
2型糖尿病に関する遺伝子検出方法
【課題】 mtDNAのハプログループに基づき、2型糖尿病の遺伝的危険度を予測する遺伝子検出方法等を提供すること、およびそれによって2型糖尿病の発症を予防すること。
【解決手段】 1289名の2型糖尿病患者と、1617名の対照者とを合わせた2906名の日本人について、ミトコンドリアゲノムのコード領域における多型に関する遺伝子型を決定することにより、10個の代表的なハプログループ(F,B,A,N9b,M7a,M7b,G1,G2,D4,およびD5)に分類した。統計解析の結果、ハプログループFが2型糖尿病に関連していること、および女性の場合には、ハプログループN9aが2型糖尿病に対する抵抗性に関係していることが明らかになった。
【解決手段】 1289名の2型糖尿病患者と、1617名の対照者とを合わせた2906名の日本人について、ミトコンドリアゲノムのコード領域における多型に関する遺伝子型を決定することにより、10個の代表的なハプログループ(F,B,A,N9b,M7a,M7b,G1,G2,D4,およびD5)に分類した。統計解析の結果、ハプログループFが2型糖尿病に関連していること、および女性の場合には、ハプログループN9aが2型糖尿病に対する抵抗性に関係していることが明らかになった。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、2型糖尿病に関する遺伝子検出方法に関し、特にヒトミトコンドリアDNAのハプログループを分類することにより2型糖尿病に関する遺伝子検出を行うものに関する。
【背景技術】
【0002】
インスリン非依存性の2型糖尿病は、先進国において高い発症率を示し、世界的規模で公衆衛生上の主要な問題の一つになっている。2型糖尿病は、複合的な要因に由来する疾患であり、そこでは様々な遺伝的または環境的な影響が相互に重要な役割を果たしている。若年発症性の成人型糖尿病(maturity-onset diabetes of the young :MODY)を引き起こす病原性突然変異ついては、既にいくつかの報告がある。
【0003】
一方、ミトコンドリアゲノムにおいて、tRNA−Leu(UUR)遺伝子中の3243A>G突然変異が、母系遺伝性糖尿病ならびに難聴の患者(非特許文献1。本明細書においては、関連文献については末尾にまとめて示す。)で検出された。更に、ミトコンドリアゲノム多型の幾つかは、日本人集団においてはインスリン抵抗性および/または高血糖と関連を持つことが報告されている(非特許文献2−4)。このように、ミトコンドリアの機能障害は、骨格筋のインシュリン抵抗性、および膵臓のベータ細胞からのインスリン分泌障害の両者に関連していると考えられている(非特許文献5)。
【0004】
ミトコンドリアゲノム中の母系遺伝性の変異は、過去二十万年以上に渡り、ホモサピエンスがアフリカからアジアおよびヨーロッパに拡張した時代に、新たな突然変異が連続的に追加された事に起因している(非特許文献6)。ミトコンドリアゲノムの突然変異率は核ゲノムの突然変異率に比べると少なくとも10倍以上高いので、ホモサピエンスに蓄積されて来た古代のミトコンドリア多型は、比較的短い歴史を持つものではあっても、現代人類の代謝特性に影響している可能性がある(非特許文献7)。世界の様々な地域で発見されたミトコンドリアハプログループが、寒冷気候と栄養条件への適応を通じて選択されて来たものであると推測することができる(非特許文献7−9)。ミトコンドリアハプログループ間で機能的差異が存在することを支持する数々の証拠が、精子運動性(非特許文献10)、敗血症後の生存率(非特許文献11)、およびパーキンソン病に対する感受性(非特許文献12)の研究により与えられている。
【0005】
これまで本発明者らは、寿命(非特許文献13)、パーキンソン病(非特許文献14、15)およびアルツハイマー病と言った加齢関連の疾病、並びに肥満(非特許文献2)、るい痩(非特許文献16)、2型糖尿病(非特許文献2)および動脈硬化症と言ったエネルギー代謝に関連する疾病に関与するミトコンドリアDNA多型(mtSNP)を同定することを目的として研究を重ねてきた。この目的のため、我々は7つの異なる群に属する672名について、16569塩基対のミトコンドリアゲノムを完全にシーケンスした。7群は、それぞれ96名から構成されており、それらは、百寿者群、パーキンソン病罹患者群、アルツハイマー病患者群、肥満の若い男性群、肥満でない若い男性群、重度の血管性病変を持つ2型糖尿病患者群、および重度の血管性病変を持たない2型糖尿病患者群であった(非特許文献17)。この調査結果から、ヒトミトコンドリアゲノム多型データベース(http://www.giib.or.jp/mtsnp/index_e.shtml)を構築した。これらのmtSNPデータに基づいて、本発明者らは、蛍光ビーズ法を用いた包括的なmtSNP分析システムを開発した。
【0006】
Poultonらは、mtDNAの16189T>C変異が2型糖尿病と関連を持つことを既に報告している(非特許文献18)。Wengらは、この多型が中国人成人においては、メタボリックシンドロームとも関連していると報告している(非特許文献19)。しかしながら、ヨーロッパ人に関する研究では、16184−16193のpoly(C)tractの遺伝変異が、2型糖尿病発症においては主要な役割を担っていないことが示されている(非特許文献20)。従って、非コード領域のみに於ける多型分析は、個々人の代謝特性とミトコンドリアゲノムに於ける機能的多型との間の相互関係については、直接的な証拠を提供し難いと思われる。
【発明の開示】
【発明が解決しようとする課題】
【0007】
上記の諸点に留意しつつ、本発明者らは、ミトコンドリアゲノムのコード領域に於ける包括的な多型分析を基づいて、日本人に於ける2型糖尿病と主要な10個のハプログループ(F,B,A,N9a,M7a,M7b,G1,G2,D4,およびD5)との関係を調べる大規模な関連解析を行った。
本発明者らの研究目的は、mtDNAのハプログループ分けに基づき、2型糖尿病の遺伝的リスクを予測し、それによって2型糖尿病の一次予防に役立てることにある。
【課題を解決するための手段】
【0008】
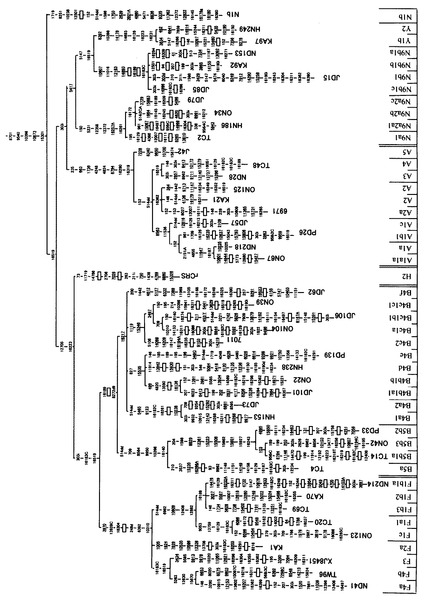
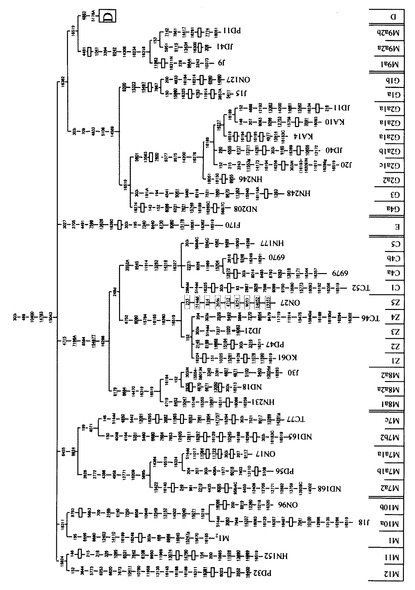
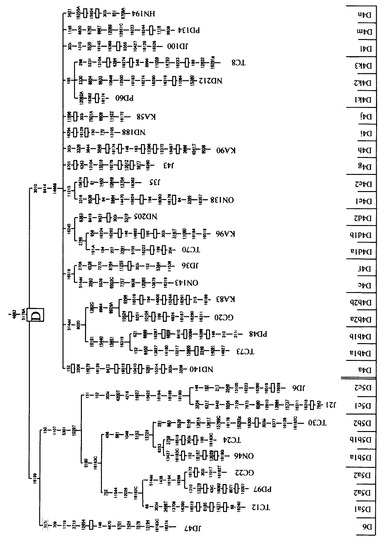
本発明者らは、長年に亘ってヒトミトコンドリアゲノムに関する研究を行っている者である。ヒトミトコンドリアDNAは、強固に連鎖した複数のmtSNPを有するハプログループに分類することができる。図3〜図5には、mtSNPとハプログループ(及びサブハプログループ)との関係を示した。このハプログループの分布は、人種間において、かなり大きな隔たりがある。個々の日本人は、主要な10個のハプログループによって、そのほとんど(約90パーセント以上)を分類することができる。各ハプログループは、数個〜20個程度のmtSNPによって特徴付けられる。例えば、ハプログループFは、8701、9540、10398、10873、15301、16519、12705、16223、16183、16519、3970、13928、16304、248、6392、10310等の位置にmtSNPを有している(その他のハプログループについても、同様の状況が存在する)。ハプログループ分類をするために、これら全てのmtSNPを調べようとするのは、大変な労力が掛かるため大規模な研究には向きにくい。しかしながら、これらのうちのいずれのmtSNPを選択・検出すれば、適切にハプログループを分類することが可能であるかについては明確には知られていなかったことに加え、選択したmtSNPを迅速かつ的確に検出する方法については開発が十分に進んでいなかった。
【0009】
そこで、本発明者らは、鋭意検討の結果、新たなmtSNPの検出法を開発し、日本人における主要な10個のハプログループ(F,B,A,N9a,M7a,M7b,G1,G2,D4及びD5)を迅速かつ的確に分類する検出法を開発した。
つまり、各ハプログループが持っている数個〜二十個程度のmtSNPのうち、適当に1個〜5個のmtSNPを選択することにより、表1に記載のmtSNPsを検出すれば、10個のハプログループへの分類が可能であることを見出した。更に、この方法を用いて、本発明者らは、ミトコンドリアゲノムのコード領域における包括的な多型分析を基にして、日本人における2型糖尿病と主要な10個のハプログループ(F,B,A,N9a,M7a,M7b,G1,G2,D4およびD5)との関係を調べるために、2906名を対象とした大規模な関連解析を行った。その結果、ハプログループFを持つ者は2型糖尿病の危険性が高いこと、ハプログループN9aを持つ者は2型糖尿病に対する抵抗性を有すること(特に、女性の場合)、および女性の場合にはハプログループAを持つ者は2型糖尿病の危険性が高いことを明らかとし、基本的には本発明を完成するに至った。
【0010】
こうして、上記課題を解決するために、本発明に係る2型糖尿病に関する遺伝子検出法は、ヒトミトコンドリアDNAにおいて、5231A、12358G、12372Aを有するハプログループN9aと、3970T、13928C、10310Aを有するハプログループFと、663G、8794Tを有するハプログループAと、12406Aを有するサブハプログループF1と、14944Tを有するサブハプログループA1a1aと、15067Cを有するサブハプログループN9a2a1と、5147Aを有するF1b1aと、5147Aを有するサブハプログループN9b1と、8856Aを有するサブハプログループB5b3と、14476Aを有するサブハプログループF1b1aと、15314Aを有するサブハプログループD4aと、4655Aを有するサブハプログループA1aと、8856Aを有するサブハプログループB5b3と、1391Cを有するサブハプログループB4b1bと、8762Cを有するサブハプログループD4mと、8383Cを有するサブハプログループD4d2と、7861Cを有するサブハプログループF4aと、15047Aを有するサブハプログループM8a1とを検出することを特徴とする。
【0011】
本明細書中において、多型の記載方法は、次の通りである。原則として、各遺伝子について、「多型が生じている位置、データベースに登録されている塩基(A:アデニン、G:グアニン、C:シトシン、T:チミン)>多型塩基」の順で記載する。例えば、「5231G>A」は、5231位のGがAとなっている多型を意味している。また、その多型が生じている遺伝子(rRNAまたはmRNA)の位置について、カッコ書きで情報を示すことがある。例えば、「5231G>A(ND2:syn)」とは、上記多型について、ND2遺伝子において、アミノ酸の変異を伴わない多型(synonymous mutation)であることを意味している。但し、挿入あるいは欠失多型については、「多型が生じている位置(欠失または挿入に関する塩基情報)」の順で記載する。例えば、「8272(9-bp deletion in non-coding region)」とは、8272位の非コード領域における9個の塩基の欠失多型であることを意味している。また、順方向(forward strand)に読んでA/G多型としてデータベースに記録されている場合であっても、逆方向(reverse strand)で読むとT/C多型となる。多くの文献について、「T/C」多型として記載されている場合には、データベース中の記録であるA/G多型と異なることがある。
【0012】
一般にミトコンドリアDNA多型(mtSNPs)或いはミトコンドリアDNAハプログループは、対象となる集団(例えば、日本人・中国人・韓国人を含む東アジア人集団、西洋人集団など)が異なると、その種類・頻度が異なる。このため、日本人以外の集団において、2型糖尿病との関係が指摘されているハプログループであっても、必ずしも日本人集団においてそのような関連が認められるわけではない。このため、従来の報告については、集団または疾患が異なる場合には、必ずしも日本人におけるハプログループおよび2型糖尿病との関連が裏付けられるわけではない。
【0013】
本発明において、mtDNAのハプログループを検出する方法としては、従来公知の方法、例えばDNAシークエンス、DNAチップ(或いはDNAマイクロアレイ)、RFLP、PCR−SSOP−Luminex法などの方法を用いることができる。これらの中でも、PCR−SSOP−Luminex法を用いることが好ましい。PCR−SSOP−Luminex法を用いることにより、数百個程度の多型であれば迅速に測定することができる。なお、同方法の詳細については後に詳述する。
【発明の効果】
【0014】
本発明によれば、mtDNAのハプログループを検出することにより、2型糖尿病の遺伝的危険度を予測することができる。この発明を用いることにより、2型糖尿病に対する予防が可能となり、高齢者の健康寿命延長・QOL向上・ねたきり防止ならびに今後の医療費削減など、医学的・社会的に大きく貢献できる。
【発明を実施するための最良の形態】
【0015】
次に、本発明の実施形態について、図面を参照しつつ説明するが、本発明の技術的範囲は、これらの実施形態によって限定されるものではなく、発明の要旨を変更することなく様々な形態で実施することができる。また、本発明の技術的範囲は、均等の範囲にまで及ぶものである。
【0016】
<研究対象>
研究対象は、互いに血縁関係の無い40歳以上の日本人2906名(男性1938名、女性968名)であった。対象者は、2002年10月から2005年3月の間に、3つの研究参加病院(岐阜県立岐阜病院、多治見病院および下呂温泉病院)の1つを外来した受診者、或いは入院患者であった。2型糖尿病患者は、絶食時血漿グルコースレベルが6.99mmol/L(126mg/dL)以上、またはヘモグロビンAlc(Glycosylated hemoglobin)濃度が6.5%以上を示す者、或いは糖尿病に対する薬物治療下にあった。2型糖尿病は、世界保健機構により承認された基準に従って定義した。これらの基準に基づいて、研究対象集団中の1289名の被験者(男性890名、女性399名)が、2型糖尿病患者であると診断された。残りの1617名(男性1048名、女性569名)は対照群とした。対照群は、研究参加病院に例年の健康チェックのために外来受診した者である。対照群は、絶食時血漿グルコースレベルが6.11mmol/L(110mg/dL)未満であり、ヘモグロビンAlcが5.6%未満を示し、かつ2型糖尿病の既往歴ならびに2型糖尿病に対する薬物治療を受けた経験が無い者であった。
研究プロトコルは、岐阜県国際バイオ研究所のヒト研究に関する倫理委員会により承認され、また各参加者からは個別に書面によるインフォームドコンセントを得た。
【0017】
<ハプログループ分類のためのミトコンドリア多型の選択>
本発明者らが作製したヒトミトコンドリアゲノム多型データベース(http://www.giib.or.jp/mtsnp/index_e.shtml)および日本人の系統樹(非特許文献17)を使用して、ミトコンドリアハプログループの分類に有用である149個の多型部位を選択した。更に、本発明者らの鋭意検討に基づき、日本人集団において発見された10個の主要なハプログループ(F,B,A,N9b,M7a,M7b,G1,G2,D4,およびD5)を定義するmtSNPsを選択した。これらのmtSNPsを表1に示した。
【0018】
【表1】

【0019】
表中、左欄より順に、ハプログループ(Haplogroup)、および各ハプログループを代表する多型(Polymorphism)を示している。また表中、「Syn」はアミノ酸の変異を伴わない多型(synonymous mutation)であることを示している。
本発明者らは、上記2906名の被験者について、これらハプログループと2型糖尿病との間の関係について研究を行った。
【0020】
<多型の遺伝子タイピング>
各参加者から50mmol/L EDTA(ジナトリウム塩)を含む試験管に、7mLの静脈血を採取し、市販のキット(Genomix, Talent, Trieste, Italy)を用いてDNAを分離した。
DNAをPCRにかけ、特定のDNAを増幅した。その際に使用したプライマーを表2および表3に示した。ミトコンドリアの多型は懸濁液アレイ技術(Luminex 100; Luminex, Austin, TX)を用い、シーケンス特異性オリゴヌクレオチド試験法(G&Gサイエンス株式会社、日本)で決定した。詳細な方法については、既報のもの(非特許文献21)を基本として、適宜に増幅条件を変えて行った。なお、条件については、後に詳述する。
【0021】
また、ハプロタイピングのために使用したプライマーは、表4〜表9に示した。今回のタイピングには、最大で100種類の多型の判定が行えるものを用いた。186種類のプローブを用いるために、全体を100種類のセット(プローブセットA)と86種類のセット(プローブセットB)とに分けて、タイピングを行った。表4〜表6にはプローブセットAの詳細を、表7〜表9にはプローブセットBの詳細を示した。
【0022】
【表2】
【0023】
【表3】
【0024】
【表4】
【0025】
【表5】
【0026】
【表6】
【0027】
【表7】
【0028】
【表8】
【0029】
【表9】
【0030】
表2および表3については、左欄よりPCR法によって合成されるフラグメント番号(Fragment)、フォワードプライマー(Forward primer)のmtDNAにおける位置(Position)と塩基配列(Sequence)、リバースプライマー(Reverse primer)のmtDNAにおける位置(Position)と塩基配列、および合成されるDNAフラグメント長(Product)を示している。
表4〜表6については、第1のセット(プローブセットA)について、mtDNAにおける位置(Position)、目的(Purpose)、プローブの塩基配列(Sequence)を示している。また、目的の欄については、「a」が多型チェック用、「b」が野生型チェック用、「p」がPCR産物チェック用、「m」および「n」がマクロハプログループチェック用のものであることを示している。表7〜表9については、第2のセット(プローブセットB)について、上記と同じ意味である。
表4〜表9に示したプローブセットは、前述の10個のハプログループを分類するmtSNPに加えて、更に細かいサブハプログループを検出するためのプローブを含んでいる。
【0031】
PCR−SSOP−Luminex法
方法の詳細については、非特許文献21に記載の通りである。以下には、この方法の概要について説明する。
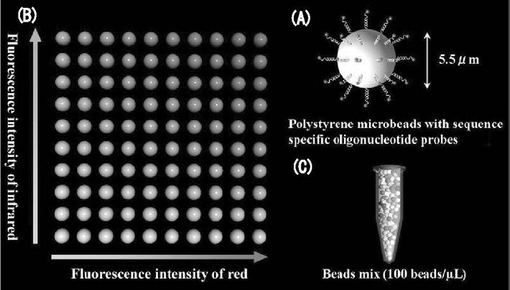
図1には、Luminex100フローサイトメトリーで検出するマイクロビーズ(本発明における固相に該当する)の微細構造と特徴を示した。マイクロビーズ(図中の符号(A))は、直径が約5.5μm程度であり、ポリスチレン製である。ビーズ表面にはカルボキシル基が存在している。このカルボキシル基と、アミノ修飾された核酸プローブとを結合することにより、ビーズ表面に核酸プローブが結合されている(本発明における核酸プローブの固相化に該当する)。各ビーズには、一種類の核酸プローブが結合されている。このマイクロビーズには、赤色色素と赤外色素との二種類の色素が割合を変化させつつ配されている(Multi-Analyte Microsphere Carboxylated: Luminex, オースチン社(Austin)、米国テキサス州)。こうして、図中の符号(B)に示すように、最大で100種類のものを混合した状態で、各ビーズの同定が行えるようになっている。
【0032】
複数種類のプローブを備えたマイクロビーズ(但し、各マイクロビーズには一種類のプローブのみ)を適当な割合で混合し、100ビーズ/μLとなるようにしたビーズミックスを調製した(図中の符号(C))。
本実施例においては、表4〜表6に記載の100種類のプローブを第1セット(セットA)とし、表7〜表9に記載の86種類のプローブを第2セット(セットB)として、2セットを用いることで、合計186種類の多型を検出可能な検出系とした。これら186種類のプローブを用いることにより、ヒトミトコンドリアDNAを少なくとも10種類のハプログループ(F,B,A,N9a,M7a,M7b,G1,G2,D4およびD5)に分類することができる。
【0033】
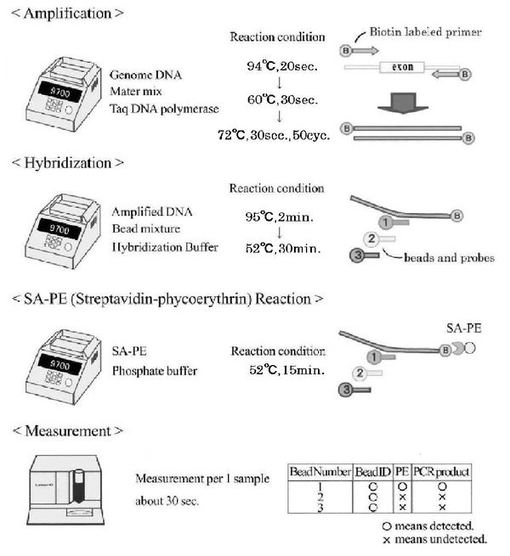
図2には、PCR−SSOP−Luminex法の手順の概要を示した。
<増幅反応(Amplification)>
目的とするDNAフラグメントを増幅するPCR反応には、5’末端をビオチン化したプライマーを用いた。mtDNAフラグメントを増幅するために28−plex PCRを行った。2mM塩化マグネシウムを含む1xPCR溶液(30mM KCl、20mM Tris−HCl、pH8.3)、2.5%DMSO、0.2mM dNTPs、0.1μMずつのプライマーセットを混合し、Taq DNAポリメラーゼ(0.625U)と1ngのゲノムDNAを加えて25μLとした。PCR反応は、95℃で10分間処理の後、94℃で20秒間の変性、60℃で30秒間のアニーリング、及び72℃で30秒間の伸長を1サイクルとし、これを50サイクル繰り返した。最後に、72℃で7分間の伸長反応を行った。機器としてGeneAmp9700サーマルサイクラー(アプライドバイオシステムズ社製)を用いた。
【0034】
<ハイブリダイゼーション(Hybridization)>
増幅したDNAを変性した後、ビーズミックスとハイブリダイズさせた。96ウエルプレートの各ウエルに、5μLの増幅反応後のPCR増幅液、5μLのビーズミックス、及び40μLのハイブリダイズ用緩衝液(1.5M TMAC、62.5mM TB(pH8.0)、0.5mM EDTA、0.125% N−ラウロイルザルコシン)を添加し、全量50μLとした。この混合液を添加した96ウエルプレートについて、95℃で2分間の変性、及び52℃で30分間のハイブリダイゼーションを行った(GeneAmp9700サーマルサイクラーを用いた。)。
図2中には、増幅したDNAを認識するプローブを有するビーズ(1)のみが、DNAと結合する様子が示されている。
【0035】
<ストレプトアビジン−フィコエリトリン反応(SA−PE Reaction)>
次に、上記ビーズミックス−DNAをSA−PEと反応させた。ハイブリッド工程の後、各ウエルに75μLのPBS−Tween(1xPBS(pH7.5)、0.01% Tween−20)を添加し、1000xgで5分間の遠心を行い、上清を取り去った後、再度100μLのPBS−Tweenを添加し、1000xgで5分間の遠心を行い、上清を取り去ることで、マイクロビーズを洗浄した。各ウエルに残ったマイクロビーズに、それぞれ70μLのSA−PE溶液(PBS−Tweenにより、市販品(G&Gサイエンス株式会社製)を100倍希釈したもの)を添加し混合した後、52℃で15分間の反応を行った(GeneAmp9700サーマルサイクラーを用いた。)。
図2中には、ビーズ(1)のプローブにのみビオチン化DNAが結合しているので、そのビオチンにSA−PEが結合する様子が示されている。
【0036】
<測定(Measurement)>
次に、反応後のサンプルはLuminex100を用いて、ビーズ種類の同定と、そのビーズにPEが結合しているか否かを判定した。測定は2種類のレーザを使用して行い、ビーズの種類は635nmレーザにより同定し、PE蛍光は532nmレーザを用いて定量した。オリゴビーズに結合したDNAは1測定あたり各々のビーズを最低50個ずつ測定し、定量されたPEの蛍光強度の中央値(MFI)を使用した。
図2中には、各ビーズ(1)〜(3)が同定され、かつビーズ(1)にのみPEが測定されたことから、ビーズ(1)に結合させたプローブが認識するDNAが増幅された様子が示されている。
【0037】
この方法に拠る遺伝子タイピングの精度を確かめるために、91個のDNAサンプルを材料に使用した。これらのDNAサンプルは、ミトコンドリアゲノムの全塩基配列が直接シーケンスによって既に決定されているものである。各例において、ルミネックスシーケンス特異性オリゴヌクレオチドハイブリッドアッセイシステム(PCR−SSOP−Luminex法)によって決定された遺伝子型は、直接シーケンスによって決定されたものと一致した。
【0038】
<統計的分析>
数量的な臨床データは、2型糖尿病患者群と対照群との間で、対応のないスチューデントt検定を用いて比較した。質的なデータは、カイ二乗検定によって比較した。
2型糖尿病と関連する多型は、交絡因子を含む多項ロジスティック回帰分析法により解析した。このとき、交絡因子については、年齢(age)、性別(gender:0=女性、1=男性)、喫煙状態(smoker:0=非喫煙、1=喫煙)、および各mtSNPの遺伝子型を独立変数とし、2型糖尿病を従属変数とした。P値、オッズ比(OR)、および95%信頼区間(CI)を計算した。特に説明しない限り、0.05未満のP値は統計上有意であると考えた。
【0039】
ハプログループを多重比較するために、ボンフェローニ(Bonferroni)の補正を適用した。我々は、10個のハプログループを調べたので、0.05を10で除して、0.005とした。こうして、0.005未満のP値を統計上有意であると見なした。
<結果>
2906名の被験者に関する基礎的データを表10に示した。
【0040】
【表10】
【0041】
表中において、最左欄は上段から順に、年齢(Age)、性別(女性/男性)(Gender)、BMI(Body mass index)、喫煙率(Smoker)、高血圧(Hypertension)、収縮期血圧(Systolic blood pressure)、拡張期血圧(Diastolic blood pressure)、高脂血症(Hypercholesterolemia)、全コレステロール(Total cholesterol)、中性脂肪(Triglycerides)、HDL−コレステロール(HDL-cholesterol)、空腹時血糖値(Fasting blood glucose)、ヘモグロビンAlc(Glycosylated hemoglobin)を意味している。また、最上段は左から順に、被験者全員(All subjects)における2型糖尿病患者群(Type 2 diabetes)の値と対照群(Controls)の値とP値(P value)、女性(Women)における2型糖尿病患者群の値と対照群の値とP値、男性(Men)における2型糖尿病患者群の値と対照群の値とP値とをそれぞれ示している。また、各値において、「±」が付してあるものは、平均値±SDで示した。喫煙率については、1日あたり10本以上の喫煙数がある者を喫煙者とした。高血圧については、収縮期血圧が140mmHg以上、または拡張期血圧が90mmHg以上、或いは両基準を満たす者、または高血圧薬を服用している者とした。高脂血症については、全コレステロール値が220mg/dL(5.72mmol/L)以上、または高脂血症剤を服用している者とした。
【0042】
年齢は2型糖尿病患者群において対照群よりも低かったが、BMI、収縮期および弛緩期血圧、トリグリセリド、絶食時血漿グルコース濃度、およびヘモグロビンAlc(HbA1c)比率は、対照群に比べると2型糖尿病患者群においては有意に高かった(P<0.05)。
年齢とHDLコレステロールは、対照群に比して2型糖尿病患者者の方が有意に低かった(P<0.05)。喫煙率、高血圧症、高コレステロール血症は、対照群に比較して2型糖尿病患者群では有意に高かった(P<0.05)。女性/男性の比率は、対照群に比して2型糖尿病患者群の方が有意に低かった(P<0.05)。
年齢、性別、喫煙状態を補正して多項ロジスティック回帰分析を行った結果を表11に示した。
【0043】
【表11】
【0044】
表中において、左欄より順に、変数(Variable)、P値(P value)、オッズ比(OR)、95%信頼区間(95%CI)を示している。また、上段より被験者全員(All subjects)における従来の危険因子(Conventional risk factors)である年齢(Age)・性別(Gender)・喫煙率(Smoking)と遺伝的危険因子(Genetic risk factors)であるハプログループF、N9aを、女性における従来の危険因子である年齢・喫煙率と遺伝的危険因子であるハプログループN9a、F、Aを、男性における従来の危険因子である年齢・喫煙率と遺伝的危険因子であるハプログループFをそれぞれ意味している。
【0045】
表に示す通り、P値が0.05未満で、ハプログループFを持つ被験者が、2型糖尿病罹患のリスクが有意に高いことが明らかとなった。これとは対照的に、ハプログループN9aを持つ被験者は、2型糖尿病に罹患するリスクが低い傾向を示した(P = 0.023, オッズ率 0.64(95% CI 0.43-0.94))。ハプログループN9aを持つ女性は、2型糖尿病罹患のリスクが有意に低かった(P = 0.0042, オッズ率 0.27(95% CI 0.10-0.62))。一方、ハプログループFおよびAを持つ女性は2型糖尿病罹患のリスクが高い傾向を示した(Fについては、P = 0.0165, オッズ率 1.78(95% CI 1.11-2.89);Aについては、P = 0.0405, オッズ率 1.67(95% CI 1.02-2.73))。ハプログループFを持つ男性は、2型糖尿病に罹患するリスクが高かった(P = 0.0482, オッズ率1.43(95% CI 1.00-2.05))。
我々は、ハプログループの多重比較について、0.005未満のP値が得られた場合に、その関連が有意であると考えた。この基準に基づくと、ハプログループFは、全ての被験者について2型糖尿病に対する感受性に有意に関与していた( P = 0.0028)。一方、女性について、ハプログループN9aは、2型糖尿病に対する抵抗性に有意に関与していた( P = 0.0042)。
表12には、ステップワイズ前向き選択法による解析結果を示した。
【0046】
【表12】
【0047】
表より、ハプログループFは全ての被験者について、2型糖尿病に対する危険因子であることが明らかとなった(P = 0.0035)。
女性の場合には、ハプログループN9aは2型糖尿病に対する防御因子であり(P = 0.0013)、ハプログループFとAは2型糖尿病に対して独立な危険因子である可能性が示された(Fについて、P = 0.0227;Aについて、P = 0.0374)。しかし男性の場合には、これらのハプログループのいずれも、2型糖尿病との関連性を示さなかった。
女性の場合、ハプログループN9a、F、およびAの2型糖尿病への抵抗性に対する総合的な寄与は0.0152(3つのハプログループの寄与率(R2)の和)であった。これはミトコンドリアゲノム多型が、特に女性においては、2型糖尿病への感受性に影響する重要な遺伝要因であることを示唆している。
表13には、mtSNPsのうち、2型糖尿病に対する保護因子或いは危険因子として関連するものを示した。
【0048】
【表13】
【0049】
表中、「*」は表1において示すように、10個のハプログループを代表させるために用いた多型マーカであることを示している。また、表中の太字で示したP値は、0.005未満であることを示している。
年齢および喫煙状態を補正した多項ロジスティック回帰分析の結果、ハプログループFを代表する3個のmtSNPである3970C>T、10310G>Aおよび13928G>C、ハプログループF1を代表するmtSNPである12406G>A、ハプログループF1b1aを代表するmtSNPである13928G>Cが、全ての被験者に関して、糖尿病と関連性を持つことが明らかになった(P = 0.0014 [10310G>A] , P = 0.0025 [12406G>A], P = 0.0028 [3970C>T], P=0.0028 [13928G>C]。これらは149 個のmtSNP の比較に対しての無修正値(ボンフェローニの補正を行っていない値)である)。個々のmtSNPについては、各ハプロタイプに対応していることから、ハプロタイプに適用した有意水準(P<0.005)で判断を行った。
【0050】
サブハプログループA1a1aを代表するmtSNPである14944C>T、サブハプログループF1b1aまたはN9b1を代表するmtSNPである5147G>A、サブハプログループB5b3を代表するmtSNPである8856G>A、およびサブハプログループF1b1aを代表するmtSNPである14476G>Aを保持する被験者は、0.05未満のP値で2型糖尿病に対する危険性が高かった(10個のハプログループにおけるサブハプログループを代表するmtSNPの有無については、表4〜表9に記載のプローブのいずれかによって測定されている。以下の記述中のサブハプログループを代表するサブハプログループについても同様である。)。
対照的に、サブハプログループN9a2a1を代表するmtSNPである15067T>C、サブハプログループN9aを代表する2個のmtSNPである5231G>Aおよび12372G>A、並びにサブハプログループD4aを代表するmtSNPである15314G>Aを保持する被験者は、0.05未満のP値で2型糖尿病に抵抗性を示した。
【0051】
女性については、年齢および喫煙状態を補正した多項ロジスティック回帰分析の結果、ハプログループN9aを代表する2個のmtSNPである5231G>Aおよび12372G>Aが、糖尿病に対する抵抗性に関係することが明らかになった( P = 0.0025 , P = 0.0035。これは149 個のmtSNP の比較に対しての無修正値である)。これに対し、サブハプログループA1a1aを代表するmtSNPである14944C>Tは、糖尿病に対する危険性が高かった(P=0.0035)。更に、サブハプログループF1を代表するmtSNPである12406G>A、ハプログループFを代表する3個のmtSNPである13928G>C、10310G>A、および3970C>T、サブハプログループA1aを代表するmtSNPである4655G>A、サブハプログループB5b3を代表するmtSNPである8856G>A、ハプログループAを代表する2個のmtSNPである663A>Gおよび8794C>Tを保持する被験者は、0.05未満のP値で2型糖尿病に対するリスクが高いことがわかった。
【0052】
これに対し、ハプログループN9aを代表するmtSNPである12358A>G、サブハプログループN9a2a1を代表するmtSNPである15067T>Cを持つ女性は、0.05未満のP値で2型糖尿病に対するリスクが低いことがわかった。
男性の場合には、年齢および喫煙状態を補正した多項ロジスティック回帰分析の結果、ハプログループFを代表するmtSNPである10310G>Aを持つと、0.05未満のP値で2型糖尿病に対するリスクが高いことがわかった。
表14に示すように、ステップワイズ前向き選択法によれば、ハプログループFを代表するmtSNPである10310G>Aが全被験者で2型糖尿病に対する危険因子であることが明らかになった(P = 0.0018)。
【0053】
【表14】
【0054】
表中、「*」は表1において示すように、10個のハプログループを代表させるために用いた多型マーカであることを示している。また、表中の太字で示したP値は、0.005未満であることを示している。
女性の場合には、ハプログループN9aを代表するmtSNPである5231G>Aが、2型糖尿病に対する防御因子であることが明らかになった(P = 0.0006)。他方、サブハプログループA1a1aを代表するmtSNPである14944C>Tは、2型糖尿病に対する危険因子であった(P = 0.0036)。7個のmtSNPs、すなわち5231G>A、14944C>T、12406G>A、8762T>C、8856G>A、12358A>G、および8383T>Cの糖尿病に対する感受性ならびに/若しくは防御性への総合的な寄与は0.0364(7個のmtSNPsの寄与率(R2)の和)であった。
【0055】
男性の場合には、サブハプログループF4aを代表するmtSNPである7861T>Cは、2型糖尿病に対する危険因子であった(P = 0.0005)。4個のmtSNPs、すなわち7861T>C、1391T>C、10310G>A、および15047G>Aの2型糖尿病に対する感受性への総合的な寄与は0.0101(4個のmtSNPsの寄与率(R2)の和)であった。
これらの結果は、上記のmtSNPsが、特に女性の場合には2型糖尿病に対する感受性に影響する重要な遺伝因子であることを示している。
【0056】
<考察>
我々は、2906名の被験者に基づく大規模な関連解析を行い、2型糖尿病と10個の主要なミトコンドリアハプログループとの間の関係を調査した。これらのハプログループのうち、ハプログループFは全ての被験者で2型糖尿病のリスクに有意に関係していた。女性の場合には、ハプログループN9aは、2型糖尿病に対する抵抗性に有意に関係していた。一方、ハプログループFおよびハプログループAは、2型糖尿病に対するリスクに関係していた。男性の場合には、ハプログループFは、2型糖尿病のリスクと関係していた。
【0057】
我々は、ミトコンドリアDNA配列の解析から、日本人集団がマクロハプログループNとMとに分割されることを報告した(非特許文献17)。マクロハプログループMは、10個の強固なmtSNPsがリンクしたマクロハプログループNとは異なっている。マクロハプログループNには、4個の主要なハプログループ、すなわちF、B、A、およびN9aが含まれる。他方、マクロハプログループMには、6個の主要なハプログループ、すなわちM7a、M7b、G1、G2、D4およびD5が含まれる。こうして、日本人集団は、これら10個の主なハプログループに分類される。
【0058】
ハプログループFに特異なmtSNPに、13928G>Cトランスバージョンは、潜在的な機能的多型の1つである。このmtSNPは、NADH脱水素酵素サブユニット5遺伝子産物(ND5)中で、S531T置換を起こすことが知られている。ND5はmtDNAにコードされたサブユニットの7個の内の一つであるが、これらのサブユニットは呼吸鎖複合体I(NADH:ユビキノン酸化還元酵素、EC1.6.5.3)の約41個のポリペプチドに含まれている。複合体Iは、NADHから電子を受け取り、それをユビキノン(補酵素Q10)に転送する。その際に得られたエネルギーを用いて、ミトコンドリア内膜を通してプロトンを輸送する。
【0059】
ND5は、複合体Iの機能には欠かすことができない。その理由は、ND5遺伝子中の様々なミスセンス変異が、レーバー遺伝性視神経萎縮症(LHON)(非特許文献22)、リー症候群(非特許文献25)、またはミトコンドリア性ミオパチー、脳症、乳酸アシドーシスおよび脳卒中様症状から成る症候群(MELAS)(非特許文献12)に罹患する患者において報告されているからである。従って、13928G>Cによって惹き起こされるS531T置換が、ND5サブユニットおよび複合体Iの機能に影響を及ぼすと考えることができる。13928G>C(ND5:S531T)がミトコンドリア機能に及ぼす具体的な作用については、このハプログループを伴うmtDNAを持つ細胞質雑種細胞系統を使う実験によって確認することができ得る。
【0060】
ハプログループFを持つ個人の代謝特性は、おそらくインスリン分泌ならびに/あるいはインスリン抵抗性に影響を与えることによって、2型糖尿病に対する抵抗性を得るものと思われる。ハプログループFに属するサブハプログループ(F1a、F1b、F1c、F2a、F3、F4a、およびF4b)の内、サブハプログループF1bは16189G>C多型によって定義される。この多型はハプログループBにおいても検出されること、およびハプログループBは本研究では糖尿病に関係しなかったことから、16189T>C多型は2型糖尿病とは関係しないと判断される。
【0061】
ハプログループN9aに特有のmtSNPsは、ほとんど同義突然変異である。これらの同義突然変異のうち、5231G>Aおよび12372G>Aは、本研究の遺伝子タイピングに使用した。このハプログループに於ける機能的な多型として考えられる候補は、150C>Tおよび338C>Tであるが、これらはミトコンドリアゲノムの非コード領域にある。150C>T置換は、イタリアの百寿者で最初に報告された(非特許文献23)。また我々は、この置換がフィンランドと日本の両国で共に健康的な長寿者と関連していることを報告している(非特許文献24)。このため、150C>Tは、女性の場合には2型糖尿病に対する抵抗性を授与するものと考えられる。
【0062】
ハプログループN9aに特異なmtSNPsのうちでは、12358A>GがND5サブユニットにT8A置換(8位のスレオニンをアラニンに置換する)を惹き起こすことから、潜在的な機能的な多型であると考えられる。12358A>Gによって惹き起こされたT8A置換が、ND5サブユニットおよび複合体Iの機能に影響を与えるかも知れない。12358A>G(ND5:T8A)がミトコンドリア機能に与える具体的な作用については、細胞実験によって調査することができ得る。150C>Tと12358A>Gの両者を備えたハプログループN9aを持つ女性の代謝特性は、2型糖尿病に対する抵抗性の機構を理解するために更なる調査が望まれる。
【0063】
本研究において、女性の場合には、ハプログループAが2型糖尿病への感受性と関連を持つことを見出した。ハプログループAは8794C>T突然変異を特徴とするが、この変異はATPaseサブユニット6においてH90Y置換(90位のヒスチジンをチロシンに置換する)を惹き起こす。FoF1−ATPaseは、核DNAにコードされた10−16個のサブユニットおよびmtDNAにコードされた2個のサブユニット(ATPase6およびATPase8)から構成される。ATP6遺伝子は、サブユニットaをコードする。サブユニットaは、ATPaseのFoセクタを横断してプロトンを輸送する際に重要な役割を果たす。L156R置換を惹き起こす8993T>Gトランスバージョン、およびL156P置換を惹き起こす8993T>Cトランジションは、NARP(神経障害、運動失調、および色素視神経網膜炎)症候群(非特許文献25、26)の原因となる病原性突然変異である。従って、8794C>Tによって惹き起こされたH90Y置換がATP6サブユニットとFoF1−ATPase(複合体V)の機能に影響を与える可能性がある。
【0064】
ミトコンドリア機能および代謝に対する8794C>T(ATP6:H90Y)の実際の効果については、この多型と2型糖尿病に対する感受性との関係を理解するために更なる調査が望まれる。ミトコンドリアには組換え機構が存在しないので、各ハプログループあるいはサブハプログループに於けるmtSNPsの全ては、相互に強く連鎖している。従って、本研究で調べたmtSNPsのいくつかが連鎖不均衡にあって、2型糖尿病に対する感受性若しくは抵抗性を実際に担う他のミトコンドリア遺伝子の機能的な多型にそのmtSNPが連鎖している可能性がある。女性の場合、ハプログループN9aは2型糖尿病に対する抵抗性に関係していたが、この関係は男性の場合には検出されなかった。
【0065】
遺伝的変異と肥満との関係は、男性の場合に比べると女性の方が強いという考えを支持する他の証拠がある(非特許文献27、28)。Comuzzieらは、性相互作用により遺伝子型を導入し、脂肪重量をメンデル混合モデルにて分析したところ、男性では脂肪重量の分散値全体の37%を主な遺伝子が担い、女性では43%を担っていた(非特許文献28)。これまでに、核遺伝子多型と糖尿病との間の関係についての関連解析がいくつか認められる。グルコキナーゼをコードする遺伝子の多型(非特許文献29)、インスリン受容体基質1(非特許文献30)、ペルオキシソーム増殖因子活性性受容体γ(非特許文献31)、カルパイン−10(非特許文献32)、およびアディポネクチン(非特許文献33)は、糖尿病に関連するものと報告されている。
しかしながら、ミトコンドリアDNA多型と2型糖尿病との関係に対する性別の果たす影響に関しては、今までのところほとんど報告が見られない。本明細書が、初めての報告である。
【0066】
このように、本実施形態によれば、mtDNAのハプログループを検出することにより、2型糖尿病の遺伝的危険度を予測することができる。このことにより、2型糖尿病の予防が可能となり、高齢者の健康寿命延長・QOL向上・ねたきり防止ならびに今後の医療費削減など、医学的・社会的に大きく貢献できる。
【0067】
【非特許文献1】van den Ouweland JM, Lemkes HH, Ruitenbeek W, Sandkuijl LA, de Vijlder MF, Struyvenberg PA, van de Kamp JJ, Maassen JA: Mutation in mitochondrial tRNA(Leu)(UUR) gene in a large pedigree with maternally transmitted type II diabetes mellitus and deafness. Nat Genet 1:368-371, 1992
【非特許文献2】Guo LJ, Oshida Y, Fuku N, Takeyasu T, Fujita Y, Kurata M, Sato Y, Ito M, Tanaka M: Mitochondrial genome polymorphisms associated with type-2 diabetes or obesity. Mitochondrion 5:15-33, 2005
【非特許文献3】Kokaze A, Ishikawa M, Matsunaga N, Yoshida M, Makita R, Satoh M, Teruya K, Sekiguchi K, Masuda Y, Harada M, Uchida Y, Takashima Y: Longevity-associated mitochondrial DNA 5178 C/A polymorphism is associated with fasting plasma glucose levels and glucose tolerance in Japanese men. Mitochondrion 5:418-425, 2005
【非特許文献4】Okura T, Koda M, Ando F, Niino N, Tanaka M, Shimokata H: Association of the mitochondrial DNA 15497G/A polymorphism with obesity in a middle-aged and elderly Japanese population. Hum Genet 113:432-436, 2003
【非特許文献5】Lowell BB, Shulman GI: Mitochondrial dysfunction and type 2 diabetes. Science 307:384-387, 2005
【非特許文献6】Macaulay V, Hill C, Achilli A, Rengo C, Clarke D, Meehan W, Blackburn J, Semino O, Scozzari R, Cruciani F, Taha A, Shaari NK, Raja JM, Ismail P, Zainuddin Z, Goodwin W, Bulbeck D, Bandelt HJ, Oppenheimer S, Torroni A, Richards M: Single, rapid coastal settlement of Asia revealed by analysis of complete mitochondrial genomes. Science 308:1034-1036, 2005
【非特許文献7】Wallace DC: A mitochondrial paradigm of metabolic and degenerative diseases, aging, and cancer: a dawn for evolutionary medicine. Annu Rev Genet 39:359-407, 2005
【非特許文献8】Mishmar D, Ruiz-Pesini E, Golik P, Macaulay V, Clark AG, Hosseini S, Brandon M, Easley K, Chen E, Brown MD, Sukernik RI, Olckers A, Wallace DC: Natural selection shaped regional mtDNA variation in humans. Proc Natl Acad Sci U S A 100:171-176, 2003
【非特許文献9】Ruiz-Pesini E, Mishmar D, Brandon M, Procaccio V, Wallace DC: Effects of purifying and adaptive selection on regional variation in human mtDNA. Science 303:223-226, 2004
【非特許文献10】Montiel-Sosa F, Ruiz-Pesini E, Enriquez JA, Marcuello A, Diez-Sanchez C, Montoya J, Wallace DC, Lopez-Perez MJ: Differences of sperm motility in mitochondrial DNA haplogroup U sublineages. Gene 368C:21-27, 2006
【非特許文献11】Baudouin SV, Saunders D, Tiangyou W, Elson JL, Poynter J, Pyle A, Keers S, Turnbull DM, Howell N, Chinnery PF: Mitochondrial DNA and survival after sepsis: a prospective study. Lancet 366:2118-2121, 2005
【非特許文献12】Pyle A, Foltynie T, Tiangyou W, Lambert C, Keers SM, Allcock LM, Davison J, Lewis SJ, Perry RH, Barker R, Burn DJ, Chinnery PF: Mitochondrial DNA haplogroup cluster UKJT reduces the risk of PD. Ann Neurol 57:564-567, 2005
【非特許文献13】Tanaka M, Gong JS, Zhang J, Yoneda M, Yagi K: Mitochondrial genotype associated with longevity. Lancet 351:185-186, 1998
【非特許文献14】Ikebe S, Tanaka M, Ozawa T: Point mutations of mitochondrial genome in Parkinson's disease. Brain Res Mol Brain Res 28:281-295, 1995
【非特許文献15】Tanaka M: Mitochondrial genotypes and cytochrome b variants associated with longevity or Parkinson's disease. J Neurol 249 Suppl 2:II11-18, 2002
【非特許文献16】Fuku N, Oshida Y, Takeyasu T, Guo LJ, Kurata M, Yamada Y, Sato Y, Tanaka M: Mitochondrial ATPase subunit 6 and cytochrome b gene polymorphisms in young obese adults. Biochem Biophys Res Commun 290:1199-1205, 2002
【非特許文献17】Tanaka M, Cabrera VM, Gonzalez AM, Larruga JM, Takeyasu T, Fuku N, Guo LJ, Hirose R, Fujita Y, Kurata M, Shinoda K, Umetsu K, Yamada Y, Oshida Y, Sato Y, Hattori N, Mizuno Y, Arai Y, Hirose N, Ohta S, Ogawa O, Tanaka Y, Kawamori R, Shamoto-Nagai M, Maruyama W, Shimokata H, Suzuki R, Shimodaira H: Mitochondrial genome variation in eastern Asia and the peopling of Japan. Genome Res 14:1832-1850, 2004
【非特許文献18】Poulton J, Brown MS, Cooper A, Marchington DR, Phillips DI: A common mitochondrial DNA variant is associated with insulin resistance in adult life. Diabetologia 41:54-58, 1998
【非特許文献19】Weng SW, Liou CW, Lin TK, Wei YH, Lee CF, Eng HL, Chen SD, Liu RT, Chen JF, Chen IY, Chen MH, Wang PW: Association of mitochondrial DNA 16189 variant (T->C transition) with metabolic syndrome in Chinese adults. J Clin Endocrinol Metab 90:5037-5040, 2005
【非特許文献20】Chinnery PF, Elliott HR, Patel S, Lambert C, Keers SM, Durham SE, McCarthy MI, Hitman GA, Hattersley AT, Walker M: Role of the mitochondrial DNA 16184-16193 poly-C tract in type 2 diabetes. Lancet 366:1650-1651, 2005
【非特許文献21】Itoh Y, Mizuki N, Shimada T, Azuma F, Itakura M, Kashiwase K, Kikkawa E, Kulski JK, Satake M, Inoko H: High-throughput DNA typing of HLA-A, -B, -C, and -DRB1 loci by a PCR-SSOP-Luminex method in the Japanese population. Immunogenetics 57:717-729, 2005
【非特許文献22】Howell N, Halvorson S, Burns J, McCullough DA, Paulton J: When does bilateral optic atrophy become Leber hereditary optic neuropathy? Am J Hum Genet 53:959-963, 1993
【非特許文献23】Zhang J, Asin-Cayuela J, Fish J, Michikawa Y, Bonafe M, Olivieri F, Passarino G, De Benedictis G, Franceschi C, Attardi G: Strikingly higher frequency in centenarians and twins of mtDNA mutation causing remodeling of replication origin in leukocytes. Proc Natl Acad Sci U S A 100:1116-1121, 2003
【非特許文献24】Niemi AK, Moilanen JS, Tanaka M, Hervonen A, Hurme M, Lehtimaki T, Arai Y, Hirose N, Majamaa K: A combination of three common inherited mitochondrial DNA polymorphisms promotes longevity in Finnish and Japanese subjects. Eur J Hum Genet 13:166-170, 2005
【非特許文献25】Holt IJ, Harding AE, Petty RK, Morgan-Hughes JA: A new mitochondrial disease associated with mitochondrial DNA heteroplasmy. Am J Hum Genet 46:428-433, 1990
【非特許文献26】Tanaka M, Borgeld HJ, Zhang J, Muramatsu S, Gong JS, Yoneda M, Maruyama W, Naoi M, Ibi T, Sahashi K, Shamoto M, Fuku N, Kurata M, Yamada Y, Nishizawa K, Akao Y, Ohishi N, Miyabayashi S, Umemoto H, Muramatsu T, Furukawa K, Kikuchi A, Nakano I, Ozawa K, Yagi K: Gene therapy for mitochondrial disease by delivering restriction endonuclease SmaI into mitochondria. J Biomed Sci 9:534-541, 2002
【非特許文献27】Borecki IB, Bonney GE, Rice T, Bouchard C, Rao DC: Influence of genotype-dependent effects of covariates on the outcome of segregation analysis of the body mass index. Am J Hum Genet 53:676-687, 1993
【非特許文献28】Comuzzie AG, Blangero J, Mahaney MC, Mitchell BD, Hixson JE, Samollow PB, Stern MP, MacCluer JW: Major gene with sex-specific effects influences fat mass in Mexican Americans. Genet Epidemiol 12:475-488, 1995
【非特許文献29】Vionnet N, Stoffel M, Takeda J, Yasuda K, Bell GI, Zouali H, Lesage S, Velho G, Iris F, Passa P, et al.: Nonsense mutation in the glucokinase gene causes early-onset non-insulin-dependent diabetes mellitus. Nature 356:721-722, 1992
【非特許文献30】Almind K, Bjorbaek C, Vestergaard H, Hansen T, Echwald S, Pedersen O: Aminoacid polymorphisms of insulin receptor substrate-1 in non-insulin-dependent diabetes mellitus. Lancet 342:828-832, 1993
【非特許文献31】Altshuler D, Hirschhorn JN, Klannemark M, Lindgren CM, Vohl MC, Nemesh J, Lane CR, Schaffner SF, Bolk S, Brewer C, Tuomi T, Gaudet D, Hudson TJ, Daly M, Groop L, Lander ES: The common PPARgamma Pro12Ala polymorphism is associated with decreased risk of type 2 diabetes. Nat Genet 26:76-80, 2000
【非特許文献32】Horikawa Y, Oda N, Cox NJ, Li X, Orho-Melander M, Hara M, Hinokio Y, Lindner TH, Mashima H, Schwarz PE, del Bosque-Plata L, Horikawa Y, Oda Y, Yoshiuchi I, Colilla S, Polonsky KS, Wei S, Concannon P, Iwasaki N, Schulze J, Baier LJ, Bogardus C, Groop L, Boerwinkle E, Hanis CL, Bell GI: Genetic variation in the gene encoding calpain-10 is associated with type 2 diabetes mellitus. Nat Genet 26:163-175, 2000
【非特許文献33】Hara K, Boutin P, Mori Y, Tobe K, Dina C, Yasuda K, Yamauchi T, Otabe S, Okada T, Eto K, Kadowaki H, Hagura R, Akanuma Y, Yazaki Y, Nagai R, Taniyama M, Matsubara K, Yoda M, Nakano Y, Tomita M, Kimura S, Ito C, Froguel P, Kadowaki T: Genetic variation in the gene encoding adiponectin is associated with an increased risk of type 2 diabetes in the Japanese population. Diabetes 51:536-540, 2002
【図面の簡単な説明】
【0068】
【図1】Luminex100で検出するマイクロビーズの微細構造と特徴を示す図である。
【図2】PCR−SSOP−Luminex法の手順の概要を示す図である。
【図3】mtSNPに基づくハプログループ分類を示す図(1)である。図の下端には、ハプログループ及びサブハプログループを示している(図4及び図4においても同じである)。
【図4】mtSNPに基づくハプログループ分類を示す図(2)である。
【図5】mtSNPに基づくハプログループ分類を示す図(3)である。
【技術分野】
【0001】
本発明は、2型糖尿病に関する遺伝子検出方法に関し、特にヒトミトコンドリアDNAのハプログループを分類することにより2型糖尿病に関する遺伝子検出を行うものに関する。
【背景技術】
【0002】
インスリン非依存性の2型糖尿病は、先進国において高い発症率を示し、世界的規模で公衆衛生上の主要な問題の一つになっている。2型糖尿病は、複合的な要因に由来する疾患であり、そこでは様々な遺伝的または環境的な影響が相互に重要な役割を果たしている。若年発症性の成人型糖尿病(maturity-onset diabetes of the young :MODY)を引き起こす病原性突然変異ついては、既にいくつかの報告がある。
【0003】
一方、ミトコンドリアゲノムにおいて、tRNA−Leu(UUR)遺伝子中の3243A>G突然変異が、母系遺伝性糖尿病ならびに難聴の患者(非特許文献1。本明細書においては、関連文献については末尾にまとめて示す。)で検出された。更に、ミトコンドリアゲノム多型の幾つかは、日本人集団においてはインスリン抵抗性および/または高血糖と関連を持つことが報告されている(非特許文献2−4)。このように、ミトコンドリアの機能障害は、骨格筋のインシュリン抵抗性、および膵臓のベータ細胞からのインスリン分泌障害の両者に関連していると考えられている(非特許文献5)。
【0004】
ミトコンドリアゲノム中の母系遺伝性の変異は、過去二十万年以上に渡り、ホモサピエンスがアフリカからアジアおよびヨーロッパに拡張した時代に、新たな突然変異が連続的に追加された事に起因している(非特許文献6)。ミトコンドリアゲノムの突然変異率は核ゲノムの突然変異率に比べると少なくとも10倍以上高いので、ホモサピエンスに蓄積されて来た古代のミトコンドリア多型は、比較的短い歴史を持つものではあっても、現代人類の代謝特性に影響している可能性がある(非特許文献7)。世界の様々な地域で発見されたミトコンドリアハプログループが、寒冷気候と栄養条件への適応を通じて選択されて来たものであると推測することができる(非特許文献7−9)。ミトコンドリアハプログループ間で機能的差異が存在することを支持する数々の証拠が、精子運動性(非特許文献10)、敗血症後の生存率(非特許文献11)、およびパーキンソン病に対する感受性(非特許文献12)の研究により与えられている。
【0005】
これまで本発明者らは、寿命(非特許文献13)、パーキンソン病(非特許文献14、15)およびアルツハイマー病と言った加齢関連の疾病、並びに肥満(非特許文献2)、るい痩(非特許文献16)、2型糖尿病(非特許文献2)および動脈硬化症と言ったエネルギー代謝に関連する疾病に関与するミトコンドリアDNA多型(mtSNP)を同定することを目的として研究を重ねてきた。この目的のため、我々は7つの異なる群に属する672名について、16569塩基対のミトコンドリアゲノムを完全にシーケンスした。7群は、それぞれ96名から構成されており、それらは、百寿者群、パーキンソン病罹患者群、アルツハイマー病患者群、肥満の若い男性群、肥満でない若い男性群、重度の血管性病変を持つ2型糖尿病患者群、および重度の血管性病変を持たない2型糖尿病患者群であった(非特許文献17)。この調査結果から、ヒトミトコンドリアゲノム多型データベース(http://www.giib.or.jp/mtsnp/index_e.shtml)を構築した。これらのmtSNPデータに基づいて、本発明者らは、蛍光ビーズ法を用いた包括的なmtSNP分析システムを開発した。
【0006】
Poultonらは、mtDNAの16189T>C変異が2型糖尿病と関連を持つことを既に報告している(非特許文献18)。Wengらは、この多型が中国人成人においては、メタボリックシンドロームとも関連していると報告している(非特許文献19)。しかしながら、ヨーロッパ人に関する研究では、16184−16193のpoly(C)tractの遺伝変異が、2型糖尿病発症においては主要な役割を担っていないことが示されている(非特許文献20)。従って、非コード領域のみに於ける多型分析は、個々人の代謝特性とミトコンドリアゲノムに於ける機能的多型との間の相互関係については、直接的な証拠を提供し難いと思われる。
【発明の開示】
【発明が解決しようとする課題】
【0007】
上記の諸点に留意しつつ、本発明者らは、ミトコンドリアゲノムのコード領域に於ける包括的な多型分析を基づいて、日本人に於ける2型糖尿病と主要な10個のハプログループ(F,B,A,N9a,M7a,M7b,G1,G2,D4,およびD5)との関係を調べる大規模な関連解析を行った。
本発明者らの研究目的は、mtDNAのハプログループ分けに基づき、2型糖尿病の遺伝的リスクを予測し、それによって2型糖尿病の一次予防に役立てることにある。
【課題を解決するための手段】
【0008】
本発明者らは、長年に亘ってヒトミトコンドリアゲノムに関する研究を行っている者である。ヒトミトコンドリアDNAは、強固に連鎖した複数のmtSNPを有するハプログループに分類することができる。図3〜図5には、mtSNPとハプログループ(及びサブハプログループ)との関係を示した。このハプログループの分布は、人種間において、かなり大きな隔たりがある。個々の日本人は、主要な10個のハプログループによって、そのほとんど(約90パーセント以上)を分類することができる。各ハプログループは、数個〜20個程度のmtSNPによって特徴付けられる。例えば、ハプログループFは、8701、9540、10398、10873、15301、16519、12705、16223、16183、16519、3970、13928、16304、248、6392、10310等の位置にmtSNPを有している(その他のハプログループについても、同様の状況が存在する)。ハプログループ分類をするために、これら全てのmtSNPを調べようとするのは、大変な労力が掛かるため大規模な研究には向きにくい。しかしながら、これらのうちのいずれのmtSNPを選択・検出すれば、適切にハプログループを分類することが可能であるかについては明確には知られていなかったことに加え、選択したmtSNPを迅速かつ的確に検出する方法については開発が十分に進んでいなかった。
【0009】
そこで、本発明者らは、鋭意検討の結果、新たなmtSNPの検出法を開発し、日本人における主要な10個のハプログループ(F,B,A,N9a,M7a,M7b,G1,G2,D4及びD5)を迅速かつ的確に分類する検出法を開発した。
つまり、各ハプログループが持っている数個〜二十個程度のmtSNPのうち、適当に1個〜5個のmtSNPを選択することにより、表1に記載のmtSNPsを検出すれば、10個のハプログループへの分類が可能であることを見出した。更に、この方法を用いて、本発明者らは、ミトコンドリアゲノムのコード領域における包括的な多型分析を基にして、日本人における2型糖尿病と主要な10個のハプログループ(F,B,A,N9a,M7a,M7b,G1,G2,D4およびD5)との関係を調べるために、2906名を対象とした大規模な関連解析を行った。その結果、ハプログループFを持つ者は2型糖尿病の危険性が高いこと、ハプログループN9aを持つ者は2型糖尿病に対する抵抗性を有すること(特に、女性の場合)、および女性の場合にはハプログループAを持つ者は2型糖尿病の危険性が高いことを明らかとし、基本的には本発明を完成するに至った。
【0010】
こうして、上記課題を解決するために、本発明に係る2型糖尿病に関する遺伝子検出法は、ヒトミトコンドリアDNAにおいて、5231A、12358G、12372Aを有するハプログループN9aと、3970T、13928C、10310Aを有するハプログループFと、663G、8794Tを有するハプログループAと、12406Aを有するサブハプログループF1と、14944Tを有するサブハプログループA1a1aと、15067Cを有するサブハプログループN9a2a1と、5147Aを有するF1b1aと、5147Aを有するサブハプログループN9b1と、8856Aを有するサブハプログループB5b3と、14476Aを有するサブハプログループF1b1aと、15314Aを有するサブハプログループD4aと、4655Aを有するサブハプログループA1aと、8856Aを有するサブハプログループB5b3と、1391Cを有するサブハプログループB4b1bと、8762Cを有するサブハプログループD4mと、8383Cを有するサブハプログループD4d2と、7861Cを有するサブハプログループF4aと、15047Aを有するサブハプログループM8a1とを検出することを特徴とする。
【0011】
本明細書中において、多型の記載方法は、次の通りである。原則として、各遺伝子について、「多型が生じている位置、データベースに登録されている塩基(A:アデニン、G:グアニン、C:シトシン、T:チミン)>多型塩基」の順で記載する。例えば、「5231G>A」は、5231位のGがAとなっている多型を意味している。また、その多型が生じている遺伝子(rRNAまたはmRNA)の位置について、カッコ書きで情報を示すことがある。例えば、「5231G>A(ND2:syn)」とは、上記多型について、ND2遺伝子において、アミノ酸の変異を伴わない多型(synonymous mutation)であることを意味している。但し、挿入あるいは欠失多型については、「多型が生じている位置(欠失または挿入に関する塩基情報)」の順で記載する。例えば、「8272(9-bp deletion in non-coding region)」とは、8272位の非コード領域における9個の塩基の欠失多型であることを意味している。また、順方向(forward strand)に読んでA/G多型としてデータベースに記録されている場合であっても、逆方向(reverse strand)で読むとT/C多型となる。多くの文献について、「T/C」多型として記載されている場合には、データベース中の記録であるA/G多型と異なることがある。
【0012】
一般にミトコンドリアDNA多型(mtSNPs)或いはミトコンドリアDNAハプログループは、対象となる集団(例えば、日本人・中国人・韓国人を含む東アジア人集団、西洋人集団など)が異なると、その種類・頻度が異なる。このため、日本人以外の集団において、2型糖尿病との関係が指摘されているハプログループであっても、必ずしも日本人集団においてそのような関連が認められるわけではない。このため、従来の報告については、集団または疾患が異なる場合には、必ずしも日本人におけるハプログループおよび2型糖尿病との関連が裏付けられるわけではない。
【0013】
本発明において、mtDNAのハプログループを検出する方法としては、従来公知の方法、例えばDNAシークエンス、DNAチップ(或いはDNAマイクロアレイ)、RFLP、PCR−SSOP−Luminex法などの方法を用いることができる。これらの中でも、PCR−SSOP−Luminex法を用いることが好ましい。PCR−SSOP−Luminex法を用いることにより、数百個程度の多型であれば迅速に測定することができる。なお、同方法の詳細については後に詳述する。
【発明の効果】
【0014】
本発明によれば、mtDNAのハプログループを検出することにより、2型糖尿病の遺伝的危険度を予測することができる。この発明を用いることにより、2型糖尿病に対する予防が可能となり、高齢者の健康寿命延長・QOL向上・ねたきり防止ならびに今後の医療費削減など、医学的・社会的に大きく貢献できる。
【発明を実施するための最良の形態】
【0015】
次に、本発明の実施形態について、図面を参照しつつ説明するが、本発明の技術的範囲は、これらの実施形態によって限定されるものではなく、発明の要旨を変更することなく様々な形態で実施することができる。また、本発明の技術的範囲は、均等の範囲にまで及ぶものである。
【0016】
<研究対象>
研究対象は、互いに血縁関係の無い40歳以上の日本人2906名(男性1938名、女性968名)であった。対象者は、2002年10月から2005年3月の間に、3つの研究参加病院(岐阜県立岐阜病院、多治見病院および下呂温泉病院)の1つを外来した受診者、或いは入院患者であった。2型糖尿病患者は、絶食時血漿グルコースレベルが6.99mmol/L(126mg/dL)以上、またはヘモグロビンAlc(Glycosylated hemoglobin)濃度が6.5%以上を示す者、或いは糖尿病に対する薬物治療下にあった。2型糖尿病は、世界保健機構により承認された基準に従って定義した。これらの基準に基づいて、研究対象集団中の1289名の被験者(男性890名、女性399名)が、2型糖尿病患者であると診断された。残りの1617名(男性1048名、女性569名)は対照群とした。対照群は、研究参加病院に例年の健康チェックのために外来受診した者である。対照群は、絶食時血漿グルコースレベルが6.11mmol/L(110mg/dL)未満であり、ヘモグロビンAlcが5.6%未満を示し、かつ2型糖尿病の既往歴ならびに2型糖尿病に対する薬物治療を受けた経験が無い者であった。
研究プロトコルは、岐阜県国際バイオ研究所のヒト研究に関する倫理委員会により承認され、また各参加者からは個別に書面によるインフォームドコンセントを得た。
【0017】
<ハプログループ分類のためのミトコンドリア多型の選択>
本発明者らが作製したヒトミトコンドリアゲノム多型データベース(http://www.giib.or.jp/mtsnp/index_e.shtml)および日本人の系統樹(非特許文献17)を使用して、ミトコンドリアハプログループの分類に有用である149個の多型部位を選択した。更に、本発明者らの鋭意検討に基づき、日本人集団において発見された10個の主要なハプログループ(F,B,A,N9b,M7a,M7b,G1,G2,D4,およびD5)を定義するmtSNPsを選択した。これらのmtSNPsを表1に示した。
【0018】
【表1】
【0019】
表中、左欄より順に、ハプログループ(Haplogroup)、および各ハプログループを代表する多型(Polymorphism)を示している。また表中、「Syn」はアミノ酸の変異を伴わない多型(synonymous mutation)であることを示している。
本発明者らは、上記2906名の被験者について、これらハプログループと2型糖尿病との間の関係について研究を行った。
【0020】
<多型の遺伝子タイピング>
各参加者から50mmol/L EDTA(ジナトリウム塩)を含む試験管に、7mLの静脈血を採取し、市販のキット(Genomix, Talent, Trieste, Italy)を用いてDNAを分離した。
DNAをPCRにかけ、特定のDNAを増幅した。その際に使用したプライマーを表2および表3に示した。ミトコンドリアの多型は懸濁液アレイ技術(Luminex 100; Luminex, Austin, TX)を用い、シーケンス特異性オリゴヌクレオチド試験法(G&Gサイエンス株式会社、日本)で決定した。詳細な方法については、既報のもの(非特許文献21)を基本として、適宜に増幅条件を変えて行った。なお、条件については、後に詳述する。
【0021】
また、ハプロタイピングのために使用したプライマーは、表4〜表9に示した。今回のタイピングには、最大で100種類の多型の判定が行えるものを用いた。186種類のプローブを用いるために、全体を100種類のセット(プローブセットA)と86種類のセット(プローブセットB)とに分けて、タイピングを行った。表4〜表6にはプローブセットAの詳細を、表7〜表9にはプローブセットBの詳細を示した。
【0022】
【表2】
【0023】
【表3】
【0024】
【表4】
【0025】
【表5】
【0026】
【表6】
【0027】
【表7】
【0028】
【表8】
【0029】
【表9】
【0030】
表2および表3については、左欄よりPCR法によって合成されるフラグメント番号(Fragment)、フォワードプライマー(Forward primer)のmtDNAにおける位置(Position)と塩基配列(Sequence)、リバースプライマー(Reverse primer)のmtDNAにおける位置(Position)と塩基配列、および合成されるDNAフラグメント長(Product)を示している。
表4〜表6については、第1のセット(プローブセットA)について、mtDNAにおける位置(Position)、目的(Purpose)、プローブの塩基配列(Sequence)を示している。また、目的の欄については、「a」が多型チェック用、「b」が野生型チェック用、「p」がPCR産物チェック用、「m」および「n」がマクロハプログループチェック用のものであることを示している。表7〜表9については、第2のセット(プローブセットB)について、上記と同じ意味である。
表4〜表9に示したプローブセットは、前述の10個のハプログループを分類するmtSNPに加えて、更に細かいサブハプログループを検出するためのプローブを含んでいる。
【0031】
PCR−SSOP−Luminex法
方法の詳細については、非特許文献21に記載の通りである。以下には、この方法の概要について説明する。
図1には、Luminex100フローサイトメトリーで検出するマイクロビーズ(本発明における固相に該当する)の微細構造と特徴を示した。マイクロビーズ(図中の符号(A))は、直径が約5.5μm程度であり、ポリスチレン製である。ビーズ表面にはカルボキシル基が存在している。このカルボキシル基と、アミノ修飾された核酸プローブとを結合することにより、ビーズ表面に核酸プローブが結合されている(本発明における核酸プローブの固相化に該当する)。各ビーズには、一種類の核酸プローブが結合されている。このマイクロビーズには、赤色色素と赤外色素との二種類の色素が割合を変化させつつ配されている(Multi-Analyte Microsphere Carboxylated: Luminex, オースチン社(Austin)、米国テキサス州)。こうして、図中の符号(B)に示すように、最大で100種類のものを混合した状態で、各ビーズの同定が行えるようになっている。
【0032】
複数種類のプローブを備えたマイクロビーズ(但し、各マイクロビーズには一種類のプローブのみ)を適当な割合で混合し、100ビーズ/μLとなるようにしたビーズミックスを調製した(図中の符号(C))。
本実施例においては、表4〜表6に記載の100種類のプローブを第1セット(セットA)とし、表7〜表9に記載の86種類のプローブを第2セット(セットB)として、2セットを用いることで、合計186種類の多型を検出可能な検出系とした。これら186種類のプローブを用いることにより、ヒトミトコンドリアDNAを少なくとも10種類のハプログループ(F,B,A,N9a,M7a,M7b,G1,G2,D4およびD5)に分類することができる。
【0033】
図2には、PCR−SSOP−Luminex法の手順の概要を示した。
<増幅反応(Amplification)>
目的とするDNAフラグメントを増幅するPCR反応には、5’末端をビオチン化したプライマーを用いた。mtDNAフラグメントを増幅するために28−plex PCRを行った。2mM塩化マグネシウムを含む1xPCR溶液(30mM KCl、20mM Tris−HCl、pH8.3)、2.5%DMSO、0.2mM dNTPs、0.1μMずつのプライマーセットを混合し、Taq DNAポリメラーゼ(0.625U)と1ngのゲノムDNAを加えて25μLとした。PCR反応は、95℃で10分間処理の後、94℃で20秒間の変性、60℃で30秒間のアニーリング、及び72℃で30秒間の伸長を1サイクルとし、これを50サイクル繰り返した。最後に、72℃で7分間の伸長反応を行った。機器としてGeneAmp9700サーマルサイクラー(アプライドバイオシステムズ社製)を用いた。
【0034】
<ハイブリダイゼーション(Hybridization)>
増幅したDNAを変性した後、ビーズミックスとハイブリダイズさせた。96ウエルプレートの各ウエルに、5μLの増幅反応後のPCR増幅液、5μLのビーズミックス、及び40μLのハイブリダイズ用緩衝液(1.5M TMAC、62.5mM TB(pH8.0)、0.5mM EDTA、0.125% N−ラウロイルザルコシン)を添加し、全量50μLとした。この混合液を添加した96ウエルプレートについて、95℃で2分間の変性、及び52℃で30分間のハイブリダイゼーションを行った(GeneAmp9700サーマルサイクラーを用いた。)。
図2中には、増幅したDNAを認識するプローブを有するビーズ(1)のみが、DNAと結合する様子が示されている。
【0035】
<ストレプトアビジン−フィコエリトリン反応(SA−PE Reaction)>
次に、上記ビーズミックス−DNAをSA−PEと反応させた。ハイブリッド工程の後、各ウエルに75μLのPBS−Tween(1xPBS(pH7.5)、0.01% Tween−20)を添加し、1000xgで5分間の遠心を行い、上清を取り去った後、再度100μLのPBS−Tweenを添加し、1000xgで5分間の遠心を行い、上清を取り去ることで、マイクロビーズを洗浄した。各ウエルに残ったマイクロビーズに、それぞれ70μLのSA−PE溶液(PBS−Tweenにより、市販品(G&Gサイエンス株式会社製)を100倍希釈したもの)を添加し混合した後、52℃で15分間の反応を行った(GeneAmp9700サーマルサイクラーを用いた。)。
図2中には、ビーズ(1)のプローブにのみビオチン化DNAが結合しているので、そのビオチンにSA−PEが結合する様子が示されている。
【0036】
<測定(Measurement)>
次に、反応後のサンプルはLuminex100を用いて、ビーズ種類の同定と、そのビーズにPEが結合しているか否かを判定した。測定は2種類のレーザを使用して行い、ビーズの種類は635nmレーザにより同定し、PE蛍光は532nmレーザを用いて定量した。オリゴビーズに結合したDNAは1測定あたり各々のビーズを最低50個ずつ測定し、定量されたPEの蛍光強度の中央値(MFI)を使用した。
図2中には、各ビーズ(1)〜(3)が同定され、かつビーズ(1)にのみPEが測定されたことから、ビーズ(1)に結合させたプローブが認識するDNAが増幅された様子が示されている。
【0037】
この方法に拠る遺伝子タイピングの精度を確かめるために、91個のDNAサンプルを材料に使用した。これらのDNAサンプルは、ミトコンドリアゲノムの全塩基配列が直接シーケンスによって既に決定されているものである。各例において、ルミネックスシーケンス特異性オリゴヌクレオチドハイブリッドアッセイシステム(PCR−SSOP−Luminex法)によって決定された遺伝子型は、直接シーケンスによって決定されたものと一致した。
【0038】
<統計的分析>
数量的な臨床データは、2型糖尿病患者群と対照群との間で、対応のないスチューデントt検定を用いて比較した。質的なデータは、カイ二乗検定によって比較した。
2型糖尿病と関連する多型は、交絡因子を含む多項ロジスティック回帰分析法により解析した。このとき、交絡因子については、年齢(age)、性別(gender:0=女性、1=男性)、喫煙状態(smoker:0=非喫煙、1=喫煙)、および各mtSNPの遺伝子型を独立変数とし、2型糖尿病を従属変数とした。P値、オッズ比(OR)、および95%信頼区間(CI)を計算した。特に説明しない限り、0.05未満のP値は統計上有意であると考えた。
【0039】
ハプログループを多重比較するために、ボンフェローニ(Bonferroni)の補正を適用した。我々は、10個のハプログループを調べたので、0.05を10で除して、0.005とした。こうして、0.005未満のP値を統計上有意であると見なした。
<結果>
2906名の被験者に関する基礎的データを表10に示した。
【0040】
【表10】
【0041】
表中において、最左欄は上段から順に、年齢(Age)、性別(女性/男性)(Gender)、BMI(Body mass index)、喫煙率(Smoker)、高血圧(Hypertension)、収縮期血圧(Systolic blood pressure)、拡張期血圧(Diastolic blood pressure)、高脂血症(Hypercholesterolemia)、全コレステロール(Total cholesterol)、中性脂肪(Triglycerides)、HDL−コレステロール(HDL-cholesterol)、空腹時血糖値(Fasting blood glucose)、ヘモグロビンAlc(Glycosylated hemoglobin)を意味している。また、最上段は左から順に、被験者全員(All subjects)における2型糖尿病患者群(Type 2 diabetes)の値と対照群(Controls)の値とP値(P value)、女性(Women)における2型糖尿病患者群の値と対照群の値とP値、男性(Men)における2型糖尿病患者群の値と対照群の値とP値とをそれぞれ示している。また、各値において、「±」が付してあるものは、平均値±SDで示した。喫煙率については、1日あたり10本以上の喫煙数がある者を喫煙者とした。高血圧については、収縮期血圧が140mmHg以上、または拡張期血圧が90mmHg以上、或いは両基準を満たす者、または高血圧薬を服用している者とした。高脂血症については、全コレステロール値が220mg/dL(5.72mmol/L)以上、または高脂血症剤を服用している者とした。
【0042】
年齢は2型糖尿病患者群において対照群よりも低かったが、BMI、収縮期および弛緩期血圧、トリグリセリド、絶食時血漿グルコース濃度、およびヘモグロビンAlc(HbA1c)比率は、対照群に比べると2型糖尿病患者群においては有意に高かった(P<0.05)。
年齢とHDLコレステロールは、対照群に比して2型糖尿病患者者の方が有意に低かった(P<0.05)。喫煙率、高血圧症、高コレステロール血症は、対照群に比較して2型糖尿病患者群では有意に高かった(P<0.05)。女性/男性の比率は、対照群に比して2型糖尿病患者群の方が有意に低かった(P<0.05)。
年齢、性別、喫煙状態を補正して多項ロジスティック回帰分析を行った結果を表11に示した。
【0043】
【表11】
【0044】
表中において、左欄より順に、変数(Variable)、P値(P value)、オッズ比(OR)、95%信頼区間(95%CI)を示している。また、上段より被験者全員(All subjects)における従来の危険因子(Conventional risk factors)である年齢(Age)・性別(Gender)・喫煙率(Smoking)と遺伝的危険因子(Genetic risk factors)であるハプログループF、N9aを、女性における従来の危険因子である年齢・喫煙率と遺伝的危険因子であるハプログループN9a、F、Aを、男性における従来の危険因子である年齢・喫煙率と遺伝的危険因子であるハプログループFをそれぞれ意味している。
【0045】
表に示す通り、P値が0.05未満で、ハプログループFを持つ被験者が、2型糖尿病罹患のリスクが有意に高いことが明らかとなった。これとは対照的に、ハプログループN9aを持つ被験者は、2型糖尿病に罹患するリスクが低い傾向を示した(P = 0.023, オッズ率 0.64(95% CI 0.43-0.94))。ハプログループN9aを持つ女性は、2型糖尿病罹患のリスクが有意に低かった(P = 0.0042, オッズ率 0.27(95% CI 0.10-0.62))。一方、ハプログループFおよびAを持つ女性は2型糖尿病罹患のリスクが高い傾向を示した(Fについては、P = 0.0165, オッズ率 1.78(95% CI 1.11-2.89);Aについては、P = 0.0405, オッズ率 1.67(95% CI 1.02-2.73))。ハプログループFを持つ男性は、2型糖尿病に罹患するリスクが高かった(P = 0.0482, オッズ率1.43(95% CI 1.00-2.05))。
我々は、ハプログループの多重比較について、0.005未満のP値が得られた場合に、その関連が有意であると考えた。この基準に基づくと、ハプログループFは、全ての被験者について2型糖尿病に対する感受性に有意に関与していた( P = 0.0028)。一方、女性について、ハプログループN9aは、2型糖尿病に対する抵抗性に有意に関与していた( P = 0.0042)。
表12には、ステップワイズ前向き選択法による解析結果を示した。
【0046】
【表12】
【0047】
表より、ハプログループFは全ての被験者について、2型糖尿病に対する危険因子であることが明らかとなった(P = 0.0035)。
女性の場合には、ハプログループN9aは2型糖尿病に対する防御因子であり(P = 0.0013)、ハプログループFとAは2型糖尿病に対して独立な危険因子である可能性が示された(Fについて、P = 0.0227;Aについて、P = 0.0374)。しかし男性の場合には、これらのハプログループのいずれも、2型糖尿病との関連性を示さなかった。
女性の場合、ハプログループN9a、F、およびAの2型糖尿病への抵抗性に対する総合的な寄与は0.0152(3つのハプログループの寄与率(R2)の和)であった。これはミトコンドリアゲノム多型が、特に女性においては、2型糖尿病への感受性に影響する重要な遺伝要因であることを示唆している。
表13には、mtSNPsのうち、2型糖尿病に対する保護因子或いは危険因子として関連するものを示した。
【0048】
【表13】
【0049】
表中、「*」は表1において示すように、10個のハプログループを代表させるために用いた多型マーカであることを示している。また、表中の太字で示したP値は、0.005未満であることを示している。
年齢および喫煙状態を補正した多項ロジスティック回帰分析の結果、ハプログループFを代表する3個のmtSNPである3970C>T、10310G>Aおよび13928G>C、ハプログループF1を代表するmtSNPである12406G>A、ハプログループF1b1aを代表するmtSNPである13928G>Cが、全ての被験者に関して、糖尿病と関連性を持つことが明らかになった(P = 0.0014 [10310G>A] , P = 0.0025 [12406G>A], P = 0.0028 [3970C>T], P=0.0028 [13928G>C]。これらは149 個のmtSNP の比較に対しての無修正値(ボンフェローニの補正を行っていない値)である)。個々のmtSNPについては、各ハプロタイプに対応していることから、ハプロタイプに適用した有意水準(P<0.005)で判断を行った。
【0050】
サブハプログループA1a1aを代表するmtSNPである14944C>T、サブハプログループF1b1aまたはN9b1を代表するmtSNPである5147G>A、サブハプログループB5b3を代表するmtSNPである8856G>A、およびサブハプログループF1b1aを代表するmtSNPである14476G>Aを保持する被験者は、0.05未満のP値で2型糖尿病に対する危険性が高かった(10個のハプログループにおけるサブハプログループを代表するmtSNPの有無については、表4〜表9に記載のプローブのいずれかによって測定されている。以下の記述中のサブハプログループを代表するサブハプログループについても同様である。)。
対照的に、サブハプログループN9a2a1を代表するmtSNPである15067T>C、サブハプログループN9aを代表する2個のmtSNPである5231G>Aおよび12372G>A、並びにサブハプログループD4aを代表するmtSNPである15314G>Aを保持する被験者は、0.05未満のP値で2型糖尿病に抵抗性を示した。
【0051】
女性については、年齢および喫煙状態を補正した多項ロジスティック回帰分析の結果、ハプログループN9aを代表する2個のmtSNPである5231G>Aおよび12372G>Aが、糖尿病に対する抵抗性に関係することが明らかになった( P = 0.0025 , P = 0.0035。これは149 個のmtSNP の比較に対しての無修正値である)。これに対し、サブハプログループA1a1aを代表するmtSNPである14944C>Tは、糖尿病に対する危険性が高かった(P=0.0035)。更に、サブハプログループF1を代表するmtSNPである12406G>A、ハプログループFを代表する3個のmtSNPである13928G>C、10310G>A、および3970C>T、サブハプログループA1aを代表するmtSNPである4655G>A、サブハプログループB5b3を代表するmtSNPである8856G>A、ハプログループAを代表する2個のmtSNPである663A>Gおよび8794C>Tを保持する被験者は、0.05未満のP値で2型糖尿病に対するリスクが高いことがわかった。
【0052】
これに対し、ハプログループN9aを代表するmtSNPである12358A>G、サブハプログループN9a2a1を代表するmtSNPである15067T>Cを持つ女性は、0.05未満のP値で2型糖尿病に対するリスクが低いことがわかった。
男性の場合には、年齢および喫煙状態を補正した多項ロジスティック回帰分析の結果、ハプログループFを代表するmtSNPである10310G>Aを持つと、0.05未満のP値で2型糖尿病に対するリスクが高いことがわかった。
表14に示すように、ステップワイズ前向き選択法によれば、ハプログループFを代表するmtSNPである10310G>Aが全被験者で2型糖尿病に対する危険因子であることが明らかになった(P = 0.0018)。
【0053】
【表14】
【0054】
表中、「*」は表1において示すように、10個のハプログループを代表させるために用いた多型マーカであることを示している。また、表中の太字で示したP値は、0.005未満であることを示している。
女性の場合には、ハプログループN9aを代表するmtSNPである5231G>Aが、2型糖尿病に対する防御因子であることが明らかになった(P = 0.0006)。他方、サブハプログループA1a1aを代表するmtSNPである14944C>Tは、2型糖尿病に対する危険因子であった(P = 0.0036)。7個のmtSNPs、すなわち5231G>A、14944C>T、12406G>A、8762T>C、8856G>A、12358A>G、および8383T>Cの糖尿病に対する感受性ならびに/若しくは防御性への総合的な寄与は0.0364(7個のmtSNPsの寄与率(R2)の和)であった。
【0055】
男性の場合には、サブハプログループF4aを代表するmtSNPである7861T>Cは、2型糖尿病に対する危険因子であった(P = 0.0005)。4個のmtSNPs、すなわち7861T>C、1391T>C、10310G>A、および15047G>Aの2型糖尿病に対する感受性への総合的な寄与は0.0101(4個のmtSNPsの寄与率(R2)の和)であった。
これらの結果は、上記のmtSNPsが、特に女性の場合には2型糖尿病に対する感受性に影響する重要な遺伝因子であることを示している。
【0056】
<考察>
我々は、2906名の被験者に基づく大規模な関連解析を行い、2型糖尿病と10個の主要なミトコンドリアハプログループとの間の関係を調査した。これらのハプログループのうち、ハプログループFは全ての被験者で2型糖尿病のリスクに有意に関係していた。女性の場合には、ハプログループN9aは、2型糖尿病に対する抵抗性に有意に関係していた。一方、ハプログループFおよびハプログループAは、2型糖尿病に対するリスクに関係していた。男性の場合には、ハプログループFは、2型糖尿病のリスクと関係していた。
【0057】
我々は、ミトコンドリアDNA配列の解析から、日本人集団がマクロハプログループNとMとに分割されることを報告した(非特許文献17)。マクロハプログループMは、10個の強固なmtSNPsがリンクしたマクロハプログループNとは異なっている。マクロハプログループNには、4個の主要なハプログループ、すなわちF、B、A、およびN9aが含まれる。他方、マクロハプログループMには、6個の主要なハプログループ、すなわちM7a、M7b、G1、G2、D4およびD5が含まれる。こうして、日本人集団は、これら10個の主なハプログループに分類される。
【0058】
ハプログループFに特異なmtSNPに、13928G>Cトランスバージョンは、潜在的な機能的多型の1つである。このmtSNPは、NADH脱水素酵素サブユニット5遺伝子産物(ND5)中で、S531T置換を起こすことが知られている。ND5はmtDNAにコードされたサブユニットの7個の内の一つであるが、これらのサブユニットは呼吸鎖複合体I(NADH:ユビキノン酸化還元酵素、EC1.6.5.3)の約41個のポリペプチドに含まれている。複合体Iは、NADHから電子を受け取り、それをユビキノン(補酵素Q10)に転送する。その際に得られたエネルギーを用いて、ミトコンドリア内膜を通してプロトンを輸送する。
【0059】
ND5は、複合体Iの機能には欠かすことができない。その理由は、ND5遺伝子中の様々なミスセンス変異が、レーバー遺伝性視神経萎縮症(LHON)(非特許文献22)、リー症候群(非特許文献25)、またはミトコンドリア性ミオパチー、脳症、乳酸アシドーシスおよび脳卒中様症状から成る症候群(MELAS)(非特許文献12)に罹患する患者において報告されているからである。従って、13928G>Cによって惹き起こされるS531T置換が、ND5サブユニットおよび複合体Iの機能に影響を及ぼすと考えることができる。13928G>C(ND5:S531T)がミトコンドリア機能に及ぼす具体的な作用については、このハプログループを伴うmtDNAを持つ細胞質雑種細胞系統を使う実験によって確認することができ得る。
【0060】
ハプログループFを持つ個人の代謝特性は、おそらくインスリン分泌ならびに/あるいはインスリン抵抗性に影響を与えることによって、2型糖尿病に対する抵抗性を得るものと思われる。ハプログループFに属するサブハプログループ(F1a、F1b、F1c、F2a、F3、F4a、およびF4b)の内、サブハプログループF1bは16189G>C多型によって定義される。この多型はハプログループBにおいても検出されること、およびハプログループBは本研究では糖尿病に関係しなかったことから、16189T>C多型は2型糖尿病とは関係しないと判断される。
【0061】
ハプログループN9aに特有のmtSNPsは、ほとんど同義突然変異である。これらの同義突然変異のうち、5231G>Aおよび12372G>Aは、本研究の遺伝子タイピングに使用した。このハプログループに於ける機能的な多型として考えられる候補は、150C>Tおよび338C>Tであるが、これらはミトコンドリアゲノムの非コード領域にある。150C>T置換は、イタリアの百寿者で最初に報告された(非特許文献23)。また我々は、この置換がフィンランドと日本の両国で共に健康的な長寿者と関連していることを報告している(非特許文献24)。このため、150C>Tは、女性の場合には2型糖尿病に対する抵抗性を授与するものと考えられる。
【0062】
ハプログループN9aに特異なmtSNPsのうちでは、12358A>GがND5サブユニットにT8A置換(8位のスレオニンをアラニンに置換する)を惹き起こすことから、潜在的な機能的な多型であると考えられる。12358A>Gによって惹き起こされたT8A置換が、ND5サブユニットおよび複合体Iの機能に影響を与えるかも知れない。12358A>G(ND5:T8A)がミトコンドリア機能に与える具体的な作用については、細胞実験によって調査することができ得る。150C>Tと12358A>Gの両者を備えたハプログループN9aを持つ女性の代謝特性は、2型糖尿病に対する抵抗性の機構を理解するために更なる調査が望まれる。
【0063】
本研究において、女性の場合には、ハプログループAが2型糖尿病への感受性と関連を持つことを見出した。ハプログループAは8794C>T突然変異を特徴とするが、この変異はATPaseサブユニット6においてH90Y置換(90位のヒスチジンをチロシンに置換する)を惹き起こす。FoF1−ATPaseは、核DNAにコードされた10−16個のサブユニットおよびmtDNAにコードされた2個のサブユニット(ATPase6およびATPase8)から構成される。ATP6遺伝子は、サブユニットaをコードする。サブユニットaは、ATPaseのFoセクタを横断してプロトンを輸送する際に重要な役割を果たす。L156R置換を惹き起こす8993T>Gトランスバージョン、およびL156P置換を惹き起こす8993T>Cトランジションは、NARP(神経障害、運動失調、および色素視神経網膜炎)症候群(非特許文献25、26)の原因となる病原性突然変異である。従って、8794C>Tによって惹き起こされたH90Y置換がATP6サブユニットとFoF1−ATPase(複合体V)の機能に影響を与える可能性がある。
【0064】
ミトコンドリア機能および代謝に対する8794C>T(ATP6:H90Y)の実際の効果については、この多型と2型糖尿病に対する感受性との関係を理解するために更なる調査が望まれる。ミトコンドリアには組換え機構が存在しないので、各ハプログループあるいはサブハプログループに於けるmtSNPsの全ては、相互に強く連鎖している。従って、本研究で調べたmtSNPsのいくつかが連鎖不均衡にあって、2型糖尿病に対する感受性若しくは抵抗性を実際に担う他のミトコンドリア遺伝子の機能的な多型にそのmtSNPが連鎖している可能性がある。女性の場合、ハプログループN9aは2型糖尿病に対する抵抗性に関係していたが、この関係は男性の場合には検出されなかった。
【0065】
遺伝的変異と肥満との関係は、男性の場合に比べると女性の方が強いという考えを支持する他の証拠がある(非特許文献27、28)。Comuzzieらは、性相互作用により遺伝子型を導入し、脂肪重量をメンデル混合モデルにて分析したところ、男性では脂肪重量の分散値全体の37%を主な遺伝子が担い、女性では43%を担っていた(非特許文献28)。これまでに、核遺伝子多型と糖尿病との間の関係についての関連解析がいくつか認められる。グルコキナーゼをコードする遺伝子の多型(非特許文献29)、インスリン受容体基質1(非特許文献30)、ペルオキシソーム増殖因子活性性受容体γ(非特許文献31)、カルパイン−10(非特許文献32)、およびアディポネクチン(非特許文献33)は、糖尿病に関連するものと報告されている。
しかしながら、ミトコンドリアDNA多型と2型糖尿病との関係に対する性別の果たす影響に関しては、今までのところほとんど報告が見られない。本明細書が、初めての報告である。
【0066】
このように、本実施形態によれば、mtDNAのハプログループを検出することにより、2型糖尿病の遺伝的危険度を予測することができる。このことにより、2型糖尿病の予防が可能となり、高齢者の健康寿命延長・QOL向上・ねたきり防止ならびに今後の医療費削減など、医学的・社会的に大きく貢献できる。
【0067】
【非特許文献1】van den Ouweland JM, Lemkes HH, Ruitenbeek W, Sandkuijl LA, de Vijlder MF, Struyvenberg PA, van de Kamp JJ, Maassen JA: Mutation in mitochondrial tRNA(Leu)(UUR) gene in a large pedigree with maternally transmitted type II diabetes mellitus and deafness. Nat Genet 1:368-371, 1992
【非特許文献2】Guo LJ, Oshida Y, Fuku N, Takeyasu T, Fujita Y, Kurata M, Sato Y, Ito M, Tanaka M: Mitochondrial genome polymorphisms associated with type-2 diabetes or obesity. Mitochondrion 5:15-33, 2005
【非特許文献3】Kokaze A, Ishikawa M, Matsunaga N, Yoshida M, Makita R, Satoh M, Teruya K, Sekiguchi K, Masuda Y, Harada M, Uchida Y, Takashima Y: Longevity-associated mitochondrial DNA 5178 C/A polymorphism is associated with fasting plasma glucose levels and glucose tolerance in Japanese men. Mitochondrion 5:418-425, 2005
【非特許文献4】Okura T, Koda M, Ando F, Niino N, Tanaka M, Shimokata H: Association of the mitochondrial DNA 15497G/A polymorphism with obesity in a middle-aged and elderly Japanese population. Hum Genet 113:432-436, 2003
【非特許文献5】Lowell BB, Shulman GI: Mitochondrial dysfunction and type 2 diabetes. Science 307:384-387, 2005
【非特許文献6】Macaulay V, Hill C, Achilli A, Rengo C, Clarke D, Meehan W, Blackburn J, Semino O, Scozzari R, Cruciani F, Taha A, Shaari NK, Raja JM, Ismail P, Zainuddin Z, Goodwin W, Bulbeck D, Bandelt HJ, Oppenheimer S, Torroni A, Richards M: Single, rapid coastal settlement of Asia revealed by analysis of complete mitochondrial genomes. Science 308:1034-1036, 2005
【非特許文献7】Wallace DC: A mitochondrial paradigm of metabolic and degenerative diseases, aging, and cancer: a dawn for evolutionary medicine. Annu Rev Genet 39:359-407, 2005
【非特許文献8】Mishmar D, Ruiz-Pesini E, Golik P, Macaulay V, Clark AG, Hosseini S, Brandon M, Easley K, Chen E, Brown MD, Sukernik RI, Olckers A, Wallace DC: Natural selection shaped regional mtDNA variation in humans. Proc Natl Acad Sci U S A 100:171-176, 2003
【非特許文献9】Ruiz-Pesini E, Mishmar D, Brandon M, Procaccio V, Wallace DC: Effects of purifying and adaptive selection on regional variation in human mtDNA. Science 303:223-226, 2004
【非特許文献10】Montiel-Sosa F, Ruiz-Pesini E, Enriquez JA, Marcuello A, Diez-Sanchez C, Montoya J, Wallace DC, Lopez-Perez MJ: Differences of sperm motility in mitochondrial DNA haplogroup U sublineages. Gene 368C:21-27, 2006
【非特許文献11】Baudouin SV, Saunders D, Tiangyou W, Elson JL, Poynter J, Pyle A, Keers S, Turnbull DM, Howell N, Chinnery PF: Mitochondrial DNA and survival after sepsis: a prospective study. Lancet 366:2118-2121, 2005
【非特許文献12】Pyle A, Foltynie T, Tiangyou W, Lambert C, Keers SM, Allcock LM, Davison J, Lewis SJ, Perry RH, Barker R, Burn DJ, Chinnery PF: Mitochondrial DNA haplogroup cluster UKJT reduces the risk of PD. Ann Neurol 57:564-567, 2005
【非特許文献13】Tanaka M, Gong JS, Zhang J, Yoneda M, Yagi K: Mitochondrial genotype associated with longevity. Lancet 351:185-186, 1998
【非特許文献14】Ikebe S, Tanaka M, Ozawa T: Point mutations of mitochondrial genome in Parkinson's disease. Brain Res Mol Brain Res 28:281-295, 1995
【非特許文献15】Tanaka M: Mitochondrial genotypes and cytochrome b variants associated with longevity or Parkinson's disease. J Neurol 249 Suppl 2:II11-18, 2002
【非特許文献16】Fuku N, Oshida Y, Takeyasu T, Guo LJ, Kurata M, Yamada Y, Sato Y, Tanaka M: Mitochondrial ATPase subunit 6 and cytochrome b gene polymorphisms in young obese adults. Biochem Biophys Res Commun 290:1199-1205, 2002
【非特許文献17】Tanaka M, Cabrera VM, Gonzalez AM, Larruga JM, Takeyasu T, Fuku N, Guo LJ, Hirose R, Fujita Y, Kurata M, Shinoda K, Umetsu K, Yamada Y, Oshida Y, Sato Y, Hattori N, Mizuno Y, Arai Y, Hirose N, Ohta S, Ogawa O, Tanaka Y, Kawamori R, Shamoto-Nagai M, Maruyama W, Shimokata H, Suzuki R, Shimodaira H: Mitochondrial genome variation in eastern Asia and the peopling of Japan. Genome Res 14:1832-1850, 2004
【非特許文献18】Poulton J, Brown MS, Cooper A, Marchington DR, Phillips DI: A common mitochondrial DNA variant is associated with insulin resistance in adult life. Diabetologia 41:54-58, 1998
【非特許文献19】Weng SW, Liou CW, Lin TK, Wei YH, Lee CF, Eng HL, Chen SD, Liu RT, Chen JF, Chen IY, Chen MH, Wang PW: Association of mitochondrial DNA 16189 variant (T->C transition) with metabolic syndrome in Chinese adults. J Clin Endocrinol Metab 90:5037-5040, 2005
【非特許文献20】Chinnery PF, Elliott HR, Patel S, Lambert C, Keers SM, Durham SE, McCarthy MI, Hitman GA, Hattersley AT, Walker M: Role of the mitochondrial DNA 16184-16193 poly-C tract in type 2 diabetes. Lancet 366:1650-1651, 2005
【非特許文献21】Itoh Y, Mizuki N, Shimada T, Azuma F, Itakura M, Kashiwase K, Kikkawa E, Kulski JK, Satake M, Inoko H: High-throughput DNA typing of HLA-A, -B, -C, and -DRB1 loci by a PCR-SSOP-Luminex method in the Japanese population. Immunogenetics 57:717-729, 2005
【非特許文献22】Howell N, Halvorson S, Burns J, McCullough DA, Paulton J: When does bilateral optic atrophy become Leber hereditary optic neuropathy? Am J Hum Genet 53:959-963, 1993
【非特許文献23】Zhang J, Asin-Cayuela J, Fish J, Michikawa Y, Bonafe M, Olivieri F, Passarino G, De Benedictis G, Franceschi C, Attardi G: Strikingly higher frequency in centenarians and twins of mtDNA mutation causing remodeling of replication origin in leukocytes. Proc Natl Acad Sci U S A 100:1116-1121, 2003
【非特許文献24】Niemi AK, Moilanen JS, Tanaka M, Hervonen A, Hurme M, Lehtimaki T, Arai Y, Hirose N, Majamaa K: A combination of three common inherited mitochondrial DNA polymorphisms promotes longevity in Finnish and Japanese subjects. Eur J Hum Genet 13:166-170, 2005
【非特許文献25】Holt IJ, Harding AE, Petty RK, Morgan-Hughes JA: A new mitochondrial disease associated with mitochondrial DNA heteroplasmy. Am J Hum Genet 46:428-433, 1990
【非特許文献26】Tanaka M, Borgeld HJ, Zhang J, Muramatsu S, Gong JS, Yoneda M, Maruyama W, Naoi M, Ibi T, Sahashi K, Shamoto M, Fuku N, Kurata M, Yamada Y, Nishizawa K, Akao Y, Ohishi N, Miyabayashi S, Umemoto H, Muramatsu T, Furukawa K, Kikuchi A, Nakano I, Ozawa K, Yagi K: Gene therapy for mitochondrial disease by delivering restriction endonuclease SmaI into mitochondria. J Biomed Sci 9:534-541, 2002
【非特許文献27】Borecki IB, Bonney GE, Rice T, Bouchard C, Rao DC: Influence of genotype-dependent effects of covariates on the outcome of segregation analysis of the body mass index. Am J Hum Genet 53:676-687, 1993
【非特許文献28】Comuzzie AG, Blangero J, Mahaney MC, Mitchell BD, Hixson JE, Samollow PB, Stern MP, MacCluer JW: Major gene with sex-specific effects influences fat mass in Mexican Americans. Genet Epidemiol 12:475-488, 1995
【非特許文献29】Vionnet N, Stoffel M, Takeda J, Yasuda K, Bell GI, Zouali H, Lesage S, Velho G, Iris F, Passa P, et al.: Nonsense mutation in the glucokinase gene causes early-onset non-insulin-dependent diabetes mellitus. Nature 356:721-722, 1992
【非特許文献30】Almind K, Bjorbaek C, Vestergaard H, Hansen T, Echwald S, Pedersen O: Aminoacid polymorphisms of insulin receptor substrate-1 in non-insulin-dependent diabetes mellitus. Lancet 342:828-832, 1993
【非特許文献31】Altshuler D, Hirschhorn JN, Klannemark M, Lindgren CM, Vohl MC, Nemesh J, Lane CR, Schaffner SF, Bolk S, Brewer C, Tuomi T, Gaudet D, Hudson TJ, Daly M, Groop L, Lander ES: The common PPARgamma Pro12Ala polymorphism is associated with decreased risk of type 2 diabetes. Nat Genet 26:76-80, 2000
【非特許文献32】Horikawa Y, Oda N, Cox NJ, Li X, Orho-Melander M, Hara M, Hinokio Y, Lindner TH, Mashima H, Schwarz PE, del Bosque-Plata L, Horikawa Y, Oda Y, Yoshiuchi I, Colilla S, Polonsky KS, Wei S, Concannon P, Iwasaki N, Schulze J, Baier LJ, Bogardus C, Groop L, Boerwinkle E, Hanis CL, Bell GI: Genetic variation in the gene encoding calpain-10 is associated with type 2 diabetes mellitus. Nat Genet 26:163-175, 2000
【非特許文献33】Hara K, Boutin P, Mori Y, Tobe K, Dina C, Yasuda K, Yamauchi T, Otabe S, Okada T, Eto K, Kadowaki H, Hagura R, Akanuma Y, Yazaki Y, Nagai R, Taniyama M, Matsubara K, Yoda M, Nakano Y, Tomita M, Kimura S, Ito C, Froguel P, Kadowaki T: Genetic variation in the gene encoding adiponectin is associated with an increased risk of type 2 diabetes in the Japanese population. Diabetes 51:536-540, 2002
【図面の簡単な説明】
【0068】
【図1】Luminex100で検出するマイクロビーズの微細構造と特徴を示す図である。
【図2】PCR−SSOP−Luminex法の手順の概要を示す図である。
【図3】mtSNPに基づくハプログループ分類を示す図(1)である。図の下端には、ハプログループ及びサブハプログループを示している(図4及び図4においても同じである)。
【図4】mtSNPに基づくハプログループ分類を示す図(2)である。
【図5】mtSNPに基づくハプログループ分類を示す図(3)である。
【特許請求の範囲】
【請求項1】
ヒトミトコンドリアDNAにおいて、5231A、12358G、12372Aを有するハプログループN9aと、3970T、13928C、10310Aを有するハプログループFと、663G、8794Tを有するハプログループAと、12406Aを有するサブハプログループF1と、14944Tを有するサブハプログループA1a1aと、15067Cを有するサブハプログループN9a2a1と、5147Aを有するF1b1aと、5147Aを有するサブハプログループN9b1と、8856Aを有するサブハプログループB5b3と、14476Aを有するサブハプログループF1b1aと、15314Aを有するサブハプログループD4aと、4655Aを有するサブハプログループA1aと、8856Aを有するサブハプログループB5b3と、1391Cを有するサブハプログループB4b1bと、8762Cを有するサブハプログループD4mと、8383Cを有するサブハプログループD4d2と、7861Cを有するサブハプログループF4aと、15047Aを有するサブハプログループM8a1とを検出することを特徴とする2型糖尿病に関する遺伝子検出法。
【請求項2】
前記ハプログループの検出方法が、PCR−SSOP−Luminex法であることを特徴とする請求項1に記載の遺伝子検出方法。
【請求項1】
ヒトミトコンドリアDNAにおいて、5231A、12358G、12372Aを有するハプログループN9aと、3970T、13928C、10310Aを有するハプログループFと、663G、8794Tを有するハプログループAと、12406Aを有するサブハプログループF1と、14944Tを有するサブハプログループA1a1aと、15067Cを有するサブハプログループN9a2a1と、5147Aを有するF1b1aと、5147Aを有するサブハプログループN9b1と、8856Aを有するサブハプログループB5b3と、14476Aを有するサブハプログループF1b1aと、15314Aを有するサブハプログループD4aと、4655Aを有するサブハプログループA1aと、8856Aを有するサブハプログループB5b3と、1391Cを有するサブハプログループB4b1bと、8762Cを有するサブハプログループD4mと、8383Cを有するサブハプログループD4d2と、7861Cを有するサブハプログループF4aと、15047Aを有するサブハプログループM8a1とを検出することを特徴とする2型糖尿病に関する遺伝子検出法。
【請求項2】
前記ハプログループの検出方法が、PCR−SSOP−Luminex法であることを特徴とする請求項1に記載の遺伝子検出方法。
【図3】

【図4】
【図5】
【図1】
【図2】
【図4】
【図5】
【図1】
【図2】
【公開番号】特開2007−330148(P2007−330148A)
【公開日】平成19年12月27日(2007.12.27)
【国際特許分類】
【出願番号】特願2006−165293(P2006−165293)
【出願日】平成18年6月14日(2006.6.14)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2005年(平成17年)12月14日 日本ミトコンドリア学会発行の「国際学術集会 ミトコンドリアと生命2006」に発表
【出願人】(597144060)
【出願人】(506023806)
【出願人】(399077674)G&Gサイエンス株式会社 (21)
【出願人】(597112472)財団法人岐阜県研究開発財団 (25)
【Fターム(参考)】
【公開日】平成19年12月27日(2007.12.27)
【国際特許分類】
【出願日】平成18年6月14日(2006.6.14)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 2005年(平成17年)12月14日 日本ミトコンドリア学会発行の「国際学術集会 ミトコンドリアと生命2006」に発表
【出願人】(597144060)
【出願人】(506023806)
【出願人】(399077674)G&Gサイエンス株式会社 (21)
【出願人】(597112472)財団法人岐阜県研究開発財団 (25)
【Fターム(参考)】
[ Back to top ]