
花色変異体植物の作出方法
【課題】電離放射線照射による花色変異体植物の獲得頻度増大・花色変異幅拡大に寄与する新しい手段を提供する。
【解決手段】電離放射線照射によって花色変異体植物を作出する方法において、電離放射線の照射前に、植物をストレス処理することによって、電離放射線照射のみによる花色変異体植物の獲得頻度に比べ、2倍以上の頻度で花色変異体植物を獲得する。
【解決手段】電離放射線照射によって花色変異体植物を作出する方法において、電離放射線の照射前に、植物をストレス処理することによって、電離放射線照射のみによる花色変異体植物の獲得頻度に比べ、2倍以上の頻度で花色変異体植物を獲得する。
【発明の詳細な説明】
【技術分野】
【0001】
本願発明は、園芸植物等における花色変異体を効率よく作出するための方法に関するものである。
【背景技術】
【0002】
電離放射線は数十年にわたって新しい植物品種を作り出すために使われてきた。電離放射線照射による植物の突然変異の頻度を高めるためには、植物が生存し得る限り線量を上げてやればよい。また、酸素濃度、水分含量及び温度などの外的要因により、植物の放射線感受性を変えられることが知られており(非特許文献1)、その要因をコントロールすることで変異頻度を高めることも可能であろう。しかしながら、望ましい突然変異を起こすために線量を上げたり放射線感受性を高めたりすれば同時に生育不良等を引き起こしうる望まない変異も同時に増え、結果的に、望ましい変異のみを有する個体獲得は困難となる。よって、実際に電離放射線照射による植物の突然変異育種においては、できるだけ望ましい変異が起きる頻度を高めつつも不要な変異の誘発を避けるために、照射線量は植物と照射の条件に応じた、一定の低いレベルに留める必要がある。ただし、その場合には望ましい変異のみを持つ個体の獲得頻度は低いため、非常に多数の植物に照射し多数の個体から望む個体を選抜する必要があった。よって、効率的な突然変異個体獲得のためには、不要な変異の頻度を高めずに望まれる変異のみを誘発する手法が望まれている。
【0003】
永富ら(非特許文献2、3)は、キクにおける炭素イオンビームの変異効果を調査し、葉培養物に照射するよりも花弁培養物に照射した際に花色変異体が高い頻度で得られたと報告しており、植物の照射部位選択によって変異頻度が変わりうることを示した。
【0004】
なお、花色は園芸植物において最も重要な形質の1つであるが、この花色に関連するアントシアニン色素の生合成に関与する遺伝子の発現は、強光、紫外線、低温及び糖などのストレスによって活性化されることが知られている(非特許文献4、5、6、7)。ただし、このような色素の蓄積は植物体の遺伝形質に影響を及ぼすものではなく、たとえストレス暴露により花色を変化させたとしても、その形質が次世代の植物体に継続されることはない。
【非特許文献1】van Harten (1998) Mutation Breeding, Theory and Practical Applications (Cambridge University Press).
【非特許文献2】Nagatomi S, Tanaka A, Watanabe H, Tano S (1997) Chrysanthemum mutants regenerated from in vitro explants irradiated with 12C5+ion beam. Institute of Radiation Breeding, Technical News No.60.
【非特許文献3】Nagatomi S, Tanaka A, Kato A, Yamaguchi H, Watanabe H, Tano (1998) Mutation induction through ion beam irradiations in rice and chrysanthemum. JAERI-Review 98-016 / TIARA Annual Report 1997: 41-43.
【非特許文献4】Dubos C, Le Gourrierec J, Baudry A, Huep G, LanetE, Debeaujon I, Routaboul J M, Alboresi A, Weisshaar B, Lapiniec L (2008) MYBL2 is a new regulator of flavonoid biosynthesis in Arabidopsis thaliana. Plant J 55: 940-953.
【非特許文献5】Hase Y, Trung K H, Matsunaga T, Tanaka A (2006) A mutation in the uvi4 gene promotes progression of endo-reduplication and confers increased tolerance towards ultraviolet B light. Plant J 46: 317-326.
【非特許文献6】Olsen K M, Slimestad R, Lea U S, Brede C, Lovdal T, Ruoff P, Verhuel M, Lillo C (2009) Temperature and nitrogen effects on regulators and products of the flavonoid pathway: experimental and kinetic model studies. Plant Cell Environ 32: 286-299.
【非特許文献7】Tsukaya H, Ohshima T, Naito S, Chino M, Komeda Y (1991) Sugar-dependent expression of the CHS-A gene for chalcone synthase from petunia in transgenic Arabidopsis. Plant Physiol 97: 1414-1421.
【発明の概要】
【発明が解決しようとする課題】
【0005】
園芸植物等としての花色変異体植物の需要は大きい。これまで花色変異体植物の作出について様々な試みがなされているが、所望の変異体獲得の効率性については改善すべき点が残されている。
【0006】
本願発明は、電離放射線照射による花色変異体植物の獲得頻度増大に寄与する新しい手段を提供することを課題としている。
【課題を解決するための手段】
【0007】
本願発明者らは、花色変異体植物の獲得頻度を向上させる手段について検討した結果、植物へのストレス処理を施した状態で電離放射線を照射することにより、花色変異体植物の獲得頻度が大きく上昇し変異幅も拡がることを見出した。
【0008】
本願発明は、上記の新規知見に基づいて完成されたものである。
【0009】
すなわち、本願発明は、電離放射線照射によって花色変異体植物を作出する方法において、電離放射線の照射前に、植物をストレス処理することを特徴とする方法である。
【0010】
この花色変異体植物の作出方法においては、電離放射線照射のみによる花色変異体植物の獲得頻度に比べ、2倍以上の頻度で花色変異体植物が獲得されることを別の特徴とする。
【0011】
またこの作出方法においては、葉緑素変異体の出現頻度が、電離放射線照射のみによる当該変異体の出現頻度と同程度であることをさらに別の特徴としている。
【0012】
さらにこの作出方法においては、例えばストレス処理が植物への高濃度糖処理であるか、あるいは植物の低温での高濃度糖処理であることを具体的な実施形態としている。
【0013】
すなわち、本願発明は、以下の発明を包含する。
[1]電離放射線照射によって花色変異体植物を作出する方法において、電離放射線の照射前に、植物をストレス処理することを特徴とする方法。
[2]ストレス処理が、植物の高濃度糖処理または植物の低温処理と高濃度糖処理の組み合わせである[1]記載の方法。
[3]電離放射線がイオンビームまたはエックス線である[1]または[2]記載の方法。
【発明の効果】
【0014】
本願発明により、電離放射線による突然変異を利用した植物の花色変異を得る効率を高めることができる。本願発明で用いるストレス処理は、植物の放射線感受性を高めるものではなく、また、葉緑素変異株の獲得頻度が変化しないことから、花色の変異誘発に特異的に作用すると考えられる。従って、本発明によって、花色に関する植物育種において、目的とする花色変異のみを有する個体を獲得する効率を高めることが可能となる。
【図面の簡単な説明】
【0015】
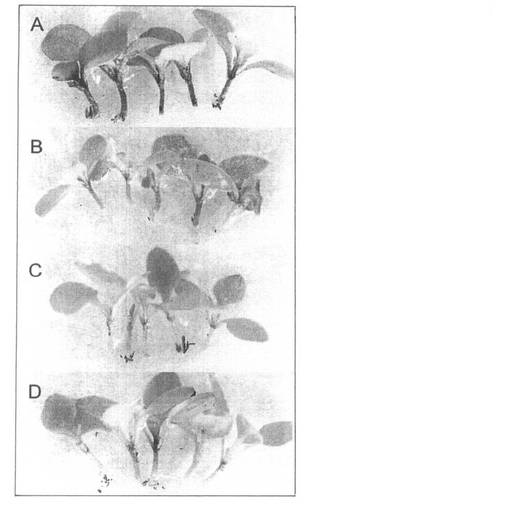
【図1】実施例1で確認したペチュニア幼苗における色素蓄積に対するストレス処理の効果。播種後8日目の幼苗を3%蔗糖(A)、15 kJ m-2の紫外線(B)、4℃・24時間(C)で処理した。写真はストレス処理後4日目の幼苗を示す。(D)には無処理の対照植物を示した。
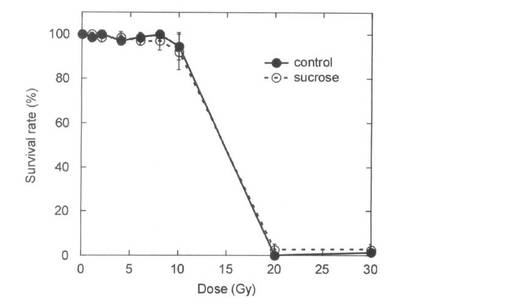
【図2】実施例1で確認した320 MeV炭素イオンビームを照射した播種後10日目のペチュニア幼苗の生存率の線量反応。終濃度3%となる蔗糖溶液(白丸)または水のみ(黒丸)を照射2日前に加えた。生存率は照射1ヶ月後に調査した。データは平均±標準偏差を示す。
【図3】ペチュニアの親系統ならびに実施例1で得られた花色変異体。(A)バイオレットの花色を持つ親系統。(B−H)花色変異体。(B)マゼンタ、(C)パープル、(D)パープルヴェイン、(E)ライトピンク、(F)ホワイト、(G)ブルーピコティー、(H)バーガンディー。
【図4】実施例2で得られたキク品種リネカーの純白変異体(左)と、実施例2の対照試験区で得られた微量のアントシアニンを持つ白色変異体(右)。
【図5】実施例2で得られた純白変異体から樹立された純白リネカー株。
【発明を実施するための形態】
【0016】
本願発明の花色変異体植物作出方法は、電離放射線照射によって花色変異体植物を作出する方法において、電離放射線を照射する前に、植物をストレス処理することを特徴とする。
【0017】
本願発明が対象とする「植物」は花冠を有する植物全般であり、花冠を有する草本植物および木本植物が含まれる。また、これまで電離放射線照射による花色変異体植物の作出が試みられてきた植物は、全て本願発明の対象植物となる。なお、以下の発明において、特に成長した個体を意味する場合には「植物体」と記載することがある。また本願発明における「花色変異体植物」とは、野生型と異なる花冠色を有するような突然変異体植物であり、特に、観賞用の園芸植物として有用な花色を有する植物である。
【0018】
ストレス処理と電離放射線照射は、花弁など植物の特定の部位に限らず、様々な部位・成長段階の材料を用いて行うことができる。具体的には、植物体の全体あるいはその一部、または植物体への成長能を有する植物組織を対象として行うことができる。この場合の植物組織は、吸水させた種子、発芽直後の種子、幼苗、培養組織等である。培養組織としては、芽、根、葉、花弁、不定芽、不定胚、管状花、カルスなど、組織培養による植物体成長が可能な全ての組織を対象とすることができる。また、ストレス処理と電離放射線照射は、色素が蓄積した状態の植物に電離放射線を照射することを前提として、ストレス処理に連続して電離放射線を照射することもでき、あるいは異なる成長段階にある植物に対して両者を時間的に分離して行うこともできる(例えば、種子にストレス処理を行い、幼苗に対して電離放射線照射を行うなど)。
【0019】
ストレスの種類は、「色素の蓄積を促進させるストレス」として公知のものを使用することができる。例えば、アントシアニン色素の蓄積を促進する強光、紫外線、低温及び糖などであり、これらのストレス種と色素蓄積の公知の関係(非特許文献4、5、6、7)から、それぞれに適宜なストレス強度、その暴露時間等を設定することができる。具体例として、糖(蔗糖、グルトース、フルクトース、マンニトールなど)の場合には、例えば組織培養の培地に、1%〜10%、好ましくは2%〜6%、特に好ましくは2.5%〜5%程度の濃度の糖を添加する。また、低温ストレスの場合には、植物を0℃〜10℃、好ましくは2℃〜6℃、さらに好ましくは3℃〜5℃程度の温度条件下で維持する。さらに、これらの高濃度糖ストレスや低温ストレスは、0.5〜10日、好ましくは1日〜5日程度の期間で植物に暴露する。
【0020】
このようにしてストレスに暴露させた後、植物に電離放射線を照射する。電離放射線は、電離作用を有する高エネルギーの電磁波または粒子線であり、例えば、イオンビーム、エックス線、ガンマ線、電子線、中性子線などである。電離放射線としてイオンビームを使用する場合は、任意の線種を採用することができ、例えば、10〜5000MeVのヘリウムイオン、炭素イオン、ネオンイオン、アルゴンイオンなどを使用することができる。また炭素イオンビームを使用する場合は、320MeVの炭素イオンビームが好ましい。
【0021】
また、イオンビームは、0.1〜100Gyの線量で照射するが、最適な照射線量は対象とする植物種により異なる。植物種と照射線量の関係は、従来のイオンビーム照射による花色変異体作出例に準じて適宜に決定することができる。例えば、ナス科植物(例えば、ペチュニア)の場合には0.1〜20Gy、好ましくは1〜15Gy、さらに好ましくは6〜12Gy程度である。
【0022】
また電離放射線としてエックス線を照射する場合の線量は0.1〜10kRの範囲であり、例えばキク科植物(例えば、キク品種リネカー)の場合には、0.1〜10kR、好ましくは0.3〜5kR、さらに好ましくは0.5〜3kR程度である。
【0023】
以上のとおりのストレス処理と電離放射線照射によって、後記実施例に例示したように、電離放射線照射のみに比較して2倍以上の高い頻度で花色変異体植物を作出することができる。また、葉緑素変異体などの出現頻度を上昇させることがない。さらに、得られた花色変異体植物の次世代においても変異花色は安定に保存される。
【実施例】
【0024】
以下、実施例を示して本願発明をさらに詳細かつ具体的に説明するが、本願発明は以下の例に限定されるものではない。
(実施例1) ペチュニアの花色変異体作出
ペチュニア(花色:バイオレット)の花色変異体の出現頻度に対するストレス処理の効果を調査した。ストレス(蔗糖)処理及び無処理の幼苗に炭素イオンビームを照射し、M2世代(照射次世代)において花色変異体の出現頻度を比較した。葉緑素変異体の出現頻度を全般的な変異頻度の内部標準として調査した。
【0025】
1.材料及び方法
植物材料として、バイオレット色の花を持つペチュニア(Petunia hybrida)のBBss11系統(キリンアグリバイオ株式会社)を用いた。BBss11系統は10世代以上の自殖によって遺伝的に固定されている。
種子は、培養土(プラグメイト、ホーネンアグリ株式会社)を入れたプラスチックシャーレに播種した。シャーレは23℃、16時間明期/8時間暗期、照度約4,000lx(45.6μmol m-2s-1光合成光量子束密度)の白色光の育成室に置き、蒸留水を与えて常に湿っている状態を保った。ストレス処理群は以下のとおりとした。蔗糖処理群は、播種後8日目に20%蔗糖溶液を1ml加えた。蔗糖の終濃度は約3%と見積もられる。紫外線処理群は、播種後8日目の幼苗を15 kJ/m2のUVBに暴露した。低温処理群は、播種後8日目の幼苗を4℃に24時間または48時間置いた。
【0026】
電離放射線照射として、播種後10日目の幼苗に対しAVFサイクロトロン(日本原子力研究開発機構)で発生させた320MeV炭素イオンを照射した。炭素イオンの線エネルギー付与はIRACMコード(Tanaka et al 1997)によって76 keV/μmと計算された。照射の翌日、幼苗をプラグトレイまたはポットに移植し、キリンアグリバイオまたは日本原子力研究開発機構の温室で育成した。放射線感受性を調査するために、蔗糖溶液または水のみを与えた幼苗に0から30Gyの炭素イオンビームを照射した。各線量について25個体の3反復を用い、照射1ヵ月後に生存率を調査した。変異体選抜のために、8Gyを照射した幼苗を温室で育成し、M1植物から個体別にM2種子を採種した(M2系統)。M2系統の種子を土の上で発芽させ、葉緑素変異体の頻度を調査した。各M2系統について5から20粒、多くの場合10粒以上を播種した。発芽した幼苗は温室で育成し、花色変異体の頻度を調査した。
【0027】
2.結果
色素の生合成を促進するのに最適なストレス条件を見つけるために、ペチュニア幼苗において蔗糖、紫外線及び低温処理の効果を調査した。3%蔗糖で処理した幼苗では、蔗糖添加後2日後に色素蓄積の増加が観察され、4日後に対照区の幼苗との差が明確になった(図1)。紫外線も色素の蓄積を促進したが、蔗糖処理に比べて効果は著しく低かった。さらに紫外線は葉に損傷を与えた。低温処理は試験した条件では効果が無かった。これらの観察から、2日間の蔗糖処理がペチュニア幼苗における色素蓄積に関わる遺伝子を活性化するに最適かつ十分であると考えられた。
【0028】
蔗糖処理の変異率に対する効果を調査するのに適当な線量を決定するために、播種後10日目の幼苗に0から30Gyの炭素イオンビームを照射した。照射した幼苗をポットへ移植する際、蔗糖処理した幼苗は無処理の幼苗に比べて根の伸長が抑制されていた。図2は照射1ヶ月後の蔗糖処理及び無処理の幼苗の生存率の線量反応を示す。植物の生育は線量の増加に伴って抑制されたが、生存率は10Gyまで低下しなかった。蔗糖処理は幼苗の生存には影響しないと考えられた。発明者らの経験から、イオンビームによる変異誘発においては、好ましくない変異の誘発を抑えるために、線量反応曲線の肩もしくはそれ以下に相当する線量が適していると考えられる。従って、本実験においては8Gyが変異誘発に適当であると決定した。
【0029】
蔗糖処理及び無処理のペチュニア幼苗に対し炭素イオンを8Gy照射し、個々のM1植物からM2種子を採種した。変異体の選抜は異なる場所における3つの独立した試験として行い、その結果を表1にまとめた。試験1では、おそらく使用したポットが小さかったために、20%以下のM1植物からしかM2種子は採種されなかった。試験2では、より大きなポットを使用してM1植物を育成し、M2種子はM1植物の約半数から得られた。非照射植物の場合は、83%の植物から次世代種子が得られた。試験3では、さらに大きなポットを使用し、90%以上のM1植物からM2種子を得た。
【0030】
【表1】

【0031】
M2世代において、葉緑素変異体は蔗糖処理及び無処理の両者から得られた。試験1では、葉緑素変異体の頻度は蔗糖処理区で0.56%、無処理区で0.48%であった。試験2では、葉緑素変異体の頻度は蔗糖処理区で0.39%、無処理区で0.33%であった。試験3では、葉緑素変異体の頻度は蔗糖処理区で0.55%、無処理区で0.43%であった。各試験において2つの処理区間で葉緑素変異体の頻度に有意な差は無かった。非照射区においては、葉緑素変異体は得られなかった。試験1で葉緑素変異体が得られた3つのM2系統では、4/16(= 25%)、1/7(=14%)、3/20(=15%)(変異体数/植物体総数)の割合で分離した。この結果は、これらの葉緑素変異が単一劣性であることを示唆している。また、播種後10日目のペチュニア幼苗において生殖系列につながる細胞は1つまたは2つであると予測される。花色変異体の頻度は蔗糖処理区で1.52%、1.20%及び1.26%、無処理区で0.56%、0.58%および0.47%であった。蔗糖処理は3つの独立した試験において花色変異体の頻度を2倍以上増加させた。また、パープルヴェイン、ホワイト、ブルーピコティーおよびバーガンディー(図3)は蔗糖処理区のみで得られ、蔗糖処理区で花色変異幅が広かった。これらの結果は、蔗糖処理がイオンビーム照射によって得られる花色変異体の獲得頻度を特異的に増加させ、さらに得られる花色変異の幅を拡大することを示している。
【0032】
(実施例2)キクの花色変異体作出
キク品種リネカー(キリンアグリバイオ株式会社、花色:桃色)の花色変異体を作成した。
なお、この実施例の組織培養で使用した「KS2培地」は、2%蔗糖、BA 2mg/l、NAA 0.2mg/lを含むMS培地であり、「再生KS培地」は、2%蔗糖、BA1mg/l、IAA0.2mg/lを含むMS培地(但し、NH4NO3が300mg/l)である。
【0033】
1.方法
試験区1:非照射区(組織培養のみ)
品種「リネカー」の管状花30個をKS2培地にて、25℃暗黒下で7日間培養した。
次に、再生KS2培地に培養組織を移植し、再生した植物体を育成し、開花検定を行った。
試験区2:エックス線照射区
品種「リネカー」の管状花30個をKS2培地にて、25℃暗黒下で7日間培養した。
線量1kRのエックス線を照射した後、再生KS2培地に培養組織を移植した。再生した植物体を育成し、開花検定を行った。
試験区3:高蔗糖+低温前培養+エックス線照射処理区
品種「リネカー」の管状花30個をKS2培地(但し、蔗糖3%)にて、25℃暗黒下で4日間、さらに、4℃暗黒下で3日間培養した後、線量1kRのエックス線を照射した。
次いで、再生KS2培地に培養組織を移植し、再生した植物体を育成し、開花検定を行った。
【0034】
2.結果
結果は表2および図4−5に示したとおりである。
【0035】
【表2】
【0036】
試験区1(非照射区)では、白色変異株を1個体得たが、微量のアントシアニン蓄積が認められ、純白ではなかった(図4右)。試験区2(エックス線照射区)では、白色変異株を2個体得たがこれらにも微量のアントシアニン蓄積が認められ純白ではなかった。
【0037】
これに対して試験区3(高濃度蔗糖処理+低温前培養処理+エックス線照射区)では、白色変異株を12個体得た。このうち1個体がアントシアニンを持たない純白個体であった(図4左)。生育もよく、栄養繁殖によって増殖させた植物体においても純白形質が保存された(図5)。
【技術分野】
【0001】
本願発明は、園芸植物等における花色変異体を効率よく作出するための方法に関するものである。
【背景技術】
【0002】
電離放射線は数十年にわたって新しい植物品種を作り出すために使われてきた。電離放射線照射による植物の突然変異の頻度を高めるためには、植物が生存し得る限り線量を上げてやればよい。また、酸素濃度、水分含量及び温度などの外的要因により、植物の放射線感受性を変えられることが知られており(非特許文献1)、その要因をコントロールすることで変異頻度を高めることも可能であろう。しかしながら、望ましい突然変異を起こすために線量を上げたり放射線感受性を高めたりすれば同時に生育不良等を引き起こしうる望まない変異も同時に増え、結果的に、望ましい変異のみを有する個体獲得は困難となる。よって、実際に電離放射線照射による植物の突然変異育種においては、できるだけ望ましい変異が起きる頻度を高めつつも不要な変異の誘発を避けるために、照射線量は植物と照射の条件に応じた、一定の低いレベルに留める必要がある。ただし、その場合には望ましい変異のみを持つ個体の獲得頻度は低いため、非常に多数の植物に照射し多数の個体から望む個体を選抜する必要があった。よって、効率的な突然変異個体獲得のためには、不要な変異の頻度を高めずに望まれる変異のみを誘発する手法が望まれている。
【0003】
永富ら(非特許文献2、3)は、キクにおける炭素イオンビームの変異効果を調査し、葉培養物に照射するよりも花弁培養物に照射した際に花色変異体が高い頻度で得られたと報告しており、植物の照射部位選択によって変異頻度が変わりうることを示した。
【0004】
なお、花色は園芸植物において最も重要な形質の1つであるが、この花色に関連するアントシアニン色素の生合成に関与する遺伝子の発現は、強光、紫外線、低温及び糖などのストレスによって活性化されることが知られている(非特許文献4、5、6、7)。ただし、このような色素の蓄積は植物体の遺伝形質に影響を及ぼすものではなく、たとえストレス暴露により花色を変化させたとしても、その形質が次世代の植物体に継続されることはない。
【非特許文献1】van Harten (1998) Mutation Breeding, Theory and Practical Applications (Cambridge University Press).
【非特許文献2】Nagatomi S, Tanaka A, Watanabe H, Tano S (1997) Chrysanthemum mutants regenerated from in vitro explants irradiated with 12C5+ion beam. Institute of Radiation Breeding, Technical News No.60.
【非特許文献3】Nagatomi S, Tanaka A, Kato A, Yamaguchi H, Watanabe H, Tano (1998) Mutation induction through ion beam irradiations in rice and chrysanthemum. JAERI-Review 98-016 / TIARA Annual Report 1997: 41-43.
【非特許文献4】Dubos C, Le Gourrierec J, Baudry A, Huep G, LanetE, Debeaujon I, Routaboul J M, Alboresi A, Weisshaar B, Lapiniec L (2008) MYBL2 is a new regulator of flavonoid biosynthesis in Arabidopsis thaliana. Plant J 55: 940-953.
【非特許文献5】Hase Y, Trung K H, Matsunaga T, Tanaka A (2006) A mutation in the uvi4 gene promotes progression of endo-reduplication and confers increased tolerance towards ultraviolet B light. Plant J 46: 317-326.
【非特許文献6】Olsen K M, Slimestad R, Lea U S, Brede C, Lovdal T, Ruoff P, Verhuel M, Lillo C (2009) Temperature and nitrogen effects on regulators and products of the flavonoid pathway: experimental and kinetic model studies. Plant Cell Environ 32: 286-299.
【非特許文献7】Tsukaya H, Ohshima T, Naito S, Chino M, Komeda Y (1991) Sugar-dependent expression of the CHS-A gene for chalcone synthase from petunia in transgenic Arabidopsis. Plant Physiol 97: 1414-1421.
【発明の概要】
【発明が解決しようとする課題】
【0005】
園芸植物等としての花色変異体植物の需要は大きい。これまで花色変異体植物の作出について様々な試みがなされているが、所望の変異体獲得の効率性については改善すべき点が残されている。
【0006】
本願発明は、電離放射線照射による花色変異体植物の獲得頻度増大に寄与する新しい手段を提供することを課題としている。
【課題を解決するための手段】
【0007】
本願発明者らは、花色変異体植物の獲得頻度を向上させる手段について検討した結果、植物へのストレス処理を施した状態で電離放射線を照射することにより、花色変異体植物の獲得頻度が大きく上昇し変異幅も拡がることを見出した。
【0008】
本願発明は、上記の新規知見に基づいて完成されたものである。
【0009】
すなわち、本願発明は、電離放射線照射によって花色変異体植物を作出する方法において、電離放射線の照射前に、植物をストレス処理することを特徴とする方法である。
【0010】
この花色変異体植物の作出方法においては、電離放射線照射のみによる花色変異体植物の獲得頻度に比べ、2倍以上の頻度で花色変異体植物が獲得されることを別の特徴とする。
【0011】
またこの作出方法においては、葉緑素変異体の出現頻度が、電離放射線照射のみによる当該変異体の出現頻度と同程度であることをさらに別の特徴としている。
【0012】
さらにこの作出方法においては、例えばストレス処理が植物への高濃度糖処理であるか、あるいは植物の低温での高濃度糖処理であることを具体的な実施形態としている。
【0013】
すなわち、本願発明は、以下の発明を包含する。
[1]電離放射線照射によって花色変異体植物を作出する方法において、電離放射線の照射前に、植物をストレス処理することを特徴とする方法。
[2]ストレス処理が、植物の高濃度糖処理または植物の低温処理と高濃度糖処理の組み合わせである[1]記載の方法。
[3]電離放射線がイオンビームまたはエックス線である[1]または[2]記載の方法。
【発明の効果】
【0014】
本願発明により、電離放射線による突然変異を利用した植物の花色変異を得る効率を高めることができる。本願発明で用いるストレス処理は、植物の放射線感受性を高めるものではなく、また、葉緑素変異株の獲得頻度が変化しないことから、花色の変異誘発に特異的に作用すると考えられる。従って、本発明によって、花色に関する植物育種において、目的とする花色変異のみを有する個体を獲得する効率を高めることが可能となる。
【図面の簡単な説明】
【0015】
【図1】実施例1で確認したペチュニア幼苗における色素蓄積に対するストレス処理の効果。播種後8日目の幼苗を3%蔗糖(A)、15 kJ m-2の紫外線(B)、4℃・24時間(C)で処理した。写真はストレス処理後4日目の幼苗を示す。(D)には無処理の対照植物を示した。
【図2】実施例1で確認した320 MeV炭素イオンビームを照射した播種後10日目のペチュニア幼苗の生存率の線量反応。終濃度3%となる蔗糖溶液(白丸)または水のみ(黒丸)を照射2日前に加えた。生存率は照射1ヶ月後に調査した。データは平均±標準偏差を示す。
【図3】ペチュニアの親系統ならびに実施例1で得られた花色変異体。(A)バイオレットの花色を持つ親系統。(B−H)花色変異体。(B)マゼンタ、(C)パープル、(D)パープルヴェイン、(E)ライトピンク、(F)ホワイト、(G)ブルーピコティー、(H)バーガンディー。
【図4】実施例2で得られたキク品種リネカーの純白変異体(左)と、実施例2の対照試験区で得られた微量のアントシアニンを持つ白色変異体(右)。
【図5】実施例2で得られた純白変異体から樹立された純白リネカー株。
【発明を実施するための形態】
【0016】
本願発明の花色変異体植物作出方法は、電離放射線照射によって花色変異体植物を作出する方法において、電離放射線を照射する前に、植物をストレス処理することを特徴とする。
【0017】
本願発明が対象とする「植物」は花冠を有する植物全般であり、花冠を有する草本植物および木本植物が含まれる。また、これまで電離放射線照射による花色変異体植物の作出が試みられてきた植物は、全て本願発明の対象植物となる。なお、以下の発明において、特に成長した個体を意味する場合には「植物体」と記載することがある。また本願発明における「花色変異体植物」とは、野生型と異なる花冠色を有するような突然変異体植物であり、特に、観賞用の園芸植物として有用な花色を有する植物である。
【0018】
ストレス処理と電離放射線照射は、花弁など植物の特定の部位に限らず、様々な部位・成長段階の材料を用いて行うことができる。具体的には、植物体の全体あるいはその一部、または植物体への成長能を有する植物組織を対象として行うことができる。この場合の植物組織は、吸水させた種子、発芽直後の種子、幼苗、培養組織等である。培養組織としては、芽、根、葉、花弁、不定芽、不定胚、管状花、カルスなど、組織培養による植物体成長が可能な全ての組織を対象とすることができる。また、ストレス処理と電離放射線照射は、色素が蓄積した状態の植物に電離放射線を照射することを前提として、ストレス処理に連続して電離放射線を照射することもでき、あるいは異なる成長段階にある植物に対して両者を時間的に分離して行うこともできる(例えば、種子にストレス処理を行い、幼苗に対して電離放射線照射を行うなど)。
【0019】
ストレスの種類は、「色素の蓄積を促進させるストレス」として公知のものを使用することができる。例えば、アントシアニン色素の蓄積を促進する強光、紫外線、低温及び糖などであり、これらのストレス種と色素蓄積の公知の関係(非特許文献4、5、6、7)から、それぞれに適宜なストレス強度、その暴露時間等を設定することができる。具体例として、糖(蔗糖、グルトース、フルクトース、マンニトールなど)の場合には、例えば組織培養の培地に、1%〜10%、好ましくは2%〜6%、特に好ましくは2.5%〜5%程度の濃度の糖を添加する。また、低温ストレスの場合には、植物を0℃〜10℃、好ましくは2℃〜6℃、さらに好ましくは3℃〜5℃程度の温度条件下で維持する。さらに、これらの高濃度糖ストレスや低温ストレスは、0.5〜10日、好ましくは1日〜5日程度の期間で植物に暴露する。
【0020】
このようにしてストレスに暴露させた後、植物に電離放射線を照射する。電離放射線は、電離作用を有する高エネルギーの電磁波または粒子線であり、例えば、イオンビーム、エックス線、ガンマ線、電子線、中性子線などである。電離放射線としてイオンビームを使用する場合は、任意の線種を採用することができ、例えば、10〜5000MeVのヘリウムイオン、炭素イオン、ネオンイオン、アルゴンイオンなどを使用することができる。また炭素イオンビームを使用する場合は、320MeVの炭素イオンビームが好ましい。
【0021】
また、イオンビームは、0.1〜100Gyの線量で照射するが、最適な照射線量は対象とする植物種により異なる。植物種と照射線量の関係は、従来のイオンビーム照射による花色変異体作出例に準じて適宜に決定することができる。例えば、ナス科植物(例えば、ペチュニア)の場合には0.1〜20Gy、好ましくは1〜15Gy、さらに好ましくは6〜12Gy程度である。
【0022】
また電離放射線としてエックス線を照射する場合の線量は0.1〜10kRの範囲であり、例えばキク科植物(例えば、キク品種リネカー)の場合には、0.1〜10kR、好ましくは0.3〜5kR、さらに好ましくは0.5〜3kR程度である。
【0023】
以上のとおりのストレス処理と電離放射線照射によって、後記実施例に例示したように、電離放射線照射のみに比較して2倍以上の高い頻度で花色変異体植物を作出することができる。また、葉緑素変異体などの出現頻度を上昇させることがない。さらに、得られた花色変異体植物の次世代においても変異花色は安定に保存される。
【実施例】
【0024】
以下、実施例を示して本願発明をさらに詳細かつ具体的に説明するが、本願発明は以下の例に限定されるものではない。
(実施例1) ペチュニアの花色変異体作出
ペチュニア(花色:バイオレット)の花色変異体の出現頻度に対するストレス処理の効果を調査した。ストレス(蔗糖)処理及び無処理の幼苗に炭素イオンビームを照射し、M2世代(照射次世代)において花色変異体の出現頻度を比較した。葉緑素変異体の出現頻度を全般的な変異頻度の内部標準として調査した。
【0025】
1.材料及び方法
植物材料として、バイオレット色の花を持つペチュニア(Petunia hybrida)のBBss11系統(キリンアグリバイオ株式会社)を用いた。BBss11系統は10世代以上の自殖によって遺伝的に固定されている。
種子は、培養土(プラグメイト、ホーネンアグリ株式会社)を入れたプラスチックシャーレに播種した。シャーレは23℃、16時間明期/8時間暗期、照度約4,000lx(45.6μmol m-2s-1光合成光量子束密度)の白色光の育成室に置き、蒸留水を与えて常に湿っている状態を保った。ストレス処理群は以下のとおりとした。蔗糖処理群は、播種後8日目に20%蔗糖溶液を1ml加えた。蔗糖の終濃度は約3%と見積もられる。紫外線処理群は、播種後8日目の幼苗を15 kJ/m2のUVBに暴露した。低温処理群は、播種後8日目の幼苗を4℃に24時間または48時間置いた。
【0026】
電離放射線照射として、播種後10日目の幼苗に対しAVFサイクロトロン(日本原子力研究開発機構)で発生させた320MeV炭素イオンを照射した。炭素イオンの線エネルギー付与はIRACMコード(Tanaka et al 1997)によって76 keV/μmと計算された。照射の翌日、幼苗をプラグトレイまたはポットに移植し、キリンアグリバイオまたは日本原子力研究開発機構の温室で育成した。放射線感受性を調査するために、蔗糖溶液または水のみを与えた幼苗に0から30Gyの炭素イオンビームを照射した。各線量について25個体の3反復を用い、照射1ヵ月後に生存率を調査した。変異体選抜のために、8Gyを照射した幼苗を温室で育成し、M1植物から個体別にM2種子を採種した(M2系統)。M2系統の種子を土の上で発芽させ、葉緑素変異体の頻度を調査した。各M2系統について5から20粒、多くの場合10粒以上を播種した。発芽した幼苗は温室で育成し、花色変異体の頻度を調査した。
【0027】
2.結果
色素の生合成を促進するのに最適なストレス条件を見つけるために、ペチュニア幼苗において蔗糖、紫外線及び低温処理の効果を調査した。3%蔗糖で処理した幼苗では、蔗糖添加後2日後に色素蓄積の増加が観察され、4日後に対照区の幼苗との差が明確になった(図1)。紫外線も色素の蓄積を促進したが、蔗糖処理に比べて効果は著しく低かった。さらに紫外線は葉に損傷を与えた。低温処理は試験した条件では効果が無かった。これらの観察から、2日間の蔗糖処理がペチュニア幼苗における色素蓄積に関わる遺伝子を活性化するに最適かつ十分であると考えられた。
【0028】
蔗糖処理の変異率に対する効果を調査するのに適当な線量を決定するために、播種後10日目の幼苗に0から30Gyの炭素イオンビームを照射した。照射した幼苗をポットへ移植する際、蔗糖処理した幼苗は無処理の幼苗に比べて根の伸長が抑制されていた。図2は照射1ヶ月後の蔗糖処理及び無処理の幼苗の生存率の線量反応を示す。植物の生育は線量の増加に伴って抑制されたが、生存率は10Gyまで低下しなかった。蔗糖処理は幼苗の生存には影響しないと考えられた。発明者らの経験から、イオンビームによる変異誘発においては、好ましくない変異の誘発を抑えるために、線量反応曲線の肩もしくはそれ以下に相当する線量が適していると考えられる。従って、本実験においては8Gyが変異誘発に適当であると決定した。
【0029】
蔗糖処理及び無処理のペチュニア幼苗に対し炭素イオンを8Gy照射し、個々のM1植物からM2種子を採種した。変異体の選抜は異なる場所における3つの独立した試験として行い、その結果を表1にまとめた。試験1では、おそらく使用したポットが小さかったために、20%以下のM1植物からしかM2種子は採種されなかった。試験2では、より大きなポットを使用してM1植物を育成し、M2種子はM1植物の約半数から得られた。非照射植物の場合は、83%の植物から次世代種子が得られた。試験3では、さらに大きなポットを使用し、90%以上のM1植物からM2種子を得た。
【0030】
【表1】
【0031】
M2世代において、葉緑素変異体は蔗糖処理及び無処理の両者から得られた。試験1では、葉緑素変異体の頻度は蔗糖処理区で0.56%、無処理区で0.48%であった。試験2では、葉緑素変異体の頻度は蔗糖処理区で0.39%、無処理区で0.33%であった。試験3では、葉緑素変異体の頻度は蔗糖処理区で0.55%、無処理区で0.43%であった。各試験において2つの処理区間で葉緑素変異体の頻度に有意な差は無かった。非照射区においては、葉緑素変異体は得られなかった。試験1で葉緑素変異体が得られた3つのM2系統では、4/16(= 25%)、1/7(=14%)、3/20(=15%)(変異体数/植物体総数)の割合で分離した。この結果は、これらの葉緑素変異が単一劣性であることを示唆している。また、播種後10日目のペチュニア幼苗において生殖系列につながる細胞は1つまたは2つであると予測される。花色変異体の頻度は蔗糖処理区で1.52%、1.20%及び1.26%、無処理区で0.56%、0.58%および0.47%であった。蔗糖処理は3つの独立した試験において花色変異体の頻度を2倍以上増加させた。また、パープルヴェイン、ホワイト、ブルーピコティーおよびバーガンディー(図3)は蔗糖処理区のみで得られ、蔗糖処理区で花色変異幅が広かった。これらの結果は、蔗糖処理がイオンビーム照射によって得られる花色変異体の獲得頻度を特異的に増加させ、さらに得られる花色変異の幅を拡大することを示している。
【0032】
(実施例2)キクの花色変異体作出
キク品種リネカー(キリンアグリバイオ株式会社、花色:桃色)の花色変異体を作成した。
なお、この実施例の組織培養で使用した「KS2培地」は、2%蔗糖、BA 2mg/l、NAA 0.2mg/lを含むMS培地であり、「再生KS培地」は、2%蔗糖、BA1mg/l、IAA0.2mg/lを含むMS培地(但し、NH4NO3が300mg/l)である。
【0033】
1.方法
試験区1:非照射区(組織培養のみ)
品種「リネカー」の管状花30個をKS2培地にて、25℃暗黒下で7日間培養した。
次に、再生KS2培地に培養組織を移植し、再生した植物体を育成し、開花検定を行った。
試験区2:エックス線照射区
品種「リネカー」の管状花30個をKS2培地にて、25℃暗黒下で7日間培養した。
線量1kRのエックス線を照射した後、再生KS2培地に培養組織を移植した。再生した植物体を育成し、開花検定を行った。
試験区3:高蔗糖+低温前培養+エックス線照射処理区
品種「リネカー」の管状花30個をKS2培地(但し、蔗糖3%)にて、25℃暗黒下で4日間、さらに、4℃暗黒下で3日間培養した後、線量1kRのエックス線を照射した。
次いで、再生KS2培地に培養組織を移植し、再生した植物体を育成し、開花検定を行った。
【0034】
2.結果
結果は表2および図4−5に示したとおりである。
【0035】
【表2】
【0036】
試験区1(非照射区)では、白色変異株を1個体得たが、微量のアントシアニン蓄積が認められ、純白ではなかった(図4右)。試験区2(エックス線照射区)では、白色変異株を2個体得たがこれらにも微量のアントシアニン蓄積が認められ純白ではなかった。
【0037】
これに対して試験区3(高濃度蔗糖処理+低温前培養処理+エックス線照射区)では、白色変異株を12個体得た。このうち1個体がアントシアニンを持たない純白個体であった(図4左)。生育もよく、栄養繁殖によって増殖させた植物体においても純白形質が保存された(図5)。
【特許請求の範囲】
【請求項1】
電離放射線照射によって花色変異体植物を作出する方法において、電離放射線の照射前に、植物をストレス処理することを特徴とする方法。
【請求項2】
ストレス処理が、植物の高濃度糖処理または植物の低温処理と高濃度糖処理の組み合わせである請求項1記載の方法。
【請求項3】
電離放射線がイオンビームまたはエックス線である請求項1または請求項2記載の方法。
【請求項1】
電離放射線照射によって花色変異体植物を作出する方法において、電離放射線の照射前に、植物をストレス処理することを特徴とする方法。
【請求項2】
ストレス処理が、植物の高濃度糖処理または植物の低温処理と高濃度糖処理の組み合わせである請求項1記載の方法。
【請求項3】
電離放射線がイオンビームまたはエックス線である請求項1または請求項2記載の方法。
【図1】

【図2】
【図3】

【図4】

【図5】

【図2】
【図3】
【図4】
【図5】
【公開番号】特開2011−30566(P2011−30566A)
【公開日】平成23年2月17日(2011.2.17)
【国際特許分類】
【出願番号】特願2010−152796(P2010−152796)
【出願日】平成22年7月5日(2010.7.5)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 平成22年3月10日 The Japanese Society for Plant Cell and Molecular Biology発行の「Plant Biotechnology」(Volume 27,Number 1)に発表
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成20年度生物系特定産業技術研究支援センター「生物系産業創出のための異分野融合研究支援事業」産業技術力強化法第19条の適用を受ける特許出願
【出願人】(505374783)独立行政法人 日本原子力研究開発機構 (727)
【出願人】(000253503)キリンホールディングス株式会社 (247)
【Fターム(参考)】
【公開日】平成23年2月17日(2011.2.17)
【国際特許分類】
【出願日】平成22年7月5日(2010.7.5)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 平成22年3月10日 The Japanese Society for Plant Cell and Molecular Biology発行の「Plant Biotechnology」(Volume 27,Number 1)に発表
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成20年度生物系特定産業技術研究支援センター「生物系産業創出のための異分野融合研究支援事業」産業技術力強化法第19条の適用を受ける特許出願
【出願人】(505374783)独立行政法人 日本原子力研究開発機構 (727)
【出願人】(000253503)キリンホールディングス株式会社 (247)
【Fターム(参考)】
[ Back to top ]