
経口投与用の低分子干渉リボ核酸(siRNA)
【課題】 少なくとも15ヌクレオチドにわたって互いに相補的である2本の別個のRNA鎖を含む経口投与用の低分子干渉リボ核酸(siRNA)であって、ここで各鎖は49ヌクレオチド以下であり、そしてここでその鎖の少なくとも1本は少なくとも一つの化学的修飾を含有するsiRNAを提供すること。
【解決手段】 少なくとも15ヌクレオチドにわたって互いに相補的である2本の別個のRNA鎖を含む経口投与用の低分子干渉リボ核酸(siRNA)であって、ここで各鎖は49ヌクレオチド以下であり、そしてここでその鎖の少なくとも1本は少なくとも一つの化学的修飾を含有するsiRNAは、血管形成障害を処置するために使用することができること見いだした。
【解決手段】 少なくとも15ヌクレオチドにわたって互いに相補的である2本の別個のRNA鎖を含む経口投与用の低分子干渉リボ核酸(siRNA)であって、ここで各鎖は49ヌクレオチド以下であり、そしてここでその鎖の少なくとも1本は少なくとも一つの化学的修飾を含有するsiRNAは、血管形成障害を処置するために使用することができること見いだした。
【発明の詳細な説明】
【技術分野】
【0001】
背景
RNA干渉は当初植物において転写後遺伝子サイレンシング(PTGS)として見出された、二本鎖RNA(dsRNA)が引き金となる高度に保存されたメカニズムであり、そしてdsRNAに相同な遺伝子の転写を下方調節できる1。dsRNAは最初にダイサーにより低分子干渉RNA(siRNA)と称される21−23ヌクレオチドの短い二重鎖にプロセシングされる2。それはRNA誘起サイレンシング複合体(RISC)に組み込まれているので、RISCの構成成分であるアルゴノート2により標的mRNAを相同性の領域の中央で切断することにより遺伝子サイレンシングを媒介することができる3。Elbashir et al(2001)4では、合成siRNAの直接導入によりショウジョウバエのみならず哺乳動物細胞においてもRNA干渉遺伝子サイレンシングが媒介されることが実証された。それ以来siRNA媒介遺伝子サイレンシングは標的同定および標的バリデーション研究の双方で、強力でそして広く用いられる分子生物学的手段になってきている。動物実験での遺伝子サイレンシングのためのsiRNAの使用は限られた量の動物モデルでしか記載されていない。未修飾siRNAは眼において局所的に5、中枢神経系において髄腔内または小脳内に6、および呼吸器系ウイルスの阻止のために鼻内に7送達された。未修飾siRNAの静脈内水力学的尾静脈注射もまた研究されている。この研究法により、主に肝臓への急速な送達が可能になる8。未修飾siRNAの全身投与に関しては非常に限られた数の研究しか報告されていない。Duxbury et al9は接着斑キナーゼを標的とする未修飾siRNAを正所性腫瘍異種移植片マウスモデルに静脈内投与し、そして腫瘍成長阻害およびゲムシタビンに対する化学増感を観察した。Soutscheck et alはアポリポタンパク質Bを内在的にサイレンシングするための高度に化学的に修飾されたsiRNAの全身使用を報告した。ほとんどの抗ApoB siRNAの高用量の50mg/kgでの腹腔内投与によりApoBタンパク質レベルおよびリポタンパク質濃度が低下した10。これらの実例にもかかわらず、全身送達時のsiRNAのインビボ使用は、このテクノロジーを標的バリデーションまたは治療適用に広く適用できるようにするために、改良を必要とする。実際、未修飾siRNAは、主に血流で豊富なヌクレアーゼによる酵素消化に供される。siRNAの薬理学的特性を改善するために、いくつかのグループはこれらの試薬の化学的修飾を調査した。記載された研究法はその中でも非常に様々で、そして系統的な研究は依然実施されていないが、結果の概要により化学的修飾に対するsiRNAの寛容性を決定することが可能になる。ホスホロチオエート11またはボラノホスフェート12、2’−O−メチル13、2’−O−アリル14、2’−メトキシエチル(MOE)および2’−デオキシフルオロヌクレオチド15またはロックされた核酸(Locked Nucleic Acid)(LNA)16のようないくつかの化学が調査されている。これらの研究により、修飾に関する寛容性は化学依存性のみならず、位置依存性でもあることが強調された。
【発明の概要】
【0002】
本発明は改善された薬理学的特性を有する最小限に修飾されたsiRNAを提供する。最小限に修飾されたsiRNAは、3’エキソヌクレアーゼ消化を防御するために各鎖の3’末端で修飾された19塩基対二本鎖RNAであり:21ヌクレオチドsiRNAの3’−ジデオキシヌクレオチドオーバーハングが一般的な3’−ヒドロキシプロピルホスホジエステル部分により置き換えられており、そして各鎖の3’末端で最初に塩基対形成している2個のヌクレオチドの修飾が血清安定性をさらに強化する。成体マウスに腹腔内または経口適用した場合、修飾されたsiRNAは成長因子誘起の血管形成モデルにおいてより高度な効力を表し、それは血清安定性の増大に相関する。
【0003】
発明の要旨
一つの態様では、本発明は経口投与用の低分子干渉リボ核酸(siRNA)を提供し、該siRNAは少なくとも15ヌクレオチドにわたって互いに相補的である2本の別個のRNA鎖を含み、ここで各鎖は49ヌクレオチド以下であり、そしてここでその鎖の少なくとも1本は少なくとも一つの化学的修飾を含有する。
【0004】
一つの実施態様では、siRNAは少なくとも1個の修飾されたヌクレオチドを含む。
別の実施態様では、siRNAは少なくとも1個の3’末端キャップを含む。
別の実施態様では、該修飾されたヌクレオチドは2’アルコキシリボヌクレオチド、2’アルコキシアルコキシリボヌクレオチド、ロックされた核酸リボヌクレオチド(LNA)、2’−フルオロリボヌクレオチド、モルホリノヌクレオチドの中から選択される。
別の実施態様では、該修飾されたヌクレオチドはホスホロチオエート、ホスホロジチオエート、ホスホロアミダート、ボラノホスホノエートおよびアミド結合の中から選択される修飾されたヌクレオシド内結合を有するヌクレオチドの中から選択される。
別の実施態様では、該2本のRNA鎖は互いに完全に相補的である。
別の実施態様では、該siRNAは5’末端または3’末端の少なくとも一つに1から6ヌクレオチドオーバーハングを含む。
【0005】
別の実施態様では、siRNAは少なくとも1個の3’キャップを含有し、それは3’炭素を介して3’末端に抱合された化学的部分であり、かつ式I:
【化1】

[式I]
(式中、
XはOまたはSであり、
R1およびR2は独立してOH、NH2、SH、アルキル、アリール、アルキル−アリール、アリール−アルキルであり、ここでアルキル、アリール、アルキル−アリール、アリール−アルキルをさらなるヘテロ原子および官能基、好ましくはN、OもしくはSの群から選択されるヘテロ原子またはOH、NH2、SH、カルボン酸もしくはエステルの群から選択される官能基により置換することができ;
またはR1およびR2は式Y−Zのものでよく、ここでYはO、N、Sであり、そしてZはH、アルキル、アリール、アルキル−アリール、アリール−アルキルであり、ここでアルキル、アリール、アルキル−アリール、アリール−アルキルをさらなるヘテロ原子、好ましくはN、OもしくはSの群から選択されるヘテロ原子により置換することができる)
の化合物の中から選択される。
【0006】
別の実施態様では、siRNAは少なくとも15ヌクレオチドにわたってVEGFR−1、VEGFR−2、VEGFR3、Tie2、bFGFR、IL8RA、IL8RB、FasまたはIGF2RのmRNAまたはプレmRNAに相補的である少なくとも1本の鎖を含有する。
別の実施態様では、siRNAは配列番号:1−900から選択される配列を含む少なくとも1本の鎖を含有する。
別の実施態様では、siRNAは配列番号:901−930からなる群から選ばれる。
別の実施態様では、siRNAは標準的な胃酸アッセイにおいて、同一ヌクレオチド配列を有する未修飾siRNAよりも大きな安定性を有する。
別の実施態様では、siRNAは標準的な胃酸アッセイにおいて、30分暴露後に50%以上である安定性を有する。
【0007】
別の実施態様では、siRNAは標準的な血清アッセイにおいて未修飾siRNAよりも大きな安定性を有する。
別の実施態様では、siRNAは標準的な血清アッセイにおいて30分暴露後に50%以上である安定性を有する。
別の実施態様では、siRNAは標準的な腸洗浄液アッセイにおいて未修飾siRNAよりも大きな安定性を有する。
別の実施態様では、siRNAは同一ヌクレオチド配列の未修飾siRNAと比較して経口バイオアベイラビリティーが強化されている。
【0008】
一つの態様では、本発明は一つまたはそれより多い前記の特性を有するsiRNAを含む医薬組成物を提供する。
別の態様では、本発明は医薬品として使用するための任意の一つまたはそれより多い前記の特性を有するsiRNAを提供する。
別の態様では、本発明は血管形成障害の処置用医薬品の製造における任意の一つまたはそれより多い前記の特性を有するsiRNAの使用を提供する。
別の態様では、本発明はインビトロで血管形成過程を阻害するための任意の一つまたはそれより多い前記の特性を有するsiRNAの使用を提供する。
【図面の簡単な説明】
【0009】
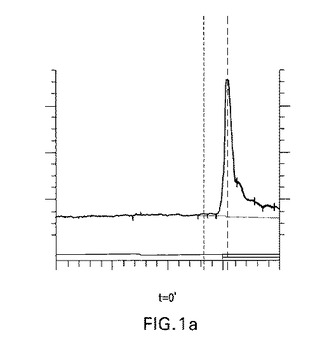
【図1a】図1a、1b、1c、1dおよび1e:未修飾siRNA pGl3−siRNA(マウス血清中の野生型siRNA)の代謝分解;a−c)マウス血清中0分、30分および180分インキュベートした後の未修飾siRNAのイオン交換−HPLC分析;37℃で30分インキュベートした後のイオン交換HPLCにおける主要なピークを単離し、そしてLC−MSに再注入した、d)検出された分子量およびその割り当ての表;e)ESI−MSスペクトル。
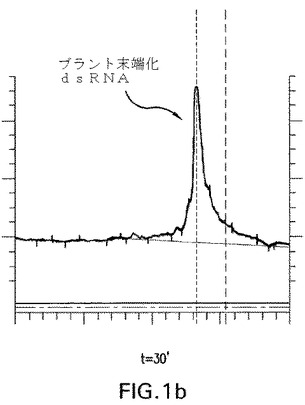
【図1b】図1a、1b、1c、1dおよび1e:未修飾siRNA pGl3−siRNA(マウス血清中の野生型siRNA)の代謝分解;a−c)マウス血清中0分、30分および180分インキュベートした後の未修飾siRNAのイオン交換−HPLC分析;37℃で30分インキュベートした後のイオン交換HPLCにおける主要なピークを単離し、そしてLC−MSに再注入した、d)検出された分子量およびその割り当ての表;e)ESI−MSスペクトル。
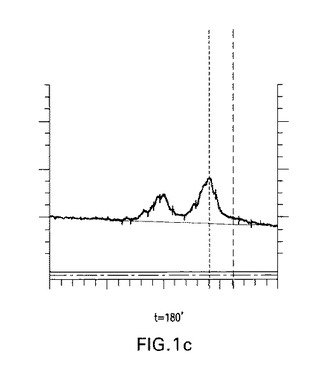
【図1c】図1a、1b、1c、1dおよび1e:未修飾siRNA pGl3−siRNA(マウス血清中の野生型siRNA)の代謝分解;a−c)マウス血清中0分、30分および180分インキュベートした後の未修飾siRNAのイオン交換−HPLC分析;37℃で30分インキュベートした後のイオン交換HPLCにおける主要なピークを単離し、そしてLC−MSに再注入した、d)検出された分子量およびその割り当ての表;e)ESI−MSスペクトル。
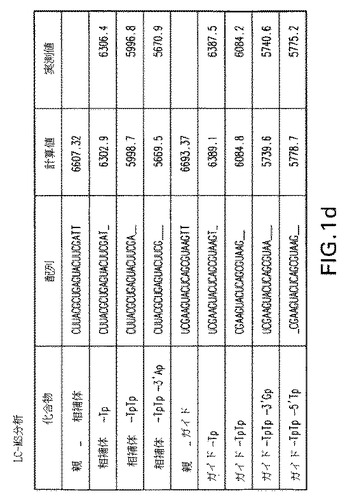
【図1d】図1a、1b、1c、1dおよび1e:未修飾siRNA pGl3−siRNA(マウス血清中の野生型siRNA)の代謝分解;a−c)マウス血清中0分、30分および180分インキュベートした後の未修飾siRNAのイオン交換−HPLC分析;37℃で30分インキュベートした後のイオン交換HPLCにおける主要なピークを単離し、そしてLC−MSに再注入した、d)検出された分子量およびその割り当ての表;e)ESI−MSスペクトル。
【図1e】図1a、1b、1c、1dおよび1e:未修飾siRNA pGl3−siRNA(マウス血清中の野生型siRNA)の代謝分解;a−c)マウス血清中0分、30分および180分インキュベートした後の未修飾siRNAのイオン交換−HPLC分析;37℃で30分インキュベートした後のイオン交換HPLCにおける主要なピークを単離し、そしてLC−MSに再注入した、d)検出された分子量およびその割り当ての表;e)ESI−MSスペクトル。
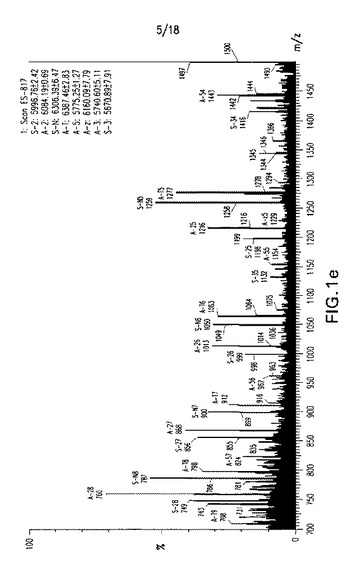
【図2】図2:四つの二本鎖RNA型式の説明:野生型(または未修飾)siRNA、MOE o/h siRNA、C3−siRNAおよびC3−MOE siRNA。
【0010】
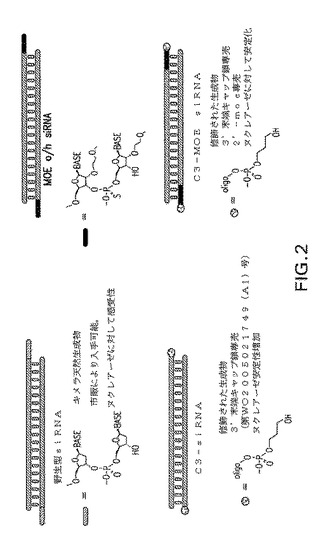
【図3】図3:マウス胃酸中三つの異なる型式でのsiRNAの安定性。試料をマウス胃酸中37℃で2μMの濃度でインキュベートした。親化合物のバンドを定量することにより親化合物の消失を2−6時間にわたって追跡した。レーン1−7:胃酸中t=0、5、10、15、30、60および120分の野生型siRNAレーン8:dsRNAラダー(30、21、19、16、13、10塩基対)レーン9−15:胃酸中t=0、5、10、15、30、60および120分のC3 siRNAレーン16:dsRNAラダー(30、21、19、16、13、10塩基対)レーン17−24:胃酸中t=0、5、10、15、30、60および120分のC3−MOE siRNA
【0011】
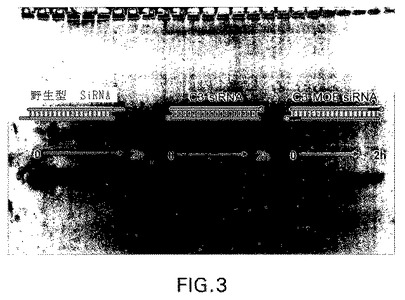
【図4】図4:腸洗浄液中四つの異なる型式でのsiRNAの安定性。試料を肝ミクロソーム中37℃で5μMの濃度でインキュベートした。(左から右に)レーン1:dsRNAラダー(30、21、19、16、13、10塩基対)レーン2−7:腸洗浄液中t=0、15、30、60、180および360分の野生型siRNAレーン8−13:腸洗浄液中t=0、15、30、60、180および360分のmoe o/h siRNAレーン14−19:腸洗浄液中t=0、15、30、60、180および360分のC3 siRNAレーン20−25:腸洗浄液中t=0、15、30、60、180および360分のC3−MOE siRNA
【0012】
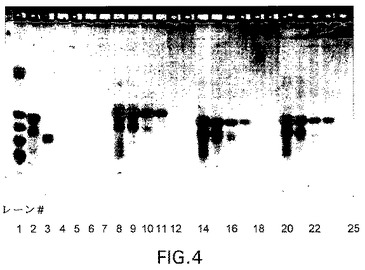
【図5】図5:肝ミクロソーム中四つの異なる型式でのsiRNAの安定性。試料をラット腸洗浄液からの腸液中37℃で2μMの濃度でインキュベートした。(左から右に)レーン1:ds
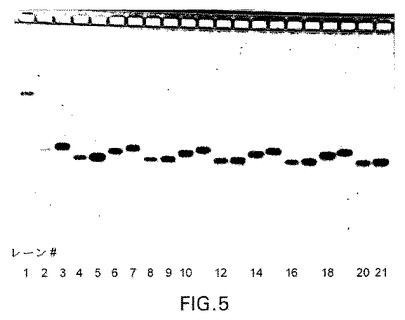
【図6】図6:マウス血清中四つの異なる型式でのsiRNAの安定性。試料をマウス血清中37℃で2μMの濃度でインキュベートした。親化合物のバンドを定量することにより親化合物の消失を6時間にわたって追跡した。(左から右に)レーン1:dsRNAラダー(30、21、19、16、13、10塩基対)RNAラダー(30、21、19、16、13、10塩基対)レーン2:野生型siRNA未処理レーン3:moe o/h siRNA未処理レーン4:C3 siRNA未処理レーン5:C3−MOE siRNA未処理レーン6−9:肝ミクロソーム中2−5と同一、t=0レーン10−13:肝ミクロソーム中2−5と同一、t=60分レーン14−17:上澄S12中2−5と同一、t=0レーン18−21:上澄S12中2−5と同一、t=60分レーン2−7:マウス血清中t=0、15、30、60、180および360分の野生型siRNAレーン8−13:マウス血清中t=0、15、30、60、180および360分のmoe o/h siRNAレーン14−19:マウス血清中t=0、15、30、60、180および360分のC3 siRNAレーン20−25:マウス血清中t=0、15、30、60、180および360分のC3−MOE siRNA
【0013】
【図7】図7:抗VEGFR2 siRNA(2個の独立した配列)の三つの型式の細胞内(in cellulo)での特徴付け。野生型siRNA、C3−siRNAおよびC3−MOE siRNAを3濃度(1、5、10nM)でMS1細胞にトランスフェクトした。FACSによりVEGFR2細胞表面レベルを測定することによりサイレンシング効力を評価した。
【0014】
【図8a】図8aおよび8b:成長因子誘起血管形成「寒天チャンバー(Agar Chamber)」マウスモデルにおける野生型siRNA、C3−siRNAおよびC3−Moe siRNAのインビボ試験。図8aは1、5および25μg/マウス/日の対照、未修飾VEGFR2 siRNAおよびC3修飾VEGFR2 siRNAの結果を示す。図8bは0.2、1および5μg/マウス/日での対照、C3修飾VEGFR2 siRNAおよびC3−MOE VEGFR2 siRNAを示す。各例で2つの抗VEGFR2 siRNAのプールを毎日3日間腹腔内に与えた。
【図8b】図8aおよび8b:成長因子誘起血管形成「寒天チャンバー(Agar Chamber)」マウスモデルにおける野生型siRNA、C3−siRNAおよびC3−Moe siRNAのインビボ試験。図8aは1、5および25μg/マウス/日の対照、未修飾VEGFR2 siRNAおよびC3修飾VEGFR2 siRNAの結果を示す。図8bは0.2、1および5μg/マウス/日での対照、C3修飾VEGFR2 siRNAおよびC3−MOE VEGFR2 siRNAを示す。各例で2つの抗VEGFR2 siRNAのプールを毎日3日間腹腔内に与えた。
【0015】
【図9a】図9:B16同種移植片メラノーマ腫瘍マウスモデルに5および20μg/マウス/日で腹腔内(i.p.)に与えられた抗VEGFR2 C3−MOE siRNAのインビボ試験。図9aは、修飾されたVEGFR2 siRNAでのi.p.処置が腫瘍発達を有意に低下させることを示す。図9bもまたVEGFR2 siRNAの20μg/マウスのi.p.注射が有意な腫瘍成長の阻害を招くことを示す。
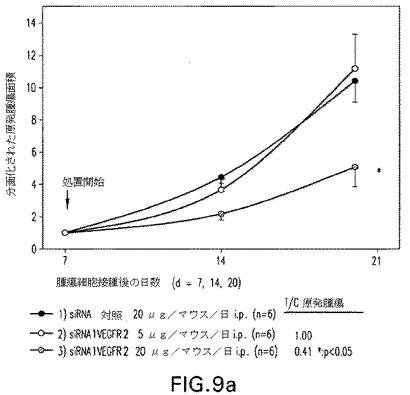
【図9b】図9:B16同種移植片メラノーマ腫瘍マウスモデルに5および20μg/マウス/日で腹腔内(i.p.)に与えられた抗VEGFR2 C3−MOE siRNAのインビボ試験。図9aは、修飾されたVEGFR2 siRNAでのi.p.処置が腫瘍発達を有意に低下させることを示す。図9bもまたVEGFR2 siRNAの20μg/マウスのi.p.注射が有意な腫瘍成長の阻害を招くことを示す。
【0016】
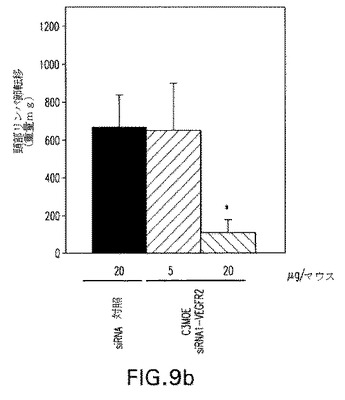
【図10】図10:成長因子誘起血管形成マウスモデルにおけるC3−MOE siRNAのインビボ試験。抗VEGFR2 siRNAを20μg/マウス/日を毎日3日間経口的に与えた。
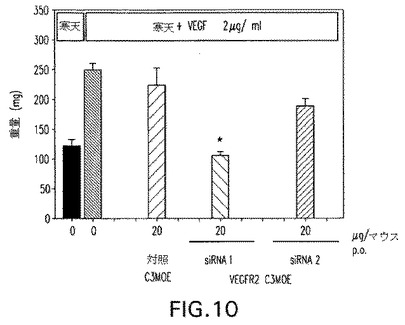
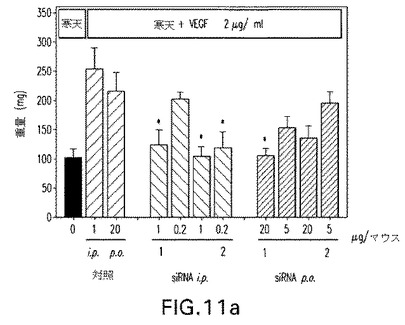
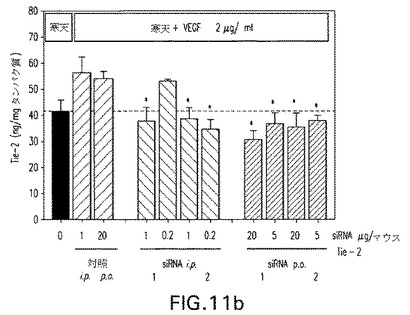
【図11a】図11:成長因子誘起血管形成マウスモデルにおけるC3−MOE siRNAのインビボ試験。抗Tie2 siRNAを毎日腹腔内(1および0.2μg/マウス/日)または経口的に(20および5μg/マウス/日)3日間経口与えた。図11a:切除組織の重量;図11b:Tie2タンパク質ノックダウン。
【図11b】図11:成長因子誘起血管形成マウスモデルにおけるC3−MOE siRNAのインビボ試験。抗Tie2 siRNAを毎日腹腔内(1および0.2μg/マウス/日)または経口的に(20および5μg/マウス/日)3日間経口与えた。図11a:切除組織の重量;図11b:Tie2タンパク質ノックダウン。
【発明を実施するための形態】
【0017】
発明の詳細な説明
本発明は哺乳動物における血管形成障害を処置するための組成物および方法に関する。特に本発明は哺乳動物への経口投与時に血管形成障害を処置するために使用することができる低分子干渉RNA(「siRNA」)に関する。
血管形成は以下の標的/遺伝子を含む血管内皮細胞を標的とする:VEGFR−1(GenBank受入番号AF06365);VEGFR−2(GenBank受入番号AF063658);VEGFR−3(GenBank受入番号(NM_002020);Tie2(TEK)(GenBank受入番号NM_000459);bFGFR(GenBank受入番号M60485);IL8RA(GenBank受入番号L19591);IL8RB(GenBank受入番号L19593);Fas(GenBank受入番号X89101);IGF2R(GenBank受入番号NM_000876)。
【0018】
本発明によるsiRNA分子はRNA干渉(「RNAi」)を媒介する。「RNAi」なる用語は当分野において周知であり、そして一般的に標的遺伝子に相補的である領域を有するsiRNAによる、細胞における一つまたはそれより多い標的遺伝子の阻害を意味すると理解される。RNAiを媒介するその能力に関してsiRNAを試験するための種々のアッセイが当分野において公知である(例えばElbashir et al., Methods 26:199−213(2002)参照)。本発明によるsiRNAの遺伝子発現に及ぼす影響は、本発明によるRNA分子で処理されていない細胞と比較した場合、典型的には少なくとも10%、33%、50%、90%、95%または99%まで阻害される標的遺伝子の発現に至る。
【0019】
本発明による「siRNA」または「低分子干渉リボ核酸」は以下の態様を含む当分野において公知の意味を有する。siRNAは生理学的条件下で相補的領域に沿ってハイブリダイズする2本の鎖のリボヌクレオチドからなる。鎖は別個であるが、特定の実施態様では、分子リンカーにより結合し得る。個々のリボヌクレオチドは未修飾天然発生リボヌクレオチド、未修飾天然発生デオキシリボヌクレオチドでよいか、またはそれらは本明細書の他の部分で記載されるような化学的に修飾された、もしくは合成性でよい。
【0020】
本発明によるsiRNA分子は標的遺伝子のmRNAの領域と実質的に同一である二本鎖領域を含む。標的遺伝子の対応する配列に対して100%同一性を有する領域が適当である。この状態は「完全に相補的」と称される。しかしながらその領域はまた、標的化されるmRNAの領域の長さに依存して、標的遺伝子の対応する領域と比較して1、2または3個の誤対合を含有してもよく、そしてそのようなものは完全に相補的ではないかもしれない。一つの実施態様では、本発明のRNA分子は1個の所定の遺伝子を特異的に標的化する。望ましいmRNAのみを標的化するために、siRNA試薬は標的mRNAに対して100%相同性、および細胞または生物に存在する全てのその他の遺伝子に対して少なくとも2個の誤対合したヌクレオチドを有し得る。具体的な標的配列の発現を有効に阻害するために、十分な配列同一性を有するsiRNAを分析および同定するための方法が当分野において公知である。当分野において公知の配列比較およびアラインメントアルゴリズムにより配列同一性を最適化し(Gribskov and Devereux, Sequence Analysis Primer, Stockton Press,(1991)およびそこに引用された参照文献を参照のこと)、そして例えばBESTFITソフトウェアプログラムでデフォルトパラメーターを用いて実行されるようなSmith−Watermanアルゴリズム(例えばUniversity of Wisconsin Genetic Computing Group)によりヌクレオチド配列間の差異パーセントを計算することができる。
【0021】
RNAi試薬の有効性に影響を及ぼす別の因子は標的遺伝子の標的領域である。RNAi試薬による阻害に有効な標的遺伝子の領域を実験方法により決定することができる。適当なmRNA標的領域はコード化領域であろう。5’−UTR、3’−UTRおよびスプライスジャンクションのような未翻訳領域もまた適当である。例えばElbashir S.M. et al, EMBO J., 20:6877−6888(2001)に記載されるようなトランスフェクションアッセイをこの目的のために実施することができる。当分野では当業者に周知である多くのその他の適当なアッセイおよび方法が存在する。
【0022】
本発明にしたがって、標的に相補的なsiRNAの領域の長さは10から100ヌクレオチド、12から25ヌクレオチド、14から22ヌクレオチドまたは15、16、17もしくは18ヌクレオチドでよい。対応する標的領域に対する誤対合が存在する場合、相補的領域の長さは一般的に幾分長くなることが要求される。
siRNAはオーバーハングした末端(標的に対して相補的であってもなくてもよい)、または標的配列にではなくそれ自体に相補的なさらなるヌクレオチドを担持し得るので、siRNAの各別個の鎖の全体の長さは10から100ヌクレオチド、15から49ヌクレオチド、17から30ヌクレオチドまたは19から25ヌクレオチドでよい。
【0023】
「各鎖が49ヌクレオチド以下である」なる語句は、全ての修飾または未修飾ヌクレオチドを含むが、鎖の3’または5’末端に付加され得る任意の化学的部分を含まない、鎖の連続したヌクレオチドの全数を意味する。鎖に挿入された短い化学的部分は計数されないが、2本の別個の鎖を結合するために設計された化学的リンカーは連続したヌクレオチドを創成するとは考えられない。
「5’末端または3’末端の少なくとも一つにおける1から6ヌクレオチドオーバーハング」なる語句は生理学的条件下で2本の別個の鎖から形成される相補的siRNAの構造を指す。末端ヌクレオチドがsiRNAの二本鎖領域の一部である場合、siRNAはブラント末端と考えられる。一つまたはそれより多いヌクレオチドが末端で対形成されない場合、オーバーハングが創成される。オーバーハングの長さはオーバーハングするヌクレオチドの数により測定される。オーバーハングするヌクレオチドはいずれかの鎖の5’末端または3’末端のいずれかでよい。
【0024】
本発明によるsiRNAは少なくとも1本の鎖に少なくとも1個の修飾されたヌクレオチドを含むことにより、インビボで経口送達に適当な高い安定性を付与する。故に本発明によるsiRNAは少なくとも1個の修飾されたまたは非天然リボヌクレオチドを含有する。多くの公知の化学的修飾についての冗長な記載は公開PCT特許出願第WO200370918号に示され、そして本明細書で繰り返さない。経口送達のための適当な修飾をさらに具体的に本明細書の実施例および記載にて示す。適当な修飾には、限定するものではないが糖部分に対する修飾(すなわち例えば2’−O−(2−メトキシエチル)または2’−MOEのような糖部分の2’位置(Martin et al., Helv. Chim. Acta, 78:486−504(1995))すなわちアルコキシアルコキシ基)または塩基部分(すなわち代替ヌクレオチド鎖の別の具体的な塩基と対形成する能力を維持する非天然または修飾された塩基)が含まれる。その他の修飾には、限定するものではないがリン酸エステル基の置換(隣接するリボヌクレオチドを例えばホスホロチオエート、キラルホスホロチオエートまたはホスホロジチオエートと連結させる)を含むいわゆる「バックボーン」修飾が含まれる。最後に、時に本明細書では3’キャップまたは5’キャップと称される末端修飾が重要である。表1にて説明されるようにキャップは、siRNAに安定性を付与することが見出されている「T−T」のようなさらなるヌクレオチドの単純な付加からなってよい。キャップは当業者に公知である錯体化学からなってよい。
【0025】
以下の実施例で用いられる一つの実施態様では、3’キャップは3’炭素を介して3’末端に抱合された化学的部分であり、かつ式I:
【化2】
[式I]
(式中、
XはOまたはSであり、
R1およびR2は独立してOH、NH2、SH、アルキル、アリール、アルキル−アリール、アリール−アルキルであり、ここでアルキル、アリール、アルキル−アリール、アリール−アルキルをさらなるヘテロ原子および官能基、好ましくはN、OもしくはSの群から選択されるヘテロ原子またはOH、NH2、SH、カルボン酸もしくはエステルの群から選択される官能基により置換することができ;
またはR1およびR2は式Y−Zのものでよく、ここでYはO、N、Sであり、そしてZはH、アルキル、アリール、アルキル−アリール、アリール−アルキルであり、ここでアルキル、アリール、アルキル−アリール、アリール−アルキルをさらなるヘテロ原子、好ましくはN、OもしくはSの群から選択されるヘテロ原子により置換することができる)
の化合物の中から選択される。
【0026】
糖部分での修飾の実例には2’アルコキシリボヌクレオチド、2’アルコキシアルコキシリボヌクレオチド、ロックされた核酸リボヌクレオチド(LNA)、2’−フルオロリボヌクレオチド、モルホリノヌクレオチドが挙げられる。
ヌクレオシド内結合を修飾することもできる。ヌクレオシド内結合の実例にはホスホロチオエート、ホスホロジチオエート、ホスホロアミダートおよびアミド結合が挙げられる。
R1はOHでよい。
R1およびR2は一緒に1から24個のC原子、1から12個のC原子、2から10個のC原子、1から8個または2から6個のC原子を含み得る。別の実施態様では、R1およびR2は独立してOH、低級アルキル、低級アリール、低級アルキル−アリール、低級アリール−アルキルであり、ここで低級アルキル、低級アリール、低級アルキル−アリール、低級アリール−アルキルを前記で定義されるようなさらなるヘテロ原子および官能基により置換することができる。別の実施態様では、R1およびR2は双方ともOHではない。
【0027】
有機ラジカルまたは化合物に関連する「低級」なる用語は7個以下の炭素原子、好ましくは1−4個の炭素原子を含む分岐しているかまたは分岐していなくてよい化合物またはラジカルを意味する。低級アルキルは、例えばメチル、エチル、n−プロピル、イソプロピル、n−ブチル、sec−ブチル、tert−ブチル、n−ペンチルおよび分岐したペンチル、n−ヘキシルおよび分岐したヘキシルを表す。
アルコキシの実例にはO−Met、O−Eth、O−prop、O−but、O−pent、O−hexが挙げられる。
【0028】
少なくとも1個の修飾されたまたは非天然リボヌクレオチドを含有するsiRNAを含むsiRNAの合成のための方法は周知であり、そして当業者に容易に利用可能である。例えば種々の合成化学が公開PCT特許出願第WO2005021749号および第WO200370918号(双方共に出典明示により本明細書の一部とする)に示されている。反応を溶液中または好ましくは固相でポリマーに支持された試薬を使用することにより実施でき、続いてRNAiを媒介することができるsiRNA分子が形成される条件下で合成されたRNA鎖を組み合わせる。
【0029】
本発明は経口送達に適当である少なくとも1個の修飾されたヌクレオチドを含有するするsiRNAを提供する。機能的な表現ではこれは、siRNAは経口投与時に適当な薬物動態および体内分布を有し、関連する標的組織への送達を達成するであろうということを意味する。とりわけこれには血清安定性、免疫応答の欠如、および薬物様の挙動が必要とされる。本明細書の別の部分で開示される標準的な胃酸アッセイおよび標準的な血清アッセイに基づいてsiRNAのこれらの様相の多くを予想することができる。
【0030】
別の態様では、本発明は、RNAiにより少なくとも1個の標的遺伝子を阻害することができる本発明によるsiRNAを細胞に導入することを含む標的遺伝子の阻害のための方法を提供する。また、各々別の標的領域に特異的である1を超える種のsiRNAを同時にまたは逐次的に細胞に導入することができる。
【0031】
本発明は任意の型の標的遺伝子またはヌクレオチド配列に限定されるものではない。例えば標的遺伝子は細胞遺伝子、内因性遺伝子、病原体関連遺伝子、ウイルス遺伝子または発癌遺伝子でよい。実施例のいくつかは、経口送達された本発明のsiRNAが脈管形成、血管新生または血管形成の部位に蓄積され得ることを強調しているので、血管形成遺伝子は本発明に特に重要である。本発明に特に興味深いこれらの部位での血管形成遺伝子の最新のリストはAngioDB:血管形成および血管形成関連分子Tae−Kwon Sohn、Eun−Joung Moon1、Seok−Ki Lee1、Hwan−Gue Cho2およびKyu−Won Kim3のデータベース、Nucleic Acids Research, 30(1):369−371(2002)およびオンラインhttp://angiodb.snu.ac.kr/に列挙される。特に重要な遺伝子は詳細に分析されており、そして本明細書の別の部分に示されている。
【0032】
別の態様では、本発明はまた細胞における標的遺伝子の発現を阻害するための試薬を含むキットを提供し、ここで該キットは本発明によるdsRNAを含む。キットはインビトロまたはインビボで本発明によるdsRNAの導入を実施して、試料または対象を試験するのに必要な少なくとも一つの試薬を含む。好ましい実施態様では、かかるキットはまたキットの構成成分が使用される手順を詳記する説明書をも含む。
【0033】
「血管形成障害の処置」とはこの開示で使用される際には、血管新生、脈管形成および/または血管形成の生理学的および病理学的過程を伴う疾患の処置のための医薬組成物における本発明の修飾されたsiRNAの使用を意味する。このようにこれらの医薬組成物は、限定するものではないが癌腫瘍成長および転移、新生物、眼血管新生(黄斑変性症、糖尿病性網膜症、虚血性網膜症、未熟児網膜症、脈絡膜血管新生を含む)、関節リウマチ、骨関節炎、慢性喘息、敗血症性ショック、炎症性疾患、滑膜炎、骨および軟骨破壊、パンヌス成長、骨棘形成、骨髄炎、乾癬、肥満、血管腫、カポジ肉腫、アテローム性動脈硬化症(アテローム性動脈硬化巣断裂を含む)、子宮内膜症、疣贅、多毛症、ケロイド瘢痕、アレルギー性浮腫、機能不全性子宮出血、卵胞嚢胞、卵巣過剰刺激、子宮内膜症、骨髄炎、炎症性および感染性過程(肝炎、肺炎、糸球体腎炎)、喘息、鼻ポリープ、移植、肝臓再生、白質軟化症、甲状腺炎、甲状腺肥大、リンパ球増殖性障害、血液学的悪性腫瘍、血管奇形および子癇前症を含む血管新生、脈管形成または血管形成の阻害を必要とする疾患、症状および障害を処置するために有用である。
【0034】
本明細書で使用される際には、「処置」とは、疾患、障害もしくは症状の過程を阻害もしくは低減させるため、疾患、障害もしくは症状の病徴を阻害もしくは低減させるため、または疾患、障害もしくは症状の発症もしくはさらなる発達を予防的に防御するために取る行動を意味する。「処置する」とはその認識動詞である。
【0035】
本発明の治療薬の有効用量は疾患状態を処置するために必要とされるその用量である。有効用量は疾患の型、使用される組成物、投与の経路、処置されている哺乳動物の型、考慮される具体的な哺乳動物の身体的特徴、併用薬、および医学の分野において当業者に認識されるその他の因子に依存する。一般的に効力に依存して0.1mg/kgと100mg/kg体重/日の間の量のsiRNAが投与される。本発明の核酸分子およびその処方を経口的、局所的、非経口的に、吸入もしくはスプレーにより、または直腸に従来の無毒性の薬学的に許容される担体、アジュバントおよび/またはベヒクルを含有する投薬単位処方で投与できる。非経口的なる用語は、本明細書で使用される際には、経皮、皮下、血管内(例えば静脈内)、筋肉内、腹腔内もしくはくも膜下腔内注射、または注入技術等を含む。加えて、本発明の核酸分子および薬学的に許容される担体を含む医薬用処方が提供される。一つまたはそれより多い本発明の核酸分子は、一つまたはそれより多い無毒性の薬学的に許容される担体および/または希釈剤および/またはアジュバント、ならびに所望によりその他の活性成分を伴って存在できる。本発明の核酸分子を含有する医薬組成物を経口使用に適当な形態、例えば錠剤、トローチ、ロゼンジ、水性もしくは油性懸濁液、分散性粉末もしくは顆粒、エマルジョン、硬質もしくは軟質カプセル、またはシロップもしくはエリキシルにできる。
【0036】
経口使用を意図される組成物を、医薬組成物の製造に関して当分野において公知の任意の方法にしたがって調製することができ、そしてかかる組成物は医薬的に洗練され、そして口当たりのよい調製物を提供するために一つまたはそれより多いかかる甘味剤、着香剤、着色剤または保存剤を含有できる。錠剤は錠剤の製造に適当である無毒性の薬学的に許容される賦形剤と混合された活性成分を含有する。これらの賦形剤は例えば炭酸カルシウム、炭酸ナトリウム、ラクトース、リン酸カルシウムまたはリン酸ナトリウムのような不活性希釈剤;顆粒化および崩壊剤、例えばトウモロコシデンプンまたはアルギン酸;結合剤、例えばデンプン、ゼラチンまたはアラビアゴム;および滑沢剤、例えばステアリン酸マグネシウム、ステアリン酸またはタルクでよい。錠剤はコーティングされていなくてよいか、またはそれらを公知の技術によりコーティングすることができる。経口使用のための処方を、活性成分が不活性固体希釈剤、例えば炭酸カルシウム、リン酸カルシウムもしくはカオリンと混合される硬質ゼラチンカプセルとして、または活性成分が水もしくは油性溶媒、例えば落花生油、液体パフィンもしくはオリーブ油と混合される軟質ゼラチンカプセルとして提示することもできる。水性懸濁液は水性懸濁液の製造に適当な賦形剤との混合物中に活性成分を含有する。
【0037】
本発明の組成物の経口投与は胃または腸に直接的に、最も重要には患者による嚥下制御投薬形態によるが、またかかる送達のその他の機械的および補助的手段による物質を投与するための全ての標準的な技術を含む。
【0038】
約0.1mgから約140mg/kg体重/日のオーダーの投薬量レベルが前記で指示された症状の処置において有用である(約0.5mgから約7g/対象/日)。単回投薬形態を生成するための担体材料と組み合わすことができる活性成分の量は、処置される宿主および投与の特定の様式に依存して異なる。一般的に投薬単位形態は約1mgから約500mgの間の活性成分を含有する。任意の特定の対象に関する具体的な用量レベルは用いられる具体的な化合物の活性、年齢、体重、一般健康状態、性別、食事、投与の時間、投与の経路、ならびに排出の速度、薬物の組み合わせおよび治療を行っている特定の疾患の重篤度を含む種々の因子に依存することは理解される。
【0039】
本発明の治療薬の治療効果をその他の薬剤との組み合わせにより強化することができる。典型的にはかかるその他の薬剤には血管形成障害のような類似の疾患の処置に使用されることが知られている薬剤が含まれよう。これに代えて、かかる薬剤を使用して本発明の治療薬により引き起こされる副作用または望ましくない影響を低減させることができる。
【0040】
本発明のsiRNAはまた重要な調査用途をも有する。かかる研究の一つにはインビトロでの血管形成過程への調査が含まれる。「インビトロでの血管形成過程」とは動物全身を用いない血管形成または脈管形成を研究するための任意の過程を意味する。このように血管形成のマーカーまたは指標を使用して血管形成過程の工程を研究するインビトロまたはエキソビボ方法およびアッセイはここに含まれる。
【0041】
RNA鎖ヌクレオチド配列
表1で同定されるsiRNA鎖配列は以下の標的に対する適当なsiRNA配列として同定されている:VEGFR−1(GenBank受入番号AF06365);VEGFR−2(GenBank受入番号AF063658);VEGFR−3(GenBank受入番号(NM_002020);Tie2(TEK)(GenBank受入番号NM_000459);bFGFR(GenBank受入番号M60485);IL8RA(GenBank受入番号L19591);IL8RB(GenBank受入番号L19593);Fas(GenBank受入番号X89101);IGF2R(GenBank受入番号NM_000876)。
【0042】
表1:ヒトVEGFR−1、VEGFR−2、VEGFR−3、Tie2、bFGFR、IL8RA、IL8RB、Fas、IGF2Rに対するsiRNA
【表1】
【表2】
【表3】
【表4】
【表5】
【表6】
【表7】
【表8】
【表9】
【表10】
【表11】
【表12】
【表13】
【表14】
【表15】
【表16】
【表17】
【表18】
【表19】
【表20】
【表21】
【表22】
【表23】
【表24】
【表25】
【表26】
【表27】
【表28】
【表29】
【表30】
【表31】
【0043】
RNA鎖ヌクレオチドの化学的修飾
本発明によるsiRNAは少なくとも1本のRNA鎖に少なくとも1個の修飾されたヌクレオチドを含み得る。様々な可能性のある修飾されたヌクレオチドを本明細書の別の部分で開示する。本発明による使用のための有用な修飾および修飾の組み合わせを表2に示す:
【0044】
表2:化学的修飾および配列構造
【表32】
N=任意の未修飾RNAヌクレオチド
n=未修飾DNAヌクレオチド
Np=修飾されたRNAヌクレオチド
s=ホスホロチオエートヌクレオシド内結合の同定
o/h=オーバーハング
【0045】
siRNA鎖の3’末端の3’位置に付加された以下の修飾は時に「3’末端キャップ」と称され、本発明の有用な実施態様としても認識され、そして本発明によるいずれかのsiRNAと共に使用され得る:
【0046】
【化3】
【0047】
本発明による活性を有する具体的な化合物には以下の表3に示されるものが含まれる:
表3:実施例で使用されるsiRNAの配列および化学
【表33】
【表34】
【表35】
【実施例】
【0048】
実施例
以下の実施例は本発明の態様を説明し、そして以下に列挙される請求の範囲に含まれる実施態様を限定することを意図されるものではない。さらに下の結果および考察のセクションは、以下のプロトコールにしたがって行われ、そして以下の材料を用いる実験に言及する。具体的に記載されていない材料およびプロトコールは当業者に日常的に利用可能であると考えられる。
【0049】
実施例1
siRNAの調製
標準的な2’−O−TOMホスホアミダイトテクノロジーにより一本鎖siRNA誘導体を合成し、そしてOasis(登録商標)HLB抽出プレート(Waters)により精製した。センス−およびアンチセンス鎖siRNAをハイブリダイゼーションバッファー(100mM酢酸カリウム、2mM酢酸マグネシウム、30mM Hepes、pH7.6)中で混合し、90°Cで3分間熱変性し、そして30°Cで60分間アニーリングした。siRNA二重鎖の100μM原液を−20°Cで保存した。
【0050】
実施例2
血清中でのインキュベーションおよびIE−HPLC(LC−MS)による分析
標準的な血清アッセイでは、20μMの各siRNA6μlを血清またはCSF 54μlと混合し、そしてインキュベーター中37°Cで加熱した、冷却した混合物50μlを分析用DNA−pac PA−100カラム(Dionex)に負荷し、そして1:10アセトニトリル:バッファー(20mM酢酸ナトリウム、1mM酢酸マグネシウム、pH6.5)溶液中NaClグラジエント(30分で0−0.6M)で分析した。
LC−MS分析用に各siRNA 100μl(20μMまたは50μM)を滅菌ウシ胎仔血清(GIBCO)900μlと混合し、37°Cでインキュベートし、そして以前に指示されたように(NaClグラジエントを除く:9分で0M−0.36M/12分で0.36M−0.6M)HPLCにより分離した。分解生成物をNAPカラムで脱塩し、そしてLC−ESI−−MSにより分析した。
【0051】
実施例3
胃酸中でのインキュベーション
標準的な胃酸アッセイを準備するために、体重18から20g(6から8週齢)のFVBおよびC57BL6マウスをCharles River Laboratories(Les Oncins, France)から入手した。CO2を用いて動物を屠殺し、そして次に胃を即座に回収した。胃液および胃の内容物を収集し、そしてプールし、次いで遠心ろ過装置(Ultrafree MC, Millipores)に負荷した。製造者の推奨にしたがってフィルターユニットを10分間回転させた。マウス胃液に相当するろ液を回収し、等分し、そしてさらなる実験の前に凍結した。
各アッセイ用に、20μM siRNA溶液を9倍容量の前記されたような胃酸で希釈し、そして37°Cで0、5、10、15、30、60および120分間インキュベートした。
【0052】
実施例4
腸洗浄液中のインキュベーション
標準的な腸洗浄液アッセイを準備するために、雄Wistarラットを絶食させ、イソフルランで麻酔した。10ml生理食塩水(0.5ml/分)、続いて水20ml(1ml/分)で小腸(十二指腸、空腸、回腸)を原位置で灌流することにより腸洗浄液を入手した。収集された流出液を遠心し(3000×g、15分、22°C)、そして上澄を1.2μmフィルターに通し、そして−20°Cで保存した。
各アッセイ用に、20μM siRNA溶液を9倍容量の腸洗浄液で希釈し、そして37°Cで0、15、30、60、180および360分間インキュベートした。
【0053】
実施例5
マウス肝ミクロソーム中のインキュベーション
標準的な肝ミクロソームアッセイでは、250μM siRNA溶液10μlに、タンパク質20mg/mlのマウス肝ミクロソーム(GEntest 452701 Charge 11)25μl、100mMリン酸バッファー(pH7.4)365μl、UDPGAコファクター(水中24mM)50μl、NADPH 50μlを加えた。t=0分およびt=60分に凍結することによりインキュベーションをクエンチした。
【0054】
実施例6
ラットS12上澄中のインキュベーション
標準的なラットS12上澄アッセイ用に250μM siRNA溶液10μlをタンパク質29.9mg/mlのラット肝臓S12 17μl、100mMリン酸バッファー(pH7.4)373μl、UDPGAコファクター(水中24mM)50μl、NADPH 50μlに加えた。t=0分およびt=60分に凍結することによりインキュベーションをクエンチした。
【0055】
実施例7
マウス血清中のインキュベーション
標準的なマウス血清中インキュベーション用に20μM siRNA溶液を9倍容量のネズミ血清(Harlanヌードマウス)で希釈し、そして37°Cで0、15、30、60、180および360分間インキュベートした。
【0056】
実施例8
ゲル電気泳動安定性アッセイ
振盪およびドライアイス上で衝撃凍結(shock-frozen)の直後にインキュベーション溶液の10μlアリコートを取り、混合物を37°Cでインキュベートし、そしてアリコートを種々の時点で衝撃凍結した。負荷バッファー(Elchrom Sc., Cham, Switzerland)30μl(各々15μl)中でアリコートを解凍し、そしてSF50ゲル(Elchrom Sc., Cham, Switzerland)上、120V、8°Cで240分間分離した。バンドをSYBR Gold(Molecular Probes)で染色し、そしてBIORAD ChemiDoc(商標)XRSシステムで写真を撮った。
【0057】
実施例9
細胞培養
1.5%ゼラチンコートした培養皿でL−グルタミンおよび10%熱不活化FCS(AMIMED, Switzerland)を補充したDMEM高グルコース(4.5g/l)中でマウス不死化内皮細胞系MS1(ATCC CRL−2279)を成長させた。HiPerfect(QIAGEN)を用いて製造者の手順にしたがって24ウェル型式でMS1細胞をsiRNAでトランスフェクトした(4検体ずつ、siRNA最終濃度は10nMまたは指示されたとおりであった)。
【0058】
実施例10
FACS分析
トランスフェクトされていない、およびsiRNAトランスフェクトされたMS1細胞をFACSによりVEGFR2レベルに関して分析した。簡単には、2検体ずつまたは3検体ずつのウェルからの細胞をトリプシン処理し、各条件に関してプールし、次いでPBS+10%FCSで2回洗浄し、そして氷上で10分間インキュベートした後にRPE抱合抗VEGFR2 Ab(1μg/106セル;Avas 12α1、BD Pharmingen)を添加した。RPE標識したアイソタイプIgG2αをFACS対照(BD Pharmingen)として使用した。Cell Quest Software(Becton−Dickinson)を用いてFACScaliburでFACS獲得および分析を実施した。
【0059】
実施例11
動物研究
雌FVBマウス(6から8週齢)をCharles River Laboratories(Les Oncins, France)から入手した。耳のマーキングによりマウスを識別し、そして通常の条件下で群を維持し(ケージあたり動物6匹)、そして毎日観察した。処置群あたり6匹のマウスを使用し、そして動物保護のためのスイス動物保護法を厳密に順守して全ての動物実験を実施した。
【0060】
参照チャンバーモデルは出版物に記載されている(例えばWood J, Bold G, Buchdunger E, et al. PTK787/ZK222584、血管内皮成長因子受容体チロシンキナーゼの新規のおよび強力な阻害剤は経口投与後に血管内皮成長因子誘起応答および腫瘍成長を損なう。Cancer Res 60:2178−89(2000))。簡単には、ペルフルオロ−アルコキシ−Teflon(Teflon(登録商標)−PFA、直径21mm×8mm、550μl容量)から作られた多孔性組織チャンバーを、3μg/ml組換えヒトVEGFおよび指示されたようなsiRNAを補充したまたは補充していない0.8%寒天(BBL(登録商標)Nr. 11849、Becton Dickinson, Meylan, France)および20U/mlヘパリン(Novo Nordisk A/S, Bagsvaerd, Denmark)で充填した。充填手順の前に溶液を42℃で維持した。3%イソフルラン(Forene(登録商標)、 Abbott AG, Cham, Switzerland)吸入を用いてマウスを麻酔した。皮下移植用に、尾の付け根に皮膚の小切開を作成して移植用外套針の挿入を可能にした。動物の背の小切開を通して無菌条件下でチャンバーを移植した。創傷クリップ(Autoclip 9mm Clay Adams)により皮膚切開を閉じた。必要とされる用量に依存して、siRNAを「注射用の品質等級」の0.9%生理食塩水で希釈し、次いで動物にi.p.(200μl/用量)または経管によるp.o.(100μl/用量)のいずれかで送達した。チャンバー移植の2から4時間前にマウスに最初の投与を行い;次いで毎日2日間処置した。特記しない場合、移植後3日にマウスを屠殺し、チャンバーを切除し、そして各移植片の周囲に形成された血管新生化された線維性組織を注意深く除去した。マウスの一般状態をモニタリングするために体重を用いた。一元配置分散分析、続いてダネット検定を用いて統計分析を行った。
【0061】
実施例12
B16メラノーマ異種移植片モデル
抗血管形成治療に応答することが以前に同定されている同系B16/BL6ネズミメラノーマモデル(例えばLaMontagne K, Littlewood−Evans A, Schnell C, O’Reilly T, Wyder L, Sanchez T, Probst B, Butler J, Wood A, Liau G, Billy E, Theuer A, Hla T, Wood J. 、FTY720によるスフィンゴシン−1−リン酸受容体の拮抗作用が血管形成および腫瘍脈管化を阻害する。Cancer Res. 66(1):221−31(2006 Jan 1))を使用して標準的なまたは修飾されたsiRNAの抗腫瘍活性を評価した。腫瘍細胞(1μl、5×104/μl)を同系雌C57BL/6マウスの両耳の背側耳介に皮内注射した。原発腫瘍面積(mm2)の測定を腫瘍細胞接種後7、14および21日にコンピューター支援画像分析ソフトウェア(KS−400 3.0撮像システム、Zeiss)および具体的に設計されたマクロを用いて実施した。7日から21日まで「注射用の品質等級」の0.9%生理食塩水で希釈したsiRNAを、i.p.(200μl/用量)または経管によるp.o.(100μl/用量)のいずれかでマウスに1日1回投与した。21日にマウスを屠殺し、そして頭部リンパ節転移を重量測定し、そして次に凍結した。
これらの結果では、用いる実際のsiRNA配列および化学は表3を参照して決定できる。
【0062】
マウス血清中野生型siRNAを双方の3’末端から分解する
ヌクレアーゼによるオリゴヌクレオチド分解は大部分が3’エキソヌクレオチド分解性である。その終端での芳香族または親油性残基の導入によるアンチセンスオリゴヌクレオチドの修飾はそのヌクレオチド分解を遅延させる17。この代謝経路もまたsiRNAに優勢であるかどうかを検証するために、未修飾siRNA(野生型siRNA)をマウス血清中37°Cで3時間までインキュベートした。
用いられた未修飾siRNA配列はpGl3−siRNAであった(表3参照)。
混合物を強陰イオン交換HPLCで、t=0分、t=30分、t=180分で分析した。
【0063】
図1a、1bおよび1cで示されるように、t=30分でブラント末端化siRNAに相当する明確なピークが観察された。t=3時間までに実質的な分解が観察された。図1dおよび1eはHPLC−ESI−MS分析により同定された代謝物を説明する。この分析により双方の鎖の3’オーバーハングおよび3’末端の最初に塩基対形成するリボヌクレオチドの喪失に相当するいくつかの代謝物の存在が明らかにされる。ガイド鎖の5’末端リボヌクレオチドの消化もまた観察された。
【0064】
図1は血清中の未修飾siRNAの分解経路を示唆する。DNAオーバーハングがもしかすると3’エキソヌクレアーゼにより最初に消化される。LC−MSでは、双方の鎖の最初に塩基対形成する3’リボヌクレオチドおよびまたガイド鎖の最初に5’塩基対形成するリボヌクレオチドの喪失に相当するさらなる代謝物もまた検出された。
【0065】
3’修飾siRNAは胃腸管を通して安定である
2’−メトキシエチルリボヌクレオチドオーバーハング(MOE o/h siRNA)を有するsiRNA、ヒドロキシプロポキシホスホジエステル部分でキャップされたブラント末端化siRNA(C3−siRNA)および各鎖の3’末端で2個の最初に塩基対形成するヌクレオチドが2’−メトキシエチルリボヌクレオチド残基により修飾されたヒドロキシプロポキシホスホジエステル3’キャップsiRNA(C3−MOE siRNA)を合成した。これらの化合物を図2で図式的に説明する。
最初にsiRNAをマウス胃酸中2時間インキュベートした(図3)。C3 siRNAおよびC3−MOE sRNA場合、分解は観察されなかったが、野生型siRNAの分解は30分後に観察された。
【0066】
ラットの腸洗浄液から得られた腸液中の安定性により15分後にほぼ完了する野生型siRNAの分解が明らかにされたが、MOE o/h siRNA、C3−siRNAおよびC3−Moe siRNAの親化合物は60分間観察された(図4)。
肝臓中の安定性を肝ミクロソームアッセイおよびS12アッセイ(代表的な肝臓細胞質酵素活性)を用いて評価した。結果を図5に示す。双方の場合で、インキュベーションの60分後に分解は観察されなかった。
【0067】
最後に、siRNAをマウス血清中2μMで、37°Cで6時間までインキュベートすることにより試験した(図6の結果)。親化合物の安定性をゲル電気泳動により追跡した。修飾されたsiRNA(C3 siRNA、MOE o/h siRNAのC3−MOE siRNA)の場合、有意な分解は観察されなかったが、野生型siRNAでは観察された。
この研究は野生型(未修飾)siRNAがマウス胃酸中およびマウス血清中で代謝されることを示している。3’末端修飾されたsiRNAの場合、胃腸管で分解は観察されなかった。したがって3’修飾されたsiRNAは野生型siRNAよりも高い経口バイオアベイラビリティーを有している可能性がある。
【0068】
全身送達された3’修飾されたsiRNAはインビボ成長因子誘起血管形成モデルにおいてさらに活性である18
最初に、修飾されたsiRNA(C3−siRNAおよびC3−MOE siRNA)が標的遺伝子を下方調節する能力を、細胞内(in cellulo)で抗VEGFR2 siRNAでトランスフェクトされたMS1細胞のVEGFR2表面レベルを測定することにより検査した。
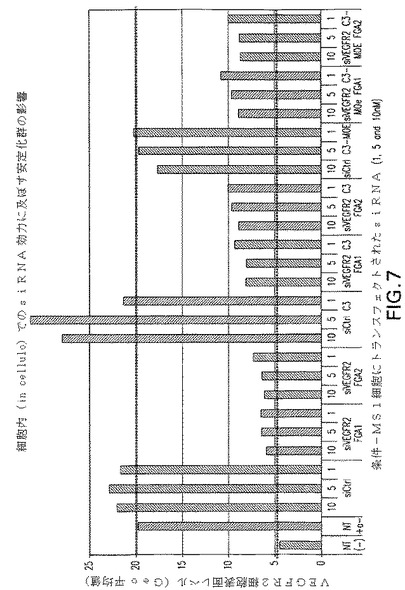
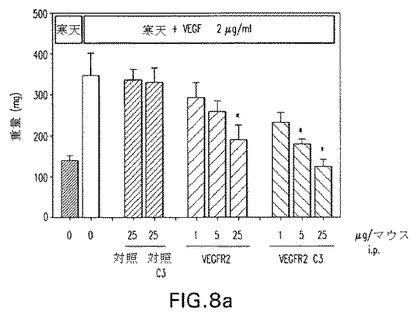
野生型siRNAとして二つの抗VEGFR2 siRNA、C3−siRNAおよびC3−MOE siRNAのプールを腹腔内投与した。結果を図7に示す。プールされた野生型siRNAは25μg/マウス/日の高用量でVEGF誘起の脈管化を有意に低減させた。C3−siRNAでは5倍低い用量で同一レベルの阻害が観察された。C3−MOE siRNAプールの場合、脈管化された組織重量の有意な低下が最低の0.2μg/マウス/日を含む全被験用量で観察された。
【0069】
図8aおよび8bは、腹腔内に与えられた場合、VEGFR2−C3およびC3−MOE siRNAの双方がマ1μg/マウス/日の用量を下回って活性であった。
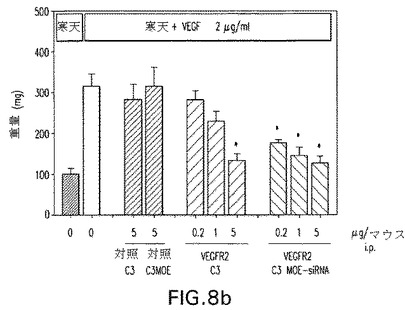
B16同種移植片メラノーマ腫瘍マウスモデルに腹腔内に与えられた(i.p.)抗VEGFR2 C3−MOE siRNAのインビボ試験。図9aは、修飾されたVEGFR2−C3−MOE−siRNAでのi.p.処置が腫瘍発達を有意に低減させることを示している。図9bもまたVEGFR2−C3−MOE−siRNAの20μg/マウスのi.p.注射の結果、腫瘍成長の有意な阻害に至ることを示している。
【0070】
血管形成障害の処置のためのsiRNAの経口送達
図10は20μg/マウス/日の用量で経口的に与えられたVEGFR2−C3−MOE−siRNA1は脈管化重量を基底レベル(例えば成長因子誘導を伴わない重量)まで低下させたことを示している。用いられた実際のsiRNA配列は表3で言及される。
抗Tie2 C3−MOE siRNAもまた成長因子誘起血管形成モデルにおいて腹腔内および経口送達の双方で試験した。図11aおよび11bは経口的に与えられた、Tie2で志向される双方のC3−MOE siRNAは20μg/マウス/日で活性であったことを示している。用いられた実際のsiRNA配列は表3を参照して決定できる。
データにより、さらなる内部修飾を伴うかまたは伴わない3’末端修飾されたsiRNAは経口投与時に妥当な用量で治療効果を実証できることが示される。
【0071】
参照文献
1. a)Y. Tomari et al. Genes and Development 19(2005), 517;b)P. Shankar et al. JAMA 11(2005), 1367;c)Y. Dorsett et al. Nature Reviews 3(2004), 318
2. a)P.D. Zamore et al. Cell 101,(2000), 25;b)S.M. Hammond et al. Nature 404(2000), 293
3. a)G. Meister et al. Molecular Cell 15(2004), 185.
4. S.M. Elbashir et al. Genes Dev. 15(2001), 188.
5. S.J. Reich et al. Molecular Vision 9(2003), 210.
6. a)Dorn et al. Nucleic Acids Research 32(2004), e49;b)D. R.Thakker et al. PNAS 101(2004), 17270;c)D.R. Thakker et al. Molecular Psychiatry 10(2005), 714
7. V. Bitko et al. Nature Medicine 11(2005), 50.
8. E. Song et al. Nature Medicine 9(2003), 347.
9. D.A.Braasch et al. Biochemistry 42(2003), 7967.
10. Harborth, Antisense Nucleic Acid Drug Devt, 2003
11. A.H.S. Hall et al. Nucleic Acids Research 32(2004), 5991.
12. M. Amarzguioui et al. Nucleic Acids Research 31(2003), 589.
13. F. Czauderna et al. Nucleic Acids Research 31(2003), 2705.
14. T. Prakash et al. Journal of Medicinal Chemistry 48(2005), 4247.
15. J. Elmen et al. Nucleic Acids Research 33(2005), 439.
16. A.S. Boutorin, L.V. Guskova, E.M. Ivanova, N.D. Kobetz, V.F. Zafytova, A.S. Ryte, L.V. Yurchenko and V.V. Vlassov FEBS Lett. 254(1989), p. 129
17. J. Wood et al. Cancer Research 60(2000), 2178.
18. K. LaMontagne et al. Cancer Res. 66(2006), 221.
【技術分野】
【0001】
背景
RNA干渉は当初植物において転写後遺伝子サイレンシング(PTGS)として見出された、二本鎖RNA(dsRNA)が引き金となる高度に保存されたメカニズムであり、そしてdsRNAに相同な遺伝子の転写を下方調節できる1。dsRNAは最初にダイサーにより低分子干渉RNA(siRNA)と称される21−23ヌクレオチドの短い二重鎖にプロセシングされる2。それはRNA誘起サイレンシング複合体(RISC)に組み込まれているので、RISCの構成成分であるアルゴノート2により標的mRNAを相同性の領域の中央で切断することにより遺伝子サイレンシングを媒介することができる3。Elbashir et al(2001)4では、合成siRNAの直接導入によりショウジョウバエのみならず哺乳動物細胞においてもRNA干渉遺伝子サイレンシングが媒介されることが実証された。それ以来siRNA媒介遺伝子サイレンシングは標的同定および標的バリデーション研究の双方で、強力でそして広く用いられる分子生物学的手段になってきている。動物実験での遺伝子サイレンシングのためのsiRNAの使用は限られた量の動物モデルでしか記載されていない。未修飾siRNAは眼において局所的に5、中枢神経系において髄腔内または小脳内に6、および呼吸器系ウイルスの阻止のために鼻内に7送達された。未修飾siRNAの静脈内水力学的尾静脈注射もまた研究されている。この研究法により、主に肝臓への急速な送達が可能になる8。未修飾siRNAの全身投与に関しては非常に限られた数の研究しか報告されていない。Duxbury et al9は接着斑キナーゼを標的とする未修飾siRNAを正所性腫瘍異種移植片マウスモデルに静脈内投与し、そして腫瘍成長阻害およびゲムシタビンに対する化学増感を観察した。Soutscheck et alはアポリポタンパク質Bを内在的にサイレンシングするための高度に化学的に修飾されたsiRNAの全身使用を報告した。ほとんどの抗ApoB siRNAの高用量の50mg/kgでの腹腔内投与によりApoBタンパク質レベルおよびリポタンパク質濃度が低下した10。これらの実例にもかかわらず、全身送達時のsiRNAのインビボ使用は、このテクノロジーを標的バリデーションまたは治療適用に広く適用できるようにするために、改良を必要とする。実際、未修飾siRNAは、主に血流で豊富なヌクレアーゼによる酵素消化に供される。siRNAの薬理学的特性を改善するために、いくつかのグループはこれらの試薬の化学的修飾を調査した。記載された研究法はその中でも非常に様々で、そして系統的な研究は依然実施されていないが、結果の概要により化学的修飾に対するsiRNAの寛容性を決定することが可能になる。ホスホロチオエート11またはボラノホスフェート12、2’−O−メチル13、2’−O−アリル14、2’−メトキシエチル(MOE)および2’−デオキシフルオロヌクレオチド15またはロックされた核酸(Locked Nucleic Acid)(LNA)16のようないくつかの化学が調査されている。これらの研究により、修飾に関する寛容性は化学依存性のみならず、位置依存性でもあることが強調された。
【発明の概要】
【0002】
本発明は改善された薬理学的特性を有する最小限に修飾されたsiRNAを提供する。最小限に修飾されたsiRNAは、3’エキソヌクレアーゼ消化を防御するために各鎖の3’末端で修飾された19塩基対二本鎖RNAであり:21ヌクレオチドsiRNAの3’−ジデオキシヌクレオチドオーバーハングが一般的な3’−ヒドロキシプロピルホスホジエステル部分により置き換えられており、そして各鎖の3’末端で最初に塩基対形成している2個のヌクレオチドの修飾が血清安定性をさらに強化する。成体マウスに腹腔内または経口適用した場合、修飾されたsiRNAは成長因子誘起の血管形成モデルにおいてより高度な効力を表し、それは血清安定性の増大に相関する。
【0003】
発明の要旨
一つの態様では、本発明は経口投与用の低分子干渉リボ核酸(siRNA)を提供し、該siRNAは少なくとも15ヌクレオチドにわたって互いに相補的である2本の別個のRNA鎖を含み、ここで各鎖は49ヌクレオチド以下であり、そしてここでその鎖の少なくとも1本は少なくとも一つの化学的修飾を含有する。
【0004】
一つの実施態様では、siRNAは少なくとも1個の修飾されたヌクレオチドを含む。
別の実施態様では、siRNAは少なくとも1個の3’末端キャップを含む。
別の実施態様では、該修飾されたヌクレオチドは2’アルコキシリボヌクレオチド、2’アルコキシアルコキシリボヌクレオチド、ロックされた核酸リボヌクレオチド(LNA)、2’−フルオロリボヌクレオチド、モルホリノヌクレオチドの中から選択される。
別の実施態様では、該修飾されたヌクレオチドはホスホロチオエート、ホスホロジチオエート、ホスホロアミダート、ボラノホスホノエートおよびアミド結合の中から選択される修飾されたヌクレオシド内結合を有するヌクレオチドの中から選択される。
別の実施態様では、該2本のRNA鎖は互いに完全に相補的である。
別の実施態様では、該siRNAは5’末端または3’末端の少なくとも一つに1から6ヌクレオチドオーバーハングを含む。
【0005】
別の実施態様では、siRNAは少なくとも1個の3’キャップを含有し、それは3’炭素を介して3’末端に抱合された化学的部分であり、かつ式I:
【化1】
[式I]
(式中、
XはOまたはSであり、
R1およびR2は独立してOH、NH2、SH、アルキル、アリール、アルキル−アリール、アリール−アルキルであり、ここでアルキル、アリール、アルキル−アリール、アリール−アルキルをさらなるヘテロ原子および官能基、好ましくはN、OもしくはSの群から選択されるヘテロ原子またはOH、NH2、SH、カルボン酸もしくはエステルの群から選択される官能基により置換することができ;
またはR1およびR2は式Y−Zのものでよく、ここでYはO、N、Sであり、そしてZはH、アルキル、アリール、アルキル−アリール、アリール−アルキルであり、ここでアルキル、アリール、アルキル−アリール、アリール−アルキルをさらなるヘテロ原子、好ましくはN、OもしくはSの群から選択されるヘテロ原子により置換することができる)
の化合物の中から選択される。
【0006】
別の実施態様では、siRNAは少なくとも15ヌクレオチドにわたってVEGFR−1、VEGFR−2、VEGFR3、Tie2、bFGFR、IL8RA、IL8RB、FasまたはIGF2RのmRNAまたはプレmRNAに相補的である少なくとも1本の鎖を含有する。
別の実施態様では、siRNAは配列番号:1−900から選択される配列を含む少なくとも1本の鎖を含有する。
別の実施態様では、siRNAは配列番号:901−930からなる群から選ばれる。
別の実施態様では、siRNAは標準的な胃酸アッセイにおいて、同一ヌクレオチド配列を有する未修飾siRNAよりも大きな安定性を有する。
別の実施態様では、siRNAは標準的な胃酸アッセイにおいて、30分暴露後に50%以上である安定性を有する。
【0007】
別の実施態様では、siRNAは標準的な血清アッセイにおいて未修飾siRNAよりも大きな安定性を有する。
別の実施態様では、siRNAは標準的な血清アッセイにおいて30分暴露後に50%以上である安定性を有する。
別の実施態様では、siRNAは標準的な腸洗浄液アッセイにおいて未修飾siRNAよりも大きな安定性を有する。
別の実施態様では、siRNAは同一ヌクレオチド配列の未修飾siRNAと比較して経口バイオアベイラビリティーが強化されている。
【0008】
一つの態様では、本発明は一つまたはそれより多い前記の特性を有するsiRNAを含む医薬組成物を提供する。
別の態様では、本発明は医薬品として使用するための任意の一つまたはそれより多い前記の特性を有するsiRNAを提供する。
別の態様では、本発明は血管形成障害の処置用医薬品の製造における任意の一つまたはそれより多い前記の特性を有するsiRNAの使用を提供する。
別の態様では、本発明はインビトロで血管形成過程を阻害するための任意の一つまたはそれより多い前記の特性を有するsiRNAの使用を提供する。
【図面の簡単な説明】
【0009】
【図1a】図1a、1b、1c、1dおよび1e:未修飾siRNA pGl3−siRNA(マウス血清中の野生型siRNA)の代謝分解;a−c)マウス血清中0分、30分および180分インキュベートした後の未修飾siRNAのイオン交換−HPLC分析;37℃で30分インキュベートした後のイオン交換HPLCにおける主要なピークを単離し、そしてLC−MSに再注入した、d)検出された分子量およびその割り当ての表;e)ESI−MSスペクトル。
【図1b】図1a、1b、1c、1dおよび1e:未修飾siRNA pGl3−siRNA(マウス血清中の野生型siRNA)の代謝分解;a−c)マウス血清中0分、30分および180分インキュベートした後の未修飾siRNAのイオン交換−HPLC分析;37℃で30分インキュベートした後のイオン交換HPLCにおける主要なピークを単離し、そしてLC−MSに再注入した、d)検出された分子量およびその割り当ての表;e)ESI−MSスペクトル。
【図1c】図1a、1b、1c、1dおよび1e:未修飾siRNA pGl3−siRNA(マウス血清中の野生型siRNA)の代謝分解;a−c)マウス血清中0分、30分および180分インキュベートした後の未修飾siRNAのイオン交換−HPLC分析;37℃で30分インキュベートした後のイオン交換HPLCにおける主要なピークを単離し、そしてLC−MSに再注入した、d)検出された分子量およびその割り当ての表;e)ESI−MSスペクトル。
【図1d】図1a、1b、1c、1dおよび1e:未修飾siRNA pGl3−siRNA(マウス血清中の野生型siRNA)の代謝分解;a−c)マウス血清中0分、30分および180分インキュベートした後の未修飾siRNAのイオン交換−HPLC分析;37℃で30分インキュベートした後のイオン交換HPLCにおける主要なピークを単離し、そしてLC−MSに再注入した、d)検出された分子量およびその割り当ての表;e)ESI−MSスペクトル。
【図1e】図1a、1b、1c、1dおよび1e:未修飾siRNA pGl3−siRNA(マウス血清中の野生型siRNA)の代謝分解;a−c)マウス血清中0分、30分および180分インキュベートした後の未修飾siRNAのイオン交換−HPLC分析;37℃で30分インキュベートした後のイオン交換HPLCにおける主要なピークを単離し、そしてLC−MSに再注入した、d)検出された分子量およびその割り当ての表;e)ESI−MSスペクトル。
【図2】図2:四つの二本鎖RNA型式の説明:野生型(または未修飾)siRNA、MOE o/h siRNA、C3−siRNAおよびC3−MOE siRNA。
【0010】
【図3】図3:マウス胃酸中三つの異なる型式でのsiRNAの安定性。試料をマウス胃酸中37℃で2μMの濃度でインキュベートした。親化合物のバンドを定量することにより親化合物の消失を2−6時間にわたって追跡した。レーン1−7:胃酸中t=0、5、10、15、30、60および120分の野生型siRNAレーン8:dsRNAラダー(30、21、19、16、13、10塩基対)レーン9−15:胃酸中t=0、5、10、15、30、60および120分のC3 siRNAレーン16:dsRNAラダー(30、21、19、16、13、10塩基対)レーン17−24:胃酸中t=0、5、10、15、30、60および120分のC3−MOE siRNA
【0011】
【図4】図4:腸洗浄液中四つの異なる型式でのsiRNAの安定性。試料を肝ミクロソーム中37℃で5μMの濃度でインキュベートした。(左から右に)レーン1:dsRNAラダー(30、21、19、16、13、10塩基対)レーン2−7:腸洗浄液中t=0、15、30、60、180および360分の野生型siRNAレーン8−13:腸洗浄液中t=0、15、30、60、180および360分のmoe o/h siRNAレーン14−19:腸洗浄液中t=0、15、30、60、180および360分のC3 siRNAレーン20−25:腸洗浄液中t=0、15、30、60、180および360分のC3−MOE siRNA
【0012】
【図5】図5:肝ミクロソーム中四つの異なる型式でのsiRNAの安定性。試料をラット腸洗浄液からの腸液中37℃で2μMの濃度でインキュベートした。(左から右に)レーン1:ds
【図6】図6:マウス血清中四つの異なる型式でのsiRNAの安定性。試料をマウス血清中37℃で2μMの濃度でインキュベートした。親化合物のバンドを定量することにより親化合物の消失を6時間にわたって追跡した。(左から右に)レーン1:dsRNAラダー(30、21、19、16、13、10塩基対)RNAラダー(30、21、19、16、13、10塩基対)レーン2:野生型siRNA未処理レーン3:moe o/h siRNA未処理レーン4:C3 siRNA未処理レーン5:C3−MOE siRNA未処理レーン6−9:肝ミクロソーム中2−5と同一、t=0レーン10−13:肝ミクロソーム中2−5と同一、t=60分レーン14−17:上澄S12中2−5と同一、t=0レーン18−21:上澄S12中2−5と同一、t=60分レーン2−7:マウス血清中t=0、15、30、60、180および360分の野生型siRNAレーン8−13:マウス血清中t=0、15、30、60、180および360分のmoe o/h siRNAレーン14−19:マウス血清中t=0、15、30、60、180および360分のC3 siRNAレーン20−25:マウス血清中t=0、15、30、60、180および360分のC3−MOE siRNA
【0013】
【図7】図7:抗VEGFR2 siRNA(2個の独立した配列)の三つの型式の細胞内(in cellulo)での特徴付け。野生型siRNA、C3−siRNAおよびC3−MOE siRNAを3濃度(1、5、10nM)でMS1細胞にトランスフェクトした。FACSによりVEGFR2細胞表面レベルを測定することによりサイレンシング効力を評価した。
【0014】
【図8a】図8aおよび8b:成長因子誘起血管形成「寒天チャンバー(Agar Chamber)」マウスモデルにおける野生型siRNA、C3−siRNAおよびC3−Moe siRNAのインビボ試験。図8aは1、5および25μg/マウス/日の対照、未修飾VEGFR2 siRNAおよびC3修飾VEGFR2 siRNAの結果を示す。図8bは0.2、1および5μg/マウス/日での対照、C3修飾VEGFR2 siRNAおよびC3−MOE VEGFR2 siRNAを示す。各例で2つの抗VEGFR2 siRNAのプールを毎日3日間腹腔内に与えた。
【図8b】図8aおよび8b:成長因子誘起血管形成「寒天チャンバー(Agar Chamber)」マウスモデルにおける野生型siRNA、C3−siRNAおよびC3−Moe siRNAのインビボ試験。図8aは1、5および25μg/マウス/日の対照、未修飾VEGFR2 siRNAおよびC3修飾VEGFR2 siRNAの結果を示す。図8bは0.2、1および5μg/マウス/日での対照、C3修飾VEGFR2 siRNAおよびC3−MOE VEGFR2 siRNAを示す。各例で2つの抗VEGFR2 siRNAのプールを毎日3日間腹腔内に与えた。
【0015】
【図9a】図9:B16同種移植片メラノーマ腫瘍マウスモデルに5および20μg/マウス/日で腹腔内(i.p.)に与えられた抗VEGFR2 C3−MOE siRNAのインビボ試験。図9aは、修飾されたVEGFR2 siRNAでのi.p.処置が腫瘍発達を有意に低下させることを示す。図9bもまたVEGFR2 siRNAの20μg/マウスのi.p.注射が有意な腫瘍成長の阻害を招くことを示す。
【図9b】図9:B16同種移植片メラノーマ腫瘍マウスモデルに5および20μg/マウス/日で腹腔内(i.p.)に与えられた抗VEGFR2 C3−MOE siRNAのインビボ試験。図9aは、修飾されたVEGFR2 siRNAでのi.p.処置が腫瘍発達を有意に低下させることを示す。図9bもまたVEGFR2 siRNAの20μg/マウスのi.p.注射が有意な腫瘍成長の阻害を招くことを示す。
【0016】
【図10】図10:成長因子誘起血管形成マウスモデルにおけるC3−MOE siRNAのインビボ試験。抗VEGFR2 siRNAを20μg/マウス/日を毎日3日間経口的に与えた。
【図11a】図11:成長因子誘起血管形成マウスモデルにおけるC3−MOE siRNAのインビボ試験。抗Tie2 siRNAを毎日腹腔内(1および0.2μg/マウス/日)または経口的に(20および5μg/マウス/日)3日間経口与えた。図11a:切除組織の重量;図11b:Tie2タンパク質ノックダウン。
【図11b】図11:成長因子誘起血管形成マウスモデルにおけるC3−MOE siRNAのインビボ試験。抗Tie2 siRNAを毎日腹腔内(1および0.2μg/マウス/日)または経口的に(20および5μg/マウス/日)3日間経口与えた。図11a:切除組織の重量;図11b:Tie2タンパク質ノックダウン。
【発明を実施するための形態】
【0017】
発明の詳細な説明
本発明は哺乳動物における血管形成障害を処置するための組成物および方法に関する。特に本発明は哺乳動物への経口投与時に血管形成障害を処置するために使用することができる低分子干渉RNA(「siRNA」)に関する。
血管形成は以下の標的/遺伝子を含む血管内皮細胞を標的とする:VEGFR−1(GenBank受入番号AF06365);VEGFR−2(GenBank受入番号AF063658);VEGFR−3(GenBank受入番号(NM_002020);Tie2(TEK)(GenBank受入番号NM_000459);bFGFR(GenBank受入番号M60485);IL8RA(GenBank受入番号L19591);IL8RB(GenBank受入番号L19593);Fas(GenBank受入番号X89101);IGF2R(GenBank受入番号NM_000876)。
【0018】
本発明によるsiRNA分子はRNA干渉(「RNAi」)を媒介する。「RNAi」なる用語は当分野において周知であり、そして一般的に標的遺伝子に相補的である領域を有するsiRNAによる、細胞における一つまたはそれより多い標的遺伝子の阻害を意味すると理解される。RNAiを媒介するその能力に関してsiRNAを試験するための種々のアッセイが当分野において公知である(例えばElbashir et al., Methods 26:199−213(2002)参照)。本発明によるsiRNAの遺伝子発現に及ぼす影響は、本発明によるRNA分子で処理されていない細胞と比較した場合、典型的には少なくとも10%、33%、50%、90%、95%または99%まで阻害される標的遺伝子の発現に至る。
【0019】
本発明による「siRNA」または「低分子干渉リボ核酸」は以下の態様を含む当分野において公知の意味を有する。siRNAは生理学的条件下で相補的領域に沿ってハイブリダイズする2本の鎖のリボヌクレオチドからなる。鎖は別個であるが、特定の実施態様では、分子リンカーにより結合し得る。個々のリボヌクレオチドは未修飾天然発生リボヌクレオチド、未修飾天然発生デオキシリボヌクレオチドでよいか、またはそれらは本明細書の他の部分で記載されるような化学的に修飾された、もしくは合成性でよい。
【0020】
本発明によるsiRNA分子は標的遺伝子のmRNAの領域と実質的に同一である二本鎖領域を含む。標的遺伝子の対応する配列に対して100%同一性を有する領域が適当である。この状態は「完全に相補的」と称される。しかしながらその領域はまた、標的化されるmRNAの領域の長さに依存して、標的遺伝子の対応する領域と比較して1、2または3個の誤対合を含有してもよく、そしてそのようなものは完全に相補的ではないかもしれない。一つの実施態様では、本発明のRNA分子は1個の所定の遺伝子を特異的に標的化する。望ましいmRNAのみを標的化するために、siRNA試薬は標的mRNAに対して100%相同性、および細胞または生物に存在する全てのその他の遺伝子に対して少なくとも2個の誤対合したヌクレオチドを有し得る。具体的な標的配列の発現を有効に阻害するために、十分な配列同一性を有するsiRNAを分析および同定するための方法が当分野において公知である。当分野において公知の配列比較およびアラインメントアルゴリズムにより配列同一性を最適化し(Gribskov and Devereux, Sequence Analysis Primer, Stockton Press,(1991)およびそこに引用された参照文献を参照のこと)、そして例えばBESTFITソフトウェアプログラムでデフォルトパラメーターを用いて実行されるようなSmith−Watermanアルゴリズム(例えばUniversity of Wisconsin Genetic Computing Group)によりヌクレオチド配列間の差異パーセントを計算することができる。
【0021】
RNAi試薬の有効性に影響を及ぼす別の因子は標的遺伝子の標的領域である。RNAi試薬による阻害に有効な標的遺伝子の領域を実験方法により決定することができる。適当なmRNA標的領域はコード化領域であろう。5’−UTR、3’−UTRおよびスプライスジャンクションのような未翻訳領域もまた適当である。例えばElbashir S.M. et al, EMBO J., 20:6877−6888(2001)に記載されるようなトランスフェクションアッセイをこの目的のために実施することができる。当分野では当業者に周知である多くのその他の適当なアッセイおよび方法が存在する。
【0022】
本発明にしたがって、標的に相補的なsiRNAの領域の長さは10から100ヌクレオチド、12から25ヌクレオチド、14から22ヌクレオチドまたは15、16、17もしくは18ヌクレオチドでよい。対応する標的領域に対する誤対合が存在する場合、相補的領域の長さは一般的に幾分長くなることが要求される。
siRNAはオーバーハングした末端(標的に対して相補的であってもなくてもよい)、または標的配列にではなくそれ自体に相補的なさらなるヌクレオチドを担持し得るので、siRNAの各別個の鎖の全体の長さは10から100ヌクレオチド、15から49ヌクレオチド、17から30ヌクレオチドまたは19から25ヌクレオチドでよい。
【0023】
「各鎖が49ヌクレオチド以下である」なる語句は、全ての修飾または未修飾ヌクレオチドを含むが、鎖の3’または5’末端に付加され得る任意の化学的部分を含まない、鎖の連続したヌクレオチドの全数を意味する。鎖に挿入された短い化学的部分は計数されないが、2本の別個の鎖を結合するために設計された化学的リンカーは連続したヌクレオチドを創成するとは考えられない。
「5’末端または3’末端の少なくとも一つにおける1から6ヌクレオチドオーバーハング」なる語句は生理学的条件下で2本の別個の鎖から形成される相補的siRNAの構造を指す。末端ヌクレオチドがsiRNAの二本鎖領域の一部である場合、siRNAはブラント末端と考えられる。一つまたはそれより多いヌクレオチドが末端で対形成されない場合、オーバーハングが創成される。オーバーハングの長さはオーバーハングするヌクレオチドの数により測定される。オーバーハングするヌクレオチドはいずれかの鎖の5’末端または3’末端のいずれかでよい。
【0024】
本発明によるsiRNAは少なくとも1本の鎖に少なくとも1個の修飾されたヌクレオチドを含むことにより、インビボで経口送達に適当な高い安定性を付与する。故に本発明によるsiRNAは少なくとも1個の修飾されたまたは非天然リボヌクレオチドを含有する。多くの公知の化学的修飾についての冗長な記載は公開PCT特許出願第WO200370918号に示され、そして本明細書で繰り返さない。経口送達のための適当な修飾をさらに具体的に本明細書の実施例および記載にて示す。適当な修飾には、限定するものではないが糖部分に対する修飾(すなわち例えば2’−O−(2−メトキシエチル)または2’−MOEのような糖部分の2’位置(Martin et al., Helv. Chim. Acta, 78:486−504(1995))すなわちアルコキシアルコキシ基)または塩基部分(すなわち代替ヌクレオチド鎖の別の具体的な塩基と対形成する能力を維持する非天然または修飾された塩基)が含まれる。その他の修飾には、限定するものではないがリン酸エステル基の置換(隣接するリボヌクレオチドを例えばホスホロチオエート、キラルホスホロチオエートまたはホスホロジチオエートと連結させる)を含むいわゆる「バックボーン」修飾が含まれる。最後に、時に本明細書では3’キャップまたは5’キャップと称される末端修飾が重要である。表1にて説明されるようにキャップは、siRNAに安定性を付与することが見出されている「T−T」のようなさらなるヌクレオチドの単純な付加からなってよい。キャップは当業者に公知である錯体化学からなってよい。
【0025】
以下の実施例で用いられる一つの実施態様では、3’キャップは3’炭素を介して3’末端に抱合された化学的部分であり、かつ式I:
【化2】
[式I]
(式中、
XはOまたはSであり、
R1およびR2は独立してOH、NH2、SH、アルキル、アリール、アルキル−アリール、アリール−アルキルであり、ここでアルキル、アリール、アルキル−アリール、アリール−アルキルをさらなるヘテロ原子および官能基、好ましくはN、OもしくはSの群から選択されるヘテロ原子またはOH、NH2、SH、カルボン酸もしくはエステルの群から選択される官能基により置換することができ;
またはR1およびR2は式Y−Zのものでよく、ここでYはO、N、Sであり、そしてZはH、アルキル、アリール、アルキル−アリール、アリール−アルキルであり、ここでアルキル、アリール、アルキル−アリール、アリール−アルキルをさらなるヘテロ原子、好ましくはN、OもしくはSの群から選択されるヘテロ原子により置換することができる)
の化合物の中から選択される。
【0026】
糖部分での修飾の実例には2’アルコキシリボヌクレオチド、2’アルコキシアルコキシリボヌクレオチド、ロックされた核酸リボヌクレオチド(LNA)、2’−フルオロリボヌクレオチド、モルホリノヌクレオチドが挙げられる。
ヌクレオシド内結合を修飾することもできる。ヌクレオシド内結合の実例にはホスホロチオエート、ホスホロジチオエート、ホスホロアミダートおよびアミド結合が挙げられる。
R1はOHでよい。
R1およびR2は一緒に1から24個のC原子、1から12個のC原子、2から10個のC原子、1から8個または2から6個のC原子を含み得る。別の実施態様では、R1およびR2は独立してOH、低級アルキル、低級アリール、低級アルキル−アリール、低級アリール−アルキルであり、ここで低級アルキル、低級アリール、低級アルキル−アリール、低級アリール−アルキルを前記で定義されるようなさらなるヘテロ原子および官能基により置換することができる。別の実施態様では、R1およびR2は双方ともOHではない。
【0027】
有機ラジカルまたは化合物に関連する「低級」なる用語は7個以下の炭素原子、好ましくは1−4個の炭素原子を含む分岐しているかまたは分岐していなくてよい化合物またはラジカルを意味する。低級アルキルは、例えばメチル、エチル、n−プロピル、イソプロピル、n−ブチル、sec−ブチル、tert−ブチル、n−ペンチルおよび分岐したペンチル、n−ヘキシルおよび分岐したヘキシルを表す。
アルコキシの実例にはO−Met、O−Eth、O−prop、O−but、O−pent、O−hexが挙げられる。
【0028】
少なくとも1個の修飾されたまたは非天然リボヌクレオチドを含有するsiRNAを含むsiRNAの合成のための方法は周知であり、そして当業者に容易に利用可能である。例えば種々の合成化学が公開PCT特許出願第WO2005021749号および第WO200370918号(双方共に出典明示により本明細書の一部とする)に示されている。反応を溶液中または好ましくは固相でポリマーに支持された試薬を使用することにより実施でき、続いてRNAiを媒介することができるsiRNA分子が形成される条件下で合成されたRNA鎖を組み合わせる。
【0029】
本発明は経口送達に適当である少なくとも1個の修飾されたヌクレオチドを含有するするsiRNAを提供する。機能的な表現ではこれは、siRNAは経口投与時に適当な薬物動態および体内分布を有し、関連する標的組織への送達を達成するであろうということを意味する。とりわけこれには血清安定性、免疫応答の欠如、および薬物様の挙動が必要とされる。本明細書の別の部分で開示される標準的な胃酸アッセイおよび標準的な血清アッセイに基づいてsiRNAのこれらの様相の多くを予想することができる。
【0030】
別の態様では、本発明は、RNAiにより少なくとも1個の標的遺伝子を阻害することができる本発明によるsiRNAを細胞に導入することを含む標的遺伝子の阻害のための方法を提供する。また、各々別の標的領域に特異的である1を超える種のsiRNAを同時にまたは逐次的に細胞に導入することができる。
【0031】
本発明は任意の型の標的遺伝子またはヌクレオチド配列に限定されるものではない。例えば標的遺伝子は細胞遺伝子、内因性遺伝子、病原体関連遺伝子、ウイルス遺伝子または発癌遺伝子でよい。実施例のいくつかは、経口送達された本発明のsiRNAが脈管形成、血管新生または血管形成の部位に蓄積され得ることを強調しているので、血管形成遺伝子は本発明に特に重要である。本発明に特に興味深いこれらの部位での血管形成遺伝子の最新のリストはAngioDB:血管形成および血管形成関連分子Tae−Kwon Sohn、Eun−Joung Moon1、Seok−Ki Lee1、Hwan−Gue Cho2およびKyu−Won Kim3のデータベース、Nucleic Acids Research, 30(1):369−371(2002)およびオンラインhttp://angiodb.snu.ac.kr/に列挙される。特に重要な遺伝子は詳細に分析されており、そして本明細書の別の部分に示されている。
【0032】
別の態様では、本発明はまた細胞における標的遺伝子の発現を阻害するための試薬を含むキットを提供し、ここで該キットは本発明によるdsRNAを含む。キットはインビトロまたはインビボで本発明によるdsRNAの導入を実施して、試料または対象を試験するのに必要な少なくとも一つの試薬を含む。好ましい実施態様では、かかるキットはまたキットの構成成分が使用される手順を詳記する説明書をも含む。
【0033】
「血管形成障害の処置」とはこの開示で使用される際には、血管新生、脈管形成および/または血管形成の生理学的および病理学的過程を伴う疾患の処置のための医薬組成物における本発明の修飾されたsiRNAの使用を意味する。このようにこれらの医薬組成物は、限定するものではないが癌腫瘍成長および転移、新生物、眼血管新生(黄斑変性症、糖尿病性網膜症、虚血性網膜症、未熟児網膜症、脈絡膜血管新生を含む)、関節リウマチ、骨関節炎、慢性喘息、敗血症性ショック、炎症性疾患、滑膜炎、骨および軟骨破壊、パンヌス成長、骨棘形成、骨髄炎、乾癬、肥満、血管腫、カポジ肉腫、アテローム性動脈硬化症(アテローム性動脈硬化巣断裂を含む)、子宮内膜症、疣贅、多毛症、ケロイド瘢痕、アレルギー性浮腫、機能不全性子宮出血、卵胞嚢胞、卵巣過剰刺激、子宮内膜症、骨髄炎、炎症性および感染性過程(肝炎、肺炎、糸球体腎炎)、喘息、鼻ポリープ、移植、肝臓再生、白質軟化症、甲状腺炎、甲状腺肥大、リンパ球増殖性障害、血液学的悪性腫瘍、血管奇形および子癇前症を含む血管新生、脈管形成または血管形成の阻害を必要とする疾患、症状および障害を処置するために有用である。
【0034】
本明細書で使用される際には、「処置」とは、疾患、障害もしくは症状の過程を阻害もしくは低減させるため、疾患、障害もしくは症状の病徴を阻害もしくは低減させるため、または疾患、障害もしくは症状の発症もしくはさらなる発達を予防的に防御するために取る行動を意味する。「処置する」とはその認識動詞である。
【0035】
本発明の治療薬の有効用量は疾患状態を処置するために必要とされるその用量である。有効用量は疾患の型、使用される組成物、投与の経路、処置されている哺乳動物の型、考慮される具体的な哺乳動物の身体的特徴、併用薬、および医学の分野において当業者に認識されるその他の因子に依存する。一般的に効力に依存して0.1mg/kgと100mg/kg体重/日の間の量のsiRNAが投与される。本発明の核酸分子およびその処方を経口的、局所的、非経口的に、吸入もしくはスプレーにより、または直腸に従来の無毒性の薬学的に許容される担体、アジュバントおよび/またはベヒクルを含有する投薬単位処方で投与できる。非経口的なる用語は、本明細書で使用される際には、経皮、皮下、血管内(例えば静脈内)、筋肉内、腹腔内もしくはくも膜下腔内注射、または注入技術等を含む。加えて、本発明の核酸分子および薬学的に許容される担体を含む医薬用処方が提供される。一つまたはそれより多い本発明の核酸分子は、一つまたはそれより多い無毒性の薬学的に許容される担体および/または希釈剤および/またはアジュバント、ならびに所望によりその他の活性成分を伴って存在できる。本発明の核酸分子を含有する医薬組成物を経口使用に適当な形態、例えば錠剤、トローチ、ロゼンジ、水性もしくは油性懸濁液、分散性粉末もしくは顆粒、エマルジョン、硬質もしくは軟質カプセル、またはシロップもしくはエリキシルにできる。
【0036】
経口使用を意図される組成物を、医薬組成物の製造に関して当分野において公知の任意の方法にしたがって調製することができ、そしてかかる組成物は医薬的に洗練され、そして口当たりのよい調製物を提供するために一つまたはそれより多いかかる甘味剤、着香剤、着色剤または保存剤を含有できる。錠剤は錠剤の製造に適当である無毒性の薬学的に許容される賦形剤と混合された活性成分を含有する。これらの賦形剤は例えば炭酸カルシウム、炭酸ナトリウム、ラクトース、リン酸カルシウムまたはリン酸ナトリウムのような不活性希釈剤;顆粒化および崩壊剤、例えばトウモロコシデンプンまたはアルギン酸;結合剤、例えばデンプン、ゼラチンまたはアラビアゴム;および滑沢剤、例えばステアリン酸マグネシウム、ステアリン酸またはタルクでよい。錠剤はコーティングされていなくてよいか、またはそれらを公知の技術によりコーティングすることができる。経口使用のための処方を、活性成分が不活性固体希釈剤、例えば炭酸カルシウム、リン酸カルシウムもしくはカオリンと混合される硬質ゼラチンカプセルとして、または活性成分が水もしくは油性溶媒、例えば落花生油、液体パフィンもしくはオリーブ油と混合される軟質ゼラチンカプセルとして提示することもできる。水性懸濁液は水性懸濁液の製造に適当な賦形剤との混合物中に活性成分を含有する。
【0037】
本発明の組成物の経口投与は胃または腸に直接的に、最も重要には患者による嚥下制御投薬形態によるが、またかかる送達のその他の機械的および補助的手段による物質を投与するための全ての標準的な技術を含む。
【0038】
約0.1mgから約140mg/kg体重/日のオーダーの投薬量レベルが前記で指示された症状の処置において有用である(約0.5mgから約7g/対象/日)。単回投薬形態を生成するための担体材料と組み合わすことができる活性成分の量は、処置される宿主および投与の特定の様式に依存して異なる。一般的に投薬単位形態は約1mgから約500mgの間の活性成分を含有する。任意の特定の対象に関する具体的な用量レベルは用いられる具体的な化合物の活性、年齢、体重、一般健康状態、性別、食事、投与の時間、投与の経路、ならびに排出の速度、薬物の組み合わせおよび治療を行っている特定の疾患の重篤度を含む種々の因子に依存することは理解される。
【0039】
本発明の治療薬の治療効果をその他の薬剤との組み合わせにより強化することができる。典型的にはかかるその他の薬剤には血管形成障害のような類似の疾患の処置に使用されることが知られている薬剤が含まれよう。これに代えて、かかる薬剤を使用して本発明の治療薬により引き起こされる副作用または望ましくない影響を低減させることができる。
【0040】
本発明のsiRNAはまた重要な調査用途をも有する。かかる研究の一つにはインビトロでの血管形成過程への調査が含まれる。「インビトロでの血管形成過程」とは動物全身を用いない血管形成または脈管形成を研究するための任意の過程を意味する。このように血管形成のマーカーまたは指標を使用して血管形成過程の工程を研究するインビトロまたはエキソビボ方法およびアッセイはここに含まれる。
【0041】
RNA鎖ヌクレオチド配列
表1で同定されるsiRNA鎖配列は以下の標的に対する適当なsiRNA配列として同定されている:VEGFR−1(GenBank受入番号AF06365);VEGFR−2(GenBank受入番号AF063658);VEGFR−3(GenBank受入番号(NM_002020);Tie2(TEK)(GenBank受入番号NM_000459);bFGFR(GenBank受入番号M60485);IL8RA(GenBank受入番号L19591);IL8RB(GenBank受入番号L19593);Fas(GenBank受入番号X89101);IGF2R(GenBank受入番号NM_000876)。
【0042】
表1:ヒトVEGFR−1、VEGFR−2、VEGFR−3、Tie2、bFGFR、IL8RA、IL8RB、Fas、IGF2Rに対するsiRNA
【表1】
【表2】
【表3】
【表4】
【表5】
【表6】
【表7】
【表8】
【表9】
【表10】
【表11】
【表12】
【表13】
【表14】
【表15】
【表16】
【表17】
【表18】
【表19】
【表20】
【表21】
【表22】
【表23】
【表24】
【表25】
【表26】
【表27】
【表28】
【表29】
【表30】
【表31】
【0043】
RNA鎖ヌクレオチドの化学的修飾
本発明によるsiRNAは少なくとも1本のRNA鎖に少なくとも1個の修飾されたヌクレオチドを含み得る。様々な可能性のある修飾されたヌクレオチドを本明細書の別の部分で開示する。本発明による使用のための有用な修飾および修飾の組み合わせを表2に示す:
【0044】
表2:化学的修飾および配列構造
【表32】
N=任意の未修飾RNAヌクレオチド
n=未修飾DNAヌクレオチド
Np=修飾されたRNAヌクレオチド
s=ホスホロチオエートヌクレオシド内結合の同定
o/h=オーバーハング
【0045】
siRNA鎖の3’末端の3’位置に付加された以下の修飾は時に「3’末端キャップ」と称され、本発明の有用な実施態様としても認識され、そして本発明によるいずれかのsiRNAと共に使用され得る:
【0046】
【化3】
【0047】
本発明による活性を有する具体的な化合物には以下の表3に示されるものが含まれる:
表3:実施例で使用されるsiRNAの配列および化学
【表33】
【表34】
【表35】
【実施例】
【0048】
実施例
以下の実施例は本発明の態様を説明し、そして以下に列挙される請求の範囲に含まれる実施態様を限定することを意図されるものではない。さらに下の結果および考察のセクションは、以下のプロトコールにしたがって行われ、そして以下の材料を用いる実験に言及する。具体的に記載されていない材料およびプロトコールは当業者に日常的に利用可能であると考えられる。
【0049】
実施例1
siRNAの調製
標準的な2’−O−TOMホスホアミダイトテクノロジーにより一本鎖siRNA誘導体を合成し、そしてOasis(登録商標)HLB抽出プレート(Waters)により精製した。センス−およびアンチセンス鎖siRNAをハイブリダイゼーションバッファー(100mM酢酸カリウム、2mM酢酸マグネシウム、30mM Hepes、pH7.6)中で混合し、90°Cで3分間熱変性し、そして30°Cで60分間アニーリングした。siRNA二重鎖の100μM原液を−20°Cで保存した。
【0050】
実施例2
血清中でのインキュベーションおよびIE−HPLC(LC−MS)による分析
標準的な血清アッセイでは、20μMの各siRNA6μlを血清またはCSF 54μlと混合し、そしてインキュベーター中37°Cで加熱した、冷却した混合物50μlを分析用DNA−pac PA−100カラム(Dionex)に負荷し、そして1:10アセトニトリル:バッファー(20mM酢酸ナトリウム、1mM酢酸マグネシウム、pH6.5)溶液中NaClグラジエント(30分で0−0.6M)で分析した。
LC−MS分析用に各siRNA 100μl(20μMまたは50μM)を滅菌ウシ胎仔血清(GIBCO)900μlと混合し、37°Cでインキュベートし、そして以前に指示されたように(NaClグラジエントを除く:9分で0M−0.36M/12分で0.36M−0.6M)HPLCにより分離した。分解生成物をNAPカラムで脱塩し、そしてLC−ESI−−MSにより分析した。
【0051】
実施例3
胃酸中でのインキュベーション
標準的な胃酸アッセイを準備するために、体重18から20g(6から8週齢)のFVBおよびC57BL6マウスをCharles River Laboratories(Les Oncins, France)から入手した。CO2を用いて動物を屠殺し、そして次に胃を即座に回収した。胃液および胃の内容物を収集し、そしてプールし、次いで遠心ろ過装置(Ultrafree MC, Millipores)に負荷した。製造者の推奨にしたがってフィルターユニットを10分間回転させた。マウス胃液に相当するろ液を回収し、等分し、そしてさらなる実験の前に凍結した。
各アッセイ用に、20μM siRNA溶液を9倍容量の前記されたような胃酸で希釈し、そして37°Cで0、5、10、15、30、60および120分間インキュベートした。
【0052】
実施例4
腸洗浄液中のインキュベーション
標準的な腸洗浄液アッセイを準備するために、雄Wistarラットを絶食させ、イソフルランで麻酔した。10ml生理食塩水(0.5ml/分)、続いて水20ml(1ml/分)で小腸(十二指腸、空腸、回腸)を原位置で灌流することにより腸洗浄液を入手した。収集された流出液を遠心し(3000×g、15分、22°C)、そして上澄を1.2μmフィルターに通し、そして−20°Cで保存した。
各アッセイ用に、20μM siRNA溶液を9倍容量の腸洗浄液で希釈し、そして37°Cで0、15、30、60、180および360分間インキュベートした。
【0053】
実施例5
マウス肝ミクロソーム中のインキュベーション
標準的な肝ミクロソームアッセイでは、250μM siRNA溶液10μlに、タンパク質20mg/mlのマウス肝ミクロソーム(GEntest 452701 Charge 11)25μl、100mMリン酸バッファー(pH7.4)365μl、UDPGAコファクター(水中24mM)50μl、NADPH 50μlを加えた。t=0分およびt=60分に凍結することによりインキュベーションをクエンチした。
【0054】
実施例6
ラットS12上澄中のインキュベーション
標準的なラットS12上澄アッセイ用に250μM siRNA溶液10μlをタンパク質29.9mg/mlのラット肝臓S12 17μl、100mMリン酸バッファー(pH7.4)373μl、UDPGAコファクター(水中24mM)50μl、NADPH 50μlに加えた。t=0分およびt=60分に凍結することによりインキュベーションをクエンチした。
【0055】
実施例7
マウス血清中のインキュベーション
標準的なマウス血清中インキュベーション用に20μM siRNA溶液を9倍容量のネズミ血清(Harlanヌードマウス)で希釈し、そして37°Cで0、15、30、60、180および360分間インキュベートした。
【0056】
実施例8
ゲル電気泳動安定性アッセイ
振盪およびドライアイス上で衝撃凍結(shock-frozen)の直後にインキュベーション溶液の10μlアリコートを取り、混合物を37°Cでインキュベートし、そしてアリコートを種々の時点で衝撃凍結した。負荷バッファー(Elchrom Sc., Cham, Switzerland)30μl(各々15μl)中でアリコートを解凍し、そしてSF50ゲル(Elchrom Sc., Cham, Switzerland)上、120V、8°Cで240分間分離した。バンドをSYBR Gold(Molecular Probes)で染色し、そしてBIORAD ChemiDoc(商標)XRSシステムで写真を撮った。
【0057】
実施例9
細胞培養
1.5%ゼラチンコートした培養皿でL−グルタミンおよび10%熱不活化FCS(AMIMED, Switzerland)を補充したDMEM高グルコース(4.5g/l)中でマウス不死化内皮細胞系MS1(ATCC CRL−2279)を成長させた。HiPerfect(QIAGEN)を用いて製造者の手順にしたがって24ウェル型式でMS1細胞をsiRNAでトランスフェクトした(4検体ずつ、siRNA最終濃度は10nMまたは指示されたとおりであった)。
【0058】
実施例10
FACS分析
トランスフェクトされていない、およびsiRNAトランスフェクトされたMS1細胞をFACSによりVEGFR2レベルに関して分析した。簡単には、2検体ずつまたは3検体ずつのウェルからの細胞をトリプシン処理し、各条件に関してプールし、次いでPBS+10%FCSで2回洗浄し、そして氷上で10分間インキュベートした後にRPE抱合抗VEGFR2 Ab(1μg/106セル;Avas 12α1、BD Pharmingen)を添加した。RPE標識したアイソタイプIgG2αをFACS対照(BD Pharmingen)として使用した。Cell Quest Software(Becton−Dickinson)を用いてFACScaliburでFACS獲得および分析を実施した。
【0059】
実施例11
動物研究
雌FVBマウス(6から8週齢)をCharles River Laboratories(Les Oncins, France)から入手した。耳のマーキングによりマウスを識別し、そして通常の条件下で群を維持し(ケージあたり動物6匹)、そして毎日観察した。処置群あたり6匹のマウスを使用し、そして動物保護のためのスイス動物保護法を厳密に順守して全ての動物実験を実施した。
【0060】
参照チャンバーモデルは出版物に記載されている(例えばWood J, Bold G, Buchdunger E, et al. PTK787/ZK222584、血管内皮成長因子受容体チロシンキナーゼの新規のおよび強力な阻害剤は経口投与後に血管内皮成長因子誘起応答および腫瘍成長を損なう。Cancer Res 60:2178−89(2000))。簡単には、ペルフルオロ−アルコキシ−Teflon(Teflon(登録商標)−PFA、直径21mm×8mm、550μl容量)から作られた多孔性組織チャンバーを、3μg/ml組換えヒトVEGFおよび指示されたようなsiRNAを補充したまたは補充していない0.8%寒天(BBL(登録商標)Nr. 11849、Becton Dickinson, Meylan, France)および20U/mlヘパリン(Novo Nordisk A/S, Bagsvaerd, Denmark)で充填した。充填手順の前に溶液を42℃で維持した。3%イソフルラン(Forene(登録商標)、 Abbott AG, Cham, Switzerland)吸入を用いてマウスを麻酔した。皮下移植用に、尾の付け根に皮膚の小切開を作成して移植用外套針の挿入を可能にした。動物の背の小切開を通して無菌条件下でチャンバーを移植した。創傷クリップ(Autoclip 9mm Clay Adams)により皮膚切開を閉じた。必要とされる用量に依存して、siRNAを「注射用の品質等級」の0.9%生理食塩水で希釈し、次いで動物にi.p.(200μl/用量)または経管によるp.o.(100μl/用量)のいずれかで送達した。チャンバー移植の2から4時間前にマウスに最初の投与を行い;次いで毎日2日間処置した。特記しない場合、移植後3日にマウスを屠殺し、チャンバーを切除し、そして各移植片の周囲に形成された血管新生化された線維性組織を注意深く除去した。マウスの一般状態をモニタリングするために体重を用いた。一元配置分散分析、続いてダネット検定を用いて統計分析を行った。
【0061】
実施例12
B16メラノーマ異種移植片モデル
抗血管形成治療に応答することが以前に同定されている同系B16/BL6ネズミメラノーマモデル(例えばLaMontagne K, Littlewood−Evans A, Schnell C, O’Reilly T, Wyder L, Sanchez T, Probst B, Butler J, Wood A, Liau G, Billy E, Theuer A, Hla T, Wood J. 、FTY720によるスフィンゴシン−1−リン酸受容体の拮抗作用が血管形成および腫瘍脈管化を阻害する。Cancer Res. 66(1):221−31(2006 Jan 1))を使用して標準的なまたは修飾されたsiRNAの抗腫瘍活性を評価した。腫瘍細胞(1μl、5×104/μl)を同系雌C57BL/6マウスの両耳の背側耳介に皮内注射した。原発腫瘍面積(mm2)の測定を腫瘍細胞接種後7、14および21日にコンピューター支援画像分析ソフトウェア(KS−400 3.0撮像システム、Zeiss)および具体的に設計されたマクロを用いて実施した。7日から21日まで「注射用の品質等級」の0.9%生理食塩水で希釈したsiRNAを、i.p.(200μl/用量)または経管によるp.o.(100μl/用量)のいずれかでマウスに1日1回投与した。21日にマウスを屠殺し、そして頭部リンパ節転移を重量測定し、そして次に凍結した。
これらの結果では、用いる実際のsiRNA配列および化学は表3を参照して決定できる。
【0062】
マウス血清中野生型siRNAを双方の3’末端から分解する
ヌクレアーゼによるオリゴヌクレオチド分解は大部分が3’エキソヌクレオチド分解性である。その終端での芳香族または親油性残基の導入によるアンチセンスオリゴヌクレオチドの修飾はそのヌクレオチド分解を遅延させる17。この代謝経路もまたsiRNAに優勢であるかどうかを検証するために、未修飾siRNA(野生型siRNA)をマウス血清中37°Cで3時間までインキュベートした。
用いられた未修飾siRNA配列はpGl3−siRNAであった(表3参照)。
混合物を強陰イオン交換HPLCで、t=0分、t=30分、t=180分で分析した。
【0063】
図1a、1bおよび1cで示されるように、t=30分でブラント末端化siRNAに相当する明確なピークが観察された。t=3時間までに実質的な分解が観察された。図1dおよび1eはHPLC−ESI−MS分析により同定された代謝物を説明する。この分析により双方の鎖の3’オーバーハングおよび3’末端の最初に塩基対形成するリボヌクレオチドの喪失に相当するいくつかの代謝物の存在が明らかにされる。ガイド鎖の5’末端リボヌクレオチドの消化もまた観察された。
【0064】
図1は血清中の未修飾siRNAの分解経路を示唆する。DNAオーバーハングがもしかすると3’エキソヌクレアーゼにより最初に消化される。LC−MSでは、双方の鎖の最初に塩基対形成する3’リボヌクレオチドおよびまたガイド鎖の最初に5’塩基対形成するリボヌクレオチドの喪失に相当するさらなる代謝物もまた検出された。
【0065】
3’修飾siRNAは胃腸管を通して安定である
2’−メトキシエチルリボヌクレオチドオーバーハング(MOE o/h siRNA)を有するsiRNA、ヒドロキシプロポキシホスホジエステル部分でキャップされたブラント末端化siRNA(C3−siRNA)および各鎖の3’末端で2個の最初に塩基対形成するヌクレオチドが2’−メトキシエチルリボヌクレオチド残基により修飾されたヒドロキシプロポキシホスホジエステル3’キャップsiRNA(C3−MOE siRNA)を合成した。これらの化合物を図2で図式的に説明する。
最初にsiRNAをマウス胃酸中2時間インキュベートした(図3)。C3 siRNAおよびC3−MOE sRNA場合、分解は観察されなかったが、野生型siRNAの分解は30分後に観察された。
【0066】
ラットの腸洗浄液から得られた腸液中の安定性により15分後にほぼ完了する野生型siRNAの分解が明らかにされたが、MOE o/h siRNA、C3−siRNAおよびC3−Moe siRNAの親化合物は60分間観察された(図4)。
肝臓中の安定性を肝ミクロソームアッセイおよびS12アッセイ(代表的な肝臓細胞質酵素活性)を用いて評価した。結果を図5に示す。双方の場合で、インキュベーションの60分後に分解は観察されなかった。
【0067】
最後に、siRNAをマウス血清中2μMで、37°Cで6時間までインキュベートすることにより試験した(図6の結果)。親化合物の安定性をゲル電気泳動により追跡した。修飾されたsiRNA(C3 siRNA、MOE o/h siRNAのC3−MOE siRNA)の場合、有意な分解は観察されなかったが、野生型siRNAでは観察された。
この研究は野生型(未修飾)siRNAがマウス胃酸中およびマウス血清中で代謝されることを示している。3’末端修飾されたsiRNAの場合、胃腸管で分解は観察されなかった。したがって3’修飾されたsiRNAは野生型siRNAよりも高い経口バイオアベイラビリティーを有している可能性がある。
【0068】
全身送達された3’修飾されたsiRNAはインビボ成長因子誘起血管形成モデルにおいてさらに活性である18
最初に、修飾されたsiRNA(C3−siRNAおよびC3−MOE siRNA)が標的遺伝子を下方調節する能力を、細胞内(in cellulo)で抗VEGFR2 siRNAでトランスフェクトされたMS1細胞のVEGFR2表面レベルを測定することにより検査した。
野生型siRNAとして二つの抗VEGFR2 siRNA、C3−siRNAおよびC3−MOE siRNAのプールを腹腔内投与した。結果を図7に示す。プールされた野生型siRNAは25μg/マウス/日の高用量でVEGF誘起の脈管化を有意に低減させた。C3−siRNAでは5倍低い用量で同一レベルの阻害が観察された。C3−MOE siRNAプールの場合、脈管化された組織重量の有意な低下が最低の0.2μg/マウス/日を含む全被験用量で観察された。
【0069】
図8aおよび8bは、腹腔内に与えられた場合、VEGFR2−C3およびC3−MOE siRNAの双方がマ1μg/マウス/日の用量を下回って活性であった。
B16同種移植片メラノーマ腫瘍マウスモデルに腹腔内に与えられた(i.p.)抗VEGFR2 C3−MOE siRNAのインビボ試験。図9aは、修飾されたVEGFR2−C3−MOE−siRNAでのi.p.処置が腫瘍発達を有意に低減させることを示している。図9bもまたVEGFR2−C3−MOE−siRNAの20μg/マウスのi.p.注射の結果、腫瘍成長の有意な阻害に至ることを示している。
【0070】
血管形成障害の処置のためのsiRNAの経口送達
図10は20μg/マウス/日の用量で経口的に与えられたVEGFR2−C3−MOE−siRNA1は脈管化重量を基底レベル(例えば成長因子誘導を伴わない重量)まで低下させたことを示している。用いられた実際のsiRNA配列は表3で言及される。
抗Tie2 C3−MOE siRNAもまた成長因子誘起血管形成モデルにおいて腹腔内および経口送達の双方で試験した。図11aおよび11bは経口的に与えられた、Tie2で志向される双方のC3−MOE siRNAは20μg/マウス/日で活性であったことを示している。用いられた実際のsiRNA配列は表3を参照して決定できる。
データにより、さらなる内部修飾を伴うかまたは伴わない3’末端修飾されたsiRNAは経口投与時に妥当な用量で治療効果を実証できることが示される。
【0071】
参照文献
1. a)Y. Tomari et al. Genes and Development 19(2005), 517;b)P. Shankar et al. JAMA 11(2005), 1367;c)Y. Dorsett et al. Nature Reviews 3(2004), 318
2. a)P.D. Zamore et al. Cell 101,(2000), 25;b)S.M. Hammond et al. Nature 404(2000), 293
3. a)G. Meister et al. Molecular Cell 15(2004), 185.
4. S.M. Elbashir et al. Genes Dev. 15(2001), 188.
5. S.J. Reich et al. Molecular Vision 9(2003), 210.
6. a)Dorn et al. Nucleic Acids Research 32(2004), e49;b)D. R.Thakker et al. PNAS 101(2004), 17270;c)D.R. Thakker et al. Molecular Psychiatry 10(2005), 714
7. V. Bitko et al. Nature Medicine 11(2005), 50.
8. E. Song et al. Nature Medicine 9(2003), 347.
9. D.A.Braasch et al. Biochemistry 42(2003), 7967.
10. Harborth, Antisense Nucleic Acid Drug Devt, 2003
11. A.H.S. Hall et al. Nucleic Acids Research 32(2004), 5991.
12. M. Amarzguioui et al. Nucleic Acids Research 31(2003), 589.
13. F. Czauderna et al. Nucleic Acids Research 31(2003), 2705.
14. T. Prakash et al. Journal of Medicinal Chemistry 48(2005), 4247.
15. J. Elmen et al. Nucleic Acids Research 33(2005), 439.
16. A.S. Boutorin, L.V. Guskova, E.M. Ivanova, N.D. Kobetz, V.F. Zafytova, A.S. Ryte, L.V. Yurchenko and V.V. Vlassov FEBS Lett. 254(1989), p. 129
17. J. Wood et al. Cancer Research 60(2000), 2178.
18. K. LaMontagne et al. Cancer Res. 66(2006), 221.
【特許請求の範囲】
【請求項1】
少なくとも15ヌクレオチドにわたって互いに相補的である2本の別個のRNA鎖を含み、ここで各鎖は49ヌクレオチド以下であり、そしてここで少なくとも1つの鎖の3’末端は3’炭素での修飾を含み、そしてここで該修飾は
【化1】
である、低分子干渉リボ核酸(siRNA)。
【請求項2】
少なくとも15ヌクレオチドにわたって互いに相補的である2本の別個のRNA鎖を含み、ここで各鎖は49ヌクレオチド以下であり、そしてここで各鎖の3’末端は3’炭素での修飾を含み、そしてここで該修飾は
【化2】
である、siRNA。
【請求項3】
該各鎖の3’末端で最初に塩基対形成している2個のヌクレオチドが修飾されている、請求項1に記載のsiRNA。
【請求項4】
該各鎖の3’末端で最初に塩基対形成している2個のヌクレオチドが2’−メトキシエチルリボヌクレオチド残基である、請求項1に記載のsiRNA。
【請求項5】
該2本の鎖が少なくとも19ヌクレオチドにわたって互いに相補的である、請求項1に記載のsiRNA。
【請求項6】
各鎖が19ヌクレオチドである、請求項5に記載のsiRNA。
【請求項7】
siRNAの両方の末端がブラント末端である、請求項1に記載のsiRNA。
【請求項8】
各鎖が19ヌクレオチドである、請求項1に記載のsiRNA。
【請求項9】
該2本の鎖が19ヌクレオチドにわたって互いに完全に相補的であり、siRNAがブラント末端である、請求項1に記載のsiRNA。
【請求項10】
少なくとも1つのさらなるヌクレオチドが修飾されている、請求項3に記載のsiRNA。
【請求項11】
標準的な胃酸アッセイにおいて、同一ヌクレオチド配列を有する未修飾siRNAよりも大きな安定性を有する、請求項1に記載のsiRNA。
【請求項12】
標準的な胃酸アッセイにおいて、30分暴露後に50%以上である安定性を有する、請求項1に記載のsiRNA。
【請求項13】
標準的な血清アッセイにおいて、同一ヌクレオチド配列を有する未修飾siRNAよりも大きな安定性を有する、請求項1に記載のsiRNA。
【請求項14】
標準的な血清アッセイにおいて、30分暴露後に50%以上である安定性を有する、請求項1に記載のsiRNA。
【請求項15】
標準的な腸洗浄液アッセイにおいて、同一ヌクレオチド配列を有する未修飾siRNAよりも大きな安定性を有する、請求項1に記載のsiRNA。
【請求項16】
同一ヌクレオチド配列の未修飾siRNAと比較してバイオアベイラビリティーが強化されている、請求項1に記載のsiRNA。
【請求項17】
請求項1に記載のsiRNAおよび薬学的に許容される担体を含む医薬組成物。
【請求項18】
医薬として使用するための、請求項1に記載のsiRNA。
【請求項19】
該各鎖の3’末端で最初に塩基対形成している2個のヌクレオチドが修飾されており、各修飾されたヌクレオチドは、アミド結合であるヌクレオシド内結合を有する、請求項1に記載のsiRNA。
【請求項20】
非経口的に投与される医薬として使用するための、請求項1に記載のsiRNA。
【請求項21】
該各鎖の3’末端で最初に塩基対形成している2個のヌクレオチドが修飾されており、各修飾されたヌクレオチドは、ホスホロチオエート、ホスホロジチオエート、ホスホロアミダート、ボラノホスホノエートおよびアミド結合の中から選択される修飾されたヌクレオシド内結合を有するヌクレオチドの中から選択される、請求項1に記載のsiRNA。
【請求項22】
siRNAの1個の末端がブラント末端である、請求項1に記載のsiRNA。
【請求項23】
5’末端または3’末端の少なくとも一つに1から6ヌクレオチドオーバーハングを含む、請求項1に記載のsiRNA。
【請求項24】
経口的、局所的、非経口的、吸入もしくはスプレーにより、または直腸、経皮、皮下、血管内、静脈内、筋肉内、腹腔内もしくはくも膜下腔内、または注入技術的に投与される医薬として使用するための、請求項1に記載のsiRNA。
【請求項1】
少なくとも15ヌクレオチドにわたって互いに相補的である2本の別個のRNA鎖を含み、ここで各鎖は49ヌクレオチド以下であり、そしてここで少なくとも1つの鎖の3’末端は3’炭素での修飾を含み、そしてここで該修飾は
【化1】
である、低分子干渉リボ核酸(siRNA)。
【請求項2】
少なくとも15ヌクレオチドにわたって互いに相補的である2本の別個のRNA鎖を含み、ここで各鎖は49ヌクレオチド以下であり、そしてここで各鎖の3’末端は3’炭素での修飾を含み、そしてここで該修飾は
【化2】
である、siRNA。
【請求項3】
該各鎖の3’末端で最初に塩基対形成している2個のヌクレオチドが修飾されている、請求項1に記載のsiRNA。
【請求項4】
該各鎖の3’末端で最初に塩基対形成している2個のヌクレオチドが2’−メトキシエチルリボヌクレオチド残基である、請求項1に記載のsiRNA。
【請求項5】
該2本の鎖が少なくとも19ヌクレオチドにわたって互いに相補的である、請求項1に記載のsiRNA。
【請求項6】
各鎖が19ヌクレオチドである、請求項5に記載のsiRNA。
【請求項7】
siRNAの両方の末端がブラント末端である、請求項1に記載のsiRNA。
【請求項8】
各鎖が19ヌクレオチドである、請求項1に記載のsiRNA。
【請求項9】
該2本の鎖が19ヌクレオチドにわたって互いに完全に相補的であり、siRNAがブラント末端である、請求項1に記載のsiRNA。
【請求項10】
少なくとも1つのさらなるヌクレオチドが修飾されている、請求項3に記載のsiRNA。
【請求項11】
標準的な胃酸アッセイにおいて、同一ヌクレオチド配列を有する未修飾siRNAよりも大きな安定性を有する、請求項1に記載のsiRNA。
【請求項12】
標準的な胃酸アッセイにおいて、30分暴露後に50%以上である安定性を有する、請求項1に記載のsiRNA。
【請求項13】
標準的な血清アッセイにおいて、同一ヌクレオチド配列を有する未修飾siRNAよりも大きな安定性を有する、請求項1に記載のsiRNA。
【請求項14】
標準的な血清アッセイにおいて、30分暴露後に50%以上である安定性を有する、請求項1に記載のsiRNA。
【請求項15】
標準的な腸洗浄液アッセイにおいて、同一ヌクレオチド配列を有する未修飾siRNAよりも大きな安定性を有する、請求項1に記載のsiRNA。
【請求項16】
同一ヌクレオチド配列の未修飾siRNAと比較してバイオアベイラビリティーが強化されている、請求項1に記載のsiRNA。
【請求項17】
請求項1に記載のsiRNAおよび薬学的に許容される担体を含む医薬組成物。
【請求項18】
医薬として使用するための、請求項1に記載のsiRNA。
【請求項19】
該各鎖の3’末端で最初に塩基対形成している2個のヌクレオチドが修飾されており、各修飾されたヌクレオチドは、アミド結合であるヌクレオシド内結合を有する、請求項1に記載のsiRNA。
【請求項20】
非経口的に投与される医薬として使用するための、請求項1に記載のsiRNA。
【請求項21】
該各鎖の3’末端で最初に塩基対形成している2個のヌクレオチドが修飾されており、各修飾されたヌクレオチドは、ホスホロチオエート、ホスホロジチオエート、ホスホロアミダート、ボラノホスホノエートおよびアミド結合の中から選択される修飾されたヌクレオシド内結合を有するヌクレオチドの中から選択される、請求項1に記載のsiRNA。
【請求項22】
siRNAの1個の末端がブラント末端である、請求項1に記載のsiRNA。
【請求項23】
5’末端または3’末端の少なくとも一つに1から6ヌクレオチドオーバーハングを含む、請求項1に記載のsiRNA。
【請求項24】
経口的、局所的、非経口的、吸入もしくはスプレーにより、または直腸、経皮、皮下、血管内、静脈内、筋肉内、腹腔内もしくはくも膜下腔内、または注入技術的に投与される医薬として使用するための、請求項1に記載のsiRNA。
【図1a】

【図1b】
【図1c】
【図1d】
【図1e】
【図2】
【図3】
【図4】
【図5】
【図6】

【図7】
【図8a】
【図8b】
【図9a】
【図9b】
【図10】
【図11a】
【図11b】
【図1b】
【図1c】
【図1d】
【図1e】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8a】
【図8b】
【図9a】
【図9b】
【図10】
【図11a】
【図11b】
【公開番号】特開2013−5812(P2013−5812A)
【公開日】平成25年1月10日(2013.1.10)
【国際特許分類】
【出願番号】特願2012−200083(P2012−200083)
【出願日】平成24年9月12日(2012.9.12)
【分割の表示】特願2009−508205(P2009−508205)の分割
【原出願日】平成19年5月2日(2007.5.2)
【出願人】(504389991)ノバルティス アーゲー (806)
【Fターム(参考)】
【公開日】平成25年1月10日(2013.1.10)
【国際特許分類】
【出願日】平成24年9月12日(2012.9.12)
【分割の表示】特願2009−508205(P2009−508205)の分割
【原出願日】平成19年5月2日(2007.5.2)
【出願人】(504389991)ノバルティス アーゲー (806)
【Fターム(参考)】
[ Back to top ]