
HER2に結合する化合物
【課題】ヒトerbB2遺伝子産物(HER2とも称されるErbB2、又はc-ErbB-2)に結合する新規な化合物、及び新規な化合物を含有する製薬組成物を提供する。
【解決手段】ErbB2に結合する非天然発生アミノ酸配列からなる化合物である。化合物は、約5〜約50のアミノ酸残基の非自然発生アミノ酸配列からなるペプチドリガンドであり、更に免疫グロブリン定常領域配列としてCH3ドメイン、ヒンジ領域を含むペプチドリガンドからなる。
【解決手段】ErbB2に結合する非天然発生アミノ酸配列からなる化合物である。化合物は、約5〜約50のアミノ酸残基の非自然発生アミノ酸配列からなるペプチドリガンドであり、更に免疫グロブリン定常領域配列としてCH3ドメイン、ヒンジ領域を含むペプチドリガンドからなる。
【発明の詳細な説明】
【発明の概要】
【0001】
(発明の背景)
(発明の分野)
本発明はヒトerbB2遺伝子産物(HER2としても知られているErbB2、又はc-ErbB-2)に結合する新規な化合物に関する。特定の態様では、本発明は、本発明の新規な化合物を利用するErbB2の発現により特徴付けられる疾患の治療に関する。また本発明は、その新規な化合物を含有する組成物、例えば製薬用組成物、並びに研究、診断、治療及び予防法におけるそれらの使用に関する。
【0002】
(関連する開示の記載)
ファージディスプレイは、テンプレート配列内の特定のアミノ酸残基の無作為化を通して又は天然ペプチドライブラリーの生成によって、ペプチド及びタンパク質変異体の産生する手段を提供する(Lowman, H. (1998) Methods Mol. Biol. 87:249-264;Lowman(1997)Annu. Rev. Biophys. Biomol. Struct. 26:401-424)。予め決められた標的分子に結合するディスプレイされたタンパク質又はタンパク質の同定と単離は、固定された標的分子上の非結合又は弱く結合している変異体に対してディスプレイファージを富ませることにより、達成することができる(上掲のLowman, H.(1998))。突然変異誘発と選択の連続の実施により、細胞レセプターに対して高親和性を持つペプチドリガンド又はタンパク質変異体を生成することができる(Lowman, H.(1998)上掲)。上記技術は、例えば細胞標的に帰着するペプチドモチーフを同定するため(Arapら, (1998)Science 279:377-380)、又は天然のタンパク質結合リガンドから親和性が改善され又は成熟したペプチドリガンドを産生するために(Lowmanら, (1991)Biochemistry 30:10832-10838)使用されている。親和性又は特異性が改善されたタンパク質の例には、ヒト成長ホルモン、Znフィンガー、プロテアーゼインヒビター、心房性ナトリウム利尿ペプチド、及び抗体が含まれる(Wells, J. 及びLowman H.(1992) Curr. Opin. Struct. Biol. 2:597-604;Clackson, T.及びWells, J.(1994)Trends Biotechnol. 12:173-184;Lowmanら, (1991)Biochemistry 30(10):832-838;Lowmanら.及びWells J.(1993)J. Mol. Biol. 234:564-578;Dennis M.及びLazarus R.(1994)J Biol. Chem. 269(22):137-144)。
【0003】
インビボでのファージ選択を利用して、ファージディスプレイは、脳及び腎臓のような種々の器官に選択的局在化を媒介可能なペプチドを同定し単離するために(Pasqualini及びRuoslohti (1996) Nature 380:364-366)、並びにαVβ3又はαVβ5インテグリンを担持する特定の腫瘍タイプに帰着するペプチドを同定するために使用されている(Arapら, (1998) Science 279:377-380)。米国特許第5627263号には、α5β1インテグリンにより認識され、これと選択的に結合するペプチドが記載されている。一価のファージディスプレイにより生成される構造的に限定されたペプチドライブラリーを使用して、インシュリン様成長因子1結合タンパク質(IGFBPs)に特異的に結合する14のアミノ酸のペプチドが単離されている(Lowmanら, (1998) Biochemistry, 37:8870-8878)。そのペプチドはヘリックス構造を有し、IGFBPsに結合してインビトロでインシュリン様成長因子α(IGF-1)活性を作用させる(上掲のLowmanら,(1998))。
新規なタンパク質に基づく治療が臨床に入っているので、特定の細胞レセプターとそれらのリガンド、特に種々のヒト悪性腫瘍の病因に関連しているものは、近年、科学界において多くの注目を集めている。例えば、上皮成長因子レセプター(EGFR)に関連した185kdの膜貫通糖タンパク質レセプター(p185HER2)をコードするヒトerbB2遺伝子(her2としても知られている、又はc-erbB-2)は、ヒトの乳癌の約25%〜30%で過剰発現していることが見出されている(Slamonら, (1987) Science 235:177-182;Slamonら, (1989) Science 244:707-712)。いくつかの証拠情報は、ErbB2を過剰発現する腫瘍の病原性及び臨床的病原力におけるErbB2の直接的な役割を裏付けている。非新生物細胞へerbB2を導入すると、その悪性形質転換を引き起こすことが示されている(Hudziakら, (1987)Proc. Natl. Acad. Sci. USA 84:7159-7163;DiFioreら, (1987)Science 237:78-182)。HER2を発現するトランスジェニックマウスは、乳房腫瘍を発生させることが見出されている(Guyら, (1992) Proc. Natl. Acad. Sci. USA)。
【0004】
ErbB2過剰発現は、特に腋窩リンパ節に関与する原疾患を持つ患者における、悪い予後のプレディクターであると一般的に考えられている(上掲のSlamonら, (1987)及び(1989);Ravdin及びChamness, Gene (1995)159:19-27;及びHynesとStern, (1994) Biochim. Biophys. Acta 1198:165-184)。また過剰発現は、CMF(シクロホスファミド、メトトレキセート及びフルオロウラシル)及びアントラサイクリン(Baselgaら, (1997) Oncology 11(3 Suppl 1):43-48)を含む、ホルモン療法及び化学療法に対する感受性及び/又は耐性に関連している。
組換えヒト化抗ErbB2モノクローナル抗体(rhuMAb HER2又はハーセプチン(HERCEPTINR)と称されるマウス抗ErbB2抗体4D5のヒト化体)は、広範な抗癌治療を前に受けたことのあるErbB2過剰発現転移性乳癌を持つ患者において治療的に活性であった(Baselgaら, (1996) J. Clin. Oncol. 14:737-744)。
【0005】
(発明の概要)
本発明は、ヒトerbB2遺伝子産物(ErbB2)に結合する化合物を提供する。本発明の化合物(ここではペプチドリガンドとも呼ぶ)は、例えばペプチド又はペプチド誘導体、例えばペプチド模倣物及びペプチド類似物である。好ましいペプチドリガンドには、線形又は環状のペプチド、好ましくは次の核となる式:
-Cys-Xaa-Xaa-Xaa-Xaa-Cys-
を含む環状ペプチド化合物、好ましくは次の一般式:
Cys-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Cys-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Cys-Cys-Xaa-Xaa-Xaa-Cys-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Cys (配列番号:70)
又は次の一般式:
Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Cys-Xaa-Xaa-Xaa-Xaa-Cys-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa
を有するものが含まれ、また上記の一般構造のダイマー及び他の組合せも含まれる。本発明のペプチドリガンドの中で好ましいものは、次の核となる式:
-Cys-Cys/Ile-Gly-Pro-Gly-Cys (配列番号:71)
を含むペプチドである。このような化合物の特定の例には、約10〜60のアミノ酸残基の環状ペプチドで、場合によってはN-もしくはC-末端、あるいは双方が修飾されたもの、例えばそれらのエステル、アミド、塩及び他の誘導体が含まれる。本発明の好ましい態様では、化合物は、好ましくはErbB2に結合する非天然発生アミノ酸配列である。好ましくは、化合物は、約5〜約50のアミノ酸残基の非自然発生アミノ酸配列である。好ましい化合物は、上記の一般式を有し、約20〜約30のアミノ酸残基を有する環状ペプチドで、そのダイマー及び他の組合せを含む。
【0006】
特定の態様において、本発明はペプチドリガンドと他のペプチドリガンド又は他のタンパク質、特に血清タンパク質又はペプチドとの組合せに関する。この組合せは、ペプチドリガンドのその標的分子に対する親和性又は親和力を、例えば種々の多量体化ドメインを使用して増加させ;例えばZプロテイン融合体としてペプチドリガンドを発現させることにより、ペプチドリガンドの安定性を増加させるか、又はその回収と精製を容易にし;例えば血清の半減期を増加又は減少させることにより、血清アルブミン、免疫グロブリン、アポリポプロテイン又はトランスフェリン等の細胞質タンパク質にペプチドリガンドを融合させることにより(そのような融合体は組換え宿主細胞において簡便に、又は二官能性架橋剤を使用することにより生成する)、及び機能性Fcドメイン又は細胞障害又は酵素部分等の付加的機能を導入することにより、ペプチドリガンドのインビボ使用を含む本発明の態様のペプチドリガンドの治療効果を改善することを含む、種々の目的をもって調製される。
本発明の好ましい態様では、本発明の化合物は多量体化ドメインに結合する。多量体化ドメインは、好ましくは免疫グロブリン配列、又は例えばロイシンジッパー配列である。本発明のこの態様では、免疫グロブリン配列は、好ましくは免疫グロブリン定常領域配列、特に免疫グロブリン重鎖の定常領域である。本発明の好ましい態様では、多量体化ドメインは一又は複数のコンパニオン多量体化ドメインと対になり、ホモ-及びヘテロ-多量体組成をもたらす。本発明のこの側面の好ましい態様はホモ-及びヘテロ-多量体、特にホモ-及びヘテロダイマーにあり、そこでは、多量体化ドメインが、対となって機能的免疫グロブリンFcドメインを提供する免疫グロブリン重鎖定常領域である。よって、好ましい態様では、本発明は、機能的免疫グロブリンFcドメインと結合するエフェクター機能を有する機能的免疫グロブリンFcドメインとErbB2担持細胞型を標的とするように機能する本発明の化合物を含有するハイブリッド分子を提供する。
【0007】
本発明のこの態様によるハイブリッド分子は、場合によってはさらなる機能的部分、例えば酵素部分及び細胞障害性部分を含む。例えば、付加的な機能的ドメインは、ハイブリッド分子に共有結合的に結合し、プロドラッグをそのより活性な形態に転換させるようにプロドラッグに作用可能な酵素であってよい。本発明の所定の好ましい態様による任意成分の機能的ドメインは、例えばハイブリッド分子に共有結合によって結合する細胞障害剤であってもよい。好ましい細胞障害剤には、例えば化学治療剤、毒素及び放射性アイソトープが含まれる。ハイブリッド分子を含む化合物とそれらを含有する組成物は、ヒトerbB2遺伝子産物への結合又は検出と、場合によってはErbB2担持細胞型への酵素又は細胞障害剤等の機能的部分の送達に使用される。
一実施態様では、本発明の化合物、例えば上述したハイブリッド分子はポリペプチドであり、本発明は、本発明のポリペプチドをコードする単離された核酸、好ましくはDNAを含有する物質の組成物を含む。この態様では、本発明は、DNA分子に作用可能に結合した発現コントロール配列、DNA分子を含有する発現ベクター、好ましくはプラスミドで、ここでコントロール配列がベクターで形質転換された宿主細胞により認識されるもの、及びベクターで形質転換された宿主細胞を更に含む。好ましい態様では、核酸が本発明のペプチド化合物と免疫グロブリン定常領域ドメイン配列を含有するハイブリッド分子をコードする。本発明のこの態様に係る核酸分子はハイブリッド分子をコードし、本発明のペプチド化合物をコードする核酸は免疫グロブリン定常ドメインに配列に作用可能に結合している(DNA配列が隣接し読み枠にあるという意味)。場合によっては、DNA配列は任意成分のリンカードメインアミノ酸配列をコードする核酸配列によって結合している。
【0008】
本発明の組成物は、本発明のアミノ酸配列の任意のものをコードする核酸配列を単離又は合成し、適切な宿主中で核酸配列を発現可能な適切な発現ベクター内に核酸配列をライゲーションし、核酸配列がライゲーションされた発現ベクターで宿主を形質転換し、核酸配列の発現に適した条件下で宿主を培養する工程を含む方法により作成され、それにより、選択された核酸配列によりコードされるタンパク質は宿主により発現される。好ましくは、ポリペプチドはついで宿主細胞培養物から回収される。この方法では、ライゲーション工程は、核酸配列が適切な分泌シグナルに作用可能に結合するように適切な発現ベクターに核酸をライゲーションすることをさらに意図しており、それにより、アミノ酸配列は宿主により分泌される。
本発明は、本発明の化合物を含有する、特にerbB2遺伝子発現に関連した病気又は疾患の検出及び治療のための製薬用組成物を含む組成物を含む。例えば、癌の検出及び治療用のキット及び製造品が提供される。キット及び製造品は、好ましくは:
(a)容器;
(b)前記容器上の又は該容器に付随するラベル;及び
(c)前記容器内に収容された本発明の化合物を含む組成物を含み、ある実施態様では、当該組成物は、erbB2遺伝子発現を伴う病気又は疾患、例えば癌の治療に有効である。好ましくは、前記容器上のラベルは、当該組成物が癌の治療に使用できることを表示しており、当該組成物中の化合物がヒトerbB2遺伝子産物に結合するここで記載された化合物を含む。キットは、場合によっては、付属品、例えば製薬的に許容されるバッファーを収容した第2の容器及び疾患の治療のために組成物を使用するための指示を含む。
【0009】
種々の病気及び疾患、例えば癌、特にErbB2に関連していることを特徴とするものの治療に有用な方法が開示される。よって、本発明は、治療的有効量の本発明の化合物を宿主に投与することを含む、その必要とする宿主におけるErbB2媒介性又は関連性の病気又は疾患の治療方法を提供する。さらに本発明は、限定するものではないが、化合物の非経口、経口、直腸及び肺投与に適切なものを含む、本発明の化合物の種々の投与形態を提供する。本発明の好ましい態様では、治療的投与形態は静脈内送達に適するように提供され、本発明は、ErbB2媒介性又は関連性のプロセス又は事象、例えば癌に関連した病気又は疾患の治療のために提供される。
【0010】
(好ましい実施態様の詳細な説明)
定義
「HER2」、「ErbB2」、「c-Erb-B2」という用語は互換的に使用される。特に別の定義を示さない限り、ここで使用される「ErbB2」、「c-Erb-B2」及び「HER2」という用語はヒトタンパク質を意味し、「her2」、「erbB2」及び「c-erb-B2」はヒト遺伝子を示す。ヒトerbB2遺伝子及びErbB2タンパク質は、例えばSembaら, (1985) PNAS(USA)82:6497-6501及びYamamotoら, (1986) Nature 319:230-234(Genebank受託番号 X03363)に記載されている。ErbB2は4つのドメインを含んでいる(ドメイン1-4)。
本発明における「ペプチドリガンド」なる用語は、その由来にかかわらず、ErbB2に結合するように機能するアミノ酸配列を指すことを意味する。本発明のペプチドリガンドは、一般的に束縛されている(つまり、例えばβターン又はβプリーツシートを開始するアミノ酸の存在、又はジスルフィド結合したCys残基の存在により環化したような構造のいくつかのエレメントを有している)か、又は束縛されていない(例えば、線形)、約50アミノ酸残基未満、好ましくは約40アミノ酸残基未満のアミノ酸配列である。約40アミノ酸残基未満のペプチドリガンドでは、約10〜約30アミノ酸残基のペプチドリガンド、特に約20〜約30アミノ酸残基のペプチドリガンドが好ましい。しかしながら、現在の開示を読むと、当業者であれば、本発明のペプチドリガンドを区別するのは特定のペプチドリガンドの長さではなく、ErbB2に結合するその能力であることを認識するであろう。よって、例えば5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29及び30アミノ酸残基のペプチドリガンドが、等しく有望な本発明のペプチドリガンドである。
【0011】
本発明のペプチドリガンドは、ペプチドリガンドがインビトロで、好ましくはインビボでErbB2、例えばErbB2担持細胞に「帰着する(homes)」、「結合する」又は「標的とする」ならば、十分な親和性と特異性を持ってErbB2に結合するであろう(例えば、Pasqualini及びRuoslahti(1996) Nature, 380:364-366、及びArapら,(1998) Science 279:377-380)における「帰着する」、「帰着」及び「結合する」という用語の使用を参照されたい)。一般的に、ペプチドリガンドは、Jonesら, (1999) FEBS Letters 447:227-231により記載されたものと同様なErbB基質を利用する競合アッセイ等のインビトロアッセイにより定量される場合に約1μM未満、好ましくは約100nM未満、特に約10nM未満の親和力でErbB2に結合する。しかしながら、ErbB2に対して約1nM未満、好ましくは約1pM〜1nMの親和力を有するペプチドリガンドが、等しく有望な本発明のペプチドリガンドである。一般に、上述したようなErbB2に結合するペプチドリガンドは、固定化されたErbB2分子に結合させるなどのここに記載された当該技術の多くの標準的な技術の任意のものにより単離され同定され得る。
上述したように、ペプチドリガンドは自然に生じた、また非自然的に生じたアミノ酸残基を含有していてもよい上述したアミノ酸配列である。よって、特定のアミノ酸又はペプチドの構造を模倣した非アミノ酸化学構造を含みうる、いわゆる「ペプチド模倣物」及び「ペプチド類似物」も本発明のペプチドリガンドとできる。このような模倣物及び類似物は、類似の物理的特徴、例えば大きさ、電荷又はそれらの対のペプチドに見出されるような適切な空間的配向性で存在する疎水性を示すものとして一般に特徴付けられる。ペプチドの模倣化合物の特定の例は、一又は複数のアミノ酸のアミド結合が、例えば炭素-炭素結合又は当該技術でよく知られている他の結合で置換された化合物である(例えば、Sawyer, Peptide Based Drug Design pp.378-422(ACS, Washington DC 1995)を参照のこと)。
【0012】
従って、本発明における「アミノ酸」なる用語は最も広い意味に使用され、自然に生じるLα-アミノ酸又は残基を含むことを意味する。自然に生じるアミノ酸に対して一般的に使用されている1文字及び3文字略号がここでも使用される(Lehninger, A.L., Biochemistry, 第2版, pp.71-92,(1975) Worth Publishers, New York)。この用語には、D-アミノ酸、並びに化学的に修飾されたアミノ酸、例えばアミノ酸類似物、ノルロイシン等のタンパク質に通常導入されない自然に生じるアミノ酸、及びアミノ酸に特徴的な当該分野において公知の特性を有する化学的に合成された化合物が含まれる。例えば、天然Phe又はProと同じペプチド化合物の配座制限を許容するフェニルアラニン又はプロリンの類似物又は模倣物が、アミノ酸の定義に含まれる。このような類似物及び模倣物はここではアミノ酸の「機能的等価物」と称する。アミノ酸の他の例は、出典を明示してここに取り込まれるRoberts及びVellaccio(The Peptides:Analysis, Synthesis, Biology)Eds. Gross及びMeiehofer, Vol.5 p341, Academic Press, Inc, N.Y. 1983に列挙されている。
例えば、標準的な固相合成法により合成されたペプチドリガンドは、遺伝子によりコードされるアミノ酸に限定されない。遺伝子コードによりコードされない一般的に遭遇するアミノ酸には、例えば国際公開第90/01940号に記載されているもの、例えばGlu及びAspについての2-アミノアジピン酸(Aad);Glu及びAspについての2-アミノピメリン酸(Apm);Met、Leu、及び他の脂肪族アミノ酸についての2-アミノブチル酸(Abu);Met、Leu、及び他の脂肪族アミノ酸についての2-アミノヘプタン酸(Ahe);Glyについての2-アミノイソブチル酸(Aib);Val、及びLeu及びIleについてのシクロヘキシルアラニン(Cha);Arg及びLysについてのホモアルギニン(Har);Lys、Arg及びHisについての2,3-ジアミノプロピオン酸(Dpr);Gly、Pro及びAlaについてのN-エチルグリシン(EtGly);Gly、Pro及びAlaについてのN-エチルグリシン(EtGly);Asn、及びGlnについてのN-エチルアスパラギン(EtAsn);Lysについてのヒドロキシルリジン(Hyl);Lysについてのアロヒドロキシルリジン(AHyl);Pro、Ser、及びThrについての3-(及び4)ヒドロキシプロリン(3Hyp、4Hyp);Ile、Leu、及びValについてのアロ-イソロイシン(AIle);Alaについてのρ-アミジノフェニルアラニン;Gly、Pro、及びAlaについてのN-メチルグリシン(MeGly、サルコシン);IleについてのN-メチルイソロイシイン(MeIle);Met及び他の脂肪族アミノ酸についてのノルバリン(Nva);Met及び他の脂肪族アミノ酸についてのノルロイシン(Nle);Lys、Arg及びHisについてのオルニチン(Orn);Thr、Asn及びGlnについてのシトルリン(Cit)及びメチオニンスルホキシド(MSO);PheについてのN-メチルフェニルアラニン(MePhe)、トリメチルフェニルアラニン、ハロ(F、Cl、Br、及びI)フェニルアラニン、トリフルオロフェニルアラニンが含まれる。
【0013】
本発明の保存的アミノ酸置換を、「例示的置換」及び「好ましい置換」と題して表1に示す。好ましい置換がHER2結合性を低減又は変化させない場合、表1に例示的置換と名前を付け、又はさらにここで記載する、より実質的な変化が導入され、生成物がHER2結合について試験される。

【0014】
本発明のペプチドリガンドは、好ましくは自然に生じないアミノ酸配列である。自然に生じないとは、特定のペプチドリガンドのアミノ酸配列が天然に見出されないことを意味する。本発明において、「非天然発生」とは、非天然の又は非天然発生のアミノ酸配列に相当するペプチドリガンドを指すために使用される。この多様なペプチドリガンドは、当業者によく知られている種々の技術を使用して生成又は選択されうる。例えば、束縛されているか又は束縛されていないペプチドライブラリーが無作為に作製され、当該分野で標準的な技術を利用してファージにディスプレイする(例えば、Lowmanら, (1998) Biochemistry 37:8870-8878)。
少なくとも3つの異なる種のペプチドリガンドが、ErbB2に結合する機能に基づいて区別されうる。これらは、ここでは「中性」、「アゴニスト」及び「アンタゴニスト」ペプチドリガンドと称される。一般に、中性ペプチドリガンドは、上述したようにErbB2に結合するように機能する。中性ペプチドリガンドは、ErbB2を担持する特定の細胞型の、例えば細胞障害剤又は酵素でのターゲティングが望まれる本発明の態様において好ましい。
一般に、ErbB2の結合に加えて「アゴニスト」ペプチドリガンドは、ErbB2担持細胞に対して直接の効果を有している。好ましくは、アゴニストペプチドリガンドは記載されたようにErbB2に結合し、さらにErbB2-ErbB3タンパク質複合体及び/又はErbB2-ErbB4タンパク質複合体に関連した事象、例えば細胞内キナーゼドメインをしてErbB2レセプター複合体のチロシン残基をホスホリル化させる能力を開始又は媒介する。また、アゴニストペプチドリガンドの結合は、ErbB2レセプターの二量体化に関連している場合がある。ErbB2レセプター媒介性ホスホリル化又はErbB2レセプター二量体化を誘発する能力は、チロシンホスホリル化アッセイ及びSDS PAGE等の当該分野で標準的な技術を使用して定量することができる。
【0015】
これに対して「アンタゴニスト」ペプチドリガンドは、ErbB2-ErbB3タンパク質複合体及び/又はErbB2-ErbB4タンパク質複合体に結合する天然リガンドに関連した活性、例えば天然リガンドの結合により誘発される細胞反応を、例えばErbB2-ErbB3タンパク質複合体及び/又はErbB2-ErbB4タンパク質複合体を天然又は自然に発生するリガンドに結合させるか又はこれをブロックすることにより、低減するように作用する。
しかしながら、上述の分類に限定されるものではない。例えば、ある実施態様では、ペプチドリガンドの結合が、ErbB2レセプターの内在化、又はプログラム細胞死、すなわちアポトーシスの誘発等の関連した細胞事象の誘発の原因になる。ErbB2又はErbB2レセプター複合体の内在化を開始するペプチドリガンドは、ここで以下に記載する細胞障害剤を細胞内送達することを必要とする本発明の実施態様において特に有用である。
「アポトーシスを誘発する」又は「アポトーシスを誘発可能な」という用語は、アネキシンVの結合、DNAの断片化、細胞収縮、小胞体の拡張、細胞断片化、及び/又は膜小胞の形成(アポトーシス体と呼ばれる)により決定されるプログラム細胞死を誘発する化合物の能力を意味する。細胞は、ErbB2レセプターを発現又は過剰発現するものである。好ましくは「細胞」は、腫瘍細胞、例えば乳房、卵巣、胃、子宮体、唾液腺、肺、腎臓、大腸、甲状腺、膵臓又は膀胱細胞である。インビトロでは、細胞はSKBR3、BT474、Calu3細胞、MDA-MB-453、MDA-MB-361又はSKOV3細胞でありうる。アポトーシスに伴う細胞事象を評価するために種々の方法が利用できる。例えば、ホスファチジルセリン(PS)転位置をアネキシン結合により測定することができ;DNA断片化はDNAラダーリングにより評価することができ;DNA断片化に伴う細胞核/染色質凝結は低二倍体細胞の何らかの増加により評価することができる。好ましくは、アポトーシスを誘発する化合物は、BT474細胞を使用するアネキシン結合アッセイにおいて、未処理細胞の約2〜50倍、好ましくは約5〜50倍、最も好ましくは約10〜50倍のアネキシン結合を誘発するという結果を生じるものである。
【0016】
ここで使用される「ヘレグリン」(HRG)は、ErbB2-ErbB3及びErbB2-ErbB4タンパク質複合体を活性化する(すなわち、そこに結合する際に複合体のチロシン残基のリン酸化を誘発する)ポリペプチドを意味する。この用語に包含される種々のポリペプチドは、例えば、Jonesら, (1990) FEBS Letters 447:227-231;Holmesら,(1992) Science, 256:1205-1210;国際公開92/20798号;Wenら, (1994) Mol. Cell. Biol., 14(3):1909-1919;及びMarchionniら, (1993) Nature, 362:312-318に開示されている。この用語には、自然に生じたHRGポリペプチドの変異体及び/又は生物学的に活性な断片、例えばそのEGF様ドメイン断片(例えばHRGβ1177−244)が含まれる。
「ErbB2-ErbB3タンパク質複合体」と「ErbB2-ErbB4タンパク質複合体」は、それぞれErbB2レセプターと、ErbB3レセプター又はErbB4レセプターの非共有的に結合したオリゴマーである。Sliwkowskiら, (1994) J. Biol. Chem., 269(20):14661-14665に記載されているように、これらのレセプターの両方を発現する細胞に複合体が形成され、免疫沈降法により単離され、SDS-PAGEにより分析される。
本発明の特定の態様において使用されるところの「多量体化ドメイン」なる用語は、直接又は「リンカードメイン」を介してペプチドリガンドが結合する分子の一部を指すことを意味する。多量体化ドメインは、好ましい実施態様では、2又はそれ以上の多量体化ドメインの相互作用を容易にするアミノ酸ドメインである。多量体化ドメインは2又はそれ以上の多量体化ドメインの間の相互作用を促進するが、本発明では、多量体化ドメインに結合するペプチドリガンドが多量体の一部として存在している必要性はない。
【0017】
本発明の好ましい態様では、多量体化ドメインは、2又はそれ以上の多量体化ドメインの安定した相互作用を促進させるポリペプチドである。例えば、限定するものではないが、多量体化ドメインは免疫グロブリン配列、例えば免疫グロブリン定常領域、ロイシンジッパー、疎水性領域、親水性領域、2又はそれ以上の多量体化ドメイン間に分子間ジスルフィド結合を形成する遊離チオールを含有するポリペプチド、又は例えば米国特許第5731168号に記載されている「キャビティ中突起(protuberance-into-cavity)」ドメインであってよい。前記特許では、突起は、第1ポリペプチドの界面からの小さなアミノ酸側鎖を、より大きな側鎖(例えばチロシン又はトリプトファン)で置換することにより形成される。突起に対して同一又はより小さなサイズの相補的キャビティは、大きなアミノ酸側鎖を小さなもの(例えばアラニン又はスレオニン)で置換することにより、第2ポリペプチドの界面に任意につくり出される。
よって、好ましい態様では、多量体化ドメインは、2又はそれ以上の多量体化ドメインの安定した相互作用を促進するか、これを可能にし、モノマー性多量体ドメインからのダイマーと他の多量体の形成を促進させるか、これを可能にする分子の一部を提供する。好ましくは、本発明のこの態様では、多量体化ドメインは免疫グロブリンの定常領域ドメインである。免疫グロブリンの定常ドメインは、本発明の化合物のインビボ循環半減期を改善し、場合によっては、当業者をして、以下に記載する「エフェクター機能」を本発明の所定の態様に導入することを可能にするという利点をもたらす。
【0018】
本明細書と特許請求の範囲を通して、免疫グロブリン重鎖中の残基の番号付けは、ここに出典を明示して取り込まれるKabat等, Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991)のEUインデックスのものである。「KabatのEUインデックス」とはヒトIgG1EU抗体の残基番号付けを指す。
「抗体」(Ab)と「免疫グロブリン」(Ig)は同じ構造的特徴を有する糖タンパク質である。抗体は特定の抗原に対して結合特異性を示すものであるが、免疫グロブリンは、抗体と抗原特異性を欠く他の抗体様分子の両方を含む。後者の種類のポリペプチドは、例えばリンパ系により低レベルで、骨髄腫により増加したレベルで産生される。
「抗体」及び「免疫グロブリン」は、通常は、2つの同一の軽(L)鎖及び2つの同一の重(H)鎖からなる、約150000ダルトンのヘテロテトラマー糖タンパク質である。各軽鎖は一つの共有ジスルフィド結合により重鎖に結合しており、ジスルフィド結合の数は、異なった免疫グロブリンアイソタイプの重鎖間で変化する。また各重鎖と軽鎖は、規則的に離間した鎖間ジスルフィド結合を有している。各重鎖は、カルボキシ(C)末端定常ドメインが続くアミノ(N)末端可変ドメイン(VH)を有している。各軽鎖は可変N-末端ドメイン(VL)及びC-末端定常ドメインを有し;軽鎖の定常ドメイン(CL)は重鎖の最初の定常ドメイン(CH1)と整列し、軽鎖の可変ドメインは重鎖の可変ドメインと整列している。免疫グロブリンポリペプチド鎖のドメインの定義に従って、軽(L)鎖は2つのコンホメーション的に類似したドメインVL及びCLを有し;重鎖は4つのドメイン(VH、CH1、CH2、及びCH3)を有しており、これらの各々は1つの鎖間ジスルフィド架橋を持つ。
【0019】
重鎖の定常(C)ドメインのアミノ酸配列に依存して、免疫グロブリンは異なるクラスに分けることができる。免疫グロブリンの5つの主要なクラス:IgA、IgD、IgE、IgG及びIgMがある。免疫グロブリンクラスはさらにサブクラス(アイソタイプ)、例えばIgG1、IgG2、IgG3、IgG4、IgA1及びIgA2に分割することができる。免疫グロブリンの異なるクラスに対応する重鎖定常ドメインは、各々α、δ、ε、γ、及びμドメインである。任意の脊椎動物種からの抗体の軽鎖には、その定常ドメインのアミノ酸配列に基づいて、カッパ(κ)又はラムダ(λ)と呼ばれる2つの区別される型の一つが割り当てられる。配列の研究により、IgMのμ鎖は5つのドメインVH、CHμ1、CHμ2、CHμ3、及びCHμ4を含むことが分かっている。IgE(ε)の重鎖も5つのドメインを含む。
異なるクラスの免疫グロブリンのサブユニット構造及び三次元的立体配置はよく知られている。これらのうちIgA及びIgMはポリマーであり、各サブユニットは2つの軽鎖と2つの重鎖を含む。IgG(γ)の重鎖はヒンジ領域として知られるCHγ1及びCHγ2ドメイン間にある所定長さのポリペプチド鎖を含む。IgAのα鎖はO-結合グリコシル化部位を含むヒンジ領域を有し、μ及びε鎖はγ及びα鎖のヒンジ領域に類似する配列は持たないが、それらは他のものが持たない第4の定常ドメインを含む。免疫グロブリン鎖のドメイン組成は次のように要約することができる。
軽鎖λ=Vλ Cλ
κ=Vκ Cκ
重鎖 IgG(γ)=VH CHγ1 ヒンジ CHγ2 CHγ3
IgM(μ)=VH CHμ1 CHμ2 CHμ3 CHμ4
IgA(α)=VH CHα1 ヒンジ CHα2 CHα3
IgE(ε)=VH CHε1 CHε2 CHε3 CHε4
IgD(δ)=VH CHδ1 ヒンジ CHδ2 CHδ3
【0020】
ヒトIgG Fc領域の「CH2ドメイン」(「Cγ2」ドメインとも呼ばれる)は通常約アミノ酸231から約アミノ酸340まで延びる。CH2ドメインは、他のドメインと密接には対をなさないという点で独特である。むしろ、二つのN-結合分岐炭水化物鎖が未変性の天然IgG分子の2つのCH2ドメインの間に挿入されている。
「CH3ドメイン」は、Fc領域における残基C-末端からCH2ドメインまでの伸展(すなわち、IgGのおよそアミノ酸残基341からおよそアミノ酸残基447まで)を含む。
「ヒンジ領域」はヒトIgG1のGlu216からPro230の伸展として一般に定義されている(Burton, Molec. Immunol. 22:161-206 (1985))。他のIgGアイソタイプのヒンジ領域は、重鎖間S-S結合を形成する最初と最後のシステイン残基を同じ位置に配することにより、IgG1配列と整列させられうる。
Fc領域の「低ヒンジ領域」は、通常は、C-末端からヒンジ領域、すなわちFc領域の233から239残基までの残基の伸展として定義されている。
抗体のパパイン消化は、「Fab」断片又は領域と呼ばれる2つの同一の抗原結合断片を生成し、各々が単一の抗原結合部位と残りの「Fc」断片又は領域を持つ。免疫グロブリン重鎖のFc領域の境界は変化するかも知れないが、ヒトIgG重鎖Fc領域は、通常はCys226またはPro230の位置のアミノ酸残基からカルボキシル末端まで伸展すると定義される。免疫グロブリンのFc領域は、例えば図1に示すように、2つの定常ドメイン、CH2及びCH3を含む。「天然Fc領域配列」は天然に見出されるFc領域のアミノ酸配列と同一のアミノ酸配列を含む。天然ヒトFc領域配列を図2及び3に示し、ヒトIgG1Fc領域(非-A及びAアロタイプ);ヒトIgG2Fc領域;ヒトIgG3Fc領域;及びヒトIgG4Fc領域、並びに自然に生じるその変異体を含むが、それらに限定されるものではない。天然マウスFc領域配列を図2Aに示す。
【0021】
ペプシン処理により、2つの抗原結合部位を有し、さらに抗原を架橋させ得るF(ab')2断片が得られる。またFab断片は、軽鎖の定常ドメインと重鎖の第1定常ドメイン(CH1)を有する。Fab'断片は、抗体ヒンジ領域からの一又は複数のシステインを含む重鎖CH1ドメインのカルボキシ末端に数個の残基が付加している点でFab断片とは異なる。Fab'-SHは、定常ドメインのシステイン残基が遊離チオール基を担持しているFab'に対するここでの命名である。F(ab')2抗体フラグメントは、間にヒンジシステインを有するFab'断片の対として生産された。抗体断片の他の化学結合も知られている。
「機能的なFc領域」は、天然のFc領域の「エフェクター機能」を保持する。例示的な「エフェクター機能」は、C1q結合;補体依存性細胞障害活性;Fcレセプター結合、抗体依存性細胞媒介細胞障害活性(ADCC);貪食作用、細胞表面レセプター(すなわち、B細胞レセプター;BCR)の低減下を含む。このようなエフェクターの機能は、一般に、Fc領域が結合ドメイン(例えばペプチドリガンド)に結合するのに必要とされており、当該技術で公知の様々なアッセイを用いることで評価される。
親又は天然Fc領域に適当なアミノ酸修飾を導入することにより、(a)ヒトエフェクター細胞の存在下で抗体依存性細胞媒介細胞傷害(ADCC)をより効果的に媒介する、及び/又は(b)親ポリペプチドよりも、より強い親和性でFcガンマレセプター(FcγR)と結合するような、Fc領域の変異体を作製することができる。そのようなFc領域の変異体は、一般に、少なくとも1つのアミノ酸修飾をFc領域に含むであろう。Fc領域の変異体は、2,3,4,5個所等の置換を含み得る。
【0022】
いくつかの抗体のエフェクター機能は、抗体のFc領域に結合するFcレセプター(FcRs)によって媒介される。FcRsは、免疫グロブリンアイソタイプに対する特異性により定義され;IgG抗体に対するFcレセプターはFcγRと、IgEに対してはFcεRと、IgAに対してはFcαRなどのように呼ばれている。FcγRの3つのサブタイプは同定されており:FcγRI(CD64),FcγRII(CD32)及びFcγRIII(CD16)である。それぞれのFcγRサブクラスは、2または3つの遺伝子によってコードされており、またRNAスプライシングにより複数の転写産物が生じるため、FcγRアイソフォームには広い多様性が存在する。これらの異なるFcRサブタイプは異なる型の細胞上で発現されている(Ravetch and Kinet, Annu.Rev.immunol. 9:457-492 (1991))。例えばヒトにおいて、FcγRIIIBは好中球上にのみ見出され、FcγRIIIAはマクロファージ、単球、ナチュラルキラー細胞(NK細胞)、ある種のT細胞上に見出される。特に、FcγRIIIAは、ADCCに関係する細胞型であるNK細胞上に存在する唯一のFcγRである。
FcγRに対するヒトおよびマウス抗体上の結合部位は、これまでに低ヒンジ領域(233-239残基:Kabatら, Sequences of Proteins of Immunological Interest, 5th ED. Public health Service, National Institutes of Health, Bethesda, MD. (1991) におけるようなEUインデックス番号付け)にマッピングされている。Woofら Molec. Immunol. 23:319-330(1986); Duncanら Nature 332:563 (1988); Canfield及びMorrison, J. Exp. Med. 173:1483-1491 (1991); Canppelら, Proc.Natl.Acad.Sci USA 88:9036-9040 (1991)。
「C1q」は免疫グロブリンのFc領域に対する結合部位を含むポリペプチドである。C1qは2つのセリンプロテアーゼC1r及びC1sと共に、補体依存性細胞障害(CDC)経路の最初の成分である複合体C1を形成する。
【0023】
「抗体依存性細胞媒介細胞障害」及び「ADCC」は、細胞が媒介する反応であって、FcRsを発現する非特異的障害細胞(例えば、ナチュラルキラー(NK)細胞、好中球、及びマクロファージ)が、標的細胞上に結合した抗体を認識して、続いて標的細胞の溶解を引き起こす反応を意味する。ADCCを媒介する主要な細胞である、NK細胞はFcγRIIIのみを発現するのに対し、単球は、FcγRI、FcγRII及びFcγRIIIを発現する。造血細胞におけるFcRの発現は、Ravetch及びKinet, Annu. Rev. Immunol 9:457-92 (1991) の464頁の表3に要約されている。
ここで使用される場合、「サルベージレセプター結合リガンド」という用語は、IgG分子のインビボでの血清半減期を増加させる原因となるIg分子(例えばIgG1、IgG2、IgG3又はIgG4)のFc領域のリガンドを意味する(1998年4月14日に公開された米国特許第5739277号)。
「治療」とは、治療的処置及び予防又は防止手段の両方を指す。治療の必要があるものには、既に羅患しているもの、並びに疾患が予防されるべきものが含まれる。
治療の目的とされる「哺乳動物」とは、ヒト、家庭又は農場用動物、及び動物園、スポーツ又はペット用動物、例えばイヌ、ウマ、ネコ、ウシ等を含む、哺乳動物に分類されるあらゆる動物を意味する。好ましくは哺乳動物はヒトである。
「疾患」は本発明のペプチドリガンドを含有する組成物で治療することで恩恵を得るあらゆる症状のことである。これには、問題の疾患に哺乳動物を罹患させる素因になる病理状態を含む、慢性及び急性の疾患又は病気が含まれる。ここで治療される疾患の例は、これに限定されるものではないが、良性及び悪性の腫瘍;白血病及びリンパ悪性腫瘍;ニューロン、神経膠、星状細胞、視床下部及び他の腺、マクロファージ、上皮、ストローマ及び割腔の疾患;及び炎症、血管形成及び免疫性疾患が含まれる。
【0024】
「癌」及び「癌性」という用語は、典型的には調節されない細胞成長により特徴付けられる、哺乳動物における生理学的状態を意味するか記述するものである。癌の例には、これらに限定されるものではないが、癌腫、リンパ腫、芽細胞腫、肉腫、及び白血病が含まれる。このような癌のより特定な例には、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、神経膠芽細胞腫、子宮頸癌、卵巣癌、肝臓癌、膀胱癌、肝腫瘍、乳癌、大腸癌、結腸直腸癌、子宮内膜癌、唾液腺癌、腎臓癌、肝臓癌、前立腺癌、陰門癌、甲状腺癌、肝癌及び様々な種類の頭部及び頸部の癌が含まれる。
ここで使用される場合、「非経口」なる用語は、腸以外によって、特に静脈内(i.v.)、動脈内(i.a.)、腹膜内(i.p.)、筋肉内(i.m.)、心室内及び皮下(s.c.)経路で本発明の化合物を導入することを意味する。
【0025】
(発明の実施形態)
ペプチドリガンド
本発明のペプチドリガンドはErbB2に結合し、次の一般式:
Xaa(1−14) -Cys-Xaa16-Gly-Pro-Gly-Cys-Xaa(21−27) (配列番号:90)
を有するペプチドリガンドを用いたインビトロアッセイにおいてErbB2への結合に対して競合する能力により同定することができ、ここで、Xaa(1−14)は存在しないか、又は1〜14のアミノ酸であり;Xaa16はMet、Thr、Cys及びIleからなる群から選択され、好ましくはIleであり;Xaa(21−27)は存在しないか、又は1〜7のアミノ酸である。好ましくは、ペプチドリガンドは、次の一般式:
Xaa(1−3)-Glu-Xaa5-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Xaa14-Xaa15-Leu-Xaa(17−20) (配列番号:91)
を有するペプチドリガンドと上述のようにErbB2との結合に対して競合するであろう。ここで、Xaa(1−3)及びXaa(17−20)は、独立して、存在しないか、又はそれぞれ1〜3及び1〜4のアミノ酸であり;Xaa1、Xaa2及びXaa3はアミノ酸である。特定の実施態様では、本発明のペプチドリガンドは、ここに記載する配列番号:1−配列番号:44、及び配列番号:57−配列番号:69で表されるペプチドリガンドの何れかと競合し、好ましくはErbB2との結合に対して配列番号:14のものと競合するであろう。
以上から理解されるように、「競合する」及び「競合する能力」という用語は相対的な用語である。よって、本発明のペプチドリガンドを記述するために用いられる場合、この用語は、ここで記載したような標準的な競合アッセイにおいて、50μMで存在する場合、好ましくは1μM、より好ましくは100μMで存在する場合、好ましくは1nM又はそれ未満で存在する場合の、例えば配列番号:14の結合を50%阻害するペプチドリガンドを意味する。このようなペプチドリガンドは、実施例のセクションに記載するもののような標準的な競合アッセイで測定されるときに約1μM未満、好ましくは約100nM未満、より好ましくは約10nM未満の親和性でErbB2に一般的に結合する。しかしながら、ErbB2に対して約1nM未満、好ましくは約1pM〜1nMの親和性を有するペプチドリガンドも等しく本発明のペプチドリガンドであるようである。
【0026】
化合物が上述したペプチドリガンドと競合するか、競合する能力を有しているか否かを決定するためのインビトロアッセイシステムに対しては、当業者であれば多数の標準的な競合アッセイの任意のものを使用することができる。このような手順には、限定するものではないが、数例を挙げると、ラジオイムノアッセイ、酵素イムノアッセイ(EIA)、好ましくは酵素結合イムノアッセイ(ELISA)、「サンドウィッチ」イムノアッセイ、免疫放射定量アッセイ、蛍光イムノアッセイ、免疫電気泳動アッセイ等の技術を用いる競合アッセイシステムを含む。
例として、限定するものではないが、ELISAアッセイを行うことができ、そこでは、マクロタイタープレート(例えば、Nunc MaxisorpTM96-ウェルプレート、Inter Med, Denmark)を、50mMの重炭酸アンモノウム、pH9.3にHER2-ECDが入ったものを5μg/ml使用し、4℃で終夜被覆する。ウェルは25℃で1時間、BSAを1%含有するPBS(PBS-BSA)を使用してブロックすることができる。候補ペプチドリガンドをPBS-BSAで希釈し、ここに記載されたようにZプロテイン融合体(1.1.FI-Z)として生成され、ビオチン化(1.1.FI-Zb)されたペプチド1.1.FI(配列番号:14)の、固定化されたHER2-ECDへの結合をブロックする能力を試験した。約1時間のインキュベーションに続いて、プレートをPBS-TWEENで洗浄し、ストレプトアビジン-HRPを約30分間で添加する。プレートをPBS-TWEENで再び洗浄し、結合したHRPをABTS/H2O2基質を使用してアッセイする。405nmの吸光度の変化を評価する。吸光度の減少を試料濃度に対してプロットし、各候補ペプチドリガンドに対するIC50を決定する。
先に見られたように、本発明の好ましいペプチドリガンドは約10〜30アミノ酸残基、好ましくは約20アミノ酸残基の自然には生じないペプチドリガンドである。上述した特に好ましいペプチドリガンドは自然に生じるアミノ酸からなり、当業者によく知られている標準的な組換え及び合成技術を使用して産生することができる。
【0027】
好ましいペプチドリガンドは次の一般式:
Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Cys-Xaa-Xaa-Xaa-Xaa-Cys-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa
を有する。些かより特定的には、好ましいペプチドリガンドは次の式:
Xaa(1−7) -Cys-Xaa9-Gly-Pro-Gly-Cys-Xaa(14−20) (配列番号:92)
を有し、ここで、Xaa(1−7)及びXaa(14−20)は、独立して、存在しないか、又は1〜7のアミノ酸であり;Xaa9はMet、Ile及びThrからなる群から選択されるアミノ酸、好ましくはIleである。些かより特定的には、好ましいペプチドリガンドは次の式:
Xaa(1−5) -Trp-Xaa7-Cys-Xaa9-Gly-Pro-Gly-Cys-Xaa(14−20) (配列番号:93)
を有し、ここで、Xaa(1−5)は、存在しないか、又は1〜5のアミノ酸であり;Xaa7はAsp、Gly、Glu及びHisからなる群から選択され、好ましくはGlyである。本発明のこの態様においてより好ましいものは、次の式:
Xaa(1−3) -Glu-Xaa5-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Xaa14-Xaa15--Leu-Xaa(17−20) (配列番号:91)
を有するペプチドリガンドであり、ここで、Xaa(1−3)-は存在しないか、又は1〜3のアミノ酸であり、Xaa5は一つのアミノ酸であり、Xaa13は一つのアミノ酸であり、Xaa14は一つのアミノ酸であり、-Xaa(17−20)は存在しないか、又は1〜4のアミノ酸である。本発明のこの態様における例示的なペプチドリガンドは、X(1−3)-は存在しないか、又はGln-Arg-Asn-、Leu-Ser-Pro-、Glu-Asn-Trp-、Ala-Ser-His-、Lys-Leu-Asn-、Thr-Gln-Ala-、Ala-Pro-Arg-、Gln-Val-Tyr-、Arg-Thr-Glu-、Phe-Ala-Gly-、Thr-Ala-Arg-、Arg-Pro-His-、Asn-Val-Cys-、Cys-Ile-Asp-、Tyr-Glu-Trp-、Arg-Trp-Asp-、His-Trp-Met-、Asn-Trp-Pro-、Phe-Asn-Trp-、Phe-Ser-Gly-、Gly-Gly-Trp-、Leu-Trp-Phe-、Gly-Ile-Pro-、Trp-Trp-Thr-、Leu-Gly-Trp-、Ser-Pro-Trp-、Arg-Gly-Trp-、Tyr-Glu-Phe-、Tyr-Glu-Gly-、Tyr-Glu-Val-、Tyr-Ser-Phe-、Tyr-Asp-Phe-、Asp-Glu-Val-、Ser-Glu-Val-、Phe-Glu-Phe-、及びHis-Asp-Val-からなる群から選択されるペプチドリガンドであり、Xaa5はAla、Thr、Met、Val、Arg、Glu、Asp、Ser、Gln、Pro、Gly、Phe及びLysからなる群から選択され;Xaa14はGlu、Lys、Arg、Asp、Ser、Ala、Asn、Thr、Gly、Pro、Val及びGlnからなる群から選択され;Xaa15はMet、Phe、Ala、Cys、Gln、Glu、Trp、Leu、Val、Tyr、Ser及びAsnからなる群から選択され;-X(17−20)は次の例示的配列:
-Phe-Gly-Ala-His (配列番号:94)
-Phe-Asp-Ala-His (配列番号:95)
-Leu-Glu-Ala-His (配列番号:96)
-Phe-Glu-Gly-His (配列番号:97)
-Phe-Gly-Ala-Leu (配列番号:98)
-Phe-Glu-Ala-Tyr (配列番号:99)
-Phe-Ala-Gly-His (配列番号:100)及び
-Phe-Glu-Ala-Phe (配列番号:101)
を有する4つのアミノ酸のペプチドである。
好ましくは、-Xaa(17−20)は、次の式:
-Xaa17 -Xaa18-Cys-Xaa20
を有する4つのアミノ酸のペプチドである。
【0028】
本発明のこの態様における例示的ペプチドリガンドは、-Xaa(17−20)が次の式:
-Gln-Ala-Cys-Met (配列番号:102)
-Leu-Gln-Cys-Trp (配列番号:103)
-Met-Ser-Cys-Val (配列番号:104)
-Leu-Arg-Cys-Ile (配列番号:105)
-Gln-Ala-Cys-Leu (配列番号:106)
-Leu-Ser-Cys-Leu (配列番号:107)
-Ile-Gly-Cys-Leu (配列番号:108)
-Leu-Ala-Cys-Leu (配列番号:109)
-Leu-Ser-Cys-Ile (配列番号:110)
-Met-Asn-Cys-Leu (配列番号:111)
-Leu-Arg-Cys-Leu (配列番号:112)
-Leu-Lys-Cys-Leu (配列番号:113)
-Leu-Gly-Cys-Leu (配列番号:114)
-Leu-Asn-Cys-Ile (配列番号:115)
-Met-Gly-Cys-Leu (配列番号:116)及び
-Met-Ala-Cys-Leu (配列番号:117)
からなる群から選択されるペプチドリガンドである。
【0029】
本発明のこの態様における他の実施態様では、-Xaa(17−20)が次の式:
-Cys-Xaa18 -Xaa19-Cys
を有する4つのアミノ酸ペプチドであり、Xaa18が一つのアミノ酸であり、Xaa19が一つのアミノ酸であるペプチドリガンドである。本発明のこの態様では、例示的ペプチドリガンドは:
-Cys-Ala-Trp-Cys (配列番号:118)
-Cys-Ser-Trp-Cys (配列番号:119)
-Cys-Glu-Pro-Cys (配列番号:120)
-Cys-Asp-Trp-Cys (配列番号:121)
-Cys-Glu-Trp-Cys (配列番号:122)
-Cys-Asn-Trp-Cys (配列番号:123)及び
-Cys-Gly-Trp-Cys (配列番号:124)
からなる群から選択される-Xaa(17−20)を含むものである。
【0030】
本発明のこの態様における例示的ペプチドリガンドは、
-Gln-Arg-Asn-Glu-Ala-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Met-Leu-Cys-Ala-Trp-Cys (配列番号:1)
-Leu-Ser-Pro-Glu-Thr-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Met-Leu-Cys-Ser-Trp-Cys (配列番号:2)
-Glu-Asn-Trp-Glu-Met-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Lys-Phe-Leu-Cys-Glu-Pro-Cys (配列番号:3)
-Glu-Val-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Lys-Ala-Leu-Cys-Asp-Trp-Cys (配列番号:4)
-Glu-Arg-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Arg-Met-Leu-Cys-Glu-Trp-Cys (配列番号:5)
-Glu-Val-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Asp-Met-Leu-Cys-Asn-Trp-Cys (配列番号:6)
-Glu-Val-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ser-Met-Leu-Cys-Gly-Trp-Cys (配列番号:7)
-Xaa-Xaa-Xaa-Glu-Xaa-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Xaa-Met-Leu-Cys-Xaa-Trp-Cys (配列番号:8)
-Ala-Ser-His-Glu-Val-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Lys-Cys-Leu-Gln-Ala-Cys-Met (配列番号:9)
-Lys-Leu-Asn-Glu-Glu-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Cys-Leu-Leu-Gln-Cys-Trp (配列番号:10)
-Lys-Leu-Asn-Glu-Asp-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Cys-Leu-Leu-Xaa-Cys-Xaa (配列番号:11)
-Thr-Gln-Ala-Glu-Arg-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Cys-Leu-Met-Ser-Cys-Val (配列番号:12)
-Ala-Pro-Arg-Glu-Val-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Cys-Leu-Leu-Arg-Cys-Ile (配列番号:13)
-Gln-Val-Tyr-Glu-Ser-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Cys-Leu-Gln-Ala-Cys-Leu (配列番号:14)
-Arg-Thr-Glu-Glu-Gln-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Arg-Cys-Leu-Leu-Ser-Cys-Leu (配列番号:15)
-Phe-Ala-Gly-Glu-Ser-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Cys-Leu-Ile-Gly-Cys-Leu (配列番号:16)
-Thr-Ala-Arg-Glu-Val-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Asn-Cys-Leu-Leu-Ala-Cys-Leu (配列番号:17)
-Arg-Pro-His-Glu-Pro-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ser-Cys-Leu-Leu-Ser-Cys-Ile (配列番号:18)
-Glu-Val-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Cys-Leu-Met-Asn-Cys-Leu (配列番号:19)
-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Cys-Leu-Leu-Arg-Cys-Leu (配列番号:20)
-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Gly-Cys-Leu-Leu-Lys-Cys-Leu (配列番号:21)
-Glu-Pro-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Cys-Leu-Leu-Gly-Cys-Leu (配列番号:22)
-Glu-Glu-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Cys-Leu-Leu-Asn-Cys-Ile (配列番号:23)
-Glu-Gln-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Lys-Cys-Leu-Met-Gly-Cys-Leu (配列番号:24)
-Glu-Gln-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Gly-Cys-Leu-Leu-Arg-Cys-Leu (配列番号:25)
-Glu-Ala-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Gly-Cys-Leu-Met-Ala-Cys-Leu (配列番号:26)
-Xaa-Xaa-Xaa-Glu-Xaa-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Xaa-Cys-Leu-Leu-Xaa-Cys-Leu (配列番号:27)
-Asn-Val-Cys-Glu-Phe-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Gln-Leu-Cys (配列番号:28)
-Cys-Ile-Asp-Glu-Thr-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Glu-Leu-Arg-Cys-Lys-Arg (配列番号:29)
-Tyr-Glu-Trp-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Pro-Ala-Leu-Gly-Phe-Gly-Tyr (配列番号:30)
-Arg-Trp-Asp-Glu-Glu-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Trp-Leu-Val-Val-Arg-Lys (配列番号:31)
-His-Trp-Met-Glu-Arg-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Gly-Phe-Leu (配列番号:32)
-Asn-Trp-Pro-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Lys-Leu-Leu (配列番号:33)
-Phe-Asn-Trp-Glu-Lys-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Arg-Thr-Leu (配列番号:34)
-Phe-Ser-Gly-Glu-Arg-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Gln-Val-Leu (配列番号:35)
-Gly-Gly-Trp-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Arg-Tyr-Leu (配列番号:36)
-Leu-Trp-Phe-Glu-Arg-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Thr-Xaa-Leu (配列番号:37)
-Gly-Ile-Pro-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Trp-Leu (配列番号:38)
-Trp-Trp-Thr-Glu-Arg-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ser-Met-Leu (配列番号:39)
-Xaa-Cys-Xaa-Glu-Arg-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ser-Met-Leu (配列番号:40)
-Leu-Gly-Trp-Glu-Arg-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Arg-Ala-Leu (配列番号:41)
-Ser-Pro-Trp-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Arg-Trp-Leu (配列番号:42)
-Arg-Gly-Trp-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ser-Phe-Leu (配列番号:43)
-Xaa-Xaa-Trp-Glu-Xaa-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Xaa-Xaa-Leu-Xaa-Xaa-Xaa-Xaa (配列番号:44)
-Tyr-Glu-Val-Glu-Ala-Trp-Asp-Cys-Met-Gly-Pro-Gly-Cys-Ala-Asn-Leu-Phe-Glu-Ala-His (配列番号:57)
-Tyr-Glu-Phe-Glu-Gly-Trp-Asp-Cys-Met-Gly-Pro-Gly-Cys-Ala-Ser-Val-Phe-Gly-Ala-His (配列番号:58)
-Tyr-Glu-Gly-Glu-Ser-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Ser-Leu-Phe-Asp-Ala-His (配列番号:59)
-Tyr-Glu-Val-Glu-Val-Trp-Glu-Cys-Ile-Gly-Pro-Gly-Cys-Gly-Tyr-Leu-Phe-Gly-Ala-His (配列番号:60)
-Tyr-Glu-Val-Glu-Gly-Trp-Gly-Cys-Met-Gly-Pro-Gly-Cys-Ala-Phe-Leu-Leu-Glu-Ala-His (配列番号:61)
-Tyr-Ser-Phe-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Tyr-Leu-Phe-Glu-Gly-His (配列番号:62)
-Tyr-Asp-Phe-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Gly-Asn-Leu-Leu-Glu-Ala-His (配列番号:63)
-Tyr-Asp-Phe-Glu-Gly-Trp-Asp-Cys-Thr-Gly-Pro-Gly-Cys-Ala-Tyr-Leu-Phe-Glu-Gly-His (配列番号:64)
-Asp-Glu-Val-Glu-Ser-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Tyr-Leu-Phe-Gly-Ala-Leu (配列番号:65)
-Ser-Glu-Val-Glu-Val-Trp-His-Cys-Ile-Gly-Pro-Gly-Cys-Val-Tyr-Leu-Phe-Glu-Ala-Tyr (配列番号:66)
-Phe-Glu-Phe-Glu-Gly-Trp-Glu-Cys-Met-Gly-Pro-Gly-Cys-Ala-Glu-Leu-Phe-Ala-Gly-His (配列番号:67)
-His-Asp-Val-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Asp-Leu-Phe-Glu-Ala-Phe (配列番号:68)
-Tyr-Glu-Phe-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Tyr-Leu-Phe-Glu-Ala-His (配列番号:69)
と、上述した例示的アミノ酸配列の保存的アミノ酸置換を含む。
【0031】
さらに好ましいペプチド化合物は次の一般式:
Cys-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Cys-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Cys-Cys-Xaa-Xaa-Xaa-Cys-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Cys (配列番号:125)
を有する。些かより特定的には、次の式:
Xaa(1−10)-Glu-Xaa12-Trp-Xaa14-Cys-Cys-Gly-Pro-Gly-Cys-Xaa21-Xaa22-Xaa23-Xaa(24−27) (配列番号:126)
を有し、ここでXaa(1−10)は存在しないか、又は1〜10のアミノ酸であり、Xaa12は一つのアミノ酸であり、Xaa14は一つのアミノ酸であり、Xaa21は一つのアミノ酸であり、Xaa22は一つのアミノ酸であり、Xaa23はVal及びLeuからなる群から選択されるアミノ酸であり、Xaa(24−27)は存在しないか、又は1〜4のアミノ酸であるものである。本発明のこの態様において、好ましくは、Xaa12はVal、Leu、Ser、Trpからなる群から選択されるアミノ酸であり;Xaa14はGln、Glu、Asp及びHisからなる群から選択されるアミノ酸であり;Xaa21はGly及びGluからなる群から選択されるアミノ酸であり;Xaa22はTrp、Leu及びPheからなる群から選択されるアミノ酸であり;Xaa23はValである。
本発明のこの態様において好ましいペプチドリガンドは、Xaa(1−10)が次の式:
Cys-Xaa(2−7)-Cys-Xaa9-Gly-
を有する10のアミノ酸であり、Xaa(24−27)が次の式:
Xaa(24−26)-Cys
を有する4つのアミノ酸であり、Xaa(2−7)が6つのアミノ酸であり、Xaa(24−26)が3つのアミノ酸であるものを含む。
【0032】
本発明のこの態様において好ましいものは、Xaa(1−10)が次の式:
Cys-Xaa2-Trp-Val-Xaa5−Xaa6-Xaa7-Cys-Xaa9-Gly- (配列番号:127)
を有し、ここでXaa2がAla及びSerからなる群から選択されるアミノ酸であり、Xaa5がSer、Leu、Ala、Arg及びValからなる群から選択されるアミノ酸であり、Xaa6がPhe、Val及びLeuからなる群から選択されるアミノ酸であり、Xaa7がAsp、Gln、Tyr、Trp、Leu及びHisからなる群から選択されるアミノ酸であり、Xaa9がGly、Phe及びLeuからなる群から選択されるアミノ酸であるペプチドリガンドである。
この群のペプチド中で好ましいものは、Xaa(1−10)が次の式:
Cys-Ala-Trp-Val-Leu-Xaa6-Xaa7-Cys-Gly-Gly- (配列番号:128)
を有し、Xaa(24−26)が次の式:
Xaa24−Xaa25-Xaa26
を有し、ここでXaa24がTrp、Val、Gly及びAlaからなる群から選択されるアミノ酸、好ましくはValであり;Xaa25がAsn、Lys、Asp、Glu及びHisからなる群から選択されるアミノ酸、好ましくはAsnであり、Xaa26がAla、Ser及びValからなる群から選択されるアミノ酸、好ましくはAlaであるペプチドリガンドである。
【0033】
例として、限定するものではないが、次のもの:
Cys-Ser-Trp-Val-Leu-Val-Gln-Cys-Gly-Gly-Glu-Trp-Trp-His-Cys-Cys-Gly-Leu-Gly-Cys-Gly-Leu-Val-Val-Asn-Ala-Cys (配列番号:45)
Cys-Ala-Trp-Val-Ser-Phe-Glu-Cys-Gly-Gly-Glu-Val-Trp-His-Cys-Cys-Gly-Leu-Gly-Cys-Gly-Trp-Val-Trp-Lys-Ala-Cys (配列番号:46)
Cys-Ala-Trp-Val-Leu-Val-Gln-Cys-Gly-Gly-Glu-Trp-Trp-His-Cys-Cys-Gly-Pro-Gly-Cys-Glu-Phe-Val-Val-Asp-Ala-Cys (配列番号:47)
Cys-Ala-Trp-Val-Ala-Val-Tyr-Cys-Gly-Gly-Glu-Leu-Trp-His-Cys-Cys-Gly-Pro-Gly-Cys-Gly-Phe-Val-Val-Asp-Ser-Cys (配列番号:48)
Cys-Ala-Trp-Val-Arg-Val-Trp-Cys-Phe-Gly-Glu-Trp-Trp-Asp-Cys-Cys-Gly-Leu-Gly-Cys-Gly-Trp-Val-Val-Asn-Val-Cys (配列番号:49)
Cys-Ala-Trp-Val-Arg-Val-Leu-Cys-Gly-Gly-Glu-Trp-Trp-His-Cys-Cys-Gly-Leu-Gly-Cys-Gly-Trp-Val-Val-Glu-Ala-Cys (配列番号:50)
Cys-Ser-Trp-Val-Ser-Val-Leu-Cys-Gly-Gly-Glu-Trp-Trp-Gln-Cys-Cys-Gly-Pro-Gly-Cys-Gly-Leu-Val-Val-Asn-Ala-Cys (配列番号:51)
Cys-Ser-Trp-Val-Ser-Leu-Gln-Cys-Gly-Gly-Glu-Trp-Trp-His-Cys-Cys-Gly-Gly-Gly-Cys-Gly-Trp-Val-Val-Asn-Val-Cys (配列番号:52)
Cys-Ser-Trp-Val-Leu-Leu-His-Cys-Gly-Gly-Glu-Trp-Trp-His-Cys-Cys-Gly-Gly-Gly-Cys-Gly-Trp-Val-Gly-Glu-Ala-Cys (配列番号:53)
Cys-Ser-Trp-Val-Leu-Leu-Glu-Cys-Gly-Gly-Glu-Leu-Trp-Glu-Cys-Cys-Gly-Leu-Gly-Cys-Gly-Trp-Val-Ala-Asp-Ala-Cys (配列番号:54)
Cys-Ser-Trp-Val-Val-Phe-Glu-Cys-Leu-Gly-Glu-Ser-Trp-His-Cys-Cys-Gly-Gly-Gly-Cys-Gly-Trp-Val-Val-His-Ala-Cys (配列番号:55)
Cys-Ala-Trp-Val-Ser-Val-Glu-Cys-Gly-Gly-Glu-Trp-Trp-His-Cys-Cys-Gly-Pro-Gly-Cys-Gly-Trp-Val-Val-Asp-Ala-Cys (配列番号:56)
が本発明の適切なペプチドリガンドである。
【0034】
ペプチドリガンドの組み合わせ
A.多量体化ドメイン
本発明の好適な実施態様では、ペプチドリガンドは多量体化ドメインと組み合わされる。本発明のこの側面においては、少なくとも二つの別のドメインを有するハイブリッド分子が提供される。各分子はペプチドリガンドドメインと多量体化ドメインを有している。本発明では、ペプチドリガンドドメインは任意には柔軟なリンカードメインを介して免疫グロブリンFc領域のような多量体化ドメインに結合されている。
本発明のハイブリッド分子は適切な多量体化ドメインにペプチドリガンドを組み合わせることにより構築される。通常は、本発明のハイブリッド分子を調製する場合、ペプチドリガンドをコード化する核酸は多量体化ドメイン配列をコード化する核酸に作用可能に結合される。典型的には、作成物は、ペプチドリガンドのC末端が多量体化ドメインのN末端又はC末端、好ましくはC末端に結合されている融合タンパク質をコードしている。しかし、ペプチドリガンドのN末端が多量体化ドメインのN末端又はC末端に結合されている融合体もまた可能である。
好適な多量体化ドメインは免疫グロブリンの定常領域配列である。典型的には、そのような融合体では、コードされたハイブリッド分子は免疫グロブリン重鎖の定常領域の少なくとも機能的に活性なヒンジ、CH2及びCH3ドメインを保持しているであろう。融合体はまた例えば定常ドメインのFc部分のC末端、又は重鎖のCH1の直ぐ近くのN末端又は軽鎖の対応する領域に対して生成される。
【0035】
免疫グロブリン定常ドメインへのペプチドリガンドの融合がなされる正確なアミノ酸部位は重要ではない;特定の部位はよく知られており、生物学的活性、分泌又は結合特性を最適化するために選択することができる。この点に関して、当業者は文献(米国特許第5116964号、同第5714147号及び同第5336603号;Caponら、(1989) Nature 337:525-531; Trauneckerら, (1989) Nature 339:68-70;及びByrnら, (1990) Nature 344:667-670; Watsonら, (1990) J. Cell. Biol. 110:2221-2229; Watsonら, (1991) Nature 349:164-167; Aruffoら, (1990) Cell 61:1303-1313; Linsleyら, (1991) J. Exp. Med. 173:721-730; Lisleyら, J. Exp. Med. 174:561-569; Stamenkovicら, Cell 66:1133-1144; Ashkenaziら, (1991) Proc. Natl. Acad. Sci. USA 88:10535-10539; Lesslaierら, (1991) Eur. J. Immunol. 27:2883-2886; 及びPeppelら, (1991) J. Exp. Med. 174:1483-1489; Mohlerら, (1993) J. Immunol. 151:1548-1561; Bennettら, (1991) J. Biol. Chem. 266:23060-23067; Kurschnerら, (1992) J. Biol. Chem. 267:9354-9360; Chalupnyら, (1992) PNAS USA 89:10360-10364; Ridgway及びGorman, (1991) J. Cell. Biol. 115, Abstract No. 1448)に記載された様々なイムノアドヘシンの構築を参考にすることができる。
特定の側面では、免疫グロブリンタイプの多量体化ドメインは機能性Fcドメインを有するダイマーのような多量体を提供するように選択される。好適な側面では、多量体化ドメインは天然の免疫グロブリンFc領域に伴うエフェクター機能を有しているFcドメインを提供するように選択される。従って、ペプチドリガンドは、特定の側面では、特定のエフェクター機能又は機能群に対して選択された機能性Fcドメインを有する多量体を提供する免疫グロブリン重鎖定常ドメインに結合される。この場合、免疫グロブリン鎖−ペプチドリガンド配列をコードするDNAは典型的には第二のペプチドリガンド−免疫グロブリン重鎖融合タンパク質をコードするDNAと同時発現される。分泌の際、ハイブリッド重鎖は二つのジスルフィド結合免疫グロブリン重鎖を有する免疫グロブリン様構造を提供するように共有結合的に結合される。当業者であれば、エフェクター機能には、例えばC1q結合;補体依存性細胞毒素;Fcレセプター結合;抗体依存細胞性細胞毒素(ADCC);ファゴサイトーシス;及びサルベージレセプター結合リガンドの導入による半減期の延長と細胞表面レセプター(例えばB細胞レセプター;BCR)のダウンレギュレーションで、例えば1998年4月14日に発行された米国特許第5739277号にされたようなものが含まれることを認識するであろう。
【0036】
好ましくは、Fc領域はヒトFc領域、例えば天然配列ヒトFc領域ヒトIgG1(A及び非Aアロタイプ)、IgG2、IgG3又はIgG4のFc領域である。そのような配列は図2及び3に示されている。また、親Fc領域に適当なアミノ酸配列の修飾を導入することにより、(a)ヒトエフェクター細胞の存在下で抗体依存細胞性細胞毒素を媒介し、及び/又は(b)天然配列よりも良好な親和性でFcガンマレセプター(Fc(R))に結合する変異体Fc領域をつくり出すことができる。そのようなFc領域変異体は一般にFc領域に少なくとも一つのアミノ酸の修飾を含んでいる。
好適な実施態様では、ペプチドリガンド配列は免疫グロブリンG1(IgG1)のFc領域のN末端に融合している。ペプチドリガンド配列に重鎖定常領域の全体を融合させることが可能である。しかし、より好ましくは、IgGのFcを化学的に定めるパパイン切断部位(すなわち、重鎖定常領域の最初の残基を114であるとして、残基216)、又は他の免疫グロブリンの類似部位の丁度上流のヒンジ領域で始まる配列が融合に使用される。特定の好適な実施態様では、ペプチドリガンドアミノ酸配列はIgG重鎖の(a)ヒンジ領域(又は他のリンカードメイン)及びCH2及びCH3又は(b)CH1、ヒンジ、CH2及びCH3ドメインに融合している。更に好適な実施態様では、ペプチドリガンドアミノ酸配列はIgGの(a)ヒンジ領域(又は他のリンカードメイン)及び(b)CH3ドメインに融合している(例えば、Huら, (1996) Cancer Res. 56:3055-3061に記載された構築物を参照されたい)。
【0037】
この実施態様の特定の側面では、ペプチドリガンドと多量体化ドメインを含むハイブリッド分子が多量体、例えばホモダイマー、又はヘテロダイマーあるいはヘテロテトラマーとしてでさえ構築される。ホモダイマーはペプチドリガンドと多量体化ドメインを含む二つのモノマーの対合又は架橋から生じる。しかし、二つの同一のモノマー対は必須ではない。本発明の特定の側面では、免疫グロブリン定常ドメインのような多量体化ドメインとペプチドリガンドを含むここに記載したハイブリッド分子は免疫グロブリンの一つのアームを含む対の他方の免疫グロブリンと対合しうる。本発明の範囲に入る様々な例示的構築ハイブリッド分子は以下に概略的に模式化される:
(a)ACH
(b)ACH−ACH
(c)ACH−VHCH−VLCL
(d)ACH−VHCH
ここで、Aはそれぞれ同一の又は異なるペプチドリガンドを表し;
VLは免疫グロブリン軽鎖可変ドメインであり;
VHは免疫グロブリン重鎖可変ドメインであり;
CLは免疫グロブリン軽鎖定常ドメインであり、及び
CHは免疫グロブリン重鎖定常ドメインである。
簡略にするために、上記の構造はキーとなる特徴のみを示している;上記構造は以下に記載されるような多量体化ドメインとペプチドリガンドドメインの間に任意のリンカードメインを示していない;上記構造は免疫グロブリンのジョイニング、ヒンジ又は他のドメインを示していないし、ジスルフィド結合も示していない。しかし、そのようなドメインが結合活性に対して必要とされる場合、それらは免疫グロブリン分子に占める通常の位置に存在する構造とされている。
【0038】
免疫グロブリン軽鎖の存在は本発明のハイブリッド分子に必要とされていないけれども、免疫グロブリン軽鎖はペプチドリガンド−免疫グロブリン重鎖融合ポリペプチドに共有結合的に結合するか、ペプチドリガンドに直接融合するかして、存在しているかもしれない。前者の場合、免疫グロブリン軽鎖をコードしているDNAは典型的にはペプチドリガンド−免疫グロブリン重鎖融合タンパク質と同時発現される。分泌の際、ハイブリッド重鎖及び軽鎖は二つのジスルフィド結合免疫グロブリン重鎖−軽鎖対を含む免疫グロブリン様構造を提供するように共有結合的に結合する。
ここに記載されたハイブリッド分子は免疫グロブリンcDNA配列に読み枠を一致させてペプチドリガンド部分をコードするcDNA配列を融合させることにより最も簡便に構築される。しかし、ゲノム免疫グロブリン断片への融合体もまた使用できる(Aruffoら, (1990), Cell 61:1303-1313;及びStamenkovicら, (1991), Cell 66:1133-1144)。後者のタイプの融合体は発現のためにIg調節配列の存在を必要とする。IgG重鎖定常領域をコードするcDNAは、ハイブリダイゼーション又はポリメラーゼ連鎖反応(PCR)法によって脾臓又は末梢血リンパ球から取り出されたcDNAライブラリーから発表された配列に基づいて単離できる。ハイブリッド分子のペプチドリガンドと免疫グロブリン部分をコードしているcDNAは、選択された宿主細胞において効果的な発現をさせるプラスミドベクターにタンデムに挿入される。
【0039】
あるいは、特にペプチドリガンドが例えば標準的な固相合成法によって合成される実施態様では、ペプチドリガンドは当業者によく知られた様々な手段の任意のものによって多量体化ドメインに結合されうる。共有結合的接合が典型的には最も簡便であるが、他の形態の接合も用途に応じて用いることができる。共有結合的な接合の好適な形態の例には、多量体化ドメインのアミノ酸側鎖と活性化された化学基を持つ分子の反応により生じる結合が含まれ、様々な二官能性タンパク質カップリング剤、例えばN-スクシンイミジル-3-(2-ピリジルチオール)プロピオナート(SPDP)、イミノチオラン(IT)、イミドエステル類の二官能性誘導体(例えばジメチルアジピミダートHCL)、活性エステル類(例えば、スベリン酸ジスクシンイミジル)、アルデヒド類(例えば、グルタルアルデヒド)、ビスアジド化合物(例えば、ビス(p-アジドベンゾイル)ヘキサンジアミン)、ビス-ジアゾニウム誘導体(例えば、ビス-(p-ジアゾニウムベンゾイル)エチレンジアミン)、ジイソシアネート(例えば、トリエン-2,6-ジイソシアネート)、及び二活性フッ素化合物(例えば、1,5-ジフルオロ-2,4-ジニトロベンゼン)を使用して作製することができる。
本発明は、HER2を有する細胞型のペプチドリガンド経由インビボでの特定のターゲティング及び本発明のこの側面によれば、Fcエフェクター機能、例えばFcレセプター結合、補体結合、プロテインA結合、半減期の増加又は分子までの血液脳関門の通過の付与を含む。
【0040】
B.ペプチドリガンド融合体
本発明では、ペプチドリガンドドメインは、任意には、例えば他のペプチドリガンドドメインに、以下に記載するように直接又は柔軟なペプチドリンカーを経由して結合される。本発明では、リンカードメインは以下に更に詳細に記載するように二以上のペプチドリガンドドメインの間に空間的ブリッジを提供する任意の分子群である。本発明のこの側面では、ペプチドリガンドは例えば融合タンパク質におけるように互いに結合される。本発明のこの側面のハイブリッド分子は例えば二以上のレセプターを架橋させるのに有用である。
【0041】
C.リンカードメイン
本発明では、ペプチドリガンドドメインは、任意には、例えば他のペプチドリガンドドメインに又は柔軟なペプチドリンカーを経由して多量体化ドメインに結合される。本発明のハイブリッド分子のリンカー成分はハイブリッド分子の機能に必ずしも関与しないが寄与しうる。従って、本発明では、リンカードメインは、二以上のペプチドリガンドドメイン又はペプチドリガンドドメインと多量体化ドメインの間に空間的ブリッジを提供する任意の分子群である。
リンカードメインは可変の長さ及び構造とできる。一般には、重要であるのは、リンカードメインにより引き起こされる配向であり、その化学構造ではない。リンカードメインは、好ましくは、配位ErbB2分子に対する空間的/立体配置的制約を実質的に生じないで、ハイブリッド分子のペプチドリガンドドメインを結合させない。従って、リンカードメインの長さは、ハイブリッド分子の例えばペプチドリガンドと多量体化ドメインの二つの機能の性質に依存する。
当業者であれば、原子の様々な組み合わせが様々な結合間の既知の距離に基づいて可変長さの分子を提供することが分かるであろう(Morrison及びBoyd, Organic Chemistry, 3版, Allyn及びBacon, Inc., Boston, MA (1977))。例えば、リンカードメインは可変長さのポリペプチドでありうる。ポリペプチドのアミノ酸組成がリンカーの性質と長さを決定する。例示的なリンカードメインは2から10のアミノ酸、好ましくは約6のアミノ酸、例えばGly-Gly-Gly-Ser-Gly-Gly(配列番号:129)、Gly-Gly-Gly-Ser-Ser-Gly(配列番号:130)、及びGly-Gly-Gly-Arg-Gly-Gly(配列番号:131)を含む。
【0042】
D.他の二重特異的組み合わせ
本発明のある側面では、少なくとも一つのペプチドリガンドドメインを含む二又は二重特異的組成物が考えられる。例えば、二重特異性抗体組成物はロイシンジッパーを使用して生産されている(Kostelnyら, (1992) J.Immunol., 148(5):1547-1553)。Fos及びJunタンパク質からのロイシンジッパーペプチドを遺伝子融合により二つの異なった抗体のFab'部分に結合させる。抗体ホモダイマーをヒンジ領域で還元してモノマーを形成し、ついで再び酸化して抗体ヘテロダイマーを形成する。この方法はまた抗体ヘテロダイマーの結合ドメインの代わりにペプチドリガンドを使用するペプチドリガンドホモダイマー及びヘテロダイマーの生産に対して使用することができる。
本発明はここに記載されたペプチドをコードする単離された核酸、好ましくはDNAを含む物質の組成物を包含する。本発明のペプチドをコードするDNAは当該分野で知られている様々な方法によって調製することができる。これらの方法は、限定されるものではないが、出典明示により開示の全体がここに取り込まれるEngelsら, (1989) Agnew. Chem. Int. Ed. Engl., 28:716-734に記載された任意の方法、例えばトリエステル、亜リン酸、ホスホラミダイト及びH-ホスホネート法による化学合成を含む。一実施態様では、発現宿主細胞により好まれるコドンはコード化DNAの設計に使用される。あるいは、ペプチドをコードしているDNAを、組換えDNA法、例えば部位特異的突然変異誘発(Kunkelら, (1991) Methods Enzymol. 204:125-139; Carter, P.ら, (186) Nucl. Acids. Res. 13:4331; Zoller, M.J.ら, (1982) Nucl. Acids Res. 10:6487)、カセット突然変異誘発(Wells. J.A.ら, (1985) Gene 34:315)、制限選択突然変異誘発(Wells, J.A.ら,(1986)Philos. Trans. R. Soc. London SerA 317, 415)等々を使用して一又は複数の変異体をコードするように変更させることができる。
【0043】
本発明は更に作用可能に結合した発現コントロール配列、及びベクターで形質転換した宿主細胞によりコントロール配列が認識されるDNA分子を含むプラスミドのような発現ベクターを更に含む。一般には、プラスミドベクターは宿主細胞と適合性がある種から取り出された複製及びコントロール配列を含む。ベクターは通常は複製部位並びに形質転換細胞において表現型の選択を提供することができるタンパク質をコードする配列を有する。
DNAを発現するために適切な宿主細胞は、原核生物、酵母、又は高等真核生物細胞を含む。適切な原核生物は、限定するものではないが、真正細菌、例えばグラム陰性又はグラム陽性生物体、例えば大腸菌(E.coli)のような腸内菌科を含む。様々な大腸菌株が公的に利用でき、例えば大腸菌K12株MM294(ATCC31446);大腸菌X1776(ATCC31537);大腸菌W3110(ATCC27325)及びK5 772(ATCC53635)である。
原核生物に加えて、真核生物体、例えば酵母菌、あるいは多細胞生物体由来の細胞を宿主細胞として使用することができる。一般的なパン酵母又はサッカロミセス・セレヴィシアのような酵母宿主細胞中での発現に対しては、好適なベクターには、2ミクロンプラスミドに基づくエピソーム的に複製するベクター、組込み型ベクター、及び酵母人工染色体(YAC)ベクターが含まれる。発現にまた好適な宿主細胞は多細胞生物体から由来する。無脊椎動物細胞の例には、ショウジョウバエS2及びスポドプテラ(Spodoptera)SF9のような昆虫細胞並びに植物細胞が含まれる。SF9細胞のような昆虫宿主細胞での発現に対しては、好適なベクターにはバキュロウイルスベクターが含まれる。植物宿主細胞、特にタバコのような双子葉植物宿主における発現に対しては、好適な発現ベクターにはアグロバクテリウムツメファシエンスのTiプラスミドに由来するベクターが含まれる。
【0044】
有用な哺乳動物宿主細胞の例には、SV40によって形質転換されたサル腎臓CV1株 (COS-7, ATCC CRL 1651);ヒト胚腎臓株(293又は懸濁培養での増殖のためにサブクローン化された293細胞、Grahamら, (1997) J. Gen Virol., 36:59);ハムスター乳児腎細胞(BHK, ATCC CCL 10);チャイニーズハムスター卵巣細胞/-DHFR(CHO, Urlaub及びChasin, (1980) Proc. Natl. Acad. Sci. USA, 77:4216);マウスのセルトリ細胞(TM4, Mather, (1980) Biol. Reprod., 23:243-251);サルの腎細胞 (CVI ATCC CCL 70);アフリカミドリザルの腎細胞(VERO-76, ATCC CRL-1587);ヒト子宮頸癌細胞 (HELA, ATCC CCL 2); イヌ腎細胞 (MDCK, ATCC CCL 34);バッファローラット肝細胞 (BRL 3A, ATCC CRL 1442);ヒト肺細胞 (W138, ATCC CCL 75);ヒト肝細胞 (Hep G2, HB 8065);マウス乳房腫瘍(MMT 060562, ATTC CCL51);TRI細胞(Motherら, (1982) Annals N.Y. Acad. Sci., 383:44-68);MRC5細胞;FS4細胞;及びヒト肝癌株(HepG2)が含まれる。
原核生物宿主中での発現に対しては、好適なベクターにはpBR322(ATCC第37017)、phGH107(ATCC第40011)、pBO475、pS0132、pRIT5、pRIT20又はpRIT30シリーズの任意のベクター(Nilsson及びAbrahmsen, (1990) Meth. Enzymol., 185:144-161)、pRIT2T、pKK233-2、pDR540及びpPL-ラムダが含まれる。本発明の発現ベクターを含む原核生物宿主細胞には、大腸菌K12株294(ATCC第31446)、大腸菌株JM101(Messingら, (1981) Nucl. Acid Res., 9:309);大腸菌株B、大腸菌χ1776(ATCC第31537)、大腸菌c600(Appleyard, Genetics, 39: 440(1954))、大腸菌W3110(F-、ガンマ-、原栄養菌株、ATCC第27325)、大腸菌株27C7(W3110、tonA、phoA、E15、(argF-lac)169、ptr3、degP41、ompT、kanχ)(米国特許第5288931号、ATCC第55244)、枯草菌、サルモネラ菌、セラチア・マルセッセンス及びシュードモナス種が含まれる。
【0045】
哺乳動物宿主細胞における発現に対しては、有用なベクターには、SV40由来のベクター、サイトメガロウイルス由来のベクター、例えばpRK5及びpRK7を含むpRKベクター(Suvaら, (1987) Science, 237:893-896; EP307247(3/15/89)、EP278776(8/17/88))、ワクシニアウイルス又は他のポックスウイルス由来のベクター、及びモロニーのマウス白血病ウイルス(MoMLV)由来のベクターのようなレトロウイルスベクターが含まれる。
場合によっては、対象のペプチドをコードしているDNAは宿主細胞による発現産物の培地中への分泌を生じさせる分泌リーダー配列に作用可能に結合される。分泌リーダー配列の例には、stII、エコチン(ecotin)、lamB、ヘルペスGD、lpp、アルカリホスファターゼ、インベルターゼ、及びアルファ因子が含まれる。ここでの使用にまた適しているのはプロテインAの36アミノ酸リーダー配列である(Abrahmsenら, (1985) EMBO J., 4:3901)。
宿主細胞はこの発明の上述の発現又はクローニングベクターで形質移入され、好ましくは形質転換され、プロモーターを誘導し、形質転換体を選択し、又は所望の配列をコードする遺伝子を増幅するのに適するように変性された一般的な培養液中で培養される。
形質移入は、コード化配列が実際に発現されようとされまいと、宿主細胞による発現ベクターの取り込みを意味する。数多くの形質移入の方法が当業者に知られており、例えばCaPO4沈殿法及びエレクトロポレーション法である。このベクターの作用の任意の兆しが宿主細胞内で生じた場合に一般に成功した形質移入が認められる。
【0046】
形質転換は、染色体外のエレメントとしてであろうと染色体構成成分によってであろうと、DNAが複製可能であるように生物体中にDNAを導入することを意味する。用いられる宿主細胞に応じて、そのような細胞に対して適した標準的な方法を用いて形質転換はなされる。Sambrookら, Molecular Cloning (2版), Cold Spring Harbor Laboratory, NY (1989)のセクション1.82に記載された塩化カルシウムを用いるカルシウム処理が、原核生物又は実質的な細胞壁障壁を含む他の細胞に対して用いられる。アグロバクテリウム・トゥメファシエンスによる感染が、Shawら, (1983) Gene, 23:315及び1989年6月29日公開の国際公開第89/05859に記載されているように、ある種の植物細胞の形質転換に用いられる。そのような細胞壁のない哺乳動物細胞に対しては、上掲のSambrookらのセクション16.30−16.37に記載されたリン酸カルシウム沈殿法が好ましい。哺乳動物宿主系の形質転換の一般的な側面は1983年8月16日に発行された米国特許第4399216号にAxelによって記載されている。酵母菌への形質転換は典型的には、Van Solingenら, (1977) J. Bact.,130:946及びHsiaoら, (1979) Proc. Natl. Acad. Sci. (USA), 76:3829の方法に従って実施される。しかし、核注入、エレクトロポレーション、あるいは原形質融合のような他の細胞にDNAを導入する方法を使用することもできる。
他の好適なベクターは上述のベクターの関連した形質を組み合わせることによって標準的な方法を使用して構築することができる。関連する形質には、プロモーター、リボソーム結合部位、対象遺伝子又は遺伝子融合体(プロテインAのZドメイン及び対象遺伝子及びリンカー)、抗生物質耐性マーカー、及び適切な複製起点が含まれる。
【0047】
上記の方法の変形例は、ベクターにおいて、所望のペプチドをコードする遺伝子が他のタンパク質又は他のタンパク質の断片をコードする遺伝子に結合している遺伝子融合体の使用を考える。これは、所望のペプチドが他のタンパク質又はペプチドとの融合体として宿主細胞によって産生される結果となる。「他の」タンパク質又はペプチドはしばしば細胞によって分泌されうるタンパク質又はペプチドであり、培地から所望のペプチドを単離し精製することを可能にし、所望のペプチドが細胞内に残る場合に生じる宿主細胞を破壊する必要性を排除する。あるいは、融合タンパク質は細胞内に発現されうる。高度に発現される融合タンパク質を使用するのが有用である。
遺伝子融合体の使用は、必須ではないが、昆虫細胞における異種性ペプチドの発現並びにその遺伝子産物の続く精製を容易にしうる。プロテインAの結合、又はより詳細にはプロテインAのIgGへの結合が融合タンパク質の精製のための「アフィニティーハンドル」を提供するので、ZドメインプロテインA融合体がしばしば使用される。例えば、所望のペプチドリガンドをコードするDNA配列を、Zドメインとして知られているプロテインAのコンセンサスドメインの遺伝子と部位特異的突然変異誘発によって融合させることができる(Nilssonら, (1987) Protein Engineering 1:107-113)。発現及び分泌後に、融合タンパク質を酵素的に切断して、酵素混合物から精製できる遊離のペプチドを生じさせることができる(例えばVaradarajanら, (1985) Proc. Natl. Acad. Sci USA 82:5681-5684; Castellanos-Serraら, (1996) FEBS Letters 378:171-176; Nilssonら, (1996) J. Biotechnol. 48:241-250を参照されたい)。
融合タンパク質は、薬品、例えばメチオニンで切断する臭化シアノゲン、又はAsn及びGly残基の間で切断するヒドロキシルアミンを使用して切断できる。標準的な組換体DNA法を使用して、これらのアミノ酸をコードするヌクレオチド塩基対を所望のペプチドをコードする遺伝子の5'末端の直ぐ前に挿入してもよい。
【0048】
あるいは、融合タンパク質のタンパク分解的切断を用いることができる。Cater, Protein Purification: From Molecular Mechanisms to Large-Scale Processes, Ladischら編, (American Chemical Society Symposium Series No.427, 1990), Ch 13, p181-193。
プロテアーゼ、例えばXa因子、トロンビン、及びサブチリシン又はその変異体、及び多くの他のものが融合タンパク質を切断するために首尾良く使用された。約30未満のアミノ酸のペプチドリガンドの生産に対して本発明において好適なものは、Arg及びLys残基に高度に特異的なプロテアーゼトリプシンである。トリプシン切断は一般にNilssonら, (1996) J. Biotech. 48:241及びSmithら, Methods Mol. Biol. 32:289にいて検討されている。典型的には、使用されるプロテアーゼによる切断に受け入れられるペプチドリンカーは「他の」タンパク質(例えば、プロテインAのZドメイン)と所望のペプチドの間に挿入される。組換体DNA法を使用して、リンカーをコードするヌクレオチド塩基対を他のタンパク質をコードする遺伝子又は遺伝子断片の間に挿入する。ついで、正しいリンカーを含む部分的に精製された融合タンパク質のタンパク分解的切断は天然融合タンパク質又は還元されもしくは変性された融合タンパク質の何れかで実施できる。
ペプチドは融合タンパク質として発現されるときに正しく折り畳まれても折り畳まれなくてもよい。また、切断部位を含む特異的ペプチドリンカーはプロテアーゼに接近可能でも可能でなくてもよい。これらの因子は、融合タンパク質が変性され、再び折り畳まれなければならないかを、そして、しかりであれば、これらの手順が切断の前又は後の何れに使用されるかを決定する。
【0049】
変性と再折り畳みが必要な場合、典型的にはペプチドはグアニジンHClのようなカオトロープで処理し、ついで、例えば還元され酸化されたジチオスレイトール又はグルタチオンを適切な比、pH、及び温度で含むレドックスバッファーで、ペプチドがその天然構造まで再折り畳みされるように処理する。
この明細書において言及される宿主細胞はインビトロ培養中の細胞並びに宿主動物内である細胞を包含する。
本発明の環化実施態様では、組換え的に産生されたペプチドは上述したような細胞内ジスルフィド結合の形成により環化できる。
一般的に言って、本発明のペプチドリガンドとハイブリッド分子は、例えば天然又は変異体ヘルグリン分子又はErbB2に対する抗体が使用されるのと同じ用途に使用することができる。もちろん、本発明のいくつかのペプチドリガンド又はハイブリッド分子は他の用途よりも特定の用途により適している場合がある。当業者であれば、ペプチドリガンド又はハイブリッド分子の生物活性を決定する一又は複数の一般的な生物学的アッセイを使用することによって与えられた用途にどの分子が適しているかを容易に確認できるであろう。
一つの例として、ヘレグリンは様々な障害、例えば神経系、筋肉組織及び上皮に影響を及ぼす障害及び疾患の治療に有用である。例えば、本発明のアゴニストペプチドリガンド又はハイブリッド分子はヘレグリン又はヘレグリン変異体が使用されるのと同じようにしてインビボでニューロンの発生、維持及び/又は再生を促進するのに使用される。本発明のこの側面に係るペプチドリガンド又はハイブリッド分子での治療を受け入れることができる疾患又は障害には、例えば外傷、手術、脳卒中、虚血、感染、代謝異常栄養不足、悪性又は毒物に伴う中枢神経系の損傷が含まれる。これらに限定されるものではないがヒト神経変性疾病又は障害、例えばアルツハイマー病、パーキンソン病、てんかん、多発性硬化症、ハンチントン舞踏病、ダウン症候群、神経性難聴及びメニエール病を治療することができる。同様に、特定のペプチドリガンドは神経障害、例えば糖尿病のような全身性疾患又は障害に伴う末梢神経障害の治療における使用に適している。
【0050】
ErbB2への結合の際に上述のようなリン酸化事象を開始させるアゴニストペプチドリガンドは特に本発明の前述の態様に有用である。本発明の特定のアゴニストペプチドリガンドは細胞の生存を亢進することができ、すなわち、特定のアゴニストペプチドリガンドに暴露されていなかった細胞の生存期間に対してインビボかインビトロでErbB2保有細胞の生存期間を増加させる。好適な実施態様では、アゴニストペプチドリガンドは、例えば細胞による3H-チミジンの取り込みを測定することによって定量されるErbB2保有細胞型の増殖をインビボかインビトロの何れかで生じさせ又は亢進する。増殖には、表現型の変化についてErbB2保有細胞集団をスクリーニングすることによって決定されるErbB2細胞の分化を伴いうる。
本発明の他の側面では、ある種のペプチドリガンド及びハイブリッド分子、特にErbB2に結合するがアゴニスト関連応答を誘発しないものは、以下に十分に説明されるように、例えば腫瘍細胞浸潤及び転移を阻害するのに有用である。例えば、ErbB2レセプターを発現する腫瘍(特にErbB2を過剰発現するもの)は細胞毒性剤に結合したペプチドリガンド又はハイブリッド分子を使用して治療することができ、又はErbB2を発現する細胞にプロドラッグをターゲティングする際に使用される。
【0051】
A.エフェクター機能の設計
例えば、本発明のハイブリッド分子の多量体化ドメインをエフェクター機能について改変し、例えば癌の治療における抗体の効能を増強することが望ましい。例えば、システイン残基を免疫グロブリンFc領域に導入して、この領域における鎖間ジスルフィド結合の形成を可能にする。このようにして産生されたホモ又はヘテロダイマーハイブリッド分子は改善されたインターナリゼーション能力及び/又は増加した補体媒介細胞死滅及び抗体依存性細胞障害活性(ADCC)を有しうる。Caronら, (1992) J. Exp. Med. 176:1191-1195及びShopes, B. (1992) J. Immunol. 148:2918-2922を参照されたい。抗腫瘍活性が高められたホモダイマーハイブリッド分子は、Wolffら, (1993) Cancer Research 53:2560-2565に記載されているようなヘテロ二官能性架橋剤を使用して調製することもできる。
別法として、二重Fc領域を有し、よって増強された補体溶解及びADCC能を有しうるヘテロダイマーハイブリッド分子を設計することができる。Stevensonら (1989) Anti-Cancer Drug Design 3:219-230を参照されたい。
【0052】
B.コンジュゲート
また本発明は、細胞障害剤、例えば化学療法剤、例えばタンパク質毒素もしくは細胞障害性薬物もしくは毒素(例えば、細菌、真菌、植物又は動物由来の酵素活性毒又はそれらの断片)、又は放射性アイソトープ(すなわち、放射性コンジュゲート)に結合したここに記載の任意のハイブリッド分子又はペプチドリガンドを含有するコンジュゲートにも関する。
このような免疫コンジュゲートの生成に有用な化学療法剤は上述した。化学療法剤は、悪性腫瘍、リンパ腫、芽細胞腫、肉腫及び白血病を含む癌の治療に有用な化合物である。化学療法剤の例には、カリケアマイシンのような微生物から単離された抗生物質(Leeら, (1987) J. Am. Chem. Soc. 109:3464-3466; Hinmanら, (1993) Cancer Res. 53:3336-3342)、メイタンシノイド(例えば、Liuら, (1996) Proc. Natl. Acad. Sci. USA 93:8618-8323に記載されているもの)、アドリアマイシン、ドキソルビシン、5-フルオロウラシル、シトシンアラビノシド、シクロホスファミド、チオテパ、ブスルファン、サイトキシン、タキソール、メトトレキセート、シスプラチン、メルファラン、ビンブラスチン、ブレオマイシン、エトポシド、イホスファミド、マイトマイシンC、ミトキサントロン、ビンクリスチン、ビノレルビン、カルボプラチン、テニポシド、ダウノマイシン、カルミノマイシン(Carminomydcin)、アミノプテリン、ダクチノマイシン、マイトマイシン、ニコチンアミド、エスペラミシン、メルファラン、及び他の関連したナイトロジェンマスタード、及びエンドクリン治療薬(例えば、ジエチルスチルベストロール、タモキシフェン、LHRH-拮抗薬、プロゲスチン、抗プロゲスチン等々)が含まれる。
【0053】
使用可能な酵素活性毒及びその断片には、ジフテリアA鎖、ジフテリア毒素の非結合性活性断片、外毒素A鎖(シュードモナス・アエルギノーサ(Pseudomonas aeruginosa)由来)、リシンA鎖、アブリンA鎖、モデシン(modeccin)A鎖、アルファ-サルシン(sarcin)、アレウライツ・フォルディイ(Aleurites fordii)プロテイン、ジアンシン(dianthin)プロテイン、フィトラッカ・アメリカーナ(Phytolaca americana)プロテイン(PAPI、PAPII及びPAP-S)、モモルディカ・キャランティア(momordica charantia)インヒビター、クルシン(curcin)、クロチン、サパオナリア(sapaonaria)オフィシナリスインヒビター、ゲロニン(gelonin)、マイトゲリン(mitogellin)、レストリクトシン(restrictocin)、フェノマイシン、エノマイシン及びトリコセセンス(tricothecenes)が含まれる。
種々の放射性核種も放射性コンジュゲートペプチドリガンド又はハイブリッド分子の生産に利用できる。例としては212Bi、131I、131In、90Y及び186Reが含まれる。炭素-14標識された1-イソチオシアナトベンジル-3-メチルジエチレントリアミン五酢酸(MX−DTPA)はハイブリッド分子へ放射性ヌクレオチドを結合させるためのキレート剤の例である。
ペプチドリガンド又はハイブリッド分子と細胞障害剤のコンジュゲートは、様々な二官能性タンパク質カップリング剤、例えばN-スクシンイミジル-3-(2-ピリジルチオール)プロピオナート(SPDP)、イミノチオラン(IT)、イミドエステル類の二官能性誘導体(例えばジメチルアジピミダートHCL)、活性エステル類(例えば、スベリン酸ジスクシンイミジル)、アルデヒド類(例えば、グルタルアルデヒド)、ビスアジド化合物(例えば、ビス(p-アジドベンゾイル)ヘキサンジアミン)、ビス-ジアゾニウム誘導体(例えば、ビス-(p-ジアゾニウムベンゾイル)エチレンジアミン)、ジイソシアネート(例えば、トリエン-2,6-ジイソシアネート)、及び二活性フッ素化合物(例えば、1,5-ジフルオロ-2,4-ジニトロベンゼン)を使用して作製することができる。例えば、リシン免疫毒素は、Vitettaら, (1987) Science 238:1098に記載されているようにして調製することができる。
他の実施態様では、腫瘍の事前ターゲティングに利用するために、「レセプター」(例えばストレプトアビジン)にペプチドリガンド又はハイブリッド分子が結合され、ペプチドリガンド又はハイブリッド分子-レセプターコンジュゲートが患者に投与され、続いてキレート剤を使用し、循環から未結合コンジュゲートが除去され、細胞障害剤(例えば放射性ヌクレオチド)に結合された「リガンド」(例えばアビジン)が投与される。
【0054】
C.リポソーム
ここで開示されているハイブリッド分子又はペプチドリガンドはまたリポソームとして処方することもできる。ハイブリッド分子を含むリポソームは、例えばEpsteinら, (1985) Proc. Natl. Acad. Sci. USA, 82:3688;Hwangら, (1980) Proc. Natl. Acad. Sci. USA 77:4030;及び米国特許第4485045号及び同4544545号に記載されているように、当該分野において知られている方法により調製される。循環時間が増したリポソームは米国特許第5013556号に開示されている。
特に有用なリポソームは、ホスファチジルコリン、コレステロール及びPEG-誘導体化ホスファチジルエタノールアミン(PEG-PE)を含有する脂質組成物を用いた逆相蒸発法により作製することができる。リポソームは孔径が定められたフィルターを通して押し出され、所望の直径を有するリポソームが得られる。例えば、ここに記載されたような免疫グロブリン定常ドメインを有するハイブリッド分子は、ジスルフィド交換反応を介して、Martinら,J. Biol. Chem. 257:286-288(1982)に記載されているようにしてリポソームに結合させることができる。場合によっては、化学療法剤はリポソーム内に包含される。Gabizonら, J. National Cancer Inst. 81(19)1484(1989)を参照されたい。
【0055】
D.酵素媒介性プロドラッグ治療法
また、本発明のペプチドリガンド又はハイブリッド分子は、プロドラッグ(例えばペプチジル化学療法剤、国際公開81/01145号を参照)を活性な抗癌剤に転化させるプロドラッグ活性化酵素にペプチドリガンド又はハイブリッド分子を結合させることにより、抗体依存性酵素媒介性プロドラッグ治療法(ADEPT)の抗体のようにして使用することもできる。例えば国際公開88/07378号及び米国特許第4975278号を参照されたい。
ADEPT型治療法に有用なコンジュゲートの酵素成分には、より活性な細胞毒形態に転化するように、プロドラッグに作用し得る任意の酵素が含まれる。限定するものではないが、この発明の方法に有用な酵素には、ホスファート含有プロドラッグを遊離の薬剤に転化するのに有用なアルカリ性ホスファターゼ;スルファート含有プロドラッグを遊離の薬剤に転化するのに有用なアリールスルファターゼ;非毒性5-フルオロシトシンを抗癌剤5-フルオロウラシルに転化するのに有用なシトシンデアミナーゼ;プロテアーゼ、例えばセラチアプロテアーゼ、サーモリシン、サブチリシン、カルボキシペプチダーゼ及びカテプシン(例えば、カテプシンB及びL)で、ペプチド含有プロドラッグを遊離の薬剤に転化するのに有用なもの;D-アミノ酸置換基を含むプロドラッグの転化に有用なD-アラニルカルボキシペプチダーゼ;炭水化物切断酵素、例えばグリコシル化プロドラッグを遊離の薬剤に転化するのに有用なβガラクトシダーゼ及びノイラミニダーゼ;βラクタムで誘導体化された薬剤を遊離の薬剤に転化させるのに有用なβラクタマーゼ;及びペニシリンアミダーゼ、例えばそれぞれフェノキシアセチル又はフェニルアセチル基で、それらのアミン性窒素において誘導体化された薬剤を遊離の薬剤に転化するのに有用なペニシリンVアミダーゼ又はペニシリンGアミダーゼが含まれる。あるいは、「アブザイム」としてもまた当該分野で知られている酵素活性を有する抗体を、遊離の活性薬剤に本発明のプロドラッグを転化させるために使用することもできる(例えば、Massey, (1987) Nature 328:457-458を参照)。ペプチドリガンド/ハイブリッド分子-アブザイムコンジュゲートは、ここで記載されているようにして、腫瘍細胞集団にアブザイムを送達するために調製することができる。
酵素は、当該分野においてよく知られている技術、例えば上で検討したヘテロ二官能性架橋試薬を使用することにより、ハイブリッド分子に共有的に結合させることができる。あるいは、本発明のハイブリッド分子の少なくともペプチドリガンド部分を含む融合タンパク質を、当該分野においてよく知られている組換えDNA技術を使用して作成することができる(Neubergerら, (1984) Nature 312:604-608参照)。
【0056】
E.医薬組成物
本発明のペプチドリガンドを含有する医薬組成物は、腸管外、局所、経口又は局部的(エアロゾル又は経皮など)又はその任意の組み合わせを含む任意の適切な形で投与されうる。好適な治療法にはまた静脈内ボーラス注射による最初の投与に続いて一又は複数のインターバルで繰り返し投与を行うことが含まれる。
本発明の化合物の医薬組成物は、所定の純度を持つ化合物を含むペプチドリガンドを、任意成分の製薬的に許容される担体、賦形剤又は安定剤と混合することにより(Remington's Pharmaceutical sciences 16th edition, Osol, A. 編, [1980])、凍結乾燥製剤又は水溶液の形態で調製され保存される。許容される担体、賦形剤又は安定化剤は、用いられる用量及び濃度で受容者に非毒性であり、リン酸塩、クエン酸塩、及び他の有機酸等のバッファー;アスコルビン酸及びメチオニンを含む酸化防止剤;防腐剤(オクタデシルジメチルベンジルアンモニウムクロリド;ヘキサメトニウムクロリド;ベンザルコニウムクロリド、ベンズエトニウムクロリド;フェノール、ブチル又はベンジルアルコール;メチル又はプロピルパラベン等のアルキルパラベン;カテコール;レゾルシノール;シクロヘキサノール;3-ペンタノール;及びm-クレゾール等);低分子量(約10残基未満)ポリペプチド;血清アルブミン、ゼラチン、又は免疫グロブリン等のタンパク質;ポリビニルピロリドン等の親水性ポリマー;グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン又はリシン等のアミノ酸;グルコース、マンノース、又はデキストリンを含む単糖類、二糖類、及び他の炭水化物;EDTA等のキレート化剤;スクロース、マンニトール、トレハロース又はソルビトール等の糖;ナトリウム等の塩形成対イオン;金属錯体(例えば、Zn-タンパク質複合体);及び/又はTWEENTM、PLURONICSTM又はポリエチレングリコール(PEG)のような非イオン性界面活性剤を含む。
ここでの組成物は、治療される特定の徴候に必要ならば一以上の活性化合物も含んでよく、好ましくは互いに悪影響を与えない相補的活性を持つものである。例えば、一つの製剤にEGFR、ErbB2(例えばErbB2上の異なるリガンドに結合する抗体)、ErbB3、ErbB4、又は血管内皮因子(VEGF)に結合する抗体のような分子を更に提供することが望ましい。あるいは、又は加えて、組成物は、細胞毒性剤、サイトカイン、成長抑制剤及び/又は心臓保護剤を含有することもできる。そのような分子は好適には意図される目的に対して効果的な量で組み合わされて存在する。
【0057】
F.疾患の診断と予後
本発明における予後及び診断のための疾患は、好ましくは、ErbB2レセプターの過剰発現により特徴づけられる良性又は悪性腫瘍、例えば、癌、例えば乳癌、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、グリア芽細胞腫、子宮頸管癌、卵巣癌、肝臓癌、膀胱癌、肝細胞腫(hepatoma)、大腸癌、結腸直腸癌、子宮体癌、唾液腺癌、腎臓癌、肝臓癌、前立腺癌、産卵口癌、甲状腺癌、肝癌(hepatic carcinoma)及び様々な種類の頭部及び頸部の癌が含まれる。従って、本発明の一実施態様は、試料中のErb2の検出及び/又は測定と、そのような検出又は測定の診断、段階決定、重症度の決定、及び疾患の予後一般への使用に関する。
特定の実施態様では、記載された診断方法は治療の進行に従って使用することができる。Erb2保持細胞の量の増加又は減少に至る治療処置を受けている患者では、試料中のErb2保持細胞の量が治療の成功又は失敗の有用な目安となりうる。しかして、本発明は、Erb−2発現細胞を含むと疑われる組織試料において発現されるErb2の量を適当な時間間隔で測定することを含む、患者における治療の効果をモニターする方法を提供する。Erb2の全量が、疾患と治療に依存して、正常な被験者、疾患の発症前又は疾患の緩解時の患者、又は治療開始前の患者の同様な試料中のErb2の量でありうる「ベースライン」又は「コントロール」値と比較される。当業者であれば、過度の実験を行うことなく特定の状況に使用される好適なベースラインが容易に分かるであろう。
本発明の化合物を使用する試料中のErb2の測定の実施においては分析物の測定に対して当該分野で知られている任意の方法を使用できる。そのような方法には、限定されるものではなく、少しだけ挙げると、競合及び非競合アッセイシステムで、ラジオイムノアッセイのような技術を使用するもの、酵素免疫アッセイ(EIA)、好ましくは酵素結合免疫吸着検定法(エライザ:ELISA)、「サンドウィッチ」イムノアッセイ、沈降反応、ゲル拡散反応、免疫拡散アッセイ、凝集アッセイ、補体結合アッセイ、及び免疫電気泳動アッセイが含まれる。好適なイムノアッセイ法の例については、米国特許第4845026号(1989年7月4日)及び同第5006459号(1991年4月9日)を参照されたい。
【0058】
診断及び予後の応用に対しては、本発明の化合物、典型的にはここに上述したハイブリッド分子は検出可能な部分で標識され、上述のような試料中のErb−2を検出するために使用される。多数の標識が利用可能であり、それらは好適には次の範疇にグループ分けされる:
(a)放射性同位体、例えば、35S、14C、125I、3H及び131I等。ハイブリッド分子は、例えばCurrent Protocols in Immunology, Volumes 1 and 2, Coligen等, 編, Wiley-Interscience, New York, Pubs. (1991)に記載された技術を用いて放射性同位体で標識でき、放射活性はシンチレーションカウンティングを使用して測定できる。
(b)蛍光標識、例えば希土類キレート(ユーロピウムキレート)又はフルオレセインとその誘導体、ローダミンとその誘導体、ダンシル、リサミン(Lissamine)、フィコエリトリン及びテキサスレッドが使用できる。蛍光標識は、例えば上掲のCurrent Protocols in Immunologyに開示された技術を用いてハイブリッド分子に結合させることができる。蛍光は蛍光定量計を用いて定量できる。
(c)様々な酵素−基質標識が利用でき、米国特許第4275149号は、それらの幾つかの概説を提供している。酵素は好ましくは種々の技術を用いて測定可能な色素原基質の化学変換を触媒する。例えば、酵素は基質における色変化を触媒し、それは分光学的に測定可能である。あるいは、酵素は基質の蛍光又は化学発光を変化させることもある。蛍光変化を定量化する技術は上述した。化学発光基質は化学反応によって電子的に励起され、ついで(例えば化学発光計を用いて)測定可能な光を放出するか、又は蛍光受容体にエネルギーを供与する。酵素標識の例には、ルシフェラーゼ(例えば、ホタルルシフェラーゼ及び細菌ルシフェラーゼ;米国特許第4737456号)、ルシフェリン、2,3-ジヒドロフタラジンジオン、リンゴ酸塩デヒドロゲナーゼ、ウレアーゼ、西洋ワサビペルオキシダーゼ(HRPO)等のペルオキシダーゼ、アルカリホスファターゼ、β-ガラクトシダーゼ、グルコアミラーゼ、リソザイム、糖類オキシダーゼ(例えば、グルコースオキシダーゼ、ガラクトースオキシダーゼ、及びグルコース-6-ホスフェートデヒドロゲナーゼ)、ヘテロ環オキシダーゼ(ウリカーゼ及びキサンチンオキシダーゼ等)、ラクトペルオキシダーゼ、ミクロペルオキシダーゼ等が含まれる。酵素を抗体に結合させる技術は、O'Sullivanら, Methods for Preparation of Enzyme-Antibody Conjugates for use in Enzyme Immunoassay, in Methods in Enzym. (J. Langone & H. Van Vunakis編)Academic press, New York, 73: 147-166 (1981)に記載されている。
【0059】
酵素−基質の組み合わせは、例えば以下を含む:
(i)西洋ワサビペルオキシダーゼ(HRPO)と基質としてのハイドロジェンペルオキシダーゼで、ハイドロジェンペルオキシダーゼが染料前駆物質(例えば、オルトフェニレンジアミン(OPD)又は3,3',5,5'-テトラメチルベンジジンヒドロクロリド(TMB))を酸化する;
(ii)アルカリホスファターゼ(AP)と色素原基質としてのパラ-ニトロフェニルホスフェート;
(iii)β-D-ガラクトシダーゼ(β-D-Gal)と色素原基質(例えば、p-ニトロフェニル-β-D-ガラクトシダーゼ)又は蛍光原基質4-メチルウンベリフェリル-β-D-ガラクトシダーゼ。
当業者には、多くの他の酵素−基質の組み合わせが利用可能である。これらの一般的な概説については、米国特許第4275149号及び第4318980号を参照されたい。
本発明のアッセイでは、好ましくは、ハイブリッド分子は、固相の支持体又は坦体に結合している。「固相の支持体又は坦体」という用語により、抗原又は抗体と結合可能な任意の支持体が意図される。周知の支持体又は坦体には、ガラス、ポリスチレン、ポリプロピレン、ポリエチレン、デキストラン、ナイロン、アミロース、天然又は修飾されたセルロース、ポリアクリルアミド、アガロース、及び磁鉄鉱が含まれる。坦体の性質は、本発明の目的に対して、ある程度可溶性であるか、不溶性であり得る。支持材料は、結合された分子が抗原又は抗体と結合可能である限り、実質的には、任意の可能な立体構造をとってもよい。従って、支持体の立体構造は、ビーズのような球状、又は試験管の内面、又はロッドの外表面のような円筒状であってもよい。あるいは、表面はシート、テストストリップ等々のような平坦面でもよい。好ましい支持体には、ポリスチレンビーズが含まれる。当業者であれば、抗体又は抗原を結合させるための他の多くの好適な坦体が分かり、日常的な実験を行うことにより、同一性について確認を行うことができるであろう。
【0060】
好ましい実施態様では、サンドイッチイムノアッセイが利用される、すなわち、Erb2は、一次抗体又はハイブリッド分子のErb2抗原に対する結合、及び二次抗体又はハイブリッド分子のErb2に対する結合を含み、一次及び二次抗体の両方、又はハイブリッド分子による免疫特異的なErb2への結合を検出又は測定することを含む方法によって検出され又は測定される。特別の実施態様では、一次又は二次抗体のどちらかがモノクローナル抗体であり、一次又は二次のどちらか又はその両方が、本発明のハイブリッド分子である。この実施態様では、ハイブリッド分子のペプチドリガンド部分は、好ましくは、一次抗体のリガンド部分とは異なる部位に結合する(例えば、抗原への結合に対する、抗体とハイブリッド分子間における競合阻害の欠如に反映される)。他の特定の実施態様では、一次又は二次抗体はポリクローナル抗体である。
また、本発明中のアッセイを実施するための成分を含む一又は複数の容器又はバイアルを含むキットも本発明の範囲内である。例えば、そのようなキットは、腫瘍のバイオプシーのような免疫組織化学的サンプル分析に必要とされる薬剤を含み得る。薬剤には、例えば、ハイブリッド分子又は抗体のような一又は複数の結合相手が含まれる。組織学的なアッセイに対しては、キットは、色素生産性の基質並びに発色したとき酵素反応を停止する薬剤を含む。キットに含まれる基質は本発明のハイブリッド分子の一つに結合した酵素に適したものである。これらは、当該技術分野において周知である。また、キットは、例えば、既知量の精製Erb2などの標準物質を含む。
以下の実施例は、例示のために提供されるもので、限定するものではない。明細書中の全ての文献の開示は、ここに明示的に出典明示により取り込まれる。
【0061】
実施例
実施例I
HER2結合ペプチドの同定及び成熟
方法
ファージライブラリー− ランダム配列の多価ペプチドファージライブラリー(表1)は、テンプレートであるpB2479.g8の大規模なクンケル突然変異導入法を用いることにより構築された。このファージミドは、Tacプロモーター、malシグナル配列、3つのストップコドン、及びgVIIIと融合されたリンカー配列を含み、LacIq及びアンピシリン耐性遺伝子を有する。従って、このペプチドライブラリーは、IPTGでコピー数を制御することが可能なpVIII融合体として、多価的にファージ上で発現される。各ライブラリーは、109クローンを越える多様性を有する。
【0062】
【0063】
選択条件− HER2-ECDは、50mM炭酸水素アンモニウム、pH9.3中にて、5μg/mlを使用し、4℃で一晩反応させ、マキシソープ(maxisorp)プレートに直接固定化された。ウェルは、1%BSAを含むPBS(PBS−BSA)を用いて、25℃で1時間ブロッキングした。上述のライブラリーから得られたファージを、表1に示すような7つのグループにプールした。各プールからのファージは、PBS−BSA にHER2-ECDを含むウェル中で、25℃で3時間反応させた;通常、約5 x 1010のファージが各実験ラウンドの開始時に添加された。結合しなかったファージは、0.05%のTween20を含むPBS(PBS-Tween)による繰り返しの洗浄により除去された;残存するファージは、500mM KCl、10mM HCl、pH2により溶出された。その後、溶出されたファージは、XL1-Blue細胞中でVCSM13ヘルパーファージ(Stratagene)を用い、37℃で一晩増幅させた。IPTG(10μM)は、最初のファージライブラリー及び増幅中の第1回目のラウンドに、添加された;その後のラウンドは、ペプチド-pVIII発現産生のためのtacプロモーターの基本発現に依存した。濃縮度は、BSAがコートされているウェルに結合するファージ数に対して、標的がコートされているウェルに結合するファージ数を希釈することにより、観察した。
HER2ファージ結合アッセイ− Erb2の固定化された細胞外ドメイン(HER2-ECD、Hudziak等、(1991) J. Biol. Chem. 266:24109-15)に対する、表面にペプチド1.1FI(表1)の単一コピーを表示するファージの結合阻害は、ファージELISAを使用して評価した。HER2-ECDは、50mM 炭酸水素アンモニウム、pH9.3中にて、5μg/mlになるように使用して、4℃で一晩反応させ、直接、マキシソープ・プレート(Nunc)に固定化させた。ウェルは、1%BSAを含むPBS(PBS−BSA)を用いて、25℃で1時間ブロッキングした。PBSによる1.1FI−Fcの希釈物は、1.1Fc−Fcを表示するファージの固定化HER2-ECDへの結合に対する阻害能についてテストされた。マイクロタイタープレートは、0.05%のTween20を含むPBS(PBS-Tween)により洗浄され、HER2-ECDに結合したファージは、抗gVIII/HRPモノクローナル抗体(Amersham Pharmacia Biotech)により検出された。結合したHRPの量は、ABTS/H2O2基質を使用し、405nmにおける変化を検出することにより測定された。
【0064】
結果
HER2-ECDへ結合する多価ペプチド-ファージの選択− 多価のペプチドライブラリーは、固定化HER2-ECDに対して7つのプール(表1)に分類された。多価のファージディスプレイ(Scott等 (1990) Science 249:386-390)が、アビディティー効果を介して結合を促進するために使用された。濃縮度、つまり、HER2-ECDでコートされたウェルから溶出されたファージの数をBSAでコートされたウェルから溶出されたファージの数で除した数は、ラウンド3及びラウンド4について図4に示されている。各プールにおけるランダムクローンから得たDNAは、配列確認され、その予想されるペプチド配列が表2に示されている。プール1は、単一クローンとして得られたが、他のプールは、複数の配列を含んでいた。
ファージ結合アッセイを用いたところ、ファージクローン1.1、5.1及び7.1は、固定化HER2-ECDに結合するが、固定化BSA又は、近縁関係にあるHER3又はHER4には結合しないことが見いだされ、これらのファージクローンがHER2-ECDを特異的に認識することを示す(図5)。これらのクローンの配列は、更に高い親和性を持つクローンの再選択のために、部分的ランダム化に供された。
【0065】
表2
【0066】
実施例II
方法
一価ファージの部分的又は完全なランダム化− ファージ表面に、gIIIによってコードされる尾タンパク質に対しリンカー配列を介して融合されたペプチドの単一コピーを表示する一価ライブラリーが、類似のファージミド、pB2479.g3に対する一本鎖鋳型指向性突然変異(Kunkel等, (1991) Met. Enz. 204:125-139)を用いて構築された。このベクターにおいて、gVIIIに対するコード配列は、gIIIによって置換され、さらに、CMPγ遺伝子は、AMPγ遺伝子の唯一のhincII部位に挿入された。薬剤耐性の変化は、弱い親和性ながらも、アビディティー効果を介して集団化を引き起こす(Cwirlaら, (1990) Proc. Natl. Acad. Sci.USA 87:6378-6382)多価クローンの関連クローンによるコンタミネーションを除去するように設計された。部分的にランダム化されたライブラリーは、各アミノ酸位置において50%の変異率を許容しながらも、初期の多価ライブラリーから同定されたペプチド配列への偏りを維持するように設計された。この突然変異率は、70-10-10-10の塩基の混合物によりオリゴを合成することにより達成される(この場合、オリゴのドープ領域の各塩基は、野生型配列に寄与する塩基の70%を含む混合物を用いて、関連づけられ、他の3塩基の各塩基は、野生型の配列塩基の10%により関連づけられる)。これに対して、ライブラリーにおける完全なランダム化は、特定のコドンに関しNNSを用いてオリゴを合成することにより、配列の他の部分を一定に保ちながら、得ることができた。
【0067】
結果
ペプチドファージの部分的なランダム化− 作製された実際のライブラリーが、およそ109クローン(潜在的な多様性のほんの一部の多様性)しか含まないのに対し、設計された最初のペプチドライブラリーは、2020(1026)以上の潜在的な多様性をコードしていた。サーチ範囲を狭め、最初に選択したペプチドの範囲内におけるペプチド多様性を更に探索するために、部分的ランダム化技術を用いた。この技術は、各アミノ酸位置に50%の突然変異率(アミノ酸レベルにおいて)を導入する間、野生型配列へ向かう偏りを維持する;従って、平均して、20アミノ酸のペプチドを表示するファージは、10のランダム突然変異を必要とするであろう。その上、更なる親和性の改善が予想されるため、このようなライブラリーを一価ファージ(gIII)(Bassら, (1990) Protein: Struct. Funct. Genet. 8:309;Lowmanら, (1991) Biochemistry 30:10832)について構築することにした。
クローン1.1、5.1、及び7.1の一価の部分的ランダム化ライブラリーが構築され、HER2に対して4ラウンドの分類がなされた。10000倍より高い濃縮度が、ライブラリー1.1及び7.1について観察された;しかし、ライブラリー5.1の濃縮は観察されなかった。再び、ランダムクローンが選択され、配列決定が行われた;ライブラリー1.1及び7.1のクローンから選択された予想されるペプチド配列が、図15に示される。ライブラリー1.1及び7.1における幾つかのアミノ酸位置は、100%保持されており、さらに、複数のコドンがこれらの多くの位置において観察された。続いて行われた部分的ランダム化において、強固に保持された残基は、直接的な接触を通じた結合又は構造的な理由の何れかに対して重要となりうる。
【0068】
HER2結合配列の完全成熟− クラス1配列の親和性の成熟を完成させるために、100%保存された位置は固定化し、残りの位置は完全にランダムである、第三のライブラリーセットが構築された。クラス1配列のジスルフィドループに隣接するアミノ酸残基の役割は、アミノ及びカルボキシ末端の何れかを欠失するライブラリーを構築することにより解明された。よって、3つの一価ライブラリーが図16に示されるようにして構築された。500倍よりも高い濃縮度が、ラウンド4における各ライブラリーによって観察された;ランダムクローンの配列は、図16に示されている。10の位置において完全にランダム化されたこれらのライブラリーは、完全にはほど遠いが、全長及びアミノ末端を欠失したライブラリーを比較することにより、クラス1配列のC末端領域における付加的なジスルフィドへの選択性が示された。両ライブラリーにおいて、付加的なジスルフィド結合は、2箇所に見いだされる。3つ目のライブラリーにおいて、C末端位置の欠失は、これらの位置における付加的なジスルフィド結合の選択を防止する。付加的なジスルフィド結合の選択とは別に、他のランダム化された位置において選択された残基は非常に多様性を有している。
【0069】
実施例III
方法
ペプチド合成− ペプチドは、PEG-ポリスチレン樹脂を用い(Bodanszkyら, (1984) Int. J Peptide and Protein Res. 23(1):111)、0.25mmolスケールにて、手動又は自動化されたFmocに基づく固相合成により合成された。側鎖の保護基が除去され、ペプチドは95%のトリフルオロ酢酸(TFA)及び5%のトリイソプロピルシランによって樹脂から切断された。酢酸による飽和ヨウ素溶液がジスルフィド結合を酸化するために添加された。ペプチドは、0.1%のTFAを含んだ水/アセトニトリルの濃度勾配を用い、逆相HPLCによって精製された。ペプチドは、分析的なHPLCによると、純度>95%であり、同一性については、マススペクトロメトリーによって確認された。
ペプチド-Z融合物の生成− HER2-ECDへの結合のために選択されたファージペプチドは、プロテインAのZタンパク質との融合体として大腸菌(27C7株)中で発現され、分泌された。オリゴは、ファージ-ペプチド配列に対するコード化配列をstIIシグナル配列とZドメインを持つプラスミド(pZCT)中のZドメインとの間に挿入するように設計された。細胞はリン酸を制限した培地中で増殖され、ペプチド-Z融合体は、記載されているように、IgGアフィニティーカラムを使用して(Dennis等, (1993) Proteins: Struct. Funct. Genetics 15:312-321)、培地から精製された。IgGでアフィニティー精製されたペプチド-Z融合体は、NHS-LC-ビオチン(Pierce)によってビオチン化され、マススペクトロメトリーによって性質決定された。
多量体を形成するペプチド-Z融合体は、PBSにてSuperdex75カラムのサイズ排除クロマトフラフィーにより、単量体及びダイマーを含む画分へ分離し得る。
【0070】
精製されたペプチド-Z融合体からのZドメインの除去− 精製されたペプチド-Z融合体からのZドメインの除去は、トリプシンによる融合体の切断により行うことができる。トリプシン(1%, w/w)は、100mM Tris pH8.0、及び10mM CaCl2中にて37℃、4時間、添加された。Zドメインが除去されたペプチドは、0.1%のTFAを含んだ20から50%のアセトニトリル濃度勾配を用い、C18逆相HPLCによって精製された。
HER2-Fc発現ベクターの構築− ベクターpVL1393(Pharmigen)を基礎とする組換体転移ベクターの構築には、標準的な組換えDNA技術が用いられた(Sambrook, J., Fritsch, E. F., 及びManiatis, T. (1989) Molecular Cloning: A Laboratory Manual, second Ed., Cold Spring Harbor Laboratory Press, New York; O’Reilly, D. R., Miller, L. K., 及びLuckow, V. A. (1994) Baculovirus Expression Vectors: A Lboratory Mnual, Oxford University Press, New York)。pVL1393由来プラスミドpbPH.Hisは、NcoIとSmaIによって直線状にされ、エビのアルカリホスファターゼで処理された(Dweyer, M. A.ら, (1999) J. Biol. Chem. 274:9738-9743)。ヒトIgG1のヒトFc部分は、他のpVL1393由来プラスミドpVL1393.IgGから、NdeIで切断し、続いてKlenowで処理し、更にNcoIで切断することで得られる700塩基対の断片として得られた。MIP.5シグナル配列は、大腸菌で切断されたPCR断片としてFc配列の前に導入され、その断片には、AscI部位が含まれる。AscI部位は、予想されるシグナル配列切断部位の後に生じる。ライゲーションの後、コンピテント化された大腸菌XL-1 Blueが形質転換され、DNAの配列決定により正しい組換体プラスミド(pVL1393.MIP.5sig.Fc)を持つ大腸菌が選択された。その後、pVL1393.MIP.5sig.Fcは、AscI及びStuIにより直線化され、エビのアルカリホスファターゼにより処理された。直線化されたベクターは、その後、整合する末端を持つ合成DNA片とライゲーションされた。合成DNA挿入断片は、以下の2オリゴをアニールすることにより生成された:5'-CGC GCC CAG GTG TAC GAG TCC TGG GGA TGC ATC GGC CCC GGC TGC GCC TGC CTG CAG GCC TGC CTG GGA GGC GGG AGC TCC GGC-3'(配列番号:159)及びGGGSSG(配列番号:130)リンカーを含むペプチド配列1.1FIに対するコード化配列である5'-GCC GGA GCT CCC GCC TCC CAG GCA GGC CTG CAG GCA GGC GCA GCC GGG GCC GAT GCA TCC CCA GGA CTC GTA CAC CTG GG-3'(配列番号:160)。
ライゲーションの後、コンピテント化された大腸菌XL-1 Blueが形質転換され、ローダミンダイターミネーター法及びApplied Biosystems ABI Model 373自動DNAシークエンサーを用いたDNAの配列決定により正しい組換体プラスミド(pVL1393.MIP.5.1.1FI-Fc)を持つ大腸菌が選択された。組換体転移ベクターは、キアゲンのMini-Prepを使用して精製され、組換体バキュロウィルスの構築のために使用された。
【0071】
組換体バキュロウィルスは、sf9細胞に対する転移ベクターと直線化された野生型バキュロウィルスDNA(Autographa californica nuclear polyhedrosis virus (AcNpV)、Pharmingen)との共形質移入により生産された。組換体バキュロウィルスの最初の増幅により、検出可能なタンパク質の発現を達成することができた。次いで、プラーク精製及びウイルスストック希釈が、プラークアッセイにおいて実施された。先に記載されたような標準的な方法が用いられた(O'Reilly, D. R., Miller, L. K., 及びLuckow, V. A. (1994) Baculovirus Expression Vectors: A Laboratory Manual, Oxford University Press, New York)。
細胞培養− スポデプテラ・フルギペルダ(Spodeptera frugiperda)(Sf9)昆虫細胞(ATCC CRL 1711)の接着培養は、GRACE's(JRH Biosciences, #51942-78p)、グルタミン、ストレプトマイシン/ペニシリン、及び10%ウシ胎仔血清(56℃、30分間、熱により不活性化されたもの)が添加されたヒンクスのTNF-FH昆虫用培地中、28℃で培養された。培養細胞は3日毎に継代された。High5細胞のスピナーによる培養細胞は、0.5の感染多重度にて感染され、感染6時間後に回収された。浮遊培養は、スピナーフラスコ中、28℃でESP-921タンパクフリーの昆虫細胞培地(Expression Systems LLC、#96-001)を用いて維持された。培養細胞は、最初の細胞密度が106細胞/mlとなるように3日毎に継代された。
ペプチド-Fcの精製− 至適化された感染方法に従い、High5細胞は、4℃、800 x gにて10分間遠心することにより除去された。細胞が除かれた上清(0.5 L)は、0.45μのNalgene社のフィルターを用いてろ過され、25℃にてPBS(リン酸緩衝生理食塩水)で平衡化されたHi-Trap プロテインAカラム(Amersham Pharmacia Biotech)に添加された。20mlのPBSで洗浄後、カラムは0.2N HOAc、3mlで溶出され、ペプチド-Fcを含む画分が凍結乾燥され、4℃にて保存された。
SDS-PAGE− サンプルが、Laemmli(Laemmli, U. K. (1970) Nature 227:680-685)の方法により、プレステインタンパク質分子量マーカー(SeaBlue、Novex)を用い、還元又は非還元状態で4−20%のトリス-グリシンSDS-PAGE(Novex)にて解析された。
【0072】
タンパク質の配列決定− 感染されたSf9細胞の上清から精製された1.1FI−Fcを、SDS-PAGEにかけ、その後PVDF膜に移した。ミリポアImmobilon-PSQ膜上への電気的なブロットは、BioRad Trans-Blot Transferセルにより、250mAの定電流で1時間行われた(Matsudaira, P. (1987) J. Biol. Chem. 262:10035-10038)。PVDF膜は、50%メタノール中の0.1%クマジーブルーで0.5分間染色し、10%酢酸、50%メタノールで2から3分間脱染色した。膜は、水で完全に洗浄した後、−20℃で保存する前に乾燥させた。50kD付近の1.1FI−Fcのバンドを切り出し、オンラインPTH分析装置を備えた494型Applied Biosystems シークエンサーを用いて最初の11残基を配列決定した。Nelson Analytical 760 インターフェースにより、Justice Innovation Softwareを用いてピーク面積を積算した。配列翻訳は、DEC alphaにより実行した(Henzel, W. J., Rodringuez, H., 及びWatanabe, C. (1987) J. Chromatog. 404:41-52)。
HER2競合ELISA− HER2-ECDに対する1.1FI−Fcへの結合は、競合ELISAにより測定した。サンプルをPBS-BSAにて希釈し、上述した方法でマイクロタイタープレートに固定化されたHER2-ECDに対する40nMのビオチン化1.1FI−Zの結合を阻害する能力を試験した。1時間のインキュベーションの後、プレートをPBS-Tweenにて洗浄し、ストレプタビジン/HRP結合体(Spreptavidin-POD, Roche Molecular Biochemicals)を30分間添加した。プレートを、再度PBS-Tweenで洗浄し、結合されたHRPをABTS/H2O2基質(Kirkegaard & Perry Laboratories)を用いてアッセイし、405 nmにおける吸収を測定した。405 nmにおける吸収値を、ウェルに元々添加されていた1.1FI−Fcの濃度に対してプロットした。非線形回帰分析により(Marquardt, J. Soc.Indust. Appl. Math. 11:431-441 (1963))、シグモイド曲線は、4つの変数の方程式にフィットさせられた;このアッセイにおいて、最大値の半数のシグナルを与えるのに必要な1.1FI−Fcの濃度は、この曲線より計算され、IC50値と呼ぶことにした。
【0073】
細胞の結合− HER2(BT 474 [3+], MDA 361[2+], 及びBaF3 [0+])を発現する細胞をトリプシンで剥がし、洗浄用緩衝液(0.5%BSA、1mM NaOHを含むPBS)で2回洗浄し、各々0.5x106細胞を含む画分に分けられた。細胞は、0、0.5又は5μMの1.1FI−Zbを含む洗浄用緩衝液に懸濁し、洗浄用緩衝液で2回洗浄し、ストレプタビジン-PEを含む洗浄用緩衝液に懸濁され、更に2回洗浄用緩衝液で洗浄し、FACS分析にかけた。
HER2-ECDに結合するペプチドの性質決定− ペプチドHER201は、部分的にランダム化されたファージ-クローン1.1.2(図15)に一致するように合成された。更にまた、完全にランダム化された全長ライブラリーに由来する代表的なクローンである1.1.FI(図16)に一致するペプチドHER212も合成された。このペプチドに存在する2つのジスルフィドに起因して、1-2及び3-4のジスルフィド配列を誘導するために、直交性の保護基が、システインのために使用された。ジスルフィド結合が選択の段階中に誘導されたため、この配列が考えられた。
また、前記ペプチド及びライブラリー7.1(図15)(7.1c)のコンセンサス配列に由来するペプチド配列が、プロテインAのZドメインタンパク質との融合体(1.1.FI-Z及び7.1c-Zと呼ぶ)として発現され、システインの酸化がファージ生産の段階と同様に生じ得るような大腸菌から分泌された。1.1FI-Zの調製物は、ビオチン化され(1.1.FI-Zb)、全タンパク質に基づいた場合2nMのEC50で、固定化HER2に結合し(Fig.6)、競合ELISAに対する試薬として役立った。
組換体ペプチド融合体である1.1.FI-Z及び7.1c-ZペプチドHER201、HER212は、固定化されたHER2に対する1.1.FI-Zbの結合を阻害する能力に関しテストされた(図7)。HER201及び7.1c-Zは、それぞれ300及び1,100 nMのIC50を示した。興味深いことに、HER212及び1.1FI-Z、これらは同じファージペプチド配列に由来するが、劇的に異なるIC50を示した;各々、9μMと50 nMであった。一つの説明として挙げられることは、他のジスルフィド配列がHER212の合成中に強制的に生じた可能性である。更に、1.1.FI-Z(Fig.8B)(レーンS)のSDS-PAGE解析によると、分子間のジスルフィド結合による多量体化に起因するらしき不均一な混合物であることが明らかにされた。マススペクトロ分析によると、HER212はモノマーでのみ存在するのに対し、ダイマー1.1FI-Zの存在が確認された。
【0074】
1.1.FI-Z調製物は、更にSuperdex75を用いてモノマーとダイマーの画分に精製された(図8A及び8B)。驚くべきことに、HER2結合は、HER2の競合ELISAでアッセイした場合、モノマーの画分よりはむしろダイマーの画分に一致した(図8B及び8C)。
更に、1.1.FI-Zは、Zドメインを除去するためにトリプシンにより切断され、逆相HPLCにかけられた。HPLCから得られる画分は、HER2競合ELISAにおける競合能に関し、テストされた。活性画分は、HER2の配列に一致する質量を有したが、ダイマーを含むことも見いだされた。この画分は、HER2の競合ELISAにおいて、タンパク質濃度に基づき3nMのIC50を示した。
ダイマー形成を促進するため、1.1FIのアミノ酸配列はZドメインと融合する前に二度繰り返されるような発現ベクターが構築された(図9)。この「一本鎖ダイマー」((FI)2-Zと称する)も、ダイマーを生産するが、生産される主な種はモノマーであった(図9B)。更に、Superdex75カラムで分画し(図9A)、HER2の競合ELISAでアッセイした場合、活性のある種は、モノマーとして同定された(図9B及び9C)。(FI)2-Zは、1.1FI-Zダイマー(IC50=20 nM)と同等の親和性を示す。
HER2上の結合部位− クラス1及びクラス7の配列は、HER2上の同一、それとも独立の部位に結合されるのかどうか検討するために、固定化HER2に対する、クラス1及びクラス7のファージクローンの結合がHER201又は7.1c-Zの存在下において調べられた。図10A及び図10Bより、HER201及び7.1c-Zはクラス1及びクラス7のファージ両方の結合を阻害するようであり、このことは、両クラスはHER2の同一の部位に結合することを示すものである。これら2つのクラスにおいて得られ配列をつぶさに見ると、コアの相同領域が存在することが分かる(図11)。
【0075】
また、クラス1及びクラス7ペプチドの結合部位は、HER2に対する既知のモノクローナル抗体が、固定化HER2に対する1.1.FI-ファージへの結合と、直接競合しないHER2上の新規な部位のようである(図12)。固定化HER2に対する1.1.FI-ファージの結合を阻害するために、種々のモノクローナル抗体の濃度を増加させても影響はなかった。HER2に対するこのモノクローナル抗体の親和性は、100pM以下であるため、3H4の非常に高濃度(>100nM)な状態における結合の減少は、間接的な効果を介した結果であると考えられる。
細胞の結合− 1.1.FI-Zbは、細胞上に発現されたHER2を認識する能力について調べられた。それぞれ異なる量にてHER2を細胞上に発現する3つのタイプの細胞が、調べられた;BT 474 [3+](3 x 106 レセプター/細胞)、MDA 361[2+](4 x 105 レセプター/細胞)及びHER2を発現しないコントロールの細胞株、BaF3である。FACS分析によると、1.1.FI-Zbは、レセプターとなるHER2の細胞あたりの数、及び用いた1.1.FI-Zbの濃度に比例して細胞に結合することが見いだされた(図13)。
ペプチド-Fc融合体− 1.1.FI配列をコードするDNAが、リンカーを介してIgG1のFc領域と融合され、バキュロウィルス中において発現された。その結果得られたタンパク質は、プロテインAカラムにより精製され、酸化及び還元状態下、SDS-PAGEにより性質決定された(図14)。これら2つの状態において、明らかな分子量のシフトが観察されており、このことは、Fcの二量体化を示すものである。この分子は、固定化されたHER2に対する1.1FIファージの結合を阻害する能力に関してテストした場合、3 nMのIC50を示した。HER2の競合ELISAによりテストした場合、1.1FI-Fcは1.1FI-Zと同様なIC50を示したが、このことは、1.1FI配列がFcに融合された場合HER2と結合することが、十分可能であったことを示す。
【図面の簡単な説明】
【0076】
【図1】天然IgG及びパパイン及びペプシン消化により得られたそれらの断片の概略図である。ジスルフィド結合は、例えばCH1とCLドメインの間の-S-S-により表される。この図において、Vは可変ドメインであり;Cは定常ドメインであり;Lは軽鎖の略であり、Hは重鎖の略である。
【図2A】図2Aは天然IgG Fc領域アミノ酸配列の整列を示す。ヒト(hum)IgG Fc領域配列を示しており、humIgG(非-A及びAアロタイプ)(それぞれ、配列番号:72及び73)、humIgG2(配列番号:74)、humIgG3(配列番号:75)及びhumIgG4(配列番号:76)が示されている。ヒトIgG1配列は非-Aアロタイプであり、この配列とAアロタイプとの差異(356及び358位置;EU番号方式)を、ヒトIgG1配列の下に示す。また、天然マウス(mur)IgG Fc領域配列を示し、murIgG1(配列番号:77)、murIgG2A(配列番号:78)、murIgG2B(配列番号:79)及びmurIgG3(配列番号:80)が示されている。
【図2B】図2Bは図2AのFc領域配列間のパーセント同一性を示す。
【図3】星印が付された配列間に差異を有する、天然ヒトIgG Fc領域配列、humIgG1(非-A及びAアロタイプ)(配列番号:72及び73)、humIgG2(配列番号:74)、humIgG3(配列番号:75)及びhumIgG4(配列番号:76)の整列を示す。
【図4】HER2細胞外ドメイン(ECD)に結合する多価ペプチドファージの富化を示す。選択の3及び4回に対して、HER2で被覆されたウェル対BSAで被覆したウェルから溶出したファージの比率が示されている。
【図5】ファージ結合アッセイの結果を示す。ファージクローン1.1,5.1及び7.1は固定化されたHER2-ECDに結合したが、BSA、又はHER2-ECDを特異的に認識することが示されている密接に関連したHER3又はHER4には結合しなかった。
【図6】固定化されたHER2に結合するペプチド1.1.FIZbのEC50の定量を示す。
【図7】固定化されたHER2-ECDへの結合に対するペプチド1.1.FI-Zbとの、合成ペプチドHER201及びHER212並びに組換えペプチド1.1.FI-Z及び7.1c-Zの競合アッセイの結果を示す。
【図8】図8Aはモノマー(M)及びダイマー(D)含有画分にSuperdex75を使用して精製した1.1.FI-Z(配列番号:81)の調製物を示す。図8Bは、出発物質とペプチド1.1.FI-ZのSUPERDEXTM75カラムからの画分のSDS PAGE分析を示す。図8Cは、競合アッセイを使用してアッセイした場合に、1.1.FI-Zのダイマー画分と符合するHER2結合活性を示す。
【図9】図9Aはモノマー及びダイマー含有画分にSUPERDEXTM75を使用して精製した(1.1FI)2-Z(配列番号:82)(1.1FIを2回繰り返して、プロテインAのZドメインに融合させたもの)調製物を示す。図9Bはペプチド(1.1FI)2-ZのSUPERDEXTM75カラムからの画分のSDS PAGE分析を示す。図9Cは、競合アッセイを使用してアッセイした場合に、(1.1FI)2-Zのダイマー画分と符合するHER2結合活性を示す。
【図10】図10A及び10Bは、ペプチドHER201と7.1c-Zがクラス1とクラス7のファージの結合をブロックすることを示しており、双方のクラスのペプチドがHER2の同じ部位に結合することを示唆している。
【図11】2つのクラスのペプチド間の核の相同性領域を表すクラス1及びクラス7の配列の例を示す。配列7.1c(配列番号:83)、1.1.2(配列番号:84)、1.1.Fc(配列番号:85)、1.1.FA(配列番号:86)、1.1.FB(配列番号:87)、1.1.FH(配列番号:88)、1.1.CF(配列番号:89)。
【図12】HER2-ECDへの結合に対する、HER2に対する既知のモノクローナル抗体とのクラス1及びクラス7のペプチドの競合アッセイの結果を示す。該ペプチドはHER2-ECDへの結合に対して、モノクローナル抗体と競合せず、既知のモノクローナル抗体により認識されるものとは異なるHER2-ECD部位に該ペプチドが結合することを示唆している。
【図13】ペプチド1.1.FI-Zbと異なるレベルでHER2を発現する細胞(MDA361[4x105レセプター/細胞]及びBT474[3x106レセプター/細胞])、又はHER2を発現しない細胞(BaF3)を使用して行われた結合アッセイの結果を示す。1.1.FI-Zbは、細胞で発現したHER2レセプターの数及び1.1.FI-Zbペプチドの濃度に比例して細胞に結合することが見出された。
【図14】ヒトIgG1のヒンジ、CH1及びCH2ドメインにリンカーを介して融合したペプチド1.1.FI(1.1FI-Fc)の、酸化及び還元条件下でのSDS PAGE分析の結果を示す。2つの条件下で分子量がシフトすることは、Ig定常領域の二量体化とFcドメインの生成を示唆している。
【図15】ライブラリー1.1及び7.1.のクローンからの推定ペプチド配列を示す。配列1.1(配列番号:57);1.1.8(配列番号:58);1.1.9(配列番号:59);1.1.10(配列番号:60);1.1.1(配列番号:61);1.1.2(配列番号:62);1.1.6(配列番号:63);1.1.11(配列番号:64);1.1.3(配列番号:65);1.1.4(配列番号:66);1.1.5(配列番号:67);1.1.7(配列番号:68);コンセンサス(配列番号:69);7.1(配列番号:45);7.1.1(配列番号:46);7.1.9(配列番号:47);7.1.6(配列番号:48);7.1.5(配列番号:49);7.1.3(配列番号:50);7.1.4(配列番号:51);7.1.2(配列番号:52);7.1.7(配列番号:53);7.1.8(配列番号:54);7.1.11(配列番号:55);コンセンサス(配列番号:56)。
【図16】第4の選択回による各ライブラリーのランダムクローンからの配列を示す。配列1.1(配列番号:57);コンセンサス(配列番号:58);1.1.FC(配列番号:30);1.1.FN(配列番号:31);1.1.FA(配列番号:1);1.1.FM(配列番号:2);1.1.FE(配列番号:3);1.1.FH(配列番号:29);1.1.FB(配列番号:12);1.1.FF(配列番号:13);1.1.FI(配列番号:14);1.1.FK(配列番号:15);1.1.FD(配列番号:16);1.1.FQ(配列番号:17);1.1.FS(配列番号:18);1.1.FX(配列番号:9);1.1.FY(配列番号:10);1.1.FZ(配列番号:11);1.1.NC(配列番号:4);1.1.NB(配列番号:5);1.1.NM(配列番号:6);1.1.NQ(配列番号:7);1.1.NA(配列番号:19);1.1.NG(配列番号:20);1.1.NH(配列番号:21);1.1.NE(配列番号:22);1.1.NJ(配列番号:23);1.1.NF(配列番号:24);1.1.NK(配列番号:25);1.1.NR(配列番号:26);1.1.CF(配列番号:28);1.1.CA(配列番号:32);1.1.CB(配列番号:33);1.1.CC(配列番号:34);1.1.CD(配列番号:35);1.1.CE(配列番号:36);1.1.CG(配列番号:37);1.1.CH(配列番号:38);1.1.CI(配列番号:39);1.1.CJ(配列番号:40);1.1.CL(配列番号:41);1.1.CM(配列番号:42);1.1.CP(配列番号:43)。
【図17】固定化されたHER2-ECDへの結合に対する、ペプチド1.1.FI-Zbとの、1.1FI-Fc及び1.1FI-Zの競合アッセイの結果を示す。
【発明の概要】
【0001】
(発明の背景)
(発明の分野)
本発明はヒトerbB2遺伝子産物(HER2としても知られているErbB2、又はc-ErbB-2)に結合する新規な化合物に関する。特定の態様では、本発明は、本発明の新規な化合物を利用するErbB2の発現により特徴付けられる疾患の治療に関する。また本発明は、その新規な化合物を含有する組成物、例えば製薬用組成物、並びに研究、診断、治療及び予防法におけるそれらの使用に関する。
【0002】
(関連する開示の記載)
ファージディスプレイは、テンプレート配列内の特定のアミノ酸残基の無作為化を通して又は天然ペプチドライブラリーの生成によって、ペプチド及びタンパク質変異体の産生する手段を提供する(Lowman, H. (1998) Methods Mol. Biol. 87:249-264;Lowman(1997)Annu. Rev. Biophys. Biomol. Struct. 26:401-424)。予め決められた標的分子に結合するディスプレイされたタンパク質又はタンパク質の同定と単離は、固定された標的分子上の非結合又は弱く結合している変異体に対してディスプレイファージを富ませることにより、達成することができる(上掲のLowman, H.(1998))。突然変異誘発と選択の連続の実施により、細胞レセプターに対して高親和性を持つペプチドリガンド又はタンパク質変異体を生成することができる(Lowman, H.(1998)上掲)。上記技術は、例えば細胞標的に帰着するペプチドモチーフを同定するため(Arapら, (1998)Science 279:377-380)、又は天然のタンパク質結合リガンドから親和性が改善され又は成熟したペプチドリガンドを産生するために(Lowmanら, (1991)Biochemistry 30:10832-10838)使用されている。親和性又は特異性が改善されたタンパク質の例には、ヒト成長ホルモン、Znフィンガー、プロテアーゼインヒビター、心房性ナトリウム利尿ペプチド、及び抗体が含まれる(Wells, J. 及びLowman H.(1992) Curr. Opin. Struct. Biol. 2:597-604;Clackson, T.及びWells, J.(1994)Trends Biotechnol. 12:173-184;Lowmanら, (1991)Biochemistry 30(10):832-838;Lowmanら.及びWells J.(1993)J. Mol. Biol. 234:564-578;Dennis M.及びLazarus R.(1994)J Biol. Chem. 269(22):137-144)。
【0003】
インビボでのファージ選択を利用して、ファージディスプレイは、脳及び腎臓のような種々の器官に選択的局在化を媒介可能なペプチドを同定し単離するために(Pasqualini及びRuoslohti (1996) Nature 380:364-366)、並びにαVβ3又はαVβ5インテグリンを担持する特定の腫瘍タイプに帰着するペプチドを同定するために使用されている(Arapら, (1998) Science 279:377-380)。米国特許第5627263号には、α5β1インテグリンにより認識され、これと選択的に結合するペプチドが記載されている。一価のファージディスプレイにより生成される構造的に限定されたペプチドライブラリーを使用して、インシュリン様成長因子1結合タンパク質(IGFBPs)に特異的に結合する14のアミノ酸のペプチドが単離されている(Lowmanら, (1998) Biochemistry, 37:8870-8878)。そのペプチドはヘリックス構造を有し、IGFBPsに結合してインビトロでインシュリン様成長因子α(IGF-1)活性を作用させる(上掲のLowmanら,(1998))。
新規なタンパク質に基づく治療が臨床に入っているので、特定の細胞レセプターとそれらのリガンド、特に種々のヒト悪性腫瘍の病因に関連しているものは、近年、科学界において多くの注目を集めている。例えば、上皮成長因子レセプター(EGFR)に関連した185kdの膜貫通糖タンパク質レセプター(p185HER2)をコードするヒトerbB2遺伝子(her2としても知られている、又はc-erbB-2)は、ヒトの乳癌の約25%〜30%で過剰発現していることが見出されている(Slamonら, (1987) Science 235:177-182;Slamonら, (1989) Science 244:707-712)。いくつかの証拠情報は、ErbB2を過剰発現する腫瘍の病原性及び臨床的病原力におけるErbB2の直接的な役割を裏付けている。非新生物細胞へerbB2を導入すると、その悪性形質転換を引き起こすことが示されている(Hudziakら, (1987)Proc. Natl. Acad. Sci. USA 84:7159-7163;DiFioreら, (1987)Science 237:78-182)。HER2を発現するトランスジェニックマウスは、乳房腫瘍を発生させることが見出されている(Guyら, (1992) Proc. Natl. Acad. Sci. USA)。
【0004】
ErbB2過剰発現は、特に腋窩リンパ節に関与する原疾患を持つ患者における、悪い予後のプレディクターであると一般的に考えられている(上掲のSlamonら, (1987)及び(1989);Ravdin及びChamness, Gene (1995)159:19-27;及びHynesとStern, (1994) Biochim. Biophys. Acta 1198:165-184)。また過剰発現は、CMF(シクロホスファミド、メトトレキセート及びフルオロウラシル)及びアントラサイクリン(Baselgaら, (1997) Oncology 11(3 Suppl 1):43-48)を含む、ホルモン療法及び化学療法に対する感受性及び/又は耐性に関連している。
組換えヒト化抗ErbB2モノクローナル抗体(rhuMAb HER2又はハーセプチン(HERCEPTINR)と称されるマウス抗ErbB2抗体4D5のヒト化体)は、広範な抗癌治療を前に受けたことのあるErbB2過剰発現転移性乳癌を持つ患者において治療的に活性であった(Baselgaら, (1996) J. Clin. Oncol. 14:737-744)。
【0005】
(発明の概要)
本発明は、ヒトerbB2遺伝子産物(ErbB2)に結合する化合物を提供する。本発明の化合物(ここではペプチドリガンドとも呼ぶ)は、例えばペプチド又はペプチド誘導体、例えばペプチド模倣物及びペプチド類似物である。好ましいペプチドリガンドには、線形又は環状のペプチド、好ましくは次の核となる式:
-Cys-Xaa-Xaa-Xaa-Xaa-Cys-
を含む環状ペプチド化合物、好ましくは次の一般式:
Cys-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Cys-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Cys-Cys-Xaa-Xaa-Xaa-Cys-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Cys (配列番号:70)
又は次の一般式:
Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Cys-Xaa-Xaa-Xaa-Xaa-Cys-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa
を有するものが含まれ、また上記の一般構造のダイマー及び他の組合せも含まれる。本発明のペプチドリガンドの中で好ましいものは、次の核となる式:
-Cys-Cys/Ile-Gly-Pro-Gly-Cys (配列番号:71)
を含むペプチドである。このような化合物の特定の例には、約10〜60のアミノ酸残基の環状ペプチドで、場合によってはN-もしくはC-末端、あるいは双方が修飾されたもの、例えばそれらのエステル、アミド、塩及び他の誘導体が含まれる。本発明の好ましい態様では、化合物は、好ましくはErbB2に結合する非天然発生アミノ酸配列である。好ましくは、化合物は、約5〜約50のアミノ酸残基の非自然発生アミノ酸配列である。好ましい化合物は、上記の一般式を有し、約20〜約30のアミノ酸残基を有する環状ペプチドで、そのダイマー及び他の組合せを含む。
【0006】
特定の態様において、本発明はペプチドリガンドと他のペプチドリガンド又は他のタンパク質、特に血清タンパク質又はペプチドとの組合せに関する。この組合せは、ペプチドリガンドのその標的分子に対する親和性又は親和力を、例えば種々の多量体化ドメインを使用して増加させ;例えばZプロテイン融合体としてペプチドリガンドを発現させることにより、ペプチドリガンドの安定性を増加させるか、又はその回収と精製を容易にし;例えば血清の半減期を増加又は減少させることにより、血清アルブミン、免疫グロブリン、アポリポプロテイン又はトランスフェリン等の細胞質タンパク質にペプチドリガンドを融合させることにより(そのような融合体は組換え宿主細胞において簡便に、又は二官能性架橋剤を使用することにより生成する)、及び機能性Fcドメイン又は細胞障害又は酵素部分等の付加的機能を導入することにより、ペプチドリガンドのインビボ使用を含む本発明の態様のペプチドリガンドの治療効果を改善することを含む、種々の目的をもって調製される。
本発明の好ましい態様では、本発明の化合物は多量体化ドメインに結合する。多量体化ドメインは、好ましくは免疫グロブリン配列、又は例えばロイシンジッパー配列である。本発明のこの態様では、免疫グロブリン配列は、好ましくは免疫グロブリン定常領域配列、特に免疫グロブリン重鎖の定常領域である。本発明の好ましい態様では、多量体化ドメインは一又は複数のコンパニオン多量体化ドメインと対になり、ホモ-及びヘテロ-多量体組成をもたらす。本発明のこの側面の好ましい態様はホモ-及びヘテロ-多量体、特にホモ-及びヘテロダイマーにあり、そこでは、多量体化ドメインが、対となって機能的免疫グロブリンFcドメインを提供する免疫グロブリン重鎖定常領域である。よって、好ましい態様では、本発明は、機能的免疫グロブリンFcドメインと結合するエフェクター機能を有する機能的免疫グロブリンFcドメインとErbB2担持細胞型を標的とするように機能する本発明の化合物を含有するハイブリッド分子を提供する。
【0007】
本発明のこの態様によるハイブリッド分子は、場合によってはさらなる機能的部分、例えば酵素部分及び細胞障害性部分を含む。例えば、付加的な機能的ドメインは、ハイブリッド分子に共有結合的に結合し、プロドラッグをそのより活性な形態に転換させるようにプロドラッグに作用可能な酵素であってよい。本発明の所定の好ましい態様による任意成分の機能的ドメインは、例えばハイブリッド分子に共有結合によって結合する細胞障害剤であってもよい。好ましい細胞障害剤には、例えば化学治療剤、毒素及び放射性アイソトープが含まれる。ハイブリッド分子を含む化合物とそれらを含有する組成物は、ヒトerbB2遺伝子産物への結合又は検出と、場合によってはErbB2担持細胞型への酵素又は細胞障害剤等の機能的部分の送達に使用される。
一実施態様では、本発明の化合物、例えば上述したハイブリッド分子はポリペプチドであり、本発明は、本発明のポリペプチドをコードする単離された核酸、好ましくはDNAを含有する物質の組成物を含む。この態様では、本発明は、DNA分子に作用可能に結合した発現コントロール配列、DNA分子を含有する発現ベクター、好ましくはプラスミドで、ここでコントロール配列がベクターで形質転換された宿主細胞により認識されるもの、及びベクターで形質転換された宿主細胞を更に含む。好ましい態様では、核酸が本発明のペプチド化合物と免疫グロブリン定常領域ドメイン配列を含有するハイブリッド分子をコードする。本発明のこの態様に係る核酸分子はハイブリッド分子をコードし、本発明のペプチド化合物をコードする核酸は免疫グロブリン定常ドメインに配列に作用可能に結合している(DNA配列が隣接し読み枠にあるという意味)。場合によっては、DNA配列は任意成分のリンカードメインアミノ酸配列をコードする核酸配列によって結合している。
【0008】
本発明の組成物は、本発明のアミノ酸配列の任意のものをコードする核酸配列を単離又は合成し、適切な宿主中で核酸配列を発現可能な適切な発現ベクター内に核酸配列をライゲーションし、核酸配列がライゲーションされた発現ベクターで宿主を形質転換し、核酸配列の発現に適した条件下で宿主を培養する工程を含む方法により作成され、それにより、選択された核酸配列によりコードされるタンパク質は宿主により発現される。好ましくは、ポリペプチドはついで宿主細胞培養物から回収される。この方法では、ライゲーション工程は、核酸配列が適切な分泌シグナルに作用可能に結合するように適切な発現ベクターに核酸をライゲーションすることをさらに意図しており、それにより、アミノ酸配列は宿主により分泌される。
本発明は、本発明の化合物を含有する、特にerbB2遺伝子発現に関連した病気又は疾患の検出及び治療のための製薬用組成物を含む組成物を含む。例えば、癌の検出及び治療用のキット及び製造品が提供される。キット及び製造品は、好ましくは:
(a)容器;
(b)前記容器上の又は該容器に付随するラベル;及び
(c)前記容器内に収容された本発明の化合物を含む組成物を含み、ある実施態様では、当該組成物は、erbB2遺伝子発現を伴う病気又は疾患、例えば癌の治療に有効である。好ましくは、前記容器上のラベルは、当該組成物が癌の治療に使用できることを表示しており、当該組成物中の化合物がヒトerbB2遺伝子産物に結合するここで記載された化合物を含む。キットは、場合によっては、付属品、例えば製薬的に許容されるバッファーを収容した第2の容器及び疾患の治療のために組成物を使用するための指示を含む。
【0009】
種々の病気及び疾患、例えば癌、特にErbB2に関連していることを特徴とするものの治療に有用な方法が開示される。よって、本発明は、治療的有効量の本発明の化合物を宿主に投与することを含む、その必要とする宿主におけるErbB2媒介性又は関連性の病気又は疾患の治療方法を提供する。さらに本発明は、限定するものではないが、化合物の非経口、経口、直腸及び肺投与に適切なものを含む、本発明の化合物の種々の投与形態を提供する。本発明の好ましい態様では、治療的投与形態は静脈内送達に適するように提供され、本発明は、ErbB2媒介性又は関連性のプロセス又は事象、例えば癌に関連した病気又は疾患の治療のために提供される。
【0010】
(好ましい実施態様の詳細な説明)
定義
「HER2」、「ErbB2」、「c-Erb-B2」という用語は互換的に使用される。特に別の定義を示さない限り、ここで使用される「ErbB2」、「c-Erb-B2」及び「HER2」という用語はヒトタンパク質を意味し、「her2」、「erbB2」及び「c-erb-B2」はヒト遺伝子を示す。ヒトerbB2遺伝子及びErbB2タンパク質は、例えばSembaら, (1985) PNAS(USA)82:6497-6501及びYamamotoら, (1986) Nature 319:230-234(Genebank受託番号 X03363)に記載されている。ErbB2は4つのドメインを含んでいる(ドメイン1-4)。
本発明における「ペプチドリガンド」なる用語は、その由来にかかわらず、ErbB2に結合するように機能するアミノ酸配列を指すことを意味する。本発明のペプチドリガンドは、一般的に束縛されている(つまり、例えばβターン又はβプリーツシートを開始するアミノ酸の存在、又はジスルフィド結合したCys残基の存在により環化したような構造のいくつかのエレメントを有している)か、又は束縛されていない(例えば、線形)、約50アミノ酸残基未満、好ましくは約40アミノ酸残基未満のアミノ酸配列である。約40アミノ酸残基未満のペプチドリガンドでは、約10〜約30アミノ酸残基のペプチドリガンド、特に約20〜約30アミノ酸残基のペプチドリガンドが好ましい。しかしながら、現在の開示を読むと、当業者であれば、本発明のペプチドリガンドを区別するのは特定のペプチドリガンドの長さではなく、ErbB2に結合するその能力であることを認識するであろう。よって、例えば5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29及び30アミノ酸残基のペプチドリガンドが、等しく有望な本発明のペプチドリガンドである。
【0011】
本発明のペプチドリガンドは、ペプチドリガンドがインビトロで、好ましくはインビボでErbB2、例えばErbB2担持細胞に「帰着する(homes)」、「結合する」又は「標的とする」ならば、十分な親和性と特異性を持ってErbB2に結合するであろう(例えば、Pasqualini及びRuoslahti(1996) Nature, 380:364-366、及びArapら,(1998) Science 279:377-380)における「帰着する」、「帰着」及び「結合する」という用語の使用を参照されたい)。一般的に、ペプチドリガンドは、Jonesら, (1999) FEBS Letters 447:227-231により記載されたものと同様なErbB基質を利用する競合アッセイ等のインビトロアッセイにより定量される場合に約1μM未満、好ましくは約100nM未満、特に約10nM未満の親和力でErbB2に結合する。しかしながら、ErbB2に対して約1nM未満、好ましくは約1pM〜1nMの親和力を有するペプチドリガンドが、等しく有望な本発明のペプチドリガンドである。一般に、上述したようなErbB2に結合するペプチドリガンドは、固定化されたErbB2分子に結合させるなどのここに記載された当該技術の多くの標準的な技術の任意のものにより単離され同定され得る。
上述したように、ペプチドリガンドは自然に生じた、また非自然的に生じたアミノ酸残基を含有していてもよい上述したアミノ酸配列である。よって、特定のアミノ酸又はペプチドの構造を模倣した非アミノ酸化学構造を含みうる、いわゆる「ペプチド模倣物」及び「ペプチド類似物」も本発明のペプチドリガンドとできる。このような模倣物及び類似物は、類似の物理的特徴、例えば大きさ、電荷又はそれらの対のペプチドに見出されるような適切な空間的配向性で存在する疎水性を示すものとして一般に特徴付けられる。ペプチドの模倣化合物の特定の例は、一又は複数のアミノ酸のアミド結合が、例えば炭素-炭素結合又は当該技術でよく知られている他の結合で置換された化合物である(例えば、Sawyer, Peptide Based Drug Design pp.378-422(ACS, Washington DC 1995)を参照のこと)。
【0012】
従って、本発明における「アミノ酸」なる用語は最も広い意味に使用され、自然に生じるLα-アミノ酸又は残基を含むことを意味する。自然に生じるアミノ酸に対して一般的に使用されている1文字及び3文字略号がここでも使用される(Lehninger, A.L., Biochemistry, 第2版, pp.71-92,(1975) Worth Publishers, New York)。この用語には、D-アミノ酸、並びに化学的に修飾されたアミノ酸、例えばアミノ酸類似物、ノルロイシン等のタンパク質に通常導入されない自然に生じるアミノ酸、及びアミノ酸に特徴的な当該分野において公知の特性を有する化学的に合成された化合物が含まれる。例えば、天然Phe又はProと同じペプチド化合物の配座制限を許容するフェニルアラニン又はプロリンの類似物又は模倣物が、アミノ酸の定義に含まれる。このような類似物及び模倣物はここではアミノ酸の「機能的等価物」と称する。アミノ酸の他の例は、出典を明示してここに取り込まれるRoberts及びVellaccio(The Peptides:Analysis, Synthesis, Biology)Eds. Gross及びMeiehofer, Vol.5 p341, Academic Press, Inc, N.Y. 1983に列挙されている。
例えば、標準的な固相合成法により合成されたペプチドリガンドは、遺伝子によりコードされるアミノ酸に限定されない。遺伝子コードによりコードされない一般的に遭遇するアミノ酸には、例えば国際公開第90/01940号に記載されているもの、例えばGlu及びAspについての2-アミノアジピン酸(Aad);Glu及びAspについての2-アミノピメリン酸(Apm);Met、Leu、及び他の脂肪族アミノ酸についての2-アミノブチル酸(Abu);Met、Leu、及び他の脂肪族アミノ酸についての2-アミノヘプタン酸(Ahe);Glyについての2-アミノイソブチル酸(Aib);Val、及びLeu及びIleについてのシクロヘキシルアラニン(Cha);Arg及びLysについてのホモアルギニン(Har);Lys、Arg及びHisについての2,3-ジアミノプロピオン酸(Dpr);Gly、Pro及びAlaについてのN-エチルグリシン(EtGly);Gly、Pro及びAlaについてのN-エチルグリシン(EtGly);Asn、及びGlnについてのN-エチルアスパラギン(EtAsn);Lysについてのヒドロキシルリジン(Hyl);Lysについてのアロヒドロキシルリジン(AHyl);Pro、Ser、及びThrについての3-(及び4)ヒドロキシプロリン(3Hyp、4Hyp);Ile、Leu、及びValについてのアロ-イソロイシン(AIle);Alaについてのρ-アミジノフェニルアラニン;Gly、Pro、及びAlaについてのN-メチルグリシン(MeGly、サルコシン);IleについてのN-メチルイソロイシイン(MeIle);Met及び他の脂肪族アミノ酸についてのノルバリン(Nva);Met及び他の脂肪族アミノ酸についてのノルロイシン(Nle);Lys、Arg及びHisについてのオルニチン(Orn);Thr、Asn及びGlnについてのシトルリン(Cit)及びメチオニンスルホキシド(MSO);PheについてのN-メチルフェニルアラニン(MePhe)、トリメチルフェニルアラニン、ハロ(F、Cl、Br、及びI)フェニルアラニン、トリフルオロフェニルアラニンが含まれる。
【0013】
本発明の保存的アミノ酸置換を、「例示的置換」及び「好ましい置換」と題して表1に示す。好ましい置換がHER2結合性を低減又は変化させない場合、表1に例示的置換と名前を付け、又はさらにここで記載する、より実質的な変化が導入され、生成物がHER2結合について試験される。
【0014】
本発明のペプチドリガンドは、好ましくは自然に生じないアミノ酸配列である。自然に生じないとは、特定のペプチドリガンドのアミノ酸配列が天然に見出されないことを意味する。本発明において、「非天然発生」とは、非天然の又は非天然発生のアミノ酸配列に相当するペプチドリガンドを指すために使用される。この多様なペプチドリガンドは、当業者によく知られている種々の技術を使用して生成又は選択されうる。例えば、束縛されているか又は束縛されていないペプチドライブラリーが無作為に作製され、当該分野で標準的な技術を利用してファージにディスプレイする(例えば、Lowmanら, (1998) Biochemistry 37:8870-8878)。
少なくとも3つの異なる種のペプチドリガンドが、ErbB2に結合する機能に基づいて区別されうる。これらは、ここでは「中性」、「アゴニスト」及び「アンタゴニスト」ペプチドリガンドと称される。一般に、中性ペプチドリガンドは、上述したようにErbB2に結合するように機能する。中性ペプチドリガンドは、ErbB2を担持する特定の細胞型の、例えば細胞障害剤又は酵素でのターゲティングが望まれる本発明の態様において好ましい。
一般に、ErbB2の結合に加えて「アゴニスト」ペプチドリガンドは、ErbB2担持細胞に対して直接の効果を有している。好ましくは、アゴニストペプチドリガンドは記載されたようにErbB2に結合し、さらにErbB2-ErbB3タンパク質複合体及び/又はErbB2-ErbB4タンパク質複合体に関連した事象、例えば細胞内キナーゼドメインをしてErbB2レセプター複合体のチロシン残基をホスホリル化させる能力を開始又は媒介する。また、アゴニストペプチドリガンドの結合は、ErbB2レセプターの二量体化に関連している場合がある。ErbB2レセプター媒介性ホスホリル化又はErbB2レセプター二量体化を誘発する能力は、チロシンホスホリル化アッセイ及びSDS PAGE等の当該分野で標準的な技術を使用して定量することができる。
【0015】
これに対して「アンタゴニスト」ペプチドリガンドは、ErbB2-ErbB3タンパク質複合体及び/又はErbB2-ErbB4タンパク質複合体に結合する天然リガンドに関連した活性、例えば天然リガンドの結合により誘発される細胞反応を、例えばErbB2-ErbB3タンパク質複合体及び/又はErbB2-ErbB4タンパク質複合体を天然又は自然に発生するリガンドに結合させるか又はこれをブロックすることにより、低減するように作用する。
しかしながら、上述の分類に限定されるものではない。例えば、ある実施態様では、ペプチドリガンドの結合が、ErbB2レセプターの内在化、又はプログラム細胞死、すなわちアポトーシスの誘発等の関連した細胞事象の誘発の原因になる。ErbB2又はErbB2レセプター複合体の内在化を開始するペプチドリガンドは、ここで以下に記載する細胞障害剤を細胞内送達することを必要とする本発明の実施態様において特に有用である。
「アポトーシスを誘発する」又は「アポトーシスを誘発可能な」という用語は、アネキシンVの結合、DNAの断片化、細胞収縮、小胞体の拡張、細胞断片化、及び/又は膜小胞の形成(アポトーシス体と呼ばれる)により決定されるプログラム細胞死を誘発する化合物の能力を意味する。細胞は、ErbB2レセプターを発現又は過剰発現するものである。好ましくは「細胞」は、腫瘍細胞、例えば乳房、卵巣、胃、子宮体、唾液腺、肺、腎臓、大腸、甲状腺、膵臓又は膀胱細胞である。インビトロでは、細胞はSKBR3、BT474、Calu3細胞、MDA-MB-453、MDA-MB-361又はSKOV3細胞でありうる。アポトーシスに伴う細胞事象を評価するために種々の方法が利用できる。例えば、ホスファチジルセリン(PS)転位置をアネキシン結合により測定することができ;DNA断片化はDNAラダーリングにより評価することができ;DNA断片化に伴う細胞核/染色質凝結は低二倍体細胞の何らかの増加により評価することができる。好ましくは、アポトーシスを誘発する化合物は、BT474細胞を使用するアネキシン結合アッセイにおいて、未処理細胞の約2〜50倍、好ましくは約5〜50倍、最も好ましくは約10〜50倍のアネキシン結合を誘発するという結果を生じるものである。
【0016】
ここで使用される「ヘレグリン」(HRG)は、ErbB2-ErbB3及びErbB2-ErbB4タンパク質複合体を活性化する(すなわち、そこに結合する際に複合体のチロシン残基のリン酸化を誘発する)ポリペプチドを意味する。この用語に包含される種々のポリペプチドは、例えば、Jonesら, (1990) FEBS Letters 447:227-231;Holmesら,(1992) Science, 256:1205-1210;国際公開92/20798号;Wenら, (1994) Mol. Cell. Biol., 14(3):1909-1919;及びMarchionniら, (1993) Nature, 362:312-318に開示されている。この用語には、自然に生じたHRGポリペプチドの変異体及び/又は生物学的に活性な断片、例えばそのEGF様ドメイン断片(例えばHRGβ1177−244)が含まれる。
「ErbB2-ErbB3タンパク質複合体」と「ErbB2-ErbB4タンパク質複合体」は、それぞれErbB2レセプターと、ErbB3レセプター又はErbB4レセプターの非共有的に結合したオリゴマーである。Sliwkowskiら, (1994) J. Biol. Chem., 269(20):14661-14665に記載されているように、これらのレセプターの両方を発現する細胞に複合体が形成され、免疫沈降法により単離され、SDS-PAGEにより分析される。
本発明の特定の態様において使用されるところの「多量体化ドメイン」なる用語は、直接又は「リンカードメイン」を介してペプチドリガンドが結合する分子の一部を指すことを意味する。多量体化ドメインは、好ましい実施態様では、2又はそれ以上の多量体化ドメインの相互作用を容易にするアミノ酸ドメインである。多量体化ドメインは2又はそれ以上の多量体化ドメインの間の相互作用を促進するが、本発明では、多量体化ドメインに結合するペプチドリガンドが多量体の一部として存在している必要性はない。
【0017】
本発明の好ましい態様では、多量体化ドメインは、2又はそれ以上の多量体化ドメインの安定した相互作用を促進させるポリペプチドである。例えば、限定するものではないが、多量体化ドメインは免疫グロブリン配列、例えば免疫グロブリン定常領域、ロイシンジッパー、疎水性領域、親水性領域、2又はそれ以上の多量体化ドメイン間に分子間ジスルフィド結合を形成する遊離チオールを含有するポリペプチド、又は例えば米国特許第5731168号に記載されている「キャビティ中突起(protuberance-into-cavity)」ドメインであってよい。前記特許では、突起は、第1ポリペプチドの界面からの小さなアミノ酸側鎖を、より大きな側鎖(例えばチロシン又はトリプトファン)で置換することにより形成される。突起に対して同一又はより小さなサイズの相補的キャビティは、大きなアミノ酸側鎖を小さなもの(例えばアラニン又はスレオニン)で置換することにより、第2ポリペプチドの界面に任意につくり出される。
よって、好ましい態様では、多量体化ドメインは、2又はそれ以上の多量体化ドメインの安定した相互作用を促進するか、これを可能にし、モノマー性多量体ドメインからのダイマーと他の多量体の形成を促進させるか、これを可能にする分子の一部を提供する。好ましくは、本発明のこの態様では、多量体化ドメインは免疫グロブリンの定常領域ドメインである。免疫グロブリンの定常ドメインは、本発明の化合物のインビボ循環半減期を改善し、場合によっては、当業者をして、以下に記載する「エフェクター機能」を本発明の所定の態様に導入することを可能にするという利点をもたらす。
【0018】
本明細書と特許請求の範囲を通して、免疫グロブリン重鎖中の残基の番号付けは、ここに出典を明示して取り込まれるKabat等, Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991)のEUインデックスのものである。「KabatのEUインデックス」とはヒトIgG1EU抗体の残基番号付けを指す。
「抗体」(Ab)と「免疫グロブリン」(Ig)は同じ構造的特徴を有する糖タンパク質である。抗体は特定の抗原に対して結合特異性を示すものであるが、免疫グロブリンは、抗体と抗原特異性を欠く他の抗体様分子の両方を含む。後者の種類のポリペプチドは、例えばリンパ系により低レベルで、骨髄腫により増加したレベルで産生される。
「抗体」及び「免疫グロブリン」は、通常は、2つの同一の軽(L)鎖及び2つの同一の重(H)鎖からなる、約150000ダルトンのヘテロテトラマー糖タンパク質である。各軽鎖は一つの共有ジスルフィド結合により重鎖に結合しており、ジスルフィド結合の数は、異なった免疫グロブリンアイソタイプの重鎖間で変化する。また各重鎖と軽鎖は、規則的に離間した鎖間ジスルフィド結合を有している。各重鎖は、カルボキシ(C)末端定常ドメインが続くアミノ(N)末端可変ドメイン(VH)を有している。各軽鎖は可変N-末端ドメイン(VL)及びC-末端定常ドメインを有し;軽鎖の定常ドメイン(CL)は重鎖の最初の定常ドメイン(CH1)と整列し、軽鎖の可変ドメインは重鎖の可変ドメインと整列している。免疫グロブリンポリペプチド鎖のドメインの定義に従って、軽(L)鎖は2つのコンホメーション的に類似したドメインVL及びCLを有し;重鎖は4つのドメイン(VH、CH1、CH2、及びCH3)を有しており、これらの各々は1つの鎖間ジスルフィド架橋を持つ。
【0019】
重鎖の定常(C)ドメインのアミノ酸配列に依存して、免疫グロブリンは異なるクラスに分けることができる。免疫グロブリンの5つの主要なクラス:IgA、IgD、IgE、IgG及びIgMがある。免疫グロブリンクラスはさらにサブクラス(アイソタイプ)、例えばIgG1、IgG2、IgG3、IgG4、IgA1及びIgA2に分割することができる。免疫グロブリンの異なるクラスに対応する重鎖定常ドメインは、各々α、δ、ε、γ、及びμドメインである。任意の脊椎動物種からの抗体の軽鎖には、その定常ドメインのアミノ酸配列に基づいて、カッパ(κ)又はラムダ(λ)と呼ばれる2つの区別される型の一つが割り当てられる。配列の研究により、IgMのμ鎖は5つのドメインVH、CHμ1、CHμ2、CHμ3、及びCHμ4を含むことが分かっている。IgE(ε)の重鎖も5つのドメインを含む。
異なるクラスの免疫グロブリンのサブユニット構造及び三次元的立体配置はよく知られている。これらのうちIgA及びIgMはポリマーであり、各サブユニットは2つの軽鎖と2つの重鎖を含む。IgG(γ)の重鎖はヒンジ領域として知られるCHγ1及びCHγ2ドメイン間にある所定長さのポリペプチド鎖を含む。IgAのα鎖はO-結合グリコシル化部位を含むヒンジ領域を有し、μ及びε鎖はγ及びα鎖のヒンジ領域に類似する配列は持たないが、それらは他のものが持たない第4の定常ドメインを含む。免疫グロブリン鎖のドメイン組成は次のように要約することができる。
軽鎖λ=Vλ Cλ
κ=Vκ Cκ
重鎖 IgG(γ)=VH CHγ1 ヒンジ CHγ2 CHγ3
IgM(μ)=VH CHμ1 CHμ2 CHμ3 CHμ4
IgA(α)=VH CHα1 ヒンジ CHα2 CHα3
IgE(ε)=VH CHε1 CHε2 CHε3 CHε4
IgD(δ)=VH CHδ1 ヒンジ CHδ2 CHδ3
【0020】
ヒトIgG Fc領域の「CH2ドメイン」(「Cγ2」ドメインとも呼ばれる)は通常約アミノ酸231から約アミノ酸340まで延びる。CH2ドメインは、他のドメインと密接には対をなさないという点で独特である。むしろ、二つのN-結合分岐炭水化物鎖が未変性の天然IgG分子の2つのCH2ドメインの間に挿入されている。
「CH3ドメイン」は、Fc領域における残基C-末端からCH2ドメインまでの伸展(すなわち、IgGのおよそアミノ酸残基341からおよそアミノ酸残基447まで)を含む。
「ヒンジ領域」はヒトIgG1のGlu216からPro230の伸展として一般に定義されている(Burton, Molec. Immunol. 22:161-206 (1985))。他のIgGアイソタイプのヒンジ領域は、重鎖間S-S結合を形成する最初と最後のシステイン残基を同じ位置に配することにより、IgG1配列と整列させられうる。
Fc領域の「低ヒンジ領域」は、通常は、C-末端からヒンジ領域、すなわちFc領域の233から239残基までの残基の伸展として定義されている。
抗体のパパイン消化は、「Fab」断片又は領域と呼ばれる2つの同一の抗原結合断片を生成し、各々が単一の抗原結合部位と残りの「Fc」断片又は領域を持つ。免疫グロブリン重鎖のFc領域の境界は変化するかも知れないが、ヒトIgG重鎖Fc領域は、通常はCys226またはPro230の位置のアミノ酸残基からカルボキシル末端まで伸展すると定義される。免疫グロブリンのFc領域は、例えば図1に示すように、2つの定常ドメイン、CH2及びCH3を含む。「天然Fc領域配列」は天然に見出されるFc領域のアミノ酸配列と同一のアミノ酸配列を含む。天然ヒトFc領域配列を図2及び3に示し、ヒトIgG1Fc領域(非-A及びAアロタイプ);ヒトIgG2Fc領域;ヒトIgG3Fc領域;及びヒトIgG4Fc領域、並びに自然に生じるその変異体を含むが、それらに限定されるものではない。天然マウスFc領域配列を図2Aに示す。
【0021】
ペプシン処理により、2つの抗原結合部位を有し、さらに抗原を架橋させ得るF(ab')2断片が得られる。またFab断片は、軽鎖の定常ドメインと重鎖の第1定常ドメイン(CH1)を有する。Fab'断片は、抗体ヒンジ領域からの一又は複数のシステインを含む重鎖CH1ドメインのカルボキシ末端に数個の残基が付加している点でFab断片とは異なる。Fab'-SHは、定常ドメインのシステイン残基が遊離チオール基を担持しているFab'に対するここでの命名である。F(ab')2抗体フラグメントは、間にヒンジシステインを有するFab'断片の対として生産された。抗体断片の他の化学結合も知られている。
「機能的なFc領域」は、天然のFc領域の「エフェクター機能」を保持する。例示的な「エフェクター機能」は、C1q結合;補体依存性細胞障害活性;Fcレセプター結合、抗体依存性細胞媒介細胞障害活性(ADCC);貪食作用、細胞表面レセプター(すなわち、B細胞レセプター;BCR)の低減下を含む。このようなエフェクターの機能は、一般に、Fc領域が結合ドメイン(例えばペプチドリガンド)に結合するのに必要とされており、当該技術で公知の様々なアッセイを用いることで評価される。
親又は天然Fc領域に適当なアミノ酸修飾を導入することにより、(a)ヒトエフェクター細胞の存在下で抗体依存性細胞媒介細胞傷害(ADCC)をより効果的に媒介する、及び/又は(b)親ポリペプチドよりも、より強い親和性でFcガンマレセプター(FcγR)と結合するような、Fc領域の変異体を作製することができる。そのようなFc領域の変異体は、一般に、少なくとも1つのアミノ酸修飾をFc領域に含むであろう。Fc領域の変異体は、2,3,4,5個所等の置換を含み得る。
【0022】
いくつかの抗体のエフェクター機能は、抗体のFc領域に結合するFcレセプター(FcRs)によって媒介される。FcRsは、免疫グロブリンアイソタイプに対する特異性により定義され;IgG抗体に対するFcレセプターはFcγRと、IgEに対してはFcεRと、IgAに対してはFcαRなどのように呼ばれている。FcγRの3つのサブタイプは同定されており:FcγRI(CD64),FcγRII(CD32)及びFcγRIII(CD16)である。それぞれのFcγRサブクラスは、2または3つの遺伝子によってコードされており、またRNAスプライシングにより複数の転写産物が生じるため、FcγRアイソフォームには広い多様性が存在する。これらの異なるFcRサブタイプは異なる型の細胞上で発現されている(Ravetch and Kinet, Annu.Rev.immunol. 9:457-492 (1991))。例えばヒトにおいて、FcγRIIIBは好中球上にのみ見出され、FcγRIIIAはマクロファージ、単球、ナチュラルキラー細胞(NK細胞)、ある種のT細胞上に見出される。特に、FcγRIIIAは、ADCCに関係する細胞型であるNK細胞上に存在する唯一のFcγRである。
FcγRに対するヒトおよびマウス抗体上の結合部位は、これまでに低ヒンジ領域(233-239残基:Kabatら, Sequences of Proteins of Immunological Interest, 5th ED. Public health Service, National Institutes of Health, Bethesda, MD. (1991) におけるようなEUインデックス番号付け)にマッピングされている。Woofら Molec. Immunol. 23:319-330(1986); Duncanら Nature 332:563 (1988); Canfield及びMorrison, J. Exp. Med. 173:1483-1491 (1991); Canppelら, Proc.Natl.Acad.Sci USA 88:9036-9040 (1991)。
「C1q」は免疫グロブリンのFc領域に対する結合部位を含むポリペプチドである。C1qは2つのセリンプロテアーゼC1r及びC1sと共に、補体依存性細胞障害(CDC)経路の最初の成分である複合体C1を形成する。
【0023】
「抗体依存性細胞媒介細胞障害」及び「ADCC」は、細胞が媒介する反応であって、FcRsを発現する非特異的障害細胞(例えば、ナチュラルキラー(NK)細胞、好中球、及びマクロファージ)が、標的細胞上に結合した抗体を認識して、続いて標的細胞の溶解を引き起こす反応を意味する。ADCCを媒介する主要な細胞である、NK細胞はFcγRIIIのみを発現するのに対し、単球は、FcγRI、FcγRII及びFcγRIIIを発現する。造血細胞におけるFcRの発現は、Ravetch及びKinet, Annu. Rev. Immunol 9:457-92 (1991) の464頁の表3に要約されている。
ここで使用される場合、「サルベージレセプター結合リガンド」という用語は、IgG分子のインビボでの血清半減期を増加させる原因となるIg分子(例えばIgG1、IgG2、IgG3又はIgG4)のFc領域のリガンドを意味する(1998年4月14日に公開された米国特許第5739277号)。
「治療」とは、治療的処置及び予防又は防止手段の両方を指す。治療の必要があるものには、既に羅患しているもの、並びに疾患が予防されるべきものが含まれる。
治療の目的とされる「哺乳動物」とは、ヒト、家庭又は農場用動物、及び動物園、スポーツ又はペット用動物、例えばイヌ、ウマ、ネコ、ウシ等を含む、哺乳動物に分類されるあらゆる動物を意味する。好ましくは哺乳動物はヒトである。
「疾患」は本発明のペプチドリガンドを含有する組成物で治療することで恩恵を得るあらゆる症状のことである。これには、問題の疾患に哺乳動物を罹患させる素因になる病理状態を含む、慢性及び急性の疾患又は病気が含まれる。ここで治療される疾患の例は、これに限定されるものではないが、良性及び悪性の腫瘍;白血病及びリンパ悪性腫瘍;ニューロン、神経膠、星状細胞、視床下部及び他の腺、マクロファージ、上皮、ストローマ及び割腔の疾患;及び炎症、血管形成及び免疫性疾患が含まれる。
【0024】
「癌」及び「癌性」という用語は、典型的には調節されない細胞成長により特徴付けられる、哺乳動物における生理学的状態を意味するか記述するものである。癌の例には、これらに限定されるものではないが、癌腫、リンパ腫、芽細胞腫、肉腫、及び白血病が含まれる。このような癌のより特定な例には、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、神経膠芽細胞腫、子宮頸癌、卵巣癌、肝臓癌、膀胱癌、肝腫瘍、乳癌、大腸癌、結腸直腸癌、子宮内膜癌、唾液腺癌、腎臓癌、肝臓癌、前立腺癌、陰門癌、甲状腺癌、肝癌及び様々な種類の頭部及び頸部の癌が含まれる。
ここで使用される場合、「非経口」なる用語は、腸以外によって、特に静脈内(i.v.)、動脈内(i.a.)、腹膜内(i.p.)、筋肉内(i.m.)、心室内及び皮下(s.c.)経路で本発明の化合物を導入することを意味する。
【0025】
(発明の実施形態)
ペプチドリガンド
本発明のペプチドリガンドはErbB2に結合し、次の一般式:
Xaa(1−14) -Cys-Xaa16-Gly-Pro-Gly-Cys-Xaa(21−27) (配列番号:90)
を有するペプチドリガンドを用いたインビトロアッセイにおいてErbB2への結合に対して競合する能力により同定することができ、ここで、Xaa(1−14)は存在しないか、又は1〜14のアミノ酸であり;Xaa16はMet、Thr、Cys及びIleからなる群から選択され、好ましくはIleであり;Xaa(21−27)は存在しないか、又は1〜7のアミノ酸である。好ましくは、ペプチドリガンドは、次の一般式:
Xaa(1−3)-Glu-Xaa5-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Xaa14-Xaa15-Leu-Xaa(17−20) (配列番号:91)
を有するペプチドリガンドと上述のようにErbB2との結合に対して競合するであろう。ここで、Xaa(1−3)及びXaa(17−20)は、独立して、存在しないか、又はそれぞれ1〜3及び1〜4のアミノ酸であり;Xaa1、Xaa2及びXaa3はアミノ酸である。特定の実施態様では、本発明のペプチドリガンドは、ここに記載する配列番号:1−配列番号:44、及び配列番号:57−配列番号:69で表されるペプチドリガンドの何れかと競合し、好ましくはErbB2との結合に対して配列番号:14のものと競合するであろう。
以上から理解されるように、「競合する」及び「競合する能力」という用語は相対的な用語である。よって、本発明のペプチドリガンドを記述するために用いられる場合、この用語は、ここで記載したような標準的な競合アッセイにおいて、50μMで存在する場合、好ましくは1μM、より好ましくは100μMで存在する場合、好ましくは1nM又はそれ未満で存在する場合の、例えば配列番号:14の結合を50%阻害するペプチドリガンドを意味する。このようなペプチドリガンドは、実施例のセクションに記載するもののような標準的な競合アッセイで測定されるときに約1μM未満、好ましくは約100nM未満、より好ましくは約10nM未満の親和性でErbB2に一般的に結合する。しかしながら、ErbB2に対して約1nM未満、好ましくは約1pM〜1nMの親和性を有するペプチドリガンドも等しく本発明のペプチドリガンドであるようである。
【0026】
化合物が上述したペプチドリガンドと競合するか、競合する能力を有しているか否かを決定するためのインビトロアッセイシステムに対しては、当業者であれば多数の標準的な競合アッセイの任意のものを使用することができる。このような手順には、限定するものではないが、数例を挙げると、ラジオイムノアッセイ、酵素イムノアッセイ(EIA)、好ましくは酵素結合イムノアッセイ(ELISA)、「サンドウィッチ」イムノアッセイ、免疫放射定量アッセイ、蛍光イムノアッセイ、免疫電気泳動アッセイ等の技術を用いる競合アッセイシステムを含む。
例として、限定するものではないが、ELISAアッセイを行うことができ、そこでは、マクロタイタープレート(例えば、Nunc MaxisorpTM96-ウェルプレート、Inter Med, Denmark)を、50mMの重炭酸アンモノウム、pH9.3にHER2-ECDが入ったものを5μg/ml使用し、4℃で終夜被覆する。ウェルは25℃で1時間、BSAを1%含有するPBS(PBS-BSA)を使用してブロックすることができる。候補ペプチドリガンドをPBS-BSAで希釈し、ここに記載されたようにZプロテイン融合体(1.1.FI-Z)として生成され、ビオチン化(1.1.FI-Zb)されたペプチド1.1.FI(配列番号:14)の、固定化されたHER2-ECDへの結合をブロックする能力を試験した。約1時間のインキュベーションに続いて、プレートをPBS-TWEENで洗浄し、ストレプトアビジン-HRPを約30分間で添加する。プレートをPBS-TWEENで再び洗浄し、結合したHRPをABTS/H2O2基質を使用してアッセイする。405nmの吸光度の変化を評価する。吸光度の減少を試料濃度に対してプロットし、各候補ペプチドリガンドに対するIC50を決定する。
先に見られたように、本発明の好ましいペプチドリガンドは約10〜30アミノ酸残基、好ましくは約20アミノ酸残基の自然には生じないペプチドリガンドである。上述した特に好ましいペプチドリガンドは自然に生じるアミノ酸からなり、当業者によく知られている標準的な組換え及び合成技術を使用して産生することができる。
【0027】
好ましいペプチドリガンドは次の一般式:
Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Cys-Xaa-Xaa-Xaa-Xaa-Cys-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa
を有する。些かより特定的には、好ましいペプチドリガンドは次の式:
Xaa(1−7) -Cys-Xaa9-Gly-Pro-Gly-Cys-Xaa(14−20) (配列番号:92)
を有し、ここで、Xaa(1−7)及びXaa(14−20)は、独立して、存在しないか、又は1〜7のアミノ酸であり;Xaa9はMet、Ile及びThrからなる群から選択されるアミノ酸、好ましくはIleである。些かより特定的には、好ましいペプチドリガンドは次の式:
Xaa(1−5) -Trp-Xaa7-Cys-Xaa9-Gly-Pro-Gly-Cys-Xaa(14−20) (配列番号:93)
を有し、ここで、Xaa(1−5)は、存在しないか、又は1〜5のアミノ酸であり;Xaa7はAsp、Gly、Glu及びHisからなる群から選択され、好ましくはGlyである。本発明のこの態様においてより好ましいものは、次の式:
Xaa(1−3) -Glu-Xaa5-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Xaa14-Xaa15--Leu-Xaa(17−20) (配列番号:91)
を有するペプチドリガンドであり、ここで、Xaa(1−3)-は存在しないか、又は1〜3のアミノ酸であり、Xaa5は一つのアミノ酸であり、Xaa13は一つのアミノ酸であり、Xaa14は一つのアミノ酸であり、-Xaa(17−20)は存在しないか、又は1〜4のアミノ酸である。本発明のこの態様における例示的なペプチドリガンドは、X(1−3)-は存在しないか、又はGln-Arg-Asn-、Leu-Ser-Pro-、Glu-Asn-Trp-、Ala-Ser-His-、Lys-Leu-Asn-、Thr-Gln-Ala-、Ala-Pro-Arg-、Gln-Val-Tyr-、Arg-Thr-Glu-、Phe-Ala-Gly-、Thr-Ala-Arg-、Arg-Pro-His-、Asn-Val-Cys-、Cys-Ile-Asp-、Tyr-Glu-Trp-、Arg-Trp-Asp-、His-Trp-Met-、Asn-Trp-Pro-、Phe-Asn-Trp-、Phe-Ser-Gly-、Gly-Gly-Trp-、Leu-Trp-Phe-、Gly-Ile-Pro-、Trp-Trp-Thr-、Leu-Gly-Trp-、Ser-Pro-Trp-、Arg-Gly-Trp-、Tyr-Glu-Phe-、Tyr-Glu-Gly-、Tyr-Glu-Val-、Tyr-Ser-Phe-、Tyr-Asp-Phe-、Asp-Glu-Val-、Ser-Glu-Val-、Phe-Glu-Phe-、及びHis-Asp-Val-からなる群から選択されるペプチドリガンドであり、Xaa5はAla、Thr、Met、Val、Arg、Glu、Asp、Ser、Gln、Pro、Gly、Phe及びLysからなる群から選択され;Xaa14はGlu、Lys、Arg、Asp、Ser、Ala、Asn、Thr、Gly、Pro、Val及びGlnからなる群から選択され;Xaa15はMet、Phe、Ala、Cys、Gln、Glu、Trp、Leu、Val、Tyr、Ser及びAsnからなる群から選択され;-X(17−20)は次の例示的配列:
-Phe-Gly-Ala-His (配列番号:94)
-Phe-Asp-Ala-His (配列番号:95)
-Leu-Glu-Ala-His (配列番号:96)
-Phe-Glu-Gly-His (配列番号:97)
-Phe-Gly-Ala-Leu (配列番号:98)
-Phe-Glu-Ala-Tyr (配列番号:99)
-Phe-Ala-Gly-His (配列番号:100)及び
-Phe-Glu-Ala-Phe (配列番号:101)
を有する4つのアミノ酸のペプチドである。
好ましくは、-Xaa(17−20)は、次の式:
-Xaa17 -Xaa18-Cys-Xaa20
を有する4つのアミノ酸のペプチドである。
【0028】
本発明のこの態様における例示的ペプチドリガンドは、-Xaa(17−20)が次の式:
-Gln-Ala-Cys-Met (配列番号:102)
-Leu-Gln-Cys-Trp (配列番号:103)
-Met-Ser-Cys-Val (配列番号:104)
-Leu-Arg-Cys-Ile (配列番号:105)
-Gln-Ala-Cys-Leu (配列番号:106)
-Leu-Ser-Cys-Leu (配列番号:107)
-Ile-Gly-Cys-Leu (配列番号:108)
-Leu-Ala-Cys-Leu (配列番号:109)
-Leu-Ser-Cys-Ile (配列番号:110)
-Met-Asn-Cys-Leu (配列番号:111)
-Leu-Arg-Cys-Leu (配列番号:112)
-Leu-Lys-Cys-Leu (配列番号:113)
-Leu-Gly-Cys-Leu (配列番号:114)
-Leu-Asn-Cys-Ile (配列番号:115)
-Met-Gly-Cys-Leu (配列番号:116)及び
-Met-Ala-Cys-Leu (配列番号:117)
からなる群から選択されるペプチドリガンドである。
【0029】
本発明のこの態様における他の実施態様では、-Xaa(17−20)が次の式:
-Cys-Xaa18 -Xaa19-Cys
を有する4つのアミノ酸ペプチドであり、Xaa18が一つのアミノ酸であり、Xaa19が一つのアミノ酸であるペプチドリガンドである。本発明のこの態様では、例示的ペプチドリガンドは:
-Cys-Ala-Trp-Cys (配列番号:118)
-Cys-Ser-Trp-Cys (配列番号:119)
-Cys-Glu-Pro-Cys (配列番号:120)
-Cys-Asp-Trp-Cys (配列番号:121)
-Cys-Glu-Trp-Cys (配列番号:122)
-Cys-Asn-Trp-Cys (配列番号:123)及び
-Cys-Gly-Trp-Cys (配列番号:124)
からなる群から選択される-Xaa(17−20)を含むものである。
【0030】
本発明のこの態様における例示的ペプチドリガンドは、
-Gln-Arg-Asn-Glu-Ala-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Met-Leu-Cys-Ala-Trp-Cys (配列番号:1)
-Leu-Ser-Pro-Glu-Thr-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Met-Leu-Cys-Ser-Trp-Cys (配列番号:2)
-Glu-Asn-Trp-Glu-Met-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Lys-Phe-Leu-Cys-Glu-Pro-Cys (配列番号:3)
-Glu-Val-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Lys-Ala-Leu-Cys-Asp-Trp-Cys (配列番号:4)
-Glu-Arg-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Arg-Met-Leu-Cys-Glu-Trp-Cys (配列番号:5)
-Glu-Val-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Asp-Met-Leu-Cys-Asn-Trp-Cys (配列番号:6)
-Glu-Val-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ser-Met-Leu-Cys-Gly-Trp-Cys (配列番号:7)
-Xaa-Xaa-Xaa-Glu-Xaa-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Xaa-Met-Leu-Cys-Xaa-Trp-Cys (配列番号:8)
-Ala-Ser-His-Glu-Val-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Lys-Cys-Leu-Gln-Ala-Cys-Met (配列番号:9)
-Lys-Leu-Asn-Glu-Glu-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Cys-Leu-Leu-Gln-Cys-Trp (配列番号:10)
-Lys-Leu-Asn-Glu-Asp-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Cys-Leu-Leu-Xaa-Cys-Xaa (配列番号:11)
-Thr-Gln-Ala-Glu-Arg-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Cys-Leu-Met-Ser-Cys-Val (配列番号:12)
-Ala-Pro-Arg-Glu-Val-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Cys-Leu-Leu-Arg-Cys-Ile (配列番号:13)
-Gln-Val-Tyr-Glu-Ser-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Cys-Leu-Gln-Ala-Cys-Leu (配列番号:14)
-Arg-Thr-Glu-Glu-Gln-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Arg-Cys-Leu-Leu-Ser-Cys-Leu (配列番号:15)
-Phe-Ala-Gly-Glu-Ser-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Cys-Leu-Ile-Gly-Cys-Leu (配列番号:16)
-Thr-Ala-Arg-Glu-Val-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Asn-Cys-Leu-Leu-Ala-Cys-Leu (配列番号:17)
-Arg-Pro-His-Glu-Pro-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ser-Cys-Leu-Leu-Ser-Cys-Ile (配列番号:18)
-Glu-Val-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Cys-Leu-Met-Asn-Cys-Leu (配列番号:19)
-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Cys-Leu-Leu-Arg-Cys-Leu (配列番号:20)
-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Gly-Cys-Leu-Leu-Lys-Cys-Leu (配列番号:21)
-Glu-Pro-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Cys-Leu-Leu-Gly-Cys-Leu (配列番号:22)
-Glu-Glu-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Cys-Leu-Leu-Asn-Cys-Ile (配列番号:23)
-Glu-Gln-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Lys-Cys-Leu-Met-Gly-Cys-Leu (配列番号:24)
-Glu-Gln-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Gly-Cys-Leu-Leu-Arg-Cys-Leu (配列番号:25)
-Glu-Ala-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Gly-Cys-Leu-Met-Ala-Cys-Leu (配列番号:26)
-Xaa-Xaa-Xaa-Glu-Xaa-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Xaa-Cys-Leu-Leu-Xaa-Cys-Leu (配列番号:27)
-Asn-Val-Cys-Glu-Phe-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Gln-Leu-Cys (配列番号:28)
-Cys-Ile-Asp-Glu-Thr-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Glu-Leu-Arg-Cys-Lys-Arg (配列番号:29)
-Tyr-Glu-Trp-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Pro-Ala-Leu-Gly-Phe-Gly-Tyr (配列番号:30)
-Arg-Trp-Asp-Glu-Glu-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Trp-Leu-Val-Val-Arg-Lys (配列番号:31)
-His-Trp-Met-Glu-Arg-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Gly-Phe-Leu (配列番号:32)
-Asn-Trp-Pro-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Lys-Leu-Leu (配列番号:33)
-Phe-Asn-Trp-Glu-Lys-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Arg-Thr-Leu (配列番号:34)
-Phe-Ser-Gly-Glu-Arg-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Gln-Val-Leu (配列番号:35)
-Gly-Gly-Trp-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Arg-Tyr-Leu (配列番号:36)
-Leu-Trp-Phe-Glu-Arg-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Thr-Xaa-Leu (配列番号:37)
-Gly-Ile-Pro-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Glu-Trp-Leu (配列番号:38)
-Trp-Trp-Thr-Glu-Arg-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ser-Met-Leu (配列番号:39)
-Xaa-Cys-Xaa-Glu-Arg-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ser-Met-Leu (配列番号:40)
-Leu-Gly-Trp-Glu-Arg-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Arg-Ala-Leu (配列番号:41)
-Ser-Pro-Trp-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Arg-Trp-Leu (配列番号:42)
-Arg-Gly-Trp-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ser-Phe-Leu (配列番号:43)
-Xaa-Xaa-Trp-Glu-Xaa-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Xaa-Xaa-Leu-Xaa-Xaa-Xaa-Xaa (配列番号:44)
-Tyr-Glu-Val-Glu-Ala-Trp-Asp-Cys-Met-Gly-Pro-Gly-Cys-Ala-Asn-Leu-Phe-Glu-Ala-His (配列番号:57)
-Tyr-Glu-Phe-Glu-Gly-Trp-Asp-Cys-Met-Gly-Pro-Gly-Cys-Ala-Ser-Val-Phe-Gly-Ala-His (配列番号:58)
-Tyr-Glu-Gly-Glu-Ser-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Ser-Leu-Phe-Asp-Ala-His (配列番号:59)
-Tyr-Glu-Val-Glu-Val-Trp-Glu-Cys-Ile-Gly-Pro-Gly-Cys-Gly-Tyr-Leu-Phe-Gly-Ala-His (配列番号:60)
-Tyr-Glu-Val-Glu-Gly-Trp-Gly-Cys-Met-Gly-Pro-Gly-Cys-Ala-Phe-Leu-Leu-Glu-Ala-His (配列番号:61)
-Tyr-Ser-Phe-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Tyr-Leu-Phe-Glu-Gly-His (配列番号:62)
-Tyr-Asp-Phe-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Gly-Asn-Leu-Leu-Glu-Ala-His (配列番号:63)
-Tyr-Asp-Phe-Glu-Gly-Trp-Asp-Cys-Thr-Gly-Pro-Gly-Cys-Ala-Tyr-Leu-Phe-Glu-Gly-His (配列番号:64)
-Asp-Glu-Val-Glu-Ser-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Tyr-Leu-Phe-Gly-Ala-Leu (配列番号:65)
-Ser-Glu-Val-Glu-Val-Trp-His-Cys-Ile-Gly-Pro-Gly-Cys-Val-Tyr-Leu-Phe-Glu-Ala-Tyr (配列番号:66)
-Phe-Glu-Phe-Glu-Gly-Trp-Glu-Cys-Met-Gly-Pro-Gly-Cys-Ala-Glu-Leu-Phe-Ala-Gly-His (配列番号:67)
-His-Asp-Val-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Asp-Leu-Phe-Glu-Ala-Phe (配列番号:68)
-Tyr-Glu-Phe-Glu-Gly-Trp-Gly-Cys-Ile-Gly-Pro-Gly-Cys-Ala-Tyr-Leu-Phe-Glu-Ala-His (配列番号:69)
と、上述した例示的アミノ酸配列の保存的アミノ酸置換を含む。
【0031】
さらに好ましいペプチド化合物は次の一般式:
Cys-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Cys-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Cys-Cys-Xaa-Xaa-Xaa-Cys-Xaa-Xaa-Xaa-Xaa-Xaa-Xaa-Cys (配列番号:125)
を有する。些かより特定的には、次の式:
Xaa(1−10)-Glu-Xaa12-Trp-Xaa14-Cys-Cys-Gly-Pro-Gly-Cys-Xaa21-Xaa22-Xaa23-Xaa(24−27) (配列番号:126)
を有し、ここでXaa(1−10)は存在しないか、又は1〜10のアミノ酸であり、Xaa12は一つのアミノ酸であり、Xaa14は一つのアミノ酸であり、Xaa21は一つのアミノ酸であり、Xaa22は一つのアミノ酸であり、Xaa23はVal及びLeuからなる群から選択されるアミノ酸であり、Xaa(24−27)は存在しないか、又は1〜4のアミノ酸であるものである。本発明のこの態様において、好ましくは、Xaa12はVal、Leu、Ser、Trpからなる群から選択されるアミノ酸であり;Xaa14はGln、Glu、Asp及びHisからなる群から選択されるアミノ酸であり;Xaa21はGly及びGluからなる群から選択されるアミノ酸であり;Xaa22はTrp、Leu及びPheからなる群から選択されるアミノ酸であり;Xaa23はValである。
本発明のこの態様において好ましいペプチドリガンドは、Xaa(1−10)が次の式:
Cys-Xaa(2−7)-Cys-Xaa9-Gly-
を有する10のアミノ酸であり、Xaa(24−27)が次の式:
Xaa(24−26)-Cys
を有する4つのアミノ酸であり、Xaa(2−7)が6つのアミノ酸であり、Xaa(24−26)が3つのアミノ酸であるものを含む。
【0032】
本発明のこの態様において好ましいものは、Xaa(1−10)が次の式:
Cys-Xaa2-Trp-Val-Xaa5−Xaa6-Xaa7-Cys-Xaa9-Gly- (配列番号:127)
を有し、ここでXaa2がAla及びSerからなる群から選択されるアミノ酸であり、Xaa5がSer、Leu、Ala、Arg及びValからなる群から選択されるアミノ酸であり、Xaa6がPhe、Val及びLeuからなる群から選択されるアミノ酸であり、Xaa7がAsp、Gln、Tyr、Trp、Leu及びHisからなる群から選択されるアミノ酸であり、Xaa9がGly、Phe及びLeuからなる群から選択されるアミノ酸であるペプチドリガンドである。
この群のペプチド中で好ましいものは、Xaa(1−10)が次の式:
Cys-Ala-Trp-Val-Leu-Xaa6-Xaa7-Cys-Gly-Gly- (配列番号:128)
を有し、Xaa(24−26)が次の式:
Xaa24−Xaa25-Xaa26
を有し、ここでXaa24がTrp、Val、Gly及びAlaからなる群から選択されるアミノ酸、好ましくはValであり;Xaa25がAsn、Lys、Asp、Glu及びHisからなる群から選択されるアミノ酸、好ましくはAsnであり、Xaa26がAla、Ser及びValからなる群から選択されるアミノ酸、好ましくはAlaであるペプチドリガンドである。
【0033】
例として、限定するものではないが、次のもの:
Cys-Ser-Trp-Val-Leu-Val-Gln-Cys-Gly-Gly-Glu-Trp-Trp-His-Cys-Cys-Gly-Leu-Gly-Cys-Gly-Leu-Val-Val-Asn-Ala-Cys (配列番号:45)
Cys-Ala-Trp-Val-Ser-Phe-Glu-Cys-Gly-Gly-Glu-Val-Trp-His-Cys-Cys-Gly-Leu-Gly-Cys-Gly-Trp-Val-Trp-Lys-Ala-Cys (配列番号:46)
Cys-Ala-Trp-Val-Leu-Val-Gln-Cys-Gly-Gly-Glu-Trp-Trp-His-Cys-Cys-Gly-Pro-Gly-Cys-Glu-Phe-Val-Val-Asp-Ala-Cys (配列番号:47)
Cys-Ala-Trp-Val-Ala-Val-Tyr-Cys-Gly-Gly-Glu-Leu-Trp-His-Cys-Cys-Gly-Pro-Gly-Cys-Gly-Phe-Val-Val-Asp-Ser-Cys (配列番号:48)
Cys-Ala-Trp-Val-Arg-Val-Trp-Cys-Phe-Gly-Glu-Trp-Trp-Asp-Cys-Cys-Gly-Leu-Gly-Cys-Gly-Trp-Val-Val-Asn-Val-Cys (配列番号:49)
Cys-Ala-Trp-Val-Arg-Val-Leu-Cys-Gly-Gly-Glu-Trp-Trp-His-Cys-Cys-Gly-Leu-Gly-Cys-Gly-Trp-Val-Val-Glu-Ala-Cys (配列番号:50)
Cys-Ser-Trp-Val-Ser-Val-Leu-Cys-Gly-Gly-Glu-Trp-Trp-Gln-Cys-Cys-Gly-Pro-Gly-Cys-Gly-Leu-Val-Val-Asn-Ala-Cys (配列番号:51)
Cys-Ser-Trp-Val-Ser-Leu-Gln-Cys-Gly-Gly-Glu-Trp-Trp-His-Cys-Cys-Gly-Gly-Gly-Cys-Gly-Trp-Val-Val-Asn-Val-Cys (配列番号:52)
Cys-Ser-Trp-Val-Leu-Leu-His-Cys-Gly-Gly-Glu-Trp-Trp-His-Cys-Cys-Gly-Gly-Gly-Cys-Gly-Trp-Val-Gly-Glu-Ala-Cys (配列番号:53)
Cys-Ser-Trp-Val-Leu-Leu-Glu-Cys-Gly-Gly-Glu-Leu-Trp-Glu-Cys-Cys-Gly-Leu-Gly-Cys-Gly-Trp-Val-Ala-Asp-Ala-Cys (配列番号:54)
Cys-Ser-Trp-Val-Val-Phe-Glu-Cys-Leu-Gly-Glu-Ser-Trp-His-Cys-Cys-Gly-Gly-Gly-Cys-Gly-Trp-Val-Val-His-Ala-Cys (配列番号:55)
Cys-Ala-Trp-Val-Ser-Val-Glu-Cys-Gly-Gly-Glu-Trp-Trp-His-Cys-Cys-Gly-Pro-Gly-Cys-Gly-Trp-Val-Val-Asp-Ala-Cys (配列番号:56)
が本発明の適切なペプチドリガンドである。
【0034】
ペプチドリガンドの組み合わせ
A.多量体化ドメイン
本発明の好適な実施態様では、ペプチドリガンドは多量体化ドメインと組み合わされる。本発明のこの側面においては、少なくとも二つの別のドメインを有するハイブリッド分子が提供される。各分子はペプチドリガンドドメインと多量体化ドメインを有している。本発明では、ペプチドリガンドドメインは任意には柔軟なリンカードメインを介して免疫グロブリンFc領域のような多量体化ドメインに結合されている。
本発明のハイブリッド分子は適切な多量体化ドメインにペプチドリガンドを組み合わせることにより構築される。通常は、本発明のハイブリッド分子を調製する場合、ペプチドリガンドをコード化する核酸は多量体化ドメイン配列をコード化する核酸に作用可能に結合される。典型的には、作成物は、ペプチドリガンドのC末端が多量体化ドメインのN末端又はC末端、好ましくはC末端に結合されている融合タンパク質をコードしている。しかし、ペプチドリガンドのN末端が多量体化ドメインのN末端又はC末端に結合されている融合体もまた可能である。
好適な多量体化ドメインは免疫グロブリンの定常領域配列である。典型的には、そのような融合体では、コードされたハイブリッド分子は免疫グロブリン重鎖の定常領域の少なくとも機能的に活性なヒンジ、CH2及びCH3ドメインを保持しているであろう。融合体はまた例えば定常ドメインのFc部分のC末端、又は重鎖のCH1の直ぐ近くのN末端又は軽鎖の対応する領域に対して生成される。
【0035】
免疫グロブリン定常ドメインへのペプチドリガンドの融合がなされる正確なアミノ酸部位は重要ではない;特定の部位はよく知られており、生物学的活性、分泌又は結合特性を最適化するために選択することができる。この点に関して、当業者は文献(米国特許第5116964号、同第5714147号及び同第5336603号;Caponら、(1989) Nature 337:525-531; Trauneckerら, (1989) Nature 339:68-70;及びByrnら, (1990) Nature 344:667-670; Watsonら, (1990) J. Cell. Biol. 110:2221-2229; Watsonら, (1991) Nature 349:164-167; Aruffoら, (1990) Cell 61:1303-1313; Linsleyら, (1991) J. Exp. Med. 173:721-730; Lisleyら, J. Exp. Med. 174:561-569; Stamenkovicら, Cell 66:1133-1144; Ashkenaziら, (1991) Proc. Natl. Acad. Sci. USA 88:10535-10539; Lesslaierら, (1991) Eur. J. Immunol. 27:2883-2886; 及びPeppelら, (1991) J. Exp. Med. 174:1483-1489; Mohlerら, (1993) J. Immunol. 151:1548-1561; Bennettら, (1991) J. Biol. Chem. 266:23060-23067; Kurschnerら, (1992) J. Biol. Chem. 267:9354-9360; Chalupnyら, (1992) PNAS USA 89:10360-10364; Ridgway及びGorman, (1991) J. Cell. Biol. 115, Abstract No. 1448)に記載された様々なイムノアドヘシンの構築を参考にすることができる。
特定の側面では、免疫グロブリンタイプの多量体化ドメインは機能性Fcドメインを有するダイマーのような多量体を提供するように選択される。好適な側面では、多量体化ドメインは天然の免疫グロブリンFc領域に伴うエフェクター機能を有しているFcドメインを提供するように選択される。従って、ペプチドリガンドは、特定の側面では、特定のエフェクター機能又は機能群に対して選択された機能性Fcドメインを有する多量体を提供する免疫グロブリン重鎖定常ドメインに結合される。この場合、免疫グロブリン鎖−ペプチドリガンド配列をコードするDNAは典型的には第二のペプチドリガンド−免疫グロブリン重鎖融合タンパク質をコードするDNAと同時発現される。分泌の際、ハイブリッド重鎖は二つのジスルフィド結合免疫グロブリン重鎖を有する免疫グロブリン様構造を提供するように共有結合的に結合される。当業者であれば、エフェクター機能には、例えばC1q結合;補体依存性細胞毒素;Fcレセプター結合;抗体依存細胞性細胞毒素(ADCC);ファゴサイトーシス;及びサルベージレセプター結合リガンドの導入による半減期の延長と細胞表面レセプター(例えばB細胞レセプター;BCR)のダウンレギュレーションで、例えば1998年4月14日に発行された米国特許第5739277号にされたようなものが含まれることを認識するであろう。
【0036】
好ましくは、Fc領域はヒトFc領域、例えば天然配列ヒトFc領域ヒトIgG1(A及び非Aアロタイプ)、IgG2、IgG3又はIgG4のFc領域である。そのような配列は図2及び3に示されている。また、親Fc領域に適当なアミノ酸配列の修飾を導入することにより、(a)ヒトエフェクター細胞の存在下で抗体依存細胞性細胞毒素を媒介し、及び/又は(b)天然配列よりも良好な親和性でFcガンマレセプター(Fc(R))に結合する変異体Fc領域をつくり出すことができる。そのようなFc領域変異体は一般にFc領域に少なくとも一つのアミノ酸の修飾を含んでいる。
好適な実施態様では、ペプチドリガンド配列は免疫グロブリンG1(IgG1)のFc領域のN末端に融合している。ペプチドリガンド配列に重鎖定常領域の全体を融合させることが可能である。しかし、より好ましくは、IgGのFcを化学的に定めるパパイン切断部位(すなわち、重鎖定常領域の最初の残基を114であるとして、残基216)、又は他の免疫グロブリンの類似部位の丁度上流のヒンジ領域で始まる配列が融合に使用される。特定の好適な実施態様では、ペプチドリガンドアミノ酸配列はIgG重鎖の(a)ヒンジ領域(又は他のリンカードメイン)及びCH2及びCH3又は(b)CH1、ヒンジ、CH2及びCH3ドメインに融合している。更に好適な実施態様では、ペプチドリガンドアミノ酸配列はIgGの(a)ヒンジ領域(又は他のリンカードメイン)及び(b)CH3ドメインに融合している(例えば、Huら, (1996) Cancer Res. 56:3055-3061に記載された構築物を参照されたい)。
【0037】
この実施態様の特定の側面では、ペプチドリガンドと多量体化ドメインを含むハイブリッド分子が多量体、例えばホモダイマー、又はヘテロダイマーあるいはヘテロテトラマーとしてでさえ構築される。ホモダイマーはペプチドリガンドと多量体化ドメインを含む二つのモノマーの対合又は架橋から生じる。しかし、二つの同一のモノマー対は必須ではない。本発明の特定の側面では、免疫グロブリン定常ドメインのような多量体化ドメインとペプチドリガンドを含むここに記載したハイブリッド分子は免疫グロブリンの一つのアームを含む対の他方の免疫グロブリンと対合しうる。本発明の範囲に入る様々な例示的構築ハイブリッド分子は以下に概略的に模式化される:
(a)ACH
(b)ACH−ACH
(c)ACH−VHCH−VLCL
(d)ACH−VHCH
ここで、Aはそれぞれ同一の又は異なるペプチドリガンドを表し;
VLは免疫グロブリン軽鎖可変ドメインであり;
VHは免疫グロブリン重鎖可変ドメインであり;
CLは免疫グロブリン軽鎖定常ドメインであり、及び
CHは免疫グロブリン重鎖定常ドメインである。
簡略にするために、上記の構造はキーとなる特徴のみを示している;上記構造は以下に記載されるような多量体化ドメインとペプチドリガンドドメインの間に任意のリンカードメインを示していない;上記構造は免疫グロブリンのジョイニング、ヒンジ又は他のドメインを示していないし、ジスルフィド結合も示していない。しかし、そのようなドメインが結合活性に対して必要とされる場合、それらは免疫グロブリン分子に占める通常の位置に存在する構造とされている。
【0038】
免疫グロブリン軽鎖の存在は本発明のハイブリッド分子に必要とされていないけれども、免疫グロブリン軽鎖はペプチドリガンド−免疫グロブリン重鎖融合ポリペプチドに共有結合的に結合するか、ペプチドリガンドに直接融合するかして、存在しているかもしれない。前者の場合、免疫グロブリン軽鎖をコードしているDNAは典型的にはペプチドリガンド−免疫グロブリン重鎖融合タンパク質と同時発現される。分泌の際、ハイブリッド重鎖及び軽鎖は二つのジスルフィド結合免疫グロブリン重鎖−軽鎖対を含む免疫グロブリン様構造を提供するように共有結合的に結合する。
ここに記載されたハイブリッド分子は免疫グロブリンcDNA配列に読み枠を一致させてペプチドリガンド部分をコードするcDNA配列を融合させることにより最も簡便に構築される。しかし、ゲノム免疫グロブリン断片への融合体もまた使用できる(Aruffoら, (1990), Cell 61:1303-1313;及びStamenkovicら, (1991), Cell 66:1133-1144)。後者のタイプの融合体は発現のためにIg調節配列の存在を必要とする。IgG重鎖定常領域をコードするcDNAは、ハイブリダイゼーション又はポリメラーゼ連鎖反応(PCR)法によって脾臓又は末梢血リンパ球から取り出されたcDNAライブラリーから発表された配列に基づいて単離できる。ハイブリッド分子のペプチドリガンドと免疫グロブリン部分をコードしているcDNAは、選択された宿主細胞において効果的な発現をさせるプラスミドベクターにタンデムに挿入される。
【0039】
あるいは、特にペプチドリガンドが例えば標準的な固相合成法によって合成される実施態様では、ペプチドリガンドは当業者によく知られた様々な手段の任意のものによって多量体化ドメインに結合されうる。共有結合的接合が典型的には最も簡便であるが、他の形態の接合も用途に応じて用いることができる。共有結合的な接合の好適な形態の例には、多量体化ドメインのアミノ酸側鎖と活性化された化学基を持つ分子の反応により生じる結合が含まれ、様々な二官能性タンパク質カップリング剤、例えばN-スクシンイミジル-3-(2-ピリジルチオール)プロピオナート(SPDP)、イミノチオラン(IT)、イミドエステル類の二官能性誘導体(例えばジメチルアジピミダートHCL)、活性エステル類(例えば、スベリン酸ジスクシンイミジル)、アルデヒド類(例えば、グルタルアルデヒド)、ビスアジド化合物(例えば、ビス(p-アジドベンゾイル)ヘキサンジアミン)、ビス-ジアゾニウム誘導体(例えば、ビス-(p-ジアゾニウムベンゾイル)エチレンジアミン)、ジイソシアネート(例えば、トリエン-2,6-ジイソシアネート)、及び二活性フッ素化合物(例えば、1,5-ジフルオロ-2,4-ジニトロベンゼン)を使用して作製することができる。
本発明は、HER2を有する細胞型のペプチドリガンド経由インビボでの特定のターゲティング及び本発明のこの側面によれば、Fcエフェクター機能、例えばFcレセプター結合、補体結合、プロテインA結合、半減期の増加又は分子までの血液脳関門の通過の付与を含む。
【0040】
B.ペプチドリガンド融合体
本発明では、ペプチドリガンドドメインは、任意には、例えば他のペプチドリガンドドメインに、以下に記載するように直接又は柔軟なペプチドリンカーを経由して結合される。本発明では、リンカードメインは以下に更に詳細に記載するように二以上のペプチドリガンドドメインの間に空間的ブリッジを提供する任意の分子群である。本発明のこの側面では、ペプチドリガンドは例えば融合タンパク質におけるように互いに結合される。本発明のこの側面のハイブリッド分子は例えば二以上のレセプターを架橋させるのに有用である。
【0041】
C.リンカードメイン
本発明では、ペプチドリガンドドメインは、任意には、例えば他のペプチドリガンドドメインに又は柔軟なペプチドリンカーを経由して多量体化ドメインに結合される。本発明のハイブリッド分子のリンカー成分はハイブリッド分子の機能に必ずしも関与しないが寄与しうる。従って、本発明では、リンカードメインは、二以上のペプチドリガンドドメイン又はペプチドリガンドドメインと多量体化ドメインの間に空間的ブリッジを提供する任意の分子群である。
リンカードメインは可変の長さ及び構造とできる。一般には、重要であるのは、リンカードメインにより引き起こされる配向であり、その化学構造ではない。リンカードメインは、好ましくは、配位ErbB2分子に対する空間的/立体配置的制約を実質的に生じないで、ハイブリッド分子のペプチドリガンドドメインを結合させない。従って、リンカードメインの長さは、ハイブリッド分子の例えばペプチドリガンドと多量体化ドメインの二つの機能の性質に依存する。
当業者であれば、原子の様々な組み合わせが様々な結合間の既知の距離に基づいて可変長さの分子を提供することが分かるであろう(Morrison及びBoyd, Organic Chemistry, 3版, Allyn及びBacon, Inc., Boston, MA (1977))。例えば、リンカードメインは可変長さのポリペプチドでありうる。ポリペプチドのアミノ酸組成がリンカーの性質と長さを決定する。例示的なリンカードメインは2から10のアミノ酸、好ましくは約6のアミノ酸、例えばGly-Gly-Gly-Ser-Gly-Gly(配列番号:129)、Gly-Gly-Gly-Ser-Ser-Gly(配列番号:130)、及びGly-Gly-Gly-Arg-Gly-Gly(配列番号:131)を含む。
【0042】
D.他の二重特異的組み合わせ
本発明のある側面では、少なくとも一つのペプチドリガンドドメインを含む二又は二重特異的組成物が考えられる。例えば、二重特異性抗体組成物はロイシンジッパーを使用して生産されている(Kostelnyら, (1992) J.Immunol., 148(5):1547-1553)。Fos及びJunタンパク質からのロイシンジッパーペプチドを遺伝子融合により二つの異なった抗体のFab'部分に結合させる。抗体ホモダイマーをヒンジ領域で還元してモノマーを形成し、ついで再び酸化して抗体ヘテロダイマーを形成する。この方法はまた抗体ヘテロダイマーの結合ドメインの代わりにペプチドリガンドを使用するペプチドリガンドホモダイマー及びヘテロダイマーの生産に対して使用することができる。
本発明はここに記載されたペプチドをコードする単離された核酸、好ましくはDNAを含む物質の組成物を包含する。本発明のペプチドをコードするDNAは当該分野で知られている様々な方法によって調製することができる。これらの方法は、限定されるものではないが、出典明示により開示の全体がここに取り込まれるEngelsら, (1989) Agnew. Chem. Int. Ed. Engl., 28:716-734に記載された任意の方法、例えばトリエステル、亜リン酸、ホスホラミダイト及びH-ホスホネート法による化学合成を含む。一実施態様では、発現宿主細胞により好まれるコドンはコード化DNAの設計に使用される。あるいは、ペプチドをコードしているDNAを、組換えDNA法、例えば部位特異的突然変異誘発(Kunkelら, (1991) Methods Enzymol. 204:125-139; Carter, P.ら, (186) Nucl. Acids. Res. 13:4331; Zoller, M.J.ら, (1982) Nucl. Acids Res. 10:6487)、カセット突然変異誘発(Wells. J.A.ら, (1985) Gene 34:315)、制限選択突然変異誘発(Wells, J.A.ら,(1986)Philos. Trans. R. Soc. London SerA 317, 415)等々を使用して一又は複数の変異体をコードするように変更させることができる。
【0043】
本発明は更に作用可能に結合した発現コントロール配列、及びベクターで形質転換した宿主細胞によりコントロール配列が認識されるDNA分子を含むプラスミドのような発現ベクターを更に含む。一般には、プラスミドベクターは宿主細胞と適合性がある種から取り出された複製及びコントロール配列を含む。ベクターは通常は複製部位並びに形質転換細胞において表現型の選択を提供することができるタンパク質をコードする配列を有する。
DNAを発現するために適切な宿主細胞は、原核生物、酵母、又は高等真核生物細胞を含む。適切な原核生物は、限定するものではないが、真正細菌、例えばグラム陰性又はグラム陽性生物体、例えば大腸菌(E.coli)のような腸内菌科を含む。様々な大腸菌株が公的に利用でき、例えば大腸菌K12株MM294(ATCC31446);大腸菌X1776(ATCC31537);大腸菌W3110(ATCC27325)及びK5 772(ATCC53635)である。
原核生物に加えて、真核生物体、例えば酵母菌、あるいは多細胞生物体由来の細胞を宿主細胞として使用することができる。一般的なパン酵母又はサッカロミセス・セレヴィシアのような酵母宿主細胞中での発現に対しては、好適なベクターには、2ミクロンプラスミドに基づくエピソーム的に複製するベクター、組込み型ベクター、及び酵母人工染色体(YAC)ベクターが含まれる。発現にまた好適な宿主細胞は多細胞生物体から由来する。無脊椎動物細胞の例には、ショウジョウバエS2及びスポドプテラ(Spodoptera)SF9のような昆虫細胞並びに植物細胞が含まれる。SF9細胞のような昆虫宿主細胞での発現に対しては、好適なベクターにはバキュロウイルスベクターが含まれる。植物宿主細胞、特にタバコのような双子葉植物宿主における発現に対しては、好適な発現ベクターにはアグロバクテリウムツメファシエンスのTiプラスミドに由来するベクターが含まれる。
【0044】
有用な哺乳動物宿主細胞の例には、SV40によって形質転換されたサル腎臓CV1株 (COS-7, ATCC CRL 1651);ヒト胚腎臓株(293又は懸濁培養での増殖のためにサブクローン化された293細胞、Grahamら, (1997) J. Gen Virol., 36:59);ハムスター乳児腎細胞(BHK, ATCC CCL 10);チャイニーズハムスター卵巣細胞/-DHFR(CHO, Urlaub及びChasin, (1980) Proc. Natl. Acad. Sci. USA, 77:4216);マウスのセルトリ細胞(TM4, Mather, (1980) Biol. Reprod., 23:243-251);サルの腎細胞 (CVI ATCC CCL 70);アフリカミドリザルの腎細胞(VERO-76, ATCC CRL-1587);ヒト子宮頸癌細胞 (HELA, ATCC CCL 2); イヌ腎細胞 (MDCK, ATCC CCL 34);バッファローラット肝細胞 (BRL 3A, ATCC CRL 1442);ヒト肺細胞 (W138, ATCC CCL 75);ヒト肝細胞 (Hep G2, HB 8065);マウス乳房腫瘍(MMT 060562, ATTC CCL51);TRI細胞(Motherら, (1982) Annals N.Y. Acad. Sci., 383:44-68);MRC5細胞;FS4細胞;及びヒト肝癌株(HepG2)が含まれる。
原核生物宿主中での発現に対しては、好適なベクターにはpBR322(ATCC第37017)、phGH107(ATCC第40011)、pBO475、pS0132、pRIT5、pRIT20又はpRIT30シリーズの任意のベクター(Nilsson及びAbrahmsen, (1990) Meth. Enzymol., 185:144-161)、pRIT2T、pKK233-2、pDR540及びpPL-ラムダが含まれる。本発明の発現ベクターを含む原核生物宿主細胞には、大腸菌K12株294(ATCC第31446)、大腸菌株JM101(Messingら, (1981) Nucl. Acid Res., 9:309);大腸菌株B、大腸菌χ1776(ATCC第31537)、大腸菌c600(Appleyard, Genetics, 39: 440(1954))、大腸菌W3110(F-、ガンマ-、原栄養菌株、ATCC第27325)、大腸菌株27C7(W3110、tonA、phoA、E15、(argF-lac)169、ptr3、degP41、ompT、kanχ)(米国特許第5288931号、ATCC第55244)、枯草菌、サルモネラ菌、セラチア・マルセッセンス及びシュードモナス種が含まれる。
【0045】
哺乳動物宿主細胞における発現に対しては、有用なベクターには、SV40由来のベクター、サイトメガロウイルス由来のベクター、例えばpRK5及びpRK7を含むpRKベクター(Suvaら, (1987) Science, 237:893-896; EP307247(3/15/89)、EP278776(8/17/88))、ワクシニアウイルス又は他のポックスウイルス由来のベクター、及びモロニーのマウス白血病ウイルス(MoMLV)由来のベクターのようなレトロウイルスベクターが含まれる。
場合によっては、対象のペプチドをコードしているDNAは宿主細胞による発現産物の培地中への分泌を生じさせる分泌リーダー配列に作用可能に結合される。分泌リーダー配列の例には、stII、エコチン(ecotin)、lamB、ヘルペスGD、lpp、アルカリホスファターゼ、インベルターゼ、及びアルファ因子が含まれる。ここでの使用にまた適しているのはプロテインAの36アミノ酸リーダー配列である(Abrahmsenら, (1985) EMBO J., 4:3901)。
宿主細胞はこの発明の上述の発現又はクローニングベクターで形質移入され、好ましくは形質転換され、プロモーターを誘導し、形質転換体を選択し、又は所望の配列をコードする遺伝子を増幅するのに適するように変性された一般的な培養液中で培養される。
形質移入は、コード化配列が実際に発現されようとされまいと、宿主細胞による発現ベクターの取り込みを意味する。数多くの形質移入の方法が当業者に知られており、例えばCaPO4沈殿法及びエレクトロポレーション法である。このベクターの作用の任意の兆しが宿主細胞内で生じた場合に一般に成功した形質移入が認められる。
【0046】
形質転換は、染色体外のエレメントとしてであろうと染色体構成成分によってであろうと、DNAが複製可能であるように生物体中にDNAを導入することを意味する。用いられる宿主細胞に応じて、そのような細胞に対して適した標準的な方法を用いて形質転換はなされる。Sambrookら, Molecular Cloning (2版), Cold Spring Harbor Laboratory, NY (1989)のセクション1.82に記載された塩化カルシウムを用いるカルシウム処理が、原核生物又は実質的な細胞壁障壁を含む他の細胞に対して用いられる。アグロバクテリウム・トゥメファシエンスによる感染が、Shawら, (1983) Gene, 23:315及び1989年6月29日公開の国際公開第89/05859に記載されているように、ある種の植物細胞の形質転換に用いられる。そのような細胞壁のない哺乳動物細胞に対しては、上掲のSambrookらのセクション16.30−16.37に記載されたリン酸カルシウム沈殿法が好ましい。哺乳動物宿主系の形質転換の一般的な側面は1983年8月16日に発行された米国特許第4399216号にAxelによって記載されている。酵母菌への形質転換は典型的には、Van Solingenら, (1977) J. Bact.,130:946及びHsiaoら, (1979) Proc. Natl. Acad. Sci. (USA), 76:3829の方法に従って実施される。しかし、核注入、エレクトロポレーション、あるいは原形質融合のような他の細胞にDNAを導入する方法を使用することもできる。
他の好適なベクターは上述のベクターの関連した形質を組み合わせることによって標準的な方法を使用して構築することができる。関連する形質には、プロモーター、リボソーム結合部位、対象遺伝子又は遺伝子融合体(プロテインAのZドメイン及び対象遺伝子及びリンカー)、抗生物質耐性マーカー、及び適切な複製起点が含まれる。
【0047】
上記の方法の変形例は、ベクターにおいて、所望のペプチドをコードする遺伝子が他のタンパク質又は他のタンパク質の断片をコードする遺伝子に結合している遺伝子融合体の使用を考える。これは、所望のペプチドが他のタンパク質又はペプチドとの融合体として宿主細胞によって産生される結果となる。「他の」タンパク質又はペプチドはしばしば細胞によって分泌されうるタンパク質又はペプチドであり、培地から所望のペプチドを単離し精製することを可能にし、所望のペプチドが細胞内に残る場合に生じる宿主細胞を破壊する必要性を排除する。あるいは、融合タンパク質は細胞内に発現されうる。高度に発現される融合タンパク質を使用するのが有用である。
遺伝子融合体の使用は、必須ではないが、昆虫細胞における異種性ペプチドの発現並びにその遺伝子産物の続く精製を容易にしうる。プロテインAの結合、又はより詳細にはプロテインAのIgGへの結合が融合タンパク質の精製のための「アフィニティーハンドル」を提供するので、ZドメインプロテインA融合体がしばしば使用される。例えば、所望のペプチドリガンドをコードするDNA配列を、Zドメインとして知られているプロテインAのコンセンサスドメインの遺伝子と部位特異的突然変異誘発によって融合させることができる(Nilssonら, (1987) Protein Engineering 1:107-113)。発現及び分泌後に、融合タンパク質を酵素的に切断して、酵素混合物から精製できる遊離のペプチドを生じさせることができる(例えばVaradarajanら, (1985) Proc. Natl. Acad. Sci USA 82:5681-5684; Castellanos-Serraら, (1996) FEBS Letters 378:171-176; Nilssonら, (1996) J. Biotechnol. 48:241-250を参照されたい)。
融合タンパク質は、薬品、例えばメチオニンで切断する臭化シアノゲン、又はAsn及びGly残基の間で切断するヒドロキシルアミンを使用して切断できる。標準的な組換体DNA法を使用して、これらのアミノ酸をコードするヌクレオチド塩基対を所望のペプチドをコードする遺伝子の5'末端の直ぐ前に挿入してもよい。
【0048】
あるいは、融合タンパク質のタンパク分解的切断を用いることができる。Cater, Protein Purification: From Molecular Mechanisms to Large-Scale Processes, Ladischら編, (American Chemical Society Symposium Series No.427, 1990), Ch 13, p181-193。
プロテアーゼ、例えばXa因子、トロンビン、及びサブチリシン又はその変異体、及び多くの他のものが融合タンパク質を切断するために首尾良く使用された。約30未満のアミノ酸のペプチドリガンドの生産に対して本発明において好適なものは、Arg及びLys残基に高度に特異的なプロテアーゼトリプシンである。トリプシン切断は一般にNilssonら, (1996) J. Biotech. 48:241及びSmithら, Methods Mol. Biol. 32:289にいて検討されている。典型的には、使用されるプロテアーゼによる切断に受け入れられるペプチドリンカーは「他の」タンパク質(例えば、プロテインAのZドメイン)と所望のペプチドの間に挿入される。組換体DNA法を使用して、リンカーをコードするヌクレオチド塩基対を他のタンパク質をコードする遺伝子又は遺伝子断片の間に挿入する。ついで、正しいリンカーを含む部分的に精製された融合タンパク質のタンパク分解的切断は天然融合タンパク質又は還元されもしくは変性された融合タンパク質の何れかで実施できる。
ペプチドは融合タンパク質として発現されるときに正しく折り畳まれても折り畳まれなくてもよい。また、切断部位を含む特異的ペプチドリンカーはプロテアーゼに接近可能でも可能でなくてもよい。これらの因子は、融合タンパク質が変性され、再び折り畳まれなければならないかを、そして、しかりであれば、これらの手順が切断の前又は後の何れに使用されるかを決定する。
【0049】
変性と再折り畳みが必要な場合、典型的にはペプチドはグアニジンHClのようなカオトロープで処理し、ついで、例えば還元され酸化されたジチオスレイトール又はグルタチオンを適切な比、pH、及び温度で含むレドックスバッファーで、ペプチドがその天然構造まで再折り畳みされるように処理する。
この明細書において言及される宿主細胞はインビトロ培養中の細胞並びに宿主動物内である細胞を包含する。
本発明の環化実施態様では、組換え的に産生されたペプチドは上述したような細胞内ジスルフィド結合の形成により環化できる。
一般的に言って、本発明のペプチドリガンドとハイブリッド分子は、例えば天然又は変異体ヘルグリン分子又はErbB2に対する抗体が使用されるのと同じ用途に使用することができる。もちろん、本発明のいくつかのペプチドリガンド又はハイブリッド分子は他の用途よりも特定の用途により適している場合がある。当業者であれば、ペプチドリガンド又はハイブリッド分子の生物活性を決定する一又は複数の一般的な生物学的アッセイを使用することによって与えられた用途にどの分子が適しているかを容易に確認できるであろう。
一つの例として、ヘレグリンは様々な障害、例えば神経系、筋肉組織及び上皮に影響を及ぼす障害及び疾患の治療に有用である。例えば、本発明のアゴニストペプチドリガンド又はハイブリッド分子はヘレグリン又はヘレグリン変異体が使用されるのと同じようにしてインビボでニューロンの発生、維持及び/又は再生を促進するのに使用される。本発明のこの側面に係るペプチドリガンド又はハイブリッド分子での治療を受け入れることができる疾患又は障害には、例えば外傷、手術、脳卒中、虚血、感染、代謝異常栄養不足、悪性又は毒物に伴う中枢神経系の損傷が含まれる。これらに限定されるものではないがヒト神経変性疾病又は障害、例えばアルツハイマー病、パーキンソン病、てんかん、多発性硬化症、ハンチントン舞踏病、ダウン症候群、神経性難聴及びメニエール病を治療することができる。同様に、特定のペプチドリガンドは神経障害、例えば糖尿病のような全身性疾患又は障害に伴う末梢神経障害の治療における使用に適している。
【0050】
ErbB2への結合の際に上述のようなリン酸化事象を開始させるアゴニストペプチドリガンドは特に本発明の前述の態様に有用である。本発明の特定のアゴニストペプチドリガンドは細胞の生存を亢進することができ、すなわち、特定のアゴニストペプチドリガンドに暴露されていなかった細胞の生存期間に対してインビボかインビトロでErbB2保有細胞の生存期間を増加させる。好適な実施態様では、アゴニストペプチドリガンドは、例えば細胞による3H-チミジンの取り込みを測定することによって定量されるErbB2保有細胞型の増殖をインビボかインビトロの何れかで生じさせ又は亢進する。増殖には、表現型の変化についてErbB2保有細胞集団をスクリーニングすることによって決定されるErbB2細胞の分化を伴いうる。
本発明の他の側面では、ある種のペプチドリガンド及びハイブリッド分子、特にErbB2に結合するがアゴニスト関連応答を誘発しないものは、以下に十分に説明されるように、例えば腫瘍細胞浸潤及び転移を阻害するのに有用である。例えば、ErbB2レセプターを発現する腫瘍(特にErbB2を過剰発現するもの)は細胞毒性剤に結合したペプチドリガンド又はハイブリッド分子を使用して治療することができ、又はErbB2を発現する細胞にプロドラッグをターゲティングする際に使用される。
【0051】
A.エフェクター機能の設計
例えば、本発明のハイブリッド分子の多量体化ドメインをエフェクター機能について改変し、例えば癌の治療における抗体の効能を増強することが望ましい。例えば、システイン残基を免疫グロブリンFc領域に導入して、この領域における鎖間ジスルフィド結合の形成を可能にする。このようにして産生されたホモ又はヘテロダイマーハイブリッド分子は改善されたインターナリゼーション能力及び/又は増加した補体媒介細胞死滅及び抗体依存性細胞障害活性(ADCC)を有しうる。Caronら, (1992) J. Exp. Med. 176:1191-1195及びShopes, B. (1992) J. Immunol. 148:2918-2922を参照されたい。抗腫瘍活性が高められたホモダイマーハイブリッド分子は、Wolffら, (1993) Cancer Research 53:2560-2565に記載されているようなヘテロ二官能性架橋剤を使用して調製することもできる。
別法として、二重Fc領域を有し、よって増強された補体溶解及びADCC能を有しうるヘテロダイマーハイブリッド分子を設計することができる。Stevensonら (1989) Anti-Cancer Drug Design 3:219-230を参照されたい。
【0052】
B.コンジュゲート
また本発明は、細胞障害剤、例えば化学療法剤、例えばタンパク質毒素もしくは細胞障害性薬物もしくは毒素(例えば、細菌、真菌、植物又は動物由来の酵素活性毒又はそれらの断片)、又は放射性アイソトープ(すなわち、放射性コンジュゲート)に結合したここに記載の任意のハイブリッド分子又はペプチドリガンドを含有するコンジュゲートにも関する。
このような免疫コンジュゲートの生成に有用な化学療法剤は上述した。化学療法剤は、悪性腫瘍、リンパ腫、芽細胞腫、肉腫及び白血病を含む癌の治療に有用な化合物である。化学療法剤の例には、カリケアマイシンのような微生物から単離された抗生物質(Leeら, (1987) J. Am. Chem. Soc. 109:3464-3466; Hinmanら, (1993) Cancer Res. 53:3336-3342)、メイタンシノイド(例えば、Liuら, (1996) Proc. Natl. Acad. Sci. USA 93:8618-8323に記載されているもの)、アドリアマイシン、ドキソルビシン、5-フルオロウラシル、シトシンアラビノシド、シクロホスファミド、チオテパ、ブスルファン、サイトキシン、タキソール、メトトレキセート、シスプラチン、メルファラン、ビンブラスチン、ブレオマイシン、エトポシド、イホスファミド、マイトマイシンC、ミトキサントロン、ビンクリスチン、ビノレルビン、カルボプラチン、テニポシド、ダウノマイシン、カルミノマイシン(Carminomydcin)、アミノプテリン、ダクチノマイシン、マイトマイシン、ニコチンアミド、エスペラミシン、メルファラン、及び他の関連したナイトロジェンマスタード、及びエンドクリン治療薬(例えば、ジエチルスチルベストロール、タモキシフェン、LHRH-拮抗薬、プロゲスチン、抗プロゲスチン等々)が含まれる。
【0053】
使用可能な酵素活性毒及びその断片には、ジフテリアA鎖、ジフテリア毒素の非結合性活性断片、外毒素A鎖(シュードモナス・アエルギノーサ(Pseudomonas aeruginosa)由来)、リシンA鎖、アブリンA鎖、モデシン(modeccin)A鎖、アルファ-サルシン(sarcin)、アレウライツ・フォルディイ(Aleurites fordii)プロテイン、ジアンシン(dianthin)プロテイン、フィトラッカ・アメリカーナ(Phytolaca americana)プロテイン(PAPI、PAPII及びPAP-S)、モモルディカ・キャランティア(momordica charantia)インヒビター、クルシン(curcin)、クロチン、サパオナリア(sapaonaria)オフィシナリスインヒビター、ゲロニン(gelonin)、マイトゲリン(mitogellin)、レストリクトシン(restrictocin)、フェノマイシン、エノマイシン及びトリコセセンス(tricothecenes)が含まれる。
種々の放射性核種も放射性コンジュゲートペプチドリガンド又はハイブリッド分子の生産に利用できる。例としては212Bi、131I、131In、90Y及び186Reが含まれる。炭素-14標識された1-イソチオシアナトベンジル-3-メチルジエチレントリアミン五酢酸(MX−DTPA)はハイブリッド分子へ放射性ヌクレオチドを結合させるためのキレート剤の例である。
ペプチドリガンド又はハイブリッド分子と細胞障害剤のコンジュゲートは、様々な二官能性タンパク質カップリング剤、例えばN-スクシンイミジル-3-(2-ピリジルチオール)プロピオナート(SPDP)、イミノチオラン(IT)、イミドエステル類の二官能性誘導体(例えばジメチルアジピミダートHCL)、活性エステル類(例えば、スベリン酸ジスクシンイミジル)、アルデヒド類(例えば、グルタルアルデヒド)、ビスアジド化合物(例えば、ビス(p-アジドベンゾイル)ヘキサンジアミン)、ビス-ジアゾニウム誘導体(例えば、ビス-(p-ジアゾニウムベンゾイル)エチレンジアミン)、ジイソシアネート(例えば、トリエン-2,6-ジイソシアネート)、及び二活性フッ素化合物(例えば、1,5-ジフルオロ-2,4-ジニトロベンゼン)を使用して作製することができる。例えば、リシン免疫毒素は、Vitettaら, (1987) Science 238:1098に記載されているようにして調製することができる。
他の実施態様では、腫瘍の事前ターゲティングに利用するために、「レセプター」(例えばストレプトアビジン)にペプチドリガンド又はハイブリッド分子が結合され、ペプチドリガンド又はハイブリッド分子-レセプターコンジュゲートが患者に投与され、続いてキレート剤を使用し、循環から未結合コンジュゲートが除去され、細胞障害剤(例えば放射性ヌクレオチド)に結合された「リガンド」(例えばアビジン)が投与される。
【0054】
C.リポソーム
ここで開示されているハイブリッド分子又はペプチドリガンドはまたリポソームとして処方することもできる。ハイブリッド分子を含むリポソームは、例えばEpsteinら, (1985) Proc. Natl. Acad. Sci. USA, 82:3688;Hwangら, (1980) Proc. Natl. Acad. Sci. USA 77:4030;及び米国特許第4485045号及び同4544545号に記載されているように、当該分野において知られている方法により調製される。循環時間が増したリポソームは米国特許第5013556号に開示されている。
特に有用なリポソームは、ホスファチジルコリン、コレステロール及びPEG-誘導体化ホスファチジルエタノールアミン(PEG-PE)を含有する脂質組成物を用いた逆相蒸発法により作製することができる。リポソームは孔径が定められたフィルターを通して押し出され、所望の直径を有するリポソームが得られる。例えば、ここに記載されたような免疫グロブリン定常ドメインを有するハイブリッド分子は、ジスルフィド交換反応を介して、Martinら,J. Biol. Chem. 257:286-288(1982)に記載されているようにしてリポソームに結合させることができる。場合によっては、化学療法剤はリポソーム内に包含される。Gabizonら, J. National Cancer Inst. 81(19)1484(1989)を参照されたい。
【0055】
D.酵素媒介性プロドラッグ治療法
また、本発明のペプチドリガンド又はハイブリッド分子は、プロドラッグ(例えばペプチジル化学療法剤、国際公開81/01145号を参照)を活性な抗癌剤に転化させるプロドラッグ活性化酵素にペプチドリガンド又はハイブリッド分子を結合させることにより、抗体依存性酵素媒介性プロドラッグ治療法(ADEPT)の抗体のようにして使用することもできる。例えば国際公開88/07378号及び米国特許第4975278号を参照されたい。
ADEPT型治療法に有用なコンジュゲートの酵素成分には、より活性な細胞毒形態に転化するように、プロドラッグに作用し得る任意の酵素が含まれる。限定するものではないが、この発明の方法に有用な酵素には、ホスファート含有プロドラッグを遊離の薬剤に転化するのに有用なアルカリ性ホスファターゼ;スルファート含有プロドラッグを遊離の薬剤に転化するのに有用なアリールスルファターゼ;非毒性5-フルオロシトシンを抗癌剤5-フルオロウラシルに転化するのに有用なシトシンデアミナーゼ;プロテアーゼ、例えばセラチアプロテアーゼ、サーモリシン、サブチリシン、カルボキシペプチダーゼ及びカテプシン(例えば、カテプシンB及びL)で、ペプチド含有プロドラッグを遊離の薬剤に転化するのに有用なもの;D-アミノ酸置換基を含むプロドラッグの転化に有用なD-アラニルカルボキシペプチダーゼ;炭水化物切断酵素、例えばグリコシル化プロドラッグを遊離の薬剤に転化するのに有用なβガラクトシダーゼ及びノイラミニダーゼ;βラクタムで誘導体化された薬剤を遊離の薬剤に転化させるのに有用なβラクタマーゼ;及びペニシリンアミダーゼ、例えばそれぞれフェノキシアセチル又はフェニルアセチル基で、それらのアミン性窒素において誘導体化された薬剤を遊離の薬剤に転化するのに有用なペニシリンVアミダーゼ又はペニシリンGアミダーゼが含まれる。あるいは、「アブザイム」としてもまた当該分野で知られている酵素活性を有する抗体を、遊離の活性薬剤に本発明のプロドラッグを転化させるために使用することもできる(例えば、Massey, (1987) Nature 328:457-458を参照)。ペプチドリガンド/ハイブリッド分子-アブザイムコンジュゲートは、ここで記載されているようにして、腫瘍細胞集団にアブザイムを送達するために調製することができる。
酵素は、当該分野においてよく知られている技術、例えば上で検討したヘテロ二官能性架橋試薬を使用することにより、ハイブリッド分子に共有的に結合させることができる。あるいは、本発明のハイブリッド分子の少なくともペプチドリガンド部分を含む融合タンパク質を、当該分野においてよく知られている組換えDNA技術を使用して作成することができる(Neubergerら, (1984) Nature 312:604-608参照)。
【0056】
E.医薬組成物
本発明のペプチドリガンドを含有する医薬組成物は、腸管外、局所、経口又は局部的(エアロゾル又は経皮など)又はその任意の組み合わせを含む任意の適切な形で投与されうる。好適な治療法にはまた静脈内ボーラス注射による最初の投与に続いて一又は複数のインターバルで繰り返し投与を行うことが含まれる。
本発明の化合物の医薬組成物は、所定の純度を持つ化合物を含むペプチドリガンドを、任意成分の製薬的に許容される担体、賦形剤又は安定剤と混合することにより(Remington's Pharmaceutical sciences 16th edition, Osol, A. 編, [1980])、凍結乾燥製剤又は水溶液の形態で調製され保存される。許容される担体、賦形剤又は安定化剤は、用いられる用量及び濃度で受容者に非毒性であり、リン酸塩、クエン酸塩、及び他の有機酸等のバッファー;アスコルビン酸及びメチオニンを含む酸化防止剤;防腐剤(オクタデシルジメチルベンジルアンモニウムクロリド;ヘキサメトニウムクロリド;ベンザルコニウムクロリド、ベンズエトニウムクロリド;フェノール、ブチル又はベンジルアルコール;メチル又はプロピルパラベン等のアルキルパラベン;カテコール;レゾルシノール;シクロヘキサノール;3-ペンタノール;及びm-クレゾール等);低分子量(約10残基未満)ポリペプチド;血清アルブミン、ゼラチン、又は免疫グロブリン等のタンパク質;ポリビニルピロリドン等の親水性ポリマー;グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン又はリシン等のアミノ酸;グルコース、マンノース、又はデキストリンを含む単糖類、二糖類、及び他の炭水化物;EDTA等のキレート化剤;スクロース、マンニトール、トレハロース又はソルビトール等の糖;ナトリウム等の塩形成対イオン;金属錯体(例えば、Zn-タンパク質複合体);及び/又はTWEENTM、PLURONICSTM又はポリエチレングリコール(PEG)のような非イオン性界面活性剤を含む。
ここでの組成物は、治療される特定の徴候に必要ならば一以上の活性化合物も含んでよく、好ましくは互いに悪影響を与えない相補的活性を持つものである。例えば、一つの製剤にEGFR、ErbB2(例えばErbB2上の異なるリガンドに結合する抗体)、ErbB3、ErbB4、又は血管内皮因子(VEGF)に結合する抗体のような分子を更に提供することが望ましい。あるいは、又は加えて、組成物は、細胞毒性剤、サイトカイン、成長抑制剤及び/又は心臓保護剤を含有することもできる。そのような分子は好適には意図される目的に対して効果的な量で組み合わされて存在する。
【0057】
F.疾患の診断と予後
本発明における予後及び診断のための疾患は、好ましくは、ErbB2レセプターの過剰発現により特徴づけられる良性又は悪性腫瘍、例えば、癌、例えば乳癌、扁平上皮細胞癌、小細胞肺癌、非小細胞肺癌、胃腸癌、膵臓癌、グリア芽細胞腫、子宮頸管癌、卵巣癌、肝臓癌、膀胱癌、肝細胞腫(hepatoma)、大腸癌、結腸直腸癌、子宮体癌、唾液腺癌、腎臓癌、肝臓癌、前立腺癌、産卵口癌、甲状腺癌、肝癌(hepatic carcinoma)及び様々な種類の頭部及び頸部の癌が含まれる。従って、本発明の一実施態様は、試料中のErb2の検出及び/又は測定と、そのような検出又は測定の診断、段階決定、重症度の決定、及び疾患の予後一般への使用に関する。
特定の実施態様では、記載された診断方法は治療の進行に従って使用することができる。Erb2保持細胞の量の増加又は減少に至る治療処置を受けている患者では、試料中のErb2保持細胞の量が治療の成功又は失敗の有用な目安となりうる。しかして、本発明は、Erb−2発現細胞を含むと疑われる組織試料において発現されるErb2の量を適当な時間間隔で測定することを含む、患者における治療の効果をモニターする方法を提供する。Erb2の全量が、疾患と治療に依存して、正常な被験者、疾患の発症前又は疾患の緩解時の患者、又は治療開始前の患者の同様な試料中のErb2の量でありうる「ベースライン」又は「コントロール」値と比較される。当業者であれば、過度の実験を行うことなく特定の状況に使用される好適なベースラインが容易に分かるであろう。
本発明の化合物を使用する試料中のErb2の測定の実施においては分析物の測定に対して当該分野で知られている任意の方法を使用できる。そのような方法には、限定されるものではなく、少しだけ挙げると、競合及び非競合アッセイシステムで、ラジオイムノアッセイのような技術を使用するもの、酵素免疫アッセイ(EIA)、好ましくは酵素結合免疫吸着検定法(エライザ:ELISA)、「サンドウィッチ」イムノアッセイ、沈降反応、ゲル拡散反応、免疫拡散アッセイ、凝集アッセイ、補体結合アッセイ、及び免疫電気泳動アッセイが含まれる。好適なイムノアッセイ法の例については、米国特許第4845026号(1989年7月4日)及び同第5006459号(1991年4月9日)を参照されたい。
【0058】
診断及び予後の応用に対しては、本発明の化合物、典型的にはここに上述したハイブリッド分子は検出可能な部分で標識され、上述のような試料中のErb−2を検出するために使用される。多数の標識が利用可能であり、それらは好適には次の範疇にグループ分けされる:
(a)放射性同位体、例えば、35S、14C、125I、3H及び131I等。ハイブリッド分子は、例えばCurrent Protocols in Immunology, Volumes 1 and 2, Coligen等, 編, Wiley-Interscience, New York, Pubs. (1991)に記載された技術を用いて放射性同位体で標識でき、放射活性はシンチレーションカウンティングを使用して測定できる。
(b)蛍光標識、例えば希土類キレート(ユーロピウムキレート)又はフルオレセインとその誘導体、ローダミンとその誘導体、ダンシル、リサミン(Lissamine)、フィコエリトリン及びテキサスレッドが使用できる。蛍光標識は、例えば上掲のCurrent Protocols in Immunologyに開示された技術を用いてハイブリッド分子に結合させることができる。蛍光は蛍光定量計を用いて定量できる。
(c)様々な酵素−基質標識が利用でき、米国特許第4275149号は、それらの幾つかの概説を提供している。酵素は好ましくは種々の技術を用いて測定可能な色素原基質の化学変換を触媒する。例えば、酵素は基質における色変化を触媒し、それは分光学的に測定可能である。あるいは、酵素は基質の蛍光又は化学発光を変化させることもある。蛍光変化を定量化する技術は上述した。化学発光基質は化学反応によって電子的に励起され、ついで(例えば化学発光計を用いて)測定可能な光を放出するか、又は蛍光受容体にエネルギーを供与する。酵素標識の例には、ルシフェラーゼ(例えば、ホタルルシフェラーゼ及び細菌ルシフェラーゼ;米国特許第4737456号)、ルシフェリン、2,3-ジヒドロフタラジンジオン、リンゴ酸塩デヒドロゲナーゼ、ウレアーゼ、西洋ワサビペルオキシダーゼ(HRPO)等のペルオキシダーゼ、アルカリホスファターゼ、β-ガラクトシダーゼ、グルコアミラーゼ、リソザイム、糖類オキシダーゼ(例えば、グルコースオキシダーゼ、ガラクトースオキシダーゼ、及びグルコース-6-ホスフェートデヒドロゲナーゼ)、ヘテロ環オキシダーゼ(ウリカーゼ及びキサンチンオキシダーゼ等)、ラクトペルオキシダーゼ、ミクロペルオキシダーゼ等が含まれる。酵素を抗体に結合させる技術は、O'Sullivanら, Methods for Preparation of Enzyme-Antibody Conjugates for use in Enzyme Immunoassay, in Methods in Enzym. (J. Langone & H. Van Vunakis編)Academic press, New York, 73: 147-166 (1981)に記載されている。
【0059】
酵素−基質の組み合わせは、例えば以下を含む:
(i)西洋ワサビペルオキシダーゼ(HRPO)と基質としてのハイドロジェンペルオキシダーゼで、ハイドロジェンペルオキシダーゼが染料前駆物質(例えば、オルトフェニレンジアミン(OPD)又は3,3',5,5'-テトラメチルベンジジンヒドロクロリド(TMB))を酸化する;
(ii)アルカリホスファターゼ(AP)と色素原基質としてのパラ-ニトロフェニルホスフェート;
(iii)β-D-ガラクトシダーゼ(β-D-Gal)と色素原基質(例えば、p-ニトロフェニル-β-D-ガラクトシダーゼ)又は蛍光原基質4-メチルウンベリフェリル-β-D-ガラクトシダーゼ。
当業者には、多くの他の酵素−基質の組み合わせが利用可能である。これらの一般的な概説については、米国特許第4275149号及び第4318980号を参照されたい。
本発明のアッセイでは、好ましくは、ハイブリッド分子は、固相の支持体又は坦体に結合している。「固相の支持体又は坦体」という用語により、抗原又は抗体と結合可能な任意の支持体が意図される。周知の支持体又は坦体には、ガラス、ポリスチレン、ポリプロピレン、ポリエチレン、デキストラン、ナイロン、アミロース、天然又は修飾されたセルロース、ポリアクリルアミド、アガロース、及び磁鉄鉱が含まれる。坦体の性質は、本発明の目的に対して、ある程度可溶性であるか、不溶性であり得る。支持材料は、結合された分子が抗原又は抗体と結合可能である限り、実質的には、任意の可能な立体構造をとってもよい。従って、支持体の立体構造は、ビーズのような球状、又は試験管の内面、又はロッドの外表面のような円筒状であってもよい。あるいは、表面はシート、テストストリップ等々のような平坦面でもよい。好ましい支持体には、ポリスチレンビーズが含まれる。当業者であれば、抗体又は抗原を結合させるための他の多くの好適な坦体が分かり、日常的な実験を行うことにより、同一性について確認を行うことができるであろう。
【0060】
好ましい実施態様では、サンドイッチイムノアッセイが利用される、すなわち、Erb2は、一次抗体又はハイブリッド分子のErb2抗原に対する結合、及び二次抗体又はハイブリッド分子のErb2に対する結合を含み、一次及び二次抗体の両方、又はハイブリッド分子による免疫特異的なErb2への結合を検出又は測定することを含む方法によって検出され又は測定される。特別の実施態様では、一次又は二次抗体のどちらかがモノクローナル抗体であり、一次又は二次のどちらか又はその両方が、本発明のハイブリッド分子である。この実施態様では、ハイブリッド分子のペプチドリガンド部分は、好ましくは、一次抗体のリガンド部分とは異なる部位に結合する(例えば、抗原への結合に対する、抗体とハイブリッド分子間における競合阻害の欠如に反映される)。他の特定の実施態様では、一次又は二次抗体はポリクローナル抗体である。
また、本発明中のアッセイを実施するための成分を含む一又は複数の容器又はバイアルを含むキットも本発明の範囲内である。例えば、そのようなキットは、腫瘍のバイオプシーのような免疫組織化学的サンプル分析に必要とされる薬剤を含み得る。薬剤には、例えば、ハイブリッド分子又は抗体のような一又は複数の結合相手が含まれる。組織学的なアッセイに対しては、キットは、色素生産性の基質並びに発色したとき酵素反応を停止する薬剤を含む。キットに含まれる基質は本発明のハイブリッド分子の一つに結合した酵素に適したものである。これらは、当該技術分野において周知である。また、キットは、例えば、既知量の精製Erb2などの標準物質を含む。
以下の実施例は、例示のために提供されるもので、限定するものではない。明細書中の全ての文献の開示は、ここに明示的に出典明示により取り込まれる。
【0061】
実施例
実施例I
HER2結合ペプチドの同定及び成熟
方法
ファージライブラリー− ランダム配列の多価ペプチドファージライブラリー(表1)は、テンプレートであるpB2479.g8の大規模なクンケル突然変異導入法を用いることにより構築された。このファージミドは、Tacプロモーター、malシグナル配列、3つのストップコドン、及びgVIIIと融合されたリンカー配列を含み、LacIq及びアンピシリン耐性遺伝子を有する。従って、このペプチドライブラリーは、IPTGでコピー数を制御することが可能なpVIII融合体として、多価的にファージ上で発現される。各ライブラリーは、109クローンを越える多様性を有する。
【0062】
【0063】
選択条件− HER2-ECDは、50mM炭酸水素アンモニウム、pH9.3中にて、5μg/mlを使用し、4℃で一晩反応させ、マキシソープ(maxisorp)プレートに直接固定化された。ウェルは、1%BSAを含むPBS(PBS−BSA)を用いて、25℃で1時間ブロッキングした。上述のライブラリーから得られたファージを、表1に示すような7つのグループにプールした。各プールからのファージは、PBS−BSA にHER2-ECDを含むウェル中で、25℃で3時間反応させた;通常、約5 x 1010のファージが各実験ラウンドの開始時に添加された。結合しなかったファージは、0.05%のTween20を含むPBS(PBS-Tween)による繰り返しの洗浄により除去された;残存するファージは、500mM KCl、10mM HCl、pH2により溶出された。その後、溶出されたファージは、XL1-Blue細胞中でVCSM13ヘルパーファージ(Stratagene)を用い、37℃で一晩増幅させた。IPTG(10μM)は、最初のファージライブラリー及び増幅中の第1回目のラウンドに、添加された;その後のラウンドは、ペプチド-pVIII発現産生のためのtacプロモーターの基本発現に依存した。濃縮度は、BSAがコートされているウェルに結合するファージ数に対して、標的がコートされているウェルに結合するファージ数を希釈することにより、観察した。
HER2ファージ結合アッセイ− Erb2の固定化された細胞外ドメイン(HER2-ECD、Hudziak等、(1991) J. Biol. Chem. 266:24109-15)に対する、表面にペプチド1.1FI(表1)の単一コピーを表示するファージの結合阻害は、ファージELISAを使用して評価した。HER2-ECDは、50mM 炭酸水素アンモニウム、pH9.3中にて、5μg/mlになるように使用して、4℃で一晩反応させ、直接、マキシソープ・プレート(Nunc)に固定化させた。ウェルは、1%BSAを含むPBS(PBS−BSA)を用いて、25℃で1時間ブロッキングした。PBSによる1.1FI−Fcの希釈物は、1.1Fc−Fcを表示するファージの固定化HER2-ECDへの結合に対する阻害能についてテストされた。マイクロタイタープレートは、0.05%のTween20を含むPBS(PBS-Tween)により洗浄され、HER2-ECDに結合したファージは、抗gVIII/HRPモノクローナル抗体(Amersham Pharmacia Biotech)により検出された。結合したHRPの量は、ABTS/H2O2基質を使用し、405nmにおける変化を検出することにより測定された。
【0064】
結果
HER2-ECDへ結合する多価ペプチド-ファージの選択− 多価のペプチドライブラリーは、固定化HER2-ECDに対して7つのプール(表1)に分類された。多価のファージディスプレイ(Scott等 (1990) Science 249:386-390)が、アビディティー効果を介して結合を促進するために使用された。濃縮度、つまり、HER2-ECDでコートされたウェルから溶出されたファージの数をBSAでコートされたウェルから溶出されたファージの数で除した数は、ラウンド3及びラウンド4について図4に示されている。各プールにおけるランダムクローンから得たDNAは、配列確認され、その予想されるペプチド配列が表2に示されている。プール1は、単一クローンとして得られたが、他のプールは、複数の配列を含んでいた。
ファージ結合アッセイを用いたところ、ファージクローン1.1、5.1及び7.1は、固定化HER2-ECDに結合するが、固定化BSA又は、近縁関係にあるHER3又はHER4には結合しないことが見いだされ、これらのファージクローンがHER2-ECDを特異的に認識することを示す(図5)。これらのクローンの配列は、更に高い親和性を持つクローンの再選択のために、部分的ランダム化に供された。
【0065】
表2
【0066】
実施例II
方法
一価ファージの部分的又は完全なランダム化− ファージ表面に、gIIIによってコードされる尾タンパク質に対しリンカー配列を介して融合されたペプチドの単一コピーを表示する一価ライブラリーが、類似のファージミド、pB2479.g3に対する一本鎖鋳型指向性突然変異(Kunkel等, (1991) Met. Enz. 204:125-139)を用いて構築された。このベクターにおいて、gVIIIに対するコード配列は、gIIIによって置換され、さらに、CMPγ遺伝子は、AMPγ遺伝子の唯一のhincII部位に挿入された。薬剤耐性の変化は、弱い親和性ながらも、アビディティー効果を介して集団化を引き起こす(Cwirlaら, (1990) Proc. Natl. Acad. Sci.USA 87:6378-6382)多価クローンの関連クローンによるコンタミネーションを除去するように設計された。部分的にランダム化されたライブラリーは、各アミノ酸位置において50%の変異率を許容しながらも、初期の多価ライブラリーから同定されたペプチド配列への偏りを維持するように設計された。この突然変異率は、70-10-10-10の塩基の混合物によりオリゴを合成することにより達成される(この場合、オリゴのドープ領域の各塩基は、野生型配列に寄与する塩基の70%を含む混合物を用いて、関連づけられ、他の3塩基の各塩基は、野生型の配列塩基の10%により関連づけられる)。これに対して、ライブラリーにおける完全なランダム化は、特定のコドンに関しNNSを用いてオリゴを合成することにより、配列の他の部分を一定に保ちながら、得ることができた。
【0067】
結果
ペプチドファージの部分的なランダム化− 作製された実際のライブラリーが、およそ109クローン(潜在的な多様性のほんの一部の多様性)しか含まないのに対し、設計された最初のペプチドライブラリーは、2020(1026)以上の潜在的な多様性をコードしていた。サーチ範囲を狭め、最初に選択したペプチドの範囲内におけるペプチド多様性を更に探索するために、部分的ランダム化技術を用いた。この技術は、各アミノ酸位置に50%の突然変異率(アミノ酸レベルにおいて)を導入する間、野生型配列へ向かう偏りを維持する;従って、平均して、20アミノ酸のペプチドを表示するファージは、10のランダム突然変異を必要とするであろう。その上、更なる親和性の改善が予想されるため、このようなライブラリーを一価ファージ(gIII)(Bassら, (1990) Protein: Struct. Funct. Genet. 8:309;Lowmanら, (1991) Biochemistry 30:10832)について構築することにした。
クローン1.1、5.1、及び7.1の一価の部分的ランダム化ライブラリーが構築され、HER2に対して4ラウンドの分類がなされた。10000倍より高い濃縮度が、ライブラリー1.1及び7.1について観察された;しかし、ライブラリー5.1の濃縮は観察されなかった。再び、ランダムクローンが選択され、配列決定が行われた;ライブラリー1.1及び7.1のクローンから選択された予想されるペプチド配列が、図15に示される。ライブラリー1.1及び7.1における幾つかのアミノ酸位置は、100%保持されており、さらに、複数のコドンがこれらの多くの位置において観察された。続いて行われた部分的ランダム化において、強固に保持された残基は、直接的な接触を通じた結合又は構造的な理由の何れかに対して重要となりうる。
【0068】
HER2結合配列の完全成熟− クラス1配列の親和性の成熟を完成させるために、100%保存された位置は固定化し、残りの位置は完全にランダムである、第三のライブラリーセットが構築された。クラス1配列のジスルフィドループに隣接するアミノ酸残基の役割は、アミノ及びカルボキシ末端の何れかを欠失するライブラリーを構築することにより解明された。よって、3つの一価ライブラリーが図16に示されるようにして構築された。500倍よりも高い濃縮度が、ラウンド4における各ライブラリーによって観察された;ランダムクローンの配列は、図16に示されている。10の位置において完全にランダム化されたこれらのライブラリーは、完全にはほど遠いが、全長及びアミノ末端を欠失したライブラリーを比較することにより、クラス1配列のC末端領域における付加的なジスルフィドへの選択性が示された。両ライブラリーにおいて、付加的なジスルフィド結合は、2箇所に見いだされる。3つ目のライブラリーにおいて、C末端位置の欠失は、これらの位置における付加的なジスルフィド結合の選択を防止する。付加的なジスルフィド結合の選択とは別に、他のランダム化された位置において選択された残基は非常に多様性を有している。
【0069】
実施例III
方法
ペプチド合成− ペプチドは、PEG-ポリスチレン樹脂を用い(Bodanszkyら, (1984) Int. J Peptide and Protein Res. 23(1):111)、0.25mmolスケールにて、手動又は自動化されたFmocに基づく固相合成により合成された。側鎖の保護基が除去され、ペプチドは95%のトリフルオロ酢酸(TFA)及び5%のトリイソプロピルシランによって樹脂から切断された。酢酸による飽和ヨウ素溶液がジスルフィド結合を酸化するために添加された。ペプチドは、0.1%のTFAを含んだ水/アセトニトリルの濃度勾配を用い、逆相HPLCによって精製された。ペプチドは、分析的なHPLCによると、純度>95%であり、同一性については、マススペクトロメトリーによって確認された。
ペプチド-Z融合物の生成− HER2-ECDへの結合のために選択されたファージペプチドは、プロテインAのZタンパク質との融合体として大腸菌(27C7株)中で発現され、分泌された。オリゴは、ファージ-ペプチド配列に対するコード化配列をstIIシグナル配列とZドメインを持つプラスミド(pZCT)中のZドメインとの間に挿入するように設計された。細胞はリン酸を制限した培地中で増殖され、ペプチド-Z融合体は、記載されているように、IgGアフィニティーカラムを使用して(Dennis等, (1993) Proteins: Struct. Funct. Genetics 15:312-321)、培地から精製された。IgGでアフィニティー精製されたペプチド-Z融合体は、NHS-LC-ビオチン(Pierce)によってビオチン化され、マススペクトロメトリーによって性質決定された。
多量体を形成するペプチド-Z融合体は、PBSにてSuperdex75カラムのサイズ排除クロマトフラフィーにより、単量体及びダイマーを含む画分へ分離し得る。
【0070】
精製されたペプチド-Z融合体からのZドメインの除去− 精製されたペプチド-Z融合体からのZドメインの除去は、トリプシンによる融合体の切断により行うことができる。トリプシン(1%, w/w)は、100mM Tris pH8.0、及び10mM CaCl2中にて37℃、4時間、添加された。Zドメインが除去されたペプチドは、0.1%のTFAを含んだ20から50%のアセトニトリル濃度勾配を用い、C18逆相HPLCによって精製された。
HER2-Fc発現ベクターの構築− ベクターpVL1393(Pharmigen)を基礎とする組換体転移ベクターの構築には、標準的な組換えDNA技術が用いられた(Sambrook, J., Fritsch, E. F., 及びManiatis, T. (1989) Molecular Cloning: A Laboratory Manual, second Ed., Cold Spring Harbor Laboratory Press, New York; O’Reilly, D. R., Miller, L. K., 及びLuckow, V. A. (1994) Baculovirus Expression Vectors: A Lboratory Mnual, Oxford University Press, New York)。pVL1393由来プラスミドpbPH.Hisは、NcoIとSmaIによって直線状にされ、エビのアルカリホスファターゼで処理された(Dweyer, M. A.ら, (1999) J. Biol. Chem. 274:9738-9743)。ヒトIgG1のヒトFc部分は、他のpVL1393由来プラスミドpVL1393.IgGから、NdeIで切断し、続いてKlenowで処理し、更にNcoIで切断することで得られる700塩基対の断片として得られた。MIP.5シグナル配列は、大腸菌で切断されたPCR断片としてFc配列の前に導入され、その断片には、AscI部位が含まれる。AscI部位は、予想されるシグナル配列切断部位の後に生じる。ライゲーションの後、コンピテント化された大腸菌XL-1 Blueが形質転換され、DNAの配列決定により正しい組換体プラスミド(pVL1393.MIP.5sig.Fc)を持つ大腸菌が選択された。その後、pVL1393.MIP.5sig.Fcは、AscI及びStuIにより直線化され、エビのアルカリホスファターゼにより処理された。直線化されたベクターは、その後、整合する末端を持つ合成DNA片とライゲーションされた。合成DNA挿入断片は、以下の2オリゴをアニールすることにより生成された:5'-CGC GCC CAG GTG TAC GAG TCC TGG GGA TGC ATC GGC CCC GGC TGC GCC TGC CTG CAG GCC TGC CTG GGA GGC GGG AGC TCC GGC-3'(配列番号:159)及びGGGSSG(配列番号:130)リンカーを含むペプチド配列1.1FIに対するコード化配列である5'-GCC GGA GCT CCC GCC TCC CAG GCA GGC CTG CAG GCA GGC GCA GCC GGG GCC GAT GCA TCC CCA GGA CTC GTA CAC CTG GG-3'(配列番号:160)。
ライゲーションの後、コンピテント化された大腸菌XL-1 Blueが形質転換され、ローダミンダイターミネーター法及びApplied Biosystems ABI Model 373自動DNAシークエンサーを用いたDNAの配列決定により正しい組換体プラスミド(pVL1393.MIP.5.1.1FI-Fc)を持つ大腸菌が選択された。組換体転移ベクターは、キアゲンのMini-Prepを使用して精製され、組換体バキュロウィルスの構築のために使用された。
【0071】
組換体バキュロウィルスは、sf9細胞に対する転移ベクターと直線化された野生型バキュロウィルスDNA(Autographa californica nuclear polyhedrosis virus (AcNpV)、Pharmingen)との共形質移入により生産された。組換体バキュロウィルスの最初の増幅により、検出可能なタンパク質の発現を達成することができた。次いで、プラーク精製及びウイルスストック希釈が、プラークアッセイにおいて実施された。先に記載されたような標準的な方法が用いられた(O'Reilly, D. R., Miller, L. K., 及びLuckow, V. A. (1994) Baculovirus Expression Vectors: A Laboratory Manual, Oxford University Press, New York)。
細胞培養− スポデプテラ・フルギペルダ(Spodeptera frugiperda)(Sf9)昆虫細胞(ATCC CRL 1711)の接着培養は、GRACE's(JRH Biosciences, #51942-78p)、グルタミン、ストレプトマイシン/ペニシリン、及び10%ウシ胎仔血清(56℃、30分間、熱により不活性化されたもの)が添加されたヒンクスのTNF-FH昆虫用培地中、28℃で培養された。培養細胞は3日毎に継代された。High5細胞のスピナーによる培養細胞は、0.5の感染多重度にて感染され、感染6時間後に回収された。浮遊培養は、スピナーフラスコ中、28℃でESP-921タンパクフリーの昆虫細胞培地(Expression Systems LLC、#96-001)を用いて維持された。培養細胞は、最初の細胞密度が106細胞/mlとなるように3日毎に継代された。
ペプチド-Fcの精製− 至適化された感染方法に従い、High5細胞は、4℃、800 x gにて10分間遠心することにより除去された。細胞が除かれた上清(0.5 L)は、0.45μのNalgene社のフィルターを用いてろ過され、25℃にてPBS(リン酸緩衝生理食塩水)で平衡化されたHi-Trap プロテインAカラム(Amersham Pharmacia Biotech)に添加された。20mlのPBSで洗浄後、カラムは0.2N HOAc、3mlで溶出され、ペプチド-Fcを含む画分が凍結乾燥され、4℃にて保存された。
SDS-PAGE− サンプルが、Laemmli(Laemmli, U. K. (1970) Nature 227:680-685)の方法により、プレステインタンパク質分子量マーカー(SeaBlue、Novex)を用い、還元又は非還元状態で4−20%のトリス-グリシンSDS-PAGE(Novex)にて解析された。
【0072】
タンパク質の配列決定− 感染されたSf9細胞の上清から精製された1.1FI−Fcを、SDS-PAGEにかけ、その後PVDF膜に移した。ミリポアImmobilon-PSQ膜上への電気的なブロットは、BioRad Trans-Blot Transferセルにより、250mAの定電流で1時間行われた(Matsudaira, P. (1987) J. Biol. Chem. 262:10035-10038)。PVDF膜は、50%メタノール中の0.1%クマジーブルーで0.5分間染色し、10%酢酸、50%メタノールで2から3分間脱染色した。膜は、水で完全に洗浄した後、−20℃で保存する前に乾燥させた。50kD付近の1.1FI−Fcのバンドを切り出し、オンラインPTH分析装置を備えた494型Applied Biosystems シークエンサーを用いて最初の11残基を配列決定した。Nelson Analytical 760 インターフェースにより、Justice Innovation Softwareを用いてピーク面積を積算した。配列翻訳は、DEC alphaにより実行した(Henzel, W. J., Rodringuez, H., 及びWatanabe, C. (1987) J. Chromatog. 404:41-52)。
HER2競合ELISA− HER2-ECDに対する1.1FI−Fcへの結合は、競合ELISAにより測定した。サンプルをPBS-BSAにて希釈し、上述した方法でマイクロタイタープレートに固定化されたHER2-ECDに対する40nMのビオチン化1.1FI−Zの結合を阻害する能力を試験した。1時間のインキュベーションの後、プレートをPBS-Tweenにて洗浄し、ストレプタビジン/HRP結合体(Spreptavidin-POD, Roche Molecular Biochemicals)を30分間添加した。プレートを、再度PBS-Tweenで洗浄し、結合されたHRPをABTS/H2O2基質(Kirkegaard & Perry Laboratories)を用いてアッセイし、405 nmにおける吸収を測定した。405 nmにおける吸収値を、ウェルに元々添加されていた1.1FI−Fcの濃度に対してプロットした。非線形回帰分析により(Marquardt, J. Soc.Indust. Appl. Math. 11:431-441 (1963))、シグモイド曲線は、4つの変数の方程式にフィットさせられた;このアッセイにおいて、最大値の半数のシグナルを与えるのに必要な1.1FI−Fcの濃度は、この曲線より計算され、IC50値と呼ぶことにした。
【0073】
細胞の結合− HER2(BT 474 [3+], MDA 361[2+], 及びBaF3 [0+])を発現する細胞をトリプシンで剥がし、洗浄用緩衝液(0.5%BSA、1mM NaOHを含むPBS)で2回洗浄し、各々0.5x106細胞を含む画分に分けられた。細胞は、0、0.5又は5μMの1.1FI−Zbを含む洗浄用緩衝液に懸濁し、洗浄用緩衝液で2回洗浄し、ストレプタビジン-PEを含む洗浄用緩衝液に懸濁され、更に2回洗浄用緩衝液で洗浄し、FACS分析にかけた。
HER2-ECDに結合するペプチドの性質決定− ペプチドHER201は、部分的にランダム化されたファージ-クローン1.1.2(図15)に一致するように合成された。更にまた、完全にランダム化された全長ライブラリーに由来する代表的なクローンである1.1.FI(図16)に一致するペプチドHER212も合成された。このペプチドに存在する2つのジスルフィドに起因して、1-2及び3-4のジスルフィド配列を誘導するために、直交性の保護基が、システインのために使用された。ジスルフィド結合が選択の段階中に誘導されたため、この配列が考えられた。
また、前記ペプチド及びライブラリー7.1(図15)(7.1c)のコンセンサス配列に由来するペプチド配列が、プロテインAのZドメインタンパク質との融合体(1.1.FI-Z及び7.1c-Zと呼ぶ)として発現され、システインの酸化がファージ生産の段階と同様に生じ得るような大腸菌から分泌された。1.1FI-Zの調製物は、ビオチン化され(1.1.FI-Zb)、全タンパク質に基づいた場合2nMのEC50で、固定化HER2に結合し(Fig.6)、競合ELISAに対する試薬として役立った。
組換体ペプチド融合体である1.1.FI-Z及び7.1c-ZペプチドHER201、HER212は、固定化されたHER2に対する1.1.FI-Zbの結合を阻害する能力に関しテストされた(図7)。HER201及び7.1c-Zは、それぞれ300及び1,100 nMのIC50を示した。興味深いことに、HER212及び1.1FI-Z、これらは同じファージペプチド配列に由来するが、劇的に異なるIC50を示した;各々、9μMと50 nMであった。一つの説明として挙げられることは、他のジスルフィド配列がHER212の合成中に強制的に生じた可能性である。更に、1.1.FI-Z(Fig.8B)(レーンS)のSDS-PAGE解析によると、分子間のジスルフィド結合による多量体化に起因するらしき不均一な混合物であることが明らかにされた。マススペクトロ分析によると、HER212はモノマーでのみ存在するのに対し、ダイマー1.1FI-Zの存在が確認された。
【0074】
1.1.FI-Z調製物は、更にSuperdex75を用いてモノマーとダイマーの画分に精製された(図8A及び8B)。驚くべきことに、HER2結合は、HER2の競合ELISAでアッセイした場合、モノマーの画分よりはむしろダイマーの画分に一致した(図8B及び8C)。
更に、1.1.FI-Zは、Zドメインを除去するためにトリプシンにより切断され、逆相HPLCにかけられた。HPLCから得られる画分は、HER2競合ELISAにおける競合能に関し、テストされた。活性画分は、HER2の配列に一致する質量を有したが、ダイマーを含むことも見いだされた。この画分は、HER2の競合ELISAにおいて、タンパク質濃度に基づき3nMのIC50を示した。
ダイマー形成を促進するため、1.1FIのアミノ酸配列はZドメインと融合する前に二度繰り返されるような発現ベクターが構築された(図9)。この「一本鎖ダイマー」((FI)2-Zと称する)も、ダイマーを生産するが、生産される主な種はモノマーであった(図9B)。更に、Superdex75カラムで分画し(図9A)、HER2の競合ELISAでアッセイした場合、活性のある種は、モノマーとして同定された(図9B及び9C)。(FI)2-Zは、1.1FI-Zダイマー(IC50=20 nM)と同等の親和性を示す。
HER2上の結合部位− クラス1及びクラス7の配列は、HER2上の同一、それとも独立の部位に結合されるのかどうか検討するために、固定化HER2に対する、クラス1及びクラス7のファージクローンの結合がHER201又は7.1c-Zの存在下において調べられた。図10A及び図10Bより、HER201及び7.1c-Zはクラス1及びクラス7のファージ両方の結合を阻害するようであり、このことは、両クラスはHER2の同一の部位に結合することを示すものである。これら2つのクラスにおいて得られ配列をつぶさに見ると、コアの相同領域が存在することが分かる(図11)。
【0075】
また、クラス1及びクラス7ペプチドの結合部位は、HER2に対する既知のモノクローナル抗体が、固定化HER2に対する1.1.FI-ファージへの結合と、直接競合しないHER2上の新規な部位のようである(図12)。固定化HER2に対する1.1.FI-ファージの結合を阻害するために、種々のモノクローナル抗体の濃度を増加させても影響はなかった。HER2に対するこのモノクローナル抗体の親和性は、100pM以下であるため、3H4の非常に高濃度(>100nM)な状態における結合の減少は、間接的な効果を介した結果であると考えられる。
細胞の結合− 1.1.FI-Zbは、細胞上に発現されたHER2を認識する能力について調べられた。それぞれ異なる量にてHER2を細胞上に発現する3つのタイプの細胞が、調べられた;BT 474 [3+](3 x 106 レセプター/細胞)、MDA 361[2+](4 x 105 レセプター/細胞)及びHER2を発現しないコントロールの細胞株、BaF3である。FACS分析によると、1.1.FI-Zbは、レセプターとなるHER2の細胞あたりの数、及び用いた1.1.FI-Zbの濃度に比例して細胞に結合することが見いだされた(図13)。
ペプチド-Fc融合体− 1.1.FI配列をコードするDNAが、リンカーを介してIgG1のFc領域と融合され、バキュロウィルス中において発現された。その結果得られたタンパク質は、プロテインAカラムにより精製され、酸化及び還元状態下、SDS-PAGEにより性質決定された(図14)。これら2つの状態において、明らかな分子量のシフトが観察されており、このことは、Fcの二量体化を示すものである。この分子は、固定化されたHER2に対する1.1FIファージの結合を阻害する能力に関してテストした場合、3 nMのIC50を示した。HER2の競合ELISAによりテストした場合、1.1FI-Fcは1.1FI-Zと同様なIC50を示したが、このことは、1.1FI配列がFcに融合された場合HER2と結合することが、十分可能であったことを示す。
【図面の簡単な説明】
【0076】
【図1】天然IgG及びパパイン及びペプシン消化により得られたそれらの断片の概略図である。ジスルフィド結合は、例えばCH1とCLドメインの間の-S-S-により表される。この図において、Vは可変ドメインであり;Cは定常ドメインであり;Lは軽鎖の略であり、Hは重鎖の略である。
【図2A】図2Aは天然IgG Fc領域アミノ酸配列の整列を示す。ヒト(hum)IgG Fc領域配列を示しており、humIgG(非-A及びAアロタイプ)(それぞれ、配列番号:72及び73)、humIgG2(配列番号:74)、humIgG3(配列番号:75)及びhumIgG4(配列番号:76)が示されている。ヒトIgG1配列は非-Aアロタイプであり、この配列とAアロタイプとの差異(356及び358位置;EU番号方式)を、ヒトIgG1配列の下に示す。また、天然マウス(mur)IgG Fc領域配列を示し、murIgG1(配列番号:77)、murIgG2A(配列番号:78)、murIgG2B(配列番号:79)及びmurIgG3(配列番号:80)が示されている。
【図2B】図2Bは図2AのFc領域配列間のパーセント同一性を示す。
【図3】星印が付された配列間に差異を有する、天然ヒトIgG Fc領域配列、humIgG1(非-A及びAアロタイプ)(配列番号:72及び73)、humIgG2(配列番号:74)、humIgG3(配列番号:75)及びhumIgG4(配列番号:76)の整列を示す。
【図4】HER2細胞外ドメイン(ECD)に結合する多価ペプチドファージの富化を示す。選択の3及び4回に対して、HER2で被覆されたウェル対BSAで被覆したウェルから溶出したファージの比率が示されている。
【図5】ファージ結合アッセイの結果を示す。ファージクローン1.1,5.1及び7.1は固定化されたHER2-ECDに結合したが、BSA、又はHER2-ECDを特異的に認識することが示されている密接に関連したHER3又はHER4には結合しなかった。
【図6】固定化されたHER2に結合するペプチド1.1.FIZbのEC50の定量を示す。
【図7】固定化されたHER2-ECDへの結合に対するペプチド1.1.FI-Zbとの、合成ペプチドHER201及びHER212並びに組換えペプチド1.1.FI-Z及び7.1c-Zの競合アッセイの結果を示す。
【図8】図8Aはモノマー(M)及びダイマー(D)含有画分にSuperdex75を使用して精製した1.1.FI-Z(配列番号:81)の調製物を示す。図8Bは、出発物質とペプチド1.1.FI-ZのSUPERDEXTM75カラムからの画分のSDS PAGE分析を示す。図8Cは、競合アッセイを使用してアッセイした場合に、1.1.FI-Zのダイマー画分と符合するHER2結合活性を示す。
【図9】図9Aはモノマー及びダイマー含有画分にSUPERDEXTM75を使用して精製した(1.1FI)2-Z(配列番号:82)(1.1FIを2回繰り返して、プロテインAのZドメインに融合させたもの)調製物を示す。図9Bはペプチド(1.1FI)2-ZのSUPERDEXTM75カラムからの画分のSDS PAGE分析を示す。図9Cは、競合アッセイを使用してアッセイした場合に、(1.1FI)2-Zのダイマー画分と符合するHER2結合活性を示す。
【図10】図10A及び10Bは、ペプチドHER201と7.1c-Zがクラス1とクラス7のファージの結合をブロックすることを示しており、双方のクラスのペプチドがHER2の同じ部位に結合することを示唆している。
【図11】2つのクラスのペプチド間の核の相同性領域を表すクラス1及びクラス7の配列の例を示す。配列7.1c(配列番号:83)、1.1.2(配列番号:84)、1.1.Fc(配列番号:85)、1.1.FA(配列番号:86)、1.1.FB(配列番号:87)、1.1.FH(配列番号:88)、1.1.CF(配列番号:89)。
【図12】HER2-ECDへの結合に対する、HER2に対する既知のモノクローナル抗体とのクラス1及びクラス7のペプチドの競合アッセイの結果を示す。該ペプチドはHER2-ECDへの結合に対して、モノクローナル抗体と競合せず、既知のモノクローナル抗体により認識されるものとは異なるHER2-ECD部位に該ペプチドが結合することを示唆している。
【図13】ペプチド1.1.FI-Zbと異なるレベルでHER2を発現する細胞(MDA361[4x105レセプター/細胞]及びBT474[3x106レセプター/細胞])、又はHER2を発現しない細胞(BaF3)を使用して行われた結合アッセイの結果を示す。1.1.FI-Zbは、細胞で発現したHER2レセプターの数及び1.1.FI-Zbペプチドの濃度に比例して細胞に結合することが見出された。
【図14】ヒトIgG1のヒンジ、CH1及びCH2ドメインにリンカーを介して融合したペプチド1.1.FI(1.1FI-Fc)の、酸化及び還元条件下でのSDS PAGE分析の結果を示す。2つの条件下で分子量がシフトすることは、Ig定常領域の二量体化とFcドメインの生成を示唆している。
【図15】ライブラリー1.1及び7.1.のクローンからの推定ペプチド配列を示す。配列1.1(配列番号:57);1.1.8(配列番号:58);1.1.9(配列番号:59);1.1.10(配列番号:60);1.1.1(配列番号:61);1.1.2(配列番号:62);1.1.6(配列番号:63);1.1.11(配列番号:64);1.1.3(配列番号:65);1.1.4(配列番号:66);1.1.5(配列番号:67);1.1.7(配列番号:68);コンセンサス(配列番号:69);7.1(配列番号:45);7.1.1(配列番号:46);7.1.9(配列番号:47);7.1.6(配列番号:48);7.1.5(配列番号:49);7.1.3(配列番号:50);7.1.4(配列番号:51);7.1.2(配列番号:52);7.1.7(配列番号:53);7.1.8(配列番号:54);7.1.11(配列番号:55);コンセンサス(配列番号:56)。
【図16】第4の選択回による各ライブラリーのランダムクローンからの配列を示す。配列1.1(配列番号:57);コンセンサス(配列番号:58);1.1.FC(配列番号:30);1.1.FN(配列番号:31);1.1.FA(配列番号:1);1.1.FM(配列番号:2);1.1.FE(配列番号:3);1.1.FH(配列番号:29);1.1.FB(配列番号:12);1.1.FF(配列番号:13);1.1.FI(配列番号:14);1.1.FK(配列番号:15);1.1.FD(配列番号:16);1.1.FQ(配列番号:17);1.1.FS(配列番号:18);1.1.FX(配列番号:9);1.1.FY(配列番号:10);1.1.FZ(配列番号:11);1.1.NC(配列番号:4);1.1.NB(配列番号:5);1.1.NM(配列番号:6);1.1.NQ(配列番号:7);1.1.NA(配列番号:19);1.1.NG(配列番号:20);1.1.NH(配列番号:21);1.1.NE(配列番号:22);1.1.NJ(配列番号:23);1.1.NF(配列番号:24);1.1.NK(配列番号:25);1.1.NR(配列番号:26);1.1.CF(配列番号:28);1.1.CA(配列番号:32);1.1.CB(配列番号:33);1.1.CC(配列番号:34);1.1.CD(配列番号:35);1.1.CE(配列番号:36);1.1.CG(配列番号:37);1.1.CH(配列番号:38);1.1.CI(配列番号:39);1.1.CJ(配列番号:40);1.1.CL(配列番号:41);1.1.CM(配列番号:42);1.1.CP(配列番号:43)。
【図17】固定化されたHER2-ECDへの結合に対する、ペプチド1.1.FI-Zbとの、1.1FI-Fc及び1.1FI-Zの競合アッセイの結果を示す。
【特許請求の範囲】
【請求項1】
インビトロアッセイにおいて、次の式:
Xaa(1−14)−Cys−Xaa16−Gly−Pro−Gly−Cys−Xaa(21−27)(配列番号:90)
を有し、ここで、Xaa(1−14)がないか、1から14のアミノ酸であり、Xaa16がMet、Thr、Cys及びIleからなる群から選択されるアミノ酸であり、Xaa(21−27)はないか、1から7のアミノ酸であるペプチドリガンドと、HER2への結合について競合する、非天然発生ペプチドリガンド。
【請求項2】
次の式:
Xaa(1−7)−Cys−Xaa9−Gly−Pro−Gly−Cys−Xaa(14−20)(配列番号:92)
を有し、ここで、Xaa(1−7)がないか、1から7のアミノ酸であり、Xaa9がMet、Ile、Thr又はCysからなる群から選択されるアミノ酸であり、Xaa(14−20)はないか、1から7のアミノ酸である、請求項1に記載のペプチドリガンド。
【請求項3】
次の式:
Xaa(1−7)−Cys−Ile−Gly−Pro−Gly−Cys−Xaa(14−20)(配列番号:161)
を有する請求項2に記載のペプチドリガンド。
【請求項4】
式A−Aを有するペプチドリガンドであって、ここでAが請求項3に記載のペプチドリガンドであり、「−」が任意成分のリンカードメインであるペプチドリガンド。
【請求項5】
次の式:
Xaa(1−5)−Trp−Gly−Cys−Ile−Gly−Pro−Gly−Cys−Xaa(14−20)(配列番号:93)
を有し、ここで、Xaa(1−5)がないか、1から5のアミノ酸である、請求項3に記載のペプチドリガンド。
【請求項6】
次の式:
Xaa(1−3)−Glu−Xaa5−Trp−Gly−Cys−Ile−Gly−Pro−Gly−Cys−Xaa14−Xaa15−Leu−Xaa(17−20)(配列番号:87)
を有し、ここで、Xaa(1−3)がないか、1から3のアミノ酸であり、Xaa5がアミノ酸であり、
Xaa5がアミノ酸であり、
Xaa14がアミノ酸であり、
Xaa19がアミノ酸であり、
−Xaa(17−20)がないか、1から4のアミノ酸である、請求項5に記載のペプチドリガンド。
【請求項7】
X(1−3)−がないか、
Gln−Arg−Asn−
Leu−Ser−Pro−
Glu−Asn−Trp−
Ala−Ser−His−
Lys−Leu−Asn−
Thr−Gln−Ala−
Ala−Pro−Arg−
Gln−Val−Tyr−
Arg−Thr−Glu−
Phe−Ala−Gly−
Thr−Ala−Arg−
Arg−Pro−His−
Asn−Val−Cys−
Cys−Ile−Asp−
Tyr−Glu−Trp−
Arg−Trp−Asp−
His−Trp−Met−
Asn−Trp−Pro−
Phe−Asn−Trp−
Phe−Ser−Gly−
Gly−Gly−Trp−
Leu−Trp−Phe−
Gly−Ile−Pro−
Trp−Trp−Thr−
Leu−Gly−Trp−
Ser−Pro−Trp−
Arg−Gly−Trp−
からなる群から選択され、
Xaa5は、Ala、Thr、Met、Val、Arg、Glu、Asp、Ser、Gln、Pro、Gly、Phe及びLysからなる群から選択され、
Xaa14は、Glu、Lys、Arg、Asp、Ser、Ala、Asn、Gly、Pro及びGlnからなる群から選択され、
Xaa15は、Met、Phe、Ala、Met、Cys、Gln、Glu、Ala、Trp、Phe、Leu、Val、Tyr、Trpからなる群から選択され、
−Xaa(17−20)は、次の式:
−Xaa17−Xaa18−Cys−Xaa20
を有する4のアミノ酸のペプチドである、請求項6に記載のペプチドリガンド。
【請求項8】
−Xaa(17−20)が、
−Gln−Ala−Cys−Met(配列番号:102)
−Leu−Gln−Cys−Trp(配列番号:103)
−Met−Ser−Cys−Val(配列番号:104)
−Leu−Arg−Cys−Ile(配列番号:105)
−Gln−Ala−Cys−Leu(配列番号:106)
−Leu−Ser−Cys−Leu(配列番号:107)
−Ile−Gly−Cys−Leu(配列番号:108)
−Leu−Ala−Cys−Leu(配列番号:109)
−Leu−Ser−Cys−Ile(配列番号:110)
−Met−Asn−Cys−Leu(配列番号:111)
−Leu−Arg−Cys−Leu(配列番号:112)
−Leu−Lys−Cys−Leu(配列番号:113)
−Leu−Gly−Cys−Leu(配列番号:114)
−Leu−Asn−Cys−Ile(配列番号:115)
−Met−Gly−Cys−Leu(配列番号:116)及び
−Met−Ala−Cys−Leu(配列番号:117)
からなる群から選択される請求項7に記載のペプチドリガンド。
【請求項9】
−Xaa(17−20)が、次の式:
−Cys−Xaa18−Xaa19−Cys
を有する4のアミノ酸であり、ここでXaa18はアミノ酸であり、Xaa19はアミノ酸である、請求項6に記載のペプチドリガンド。
【請求項10】
−Xaa(17−20)が、
−Cys−Ala−Trp−Cys(配列番号:118)
−Cys−Ser−Trp−Cys(配列番号:119)
−Cys−Glu−Pro−Cys(配列番号:120)
−Cys−Asp−Trp−Cys(配列番号:121)
−Cys−Glu−Trp−Cys(配列番号:122)
−Cys−Asn−Trp−Cys(配列番号:123)及び
−Cys−Gly−Trp−Cys(配列番号:124)
からなる群から選択される請求項9に記載のペプチドリガンド。
【請求項11】
次の式:
Xaa(1−10)−Glu−Xaa12−Trp−Xaa14−Cys−Cys−Gly−Pro−Gly−Cys−Xaa21−Xaa22−Xaa23−Xaa(24−27)(配列番号:126)
を有し、ここで、Xaa(1−10)がないか、1から10のアミノ酸であり、
Xaa12はアミノ酸であり;
Xaa14はアミノ酸であり;
Xaa21はアミノ酸であり;
Xaa22はアミノ酸であり;
Xaa23はVal及びLeuからなる群から選択されるアミノ酸であり、
Xaa(24−27)がないか、1から4のアミノ酸である、請求項1に記載のペプチドリガンド。
【請求項12】
Xaa(1−10)が、次の式:
−Cys−Xaa(2−7)−Cys−Xaa9−Gly−
を有する10のアミノ酸のペプチドであり、
Xaa(24−27)が、次の式:
Xaa(24−26)−Cys
を有する4のアミノ酸のペプチドであり、ここで、Xaa(2−7)が6のアミノ酸ペプチドであり、Xaa(24−26)が3のアミノ酸のペプチドである請求項11に記載のペプチドリガンド。
【請求項13】
式B−Bを有するペプチドリガンドであって、Bが請求項11に記載のペプチドリガンドであり、ここで「−」は任意成分のリンカードメインであるペプチドリガンド。
【請求項14】
式A−Bを有するペプチドリガンドであって、Aが請求項2に記載のペプチドリガンドで、Bが請求項11に記載のペプチドリガンドであり、ここで「−」は任意成分のリンカードメインであるペプチドリガンド。
【請求項15】
Xaa(1−10)が次の式:
Cys−Xaa2−Trp−Val−Xaa5−Xaa6−Xaa7−Cys−Xaa9−Gly− (配列番号:127)
を有している請求項12に記載のペプチドリガンド。
【請求項16】
Xaa(1−10)が次の式:
Cys−Xaa2−Trp−Val−Xaa5−Xaa6−Xaa7−Cys−Xaa9−Gly− (配列番号:127)
を有し、ここで、Xaa2は、Ala及びSerからなる群から選択されるアミノ酸であり;
Xaa5は、Ser、Leu、Ala、Arg及びValからなる群から選択されるアミノ酸であり;
Xaa6は、Phe、Val及びLeuからなる群から選択されるアミノ酸であり、
Xaa7は、Asp、Gln、Tyr、Trp、Leu及びHisからなる群から選択されるアミノ酸であり;
Xaa9は、Gly、Phe及びLeuからなる群から選択されるアミノ酸であり、
る、請求項15に記載のペプチドリガンド。
【請求項17】
Xaa(1−10)が次の式:
Cys−Ser−Trp−Val−Leu−Xaa6−Xaa7−Cys−Gly−Gly− (配列番号:162)
を有している請求項16に記載のペプチドリガンド。
【請求項18】
Xaa(24−26)が次の式:
Xaa24−Xaa25−Xaa26
を有し、ここで、Xaa24は、Trp、Val、Gly及びAlaからなる群から選択されるアミノ酸であり;
Xaa25は、Asn、Lys、Asp、Glu及びHisからなる群から選択されるアミノ酸であり;
Xaa26は、Ala、Ser及びValからなる群から選択されるアミノ酸である、請求項17に記載のペプチドリガンド。
【請求項19】
Xaa(24−26)が次の式:
Xaa24−Xaa25−Xaa26
を有し、ここで、Xaa24は、Trp、Val、Gly及びAlaからなる群から選択されるアミノ酸であり;
Xaa25は、Asn、Lys、Asp、Glu及びHisからなる群から選択されるアミノ酸であり;
Xaa26は、Ala、Ser及びValからなる群から選択されるアミノ酸である、請求項17に記載のペプチドリガンド。
【請求項20】
Xaa24がValであり;
Xaa25がAsnであり、
Xaa26がAlaである、請求項19に記載のペプチドリガンド。
【請求項21】
(a)請求項1、4、11又は13の何れか1項に記載のペプチドリガンドと(b)免疫グロブリン定常領域配列を含むポリペプチド。
【請求項22】
リンカー配列を更に含んでなる請求項21に記載のポリペプチド。
【請求項23】
免疫グロブリン定常領域配列が、IgG重鎖の定常ドメインである請求項20又は21に記載のポリペプチド。
【請求項24】
上記定常領域配列が、免疫グロブリン重鎖のCH3ドメインを含む請求項22に記載のポリペプチド。
【請求項25】
免疫グロブリン重鎖のヒンジ領域を更に含む請求項24に記載のポリペプチド。
【請求項26】
上記定常領域配列が、免疫グロブリン重鎖の定常領域の少なくとも機能的に活性なヒンジ、CH2及びCH3ドメインを含む請求項25に記載のポリペプチド。
【請求項27】
更なる機能部分を更に含んでなる請求項21に記載のポリペプチド。
【請求項28】
更なる機能部分が細胞毒性剤と酵素からなる群から選択される請求項27に記載のポリペプチド。
【請求項29】
更なる機能部分が細胞毒性剤である請求項27に記載のポリペプチド。
【請求項30】
請求項1に記載のペプチドリガンドと製薬的に許容可能な賦形剤を含有する製薬組成物。
【請求項31】
請求項21に記載のポリペプチドと製薬的に許容可能な賦形剤を含有する製薬組成物。
【請求項32】
請求項28に記載のポリペプチドと製薬的に許容可能な賦形剤を含有する製薬組成物。
【請求項33】
請求項1に記載のペプチドリガンドをコードする単離されたDNA分子。
【請求項34】
DNA分子に作用可能に結合した発現コントロール配列を更に含んでなる請求項33に記載のDNA分子。
【請求項35】
コントロール配列がベクターで形質転換された宿主細胞によって認識される、請求項33のDNA分子を含んでなる発現ベクター。
【請求項36】
請求項35に記載のベクターで形質転換された宿主細胞。
【請求項37】
宿主細胞中でペプチドリガンドをコードするDNA分子を発現させる方法において、ペプチドリガンドの発現に適した条件下で請求項36に記載の宿主細胞を培養することを含んでなる方法。
【請求項38】
培地からペプチドリガンドを回収することを更に含む請求項37に記載の方法。
【請求項1】
インビトロアッセイにおいて、次の式:
Xaa(1−14)−Cys−Xaa16−Gly−Pro−Gly−Cys−Xaa(21−27)(配列番号:90)
を有し、ここで、Xaa(1−14)がないか、1から14のアミノ酸であり、Xaa16がMet、Thr、Cys及びIleからなる群から選択されるアミノ酸であり、Xaa(21−27)はないか、1から7のアミノ酸であるペプチドリガンドと、HER2への結合について競合する、非天然発生ペプチドリガンド。
【請求項2】
次の式:
Xaa(1−7)−Cys−Xaa9−Gly−Pro−Gly−Cys−Xaa(14−20)(配列番号:92)
を有し、ここで、Xaa(1−7)がないか、1から7のアミノ酸であり、Xaa9がMet、Ile、Thr又はCysからなる群から選択されるアミノ酸であり、Xaa(14−20)はないか、1から7のアミノ酸である、請求項1に記載のペプチドリガンド。
【請求項3】
次の式:
Xaa(1−7)−Cys−Ile−Gly−Pro−Gly−Cys−Xaa(14−20)(配列番号:161)
を有する請求項2に記載のペプチドリガンド。
【請求項4】
式A−Aを有するペプチドリガンドであって、ここでAが請求項3に記載のペプチドリガンドであり、「−」が任意成分のリンカードメインであるペプチドリガンド。
【請求項5】
次の式:
Xaa(1−5)−Trp−Gly−Cys−Ile−Gly−Pro−Gly−Cys−Xaa(14−20)(配列番号:93)
を有し、ここで、Xaa(1−5)がないか、1から5のアミノ酸である、請求項3に記載のペプチドリガンド。
【請求項6】
次の式:
Xaa(1−3)−Glu−Xaa5−Trp−Gly−Cys−Ile−Gly−Pro−Gly−Cys−Xaa14−Xaa15−Leu−Xaa(17−20)(配列番号:87)
を有し、ここで、Xaa(1−3)がないか、1から3のアミノ酸であり、Xaa5がアミノ酸であり、
Xaa5がアミノ酸であり、
Xaa14がアミノ酸であり、
Xaa19がアミノ酸であり、
−Xaa(17−20)がないか、1から4のアミノ酸である、請求項5に記載のペプチドリガンド。
【請求項7】
X(1−3)−がないか、
Gln−Arg−Asn−
Leu−Ser−Pro−
Glu−Asn−Trp−
Ala−Ser−His−
Lys−Leu−Asn−
Thr−Gln−Ala−
Ala−Pro−Arg−
Gln−Val−Tyr−
Arg−Thr−Glu−
Phe−Ala−Gly−
Thr−Ala−Arg−
Arg−Pro−His−
Asn−Val−Cys−
Cys−Ile−Asp−
Tyr−Glu−Trp−
Arg−Trp−Asp−
His−Trp−Met−
Asn−Trp−Pro−
Phe−Asn−Trp−
Phe−Ser−Gly−
Gly−Gly−Trp−
Leu−Trp−Phe−
Gly−Ile−Pro−
Trp−Trp−Thr−
Leu−Gly−Trp−
Ser−Pro−Trp−
Arg−Gly−Trp−
からなる群から選択され、
Xaa5は、Ala、Thr、Met、Val、Arg、Glu、Asp、Ser、Gln、Pro、Gly、Phe及びLysからなる群から選択され、
Xaa14は、Glu、Lys、Arg、Asp、Ser、Ala、Asn、Gly、Pro及びGlnからなる群から選択され、
Xaa15は、Met、Phe、Ala、Met、Cys、Gln、Glu、Ala、Trp、Phe、Leu、Val、Tyr、Trpからなる群から選択され、
−Xaa(17−20)は、次の式:
−Xaa17−Xaa18−Cys−Xaa20
を有する4のアミノ酸のペプチドである、請求項6に記載のペプチドリガンド。
【請求項8】
−Xaa(17−20)が、
−Gln−Ala−Cys−Met(配列番号:102)
−Leu−Gln−Cys−Trp(配列番号:103)
−Met−Ser−Cys−Val(配列番号:104)
−Leu−Arg−Cys−Ile(配列番号:105)
−Gln−Ala−Cys−Leu(配列番号:106)
−Leu−Ser−Cys−Leu(配列番号:107)
−Ile−Gly−Cys−Leu(配列番号:108)
−Leu−Ala−Cys−Leu(配列番号:109)
−Leu−Ser−Cys−Ile(配列番号:110)
−Met−Asn−Cys−Leu(配列番号:111)
−Leu−Arg−Cys−Leu(配列番号:112)
−Leu−Lys−Cys−Leu(配列番号:113)
−Leu−Gly−Cys−Leu(配列番号:114)
−Leu−Asn−Cys−Ile(配列番号:115)
−Met−Gly−Cys−Leu(配列番号:116)及び
−Met−Ala−Cys−Leu(配列番号:117)
からなる群から選択される請求項7に記載のペプチドリガンド。
【請求項9】
−Xaa(17−20)が、次の式:
−Cys−Xaa18−Xaa19−Cys
を有する4のアミノ酸であり、ここでXaa18はアミノ酸であり、Xaa19はアミノ酸である、請求項6に記載のペプチドリガンド。
【請求項10】
−Xaa(17−20)が、
−Cys−Ala−Trp−Cys(配列番号:118)
−Cys−Ser−Trp−Cys(配列番号:119)
−Cys−Glu−Pro−Cys(配列番号:120)
−Cys−Asp−Trp−Cys(配列番号:121)
−Cys−Glu−Trp−Cys(配列番号:122)
−Cys−Asn−Trp−Cys(配列番号:123)及び
−Cys−Gly−Trp−Cys(配列番号:124)
からなる群から選択される請求項9に記載のペプチドリガンド。
【請求項11】
次の式:
Xaa(1−10)−Glu−Xaa12−Trp−Xaa14−Cys−Cys−Gly−Pro−Gly−Cys−Xaa21−Xaa22−Xaa23−Xaa(24−27)(配列番号:126)
を有し、ここで、Xaa(1−10)がないか、1から10のアミノ酸であり、
Xaa12はアミノ酸であり;
Xaa14はアミノ酸であり;
Xaa21はアミノ酸であり;
Xaa22はアミノ酸であり;
Xaa23はVal及びLeuからなる群から選択されるアミノ酸であり、
Xaa(24−27)がないか、1から4のアミノ酸である、請求項1に記載のペプチドリガンド。
【請求項12】
Xaa(1−10)が、次の式:
−Cys−Xaa(2−7)−Cys−Xaa9−Gly−
を有する10のアミノ酸のペプチドであり、
Xaa(24−27)が、次の式:
Xaa(24−26)−Cys
を有する4のアミノ酸のペプチドであり、ここで、Xaa(2−7)が6のアミノ酸ペプチドであり、Xaa(24−26)が3のアミノ酸のペプチドである請求項11に記載のペプチドリガンド。
【請求項13】
式B−Bを有するペプチドリガンドであって、Bが請求項11に記載のペプチドリガンドであり、ここで「−」は任意成分のリンカードメインであるペプチドリガンド。
【請求項14】
式A−Bを有するペプチドリガンドであって、Aが請求項2に記載のペプチドリガンドで、Bが請求項11に記載のペプチドリガンドであり、ここで「−」は任意成分のリンカードメインであるペプチドリガンド。
【請求項15】
Xaa(1−10)が次の式:
Cys−Xaa2−Trp−Val−Xaa5−Xaa6−Xaa7−Cys−Xaa9−Gly− (配列番号:127)
を有している請求項12に記載のペプチドリガンド。
【請求項16】
Xaa(1−10)が次の式:
Cys−Xaa2−Trp−Val−Xaa5−Xaa6−Xaa7−Cys−Xaa9−Gly− (配列番号:127)
を有し、ここで、Xaa2は、Ala及びSerからなる群から選択されるアミノ酸であり;
Xaa5は、Ser、Leu、Ala、Arg及びValからなる群から選択されるアミノ酸であり;
Xaa6は、Phe、Val及びLeuからなる群から選択されるアミノ酸であり、
Xaa7は、Asp、Gln、Tyr、Trp、Leu及びHisからなる群から選択されるアミノ酸であり;
Xaa9は、Gly、Phe及びLeuからなる群から選択されるアミノ酸であり、
る、請求項15に記載のペプチドリガンド。
【請求項17】
Xaa(1−10)が次の式:
Cys−Ser−Trp−Val−Leu−Xaa6−Xaa7−Cys−Gly−Gly− (配列番号:162)
を有している請求項16に記載のペプチドリガンド。
【請求項18】
Xaa(24−26)が次の式:
Xaa24−Xaa25−Xaa26
を有し、ここで、Xaa24は、Trp、Val、Gly及びAlaからなる群から選択されるアミノ酸であり;
Xaa25は、Asn、Lys、Asp、Glu及びHisからなる群から選択されるアミノ酸であり;
Xaa26は、Ala、Ser及びValからなる群から選択されるアミノ酸である、請求項17に記載のペプチドリガンド。
【請求項19】
Xaa(24−26)が次の式:
Xaa24−Xaa25−Xaa26
を有し、ここで、Xaa24は、Trp、Val、Gly及びAlaからなる群から選択されるアミノ酸であり;
Xaa25は、Asn、Lys、Asp、Glu及びHisからなる群から選択されるアミノ酸であり;
Xaa26は、Ala、Ser及びValからなる群から選択されるアミノ酸である、請求項17に記載のペプチドリガンド。
【請求項20】
Xaa24がValであり;
Xaa25がAsnであり、
Xaa26がAlaである、請求項19に記載のペプチドリガンド。
【請求項21】
(a)請求項1、4、11又は13の何れか1項に記載のペプチドリガンドと(b)免疫グロブリン定常領域配列を含むポリペプチド。
【請求項22】
リンカー配列を更に含んでなる請求項21に記載のポリペプチド。
【請求項23】
免疫グロブリン定常領域配列が、IgG重鎖の定常ドメインである請求項20又は21に記載のポリペプチド。
【請求項24】
上記定常領域配列が、免疫グロブリン重鎖のCH3ドメインを含む請求項22に記載のポリペプチド。
【請求項25】
免疫グロブリン重鎖のヒンジ領域を更に含む請求項24に記載のポリペプチド。
【請求項26】
上記定常領域配列が、免疫グロブリン重鎖の定常領域の少なくとも機能的に活性なヒンジ、CH2及びCH3ドメインを含む請求項25に記載のポリペプチド。
【請求項27】
更なる機能部分を更に含んでなる請求項21に記載のポリペプチド。
【請求項28】
更なる機能部分が細胞毒性剤と酵素からなる群から選択される請求項27に記載のポリペプチド。
【請求項29】
更なる機能部分が細胞毒性剤である請求項27に記載のポリペプチド。
【請求項30】
請求項1に記載のペプチドリガンドと製薬的に許容可能な賦形剤を含有する製薬組成物。
【請求項31】
請求項21に記載のポリペプチドと製薬的に許容可能な賦形剤を含有する製薬組成物。
【請求項32】
請求項28に記載のポリペプチドと製薬的に許容可能な賦形剤を含有する製薬組成物。
【請求項33】
請求項1に記載のペプチドリガンドをコードする単離されたDNA分子。
【請求項34】
DNA分子に作用可能に結合した発現コントロール配列を更に含んでなる請求項33に記載のDNA分子。
【請求項35】
コントロール配列がベクターで形質転換された宿主細胞によって認識される、請求項33のDNA分子を含んでなる発現ベクター。
【請求項36】
請求項35に記載のベクターで形質転換された宿主細胞。
【請求項37】
宿主細胞中でペプチドリガンドをコードするDNA分子を発現させる方法において、ペプチドリガンドの発現に適した条件下で請求項36に記載の宿主細胞を培養することを含んでなる方法。
【請求項38】
培地からペプチドリガンドを回収することを更に含む請求項37に記載の方法。
【図1】

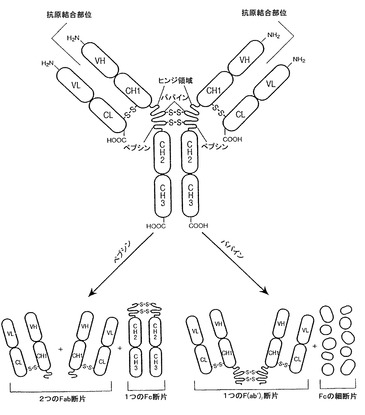
【図2A】

【図2B】

【図3】
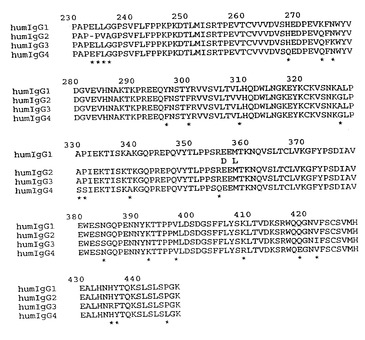
【図4】
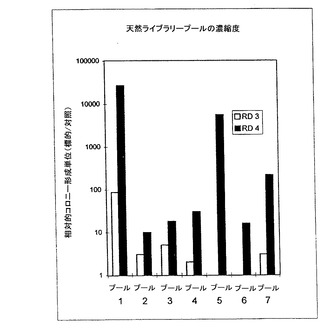
【図5】
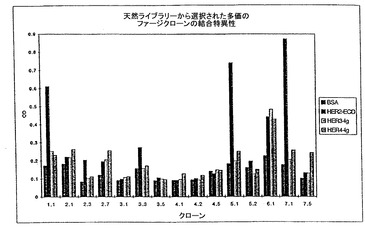
【図6】
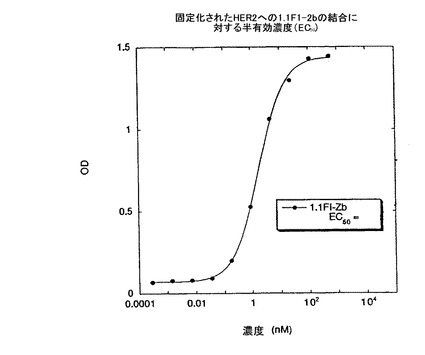
【図7】
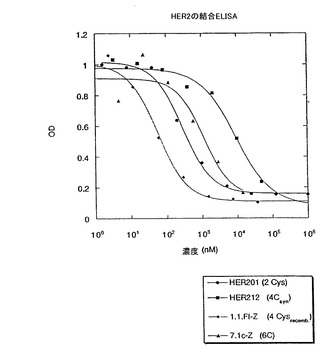
【図8】
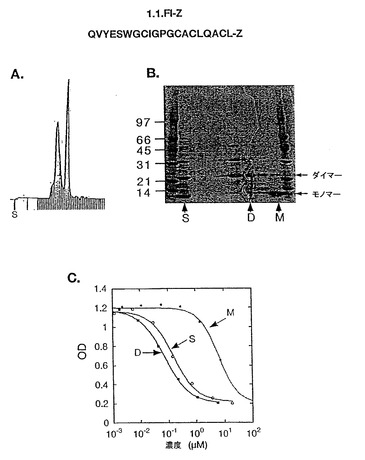
【図9】
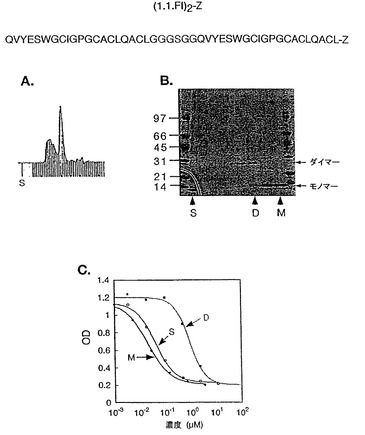
【図10】
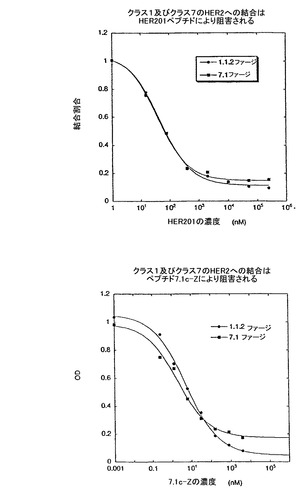
【図11】

【図12】
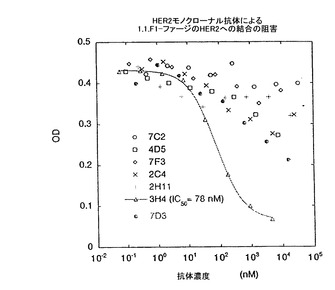
【図13】
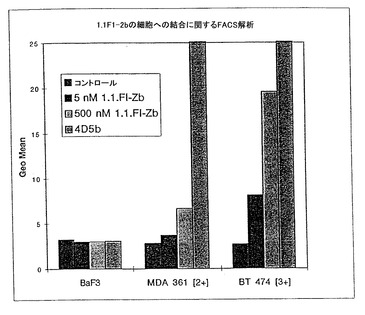
【図14】
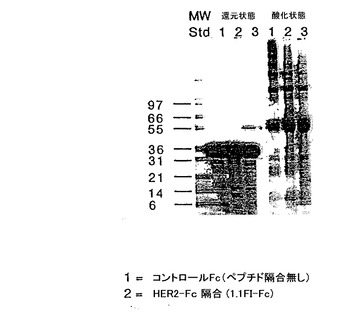
【図15】

【図16】

【図17】
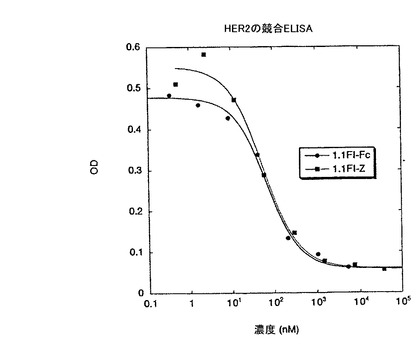
【図2A】
【図2B】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【公開番号】特開2010−4885(P2010−4885A)
【公開日】平成22年1月14日(2010.1.14)
【国際特許分類】
【外国語出願】
【出願番号】特願2009−171647(P2009−171647)
【出願日】平成21年7月22日(2009.7.22)
【分割の表示】特願2001−507269(P2001−507269)の分割
【原出願日】平成12年6月30日(2000.6.30)
【出願人】(596168317)ジェネンテック・インコーポレーテッド (372)
【氏名又は名称原語表記】GENENTECH,INC.
【Fターム(参考)】
【公開日】平成22年1月14日(2010.1.14)
【国際特許分類】
【出願番号】特願2009−171647(P2009−171647)
【出願日】平成21年7月22日(2009.7.22)
【分割の表示】特願2001−507269(P2001−507269)の分割
【原出願日】平成12年6月30日(2000.6.30)
【出願人】(596168317)ジェネンテック・インコーポレーテッド (372)
【氏名又は名称原語表記】GENENTECH,INC.
【Fターム(参考)】
[ Back to top ]