮で 発現していることが示されている。外来性ガレクチン‐3及び‐7は、マウス動物モデルにおいて角膜傷の再上皮化を促進させ、角膜上皮細胞の輸送に関与している可能性がある(Cao et al, 2002, J Biol Chem. 277:42299-4230及び2003, Arch Ophthalmol, 121: 82-86)。
【0068】
本明細書の例にある予備的な研究では、2つの異なったモデルの角膜傷治癒が用いられ、創傷の再上皮化は、以前に、ガレクチン‐3欠損(gal3-/-)マウスでは野生型(gal3+/+)マウスに比べて著しく遅いことが発見されている。一方、角膜上皮創傷閉塞率は、ガレクチン‐1欠損マウスと野生型と比べて差は見られなかった。外来性ガレクチン‐3及びガレクチン‐7は角膜アルカリ熱傷創傷の再上皮化を、濃度依存的に促進させた。該作用は、競合する糖、即ちβ‐ラクトースにより抑制されたが、関係のない二糖であるショ糖には抑制されなかった(Cao et al, 2002, J Biol Chem. 277:42299-4230及び2003, Arch Ophthalmol, 121: 82-86)。
【実施例】
【0069】
<材料と方法>
以下の材料と方法は、本明細書全般の例で使用される。
【0070】
<治療濃度のガレクチン‐1、‐3、‐7及び‐8を含む人工涙液溶液>
標準的な人工涙液溶液は動物研究に基づくガレクチン濃度で処方されている。該溶液は緩衝液で処理され、pHは中性、等張でCMC及びHMPC等の粘度を増すための増粘剤を含む。理想的な対照溶液は、ガレクチン以外全ての成分を含み、作成される。保存料が上皮損傷を悪化させる為、該溶液は保存用ではない。従って、新しい溶液はサンプルごとに調整され、投与時までに冷凍される。種々の処方には、対照人工涙液を含む。この対照人工涙液は、10μg/mlの各ガレクチン‐1、‐3、‐7若しくは‐8、又は対の組み合わせを含む人工涙液、及び、20μg/mlのガレクチン‐1、‐3、‐7若しくは‐8、又は対の組み合わせを含む人工涙液を含み、さらに、これら人工涙液に0.1Mのβ‐ラクトース及び二糖を含む人工涙液を含む。尚、二糖はガレクチン阻害剤として働く。
【0071】
<DES動物モデル:正常マウスにおいてスコポラミンより誘発される涙液不足>
スコポラミンは抗コリン作用薬で、ムスカリン性コリン受容体を標的とする。経皮スコポラミン(scop)パッチは、ヒトにおいて抗嘔気目的で及び動揺病治療で一般的に使われている。公表された研究(Dursun et al., 2002, Invest Ophthalmol & Vis Sci. 43:632-638)によると、この方法を用いたscop治療により、対照群の涙液生産の20%まで減少させるという、顕著で統計学的に有意な結果が得られた。涙液中のフルオレセインのクリアランス、及び角膜におけるカルボキシフルオレセイン取り込みは、どちらも約3倍となった。更に該動物を送風フード内に置くことにより、これらのパラメーターは一層劇的に増大した。更に、scop+送風フード処理したマウスでは、結膜杯細胞濃度が対照と比べて90%以上も減少した。径皮パッチ送達系により制御され継続的な薬剤送達が可能となるため、涙液生産の減少は24時間以上も続いた。一方、局所的アトロピン投与では、涙液の減少は数時間しか持続しなかった。新しいスコポラミンパッチの再投与により、効果は更に持続する。
【0072】
該例では、C57BL/6系マウス(Charles River Laboratories, Wilmington, MA)が用いられる。該マウスは6から8週齢で性混合である。無麻酔のマウスを手で捕まえ、1インチ部分の尾部中部を電気かみそりで剃る。経皮scopパッチ(Novartis, Summit, NJ)を4つに切り、4分の1を脱毛した尾部中部に投与する。パッチは48時間ごとに張り替えられ、安定した薬剤送達が維持される。動物を1時間、1日3回、実験の間毎日送風フード内に置く。
【0073】
<MRL/lpr自己免疫性マウス>
本実験では、MRL/Mp‐lpr/lpr(MRL/lpr)系のマウス(Jackson Laboratory, Bar Harbor, ME)が用いる。該マウスは類遺伝子亜系で、自己免疫性疾患を発病し、16週齢以内に涙腺炎症性病変を引き起こす。病理学及び組織学的評価に基づき、該マウスはヒトシューグレン症候群(Jabs et al., 1991, Invest Ophthamol Vis Sci. 32:371-380及び2001, Curr Eye Res. 16:909-916)のモデルとして提唱された。本明細書中例に用いられるマウスは、少なくとも16週齢以上で性混合のものである。
【0074】
<動物における涙液生産、涙液クリアランス及び角膜染色の測定>
水溶性類液生産は、フェノールレッドを含侵させた綿糸(Zone-quick, Oasis, Glendora, CA)を用いて測定した。該糸をピンセットで持ち、外眼角内の眼表面に1分間当てる。綿糸上の目盛りを使って、何ミリ綿糸が濡れたかで水性の涙液生産を測定する。
【0075】
涙液破砕時間(TBUT)は、1%ナトリウムフルオレセイン2μLを滴剤として用いて、これを眼表面に投与して測定した。涙液層を隙灯生体顕微鏡で観察し、涙液層が拡散するまでの時間を記録する。
【0076】
フルオレセイン染色はTBUT測定直後に行い、1%ナトリウムフルオレセイン2μLを滴剤として用い、これを眼表面に投与する。投与5分後、隙灯生体顕微鏡を用い、角膜におけるフルオレセイン染色を評価する。フルオレセイン染色は、臨床では角膜表面損傷を判定する為に一般的に用いられている。眼乾燥症の動物の角膜表面はフルオレセイン染色するが、正常の角膜表面は染色しないとされている。フルオレセイン染色の程度は、眼乾燥臨床治験でも使われた尺度と同様の標準F1評価尺度を用いて記述する。
【0077】
<同等の治療>
治療及び測定計画は、短期的効果(ガレクエン治療後2時間)及び長期的効果(ガレクエン治療後14時間まで)が測定できるようにされている。ガレクチンは、1日4回の治療計画に長期的効果があると期待される。治療計画、測定計画又は投与量の変更は、角膜に有益な効果をもたらすために必要なものであれば、本明細書に例と同等のものとする。例えば、20μL/mLのガレクチン治療で反応が見られ、10μL/mLで反応が見られない場合は、追加例はより高い投与量(50μg/mL)を備える。
【0078】
<ガレクチン‐1、‐3、ガレクチン‐7及びガレクチン‐8の製剤>
本発明のガレクチンは、当業者が入手可能である。入手可能なガレクチンには、単離されたタンパク質だけでなく、遺伝子組み換えや合成により生産されたタンパク質が含まれ、固相方法によるタンパク質が例に挙げられる。本発明によると、ガレクチン‐3、ガレクチン‐7又はガレクチン‐8をコードするポリヌクレオチド配列は、組み換えDNA分子として用いられることが可能で、該組換え分子は適当な宿主細胞内で本発明のガレクチン発現を指示する。参考文献(Cherayil et al., supra、Madsen et al., supra 及びHadri et al., supra)には、ヒトガレクチン‐1、‐3、‐7及び‐8夫々のクローニング方法が詳細に記載されている。生物学的に活性を持つガレクチン‐1、ガレクチン‐3、ガレクチン‐7、ガレクチン‐8を発現するために、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7、ガレクチン‐8をエンコードするヌクレオチド配列又は機能的に同等な配列が適当な発現ベクターに挿入される。発現ベクターには、挿入したコードされた配列の転写及び翻訳に必要な要素を含むものとする。当業者に公知の方法を用いて、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7若しくはガレクチン‐8をコードする配列、並びに適当な転写又は翻訳制御を含む発現ベクターを構築してもよい。該方法には、生体外遺伝子組み換え技術、合成技術、並びに生体内組み換え若しくは遺伝子組み換えが含まれる。欠失、付加、又は置換は、当業者に公知の任意の技術を用いて導入することができる。例えば、PCRによる突然変異誘導が挙げられる。これらの技術は、参考文献(Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Press, Plainview, NY, 1989及びAusubel et al, Current Protocols in Molecular Biology, John Wiley & Sons, New York, NY, 1989))に記載されている。様々な発現ベクター/宿主系に導入され、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7又はガレクチン‐8を発現させることが可能である。これらの発現ベクター/宿主系には、組換えバクテリオファージ、プラスミド又はコスミドDNA発現ベクターにより形質転換した細菌等の微生物、酵母発現ベクターにより形質転換した酵母、ウイルス発現ベクター(例えば、バキュロウイルス)に感染した昆虫細胞系、多種の発現ベクター(例えば、カリフラワーモザイクウイルス(CaMV)、タバコモザイクウイルス(TMV))若しくは細菌発現ベクター(例えば、Ti、pBR322又はpET25bプラスミド)により形質転換した植物細胞系、又は動物細胞系が含まれるが、限定されるものではない。或いは、本発明のガレクチンは、化学的手法を用いてガレクチン‐1、ガレクチン‐3、ガレクチン‐7又はガレクチン‐8アミノ酸配列を全体又は一部を合成してもよい。例えば、ペプチド合成は、多様な固相法(Roberge et al., Science 269:202, 1995)を用いて行うことが可能であり、自動合成も実施可能であり、例えば、431Aペプチド合成機(Applied Biosystems of Foster City, CAから入手可能)を用い、機器添付の取扱説明書に従いペプチド合成することができる。
【0079】
<医薬組成物>
本発明の特徴の一つは医薬組成物が提供されていることである。該組成物は、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7及び/又はガレクチン‐8を備え、薬学的に許容可能な担体を任意に備える。実施例によっては、これらの組成は、一又はそれ以上の追加治療剤を任意に備える。実施例によっては、治療剤又は薬剤は以下の群から選択される。該薬剤は増殖因子、抗炎症剤、血管収縮剤、コラゲナーゼ阻害剤、局所ステロイド、マトリックスメタロプロテナーゼ阻害剤、アスコルビン酸、アンジオテンシンII、アンジオテンシンIII、カルレティキュリン、テトラサイクリン、フィブロネクチン、コラーゲン、トロンボスポンジン、形質転換増殖因子(TGF)、ケラチノサイト増殖因子(KGF)、線維眼細胞増殖因子(FGF)、インシュリン様増殖因子(IGF)、上皮増殖因子(EGF)、血小板由来増殖因子(PDGF)、ニューレギュリン(NDF)、肝細胞増殖因子(HGF)ビオチン等のビタミンB及びニアルロン酸からなる。
【0080】
本明細書で用いられている「薬学的に許容可能な担体」は、所望の投与形態により、ありとあらゆる溶媒、希釈剤又は他の液体媒体、分散若しくは懸濁酸、表面活性剤、等張剤、増粘若しくは乳化剤、防腐剤、固体結合剤並びに潤滑剤等からなる。参考文献(Remington's Pharmaceutical Sciences Ed. by Gennaro, Mack Publishing, Easton, PA, 1995)には、医薬品組成物の処方に用いられる多種多様の担体が開示されており、公知の調剤方法が記載されている。薬学的に許容可能な担体の物質例としては、ブドウ糖及びショ糖等の糖類、コーンスターチ及びジャガイモ澱粉等の澱粉、カルボキシメチルセルロースナトリウム、エチルセルロース及び酢酸セルロース等のセルロース並びにその派生物、粉末トラガカント、麦芽、ゼラチン、ココアバター及び座薬ワックス等の賦形剤、ピーナッツ油、綿実油、紅花油、ゴマ油、オリーブ油、コーン油及び大豆油等の油、プロピレングリコール等のグリコール、オレイン酸エチル及びウラリル酸エチル等のエステル、寒天、水酸化マグネシウム及び水酸化アルミニウム等の緩衝剤、アルギン酸、pyrogen-free water、等張食塩水、リンガー溶液、エチルアルコール並びにリン酸緩衝液が挙げられ、また、ラウリルシュウ酸ナトリウム及びステアリン酸マグネシウムであるような非毒性相溶性の滑剤、さらに、着色剤、解除剤、コーティング剤、甘味、香料添加及び香料剤、並びに保存料及び抗酸化剤も、策定者の判断により組成物に含有可能である。
【0081】
<治療に有効な投与量>
他の特徴としては、本発明の治療法によると、眼と医薬品組成物を接触させることによる眼乾燥の治療があげられ、以下に記載する。従って、本発明は眼乾燥の治療方法を提供し、治療に有効量の医薬品組成物を、治療を必要とする対象に投与する過程を備える。該組成は、ガレクチン‐3、ガレクチン‐7及び/又はガレクチン‐8を含む有効成分を備え、投与量及び投与回数は、所望の結果に応じて決められる。当然のことながら、これには本発明の医薬品を治療的処置として投与する過程を含み、この過程により角膜及び結膜表面における涙液層の拡散が促進される。或いは、眼乾燥に伴う合併症を最小限に抑える為の予防手段(例えば、手術中及び/又は手術後、或いは抗ヒスタミン剤と併用した炎症症状の治療中及び/又は治療後の傷洗浄液として)として投与する過程を含む。本発明の実施例によっては、「治療に有効な量」の医薬品組成物とは、眼乾燥治療に有効な量である。本発明の方法によると、該組成物は、眼乾燥治療を促進させるために有効な任意の量を任意の投与経路により投与可能である。従って、本明細書にある「眼乾燥治療を促進させるために有効な量」との表現は、涙液層を促進させる為に十分な量の組成物に言及する。的確な投与量は、個々の医師により、治療する患者を考慮し選択される。投与量と投与は、十分量の有効成分を提供する為に、又は所望の効果を維持するために調節される。考慮すべき更なる要因としては症状の重症度が挙げられ、例えば眼乾燥の程度、症歴、患者の体重、年齢、性別、食習慣、投与時間及び回数、併用する薬剤、反応特異性、並びに治療に対する耐性/反応が挙げられる。長時間作用性医薬品組成物は、1日に数回、毎日、3から4日間、毎週又は2週間に一度、組成物の半減期及びクリアランス時間により投与可能である。
【0082】
本発明の有効成分は、好適には、投与のし易さ及び投与の均一性を考慮して投与単位形態として処方されるものとする。「投与単位形態」との表現は、本明細書では、物理的に独立した単位に言及し、治療対象の患者に適当な有効成分の単位である。しかしながら、本発明組成の1日当たりの使用量は、主治医により医学的に妥当な判断内で決定されるものとする。有効成分の治療に有効な投与量は、先ずは細胞培養試験又は動物モデルを用いて概算され、一般的にはマウス、ウサギ、イヌ又は豚が用いられる。動物モデルは所望の濃度範囲及び投与経路を特定する際にも用いられる。直接投与が投与経路と考えられるが、得られた情報は、ヒトへの投与に有用な投与量及び追加投与経路を特定する為に利用される。治療に有効な投与量とは、症状や状態を回復させる有効成分量のに言及する。有効成分の治療的有効性及び毒性は、培養細胞又は実験動物を用いた標準的な薬学手法により特定可能であり、例えば、ED50(投与量は集団の50%に治療的有効)及びLD50(投与量は集団の50%で致死)が挙げられる。毒性用量と治療有効性用量の比が治療指数とされ、例えばLD50/ED50の比率で表される。大きい治療指数を示す医薬品組成物が好適である。細胞培養解析及び動物実験により得られたデータはヒトに用いられる一連の処方に利用される。
【0083】
<医薬品組成物の投与>
薬学的に許容な適当な担体と共に所望の投与量に処方後、本発明の医薬品組成物は、ヒト及び他の哺乳類に、眼等(粉末、軟膏又は点滴薬により)局所的に、例えば眼に直接投与可能である。代替及び付加経路としては、経口、直腸、非経口、大槽内、膣内、腹腔内、口腔内又は経鼻的経路等が挙げられ、治療対象の症状の重症度により想定される。
【0084】
眼投与のための液状剤形は、緩衝剤及び可溶化剤、水等所望の希釈剤、チモゾール等の保存剤、並びに溶液調製の為の一以上の生重合体又は重合体を含む。生重合体又は重合体には、ポリエチレングリコール、メチルセルロース水酸化プロプル、ヒアルロン酸ナトリウム、ポリアクリル酸ナトリウム又はタマリンドゴム等がある。
【0085】
経口投与のための液状剤形には、限定されるものではないが、薬学的に許容可能な乳剤、微小乳剤、液剤、懸濁剤、シロップ及びエリキシル剤が含まれる。これらの有効成分に加えて、液体剤形は不活性希釈剤を含み、該技術分野で一般的に用いられているものとしては、例えば、水若しくは他の溶媒、可溶化剤又は乳化剤{例えば、エチルアルコール、イソプロピルアルコール、炭酸エステル、酢酸エチル、ベンジルアルコール、安息香酸ベンジル、プロピレングリコール、1,3−ブチレングリコール、ジメチルホルムアミド、油(特に綿実油、落花生油、コーン油、オリーブ油、ひまし油及びゴマ油)、グリセロール及びテトラヒドロフリフリルアルコール等}、ポリエチレングリコール、ソルビタン脂肪酸エステル、並びにこれらの混合物が挙げられる。不活性希釈剤の他には、経口組成物は湿潤剤等の補助剤、乳化剤及び懸濁剤、甘味剤、香料添加剤、並びに香料剤を含むことも可能である。
【0086】
発明の医薬品組成物の局所投与又は経皮投与剤形は、軟膏、ペースト、クリーム、ローション、ゲル、粉末、水溶液、スプレー、吸入剤又はパッチを含む。該有効成分は、無菌状態で混合され、薬学的に許容可能な担体、並びに必要に応じて保存料若しくは緩衝剤が加えられる。例えば、眼又は皮膚感染は水溶性点滴剤、ミスト、乳剤又はクリームにより治療される。薬剤投与は、治療にも予防にもなりうる。予防的処方は潜在性創傷又は創傷の原因に適用され、コンタクトレンズ、コンタクトレンズ洗浄及び水洗液、コンタクトレンズ保存又は運搬容器、コンタクトレンズ取り扱い器具、目薬、手術用洗浄液、点耳薬、眼帯及び目周辺用化粧品(クリーム、ローション、マスカラ、アイライナー及びアイシャドウ等を含む)に用いられる。本発明は、眼科機器、手術用機器、聴覚機器又は開示の組成物を含む製品(ガーゼ、包帯又はストリップ)、並びにこれらの機器若しくは製品を作るための若しくは使うための方法を含む。これらの機器は開示の組成物により被膜し、組成物を浸透させ、結合させ、又は他の方法で処理しても良い。
【0087】
該軟膏、ペースト、クリーム及びゲルは本発明の有効成分だけでなく、動物性若しくは植物性脂肪等の賦形剤、油、ワックス、パラフィン、スターチ、トラガカント、セルロース派生物、ポリエチレングリコール、シリコン、ベントナイト、ケイ酸、タルク並びに酸化亜鉛又はこれらの混合物を含むことが可能である。
【0088】
粉末及びスプレーは、本発明の薬剤に加えて、タルク、ケイ酸、水酸化アルミニウム、ケイ酸カルシウム、ポリアミン粉末又は該薬物の混合物等の賦形剤を含むことが可能である。スプレーは更にクロロフルオロ炭化水素等の噴霧剤を含む。
【0089】
経皮パッチは、体への有効成分供給を制御できるという点で更に優れている。そのような剤形は、化合物を適当な溶剤に溶解又は分注し作られる。吸収促進剤を用いて、化合物が皮膚上に良く行渡らせることも可能である。該速度は、速度調整膜を用いることで、又は、化合物をポリマーマトリックス若しくはゲルに分散させることで制御可能である。
【0090】
注射製剤は、例えば無菌の水溶性又は油性の懸濁液が既知の技術に基づき、分散剤又は湿潤剤及び懸濁剤を用いて処方することができる。該無菌注射製剤は、非毒性で非経口的に許容可能な希釈又は溶媒(例えば1,3−ブタンジオール水溶液等)含有の無菌の注射剤、懸濁剤又は乳剤でもよい。許容可能な溶媒又は溶媒として使用されるものには、水、リンガー水溶液、U.S.P.及び等張塩化ナトリウム溶液がある。更に、通常は無菌の、不揮発性油が溶剤又は懸濁剤として用いられている。この用途には、合成モノ‐又はジグリセリドを含む、どの製品の不揮発性油も用いることが可能である。また、オレイン酸等の脂肪酸が注射製剤に用いられている。注射製剤は滅菌可能で、例えば、細菌保持フィルターを用いた濾過により、又は、事前に滅菌水若しくは他の滅菌注射溶剤に溶解または拡散させるなどして混入した無菌固体組成形状の無菌剤により、滅菌することができる。有効成分の効果を持続させるためには、皮下又は筋肉内注射による有効成分の吸収を遅らせることが好適である。非経口投与による有効成分の吸収を遅らせるために、有効成分を油溶媒に溶解又は懸濁することも可能である。デポー注射形態は、ポリラクチド-ポリグリコライド(polylactide-polyglycolide)であるような生分解性重合体に有効成分を含んだマイクロカプセル剤を形成することにより製造される。有効成分と重合体の割合、及び用いられる重合体の性質により、放出される有効成分の割合は制御可能である。生分解性重合体には、poly(orthoesters)及びpoly(anhydrides)が例として挙げられる。デポー注射製剤は有効成分をリポソーム又はマイクロエマルジョンに封入し調整することも可能で、これらは体組織と適合性がある。
【0091】
直腸又は膣内投与用組成物は好適には座薬で、本発明の有効成分を適当な非刺激性の賦形剤又は担体と混合し調整することが可能である。該賦形剤又は担体には、ココアバター、ポリエチレングリコール又は座薬ワックス等が挙げられ、これらは室温では固体であるが体温で液体になるため、直腸又は膣腔で溶け該有効成分を放出する。
【0092】
経口投与用固体剤形はカプセル剤、錠剤、丸剤、散剤及び顆粒剤を含む。該固体剤形には、有効成分は、少なくとも一の非活性で薬学的許容可能な賦形剤又は担体(クエン酸ナトリウム又は第二リン酸カルシウム等)並びに/又は、
a)注入剤若しくは増量剤(スターチ、ショ糖、ブドウ糖、マニトール及びケイ酸等)、
b)結合剤(例えば、カルボキシメチルセルロース、アルギン酸塩、ゼラチン、ポリビニルピロリジノン、ショ糖及びアカシア等)、
c)保湿剤(グリセロール等)、d)崩壊剤(寒天、炭酸カルシウム、ジャガイモ若しくはタピオカ澱粉、アルギン酸、特定のケイ酸並びに炭酸ナトリウム等)、
e)溶液緩染剤(パラフィン等)、
f)吸収促進剤(第四アンモニア化合物等)、
g)湿潤剤(例えば、セチルアルコール及びモノステアレートグリセロール等)、
h)吸収剤(カオリン及びベントナイト粘土等)、
i)潤滑剤(タルク、ステアリン酸カルシウム、ステアリン酸マグネシウム、固体ポリエチレングリコール、ラウリル硫酸ナトリウム)
及びこれらの混合物と混合されている
【0093】
同類の固体組成物を軟又は硬ゼラチンカプセル状の注入剤として使用することも可能で、高分子ポリエチレングリコール等は言うまでもなく乳糖等の賦形剤が用いられる。固体剤形は、錠剤、糖衣錠、カプセル剤、丸剤及び顆粒剤が、コーティング剤又はシェル剤(腸溶コーティング、放出制御コーティング及び他の薬剤処方技術として公知のコーティング)を用いて調剤される。前記固形剤形状では、有効成分はショ糖又はスターチ等の少なくとも一の非活性希釈剤と混合される。前記剤形は、通常行われている様に、非活性希釈剤以外の追加物質を備えることも可能で、例えば、錠剤潤滑剤並びにステアリン酸マグネシウム及びマイクロクリスタリンセルロース等の他の錠剤補助剤が挙げられる。カプセル剤、錠剤及び丸剤の場合は、剤形は緩衝剤を備えることも可能である。任意に乳白剤を含むことも可能である。乳白剤は腸管の特定部位だけで、又は腸管の特定部位選択的に遅発的に有効成分を放出する。使用可能な包埋組成の例には、ポリマー物質及びワックスが挙げられる。
【0094】
<医薬品組成物の使用>
上述の記載及び例により更に詳細に記載の如く、ガレクチン‐3、ガレクチン‐7及びガレクチン‐8は、涙液層の拡散促進に有用であり、これは該ガレクチンが分泌性ムチンのオリゴ糖鎖を膜貫通型ムチン(又は他の糖タンパク質)と結合させることでなされる。一般的には、ガレクチンは臨床的に眼乾燥の治癒を促進するのに有効と考えられている。
【0095】
一般的には、ガレクチンは、皮膚上皮に限定されずあらゆる皮膚上皮組織、角膜上皮、消化器管の内張り、肺上皮、並びに腎管、血管上皮、子宮及び膣上皮、尿道上皮、又は呼吸器上皮の内表皮に関連した治癒の促進に有用であることが本明細書に示されている。本発明は多種多様の実施例を網羅し、種々の上皮創傷を治癒する。上皮創傷には、外科創傷、切除損傷、疱疹、潰瘍、病変部、剥離、糜爛、裂傷、おでき、傷口、痛み、並びに熱又は化学物質暴露による熱傷が挙げられるが、限定されるものではない。該創傷は健常者のものである場合もあるが、異常な損傷治癒を誘発する症状(糖尿病、角膜ジストロフィー、尿道症、栄養失調、ビタミン欠乏、肥満、感染症、免疫抑制、並びに、ステロイド、放射線治療、非ステロイド系抗炎症薬(NSAID)、抗新生物治療薬及び代謝拮抗物による全身治療に伴う合併症)を持つ患者の創傷でもよい。
【0096】
ガレクチン‐1、‐3及び‐7及び/又は‐8は、本明細書では、例えば、皮膚の損失の後で起こる皮膚の回復の促進に役立つ。或いは、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7及び/又はガレクチン‐8について、本明細書では、創傷を起こしている層に対する植皮の密着性を増強すること、及び創傷を起こしている層で再度上皮形成を促すことが示された。適切な植皮は、自己移植片、人工皮膚、同種移植片、自己植皮片、自己上皮植皮片、無血管性植皮片、ブレア‐ブラウン植皮(Blair‐Brown graft)、骨移植、胚胎組織移植片、真皮移植片、遅延移植片、皮膚移植片、表皮移植片、筋膜移植片、全層植皮、異種移植片、同種移植片、増植性移植片、表層移植片、網状移植片、粘膜移植片、オレア‐ティールシュ植皮片(Ollier-Thiersch graft)、大網移植片、切り張り移植片、茎状移植片、穿通性植皮片、中間層皮膚移植片、分層植皮片を含むが、これらに限定されない。
【0097】
ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、本明細書では、疱疹状皮膚炎を治療するのに役立つ。この疱疹状皮膚炎は、真皮と上皮の接合部で疱疹を形成する。ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、本明細書では、表皮水疱症を治療することに、また、上皮とその下の真皮との密着性の欠損を治療することに役立つ。尚、また、この密着性の欠損は頻発することとなる。ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、開いて痛みを伴う水膨れを治療することに役立つ。これらの治療は、ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8が、これら損傷部の上皮再形成を促進ことによりなしえる。ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、さらに上皮内の細胞間の接着の損失を伴う天疱瘡を治療すること、或いは、真皮と上皮の接合部の細胞間の接着の損失を伴う接合部類天疱瘡を治療することに役立つ。ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、様々な潰瘍を治療することに使用される。前記潰瘍には、糖尿病(性)潰瘍、皮膚潰瘍、褥瘡性潰瘍、動脈潰瘍、及び様々なうっ血性潰瘍が含まれるが、これらに限定されない。
【0098】
本発明は、角膜組織修復を促進するための方法も含むものとする。前記方法は、角膜潰瘍、熱、放射線、泡性角膜炎、角膜潰瘍又は断裂、近視矯正のための光屈折率手術、異物性及び無菌性角膜浸潤、酸又はアルカリ(例えば、フッ化水素酸、ギ酸、無水アンモニア、セメント、フェノール類、或いは他の化学薬剤(黄燐、元素金属、硝酸塩炭化水素、タール))の暴露による化学熱傷、神経栄養角膜症、糖尿病性の角膜症、Thygeson's点状表層角膜炎等の角膜症、ウイルス性角膜炎(メタヘルペス性(metaherpetic)角膜炎、ヘルペス性角膜炎)及び細菌性角膜炎等の角膜炎、格子状ジストロフィー等の角膜ジストロフィー、上皮基底膜ジストロフィー(EBMD)及びフックス内皮ジストロフィーにより引き起こされる角膜上皮欠損を治療することを含む。
【0099】
本明細書における方法により、ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、胃腸の潰瘍を治療することに有用であり、粘膜内層の治癒を助け、また腺性粘膜及び十二指腸粘膜内層をより早く再生させることができる。クローン病及び潰瘍性大腸炎等の炎症性腸疾患は、小腸及び大腸それぞれの粘膜表面の破壊を引き起こす。従って、ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、その粘膜表面を再形成することを促進することにより使用されることができので、より早い治癒を助け、炎症性腸疾患の進行を遅らせることができる。ガレクチン‐1、‐3、‐7及びガレクチン‐8は、ムチンを固定し、ムチンと上皮の先端面を結合させることにより、有害な物質(例えば摂取される有害物質または手術に伴い生じる有害物質)から胃腸管を保護することができる。ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、腸毒性の副作用を減らすために使用される。前記腸毒性は、細菌感染、ウイルス感染、放射線治療、化学療法、或いは他の治療により引き起こされる。ガレクチン‐8、‐3及び/又はガレクチン‐7は、例えば、粘膜炎、食道炎或いは胃炎を、予防的に或いは治療的に予防或いは緩和する。(例えば、経口、食道、腸管、結腸、直腸、肛門潰瘍に関連する損傷が治癒される。)
【0100】
ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は尿路上皮の治癒を促進するのに有益である。尿路上皮細胞を含む組織層は、多くの手段により損傷している。該手段とは、カテーテル法、外科手術、或いはバクテリア感染(例えば、淋病といった、性感染症を引き起こす媒介物による感染)を含む。本発明はまた女性生殖系の組織の治癒の促進方法も包含する。該方法は有効量のガレクチン‐8、‐3及び又はガレクチン‐7を投与することを含む。女性生殖系の組織の損傷は、カンジダ菌、トリコモナス、ガードネレラ、淋病、クラミジア、マイコプラズマ感染症及びその他の性感染症といった感染を含む広範囲の病気により引き起こされる。
【0101】
ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は腎上皮細胞の治療を促進する方法により有益とされる。そしてそれ故に、急性又は慢性腎不全や末期の腎臓病などの腎疾患や病状の緩和や治療に有益でありうる。ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は乳房組織の治療を促進する方法により有益とされる。そしてそれ故に、外科手術、外傷又は癌によって受けた乳房組織の治癒の促進に有益でありうる。ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、脳組織の損傷の治癒や緩和を促進する本明細書に示された方法により有益とされる。該脳組織の損傷とは、外傷、外科手術又は化学薬品によるものである。
【0102】
ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は予防的に投与されて、様々な病理状態により引き起こされる肺への損傷を軽減又は回避することが可能である。例えば、ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は肺胞や細気管支上皮の治療を促進し、急性又は慢性肺損傷を回避、減弱又は治療することが可能である。肺胞の進行性消失をもたらす肺気腫や、気道障害、すなわち煙の吸入や火傷に起因し、細気管支上皮や肺胞の壊死の原因となるものは、ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8を用いて効果的に治療することができる。該ガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、化学療法、放射線治療、肺癌、ぜんそく、黒色肺及びその他の肺損傷状況に起因する損傷を治療することができる。
【0103】
当然のことながら、本発明に包含される治療方法は、ヒトの創傷の治療に限定されるものではなく、任意の哺乳類の創傷の治療に用いられてもよい。該哺乳類は、ウシ、イヌ、ネコ、ヤギ、ヒツジ、ブタ、ネズミ及びウマの種を含むが、これらに限定されない。ある種の創傷を治療する場合、使用されたガレクチン‐1、‐3、‐7及び/又はガレクチン‐8は、種に自然に発生するガレクチン‐1、‐3、‐7及び/又はガレクチン‐8のアミノ酸配列と実質的に同一であるアミノ酸配列を持つことが好まれるが、これに限定されない。
【0104】
これらの例の中に述べられてきた全ての動物は、眼科研究での動物の使用の、眼科と視覚に関する研究会議の決議、またNIHの実験動物の管理と使用に関する指針による勧告に準拠している。
【0105】
<実施例1:損傷に伴う角膜上皮の移動におけるガレクチン−3の上方制御>
ガレクチン−3の発現レベルが、損傷に伴った治癒角膜の上皮において変わるかどうかを決定するために、生体内において部分的に治癒することができる程度の2mmのエクシマレーザー切除と切除創傷をもつマウスの角膜を、ラットの抗ヒトガレクチン3-mAb M3/38(アメリカ培養コレクション、Rockville, MD)により、免疫染色した。角膜上皮は、層鱗状上皮の原型である。マウスにおいて、角膜上皮は全体の角膜の厚さの20〜25%を構成し、5〜6層の細胞からなる。上皮基底膜の下面には角膜実質を備え、マウスの場合、該角膜実質は全体の角膜の厚さの70〜80%である。切除創傷は、角膜実質に傷をつけずに上皮を除去する。反対に、エクシマレーザー処置は一般的に近視の矯正に用いられ、上皮だけでなくその下面に位置する角膜実質も除去する。
【0106】
スイス・ウェブスターマウス(Taconic Laboratory Animal Service, Germantown, NY)を、1.25%のアバーティン(0.2ml/10kg体重)を用いて筋肉注射によって麻酔した。該アバーティンは、2,2,2トリブロモエタノール2.5gと、2−メチル−2ブタノール(Aldrich, Milwaukee, WI)5mlと、蒸留水195mlを混合して調製した。プロパラカイン点眼薬(Alcon Labs Fort Worth, TXより入手可能なALCAINE(登録商標))を角膜の局所麻酔として用いた。第1群マウスの右目に、APEX PLUS(登録商標)エクシマレーザー(Summit Technology of Waltham、MA)を用いて、経上皮エクシマレーザー切除術を行った(2mmの光学部、42から44μmの切除の深さ、PTKモード)。第2群のマウスの右目に、アルジャー・ブラッシュ(Alger brush (Alger Equipment Company of Lago Vista,TX))を用いて、2mmの切除創傷を作製した。
【0107】
手術に伴い、全ての動物に鎮痛薬として、ブプレノルフィン(0.3mg/mlを0.2ml、Reckitt&Colman Pharmaceuticals, Richmond, VAより入手可能なBUPRENEX(登録商標))を用いて筋肉注射を行った。抗生物質軟膏(Pharmaderm, Melville, NY から入手可能なVETROPOLYCIN(登録商標))を塗ると、角膜は、生体内で16〜18時間後に部分的に治癒された。治癒期間の終了時、上記のように動物に麻酔をかけた後、頸部脱臼させて解剖を行った。解剖後、その眼をホルマリンで2時間固定した後、パラフィンろうに包埋した。組織断片(厚さ5μm)は眼球軸に対して平行な位置で切断した。その断片はキシレンによる処置と、段階的なエタノール溶液(100%、70%、30%)を用いて再水和することによって脱パラフィン化した。免疫染色については、組織断片は、連続して3%のH2O2(37℃、10分)、2.5%の標準的なヤギ血清で培養した。それぞれ、H2O2は内因性のペルオキシダーゼ活性を阻害し、ヤギ血清は非特異的結合を阻害する。その断片は、連続してmAb M3/38(不希釈のハイブリドーマ流動体、1時間)で培養し、抗ラット免疫グロブリン(IgG)を1時間ビオチン化し(1:200、Vector Labs、Burlingame、CA)、アビジンDとビオチンペルオキシダーゼ(Vector Lab)とジアミノベンジジン(DAB)―H2O2試薬(Kirkegaard&Perry Labs, Gaithersburg, MD)との複合体を20時間で新たに調製した。負対照として、関連のないmAbまたは培地のみを用いて断片を処置した。
【0108】
図9に、免疫組織化学的染色したパラフィン断片における標準的な角膜(図9AとB)と治癒角膜(図9CとD)を示す。この図9には、角膜創傷治癒モデルのどちらにおいても、移動性の治癒角膜上皮が、標準的な表皮よりもmAb M3/38によって濃く染色され、詳細には基底細胞層と中間細胞層が濃く染色されていることが示されている。治癒角膜上皮と標準的な角膜上皮のどちらも、免疫染色は細胞−マトリックスの接着部位においてさらに濃く染色されている。標準的な角膜の間質細胞は、mAb M3/38と反応しない一方、治癒角膜の下の前部角膜実質に位置する細胞は、特に移動性上皮の下の領域において、確実にガレクチン−3を発現する。
【0109】
角膜上皮におけるガレクチン−3の免疫活性は、血清を含まないイーグルの最小必須培地において、器官培養を16〜18時間することにより角膜が治癒された場合と類似している。該イーグルの最小必須培地には、非必須アミノ酸、L−グルタミン、抗生物質、0.4%のウシ血清アルブミン(BSA)が含まれる。しかしながら、前部角膜実質は、ガレクチン−3を発現する細胞なしに生体内で治癒され、これにより、生体内で治癒を行う可能性がある角膜実質で見られるガレクチン−3陽性の細胞は、ケラトサイトではなく白血球である可能性が高いことが示唆された。
【0110】
ガレクチン−3の糖質認識領域が、損傷に伴う角膜上皮層の移動に関与するかどうかを決定するために、2mmのエクシマレーザー切除と切除創傷をもつ角膜を、二糖のβ−ラクトースとスクロースの存在下または非存在下で、器官培養を行うことで治癒させた。β−ラクトースがガラクトースを含み、ガレクチンと結合する一方で、スクロースはガラクトースをもたず、ガレクチンと結合しなかった。これらの実験によると、角膜創傷の再上皮形成率は、β−ラクトースの存在下では著しく遅く、一方スクロースは何も影響しなかった。図10で示すとおり、異なる群間の治癒率(少なくとも2つの実験による平均±標準誤差)は、mm2/hで以下のように示される。培地のみは0.088±0.003(N=29)、培地とβ−ラクトースは0.063±0.003(N=19)、培地とスクロースは0.084±0.004(N=10)であった。
【0111】
<実施例2:野生型マウスとガレクチン−3欠損マウスにおける角膜上皮創傷閉塞>
角膜創傷の再上皮形成が、ガレクチン−3欠損マウスで低下するかどうかを決定するために、角膜創傷治癒の4つの異なるモデルを用いた。ガレクチン−3欠損マウス(gal-3-/-)は、Hsu et al., Am J. Pathol 156:1073,2000に記載されたガレクチン−3遺伝子の遮断を対象として生産された。特に、CRDをコードする領域は、ネオマイシン抵抗性遺伝子によって遮断された。これは、0.5kbイントロン4−エクソンの5断片を抗生物質抵抗性遺伝子(neo)に置換することを含む。不活性化されたガレクチン−3遺伝子が、サザンブロット法及びウエスタンブロット法によって確認された。
【0112】
簡潔に説明すると、エクシマレーザー切除術を行った角膜(実施例1で記載したように)又はアルカリによる火傷は、生体内又は生体外で(実施例1で記載したように)部分的に治癒された。アルカリ損傷については、2mmのフィルターディスク(Whatman 50, Whatman International, Maidstone, UK)を、トレフィンを用いることにより作製し、0.5Nの水酸化ナトリウムに浸し、それを第2群のマウスの右目の角膜表面に30秒おいた。その後、該眼を十分なPBSで洗浄した。治癒期間の終了時、創傷範囲をメチレンブルーで染色することにより視覚化した。その後、染色された創傷を標準距離で撮影し、染色された創傷の映像から創傷範囲の輪郭を紙に写しとった。SIGMASCAN(登録商標)のソフトウェア(SPSS Science of Chicago, IL)を用いて、これらの輪郭をデジタル化して定量化した。創傷治癒の異なるモデルのgal-3+/+マウスの創傷閉塞率の分析では、gal-3+/+のmm2/hで示される創傷閉塞率は、エクシマレーザーによって損傷した角膜の方がアルカリによる火傷によって損傷した角膜に比べて、低いことが明らかになった。また、損傷方法に関らず、創傷閉塞率は、生体内で治癒する角膜の方が器官培養で治癒する角膜に比べて高い。図11に示すように、gal-3+/+群間の創傷閉塞率は、エクシマレーザー/生体内群は0.076±0.003mm2/h、エクシマレーザー/生体外群は0.050±0.003mm2/h、アルカリによる火傷/生体内群は0.182±0.003mm2/h、アルカリによる火傷/生体外群は0.106±0.005mm2/hである。それぞれの群の値は、少なくとも2つの実験(各群N=9以上)の平均±標準誤差で示されている。gal-3+/+群とgal-3-/-群の創傷閉塞率を比較すると、角膜がエクシマレーザーによる損傷を受けたかアルカリ処置による損傷を受けたかに関らず、また角膜が生体内又は生体外で治癒されたかに関らず、mm2/hで表される角膜上皮創傷閉塞率は、gal-3-/-群のマウスがgal-3+/+群のマウスと比較して著しく低いことが明らかになった。異なるgal-3-/-群における創傷閉塞率は、エクシマレーザー/生体内群は0.060±0.004mm2/h、エクシマレーザー/生体外群は0.036±0.005mm2/h、アルカリによる火傷/生体内群は0.150±0.008mm2、アルカリによる火傷/生体外群は0.081±0.004mm2/hである。この場合も、すべての値が少なくとも2つの実験(各群N=8以上)の平均±標準誤差で示されている。
【0113】
<実施例3:損傷に伴うガレクチン−3欠損マウスの角膜上皮の移動における遺伝子の発現パターン>
角膜上皮創傷の再上皮形成が、なぜgal-3-/-マウスで乱れるのかを理解する試みとして、gal-3+/+とgal-3-/-の角膜の治癒における遺伝子発現パターンを、cDNAのマイクロアレイを用いて比較し、結果をさらに半定量的RT−PCRによって確かめた。経上皮エクシマレーザー切除(半径2mm)は、gal-3+/+とgal-3-/-のマウスの右目において作製した(実施例1のように)。角膜は、部分的に20〜24時間で生体内で治癒された。治癒期間の終了時、動物を解剖し、角膜を摘出してすぐに液体窒素に入れ、SMART(登録商標)のcDNA技術を用いた遺伝子発現分析のため、Clontech Labolatories, Palo Alto, CAに発送した。簡潔に説明すると、ATLAS(登録商標)のピュアトータルRNAラベリングシステムで定める試薬を用いて全RNAを分離した。30のgal-3+/+とgal-3-/-の角膜から生産されるRNAは、それぞれ3.5μgと2.6μgであった。gal-3+/+とgal-3-/-のマウスの角膜のRNA作製のA260:A280比率は、それぞれ1.48と1.37であった。リボソームRNAの28S:18S比率は、どちらの作製においても1.8であった。これにより、RNA作製量は十分であるということが確かとなった。プローブ作製については、第1のらせん構造cDNAは、RNAを175ng、修飾オリゴ(dT)プライマー(CDSプライマー)、POWERSCRIPT(登録商標)逆転写酵素と、SMART(登録商標)IIオリゴヌクレオチドを用いて合成した。対照群は、逆転写酵素なしにサンプルを培養した。長距離(LD)−PCRによってcDNAを増幅させた。増幅サイクルの最適数を決定するために、試薬製品の一定分量を15、18、21、24サイクル時に集め、アガロースゲル電気泳動によって分析した。最適数の増幅サイクル(例えば23サイクル)を用いて増幅した2重らせんcDNAの生産量は1〜1.6μgであった。増幅したcDNA(500ng)は、クレノウ酵素と33P−αATPを、ATLAS(登録商標)のマイクロアレイ(Clontech)に用いるSMART(登録商標)cDNAプローブ合成用の取扱説明書を用いて、放射性同位体でラベル付けした。ラベルしたプローブをNUCLEOSPIN(登録商標)フィルターによる濾過によって精製し、マウスの1.2k-I ATLAS (登録商標)ナイロンのcDNAマイクロアレイ(Clonetech)とハイブリッド形成した。これは、1200までのマウス遺伝子からなる広域スペクトルアレイである。ハイブリダイゼーションに伴い、その膜を放射線画像解析装置の画面にさらし、結果をATLAS IMAGE (登録商標) 2.0 ソフトウェア (Clonetech)で分析した。データは半定量的RT−PCRによって確認した。
【0114】
RT−PCRとして、全RNAと第1のらせん構造cDNAは、前述した手順によって、gal-3+/+治癒角膜とgal-3-/-の治癒角膜により調製した。PCR増幅は、cDNAを14ng、Clonetechから購入した遺伝子特異性の特別プライマー、及びADVANTAGE (登録商標) 2 PCRキット(Clonetech)によるその他の試薬を用いて50μl量中で行った。アニーリングの温度は68℃とし、試薬はPCR増幅のサイクル数の変化に従った。ハウスキーピング遺伝子の分析として、増幅生成物の一定分量5μlを5サイクルごと及び18サイクル開始時に集める一方、異なる発現遺伝子反応の増幅生成物を1つおきのサイクルごと及び28サイクル開始時に集めた。様々なサイクルで集められた増幅生成物は、1.5%アガロース/エチジウムブロマイドゲルで電気泳動することにより分析した(図12)。これらの実験から、gal-3+/+マウスの治癒角膜と比較して、gal-3-/-マウスの治癒角膜は、ガレクチンー7のmRNA転写産物、その他のガラクトース結合タンパク質及びトロイド様タンパク質(TLL)、メタロプロテイナーゼが顕著に少ない量であることが明らかになった。全体的には、gal-3+/+の治癒角膜と比較して、gal-3-/-の治癒角膜は、ガレクチン−7を約1/12、TLL遺伝子転写産物を約1/14(データ示さず)含んでいた(図12)。様々なハウスキーピング遺伝子のmRNA転写産物の発現レベルは、マイクロアレイ技術(図12)と半定量的RT−PCR(図12、GAPDHはD−グリセルアルデヒド−3−リン酸脱水素酵素、RPS29はリボソームタンパク質S29、ODCはオルニチンデカルボキシラーゼである)によって検出した結果、gal-3+/+治癒とgal-3-/-治癒の両方が類似していた。
【0115】
ガレクチン−7タンパク質の発現レベルがgal-3-/-マウスの治癒角膜でも減少するかどうかを確認するために、gal-3+/+治癒角膜とgal-3-/-の治癒角膜の界面活性剤抽出物を用いたウエスタンブロット分析(図13A)と、gal-3+/+とgal-3-/-マウスの角膜由来のパラフィン断片を用いた抗ガレクチン−7のポリクローナル抗体の免疫組織化学的研究(図13B)を行った。この免疫活性は、激しい(+++)、中程度(++)、弱い(+)、又は負(−)で級付けをした。gal-3-/-治癒角膜上皮の移動が、gal-3+/+治癒角膜上皮の移動と比較して、著しくガレクチン−7の免疫活性が少ないことを発見した(gal-3+/+:+++36/42、++5/42、+1/42以下、gal-3-/-:+++3/42、++26/42、+13/42以下)。また、細胞培養で成長したgal-3-/-マウスの胎児線維芽細胞(MEF)は、gal-3+/+のMEF培養と比較して減少量を示した(図13C)。
【0116】
<実施例4:外因性のガレクチン−3が、野生型マウスとガレクチン−3欠損マウス内で角膜創の上皮再形成を促進する>
角膜上皮創閉塞速度がガレクチン−3欠損マウス(gal-3-/-マウス)内で変動したが(実施例2)、器官培養において、角膜を治療する上皮再形成を外因性ガレクチンが促進させるか否かという点は興味深いところである。本研究において、アルカリによる火傷を負ったgal-3+/+マウスとgal-3-/-マウスの角膜が、無血清培地内で培養された。尚、この培養は、様々な量の組み換え型ガレクチン−3の存在下及び不存在下で行なわれている。
【0117】
上記の如く(Yang et al., Biochemistry 37:4086,1998)、大腸菌内で、組み換え型全長ヒトガレクチン−3が作り出されるとともに精製された。実施例2において説明されたように、アルカリが含浸されたフィルターディスクを用いて、麻酔されたマウスの両目にアルカリ火傷創(2mm直径)が作られた。この創傷の後、マウスは殺傷され、眼球が摘出され、外因性ガレクチン−3の存在下或いは不存在下で、眼球の培養が18時間から20時間行なわれた。
マウスの左目は対象として用いられるものであるが、無血清培地でのみ培養された。右目は、様々なテスト試薬を含む無血清培地内で培養された。このテスト試薬には、以下のものが含まれる。
(i)ガレクチン−3(5μg/mLから20μg/mL)
(ii)ガレクチン−3(10μg/mL)と0.1Mのβ−ラクトース
(iii)ガレクチン−3(10μg/mL)と0.1Mのスクロース
(iv)0.1Mのβ−ラクトース
(v)0.1Mのスクロース
治癒期間の終わりに、実施例2で説明した如く、SIGMASCAN(登録商標)ソフトウェア(イリノイ州シカゴのSPSS Science社製)を用いて、残存する創傷領域が染色され、撮像され、定量化された。各グループは、最低3つの眼球を含み、全ての実験は、少なくとも2回行なわれた。
【0118】
外因性ガレクチン−3は、gal-3-/-マウスの角膜創傷の上皮再形成速度に何ら盈虚を与えなかった(図14A参照)。しかし、外因性ガレクチンは、gal-3+/+マウスの創傷閉塞速度を促進した。この創傷閉塞速度は、10μg/mLから20μg/mLの濃度に依存する(0及び5μg/mLの濃度では、0.090±0.010mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は7以上)であり、10μg/mLでは、0.129±0.010 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は7以上)であり、20μg/mLでは、0.154±0.004 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は7以上)である。
図15に示す如く、gal-3+/+マウスの角膜上皮創傷閉塞に対するガレクチン−3の促進効果は、β−ラクトースにより特に抑制されたがスクロースによっては抑制されなかった(10μg/mLのガレクチン−3に対して、0.127±0.010 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は7以上)であり、ガレクチン−3(10μg/mL)と0.1Mのβ−ラクトースに対して、0.103±0.014 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は7以上)であり、ガレクチン−3(10μg/mL)と0.1Mのスクロースに対して、0.130±0.003 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は7以上)である)。
【0119】
<実施例5:外因性ガレクチン−7が、野生型マウスとガレクチン−3欠損マウスの角膜創傷の上皮再形成を促す>
別個の研究において、cDNAマイクロアレイ(例えば、実施例3に示すような)を用いて、gal-3+/+マウスの正常角膜と治癒角膜の遺伝子発現パターンの比較が行なわれ、治癒角膜においては、ガレクチン−7の発現が著しく向上することが明らかとなった。実施例3で説明した研究と関連して、これらの発見は、gal-3-/-マウスの治癒角膜において、ガレクチン−7の発現が低下することを示している。これに起因して、これらの発見は、外因性ガレクチン−7が器官培養において治癒角膜の上皮再形成を促すか否かを判定する実験設計に繋がっている。
この研究において、アルカリ火傷創を負ったgal-3-/-マウスの角膜が、無血清培地で培養された。この培養は、様々な量の組み換え型ガレクチン−7の存在下及び不存在下で行なわれている。
【0120】
cDNA(ヴァージニア州のマナサスにあるAmerican Type Culture Collection社からESTクローンとして入手可能である)をpET25bプラスミド(ウィスコンシン州マジソンにあるNovagen社から入手可能である)内にクローン化することにより、大腸菌内に組み換え型全長ヒトガレクチン−7が作り出された。
実施例2において説明されたように、アルカリが含浸されたフィルターディスクを用いて、麻酔されたマウスの両目にアルカリ火傷創(2mm直径)が作られた。この創傷の後、マウスは殺傷され、眼球が摘出され、外因性ガレクチン−7の存在下或いは不存在下で、眼球の培養が18時間から20時間行なわれた。
マウスの左目は対象として用いられるものであるが、無血清培地でのみ培養された。右目は、様々なテスト試薬を含む無血清培地内で培養された。このテスト試薬には、以下のものが含まれる。
(i)ガレクチン−7(20μg/mL)
(ii)ガレクチン−7(20μg/mL)と0.1Mのβ−ラクトース
(iii)ガレクチン−7(20μg/mL)と0.1Mのスクロース
治癒期間の終わりに、実施例2で説明した如く、SIGMASCAN(登録商標)ソフトウェア(イリノイ州シカゴのSPSS Science社製)を用いて、残存する創傷領域が染色され、撮像され、定量化された。各グループは、最低6つの眼球を含み、全ての実験は、少なくとも2回行なわれた。
【0121】
図16に示す如く、外因性ガレクチン−7は、創傷閉塞速度を促進させた(培地単独の場合は、0.036±0.006 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は10以上)であり、ガレクチン−7(20μg/mL)の場合は、0.072±0.004 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は10以上)である。
図16に示す如く、角膜上皮創傷閉塞に対するガレクチン−7の促進効果は、β−ラクトースにより特に抑制されたが、スクロースによっては抑制されなかった(20μg/mLのガレクチン−7に対して、0.072±0.004 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は9以上)であり、ガレクチン−7(20μg/mL)と0.1Mのβ−ラクトースに対して、0.050±0.004 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は9以上)であり、ガレクチン−7(20μg/mL)と0.1Mのスクロースに対して、0.079±0.007 mm2/h(少なくとも2回の実験の平均値±標準偏差:サンプル数は9以上)である)。
図16に示す如く、外因性ガレクチン−7が、gal-3-/-マウスに代えて、gal-3+/+マウスの損傷した角膜に添加されると、損傷閉塞速度が更に高まった(0.094±0.003 gal-3+/+ mm2/h)。
【0122】
<実施例6:野生型マウスとガレクチン−3欠損マウス内での皮膚上皮創閉塞>
gal-3+/+マウスとgal-3-/-マウスは、1.25%のトリブロモエタノール(Avertin)(0.2mL/10g体重)の腹腔内投与により、麻酔された。レーザ処置の前に、剃刀刃を用いて、背部領域の体毛が剃られた。上皮を透過する6mmの背部の皮膚の創傷が、エキシマレーザ(マサチューセッツ州のワルシャムにあるSummit Technology社製)を用いて作られた。外科的処置の後、抗菌性軟膏が、創傷表面に塗布され、ブプノルフィン(2 mg/kg体重)が、皮下に投与され、術後の痛感を最小限化させている。当該創傷は、部分的に生体内治癒が可能であり、術後、24、48及び72時間で試験が行なわれている。
治癒期間の終わりに、マウスは、1.25%のトリブロモエタノール(Avertin)(0.2mL/10g体重)の腹腔内投与により、再度麻酔され、Sigma Scanソフトウェアを用いて撮像され、その後定量化される。
実験動物の2つのグループ(即ち、gal-3+/+マウスとgal-3-/-マウス)の間で創傷閉塞速度が比較される。実験動物は、二酸化炭素吸入或いはペントバービタルの過投与により、その後殺傷される。
【0123】
<実施例7:皮膚創傷の上皮再形成に対する外因性ガレクチン−3の効果>
実験動物(マウス:57BL/6及び129混合遺伝背景;齢:6週から8週;性別:混合)が、1.25%のトリブロモエタノール(Avertin)(0.2mL/10g体重)の腹腔内投与により麻酔された。
レーザ処置の前に、剃刀刃を用いて、背部領域の体毛が剃られた。上皮を透過する6mmの背部の2つの皮膚上の創傷(各側1つの創傷)が、エキシマレーザー(マサチューセッツ州のワルシャムにあるSummit Technology社製)を用いて作られた。
外科的処置の後、抗菌性軟膏が、創傷表面に塗布され、ブプノルフィン(2 mg/kg体重)が、皮下に投与され、術後の痛感を最小限化させている。当該創傷は、部分的に生体内治癒が可能であり、術後、4時間から6時間おきにガレクチン−3を含有する軟膏が右側の創傷に塗布され、キャリアのみが左側の創傷に塗布された。尚、左側の創傷は対象としての役割を担う。
治癒期間の終わり(24時間から48時間後)に、実験動物は、1.25%のトリブロモエタノール(Avertin)(0.2mL/10g体重)の腹腔内投与により、麻酔され、Sigma Scanソフトウェアを用いて撮像され、その後定量化される。
実験動物の2つのグループ(即ち、ガレクチン−3を用いて治療されたものと対象)の間で創傷閉塞速度が比較される。実験動物は、二酸化炭素吸入或いはペントバービタルの過投与により、その後殺傷される。
【0124】
<実施例8:皮膚創傷の上皮再形成に対する外因性ガレクチン−7の効果>
実験動物(マウス:57BL/6及び129混合遺伝背景;齢:6週から8週;性別:混合)が、1.25%のトリブロモエタノール(Avertin)(0.2mL/10g体重)の腹腔内投与により麻酔された。
レーザ処置の前に、剃刀刃を用いて、背部領域の体毛が剃られた。上皮を透過する6mmの背部の2つの皮膚上の創傷(各側1つの創傷)が、エキシマレーザー(マサチューセッツ州のワルシャムにあるSummit Technology社製)を用いて作られた。
外科的処置の後、抗菌性軟膏が、創傷表面に塗布され、ブプノルフィン(2 mg/kg体重)が、皮下に投与され、術後の痛感を最小限化させている。当該創傷は、部分的に生体内治癒が可能であり、術後、4時間から6時間おきにガレクチン−7を含有する軟膏が右側の創傷に塗布され、キャリアのみが左側の創傷に塗布された。尚、左側の創傷は対象としての役割を担う。
治癒期間の終わり(24時間から48時間後)に、実験動物は、1.25%のトリブロモエタノール(Avertin)(0.2mL/10g体重)の腹腔内投与により、麻酔され、Sigma Scanソフトウェアを用いて撮像され、その後定量化される。
実験動物の2つのグループ(即ち、ガレクチン−7を用いて治療されたものと対象)の間で創傷閉塞速度が比較される。実験動物は、二酸化炭素吸入或いはペントバービタルの過投与により、その後殺傷される。
【0125】
<実施例9:アルビノウサギのドライアイ症状に対する外因性ガレクチン−8或いはガレクチン−1の影響>
実験動物(ウサギ:アルビノ;齢:6週から8週;性別:混合)が、1.25%のトリブロモエタノール(Avertin)(0.2mL/10g体重)の腹腔内投与により麻酔された。1.0%の硫酸アトロピンの日常的な繰り返し滴下(Burgalassi, S., et al., Ophthalm Res 31:229-235, 1997)により、各ウサギにドライアイ症状を引き起こす。Schirmer I test及び蛍光染色後の角膜の試験により、ドライアイ症候群の評価が各実験動物に対してなされる。
【0126】
4時間から6時間おきに、ガレクチン−8の溶液を含有する点眼薬が、各実験動物の右眼に投与される。また、キャリアのみが左目に投与される。尚、左目は対象としての役割を担う。
治癒期間の終わり(24時間から48時間後)に、実験動物は、1.25%のトリブロモエタノール(Avertin)(0.2mL/10g体重)の腹腔内投与により、麻酔され、眼領域がSigma Scanソフトウェアを用いて撮像され、その後定量化される。
眼表面及びドライアイの症状が2つの眼のグループ(即ち、ガレクチン−8により治療された右目と対象となる左目)の間で評価され、これらの表面が比較される。
実験動物は、二酸化炭素吸入或いはペントバービタルの過投与により、その後殺傷され、標準的な毒学的基準により解析がなされた。
【0127】
ヒトと他の脊椎動物(図20参照)の多種のガレクチン−8タンパク質(図18及び配列識別番号:4及び5)のアミノ酸配列は略同一であり、このことは、他の種の哺乳類に当てはまる。同様に、ヒトと他の哺乳類の多種のガレクチン−1タンパク質(図19及び配列識別番号:6)のアミノ酸配列は略同一である。保存部位及び保存変化が少ない部位は、残基が変化し他の機能性ガレクチン−8及び/又は機能性ガレクチン−1タンパク質を得られ、これらタンパク質が、ドライアイ症状及び他の眼症状を治療する能力を備える。
【0128】
<実施例10:スコポラミンモデル:DESに対するガレクチン治療並びにスコポラミン治療前のガレクチン溶液を用いた前治療の効果>
溶液1mL当たり0, 10又は20μgのいずれかの量のガレクチン−1、ガレクチン−3、ガレクチン−7及びガレクチン−8のうち1つ或いは2つを含有する涙液溶液が、以下に示す治療スケジュールに則って投与される。
10μLの滴下量を以って、各実験動物の一方の眼に対する治療が各時点で行なわれる。スコポラミンパッチモデルのグループに対して、10μg/mLのガレクチン溶液が、1日当たり4回、一方の目に投与される。この投与は、スコポラミンパッチの使用直後に開始されるものである。
マウスで構成される治療グループが使用される。治療グループ及び眼は、ランダム化並びにコード化され、無作為に測定が行われる。各実験動物の一方の眼が治療溶液を投与され、他方の眼は対象滴を投与される。測定は、各時点で相互に行われる。
【0129】
<実施例11:スコポラミンモデル:βラクトース、ガレクチン阻害剤とともに投与されるガレクチンの混合治療>
実施例1に示すものと同一の手順が実行された。尚、0.1 Mのβ−ラクトースとともに10μg/mLのガレクチン溶液が用いられ、ガレクチンの効果が、競合するに糖類により抑制されるか否かを判断する。
【0130】
【表3】
【0131】
<実施例12:DESにおけるガレクチン−1、ガレクチン−3、ガレクチン−7及びガレクチン−8の役割:マウスの自己免疫MRL/lpr染色>
治療グループ及び眼は、ランダム化並びにコード化され、無作為に測定が行われる。各実験動物の一方の眼が治療溶液を投与され、他方の眼は対象滴を投与される。測定は、各時点で相互に行われる。
【0132】
【表4】
【0133】
<実施例13:ドライアイ症状を有する被験体に対するガレクチン−3による治療効果>
ドライアイ症状に対するガレクチンの効果を判断するために、インターロイキン−1を用いた動物モデルシステムが用いられ、4つのマウスグループが使用された。各グループは、5つの実験動物からなり、以下のように処置を施された。
3つのグループに対して、第0日目にインターロイキン−1(IL-1)が被検体に注入された。この注入は、シェーグレン症候群のマウスモデル用に創られた標準的手順にしたがって行われた(Zoukhri, D. et al., Invest Ophthalmol Vis Sci 42 (5): 925-932)。対象被検体は当該注入を受けておらず、点眼薬を用いた更なる治療を施されていない。IL-1の注入を受けた被検体は、その後、点眼薬の投与による治療を受けた。この点眼薬には、ガレクチン−3が含まれ、点眼量は、75μg/mL又は150μg/mLであり、1日当たり4回の点眼が行なわれる。この点眼は、第1日目から行なわれる。1つの被検体グループは、同様にバッファのみ(ガレクチンなし)を1日当たり4回投与された。ドライアイの程度は、上述の如く(Zoukhri et al.)、フルオレセインのクリアランスによって、各被検体ごとに評価された。
【0134】
第1日目の治療初期において、涙液分泌液が、フルオレセインのクリアランスにより測定され、75μg/mL或いは150μg/mLの投与量のガレクチン−3それぞれにより治療された被検体において、対象の被検体と比して、改善が見られた。第2日目までに、ガレクチン治療を施された被検体の症状は完全に改善された。実際に、第2日目において、幾分良好な涙液クリアランスが、低投与量75μg/mLの被検体で観察されたことから、これより少ない投与量が最適な投与量である可能性が示されている。図21において、棒グラフの棒(第2日目の治療された被検体に対する)上のアスタリスクは、ドライアイ症状を有する対象の被検体グループであって、バッファ賦形剤のみを投与されたものからのデータと比してこれらのデータが統計学的に有意であることを示している(p<0.05)。結論として、ガレクチン−3は、ドライアイを治療するとともに、1日から2日の治療期間内でこの症状を改善する能力を備えるということができる。哺乳類間の高程度の同一性並びにガレクチンファミリーの異なる型間でのガレクチン機能の同一性及び特異的アミノ酸コンセンサス配列の同一性の存在により、ガレクチン−1、ガレクチン−3、ガレクチン−7及びガレクチン−8のいずれか或いはこれらの組み合わせが、同様にドライアイ及びこれに関連する症状の治療に有効であることが予測できる。
【0135】
<結論>
本明細書では、ガレクチン−3及びガレクチン−7が角膜創傷の再上皮化の役割を担うことが実証された。実施例1では、免疫組織化学の研究により、損傷後にガレクチン−3が角膜上皮の細胞間基質接着部位で高密度存在することが明らかになった。そこは細胞基質の相互作用に影響を与え、細胞移動にも理想的な部位である。実施例2では、角膜創傷の再上皮化の進度がガレクチン−3欠損マウスにおいては、野生型マウスよりも著しく遅れることが示された。実施例3では、損傷後にガレクチン−7の発現がガレクチン−3欠損マウスにおいては、野生型マウスよりも著しく減少することが確認された。実施例4、5では、外因性の組み換え型ガレクチン−3とガレクチン−7が、gal3+/+マウスにおいて角膜創傷の再上皮化を刺激することが示された。実施例6−8では、創傷の再上皮化に際するガレクチンの効果の測定手段を明示した。実施例9−12においては、眼乾燥症候群におけるガレクチンの効果の測定手段を明らかにした。実施例13では、ガレクチン−3の眼乾燥への即効性治療が示された。さらに、実施例1においては、角膜上皮創閉塞速度におけるガレクチン−3の刺激性効果が、ガラクトシド化学構造の競合二糖(β−ラクトース)によって抑制されることが実証された。同時に、科学的に関連性のない二糖(スクロース)の影響を受けないことも示された。本結果として、糖鎖認識部位は角膜に対する外因性ガレクチンの有用な効果に直接関与していることが示された。
【0136】
ガレクチン1、3、7及びガレクチン8が眼乾燥症候群の治療に影響するというメカニズムに関して、いかなる特定の理論にも結びつけるという意図ではないが、以下を提案する。
【0137】
上記の通り、ガレクチン−3は糖鎖ポリラクトサミンを包含する補完的複合等質と結合して、細胞同士の、又は細胞基質の相互作用の仲介をすると考えられている。糖鎖ポリラクトサミンは多数の細胞外基質(ECM)において、又はイソ型のフィブロネクチン、ラミニン及び、インテグリンのような細胞表面分子において見られる。(Liu, Clin. Immunol. 97:79, 2000 及び Perillo, supra) しかしながら、外因性のガレクチン−3がgal3-/-マウスにおいては創傷再上皮化を促進しないという本明細書の所見(例4参照)は、細胞内のガレクチン−3が創傷治癒の過程で有意に貢献しているということを示唆している。細胞内のガレクチン−3は、特定の細胞表面、又は/あるいは、細胞外基質(ECM)受容体の発現において最も影響を与えており、結果として、細胞基質の相互作用や細胞移動にも影響を与えている。この考えはすでに発表された研究と一致しており、その研究ではガレクチン−3は乳癌細胞株内で安定的に過剰発現した。親細胞株はガレクチンをほとんど、あるいはまったく発現しなかった。結果として、α4β7、α6β1インテグリンが増加し、ラミニン、フィブロネクチン、及びビトロネクチンを含む様々な細胞外基質(ECM)分子との接着が強固になった。(Warfield, supra及びMattarese, supra) 別の研究においては(Dudas et al, Gastroenterology 118:1553, 2000)発現ガレクチン−3を形質移入された結腸癌細胞株が、レクチン本体の主要な結合基である特定ムチン、MUC2を増加させた。(Bresalier et al, Cancer Research 56:4354,1996) gal3+/+マウスにおいて創傷再上皮化速度に対する外因性ガレクチン−3の刺激性効果がラクロース阻害であるという事実は、以下の興味深い可能性を示唆している。細胞内ガレクチン−3は、実際にレクチン本体の細胞外基質(ECM)受容体や細胞表面として機能するタンパク質糖鎖付加を制御しているのかもしれない。細胞内ガレクチン−3が核基質に働きかけ、複雑な生物学的過程に影響を与える可能性を有しているということはまた、以下の所見を示唆している。特定の状況下では、レクチンは核内でリボ核タンパク質複合体と結合したり、RNA前駆体のスプライシング因子として作用したりすることが可能である。(Dagher et al., Proc. Natl. Acad. ScL USA 92:1213, 1995). 同様に、ワング(Wang)などによって、ガレクチン−3は前立腺癌細胞において核基質と結合し、又、一本鎖DNAとRNAの双方と結びつくことが実証された。(Wang et al., Biochem. Biophys. Res. Commun. 217:292, 1995).
【0138】
マウスの相補DNAマイクロアレイを用いたgal3+/+及びgal3-/-マウスの角膜治癒の遺伝子発現パターンの分析により、発現したgal3-/-マウスの角膜治癒は野生タイプのマウスよりもガレクチン−7を著しく減少させることが明らかになった。(例3、5参照)ガレクチン−7は1994年に初めて報告され(Barondes, supra)、ガレクチン−3ほど十分に特徴づけられていない。ガレクチン−3とは異なり、ガレクチン−7には顕著な組織特異性がある。動物の成体において、ガレクチン−7の発現は上皮に限られており、上皮は重層化しているか、重層化することになる。(Timmons et al., supra). タンパク質は細胞基質や細胞間の相互作用、及びアポトーシスに関与していると考えられている。(Leonidas, Biochemistry 37:13930, 1998 及び Bernerd et al., Proc. Natl. Acad. Sci. USA 96:11329, 1999). 一般的に、ガレクチン−7の発現とケラチノサイト増殖との間には逆相関が存在しており、ガレクチン−7の発現はSV40ウィルスに形質転換したケラチノサイトで抑制され、表皮性腫瘍に由来する株細胞でも同様に抑制される。外因性ガレクチン−3はgal3-/-角膜では創傷再上皮化を刺激せず、gal3-/-角膜の治癒はガレクチン−7の量を減少させる。この発見により、ガレクチン−3がガレクチン−7を調節することで創傷再上皮化に少なくとも部分的には影響を与えるということが示唆されている。実際に、ガレクチン−3と異なり、ガレクチン−7はラクトースを抑制してgal3-/-角膜で創傷の再上皮化を促進することが発見されている。また、gal3-/-マウスの胚繊維芽細胞は発現してガレクチン−7の量を減少させた。
【0139】
関与するメカニズムにかかわらず、ガレクチン−3とガレクチン−7の双方が角膜創傷の再上皮化を刺激するという所見は、上皮創傷の治療やとりわけ上皮創傷の非治癒に広範な意味を持つ。今日では、角膜の持続性上皮欠損の治療が主要な臨床問題となっている。さらに、移植後の創傷、高齢者の慢性創傷、褥瘡、及び皮膚の静脈うっ血性潰瘍への効果的な治療の必要性が高まっている。細胞増殖を刺激するとして公知の多数の成長因子(例えば、上皮成長細胞因子(EGF)、トランスフォーミング増殖因子(TGF)、繊維芽細胞増殖因子(FGF)、KGF、肝細胞増殖因子(HGF))が角膜での有用性を検査されており、皮膚の上皮創傷治癒においては、全体的に失望させる結果に終わっている。(Eaglstein, Surg. Clin. North Am. 77:689, 1997; Singer 及び Clark, N. Engl. J. Med. 341:738, 1999; Zieske 及び Gipson, pp. 364-372 in "Principle 及び Practice of Ophthalmology" Ed. by D. M. Albert 及び F. A. Jakobiec, W.B. Saunders Company, Philadelphia, PA, 2000; Schultz et al., Eye 8:184, 1994; Kandarakiset al., Am. J. Ophthalmol. 98:411, 1984; and Singh 及び Foster, Am. J. Ophthalmol. 103:802, 1987). 成長因子を用いる上記研究の大部分で、創傷の再上皮化の加速度範囲は今日の研究でガレクチンによって確認される範囲より著しく小さかった。また、上皮細胞成長因子(EGF)のような成長因子として処遇される角膜上皮は過形成のものであり、(Singh 及び Foster, Cornea 8:45, 1989) 明らかに望ましくない状態である。この観点においては、上皮細胞でレクチンの細胞有糸分裂の誘発性は観察されなかったため、臨床的な可能性はガレクチン−3とガレクチン−7の方が成長因子よりも魅力的である。ここ10年以上に渡って、近視の矯正のために角膜特性を修正するエキシマレーザー角膜切除術の可能性が現実味を帯びてきている。上記のような施術が毎週数千も行われ、近視の人々はメガネやコンタクトレンズから解放されている。世界の成人人口の25%−30%が近視であるという事実に着目すれば、年間約50万もの施術がアメリカだけで行われると想定されるだろう。エキシマレーザー手術のあとに、上皮治癒で遅延が生じる場合もある。上記のような遅延は可能な限り避けなければならず、そうでないと角膜が術後濁度や感染性角膜炎、及び角膜腫瘍に罹ってしまう危険性がある。やはり、ガレクチンに基づく治療はそのような症例で創傷の上皮化を促進する。
【0140】
<他の実施例>
本発明の他の具体例については、本明細書で開示された本発明の実施や規格を考慮すれば、同業者には明白である。規格や諸例については例示しただけに過ぎず、本発明の正確な範囲は以下の請求項で示されている。
【図面の簡単な説明】
【0141】
【図1】アミノ酸配列とヒトのガレクチン‐3を示す。(ジェンバンク受入番号BAA22164、配列番号1)
【図2】アミノ酸配列とヒトのガレクチン‐7を示す。(ジェンバンク受入番号I55469、配列番号2)
【図3】ヒトのガレクチン3(配列番号1)と、ウサギのガレクチン‐3のアミノ酸配列(ジェンバンク受入番号JC4300)、ニワトリのガレクチン‐3のアミノ酸配列(ジェンバンク受入番号AAB02856)及びハムスターのガレクチン‐3のアミノ酸配列(ジェンバンク受入番号CAA55479)をクラスタルWを用いて縦に配列したものを示す。図中の最初(上部)の配列はヒトのガレクチン‐3(配列番号1)のアミノ酸1から250、図中の2番目の配列はハムスターのガレクチン‐3のアミノ酸1から245、図中の3番目の配列はウサギのガレクチン‐3のアミノ酸1から242、図中の4番目(下部)の配列はニワトリのガレクチン‐3のアミノ酸1から262である。
【図4】ヒトのガレクチン‐7(配列番号2)とラットのガレクチン‐7(ジェンバンク受入番号P97590)及びマウスのガレクチン‐7(ジェンバンク受入番号O54974)をクラスタルWを用いて縦に配列したものを示す。図中の最初(上部)の配列はラットのガレクチン‐7のアミノ酸1から136、図中の2番目の配列はマウスのガレクチン‐7のアミノ酸1から136、図中3番目(下部)の配列はヒトのガレクチン‐7(配列番号2)のアミノ酸1から136である。
【図5】ヒトのガレクチン‐3(配列番号1)をPROSITEでスキャンした結果の一覧である。
【図6】ヒトのガレクチン‐7(配列番号2)をPROSITEでスキャンした結果の一覧である。
【図7】ヒトのガレクチン‐3のガラクトシド結合領域と、PFAM中の隠れマルコフモデル(HMM)由来の共通アミノ酸配列(PF00337)を縦に配列したものを示す。上部の配列は共通アミノ酸配列(PF00337、配列番号3)であり、また下部のアミノ酸配列は配列番号1のアミノ酸117から247に対応する。
【図8】ヒトのガレクチン‐7のガラクトシド結合領域と、PFAM中の隠れマルコフモデル(HMM)由来の共通アミノ酸配列(PF00337)を縦に配列したものを示す。上部の配列は共通アミノ酸配列(PF00337、配列番号3)であり、また下部のアミノ酸配列は配列番号2のアミノ酸5から135に対応する。
【図9】生体内で治癒可能な程度で角膜を2ミリ剥離し、或いは角膜にエクシマーレーザー創傷を与え、それからパラフィン切片中でガレクチン‐3の免疫反応性を分析した一連の写真を含む。(A)は、メマトキシリンとエオシンで色づけした(i)正常な角膜、(ii)剥離直後の角膜、(iii)エクシマーレーザー創傷を与えた直後の角膜である。(B)は、免疫組織学上色づけした(i)通常のgal-3+/+角膜、(ii)エクシマーレーザー創傷後の治癒gal-3+/+角膜である。免疫組織学上色づけした(iii)通常のgal-3-/-角膜と(iv)エクシマーレーザー創傷後の治癒gal-3-/-角膜である。濃い色は陽性免疫染色を示す。WE、(傷の端(wound edge))、LE(移動した上皮の先端(leading edge of migrating epithelium))、すなわち矢印(上皮)、矢頭(白血球/間質細胞)である。
【図10】創傷を受けた角膜上皮の治癒率に及ぼされるβ‐ラクトース(Lac)とサッカロース(Suc)の影響を図示するグラフである。
【図11】野生型(gal-3+/+)マウス及びガレクチン‐3欠損(gal-3-/-)マウスに、エクシマーレーザー或いはアルカリ処理を用いて、体内又は体外で治癒可能な程度の創傷を与えた角膜上皮の治癒率を図示している。
【図12】ガレクチン‐7の遺伝子発現とハウスキーピング遺伝子の選択における野生型(gal-3+/+)とガレクチン‐3欠損(gal-3-/-)マウス間での差を図示している。(GAPDHはD‐グリセルアルデヒド3リン酸デヒドロゲナーゼ、RPS29はリボソームたんぱく質S29、ODCはオルニチンデカルボキシラーゼである。)該差は、cDNAマイクロアレイ及び半定量PCRにより決定される。
【図13】ガレクチン‐7の発現における野生型(gal-3+/+)マウスとガレクチン‐3欠損(gal-3-/-)マウス間での差を図示している。該差は、(A)ウェスタンブロット分析、(B)免疫組織化学分析、(C)マウスの胚前脳部分を用いることにより決定される。
【図14】(図14Aの)ガレクチン‐3欠損(gal-3-/-)マウス及び(図14Bの)野生型(gal-3+/+)マウスの創傷を受けた角膜上皮の治癒率に及ぼされる外因性ガレクチン‐3の影響を図示するグラフである。
【図15】外因性ガレクチン‐3の存在下で、野生型(gal-3+/+)マウスの創傷を受けた角膜上皮の治癒率に及ぼされるβ‐ラクトース(Lac)とサッカロース(Suc)の影響を図示するグラフである。
【図16】(A)は外因性ガレクチン‐7が単独で、もしくはβ‐ラクトース(Lac)と共に、或いはサッカロース(Suc)と共に用いられた場合の、野生型(gal-3+/+)マウスの創傷を受けた角膜上皮の治癒率に及ぼされる外因性ガレクチン‐7の影響を図示したグラフである。また(B)は、野生型(gal-3+/+)マウス及びガレクチン‐3欠損(gal-3-/-)マウスの創傷を受けた角膜上皮の治癒率に及ぼされる外因性ガレクチン‐7の影響を比較するグラフである。
【図17】ヒトのガレクチン‐1(配列番号6)のアミノ酸配列を図示する。
【図18】ヒトのガレクチン‐8(配列番号4)の短形のアミノ酸配列を図示する。
【図19】ヒトのガレクチン‐1(配列番号6)、ラットのガレクチン‐1、マウスのガレクチン‐1及びハムスターのガレクチン‐1それぞれのアミノ酸配列を示す。しかもこれらは非常に高率な同一性を共有しているので、ガレクチン‐1配列は哺乳類種の間で強力に保存されている。黒い部分は残基の位置を表し、該位置は全ての哺乳類の配列において同一ではない。ラットとヒトは90.4%が同一で、マウスとヒトは88.2%同一で、またハムスターとヒトは91.2%同一である。また、図は非同一の位置にある残基の大部分が保存的な変化をすることを示す。例えば、アルギニンはポジション19でリシンに置換され、ロイシンはポジション24でバリンに、セリンはポジション73でトレオニンに、そしてアスパラギン酸はポジション136でグルタミン酸に置換される。
【図20】ヒトのガレクチン‐8(配列番号4)の短形(316アミノ酸)のそれぞれのアミノ酸配列と、マウスのガレクチン‐8、ラットのガレクチン‐8、ニワトリのガレクチン‐8及びカエルのガレクチン‐8の短形の対応するアミノ酸配列を示す。該アミノ酸配列は、強調部分と共に配列され、ヒトとラットガレクチン‐8の配列に匹敵する非ヒトアミノ酸配列が相違していることを表す。3つの哺乳類のガレクチン‐8アミノ酸配列は、非常に高率の同一性を共有し、また78.2%同一であって、非同一部分(21.8%)の大部分においてアミノ酸変化が保存されている。該アミノ酸変化は、マウス及びラットの類似のポジションにあるチロシンに匹敵するヒトのポジション20にあるフェニルアラニン、ヒトのポジション22にあるグリシンとラットのセリン、ヒトのポジション26にあるグリシンとげっ歯類配列中のアスパラギン酸塩である。さらに、脊椎動物種は43.0%のかなりの同一性を共有しているので、ガレクチン-8配列は哺乳類種の間で高い同一性を持ち、脊椎動物種間両方で保存されている。
【図21】マウスモデル中の眼乾燥症候群にガレクチンを投与した影響を示す棒グラフである。該症候群は時間の関数としての0日目にインターロイキン-1α(図では「IL-1」と示される)を注入して引き起こされたものである。ガレクチン-3は1日目と2日目に、一日4回、目薬を用いて1グループにつき5つの被験物それぞれに投与した。該目薬は75μg/mlまたは150μg/mlの濃度である。対照群の被験物にはIL-1を注入せず、目薬を投与しなかった。「緩衝液」グループ中の被験物には一日4回キャリア媒体のみの治療薬を投与した。Zoukhri,D.et.al.,Invest Ophthalmol Vis Sci 42(5):925−932に述べられているように、フルオレセインクリアランスにより涙の動態が評価される。低い値はより良く涙が流れることを指し示す。2日目のp<0.05の統計学上の有意差を持って対照群のデータと治療を施したグループのデータを比較すると、ガレクチン治療により、ガレクチンを投与されていない制御動物と比較して、涙の動態を正常なグループの涙の動態に回復したことがデータに示している。
【特許請求の範囲】
【請求項1】
眼乾燥の治療を必要とする哺乳類の眼乾燥を治療する方法であって、治療に有効な量のガレクチンタンパク質を哺乳類に投与することを特徴とする治療方法。
【請求項2】
投与前に、眼乾燥の予防を必要とする前記哺乳類を特定する工程を備えることを特徴とする請求項1に記載の治療方法。
【請求項3】
前記治療又は予防を必要とする前記哺乳類が、眼上皮創傷である哺乳類、抗ヒスタミン剤の使用前である哺乳類、抗炎症剤の使用前である哺乳類、及びエキシマーレーザー治療を施す前の哺乳類のうち少なくとも1つの哺乳類であることを特徴とする請求項1に記載の方法。
【請求項4】
前記哺乳類がヒトであることを特徴とする請求項1に記載の方法。
【請求項5】
前記ガレクチンタンパク質が、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7及びガレクチン‐8から選択されるタンパク質のうち少なくとも一つであることを特徴とする請求項1に記載の方法。
【請求項6】
前記眼乾燥が、持続性症候群であることを特徴とする請求項5記載の方法。
【請求項7】
前記眼乾燥が、上皮びらんを引き起こすことを特徴とする請求項5記載の方法。
【請求項8】
前記上皮びらんが、角膜創傷を引き起こすことを特徴とする請求項5記載の方法。
【請求項9】
前記ガレクチン‐8タンパク質が、配列番号4又は5のアミノ酸配列を備えることを特徴とする請求項5記載の方法。
【請求項10】
前記ガレクチン‐8タンパク質が、配列番号4又は5のアミノ酸配列と実質的に同一なアミノ酸配列であることを特徴とする請求項5記載の方法。
【請求項11】
前記実質的に同一であることが、配列番号4又は5のアミノ酸配列と少なくとも60%の同一性、少なくとも70%の同一性、少なくとも80%の同一性、又は、少なくとも90%の同一性を有するものとすることを特徴とする請求項10記載の方法。
【請求項12】
前記ガレクチン‐8タンパク質が、ガレクチン‐8‐N末端領域とガレクチン‐8プロリン、グリシン及びチロシンが豊富な領域、並びにガレクチン‐8ガラクトシド結合領域とを備えることを特徴とする請求項5記載の方法。
【請求項13】
前記ガレクチン‐3タンパク質が、ガレクチン‐3プロリン、グリシン、及びチロシンが豊富な領域、及びガレクチン‐3ガラクトシド結合領域を備えることを特徴とする請求項5記載の方法。
【請求項14】
前記ガレクチン‐7タンパク質が、ガレクチン‐7‐N末端領域とガレクチン‐7プロリン、グリシン及びチロシンが豊富な領域、並びにガレクチン‐7ガラクトシド結合領域とを備えることを特徴とする請求項5記載の方法。
【請求項15】
前記ガレクチン‐1タンパク質が、ガレクチン‐1‐N末端領域とガレクチン‐1プロリン、グリシン及びチロシンが豊富な領域、並びにガレクチン‐1ガラクトシド結合領域とを備えることを特徴とする請求項5記載の方法。
【請求項16】
前記ガレクチン‐7タンパク質が、配列番号2のアミノ酸配列を備えることを特徴とする請求項5記載の方法。
【請求項17】
前記ガレクチン‐7タンパク質が、配列番号2のアミノ酸配列と実質的に同一なアミノ酸配列を備えることを特徴とする請求項5記載の方法。
【請求項18】
前記実質的に同一であることが、配列番号2のアミノ酸配列と少なくとも70%同一性、少なくとも80%同一性、又は、少なくとも90%の同一性を有するものとすることを特徴とする請求項17記載の方法。
【請求項19】
前記ガレクチン‐3タンパク質が、配列番号1のアミノ酸配列を備えることを特徴とする請求項5記載の方法。
【請求項20】
前記ガレクチン‐3タンパク質が、配列番号1のアミノ酸配列と実質的に同一なアミノ酸配列を備えることを特徴とする請求項5記載の方法。
【請求項21】
前記実質的に同一であることが、配列番号1のアミノ酸配列と少なくとも70%同一性、少なくとも80%同一性、又は、少なくとも90%の同一性を有するものとすることを特徴とする請求項20記載の方法。
【請求項22】
前記ガレクチン‐3タンパク質が、ガレクチン‐3ガラクトシド結合領域を備えることを特徴とする請求項5記載の方法。
【請求項23】
前記ガレクチン‐1タンパク質が、配列番号6のアミノ酸配列を備えることを特徴とする請求項5記載の方法。
【請求項24】
前記ガレクチン‐1タンパク質が、配列番号6のアミノ酸配列と実質的に同一なアミノ酸配列を備えることを特徴とする請求項5記載の方法。
【請求項25】
前記実質的に同一が、配列番号6のアミノ酸配列と少なくとも60%同一性、少なくとも70%同一性、少なくとも80%同一性、又は、少なくとも90%の同一性を有するものとすることを特徴とする請求項24記載の方法。
【請求項26】
前記ガレクチン‐1タンパク質が、ガレクチン‐1ガラクトシド結合領域を備えることを特徴とする請求項5記載の方法。
【請求項27】
眼乾燥の治療の促進効果がある医薬品組成物であり、前記組成物は、薬学的に適当な担体又は希釈剤、並びにガレクチン‐1、ガレクチン‐3、ガレクチン‐7及びガレクチン‐8タンパク質のうち少なくとも1種のガレクチンを備え、該ガレクチンは結膜及び/又は角膜上皮の完全性を促進させる十分量である医薬品組成物。
【請求項28】
前記眼乾燥が、再発性上皮びらんを伴う眼乾燥症であることを特徴とする請求項27の医薬品組成物。
【請求項29】
前記眼乾燥症が、創傷を引き起こし、該創傷は角膜創傷であることを特徴とする請求項28の医薬品組成物。
【請求項30】
前記眼乾燥が、エクシマーレーザー角膜切除術により引き起こされることを特徴とする請求項28の医薬品組成物。
【請求項31】
前記ガレクチン‐8タンパク質が、配列番号4又は5のアミノ酸配列を備えるタンパク質であることを特徴とする請求項27の医薬品組成物。
【請求項32】
前記ガレクチン‐8タンパク質が、配列番号4又は5のアミノ酸配列と実質的に同一なアミノ酸配列を備えるタンパク質であることを特徴とする請求項27の医薬品組成物。
【請求項33】
前記ガレクチン‐8タンパク質が、ガレクチン‐8N末端領域、並びにガレクチン‐8プロリン、グリシン、及びチロシンが豊富な領域を備えるタンパク質であることを特徴とする請求項27の医薬品組成物。
【請求項34】
前記ガレクチン‐8タンパク質が、ガレクチン‐8プロリン、グリシン及びチロシンが豊富な領域、並びにガレクチン‐8ガラクトシド結合領域を備えるタンパク質であることを特徴とする請求項27の医薬品組成物。
【請求項35】
前記ガレクチン‐8タンパク質が、ガレクチン‐8ガラクトシド結合領域を備えるタンパク質であることを特徴とする請求項27の医薬品組成物。
【請求項36】
前記実質的に同一であることが、配列番号4又は5のアミノ酸配列と少なくとも60%の同一性、少なくとも70%の同一性、少なくとも80%の同一性、又は少なくとも90%の同一性を有するものとすることを特徴とする請求項32の医薬品組成物。
【請求項37】
前記ガレクチン‐7タンパク質が、配列番号2のアミノ酸配列を備えることを特徴とする請求項27の医薬品組成物。
【請求項38】
前記ガレクチン‐7タンパク質が、配列番号2のアミノ酸配列と実質的に同一なアミノ酸配列を備えることを特徴とする請求項27の医薬品組成物。
【請求項39】
前記ガレクチン‐7タンパク質はガレクチン‐7ガラクトシド結合領域を備えることを特徴とする請求項27の医薬品組成物。
【請求項40】
前記実質的に同一であることが、配列番号2のアミノ酸配列と少なくとも60%の同一性、少なくとも70%の同一性、少なくとも80%の同一性、又は少なくとも90%の同一性を有するものとすることを特徴とする請求項38の医薬品組成物。
【請求項41】
前記ガレクチン‐3タンパク質が、配列番号1のアミノ酸配列を備えることを特徴とする請求項27の医薬品組成物。
【請求項42】
前記ガレクチン‐3タンパク質が、配列番号1のアミノ酸配列と実質的に同一なアミノ酸配列を備えることを特徴とする請求項27の医薬品組成物。
【請求項43】
前記実質的に同一であることが、配列番号1のアミノ酸配列と少なくとも60%の同一性、少なくとも70%の同一性、少なくとも80%の同一性、又は少なくとも90%の同一性を有するものとすることを特徴とする請求項42の医薬品組成物。
【請求項44】
前記ガレクチン‐3タンパク質が、ガレクチン‐3ガラクトシド結合領域を備えるタンパク質であることを特徴とする請求項27の医薬品組成物。
【請求項45】
前記ガレクチン‐3タンパク質が、ガレクチン‐3N末端領域、並びにガレクチン‐3プロリン、グリシン及びチロシンが豊富な領域を備えることを特徴とする請求項27の医薬品組成物。
【請求項46】
前記ガレクチン‐3タンパク質が、ガレクチン‐3プロリン、グリシン、及びチロシンが豊富な領域、並びに、ガレクチン‐3ガラクトシド結合領域を備えることを特徴とする請求項27の医薬品組成物。
【請求項47】
前記ガレクチン‐1タンパク質が、配列番号6のアミノ酸配列を備えることを特徴とする請求項27の医薬品組成物。
【請求項48】
前記ガレクチン‐1タンパク質が、配列番号6のアミノ酸配列と実質的に同一なアミノ酸配列を備えることを特徴とする請求項27の医薬品組成物。
【請求項49】
前記実質的に同一であることが、配列番号6のアミノ酸配列と少なくとも60%の同一性、少なくとも70%の同一性、少なくとも80%の同一性、又は少なくとも90%の同一性を有するものとすることを特徴とする請求項48の医薬品組成物。
【請求項50】
前記ガレクチン‐1タンパク質が、ガレクチン‐1N末端領域並びにガレクチン‐1プロリン、グリシン、及びチロシンが豊富な領域を備えることを特徴とする請求項27の医薬品組成物。
【請求項51】
前記ガレクチン‐1タンパク質が、ガレクチン‐1プロリン、グリシン、及びチロシンが豊富な領域、並びに、ガレクチン‐1ガラクトシド結合領域を備えることを特徴とする請求項27の医薬品組成物。
【請求項52】
前記ガレクチン‐1タンパク質が、ガレクチン‐1ガラクトシド結合領域を備えることを特徴とする請求項27の医薬品組成物。
【請求項53】
眼乾燥症の治療を必要とする哺乳類の眼乾燥を治療する方法であって、
治療に有効量の物質を哺乳類に投与する工程を備え、該物質は角膜表面又は結膜表面における涙液層の拡散に影響を与えることを特徴とする治療方法。
【請求項54】
前記物質が、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7及びガレクチン‐8からなる群から選択される少なくとも一のタンパク質であることを特徴とする請求項53記載の方法。
【請求項55】
眼乾燥症の治療を必要とする哺乳類の眼乾燥を治療する方法であって、
治療に有効量の物質を、眼乾燥症に苦しむ哺乳類に投与する工程を備え、
該物質は、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7及びガレクチン‐8からなる群から選択される少なくとも一のタンパク質の発現に影響を与えることを特徴とする治療方法。
【請求項56】
容器と、
薬学的に適切な担体又は希釈剤に含まれるガレクチン‐1、ガレクチン‐3、ガレクチン‐7及びガレクチン‐8からなる群から選択される少なくとも一のタンパク質、並びに使用説明書を備える眼乾燥を治療するためのキット。
【請求項57】
前記タンパク質が、一回分の投与量であることを特徴とする請求項56に記載のキット。
【請求項1】
眼乾燥の治療を必要とする哺乳類の眼乾燥を治療する方法であって、治療に有効な量のガレクチンタンパク質を哺乳類に投与することを特徴とする治療方法。
【請求項2】
投与前に、眼乾燥の予防を必要とする前記哺乳類を特定する工程を備えることを特徴とする請求項1に記載の治療方法。
【請求項3】
前記治療又は予防を必要とする前記哺乳類が、眼上皮創傷である哺乳類、抗ヒスタミン剤の使用前である哺乳類、抗炎症剤の使用前である哺乳類、及びエキシマーレーザー治療を施す前の哺乳類のうち少なくとも1つの哺乳類であることを特徴とする請求項1に記載の方法。
【請求項4】
前記哺乳類がヒトであることを特徴とする請求項1に記載の方法。
【請求項5】
前記ガレクチンタンパク質が、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7及びガレクチン‐8から選択されるタンパク質のうち少なくとも一つであることを特徴とする請求項1に記載の方法。
【請求項6】
前記眼乾燥が、持続性症候群であることを特徴とする請求項5記載の方法。
【請求項7】
前記眼乾燥が、上皮びらんを引き起こすことを特徴とする請求項5記載の方法。
【請求項8】
前記上皮びらんが、角膜創傷を引き起こすことを特徴とする請求項5記載の方法。
【請求項9】
前記ガレクチン‐8タンパク質が、配列番号4又は5のアミノ酸配列を備えることを特徴とする請求項5記載の方法。
【請求項10】
前記ガレクチン‐8タンパク質が、配列番号4又は5のアミノ酸配列と実質的に同一なアミノ酸配列であることを特徴とする請求項5記載の方法。
【請求項11】
前記実質的に同一であることが、配列番号4又は5のアミノ酸配列と少なくとも60%の同一性、少なくとも70%の同一性、少なくとも80%の同一性、又は、少なくとも90%の同一性を有するものとすることを特徴とする請求項10記載の方法。
【請求項12】
前記ガレクチン‐8タンパク質が、ガレクチン‐8‐N末端領域とガレクチン‐8プロリン、グリシン及びチロシンが豊富な領域、並びにガレクチン‐8ガラクトシド結合領域とを備えることを特徴とする請求項5記載の方法。
【請求項13】
前記ガレクチン‐3タンパク質が、ガレクチン‐3プロリン、グリシン、及びチロシンが豊富な領域、及びガレクチン‐3ガラクトシド結合領域を備えることを特徴とする請求項5記載の方法。
【請求項14】
前記ガレクチン‐7タンパク質が、ガレクチン‐7‐N末端領域とガレクチン‐7プロリン、グリシン及びチロシンが豊富な領域、並びにガレクチン‐7ガラクトシド結合領域とを備えることを特徴とする請求項5記載の方法。
【請求項15】
前記ガレクチン‐1タンパク質が、ガレクチン‐1‐N末端領域とガレクチン‐1プロリン、グリシン及びチロシンが豊富な領域、並びにガレクチン‐1ガラクトシド結合領域とを備えることを特徴とする請求項5記載の方法。
【請求項16】
前記ガレクチン‐7タンパク質が、配列番号2のアミノ酸配列を備えることを特徴とする請求項5記載の方法。
【請求項17】
前記ガレクチン‐7タンパク質が、配列番号2のアミノ酸配列と実質的に同一なアミノ酸配列を備えることを特徴とする請求項5記載の方法。
【請求項18】
前記実質的に同一であることが、配列番号2のアミノ酸配列と少なくとも70%同一性、少なくとも80%同一性、又は、少なくとも90%の同一性を有するものとすることを特徴とする請求項17記載の方法。
【請求項19】
前記ガレクチン‐3タンパク質が、配列番号1のアミノ酸配列を備えることを特徴とする請求項5記載の方法。
【請求項20】
前記ガレクチン‐3タンパク質が、配列番号1のアミノ酸配列と実質的に同一なアミノ酸配列を備えることを特徴とする請求項5記載の方法。
【請求項21】
前記実質的に同一であることが、配列番号1のアミノ酸配列と少なくとも70%同一性、少なくとも80%同一性、又は、少なくとも90%の同一性を有するものとすることを特徴とする請求項20記載の方法。
【請求項22】
前記ガレクチン‐3タンパク質が、ガレクチン‐3ガラクトシド結合領域を備えることを特徴とする請求項5記載の方法。
【請求項23】
前記ガレクチン‐1タンパク質が、配列番号6のアミノ酸配列を備えることを特徴とする請求項5記載の方法。
【請求項24】
前記ガレクチン‐1タンパク質が、配列番号6のアミノ酸配列と実質的に同一なアミノ酸配列を備えることを特徴とする請求項5記載の方法。
【請求項25】
前記実質的に同一が、配列番号6のアミノ酸配列と少なくとも60%同一性、少なくとも70%同一性、少なくとも80%同一性、又は、少なくとも90%の同一性を有するものとすることを特徴とする請求項24記載の方法。
【請求項26】
前記ガレクチン‐1タンパク質が、ガレクチン‐1ガラクトシド結合領域を備えることを特徴とする請求項5記載の方法。
【請求項27】
眼乾燥の治療の促進効果がある医薬品組成物であり、前記組成物は、薬学的に適当な担体又は希釈剤、並びにガレクチン‐1、ガレクチン‐3、ガレクチン‐7及びガレクチン‐8タンパク質のうち少なくとも1種のガレクチンを備え、該ガレクチンは結膜及び/又は角膜上皮の完全性を促進させる十分量である医薬品組成物。
【請求項28】
前記眼乾燥が、再発性上皮びらんを伴う眼乾燥症であることを特徴とする請求項27の医薬品組成物。
【請求項29】
前記眼乾燥症が、創傷を引き起こし、該創傷は角膜創傷であることを特徴とする請求項28の医薬品組成物。
【請求項30】
前記眼乾燥が、エクシマーレーザー角膜切除術により引き起こされることを特徴とする請求項28の医薬品組成物。
【請求項31】
前記ガレクチン‐8タンパク質が、配列番号4又は5のアミノ酸配列を備えるタンパク質であることを特徴とする請求項27の医薬品組成物。
【請求項32】
前記ガレクチン‐8タンパク質が、配列番号4又は5のアミノ酸配列と実質的に同一なアミノ酸配列を備えるタンパク質であることを特徴とする請求項27の医薬品組成物。
【請求項33】
前記ガレクチン‐8タンパク質が、ガレクチン‐8N末端領域、並びにガレクチン‐8プロリン、グリシン、及びチロシンが豊富な領域を備えるタンパク質であることを特徴とする請求項27の医薬品組成物。
【請求項34】
前記ガレクチン‐8タンパク質が、ガレクチン‐8プロリン、グリシン及びチロシンが豊富な領域、並びにガレクチン‐8ガラクトシド結合領域を備えるタンパク質であることを特徴とする請求項27の医薬品組成物。
【請求項35】
前記ガレクチン‐8タンパク質が、ガレクチン‐8ガラクトシド結合領域を備えるタンパク質であることを特徴とする請求項27の医薬品組成物。
【請求項36】
前記実質的に同一であることが、配列番号4又は5のアミノ酸配列と少なくとも60%の同一性、少なくとも70%の同一性、少なくとも80%の同一性、又は少なくとも90%の同一性を有するものとすることを特徴とする請求項32の医薬品組成物。
【請求項37】
前記ガレクチン‐7タンパク質が、配列番号2のアミノ酸配列を備えることを特徴とする請求項27の医薬品組成物。
【請求項38】
前記ガレクチン‐7タンパク質が、配列番号2のアミノ酸配列と実質的に同一なアミノ酸配列を備えることを特徴とする請求項27の医薬品組成物。
【請求項39】
前記ガレクチン‐7タンパク質はガレクチン‐7ガラクトシド結合領域を備えることを特徴とする請求項27の医薬品組成物。
【請求項40】
前記実質的に同一であることが、配列番号2のアミノ酸配列と少なくとも60%の同一性、少なくとも70%の同一性、少なくとも80%の同一性、又は少なくとも90%の同一性を有するものとすることを特徴とする請求項38の医薬品組成物。
【請求項41】
前記ガレクチン‐3タンパク質が、配列番号1のアミノ酸配列を備えることを特徴とする請求項27の医薬品組成物。
【請求項42】
前記ガレクチン‐3タンパク質が、配列番号1のアミノ酸配列と実質的に同一なアミノ酸配列を備えることを特徴とする請求項27の医薬品組成物。
【請求項43】
前記実質的に同一であることが、配列番号1のアミノ酸配列と少なくとも60%の同一性、少なくとも70%の同一性、少なくとも80%の同一性、又は少なくとも90%の同一性を有するものとすることを特徴とする請求項42の医薬品組成物。
【請求項44】
前記ガレクチン‐3タンパク質が、ガレクチン‐3ガラクトシド結合領域を備えるタンパク質であることを特徴とする請求項27の医薬品組成物。
【請求項45】
前記ガレクチン‐3タンパク質が、ガレクチン‐3N末端領域、並びにガレクチン‐3プロリン、グリシン及びチロシンが豊富な領域を備えることを特徴とする請求項27の医薬品組成物。
【請求項46】
前記ガレクチン‐3タンパク質が、ガレクチン‐3プロリン、グリシン、及びチロシンが豊富な領域、並びに、ガレクチン‐3ガラクトシド結合領域を備えることを特徴とする請求項27の医薬品組成物。
【請求項47】
前記ガレクチン‐1タンパク質が、配列番号6のアミノ酸配列を備えることを特徴とする請求項27の医薬品組成物。
【請求項48】
前記ガレクチン‐1タンパク質が、配列番号6のアミノ酸配列と実質的に同一なアミノ酸配列を備えることを特徴とする請求項27の医薬品組成物。
【請求項49】
前記実質的に同一であることが、配列番号6のアミノ酸配列と少なくとも60%の同一性、少なくとも70%の同一性、少なくとも80%の同一性、又は少なくとも90%の同一性を有するものとすることを特徴とする請求項48の医薬品組成物。
【請求項50】
前記ガレクチン‐1タンパク質が、ガレクチン‐1N末端領域並びにガレクチン‐1プロリン、グリシン、及びチロシンが豊富な領域を備えることを特徴とする請求項27の医薬品組成物。
【請求項51】
前記ガレクチン‐1タンパク質が、ガレクチン‐1プロリン、グリシン、及びチロシンが豊富な領域、並びに、ガレクチン‐1ガラクトシド結合領域を備えることを特徴とする請求項27の医薬品組成物。
【請求項52】
前記ガレクチン‐1タンパク質が、ガレクチン‐1ガラクトシド結合領域を備えることを特徴とする請求項27の医薬品組成物。
【請求項53】
眼乾燥症の治療を必要とする哺乳類の眼乾燥を治療する方法であって、
治療に有効量の物質を哺乳類に投与する工程を備え、該物質は角膜表面又は結膜表面における涙液層の拡散に影響を与えることを特徴とする治療方法。
【請求項54】
前記物質が、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7及びガレクチン‐8からなる群から選択される少なくとも一のタンパク質であることを特徴とする請求項53記載の方法。
【請求項55】
眼乾燥症の治療を必要とする哺乳類の眼乾燥を治療する方法であって、
治療に有効量の物質を、眼乾燥症に苦しむ哺乳類に投与する工程を備え、
該物質は、ガレクチン‐1、ガレクチン‐3、ガレクチン‐7及びガレクチン‐8からなる群から選択される少なくとも一のタンパク質の発現に影響を与えることを特徴とする治療方法。
【請求項56】
容器と、
薬学的に適切な担体又は希釈剤に含まれるガレクチン‐1、ガレクチン‐3、ガレクチン‐7及びガレクチン‐8からなる群から選択される少なくとも一のタンパク質、並びに使用説明書を備える眼乾燥を治療するためのキット。
【請求項57】
前記タンパク質が、一回分の投与量であることを特徴とする請求項56に記載のキット。
【図1】


【図2】

【図3】

【図3】

【図4】

【図5】

【図6】

【図7】

【図8】

【図9A】

【図9B】

【図10】
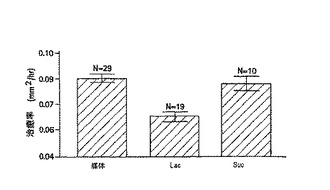
【図11】
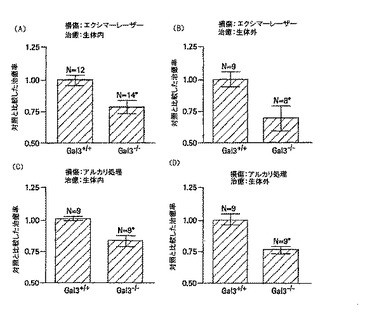
【図12】
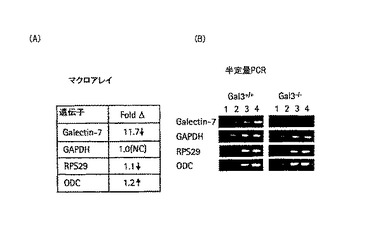
【図13】
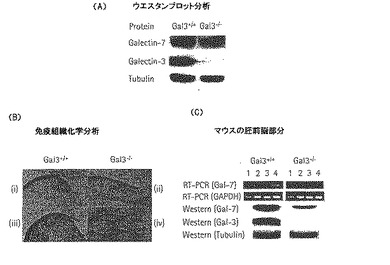
【図14】

【図15】

【図16】
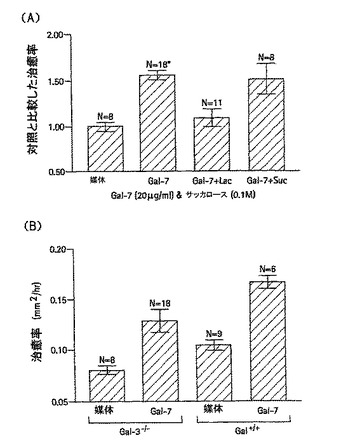
【図17】

【図18】

【図19】
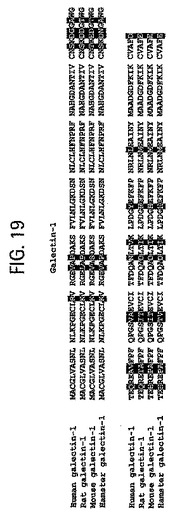
【図20】

【図21】
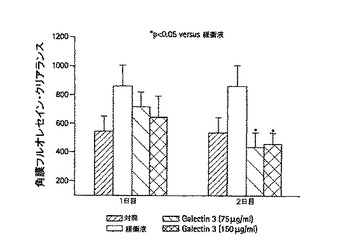
【図2】
【図3】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9A】
【図9B】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【公表番号】特表2008−536857(P2008−536857A)
【公表日】平成20年9月11日(2008.9.11)
【国際特許分類】
【出願番号】特願2008−506673(P2008−506673)
【出願日】平成18年4月13日(2006.4.13)
【国際出願番号】PCT/US2006/013785
【国際公開番号】WO2006/113311
【国際公開日】平成18年10月26日(2006.10.26)
【出願人】(507333649)タフツ ユニバーシティー (4)
【Fターム(参考)】
【公表日】平成20年9月11日(2008.9.11)
【国際特許分類】
【出願日】平成18年4月13日(2006.4.13)
【国際出願番号】PCT/US2006/013785
【国際公開番号】WO2006/113311
【国際公開日】平成18年10月26日(2006.10.26)
【出願人】(507333649)タフツ ユニバーシティー (4)
【Fターム(参考)】
[ Back to top ]