
細胞ベースの治療に関連する材料および方法
本発明は、組織再生および加齢性組織変化についての細胞変性と関連した疾患状態の処置のために使用され得る新規な細胞集団の提供に関する。上記細胞集団は、Thy1.1細胞マーカーに対して陽性もしくは陰性であることによって特徴付けられる成体幹細胞/前駆細胞に由来する。一実施形態において、本発明は、成体組織に由来する単離された細胞集団を提供し、ここで該細胞は、血清の存在下でマトリゲルなしの培養システムにおいて増殖でき;該細胞は、Pdx−1陽性であることで特徴付けられる。
【発明の詳細な説明】
【技術分野】
【0001】
(関連出願への相互参照)
この出願は、2008年5月9日に出願された、米国仮特許出願第61/052,098号と同時係属中であり、かつ少なくとも1人の発明者と共有され、かつそれへの優先権を主張する。先行する出願は、その全体が参考として本明細書に援用される。
【0002】
(発明の分野)
本発明は、組織再生、および細胞変性もしくは加齢性組織変化(age related tissue change)と関連する疾患状態の処置のために使用され得る新規な細胞集団の提供に関する。特に(しかし網羅的ではない)、本発明は、膵臓細胞タイプおよび肝細胞タイプの両方に分化するための両能性を示す新規な細胞集団の決定から生じる材料および方法を提供する。
【背景技術】
【0003】
(発明の背景)
幹細胞は、培養において長期間増殖する能力を有し、特定の細胞タイプになるために誘導され得る分化していない細胞である。幹細胞は、主に胚もしくは生体から単離され得るが、これらは、異なる特徴および機能を有するようである。
【0004】
幹細胞は、多くの哺乳動物種の胚(胚性/胚幹細胞−ESC)から単離されてきた。マウス由来のものは、20年間にわたって集中した研究の対象であり、単離されかつ1998年以来研究されてきたヒトESCの単離のための方法を可能にした。
【0005】
ESCは、代表的には、胚盤胞(embryonic blastocyst)に由来し、ここで胚盤胞は、インビボで全てのその後の発生細胞タイプを生じるように続く。成体幹細胞(ASC)は、他方で、多くの組織に存在し、そこでは、それらは、傷害および、または消耗(wear and tear)を介して失われた細胞の置換を可能にする。これら細胞は、疾患(例えば、糖尿病およびパーキンソン病)の処置のための細胞ベースの治療を提供する能力を有する。上記疾患では、特定の病理をもたらす細胞損失および損傷が存在する。これら細胞はまた、薬物のスクリーニング、毒物学的調査、発生プログラミングの調査、加齢性組織損失および変性の処置のための優れた能力を有する。これら細胞はまた、外科手術的除去後の、傷害後の、もしくは美容外科手順の一部としての組織再生の能力を有する。
【0006】
ASCは、成体組織もしくは器官中の分化した細胞の中で見いだされる未分化の細胞であり、ASCは、分化しておらず、そしてそれ自体を自分で補充する(self renew)し得、組織もしくは器官の主要な細胞タイプを得るように分化する能力を維持する。ASCは、組織修復を維持しかつもたらすと考えられる。成熟組織における成体幹細胞の起源は未知であり、それらの可塑性の程度は、未だ決定されていない。移植におけるそれらの用途は、広く知られている。骨髄由来のASCは、30年間にわたって移植で使用されてきた。成体非HSCの使用、それらの効力、可塑性および長期追跡での安全性は、未だ証明されていない。非間質性ASCの報告は、現場で討議されたままであるが、神経幹細胞は、今では確立されており、真正な(bone fide)ASCタイプとして受け入れられている。
【0007】
ESCの証明された多能性特徴およびESCを多数増殖させる能力は、ESCを細胞ベースの治療の魅力的な候補にしている。この状況においてASCの使用には厳しい制限が存在する。ここでこのような細胞は、非常に稀であると考えられておりかつそれらの増殖条件は、可能性のある治療に十分な数で適切な細胞を生成するほど十分に明確でない。
【0008】
しかし、ASCは、「自己」に由来し得るという点で、決定的な利点を確かに有しているので、いかなる患者でも、彼ら自身の細胞を受容し、顕著に高まった癌のリスクを含め、拒絶を防止するために免疫抑制の有害な副作用に苦しむ必要はない。制限された有効性/可塑性はまた、以上型の細胞分化が制限され、新生物のリスクが低減されるという点で、高められた安全性因子であると考えられる。
【0009】
肝臓および膵臓の共通する発生起源は、これら器官が、共通する幹細胞/前駆細胞集団を共有し得ることを示唆する。この過程の裏付けにおいてかなりの間接的証拠が存在する。外植片実験から、Pdx−1を発現する腹側内胚葉は、心臓中胚葉への近接によって、肝臓系統に流用されることが実証されている(非特許文献1)。肝細胞特性を有する細胞(Deutsch et al., 2001)はまた、マウス膵臓内の管領域において観察された。銅欠乏性食餌を与えられたマウスは、膵臓損傷、腺房細胞の損失を受け、4〜6週間後には、肝円形様細胞(hepatic oval−like cell)が観察される(非特許文献2;非特許文献3)。さらに、脾臓に移植された、銅欠乏性食餌に供されたラットから単離した膵臓細胞は、実質構造へと組み込まれかつ成熟肝臓特異的タンパク質を発現することによって、形態的にも機能的にも肝細胞への分化を示した(Dabevaら,1997)。フマリルアセトアセテートヒドロラーゼが欠損し、後にチロシン血症を伴う共通遺伝子のレシピエントに移植した野生型マウス膵臓細胞の懸濁物はまた、肝機能の正常化とともに、ドナー由来細胞によって生合成レスキューを生じることが報告された(Wangら,2001)。さらに、成体マウス膵臓におけるKGFのトランスジェニック過剰発現は、膵島内で肝細胞の出現および付随して膵管増殖を生じる(Krakowskiら,1999b;Krakowskiら,1999a)。
【0010】
いくつかの推定される膵臓前駆細胞が、膵管細胞および島細胞の両方から特徴付けられた(Abrahamら,2004;Corneliusら,1997;Lechnerら,2002;Ramiyaら,2000)。これら細胞集団のうちの一方は、インビトロでの分化後に、αフェトプロテインおよびc−Met(代表的には、肝細胞によって発現されるタンパク質)を発現することが観察された(Zulewskiら,2001)。(Zulewskiら, 2001)より近年になってからは、ヒト膵管上皮から単離された間葉系幹細胞集団は、膵臓分化、肝臓分化および中胚葉分化の能力を有することが報告された(Seebergerら,2006)。これら細胞は、Gata 4、アルブミンおよびTAT(チロシンアミノトランスフェラーゼ)を発現するように誘導され得るが、機能的評価は未だ報告されていない。
【0011】
本発明者は、以前に、さらなる細胞タイプである、STZ糖尿病モデルにおいて多能性および機能的効力を示す、成体ラット膵管から単離された膵臓由来開通細胞(Pancreatic Derived Pathfinder Cell)(PDPC)を決定した(Shiels 2004;および特許文献1(本明細書に参考として援用される))。成体ラット肝臓において、推定される幹細胞の集団は、化学的傷害、もしくは部分的肝切除術による誘導後に誘導され得る(Petersenら,1998;Petersenら,2003)(Petersenら, 1998; Petersenら, 2003)。
【先行技術文献】
【特許文献】
【0012】
【特許文献1】国際公開第2006/120476号
【非特許文献】
【0013】
【非特許文献1】Deutsch,G., J.Jung, M.Zheng, J.Lora and K.S.Zaret: A bipotential precursor population for pancreas and liver within the embryonic endoderm. Development Dev.(2001)128、871−881
【非特許文献2】Rao,M.S., V.Subbarao and J.K.Reddy: Induction of hepatocytes in the pancreas of copper−depleted rats following copper repletion. Cell Differ. Dev.(1986)18、109−117
【非特許文献3】Rao,M.S. and J.K.Reddy: Hepatic transdifferentiation in the pancreas. Semin.Cell Biol. Dev.(1995)6、151−156
【非特許文献4】Dabeva,M.D., S.G.Hwang, S.R.Vasa, E.Hurston, P.M.Novikoff, D.C.Hixson, S.Gupta and D.A.Shafritz: Differentiation of pancreatic epithelial progenitor cells into hepatocytes following transplantation into rat liver. Proc.Natl.Acad.Sci.U.S.A Dev.(1997)94、7356−7361
【発明の概要】
【課題を解決するための手段】
【0014】
(発明の要旨)
細胞変性と特に関連する疾患を処置するために、細胞ベースの治療において使用され得る細胞集団を提供することが継続して必要である。糖尿病は、このような疾患の一例である。膵臓におけるインスリンは、インスリン分泌β細胞によって作られる。インビボでは、β細胞ターンオーバーは、生涯全体を通じて起こると考えられているが、置換細胞の起源に関しては、論争がある。従って、糖尿病は、完全に機能するネイティブ細胞に類似した細胞の提供(特に、自己に由来する細胞)が処置の重要な選択肢を提供する疾患の一例である。糖尿病の場合には、インスリンを生成し得るという点で、β細胞に類似の細胞の提供は、上記疾患にとって重大な細胞ベースの治療を提供する。
【0015】
この疾患および他の疾患を考慮に入れて、本発明者は、成体膵管から肝細胞/前駆細胞集団に由来する細胞の亜集団を単離した。
【0016】
いくつかの実施形態において、本明細書で提供される細胞の亜集団は、Pdx−1(HUMAN: NP_000200.1 GI:4557673; RAT: NP_074043.3 GI:50838802)陽性である。Pdx−1発現の存在は、細胞が非β細胞由来インスリンの供給源としての能力を有することを示す。
【0017】
いくつかの実施形態において、本明細書で提供される成体幹細胞/前駆細胞の亜集団は、Thy1.1(CD90)(HUMAN: NP_006279)の存在もしくは非存在によって特徴付けられる。Thy1.1陽性亜集団およびThy1.1陰性亜集団の特性は、以下で議論される。
【0018】
(1.Thy1.1陽性細胞)
維持(非分化)培地中で、Thy1.1陽性細胞亜集団は、Pdx−1陽性、インスリン陰性およびグルカゴン陰性である。Thy1.1陽性細胞集団は、膵臓分化培地中に置かれた場合、最初に、線維芽細胞様形態を示し、次いで、親細胞層から最後には分離する絡み合った細胞クラスタを形成する。得られた分化した細胞クラスタは、Pdx−1、インスリンおよびグルカゴンについて陽性である(図4、パネルB)。
【0019】
本明細書に記載されるThy1.1陽性細胞亜集団に関する驚くべき決定は、それらが、少なくとも両能性のある(bipotent)ことである。具体的には、適切な分化培地とともに提供される場合、Thy1.1陽性細胞は、膵臓細胞タイプもしくは肝細胞タイプのいずれかに分化し得る。
【0020】
(2.Thy1.1陰性細胞)
非分化状態において、Thy1.1陰性細胞集団は、Pdx−1について陽性であるが、インスリンおよびグルカゴンについては陰性である。分化培地中で増殖される場合、Thy1.1陰性細胞は、形態学的変化を示さなかったが、インスリン転写が検出された。よって、Thy1.1陰性集団は、非β細胞由来インスリンの新規な供給源を提供する。
【0021】
よって、その最も一般には、本発明は、多能性成体幹細胞集団に由来する細胞の新規な亜集団を使用して、細胞ベースの治療に基づく加齢の疾患および状態を処置するための材料および方法を提供する。上記亜集団は、Pdx−1陽性である。
【0022】
このような細胞集団は、糖尿病のような疾患およびパーキンソン病のような神経変性障害の細胞ベースの治療に対する能力を提供する。
【0023】
成体幹細胞集団(前駆細胞としても公知)は、成体膵臓組織(例えば、ヒト、ラット、マウス、霊長類、ブタなど)に由来し得る。上記成体組織は、好ましくは、膵臓組織であるが、乳房、骨髄、心臓、肝臓もしくは腎臓のような他の器官に由来する組織であってもよい。
【0024】
本発明の一実施形態において、成体幹細胞は成体ラット膵臓に由来し、これは、1977年のブダペスト条約に従って、University of GlasgowのThe University Courtによって、Cell Cultures,Porton Down,Salisbury Wiltshire,UK,SP4 0JGのThe European Collectionにおいて、2005年5月12日にECACC番号Q6203の下で寄託された。これら細胞は、本明細書中以降、PDPC(膵臓由来前駆細胞)として知られる。
【0025】
上記のように、本発明者はまた、マーカーThy1.1(CD90)(HUMAN: NP_006279)を参照して、2つのさらなる亜集団を決定した。これら2つの亜集団は異なるが、等しく重要な特性を有する。特に、Thy1.1陽性細胞は、膵臓細胞タイプおよび肝細胞タイプの両方への分化能力を示す。いくつかの実施形態において、本明細書で記載される細胞亜集団は、インスリンの非β細胞供給源を提供する。いくつかの実施形態において、本明細書で記載される細胞亜集団は、試験物質の毒性を評価するための細胞タイプを提供する(例えば、毒物学試験のために)。
【0026】
よって、第1の局面において、成体組織から発生する幹細胞/前駆細胞の集団を提供する。ここで上記細胞は、血清の存在下でマトリゲルなしの培養システムにおいて増殖でき、上記細胞は、Thy1.1陽性である。いくつかの実施形態において、Thy1.1陽性細胞はまた、Pdx−1陽性である。いくつかの実施形態において、Thy1.1陽性細胞は、ネスチン(RAT:NP_037119.1 GI:6981262; HUMAN: NP_006608.1 GI:38176300)陽性である。
【0027】
いくつかの実施形態において、上記集団中のネスチン陽性細胞のパーセンテージは、フローサイトメトリー分析によって決定される場合、50%未満(例えば、40%、30%、20%、10%、5%、2%もしくは1%未満)である。いくつかの実施形態において、細胞は、例えば、PCRによって、ネスチン核酸の発現によって、ネスチン陽性細胞として区別可能である。
【0028】
細胞集団は、Thy1.1陽性(CD90陽性)(RAT:LOCUS:P01830 GI135832; NP_006279 GI:19923362)として見なされ得る。ここでThy1.1陽性細胞のパーセンテージは、50%より大きく、好ましくは、60%より大きく、より好ましくは、70%、80%、90%、もしくは95%より大きい。いくつかの実施形態において、Thy1.1陽性細胞集団は、純度が98%より高い。Thy1.1陽性細胞のパーセンテージは、例えば、フローサイトメトリーによって、もしくはPCRによって決定され得る。
【0029】
いくつかの実施形態において、本発明に従うThy1.1陽性細胞集団は、Pdx−1、CD49f、CD147、CD44、c−Met、およびネスチンのうちの1種以上の発現について陽性である。いくつかの実施形態において、Thy1.1陽性細胞集団は、CD24、CD45、CD31、c−kit、およびCK19のうちの1種以上の発現について陰性である。いくつかの実施形態において、本発明に従うThy1.1陽性細胞集団は、以下の細胞表面マーカープロフィールを有する:
Thy1.1(CD90) 陽性
Pdx−1 陽性
CD49f 約95%+
CD24 陰性
CD147 約90%+
CD45 陰性
CD44 約85%+
CD71 低い
CD31 陰性
C−KIT 陰性
CK19 陰性
c−Met 陽性
ネスチン 陽性
(低い=細胞のうちの約5%以下が上記マーカーを発現する)。
【0030】
本発明はまた、本発明のこの局面に従う細胞集団とともに、薬学的に受容可能なキャリアを含む薬学的組成物を提供する。
【0031】
本発明の第2の局面において、成体哺乳動物組織から単離された両能性のある幹細胞集団を生成するための方法が提供され、上記方法は、上記成体哺乳動物組織を培養する工程;発生する細胞集団単層を得る工程;およびThy1.1について陽性である細胞を含む亜集団を単離する工程を包含する。いくつかの実施形態において、上記亜集団のうちの少なくとも50%、60%、70%、80%、90%、95%が、Thy1.1陽性である。
【0032】
上記方法はまた、上記で提供されるプロフィール(例えば、Pdx−1、CD49f、CD147、CD44、c−Met、およびネスチンのうちの1種以上について陽性ならびに/またはCD24、CD45、CD31、c−kit、およびCK19のうちの1種以上について陰性)中に提供される1種以上の他の細胞表面マーカーと組み合わせて、Thy1.1について陽性樽細胞を単離する工程を包含し得る。
【0033】
いくつかの実施形態において、上記成体哺乳動物組織は、膵臓であり、例えば、膵管由来である。いくつかの実施形態において、上記成体哺乳動物組織は、乳房、肝臓もしくは腎臓である。いくつかの実施形態において、上記成体哺乳動物組織は、ヒト組織である。
【0034】
発生する細胞集団単層を得るために成体哺乳動物組織を得る代わりに、方法は、既に単離された成体幹細胞(例えば、2005年5月12日に、アクセッション番号Q6203の下でECACCに寄託されたもの)を得ることを包含し得る。
【0035】
第3の局面において、培養において幹細胞の集団を生成するための方法が提供され、上記方法は、本発明に従うThy1.1陽性細胞集団を、肝臓系統分化に適した培地中で培養する工程を包含する。一例として(他は、当業者に公知である)、上記培養培地は、無血清FGF−4含有分化培地であり得る。
【0036】
第4の局面において、培養において膵臓細胞の集団を生成するための方法が提供され、上記方法は、本発明に従うThy1.1陽性細胞集団を、膵臓系統分化に適した培地中で培養する工程を包含する。
【0037】
本発明は、本明細書で記載される方法から得られるもしくは得ることができる細胞および細胞集団に及ぶ。
【0038】
本発明の第5の局面において、細胞変性もしくは加齢性組織変化と関連する疾患状態を処置するための方法が提供され、上記方法は、本発明に従うThy1.1陽性成体幹細胞集団もしくは上記Thy1.1陽性細胞集団を含む薬学的組成物を、上記疾患もしくは加齢性状態を有する患者に投与する工程を包含する。
【0039】
Thy1.1陽性細胞集団は、静脈内に投与され得るか、または疾患部位に移植され得る。
【0040】
いくつかの実施形態において、上記疾患は、膵臓細胞、神経細胞、心血管細胞(例えば、心筋細胞)、上皮細胞、肝細胞、もしくは腎細胞の変性と関連する。
【0041】
上記処置されるべき疾患状態としては、糖尿病(I型およびII型)、肝臓疾患、腎臓疾患、眼の疾患、パーキンソン病および心血管疾患ならびに身体の器官および組織の加齢性変性状態が挙げられ得る。本発明のこの局面はまた、美容外科手術の形態として(例えば、組織の細胞再生および加齢の形態を妨げるため)使用され得る。
【0042】
いくつかの実施形態において、上記細胞のドナーおよび上記レシピエントは、同じ種である(例えば、両方ともヒト)。いくつかの実施形態において、上記細胞のドナーおよび上記レシピエントは、異なる種である。よって、本発明の一実施形態は、ヒト患者の処置における、ラットに由来する成体幹細胞集団の使用を包含する。
【0043】
本発明の第6の局面において、指定された分化した細胞集団(例えば、膵臓細胞もしくは肝細胞)を生成するための方法が提供され、上記方法は、成体幹細胞集団を提供する工程;Pdx−1および/もしくはThy1.1マーカー、ならびに必要に応じて、Thy1.1亜集団細胞表面マーカープロフィールに関連して本明細書で同定される1種以上の他のマーカーを使用して、細胞亜集団を選択する工程;ならびに細胞分化の助けとなる条件下で上記細胞亜集団を培養する工程を包含する。
【0044】
本発明は、美容外科手術を含め、医療処置の方法において使用するための本発明の第1の局面に従う細胞集団をさらに提供する。上記方法は、細胞損失もしくは変性(例えば、糖尿病もしくはパーキンソン病)と関連する疾患状態もしくは加齢の状態を処置し得る。
【0045】
本発明の第6の局面において、成体組織から発生する細胞集団が提供され、ここで上記細胞は、血清の存在下でマトリゲルなしの培養システムにおいて増殖でき、上記細胞は、Thy1.1陰性である。
【0046】
いくつかの実施形態において、上記細胞集団は、Pdx−1陽性である。上記細胞集団はまた、ネスチン陽性であり得る。
【0047】
本発明はまた、成体幹Thy1.1陰性細胞集団を、薬学的に受容可能なキャリアとともに含む薬学的組成物を提供する。
【0048】
本発明の第7の局面において、成体哺乳動物組織から単離された幹細胞集団を生成するための方法が提供され、上記方法は、上記成体哺乳動物組織を培養する工程;発生する細胞集団単層を得る工程;およびThy1.1について陰性の細胞の亜集団を単離する工程を包含する。一実施形態において、上記方法は、Pdx−1陽性および/もしくはネスチン陽性でもある細胞の亜集団を単離する工程をさらに包含する。
【0049】
一実施形態において、Thy1.1陰性細胞の亜集団は、CD49f、CD24、CD147、CD44、c−Metのうちの1種以上の発現について陽性であり、そして/またはCD31、c−kit、およびck7のうちの1種以上の発現について陰性である。一実施形態において、以下の細胞表面マーカープロフィールであるThy1.1陰性細胞の亜集団が単離される:
Thy1.1(CD90) 陰性
Pdx−1 陽性
CD49f 約95%+
CD24 約80%+
CD147 約80%+
CD45 陰性
CD44 約60%+
CD71 低い
CD31 陰性
c−KIT 陰性
ck7 陰性
CK19 弱陽性
c−Met 陽性
(低い=細胞のうちの5%以下がそのマーカーを発現する)。
【0050】
発生する細胞集団単層を得るために成体哺乳動物組織を得る代わりに、上記方法は、既に単離された成体幹細胞/前駆細胞(例えば、2005年5月12日にアクセッション番号Q6203の下でECACCに寄託されたもの)を得る工程を包含し得る。
【0051】
本発明の第8の局面において、糖尿病もしくはインスリン生成の低下と関連する疾患状態を処置するための方法が提供され、上記方法は、本発明に従うThy1.1陰性細胞集団、もしくは上記Thy1.1陰性細胞集団を含む薬学的組成物を、上記疾患もしくは加齢性状態を有する患者に投与する工程を包含する。
【0052】
Thy1.1陰性細胞集団は、静脈内投与され得るか、または上記疾患部位に移植され得る。
【0053】
いくつかの実施形態において、上記細胞のドナーおよび上記レシピエントは、同じ種(例えば、ヒト)である。いくつかの実施形態において、上記細胞および上記レシピエントは、異なる種である。よって、本発明の一実施形態は、ヒト患者の処置における成体ラット幹細胞/前駆細胞の使用を包含する。
【0054】
本発明の第9の局面において、インスリンを生成することができる指定された分化した細胞集団を生成するための方法が提供され、上記方法は、成体幹細胞集団を提供する工程;Pdx−1について陽性でありかつThy1.1マーカーについて陰性である成体幹細胞亜集団を選択する工程;ならびに細胞分化の助けとなる条件下で、上記細胞亜集団を培養する工程を包含する。上記方法は、Thy1.1細胞表面マーカープロフィールの一部として上記で同定された1種以上のさらなるマーカーに基づいて、上記成体幹細胞集団を選択する工程をさらに包含し得る。
【0055】
本発明は、医療処置の方法において使用するための、本発明に従う成体幹細胞Thy1.1陰性細胞集団をさらに提供する。特に、上記方法は、糖尿病を処置することであり得る。
【0056】
インスリンを生成するための方法もまた、本明細書で提供される。インスリンを生成するための方法は、本明細書に記載されるThy1.1陰性のPdx−1陽性細胞の集団(例えば、成体組織(例えば、成体膵臓組織)に由来するThy1.1陰性のPdx−1陽性細胞の集団)を、インスリンが生成される条件下で培養する工程を包含し得る。上記方法は、上記培養物からインスリンを単離する工程をさらに包含し得る。いくつかの実施形態において、上記Thy1.1陰性細胞は、CD49f、CD24、CD147、CD44、c−Metのうちの1種以上の発現について陽性であり、そして/またはCD31、c−kit、およびck7のうちの1種以上の発現について陰性である。
【0057】
別の実施形態において、インスリンを生成するための方法は、本明細書に記載されるThy1.1陽性のPdx−1陽性細胞の集団(例えば、成体組織(例えば、成体膵臓組織)に由来する分化したThy1.1陽性のPdx−1陽性細胞の集団)を、インスリンが生成される条件下で培養する工程を包含する。上記方法は、インスリンを上記培養物から単離する工程をさらに包含し得る。いくつかの実施形態において、Thy1.1陽性細胞集団は、Pdx−1、CD49f、CD147、CD44、c−Met、およびネスチンのうちの1種以上の発現について陽性であり、そして/またはCD24、CD45、CD31、c−kit、およびCK19のうちの1種以上の発現について陰性である。
【0058】
本発明の局面および実施形態は、ここで添付の図面を参照しながら、例示によって示される。さらなる局面および実施形態は、当業者に明らかである。本文において言及される全ての文書は、本明細書に参考として援用される。
【図面の簡単な説明】
【0059】
【図1】図1は、未分化のThy1.1陽性集団およびThy1.1陰性集団のインビトロでの形態である。Thy1.1陽性PDPC集団およびThy1.1陰性PDPC集団のインビトロでの肝臓分化および膵臓分化(図1A〜C):形態学的変化は、分化の間にThy1.1陰性集団においてほとんど認められない。(図1A)未分化のThy1.1陰性PDPC。(図1B)膵臓分化誘導後28日目のThy1.1陰性PDPC。(図1C)肝臓分化誘導後28日目のThy1.1陰性PDPC。(図1D-I):Thy1.1陽性PDPCの肝臓分化の誘導に際して、顕著な形態学的変化が認められた。(図1D)未分化のThy1.1陽性PDPC(線維芽細胞様形態)。(図1E)肝臓分化誘導後14日目のThy1陽性PDPC。(図1F)肝臓分化誘導後28日目のThy1.1陽性PDPC−管腔構造が優勢な上皮形態が存在する。(図1G)肝臓誘導後28日目のthy1陽性PDPCの立方形の形態。(図1H)膵臓誘導後14日目のThy1.1陽性PDPC。(図1I)膵臓誘導後28日目のThy1.1陽性PDPC−島様クラスタが形成され、その後、培地へと分離する。
【図2】図2は、フローサイトメトリーによってMACSソートしたPDPCの細胞表面特徴付けである。(図2A)Thy1(CD90)陽性集団は、CD24、CD31、CD45、c−kitの発現について陰性であり、CD71については低いが、CD147、CD44およびCD49fの発現については陽性である。(図2B)Thy1(CD90)陰性集団は、CD31、CD45、c−kitの発現については陰性であり、CD71については低いが、CD24、CD147、CD44およびCD49fの発現については陽性である。
【図3】図3は、Thy1.1ソートしたPDPC集団の免疫細胞化学である。陽性コントロールは、アルブミン(図3A)、サイトケラチン7(図3E)、ビメンチン(図3I)およびサイトケラチン19(図3M)について示される。上記未分化のThy1.1陰性PDPC集団は、アルブミン(図3B)、サイトケラチン7(図3F)、ビメンチン(図3J)について陰性染色を示すが、サイトケラチン19(図3N)については弱い染色を示す。上記未分化のThy1.1陽性集団は、アルブミン(図3C)、サイトケラチン7(図3G)、ビメンチン(図3K)およびサイトケラチン19(図3O)について陰性であった。しかし、肝臓分化培地中での14日目までに、Thy1.1陽性PDPCは、アルブミンについて陽性であり(図3D)、管腔様領域においては、ビメンチン(図3L)およびサイトケラチン19(図3P)について陽性であったが、サイトケラチン7は、全体を通じて陰性のままであった(図3H)。
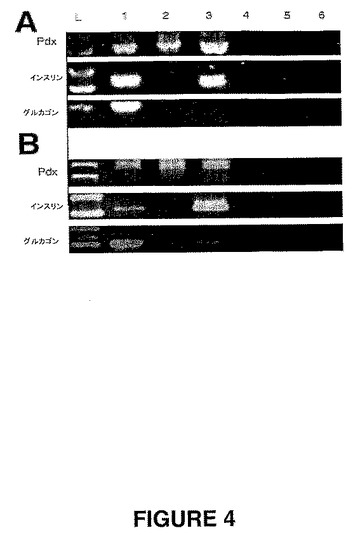
【図4】図4は、膵臓分化培地中のThy1.1陽性PDPC(図4A)およびThy1.1陰性PDPC(図4B)のRT−PCR。Thy1.1陰性PDPCおよびThy1.1陽性PDPCを、膵臓分化培地中で28日間にわたって培養した。列:(1)陽性コントロール、(2)未分化のPDPC、(3)28日目の分化したPDPC、(4〜6) サンプル1〜3のRTなし(no RT)コントロール。
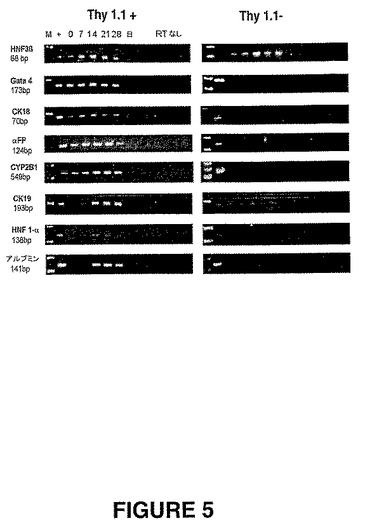
【図5】図5は、肝臓分化培地中のThy1.1陽性PDPCおよびThy1.1陰性PDPCのRT−PCRである。Thy1.1陽性PDPC(図5A)およびThy1.1陰性PDPC(図5B)を、肝臓分化培地中で28日間培養した。列:(1)陽性コントロール、(2)未分化のPDPC、(3)7日目、(4)14日目、(5)21日目、(6)28日目、(7〜13)サンプル1〜6のRTなしコントロール。アルブミン、CK19およびHNF1αの発現は、上記Thy1.1陽性集団において誘導されたが、上記Thy1.1陰性集団においては認められなかった。
【図6】図6は、未分化のThy1.1陽性PDPC集団(図6B)もしくはThy1.1陰性PDPC集団(図6C)がグリコーゲンを貯蔵しないことを示す。FGF−4含有培地中で培養した後のThy1.1陽性PDPCは、グリコーゲンを生成および貯蔵する(図6DおよびE)。グリコーゲン貯蔵は、過ヨウ素酸−シッフによって染色される場合に、赤紫色の染色の蓄積として認められる。FGF−4含有肝臓分化培地中で培養した後のThy1.1陰性PDPCは、染色を示さない(図6F)。陽性コントロール(図6A)。
【発明を実施するための形態】
【0060】
(詳細な説明)
(材料および方法)
(ラットPDPCの単離および維持培養)
膵管を、切開することによって12ヶ月齢のAlbino Swiss(Glasgow)ラットから単離し、CMRL培地中に播種する前に切り刻んだ。上記PDP細胞は、培養において約5週間後にコンフルエントな単層として発生した。次いで、これらを採取し、PBS中で洗浄した。PDPCを、0.2μm フィルターキャップ付きのT75培養フラスコ(Corning,UK)において、37℃で5% CO2雰囲気中、20mlのCMRL 1066培地(Invitrogen,Paisley,UK.)(10% ウシ胎仔血清(Sigma,Poole,UK)、2mM glutamax、1.25μg/ml アンホテリシンB、および100u/ml ペニシリン、100μg/ml ストレプトマイシン(全てInvitrogen,Paisley,UK)を補充)中の培養において維持した。コンフルエント未満の培養物を、ピペットでの培養培地の完全な除去、ならびに10mlのカルシウムおよびマグネシウム非含有ハンクス平衡塩類溶液(HBSS)(Cambrex Bio−Science,Wokingham,UK)を上記フラスコに室温で5分間添加することによる、接着性細胞の洗浄することによって継代した。上記HBSSをピペットで上記フラスコから除去した後、2mlのトリプシン−Versene溶液(200mg/L Versene、500mg/L トリプシン)を、上記フラスコに添加した。上記フラスコを、上記細胞単層の分離が確認されうるまで、顕微鏡で定期的に観察した。次いで、細胞を、ピペットで取り出し、望ましい場合、1/5〜1/10の密度で20mlの新鮮な培養培地を添加して上記のように再培養した。
【0061】
PDPCを、0.2μm フィルターキャップ付きのT75において、37℃で5% CO2雰囲気中、CMRL 1066培地(Invitrogen,Paisley,UK)(5% ウシ胎仔血清(Sigma,Poole,UK)、2mM Glutamax、1.25μg/ml アンホテリシンB、および100u/ml ペニシリン/ストレプトマイシン(全てInvitrogen,Paisley)を補充)中で長期間維持した。PDPCを、37℃で加湿5% CO2雰囲気中、単層として増殖させ、90% コンフルエントになったときに、トリプシン−EDTA(Invitrogen)を用いて継代した。細胞を計数し、3300細胞/cm2の密度で再度プレートした。
【0062】
(磁性活性化細胞ソーティング)
磁性活性化細胞ソーティング(MACS)を、製造業者のプロトコル(Dynabeads ヤギ抗マウスIgG(Dynal Biotech))に従って、106標的細胞あたり1μgの一次抗体(マウス抗ラットThy1.1(CD90)(Serotec))を使用して、4℃において20分間にわたって、単離および除去のために行った。ソートした細胞集団を、維持培養培地中で再懸濁し、組織培養フラスコに再度プレートした。MACSを、実験において使用する前に、各陽性ソートした集団および陰性ソートした集団に対して2回行った。全てのソートした集団を、後の分化実験の前に、蛍光活性化フローサイトメトリーでチェックした。
【0063】
(細胞表面抗原のフローサイトメトリー評価)
細胞を、HBSS中の0.5% BSAにおいて再懸濁した。次いで、それらを1000×rpmで10分間にわたって遠心分離し、得られた細胞ペレットを、HBBS中に再懸濁する。トリパンブルー(Invitrogen,Paisley,UK)で生存性係数した後、1×106細胞/mlを、100μl 一次抗体で標識した。使用した一次抗体は、CD90に対するもの(Serotec MCA47R,1:75)、CD44に対するもの(Serotec MCA643,1:10)、CD49fに対するもの(Serotec MCA2034,1:50)、CD147に対するもの(Serotec MCA729,1:10)、c−KITに対するもの(Santa Cruz SC−19983,1:20)、CD71に対するもの(Serotec MCA 155FT,1:10)、CD24に対するもの(BD Biosciences 551133,1:50)、CD45に対するもの(BD Biosciences 554875,1:50)、CD31に対するもの(Serotec MCA1334GA,1:50)およびCD34に対するもの(Santa Cruz sc−7324,1:50)であった。2次抗体を、0.2% BSA/PBS中、遮光して、4℃において45分間にわたって添加した。次いで、上記細胞を洗浄し、0.2% BSA中のウサギ抗マウス免疫グロブリン(Dako Cytomation,Ely,UK 1:20)の100μl FITC結合体化Fab2フラグメントを遮光して4℃で45分間にわたって添加することによって標識する前に、1000×rpmで、0.2% BSA/PBS中、3回遠心分離した。アイソタイプFITCコントロールもまた行った。前述のように、3回洗浄し、遠心分離した後、得られた細胞ペレットを、1mlのHBSS中で再懸濁し、上記細胞を、Beckman Coulter XLフローサイトメーター(Beckman Coulter,High Wycombe,UK)を使用して分析した。
【0064】
(分化実験)
膵臓分化のために、Thy1.1陽性細胞集団およびThy1.1陰性細胞集団(継代30)を、6600細胞/cm2の細胞密度においてプレートした。24時間後、維持培地を除去し、単層をHBSSで3回洗浄した。引き続き、細胞をDMEM:F12(Lonza)(1×ITS、1.25μg/ml アンホテリシンB、および100μ/ml ペニシリン/ストレプトマイシン(全てInvitrogen,UK)、ニコチンアミド 10mM(Sigma)、KGF 10ng/ml(Sigma)および0.2% BSA(Sigma)を補充)中で培養した。
【0065】
肝性分化のために、細胞を、T75および6ウェルプレート中、6600細胞/cm2においてプレートし、24時間維持で、チャンバースライド(Nunc)中2500細胞/cm2において、HBSS、DMEM:F12(Lonza)(線維芽細胞増殖因子−4 10ng/ml(Sigma)、1×ITS、100μ/ml ペニシリン/ストレプトマイシン(Invitrogen)および0.2% ウシ血清アルブミン(Sigma)を補充)で3回洗浄した後に、置換した。培地好感を1週間に3回行い、細胞を、膵臓分化については0日目および28日目に、および肝臓分化については0日目、7日目、14日目、21日目および28日目に、未分化のThy1.1陽性細胞およびThy1.1陰性細胞からのRNA抽出のために採取した。チャンバースライドにおいて肝臓分化を受けている細胞を、PBSで3回洗浄し、10〜14日の間で4% パラホルムアルデヒドで室温において15分間固定した。未分化のThy1.1陽性集団およびThy1.1陰性集団を、同時にチャンバースライド中でも増殖させ、上記のように、90%コンフルエントにおいて固定した。
【0066】
(免疫蛍光)
細胞内タンパク質の染色のために、細胞を上記のように固定した。細胞をPBS中で3回洗浄し、0.1% Triton X−100(Sigma−Aldrich)で10分間にわたって透過性にした。スライドを、ロバ血清とともに20分間にわたってインキュベートし、0.5%BSA/PBS中で希釈した以前に最適化した、ラットアルブミンに対する一次抗体(Abcam ab14255,1:100)、CK19に対する一次抗体(Biodesign Int. M08029M,1:100)、CK7に対する一次抗体(Chemicon MAB3226,1:100)、CK18に対する一次抗体(Sigma F−4772)およびビメンチンに対する一次抗体(Abcam ab8979,1:50)と1時間にわたってインキュベートした。スライドを、PBS中で3回洗浄し、続いて、適切なFITC標識2次抗体(Abcam ab6749もしくはDako Cytomation F0313,Ely,UK)で洗浄した。上記一次抗体の省略を、陰性コントロールとして行った。ラット肝臓の凍結切片を、陽性コントロールとして使用した。スライドを、Vectashield中にマウントする前に3回洗浄し、そして蛍光顕微鏡によって可視化し、撮影した。
【0067】
(RT−PCR)
総RNAを、製造業者の説明書に従ってTrizol(登録商標)を使用することによって抽出し、GeneQuant分析器によって定量した。サンプルを、DNAse処理(Ambion)し、cDNAへの逆転写を、製造業者の説明書に従って、SuperScript II逆転写酵素(Invitrogen)を使用して行った。RTなし陰性コントロールを、全てのサンプルについて行った。RT−PCRを、Taqポリメラーゼ(Invitrogen)を使用して行った。ハウスキーピング遺伝子であるBactinを使用して、テンプレートの品質を評価した。全てのPCR反応を、Peltier Thermal Cycler −200を使用して行った。ネスト化PCRを、膵臓分化実験について行った。
【0068】
以下の特異的オリゴヌクレオチドプライマーに、PDX 1、インスリンIIおよびグルカゴンを使用した(PDX−1,順方向 5−cggccacacagctctacaagg−3(配列番号1)、逆方向 5−ctccggttctgctgcgtatgc−3(配列番号2)、ネスト化逆方向 5−ttccaggcccccagtctcgg−3(配列番号3)(305bp)、インスリン,順方向 5−atggccctgtggatccgctt−3(配列番号4);逆方向 5−tgccaaggtctgaaggtcac−3(配列番号5);ネスト化順方向 5−cctgctcatcctctgggagcc−3(配列番号6)(209bp);グルカゴン,順方向 5−gaccgtttacgtggctgg−3(配列番号7);逆方向 5−cggttcctcttggtgttcatcaag−3(配列番号8);ネスト化順方向 5−acaaggcagctggcagcatgc−3(配列番号9)(210bp)。ラット膵臓総RNAを逆転写し、陽性コントロールとして使用した。以下の特異的オリゴヌクレオチドプライマーを、肝臓分化実験のために使用した:アルブミン(141bp) 順方向 5-ctgggagtgtgcagatatcagagt−3(配列番号10),逆方向 5−gagaaggtcaccaagtgctgtagt−3(配列番号11)、HNF3β(63bp) 順方向 5−cctactcgtacatctcgctcatca−3(配列番号12),逆方向 −cgctcagcgtcagcatctt(配列番号13)、HNF1(138bp)α 順方向 5−agctgctcctccatcatcaga−3(配列番号14),逆方向 5−tgttccaagcattaagttttctattctaa−3(配列番号15)、Gata4(173bp) 順方向 5−catgcttgcagttgtgctag−3(配列番号16),逆方向 5−attctctgctacggccagta−3(配列番号17)、α−フェトプロテイン(124bp) 順方向 5−gtcctttcttcctcctggagat−3(配列番号18),逆方向 5−ctgtcactgctgatttctctgg−3(配列番号19)、CYP2B1(549bp) 順方向 5−gagttcttctctgggttcctg−3(配列番号20),逆方向 5−actgtgggtcatggagagct−3(配列番号21)、CK19(193bp) 順方向 5−agtaacgtgcgtgctgacac−3(配列番号22),逆方向 5−agtcgcactggtagcaaggt−3(配列番号23)、CK18(70bp) 順方向 5ggacctcagcaagatcatggc−3(配列番号24),逆方向 5 ccacgatcttacgggtagttg−3(配列番号25)。次いで、上記PCR産物に、アガロースゲル電気泳動をかけて、エチジウムブロミド染色で可視化した。ラット肝臓組織を、陽性コントロールとして使用した。
【0069】
(過ヨウ素酸シッフ染色)
グリコーゲン貯蔵についての過ヨウ素酸シッフ染色を、未分化のThy1.1陽性細胞およびThy1.1陰性細胞、ならびに肝臓分化21日目のThy1.1陽性集団およびThy1.1陰性集団に対して行った。ヒト肝臓切片を、陽性コントロールとして使用した。細胞を、4% パラホルムアルデヒド中、室温で10分間にわたって固定した。細胞をPBS中で3回洗浄し、0.1% Triton X−100(Sigma−Aldrich)で10分間にわたって透過性にし、PBSで2回およびddH2Oで1回洗浄した。細胞を、過ヨウ素酸溶液(1g/dL)に室温で5分間浸漬した。ウェルを、蒸留水で3回すすいだ。細胞を、シッフ試薬中に室温で15分間浸漬した。細胞を、水道の流水で5分間洗浄した。細胞をヘマトキシリン溶液で90秒間、対比染色した。水道の流水で15〜30秒間、細胞をすすぐ。
【0070】
(結果)
(Thy1.1陽性集団およびThy1.1陰性集団の特徴付け)
PDPCのMACソーティングを使用して、それぞれ、Thy1.1陽性細胞を発現する集団を98.5%超で、およびThy1.1陰性細胞を発現する集団を98.7%純度で単離した。次いで、これら集団を培養し、いかなる分化実験もしくは特徴付け実験の前にも、10〜12日ごとに定期的にフローサイトメトリーによって再評価した。
【0071】
表現型的には、Thy1.1陽性集団およびThy1.1陰性集団は、形態学において別個の差異を示した:上記Thy1.1陽性集団は、線維芽細胞様形態を示した(図1,パネルA)一方で、Thy1.1陰性集団は、より上皮様の形態を示した(図1,パネルD)。
【0072】
いくつかの差異がまた、Thy1.1陽性細胞集団と、Thy1.1陰性細胞集団との間で、細胞表面マーカーの発現において認められた。両方の細胞系統は、CD147、CD44およびCD49fを発現した。両方とも、CD71は低く、造血(haematopoetic)マーカーであるCD31、CD34、CD45およびc−kitを発現しなかった。上記Thy1.1陽性ソートした細胞集団とは対照的に、上記Thy1.1陰性集団は、CD24について陽性であった(図2aおよび2b)。上記亜集団はまた、肝性マーカー、胆嚢マーカーおよび間葉性マーカーであるアルブミン、ビメンチン、CK7およびCK19での免疫細胞化学によって評価した(図3)。
【0073】
両方のThy1.1ソートした集団は、アルブミン、CK7およびビメンチンについて陰性であった(図3−パネルB、C、F、G、JおよびK)。上記Thy1.1陽性集団は、CK19についても陰性であった(図3,パネルO)のに対して、上記Thy1.1陰性細胞は、弱陽性であった(図3,パネルN)。両方の集団は、RT PCRによって、c−Metおよびネスチンについて陽性であった(データは示さず)。
【0074】
まとめると、上記Thy1.1陰性細胞集団は、以下の細胞表面マーカープロフィールを発現する:
Thy1.1(CD90) 陰性
Pdx−1 陽性
CD49f 約95%+
CD24 約80%+
CD147 約80%+
CD45 陰性
CD44 約60%+
CD71 低い
CD31 陰性
c−Kit 陰性
ck7 陰性
CK19 弱陽性
c−Met 陽性
上記Thy1.1陽性細胞は、以下の細胞表面マーカープロフィールを発現する:
Thy1.1(CD90) 陽性
Pdx−1 陽性
CD49f 約95%+
CD24 陰性
CD147 約90%+
CD45 陰性
CD44 約85%+
CD71 低い
CD31 陰性
C−KIT 陰性
CK19 陰性
c−Met 陽性
ネスチン 陽性。
【0075】
(Thy1.1陽性集団およびThy1.1陰性集団の分化能)
(膵臓分化)
Thy1.1陽性集団およびThy1.1陰性集団は、膵臓分化培地において顕著に異なる形態学的変化を示した。上記Thy1.1陽性集団は、形態において最初に線維芽細胞様を示し、14〜21日目までに絡み合った細胞クラスタを形成し、28日目までに島様の球形クラスタへと形成された。これは、最終的に、親細胞層から分離した(図1,パネルD、H、I)。対照的に、上記Thy−1.1陰性細胞は、小さな上皮様形態を有する単層のままであり、三次元構造へと発展しなかった(図1,パネルAおよびB)。
【0076】
未分化のThy1.1陽性細胞のRT−PCR分析は、陽性Pdx−1発現を示したが、インスリンの発現もグルカゴンの発現も示さなかった(図4−B)。分化した細胞クラスタは、3種のマーカー全ての転写発現について全て陽性であった(図4 パネルB)。
【0077】
しかし、Thy1.1陰性細胞は、維持培地もしくは分化培地のいずれかで増殖させた場合に、Pdx−1を発現した。著しいことには、分化培地中で増殖させた場合、形態学的な変化を示さないにも拘わらず、上記Thy1.1陰性集団においてインスリン転写が検出された。グルカゴンは、上記未分化のThy−1.1陰性細胞において発現せず、分化後にインビトロで誘導もされなかった(図4 パネルA)。
【0078】
(肝臓分化)
Thy−1.1陽性PDPCおよびThy−1.1陰性PDPCを、血清非含有FGF4含有培地中で培養して、肝性の能力を評価した。Thy−1.1陽性細胞は、線維芽細胞様から上皮/立方形形態へと形態学的変化を示した。さらに、28日目までには、平らになった上皮との培養全体を通じて、管腔構造が明らかになった。上記膵臓分化プレートにおいて認められるものに類似したときおりの三次元島様構造が、肝臓分化プレートにおいても認められた。上記Thy−1.1陰性集団は、単層のままであり、三次元構造も管腔様構造も明らかでなく、顕著な形態学的変化もなかった(図1A,C)。
【0079】
本発明者は、RT−PCRによって28日間にわたって、上記2つの集団の分化をさらに試験した。RT−PCRを、内胚葉特異的遺伝子であるHNF3βおよびGATA 4、初期肝臓マーカーであるαフェトプロテインおよびCK18、成熟肝臓マーカーであるHNF1α、アルブミンおよびシトクロームP450酵素CYP2B1について行った。未分化のThy1.1陰性細胞は、RT−PCRによって、HNF3−βおよびCK19を発現したが、アルブミン、CK18、HNF1α、CY2B1、Gata4およびαフェトプロテインを発現しなかった(図5,パネルB)。他の初期肝臓マーカーもしくは成熟肝臓マーカー、またはCY2B1のいずれも、上記Thy1.1陰性集団において誘導されなかった。興味深いことには、未分化のThy1.1陽性PDPCは、初期内胚葉マーカーであるHNF3−β、GATA 4およびαフェトプロテインを発現したが、肝臓分化の14日目までに、肝細胞分化の後期マーカーであるHNF1−αおよびアルブミンを発現しなかった(図5 パネルA)。CK 19(胆嚢細胞によって正常に発現される)はまた、培養における上記管腔構造の出現と一致して、14〜28日目の間に誘導された。アルブミン発現の誘導は、免疫細胞化学によって確認された。未分化の細胞は、アルブミン内容物について陰性に染色される(図3,パネルA)一方で、14日目の分化した細胞は、アルブミン染色について強く陽性であった(図3,パネルD)。興味深いことには、10日目および14日目の時点で分化した細胞は、胆嚢マーカーであるCK19およびCK7について陰性に染色された(図3,パネルNおよびG)が、上記管腔様構造のみにおいてビメンチンについては陽性に染色された(図3,パネルL)。CK 18はまた、未分化のThy1.1陽性細胞において、および上記28日間の分化期間全体を通じて発現した。CYP2B1は、未分化のThy1.1陽性細胞において、および上記分化期間全体を通じて存在した。
【0080】
未分化のThy1.1陰性細胞は、免疫細胞化学によれば、CK7、ビメンチンおよびアルブミンの発現について陰性であり、CK19について弱陽性であった。上記Thy1.1陽性集団は、CK19について陰性であった。
【0081】
(グリコーゲン貯蔵についての過ヨウ素酸シッフ(PAS)染色)
貯蔵グリコーゲンの存在は、PAS染色によって決定される場合、Thy−1.1陽性PDPCでもThy−1.1陰性PDPCでも認められず、21日目の分化したThy−1.1陰性細胞においても認められなかった。しかし、グリコーゲン貯蔵を示す、PASでの陽性染色は、21日目までに上記Thy1.1陽性の分化した細胞において認められた(図6)。
【0082】
(糖尿病の動物モデルにおける細胞亜集団の試験)
本明細書に記載の細胞亜集団(例えば、Thy1.1陽性細胞集団もしくはThy1.1陰性細胞集団)は、糖尿病のような疾患の動物モデルにおいて使用されうる。一実施形態において、細胞の亜集団は、ストレプトゾトシン(STZ)誘導性の糖尿病の齧歯類和合(rodent concordant)異種移植片モデルにおいて使用される。C57BL/6マウスを、0日目にSTZを注射することによって糖尿病にする一方で、750,000個の細胞(例えば、Thy1.1陽性細胞)を、3日目に、処置した動物の尾静脈に注射する。コントロール動物に、生理食塩水の注射を与えるか、または等数のC57BL/6骨髄細胞を与える。3日ごとに血糖をモニターする。血糖の安定化および/もしくはコントロールに対して増大した生存は、上記投与した細胞が、上記動物においてインスリン生成をもたらすことを示す。
【0083】
(考察)
本明細書で提供されるのは、Thy1.1陽性PDPC亜集団およびThy1.1陰性PDPC亜集団のインビトロ培養、選択および特徴付けに関する詳細である。さらに、膵臓系統および肝臓系統への分化、ならびにマーカーThy1.1(これは、インビトロで系統への両能性を示す)を使用してソートした細胞集団の提供に関するそれらの効力について詳細が開示される。
【0084】
Thy1.1は、機能が明らかには理解されていない細胞表面タンパク質である。しかし、細胞認識(Gunterら,1984;Williams,1985)、細胞接着(Gunterら, 1984; Williams, 1985)(Heら,1991;Hueberら,1992)およびシグナル伝達(Kroczekら,1986)に関与していることが示唆された。Thy1.1発現は種々の幹細胞集団において認められ、顕著には、成体ラット肝臓における円形細胞集団は、Thy1.1が、潜在的に損傷後の修復細胞として、細胞が間質組織を認識しかつこれに接着することを可能にし得るという仮説をもたらした(Massonら,2006;Petersenら,1998;Terraceら,2007)。Thy1.1はまた、ヒト、マウスおよびラットにおける胎児肝臓の幹細胞、臍帯血および間葉性幹細胞上で発現される。Thy1.1陽性集団内のより大きなインビトロ効力における本知見は、これら観察と一致している。これら観察はまた、このような細胞のさらなる使用を可能にするために、単離および精製のための方法もまた、示す。
【0085】
以前の研究から、多くの異なる細胞タイプ(骨髄由来MSC、MAPC、子宮内膜および膵臓由来のMSCを含む)の肝臓分化が示された(Jiangら,2002;Mengら,2007;Schwartzら,2002;Seebergerら,2006)。PDPCの(Jiangら, 2002; Mengら, 2007; Schwartzら, 2002; Seebergerら, 2006)Thy1.1陽性亜集団は、形態学的表現型を共有し、これら集団とともに多くの細胞表面マーカーを発現する(CD44+、CD24−、CD45−、CD31−およびCD34を含む)。しかし、このこととは対照的に、Thy1.1陽性PDPCは、GATA4、HNF3−βおよびαフェトプロテインを発現する別個の細胞タイプであるようであり、これは、これら他の細胞タイプのうちのいずれかについて発現される場合に記載されていない。
【0086】
HNF3−βは、内胚葉能力において重要な役割を果たすと考えられている完全な兄はいようのマーカーである(Gualdiら 1996)一方で、GATA4は、腹側前腸内胚葉発生および初期肝臓遺伝子発現について必要とされる転写因子である(Gualdiら,1996;Rossiら,2001)。HNF3−βは、上記アルブミンエンハンサーへのGATA4の結合のその後の促進とともに、アルブミンエンハンサーの前後に位置しているヌクレオソームに指向されることが示された(McPhersonら,1996;Cirillo and Zaret,1999)。GATA4−/−胚およびHNF3−β−/−胚の両方は、前腸形態発生の欠損を示す(Duncanら,1997)。従って、未分化のPDPCにおけるHNF3−βおよびGATA4の発現、ならびにその後の肝臓特異的遺伝子(例えば、アルブミンおよびHNF1−α)の発現のFGF刺激誘導は、HNF 3−βおよびGATA 4が、これら細胞が肝臓へと発達する結果(hepatic fate)になる可能性を制御するように協同して作用するという提唱と一致する。さらに、分化の21日後の上記Thy1.1陽性集団におけるPAS染色の存在は、より成熟した肝細胞の機能的特徴を示した。このことは、HNF1−αの発現と一致する。HNF1−αは、遺伝子産物が成熟肝機能(炭水化物貯蔵および合成、ならびに脂質代謝を含む)と関連する遺伝子に結合することが公知である(Odomら,2004)。
【0087】
未分化のThy1.1陽性PDPCは、AFPを発現する。この観察は、初期腹側前腸内胚葉における肝形成の前に、ネスチン陽性島由来の前駆細胞におけるAFP発現、ならびに低レベルのAFPおよびTTR発現を記載する報告と一致する。この発現は、その後、心臓の中胚葉シグナル伝達から分離された内胚葉において失われる(Gualdiら,1996;Jungら,1999;Zulewskiら,2001)。このことは、腹側前腸内胚葉の標準的な(default)膵臓となる運命の特徴であることもまた、示唆された(Deuschら)。前記Thy1.1陽性PDPC集団におけるAFPの発現(これは、膵臓系統および肝臓系統の両方への能力を示す)は、この知見と矛盾しない(Deutschら,2001)。
【0088】
重大なことには、上記未分化のThy1.1陰性集団は、HNF3βを発現するが、GATA 4も、αフェトプロテインも発現せず、上記分化実験の間に誘導されなかった。肝臓の能力の証拠は、上記Thy1.1陰性集団において認められなかった。このことは、この未分化の集団内でのPdx−1発現、およびGata4発現の非存在と一致する。ビメンチンは、未分化のThy1.1陽性集団においても、Thy1.1陰性集団においても発現されなかったが、肝臓分化の間に管様構造を形成する細胞において発現された。ビメンチンは、間葉マーカーを表すと考えられる。しかし、Massonらは、肝門構造(portal structure)におけるThy1.1およびビメンチンの発現を観察し、組織切片内の上皮細胞における、および胎児肝臓上皮細胞の培養物におけるビメンチン発現も示した(Massonら,2006)。(Massonら, 2006)
上記膵臓分化に関するデータは、興味をそそるものである。島様クラスタの形態学的な証拠は、上記Thy1.1陰性集団において認められなかった。対照的に、Thy1.1陽性PDPCは、三次元島様構造およびPDX−1、インスリンおよびグルカゴンの転写発現を生じる特徴的な形態学的変化とともに、容易に膵臓系統へと誘導できた。
【0089】
両方の集団におけるPdx−1転写発現の検出は、それらがインスリン生成細胞になる可能性を示す。しかし、顕著なことには、Thy1.1陰性細胞は、分化培地中で増殖される場合、形態学的屁かを示さないにも拘わらず、インスリンを発現した。グルカゴンは、上記未分化のThy−1.1陰性細胞において発現されず、分化後にインビトロで誘導もされなかった(図4 パネル)。
【0090】
膵臓前駆細胞/幹細胞の種々の異なる候補集団が、以前から記載されてきた(ネスチンもしくは他の神経幹細胞マーカーを発現する島前駆細胞を含む)(Abrahamら,2004;Corneliusら,1997;Lechnerら,2002;Ramiyaら,2000)。別の集団は、PDX−1(インスリン生成細胞についての公知のマーカー)を発現することが示された。そしてこれら細胞は、適切な条件下で、インビトロで導管および内分泌腺分化の両方を刺激し得る(Bonner−Weirら,2000;Otonkoskiら,1993)。さらに、膵管の上皮細胞が、新たな島および腺房の増殖および形成の能力がある前駆細胞へ分化する能力を有するという証拠が存在し(Bonner−Weirら,2004)、もっとも近年では、(Bonner−Weirら, 2004)CK19+ 非ナイン分泌性膵臓上皮細胞(NEPC)が、胎児膵臓組織の存在下にある場合、インビボでインスリン生成細胞へと分化するように部分的に誘導されることが報告された(Haoら,2006)。
【0091】
これら細胞が果たす正確な生理学的役割が探究されてきた。Dorらは、新たな島細胞生成よりむしろβ細胞複製が、優勢な機構である(これによって、膵臓内分泌組織が、ほぼ完全な膵切除術後に再生する)という議論の代わりに、導管細胞もしくは前駆細胞からの再生が生じるという見方を吟味してきた(Dorら,2004)が、この解釈は、未だ論議の的である(Bonner−Weir and Weir,2005)。本知見は、このようなプロセスを潜在的に促進する場合に、インスリンを生成し得る非β細胞の役割と一致する。これら細胞が移植治療の代替のインスリン生成細胞供給源を提供することは明らかである。
【0092】
本発明者は、無血清FGF−4含有分化プロトコルの使用によって、肝臓系統分化を示す、形態学的発現変化および遺伝子発現変化の両方の過程を観察した。膵臓分化および肝臓分化についての胚性腹側内胚葉の潜在的な両能性は、腹側内胚葉が心臓中胚葉への近接によって肝臓系統へ分化した外植片実験において調査された。心臓中胚葉によって分泌される誘導因子(例えば、FGF−1、FGF−2およびFGF−4)の非存在は、腹側内胚葉の標準的な膵臓経路が継続することを可能にし(Deutschら,2001)、そして一般的なFGFシグナル伝達アンタゴニストは、インビトロでの肝発生(Jungら,1999)およびFGF−4(Zhuら,1999)を阻害する。本データは、この概念と完全に一致する。
【0093】
本発明者は、両能性のある内胚葉前駆体の集団と一致して、インビトロで、効力およびシグナル伝達に対する転写応答を示すPDPCの単離および特徴付けを示した。前述で、マウスストレプトゾトシン誘導性糖尿病モデルにおける、ソートしていないPDPC集団の投与は、分化およびラットインスリンの生成と同時に、マウス膵臓再生の刺激を実証した(Shiels 2005およびWO 2006/120476(ともに、本明細書に参考として援用される))。
【0094】
参考文献のリスト
【0095】
【化1】

【0096】
【化2】
【0097】
【化3】
【0098】
【化4】
【0099】
【化5】
【0100】
【化6】
【0101】
【化7】
【図1A】
【図1B】
【図1C】
【図1D】
【図1E】
【図1F】
【図1G】
【図1H】
【図1I】
【図2A】
【図2B】
【図3A】
【図3B】
【図3C】
【図3D】
【図3E】
【図3F】
【図3G】
【図3H】
【図3I】
【図3J】
【図3K】
【図3L】
【図3M】
【図3N】
【図3O】
【図3P】
【技術分野】
【0001】
(関連出願への相互参照)
この出願は、2008年5月9日に出願された、米国仮特許出願第61/052,098号と同時係属中であり、かつ少なくとも1人の発明者と共有され、かつそれへの優先権を主張する。先行する出願は、その全体が参考として本明細書に援用される。
【0002】
(発明の分野)
本発明は、組織再生、および細胞変性もしくは加齢性組織変化(age related tissue change)と関連する疾患状態の処置のために使用され得る新規な細胞集団の提供に関する。特に(しかし網羅的ではない)、本発明は、膵臓細胞タイプおよび肝細胞タイプの両方に分化するための両能性を示す新規な細胞集団の決定から生じる材料および方法を提供する。
【背景技術】
【0003】
(発明の背景)
幹細胞は、培養において長期間増殖する能力を有し、特定の細胞タイプになるために誘導され得る分化していない細胞である。幹細胞は、主に胚もしくは生体から単離され得るが、これらは、異なる特徴および機能を有するようである。
【0004】
幹細胞は、多くの哺乳動物種の胚(胚性/胚幹細胞−ESC)から単離されてきた。マウス由来のものは、20年間にわたって集中した研究の対象であり、単離されかつ1998年以来研究されてきたヒトESCの単離のための方法を可能にした。
【0005】
ESCは、代表的には、胚盤胞(embryonic blastocyst)に由来し、ここで胚盤胞は、インビボで全てのその後の発生細胞タイプを生じるように続く。成体幹細胞(ASC)は、他方で、多くの組織に存在し、そこでは、それらは、傷害および、または消耗(wear and tear)を介して失われた細胞の置換を可能にする。これら細胞は、疾患(例えば、糖尿病およびパーキンソン病)の処置のための細胞ベースの治療を提供する能力を有する。上記疾患では、特定の病理をもたらす細胞損失および損傷が存在する。これら細胞はまた、薬物のスクリーニング、毒物学的調査、発生プログラミングの調査、加齢性組織損失および変性の処置のための優れた能力を有する。これら細胞はまた、外科手術的除去後の、傷害後の、もしくは美容外科手順の一部としての組織再生の能力を有する。
【0006】
ASCは、成体組織もしくは器官中の分化した細胞の中で見いだされる未分化の細胞であり、ASCは、分化しておらず、そしてそれ自体を自分で補充する(self renew)し得、組織もしくは器官の主要な細胞タイプを得るように分化する能力を維持する。ASCは、組織修復を維持しかつもたらすと考えられる。成熟組織における成体幹細胞の起源は未知であり、それらの可塑性の程度は、未だ決定されていない。移植におけるそれらの用途は、広く知られている。骨髄由来のASCは、30年間にわたって移植で使用されてきた。成体非HSCの使用、それらの効力、可塑性および長期追跡での安全性は、未だ証明されていない。非間質性ASCの報告は、現場で討議されたままであるが、神経幹細胞は、今では確立されており、真正な(bone fide)ASCタイプとして受け入れられている。
【0007】
ESCの証明された多能性特徴およびESCを多数増殖させる能力は、ESCを細胞ベースの治療の魅力的な候補にしている。この状況においてASCの使用には厳しい制限が存在する。ここでこのような細胞は、非常に稀であると考えられておりかつそれらの増殖条件は、可能性のある治療に十分な数で適切な細胞を生成するほど十分に明確でない。
【0008】
しかし、ASCは、「自己」に由来し得るという点で、決定的な利点を確かに有しているので、いかなる患者でも、彼ら自身の細胞を受容し、顕著に高まった癌のリスクを含め、拒絶を防止するために免疫抑制の有害な副作用に苦しむ必要はない。制限された有効性/可塑性はまた、以上型の細胞分化が制限され、新生物のリスクが低減されるという点で、高められた安全性因子であると考えられる。
【0009】
肝臓および膵臓の共通する発生起源は、これら器官が、共通する幹細胞/前駆細胞集団を共有し得ることを示唆する。この過程の裏付けにおいてかなりの間接的証拠が存在する。外植片実験から、Pdx−1を発現する腹側内胚葉は、心臓中胚葉への近接によって、肝臓系統に流用されることが実証されている(非特許文献1)。肝細胞特性を有する細胞(Deutsch et al., 2001)はまた、マウス膵臓内の管領域において観察された。銅欠乏性食餌を与えられたマウスは、膵臓損傷、腺房細胞の損失を受け、4〜6週間後には、肝円形様細胞(hepatic oval−like cell)が観察される(非特許文献2;非特許文献3)。さらに、脾臓に移植された、銅欠乏性食餌に供されたラットから単離した膵臓細胞は、実質構造へと組み込まれかつ成熟肝臓特異的タンパク質を発現することによって、形態的にも機能的にも肝細胞への分化を示した(Dabevaら,1997)。フマリルアセトアセテートヒドロラーゼが欠損し、後にチロシン血症を伴う共通遺伝子のレシピエントに移植した野生型マウス膵臓細胞の懸濁物はまた、肝機能の正常化とともに、ドナー由来細胞によって生合成レスキューを生じることが報告された(Wangら,2001)。さらに、成体マウス膵臓におけるKGFのトランスジェニック過剰発現は、膵島内で肝細胞の出現および付随して膵管増殖を生じる(Krakowskiら,1999b;Krakowskiら,1999a)。
【0010】
いくつかの推定される膵臓前駆細胞が、膵管細胞および島細胞の両方から特徴付けられた(Abrahamら,2004;Corneliusら,1997;Lechnerら,2002;Ramiyaら,2000)。これら細胞集団のうちの一方は、インビトロでの分化後に、αフェトプロテインおよびc−Met(代表的には、肝細胞によって発現されるタンパク質)を発現することが観察された(Zulewskiら,2001)。(Zulewskiら, 2001)より近年になってからは、ヒト膵管上皮から単離された間葉系幹細胞集団は、膵臓分化、肝臓分化および中胚葉分化の能力を有することが報告された(Seebergerら,2006)。これら細胞は、Gata 4、アルブミンおよびTAT(チロシンアミノトランスフェラーゼ)を発現するように誘導され得るが、機能的評価は未だ報告されていない。
【0011】
本発明者は、以前に、さらなる細胞タイプである、STZ糖尿病モデルにおいて多能性および機能的効力を示す、成体ラット膵管から単離された膵臓由来開通細胞(Pancreatic Derived Pathfinder Cell)(PDPC)を決定した(Shiels 2004;および特許文献1(本明細書に参考として援用される))。成体ラット肝臓において、推定される幹細胞の集団は、化学的傷害、もしくは部分的肝切除術による誘導後に誘導され得る(Petersenら,1998;Petersenら,2003)(Petersenら, 1998; Petersenら, 2003)。
【先行技術文献】
【特許文献】
【0012】
【特許文献1】国際公開第2006/120476号
【非特許文献】
【0013】
【非特許文献1】Deutsch,G., J.Jung, M.Zheng, J.Lora and K.S.Zaret: A bipotential precursor population for pancreas and liver within the embryonic endoderm. Development Dev.(2001)128、871−881
【非特許文献2】Rao,M.S., V.Subbarao and J.K.Reddy: Induction of hepatocytes in the pancreas of copper−depleted rats following copper repletion. Cell Differ. Dev.(1986)18、109−117
【非特許文献3】Rao,M.S. and J.K.Reddy: Hepatic transdifferentiation in the pancreas. Semin.Cell Biol. Dev.(1995)6、151−156
【非特許文献4】Dabeva,M.D., S.G.Hwang, S.R.Vasa, E.Hurston, P.M.Novikoff, D.C.Hixson, S.Gupta and D.A.Shafritz: Differentiation of pancreatic epithelial progenitor cells into hepatocytes following transplantation into rat liver. Proc.Natl.Acad.Sci.U.S.A Dev.(1997)94、7356−7361
【発明の概要】
【課題を解決するための手段】
【0014】
(発明の要旨)
細胞変性と特に関連する疾患を処置するために、細胞ベースの治療において使用され得る細胞集団を提供することが継続して必要である。糖尿病は、このような疾患の一例である。膵臓におけるインスリンは、インスリン分泌β細胞によって作られる。インビボでは、β細胞ターンオーバーは、生涯全体を通じて起こると考えられているが、置換細胞の起源に関しては、論争がある。従って、糖尿病は、完全に機能するネイティブ細胞に類似した細胞の提供(特に、自己に由来する細胞)が処置の重要な選択肢を提供する疾患の一例である。糖尿病の場合には、インスリンを生成し得るという点で、β細胞に類似の細胞の提供は、上記疾患にとって重大な細胞ベースの治療を提供する。
【0015】
この疾患および他の疾患を考慮に入れて、本発明者は、成体膵管から肝細胞/前駆細胞集団に由来する細胞の亜集団を単離した。
【0016】
いくつかの実施形態において、本明細書で提供される細胞の亜集団は、Pdx−1(HUMAN: NP_000200.1 GI:4557673; RAT: NP_074043.3 GI:50838802)陽性である。Pdx−1発現の存在は、細胞が非β細胞由来インスリンの供給源としての能力を有することを示す。
【0017】
いくつかの実施形態において、本明細書で提供される成体幹細胞/前駆細胞の亜集団は、Thy1.1(CD90)(HUMAN: NP_006279)の存在もしくは非存在によって特徴付けられる。Thy1.1陽性亜集団およびThy1.1陰性亜集団の特性は、以下で議論される。
【0018】
(1.Thy1.1陽性細胞)
維持(非分化)培地中で、Thy1.1陽性細胞亜集団は、Pdx−1陽性、インスリン陰性およびグルカゴン陰性である。Thy1.1陽性細胞集団は、膵臓分化培地中に置かれた場合、最初に、線維芽細胞様形態を示し、次いで、親細胞層から最後には分離する絡み合った細胞クラスタを形成する。得られた分化した細胞クラスタは、Pdx−1、インスリンおよびグルカゴンについて陽性である(図4、パネルB)。
【0019】
本明細書に記載されるThy1.1陽性細胞亜集団に関する驚くべき決定は、それらが、少なくとも両能性のある(bipotent)ことである。具体的には、適切な分化培地とともに提供される場合、Thy1.1陽性細胞は、膵臓細胞タイプもしくは肝細胞タイプのいずれかに分化し得る。
【0020】
(2.Thy1.1陰性細胞)
非分化状態において、Thy1.1陰性細胞集団は、Pdx−1について陽性であるが、インスリンおよびグルカゴンについては陰性である。分化培地中で増殖される場合、Thy1.1陰性細胞は、形態学的変化を示さなかったが、インスリン転写が検出された。よって、Thy1.1陰性集団は、非β細胞由来インスリンの新規な供給源を提供する。
【0021】
よって、その最も一般には、本発明は、多能性成体幹細胞集団に由来する細胞の新規な亜集団を使用して、細胞ベースの治療に基づく加齢の疾患および状態を処置するための材料および方法を提供する。上記亜集団は、Pdx−1陽性である。
【0022】
このような細胞集団は、糖尿病のような疾患およびパーキンソン病のような神経変性障害の細胞ベースの治療に対する能力を提供する。
【0023】
成体幹細胞集団(前駆細胞としても公知)は、成体膵臓組織(例えば、ヒト、ラット、マウス、霊長類、ブタなど)に由来し得る。上記成体組織は、好ましくは、膵臓組織であるが、乳房、骨髄、心臓、肝臓もしくは腎臓のような他の器官に由来する組織であってもよい。
【0024】
本発明の一実施形態において、成体幹細胞は成体ラット膵臓に由来し、これは、1977年のブダペスト条約に従って、University of GlasgowのThe University Courtによって、Cell Cultures,Porton Down,Salisbury Wiltshire,UK,SP4 0JGのThe European Collectionにおいて、2005年5月12日にECACC番号Q6203の下で寄託された。これら細胞は、本明細書中以降、PDPC(膵臓由来前駆細胞)として知られる。
【0025】
上記のように、本発明者はまた、マーカーThy1.1(CD90)(HUMAN: NP_006279)を参照して、2つのさらなる亜集団を決定した。これら2つの亜集団は異なるが、等しく重要な特性を有する。特に、Thy1.1陽性細胞は、膵臓細胞タイプおよび肝細胞タイプの両方への分化能力を示す。いくつかの実施形態において、本明細書で記載される細胞亜集団は、インスリンの非β細胞供給源を提供する。いくつかの実施形態において、本明細書で記載される細胞亜集団は、試験物質の毒性を評価するための細胞タイプを提供する(例えば、毒物学試験のために)。
【0026】
よって、第1の局面において、成体組織から発生する幹細胞/前駆細胞の集団を提供する。ここで上記細胞は、血清の存在下でマトリゲルなしの培養システムにおいて増殖でき、上記細胞は、Thy1.1陽性である。いくつかの実施形態において、Thy1.1陽性細胞はまた、Pdx−1陽性である。いくつかの実施形態において、Thy1.1陽性細胞は、ネスチン(RAT:NP_037119.1 GI:6981262; HUMAN: NP_006608.1 GI:38176300)陽性である。
【0027】
いくつかの実施形態において、上記集団中のネスチン陽性細胞のパーセンテージは、フローサイトメトリー分析によって決定される場合、50%未満(例えば、40%、30%、20%、10%、5%、2%もしくは1%未満)である。いくつかの実施形態において、細胞は、例えば、PCRによって、ネスチン核酸の発現によって、ネスチン陽性細胞として区別可能である。
【0028】
細胞集団は、Thy1.1陽性(CD90陽性)(RAT:LOCUS:P01830 GI135832; NP_006279 GI:19923362)として見なされ得る。ここでThy1.1陽性細胞のパーセンテージは、50%より大きく、好ましくは、60%より大きく、より好ましくは、70%、80%、90%、もしくは95%より大きい。いくつかの実施形態において、Thy1.1陽性細胞集団は、純度が98%より高い。Thy1.1陽性細胞のパーセンテージは、例えば、フローサイトメトリーによって、もしくはPCRによって決定され得る。
【0029】
いくつかの実施形態において、本発明に従うThy1.1陽性細胞集団は、Pdx−1、CD49f、CD147、CD44、c−Met、およびネスチンのうちの1種以上の発現について陽性である。いくつかの実施形態において、Thy1.1陽性細胞集団は、CD24、CD45、CD31、c−kit、およびCK19のうちの1種以上の発現について陰性である。いくつかの実施形態において、本発明に従うThy1.1陽性細胞集団は、以下の細胞表面マーカープロフィールを有する:
Thy1.1(CD90) 陽性
Pdx−1 陽性
CD49f 約95%+
CD24 陰性
CD147 約90%+
CD45 陰性
CD44 約85%+
CD71 低い
CD31 陰性
C−KIT 陰性
CK19 陰性
c−Met 陽性
ネスチン 陽性
(低い=細胞のうちの約5%以下が上記マーカーを発現する)。
【0030】
本発明はまた、本発明のこの局面に従う細胞集団とともに、薬学的に受容可能なキャリアを含む薬学的組成物を提供する。
【0031】
本発明の第2の局面において、成体哺乳動物組織から単離された両能性のある幹細胞集団を生成するための方法が提供され、上記方法は、上記成体哺乳動物組織を培養する工程;発生する細胞集団単層を得る工程;およびThy1.1について陽性である細胞を含む亜集団を単離する工程を包含する。いくつかの実施形態において、上記亜集団のうちの少なくとも50%、60%、70%、80%、90%、95%が、Thy1.1陽性である。
【0032】
上記方法はまた、上記で提供されるプロフィール(例えば、Pdx−1、CD49f、CD147、CD44、c−Met、およびネスチンのうちの1種以上について陽性ならびに/またはCD24、CD45、CD31、c−kit、およびCK19のうちの1種以上について陰性)中に提供される1種以上の他の細胞表面マーカーと組み合わせて、Thy1.1について陽性樽細胞を単離する工程を包含し得る。
【0033】
いくつかの実施形態において、上記成体哺乳動物組織は、膵臓であり、例えば、膵管由来である。いくつかの実施形態において、上記成体哺乳動物組織は、乳房、肝臓もしくは腎臓である。いくつかの実施形態において、上記成体哺乳動物組織は、ヒト組織である。
【0034】
発生する細胞集団単層を得るために成体哺乳動物組織を得る代わりに、方法は、既に単離された成体幹細胞(例えば、2005年5月12日に、アクセッション番号Q6203の下でECACCに寄託されたもの)を得ることを包含し得る。
【0035】
第3の局面において、培養において幹細胞の集団を生成するための方法が提供され、上記方法は、本発明に従うThy1.1陽性細胞集団を、肝臓系統分化に適した培地中で培養する工程を包含する。一例として(他は、当業者に公知である)、上記培養培地は、無血清FGF−4含有分化培地であり得る。
【0036】
第4の局面において、培養において膵臓細胞の集団を生成するための方法が提供され、上記方法は、本発明に従うThy1.1陽性細胞集団を、膵臓系統分化に適した培地中で培養する工程を包含する。
【0037】
本発明は、本明細書で記載される方法から得られるもしくは得ることができる細胞および細胞集団に及ぶ。
【0038】
本発明の第5の局面において、細胞変性もしくは加齢性組織変化と関連する疾患状態を処置するための方法が提供され、上記方法は、本発明に従うThy1.1陽性成体幹細胞集団もしくは上記Thy1.1陽性細胞集団を含む薬学的組成物を、上記疾患もしくは加齢性状態を有する患者に投与する工程を包含する。
【0039】
Thy1.1陽性細胞集団は、静脈内に投与され得るか、または疾患部位に移植され得る。
【0040】
いくつかの実施形態において、上記疾患は、膵臓細胞、神経細胞、心血管細胞(例えば、心筋細胞)、上皮細胞、肝細胞、もしくは腎細胞の変性と関連する。
【0041】
上記処置されるべき疾患状態としては、糖尿病(I型およびII型)、肝臓疾患、腎臓疾患、眼の疾患、パーキンソン病および心血管疾患ならびに身体の器官および組織の加齢性変性状態が挙げられ得る。本発明のこの局面はまた、美容外科手術の形態として(例えば、組織の細胞再生および加齢の形態を妨げるため)使用され得る。
【0042】
いくつかの実施形態において、上記細胞のドナーおよび上記レシピエントは、同じ種である(例えば、両方ともヒト)。いくつかの実施形態において、上記細胞のドナーおよび上記レシピエントは、異なる種である。よって、本発明の一実施形態は、ヒト患者の処置における、ラットに由来する成体幹細胞集団の使用を包含する。
【0043】
本発明の第6の局面において、指定された分化した細胞集団(例えば、膵臓細胞もしくは肝細胞)を生成するための方法が提供され、上記方法は、成体幹細胞集団を提供する工程;Pdx−1および/もしくはThy1.1マーカー、ならびに必要に応じて、Thy1.1亜集団細胞表面マーカープロフィールに関連して本明細書で同定される1種以上の他のマーカーを使用して、細胞亜集団を選択する工程;ならびに細胞分化の助けとなる条件下で上記細胞亜集団を培養する工程を包含する。
【0044】
本発明は、美容外科手術を含め、医療処置の方法において使用するための本発明の第1の局面に従う細胞集団をさらに提供する。上記方法は、細胞損失もしくは変性(例えば、糖尿病もしくはパーキンソン病)と関連する疾患状態もしくは加齢の状態を処置し得る。
【0045】
本発明の第6の局面において、成体組織から発生する細胞集団が提供され、ここで上記細胞は、血清の存在下でマトリゲルなしの培養システムにおいて増殖でき、上記細胞は、Thy1.1陰性である。
【0046】
いくつかの実施形態において、上記細胞集団は、Pdx−1陽性である。上記細胞集団はまた、ネスチン陽性であり得る。
【0047】
本発明はまた、成体幹Thy1.1陰性細胞集団を、薬学的に受容可能なキャリアとともに含む薬学的組成物を提供する。
【0048】
本発明の第7の局面において、成体哺乳動物組織から単離された幹細胞集団を生成するための方法が提供され、上記方法は、上記成体哺乳動物組織を培養する工程;発生する細胞集団単層を得る工程;およびThy1.1について陰性の細胞の亜集団を単離する工程を包含する。一実施形態において、上記方法は、Pdx−1陽性および/もしくはネスチン陽性でもある細胞の亜集団を単離する工程をさらに包含する。
【0049】
一実施形態において、Thy1.1陰性細胞の亜集団は、CD49f、CD24、CD147、CD44、c−Metのうちの1種以上の発現について陽性であり、そして/またはCD31、c−kit、およびck7のうちの1種以上の発現について陰性である。一実施形態において、以下の細胞表面マーカープロフィールであるThy1.1陰性細胞の亜集団が単離される:
Thy1.1(CD90) 陰性
Pdx−1 陽性
CD49f 約95%+
CD24 約80%+
CD147 約80%+
CD45 陰性
CD44 約60%+
CD71 低い
CD31 陰性
c−KIT 陰性
ck7 陰性
CK19 弱陽性
c−Met 陽性
(低い=細胞のうちの5%以下がそのマーカーを発現する)。
【0050】
発生する細胞集団単層を得るために成体哺乳動物組織を得る代わりに、上記方法は、既に単離された成体幹細胞/前駆細胞(例えば、2005年5月12日にアクセッション番号Q6203の下でECACCに寄託されたもの)を得る工程を包含し得る。
【0051】
本発明の第8の局面において、糖尿病もしくはインスリン生成の低下と関連する疾患状態を処置するための方法が提供され、上記方法は、本発明に従うThy1.1陰性細胞集団、もしくは上記Thy1.1陰性細胞集団を含む薬学的組成物を、上記疾患もしくは加齢性状態を有する患者に投与する工程を包含する。
【0052】
Thy1.1陰性細胞集団は、静脈内投与され得るか、または上記疾患部位に移植され得る。
【0053】
いくつかの実施形態において、上記細胞のドナーおよび上記レシピエントは、同じ種(例えば、ヒト)である。いくつかの実施形態において、上記細胞および上記レシピエントは、異なる種である。よって、本発明の一実施形態は、ヒト患者の処置における成体ラット幹細胞/前駆細胞の使用を包含する。
【0054】
本発明の第9の局面において、インスリンを生成することができる指定された分化した細胞集団を生成するための方法が提供され、上記方法は、成体幹細胞集団を提供する工程;Pdx−1について陽性でありかつThy1.1マーカーについて陰性である成体幹細胞亜集団を選択する工程;ならびに細胞分化の助けとなる条件下で、上記細胞亜集団を培養する工程を包含する。上記方法は、Thy1.1細胞表面マーカープロフィールの一部として上記で同定された1種以上のさらなるマーカーに基づいて、上記成体幹細胞集団を選択する工程をさらに包含し得る。
【0055】
本発明は、医療処置の方法において使用するための、本発明に従う成体幹細胞Thy1.1陰性細胞集団をさらに提供する。特に、上記方法は、糖尿病を処置することであり得る。
【0056】
インスリンを生成するための方法もまた、本明細書で提供される。インスリンを生成するための方法は、本明細書に記載されるThy1.1陰性のPdx−1陽性細胞の集団(例えば、成体組織(例えば、成体膵臓組織)に由来するThy1.1陰性のPdx−1陽性細胞の集団)を、インスリンが生成される条件下で培養する工程を包含し得る。上記方法は、上記培養物からインスリンを単離する工程をさらに包含し得る。いくつかの実施形態において、上記Thy1.1陰性細胞は、CD49f、CD24、CD147、CD44、c−Metのうちの1種以上の発現について陽性であり、そして/またはCD31、c−kit、およびck7のうちの1種以上の発現について陰性である。
【0057】
別の実施形態において、インスリンを生成するための方法は、本明細書に記載されるThy1.1陽性のPdx−1陽性細胞の集団(例えば、成体組織(例えば、成体膵臓組織)に由来する分化したThy1.1陽性のPdx−1陽性細胞の集団)を、インスリンが生成される条件下で培養する工程を包含する。上記方法は、インスリンを上記培養物から単離する工程をさらに包含し得る。いくつかの実施形態において、Thy1.1陽性細胞集団は、Pdx−1、CD49f、CD147、CD44、c−Met、およびネスチンのうちの1種以上の発現について陽性であり、そして/またはCD24、CD45、CD31、c−kit、およびCK19のうちの1種以上の発現について陰性である。
【0058】
本発明の局面および実施形態は、ここで添付の図面を参照しながら、例示によって示される。さらなる局面および実施形態は、当業者に明らかである。本文において言及される全ての文書は、本明細書に参考として援用される。
【図面の簡単な説明】
【0059】
【図1】図1は、未分化のThy1.1陽性集団およびThy1.1陰性集団のインビトロでの形態である。Thy1.1陽性PDPC集団およびThy1.1陰性PDPC集団のインビトロでの肝臓分化および膵臓分化(図1A〜C):形態学的変化は、分化の間にThy1.1陰性集団においてほとんど認められない。(図1A)未分化のThy1.1陰性PDPC。(図1B)膵臓分化誘導後28日目のThy1.1陰性PDPC。(図1C)肝臓分化誘導後28日目のThy1.1陰性PDPC。(図1D-I):Thy1.1陽性PDPCの肝臓分化の誘導に際して、顕著な形態学的変化が認められた。(図1D)未分化のThy1.1陽性PDPC(線維芽細胞様形態)。(図1E)肝臓分化誘導後14日目のThy1陽性PDPC。(図1F)肝臓分化誘導後28日目のThy1.1陽性PDPC−管腔構造が優勢な上皮形態が存在する。(図1G)肝臓誘導後28日目のthy1陽性PDPCの立方形の形態。(図1H)膵臓誘導後14日目のThy1.1陽性PDPC。(図1I)膵臓誘導後28日目のThy1.1陽性PDPC−島様クラスタが形成され、その後、培地へと分離する。
【図2】図2は、フローサイトメトリーによってMACSソートしたPDPCの細胞表面特徴付けである。(図2A)Thy1(CD90)陽性集団は、CD24、CD31、CD45、c−kitの発現について陰性であり、CD71については低いが、CD147、CD44およびCD49fの発現については陽性である。(図2B)Thy1(CD90)陰性集団は、CD31、CD45、c−kitの発現については陰性であり、CD71については低いが、CD24、CD147、CD44およびCD49fの発現については陽性である。
【図3】図3は、Thy1.1ソートしたPDPC集団の免疫細胞化学である。陽性コントロールは、アルブミン(図3A)、サイトケラチン7(図3E)、ビメンチン(図3I)およびサイトケラチン19(図3M)について示される。上記未分化のThy1.1陰性PDPC集団は、アルブミン(図3B)、サイトケラチン7(図3F)、ビメンチン(図3J)について陰性染色を示すが、サイトケラチン19(図3N)については弱い染色を示す。上記未分化のThy1.1陽性集団は、アルブミン(図3C)、サイトケラチン7(図3G)、ビメンチン(図3K)およびサイトケラチン19(図3O)について陰性であった。しかし、肝臓分化培地中での14日目までに、Thy1.1陽性PDPCは、アルブミンについて陽性であり(図3D)、管腔様領域においては、ビメンチン(図3L)およびサイトケラチン19(図3P)について陽性であったが、サイトケラチン7は、全体を通じて陰性のままであった(図3H)。
【図4】図4は、膵臓分化培地中のThy1.1陽性PDPC(図4A)およびThy1.1陰性PDPC(図4B)のRT−PCR。Thy1.1陰性PDPCおよびThy1.1陽性PDPCを、膵臓分化培地中で28日間にわたって培養した。列:(1)陽性コントロール、(2)未分化のPDPC、(3)28日目の分化したPDPC、(4〜6) サンプル1〜3のRTなし(no RT)コントロール。
【図5】図5は、肝臓分化培地中のThy1.1陽性PDPCおよびThy1.1陰性PDPCのRT−PCRである。Thy1.1陽性PDPC(図5A)およびThy1.1陰性PDPC(図5B)を、肝臓分化培地中で28日間培養した。列:(1)陽性コントロール、(2)未分化のPDPC、(3)7日目、(4)14日目、(5)21日目、(6)28日目、(7〜13)サンプル1〜6のRTなしコントロール。アルブミン、CK19およびHNF1αの発現は、上記Thy1.1陽性集団において誘導されたが、上記Thy1.1陰性集団においては認められなかった。
【図6】図6は、未分化のThy1.1陽性PDPC集団(図6B)もしくはThy1.1陰性PDPC集団(図6C)がグリコーゲンを貯蔵しないことを示す。FGF−4含有培地中で培養した後のThy1.1陽性PDPCは、グリコーゲンを生成および貯蔵する(図6DおよびE)。グリコーゲン貯蔵は、過ヨウ素酸−シッフによって染色される場合に、赤紫色の染色の蓄積として認められる。FGF−4含有肝臓分化培地中で培養した後のThy1.1陰性PDPCは、染色を示さない(図6F)。陽性コントロール(図6A)。
【発明を実施するための形態】
【0060】
(詳細な説明)
(材料および方法)
(ラットPDPCの単離および維持培養)
膵管を、切開することによって12ヶ月齢のAlbino Swiss(Glasgow)ラットから単離し、CMRL培地中に播種する前に切り刻んだ。上記PDP細胞は、培養において約5週間後にコンフルエントな単層として発生した。次いで、これらを採取し、PBS中で洗浄した。PDPCを、0.2μm フィルターキャップ付きのT75培養フラスコ(Corning,UK)において、37℃で5% CO2雰囲気中、20mlのCMRL 1066培地(Invitrogen,Paisley,UK.)(10% ウシ胎仔血清(Sigma,Poole,UK)、2mM glutamax、1.25μg/ml アンホテリシンB、および100u/ml ペニシリン、100μg/ml ストレプトマイシン(全てInvitrogen,Paisley,UK)を補充)中の培養において維持した。コンフルエント未満の培養物を、ピペットでの培養培地の完全な除去、ならびに10mlのカルシウムおよびマグネシウム非含有ハンクス平衡塩類溶液(HBSS)(Cambrex Bio−Science,Wokingham,UK)を上記フラスコに室温で5分間添加することによる、接着性細胞の洗浄することによって継代した。上記HBSSをピペットで上記フラスコから除去した後、2mlのトリプシン−Versene溶液(200mg/L Versene、500mg/L トリプシン)を、上記フラスコに添加した。上記フラスコを、上記細胞単層の分離が確認されうるまで、顕微鏡で定期的に観察した。次いで、細胞を、ピペットで取り出し、望ましい場合、1/5〜1/10の密度で20mlの新鮮な培養培地を添加して上記のように再培養した。
【0061】
PDPCを、0.2μm フィルターキャップ付きのT75において、37℃で5% CO2雰囲気中、CMRL 1066培地(Invitrogen,Paisley,UK)(5% ウシ胎仔血清(Sigma,Poole,UK)、2mM Glutamax、1.25μg/ml アンホテリシンB、および100u/ml ペニシリン/ストレプトマイシン(全てInvitrogen,Paisley)を補充)中で長期間維持した。PDPCを、37℃で加湿5% CO2雰囲気中、単層として増殖させ、90% コンフルエントになったときに、トリプシン−EDTA(Invitrogen)を用いて継代した。細胞を計数し、3300細胞/cm2の密度で再度プレートした。
【0062】
(磁性活性化細胞ソーティング)
磁性活性化細胞ソーティング(MACS)を、製造業者のプロトコル(Dynabeads ヤギ抗マウスIgG(Dynal Biotech))に従って、106標的細胞あたり1μgの一次抗体(マウス抗ラットThy1.1(CD90)(Serotec))を使用して、4℃において20分間にわたって、単離および除去のために行った。ソートした細胞集団を、維持培養培地中で再懸濁し、組織培養フラスコに再度プレートした。MACSを、実験において使用する前に、各陽性ソートした集団および陰性ソートした集団に対して2回行った。全てのソートした集団を、後の分化実験の前に、蛍光活性化フローサイトメトリーでチェックした。
【0063】
(細胞表面抗原のフローサイトメトリー評価)
細胞を、HBSS中の0.5% BSAにおいて再懸濁した。次いで、それらを1000×rpmで10分間にわたって遠心分離し、得られた細胞ペレットを、HBBS中に再懸濁する。トリパンブルー(Invitrogen,Paisley,UK)で生存性係数した後、1×106細胞/mlを、100μl 一次抗体で標識した。使用した一次抗体は、CD90に対するもの(Serotec MCA47R,1:75)、CD44に対するもの(Serotec MCA643,1:10)、CD49fに対するもの(Serotec MCA2034,1:50)、CD147に対するもの(Serotec MCA729,1:10)、c−KITに対するもの(Santa Cruz SC−19983,1:20)、CD71に対するもの(Serotec MCA 155FT,1:10)、CD24に対するもの(BD Biosciences 551133,1:50)、CD45に対するもの(BD Biosciences 554875,1:50)、CD31に対するもの(Serotec MCA1334GA,1:50)およびCD34に対するもの(Santa Cruz sc−7324,1:50)であった。2次抗体を、0.2% BSA/PBS中、遮光して、4℃において45分間にわたって添加した。次いで、上記細胞を洗浄し、0.2% BSA中のウサギ抗マウス免疫グロブリン(Dako Cytomation,Ely,UK 1:20)の100μl FITC結合体化Fab2フラグメントを遮光して4℃で45分間にわたって添加することによって標識する前に、1000×rpmで、0.2% BSA/PBS中、3回遠心分離した。アイソタイプFITCコントロールもまた行った。前述のように、3回洗浄し、遠心分離した後、得られた細胞ペレットを、1mlのHBSS中で再懸濁し、上記細胞を、Beckman Coulter XLフローサイトメーター(Beckman Coulter,High Wycombe,UK)を使用して分析した。
【0064】
(分化実験)
膵臓分化のために、Thy1.1陽性細胞集団およびThy1.1陰性細胞集団(継代30)を、6600細胞/cm2の細胞密度においてプレートした。24時間後、維持培地を除去し、単層をHBSSで3回洗浄した。引き続き、細胞をDMEM:F12(Lonza)(1×ITS、1.25μg/ml アンホテリシンB、および100μ/ml ペニシリン/ストレプトマイシン(全てInvitrogen,UK)、ニコチンアミド 10mM(Sigma)、KGF 10ng/ml(Sigma)および0.2% BSA(Sigma)を補充)中で培養した。
【0065】
肝性分化のために、細胞を、T75および6ウェルプレート中、6600細胞/cm2においてプレートし、24時間維持で、チャンバースライド(Nunc)中2500細胞/cm2において、HBSS、DMEM:F12(Lonza)(線維芽細胞増殖因子−4 10ng/ml(Sigma)、1×ITS、100μ/ml ペニシリン/ストレプトマイシン(Invitrogen)および0.2% ウシ血清アルブミン(Sigma)を補充)で3回洗浄した後に、置換した。培地好感を1週間に3回行い、細胞を、膵臓分化については0日目および28日目に、および肝臓分化については0日目、7日目、14日目、21日目および28日目に、未分化のThy1.1陽性細胞およびThy1.1陰性細胞からのRNA抽出のために採取した。チャンバースライドにおいて肝臓分化を受けている細胞を、PBSで3回洗浄し、10〜14日の間で4% パラホルムアルデヒドで室温において15分間固定した。未分化のThy1.1陽性集団およびThy1.1陰性集団を、同時にチャンバースライド中でも増殖させ、上記のように、90%コンフルエントにおいて固定した。
【0066】
(免疫蛍光)
細胞内タンパク質の染色のために、細胞を上記のように固定した。細胞をPBS中で3回洗浄し、0.1% Triton X−100(Sigma−Aldrich)で10分間にわたって透過性にした。スライドを、ロバ血清とともに20分間にわたってインキュベートし、0.5%BSA/PBS中で希釈した以前に最適化した、ラットアルブミンに対する一次抗体(Abcam ab14255,1:100)、CK19に対する一次抗体(Biodesign Int. M08029M,1:100)、CK7に対する一次抗体(Chemicon MAB3226,1:100)、CK18に対する一次抗体(Sigma F−4772)およびビメンチンに対する一次抗体(Abcam ab8979,1:50)と1時間にわたってインキュベートした。スライドを、PBS中で3回洗浄し、続いて、適切なFITC標識2次抗体(Abcam ab6749もしくはDako Cytomation F0313,Ely,UK)で洗浄した。上記一次抗体の省略を、陰性コントロールとして行った。ラット肝臓の凍結切片を、陽性コントロールとして使用した。スライドを、Vectashield中にマウントする前に3回洗浄し、そして蛍光顕微鏡によって可視化し、撮影した。
【0067】
(RT−PCR)
総RNAを、製造業者の説明書に従ってTrizol(登録商標)を使用することによって抽出し、GeneQuant分析器によって定量した。サンプルを、DNAse処理(Ambion)し、cDNAへの逆転写を、製造業者の説明書に従って、SuperScript II逆転写酵素(Invitrogen)を使用して行った。RTなし陰性コントロールを、全てのサンプルについて行った。RT−PCRを、Taqポリメラーゼ(Invitrogen)を使用して行った。ハウスキーピング遺伝子であるBactinを使用して、テンプレートの品質を評価した。全てのPCR反応を、Peltier Thermal Cycler −200を使用して行った。ネスト化PCRを、膵臓分化実験について行った。
【0068】
以下の特異的オリゴヌクレオチドプライマーに、PDX 1、インスリンIIおよびグルカゴンを使用した(PDX−1,順方向 5−cggccacacagctctacaagg−3(配列番号1)、逆方向 5−ctccggttctgctgcgtatgc−3(配列番号2)、ネスト化逆方向 5−ttccaggcccccagtctcgg−3(配列番号3)(305bp)、インスリン,順方向 5−atggccctgtggatccgctt−3(配列番号4);逆方向 5−tgccaaggtctgaaggtcac−3(配列番号5);ネスト化順方向 5−cctgctcatcctctgggagcc−3(配列番号6)(209bp);グルカゴン,順方向 5−gaccgtttacgtggctgg−3(配列番号7);逆方向 5−cggttcctcttggtgttcatcaag−3(配列番号8);ネスト化順方向 5−acaaggcagctggcagcatgc−3(配列番号9)(210bp)。ラット膵臓総RNAを逆転写し、陽性コントロールとして使用した。以下の特異的オリゴヌクレオチドプライマーを、肝臓分化実験のために使用した:アルブミン(141bp) 順方向 5-ctgggagtgtgcagatatcagagt−3(配列番号10),逆方向 5−gagaaggtcaccaagtgctgtagt−3(配列番号11)、HNF3β(63bp) 順方向 5−cctactcgtacatctcgctcatca−3(配列番号12),逆方向 −cgctcagcgtcagcatctt(配列番号13)、HNF1(138bp)α 順方向 5−agctgctcctccatcatcaga−3(配列番号14),逆方向 5−tgttccaagcattaagttttctattctaa−3(配列番号15)、Gata4(173bp) 順方向 5−catgcttgcagttgtgctag−3(配列番号16),逆方向 5−attctctgctacggccagta−3(配列番号17)、α−フェトプロテイン(124bp) 順方向 5−gtcctttcttcctcctggagat−3(配列番号18),逆方向 5−ctgtcactgctgatttctctgg−3(配列番号19)、CYP2B1(549bp) 順方向 5−gagttcttctctgggttcctg−3(配列番号20),逆方向 5−actgtgggtcatggagagct−3(配列番号21)、CK19(193bp) 順方向 5−agtaacgtgcgtgctgacac−3(配列番号22),逆方向 5−agtcgcactggtagcaaggt−3(配列番号23)、CK18(70bp) 順方向 5ggacctcagcaagatcatggc−3(配列番号24),逆方向 5 ccacgatcttacgggtagttg−3(配列番号25)。次いで、上記PCR産物に、アガロースゲル電気泳動をかけて、エチジウムブロミド染色で可視化した。ラット肝臓組織を、陽性コントロールとして使用した。
【0069】
(過ヨウ素酸シッフ染色)
グリコーゲン貯蔵についての過ヨウ素酸シッフ染色を、未分化のThy1.1陽性細胞およびThy1.1陰性細胞、ならびに肝臓分化21日目のThy1.1陽性集団およびThy1.1陰性集団に対して行った。ヒト肝臓切片を、陽性コントロールとして使用した。細胞を、4% パラホルムアルデヒド中、室温で10分間にわたって固定した。細胞をPBS中で3回洗浄し、0.1% Triton X−100(Sigma−Aldrich)で10分間にわたって透過性にし、PBSで2回およびddH2Oで1回洗浄した。細胞を、過ヨウ素酸溶液(1g/dL)に室温で5分間浸漬した。ウェルを、蒸留水で3回すすいだ。細胞を、シッフ試薬中に室温で15分間浸漬した。細胞を、水道の流水で5分間洗浄した。細胞をヘマトキシリン溶液で90秒間、対比染色した。水道の流水で15〜30秒間、細胞をすすぐ。
【0070】
(結果)
(Thy1.1陽性集団およびThy1.1陰性集団の特徴付け)
PDPCのMACソーティングを使用して、それぞれ、Thy1.1陽性細胞を発現する集団を98.5%超で、およびThy1.1陰性細胞を発現する集団を98.7%純度で単離した。次いで、これら集団を培養し、いかなる分化実験もしくは特徴付け実験の前にも、10〜12日ごとに定期的にフローサイトメトリーによって再評価した。
【0071】
表現型的には、Thy1.1陽性集団およびThy1.1陰性集団は、形態学において別個の差異を示した:上記Thy1.1陽性集団は、線維芽細胞様形態を示した(図1,パネルA)一方で、Thy1.1陰性集団は、より上皮様の形態を示した(図1,パネルD)。
【0072】
いくつかの差異がまた、Thy1.1陽性細胞集団と、Thy1.1陰性細胞集団との間で、細胞表面マーカーの発現において認められた。両方の細胞系統は、CD147、CD44およびCD49fを発現した。両方とも、CD71は低く、造血(haematopoetic)マーカーであるCD31、CD34、CD45およびc−kitを発現しなかった。上記Thy1.1陽性ソートした細胞集団とは対照的に、上記Thy1.1陰性集団は、CD24について陽性であった(図2aおよび2b)。上記亜集団はまた、肝性マーカー、胆嚢マーカーおよび間葉性マーカーであるアルブミン、ビメンチン、CK7およびCK19での免疫細胞化学によって評価した(図3)。
【0073】
両方のThy1.1ソートした集団は、アルブミン、CK7およびビメンチンについて陰性であった(図3−パネルB、C、F、G、JおよびK)。上記Thy1.1陽性集団は、CK19についても陰性であった(図3,パネルO)のに対して、上記Thy1.1陰性細胞は、弱陽性であった(図3,パネルN)。両方の集団は、RT PCRによって、c−Metおよびネスチンについて陽性であった(データは示さず)。
【0074】
まとめると、上記Thy1.1陰性細胞集団は、以下の細胞表面マーカープロフィールを発現する:
Thy1.1(CD90) 陰性
Pdx−1 陽性
CD49f 約95%+
CD24 約80%+
CD147 約80%+
CD45 陰性
CD44 約60%+
CD71 低い
CD31 陰性
c−Kit 陰性
ck7 陰性
CK19 弱陽性
c−Met 陽性
上記Thy1.1陽性細胞は、以下の細胞表面マーカープロフィールを発現する:
Thy1.1(CD90) 陽性
Pdx−1 陽性
CD49f 約95%+
CD24 陰性
CD147 約90%+
CD45 陰性
CD44 約85%+
CD71 低い
CD31 陰性
C−KIT 陰性
CK19 陰性
c−Met 陽性
ネスチン 陽性。
【0075】
(Thy1.1陽性集団およびThy1.1陰性集団の分化能)
(膵臓分化)
Thy1.1陽性集団およびThy1.1陰性集団は、膵臓分化培地において顕著に異なる形態学的変化を示した。上記Thy1.1陽性集団は、形態において最初に線維芽細胞様を示し、14〜21日目までに絡み合った細胞クラスタを形成し、28日目までに島様の球形クラスタへと形成された。これは、最終的に、親細胞層から分離した(図1,パネルD、H、I)。対照的に、上記Thy−1.1陰性細胞は、小さな上皮様形態を有する単層のままであり、三次元構造へと発展しなかった(図1,パネルAおよびB)。
【0076】
未分化のThy1.1陽性細胞のRT−PCR分析は、陽性Pdx−1発現を示したが、インスリンの発現もグルカゴンの発現も示さなかった(図4−B)。分化した細胞クラスタは、3種のマーカー全ての転写発現について全て陽性であった(図4 パネルB)。
【0077】
しかし、Thy1.1陰性細胞は、維持培地もしくは分化培地のいずれかで増殖させた場合に、Pdx−1を発現した。著しいことには、分化培地中で増殖させた場合、形態学的な変化を示さないにも拘わらず、上記Thy1.1陰性集団においてインスリン転写が検出された。グルカゴンは、上記未分化のThy−1.1陰性細胞において発現せず、分化後にインビトロで誘導もされなかった(図4 パネルA)。
【0078】
(肝臓分化)
Thy−1.1陽性PDPCおよびThy−1.1陰性PDPCを、血清非含有FGF4含有培地中で培養して、肝性の能力を評価した。Thy−1.1陽性細胞は、線維芽細胞様から上皮/立方形形態へと形態学的変化を示した。さらに、28日目までには、平らになった上皮との培養全体を通じて、管腔構造が明らかになった。上記膵臓分化プレートにおいて認められるものに類似したときおりの三次元島様構造が、肝臓分化プレートにおいても認められた。上記Thy−1.1陰性集団は、単層のままであり、三次元構造も管腔様構造も明らかでなく、顕著な形態学的変化もなかった(図1A,C)。
【0079】
本発明者は、RT−PCRによって28日間にわたって、上記2つの集団の分化をさらに試験した。RT−PCRを、内胚葉特異的遺伝子であるHNF3βおよびGATA 4、初期肝臓マーカーであるαフェトプロテインおよびCK18、成熟肝臓マーカーであるHNF1α、アルブミンおよびシトクロームP450酵素CYP2B1について行った。未分化のThy1.1陰性細胞は、RT−PCRによって、HNF3−βおよびCK19を発現したが、アルブミン、CK18、HNF1α、CY2B1、Gata4およびαフェトプロテインを発現しなかった(図5,パネルB)。他の初期肝臓マーカーもしくは成熟肝臓マーカー、またはCY2B1のいずれも、上記Thy1.1陰性集団において誘導されなかった。興味深いことには、未分化のThy1.1陽性PDPCは、初期内胚葉マーカーであるHNF3−β、GATA 4およびαフェトプロテインを発現したが、肝臓分化の14日目までに、肝細胞分化の後期マーカーであるHNF1−αおよびアルブミンを発現しなかった(図5 パネルA)。CK 19(胆嚢細胞によって正常に発現される)はまた、培養における上記管腔構造の出現と一致して、14〜28日目の間に誘導された。アルブミン発現の誘導は、免疫細胞化学によって確認された。未分化の細胞は、アルブミン内容物について陰性に染色される(図3,パネルA)一方で、14日目の分化した細胞は、アルブミン染色について強く陽性であった(図3,パネルD)。興味深いことには、10日目および14日目の時点で分化した細胞は、胆嚢マーカーであるCK19およびCK7について陰性に染色された(図3,パネルNおよびG)が、上記管腔様構造のみにおいてビメンチンについては陽性に染色された(図3,パネルL)。CK 18はまた、未分化のThy1.1陽性細胞において、および上記28日間の分化期間全体を通じて発現した。CYP2B1は、未分化のThy1.1陽性細胞において、および上記分化期間全体を通じて存在した。
【0080】
未分化のThy1.1陰性細胞は、免疫細胞化学によれば、CK7、ビメンチンおよびアルブミンの発現について陰性であり、CK19について弱陽性であった。上記Thy1.1陽性集団は、CK19について陰性であった。
【0081】
(グリコーゲン貯蔵についての過ヨウ素酸シッフ(PAS)染色)
貯蔵グリコーゲンの存在は、PAS染色によって決定される場合、Thy−1.1陽性PDPCでもThy−1.1陰性PDPCでも認められず、21日目の分化したThy−1.1陰性細胞においても認められなかった。しかし、グリコーゲン貯蔵を示す、PASでの陽性染色は、21日目までに上記Thy1.1陽性の分化した細胞において認められた(図6)。
【0082】
(糖尿病の動物モデルにおける細胞亜集団の試験)
本明細書に記載の細胞亜集団(例えば、Thy1.1陽性細胞集団もしくはThy1.1陰性細胞集団)は、糖尿病のような疾患の動物モデルにおいて使用されうる。一実施形態において、細胞の亜集団は、ストレプトゾトシン(STZ)誘導性の糖尿病の齧歯類和合(rodent concordant)異種移植片モデルにおいて使用される。C57BL/6マウスを、0日目にSTZを注射することによって糖尿病にする一方で、750,000個の細胞(例えば、Thy1.1陽性細胞)を、3日目に、処置した動物の尾静脈に注射する。コントロール動物に、生理食塩水の注射を与えるか、または等数のC57BL/6骨髄細胞を与える。3日ごとに血糖をモニターする。血糖の安定化および/もしくはコントロールに対して増大した生存は、上記投与した細胞が、上記動物においてインスリン生成をもたらすことを示す。
【0083】
(考察)
本明細書で提供されるのは、Thy1.1陽性PDPC亜集団およびThy1.1陰性PDPC亜集団のインビトロ培養、選択および特徴付けに関する詳細である。さらに、膵臓系統および肝臓系統への分化、ならびにマーカーThy1.1(これは、インビトロで系統への両能性を示す)を使用してソートした細胞集団の提供に関するそれらの効力について詳細が開示される。
【0084】
Thy1.1は、機能が明らかには理解されていない細胞表面タンパク質である。しかし、細胞認識(Gunterら,1984;Williams,1985)、細胞接着(Gunterら, 1984; Williams, 1985)(Heら,1991;Hueberら,1992)およびシグナル伝達(Kroczekら,1986)に関与していることが示唆された。Thy1.1発現は種々の幹細胞集団において認められ、顕著には、成体ラット肝臓における円形細胞集団は、Thy1.1が、潜在的に損傷後の修復細胞として、細胞が間質組織を認識しかつこれに接着することを可能にし得るという仮説をもたらした(Massonら,2006;Petersenら,1998;Terraceら,2007)。Thy1.1はまた、ヒト、マウスおよびラットにおける胎児肝臓の幹細胞、臍帯血および間葉性幹細胞上で発現される。Thy1.1陽性集団内のより大きなインビトロ効力における本知見は、これら観察と一致している。これら観察はまた、このような細胞のさらなる使用を可能にするために、単離および精製のための方法もまた、示す。
【0085】
以前の研究から、多くの異なる細胞タイプ(骨髄由来MSC、MAPC、子宮内膜および膵臓由来のMSCを含む)の肝臓分化が示された(Jiangら,2002;Mengら,2007;Schwartzら,2002;Seebergerら,2006)。PDPCの(Jiangら, 2002; Mengら, 2007; Schwartzら, 2002; Seebergerら, 2006)Thy1.1陽性亜集団は、形態学的表現型を共有し、これら集団とともに多くの細胞表面マーカーを発現する(CD44+、CD24−、CD45−、CD31−およびCD34を含む)。しかし、このこととは対照的に、Thy1.1陽性PDPCは、GATA4、HNF3−βおよびαフェトプロテインを発現する別個の細胞タイプであるようであり、これは、これら他の細胞タイプのうちのいずれかについて発現される場合に記載されていない。
【0086】
HNF3−βは、内胚葉能力において重要な役割を果たすと考えられている完全な兄はいようのマーカーである(Gualdiら 1996)一方で、GATA4は、腹側前腸内胚葉発生および初期肝臓遺伝子発現について必要とされる転写因子である(Gualdiら,1996;Rossiら,2001)。HNF3−βは、上記アルブミンエンハンサーへのGATA4の結合のその後の促進とともに、アルブミンエンハンサーの前後に位置しているヌクレオソームに指向されることが示された(McPhersonら,1996;Cirillo and Zaret,1999)。GATA4−/−胚およびHNF3−β−/−胚の両方は、前腸形態発生の欠損を示す(Duncanら,1997)。従って、未分化のPDPCにおけるHNF3−βおよびGATA4の発現、ならびにその後の肝臓特異的遺伝子(例えば、アルブミンおよびHNF1−α)の発現のFGF刺激誘導は、HNF 3−βおよびGATA 4が、これら細胞が肝臓へと発達する結果(hepatic fate)になる可能性を制御するように協同して作用するという提唱と一致する。さらに、分化の21日後の上記Thy1.1陽性集団におけるPAS染色の存在は、より成熟した肝細胞の機能的特徴を示した。このことは、HNF1−αの発現と一致する。HNF1−αは、遺伝子産物が成熟肝機能(炭水化物貯蔵および合成、ならびに脂質代謝を含む)と関連する遺伝子に結合することが公知である(Odomら,2004)。
【0087】
未分化のThy1.1陽性PDPCは、AFPを発現する。この観察は、初期腹側前腸内胚葉における肝形成の前に、ネスチン陽性島由来の前駆細胞におけるAFP発現、ならびに低レベルのAFPおよびTTR発現を記載する報告と一致する。この発現は、その後、心臓の中胚葉シグナル伝達から分離された内胚葉において失われる(Gualdiら,1996;Jungら,1999;Zulewskiら,2001)。このことは、腹側前腸内胚葉の標準的な(default)膵臓となる運命の特徴であることもまた、示唆された(Deuschら)。前記Thy1.1陽性PDPC集団におけるAFPの発現(これは、膵臓系統および肝臓系統の両方への能力を示す)は、この知見と矛盾しない(Deutschら,2001)。
【0088】
重大なことには、上記未分化のThy1.1陰性集団は、HNF3βを発現するが、GATA 4も、αフェトプロテインも発現せず、上記分化実験の間に誘導されなかった。肝臓の能力の証拠は、上記Thy1.1陰性集団において認められなかった。このことは、この未分化の集団内でのPdx−1発現、およびGata4発現の非存在と一致する。ビメンチンは、未分化のThy1.1陽性集団においても、Thy1.1陰性集団においても発現されなかったが、肝臓分化の間に管様構造を形成する細胞において発現された。ビメンチンは、間葉マーカーを表すと考えられる。しかし、Massonらは、肝門構造(portal structure)におけるThy1.1およびビメンチンの発現を観察し、組織切片内の上皮細胞における、および胎児肝臓上皮細胞の培養物におけるビメンチン発現も示した(Massonら,2006)。(Massonら, 2006)
上記膵臓分化に関するデータは、興味をそそるものである。島様クラスタの形態学的な証拠は、上記Thy1.1陰性集団において認められなかった。対照的に、Thy1.1陽性PDPCは、三次元島様構造およびPDX−1、インスリンおよびグルカゴンの転写発現を生じる特徴的な形態学的変化とともに、容易に膵臓系統へと誘導できた。
【0089】
両方の集団におけるPdx−1転写発現の検出は、それらがインスリン生成細胞になる可能性を示す。しかし、顕著なことには、Thy1.1陰性細胞は、分化培地中で増殖される場合、形態学的屁かを示さないにも拘わらず、インスリンを発現した。グルカゴンは、上記未分化のThy−1.1陰性細胞において発現されず、分化後にインビトロで誘導もされなかった(図4 パネル)。
【0090】
膵臓前駆細胞/幹細胞の種々の異なる候補集団が、以前から記載されてきた(ネスチンもしくは他の神経幹細胞マーカーを発現する島前駆細胞を含む)(Abrahamら,2004;Corneliusら,1997;Lechnerら,2002;Ramiyaら,2000)。別の集団は、PDX−1(インスリン生成細胞についての公知のマーカー)を発現することが示された。そしてこれら細胞は、適切な条件下で、インビトロで導管および内分泌腺分化の両方を刺激し得る(Bonner−Weirら,2000;Otonkoskiら,1993)。さらに、膵管の上皮細胞が、新たな島および腺房の増殖および形成の能力がある前駆細胞へ分化する能力を有するという証拠が存在し(Bonner−Weirら,2004)、もっとも近年では、(Bonner−Weirら, 2004)CK19+ 非ナイン分泌性膵臓上皮細胞(NEPC)が、胎児膵臓組織の存在下にある場合、インビボでインスリン生成細胞へと分化するように部分的に誘導されることが報告された(Haoら,2006)。
【0091】
これら細胞が果たす正確な生理学的役割が探究されてきた。Dorらは、新たな島細胞生成よりむしろβ細胞複製が、優勢な機構である(これによって、膵臓内分泌組織が、ほぼ完全な膵切除術後に再生する)という議論の代わりに、導管細胞もしくは前駆細胞からの再生が生じるという見方を吟味してきた(Dorら,2004)が、この解釈は、未だ論議の的である(Bonner−Weir and Weir,2005)。本知見は、このようなプロセスを潜在的に促進する場合に、インスリンを生成し得る非β細胞の役割と一致する。これら細胞が移植治療の代替のインスリン生成細胞供給源を提供することは明らかである。
【0092】
本発明者は、無血清FGF−4含有分化プロトコルの使用によって、肝臓系統分化を示す、形態学的発現変化および遺伝子発現変化の両方の過程を観察した。膵臓分化および肝臓分化についての胚性腹側内胚葉の潜在的な両能性は、腹側内胚葉が心臓中胚葉への近接によって肝臓系統へ分化した外植片実験において調査された。心臓中胚葉によって分泌される誘導因子(例えば、FGF−1、FGF−2およびFGF−4)の非存在は、腹側内胚葉の標準的な膵臓経路が継続することを可能にし(Deutschら,2001)、そして一般的なFGFシグナル伝達アンタゴニストは、インビトロでの肝発生(Jungら,1999)およびFGF−4(Zhuら,1999)を阻害する。本データは、この概念と完全に一致する。
【0093】
本発明者は、両能性のある内胚葉前駆体の集団と一致して、インビトロで、効力およびシグナル伝達に対する転写応答を示すPDPCの単離および特徴付けを示した。前述で、マウスストレプトゾトシン誘導性糖尿病モデルにおける、ソートしていないPDPC集団の投与は、分化およびラットインスリンの生成と同時に、マウス膵臓再生の刺激を実証した(Shiels 2005およびWO 2006/120476(ともに、本明細書に参考として援用される))。
【0094】
参考文献のリスト
【0095】
【化1】
【0096】
【化2】
【0097】
【化3】
【0098】
【化4】
【0099】
【化5】
【0100】
【化6】
【0101】
【化7】
【図1A】
【図1B】
【図1C】
【図1D】
【図1E】
【図1F】
【図1G】
【図1H】
【図1I】
【図2A】
【図2B】
【図3A】
【図3B】
【図3C】
【図3D】
【図3E】
【図3F】
【図3G】
【図3H】
【図3I】
【図3J】
【図3K】
【図3L】
【図3M】
【図3N】
【図3O】
【図3P】
【特許請求の範囲】
【請求項1】
成体組織に由来する単離された細胞集団であって、ここで該細胞は、血清の存在下でマトリゲルなしの培養システムにおいて増殖でき;該細胞は、Pdx−1陽性であることで特徴付けられる、単離された細胞集団。
【請求項2】
前記細胞は、ネスチン陰性である、請求項1に記載の単離された細胞集団。
【請求項3】
前記細胞は、ネスチン陽性である、請求項1に記載の単離された細胞集団。
【請求項4】
前記成体組織は、膵臓組織、骨髄組織、心臓組織、乳房組織、肝臓組織もしくは腎臓組織である、上記請求項のいずれか1項に記載の単離された細胞集団。
【請求項5】
前記成体組織は、ヒトのものである、請求項3に記載の単離された細胞集団。
【請求項6】
前記成体組織は、膵臓組織である、請求項4に記載の単離された細胞集団。
【請求項7】
前記細胞は、2005年5月12日にECACCにおいてアクセッション番号Q6203の下で寄託された細胞に由来する、請求項6に記載の単離された細胞集団。
【請求項8】
前記細胞は、細胞表面マーカーThy1.1について陽性である、上記請求項のいずれか1項に記載の単離された細胞集団。
【請求項9】
前記細胞は、細胞表面マーカーThy1.1について陰性である、請求項1〜7のいずれか1項に記載の単離された細胞集団。
【請求項10】
上記請求項のいずれか1項に記載の単離された細胞集団および薬学的に受容可能なキャリアを含む、薬学的組成物。
【請求項11】
成体哺乳動物組織から両能性のある幹細胞集団を単離する方法であって、該方法は、
該成体哺乳動物組織を培養する工程;
発生する細胞集団単層を単離する工程;および
細胞表面マーカーThy1.1について陽性である細胞をさらに単離し、それによって、両能性のある幹細胞集団を提供する工程、
を包含する、方法。
【請求項12】
前記単離された細胞は、CD147、CD44、CD49F、C−Metおよびネスチンからなる群より選択される1種以上の細胞表面マーカーについても陽性である、請求項11に記載の方法。
【請求項13】
前記成体哺乳動物組織は、ヒトのものである、請求項11または12に記載の方法。
【請求項14】
前記成体哺乳動物組織は、膵臓、乳房、肝臓もしくは腎臓からなる群より選択される、請求項11〜13のいずれか1項に記載の方法。
【請求項15】
前記組織は、CMRL−1066培地(sigma−C−0422)中で培養される、請求項11〜14のいずれか1項に記載の方法。
【請求項16】
前記組織は、管全体(whole duct)を含む成体膵管に由来する、請求項11〜15のいずれか1項に記載の方法。
【請求項17】
前記管組織は、前記培養工程を補助するために切り刻まれる、請求項16に記載の方法。
【請求項18】
両能性のある幹細胞集団を単離する方法であって、該方法は、2005年5月12日にECACCにおいてアクセッション番号Q6203の下で寄託されたような細胞集団を得る工程;および細胞表面マーカーThy1.1について陽性である細胞の亜集団を単離する工程、を包含する、方法。
【請求項19】
請求項11〜18のいずれか1項に記載の方法によって得ることができる、成体細胞集団。
【請求項20】
細胞変性もしくは加齢性組織変化と関連する疾患状態を処置する方法であって、該方法は、請求項1〜9のいずれか1項に記載の細胞集団もしくは請求項10に記載の薬学的組成物を、該疾患を有する患者に投与する工程を包含する、方法。
【請求項21】
前記細胞は、前記患者に静脈内投与される、請求項20に記載の方法。
【請求項22】
前記細胞は、前記疾患部位もしくは加齢性変性の部位に移植される、請求項20に記載の方法。
【請求項23】
前記疾患は、膵臓細胞、ニューロン細胞、心血管細胞、上皮細胞、肝細胞、筋細胞、網膜細胞、毛包細胞もしくは腎細胞の変性と関連する、請求項20〜22のいずれか1項に記載の方法。
【請求項24】
前記疾患は、糖尿病(I型もしくはII型)、パーキンソン病、アルツハイマー病、腎疾患、眼の疾患、肝疾患もしくは心血管疾患である、請求項23に記載の方法。
【請求項25】
前記細胞のドナーおよび前記患者は、同じ種である、請求項20〜24のいずれか1項に記載の方法。
【請求項26】
前記患者はヒトである、請求項25に記載の方法。
【請求項27】
請求項9に記載の成体幹細胞集団を、成体哺乳動物組織から単離する方法であって、該方法は、
該成体哺乳動物組織を培養する工程;
発生する細胞集団単層を単離する工程;および
細胞表面マーカーThy1.1について陰性である細胞をさらに単離する工程、
を包含する、方法。
【請求項28】
前記単離された細胞は、CD147、CD49f、CD44、CK19、C−Metおよびネスチンからなる群より選択される1種以上の細胞表面マーカーについて陽性である、請求項27に記載の方法。
【請求項29】
前記成体哺乳動物組織は、ヒトのものである、請求項27または28に記載の方法。
【請求項30】
前記成体哺乳動物組織は、膵臓、乳房、肝臓もしくは腎臓からなる群より選択される、請求項27〜29のいずれか1項に記載の方法。
【請求項31】
請求項9に記載の成体幹細胞集団を単離する方法であって、該方法は、2005年5月12日にECACCにおいてアクセッション番号Q6203の下で寄託されたような細胞集団を得る工程;および細胞表面マーカーThy1.1について陰性である細胞の亜集団を単離する工程を包含する、方法。
【請求項32】
処置方法において使用するための、請求項8に記載の単離された成体幹細胞集団。
【請求項33】
前記処置方法は、膵臓細胞もしくは肝細胞の変性と関連する疾患のためである、請求項32に記載の単離された幹細胞集団。
【請求項34】
処置方法において使用するための、請求項9に記載の単離された幹細胞集団。
【請求項35】
前記処置方法は、糖尿病のためである、請求項34に記載の単離された幹細胞集団。
【請求項36】
前記処置方法は、前記細胞を疾患部位に移植する工程を包含する、請求項32〜35のいずれか1項に記載の単離された幹細胞集団。
【請求項1】
成体組織に由来する単離された細胞集団であって、ここで該細胞は、血清の存在下でマトリゲルなしの培養システムにおいて増殖でき;該細胞は、Pdx−1陽性であることで特徴付けられる、単離された細胞集団。
【請求項2】
前記細胞は、ネスチン陰性である、請求項1に記載の単離された細胞集団。
【請求項3】
前記細胞は、ネスチン陽性である、請求項1に記載の単離された細胞集団。
【請求項4】
前記成体組織は、膵臓組織、骨髄組織、心臓組織、乳房組織、肝臓組織もしくは腎臓組織である、上記請求項のいずれか1項に記載の単離された細胞集団。
【請求項5】
前記成体組織は、ヒトのものである、請求項3に記載の単離された細胞集団。
【請求項6】
前記成体組織は、膵臓組織である、請求項4に記載の単離された細胞集団。
【請求項7】
前記細胞は、2005年5月12日にECACCにおいてアクセッション番号Q6203の下で寄託された細胞に由来する、請求項6に記載の単離された細胞集団。
【請求項8】
前記細胞は、細胞表面マーカーThy1.1について陽性である、上記請求項のいずれか1項に記載の単離された細胞集団。
【請求項9】
前記細胞は、細胞表面マーカーThy1.1について陰性である、請求項1〜7のいずれか1項に記載の単離された細胞集団。
【請求項10】
上記請求項のいずれか1項に記載の単離された細胞集団および薬学的に受容可能なキャリアを含む、薬学的組成物。
【請求項11】
成体哺乳動物組織から両能性のある幹細胞集団を単離する方法であって、該方法は、
該成体哺乳動物組織を培養する工程;
発生する細胞集団単層を単離する工程;および
細胞表面マーカーThy1.1について陽性である細胞をさらに単離し、それによって、両能性のある幹細胞集団を提供する工程、
を包含する、方法。
【請求項12】
前記単離された細胞は、CD147、CD44、CD49F、C−Metおよびネスチンからなる群より選択される1種以上の細胞表面マーカーについても陽性である、請求項11に記載の方法。
【請求項13】
前記成体哺乳動物組織は、ヒトのものである、請求項11または12に記載の方法。
【請求項14】
前記成体哺乳動物組織は、膵臓、乳房、肝臓もしくは腎臓からなる群より選択される、請求項11〜13のいずれか1項に記載の方法。
【請求項15】
前記組織は、CMRL−1066培地(sigma−C−0422)中で培養される、請求項11〜14のいずれか1項に記載の方法。
【請求項16】
前記組織は、管全体(whole duct)を含む成体膵管に由来する、請求項11〜15のいずれか1項に記載の方法。
【請求項17】
前記管組織は、前記培養工程を補助するために切り刻まれる、請求項16に記載の方法。
【請求項18】
両能性のある幹細胞集団を単離する方法であって、該方法は、2005年5月12日にECACCにおいてアクセッション番号Q6203の下で寄託されたような細胞集団を得る工程;および細胞表面マーカーThy1.1について陽性である細胞の亜集団を単離する工程、を包含する、方法。
【請求項19】
請求項11〜18のいずれか1項に記載の方法によって得ることができる、成体細胞集団。
【請求項20】
細胞変性もしくは加齢性組織変化と関連する疾患状態を処置する方法であって、該方法は、請求項1〜9のいずれか1項に記載の細胞集団もしくは請求項10に記載の薬学的組成物を、該疾患を有する患者に投与する工程を包含する、方法。
【請求項21】
前記細胞は、前記患者に静脈内投与される、請求項20に記載の方法。
【請求項22】
前記細胞は、前記疾患部位もしくは加齢性変性の部位に移植される、請求項20に記載の方法。
【請求項23】
前記疾患は、膵臓細胞、ニューロン細胞、心血管細胞、上皮細胞、肝細胞、筋細胞、網膜細胞、毛包細胞もしくは腎細胞の変性と関連する、請求項20〜22のいずれか1項に記載の方法。
【請求項24】
前記疾患は、糖尿病(I型もしくはII型)、パーキンソン病、アルツハイマー病、腎疾患、眼の疾患、肝疾患もしくは心血管疾患である、請求項23に記載の方法。
【請求項25】
前記細胞のドナーおよび前記患者は、同じ種である、請求項20〜24のいずれか1項に記載の方法。
【請求項26】
前記患者はヒトである、請求項25に記載の方法。
【請求項27】
請求項9に記載の成体幹細胞集団を、成体哺乳動物組織から単離する方法であって、該方法は、
該成体哺乳動物組織を培養する工程;
発生する細胞集団単層を単離する工程;および
細胞表面マーカーThy1.1について陰性である細胞をさらに単離する工程、
を包含する、方法。
【請求項28】
前記単離された細胞は、CD147、CD49f、CD44、CK19、C−Metおよびネスチンからなる群より選択される1種以上の細胞表面マーカーについて陽性である、請求項27に記載の方法。
【請求項29】
前記成体哺乳動物組織は、ヒトのものである、請求項27または28に記載の方法。
【請求項30】
前記成体哺乳動物組織は、膵臓、乳房、肝臓もしくは腎臓からなる群より選択される、請求項27〜29のいずれか1項に記載の方法。
【請求項31】
請求項9に記載の成体幹細胞集団を単離する方法であって、該方法は、2005年5月12日にECACCにおいてアクセッション番号Q6203の下で寄託されたような細胞集団を得る工程;および細胞表面マーカーThy1.1について陰性である細胞の亜集団を単離する工程を包含する、方法。
【請求項32】
処置方法において使用するための、請求項8に記載の単離された成体幹細胞集団。
【請求項33】
前記処置方法は、膵臓細胞もしくは肝細胞の変性と関連する疾患のためである、請求項32に記載の単離された幹細胞集団。
【請求項34】
処置方法において使用するための、請求項9に記載の単離された幹細胞集団。
【請求項35】
前記処置方法は、糖尿病のためである、請求項34に記載の単離された幹細胞集団。
【請求項36】
前記処置方法は、前記細胞を疾患部位に移植する工程を包含する、請求項32〜35のいずれか1項に記載の単離された幹細胞集団。
【図4】

【図5】
【図6】

【図5】
【図6】
【公表番号】特表2011−519574(P2011−519574A)
【公表日】平成23年7月14日(2011.7.14)
【国際特許分類】
【出願番号】特願2011−507987(P2011−507987)
【出願日】平成21年5月8日(2009.5.8)
【国際出願番号】PCT/GB2009/001149
【国際公開番号】WO2009/136168
【国際公開日】平成21年11月12日(2009.11.12)
【出願人】(506277591)ザ ユニバーシティ コート オブ ザ ユニバーシティ オブ グラスゴー (1)
【Fターム(参考)】
【公表日】平成23年7月14日(2011.7.14)
【国際特許分類】
【出願日】平成21年5月8日(2009.5.8)
【国際出願番号】PCT/GB2009/001149
【国際公開番号】WO2009/136168
【国際公開日】平成21年11月12日(2009.11.12)
【出願人】(506277591)ザ ユニバーシティ コート オブ ザ ユニバーシティ オブ グラスゴー (1)
【Fターム(参考)】
[ Back to top ]