
視神経症に対する感受性を診断または予測するための方法
緑内障およびレーベル病を含む視神経症に連関する遺伝子多型のセットを開示する。これらの多型は視神経症に対する感受性を診断し予測するのに有用である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、視神経症に連関する遺伝子多型のセットに関する。
【背景技術】
【0002】
緑内障は世界的に失明の主原因であり、およそ6700万人が何らかの形態の緑内障を患っていると見積られている。大多数の症例は原発開放隅角緑内障(POAG)の後期成人発症(通例、40歳超)として起こる。これは、最も一般的な形態の緑内障であり、40歳を超えた白人人口の約2%および黒人人口の7%を冒す。POAGは、通常は眼内圧(IOP)の上昇と関連する視神経乳頭の陥凹に対応した特徴的な視野変化をもたらす。正常眼圧緑内障(NTG)は、典型的な緑内障性視神経乳頭陥凹および視野喪失が存在するものの、常に21mmHgを超える上昇したIOPの証拠がない開放隅角緑内障の一形態である。日本の場合、緑内障の有病率は40歳超の約3.5%(POAG 0.58%およびNTG 2.04%)である。日本人集団におけるNTGの有病率は他の集団と比較して高い。眼内圧(IOP)は緑内障の主要リスク因子であるが、緑内障は特有の視野喪失と関連する進行性視神経症を特徴とする多因子性障害であり、複数の遺伝子および環境的影響の相互作用によってもたらされる。
【0003】
緑内障を発病させるリスク因子には、高いIOP、年齢、人種、家族歴あり、近視、糖尿病または高血圧の存在、および遺伝的因子が含まれる。緑内障性視神経症の正確な病理発生過程はまだ不明であるが、上昇したIOPが主要リスク因子であることは、広く受け入れられている。現在行なわれている緑内障の処置はIOPを低下させる介入からなる。しかし、緑内障、NTGまたは末期POAGを持つ一部の患者においては、IOPの低下では疾患の進行が予防されないことから、上昇したIOP以外の因子が緑内障の発病または進行に関与しうることが示される。
【0004】
POAGおよびNTGは不均質な病態群であり、これらはおそらく、視神経乳頭に観察されるニューロン喪失パターンをもたらす異なる多因子性病因を持っている。緑内障と、低体血圧、血圧の夜間低下、高血圧、片頭痛、血管痙攣、および糖尿病を含む多くの全身性血管疾患の存在との関連が報告されている。NTG患者における視神経乳頭出血の存在は、NTGの発病および進行に血管不全が深く関与することを示唆している。POAGを持つ患者は高い割合で、併存障害に対する多種多様な投薬を受ける。特に、全身性高血圧は最もよく見られる障害であり、集団全体の48%に見いだされた。
【0005】
萎縮期のレーベル遺伝性視神経症(LHON)および優性遺伝性視神経萎縮(DAO)を持つ患者に、緑内障様の形態変化が報告されている。最近、本発明者らは、11778突然変異を保持する15人の日本人LHON患者の萎縮期において、ハイデルベルグ網膜断層撮影(HRT)を使った定量的解析により、視神経乳頭陥凹を報告した(Mashima Yら,Arch Clin Exp Ophthalmol 2003;241:75-80,この引用文献の内容は参照により本明細書に組み込まれる)。LHONは主に若年成人男性を冒す母系遺伝性の眼疾患である。患者の約70%は男性だった。この疾患は通常、重度で永続的な視覚喪失を引き起こし、視力は0.1未満になる。視野欠損は中心暗点または盲点中心暗点として存在する。今までに20を超えるミトコンドリアDNA(mtDNA)の点突然変異が世界中のLHON患者に報告されており(Brown MDら,Clin Neurosci 1994;2:138-145,この引用文献の内容は参照により本明細書に組み込まれる)、LHON患者の80%超が、ヌクレオチド位置3460、11778、または14484にある三つのmtDNA突然変異のうちの一つを保有する(Mackey DAら,Am J Hum Genet 1996;59:481-485,この引用文献の内容は参照により本明細書に組み込まれる)。mtDNAヌクレオチド位置3460、11778および14484の三つの突然変異についてNTG患者を調べたが、突然変異も、骨格筋試料における呼吸鎖活性の欠損も、検出されなかった(Brierley EJら,Arch Ophthalmol 114:142-146およびOpial Dら,Graefes Arch Clin Exp Ophthalmol 239:437-440,これらの引用文献の内容は参照により本明細書に組み込まれる)。
【0006】
これらのmtDNA突然変異の一つを持つLHON患者の間に見られる主な相違は、その臨床経過にある。3460突然変異および14484突然変異は、4%〜7%の視覚回復率しか示さない11778突然変異よりも良好な視覚予後と関連する(Oostra RJら,J med Genet 1994;31:280-286、Riordan-Eva Pら,Brain 1995;118:319-337、Mashima Yら,Curr Eye Res 1998;17:403-408,これらの引用文献の内容は参照により本明細書に組み込まれる)。しかし、11778突然変異を持ち発症年齢が十代前半である一部の患者では、視覚回復が記録されている(Stone EMら,J clin Meuro-Ophthalmol 1992;12:10-14、Zhu Dら,Am J Med Genet 1992;42:173-179、Salmaggi Aら,Intern J Neuroscience 1994;77:261-266、Oostra RJら,Clin Genet 1997;51:388-393、Mashima Yら,Jpn J Ophthalmol 2002;46:660-667,これらの引用文献の内容は参照により本明細書に組み込まれる)。視覚の回復は、11778突然変異を持つ患者であっても、視覚悪化が若年時に始まる場合の方が、可能性が高くなるようである。
【0007】
発症時年齢、男性好発、不完全浸透、および視覚回復を含むLHON患者の臨床的ばらつきは、おそらくこの疾患が、環境ストレス要因、X染色体遺伝子座、および他のmtDNA突然変異の関与が考えられる多遺伝子性機序または多因子性機序によるものであることを示唆している(Man PYWら,J Med Genet 2002;39:162-169、この引用文献の内容は参照により本明細書に組み込まれる)。しかし、X染色体上の関連遺伝子座を同定しようとする試みは不成功に終わっている(Chalmers RMら,Am J Hum Genet 1996;59:103-108およびPegoraro Eら,Am J Med Genet 2003;119A:37-40,これらの引用文献の内容は参照により本明細書に組み込まれる)。いわゆる「二次LHON突然変異」は、欧州人LHON患者に非罹患欧州人よりも高い頻度で見いだされ、欧州人ハプロタイプJに連関する多型である。これらの多型は強い独立リスク因子ではない(Brown MDら,Am J Hum Genet 1997;60:381-387およびTorroni Aら,Am J Hum Genet 1997;60:1107-1121,これらの引用文献の内容は参照により本明細書に組み込まれる)。
【0008】
したがって、一次突然変異がLHONにおける主要リスク因子であるが、その病原性表現型を増強または調整する付加的な病因因子が必要であるらしい。アルコールおよび/またはタバコの多用がLHON家系における視神経症のリスクを増加させることは、かなりの証拠が示している(Smith PRら,Q J Med 1993;86:657-660、Chalmers RMら,Brain 1996;119:1481-1486およびTsao Kら,Br J Ophthalmol 1999;83:577-581,これらの引用文献の内容は参照により本明細書に組み込まれる)。ただし、この関連性を見いださなかった研究も一つある。考えうる二次的遺伝子相互作用は複雑であり、確立されていない(Kerrison JBら,Am J Ophthalmol 2000;130:803-812,この引用文献の内容は参照により本明細書に組み込まれる)。
【0009】
酸化ストレスはmtDNAの突然変異と関連する多くの障害に関係づけられている。最近行なわれた動物モデルでの研究では、LHONの病理発生過程における有望な因子として、反応性酸素種(ROS)が同定された(Qi Xら,Invest Ophthalmol Vis Sci 2003;44:1088-1096,この引用文献の内容は参照により本明細書に組み込まれる)。さらにまた、mtDNA LHON病原性突然変異により、細胞は、インビトロでFas依存性アポトーシス死を起こしやすくなることが見いだされた(Danielson SRら,J Biol Chem 2002;277:5810-5815,この引用文献の内容は参照により本明細書に組み込まれる)。これらの知見により、LHONの発病に関与するいくつかの核修飾因子遺伝子が存在するに違いないことが暗示された。
【発明の開示】
【課題を解決するための手段】
【0010】
本発明者らは、いくつかの既知SNPおよび未知SNPが緑内障およびレーベル病を含む視神経症の発症に連関することを明らかにし、本発明を完成した。
【0011】
したがって本発明は、
(1)エンドセリン-1遺伝子のコドン198におけるAAG→AAT置換(Lys198Asn);
(2)エンドセリン-1遺伝子プロモーター領域の-1370T>G多型;
(3)エンドセリン-1遺伝子のエクソン1におけるA138挿入/欠失(A138I/D)多型;
(4)エンドセリン受容体A遺伝子の3'非コード領域における+70C>G多型;
(5)エンドセリン受容体A遺伝子の+1222C>T多型;
(6)エンドセリン受容体A遺伝子のエクソン6中のコドン323におけるCAC→CAT置換(His323His);
(7)エンドセリン受容体A遺伝子プロモーター領域の-231A>G多型;
(8)エンドセリン受容体B遺伝子のエクソン4中のコドン277におけるCTG→CTA置換;
(9)ミトコンドリア遺伝子の9099C>A多型;
(10)ミトコンドリア遺伝子の9101T>G多型;
(11)ミトコンドリア遺伝子の9101T>C多型;
(12)ミトコンドリア遺伝子の9804G>A多型;
(13)ミトコンドリア遺伝子の11778G>A多型;
(14)アンギオテンシンII1型受容体遺伝子プロモーター領域の-713T>G多型;
(16)アンギオテンシンII2型受容体遺伝子の3123C>A多型;
(25)パラオキソナーゼ1遺伝子のコドン192におけるCAA→CGA置換(Gln192Arg);
(26)パラオキソナーゼ1遺伝子のコドン55におけるTTG→ATG置換(Leu55Met);
(27)ノエリン(Noelin)2遺伝子のコドン144におけるCGG→CAG置換(Arg144Gln);
(32)β1アドレナリン作動性受容体遺伝子のコドン389におけるGGA→CGA置換(Gly389Arg);
(35)ミオシリン遺伝子の1105T>C多型(Phe369Leu);
(36)オプチニューリン遺伝子の412G>A多型;
(37)E-セレクチン遺伝子の1402C>T多型;
(38)腫瘍壊死因子α遺伝子プロモーター領域の-857C>T多型とオプチニューリン遺伝子の412G>A多型との組み合わせ;
(39)腫瘍壊死因子α遺伝子プロモーター領域の-863C>A多型とオプチニューリン遺伝子の603T>A多型との組み合わせ;
(40)TP53遺伝子のコドン72におけるCGC→CCC置換(Arg72Pro);
(41)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン113におけるTAC→CAC置換(Tyr113His);
(42)熱ショックタンパク質70-1遺伝子プロモーター領域の-110A>C多型;
(43)エンドセリン変換酵素遺伝子プロモーター領域の-338C>A多型;
(44)CD95遺伝子プロモーター領域の-670A>G多型;
(45)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン119におけるAAG→AAA置換(Lys119Lys);
(47)β2アドレナリン作動性受容体遺伝子のコドン16におけるGGA→AGA置換(Gly16Arg);および
(48)β2アドレナリン作動性受容体遺伝子のコドン27におけるCAA→GAA置換(Gln27Glu)
からなる群より選択される少なくとも一つの多型を含む、視神経症と関連する遺伝子多型のセットを提供する。
【0012】
さらに本発明は、ヒト被験者における視神経症に対する感受性を診断または予測するための方法であって、
i)被験者から生物学的試料を取得する工程、
ii)上に定義した多型のセットに関して、試料の遺伝子型を決定する工程、および
iii)被験者における視神経症に対する感受性を遺伝子型に基づいて診断または予測する工程
を含む方法も提供する。
【0013】
本発明によれば、視神経症は、好ましくは、緑内障またはレーベル病であることができる。多型(1)〜(39)および(42)〜(48)は、特に緑内障に使用することができる。それらのうち、多型(1)、(2)、(5)〜(7)、(16)、(26)、(32)、(43)および(45)は、特に正常眼圧緑内障に使用することができ、多型(4)、(14)、(25)、(35)、(36)、(38)、(42)、(44)、(47)〜(48)は、特に原発開放隅角緑内障に使用することができる。多型(40)および(41)は、特にレーベル病に使用することができる。
【0014】
本発明によれば、多型のセットは、視神経症と関連することが知られている少なくとも一つの他の多型を、さらに含みうる。
【0015】
本発明のもう一つの態様として、上に定義した遺伝子多型のセットに関して遺伝子型を決定するのに適したプライマーセットおよび/またはプローブを含む、ヒト被験者における視神経症に対する感受性を診断または予測するためのキットが提供される。
【0016】
本発明のさらなる態様として、ミトコンドリア遺伝子、ミオシリン遺伝子およびノエリン2遺伝子内に新たに同定されたSNPを提供する。したがって本発明は、これらのSNPを含むヌクレオチド断片を包含する。一般に、前記SNPに関して遺伝子型を決定するには、そのSNPを含有する90個以上の連続するヌクレオチドの配列が必要だろう。すなわち、配列:

のセグメントからなる単離ポリヌクレオチドであって、
前記セグメントは少なくとも90個の連続するヌクレオチドを含んでなり、その少なくとも90個の連続するヌクレオチドは上記配列の9099位を含み、上記配列の9099位がAである、単離ポリヌクレオチド、もしくは上記セグメントに対して完全に相補的な単離ポリヌクレオチド;または、
前記セグメントは少なくとも90個の連続するヌクレオチドを含んでなり、その少なくとも90個の連続するヌクレオチドは上記配列の9101位を含み、上記配列の9101位がGである、単離ポリヌクレオチド;または、
上記セグメントのいずれか一方に対して完全に相補的な単離ポリヌクレオチドが提供される。
【0017】
さらに本発明は、配列:
のセグメントからなる単離ポリヌクレオチドであって、前記セグメントは少なくとも90個の連続するヌクレオチドを含んでなり、その少なくとも90個の連続するヌクレオチドは上記配列の下線部のヌクレオチドに相当するコドン369を含み、コドン369は、それがLeuをコードするように置換されている、単離ポリヌクレオチド、または上記セグメントに対して完全に相補的な単離ポリヌクレオチドを提供する。
【0018】
さらに本発明は、配列:
のセグメントからなる単離ポリヌクレオチドであって、前記セグメントは少なくとも90個の連続するヌクレオチドを含んでなり、その少なくとも90個の連続するヌクレオチドは上記配列の下線部のヌクレオチドに相当するコドン144を含み、コドン144は、それがGlnをコードするように置換されている、単離ポリヌクレオチド、または上記セグメントに対して完全に相補的な単離ポリヌクレオチドを提供する。
【発明を実施するための最良の形態】
【0019】
本明細書および特許請求の範囲において、「遺伝子多型」とは、ある遺伝子座における個体間のゲノム多様性を意味する。遺伝子多型は「一塩基多型」または「SNP」と呼ばれる一塩基置換であってもよいし、複数のヌクレオチドからなるものであってもよい。遺伝子多型は、その個体の表現型に影響を及ぼすものであっても、影響を及ぼさないものであってもよい。さらに、ある個体のヌクレオチド配列が、対応する野生型配列とは異なる(すなわち野生型配列上に挿入、欠失または置換を持つ)場合、そのヌクレオチド配列を「遺伝子突然変異体」と呼ぶが、その遺伝子突然変異体も本明細書にいう「多型変異体」に含まれる。
【0020】
本明細書および特許請求の範囲において、「9099C>A」または「C9099A」などの表現は、その遺伝子が9099位に多型を持つこと、すなわちその遺伝子には二つの対立遺伝子があり、一方は9099位にシトシン、すなわちCを持ち、他方は9099位にアデニン、すなわちAを持つこと(二対立遺伝子性)を意味する。これは、高頻度の対立遺伝子がその位置にCを持ち、低頻度の対立遺伝子がその位置にAを持つことを、必ずしも意味しない。
【0021】
「Gln192Arg」などの表現は、アミノ酸配列をコードする遺伝子における塩基置換によるアミノ酸置換を表す。例えば、Gln192Argは、コドン192(すなわちアミノ酸番号192)のグリシンが、アルギニン、すなわちArgで置き換えられることを表す。これは、そのタンパク質に、コドン192のアミノ酸がGlnまたはArgである多型変異体が存在することも意味する。
【0022】
本発明によれば、遺伝子多型に関する遺伝子型の決定は、単一の多型ごとに行なうか、または複数の多型もしくは全ての多型を同時に決定することができる。
【0023】
本発明において、ヒト被験者における視神経症に対する感受性を診断または予測するための方法は、本願で新たに報告する視神経症との関係を持つ遺伝子多型のセットに関して、遺伝子型を決定することを含む。本発明によって視神経症に連関すると同定された遺伝子多型に加えて、視神経症に連関することが既に明らかにされていた他の任意の多型を一緒に検出することができる。視神経症に連関する複数の遺伝子多型を使用することにより、診断確率を改善することができる。
【0024】
本発明によれば、遺伝子多型に関して遺伝子型を決定するために使用する方法は限定されず、当技術分野で知られている任意の方法を使用することができる。遺伝子多型に関して遺伝子型を決定するための代表的方法には、ポリメラーゼ連鎖反応・制限断片長多型(PCR-RFLP)解析、ポリメラーゼ連鎖反応後の一本鎖高次構造多型(PCR-SSCP)解析、ASOハイブリダイゼーション解析、ダイレクトシークエンス解析、ARMS解析、DGGE解析、RNaseA切断解析、化学的制限切断解析(chemical restriction analysis)、DPL解析、TaqMan(登録商標)PCR解析、Invader(登録商標)アッセイ、MALDI-TOF/MS解析、TDI解析、一塩基伸長アッセイ、WAVEアッセイ、一分子蛍光検出アッセイなどがある。本発明によれば、検出方法はこれらの方法の一つであってもよいし、二つ以上の組み合わせであってもよい。
【0025】
本発明によれば、遺伝子多型の検出に用いるべき生物学的試料は特に限定されず、毛髪、血液、唾液、リンパ液、気道粘膜、培養細胞および尿であることができる。
【0026】
本明細書および特許請求の範囲において、「視神経症に対する感受性を診断または予測すること」には、視神経症の発症を診断することだけでなく、疾患の発症を早め、疾患の進行を加速するリスク因子を決定することも含まれる。
【0027】
本発明によれば、上に同定した遺伝子多型ならびにタンパク質多型を検出するためのキットも提供される。このキットは、上に同定した遺伝子多型を検出するために特に設計されたプライマーおよび/またはプローブ、上に同定したタンパク質多型を検出するための抗体を含む。本発明によれば、このキットは、視神経症に対する感受性を診断または予測するために使用することができる。
【0028】
本明細書および特許請求の範囲において、用語「プライマー」は、標的ヌクレオチド配列の一部に相補的であって、標的ヌクレオチド配列にハイブリダイズさせるために使用される、特異的オリゴヌクレオチドを意味する。プライマーは、DNAポリメラーゼ、RNAポリメラーゼまたは逆転写酵素のいずれかによって触媒されるヌクレオチド重合の開始点として役立つ。
【0029】
用語「プローブ」は、試料中に存在する特異的ポリヌクレオチド配列を同定するために、または遺伝子修飾過程において標的DNAもしくはRNAを確認するために、使用することができる所定の核酸セグメントであって、同定されるべき特異的ポリヌクレオチド配列に相補的なヌクレオチド配列を含むものを意味する。
【0030】
本発明によれば、プライマーおよびプローブは、標的とする配列に基づいて、それらが他のいくつかの遺伝子と同一でない限り(すなわち反復配列でもパリンドローム配列でもないことが必要である)、標的とする多型が予想される位置および/またはその位置の周辺配列に対して、それらが相補的になるように、設計することができる。
【0031】
本発明によれば、視神経症、特に緑内障およびレーベル病に連関する遺伝子多型が同定される。これらの知見に基づいて、個体から取得した生物学的試料の遺伝子多型に関する遺伝子型を決定し、そのようにして取得した遺伝子型に基づいて、疾患の発症または疾患の発症に関して予測されるリスクを決定することができる。
【0032】
上に同定した多型(1)〜(48)に加えて、視神経症と高度に関連することが当技術分野で既に知られていた他のいくつかの遺伝子多型に関する遺伝子型も、診断または予測の信頼性を改善するために決定することができる。
【0033】
例えば、ミオシリン遺伝子およびオプチニューリン遺伝子中の2タイプの遺伝子多型は原発開放隅角緑内障の発症と関連することが、本発明者らによって明らかにされている。これら二つの遺伝子に加えて、突然変異を含む他の四つの遺伝子多型が原発開放隅角緑内障と関連すると既に同定されていた。本発明の遺伝子多型に関するリスク遺伝子型と当技術分野で既に知られていた遺伝子多型に関するリスク遺伝子型をどちらも持つ被験者は、ほぼ100%が、緑内障を発病しうる。すなわち、これら遺伝子多型のセットは、発症前検査に有用であるだろう。
【0034】
いくつかのSNPに関して、本発明者らは、人種または性別などの特定の群で、視神経症との相関を確認した。したがって、指定された群における視神経症のリスクを診断または予測するには、それらのSNPを好ましく使用することができる。
【0035】
さらに、多型のセットに関する遺伝子型の統計学的解析により、予測発症年齢、生活習慣病との予測的関連性、症状因子との予測的関連性などの有用な情報が得られうる。さらに、その情報に基づいて、いくつかの医学的処置の効果を予測することもできるだろう。
【0036】
本発明によれば、視神経症に対する感受性の予測を、遺伝子型に基づいて疾患の発症前に行なうことができ、被験者はリスク因子を取り除く方法についての助言(例えば生活習慣を改善するようにという助言、または環境を変えるようにという助言)を受けることができる。さらに、リスク遺伝子の減少などの早期処置を受けることもできるだろう。適切な処置を早期に開始することができる。結果として、これらの「オーダーメード処置」は、視覚喪失のリスクを低下させることができる。
【0037】
例えば、ある被験者が視神経症の発症に関する高リスクに連関する遺伝子型を持つ場合、低い発症リスクに連関する遺伝子型をその被験者に導入し、それを発現させることによって、発症リスクの低下または症状の軽減を期待することができる。さらに、高リスク対立遺伝子の発現を阻害するために、視神経症の発症に関して高いリスクを持つ対立遺伝子のmRNAに対するアンチセンス、またはRNAi法を使用することもできる。
【0038】
もう一つの態様として、本発明に示される多型のセットに関する遺伝子型決定に基づいて、視神経症の遺伝的病因を明らかにすることができ、そのようにして得られた病因は新規医薬剤の開発に役立ちうる。
【0039】
さらに、本発明によって得られた視神経症と関連する遺伝子型情報と、生活習慣病などと関連する他の遺伝子型情報とを組み合わせることにより、加齢関連生活習慣関連疾患の総合的リスクを予測し、生活の質を高めるために使用することができる。
【0040】
下記の実施例を使って本発明をさらに例示する。しかし、これらの実施例は単に例示が目的であって、本発明の範囲を限定しようとするものではないと、明確に理解すべきである。
【実施例1】
【0041】
レーベル遺伝性視神経症におけるTP53およびEPHX1の遺伝子変異体ならびにそれらの発症時年齢との関係
目的:酸化ストレスおよびアポトーシスに関する遺伝子の遺伝子多型がレーベル遺伝性視神経症(LHON)を持つ患者における臨床的ばらつきを引き起こすかどうかを決定すること。
【0042】
材料および方法
患者
本発明者らは、ホモプラスミー状態の11778突然変異を保有する血縁関係にない86人の日本人LHON患者を研究した。彼らのmtDNA突然変異を、ポリメラーゼ連鎖反応後の制限酵素アッセイによって確認したところ、一貫してMaeIII部位が得られることが明らかになった(Mashima Yら,Curr Eye Res 1998;17:403-408,この引用文献の内容は参照により本明細書に組み込まれる)。
【0043】
86人のLHON患者における視覚喪失の平均発症時年齢は25.1±13.0歳で、3歳〜65歳の範囲だった。
【0044】
ゲノムDNA抽出および遺伝子型判定
SDS-プロテイナーゼKおよびフェノール/クロロホルム抽出法により、末梢血白血球からDNAを抽出した。酸化ストレス関連遺伝子、ミクロソームエポキシドヒドロラーゼ(EPHX1)(Kimura Kら,Am J Ophthalmol 2000;130:769-773,この引用文献の内容は参照により本明細書に組み込まれる)、およびアポトーシス関連遺伝子TP53中のArg72Pro(Ara Sら,Nucleic Acids Res 1990;18:4961,この引用文献の内容は参照により本明細書に組み込まれる)における多型を調べた。
【0045】
ポリメラーゼ連鎖反応-制限断片長多型(PCR-RFLP)技術を使って各多型を同定した(表1)。
【0046】
[表1]
【0047】
結果
発症時年齢と多型との関連を表2-1および表2-2に記載した。
【0048】
[表2−1]
レーベル遺伝性視神経症における発症時年齢とTP53(Arg72Pro)遺伝子多型およびEPHX1(Tyr113His)遺伝子多型との関連
【0049】
[表2−2]
レーベル遺伝性視神経症における発症時年齢とTP53(Arg/Arg)遺伝子多型およびEPHX1(His/His)遺伝子多型との関連
第1群:TP53のコドン72にArg/Argを持ち、EPHX1のコドン113にHis/Hisを持つ患者。
第2群:TP53のコドン72にArg/Argを持つがEPHX1のコドン113にHis/Hisを持たない患者またはEPHX1のコドン113にHis/Hisを持つがTP53のコドン72にArg/Argを持たない患者。
第3群:第1群および第2群以外の患者。
【0050】
表2-1に示すように、TP53中のコドン72遺伝子型およびEPHX1中のコドン113遺伝子型は、レーベル遺伝性視神経症の発症時年齢の若さと有意に関連した。
【0051】
表2-2に示すように、TP53中のコドン72遺伝子型とEPHX1中のコドン113遺伝子型との併存は、レーベル遺伝性視神経症の発症時年齢の若さと有意に関連した。
【0052】
これらの結果から、TP53におけるArg/Argホモ接合体およびEPHX1におけるHis/Hisホモ接合体を検出することにより、レーベル遺伝性視神経症の早期診断および早期処置が可能になることが示された。
【0053】
これらの結果は、コドン72多型がミトコンドリア機能障害と相互作用して疾患発現に影響を及ぼしうることも示した。p53のコドン72における多型と相関するアポトーシス応答には個体変動が存在しうる。Bonafeら(Biochem Biophys Res Commun 2002;299:539-541)は、Arg/Arg遺伝子型を保有する健常被験者に由来する培養細胞が、細胞毒性薬シトシンアラビノシドに応答して、Arg/Pro被験者に由来する細胞よりも大規模なアポトーシスを起こすことを報告した。したがって、p53遺伝子に天然に存在する遺伝的ばらつきは、化学療法薬に対する細胞のインビボ感受性の個体差に関する説明の一つになるだろう。Dumountら(Nat Genet 2003;33:357-365)は、Arg72変異体がPro72変異体よりも効率よくアポトーシスを誘発することを報告した。この効率増大の基礎にある少なくとも一つの機序は、腫瘍細胞中のミトコンドリアへのArg72変異体の局在の強化である。合成p53阻害剤は、ミトコンドリア損傷および酸化ストレスが惹起するアポトーシスによってニューロンが死ぬLHONの処置に、著しく有効であるかもしれない。
【0054】
標的とする多型を含有するEPHX1遺伝子およびTP53遺伝子の部分ヌクレオチド配列は以下のとおりである。
EPHX1 Tyr113Hisコドン113(下線部)(TAC→CAC変化)
【0055】
TP53コドン72(下線部):CGC(Arg)→CCC(Pro)
【実施例2】
【0056】
原発開放隅角緑内障および正常眼圧緑内障における、レーベル遺伝性視神経症と関係するミトコンドリアDNA突然変異
材料および方法
患者
日本国内の7施設で合計651の血液試料を集めた。POAG患者が201人、NTG患者が232人、正常対照が218人であり、どの被験者もこの研究における他の被験者とは血縁関係になかった。
【0057】
検査時の平均年齢は、POAGでは61.2±16.0歳、NTGでは58.8±13.6歳、対照被験者では70.6±10.9歳だった。対照被験者の平均年齢はPOAG患者(P<0.001)およびNTG患者(P<0.001)の平均年齢より有意に高かった。本発明者らは、対照被験者の一部が最終的には緑内障を発病してしまう確率を低下させるために、より高齢の対照被験者を意図的に選択した。男性はPOAG群では112人(55.7%)、NTG群では108人(46.6%)、対照群では89人(40.8%)だった。
【0058】
正常開放隅角、ゴールドマン視野測定またはハンフリー視野測定で典型的な緑内障性視野喪失を伴う0.7より大きい陥凹/乳頭径比を持ち、視神経損傷の原因となりうる眼障害、鼻障害、神経障害または全身性障害を持たない場合に、その患者はPOAGを有するとみなした。NTGを有する患者は21mmHg以下のIOPを持っていた。剥脱性緑内障、色素性緑内障、およびコルチコステロイド誘発性緑内障を持つ患者は除外した。
【0059】
218の対照試料は、白内障を除く既知の眼異常を持たない日本人被験者から取得した。これらの被験者は40歳を超えていて、21mmHg未満のIOPを持ち、正常な視神経乳頭を持ち、緑内障の家族歴はなかった。
【0060】
Invader(登録商標)アッセイによるmtDNA突然変異の検出
標準的なフェノール-クロロホルム抽出法により、末梢血リンパ球からゲノムDNAを単離した。
【0061】
Invader(登録商標)アッセイによって六つのmtDNA突然変異(G3460A、T9101C、G9804A、G11778A、T14484C、およびT14498C)を検出するために使用した一次プローブ(野生型プローブおよび突然変異型プローブ)ならびにInvader(登録商標)オリゴヌクレオチド(Invader(登録商標)プローブ)を、表3に示す。
【0062】
[表3]
【0063】
Invader(登録商標)アッセイFRET検出256穴プレート(Third Wave Technologies, Inc.,ウィスコンシン州マディソン)は、個々のウェルのそれぞれに乾燥されたInvader(登録商標)アッセイの一般成分(Cleavase(登録商標)酵素VIII、FRETプローブ、MOPSバッファー、およびポリエチレングリコール)を含有している。二重(biplex)形式のInvader(登録商標)アッセイにより、一つのウェルで二つのDNA配列を同時に検出することができた。
【0064】
詳細な方法は先に記述されている。簡単に述べると、一次プローブ/Invader(登録商標)/混合物8μlおよび全DNA(10ng)試料を96穴プレートの各ウェルに加え、95℃で10分間インキュベートすることによって変性させた。鉱油(Sigma,ミズーリ州セントルイス)15μlを全ての反応ウェルに重層し、プレートをPTC-100サーマルサイクラー(MJ Research,マサチューセッツ州ウォルサム)にて等温的に63℃で2時間インキュベートした後、蛍光測定まで4℃で保存した。蛍光強度は、CytoFlour 4000蛍光プレートリーダー(Applied Biosystems,カリフォルニア州フォスターシティ)を使って、FAM色素については励起485nm/20nm(波長/バンド幅)および放射530nm/25nmで、またRedmond RED(RED)色素については励起560nm/20nmおよび放射620nm/40nmで測定した。各試料を同じプレートで二重に試験し、各プレートで2回の蛍光測定を行なった。そのようにして、各試料につき四つの測定値を取得し、それらを平均した。
【0065】
ダイレクトDNAシークエンス
ダイレクトシークエンス法によって突然変異を検出するために、PCR産物をまずQIAquick PCR精製キット(QIAGEN,米国カリフォルニア州バレンシア)で精製することにより、未反応のプライマーおよび前駆体を除去した。次に、ABI PRISM BigDye Terminator(v.3.1)サイクルシークエンスキットを、製造者のプロトコール(Applied Biosystems)に従って使用することにより、シークエンス反応を行なった。ABI PRISM 310遺伝子解析装置によってデータを収集し、ABI PRISMシークエンス解析プログラム(v.3.7)によって解析した。
【0066】
[表4]
【0067】
結果
合計651人の日本人被験者を調べた。ヌクレオチド置換が一次プローブ内またはInvaderプローブ内にある場合、検査した症例は、Invaderアッセイでは、どちらのプローブにも反応を示さなかった。そのような症例において、ダイレクトシークエンス解析を行なったところ、9099、9101、9102、9797、および9815のヌクレオチド位置に一塩基多型(SNP)が見いだされた。
【0068】
表5に示すように、女性5人および男性2人を含む7人の患者が、mtDNAに五つの突然変異を保持しており、LHONを発病していなかった。2人の患者(症例1および症例2)は、LHONと関連づけられていない新規なアミノ酸変化を保持しており、5人の患者(症例3〜7)は、LHON突然変異を保持していた。
【0069】
これらのmtDNA突然変異は正常対照には検出されなかった。
【0070】
[表5]
【0071】
上述のように、本発明者らは、緑内障患者に二つの新規mtDNA突然変異を含む五つのmtDNA突然変異を見いだした。これらの結果から、mtDNA突然変異は緑内障を発病または進行させるリスク因子の一つであること、そしてmtDNA突然変異の検出により、緑内障の早期診断および早期処置が可能になることが示された。
【0072】
標的とする突然変異/多型を含有するミトコンドリア遺伝子の部分ヌクレオチド配列は以下のとおりである。
C9099A,T9101G(下線部)
【0073】
G9804A(下線部)
【0074】
G11778A(下線部)
【実施例3】
【0075】
レニン-アンギオテンシン-アルドステロン系の遺伝子多型は原発開放隅角緑内障および正常眼圧緑内障を発病するリスクと関連する
目的:緑内障の病理発生過程には複数の環境的因子および遺伝的因子が関与しうる。緑内障の遺伝的リスクを予測するために、レニン-アンギオテンシン-アルドステロン(R-A-A)系の遺伝子多型における関連研究を行なった。
【0076】
材料および方法
患者および対照研究被験者
日本国内の7施設で合計551の血液試料を集めた。POAG患者が162人、NTG患者が193人、正常被験者が196人であり、どの被験者もこの研究における他の被験者とは血縁関係になかった。
【0077】
検査時の平均年齢は、NTGでは58.8±13.7歳、POAGでは62.0±15.4歳、および正常被験者では71.2±10.4歳だった。正常対照被験者の平均年齢は、NTG患者(P<0.001)またはPOAG患者(p<0.001)より、それぞれ有意に高い。これにより、一部が最終的には緑内障を発病してしまう可能性を低減することができた。NTG患者127人のうち66人(34.2%)およびPOAG患者113人のうち49人(30.2%)に、家族歴が記録された。男性患者はNTGでは89人(46.1%)、POAGでは87人(53.7%)、正常被験者では77人(39.3%)だった。
【0078】
196の日本人対照試料は、白内障を除く既知の眼異常を持たない個体から取得した。これらの被験者は40歳を超えていて、21mmHg未満のIOPを持ち、緑内障性乳頭変化を持たず、緑内障の家族歴はなかった。
【0079】
遺伝子型判定
R-A-A系中の七つの遺伝子および10個の多型を、緑内障を持つ被験者または正常日本人対照のそれぞれについて決定した。レニン(REN)I8-83G>A(Frossard PMら,Hypertens Res 1998;21:221-225,この引用文献の内容は参照により本明細書に組み込まれる)、アンギオテンシンII1型受容体(AT1R)1166A>C、-521C>T、-713T>G(Nalogowska-Glosnicka Kら,Med Sci Monit 2000;6:523-529およびErdmann Jら,Ann Hum Genet 1999;63:369-374,これらの引用文献の内容は参照により本明細書に組み込まれる)、アンギオテンシンII2型受容体(AT2R)3123C>A(Katsuya Tら,Mol Cell Endocrinol 1997;127:221-228,この引用文献の内容は参照により本明細書に組み込まれる)、シトクロムP45011B1(CYP11B1)-344T>C(Tsujita Yら,Hypertens Res 2001;24:105-109,この引用文献の内容は参照により本明細書に組み込まれる)、およびキマーゼ(CYM)3123C>Aは、ポリメラーゼ連鎖反応-制限断片長多型(PCR-RFLP)を使って同定した。アンギオテンシン変換酵素(ACE)挿入/欠失(I/D)はPCRおよびアガロースゲル電気泳動だけで決定した。ACE/ID多型の偽決定を避けるために、Lindpaintnerらのプロトコール(N Engl J Med 1995;332:706-711,この引用文献の内容は参照により本明細書に組み込まれる)に従って、I対立遺伝子特異的な増幅を行なった。ゲノムDNAは、フェノール-クロロホルム抽出により、末梢血リンパ球から単離した。使用したプライマーセットおよび制限酵素を表6に記載した。
【0080】
[表6]
【0081】
アンギオテンシノゲン(AGT)T174M、M235Tの遺伝子型はInvaderアッセイ(登録商標)を使って決定した(Lyamichev Vら,Nat Biotechnol 1999;17:292-296,この引用文献の内容は参照により本明細書に組み込まれる)。
【0082】
結果
日本人集団におけるR-A-A系の遺伝子型分布
R-A-A系中の10個の多型のうちの二つ、すなわちPOAGに関してAT1R/-713T>Gと、NTGに関してAT2/3123C>Aは、遺伝子型の頻度に有意差を示した(表7)。3123C>A多型はNTGを持つ女性患者とだけ関連した。
【0083】
AT1R/-713T>G多型におけるホモ接合性G遺伝子型(GG)の頻度は、POAG患者(4.2%)では対照(0.5%)よりも有意に高かった(TT+TG対GGについてp=0.04)。AT2R/3123C>A多型におけるCA+AA遺伝子型の頻度は、NTGを持つ女性患者(70.8%)では女性対照(55.0%)よりも有意に高かった(CC対CA+AAについてp=0.011)。
【0084】
[表7]
緑内障(POAGおよびNTG)とレニン-アンギオテンシン-アルドステロン系の遺伝子多型との関連
【0085】
POAG患者におけるAT1R中の二つのプロモーター多型間の関連
ホモ接合性-521Tとホモ接合性-713Gとを合わせ持つPOAG保有者の頻度(4.2%)は、正常者(0%)より有意に高かった(p=0.011)(表8-1)。POAG患者だけがこの遺伝子型を持ち、NTG被験者も正常被験者もこの遺伝子型を持たなかった。
【0086】
[表8−1]
AT1R -521T対立遺伝子および-713G対立遺伝子の遺伝子型の分布
A:二つの-521対立遺伝子および二つの-713G対立遺伝子を持つ被験者。
B:A群の基準を満たさない被験者。
【0087】
これらの結果から、レニン-アンギオテンシン-アルドステロン系の遺伝子多型は緑内障の発病に関する重要な遺伝的リスク因子の一つであることが示された。AT1R/-731T>G多型の検出により、POAGの早期診断および早期処置が可能になる。特に、AT1R遺伝子中にホモ接合性-521Tとホモ接合性-713Gとを合わせ持つ特異な遺伝子型は、POAGの早期診断に有用である。AT2R/3123C>A多型の検出により、NTGを持つ女性患者の早期診断および早期処置が可能になる。
【0088】
AT2R 3123C>A多型およびACE I/D多型を持つNTG患者の臨床特性
緑内障患者に関して記録された臨床特徴は、診断時の年齢、無処置時最大IOP(診断時のIOPと定義される)、および初診時の視野欠損(診断時の視野欠損と定義される)だった。視野欠損の重症度は1〜5にスコア化した。異なる視野計を使って取得したデータを、以下に定義する5点尺度を使って集約した。1=変化なし;2=早期欠損;3=中等度欠損;4=重度欠損;および5=光覚のみまたは視覚なし。視野欠損は、ゴールドマン視野測定の結果に基づくKozakiの分類に従って、またはハンフリー視野解析機に用いられる分類に従って、早期、中等度、または重度と判定した。日本では前者の分類が最も広く用いられている。
【0089】
AT2R遺伝子型を持つ男性緑内障患者との間に、視野スコア臨床特性の有意な関連が検出された。C遺伝子型を持つ男性POAG患者における視野スコアは、A遺伝子型を持つ者より悪かった(P=0.04,表8-2)。C/Cを持つ女性緑内障患者およびC/A+A/Aを持つ女性緑内障患者との間に、臨床特性(年齢、IOP、および視野スコア)の有意な関連は検出されなかった。視野スコアは、C/C遺伝子型を持つNTG患者では、C/A+A/A遺伝子型を持つNTG患者より悪い傾向があった(P=0.165)。
【0090】
しかし、ACE挿入/欠失多型と組み合わせた場合、AT2R遺伝子中にC/Cを保有すると共にACE遺伝子中にID+DDを保有する女性NTG患者は、他の三つの複合遺伝子型よりも視野スコアが有意に悪かった(P=0.012;表8-3,図1)。
【0091】
[表8−2]
AT2R遺伝子型による男性緑内障患者の臨床特性の比較
【0092】
[表8−3]
【0093】
標的とする多型を含有するAT1R遺伝子およびAT2R遺伝子の部分ヌクレオチド配列は以下のとおりである。
AT1R -713(下線部の「t」) T>G
【0094】
AT2R 3123(下線部の「c」)C>A,下線部のオリゴヌクレオチド配列をプライマーに使用した
【0095】
(GenBankアクセッション番号AY536522によれば、AT2R 3123はヌクレオチド番号4926に対応する)
【実施例4】
【0096】
正常眼圧緑内障を発病するリスクと関連するエンドセリン遺伝子の遺伝子多型
方法
患者
合計605の血液試料を集めた。POAG患者が178人、NTG患者が214人、正常対照が213人であり、どの被験者もこの研究における他の被験者とは血縁関係になかった。正常開放隅角、ゴールドマン視野測定またはハンフリー視野測定で典型的な緑内障性視野喪失を伴う0.7より大きい陥凹/乳頭径比を持ち、視神経損傷の原因となりうる眼障害、鼻障害、神経障害または全身性障害を持たない場合に、その患者はPOAGを有するとみなした。NTGを有する患者は21mmHg以下のIOPを持っていた。剥脱性緑内障、色素性緑内障、およびコルチコステロイド誘発性緑内障を持つ患者は除外した。対照試料は、白内障を除く既知の眼異常を持たない日本人被験者から取得した。これらの被験者は21mmHg未満のIOPを持ち、正常な視神経乳頭を持ち、緑内障の家族歴はなかった。
【0097】
Invaderアッセイによるエンドセリン(ET)遺伝子のG/T多型の検出
標準的なフェノール-クロロホルム抽出法によって末梢血リンパ球からDNAを単離し、ET遺伝子のエクソン5中のコドン198におけるG/T多型(Lys/Lys、Lys/AsnおよびAsn/Asn)を、Invader(登録商標)アッセイによって決定した。ET遺伝子のG/T多型を検出するために使用した一次プローブ(野生型プローブおよび突然変異型プローブ)ならびにInvader(登録商標)オリゴヌクレオチド(Invader(登録商標)プローブ)を表9に示す。
【0098】
[表9]
【0099】
Invader(登録商標)アッセイFRET検出96穴プレート(Third Wave Technologies, Inc.,ウィスコンシン州マディソン)は、個々のウェルのそれぞれに乾燥されたInvader(登録商標)アッセイの一般成分(Cleavase(登録商標)酵素VIII、FRETプローブ、MOPSバッファー、およびポリエチレングリコール)を含有している。簡単に述べると、一次プローブ/Invader(登録商標)/混合物8μlおよび全DNA(10ng)試料を96穴プレートの各ウェルに加え、95℃で10分間インキュベートすることによって変性させた。鉱油(Sigma,ミズーリ州セントルイス)15μlを全ての反応ウェルに重層し、プレートをPTC-100サーマルサイクラー(MJ Research,マサチューセッツ州ウォルサム)にて等温的に63℃で2時間インキュベートした後、蛍光測定まで4℃で保存した。蛍光強度は、CytoFlour 4000蛍光プレートリーダー(Applied Biosystems,カリフォルニア州フォスターシティ)を使って、FAM色素については励起485nm/20nm(波長/バンド幅)および放射530nm/25nmで、またRedmond RED(RED)色素については励起560nm/20nmおよび放射620nm/40nmで測定した。各試料を同じプレートで二重に試験し、各プレートで2回の蛍光測定を行なった。そのようにして、各試料につき四つの測定値を取得し、それらを平均した。
【0100】
結果
ET遺伝子のエクソン5中のコドン198におけるG/T多型(Lys/Lys、Lys/AsnおよびAsn/Asn)の遺伝子型頻度を表10に示す。
【0101】
[表10]
ET遺伝子のエクソン5中のコドン198における遺伝子型頻度
【0102】
これらの結果から、エクソン5中のコドン198におけるET-1遺伝子のLys/Lysホモ接合体は、NTGを発病または進行させるリスク因子の一つであり、Lys/Lysホモ接合体の検出により、NTGの早期診断および早期処置が可能になる。
【0103】
コドン198を含むEDN1の部分配列は以下のとおりである。
EDN1 コドン198(下線部):aag(Lys)→aat(Asn)
【実施例5】
【0104】
変性高速液体クロマトグラフィーによって検出される、原発開放隅角緑内障を持つ日本人患者における新規MYOC遺伝子突然変異Phe369Leu
目的:変性高速液体クロマトグラフィー(DHPLC)を使って原発開放隅角緑内障(POAG)を持つ日本人患者におけるMYOC遺伝子中の突然変異についてスクリーニングすること。
【0105】
材料および方法
患者
日本の医療施設7ヶ所でPOAG患者171人および正常被験者100人から血液試料を集めた。被験者らは血縁関係になく、検査時の平均年齢は、POAGを持つ患者では55.1±16.0歳(±標準偏差)、正常被験者では70.5±10.6歳だった。本発明者らは、対照被験者の一部が最終的には緑内障を発病してしまう確率を低下させるために、より高齢の対照被験者を意図的に選択した。
【0106】
55人のPOAG患者(32.2%)では問診によって詳細な家族歴が得られた。男性はPOAG患者では91人(53.2%)、正常被験者では41人(41.0%)だった。
【0107】
DNA抽出およびPCR条件
標準的方法により末梢血リンパ球からゲノムDNAを単離した。ポリメラーゼ連鎖反応(PCR)により、表11に記載するプライマーセットを使って、MYOC遺伝子の七つのエクソン領域を増幅した。患者のハイスループット解析を行なう場合は、3人の患者から得た試料をプールした。PCR反応はサーマルサイクラー(iCyceler;BioRad,カリフォルニア州ハーキュリーズ)を使って25μlの総液量で行なった。PCR条件は以下のとおりとした:95℃で9分間変性;次に、95℃で1分、58℃で30秒(表1)、そして72℃で1.5分を35サイクル;次に、72℃で7分間の最終伸長工程を行なった。ヘテロ二重鎖形成のために、各PCR産物(25μl)を95℃で5分間変性し、25℃まで徐冷した。
【0108】
数個の試料を解析する場合は、96穴プレート(96穴マルチプレートMLP-9601;MJ Research,マサチューセッツ州ウォルサム)でのPCRによって、七つのエクソン領域のそれぞれを同時に増幅した。各患者につき七つのウェルを使用した。58℃という単一のアニーリング温度を使った場合に有効であるように、プライマーセットを設計した(表11)。
【0109】
[表11]
プライマー配列、産物サイズ、ならびにPCRアニーリング温度およびDHPLC解析温度
【0110】
変性HPLC解析
ハイスループット解析の場合、3人の患者から得られたPCR産物25μlを、DHPLC解析用のWAVE(登録商標)システム(Transgenomic,ネブラスカ州オマハ)を使って、解析用クロマトグラフに自動注入した。DHPLC融解温度を表1に記載する。小数の試料を解析する場合は、96穴プレートPCR後に、2〜3の融解温度で各ウェルを自動的に解析するようにプログラムされたWAVE(登録商標)システムに、プレートを入れた。一個体の試料を解析するには、約3時間あれば十分だった。
【0111】
ハイスループットプロトコールでプール試料に異常なクロマトグラフパターンが検出された場合は、その試料をWAVE(登録商標)システムで個別に再解析した。次に、異常なクロマトグラフパターンを示したPCR産物をシークエンスした。
【0112】
ダイレクトDNAシークエンス
ダイレクトシークエンスのために、PCR産物をQIA Quick PCR精製キット(Qiagen,カリフォルニア州バレンシア)で精製することにより、未使用のプライマーおよび前駆体を除去した。BigDyeケミストリーを製造者の推奨するプロトコール(Applied Biosystems,カリフォルニア州フォスターシティ)に従って使用することにより、ABI310自動シークエンサーで、同じフォワードおよびリバースPCR増幅プライマーを使って、PCR産物をダイレクトシークエンスした。
【0113】
結果
患者171人でのDNAプールのスクリーニング
エクソン3C領域における四つのDHPLC記録線パターンを図2に示した。一番上のパターン(A)が正常な外観を持つのに対して、中央のパターン(B)は幅広いショルダーを示し、下側のパターン(CおよびD)はこの領域における配列変異を示す特徴的な二重ピークパターンを持っていた。試料B、CおよびDのシークエンス解析により、Thr448Pro、Pro481Ser、およびAla488Ala突然変異が明らかになった(表12)。
【0114】
4つの緑内障原因突然変異が、POAGを持つ患者171人のうち5人(2.9%)に同定された。さらに、八つの多型および五つの同義コドン変化が同定された(表12)。エクソン3中に検出される新規ミスセンス突然変異Phe369Leu(図3)は、100人の正常日本人被験者には存在しなかった。他の三つのミスセンス突然変異、Ile360Asn、Ala363Thr、およびThr448Proは、POAGを持つ日本人患者で報告されている
【0115】
[表12]
POAGを持つ患者および対照におけるMYOC突然変異および多型
【0116】
プレートPCRとそれに続くDHPLCによる個々の患者のスクリーニング
POAGを持つ患者から得たDHPLC記録線を図4に示す。エクソン3B領域には、配列変異を示す異常記録線を見ることができ、これはダイレクトシークエンス法でPhe369Leu突然変異を表すことがわかった。
【0117】
標的とする多型を含有するMYOCエクソン3遺伝子の部分ヌクレオチド配列は以下のとおりである。
MYOC エクソン3,コドン369(下線部)TTC(Phe)→CTC(Leu)
【0118】
MYOCエクソン1〜3のヌクレオチド配列はGenBankからアクセッション番号AB006686-AB006688として入手することができる。
【実施例6】
【0119】
オプチニューリン遺伝子中の変異体および緑内障を持つ日本人患者におけるそれらと腫瘍壊死因子α多型との関連
目的:オプチニューリン(OPTN)遺伝子中の配列変異および緑内障を持つ日本人患者におけるそれらとTNF-α多型との関連を調べること。
【0120】
被験者および方法
患者および対照被験者
日本国内の7施設で合計629の血液試料を集めた。POAG患者が194人、NTG患者が217人、正常対照が218人であり、どの被験者もこの研究における他の被験者とは血縁関係になかった。診断時の年齢が35歳未満の患者および-5.5Dを超える近視を持つ患者は除外した。MYOC突然変異を持つPOAG患者も除外した。
【0121】
DNA抽出およびPCR条件
フェノール-クロロホルム抽出によって末梢血リンパ球からゲノムDNAを単離した。ポリメラーゼ連鎖反応(PCR)により、表13に記載するプライマーセットを使って、OPTN遺伝子の13のエクソンコード領域を増幅した。融解温度が高いドメイン中の突然変異をDHPLC解析によって検出するために、フォワードプライマーの一部に20塩基GCクランプを取り付けた(Narayanaswami Gら,Genet Test. 2001;5:9-16)。ハイスループット解析では、3人の患者から得た試料をプールした。PCRは、サーマルサイクラー(iCyceler;BioRad,カリフォルニア州ハーキュリーズ)を使って、ゲノムDNA 45ng、GeneAmp 10×PCRバッファーII 2μl、濃度2.0mMの各dNTPを含むGeneAmp dNTPミックス2μl、25mM MgCl2溶液2.4μl、各プライマー4pmol、およびAmpliTaq Gold DNAポリメラーゼ(Applied Biosystems,カリフォルニア州フォスターシティ)0.1Uを含有する20μlの総液量で行なった。PCR条件は以下のとおりとした:95℃で9分間変性した後、95℃で1分、55〜60℃で30秒(表13)、および72℃で1分30秒を35サイクル、そして72℃で7分間の最終伸長工程。
【0122】
[表13]
プライマー配列、PCR産物サイズ、ならびにPCRアニーリング温度およびDHPLC解析温度
【0123】
変性HPLC解析
WAVE(登録商標)システム(Transgenomic,ネブラスカ州オマハ)を使ってDHPLC解析を行なった。ヘテロ二重鎖形成のために、各PCRの産物(20μl)を95℃で5分間変性し、25℃まで徐冷した。混合した3試料から得られるアニールしたPCR産物を、DNASep(登録商標)カートリッジ(Transgenomic,ネブラスカ州オマハ)に自動注入した。
【0124】
バッファーA(Transgenomic,ネブラスカ州オマハ)は0.1M酢酸トリエチルアンモニウム(TEAA)から構成され、バッファーBは0.1M TEAAおよび25%アセトニトリルから構成された。流速0.9ml/分、2%/分ずつ増加する4.5分間のバッファーB勾配で、解析を行なった。カートリッジからのDNA断片の溶出は260nmでの吸光度によって検出した。解析に使用する温度は、DNA断片の配列に応じて選択した。WAVEMAKERソフトウェア(v.4.1,Transgenomic,ネブラスカ州オマハ)により、さまざまな温度におけるDNA断片の融解挙動が予測された。DNA断片内の予測融解ドメインが、DHPLC解析の温度を決定した(表13)。異常なクロマトグラフパターンが3試料のプール中に検出された場合は、それら3試料のそれぞれを、WAVE(登録商標)システムで個別に再解析した。次に、異常なクロマトグラフパターンを示したPCR産物をシークエンスした。異常なクロマトグラフパターンと塩基変化との相関がダイレクトシークエンス解析によって確認された後は、既知の異常クロマトグラフパターンのいずれかがDHPLC解析で観察された場合に、シークエンス解析をさらに行なうことはしなかった。
【0125】
ダイレクトDNAシークエンス
ダイレクトシークエンスによって突然変異を検出するために、PCR産物をまずQIAquick PCR精製キット(QIAGEN,米国カリフォルニア州バレンシア)で精製することにより、未反応のプライマーおよび前駆体を除去した。次に、ABI PRISM BigDye Terminator(v.3.1)サイクルシークエンスキットを製造者のプロトコール(Applied Biosystems)に従って使用することにより、シークエンス反応を行なった。ABI PRISM 310遺伝子解析装置によってデータを収集し、ABI PRISMシークエンス解析プログラム(v.3.7)によって解析した。
【0126】
OPTN c.412G>A(Thr34Thr)多型の遺伝子型判定
OPTN遺伝子のエクソン4中のc.412位におけるG→A置換を、制限酵素HpyCH4IV(New England BioLabs,マサチューセッツ州ベバリー)と、DHPLC解析に関して表13に挙げたプライマーと同じプライマーとを使って検出した。G対立遺伝子配列はHpyCH4IVによって二つの断片(188bp+129bp)に切断されたが、A対立遺伝子配列は無傷のまま(317bp)だった。制限酵素アッセイおよびDHPLCのクロマトグラフパターンによって多型を確認した。
【0127】
OPTN c.603T>A(Met98Lys)多型の遺伝子型判定
OPTN遺伝子のエクソン5中のc.603位におけるT→A置換は、DHPLC解析用のプライマーと同じプライマー(表13)を使って、制限酵素StuI(タカラ,日本国滋賀県)により、検出した。A対立遺伝子配列はStuIによって二つの断片(175bp+102bp)に切断されたが、T対立遺伝子配列は無傷のまま(277bp)だった。制限酵素アッセイおよびDHPLCのクロマトグラフパターンによって多型を確認した。
【0128】
OPTN c.1944G>A(Arg545Gln)多型の遺伝子型判定
OPTN遺伝子のエクソン16中のc.1944位におけるG→A置換を、ビー・エム・エルR&Dセンター研究部門(日本国埼玉県)によって提供されたInvaderアッセイで解析した。Invader(登録商標)アッセイおよびDHPLCのクロマトグラフパターンによって多型を確認した。
【0129】
TNF-α -308G>A多型の遺伝子型判定
TNF-αプロモーター領域中の308G>A多型の遺伝子型判定は、制限酵素NcoI(New England BioLabs,マサチューセッツ州ベバリー)と、フォワードプライマー5'-AGGCAATAGGTTTTGAGGGCCAT-3'およびリバースプライマー5'-GTAGTGGGCCCTGCACCTTCT-3'とを使って行なった。フォワードプライマーは制限酵素の使用を可能にする一塩基ミスマッチ(太字および下線部)を含有した。G対立遺伝子配列はNcoIによって二つの断片(192bp+20bp)に切断されたが、A対立遺伝子配列は無傷のまま(212bp)だった。
【0130】
TNF-α -857C>T多型の遺伝子型判定
TNF-αプロモーター領域中の857C>T多型の遺伝子型判定は、制限酵素HincII(タカラ,日本国滋賀県)と、フォワードプライマー5'-AAGTCGAGTATGGGGACCCCCCGTTAA-3'およびリバースプライマー5'-CCCCAGTGTGTGGCCATATCTTCTT-3'とを使って行なった。フォワードプライマーは制限酵素の使用を可能にする一塩基ミスマッチ(太字および下線部)を含有した。C対立遺伝子配列はHincIIによって二つの断片(106bp+25bp)に切断されたが、T対立遺伝子配列は無傷のまま(131bp)だった。-857T対立遺伝子の転写活性は-857C対立遺伝子の転写活性よりも有意に高かった。
【0131】
TNF-α -863C>A多型の遺伝子型判定
TNF-αプロモーター領域中の863C>A多型の遺伝子型判定は、制限酵素EcoNI(New England BioLabs,マサチューセッツ州ベバリー)と、フォワードプライマー5'-GCTGAGAAGATGAAGGAAAAGTC-3'およびリバースプライマー5'-CCTCTACATGGCCCTGTCCT-3'とを使って行なった。リバースプライマーは制限酵素の使用を可能にする一塩基ミスマッチ(太字および下線部)を含有した。C対立遺伝子配列はEcoNIによって二つの断片(183bp+23bp)に切断されたが、A対立遺伝子配列は無傷のまま(206bp)だった。-863A対立遺伝子の転写活性は-863C対立遺伝子の転写活性よりも有意に高かった。
【0132】
統計解析
患者および対照における遺伝子型および対立遺伝子の頻度を、カイ二乗検定およびフィッシャーの正確検定で比較した。オッズ比および95%信頼区間(CI)も計算した。観察された頻度についてハーディー・ワインベルグ平衡も計算した。これら二つの群間での臨床特性の比較は、適宜、マン・ホイットニーのU検定またはスチューデントの対応のないt検定を使って行なった。分散分析(ANOVA)を行なうための正規分布を得るために、POAG診断時のIOP、NTGおよびPOAG診断時の視野スコアである歪んだ分布の臨床データに対して、対数変換を行なった。一元配置ANOVAを使って、TNF-α/-857C>T遺伝子型およびオプチニューリン/412G>A遺伝子型、またはTNF-α/-863C>A遺伝子型およびオプチニューリン/603T>A遺伝子型の四つの異なる組み合わせを持つ患者間で、三つの臨床特性を比較した(表17参照)。
【0133】
統計解析はSPSSプログラム(SPSS Inc.,米国シカゴ)で行なった。0.05未満のP値を有意であるとみなした。
【0134】
結果
日本人被験者におけるOPTN変異体
合計629人の日本人被験者を研究した。結果を表14に示す。
【0135】
[表14]
緑内障患者および対照被験者に観察されるOPTN変異体
【0136】
緑内障患者および対照被験者に17の配列変化が同定された。これらのうち、3個はミスセンス変化であり、1個は一アミノ酸残基の欠失であり、4個は同義コドン変化であり、9個は非コード配列中の変化だった。疾患原因突然変異候補の一つ、His26Aspは、1人のPOAG発端者に同定され、218人の正常日本人対照には存在しなかった。彼女の兄弟(55歳)はこの突然変異を保持し、NTGと診断された。彼女の兄弟の娘(23歳)もこの突然変異を持ち、陥凹/乳頭径比が0.7で視神経乳頭の陥凹を示し、ハンフリー視野測定では視野欠損の徴候はなかった。
【0137】
Leu47の欠失(3bp欠失,CTC)が1人の対照に見つかった。Met98Lysは33人のPOAG患者、48人のNTG患者、および36人の対照に同定され、Arg545Glnは11人のPOAG患者、15人のNTG患者、および11人の対照に同定された。
【0138】
4個の同義ヌクレオチド置換、c.412G>A(Thr34Thr)、c.421G>A(Pro37Pro)、c.457C>T(Thr49Thr)、およびc.2023C>T(His571His)が見つかった。Thr34Thr置換は69人(35.6%)のPOAG患者、69人(31.8%)のNTG患者、および52人(23.9%)の対照に存在し、Pro37Proは1人のNTG患者に見いだされた。Thr49Thrは1人のPOAG患者に同定され、His571Hisは2人の対照に存在した。
【0139】
日本人被験者におけるOPTN変異体の分布
Thr34Thr(c.412G>A)多型はPOAGおよびNTGと関連した(表15)。有意な関連がPOAGを持つ患者に見いだされた(遺伝子型頻度:G/G対G/A+A/AでP=0.009、対立遺伝子頻度でP=0.003)。Met98Lys(c.603T>A)多型またはArg545Gln(c.1944G>A)多型については、遺伝子型頻度にも対立遺伝子頻度にも、緑内障患者と対照との間に有意差は検出されなかった。しかしMet98Lys多型は、POAGよりもNTGと関連する傾向が高かった。観察された遺伝子型頻度はハーディー・ワインベルグ平衡によって予測されるものと一致した。
【0140】
[表15]
【0141】
緑内障患者の三つの臨床特性、すなわち診断時年齢、診断時IOP、および診断時視野スコアを、c.412G>A(Thr34Thr)多型またはc.603T>A(Met98Lys)多型との関連について調べた(表16)。緑内障患者は、c.412G>A多型について臨床特性との関連を示さなかった。G/A+A/A遺伝子型を持つPOAG患者(すなわち412A保有者)は、G/G遺伝子型を持つ者(すなわち非412A保有者)よりも進行した視野スコアを持つ傾向があった(P=0.093)。603T>A多型を持つPOAG患者は、診断時年齢と弱い関連を示した(P=0.046)。
【0142】
[表16]
OPTN遺伝子型による緑内障患者の臨床特性の比較
【0143】
緑内障患者におけるOPTN多型とTNF-α多型との関連
三つの多型、308G>A、-857C>Tまたは863C>Aに関して、患者と対照との間に、遺伝子型頻度または対立遺伝子頻度の有意差は認められなかった。また、緑内障患者はこれら三つの多型について臨床特性との関連を示さなかった(データ未掲載)。観察された遺伝子型頻度は、ハーディー・ワインベルグ平衡によって予測されるものと一致した。
【0144】
しかし、TNF-α遺伝子中にC/T+T/T遺伝子型を持つ個体(すなわち-857T保有者)のうち、OPTN遺伝子がG/A+A/A遺伝子型(すなわち412A保有者)であるのは、対照では21.6%だったのに対して、POAG患者では44.1%だった(表17)。この頻度の差は有意だった(P=0.006)。TNF-α遺伝子にC/A+A/A遺伝子型を持つ個体(すなわち-863A保有者)のうち、OPTN遺伝子に603Aを持つ個体(すなわちLys98保有者)は、POAGおよびNTGと有意に関連した(それぞれP=0.008および0.027)。
【0145】
[表17]
TNF-α遺伝子型(-857C>Tおよび-863C>A)によるオプチニューリン遺伝子型(c.412G>Aおよびc.603T>A)の分布
【0146】
これら複合遺伝子型の臨床特性、例えば診断時年齢、診断時IOP、および診断時視野スコアを、表18に示す。TNF-α/-857T保有者でありかつオプチニューリン/412A保有者であるPOAG患者は、TNF-α/-857T保有者でありかつ非オプチニューリン/412A保有者である者よりも有意に悪い(P=0.020)視野スコアを持っていた。しかし、一元配置ANOVAでは、-857T>A多型とc.412G>A多型とを組み合わせたの四つの遺伝子型(表6)間で、POAG患者の三つの臨床特性に有意差はなかった:診断時年齢に関してP=0.823、診断時IOPに関してP=0.692、診断時視野スコアに関してP=0.152。
【0147】
TNF-α/-863A保有者でありかつ オプチニューリン/603A保有者であるPOAG患者は、TNF-α/-863A保有者でありかつ非オプチニューリン/603A保有者である者よりも有意に悪い(P=0.026)視野スコアを持っていた。しかし、-863C>A多型と-603T>A多型とを組み合わせた四つの遺伝子型(表6)間で、POAG患者の視野スコアに有意差はなかった(一元配置ANOVA:P=0.200)。
【0148】
[表18]
TNF-α遺伝子型(-857Tおよび-863A)およびオプチニューリン遺伝子型(412Aおよび603A)による緑内障患者の臨床特性の比較
【0149】
標的とする多型412G>A(下線部)を含むOPTNエクソン4の部分ヌクレオチド配列
【0150】
OPTN遺伝子の配列、GenBankアクセッション番号AF423071
【0151】
標的とする多型位置を含むTNF-α遺伝子の部分ヌクレオチド配列は以下のとおりである。
TNF-α -863C>A;-857C>T(下線部)
【実施例7】
【0152】
経口アンギオテンシンII受容体遮断薬が正常被験者のIOPに及ぼす効果、ならびにAT1R遺伝子およびAT2R遺伝子中のSNPとのその関連
実施例7-1
方法
X染色体上のアンギオテンシンII受容体2遺伝子(AT2R)のヌクレオチド番号3123における多型(CまたはA)と、アンギオテンシンII受容体遮断薬カンデサルタンシレキセチルの効果との関係を調べた。この研究は全身性疾患および眼疾患を持たない20人の健常ボランティア(男性13人および女性8人)で行なわれた。多型位置において、彼らのうち男性9人はC遺伝子型、男性4人はA遺伝子型、女性4人はCC遺伝子型、女性4人はCA遺伝子型を持っていた。各被験者にカンデサルタンシレキセチルを経口投与し、投与の1〜24時間後にIOPを記録した。
【0153】
結果
薬物投与後1〜24時間の眼内圧の変化を表19に示す。
[表19]
【0154】
男性の場合、AT2R遺伝子のヌクレオチド3123にA遺伝子型を持つ人の75%では、カンデサルタンシレキセチルを経口投与しても、IOPがほとんど低下せず、一方、C遺伝子型を持つ人は100%が、IOPを効果的に低下させた。女性の場合、カンデサルタンシレキセチルの経口投与により、CC遺伝子型を持つ人の100%で、IOPが効果的に低下した。
【0155】
この結果は、AT2(AGTR2)のヌクレオチド3123における遺伝子多型がカンデサルタンシレキセチルの効果と関連することを示唆している。
【0156】
実施例7-2
方法
この研究は全身性疾患および眼疾患を持たない20人の健常ボランティア(23〜28歳の男性13人および女性7人)で行なわれた。各被験者には、無作為化クロスオーバー方式で、朝(10:00)に、12mgの経口カンデサルタンシレキセチル(Blopress(登録商標),武田,日本)または偽薬を与えた。
【0157】
ベースライン心拍数、ベースライン収縮期/拡張期動脈圧(SBP/DBP)、およびベースラインIOPを記録した。次に、被験者に経口カンデサルタンシレキセチルまたは偽薬を与え、6時間までの毎時間と24時間後に測定を繰り返した。1ヶ月後に、各被験者にもう一方の処置を施した。右眼だけを測定し、解析した。
【0158】
眼灌流圧(OPP)は、組織に入る動脈内の圧力と、それを離れる静脈内の圧力との差と定義される。OPPは、平均血圧(BPm)およびIOPを使って以下の式で近似することができる。
OPP=2/3×BPm−IOP
[式中、BPm=DBP+1/3×(SBP-DBP)]
【0159】
ATR1およびATR2中の多型を20人の被験者で検索し、IOPの変化と相関させた。この研究はヘルシンキ宣言の精神に則って行なわれた。研究の性質および考えうる帰結を説明した上で、文書によるインフォームド・コンセントを得た。この研究は、適用可能な場合には、DNA解析に関する施設内人体実験委員会による承認を得た。
【0160】
統計解析
ARB投与後の結果の統計解析は、反復測定ANOVA検定を使って、StatView(SAS Institute,米国)で行なった。各IOP値の統計解析にはボンフェローニ補正ANOVA検定を使用し、P値<0.0004を統計的に有意であるとみなした。
【0161】
結果
経口カンデサルタンシレキセチル投与または偽薬投与後のIOPの変化を図5Aに示す。偽薬の投与を受けた被験者のIOPに有意な変化はなかった。これに対し、経口カンデサルタンシレキセチル投与の1時間後には早くもIOPが有意に低下しており、5時間にわたって偽薬より低い状態を保った(P<0.0001)。カンデサルタンシレキセチルは灌流圧に有意な影響を及ぼさなかった(図5B)。カンデサルタンシレキセチルまたは偽薬の単回経口投与後にSBP、DBP、および心拍数の有意な変化は検出されなかった(データ未掲載)。
【0162】
被験者20人のそれぞれにおける経口カンデサルタンシレキセチル投与後のIOP変化を図5Cに示す。この20人の被験者では、カンデサルタンシレキセチルの効果とATR1遺伝子中の3個のSNPとの間に、有意な関連はなかった(表19-2)。しかし、ATR2遺伝子型については、A遺伝子型を持つ男性4人が2.3±0.5mmHgのIOP低下を示し、これは偽薬の投与を受けた被験者と同じ値であり、C遺伝子型を持つ男性9人(5.0±1.1mmHg,P=0.014)よりも有意に少ない低下量だった。この研究ではAA遺伝子型を持つ女性はいなかった。
【0163】
[表19−2]
アンギオテンシンII受容体遺伝子の遺伝子型と関連するアンギオテンシンII受容体遮断薬の眼内圧に対する効果
【実施例8】
【0164】
緑内障とエンドセリン-1およびエンドセリンA型受容体の遺伝子多型との関連
目的:強力な血管収縮因子エンドセリン1(ET-1)は眼内圧および眼血管張力の調節に影響を及ぼしうる。したがってET-1およびその受容体は緑内障の発病の一因になりうる。本発明者らは、ET-1(EDN1)ならびにその受容体ETA(EDNRA)およびETB(EDNRB)の遺伝子多型が緑内障表現型および臨床特性と関連するかどうかを調べた。
【0165】
方法
研究対象集団:
この研究では、日本の医療施設7ヶ所から採用した合計650人の日本人被験者(正常対照224人、POAG患者176人、およびNTG患者250人)を調べた。どの被験者も血縁関係にはなかった。OAG診断時の平均年齢(±標準偏差)は57.2±12.8歳だった。OAG被験者はPOAG患者とNTG患者に分割され、その診断時年齢はそれぞれ58.8±12.2歳および56.1±13.2歳だった(表1)。検査時の平均年齢は対照では70.0±11.2歳だった。本発明者らは、対照の一部が後に緑内障を発病する可能性が低下させるために、より高齢の対照被験者を意図的に選択した。
【0166】
眼科検査には、細隙灯生体顕微鏡検査、視神経乳頭検査、ゴールドマン圧平眼圧測定によるIOP測定、および隅角鏡検査を含めた。視野はハンフリー自動視野測定(プログラム30-2)またはゴールドマン視野測定で評価した。視野欠損の重症度を1〜5にスコア化した。2タイプの視野測定によって取得したデータを5点尺度(1=変化なし;2=早期欠損;3=中等度欠損;4=重度欠損;および5=光覚のみまたは視覚なし)を使って集約した。この重症度尺度は、ゴールドマン視野測定に基づくKozakiの分類(今まで日本では最も広く用いられてきたもの)に従うか、ハンフリー視野解析機用に確立された分類によった。
【0167】
以下の基準を全て満たす場合にPOAGと診断した:最大IOPが21mmHgを超えたこと;隅角鏡検査での開放隅角;視野変化を伴う典型的な緑内障性乳頭陥凹;および視神経損傷を引き起こす可能性がある他の眼障害、鼻障害、神経障害または全身性障害が存在しないこと。本発明者らは、明確な原因(例えば外傷、ぶどう膜炎、ステロイド投与、または剥脱性緑内障、色素性緑内障、もしくは血管新生緑内障)に続発するIOPの上昇を持つ患者を除外した。MYOC突然変異を持つPOAG患者およびJOAG患者も除外した。NTGは、追跡期間中は常にIOPが21mmHgを超えなかったという点を除いてPOAGと同じ基準によって診断した。正常対照被験者は20mmHg未満のIOPを持ち、緑内障性乳頭変化がなく、緑内障の家族歴を持っていなかった。
【0168】
DNA抽出および多型の遺伝子型判定
標準的方法によって末梢血リンパ球からゲノムDNAを単離した。参加者全員に9個の一塩基多型(SNP)、すなわちEDN1について4個(T-1370G、+138/ex1 del/ins、G8002A、K198N)、EDNRAについて4個(G-231A、H323H、C+70G、C+1222T)、およびEDNRBについて1個(L277L)を検出した。これらの多型は、http://genecanvas.idf.inserm.fr/に列挙されている。本発明者らは、SNPのハイスループット遺伝子型判定のために最近開発されたInvader(登録商標)アッセイ(Thrid Wave Technologies, Inc.,ウィスコンシン州マディソン)を使って、これらのSNPを遺伝子型判定した(Lyamichev Vら,Nat Biotechnol 1999;17:292-296,この引用文献の内容は参照により本明細書に組み込まれる)。
【0169】
この研究で使用した一次プローブおよびInvader(登録商標)プローブのオリゴヌクレオチド配列を表20に記載する。
【0170】
[表20]
【0171】
統計解析
正常対照における遺伝子型分布と、OAG患者、POAG患者、およびNTG患者における遺伝子型分布との比較は、χ2解析によって行なった。臨床特性(診断時年齢、無処置時最大IOP、および診断時視野スコア)と遺伝子型との関連はマン・ホイットニーのU検定によって評価した。統計解析はWindows用SPSS(バージョン12.0;SPSS Inc.,イリノイ州シカゴ)で行なった。p<0.05の値を有意であるとみなした。
【0172】
結果
この研究で得られた遺伝子型頻度および対立遺伝子頻度を表21に示す。分布はハーディー・ワインベルグ平衡と一致した。EDN1/+138/ex1 del/ins多型の場合、del/del遺伝子型およびdel/ins+ins/ins遺伝子型の頻度は、それぞれ、対照被験者で65.2%および34.8%だったのに対して、OAG患者全体では74.2%および25.8%(p=0.016)、POAG患者では74.4%および25.6%(p=0.047)、NTG患者では74.0%および26.0%(p=0.037)だった。EDN1/K198N多型の場合、OAG患者の53.2%はKK遺伝子型を持つことがわかった。これは対照被験者における43.8%という優勢率よりも有意に高かった(p=0.022)。OAG患者をPOAG患者とNTG患者とに分割した場合、NTG患者におけるKK遺伝子型の頻度は対照よりもはるかに高く(p=0.008)、POAG患者における遺伝子型頻度分布および対立遺伝子頻度分布は、対照における分布と統計的に相違しなかった。性差が認められた。具体的には、KK遺伝子型は、女性NTG患者の間では(女性対照に対してp=0.010)、男性NTG患者の場合よりも(男性対照に対してp=0.251;表22)、有意に優勢だった。イントロン4領域におけるEDN1/G8002Aの多型は一試料を除いてEDN1/K198Nと高度に共在した(データ未掲載)。
【0173】
EDNRA/C+1222T遺伝子型(CC対CT+TT)の頻度はOAG患者と対照の間でわずかに異なった(p=0.036)。他の多型に関する遺伝子型の分布はどの患者群と対照との間にも有意差を示さなかった。
【0174】
患者の特徴を各多型の優性モデルおよび劣性モデルで調べ、データを有意差と共に表23に示す。OAG患者全体およびPOAG患者では、どの特徴も遺伝子型群間に有意差を示さなかった。しかし、NTG患者では、EDNRA/G-231AのAA群が、GG+GA群よりも悪い診断時視野スコアを持っていた(3.0±0.8対2.7±0.6,p=0.043)。また、NTG患者の間で、EDNRA/C+70GのGG群は、診断時視野スコアが、CC+CG群よりも有意に悪いこともわかった(3.0±0.7対2.7±0.7,p=0.014)。NTG患者では、ENDRA/H323HのTT群における無処置時最大IOPが、CC+CT群よりも統計的に高かった(17.2±2.2対16.6±2.3,p=0.040)。NTG患者における他の多型は、遺伝子型群間で、特性に有意差を示さなかった。
【0175】
[表21]
対照被験者および緑内障患者におけるEDN1、EDNRAおよびEDNRB多型の遺伝子型頻度および対立遺伝子頻度
データはn(%)である。
* P<0.05(χ2検定)
遺伝子型分布は、OAG患者と対照との間で、EDN1/+138/ex1 del/ins多型(p=0.016)およびEDN1/K198N多型(p=0.022)に関して有意差を示し、EDNRA/C+1222T多型(p=0.036)に関してわずかな差を示した。OAG群をPOAGおよびNTGに分割すると、NTG患者におけるEDN1/K198N多型のKK遺伝子型の頻度は、対照よりもはるかに高くなった(p=0.008)。
【0176】
[表22]
【0177】
[表23]
【0178】
男性被験者では、以下の相関関係が確認された:
1)エンドセリン-1遺伝子のエクソン1におけるA138挿入/欠失(A138I/D)多型はPOAGともNTGとも関連する(表24)
2)エンドセリン受容体A遺伝子のプロモーター領域の-231A>G多型は、NTG、特に15mmHg未満の眼内圧を持つ患者と関連する(表25)。
3)エンドセリン受容体A遺伝子のエクソン6中のコドン番号233におけるCAC→CAT置換(His323His)は、NTG、特に15mmHg未満の眼内圧を持つ患者と関連する(表26)。
4)エンドセリン受容体B遺伝子のエクソン4中のコドン番号277におけるCTG→CTA置換はPOAGともNTGとも関連する(表27)。
【0179】
女性患者では、以下の相関関係が確認された:
1)エンドセリン-1遺伝子のコドン番号198におけるAAG→AAT置換(Lys198Asn)はNTGと関連する(表28)。
2)エンドセリン-1遺伝子プロモーター領域の-1370T>G多型はNTGと関連する(表29)。
3)エンドセリン受容体Aの3'非コード領域における+70C>G(停止コドンから70塩基)多型はPOAGと関連する(表30)。
4)エンドセリン受容体Aの3'非コード領域における+1222C>T(停止コドンから1222塩基)多型はNTG(眼内圧が16mmHg〜21mmHgであるもの)と関連する(表31)。
【0180】
[表24]
エンドセリンA138I/D(男性)
【0181】
[表25]
エンドセリン受容体A -231A/G(男性)
H-NTG:16mmHg〜21mmHgの眼内圧を持つNTG患者。
L-NTG:15mmHg以下の最大眼内圧を持つNTG患者。
【0182】
[表26]
エンドセリン受容体A H323H C>T His323His(男性)
H-NTG:16mmHg〜21mmHgの眼内圧を持つNTG患者。
L-NTG:15mmHg以下の最大眼内圧を持つMTG患者。
【0183】
[表27]
エンドセリン受容体B L277L G>A Leu277Leu(男性)
【0184】
[表28]
エンドセリン Lys198Asn G>TまたはK198N(女性)
【0185】
[表29]
エンドセリン -1370T>G(女性)
【0186】
[表30]
エンドセリン受容体A +70C>G(女性)
【0187】
[表31]
エンドセリン受容体A +1222C>T(女性)
H-NTG:16mmHg〜21mmHgの眼内圧を持つNTG患者。
L-NTG:15mmHg以下の最大眼内圧を持つMTG患者。
【0188】
標的とする多型を含むエンドセリン-1(EDN1)ならびにエンドセリン受容体A(EDNRA)およびエンドセリン受容体B(EDNRB)の部分ヌクレオチド配列を以下に示す。
【0189】
EDN1 -1370(下線部)T>G
【0190】
END1 +138(下線部)ins/del(それぞれ3743-3745にあるaのそれぞれ一つ)
【0191】
EDNRA +70(下線部)C>G
【0192】
EDNRA +1222(下線部)C>T
【0193】
EDNRAコドン番号323(下線部)(T>C)His323His
【0194】
EDNRA -231(下線部)A>G
【0195】
EDNRBコドン番号277(下線部)Leu277Leu(CTG→CTA)
【実施例9】
【0196】
β1アドレナリン作動性受容体(ADRB1)の遺伝子多型と緑内障との関連
方法
PCR-RFLP技術を使って、POAG患者、NTG患者および正常(対照)被験者の間で、ADRB1の遺伝子多型と緑内障との関連を調べた(表32-1)。
[表32−1]
プライマー配列
【0197】
結果
表32-2に示すように、ADRB1中のGly389Argの多型はNTGと関連する(表32-2)。
【0198】
[表32−2]
β1-アドレナリン受容体Gly389Arg
【0199】
標的とする多型を含むβ1-アドレナリン受容体の部分ヌクレオチド配列。
B1ARコドン389(下線部)GGA(Gly)→CGA(Arg)Gly389Arg
【実施例10】
【0200】
E-セレクチンの遺伝子多型と緑内障との相関関係
方法
Invader(登録商標)法を使って、POAGを持つ被験者、NTGを持つ被験者および正常被験者の間で、E-セレクチン遺伝子多型と緑内障との関係を調べた。
【0201】
SELE遺伝子のC/T多型を検出するために使用したInvader(登録商標)オリゴヌクレオチド(Invader(登録商標)プローブ)を表33-1に示す。
[表33−1]
【0202】
結果
E-セレクチン遺伝子の1402C>T多型は、POAGともNTGとも関連することが確認された(表33-2)。
[表33−2]
E-セレクチン1402C>T
【0203】
標的とする多型を含むE-セレクチンの部分ヌクレオチド配列は以下のとおりである:
SELE 番号1402(下線部)C>T
【実施例11】
【0204】
パラオキソナーゼ1遺伝子多型は開放隅角緑内障の臨床特徴と関連する
目的:低密度リポタンパク質(LDL)の酸化誘導体は内皮にとって有害である。内皮機能障害は、開放隅角緑内障(OAG)の病理発生過程に関与することが知られている。高密度リポタンパク質(HDL)はLDLの酸化的修飾を防止する。本発明者らは、HDL関連抗酸化酵素であるパラオキソナーゼ1(PON1)、PON2、および血小板活性化因子アセチルヒドロラーゼ(PAF-AH)遺伝子中の多型が、日本人集団においてOAGと関連するかどうかを調べた。
【0205】
材料および方法
患者および対照被験者
日本の施設7ヶ所で698の血液試料を集めた。被験者には、POAG患者190人、NTG患者268人、および正常対照240人を含めた。どの被験者も互いに血縁関係にはなかった。
【0206】
血液試料採取時の年齢(平均±SD)は、POAG患者で65.3±11.9歳、NTG患者で58.8±13.4歳、正常被験者で69.7±11.2歳だった。正常対照被験者はPOAG患者(p<0.001)またはNTG患者(p<0.001)より有意に高齢だった。これにより、対照被験者が最終的に緑内障を発病する可能性が低減されるだろう。
【0207】
緑内障患者に関して記録された臨床特徴は、診断時年齢、診断時IOP、および診断時視野欠損だった。視野欠損の重症度は1〜5にスコア化した。異なる視野計を使って取得したデータを、以下に定義する5点尺度を使って集約した。1=変化なし;2=早期欠損;3=中等度欠損;4=重度欠損;および5=光覚のみまたは視覚なし。視野欠損は、ゴールドマン視野測定の結果に基づくKozakiの分類に従って、またはハンフリー視野解析機に用いられる分類によって、早期、中等度、または重度と判定した。日本では今まで前者の分類が最も広く用いられてきた。
【0208】
ゴールドマン圧平眼圧測定によるIOP測定、ハンフリー視野測定(30-2)またはゴールドマン視野測定、隅角鏡検査、および眼底撮影を含む視神経乳頭検査などといった一連の眼科検査を、全ての患者に行なった。緑内障患者は全員、以下の基準に従って診断された:緑内障性陥凹(陥凹/乳頭径比>0.7)および神経網膜辺縁部の喪失を伴う典型的視神経乳頭損傷の存在、緑内障性陥凹と矛盾しない再現性のある視野欠損、ならびに隅角鏡検査での開放隅角。OAG患者のうち、追跡期間中どの時点でもIOP>21mmHgである場合は、POAGと診断した。剥脱性緑内障、色素性緑内障、およびコルチコステロイド誘発性緑内障を持つ患者は除外した。OAG患者のうち、無処置時最大IOPが3回のベースライン測定値および毎日の試験値(6時から24時まで3時間ごと)を含めて常に一貫して21mmHg以下であり、診断後の投薬時または無投薬時の最大IOPが追跡期間中常に一貫して22mmHgであり、緑内障性視神経症の二次的原因、例えば外傷後のIOP上昇を過去に経験していること、ステロイド投与の期間、またはぶどう膜炎などが存在しなかった場合に、NTGと診断した。
【0209】
対照被験者は、白内障を除く既知の眼異常を持たない日本人個体から採用した。これらの被験者は196人に達し、40歳より高齢で、20mmHg未満のIOPを持ち、緑内障性乳頭変化がなく、緑内障の家族歴もなかった。
【0210】
遺伝子型判定
標準的方法によって末梢血リンパ球からゲノムDNAを単離した。次に、全ての参加者に4個のSNP(PON1について2個(L55M、Q192R)、PON2について1個(Cys311Ser,C311S)、およびPAF-AHについて1個(V279F))を検出した。
【0211】
SNPのハイスループット遺伝子型判定のために最近開発されたInvader(登録商標)アッセイ(Third Wave Technologies, Inc,米国ウィスコンシン州マディソン)を使って、これらのSNPを遺伝子型判定した。この研究で使用した一次プローブおよびInvader(登録商標)プローブのオリゴヌクレオチド配列を表34に挙げた。
【0212】
[表34]
【0213】
統計解析
カイ二乗解析によってハーディー・ワインベルグ平衡を評価した。遺伝子型の頻度および対立遺伝子の頻度を、カイ二乗解析により、症例と対照とで比較した。三つの臨床変量と遺伝子型の間の関連を確認するために、ロジスティック回帰モデルで多変量解析を行なった。PON1遺伝子におけるQ192Rの遺伝子型群間のIOPの比較は、クラスカル・ワリス検定によって行なった。統計解析はSPSS(バージョン12.0;SPSS,イリノイ州シカゴ)で行なった。p<0.05の値は有意性を示すとみなした。
【0214】
結果
緑内障患者および対象における4個のSNPに関する遺伝子型の分布を表35に示す。NTGを持つ患者ではPON1遺伝子のL55M多型が有意に異なる遺伝子型頻度を持っていた。
【0215】
PON2遺伝子およびPAF-AH遺伝子中の多型に関する遺伝子型の分布は、どの患者群と対照との間にも有意差を示さなかった(表35)。また、4個のSNPの対立遺伝子頻度にも有意差はなかった。
【0216】
[表35]
【0217】
OAG集団におけるPON1遺伝子中の二つの多型を組み合わせたものの分布を表36に示す。明瞭に示されるように、55位のメチオニン(M)(M対立遺伝子)が192位のアルギニン(R)(R対立遺伝子)を伴うことはほとんどなかった。解析により、55位にロイシン(L)をもたらす多型と192位にアルギン(R)をもたらす多型との間に連鎖不平衡が確認された(p<0.001)。
【0218】
[表36]
【0219】
患者の特性を各多型について優性モデルおよび劣性モデルで調べた。劣性モデルでは、どの多型についても、OAGを持つ患者における三つの特性に有意差が見られなかった。PON1多型の優性モデルによる有意差を表37および表38に示す。OAG患者におけるPON1遺伝子中のL55M多型について、LL群(非55M保有者)は診断時の年齢がLM+MM群(55M保有者)より有意に低かった(56.8±12.8歳対60.1±11.4歳,p=0.028)(表37)。この関連はPOAG患者には観察されなかったが、NTG患者には観察された(55.6±13.1歳対63.7±9.6,p=0.001)。
【0220】
Q192R多型に関して、診断時の無処置時最大IOPは、QR+RR群を持つOAG患者(192R保有者)(21.5±7.4mmHg)の方が、QQ群(非192R保有者)の場合(18.7±5.3mmHg,P=0.006,表38)よりも有意に高かった。無処置時最大IOPは、POAG患者(POAGについて27.5±7.0mmHg対24.0±4.9,p=0.049)でも、NTG患者(NTGについて15.8±2.8mmHg対16.7±2.4,p=0.030)でも、192R保有者の方が非保有者より高かった。
【0221】
[表37]
PON1遺伝子中のL55Mの遺伝子型によるNTG患者の臨床特性
【0222】
[表38]
PON1遺伝子中のQ192Rの遺伝子型による緑内障患者の臨床特性
【0223】
PON1遺伝子中のGly192Arg(Q192R)多型はPOAGと関連した(表39)。Leu55Met多型はNTG(特に15mmHg未満のもの)と関連した(表40)。
【0224】
[表39]
PON1 Gln192Arg(Q192R)
【0225】
[表40]
PON1 Leu55Met(L55M)
H-NTG:16mmHg〜21mmHgの眼内圧を持つNTG患者。
L-NTG:15mmHg以下の最大眼内圧を持つMTG患者。
【0226】
結論:PON遺伝子多型はOAGを持つ日本人患者、特にNTGを持つ日本人患者の特徴に影響を及ぼしうる。
【0227】
標的とする多型を含有するパラオキソナーゼ1遺伝子の部分ヌクレオチド配列は以下のとおりである。
PON1コドン55(下線部)TTG(Leu)→ATG(Met)(Leu55Met)
および
PON1コドン192(下線部)CAA(Gln)→CGA(Arg)(Gln192Arg)
【実施例12】
【0228】
開放隅角緑内障の病因におけるノエリン2遺伝子の評価
目的:開放隅角緑内障を持つ日本人患者におけるノエリン2遺伝子中の突然変異について、変性高速液体クロマトグラフィー(DHPLC)を使ってスクリニーングすること。
【0229】
方法
被験者
日本国内の8施設で合計616の血液試料を集めた。POAG患者276人、NTG患者340人、および正常被験者300人であり、どの被験者もこの研究における他の被験者とは血縁関係になかった。
【0230】
DNA抽出およびPCR条件
血液試料の解析は全て慶応大学で行なわれた。フェノール-クロロホルム抽出によって末梢血リンパ球からゲノムDNAを単離した。ポリメラーゼ連鎖反応(PCR)により、表41に挙げるプライマーセットを使って、ノエリン2遺伝子の六つのエクソンコード領域を増幅した。
【0231】
[表41]
プライマー配列、PCR産物サイズ、ならびにPCRアニーリング温度およびDHPLC解析温度
【0232】
ハイスループット解析では、3人の患者からの試料をプールした。PCRは、サーマルサイクラー(iCycler,Bio-Rad,カリフォルニア州ハーキュリーズ)を使って、ゲノムDNA 45ng、GeneAmp 10×PCRバッファーII 2μl、濃度2.0mMの各dNTPを含むGeneAmp dNTPミックス2μl、25mM MgCl2溶液2.4μl、各プライマー4pmol、およびAmpliTaq Gold DNAポリメメラーゼ(Applied Biosystems,カリフォルニア州フォスターシティ)0.1Uを含有する20μlの総液量で行なった。PCR条件は以下のとおりとした:95℃で9分間変性した後、95℃で1分、65〜67℃で30秒(表1)、および72℃で1分30秒を35サイクル、そして72℃で7分間の最終伸長工程。
【0233】
変性HPLC解析
ハイスループット解析の場合、3人の患者から得られたPCR産物25μlを、DHPLC解析用のWAVE(登録商標)システム(Transgenomic,ネブラスカ州オマハ)を使って、解析用クロマトグラフに自動注入した。DHPLC融解温度を表41に記載する。
【0234】
ハイスループットプロトコールでプール試料に異常なクロマトグラフパターンが検出された場合は、その試料をWAVE(登録商標)システムで個別に再解析した。次に、異常なクロマトグラフパターンを示したPCR産物をシークエンスした。
【0235】
ダイレクトDNAシークエンス
ダイレクトシークエンスのために、PCR産物をQIA Quick PCR精製キット(Qiagen,カリフォルニア州バレンシア)で精製することにより、未使用のプライマーおよび前駆体を除去した。BigDyeケミストリーを製造者の推奨するプロトコール(Applied Biosystems,カリフォルニア州フォスターシティ)に従って使用することにより、ABI310自動シークエンサーで、同じフォワードおよびリバースPCR増幅プライマーを使って、PCR産物をダイレクトシークエンスした。
【0236】
ミオシリン遺伝子のスクリーニング
ノエリン2遺伝子中に突然変異を保持する緑内障患者2人について、DHPLCにより、ミオシリン遺伝子のスクリーニングを行なった。
【0237】
ノエリン2 c.462G>A(Arg144Gln)多型の遺伝子型判定
ノエリン2遺伝子のエクソン4中のc.462位におけるG→A置換を、制限酵素BstU1を使って検出した。G対立遺伝子配列はBstU1によって二つの断片(140bp+200bp)に切断されたが、A対立遺伝子配列は無傷のまま(344bp)だった。
【0238】
制限酵素アッセイおよびDHPLCのクロマトグラフパターンによって多型を確認した。
【0239】
統計解析
患者および対照における遺伝子型および対立遺伝子の頻度をカイ二乗検定またはフィッシャーの正確検定で比較した。観察された頻度についてハーディー・ワインベルグ平衡も計算した。統計解析はSPSSプログラム(SPSS Inc.,米国シカゴ)で行なった。0.05未満のP値を有意であるとみなした。
【0240】
結果
日本人被験者におけるノエリン2変異体
合計616人の日本人被験者を調べた。その結果を表42に記載する。緑内障患者および対照被験者中の配列変化を同定した。これらのうち二つはミスセンス変化であり、七つは同義コドン変化であり、一つはイントロン配列中の変化だった。疾患原因突然変異候補の一つ、Arg144Glnは、一人のPOAG発端者および一人のPOAG発端者に同定され、300人の正常日本人対照には存在しなかった。Arg106Gln(P=0.30)、Ala226Ala(P=0.30)、およびArg427Arg(P=0.30)については、緑内障患者と対照との間に有意差は検出されなかった。
【0241】
Arg144Glnを持つNTG患者はミオシリン遺伝子中のArg76Lys変化を保持していた。
【0242】
エクソン4中の緑内障原因突然変異候補Arg144Glnは616人の日本人緑内障患者のうち2人(0.3%)に同定された。
【0243】
[表42]
緑内障患者および対照被験者に観察されたOLFM2変異体
【0244】
標的とする多型を含むノエリン2の部分ヌクレオチド配列は以下のとおりである。
Noelin2コドン144(下線部)CGG(Arg)→CAG(Gln):(GG:200bp+144bp,GA:344bp+200bp+144bp,AA:344bp)(BstUI)
コドン140(下線部)Lys140Lys(AAG>AAA)
コドン152(下線部)Glu152Glu(GAG>CAA)
79841-79862,80164-80184;コドン144における多型を検出するためのプライマー。
79916-80131,コード領域。
【実施例13】
【0245】
緑内障の病因における熱ショックタンパク質70-1(HSP70-1)遺伝子の評価
POAG患者、NTG患者および対照被験者の間で、Invaderアッセイを使って、緑内障とHSP70-1の遺伝子多型(Biogerontology 4:215-220,2003およびHum Genet 114:236-241,2004)との関連を調べた。
【0246】
HSP70-1遺伝子の多型を検出するために使用した一次プローブ(野生型プローブおよび突然変異型プローブ)ならびにInvader(登録商標)オリゴヌクレオチド(Invader(登録商標)プローブ)を表43に示す。
[表43]
HSP70-1のオリゴヌクレオチド配列
【0247】
結果
表44に示すように、HSP70-1における-110A>Cの多型は緑内障、特にPOAGと関連する。
[表44]
【0248】
標的とする配列を含むHSP70-1の部分ヌクレオチド配列は以下のとおりである。
HSP70-1 -110A>C(以下の配列はC対立遺伝子である)
【実施例14】
【0249】
緑内障の病因におけるエンドセリン変換酵素1(ECE1)遺伝子の評価
POAG患者およびNTG患者で、Invaderアッセイを使って、緑内障とECE1の遺伝子多型との関連を調べた。
【0250】
ECE1遺伝子の多型を検出するために使用した一次プローブ(野生型プローブおよび突然変異型プローブ)ならびにInvader(登録商標)オリゴヌクレオチド(Invader(登録商標)プローブ)を表45に示す。
[表45]
【0251】
結果
表46に示すように、ECE1における-338C>Aの多型は、NTGにおける高いIOPと関連した。
[表46]
【0252】
標的とする多型を含むECE-1の部分ヌクレオチド配列を以下に示す。
ECE1 -338C>A(下線部)
【実施例15】
【0253】
開放隅角緑内障の病因におけるCD50遺伝子の評価
ポリメラーゼ連鎖反応-制限断片長多型(PCR-RFLP)技術を使ってCD50遺伝子の多型を同定した(表47)。
[表47]
【0254】
結果
表48に示すように、CD95におけるA-670Gの多型は緑内障、特にPOAGと関連する。
[表48]
【実施例16】
【0255】
緑内障の病因におけるEPHX1遺伝子の評価
Invaderアッセイを使って、POAG患者、NTG患者および対照被験者の間で、緑内障とEPHX1の遺伝子多型との関連を調べた。
【0256】
ECE1遺伝子の多型を検出するために使用した一次プローブ(野生型および突然変異型プローブ)ならびにInvader(登録商標)オリゴヌクレオチド(Invader(登録商標)プローブ)を表49に示す。
[表49]
【0257】
結果
表50に示すように、コドン119LysにおけるG>Aの多型は緑内障、特にNTGと関連した。
[表50]
【0258】
標的とする多型を含むEPHX1の部分ヌクレオチド配列は以下のとおりである。
【実施例17】
【0259】
緑内障の病因におけるβ2アドレナリン作動性受容体(ADRB2)遺伝子の評価
Invader アッセイを使って、開放隅角緑内障患者(POAG患者およびNTG患者)で、緑内障とADRB2の遺伝子多型との関連を調べた。
【0260】
ADRB2遺伝子の多型を検出するために使用した一次プローブ(野生型プローブおよび突然変異型プローブ)ならびにInvader(登録商標)オリゴヌクレオチド(Invader(登録商標)プローブ)を表51に示す。
[表51]
【0261】
結果
表52に示すように、ADRB2におけるGly16Arg(G46A)の多型はPOAGの早期発症と関連する。
[表52]
ADRB2遺伝子におけるGln16Argの遺伝子型による緑内障患者の臨床特性
【0262】
表53に示すように、Gln27Glu(C79G)の多型はOAG、特にPOAGにおける高い眼内圧(IOP)と関連する。
[表53]
ADRB2遺伝子におけるGln27Gluの遺伝子型による緑内障患者の臨床特性
【0263】
標的とする多型を含有するADRB2遺伝子の部分ヌクレオチド配列は以下のとおりである。
ADRB2コドン番号Gly16Arg(GGA>AGA):Gln27Glu(CAA>GAA)(下線部)
【図面の簡単な説明】
【0264】
【図1】NTG患者の臨床特性とAT2R 3123C>A多型およびACE I/D多型との相関を表す。
【図2】MYOC遺伝子のエクソン3CにおけるDHPLC記録線パターンを表す。
【図3】MYOC遺伝子のエクソン3中に検出された新規ミスセンス突然変異Phe369Leuを表す。
【図4】POAGを持つ患者に由来するMYOC遺伝子のDHPLC記録線を表す。
【図5A】経口カンデサルタンシレキセチル投与後または偽薬投与後のIOPを表す。
【図5B】経口カンデサルタンシレキセチル投与後または偽薬投与後の眼灌流圧を表す。
【図5C】被験者15人のそれぞれにおける経口カンデサルタンシレキセチル投与後のIOPを表す。
【技術分野】
【0001】
本発明は、視神経症に連関する遺伝子多型のセットに関する。
【背景技術】
【0002】
緑内障は世界的に失明の主原因であり、およそ6700万人が何らかの形態の緑内障を患っていると見積られている。大多数の症例は原発開放隅角緑内障(POAG)の後期成人発症(通例、40歳超)として起こる。これは、最も一般的な形態の緑内障であり、40歳を超えた白人人口の約2%および黒人人口の7%を冒す。POAGは、通常は眼内圧(IOP)の上昇と関連する視神経乳頭の陥凹に対応した特徴的な視野変化をもたらす。正常眼圧緑内障(NTG)は、典型的な緑内障性視神経乳頭陥凹および視野喪失が存在するものの、常に21mmHgを超える上昇したIOPの証拠がない開放隅角緑内障の一形態である。日本の場合、緑内障の有病率は40歳超の約3.5%(POAG 0.58%およびNTG 2.04%)である。日本人集団におけるNTGの有病率は他の集団と比較して高い。眼内圧(IOP)は緑内障の主要リスク因子であるが、緑内障は特有の視野喪失と関連する進行性視神経症を特徴とする多因子性障害であり、複数の遺伝子および環境的影響の相互作用によってもたらされる。
【0003】
緑内障を発病させるリスク因子には、高いIOP、年齢、人種、家族歴あり、近視、糖尿病または高血圧の存在、および遺伝的因子が含まれる。緑内障性視神経症の正確な病理発生過程はまだ不明であるが、上昇したIOPが主要リスク因子であることは、広く受け入れられている。現在行なわれている緑内障の処置はIOPを低下させる介入からなる。しかし、緑内障、NTGまたは末期POAGを持つ一部の患者においては、IOPの低下では疾患の進行が予防されないことから、上昇したIOP以外の因子が緑内障の発病または進行に関与しうることが示される。
【0004】
POAGおよびNTGは不均質な病態群であり、これらはおそらく、視神経乳頭に観察されるニューロン喪失パターンをもたらす異なる多因子性病因を持っている。緑内障と、低体血圧、血圧の夜間低下、高血圧、片頭痛、血管痙攣、および糖尿病を含む多くの全身性血管疾患の存在との関連が報告されている。NTG患者における視神経乳頭出血の存在は、NTGの発病および進行に血管不全が深く関与することを示唆している。POAGを持つ患者は高い割合で、併存障害に対する多種多様な投薬を受ける。特に、全身性高血圧は最もよく見られる障害であり、集団全体の48%に見いだされた。
【0005】
萎縮期のレーベル遺伝性視神経症(LHON)および優性遺伝性視神経萎縮(DAO)を持つ患者に、緑内障様の形態変化が報告されている。最近、本発明者らは、11778突然変異を保持する15人の日本人LHON患者の萎縮期において、ハイデルベルグ網膜断層撮影(HRT)を使った定量的解析により、視神経乳頭陥凹を報告した(Mashima Yら,Arch Clin Exp Ophthalmol 2003;241:75-80,この引用文献の内容は参照により本明細書に組み込まれる)。LHONは主に若年成人男性を冒す母系遺伝性の眼疾患である。患者の約70%は男性だった。この疾患は通常、重度で永続的な視覚喪失を引き起こし、視力は0.1未満になる。視野欠損は中心暗点または盲点中心暗点として存在する。今までに20を超えるミトコンドリアDNA(mtDNA)の点突然変異が世界中のLHON患者に報告されており(Brown MDら,Clin Neurosci 1994;2:138-145,この引用文献の内容は参照により本明細書に組み込まれる)、LHON患者の80%超が、ヌクレオチド位置3460、11778、または14484にある三つのmtDNA突然変異のうちの一つを保有する(Mackey DAら,Am J Hum Genet 1996;59:481-485,この引用文献の内容は参照により本明細書に組み込まれる)。mtDNAヌクレオチド位置3460、11778および14484の三つの突然変異についてNTG患者を調べたが、突然変異も、骨格筋試料における呼吸鎖活性の欠損も、検出されなかった(Brierley EJら,Arch Ophthalmol 114:142-146およびOpial Dら,Graefes Arch Clin Exp Ophthalmol 239:437-440,これらの引用文献の内容は参照により本明細書に組み込まれる)。
【0006】
これらのmtDNA突然変異の一つを持つLHON患者の間に見られる主な相違は、その臨床経過にある。3460突然変異および14484突然変異は、4%〜7%の視覚回復率しか示さない11778突然変異よりも良好な視覚予後と関連する(Oostra RJら,J med Genet 1994;31:280-286、Riordan-Eva Pら,Brain 1995;118:319-337、Mashima Yら,Curr Eye Res 1998;17:403-408,これらの引用文献の内容は参照により本明細書に組み込まれる)。しかし、11778突然変異を持ち発症年齢が十代前半である一部の患者では、視覚回復が記録されている(Stone EMら,J clin Meuro-Ophthalmol 1992;12:10-14、Zhu Dら,Am J Med Genet 1992;42:173-179、Salmaggi Aら,Intern J Neuroscience 1994;77:261-266、Oostra RJら,Clin Genet 1997;51:388-393、Mashima Yら,Jpn J Ophthalmol 2002;46:660-667,これらの引用文献の内容は参照により本明細書に組み込まれる)。視覚の回復は、11778突然変異を持つ患者であっても、視覚悪化が若年時に始まる場合の方が、可能性が高くなるようである。
【0007】
発症時年齢、男性好発、不完全浸透、および視覚回復を含むLHON患者の臨床的ばらつきは、おそらくこの疾患が、環境ストレス要因、X染色体遺伝子座、および他のmtDNA突然変異の関与が考えられる多遺伝子性機序または多因子性機序によるものであることを示唆している(Man PYWら,J Med Genet 2002;39:162-169、この引用文献の内容は参照により本明細書に組み込まれる)。しかし、X染色体上の関連遺伝子座を同定しようとする試みは不成功に終わっている(Chalmers RMら,Am J Hum Genet 1996;59:103-108およびPegoraro Eら,Am J Med Genet 2003;119A:37-40,これらの引用文献の内容は参照により本明細書に組み込まれる)。いわゆる「二次LHON突然変異」は、欧州人LHON患者に非罹患欧州人よりも高い頻度で見いだされ、欧州人ハプロタイプJに連関する多型である。これらの多型は強い独立リスク因子ではない(Brown MDら,Am J Hum Genet 1997;60:381-387およびTorroni Aら,Am J Hum Genet 1997;60:1107-1121,これらの引用文献の内容は参照により本明細書に組み込まれる)。
【0008】
したがって、一次突然変異がLHONにおける主要リスク因子であるが、その病原性表現型を増強または調整する付加的な病因因子が必要であるらしい。アルコールおよび/またはタバコの多用がLHON家系における視神経症のリスクを増加させることは、かなりの証拠が示している(Smith PRら,Q J Med 1993;86:657-660、Chalmers RMら,Brain 1996;119:1481-1486およびTsao Kら,Br J Ophthalmol 1999;83:577-581,これらの引用文献の内容は参照により本明細書に組み込まれる)。ただし、この関連性を見いださなかった研究も一つある。考えうる二次的遺伝子相互作用は複雑であり、確立されていない(Kerrison JBら,Am J Ophthalmol 2000;130:803-812,この引用文献の内容は参照により本明細書に組み込まれる)。
【0009】
酸化ストレスはmtDNAの突然変異と関連する多くの障害に関係づけられている。最近行なわれた動物モデルでの研究では、LHONの病理発生過程における有望な因子として、反応性酸素種(ROS)が同定された(Qi Xら,Invest Ophthalmol Vis Sci 2003;44:1088-1096,この引用文献の内容は参照により本明細書に組み込まれる)。さらにまた、mtDNA LHON病原性突然変異により、細胞は、インビトロでFas依存性アポトーシス死を起こしやすくなることが見いだされた(Danielson SRら,J Biol Chem 2002;277:5810-5815,この引用文献の内容は参照により本明細書に組み込まれる)。これらの知見により、LHONの発病に関与するいくつかの核修飾因子遺伝子が存在するに違いないことが暗示された。
【発明の開示】
【課題を解決するための手段】
【0010】
本発明者らは、いくつかの既知SNPおよび未知SNPが緑内障およびレーベル病を含む視神経症の発症に連関することを明らかにし、本発明を完成した。
【0011】
したがって本発明は、
(1)エンドセリン-1遺伝子のコドン198におけるAAG→AAT置換(Lys198Asn);
(2)エンドセリン-1遺伝子プロモーター領域の-1370T>G多型;
(3)エンドセリン-1遺伝子のエクソン1におけるA138挿入/欠失(A138I/D)多型;
(4)エンドセリン受容体A遺伝子の3'非コード領域における+70C>G多型;
(5)エンドセリン受容体A遺伝子の+1222C>T多型;
(6)エンドセリン受容体A遺伝子のエクソン6中のコドン323におけるCAC→CAT置換(His323His);
(7)エンドセリン受容体A遺伝子プロモーター領域の-231A>G多型;
(8)エンドセリン受容体B遺伝子のエクソン4中のコドン277におけるCTG→CTA置換;
(9)ミトコンドリア遺伝子の9099C>A多型;
(10)ミトコンドリア遺伝子の9101T>G多型;
(11)ミトコンドリア遺伝子の9101T>C多型;
(12)ミトコンドリア遺伝子の9804G>A多型;
(13)ミトコンドリア遺伝子の11778G>A多型;
(14)アンギオテンシンII1型受容体遺伝子プロモーター領域の-713T>G多型;
(16)アンギオテンシンII2型受容体遺伝子の3123C>A多型;
(25)パラオキソナーゼ1遺伝子のコドン192におけるCAA→CGA置換(Gln192Arg);
(26)パラオキソナーゼ1遺伝子のコドン55におけるTTG→ATG置換(Leu55Met);
(27)ノエリン(Noelin)2遺伝子のコドン144におけるCGG→CAG置換(Arg144Gln);
(32)β1アドレナリン作動性受容体遺伝子のコドン389におけるGGA→CGA置換(Gly389Arg);
(35)ミオシリン遺伝子の1105T>C多型(Phe369Leu);
(36)オプチニューリン遺伝子の412G>A多型;
(37)E-セレクチン遺伝子の1402C>T多型;
(38)腫瘍壊死因子α遺伝子プロモーター領域の-857C>T多型とオプチニューリン遺伝子の412G>A多型との組み合わせ;
(39)腫瘍壊死因子α遺伝子プロモーター領域の-863C>A多型とオプチニューリン遺伝子の603T>A多型との組み合わせ;
(40)TP53遺伝子のコドン72におけるCGC→CCC置換(Arg72Pro);
(41)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン113におけるTAC→CAC置換(Tyr113His);
(42)熱ショックタンパク質70-1遺伝子プロモーター領域の-110A>C多型;
(43)エンドセリン変換酵素遺伝子プロモーター領域の-338C>A多型;
(44)CD95遺伝子プロモーター領域の-670A>G多型;
(45)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン119におけるAAG→AAA置換(Lys119Lys);
(47)β2アドレナリン作動性受容体遺伝子のコドン16におけるGGA→AGA置換(Gly16Arg);および
(48)β2アドレナリン作動性受容体遺伝子のコドン27におけるCAA→GAA置換(Gln27Glu)
からなる群より選択される少なくとも一つの多型を含む、視神経症と関連する遺伝子多型のセットを提供する。
【0012】
さらに本発明は、ヒト被験者における視神経症に対する感受性を診断または予測するための方法であって、
i)被験者から生物学的試料を取得する工程、
ii)上に定義した多型のセットに関して、試料の遺伝子型を決定する工程、および
iii)被験者における視神経症に対する感受性を遺伝子型に基づいて診断または予測する工程
を含む方法も提供する。
【0013】
本発明によれば、視神経症は、好ましくは、緑内障またはレーベル病であることができる。多型(1)〜(39)および(42)〜(48)は、特に緑内障に使用することができる。それらのうち、多型(1)、(2)、(5)〜(7)、(16)、(26)、(32)、(43)および(45)は、特に正常眼圧緑内障に使用することができ、多型(4)、(14)、(25)、(35)、(36)、(38)、(42)、(44)、(47)〜(48)は、特に原発開放隅角緑内障に使用することができる。多型(40)および(41)は、特にレーベル病に使用することができる。
【0014】
本発明によれば、多型のセットは、視神経症と関連することが知られている少なくとも一つの他の多型を、さらに含みうる。
【0015】
本発明のもう一つの態様として、上に定義した遺伝子多型のセットに関して遺伝子型を決定するのに適したプライマーセットおよび/またはプローブを含む、ヒト被験者における視神経症に対する感受性を診断または予測するためのキットが提供される。
【0016】
本発明のさらなる態様として、ミトコンドリア遺伝子、ミオシリン遺伝子およびノエリン2遺伝子内に新たに同定されたSNPを提供する。したがって本発明は、これらのSNPを含むヌクレオチド断片を包含する。一般に、前記SNPに関して遺伝子型を決定するには、そのSNPを含有する90個以上の連続するヌクレオチドの配列が必要だろう。すなわち、配列:
のセグメントからなる単離ポリヌクレオチドであって、
前記セグメントは少なくとも90個の連続するヌクレオチドを含んでなり、その少なくとも90個の連続するヌクレオチドは上記配列の9099位を含み、上記配列の9099位がAである、単離ポリヌクレオチド、もしくは上記セグメントに対して完全に相補的な単離ポリヌクレオチド;または、
前記セグメントは少なくとも90個の連続するヌクレオチドを含んでなり、その少なくとも90個の連続するヌクレオチドは上記配列の9101位を含み、上記配列の9101位がGである、単離ポリヌクレオチド;または、
上記セグメントのいずれか一方に対して完全に相補的な単離ポリヌクレオチドが提供される。
【0017】
さらに本発明は、配列:
のセグメントからなる単離ポリヌクレオチドであって、前記セグメントは少なくとも90個の連続するヌクレオチドを含んでなり、その少なくとも90個の連続するヌクレオチドは上記配列の下線部のヌクレオチドに相当するコドン369を含み、コドン369は、それがLeuをコードするように置換されている、単離ポリヌクレオチド、または上記セグメントに対して完全に相補的な単離ポリヌクレオチドを提供する。
【0018】
さらに本発明は、配列:
のセグメントからなる単離ポリヌクレオチドであって、前記セグメントは少なくとも90個の連続するヌクレオチドを含んでなり、その少なくとも90個の連続するヌクレオチドは上記配列の下線部のヌクレオチドに相当するコドン144を含み、コドン144は、それがGlnをコードするように置換されている、単離ポリヌクレオチド、または上記セグメントに対して完全に相補的な単離ポリヌクレオチドを提供する。
【発明を実施するための最良の形態】
【0019】
本明細書および特許請求の範囲において、「遺伝子多型」とは、ある遺伝子座における個体間のゲノム多様性を意味する。遺伝子多型は「一塩基多型」または「SNP」と呼ばれる一塩基置換であってもよいし、複数のヌクレオチドからなるものであってもよい。遺伝子多型は、その個体の表現型に影響を及ぼすものであっても、影響を及ぼさないものであってもよい。さらに、ある個体のヌクレオチド配列が、対応する野生型配列とは異なる(すなわち野生型配列上に挿入、欠失または置換を持つ)場合、そのヌクレオチド配列を「遺伝子突然変異体」と呼ぶが、その遺伝子突然変異体も本明細書にいう「多型変異体」に含まれる。
【0020】
本明細書および特許請求の範囲において、「9099C>A」または「C9099A」などの表現は、その遺伝子が9099位に多型を持つこと、すなわちその遺伝子には二つの対立遺伝子があり、一方は9099位にシトシン、すなわちCを持ち、他方は9099位にアデニン、すなわちAを持つこと(二対立遺伝子性)を意味する。これは、高頻度の対立遺伝子がその位置にCを持ち、低頻度の対立遺伝子がその位置にAを持つことを、必ずしも意味しない。
【0021】
「Gln192Arg」などの表現は、アミノ酸配列をコードする遺伝子における塩基置換によるアミノ酸置換を表す。例えば、Gln192Argは、コドン192(すなわちアミノ酸番号192)のグリシンが、アルギニン、すなわちArgで置き換えられることを表す。これは、そのタンパク質に、コドン192のアミノ酸がGlnまたはArgである多型変異体が存在することも意味する。
【0022】
本発明によれば、遺伝子多型に関する遺伝子型の決定は、単一の多型ごとに行なうか、または複数の多型もしくは全ての多型を同時に決定することができる。
【0023】
本発明において、ヒト被験者における視神経症に対する感受性を診断または予測するための方法は、本願で新たに報告する視神経症との関係を持つ遺伝子多型のセットに関して、遺伝子型を決定することを含む。本発明によって視神経症に連関すると同定された遺伝子多型に加えて、視神経症に連関することが既に明らかにされていた他の任意の多型を一緒に検出することができる。視神経症に連関する複数の遺伝子多型を使用することにより、診断確率を改善することができる。
【0024】
本発明によれば、遺伝子多型に関して遺伝子型を決定するために使用する方法は限定されず、当技術分野で知られている任意の方法を使用することができる。遺伝子多型に関して遺伝子型を決定するための代表的方法には、ポリメラーゼ連鎖反応・制限断片長多型(PCR-RFLP)解析、ポリメラーゼ連鎖反応後の一本鎖高次構造多型(PCR-SSCP)解析、ASOハイブリダイゼーション解析、ダイレクトシークエンス解析、ARMS解析、DGGE解析、RNaseA切断解析、化学的制限切断解析(chemical restriction analysis)、DPL解析、TaqMan(登録商標)PCR解析、Invader(登録商標)アッセイ、MALDI-TOF/MS解析、TDI解析、一塩基伸長アッセイ、WAVEアッセイ、一分子蛍光検出アッセイなどがある。本発明によれば、検出方法はこれらの方法の一つであってもよいし、二つ以上の組み合わせであってもよい。
【0025】
本発明によれば、遺伝子多型の検出に用いるべき生物学的試料は特に限定されず、毛髪、血液、唾液、リンパ液、気道粘膜、培養細胞および尿であることができる。
【0026】
本明細書および特許請求の範囲において、「視神経症に対する感受性を診断または予測すること」には、視神経症の発症を診断することだけでなく、疾患の発症を早め、疾患の進行を加速するリスク因子を決定することも含まれる。
【0027】
本発明によれば、上に同定した遺伝子多型ならびにタンパク質多型を検出するためのキットも提供される。このキットは、上に同定した遺伝子多型を検出するために特に設計されたプライマーおよび/またはプローブ、上に同定したタンパク質多型を検出するための抗体を含む。本発明によれば、このキットは、視神経症に対する感受性を診断または予測するために使用することができる。
【0028】
本明細書および特許請求の範囲において、用語「プライマー」は、標的ヌクレオチド配列の一部に相補的であって、標的ヌクレオチド配列にハイブリダイズさせるために使用される、特異的オリゴヌクレオチドを意味する。プライマーは、DNAポリメラーゼ、RNAポリメラーゼまたは逆転写酵素のいずれかによって触媒されるヌクレオチド重合の開始点として役立つ。
【0029】
用語「プローブ」は、試料中に存在する特異的ポリヌクレオチド配列を同定するために、または遺伝子修飾過程において標的DNAもしくはRNAを確認するために、使用することができる所定の核酸セグメントであって、同定されるべき特異的ポリヌクレオチド配列に相補的なヌクレオチド配列を含むものを意味する。
【0030】
本発明によれば、プライマーおよびプローブは、標的とする配列に基づいて、それらが他のいくつかの遺伝子と同一でない限り(すなわち反復配列でもパリンドローム配列でもないことが必要である)、標的とする多型が予想される位置および/またはその位置の周辺配列に対して、それらが相補的になるように、設計することができる。
【0031】
本発明によれば、視神経症、特に緑内障およびレーベル病に連関する遺伝子多型が同定される。これらの知見に基づいて、個体から取得した生物学的試料の遺伝子多型に関する遺伝子型を決定し、そのようにして取得した遺伝子型に基づいて、疾患の発症または疾患の発症に関して予測されるリスクを決定することができる。
【0032】
上に同定した多型(1)〜(48)に加えて、視神経症と高度に関連することが当技術分野で既に知られていた他のいくつかの遺伝子多型に関する遺伝子型も、診断または予測の信頼性を改善するために決定することができる。
【0033】
例えば、ミオシリン遺伝子およびオプチニューリン遺伝子中の2タイプの遺伝子多型は原発開放隅角緑内障の発症と関連することが、本発明者らによって明らかにされている。これら二つの遺伝子に加えて、突然変異を含む他の四つの遺伝子多型が原発開放隅角緑内障と関連すると既に同定されていた。本発明の遺伝子多型に関するリスク遺伝子型と当技術分野で既に知られていた遺伝子多型に関するリスク遺伝子型をどちらも持つ被験者は、ほぼ100%が、緑内障を発病しうる。すなわち、これら遺伝子多型のセットは、発症前検査に有用であるだろう。
【0034】
いくつかのSNPに関して、本発明者らは、人種または性別などの特定の群で、視神経症との相関を確認した。したがって、指定された群における視神経症のリスクを診断または予測するには、それらのSNPを好ましく使用することができる。
【0035】
さらに、多型のセットに関する遺伝子型の統計学的解析により、予測発症年齢、生活習慣病との予測的関連性、症状因子との予測的関連性などの有用な情報が得られうる。さらに、その情報に基づいて、いくつかの医学的処置の効果を予測することもできるだろう。
【0036】
本発明によれば、視神経症に対する感受性の予測を、遺伝子型に基づいて疾患の発症前に行なうことができ、被験者はリスク因子を取り除く方法についての助言(例えば生活習慣を改善するようにという助言、または環境を変えるようにという助言)を受けることができる。さらに、リスク遺伝子の減少などの早期処置を受けることもできるだろう。適切な処置を早期に開始することができる。結果として、これらの「オーダーメード処置」は、視覚喪失のリスクを低下させることができる。
【0037】
例えば、ある被験者が視神経症の発症に関する高リスクに連関する遺伝子型を持つ場合、低い発症リスクに連関する遺伝子型をその被験者に導入し、それを発現させることによって、発症リスクの低下または症状の軽減を期待することができる。さらに、高リスク対立遺伝子の発現を阻害するために、視神経症の発症に関して高いリスクを持つ対立遺伝子のmRNAに対するアンチセンス、またはRNAi法を使用することもできる。
【0038】
もう一つの態様として、本発明に示される多型のセットに関する遺伝子型決定に基づいて、視神経症の遺伝的病因を明らかにすることができ、そのようにして得られた病因は新規医薬剤の開発に役立ちうる。
【0039】
さらに、本発明によって得られた視神経症と関連する遺伝子型情報と、生活習慣病などと関連する他の遺伝子型情報とを組み合わせることにより、加齢関連生活習慣関連疾患の総合的リスクを予測し、生活の質を高めるために使用することができる。
【0040】
下記の実施例を使って本発明をさらに例示する。しかし、これらの実施例は単に例示が目的であって、本発明の範囲を限定しようとするものではないと、明確に理解すべきである。
【実施例1】
【0041】
レーベル遺伝性視神経症におけるTP53およびEPHX1の遺伝子変異体ならびにそれらの発症時年齢との関係
目的:酸化ストレスおよびアポトーシスに関する遺伝子の遺伝子多型がレーベル遺伝性視神経症(LHON)を持つ患者における臨床的ばらつきを引き起こすかどうかを決定すること。
【0042】
材料および方法
患者
本発明者らは、ホモプラスミー状態の11778突然変異を保有する血縁関係にない86人の日本人LHON患者を研究した。彼らのmtDNA突然変異を、ポリメラーゼ連鎖反応後の制限酵素アッセイによって確認したところ、一貫してMaeIII部位が得られることが明らかになった(Mashima Yら,Curr Eye Res 1998;17:403-408,この引用文献の内容は参照により本明細書に組み込まれる)。
【0043】
86人のLHON患者における視覚喪失の平均発症時年齢は25.1±13.0歳で、3歳〜65歳の範囲だった。
【0044】
ゲノムDNA抽出および遺伝子型判定
SDS-プロテイナーゼKおよびフェノール/クロロホルム抽出法により、末梢血白血球からDNAを抽出した。酸化ストレス関連遺伝子、ミクロソームエポキシドヒドロラーゼ(EPHX1)(Kimura Kら,Am J Ophthalmol 2000;130:769-773,この引用文献の内容は参照により本明細書に組み込まれる)、およびアポトーシス関連遺伝子TP53中のArg72Pro(Ara Sら,Nucleic Acids Res 1990;18:4961,この引用文献の内容は参照により本明細書に組み込まれる)における多型を調べた。
【0045】
ポリメラーゼ連鎖反応-制限断片長多型(PCR-RFLP)技術を使って各多型を同定した(表1)。
【0046】
[表1]
【0047】
結果
発症時年齢と多型との関連を表2-1および表2-2に記載した。
【0048】
[表2−1]
レーベル遺伝性視神経症における発症時年齢とTP53(Arg72Pro)遺伝子多型およびEPHX1(Tyr113His)遺伝子多型との関連
【0049】
[表2−2]
レーベル遺伝性視神経症における発症時年齢とTP53(Arg/Arg)遺伝子多型およびEPHX1(His/His)遺伝子多型との関連
第1群:TP53のコドン72にArg/Argを持ち、EPHX1のコドン113にHis/Hisを持つ患者。
第2群:TP53のコドン72にArg/Argを持つがEPHX1のコドン113にHis/Hisを持たない患者またはEPHX1のコドン113にHis/Hisを持つがTP53のコドン72にArg/Argを持たない患者。
第3群:第1群および第2群以外の患者。
【0050】
表2-1に示すように、TP53中のコドン72遺伝子型およびEPHX1中のコドン113遺伝子型は、レーベル遺伝性視神経症の発症時年齢の若さと有意に関連した。
【0051】
表2-2に示すように、TP53中のコドン72遺伝子型とEPHX1中のコドン113遺伝子型との併存は、レーベル遺伝性視神経症の発症時年齢の若さと有意に関連した。
【0052】
これらの結果から、TP53におけるArg/Argホモ接合体およびEPHX1におけるHis/Hisホモ接合体を検出することにより、レーベル遺伝性視神経症の早期診断および早期処置が可能になることが示された。
【0053】
これらの結果は、コドン72多型がミトコンドリア機能障害と相互作用して疾患発現に影響を及ぼしうることも示した。p53のコドン72における多型と相関するアポトーシス応答には個体変動が存在しうる。Bonafeら(Biochem Biophys Res Commun 2002;299:539-541)は、Arg/Arg遺伝子型を保有する健常被験者に由来する培養細胞が、細胞毒性薬シトシンアラビノシドに応答して、Arg/Pro被験者に由来する細胞よりも大規模なアポトーシスを起こすことを報告した。したがって、p53遺伝子に天然に存在する遺伝的ばらつきは、化学療法薬に対する細胞のインビボ感受性の個体差に関する説明の一つになるだろう。Dumountら(Nat Genet 2003;33:357-365)は、Arg72変異体がPro72変異体よりも効率よくアポトーシスを誘発することを報告した。この効率増大の基礎にある少なくとも一つの機序は、腫瘍細胞中のミトコンドリアへのArg72変異体の局在の強化である。合成p53阻害剤は、ミトコンドリア損傷および酸化ストレスが惹起するアポトーシスによってニューロンが死ぬLHONの処置に、著しく有効であるかもしれない。
【0054】
標的とする多型を含有するEPHX1遺伝子およびTP53遺伝子の部分ヌクレオチド配列は以下のとおりである。
EPHX1 Tyr113Hisコドン113(下線部)(TAC→CAC変化)
【0055】
TP53コドン72(下線部):CGC(Arg)→CCC(Pro)
【実施例2】
【0056】
原発開放隅角緑内障および正常眼圧緑内障における、レーベル遺伝性視神経症と関係するミトコンドリアDNA突然変異
材料および方法
患者
日本国内の7施設で合計651の血液試料を集めた。POAG患者が201人、NTG患者が232人、正常対照が218人であり、どの被験者もこの研究における他の被験者とは血縁関係になかった。
【0057】
検査時の平均年齢は、POAGでは61.2±16.0歳、NTGでは58.8±13.6歳、対照被験者では70.6±10.9歳だった。対照被験者の平均年齢はPOAG患者(P<0.001)およびNTG患者(P<0.001)の平均年齢より有意に高かった。本発明者らは、対照被験者の一部が最終的には緑内障を発病してしまう確率を低下させるために、より高齢の対照被験者を意図的に選択した。男性はPOAG群では112人(55.7%)、NTG群では108人(46.6%)、対照群では89人(40.8%)だった。
【0058】
正常開放隅角、ゴールドマン視野測定またはハンフリー視野測定で典型的な緑内障性視野喪失を伴う0.7より大きい陥凹/乳頭径比を持ち、視神経損傷の原因となりうる眼障害、鼻障害、神経障害または全身性障害を持たない場合に、その患者はPOAGを有するとみなした。NTGを有する患者は21mmHg以下のIOPを持っていた。剥脱性緑内障、色素性緑内障、およびコルチコステロイド誘発性緑内障を持つ患者は除外した。
【0059】
218の対照試料は、白内障を除く既知の眼異常を持たない日本人被験者から取得した。これらの被験者は40歳を超えていて、21mmHg未満のIOPを持ち、正常な視神経乳頭を持ち、緑内障の家族歴はなかった。
【0060】
Invader(登録商標)アッセイによるmtDNA突然変異の検出
標準的なフェノール-クロロホルム抽出法により、末梢血リンパ球からゲノムDNAを単離した。
【0061】
Invader(登録商標)アッセイによって六つのmtDNA突然変異(G3460A、T9101C、G9804A、G11778A、T14484C、およびT14498C)を検出するために使用した一次プローブ(野生型プローブおよび突然変異型プローブ)ならびにInvader(登録商標)オリゴヌクレオチド(Invader(登録商標)プローブ)を、表3に示す。
【0062】
[表3]
【0063】
Invader(登録商標)アッセイFRET検出256穴プレート(Third Wave Technologies, Inc.,ウィスコンシン州マディソン)は、個々のウェルのそれぞれに乾燥されたInvader(登録商標)アッセイの一般成分(Cleavase(登録商標)酵素VIII、FRETプローブ、MOPSバッファー、およびポリエチレングリコール)を含有している。二重(biplex)形式のInvader(登録商標)アッセイにより、一つのウェルで二つのDNA配列を同時に検出することができた。
【0064】
詳細な方法は先に記述されている。簡単に述べると、一次プローブ/Invader(登録商標)/混合物8μlおよび全DNA(10ng)試料を96穴プレートの各ウェルに加え、95℃で10分間インキュベートすることによって変性させた。鉱油(Sigma,ミズーリ州セントルイス)15μlを全ての反応ウェルに重層し、プレートをPTC-100サーマルサイクラー(MJ Research,マサチューセッツ州ウォルサム)にて等温的に63℃で2時間インキュベートした後、蛍光測定まで4℃で保存した。蛍光強度は、CytoFlour 4000蛍光プレートリーダー(Applied Biosystems,カリフォルニア州フォスターシティ)を使って、FAM色素については励起485nm/20nm(波長/バンド幅)および放射530nm/25nmで、またRedmond RED(RED)色素については励起560nm/20nmおよび放射620nm/40nmで測定した。各試料を同じプレートで二重に試験し、各プレートで2回の蛍光測定を行なった。そのようにして、各試料につき四つの測定値を取得し、それらを平均した。
【0065】
ダイレクトDNAシークエンス
ダイレクトシークエンス法によって突然変異を検出するために、PCR産物をまずQIAquick PCR精製キット(QIAGEN,米国カリフォルニア州バレンシア)で精製することにより、未反応のプライマーおよび前駆体を除去した。次に、ABI PRISM BigDye Terminator(v.3.1)サイクルシークエンスキットを、製造者のプロトコール(Applied Biosystems)に従って使用することにより、シークエンス反応を行なった。ABI PRISM 310遺伝子解析装置によってデータを収集し、ABI PRISMシークエンス解析プログラム(v.3.7)によって解析した。
【0066】
[表4]
【0067】
結果
合計651人の日本人被験者を調べた。ヌクレオチド置換が一次プローブ内またはInvaderプローブ内にある場合、検査した症例は、Invaderアッセイでは、どちらのプローブにも反応を示さなかった。そのような症例において、ダイレクトシークエンス解析を行なったところ、9099、9101、9102、9797、および9815のヌクレオチド位置に一塩基多型(SNP)が見いだされた。
【0068】
表5に示すように、女性5人および男性2人を含む7人の患者が、mtDNAに五つの突然変異を保持しており、LHONを発病していなかった。2人の患者(症例1および症例2)は、LHONと関連づけられていない新規なアミノ酸変化を保持しており、5人の患者(症例3〜7)は、LHON突然変異を保持していた。
【0069】
これらのmtDNA突然変異は正常対照には検出されなかった。
【0070】
[表5]
【0071】
上述のように、本発明者らは、緑内障患者に二つの新規mtDNA突然変異を含む五つのmtDNA突然変異を見いだした。これらの結果から、mtDNA突然変異は緑内障を発病または進行させるリスク因子の一つであること、そしてmtDNA突然変異の検出により、緑内障の早期診断および早期処置が可能になることが示された。
【0072】
標的とする突然変異/多型を含有するミトコンドリア遺伝子の部分ヌクレオチド配列は以下のとおりである。
C9099A,T9101G(下線部)
【0073】
G9804A(下線部)
【0074】
G11778A(下線部)
【実施例3】
【0075】
レニン-アンギオテンシン-アルドステロン系の遺伝子多型は原発開放隅角緑内障および正常眼圧緑内障を発病するリスクと関連する
目的:緑内障の病理発生過程には複数の環境的因子および遺伝的因子が関与しうる。緑内障の遺伝的リスクを予測するために、レニン-アンギオテンシン-アルドステロン(R-A-A)系の遺伝子多型における関連研究を行なった。
【0076】
材料および方法
患者および対照研究被験者
日本国内の7施設で合計551の血液試料を集めた。POAG患者が162人、NTG患者が193人、正常被験者が196人であり、どの被験者もこの研究における他の被験者とは血縁関係になかった。
【0077】
検査時の平均年齢は、NTGでは58.8±13.7歳、POAGでは62.0±15.4歳、および正常被験者では71.2±10.4歳だった。正常対照被験者の平均年齢は、NTG患者(P<0.001)またはPOAG患者(p<0.001)より、それぞれ有意に高い。これにより、一部が最終的には緑内障を発病してしまう可能性を低減することができた。NTG患者127人のうち66人(34.2%)およびPOAG患者113人のうち49人(30.2%)に、家族歴が記録された。男性患者はNTGでは89人(46.1%)、POAGでは87人(53.7%)、正常被験者では77人(39.3%)だった。
【0078】
196の日本人対照試料は、白内障を除く既知の眼異常を持たない個体から取得した。これらの被験者は40歳を超えていて、21mmHg未満のIOPを持ち、緑内障性乳頭変化を持たず、緑内障の家族歴はなかった。
【0079】
遺伝子型判定
R-A-A系中の七つの遺伝子および10個の多型を、緑内障を持つ被験者または正常日本人対照のそれぞれについて決定した。レニン(REN)I8-83G>A(Frossard PMら,Hypertens Res 1998;21:221-225,この引用文献の内容は参照により本明細書に組み込まれる)、アンギオテンシンII1型受容体(AT1R)1166A>C、-521C>T、-713T>G(Nalogowska-Glosnicka Kら,Med Sci Monit 2000;6:523-529およびErdmann Jら,Ann Hum Genet 1999;63:369-374,これらの引用文献の内容は参照により本明細書に組み込まれる)、アンギオテンシンII2型受容体(AT2R)3123C>A(Katsuya Tら,Mol Cell Endocrinol 1997;127:221-228,この引用文献の内容は参照により本明細書に組み込まれる)、シトクロムP45011B1(CYP11B1)-344T>C(Tsujita Yら,Hypertens Res 2001;24:105-109,この引用文献の内容は参照により本明細書に組み込まれる)、およびキマーゼ(CYM)3123C>Aは、ポリメラーゼ連鎖反応-制限断片長多型(PCR-RFLP)を使って同定した。アンギオテンシン変換酵素(ACE)挿入/欠失(I/D)はPCRおよびアガロースゲル電気泳動だけで決定した。ACE/ID多型の偽決定を避けるために、Lindpaintnerらのプロトコール(N Engl J Med 1995;332:706-711,この引用文献の内容は参照により本明細書に組み込まれる)に従って、I対立遺伝子特異的な増幅を行なった。ゲノムDNAは、フェノール-クロロホルム抽出により、末梢血リンパ球から単離した。使用したプライマーセットおよび制限酵素を表6に記載した。
【0080】
[表6]
【0081】
アンギオテンシノゲン(AGT)T174M、M235Tの遺伝子型はInvaderアッセイ(登録商標)を使って決定した(Lyamichev Vら,Nat Biotechnol 1999;17:292-296,この引用文献の内容は参照により本明細書に組み込まれる)。
【0082】
結果
日本人集団におけるR-A-A系の遺伝子型分布
R-A-A系中の10個の多型のうちの二つ、すなわちPOAGに関してAT1R/-713T>Gと、NTGに関してAT2/3123C>Aは、遺伝子型の頻度に有意差を示した(表7)。3123C>A多型はNTGを持つ女性患者とだけ関連した。
【0083】
AT1R/-713T>G多型におけるホモ接合性G遺伝子型(GG)の頻度は、POAG患者(4.2%)では対照(0.5%)よりも有意に高かった(TT+TG対GGについてp=0.04)。AT2R/3123C>A多型におけるCA+AA遺伝子型の頻度は、NTGを持つ女性患者(70.8%)では女性対照(55.0%)よりも有意に高かった(CC対CA+AAについてp=0.011)。
【0084】
[表7]
緑内障(POAGおよびNTG)とレニン-アンギオテンシン-アルドステロン系の遺伝子多型との関連
【0085】
POAG患者におけるAT1R中の二つのプロモーター多型間の関連
ホモ接合性-521Tとホモ接合性-713Gとを合わせ持つPOAG保有者の頻度(4.2%)は、正常者(0%)より有意に高かった(p=0.011)(表8-1)。POAG患者だけがこの遺伝子型を持ち、NTG被験者も正常被験者もこの遺伝子型を持たなかった。
【0086】
[表8−1]
AT1R -521T対立遺伝子および-713G対立遺伝子の遺伝子型の分布
A:二つの-521対立遺伝子および二つの-713G対立遺伝子を持つ被験者。
B:A群の基準を満たさない被験者。
【0087】
これらの結果から、レニン-アンギオテンシン-アルドステロン系の遺伝子多型は緑内障の発病に関する重要な遺伝的リスク因子の一つであることが示された。AT1R/-731T>G多型の検出により、POAGの早期診断および早期処置が可能になる。特に、AT1R遺伝子中にホモ接合性-521Tとホモ接合性-713Gとを合わせ持つ特異な遺伝子型は、POAGの早期診断に有用である。AT2R/3123C>A多型の検出により、NTGを持つ女性患者の早期診断および早期処置が可能になる。
【0088】
AT2R 3123C>A多型およびACE I/D多型を持つNTG患者の臨床特性
緑内障患者に関して記録された臨床特徴は、診断時の年齢、無処置時最大IOP(診断時のIOPと定義される)、および初診時の視野欠損(診断時の視野欠損と定義される)だった。視野欠損の重症度は1〜5にスコア化した。異なる視野計を使って取得したデータを、以下に定義する5点尺度を使って集約した。1=変化なし;2=早期欠損;3=中等度欠損;4=重度欠損;および5=光覚のみまたは視覚なし。視野欠損は、ゴールドマン視野測定の結果に基づくKozakiの分類に従って、またはハンフリー視野解析機に用いられる分類に従って、早期、中等度、または重度と判定した。日本では前者の分類が最も広く用いられている。
【0089】
AT2R遺伝子型を持つ男性緑内障患者との間に、視野スコア臨床特性の有意な関連が検出された。C遺伝子型を持つ男性POAG患者における視野スコアは、A遺伝子型を持つ者より悪かった(P=0.04,表8-2)。C/Cを持つ女性緑内障患者およびC/A+A/Aを持つ女性緑内障患者との間に、臨床特性(年齢、IOP、および視野スコア)の有意な関連は検出されなかった。視野スコアは、C/C遺伝子型を持つNTG患者では、C/A+A/A遺伝子型を持つNTG患者より悪い傾向があった(P=0.165)。
【0090】
しかし、ACE挿入/欠失多型と組み合わせた場合、AT2R遺伝子中にC/Cを保有すると共にACE遺伝子中にID+DDを保有する女性NTG患者は、他の三つの複合遺伝子型よりも視野スコアが有意に悪かった(P=0.012;表8-3,図1)。
【0091】
[表8−2]
AT2R遺伝子型による男性緑内障患者の臨床特性の比較
【0092】
[表8−3]
【0093】
標的とする多型を含有するAT1R遺伝子およびAT2R遺伝子の部分ヌクレオチド配列は以下のとおりである。
AT1R -713(下線部の「t」) T>G
【0094】
AT2R 3123(下線部の「c」)C>A,下線部のオリゴヌクレオチド配列をプライマーに使用した
【0095】
(GenBankアクセッション番号AY536522によれば、AT2R 3123はヌクレオチド番号4926に対応する)
【実施例4】
【0096】
正常眼圧緑内障を発病するリスクと関連するエンドセリン遺伝子の遺伝子多型
方法
患者
合計605の血液試料を集めた。POAG患者が178人、NTG患者が214人、正常対照が213人であり、どの被験者もこの研究における他の被験者とは血縁関係になかった。正常開放隅角、ゴールドマン視野測定またはハンフリー視野測定で典型的な緑内障性視野喪失を伴う0.7より大きい陥凹/乳頭径比を持ち、視神経損傷の原因となりうる眼障害、鼻障害、神経障害または全身性障害を持たない場合に、その患者はPOAGを有するとみなした。NTGを有する患者は21mmHg以下のIOPを持っていた。剥脱性緑内障、色素性緑内障、およびコルチコステロイド誘発性緑内障を持つ患者は除外した。対照試料は、白内障を除く既知の眼異常を持たない日本人被験者から取得した。これらの被験者は21mmHg未満のIOPを持ち、正常な視神経乳頭を持ち、緑内障の家族歴はなかった。
【0097】
Invaderアッセイによるエンドセリン(ET)遺伝子のG/T多型の検出
標準的なフェノール-クロロホルム抽出法によって末梢血リンパ球からDNAを単離し、ET遺伝子のエクソン5中のコドン198におけるG/T多型(Lys/Lys、Lys/AsnおよびAsn/Asn)を、Invader(登録商標)アッセイによって決定した。ET遺伝子のG/T多型を検出するために使用した一次プローブ(野生型プローブおよび突然変異型プローブ)ならびにInvader(登録商標)オリゴヌクレオチド(Invader(登録商標)プローブ)を表9に示す。
【0098】
[表9]
【0099】
Invader(登録商標)アッセイFRET検出96穴プレート(Third Wave Technologies, Inc.,ウィスコンシン州マディソン)は、個々のウェルのそれぞれに乾燥されたInvader(登録商標)アッセイの一般成分(Cleavase(登録商標)酵素VIII、FRETプローブ、MOPSバッファー、およびポリエチレングリコール)を含有している。簡単に述べると、一次プローブ/Invader(登録商標)/混合物8μlおよび全DNA(10ng)試料を96穴プレートの各ウェルに加え、95℃で10分間インキュベートすることによって変性させた。鉱油(Sigma,ミズーリ州セントルイス)15μlを全ての反応ウェルに重層し、プレートをPTC-100サーマルサイクラー(MJ Research,マサチューセッツ州ウォルサム)にて等温的に63℃で2時間インキュベートした後、蛍光測定まで4℃で保存した。蛍光強度は、CytoFlour 4000蛍光プレートリーダー(Applied Biosystems,カリフォルニア州フォスターシティ)を使って、FAM色素については励起485nm/20nm(波長/バンド幅)および放射530nm/25nmで、またRedmond RED(RED)色素については励起560nm/20nmおよび放射620nm/40nmで測定した。各試料を同じプレートで二重に試験し、各プレートで2回の蛍光測定を行なった。そのようにして、各試料につき四つの測定値を取得し、それらを平均した。
【0100】
結果
ET遺伝子のエクソン5中のコドン198におけるG/T多型(Lys/Lys、Lys/AsnおよびAsn/Asn)の遺伝子型頻度を表10に示す。
【0101】
[表10]
ET遺伝子のエクソン5中のコドン198における遺伝子型頻度
【0102】
これらの結果から、エクソン5中のコドン198におけるET-1遺伝子のLys/Lysホモ接合体は、NTGを発病または進行させるリスク因子の一つであり、Lys/Lysホモ接合体の検出により、NTGの早期診断および早期処置が可能になる。
【0103】
コドン198を含むEDN1の部分配列は以下のとおりである。
EDN1 コドン198(下線部):aag(Lys)→aat(Asn)
【実施例5】
【0104】
変性高速液体クロマトグラフィーによって検出される、原発開放隅角緑内障を持つ日本人患者における新規MYOC遺伝子突然変異Phe369Leu
目的:変性高速液体クロマトグラフィー(DHPLC)を使って原発開放隅角緑内障(POAG)を持つ日本人患者におけるMYOC遺伝子中の突然変異についてスクリーニングすること。
【0105】
材料および方法
患者
日本の医療施設7ヶ所でPOAG患者171人および正常被験者100人から血液試料を集めた。被験者らは血縁関係になく、検査時の平均年齢は、POAGを持つ患者では55.1±16.0歳(±標準偏差)、正常被験者では70.5±10.6歳だった。本発明者らは、対照被験者の一部が最終的には緑内障を発病してしまう確率を低下させるために、より高齢の対照被験者を意図的に選択した。
【0106】
55人のPOAG患者(32.2%)では問診によって詳細な家族歴が得られた。男性はPOAG患者では91人(53.2%)、正常被験者では41人(41.0%)だった。
【0107】
DNA抽出およびPCR条件
標準的方法により末梢血リンパ球からゲノムDNAを単離した。ポリメラーゼ連鎖反応(PCR)により、表11に記載するプライマーセットを使って、MYOC遺伝子の七つのエクソン領域を増幅した。患者のハイスループット解析を行なう場合は、3人の患者から得た試料をプールした。PCR反応はサーマルサイクラー(iCyceler;BioRad,カリフォルニア州ハーキュリーズ)を使って25μlの総液量で行なった。PCR条件は以下のとおりとした:95℃で9分間変性;次に、95℃で1分、58℃で30秒(表1)、そして72℃で1.5分を35サイクル;次に、72℃で7分間の最終伸長工程を行なった。ヘテロ二重鎖形成のために、各PCR産物(25μl)を95℃で5分間変性し、25℃まで徐冷した。
【0108】
数個の試料を解析する場合は、96穴プレート(96穴マルチプレートMLP-9601;MJ Research,マサチューセッツ州ウォルサム)でのPCRによって、七つのエクソン領域のそれぞれを同時に増幅した。各患者につき七つのウェルを使用した。58℃という単一のアニーリング温度を使った場合に有効であるように、プライマーセットを設計した(表11)。
【0109】
[表11]
プライマー配列、産物サイズ、ならびにPCRアニーリング温度およびDHPLC解析温度
【0110】
変性HPLC解析
ハイスループット解析の場合、3人の患者から得られたPCR産物25μlを、DHPLC解析用のWAVE(登録商標)システム(Transgenomic,ネブラスカ州オマハ)を使って、解析用クロマトグラフに自動注入した。DHPLC融解温度を表1に記載する。小数の試料を解析する場合は、96穴プレートPCR後に、2〜3の融解温度で各ウェルを自動的に解析するようにプログラムされたWAVE(登録商標)システムに、プレートを入れた。一個体の試料を解析するには、約3時間あれば十分だった。
【0111】
ハイスループットプロトコールでプール試料に異常なクロマトグラフパターンが検出された場合は、その試料をWAVE(登録商標)システムで個別に再解析した。次に、異常なクロマトグラフパターンを示したPCR産物をシークエンスした。
【0112】
ダイレクトDNAシークエンス
ダイレクトシークエンスのために、PCR産物をQIA Quick PCR精製キット(Qiagen,カリフォルニア州バレンシア)で精製することにより、未使用のプライマーおよび前駆体を除去した。BigDyeケミストリーを製造者の推奨するプロトコール(Applied Biosystems,カリフォルニア州フォスターシティ)に従って使用することにより、ABI310自動シークエンサーで、同じフォワードおよびリバースPCR増幅プライマーを使って、PCR産物をダイレクトシークエンスした。
【0113】
結果
患者171人でのDNAプールのスクリーニング
エクソン3C領域における四つのDHPLC記録線パターンを図2に示した。一番上のパターン(A)が正常な外観を持つのに対して、中央のパターン(B)は幅広いショルダーを示し、下側のパターン(CおよびD)はこの領域における配列変異を示す特徴的な二重ピークパターンを持っていた。試料B、CおよびDのシークエンス解析により、Thr448Pro、Pro481Ser、およびAla488Ala突然変異が明らかになった(表12)。
【0114】
4つの緑内障原因突然変異が、POAGを持つ患者171人のうち5人(2.9%)に同定された。さらに、八つの多型および五つの同義コドン変化が同定された(表12)。エクソン3中に検出される新規ミスセンス突然変異Phe369Leu(図3)は、100人の正常日本人被験者には存在しなかった。他の三つのミスセンス突然変異、Ile360Asn、Ala363Thr、およびThr448Proは、POAGを持つ日本人患者で報告されている
【0115】
[表12]
POAGを持つ患者および対照におけるMYOC突然変異および多型
【0116】
プレートPCRとそれに続くDHPLCによる個々の患者のスクリーニング
POAGを持つ患者から得たDHPLC記録線を図4に示す。エクソン3B領域には、配列変異を示す異常記録線を見ることができ、これはダイレクトシークエンス法でPhe369Leu突然変異を表すことがわかった。
【0117】
標的とする多型を含有するMYOCエクソン3遺伝子の部分ヌクレオチド配列は以下のとおりである。
MYOC エクソン3,コドン369(下線部)TTC(Phe)→CTC(Leu)
【0118】
MYOCエクソン1〜3のヌクレオチド配列はGenBankからアクセッション番号AB006686-AB006688として入手することができる。
【実施例6】
【0119】
オプチニューリン遺伝子中の変異体および緑内障を持つ日本人患者におけるそれらと腫瘍壊死因子α多型との関連
目的:オプチニューリン(OPTN)遺伝子中の配列変異および緑内障を持つ日本人患者におけるそれらとTNF-α多型との関連を調べること。
【0120】
被験者および方法
患者および対照被験者
日本国内の7施設で合計629の血液試料を集めた。POAG患者が194人、NTG患者が217人、正常対照が218人であり、どの被験者もこの研究における他の被験者とは血縁関係になかった。診断時の年齢が35歳未満の患者および-5.5Dを超える近視を持つ患者は除外した。MYOC突然変異を持つPOAG患者も除外した。
【0121】
DNA抽出およびPCR条件
フェノール-クロロホルム抽出によって末梢血リンパ球からゲノムDNAを単離した。ポリメラーゼ連鎖反応(PCR)により、表13に記載するプライマーセットを使って、OPTN遺伝子の13のエクソンコード領域を増幅した。融解温度が高いドメイン中の突然変異をDHPLC解析によって検出するために、フォワードプライマーの一部に20塩基GCクランプを取り付けた(Narayanaswami Gら,Genet Test. 2001;5:9-16)。ハイスループット解析では、3人の患者から得た試料をプールした。PCRは、サーマルサイクラー(iCyceler;BioRad,カリフォルニア州ハーキュリーズ)を使って、ゲノムDNA 45ng、GeneAmp 10×PCRバッファーII 2μl、濃度2.0mMの各dNTPを含むGeneAmp dNTPミックス2μl、25mM MgCl2溶液2.4μl、各プライマー4pmol、およびAmpliTaq Gold DNAポリメラーゼ(Applied Biosystems,カリフォルニア州フォスターシティ)0.1Uを含有する20μlの総液量で行なった。PCR条件は以下のとおりとした:95℃で9分間変性した後、95℃で1分、55〜60℃で30秒(表13)、および72℃で1分30秒を35サイクル、そして72℃で7分間の最終伸長工程。
【0122】
[表13]
プライマー配列、PCR産物サイズ、ならびにPCRアニーリング温度およびDHPLC解析温度
【0123】
変性HPLC解析
WAVE(登録商標)システム(Transgenomic,ネブラスカ州オマハ)を使ってDHPLC解析を行なった。ヘテロ二重鎖形成のために、各PCRの産物(20μl)を95℃で5分間変性し、25℃まで徐冷した。混合した3試料から得られるアニールしたPCR産物を、DNASep(登録商標)カートリッジ(Transgenomic,ネブラスカ州オマハ)に自動注入した。
【0124】
バッファーA(Transgenomic,ネブラスカ州オマハ)は0.1M酢酸トリエチルアンモニウム(TEAA)から構成され、バッファーBは0.1M TEAAおよび25%アセトニトリルから構成された。流速0.9ml/分、2%/分ずつ増加する4.5分間のバッファーB勾配で、解析を行なった。カートリッジからのDNA断片の溶出は260nmでの吸光度によって検出した。解析に使用する温度は、DNA断片の配列に応じて選択した。WAVEMAKERソフトウェア(v.4.1,Transgenomic,ネブラスカ州オマハ)により、さまざまな温度におけるDNA断片の融解挙動が予測された。DNA断片内の予測融解ドメインが、DHPLC解析の温度を決定した(表13)。異常なクロマトグラフパターンが3試料のプール中に検出された場合は、それら3試料のそれぞれを、WAVE(登録商標)システムで個別に再解析した。次に、異常なクロマトグラフパターンを示したPCR産物をシークエンスした。異常なクロマトグラフパターンと塩基変化との相関がダイレクトシークエンス解析によって確認された後は、既知の異常クロマトグラフパターンのいずれかがDHPLC解析で観察された場合に、シークエンス解析をさらに行なうことはしなかった。
【0125】
ダイレクトDNAシークエンス
ダイレクトシークエンスによって突然変異を検出するために、PCR産物をまずQIAquick PCR精製キット(QIAGEN,米国カリフォルニア州バレンシア)で精製することにより、未反応のプライマーおよび前駆体を除去した。次に、ABI PRISM BigDye Terminator(v.3.1)サイクルシークエンスキットを製造者のプロトコール(Applied Biosystems)に従って使用することにより、シークエンス反応を行なった。ABI PRISM 310遺伝子解析装置によってデータを収集し、ABI PRISMシークエンス解析プログラム(v.3.7)によって解析した。
【0126】
OPTN c.412G>A(Thr34Thr)多型の遺伝子型判定
OPTN遺伝子のエクソン4中のc.412位におけるG→A置換を、制限酵素HpyCH4IV(New England BioLabs,マサチューセッツ州ベバリー)と、DHPLC解析に関して表13に挙げたプライマーと同じプライマーとを使って検出した。G対立遺伝子配列はHpyCH4IVによって二つの断片(188bp+129bp)に切断されたが、A対立遺伝子配列は無傷のまま(317bp)だった。制限酵素アッセイおよびDHPLCのクロマトグラフパターンによって多型を確認した。
【0127】
OPTN c.603T>A(Met98Lys)多型の遺伝子型判定
OPTN遺伝子のエクソン5中のc.603位におけるT→A置換は、DHPLC解析用のプライマーと同じプライマー(表13)を使って、制限酵素StuI(タカラ,日本国滋賀県)により、検出した。A対立遺伝子配列はStuIによって二つの断片(175bp+102bp)に切断されたが、T対立遺伝子配列は無傷のまま(277bp)だった。制限酵素アッセイおよびDHPLCのクロマトグラフパターンによって多型を確認した。
【0128】
OPTN c.1944G>A(Arg545Gln)多型の遺伝子型判定
OPTN遺伝子のエクソン16中のc.1944位におけるG→A置換を、ビー・エム・エルR&Dセンター研究部門(日本国埼玉県)によって提供されたInvaderアッセイで解析した。Invader(登録商標)アッセイおよびDHPLCのクロマトグラフパターンによって多型を確認した。
【0129】
TNF-α -308G>A多型の遺伝子型判定
TNF-αプロモーター領域中の308G>A多型の遺伝子型判定は、制限酵素NcoI(New England BioLabs,マサチューセッツ州ベバリー)と、フォワードプライマー5'-AGGCAATAGGTTTTGAGGGCCAT-3'およびリバースプライマー5'-GTAGTGGGCCCTGCACCTTCT-3'とを使って行なった。フォワードプライマーは制限酵素の使用を可能にする一塩基ミスマッチ(太字および下線部)を含有した。G対立遺伝子配列はNcoIによって二つの断片(192bp+20bp)に切断されたが、A対立遺伝子配列は無傷のまま(212bp)だった。
【0130】
TNF-α -857C>T多型の遺伝子型判定
TNF-αプロモーター領域中の857C>T多型の遺伝子型判定は、制限酵素HincII(タカラ,日本国滋賀県)と、フォワードプライマー5'-AAGTCGAGTATGGGGACCCCCCGTTAA-3'およびリバースプライマー5'-CCCCAGTGTGTGGCCATATCTTCTT-3'とを使って行なった。フォワードプライマーは制限酵素の使用を可能にする一塩基ミスマッチ(太字および下線部)を含有した。C対立遺伝子配列はHincIIによって二つの断片(106bp+25bp)に切断されたが、T対立遺伝子配列は無傷のまま(131bp)だった。-857T対立遺伝子の転写活性は-857C対立遺伝子の転写活性よりも有意に高かった。
【0131】
TNF-α -863C>A多型の遺伝子型判定
TNF-αプロモーター領域中の863C>A多型の遺伝子型判定は、制限酵素EcoNI(New England BioLabs,マサチューセッツ州ベバリー)と、フォワードプライマー5'-GCTGAGAAGATGAAGGAAAAGTC-3'およびリバースプライマー5'-CCTCTACATGGCCCTGTCCT-3'とを使って行なった。リバースプライマーは制限酵素の使用を可能にする一塩基ミスマッチ(太字および下線部)を含有した。C対立遺伝子配列はEcoNIによって二つの断片(183bp+23bp)に切断されたが、A対立遺伝子配列は無傷のまま(206bp)だった。-863A対立遺伝子の転写活性は-863C対立遺伝子の転写活性よりも有意に高かった。
【0132】
統計解析
患者および対照における遺伝子型および対立遺伝子の頻度を、カイ二乗検定およびフィッシャーの正確検定で比較した。オッズ比および95%信頼区間(CI)も計算した。観察された頻度についてハーディー・ワインベルグ平衡も計算した。これら二つの群間での臨床特性の比較は、適宜、マン・ホイットニーのU検定またはスチューデントの対応のないt検定を使って行なった。分散分析(ANOVA)を行なうための正規分布を得るために、POAG診断時のIOP、NTGおよびPOAG診断時の視野スコアである歪んだ分布の臨床データに対して、対数変換を行なった。一元配置ANOVAを使って、TNF-α/-857C>T遺伝子型およびオプチニューリン/412G>A遺伝子型、またはTNF-α/-863C>A遺伝子型およびオプチニューリン/603T>A遺伝子型の四つの異なる組み合わせを持つ患者間で、三つの臨床特性を比較した(表17参照)。
【0133】
統計解析はSPSSプログラム(SPSS Inc.,米国シカゴ)で行なった。0.05未満のP値を有意であるとみなした。
【0134】
結果
日本人被験者におけるOPTN変異体
合計629人の日本人被験者を研究した。結果を表14に示す。
【0135】
[表14]
緑内障患者および対照被験者に観察されるOPTN変異体
【0136】
緑内障患者および対照被験者に17の配列変化が同定された。これらのうち、3個はミスセンス変化であり、1個は一アミノ酸残基の欠失であり、4個は同義コドン変化であり、9個は非コード配列中の変化だった。疾患原因突然変異候補の一つ、His26Aspは、1人のPOAG発端者に同定され、218人の正常日本人対照には存在しなかった。彼女の兄弟(55歳)はこの突然変異を保持し、NTGと診断された。彼女の兄弟の娘(23歳)もこの突然変異を持ち、陥凹/乳頭径比が0.7で視神経乳頭の陥凹を示し、ハンフリー視野測定では視野欠損の徴候はなかった。
【0137】
Leu47の欠失(3bp欠失,CTC)が1人の対照に見つかった。Met98Lysは33人のPOAG患者、48人のNTG患者、および36人の対照に同定され、Arg545Glnは11人のPOAG患者、15人のNTG患者、および11人の対照に同定された。
【0138】
4個の同義ヌクレオチド置換、c.412G>A(Thr34Thr)、c.421G>A(Pro37Pro)、c.457C>T(Thr49Thr)、およびc.2023C>T(His571His)が見つかった。Thr34Thr置換は69人(35.6%)のPOAG患者、69人(31.8%)のNTG患者、および52人(23.9%)の対照に存在し、Pro37Proは1人のNTG患者に見いだされた。Thr49Thrは1人のPOAG患者に同定され、His571Hisは2人の対照に存在した。
【0139】
日本人被験者におけるOPTN変異体の分布
Thr34Thr(c.412G>A)多型はPOAGおよびNTGと関連した(表15)。有意な関連がPOAGを持つ患者に見いだされた(遺伝子型頻度:G/G対G/A+A/AでP=0.009、対立遺伝子頻度でP=0.003)。Met98Lys(c.603T>A)多型またはArg545Gln(c.1944G>A)多型については、遺伝子型頻度にも対立遺伝子頻度にも、緑内障患者と対照との間に有意差は検出されなかった。しかしMet98Lys多型は、POAGよりもNTGと関連する傾向が高かった。観察された遺伝子型頻度はハーディー・ワインベルグ平衡によって予測されるものと一致した。
【0140】
[表15]
【0141】
緑内障患者の三つの臨床特性、すなわち診断時年齢、診断時IOP、および診断時視野スコアを、c.412G>A(Thr34Thr)多型またはc.603T>A(Met98Lys)多型との関連について調べた(表16)。緑内障患者は、c.412G>A多型について臨床特性との関連を示さなかった。G/A+A/A遺伝子型を持つPOAG患者(すなわち412A保有者)は、G/G遺伝子型を持つ者(すなわち非412A保有者)よりも進行した視野スコアを持つ傾向があった(P=0.093)。603T>A多型を持つPOAG患者は、診断時年齢と弱い関連を示した(P=0.046)。
【0142】
[表16]
OPTN遺伝子型による緑内障患者の臨床特性の比較
【0143】
緑内障患者におけるOPTN多型とTNF-α多型との関連
三つの多型、308G>A、-857C>Tまたは863C>Aに関して、患者と対照との間に、遺伝子型頻度または対立遺伝子頻度の有意差は認められなかった。また、緑内障患者はこれら三つの多型について臨床特性との関連を示さなかった(データ未掲載)。観察された遺伝子型頻度は、ハーディー・ワインベルグ平衡によって予測されるものと一致した。
【0144】
しかし、TNF-α遺伝子中にC/T+T/T遺伝子型を持つ個体(すなわち-857T保有者)のうち、OPTN遺伝子がG/A+A/A遺伝子型(すなわち412A保有者)であるのは、対照では21.6%だったのに対して、POAG患者では44.1%だった(表17)。この頻度の差は有意だった(P=0.006)。TNF-α遺伝子にC/A+A/A遺伝子型を持つ個体(すなわち-863A保有者)のうち、OPTN遺伝子に603Aを持つ個体(すなわちLys98保有者)は、POAGおよびNTGと有意に関連した(それぞれP=0.008および0.027)。
【0145】
[表17]
TNF-α遺伝子型(-857C>Tおよび-863C>A)によるオプチニューリン遺伝子型(c.412G>Aおよびc.603T>A)の分布
【0146】
これら複合遺伝子型の臨床特性、例えば診断時年齢、診断時IOP、および診断時視野スコアを、表18に示す。TNF-α/-857T保有者でありかつオプチニューリン/412A保有者であるPOAG患者は、TNF-α/-857T保有者でありかつ非オプチニューリン/412A保有者である者よりも有意に悪い(P=0.020)視野スコアを持っていた。しかし、一元配置ANOVAでは、-857T>A多型とc.412G>A多型とを組み合わせたの四つの遺伝子型(表6)間で、POAG患者の三つの臨床特性に有意差はなかった:診断時年齢に関してP=0.823、診断時IOPに関してP=0.692、診断時視野スコアに関してP=0.152。
【0147】
TNF-α/-863A保有者でありかつ オプチニューリン/603A保有者であるPOAG患者は、TNF-α/-863A保有者でありかつ非オプチニューリン/603A保有者である者よりも有意に悪い(P=0.026)視野スコアを持っていた。しかし、-863C>A多型と-603T>A多型とを組み合わせた四つの遺伝子型(表6)間で、POAG患者の視野スコアに有意差はなかった(一元配置ANOVA:P=0.200)。
【0148】
[表18]
TNF-α遺伝子型(-857Tおよび-863A)およびオプチニューリン遺伝子型(412Aおよび603A)による緑内障患者の臨床特性の比較
【0149】
標的とする多型412G>A(下線部)を含むOPTNエクソン4の部分ヌクレオチド配列
【0150】
OPTN遺伝子の配列、GenBankアクセッション番号AF423071
【0151】
標的とする多型位置を含むTNF-α遺伝子の部分ヌクレオチド配列は以下のとおりである。
TNF-α -863C>A;-857C>T(下線部)
【実施例7】
【0152】
経口アンギオテンシンII受容体遮断薬が正常被験者のIOPに及ぼす効果、ならびにAT1R遺伝子およびAT2R遺伝子中のSNPとのその関連
実施例7-1
方法
X染色体上のアンギオテンシンII受容体2遺伝子(AT2R)のヌクレオチド番号3123における多型(CまたはA)と、アンギオテンシンII受容体遮断薬カンデサルタンシレキセチルの効果との関係を調べた。この研究は全身性疾患および眼疾患を持たない20人の健常ボランティア(男性13人および女性8人)で行なわれた。多型位置において、彼らのうち男性9人はC遺伝子型、男性4人はA遺伝子型、女性4人はCC遺伝子型、女性4人はCA遺伝子型を持っていた。各被験者にカンデサルタンシレキセチルを経口投与し、投与の1〜24時間後にIOPを記録した。
【0153】
結果
薬物投与後1〜24時間の眼内圧の変化を表19に示す。
[表19]
【0154】
男性の場合、AT2R遺伝子のヌクレオチド3123にA遺伝子型を持つ人の75%では、カンデサルタンシレキセチルを経口投与しても、IOPがほとんど低下せず、一方、C遺伝子型を持つ人は100%が、IOPを効果的に低下させた。女性の場合、カンデサルタンシレキセチルの経口投与により、CC遺伝子型を持つ人の100%で、IOPが効果的に低下した。
【0155】
この結果は、AT2(AGTR2)のヌクレオチド3123における遺伝子多型がカンデサルタンシレキセチルの効果と関連することを示唆している。
【0156】
実施例7-2
方法
この研究は全身性疾患および眼疾患を持たない20人の健常ボランティア(23〜28歳の男性13人および女性7人)で行なわれた。各被験者には、無作為化クロスオーバー方式で、朝(10:00)に、12mgの経口カンデサルタンシレキセチル(Blopress(登録商標),武田,日本)または偽薬を与えた。
【0157】
ベースライン心拍数、ベースライン収縮期/拡張期動脈圧(SBP/DBP)、およびベースラインIOPを記録した。次に、被験者に経口カンデサルタンシレキセチルまたは偽薬を与え、6時間までの毎時間と24時間後に測定を繰り返した。1ヶ月後に、各被験者にもう一方の処置を施した。右眼だけを測定し、解析した。
【0158】
眼灌流圧(OPP)は、組織に入る動脈内の圧力と、それを離れる静脈内の圧力との差と定義される。OPPは、平均血圧(BPm)およびIOPを使って以下の式で近似することができる。
OPP=2/3×BPm−IOP
[式中、BPm=DBP+1/3×(SBP-DBP)]
【0159】
ATR1およびATR2中の多型を20人の被験者で検索し、IOPの変化と相関させた。この研究はヘルシンキ宣言の精神に則って行なわれた。研究の性質および考えうる帰結を説明した上で、文書によるインフォームド・コンセントを得た。この研究は、適用可能な場合には、DNA解析に関する施設内人体実験委員会による承認を得た。
【0160】
統計解析
ARB投与後の結果の統計解析は、反復測定ANOVA検定を使って、StatView(SAS Institute,米国)で行なった。各IOP値の統計解析にはボンフェローニ補正ANOVA検定を使用し、P値<0.0004を統計的に有意であるとみなした。
【0161】
結果
経口カンデサルタンシレキセチル投与または偽薬投与後のIOPの変化を図5Aに示す。偽薬の投与を受けた被験者のIOPに有意な変化はなかった。これに対し、経口カンデサルタンシレキセチル投与の1時間後には早くもIOPが有意に低下しており、5時間にわたって偽薬より低い状態を保った(P<0.0001)。カンデサルタンシレキセチルは灌流圧に有意な影響を及ぼさなかった(図5B)。カンデサルタンシレキセチルまたは偽薬の単回経口投与後にSBP、DBP、および心拍数の有意な変化は検出されなかった(データ未掲載)。
【0162】
被験者20人のそれぞれにおける経口カンデサルタンシレキセチル投与後のIOP変化を図5Cに示す。この20人の被験者では、カンデサルタンシレキセチルの効果とATR1遺伝子中の3個のSNPとの間に、有意な関連はなかった(表19-2)。しかし、ATR2遺伝子型については、A遺伝子型を持つ男性4人が2.3±0.5mmHgのIOP低下を示し、これは偽薬の投与を受けた被験者と同じ値であり、C遺伝子型を持つ男性9人(5.0±1.1mmHg,P=0.014)よりも有意に少ない低下量だった。この研究ではAA遺伝子型を持つ女性はいなかった。
【0163】
[表19−2]
アンギオテンシンII受容体遺伝子の遺伝子型と関連するアンギオテンシンII受容体遮断薬の眼内圧に対する効果
【実施例8】
【0164】
緑内障とエンドセリン-1およびエンドセリンA型受容体の遺伝子多型との関連
目的:強力な血管収縮因子エンドセリン1(ET-1)は眼内圧および眼血管張力の調節に影響を及ぼしうる。したがってET-1およびその受容体は緑内障の発病の一因になりうる。本発明者らは、ET-1(EDN1)ならびにその受容体ETA(EDNRA)およびETB(EDNRB)の遺伝子多型が緑内障表現型および臨床特性と関連するかどうかを調べた。
【0165】
方法
研究対象集団:
この研究では、日本の医療施設7ヶ所から採用した合計650人の日本人被験者(正常対照224人、POAG患者176人、およびNTG患者250人)を調べた。どの被験者も血縁関係にはなかった。OAG診断時の平均年齢(±標準偏差)は57.2±12.8歳だった。OAG被験者はPOAG患者とNTG患者に分割され、その診断時年齢はそれぞれ58.8±12.2歳および56.1±13.2歳だった(表1)。検査時の平均年齢は対照では70.0±11.2歳だった。本発明者らは、対照の一部が後に緑内障を発病する可能性が低下させるために、より高齢の対照被験者を意図的に選択した。
【0166】
眼科検査には、細隙灯生体顕微鏡検査、視神経乳頭検査、ゴールドマン圧平眼圧測定によるIOP測定、および隅角鏡検査を含めた。視野はハンフリー自動視野測定(プログラム30-2)またはゴールドマン視野測定で評価した。視野欠損の重症度を1〜5にスコア化した。2タイプの視野測定によって取得したデータを5点尺度(1=変化なし;2=早期欠損;3=中等度欠損;4=重度欠損;および5=光覚のみまたは視覚なし)を使って集約した。この重症度尺度は、ゴールドマン視野測定に基づくKozakiの分類(今まで日本では最も広く用いられてきたもの)に従うか、ハンフリー視野解析機用に確立された分類によった。
【0167】
以下の基準を全て満たす場合にPOAGと診断した:最大IOPが21mmHgを超えたこと;隅角鏡検査での開放隅角;視野変化を伴う典型的な緑内障性乳頭陥凹;および視神経損傷を引き起こす可能性がある他の眼障害、鼻障害、神経障害または全身性障害が存在しないこと。本発明者らは、明確な原因(例えば外傷、ぶどう膜炎、ステロイド投与、または剥脱性緑内障、色素性緑内障、もしくは血管新生緑内障)に続発するIOPの上昇を持つ患者を除外した。MYOC突然変異を持つPOAG患者およびJOAG患者も除外した。NTGは、追跡期間中は常にIOPが21mmHgを超えなかったという点を除いてPOAGと同じ基準によって診断した。正常対照被験者は20mmHg未満のIOPを持ち、緑内障性乳頭変化がなく、緑内障の家族歴を持っていなかった。
【0168】
DNA抽出および多型の遺伝子型判定
標準的方法によって末梢血リンパ球からゲノムDNAを単離した。参加者全員に9個の一塩基多型(SNP)、すなわちEDN1について4個(T-1370G、+138/ex1 del/ins、G8002A、K198N)、EDNRAについて4個(G-231A、H323H、C+70G、C+1222T)、およびEDNRBについて1個(L277L)を検出した。これらの多型は、http://genecanvas.idf.inserm.fr/に列挙されている。本発明者らは、SNPのハイスループット遺伝子型判定のために最近開発されたInvader(登録商標)アッセイ(Thrid Wave Technologies, Inc.,ウィスコンシン州マディソン)を使って、これらのSNPを遺伝子型判定した(Lyamichev Vら,Nat Biotechnol 1999;17:292-296,この引用文献の内容は参照により本明細書に組み込まれる)。
【0169】
この研究で使用した一次プローブおよびInvader(登録商標)プローブのオリゴヌクレオチド配列を表20に記載する。
【0170】
[表20]
【0171】
統計解析
正常対照における遺伝子型分布と、OAG患者、POAG患者、およびNTG患者における遺伝子型分布との比較は、χ2解析によって行なった。臨床特性(診断時年齢、無処置時最大IOP、および診断時視野スコア)と遺伝子型との関連はマン・ホイットニーのU検定によって評価した。統計解析はWindows用SPSS(バージョン12.0;SPSS Inc.,イリノイ州シカゴ)で行なった。p<0.05の値を有意であるとみなした。
【0172】
結果
この研究で得られた遺伝子型頻度および対立遺伝子頻度を表21に示す。分布はハーディー・ワインベルグ平衡と一致した。EDN1/+138/ex1 del/ins多型の場合、del/del遺伝子型およびdel/ins+ins/ins遺伝子型の頻度は、それぞれ、対照被験者で65.2%および34.8%だったのに対して、OAG患者全体では74.2%および25.8%(p=0.016)、POAG患者では74.4%および25.6%(p=0.047)、NTG患者では74.0%および26.0%(p=0.037)だった。EDN1/K198N多型の場合、OAG患者の53.2%はKK遺伝子型を持つことがわかった。これは対照被験者における43.8%という優勢率よりも有意に高かった(p=0.022)。OAG患者をPOAG患者とNTG患者とに分割した場合、NTG患者におけるKK遺伝子型の頻度は対照よりもはるかに高く(p=0.008)、POAG患者における遺伝子型頻度分布および対立遺伝子頻度分布は、対照における分布と統計的に相違しなかった。性差が認められた。具体的には、KK遺伝子型は、女性NTG患者の間では(女性対照に対してp=0.010)、男性NTG患者の場合よりも(男性対照に対してp=0.251;表22)、有意に優勢だった。イントロン4領域におけるEDN1/G8002Aの多型は一試料を除いてEDN1/K198Nと高度に共在した(データ未掲載)。
【0173】
EDNRA/C+1222T遺伝子型(CC対CT+TT)の頻度はOAG患者と対照の間でわずかに異なった(p=0.036)。他の多型に関する遺伝子型の分布はどの患者群と対照との間にも有意差を示さなかった。
【0174】
患者の特徴を各多型の優性モデルおよび劣性モデルで調べ、データを有意差と共に表23に示す。OAG患者全体およびPOAG患者では、どの特徴も遺伝子型群間に有意差を示さなかった。しかし、NTG患者では、EDNRA/G-231AのAA群が、GG+GA群よりも悪い診断時視野スコアを持っていた(3.0±0.8対2.7±0.6,p=0.043)。また、NTG患者の間で、EDNRA/C+70GのGG群は、診断時視野スコアが、CC+CG群よりも有意に悪いこともわかった(3.0±0.7対2.7±0.7,p=0.014)。NTG患者では、ENDRA/H323HのTT群における無処置時最大IOPが、CC+CT群よりも統計的に高かった(17.2±2.2対16.6±2.3,p=0.040)。NTG患者における他の多型は、遺伝子型群間で、特性に有意差を示さなかった。
【0175】
[表21]
対照被験者および緑内障患者におけるEDN1、EDNRAおよびEDNRB多型の遺伝子型頻度および対立遺伝子頻度
データはn(%)である。
* P<0.05(χ2検定)
遺伝子型分布は、OAG患者と対照との間で、EDN1/+138/ex1 del/ins多型(p=0.016)およびEDN1/K198N多型(p=0.022)に関して有意差を示し、EDNRA/C+1222T多型(p=0.036)に関してわずかな差を示した。OAG群をPOAGおよびNTGに分割すると、NTG患者におけるEDN1/K198N多型のKK遺伝子型の頻度は、対照よりもはるかに高くなった(p=0.008)。
【0176】
[表22]
【0177】
[表23]
【0178】
男性被験者では、以下の相関関係が確認された:
1)エンドセリン-1遺伝子のエクソン1におけるA138挿入/欠失(A138I/D)多型はPOAGともNTGとも関連する(表24)
2)エンドセリン受容体A遺伝子のプロモーター領域の-231A>G多型は、NTG、特に15mmHg未満の眼内圧を持つ患者と関連する(表25)。
3)エンドセリン受容体A遺伝子のエクソン6中のコドン番号233におけるCAC→CAT置換(His323His)は、NTG、特に15mmHg未満の眼内圧を持つ患者と関連する(表26)。
4)エンドセリン受容体B遺伝子のエクソン4中のコドン番号277におけるCTG→CTA置換はPOAGともNTGとも関連する(表27)。
【0179】
女性患者では、以下の相関関係が確認された:
1)エンドセリン-1遺伝子のコドン番号198におけるAAG→AAT置換(Lys198Asn)はNTGと関連する(表28)。
2)エンドセリン-1遺伝子プロモーター領域の-1370T>G多型はNTGと関連する(表29)。
3)エンドセリン受容体Aの3'非コード領域における+70C>G(停止コドンから70塩基)多型はPOAGと関連する(表30)。
4)エンドセリン受容体Aの3'非コード領域における+1222C>T(停止コドンから1222塩基)多型はNTG(眼内圧が16mmHg〜21mmHgであるもの)と関連する(表31)。
【0180】
[表24]
エンドセリンA138I/D(男性)
【0181】
[表25]
エンドセリン受容体A -231A/G(男性)
H-NTG:16mmHg〜21mmHgの眼内圧を持つNTG患者。
L-NTG:15mmHg以下の最大眼内圧を持つNTG患者。
【0182】
[表26]
エンドセリン受容体A H323H C>T His323His(男性)
H-NTG:16mmHg〜21mmHgの眼内圧を持つNTG患者。
L-NTG:15mmHg以下の最大眼内圧を持つMTG患者。
【0183】
[表27]
エンドセリン受容体B L277L G>A Leu277Leu(男性)
【0184】
[表28]
エンドセリン Lys198Asn G>TまたはK198N(女性)
【0185】
[表29]
エンドセリン -1370T>G(女性)
【0186】
[表30]
エンドセリン受容体A +70C>G(女性)
【0187】
[表31]
エンドセリン受容体A +1222C>T(女性)
H-NTG:16mmHg〜21mmHgの眼内圧を持つNTG患者。
L-NTG:15mmHg以下の最大眼内圧を持つMTG患者。
【0188】
標的とする多型を含むエンドセリン-1(EDN1)ならびにエンドセリン受容体A(EDNRA)およびエンドセリン受容体B(EDNRB)の部分ヌクレオチド配列を以下に示す。
【0189】
EDN1 -1370(下線部)T>G
【0190】
END1 +138(下線部)ins/del(それぞれ3743-3745にあるaのそれぞれ一つ)
【0191】
EDNRA +70(下線部)C>G
【0192】
EDNRA +1222(下線部)C>T
【0193】
EDNRAコドン番号323(下線部)(T>C)His323His
【0194】
EDNRA -231(下線部)A>G
【0195】
EDNRBコドン番号277(下線部)Leu277Leu(CTG→CTA)
【実施例9】
【0196】
β1アドレナリン作動性受容体(ADRB1)の遺伝子多型と緑内障との関連
方法
PCR-RFLP技術を使って、POAG患者、NTG患者および正常(対照)被験者の間で、ADRB1の遺伝子多型と緑内障との関連を調べた(表32-1)。
[表32−1]
プライマー配列
【0197】
結果
表32-2に示すように、ADRB1中のGly389Argの多型はNTGと関連する(表32-2)。
【0198】
[表32−2]
β1-アドレナリン受容体Gly389Arg
【0199】
標的とする多型を含むβ1-アドレナリン受容体の部分ヌクレオチド配列。
B1ARコドン389(下線部)GGA(Gly)→CGA(Arg)Gly389Arg
【実施例10】
【0200】
E-セレクチンの遺伝子多型と緑内障との相関関係
方法
Invader(登録商標)法を使って、POAGを持つ被験者、NTGを持つ被験者および正常被験者の間で、E-セレクチン遺伝子多型と緑内障との関係を調べた。
【0201】
SELE遺伝子のC/T多型を検出するために使用したInvader(登録商標)オリゴヌクレオチド(Invader(登録商標)プローブ)を表33-1に示す。
[表33−1]
【0202】
結果
E-セレクチン遺伝子の1402C>T多型は、POAGともNTGとも関連することが確認された(表33-2)。
[表33−2]
E-セレクチン1402C>T
【0203】
標的とする多型を含むE-セレクチンの部分ヌクレオチド配列は以下のとおりである:
SELE 番号1402(下線部)C>T
【実施例11】
【0204】
パラオキソナーゼ1遺伝子多型は開放隅角緑内障の臨床特徴と関連する
目的:低密度リポタンパク質(LDL)の酸化誘導体は内皮にとって有害である。内皮機能障害は、開放隅角緑内障(OAG)の病理発生過程に関与することが知られている。高密度リポタンパク質(HDL)はLDLの酸化的修飾を防止する。本発明者らは、HDL関連抗酸化酵素であるパラオキソナーゼ1(PON1)、PON2、および血小板活性化因子アセチルヒドロラーゼ(PAF-AH)遺伝子中の多型が、日本人集団においてOAGと関連するかどうかを調べた。
【0205】
材料および方法
患者および対照被験者
日本の施設7ヶ所で698の血液試料を集めた。被験者には、POAG患者190人、NTG患者268人、および正常対照240人を含めた。どの被験者も互いに血縁関係にはなかった。
【0206】
血液試料採取時の年齢(平均±SD)は、POAG患者で65.3±11.9歳、NTG患者で58.8±13.4歳、正常被験者で69.7±11.2歳だった。正常対照被験者はPOAG患者(p<0.001)またはNTG患者(p<0.001)より有意に高齢だった。これにより、対照被験者が最終的に緑内障を発病する可能性が低減されるだろう。
【0207】
緑内障患者に関して記録された臨床特徴は、診断時年齢、診断時IOP、および診断時視野欠損だった。視野欠損の重症度は1〜5にスコア化した。異なる視野計を使って取得したデータを、以下に定義する5点尺度を使って集約した。1=変化なし;2=早期欠損;3=中等度欠損;4=重度欠損;および5=光覚のみまたは視覚なし。視野欠損は、ゴールドマン視野測定の結果に基づくKozakiの分類に従って、またはハンフリー視野解析機に用いられる分類によって、早期、中等度、または重度と判定した。日本では今まで前者の分類が最も広く用いられてきた。
【0208】
ゴールドマン圧平眼圧測定によるIOP測定、ハンフリー視野測定(30-2)またはゴールドマン視野測定、隅角鏡検査、および眼底撮影を含む視神経乳頭検査などといった一連の眼科検査を、全ての患者に行なった。緑内障患者は全員、以下の基準に従って診断された:緑内障性陥凹(陥凹/乳頭径比>0.7)および神経網膜辺縁部の喪失を伴う典型的視神経乳頭損傷の存在、緑内障性陥凹と矛盾しない再現性のある視野欠損、ならびに隅角鏡検査での開放隅角。OAG患者のうち、追跡期間中どの時点でもIOP>21mmHgである場合は、POAGと診断した。剥脱性緑内障、色素性緑内障、およびコルチコステロイド誘発性緑内障を持つ患者は除外した。OAG患者のうち、無処置時最大IOPが3回のベースライン測定値および毎日の試験値(6時から24時まで3時間ごと)を含めて常に一貫して21mmHg以下であり、診断後の投薬時または無投薬時の最大IOPが追跡期間中常に一貫して22mmHgであり、緑内障性視神経症の二次的原因、例えば外傷後のIOP上昇を過去に経験していること、ステロイド投与の期間、またはぶどう膜炎などが存在しなかった場合に、NTGと診断した。
【0209】
対照被験者は、白内障を除く既知の眼異常を持たない日本人個体から採用した。これらの被験者は196人に達し、40歳より高齢で、20mmHg未満のIOPを持ち、緑内障性乳頭変化がなく、緑内障の家族歴もなかった。
【0210】
遺伝子型判定
標準的方法によって末梢血リンパ球からゲノムDNAを単離した。次に、全ての参加者に4個のSNP(PON1について2個(L55M、Q192R)、PON2について1個(Cys311Ser,C311S)、およびPAF-AHについて1個(V279F))を検出した。
【0211】
SNPのハイスループット遺伝子型判定のために最近開発されたInvader(登録商標)アッセイ(Third Wave Technologies, Inc,米国ウィスコンシン州マディソン)を使って、これらのSNPを遺伝子型判定した。この研究で使用した一次プローブおよびInvader(登録商標)プローブのオリゴヌクレオチド配列を表34に挙げた。
【0212】
[表34]
【0213】
統計解析
カイ二乗解析によってハーディー・ワインベルグ平衡を評価した。遺伝子型の頻度および対立遺伝子の頻度を、カイ二乗解析により、症例と対照とで比較した。三つの臨床変量と遺伝子型の間の関連を確認するために、ロジスティック回帰モデルで多変量解析を行なった。PON1遺伝子におけるQ192Rの遺伝子型群間のIOPの比較は、クラスカル・ワリス検定によって行なった。統計解析はSPSS(バージョン12.0;SPSS,イリノイ州シカゴ)で行なった。p<0.05の値は有意性を示すとみなした。
【0214】
結果
緑内障患者および対象における4個のSNPに関する遺伝子型の分布を表35に示す。NTGを持つ患者ではPON1遺伝子のL55M多型が有意に異なる遺伝子型頻度を持っていた。
【0215】
PON2遺伝子およびPAF-AH遺伝子中の多型に関する遺伝子型の分布は、どの患者群と対照との間にも有意差を示さなかった(表35)。また、4個のSNPの対立遺伝子頻度にも有意差はなかった。
【0216】
[表35]
【0217】
OAG集団におけるPON1遺伝子中の二つの多型を組み合わせたものの分布を表36に示す。明瞭に示されるように、55位のメチオニン(M)(M対立遺伝子)が192位のアルギニン(R)(R対立遺伝子)を伴うことはほとんどなかった。解析により、55位にロイシン(L)をもたらす多型と192位にアルギン(R)をもたらす多型との間に連鎖不平衡が確認された(p<0.001)。
【0218】
[表36]
【0219】
患者の特性を各多型について優性モデルおよび劣性モデルで調べた。劣性モデルでは、どの多型についても、OAGを持つ患者における三つの特性に有意差が見られなかった。PON1多型の優性モデルによる有意差を表37および表38に示す。OAG患者におけるPON1遺伝子中のL55M多型について、LL群(非55M保有者)は診断時の年齢がLM+MM群(55M保有者)より有意に低かった(56.8±12.8歳対60.1±11.4歳,p=0.028)(表37)。この関連はPOAG患者には観察されなかったが、NTG患者には観察された(55.6±13.1歳対63.7±9.6,p=0.001)。
【0220】
Q192R多型に関して、診断時の無処置時最大IOPは、QR+RR群を持つOAG患者(192R保有者)(21.5±7.4mmHg)の方が、QQ群(非192R保有者)の場合(18.7±5.3mmHg,P=0.006,表38)よりも有意に高かった。無処置時最大IOPは、POAG患者(POAGについて27.5±7.0mmHg対24.0±4.9,p=0.049)でも、NTG患者(NTGについて15.8±2.8mmHg対16.7±2.4,p=0.030)でも、192R保有者の方が非保有者より高かった。
【0221】
[表37]
PON1遺伝子中のL55Mの遺伝子型によるNTG患者の臨床特性
【0222】
[表38]
PON1遺伝子中のQ192Rの遺伝子型による緑内障患者の臨床特性
【0223】
PON1遺伝子中のGly192Arg(Q192R)多型はPOAGと関連した(表39)。Leu55Met多型はNTG(特に15mmHg未満のもの)と関連した(表40)。
【0224】
[表39]
PON1 Gln192Arg(Q192R)
【0225】
[表40]
PON1 Leu55Met(L55M)
H-NTG:16mmHg〜21mmHgの眼内圧を持つNTG患者。
L-NTG:15mmHg以下の最大眼内圧を持つMTG患者。
【0226】
結論:PON遺伝子多型はOAGを持つ日本人患者、特にNTGを持つ日本人患者の特徴に影響を及ぼしうる。
【0227】
標的とする多型を含有するパラオキソナーゼ1遺伝子の部分ヌクレオチド配列は以下のとおりである。
PON1コドン55(下線部)TTG(Leu)→ATG(Met)(Leu55Met)
および
PON1コドン192(下線部)CAA(Gln)→CGA(Arg)(Gln192Arg)
【実施例12】
【0228】
開放隅角緑内障の病因におけるノエリン2遺伝子の評価
目的:開放隅角緑内障を持つ日本人患者におけるノエリン2遺伝子中の突然変異について、変性高速液体クロマトグラフィー(DHPLC)を使ってスクリニーングすること。
【0229】
方法
被験者
日本国内の8施設で合計616の血液試料を集めた。POAG患者276人、NTG患者340人、および正常被験者300人であり、どの被験者もこの研究における他の被験者とは血縁関係になかった。
【0230】
DNA抽出およびPCR条件
血液試料の解析は全て慶応大学で行なわれた。フェノール-クロロホルム抽出によって末梢血リンパ球からゲノムDNAを単離した。ポリメラーゼ連鎖反応(PCR)により、表41に挙げるプライマーセットを使って、ノエリン2遺伝子の六つのエクソンコード領域を増幅した。
【0231】
[表41]
プライマー配列、PCR産物サイズ、ならびにPCRアニーリング温度およびDHPLC解析温度
【0232】
ハイスループット解析では、3人の患者からの試料をプールした。PCRは、サーマルサイクラー(iCycler,Bio-Rad,カリフォルニア州ハーキュリーズ)を使って、ゲノムDNA 45ng、GeneAmp 10×PCRバッファーII 2μl、濃度2.0mMの各dNTPを含むGeneAmp dNTPミックス2μl、25mM MgCl2溶液2.4μl、各プライマー4pmol、およびAmpliTaq Gold DNAポリメメラーゼ(Applied Biosystems,カリフォルニア州フォスターシティ)0.1Uを含有する20μlの総液量で行なった。PCR条件は以下のとおりとした:95℃で9分間変性した後、95℃で1分、65〜67℃で30秒(表1)、および72℃で1分30秒を35サイクル、そして72℃で7分間の最終伸長工程。
【0233】
変性HPLC解析
ハイスループット解析の場合、3人の患者から得られたPCR産物25μlを、DHPLC解析用のWAVE(登録商標)システム(Transgenomic,ネブラスカ州オマハ)を使って、解析用クロマトグラフに自動注入した。DHPLC融解温度を表41に記載する。
【0234】
ハイスループットプロトコールでプール試料に異常なクロマトグラフパターンが検出された場合は、その試料をWAVE(登録商標)システムで個別に再解析した。次に、異常なクロマトグラフパターンを示したPCR産物をシークエンスした。
【0235】
ダイレクトDNAシークエンス
ダイレクトシークエンスのために、PCR産物をQIA Quick PCR精製キット(Qiagen,カリフォルニア州バレンシア)で精製することにより、未使用のプライマーおよび前駆体を除去した。BigDyeケミストリーを製造者の推奨するプロトコール(Applied Biosystems,カリフォルニア州フォスターシティ)に従って使用することにより、ABI310自動シークエンサーで、同じフォワードおよびリバースPCR増幅プライマーを使って、PCR産物をダイレクトシークエンスした。
【0236】
ミオシリン遺伝子のスクリーニング
ノエリン2遺伝子中に突然変異を保持する緑内障患者2人について、DHPLCにより、ミオシリン遺伝子のスクリーニングを行なった。
【0237】
ノエリン2 c.462G>A(Arg144Gln)多型の遺伝子型判定
ノエリン2遺伝子のエクソン4中のc.462位におけるG→A置換を、制限酵素BstU1を使って検出した。G対立遺伝子配列はBstU1によって二つの断片(140bp+200bp)に切断されたが、A対立遺伝子配列は無傷のまま(344bp)だった。
【0238】
制限酵素アッセイおよびDHPLCのクロマトグラフパターンによって多型を確認した。
【0239】
統計解析
患者および対照における遺伝子型および対立遺伝子の頻度をカイ二乗検定またはフィッシャーの正確検定で比較した。観察された頻度についてハーディー・ワインベルグ平衡も計算した。統計解析はSPSSプログラム(SPSS Inc.,米国シカゴ)で行なった。0.05未満のP値を有意であるとみなした。
【0240】
結果
日本人被験者におけるノエリン2変異体
合計616人の日本人被験者を調べた。その結果を表42に記載する。緑内障患者および対照被験者中の配列変化を同定した。これらのうち二つはミスセンス変化であり、七つは同義コドン変化であり、一つはイントロン配列中の変化だった。疾患原因突然変異候補の一つ、Arg144Glnは、一人のPOAG発端者および一人のPOAG発端者に同定され、300人の正常日本人対照には存在しなかった。Arg106Gln(P=0.30)、Ala226Ala(P=0.30)、およびArg427Arg(P=0.30)については、緑内障患者と対照との間に有意差は検出されなかった。
【0241】
Arg144Glnを持つNTG患者はミオシリン遺伝子中のArg76Lys変化を保持していた。
【0242】
エクソン4中の緑内障原因突然変異候補Arg144Glnは616人の日本人緑内障患者のうち2人(0.3%)に同定された。
【0243】
[表42]
緑内障患者および対照被験者に観察されたOLFM2変異体
【0244】
標的とする多型を含むノエリン2の部分ヌクレオチド配列は以下のとおりである。
Noelin2コドン144(下線部)CGG(Arg)→CAG(Gln):(GG:200bp+144bp,GA:344bp+200bp+144bp,AA:344bp)(BstUI)
コドン140(下線部)Lys140Lys(AAG>AAA)
コドン152(下線部)Glu152Glu(GAG>CAA)
79841-79862,80164-80184;コドン144における多型を検出するためのプライマー。
79916-80131,コード領域。
【実施例13】
【0245】
緑内障の病因における熱ショックタンパク質70-1(HSP70-1)遺伝子の評価
POAG患者、NTG患者および対照被験者の間で、Invaderアッセイを使って、緑内障とHSP70-1の遺伝子多型(Biogerontology 4:215-220,2003およびHum Genet 114:236-241,2004)との関連を調べた。
【0246】
HSP70-1遺伝子の多型を検出するために使用した一次プローブ(野生型プローブおよび突然変異型プローブ)ならびにInvader(登録商標)オリゴヌクレオチド(Invader(登録商標)プローブ)を表43に示す。
[表43]
HSP70-1のオリゴヌクレオチド配列
【0247】
結果
表44に示すように、HSP70-1における-110A>Cの多型は緑内障、特にPOAGと関連する。
[表44]
【0248】
標的とする配列を含むHSP70-1の部分ヌクレオチド配列は以下のとおりである。
HSP70-1 -110A>C(以下の配列はC対立遺伝子である)
【実施例14】
【0249】
緑内障の病因におけるエンドセリン変換酵素1(ECE1)遺伝子の評価
POAG患者およびNTG患者で、Invaderアッセイを使って、緑内障とECE1の遺伝子多型との関連を調べた。
【0250】
ECE1遺伝子の多型を検出するために使用した一次プローブ(野生型プローブおよび突然変異型プローブ)ならびにInvader(登録商標)オリゴヌクレオチド(Invader(登録商標)プローブ)を表45に示す。
[表45]
【0251】
結果
表46に示すように、ECE1における-338C>Aの多型は、NTGにおける高いIOPと関連した。
[表46]
【0252】
標的とする多型を含むECE-1の部分ヌクレオチド配列を以下に示す。
ECE1 -338C>A(下線部)
【実施例15】
【0253】
開放隅角緑内障の病因におけるCD50遺伝子の評価
ポリメラーゼ連鎖反応-制限断片長多型(PCR-RFLP)技術を使ってCD50遺伝子の多型を同定した(表47)。
[表47]
【0254】
結果
表48に示すように、CD95におけるA-670Gの多型は緑内障、特にPOAGと関連する。
[表48]
【実施例16】
【0255】
緑内障の病因におけるEPHX1遺伝子の評価
Invaderアッセイを使って、POAG患者、NTG患者および対照被験者の間で、緑内障とEPHX1の遺伝子多型との関連を調べた。
【0256】
ECE1遺伝子の多型を検出するために使用した一次プローブ(野生型および突然変異型プローブ)ならびにInvader(登録商標)オリゴヌクレオチド(Invader(登録商標)プローブ)を表49に示す。
[表49]
【0257】
結果
表50に示すように、コドン119LysにおけるG>Aの多型は緑内障、特にNTGと関連した。
[表50]
【0258】
標的とする多型を含むEPHX1の部分ヌクレオチド配列は以下のとおりである。
【実施例17】
【0259】
緑内障の病因におけるβ2アドレナリン作動性受容体(ADRB2)遺伝子の評価
Invader アッセイを使って、開放隅角緑内障患者(POAG患者およびNTG患者)で、緑内障とADRB2の遺伝子多型との関連を調べた。
【0260】
ADRB2遺伝子の多型を検出するために使用した一次プローブ(野生型プローブおよび突然変異型プローブ)ならびにInvader(登録商標)オリゴヌクレオチド(Invader(登録商標)プローブ)を表51に示す。
[表51]
【0261】
結果
表52に示すように、ADRB2におけるGly16Arg(G46A)の多型はPOAGの早期発症と関連する。
[表52]
ADRB2遺伝子におけるGln16Argの遺伝子型による緑内障患者の臨床特性
【0262】
表53に示すように、Gln27Glu(C79G)の多型はOAG、特にPOAGにおける高い眼内圧(IOP)と関連する。
[表53]
ADRB2遺伝子におけるGln27Gluの遺伝子型による緑内障患者の臨床特性
【0263】
標的とする多型を含有するADRB2遺伝子の部分ヌクレオチド配列は以下のとおりである。
ADRB2コドン番号Gly16Arg(GGA>AGA):Gln27Glu(CAA>GAA)(下線部)
【図面の簡単な説明】
【0264】
【図1】NTG患者の臨床特性とAT2R 3123C>A多型およびACE I/D多型との相関を表す。
【図2】MYOC遺伝子のエクソン3CにおけるDHPLC記録線パターンを表す。
【図3】MYOC遺伝子のエクソン3中に検出された新規ミスセンス突然変異Phe369Leuを表す。
【図4】POAGを持つ患者に由来するMYOC遺伝子のDHPLC記録線を表す。
【図5A】経口カンデサルタンシレキセチル投与後または偽薬投与後のIOPを表す。
【図5B】経口カンデサルタンシレキセチル投与後または偽薬投与後の眼灌流圧を表す。
【図5C】被験者15人のそれぞれにおける経口カンデサルタンシレキセチル投与後のIOPを表す。
【特許請求の範囲】
【請求項1】
(1)エンドセリン-1遺伝子のコドン198におけるAAG→AAT置換(Lys198Asn);
(2)エンドセリン-1遺伝子プロモーター領域の-1370T>G多型;
(3)エンドセリン-1遺伝子のエクソン1におけるA138挿入/欠失(A138I/D)多型;
(4)エンドセリン受容体A遺伝子の3'非コード領域における+70C>G多型;
(5)エンドセリン受容体A遺伝子の+1222C>T多型;
(6)エンドセリン受容体A遺伝子のエクソン6中のコドン323におけるCAC→CAT置換(His323His);
(7)エンドセリン受容体A遺伝子プロモーター領域の-231A>G多型;
(8)エンドセリン受容体B遺伝子のエクソン4中のコドン277におけるCTG→CTA置換;
(9)ミトコンドリア遺伝子の9099C>A多型;
(10)ミトコンドリア遺伝子の9101T>G多型;
(11)ミトコンドリア遺伝子の9101T>C多型;
(12)ミトコンドリア遺伝子の9804G>A多型;
(13)ミトコンドリア遺伝子の11778G>A多型;
(14)アンギオテンシンII1型受容体遺伝子プロモーター領域の-713T>G多型;
(16)アンギオテンシンII2型受容体遺伝子の3123C>A多型;
(25)パラオキソナーゼ1遺伝子のコドン192におけるCAA→CGA置換(Gln192Arg);
(26)パラオキソナーゼ1遺伝子のコドン55におけるTTG→ATG置換(Leu55Met);
(27)ノエリン2遺伝子のコドン144におけるCGG→CAG置換(Arg144Gln);
(32)β1アドレナリン作動性受容体遺伝子のコドン389におけるGGA→CGA置換(Gly389Arg);
(35)ミオシリン遺伝子の1105T>C多型(Phe369Leu);
(36)オプチニューリン遺伝子の412G>A多型;
(37)E-セレクチン遺伝子の1402C>T多型;
(38)腫瘍壊死因子α遺伝子プロモーター領域の-857C>T多型とオプチニューリン遺伝子の412G>A多型との組み合わせ;
(39)腫瘍壊死因子α遺伝子プロモーター領域の-863C>A多型とオプチニューリン遺伝子の603T>A多型との組み合わせ;
(40)TP53遺伝子のコドン72におけるCGC→CCC置換(Arg72Pro);
(41)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン113におけるTAC→CAC置換(Tyr113His);
(42)熱ショックタンパク質70-1遺伝子プロモーター領域の-110A>C多型;
(43)エンドセリン変換酵素遺伝子プロモーター領域の-338C>A多型;
(44)CD95遺伝子プロモーター領域の-670A>G多型;
(45)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン119におけるAAG→AAA置換(Lys119Lys);
(47)β2アドレナリン作動性受容体遺伝子のコドン16におけるGGA→AGA置換(Gly16Arg);および
(48)β2アドレナリン作動性受容体遺伝子のコドン27におけるCAA→GAA置換(Gln27Glu)
からなる群より選択される少なくとも一つの多型を含む、視神経症と関連する遺伝子多型のセット。
【請求項2】
ヒト被験者における視神経症に対する感受性を診断または予測するための方法であって、
i)被験者から生物学的試料を取得する工程、
ii)請求項1に記載した多型のセットに関して、試料の遺伝子型を決定する工程、および
iii)被験者における視神経症に対する感受性を遺伝子型に基づいて診断または予測する工程
を含む方法。
【請求項3】
視神経症が緑内障またはレーベル病である、請求項2に記載の方法。
【請求項4】
多型のセットが、視神経症と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項2に記載の方法。
【請求項5】
ヒト被験者における緑内障に対する感受性を診断または予測するための方法であって、
i)被験者から生物学的試料を取得する工程、
ii)(1)エンドセリン-1遺伝子のコドン198におけるAAG→AAT置換(Lys198Asn);
(2)エンドセリン-1遺伝子プロモーター領域の-1370T>G多型;
(3)エンドセリン-1遺伝子のエクソン1におけるA138挿入/欠失(A138I/D)多型;
(4)エンドセリン受容体A遺伝子の3'非コード領域における+70C>G多型;
(5)エンドセリン受容体A遺伝子の+1222C>T多型;
(6)エンドセリン受容体A遺伝子のエクソン6中のコドン323におけるCAC→CAT置換(His323His);
(7)エンドセリン受容体A遺伝子プロモーター領域の-231A>G多型;
(8)エンドセリン受容体B遺伝子のエクソン4中のコドン277におけるCTG→CTA置換;
(9)ミトコンドリア遺伝子の9099C>A多型;
(10)ミトコンドリア遺伝子の9101T>G多型;
(11)ミトコンドリア遺伝子の9101T>C多型;
(12)ミトコンドリア遺伝子の9804G>A多型;
(13)ミトコンドリア遺伝子の11778G>A多型;
(14)アンギオテンシンII1型受容体遺伝子プロモーター領域の-713T>G多型;
(16)アンギオテンシンII2型受容体遺伝子の3123C>A多型;
(25)パラオキソナーゼ1遺伝子のコドン192におけるCAA→CGA置換(Gln192Arg);
(26)パラオキソナーゼ1遺伝子のコドン55におけるTTG→ATG置換(Leu55Met);
(27)ノエリン2遺伝子のコドン144におけるCGG→CAG置換(Arg144Gln);
(32)β1アドレナリン作動性受容体遺伝子のコドン389におけるGGA→CGA置換(Gly389Arg);
(35)ミオシリン遺伝子の1105T>C多型(Phe369Leu);
(36)オプチニューリン遺伝子の412G>A多型;
(37)E-セレクチン遺伝子の1402C>T多型;
(38)腫瘍壊死因子α遺伝子プロモーター領域の-857C>T多型とオプチニューリン遺伝子の412G>A多型との組み合わせ;
(39)腫瘍壊死因子α遺伝子プロモーター領域の-863C>A多型とオプチニューリン遺伝子の603T>A多型との組み合わせ;
(42)熱ショックタンパク質70-1遺伝子プロモーター領域の-110A>C多型;
(43)エンドセリン変換酵素遺伝子プロモーター領域の-338C>A多型;
(44)CD95遺伝子プロモーター領域の-670A>G多型;
(45)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン119におけるAAG→AAA置換(Lys119Lys);
(47)β2アドレナリン作動性受容体遺伝子のコドン16におけるGGA→AGA置換(Gly16Arg);および
(48)β2アドレナリン作動性受容体遺伝子のコドン27におけるCAA→GAA置換(Gln27Glu)
からなる群より選択される少なくとも一つの多型を含む多型のセットに関して、試料の遺伝子型を決定する工程、ならびに
iii)被験者における緑内障に対する感受性を遺伝子型に基づいて診断または予測する工程
を含む方法。
【請求項6】
多型のセットが、緑内障と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項5に記載の方法。
【請求項7】
少なくとも一つの遺伝子多型が、
(1)エンドセリン-1遺伝子のコドン198におけるAAG→AAT置換(Lys198Asn);
(2)エンドセリン-1遺伝子プロモーター領域の-1370T>G多型;
(5)エンドセリン受容体A遺伝子の+1222C>T多型;
(6)エンドセリン受容体A遺伝子のエクソン6中のコドン323におけるCAC→CAT置換(His323His);
(7)エンドセリン受容体A遺伝子プロモーター領域の-231A>G多型;
(16)アンギオテンシンII2型受容体遺伝子の3123C>A多型;
(26)パラオキソナーゼ1遺伝子のコドン55におけるTTG→ATG置換(Leu55Met);
(32)β1アドレナリン作動性受容体遺伝子のコドン389におけるGGA→CGA置換(Gly389Arg);
(43)エンドセリン変換酵素遺伝子プロモーター領域の-338C>A多型;
(45)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン119におけるAAG→AAA置換(Lys119Lys);
からなる群より選択され、かつ緑内障が正常眼圧緑内障である、請求項5に記載の方法。
【請求項8】
多型のセットが、正常眼圧緑内障と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項7に記載の方法。
【請求項9】
少なくとも一つの遺伝子多型が、
(4)エンドセリン受容体A遺伝子の3'非コード領域における+70C>G多型;
(14)アンギオテンシンII1型受容体遺伝子プロモーター領域の-713T>G多型;
(25)パラオキソナーゼ1遺伝子のコドン192におけるCAA→CGA置換(Gln192Arg);
(35)ミオシリン遺伝子の1105T>C多型(Phe369Leu);
(36)オプチニューリン遺伝子の412G>A多型;
(38)腫瘍壊死因子α遺伝子プロモーター領域の-857C>T多型とオプチニューリン遺伝子の412G>A多型との組み合わせ;
(42)熱ショックタンパク質70-1遺伝子プロモーター領域の-110A>C多型;
(44)CD95遺伝子プロモーター領域の-670A>G多型;
(47)β2アドレナリン作動性受容体遺伝子のコドン16におけるGGA→AGA置換(Gly16Arg);および
(48)β2アドレナリン作動性受容体遺伝子のコドン27におけるCAA→GAA置換(Gln27Glu)
からなる群より選択され、かつ緑内障が原発開放隅角緑内障である、請求項5に記載の方法。
【請求項10】
多型のセットが、原発開放隅角緑内障と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項9に記載の方法。
【請求項11】
ヒト被験者におけるレーベル病に対する感受性を診断または予測するための方法であって、
i)被験者から生物学的試料を取得する工程、
ii)(40)TP53遺伝子のコドン72におけるCGC→CCC置換(Arg72Pro);および
(41)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン113におけるTAC→CAC置換(Tyr113His)
からなる群より選択される少なくとも一つの多型を含む多型のセットに関して、試料の遺伝子型を決定する工程、ならびに
iii)被験者におけるレーベル病に対する感受性を遺伝子型に基づいて診断または予測する工程
を含む方法。
【請求項12】
多型のセットが、レーベル病と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項11に記載の方法。
【請求項13】
遺伝子型が、ポリメラーゼ連鎖反応・制限断片長多型(PCR-RFLP)解析、ポリメラーゼ連鎖反応後の一本鎖高次構造多型(PCR-SSCP)解析、ASOハイブリダイゼーション解析、ダイレクトシークエンス解析、ARMS解析、DGGE解析、RNaseA切断解析、化学的制限切断解析、DPL解析、TaqMan(登録商標)PCR解析、Invader(登録商標)アッセイ、MALDI-TOF/MS解析、TDI解析、一塩基伸長アッセイ、WAVEアッセイおよび一分子蛍光検出アッセイ、ならびにそれらの組み合わせからなる群より選択される方法によって決定される、請求項2〜12のいずれか一項に記載の方法。
【請求項14】
(1)エンドセリン-1遺伝子のコドン198におけるAAG→AAT置換(Lys198Asn);
(2)エンドセリン-1遺伝子プロモーター領域の-1370T>G多型;
(3)エンドセリン-1遺伝子のエクソン1におけるA138挿入/欠失(A138I/D)多型;
(4)エンドセリン受容体A遺伝子の3'非コード領域における+70C>G多型;
(5)エンドセリン受容体A遺伝子の+1222C>T多型;
(6)エンドセリン受容体A遺伝子のエクソン6中のコドン323におけるCAC→CAT置換(His323His);
(7)エンドセリン受容体A遺伝子プロモーター領域の-231A>G多型;
(8)エンドセリン受容体B遺伝子のエクソン4中のコドン277におけるCTG→CTA置換;
(9)ミトコンドリア遺伝子の9099C>A多型;
(10)ミトコンドリア遺伝子の9101T>G多型;
(11)ミトコンドリア遺伝子の9101T>C多型;
(12)ミトコンドリア遺伝子の9804G>A多型;
(13)ミトコンドリア遺伝子の11778G>A多型;
(14)アンギオテンシンII1型受容体遺伝子プロモーター領域の-713T>G多型;
(16)アンギオテンシンII2型受容体遺伝子の3123C>A多型;
(25)パラオキソナーゼ1遺伝子のコドン192におけるCAA→CGA置換(Gln192Arg);
(26)パラオキソナーゼ1遺伝子のコドン55におけるTTG→ATG置換(Leu55Met);
(27)ノエリン2遺伝子のコドン144におけるCGG→CAG置換(Arg144Gln);
(32)β1アドレナリン作動性受容体遺伝子のコドン389におけるGGA→CGA置換(Gly389Arg);
(35)ミオシリン遺伝子の1105T>C多型(Phe369Leu);
(36)オプチニューリン遺伝子の412G>A多型;
(37)E-セレクチン遺伝子の1402C>T多型;
(38)腫瘍壊死因子α遺伝子プロモーター領域の-857C>T多型とオプチニューリン遺伝子の412G>A多型との組み合わせ;
(39)腫瘍壊死因子α遺伝子プロモーター領域の-863C>A多型とオプチニューリン遺伝子の603T>A多型との組み合わせ;
(40)TP53遺伝子のコドン72におけるCGC→CCC置換(Arg72Pro);
(41)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン113におけるTAC→CAC置換(Tyr113His);
(42)熱ショックタンパク質70-1遺伝子プロモーター領域の-110A>C多型;
(43)エンドセリン変換酵素遺伝子プロモーター領域の-338C>A多型;
(44)CD95遺伝子プロモーター領域の-670A>G多型;
(45)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン119におけるAAG→AAA置換(Lys119Lys);
(47)β2アドレナリン作動性受容体遺伝子のコドン16におけるGGA→AGA置換(Gly16Arg);および
(48)β2アドレナリン作動性受容体遺伝子のコドン27におけるCAA→GAA置換(Gln27Glu)
からなる群より選択される少なくとも一つの遺伝子多型を含む遺伝子多型のセットに関して遺伝子型を決定するのに適したプライマーセットおよび/またはプローブを含む、ヒト被験者における視神経症に対する感受性を診断または予測するためのキット。
【請求項15】
視神経症が緑内障またはレーベル病である、請求項14に記載のキット。
【請求項16】
遺伝子多型のセットが、視神経症と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項14に記載のキット
【請求項17】
(1)エンドセリン-1遺伝子のコドン198におけるAAG→AAT置換(Lys198Asn);
(2)エンドセリン-1遺伝子プロモーター領域の-1370T>G多型;
(3)エンドセリン-1遺伝子のエクソン1におけるA138挿入/欠失(A138I/D)多型;
(4)エンドセリン受容体A遺伝子の3'非コード領域における+70C>G多型;
(5)エンドセリン受容体A遺伝子の+1222C>T多型;
(6)エンドセリン受容体A遺伝子のエクソン6中のコドン323におけるCAC→CAT置換(His323His);
(7)エンドセリン受容体A遺伝子プロモーター領域の-231A>G多型;
(8)エンドセリン受容体B遺伝子のエクソン4中のコドン277におけるCTG→CTA置換;
(9)ミトコンドリア遺伝子の9099C>A多型;
(10)ミトコンドリア遺伝子の9101T>G多型;
(11)ミトコンドリア遺伝子の9101T>C多型;
(12)ミトコンドリア遺伝子の9804G>A多型;
(13)ミトコンドリア遺伝子の11778G>A多型;
(14)アンギオテンシンII1型受容体遺伝子プロモーター領域の-713T>G多型;
(16)アンギオテンシンII2型受容体遺伝子の3123C>A多型;
(25)パラオキソナーゼ1遺伝子のコドン192におけるCAA→CGA置換(Gln192Arg);
(26)パラオキソナーゼ1遺伝子のコドン55におけるTTG→ATG置換(Leu55Met);
(27)ノエリン2遺伝子のコドン144におけるCGG→CAG置換(Arg144Gln);
(32)β1アドレナリン作動性受容体遺伝子のコドン389におけるGGA→CGA置換(Gly389Arg);
(35)ミオシリン遺伝子の1105T>C多型(Phe369Leu);
(36)オプチニューリン遺伝子の412G>A多型;
(37)E-セレクチン遺伝子の1402C>T多型;
(38)腫瘍壊死因子α遺伝子プロモーター領域の-857C>T多型とオプチニューリン遺伝子の412G>A多型との組み合わせ;
(39)腫瘍壊死因子α遺伝子プロモーター領域の-863C>A多型とオプチニューリン遺伝子の603T>A多型との組み合わせ;
(42)熱ショックタンパク質70-1遺伝子プロモーター領域の-110A>C多型;
(43)エンドセリン変換酵素遺伝子プロモーター領域の-338C>A多型;
(44)CD95遺伝子プロモーター領域の-670A>G多型;
(45)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン119におけるAAG→AAA置換(Lys119Lys);
(47)β2アドレナリン作動性受容体遺伝子のコドン16におけるGGA→AGA置換(Gly16Arg);および
(48)β2アドレナリン作動性受容体遺伝子のコドン27におけるCAA→GAA置換(Gln27Glu)
からなる群より選択される少なくとも一つの遺伝子多型を含む遺伝子多型のセットに関して遺伝子型を決定するのに適したプライマーセットおよび/またはプローブを含む、ヒト被験者における緑内障に対する感受性を診断または予測するためのキット。
【請求項18】
遺伝子多型のセットが、視神経症と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項17に記載のキット。
【請求項19】
(1)エンドセリン-1遺伝子のコドン198におけるAAG→AAT置換(Lys198Asn);
(2)エンドセリン-1遺伝子プロモーター領域の-1370T>G多型;
(5)エンドセリン受容体A遺伝子の+1222C>T多型;
(6)エンドセリン受容体A遺伝子のエクソン6中のコドン323におけるCAC→CAT置換(His323His);
(7)エンドセリン受容体A遺伝子プロモーター領域の-231A>G多型;
(16)アンギオテンシンII2型受容体遺伝子の3123C>A多型;
(26)パラオキソナーゼ1遺伝子のコドン55におけるTTG→ATG置換(Leu55Met);
(32)β1アドレナリン作動性受容体遺伝子のコドン389におけるGGA→CGA置換(Gly389Arg);
(43)エンドセリン変換酵素遺伝子プロモーター領域の-338C>A多型;
(45)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン119におけるAAG→AAA置換(Lys119Lys)
からなる群より選択される少なくとも一つの遺伝子多型を含む遺伝子多型のセットに関して遺伝子型を決定するのに適したプライマーセットおよび/またはプローブを含む、ヒト被験者における正常眼圧緑内障に対する感受性を診断または予測するためのキット。
【請求項20】
遺伝子多型のセットが、正常眼圧緑内障と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項19に記載のキット。
【請求項21】
(4)エンドセリン受容体A遺伝子の3'非コード領域における+70C>G多型;
(14)アンギオテンシンII1型受容体遺伝子プロモーター領域の-713T>G多型;
(25)パラオキソナーゼ1遺伝子のコドン192におけるCAA→CGA置換(Gln192Arg);
(35)ミオシリン遺伝子の1105T>C多型(Phe369Leu);
(36)オプチニューリン遺伝子の412G>A多型;
(38)腫瘍壊死因子α遺伝子プロモーター領域の-857C>T多型とオプチニューリン遺伝子の412G>A多型との組み合わせ;
(42)熱ショックタンパク質70-1遺伝子プロモーター領域の-110A>C多型;
(44)CD95遺伝子プロモーター領域の-670A>G多型;
(47)β2アドレナリン作動性受容体遺伝子のコドン16におけるGGA→AGA置換(Gly16Arg);および
(48)β2アドレナリン作動性受容体遺伝子のコドン27におけるCAA→GAA置換(Gln27Glu)
からなる群より選択される少なくとも一つの遺伝子多型を含む遺伝子多型のセットに関して遺伝子型を決定するのに適したプライマーセットおよび/またはプローブを含む、ヒト被験者における原発開放隅角緑内障に対する感受性を診断または予測するためのキット。
【請求項22】
遺伝子多型のセットが、原発開放隅角緑内障と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項21に記載のキット。
【請求項23】
(40)TP53遺伝子のコドン72におけるCGC→CCC置換(Arg72Pro);
(41)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン113におけるTAC→CAC置換(Tyr113His)
からなる群より選択される少なくとも一つの遺伝子多型を含む遺伝子多型のセットに関して遺伝子型を決定するのに適したプライマーセットおよび/またはプローブを含む、ヒト被験者におけるレーベル病に対する感受性を診断または予測するためのキット。
【請求項24】
遺伝子多型のセットが、レーベル病と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項23に記載のキット。
【請求項25】
配列:
のセグメントからなる単離ポリヌクレオチドであって、
セグメントは少なくとも90個の連続するヌクレオチドを含んでなり、その少なくとも90個の連続するヌクレオチドは上記配列の9099位を含み、上記配列の9099位がAである、単離ポリヌクレオチド、または上記セグメントに対して完全に相補的な単離ポリヌクレオチド。
【請求項26】
請求項25に記載の配列のセグメントからなる単離ポリヌクレオチドであって、セグメントは少なくとも90個の連続するヌクレオチドを含んでなり、その少なくとも90個の連続するヌクレオチドは上記配列の9101位を含み、上記配列の9101位がGである、単離ポリヌクレオチド、または上記セグメントに対して完全に相補的な単離ポリヌクレオチド。
【請求項27】
配列:
のセグメントからなる単離ポリヌクレオチドであって、セグメントは少なくとも90個の連続するヌクレオチドを含んでなり、その少なくとも90個の連続するヌクレオチドは上記配列の下線部のヌクレオチドに相当するコドン369を含み、コドン369は、それがLeuをコードするように置換されている、単離ポリヌクレオチド、または上記セグメントに対して完全に相補的な単離ポリヌクレオチド。
【請求項28】
配列:
のセグメントからなる単離ポリヌクレオチドであって、セグメントは少なくとも90個の連続するヌクレオチドを含んでなり、その少なくとも90個の連続するヌクレオチドは上記配列の下線部のヌクレオチドに相当するコドン144を含み、コドン144は、それがGlnをコードするように置換されている、単離ポリヌクレオチド、または上記セグメントに対して完全に相補的な単離ポリヌクレオチド。
【請求項29】
ミオシリン遺伝子中に異常を持つ患者の緑内障を処置するための方法であって、患者における異常ミオシリン遺伝子の発現を抑制することを含む方法。
【請求項30】
抑制がRNA干渉法を使って行なわれる、請求項29に記載の方法。
【請求項31】
薬物による処置に対する被験者の応答を予測するための方法であって、
視神経症と関連する少なくとも一つの遺伝子多型に関して遺伝子型を決定する工程、および
遺伝子型に基づいて患者の応答を予測する工程、
を含む方法。
【請求項32】
視神経症が緑内障またはレーベル病である、請求項31に記載の方法。
【請求項33】
視神経症が緑内障である、請求項31に記載の方法。
【請求項34】
少なくとも一つの遺伝子多型がアンギオテンシンII2型受容体遺伝子の3123C>A多型である、請求項31に記載の方法。
【請求項35】
薬物がアンギオテンシン受容体IIアンタゴニストである、請求項31に記載の方法。
【請求項1】
(1)エンドセリン-1遺伝子のコドン198におけるAAG→AAT置換(Lys198Asn);
(2)エンドセリン-1遺伝子プロモーター領域の-1370T>G多型;
(3)エンドセリン-1遺伝子のエクソン1におけるA138挿入/欠失(A138I/D)多型;
(4)エンドセリン受容体A遺伝子の3'非コード領域における+70C>G多型;
(5)エンドセリン受容体A遺伝子の+1222C>T多型;
(6)エンドセリン受容体A遺伝子のエクソン6中のコドン323におけるCAC→CAT置換(His323His);
(7)エンドセリン受容体A遺伝子プロモーター領域の-231A>G多型;
(8)エンドセリン受容体B遺伝子のエクソン4中のコドン277におけるCTG→CTA置換;
(9)ミトコンドリア遺伝子の9099C>A多型;
(10)ミトコンドリア遺伝子の9101T>G多型;
(11)ミトコンドリア遺伝子の9101T>C多型;
(12)ミトコンドリア遺伝子の9804G>A多型;
(13)ミトコンドリア遺伝子の11778G>A多型;
(14)アンギオテンシンII1型受容体遺伝子プロモーター領域の-713T>G多型;
(16)アンギオテンシンII2型受容体遺伝子の3123C>A多型;
(25)パラオキソナーゼ1遺伝子のコドン192におけるCAA→CGA置換(Gln192Arg);
(26)パラオキソナーゼ1遺伝子のコドン55におけるTTG→ATG置換(Leu55Met);
(27)ノエリン2遺伝子のコドン144におけるCGG→CAG置換(Arg144Gln);
(32)β1アドレナリン作動性受容体遺伝子のコドン389におけるGGA→CGA置換(Gly389Arg);
(35)ミオシリン遺伝子の1105T>C多型(Phe369Leu);
(36)オプチニューリン遺伝子の412G>A多型;
(37)E-セレクチン遺伝子の1402C>T多型;
(38)腫瘍壊死因子α遺伝子プロモーター領域の-857C>T多型とオプチニューリン遺伝子の412G>A多型との組み合わせ;
(39)腫瘍壊死因子α遺伝子プロモーター領域の-863C>A多型とオプチニューリン遺伝子の603T>A多型との組み合わせ;
(40)TP53遺伝子のコドン72におけるCGC→CCC置換(Arg72Pro);
(41)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン113におけるTAC→CAC置換(Tyr113His);
(42)熱ショックタンパク質70-1遺伝子プロモーター領域の-110A>C多型;
(43)エンドセリン変換酵素遺伝子プロモーター領域の-338C>A多型;
(44)CD95遺伝子プロモーター領域の-670A>G多型;
(45)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン119におけるAAG→AAA置換(Lys119Lys);
(47)β2アドレナリン作動性受容体遺伝子のコドン16におけるGGA→AGA置換(Gly16Arg);および
(48)β2アドレナリン作動性受容体遺伝子のコドン27におけるCAA→GAA置換(Gln27Glu)
からなる群より選択される少なくとも一つの多型を含む、視神経症と関連する遺伝子多型のセット。
【請求項2】
ヒト被験者における視神経症に対する感受性を診断または予測するための方法であって、
i)被験者から生物学的試料を取得する工程、
ii)請求項1に記載した多型のセットに関して、試料の遺伝子型を決定する工程、および
iii)被験者における視神経症に対する感受性を遺伝子型に基づいて診断または予測する工程
を含む方法。
【請求項3】
視神経症が緑内障またはレーベル病である、請求項2に記載の方法。
【請求項4】
多型のセットが、視神経症と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項2に記載の方法。
【請求項5】
ヒト被験者における緑内障に対する感受性を診断または予測するための方法であって、
i)被験者から生物学的試料を取得する工程、
ii)(1)エンドセリン-1遺伝子のコドン198におけるAAG→AAT置換(Lys198Asn);
(2)エンドセリン-1遺伝子プロモーター領域の-1370T>G多型;
(3)エンドセリン-1遺伝子のエクソン1におけるA138挿入/欠失(A138I/D)多型;
(4)エンドセリン受容体A遺伝子の3'非コード領域における+70C>G多型;
(5)エンドセリン受容体A遺伝子の+1222C>T多型;
(6)エンドセリン受容体A遺伝子のエクソン6中のコドン323におけるCAC→CAT置換(His323His);
(7)エンドセリン受容体A遺伝子プロモーター領域の-231A>G多型;
(8)エンドセリン受容体B遺伝子のエクソン4中のコドン277におけるCTG→CTA置換;
(9)ミトコンドリア遺伝子の9099C>A多型;
(10)ミトコンドリア遺伝子の9101T>G多型;
(11)ミトコンドリア遺伝子の9101T>C多型;
(12)ミトコンドリア遺伝子の9804G>A多型;
(13)ミトコンドリア遺伝子の11778G>A多型;
(14)アンギオテンシンII1型受容体遺伝子プロモーター領域の-713T>G多型;
(16)アンギオテンシンII2型受容体遺伝子の3123C>A多型;
(25)パラオキソナーゼ1遺伝子のコドン192におけるCAA→CGA置換(Gln192Arg);
(26)パラオキソナーゼ1遺伝子のコドン55におけるTTG→ATG置換(Leu55Met);
(27)ノエリン2遺伝子のコドン144におけるCGG→CAG置換(Arg144Gln);
(32)β1アドレナリン作動性受容体遺伝子のコドン389におけるGGA→CGA置換(Gly389Arg);
(35)ミオシリン遺伝子の1105T>C多型(Phe369Leu);
(36)オプチニューリン遺伝子の412G>A多型;
(37)E-セレクチン遺伝子の1402C>T多型;
(38)腫瘍壊死因子α遺伝子プロモーター領域の-857C>T多型とオプチニューリン遺伝子の412G>A多型との組み合わせ;
(39)腫瘍壊死因子α遺伝子プロモーター領域の-863C>A多型とオプチニューリン遺伝子の603T>A多型との組み合わせ;
(42)熱ショックタンパク質70-1遺伝子プロモーター領域の-110A>C多型;
(43)エンドセリン変換酵素遺伝子プロモーター領域の-338C>A多型;
(44)CD95遺伝子プロモーター領域の-670A>G多型;
(45)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン119におけるAAG→AAA置換(Lys119Lys);
(47)β2アドレナリン作動性受容体遺伝子のコドン16におけるGGA→AGA置換(Gly16Arg);および
(48)β2アドレナリン作動性受容体遺伝子のコドン27におけるCAA→GAA置換(Gln27Glu)
からなる群より選択される少なくとも一つの多型を含む多型のセットに関して、試料の遺伝子型を決定する工程、ならびに
iii)被験者における緑内障に対する感受性を遺伝子型に基づいて診断または予測する工程
を含む方法。
【請求項6】
多型のセットが、緑内障と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項5に記載の方法。
【請求項7】
少なくとも一つの遺伝子多型が、
(1)エンドセリン-1遺伝子のコドン198におけるAAG→AAT置換(Lys198Asn);
(2)エンドセリン-1遺伝子プロモーター領域の-1370T>G多型;
(5)エンドセリン受容体A遺伝子の+1222C>T多型;
(6)エンドセリン受容体A遺伝子のエクソン6中のコドン323におけるCAC→CAT置換(His323His);
(7)エンドセリン受容体A遺伝子プロモーター領域の-231A>G多型;
(16)アンギオテンシンII2型受容体遺伝子の3123C>A多型;
(26)パラオキソナーゼ1遺伝子のコドン55におけるTTG→ATG置換(Leu55Met);
(32)β1アドレナリン作動性受容体遺伝子のコドン389におけるGGA→CGA置換(Gly389Arg);
(43)エンドセリン変換酵素遺伝子プロモーター領域の-338C>A多型;
(45)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン119におけるAAG→AAA置換(Lys119Lys);
からなる群より選択され、かつ緑内障が正常眼圧緑内障である、請求項5に記載の方法。
【請求項8】
多型のセットが、正常眼圧緑内障と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項7に記載の方法。
【請求項9】
少なくとも一つの遺伝子多型が、
(4)エンドセリン受容体A遺伝子の3'非コード領域における+70C>G多型;
(14)アンギオテンシンII1型受容体遺伝子プロモーター領域の-713T>G多型;
(25)パラオキソナーゼ1遺伝子のコドン192におけるCAA→CGA置換(Gln192Arg);
(35)ミオシリン遺伝子の1105T>C多型(Phe369Leu);
(36)オプチニューリン遺伝子の412G>A多型;
(38)腫瘍壊死因子α遺伝子プロモーター領域の-857C>T多型とオプチニューリン遺伝子の412G>A多型との組み合わせ;
(42)熱ショックタンパク質70-1遺伝子プロモーター領域の-110A>C多型;
(44)CD95遺伝子プロモーター領域の-670A>G多型;
(47)β2アドレナリン作動性受容体遺伝子のコドン16におけるGGA→AGA置換(Gly16Arg);および
(48)β2アドレナリン作動性受容体遺伝子のコドン27におけるCAA→GAA置換(Gln27Glu)
からなる群より選択され、かつ緑内障が原発開放隅角緑内障である、請求項5に記載の方法。
【請求項10】
多型のセットが、原発開放隅角緑内障と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項9に記載の方法。
【請求項11】
ヒト被験者におけるレーベル病に対する感受性を診断または予測するための方法であって、
i)被験者から生物学的試料を取得する工程、
ii)(40)TP53遺伝子のコドン72におけるCGC→CCC置換(Arg72Pro);および
(41)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン113におけるTAC→CAC置換(Tyr113His)
からなる群より選択される少なくとも一つの多型を含む多型のセットに関して、試料の遺伝子型を決定する工程、ならびに
iii)被験者におけるレーベル病に対する感受性を遺伝子型に基づいて診断または予測する工程
を含む方法。
【請求項12】
多型のセットが、レーベル病と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項11に記載の方法。
【請求項13】
遺伝子型が、ポリメラーゼ連鎖反応・制限断片長多型(PCR-RFLP)解析、ポリメラーゼ連鎖反応後の一本鎖高次構造多型(PCR-SSCP)解析、ASOハイブリダイゼーション解析、ダイレクトシークエンス解析、ARMS解析、DGGE解析、RNaseA切断解析、化学的制限切断解析、DPL解析、TaqMan(登録商標)PCR解析、Invader(登録商標)アッセイ、MALDI-TOF/MS解析、TDI解析、一塩基伸長アッセイ、WAVEアッセイおよび一分子蛍光検出アッセイ、ならびにそれらの組み合わせからなる群より選択される方法によって決定される、請求項2〜12のいずれか一項に記載の方法。
【請求項14】
(1)エンドセリン-1遺伝子のコドン198におけるAAG→AAT置換(Lys198Asn);
(2)エンドセリン-1遺伝子プロモーター領域の-1370T>G多型;
(3)エンドセリン-1遺伝子のエクソン1におけるA138挿入/欠失(A138I/D)多型;
(4)エンドセリン受容体A遺伝子の3'非コード領域における+70C>G多型;
(5)エンドセリン受容体A遺伝子の+1222C>T多型;
(6)エンドセリン受容体A遺伝子のエクソン6中のコドン323におけるCAC→CAT置換(His323His);
(7)エンドセリン受容体A遺伝子プロモーター領域の-231A>G多型;
(8)エンドセリン受容体B遺伝子のエクソン4中のコドン277におけるCTG→CTA置換;
(9)ミトコンドリア遺伝子の9099C>A多型;
(10)ミトコンドリア遺伝子の9101T>G多型;
(11)ミトコンドリア遺伝子の9101T>C多型;
(12)ミトコンドリア遺伝子の9804G>A多型;
(13)ミトコンドリア遺伝子の11778G>A多型;
(14)アンギオテンシンII1型受容体遺伝子プロモーター領域の-713T>G多型;
(16)アンギオテンシンII2型受容体遺伝子の3123C>A多型;
(25)パラオキソナーゼ1遺伝子のコドン192におけるCAA→CGA置換(Gln192Arg);
(26)パラオキソナーゼ1遺伝子のコドン55におけるTTG→ATG置換(Leu55Met);
(27)ノエリン2遺伝子のコドン144におけるCGG→CAG置換(Arg144Gln);
(32)β1アドレナリン作動性受容体遺伝子のコドン389におけるGGA→CGA置換(Gly389Arg);
(35)ミオシリン遺伝子の1105T>C多型(Phe369Leu);
(36)オプチニューリン遺伝子の412G>A多型;
(37)E-セレクチン遺伝子の1402C>T多型;
(38)腫瘍壊死因子α遺伝子プロモーター領域の-857C>T多型とオプチニューリン遺伝子の412G>A多型との組み合わせ;
(39)腫瘍壊死因子α遺伝子プロモーター領域の-863C>A多型とオプチニューリン遺伝子の603T>A多型との組み合わせ;
(40)TP53遺伝子のコドン72におけるCGC→CCC置換(Arg72Pro);
(41)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン113におけるTAC→CAC置換(Tyr113His);
(42)熱ショックタンパク質70-1遺伝子プロモーター領域の-110A>C多型;
(43)エンドセリン変換酵素遺伝子プロモーター領域の-338C>A多型;
(44)CD95遺伝子プロモーター領域の-670A>G多型;
(45)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン119におけるAAG→AAA置換(Lys119Lys);
(47)β2アドレナリン作動性受容体遺伝子のコドン16におけるGGA→AGA置換(Gly16Arg);および
(48)β2アドレナリン作動性受容体遺伝子のコドン27におけるCAA→GAA置換(Gln27Glu)
からなる群より選択される少なくとも一つの遺伝子多型を含む遺伝子多型のセットに関して遺伝子型を決定するのに適したプライマーセットおよび/またはプローブを含む、ヒト被験者における視神経症に対する感受性を診断または予測するためのキット。
【請求項15】
視神経症が緑内障またはレーベル病である、請求項14に記載のキット。
【請求項16】
遺伝子多型のセットが、視神経症と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項14に記載のキット
【請求項17】
(1)エンドセリン-1遺伝子のコドン198におけるAAG→AAT置換(Lys198Asn);
(2)エンドセリン-1遺伝子プロモーター領域の-1370T>G多型;
(3)エンドセリン-1遺伝子のエクソン1におけるA138挿入/欠失(A138I/D)多型;
(4)エンドセリン受容体A遺伝子の3'非コード領域における+70C>G多型;
(5)エンドセリン受容体A遺伝子の+1222C>T多型;
(6)エンドセリン受容体A遺伝子のエクソン6中のコドン323におけるCAC→CAT置換(His323His);
(7)エンドセリン受容体A遺伝子プロモーター領域の-231A>G多型;
(8)エンドセリン受容体B遺伝子のエクソン4中のコドン277におけるCTG→CTA置換;
(9)ミトコンドリア遺伝子の9099C>A多型;
(10)ミトコンドリア遺伝子の9101T>G多型;
(11)ミトコンドリア遺伝子の9101T>C多型;
(12)ミトコンドリア遺伝子の9804G>A多型;
(13)ミトコンドリア遺伝子の11778G>A多型;
(14)アンギオテンシンII1型受容体遺伝子プロモーター領域の-713T>G多型;
(16)アンギオテンシンII2型受容体遺伝子の3123C>A多型;
(25)パラオキソナーゼ1遺伝子のコドン192におけるCAA→CGA置換(Gln192Arg);
(26)パラオキソナーゼ1遺伝子のコドン55におけるTTG→ATG置換(Leu55Met);
(27)ノエリン2遺伝子のコドン144におけるCGG→CAG置換(Arg144Gln);
(32)β1アドレナリン作動性受容体遺伝子のコドン389におけるGGA→CGA置換(Gly389Arg);
(35)ミオシリン遺伝子の1105T>C多型(Phe369Leu);
(36)オプチニューリン遺伝子の412G>A多型;
(37)E-セレクチン遺伝子の1402C>T多型;
(38)腫瘍壊死因子α遺伝子プロモーター領域の-857C>T多型とオプチニューリン遺伝子の412G>A多型との組み合わせ;
(39)腫瘍壊死因子α遺伝子プロモーター領域の-863C>A多型とオプチニューリン遺伝子の603T>A多型との組み合わせ;
(42)熱ショックタンパク質70-1遺伝子プロモーター領域の-110A>C多型;
(43)エンドセリン変換酵素遺伝子プロモーター領域の-338C>A多型;
(44)CD95遺伝子プロモーター領域の-670A>G多型;
(45)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン119におけるAAG→AAA置換(Lys119Lys);
(47)β2アドレナリン作動性受容体遺伝子のコドン16におけるGGA→AGA置換(Gly16Arg);および
(48)β2アドレナリン作動性受容体遺伝子のコドン27におけるCAA→GAA置換(Gln27Glu)
からなる群より選択される少なくとも一つの遺伝子多型を含む遺伝子多型のセットに関して遺伝子型を決定するのに適したプライマーセットおよび/またはプローブを含む、ヒト被験者における緑内障に対する感受性を診断または予測するためのキット。
【請求項18】
遺伝子多型のセットが、視神経症と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項17に記載のキット。
【請求項19】
(1)エンドセリン-1遺伝子のコドン198におけるAAG→AAT置換(Lys198Asn);
(2)エンドセリン-1遺伝子プロモーター領域の-1370T>G多型;
(5)エンドセリン受容体A遺伝子の+1222C>T多型;
(6)エンドセリン受容体A遺伝子のエクソン6中のコドン323におけるCAC→CAT置換(His323His);
(7)エンドセリン受容体A遺伝子プロモーター領域の-231A>G多型;
(16)アンギオテンシンII2型受容体遺伝子の3123C>A多型;
(26)パラオキソナーゼ1遺伝子のコドン55におけるTTG→ATG置換(Leu55Met);
(32)β1アドレナリン作動性受容体遺伝子のコドン389におけるGGA→CGA置換(Gly389Arg);
(43)エンドセリン変換酵素遺伝子プロモーター領域の-338C>A多型;
(45)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン119におけるAAG→AAA置換(Lys119Lys)
からなる群より選択される少なくとも一つの遺伝子多型を含む遺伝子多型のセットに関して遺伝子型を決定するのに適したプライマーセットおよび/またはプローブを含む、ヒト被験者における正常眼圧緑内障に対する感受性を診断または予測するためのキット。
【請求項20】
遺伝子多型のセットが、正常眼圧緑内障と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項19に記載のキット。
【請求項21】
(4)エンドセリン受容体A遺伝子の3'非コード領域における+70C>G多型;
(14)アンギオテンシンII1型受容体遺伝子プロモーター領域の-713T>G多型;
(25)パラオキソナーゼ1遺伝子のコドン192におけるCAA→CGA置換(Gln192Arg);
(35)ミオシリン遺伝子の1105T>C多型(Phe369Leu);
(36)オプチニューリン遺伝子の412G>A多型;
(38)腫瘍壊死因子α遺伝子プロモーター領域の-857C>T多型とオプチニューリン遺伝子の412G>A多型との組み合わせ;
(42)熱ショックタンパク質70-1遺伝子プロモーター領域の-110A>C多型;
(44)CD95遺伝子プロモーター領域の-670A>G多型;
(47)β2アドレナリン作動性受容体遺伝子のコドン16におけるGGA→AGA置換(Gly16Arg);および
(48)β2アドレナリン作動性受容体遺伝子のコドン27におけるCAA→GAA置換(Gln27Glu)
からなる群より選択される少なくとも一つの遺伝子多型を含む遺伝子多型のセットに関して遺伝子型を決定するのに適したプライマーセットおよび/またはプローブを含む、ヒト被験者における原発開放隅角緑内障に対する感受性を診断または予測するためのキット。
【請求項22】
遺伝子多型のセットが、原発開放隅角緑内障と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項21に記載のキット。
【請求項23】
(40)TP53遺伝子のコドン72におけるCGC→CCC置換(Arg72Pro);
(41)ミクロソームエポキシドヒドラーゼ1遺伝子のコドン113におけるTAC→CAC置換(Tyr113His)
からなる群より選択される少なくとも一つの遺伝子多型を含む遺伝子多型のセットに関して遺伝子型を決定するのに適したプライマーセットおよび/またはプローブを含む、ヒト被験者におけるレーベル病に対する感受性を診断または予測するためのキット。
【請求項24】
遺伝子多型のセットが、レーベル病と関連することが知られている少なくとも一つの遺伝子多型をさらに含む、請求項23に記載のキット。
【請求項25】
配列:
のセグメントからなる単離ポリヌクレオチドであって、
セグメントは少なくとも90個の連続するヌクレオチドを含んでなり、その少なくとも90個の連続するヌクレオチドは上記配列の9099位を含み、上記配列の9099位がAである、単離ポリヌクレオチド、または上記セグメントに対して完全に相補的な単離ポリヌクレオチド。
【請求項26】
請求項25に記載の配列のセグメントからなる単離ポリヌクレオチドであって、セグメントは少なくとも90個の連続するヌクレオチドを含んでなり、その少なくとも90個の連続するヌクレオチドは上記配列の9101位を含み、上記配列の9101位がGである、単離ポリヌクレオチド、または上記セグメントに対して完全に相補的な単離ポリヌクレオチド。
【請求項27】
配列:
のセグメントからなる単離ポリヌクレオチドであって、セグメントは少なくとも90個の連続するヌクレオチドを含んでなり、その少なくとも90個の連続するヌクレオチドは上記配列の下線部のヌクレオチドに相当するコドン369を含み、コドン369は、それがLeuをコードするように置換されている、単離ポリヌクレオチド、または上記セグメントに対して完全に相補的な単離ポリヌクレオチド。
【請求項28】
配列:
のセグメントからなる単離ポリヌクレオチドであって、セグメントは少なくとも90個の連続するヌクレオチドを含んでなり、その少なくとも90個の連続するヌクレオチドは上記配列の下線部のヌクレオチドに相当するコドン144を含み、コドン144は、それがGlnをコードするように置換されている、単離ポリヌクレオチド、または上記セグメントに対して完全に相補的な単離ポリヌクレオチド。
【請求項29】
ミオシリン遺伝子中に異常を持つ患者の緑内障を処置するための方法であって、患者における異常ミオシリン遺伝子の発現を抑制することを含む方法。
【請求項30】
抑制がRNA干渉法を使って行なわれる、請求項29に記載の方法。
【請求項31】
薬物による処置に対する被験者の応答を予測するための方法であって、
視神経症と関連する少なくとも一つの遺伝子多型に関して遺伝子型を決定する工程、および
遺伝子型に基づいて患者の応答を予測する工程、
を含む方法。
【請求項32】
視神経症が緑内障またはレーベル病である、請求項31に記載の方法。
【請求項33】
視神経症が緑内障である、請求項31に記載の方法。
【請求項34】
少なくとも一つの遺伝子多型がアンギオテンシンII2型受容体遺伝子の3123C>A多型である、請求項31に記載の方法。
【請求項35】
薬物がアンギオテンシン受容体IIアンタゴニストである、請求項31に記載の方法。
【図1】

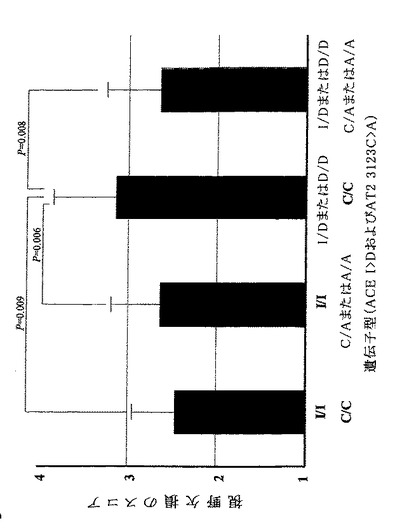
【図2】
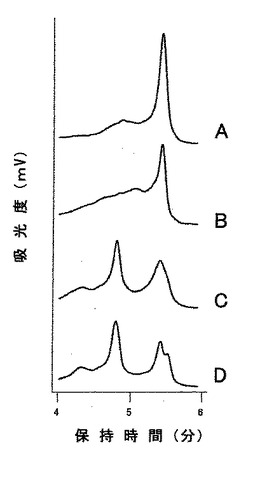
【図3】
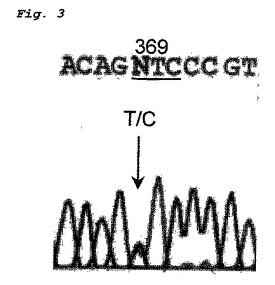
【図4】
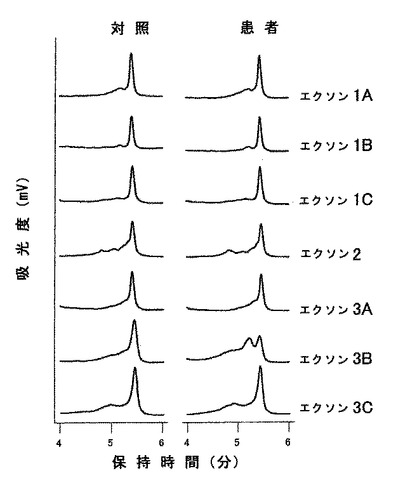
【図5】
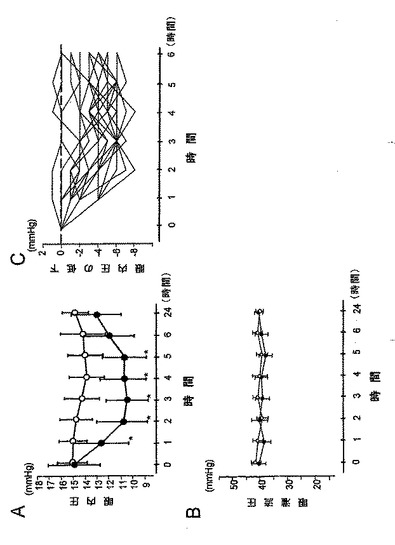
【図2】
【図3】
【図4】
【図5】
【公表番号】特表2007−529218(P2007−529218A)
【公表日】平成19年10月25日(2007.10.25)
【国際特許分類】
【出願番号】特願2007−503564(P2007−503564)
【出願日】平成17年3月18日(2005.3.18)
【国際出願番号】PCT/JP2005/005601
【国際公開番号】WO2005/090602
【国際公開日】平成17年9月29日(2005.9.29)
【出願人】(501131276)スキャンポ・アーゲー (37)
【氏名又は名称原語表記】Sucampo AG
【Fターム(参考)】
【公表日】平成19年10月25日(2007.10.25)
【国際特許分類】
【出願日】平成17年3月18日(2005.3.18)
【国際出願番号】PCT/JP2005/005601
【国際公開番号】WO2005/090602
【国際公開日】平成17年9月29日(2005.9.29)
【出願人】(501131276)スキャンポ・アーゲー (37)
【氏名又は名称原語表記】Sucampo AG
【Fターム(参考)】
[ Back to top ]