
腱及び/又は靭帯細胞の分化誘導剤

【課題】腱及び/又は靭帯細胞の分化誘導剤、並びに分化誘導剤を含む腱又は靭帯関連疾患を予防又は治療するための医薬を提供する。
【解決手段】腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化誘導剤は、(A)Mkxタンパク質、(B)前記Mkxタンパク質のアミノ酸配列において、1〜数個のアミノ酸が欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも一種の変異を有し、かつ、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する活性を有する変異体、(C)前記(A)又は(B)のタンパク質をコードする核酸、並びに、(D)前記(C)の核酸を含む発現ベクターの少なくとも一種を有効成分として含む。
【解決手段】腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化誘導剤は、(A)Mkxタンパク質、(B)前記Mkxタンパク質のアミノ酸配列において、1〜数個のアミノ酸が欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも一種の変異を有し、かつ、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する活性を有する変異体、(C)前記(A)又は(B)のタンパク質をコードする核酸、並びに、(D)前記(C)の核酸を含む発現ベクターの少なくとも一種を有効成分として含む。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、腱及び/又は靭帯細胞の分化誘導剤、分化誘導剤を用いた医薬、分化誘導剤を用いたスクリーニング方法等に関する。
【背景技術】
【0002】
腱は筋肉と骨とを結合する組織であり、靱帯は骨と骨とを結合する組織である。これらの組織は、力や運動を伝達するという重要な機能を持っている。腱や靱帯は、酷使や加齢によって損傷し、また減少するが、栄養血管が少ないため、腱の治癒力は非常に弱く、完治が極めて難しい。腱細胞の分化機構等の生物学的理解も不十分であるため、現在、再生医療等の治療法は確立していない。
【0003】
腱や靱帯は、コラーゲン原線維がクロスリンクしてできる線維でできている。腱の線維間には腱細胞が僅かに存在し、コラーゲン、プロテオグリカン等の細胞外基質の産生を行っている。腱の大部分を構成するコラーゲンは、腱に弾性力を与えるが、そのほとんどはI型コラーゲンである。プロテオグリカンは、腱においてはdecorin、fibromodulin、biglycan、lumican等が存在し、コラーゲン線維同士の結合や集合に関与している。プロテオグリカンの遺伝子欠損マウスでは、腱のコラーゲン原線維の異常が観察される。エーラスダンロス(Ehlers−Danlos)症候群に代表されるコラーゲン産生に異常を有する疾患患者に腱の障害が報告されている。腱細胞における細胞外基質の産生は、腱の形成にとって重要であることが示唆される。
【0004】
近年、腱及び靭帯細胞並びに腱及び靭帯前駆細胞で発現するbasic helix−loop−helix(bHLH)型の転写因子であるScleraxis(Scx)が腱発生に重要な因子であることが報告され、Scxノックアウトマウスでは、腱の著しい形成不全が観察された(非特許文献1)。また、腱の主成分であるI型コラーゲンを直接誘導することが示唆された(非特許文献2)。しかし、Scxノックアウトマウスは、後述の実施例に示すように、腱においてすべてのI型コラーゲンが消失するわけではない。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Cserjesi P,et al.(1995)Scleraxis: a basic helix−loop−helix protein that prefigures skeletal formation during mouse embryogenesis Development 121(4) p1099−1110
【非特許文献2】Lejard V,et al.(2007)Scleraxis and NFATc regulate the expression of the pro−alpha1(I) collagen gene in tendon fibroblasts J Biol Chem 282(24) p17665−17675
【発明の概要】
【発明が解決しようとする課題】
【0006】
腱及び靭帯細胞は、I型コラーゲン等の基質を産生する細胞であって、それぞれ、腱及び靭帯組織の形成及び維持に関与する。腱及び靭帯は、上記したように再生能の低い組織であり、腱及び靭帯疾患治療に向けての再生医療が望まれる。Scxは、腱及び靭帯細胞で発現し、腱及び靭帯の主成分であるI型コラーゲンの発現を誘導することが示唆されるものの、他のI型コラーゲンの発現調節因子の存在が示唆される等、腱発生のメカニズムには未解明な部分が多いのが現状である。腱及び靭帯細胞への分化誘導方法も、確立されていない。
【0007】
そこで、本発明の目的は、腱及び/又は靭帯細胞の分化誘導剤、並びに該分化誘導剤を含む、腱又は靭帯関連疾患を予防又は治療するための医薬を提供することにある。
【0008】
さらに、本発明の目的は、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞へ分化誘導させるために使用する物質をスクリーニングする方法を提供することにある。
【課題を解決するための手段】
【0009】
本発明者らは、腱及び靭帯細胞への分化方法の確立には、分化に重要な遺伝子を特定し、その遺伝子を導入又は誘導することが重要であると考えた。本発明者らは、後述の実施例に示すように、Mkx遺伝子を破壊すると、腱組織と腱細胞のI型コラーゲン生産量が減少し、逆に細胞内に過剰発現させると、I型コラーゲン量が増大することを発見した。Mkxは、従来、胎生期において腱に発現するホメオボックス型の転写因子として知られていたが、Mkxが腱細胞においてその分化を制御しI型コラーゲンの発現を調節する重要な因子であることが初めて明らかとなった。これらの知見をもとに、本発明者らは、以下の発明を完成させた。
【0010】
すなわち、本発明は、
(A)Mkxタンパク質、
(B)前記Mkxタンパク質のアミノ酸配列において、1〜数個のアミノ酸が欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも一種の変異を有し、かつ、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する活性を有する変異体、
(C)前記(A)又は(B)のタンパク質をコードする核酸、並びに、
(D)前記(C)の核酸を含む発現ベクター
の少なくとも一種を有効成分として含む、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化誘導剤を提供する。
【0011】
上記分化誘導剤は、
(A1)Scxタンパク質、
(B1)前記Scxタンパク質のアミノ酸配列において、1〜数個のアミノ酸が欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも一種の変異を有し、かつ、I型コラーゲンを誘導する活性を有する変異体、
(C1)前記(A1)又は(B1)のタンパク質をコードする核酸、並びに、
(D1)前記(C1)の核酸を含む発現ベクター、
の少なくとも一種をさらに含んでもよい。
【0012】
本発明は、腱及び/又は靭帯細胞への分化能を有する細胞の生育に適した条件下で、上記分化誘導剤を用いて前記細胞を培養する工程、及び前記培養された細胞の中から、腱及び/又は靭帯細胞を選択する工程を含む、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する方法を提供する。
【0013】
前記細胞は、例えば繊維芽細胞、腱前駆細胞、靱帯前駆細胞、胚性幹細胞、胚性生殖細胞、体性幹細胞及びiPS細胞からなる群から選ばれる少なくとも一種である。
【0014】
本発明は、また、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化誘導剤を含む腱又は靭帯関連疾患を予防又は治療するための医薬を提供する。
【0015】
上記腱又は靭帯関連疾患は、例えばエーラスダンロス症候群である。
【0016】
本発明は、また、上記分化誘導方法により得られる、腱及び/又は靭帯細胞を含む、腱又は靭帯関連疾患を予防又は治療するための医薬を提供する。
【0017】
上記腱又は靭帯関連疾患は、例えばエーラスダンロス症候群である。
【0018】
本発明は、また、Mkx遺伝子の機能的欠損を含む、腱又は靱帯関連疾患の研究用の遺伝子改変非ヒト動物を提供する。
【0019】
前記機能的に欠損したMkx遺伝子は、Mkx遺伝子のプロモーターの下流にレポーター遺伝子が挿入されたことにより腱細胞特異的にレポーター遺伝子を発現することが好ましい。
【0020】
前記レポーター遺伝子は、例えばVenus遺伝子である。
【0021】
本発明は、また、上記分化誘導剤を用いることを特徴とする、腱又は靭帯関連疾患の解析又は分析方法を提供する。
【0022】
本発明は、また、上記分化誘導方法により得られる腱及び/又は靭帯細胞を用いることを特徴とする、腱又は靭帯関連疾患の解析又は分析方法を提供する。
【0023】
本発明は、また、上記遺伝子改変非ヒト動物の腱及び/靱帯、それらの組織又は細胞を用いることを特徴とする、腱又は靭帯関連疾患の解析又は分析方法を提供する
【0024】
本発明は、また、被験物質の存在下又は非存在下で、上記分化誘導方法を行い、該被験物質存在下又は非存在下での腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を比較することを特徴とする、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質をスクリーニング又は評価する方法を提供する。
【0025】
上記スクリーニング又は評価する方法は、例えば被験物質の存在下で培養した細胞における、Mkxの発現量及び/又はMkxをコードする塩基配列に対するmRNAの転写量を測定すること、並びに前記被験物質を非存在下で培養した細胞内の量と比べて、前記測定した量が多い又は少ない場合に、前記被験物質を腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質として選択することを含む。
【0026】
本発明は、また、上記遺伝子改変非ヒト動物から得られる腱、その組織又はその細胞を用いることを特徴とする、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質をスクリーニング又は評価する方法を提供する。
【0027】
上記スクリーニング又は評価する方法は、例えば遺伝子改変非ヒト動物から得られる腱、その組織又はその細胞に被験物質を接触させること、及びノックインされたレポーター遺伝子の発現を上昇又は低下させる被験物質を選択することを含む。
【発明の効果】
【0028】
関節の再生等の医学において、腱や靱帯は筋・骨格をつなぐ重要な組織である。そして、Mkx遺伝子を過剰発現させると、腱の主要コンポーネントであるI型コラーゲンの発現が上昇することから、Mkxは腱の成熟過程において重要であることが示唆される。損傷した腱や靱帯の部位にMkx及びその上流・下流遺伝子等からなる医薬を外来的に導入することで、腱の治癒、運動器の再生や再建を促進することが期待される。
【0029】
エーラスダンロス症候群等の腱及び/又は靭帯関連疾患に罹患している患者から採取した細胞を用いて、本発明の分化誘導剤で自家腱及び/又は靭帯細胞を製造することもでき、結果として該疾患に対する再生医療が可能となる。
【0030】
Mkx遺伝子が腱細胞や靱帯細胞特異的に蛍光タンパク質のようなレポーター遺伝子で置き換えられている本発明の遺伝子改変非ヒト動物は、蛍光タンパクの発光を指標にMkxを調節する被験物質のスクリーニングや評価、腱損傷モデル実験等における治癒過程でのMkxの局所的な発現をモニターするためのモデル動物として有用である。
【0031】
分化誘導剤や分化誘導方法により得られる腱及び/又は靭帯細胞を用いて、腱又は靭帯関連疾患の解析又は分析方法は、腱分化の分子メカニズム理解の基礎となり、腱の障害やエーラスダンロス症候群等の腱関連疾患の治療標的発見を導くきっかけとなるかもしれない。
【0032】
Mkxは、腱の発生・成熟に重要であることから、腱・靱帯等の疾患等で、Mkxそのものの遺伝子配列又はMkxの遺伝子ネットワークに関与する上流及び下流遺伝子等の遺伝子に異常を起こしている可能性が大きい。それらの探索、診断、治療に、本発発明のスクリーニング方法が応用される。
【0033】
本発明のスクリーニング方法によって選択された物質や本発明の医薬によれば、腱又は靭帯関連疾患、特にMkxに異常を抱える疾患を、予防又は治療することができる。
【図面の簡単な説明】
【0034】
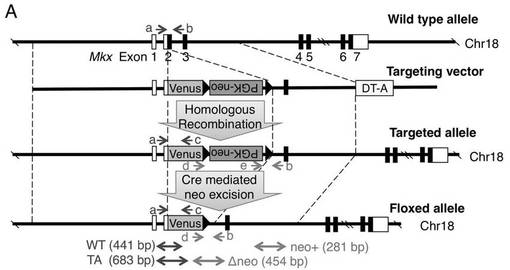
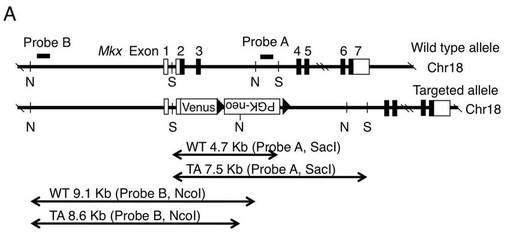
【図1A】Mkx変異マウスを作製するために設計するMkxターゲッティングコンストラクトの一例を示す。ここで、矢印a〜eはジェノタイピング用PCRプライマー、□はUTR、■は翻訳領域、DT−AはジフテリアトキシンA、WTは野生型アリル、TAはターゲッティングされたアリルを意味する。
【図1B】野生型マウスとMkx変異マウスでの図1Aのプライマーa、b及びcを用いたゲノミックPCR(図面代用写真)を示す。
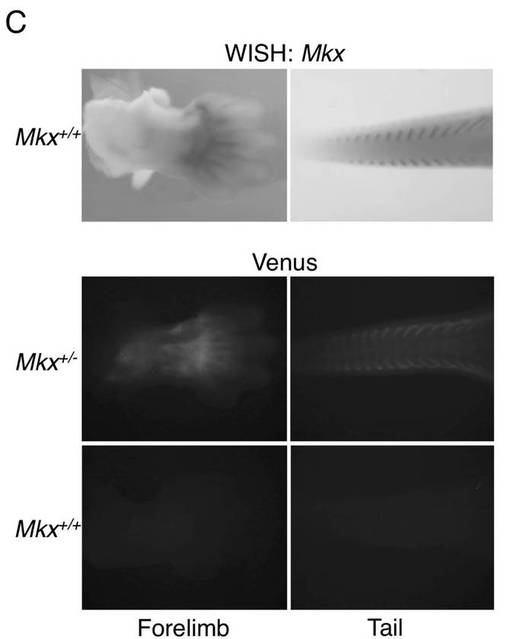
【図1C】上段は、野生型マウスの胚のE13.5前肢(Forelimb)及び尾(Tail)におけるMkxのWISH(ホールマウント in situ ハイブリダイゼーション)(図面代用写真)を示し、そして下段は、野生型マウス及びMkx変異マウスのVenus蛍光シグナルの観察(図面代用写真)を示す。
【図1D】E16.5のVenusノックインへテロマウスの胚の尾におけるミオシン重鎖抗体(MF20;red)を用いた免疫組織染色とVenus蛍光シグナルを観察した顕微鏡写真を示す。図中、○で囲んだ部分が筋肉のマーカー遺伝子であるミオシン重鎖発現細胞を示し、それ以外はVenus発現細胞の蛍光シグナルを示す。また、Barは100 μmである。
【図1E】野生型マウスとMkx変異マウスのアキレス腱におけるMkxのRT−PCR解析(図面代用写真)を示す。Gapdhは内在性コントールを示す。
【図2A】3ヶ月齢の野生型マウスにおいて、膝蓋腱(Aの矢印)、アキレス腱(Cの矢印、後肢(Eの矢印)、前肢の腱(Gの矢印)、尾腱(Iの矢印)、背中の腱(Kの矢印)及び広頸筋腱(Mの矢印)を観察した図面代用写真、並びに3ヶ月齢のMkxノックアウトマウスにおいて、膝蓋腱(Bの矢印)、アキレス腱(Dの矢印、後肢(Fの矢印)、前肢の腱(Hの矢印)、尾腱(Jの矢印)、背中の腱(Lの矢印)及び広頸筋腱(Nの矢印)を観察した図面代用写真を示す。Mkxノックアウトマウスでは、腱の異常が観察される。
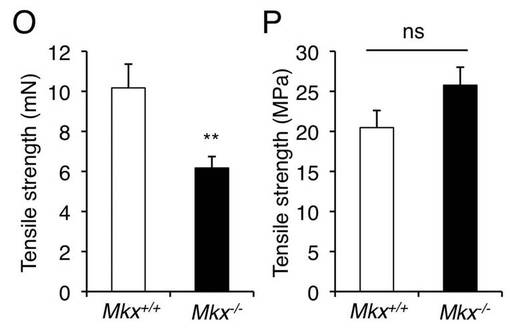
【図2B】野生型マウスとMkxノックアウトマウスのアキレス腱の引張強度を測定したグラフである。
【図2C】野生型マウスとMkxノックアウトマウスのアキレス腱のヤング率を測定したグラフである。
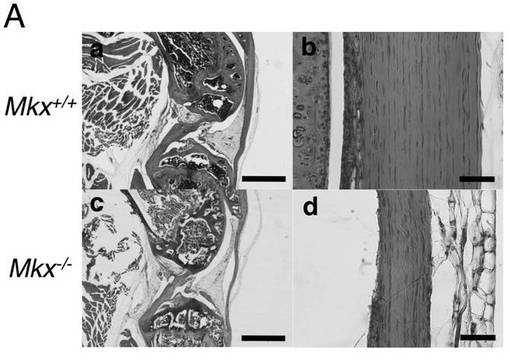
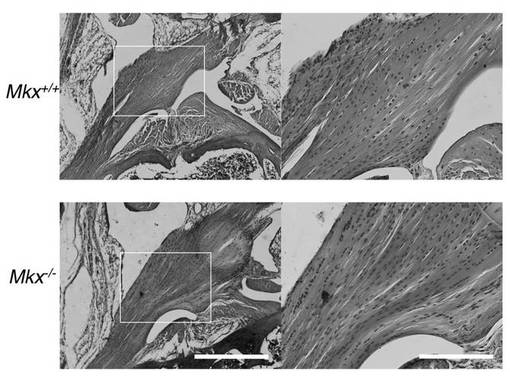
【図3A】3ヶ月齢の野生型マウスとMkxノックアウトマウスの膝蓋腱のH&E染色像(図面代用写真)を示す。a及びcのバーは1mmを意味し、b及びdのバーは50μmを意味する。Mkx欠損マウスでは、腱の低形成が起きていることがわかる。
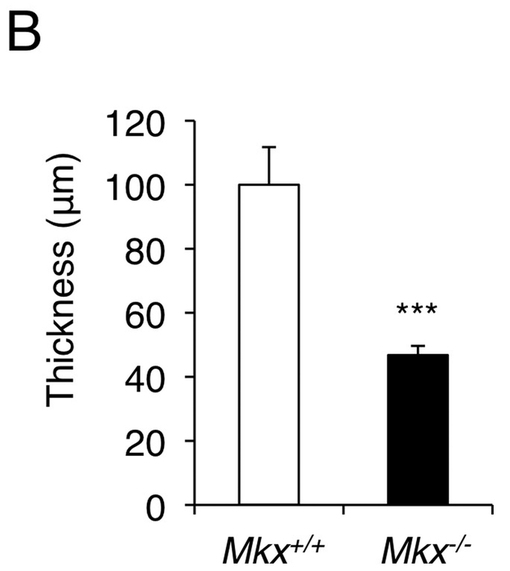
【図3B】7日齢の野生型マウスとMkxノックアウトマウスの膝蓋腱の厚さを測定したグラフである。Mkxノックアウトマウスは、膝蓋腱の厚さが有意に減少している。
【図3C】3ヶ月齢の野生型マウスとMkxノックアウトマウスの尾腱のH&E染色像(図面代用写真)を示す。a及びcのバーは200μmを意味し、b及びdのバーは50μmを意味する。△矢頭は、尾腱の線維束を示す。Mkxノックアウトマウスの尾腱の線維束は有意に細っている。
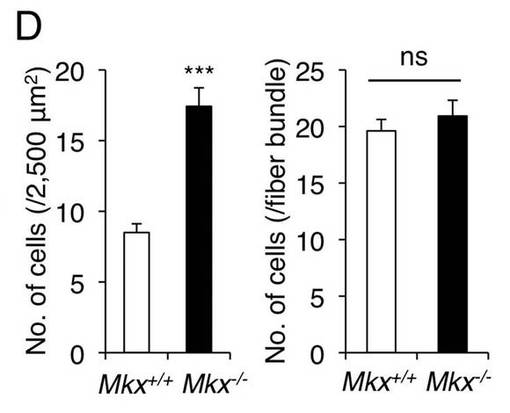
【図3D】3ヶ月齢の野生型マウスとMkxノックアウトマウスの尾腱の細胞密度を測定したグラフである。
【図3E】E18.5の野生型マウスとMkxノックアウトマウスの胚の尾腱のアザン染色像(図面代用写真)を示す。△矢頭は尾腱の線維束を示す。また、左図のバーは500μm、右図のバーは200μmを意味する。
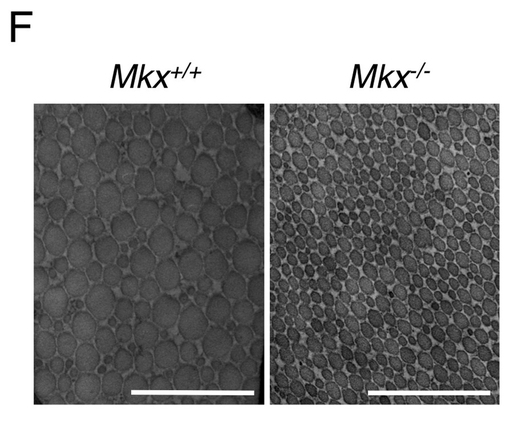
【図3F】野生型マウスとMkxノックアウトマウスのアキレス腱のコラーゲン原線維の電子顕微鏡観察像(図面代用写真)を示す。ここで、バーは200nmを意味する。
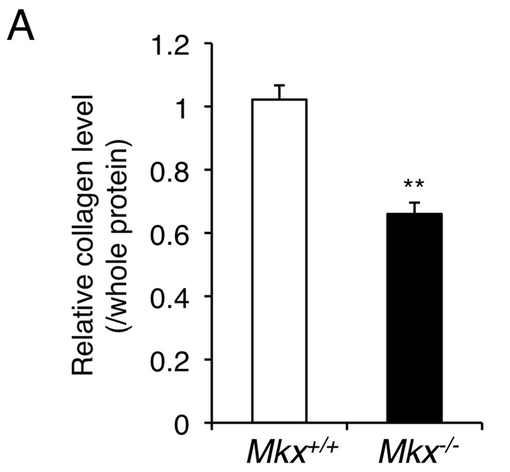
【図4A】8週齢の野生型マウスとMkxノックアウトマウスのアキレス腱のコラーゲン量の測定結果を示す。Mkx欠損腱細胞では、I型コラーゲン産生能が低下することがわかる。
【図4B】8週齢の野生型マウスとMkxノックアウトマウスのアキレス腱のI型コラーゲン(Col1)とβ−actinのウエスタンブロット解析の結果(図面代用写真)を示す。
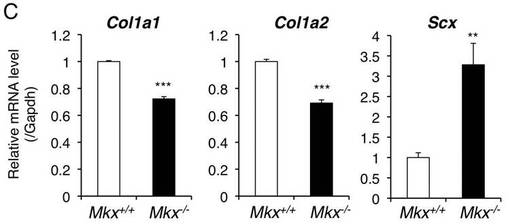
【図4C】8週齢の野生型マウスとMkxノックアウトマウスのアキレス腱のCol1a1、Col1a2及びScxのreal time PCR解析結果を示す。
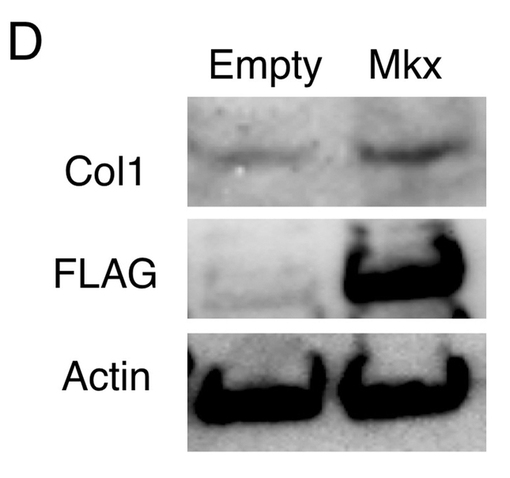
【図4D】FLAG−Mkx発現ベクター(Mkx)又は空ベクター(Empty)をトランスフェクションしたC3H10T1/2細胞のI型コラーゲン(Col1)、FLAG及びβ−actinのウエスタンブロットの結果(図面代用写真)を示す。
【図4E】腱分化ネットワークの模式図を示す。
【図5A】Mkx変異ES又はマウスのサザンブロット解析のために、野生型(WT)及びターゲッティングされたアリル(TA)と各プローブの位置を示す。ここで、NはNcoIを意味し、SはSacIを意味する。
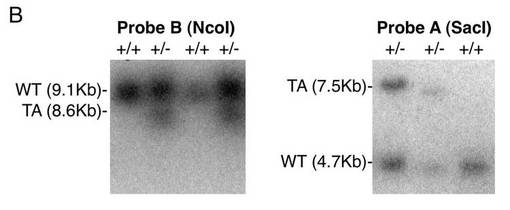
【図5B】組換え胚性幹細胞のサザンブロット解析結果(図面代用写真)を示す。ここで、TAはターゲッティングされたアリルを意味し、WTは野生型アリルを意味する。
【図5C】野生型マウス及びMkx変異マウスのサザンブロット解析結果(図面代用写真)を示す。
【図6A】野生型マウス(成体)の筋肉、アキレス腱及び尾腱におけるMkxのRT−PCR解析(図面代用写真)を示す。Mkxヘテロマウス(成体)のVenusの発現は、Mkx発現とリンクすることがわかる。Gapdhは内在性コントロールを示す。
【図6B】Mkxヘテロマウス及び野生型マウスのアキレス腱(B−E)、尾腱(F−I)及び体幹の腱(J−M)におけるVenusの発現と外観写真を示す。
【図7】出生後の各ステージにおけるMkx+/+,Mkx+/−及びMkx−/−マウスの体重を比較したグラフを示す。野生型マウスとMkxノックアウトマウスとで体重の差は見られない。
【図8】3ヶ月齢の野生型マウス(WT)のアキレス腱(A)、尾腱(C)、膝蓋腱(E)及び体幹の腱(G)の外観写真と、3ヶ月齢のネオマイシン耐性遺伝子を除去したMkxノックアウトマウス(KO)のアキレス腱(B)、尾腱(D)、膝蓋腱(F)及び体幹の腱(H)の外観写真を示す。腱の異常は、ネオマイシン耐性遺伝子を除去させたノックアウトマウスにおいても起きている。
【図9】野生型マウス及びMkxノックアウトマウスの膝十字靭帯のH&E染色像(図面代用写真)を示す。左図のバーは500μm、右図のバーは200μmを意味する。膝十字靭帯は、Mkxノックアウトマウスにおいても影響はない。
【発明を実施するための形態】
【0035】
本発明の腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化誘導剤は、以下の(A)から(D)の少なくとも一種を有効成分として含む。
(A)Mkxタンパク質、
(B)前記Mkxタンパク質のアミノ酸配列において、1〜数個のアミノ酸が欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも一種の変異を有し、かつ、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する活性を有する変異体、
(C)前記(A)又は(B)のタンパク質をコードする核酸、並びに、
(D)前記(C)の核酸を含む発現ベクター
【0036】
Mkx(Mohawk、別名 Irxl1)とは、胎生期において、腱に発現するホメオボックス型の転写因子である。Mkxは、不定型ホメオボックス遺伝子のTALEスーパークラスの中の新しく分類されたクラスに属する唯一の遺伝子である(Anderson DM,et al.(2006).Dev Dyn 235(3) p792−801)。Mkxは、腱、筋肉、軟骨の前駆細胞、オスの生殖腺、及び腎臓の尿管芽先端で発現することが報告されている(Dev Dyn 235(3) p792−801等)。本発明者らも、E9.5、E10.5及びE11.5のマウス胚を用いた1520個の転写因子及び転写コファクターのホールマウントin situハイブリダイゼーションデータベース(EMBRYS)の構築過程で、腱で発現する転写因子として同定していた(Yokoyama S,et al.2009 Dev Cell 17(6) p836−848)。
【0037】
本発明では、細胞から腱細胞及び/又は靱帯細胞を作製、再生、増強等する分化誘導剤としてMkxタンパク質を用いる。
【0038】
本発明の分化誘導剤を実験及び研究用に供する場合は、Mkxタンパク質の由来は問わない。例えば、ヒト、マウス、サル、チンパンジー、ブタ、イヌ等の哺乳動物が挙げられる。本発明の分化誘導剤をヒトの腱細胞及び/又は靱帯細胞を再生等の医療行為に使用する場合は、ヒトのMkxタンパク質を用いることが好ましい。
【0039】
ヒトMkxタンパク質に関する情報は、NCBIからアクセッション番号NP_775847として入手することができ、Mkxタンパク質は、352個のアミノ酸で構成されるタンパク質である。また、ヒトMkx遺伝子に関する情報は、NCBIからアクセッション番号NM_173576として入手可能である。ヒトMkx遺伝子は、ヒト第10染色体上に存在し、全長で約3658の塩基対よりなり、7個のエキソンで構成される。マウスMkx遺伝子は、NCBIから、アクセッション番号NM_177595として入手することができる。
【0040】
本発明の分化誘導剤に用いる(A)Mkxタンパク質は、天然由来のタンパク質の他に、組み換えタンパク質でもよい。組み換えタンパク質は、常法により、Mkxタンパク質をコードする核酸を組み込んだ形質転換体を培養等することにより得ることができる。
【0041】
本発明の分化誘導剤に用いる(B)Mkxタンパク質のアミノ酸配列において、1〜数個のアミノ酸が欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも一種の変異を有し、かつ、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する活性を有する変異体は、当業者に公知の方法により容易に取得することができる。変異体が分化誘導活性を有するか否かは、ウエスタンブロット、ノーザンブロット、リアルタイムPCR等で測定することができる。
【0042】
Mkxタンパク質の変異体は、また、Mkxタンパク質のアミノ酸配列と通常、70%以上、好ましくは80%以上、さらに好ましくは90%の相同性を有するアミノ酸配列からなり、かつ、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する活性を有するものでもよい。
【0043】
本発明の分化誘導剤は、前記(A)又は(B)のタンパク質をコードする(C)核酸でもよい。核酸には、染色体DNAやcDNAが挙げられる。染色体DNAは、例えばPCRによる増幅や細胞等からDNAライブラリ−を調製し、Mkxタンパク質をコードするDNAにハイブリダイズするプローブでスクリーニングすることにより取得する。cDNAは、例えば腱細胞等からRNAを抽出し、Mkxタンパク質をコードするDNAにハイブリダイズするプライマーを用いたPCR法により取得する。
【0044】
本発明の分化誘導剤は、(D)前記(C)の核酸を含む発現ベクターでもよい。発現ベクターは、公知のベクターにMkx遺伝子を発現可能に連結することで得る。前記ベクターは、宿主中で複製可能なものであれば特に限定されず、例えば、プラスミドベクター、ウイルスベクター、ファージベクター等が挙げられる。
【0045】
前記プラスミドベクターとしては、大腸菌由来のプラスミド(例えばpBR322,pUC18,pUC119,pTrcHis,pBlueBacHis等)、枯草菌由来のプラスミド(例えばpUB110,pTP5等)、酵母由来のプラスミド(例えばYEp13,YEp24,YCp50,pYE52等)が挙げられる。ファージベクターとしては、λファージ等が挙げられる。ウイルスベクターとしては、レトロウイルス、レンチウィルス、アデノウイルス、アデノ随伴ウイルス、センダイウイルス、バキュロウイルス、ワクシニアウイルス、ポックスウイルス、ポリオウイルス、シンドビスウイルス、ジャガイモエックスウイルス等が挙げられる。
【0046】
Mkx遺伝子をベクターへ挿入するには、DNAを制限酵素で切断し、ベクターDNAの適当な制限酵素部位に挿入する方法又はリコンビナーゼやGatwayシステム等を用いたクローニング法等が採用される。その際、Mkx遺伝子の機能を発揮できるようにベクターに組み込む必要がある。そのために、ベクターには、Mkx遺伝子の他に、プロモーター、エンハンサー等のシスエレメント、スプライシングシグナル、ポリA付加シグナル、選択マーカー、リボソーム結合配列(SD配列)等を含有させるとよい。選択マーカーとしては、例えばアンピシリン耐性遺伝子、カナマイシン耐性遺伝子、クロラムフェニコール耐性遺伝子、ネオマイシン(neo)耐性遺伝子、ジヒドロ葉酸還元酵素遺伝子、等を挙げることができる。
【0047】
発現ベクターは、Mkx遺伝子を直接発現させるものでも、例えばFLAGタグやMycタグ等の他のタンパク質又はペプチドとの融合タンパク質として発現させるものでもよい。
【0048】
本発明の分化誘導剤は、さらに、以下の(A1)〜(D1)の少なくとも一種を含んでいてもよい。
(A1)Scxタンパク質、
(B1)前記Scxタンパク質のアミノ酸配列において、1〜数個のアミノ酸が欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも一種の変異を有し、かつ、I型コラーゲンを誘導する活性を有する変異体、
(C1)前記(A1)又は(B1)のタンパク質をコードする核酸、並びに
(D1)前記(C1)の核酸を含む発現ベクター。
【0049】
Scxタンパク質は、前述のとおり、腱の主成分であるI型コラーゲンを直接誘導することが示唆されている(非特許文献1)。Scxノックアウトマウスは、腱を部分的に消失する。一方、Mkxノックアウトマウスは、実施例に示すように、腱の低形成は認められるが、腱細胞数は変わらない。このことから、Scx腱細胞分化の初期段階から必須の因子であり、Mkxは、後期分化・成熟に重要であることが予想される。また、腱細胞の分化とそこでのI型コラーゲンの産生には、Scx遺伝子による初期分化と、Mkxによる成熟の両者の協同作業が重要といえる。したがって、本発明の分化誘導剤に(A1)〜(D1)のScx関連物質を含めることは、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化をより効率的に行う点で好ましい。
【0050】
Scxタンパク質に関する情報は、NCBIからアクセッション番号NP_001008272として入手することができる。また、Scx遺伝子に関する情報は、NCBIからアクセッション番号NM_001008271として入手可能である。
【0051】
(A1)〜(D1)の作製方法は、タンパク質及び遺伝子がMkxと異なる以外は(A)〜(D)で述べた内容と同じである。
【0052】
本発明は、また、腱及び/又は靭帯細胞への分化能を有する細胞の生育に適した条件下で、上記分化誘導剤を用いて前記細胞を培養する工程、及び前記培養された細胞の中から、腱及び/又は靭帯細胞を選択する工程を含む、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する方法を提供する。
【0053】
前記腱及び/又は靭帯細胞への分化能を有する細胞は、例えば、繊維芽細胞(C3H10T1/2等)、腱前駆細胞、靱帯前駆細胞、胚性幹細胞、胚性生殖細胞、体性幹細胞、iPS細胞等が挙げられる。
【0054】
以下に、分化誘導方法の一態様を説明する。まず、腱及び/又は靭帯細胞への分化能を有する細胞に分化誘導剤を導入して、それを培養する。具体的には、C3H10T1/2細胞をDMEM培地等で培養して得られる50%コンフルエントのC3H10T1/2細胞に、Mkx発現ベクターをトランスフェクションする。トランスフェクションは、公知の方法で行え、またトランスフェクション以外の導入法として例えばウィルスベクターによる導入法、エレクトロポレーション法、リン酸カルシウム沈殿法、マイクロインジェクション法、リポソーム法等が挙げられる。
【0055】
培養方法は、分化誘導方法に適したものであれば、特に制限されない。例えば、平面培養法、浮遊凝集培養法、懸濁培養法、フィーダー細胞との共培養法、旋回培養法、軟寒天培養法、マイクロキャリア培養法等が挙げられる。
【0056】
培養後、回収した細胞を細胞溶解バッファーで溶解して全可溶型タンパク質を取得する。全可溶型タンパク質をウエスタンブロット等にかけ、I型コラーゲンの存在を確認することが好ましい。
【0057】
上記分化誘導剤を用いてMkx遺伝子を過剰発現させると、腱の主要コンポーネントであるI型コラーゲンの発現が上昇する。損傷した腱部位に分化誘導剤や分化誘導剤を用いて製造した腱細胞及び/又は靱帯細胞を導入すれば、腱の治癒、運動器の再生や再建を促進することが期待できる。そこで、本発明は、上記分化誘導剤、又は上記分化誘導剤を用いて製造した腱及び/又は靭帯細胞を含む、腱又は靭帯関連疾患を予防又は治療するための医薬を提供する。
【0058】
本発明の分化誘導剤は、遺伝子治療用にも好適である。遺伝子治療の腱又は靭帯関連疾患の候補は、エーラスダンロス症候群がある。エーラスダンロス症候群は、ヒトの皮膚や組織を形成するコラーゲン等の結合組織成分が先天性代謝異常を示し、その結果、靱帯や関節の異常な可動性亢進等を引き起こす疾患である。現在、根本的治療法は見つかっていない。エーラスダンロス症候群で腱及び/又は靭帯が罹患している患者から採取した細胞を用いて、分化誘導剤で自家腱及び/又は靭帯細胞を製造することにより、該疾患に対する再生医療が期待される。
【0059】
上記医薬の投与形態は、特に限定されないが、好ましくは損傷した腱及び/又は靱帯部位への直接投与である。具体的には、皮下注射剤、筋肉注射剤等の注射剤、点滴剤、経皮投与剤、軟膏剤等の外用薬剤、細胞の単層又は複層のシート等が挙げられる。
【0060】
これらの剤形に適用される助剤や製剤技術には当業分野に公知のものを使用可能である。例えば、注射剤であれば、適当な緩衝剤、安定剤、保存剤、無痛化剤、等張化剤、溶解補助剤等が挙げられる。
【0061】
本発明の医薬は、ヒトを含む動物に対して、薬理上必要量が投与される。薬理上必要量は、剤形、投与方法、患者や実験動物の症状、治療部位、患者や実験動物の年齢、性別、体重等を考慮して、医師等の判断により適宜決められる。
【0062】
本発明は、また、Mkx遺伝子の機能的欠損を含む、腱又は靱帯関連疾患の研究用の遺伝子改変非ヒト動物を提供する。「Mkx遺伝子の機能的欠損」とは、本明細書において、Mkx遺伝子が本来有する正常な機能を十分に発揮しないか、全く発揮しない状態を意味する。
【0063】
遺伝子改変非ヒト動物には、Mkx欠損遺伝子のヘテロ接合体(Mkx+/−と表示することがある)とホモ接合体(Mkx−/−と表示することがある)の二種類がある。本明細書中、「Mkx変異」(例えばMkx変異マウス)の記載は、両者を含む意味で使用する。
【0064】
ノックアウトマウスのようなホモ接合体は、遺伝子改変非ヒト動物の腱及び/又は靭帯が野生型動物と比べて有意に低形成を示す。具体的には、Mkxノックアウトマウスは、全身の腱が低形成になり、尾腱の線維束が細くなるにもかかわらず、腱細胞の数は野生型と同程度である。さらに、Mkxノックアウトマウスの腱は、コラーゲン原線維の径が小さく、I型コラーゲンの量が低下する。以上の結果から、Mkxは、腱細胞において、I型コラーゲンの産生能に寄与し、腱細胞の分化に重要な役割を担っていることが示唆される。この知見に基づく本発明のMkxノックアウトマウスは、Mkxの腱及び/又は靭帯細胞への分化誘導への研究や腱又は靭帯関連疾患を予防又は治療するための医薬の探索に有用である。
【0065】
一方、ヘテロマウスのようなヘテロ接合体は、腱形成に異常が見られない。ヘテロマウスは、腱に異常がなく、腱や靭帯などMkxが発現する細胞で蛍光タンパク質のVenusが発現するので、Mkxがどこで発現しているか、Mkxがどのような物質や遺伝子で発現が誘導されるか等の解析や、腱組織がどのようにできるかの観察、腱細胞を分離して細胞の性状を調査する等に有用である。
【0066】
非ヒト動物の例は、マウス、ラット、ハムスター、モルモット、チンパンジー、サル、ウサギ等の実験動物、ブタ、ウシ、ヒツジ等の家畜類、イヌ、ネコ等の愛玩動物であり、好ましくはマウスである。
【0067】
Mkx遺伝子を機能的に欠損させる方法は、当業分野で公知の方法のいずれも採用することができる。具体的には、Mkx遺伝子を含むゲノムDNAのエキソンを含む領域をターゲッティングベクターで相同組み換えする方法が例示される。
【0068】
Mkx遺伝子を機能的に欠損させる際に、Mkx遺伝子のプロモーターの下流にレポーター遺伝子が挿入されたことにより腱細胞特異的にレポーター遺伝子を発現させることが好ましい。ヘテロ接合体のMkxヘテロマウスのMkx欠損部位を蛍光タンパク質等のレポーター遺伝子で置き換えると、レポーター遺伝子が腱細胞特異的に発現するとともに、正常な腱細胞もまた視認できる(図1D)。このようなMkxヘテロマウスは、Mkxの腱及び/又は靭帯細胞への分化誘導への研究や、腱又は靭帯関連疾患を予防又は治療するための医薬の探索に極めて有用である。
【0069】
前記レポーター遺伝子には、Venus、BFP、CFP、dsRed、EBFP、ECFP、EGFP、EYFP、FlAsH、GFP、HcRed1、mRFP1、mPlum、PhiYFP、RFP、YFP等の蛍光タンパク質、ルシフェラーゼ遺伝子、カタラーゼ遺伝子、LacZ遺伝子等を使用することができる。
【0070】
さらに、Mkx遺伝子の下流に、ネオマイシン耐性遺伝子等の選択マーカーを挿入することが好ましい。
【0071】
上記選択マーカーの両端をloxP配列で挟んでおくことがより好ましい。こうすると、Cre−loxPシステムを用いて、選択マーカーの除去が可能である。
【0072】
遺伝子改変非ヒト動物は、それ自体公知の方法により製造することができる。具体的には、
1.Mkx遺伝子の機能的欠損を含む胚性幹細胞クローンを作製する工程、
2.上記胚性幹細胞クローンを胚にインジェクションする工程、
3.キメラ胚を動物の子宮に移して、キメラ動物を作る工程、
4.キメラ動物を交配させ、ヘテロ接合体を得る工程、及び
5.ヘテロ接合体同士を交配させ、ホモ接合体を得る工程、
を含む。
【0073】
Mkxヘテロマウス及びMkxノックアウトマウスを例に、遺伝子改変非ヒト動物をより具体的に説明する。ゲノムDNAのMkx遺伝子を欠損させるために、胚性幹細胞での相同組み換えにおいてMkx遺伝子の部分領域(エキソンの一部を含む)を、例えば蛍光タンパク質遺伝子とポジティブ選択マーカー(例えばネオマイシン耐性遺伝子、ピューロマイシン耐性遺伝子等)とのカセットに置き換わるようなターゲッティングベクターを設計する(図1A)。また、ポジティブ選択マーカー遺伝子の両側にCre/loxPシステムで用いられるloxP配列のようなリコンビナーゼ標的配列を加えると、Cre酵素を介したマーカー遺伝子の切除が可能である。ネガティブ選択用にジフテリアトキシンA(DT−A)遺伝子、チミジンキナーゼ(TK)遺伝子等を組み込む。
【0074】
上記のターゲッティングベクターを、胚性幹細胞クローンや、初代培養細胞株等の体細胞へ導入する。胚性幹細胞クローンは、マウス、ヒト、ハムスター、ブタ等の動物の胚盤胞から分離した内部細胞塊をフィーダー細胞上で培養することにより樹立することができる。市販の胚性幹細胞クローンを使用することができ、例えばTT2、ES−D3、SCC−PSA1等が挙げられる。
【0075】
ターゲッティングベクターの組み込まれた胚性幹細胞等を、ネオマイシン耐性選択ならG418抗生物質を含む培地といった適宜の選択培地で培養後、組換え胚性幹細胞クローンを分離する。
【0076】
適切に組み込まれた組換え胚性幹細胞クローンのスクリーニングを、サザンブロットやPCT法により行う。図5Aに、サザンブロット解析のための野生型(WT)及びターゲッティングされたアリル(TA)と各プローブの位置を示す。
【0077】
相同組み換えの確認された胚性幹細胞クローンを、マイクロインジェクション法、凝集法等で8細胞期等のマウス胚に導入してキメラ胚を作る。
【0078】
得られたキメラ胚を動物の子宮に移してキメラ動物を作る。得られたキメラ動物を野生型動物(例えばC57BL/6マウス)と掛け合わせる。生殖細胞への伝達をゲノミックPCRやサザンブロット解析により確認する。LoxP配列に挟まれたネオマイシン耐性遺伝子カセットを、Meox−Creトランスジェニック動物との掛け合わせにより除去する。Creを介したネオマイシン耐性遺伝子除去をゲノミックPCRで確認する。キメラ動物を野生型動物と交配させ、Mkxヘテロ接合体を得る。
【0079】
Mkxヘテロ接合体同士を交配させて、ホモ接合体(Mkxノックアウトマウス)を得る。
【0080】
本発明は、また、上記分化誘導剤、上記分化誘導方法により得られる腱及び/又は靭帯細胞、若しくは遺伝子改変非ヒト動物の腱及び/靱帯、それらの組織又は細胞を用いることを特徴とする、腱又は靭帯関連疾患を解析又は分析する方法を提供する。分析や解析の対象は、細胞に分化誘導剤を作用させ分化させて得られるMkxヘテロマウスの胚とその組織、トランスジェニック非ヒト動物からを得られた腱及び/靱帯、それらの組織又は細胞等である。分析や解析の手法を以下に例示する。
【0081】
ホールマウントin situハイブリダイゼーション(WISH)は、マウス胚での遺伝子発現パターンの解析に有効である。WISHは、常法を用いることができ、例えばYokoyama S, et al.(2008)Dynamic gene expression of Lin−28 during embryonic development in mouse and chicken. Gene Expr Patterns 8(3):155−160に記載の方法に従う。これに用いるプローブの合成方法の詳細は、EMBRYSウェブサイト(http://embrys.jp/html/MainMenu.html?)にて閲覧可能である。
【0082】
Venus等の蛍光タンパク質は、Mkx遺伝子の各組織での発現パターンを観察するのに有効である。
【0083】
免疫組織染色は、マウス胚や成体マウスの各組織の観察に有効である。各組織に特異的なマーカーとして、筋肉ではミオシン重鎖、血管ではPECAM1等がある。これらのマーカー遺伝子の発現を、マーカー特異的ポリクローナル又はモノクローナル抗体を用いて免疫組織染色する。まず、マウス胚をパラホルムアルデヒドに固定した組織を凍結組織切片作製用包埋剤に包埋して凍結する。検体を薄切りして風乾後、ブロッキング剤にて1時間ブロッキングする。その切片を抗ミオシン重鎖抗体(MF20,DSHB)でインキュベートし、さらに、二次抗体と共にインキュベートする。VenusノックインMkxへテロマウスでは、ミオシン重鎖抗体(MF20)の赤い染色とVenusの緑色シグナルの両方が観察される。
【0084】
Mkxノックアウトマウスの腱の異常を調査するために、腱及び靱帯の組織学的観察を行うことが有効である。その手法として、以下に電子顕微鏡観察、H&E染色及びアザン染色を例示する。
【0085】
電子顕微鏡を用いると、Mkxノックアウトマウスの腱におけるコラーゲン原線維を観察できる。まず、アキレス腱断片を採取し、緩衝液にて固定する。緩衝液及び生理食塩水で洗浄し、エタノールにて脱水後、樹脂に包埋して極薄切りにする。得られた切片を銅の格子上に置き、透過型電子顕微鏡にて観察する。
【0086】
H&E染色から、野生型マウスやMkxノックアウトマウスの尾腱の細胞密度を求めることができる。まず、成体の膝、尾等の組織を採取し、パラホルムアルデヒド等に固定する。組織を、脱水後、パラフィン包埋し、薄切りし、ヘマトキシリン・エオシン(H&E)で染色する。
【0087】
アザン染色は、コラーゲンを検出する。まず、マウスの尾を採取し、4%パラホルムアルデヒドに固定する。組織を、脱水後、パラフィン包埋し、薄切りし、アゾカルミン−アニリンブルー(アザン)で染色する。
【0088】
Mkx欠損した腱の力学的特性を調べるには、例えば筋腱ジャンクションから踵結節までの全腱を用いて引張試験を行う。張力の決定は、Gentleman E, et al.(2003) Mechanical characterization of collagen fibers and scaffolds for tissue engineering. Biomaterials 24(21):3805−3813で報告された方法に従って、単軸の材料テストシステム(Autograph AGS−G, Shimadzu Corp. Ltd.)を使用するとよい。それぞれの検体の最初の長さと断面積を、デジタルカリパスを使用して、H&E染色した切片の顕微鏡検査によって測定する。データ収集には、例えばTrapeziumソフトウェア(登録商標、Shimadzu社)を用いる。各検体の応力−歪み曲線は、load−displacementカーブから作られ、ヤング率はそれぞれの応力−歪み曲線と腱断面積から算出される。
【0089】
本発明は、また、被験物質の存在下又は非存在下で、上記分化誘導方法を行い、該被験物質存在下又は非存在下での腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を比較することを特徴とする、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質をスクリーニング又は評価する方法を提供する。
【0090】
好ましくは、被験物質の存在下で培養した細胞における、Mkxの発現量及び/又はMkxをコードする塩基配列に対するmRNAの転写量を測定すること、並びに前記被験物質を非存在下で培養した細胞内の量と比べることを含む。測定量が多いか又は少ない場合に、前記被験物質を腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質として選択する。
【0091】
上記培養培地は、用いる細胞に応じて適宜決められる。例えば、MEM培地、DMEM培地、RPM1640培地、199培地等である。培養条件も、細胞に応じて適宜、決められる。
【0092】
本発明は、また、上記遺伝子改変非ヒト動物から得られる腱、その組織又はその細胞を用いることを特徴とする、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質をスクリーニング又は評価する方法を提供する。好ましくは、遺伝子改変非ヒト動物から採取した腱、その組織又はその細胞に被験物質を接触させること、及びノックインされた蛍光タンパク質等のレポーター遺伝子の発現を上昇又は低下させる被験物質を選択することを含む。
【0093】
上記スクリーニング又は評価する方法に供する被験物質は、特に制限されない。例えば、天然又は人工の有機化合物や無機化合物、核酸、ペプチド、タンパク質、糖質、脂質、細胞、植物、動物等の抽出物等である。
【0094】
被験物質と分化誘導させる細胞や遺伝子改変非ヒト動物の組織等との接触は、特に制限されないが、添加、トランスフェクション、ウィルスベクターによる導入等である。
【0095】
Mkxの発現量及び/又はMkxをコードする塩基配列に対するmRNAの転写量、レポーター遺伝子の発現の上昇又は低下の測定は、常法に基づく、例えばノーザンブロット法、リアルタイムPCR、RT−PCR、ウエスタンブロット法、免疫学的手法、 等である。
【実施例】
【0096】
以下に、実施例を用いて本発明をより詳細に説明する。しかし、本発明は、以下の実施例に限定されるものではない。
〔実施例1〕(分化誘導剤の作製と評価)
Mkxタンパク質をコードする核酸を含む発現ベクターからなる分化誘導剤を、C3H10T1/2マウス線維芽細胞に導入してMkxを過剰発現させることにより、分化誘導剤の効果を調べた。
【0097】
まず、マウスMkx遺伝子(NCBIアクセッション番号NM_177595)の全タンパク質コード領域をp3XFLAG−CMVTM−7.1ベクター(SIGMA社)に組み込むことにより、FLAG−Mkx発現ベクターを作製した。また、FLAG−Mkx遺伝子を組み込まない空ベクターを陰性対照として用いた。
【0098】
C3H10T1/2細胞(ATDCから入手)を、10%FBSを含むDMEM培地で培養した。6ウエルプレートにて培養した50%コンフルエントのC3H10T1/2細胞に、トランスフェクション試薬(製品名、FugeneHD、Roche社)10μl、及び上記FLAG−Mkx発現ベクター又は空ベクター1μgをトランスフェクションした。
【0099】
48時間培養後、細胞を回収した。RIPAバッファー(50mM Tris−HCl, 150mM NaCl,0.5%DOC,0.1%SDS,1%NP40;pH 8.0)で細胞を溶解して、C3H10T1/2細胞中の全可溶型タンパク質を取得し、以下のウエスタンブロット解析に供した。
【0100】
取得した全可溶型タンパク質をSDS−PAGEにより分離し、PVDF膜にセミドライ法でブロットした。メンブレンをブロッキング剤(製品名Blocking One、Nacalai Tesque社)にて30分間ブロッキングし、抗I型コラーゲン抗体又は抗β−アクチン抗体と共に4℃、一晩インキュベートした。
【0101】
洗浄後、西洋ワサビペルオキダーゼ(HRP)標識抗ウサギイムノグロブリンG(IgG)抗体(A6154,SIGMA社)、又はHRP標識抗マウスIgG抗体(A2304,SIGMA社)と共に1時間インキュベートした。抗原−抗体の発色には、化学発光試薬(製品名Chemi−Lumi One、ナカライテスク社)を用いた。
【0102】
図4Dに、FLAG−Mkx発現ベクター又は空ベクターをトランスフェクションしたC3H10T1/2細胞のI型コラーゲン(Col1)、FLAG及びβ−actinのウエスタンブロットの結果を示す。図4Dに示すとおり、C3H10T1/2細胞内にMkxを過剰発現させると、I型コラーゲン量が増大した。
【0103】
〔実施例2〕(Mkx変異マウスの作製と評価)
マウスの染色体上のMkx遺伝子を機能的に欠損させるために、図1Aに、胚性細胞での相同組み換えにおいてMkxの翻訳スタートからエキソン2の終わりまでをMkx遺伝子がVenus遺伝子とネオマイシン耐性遺伝子とのカセットで置き換わるようなターゲッティングベクターを設計した。
【0104】
図1Aのターゲッティングコンストラクトをもとに、C57BL/6ゲノムBACクローン(BACPAC Resource Center社)を鋳型にしたPCRを行うことによって、Mkx遺伝子の5‘及び3’側の隣接領域を増幅した。これらの相同組み換え領域を、Venus遺伝子とポジティブ選択用のネオマイシン耐性遺伝子とネガティブ選択用のジフテリアトキシンA(DT−A)遺伝子を含むpBluescript由来のベクターに組み込むことにより、ターゲッティングベクターを作製した。
【0105】
上記のターゲッティングベクターを制限酵素NotIで直鎖化し、TT2F胚性幹細胞へエレクトロポレーション法により導入した。ターゲッティングベクターの組み込まれたTT2F胚性幹細胞を、G418を含む培地で培養して、組換え胚性幹細胞クローンを分離した。
【0106】
適切に組み込まれた組換え胚性幹細胞クローンのスクリーニングを、5‘プローブ、3’プローブ及びneoプローブを用いたサザンブロット法により行った。
【0107】
図5Aに、サザンブロット解析のための、野生型(WT)及びターゲッティングされたアリル(TA)と各プローブの位置を示す。図5Bに、組換え胚性幹細胞のサザンブロットの結果を示す。図5Cに、野生型マウス及びMkx変異マウスのサザンブロットの結果を示す。
【0108】
相同組み換えの確認された2つの胚性幹細胞クローンを8細胞期のマウス胚にインジェクションした。得られたキメラマウスをC57BL/6マウスと掛け合わせ、生殖細胞への伝達をゲノミックPCRにより確認した。LoxP配列に挟まれたネオマイシン耐性遺伝子カセットを、Meox−Creトランスジェニックマウスとの掛け合わせにより除去し、Creを介したネオマイシン耐性遺伝子除去の確認をゲノミックPCRにより行った。ジェノタイピングに使用したPCRプライマーの配列を表1に記載する。
【0109】
【表1】

【0110】
図1Bに、野生型マウスとMkx変異マウスのプライマーa、b及びcを用いたゲノミックPCR結果を示す。
【0111】
ターゲッティングベクターを用いて作製されたVenusノックインMkxヘテロマウスは、Mkxの発現が陽性であり、かつ、ノックインしたVenus遺伝子の発現もまた予想される。そこで、マウス胚の蛍光シグナルの観察を行った。Mkxプローブを用いたホールマウントin situハイブリダイゼーションを行った。
【0112】
図1Cに、E13.5前肢(Forelimb)及び尾(Tail)における野生型マウスとMkxヘテロマウスの胚のMkxのホールマウントin situ ハイブリダイゼーションを示す。VenusノックインMkxヘテロマウスの前肢と尾では、Mkxの発現と同様の場所で、Venusの発現が観察された。
【0113】
Mkxヘテロマウス胚の免疫組織染色のために、E16.5マウス胚の尾を採取し、4%パラホルムアルデヒドに4℃で2時間固定した。組織を、凍結組織切片作製用包埋剤(製品名O.C.T.compound、Sakura Finetek社)に包埋し、液体窒素で直ちに凍結した。検体を10μmに薄切りし、その凍結切片を風乾後、Blocking One(Nacalai Tesque社)にて1時間ブロッキングした。その切片を抗ミオシン重鎖抗体(MF20,DSHB)と共に4℃、一晩インキュベートした。洗浄後、二次抗体蛍光試薬(製品名Molecular Probes Alexa 594)と共に1時間インキュベートした。
【0114】
図1Dに、E16.5のVenusノックインMkxへテロマウスの胚の尾における抗ミオシン重鎖抗体(MF20;red)を用いた免疫組織染色とVenus蛍光シグナルを観察した顕微鏡写真を示す。図中、○で囲んだ部分はMF20免疫組織染色(実際は赤色に呈色)を示し、それ以外はVenus蛍光シグナル(実際は緑色に呈色)を示す。VenusノックインMkxヘテロマウスの胚は、E16.5の尾において腱特異的なVenusの発現を示した。
【0115】
Mkxヘテロマウス胚の解析に加え、その成体マウスにおけるVenusの発現についても調査した。成体の腱でのMkxの発現は明らかではなかったので、成体の腱でのMkxの発現をRT−PCRにより調査した。図6Aに、野生型マウス(成体)の筋肉、アキレス腱及び尾腱におけるMkx及びGapdhのRT−PCR解析を示す。図6Aから、Mkxがアキレス腱及び尾腱で強く発現していることを確認した。
【0116】
図6Bに、Mkxヘテロマウス及び野生型マウスのアキレス腱(B−E)、尾腱(F−I)及び体幹の腱(J−M)におけるVenusの発現と外観写真を示す。図6Bから、MkxヘテロマウスにおけるVenusは、アキレス腱、尾腱及び体幹の腱において特異的発現を示した。
【0117】
Mkx遺伝子の欠損を確認するために、Mkx変異マウスを用いたRT−PCRによる発現解析を行った。核酸抽出薬(製品名ISOGEN、Nippongene社)を用いて全RNAを分離した。分離した全RNAから、cDNA合成キット(製品名Ready−To−Go You−Prime First−Strand Beads、GE Healthcare社)を用いて逆転写し、そのcDNAを鋳型にDNAポリメラーゼ(製品名Go−Taq(登録商標)、Promega社)を用いてPCRした。プライマー配列を表2に示す。Gapdhは、mRNAのコントロールとして用いた。
【0118】
【表2】
【0119】
図1Eに、野生型マウスとMkx変異マウスのアキレス腱におけるMkx及びGapdhのRT−PCRの解析結果を示す。図1Eから、Mkxノックアウトマウス(成体)のアキレス腱において、Mkxの発現が完全に消失していることがわかった。
【0120】
以上の結果から、Mkx変異マウス(VenusノックインMkxヘテロマウス及びMkxノックアウトマウス)は、Mkxの機能解析だけでなく、腱の生物学的解析やMkxの発現解析に有用であることが示された。
【0121】
〔実施例3〕(Mkxノックアウトマウスの作製と評価)
実施例2で作製したMkxヘテロマウス同士を掛け合わせることにより、Mkxノックアウトマウスを作製した。産まれた各遺伝子型の新生仔マウスの数と比を表3に示す。
【0122】
【表3】
【0123】
ヘテロマウス同士の掛け合わせによって産まれた206匹の新生仔マウスのジェノタイピングの結果は、メンデルの法則に従った。
【0124】
Mkxヘテロマウス及びMkxホモマウス(ノックアウトマウス)は、問題なく生存、生殖し、重量測定の結果も野生型マウスと変わらなかった(図7)。
【0125】
Mkxヘテロマウスでは、腱の異常は見られなかった(図6B)。腱形成におけるMkxの役割を調査するために、Mkxノックアウトマウスで腱の異常が観察されるか否かを調べた。
【0126】
図2Aに、野生型マウスにおいて、膝蓋腱(Aの矢印)、アキレス腱(Cの矢印、後肢(Eの矢印)、前肢の腱(Gの矢印)、尾腱(Iの矢印)、背中の腱(Kの矢印)及び広頸筋腱(Mの矢印)を観察した写真、並びにMkxノックアウトマウスにおいて、膝蓋腱(Bの矢印)、アキレス腱(Dの矢印、後肢(Fの矢印)、前肢の腱(Hの矢印)、尾腱(Jの矢印)、背中の腱(Lの矢印)及び広頸筋腱(Nの矢印)を観察した写真を示す。Mkxノックアウトマウスでは、膝蓋腱、アキレス腱、尾腱、四肢の腱、体幹の腱及び広頸筋腱が、野生型マウスに比べて低形成であり、白色が薄くなっていた。この腱の異常は、7日齢から8ヶ月齢までのMkxノックアウトマウスとネオマイシン耐性遺伝子を除去したマウス(図8)においても観察された。
【0127】
Mkxが欠損した腱の力学的特性を調べるために、筋腱ジャンクションから踵結節までの全腱を用いて引張試験を行った。張力の決定は、Gentleman E, et al. (2003) Mechanical characterization of collagen fibers and scaffolds for tissue engineering. Biomaterials 24(21):3805−3813で報告された方法に従って、単軸の材料テストシステム(Autograph AGS−G、島津社)を使用して行った。検体を、0.5 mm/secの重圧率で引張した。
【0128】
それぞれの検体の最初の長さと断面積を、デジタルカリパスを使用して、H&E染色した切片の顕微鏡検査によって測定した。データ収集はTrapezium(登録商標、島津社)ソフトウェアを用いて行った。各検体の応力−歪み曲線は、load−displacementカーブから作られ、ヤング率はそれぞれの応力−歪み曲線と腱断面積から算出した。
【0129】
図2Bに、野生型マウスとMkxノックアウトマウスのアキレス腱の引張強度を示す。Mkxノックアウトマウスのアキレス腱は、引張強度が低く、機能低下が見受けられた。図2Cに、野生型マウスとMkxノックアウトマウスのアキレス腱のヤング率を示す。単位面積あたりの引張強度及び単位面積あたりの弾力性を示すヤング率の値は、野生型マウスとMkxノックアウトマウスでほぼ同程度であった。以上の結果から、Mkxノックアウトマウスのアキレス腱における引張強度の低下は、腱が細くなることに起因していることが示唆された。
【0130】
Mkxノックアウトマウスの腱の異常をさらに詳細に調査するために、組織学的解析を行った。成体マウスの膝又は尾を採取し、4%パラホルムアルデヒドに4℃で一晩固定した。組織を、脱水後、パラフィン包埋し、薄切りし、ヘマトキシリン・エオシン(H&E)で染色した。
【0131】
図3Aに、3ヶ月齢の野生型マウスとMkxノックアウトマウスの膝蓋腱のH&E染色像を示す。H&E染色の結果、3ヶ月齢のMkxノックアウトマウスの膝蓋腱は、野生型マウスに比べて薄いことが分かった。
【0132】
図3Bに、7日齢の野生型マウスとMkxノックアウトマウスの膝蓋腱の厚さを示す。7日齢のノックアウトマウスにおいても、膝蓋腱が薄く、野生型マウスの腱の約半分の厚さになっていた(図3B)。
【0133】
図9に、野生型マウス及びMkxノックアウトマウスの膝十字靭帯のH&E染色像を示す。腱と同様の組成を持つ組織である靭帯の1つである膝十字靭帯においては、野生型マウスとMkxノックアウトマウスとの間に差は見られなかった。
【0134】
図3Cに、3ヶ月齢の野生型マウスとMkxノックアウトマウスの尾腱のH&E染色像を示す。Mkxノックアウトマウスの尾腱でも、腱線維束の径の縮小(図3Cの△矢頭)が見られた。
【0135】
図3Dに、3ヶ月齢の野生型マウスとMkxノックアウトマウスの尾腱の細胞密度を示す。Mkxノックアウトマウスの尾腱では、細胞密度が高く、1つの尾腱線維束における細胞数は野生型マウスと同程度であった。このことから、Mkxノックアウト腱細胞は、機能的に不完全であることが示唆される。
【0136】
Mkxノックアウトマウスにおける腱の形成不全が胎生期においても観察されるか否かを調査するため、E18.5のMkx欠損胚の尾腱のアザン染色を行った。具体的には、尾を採取し、4%パラホルムアルデヒドに4℃で一晩固定した。組織を、脱水後、パラフィン包埋し、薄切りし、アゾカルミン−アニリンブルー(アザン)で染色した。
【0137】
図3Eに、E18.5の野生型マウスとMkxノックアウトマウスの胚の尾腱のアザン染色像を示す。図3Eからは、尾腱の大きさは、野生型マウスと違いが見られない。Mkxノックアウト胚では、コラーゲンを検出するアニリンブルーの染色の密度が低い(△矢頭部)ことが分かった。
【0138】
Mkxノックアウトマウスの腱におけるコラーゲン原線維について解析するため、アキレス腱を電子顕微鏡で観察した。約1mm3のアキレス腱断片を採取し、4%パラホルムアルデヒドと2.5%グルタルアルデヒドを含む0.1Mカコジル酸バッファーにて一晩固定した。0.1Mカコジル酸バッファー及び生理食塩水で洗浄し、エタノールにて脱水後、エポキシ樹脂に包埋して極薄切りにした。得られた切片を銅の格子上に置き、水性ウラニルアセテートとクエン酸鉛と対比させて、透過型電子顕微鏡(H−7100,日立社)にて観察した。
【0139】
図3Fに、野生型マウスとMkxノックアウトマウスのアキレス腱のコラーゲン原線維の電子顕微鏡観察像を示す。ノックアウトマウスの腱におけるコラーゲン原線維の径は、野生型マウスと比べて均一に小さいことが分かった。これらの結果から、Mkxノックアウトマウスの腱では、コラーゲンが減少していること、さらにはMkxが腱の分化に重要であることが示唆された。
【0140】
3ヶ月齢のMkxノックアウトマウスの尾腱の細胞密度(図3D)に示されるように、Mkxノックアウトマウスでは、腱の量が減少しているにもかかわらず、腱細胞の数は野生型マウスと変わらなかった。これは、Mkxノックアウト腱細胞において、細胞外基質の産生能が低下していることを示唆する。また、図3Eに示すE18.5のMkxノックアウトマウスの胚の尾腱のアザン染色像、並びに図3Fに示すMkxノックアウトマウスのアキレス腱のコラーゲン原線維の微細構造解析から、Mkxノックアウトマウスの腱ではコラーゲン産生能が減少していると考えられる。
【0141】
Mkxノックアウトマウスの腱細胞のコラーゲン産生能を調査するため、8週齢の野生型マウス及びMkxノックアウトマウスの腱のコラーゲン量を測定した。コラーゲンの定量は、市販のコラーゲン定量キット(製品名Sircol Soluble Collagen Assay、Biocolor社)の説明書に従った。結果を図4Aに示す。ノックアウトマウスの腱のコラーゲン量は野生型マウスと比べて少ないことが分かった。
【0142】
図4Bに8週齢の野生型マウスとMkxノックアウトマウスのアキレス腱のI型コラーゲン(Col1)とβ−actinのウエスタンブロット解析を示す。腱の細胞外基質の主要成分であるI型コラーゲンの発現量が示すように、Mkxノックアウトマウスでのタンパク質及びmRNAの両方とも減少していた。
【0143】
8週齢の野生型マウスとMkxノックアウトマウスのアキレス腱のCol1a1、Col1a2及びScxのreal time PCRを、SYBR Green PCR Master Mix(Applied Biosystems社)を用いて行った。プライマー配列を表4に示す。Gapdhは、mRNAのコントロールとして用いた。
【0144】
【表4】
【0145】
delta−delta CT法により測定した相対発現量のグラフを図4Cに示す。図4Cでは、I型コラーゲンの発現を誘導することが示唆されている転写因子であるScxの発現は減少していなかった。これから、MkxノックアウトマウスにおけるI型コラーゲンの減少は、Scxの減少ではないことが示唆された。
【0146】
以上の結果から、Mkxは、腱細胞においてI型コラーゲンの発現を調節する重要な因子であることが実証された。
【技術分野】
【0001】
本発明は、腱及び/又は靭帯細胞の分化誘導剤、分化誘導剤を用いた医薬、分化誘導剤を用いたスクリーニング方法等に関する。
【背景技術】
【0002】
腱は筋肉と骨とを結合する組織であり、靱帯は骨と骨とを結合する組織である。これらの組織は、力や運動を伝達するという重要な機能を持っている。腱や靱帯は、酷使や加齢によって損傷し、また減少するが、栄養血管が少ないため、腱の治癒力は非常に弱く、完治が極めて難しい。腱細胞の分化機構等の生物学的理解も不十分であるため、現在、再生医療等の治療法は確立していない。
【0003】
腱や靱帯は、コラーゲン原線維がクロスリンクしてできる線維でできている。腱の線維間には腱細胞が僅かに存在し、コラーゲン、プロテオグリカン等の細胞外基質の産生を行っている。腱の大部分を構成するコラーゲンは、腱に弾性力を与えるが、そのほとんどはI型コラーゲンである。プロテオグリカンは、腱においてはdecorin、fibromodulin、biglycan、lumican等が存在し、コラーゲン線維同士の結合や集合に関与している。プロテオグリカンの遺伝子欠損マウスでは、腱のコラーゲン原線維の異常が観察される。エーラスダンロス(Ehlers−Danlos)症候群に代表されるコラーゲン産生に異常を有する疾患患者に腱の障害が報告されている。腱細胞における細胞外基質の産生は、腱の形成にとって重要であることが示唆される。
【0004】
近年、腱及び靭帯細胞並びに腱及び靭帯前駆細胞で発現するbasic helix−loop−helix(bHLH)型の転写因子であるScleraxis(Scx)が腱発生に重要な因子であることが報告され、Scxノックアウトマウスでは、腱の著しい形成不全が観察された(非特許文献1)。また、腱の主成分であるI型コラーゲンを直接誘導することが示唆された(非特許文献2)。しかし、Scxノックアウトマウスは、後述の実施例に示すように、腱においてすべてのI型コラーゲンが消失するわけではない。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Cserjesi P,et al.(1995)Scleraxis: a basic helix−loop−helix protein that prefigures skeletal formation during mouse embryogenesis Development 121(4) p1099−1110
【非特許文献2】Lejard V,et al.(2007)Scleraxis and NFATc regulate the expression of the pro−alpha1(I) collagen gene in tendon fibroblasts J Biol Chem 282(24) p17665−17675
【発明の概要】
【発明が解決しようとする課題】
【0006】
腱及び靭帯細胞は、I型コラーゲン等の基質を産生する細胞であって、それぞれ、腱及び靭帯組織の形成及び維持に関与する。腱及び靭帯は、上記したように再生能の低い組織であり、腱及び靭帯疾患治療に向けての再生医療が望まれる。Scxは、腱及び靭帯細胞で発現し、腱及び靭帯の主成分であるI型コラーゲンの発現を誘導することが示唆されるものの、他のI型コラーゲンの発現調節因子の存在が示唆される等、腱発生のメカニズムには未解明な部分が多いのが現状である。腱及び靭帯細胞への分化誘導方法も、確立されていない。
【0007】
そこで、本発明の目的は、腱及び/又は靭帯細胞の分化誘導剤、並びに該分化誘導剤を含む、腱又は靭帯関連疾患を予防又は治療するための医薬を提供することにある。
【0008】
さらに、本発明の目的は、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞へ分化誘導させるために使用する物質をスクリーニングする方法を提供することにある。
【課題を解決するための手段】
【0009】
本発明者らは、腱及び靭帯細胞への分化方法の確立には、分化に重要な遺伝子を特定し、その遺伝子を導入又は誘導することが重要であると考えた。本発明者らは、後述の実施例に示すように、Mkx遺伝子を破壊すると、腱組織と腱細胞のI型コラーゲン生産量が減少し、逆に細胞内に過剰発現させると、I型コラーゲン量が増大することを発見した。Mkxは、従来、胎生期において腱に発現するホメオボックス型の転写因子として知られていたが、Mkxが腱細胞においてその分化を制御しI型コラーゲンの発現を調節する重要な因子であることが初めて明らかとなった。これらの知見をもとに、本発明者らは、以下の発明を完成させた。
【0010】
すなわち、本発明は、
(A)Mkxタンパク質、
(B)前記Mkxタンパク質のアミノ酸配列において、1〜数個のアミノ酸が欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも一種の変異を有し、かつ、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する活性を有する変異体、
(C)前記(A)又は(B)のタンパク質をコードする核酸、並びに、
(D)前記(C)の核酸を含む発現ベクター
の少なくとも一種を有効成分として含む、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化誘導剤を提供する。
【0011】
上記分化誘導剤は、
(A1)Scxタンパク質、
(B1)前記Scxタンパク質のアミノ酸配列において、1〜数個のアミノ酸が欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも一種の変異を有し、かつ、I型コラーゲンを誘導する活性を有する変異体、
(C1)前記(A1)又は(B1)のタンパク質をコードする核酸、並びに、
(D1)前記(C1)の核酸を含む発現ベクター、
の少なくとも一種をさらに含んでもよい。
【0012】
本発明は、腱及び/又は靭帯細胞への分化能を有する細胞の生育に適した条件下で、上記分化誘導剤を用いて前記細胞を培養する工程、及び前記培養された細胞の中から、腱及び/又は靭帯細胞を選択する工程を含む、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する方法を提供する。
【0013】
前記細胞は、例えば繊維芽細胞、腱前駆細胞、靱帯前駆細胞、胚性幹細胞、胚性生殖細胞、体性幹細胞及びiPS細胞からなる群から選ばれる少なくとも一種である。
【0014】
本発明は、また、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化誘導剤を含む腱又は靭帯関連疾患を予防又は治療するための医薬を提供する。
【0015】
上記腱又は靭帯関連疾患は、例えばエーラスダンロス症候群である。
【0016】
本発明は、また、上記分化誘導方法により得られる、腱及び/又は靭帯細胞を含む、腱又は靭帯関連疾患を予防又は治療するための医薬を提供する。
【0017】
上記腱又は靭帯関連疾患は、例えばエーラスダンロス症候群である。
【0018】
本発明は、また、Mkx遺伝子の機能的欠損を含む、腱又は靱帯関連疾患の研究用の遺伝子改変非ヒト動物を提供する。
【0019】
前記機能的に欠損したMkx遺伝子は、Mkx遺伝子のプロモーターの下流にレポーター遺伝子が挿入されたことにより腱細胞特異的にレポーター遺伝子を発現することが好ましい。
【0020】
前記レポーター遺伝子は、例えばVenus遺伝子である。
【0021】
本発明は、また、上記分化誘導剤を用いることを特徴とする、腱又は靭帯関連疾患の解析又は分析方法を提供する。
【0022】
本発明は、また、上記分化誘導方法により得られる腱及び/又は靭帯細胞を用いることを特徴とする、腱又は靭帯関連疾患の解析又は分析方法を提供する。
【0023】
本発明は、また、上記遺伝子改変非ヒト動物の腱及び/靱帯、それらの組織又は細胞を用いることを特徴とする、腱又は靭帯関連疾患の解析又は分析方法を提供する
【0024】
本発明は、また、被験物質の存在下又は非存在下で、上記分化誘導方法を行い、該被験物質存在下又は非存在下での腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を比較することを特徴とする、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質をスクリーニング又は評価する方法を提供する。
【0025】
上記スクリーニング又は評価する方法は、例えば被験物質の存在下で培養した細胞における、Mkxの発現量及び/又はMkxをコードする塩基配列に対するmRNAの転写量を測定すること、並びに前記被験物質を非存在下で培養した細胞内の量と比べて、前記測定した量が多い又は少ない場合に、前記被験物質を腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質として選択することを含む。
【0026】
本発明は、また、上記遺伝子改変非ヒト動物から得られる腱、その組織又はその細胞を用いることを特徴とする、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質をスクリーニング又は評価する方法を提供する。
【0027】
上記スクリーニング又は評価する方法は、例えば遺伝子改変非ヒト動物から得られる腱、その組織又はその細胞に被験物質を接触させること、及びノックインされたレポーター遺伝子の発現を上昇又は低下させる被験物質を選択することを含む。
【発明の効果】
【0028】
関節の再生等の医学において、腱や靱帯は筋・骨格をつなぐ重要な組織である。そして、Mkx遺伝子を過剰発現させると、腱の主要コンポーネントであるI型コラーゲンの発現が上昇することから、Mkxは腱の成熟過程において重要であることが示唆される。損傷した腱や靱帯の部位にMkx及びその上流・下流遺伝子等からなる医薬を外来的に導入することで、腱の治癒、運動器の再生や再建を促進することが期待される。
【0029】
エーラスダンロス症候群等の腱及び/又は靭帯関連疾患に罹患している患者から採取した細胞を用いて、本発明の分化誘導剤で自家腱及び/又は靭帯細胞を製造することもでき、結果として該疾患に対する再生医療が可能となる。
【0030】
Mkx遺伝子が腱細胞や靱帯細胞特異的に蛍光タンパク質のようなレポーター遺伝子で置き換えられている本発明の遺伝子改変非ヒト動物は、蛍光タンパクの発光を指標にMkxを調節する被験物質のスクリーニングや評価、腱損傷モデル実験等における治癒過程でのMkxの局所的な発現をモニターするためのモデル動物として有用である。
【0031】
分化誘導剤や分化誘導方法により得られる腱及び/又は靭帯細胞を用いて、腱又は靭帯関連疾患の解析又は分析方法は、腱分化の分子メカニズム理解の基礎となり、腱の障害やエーラスダンロス症候群等の腱関連疾患の治療標的発見を導くきっかけとなるかもしれない。
【0032】
Mkxは、腱の発生・成熟に重要であることから、腱・靱帯等の疾患等で、Mkxそのものの遺伝子配列又はMkxの遺伝子ネットワークに関与する上流及び下流遺伝子等の遺伝子に異常を起こしている可能性が大きい。それらの探索、診断、治療に、本発発明のスクリーニング方法が応用される。
【0033】
本発明のスクリーニング方法によって選択された物質や本発明の医薬によれば、腱又は靭帯関連疾患、特にMkxに異常を抱える疾患を、予防又は治療することができる。
【図面の簡単な説明】
【0034】
【図1A】Mkx変異マウスを作製するために設計するMkxターゲッティングコンストラクトの一例を示す。ここで、矢印a〜eはジェノタイピング用PCRプライマー、□はUTR、■は翻訳領域、DT−AはジフテリアトキシンA、WTは野生型アリル、TAはターゲッティングされたアリルを意味する。
【図1B】野生型マウスとMkx変異マウスでの図1Aのプライマーa、b及びcを用いたゲノミックPCR(図面代用写真)を示す。
【図1C】上段は、野生型マウスの胚のE13.5前肢(Forelimb)及び尾(Tail)におけるMkxのWISH(ホールマウント in situ ハイブリダイゼーション)(図面代用写真)を示し、そして下段は、野生型マウス及びMkx変異マウスのVenus蛍光シグナルの観察(図面代用写真)を示す。
【図1D】E16.5のVenusノックインへテロマウスの胚の尾におけるミオシン重鎖抗体(MF20;red)を用いた免疫組織染色とVenus蛍光シグナルを観察した顕微鏡写真を示す。図中、○で囲んだ部分が筋肉のマーカー遺伝子であるミオシン重鎖発現細胞を示し、それ以外はVenus発現細胞の蛍光シグナルを示す。また、Barは100 μmである。
【図1E】野生型マウスとMkx変異マウスのアキレス腱におけるMkxのRT−PCR解析(図面代用写真)を示す。Gapdhは内在性コントールを示す。
【図2A】3ヶ月齢の野生型マウスにおいて、膝蓋腱(Aの矢印)、アキレス腱(Cの矢印、後肢(Eの矢印)、前肢の腱(Gの矢印)、尾腱(Iの矢印)、背中の腱(Kの矢印)及び広頸筋腱(Mの矢印)を観察した図面代用写真、並びに3ヶ月齢のMkxノックアウトマウスにおいて、膝蓋腱(Bの矢印)、アキレス腱(Dの矢印、後肢(Fの矢印)、前肢の腱(Hの矢印)、尾腱(Jの矢印)、背中の腱(Lの矢印)及び広頸筋腱(Nの矢印)を観察した図面代用写真を示す。Mkxノックアウトマウスでは、腱の異常が観察される。
【図2B】野生型マウスとMkxノックアウトマウスのアキレス腱の引張強度を測定したグラフである。
【図2C】野生型マウスとMkxノックアウトマウスのアキレス腱のヤング率を測定したグラフである。
【図3A】3ヶ月齢の野生型マウスとMkxノックアウトマウスの膝蓋腱のH&E染色像(図面代用写真)を示す。a及びcのバーは1mmを意味し、b及びdのバーは50μmを意味する。Mkx欠損マウスでは、腱の低形成が起きていることがわかる。
【図3B】7日齢の野生型マウスとMkxノックアウトマウスの膝蓋腱の厚さを測定したグラフである。Mkxノックアウトマウスは、膝蓋腱の厚さが有意に減少している。
【図3C】3ヶ月齢の野生型マウスとMkxノックアウトマウスの尾腱のH&E染色像(図面代用写真)を示す。a及びcのバーは200μmを意味し、b及びdのバーは50μmを意味する。△矢頭は、尾腱の線維束を示す。Mkxノックアウトマウスの尾腱の線維束は有意に細っている。
【図3D】3ヶ月齢の野生型マウスとMkxノックアウトマウスの尾腱の細胞密度を測定したグラフである。
【図3E】E18.5の野生型マウスとMkxノックアウトマウスの胚の尾腱のアザン染色像(図面代用写真)を示す。△矢頭は尾腱の線維束を示す。また、左図のバーは500μm、右図のバーは200μmを意味する。
【図3F】野生型マウスとMkxノックアウトマウスのアキレス腱のコラーゲン原線維の電子顕微鏡観察像(図面代用写真)を示す。ここで、バーは200nmを意味する。
【図4A】8週齢の野生型マウスとMkxノックアウトマウスのアキレス腱のコラーゲン量の測定結果を示す。Mkx欠損腱細胞では、I型コラーゲン産生能が低下することがわかる。
【図4B】8週齢の野生型マウスとMkxノックアウトマウスのアキレス腱のI型コラーゲン(Col1)とβ−actinのウエスタンブロット解析の結果(図面代用写真)を示す。
【図4C】8週齢の野生型マウスとMkxノックアウトマウスのアキレス腱のCol1a1、Col1a2及びScxのreal time PCR解析結果を示す。
【図4D】FLAG−Mkx発現ベクター(Mkx)又は空ベクター(Empty)をトランスフェクションしたC3H10T1/2細胞のI型コラーゲン(Col1)、FLAG及びβ−actinのウエスタンブロットの結果(図面代用写真)を示す。
【図4E】腱分化ネットワークの模式図を示す。
【図5A】Mkx変異ES又はマウスのサザンブロット解析のために、野生型(WT)及びターゲッティングされたアリル(TA)と各プローブの位置を示す。ここで、NはNcoIを意味し、SはSacIを意味する。
【図5B】組換え胚性幹細胞のサザンブロット解析結果(図面代用写真)を示す。ここで、TAはターゲッティングされたアリルを意味し、WTは野生型アリルを意味する。
【図5C】野生型マウス及びMkx変異マウスのサザンブロット解析結果(図面代用写真)を示す。
【図6A】野生型マウス(成体)の筋肉、アキレス腱及び尾腱におけるMkxのRT−PCR解析(図面代用写真)を示す。Mkxヘテロマウス(成体)のVenusの発現は、Mkx発現とリンクすることがわかる。Gapdhは内在性コントロールを示す。
【図6B】Mkxヘテロマウス及び野生型マウスのアキレス腱(B−E)、尾腱(F−I)及び体幹の腱(J−M)におけるVenusの発現と外観写真を示す。
【図7】出生後の各ステージにおけるMkx+/+,Mkx+/−及びMkx−/−マウスの体重を比較したグラフを示す。野生型マウスとMkxノックアウトマウスとで体重の差は見られない。
【図8】3ヶ月齢の野生型マウス(WT)のアキレス腱(A)、尾腱(C)、膝蓋腱(E)及び体幹の腱(G)の外観写真と、3ヶ月齢のネオマイシン耐性遺伝子を除去したMkxノックアウトマウス(KO)のアキレス腱(B)、尾腱(D)、膝蓋腱(F)及び体幹の腱(H)の外観写真を示す。腱の異常は、ネオマイシン耐性遺伝子を除去させたノックアウトマウスにおいても起きている。
【図9】野生型マウス及びMkxノックアウトマウスの膝十字靭帯のH&E染色像(図面代用写真)を示す。左図のバーは500μm、右図のバーは200μmを意味する。膝十字靭帯は、Mkxノックアウトマウスにおいても影響はない。
【発明を実施するための形態】
【0035】
本発明の腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化誘導剤は、以下の(A)から(D)の少なくとも一種を有効成分として含む。
(A)Mkxタンパク質、
(B)前記Mkxタンパク質のアミノ酸配列において、1〜数個のアミノ酸が欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも一種の変異を有し、かつ、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する活性を有する変異体、
(C)前記(A)又は(B)のタンパク質をコードする核酸、並びに、
(D)前記(C)の核酸を含む発現ベクター
【0036】
Mkx(Mohawk、別名 Irxl1)とは、胎生期において、腱に発現するホメオボックス型の転写因子である。Mkxは、不定型ホメオボックス遺伝子のTALEスーパークラスの中の新しく分類されたクラスに属する唯一の遺伝子である(Anderson DM,et al.(2006).Dev Dyn 235(3) p792−801)。Mkxは、腱、筋肉、軟骨の前駆細胞、オスの生殖腺、及び腎臓の尿管芽先端で発現することが報告されている(Dev Dyn 235(3) p792−801等)。本発明者らも、E9.5、E10.5及びE11.5のマウス胚を用いた1520個の転写因子及び転写コファクターのホールマウントin situハイブリダイゼーションデータベース(EMBRYS)の構築過程で、腱で発現する転写因子として同定していた(Yokoyama S,et al.2009 Dev Cell 17(6) p836−848)。
【0037】
本発明では、細胞から腱細胞及び/又は靱帯細胞を作製、再生、増強等する分化誘導剤としてMkxタンパク質を用いる。
【0038】
本発明の分化誘導剤を実験及び研究用に供する場合は、Mkxタンパク質の由来は問わない。例えば、ヒト、マウス、サル、チンパンジー、ブタ、イヌ等の哺乳動物が挙げられる。本発明の分化誘導剤をヒトの腱細胞及び/又は靱帯細胞を再生等の医療行為に使用する場合は、ヒトのMkxタンパク質を用いることが好ましい。
【0039】
ヒトMkxタンパク質に関する情報は、NCBIからアクセッション番号NP_775847として入手することができ、Mkxタンパク質は、352個のアミノ酸で構成されるタンパク質である。また、ヒトMkx遺伝子に関する情報は、NCBIからアクセッション番号NM_173576として入手可能である。ヒトMkx遺伝子は、ヒト第10染色体上に存在し、全長で約3658の塩基対よりなり、7個のエキソンで構成される。マウスMkx遺伝子は、NCBIから、アクセッション番号NM_177595として入手することができる。
【0040】
本発明の分化誘導剤に用いる(A)Mkxタンパク質は、天然由来のタンパク質の他に、組み換えタンパク質でもよい。組み換えタンパク質は、常法により、Mkxタンパク質をコードする核酸を組み込んだ形質転換体を培養等することにより得ることができる。
【0041】
本発明の分化誘導剤に用いる(B)Mkxタンパク質のアミノ酸配列において、1〜数個のアミノ酸が欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも一種の変異を有し、かつ、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する活性を有する変異体は、当業者に公知の方法により容易に取得することができる。変異体が分化誘導活性を有するか否かは、ウエスタンブロット、ノーザンブロット、リアルタイムPCR等で測定することができる。
【0042】
Mkxタンパク質の変異体は、また、Mkxタンパク質のアミノ酸配列と通常、70%以上、好ましくは80%以上、さらに好ましくは90%の相同性を有するアミノ酸配列からなり、かつ、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する活性を有するものでもよい。
【0043】
本発明の分化誘導剤は、前記(A)又は(B)のタンパク質をコードする(C)核酸でもよい。核酸には、染色体DNAやcDNAが挙げられる。染色体DNAは、例えばPCRによる増幅や細胞等からDNAライブラリ−を調製し、Mkxタンパク質をコードするDNAにハイブリダイズするプローブでスクリーニングすることにより取得する。cDNAは、例えば腱細胞等からRNAを抽出し、Mkxタンパク質をコードするDNAにハイブリダイズするプライマーを用いたPCR法により取得する。
【0044】
本発明の分化誘導剤は、(D)前記(C)の核酸を含む発現ベクターでもよい。発現ベクターは、公知のベクターにMkx遺伝子を発現可能に連結することで得る。前記ベクターは、宿主中で複製可能なものであれば特に限定されず、例えば、プラスミドベクター、ウイルスベクター、ファージベクター等が挙げられる。
【0045】
前記プラスミドベクターとしては、大腸菌由来のプラスミド(例えばpBR322,pUC18,pUC119,pTrcHis,pBlueBacHis等)、枯草菌由来のプラスミド(例えばpUB110,pTP5等)、酵母由来のプラスミド(例えばYEp13,YEp24,YCp50,pYE52等)が挙げられる。ファージベクターとしては、λファージ等が挙げられる。ウイルスベクターとしては、レトロウイルス、レンチウィルス、アデノウイルス、アデノ随伴ウイルス、センダイウイルス、バキュロウイルス、ワクシニアウイルス、ポックスウイルス、ポリオウイルス、シンドビスウイルス、ジャガイモエックスウイルス等が挙げられる。
【0046】
Mkx遺伝子をベクターへ挿入するには、DNAを制限酵素で切断し、ベクターDNAの適当な制限酵素部位に挿入する方法又はリコンビナーゼやGatwayシステム等を用いたクローニング法等が採用される。その際、Mkx遺伝子の機能を発揮できるようにベクターに組み込む必要がある。そのために、ベクターには、Mkx遺伝子の他に、プロモーター、エンハンサー等のシスエレメント、スプライシングシグナル、ポリA付加シグナル、選択マーカー、リボソーム結合配列(SD配列)等を含有させるとよい。選択マーカーとしては、例えばアンピシリン耐性遺伝子、カナマイシン耐性遺伝子、クロラムフェニコール耐性遺伝子、ネオマイシン(neo)耐性遺伝子、ジヒドロ葉酸還元酵素遺伝子、等を挙げることができる。
【0047】
発現ベクターは、Mkx遺伝子を直接発現させるものでも、例えばFLAGタグやMycタグ等の他のタンパク質又はペプチドとの融合タンパク質として発現させるものでもよい。
【0048】
本発明の分化誘導剤は、さらに、以下の(A1)〜(D1)の少なくとも一種を含んでいてもよい。
(A1)Scxタンパク質、
(B1)前記Scxタンパク質のアミノ酸配列において、1〜数個のアミノ酸が欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも一種の変異を有し、かつ、I型コラーゲンを誘導する活性を有する変異体、
(C1)前記(A1)又は(B1)のタンパク質をコードする核酸、並びに
(D1)前記(C1)の核酸を含む発現ベクター。
【0049】
Scxタンパク質は、前述のとおり、腱の主成分であるI型コラーゲンを直接誘導することが示唆されている(非特許文献1)。Scxノックアウトマウスは、腱を部分的に消失する。一方、Mkxノックアウトマウスは、実施例に示すように、腱の低形成は認められるが、腱細胞数は変わらない。このことから、Scx腱細胞分化の初期段階から必須の因子であり、Mkxは、後期分化・成熟に重要であることが予想される。また、腱細胞の分化とそこでのI型コラーゲンの産生には、Scx遺伝子による初期分化と、Mkxによる成熟の両者の協同作業が重要といえる。したがって、本発明の分化誘導剤に(A1)〜(D1)のScx関連物質を含めることは、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化をより効率的に行う点で好ましい。
【0050】
Scxタンパク質に関する情報は、NCBIからアクセッション番号NP_001008272として入手することができる。また、Scx遺伝子に関する情報は、NCBIからアクセッション番号NM_001008271として入手可能である。
【0051】
(A1)〜(D1)の作製方法は、タンパク質及び遺伝子がMkxと異なる以外は(A)〜(D)で述べた内容と同じである。
【0052】
本発明は、また、腱及び/又は靭帯細胞への分化能を有する細胞の生育に適した条件下で、上記分化誘導剤を用いて前記細胞を培養する工程、及び前記培養された細胞の中から、腱及び/又は靭帯細胞を選択する工程を含む、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する方法を提供する。
【0053】
前記腱及び/又は靭帯細胞への分化能を有する細胞は、例えば、繊維芽細胞(C3H10T1/2等)、腱前駆細胞、靱帯前駆細胞、胚性幹細胞、胚性生殖細胞、体性幹細胞、iPS細胞等が挙げられる。
【0054】
以下に、分化誘導方法の一態様を説明する。まず、腱及び/又は靭帯細胞への分化能を有する細胞に分化誘導剤を導入して、それを培養する。具体的には、C3H10T1/2細胞をDMEM培地等で培養して得られる50%コンフルエントのC3H10T1/2細胞に、Mkx発現ベクターをトランスフェクションする。トランスフェクションは、公知の方法で行え、またトランスフェクション以外の導入法として例えばウィルスベクターによる導入法、エレクトロポレーション法、リン酸カルシウム沈殿法、マイクロインジェクション法、リポソーム法等が挙げられる。
【0055】
培養方法は、分化誘導方法に適したものであれば、特に制限されない。例えば、平面培養法、浮遊凝集培養法、懸濁培養法、フィーダー細胞との共培養法、旋回培養法、軟寒天培養法、マイクロキャリア培養法等が挙げられる。
【0056】
培養後、回収した細胞を細胞溶解バッファーで溶解して全可溶型タンパク質を取得する。全可溶型タンパク質をウエスタンブロット等にかけ、I型コラーゲンの存在を確認することが好ましい。
【0057】
上記分化誘導剤を用いてMkx遺伝子を過剰発現させると、腱の主要コンポーネントであるI型コラーゲンの発現が上昇する。損傷した腱部位に分化誘導剤や分化誘導剤を用いて製造した腱細胞及び/又は靱帯細胞を導入すれば、腱の治癒、運動器の再生や再建を促進することが期待できる。そこで、本発明は、上記分化誘導剤、又は上記分化誘導剤を用いて製造した腱及び/又は靭帯細胞を含む、腱又は靭帯関連疾患を予防又は治療するための医薬を提供する。
【0058】
本発明の分化誘導剤は、遺伝子治療用にも好適である。遺伝子治療の腱又は靭帯関連疾患の候補は、エーラスダンロス症候群がある。エーラスダンロス症候群は、ヒトの皮膚や組織を形成するコラーゲン等の結合組織成分が先天性代謝異常を示し、その結果、靱帯や関節の異常な可動性亢進等を引き起こす疾患である。現在、根本的治療法は見つかっていない。エーラスダンロス症候群で腱及び/又は靭帯が罹患している患者から採取した細胞を用いて、分化誘導剤で自家腱及び/又は靭帯細胞を製造することにより、該疾患に対する再生医療が期待される。
【0059】
上記医薬の投与形態は、特に限定されないが、好ましくは損傷した腱及び/又は靱帯部位への直接投与である。具体的には、皮下注射剤、筋肉注射剤等の注射剤、点滴剤、経皮投与剤、軟膏剤等の外用薬剤、細胞の単層又は複層のシート等が挙げられる。
【0060】
これらの剤形に適用される助剤や製剤技術には当業分野に公知のものを使用可能である。例えば、注射剤であれば、適当な緩衝剤、安定剤、保存剤、無痛化剤、等張化剤、溶解補助剤等が挙げられる。
【0061】
本発明の医薬は、ヒトを含む動物に対して、薬理上必要量が投与される。薬理上必要量は、剤形、投与方法、患者や実験動物の症状、治療部位、患者や実験動物の年齢、性別、体重等を考慮して、医師等の判断により適宜決められる。
【0062】
本発明は、また、Mkx遺伝子の機能的欠損を含む、腱又は靱帯関連疾患の研究用の遺伝子改変非ヒト動物を提供する。「Mkx遺伝子の機能的欠損」とは、本明細書において、Mkx遺伝子が本来有する正常な機能を十分に発揮しないか、全く発揮しない状態を意味する。
【0063】
遺伝子改変非ヒト動物には、Mkx欠損遺伝子のヘテロ接合体(Mkx+/−と表示することがある)とホモ接合体(Mkx−/−と表示することがある)の二種類がある。本明細書中、「Mkx変異」(例えばMkx変異マウス)の記載は、両者を含む意味で使用する。
【0064】
ノックアウトマウスのようなホモ接合体は、遺伝子改変非ヒト動物の腱及び/又は靭帯が野生型動物と比べて有意に低形成を示す。具体的には、Mkxノックアウトマウスは、全身の腱が低形成になり、尾腱の線維束が細くなるにもかかわらず、腱細胞の数は野生型と同程度である。さらに、Mkxノックアウトマウスの腱は、コラーゲン原線維の径が小さく、I型コラーゲンの量が低下する。以上の結果から、Mkxは、腱細胞において、I型コラーゲンの産生能に寄与し、腱細胞の分化に重要な役割を担っていることが示唆される。この知見に基づく本発明のMkxノックアウトマウスは、Mkxの腱及び/又は靭帯細胞への分化誘導への研究や腱又は靭帯関連疾患を予防又は治療するための医薬の探索に有用である。
【0065】
一方、ヘテロマウスのようなヘテロ接合体は、腱形成に異常が見られない。ヘテロマウスは、腱に異常がなく、腱や靭帯などMkxが発現する細胞で蛍光タンパク質のVenusが発現するので、Mkxがどこで発現しているか、Mkxがどのような物質や遺伝子で発現が誘導されるか等の解析や、腱組織がどのようにできるかの観察、腱細胞を分離して細胞の性状を調査する等に有用である。
【0066】
非ヒト動物の例は、マウス、ラット、ハムスター、モルモット、チンパンジー、サル、ウサギ等の実験動物、ブタ、ウシ、ヒツジ等の家畜類、イヌ、ネコ等の愛玩動物であり、好ましくはマウスである。
【0067】
Mkx遺伝子を機能的に欠損させる方法は、当業分野で公知の方法のいずれも採用することができる。具体的には、Mkx遺伝子を含むゲノムDNAのエキソンを含む領域をターゲッティングベクターで相同組み換えする方法が例示される。
【0068】
Mkx遺伝子を機能的に欠損させる際に、Mkx遺伝子のプロモーターの下流にレポーター遺伝子が挿入されたことにより腱細胞特異的にレポーター遺伝子を発現させることが好ましい。ヘテロ接合体のMkxヘテロマウスのMkx欠損部位を蛍光タンパク質等のレポーター遺伝子で置き換えると、レポーター遺伝子が腱細胞特異的に発現するとともに、正常な腱細胞もまた視認できる(図1D)。このようなMkxヘテロマウスは、Mkxの腱及び/又は靭帯細胞への分化誘導への研究や、腱又は靭帯関連疾患を予防又は治療するための医薬の探索に極めて有用である。
【0069】
前記レポーター遺伝子には、Venus、BFP、CFP、dsRed、EBFP、ECFP、EGFP、EYFP、FlAsH、GFP、HcRed1、mRFP1、mPlum、PhiYFP、RFP、YFP等の蛍光タンパク質、ルシフェラーゼ遺伝子、カタラーゼ遺伝子、LacZ遺伝子等を使用することができる。
【0070】
さらに、Mkx遺伝子の下流に、ネオマイシン耐性遺伝子等の選択マーカーを挿入することが好ましい。
【0071】
上記選択マーカーの両端をloxP配列で挟んでおくことがより好ましい。こうすると、Cre−loxPシステムを用いて、選択マーカーの除去が可能である。
【0072】
遺伝子改変非ヒト動物は、それ自体公知の方法により製造することができる。具体的には、
1.Mkx遺伝子の機能的欠損を含む胚性幹細胞クローンを作製する工程、
2.上記胚性幹細胞クローンを胚にインジェクションする工程、
3.キメラ胚を動物の子宮に移して、キメラ動物を作る工程、
4.キメラ動物を交配させ、ヘテロ接合体を得る工程、及び
5.ヘテロ接合体同士を交配させ、ホモ接合体を得る工程、
を含む。
【0073】
Mkxヘテロマウス及びMkxノックアウトマウスを例に、遺伝子改変非ヒト動物をより具体的に説明する。ゲノムDNAのMkx遺伝子を欠損させるために、胚性幹細胞での相同組み換えにおいてMkx遺伝子の部分領域(エキソンの一部を含む)を、例えば蛍光タンパク質遺伝子とポジティブ選択マーカー(例えばネオマイシン耐性遺伝子、ピューロマイシン耐性遺伝子等)とのカセットに置き換わるようなターゲッティングベクターを設計する(図1A)。また、ポジティブ選択マーカー遺伝子の両側にCre/loxPシステムで用いられるloxP配列のようなリコンビナーゼ標的配列を加えると、Cre酵素を介したマーカー遺伝子の切除が可能である。ネガティブ選択用にジフテリアトキシンA(DT−A)遺伝子、チミジンキナーゼ(TK)遺伝子等を組み込む。
【0074】
上記のターゲッティングベクターを、胚性幹細胞クローンや、初代培養細胞株等の体細胞へ導入する。胚性幹細胞クローンは、マウス、ヒト、ハムスター、ブタ等の動物の胚盤胞から分離した内部細胞塊をフィーダー細胞上で培養することにより樹立することができる。市販の胚性幹細胞クローンを使用することができ、例えばTT2、ES−D3、SCC−PSA1等が挙げられる。
【0075】
ターゲッティングベクターの組み込まれた胚性幹細胞等を、ネオマイシン耐性選択ならG418抗生物質を含む培地といった適宜の選択培地で培養後、組換え胚性幹細胞クローンを分離する。
【0076】
適切に組み込まれた組換え胚性幹細胞クローンのスクリーニングを、サザンブロットやPCT法により行う。図5Aに、サザンブロット解析のための野生型(WT)及びターゲッティングされたアリル(TA)と各プローブの位置を示す。
【0077】
相同組み換えの確認された胚性幹細胞クローンを、マイクロインジェクション法、凝集法等で8細胞期等のマウス胚に導入してキメラ胚を作る。
【0078】
得られたキメラ胚を動物の子宮に移してキメラ動物を作る。得られたキメラ動物を野生型動物(例えばC57BL/6マウス)と掛け合わせる。生殖細胞への伝達をゲノミックPCRやサザンブロット解析により確認する。LoxP配列に挟まれたネオマイシン耐性遺伝子カセットを、Meox−Creトランスジェニック動物との掛け合わせにより除去する。Creを介したネオマイシン耐性遺伝子除去をゲノミックPCRで確認する。キメラ動物を野生型動物と交配させ、Mkxヘテロ接合体を得る。
【0079】
Mkxヘテロ接合体同士を交配させて、ホモ接合体(Mkxノックアウトマウス)を得る。
【0080】
本発明は、また、上記分化誘導剤、上記分化誘導方法により得られる腱及び/又は靭帯細胞、若しくは遺伝子改変非ヒト動物の腱及び/靱帯、それらの組織又は細胞を用いることを特徴とする、腱又は靭帯関連疾患を解析又は分析する方法を提供する。分析や解析の対象は、細胞に分化誘導剤を作用させ分化させて得られるMkxヘテロマウスの胚とその組織、トランスジェニック非ヒト動物からを得られた腱及び/靱帯、それらの組織又は細胞等である。分析や解析の手法を以下に例示する。
【0081】
ホールマウントin situハイブリダイゼーション(WISH)は、マウス胚での遺伝子発現パターンの解析に有効である。WISHは、常法を用いることができ、例えばYokoyama S, et al.(2008)Dynamic gene expression of Lin−28 during embryonic development in mouse and chicken. Gene Expr Patterns 8(3):155−160に記載の方法に従う。これに用いるプローブの合成方法の詳細は、EMBRYSウェブサイト(http://embrys.jp/html/MainMenu.html?)にて閲覧可能である。
【0082】
Venus等の蛍光タンパク質は、Mkx遺伝子の各組織での発現パターンを観察するのに有効である。
【0083】
免疫組織染色は、マウス胚や成体マウスの各組織の観察に有効である。各組織に特異的なマーカーとして、筋肉ではミオシン重鎖、血管ではPECAM1等がある。これらのマーカー遺伝子の発現を、マーカー特異的ポリクローナル又はモノクローナル抗体を用いて免疫組織染色する。まず、マウス胚をパラホルムアルデヒドに固定した組織を凍結組織切片作製用包埋剤に包埋して凍結する。検体を薄切りして風乾後、ブロッキング剤にて1時間ブロッキングする。その切片を抗ミオシン重鎖抗体(MF20,DSHB)でインキュベートし、さらに、二次抗体と共にインキュベートする。VenusノックインMkxへテロマウスでは、ミオシン重鎖抗体(MF20)の赤い染色とVenusの緑色シグナルの両方が観察される。
【0084】
Mkxノックアウトマウスの腱の異常を調査するために、腱及び靱帯の組織学的観察を行うことが有効である。その手法として、以下に電子顕微鏡観察、H&E染色及びアザン染色を例示する。
【0085】
電子顕微鏡を用いると、Mkxノックアウトマウスの腱におけるコラーゲン原線維を観察できる。まず、アキレス腱断片を採取し、緩衝液にて固定する。緩衝液及び生理食塩水で洗浄し、エタノールにて脱水後、樹脂に包埋して極薄切りにする。得られた切片を銅の格子上に置き、透過型電子顕微鏡にて観察する。
【0086】
H&E染色から、野生型マウスやMkxノックアウトマウスの尾腱の細胞密度を求めることができる。まず、成体の膝、尾等の組織を採取し、パラホルムアルデヒド等に固定する。組織を、脱水後、パラフィン包埋し、薄切りし、ヘマトキシリン・エオシン(H&E)で染色する。
【0087】
アザン染色は、コラーゲンを検出する。まず、マウスの尾を採取し、4%パラホルムアルデヒドに固定する。組織を、脱水後、パラフィン包埋し、薄切りし、アゾカルミン−アニリンブルー(アザン)で染色する。
【0088】
Mkx欠損した腱の力学的特性を調べるには、例えば筋腱ジャンクションから踵結節までの全腱を用いて引張試験を行う。張力の決定は、Gentleman E, et al.(2003) Mechanical characterization of collagen fibers and scaffolds for tissue engineering. Biomaterials 24(21):3805−3813で報告された方法に従って、単軸の材料テストシステム(Autograph AGS−G, Shimadzu Corp. Ltd.)を使用するとよい。それぞれの検体の最初の長さと断面積を、デジタルカリパスを使用して、H&E染色した切片の顕微鏡検査によって測定する。データ収集には、例えばTrapeziumソフトウェア(登録商標、Shimadzu社)を用いる。各検体の応力−歪み曲線は、load−displacementカーブから作られ、ヤング率はそれぞれの応力−歪み曲線と腱断面積から算出される。
【0089】
本発明は、また、被験物質の存在下又は非存在下で、上記分化誘導方法を行い、該被験物質存在下又は非存在下での腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を比較することを特徴とする、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質をスクリーニング又は評価する方法を提供する。
【0090】
好ましくは、被験物質の存在下で培養した細胞における、Mkxの発現量及び/又はMkxをコードする塩基配列に対するmRNAの転写量を測定すること、並びに前記被験物質を非存在下で培養した細胞内の量と比べることを含む。測定量が多いか又は少ない場合に、前記被験物質を腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質として選択する。
【0091】
上記培養培地は、用いる細胞に応じて適宜決められる。例えば、MEM培地、DMEM培地、RPM1640培地、199培地等である。培養条件も、細胞に応じて適宜、決められる。
【0092】
本発明は、また、上記遺伝子改変非ヒト動物から得られる腱、その組織又はその細胞を用いることを特徴とする、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質をスクリーニング又は評価する方法を提供する。好ましくは、遺伝子改変非ヒト動物から採取した腱、その組織又はその細胞に被験物質を接触させること、及びノックインされた蛍光タンパク質等のレポーター遺伝子の発現を上昇又は低下させる被験物質を選択することを含む。
【0093】
上記スクリーニング又は評価する方法に供する被験物質は、特に制限されない。例えば、天然又は人工の有機化合物や無機化合物、核酸、ペプチド、タンパク質、糖質、脂質、細胞、植物、動物等の抽出物等である。
【0094】
被験物質と分化誘導させる細胞や遺伝子改変非ヒト動物の組織等との接触は、特に制限されないが、添加、トランスフェクション、ウィルスベクターによる導入等である。
【0095】
Mkxの発現量及び/又はMkxをコードする塩基配列に対するmRNAの転写量、レポーター遺伝子の発現の上昇又は低下の測定は、常法に基づく、例えばノーザンブロット法、リアルタイムPCR、RT−PCR、ウエスタンブロット法、免疫学的手法、 等である。
【実施例】
【0096】
以下に、実施例を用いて本発明をより詳細に説明する。しかし、本発明は、以下の実施例に限定されるものではない。
〔実施例1〕(分化誘導剤の作製と評価)
Mkxタンパク質をコードする核酸を含む発現ベクターからなる分化誘導剤を、C3H10T1/2マウス線維芽細胞に導入してMkxを過剰発現させることにより、分化誘導剤の効果を調べた。
【0097】
まず、マウスMkx遺伝子(NCBIアクセッション番号NM_177595)の全タンパク質コード領域をp3XFLAG−CMVTM−7.1ベクター(SIGMA社)に組み込むことにより、FLAG−Mkx発現ベクターを作製した。また、FLAG−Mkx遺伝子を組み込まない空ベクターを陰性対照として用いた。
【0098】
C3H10T1/2細胞(ATDCから入手)を、10%FBSを含むDMEM培地で培養した。6ウエルプレートにて培養した50%コンフルエントのC3H10T1/2細胞に、トランスフェクション試薬(製品名、FugeneHD、Roche社)10μl、及び上記FLAG−Mkx発現ベクター又は空ベクター1μgをトランスフェクションした。
【0099】
48時間培養後、細胞を回収した。RIPAバッファー(50mM Tris−HCl, 150mM NaCl,0.5%DOC,0.1%SDS,1%NP40;pH 8.0)で細胞を溶解して、C3H10T1/2細胞中の全可溶型タンパク質を取得し、以下のウエスタンブロット解析に供した。
【0100】
取得した全可溶型タンパク質をSDS−PAGEにより分離し、PVDF膜にセミドライ法でブロットした。メンブレンをブロッキング剤(製品名Blocking One、Nacalai Tesque社)にて30分間ブロッキングし、抗I型コラーゲン抗体又は抗β−アクチン抗体と共に4℃、一晩インキュベートした。
【0101】
洗浄後、西洋ワサビペルオキダーゼ(HRP)標識抗ウサギイムノグロブリンG(IgG)抗体(A6154,SIGMA社)、又はHRP標識抗マウスIgG抗体(A2304,SIGMA社)と共に1時間インキュベートした。抗原−抗体の発色には、化学発光試薬(製品名Chemi−Lumi One、ナカライテスク社)を用いた。
【0102】
図4Dに、FLAG−Mkx発現ベクター又は空ベクターをトランスフェクションしたC3H10T1/2細胞のI型コラーゲン(Col1)、FLAG及びβ−actinのウエスタンブロットの結果を示す。図4Dに示すとおり、C3H10T1/2細胞内にMkxを過剰発現させると、I型コラーゲン量が増大した。
【0103】
〔実施例2〕(Mkx変異マウスの作製と評価)
マウスの染色体上のMkx遺伝子を機能的に欠損させるために、図1Aに、胚性細胞での相同組み換えにおいてMkxの翻訳スタートからエキソン2の終わりまでをMkx遺伝子がVenus遺伝子とネオマイシン耐性遺伝子とのカセットで置き換わるようなターゲッティングベクターを設計した。
【0104】
図1Aのターゲッティングコンストラクトをもとに、C57BL/6ゲノムBACクローン(BACPAC Resource Center社)を鋳型にしたPCRを行うことによって、Mkx遺伝子の5‘及び3’側の隣接領域を増幅した。これらの相同組み換え領域を、Venus遺伝子とポジティブ選択用のネオマイシン耐性遺伝子とネガティブ選択用のジフテリアトキシンA(DT−A)遺伝子を含むpBluescript由来のベクターに組み込むことにより、ターゲッティングベクターを作製した。
【0105】
上記のターゲッティングベクターを制限酵素NotIで直鎖化し、TT2F胚性幹細胞へエレクトロポレーション法により導入した。ターゲッティングベクターの組み込まれたTT2F胚性幹細胞を、G418を含む培地で培養して、組換え胚性幹細胞クローンを分離した。
【0106】
適切に組み込まれた組換え胚性幹細胞クローンのスクリーニングを、5‘プローブ、3’プローブ及びneoプローブを用いたサザンブロット法により行った。
【0107】
図5Aに、サザンブロット解析のための、野生型(WT)及びターゲッティングされたアリル(TA)と各プローブの位置を示す。図5Bに、組換え胚性幹細胞のサザンブロットの結果を示す。図5Cに、野生型マウス及びMkx変異マウスのサザンブロットの結果を示す。
【0108】
相同組み換えの確認された2つの胚性幹細胞クローンを8細胞期のマウス胚にインジェクションした。得られたキメラマウスをC57BL/6マウスと掛け合わせ、生殖細胞への伝達をゲノミックPCRにより確認した。LoxP配列に挟まれたネオマイシン耐性遺伝子カセットを、Meox−Creトランスジェニックマウスとの掛け合わせにより除去し、Creを介したネオマイシン耐性遺伝子除去の確認をゲノミックPCRにより行った。ジェノタイピングに使用したPCRプライマーの配列を表1に記載する。
【0109】
【表1】
【0110】
図1Bに、野生型マウスとMkx変異マウスのプライマーa、b及びcを用いたゲノミックPCR結果を示す。
【0111】
ターゲッティングベクターを用いて作製されたVenusノックインMkxヘテロマウスは、Mkxの発現が陽性であり、かつ、ノックインしたVenus遺伝子の発現もまた予想される。そこで、マウス胚の蛍光シグナルの観察を行った。Mkxプローブを用いたホールマウントin situハイブリダイゼーションを行った。
【0112】
図1Cに、E13.5前肢(Forelimb)及び尾(Tail)における野生型マウスとMkxヘテロマウスの胚のMkxのホールマウントin situ ハイブリダイゼーションを示す。VenusノックインMkxヘテロマウスの前肢と尾では、Mkxの発現と同様の場所で、Venusの発現が観察された。
【0113】
Mkxヘテロマウス胚の免疫組織染色のために、E16.5マウス胚の尾を採取し、4%パラホルムアルデヒドに4℃で2時間固定した。組織を、凍結組織切片作製用包埋剤(製品名O.C.T.compound、Sakura Finetek社)に包埋し、液体窒素で直ちに凍結した。検体を10μmに薄切りし、その凍結切片を風乾後、Blocking One(Nacalai Tesque社)にて1時間ブロッキングした。その切片を抗ミオシン重鎖抗体(MF20,DSHB)と共に4℃、一晩インキュベートした。洗浄後、二次抗体蛍光試薬(製品名Molecular Probes Alexa 594)と共に1時間インキュベートした。
【0114】
図1Dに、E16.5のVenusノックインMkxへテロマウスの胚の尾における抗ミオシン重鎖抗体(MF20;red)を用いた免疫組織染色とVenus蛍光シグナルを観察した顕微鏡写真を示す。図中、○で囲んだ部分はMF20免疫組織染色(実際は赤色に呈色)を示し、それ以外はVenus蛍光シグナル(実際は緑色に呈色)を示す。VenusノックインMkxヘテロマウスの胚は、E16.5の尾において腱特異的なVenusの発現を示した。
【0115】
Mkxヘテロマウス胚の解析に加え、その成体マウスにおけるVenusの発現についても調査した。成体の腱でのMkxの発現は明らかではなかったので、成体の腱でのMkxの発現をRT−PCRにより調査した。図6Aに、野生型マウス(成体)の筋肉、アキレス腱及び尾腱におけるMkx及びGapdhのRT−PCR解析を示す。図6Aから、Mkxがアキレス腱及び尾腱で強く発現していることを確認した。
【0116】
図6Bに、Mkxヘテロマウス及び野生型マウスのアキレス腱(B−E)、尾腱(F−I)及び体幹の腱(J−M)におけるVenusの発現と外観写真を示す。図6Bから、MkxヘテロマウスにおけるVenusは、アキレス腱、尾腱及び体幹の腱において特異的発現を示した。
【0117】
Mkx遺伝子の欠損を確認するために、Mkx変異マウスを用いたRT−PCRによる発現解析を行った。核酸抽出薬(製品名ISOGEN、Nippongene社)を用いて全RNAを分離した。分離した全RNAから、cDNA合成キット(製品名Ready−To−Go You−Prime First−Strand Beads、GE Healthcare社)を用いて逆転写し、そのcDNAを鋳型にDNAポリメラーゼ(製品名Go−Taq(登録商標)、Promega社)を用いてPCRした。プライマー配列を表2に示す。Gapdhは、mRNAのコントロールとして用いた。
【0118】
【表2】
【0119】
図1Eに、野生型マウスとMkx変異マウスのアキレス腱におけるMkx及びGapdhのRT−PCRの解析結果を示す。図1Eから、Mkxノックアウトマウス(成体)のアキレス腱において、Mkxの発現が完全に消失していることがわかった。
【0120】
以上の結果から、Mkx変異マウス(VenusノックインMkxヘテロマウス及びMkxノックアウトマウス)は、Mkxの機能解析だけでなく、腱の生物学的解析やMkxの発現解析に有用であることが示された。
【0121】
〔実施例3〕(Mkxノックアウトマウスの作製と評価)
実施例2で作製したMkxヘテロマウス同士を掛け合わせることにより、Mkxノックアウトマウスを作製した。産まれた各遺伝子型の新生仔マウスの数と比を表3に示す。
【0122】
【表3】
【0123】
ヘテロマウス同士の掛け合わせによって産まれた206匹の新生仔マウスのジェノタイピングの結果は、メンデルの法則に従った。
【0124】
Mkxヘテロマウス及びMkxホモマウス(ノックアウトマウス)は、問題なく生存、生殖し、重量測定の結果も野生型マウスと変わらなかった(図7)。
【0125】
Mkxヘテロマウスでは、腱の異常は見られなかった(図6B)。腱形成におけるMkxの役割を調査するために、Mkxノックアウトマウスで腱の異常が観察されるか否かを調べた。
【0126】
図2Aに、野生型マウスにおいて、膝蓋腱(Aの矢印)、アキレス腱(Cの矢印、後肢(Eの矢印)、前肢の腱(Gの矢印)、尾腱(Iの矢印)、背中の腱(Kの矢印)及び広頸筋腱(Mの矢印)を観察した写真、並びにMkxノックアウトマウスにおいて、膝蓋腱(Bの矢印)、アキレス腱(Dの矢印、後肢(Fの矢印)、前肢の腱(Hの矢印)、尾腱(Jの矢印)、背中の腱(Lの矢印)及び広頸筋腱(Nの矢印)を観察した写真を示す。Mkxノックアウトマウスでは、膝蓋腱、アキレス腱、尾腱、四肢の腱、体幹の腱及び広頸筋腱が、野生型マウスに比べて低形成であり、白色が薄くなっていた。この腱の異常は、7日齢から8ヶ月齢までのMkxノックアウトマウスとネオマイシン耐性遺伝子を除去したマウス(図8)においても観察された。
【0127】
Mkxが欠損した腱の力学的特性を調べるために、筋腱ジャンクションから踵結節までの全腱を用いて引張試験を行った。張力の決定は、Gentleman E, et al. (2003) Mechanical characterization of collagen fibers and scaffolds for tissue engineering. Biomaterials 24(21):3805−3813で報告された方法に従って、単軸の材料テストシステム(Autograph AGS−G、島津社)を使用して行った。検体を、0.5 mm/secの重圧率で引張した。
【0128】
それぞれの検体の最初の長さと断面積を、デジタルカリパスを使用して、H&E染色した切片の顕微鏡検査によって測定した。データ収集はTrapezium(登録商標、島津社)ソフトウェアを用いて行った。各検体の応力−歪み曲線は、load−displacementカーブから作られ、ヤング率はそれぞれの応力−歪み曲線と腱断面積から算出した。
【0129】
図2Bに、野生型マウスとMkxノックアウトマウスのアキレス腱の引張強度を示す。Mkxノックアウトマウスのアキレス腱は、引張強度が低く、機能低下が見受けられた。図2Cに、野生型マウスとMkxノックアウトマウスのアキレス腱のヤング率を示す。単位面積あたりの引張強度及び単位面積あたりの弾力性を示すヤング率の値は、野生型マウスとMkxノックアウトマウスでほぼ同程度であった。以上の結果から、Mkxノックアウトマウスのアキレス腱における引張強度の低下は、腱が細くなることに起因していることが示唆された。
【0130】
Mkxノックアウトマウスの腱の異常をさらに詳細に調査するために、組織学的解析を行った。成体マウスの膝又は尾を採取し、4%パラホルムアルデヒドに4℃で一晩固定した。組織を、脱水後、パラフィン包埋し、薄切りし、ヘマトキシリン・エオシン(H&E)で染色した。
【0131】
図3Aに、3ヶ月齢の野生型マウスとMkxノックアウトマウスの膝蓋腱のH&E染色像を示す。H&E染色の結果、3ヶ月齢のMkxノックアウトマウスの膝蓋腱は、野生型マウスに比べて薄いことが分かった。
【0132】
図3Bに、7日齢の野生型マウスとMkxノックアウトマウスの膝蓋腱の厚さを示す。7日齢のノックアウトマウスにおいても、膝蓋腱が薄く、野生型マウスの腱の約半分の厚さになっていた(図3B)。
【0133】
図9に、野生型マウス及びMkxノックアウトマウスの膝十字靭帯のH&E染色像を示す。腱と同様の組成を持つ組織である靭帯の1つである膝十字靭帯においては、野生型マウスとMkxノックアウトマウスとの間に差は見られなかった。
【0134】
図3Cに、3ヶ月齢の野生型マウスとMkxノックアウトマウスの尾腱のH&E染色像を示す。Mkxノックアウトマウスの尾腱でも、腱線維束の径の縮小(図3Cの△矢頭)が見られた。
【0135】
図3Dに、3ヶ月齢の野生型マウスとMkxノックアウトマウスの尾腱の細胞密度を示す。Mkxノックアウトマウスの尾腱では、細胞密度が高く、1つの尾腱線維束における細胞数は野生型マウスと同程度であった。このことから、Mkxノックアウト腱細胞は、機能的に不完全であることが示唆される。
【0136】
Mkxノックアウトマウスにおける腱の形成不全が胎生期においても観察されるか否かを調査するため、E18.5のMkx欠損胚の尾腱のアザン染色を行った。具体的には、尾を採取し、4%パラホルムアルデヒドに4℃で一晩固定した。組織を、脱水後、パラフィン包埋し、薄切りし、アゾカルミン−アニリンブルー(アザン)で染色した。
【0137】
図3Eに、E18.5の野生型マウスとMkxノックアウトマウスの胚の尾腱のアザン染色像を示す。図3Eからは、尾腱の大きさは、野生型マウスと違いが見られない。Mkxノックアウト胚では、コラーゲンを検出するアニリンブルーの染色の密度が低い(△矢頭部)ことが分かった。
【0138】
Mkxノックアウトマウスの腱におけるコラーゲン原線維について解析するため、アキレス腱を電子顕微鏡で観察した。約1mm3のアキレス腱断片を採取し、4%パラホルムアルデヒドと2.5%グルタルアルデヒドを含む0.1Mカコジル酸バッファーにて一晩固定した。0.1Mカコジル酸バッファー及び生理食塩水で洗浄し、エタノールにて脱水後、エポキシ樹脂に包埋して極薄切りにした。得られた切片を銅の格子上に置き、水性ウラニルアセテートとクエン酸鉛と対比させて、透過型電子顕微鏡(H−7100,日立社)にて観察した。
【0139】
図3Fに、野生型マウスとMkxノックアウトマウスのアキレス腱のコラーゲン原線維の電子顕微鏡観察像を示す。ノックアウトマウスの腱におけるコラーゲン原線維の径は、野生型マウスと比べて均一に小さいことが分かった。これらの結果から、Mkxノックアウトマウスの腱では、コラーゲンが減少していること、さらにはMkxが腱の分化に重要であることが示唆された。
【0140】
3ヶ月齢のMkxノックアウトマウスの尾腱の細胞密度(図3D)に示されるように、Mkxノックアウトマウスでは、腱の量が減少しているにもかかわらず、腱細胞の数は野生型マウスと変わらなかった。これは、Mkxノックアウト腱細胞において、細胞外基質の産生能が低下していることを示唆する。また、図3Eに示すE18.5のMkxノックアウトマウスの胚の尾腱のアザン染色像、並びに図3Fに示すMkxノックアウトマウスのアキレス腱のコラーゲン原線維の微細構造解析から、Mkxノックアウトマウスの腱ではコラーゲン産生能が減少していると考えられる。
【0141】
Mkxノックアウトマウスの腱細胞のコラーゲン産生能を調査するため、8週齢の野生型マウス及びMkxノックアウトマウスの腱のコラーゲン量を測定した。コラーゲンの定量は、市販のコラーゲン定量キット(製品名Sircol Soluble Collagen Assay、Biocolor社)の説明書に従った。結果を図4Aに示す。ノックアウトマウスの腱のコラーゲン量は野生型マウスと比べて少ないことが分かった。
【0142】
図4Bに8週齢の野生型マウスとMkxノックアウトマウスのアキレス腱のI型コラーゲン(Col1)とβ−actinのウエスタンブロット解析を示す。腱の細胞外基質の主要成分であるI型コラーゲンの発現量が示すように、Mkxノックアウトマウスでのタンパク質及びmRNAの両方とも減少していた。
【0143】
8週齢の野生型マウスとMkxノックアウトマウスのアキレス腱のCol1a1、Col1a2及びScxのreal time PCRを、SYBR Green PCR Master Mix(Applied Biosystems社)を用いて行った。プライマー配列を表4に示す。Gapdhは、mRNAのコントロールとして用いた。
【0144】
【表4】
【0145】
delta−delta CT法により測定した相対発現量のグラフを図4Cに示す。図4Cでは、I型コラーゲンの発現を誘導することが示唆されている転写因子であるScxの発現は減少していなかった。これから、MkxノックアウトマウスにおけるI型コラーゲンの減少は、Scxの減少ではないことが示唆された。
【0146】
以上の結果から、Mkxは、腱細胞においてI型コラーゲンの発現を調節する重要な因子であることが実証された。
【特許請求の範囲】
【請求項1】
(A)Mkxタンパク質、
(B)前記Mkxタンパク質のアミノ酸配列において、1〜数個のアミノ酸が欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも一種の変異を有し、かつ、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する活性を有する変異体、
(C)前記(A)又は(B)のタンパク質をコードする核酸、並びに、
(D)前記(C)の核酸を含む発現ベクター
の少なくとも一種を有効成分として含む、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化誘導剤。
【請求項2】
(A1)Scxタンパク質、
(B1)前記Scxタンパク質のアミノ酸配列において、1〜数個のアミノ酸が欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも一種の変異を有し、かつ、I型コラーゲンを誘導する活性を有する変異体、
(C1)前記(A1)又は(B1)のタンパク質をコードする核酸、並びに、
(D1)前記(C1)の核酸を含む発現ベクター、
の少なくとも一種を含む、請求項1に記載の腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化誘導剤。
【請求項3】
腱及び/又は靭帯細胞への分化能を有する細胞の生育に適した条件下で、請求項1又は2に記載の分化誘導剤を用いて前記細胞を培養する工程、及び前記培養された細胞の中から、腱及び/又は靭帯細胞を選択する工程を含む、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する方法。
【請求項4】
前記腱及び/又は靭帯細胞への分化能を有する細胞が、繊維芽細胞、腱前駆細胞、靱帯前駆細胞、胚性幹細胞、胚性生殖細胞、体性幹細胞及びiPS細胞からなる群から選ばれる少なくとも一種である、請求項3に記載の腱及び/又は靭帯細胞への分化能を有する細胞を腱及び/又は靭帯細胞に分化誘導する方法。
【請求項5】
請求項1又は2に記載の腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化誘導剤を含む、腱又は靭帯関連疾患を予防又は治療するための医薬。
【請求項6】
腱又は靭帯関連疾患が、エーラスダンロス症候群である、請求項5に記載の腱又は靭帯関連疾患を予防又は治療するための医薬。
【請求項7】
請求項3又は4に記載の分化誘導方法により得られる腱及び/又は靭帯細胞を含む、腱又は靭帯関連疾患を予防又は治療するための医薬。
【請求項8】
Mkx遺伝子の機能的欠損を含む、腱又は靱帯関連疾患の研究用の遺伝子改変非ヒト動物。
【請求項9】
前記機能的に欠損したMkx遺伝子は、Mkx遺伝子のプロモーターの下流にレポーター遺伝子が挿入されたことにより腱細胞特異的にレポーター遺伝子を発現する、請求項8に記載の腱又は靱帯関連疾患の研究用の遺伝子改変非ヒト動物。
【請求項10】
前記レポーター遺伝子が、Venus遺伝子である、請求項9に記載の腱又は靱帯関連疾患の研究用の遺伝子改変非ヒト動物。
【請求項11】
請求項1又は2に記載の分化誘導剤を用いることを特徴とする、腱又は靭帯関連疾患の解析又は分析方法。
【請求項12】
請求項3又は4に記載の分化誘導方法により得られる腱及び/又は靭帯細胞を用いることを特徴とする、腱又は靭帯関連疾患の解析又は分析方法。
【請求項13】
請求項8、9又は10に記載の遺伝子改変非ヒト動物の腱及び/靱帯、それらの組織又は細胞を用いることを特徴とする、腱又は靭帯関連疾患の解析又は分析方法。
【請求項14】
被験物質の存在下又は非存在下で、請求項3又は4に記載の分化誘導方法を行い、該被験物質存在下又は非存在下での腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を比較することを特徴とする、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質をスクリーニング又は評価する方法。
【請求項15】
被験物質の存在下で培養した細胞における、Mkxの発現量及び/又はMkxをコードする塩基配列に対するmRNAの転写量を測定すること、並びに前記被験物質を非存在下で培養した細胞内の量と比べて、前記測定した量が多い又は少ない場合に、前記被験物質を腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質として選択することを含む、請求項14に記載の腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質をスクリーニング又は評価する方法。
【請求項16】
請求項8、9又は10記載の遺伝子改変非ヒト動物から得られる腱、その組織又はその細胞を用いることを特徴とする、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質をスクリーニング又は評価する方法。
【請求項17】
遺伝子改変非ヒト動物から得られる腱、その組織又はその細胞に被験物質を接触させること、及びノックインされたレポーター遺伝子の発現を上昇又は低下させる被験物質を選択することを含む、請求項16に記載の腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質をスクリーニング又は評価する方法。
【請求項1】
(A)Mkxタンパク質、
(B)前記Mkxタンパク質のアミノ酸配列において、1〜数個のアミノ酸が欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも一種の変異を有し、かつ、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する活性を有する変異体、
(C)前記(A)又は(B)のタンパク質をコードする核酸、並びに、
(D)前記(C)の核酸を含む発現ベクター
の少なくとも一種を有効成分として含む、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化誘導剤。
【請求項2】
(A1)Scxタンパク質、
(B1)前記Scxタンパク質のアミノ酸配列において、1〜数個のアミノ酸が欠失、置換、逆位、付加及び挿入からなる群から選ばれる少なくとも一種の変異を有し、かつ、I型コラーゲンを誘導する活性を有する変異体、
(C1)前記(A1)又は(B1)のタンパク質をコードする核酸、並びに、
(D1)前記(C1)の核酸を含む発現ベクター、
の少なくとも一種を含む、請求項1に記載の腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化誘導剤。
【請求項3】
腱及び/又は靭帯細胞への分化能を有する細胞の生育に適した条件下で、請求項1又は2に記載の分化誘導剤を用いて前記細胞を培養する工程、及び前記培養された細胞の中から、腱及び/又は靭帯細胞を選択する工程を含む、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を誘導する方法。
【請求項4】
前記腱及び/又は靭帯細胞への分化能を有する細胞が、繊維芽細胞、腱前駆細胞、靱帯前駆細胞、胚性幹細胞、胚性生殖細胞、体性幹細胞及びiPS細胞からなる群から選ばれる少なくとも一種である、請求項3に記載の腱及び/又は靭帯細胞への分化能を有する細胞を腱及び/又は靭帯細胞に分化誘導する方法。
【請求項5】
請求項1又は2に記載の腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化誘導剤を含む、腱又は靭帯関連疾患を予防又は治療するための医薬。
【請求項6】
腱又は靭帯関連疾患が、エーラスダンロス症候群である、請求項5に記載の腱又は靭帯関連疾患を予防又は治療するための医薬。
【請求項7】
請求項3又は4に記載の分化誘導方法により得られる腱及び/又は靭帯細胞を含む、腱又は靭帯関連疾患を予防又は治療するための医薬。
【請求項8】
Mkx遺伝子の機能的欠損を含む、腱又は靱帯関連疾患の研究用の遺伝子改変非ヒト動物。
【請求項9】
前記機能的に欠損したMkx遺伝子は、Mkx遺伝子のプロモーターの下流にレポーター遺伝子が挿入されたことにより腱細胞特異的にレポーター遺伝子を発現する、請求項8に記載の腱又は靱帯関連疾患の研究用の遺伝子改変非ヒト動物。
【請求項10】
前記レポーター遺伝子が、Venus遺伝子である、請求項9に記載の腱又は靱帯関連疾患の研究用の遺伝子改変非ヒト動物。
【請求項11】
請求項1又は2に記載の分化誘導剤を用いることを特徴とする、腱又は靭帯関連疾患の解析又は分析方法。
【請求項12】
請求項3又は4に記載の分化誘導方法により得られる腱及び/又は靭帯細胞を用いることを特徴とする、腱又は靭帯関連疾患の解析又は分析方法。
【請求項13】
請求項8、9又は10に記載の遺伝子改変非ヒト動物の腱及び/靱帯、それらの組織又は細胞を用いることを特徴とする、腱又は靭帯関連疾患の解析又は分析方法。
【請求項14】
被験物質の存在下又は非存在下で、請求項3又は4に記載の分化誘導方法を行い、該被験物質存在下又は非存在下での腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化を比較することを特徴とする、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質をスクリーニング又は評価する方法。
【請求項15】
被験物質の存在下で培養した細胞における、Mkxの発現量及び/又はMkxをコードする塩基配列に対するmRNAの転写量を測定すること、並びに前記被験物質を非存在下で培養した細胞内の量と比べて、前記測定した量が多い又は少ない場合に、前記被験物質を腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質として選択することを含む、請求項14に記載の腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質をスクリーニング又は評価する方法。
【請求項16】
請求項8、9又は10記載の遺伝子改変非ヒト動物から得られる腱、その組織又はその細胞を用いることを特徴とする、腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質をスクリーニング又は評価する方法。
【請求項17】
遺伝子改変非ヒト動物から得られる腱、その組織又はその細胞に被験物質を接触させること、及びノックインされたレポーター遺伝子の発現を上昇又は低下させる被験物質を選択することを含む、請求項16に記載の腱及び/又は靭帯細胞への分化能を有する細胞から腱及び/又は靭帯細胞への分化調節に関連する物質をスクリーニング又は評価する方法。
【図1A】

【図1B】

【図1C】
【図1D】

【図1E】

【図2A】

【図2B】
【図2C】

【図3A】
【図3B】
【図3C】

【図3D】
【図3E】

【図3F】
【図4A】
【図4B】

【図4C】
【図4D】
【図4E】

【図5A】
【図5B】
【図5C】

【図6A】
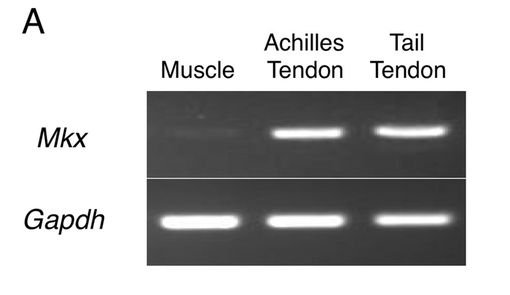
【図6B】
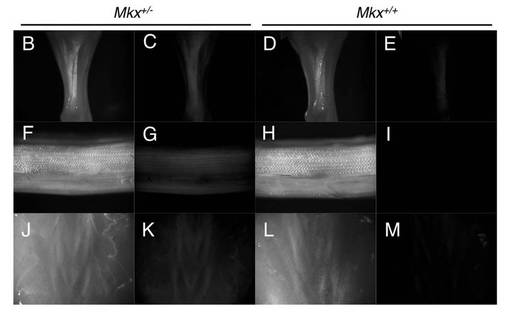
【図7】
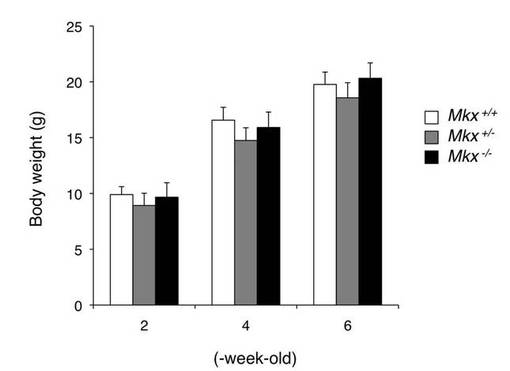
【図8】
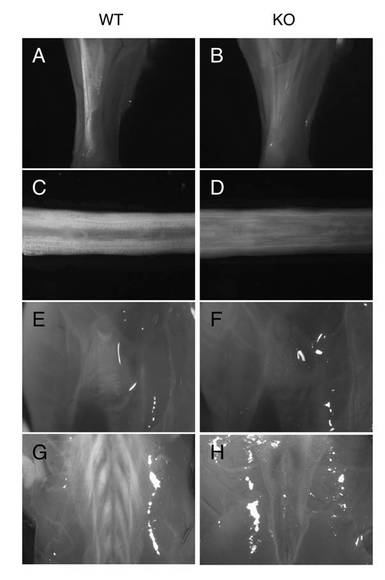
【図9】
【図1B】
【図1C】
【図1D】
【図1E】
【図2A】
【図2B】
【図2C】
【図3A】
【図3B】
【図3C】
【図3D】
【図3E】
【図3F】
【図4A】
【図4B】
【図4C】
【図4D】
【図4E】
【図5A】
【図5B】
【図5C】
【図6A】
【図6B】
【図7】
【図8】
【図9】
【公開番号】特開2011−205964(P2011−205964A)
【公開日】平成23年10月20日(2011.10.20)
【国際特許分類】
【出願番号】特願2010−76625(P2010−76625)
【出願日】平成22年3月30日(2010.3.30)
【出願人】(803000056)財団法人ヒューマンサイエンス振興財団 (341)
【Fターム(参考)】
【公開日】平成23年10月20日(2011.10.20)
【国際特許分類】
【出願日】平成22年3月30日(2010.3.30)
【出願人】(803000056)財団法人ヒューマンサイエンス振興財団 (341)
【Fターム(参考)】
[ Back to top ]