
RNAの修飾とRNAからDNAを調製する方法
【課題】 本発明は一つ又は複数のRNA分子の5’末端の塩基配列情報を得るためのそれら分子の修飾に関する。
【解決手段】 RNA分子の末端を修飾する手段と、そのような修飾されたRNA分子を、修飾されたRNA分子に由来する任意のcDNA、すべてのRNA、又は一部のRNAのクローニング、検出、塩基配列決定、増幅を目的としたcDNAの合成に用いる手段を開示する。さらに本発明は、一分子検出およびハイスループット塩基配列決定を目的とした核酸分子の増幅と同定に関係するものである。加えて、 「Identifier配列」によってその由来を印付けられた各々の分子を含むプールサンプルを調製する手段を提供する。本発明は、生物学的システムとそこで発現する遺伝子の解析研究を促進する。
【解決手段】 RNA分子の末端を修飾する手段と、そのような修飾されたRNA分子を、修飾されたRNA分子に由来する任意のcDNA、すべてのRNA、又は一部のRNAのクローニング、検出、塩基配列決定、増幅を目的としたcDNAの合成に用いる手段を開示する。さらに本発明は、一分子検出およびハイスループット塩基配列決定を目的とした核酸分子の増幅と同定に関係するものである。加えて、 「Identifier配列」によってその由来を印付けられた各々の分子を含むプールサンプルを調製する手段を提供する。本発明は、生物学的システムとそこで発現する遺伝子の解析研究を促進する。
【発明の詳細な説明】
【技術分野】
【0001】
ゲノムは、生物の発生や恒常性の維持に必要不可欠な情報を含んでいる。生物学的現象を理解するためには、ある時点の細胞や組織で遺伝情報がどのように利用されているかを知ることが必要である。ヒト、動物または植物において、遺伝情報や関係した制御パスウェイが誤った形で使われることが疾病の原因となる例が数多く知られている。RNAの発現は、個々の細胞、組織又は生命体によって非常に多様である。そのため、限られた量の細胞からRNAとDNA分子を取り出して調製できる新しい方法の開発が期待されており、このような方法が開発されれば、組織の中の個々の細胞のRNA発現、プロモーター利用及び発現しているゲノム情報を解析することができる。ライフサイエンスの新しい方向は、このような要求に取り組んでいる。個々のDNAおよびRNA分子を捕捉、解析し、遺伝子ネットワーク研究のような生物学的システム全体を理解するための技術の開発が進められている。
【背景技術】
【0002】
[遺伝子発現解析]
発現プロファイリングや転写物の注釈付けには、種々の方法が用いられている。簡単に言うと、最近の大規模発現研究にはマイクロアレイのようなin situハイブリダイゼーションによる方法、あるいはSAGE、CAGE、MPSSなどの短いタグの高速塩基配列による方法が用いられている。このような研究では、個々の遺伝子の発現レベルを調べるために、さらにノーザンブロッティングやRT−PCRなどの従来の方法が組み合わせて用いられる。
【0003】
大規模な発現プロファイリングには、通常、DNAマイクロアレイ(Jordan B., DNA Microarrays: Gene Expression Applications,Springer-Verlag,Berlin Heidelberg New York、 2001: Schena A,DNA Microarrays,A Practical Approach,Oxford University Press,Oxford 1999、これらは、参照することによって本明細書に組み込まれるものとする) と呼ばれる技術が用いられている。この実験では、個々の遺伝子や転写物に対応するプローブを支持体上に固定し、そこに種々のDNA分子をハイブリダイズさせて行なわれる。ポジティブ信号は、支持体上に固定されたプローブとサンプル中の分子が反応した場合に得られる。これらの実験は、多数の遺伝子又は転写物を同時並行的に解析できるが、その反面、解析の対象が、他の実験や手段によって同定された遺伝子や転写物に限られるという制限がある。このような実験や手段は、cDNAライブラリーや部分的に塩基配列がわかっているタグ、およびコンピューターによる予測結果を含む。将来的には、タイリングアレイによって、ゲノムの塩基配列が判っている生物の偏りのない発現プロファイリングが可能となる(Kapranov P. et al.,Science 296,916−919,2002、本文献は、参照することによって本明細書に組み込まれるものとする)であろうが、しかし、実験から得られる転写物の性質についてはタイリングアレイでの解釈は困難である。
【0004】
DNAマイクロアレイによる解析が持つ問題を解決するために、複数のRNAサンプルより得られる部分シーケンスやタグをベースとした発現プロファイリングや遺伝子探索が行われている。いわゆるSAGE(Serial Analysis of Gene Expression)法は、mRNAの塩基配列から効率的に情報を得る方法として知られている(Velculescu V.E. et at.,Science 270,484−487,1995、本文献は、参照することによって本明細書に組み込まれるものとする)。タグの塩基配列決定をハイスループットに行うために、この方法では、複数のRNAサンプルの3’末端の塩基配列情報を含む短いDNAフラグメント(当初は約10塩基対)をつなぎ合わせてコンカテマーを作る。このコンカテマーの一回の塩基配列解析により、DNAコンカテマーに含まれる異なるRNA分子から得られるたくさんのタグの塩基配列を決定することができる。最近、SAGE法の一つであるLong SAGE法が発表され、長いタグをクローニングすることが可能となった(Saha S. et al.,Nat. Biotechnol.20,508−12,2002、米国特許出願20030008290号、米国特許出願20030049653号、これらは、参照することによって本明細書に組み込まれるものとする)。この概念は「Super SAGE」と呼ばれる方法に発展し、25塩基対程度のタグの塩基配列を得ることができる(Matsumura,H et al.,Cell.Microbiol.7,11−18,2005、本文献は、参照することによって本明細書に組み込まれるものとする)。SAGE法は現在、生物、組織、特定の細胞などの遺伝子発現を解析する重要な手法として広く用いられており、このタグの情報は例えばhttp://cgap.nci.nih.gov/SAGEで公開されている。最近のSAGE研究の発展に関する情報は、Wang,S.M.:SAGE:Current Technologies and Applications,Horizon Bioscience,Norwich(2005)(本文献は、参照することによって本明細書に組み込まれるものとする)で参照できる。
【0005】
米国特許第6,352,828号、米国特許第6,306,597号、米国特許第6,280,935号、米国特許第6,265,163号、米国特許第5,695,934号(これらは、参照することによって本明細書に組み込まれるものとする)では、短いタグ配列を大規模に塩基配列決定する別の方法が開示されている。Massively Parallerl Signature Sequencing(MPSS)と呼ばれるこの方法は、Brennerらによれば(Brenner S.,et al.,Nat. Biotechnol. 18,630−634,2000、Brenner S.,et al.,Proc. Natl. Acad. Sci. USA 97,1655−1670,2000、これらは、参照することによって本明細書に組み込まれるものとする)、単層のビーズ上で異なる酵素を使ってサイクル反応を行ない、転写物の3’末端の短い塩基配列を超並列に得る方法である。WO03/091416(本文献は、参照することによって本明細書に組み込まれるものとする)では、この方法の改良が開示されており、転写物の5’末端の短い塩基配列を決定することが出来るようになった。しかし、その初期の様態では、MPSSはシグニチャータグの解読塩基長の短さのため限定された方法である。
【0006】
3’末端塩基配列情報から5’末端塩基配列情報へのシフトは、それが技術的にはより難度が高いものであるが、発現情報と転写の調製原理を結びつけるために不可欠なものである。遺伝子発現の制御をする一般的な調節因子は転写物の5’末端近くに存在し、その遺伝子のプロモーターと呼ばれている。選択的プロモーターの利用や、RNAプロセッシング、スプライシングによる一次転写物の再構成により、高等動物では大部分の転写物のプロモーター領域を3’末端の情報から特定することは出来ない。従って、転写物の5’末端から特異的に配列タグを得る新しいアプローチが開発されてきたのである。そのようなアプローチはPCT/JP03/07514、Shiraki T.et al.,Prog.Natl.Acad.Sci.USA 100,15776−15781(2003)、Kozdius R.et al.,Nature Method 3,211−222(2006)、米国特許出願番号20050250100号(これらは、参照することによって本明細書に組み込まれるものとする)に開示されている。この方法はCAGE(Cap Analysis Gene Expression)法とよばれ、転写物の5’末端から特異的に得たタグ配列をSAGE法と同様にコンカテマーにしてクローニングする方法である。この方法により得られるCAGEタグは、転写物の検出や発現プロファイリングだけでなく、転写開始点の情報を得るために利用され、転写調節のメカニズム解析や転写物の高度な注釈付けを行なうことができる。同様の5’末端の塩基配列情報をコンカテマーにしてクローニングするアプローチが、最近いくつもの研究室より発表されている。例えば、Hwang B.J. et al.,Proc.Natl.Acad.Sci.USA 101,1650−1655(2004);Hashimoto S.et al.,Nat.Biotechnol.22,1146−1149(2004);Zhang Z.and Dietrich F.S.,Nuc. Acids Res.33,2838−2851(2005);Wei C.L. et al.Proc.Natl.Acad.Sci.USA 101,11701−11706(2004)(これらの文献は、参照することによって本明細書に組み込まれるものとする)などが挙げられる。これらのアプローチは、すべて5’末端を捕獲する方法に特徴がある。例えば、Cap−Trapper又はOligo−Cappingなどの方法である。5’末端タグの意義に関する詳細は、Harbers M.and Carninci P.,Nature Methods.2,495−502(2005)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。
【0007】
前述のアプローチは、タグの塩基配列決定の処理量の点で今もなお制限がある。さらに、それらのアプローチは、多くの操作を必要とするためコンカテマーより得られる塩基配列情報に間違いが起きる可能性がある。特に、増幅のステップにおいて、個々のDNA断片の異なる増幅効率に起因するタグ頻度のバイアスや人為的な影響が発生する可能性がある。これらの制限を解決するために、不必要に複雑な操作やクローニングステップを省くためのDNAおよびRNA分子の直接的捕獲と、よりハイスループットなデータ収集とを目的としたさらなる開発が必要とされている。
【0008】
[塩基配列決定技術]
遺伝コードの読み取りと解読はライフサイエンスの最も大きな発展の一つである。塩基配列決定技術は遺伝情報を得るための重要なキーとなってきている。当業者が知り得る塩基配列情報を得る手段としては、Sanger法やMaxim−Gilbert法のようなSambrook J. and Russuell D.W.,Molecular Cloning,A Laboratory Manual,Cold Spring Harbor Laboratory Press,New York(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載された方法が挙げられるが、これに限定するものではない。近年、ハイスループット塩基配列決定の新しい技術が開発された。電気泳動、ハイブリダイゼーションによる塩基配列決定、合成による塩基配列決定、MPSS塩基配列決定、非酵素的一分子塩基配列決定などの利用が挙げられる。例えば、ハイブリダイゼーションによる塩基配列決定では、決められた塩基配列のオリゴヌクレオチドセットが載っている高密度マイクロアレイを用い、ハイブリダイゼーションのパターンによって未知配列の新たな塩基配列決定を行うことができる。例えば、Perlegenは、このような方法をヒトゲノム中の点突然変異解析に用いた。また、合成による塩基配列決定方法は、鋳型DNAに沿ってDNA分子が伸長する間にポリメラーゼがヌクレオチドを結合させることによって行われる。表面上で決められた位置に個々のヌクレオチドが結合するのをモニターし、さらにそれぞれの位置で引き続き結合するヌクレオチドを順番に見ることで、それぞれの位置にある核酸分子の塩基配列を決定する。非常に大量の伸長反応がひとつの反応器のなかで起こるため、例えばビーズアレイのガラススライド表面上では、使用する機器によって100、000から1、000、000種のサンプルを、一反応で並行的に塩基配列決定にかけることができる。とりわけ興味深いのは、本発明のDNAの一分子検出と塩基配列決定への応用であり、最近、Metzker M.L.,Genome Res.15,1767−1776(2005); Kling J.,Nature Biotechnology 23,1333−1335(2005); Shendure J. et al.,Nature Review Genetics 5,335−344(2004)(これらの文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。これらアプローチのうちいくつかは、454 Life Sciences(http://www.454.com/)、 Helicos BioScience(http://www.helicosbio.com/)、Solexa(http://www.solexa.com/Company/overview.htm)、Visigen(http://www.visigenbio.com/index.html)、GeneoVoxx GmbH(http://www.genovoxx.de/)(これらは、参照することによって本明細書に組み込まれるものとする)のような企業により商業的に利用されている。このような新しい技術は、現在の塩基配列決定速度を飛躍的にあげるであろう。使用する方法によっては、20から25塩基対の非常に短いタグ配列を、1ランあたり1,000,000種以上読むことができるという超ハイスループットの機器(例えば、Helicos、Solexaなど)もある。また、454Life Sciences社の機器ははるかに長い100塩基対以上の解読長を1ランあたり200,000から300,000種解読することができる。
【0009】
[完全長cDNA分子の単離]
生物サンプルのRNAを解析では、完全長RNA分子を単離し、そのRNAからcDNAを調製することにより大きな利益を得ることができる。特に、前述のようにRNAの5’末端の塩基配列情報は、データの幅広い解析を可能とする。さらに、完全長cDNA技術はRNAのプロセッシングやスプライシングに関係したプロセスを解析するためには、必須である。最近の種々の実験より得られたデータは、高等動物の複雑性がゲノム情報の選択的な利用によるものであることを示している。転写情報における組み合わせの再構成とプロセッシングのメカニズムは、遺伝プールの拡大と多様化に重要である(Zavolan M.,et al.,Genome Res. 13,2003,1290−1300、本文献は、参照することによって本明細書に組み込まれるものとする)。
【0010】
完全長cDNAを調製する種々の方法は、Das,M.,et al.,Physiol Genomics 6,57−80(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。これらの中から、Cap−Trapper法とOligo−Capping法は、当業者が知り得る他の方法の中でも最も頻繁に使用されている。これらのアプローチは、ゲノム、転写物の構造、タンパク質配列の解析の手段として利用されてきた。さらに、完全長cDNAだけがそれにコードされたタンパク質へとつながる手段であり、実験において必要とされるとき、あるいは産業応用に際して、機能的ベクターを使ってタンパク質を発現させることができる。
【0011】
Cap−Trapper法(Carninci P.and Hayashizaki Y.,Methods Enzymol. 303,19−44,1999;米国特許第5,962,272号、米国特許第6,022,715号、これらの文献は、参照することによって本明細書に組み込まれるものとする)では、Cap構造のジオール基を化学的にビオチン化し、mRNAとcDNAの複合体をストレプトアビジンでコートしたビーズで捕獲する。残っているtRNA、およびRNAタンパク質などの一本鎖RNAと部分的にDNA−RNAがハイブリダイズしている複合体は、RNaseI分解によって破壊されるが、mRNAとcDNAの複合体は分解されない。その後、濃縮された完全長cDNAはRNAの加水分解によってビーズより遊離する。
【0012】
代替法となるOligo−Capping法では、まずmRNAの5’末端を修飾する(Maruyama K.and Sugano S.,Gene 138,171−174,1994;Suzuki Y.Sugano S.,Methods Mol Biol.221,73−91,2003、これらの文献は、参照することによって本明細書に組み込まれるものとする)。最初のステップで、CapされていないmRNAと切断されているmRNAより脱リン酸化酵素によってリン酸基を除去し、次にタバコ酸ピロホスファターゼ処理によってmRNAのCap構造を取り除く。この処理により、完全長mRNAのみが、リン酸化された5’末端を持つことになる。そのため、RNAリガーゼによりリン酸化された完全長mRNAの5’末端にのみ、オリゴヌクレオチドを結合させることができる。mRNAの5’末端に結合したオリゴヌクレオチドは、例えば第二鎖cDNAの合成など、その後mRNAに由来するcDNAの操作に利用することができる。
【0013】
完全長cDNAを選択するその他の方法には、Cap−binding proteinを利用する方法(Edery,I.,et al.,Mol. Cell Biol. 15,3363−3371,1995、本文献は、参照することによって本明細書に組み込まれるものとする)、Cap構造に対する抗体を利用する方法(Theissen,H.,et al.,Embo J 5,3209−3217(1986)、本文献は、参照することによって本明細書に組み込まれるものとする)、Cap構造にオリゴヌクレオチドを結合させる方法(米国特許第5,962,272号、米国特許第6,022,715号、これらの文献は、参照することによって本明細書に組み込まれるものとする)、Clontech社(http://www.clontech.com/clontech/smart/index.shtml、ここに記載された情報は、参照することによって本明細書に組み込まれるものとする)のSMART(商標)法などがある。しかし、Cap−Switching法(Zhu Y. et al.,Biotechniques 30,892−897,2001、本文献は、参照することによって本明細書に組み込まれるものとする)を使うSMART(商標)法は、mRNAの5’末端にGGGという3つのヌクレオチドを追加するので、タグ部分の情報の長さを短縮させることになり、特に、塩基配列決定の場合には、解読長が非常に短いものとなってしまうため、5’末端タグのクローニングには好ましくない。
【発明の開示】
【0014】
本発明は、一つ又は複数のRNA分子の5’末端の塩基配列情報を得るためのRNA分子の修飾に関するものである。本発明は、RNA分子の修飾に関するものであり、RNA分子に加えられた情報は解析および操作に用いられる。
【0015】
本発明は、RNAおよびDNA分子、又はそれらの断片を取り出し一分子検出と塩基配列決定を行う手段を提供する。本発明は、新しい一分子検出のアプローチに必要な個々の分子を捕獲する手段を提供する。本発明は、RNA分子およびそれに由来するDNA分子を、それら分子の特定の領域の塩基配列情報を得るために修飾する。従って、本発明は新しいハイスループット塩基配列決定方法とその応用手段、例えば発現プロファイリング、転写物特性解析、ゲノム注釈付、さらに解析するためのクローニング、その他従来の方法などへの応用手段を提供する。特に、本発明は重要な研究に手段を提供する。重要な研究とは、例えば5’末端特異的塩基配列に基づいた発現プロファイリングであり、これは法医学、診断、創薬、ライフサイエンスを含むが、これに限定されない商業的応用、試薬、サービスには不可欠な要素である。
【0016】
一つの実施態様では、本発明は、天然又は人工的な一つ又は複数のRNA分子をcDNAに転写的に転換することに関係する。よって、本発明は一本鎖DNA分子の合成と調製に関係するものであり、検出と解析を目的とした核酸分子の断片の単離方法に関係するものである。さらに、本発明は単一又は複数の種類の核酸分子を含むRNAサンプルのDNA分子への転換に関係するものである。
【0017】
別の実施態様において、本発明は核酸分子を直線状一本鎖DNAの形に調製するために核酸分子を操作する方法に関係するものであり、また、RNAから複写された直線状一本鎖DNAの操作、調製に関係するものである。
【0018】
別の実施態様において、本発明はRNA分子又はDNA分子の末端に官能基を付加する手段を提供する。よって本発明は官能基を用いて、操作と解析のためにRNAとDNA分子を捕獲する手段を提供する。この実施態様において、本発明は解析、検出、塩基配列決定を目的とした核酸一分子の単離に関係するものであり、一分子検出とハイスループット塩基配列決定の鋳型調製の手段を提供する。
【0019】
別の実施態様において、本発明は一本鎖DNA分子を直接的にその塩基配列情報を得るために用いる方法に関係するものであり、また一本鎖DNA断片の特定した領域の塩基配列情報を得ることに関係するものである。ある実施態様において、本発明によって調製されたDNA断片から得られた5’末端特異的塩基配列情報は、RNA分子の5’末端塩基配列と関係するものである。このように本発明は、RNA分子の塩基配列情報の取得に関係するものである。
【0020】
別の実施態様において、本発明は、RNAに由来するDNA分子の反対側の末端を修飾する手段を提供する。この場合、RNAの5’末端に対応する末端およびRNAの3’末端に対応する末端への修飾は、官能基または後のDNA分子の操作、検出及び解析のための機能を持った基の導入である。一つの実施態様においては、RNAの3’末端に対応する末端に官能基が導入され、それを表面上に結合させることによりcDNAを捕獲する。一つの実施態様においては、本発明に沿って調製されたDNA断片より得られた3’末端特異的な塩基配列情報は、RNAまたはDNA分子の末端塩基配列に関係する。別の実施態様においては、cDNA分子がLAMP法またはローリングサークル法によって増幅され得るようにcDNA分子の両末端が修飾される。
【0021】
別の実施態様において、本発明は、RNAの5’末端にある「Identifier配列」と呼ばれる、規定された塩基配列の領域を導入する手段を提供する。そのようなIdentifier配列は、分子の由来を特定する。従って、Identifier配列の導入により、由来の異なるサンプルからなるサンプルプールを解析することが可能となる。そのような異なる由来は、用いられる異なる細胞系、組織、器官または異なる発生段階、実験上の多様な時点などに関係する。別の実施態様において、Identifier配列は、本発明に従って調製された分子から得られる塩基配列の一部分である。別の実施態様において、Identifier配列は、この配列を持つ分子を検出、解析または塩基配列決定を目的として表面上の特定の位置に捕獲するために用いられる。別の実施態様において、Identifier配列は、プールされたサンプルの複数の分子の中から一つの分子の塩基配列決定をプライミングするために用いられる。前述の実施態様と同様に、本発明は、Identifier配列をRNAの5’末端だけでなくRNAの3’末端に相当するcDNAの領域に導入する手段を提供する。Identifier配列の利用は、RNAの5’末端に限定されることなく、実験的な必要性に応じてRNAに由来するcDNAのいずれの末端でも用いることができる。
【0022】
本発明は、統計的解析を含むコンピューターによる注釈付や参照情報に対するアライメントによる注釈付、およびゲノム配列に対するマッピングなどを目的とし、本発明に従って得られたDNA断片のある領域の塩基配列決定に関係するものである。従って、本発明は、遺伝子探索、遺伝子同定、遺伝子発現プロファイル、およびそれらの注釈付の手段に関係するものである。
【0023】
別の実施態様において、本発明は、本発明に沿って得られたDNA断片の塩基配列決定に関係し、それらの配列がゲノムの領域に関するものである場合には、その配列のコンピューターを用いた注釈付を可能にする。またIdentifier配列の解読、塩基配列の統計的解析を可能とする。よって本発明は、転写開始位置に関連して遺伝的要素の特性解析に関係するものである。
【0024】
さらに別の実施態様において、本発明は核酸の末端より得られるハイブリダイゼーションプローブの調製に関係するものであり、その領域はin situハイブリダイゼーションを用いて解析される。好ましい実施態様においては、そのin situハイブリダイゼーションの実験は、タイリングアレイを用いて行われる。
【0025】
また実施態様において、本発明は、本発明に従って得られたDNA断片の塩基配列情報が増幅される方法を用いた、核酸分子の完全長クローニングに関係するものである。ゲノム断片と同様に転写された領域のクローニングや増幅も本発明に含まれる。そのような断片は、プロモーター領域を含んでも良い。
【0026】
従って、本発明は、例えば生物学的サンプルの特性解析のような必要に応じて、核酸分子やその短い断片を解析する手段を提供する。さらに、本発明は、迅速で効率的なRNAやDNA断片の解析および塩基配列解析を行い、それら断片の一分子検出のような分析的検定を行う手段を提供する。よって、本発明は、特にハイスループット塩基配列決定や固体支持体上でのRNA分子又はDNA分子の並列的検出に適している。
【0027】
特にある実施態様では、本発明は環状一本鎖DNAに転換可能なDNAの修飾による双方向的な鋳型構築に関係するものである。RCA反応により環状一本鎖DNA分子の増幅を行った後、双方向性の直線状一本鎖DNA分子を得て、固体支持体上の規定された位置に直接結合させることができる。本発明は、同じ鋳型から複数の解読塩基配列を得ることを可能とするもので、規定された位置により種々の解読配列を同じ鋳型に関連付けることができる。双方向的直線状一本鎖DNA分子を鋳型として用いることにより、修飾されたDNA分子の両鎖又はRNA分子の両端の配列情報を同じ鋳型から得ることができる。
【0028】
本発明は、ライフサイエンス研究や診断に利用する分析的検定をデザインし実行する手段を提供する。それゆえ、本発明は生物学的システムの解析又は診断の手段に関係するものである。
【0029】
本発明はまた、実験的な条件を充足させるため必要とされる本発明の部分的、又は全部の実行のための試薬やキットの製造およびデザインの手段を提供する。
【発明を実施するための最良の形態】
【0030】
本発明は、直線状と環状の核酸分子の形状である二本鎖核酸と一本鎖核酸の処理を行う方法を含む。二本鎖DNAとは、デオキシヌクレオチドから構成される二つの分子から成り、二つの分子が、実質的に互いに相補的な配列を持ち二量体を形成するように対合する任意の核酸分子を含む。二つのポリマーは、デオキシヌクレオチドの対応する塩基対の間で特有の水素結合によって結合している。二つもしくはそれ以上のデオキシリボヌクレオチドにより形成される一つのポリマー鎖で構成され、対合する相補的なDNA分子がない任意のDNA分子も、本発明の目的によれば一本鎖DNA分子とみなされる。そのような分子が二次元構造を形成し、二本鎖DNAの部分を含んでいる場合も同様である。互換性をもって用いられる「核酸分子」や「ポリヌクレオチド」の語句は一本鎖であるか二本鎖であるか、コーディングがノンコーディングか、相補的かそうでないか、センスかアンチセンスかによらずRNAやDNAを含み、またハイブリッド配列も含む。特に、ゲノムDNAや相補的DNAを含み、それらは転写されることもされないこともあり、スプライスされることもされないこともあり、不完全なスプライシング又はプロセッシングを受けることもあり、その由来にはよらず、生物学的材料からクローニングされることもあり、あるいは合成によっても得られる。本発明の目的において、RNAとは、一本鎖の核酸分子であり、たとえ二次構造をとって二本鎖RNAを形成している部分があったとしても同様である。特に、本発明の目的においてRNAはリボヌクレオチドで構成される任意の形状の核酸分子を含み、特定の塩基配列や由来とは関係ない。従って、RNAはin vivo又はin vitroで人工システムによって転写されても良いし、翻訳されなくても、スプライスされてもされなくても、不完全にスプライス又はプロセッシングを受けても良く、天然由来か人工的にデザインされた鋳型から生成されたかどうかは関係ない。それらは合成によって得られたmRNA、tRNA、rRNA、miRNA、siRNA、RNAi、又はそれらの任意の混合物も含む。RNAは生物学的サンプル、あるいはより具体的には、血液や血清のような生物学的由来の液体から生成されても良い。例えば、ウイルスRNA又は個々の人の血液にあり得る他の寄生生物を含んでも良い。あるいは、RNAは、解剖された組織よりフローソートされた細胞を含む精製細胞から得ても良く、それら細胞はセルソートのための蛍光を発する抗体でラベルされていても良い。あるいは、当業者が知り得る手段を用いて、緑色蛍光タンパク質(GFP)のようなマーカーのトランジェニック発現によりラベルされても良い。他の方法として、これらの細胞は形態に基づいて選ばれることもでき、あるいはレーザーによるマイクロダイゼクションによって選ばれることも出来る。さらに正確には、「DNA」、「RNA」、「核酸」、「塩基配列」の表現は核酸材料そのものを含み、また塩基配列情報やベクター、ファージミド、又はその他の特定の核酸分子に限定されるものではない。「核酸」という語句は、また、本明細書では天然に存在し得る核酸、人工的に合成された又は調製された核酸、当業者の知り得る方法によってあるいは天然に発生する事象によって、少なくとも一つ又はそれ以上の修飾が導入された修飾核酸を含む。同様に、本発明の「タグ」または「Identifier配列」とは本発明によって調製される核酸分子の任意の領域でも良く、ここでいう「タグ」または「Identifier配列」とは、任意の核酸フラグメントを含み、由来が天然の事象により発生したものでも良く、人工的に合成又は調製されたものでも良く、また天然の事象によりあるいは当業者が知りえる方法により少なくともひとつの修飾が導入された核酸でも良い。さらに、「タグ」または「Identifier配列」は、特定の塩基配列情報や構成に関係するものではない。本発明においては、「純度」、「濃縮された」、「精製」、「濃縮」、又は「選択」という語句は、置換可能であり、生成物の絶対的な純度や濃縮を必要とするものではなく、むしろ、相対的な意味で用いられる。本発明において、「特異的」、「好ましい」、「優先的」という語句は、置換可能であり、DNA又はRNAのハイブリダイゼーションにおいて、絶対的な特異性や酵素の基質に対する特異性、反応を意味するものではなく、むしろ、相対的な意味で用いられ、酵素がその基質と関係したあるいは無関係の他の物質に対して低い、あるいは比較してより低い親和性を持つという意味も含まれる。同様に、酵素、又は酵素活性という語句は、本発明においては要素の機能や活性を表し、要素の絶対的な純度を必要とするものではない。従って、酵素若しくは同等の、又は関連した若しくは関連しない機能を持つ要素を含む混合物も本発明の対象となる。同様に、本発明におけるDNA分子又はRNA分子は、ハイブリダイゼーションプローブとして機能することもでき、本発明の目的のために相補的配列を含んでも良い。相補配列を含むDNA又はRNAは、関連する核酸分子を検出するためのプローブとして実験に使用することもでき、さらにそのようなプローブとターゲット分子は、異なる位置に突然変異を人工的又は自然発生的に導入することにより区別する実験に使用することもできる。「生物学的サンプル」とは、微生物、動物、植物、宿主に依存して複製するウイルスやプリオンなどの感染性粒子など、生物体から得られるあらゆる材料を含む。このような「生物学的サンプル」は、研究、開発、診断、治療などを目的として、患者、動物、植物、感染粒子から得られたあらゆる材料を含む。従って、本発明は特定の核酸分子又はその由来に限定されることはなく、本発明は、あらゆる核酸の操作、利用、応用のために普遍的な手段を与える。本発明に適用される核酸分子は、当業者が知りえる任意の方法によって調製、入手することができ、例えばSambrook J. and Russuell D.W. (Molecular Cloning,A Laboratory Manual,Cold Spring Harbor Laboratory Press,New York,2001、本文献は、参照することによって本明細書に組み込まれるものとする)らの方法により得ることができるが、これに限定されるものではない。
【0031】
本発明は、核酸分子より検出と解析のためにフラグメントを単離する方法に関するものである。この核酸分子の解析は、核酸分子の全体的又は部分的な塩基配列情報を得ることを含むが、これに限定されるものではない。当業者が知り得る塩基配列情報を得る方法は種々あり、例えばSambrook J. and Russuell D.W.,Molecular Cloning,A Laboratory Manual,Cold Spring Harbor Laboratory Press,New York(2001)に記載されており、また従来技術として記載した新しく発展してきたハイスループット技術が含まれるが、これに限定されるものではない。本発明は、ある特有の塩基配列決定方法や技術への利用に限定されず、実験の条件に最も適した形やあるいは分野の新しい発展に適応した形で解析や検出をおこなうためにRNAやDNAを操作する汎用的な技術を提供する。
【0032】
塩基配列決定反応を含む解析、検出又は操作のために、核酸分子は固体支持体上に結合されても良い。固体支持体は、要素が直接又は間接的に結合できる任意の固体材料でも良い。そのような材料はアクリルアミド、アガロース、セルロース、ニトロセルロース、ガラス、金、ポリスチレン、ポリエチレンビニルアセテート、ポリプロピレン、ポリメタクリレート、ポリエチレン、ポリエチレンオキサイド、ポリシリケート、テフロン、フッ化炭素、ナイロン、シリコンゴム、ポリアンハイドライド、ポリグリコール酸、ポリ酢酸、ポリオルトエステル、機能性シラン、ポリプロピルフマル酸、コラーゲン、グリコサミノグリカン、ポリアミノ酸、またはそれらの混合物が含まれるが、これに限定するものではない。固体支持体はさらに、薄いフィルム、膜、ビン、皿、繊維、編まれた繊維、チューブ状、粒子状、ビーズ状、微粒子状などに成形されたポリマー、またはそれらの組み合わせを含む。
【0033】
本発明は、ひとつ又はそれ以上の核酸分子を含むサンプルの転換に関するものである。そのような核酸分子又は核酸分子の混合物は、DNAに転換される。本発明を実施するために、核酸分子は、自然にあるゲノムDNA、RNAサンプル、人工的に作られたDNAライブラリーから得ることができ、またそれらの任意の混合物でも良い。本発明は、単独又は任意の複数の核酸分子への適用に限定されるものではなく、核酸分子が単独であっても、複数であっても良く、自然発生的におきるか、あるいは既存のライブラリーから得られるか、あるいは人工的に作製されるかによらない。さらに、本発明は、その由来や性質に関わらず、あらゆる核酸分子を処理することができる。従って、自然事象によって得られる核酸分子でも完全長分子であっても良く、それらの断片でも良い。さらに、それらの核酸分子の断片は、ランダムなプロセスによって、あるいはある特定の塩基配列に作用する酵素を用いて核酸分子のターゲットを絞った方法によって、あるいは転写領域の中のエクソン、イントロンなどの核酸分子の構造をもとに断片化する方法によって調製することができるが、これに限定されるものではない。従って、本発明は、特定の出発材料に限定されるものではない。
【0034】
本発明は、プールされたRNA分子の中のRNA分子、又は一つのRNA分子の5’末端に官能基および塩基配列情報を導入するために、一つ又は複数のRNA分子を修飾することに関係するものである。そのような官能基は、1、3、1〜5、5〜10、10〜15、15〜25、25〜35、35〜45または45以上のヌクレオチドを含んでも良い。よって、本発明は、RNA分子の操作および解析や修飾されたRNAの調製や解析に用いられるRNA分子に情報を付加するために行うRNAの修飾に関係するものである。
【0035】
当業者が知り得る酵素的又は化学的なRNAの修飾方法は、種々ある。好ましくは、本発明を実施するためには、RNAの酵素反応による修飾が望ましく、そうすることにより異なる酵素反応を用いてRNAのグループ中において標的とする特定のRNAの修飾を行うことができる。さらに好ましくは、トータルRNAの中のmRNA分子が優先的に修飾を受け選択的な濃縮が可能となる。しかしながら、本発明は、mRNAの解析に限定されることなく、解析と検出のためにRNAを捕獲する汎用的な手段を提供する。miRNAやほかの短いRNA分子のような全く新しいRNAの種類を特定の修飾と解析の対象とすることができる最近の研究の結果がAlvarez−Garcia I.and Miska EA.,Development 135,4653−4662(2005)(本文献は、参照することによって本明細書に組み込まれるものとする)に示されている。
【0036】
本発明を実施するために、標的とするRNAは3つの概念的に異なるステップの対象とされる。(1)非完全長mRNA分子のマスキング、(2)分子に含まれるCap構造の反応分子への転換、(3)標的とするRNAの5’末端への標的RNA分子の付加の3つのステップである。mRNAの5’末端へRNAオリゴヌクレオチドを付加する標準的な方法は、Oligo−Capping法(Maruyama K. and Sugano S.,Gene 138,171−174,1994;Suzuki Y. and Sugano S,Methods Mol Biol.221,73−91,2003、これらの文献は、参照することによって本明細書に組み込まれるものとする)と呼ばれるものおよびその改変法がある。生きた生物からのRNA調製は、完全長mRNAの5’末端にあるCap構造により特徴付けられるRNA種を含む。このCap構造によって、Cap構造を欠き、そのかわりに自由なリン酸基を5’末端に含んでいる切断されたRNA種から区別することができる。Oligo−Capping法は、完全長mRNAの独特な性質を選択的な濃縮に用いるものである。Oligo−Capping法は、プールされたRNA中においてmRNAを特異的に修飾するいくつもの酵素的ステップを含む。最初の酵素反応により、Cap構造を持たない切断されたmRNAや小さいRNA、tRNA、rRNAなどがホスファターゼにより5’末端を脱リン酸化される。次に第二の反応により、Cap構造を持つmRNAからタバコ酸ピロホスファターゼ(TAP)によりCap構造を除去する。この処理により、完全長mRNAのみが5’末端にリン酸基を持つことになる。よって、第三の酵素反応においてRNAリガーゼは、完全長mRNAのリン酸化された5’末端にのみ、オリゴヌクレオチドを付加することができる。
【0037】
第一の反応ステップでは、RNAの5’末端からリン酸基を除去することが可能な任意のホスファターゼを使用することができる。さらに具体的には、ホスファターゼはバクテリアアルカリホスファターゼ(BAP)、仔ウシ腸アルカリホスファターゼ(CIAP)、エビアルカリホスファターゼ(SAP)、Antarcticホスファターゼの中より選ばれる。同様に本発明を実施するために異なるホスファターゼが用いられても良く、また最も一般的には、タバコ酸ホスファターゼ(TAP)がCap構造の除去に用いられる。RNAのライゲーションステップでは、リン酸化RNAにDNAおよびRNAオリゴヌクレオチドを連結できる任意のRNAリガーゼを使用することができる。最も一般的には、T4 RNAリガーゼまたはサーモファージ一本鎖DNAリガーゼがこの反応に用いられる。サーモファージ一本鎖DNAリガーゼは商業的に入手可能であり、一本鎖DNAとRNAのどちらにも使用できる(この酵素のより詳細な情報は、製品情報に記載されている。http://www.prokaria.com/upload/files/Thermophage−ssDNA−ligase−version−4−2.pdf(本文献は、参照することによって本明細書に組み込まれるものとする)。よって、この酵素はRNAに直接DNAを連結させるために好ましい。このように本発明は、RNAに直接DNAを連結し、デオキシリボヌクレオチドまたはDNAオリゴヌクレオチドとリボヌクレオチドまたはRNAまたはRNAオリゴヌクレオチドを含む直線的ヘテロポリマーを調製することができる。
【0038】
当業者が知り得る本発明を実施するために用いられるOligo−Capping法の改変法は種々ある。最も好ましくは、本発明は、特許出願2006−106770号(本文献は、参照することによって本明細書に組み込まれるものとする)に開示されているような一つの反応容器中ですべての酵素反応が行える方法を利用する。簡単に言うと、第一の反応ステップでAntarcticホスファターゼのような熱処理によって不活性化される酵素を用いる。一連の反応の中の第一の酵素を不活性化した後、新しく成分を添加することにより、バッファー条件をTAP反応に適した状態に変更する。TAPは、再度熱処理によって不活性化される。従って、追加成分とオリゴヌクレオチドの添加で再度バッファー条件を変更するだけで、最終ステップとしてリン酸化RNAにオリゴヌクレオチドを連結することができる(図1参照)。
【0039】
前述した中で修飾反応は、プールされたRNAの中のmRNA分子に対して、後の操作のために行われる。しかしながら、本発明は、mRNAの修飾に限定されるものではない。異なる例では、Cap構造を欠き、リン酸化されたすべてのRNA分子が、RNAの5’末端にオリゴヌクレオチドを連結することによって直接修飾される。この例では、本発明は切断された(truncated)RNA分子と非mRNA分子の選択的な修飾を可能とする。本発明の他の例では、Cap構造を欠くRNA分子は、第一の酵素反応において修飾される。ある例では、Cap構造を欠いたRNA分子のみが本発明によって操作のために修飾される。他の例では、第一の反応の後に異なるRNA分子を段階的に修飾する他のステップを行う。従って、第二の酵素反応では、完全長mRNA分子のCap構造は、TAP酵素反応によって除去され、完全長mRNA分子の5’末端にリン酸基を与える。最後の反応ステップでは、同じ又は異なる塩基配列のオリゴヌクレオチドが完全長mRNA分子に連結される。この実施態様では、本発明は種々のRNA分子のプールの中にある異なるRNA種に異なるオリゴヌクレオチドを付加する手段を提供する。
【0040】
上記の、あるいは図1に示した一連の操作に続いて、第一の反応では、RNAリガーゼが一つ又は複数のRNA分子に異なる種類のオリゴヌクレオチドを連結するために用いられる。本発明によれば、オリゴヌクレオチドは、デオキシリボヌクレオチド(DNAオリゴヌクレオチド)、又はリボヌクレオチド(RNAヌクレオチド)から構成されるホモポリマーでもよく、またデオキシヌクレオチドとリボヌクレオチドから構成されるヘテロポリマー(DNA/RNAハイブリッドオリゴヌクレオチド)でも良く、あるいは、官能基をもった任意の種類のホモポリマー又はヘテロポリマーとなることもできる(図2参照)。本発明は、特有の核酸分子の利用に限定されず、修飾されたRNAが本発明の異なる実施態様で使用される方法に応じた、違うタイプのオリゴヌクレオチドを用いることができる。当業者が知り得る多くのDNAとRNAの修飾がある。例えば、異なる修飾のオリゴヌクレオチドを調製するためのオリゴヌクレオチドの情報は、MWG Biotechのウェブサイトhttp://www.mwg−biotech.com/html/s_synthetic_acids/s_modifications.shtml(ここに示された情報は、参照することによって本明細書に組み込まれるものとする)に記載されている。MWG Biotechは、異なる位置に官能基としてビオチンやジゴキシゲニンを持つオリゴヌクレオチドを提供している。さらに、修飾されたオリゴヌクレオチドは、5'Aminolink C3/C5/C6/C12、3'Aminolink C3/C6/C7、3'Aminolink C3/C6/C7、Amino(C2/C6)−dT、Amino C6−dC、Spacer C3/C9 (TEG)、Spacer C12/C18 (HEG)、reduced Thiol modifierのようなクロスリンクのための反応基など、ひとつ又はそれ以上の官能基を得ることができる。RNAオリゴヌクレオチドは、例えばInvitrogen(http://www.invitrogen.com/content.cfm?pageid=9900、ここに示された情報は、参照することによって本明細書に組み込まれるものとする)、またはOperon(http://www.operon.com/、ここに示された情報は、参照することによって本明細書に組み込まれるものとする)から入手することができる。このように、RNAの5’末端に付加されるオリゴヌクレオチドを異なる機能を持つようにデザインすることができる。一つの例では、オリゴヌクレオチドは、10から25ヌクレオチドを持つ。別の例では、オリゴヌクレオチドは、25から50ヌクレオチドを持つ。また別の例では、オリゴヌクレオチドは、50から100ヌクレオチドを持つ。また別の例では、オリゴヌクレオチドは、100以上のヌクレオチドを持つ。オリゴヌクレオチドは、化学合成によって、あるいは酵素的反応によって得ることができる。当業者が知り得るDNA依存性RNAポリメラーゼはいくつもあり、例えば鋳型DNAからRNA分子を調製できるT3 RNAポリメラーゼ、T7 RNAポリメラーゼ、SP6 RNAポリメラーゼなどがある。RNAポリメラーゼを用いたRNA調製の一つの例は、Fermentas(http://www.fermentas.com/techinfo/modifyingenzymes/protocols/p_synthstrspecrna.htm、ここに示された情報は、参照することによって本明細書に組み込まれるものとする)に記載されている。さらに、オリゴヌクレオチドは、リボゾームRNAのような天然のものでも良い。任意の生物から得られる全RNAの調製では、得られる大部分のRNAはrRNAであり、例えば、仔ウシ肝臓より得られる18S+28SリボゾームRNAは、Sigma−Aldrich(St. Louis,USA,catalog number R0889)より商業的に入手可能である。RNAオリゴヌクレオチド、DNAオリゴヌクレオチド、あるいは修飾されたオリゴヌクレオチドは、前述のようにRNAリガーゼによって直接RNA分子へ連結することができる。このように、反応条件は特定のリガーゼ、特定のオリゴヌクレオチドの修飾、特定の塩基配列のオリゴヌクレオチド、又は特定のオリゴヌクレオチド自体の利用に制限されるものではない。
【0041】
最も一般的には、オリゴヌクレオチドは、RNAの5’末端にプライミング位置を導入するための「プライマー」として機能するようにデザインされている。プライマーは5、6、5〜10、10〜15、15〜20、20〜30、30〜40、40〜50、または50以上のヌクレオチドからなっても良い。相補的核酸鎖合成の後、新しく合成された第二鎖は、3’末端にRNAの5’末端に連結されたオリゴヌクレオチドと相補的な配列を持つ。RNAの5’末端に付加されたオリゴヌクレオチドと全く同じ、又は部分的に同じ配列を持つオリゴヌクレオチドは、修飾されたmRNAと全体的、又は部分的に同じ配列を持つ核酸分子の合成をプライミングするのに用いられる。第二鎖のプライミングは、例えば、二本鎖又は一本鎖DNAの調製、DNA又はRNAの増幅、塩基配列決定に用いられる。DNAポリメラーゼによる第二鎖DNA合成の別の方法は、Sambrook J.and Russuell D.W.,Molecular Cloning,A Laboratory Manual,Cold Spring Harbor Laboratory Press,New York,2001、本文献は、参照することによって本明細書に組み込まれるものとする)などの標準的な教科書に記載されている。そのようなDNAポリメラーゼは、DNAポリメラーゼのクレノー断片、T4またはT7 DNAポリメラーゼ、DNAポリメラーゼI、Taqポリメラーゼ、Tfl DNAポリメラーゼ、Tth DNAポリメラーゼ、Tli DNAポリメラーゼ、または他の既知のDNAポリメラーゼを含むが、これに限定されない。例えば、直線状一本鎖DNAの調製には、当業者が知り得る様々な技術が発展してきた。いくつかの方法では、DNAポリメラーゼを用いてDNA又はRNAの鋳型より一本鎖DNAを合成する。特に、一本鎖DNAの合成は、非対象PCRと呼ばれる方法によって達成することができ、この方法では二つのプライマーが異なる濃度で使われる。律速となるプライマーが消費されたあと、反応は、二本鎖DNAの指数関数的増幅から律速となるプライマーより多量に入れられた片鎖のプライマーによる直線的増幅に切り替わる。他の方法では、ラムダエクソヌクレアーゼがリン酸化された5’末端をもつ二本鎖DNAの片鎖を分解するために用いられる。そのような鋳型は、二つのプライマーのうち、片方だけがリン酸化された5’末端を持つようにPCRで調製される。ラムダエクソヌクレアーゼは、また「Strandase(商標)」という名称で呼ばれており、Novagen、 (Madison,USA)から商業的に入手可能であり、「Strandase(商標) ssDNA Preparation Kit」、カタログ番号69202(これは、参照することによって本明細書に組み込まれるものとする)に説明が記載されている。同様に、ラムダエクソヌクレアーゼはEpicentre、(Madison,USA)(カタログ番号LE035HおよびLE032K)より入手可能である。数多くの直線状一本鎖DNAの応用のため、一本鎖DNAは、二つのプライマーのうち片方が特にタグづけられたプライマーセットを用いるPCRによって調製される。官能基を持った鎖と鋳型DNAからの二次的な望まれない鎖を分けるために、ビオチンラベルが最も頻繁に用いられているが、これに限定されるものではない。この方法は、目的とする鎖を固体支持体や基盤上に結合させて用いる場合には、特に有用である。固定された一本鎖DNAは、支持体上で直接精製され、一本鎖DNAの分離及び調製に応じた検出検定やDNA塩基配列決定のための鋳型調製に用いられる。そのような応用は、例えば、DASH SNP検出システムと呼ばれるようなゲノムDNA中のSNPの特性解析などを含むが、これに限定されるものではない。この方法は、米国特許出願2001046670号(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。S. Stahl et al.(Stahl,S.et al,Nucleic Acid Research 16,3025−3038(1988)(本文献は、参照することによって本明細書に組み込まれるものとする)には、ビオチン化されたDNAの異なる応用が記載されている。例えば、固相での塩基配列決定などである。他の実施態様として、Guatelli J.C. et al.,Proc. Natl. Acad. Sci. USA 87、1874−1878(1990、本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている方法によれば、逆転写酵素、RNase H、DNA依存性RNAポリメラーゼの活性を組み合わせた反応サイクルにより、修飾されたRNA分子を増幅することができる。
【0042】
他の実施態様において、RNAの5’末端に付加されたオリゴヌクレオチドは、DNA又はRNA分子の操作を可能にするための配列情報を持つようにデザインされている。当業者が知り得る特定の塩基配列や認識部位への結合に依存する酵素活性は数多くある。それらの酵素の多くは、異なる業者から商業的に入手可能である。例えば、FERMENTAS UAB(Vilnius,Lithuania)、New England Biolabs Inc.(Beverly,USA)、Promega(Madison,USA)、Takara(Tokyo,Japan)、 Roche(Mannheim,Germany)、GE Biosciences (Cardiff,United Kingdom)などが含まれるが、これに限定されるものではない。通常、制限酵素はDNA分子を認識部位や認識部位の近傍の規定された位置で分解するために用いられる。一つの例では、オリゴヌクレオチドによって導入されRNAの5’末端に付加された認識部位は、クラスII制限酵素の認識部位である。これらの酵素は、認識配列の外側で切断するもので、例えば、クラスII制限酵素であるMmeIは認識部位から20/18塩基対離れた部位を切断する。それゆえ、MmeIは、通常、例えば、前述のLongSAGE、5’−SAGE、またはCAGE法などの短い塩基配列決定用タグの分離に用いられる。他の応用では、当業者が知るようにDNAの組み換えやクローニングのために制限酵素を利用することができる。より詳しくは、Sambrook J. and Russuell D.W.,Molecular Cloning,A Laboratory Manual,Cold Spring Harbor Laboratory Press,New York(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。さらに、修飾された又はデザインされた制限酵素は、鎖特異的ニッキング酵素として機能することができる。そのような制限酵素は、二本鎖DNAを基質として認識配列の中で一方のDNA鎖のみを切断する。そのような酵素は、商業的に入手可能であるN Bpu 10I(FERMENTAS UAB,Vilnius,Lithuania)、N.Bbv C IA、N.Bst NB I、N. Alw I(New England Biolabs Inc,Beverly,USA)などを含むが、これに限定されるものではない。ニッキング酵素は二本鎖DNAにプライミング部位を作る点で特に興味深く、DNAの合成や塩基配列決定に用いるためプライマー伸長反応に用いることができる。二本鎖のDNA分子の中の片鎖にDNA合成のためのプライミングサイトを作るために、酵素活性によってニックを入れる一つの例が、Walker T.G. et al.,Proc. Natl. Acad. Sci. USA 89,392−396(1992)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。他の例では、オリゴヌクレオチドは、RNAポリメラーゼの認識部位を導入する。RNAポリメラーゼは、T3 RNAポリメラーゼ、T7 RNAポリメラーゼ、SP6 RNAポリメラーゼを含むが、これに限定されるものではない。これらはすべてDNA依存性RNAポリメラーゼであり、それぞれ特異性のある二本鎖プロモーターがある。プロモーター又はそれらの認識部位より始まり、これらは一本鎖又は二本鎖DNAのいずれかの鋳型から相補的なRNAの合成を5’から3’方向へ触媒する。Guatelli J.C. et al.はProc. Natl. Acad. Sci. USA 87,187401878(1990)(本文献は、参照することによって本明細書に組み込まれるものとする)にDNA依存性RNAポリメラーゼの使用例を記載している。異なる例では、オリゴヌクレオチドはDNA結合タンパク質の認識部位を含んでも良い。多くのDNA結合タンパク質は当業者の知るところであり、天然に存在するものでも、タンパク質デザインによって調製されたものでもあり得る。そのようなDNA結合タンパク質は、転写因子、ゲノムDNAの認識部位に直接又は間接的に結合する調製機能タンパク質などを含むが、これに限定されるものではない。転写因子は生命に不可欠の分子であり、ゲノム情報を利用するのに必要である。あらゆる生命体は多くの転写因子を含んでいる、例えば、Kanamori M.,787−93(2004)(本文献は、参照することによって本明細書に組み込まれるものとする)に既知のマウス転写因子すべてのデータベースが公開されている。転写因子は、異なる認識部位への親和性という点でほかのものと区別できる。この特異性は、与えられた転写因子および転写因子のグループの認識部位を含むDNA分子を濃縮するために用いることができる。しかしながら、当業者は、特異的配列よりもむしろ構造を認識するタンパク質について知るように、転写因子の結合特異性は、塩基配列への結合に限定されない。例えば、転写因子DAX−1は、Zazopoulos E. et al.,Nature 390,311−315(1997)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されているように異なるDNA構造に結合する。他の例では、DNA結合タンパク質は一本鎖DNAに特異的に結合する。一本鎖DNA結合タンパク質は、E.coliのSSB、ファージT4のGene32の産物、アデノウイルスのDBP、一本鎖DNAに対して作られた抗体、仔ウシ胸腺 UP1、またはそれらの混合物などが含まれるが、これに限定するものではない。加えて、二本鎖DNAのミスマッチに特異的に結合するタンパク質がある。このグループのタンパク質は、E.coliの主要なミスマッチ修復パスウェイに関係するMutSタンパク質ファミリー(タンパク質のファミリーについてはhttp://www.tigr.org/〜jeisen/MutS/MutS.htmlを参照、本ウェブページの内容は、参照することによって本明細書に組み込まれるものとする)が含まれるが、これに限定されるものではない。修飾されたRNAやDNAに相補配列又はその部分的配列が付着される場合と比べて、塩基配列中にミスマッチがある場合のプライマーの伸長反応にプライマーが使われる場合、MutSタンパク質やその他の遺伝子ファミリーは、ミスマッチをもつ二本鎖のDNA種を特に濃縮するように用いても良い。加えて、MutSや遺伝子ファミリーは、プライマー伸長反応を操作するあるいはブロックするために用いられても良い。MutSタンパク質のいくつかは、商業的に入手可能であり、例えばNippongeneのTaq MutS(Tokyo,Japan,コード番号316−04011)がある。異なる例では、オリゴヌクレオチドは「Identifierh配列」や「Barcode」として用いられる塩基配列の領域を含んでも良い。そのような塩基配列は、Identifier配列としてサンプルの由来の印となったり、核酸やIdentifier配列に相補的な塩基配列を持つ核酸とのハイブリダイゼーションによって、修飾されたRNAやDNAを特異的に捕獲するためのタグとして機能させることもできる。そのような塩基配列は、前述した酵素反応の任意の反応の特異的、選択的なプライミング部位として用いることもできる。Identifier配列は、DNAポリメラーゼによる第二鎖の合成や、例えばPCRによる増幅、一本鎖DNAの調製などのための選択的なプライミング部位となることもできる。このように、異なるRNA種に認識部位やIdentifier配列として異なるオリゴヌクレオチドを導入する前述の方法の組み合わせによって、本発明は、全RNAや複数のRNA分子中で、個々のRNA分子を分けて操作する手段を提供する。
【0043】
異なる実施態様において、本発明は、RNA分子又は種々のRNA分子の末端に官能基を導入する手段を提供する。多くの異なる官能基が結合分子に結合する親和性を持っている。官能基は、反応基又は化学反応において共有結合を形成するのに適した架橋リンカー、アミノ基、ビオチン、ジゴキシゲニン、抗体、抗原、タンパク質、核酸、核酸結合分子、またはそれらの組み合わせを含むが、これに限定するものではない。官能基や官能基に結合された分子は、基盤上にある結合分子と結合することができる。本発明を実施するための基盤は、アビジン、ストレプトアビジン、ジゴキシゲニン結合分子、所定の配列をもったオリゴヌクレオチド、抗体またはリガンド、又は化学的基盤など、共有結合を形成する化学反応に用いられる反応基を固定化したものから選ぶことができる。用いられる官能基がビオチンの場合、対応する基盤はアビジンまたはストレプトアビジンである。同様に、官能基がジゴキシゲニンの場合、基盤はジゴキシゲニン結合分子である(Roche Diagnostics GmbH Catalog、本文書は、参照することによって本明細書に組み込まれるものとする)。官能基がオリゴヌクレオチドの場合、基盤は官能基の配列に相補的配列を持ったオリゴヌクレオチドであり、官能基が抗原の場合は、基盤は抗体またはprotein I又はprotein Gのような抗体結合タンパク質である。従って、本発明はRNA分子へ官能基を導入する手段を提供し、官能基はオリゴヌクレオチドに付加される。修飾されたオリゴヌクレオチドは、多くの業者より商業的に入手可能である。最もよく使用されるのはビオチンラベル化オリゴヌクレオチドである。異なる修飾オリゴヌクレオチドの一つの例として、MWG Biotechのウェブサイト(http://www.mwg−biotech.com/html/s_synthetic_acids/s_modifications.shtml、ここから入手できる情報は、参照することによって本明細書に組み込まれるものとする)に記載がある。MWG Biotechはオリゴヌクレオチドの異なる位置に官能基としてビオチンやジゴキシゲニンを持つオリゴヌクレオチドを提供できる。さらに、一つ又はそれ以上の官能基を持つ修飾オリゴヌクレオチドを得ることができる。例えば、5’Aminolink C3/C5/C6/C12、3’Aminolink C3/C6/C7、3’Aminolink C3/C6/C7、Amino(C2/C6)−dT、Amino C6−dC、Spacer C3/C9(TEG)、Spacer C12/C18(HEG)、あるいはreduced Thiol modifierなどのクロスリンクのための反応基がある。RNAオリゴヌクレオチドは、例えばInvitrogenより購入でき、そのようなRNAオリゴヌクレオチドの情報はhttp://www.invitrogen.com/content.cfm?pageid=9900(ここに示された情報は、参照することによって本明細書に組み込まれるものとする)で得ることができる。また、Operonはhttp://www.operon.com/(ここに示される情報は、参照することによって本明細書に組み込まれるものとする)で役立つ情報を提供している。
【0044】
前述の実施態様において、図1〜2で示すように、RNAリガーゼはRNA分子のリン酸化された5’末端にオリゴヌクレオチドを連結するために用いられる。通常用いられるT4 リガーゼのようなRNAリガーゼは、オリゴヌクレオチドの3’末端のリボヌクレオチドに依存し、RNAにデオキシリボヌクレオチドを直接的に連結しなくても良い。さらに、RNA分子へのオリゴヌクレオチドの連結は配列特異的ではなく、任意の配列のオリゴヌクレオチドをRNA分子と組み合わせることができる。代替的な方法がClepet C. et al. in Nucleic Acids Res. 32、 e6 (2004)(本文献は、参照することによって本明細書に組み込まれるものとする)に示されており、その中でDNAリガーゼはRNAに二本鎖DNA又は部分的に二本鎖になったDNA分子を連結する(RNA−taggingと呼ばれる)。例えば、T4 DNAリガーゼは鋳型DNAへのRNA断片の連結を触媒することができる(Kleppe、 K. et al.,Proc. Natl Acad. Sci. USA,67,8-73(1970)、Fareed,G.C. et al.,J. Biol. Chem.,246、925-932(1971)(両文献は、参照することによって本明細書に組み込まれるものとする)。連結反応を仲介する鋳型DNAは、個々のRNA分子を修飾するための塩基配列特異的な連結反応に用いることができる。配列特異性は、部分的に二本鎖となったオリゴヌクレオチドによって達成できる。そのようなオリゴヌクレオチドは、RNA分子の5’末端にハイブリダイズする突出領域を持つ(図3参照)。突出領域は、4から6、又は6から8ヌクレオチドの長さでも良い。また8〜12、又は12ヌクレオチド以上の長さでも良い。当業者が知り得る、例えばShibata Y.et al.in Biotechniques,30,1250−1254(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に述べられているような連結反応において部分的に二本鎖となったオリゴヌクレオチドを用いる種々の方法がある。反応によって、突出領域は、例えば前述のShibata.et.al.で用いられているようなランダム配列や特有のRNA分子を標的とした規定の配列を持つことができる。他の例として、突出領域は、認識部位にランダム配列を含む制限酵素や認識部位の外側で切断する制限酵素を用いて酵素反応的に作っても良い。そのような酵素は、BstXI(CCANNNNN↓NTGG)、DrdI(GACNNNN↓NNGTC)、BglI(GCCNNN↓NGGC)、BoxI(GACNN↓NNGTC)、BseJI(GATNN↓NNATC)、BseLI(CCNNNNN↓NNGG)、CaiI(CAGNNN↓CTG)、CseI(GACGC(5/10)↓)、Eam1105I(GACNNN↓NNGTC)、Eco31I(GGTCTC(1/5)↓)、Eco57I(CTGAAG(16/14)↓)、Esp3I(CGTCTC(1/5)↓)、HpyF10VI(GCNNNN↓NNGC)、LguI(GCTCTTC(1/4)↓)、OliI(CACNN↓NNGTG)、PdmI(GAANN↓NNTTC)、PsyI(GACN↓NNGTC)、SfiI(GGCCNNNN↓NGGCC)、SmuI(CCGGC(4/6)↓)、Van91I(CCANNN↓NTGG)、XagI(CCTNN↓NNNAGG)、又はそれらのアイソザイムを含むが、これに限定されるものではない。これらの酵素は特に有用であり、ランダム突出領域は複数の核酸分子より調製される。例えば、cDNAライブラリーは、前述の酵素の一つの認識部位を持つリンカーが5’末端に持つcDNAインサートで構築することができる。そのようなライブラリーの分子の切断は、もとのcDNAライブラリーに存在する分子を代表するランダムな突出領域を持つ複数の分子を生成する。よって、本発明は、反応に使用されるオリゴヌクレオチドに相補的な配列とRNA分子の5’末端に相補的な配列を持つ鋳型DNAによって、個々のRNA分子の目的とされた修飾を行う手段を提供する。一つの例では、それらの配列は5〜10ヌクレオチドの長さであり、別の例では、10〜25ヌクレオチドの長さであり、また別の例では、25ヌクレオチドよりも長い。RNA分子の5’末端に相補的な領域は、実験的手法で得ることができ、例えば、複数の核酸分子の操作によって、あるいはコンピューターデザインによる組み合わせを用いた化学合成によって得ることができる。RNA分子の配列情報は、当業者が知り得る公共データベース、例えばNCBI(http://www.ncbi.nlm.nih.gov/)、EMBL(http://www.ebi.ac.uk/Databases/)、DDBJ(http://www.ddbj.nig.ac.jp/)などより入手できる。ある例では、一つの鋳型DNAは特有のRNA分子を標的として用いることができる。別の例では、ランダム配列を含む複数の鋳型DNAが用いられる。さらに別の例では、複数の鋳型DNAは、特有の配列を含む。よって、本発明は、全RNA中で、特有のRNA分子やmRNAなどのRNAを標的とした操作を行う柔軟な手段を提供する。
【0045】
RNA分子はSambrook J. and Russuell D.W.,Molecular Cloning,A Laboratory Manual,Cold Spring Harbor Laboratory Press,New York、(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載の逆転写によるDNA転写物の調製の鋳型として用いることができる。当業者が知り得る異なる反応条件や酵素修飾などを含む多くの改変された方法がある。例えば、逆転写酵素は、AMV reverse transcriptase、M−MLV reverse transcriptase、M−MLV reverse transcriptase RNase H minus、あるいはそれらの改変物などを含むが、これに限定されるものではない。任意の修飾されたRNAは、前述の又は図1〜3で示されたすべての又は部分的なステップによって得ることができ、それらを鋳型にして、逆転写酵素によってRNA鋳型からコピーDNAを合成するcDNA合成反応に用いることができる。この反応は、鋳型RNAにハイブリダイズし、DNA合成を開始するプライマーを必要とする。一つの例では、3’末端にランダム配列とそれに続く所定の配列を持ったプライマーセットが用いられる。所定の配列はDNAの後の操作に役立てられるが、逆転写反応のプライミングには必要とされない。従って、本発明は、ランダム配列のみを持つプライマーを用いることができる。そのようなランダム配列自体又は規定配列を持つオリゴヌクレオチドの一部としてのランダム配列は、4〜6ヌクレオチド、又は6〜10ヌクレオチド、又は10〜15ヌクレオチドの長さを取ることができる。ランダムプライマーを用いることにより、鋳型RNAの5’末端の配列に相補的な配列を持つDNA断片を合成することができる。ランダムプライマーは、鋳型RNAの任意の領域にハイブリダイズできるので、反応は異なる長さのDNA分子の混合物を生成する。ランダムプライミングでは、完全長cDNAの調製はできないが、長いRNAの本来の5’末端まで到達できるという有利な面があり、それを用いなければ逆転写反応の限界のため困難である。別の例では、オリゴdTプライマーが通常多くのmRNA種の3’末端にあるポリA配列にハイブリダイズすることができる。ランダムプライミングとは対照的に、オリゴdTプライミングは、RNA分子全体を反映した完全長cDNAの合成に通常用いられる。さらに、規定配列のプライマーは、逆転写酵素反応を開始するために用いられ、例えば、通常RACE法(Rapid Amplification of CDNA Ends)などの応用に用いられる。完全長cDNA合成を開始する他の方法は、WO2006/003721(本文献は、参照することによって本明細書に組み込まれるものとする)に開示されている。従って、本発明は図4と5に示されたように修飾されたRNAからDNA転写物を調製する異なる手段を提供する。
【0046】
前述の鋳型RNAからのDNAの合成は、二本鎖DNA/RNA分子を生成する。このような二本鎖DNA/RNA分子の中のRNA部分は、RNA分解酵素やバッファーのpH変更により除去される。例えば、RNase H酵素は、DNA/RNA二本鎖分子のRNAを特異的に分解するので、本発明の実施には好ましい酵素である。RNAの除去は、逆転写反応で使われたプライマーに関係なく、ランダムプライミングであっても特異的プライミングであってもオリゴdTプライミングであっても関係なく、任意の種類のcDNAに適用される。鋳型DNA/RNAからのRNAの除去の例は、Sambrook J. and Russuell D.W.,Molecular Cloning,A Laboratory Manual,Cold Spring Harbor Laboratory Press,New York、(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。任意のこのようなDNA/RNA二本鎖分子の処理は、リボヌクレオチドから構成される鋳型の大部分又は全体より作られた一本鎖DNAとしてのDNA分子を遊離する。DNA/RNAハイブリッドが二本鎖DNA領域を含む場合は、例えば、前のステップでRNA分子に付加されたオリゴヌクレオチドの一部又は全部がDNAを含む場合、RNA部分の除去により、鋳型RNAの5’末端に相当する末端に二本鎖DNA領域を含むDNA分子が調製される。よって、本発明は、全RNA又はmRNAに由来する塩基配列を持つ、あるいはそれらのRNA分子の操作により導入された塩基配列を持つ一本鎖又は部分的一本鎖DNA分子を調製する手段を提供する。
【0047】
一本鎖DNAはDNA解析や操作に重要であり、またバイオテクノロジーや分子生物学において多くの応用や技術は、一本鎖DNAの鎖特異的な調製を必要とする。そのような応用は、塩基配列決定や標識プローブの合成を含む鎖特異的DNA合成のための鋳型DNAの調製、チミン残基のウラシルへの置換、点突然変異の導入、種々のDNAまたはRNA分子の混合物からの個々のクローンの分離、検出やディファレンシャルハイブリダイゼーションのためのドライバーやテスターの調製、一塩基多型(SNPs)の検出と解析、マイクロアレイの調製を含むが、これに限定されるものではない。これらの方法や応用は分子生物学の当業者がよく知るところであり、Sambrook J. and Russuell D.W.,Molecular Cloning,A Laboratory Manual, Cold Spring Harbor Laboratory Press,New York,(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。
【0048】
本発明により得られた鋳型DNAは、初期の段階でRNAに付加されたオリゴヌクレオチドの性質によって区別でき、例えば、図2〜3に示される。よって、本発明は、図7に示されるような一本鎖DNA又は部分的一本鎖DNAの調製に用いることができる。例えば、RNAにRNAオリゴヌクレオチドが付加され、その修飾されたRNAが逆転写酵素によるcDNA合成に用いられた場合、RNA/DNAハイブリッド分子からのRNA部分の除去は、鋳型RNAに相補的配列、初期段階でRNAの3’末端に付加されたRNAオリゴヌクレオチドに相補的な配列、5’末端に逆転写反応に用いたプライマーより直接生成した配列(図7A参照)を持つ一本鎖DNAを生成する。よって、そのようなDNA分子は、RNAに付加されたオリゴヌクレオチドより直接的又は間接的に生成された既知配列の領域に並んで潜在的に未知の配列を中心領域に含む。並んでいる領域の配列は既知なので、そのような分子は塩基配列決定や操作の鋳型として直接用いることができる。例えば、DNAの3’末端に相補的配列を持っているプライマーは、塩基配列決定反応のプライマーに用いることができる。鋳型DNAの塩基配列決定を行う従来の方法は、例えばSambrook J. and Russuell D.W., Molecular Cloning、 A Laboratory Manual,Cold Spring Harbor Laboratory Press,New York,(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されているサンガー法がある。しかしながら、本発明はサンガー法による鋳型DNAの塩基配列決定に限定されるのものではなく、最近数々の塩基配列決定法が開発されており、詳細はMetzker M.L. Genome Res. 15,1767−1776(2005),Kling J.,Nature Biotechnology 23,1333−1335(2005);Shendure J. et al.,Nature Review Genetics 5,335−344(2004)(これらの文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。これらの方法のいくつかは商業的な応用の対象となっており、例えば454Life Sciences、(http://www.454.com/、Helicos BioScience(http://www.helicosbio.com/)、Solexa(http://www.solexa.com/Company/overview.htm)、Visigen、(http://www.visigenbio.com/index.html)、GeneoVoxx GmbH、(http://www.genovoxx.de/)(これらのウェブページの一部又はすべてに見出される情報は、参照することによって本明細書に組み込まれるものとする)などが挙げられる。本発明は、鋳型DNAの向かい合った末端をデザインする手段を提供するので、鋳型DNAは必要に応じた性質を持つようにデザインされることができ、例えば、前述の任意の方法又は他の新しい方法により塩基配列決定を行う必要に応じてデザインされることができる。例えば、454 Life Sciencesによる用法で塩基配列決定を行う場合には、鋳型の向かい合った末端に特有のプライマー部位が要求され、これによりエマルジョンPCRによるDNA分子のクローンの増幅が可能となる。好ましい例として、鋳型DNAは、Margulies M.et al.,Nature 437,376−80(2005)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されているように、一分子のエマルジョンPCRに適応するように5’末端と3’末端に二つの異なる塩基配列を含む。異なる例においては、DNA/RNAのハイブリッドオリゴヌクレオチドは、mRNAに付加され、逆転写によるcDNA合成に用いられる。RNA/DNAハイブリッド分子からのRNA部分の除去はRNAの5’末端に対応する長いDNA鎖の3’末端に二本鎖DNAの領域を持つ部分的な一本鎖DNA分子を生成する。さらにこのDNA分子は、鋳型RNAに相補的な配列を含み、また3’末端に初期段階でRNAに付加されたRNA/DNAオリゴヌクレオチドに相補的な配列を含み、5’末端に逆転写反応で使われたプライマーから直接生成される配列を含む(図7B参照)。この一群では、DNA分子の二本鎖領域は、DNAポリメラーゼ反応のプライマーとして機能する短いDNAストレッチを含む。従って、本発明は、他に塩基配列決定用のプライマーを添加することなく、配列決定用の鋳型を直接提供する手段を提供する。別の例では、DNAオリゴヌクレオチドがmRNAに付加され、その結果作られる修飾されたRNA分子は逆転写によるcDNA合成に用いられ、RNA/DNAハイブリッド分子からのRNA部分の除去は、RNAの5’末端に対応する長いDNA鎖の3’末端に二本鎖DNAの領域を持つ部分的な一本鎖DNA分子を生成する。さらにこのDNA分子は、鋳型RNAに相補的な配列を含み、また3’末端に初期段階でRNAに付加されたRNA/DNAオリゴヌクレオチドに相補的な配列を含み、5’末端に逆転写反応で使われたプライマーから直接生成される配列を含む(図7C参照)。これらの方法により得られた鋳型は、図7Bに記載されている鋳型とよく似ていて、二本鎖領域は鋳型RNAの5’末端で正確に止まっている。よって、DNAオリゴヌクレオチドやDNA/RNAハイブリッドオリゴヌクレオチドにより、もとのRNA分子の5’末端に直接に接しているか、もとのRNAの5’末端より1〜5ヌクレオチド、又は5〜10ヌクレオチド、又は10〜15ヌクレオチド、又は15ヌクレオチド以上離れている。図7Cに示されるDNA分子は、図7Bに記載されている鋳型と同様の方法で塩基配列決定反応に用いられる。図7Aと7Bに示されているDNA分子を調製する前述した例の改変法では、RNAに付加されるDNAオリゴヌクレオチド、又はDNA/RNAオリゴヌクレオチドは官能基を含む。この例では、本発明は、RNAの5’末端に対応する長いDNA鎖の3’末端に二本鎖DNAの領域を持つ部分的な一本鎖DNA分子を調製し、二本鎖領域の短いほうの断片は官能基を含む。分子中の長いほうの断片は、鋳型RNAに相補的な配列を含み、また3’末端に初期段階でRNAに付加されたRNA/DNAオリゴヌクレオチドに相補的な配列を含み、5’末端に逆転写反応で使われたプライマーから直接生成される配列含む(図7D参照)。このような鋳型DNAは、塩基配列決定反応で用いられ、また官能基は支持体や基盤へ鋳型を直接結合させることを可能とする。官能基や官能基に付加された分子は、基盤上の結合分子と結合することができる。そのような基盤は、官能基の性質に応じて選択することができる。例えば、官能基がアミノ基だった場合、反応基は、化学反応で基盤に共有結合するものになる。官能基がビオチンだった場合には、基盤はアビジン又はストレプトアビジンになる。官能基がジゴキシゲニンの場合、基盤はジゴキシゲニン結合分子である(Roche Diagnostics GmbH Catalog、本文献は、参照することによって本明細書に組み込まれるものとする)。官能基がオリゴヌクレオチドの場合、基盤は官能基の配列に相補的配列を持ったオリゴヌクレオチドであり、官能基が抗原の場合は、基盤は抗体またはprotein I又はprotein Gのような抗体結合タンパク質である。従って、本発明は、プライマー部位と基盤又は支持体に直接結合する性質を持ったDNA分子又は塩基配列決定用鋳型を調製する手段を提供する。本発明の好ましい例においては、そのような鋳型は新しい塩基配列決定方法に適応し、個々の分子が直接塩基配列決定用の支持体に結合できる。そのような塩基配列決定方法は、例えば454Life Sciences、(http://www.454.com/、Helicos BioScience(http://www.helicosbio.com/)、Solexa(http://www.solexa.com/Company/overview.htm)、Visigen、(http://www.visigenbio.com/index.html)、GeneoVoxx GmbH、(http://www.genovoxx.de/)(これらの一部又は全てのウェブページの情報は、参照することによって本明細書に組み込まれるものとする)などが含まれるが、これに限定するものではない。
【0049】
前述の例では、本発明はmRNAの5’末端を修飾する手段を提供する。しかしながら、本発明は5’末端の修飾に限定されるものではない。上述の逆転写反応によるcDNA合成のプライミングと図4〜5に示される例によれば、オリゴdTのような一般的なプライマーやランダムプライマー又は特定の配列をもったプライマーのセットは鋳型RNAからcDNAの第一鎖を合成するのに用いられる。cDNA合成をプライミングするプライマーは、一本鎖DNA、又はWO2006/003721(本文献は、参照することによって本明細書に組み込まれるものとする)に開示されるように部分的に一本鎖と二本鎖部分を持つDNAを含んでも良い。プライマーは、鋳型RNAの中の配列に相補的な配列を含んでも良く、cDNAの後の操作に用いられるようにデザインされた配列又はIdentifier配列として機能する配列を含んでも良い。RNAにハイブリダイズするヌクレオチドはランダム配列でも良く、特定のRNA種にプライミングするために公共のデータベースより得られた配列でも良く、またmRNAのポリA配列にハイブリダイズするようなdTの長いストレッチ配列から構成されるものでも良い。RNA分子の配列にある情報は、NCBI(http://www.ncbi.nlm.nih.gov/)、EMBL(http://www.ebi.ac.uk/Databases/)、DDBJ(http://www.ddbj.nig.ac.jp/)などの当業者が知り得る公共のデータベースを調べることにより得られ、また多くの場合、得られた情報は3’末端の特定のプライマーをデザインするのに充分である。別の例では、そのような配列情報は、スプライス部位やオープンリーディングフレームと呼ばれるコーディング領域の末端の一つのようなRNAの規定領域を標的とするプライマーのデザインに用いられる。逆転写反応のプライミングに用いられるプライマーは、RNAに相補的な配列のみを含んでもよい。この例として、そのような反応より得られたcDNAは、その5’末端にRNAの中の配列に相補的な配列を持つ。別の例では、プライマーは、RNA配列に相補的な配列とRNAの配列には関係のない配列とを含む。よって、本発明は、cDNAの5’末端につなげる新しい配列をcDNAに導入する手段を提供する。当業者が知り得る特定の配列や認識部位への結合に依存する酵素活性は多々あり、多くのそれら酵素は商業的に種々の業者より入手可能である。それらの業者は、FERMENTAS UAB(Vilnius,Lithuania)、New England Biolabs Inc.(Beverly,USA)、Promega(Madison,USA)、Takara(Tokyo,Japan)、Roche(Mannheim,Germany)、GE Biosciences(Cardiff,United Kingdom)を含むが、これに限定されるものではない。通常、制限酵素は二本鎖DNAのみを切断し、一本鎖DNAは切断しない。よって、制限酵素は、第二鎖の合成後にのみ適用することができ、例えば、クローニングなどの目的に用いられる。よって、本発明は、核酸分子の完全長クローニングに関係し、得られたDNA断片より配列情報を得ることができる。特定の例では、制限酵素は、例えばShibata Y et al. in Biotechniques. 31,1048−1049(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されているような方法によって、cDNAからポリA/Tストレッチを除去するのに用いることができる。ポリA/Tストレッチの除去の方法は、RNAの3’末端のタグ配列を得るのに特に重要であり、それらの方法は特許出願US2005/059022、US2005/0255501、WO2004/050918、米国特許第6,136,537号(これらの文献は、参照することによって本明細書に組み込まれるものとする)に開示されているが、これに限定するものではない。
【0050】
ほかの例では、逆転写反応に用いられるプライマーは官能基を含む。そのような官能基は、プライマーがランダムプライマーであるか、特定のプライマーであるか、又はオリゴdTプライマーであるかというプライマーの性質とは関係なく、cDNAに組み入れられる。よって、本発明は官能基を持った修飾された5’末端を持つcDNA断片を調製する手段を提供する。当業者が知り得るところの結合分子へ結合する親和力をもった多くの官能基がある。官能基は反応基、アミノ基、ビオチン、ジゴキシゲニン、抗体、抗原、タンパク質、核酸結合分子などを含むが、これに限定されるものではない。官能基と官能基に付加された任意の分子は、基盤上に存在する結合分子と結合することができる。本発明の目的のために、基盤は、アビジン、ストレプトアビジン、ジゴキシゲニン結合分子、抗体とそのリガンドが固定された任意の形状、および化学的基盤の中より選ぶことができる。用いられる官能基がアミノ基のような反応基の場合、反応基は化学反応で基盤に共有結合するために用いられる。官能基がビオチンだった場合には、基盤はアビジン又はストレプトアビジンになる。同様に官能基がジゴキシゲニンの場合、基盤はジゴキシゲニン結合分子である(Roche Diagnostics GmbH Catalog、本文献は、参照することによって本明細書に組み込まれるものとする)。官能基が抗原の場合は、基盤は抗体またはprotein I又はprotein Gのような抗体結合タンパク質である。修飾されたオリゴヌクレオチドは、多くの業者より商業的に入手可能であリ、ビオチンラベル化オリゴヌクレオチドが頻繁に使われている。種々の修飾オリゴヌクレオチドの調製の例は、MWG Biotech(http://www.mwg−biotech.com/html/s_synthetic_acids/s_modifications.shtml)、Invitrogen(http://www.invitrogen.com/content.cfm?pageid=9900、Operon(http://www.operon.com/)(これらのウェブページに示された情報は、参照することによって本明細書に組み込まれるものとする)のウェブページに記載されている。別の例では、オリゴヌクレオチドはDNA結合タンパク質の認識部位を持っても良い。当業者が知り得るDNA結合タンパク質は多数あり、それらは天然に存在するものでも、タンパク質デザインを使って調製されるものでも良い。そのようなDNA結合タンパク質は、転写因子、ゲノムDNAの認識部位直接又は間接的に結合する調製機能タンパク質などを含むが、これに限定されない。転写因子は生命に不可欠の分子であり、ゲノム情報を利用するのに必要である。あらゆる生命体は多くの転写因子を含んでいる、例えば、Kanamori M. et al.はBiochem Biophys Res Commun,322,787−93(2004)(本文献は、参照することによって本明細書に組み込まれるものとする)に、既知のマウス転写因子すべてのデータベースが公開されている。転写因子は異なる認識部位への親和性という点でほかのものとは区別できる。この特異性は、与えられた転写因子および転写因子のグループの認識部位を含むDNA分子を濃縮するために用いることができる。しかしながら、当業者が特異的配列よりもむしろ構造を認識するタンパク質について知るように、転写因子の結合特異性は、塩基配列への結合に限定されず、例えば、転写因子DAX−1はZazopoulos E. et al.,Nature 390,311−315(1997)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されているように異なるDNA構造に結合する。他の例では、DNA結合タンパク質は、一本鎖DNAに特異的に結合する。一本鎖DNA結合タンパク質は、E.coliのSSB、ファージT4のGene32の産物、アデノウイルスのDBP、一本鎖DNAに対して作られた抗体、仔ウシ胸腺 UP1、またはそれらの混合物などが含まれるが、これに限定するものではない。別の例では、結合タンパク質はMutS又はMutS遺伝子のファミリーであっても良い。さらに別の例では、オリゴヌクレオチドはIdentifier配列として用いられる配列の領域を含んでも良い。このような配列やIdentifier配列は、サンプルの由来の印となったり、核酸やIdentifier配列に相補的な塩基配列を持つ核酸とのハイブリダイゼーションによって、修飾されたRNAやDNAを特異的に捕獲するためのタグとして機能することもできる。また、前述した酵素反応の任意の反応の特異的、選択的なプライミング部位として用いることもできる。Identifier配列は、DNAポリメラーゼによる第二鎖の合成や、例えばPCRによる増幅、一本鎖DNAの調製、又は塩基配列反応のプライミングのための選択的なプライミング部位となることもできる。よって、先述の方法を組み合わせることによって、本発明は複数のRNAの中のRNA分子に由来するcDNAに異なるオリゴヌクレオチドを導入する手段を提供する。
【0051】
先述した任意のステップによって、本発明は、RNAの5’末端に相当する位置に官能基を導入する手段を提供する。加えて、本発明は、RNAの3’末端に相当する位置又はcDNA第一鎖の5’末端に相当する位置に官能基を導入する手段を提供する。従って、本発明は、RNAより生成されるcDNAのいずれかの末端に官能基を導入する手段を提供する。RNA又はcDNAに付加された官能基は、他の分子に対する結合親和力を持ち、また官能基は修飾されたRNA又はDNAを捕獲し表面上に結合するために用いることができる。官能基と結合分子の組み合わせ例は、化学反応により基盤上に共有結合を形成するアミノ基などの反応基、ビオチンとアビジン又はストレプトアビジンの結合、ジゴキシゲニンとジゴキシゲニン結合分子の結合、オリゴヌクレオチドの相補配列への結合、抗原の抗体への結合、抗体のprotein Iやprotein Gなどの抗体結合タンパク質への結合を含むが、これに限定されるものではない。官能基の位置によって、修飾されたRNA又はDNAが表面上へ結合される方法や方向は異なる。例えば、部分的に二本鎖となっているcDNA分子はRNAの5’末端に相当する位置の官能基によって表面に結合することができる。この例では、部分的に二本鎖となったcDNA分子は、RNAの5’末端に相当する位置からcDNA断片の塩基配列決定を開始することができる(図9A参照)。別の例では、一本鎖cDNA分子は、RNAの3’末端に対応する一本鎖DNA分子の5’末端の位置に付加された官能基の相互作用によって表面に結合する。RNAの3’末端に対応する位置でcDNAが表面に結合するとき、外側プライマーはcDNA分子の塩基配列決定を開始する位置を決定するために用いられても良い(図9B参照)。別の例においては、修飾されたRNAまたはcDNAの第一鎖に付加された塩基配列は表面上に存在するオリゴヌクレオチドと相補的な配列を含んでも良い。図9Cは、表面上に結合しプライマーの機能を持つオリゴヌクレオチドにハイブリダイゼーションによって表面上に結合する一本鎖RNAまたはcDNA分子の原理を示している。表面上のプライマーは、その配列に相補的な配列を含むか否かにより、どのRNA又はcDNA断片が表面上に結合できるかを決定する。さらに、表面上のプライマーはcDNA分子の塩基配列決定を始める位置を決定する。固相上の塩基配列決定の例はStahl,S. et al.,Nucleic Acid Research 16,3025−3038(1988)、Lindroos K.et al.,Nucleic Acid Research 29,No.13 e69(2001)(これらの文献は、参照することによって本明細書に組み込まれるものとする)に公表されている。当業者が知り得る規定された配列のオリゴヌクレオチド合成を支持体上で直接行う方法は種々あり、また支持体上にそれらオリゴヌクレオチドを結合する方法も種々ある。そのような方法は、通常マイクロアレイの作成に用いられ、より詳細にはJordan B.,DNA Microarrays: Gene Expression Applications、 Springer−Verlag、 Berlin Heidelberg New York,2001: Schena A, DNA Microarrays,A Practical Approach,Oxford University Press, Oxford(1999)(両文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。従って、本発明は、修飾された核酸分子を調製し、それらをRNAやDNAの塩基配列決定のような解析のために捕獲する手段を提供する。
【0052】
前述の実施態様において、本発明は一本鎖又は部分的一本鎖RNAおよびDNA分子を調製する手段を提供する。官能基を用いて、それらの分子を表面上に結合することができる。表面上の分子は種々のバッファーによって精製や後の操作のために洗浄され得る。よって、本発明は一本鎖DNA、部分的一本鎖DNA、またはRNAを精製する手段を提供する。そのような一本鎖DNA、部分的一本鎖DNA、またはRNAを精製する方法は新しい技術によって達成される一分子検出には必須であり、そのような技術にはMetzker M.L. Genome Res. 15,1767−1776 (2005)、 Kling J.,Nature Biotechnology 23、 1333−1335 (2005)、Shendure J. et al.,Nature Review Genetics 5、 335−344 (2004)(両文献は、参照することによって本明細書に組み込まれるものとする)が含まれるが、これに限定するものではない。従って、本発明は、塩基配列情報を直接得るために、一本鎖DNA、部分的一本鎖DNA、またはRNA分子を利用することに関する。好ましい実施態様においては、本発明は一本鎖DNA断片の規定した領域から塩基配列情報を得ることに関する。好ましい例においては、RNAの5’末端特異的塩基配列情報は、RNA分子の5’末端配列に相補的な配列を含み、本発明によって調製されるDNA断片より得られる。よって、本発明は、RNAの塩基配列情報を得ることに関する。
【0053】
先述の図1から7で示される任意のステップによって、本発明はRNAの5’末端に対応する位置にオリゴヌクレオチドを導入する手段を提供する。加えて、先述の図8に示される任意のステップによって、本発明はRNAの3’末端に相当する位置にオリゴヌクレオチドを導入する手段を提供する。このように、本発明はRNAに由来するcDNAのいずれかの末端に特定の塩基配列を導入する手段と修飾された末端を含むcDNA断片の調製手段を提供する。ある例では、本発明により導入される配列は、一本鎖DNAがその鎖中にある相補的配列の部分で二本鎖を形成し末端に閉じたループを作るようなヘアピン構造を形成することができる。よって、前述の処理によりRNAから向かい合った末端にヘアピン構造を持つように修飾されたcDNA分子を得ることができる(図10参照)。第一鎖cDNA合成がランダムプライマーを用いたかオリゴdTプライマーを用いたかによって、このような分子はRNAに由来する部分的配列を含むか、又はRNAに由来する全体の配列を含む(完全長cDNA)。ある例として、先述に従って調製されRNAの5’末端に相当する末端にヘアピン構造を持つ一本鎖cDNA分子は、直接塩基配列決定ができ、その場合ヘアピン構造は塩基配列決定反応のプライミング部位として機能する。別の例では、先述に従って調製され両端にヘアピン構造を持つ一本鎖cDNA分子は、二つのヘアピン構造を利用して増幅することができる。図11Aに示されるように、各々の末端にループ構造を持つ一本鎖DNA断片は、Notomi T. et al.,Nuc Acids Res. 8,e63(2000)(本文献は、参照することによって本明細書に組み込まれるものとする)に開示されているLoop−Mediated Isothermal Amplification (LAMP法)の鋳型となる。よって、本発明によって調製されるDNA分子は繰り返し配列が得られる方法で増幅することができ、このようなポリマーにあるループ構造は、増幅反応の伸長をプライミングするために用いることができ、又は塩基配列決定反応を推進するために用いることができる。ある例においては、増幅された断片は、25〜50塩基対、又は50〜100塩基対、又は100〜200塩基対、又は200〜300塩基対の長さをとる。別の例では、増幅されたDNA断片は300塩基対以上の長さになる。よって、本発明によって調製されるDNA分子は繰り返し配列の直線状ポリマーが得られる方法で増幅することができ、そのようなポリマーは塩基配列決定反応を推進するために用いることができる。
【0054】
好ましい例において、ループ構造を両端に持つDNA分子は、エクソヌクレアーゼ活性と鎖置換活性を持たないDNAポリメラーゼによって、ヘアピン構造の自由3’末端が伸長される第一の反応を含むステップによって、環状一本鎖DNA分子に転換され得る。このようなDNAポリメラーゼは、M−MuLV Reverse Transcriptase、H Minus M−MuLV Reverse Transcriptase、Superscript II、Superscript III、AMV Reverse Transcriptase、MonsterScript、Expand Reverse Transcriptase、又はそれらの混合物を含む任意の逆転写酵素を含むが、これに限定されるものではない。その他のDNAポリメラーゼは、DNAポリメラーゼIのクレノー断片、T4とT7のDNAポリメラーゼDNAポリメラーゼI、Taqポリメラーゼ、Tfl DNAポリメラーゼ、Tli DNAポリメラーゼ、Tth DNAポリメラーゼ、又は既知の任意のDNAポリメラーゼを含むが、これに限定されるものではない。鎖置換活性の欠如により、DNAポリメラーゼは向かい合ったヘアピン構造の5’末端に達した時に反応が停止する。第二の反応ステップでは、一本鎖DNAの開いた末端が環状DNA分子を作るように結合される。そのような結合反応は任意のDNAリガーゼによって行われ、それらDNAリガーゼは、T4 DNAリガーゼ、E. coli DNAリガーゼ、Taq DNAリガーゼを含むが、これに限定されるものではない。環状一本鎖DNA分子はローリングサークル増幅法(RCA法と呼ばれる)を用いて増幅できる。RCA反応は、DNAポリメラーゼにより環状鋳型上のオリゴヌクレオチドプライマーを等温反応で伸長させることにより遂行される。詳細は米国特許5、854、033号および米国特許6、143、495号(両文献は、参照することによって本明細書に組み込まれるものとする)に開示されている。反応生成物は直線状環状DNAで、鋳型のコピーが直列に並んでいる。反応の時間や条件によって、反応生成物は数十、数百又は数千の鋳型のコピーを一分子中に持つ。RCA反応で特に用いられるDNAポリメラーゼは、当業者に公知の等温DNA増幅に必要とされる強い鎖置換活性を持つphi29 DNAポリメラーゼがあるが、これに限定されるものではない。当業者に公知の数多くのCA法の応用や改変法がある。RCA法に関するより詳細な記載は総説Gusev、Y.et al.,American J.Pathology 159,63−69(2001)、Zhang D.et al.Clin.Chim.Acta.363,61−70(2006)(これらの文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。従って、本発明により調製されたDNA分子は、繰り返し配列の直線状ポリマーが得られる方法により増幅することができ、それらのポリマーは塩基配列決定反応を推進するのに用いられる配列を含む。
【0055】
一つの例では、RCA反応は、反応生成物が直接又は間接的に規定された位置又は検出、解析、又は塩基配列決定のポイントと呼ばれる位置に結合できるような方法で実行される。一つ例では、Nallur G.et al.,Nucleic Acid Res,29,e118(2001)に開示されているガラススライド上の信号増幅に仲介されたRCAの手段がある。この例では、RCAは、複数の核酸分子の中の個々の標的のクローン増幅を実行するためのステップを可能とし、そのステップでは各々の分子は表面上の規定された位置で増幅される。ここでは、RCA反応は、例えば従来のPCR法に見られるような増幅バイアスの危険を伴うことなく、高い並列性を持った方法で実行される。RCA反応で表面上に結合されたプライマーを使うことにより、反応生成物が並んだ基盤を得ることができ、各々の反応生成物は一分子の中に鋳型のコピーを複数含んでいる。よって、RCA反応は検出や解析の感度を大きく増幅することができ、規定の位置で信頼のおける塩基配列決定反応を実施することを可能とする。
【0056】
別の例では、RCA反応は規定の位置での検出、解析および塩基配列決定のための鋳型を調製することができ、鋳型は一つ又はそれ以上の検出ステップ、解析、又は塩基配列決定反応の対象となる。従って、本発明は鋳型の塩基配列決定を行う第一のステップと、塩基配列決定反応中に鋳型より生成された増幅産物を除去する第二のステップと、異なるプライマーによって同じ位置で同じ鋳型の塩基配列決定を再度行う第三のステップからなる手段を提供する。そのような一連の反応は、一つの鋳型より二回、三回、四回、五回、さらには五回以上回数を重ねて塩基配列決定を行い、解読配列を得るために実施されても良い。DNAの表面への共有結合的な結合は、Beier M.and Hoheisel J.D.,Nucleic Acid Research、27,1970−1977(1999)によれば、少なくとも30サイクルのハイブリダイゼーションとハイブリダイズしたDNAの除去を可能とすると記載されている。よって、本発明は一つの鋳型から一種以上の配列情報を得る手段を提供し、その異なる塩基配列決定反応は同じ位置で実行され、その位置により同じ鋳型より得られる違う解読塩基配列のつながりが規定される。
【0057】
別の例では、RCA反応は、もとのRNA分子のセンス鎖およびアンチセンス鎖の多数のコピーを含む反応生成物を調製するために用いられる。そのような反応生成物は、図11Bで示されるステップによって環状鋳型が調製される場合に得られる。環状鋳型は、RNAより得られる二本鎖cDNAの中にあるセンスとアンチセンス鎖を含み、もとのRNAの向かい合った末端に対応する位置にヘアピン構造によりつながっている。この鋳型は双方向性である。本発明の好ましい例では、ヘアピン構造はセンス、アンチセンス鎖がそれらの末端からの塩基配列決定を可能にするプライミング位置を含む。よって、RNA分子の向かい合った末端より末端配列を得る手段を提供する。このとき、RNA分子はcDNAに転換され、cDNAは最初のRNA分子配列を持つセンス、アンチセンス鎖を持つ二本鎖から成り、二本鎖cDNAにあるセンス、アンチセンス鎖は一本鎖DNAを含む環状分子を構成するヘアピン構造により連結され、環状一本鎖DNA分子はセンス、アンチセンス鎖を含む塩基配列決定のための直線状鋳型DNAを生成するためにRCAにより増幅され、DNA又はもとのRNAの向かい合う末端から得られる配列情報は、二つ又はそれ以上の連続した塩基配列決定反応が同じ位置で行われることにより得られる。この例では、本発明は、RNAの末端配列、転写物の境界、ゲノム中の転写の開始、終止点、又は任意のDNA分子の末端配列を決定するために用いることができる。別の例では、本発明は、RNA、cDNA、又はゲノムDNA内における規定された領域の末端配列を決定するために用いることができる。そのような規定された領域の境界は、それらの調製段階における特定のステップによって決定される。その断片は、生物学的由来のものでも良く、また完全なランダム切断によって生成されても良い。さらに、塩基配列決定用のプライマーは、従来のプライマーウォーキング法と同じように、鋳型の任意の領域にハイブリダイズするようにデザインされることができ、又は鋳型中の特定の部位、例えばスプライス部位などに対してデザインされても良い。例えば環状一本鎖DNA分子を形成するために、直線状二本鎖DNAの末端にヘアピン構造を持ったオリゴヌクレオチドを連結することによって、任意の二本鎖DNAを双方向性鋳型へ転換することは、本発明の範囲内である。環状一本鎖DNA分子は、RCAによって増幅され、もとのDNA分子のセンス、アンチセンス鎖を含む塩基配列決定用の直線状鋳型DNAが生成される。DNAの向かい合った末端からの配列情報は、同じ位置で行われる二つ又はそれ以上の連続した塩基配列決定反応によって得られる。この例では、本発明は、例えばエクソンの末端配列やクロマチンIPより得られたゲノム断片の末端配列、ハイパーセンシティブ部位の境界配列などを決定するために用いることができる。
【0058】
別の実施態様では、本発明は、RNAの5’末端やRNAの3’末端に相当する領域に導入されたIdentifier配列の利用に関する。先述のとおり、本発明によって調製されたcDNAの向かい合った末端にIdentifier配列、又は特定の配列を導入する手段を提供する。好ましい例では、Identifier配列は、RNAまたはcDNAの末端の近傍に位置し、RNAやcDNAそのものから配列情報をえるための同じ塩基配列決定反応より塩基配列を決定することを可能とする。Identifier配列は、1塩基対、2塩基対、3塩基対、4塩基対、5塩基対、6塩基対、6〜10塩基対、10〜15塩基対、15〜20塩基対、又は20塩基対以上の長さでも良い。好ましいIdentifier配列は、6〜12塩基対、又は25〜75塩基対の長さである。Identifier配列は任意の性質、ランダム配列であっても良い。それらはコンピューターによってデザインされても良く、生物学的サンプルからとっても良く、また人工的に作られても良い。それらは、制限酵素又はその他の酵素やタンパク質の認識部位やプライミング部位を含んでも良い。Identifier配列は、以下のすべて又は任意のいずれかの法則によってデザインされることができる。
− 塩基配列決定またはハイブリダイゼーションによる同定を行うのに充分な長さを持つ
− 同じ実験で用いる異なるIdentifier配列の塩基配列は、区別可能である。
− 同じ実験で用いる異なるIdentifier配列は、Identifier配列の塩基配列決定にエラーが起こった場合でも明確な同定を可能にするため、一つの位置以上において区別できる。
− Identifier配列は、塩基配列決定反応を妨げるような配列や構造をとる塩基配列を避ける(例えばGリッチ配列、パリンドローム配列)。
− Identifier配列は、相補的配列を安定なハイブリッドを形成するように選択されても良い。
− Identifier配列は、制限酵素や転写因子のようなタンパク質への結合や特定の操作を可能にする塩基配列を持っても良い。
− Identifier配列は、本発明を実施する際にRNAやDNAの操作を妨げる塩基配列を避ける(例えば、操作の過程で使用される制限酵素の認識部位を持たない)。
【0059】
本発明の好ましい例では、Identifier配列は、サンプルのプールの中で、サンプルの由来を印付けるために用いられ、サンプルプールのすべての構成物は同じ実験で同時に扱われる。サンプルプールのサンプルは、出来るだけ早く混合されるべきであり、好ましくは修飾されるRNAサンプルの段階で混合されているべきである。異なるIdentifier配列を持つ異なるRNAサンプルの混合によって得られるサンプルは「プールサンプル」となり、異なる形で修飾されたRNAを含む(図12B参照)。それゆえ、Identifier配列は、好ましくはRNAの5’末端近くに位置し、それゆえmRNAやトータルRNAの修飾のための最初のステップで導入される。
【0060】
一つの実施態様では、Identifier配列は、異なる生物種や組織、生物学的実験の種々の処理あるいは時間的な段階、あるいは異なる細胞種を含む多数の生物学的サンプルから得られる特定のRNAにある核酸分子を印付けるために用いられる。実験のデザインにおいてサンプルのプーリングは、種々の機能的な意味を持つ。例えば新しい塩基配列決定方法のハイスループット性を十分に活用するためにサンプルの複雑性を高めること、同時に扱うサンプルの数を減らすことにより多くのサンプルの処理を単純化すること、特定のデータ解析の形態を可能にすることなどを含むが、これに限定されるものではない。一つの好ましい実施態様において、異なるサンプルに異なるシステムエラーが起きるような個々の実験と比較し、共通の統計的解析のためにすべての操作のステップにおいて同じシステムエラーがあるようにするためサンプルはプールされる。例えば、一つの典型的な応用において、Identifier配列は、本発明によってRNAが修飾される際にRNAの5’末端近傍に付加され、その後修飾されたRNAサンプルはcDNAの調製より先に混合される。プールサンプルは、区別できるIdentifier配列を持つ修飾されたRNAサンプルの異なる種類の混合物であるように調製される。修飾されたRNAサンプルのプールは、その後データを得るために本発明に沿って一つのサンプルとして処理される。例えば、修飾されたRNAサンプルの解読配列はプールライブラリーより得られる。配列情報はこの分野で知られる任意の方法によって決定することができるが、好ましくは、各々の解読配列がIdentifier配列ともとのRNA又はcDNAサンプルに由来する配列情報の両方を含む。配列の決定後、個々の解読配列からIdentifier配列を認識し、同じIdentifier配列を持つ解読配列をその後の解析のためにグループ化するためにコンピューターにより処理されることができる。もとのRNAに関係する塩基配列は、Identifier配列とは分けて実験的なデザインの必要に沿って解析される。これらの塩基配列は、「配列タグ」と呼ばれるものやRNAやcDNAに由来する部分的な配列を含む短い解読配列に関連する。配列タグは、転写物やゲノム中の位置を同定するために用いることができ、あるいはプールサンプルの中の転写物の発現レベルを統計的に解析するために用いても良い。配列タグの利用についてより詳細な情報は、Harbers M.and Carninci P.,Nature Meth.2,495−502(2005)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。配列情報は、データベースに格納され、または解析、保管、又は参照データセットの構築の目的でコンピューターに格納される。そのようなデータベースは、例えば、塩基配列情報、異なる組織、細胞株より得られた配列タグの発現頻度、ゲノム中の転写物、遺伝子、機能要素に関する注釈付データをふくむことができる。
【0061】
別の例においては、Identifier配列は塩基配列決定によって同定されず、基盤や支持体に結合したIdentifier配列に相補的な配列を持つ核酸分子と特定のハイブリッドを形成するために用いられる(図12C参照)。この例では、Identifier配列は、表面上の規定された位置に同じ由来から生成されたサンプルをグループ化するために用いられる。その位置は、Identifier配列の性質やプールサンプルの中のRNA、DNA、サンプルの由来を明らかにするために用いられる。従って、Identifier配列の読み取りは直接塩基配列決定を必要とせず、かわりに特定のハイブリダイゼーション反応に行うことができる。
【0062】
別の例では、Identifier配列は、塩基配列決定やハイブリダイゼーションによって同定されず、支持体や基盤に結合したIdentifier配列に結合親和性をもったタンパク質に特異的に結合するために用いられる。この例では、Identifier配列は、同じ由来のサンプルを表面上の規定された位置にグループ化するために用いられる。その位置は、Identifier配列の性質やプールサンプルの中のRNA、DNA、サンプルの由来を明らかにする。従って、Identifier配列の読み取りは直接塩基配列決定やハイブリダイゼーションを必要とせず、かわりにIdentifier配列に高い親和性を持つたんぱく質に結合することによって行われる。
【0063】
本発明は、コンピューターによる注釈付を目的とし、本発明に従って得られるDNA断片の領域の塩基配列決定に関係する。注釈付は、統計的解析、参照情報に対するアライメント、およびゲノム配列へのマッピングなどを含む。よって、本発明は、遺伝子発見、遺伝子同定、遺伝子発現プロファイリング、注釈付の手段に関する。
【0064】
他の実施態様では、in situハイブリダイゼーションによる領域解析のために、核酸分子の末端からハイブリダイゼーションプローブを調製することに関する。好ましい例では、in situハイブリダイゼーション実験は、タイリングアレイを用いて行う。この実施態様では、本発明は、ハイブリダイゼーションプローブのデザインに関する。プローブはマイクロアレイ上のものを含むがこれに限定されるものではない。
【0065】
よって、本発明は、例えば生物学的サンプルの特性解析のような必要に応じて、核酸分子やそれらの短い断片を解析する手段を提供する。さらに、本発明は、RNAやDNA断片を解析的検定に利用するために効率的で迅速な操作を行う手段を提供する。この意味では、本発明は、新しいハイスループット塩基配列機器と技術を利用するための手段を提供する。
【0066】
他の実施態様では、逆転写反応に特定プライマーを用いて、本発明に従って調製された修飾されたRNAは、当業者が知るRACE法と類似のプロトコールによって実際の5’末端配列を決定するために用いられる。
【0067】
本発明は、生物学的サンプルの状態、すなわち遺伝情報や発現プロファイル、調節パスウェイやネットワークの活性などを調べるための重要な情報を得るために必要な手段を提供する。よって、本発明は、ライフサイエンスや診断の研究に用いられる解析的検定のデザインや実行に関する。本発明は、生物学的システムの解析と診断の手段を提供する。
【0068】
本発明、又はその一部は、試薬、核酸分子、およびRNAの操作やDNAの調製に必要な酵素、その他の要素を含むキットの製造に用いることができる。一つの実施態様では、キットはRNAを修飾するために必要な試薬を提供する。別の実施態様では、キットは鋳型DNAを調製するために用いる試薬を提供する。好ましい実施態様では、キットは一分子検出のための鋳型を調製するための試薬を提供する。好ましい実施態様では、キットは研究目的の試薬を提供する。さらに好ましい実施態様では、キットは診断的な検定のための試薬を提供する。
【0069】
本発明の主要なステップを詳細に説明する。ここで用いるすべての名前や省略形は、当業者に公知の意味で用いられる。
【実施例1】
【0070】
[RNAの単離]
本発明を実施するために、mRNAまたはトータルRNAのサンプルは,当業者に公知の方法、例えばSambrook J.and Russuell D.W.,Molecular Cloning,A Laboratory Manual、Cold Spring Harbor Laboratory Press,New York(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に開示されている方法により調製された。さらにCarninci P.et al.(Biotechniques 33,2002,306−309、本文献は、参照することによって本明細書に組み込まれるものとする)によれば、細胞質mRNAの分画を得る方法が開示されている。細胞質mRNAの分画を用いるのが好ましいが、本発明はこれに限定されるものではなく、他の任意のmRNAやトータルRNAの調製方法を用いても本発明を同様の方法で実施することができる。
【0071】
本発明を実施するには、トータルRNAを用いると完全長cDNAライブラリーを調製する際に行うCap−selectionとの組み合わせで十分は結果を得られるので、本発明を実施するためには、全RNA又は細胞質RNAからのmRNAの調製が好ましいが、必須ではない。mRNAの量はトータルRNAの調製の約1〜3%であり、その後、商業的に販売されているオリゴdT−セルロースマトリックスを使ったキットを用いて調製される。このような商業的キットは、本発明を実施するためにmRNAの分画を調製するのに用い、推奨される条件下で十分なmRNAを得られるMACS mRNA分離キット(Milteny)を含むがこれに限定されるものではない。本発明を実施するために、1サイクルのオリゴdT−mRNA選択で十分であり、強いmRNA精製は長いmRNAを失う可能性がある。
【0072】
本発明の実施に用いられるすべてのRNAサンプルは、RNAの純度を調べるために230、260、280nmのOD比率が計測された。230/260比が0.5より低い場合には、多糖類の除去が成功したと考えられ、260/280比が1.8より高いもしくは2.0付近のとき、効果的なタンパク質の除去が行われたと考えられた。RNAサンプルはさらに、トータルRNA調製時の28Sと18S rRNAの比率が適当であることを確認し、RNA分画の品質を調べるために、アガロースゲルの電気泳動によって解析された(rRNAのサイズは哺乳類以外の種から得られるトータルRNAではことなることがある)。
【実施例2】
【0073】
[5’末端誘導体化RNA分子のライブラリーの調製]
本実施例は、RNAオリゴヌクレオチドを用いてRNA分子の5’末端を誘導体化する典型的なプロトコールである。すべての反応は、核酸の損失を避けるために、500μlのシリコン化したマイクロチューブとシリコン化したチップを用いて行われた。
【0074】
RNAサンプルは、最初に脱リン酸化された。RNA(例えば1ngから1μg)と2μgのグリコーゲンを一緒にチューブに加え、全量5μlに調製された。反応バッファーは、通常の濃度の1/10または、5mMのBis−Tris−Propane−HCl、0.1mMのMgCl2、0.01mMのZnCl2(pH 6.0)、25°Cを用いた。グリコーゲンは、操作中にRNAがプラスチックに付着するのを避けるために加えられた。サンプルを65℃で5分間変性し、後で除去するためにリン酸基を露出させた。その後37℃で2分間置き、Anctartic phosphatase(New England Biolabs)を2.5ユニット加えた。サンプルは37℃で3時間から一晩処理された。一晩脱リン酸化を行うことにより、リン酸基の98〜99%が除去された。45℃で活性が高くなるトレハロースを最終濃度0.6Mとなるように加え、45℃で短いインキュベーションを行うこともできる。
【0075】
その後、Anctartic phosphataseは65℃で不活性化されたが、この処理の前に二価イオンをキレートする必要があった。このため、0.55μlの溶液(0.5Mの酢酸ナトリウム(pH 6.0)、10mMのEDTA、1%のβ−メルカプトエタノール及び0.1% Triton X−100)を加えた。EDTAは、二価イオンをキレートし、次のTAP処理に適した条件となった。Anctartic phosphataseも、またバッファー中のEDTAによって阻害された。不活性化は65℃で5から15分間行われた。
【0076】
次に、0.2μl(2ユニット)のタバコ酸ピロホスファターゼ(TAP)を加えdecappingを行った。TAPは、実験当たり20ユニットまで増やすことが可能であった。反応は37℃で2時間行い、続いて65℃で15分間熱不活性化を行った後、サンプルを氷上で冷却した。また、GCリッチなRNAの二次構造を解くために役立つベタインを1M加えても良い。この処理の後、TAPはATPをそれ以上分解しなかった。ATPはその後のステップにおいて必要である。その後、任意の配列の5μMの「capping RNA」オリゴヌクレオチドを加え、ライゲーションを行った。反応液6.75μlに対して2μlのRNAリガーゼ(500mMのHEPES−NaOH(pH8.0、25°C)、100mMのMgCl2、100mMのDTT)を加えた。DTTはTAPを阻害した。1mMのhexamino cobaltum chloride(HCC)を加えても良いが、これは任意であり、必須ではない。ポリエチレングリコール(PEG8000)を最終濃度が25%となるように加え、ATPの125μM、T4 RNAリガーゼ(Fermentas)10ユニットを加えた。このような条件では、最終的な混合液はライゲーションステップを阻害するものではなかった。
【0077】
2時間から一晩(16時間)20℃でライゲーション反応を行った。この時点で、RNAのCap構造はオリゴヌクレオチドに置き換えられ、これは完全長cDNA調製のような他の例で示すように、他のテストに用いられる。
【実施例3】
【0078】
[修飾RNAの調製に用いられる酵素の活性試験]
実施例2で用いたれた酵素とバッファーの活性試験を行った。
(A)Antarctic Phosphatase(New England Biolabs)の活性試験
5’リン酸化オリゴヌクレオチドを次のバッファー中、37℃で2時間脱リン酸化を行った。続いてオリゴヌクレオチドをT4リガーゼとγ−32P−ATPによってRIラベルし、PAGEによって解析した。脱リン酸化されていなければ、5’リン酸基のためにRIラベルは不可能である。
【0079】
(B)タバコ酸ピロホスファターゼ(TAP)(Epicentre)活性試験
γ−32P−ATPを2ユニットのTAPと反応バッファー中でインキュベートした。放射性ATPとインキュベーションを行う前に、TAPを15分間加熱した。
【0080】
(C)T4 RNAリガーゼ(Fermentas)の活性試験
放射性ラベル化オリゴヌクレオチドを、ラベルしていないオリゴリボヌクレオチドの存在下でインキュベートした。ライゲーションの結果、ポリアクリルアミドゲルの移動度が変化した。
【実施例4】
【0081】
[修飾RNAからの完全長cDNAの調製]
上記で調製したサンプルをmicrocon YM−100フィルターを使い、製造者(Millipore)の説明書に従って脱塩した。ライゲートされたRNAに水、逆転写酵素、プライマー(Invitrogen)を加えた。プライマー(AGA GAG AGA CCU CGA GCC UAG GUC CGA C)800ngを使い、3.3Mのソルビトール−トレハロース混合液を、最終濃度が0.5Mのソルビトール及び4%トレハロースになるように加え、逆転写反応液を調製する。RNAとプライマーの混合液を65℃で10分間加熱した後、他の試薬を準備する間氷上で保持する。11μlの2倍濃度GCバッファー(Carninci,Shiraki et al,Biotechniques,2002;32,984−985、本文献は、参照することによって本明細書に組み込まれるものとする)から構成されるプレミックスを加え、その後、10mMのdNTPを1μl及びMMLV逆転写酵素(RNaseH minus、Fermentas)を1μl加えた。GCバッファーシステムは、製造者の推奨するバッファーに置き換えた。この反応液にRNAサンプルを加え、25℃で2分間(サンプルのアニーリングのため)、42℃で30分、52℃で10分、56℃で10分インキュベートした後、反応を停止した。この方法では、もとのmRNAの5’末端までのびたcDNAを高い頻度で得ることができた。その後、精製、処理された。例えば、プロテアーゼKにより処理を行った(20μgをEDTA(最終濃度が10mM)と一緒に加え、95℃で15分インキュベートした)。このサンプルを、サイズ分画しプライマーを除去するため、C14B(Amersham−Pharmacia)で処理した。
【実施例5】
【0082】
[cDNAの第二鎖合成とPCR増幅]
cDNAをPCRにより増幅した。cDNAに最終濃度が1倍となるようにTAKARA EXバッファーを加え、さらにdNTP(最終濃度200μM)、5’オリゴヌクレオチド(配列: acc tcg agc cta ggt ccg ac)、3’オリゴヌクレオチド(配列:ca gcg tcc tca agc ggc cgc)をそれぞれ400nM、MgCl2を2.5mM、KClを最終濃度50mMとなるように加えた。成分を混合した後、94℃で5分加熱し、94℃で30秒、58℃で30秒、68℃で1.5分のサイクルを30サイクル繰り返した。
【0083】
生成された5’末端cDNAを平滑末端化し、標準的な方法によりプラスミドベクターへクローニングした(クローニングと塩基配列決定の一般的な情報についてはSambrook et al.,supra参照)。
【実施例6】
【0084】
[RACE法の実験]
Capped RNAを実施例2と同じようにRNAオリゴヌクレオチドの配列のみ下記の様に変更して調製した。実施例2のプロセスを用い、次にPCRを行い、RACEによる5’末端増幅が可能となった。500ngの肝臓トータルRNAをONE−Tube oligo−capping法で処理し、未反応のオリゴリボヌクレオチドを除去、その後ランダムプライマーを用いて逆転写反応を行った。5’末端の増幅には下記の遺伝子特異的プライマー(TTGGAGAGAGGGTTTCGACGAGTCA)と、oligo−capに相補的なプライマー(CGACTGGAGCACGAGGACACTGA)を用いた。
【実施例7】
【0085】
[454塩基配列決定の応用と他の基盤]
実施例2の方法でcDNAを調製したが、オリゴヌクレオチドのデザインと調製は、RNAの3’末端と5’末端に異なるアダプターを付けるため下記の通りとした。
アダプターA
CCATCTCATCCCTGCGTGTCCCATCTGTTCCCTCCCTGTCTCAG
アダプターB
/5BioTEG/CCUAUCCCCUGUGUGCCUUGCCUAUCCCCUGUUGCGUGUCUCAG
【0086】
アダプターBは「oligo−capping」配列として用い、アダプターAは第一鎖合成のためのオリゴランダムプライマーと一緒に用いた。第一鎖合成後、Cl−4Bスパンカラムを用いて未反応のプライマーと分離した。続いて、サンプルをエマルジョンPCRにかけた。塩基配列決定反応は454−Life Science sequencing instrumentに沿って行った(Margulies et al,Nature,2005; 437(7057):376−380、本文献は、参照することによって本明細書に組み込まれるものとする)。この結果、1ランで数十万の解読配列を得ることができた。
【実施例8】
【0087】
[5’末端塩基配列タグの応用]
実施例2とそれに続く実施例のようにcDNAを合成し、当業者が知る標準的な方法でcDNA第二鎖の合成を行った。例えば、Kodzius et al.,Nat. Methods.2006 Mar; 3(3):211−22(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。DNAをMme Iにより切断し、第二リンカーを付加、増幅、精製を行い、コンカテマーを作成した。この方法の詳細は種々に記載されているが、例えば、Kodzius et al.,Nat. Methods. 2006 Mar; 3(3):211−22(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。これらの塩基配列タグは、その後塩基配列決定、遺伝子の境界の同定(Carninci et al,Science. 2005 Sep 2; 309(5740):1559−63、本文献は、参照することによって本明細書に組み込まれるものとする)、発現プロファイリング、遺伝子のプロモーター(Harbers and Carninci,Nat. Methods,2005 Jul; 2(7): 495−502、本文献は、参照することによって本明細書に組み込まれるものとする)などに用いることができる。
【実施例9】
【0088】
[固体支持体に結合したDNAの塩基配列決定]
実施例2とその後の例にそってcDNAを合成し、当業者の知る標準的なプロトコールを用いてcDNAの第一鎖を合成した。それらは例えば、Kodzius et al.,Nat. Methods. 2006 Mar、 3(3):211−22に記載されている。続いて、核酸を米国特許出願番号20060012793号、米国特許出願番号20060012784号、米国特許出願番号20060008824号に開示された方法により固相基盤に結合した。
【図面の簡単な説明】
【0089】
【図1】図1は、RNAの酵素的修飾の概念図である。この図にあるように、RNA調製はmRNAの5’末端のCap構造の存在によって特徴づけられたRNAの種類を含んでも良い。Cap構造は、他のCap構造を欠き、自由リン酸基を5’末端に持つ切断されたRNA種から区別するために使われる。一連の酵素反応の中で、まずホスファターゼにより、切断されたRNAの自由リン酸基が除去される。次にピロホスファターゼにより完全長mRNAよりCap構造が取り除かれ、Cap構造の代わりにリン酸基が露出する。5’末端にリン酸基をもったRNA分子のみが、RNAリガーゼによってオリゴヌクレオチドと共有結合して修飾を受けることができる。オリゴヌクレオチドは斜線で示されている。
【図2】図2は、オリゴヌクレオチドの付加によるRNAの修飾を概念的に示したものである。図1で示されたその後のステップの前に、RNAリガーゼは一つ又は複数のRNA分子に異なる種類のオリゴヌクレオチドを結合させるのに用いられる。本発明によれば、オリゴヌクレオチドはデオキシリボヌクレオチドまたはDNAオリゴヌクレオチド、又はリボヌクレオチドまたはRNAヌクレオチドからなるホモポリマーであることが可能である(パネルA)。またデオキシヌクレオチドとリボヌクレオチドからなるヘテロポリマーとなることも可能である(パネルB)。あるいは、官能基(パネルC中で「F」で示されている)をもった任意の種類のホモポリマー又はヘテロポリマーとなることもできる。
【図3】図3は、RNAの5’末端へオリゴヌクレオチドを連結する代替法の概念図である。図1、2で示すように、RNAリガーゼはRNAのリン酸化された5’末端にオリゴヌクレオチドを結合させることができる。しかし、そのような反応はオリゴヌクレオチドの3’末端のリボヌクレオチドに依存することもあるが塩基配列の特異性はなく、任意のオリゴヌクレオチドがRNA分子と結合することができる。反応に配列特異性を与えるには、部分的に二本鎖になっているオリゴヌクレオチドを用い、突出領域をRNA分子の5’末端の配列とハイブリダイズさせる。反応によっては、突出領域はランダム配列でも良いし、特定のRNA分子を標的とした決められた配列でも良い。さらに、部分的に二本鎖になったオリゴヌクレオチドはDNAリガーゼを用いてRNAにオリゴヌクレオチドを連結させるのに用いることもできる。その後、適当なプライマーの結合、逆転写酵素の処理、RNAの分解を行い、cDNAを得る。下記でさらに詳細とバリエーションを述べる。
【図4】図4は、ランダムプライミングを用いたcDNA第一鎖合成の概念図である。図1から3に示されるすべてのステップ、又は部分的なステップに沿って得られた単一又は複数の種類の修飾されたRNA分子は、逆転写酵素を用いて鋳型RNAのコピーcDNAを合成する反応に用いることができる。この反応は鋳型RNAにハイブリダイズしDNA合成を開始するプライマーが必要である。この図に示されている例では、3’末端にランダム配列(図中NNNNで示す)とそれに続く確定した配列を持つプライマーセットが用いられている。任意のこのようなプライマーセットが、塩基配列および5’末端に付加したオリゴヌクレオチド(パネルAからC)から得られる官能基を持った修飾RNAを鋳型として行われるDNA合成のプライミングに用いることができる。
【図5】図5は、オリゴdTプライマーを用いたcDNA第一鎖合成の概念図である。図1から3に示されるすべてのステップ、又は部分的なステップに沿って得られた単一又は複数の種類の修飾されたRNA分子は、逆転写酵素を用いて鋳型RNAのコピーcDNAを合成する反応に用いることができる。この反応は鋳型RNAにハイブリダイズしDNA合成を開始するプライマーが必要である。この図に示されている例では、多種類のmRNAの3’末端に通常見られるポリA配列にハイブリダイズするオリゴdTプライマーが用いられている。オリゴdTプライマーは、塩基配列および5’末端に付加したオリゴヌクレオチド(パネルAからC)から得られる官能基を持った修飾RNAを鋳型として行われるDNA合成のプライミングに用いることができる。
【図6】図6は、DNA/RNAの複合体からRNA除去を示す概念図である。図4、5で示すRNAを鋳型としたDNAの合成はDNA/RNA複合体を形成する。このような任意のDNA/RNA複合体分子のRNA部分はRNA分解酵素又は反応バッファーのpHを変えることによって取り除くことができる。例えば、RNase H酵素は特にDNA/RNA複合体のRNAを分解するので本発明の実施には好ましい酵素である。このような任意のDNA/RNA複合体に対する処理はDNA鎖を一本鎖DNA(パネルA)として、又は部分的に二本鎖のDNA分子として遊離する。部分的に二本鎖のDNA分子には前のステップでRNA分子に付加されたオリゴヌクレオチド(パネルB〜D)の部分がある。図にはオリゴdTを用いて得られるcDNAが描かれているが、ここで述べるRNAの除去は逆転写反応に用いられるプライミングの種類によらず任意のcDNAにも適用できる。
【図7】図7は、本発明によって得られるDNA分子の応用の概念図である。本発明によって得られる図1〜6のいずれかに描かれている鋳型DNAは、例えば図2と3にある初期の段階でRNAに付加されたオリゴヌクレオチドの性質の点で特徴を持っている。基本的な構造と応用はパネルA〜Dに示されている。
【図8】図8は、cDNAの3’末端への官能基の導入を示した概念図である。図4と5で示すステップによって通常のオリゴdTプライマーまたはランダムプライマーが鋳型RNAからの第一鎖cDNA合成をプライミングするために必要である。そのようなプライマーは鋳型RNAの配列と相補配列領域を含んでも良く、後のcDNA操作に用いるようにデザインされた配列を含んでも良い。また、図中「F」で示されているようにプライマーに官能基が付加されていても良い。そのような官能基は、ランダムプライマー(パネルA)又はオリゴdTプライマー(パネルB)のどちらを用いるかに関わらずcDNAに導入することができる。従って、本発明は修飾された末端をもつcDNA断片を調製する手段を提供する。
【図9】図9は、修飾されたDNA断片の捕獲を示した概念図である。図1〜7のいずれかに示されたステップに従って本発明はRNA分子の5’末端に相当する位置に官能基を導入する手段を提供する。加えて、図8に示されたいずれかのステップに従って本発明はRNA分子の3’末端に相当する位置に官能基を導入する手段を提供する。本発明はRNAに由来するcDNAのいずれかの末端に官能基を導入する手段を提供する。図中「F」で示されcDNAに付加される官能基が図中の開いたクランプで示される他の分子に対して結合親和性を持っている場合、官能基は修飾されたcDNAを捕獲し表面上に結合させるために用いることができる。パネルAは官能基と表面上の結合相手の反応によって部分的二本鎖を持つcDNAが表面上に結合される原理を示している。cDNA分子の部分的に二本鎖になっている領域は、RNAの5’末端に相当するcDNA断片の末端からの塩基配列決定を可能にする。パネルBは官能基と表面上の結合相手の反応によって一本鎖cDNAが表面上に付けられる原理を示している。cDNAはRNAの3’末端に相当する位置で表面上に結合するのに対し、cDNA分子の塩基配列決定を始める位置は外部プライマーを用いて決めても良い。パネルCは表面上に結合したオリゴヌクレオチドまたはプライマーとハイブリダイゼーションすることによって一本鎖cDNA分子が表面上に結合する原理を示している。表面上のプライマーによって、cDNA断片に含まれる表面上プライマーとの配列相補性に基づいてどのcDNA断片が表面に結合しているかを決定することができる。さらに、表面上プライマーはcDNA分子の塩基配列決定の開始位置を決定する。
【図10】図10は、ヘアピン構造の導入を示す概念図である。図1〜7に記載のいずれかのステップによって、本発明はRNAの5’末端に相当する位置にオリゴヌクレオチドを導入する手段を提供する。加えて、図8に示されたいずれかのステップに従って本発明はRNA分子の3’末端に相当する位置にオリゴヌクレオチドを導入する手段を提供する。本発明は、cDNA断片に修飾された末端を与えるために、RNAに由来するcDNAのいずれかの末端に特定の配列を導入する手段を提供する。図で示されるプロセスでは、本発明により導入される配列は、一本鎖DNAがその鎖中にある相補的配列の部分で二本鎖を形成し末端に閉じたループを作ってヘアピン構造を形成することができる。ここで開示された手段を用いて、RNAに由来するDNA分子は、cDNA合成にランダムプライマー(パネルA)を用いたかオリゴdTプライマー(パネルB)を用いたかに関わらず、向かい合った末端にヘアピン構造を含むように修飾されることができる。
【図11】図11は、修飾されたDNA断片の増幅を示した概念図である。図10で示されたステップによって調製されたDNA断片は、二つの向かい合った末端にあるヘアピン構造を使って増幅することができる。パネルAに示すように、向かい合った末端にループ構造を持つ一本鎖DNA断片はLAMP法と呼ばれるLoop−Mediated Isothermal Amplification(Notomi T. et al.,Nuc. Acids Res. 8,e63,2000、本文献は、参照することによって本明細書に組み込まれるものとする)の鋳型となることができる。本発明により調製されたDNA断片は繰り返し配列を含むポリマーが得られる方法で増幅することが可能であり、このようなポリマーが含むループ構造は増幅反応の伸長をプライミングするために用いることができ、また塩基配列決定反応を推進するために用いることができる。パネルBに示されるように、各々の末端にループ構造を持つDNA断片は、エクソヌクレアーゼ活性と鎖置換活性を持たないDNAポリメラーゼによって、ヘアピン構造の自由3’末端が伸長される第一の反応を含むステップによって、環状一本鎖DNA分子に転換され得る。鎖置換活性の欠如により、DNAポリメラーゼは向かい合ったヘアピン構造の5’末端に達した時に反応が停止する。第二の反応ステップでは、一本鎖DNAの開いた末端が環状DNA分子を作るように結合される。環状一本鎖DNA分子はRCA法によって増幅することが可能である。RCA法における種々の改変や応用は当業者が知り得るものである。他のRCA法の参照文献としてGusev、 Y. et al.,American J. Pathology 159,63−69 (2001)、 Zhang D. et al. Clin. Chim. Acta,363,61−70 (2006)(これらの文献は、参照することによって本明細書に組み込まれるものとする)が挙げられる。従って、本発明によって調製されたDNA断片は直線状の繰り返し配列を含むポリマーが得られる方法で増幅することができる。そのようなポリマーは、少なくとも最初のRNAの配列のコピーを二つ含み、よって二つの配列はお互いに相補的となる。そのようなポリマーはまた、最初のRNAに由来する二つの配列のいずれかより塩基配列決定反応を推進するために用いることができる配列を含んでいる。
【図12】図12は、プールされたサンプルとidentifier配列との導入を示す概念図である。図1と2で示すいずれかのステップは完全長mRNAの5’末端にオリゴヌクレオチドを導入するために用いることができる。そのようなオリゴヌクレオチドは、Identifier配列と呼ばれる配列情報を含む領域を含んでも良く、それは修飾されたRNAおよびRNAに由来する任意のDNAの由来をプールされたサンプル中で同定するために用いられる。プールされたサンプルは、異なるIdentifier配列(パネルB)を含むRNAサンプルを混合して得られる。さらに、Identifier配列は検出と解析のために一分子を捕獲するために用いても良く、またプールサンプルの中にある選ばれたサンプルの塩基配列決定反応をプライミングするために用いても良い。そのようなIdentifier配列は、Identifier配列を塩基配列反応で決定することにより、あるいはIdentifier配列と相補的な配列(パネルC)かまたはIdentifier配列を含む分子を表面上の規定された位置で選択的に捕獲することにより、RNAに由来する個々のDNAまたはRNAの由来を同定するために用いることができる。
【技術分野】
【0001】
ゲノムは、生物の発生や恒常性の維持に必要不可欠な情報を含んでいる。生物学的現象を理解するためには、ある時点の細胞や組織で遺伝情報がどのように利用されているかを知ることが必要である。ヒト、動物または植物において、遺伝情報や関係した制御パスウェイが誤った形で使われることが疾病の原因となる例が数多く知られている。RNAの発現は、個々の細胞、組織又は生命体によって非常に多様である。そのため、限られた量の細胞からRNAとDNA分子を取り出して調製できる新しい方法の開発が期待されており、このような方法が開発されれば、組織の中の個々の細胞のRNA発現、プロモーター利用及び発現しているゲノム情報を解析することができる。ライフサイエンスの新しい方向は、このような要求に取り組んでいる。個々のDNAおよびRNA分子を捕捉、解析し、遺伝子ネットワーク研究のような生物学的システム全体を理解するための技術の開発が進められている。
【背景技術】
【0002】
[遺伝子発現解析]
発現プロファイリングや転写物の注釈付けには、種々の方法が用いられている。簡単に言うと、最近の大規模発現研究にはマイクロアレイのようなin situハイブリダイゼーションによる方法、あるいはSAGE、CAGE、MPSSなどの短いタグの高速塩基配列による方法が用いられている。このような研究では、個々の遺伝子の発現レベルを調べるために、さらにノーザンブロッティングやRT−PCRなどの従来の方法が組み合わせて用いられる。
【0003】
大規模な発現プロファイリングには、通常、DNAマイクロアレイ(Jordan B., DNA Microarrays: Gene Expression Applications,Springer-Verlag,Berlin Heidelberg New York、 2001: Schena A,DNA Microarrays,A Practical Approach,Oxford University Press,Oxford 1999、これらは、参照することによって本明細書に組み込まれるものとする) と呼ばれる技術が用いられている。この実験では、個々の遺伝子や転写物に対応するプローブを支持体上に固定し、そこに種々のDNA分子をハイブリダイズさせて行なわれる。ポジティブ信号は、支持体上に固定されたプローブとサンプル中の分子が反応した場合に得られる。これらの実験は、多数の遺伝子又は転写物を同時並行的に解析できるが、その反面、解析の対象が、他の実験や手段によって同定された遺伝子や転写物に限られるという制限がある。このような実験や手段は、cDNAライブラリーや部分的に塩基配列がわかっているタグ、およびコンピューターによる予測結果を含む。将来的には、タイリングアレイによって、ゲノムの塩基配列が判っている生物の偏りのない発現プロファイリングが可能となる(Kapranov P. et al.,Science 296,916−919,2002、本文献は、参照することによって本明細書に組み込まれるものとする)であろうが、しかし、実験から得られる転写物の性質についてはタイリングアレイでの解釈は困難である。
【0004】
DNAマイクロアレイによる解析が持つ問題を解決するために、複数のRNAサンプルより得られる部分シーケンスやタグをベースとした発現プロファイリングや遺伝子探索が行われている。いわゆるSAGE(Serial Analysis of Gene Expression)法は、mRNAの塩基配列から効率的に情報を得る方法として知られている(Velculescu V.E. et at.,Science 270,484−487,1995、本文献は、参照することによって本明細書に組み込まれるものとする)。タグの塩基配列決定をハイスループットに行うために、この方法では、複数のRNAサンプルの3’末端の塩基配列情報を含む短いDNAフラグメント(当初は約10塩基対)をつなぎ合わせてコンカテマーを作る。このコンカテマーの一回の塩基配列解析により、DNAコンカテマーに含まれる異なるRNA分子から得られるたくさんのタグの塩基配列を決定することができる。最近、SAGE法の一つであるLong SAGE法が発表され、長いタグをクローニングすることが可能となった(Saha S. et al.,Nat. Biotechnol.20,508−12,2002、米国特許出願20030008290号、米国特許出願20030049653号、これらは、参照することによって本明細書に組み込まれるものとする)。この概念は「Super SAGE」と呼ばれる方法に発展し、25塩基対程度のタグの塩基配列を得ることができる(Matsumura,H et al.,Cell.Microbiol.7,11−18,2005、本文献は、参照することによって本明細書に組み込まれるものとする)。SAGE法は現在、生物、組織、特定の細胞などの遺伝子発現を解析する重要な手法として広く用いられており、このタグの情報は例えばhttp://cgap.nci.nih.gov/SAGEで公開されている。最近のSAGE研究の発展に関する情報は、Wang,S.M.:SAGE:Current Technologies and Applications,Horizon Bioscience,Norwich(2005)(本文献は、参照することによって本明細書に組み込まれるものとする)で参照できる。
【0005】
米国特許第6,352,828号、米国特許第6,306,597号、米国特許第6,280,935号、米国特許第6,265,163号、米国特許第5,695,934号(これらは、参照することによって本明細書に組み込まれるものとする)では、短いタグ配列を大規模に塩基配列決定する別の方法が開示されている。Massively Parallerl Signature Sequencing(MPSS)と呼ばれるこの方法は、Brennerらによれば(Brenner S.,et al.,Nat. Biotechnol. 18,630−634,2000、Brenner S.,et al.,Proc. Natl. Acad. Sci. USA 97,1655−1670,2000、これらは、参照することによって本明細書に組み込まれるものとする)、単層のビーズ上で異なる酵素を使ってサイクル反応を行ない、転写物の3’末端の短い塩基配列を超並列に得る方法である。WO03/091416(本文献は、参照することによって本明細書に組み込まれるものとする)では、この方法の改良が開示されており、転写物の5’末端の短い塩基配列を決定することが出来るようになった。しかし、その初期の様態では、MPSSはシグニチャータグの解読塩基長の短さのため限定された方法である。
【0006】
3’末端塩基配列情報から5’末端塩基配列情報へのシフトは、それが技術的にはより難度が高いものであるが、発現情報と転写の調製原理を結びつけるために不可欠なものである。遺伝子発現の制御をする一般的な調節因子は転写物の5’末端近くに存在し、その遺伝子のプロモーターと呼ばれている。選択的プロモーターの利用や、RNAプロセッシング、スプライシングによる一次転写物の再構成により、高等動物では大部分の転写物のプロモーター領域を3’末端の情報から特定することは出来ない。従って、転写物の5’末端から特異的に配列タグを得る新しいアプローチが開発されてきたのである。そのようなアプローチはPCT/JP03/07514、Shiraki T.et al.,Prog.Natl.Acad.Sci.USA 100,15776−15781(2003)、Kozdius R.et al.,Nature Method 3,211−222(2006)、米国特許出願番号20050250100号(これらは、参照することによって本明細書に組み込まれるものとする)に開示されている。この方法はCAGE(Cap Analysis Gene Expression)法とよばれ、転写物の5’末端から特異的に得たタグ配列をSAGE法と同様にコンカテマーにしてクローニングする方法である。この方法により得られるCAGEタグは、転写物の検出や発現プロファイリングだけでなく、転写開始点の情報を得るために利用され、転写調節のメカニズム解析や転写物の高度な注釈付けを行なうことができる。同様の5’末端の塩基配列情報をコンカテマーにしてクローニングするアプローチが、最近いくつもの研究室より発表されている。例えば、Hwang B.J. et al.,Proc.Natl.Acad.Sci.USA 101,1650−1655(2004);Hashimoto S.et al.,Nat.Biotechnol.22,1146−1149(2004);Zhang Z.and Dietrich F.S.,Nuc. Acids Res.33,2838−2851(2005);Wei C.L. et al.Proc.Natl.Acad.Sci.USA 101,11701−11706(2004)(これらの文献は、参照することによって本明細書に組み込まれるものとする)などが挙げられる。これらのアプローチは、すべて5’末端を捕獲する方法に特徴がある。例えば、Cap−Trapper又はOligo−Cappingなどの方法である。5’末端タグの意義に関する詳細は、Harbers M.and Carninci P.,Nature Methods.2,495−502(2005)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。
【0007】
前述のアプローチは、タグの塩基配列決定の処理量の点で今もなお制限がある。さらに、それらのアプローチは、多くの操作を必要とするためコンカテマーより得られる塩基配列情報に間違いが起きる可能性がある。特に、増幅のステップにおいて、個々のDNA断片の異なる増幅効率に起因するタグ頻度のバイアスや人為的な影響が発生する可能性がある。これらの制限を解決するために、不必要に複雑な操作やクローニングステップを省くためのDNAおよびRNA分子の直接的捕獲と、よりハイスループットなデータ収集とを目的としたさらなる開発が必要とされている。
【0008】
[塩基配列決定技術]
遺伝コードの読み取りと解読はライフサイエンスの最も大きな発展の一つである。塩基配列決定技術は遺伝情報を得るための重要なキーとなってきている。当業者が知り得る塩基配列情報を得る手段としては、Sanger法やMaxim−Gilbert法のようなSambrook J. and Russuell D.W.,Molecular Cloning,A Laboratory Manual,Cold Spring Harbor Laboratory Press,New York(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載された方法が挙げられるが、これに限定するものではない。近年、ハイスループット塩基配列決定の新しい技術が開発された。電気泳動、ハイブリダイゼーションによる塩基配列決定、合成による塩基配列決定、MPSS塩基配列決定、非酵素的一分子塩基配列決定などの利用が挙げられる。例えば、ハイブリダイゼーションによる塩基配列決定では、決められた塩基配列のオリゴヌクレオチドセットが載っている高密度マイクロアレイを用い、ハイブリダイゼーションのパターンによって未知配列の新たな塩基配列決定を行うことができる。例えば、Perlegenは、このような方法をヒトゲノム中の点突然変異解析に用いた。また、合成による塩基配列決定方法は、鋳型DNAに沿ってDNA分子が伸長する間にポリメラーゼがヌクレオチドを結合させることによって行われる。表面上で決められた位置に個々のヌクレオチドが結合するのをモニターし、さらにそれぞれの位置で引き続き結合するヌクレオチドを順番に見ることで、それぞれの位置にある核酸分子の塩基配列を決定する。非常に大量の伸長反応がひとつの反応器のなかで起こるため、例えばビーズアレイのガラススライド表面上では、使用する機器によって100、000から1、000、000種のサンプルを、一反応で並行的に塩基配列決定にかけることができる。とりわけ興味深いのは、本発明のDNAの一分子検出と塩基配列決定への応用であり、最近、Metzker M.L.,Genome Res.15,1767−1776(2005); Kling J.,Nature Biotechnology 23,1333−1335(2005); Shendure J. et al.,Nature Review Genetics 5,335−344(2004)(これらの文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。これらアプローチのうちいくつかは、454 Life Sciences(http://www.454.com/)、 Helicos BioScience(http://www.helicosbio.com/)、Solexa(http://www.solexa.com/Company/overview.htm)、Visigen(http://www.visigenbio.com/index.html)、GeneoVoxx GmbH(http://www.genovoxx.de/)(これらは、参照することによって本明細書に組み込まれるものとする)のような企業により商業的に利用されている。このような新しい技術は、現在の塩基配列決定速度を飛躍的にあげるであろう。使用する方法によっては、20から25塩基対の非常に短いタグ配列を、1ランあたり1,000,000種以上読むことができるという超ハイスループットの機器(例えば、Helicos、Solexaなど)もある。また、454Life Sciences社の機器ははるかに長い100塩基対以上の解読長を1ランあたり200,000から300,000種解読することができる。
【0009】
[完全長cDNA分子の単離]
生物サンプルのRNAを解析では、完全長RNA分子を単離し、そのRNAからcDNAを調製することにより大きな利益を得ることができる。特に、前述のようにRNAの5’末端の塩基配列情報は、データの幅広い解析を可能とする。さらに、完全長cDNA技術はRNAのプロセッシングやスプライシングに関係したプロセスを解析するためには、必須である。最近の種々の実験より得られたデータは、高等動物の複雑性がゲノム情報の選択的な利用によるものであることを示している。転写情報における組み合わせの再構成とプロセッシングのメカニズムは、遺伝プールの拡大と多様化に重要である(Zavolan M.,et al.,Genome Res. 13,2003,1290−1300、本文献は、参照することによって本明細書に組み込まれるものとする)。
【0010】
完全長cDNAを調製する種々の方法は、Das,M.,et al.,Physiol Genomics 6,57−80(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。これらの中から、Cap−Trapper法とOligo−Capping法は、当業者が知り得る他の方法の中でも最も頻繁に使用されている。これらのアプローチは、ゲノム、転写物の構造、タンパク質配列の解析の手段として利用されてきた。さらに、完全長cDNAだけがそれにコードされたタンパク質へとつながる手段であり、実験において必要とされるとき、あるいは産業応用に際して、機能的ベクターを使ってタンパク質を発現させることができる。
【0011】
Cap−Trapper法(Carninci P.and Hayashizaki Y.,Methods Enzymol. 303,19−44,1999;米国特許第5,962,272号、米国特許第6,022,715号、これらの文献は、参照することによって本明細書に組み込まれるものとする)では、Cap構造のジオール基を化学的にビオチン化し、mRNAとcDNAの複合体をストレプトアビジンでコートしたビーズで捕獲する。残っているtRNA、およびRNAタンパク質などの一本鎖RNAと部分的にDNA−RNAがハイブリダイズしている複合体は、RNaseI分解によって破壊されるが、mRNAとcDNAの複合体は分解されない。その後、濃縮された完全長cDNAはRNAの加水分解によってビーズより遊離する。
【0012】
代替法となるOligo−Capping法では、まずmRNAの5’末端を修飾する(Maruyama K.and Sugano S.,Gene 138,171−174,1994;Suzuki Y.Sugano S.,Methods Mol Biol.221,73−91,2003、これらの文献は、参照することによって本明細書に組み込まれるものとする)。最初のステップで、CapされていないmRNAと切断されているmRNAより脱リン酸化酵素によってリン酸基を除去し、次にタバコ酸ピロホスファターゼ処理によってmRNAのCap構造を取り除く。この処理により、完全長mRNAのみが、リン酸化された5’末端を持つことになる。そのため、RNAリガーゼによりリン酸化された完全長mRNAの5’末端にのみ、オリゴヌクレオチドを結合させることができる。mRNAの5’末端に結合したオリゴヌクレオチドは、例えば第二鎖cDNAの合成など、その後mRNAに由来するcDNAの操作に利用することができる。
【0013】
完全長cDNAを選択するその他の方法には、Cap−binding proteinを利用する方法(Edery,I.,et al.,Mol. Cell Biol. 15,3363−3371,1995、本文献は、参照することによって本明細書に組み込まれるものとする)、Cap構造に対する抗体を利用する方法(Theissen,H.,et al.,Embo J 5,3209−3217(1986)、本文献は、参照することによって本明細書に組み込まれるものとする)、Cap構造にオリゴヌクレオチドを結合させる方法(米国特許第5,962,272号、米国特許第6,022,715号、これらの文献は、参照することによって本明細書に組み込まれるものとする)、Clontech社(http://www.clontech.com/clontech/smart/index.shtml、ここに記載された情報は、参照することによって本明細書に組み込まれるものとする)のSMART(商標)法などがある。しかし、Cap−Switching法(Zhu Y. et al.,Biotechniques 30,892−897,2001、本文献は、参照することによって本明細書に組み込まれるものとする)を使うSMART(商標)法は、mRNAの5’末端にGGGという3つのヌクレオチドを追加するので、タグ部分の情報の長さを短縮させることになり、特に、塩基配列決定の場合には、解読長が非常に短いものとなってしまうため、5’末端タグのクローニングには好ましくない。
【発明の開示】
【0014】
本発明は、一つ又は複数のRNA分子の5’末端の塩基配列情報を得るためのRNA分子の修飾に関するものである。本発明は、RNA分子の修飾に関するものであり、RNA分子に加えられた情報は解析および操作に用いられる。
【0015】
本発明は、RNAおよびDNA分子、又はそれらの断片を取り出し一分子検出と塩基配列決定を行う手段を提供する。本発明は、新しい一分子検出のアプローチに必要な個々の分子を捕獲する手段を提供する。本発明は、RNA分子およびそれに由来するDNA分子を、それら分子の特定の領域の塩基配列情報を得るために修飾する。従って、本発明は新しいハイスループット塩基配列決定方法とその応用手段、例えば発現プロファイリング、転写物特性解析、ゲノム注釈付、さらに解析するためのクローニング、その他従来の方法などへの応用手段を提供する。特に、本発明は重要な研究に手段を提供する。重要な研究とは、例えば5’末端特異的塩基配列に基づいた発現プロファイリングであり、これは法医学、診断、創薬、ライフサイエンスを含むが、これに限定されない商業的応用、試薬、サービスには不可欠な要素である。
【0016】
一つの実施態様では、本発明は、天然又は人工的な一つ又は複数のRNA分子をcDNAに転写的に転換することに関係する。よって、本発明は一本鎖DNA分子の合成と調製に関係するものであり、検出と解析を目的とした核酸分子の断片の単離方法に関係するものである。さらに、本発明は単一又は複数の種類の核酸分子を含むRNAサンプルのDNA分子への転換に関係するものである。
【0017】
別の実施態様において、本発明は核酸分子を直線状一本鎖DNAの形に調製するために核酸分子を操作する方法に関係するものであり、また、RNAから複写された直線状一本鎖DNAの操作、調製に関係するものである。
【0018】
別の実施態様において、本発明はRNA分子又はDNA分子の末端に官能基を付加する手段を提供する。よって本発明は官能基を用いて、操作と解析のためにRNAとDNA分子を捕獲する手段を提供する。この実施態様において、本発明は解析、検出、塩基配列決定を目的とした核酸一分子の単離に関係するものであり、一分子検出とハイスループット塩基配列決定の鋳型調製の手段を提供する。
【0019】
別の実施態様において、本発明は一本鎖DNA分子を直接的にその塩基配列情報を得るために用いる方法に関係するものであり、また一本鎖DNA断片の特定した領域の塩基配列情報を得ることに関係するものである。ある実施態様において、本発明によって調製されたDNA断片から得られた5’末端特異的塩基配列情報は、RNA分子の5’末端塩基配列と関係するものである。このように本発明は、RNA分子の塩基配列情報の取得に関係するものである。
【0020】
別の実施態様において、本発明は、RNAに由来するDNA分子の反対側の末端を修飾する手段を提供する。この場合、RNAの5’末端に対応する末端およびRNAの3’末端に対応する末端への修飾は、官能基または後のDNA分子の操作、検出及び解析のための機能を持った基の導入である。一つの実施態様においては、RNAの3’末端に対応する末端に官能基が導入され、それを表面上に結合させることによりcDNAを捕獲する。一つの実施態様においては、本発明に沿って調製されたDNA断片より得られた3’末端特異的な塩基配列情報は、RNAまたはDNA分子の末端塩基配列に関係する。別の実施態様においては、cDNA分子がLAMP法またはローリングサークル法によって増幅され得るようにcDNA分子の両末端が修飾される。
【0021】
別の実施態様において、本発明は、RNAの5’末端にある「Identifier配列」と呼ばれる、規定された塩基配列の領域を導入する手段を提供する。そのようなIdentifier配列は、分子の由来を特定する。従って、Identifier配列の導入により、由来の異なるサンプルからなるサンプルプールを解析することが可能となる。そのような異なる由来は、用いられる異なる細胞系、組織、器官または異なる発生段階、実験上の多様な時点などに関係する。別の実施態様において、Identifier配列は、本発明に従って調製された分子から得られる塩基配列の一部分である。別の実施態様において、Identifier配列は、この配列を持つ分子を検出、解析または塩基配列決定を目的として表面上の特定の位置に捕獲するために用いられる。別の実施態様において、Identifier配列は、プールされたサンプルの複数の分子の中から一つの分子の塩基配列決定をプライミングするために用いられる。前述の実施態様と同様に、本発明は、Identifier配列をRNAの5’末端だけでなくRNAの3’末端に相当するcDNAの領域に導入する手段を提供する。Identifier配列の利用は、RNAの5’末端に限定されることなく、実験的な必要性に応じてRNAに由来するcDNAのいずれの末端でも用いることができる。
【0022】
本発明は、統計的解析を含むコンピューターによる注釈付や参照情報に対するアライメントによる注釈付、およびゲノム配列に対するマッピングなどを目的とし、本発明に従って得られたDNA断片のある領域の塩基配列決定に関係するものである。従って、本発明は、遺伝子探索、遺伝子同定、遺伝子発現プロファイル、およびそれらの注釈付の手段に関係するものである。
【0023】
別の実施態様において、本発明は、本発明に沿って得られたDNA断片の塩基配列決定に関係し、それらの配列がゲノムの領域に関するものである場合には、その配列のコンピューターを用いた注釈付を可能にする。またIdentifier配列の解読、塩基配列の統計的解析を可能とする。よって本発明は、転写開始位置に関連して遺伝的要素の特性解析に関係するものである。
【0024】
さらに別の実施態様において、本発明は核酸の末端より得られるハイブリダイゼーションプローブの調製に関係するものであり、その領域はin situハイブリダイゼーションを用いて解析される。好ましい実施態様においては、そのin situハイブリダイゼーションの実験は、タイリングアレイを用いて行われる。
【0025】
また実施態様において、本発明は、本発明に従って得られたDNA断片の塩基配列情報が増幅される方法を用いた、核酸分子の完全長クローニングに関係するものである。ゲノム断片と同様に転写された領域のクローニングや増幅も本発明に含まれる。そのような断片は、プロモーター領域を含んでも良い。
【0026】
従って、本発明は、例えば生物学的サンプルの特性解析のような必要に応じて、核酸分子やその短い断片を解析する手段を提供する。さらに、本発明は、迅速で効率的なRNAやDNA断片の解析および塩基配列解析を行い、それら断片の一分子検出のような分析的検定を行う手段を提供する。よって、本発明は、特にハイスループット塩基配列決定や固体支持体上でのRNA分子又はDNA分子の並列的検出に適している。
【0027】
特にある実施態様では、本発明は環状一本鎖DNAに転換可能なDNAの修飾による双方向的な鋳型構築に関係するものである。RCA反応により環状一本鎖DNA分子の増幅を行った後、双方向性の直線状一本鎖DNA分子を得て、固体支持体上の規定された位置に直接結合させることができる。本発明は、同じ鋳型から複数の解読塩基配列を得ることを可能とするもので、規定された位置により種々の解読配列を同じ鋳型に関連付けることができる。双方向的直線状一本鎖DNA分子を鋳型として用いることにより、修飾されたDNA分子の両鎖又はRNA分子の両端の配列情報を同じ鋳型から得ることができる。
【0028】
本発明は、ライフサイエンス研究や診断に利用する分析的検定をデザインし実行する手段を提供する。それゆえ、本発明は生物学的システムの解析又は診断の手段に関係するものである。
【0029】
本発明はまた、実験的な条件を充足させるため必要とされる本発明の部分的、又は全部の実行のための試薬やキットの製造およびデザインの手段を提供する。
【発明を実施するための最良の形態】
【0030】
本発明は、直線状と環状の核酸分子の形状である二本鎖核酸と一本鎖核酸の処理を行う方法を含む。二本鎖DNAとは、デオキシヌクレオチドから構成される二つの分子から成り、二つの分子が、実質的に互いに相補的な配列を持ち二量体を形成するように対合する任意の核酸分子を含む。二つのポリマーは、デオキシヌクレオチドの対応する塩基対の間で特有の水素結合によって結合している。二つもしくはそれ以上のデオキシリボヌクレオチドにより形成される一つのポリマー鎖で構成され、対合する相補的なDNA分子がない任意のDNA分子も、本発明の目的によれば一本鎖DNA分子とみなされる。そのような分子が二次元構造を形成し、二本鎖DNAの部分を含んでいる場合も同様である。互換性をもって用いられる「核酸分子」や「ポリヌクレオチド」の語句は一本鎖であるか二本鎖であるか、コーディングがノンコーディングか、相補的かそうでないか、センスかアンチセンスかによらずRNAやDNAを含み、またハイブリッド配列も含む。特に、ゲノムDNAや相補的DNAを含み、それらは転写されることもされないこともあり、スプライスされることもされないこともあり、不完全なスプライシング又はプロセッシングを受けることもあり、その由来にはよらず、生物学的材料からクローニングされることもあり、あるいは合成によっても得られる。本発明の目的において、RNAとは、一本鎖の核酸分子であり、たとえ二次構造をとって二本鎖RNAを形成している部分があったとしても同様である。特に、本発明の目的においてRNAはリボヌクレオチドで構成される任意の形状の核酸分子を含み、特定の塩基配列や由来とは関係ない。従って、RNAはin vivo又はin vitroで人工システムによって転写されても良いし、翻訳されなくても、スプライスされてもされなくても、不完全にスプライス又はプロセッシングを受けても良く、天然由来か人工的にデザインされた鋳型から生成されたかどうかは関係ない。それらは合成によって得られたmRNA、tRNA、rRNA、miRNA、siRNA、RNAi、又はそれらの任意の混合物も含む。RNAは生物学的サンプル、あるいはより具体的には、血液や血清のような生物学的由来の液体から生成されても良い。例えば、ウイルスRNA又は個々の人の血液にあり得る他の寄生生物を含んでも良い。あるいは、RNAは、解剖された組織よりフローソートされた細胞を含む精製細胞から得ても良く、それら細胞はセルソートのための蛍光を発する抗体でラベルされていても良い。あるいは、当業者が知り得る手段を用いて、緑色蛍光タンパク質(GFP)のようなマーカーのトランジェニック発現によりラベルされても良い。他の方法として、これらの細胞は形態に基づいて選ばれることもでき、あるいはレーザーによるマイクロダイゼクションによって選ばれることも出来る。さらに正確には、「DNA」、「RNA」、「核酸」、「塩基配列」の表現は核酸材料そのものを含み、また塩基配列情報やベクター、ファージミド、又はその他の特定の核酸分子に限定されるものではない。「核酸」という語句は、また、本明細書では天然に存在し得る核酸、人工的に合成された又は調製された核酸、当業者の知り得る方法によってあるいは天然に発生する事象によって、少なくとも一つ又はそれ以上の修飾が導入された修飾核酸を含む。同様に、本発明の「タグ」または「Identifier配列」とは本発明によって調製される核酸分子の任意の領域でも良く、ここでいう「タグ」または「Identifier配列」とは、任意の核酸フラグメントを含み、由来が天然の事象により発生したものでも良く、人工的に合成又は調製されたものでも良く、また天然の事象によりあるいは当業者が知りえる方法により少なくともひとつの修飾が導入された核酸でも良い。さらに、「タグ」または「Identifier配列」は、特定の塩基配列情報や構成に関係するものではない。本発明においては、「純度」、「濃縮された」、「精製」、「濃縮」、又は「選択」という語句は、置換可能であり、生成物の絶対的な純度や濃縮を必要とするものではなく、むしろ、相対的な意味で用いられる。本発明において、「特異的」、「好ましい」、「優先的」という語句は、置換可能であり、DNA又はRNAのハイブリダイゼーションにおいて、絶対的な特異性や酵素の基質に対する特異性、反応を意味するものではなく、むしろ、相対的な意味で用いられ、酵素がその基質と関係したあるいは無関係の他の物質に対して低い、あるいは比較してより低い親和性を持つという意味も含まれる。同様に、酵素、又は酵素活性という語句は、本発明においては要素の機能や活性を表し、要素の絶対的な純度を必要とするものではない。従って、酵素若しくは同等の、又は関連した若しくは関連しない機能を持つ要素を含む混合物も本発明の対象となる。同様に、本発明におけるDNA分子又はRNA分子は、ハイブリダイゼーションプローブとして機能することもでき、本発明の目的のために相補的配列を含んでも良い。相補配列を含むDNA又はRNAは、関連する核酸分子を検出するためのプローブとして実験に使用することもでき、さらにそのようなプローブとターゲット分子は、異なる位置に突然変異を人工的又は自然発生的に導入することにより区別する実験に使用することもできる。「生物学的サンプル」とは、微生物、動物、植物、宿主に依存して複製するウイルスやプリオンなどの感染性粒子など、生物体から得られるあらゆる材料を含む。このような「生物学的サンプル」は、研究、開発、診断、治療などを目的として、患者、動物、植物、感染粒子から得られたあらゆる材料を含む。従って、本発明は特定の核酸分子又はその由来に限定されることはなく、本発明は、あらゆる核酸の操作、利用、応用のために普遍的な手段を与える。本発明に適用される核酸分子は、当業者が知りえる任意の方法によって調製、入手することができ、例えばSambrook J. and Russuell D.W. (Molecular Cloning,A Laboratory Manual,Cold Spring Harbor Laboratory Press,New York,2001、本文献は、参照することによって本明細書に組み込まれるものとする)らの方法により得ることができるが、これに限定されるものではない。
【0031】
本発明は、核酸分子より検出と解析のためにフラグメントを単離する方法に関するものである。この核酸分子の解析は、核酸分子の全体的又は部分的な塩基配列情報を得ることを含むが、これに限定されるものではない。当業者が知り得る塩基配列情報を得る方法は種々あり、例えばSambrook J. and Russuell D.W.,Molecular Cloning,A Laboratory Manual,Cold Spring Harbor Laboratory Press,New York(2001)に記載されており、また従来技術として記載した新しく発展してきたハイスループット技術が含まれるが、これに限定されるものではない。本発明は、ある特有の塩基配列決定方法や技術への利用に限定されず、実験の条件に最も適した形やあるいは分野の新しい発展に適応した形で解析や検出をおこなうためにRNAやDNAを操作する汎用的な技術を提供する。
【0032】
塩基配列決定反応を含む解析、検出又は操作のために、核酸分子は固体支持体上に結合されても良い。固体支持体は、要素が直接又は間接的に結合できる任意の固体材料でも良い。そのような材料はアクリルアミド、アガロース、セルロース、ニトロセルロース、ガラス、金、ポリスチレン、ポリエチレンビニルアセテート、ポリプロピレン、ポリメタクリレート、ポリエチレン、ポリエチレンオキサイド、ポリシリケート、テフロン、フッ化炭素、ナイロン、シリコンゴム、ポリアンハイドライド、ポリグリコール酸、ポリ酢酸、ポリオルトエステル、機能性シラン、ポリプロピルフマル酸、コラーゲン、グリコサミノグリカン、ポリアミノ酸、またはそれらの混合物が含まれるが、これに限定するものではない。固体支持体はさらに、薄いフィルム、膜、ビン、皿、繊維、編まれた繊維、チューブ状、粒子状、ビーズ状、微粒子状などに成形されたポリマー、またはそれらの組み合わせを含む。
【0033】
本発明は、ひとつ又はそれ以上の核酸分子を含むサンプルの転換に関するものである。そのような核酸分子又は核酸分子の混合物は、DNAに転換される。本発明を実施するために、核酸分子は、自然にあるゲノムDNA、RNAサンプル、人工的に作られたDNAライブラリーから得ることができ、またそれらの任意の混合物でも良い。本発明は、単独又は任意の複数の核酸分子への適用に限定されるものではなく、核酸分子が単独であっても、複数であっても良く、自然発生的におきるか、あるいは既存のライブラリーから得られるか、あるいは人工的に作製されるかによらない。さらに、本発明は、その由来や性質に関わらず、あらゆる核酸分子を処理することができる。従って、自然事象によって得られる核酸分子でも完全長分子であっても良く、それらの断片でも良い。さらに、それらの核酸分子の断片は、ランダムなプロセスによって、あるいはある特定の塩基配列に作用する酵素を用いて核酸分子のターゲットを絞った方法によって、あるいは転写領域の中のエクソン、イントロンなどの核酸分子の構造をもとに断片化する方法によって調製することができるが、これに限定されるものではない。従って、本発明は、特定の出発材料に限定されるものではない。
【0034】
本発明は、プールされたRNA分子の中のRNA分子、又は一つのRNA分子の5’末端に官能基および塩基配列情報を導入するために、一つ又は複数のRNA分子を修飾することに関係するものである。そのような官能基は、1、3、1〜5、5〜10、10〜15、15〜25、25〜35、35〜45または45以上のヌクレオチドを含んでも良い。よって、本発明は、RNA分子の操作および解析や修飾されたRNAの調製や解析に用いられるRNA分子に情報を付加するために行うRNAの修飾に関係するものである。
【0035】
当業者が知り得る酵素的又は化学的なRNAの修飾方法は、種々ある。好ましくは、本発明を実施するためには、RNAの酵素反応による修飾が望ましく、そうすることにより異なる酵素反応を用いてRNAのグループ中において標的とする特定のRNAの修飾を行うことができる。さらに好ましくは、トータルRNAの中のmRNA分子が優先的に修飾を受け選択的な濃縮が可能となる。しかしながら、本発明は、mRNAの解析に限定されることなく、解析と検出のためにRNAを捕獲する汎用的な手段を提供する。miRNAやほかの短いRNA分子のような全く新しいRNAの種類を特定の修飾と解析の対象とすることができる最近の研究の結果がAlvarez−Garcia I.and Miska EA.,Development 135,4653−4662(2005)(本文献は、参照することによって本明細書に組み込まれるものとする)に示されている。
【0036】
本発明を実施するために、標的とするRNAは3つの概念的に異なるステップの対象とされる。(1)非完全長mRNA分子のマスキング、(2)分子に含まれるCap構造の反応分子への転換、(3)標的とするRNAの5’末端への標的RNA分子の付加の3つのステップである。mRNAの5’末端へRNAオリゴヌクレオチドを付加する標準的な方法は、Oligo−Capping法(Maruyama K. and Sugano S.,Gene 138,171−174,1994;Suzuki Y. and Sugano S,Methods Mol Biol.221,73−91,2003、これらの文献は、参照することによって本明細書に組み込まれるものとする)と呼ばれるものおよびその改変法がある。生きた生物からのRNA調製は、完全長mRNAの5’末端にあるCap構造により特徴付けられるRNA種を含む。このCap構造によって、Cap構造を欠き、そのかわりに自由なリン酸基を5’末端に含んでいる切断されたRNA種から区別することができる。Oligo−Capping法は、完全長mRNAの独特な性質を選択的な濃縮に用いるものである。Oligo−Capping法は、プールされたRNA中においてmRNAを特異的に修飾するいくつもの酵素的ステップを含む。最初の酵素反応により、Cap構造を持たない切断されたmRNAや小さいRNA、tRNA、rRNAなどがホスファターゼにより5’末端を脱リン酸化される。次に第二の反応により、Cap構造を持つmRNAからタバコ酸ピロホスファターゼ(TAP)によりCap構造を除去する。この処理により、完全長mRNAのみが5’末端にリン酸基を持つことになる。よって、第三の酵素反応においてRNAリガーゼは、完全長mRNAのリン酸化された5’末端にのみ、オリゴヌクレオチドを付加することができる。
【0037】
第一の反応ステップでは、RNAの5’末端からリン酸基を除去することが可能な任意のホスファターゼを使用することができる。さらに具体的には、ホスファターゼはバクテリアアルカリホスファターゼ(BAP)、仔ウシ腸アルカリホスファターゼ(CIAP)、エビアルカリホスファターゼ(SAP)、Antarcticホスファターゼの中より選ばれる。同様に本発明を実施するために異なるホスファターゼが用いられても良く、また最も一般的には、タバコ酸ホスファターゼ(TAP)がCap構造の除去に用いられる。RNAのライゲーションステップでは、リン酸化RNAにDNAおよびRNAオリゴヌクレオチドを連結できる任意のRNAリガーゼを使用することができる。最も一般的には、T4 RNAリガーゼまたはサーモファージ一本鎖DNAリガーゼがこの反応に用いられる。サーモファージ一本鎖DNAリガーゼは商業的に入手可能であり、一本鎖DNAとRNAのどちらにも使用できる(この酵素のより詳細な情報は、製品情報に記載されている。http://www.prokaria.com/upload/files/Thermophage−ssDNA−ligase−version−4−2.pdf(本文献は、参照することによって本明細書に組み込まれるものとする)。よって、この酵素はRNAに直接DNAを連結させるために好ましい。このように本発明は、RNAに直接DNAを連結し、デオキシリボヌクレオチドまたはDNAオリゴヌクレオチドとリボヌクレオチドまたはRNAまたはRNAオリゴヌクレオチドを含む直線的ヘテロポリマーを調製することができる。
【0038】
当業者が知り得る本発明を実施するために用いられるOligo−Capping法の改変法は種々ある。最も好ましくは、本発明は、特許出願2006−106770号(本文献は、参照することによって本明細書に組み込まれるものとする)に開示されているような一つの反応容器中ですべての酵素反応が行える方法を利用する。簡単に言うと、第一の反応ステップでAntarcticホスファターゼのような熱処理によって不活性化される酵素を用いる。一連の反応の中の第一の酵素を不活性化した後、新しく成分を添加することにより、バッファー条件をTAP反応に適した状態に変更する。TAPは、再度熱処理によって不活性化される。従って、追加成分とオリゴヌクレオチドの添加で再度バッファー条件を変更するだけで、最終ステップとしてリン酸化RNAにオリゴヌクレオチドを連結することができる(図1参照)。
【0039】
前述した中で修飾反応は、プールされたRNAの中のmRNA分子に対して、後の操作のために行われる。しかしながら、本発明は、mRNAの修飾に限定されるものではない。異なる例では、Cap構造を欠き、リン酸化されたすべてのRNA分子が、RNAの5’末端にオリゴヌクレオチドを連結することによって直接修飾される。この例では、本発明は切断された(truncated)RNA分子と非mRNA分子の選択的な修飾を可能とする。本発明の他の例では、Cap構造を欠くRNA分子は、第一の酵素反応において修飾される。ある例では、Cap構造を欠いたRNA分子のみが本発明によって操作のために修飾される。他の例では、第一の反応の後に異なるRNA分子を段階的に修飾する他のステップを行う。従って、第二の酵素反応では、完全長mRNA分子のCap構造は、TAP酵素反応によって除去され、完全長mRNA分子の5’末端にリン酸基を与える。最後の反応ステップでは、同じ又は異なる塩基配列のオリゴヌクレオチドが完全長mRNA分子に連結される。この実施態様では、本発明は種々のRNA分子のプールの中にある異なるRNA種に異なるオリゴヌクレオチドを付加する手段を提供する。
【0040】
上記の、あるいは図1に示した一連の操作に続いて、第一の反応では、RNAリガーゼが一つ又は複数のRNA分子に異なる種類のオリゴヌクレオチドを連結するために用いられる。本発明によれば、オリゴヌクレオチドは、デオキシリボヌクレオチド(DNAオリゴヌクレオチド)、又はリボヌクレオチド(RNAヌクレオチド)から構成されるホモポリマーでもよく、またデオキシヌクレオチドとリボヌクレオチドから構成されるヘテロポリマー(DNA/RNAハイブリッドオリゴヌクレオチド)でも良く、あるいは、官能基をもった任意の種類のホモポリマー又はヘテロポリマーとなることもできる(図2参照)。本発明は、特有の核酸分子の利用に限定されず、修飾されたRNAが本発明の異なる実施態様で使用される方法に応じた、違うタイプのオリゴヌクレオチドを用いることができる。当業者が知り得る多くのDNAとRNAの修飾がある。例えば、異なる修飾のオリゴヌクレオチドを調製するためのオリゴヌクレオチドの情報は、MWG Biotechのウェブサイトhttp://www.mwg−biotech.com/html/s_synthetic_acids/s_modifications.shtml(ここに示された情報は、参照することによって本明細書に組み込まれるものとする)に記載されている。MWG Biotechは、異なる位置に官能基としてビオチンやジゴキシゲニンを持つオリゴヌクレオチドを提供している。さらに、修飾されたオリゴヌクレオチドは、5'Aminolink C3/C5/C6/C12、3'Aminolink C3/C6/C7、3'Aminolink C3/C6/C7、Amino(C2/C6)−dT、Amino C6−dC、Spacer C3/C9 (TEG)、Spacer C12/C18 (HEG)、reduced Thiol modifierのようなクロスリンクのための反応基など、ひとつ又はそれ以上の官能基を得ることができる。RNAオリゴヌクレオチドは、例えばInvitrogen(http://www.invitrogen.com/content.cfm?pageid=9900、ここに示された情報は、参照することによって本明細書に組み込まれるものとする)、またはOperon(http://www.operon.com/、ここに示された情報は、参照することによって本明細書に組み込まれるものとする)から入手することができる。このように、RNAの5’末端に付加されるオリゴヌクレオチドを異なる機能を持つようにデザインすることができる。一つの例では、オリゴヌクレオチドは、10から25ヌクレオチドを持つ。別の例では、オリゴヌクレオチドは、25から50ヌクレオチドを持つ。また別の例では、オリゴヌクレオチドは、50から100ヌクレオチドを持つ。また別の例では、オリゴヌクレオチドは、100以上のヌクレオチドを持つ。オリゴヌクレオチドは、化学合成によって、あるいは酵素的反応によって得ることができる。当業者が知り得るDNA依存性RNAポリメラーゼはいくつもあり、例えば鋳型DNAからRNA分子を調製できるT3 RNAポリメラーゼ、T7 RNAポリメラーゼ、SP6 RNAポリメラーゼなどがある。RNAポリメラーゼを用いたRNA調製の一つの例は、Fermentas(http://www.fermentas.com/techinfo/modifyingenzymes/protocols/p_synthstrspecrna.htm、ここに示された情報は、参照することによって本明細書に組み込まれるものとする)に記載されている。さらに、オリゴヌクレオチドは、リボゾームRNAのような天然のものでも良い。任意の生物から得られる全RNAの調製では、得られる大部分のRNAはrRNAであり、例えば、仔ウシ肝臓より得られる18S+28SリボゾームRNAは、Sigma−Aldrich(St. Louis,USA,catalog number R0889)より商業的に入手可能である。RNAオリゴヌクレオチド、DNAオリゴヌクレオチド、あるいは修飾されたオリゴヌクレオチドは、前述のようにRNAリガーゼによって直接RNA分子へ連結することができる。このように、反応条件は特定のリガーゼ、特定のオリゴヌクレオチドの修飾、特定の塩基配列のオリゴヌクレオチド、又は特定のオリゴヌクレオチド自体の利用に制限されるものではない。
【0041】
最も一般的には、オリゴヌクレオチドは、RNAの5’末端にプライミング位置を導入するための「プライマー」として機能するようにデザインされている。プライマーは5、6、5〜10、10〜15、15〜20、20〜30、30〜40、40〜50、または50以上のヌクレオチドからなっても良い。相補的核酸鎖合成の後、新しく合成された第二鎖は、3’末端にRNAの5’末端に連結されたオリゴヌクレオチドと相補的な配列を持つ。RNAの5’末端に付加されたオリゴヌクレオチドと全く同じ、又は部分的に同じ配列を持つオリゴヌクレオチドは、修飾されたmRNAと全体的、又は部分的に同じ配列を持つ核酸分子の合成をプライミングするのに用いられる。第二鎖のプライミングは、例えば、二本鎖又は一本鎖DNAの調製、DNA又はRNAの増幅、塩基配列決定に用いられる。DNAポリメラーゼによる第二鎖DNA合成の別の方法は、Sambrook J.and Russuell D.W.,Molecular Cloning,A Laboratory Manual,Cold Spring Harbor Laboratory Press,New York,2001、本文献は、参照することによって本明細書に組み込まれるものとする)などの標準的な教科書に記載されている。そのようなDNAポリメラーゼは、DNAポリメラーゼのクレノー断片、T4またはT7 DNAポリメラーゼ、DNAポリメラーゼI、Taqポリメラーゼ、Tfl DNAポリメラーゼ、Tth DNAポリメラーゼ、Tli DNAポリメラーゼ、または他の既知のDNAポリメラーゼを含むが、これに限定されない。例えば、直線状一本鎖DNAの調製には、当業者が知り得る様々な技術が発展してきた。いくつかの方法では、DNAポリメラーゼを用いてDNA又はRNAの鋳型より一本鎖DNAを合成する。特に、一本鎖DNAの合成は、非対象PCRと呼ばれる方法によって達成することができ、この方法では二つのプライマーが異なる濃度で使われる。律速となるプライマーが消費されたあと、反応は、二本鎖DNAの指数関数的増幅から律速となるプライマーより多量に入れられた片鎖のプライマーによる直線的増幅に切り替わる。他の方法では、ラムダエクソヌクレアーゼがリン酸化された5’末端をもつ二本鎖DNAの片鎖を分解するために用いられる。そのような鋳型は、二つのプライマーのうち、片方だけがリン酸化された5’末端を持つようにPCRで調製される。ラムダエクソヌクレアーゼは、また「Strandase(商標)」という名称で呼ばれており、Novagen、 (Madison,USA)から商業的に入手可能であり、「Strandase(商標) ssDNA Preparation Kit」、カタログ番号69202(これは、参照することによって本明細書に組み込まれるものとする)に説明が記載されている。同様に、ラムダエクソヌクレアーゼはEpicentre、(Madison,USA)(カタログ番号LE035HおよびLE032K)より入手可能である。数多くの直線状一本鎖DNAの応用のため、一本鎖DNAは、二つのプライマーのうち片方が特にタグづけられたプライマーセットを用いるPCRによって調製される。官能基を持った鎖と鋳型DNAからの二次的な望まれない鎖を分けるために、ビオチンラベルが最も頻繁に用いられているが、これに限定されるものではない。この方法は、目的とする鎖を固体支持体や基盤上に結合させて用いる場合には、特に有用である。固定された一本鎖DNAは、支持体上で直接精製され、一本鎖DNAの分離及び調製に応じた検出検定やDNA塩基配列決定のための鋳型調製に用いられる。そのような応用は、例えば、DASH SNP検出システムと呼ばれるようなゲノムDNA中のSNPの特性解析などを含むが、これに限定されるものではない。この方法は、米国特許出願2001046670号(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。S. Stahl et al.(Stahl,S.et al,Nucleic Acid Research 16,3025−3038(1988)(本文献は、参照することによって本明細書に組み込まれるものとする)には、ビオチン化されたDNAの異なる応用が記載されている。例えば、固相での塩基配列決定などである。他の実施態様として、Guatelli J.C. et al.,Proc. Natl. Acad. Sci. USA 87、1874−1878(1990、本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている方法によれば、逆転写酵素、RNase H、DNA依存性RNAポリメラーゼの活性を組み合わせた反応サイクルにより、修飾されたRNA分子を増幅することができる。
【0042】
他の実施態様において、RNAの5’末端に付加されたオリゴヌクレオチドは、DNA又はRNA分子の操作を可能にするための配列情報を持つようにデザインされている。当業者が知り得る特定の塩基配列や認識部位への結合に依存する酵素活性は数多くある。それらの酵素の多くは、異なる業者から商業的に入手可能である。例えば、FERMENTAS UAB(Vilnius,Lithuania)、New England Biolabs Inc.(Beverly,USA)、Promega(Madison,USA)、Takara(Tokyo,Japan)、 Roche(Mannheim,Germany)、GE Biosciences (Cardiff,United Kingdom)などが含まれるが、これに限定されるものではない。通常、制限酵素はDNA分子を認識部位や認識部位の近傍の規定された位置で分解するために用いられる。一つの例では、オリゴヌクレオチドによって導入されRNAの5’末端に付加された認識部位は、クラスII制限酵素の認識部位である。これらの酵素は、認識配列の外側で切断するもので、例えば、クラスII制限酵素であるMmeIは認識部位から20/18塩基対離れた部位を切断する。それゆえ、MmeIは、通常、例えば、前述のLongSAGE、5’−SAGE、またはCAGE法などの短い塩基配列決定用タグの分離に用いられる。他の応用では、当業者が知るようにDNAの組み換えやクローニングのために制限酵素を利用することができる。より詳しくは、Sambrook J. and Russuell D.W.,Molecular Cloning,A Laboratory Manual,Cold Spring Harbor Laboratory Press,New York(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。さらに、修飾された又はデザインされた制限酵素は、鎖特異的ニッキング酵素として機能することができる。そのような制限酵素は、二本鎖DNAを基質として認識配列の中で一方のDNA鎖のみを切断する。そのような酵素は、商業的に入手可能であるN Bpu 10I(FERMENTAS UAB,Vilnius,Lithuania)、N.Bbv C IA、N.Bst NB I、N. Alw I(New England Biolabs Inc,Beverly,USA)などを含むが、これに限定されるものではない。ニッキング酵素は二本鎖DNAにプライミング部位を作る点で特に興味深く、DNAの合成や塩基配列決定に用いるためプライマー伸長反応に用いることができる。二本鎖のDNA分子の中の片鎖にDNA合成のためのプライミングサイトを作るために、酵素活性によってニックを入れる一つの例が、Walker T.G. et al.,Proc. Natl. Acad. Sci. USA 89,392−396(1992)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。他の例では、オリゴヌクレオチドは、RNAポリメラーゼの認識部位を導入する。RNAポリメラーゼは、T3 RNAポリメラーゼ、T7 RNAポリメラーゼ、SP6 RNAポリメラーゼを含むが、これに限定されるものではない。これらはすべてDNA依存性RNAポリメラーゼであり、それぞれ特異性のある二本鎖プロモーターがある。プロモーター又はそれらの認識部位より始まり、これらは一本鎖又は二本鎖DNAのいずれかの鋳型から相補的なRNAの合成を5’から3’方向へ触媒する。Guatelli J.C. et al.はProc. Natl. Acad. Sci. USA 87,187401878(1990)(本文献は、参照することによって本明細書に組み込まれるものとする)にDNA依存性RNAポリメラーゼの使用例を記載している。異なる例では、オリゴヌクレオチドはDNA結合タンパク質の認識部位を含んでも良い。多くのDNA結合タンパク質は当業者の知るところであり、天然に存在するものでも、タンパク質デザインによって調製されたものでもあり得る。そのようなDNA結合タンパク質は、転写因子、ゲノムDNAの認識部位に直接又は間接的に結合する調製機能タンパク質などを含むが、これに限定されるものではない。転写因子は生命に不可欠の分子であり、ゲノム情報を利用するのに必要である。あらゆる生命体は多くの転写因子を含んでいる、例えば、Kanamori M.,787−93(2004)(本文献は、参照することによって本明細書に組み込まれるものとする)に既知のマウス転写因子すべてのデータベースが公開されている。転写因子は、異なる認識部位への親和性という点でほかのものと区別できる。この特異性は、与えられた転写因子および転写因子のグループの認識部位を含むDNA分子を濃縮するために用いることができる。しかしながら、当業者は、特異的配列よりもむしろ構造を認識するタンパク質について知るように、転写因子の結合特異性は、塩基配列への結合に限定されない。例えば、転写因子DAX−1は、Zazopoulos E. et al.,Nature 390,311−315(1997)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されているように異なるDNA構造に結合する。他の例では、DNA結合タンパク質は一本鎖DNAに特異的に結合する。一本鎖DNA結合タンパク質は、E.coliのSSB、ファージT4のGene32の産物、アデノウイルスのDBP、一本鎖DNAに対して作られた抗体、仔ウシ胸腺 UP1、またはそれらの混合物などが含まれるが、これに限定するものではない。加えて、二本鎖DNAのミスマッチに特異的に結合するタンパク質がある。このグループのタンパク質は、E.coliの主要なミスマッチ修復パスウェイに関係するMutSタンパク質ファミリー(タンパク質のファミリーについてはhttp://www.tigr.org/〜jeisen/MutS/MutS.htmlを参照、本ウェブページの内容は、参照することによって本明細書に組み込まれるものとする)が含まれるが、これに限定されるものではない。修飾されたRNAやDNAに相補配列又はその部分的配列が付着される場合と比べて、塩基配列中にミスマッチがある場合のプライマーの伸長反応にプライマーが使われる場合、MutSタンパク質やその他の遺伝子ファミリーは、ミスマッチをもつ二本鎖のDNA種を特に濃縮するように用いても良い。加えて、MutSや遺伝子ファミリーは、プライマー伸長反応を操作するあるいはブロックするために用いられても良い。MutSタンパク質のいくつかは、商業的に入手可能であり、例えばNippongeneのTaq MutS(Tokyo,Japan,コード番号316−04011)がある。異なる例では、オリゴヌクレオチドは「Identifierh配列」や「Barcode」として用いられる塩基配列の領域を含んでも良い。そのような塩基配列は、Identifier配列としてサンプルの由来の印となったり、核酸やIdentifier配列に相補的な塩基配列を持つ核酸とのハイブリダイゼーションによって、修飾されたRNAやDNAを特異的に捕獲するためのタグとして機能させることもできる。そのような塩基配列は、前述した酵素反応の任意の反応の特異的、選択的なプライミング部位として用いることもできる。Identifier配列は、DNAポリメラーゼによる第二鎖の合成や、例えばPCRによる増幅、一本鎖DNAの調製などのための選択的なプライミング部位となることもできる。このように、異なるRNA種に認識部位やIdentifier配列として異なるオリゴヌクレオチドを導入する前述の方法の組み合わせによって、本発明は、全RNAや複数のRNA分子中で、個々のRNA分子を分けて操作する手段を提供する。
【0043】
異なる実施態様において、本発明は、RNA分子又は種々のRNA分子の末端に官能基を導入する手段を提供する。多くの異なる官能基が結合分子に結合する親和性を持っている。官能基は、反応基又は化学反応において共有結合を形成するのに適した架橋リンカー、アミノ基、ビオチン、ジゴキシゲニン、抗体、抗原、タンパク質、核酸、核酸結合分子、またはそれらの組み合わせを含むが、これに限定するものではない。官能基や官能基に結合された分子は、基盤上にある結合分子と結合することができる。本発明を実施するための基盤は、アビジン、ストレプトアビジン、ジゴキシゲニン結合分子、所定の配列をもったオリゴヌクレオチド、抗体またはリガンド、又は化学的基盤など、共有結合を形成する化学反応に用いられる反応基を固定化したものから選ぶことができる。用いられる官能基がビオチンの場合、対応する基盤はアビジンまたはストレプトアビジンである。同様に、官能基がジゴキシゲニンの場合、基盤はジゴキシゲニン結合分子である(Roche Diagnostics GmbH Catalog、本文書は、参照することによって本明細書に組み込まれるものとする)。官能基がオリゴヌクレオチドの場合、基盤は官能基の配列に相補的配列を持ったオリゴヌクレオチドであり、官能基が抗原の場合は、基盤は抗体またはprotein I又はprotein Gのような抗体結合タンパク質である。従って、本発明はRNA分子へ官能基を導入する手段を提供し、官能基はオリゴヌクレオチドに付加される。修飾されたオリゴヌクレオチドは、多くの業者より商業的に入手可能である。最もよく使用されるのはビオチンラベル化オリゴヌクレオチドである。異なる修飾オリゴヌクレオチドの一つの例として、MWG Biotechのウェブサイト(http://www.mwg−biotech.com/html/s_synthetic_acids/s_modifications.shtml、ここから入手できる情報は、参照することによって本明細書に組み込まれるものとする)に記載がある。MWG Biotechはオリゴヌクレオチドの異なる位置に官能基としてビオチンやジゴキシゲニンを持つオリゴヌクレオチドを提供できる。さらに、一つ又はそれ以上の官能基を持つ修飾オリゴヌクレオチドを得ることができる。例えば、5’Aminolink C3/C5/C6/C12、3’Aminolink C3/C6/C7、3’Aminolink C3/C6/C7、Amino(C2/C6)−dT、Amino C6−dC、Spacer C3/C9(TEG)、Spacer C12/C18(HEG)、あるいはreduced Thiol modifierなどのクロスリンクのための反応基がある。RNAオリゴヌクレオチドは、例えばInvitrogenより購入でき、そのようなRNAオリゴヌクレオチドの情報はhttp://www.invitrogen.com/content.cfm?pageid=9900(ここに示された情報は、参照することによって本明細書に組み込まれるものとする)で得ることができる。また、Operonはhttp://www.operon.com/(ここに示される情報は、参照することによって本明細書に組み込まれるものとする)で役立つ情報を提供している。
【0044】
前述の実施態様において、図1〜2で示すように、RNAリガーゼはRNA分子のリン酸化された5’末端にオリゴヌクレオチドを連結するために用いられる。通常用いられるT4 リガーゼのようなRNAリガーゼは、オリゴヌクレオチドの3’末端のリボヌクレオチドに依存し、RNAにデオキシリボヌクレオチドを直接的に連結しなくても良い。さらに、RNA分子へのオリゴヌクレオチドの連結は配列特異的ではなく、任意の配列のオリゴヌクレオチドをRNA分子と組み合わせることができる。代替的な方法がClepet C. et al. in Nucleic Acids Res. 32、 e6 (2004)(本文献は、参照することによって本明細書に組み込まれるものとする)に示されており、その中でDNAリガーゼはRNAに二本鎖DNA又は部分的に二本鎖になったDNA分子を連結する(RNA−taggingと呼ばれる)。例えば、T4 DNAリガーゼは鋳型DNAへのRNA断片の連結を触媒することができる(Kleppe、 K. et al.,Proc. Natl Acad. Sci. USA,67,8-73(1970)、Fareed,G.C. et al.,J. Biol. Chem.,246、925-932(1971)(両文献は、参照することによって本明細書に組み込まれるものとする)。連結反応を仲介する鋳型DNAは、個々のRNA分子を修飾するための塩基配列特異的な連結反応に用いることができる。配列特異性は、部分的に二本鎖となったオリゴヌクレオチドによって達成できる。そのようなオリゴヌクレオチドは、RNA分子の5’末端にハイブリダイズする突出領域を持つ(図3参照)。突出領域は、4から6、又は6から8ヌクレオチドの長さでも良い。また8〜12、又は12ヌクレオチド以上の長さでも良い。当業者が知り得る、例えばShibata Y.et al.in Biotechniques,30,1250−1254(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に述べられているような連結反応において部分的に二本鎖となったオリゴヌクレオチドを用いる種々の方法がある。反応によって、突出領域は、例えば前述のShibata.et.al.で用いられているようなランダム配列や特有のRNA分子を標的とした規定の配列を持つことができる。他の例として、突出領域は、認識部位にランダム配列を含む制限酵素や認識部位の外側で切断する制限酵素を用いて酵素反応的に作っても良い。そのような酵素は、BstXI(CCANNNNN↓NTGG)、DrdI(GACNNNN↓NNGTC)、BglI(GCCNNN↓NGGC)、BoxI(GACNN↓NNGTC)、BseJI(GATNN↓NNATC)、BseLI(CCNNNNN↓NNGG)、CaiI(CAGNNN↓CTG)、CseI(GACGC(5/10)↓)、Eam1105I(GACNNN↓NNGTC)、Eco31I(GGTCTC(1/5)↓)、Eco57I(CTGAAG(16/14)↓)、Esp3I(CGTCTC(1/5)↓)、HpyF10VI(GCNNNN↓NNGC)、LguI(GCTCTTC(1/4)↓)、OliI(CACNN↓NNGTG)、PdmI(GAANN↓NNTTC)、PsyI(GACN↓NNGTC)、SfiI(GGCCNNNN↓NGGCC)、SmuI(CCGGC(4/6)↓)、Van91I(CCANNN↓NTGG)、XagI(CCTNN↓NNNAGG)、又はそれらのアイソザイムを含むが、これに限定されるものではない。これらの酵素は特に有用であり、ランダム突出領域は複数の核酸分子より調製される。例えば、cDNAライブラリーは、前述の酵素の一つの認識部位を持つリンカーが5’末端に持つcDNAインサートで構築することができる。そのようなライブラリーの分子の切断は、もとのcDNAライブラリーに存在する分子を代表するランダムな突出領域を持つ複数の分子を生成する。よって、本発明は、反応に使用されるオリゴヌクレオチドに相補的な配列とRNA分子の5’末端に相補的な配列を持つ鋳型DNAによって、個々のRNA分子の目的とされた修飾を行う手段を提供する。一つの例では、それらの配列は5〜10ヌクレオチドの長さであり、別の例では、10〜25ヌクレオチドの長さであり、また別の例では、25ヌクレオチドよりも長い。RNA分子の5’末端に相補的な領域は、実験的手法で得ることができ、例えば、複数の核酸分子の操作によって、あるいはコンピューターデザインによる組み合わせを用いた化学合成によって得ることができる。RNA分子の配列情報は、当業者が知り得る公共データベース、例えばNCBI(http://www.ncbi.nlm.nih.gov/)、EMBL(http://www.ebi.ac.uk/Databases/)、DDBJ(http://www.ddbj.nig.ac.jp/)などより入手できる。ある例では、一つの鋳型DNAは特有のRNA分子を標的として用いることができる。別の例では、ランダム配列を含む複数の鋳型DNAが用いられる。さらに別の例では、複数の鋳型DNAは、特有の配列を含む。よって、本発明は、全RNA中で、特有のRNA分子やmRNAなどのRNAを標的とした操作を行う柔軟な手段を提供する。
【0045】
RNA分子はSambrook J. and Russuell D.W.,Molecular Cloning,A Laboratory Manual,Cold Spring Harbor Laboratory Press,New York、(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載の逆転写によるDNA転写物の調製の鋳型として用いることができる。当業者が知り得る異なる反応条件や酵素修飾などを含む多くの改変された方法がある。例えば、逆転写酵素は、AMV reverse transcriptase、M−MLV reverse transcriptase、M−MLV reverse transcriptase RNase H minus、あるいはそれらの改変物などを含むが、これに限定されるものではない。任意の修飾されたRNAは、前述の又は図1〜3で示されたすべての又は部分的なステップによって得ることができ、それらを鋳型にして、逆転写酵素によってRNA鋳型からコピーDNAを合成するcDNA合成反応に用いることができる。この反応は、鋳型RNAにハイブリダイズし、DNA合成を開始するプライマーを必要とする。一つの例では、3’末端にランダム配列とそれに続く所定の配列を持ったプライマーセットが用いられる。所定の配列はDNAの後の操作に役立てられるが、逆転写反応のプライミングには必要とされない。従って、本発明は、ランダム配列のみを持つプライマーを用いることができる。そのようなランダム配列自体又は規定配列を持つオリゴヌクレオチドの一部としてのランダム配列は、4〜6ヌクレオチド、又は6〜10ヌクレオチド、又は10〜15ヌクレオチドの長さを取ることができる。ランダムプライマーを用いることにより、鋳型RNAの5’末端の配列に相補的な配列を持つDNA断片を合成することができる。ランダムプライマーは、鋳型RNAの任意の領域にハイブリダイズできるので、反応は異なる長さのDNA分子の混合物を生成する。ランダムプライミングでは、完全長cDNAの調製はできないが、長いRNAの本来の5’末端まで到達できるという有利な面があり、それを用いなければ逆転写反応の限界のため困難である。別の例では、オリゴdTプライマーが通常多くのmRNA種の3’末端にあるポリA配列にハイブリダイズすることができる。ランダムプライミングとは対照的に、オリゴdTプライミングは、RNA分子全体を反映した完全長cDNAの合成に通常用いられる。さらに、規定配列のプライマーは、逆転写酵素反応を開始するために用いられ、例えば、通常RACE法(Rapid Amplification of CDNA Ends)などの応用に用いられる。完全長cDNA合成を開始する他の方法は、WO2006/003721(本文献は、参照することによって本明細書に組み込まれるものとする)に開示されている。従って、本発明は図4と5に示されたように修飾されたRNAからDNA転写物を調製する異なる手段を提供する。
【0046】
前述の鋳型RNAからのDNAの合成は、二本鎖DNA/RNA分子を生成する。このような二本鎖DNA/RNA分子の中のRNA部分は、RNA分解酵素やバッファーのpH変更により除去される。例えば、RNase H酵素は、DNA/RNA二本鎖分子のRNAを特異的に分解するので、本発明の実施には好ましい酵素である。RNAの除去は、逆転写反応で使われたプライマーに関係なく、ランダムプライミングであっても特異的プライミングであってもオリゴdTプライミングであっても関係なく、任意の種類のcDNAに適用される。鋳型DNA/RNAからのRNAの除去の例は、Sambrook J. and Russuell D.W.,Molecular Cloning,A Laboratory Manual,Cold Spring Harbor Laboratory Press,New York、(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。任意のこのようなDNA/RNA二本鎖分子の処理は、リボヌクレオチドから構成される鋳型の大部分又は全体より作られた一本鎖DNAとしてのDNA分子を遊離する。DNA/RNAハイブリッドが二本鎖DNA領域を含む場合は、例えば、前のステップでRNA分子に付加されたオリゴヌクレオチドの一部又は全部がDNAを含む場合、RNA部分の除去により、鋳型RNAの5’末端に相当する末端に二本鎖DNA領域を含むDNA分子が調製される。よって、本発明は、全RNA又はmRNAに由来する塩基配列を持つ、あるいはそれらのRNA分子の操作により導入された塩基配列を持つ一本鎖又は部分的一本鎖DNA分子を調製する手段を提供する。
【0047】
一本鎖DNAはDNA解析や操作に重要であり、またバイオテクノロジーや分子生物学において多くの応用や技術は、一本鎖DNAの鎖特異的な調製を必要とする。そのような応用は、塩基配列決定や標識プローブの合成を含む鎖特異的DNA合成のための鋳型DNAの調製、チミン残基のウラシルへの置換、点突然変異の導入、種々のDNAまたはRNA分子の混合物からの個々のクローンの分離、検出やディファレンシャルハイブリダイゼーションのためのドライバーやテスターの調製、一塩基多型(SNPs)の検出と解析、マイクロアレイの調製を含むが、これに限定されるものではない。これらの方法や応用は分子生物学の当業者がよく知るところであり、Sambrook J. and Russuell D.W.,Molecular Cloning,A Laboratory Manual, Cold Spring Harbor Laboratory Press,New York,(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。
【0048】
本発明により得られた鋳型DNAは、初期の段階でRNAに付加されたオリゴヌクレオチドの性質によって区別でき、例えば、図2〜3に示される。よって、本発明は、図7に示されるような一本鎖DNA又は部分的一本鎖DNAの調製に用いることができる。例えば、RNAにRNAオリゴヌクレオチドが付加され、その修飾されたRNAが逆転写酵素によるcDNA合成に用いられた場合、RNA/DNAハイブリッド分子からのRNA部分の除去は、鋳型RNAに相補的配列、初期段階でRNAの3’末端に付加されたRNAオリゴヌクレオチドに相補的な配列、5’末端に逆転写反応に用いたプライマーより直接生成した配列(図7A参照)を持つ一本鎖DNAを生成する。よって、そのようなDNA分子は、RNAに付加されたオリゴヌクレオチドより直接的又は間接的に生成された既知配列の領域に並んで潜在的に未知の配列を中心領域に含む。並んでいる領域の配列は既知なので、そのような分子は塩基配列決定や操作の鋳型として直接用いることができる。例えば、DNAの3’末端に相補的配列を持っているプライマーは、塩基配列決定反応のプライマーに用いることができる。鋳型DNAの塩基配列決定を行う従来の方法は、例えばSambrook J. and Russuell D.W., Molecular Cloning、 A Laboratory Manual,Cold Spring Harbor Laboratory Press,New York,(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されているサンガー法がある。しかしながら、本発明はサンガー法による鋳型DNAの塩基配列決定に限定されるのものではなく、最近数々の塩基配列決定法が開発されており、詳細はMetzker M.L. Genome Res. 15,1767−1776(2005),Kling J.,Nature Biotechnology 23,1333−1335(2005);Shendure J. et al.,Nature Review Genetics 5,335−344(2004)(これらの文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。これらの方法のいくつかは商業的な応用の対象となっており、例えば454Life Sciences、(http://www.454.com/、Helicos BioScience(http://www.helicosbio.com/)、Solexa(http://www.solexa.com/Company/overview.htm)、Visigen、(http://www.visigenbio.com/index.html)、GeneoVoxx GmbH、(http://www.genovoxx.de/)(これらのウェブページの一部又はすべてに見出される情報は、参照することによって本明細書に組み込まれるものとする)などが挙げられる。本発明は、鋳型DNAの向かい合った末端をデザインする手段を提供するので、鋳型DNAは必要に応じた性質を持つようにデザインされることができ、例えば、前述の任意の方法又は他の新しい方法により塩基配列決定を行う必要に応じてデザインされることができる。例えば、454 Life Sciencesによる用法で塩基配列決定を行う場合には、鋳型の向かい合った末端に特有のプライマー部位が要求され、これによりエマルジョンPCRによるDNA分子のクローンの増幅が可能となる。好ましい例として、鋳型DNAは、Margulies M.et al.,Nature 437,376−80(2005)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されているように、一分子のエマルジョンPCRに適応するように5’末端と3’末端に二つの異なる塩基配列を含む。異なる例においては、DNA/RNAのハイブリッドオリゴヌクレオチドは、mRNAに付加され、逆転写によるcDNA合成に用いられる。RNA/DNAハイブリッド分子からのRNA部分の除去はRNAの5’末端に対応する長いDNA鎖の3’末端に二本鎖DNAの領域を持つ部分的な一本鎖DNA分子を生成する。さらにこのDNA分子は、鋳型RNAに相補的な配列を含み、また3’末端に初期段階でRNAに付加されたRNA/DNAオリゴヌクレオチドに相補的な配列を含み、5’末端に逆転写反応で使われたプライマーから直接生成される配列を含む(図7B参照)。この一群では、DNA分子の二本鎖領域は、DNAポリメラーゼ反応のプライマーとして機能する短いDNAストレッチを含む。従って、本発明は、他に塩基配列決定用のプライマーを添加することなく、配列決定用の鋳型を直接提供する手段を提供する。別の例では、DNAオリゴヌクレオチドがmRNAに付加され、その結果作られる修飾されたRNA分子は逆転写によるcDNA合成に用いられ、RNA/DNAハイブリッド分子からのRNA部分の除去は、RNAの5’末端に対応する長いDNA鎖の3’末端に二本鎖DNAの領域を持つ部分的な一本鎖DNA分子を生成する。さらにこのDNA分子は、鋳型RNAに相補的な配列を含み、また3’末端に初期段階でRNAに付加されたRNA/DNAオリゴヌクレオチドに相補的な配列を含み、5’末端に逆転写反応で使われたプライマーから直接生成される配列を含む(図7C参照)。これらの方法により得られた鋳型は、図7Bに記載されている鋳型とよく似ていて、二本鎖領域は鋳型RNAの5’末端で正確に止まっている。よって、DNAオリゴヌクレオチドやDNA/RNAハイブリッドオリゴヌクレオチドにより、もとのRNA分子の5’末端に直接に接しているか、もとのRNAの5’末端より1〜5ヌクレオチド、又は5〜10ヌクレオチド、又は10〜15ヌクレオチド、又は15ヌクレオチド以上離れている。図7Cに示されるDNA分子は、図7Bに記載されている鋳型と同様の方法で塩基配列決定反応に用いられる。図7Aと7Bに示されているDNA分子を調製する前述した例の改変法では、RNAに付加されるDNAオリゴヌクレオチド、又はDNA/RNAオリゴヌクレオチドは官能基を含む。この例では、本発明は、RNAの5’末端に対応する長いDNA鎖の3’末端に二本鎖DNAの領域を持つ部分的な一本鎖DNA分子を調製し、二本鎖領域の短いほうの断片は官能基を含む。分子中の長いほうの断片は、鋳型RNAに相補的な配列を含み、また3’末端に初期段階でRNAに付加されたRNA/DNAオリゴヌクレオチドに相補的な配列を含み、5’末端に逆転写反応で使われたプライマーから直接生成される配列含む(図7D参照)。このような鋳型DNAは、塩基配列決定反応で用いられ、また官能基は支持体や基盤へ鋳型を直接結合させることを可能とする。官能基や官能基に付加された分子は、基盤上の結合分子と結合することができる。そのような基盤は、官能基の性質に応じて選択することができる。例えば、官能基がアミノ基だった場合、反応基は、化学反応で基盤に共有結合するものになる。官能基がビオチンだった場合には、基盤はアビジン又はストレプトアビジンになる。官能基がジゴキシゲニンの場合、基盤はジゴキシゲニン結合分子である(Roche Diagnostics GmbH Catalog、本文献は、参照することによって本明細書に組み込まれるものとする)。官能基がオリゴヌクレオチドの場合、基盤は官能基の配列に相補的配列を持ったオリゴヌクレオチドであり、官能基が抗原の場合は、基盤は抗体またはprotein I又はprotein Gのような抗体結合タンパク質である。従って、本発明は、プライマー部位と基盤又は支持体に直接結合する性質を持ったDNA分子又は塩基配列決定用鋳型を調製する手段を提供する。本発明の好ましい例においては、そのような鋳型は新しい塩基配列決定方法に適応し、個々の分子が直接塩基配列決定用の支持体に結合できる。そのような塩基配列決定方法は、例えば454Life Sciences、(http://www.454.com/、Helicos BioScience(http://www.helicosbio.com/)、Solexa(http://www.solexa.com/Company/overview.htm)、Visigen、(http://www.visigenbio.com/index.html)、GeneoVoxx GmbH、(http://www.genovoxx.de/)(これらの一部又は全てのウェブページの情報は、参照することによって本明細書に組み込まれるものとする)などが含まれるが、これに限定するものではない。
【0049】
前述の例では、本発明はmRNAの5’末端を修飾する手段を提供する。しかしながら、本発明は5’末端の修飾に限定されるものではない。上述の逆転写反応によるcDNA合成のプライミングと図4〜5に示される例によれば、オリゴdTのような一般的なプライマーやランダムプライマー又は特定の配列をもったプライマーのセットは鋳型RNAからcDNAの第一鎖を合成するのに用いられる。cDNA合成をプライミングするプライマーは、一本鎖DNA、又はWO2006/003721(本文献は、参照することによって本明細書に組み込まれるものとする)に開示されるように部分的に一本鎖と二本鎖部分を持つDNAを含んでも良い。プライマーは、鋳型RNAの中の配列に相補的な配列を含んでも良く、cDNAの後の操作に用いられるようにデザインされた配列又はIdentifier配列として機能する配列を含んでも良い。RNAにハイブリダイズするヌクレオチドはランダム配列でも良く、特定のRNA種にプライミングするために公共のデータベースより得られた配列でも良く、またmRNAのポリA配列にハイブリダイズするようなdTの長いストレッチ配列から構成されるものでも良い。RNA分子の配列にある情報は、NCBI(http://www.ncbi.nlm.nih.gov/)、EMBL(http://www.ebi.ac.uk/Databases/)、DDBJ(http://www.ddbj.nig.ac.jp/)などの当業者が知り得る公共のデータベースを調べることにより得られ、また多くの場合、得られた情報は3’末端の特定のプライマーをデザインするのに充分である。別の例では、そのような配列情報は、スプライス部位やオープンリーディングフレームと呼ばれるコーディング領域の末端の一つのようなRNAの規定領域を標的とするプライマーのデザインに用いられる。逆転写反応のプライミングに用いられるプライマーは、RNAに相補的な配列のみを含んでもよい。この例として、そのような反応より得られたcDNAは、その5’末端にRNAの中の配列に相補的な配列を持つ。別の例では、プライマーは、RNA配列に相補的な配列とRNAの配列には関係のない配列とを含む。よって、本発明は、cDNAの5’末端につなげる新しい配列をcDNAに導入する手段を提供する。当業者が知り得る特定の配列や認識部位への結合に依存する酵素活性は多々あり、多くのそれら酵素は商業的に種々の業者より入手可能である。それらの業者は、FERMENTAS UAB(Vilnius,Lithuania)、New England Biolabs Inc.(Beverly,USA)、Promega(Madison,USA)、Takara(Tokyo,Japan)、Roche(Mannheim,Germany)、GE Biosciences(Cardiff,United Kingdom)を含むが、これに限定されるものではない。通常、制限酵素は二本鎖DNAのみを切断し、一本鎖DNAは切断しない。よって、制限酵素は、第二鎖の合成後にのみ適用することができ、例えば、クローニングなどの目的に用いられる。よって、本発明は、核酸分子の完全長クローニングに関係し、得られたDNA断片より配列情報を得ることができる。特定の例では、制限酵素は、例えばShibata Y et al. in Biotechniques. 31,1048−1049(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されているような方法によって、cDNAからポリA/Tストレッチを除去するのに用いることができる。ポリA/Tストレッチの除去の方法は、RNAの3’末端のタグ配列を得るのに特に重要であり、それらの方法は特許出願US2005/059022、US2005/0255501、WO2004/050918、米国特許第6,136,537号(これらの文献は、参照することによって本明細書に組み込まれるものとする)に開示されているが、これに限定するものではない。
【0050】
ほかの例では、逆転写反応に用いられるプライマーは官能基を含む。そのような官能基は、プライマーがランダムプライマーであるか、特定のプライマーであるか、又はオリゴdTプライマーであるかというプライマーの性質とは関係なく、cDNAに組み入れられる。よって、本発明は官能基を持った修飾された5’末端を持つcDNA断片を調製する手段を提供する。当業者が知り得るところの結合分子へ結合する親和力をもった多くの官能基がある。官能基は反応基、アミノ基、ビオチン、ジゴキシゲニン、抗体、抗原、タンパク質、核酸結合分子などを含むが、これに限定されるものではない。官能基と官能基に付加された任意の分子は、基盤上に存在する結合分子と結合することができる。本発明の目的のために、基盤は、アビジン、ストレプトアビジン、ジゴキシゲニン結合分子、抗体とそのリガンドが固定された任意の形状、および化学的基盤の中より選ぶことができる。用いられる官能基がアミノ基のような反応基の場合、反応基は化学反応で基盤に共有結合するために用いられる。官能基がビオチンだった場合には、基盤はアビジン又はストレプトアビジンになる。同様に官能基がジゴキシゲニンの場合、基盤はジゴキシゲニン結合分子である(Roche Diagnostics GmbH Catalog、本文献は、参照することによって本明細書に組み込まれるものとする)。官能基が抗原の場合は、基盤は抗体またはprotein I又はprotein Gのような抗体結合タンパク質である。修飾されたオリゴヌクレオチドは、多くの業者より商業的に入手可能であリ、ビオチンラベル化オリゴヌクレオチドが頻繁に使われている。種々の修飾オリゴヌクレオチドの調製の例は、MWG Biotech(http://www.mwg−biotech.com/html/s_synthetic_acids/s_modifications.shtml)、Invitrogen(http://www.invitrogen.com/content.cfm?pageid=9900、Operon(http://www.operon.com/)(これらのウェブページに示された情報は、参照することによって本明細書に組み込まれるものとする)のウェブページに記載されている。別の例では、オリゴヌクレオチドはDNA結合タンパク質の認識部位を持っても良い。当業者が知り得るDNA結合タンパク質は多数あり、それらは天然に存在するものでも、タンパク質デザインを使って調製されるものでも良い。そのようなDNA結合タンパク質は、転写因子、ゲノムDNAの認識部位直接又は間接的に結合する調製機能タンパク質などを含むが、これに限定されない。転写因子は生命に不可欠の分子であり、ゲノム情報を利用するのに必要である。あらゆる生命体は多くの転写因子を含んでいる、例えば、Kanamori M. et al.はBiochem Biophys Res Commun,322,787−93(2004)(本文献は、参照することによって本明細書に組み込まれるものとする)に、既知のマウス転写因子すべてのデータベースが公開されている。転写因子は異なる認識部位への親和性という点でほかのものとは区別できる。この特異性は、与えられた転写因子および転写因子のグループの認識部位を含むDNA分子を濃縮するために用いることができる。しかしながら、当業者が特異的配列よりもむしろ構造を認識するタンパク質について知るように、転写因子の結合特異性は、塩基配列への結合に限定されず、例えば、転写因子DAX−1はZazopoulos E. et al.,Nature 390,311−315(1997)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されているように異なるDNA構造に結合する。他の例では、DNA結合タンパク質は、一本鎖DNAに特異的に結合する。一本鎖DNA結合タンパク質は、E.coliのSSB、ファージT4のGene32の産物、アデノウイルスのDBP、一本鎖DNAに対して作られた抗体、仔ウシ胸腺 UP1、またはそれらの混合物などが含まれるが、これに限定するものではない。別の例では、結合タンパク質はMutS又はMutS遺伝子のファミリーであっても良い。さらに別の例では、オリゴヌクレオチドはIdentifier配列として用いられる配列の領域を含んでも良い。このような配列やIdentifier配列は、サンプルの由来の印となったり、核酸やIdentifier配列に相補的な塩基配列を持つ核酸とのハイブリダイゼーションによって、修飾されたRNAやDNAを特異的に捕獲するためのタグとして機能することもできる。また、前述した酵素反応の任意の反応の特異的、選択的なプライミング部位として用いることもできる。Identifier配列は、DNAポリメラーゼによる第二鎖の合成や、例えばPCRによる増幅、一本鎖DNAの調製、又は塩基配列反応のプライミングのための選択的なプライミング部位となることもできる。よって、先述の方法を組み合わせることによって、本発明は複数のRNAの中のRNA分子に由来するcDNAに異なるオリゴヌクレオチドを導入する手段を提供する。
【0051】
先述した任意のステップによって、本発明は、RNAの5’末端に相当する位置に官能基を導入する手段を提供する。加えて、本発明は、RNAの3’末端に相当する位置又はcDNA第一鎖の5’末端に相当する位置に官能基を導入する手段を提供する。従って、本発明は、RNAより生成されるcDNAのいずれかの末端に官能基を導入する手段を提供する。RNA又はcDNAに付加された官能基は、他の分子に対する結合親和力を持ち、また官能基は修飾されたRNA又はDNAを捕獲し表面上に結合するために用いることができる。官能基と結合分子の組み合わせ例は、化学反応により基盤上に共有結合を形成するアミノ基などの反応基、ビオチンとアビジン又はストレプトアビジンの結合、ジゴキシゲニンとジゴキシゲニン結合分子の結合、オリゴヌクレオチドの相補配列への結合、抗原の抗体への結合、抗体のprotein Iやprotein Gなどの抗体結合タンパク質への結合を含むが、これに限定されるものではない。官能基の位置によって、修飾されたRNA又はDNAが表面上へ結合される方法や方向は異なる。例えば、部分的に二本鎖となっているcDNA分子はRNAの5’末端に相当する位置の官能基によって表面に結合することができる。この例では、部分的に二本鎖となったcDNA分子は、RNAの5’末端に相当する位置からcDNA断片の塩基配列決定を開始することができる(図9A参照)。別の例では、一本鎖cDNA分子は、RNAの3’末端に対応する一本鎖DNA分子の5’末端の位置に付加された官能基の相互作用によって表面に結合する。RNAの3’末端に対応する位置でcDNAが表面に結合するとき、外側プライマーはcDNA分子の塩基配列決定を開始する位置を決定するために用いられても良い(図9B参照)。別の例においては、修飾されたRNAまたはcDNAの第一鎖に付加された塩基配列は表面上に存在するオリゴヌクレオチドと相補的な配列を含んでも良い。図9Cは、表面上に結合しプライマーの機能を持つオリゴヌクレオチドにハイブリダイゼーションによって表面上に結合する一本鎖RNAまたはcDNA分子の原理を示している。表面上のプライマーは、その配列に相補的な配列を含むか否かにより、どのRNA又はcDNA断片が表面上に結合できるかを決定する。さらに、表面上のプライマーはcDNA分子の塩基配列決定を始める位置を決定する。固相上の塩基配列決定の例はStahl,S. et al.,Nucleic Acid Research 16,3025−3038(1988)、Lindroos K.et al.,Nucleic Acid Research 29,No.13 e69(2001)(これらの文献は、参照することによって本明細書に組み込まれるものとする)に公表されている。当業者が知り得る規定された配列のオリゴヌクレオチド合成を支持体上で直接行う方法は種々あり、また支持体上にそれらオリゴヌクレオチドを結合する方法も種々ある。そのような方法は、通常マイクロアレイの作成に用いられ、より詳細にはJordan B.,DNA Microarrays: Gene Expression Applications、 Springer−Verlag、 Berlin Heidelberg New York,2001: Schena A, DNA Microarrays,A Practical Approach,Oxford University Press, Oxford(1999)(両文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。従って、本発明は、修飾された核酸分子を調製し、それらをRNAやDNAの塩基配列決定のような解析のために捕獲する手段を提供する。
【0052】
前述の実施態様において、本発明は一本鎖又は部分的一本鎖RNAおよびDNA分子を調製する手段を提供する。官能基を用いて、それらの分子を表面上に結合することができる。表面上の分子は種々のバッファーによって精製や後の操作のために洗浄され得る。よって、本発明は一本鎖DNA、部分的一本鎖DNA、またはRNAを精製する手段を提供する。そのような一本鎖DNA、部分的一本鎖DNA、またはRNAを精製する方法は新しい技術によって達成される一分子検出には必須であり、そのような技術にはMetzker M.L. Genome Res. 15,1767−1776 (2005)、 Kling J.,Nature Biotechnology 23、 1333−1335 (2005)、Shendure J. et al.,Nature Review Genetics 5、 335−344 (2004)(両文献は、参照することによって本明細書に組み込まれるものとする)が含まれるが、これに限定するものではない。従って、本発明は、塩基配列情報を直接得るために、一本鎖DNA、部分的一本鎖DNA、またはRNA分子を利用することに関する。好ましい実施態様においては、本発明は一本鎖DNA断片の規定した領域から塩基配列情報を得ることに関する。好ましい例においては、RNAの5’末端特異的塩基配列情報は、RNA分子の5’末端配列に相補的な配列を含み、本発明によって調製されるDNA断片より得られる。よって、本発明は、RNAの塩基配列情報を得ることに関する。
【0053】
先述の図1から7で示される任意のステップによって、本発明はRNAの5’末端に対応する位置にオリゴヌクレオチドを導入する手段を提供する。加えて、先述の図8に示される任意のステップによって、本発明はRNAの3’末端に相当する位置にオリゴヌクレオチドを導入する手段を提供する。このように、本発明はRNAに由来するcDNAのいずれかの末端に特定の塩基配列を導入する手段と修飾された末端を含むcDNA断片の調製手段を提供する。ある例では、本発明により導入される配列は、一本鎖DNAがその鎖中にある相補的配列の部分で二本鎖を形成し末端に閉じたループを作るようなヘアピン構造を形成することができる。よって、前述の処理によりRNAから向かい合った末端にヘアピン構造を持つように修飾されたcDNA分子を得ることができる(図10参照)。第一鎖cDNA合成がランダムプライマーを用いたかオリゴdTプライマーを用いたかによって、このような分子はRNAに由来する部分的配列を含むか、又はRNAに由来する全体の配列を含む(完全長cDNA)。ある例として、先述に従って調製されRNAの5’末端に相当する末端にヘアピン構造を持つ一本鎖cDNA分子は、直接塩基配列決定ができ、その場合ヘアピン構造は塩基配列決定反応のプライミング部位として機能する。別の例では、先述に従って調製され両端にヘアピン構造を持つ一本鎖cDNA分子は、二つのヘアピン構造を利用して増幅することができる。図11Aに示されるように、各々の末端にループ構造を持つ一本鎖DNA断片は、Notomi T. et al.,Nuc Acids Res. 8,e63(2000)(本文献は、参照することによって本明細書に組み込まれるものとする)に開示されているLoop−Mediated Isothermal Amplification (LAMP法)の鋳型となる。よって、本発明によって調製されるDNA分子は繰り返し配列が得られる方法で増幅することができ、このようなポリマーにあるループ構造は、増幅反応の伸長をプライミングするために用いることができ、又は塩基配列決定反応を推進するために用いることができる。ある例においては、増幅された断片は、25〜50塩基対、又は50〜100塩基対、又は100〜200塩基対、又は200〜300塩基対の長さをとる。別の例では、増幅されたDNA断片は300塩基対以上の長さになる。よって、本発明によって調製されるDNA分子は繰り返し配列の直線状ポリマーが得られる方法で増幅することができ、そのようなポリマーは塩基配列決定反応を推進するために用いることができる。
【0054】
好ましい例において、ループ構造を両端に持つDNA分子は、エクソヌクレアーゼ活性と鎖置換活性を持たないDNAポリメラーゼによって、ヘアピン構造の自由3’末端が伸長される第一の反応を含むステップによって、環状一本鎖DNA分子に転換され得る。このようなDNAポリメラーゼは、M−MuLV Reverse Transcriptase、H Minus M−MuLV Reverse Transcriptase、Superscript II、Superscript III、AMV Reverse Transcriptase、MonsterScript、Expand Reverse Transcriptase、又はそれらの混合物を含む任意の逆転写酵素を含むが、これに限定されるものではない。その他のDNAポリメラーゼは、DNAポリメラーゼIのクレノー断片、T4とT7のDNAポリメラーゼDNAポリメラーゼI、Taqポリメラーゼ、Tfl DNAポリメラーゼ、Tli DNAポリメラーゼ、Tth DNAポリメラーゼ、又は既知の任意のDNAポリメラーゼを含むが、これに限定されるものではない。鎖置換活性の欠如により、DNAポリメラーゼは向かい合ったヘアピン構造の5’末端に達した時に反応が停止する。第二の反応ステップでは、一本鎖DNAの開いた末端が環状DNA分子を作るように結合される。そのような結合反応は任意のDNAリガーゼによって行われ、それらDNAリガーゼは、T4 DNAリガーゼ、E. coli DNAリガーゼ、Taq DNAリガーゼを含むが、これに限定されるものではない。環状一本鎖DNA分子はローリングサークル増幅法(RCA法と呼ばれる)を用いて増幅できる。RCA反応は、DNAポリメラーゼにより環状鋳型上のオリゴヌクレオチドプライマーを等温反応で伸長させることにより遂行される。詳細は米国特許5、854、033号および米国特許6、143、495号(両文献は、参照することによって本明細書に組み込まれるものとする)に開示されている。反応生成物は直線状環状DNAで、鋳型のコピーが直列に並んでいる。反応の時間や条件によって、反応生成物は数十、数百又は数千の鋳型のコピーを一分子中に持つ。RCA反応で特に用いられるDNAポリメラーゼは、当業者に公知の等温DNA増幅に必要とされる強い鎖置換活性を持つphi29 DNAポリメラーゼがあるが、これに限定されるものではない。当業者に公知の数多くのCA法の応用や改変法がある。RCA法に関するより詳細な記載は総説Gusev、Y.et al.,American J.Pathology 159,63−69(2001)、Zhang D.et al.Clin.Chim.Acta.363,61−70(2006)(これらの文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。従って、本発明により調製されたDNA分子は、繰り返し配列の直線状ポリマーが得られる方法により増幅することができ、それらのポリマーは塩基配列決定反応を推進するのに用いられる配列を含む。
【0055】
一つの例では、RCA反応は、反応生成物が直接又は間接的に規定された位置又は検出、解析、又は塩基配列決定のポイントと呼ばれる位置に結合できるような方法で実行される。一つ例では、Nallur G.et al.,Nucleic Acid Res,29,e118(2001)に開示されているガラススライド上の信号増幅に仲介されたRCAの手段がある。この例では、RCAは、複数の核酸分子の中の個々の標的のクローン増幅を実行するためのステップを可能とし、そのステップでは各々の分子は表面上の規定された位置で増幅される。ここでは、RCA反応は、例えば従来のPCR法に見られるような増幅バイアスの危険を伴うことなく、高い並列性を持った方法で実行される。RCA反応で表面上に結合されたプライマーを使うことにより、反応生成物が並んだ基盤を得ることができ、各々の反応生成物は一分子の中に鋳型のコピーを複数含んでいる。よって、RCA反応は検出や解析の感度を大きく増幅することができ、規定の位置で信頼のおける塩基配列決定反応を実施することを可能とする。
【0056】
別の例では、RCA反応は規定の位置での検出、解析および塩基配列決定のための鋳型を調製することができ、鋳型は一つ又はそれ以上の検出ステップ、解析、又は塩基配列決定反応の対象となる。従って、本発明は鋳型の塩基配列決定を行う第一のステップと、塩基配列決定反応中に鋳型より生成された増幅産物を除去する第二のステップと、異なるプライマーによって同じ位置で同じ鋳型の塩基配列決定を再度行う第三のステップからなる手段を提供する。そのような一連の反応は、一つの鋳型より二回、三回、四回、五回、さらには五回以上回数を重ねて塩基配列決定を行い、解読配列を得るために実施されても良い。DNAの表面への共有結合的な結合は、Beier M.and Hoheisel J.D.,Nucleic Acid Research、27,1970−1977(1999)によれば、少なくとも30サイクルのハイブリダイゼーションとハイブリダイズしたDNAの除去を可能とすると記載されている。よって、本発明は一つの鋳型から一種以上の配列情報を得る手段を提供し、その異なる塩基配列決定反応は同じ位置で実行され、その位置により同じ鋳型より得られる違う解読塩基配列のつながりが規定される。
【0057】
別の例では、RCA反応は、もとのRNA分子のセンス鎖およびアンチセンス鎖の多数のコピーを含む反応生成物を調製するために用いられる。そのような反応生成物は、図11Bで示されるステップによって環状鋳型が調製される場合に得られる。環状鋳型は、RNAより得られる二本鎖cDNAの中にあるセンスとアンチセンス鎖を含み、もとのRNAの向かい合った末端に対応する位置にヘアピン構造によりつながっている。この鋳型は双方向性である。本発明の好ましい例では、ヘアピン構造はセンス、アンチセンス鎖がそれらの末端からの塩基配列決定を可能にするプライミング位置を含む。よって、RNA分子の向かい合った末端より末端配列を得る手段を提供する。このとき、RNA分子はcDNAに転換され、cDNAは最初のRNA分子配列を持つセンス、アンチセンス鎖を持つ二本鎖から成り、二本鎖cDNAにあるセンス、アンチセンス鎖は一本鎖DNAを含む環状分子を構成するヘアピン構造により連結され、環状一本鎖DNA分子はセンス、アンチセンス鎖を含む塩基配列決定のための直線状鋳型DNAを生成するためにRCAにより増幅され、DNA又はもとのRNAの向かい合う末端から得られる配列情報は、二つ又はそれ以上の連続した塩基配列決定反応が同じ位置で行われることにより得られる。この例では、本発明は、RNAの末端配列、転写物の境界、ゲノム中の転写の開始、終止点、又は任意のDNA分子の末端配列を決定するために用いることができる。別の例では、本発明は、RNA、cDNA、又はゲノムDNA内における規定された領域の末端配列を決定するために用いることができる。そのような規定された領域の境界は、それらの調製段階における特定のステップによって決定される。その断片は、生物学的由来のものでも良く、また完全なランダム切断によって生成されても良い。さらに、塩基配列決定用のプライマーは、従来のプライマーウォーキング法と同じように、鋳型の任意の領域にハイブリダイズするようにデザインされることができ、又は鋳型中の特定の部位、例えばスプライス部位などに対してデザインされても良い。例えば環状一本鎖DNA分子を形成するために、直線状二本鎖DNAの末端にヘアピン構造を持ったオリゴヌクレオチドを連結することによって、任意の二本鎖DNAを双方向性鋳型へ転換することは、本発明の範囲内である。環状一本鎖DNA分子は、RCAによって増幅され、もとのDNA分子のセンス、アンチセンス鎖を含む塩基配列決定用の直線状鋳型DNAが生成される。DNAの向かい合った末端からの配列情報は、同じ位置で行われる二つ又はそれ以上の連続した塩基配列決定反応によって得られる。この例では、本発明は、例えばエクソンの末端配列やクロマチンIPより得られたゲノム断片の末端配列、ハイパーセンシティブ部位の境界配列などを決定するために用いることができる。
【0058】
別の実施態様では、本発明は、RNAの5’末端やRNAの3’末端に相当する領域に導入されたIdentifier配列の利用に関する。先述のとおり、本発明によって調製されたcDNAの向かい合った末端にIdentifier配列、又は特定の配列を導入する手段を提供する。好ましい例では、Identifier配列は、RNAまたはcDNAの末端の近傍に位置し、RNAやcDNAそのものから配列情報をえるための同じ塩基配列決定反応より塩基配列を決定することを可能とする。Identifier配列は、1塩基対、2塩基対、3塩基対、4塩基対、5塩基対、6塩基対、6〜10塩基対、10〜15塩基対、15〜20塩基対、又は20塩基対以上の長さでも良い。好ましいIdentifier配列は、6〜12塩基対、又は25〜75塩基対の長さである。Identifier配列は任意の性質、ランダム配列であっても良い。それらはコンピューターによってデザインされても良く、生物学的サンプルからとっても良く、また人工的に作られても良い。それらは、制限酵素又はその他の酵素やタンパク質の認識部位やプライミング部位を含んでも良い。Identifier配列は、以下のすべて又は任意のいずれかの法則によってデザインされることができる。
− 塩基配列決定またはハイブリダイゼーションによる同定を行うのに充分な長さを持つ
− 同じ実験で用いる異なるIdentifier配列の塩基配列は、区別可能である。
− 同じ実験で用いる異なるIdentifier配列は、Identifier配列の塩基配列決定にエラーが起こった場合でも明確な同定を可能にするため、一つの位置以上において区別できる。
− Identifier配列は、塩基配列決定反応を妨げるような配列や構造をとる塩基配列を避ける(例えばGリッチ配列、パリンドローム配列)。
− Identifier配列は、相補的配列を安定なハイブリッドを形成するように選択されても良い。
− Identifier配列は、制限酵素や転写因子のようなタンパク質への結合や特定の操作を可能にする塩基配列を持っても良い。
− Identifier配列は、本発明を実施する際にRNAやDNAの操作を妨げる塩基配列を避ける(例えば、操作の過程で使用される制限酵素の認識部位を持たない)。
【0059】
本発明の好ましい例では、Identifier配列は、サンプルのプールの中で、サンプルの由来を印付けるために用いられ、サンプルプールのすべての構成物は同じ実験で同時に扱われる。サンプルプールのサンプルは、出来るだけ早く混合されるべきであり、好ましくは修飾されるRNAサンプルの段階で混合されているべきである。異なるIdentifier配列を持つ異なるRNAサンプルの混合によって得られるサンプルは「プールサンプル」となり、異なる形で修飾されたRNAを含む(図12B参照)。それゆえ、Identifier配列は、好ましくはRNAの5’末端近くに位置し、それゆえmRNAやトータルRNAの修飾のための最初のステップで導入される。
【0060】
一つの実施態様では、Identifier配列は、異なる生物種や組織、生物学的実験の種々の処理あるいは時間的な段階、あるいは異なる細胞種を含む多数の生物学的サンプルから得られる特定のRNAにある核酸分子を印付けるために用いられる。実験のデザインにおいてサンプルのプーリングは、種々の機能的な意味を持つ。例えば新しい塩基配列決定方法のハイスループット性を十分に活用するためにサンプルの複雑性を高めること、同時に扱うサンプルの数を減らすことにより多くのサンプルの処理を単純化すること、特定のデータ解析の形態を可能にすることなどを含むが、これに限定されるものではない。一つの好ましい実施態様において、異なるサンプルに異なるシステムエラーが起きるような個々の実験と比較し、共通の統計的解析のためにすべての操作のステップにおいて同じシステムエラーがあるようにするためサンプルはプールされる。例えば、一つの典型的な応用において、Identifier配列は、本発明によってRNAが修飾される際にRNAの5’末端近傍に付加され、その後修飾されたRNAサンプルはcDNAの調製より先に混合される。プールサンプルは、区別できるIdentifier配列を持つ修飾されたRNAサンプルの異なる種類の混合物であるように調製される。修飾されたRNAサンプルのプールは、その後データを得るために本発明に沿って一つのサンプルとして処理される。例えば、修飾されたRNAサンプルの解読配列はプールライブラリーより得られる。配列情報はこの分野で知られる任意の方法によって決定することができるが、好ましくは、各々の解読配列がIdentifier配列ともとのRNA又はcDNAサンプルに由来する配列情報の両方を含む。配列の決定後、個々の解読配列からIdentifier配列を認識し、同じIdentifier配列を持つ解読配列をその後の解析のためにグループ化するためにコンピューターにより処理されることができる。もとのRNAに関係する塩基配列は、Identifier配列とは分けて実験的なデザインの必要に沿って解析される。これらの塩基配列は、「配列タグ」と呼ばれるものやRNAやcDNAに由来する部分的な配列を含む短い解読配列に関連する。配列タグは、転写物やゲノム中の位置を同定するために用いることができ、あるいはプールサンプルの中の転写物の発現レベルを統計的に解析するために用いても良い。配列タグの利用についてより詳細な情報は、Harbers M.and Carninci P.,Nature Meth.2,495−502(2005)(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。配列情報は、データベースに格納され、または解析、保管、又は参照データセットの構築の目的でコンピューターに格納される。そのようなデータベースは、例えば、塩基配列情報、異なる組織、細胞株より得られた配列タグの発現頻度、ゲノム中の転写物、遺伝子、機能要素に関する注釈付データをふくむことができる。
【0061】
別の例においては、Identifier配列は塩基配列決定によって同定されず、基盤や支持体に結合したIdentifier配列に相補的な配列を持つ核酸分子と特定のハイブリッドを形成するために用いられる(図12C参照)。この例では、Identifier配列は、表面上の規定された位置に同じ由来から生成されたサンプルをグループ化するために用いられる。その位置は、Identifier配列の性質やプールサンプルの中のRNA、DNA、サンプルの由来を明らかにするために用いられる。従って、Identifier配列の読み取りは直接塩基配列決定を必要とせず、かわりに特定のハイブリダイゼーション反応に行うことができる。
【0062】
別の例では、Identifier配列は、塩基配列決定やハイブリダイゼーションによって同定されず、支持体や基盤に結合したIdentifier配列に結合親和性をもったタンパク質に特異的に結合するために用いられる。この例では、Identifier配列は、同じ由来のサンプルを表面上の規定された位置にグループ化するために用いられる。その位置は、Identifier配列の性質やプールサンプルの中のRNA、DNA、サンプルの由来を明らかにする。従って、Identifier配列の読み取りは直接塩基配列決定やハイブリダイゼーションを必要とせず、かわりにIdentifier配列に高い親和性を持つたんぱく質に結合することによって行われる。
【0063】
本発明は、コンピューターによる注釈付を目的とし、本発明に従って得られるDNA断片の領域の塩基配列決定に関係する。注釈付は、統計的解析、参照情報に対するアライメント、およびゲノム配列へのマッピングなどを含む。よって、本発明は、遺伝子発見、遺伝子同定、遺伝子発現プロファイリング、注釈付の手段に関する。
【0064】
他の実施態様では、in situハイブリダイゼーションによる領域解析のために、核酸分子の末端からハイブリダイゼーションプローブを調製することに関する。好ましい例では、in situハイブリダイゼーション実験は、タイリングアレイを用いて行う。この実施態様では、本発明は、ハイブリダイゼーションプローブのデザインに関する。プローブはマイクロアレイ上のものを含むがこれに限定されるものではない。
【0065】
よって、本発明は、例えば生物学的サンプルの特性解析のような必要に応じて、核酸分子やそれらの短い断片を解析する手段を提供する。さらに、本発明は、RNAやDNA断片を解析的検定に利用するために効率的で迅速な操作を行う手段を提供する。この意味では、本発明は、新しいハイスループット塩基配列機器と技術を利用するための手段を提供する。
【0066】
他の実施態様では、逆転写反応に特定プライマーを用いて、本発明に従って調製された修飾されたRNAは、当業者が知るRACE法と類似のプロトコールによって実際の5’末端配列を決定するために用いられる。
【0067】
本発明は、生物学的サンプルの状態、すなわち遺伝情報や発現プロファイル、調節パスウェイやネットワークの活性などを調べるための重要な情報を得るために必要な手段を提供する。よって、本発明は、ライフサイエンスや診断の研究に用いられる解析的検定のデザインや実行に関する。本発明は、生物学的システムの解析と診断の手段を提供する。
【0068】
本発明、又はその一部は、試薬、核酸分子、およびRNAの操作やDNAの調製に必要な酵素、その他の要素を含むキットの製造に用いることができる。一つの実施態様では、キットはRNAを修飾するために必要な試薬を提供する。別の実施態様では、キットは鋳型DNAを調製するために用いる試薬を提供する。好ましい実施態様では、キットは一分子検出のための鋳型を調製するための試薬を提供する。好ましい実施態様では、キットは研究目的の試薬を提供する。さらに好ましい実施態様では、キットは診断的な検定のための試薬を提供する。
【0069】
本発明の主要なステップを詳細に説明する。ここで用いるすべての名前や省略形は、当業者に公知の意味で用いられる。
【実施例1】
【0070】
[RNAの単離]
本発明を実施するために、mRNAまたはトータルRNAのサンプルは,当業者に公知の方法、例えばSambrook J.and Russuell D.W.,Molecular Cloning,A Laboratory Manual、Cold Spring Harbor Laboratory Press,New York(2001)(本文献は、参照することによって本明細書に組み込まれるものとする)に開示されている方法により調製された。さらにCarninci P.et al.(Biotechniques 33,2002,306−309、本文献は、参照することによって本明細書に組み込まれるものとする)によれば、細胞質mRNAの分画を得る方法が開示されている。細胞質mRNAの分画を用いるのが好ましいが、本発明はこれに限定されるものではなく、他の任意のmRNAやトータルRNAの調製方法を用いても本発明を同様の方法で実施することができる。
【0071】
本発明を実施するには、トータルRNAを用いると完全長cDNAライブラリーを調製する際に行うCap−selectionとの組み合わせで十分は結果を得られるので、本発明を実施するためには、全RNA又は細胞質RNAからのmRNAの調製が好ましいが、必須ではない。mRNAの量はトータルRNAの調製の約1〜3%であり、その後、商業的に販売されているオリゴdT−セルロースマトリックスを使ったキットを用いて調製される。このような商業的キットは、本発明を実施するためにmRNAの分画を調製するのに用い、推奨される条件下で十分なmRNAを得られるMACS mRNA分離キット(Milteny)を含むがこれに限定されるものではない。本発明を実施するために、1サイクルのオリゴdT−mRNA選択で十分であり、強いmRNA精製は長いmRNAを失う可能性がある。
【0072】
本発明の実施に用いられるすべてのRNAサンプルは、RNAの純度を調べるために230、260、280nmのOD比率が計測された。230/260比が0.5より低い場合には、多糖類の除去が成功したと考えられ、260/280比が1.8より高いもしくは2.0付近のとき、効果的なタンパク質の除去が行われたと考えられた。RNAサンプルはさらに、トータルRNA調製時の28Sと18S rRNAの比率が適当であることを確認し、RNA分画の品質を調べるために、アガロースゲルの電気泳動によって解析された(rRNAのサイズは哺乳類以外の種から得られるトータルRNAではことなることがある)。
【実施例2】
【0073】
[5’末端誘導体化RNA分子のライブラリーの調製]
本実施例は、RNAオリゴヌクレオチドを用いてRNA分子の5’末端を誘導体化する典型的なプロトコールである。すべての反応は、核酸の損失を避けるために、500μlのシリコン化したマイクロチューブとシリコン化したチップを用いて行われた。
【0074】
RNAサンプルは、最初に脱リン酸化された。RNA(例えば1ngから1μg)と2μgのグリコーゲンを一緒にチューブに加え、全量5μlに調製された。反応バッファーは、通常の濃度の1/10または、5mMのBis−Tris−Propane−HCl、0.1mMのMgCl2、0.01mMのZnCl2(pH 6.0)、25°Cを用いた。グリコーゲンは、操作中にRNAがプラスチックに付着するのを避けるために加えられた。サンプルを65℃で5分間変性し、後で除去するためにリン酸基を露出させた。その後37℃で2分間置き、Anctartic phosphatase(New England Biolabs)を2.5ユニット加えた。サンプルは37℃で3時間から一晩処理された。一晩脱リン酸化を行うことにより、リン酸基の98〜99%が除去された。45℃で活性が高くなるトレハロースを最終濃度0.6Mとなるように加え、45℃で短いインキュベーションを行うこともできる。
【0075】
その後、Anctartic phosphataseは65℃で不活性化されたが、この処理の前に二価イオンをキレートする必要があった。このため、0.55μlの溶液(0.5Mの酢酸ナトリウム(pH 6.0)、10mMのEDTA、1%のβ−メルカプトエタノール及び0.1% Triton X−100)を加えた。EDTAは、二価イオンをキレートし、次のTAP処理に適した条件となった。Anctartic phosphataseも、またバッファー中のEDTAによって阻害された。不活性化は65℃で5から15分間行われた。
【0076】
次に、0.2μl(2ユニット)のタバコ酸ピロホスファターゼ(TAP)を加えdecappingを行った。TAPは、実験当たり20ユニットまで増やすことが可能であった。反応は37℃で2時間行い、続いて65℃で15分間熱不活性化を行った後、サンプルを氷上で冷却した。また、GCリッチなRNAの二次構造を解くために役立つベタインを1M加えても良い。この処理の後、TAPはATPをそれ以上分解しなかった。ATPはその後のステップにおいて必要である。その後、任意の配列の5μMの「capping RNA」オリゴヌクレオチドを加え、ライゲーションを行った。反応液6.75μlに対して2μlのRNAリガーゼ(500mMのHEPES−NaOH(pH8.0、25°C)、100mMのMgCl2、100mMのDTT)を加えた。DTTはTAPを阻害した。1mMのhexamino cobaltum chloride(HCC)を加えても良いが、これは任意であり、必須ではない。ポリエチレングリコール(PEG8000)を最終濃度が25%となるように加え、ATPの125μM、T4 RNAリガーゼ(Fermentas)10ユニットを加えた。このような条件では、最終的な混合液はライゲーションステップを阻害するものではなかった。
【0077】
2時間から一晩(16時間)20℃でライゲーション反応を行った。この時点で、RNAのCap構造はオリゴヌクレオチドに置き換えられ、これは完全長cDNA調製のような他の例で示すように、他のテストに用いられる。
【実施例3】
【0078】
[修飾RNAの調製に用いられる酵素の活性試験]
実施例2で用いたれた酵素とバッファーの活性試験を行った。
(A)Antarctic Phosphatase(New England Biolabs)の活性試験
5’リン酸化オリゴヌクレオチドを次のバッファー中、37℃で2時間脱リン酸化を行った。続いてオリゴヌクレオチドをT4リガーゼとγ−32P−ATPによってRIラベルし、PAGEによって解析した。脱リン酸化されていなければ、5’リン酸基のためにRIラベルは不可能である。
【0079】
(B)タバコ酸ピロホスファターゼ(TAP)(Epicentre)活性試験
γ−32P−ATPを2ユニットのTAPと反応バッファー中でインキュベートした。放射性ATPとインキュベーションを行う前に、TAPを15分間加熱した。
【0080】
(C)T4 RNAリガーゼ(Fermentas)の活性試験
放射性ラベル化オリゴヌクレオチドを、ラベルしていないオリゴリボヌクレオチドの存在下でインキュベートした。ライゲーションの結果、ポリアクリルアミドゲルの移動度が変化した。
【実施例4】
【0081】
[修飾RNAからの完全長cDNAの調製]
上記で調製したサンプルをmicrocon YM−100フィルターを使い、製造者(Millipore)の説明書に従って脱塩した。ライゲートされたRNAに水、逆転写酵素、プライマー(Invitrogen)を加えた。プライマー(AGA GAG AGA CCU CGA GCC UAG GUC CGA C)800ngを使い、3.3Mのソルビトール−トレハロース混合液を、最終濃度が0.5Mのソルビトール及び4%トレハロースになるように加え、逆転写反応液を調製する。RNAとプライマーの混合液を65℃で10分間加熱した後、他の試薬を準備する間氷上で保持する。11μlの2倍濃度GCバッファー(Carninci,Shiraki et al,Biotechniques,2002;32,984−985、本文献は、参照することによって本明細書に組み込まれるものとする)から構成されるプレミックスを加え、その後、10mMのdNTPを1μl及びMMLV逆転写酵素(RNaseH minus、Fermentas)を1μl加えた。GCバッファーシステムは、製造者の推奨するバッファーに置き換えた。この反応液にRNAサンプルを加え、25℃で2分間(サンプルのアニーリングのため)、42℃で30分、52℃で10分、56℃で10分インキュベートした後、反応を停止した。この方法では、もとのmRNAの5’末端までのびたcDNAを高い頻度で得ることができた。その後、精製、処理された。例えば、プロテアーゼKにより処理を行った(20μgをEDTA(最終濃度が10mM)と一緒に加え、95℃で15分インキュベートした)。このサンプルを、サイズ分画しプライマーを除去するため、C14B(Amersham−Pharmacia)で処理した。
【実施例5】
【0082】
[cDNAの第二鎖合成とPCR増幅]
cDNAをPCRにより増幅した。cDNAに最終濃度が1倍となるようにTAKARA EXバッファーを加え、さらにdNTP(最終濃度200μM)、5’オリゴヌクレオチド(配列: acc tcg agc cta ggt ccg ac)、3’オリゴヌクレオチド(配列:ca gcg tcc tca agc ggc cgc)をそれぞれ400nM、MgCl2を2.5mM、KClを最終濃度50mMとなるように加えた。成分を混合した後、94℃で5分加熱し、94℃で30秒、58℃で30秒、68℃で1.5分のサイクルを30サイクル繰り返した。
【0083】
生成された5’末端cDNAを平滑末端化し、標準的な方法によりプラスミドベクターへクローニングした(クローニングと塩基配列決定の一般的な情報についてはSambrook et al.,supra参照)。
【実施例6】
【0084】
[RACE法の実験]
Capped RNAを実施例2と同じようにRNAオリゴヌクレオチドの配列のみ下記の様に変更して調製した。実施例2のプロセスを用い、次にPCRを行い、RACEによる5’末端増幅が可能となった。500ngの肝臓トータルRNAをONE−Tube oligo−capping法で処理し、未反応のオリゴリボヌクレオチドを除去、その後ランダムプライマーを用いて逆転写反応を行った。5’末端の増幅には下記の遺伝子特異的プライマー(TTGGAGAGAGGGTTTCGACGAGTCA)と、oligo−capに相補的なプライマー(CGACTGGAGCACGAGGACACTGA)を用いた。
【実施例7】
【0085】
[454塩基配列決定の応用と他の基盤]
実施例2の方法でcDNAを調製したが、オリゴヌクレオチドのデザインと調製は、RNAの3’末端と5’末端に異なるアダプターを付けるため下記の通りとした。
アダプターA
CCATCTCATCCCTGCGTGTCCCATCTGTTCCCTCCCTGTCTCAG
アダプターB
/5BioTEG/CCUAUCCCCUGUGUGCCUUGCCUAUCCCCUGUUGCGUGUCUCAG
【0086】
アダプターBは「oligo−capping」配列として用い、アダプターAは第一鎖合成のためのオリゴランダムプライマーと一緒に用いた。第一鎖合成後、Cl−4Bスパンカラムを用いて未反応のプライマーと分離した。続いて、サンプルをエマルジョンPCRにかけた。塩基配列決定反応は454−Life Science sequencing instrumentに沿って行った(Margulies et al,Nature,2005; 437(7057):376−380、本文献は、参照することによって本明細書に組み込まれるものとする)。この結果、1ランで数十万の解読配列を得ることができた。
【実施例8】
【0087】
[5’末端塩基配列タグの応用]
実施例2とそれに続く実施例のようにcDNAを合成し、当業者が知る標準的な方法でcDNA第二鎖の合成を行った。例えば、Kodzius et al.,Nat. Methods.2006 Mar; 3(3):211−22(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。DNAをMme Iにより切断し、第二リンカーを付加、増幅、精製を行い、コンカテマーを作成した。この方法の詳細は種々に記載されているが、例えば、Kodzius et al.,Nat. Methods. 2006 Mar; 3(3):211−22(本文献は、参照することによって本明細書に組み込まれるものとする)に記載されている。これらの塩基配列タグは、その後塩基配列決定、遺伝子の境界の同定(Carninci et al,Science. 2005 Sep 2; 309(5740):1559−63、本文献は、参照することによって本明細書に組み込まれるものとする)、発現プロファイリング、遺伝子のプロモーター(Harbers and Carninci,Nat. Methods,2005 Jul; 2(7): 495−502、本文献は、参照することによって本明細書に組み込まれるものとする)などに用いることができる。
【実施例9】
【0088】
[固体支持体に結合したDNAの塩基配列決定]
実施例2とその後の例にそってcDNAを合成し、当業者の知る標準的なプロトコールを用いてcDNAの第一鎖を合成した。それらは例えば、Kodzius et al.,Nat. Methods. 2006 Mar、 3(3):211−22に記載されている。続いて、核酸を米国特許出願番号20060012793号、米国特許出願番号20060012784号、米国特許出願番号20060008824号に開示された方法により固相基盤に結合した。
【図面の簡単な説明】
【0089】
【図1】図1は、RNAの酵素的修飾の概念図である。この図にあるように、RNA調製はmRNAの5’末端のCap構造の存在によって特徴づけられたRNAの種類を含んでも良い。Cap構造は、他のCap構造を欠き、自由リン酸基を5’末端に持つ切断されたRNA種から区別するために使われる。一連の酵素反応の中で、まずホスファターゼにより、切断されたRNAの自由リン酸基が除去される。次にピロホスファターゼにより完全長mRNAよりCap構造が取り除かれ、Cap構造の代わりにリン酸基が露出する。5’末端にリン酸基をもったRNA分子のみが、RNAリガーゼによってオリゴヌクレオチドと共有結合して修飾を受けることができる。オリゴヌクレオチドは斜線で示されている。
【図2】図2は、オリゴヌクレオチドの付加によるRNAの修飾を概念的に示したものである。図1で示されたその後のステップの前に、RNAリガーゼは一つ又は複数のRNA分子に異なる種類のオリゴヌクレオチドを結合させるのに用いられる。本発明によれば、オリゴヌクレオチドはデオキシリボヌクレオチドまたはDNAオリゴヌクレオチド、又はリボヌクレオチドまたはRNAヌクレオチドからなるホモポリマーであることが可能である(パネルA)。またデオキシヌクレオチドとリボヌクレオチドからなるヘテロポリマーとなることも可能である(パネルB)。あるいは、官能基(パネルC中で「F」で示されている)をもった任意の種類のホモポリマー又はヘテロポリマーとなることもできる。
【図3】図3は、RNAの5’末端へオリゴヌクレオチドを連結する代替法の概念図である。図1、2で示すように、RNAリガーゼはRNAのリン酸化された5’末端にオリゴヌクレオチドを結合させることができる。しかし、そのような反応はオリゴヌクレオチドの3’末端のリボヌクレオチドに依存することもあるが塩基配列の特異性はなく、任意のオリゴヌクレオチドがRNA分子と結合することができる。反応に配列特異性を与えるには、部分的に二本鎖になっているオリゴヌクレオチドを用い、突出領域をRNA分子の5’末端の配列とハイブリダイズさせる。反応によっては、突出領域はランダム配列でも良いし、特定のRNA分子を標的とした決められた配列でも良い。さらに、部分的に二本鎖になったオリゴヌクレオチドはDNAリガーゼを用いてRNAにオリゴヌクレオチドを連結させるのに用いることもできる。その後、適当なプライマーの結合、逆転写酵素の処理、RNAの分解を行い、cDNAを得る。下記でさらに詳細とバリエーションを述べる。
【図4】図4は、ランダムプライミングを用いたcDNA第一鎖合成の概念図である。図1から3に示されるすべてのステップ、又は部分的なステップに沿って得られた単一又は複数の種類の修飾されたRNA分子は、逆転写酵素を用いて鋳型RNAのコピーcDNAを合成する反応に用いることができる。この反応は鋳型RNAにハイブリダイズしDNA合成を開始するプライマーが必要である。この図に示されている例では、3’末端にランダム配列(図中NNNNで示す)とそれに続く確定した配列を持つプライマーセットが用いられている。任意のこのようなプライマーセットが、塩基配列および5’末端に付加したオリゴヌクレオチド(パネルAからC)から得られる官能基を持った修飾RNAを鋳型として行われるDNA合成のプライミングに用いることができる。
【図5】図5は、オリゴdTプライマーを用いたcDNA第一鎖合成の概念図である。図1から3に示されるすべてのステップ、又は部分的なステップに沿って得られた単一又は複数の種類の修飾されたRNA分子は、逆転写酵素を用いて鋳型RNAのコピーcDNAを合成する反応に用いることができる。この反応は鋳型RNAにハイブリダイズしDNA合成を開始するプライマーが必要である。この図に示されている例では、多種類のmRNAの3’末端に通常見られるポリA配列にハイブリダイズするオリゴdTプライマーが用いられている。オリゴdTプライマーは、塩基配列および5’末端に付加したオリゴヌクレオチド(パネルAからC)から得られる官能基を持った修飾RNAを鋳型として行われるDNA合成のプライミングに用いることができる。
【図6】図6は、DNA/RNAの複合体からRNA除去を示す概念図である。図4、5で示すRNAを鋳型としたDNAの合成はDNA/RNA複合体を形成する。このような任意のDNA/RNA複合体分子のRNA部分はRNA分解酵素又は反応バッファーのpHを変えることによって取り除くことができる。例えば、RNase H酵素は特にDNA/RNA複合体のRNAを分解するので本発明の実施には好ましい酵素である。このような任意のDNA/RNA複合体に対する処理はDNA鎖を一本鎖DNA(パネルA)として、又は部分的に二本鎖のDNA分子として遊離する。部分的に二本鎖のDNA分子には前のステップでRNA分子に付加されたオリゴヌクレオチド(パネルB〜D)の部分がある。図にはオリゴdTを用いて得られるcDNAが描かれているが、ここで述べるRNAの除去は逆転写反応に用いられるプライミングの種類によらず任意のcDNAにも適用できる。
【図7】図7は、本発明によって得られるDNA分子の応用の概念図である。本発明によって得られる図1〜6のいずれかに描かれている鋳型DNAは、例えば図2と3にある初期の段階でRNAに付加されたオリゴヌクレオチドの性質の点で特徴を持っている。基本的な構造と応用はパネルA〜Dに示されている。
【図8】図8は、cDNAの3’末端への官能基の導入を示した概念図である。図4と5で示すステップによって通常のオリゴdTプライマーまたはランダムプライマーが鋳型RNAからの第一鎖cDNA合成をプライミングするために必要である。そのようなプライマーは鋳型RNAの配列と相補配列領域を含んでも良く、後のcDNA操作に用いるようにデザインされた配列を含んでも良い。また、図中「F」で示されているようにプライマーに官能基が付加されていても良い。そのような官能基は、ランダムプライマー(パネルA)又はオリゴdTプライマー(パネルB)のどちらを用いるかに関わらずcDNAに導入することができる。従って、本発明は修飾された末端をもつcDNA断片を調製する手段を提供する。
【図9】図9は、修飾されたDNA断片の捕獲を示した概念図である。図1〜7のいずれかに示されたステップに従って本発明はRNA分子の5’末端に相当する位置に官能基を導入する手段を提供する。加えて、図8に示されたいずれかのステップに従って本発明はRNA分子の3’末端に相当する位置に官能基を導入する手段を提供する。本発明はRNAに由来するcDNAのいずれかの末端に官能基を導入する手段を提供する。図中「F」で示されcDNAに付加される官能基が図中の開いたクランプで示される他の分子に対して結合親和性を持っている場合、官能基は修飾されたcDNAを捕獲し表面上に結合させるために用いることができる。パネルAは官能基と表面上の結合相手の反応によって部分的二本鎖を持つcDNAが表面上に結合される原理を示している。cDNA分子の部分的に二本鎖になっている領域は、RNAの5’末端に相当するcDNA断片の末端からの塩基配列決定を可能にする。パネルBは官能基と表面上の結合相手の反応によって一本鎖cDNAが表面上に付けられる原理を示している。cDNAはRNAの3’末端に相当する位置で表面上に結合するのに対し、cDNA分子の塩基配列決定を始める位置は外部プライマーを用いて決めても良い。パネルCは表面上に結合したオリゴヌクレオチドまたはプライマーとハイブリダイゼーションすることによって一本鎖cDNA分子が表面上に結合する原理を示している。表面上のプライマーによって、cDNA断片に含まれる表面上プライマーとの配列相補性に基づいてどのcDNA断片が表面に結合しているかを決定することができる。さらに、表面上プライマーはcDNA分子の塩基配列決定の開始位置を決定する。
【図10】図10は、ヘアピン構造の導入を示す概念図である。図1〜7に記載のいずれかのステップによって、本発明はRNAの5’末端に相当する位置にオリゴヌクレオチドを導入する手段を提供する。加えて、図8に示されたいずれかのステップに従って本発明はRNA分子の3’末端に相当する位置にオリゴヌクレオチドを導入する手段を提供する。本発明は、cDNA断片に修飾された末端を与えるために、RNAに由来するcDNAのいずれかの末端に特定の配列を導入する手段を提供する。図で示されるプロセスでは、本発明により導入される配列は、一本鎖DNAがその鎖中にある相補的配列の部分で二本鎖を形成し末端に閉じたループを作ってヘアピン構造を形成することができる。ここで開示された手段を用いて、RNAに由来するDNA分子は、cDNA合成にランダムプライマー(パネルA)を用いたかオリゴdTプライマー(パネルB)を用いたかに関わらず、向かい合った末端にヘアピン構造を含むように修飾されることができる。
【図11】図11は、修飾されたDNA断片の増幅を示した概念図である。図10で示されたステップによって調製されたDNA断片は、二つの向かい合った末端にあるヘアピン構造を使って増幅することができる。パネルAに示すように、向かい合った末端にループ構造を持つ一本鎖DNA断片はLAMP法と呼ばれるLoop−Mediated Isothermal Amplification(Notomi T. et al.,Nuc. Acids Res. 8,e63,2000、本文献は、参照することによって本明細書に組み込まれるものとする)の鋳型となることができる。本発明により調製されたDNA断片は繰り返し配列を含むポリマーが得られる方法で増幅することが可能であり、このようなポリマーが含むループ構造は増幅反応の伸長をプライミングするために用いることができ、また塩基配列決定反応を推進するために用いることができる。パネルBに示されるように、各々の末端にループ構造を持つDNA断片は、エクソヌクレアーゼ活性と鎖置換活性を持たないDNAポリメラーゼによって、ヘアピン構造の自由3’末端が伸長される第一の反応を含むステップによって、環状一本鎖DNA分子に転換され得る。鎖置換活性の欠如により、DNAポリメラーゼは向かい合ったヘアピン構造の5’末端に達した時に反応が停止する。第二の反応ステップでは、一本鎖DNAの開いた末端が環状DNA分子を作るように結合される。環状一本鎖DNA分子はRCA法によって増幅することが可能である。RCA法における種々の改変や応用は当業者が知り得るものである。他のRCA法の参照文献としてGusev、 Y. et al.,American J. Pathology 159,63−69 (2001)、 Zhang D. et al. Clin. Chim. Acta,363,61−70 (2006)(これらの文献は、参照することによって本明細書に組み込まれるものとする)が挙げられる。従って、本発明によって調製されたDNA断片は直線状の繰り返し配列を含むポリマーが得られる方法で増幅することができる。そのようなポリマーは、少なくとも最初のRNAの配列のコピーを二つ含み、よって二つの配列はお互いに相補的となる。そのようなポリマーはまた、最初のRNAに由来する二つの配列のいずれかより塩基配列決定反応を推進するために用いることができる配列を含んでいる。
【図12】図12は、プールされたサンプルとidentifier配列との導入を示す概念図である。図1と2で示すいずれかのステップは完全長mRNAの5’末端にオリゴヌクレオチドを導入するために用いることができる。そのようなオリゴヌクレオチドは、Identifier配列と呼ばれる配列情報を含む領域を含んでも良く、それは修飾されたRNAおよびRNAに由来する任意のDNAの由来をプールされたサンプル中で同定するために用いられる。プールされたサンプルは、異なるIdentifier配列(パネルB)を含むRNAサンプルを混合して得られる。さらに、Identifier配列は検出と解析のために一分子を捕獲するために用いても良く、またプールサンプルの中にある選ばれたサンプルの塩基配列決定反応をプライミングするために用いても良い。そのようなIdentifier配列は、Identifier配列を塩基配列反応で決定することにより、あるいはIdentifier配列と相補的な配列(パネルC)かまたはIdentifier配列を含む分子を表面上の規定された位置で選択的に捕獲することにより、RNAに由来する個々のDNAまたはRNAの由来を同定するために用いることができる。
【特許請求の範囲】
【請求項1】
解析、ハイブリダイゼーション及び塩基配列決定のための鋳型RNAを調製する方法であって、
(A)サンプル中の複数のRNA分子中における第一のグループのRNA分子の5’末端からリン酸基を除去するステップと、
(B)複数のRNA分子中における第二のグループのRNA分子について5’末端にあるCap構造をリン酸基に転換するステップと、
(C)修飾されたRNA分子を生成するため、複数のRNA分子中における第二のRNAグループについて5’末端にリン酸基を持つRNA分子に共有結合により官能基を付加するステップと、
(D)第二のRNA分子のグループにおける修飾されたRNA分子を官能基を用いて固体支持体上に結合するステップと、
(E)固体支持体上に結合した修飾されたRNA分子より非修飾RNA分子を除去するステップと
を含む方法。
【請求項2】
Cap構造を持つRNA分子が完全長RNA分子である請求項1の方法。
【請求項3】
RNA分子の5’末端リン酸基をホスファターゼの酵素活性により除去する請求項1の方法。
【請求項4】
Cap構造のリン酸基への転換が、ピロホスファターゼの酵素活性により行われる請求項1の方法。
【請求項5】
官能基をリガーゼにより脱リン酸化RNAに共有結合させる請求項1の方法。
【請求項6】
解析、ハイブリダイゼーション及び塩基配列決定のための鋳型RNAを調製する方法であって、
(A)5’末端にリン酸基をもつRNA分子の5’末端に官能基を共有結合させ、サンプル中の複数のRNA分子の中で修飾されたRNA分子グループを調製するステップと、
(B)官能基により修飾されたRNAを固体支持体に結合するステップと
(C)固体支持体に結合した修飾されたRNA分子から非修飾RNA分子を除去するステップと
を含む方法。
【請求項7】
解析、ハイブリダイゼーション及び塩基配列決定のための鋳型DNAを調製する方法であって、
(A)サンプル中の複数のRNA分子中におけるRNA分子の5’末端からリン酸基を除去するステップと、
(B)複数のRNA分子中における第二のグループのRNA分子の5’末端にあるCap構造をリン酸基に転換するステップと、
(C)5’末端にリン酸基を持つRNA分子の5’末端に共有結合により官能基を付加し、複数のRNA分子中において修飾されたRNA分子を調製するステップと、
(D)複数のRNA分子中においてRNA分子に少なくとも一つのプライマーをハイブリダイズするステップと、
(E)複数のRNA分子に含まれ、プライマーに結合したRNA分子に相補的な部分配列又は全体的な配列を持つ一本鎖DNAを調製するステップと、
(F)一本鎖DNAからRNA分子を除去するステップと、
(G)官能基又はそれに由来する情報を用いて、一本鎖DNAを固体支持体に結合させるステップと
を含む方法。
【請求項8】
一本鎖DNAが逆転写酵素により鋳型RNAから調製される請求項7の方法
【請求項9】
逆転写酵素が部分的又は全体的にRNase H活性を欠いている請求項8の方法
【請求項10】
RNAの除去がアルカリ処理又はRNaseにより行われる請求項7の方法
【請求項11】
官能基が、部分的又は全体的にデオキシリボヌクレオチドを含み、前記官能基又はその一部がRNAの除去後にDNA分子に結合されたままの状態で、DNA分子中に二本鎖DNA領域を形成し、前記官能基が結合分子を含み、前記結合分子がDNA分子の固体支持体への結合に用いられる請求項7の方法。
【請求項12】
増幅、解析、ハイブリダイゼーション及び塩基配列決定のための鋳型DNAを調製する方法であって、
(A)複数のRNA分子に含まれるRNA分子のグループから5’末端のリン酸基を除去するステップと、
(B)複数のRNA分子に含まれるRNA分子のグループの5’末端のCap構造をリン酸基に転換するステップと、
(C)5’末端にリン酸基を持つRNA分子に官能基を共有結合させて複数のRNA分子の中で修飾されたRNAを調製するステップと、
(D)複数のRNA分子中において官能基を持つ少なくとも一つのプライマーをRNA分子にハイブリダイズさせるステップと、
(E)複数のRNA分子に含まれ、プライマーに結合したRNA分子に相補的な部分配列又は全体配列を持つ一本鎖DNAを調製するステップと、
(F)一本鎖DNAからRNA分子を除去するステップと、
(G)DNAポリメラーゼを用いてDNA分子中に二本鎖DNA領域を合成するステップと、
(H)DNAリガーゼを用いて部分的二本鎖DNA分子の5’末端と3’末端を連結して、一本鎖環状DNA分子を形成するステップと、
(I)ローリングサークル法を用いて一本鎖環状DNA分子を増幅し、直線状一本鎖DNAを作成するステップと、
(J)直線状一本鎖DNA分子を固体支持体に結合するステップと
を含む方法。
【請求項13】
一回又はそれ以上の塩基配列決定反応により双方向性鋳型からセンス鎖とアンチセンス鎖の塩基配列情報を得る請求項12の方法
【請求項14】
直線状一本鎖DNAの塩基配列決定反応を双方向性鋳型から固定支持体上の定義された位置において一回又は複数回行い、前記定義された位置が他の解読配列を同じ直線状一本鎖DNA分子と関連付けることに使用される請求項12の方法。
【請求項15】
解析、ハイブリダイゼーション及び塩基配列決定のための鋳型DNAを調製する方法であって、
(A)複数のRNA分子中において官能基を持つ少なくとも一つのプライマーをRNA分子にハイブリダイズさせるステップと、
(B)複数のRNA分子に含まれ、プライマーに結合したRNA分子に相補的な部分配列又は全体配列を持つ一本鎖DNAを調製し、プライマーと官能基を一本鎖DNA分子に組み込むステップと、
(C)一本鎖DNAからRNA分子を除去するステップと、
(D)官能基を用いてDNA分子を固体支持体に結合するステップと、
(E)官能基を持たないDNA分子を、官能基を持ち固体支持体に結合しているDNA分子から除去するステップと
を含む方法。
【請求項16】
プライマーが鋳型RNAにハイブリダイズしない一部の領域を含む請求項15の方法。
【請求項17】
逆転写酵素を用いて鋳型RNAから一本鎖DNAを調製する請求項15の方法。
【請求項18】
DNA分子に含まれる全体又は一部の塩基配列情報を得るために、固体支持体上で鋳型の一つ又はそれ以上の解読配列を得る方法。
【請求項19】
鋳型が固体支持体に結合した修飾RNAまたはDNAである請求項18の方法。
【請求項20】
鋳型の塩基配列決定反応を固定支持体上の定義された位置において一回又は複数回行い、前記定義された位置が他の解読配列を同じ鋳型と関連付けることに使用され、同じ鋳型より解読配列を得る請求項18の方法。
【請求項21】
官能基がIdentifier配列の領域を含み、異なるRNAグループの混合物中において一つのRNAグループをラベルする、又は異なるDNAグループの混合物中においてRNAに由来するDNA分子のグループをラベルする方法。
【請求項22】
異なるRNA又はDNAグループの混合物が、Identifier配列を導入後同一反応容器内で同時に処理される請求項21の方法。
【請求項23】
塩基配列決定反応が、異なるRNA又はDNAグループの混合物中においてRNA分子又はDNA分子のグループをIdentifier配列により同定するために用いられる請求項21または22の方法
【請求項24】
修飾されたRNAまたはDNA分子が、ハイブリダイゼーション反応に用いられる請求項21または22の方法。
【請求項25】
ハイブリダイゼーション反応が、異なるRNAグループの混合物中においてIdentifier配列によりRNA分子グループの同定のために用いられる請求項24の方法
【請求項26】
RNAまたはDNA分子が混合され、RNAまたはDNA分子のグループの中でRNAまたはDNA分子の統計的解析を可能とするために、同一反応容器内で同時に処理される請求項21〜25のいずれか1項に記載の方法。
【請求項1】
解析、ハイブリダイゼーション及び塩基配列決定のための鋳型RNAを調製する方法であって、
(A)サンプル中の複数のRNA分子中における第一のグループのRNA分子の5’末端からリン酸基を除去するステップと、
(B)複数のRNA分子中における第二のグループのRNA分子について5’末端にあるCap構造をリン酸基に転換するステップと、
(C)修飾されたRNA分子を生成するため、複数のRNA分子中における第二のRNAグループについて5’末端にリン酸基を持つRNA分子に共有結合により官能基を付加するステップと、
(D)第二のRNA分子のグループにおける修飾されたRNA分子を官能基を用いて固体支持体上に結合するステップと、
(E)固体支持体上に結合した修飾されたRNA分子より非修飾RNA分子を除去するステップと
を含む方法。
【請求項2】
Cap構造を持つRNA分子が完全長RNA分子である請求項1の方法。
【請求項3】
RNA分子の5’末端リン酸基をホスファターゼの酵素活性により除去する請求項1の方法。
【請求項4】
Cap構造のリン酸基への転換が、ピロホスファターゼの酵素活性により行われる請求項1の方法。
【請求項5】
官能基をリガーゼにより脱リン酸化RNAに共有結合させる請求項1の方法。
【請求項6】
解析、ハイブリダイゼーション及び塩基配列決定のための鋳型RNAを調製する方法であって、
(A)5’末端にリン酸基をもつRNA分子の5’末端に官能基を共有結合させ、サンプル中の複数のRNA分子の中で修飾されたRNA分子グループを調製するステップと、
(B)官能基により修飾されたRNAを固体支持体に結合するステップと
(C)固体支持体に結合した修飾されたRNA分子から非修飾RNA分子を除去するステップと
を含む方法。
【請求項7】
解析、ハイブリダイゼーション及び塩基配列決定のための鋳型DNAを調製する方法であって、
(A)サンプル中の複数のRNA分子中におけるRNA分子の5’末端からリン酸基を除去するステップと、
(B)複数のRNA分子中における第二のグループのRNA分子の5’末端にあるCap構造をリン酸基に転換するステップと、
(C)5’末端にリン酸基を持つRNA分子の5’末端に共有結合により官能基を付加し、複数のRNA分子中において修飾されたRNA分子を調製するステップと、
(D)複数のRNA分子中においてRNA分子に少なくとも一つのプライマーをハイブリダイズするステップと、
(E)複数のRNA分子に含まれ、プライマーに結合したRNA分子に相補的な部分配列又は全体的な配列を持つ一本鎖DNAを調製するステップと、
(F)一本鎖DNAからRNA分子を除去するステップと、
(G)官能基又はそれに由来する情報を用いて、一本鎖DNAを固体支持体に結合させるステップと
を含む方法。
【請求項8】
一本鎖DNAが逆転写酵素により鋳型RNAから調製される請求項7の方法
【請求項9】
逆転写酵素が部分的又は全体的にRNase H活性を欠いている請求項8の方法
【請求項10】
RNAの除去がアルカリ処理又はRNaseにより行われる請求項7の方法
【請求項11】
官能基が、部分的又は全体的にデオキシリボヌクレオチドを含み、前記官能基又はその一部がRNAの除去後にDNA分子に結合されたままの状態で、DNA分子中に二本鎖DNA領域を形成し、前記官能基が結合分子を含み、前記結合分子がDNA分子の固体支持体への結合に用いられる請求項7の方法。
【請求項12】
増幅、解析、ハイブリダイゼーション及び塩基配列決定のための鋳型DNAを調製する方法であって、
(A)複数のRNA分子に含まれるRNA分子のグループから5’末端のリン酸基を除去するステップと、
(B)複数のRNA分子に含まれるRNA分子のグループの5’末端のCap構造をリン酸基に転換するステップと、
(C)5’末端にリン酸基を持つRNA分子に官能基を共有結合させて複数のRNA分子の中で修飾されたRNAを調製するステップと、
(D)複数のRNA分子中において官能基を持つ少なくとも一つのプライマーをRNA分子にハイブリダイズさせるステップと、
(E)複数のRNA分子に含まれ、プライマーに結合したRNA分子に相補的な部分配列又は全体配列を持つ一本鎖DNAを調製するステップと、
(F)一本鎖DNAからRNA分子を除去するステップと、
(G)DNAポリメラーゼを用いてDNA分子中に二本鎖DNA領域を合成するステップと、
(H)DNAリガーゼを用いて部分的二本鎖DNA分子の5’末端と3’末端を連結して、一本鎖環状DNA分子を形成するステップと、
(I)ローリングサークル法を用いて一本鎖環状DNA分子を増幅し、直線状一本鎖DNAを作成するステップと、
(J)直線状一本鎖DNA分子を固体支持体に結合するステップと
を含む方法。
【請求項13】
一回又はそれ以上の塩基配列決定反応により双方向性鋳型からセンス鎖とアンチセンス鎖の塩基配列情報を得る請求項12の方法
【請求項14】
直線状一本鎖DNAの塩基配列決定反応を双方向性鋳型から固定支持体上の定義された位置において一回又は複数回行い、前記定義された位置が他の解読配列を同じ直線状一本鎖DNA分子と関連付けることに使用される請求項12の方法。
【請求項15】
解析、ハイブリダイゼーション及び塩基配列決定のための鋳型DNAを調製する方法であって、
(A)複数のRNA分子中において官能基を持つ少なくとも一つのプライマーをRNA分子にハイブリダイズさせるステップと、
(B)複数のRNA分子に含まれ、プライマーに結合したRNA分子に相補的な部分配列又は全体配列を持つ一本鎖DNAを調製し、プライマーと官能基を一本鎖DNA分子に組み込むステップと、
(C)一本鎖DNAからRNA分子を除去するステップと、
(D)官能基を用いてDNA分子を固体支持体に結合するステップと、
(E)官能基を持たないDNA分子を、官能基を持ち固体支持体に結合しているDNA分子から除去するステップと
を含む方法。
【請求項16】
プライマーが鋳型RNAにハイブリダイズしない一部の領域を含む請求項15の方法。
【請求項17】
逆転写酵素を用いて鋳型RNAから一本鎖DNAを調製する請求項15の方法。
【請求項18】
DNA分子に含まれる全体又は一部の塩基配列情報を得るために、固体支持体上で鋳型の一つ又はそれ以上の解読配列を得る方法。
【請求項19】
鋳型が固体支持体に結合した修飾RNAまたはDNAである請求項18の方法。
【請求項20】
鋳型の塩基配列決定反応を固定支持体上の定義された位置において一回又は複数回行い、前記定義された位置が他の解読配列を同じ鋳型と関連付けることに使用され、同じ鋳型より解読配列を得る請求項18の方法。
【請求項21】
官能基がIdentifier配列の領域を含み、異なるRNAグループの混合物中において一つのRNAグループをラベルする、又は異なるDNAグループの混合物中においてRNAに由来するDNA分子のグループをラベルする方法。
【請求項22】
異なるRNA又はDNAグループの混合物が、Identifier配列を導入後同一反応容器内で同時に処理される請求項21の方法。
【請求項23】
塩基配列決定反応が、異なるRNA又はDNAグループの混合物中においてRNA分子又はDNA分子のグループをIdentifier配列により同定するために用いられる請求項21または22の方法
【請求項24】
修飾されたRNAまたはDNA分子が、ハイブリダイゼーション反応に用いられる請求項21または22の方法。
【請求項25】
ハイブリダイゼーション反応が、異なるRNAグループの混合物中においてIdentifier配列によりRNA分子グループの同定のために用いられる請求項24の方法
【請求項26】
RNAまたはDNA分子が混合され、RNAまたはDNA分子のグループの中でRNAまたはDNA分子の統計的解析を可能とするために、同一反応容器内で同時に処理される請求項21〜25のいずれか1項に記載の方法。
【図1】


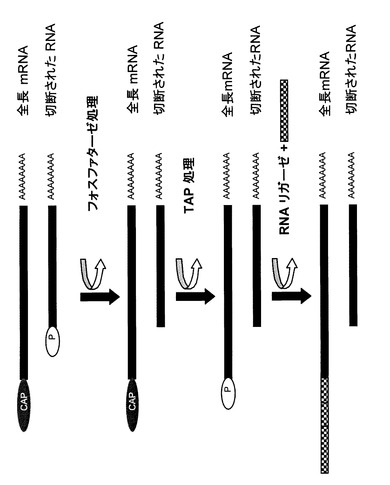
【図2】
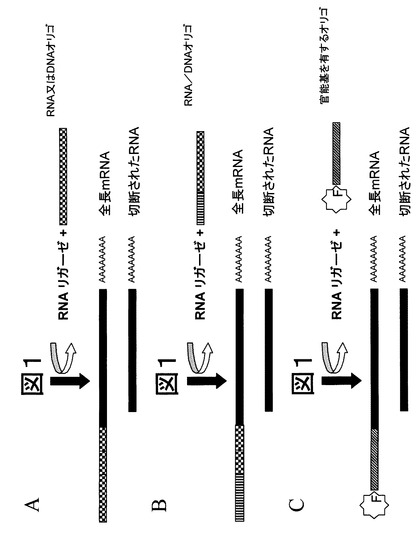
【図3】
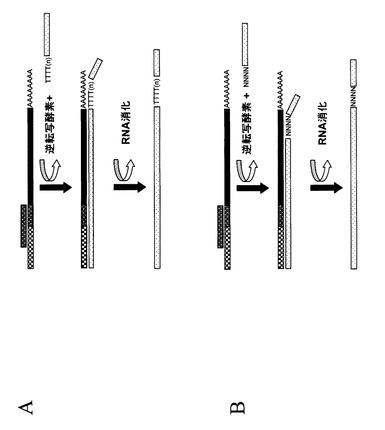
【図4】
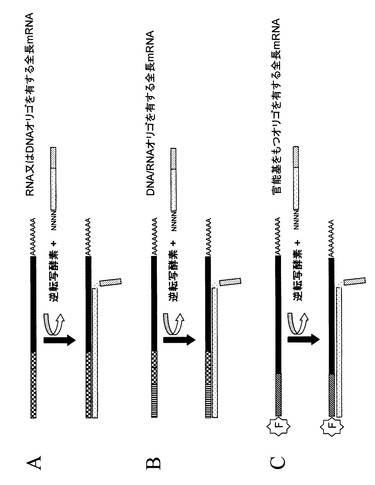
【図5】
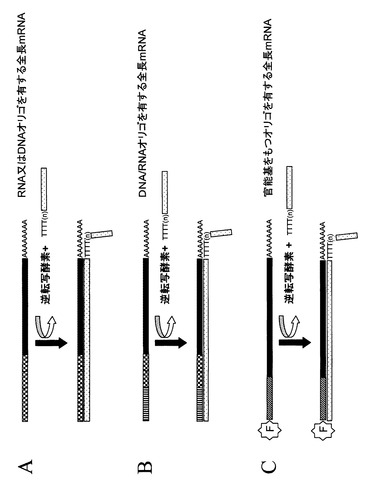
【図6】
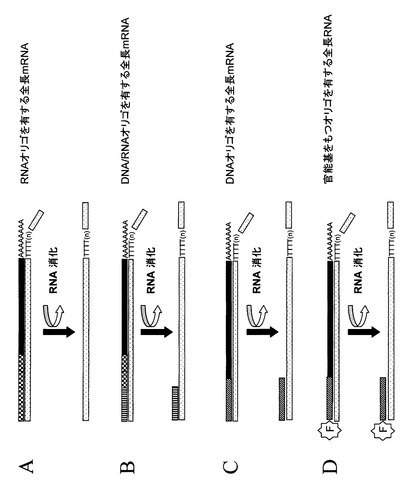
【図7】

【図8】
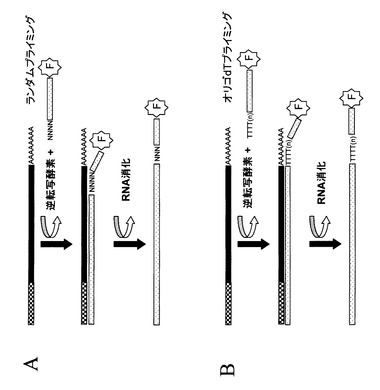
【図9−1】

【図9−2】

【図10】
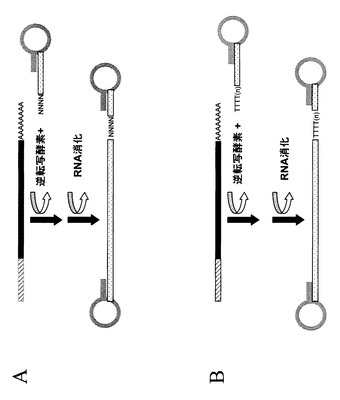
【図11】
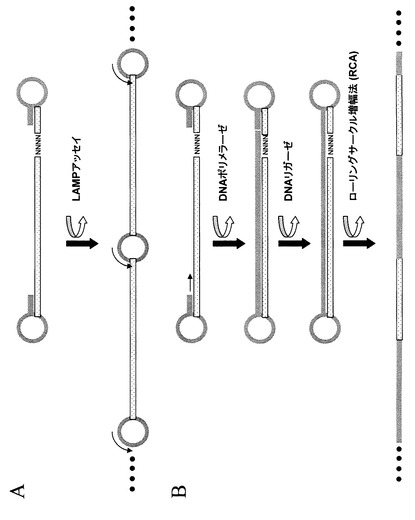
【図12】
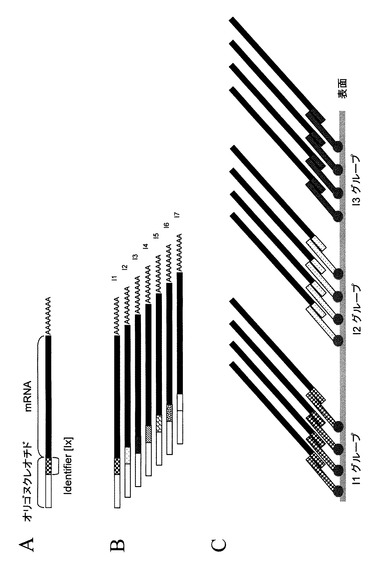
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9−1】
【図9−2】
【図10】
【図11】
【図12】
【公開番号】特開2009−268362(P2009−268362A)
【公開日】平成21年11月19日(2009.11.19)
【国際特許分類】
【外国語出願】
【出願番号】特願2008−118922(P2008−118922)
【出願日】平成20年4月30日(2008.4.30)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.テフロン
【出願人】(501293666)株式会社ダナフォーム (25)
【Fターム(参考)】
【公開日】平成21年11月19日(2009.11.19)
【国際特許分類】
【出願番号】特願2008−118922(P2008−118922)
【出願日】平成20年4月30日(2008.4.30)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.テフロン
【出願人】(501293666)株式会社ダナフォーム (25)
【Fターム(参考)】
[ Back to top ]