
Toll様受容体3アンタゴニスト
Toll様受容体3(TLR3)抗体アンタゴニスト、TLR3抗体アンタゴニスト又はそのフラグメントをコードするポリヌクレオチド、並びに上記を製造及び使用する方法を開示する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、Toll様受容体3(TLR3)抗体アンタゴニスト、TLR3抗体アンタゴニスト又はそのフラグメントをコードするポリヌクレオチド、並びに上記の製造及び使用方法に関する。
【背景技術】
【0002】
Toll様受容体(TLR)は、細菌、ウイルス、寄生虫、及び場合によりホスト由来のリガンドに応答してシグナル伝達カスケードを開始させることにより、自然免疫反応の活性化を調節し、獲得免疫の発現に影響を及ぼす(Lancaster et al.,J.Physiol.563:945〜955、2005)。細胞膜に局在するTLRであるTLR1、TLR2、TLR4及びTLR6は、細菌及び真菌のタンパク質又は脂質成分を含むリガンドを認識する。主として細胞内に存在するTLRであるTLR3、TLR7及びTLR9は、それぞれ2本鎖RNA、1本鎖RNA及び非メチル化CpG DNAに対して応答する。TLRシグナル伝達の異常調節は、多数の問題を引き起こすと考えられ、異常調節に対する治療戦略が開発されている(Hoffmanら、Nat.Rev.Drug Discov.4:879〜880,2005;Rezaei,Int.Immunopharmacol.6:863〜869,2006;Wickelgren,Science 312:184〜187,2006)。例えば、重度の敗血症及びループスに対して、それぞれTLR4、並びにTLR7及び9のアンタゴニストが臨床開発中である(Kanzlerら、Nat.Med.13:552〜559、2007)。
【0003】
TLR3シグナル伝達は、炎症又はウイルス感染時に壊死細胞から放出される二本鎖RNA、mRNA又はRNAによって活性化される。TLR3の活性化によってインターフェロン及び炎症性サイトカインの分泌が誘導され、特定の微生物感染において保護作用のある免疫細胞の活性化及び動員が誘導される。例えば、ドミナントネガティブなTLR3対立遺伝子では、小児期におけるHSV−1による一次感染時に単純ヘルペス脳炎を発症しやすくなることが示されている(Zhengら、Science 317:1522〜1527,2007)。マウスではTLR3の欠損は、コクサッキーウイルスによるチャレンジ試験における生存率の低下との関連が示されている(Richerら、PLoS One4:e4127、2009)。しかしながら、TLR3シグナル伝達の非制御又は異常調節は、西ナイルウイルス、フレボウイルス、ワクチニア、及びインフルエンザAを含む特定のウイルス感染症モデルにおける罹患率及び死亡率の一因となることが示されてきた(Wangら、Nat.Med.10:1366〜1373、2004;Gowenら、J.Immunol.177:6301〜6307、2006;Hutchensら、J.Immunol.180:483〜491、2008;LeGofficら、PloS Pathog.2:E53、2006)。
【0004】
ヒト及びマウスTLR3の細胞外ドメインの結晶構造が決定された(Bellら、Proc.Natl.Acad.Sci.(USA)、102:10976〜80、2005;Choeら、Science 309:581〜585、2005;Liuら、Science 320:379〜381,2008)。TLR3は、グリカンにより修飾されたソレノイド馬蹄様の全体形状をとり、ロイシンに富んだ繰り返し(LRR)モチーフのタンデム単位を23個有する。二本鎖RNA結合部位が、2つの別の領域にマッピングされている(Liuら、Science,320:379〜81、2008)。シグナル伝達集合体は、1つの二本鎖RNA及び2つのTLR3細胞外ドメインからなると提案された。(Leonardら、Proc.Natl.Acad.Natl.Acad.Sci.(USA)105:258〜263、2008)。
【0005】
TLR3は、例えば、敗血性ショック(Cavassaniら、J.Exp.Med.205:2609〜2621、2008)、急性肺損傷(Murrayら、Am.J.Respir.Crit.CareMed.178:1227〜1237、2008)、関節リウマチ(Kimら、Immunol.Lett.124:9〜17、2009;Brentanoら、Arth.Rheum.52:2656〜2665、2005)、喘息(Sugiuraら、Am.J.Resp.Cell Mol.Biol.40:654〜662、2009;Morishimaら、Int.Arch.Allergy Immunol.145:163〜174、2008;Stowellら、Respir.Res.10:43、2009)、クローン病及び潰瘍性大腸炎などの炎症性腸疾患(Zhouら、J.Immunol.178:4548〜4556、2007;Zhouら、Proc.Natl.Acad.Sci.(USA)104:7512〜7515、2007)、自己免疫性肝臓疾患(Langら、J.Clin.Invest.116:2456〜2463、2006)及びI型糖尿病(Dogusanら、Diabetes 57:1236〜1245、2008;Lien及びZipris、Curr.Mol.Med.9:52〜68、2009)を含む、多種多様の炎症性の免疫介在性及び自己免疫性の疾患における発病機構を促進することが示された。更に、TLR3発現の器官特異的な増加は、原発性胆汁性肝硬変における肝臓組織(Takiiら、Lab Invest.85:908〜920、2005)、関節リウマチにおける関節(Ospeltら、ArthritisRheum.58:3684〜3692、2008)、及びアレルギー性鼻炎患者における鼻粘膜(Franssonら、Respir.Res.6:100、2005)などの、局部炎症性応答が異常調節されたことにより促進される多数の発病状態と相関することが示されてきた。
【0006】
壊死状態では、内因性のmRNAを含む細胞内容物が放出されることによってサイトカイン及びケモカインの分泌、及び他の、局所炎症を誘導し、死細胞の残留物の除去を促進し、損傷を修復する因子の分泌が誘導される。壊死はしばしば炎症プロセスを継続させ、慢性炎症又は炎症悪化の一因となる(Bergsbakenら、Nature Reviews 7:99〜109、2009)。TLR3の活性化は、壊死部位においてこれらの異常な炎症プロセスに寄与し、放出されたTLR3リガンドを介して更に炎症促進性の生のフィードバックループを生じる場合がある。したがって、TLR3に対する拮抗作用は、慢性炎症若しくは炎症悪化及び/又は壊死を含むさまざまな疾患において有用であり得る。
【0007】
TLR3活性の下方制御は、腎臓細胞癌、並びに頭部及び頸部扁平上皮細胞癌が含まれる腫瘍学的な兆候に対する、新規の治療法にもなり得る(Morikawaら、Clin.Cancer Res.13:5703〜5709、2007;Priesら、Int.J.Mol.Med.21:209〜15、2008)。更には、タンパク質を低活性でコードするTLR3L423F対立遺伝子は、進行性の年齢関連の「乾燥」黄斑変性症(Yangら、N.Engl.J.Med.359:1456〜1463、2008)に対する防御と関連づけられ、この疾病においてTLR3アンタゴニストが有益であり得ることを示した。
【0008】
炎症状態にともなう病理、及び他の例えば感染症にともなう病理が、健康及び経済に及ぼす影響は多大なものである。しかしながら、多くの医学領域における進歩にも拘らず、これらの状態の多くに対する療上の選択肢及び治療法は比較的少ない。
【発明の概要】
【発明が解決しようとする課題】
【0009】
したがって、TLR3に関連する状態を治療するためにTLR3活性を抑制することが求められている。
【課題を解決するための手段】
【0010】
本発明の一つの態様は単離された抗体又はそのフラグメントであって、抗体がtoll様受容体3(TLR3)の、配列番号2のアミノ酸残基;K416、K418、L440、N441、E442、Y465、N466、K467、Y468、R488、R489、A491、K493、N515、N516、N517、H539、N541、S571、L595、及びK619に結合することを特徴とする。
【0011】
本発明の別の態様は、単離された抗体又はそのフラグメントであって、抗体がtoll様受容体3(TLR3)の、配列番号2のアミノ酸残基;S115、D116、K117、A120、K139、N140、N141、V144、K145、T166、Q167、V168、S188、E189、D192、A195、及びA219に結合することを特徴とする。
【0012】
本発明の別の態様は、単離された抗体又はそのフラグメントであって、抗体が、重鎖可変領域及び軽鎖可変領域又はそのフラグメントを有し、重鎖可変領域のChothia残基;W33、F50、D52、D54、Y56、N58、P61、E95、Y97、Y100及びD100b、並びに軽鎖可変領域のChothia残基;Q27、Y32、N92、T93、L94及びS95により、配列番号2で示されるアミノ酸配列を有するTLR3に結合することを特徴とする。
【0013】
本発明の別の態様は、単離された抗体又はそのフラグメントであって、抗体が、重鎖可変領域及び軽鎖可変領域又はそれらのフラグメントを有し、重鎖可変領域のChothia残基;N31a、Q52、R52b、S53、K54、Y56、Y97、P98、F99及びY100、並びに軽鎖可変領域のChothia残基;G29、S30、Y31、Y32、E50、D51、Y91、D92及びD93により、配列番号2に示されるアミノ酸配列を有するTLR3に結合することを特徴とする。
【0014】
本発明の別の態様は、TLR3反応性を有し、以下のa〜iの特性のうち少なくとも1つを有する、単離された抗体である;
a.Kd<10nMでヒトTLR3に結合する;
b.インビトロでのポリ(I:C)NF−κBレポーター遺伝子アッセイにおいて、1μg/mLで、ヒトTLR3の生物活性を50%超低減させる;
c.10μg/mLで、100ng/mL未満のポリ(I:C)で刺激されたBEAS−2B細胞からのIL−6又はCXCL10/IP−10の産生を60%超阻害する;
d.0.4μg/mLで、100ng/mL未満のポリ(I:C)で刺激されたBEAS−2B細胞からのIL−6又はCXCL10/IP−10の産生を50%超阻害する;
e.5μg/mLで、62.5ng/mLのポリ(I:C)で刺激されたNHBE細胞からのIL−6の産生を50%超阻害する;
f.1μg/mLで、62.5ng/mLのポリ(I:C)で刺激されたNHBE細胞からのIL−6の産生を50%超阻害する;
g.1μg/mLで、PBMC細胞による、ポリ(I:C)により誘導されたIFN−γ、IL−6又はIL−12の産生を20%超阻害する;
h.インビトロでのNF−kBレポーター遺伝子アッセイにおいて、IC50<10μg/mLで、カニクイザルのTLR3の生物活性を阻害する;
i.インビトロでのISREレポーター遺伝子アッセイにおいて、IC50<5μg/mLで、カニクイザルのTLR3の生物活性を阻害する。
【0015】
本発明の別の態様は、特定の重鎖相補性決定領域(CDR)1、2及び3のアミノ酸配列、特定の軽鎖CDR1、2及び3のアミノ酸配列、特定の重鎖可変領域(VH)のアミノ酸配列、又は特定の軽鎖可変領域(VL)のアミノ酸配列を有するモノクローナル抗体とTLR3との結合について競合する、TLR3反応性を有する単離された抗体である。
【0016】
本発明の別の態様は、重鎖及び軽鎖可変領域を有し、TLR3反応性を有する単離された抗体であって、前記抗体は特定の重鎖相補性決定領域(CDR)1、2及び3、並びに特定の軽鎖CDR 1、2及び3のアミノ酸配列を有する。
【0017】
本発明の別の態様は、重鎖及び軽鎖可変領域を有する、TLR3反応性を有する単離された抗体であって、前記抗体は特定の重鎖可変領域(VH)のアミノ酸配列、及び特定の軽鎖可変領域(VL)のアミノ酸配列を有する。
【0018】
本発明の別の態様は、重鎖及び軽鎖可変領域を有する、TLR3反応性を有する単離された抗体であって、前記抗体は特定の重鎖のアミノ酸配列、及び特定の軽鎖のアミノ酸配列を有する。
【0019】
本発明の別の態様は、単離された抗体重鎖であって、前記抗体重鎖は配列番号6、8、10、12、14、16、18、20、22、24、26、28、30、32、34、36、38、40、42、124、125、126、127、128、129、159、198、200、202、164、212、213、214、215又は216に示されるアミノ酸配列を有する。
【0020】
本発明の別の態様は、単離された抗体軽鎖であって、前記抗体軽鎖は、配列番号5、7、9、11、13、15、17、19、21、23、25、27、29、31、33、35、37、39、41、122、123、197、199、201、163、209、210、211又は225に示されるアミノ酸配列を有する。
【0021】
本発明の別の態様は、単離された抗体重鎖であって、前記抗体重鎖は配列番号102、130、131、132、133、134、135、160、204、206、208、220、166又は168に示されるアミノ酸配列を有する。
【0022】
本発明の別の態様は、単離された抗体軽鎖であって、前記抗体軽鎖は配列番号155、156、157、158、203、205、207、165、167又は227に示されるアミノ酸配列を有する。
【0023】
本発明の別の態様は抗体重鎖をコードする単離されたポリヌクレオチドであって、前記抗体重鎖は配列番号6、8、10、12、14、16、18、20、22、24、26、28、30、32、34、36、38、40、42、124、125、126、127、128、129、159、198、200、202、164、212、213、214、215又は216に示されるアミノ酸配列を有する。
【0024】
本発明の別の態様は、抗体軽鎖をコードする単離されたポリヌクレオチドであって、前記抗体軽鎖は配列番号5、7、9、11、13、15、17、19、21、23、25、27、29、31、33、35、37、39、41、122、123、197、199、201、163、209、210、211又は225に示されるアミノ酸配列を有する。
【0025】
本発明の別の態様は、抗体重鎖をコードする単離されたポリペプチドであって、前記抗体重鎖は配列番号102、130、131、132、133、134、135、160、204、206、208、220、166又は168に示されるアミノ酸配列を有する。
【0026】
本発明の別の態様は、単離されたポリペプチドであって、前記ポリペプチドは、配列番号155、156、157、158、203、205、207、165、167又は227に示されるアミノ酸配列を有する抗体軽鎖をコードする。
【0027】
本発明の別の態様は、本発明の単離された抗体及び製薬上許容され得る担体を含む製薬学的組成物である。
【0028】
本発明の別の態様は、少なくとも1つの本発明のポリヌクレオチドを含むベクターである。
【0029】
本発明の別の態様は、本発明のベクターを有するホスト細胞である。
【0030】
本発明の別の態様は、本発明のホスト細胞を培養することと、前記ホスト細胞によって産生された抗体を回収することとを含む、TLR3反応性を有する抗体の製造方法である。
【0031】
本発明の別の態様は、治療上の有効量の本発明の単離された抗体を、炎症性疾患の治療又は予防を必要とする患者に、炎症性疾患を治療又は予防するうえで充分な時間にわたって投与することを含む、炎症性疾患の治療又は予防方法である。
【0032】
本発明の別の態様は、治療上の有効量の本発明の単離された抗体を、それらを必要とする患者に、全身性の炎症状態を処置又は予防するうえで充分な時間にわたって投与することを含む、全身性炎症性疾患の処置又は予防方法である。
【0033】
本発明の別の態様は、治療上の有効量の本発明の単離された抗体を、それらを必要とする患者に、II型糖尿病を処置するうえで充分な時間にわたって投与することを含む、II型糖尿病の処置方法である。
【0034】
本発明の別の態様は、治療上の有効量の本発明の単離された抗体を、それらを必要とする患者に、高血糖を処置するうえで充分な時間にわたって投与することを含む、高血糖の処置方法である。
【0035】
本発明の別の態様は、治療上の有効量の本発明の単離された抗体を、それらを必要とする患者に、インスリン抵抗性を処置するうえで充分な時間にわたって投与することを含む、高インスリン血症の治療方法である。
【0036】
本発明の別の態様は、治療上の有効量の本発明の単離された抗体を、それらを必要とする患者に、ウイルス感染を処置又は予防するうえで充分な時間にわたって投与することを含む、ウイルス感染の処置又は予防方法である。
【図面の簡単な説明】
【0037】
【図1】NF−κBレポーター遺伝子アッセイにおける抗ヒトTLR3(huTLR3)mAbの作用を示す図。
【図2A】BEAS−2Bアッセイにおける抗huTLR3 mAbの作用(阻害率(%))を示す図。
【図2B】BEAS−2Bアッセイにおける抗huTLR3 mAbの作用(阻害率(%))を示す図。
【図3A】NHBEアッセイにおける抗huTLR3 mAbの作用を示す図。
【図3B】NHBEアッセイにおける抗huTLR3 mAbの作用を示す図。
【図4】PBMCアッセイにおける抗huTLR3 mAbの作用を示す図。
【図5A】HASMアッセイにおける抗huTLR3 mAbの作用を示す図。
【図5B】HASMアッセイにおける抗huTLR3 mAbの作用を示す図。
【図6A】TLR3変異体に対する抗huTLR3 mAbの結合を示す図。
【図6B】TLR3変異体に対する抗huTLR3 mAbの結合を示す図。
【図6C】TLR3変異体に対する抗huTLR3 mAbの結合を示す図。
【図7A】ヒトTLR3 ECD上の、mAb 15EVQ(黒)及びC1068 mAb(灰色)のエピトープ(上の画像)、並びにmAb 12QVQ/QSV(黒、下の画像)のエピトープを示す図。
【図7B】mAb 15EVQと複合体を形成させたTLR3 ECDタンパク質の局所的H/D交換撹乱マップを示す図。
【図8A】A)NF−κBレポーター遺伝子アッセイ及びB)ISREレポーター遺伝子アッセイにおけるラット/マウスの抗マウスTLR3 mAb mAb 5429(代替抗体)の作用を示す図。
【図8B】A)NF−κBレポーター遺伝子アッセイ及びB)ISREレポーター遺伝子アッセイにおけるラット/マウスの抗マウスTLR3 mAb mAb 5429(代替抗体)の作用を示す図。
【図9】MEF CXCL10/IP−10アッセイにおける代替mAb(mAb 5429、mAb c1811)の作用を示す図。
【図10】TLR3に対する代替mAbの結合の特異性を示す図。上側のパネル:アイソタイプコントロール、下側のパネル:mAbc1811。
【図11】AHRモデルにおけるpenHレベルに対する代替mAbの効果を示す図。
【図12】AHRモデルにおけるBAL液中の全好中球数に対する代替mAbの効果を示す図。
【図13】AHRモデルにおけるBAL液中のCXCL10/IP−10レベルに対する代替mAbの効果を示す図。
【図14】DSSモデルにおける組織病理学のスコアに対する代替mAbの効果を示す図。
【図15】T細胞移植モデルにおける、A)組織病理学のスコア、及びB)好中球流入に対する代替mAbの効果、を示す図。
【図16】CIAモデルにおける臨床スコアに対する代替mAbの効果を示す図。
【図17】CIAモデルにおける臨床AUCスコアに対する代替mAbの効果を示す図。
【図18】インフルエンザA/PR/8/34の経鼻投与の後のC57BL/6マウスの生存率に及ぼす代替mAbの効果を示す図。mAbの投与は1日目に始まった。
【図19】インフルエンザA/PR/8/34の投与後の臨床スコアに対する代替mAbの効果を示す図。mAbの投与は1日目に始まった。
【図20】インフルエンザA/PR/8/34の投与後に代替mAbを投与した後の14日にわたる体重変化に及ぼす代替mAbの効果を示す図。mAbの投与は1日目に始まった。
【図21】グルコースチャレンジ後の(A)野生型DIO及び(B)TLR3ノックアウトDIO動物における血中グルコース濃度に対する代替mAbの効果を示す図。
【図22】野生型DIO動物のインスリンレベルに対する代替mAbの効果を示す図。
【図23】NHBE細胞における、(A)NTHi、及び(B)ライノウイルス誘導CXCL10/IP−10及びCCL5/RANTESレベルに対するmAb 15EVQの効果を示す図。
【図24】HUVEC細胞における、(A)sICAM−1レベル及び(B)生存率に対する、mAb 15EVQの効果を示す図。
【図25】インフルエンザAの感染3日後に代替mAbを投与した後の動物の生存率を示す図。
【図26】インフルエンザAの感染3日後に代替mAbを投与した後の臨床スコアを示す図。
【図27】インフルエンザAの感染3日後に代替mAbを投与した後の動物の体重変化を示す図。
【図28】AではhuTLR3 ECDとFab 12QVQ/QSV、Fab 15EVQ及びFab c1068との四次構造の分子構造をリボン及び表面表示で示す図。TLR3 ECDの、Nで標識したN末端を薄い灰色で示し、すべてのFab分子を暗い灰色のリボン表示で示す。B.エピトープを薄い灰色で着色し、AにおけるFabについてのようにTLR3 ECD上で標識した。図28、29及び30では、判り易くするために、Fab 12QVQ/QSV、Fab c1068及びFab 15EVQの標識を、それぞれ、Fab 12、Fab 1068及びFab 15と略記する。
【図29】Fab 15EVQによる中和機構を示す図。A.2つのTLR3 ECD(薄い灰色及び暗い灰色で示す図、並び標識されたTLR3図)の一方において、Fab 15EVQエピトープをハイライト(薄い灰色)した二本鎖RNA:TLR3シグナル伝達ユニット(SU)を示す。二本鎖RNAリガンドは薄い灰色の二重らせんとして示される。B.二本鎖RNAの結合を立体的に阻害することで、SUの形成を阻害する、Fab 15EVQの結合が図示される。Fab 15EVQの結合は高親和性であり、SUの形成を阻害するか、又はすでに形成されているSUを分解する。
【図30】Fab 12QVQ/QSV及びFab c1068及びTLR3シグナル伝達ユニット(SU)のクラスター形成の機構を示す図。A.Fab 12QVQ/QSV及びFab c1068は、単一SUを結合(又は共結合)することができる。B.約76塩基対の二本鎖RNA上の2つのSUの最近接クラスター形成のモデルである。判り易くするために3つのエピトープが異なる分子中でハイライトされている。C.Fab 12QVQ/QSV及びFab c1068の結合は、抗体と、隣接するSUとの間の立体的な衝突によりSUのクラスター形成を防止する。2つの左を指す矢印は、抗体によるSUの分離の異なる度合いを定性的に表す(底部の矢印はFab 12QVQ/QSVに対するものであり、及び頂部の矢印はFab c1068に対するものである)。
【図31】代表的な抗体に対する、配列番号と、Kabatによる番号付けと、Chothiaによる番号付けの対応を示す図。CDR及びHVを灰色でハイライトしてある。
【図32】mAb 15EVQのVLと、ヒトVk1フレームワークとのアライメント結果を示す図。Chothiaによるハイパー可変ループにアンダーラインし、パラトープ残基に二重アンダーラインし、及びフレームワークの差異を灰色でハイライトしてある。特記しない限り、Vκ1遺伝子は*01対立遺伝子である。残基番号付けは連続的に行っている。
【図33】mAb 15EVQのVHと、ヒトVh5フレームワークとのアライメント結果を示す図。配列の特徴は図32のように示される。
【図34】mAb 12QVQ/QSVのVLと、ヒトVk3フレームワークとのアライメント結果を示す図。配列の特徴は図32のように示される。
【図35】mAb 15EVQ又はmAb 12QVQ/QSVのVL及びVHと、ヒトJκ、Jλ又はJhフレームワークとのアライメント結果を示す図。配列の特徴は図32のように示される。
【発明を実施するための形態】
【0038】
本明細書に引用する特許及び特許出願を含むがそれらに限定されないすべての刊行物は、あたかもその全体が記載されているのと同様に本願に参照によって組み入れるものである。
【0039】
本明細書で用いる「アンタゴニスト」なる用語は、何らかの機構によって、受容体又は細胞内メディエーターなどの別の分子の作用を部分的又は完全に阻害する分子のことを意味する。
【0040】
本明細書で用いるとき、「TLR3抗体アンタゴニスト」又は「TLR3反応性の」抗体とは、TLR3の生物活性又はTLR3受容体の活性化を直接的又は間接的に実質上相殺、低減、又は阻害する能力を有する抗体のことを言う。例えば、TLR3反応性を有する抗体は、TLR3に直接結合してTLR3の活性を中和することによって、すなわちTLR3のシグナル伝達を遮断することによって、サイトカイン及びケモカインの放出、又はNF−κBの活性化を低減させることができる。
【0041】
用語「抗体」は、本明細書において広義に用いられ、ポリクローナル抗体、マウス、ヒト、ヒト適合化、ヒト化及びキメラモノクローナル抗体を含むモノクローナル抗体、並びに抗体フラグメントを含む、免疫グロブリン又は抗体分子を含む。
【0042】
一般に抗体とは、特定の抗原に対する結合特異性を示すタンパク質又はペプチド鎖である。完全な抗体とは、2個の同一の軽鎖と2個の同一の重鎖とからなるヘテロ4量体の糖タンパク質である。通常、それぞれの軽鎖は1個の共有ジスルフィド結合によって重鎖と結合しているが、ジスルフィド結合の数は、アイソタイプの異なる免疫グロブリン重鎖間で異なる。それぞれの重鎖及び軽鎖はまた、規則的な間隔をおいた鎖内ジスルフィド架橋も有する。それぞれの重鎖は、一方の末端で可変ドメイン(可変領域)(VH)を有し、続いて多数の定常性のドメイン(定常領域)を有する。それぞれの軽鎖は一端に可変領域(VL)を有し、もう一方の端に定常領域を有する。軽鎖の定常領域は重鎖の第1の定常領域と整列し、軽鎖の可変領域は重鎖の可変領域と整列している。あらゆる脊椎動物種の抗体の軽鎖は、定常領域のアミノ酸配列に基づき、カッパー(κ)及びラムダ(λ)の2つの明確に異なるタイプの内のどちらかに分類することができる。
【0043】
免疫グロブリンは、重鎖の定常領域のアミノ酸配列に応じて、IgA、IgD、IgE、IgG及びIgMの5つの大きなクラスに分類することができる。IgA及びIgGは、IgA1、IgA2、IgG1、IgG2、IgG3、IgG4というアイソタイプに更に分類される。
【0044】
用語、「抗体フラグメント」は完全な抗体の一部を意味し、通常は完全な抗体の抗原結合領域又は可変領域を意味する。抗体フラグメントの例としては、Fab、Fab’、F(ab’)2、Fvフラグメント、二重特異性抗体、単鎖抗体分子、及び少なくとも2つの完全な抗体から形成された多特異的抗体が挙げられる。
【0045】
免疫グロブリンの軽鎖可変領域又は重鎖可変領域は、3つの「抗原結合部位」を取り囲む「フレームワーク」領域からなる。抗原結合部位は、さまざまな用語を次のように使用して定義される:(i)用語相補性決定領域(CDR)は配列可変性に基づく(Wu及びKabat、J.Exp.Med.132:211〜250、1970)。一般に、抗原結合部位は、VH(HCDR1、HCDR2、HCDR3)内に3つ及びVL(LCDR1、LCDR2、LCDR)内に3つの6つのCDRを有している(Kabatら、Sequences of Proteins of Immunological Interest、5th Ed.Public Health Service,National Institutes of Health,Bethesda,Md.,1991)。(ii)用語「超可変領域」、「HVR」、又は「HV」は、Chothia及びLesk(Chothia及びLesk,Mol.Biol.196:901〜917,1987)により定義されているように、構造が超可変性である、抗体の可変ドメイン領域を指す。一般に抗原結合部位は、VH内に3つ(H1、H2、H3)及びVL内に3つ(L1、L2、L3)の6つの超可変領域を有している。Chothia及びLeskは、構造的に保持されているHVを「カノニカル構造」と呼ぶ。(iii)Lefrancにより提案された「IMGT−CDR」(Lefrancら、Dev.Comparat.Immunol.27:55〜77,2003)は、免疫グロブリン及びT細胞受容体からのVドメインの比較に基づくものである。International ImMunoGeneTics(IMGT)データベース(http://www_imgt_org)は、これらの領域について標準化された番号及び定義を提供している。各CDR、HV、及びIMGTの表記間の対応についてはLefrancら、Dev.Comparat.Immunol.27:55〜77,2003に述べられている。(iv)抗原結合部位は特異性決定残基の使用(SDRU)(Almagro,Mol.Recognit.17:132〜143,2004)に基づいて概略説明することができる。特異性決定残基(SDR)は、抗原との接触に直接関与する免疫グロブリンのアミノ酸残基を指す。SDRUは、抗原−抗体複合体についての結晶構造解析により定義される、異なるタイプの抗原に対するSDRの数及び分布の精密な尺度である。(v)抗原結合部位を、抗原抗体複合体の結晶構造から同定される抗体パラトープ残基とも定義することもできる。
【0046】
用語「複合配列」は、本明細書で使用するとき、Kabat、Chothia又はIMGTによって個別に表記されるすべてのアミノ酸残基又は他の任意の好適な抗原結合領域の表記を含むものとして定義される抗原結合部位を意味する。
【0047】
用語「Chothia残基」は、本明細書で使用するとき、Al−Lazikani(Al−Lazikaniら、J.Mol.Biol.,273,927〜48,1997)に従って番号付けされた抗体VL残基及びVH残基である。ポリペプチド番号付けに関連して2つの最も使用される番号付けシステム、Kabat(Kabatら、Sequences of Immunological Interest,5th Ed.Public Health Service,NIH,Bethesda,MD,1991)とChothia(Chothia及びLesk,Mol.Biol.196:901〜17,1987)との間の対応を、本発明の代表的な抗体に対して図31に示す。
【0048】
「フレームワーク」又は「フレームワーク配列」は、抗原結合部位として定義されていない、可変領域の残りの配列である。フレームワークは、通常、それぞれの可変領域中で、3つの抗原結合部位に対して骨格を形成する、4つの領域、FR1、FR2、FR3、及びFR3に分割される。抗原結合部位は上記に述べたようなさまざまな用語によって定義され得るため、フレームワークの正確なアミノ酸配列は抗原結合部位がどのように定義されるかによって決まる。
【0049】
「軽鎖可変領域カッパー1(Vκ1)フレームワーク」又は「Vκ1」は、本発明で使用するとき、ヒトVκ1機能性遺伝子又はその対立遺伝子のいずれかによりコードされている、アミノ酸配列を有するフレームワークを指す。代表的な機能性ヒトVκ1遺伝子は、IGKV1−5*01、IGKV1−6*01、IGKV1−8*01、IGKV1−9*01、IGKV1−12*01、IGKV1−13*02、IGKV1−16*01、IGKV1−17*01、IGKV1−27*01、IGKV1−33*01、IGKV1−37*01、IGKV1−39*01、IGKV1D−8*01、IGKV1D−12*01、IGKV1D−13*01、IGKV1D−16*01、IGKV1D−17*01、IGKV1D−33*01、IGKV1D−37*01、IGKV1D−39*01、IGKV1D−42*01、又はIGKV1D−43*01である。免疫グロブリン遺伝子の命名法は周知されたものである。
【0050】
「軽鎖可変領域ラムダ3(Vλ3)フレームワーク」又は」「Vλ3」は、本明細書で使用するとき、ヒトVλ3機能性遺伝子又はその対立遺伝子のいずれかによりコードされたアミノ酸配列を有するフレームワークを指す。代表的な機能性ヒトVλ3遺伝子は、IGLV3−1*01、IGLV3−9*01、IGLV3−10*01、IGLV3−12*01、IGLV3−16*01、IGLV3−19*01、IGLV3−21*01、IGLV3−22*01、IGLV3−25*01、IGLV3−27*01、及びIGLV3−32*01である。
【0051】
「重鎖可変領域Vh5フレームワーク」又は「Vh5」は、本明細書で使用するとき、Vh5機能性遺伝子又はその対立遺伝子のいずれかによりコードされたアミノ酸配列を有するフレームワークを指す。代表的な機能性ヒトVh5遺伝子は、IGHV5−51*01及びIGHV5−1*01である。
【0052】
「重鎖可変領域Vh6フレームワーク」又は「Vh6」は、本明細書で使用するとき、Vh6機能性遺伝子又はその対立遺伝子のいずれかによりコードされたアミノ酸配列を有するフレームワークを指す。代表的な機能性ヒトVh6遺伝子はIGHV6−1*01である。
【0053】
「軽鎖カッパーJ領域(Jκ)フレームワーク」又は「Jκ」は、本明細書で使用するとき、ヒトJκ機能性遺伝子又はその対立遺伝子のいずれかによりコードされたアミノ酸配列を有するフレームワークを指す。代表的な機能性ヒトVκ遺伝子は、IGKJ1、IGKJ2、IGKJ3、IGKJ4、及びIGKJ5である。
【0054】
「軽鎖ラムダJ領域(Jλ)フレームワーク」又は「Jλ」は、本明細書で使用するとき、ヒトJλ機能性遺伝子又はその対立遺伝子のいずれかによりコードされたアミノ酸配列を有するフレームワークを指す。代表的な機能性ヒトJλ遺伝子は、IGLJ1、IGLJ2、IGLJ3、IGLJ4、IGLJ5、IGLJ6、及びIGLJ7である。
【0055】
「重鎖J領域(Jh)フレームワーク」又は「Jh」は、本明細書で使用するとき、ヒトJh機能性遺伝子又はその対立遺伝子のいずれかによりコードされたアミノ酸配列を有するフレームワークを指す。代表的な機能性ヒトJh遺伝子は、IGHJ1、IGHJ2、IGHJ3、IGHJ4、IGHJ5、及びIGHJ6である。
【0056】
「生殖細胞系列遺伝子」又は「抗体生殖細胞系列遺伝子」は、本明細書で使用するとき、特定の免疫グロブリンの発現のための遺伝的再配列及び変異につながる成熟プロセスを経ていない非リンパ細胞によってコードされる免疫グロブリン配列である。
【0057】
「スカフォールド」は、本明細書で使用するとき、ヒト生殖細胞系列遺伝子によってコードされる、軽鎖又は重鎖の可変領域のアミノ酸配列を指す。したがって、スカフォールドは、フレームワークと抗原結合部位の両方を包含する。
【0058】
用語「抗原」は、本明細書で使用するとき、直接又は間接に抗体を生成する能力を有する任意の分子を意味する。「抗原」の定義内には、タンパク質コーディング核酸が含まれる。
【0059】
用語「ホモログ」は、参照配列との配列同一性が40%〜100%であるタンパク質配列を意味する。ヒトTLR3のホモログには、既知のヒトTLR3配列との配列同一性が40%〜100%であるような他種由来のポリペプチドが含まれる。2つのペプチド鎖の配列の同一性(%)は、Vector NTIバージョン9.0.0(Invitrogen,Carlsbad,CA)のAlignXモジュールのデフォルト設定を使用して、配列をペアでアライメントすることによって決定することができる。「TLR3」とはヒトTLR3(huTLR3)及びそのホモログの意味である。完全長huTLR3のヌクレオチド及びアミノ酸配列をそれぞれ配列番号1及び2で示す。huTLR3の細胞外ドメイン(ECD)のヌクレオチド及びアミノ酸配列をそれぞれ配列番号3及び配列番号4に示す。
【0060】
「実質的に同一の」は、本明細書で使用するとき、比較される2つの抗体又は抗体フラグメントのアミノ酸配列が同一であるか、又は「実体のない差異」を有するということを意味する。抗体又は抗体フラグメントのアミノ酸配列における1、2、3、4、5又は6個のアミノ酸の置換が、実体のない差異として挙げられる。本明細書で開示する配列とほぼ同じアミノ酸配列も本願の一部である。特定の実施形態では、配列の同一性は、約90%、91%、92%、93%、94%、95%、96%、97%、98%、99%又はそれよりも高い。同一性(%)は、上記に述べたようにして決定することができる。比較するペプチド鎖の例としては、重鎖又は軽鎖の可変領域がある。
【0061】
用語「〜と組み合わせて」は、本明細書で使用するとき、記述の対象である薬剤が、他の薬剤と共に、混合物として一緒に、又は単独薬剤として同時に、又は単独薬剤として順次に任意の順序で、動物に投与され得ることを意味する。
【0062】
用語「炎症性状態」は、本明細書で使用するとき、大部分の場合痛み、発赤、腫張、及び組織機能の喪失を特徴とし、サイトカイン、ケモカイン、又は炎症性細胞(例えば、好中球、単球、リンパ球、マクロファージ)により一部介在される、細胞損傷に対する局所化された応答を意味する。用語「炎症性の肺状態」は、本明細書で使用するとき、肺を冒す又は肺と関連する炎症状態を意味する。
【0063】
「用語モノクローナル抗体」(mAb)は、本明細書で使用するとき、ほぼ均質な抗体の集団から得られる抗体(又は抗体フラグメント)を意味する。モノクローナル抗体は極めて特異性が高く、通常、単一の抗原決定基を標的とする。「モノクローナル」という修飾語は、抗体のほぼ均質な性質を指して言うものであり、抗体がいずれかの特定の方法によって作製される必要はない。例えば、マウスmAbは、Kohlerら、Nature 256:495〜497、1975のハイブリドーマ法によって製造することができる。ドナー抗体(通常はマウス)由来の軽鎖及び重鎖の可変領域を、アクセプター抗体(通常、例えばヒトなどの別の哺乳動物種)由来の軽鎖及び重鎖の定常領域とともに含むキメラmAbを、米国特許第4,816,567号に開示される方法によって作製することができる。非ヒトドナー免疫グロブリン(通常はマウス)に由来するCDR、及び1以上のヒト免疫グロブリンに由来するCDR以外の部分を有する、ヒト適合化mAbを、米国特許第5,225,539号に開示されるような当業者には周知の方法によって作製することができる。ヒト適合化に有用なヒトフレームワーク配列は当業者であれば関連するデータベースから選択することができる。場合により、Queenら、Proc.Natl.Acad.Sci.(USA),86:10029〜10032,1989及びHodgsonら、Bio/Technology,9:421,1991に開示されるような方法によって、結合親和性を維持するために変化させたフレームワーク支持残基を組み込むことによってヒト適合化mAbを更に改変することができる。
【0064】
ヒト以外のいかなる配列も持たない完全なヒトmAbは、例えば、Lonbergら、Nature 368:856〜859,1994、Fishwildら、Nature Biotechnology 14:845〜851,1996、及びMendezら、Nature Genetics15:146〜156,1997に参照される方法により、ヒト免疫グロブリン形質転換マウスから作製することができる。同様に、ヒトmAbは、ファージディスプレイライブラリを元に、例えば。Knappikら、J.Mol.Biol.296:57〜86 2000及びKrebsら、J.Immunol.Meth.254:67〜84 2001で参照されている方法により作製及び最適化することもできる。抗体のフラグメント、例えばFab、F(ab’)2、Fd、及びdAbフラグメントを、抗体の開裂又は遺伝子組み換え操作により生成してもよい。例えば、Fab及びF(ab’)2フラグメントを、抗体をペプシンなどの酵素により処理することにより、生成することもできる。
【0065】
用語「エピトープ」は、本明細書で使用するとき、抗体が特異的に結合する抗原の部分を意味する。エピトープは、通常、アミノ酸又は多糖類側鎖のような化学的に活性な(極性、非極性又は疎水性)部分の表面集団からなり、特定の3次元構造特性並びに特定の帯電特性を有し得る。エピトープは直線状のものであってもよく、あるいはアミノ酸の直線状の配列によってではなく、抗原の連続していないアミノ酸間の空間的な関係によって形成される不連続エピトープ(例えば立体構造エピトープ)であってもよい。立体構造エピトープには、抗原の直線状配列の異なる部分のアミノ酸同士が3次元空間で近接するような抗原の折り畳みによって生じるエピトープが含まれる。
【0066】
用語「パラトープ」は、本明細書で使用するとき、抗原が特異的に結合する抗体の一部を指す。パラトープは元来線状であるか、又は不連続であることができ、抗体のアミノ酸の線状構造よりも、隣接していないアミノ酸間の空間的な関係により形成される。「軽鎖パラトープ」及び「重鎖パラトープ」又は「軽鎖パラトープアミノ酸残基」及び「重鎖パラトープアミノ酸残基」は、それぞれ、抗原と接触する抗体の軽鎖及び重鎖残基を指す。
【0067】
用語「特異的結合」は、本明細書で使用するとき、抗体が特定の抗原と、他の抗原又はタンパク質に対するよりも高い親和性で結合することを指す。通常、抗体の結合における解離定数(KD)は10-7M以下であり、抗体が特定の抗原と結合する際のKDは、特定の抗原以外の非特異的抗原(例えば、BSA、カゼイン、又は他の任意の特定のポリペプチド)と結合する際のKDと比較して1/2以下である。本明細書では、「抗原を認識する抗体」及び「抗原に特異的な抗体」という語句を、「抗原と特異的に結合する抗体」又は「抗原特異的抗体」、例えばTLR3特異的抗体という用語と互換可能に用いる。解離定数は後述するような標準的な方法を用いて測定することができる。
【0068】
用語「TLR3の生物活性」又は「TLR3の活性化」は、本明細書で使用するとき、リガンドとTLR3との結合によって生ずる任意の活性を指す。TLR3のリガンドには、2本鎖DNA、ポリイノシンポリシチジン酸(ポリ(I:C))、及び例えば壊死細胞から放出される内因性mRNAなどの内因性mRNAが含まれる。TLR3の活性化の一例では、TLR3のリガンドに応じてNF−κBが活性化される。レポーター遺伝子アッセイを用いることで、ポリ(I:C)による受容体の誘導に基づき、NF−κBの活性化をアッセイすることができる(Alexopoulosら、Nature 413:732〜738,2001;Hackerら、EMBO J.18:6973〜6982,1999)。他の代表的なTLR3受容体の活性化は、TLR3リガンドに対する応答として、インターフェロン応答因子(IRF−3、IRF−7)の活性化をもたらす。TLR3によって介在されるIRFの活性化は、インターフェロン刺激応答配列(ISRE)によって制御されるレポーター遺伝子を用いてアッセイすることができる。別のTLR3の活性化の例では、例えばTNF−α、IL−6、IL−8、IL−12、CXCL5/IP−10及びRANTESなどの炎症性サイトカイン及びケモカインが分泌される。細胞若しくは組織からの、又は血液循環中へのサイトカイン及びケモカインの放出は、ELISA免疫アッセイなどの周知の免疫アッセイによって測定することができる。
【0069】
本明細書では以下のようにアミノ酸の慣習的な1文字及び3文字の略号を用いる。
【0070】
【表1】

【0071】
物質の組成物
本発明はTLR3の生物活性を阻害することが可能な抗体アンタゴニスト及びかかる抗体の使用法を提供する。かかるTLR3アンタゴニストは、TLR3に結合してTLR3の活性化を阻害する性質を有し得る。かかる抗体によってTLR3活性が阻害され得る機序の例としては、TLR3へのリガンドの結合のインビトロ、インビボ、若しくはインサイチューでの阻害、受容体の2量化の阻害、エンドソーム区画へのTLR3の局在化の阻害、下流のシグナル伝達経路のキナーゼ活性の阻害、又はTLR3のmRNAの転写の阻害が挙げられる。他の機序によってTLR3の活性を阻害することが可能な他の抗体アンタゴニストもまた、本発明の異なる態様及び実施形態の範囲に含まれるものである。これらのアンタゴニストは、研究用試薬、診断用試薬及び治療薬として有用である。
【0072】
抗体の多様性は、自然の系では、可変領域をコードする多数の生殖細胞系列遺伝子の使用及びさまざまな体細胞イベントにより作り出される。体細胞イベントとしては、完全なVH領域を作るための多様性(D)及び接合(J)遺伝子セグメントを持つ可変遺伝子セグメントの組換え、並びに完全なVL領域を作るための可変及び接合遺伝子セグメントの組換えがある。組み換えプロセスそれ自体は不正確であり、V(D)J接合部におけるアミノ酸の喪失又は追加を生じる可能性がある。多様性のこれらの機序は、抗原暴露に先立つB細胞の発生時に起こる。抗原刺激の後、B細胞中で発現された抗体遺伝子は、体細胞変異を受ける。生殖細胞系列遺伝子セグメント、これらのセグメントのランダムな組み換え、及びランダムなVH−VLの対形成の見積り数に基づいて,1.6×107までの異なる抗体が生成可能である(Fundamental Immunology,3rd ed.(1993),ed.Paul,RavenPress,New York,N.Y.)。抗体多様性に寄与する他のプロセス(体細胞変異などの)を考慮に入れた場合、1010以上の異なる抗体が生成可能であると考えられる(Immunoglobulin Genes,2nd ed.(1995),eds.Jonioら、Academic Press,San Diego,Calif.)。多数のプロセスが抗体多様性の生成において関与するために、同一の抗原特異性を有する独立に由来したモノクローナル抗体が同一のアミノ酸配列を有するということは極めてあり得ないことである。
【0073】
本発明は、新規の抗原結合部位及びヒト免疫グロブリン遺伝子ライブラリに由来する免疫グロブリン鎖を提供する。抗原結合部位を担持するための構造は一般に抗体の重鎖又は軽鎖若しくはそれらの部分であり、抗原結合部位は上記に述べたようにして決定された天然に生じる抗原結合部位に配置される。
【0074】
本発明は、重鎖可変領域及び軽鎖可変領域の両方を有する、TLR3反応性を有する単離された抗体又はそのフラグメントを提供するものであり、前記抗体は表1aに示されるような重鎖の相補性決定領域(CDR)のアミノ酸配列1、2及び3(HCDR1、HCDR2及びHCDR3)、及び軽鎖の相補性決定領域(CDR)のアミノ酸配列1、2及び3(LCDR1、LCDR2及びLCDR3)を有している。
【0075】
【表2】
【0076】
特定の実施形態では、本発明は、重鎖可変領域及び軽鎖可変領域の両方を有する、TLR3反応性を有する単離された抗体又はそのフラグメントを提供するものであり、前記抗体は配列番号192に示されるようなHCDR2のアミノ酸配列を有し、配列番号192のHCDR2は式(I)に示されるように定義される。
Xaa6−I−Xaa7−Xaa8−R−S−Xaa9−W−Y−N−D−Y−A−V−S−V−K−S
(I)
式中、
Xaa6は、Arg又はLysであってよく、
Xaa7は、Tyr、His又はSerであってよく、
Xaa8は、Met、Arg又はTyrであってよく、
Xaa9は、Lys又はArgであってよい。
【0077】
他の実施形態では、本発明は、重鎖可変領域及び軽鎖可変領域の両方を有する、TLR3反応性を有する単離された抗体又はそのフラグメントを提供するものであり、前記抗体は配列番号194に示されるようなHCDR2のアミノ酸配列を有し、配列番号194のHCDR2は式(III)に示されるように定義される。
I−I−Q−Xaa15−R−S−K−W−Y−N−Xaa16−Y−A−Xaa17−S−V−K−S
(III)
式中、
Xaa15は、Lys、Thr又はIleであってよく、
Xaa16は、Asn又はAspであってよく、
Xaa17は、Val又はLeuであってよい。
【0078】
他の実施形態では、本発明は、重鎖可変領域及び軽鎖可変領域の両方を有する、TLR3反応性を有する単離された抗体又はそのフラグメントを提供するものであり、前記抗体は配列番号196に示されるようなHCDR2のアミノ酸配列を有し、配列番号196のHCDR2は式(V)に示されるように定義される。
Xaa24−I−D−P−S−D−S−Y−T−N−Y−Xaa25−P−S−F−Q−G
(V)
式中、
Xaa24は、Phe又はArgであってよく、
Xaa25は、Ala又はSerであってよい。
【0079】
他の実施形態では、本発明は、重鎖可変領域及び軽鎖可変領域の両方を有する、TLR3反応性を有する単離された抗体又はそのフラグメントを提供するものであり、前記抗体は配列番号191に示されるようなLCDR3のアミノ酸配列を有し、配列番号191のLCDR3は式(II)に示されるように定義される。
Xaa1−S−Y−D−Xaa2−Xaa3−Xaa4−Xaa5−T−V
(II)
式中、
Xaa1は、Ala、Gln、Gly又はSerであってよく、
Xaa2は、Gly、Glu又はSerであってよく、
Xaa3は、Asp又はAsnであってよく、
Xaa4は、Glu又はSerであってよく、
Xaa5は、Phe、Ala又はLeuであってよい。
【0080】
他の実施形態では、本発明は、重鎖可変領域及び軽鎖可変領域の両方を有する、TLR3反応性を有する単離された抗体又はそのフラグメントを提供するものであり、前記抗体は配列番号193に示されるようなLCDR3のアミノ酸配列を有し、配列番号193のLCDR3は式(IV)に示されるように定義される。
Xaa10−S−Y−D−Xaa11−P−Xaa12−Xaa13−Xaa14−V
(IV)
式中、
Xaa10は、Gln又はSerであってよく、
Xaa11は、Thr、Glu又はAspであってよく、
Xaa12は、Val又はAsnであってよく、
Xaa13は、Tyr又はPheであってよく、
Xaa14は、Ser、Asn又はGlnであってよい。
【0081】
他の実施形態では、本発明は、重鎖可変領域及び軽鎖可変領域の両方を有する、TLR3反応性を有する単離された抗体又はそのフラグメントを提供するものであり、前記抗体は配列番号195に示されるようなLCDR3のアミノ酸配列を有し、配列番号195のLCDR3は式(VI)に示されるように定義される。
Q−Q−Xaa18−Xaa19−Xaa20−Xaa21−Xaa22−Xaa23−T
(VI)
式中、
Xaa18は、Tyr、Gly又はAlaであってよく、
Xaa19は、Gly、Glu又はAsnであってよく、
Xaa20は、Ser又はThrであってよく、
Xaa21は、Val、Ile又はLeuであってよく、
Xaa22は、Ser又はLeuであってよく、
Xaa23は、Ile、Ser、Pro又はTyrであってよい。
【0082】
本発明は、また、表1aに示されるような重鎖の相補性決定領域(CDR)のアミノ酸配列1、2及び3(HCDR1、HCDR2及びHCDR3)及び軽鎖の相補性決定領域(CDR)のアミノ酸配列1、2及び3(LCDR1、LCDR2及びLCDR3)を有する、TLR3反応性を有する単離された抗体又はそのフラグメントも提供するものである。
【0083】
抗原結合部位のアミノ酸配列が表1aのもの(配列番号49〜121及び191〜196)と大きくは異ならない抗体は本発明の範囲に含まれるものである。一般に、これには1以上のアミノ酸残基を、同様の電荷、疎水性又は立体化学的特性を有するアミノ酸によって置換することが含まれる。抗原結合部位ではなく、フレームワーク領域における更なる置換も、これらの置換が抗体の特性に悪影響を及ぼさない限りは行うことができる。置換は、例えば安定性又は親和性といった抗体の特性を向上させるために行うことができる。1、2、3、4、5又は6個の置換を抗原結合部位に行うことができる。得られる抗体が所望の特性を保持するならば、フレームワーク残基の1%、2%、3%、4%、5%、10%、15%、20%、25%、又は30%を置換することができる。
【0084】
保存的な改変では、こうした改変が行われた分子と同様の機能的及び化学的性質を有する分子が生ずる。分子の機能的及び/又は化学的特性における大幅な改変は、(1)置換領域における分子骨格の構造を例えばシート又はヘリックス立体構造として維持する、(2)標的部位における分子の電荷又は疎水性を維持する、又は(3)分子のサイズを維持する効果において、大きく異なるアミノ酸配列中の置換を選択することによって実現することができる。例えば、「保存的アミノ酸置換」では、その位置のアミノ酸残基の極性又は電荷にほとんどあるいはまったく影響しないように天然アミノ酸残基を非天然残基と置換することができる。更に、アラニン・スキャニング変異導入法についてこれまでに述べられているように(MacLennanら、Acta Physiol.Scand.Suppl.643:55〜67、1998;Sasakiら、Adv.Biophys.35:1〜24、1998)、ポリペプチド内の任意の内因性の残基をアラニンで置換することもできる。所望のアミノ酸置換(保存的又は非保存的であるかによらず)は、当業者であればこうした置換が望ましい時点で決定することができる。例えば、アミノ酸置換を用いることによってその分子の配列の重要な残基を特定したり、又は本明細書で述べる分子の親和性を増大若しくは減少させることができる。アミノ酸置換の例を表1bに示す。
【0085】
特定の実施形態においては、保存的アミノ酸置換には、生物学的システムによって合成されるのではなく一般的に化学的ペプチド合成によって組み込まれる、非天然のアミノ酸残基も含まれる。アミノ酸置換は、例えばPCR変異誘発(米国特許第4,683,195号)によって行うことができる。周知の方法を用いて、例えば、ランダムコドン(NNK)又は非ランダムコドン、例えば11個のアミノ酸(ACDEGKNRSYW)をコードするDVKコドンを用い、実施例1に示すような所望の特性を持つ変異体についてライブラリをスクリーニングすることで、変異体のライブラリを生成させることができる。表1cは、抗体の特性を改善するためにLCDR3及びHCDR2領域内で3つの親TLR3抗体アンタゴニストに対して行われた置換を示す。
【0086】
抗原結合部位の表記に応じて、本発明の抗体の抗原結合部位、次いでフレームワーク残基は、各重鎖及び軽鎖ごとに若干異なってもよい。
【0087】
【表3】
【0088】
表2a及び2bは、Kabat、Chothia及びIMGTに基づいて表記された本発明の例示的抗体の抗原結合部位の残基及びそれらの複合配列を示したものである。
【0089】
他の実施形態において、本発明は、重鎖可変領域及び軽鎖可変領域の両方を有する、TLR3反応性を有する単離された抗体又はそのフラグメントを提供するものであり、前記抗体は重鎖可変領域(VH)及び軽鎖可変領域(VL)のアミノ酸配列を有するとともに表3aに示されるようなそれぞれの単離された重鎖可変領域及び軽鎖可変領域を提供する。F17、F18及びF19は、それぞれ、ファミリー17、18及び19のコンセンサスなアミノ酸配列を有する抗体の変異体を表す(実施例1を参照)。
【0090】
【表4−1】
【0091】
【表4−2】
【0092】
【表4−3】
【0093】
実施例で説明する実施形態は、1個が重鎖に由来し、1個が軽鎖に由来する1対の可変領域を有しているが、当業者であれば代替的な実施形態は、1個の重鎖又は軽鎖の可変領域を有してもよいということを認識するであろう。単一の可変領域を用いることで、例えばTLR3に結合することが可能な2ドメイン特異的抗原結合フラグメントを形成することが可能な第2の可変ドメインについてスクリーニングを行うことができる。こうしたスクリーニングは、例えば国際公開第92/01047号に開示される、階層的に2重であるコンビナトリアルなアプローチを用いたファージディスプレイスクリーニング法によって実現することが可能である。このアプローチでは、H又はL鎖のいずれかのクローンを含む個別のコロニーを用いて他方の鎖(L又はH)をコードするクローンの完全なライブラリに感染させ、得られた2鎖特異的抗原結合ドメインを上記に述べたようなファージディスプレイ法に基づいて選択する。
【0094】
【表5】
【0095】
他の実施形態では、本発明は、表3aに示されるような可変領域のアミノ酸配列と少なくとも95%同一のアミノ酸配列を有する重鎖及び軽鎖の可変領域の両方を有し、TLR3反応性を有する単離された抗体又はそのフラグメントを提供する。
【0096】
別の態様では、本発明は、表3bに示されるような特定の重鎖及び軽鎖のアミノ酸配列を有する単離された抗体を提供する。
【0097】
本発明の別の態様は、本発明の抗体又はその補体のいずれかをコードする単離されたポリヌクレオチドである。特定のポリヌクレオチドの例を本明細書に開示するが、遺伝暗号の縮重及び特定の発現系におけるコドンの選択性を考慮すると、本発明の抗体アンタゴニストをコードする他のポリヌクレオチドも本発明の範囲内に含まれる。
【0098】
【表6】
【0099】
【表7】
【0100】
抗体アンタゴニストの例としては、IgG、IgD、IgG、IgA又はIgMアイソタイプの抗体が挙げられる。更にこうした抗体アンタゴニストは、グリコシル化、異性化、脱グリコシル化、又はポリエチレングリコール部分の付加(ペギレーション)及び脂質化などの非天然の共有結合修飾などの反応によって翻訳後修飾されてもよい。こうした修飾はインビボあるいはインビトロで行われ得る。例えば、本発明の抗体はポリエチレングリコールと結合(ペギレート)させることによって薬物動態的なプロファイルを向上させることができる。結合は当業者には周知の方法によって行うことができる。治療用抗体とPEGの結合は、機能を妨害せずに薬物動態を増強することが示されてきた(Deckertら、Int.J.Cancer 87:382〜390,2000;Knightら、Platelets 15:409〜418,2004;Leongら、Cytokine 16:106〜119,2001;Yangら、Protein Eng.16:761〜770,2003)。
【0101】
【表8】
【0102】
本発明の抗体の薬物動態的性質は、当業者には周知の技法により、Fc修飾によって向上させることも可能である。例えばIgG4アイソタイプの重鎖は、ヒンジ領域に重鎖間又は重鎖内ジスルフィド結合を形成することが可能なCys−Pro−Ser−Cys(CPSC)というモチーフを含んでいる。すなわち、CPSCモチーフ内の2個のCys残基が他の重鎖の対応するCys残基とジスルフィド結合する(重鎖間)か、あるいは特定のCPSCモチーフ内の2個のCys残基が互いにジスルフィド結合し得る(重鎖内)。生体内のイソメラーゼ酵素は、IgG4分子の重鎖間結合を重鎖内結合に変換し、またその逆反応を行うことが可能であると考えられている(Aalberse及びSchuurman、Immunology 105:9〜19、2002)。したがって、ヒンジ領域に重鎖内結合を有するこうしたIgG4分子内の重鎖:軽鎖(H:L)の各ペアは互いに共有結合していないことからH:Lモノマーに解離し、更に他のIgG4分子に由来するH:Lモノマーと再会合して、2重特異的なヘテロダイマーのIgG4分子を形成する。2重特異的なIgG抗体では、抗体分子の2個のFabの結合するエピトープが異なる。IgG4のヒンジ領域CPSCモチーフ中のSer残基をProにより置換することによって「IgG1様の挙動」が得られる。すなわち、この分子が重鎖間で安定なジスルフィド結合を形成し、それゆえ、他のIgG4分子とのH:L交換を受けにくい。一つの実施形態では、本発明の抗体は、SからPの変異と共にCPSCモチーフ中にIgG4Fcドメインを含む。CPSCモチーフの位置は通常、成熟した重鎖の残基228に見いだされるが、CDRの長さに応じて変化し得る。
【0103】
更に、本発明の抗体のFcRnサルベージ受容体以外のFc受容体への結合に影響する部位を除去することもできる。例えば、ADCC活性に関与するFc受容体結合領域を本発明の抗体において除去することができる。例えば、IgG1のヒンジ領域のLeu234/Leu235のL234A/L235Aへの変異、又はIgG4のヒンジ領域のPhe235/Leu236のP235A/L236Aへの変異によって、FcRへの結合性が最小となり、補体依存性細胞傷害活性及びADCCを介在する免疫グロブリンの能力が低減する。一実施形態では、本発明の抗体は、P235A/L236A変異を有するIgG4のFcドメインを有する。上記で特定したこれらの残基の位置は成熟した重鎖においては典型的なものであるが、CDRの長さに応じて変化し得る。P235A/L236A変異を有する抗体の例としては、配列番号218、219又は220に示される重鎖アミノ酸配列を有する抗体が挙げられる。
【0104】
完全なヒト、ヒト適合化、ヒト化、及び親和性成熟抗体分子又は抗体フラグメントは、融合タンパク質及びキメラタンパク質と同様、本発明の範囲に含まれる。抗原に対する抗体親和性は、ランダム又は部位特異的な変異誘導などの周知の方法を用いる合理的設計(rational design)又はランダムな親和性成熟によるか、あるいはファージディスプレイライブラリを用いることによって高めることができる。例えば、大部分がフレームワーク領域中に存在するバーニア残基又は「親和性決定残基」、ADRに対して置換を行うことで、抗体の親和性を調整することができる(米国特許第6,639,055号、国際公開第10/045340号)。
【0105】
安定性、選択性、交差反応性、親和性、免疫原性、又はタンパク質の所望の生物学的若しくは生物物理学的特性を高めるように改変された完全なヒト、ヒト適合化、ヒト化、親和性成熟抗体分子又は抗体フラグメントは本発明の範囲に含まれる。抗体の安定性は、(1)内在的な安定性に影響する個々のドメインのコアパッキング、(2)重鎖と軽鎖とのペアリングに影響するタンパク質/タンパク質の界面相互作用、(3)極性及び荷電残基の埋め込み、(4)極性及び荷電残基の水素結合ネットワーク、及び(5)分子内及び分子間力の中でも特に表面電荷及び極性残基の分布、などの多くの因子によって影響される。残基を不安定化させる可能性のある構造は、抗体の結晶構造に基づいて、あるいは場合によっては分子モデリングによって識別することが可能であり、抗体の安定性に対するこれらの残基の影響を、識別された残基中で変異を有する変異体を生成及び評価することにより試験することができる。抗体の安定性を高める方法の1つは、示差走査熱量測定法(DSC)によって測定される熱転移中点(Tm)を高くすることである。一般に、タンパク質のTmはその安定性との相関を示し、溶液中でのアンフォールディング及び変性のしやすさ、並びにそのタンパク質のアンフォールドのしやすさに依存する分解プロセスと負の相関を示す(Remmeleら、Biopharm.,13:36〜46,2000)。多数の研究によって、DSCにより熱安定性として測定される配合物の物理的安定性と他の方法により測定される物理的安定性のランキングの間に相関性が見出された(Guptaら、AAPS PharmSci.5E8,2003;Zhangら、J.Pharm.Sci.93:3076〜3089,2004;Maaら、Int.J.Pharm.,140:155〜168,1996;Bedu−Addoら、Pharm.Res.,21:1353〜1361,2004;Remmeleら、Pharm.Res.,15:200〜208,1997)。これらの配合物の研究は、FabのTmが対応するmAbの長期の物理的安定性と密接な関係があることを示唆している。フレームワーク中又は抗原結合部位内のアミノ酸の差は、Fabドメインの熱安定性に及ぼす著しい影響を有する可能性がある(Yasuiら、FEBS Lett.353:143〜146,1994)。
【0106】
本発明の抗体アンタゴニストは、約10-7、10-8、10-9、10-10、10-11又は10-12M以下のKdでTLR3と結合することができる。抗体などの特定の分子のTLR3に対する親和性は、任意の適当な方法によって実験的に求めることができる。こうした方法では、当業者には周知であるBiacore又はKinExa装置、ELISA又は競合的結合アッセイを使用することができる。
【0107】
特定のTLR3ホモログと所望の親和性で結合する抗体アンタゴニストは、抗体親和性成熟などの方法によって変異体又はフラグメントのライブラリから選択することができる。抗体アンタゴニストは、任意の適当な方法を用いてTLR3の生物活性の阻害に基づいて同定することができる。こうした方法では、周知の方法を用いた、本願に述べるようなレポーター遺伝子アッセイ又はサイトカイン産生を測定するアッセイを利用することができる。
【0108】
本発明の別の実施形態は、少なくとも1つの本発明のポリヌクレオチドを含むベクターである。かかるベクターは、プラスミドベクター、ウイルスベクター、バキュロウイルス発現ベクター、トランスポゾンに基づいたベクター、又は任意の手段によって特定の生物又は遺伝子的バックグラウンドに本発明のポリヌクレオチドを導入するのに適した他の任意のベクターであってよい。
【0109】
本発明の別の実施形態は、配列番号6、8、10、12、14、16、18、20、22、24、26、28、30、32、34、36、38、40、42、124、125、126、127、128、129、159、198、200、202、164、212、213、214、215又は216に示されるアミノ酸配列を有する免疫グロブリン重鎖の可変領域、又は配列番号5、7、9、11、13、15、17、19、21、23、25、27、29、31、33、35、37、39、41、122、123、197、199、201、163、209、210、211又は225に示されるアミノ酸配列を有する免疫グロブリン軽鎖の可変領域を含むポリペプチドをコードするポリヌクレオチドなどの、本発明のポリヌクレオチドのいずれかを有するホスト細胞である。
【0110】
本発明の別の実施形態は、配列番号102、130、131、132、133、134、135、160、204、206、208、220、166又は168に示されるアミノ酸配列を有する免疫グロブリン重鎖の可変領域、又は配列番号155、156、157、158、203、205、207、165、167又は227に示されるアミノ酸配列を有する免疫グロブリン軽鎖の可変領域を含むポリペプチドをコードするポリヌクレオチドを有するホスト細胞である。かかるホスト細胞は、真核細胞、細菌細胞、植物細胞又は古細菌細胞であってよい。真核細胞の例としては、哺乳動物、昆虫、鳥類又は他の動物由来のものであり得る。哺乳動物真核細胞としては、SP2/0(ATCC(American Type Culture Collection),Manassas,VA,CRL−1581)、NS0(ECACC(European Collection of Cell Cultures),Salisbury,Wiltshire,UK,ECACC No.85110503)、FO(ATCC CRL−1646)及びAg653(ATCC CRL−1580)マウス細胞株などのハイブリドーマ又はミエローマなどの不死化細胞株が挙げられる。ヒトミエローマ細胞株の一例には、U266(ATTC CRL−TIB−196)がある。他の有用な細胞株としては、CHO−K1SV(Lonza Biologics,Walkersville,MD)、CHO−K1(ATCC CRL−61)、又はDG44などの、チャイニーズハムスターの卵巣(CHO)細胞に由来するものが挙げられる。
【0111】
本発明の別の実施形態は、本発明のホスト細胞を培養することと、前記ホスト細胞によって産生された抗体を回収することとを含む、TLR3と反応性を有する抗体の製造方法である。抗体を製造して精製する方法は当該技術分野では周知のものである。
【0112】
本発明の別の実施形態は、本発明の抗体を産生するハイブリドーマ細胞株である。
【0113】
本発明の別の実施形態は、toll様受容体3(TLR3)の、配列番号2のアミノ酸残基;K416、K418、L440、N441、E442、Y465、N466、K467、Y468、R488、R489、A491、K493、N515、N516、N517、H539、N541、S571、L595、及びK619に結合する、単離された抗体又はそのフラグメントである。
【0114】
別の実施形態は、toll様受容体3(TLR3)の、配列番号2のアミノ酸残基;S115、D116、K117、A120、K139、N140、N141、V144、K145、T166、Q167、V168、S188、E189、D192、A195、及びA219に結合する、単離された抗体又はそのフラグメントである。
【0115】
幾つかの周知の方法を用いて本発明の抗体の結合エピトープを決定することができる。例えば、両方の個別の要素の構造が分かっている場合、インシリコでタンパク質−タンパク質のドッキングを行って、適合する相互作用部位を同定することができる。抗原抗体複合体において水素−重水素(H/D)交換を行うことによって抗体が結合し得る抗原の領域をマッピングすることができる。抗原のセグメント及び点変異誘導を用いることにより、抗体の結合に重要なアミノ酸の位置を特定することができる。TLR3のような大きなタンパク質では、最初にドッキング、セグメント変異誘導、又はH/D交換などによって結合部位の位置をそのタンパク質の特定の領域に限定することで点変異誘導によるマッピングが単純化される。両方の個別の要素の構造が分かっている場合、インシリコでタンパク質−タンパク質のドッキングを行って適合する相互作用の部位を同定することができる。抗体−抗原複合体の共結晶構造を使用して、エピトープ及びパラトープに寄与する残基を同定することができる。
【0116】
本発明の別の実施形態は、単離された抗体又はそのフラグメントであって、重鎖可変領域のChothia残基;W33、F50、D52、D54、Y56、N58、P61、E95、Y97、Y100及びD100b、並びに軽鎖可変領域のChothia残基;Q27、Y32、N92、T93、L94及びS95により、配列番号2で示されるアミノ酸配列を有するTLR3に結合することを特徴とする。重鎖パラトープ及び軽鎖パラトープChothia残基は、重鎖残基の配列番号216のW33、F50、D52、D55、Y57、N59、P62、E99、Y101、Y104及びD106並びに軽鎖残基の配列番号41のQ27、Y32、N92、T93、L94及びS95に対応する。
【0117】
本発明の別の実施形態は、単離された抗体又はそのフラグメントであって、重鎖可変領域のChothia残基;N31a、Q52、R52b、S53、K54、Y56、Y97、P98、F99及びY100、並びに軽鎖可変領域のChothia残基;G29、S30、Y31、Y32、E50、D51、Y91、D92及びD93により、配列番号2に示されるアミノ酸配列を有するTLR3に結合することを特徴とする。重鎖パラトープ及び軽鎖パラトープChothia残基は、重鎖残基の配列番号214のN32、Q54、R56、S57、K58、Y60、Y104、P105、F106及びY107並びに軽鎖残基の配列番号211のG28、S29、Y30、Y31、E49、D50、Y90、D91及びD92に対応する。
【0118】
例えば、パラトープ残基を好適な骨格にグラフトし、作製した骨格を完全な抗体に組立て、得られた抗体を発現させ、及びTLR3への結合又はTLR3の生物活性に及ぼす影響について抗体を試験することにより、TLR3を結合する特定のパラトープ残基を有する単離された抗体を作製することができる。代表的な骨格は、ヒト生殖細胞系列遺伝子によりコードされたヒト抗体可変領域のアミノ酸配列である。例えば、全体配列同一性、パラトープ残基間の同一性(%)、又は骨格及びmAb 15EVQ又はmAb 12QVQ/QSVなどの代表的な抗体との間の正準構造クラス同一性に基づいて、骨格を選択することができる。ヒト抗体生殖細胞系列遺伝子は、例えば、TomLinsonら、J.Mol.Biol 227:776〜798、及びInternational ImMunoGeneTics(IMGT)データベース(http_://_www_imgt_org)で開示されている。例えば、米国特許第6,300,064号で述べられているように、コンセンサスなヒトフレームワーク領域も使用することができる。例えば、国際公開第10/045340号に述べられている方法に従って、好適な骨格の選択を行うことができる。
【0119】
パラトープ残基をグラフトする骨格として使用可能な代表的なヒト生殖細胞系列遺伝子は、Vκ1、Vλ3、Vh5、Vh6、Jκ、Jλ、及びJhフレームワークによりコードされた遺伝子である。この生殖細胞系列J領域は、FR4配列を選択するために、その全体又は一部を使用される。例えば、mAb 15EVQ軽鎖パラトープ残基を、IGκJ1によりコードされたJ領域配列に直接連結されている、IGKV1−39*01によりコードされたVκ1フレームワークにグラフトすることができる。他のVκ1遺伝子由来の配列を使用することもでき、IGκJ1の代わりに、他のJκ遺伝子のFR4配列を置換することもできる。mAb 15EVQ重鎖パラトープ残基と、続いて、HCDR3及びIGHJ1によりコードされたFR4配列を構成する約11〜13個の残基、例えば12個の残基を、IGHV5−51*01によりコードされたVh5フレームワークにグラフトすることができる。11〜13個の残基は、FR3領域(「CAR」)の終端とFR4領域(大部分のJH領域に対するWGQ)の初端の間に懸り、mAb 15EVQVh由来の4個の規定されたパラトープ残基を含む。他のVh5遺伝子由来の配列を使用することもでき、IGJH1の代わりに他のJh遺伝子のFR4配列を置換することもできる。別の例では、mAb 12QVQ/QSV軽鎖パラトープ残基を、IGJL2によりコードされたJ領域配列に直接連結したIGLV3−1*01によりコードされたVλ3フレームワークにグラフトすることができる。他のVλ3及びJλ遺伝子の配列を使用することもできる。LCDR3の長さは、約9〜11個の残基、例えば、10個の残基で維持される。これらの約9〜11個の残基は、FR3領域(大部分のVラムダ骨格に対する「YYC」)の終端とFR4領域(大部分のJL領域に対する「FGG」)の初端との間に懸り、mAb 12QVQ/QSV由来の3個の規定されたパラトープ残基を含む。mAb 12QVQ/QSV重鎖パラトープ残基と、続いてHCDR3及びIGJH1によりコードされたFR4配列を構成する約9〜11個の残基、例えば10個の残基を、IGHV6−1*01によりコードされたVh6フレームワークにグラフトすることができる。これらの約9〜11個の残基は、FR3領域(「CAR」)の終端とFR4領域(大部分のJH領域に対するWGQ)の初端との間に懸り、mAb 12QVQ/QSVVhからの4個の規定されたパラトープ残基を含む。IGHJ1の代わりに他のJh遺伝子のFR4配列を置換することもできる。TLR3への結合及び得られた抗体の生物活性を、標準的な方法を用いて評価することができる。mAb 15EVQ及びmAb 12QVQ/QSV軽鎖可変領域及び重鎖可変領域と、代表的なVκ1、Vh5、Vλ3、Vh6、Jκ、Jλ又はJh遺伝子とのアライメントを、図32〜35に示す。ラトープをグラフトした、作製された抗体を、バーニア残基又は親和性決定残基の置換により更に改変して、抗体の特性、例えば上述のように親和性を改善することができる。パラトープをグラフトした抗体がTLR3への結合を保持する限り、パラトープをグラフトした抗体中のフレームワークアミノ酸配列は、mAb 15EVQ又は12QVQ/QSVフレームワーク配列と70%、75%、80%、85%、90%、95%、96%、97%、98%、又は99%同一であってもよい。
【0120】
標準的な方法を用いて、パラトープ残基に追加して抗原結合部位由来の配列をグラフトすることもできる。例えば、完全なHCDR3又はLCDR3をグラフトしてもよい。
【0121】
本発明の別の態様は、単離された抗体又はそのフラグメントであって、特定の重鎖相補性決定領域(CDR)1、2及び3のアミノ酸配列、特定の軽鎖CDR1、2及び3のアミノ酸配列、特定の重鎖可変領域(VH)のアミノ酸配列、又は特定の軽鎖可変領域(VL)のアミノ酸配列を有するモノクローナル抗体とTLR3との結合について競合する、TLR3反応性を有することを特徴とする。本発明のモノクローナル抗体の例としては、配列番号216に示されるアミノ酸配列を有する重鎖可変領域及び配列番号41に示される軽鎖可変領域のアミノ酸配列を有する単離された抗体、並びに配列番号214に示されるアミノ酸配列を有する重鎖可変領域及び配列番号211に示される軽鎖可変領域のアミノ酸配列を有する抗体が挙げられる。
【0122】
TLR3との結合についての競合は公知の方法によってインビトロでアッセイすることができる。例えば、非標識抗体の存在下でのMSD Sulfo−Tag(商標)NHS−エステル標識抗体のTLR3への結合を、ELISAによって評価することができる。本発明の抗体の例としては、mAb 12、mAb 15及びmAb c1811がある(表3aを参照)。前述の抗TLR3抗体c1068及びその誘導体(国際公開第06/060513(A2)号)、TLR3.7(eBiosciences,カタログ番号14−9039)、並びにImgenex IMG−315A(Imgenex IMG−315A、ヒトTLR3のアミノ酸55〜70のVLTHNQLRRLPAANに対して生成されたもの)は、実施例5に示されるようにmAb 12、15又はc1811によるTLR3への結合と競合しない。
【0123】
本発明の別の態様は、TLR3反応性を有し、以下のa〜iの特性のうち少なくとも1つを有する、単離された抗体である;
a.Kd<10nMでヒトTLR3に結合する;
b.インビトロでのポリ(I:C)NF−κBレポーター遺伝子アッセイにおいて、1μg/mLで、ヒトTLR3の生物活性を50%超低減させる;
c.10μg/mLで、100ng/mL未満のポリ(I:C)で刺激されたBEAS−2B細胞からのIL−6又はCXCL5/IP−10の産生を60%超阻害する;
d.0.4μg/mLで、100ng/mL未満のポリ(I:C)で刺激されたBEAS−2B細胞からのIL−6又はCXCL5/IP−10の産生を50%超阻害する;
e.5μg/mLで、62.5ng/mLのポリ(I:C)で刺激されたNHBE細胞からのIL−6の産生を50%超阻害する;
f.1μg/mLで、62.5ng/mLのポリ(I:C)で刺激されたNHBE細胞からのIL−6の産生を50%超阻害する;
g.1μg/mLで、PBMC細胞による、ポリ(I:C)により誘導されたIFN−γ、IL−6又はIL−12の産生を20%超阻害する;
h.インビトロでのNF−κBレポーター遺伝子アッセイにおいて、IC50<10μg/mLで、カニクイザルのTLR3の生物活性を阻害する;
i.インビトロでのISREレポーター遺伝子アッセイにおいて、IC50<5μg/mLで、カニクイザルのTLR3の生物活性を阻害する。
【0124】
治療方法
本発明のTLR3アンタゴニスト、例えばTLR3抗体アンタゴニストは、免疫系を調節する目的で使用することができる。何らのかの特定の理論に束縛されることも望むものではないが、本発明のアンタゴニストは、TLR3へのリガンドの結合、TLR3の2量化、TLR3の細胞内移行、又はTLR3の輸送を防止又は低減することによって免疫系を調節し得るものと考えられる。本発明の方法を用いて任意の分類に属する動物患者を治療することができる。こうした動物の例としては、ヒト、齧歯類、犬、猫、及び家畜などの哺乳動物が挙げられる。例えば本発明の抗体は、TLR3活性の中和、炎症、炎症性及び代謝性疾患の治療に有用であるだけでなく、更にこうした治療のための薬剤の調製においても有用であり、その場合、薬剤は本明細書で定義する用量で投与されるように調製される。
【0125】
一般的に、本発明のTLR3抗体アンタゴニストの投与によって予防又は治療することが可能な炎症状態、感染にともなう状態、又は免疫介在性炎症性疾患には、サイトカイン又はケモカインによって介在されるもの、及びTLR3の活性化又はTLR3経路を介したシグナル伝達に全体的又は部分的に起因する状態が含まれる。こうした炎症性状態の例としては、敗血症にともなう状態、炎症性腸疾患、自己免疫性疾患、炎症性疾患、及び感染にともなう状態が挙げられる。更に、本発明のTLR3抗体アンタゴニストの投与によって癌、心血管及び代謝性の状態、神経病及び線維症の状態を予防又は治療することも可能であると考えられる。炎症は組織を冒し、全身性の症状となり得る。罹患する組織の例としては、気道、肺、消化管、小腸、大腸、結腸、直腸、心血管系、心組織、血管、関節、骨及び滑膜組織、軟骨、上皮、内皮、肝又は脂肪組織が挙げられる。全身性の炎症状態の例としては、サイトカインストームすなわち高サイトカイン血症、全身性炎症反応症候群(SIRS)、移植片対宿主病(GVHD)、急性呼吸窮迫症候群(ARDS)、重症急性呼吸器症候群(SARS)、劇症型抗リン脂質症候群、重篤なウイルス感染、インフルエンザ、肺炎、ショック、又は敗血症がある。
【0126】
炎症は、侵入する病原体を撃退するための生物による防御反応である。炎症は、多くの細胞性及び体液性のメディエーターが関与するカスケードイベントである。一方で炎症反応の抑制はホストを免疫的に無防備状態にし、炎症を放置した場合には慢性炎症性疾患(例えば、喘息、乾癬、関節炎、リウマチ様関節炎、多発性硬化症、炎症性腸疾患など)、敗血性ショック、及び多臓器不全などの重篤な合併症を引き起こし得る。重要な点として、これらの多様な疾患状態は、サイトカイン、ケモカイン、炎症性細胞、及びこれらの細胞によって分泌される他のメディエーターなどの共通の炎症性メディエーターを共有している。
【0127】
リガンドであるポリ(I:C)、2本鎖RNA又は内因性のmRNAによってTLR3が活性化されるとシグナル伝達経路が活性化され、これにより炎症性サイトカインの合成及び分泌、マクロファージ、顆粒球、好中球及び好酸球などの炎症性細胞の活性化及び動員、細胞死、並びに組織の破壊が引き起こされる。TLR3は、IL−6、IL−8、IL−12、TNF−α、MIP−1、CXCL5/IP−10及びRANTES、並びに免疫細胞の動員及び活性化に関わる他の炎症性サイトカイン及びケモカインの分泌を誘導することにより、自己免疫疾患及び他の炎症性疾患における組織の破壊に寄与する。TLR3のリガンドである内因性のmRNAは炎症時に壊死細胞から放出され、ポジティブフィードバックループを引き起こすことによってTLR3を活性化し、炎症及び更なる組織破壊を持続させる。TLR3抗体アンタゴニストのようなTLR3アンタゴニストはサイトカインの分泌を正常化し、炎症性細胞の動員を低減し、組織破壊及び細胞死を低減させると考えられる。したがって、TLR3アンタゴニストは炎症及び広範な炎症性状態を治療するうえで治療上の可能性を有するものである。
【0128】
炎症性状態の1つの例は、全身性炎症反応症候群(SIRS)、敗血性ショック又は多臓器不全症候群(MODS)が含まれ得る、敗血症にともなう状態である。ウイルス、細菌、真菌、又は寄生虫感染によって、及び壊死細胞によって放出される2本鎖RNAは敗血症の発症に寄与し得る。特定の理論によって束縛されることを望むものではないが、TLR3アンタゴニストによる治療は、敗血症にともなう炎症状態を有する患者における生存時間を延ばすか又は局所的な炎症性イベント(例えば肺の)が広がって全身症状となることを防止することにより、あるいは内在的な抗微生物活性を強化することにより、あるいは抗微生物剤と組み合わされる場合に相乗的な活性を示すことにより、あるいは病状に寄与する局所的炎症状態を抑制することにより、あるいは上記の任意の組み合わせにより、所定の治療効果を与え得るものと考えられる。こうした介入は、患者の生存を確実にするために必要とされる更なる治療(例えば基礎となる炎症の治療又はサイトカインレベルの低減)を可能とするうえで充分なものとなり得る。敗血症はD−ガラクトサミン及びポリ(I:C)を投与することによってマウスなどの動物でモデル化することができる。こうしたモデルにおいて、D−ガラクトサミンは、敗血症感作物質として機能する肝臓毒素であり、ポリ(I:C)は2本鎖RNAを模倣してTLR3を活性化させる敗血症誘導分子である。TLR3アンタゴニストによる治療は、敗血症のマウスモデルにおいてマウスの生存率を高め得るものであることから、TLR3アンタゴニストは敗血症の治療に有用であると考えられる。
【0129】
消化管の炎症は消化管の粘膜の炎症であり、急性及び慢性の炎症状態が含まれる。急性炎症は、短時間の発症及び好中球の浸潤又は流入によって一般的に特徴付けられる。慢性炎症は、比較的長期の発症及び単核球の浸潤又は流入によって一般的に特徴付けられる。粘膜層は、腸(小腸及び大腸を含む)、直腸、胃(胃部)壁、又は口腔の粘膜であってよい。慢性の消化管炎症状態の例としては、炎症性腸疾患(IBD)、外界からの刺激によって誘導される大腸炎(例えば、化学療法、放射線療法などの投与のような治療レジメンによって引き起こされるかあるいはこれにともなう(例えば副作用として)消化管炎症(例えば大腸炎))、感染性大腸炎、虚血性大腸炎、膠原性又はリンパ性大腸炎、壊死性腸炎、慢性肉芽腫症又はセリアック病などの状態における大腸炎、食品アレルギー、胃炎、感染性胃炎又は小腸結腸炎(例えばヘリコバクター・ピロリの感染による慢性活動性胃炎)、及び感染性病原体によって引き起こされる消化管炎症の他の形態がある。
【0130】
炎症性腸疾患(IBD)には、例えば潰瘍性大腸炎(UC)及びクローン病(CD)などの、一般に病因が不明の一群の慢性炎症性疾患が含まれる。臨床的及び実験的証拠は、IBDの病因が感受性遺伝子及び環境因子が関与する多因子的なものであることを示唆している。炎症性腸疾患では、組織の傷害は、腸内の微生物叢の、抗原に対する不適切又は過剰な免疫反応によってもたらされる。炎症性腸疾患の幾つかの動物モデルが存在している。最も広く用いられているモデルは、結腸に慢性炎症及び潰瘍形成を誘発する2,4,6−トリニトロベンゼンスルホン酸/エタノール(TNBS)誘発大腸炎モデル又はオキサゾロンモデルである(Neurathら、Intern.Rev.Immunol 19:51〜62、2000)。別のモデルでは、血性下痢、体重減少、結腸の短縮、及び好中球の浸潤をともなう粘膜の潰瘍形成として発現する急性大腸炎を誘発する硫酸デキストランナトリウム(DSS)を使用する。DSS誘発大腸炎は、リンパ組織過形成、陰窩傷害(focal crypt damage)、及び上皮潰瘍形成をともなう、基底膜内への炎症性細胞の浸潤によって組織学的に特徴付けられる(Hendricksonら、Clinical Microbiology Reviews 15:79〜94、2002)。別のモデルは、ナイーブなCD45RBhigh CD4 T細胞をRAG又はSCIDマウスに養子移入することを含む。このモデルでは、ドナーのナイーブT細胞がレシピエントの腸を攻撃することにより、ヒトの炎症性腸疾患に似た慢性腸炎症及び症状を引き起こす(Read及びPowrie、Curr.Protoc.Immunol.Chapter 15 unit 15.13、2001)。これらのモデルのいずれかにおいて本発明のアンタゴニストの投与を用いることによって、これらのアンタゴニストが、炎症性腸疾患などの腸内の炎症にともなう症状を改善し、疾患の経過を変化させる潜在的な有効性を評価することができる。IBDに対しては幾つかの治療選択肢が存在し、例えば抗TNFα抗体による治療法が、クローン病を治療する目的で10年前から用いられている(Van Asscheら、Eur.J.Pharmacol.Epub Oct 2009)。しかしながら、かなりの割合の患者が現在の治療法に応答しない(Hanauerら、Lancet 359:1541〜1549、2002;Hanauerら、Gastroenterology 130:323〜333、2006)ことから、難治性の患者集団を標的とした新たな治療法が求められている。
【0131】
炎症状態の別の一例は炎症性の肺状態である。炎症性の肺状態の例としては、ウイルス、細菌、真菌、寄生虫又はプリオンによる感染にともなうものなどの感染により誘発される肺状態、アレルゲンにより誘発される肺状態、アスベスト症、珪肺症、又はベリリウム肺症などの汚染物質により誘発される肺状態、胃吸引により誘発される肺状態、嚢胞性線維症などの遺伝的素因にともなう免疫調節不全炎症状態、及び人工呼吸器傷害などの物理的外傷により誘発される肺状態が挙げられる。これらの炎症状態には更に、喘息、気腫、気管支炎、慢性閉塞性肺疾患(COPD)、サルコイドーシス、ヒスチオサイトーシス、リンパ管筋腫症、急性肺傷害、急性呼吸窮迫症候群、慢性肺疾患、気管支肺異形成症、院外感染性肺炎、院内肺炎、人工呼吸器関連肺炎、敗血症、ウイルス性肺炎、インフルエンザ感染、パラインフルエンザ感染、ロタウイルス感染、ヒトメタニューモウイルス感染、呼吸系発疹ウイルス感染、及びアスペルギルス又は他の真菌感染も含まれる。感染にともなう炎症性疾患の例としては、重症肺炎、嚢胞性線維症、気管支炎、気道増悪、及び急性呼吸窮迫症候群(ARDS)を含むウイルス又は細菌性肺炎が挙げられる。こうした感染にともなう状態は、一次ウイルス感染及び二次ウイルス感染のように複数の感染をともなう場合がある。
【0132】
喘息は、気道過敏症(AHR)、気管支収縮、喘鳴、好酸球又は好中球性炎症、粘液分泌過多、上皮下線維症、及び高いIgEレベルによって特徴付けられる肺の炎症性疾患である。喘息の患者は、最も一般的には気道の微生物感染(例えばライノウイルス、インフルエンザウイルス、ヘモフィルスインフルエンザなど)により症状の悪化である「増悪」を経験する。喘息の発作は、環境因子(例えば、回虫、昆虫、動物(例えばネコ、イヌ、ウサギ、マウス、ラット、ハムスター、モルモット及び鳥)、真菌、空気汚染物質(例えば煙草の煙)、刺激性ガス、フューム、蒸気、エアゾール、化学物質、花粉、運動、又は冷気によって誘発される。喘息は別にして、肺を冒すいくつかの慢性炎症性疾患、例えば、慢性閉塞性肺疾患(COPD)、細菌性肺炎及び嚢胞性線維症は、気道への好中球浸潤により特徴付けられ(Lindenら、Eur.Respir.J.15:973〜977,2000;Rahmanら、Clin.Immunol.115:268〜276,2005)、COPD、アレルギー性鼻炎及び嚢胞性線維症などの疾患は、気道過敏症により特徴付けられる(Fahy及びO’Byrne、Am.J.Respir.Crit.Care Med.163:822〜823,2001)。喘息及び気道炎症の一般的に用いられる動物モデルとしては、オボアルブミンチャレンジモデル及びメタコリン感作モデル(Hesselら、Eur.J.Pharmacol.293:401〜412、1995)が挙げられる。培養したヒト気管支上皮細胞、気管支線維芽細胞又は気道平滑筋細胞からのサイトカイン及びケモカインの産生の阻害をインビトロモデルとして用いることもできる。本発明のアンタゴニストをこれらのモデルのいずれかに投与することによって、喘息、気道炎症、COPDなどの症状を改善し、経過を変化させるためのこれらのアンタゴニストの使用を評価することができる。
【0133】
本発明の方法によって予防又は治療し得る他の炎症状態及び神経障害は、自己免疫疾患によって引き起こされるものである。これらの状態及び神経障害としては、多発性硬化症、全身性エリテマトーデス、並びに、アルツハイマー病、パーキンソン病、ハンチントン病、双極性疾患、及び筋萎縮性側索硬化症(ALS)を含む神経変性及び中枢神経系(CNS)疾患、原発性胆汁性硬変、原発性硬化性胆管炎、非アルコール性脂肪性肝疾患/脂肪性肝炎、線維症、C型肝炎ウイルス(HCV)、及びB型肝炎ウイルス(HBV)などの肝臓疾患、糖尿病及びインスリン抵抗性、アテローム性動脈硬化症、脳出血、脳卒中、及び心筋梗塞などの心血管疾患、関節炎、関節リウマチ、乾癬性関節炎及び若年性関節リウマチ(JRA)、骨粗鬆症、変形性関節症、膵炎、線維症、脳炎、乾癬、巨細胞動脈炎、強直性脊椎炎、自己免疫性肝炎、ヒト免疫不全ウイルス(HIV)、炎症性皮膚状態、移植、癌、アレルギー、内分泌疾患、創傷治癒、他の自己免疫性疾患、気道過敏症、並びに、細胞、ウイルス又はプリオンによって介在される感染症又は疾患が挙げられる。
【0134】
変形性関節症、関節リウマチ、怪我による関節炎を有する関節などを含む関節炎は、本発明のアンタゴニストのような抗炎症性タンパク質の治療的使用によって利するところのある一般的な炎症状態である。例えば、関節リウマチ(RA)は全身を冒す全身性疾患であり、関節炎の最も一般的な形態の1つである。関節リウマチは組織の傷害を引き起こすため、TLR3リガンドが炎症部位に存在し得る。TLR3シグナル伝達の活性化によって、炎症を起こした関節の炎症及び更なる組織傷害が持続し得る。関節リウマチの幾つかの動物モデルが当該技術分野において知られている。例えばコラーゲン誘導性関節炎(CIA)モデルでは、ヒト関節リウマチとよく似た慢性炎症性関節炎をマウスに発症させる。本発明のTLR3アンタゴニストをCIAモデルマウスに投与することによって、疾患の症状を改善して疾患の経過を変化させるためのこれらのアンタゴニストの使用を評価することができる。
【0135】
真性糖尿病すなわち糖尿病は、複数の原因因子によって誘導され、高血糖によって特徴付けられる疾患プロセスのことを指す(LeRoithら(eds.)、Diabetes Mellitus,Lippincott−Raven Publishers,Philadelphia,Pa.U.S.A.1996、及び本明細書により引用されるすべての参考文献)。制御されない高血糖は、腎障害、神経障害、網膜障害、高血圧、脳血管疾患及び冠状動脈性心疾患などの微小血管及び大血管性疾患のリスクが増大するために死亡率が増加し、若年死をともなう。したがって、グルコースホメオスタシスの制御は糖尿病の治療にとって極めて重要なアプローチである。
【0136】
糖尿病を導く欠陥に基づき、糖尿病は、患者の膵線にインスリンを産生するβ細胞が存在しない場合に発症する1型糖尿病(インスリン依存性糖尿病、IDDM)と、β細胞のインスリン分泌不全及び/又はインスリンの作用に対する抵抗性を有する患者において発症する2型糖尿病(インスリン非依存性糖尿病、NIDDM)の2つの大きなグループに分類される。
【0137】
II型糖尿病は、絶対的ではなく、相対的なインスリン欠乏をともなうインスリン抵抗性によって特徴付けられる。インスリン抵抗性の患者では、身体はこの欠陥を補償するために異常に高量のインスリンを分泌する。インスリン抵抗性を補償し、血糖値を適正に制御するために不適切な量のインスリンが存在すると、境界型糖尿病の状態が現れる。多くの患者でインスリン分泌は更に低下し、血漿グルコース濃度が上昇し、糖尿病の臨床状態がもたらされる。肥満症にともなう炎症は、インスリン抵抗性、II型糖尿病、脂質代謝異常、及び心血管疾患の発症に深く関わっていることが示唆されている。肥満脂肪はマクロファージを動員及び維持し、TNF−α及びIL−6などの炎症性サイトカイン、遊離脂肪酸及びアディポカインを過剰に産生する場合があり、インスリンシグナル伝達を妨害してインスリン抵抗性を誘発し得る。マクロファージのTLR3活性化は、こうした脂肪の炎症誘発状態に寄与し得る。インスリン抵抗性の幾つかの動物モデルが知られている。例えば、食餌誘発肥満モデル(DIO)では、動物は体重増加をともなう高血糖症及びインスリン抵抗性を発症する。DIOモデルに本発明のTLR3アンタゴニストを投与することによって、2型糖尿病にともなう合併症を改善して疾患の経過を変化させるためのこれらのアンタゴニストの使用を評価することができる。
【0138】
癌の例としては、白血病、急性白血病、急性リンパ芽球性白血病(ALL)、B細胞又はT細胞のALL、急性骨髄性白血病(AmL)、慢性骨髄性白血病(CmL)、慢性リンパ球性白血病(CLL)、有毛細胞白血病、骨髄異形成症候群(MDS)、リンパ腫、ホジキン病、悪性リンパ腫、非ホジキンリンパ腫、バーキットリンパ腫、多発性骨髄腫、カポジ肉腫、直腸結腸癌、膵臓癌、腎細胞癌、乳癌、鼻咽頭癌、悪性組織球増殖症、悪性の腫瘍随伴症候群/高カルシウム血症、固形腫瘍、腺癌、扁平上皮細胞癌、肉腫、悪性黒色腫、特に転移性黒色腫、血管腫、転移性疾患、癌関連骨吸収、及び癌関連骨痛を含む(ただしこれらに限定されない)、細胞、組織、臓器、動物、又は患者における少なくとも1つの悪性疾患が挙げられる。
【0139】
心血管疾患の例としては、心臓性昏倒症候群、心筋梗塞、鬱血性心不全、卒中、虚血性卒中、出血、動脈硬化症、アテローム性硬化症、再狭窄、糖尿病性動脈硬化疾患、高血圧、動脈高血圧、腎血管性高血圧、失神、ショック、心血管系の梅毒、心不全、肺性心、原発性肺高血圧、心臓不整脈、心房異所性収縮、心房粗動、心房細動(持続性又は痙攣性)、潅流後症候群、心肺バイパス炎症反応、多源性(chaotic又はmultifocal)心房頻拍、規則的で狭いQRS頻拍、特異的不整脈、心室細動、ヒス束不整脈、房室ブロック、脚ブロック、心筋虚血性障害、冠動脈疾患、狭心症、心筋梗塞、心筋症、拡張型うっ血性心筋症、拘束型心筋症、心臓弁疾患、心内膜炎、心臓周囲疾患、心臓腫瘍、大動脈及び抹消動脈瘤、大動脈解離、大動脈の炎症、腹部大動脈及びその分枝の閉塞、抹消血管障害、閉塞性動脈障害、抹消性アテローム硬化疾患、閉塞性血栓血管炎、機能性抹消動脈障害、レイノー現象及び障害、先端チアノーゼ、先端紅痛症、静脈疾患、静脈血栓症、静脈瘤血管、動静脈瘤、リンパ水腫、脂肪性浮腫、不安定狭心症、再潅流傷害、ポンプ後症候群、及び虚血性再潅流傷害を含む(ただしこれらに限定されない)、細胞、組織、臓器、動物又は患者における心血管疾患が挙げられる。
【0140】
代表的な神経性疾患の例としては、神経変性疾患、多発性硬化症、偏頭痛、AIDS痴呆複合症、例えば多発性硬化症及び急性横断性脊髄炎などの髄鞘脱落性疾患;例えば皮質脊髄系の病変などの錐体外路及び小脳疾患;基底核の疾患又は小脳疾患;脳幹神経節の障害;例えばハンチントン舞踏病及び老年舞踏病などの運動亢進性運動障害;例えばCNSドーパミンレセプターを遮断する薬物によって誘導される運動障害などの薬物誘導性運動障害;例えばパーキンソン病などの運動低下性運動障害;進行性核上性麻痺;小脳の構造的病変;例えば脊髄性運動失調、フリードライヒ運動失調、小脳皮質変性、多発性系変性(Mencel、Dejerine−Thomas、Shi−Drager及びMachado−Joseph)などの脊髄小脳の変性;全身性疾患(Refsum’s疾患、無β−リポタンパク血症、麻痺、毛細管拡張症及びミトコンドリア性多系疾患);例えば多発性硬化症、急性横断性脊髄炎などの髄鞘脱落性コア疾患;及び神経性筋萎縮などの運動ユニットの障害(例えば筋委縮性測索硬化症、小児性脊髄性筋委縮及び若年性脊髄性筋委縮などの前角細胞変性);アルツハイマー病;中年のダウン症候群;びまん性レビー小体病;レビー小体型の老年性認知症;ウェルニッケ‐コルサコフ症候群;慢性アルコール中毒;クロイツフェルト・ヤコブ病;亜急性硬化性全脳炎、ハラーフォルデン・シュパッツ病;ボクサー痴呆を含む(ただしこれらに限定されない)細胞、組織、臓器、動物又は患者における神経疾患が挙げられる。
【0141】
線維症状態の例としては、肝臓線維症(アルコール性肝硬変、ウイルス性肝硬変、自己免疫性肝炎を含むがこれらに限定されない);肺線維症(強皮症、特発性肺線維症を含むがこれらに限定されない);腎臓線維症(強皮症、糖尿病性腎炎、糸球体腎炎、ループス腎炎を含むがこれらに限定されない);皮膚線維症(強皮症、肥厚性及びケロイド瘢痕、火傷を含むがこれらに限定されない);骨髄線維症;神経線維腫症;線維腫;腸線維症;及び外科手術による線維化付着などが挙げられる。こうした方法では、線維症は器官特異的な線維症又は全身性線維症であり得る。器官特異的な線維症は、肺線維症、肝線維症、腎線維症、心線維症、血管線維症、皮膚線維症、眼線維症、骨髄線維症、又はその他の線維症の少なくとも1つにともない得る。肺線維症は、特発性肺線維症、薬剤誘発性肺線維症、喘息、サルコイドーシス、又は慢性閉塞性肺疾患のうち少なくとも1つにともない得る。肝臓線維症は、肝硬変、住血吸虫症、又は胆管炎のうち少なくとも1つにともない得る。肝硬変は、アルコール性肝硬変、C型肝炎後肝硬変、原発性胆汁性肝硬変の中から選択され得る。胆管炎は硬化性胆管炎である。腎線維症は、糖尿病性腎障害又はループス糸球体腎炎にともない得る。心線維症は、心筋梗塞にともない得る。血管線維症は、血管形成術後動脈再狭窄、又はアテローム性動脈硬化にともない得る。皮膚線維症は、火傷瘢痕化、肥厚性瘢痕化、ケロイド、又は腎性線維性皮膚症にともない得る。眼線維症は、後眼窩線維症、白内障手術後又は増殖性硝子体網膜症にともない得る。骨髄線維症は、特発性骨髄線維症又は薬剤誘発性骨髄線維症にともない得る。他の線維症は、ペイロニー病、デュピュイトラン拘縮又は皮膚筋炎から選択され得る。全身性線維症は、全身性硬化症及び移植片体宿主病から選択され得る。
【0142】
投与/製薬学的組成物
TLR3活性の抑制が所望される病態の治療又は予防に効果的な剤の「治療上の有効量」は、標準的な研究技術により判定され得る。例えば、喘息、クローン病、潰瘍性大腸炎又は関節リウマチなどの炎症状態の治療又は予防において有効となる薬剤の用量は、本明細書に述べるモデルのような関連する動物モデルに薬剤を投与することによって決定することができる。
【0143】
更に、場合によりインビトロアッセイを用いて最適な用量範囲を特定することができる。特定の有効用量の選択は、当業者であれば幾つかの因子の考慮に基づいて(例えば臨床試験によって)決定することができる。こうした因子には、治療又は防止しようとする疾患、関与する症状、患者の体重、患者の免疫状態、及び当業者には周知の他の因子が含まれる。製剤に使用される正確な用量は、投与経路、及び疾患の重篤度にも依存し、医師の判断及び各患者の状況に基づいて決定されなければならない。有効用量は、インビトロ又は動物モデル試験系から導出される用量反応曲線から外挿することができる。
【0144】
本発明の方法では、TLR3アンタゴニストは単独で、又は少なくとも1種類の他の分子と組み合わせて投与することができる。こうした更なる分子は、他のTLR3アンタゴニスト分子か、又はTLR3受容体のシグナル伝達によって介在されない治療効果を有する分子であってよい。抗生剤、抗ウイルス剤、緩和剤、及びサイトカインのレベル又は活性を低減する他の化合物はこうした更なる分子の例である。
【0145】
本発明の薬剤の治療上の使用のための投与方法は、薬剤をホストに送達する任意の好適な経路でよい。これらの薬剤の製薬学的組成物は、例えば、皮内、筋肉内、腹腔内、静脈内、皮下、又は鼻腔内などの非経口投与に特に有用である。
【0146】
本発明の薬剤は、有効量の薬剤を製薬上許容され得る担体中の有効成分として含有する製薬学的組成物として調製することができる。「担体」なる用語は、活性化合物が一緒に投与される希釈剤、助剤、賦形剤、又は溶媒のことを指す。こうした医薬用溶媒は、落花生油、大豆油、鉱物油、ゴマ油などの、石油、動物、植物又は合成物由来の水及び油などの液体であってよい。例えば、0.4%生理食塩水及び0.3%グリシンを使用することができる。これらの溶液は滅菌され、粒子状物質を含まないものである。これらの溶液は、従来の公知の滅菌法(例えば濾過)によって滅菌することができる。組成物は、pH調整剤及び緩衝剤、安定化剤、増粘剤、潤滑剤、及び着色剤などの、適切な生理学的状態に必要とされるような、製薬上許容できる補助物質を含有し得る。そのような製剤処方中の本発明の剤の濃度は、幅広く、すなわち、約0.5重量%未満から通常は約1重量%、又は少なくとも約1重量%から、15又は20重量%まで変化させることができ、主に、選択された特定の投与様式による、必要とされる投与量、液体の容積及び粘度などに基づいて選択される。
【0147】
したがって、筋肉内注射用の本発明の製薬学的組成物は、1mLの滅菌緩衝水、及び約1ng〜約100mg、例えば約50ng〜約30mg、又はより好ましくは約5mg〜約25mgの本発明のTLR3アンタゴニストを含むように調製することができる。同様に、静脈内注射用の本発明の製薬学的組成物は、250mLの滅菌リンゲル溶液、及び約1mg〜約30mg、好ましくは5mg〜約25mgの本発明のアンタゴニストを含むように調製することができる。非経口投与可能な組成物を調製する実際の方法は周知のものであり、例えば、「Remington’s Pharmaceutical Science」、15th ed.、Mack Publishing Company,Easton,PAにより詳細に記載されている。
【0148】
本発明の抗体アンタゴニストは凍結乾燥して保存することができ、使用の前に適切な溶媒中で還元することができる。この方法は従来の免疫グロブリン及びタンパク質の作製において有効であることが示されており、当該技術分野では周知の凍結乾燥及び還元の技術を使用することができる。
【0149】
次に以下の具体的かつ非限定的な実施例を参照して本発明を説明する。
【実施例】
【0150】
(実施例1)
抗huTLR3アンタゴニストmAbの同定及び誘導
MorphoSys Human Combinatorial Antibody Library(HuCAL(登録商標))Goldファージディスプレイライブラリ(Morphosys AG,Martinsried,Germany)をヒト抗体フラグメントの供給源として使用し、C末端にポリヒスチジンタグを有するヒトTLR3(huTLR3)(配列番号4)のアミノ酸1〜703を発現させることによって生成した精製TLR3抗原に対してパニングさせ、固定化金属アフィニティークロマトグラフィーによって精製した。アミノ酸1〜703はhuTLR3の予想される細胞外ドメイン(ECD)と一致している。多様な抗体フラグメント群を固有のものとして同定、配列決定及び確認できるさまざまな方法において、TLR3タンパク質を提示することにより、huTLR3と特異的に結合したFabフラグメント(Fab)を選別した。手法の異なるパニングにより、62種類の候補物質(V領域配列が異なる)をhTLR3のECDに結合する固有の物質として同定した。
【0151】
huTLR3のECD結合物質として同定されたこれら62種類の候補物質を、抗炎症活性の特定に関するさまざまな細胞アッセイにより中和活性についてスクリーニングした。予備活性データ(下記実施例2を参照)を用いて、重鎖CDR2(HCDR2)及び軽鎖CDR3(LCDR3)のCDR成熟のための親候補物質として、62種類の候補物質からファミリー16〜19を形成する4種類の候補物質(Fab 16〜19)を選別した。これらの親候補物質の1つ(候補物質19)でHCDR2にN結合型グリコシル化部位が認められたため、この候補物質にSer→Ala(S→A)の変異を導入してこの部位を除去した。4種類の親候補物質をCDR成熟させた後、全部で15種類の子孫候補物質(候補物質1〜15)が同定され、これを下記実施例2で述べるように更なる特性評価に供した。19種類の候補物質のそれぞれに存在する軽鎖及び重鎖の可変領域のリストを上記表3に示した。ここで各候補物質は、Fabであるか完全長の抗体鎖としてクローニングされた(実施例3)ものであるかに応じてmAb 1〜19又はFab 1〜19と呼ぶものとする。発現ベクターの設計により、すべての候補物質について、可変領域の成熟したアミノ末端は重鎖ではQVEであり、軽鎖ではDIであった。これらの末端の好ましい配列は、それぞれの生殖細胞遺伝子内に存在し、候補配列と高い相同性を有する。ファミリー17及び18では、生殖細胞系の配列はVHでQVQ、VLでSYである。ファミリー19ではこの配列はVHでEVQ、VLでDIである。SY配列はλサブグループ3に固有であり、アミノ末端残基がS又はYのいずれかであることについての異種性が報告されている。したがって、有名なλサブグループ1由来のQSVコンセンサス末端は、ファミリー17及び18のVLのDIEに対するより好適な置換になるものと考えられる。これらの改変を、ファミリー18から候補物質9、10及び12に、ファミリー19から候補物質14及び15に導入した。このプロセスでは、これらの抗体のVH及びVL領域のコドンを最適化した。それぞれ、候補物質9、10及び11の軽鎖可変領域のN末端生殖細胞変異体のアミノ酸配列を配列番号209〜211に示し、候補物質9、10、12、14及び15の重鎖可変領域のN末端生殖細胞変異体のアミノ酸配列を配列番号212〜216に示す。本明細書では、これらの候補物質のN末端変異体を候補物質/mAb/Fab 9QVQ/QSV、10QVQ/QSV、12QVQ/QSV、14EVQ又は15EVQと呼ぶ。これらのN末端生殖細胞変異体をmAbとして発現させたところ、それぞれの親候補物質と比較した場合に、TLR3への結合についても、TLR3の生物活性を阻害する能力においても何ら影響を示さなかった(データは示されていない)。
【0152】
(実施例2)
インビトロでのTLR3アンタゴニスト活性の判定
上記に述べたCDRを成熟させた15種類の候補物質を潜在的なヒトの治療物質として選択し、結合範囲及び中和活性を調べた。4種類の親Fab(Fab 16〜19)及びCDR成熟させた15種類のFab(Fab 1〜15)又はこれらの非生殖細胞V領域変異体についての、活性のアッセイ及び結果を下記に述べる。
【0153】
NF−κB及びISREシグナル伝達カスケードの阻害
熱で不活化したFBSを添加したDMEM及びGlutaMax培地(Invitrogen,Carlsbad,CA)中で293T細胞を増殖させ、pNF−κB又はISREホタルルシフェラーゼレポータープラスミド30ng、pcDNA3.1ベクター13.5ng、phRL−TK 5ng、及びFLTLR3をコードするpCDNA(配列番号2)1.5ngをトランスフェクトした。phRL−TKプラスミドは、HSV−1チミジンキナーゼプロモーターによって制御されるウミシイタケルシフェラーゼ遺伝子(Promega,Madion,WI)を有している。TLR3抗体を30〜60分インキュベートした後、ポリ(I:C)(GE Healthcare,Piscataway,NJ)を加えた。各プレートを37℃で6時間又は24時間インキュベートした後、DualGloルシフェラーゼ試薬を加えて各プレートをFLUOstarプレートリーダーで読み取った。ホタルルシフェラーゼの相対発光量(RLU)をウミシイタケルシフェラーゼのRLUで割ることによって、正規化された値(ルシフェラーゼ比)を求めた。ホタルルシフェラーゼの産生を刺激するNF−κB又はISREシグナル伝達カスケードは、TLR3アゴニストであるポリ(I:C)(1μg/mL)による刺激前に、細胞を抗TLR3抗体(0.4、2.0及び10μg/mL)とインキュベートすることによって特異的に阻害された。NF−κBアッセイの結果を図1に示す。図中、5465をポジティブコントロール(抗ヒトTLR3 mAbを中和する)として、また抗ヒト組織因子mAb(859)をヒトIgG4アイソタイプコントロールとして用いて、ホタル/ウミシイタケ比の阻害率(%)を表す。0.4〜10μg/mLのmAb濃度で50%を上回る阻害率が得られた。c1068及びTLR3.7は、10μg/mLでTLR3の生物活性の約38%及び8%を阻害した。同様の結果がISREレポーター遺伝子アッセイで得られた(データは示されていない)。
【0154】
BEAS−2B細胞におけるサイトカイン放出
BEAS−2B細胞(SV−40で形質転換した正常なヒト気管支上皮細胞)を、I型コラーゲンをコーティングした培養皿に播種し、抗ヒトTLR3抗体の存在下又は非存在下でインキュベートした後、ポリ(I:C)を加えた。処理の24時間後に上澄み液を回収し、IL−6、IL−8、CCL−2/MCP−1、CCL5/RANTES及びCXCL10/IP−10の検出用にカスタムしたマルチプレックスビーズアッセイを用いて、サイトカイン及びケモカインレベルについてアッセイした。結果を、0.4、2.0及び10μg/mLのmAbで処理した後の、サイトカイン/ケモカインの個別の阻害率(%)として図2に示す。5465はポジティブコントロールであり、859はアイソタイプコントロールである。
【0155】
NHBE細胞におけるサイトカイン放出
サイトカインの放出を、正常なヒト気管支上皮(NHBE)細胞(Lonza,Walkersville,MD)においてもアッセイした。NHBE細胞を増殖させ、コラーゲンコーティングした培養皿に移して48時間インキュベートした後、培地を除去して0.2mLの新鮮な培地を補充した。次いで細胞を抗ヒトTLR3 mAbの存在下又は非存在下で60分間インキュベートした後、ポリ(I:C)を加えた。24時間後に上澄み液を回収し、−20℃で保存するか、又は直ちにIL−6のレベルについてアッセイした。結果を、投与量0.001〜50μg/mLでのmAb処理後の、IL−6分泌の阻害率(%)として図3にグラフで示す。5465はポジティブコントロールであり、859はアイソタイプコントロールである。大部分のmAbは1μg/mLでIL−6の産生の少なくとも50%を阻害し、5μg/mLで阻害率75%を達成した。
【0156】
PBMC細胞におけるサイトカイン放出
サイトカインの放出を、ヒト末梢血単核球細胞(PBMC)においてもアッセイした。ヒトドナーから全血をヘパリン回収試験管に回収し、これにFicoll−Paque Plus溶液を下側に層を形成するようにゆっくりと加えた。試験管を遠心してFicollの直ぐ上に白色の層を形成しているPBMCを回収して播種した。この後、PBMCを抗ヒトTLR3 mAbの存在下又は非存在下でインキュベートした後、25μg/mLのポリ(I:C)を加えた。24時間後に上澄み液を回収し、Luminex法によってサイトカインのレベルを調べた。5465をポジティブコントロールとし、hIgG4をアイソタイプコントロールとして用い、mAbの単回用量(0.4μg/mL)によるIFN−γ、IL−12及びIL−6の累積阻害率(%)として、結果を図4にグラフ化して示す。
【0157】
HASM細胞におけるサイトカイン放出
簡潔に言えば、500ng/mLのポリ(I:C)及び10ng/mLのTNF−αの相乗効果のある組み合わせを添加する前に、ヒト気道平滑筋(HASM)細胞を、抗ヒトTLR3 mAbsの存在又は非存在下でインキュベートした。24時間後に上澄み液を回収し、Luminex技術を用いて、サイトカインレベルを求めた。結果を、3段階の容量(0.4、2及び10μg/mL)でmAbを用いた場合のケモカインCCL5/RANTESのレベルとして図5にグラフで示す。5465はポジティブコントロールであり、hIgG4はアイソタイプコントロールである。
【0158】
huTLR3に結合することによりサイトカイン及びケモカインの放出を低減させる本発明の抗体の能力が、ヒト細胞におけるインビトロアッセイの結果によって確認された。
【0159】
(実施例3)
完全長の抗体コンストラクト
4種類の親Fab(候補物質番号16〜19)及び15種類の子孫Fab(候補物質番号1〜15)の重鎖をS229P Fc変異を有するヒトIgG4バックグラウンド上にクローニングした。候補物質9QVQ/QSV、10QVQ/QSV、12QVQ/QSV、14EVQ又は15EVQを、F235A/L236及びS229P Fc変異を有するヒトIgG4バックグラウンド上にクローニングした。
【0160】
成熟した完全長の重鎖アミノ酸配列を以下のように配列番号90〜102及び218〜220に示す。
【0161】
【表9】
【0162】
発現のために、これらの重鎖はMAWVWTLLFLMAAAQSIQA(配列番号103)のようなN末端のリーダー配列を有してもよい。リーダー配列を有する候補物質14EVQ及び15EVQの重鎖、及び成熟型(リーダー配列を有さない)をコードするヌクレオチド配列の例を、配列番号104及び105にそれぞれ示す。同様に、発現のために、本発明の抗体の軽鎖の配列は、MGVPTQVLGLLLLWLTDARC(配列番号106)のようなN末端のリーダー配列を有してもよい。リーダー配列を有するコドン最適化された候補物質15の軽鎖、及び成熟型(リーダー配列を有さない)をコードするヌクレオチド配列の例を、配列番号107及び108にそれぞれ示す。
【0163】
(実施例4)
抗TLR3 mAbの結合の特性評価
ヒトTLR3の細胞外ドメイン(ECD)に対するmAbの結合のEC50値をELISAによって求めた。ヒトTLR3のECDタンパク質をPBSで2μg/mLに希釈し、100μLの一定分量を96穴プレート(Corning Inc.,Acton,MA)の各ウェルに分配した。4℃で一晩インキュベートした後、PBSに0.05%のTween−20(Sigma−Aldrich)を加えた洗浄緩衝液によりプレートを3回洗浄した。ウェルを2%のI−Block(Applied Biosystems,Foster City,CA)及び0.05%のTween−20/PBSからなる200μLのブロッキング溶液によりブロッキングした。室温で2時間ブロッキングした後、プレートを3回洗浄し、続いてブロッキング緩衝液に希釈した抗TLR3 mAb候補物質1〜19を連続的に添加した。
【0164】
【表10】
【0165】
抗TLR3 mAbを室温で2時間インキュベートた後3回洗浄した。この後、ブロッキング緩衝液で1:4000に希釈したペルオキシダーゼ接合ヒツジ抗ヒトIgG(GE Health care,Piscataway,NJ)を加え、室温で1時間インキュベートした後、洗浄緩衝液で3回洗浄した。結合は、TMB−S(Fitzgerald Industries International,Inc.,Concord,MA)中で10〜15分間インキュベートすることによって検出した。反応を、2N H2SO4 25μLにより停止させ、及びSPECTRA Max分光光度計(Molecular Devices Corp.,Sunnyvale,CA)を用いて、吸光度を450nmで読み取り650nmでの値を差し引いた。EC50値をGraph Pad Prismプリズムソフトウェア(Graph Pad Software,Inc.,San Diego,CA)を使用して非線形回帰によって求めた。
【0166】
mAbを2.5μg/mL〜0.6pg/mLまで4倍ずつ希釈した100μLの連続希釈液とインキュベートすることにより、huTLR3に対する結合についてEC50値を求めた(表4)。抗ヒト組織因子mAb 859及びhu IgG4κをネガティブコントロールとして含めた。
【0167】
huTLR3 ECDに対する結合親和性もBiacore解析によって求めた。データ(示されていない)は、mAb 1〜19ではhuTLR3 ECDに対するKdが10-8M未満であることを示した。
【0168】
(実施例5)
競合的エピトープ結合
抗TLR3抗体競合グループ又は「エピトープビン」を決定するためにエピトープ結合実験を行った。
【0169】
競合的ELISAを行うため、実施例1に述べるようにして作製した5μL(20μg/mL)の精製ヒトTLR3 ECDタンパク質を、MSD High Bindプレート(Meso Scale Discovery,Gaithersburg,MD)の各ウェルに室温で2時間コーティングした。150μLの5% MSDブロッカーA緩衝液(Meso Scale Discovery)を各ウェルに加えて室温で2時間インキュベートした。プレートを0.1MのHEPES緩衝液(pH 7.4)で3回洗浄した後、標識した抗TLR3 mAbと異なる競合物質の混合液を加えた。標識した抗体(10nM)を、濃度を増加させた(1nM〜2μM)非標識抗TLR3抗体とインキュベートし、次いで25μLの量の混合液を、指定されたウェルに加えた。室温で穏やかに振盪しながら2時間インキュベートした後、プレートを0.1MのHEPES緩衝液(pH 7.4)で3回洗浄した。MSD Read緩衝液Tを蒸留水で希釈し(4倍)、150μL/ウェルの量で分配してSECTOR Imager 6000によって分析した。抗体を製造者(Meso Scale Discovery)の使用説明書に従ってMSD Sulfo−Tag(商標)NHS−エステルで標識した。
【0170】
以下の抗TLR3抗体を評価した。すなわち、MorphoSysヒト組み換え抗体ライブラリより入手したmAb 1〜19(表3に示すもの);c1068(国際公開第06/060513(A2)号に述べられるもの)、c1811(マウスTLR3タンパク質で免疫性を与えられたラットから作製されるハイブリドーマにより産生されるラット抗マウスTLR3 mAb)、TLR3.7(eBiosciences,San Diego,CA,カタログ番号14−9039)、及びIMG−315A(Imgenex,San Diego,CAより入手したヒトTLR3アミノ酸55〜70(VLNLTHNQLRRLPAAN)に対して生成したもの)。mAb 9、10、12、14及び15については、変異体9QVQ/QSV、10QVQ/QSV、12QVQ/QSV、14EVQ、又は15EVQをこの実験で使用した。
【0171】
競合アッセイに基づき、抗TLR3抗体を、次の5つの異なるビンに割り当てた。ビンA:mAb 1、2、13、14EVQ、15EVQ、16、19;ビンB:mAb 3、4、5、6、7、8、9QVQ/QSV、10QVQ/QSV、11、12QVQ/QSV、17、18;ビンC:抗体Imgenex IMG−315A;ビンD:抗体TLR3.7、c1068;及びビンE:抗体c1811。
【0172】
(実施例6)
エピトープマッピング
実施例5で述べた異なるエピトープビンからの代表的な抗体を更なるエピトープマッピングのために選択した。エピトープマッピングを、TLR3セグメント交換実験、変異誘発、H/D交換、及びインシリコタンパク質−タンパク質ドッキングを含む各種のアプローチを用いて行った(The Epitope Mapping Protocols,Methods in Molecular Biology,Volume 6,GlenE.Morrised.,1996)。
【0173】
TLR3セグメント交換TLR3ヒト−マウスキメラタンパク質を用いてTLR3上の全体の抗体結合ドメインの位置を特定した。ヒトTLR3タンパク質の細胞外ドメインを3つのセグメント(GenBank受諾番号NP_003256のヒトTLR3アミノ酸配列に基づくアミノ酸番号に従ってaa 1〜209、aa 210〜436、aa 437〜708)に分割した。ヒトTLR3のアミノ酸210〜436及び437〜708を対応するマウスのアミノ酸(マウスTLR3、GenBank受諾番号NP_569054、アミノ酸211〜437及び438〜709)と置き換えることによってMT5420キメラタンパク質を作製した。ヒトアミノ酸の位置437〜708をマウスTLR3のアミノ酸(マウスTLR3、GenBank受諾番号NP_569054、アミノ酸438〜709)で置き換えることによってMT6251キメラを作製した。すべてのコンストラクトを、標準的なクローニング手法を用いてpCEP4ベクター(Life Technologies,Carslbad,CA)で生成させた。各タンパク質をHEK293細胞内でV5−His6 C末端融合タンパク質として一過性に発現させ、実施例1で述べたようにして精製した。
【0174】
mAb c1068.mAb c1068は、ヒトTLR3 ECDに高親和性で結合したが、マウスTLR3には良好に結合しなかった。c1068は、MT5420及びMT6251の両方に結合する能力を喪失したことから、結合部位がWTヒトTLR3タンパク質のアミノ酸437〜708内に局在するということが示された。
【0175】
mAb 12QVQ/QSV:mAb 12QVQ/QSVは両方のキメラに結合したが、このことはmAb 12QVQ/QSVの結合部位が配列番号2に示される配列を有するヒトTLR3タンパク質のアミノ酸1〜209内に位置していることを示している。
【0176】
インシリコタンパク質−タンパク質ドッキングmAb 15EVQ(下記を参照)の結晶構造及び発表されているヒトTLR3構造(Bellら、J.Endotoxin Res.12:375〜378,2006)を、ドッキングの開始モデルとして使用するために、CHARMm(Brooksら、J.Computat.Chem.4:187〜217,1983)でエネルギー最小化した。タンパク質ドッキングは、ZDOCK 2.1(Chen及びWeng、Proteins 51:397(Accelrys,San Diego,CA)−408、2003)と同様のZDOCKpro 1.0により6°の角度グリッドで行った。ZDOCKアルゴリズム中のエネルギー項により、ヒトTLR3における既知のN結合グリコシル化部位のAsn残基(Asn 52、70、196、252、265、275、291、398、413、507及び636)(Sunら、J.Biol.Chem.281:11144〜11151,2006)を、抗体−抗原複合体インターフェースへの参加からブロックした。2000個の最初のポーズを出力してクラスター化し、ドッキングポーズを再精密化してRDOCKで再スコアリングを行った(Liら、Proteins 53:693〜707、2003)。最初のZDOCKスコアが最も高かった200個のポーズ及びRDOCKの上位200個のポーズを視覚的に調べた。
【0177】
Fab 15EVQの結晶化を20℃で蒸気拡散法によって行った(Benvenuti及びMangani、Nature Protocols 2:1633〜51、2007)。最初のスクリーニングを、96穴プレートでHydraロボットを使用してセットアップした。実験を、0.5μLのタンパク質溶液を0.5μLのリザーバ溶液に混合した液滴で構成した。液滴を90μLのリザーバ溶液に対して平衡化した。50mMのNaClを含む20mMのTris緩衝液(pH 7.4)中のFab溶液をAmicon Ultra−5kDaセルを使用して14.3mg/mLにまで濃縮した。スクリーニングは、Wizard I & II(Emerald BioSystems,Bainbridge Island,WA)及びインハウス結晶化スクリーンによって行った。Fab 12QVQ/QSVを類似の方法で結晶化させた。
【0178】
X線回折データを収集し、Osmic(商標)VariMax(商標)共焦光学系、Saturn944CCD検出器、及びX−stream(商標)2000低温冷却システムを備えたRigaku MicroMax(商標)−007HFs微小焦点X線発生装置(Rigaku,Woodlands,TX)を使用して処理した。回折強度を、1/2°の像当たり120秒の露光時間で270°(結晶回転)にわたって検出した。X線データをD*TREKプログラム(Rigaku)により処理した。構造を、Phaser又はCNXプログラム(Accelrys,SanDiego,CA)を使用して分子置換法によって決定した。すべてのデータを用いて、原子位置及び温度因子をREFMACによりFab 15EVQに対して15〜2.2Å及びFab 12QVQ/QSVに対して50〜1.9Åの分解能範囲で精密化した。3σのカットオフレベルを用いて、(Fo−Fc)電子密度ピークで水分子を追加した。すべての結晶学的計算を、プログラムのCCP4に適したプログラムにより行った(Collaborative Computational Project,Number 4.1994.The CCP4 suite:programs for protein crystallography.Acta Crystallogr.D50:760〜763)。COOTプログラム(Emsleyら、Acta Crystallogr.D60:2126〜2132、2004)を使用して、モデル調整を行った。
【0179】
解像されたmAb 15EVQの結晶構造によって、抗体結合部位が重鎖の多数の負に帯電した残基(D52、D55、E99、D106及びD109)によって特徴付けられることが示された。したがって、mAb 15EQVとTLR3との間の認識には正に帯電した残基が関与している可能性が高いと考えられた。実施したタンパク質−タンパク質ドッキングシミュレーションは、正に帯電した残基を複数含む、TLR3上の2個の大きなパッチが、抗体に対して優れた相補性を示すことを示唆するものであった。シミュレートしたTLR3−抗TLR3抗体複合体の界面内のTLR3上の残基は、R64、K182、K416、K467、Y468、R488、R489及びK493であった。
【0180】
変異誘発実験単一及び組み合わせの変異を、TLR3 ECDの表面残基の内の、上記でmAb 12及びmAb 15EVQのエピトープを含んでいることが特定された領域に導入し、この変異タンパク質を抗体結合性について試験した。
【0181】
ヒトTLR3のアミノ酸1〜703(ECD)をコードするヌクレオチド配列(配列番号4、GenBank受諾番号NP_003256)を標準的なプロトコールを用いてクローニングした。表5aに示されるオリゴヌクレオチドを用い、製造者のプロトコールに従ってStrategene Quickchange II XLキット(Stratagene,San Diego,CA)を使用して、すべての変異体を、部位特異的な変異の誘導によって作製した。変異はDNAの配列決定によって確認した。タンパク質はCMVプロモーターの制御下で、HEK293細胞内でC末端His−タグ融合タンパク質として発現させ、実施例1で述べたようにして精製した。
【0182】
結合アッセイヒトTLR3に対するmAb 12QVQ/QSV及びmAb 15EVQの結合活性及び生成された変異体をELISAによって評価した。プロセスを促進するため、mAb 15EVQの結合が予想される部位についての変異体を、mAb 12QVQ/QSVと共にC末端Hisタグを含むTLR3 ECD変異体と、HEK細胞にコトランスフェクトすることで同時発現させた後、金属アフィニティークロマトグラフィーにより精製した。回収された試料は、TLR3変異体とmAbとの複合体であった。このアプローチは、mAb 12QVQ/QSV及びmAb 15EVQの結合部位が互いに離れており、したがって一方の部位における点変異が他方の部位のエピトープに影響する可能性が低いことから実行可能であった。これらの複合体をELISA結合アッセイで使用した。PBSに加えた20μg/mL野生型TLR3 ECD又は変異タンパク質を各ウェル当たり5μLずつ、MSD HighBindプレート(Meso Scale Discovery,Gaithersburg,MD)にコーティングした。プレートを室温で60分間インキュベートし、ブロッキングした。
【0183】
【表11】
【0184】
相補的配列を有するアンチセンスオリゴヌクレオチドを使用し、MSDBlockerA緩衝液(Meso Scale Discovery,Gaithersburg,MD)で4℃で一晩変異誘発反応を行った翌日、プレートを洗浄し、MSD Sulfo−タグ標識したmAb 15EVQを500pM〜1pMの濃度で1.5時間添加した。洗浄後、MSD Read緩衝液Tを使用して標識抗体を検出し、SECTOR Imager 6000を使用してプレートを読み取った。ヒトTLR3及び変異体に対するmAb 12QVQ/QSVの結合活性を評価するため、mAb 15EVQと同時発現させ、検出抗体を標識mAb 12QVQ/QSVとした以外はmAb 15EVQについて述べたのと同様にして結合ELISAを行った。
【0185】
mAb 12QVQ/QSV:mAb 12QVQ/QSVの結合部位は、セグメント交換実験において決定されたようにヒトTLR3タンパク質のアミノ酸1〜209内に位置が特定された。以下のTLR3変異体について評価を行った:D116R、N196A、N140A、V144A、K145E、K147E、K163E、及びQ167A。野生型TLR3及びV144A変異体は、mAb 12QVQ/QSVに対して同等の結合性を示した(図6A)。抗体はTLR3 D116R変異体には結合せず、K145E変異体に対しては大幅に低い結合親和性を示した。したがって、TLR3上で近接して配置されている残基D116及びK145は、mAb 12QVQ/QSVに対する中心的なエピトープ部位と特定された(図7A)。
【0186】
mAb 12QVQ/QSV結合エピトープのこれら2つの重要な残基は、TLR3の外部ドメインのN末端セグメントの2本鎖RNA結合部位の面の近くに配置されている(Pirherら、Nature Struct.& Mol.Biol.,15:761〜763,2008)。完全なエピトープは、隣接領域に、実施した変異分析によっては明らかにされなかった他の残基を含むと考えられる。いずれの理論に束縛されることも望むものではないが、mAb 12QVQ/QSVがそのTLR3エピトープに結合すると、TLR外部ドメインへの2本鎖RNAの結合に直接的又は間接的に干渉することにより、受容体の2量化及び下流のシグナル伝達経路の活性化が妨害されるものと考えられる。
【0187】
mAb 15EVQ:以下のTLR3変異体について評価を行った。すなわち、R64E、K182E、K416E、Y465A、K467E、R488E、R489E、N517A、D536A、D536K、Q538A、H539A、H539E、N541A、E570R、K619A、K619E、2重変異体K467E/Y468A、3重変異体T472S/R473T/N474S、及び3重変異体R488E/R489E/K493E。野生型TLR3、R64E、K182E、K416E変異体及び3重変異体T472S/R473T/N474Sは、mAb 15EVQに対して同等の結合性を示した(図6B及び表5b)。抗体は、TLR3変異体K467E、R489E、K467E/Y468A及びR488E/R489E/K493Eには結合しなかった(図6B及び6C)。残りの変異体は中間的な結合性を示し、R488Eが最も高い作用を示した。これらの変異体のすべてがmAb 12QVQ/QSVに結合した。これらの結果は、残基K467及びR489がmAb 15EVQエピトープの重要な決定因子であることを示すものである。残基R488もエピトープに寄与した。これらの残基は、TLR3の同じ表面上に近接して配置されていた(図7A)。この結果はまた、いずれもK467、R488及びR489と同じ表面上に位置する残基Y465、Y468、N517、D536、Q538、H539、N541、E570、及びK619がエピトープに寄与することを示している。この結果は、mAb 15EVQを用いたH/D交換実験によって更に支持された。図7Aは、mAb 12QVQ/QSV及び15EVQ(黒)に対する結合エピトープ部位と、C1068 mAb(灰色)に対する結合エピトープ部位とがヒトTLR3の構造上で重なっていることを示している。mAb 15EVQに対するエピトープは、残基Y465、K467、Y468、R488、R489、N517、D536、Q538、H539、N541、E570、及びK619の範囲にわたっている。
【0188】
H/D交換実験H/D交換に対しては、抗体摂動の解析に使用した手順は、以前に述べたもの(Hamuroら、J.Biomol.Techniques 14:171〜182,2003;Hornら、Biochemistry 45:8488〜8498,2006)に類似し、若干の改変を加えたものである。組換えTLR3 ECD(C末端Hisタグを付加してSf9細胞で発現させて精製したもの)を重水素化水溶液中で所定の時間インキュベートして、交換可能な水素原子を重水素と交換した。重水素化TLR3 ECDを、固定されたmAb 15EVQを含むカラムで捕捉した後、水性緩衝液で洗浄した。逆交換した(back-exchanged)TLR3 ECDタンパク質をカラムから溶出し、重水素含有フラグメントの局在性をプロテアーゼ消化及び質量分析により調べた。参照対照として、TLR3 ECD試料を、抗体カラムで捕捉した後に重水素化水と接触させてから実験試料と同様にして洗浄及び溶出した点を除いて同様に処理した。抗体と結合する領域は比較的交換から保護される部位であり、そのため、基準TLR3 ECD試料と比較して高い割合の重水素を含むものと推測された。タンパク質の約80%を特定のペプチドにマッピングすることができた。mAb 15EVQによるTLR3 ECDのH/D交換の攪乱のマップを図7Bに示す。分かりやすさのために、TLR3の、mAb 15EVQに影響される部分の周囲のセグメントのみを示した。TLR3 ECDのアミノ末端及びカルボキシル末端に延びる、タンパク質の残りの部分は大きく影響されなかった。
【0189】
H/D交換実験により、配列番号2のペプチドセグメント465YNKYLQL471、514SNNNIANINDDmL526及び529LEKL532が、mAb 15EVQとの結合によってTLR3上の交換が特に変化する領域として特定された。その特性上、H/D交換は線形マッピング法であり、ペプチド内のどの残基が抗体結合によって最も影響されるかを定義することは通常できない。しかしながら、H/D交換と変異の結果が大きく重なっていることは、図7Aに示される表面がmAb 15の結合部位であることの更なる確証を与えるものである。この結合部位は、mAb c1068について過去に述べられているもの(国際公開第06/060513(A2)号)と同じ直線状アミノ酸配列領域内にあったが、まったく重ならない表面(図7A)上に位置することが判明し、これらの抗体間で交差競合が見られないことと一致する。
【0190】
mAb 15EVQ結合エピトープは、TLR3上のC末端セグメントにおいて二本鎖RNA結合部位に空間的に近位であった(Bellら、Proc.Natl.Acad.Sci.(USA)103:8792〜8797,2006;Ranjith−Kumarら、J.Biol.Chem,282:7668〜7678,2007;Liuら、Science 320:379〜381,2008)。いずれの特定の理論に束縛されることも望むものではないが、mAb 15EVQがそのTLR3エピトープと結合すると、リガンドである2本鎖RNA分子及び/又は2量体のもう一方との立体障害を生じ、リガンドの結合及びリガンドによって誘導される受容体の2量化が妨げられるものと考えられる。
【0191】
【表12】
【0192】
(実施例7)
熱安定性が向上した変異体の作製
結晶構造に基づいた機能改変(Structure−based engineering)を行って高い熱安定性を有する抗体の変異体を作製すると同時に、生物活性を維持しつつ免疫原性を最小に抑えようと試みた。
【0193】
mAb 15EVQを機能改変用に選択した。免疫原性を最小に抑えるため、構造的考慮に基づいて有益であることが予想される生殖細胞の変異のみを行った。mAb 15EVQのVL及びVHの配列(それぞれ配列番号41及び配列番号216)を、BLASTサーチを使用してヒト生殖細胞の遺伝子と並べた。同定された最近接の生殖細胞系列配列は、VH及びVLに対してそれぞれGenBank Acc.No.AAC09093及びX59318であった。生殖細胞のVH及びVLとmAb 15EVQのVH及びVL配列との間で以下の違いが特定された。すなわち、(VH)V34I、G35S、F50R、A61S、及びQ67H;(VL)G30S、L31S、及びA34N。特定された配列の違いをmAb 15EVQの結晶構造上にマッピングし、パッキング及び界面相互作用を変化させることが予測される残基を機能改変用に選択した。抗体の結晶構造(実施例6を参照)に基づいて、潜在的に構造を不安定にし得る残基を同定した。(1)小さな囲われた空洞がV34の近くのVHのコア中で同定された。この空洞は、Ileのようなわずかに大きな側鎖が収まるだけの充分な大きさを有していた。(2)VH CDR3のE99は、水素結合ネットワークに依らずVH/VL界面に埋め込まれていた。E99の負に帯電したカルボキシレート基は概ね疎水性環境にあり、周囲の残基と主としてファンデルワールス(vds)力による接触状態にあった。荷電基を埋め込むことは通常はエネルギー的に好ましくなく、したがって不安定化作用をもたらす。(3)VHのF50は、VH/VLの界面残基である。その芳香族側鎖は嵩張るため、ペアリングに負の影響を及ぼし得る。Fvの水素結合及びvdwパッキングネットワークを計算し、Pymolにより視覚的に調べた(www://_pymol_org)。VH及びVLドメイン内の埋め込まれた空洞はCaver(Petrekら、BMC Bioinformatics、7:316、2006)によって計算した。分子のグラフィック図をすべてPymolで作製した。QuickChange II XL部位特異的変異誘発キット(Stratagene,San Diego,CA)、Change−IT多重変異部位特異的変異誘発キット(USB Corporation,Cleveland,OH)、又はQuick Change II部位特異的変異誘発キット(Stratagene,San Diego,CA)を使用し、標準的なクローニング法によって、実施例3で述べたようにして作製したFabフラグメント又はIgG4完全ヒト抗体をコードする発現ベクターに変異を導入した。反応を、それぞれの製造者の推奨に従って行った。得られたクローンの配列を、確認のために配列決定し、得られた機能改変された変異体を、改変された重鎖又は軽鎖に従ってmAb s15−1〜15−10と命名した。mAb 15−10用の重鎖がmAb 15−6からのものであることを除いて、それぞれの変異体鎖(H又はL)を野生型mAb 15EVQL又はH鎖により発現させて、抗体を作製した。配列番号のリスト化:CDRに対して、mAb 15EVQ及びその機能改変された変異体に対する軽鎖及び重鎖並びに完全長重鎖及び軽鎖の可変領域を、表6に示す。表7は、それぞれの変異体を作製するためのプライマーを示す。
【0194】
【表13】
【0195】
TLR3に対するmAb 15−1〜15−9の結合性をELISA免疫アッセイによって評価した。ヒトTLR3 ECD(100μLの2μg/mL TLR3 ECD)を黒色のMaxisorbプレート(eBioscience)に4℃で一晩結合させた。プレートを洗浄し、ブロッキングし、希釈した抗体を各ウェル当たり50μLで二つ組複製で分注した。プレートを穏やかに振盪しながら室温で2時間インキュベートした。ルミネッセンスPOD基盤(Roche Applied Science,Mannheim,Germany,カタログ番号11 582 950 001)及びヤギ抗ヒトFc:HRP(Jackson ImmunoResearch,West Grove,PA,Cat.No.109−035−098)を用いて、結合を検出し、プレートをSpectraMaxプレートリーダー(Molecular Devices,Sunnyvale,CA)で読み取った。
【0196】
DSC実験を、MicroCalの自動VP−キャピラリーDSCシステム(MicroCal’s,LLC,Northampton,MA)で行って、基準細胞と試料細胞との間の温度差を継続的に測定して出力単位に較正した。試料は10℃〜95℃まで60℃/時の加熱速度で加熱した。前走査時間は15分とし、フィルタリング時間は10秒間とした。DSC実験で使用した濃度は約0.5mg/mLである。得られたサーモグラムの分析を、MicroCal Origin 7ソフトウェア(MicroCal,LLC)を用いて行った。
【0197】
【表14】
【0198】
作製した変異体の熱安定性(Tm)をDSCによって測定した(表8)。TLR3に対する抗体の変異体の結合性は親抗体の結合性と同等であった。
【0199】
【表15】
【0200】
(実施例8)
代替抗TLR3抗体の作製
実施例1で作製したヒト化抗体がマウスTLR3に対して充分な特異性もアンタゴニスト活性も有さなかったことから、本明細書ではmAb 5429と命名するキメラアンタゴニストのラット/マウス抗マウスTLR3抗体を作製して、さまざまなインビボモデルにおいてTLR3シグナル伝達を阻害する作用について評価を行った。この代替キメラmAb 5429及びその親ラット抗マウスTLR3抗体c1811はインビトロ及びインビボでマウスのTLR3シグナル伝達を阻害し、マウスのいくつかの疾患モデルにおいて発症機構を改善した。
【0201】
以下に考察するデータは、有害な炎症の誘発及び持続化におけるTLR3の役割を示唆するものであり、例えば高サイトカイン血症、喘息及び気道炎症、炎症性腸疾患及び関節リウマチ、ウイルス感染、及びII型糖尿病などの急性及び慢性の炎症状態におけるTLR3アンタゴニスト及びTLR3抗体アンタゴニストの治療的使用の根拠を与えるものである。
【0202】
代替mAb 5429の作製
通常の方法を用いて生成させた、遺伝子組み換え型マウスTLR3の外部ドメイン(配列番号162のアミノ酸1〜703、GenBank Acc.No.NP_569054)により、CDラットに免疫性を与えた。マウスTLR3に特異的な抗体力価を示す2匹のラット由来のリンパ球を、FO骨髄腫細胞に融合した。マウスTLR3に反応性のあるモノクローナル抗体のパネルを同定し、マウスルシフェラーゼレポーター及びマウス胎児線維芽細胞アッセイにおいて、インビトロアンタゴニスト活性について試験した。ハイブリドーマ株C1811Aを更なる研究のために選択した。ハイブリドーマによって分泌されるmAb c1811から、機能性可変領域の遺伝子を配列決定した。次いで、クローニングされた重鎖及び軽鎖可変領域遺伝子をプラスミド発現ベクターにそれぞれ挿入し、mAb 5429と表示されるキメララット/Balb C muIgG1/κ mAbを常法によって作製するためのコード配列を得た。抗体を実施例3で述べたようにして発現させた。mAb 5429の重鎖及び軽鎖可変領域のアミノ酸配列を配列番号164及び配列番号163にそれぞれ示し、重鎖及び軽鎖の完全長の配列を配列番号166及び配列番号165にそれぞれ示す。mAb c1811の重鎖及び軽鎖の完全長の配列を配列番号168及び配列番号167にそれぞれ示す。
【0203】
mAb 5429の特性評価
mAb 5429を、TLR3シグナル伝達に対する中和能力について一連のインビトロアッセイにおいて特性評価した。活性のアッセイ及び結果を以下に述べる。
【0204】
マウスルシフェラーゼレポーター遺伝子アッセイ
マウスTLR3のcDNA(配列番号161、GenBank受諾番号NM_126166)をマウス脾臓cDNA(BD Biosciences,Bedford,MA)からPCRによって増幅し、標準的な方法を用いてpCEP4ベクター(Life Technologies,Carslbad,CA)にクローニングした。200μLのHEK293T細胞を、完全DMEM中、4×104細胞/ウェルの濃度で96穴白色透明底プレートに播種し、30ngのpNF−κBホタルルシフェラーゼ(Stratagene,Carslbad,CA)又は30ngのpISREホタルルシフェラーゼ(BD Biosciences,Bedford,MA)、5ngのphRL−TKコントロールウミシイタケルシフェラーゼ(Promega Corp.,Madison,WI)レポータープラスミド、1.5ngの完全長マウスTLR3をコードするpCEP4、及び13.5ngの空のpcDNA3.1ベクター(Life Technologies,Carslbad,CA)を使用して、翌日Lipofectamine 2000(Invitrogen Corp.,Carslbad,CA)を用いて全DNA量を50ng/ウェルとし、トランスフェクションに使用した。トランスフェクションの24時間後、細胞を抗マウスTLR3抗体と新鮮な無血清DMEM中で37℃で30分〜1時間インキュベートした後、0.1又は1μg/μLのポリ(I:C)を加えた。24時間後にDual−Gloルシフェラーゼアッセイシステム(Promega,Madison,WI)を使用してプレートを回収した。相対的な光量単位を、FLUOstar OPTIMA多重検出リーダーを使用し、OPTIMALソフトウェア(BMG Labtech GmbH,Germany)により測定した。標準化された値(ルシフェラーゼ比)はホタルルシフェラーゼの相対光量単位(RLU)をウミシイタケルシフェラーゼのRLUで割ることによって得た。mAb 5429、並びにその親mAb c1811及びmAb 15(表3a)が、用量依存的にポリ(I:C)誘発NF−κB及びISRE活性化を低下させたことから(図8A及び8B)、TLR3の活性を中和するこれらの抗体の能力が実証された。ISREアッセイにおいて測定されたIC50は、mAb 5249、mAb 15及びmAb c1811についてそれぞれ0.5、22、及び0.7μg/mLであった。
【0205】
マウス胚性線維芽細胞(MEF)アッセイ
C57BL/6 MEF細胞をArtis Optimusより入手した(Opti−MEF(商標)C57BL/6−0001)。200μLのMEF培地(glutamax、10%熱失活FBS、1×NEAA、及び10μg/mLゲンタマイシンを加えたDMEM)中の細胞を、96穴平底プレート(BD Falcon)に20,000細胞/ウェルで播種した。インキュベーションはすべて、37℃/5%CO2で行った。播種の24時間後に、mAb 5429又はmAb c1811を各ウェルに加えた。プレートをmAbと1時間インキュベートした後、各ウェルにポリ(I:C)を1μg/mL加えた。24時間のインキュベーションの後、上澄み液を回収した。ビーズキット(Invitrogen Corp.,Carslbad,CA)を用い、サイトカインレベルを定量して、CXCL10/IP−10を製造者のプロトコールに従って検出した。結果をGraph Pad Prismソフトウェアを使用してグラフ化した。いずれの抗体もポリ(I:C)誘発CXCL10/IP−10レベルを用量依存的に低下させたことから、これらの抗体が、内因性のTLR3を中和してTLR3シグナル伝達を阻害する能力が実証された(図9)。
【0206】
フローサイトメトリー−表面染色
各群10匹ずつのC57BL/6及びTLR3ノックアウトマウス(TLR3 KO)(C57BL/6バックグラウンド;雌、8〜12週齢、Ace Animals,Inc.)に1mLの3%チオグリコレート培地(Sigma)を腹腔内投与し、96時間後にマウスを安楽死させ、各マウスから得た腹膜を10mLの滅菌PBSで洗浄した。チオグリコレートにより腹膜に誘導されたマクロファージをPBS中に再懸濁し、細胞の生存率をTrypan Blue染色によって評価した。細胞を遠心してペレットとし、250μLのFACS緩衝液(PBS−Ca2+−Mg2+、1%熱失活FBS、0.09%アジ化ナトリウム)に再懸濁し、濡れた氷上に保持した。CD16/32試薬(eBioscience)を10μg/106細胞に10分間使用してマクロファージ上のFc受容体をブロッキングした。細胞を100μL/ウェル中、106個細胞で分配して、表面染色を行った。Alexa−Fluor 647(Molecular Probe)−接合mAb c1811及びmAb 1679(TLR3特異性を有さないことからアイソタイプコントロールとして用いられるラット抗マウスTLR3抗体)を0.25μg/106個細胞で加え、氷上、暗所で30分間インキュベートした。細胞を洗浄し、250μLのFACS緩衝液に再懸濁した。生存率染色として7−AAD(BD Biosciences,Bedford,MA)を5μL/ウェルで30分を超えないように加えた後、FACS Caliburにより試料を取得して死細胞集団を検出した。Cell Quest Proソフトウェアを使用してFACS Caliburにより、試料を採取した。収集したデータを、FCS Expressを使用してヒストグラムを作成することによって解析した。
【0207】
C57BL/6及びTLR3KOマウスから得た、チオグリコレートで誘発されたマウス腹膜マクロファージへのmAb c1811の結合を、フローサイトメトリーにより評価して結合特異性を求めた。このキメラ抗体のマウスFc領域は、非特異的結合に寄与すると予期されたので、mAb 5429はこのアッセイで使用しなかった。mAb c1811は、TLR3KOマクロファージへの結合を示さず、C57BL/6腹膜マクロファージの細胞表面への結合を上昇させたことから、TLR3に対するmAbの特異性を示唆した。(図10)mAb 5429は、mAb c1811と同一の結合領域を有し、mAb c1811と同一の結合特異性を有すると推定される。
【0208】
(実施例9)
TLR3抗体アンタゴニストはTLR3介在型の全身性炎症を防御する
モデル
ポリ(I:C)誘発全身性サイトカイン/ケモカインモデルを、TLR3介在性全身性炎症のモデルとして用いた。このモデルでは、ポリ(I:C)(PIC)を腹腔内投与することで、TLR3によって一部介在される全身性のサイトカイン及びケモカイン応答を誘導した。
【0209】
雌C57BL/6マウス(8〜10週齢)又は雌TLR3ノックアウトマウス(C57BL/6バックグラウンド;8〜10週齢、Ace Animals、Inc.)に、mAb 5429を0.5mLのPBS中、10、20又は50mg/kgで、mAb c1811を、0.5mLのPBS中、2、10又は20mg/kgで、又は0.5mLのPBSを単独(溶媒コントロール)で皮下投与した。抗体投与の24時間後、マウスに、0.1mLのPBSに加えた50μgのポリ(I:C)(Amershamカタログ番号26−4732、ロット番号IH0156)を腹腔内投与した。ポリ(I:C)チャレンジの1及び4時間後に後眼窩より血液を採取した。全血より血清を調製してLuminexによりサイトカイン及びケモカインの濃度について解析した。
【0210】
結果
腹腔内投与されたポリ(I:C)は、TLR3ノックアウト動物においてケモカイン及びサイトカイン群の産生が大幅に低減されたことによって示されるように、TLR3によって一部介在される全身性のサイトカイン及びケモカイン応答を誘導した(表9A)。TLR3依存性のポリ(I:C)誘導型メディエーターは、ポリ(I:C)ポストチャレンジ1時間では、IL−6、KC、CCL2/MCP−1及びTNF−αであり、ポリ(I:C)ポストチャレンジ4時間では、IL−1α、CCL5/RANTES及びTNF−αであった。mAb c1811及びmAb 5429は、両方ともこれらのTLR3依存性のメディエーターのレベルを著しく低下させ、抗体がインビボでTLR3シグナル伝達を低下させる能力を実証した(表9B)。表9中の値は、6匹の動物/群±SEMのpg/mLでの平均のサイトカイン又はケモカイン濃度として示したものである。これらのデータは、TLR3の拮抗作用が、サイトカインストーム又は致死的ショックなどの状態における過剰なTLR3介在サイトカイン及びケモカインレベルを低減させるうえで有用となり得ることを示唆するものである。
【0211】
【表16】
【0212】
【表17】
【0213】
(実施例10)
TLR3抗体アンタゴニストは気道過敏症を軽減する。
モデル
気道過敏症をポリ(I:C)によって誘発させた。
【0214】
雌C57BL/6マウス(12週齢)又は雌TLR3ノックアウトマウス(C57BL/6バックグラウンド;12週齢、Ace Animals,Inc.)をイソフルランで麻酔し、50μLの滅菌PBS中のポリ(I:C)を数回投与量(10〜100μg)で鼻腔内投与した。各投与の間に24時間の休止時間を置いてポリ(I:C)(又はPBS)を3回マウスに投与した。最後のポリ(I:C)(又はPBS)の投与の24時間後に、全身プレチスモグラフィー(BUXCOシステム)を用いて肺機能及びメタコリンに対する気道過敏症を測定した。マウスを全身プレチスモグラフチャンバに入れ、少なくとも5分間順応させた。ベースラインの読み取りの後、投与量を増大させた霧状化したメタコリン(Sigma,St.Louis、MO)にマウスを曝露した。霧状化メタコリンを2分間投与した後、5分間のデータ収集時間、その後、10分間の休止時間を置いた後、メタコリンの用量を増大させた後続のチャレンジを行った。空気流の抵抗の増加をエンハンストポーズ(Enhanced Pause)(Penh)として測定し、5分間の記録時間にわたった平均のPenh値として表した(BUXCOシステム)。肺機能の測定の後、マウスを安楽死させ、肺にカニューレを挿管した。肺に1mLのPBSを注入し、流出液を回収することによって気管支肺胞洗浄(BAL)を行った。肺組織を切除して凍結した。BAL液を遠心(1200rpm、10分)し、細胞を含まない上澄み液を回収して、解析を行うまでの間−80℃で保存した。細胞ペレットを200μLのPBSに再懸濁して全細胞計数及び分画別の細胞計数を行った。製造者のプロトコール及びMultiplex Immunoassayキット(Millipore,Billercia,MA)に従って多重アッセイを行った。
【0215】
結果
過去の観察により、ポリ(I:C)の鼻腔内投与はマウスの肺機能にTLR3によって介在される障害を誘発し、全身プレチスモグラフィー(BUXCO)におけるベースラインのエンハンストポーズ(PenH)測定値が高くなり、霧状化されたメタコリンに対する応答性が高くなることが示されている(国際公開第06/060513(A2)号)。肺機能のこうした障害は、肺への好中球の動員、及び肺における炎症性サイトカイン/ケモカインのレベルの上昇をともなった。この実験では、肺機能におけるポリ(I:C)誘発障害におけるmAb 1811及びmAb 5429の作用を、ポリ(I:C)チャレンジに先立ってそれぞれの抗体を50mg/kgで皮下投与することによって評価した。
【0216】
TLR3によって介在される肺機能の障害は、ポリ(I:C)チャレンジに先立って動物をTLR3抗体アンタゴニストで処置することによって大幅に軽減された。抗TLR3抗体で処置された動物ではTLR3によって介在される、ベースラインPenH及びメタコリンに対する気道の感受性の増大を防止した(図11)。更に、抗TLR3抗体で処置した動物ではTLR3によって介在される、マウスの肺への好中球の動員及び気道におけるケモカインの生成を低減した。好中球の数(図12)及びCXCL10/IP−10のレベル(図13)は、回収した気管支肺胞洗浄液(BALF)から測定した。この実験を少なくとも3回繰り返して同様の結果を得た。図11、12、及び13に示されるデータは、1つの代表的な実験からのものである。各記号は1匹のマウスからのデータポイントを表し、横棒線は群の平均を示す。この実験によって、使用したモデルにおいて、全身投与されたTLR3抗体アンタゴニストが肺に達し、TLR3によって介在される肺機能の障害、気道内への好中球の浸潤、ケモカインの生成及び気道の炎症を軽減したことが示された。したがって、TLR3アンタゴニストは、喘息、アレルギー性鼻炎、慢性閉塞性肺疾患(COPD)、及び嚢胞性線維症などの、気道過敏症によって特徴付けられる呼吸器疾患の治療又は予防に有用であり得る。
【0217】
(実施例11)
TLR3抗体アンタゴニストは炎症性腸疾患を防御する。
モデル
DSS大腸炎モデルを炎症性腸疾患のモデルとして用いた。
【0218】
雌C57BL/6マウス(8週齢未満)又は雌TLR3ノックアウトマウス(C57BL/6バックグラウンド;8週齢未満で体重16.5g〜18g、Ace Animals,Inc)にγ線照射した餌を1日目から与えた。DSS(硫酸デキストラン)(MP Biomedicals,Aurora,OH,カタログ番号160110;35〜50kDa;18〜20%イオウ,ロット番号8247J)を、オートクレーブにかけた酸性化した飲料水中で5%の最終濃度まで希釈した。DSS水を5日間投与し、その後DSS水をただの水に置き換えた。マウスには実験全体を通じて自由に水を飲ませた。すべての水ボトルの重量を毎日測定して水の消費量を記録した。0、2、及び4日目にマウスに5mg/kg(0.1mL PBSに0.1mgを加えた)のmAb 5429、マウス抗TNF−α抗体、又はコントロールとしてのPBSを腹腔内投与した。マウスを実験の全体を通じて毎日監視し、0〜4日目及び7日目に体重を測定した。マウスを実験の2日目及び7日目に安楽死させた。腹腔を開き、上行結腸を盲腸と連結する位置で切断した。結腸を採取し、10%中性緩衝ホルマリン中で固定した。結腸をパラフィンに埋め込み、薄片にスライスし、H&E染色(Qualtek Molecular Labs,Santa Barbara,CA)した。下記に述べるように獣医病理学者によって結腸の組織病理学的評価を盲検により行った(PathoMetrix,San Jose,CA)。
【0219】
組織病理学的評価
大腸、結腸、及び直腸の2つの切片を評価し、以下の変化についてスコア化した。すなわち、(i)単一の細胞の壊死、(ii)上皮の潰瘍形成、(iii)上皮の脱落、(iv)陰窩腫瘍、(v)細胞増殖、(vi)陰窩細胞増殖、(vii)粘膜固有層における肉芽組織形成、(viii)粘膜下組織における肉芽組織、(ix)粘膜下炎症性細胞の浸潤物、好中球優位性、及び(x)粘膜下浮腫。
【0220】
重篤度の単一の全体的スコアを以下の基準に基づいて与えた。すなわち、
0−所見を認めず。
1−軽度、限局性、又は時折認められる。
2−軽度、多病巣である。
3−中度、頻繁だが限定された領域に認められる。
4−重度、提供された組織の多くの領域又は広範囲に認められる。
5−極めて重度、提供された組織の大部分に拡がっている。
【0221】
結果
過去の観察によれば、TLR3ノックアウト動物は、DSSの摂取によって誘発された炎症性腸疾患のモデルにおいて、野生型マウスと比較して大幅に軽減された組織病理学的所見を示すことが実証されており(国際公開第06/60513(A2)号)、このことはTLR3シグナル伝達がこのモデルにおける病因に一定の役割を有することを示唆するものである。壊死細胞から放出される共生生物のバクテリアRNA又は哺乳類RNAは、内因性リガンドとして作用して、TLR3シグナル伝達を刺激することができ(Karikoら、Immunity 23165〜231175 2005;Karikoら、J.Biol.Chem.279:12542〜12550 2004)、それゆえ、腸中の内因性リガンドによるTLR3刺激は、DSS大腸炎モデルにおける炎症を増進させ、及び持続させ得るということが報告された。
【0222】
複合組織病理学スコア(図14)によって評価されるように、DSSに曝露した動物における疾患重篤度は、抗TLR3抗体による処置によって改善された。図14は、疾患重篤度スコアについて平均、標準偏差及び95%信頼区間を横棒線として示している。抗TLR3抗体で処置した野生型のDSS曝露動物では、非処置の野生型動物と比較してスコアの有意な低下が認められた(p<0.05)。DSSに曝露したTLR3ノックアウト動物は、DSSによって誘発される変化から防御された。抗マウスTNF−α mAbを投与したDSS曝露動物では、DSSモデルにおいて組織病理学的所見の改善は認められなかった。したがって、DSSモデルは、抗TNF−αによる治療に応答しないヒト患者集団を対象とする治療法の評価において有用であり得、抗TLR3抗体を中和することは、抗TNF−αによる治療に応答しない炎症性腸疾患の患者にとって利益となる可能性を有すると考えられる。
【0223】
モデル
T細胞移植モデルを炎症性腸疾患のモデルとして用いた。このモデルでは、免疫応答能のあるマウスから得た制御性T細胞を含まないナイーブT細胞(腸粘膜の抗原提示細胞を攻撃する)の集団を移植することによって、SCIDマウスに腸の炎症を誘発した。
【0224】
ナイーブT細胞(CD4+CD45RBhighT細胞)をSCIDレシピエントに腹腔内注射することによって慢性大腸炎を誘発した。マウスに、PBS(500μL/マウス、腹腔内投与;溶媒コントロール)、mAb 5429(0.1mg/マウス、腹腔内投与)、又は抗TNF−α抗体(0.05mg/マウス、腹腔内投与;ポジティブコントロール)のいずれかを、T細胞の移植48時間後から開始して毎週2回、8週間の実験期間全体を通じて投与した。T細胞の移植から8週間後(又はマウスが初期の体重の15%以上を失った時点)に、動物を安楽死させて結腸を取り出した。結腸を固定し、パラフィンに埋め込んでH&E染色した。組織病理学的所見(細胞浸潤、陰窩腫瘍、上皮びらん、杯細胞喪失、及び腸壁の肥厚)を盲検的に定量的に評価した。
【0225】
結果
コントロール動物と比較した場合の組織病理学的スコアの合計の有意な低下(p<0.05)(図15A)によって評価されるように、疾患重篤度は、T細胞移植を受けた動物において抗TLR3抗体による処置に応じて改善された。スコアの合計には、陰窩腫瘍、潰瘍形成、好中球流入、杯細胞喪失、異常陰窩、粘膜固有層の炎症、及び経壁併発(transmural involvement)を評価した。陰窩腫瘍、潰瘍形成、及び好中球流入では有意な低下が認められた(いずれもp<0.05)(図15B)。抗TNFα抗体を、最適な効果を与えることが知られている用量でポジティブコントロールとして用いた。
【0226】
DSS及びT細胞移植モデルという2つの広く知られた炎症性腸疾患のモデルを用いた実験により、全身投与されたTLR3抗体アンタゴニストは腸粘膜に達し、2つの異なる病因機構によって誘発された消化管の炎症を軽減させることが実証された。したがって、TLR3アンタゴニストは、抗TNFα不応性の症例及び消化管の他の免疫介在性の病態を含む炎症性腸疾患の治療に有益であると考えられる。
【0227】
(実施例12)
TLR3抗体アンタゴニストはコラーゲン誘導性関節炎を防御する。
モデル
コラーゲン誘導性関節炎(CIA)モデルを関節リウマチのモデルとして用いた。
【0228】
雄B10RIIIマウス(6〜8週齢、Jackson Labs)を1群当たり15匹(関節炎群)又は1群当たり4匹(コントロール群)の群に分けた。関節炎群をイソフルランで麻酔し、II型コラーゲン(Elastin Products)、及び結核菌(M.tuberculosis)(Difco)を添加したフロイントの完全アジュバントを0及び15日目に投与した。12日目にII型コラーゲン誘導関節炎を発症したマウスを体重により治療群にランダム化し、mAb 5429(25mg/kg)、ネガティブコントロール抗体としてCVAM(マウスに既知の特異性を示さない組換えmAb)(5mg/kg)、又は抗TNF−α抗体(5mg/kg、ポジティブコントロール)を、12、17、及び22日目(d12、d17、d22)に皮下投与(SC)した。更に、マウスのコントロール群に溶媒(PBS)又はデキサメタゾン(0.5mg/kg、Dex、基準化合物)を12〜25日目まで毎日(QD)皮下投与(SC)した。動物を12〜26日まで毎日観察した。前肢及び後肢を臨床スコアシステム(下記に示す)によって評価した。実験26日目に動物を安楽死させ、組織病理学的所見を盲検的に評価した(下記にスコアシステムを述べる)。有効性評価は、動物の体重及び臨床的な関節炎スコアに基づいて行った。すべての動物が実験終了時まで生存した。
【0229】
前肢及び後肢の臨床的スコアの判定基準
0−正常
1−後肢又は前肢関節が冒される、すなわちわずかなびまん性紅斑及び腫脹を呈する。
2−後肢又は前肢関節が冒される、すなわち軽度のびまん性紅斑及び腫脹を呈する。
3−後肢又は前肢関節が冒される、すなわち中度のびまん性紅斑及び腫脹を呈する。
4−顕著なびまん性紅斑及び腫脹、すなわち4本の指関節が冒される。
5−足全体に重篤なびまん性紅斑及び重篤な腫脹を呈し、指を曲げられない。
【0230】
II型コラーゲン誘導関節炎を有するマウス関節の組織病理学的スコアリング方法
II型コラーゲン誘導関節炎の病変を有するマウスの肢又は足首関節をスコアリングする場合、変化の重篤度及び冒された個々の関節の数を考慮する。中手骨/中足骨/指又は足根骨/脛足根骨の多数の可能性のある関節のうち、1〜3個のみの足又は足首の関節が冒されている場合には、変化の重篤度に応じて下記のパラメータの1、2又は3の最大スコアが任意に与えられる。2個よりも多い関節が関与している場合には、最も重く冒された/大多数の関節に下記の基準を適用する。
【0231】
足スコアの臨床データは、1〜15日目についてAUCによって分析し、コントロールから阻害率(%)を計算した。
【0232】
炎症
0−正常
1−冒された関節の滑膜及び関節周囲組織に炎症性細胞が最小限に浸潤している。
2−軽度の浸潤を呈している。足の場合には、冒された関節に限定された軽度の浸潤を呈している。
3−中度の浸潤を呈している。足の場合には、冒された関節に限定された中度の浸潤を呈している。
4−大部分の領域が顕著な浮腫をともなう顕著な浸潤を呈している。
5−重篤な浮腫をともなう重篤なびまん性浸潤を呈している。
【0233】
パンヌス
0−正常
1−軟骨及び肋軟骨下骨にパンヌスが最小限に浸潤している。
2−冒された関節の硬組織の周辺帯の破壊をともなう浸潤を呈している。
3−冒された関節の中度の高組織破壊をともなう中度の浸潤を呈している。
4−大部分の関節で関節構造の顕著な破壊をともなう顕著な浸潤を呈している。
5−関節構造の全体的又はほぼ全体的な破壊をともなう重篤な浸潤がすべての関節を冒している。
【0234】
軟骨の傷害
0−正常
1−トルイジンブルー染色が最小限〜軽度に損なわれている。冒された関節に明らかな軟骨細胞の損失もコラーゲン破壊も認められない。
2−トルイジンブルー染色が軽度に損傷を受けている。冒された関節に限局性の軽度(表在性)の軟骨細胞の損失及び/又はコラーゲン破壊が認められる。
3−トルイジンブルー染色が中度に損傷を受けている。冒された関節に多病巣性の中度(深部又は中間層)の軟骨細胞の損失及び/又はコラーゲン破壊が認められる。
4−トルイジンブルー染色が顕著に損傷を受けている。大部分の関節に多病巣性の顕著な(深部〜深層)軟骨細胞の損失及び/又はコラーゲン破壊が認められる。
5−トルイジンブルー染色が重度に損傷を受けている。すべての関節に多病巣性の重度(深部〜タイドマーク)の軟骨細胞の損失及び/又はコラーゲン破壊が認められる。
【0235】
骨吸収
0−正常
1−小さい範囲の吸収が認められ、最小の程度に生じている。低倍率では容易に確認できず、冒された関節に破骨細胞をほとんど認めない。
2−より多くの範囲の吸収が認められ、軽度に生じている。低倍率では容易に確認できず、冒された関節により多くの破骨細胞が認められる。
3−皮質の厚さ全体の欠損をともなわない髄様骨梁(medullary trabecular)及び皮質骨の明らかな吸収が、中度に生じている。一部の髄様骨梁が損傷し、病変が低倍率で明瞭に確認できる。冒された関節により多くの破骨細胞が認められる。
4−残存する皮質表面の断面の歪みをしばしばともなう、皮質骨の厚さ全体の欠損が、顕著な程度に生じている。髄様骨の顕著な損失、多数の破骨細胞が認められる。大部分の関節が冒されている。
5−皮質骨の厚さ全体の欠損及びすべての関節の関節構造の破壊が認められる。重度。
【0236】
結果
デキサメタゾン(Dex)及び抗マウスTNF−α抗体をポジティブコントロールとして用い、PBSを溶媒コントロールとして用い、CVAMをネガティブコントロール抗体として用いた。すべての処置は、実験の12日目の関節疾患が進行する間に開始した。溶媒で処置した疾患コントロール動物では疾患発症率は実験22日目までに100%となった。溶媒又はCVAM抗体で処置したネガティブコントロール群では臨床スコアは最も高かった。Dex(d18〜d26についてp<0.05)、5mg/kgの抗TNF−α抗体(d18〜26についてp<0.05)、又は25mg/kgのmAb 5429(d18〜d23及びd25〜d26についてp<0.05)で処置した各群では有意に低い臨床スコアが認められた(図16)。曲線下面積(AUC)として表した臨床的関節炎スコアは、25mg/kgのmAb 5429(低下率43%)、5mg/kgの抗TNF−α抗体(52%)、又はDex(69%)による処置により、溶媒コントロールと比較して有意に低下した。図17は、各群についてAUCの平均及び標準偏差を示す。
【0237】
各処置の組織病理学的作用についても評価を行った。足の骨吸収は、25mg/kgのmAb 5429(低下率47%)による処置によって溶媒コントロールと比較して有意に低下した。5mg/kgの抗TNF−α抗体で処置したポジティブコントロールマウスでは、足の炎症(33%)、軟骨傷害(38%)、及び合計した足スコア(37%)は有意に低下した。Dexによる処置によって、足の組織病理学的パラメータのすべてが有意に低下した(合計スコアにおいて73%の低下率)。
【0238】
これらのデータは、TLR3抗体アンタゴニストが、CIAモデルにおいて臨床的及び組織病理学的疾患症状を改善することを示すものであり、関節リウマチの治療におけるTLR3アンタゴニストの使用を示唆するものである。
【0239】
(実施例14)
TLR3抗体アンタゴニストは急性致死性ウイルス感染を防御する。
モデル
インフルエンザAウイルスチャレンジモデルを急性致死性ウイルス感染のモデルとして使用した。
【0240】
1、4、8及び12日目に雌C57BL/6マウス(12週齢)又は雌TLR3ノックアウトマウス(C57BL/6バックグラウンド;12週齢、ACE Animals,Inc.、1群当たり15匹のマウス)に20mg/kgのmAb 5429、又はPBS単独を皮下投与した。0日目にイソフルランでマウスを麻酔し、インフルエンザA/PR/8/34ウイルス(ATCC、Rockland、MD、ロット番号218171)を25μLのPBSに加えたもの(105.55CEID50に相当)を鼻腔内投与した。動物を体重変化及び生存率について、14日間の期間にわたって1日2回観察した。臨床的スコアリングシステムを用いて疾患の進行のレベル及びインフルエンザAウイルス治療に対するわずかな改善を評価した。
【0241】
臨床スコア
0−正常。機敏で反応性があり、目に見える病気の兆候が認められない。
1−毛皮が逆立っている。歩行のわずかな低下が認められるか、又は認められない。
2−歩行時に背中が曲がっている。歩行をいやがる。呼吸困難である。
3−毛皮が逆立っている。呼吸困難である。運動失調。震えが生じている。
4−毛皮が逆立っている。そっと突いても歩行不可能。意識不明。触れると冷たく感じる。
5−死亡。
【0242】
結果
この実験では、生存率、毎日の臨床スコア、及び体重変化を評価した。mAb 5429(20mg/kg)を投与したインフルエンザA感染野生型マウス及びmAb 5429を投与しないインフルエンザA感染TLR3ノックアウトマウスのいずれも、インフルエンザウイルスを接種したC57BL/6マウスと比較して生存率が統計的に有意に増大した(それぞれp<0.001及びp<0.01)。このことは、TLR3の阻害又は欠損がインフルエンザによって誘発される死を予防し得ることを示すものである(図18)。臨床スコアは、20mg/kgのmAb 5429を投与した群、及びTLR3ノックアウト群において有意に低下した(図19)。マウスの体重をインフルエンザウイルス投与後14日間にわたって観察した。インフルエンザAウイルスを投与したC57BL/6マウスでは体重は安定的に減少した。しかしながら、20mg/kgのmAb 5429を投与したC57BL/6マウス及びTLR3ノックアウトマウスのいずれも、インフルエンザウイルスを接種した野生型C57BL/6マウスに対して有意に大きな体重を示した(図20)。これらの結果は、TLR3抗体アンタゴニストが急性致死性インフルエンザウイルス感染モデルにおける臨床症状及び死亡率を低下させることを示すものであり、TLR3アンタゴニストが急性の感染状態にあるヒトに防御を与えることを示唆するものである。
【0243】
(実施例15)
TLR3抗体アンタゴニストは高血糖を改善し、血症インスリンを低下させる。
モデル
食餌誘発型肥満(DIO)モデルを高血糖及びインスリン抵抗性、並びに肥満のモデルとして用いた。
【0244】
C57BL/6野生型動物(約3週齢、Jackson Labs)及びTLR3ノックアウト動物(C57BL/6バックグラウンド;約3週齢、Ace Animals,Inc.)に12〜16週間持続的に高脂肪の食餌を与えた。TLR3ノックアウト及び野生型C57BL/6マウスの両方に、通常の固形飼料又は60.9% kcalの脂肪及び20.8% kcalの炭水化物からなる高脂肪の食餌(Purina Test Dietカタログ番号58126)のいずれかを与えた。マウスには、水及び食餌を自由摂取させ、12時間/12時間の明暗サイクル中に維持した。各群の各マウスの体重を毎週測定した。mAb 5429を第1週に週2回、その後は全体で7週間にわたり、週1回腹腔内投与した。空腹時に後眼窩から採取した血清試料を、示した時点でのインスリンの測定に用いた。7週目に一晩絶食させた後、1.0mg/g(体重)のグルコースを腹腔内投与することによって耐糖試験を行った。更に、空腹時のインスリン及びグルコースレベルを測定した。
【0245】
以下の式を用いて、空腹時グルコースレベル及びインスリンレベル(12)に基づきHOMA−IRを式から求めた。すなわち、HOMA−IR=((空腹時グルコース(mmol/l)×空腹時インスリン(mU/l))/22.5(Wallaceら、Diabetes Care 27:1487〜1495、2004)。空腹時血中グルコース(血糖値)(BG)をグルコースオキシダーゼアッセイによって求めた。空腹時インスリンレベルをインスリンラット/マウスELISAキット(Crystal Chem、カタログ番号90060)を使用して求めた。
【0246】
結果
高脂肪の食餌を12〜16週間与えた後、野生型DIO動物は高血糖及び高インスリンとなった。mAb 5429による処置により、野生型DIO動物では耐糖能が改善したがTLR3ノックアウトDIO動物では改善が見られなかった。グルコースチャレンジの60、90、120及び180分後に、mAb 5429で処置した動物においてコントロール(PBSのみ)と比較して有意に低い血中グルコース濃度が観察された(図21A)。mAb 5429で処置した野生型DIO動物では、mAbを投与しない野生型DIOマウスと比較してAUCの約21%の低下が観察された。mAb 5429で処置した野生型DIO動物では空腹時インスリンレベルも低下した(図22)。TLR3ノックアウトDIO動物では、mAb 5429による処置により空腹時インスリンに改善は認められなかった。インスリン抵抗性指数(Homeostatic model assessment)(HOMA)分析は、mAb 5429で処置した野生型DIO動物においてインスリン感受性の改善を示したが、TLR3ノックアウトDIO動物では改善を示さなかった。HOMA−IR値は、野生型DIO、5mg/kgの野生型DIO(mAb 5429投与)、及び20mg/kgの野生型DIO(mAb 5429投与)動物で、それぞれ14.0±9.8、8.7±4.9、9.0±3.0であった。TLR3ノックアウトDIO動物では何の効果も認められなかった。
【0247】
この実験により、TLR3抗体アンタゴニストがDIOモデルにおいて体重の損失をともなわずにインスリン抵抗性を改善し、空腹時グルコースを低下させることが示されたが、このことは、TLR3アンタゴニストが高血糖症、インスリン抵抗性、及びII型糖尿病の治療に有用であり得ることを示唆するものである。
【0248】
(実施例16)
TLR3抗体アンタゴニストは細菌及びウイルス誘発炎症反応を防御する。
試薬
細菌性増悪したCOPD患者から単離された分類不能型ヘモフィルスインフルエンザ(Nontypeable Haemophilusインフルエンザ)(HNTHi)株35をDr.T.F.Murphy(Buffalo VA Medical Center,Buffalo,NY)より入手した。ヒトライノウイルス16を、アメリカ培養細胞系統保存機関(American Type Culture Collection)(ATCC)よりTCID(50)=2.8×107/mLのものを入手した。
【0249】
NTHi刺激アッセイ
NHBE細胞(Lonza,Wakersville,MD)をMicrotest 96穴組織培養プレート(BD Biosciences,Bedford,MA)に1×105/ウェルで播いた。寒天プレート上で16〜20時間増殖させたNTHiを増殖培地に約2×108cfu/mlで再懸濁し、100μg/mlのゲンタマイシンで30分間処理してから約2×107/ウェルでNHBEを含む96穴プレートに加えた。3時間後に上澄み液を除去し、抗体(0.08〜50μg/mLの最終濃度)を添加又は非添加の新鮮な増殖培地に交換した。更に24時間インキュベートした後、細胞上澄み液中のサイトカイン及びケモカインの存在を、Luminex 100IS多重蛍光分析器及びリーダーシステム(Luminex Corporation,Austin,TX)でサイトカイン25重ABビーズキット、ヒト(IL−1β、IL−1RA、IL−2、IL−2R、IL−4、IL−5、IL−6、IL−7、IL−8、IL−10、IL12p40p70、IL−13、IL−15、IL−17、TNF−α、IFN−α、IFN−γ、GM−CSF、MIP−1α、MIP−1β、IP−10、MIG、エオタキシン、RANTES及びMCP−1を含む)によって3重にアッセイした。
【0250】
ライノウイルス刺激アッセイ
NHBE細胞をMicrotest 96穴組織培養プレート(BD Biosciences,Bedford,MA)に1×105細胞/ウェルで播いた。翌日、抗体(0.08〜50μg/mLの最終濃度)をNHBE又はBEAS−2B細胞に加えて1時間インキュベートした後、10μL/ウェルのライノウイルスを加えた。更に24時間インキュベーションした後、細胞上澄み液中のサイトカイン及びケモカインの存在について上記に述べたようにluminexアッセイによってアッセイを行った。
【0251】
結果
mAb 15EVQが、NTHiにより誘導されたIP−10/CXCL10及びRANTES/CCL5の産生を用量依存的に阻害したのに対して、コントロール抗体であるヒトIgG4(Sigma,Louis,MO)はNTHiによる刺激に対して阻害作用を示さなかった(図23A)。mAb 15EVQは、ライノウイルスにより誘導されたCXCL10/IP−10及びCCL5/RANTESの産生も阻害した(図23B)。
【0252】
(実施例17)
TLR3抗体アンタゴニストは星状細胞における炎症反応を抑制する。
方法
2人のドナーから得た正常なヒト星状細胞(Lonza,Walkersville,MD)を24穴プレートに75,000細胞/ウェルで播き、一晩接着させた。翌日、星状細胞を200ng/mLのポリ(I:C)及び/又は10μg/mLのmAb 18で24時間処理した。サイトカインをLuminexによって測定した。
【0253】
結果
表10に示すように、ポリ(I:C)により誘導されたIL−6、IL−8、IL−12、IFN−α、IFN−γ、CXCL9/MIG、CCL3/MIP−1a、CCL4、CCL5/RANTES及びCXCL10/IP−10の産生はmAb 18によって阻害された。
【0254】
【表18】
【0255】
(実施例18)
TLR3抗体アンタゴニストは上皮細胞における炎症反応を抑制する。
方法
HUVEC細胞(Lonza,Walkersville,MD)をロンザ社の推奨する血清含有増殖培地中で培養した。細胞を無血清培地(Lonza,Walkersville,MD)に再懸濁し、96穴プレートに3×105細胞/mLで播き、37℃、5%CO2で24時間インキュベートした。ポリ(I:C)(GE Healthcare、Piscataway,NJ)を濃度を増加させながら(1.5〜100μg/mL)加え、更に24時間37℃でインキュベートした。サイトカイン阻害アッセイを行うため、mAb 15EVQを細胞に異なる濃度(0〜50μg/mL)で加えて30分間インキュベートし、その後、20μg/mLのポリ(I:C)を24時間加えた。細胞上澄み液を回収し、ヒトサイトカイン30重キット及びLuminex MAP技術(Invitrogen Corp.,Carslbad,CA)を用いてサイトカインレベルを測定した。sICAM−1の発現を測定するため、HUVEC細胞を20μg/mLのポリ(I:C)及び異なる濃度のmAb 15EVQ(0.8〜50μg/mL)で処理した。細胞上澄み液をELISA(R&D Systems)によりsICAM−1の発現について分析した。細胞の生存率をCellTiterGloキット(Promega,Madison,WI)により測定した。
【0256】
結果
HUVEC細胞は、ポリ(I:C)に応答して次のサイトカインを産生した:IL−1RA、IL−2、IL−2R、IL−6、IL−7、CXCL8/IL−8、IL−12(p40/p70)、IL−15、IL−17、TNF−α、IFN−α、IFN−γ、GM−CSF、CCL3/MIP−1α、CCL4/MIP−1β、CXCL10/IP−10、CCL5/RANTES、CCL2/MCP−1、VEGF、G−CSF、FGF−basic、及びHGF(表11)。mAb 15EVQは、ポリ(I:C)により誘導される全サイトカインのレベルを用量依存的に低下させた(表12)。ポリ(I:C)により誘導されるTNF−α、CCL2/MCP−1、CCL5/RANTES、及びCXCL10/IP−10の産生を低下させるmAb 15EVQの能力は、TLR3により介在される活性の阻害が、アテローム性動脈硬化症につながり得る白血球及びT細胞の浸潤に対する防御を与え得ることを示唆するものである。更に、mAb 15EVQによるVEGFの阻害は、各種の癌及び加齢性黄斑変性などの眼疾患における脈管形成を含む、VEGFによって介在される病態におけるTLR3の遮断の潜在的な効果を示唆するものである。
【0257】
TNF−α及びIFN−γは白血球の動員において機能し、かつ活性化された内皮上の接着分子の発現を増大させる(Doukasら、Am.J.Pathol.145:137〜47、1994;Poberら、Am.J.Pathol.133:426〜33、1988)。CCL2/MCP−1、CCL5/RANTES及びCXCL10/IP−10は、単球及びT細胞の動員に関与していることが示されており、アテローム性動脈硬化症の発症に寄与する(Lundgergら、Clin.Immunol.2009)。内皮細胞によるVEGFの生成は、各種の癌において脈管形成時の異常な組織増殖又は腫瘍との関連が示されている(Livengoodら、Cell.Immunol.249:55〜62、2007)。
【0258】
【表19】
【0259】
可溶性細胞内接着分子1(sICAM−1)はタンパク質切断によって生成される、内皮細胞活性化のマーカーである。ICAM−1は白血球の遊走及び活性化において重要な役割を有し、炎症時に内皮細胞及び上皮細胞において発現が上昇して、インテグリン分子LFA−1及びMac−1を介した白血球への接着を介在する。ポリ(I:C)は内皮細胞を活性化してsICAM−1の発現を上昇させたが、この発現上昇はmAb 15EVQによる処理によって低減した(図24A)。
【0260】
【表20】
【0261】
このことは、TLR3抗体アンタゴニストが白血球の輸送を阻害し、したがって炎症性細胞の流入によって引き起こされる組織障害を阻害し得ることを示す。
【0262】
生存率アッセイを行うため、上記で述べたようにHUVECを培養、播種し、ポリ(I:C)で刺激した。mAb 15EVQはポリ(I:C)により誘導されたHUVEC細胞の生存率の低下を用量依存的に回復させた(図24B)。
【0263】
内皮細胞の活性化の下方調節は、過剰な免疫細胞の浸潤を抑制し、炎症状態において増加するサイトカインによって引き起こされる組織障害を低減させ得る。炎症及び内皮細胞におけるサイトカイン及び接着分子の過剰発現は、アテローム性動脈硬化症及び高血圧の発症の主要な寄与因子である。これらのデータは、脈管炎及び内皮機能不全をともなう血管疾患などの血管の疾患におけるTLR3アンタゴニストの使用の潜在的な有用性を探求することの根拠を与えるものである。炎症及び過剰発現したサイトカインによって引き起こされる別の疾患として、免疫抑制患者及びHIV感染患者において一般的に見られ、カポジ肉腫ヘルペスウイルス(KSHV)によって引き起こされるカポジ肉腫(KS)がある。VEGF及びサイトカインの産生はKS細胞の生存に寄与する(Livengoodら、Cell Immunol.249:55〜62、2007)。TLR3アンタゴニストは、KS及び他の腫瘍にともなう血管新生のリスクを低減するうえで有用であるばかりでなく、細胞生存率の低下を防止し、内皮バリアの完全性を保護することによって、臓器不全及び敗血症などの重篤な炎症状態にともなう潜在的に重大な状態である血管漏出を防止するうえでも有用であり得る。TLR3拮抗作用は、また、フラビウイルス科(例えば、Dengue,yellow fever)、フロビウイルス科(Ebola,Marburg)、ブニヤウイルス科(例えば、Hantavirus,Nairovirus,Phlebovirus)、及びアレナウイルス科(例えば、Lujo,Lassa,Argentine,Bolivian,Venezuelan出血熱(Sihibamiyaら、Blood 113:714〜722,2009))の多数の属により引き起こされるウイルス出血熱などの内皮細胞病変を含むウイルス感染症においても有益でもあり得る。
【0264】
(実施例20)
TLR3抗体アンタゴニストのカニクイザル及びマウスTLR3との交差反応性
実施例2で述べたようにISREレポーター遺伝子アッセイを用いて、カニクイザル又はマウスTLR3に対する活性を評価した。カニクイザル(配列番号217)及びマウスTLR3 cDNA(配列番号161)を全血から増幅し、pCEP4ベクター(Clontech)の中にクローニングし、及び上述のように発現させた。mAb 15EVQは、ヒトTLR3 NF−κB及びISREアッセイにおけるそれぞれ0.44及び0.65μg/mLのIC50と比較して、cyno NF−κB及びISREアッセイにおいてそれぞれ4.18μg/mL及び1.74μg/mLのIC50を有していた。アイソタイプコントロール抗体はこれらのアッセイでは何の作用も示さなかった。
【0265】
(実施例21)
TLR3抗体アンタゴニストの治療用投与は、急性の致命的なウイルス感染症を防御する。
実施例14は、インフルエンザA感染症に対するTLR3抗体アンタゴニストによる予防的な治療(−1、4、8、及び12日目に投与)について述べている。この実施例は、TLR3抗体アンタゴニストの治療用投与(インフルエンザA感染後臨床症状の発症3日後)が生存率の向上において効果的であるということを実証している。
【0266】
モデル
動物へのmAb 5249の投与をインフルエンザAの感染後3日後に行ったこと、及び投与した動物が8週齢であったことを除いて、インフルエンザAウイルスチャレンジモデルを、実施例14に述べたように急性の致命的なウイルス感染症のモデルとして使用した。抗マウスIgG1アイソタイプコントロールmAbはBioLegendからのものであった。インフルエンザAの感染後3、7及び11日に動物に投与した。
【0267】
この実験では、生存率、毎日の臨床スコア、及び体重変化を評価した。mAb 5249を投与したC57Bl/6マウス及びTLR3 KOマウスは、抗マウスIgG1アイソタイプコントロールmAb及びインフルエンザウイルスを接種したC57BL/6マウスと比べて、両方とも生存率の統計的に有意な増加(p<0.028及びp<0.001)を示した(図25)。抗マウスIgG1アイソタイプコントロールmAb及びインフルエンザAを投与したC57BL/6マウスと比較した場合、mAb 5249を投与したC57BL/6マウス及びTLR3 KO動物において臨床的なスコアは低下し(図26)、体重は増加した(図27)。これらの結果は、TLR3抗体アンタゴニストが急性致死性インフルエンザウイルス感染モデルにおける臨床症状及び死亡率を低下させることを示すものであり、TLR3アンタゴニストが急性の感染状態にあるヒトに防御を与えることを示唆するものである。
【0268】
(実施例22)
X線結晶法によるTLR3抗体アンタゴニストのエピトープ及びパラトープ。
ヒトTLR3細胞外ドメインを、mAb 15EVQ、mAb 12QVQ/QSV及びmAb c1068のFabsと複合体の状態で結晶化させた。
【0269】
方法
タンパク質の発現及び精製。
TLR3 ECD(配列番号2のアミノ酸1〜703)の3つのFabの発現及び精製は上述の通りであった。
【0270】
TLR3 ECDの3つのFabの四次複合体の作製
1TLR3 ECD:1.1Fabのモル比に対応させて、4mgのヒトTLR3 ECDを2.4mgのそれぞれのFabと混合し、4℃で3.5時間インキュベートした。複合体をアニオン交換クロマトグラフ法により20mMのTris(pH 8.5)、10%のグリセロール(緩衝液A)で平衡化したMonoQ5/50GLカラム(GE Healthcare,Piscataway,NJ)で精製し、20mMのTris(pH 8.5)、10%のグリセロール、1MのNaCl(緩衝液B)により溶離した。1.74mL中のほぼ2.48mgの複合体を緩衝液Aにより10mLまで希釈し、カラム上に1mL/minで充填し、B濃度の0〜40%の直線的な勾配により40カラム容積にわたって溶離した。5回の連続的な精製操作を行った。ピーク1由来の区分をプールし、Amicon−15mLのUltra−30000 MWCO及びMicrocon 30000 MWCOにより20mMのTris(pH 8.5)/27mMのNaCl/10%グリセロール中に、14.49mg/mLまで濃縮した(吸光係数:A280(1mg/mL)=1.31)。
【0271】
結晶化
Corningプレート3550(Corning Inc.,Acton,MA)を用いて等容量のタンパク質及びリザーバ溶液を、シッティングドロップ法で分注するOryx4自動タンパク質結晶化ロボット(Douglas Instruments)を用いて、自動化結晶化スクリーニングを行った。初期のスクリーニングは、Hampton Crystal Screen HT(HR2−130,Hampton Research,Aliso Viejo,CA)により実施した。いくつかの条件から、小さい結晶を使用してシードを生成し、次いでこれをMicroseed−Matrix Screening(MMS)で使用した。小さい結晶を与える初期のスクリーニング条件に基づき、何回かの精密化を行った。MMSに使用するリザーバ条件は、精密化後に小さい結晶を与えた条件に基づくものであった:18〜28%のポリエチレングリコール(PEG)3350、1MのLiCl、pH4.5及び2.0〜2.9Mの(NH4)2SO4、5%のPEG400、pH4.5、及び探索したpH及び異なる添加物。成分を1タンパク質溶液:0.25シード原料:0.75リザーバ溶液の容量比で分注することにより、Oryx4自動タンパク質結晶化ロボット(Douglas Instruments)を用いて、MMS結晶化スクリーニングを行った。〜10Åの分解能で回折する結晶は、0.1Mの酢酸Na(pH 4.5)、2.9Mの(NH4)2SO4、5%メチル−ペンタン−ジオール(MPD)及び0.1Mの酢酸Na(pH 4.5)、26%のPEG3350、1MのLiClから成長させたものであった。
【0272】
結晶の分解能を改善するための試みとして、上記の条件を有するMMSを、Hampton Additive Screen HR2−428(Hampton Research,Aliso Viejo,CA)から選択された成分を容積比:1タンパク質溶液:0.125シード原料:0.2添加物溶液:0.675リザーバ溶液で用いる、添加物スクリーニングと組み合わせた。MMSと、0.1Mの酢酸Na(pH 4.5)、28%のPEG 3350、1MのLiCl、及び30mMのGly−Gly−Glyを含有する溶液からの添加物スクリーニングとの組み合わせを適用した後、Fabと複合体形成し、約5Åの分解能で回折する、TLR3 ECDのX線品質の結晶を得た。
【0273】
TLR3 ECD四次複合体のX線データ収集
X線データを収集するために、結晶(サイズ約1.0×0.5×0.1mm3)を、合成母液(0.1Mの酢酸Na(pH 4.5)、28% PEG 3350、1MのLiCl、16%グリセロール)中に数秒浸漬し、窒素流中100Kで瞬間凍結した。X線回折データを収集し、Osmic(商標)VariMax(商標)共焦光学系、Saturn 944 CCD検出器、及びX−stream(商標)2000低温冷却システムを備えたRigaku MicroMax(商標)−007HF微小焦点X線発生装置(Rigaku,Woodlands,TX)を使用して処理した。回折強度を、1/2度の像当たり1minの露出時間、5Åの最大分解能で250°(結晶回転)にわたって検出した。X線データを、プログラムD*TREK(Pflugrath,Acta Crystallographica Section D,55:1718〜1725,1999)により処理した。この結晶は、単位セルパラメーター:a=214.90Å、b=142.08Å、c=125.04Å、及びβ=103.17の単斜晶系空間群C2に属する。この非対称ユニットには1分子の複合体を含む。X線データ統計値を表13に示す。
【0274】
【表21】
【0275】
構造決定
TLR3 ECD−Fab 15EVQ−Fab 12QVQ/QSV−Fab c1068の結晶構造を、Phaser(Read,Acta Crystallogr.Crystallogr.D.Biol.Crystallogr.57:1373〜1382,2001)を用い分子置換により決定した。探索モデルはTLR3 ECD(すべてのグルカンを取り除いた、タンパク質データバンク(PDB)構造番号1ziw,Choeら、Science 309:581〜585,2005)であり、3つのFabの高分解能結晶構造を決定した(これらのFab構造に対する結晶データ及び精密化統計値の要約としては表13を参照のこと)。Fab 12QVQ/QSVのエルボー角度は、遊離形態におけるものから著しくはずれていることが判明した。エルボー角度を約5°間隔で調整することにより、一連のFab 12QVQ/QSVモデルを生成し、その1つが電子密度とよく一致することが判明した。構造精密化をPHENIX(Adamsら、J.Synchrotron Radiat.11:53〜55,2004)により行った。TLR3 ECDに対して構造を、Fab及び13個の剛体セグメント(精密化で使用される定義:30〜60、61〜108、109〜156、157〜206、207〜257、258〜307、308〜363、364〜415、416〜464、465〜514、515〜570、571〜618、619〜687)に対する剛体ドメイン(各V又はCドメイン)として、それぞれのFab剛体に対する1つのB因子及びTLR3 ECD全体に対する単一のB因子によって精密化した。
【0276】
Fab剛体のそれぞれに対して並進/回転/回転を伴った並進(TLS)精密化を導入し、配列番号2の残基330において、TLR3 ECDを2つのTLSセグメントに分割した。グリカン密度は15個のN−グリコシル化部位の10個に対して目視可能であった。次いで、ヒトTLR3細胞外ドメイン(Choeら、Science 309:581〜585,2005,PDB構造番号1ziw)の結晶構造からの炭水化物モデルを付加した。TLR3 ECD中の短い見えないセグメント(配列番号2の残基337〜342)に対する密度が剛体の精密化後に目視可能となった。それは、細胞外構造(2a0zBellら、Proc.Acad.Sci.(USA)102:10976〜10980,2005,PDB構造番号2a0z)からの対応するセグメントにより充填されていた。TLR3 ECDのC末端は、2a0zのそれに釣り合う追加の密度を含んでいた。次いで、このセグメント(配列番号2の647〜703)を2a0wの(残基647〜687)により置き換えた。このように、TLR3 ECDモデルは、TLR3構造体1ziwと2a0zとの間のハイブリッドであり、13個の剛体セグメント(アミノ酸範囲:30〜60、61〜108、109〜156、157〜206、207〜257、258〜307、308〜363、364〜415、416〜464、465〜514、515〜570、571〜618、619〜687)として精密化されたものであった。
【0277】
Fab 12QVQ/QSVのLCDR3は、遊離形態と異なる立体配置を見掛け上とっていた。マルチスタートシミュレーションアニーリングをPHENIXの標準的なパラメータにより行った。このLCDR3のモデルを電子密度マップで目視的に検査し、「最もよく合う」立体配置を元のモデル上に接合した。精密化のプロセスを、計算開始の前に脇に置いておいた反射の5%に対してRfreeによりモニターした。最終回では、それぞれの残基に対して1つのB因子を含めた。モデル検査、並びにFab及びタンパク質−タンパク質界面における側鎖のエルボー領域の手動による再組立てを、COOT(Emsleyら、Acta Crystallogr.D.Biol.Crystallogr.60:2126〜32,2004)を用いて行った。最終のRcryst及びRfreeは、5.0Å対する全15,792個の独立の反射のそれぞれについて26.8%及び30.0%であった。精密化統計値を表13及び14に示す。
【0278】
結果
TLR3 ECD−3つのFab四次複合体の分子構造
複合体の全体的な分子構造を図28に示す。非対称単位中に1つのTLR3 ECD及びそれぞれのFabの1分子が存在する。TLR3 ECDに対する構造モデルは、huTLR3(配列番号2)の30〜687個のすべての残基を含む。3つのFabに対しては、それぞれの非結合形からのすべての残基を、溶媒イオン及び水分子を除いて含めた。TLR3 ECD分子は、全体のトポロジー(1ziwに対して0.79Åのrms、613Cα’、及び2a0zに対して1.37Å、595Cα’)において以前に報告された構造と極めて類似している。このFab構造は、方法に記載のFab 12QVQ/QSVのLCDR3、並びにTLR3 ECD/Fab界面におけるエルボー領域及び一部の側鎖を除いて、それぞれの非結合形とすべて同一である。
【0279】
【表22】
【0280】
最高の分解能シェルに対する値は()中のものである。
a Rmerge=Σ|I−<I>|/ΣI、式中、Iは測定された反射の強度であり、<I>はこの反射のすべての測定値の平均強度である。
b Rcryst=Σ||Fobs|−|Fcalc||/Σ|Fobs|、式中、Fobs及びFcalcは測定及び計算された構造因子であり、Rfreeは精密化の前に反射のランダムに選択された5%の組に対して計算されたものである。
c ラマチャンドランプロットをMolProbity(Davis,I.W.ら、Nucleic Acids Res,32:W615−9,2004)により計算した。
【0281】
エピトープ及びパラトープ
TLR3 ECDと3つのFabとの間の結合に関与する残基を、図28Bに示す。Fab 12QVQ/QSVはTLR3 ECDのN末端近くで結合した。立体配置エピトープは、TLR3 LRR 3〜7からの残基(配列番号2のアミノ酸100〜221)から構成されるものであった。Fab 12QVQ/QSVの結合は、抗原及び抗体上でそれぞれほぼ928Å2及び896Å2を埋め込んだ。Fab 12QVQ/QSVに対しては、結晶構造は、次のTLR3(配列番号2)エピトープ残基:S115、D116、K117、A120、K139、N140、N141、V144、K145、T166、Q167、V168、S188、E189、D192、A195、及びA219を同定した。Fab 12QVQ/QSVに対しては、結晶構造は、次のパラトープ残基:軽鎖(配列番号211):G28、S29、Y30、Y31、E49、D50、Y90、D91及びD92、重鎖(配列番号214):N32、Q54、R56、S57、K58、Y60、Y104、P105、F106及びY107を同定した。
【0282】
Fab 15EVQ及びFab c1068は、C末端近くのLRR 15−23(配列番号2のアミノ酸406〜635)に懸る、非折りたたみ型エピトープを結合していた。(図28)。Fab 15EVQは、抗原及び抗体上でそれぞれ1080Å2及び1064Å2を埋め込み、Fab c1068は抗原及び抗体上でそれぞれ963Å2及び914Å2を埋め込んでいた。Fab 15EVQに対するエピトープは、配列番号2で示されるTLR3の残基K416、K418、L440、N441、E442、Y465、N466、K467、Y468、R488、R489、A491、K493、N515、N516、N517、H539、N541、S571、L595及びK619を覆う。Fab 15EVQに対しては、結晶構造は、次のパラトープ残基:軽鎖(配列番号41):Q27、Y32、N92、T93、L94及びS95、重鎖(配列番号216):W33、F50、D52、D55、Y57、N59、P62、E99、Y101、Y104及びD106を同定した。
【0283】
Fab c1068に対しては、結晶構造は、TLR3(配列番号2)上の次のエピトープ残基:E446、T448、Q450、R453、R473、N474、A477、L478、P480、S498、P499、Q503、P504、R507、D523、D524、E527、E530、及びK559を同定した。Fab c1068に対しては、結晶構造は、次のパラトープ残基、軽鎖:H30、N31、Y32、N50、E66、S67、G68(glyc)、重鎖:T30、T31、Y32、W33、H35、E50、N52、N54、N55、R57、N59、V99、M102、I103、及びT104を同定した。
【0284】
中和の機構及びTLR3機能との関連
mAb 15EVQ:mAb 15EVQエピトープは、TLR3残基N517、H539及びN541を含み、これらはC末端二本鎖RNA結合部位と折りたたみする。(Bellら、Proc.Natl.Acad.Sci.USA,103:8792〜7,2006).いかなる特定の理論からも束縛されることを望むものではないが、このように、mAb 15EVQは、リガンドに対して結合するTLR3に対して競合し、及びシグナル伝達単位の形成に必要とされる、リガンドにより誘起される受容体二量体化を防止すると考えられる。(Liuら、Science 320:379〜81,2008)。図29は、mAb 15EVQに対するこの直接の競合機構を図示する。抗体濃度に依存して、この機構は、ポリ(I:C)の全阻害又は二本鎖RNAにより誘起されるTLR3活性化を生じる。
【0285】
mAb 12QVQ/QSV及びmAb c1068:図30に示すように、これらの2つの抗体は二本鎖RNAリガンドとの直接的な衝突を持たない。したがって、これらがTLR3機能をmAb 15EVQのそれに類似の機構で中和することはありえない。Fabフラグメントもリガンドから離れて配向する(図30)。構造的に、mAb 12QVQ/QSV及びFab c1068は、両方とも機能を損なわずにシグナル伝達単位(SU)に結合することができる。立体的に、mAb分子の2つのFabフラグメントが、2つのTLR3分子を1つのSU中で同時に結合し、ひいては二本鎖RNA介在型TLR3二量体化を防止し得るということはありえない。いかなる特定の理論からも束縛されることを望むものではないが、TLR3へのmAb 12QVQ/QSV又はmAb c1068の結合が、抗体と隣接するシグナル伝達単位との間の立体的な衝突により、シグナル伝達単位のクラスター形成を防止すると考えられる。二本鎖RNAへのTLR3の結合は、二本鎖RNA:TLR3複合体により定義されるシグナル伝達単位に限定されない(Liuら、Science 320:379〜81,2008)。多数のSUのクラスター形成がシグナル伝達の増強を生じる得、又は効率的なTLR3シグナル伝達がこのクラスター形成を必要とし得る。mAb 12QVQ/QSV及びmAb c1068の配置が、クラスター形成をブロックし、TLR3活性の中和を生じることができる。それゆえ、抗体の最大中和効果は、抗体結合によるSUの分離の度合いに依存する。図30に図示するように、mAb 12QVQ/QSVは、mAb c1068よりも大きい分離を引き起こし、このことがmAb 12QVQ/QSVのより大きい効能に転換する可能性がある。このことは、mAb c1068及びmAb 15EVQが、飽和濃度においてそれぞれ約50%及び100%のTLR3中和を生じることができ、mAb 12QVQ/QSVが中間の活性を呈するという観察と一致する。このように、構造及びTLR3中和を組み合わせた研究は、二本鎖RNA:TLR3シグナル伝達単位がクラスターを形成して、効率的なシグナル伝達を達成する、TLR3シグナル伝達モデルを示唆する。mAb 12QVQ/QSV及びmAb c1068は、また、リガンド結合又は受容体二量体化を妨害せずにTLR3シグナル伝達を部分的に調節することができる抗体の類も定義する。
【技術分野】
【0001】
本発明は、Toll様受容体3(TLR3)抗体アンタゴニスト、TLR3抗体アンタゴニスト又はそのフラグメントをコードするポリヌクレオチド、並びに上記の製造及び使用方法に関する。
【背景技術】
【0002】
Toll様受容体(TLR)は、細菌、ウイルス、寄生虫、及び場合によりホスト由来のリガンドに応答してシグナル伝達カスケードを開始させることにより、自然免疫反応の活性化を調節し、獲得免疫の発現に影響を及ぼす(Lancaster et al.,J.Physiol.563:945〜955、2005)。細胞膜に局在するTLRであるTLR1、TLR2、TLR4及びTLR6は、細菌及び真菌のタンパク質又は脂質成分を含むリガンドを認識する。主として細胞内に存在するTLRであるTLR3、TLR7及びTLR9は、それぞれ2本鎖RNA、1本鎖RNA及び非メチル化CpG DNAに対して応答する。TLRシグナル伝達の異常調節は、多数の問題を引き起こすと考えられ、異常調節に対する治療戦略が開発されている(Hoffmanら、Nat.Rev.Drug Discov.4:879〜880,2005;Rezaei,Int.Immunopharmacol.6:863〜869,2006;Wickelgren,Science 312:184〜187,2006)。例えば、重度の敗血症及びループスに対して、それぞれTLR4、並びにTLR7及び9のアンタゴニストが臨床開発中である(Kanzlerら、Nat.Med.13:552〜559、2007)。
【0003】
TLR3シグナル伝達は、炎症又はウイルス感染時に壊死細胞から放出される二本鎖RNA、mRNA又はRNAによって活性化される。TLR3の活性化によってインターフェロン及び炎症性サイトカインの分泌が誘導され、特定の微生物感染において保護作用のある免疫細胞の活性化及び動員が誘導される。例えば、ドミナントネガティブなTLR3対立遺伝子では、小児期におけるHSV−1による一次感染時に単純ヘルペス脳炎を発症しやすくなることが示されている(Zhengら、Science 317:1522〜1527,2007)。マウスではTLR3の欠損は、コクサッキーウイルスによるチャレンジ試験における生存率の低下との関連が示されている(Richerら、PLoS One4:e4127、2009)。しかしながら、TLR3シグナル伝達の非制御又は異常調節は、西ナイルウイルス、フレボウイルス、ワクチニア、及びインフルエンザAを含む特定のウイルス感染症モデルにおける罹患率及び死亡率の一因となることが示されてきた(Wangら、Nat.Med.10:1366〜1373、2004;Gowenら、J.Immunol.177:6301〜6307、2006;Hutchensら、J.Immunol.180:483〜491、2008;LeGofficら、PloS Pathog.2:E53、2006)。
【0004】
ヒト及びマウスTLR3の細胞外ドメインの結晶構造が決定された(Bellら、Proc.Natl.Acad.Sci.(USA)、102:10976〜80、2005;Choeら、Science 309:581〜585、2005;Liuら、Science 320:379〜381,2008)。TLR3は、グリカンにより修飾されたソレノイド馬蹄様の全体形状をとり、ロイシンに富んだ繰り返し(LRR)モチーフのタンデム単位を23個有する。二本鎖RNA結合部位が、2つの別の領域にマッピングされている(Liuら、Science,320:379〜81、2008)。シグナル伝達集合体は、1つの二本鎖RNA及び2つのTLR3細胞外ドメインからなると提案された。(Leonardら、Proc.Natl.Acad.Natl.Acad.Sci.(USA)105:258〜263、2008)。
【0005】
TLR3は、例えば、敗血性ショック(Cavassaniら、J.Exp.Med.205:2609〜2621、2008)、急性肺損傷(Murrayら、Am.J.Respir.Crit.CareMed.178:1227〜1237、2008)、関節リウマチ(Kimら、Immunol.Lett.124:9〜17、2009;Brentanoら、Arth.Rheum.52:2656〜2665、2005)、喘息(Sugiuraら、Am.J.Resp.Cell Mol.Biol.40:654〜662、2009;Morishimaら、Int.Arch.Allergy Immunol.145:163〜174、2008;Stowellら、Respir.Res.10:43、2009)、クローン病及び潰瘍性大腸炎などの炎症性腸疾患(Zhouら、J.Immunol.178:4548〜4556、2007;Zhouら、Proc.Natl.Acad.Sci.(USA)104:7512〜7515、2007)、自己免疫性肝臓疾患(Langら、J.Clin.Invest.116:2456〜2463、2006)及びI型糖尿病(Dogusanら、Diabetes 57:1236〜1245、2008;Lien及びZipris、Curr.Mol.Med.9:52〜68、2009)を含む、多種多様の炎症性の免疫介在性及び自己免疫性の疾患における発病機構を促進することが示された。更に、TLR3発現の器官特異的な増加は、原発性胆汁性肝硬変における肝臓組織(Takiiら、Lab Invest.85:908〜920、2005)、関節リウマチにおける関節(Ospeltら、ArthritisRheum.58:3684〜3692、2008)、及びアレルギー性鼻炎患者における鼻粘膜(Franssonら、Respir.Res.6:100、2005)などの、局部炎症性応答が異常調節されたことにより促進される多数の発病状態と相関することが示されてきた。
【0006】
壊死状態では、内因性のmRNAを含む細胞内容物が放出されることによってサイトカイン及びケモカインの分泌、及び他の、局所炎症を誘導し、死細胞の残留物の除去を促進し、損傷を修復する因子の分泌が誘導される。壊死はしばしば炎症プロセスを継続させ、慢性炎症又は炎症悪化の一因となる(Bergsbakenら、Nature Reviews 7:99〜109、2009)。TLR3の活性化は、壊死部位においてこれらの異常な炎症プロセスに寄与し、放出されたTLR3リガンドを介して更に炎症促進性の生のフィードバックループを生じる場合がある。したがって、TLR3に対する拮抗作用は、慢性炎症若しくは炎症悪化及び/又は壊死を含むさまざまな疾患において有用であり得る。
【0007】
TLR3活性の下方制御は、腎臓細胞癌、並びに頭部及び頸部扁平上皮細胞癌が含まれる腫瘍学的な兆候に対する、新規の治療法にもなり得る(Morikawaら、Clin.Cancer Res.13:5703〜5709、2007;Priesら、Int.J.Mol.Med.21:209〜15、2008)。更には、タンパク質を低活性でコードするTLR3L423F対立遺伝子は、進行性の年齢関連の「乾燥」黄斑変性症(Yangら、N.Engl.J.Med.359:1456〜1463、2008)に対する防御と関連づけられ、この疾病においてTLR3アンタゴニストが有益であり得ることを示した。
【0008】
炎症状態にともなう病理、及び他の例えば感染症にともなう病理が、健康及び経済に及ぼす影響は多大なものである。しかしながら、多くの医学領域における進歩にも拘らず、これらの状態の多くに対する療上の選択肢及び治療法は比較的少ない。
【発明の概要】
【発明が解決しようとする課題】
【0009】
したがって、TLR3に関連する状態を治療するためにTLR3活性を抑制することが求められている。
【課題を解決するための手段】
【0010】
本発明の一つの態様は単離された抗体又はそのフラグメントであって、抗体がtoll様受容体3(TLR3)の、配列番号2のアミノ酸残基;K416、K418、L440、N441、E442、Y465、N466、K467、Y468、R488、R489、A491、K493、N515、N516、N517、H539、N541、S571、L595、及びK619に結合することを特徴とする。
【0011】
本発明の別の態様は、単離された抗体又はそのフラグメントであって、抗体がtoll様受容体3(TLR3)の、配列番号2のアミノ酸残基;S115、D116、K117、A120、K139、N140、N141、V144、K145、T166、Q167、V168、S188、E189、D192、A195、及びA219に結合することを特徴とする。
【0012】
本発明の別の態様は、単離された抗体又はそのフラグメントであって、抗体が、重鎖可変領域及び軽鎖可変領域又はそのフラグメントを有し、重鎖可変領域のChothia残基;W33、F50、D52、D54、Y56、N58、P61、E95、Y97、Y100及びD100b、並びに軽鎖可変領域のChothia残基;Q27、Y32、N92、T93、L94及びS95により、配列番号2で示されるアミノ酸配列を有するTLR3に結合することを特徴とする。
【0013】
本発明の別の態様は、単離された抗体又はそのフラグメントであって、抗体が、重鎖可変領域及び軽鎖可変領域又はそれらのフラグメントを有し、重鎖可変領域のChothia残基;N31a、Q52、R52b、S53、K54、Y56、Y97、P98、F99及びY100、並びに軽鎖可変領域のChothia残基;G29、S30、Y31、Y32、E50、D51、Y91、D92及びD93により、配列番号2に示されるアミノ酸配列を有するTLR3に結合することを特徴とする。
【0014】
本発明の別の態様は、TLR3反応性を有し、以下のa〜iの特性のうち少なくとも1つを有する、単離された抗体である;
a.Kd<10nMでヒトTLR3に結合する;
b.インビトロでのポリ(I:C)NF−κBレポーター遺伝子アッセイにおいて、1μg/mLで、ヒトTLR3の生物活性を50%超低減させる;
c.10μg/mLで、100ng/mL未満のポリ(I:C)で刺激されたBEAS−2B細胞からのIL−6又はCXCL10/IP−10の産生を60%超阻害する;
d.0.4μg/mLで、100ng/mL未満のポリ(I:C)で刺激されたBEAS−2B細胞からのIL−6又はCXCL10/IP−10の産生を50%超阻害する;
e.5μg/mLで、62.5ng/mLのポリ(I:C)で刺激されたNHBE細胞からのIL−6の産生を50%超阻害する;
f.1μg/mLで、62.5ng/mLのポリ(I:C)で刺激されたNHBE細胞からのIL−6の産生を50%超阻害する;
g.1μg/mLで、PBMC細胞による、ポリ(I:C)により誘導されたIFN−γ、IL−6又はIL−12の産生を20%超阻害する;
h.インビトロでのNF−kBレポーター遺伝子アッセイにおいて、IC50<10μg/mLで、カニクイザルのTLR3の生物活性を阻害する;
i.インビトロでのISREレポーター遺伝子アッセイにおいて、IC50<5μg/mLで、カニクイザルのTLR3の生物活性を阻害する。
【0015】
本発明の別の態様は、特定の重鎖相補性決定領域(CDR)1、2及び3のアミノ酸配列、特定の軽鎖CDR1、2及び3のアミノ酸配列、特定の重鎖可変領域(VH)のアミノ酸配列、又は特定の軽鎖可変領域(VL)のアミノ酸配列を有するモノクローナル抗体とTLR3との結合について競合する、TLR3反応性を有する単離された抗体である。
【0016】
本発明の別の態様は、重鎖及び軽鎖可変領域を有し、TLR3反応性を有する単離された抗体であって、前記抗体は特定の重鎖相補性決定領域(CDR)1、2及び3、並びに特定の軽鎖CDR 1、2及び3のアミノ酸配列を有する。
【0017】
本発明の別の態様は、重鎖及び軽鎖可変領域を有する、TLR3反応性を有する単離された抗体であって、前記抗体は特定の重鎖可変領域(VH)のアミノ酸配列、及び特定の軽鎖可変領域(VL)のアミノ酸配列を有する。
【0018】
本発明の別の態様は、重鎖及び軽鎖可変領域を有する、TLR3反応性を有する単離された抗体であって、前記抗体は特定の重鎖のアミノ酸配列、及び特定の軽鎖のアミノ酸配列を有する。
【0019】
本発明の別の態様は、単離された抗体重鎖であって、前記抗体重鎖は配列番号6、8、10、12、14、16、18、20、22、24、26、28、30、32、34、36、38、40、42、124、125、126、127、128、129、159、198、200、202、164、212、213、214、215又は216に示されるアミノ酸配列を有する。
【0020】
本発明の別の態様は、単離された抗体軽鎖であって、前記抗体軽鎖は、配列番号5、7、9、11、13、15、17、19、21、23、25、27、29、31、33、35、37、39、41、122、123、197、199、201、163、209、210、211又は225に示されるアミノ酸配列を有する。
【0021】
本発明の別の態様は、単離された抗体重鎖であって、前記抗体重鎖は配列番号102、130、131、132、133、134、135、160、204、206、208、220、166又は168に示されるアミノ酸配列を有する。
【0022】
本発明の別の態様は、単離された抗体軽鎖であって、前記抗体軽鎖は配列番号155、156、157、158、203、205、207、165、167又は227に示されるアミノ酸配列を有する。
【0023】
本発明の別の態様は抗体重鎖をコードする単離されたポリヌクレオチドであって、前記抗体重鎖は配列番号6、8、10、12、14、16、18、20、22、24、26、28、30、32、34、36、38、40、42、124、125、126、127、128、129、159、198、200、202、164、212、213、214、215又は216に示されるアミノ酸配列を有する。
【0024】
本発明の別の態様は、抗体軽鎖をコードする単離されたポリヌクレオチドであって、前記抗体軽鎖は配列番号5、7、9、11、13、15、17、19、21、23、25、27、29、31、33、35、37、39、41、122、123、197、199、201、163、209、210、211又は225に示されるアミノ酸配列を有する。
【0025】
本発明の別の態様は、抗体重鎖をコードする単離されたポリペプチドであって、前記抗体重鎖は配列番号102、130、131、132、133、134、135、160、204、206、208、220、166又は168に示されるアミノ酸配列を有する。
【0026】
本発明の別の態様は、単離されたポリペプチドであって、前記ポリペプチドは、配列番号155、156、157、158、203、205、207、165、167又は227に示されるアミノ酸配列を有する抗体軽鎖をコードする。
【0027】
本発明の別の態様は、本発明の単離された抗体及び製薬上許容され得る担体を含む製薬学的組成物である。
【0028】
本発明の別の態様は、少なくとも1つの本発明のポリヌクレオチドを含むベクターである。
【0029】
本発明の別の態様は、本発明のベクターを有するホスト細胞である。
【0030】
本発明の別の態様は、本発明のホスト細胞を培養することと、前記ホスト細胞によって産生された抗体を回収することとを含む、TLR3反応性を有する抗体の製造方法である。
【0031】
本発明の別の態様は、治療上の有効量の本発明の単離された抗体を、炎症性疾患の治療又は予防を必要とする患者に、炎症性疾患を治療又は予防するうえで充分な時間にわたって投与することを含む、炎症性疾患の治療又は予防方法である。
【0032】
本発明の別の態様は、治療上の有効量の本発明の単離された抗体を、それらを必要とする患者に、全身性の炎症状態を処置又は予防するうえで充分な時間にわたって投与することを含む、全身性炎症性疾患の処置又は予防方法である。
【0033】
本発明の別の態様は、治療上の有効量の本発明の単離された抗体を、それらを必要とする患者に、II型糖尿病を処置するうえで充分な時間にわたって投与することを含む、II型糖尿病の処置方法である。
【0034】
本発明の別の態様は、治療上の有効量の本発明の単離された抗体を、それらを必要とする患者に、高血糖を処置するうえで充分な時間にわたって投与することを含む、高血糖の処置方法である。
【0035】
本発明の別の態様は、治療上の有効量の本発明の単離された抗体を、それらを必要とする患者に、インスリン抵抗性を処置するうえで充分な時間にわたって投与することを含む、高インスリン血症の治療方法である。
【0036】
本発明の別の態様は、治療上の有効量の本発明の単離された抗体を、それらを必要とする患者に、ウイルス感染を処置又は予防するうえで充分な時間にわたって投与することを含む、ウイルス感染の処置又は予防方法である。
【図面の簡単な説明】
【0037】
【図1】NF−κBレポーター遺伝子アッセイにおける抗ヒトTLR3(huTLR3)mAbの作用を示す図。
【図2A】BEAS−2Bアッセイにおける抗huTLR3 mAbの作用(阻害率(%))を示す図。
【図2B】BEAS−2Bアッセイにおける抗huTLR3 mAbの作用(阻害率(%))を示す図。
【図3A】NHBEアッセイにおける抗huTLR3 mAbの作用を示す図。
【図3B】NHBEアッセイにおける抗huTLR3 mAbの作用を示す図。
【図4】PBMCアッセイにおける抗huTLR3 mAbの作用を示す図。
【図5A】HASMアッセイにおける抗huTLR3 mAbの作用を示す図。
【図5B】HASMアッセイにおける抗huTLR3 mAbの作用を示す図。
【図6A】TLR3変異体に対する抗huTLR3 mAbの結合を示す図。
【図6B】TLR3変異体に対する抗huTLR3 mAbの結合を示す図。
【図6C】TLR3変異体に対する抗huTLR3 mAbの結合を示す図。
【図7A】ヒトTLR3 ECD上の、mAb 15EVQ(黒)及びC1068 mAb(灰色)のエピトープ(上の画像)、並びにmAb 12QVQ/QSV(黒、下の画像)のエピトープを示す図。
【図7B】mAb 15EVQと複合体を形成させたTLR3 ECDタンパク質の局所的H/D交換撹乱マップを示す図。
【図8A】A)NF−κBレポーター遺伝子アッセイ及びB)ISREレポーター遺伝子アッセイにおけるラット/マウスの抗マウスTLR3 mAb mAb 5429(代替抗体)の作用を示す図。
【図8B】A)NF−κBレポーター遺伝子アッセイ及びB)ISREレポーター遺伝子アッセイにおけるラット/マウスの抗マウスTLR3 mAb mAb 5429(代替抗体)の作用を示す図。
【図9】MEF CXCL10/IP−10アッセイにおける代替mAb(mAb 5429、mAb c1811)の作用を示す図。
【図10】TLR3に対する代替mAbの結合の特異性を示す図。上側のパネル:アイソタイプコントロール、下側のパネル:mAbc1811。
【図11】AHRモデルにおけるpenHレベルに対する代替mAbの効果を示す図。
【図12】AHRモデルにおけるBAL液中の全好中球数に対する代替mAbの効果を示す図。
【図13】AHRモデルにおけるBAL液中のCXCL10/IP−10レベルに対する代替mAbの効果を示す図。
【図14】DSSモデルにおける組織病理学のスコアに対する代替mAbの効果を示す図。
【図15】T細胞移植モデルにおける、A)組織病理学のスコア、及びB)好中球流入に対する代替mAbの効果、を示す図。
【図16】CIAモデルにおける臨床スコアに対する代替mAbの効果を示す図。
【図17】CIAモデルにおける臨床AUCスコアに対する代替mAbの効果を示す図。
【図18】インフルエンザA/PR/8/34の経鼻投与の後のC57BL/6マウスの生存率に及ぼす代替mAbの効果を示す図。mAbの投与は1日目に始まった。
【図19】インフルエンザA/PR/8/34の投与後の臨床スコアに対する代替mAbの効果を示す図。mAbの投与は1日目に始まった。
【図20】インフルエンザA/PR/8/34の投与後に代替mAbを投与した後の14日にわたる体重変化に及ぼす代替mAbの効果を示す図。mAbの投与は1日目に始まった。
【図21】グルコースチャレンジ後の(A)野生型DIO及び(B)TLR3ノックアウトDIO動物における血中グルコース濃度に対する代替mAbの効果を示す図。
【図22】野生型DIO動物のインスリンレベルに対する代替mAbの効果を示す図。
【図23】NHBE細胞における、(A)NTHi、及び(B)ライノウイルス誘導CXCL10/IP−10及びCCL5/RANTESレベルに対するmAb 15EVQの効果を示す図。
【図24】HUVEC細胞における、(A)sICAM−1レベル及び(B)生存率に対する、mAb 15EVQの効果を示す図。
【図25】インフルエンザAの感染3日後に代替mAbを投与した後の動物の生存率を示す図。
【図26】インフルエンザAの感染3日後に代替mAbを投与した後の臨床スコアを示す図。
【図27】インフルエンザAの感染3日後に代替mAbを投与した後の動物の体重変化を示す図。
【図28】AではhuTLR3 ECDとFab 12QVQ/QSV、Fab 15EVQ及びFab c1068との四次構造の分子構造をリボン及び表面表示で示す図。TLR3 ECDの、Nで標識したN末端を薄い灰色で示し、すべてのFab分子を暗い灰色のリボン表示で示す。B.エピトープを薄い灰色で着色し、AにおけるFabについてのようにTLR3 ECD上で標識した。図28、29及び30では、判り易くするために、Fab 12QVQ/QSV、Fab c1068及びFab 15EVQの標識を、それぞれ、Fab 12、Fab 1068及びFab 15と略記する。
【図29】Fab 15EVQによる中和機構を示す図。A.2つのTLR3 ECD(薄い灰色及び暗い灰色で示す図、並び標識されたTLR3図)の一方において、Fab 15EVQエピトープをハイライト(薄い灰色)した二本鎖RNA:TLR3シグナル伝達ユニット(SU)を示す。二本鎖RNAリガンドは薄い灰色の二重らせんとして示される。B.二本鎖RNAの結合を立体的に阻害することで、SUの形成を阻害する、Fab 15EVQの結合が図示される。Fab 15EVQの結合は高親和性であり、SUの形成を阻害するか、又はすでに形成されているSUを分解する。
【図30】Fab 12QVQ/QSV及びFab c1068及びTLR3シグナル伝達ユニット(SU)のクラスター形成の機構を示す図。A.Fab 12QVQ/QSV及びFab c1068は、単一SUを結合(又は共結合)することができる。B.約76塩基対の二本鎖RNA上の2つのSUの最近接クラスター形成のモデルである。判り易くするために3つのエピトープが異なる分子中でハイライトされている。C.Fab 12QVQ/QSV及びFab c1068の結合は、抗体と、隣接するSUとの間の立体的な衝突によりSUのクラスター形成を防止する。2つの左を指す矢印は、抗体によるSUの分離の異なる度合いを定性的に表す(底部の矢印はFab 12QVQ/QSVに対するものであり、及び頂部の矢印はFab c1068に対するものである)。
【図31】代表的な抗体に対する、配列番号と、Kabatによる番号付けと、Chothiaによる番号付けの対応を示す図。CDR及びHVを灰色でハイライトしてある。
【図32】mAb 15EVQのVLと、ヒトVk1フレームワークとのアライメント結果を示す図。Chothiaによるハイパー可変ループにアンダーラインし、パラトープ残基に二重アンダーラインし、及びフレームワークの差異を灰色でハイライトしてある。特記しない限り、Vκ1遺伝子は*01対立遺伝子である。残基番号付けは連続的に行っている。
【図33】mAb 15EVQのVHと、ヒトVh5フレームワークとのアライメント結果を示す図。配列の特徴は図32のように示される。
【図34】mAb 12QVQ/QSVのVLと、ヒトVk3フレームワークとのアライメント結果を示す図。配列の特徴は図32のように示される。
【図35】mAb 15EVQ又はmAb 12QVQ/QSVのVL及びVHと、ヒトJκ、Jλ又はJhフレームワークとのアライメント結果を示す図。配列の特徴は図32のように示される。
【発明を実施するための形態】
【0038】
本明細書に引用する特許及び特許出願を含むがそれらに限定されないすべての刊行物は、あたかもその全体が記載されているのと同様に本願に参照によって組み入れるものである。
【0039】
本明細書で用いる「アンタゴニスト」なる用語は、何らかの機構によって、受容体又は細胞内メディエーターなどの別の分子の作用を部分的又は完全に阻害する分子のことを意味する。
【0040】
本明細書で用いるとき、「TLR3抗体アンタゴニスト」又は「TLR3反応性の」抗体とは、TLR3の生物活性又はTLR3受容体の活性化を直接的又は間接的に実質上相殺、低減、又は阻害する能力を有する抗体のことを言う。例えば、TLR3反応性を有する抗体は、TLR3に直接結合してTLR3の活性を中和することによって、すなわちTLR3のシグナル伝達を遮断することによって、サイトカイン及びケモカインの放出、又はNF−κBの活性化を低減させることができる。
【0041】
用語「抗体」は、本明細書において広義に用いられ、ポリクローナル抗体、マウス、ヒト、ヒト適合化、ヒト化及びキメラモノクローナル抗体を含むモノクローナル抗体、並びに抗体フラグメントを含む、免疫グロブリン又は抗体分子を含む。
【0042】
一般に抗体とは、特定の抗原に対する結合特異性を示すタンパク質又はペプチド鎖である。完全な抗体とは、2個の同一の軽鎖と2個の同一の重鎖とからなるヘテロ4量体の糖タンパク質である。通常、それぞれの軽鎖は1個の共有ジスルフィド結合によって重鎖と結合しているが、ジスルフィド結合の数は、アイソタイプの異なる免疫グロブリン重鎖間で異なる。それぞれの重鎖及び軽鎖はまた、規則的な間隔をおいた鎖内ジスルフィド架橋も有する。それぞれの重鎖は、一方の末端で可変ドメイン(可変領域)(VH)を有し、続いて多数の定常性のドメイン(定常領域)を有する。それぞれの軽鎖は一端に可変領域(VL)を有し、もう一方の端に定常領域を有する。軽鎖の定常領域は重鎖の第1の定常領域と整列し、軽鎖の可変領域は重鎖の可変領域と整列している。あらゆる脊椎動物種の抗体の軽鎖は、定常領域のアミノ酸配列に基づき、カッパー(κ)及びラムダ(λ)の2つの明確に異なるタイプの内のどちらかに分類することができる。
【0043】
免疫グロブリンは、重鎖の定常領域のアミノ酸配列に応じて、IgA、IgD、IgE、IgG及びIgMの5つの大きなクラスに分類することができる。IgA及びIgGは、IgA1、IgA2、IgG1、IgG2、IgG3、IgG4というアイソタイプに更に分類される。
【0044】
用語、「抗体フラグメント」は完全な抗体の一部を意味し、通常は完全な抗体の抗原結合領域又は可変領域を意味する。抗体フラグメントの例としては、Fab、Fab’、F(ab’)2、Fvフラグメント、二重特異性抗体、単鎖抗体分子、及び少なくとも2つの完全な抗体から形成された多特異的抗体が挙げられる。
【0045】
免疫グロブリンの軽鎖可変領域又は重鎖可変領域は、3つの「抗原結合部位」を取り囲む「フレームワーク」領域からなる。抗原結合部位は、さまざまな用語を次のように使用して定義される:(i)用語相補性決定領域(CDR)は配列可変性に基づく(Wu及びKabat、J.Exp.Med.132:211〜250、1970)。一般に、抗原結合部位は、VH(HCDR1、HCDR2、HCDR3)内に3つ及びVL(LCDR1、LCDR2、LCDR)内に3つの6つのCDRを有している(Kabatら、Sequences of Proteins of Immunological Interest、5th Ed.Public Health Service,National Institutes of Health,Bethesda,Md.,1991)。(ii)用語「超可変領域」、「HVR」、又は「HV」は、Chothia及びLesk(Chothia及びLesk,Mol.Biol.196:901〜917,1987)により定義されているように、構造が超可変性である、抗体の可変ドメイン領域を指す。一般に抗原結合部位は、VH内に3つ(H1、H2、H3)及びVL内に3つ(L1、L2、L3)の6つの超可変領域を有している。Chothia及びLeskは、構造的に保持されているHVを「カノニカル構造」と呼ぶ。(iii)Lefrancにより提案された「IMGT−CDR」(Lefrancら、Dev.Comparat.Immunol.27:55〜77,2003)は、免疫グロブリン及びT細胞受容体からのVドメインの比較に基づくものである。International ImMunoGeneTics(IMGT)データベース(http://www_imgt_org)は、これらの領域について標準化された番号及び定義を提供している。各CDR、HV、及びIMGTの表記間の対応についてはLefrancら、Dev.Comparat.Immunol.27:55〜77,2003に述べられている。(iv)抗原結合部位は特異性決定残基の使用(SDRU)(Almagro,Mol.Recognit.17:132〜143,2004)に基づいて概略説明することができる。特異性決定残基(SDR)は、抗原との接触に直接関与する免疫グロブリンのアミノ酸残基を指す。SDRUは、抗原−抗体複合体についての結晶構造解析により定義される、異なるタイプの抗原に対するSDRの数及び分布の精密な尺度である。(v)抗原結合部位を、抗原抗体複合体の結晶構造から同定される抗体パラトープ残基とも定義することもできる。
【0046】
用語「複合配列」は、本明細書で使用するとき、Kabat、Chothia又はIMGTによって個別に表記されるすべてのアミノ酸残基又は他の任意の好適な抗原結合領域の表記を含むものとして定義される抗原結合部位を意味する。
【0047】
用語「Chothia残基」は、本明細書で使用するとき、Al−Lazikani(Al−Lazikaniら、J.Mol.Biol.,273,927〜48,1997)に従って番号付けされた抗体VL残基及びVH残基である。ポリペプチド番号付けに関連して2つの最も使用される番号付けシステム、Kabat(Kabatら、Sequences of Immunological Interest,5th Ed.Public Health Service,NIH,Bethesda,MD,1991)とChothia(Chothia及びLesk,Mol.Biol.196:901〜17,1987)との間の対応を、本発明の代表的な抗体に対して図31に示す。
【0048】
「フレームワーク」又は「フレームワーク配列」は、抗原結合部位として定義されていない、可変領域の残りの配列である。フレームワークは、通常、それぞれの可変領域中で、3つの抗原結合部位に対して骨格を形成する、4つの領域、FR1、FR2、FR3、及びFR3に分割される。抗原結合部位は上記に述べたようなさまざまな用語によって定義され得るため、フレームワークの正確なアミノ酸配列は抗原結合部位がどのように定義されるかによって決まる。
【0049】
「軽鎖可変領域カッパー1(Vκ1)フレームワーク」又は「Vκ1」は、本発明で使用するとき、ヒトVκ1機能性遺伝子又はその対立遺伝子のいずれかによりコードされている、アミノ酸配列を有するフレームワークを指す。代表的な機能性ヒトVκ1遺伝子は、IGKV1−5*01、IGKV1−6*01、IGKV1−8*01、IGKV1−9*01、IGKV1−12*01、IGKV1−13*02、IGKV1−16*01、IGKV1−17*01、IGKV1−27*01、IGKV1−33*01、IGKV1−37*01、IGKV1−39*01、IGKV1D−8*01、IGKV1D−12*01、IGKV1D−13*01、IGKV1D−16*01、IGKV1D−17*01、IGKV1D−33*01、IGKV1D−37*01、IGKV1D−39*01、IGKV1D−42*01、又はIGKV1D−43*01である。免疫グロブリン遺伝子の命名法は周知されたものである。
【0050】
「軽鎖可変領域ラムダ3(Vλ3)フレームワーク」又は」「Vλ3」は、本明細書で使用するとき、ヒトVλ3機能性遺伝子又はその対立遺伝子のいずれかによりコードされたアミノ酸配列を有するフレームワークを指す。代表的な機能性ヒトVλ3遺伝子は、IGLV3−1*01、IGLV3−9*01、IGLV3−10*01、IGLV3−12*01、IGLV3−16*01、IGLV3−19*01、IGLV3−21*01、IGLV3−22*01、IGLV3−25*01、IGLV3−27*01、及びIGLV3−32*01である。
【0051】
「重鎖可変領域Vh5フレームワーク」又は「Vh5」は、本明細書で使用するとき、Vh5機能性遺伝子又はその対立遺伝子のいずれかによりコードされたアミノ酸配列を有するフレームワークを指す。代表的な機能性ヒトVh5遺伝子は、IGHV5−51*01及びIGHV5−1*01である。
【0052】
「重鎖可変領域Vh6フレームワーク」又は「Vh6」は、本明細書で使用するとき、Vh6機能性遺伝子又はその対立遺伝子のいずれかによりコードされたアミノ酸配列を有するフレームワークを指す。代表的な機能性ヒトVh6遺伝子はIGHV6−1*01である。
【0053】
「軽鎖カッパーJ領域(Jκ)フレームワーク」又は「Jκ」は、本明細書で使用するとき、ヒトJκ機能性遺伝子又はその対立遺伝子のいずれかによりコードされたアミノ酸配列を有するフレームワークを指す。代表的な機能性ヒトVκ遺伝子は、IGKJ1、IGKJ2、IGKJ3、IGKJ4、及びIGKJ5である。
【0054】
「軽鎖ラムダJ領域(Jλ)フレームワーク」又は「Jλ」は、本明細書で使用するとき、ヒトJλ機能性遺伝子又はその対立遺伝子のいずれかによりコードされたアミノ酸配列を有するフレームワークを指す。代表的な機能性ヒトJλ遺伝子は、IGLJ1、IGLJ2、IGLJ3、IGLJ4、IGLJ5、IGLJ6、及びIGLJ7である。
【0055】
「重鎖J領域(Jh)フレームワーク」又は「Jh」は、本明細書で使用するとき、ヒトJh機能性遺伝子又はその対立遺伝子のいずれかによりコードされたアミノ酸配列を有するフレームワークを指す。代表的な機能性ヒトJh遺伝子は、IGHJ1、IGHJ2、IGHJ3、IGHJ4、IGHJ5、及びIGHJ6である。
【0056】
「生殖細胞系列遺伝子」又は「抗体生殖細胞系列遺伝子」は、本明細書で使用するとき、特定の免疫グロブリンの発現のための遺伝的再配列及び変異につながる成熟プロセスを経ていない非リンパ細胞によってコードされる免疫グロブリン配列である。
【0057】
「スカフォールド」は、本明細書で使用するとき、ヒト生殖細胞系列遺伝子によってコードされる、軽鎖又は重鎖の可変領域のアミノ酸配列を指す。したがって、スカフォールドは、フレームワークと抗原結合部位の両方を包含する。
【0058】
用語「抗原」は、本明細書で使用するとき、直接又は間接に抗体を生成する能力を有する任意の分子を意味する。「抗原」の定義内には、タンパク質コーディング核酸が含まれる。
【0059】
用語「ホモログ」は、参照配列との配列同一性が40%〜100%であるタンパク質配列を意味する。ヒトTLR3のホモログには、既知のヒトTLR3配列との配列同一性が40%〜100%であるような他種由来のポリペプチドが含まれる。2つのペプチド鎖の配列の同一性(%)は、Vector NTIバージョン9.0.0(Invitrogen,Carlsbad,CA)のAlignXモジュールのデフォルト設定を使用して、配列をペアでアライメントすることによって決定することができる。「TLR3」とはヒトTLR3(huTLR3)及びそのホモログの意味である。完全長huTLR3のヌクレオチド及びアミノ酸配列をそれぞれ配列番号1及び2で示す。huTLR3の細胞外ドメイン(ECD)のヌクレオチド及びアミノ酸配列をそれぞれ配列番号3及び配列番号4に示す。
【0060】
「実質的に同一の」は、本明細書で使用するとき、比較される2つの抗体又は抗体フラグメントのアミノ酸配列が同一であるか、又は「実体のない差異」を有するということを意味する。抗体又は抗体フラグメントのアミノ酸配列における1、2、3、4、5又は6個のアミノ酸の置換が、実体のない差異として挙げられる。本明細書で開示する配列とほぼ同じアミノ酸配列も本願の一部である。特定の実施形態では、配列の同一性は、約90%、91%、92%、93%、94%、95%、96%、97%、98%、99%又はそれよりも高い。同一性(%)は、上記に述べたようにして決定することができる。比較するペプチド鎖の例としては、重鎖又は軽鎖の可変領域がある。
【0061】
用語「〜と組み合わせて」は、本明細書で使用するとき、記述の対象である薬剤が、他の薬剤と共に、混合物として一緒に、又は単独薬剤として同時に、又は単独薬剤として順次に任意の順序で、動物に投与され得ることを意味する。
【0062】
用語「炎症性状態」は、本明細書で使用するとき、大部分の場合痛み、発赤、腫張、及び組織機能の喪失を特徴とし、サイトカイン、ケモカイン、又は炎症性細胞(例えば、好中球、単球、リンパ球、マクロファージ)により一部介在される、細胞損傷に対する局所化された応答を意味する。用語「炎症性の肺状態」は、本明細書で使用するとき、肺を冒す又は肺と関連する炎症状態を意味する。
【0063】
「用語モノクローナル抗体」(mAb)は、本明細書で使用するとき、ほぼ均質な抗体の集団から得られる抗体(又は抗体フラグメント)を意味する。モノクローナル抗体は極めて特異性が高く、通常、単一の抗原決定基を標的とする。「モノクローナル」という修飾語は、抗体のほぼ均質な性質を指して言うものであり、抗体がいずれかの特定の方法によって作製される必要はない。例えば、マウスmAbは、Kohlerら、Nature 256:495〜497、1975のハイブリドーマ法によって製造することができる。ドナー抗体(通常はマウス)由来の軽鎖及び重鎖の可変領域を、アクセプター抗体(通常、例えばヒトなどの別の哺乳動物種)由来の軽鎖及び重鎖の定常領域とともに含むキメラmAbを、米国特許第4,816,567号に開示される方法によって作製することができる。非ヒトドナー免疫グロブリン(通常はマウス)に由来するCDR、及び1以上のヒト免疫グロブリンに由来するCDR以外の部分を有する、ヒト適合化mAbを、米国特許第5,225,539号に開示されるような当業者には周知の方法によって作製することができる。ヒト適合化に有用なヒトフレームワーク配列は当業者であれば関連するデータベースから選択することができる。場合により、Queenら、Proc.Natl.Acad.Sci.(USA),86:10029〜10032,1989及びHodgsonら、Bio/Technology,9:421,1991に開示されるような方法によって、結合親和性を維持するために変化させたフレームワーク支持残基を組み込むことによってヒト適合化mAbを更に改変することができる。
【0064】
ヒト以外のいかなる配列も持たない完全なヒトmAbは、例えば、Lonbergら、Nature 368:856〜859,1994、Fishwildら、Nature Biotechnology 14:845〜851,1996、及びMendezら、Nature Genetics15:146〜156,1997に参照される方法により、ヒト免疫グロブリン形質転換マウスから作製することができる。同様に、ヒトmAbは、ファージディスプレイライブラリを元に、例えば。Knappikら、J.Mol.Biol.296:57〜86 2000及びKrebsら、J.Immunol.Meth.254:67〜84 2001で参照されている方法により作製及び最適化することもできる。抗体のフラグメント、例えばFab、F(ab’)2、Fd、及びdAbフラグメントを、抗体の開裂又は遺伝子組み換え操作により生成してもよい。例えば、Fab及びF(ab’)2フラグメントを、抗体をペプシンなどの酵素により処理することにより、生成することもできる。
【0065】
用語「エピトープ」は、本明細書で使用するとき、抗体が特異的に結合する抗原の部分を意味する。エピトープは、通常、アミノ酸又は多糖類側鎖のような化学的に活性な(極性、非極性又は疎水性)部分の表面集団からなり、特定の3次元構造特性並びに特定の帯電特性を有し得る。エピトープは直線状のものであってもよく、あるいはアミノ酸の直線状の配列によってではなく、抗原の連続していないアミノ酸間の空間的な関係によって形成される不連続エピトープ(例えば立体構造エピトープ)であってもよい。立体構造エピトープには、抗原の直線状配列の異なる部分のアミノ酸同士が3次元空間で近接するような抗原の折り畳みによって生じるエピトープが含まれる。
【0066】
用語「パラトープ」は、本明細書で使用するとき、抗原が特異的に結合する抗体の一部を指す。パラトープは元来線状であるか、又は不連続であることができ、抗体のアミノ酸の線状構造よりも、隣接していないアミノ酸間の空間的な関係により形成される。「軽鎖パラトープ」及び「重鎖パラトープ」又は「軽鎖パラトープアミノ酸残基」及び「重鎖パラトープアミノ酸残基」は、それぞれ、抗原と接触する抗体の軽鎖及び重鎖残基を指す。
【0067】
用語「特異的結合」は、本明細書で使用するとき、抗体が特定の抗原と、他の抗原又はタンパク質に対するよりも高い親和性で結合することを指す。通常、抗体の結合における解離定数(KD)は10-7M以下であり、抗体が特定の抗原と結合する際のKDは、特定の抗原以外の非特異的抗原(例えば、BSA、カゼイン、又は他の任意の特定のポリペプチド)と結合する際のKDと比較して1/2以下である。本明細書では、「抗原を認識する抗体」及び「抗原に特異的な抗体」という語句を、「抗原と特異的に結合する抗体」又は「抗原特異的抗体」、例えばTLR3特異的抗体という用語と互換可能に用いる。解離定数は後述するような標準的な方法を用いて測定することができる。
【0068】
用語「TLR3の生物活性」又は「TLR3の活性化」は、本明細書で使用するとき、リガンドとTLR3との結合によって生ずる任意の活性を指す。TLR3のリガンドには、2本鎖DNA、ポリイノシンポリシチジン酸(ポリ(I:C))、及び例えば壊死細胞から放出される内因性mRNAなどの内因性mRNAが含まれる。TLR3の活性化の一例では、TLR3のリガンドに応じてNF−κBが活性化される。レポーター遺伝子アッセイを用いることで、ポリ(I:C)による受容体の誘導に基づき、NF−κBの活性化をアッセイすることができる(Alexopoulosら、Nature 413:732〜738,2001;Hackerら、EMBO J.18:6973〜6982,1999)。他の代表的なTLR3受容体の活性化は、TLR3リガンドに対する応答として、インターフェロン応答因子(IRF−3、IRF−7)の活性化をもたらす。TLR3によって介在されるIRFの活性化は、インターフェロン刺激応答配列(ISRE)によって制御されるレポーター遺伝子を用いてアッセイすることができる。別のTLR3の活性化の例では、例えばTNF−α、IL−6、IL−8、IL−12、CXCL5/IP−10及びRANTESなどの炎症性サイトカイン及びケモカインが分泌される。細胞若しくは組織からの、又は血液循環中へのサイトカイン及びケモカインの放出は、ELISA免疫アッセイなどの周知の免疫アッセイによって測定することができる。
【0069】
本明細書では以下のようにアミノ酸の慣習的な1文字及び3文字の略号を用いる。
【0070】
【表1】
【0071】
物質の組成物
本発明はTLR3の生物活性を阻害することが可能な抗体アンタゴニスト及びかかる抗体の使用法を提供する。かかるTLR3アンタゴニストは、TLR3に結合してTLR3の活性化を阻害する性質を有し得る。かかる抗体によってTLR3活性が阻害され得る機序の例としては、TLR3へのリガンドの結合のインビトロ、インビボ、若しくはインサイチューでの阻害、受容体の2量化の阻害、エンドソーム区画へのTLR3の局在化の阻害、下流のシグナル伝達経路のキナーゼ活性の阻害、又はTLR3のmRNAの転写の阻害が挙げられる。他の機序によってTLR3の活性を阻害することが可能な他の抗体アンタゴニストもまた、本発明の異なる態様及び実施形態の範囲に含まれるものである。これらのアンタゴニストは、研究用試薬、診断用試薬及び治療薬として有用である。
【0072】
抗体の多様性は、自然の系では、可変領域をコードする多数の生殖細胞系列遺伝子の使用及びさまざまな体細胞イベントにより作り出される。体細胞イベントとしては、完全なVH領域を作るための多様性(D)及び接合(J)遺伝子セグメントを持つ可変遺伝子セグメントの組換え、並びに完全なVL領域を作るための可変及び接合遺伝子セグメントの組換えがある。組み換えプロセスそれ自体は不正確であり、V(D)J接合部におけるアミノ酸の喪失又は追加を生じる可能性がある。多様性のこれらの機序は、抗原暴露に先立つB細胞の発生時に起こる。抗原刺激の後、B細胞中で発現された抗体遺伝子は、体細胞変異を受ける。生殖細胞系列遺伝子セグメント、これらのセグメントのランダムな組み換え、及びランダムなVH−VLの対形成の見積り数に基づいて,1.6×107までの異なる抗体が生成可能である(Fundamental Immunology,3rd ed.(1993),ed.Paul,RavenPress,New York,N.Y.)。抗体多様性に寄与する他のプロセス(体細胞変異などの)を考慮に入れた場合、1010以上の異なる抗体が生成可能であると考えられる(Immunoglobulin Genes,2nd ed.(1995),eds.Jonioら、Academic Press,San Diego,Calif.)。多数のプロセスが抗体多様性の生成において関与するために、同一の抗原特異性を有する独立に由来したモノクローナル抗体が同一のアミノ酸配列を有するということは極めてあり得ないことである。
【0073】
本発明は、新規の抗原結合部位及びヒト免疫グロブリン遺伝子ライブラリに由来する免疫グロブリン鎖を提供する。抗原結合部位を担持するための構造は一般に抗体の重鎖又は軽鎖若しくはそれらの部分であり、抗原結合部位は上記に述べたようにして決定された天然に生じる抗原結合部位に配置される。
【0074】
本発明は、重鎖可変領域及び軽鎖可変領域の両方を有する、TLR3反応性を有する単離された抗体又はそのフラグメントを提供するものであり、前記抗体は表1aに示されるような重鎖の相補性決定領域(CDR)のアミノ酸配列1、2及び3(HCDR1、HCDR2及びHCDR3)、及び軽鎖の相補性決定領域(CDR)のアミノ酸配列1、2及び3(LCDR1、LCDR2及びLCDR3)を有している。
【0075】
【表2】
【0076】
特定の実施形態では、本発明は、重鎖可変領域及び軽鎖可変領域の両方を有する、TLR3反応性を有する単離された抗体又はそのフラグメントを提供するものであり、前記抗体は配列番号192に示されるようなHCDR2のアミノ酸配列を有し、配列番号192のHCDR2は式(I)に示されるように定義される。
Xaa6−I−Xaa7−Xaa8−R−S−Xaa9−W−Y−N−D−Y−A−V−S−V−K−S
(I)
式中、
Xaa6は、Arg又はLysであってよく、
Xaa7は、Tyr、His又はSerであってよく、
Xaa8は、Met、Arg又はTyrであってよく、
Xaa9は、Lys又はArgであってよい。
【0077】
他の実施形態では、本発明は、重鎖可変領域及び軽鎖可変領域の両方を有する、TLR3反応性を有する単離された抗体又はそのフラグメントを提供するものであり、前記抗体は配列番号194に示されるようなHCDR2のアミノ酸配列を有し、配列番号194のHCDR2は式(III)に示されるように定義される。
I−I−Q−Xaa15−R−S−K−W−Y−N−Xaa16−Y−A−Xaa17−S−V−K−S
(III)
式中、
Xaa15は、Lys、Thr又はIleであってよく、
Xaa16は、Asn又はAspであってよく、
Xaa17は、Val又はLeuであってよい。
【0078】
他の実施形態では、本発明は、重鎖可変領域及び軽鎖可変領域の両方を有する、TLR3反応性を有する単離された抗体又はそのフラグメントを提供するものであり、前記抗体は配列番号196に示されるようなHCDR2のアミノ酸配列を有し、配列番号196のHCDR2は式(V)に示されるように定義される。
Xaa24−I−D−P−S−D−S−Y−T−N−Y−Xaa25−P−S−F−Q−G
(V)
式中、
Xaa24は、Phe又はArgであってよく、
Xaa25は、Ala又はSerであってよい。
【0079】
他の実施形態では、本発明は、重鎖可変領域及び軽鎖可変領域の両方を有する、TLR3反応性を有する単離された抗体又はそのフラグメントを提供するものであり、前記抗体は配列番号191に示されるようなLCDR3のアミノ酸配列を有し、配列番号191のLCDR3は式(II)に示されるように定義される。
Xaa1−S−Y−D−Xaa2−Xaa3−Xaa4−Xaa5−T−V
(II)
式中、
Xaa1は、Ala、Gln、Gly又はSerであってよく、
Xaa2は、Gly、Glu又はSerであってよく、
Xaa3は、Asp又はAsnであってよく、
Xaa4は、Glu又はSerであってよく、
Xaa5は、Phe、Ala又はLeuであってよい。
【0080】
他の実施形態では、本発明は、重鎖可変領域及び軽鎖可変領域の両方を有する、TLR3反応性を有する単離された抗体又はそのフラグメントを提供するものであり、前記抗体は配列番号193に示されるようなLCDR3のアミノ酸配列を有し、配列番号193のLCDR3は式(IV)に示されるように定義される。
Xaa10−S−Y−D−Xaa11−P−Xaa12−Xaa13−Xaa14−V
(IV)
式中、
Xaa10は、Gln又はSerであってよく、
Xaa11は、Thr、Glu又はAspであってよく、
Xaa12は、Val又はAsnであってよく、
Xaa13は、Tyr又はPheであってよく、
Xaa14は、Ser、Asn又はGlnであってよい。
【0081】
他の実施形態では、本発明は、重鎖可変領域及び軽鎖可変領域の両方を有する、TLR3反応性を有する単離された抗体又はそのフラグメントを提供するものであり、前記抗体は配列番号195に示されるようなLCDR3のアミノ酸配列を有し、配列番号195のLCDR3は式(VI)に示されるように定義される。
Q−Q−Xaa18−Xaa19−Xaa20−Xaa21−Xaa22−Xaa23−T
(VI)
式中、
Xaa18は、Tyr、Gly又はAlaであってよく、
Xaa19は、Gly、Glu又はAsnであってよく、
Xaa20は、Ser又はThrであってよく、
Xaa21は、Val、Ile又はLeuであってよく、
Xaa22は、Ser又はLeuであってよく、
Xaa23は、Ile、Ser、Pro又はTyrであってよい。
【0082】
本発明は、また、表1aに示されるような重鎖の相補性決定領域(CDR)のアミノ酸配列1、2及び3(HCDR1、HCDR2及びHCDR3)及び軽鎖の相補性決定領域(CDR)のアミノ酸配列1、2及び3(LCDR1、LCDR2及びLCDR3)を有する、TLR3反応性を有する単離された抗体又はそのフラグメントも提供するものである。
【0083】
抗原結合部位のアミノ酸配列が表1aのもの(配列番号49〜121及び191〜196)と大きくは異ならない抗体は本発明の範囲に含まれるものである。一般に、これには1以上のアミノ酸残基を、同様の電荷、疎水性又は立体化学的特性を有するアミノ酸によって置換することが含まれる。抗原結合部位ではなく、フレームワーク領域における更なる置換も、これらの置換が抗体の特性に悪影響を及ぼさない限りは行うことができる。置換は、例えば安定性又は親和性といった抗体の特性を向上させるために行うことができる。1、2、3、4、5又は6個の置換を抗原結合部位に行うことができる。得られる抗体が所望の特性を保持するならば、フレームワーク残基の1%、2%、3%、4%、5%、10%、15%、20%、25%、又は30%を置換することができる。
【0084】
保存的な改変では、こうした改変が行われた分子と同様の機能的及び化学的性質を有する分子が生ずる。分子の機能的及び/又は化学的特性における大幅な改変は、(1)置換領域における分子骨格の構造を例えばシート又はヘリックス立体構造として維持する、(2)標的部位における分子の電荷又は疎水性を維持する、又は(3)分子のサイズを維持する効果において、大きく異なるアミノ酸配列中の置換を選択することによって実現することができる。例えば、「保存的アミノ酸置換」では、その位置のアミノ酸残基の極性又は電荷にほとんどあるいはまったく影響しないように天然アミノ酸残基を非天然残基と置換することができる。更に、アラニン・スキャニング変異導入法についてこれまでに述べられているように(MacLennanら、Acta Physiol.Scand.Suppl.643:55〜67、1998;Sasakiら、Adv.Biophys.35:1〜24、1998)、ポリペプチド内の任意の内因性の残基をアラニンで置換することもできる。所望のアミノ酸置換(保存的又は非保存的であるかによらず)は、当業者であればこうした置換が望ましい時点で決定することができる。例えば、アミノ酸置換を用いることによってその分子の配列の重要な残基を特定したり、又は本明細書で述べる分子の親和性を増大若しくは減少させることができる。アミノ酸置換の例を表1bに示す。
【0085】
特定の実施形態においては、保存的アミノ酸置換には、生物学的システムによって合成されるのではなく一般的に化学的ペプチド合成によって組み込まれる、非天然のアミノ酸残基も含まれる。アミノ酸置換は、例えばPCR変異誘発(米国特許第4,683,195号)によって行うことができる。周知の方法を用いて、例えば、ランダムコドン(NNK)又は非ランダムコドン、例えば11個のアミノ酸(ACDEGKNRSYW)をコードするDVKコドンを用い、実施例1に示すような所望の特性を持つ変異体についてライブラリをスクリーニングすることで、変異体のライブラリを生成させることができる。表1cは、抗体の特性を改善するためにLCDR3及びHCDR2領域内で3つの親TLR3抗体アンタゴニストに対して行われた置換を示す。
【0086】
抗原結合部位の表記に応じて、本発明の抗体の抗原結合部位、次いでフレームワーク残基は、各重鎖及び軽鎖ごとに若干異なってもよい。
【0087】
【表3】
【0088】
表2a及び2bは、Kabat、Chothia及びIMGTに基づいて表記された本発明の例示的抗体の抗原結合部位の残基及びそれらの複合配列を示したものである。
【0089】
他の実施形態において、本発明は、重鎖可変領域及び軽鎖可変領域の両方を有する、TLR3反応性を有する単離された抗体又はそのフラグメントを提供するものであり、前記抗体は重鎖可変領域(VH)及び軽鎖可変領域(VL)のアミノ酸配列を有するとともに表3aに示されるようなそれぞれの単離された重鎖可変領域及び軽鎖可変領域を提供する。F17、F18及びF19は、それぞれ、ファミリー17、18及び19のコンセンサスなアミノ酸配列を有する抗体の変異体を表す(実施例1を参照)。
【0090】
【表4−1】
【0091】
【表4−2】
【0092】
【表4−3】
【0093】
実施例で説明する実施形態は、1個が重鎖に由来し、1個が軽鎖に由来する1対の可変領域を有しているが、当業者であれば代替的な実施形態は、1個の重鎖又は軽鎖の可変領域を有してもよいということを認識するであろう。単一の可変領域を用いることで、例えばTLR3に結合することが可能な2ドメイン特異的抗原結合フラグメントを形成することが可能な第2の可変ドメインについてスクリーニングを行うことができる。こうしたスクリーニングは、例えば国際公開第92/01047号に開示される、階層的に2重であるコンビナトリアルなアプローチを用いたファージディスプレイスクリーニング法によって実現することが可能である。このアプローチでは、H又はL鎖のいずれかのクローンを含む個別のコロニーを用いて他方の鎖(L又はH)をコードするクローンの完全なライブラリに感染させ、得られた2鎖特異的抗原結合ドメインを上記に述べたようなファージディスプレイ法に基づいて選択する。
【0094】
【表5】
【0095】
他の実施形態では、本発明は、表3aに示されるような可変領域のアミノ酸配列と少なくとも95%同一のアミノ酸配列を有する重鎖及び軽鎖の可変領域の両方を有し、TLR3反応性を有する単離された抗体又はそのフラグメントを提供する。
【0096】
別の態様では、本発明は、表3bに示されるような特定の重鎖及び軽鎖のアミノ酸配列を有する単離された抗体を提供する。
【0097】
本発明の別の態様は、本発明の抗体又はその補体のいずれかをコードする単離されたポリヌクレオチドである。特定のポリヌクレオチドの例を本明細書に開示するが、遺伝暗号の縮重及び特定の発現系におけるコドンの選択性を考慮すると、本発明の抗体アンタゴニストをコードする他のポリヌクレオチドも本発明の範囲内に含まれる。
【0098】
【表6】
【0099】
【表7】
【0100】
抗体アンタゴニストの例としては、IgG、IgD、IgG、IgA又はIgMアイソタイプの抗体が挙げられる。更にこうした抗体アンタゴニストは、グリコシル化、異性化、脱グリコシル化、又はポリエチレングリコール部分の付加(ペギレーション)及び脂質化などの非天然の共有結合修飾などの反応によって翻訳後修飾されてもよい。こうした修飾はインビボあるいはインビトロで行われ得る。例えば、本発明の抗体はポリエチレングリコールと結合(ペギレート)させることによって薬物動態的なプロファイルを向上させることができる。結合は当業者には周知の方法によって行うことができる。治療用抗体とPEGの結合は、機能を妨害せずに薬物動態を増強することが示されてきた(Deckertら、Int.J.Cancer 87:382〜390,2000;Knightら、Platelets 15:409〜418,2004;Leongら、Cytokine 16:106〜119,2001;Yangら、Protein Eng.16:761〜770,2003)。
【0101】
【表8】
【0102】
本発明の抗体の薬物動態的性質は、当業者には周知の技法により、Fc修飾によって向上させることも可能である。例えばIgG4アイソタイプの重鎖は、ヒンジ領域に重鎖間又は重鎖内ジスルフィド結合を形成することが可能なCys−Pro−Ser−Cys(CPSC)というモチーフを含んでいる。すなわち、CPSCモチーフ内の2個のCys残基が他の重鎖の対応するCys残基とジスルフィド結合する(重鎖間)か、あるいは特定のCPSCモチーフ内の2個のCys残基が互いにジスルフィド結合し得る(重鎖内)。生体内のイソメラーゼ酵素は、IgG4分子の重鎖間結合を重鎖内結合に変換し、またその逆反応を行うことが可能であると考えられている(Aalberse及びSchuurman、Immunology 105:9〜19、2002)。したがって、ヒンジ領域に重鎖内結合を有するこうしたIgG4分子内の重鎖:軽鎖(H:L)の各ペアは互いに共有結合していないことからH:Lモノマーに解離し、更に他のIgG4分子に由来するH:Lモノマーと再会合して、2重特異的なヘテロダイマーのIgG4分子を形成する。2重特異的なIgG抗体では、抗体分子の2個のFabの結合するエピトープが異なる。IgG4のヒンジ領域CPSCモチーフ中のSer残基をProにより置換することによって「IgG1様の挙動」が得られる。すなわち、この分子が重鎖間で安定なジスルフィド結合を形成し、それゆえ、他のIgG4分子とのH:L交換を受けにくい。一つの実施形態では、本発明の抗体は、SからPの変異と共にCPSCモチーフ中にIgG4Fcドメインを含む。CPSCモチーフの位置は通常、成熟した重鎖の残基228に見いだされるが、CDRの長さに応じて変化し得る。
【0103】
更に、本発明の抗体のFcRnサルベージ受容体以外のFc受容体への結合に影響する部位を除去することもできる。例えば、ADCC活性に関与するFc受容体結合領域を本発明の抗体において除去することができる。例えば、IgG1のヒンジ領域のLeu234/Leu235のL234A/L235Aへの変異、又はIgG4のヒンジ領域のPhe235/Leu236のP235A/L236Aへの変異によって、FcRへの結合性が最小となり、補体依存性細胞傷害活性及びADCCを介在する免疫グロブリンの能力が低減する。一実施形態では、本発明の抗体は、P235A/L236A変異を有するIgG4のFcドメインを有する。上記で特定したこれらの残基の位置は成熟した重鎖においては典型的なものであるが、CDRの長さに応じて変化し得る。P235A/L236A変異を有する抗体の例としては、配列番号218、219又は220に示される重鎖アミノ酸配列を有する抗体が挙げられる。
【0104】
完全なヒト、ヒト適合化、ヒト化、及び親和性成熟抗体分子又は抗体フラグメントは、融合タンパク質及びキメラタンパク質と同様、本発明の範囲に含まれる。抗原に対する抗体親和性は、ランダム又は部位特異的な変異誘導などの周知の方法を用いる合理的設計(rational design)又はランダムな親和性成熟によるか、あるいはファージディスプレイライブラリを用いることによって高めることができる。例えば、大部分がフレームワーク領域中に存在するバーニア残基又は「親和性決定残基」、ADRに対して置換を行うことで、抗体の親和性を調整することができる(米国特許第6,639,055号、国際公開第10/045340号)。
【0105】
安定性、選択性、交差反応性、親和性、免疫原性、又はタンパク質の所望の生物学的若しくは生物物理学的特性を高めるように改変された完全なヒト、ヒト適合化、ヒト化、親和性成熟抗体分子又は抗体フラグメントは本発明の範囲に含まれる。抗体の安定性は、(1)内在的な安定性に影響する個々のドメインのコアパッキング、(2)重鎖と軽鎖とのペアリングに影響するタンパク質/タンパク質の界面相互作用、(3)極性及び荷電残基の埋め込み、(4)極性及び荷電残基の水素結合ネットワーク、及び(5)分子内及び分子間力の中でも特に表面電荷及び極性残基の分布、などの多くの因子によって影響される。残基を不安定化させる可能性のある構造は、抗体の結晶構造に基づいて、あるいは場合によっては分子モデリングによって識別することが可能であり、抗体の安定性に対するこれらの残基の影響を、識別された残基中で変異を有する変異体を生成及び評価することにより試験することができる。抗体の安定性を高める方法の1つは、示差走査熱量測定法(DSC)によって測定される熱転移中点(Tm)を高くすることである。一般に、タンパク質のTmはその安定性との相関を示し、溶液中でのアンフォールディング及び変性のしやすさ、並びにそのタンパク質のアンフォールドのしやすさに依存する分解プロセスと負の相関を示す(Remmeleら、Biopharm.,13:36〜46,2000)。多数の研究によって、DSCにより熱安定性として測定される配合物の物理的安定性と他の方法により測定される物理的安定性のランキングの間に相関性が見出された(Guptaら、AAPS PharmSci.5E8,2003;Zhangら、J.Pharm.Sci.93:3076〜3089,2004;Maaら、Int.J.Pharm.,140:155〜168,1996;Bedu−Addoら、Pharm.Res.,21:1353〜1361,2004;Remmeleら、Pharm.Res.,15:200〜208,1997)。これらの配合物の研究は、FabのTmが対応するmAbの長期の物理的安定性と密接な関係があることを示唆している。フレームワーク中又は抗原結合部位内のアミノ酸の差は、Fabドメインの熱安定性に及ぼす著しい影響を有する可能性がある(Yasuiら、FEBS Lett.353:143〜146,1994)。
【0106】
本発明の抗体アンタゴニストは、約10-7、10-8、10-9、10-10、10-11又は10-12M以下のKdでTLR3と結合することができる。抗体などの特定の分子のTLR3に対する親和性は、任意の適当な方法によって実験的に求めることができる。こうした方法では、当業者には周知であるBiacore又はKinExa装置、ELISA又は競合的結合アッセイを使用することができる。
【0107】
特定のTLR3ホモログと所望の親和性で結合する抗体アンタゴニストは、抗体親和性成熟などの方法によって変異体又はフラグメントのライブラリから選択することができる。抗体アンタゴニストは、任意の適当な方法を用いてTLR3の生物活性の阻害に基づいて同定することができる。こうした方法では、周知の方法を用いた、本願に述べるようなレポーター遺伝子アッセイ又はサイトカイン産生を測定するアッセイを利用することができる。
【0108】
本発明の別の実施形態は、少なくとも1つの本発明のポリヌクレオチドを含むベクターである。かかるベクターは、プラスミドベクター、ウイルスベクター、バキュロウイルス発現ベクター、トランスポゾンに基づいたベクター、又は任意の手段によって特定の生物又は遺伝子的バックグラウンドに本発明のポリヌクレオチドを導入するのに適した他の任意のベクターであってよい。
【0109】
本発明の別の実施形態は、配列番号6、8、10、12、14、16、18、20、22、24、26、28、30、32、34、36、38、40、42、124、125、126、127、128、129、159、198、200、202、164、212、213、214、215又は216に示されるアミノ酸配列を有する免疫グロブリン重鎖の可変領域、又は配列番号5、7、9、11、13、15、17、19、21、23、25、27、29、31、33、35、37、39、41、122、123、197、199、201、163、209、210、211又は225に示されるアミノ酸配列を有する免疫グロブリン軽鎖の可変領域を含むポリペプチドをコードするポリヌクレオチドなどの、本発明のポリヌクレオチドのいずれかを有するホスト細胞である。
【0110】
本発明の別の実施形態は、配列番号102、130、131、132、133、134、135、160、204、206、208、220、166又は168に示されるアミノ酸配列を有する免疫グロブリン重鎖の可変領域、又は配列番号155、156、157、158、203、205、207、165、167又は227に示されるアミノ酸配列を有する免疫グロブリン軽鎖の可変領域を含むポリペプチドをコードするポリヌクレオチドを有するホスト細胞である。かかるホスト細胞は、真核細胞、細菌細胞、植物細胞又は古細菌細胞であってよい。真核細胞の例としては、哺乳動物、昆虫、鳥類又は他の動物由来のものであり得る。哺乳動物真核細胞としては、SP2/0(ATCC(American Type Culture Collection),Manassas,VA,CRL−1581)、NS0(ECACC(European Collection of Cell Cultures),Salisbury,Wiltshire,UK,ECACC No.85110503)、FO(ATCC CRL−1646)及びAg653(ATCC CRL−1580)マウス細胞株などのハイブリドーマ又はミエローマなどの不死化細胞株が挙げられる。ヒトミエローマ細胞株の一例には、U266(ATTC CRL−TIB−196)がある。他の有用な細胞株としては、CHO−K1SV(Lonza Biologics,Walkersville,MD)、CHO−K1(ATCC CRL−61)、又はDG44などの、チャイニーズハムスターの卵巣(CHO)細胞に由来するものが挙げられる。
【0111】
本発明の別の実施形態は、本発明のホスト細胞を培養することと、前記ホスト細胞によって産生された抗体を回収することとを含む、TLR3と反応性を有する抗体の製造方法である。抗体を製造して精製する方法は当該技術分野では周知のものである。
【0112】
本発明の別の実施形態は、本発明の抗体を産生するハイブリドーマ細胞株である。
【0113】
本発明の別の実施形態は、toll様受容体3(TLR3)の、配列番号2のアミノ酸残基;K416、K418、L440、N441、E442、Y465、N466、K467、Y468、R488、R489、A491、K493、N515、N516、N517、H539、N541、S571、L595、及びK619に結合する、単離された抗体又はそのフラグメントである。
【0114】
別の実施形態は、toll様受容体3(TLR3)の、配列番号2のアミノ酸残基;S115、D116、K117、A120、K139、N140、N141、V144、K145、T166、Q167、V168、S188、E189、D192、A195、及びA219に結合する、単離された抗体又はそのフラグメントである。
【0115】
幾つかの周知の方法を用いて本発明の抗体の結合エピトープを決定することができる。例えば、両方の個別の要素の構造が分かっている場合、インシリコでタンパク質−タンパク質のドッキングを行って、適合する相互作用部位を同定することができる。抗原抗体複合体において水素−重水素(H/D)交換を行うことによって抗体が結合し得る抗原の領域をマッピングすることができる。抗原のセグメント及び点変異誘導を用いることにより、抗体の結合に重要なアミノ酸の位置を特定することができる。TLR3のような大きなタンパク質では、最初にドッキング、セグメント変異誘導、又はH/D交換などによって結合部位の位置をそのタンパク質の特定の領域に限定することで点変異誘導によるマッピングが単純化される。両方の個別の要素の構造が分かっている場合、インシリコでタンパク質−タンパク質のドッキングを行って適合する相互作用の部位を同定することができる。抗体−抗原複合体の共結晶構造を使用して、エピトープ及びパラトープに寄与する残基を同定することができる。
【0116】
本発明の別の実施形態は、単離された抗体又はそのフラグメントであって、重鎖可変領域のChothia残基;W33、F50、D52、D54、Y56、N58、P61、E95、Y97、Y100及びD100b、並びに軽鎖可変領域のChothia残基;Q27、Y32、N92、T93、L94及びS95により、配列番号2で示されるアミノ酸配列を有するTLR3に結合することを特徴とする。重鎖パラトープ及び軽鎖パラトープChothia残基は、重鎖残基の配列番号216のW33、F50、D52、D55、Y57、N59、P62、E99、Y101、Y104及びD106並びに軽鎖残基の配列番号41のQ27、Y32、N92、T93、L94及びS95に対応する。
【0117】
本発明の別の実施形態は、単離された抗体又はそのフラグメントであって、重鎖可変領域のChothia残基;N31a、Q52、R52b、S53、K54、Y56、Y97、P98、F99及びY100、並びに軽鎖可変領域のChothia残基;G29、S30、Y31、Y32、E50、D51、Y91、D92及びD93により、配列番号2に示されるアミノ酸配列を有するTLR3に結合することを特徴とする。重鎖パラトープ及び軽鎖パラトープChothia残基は、重鎖残基の配列番号214のN32、Q54、R56、S57、K58、Y60、Y104、P105、F106及びY107並びに軽鎖残基の配列番号211のG28、S29、Y30、Y31、E49、D50、Y90、D91及びD92に対応する。
【0118】
例えば、パラトープ残基を好適な骨格にグラフトし、作製した骨格を完全な抗体に組立て、得られた抗体を発現させ、及びTLR3への結合又はTLR3の生物活性に及ぼす影響について抗体を試験することにより、TLR3を結合する特定のパラトープ残基を有する単離された抗体を作製することができる。代表的な骨格は、ヒト生殖細胞系列遺伝子によりコードされたヒト抗体可変領域のアミノ酸配列である。例えば、全体配列同一性、パラトープ残基間の同一性(%)、又は骨格及びmAb 15EVQ又はmAb 12QVQ/QSVなどの代表的な抗体との間の正準構造クラス同一性に基づいて、骨格を選択することができる。ヒト抗体生殖細胞系列遺伝子は、例えば、TomLinsonら、J.Mol.Biol 227:776〜798、及びInternational ImMunoGeneTics(IMGT)データベース(http_://_www_imgt_org)で開示されている。例えば、米国特許第6,300,064号で述べられているように、コンセンサスなヒトフレームワーク領域も使用することができる。例えば、国際公開第10/045340号に述べられている方法に従って、好適な骨格の選択を行うことができる。
【0119】
パラトープ残基をグラフトする骨格として使用可能な代表的なヒト生殖細胞系列遺伝子は、Vκ1、Vλ3、Vh5、Vh6、Jκ、Jλ、及びJhフレームワークによりコードされた遺伝子である。この生殖細胞系列J領域は、FR4配列を選択するために、その全体又は一部を使用される。例えば、mAb 15EVQ軽鎖パラトープ残基を、IGκJ1によりコードされたJ領域配列に直接連結されている、IGKV1−39*01によりコードされたVκ1フレームワークにグラフトすることができる。他のVκ1遺伝子由来の配列を使用することもでき、IGκJ1の代わりに、他のJκ遺伝子のFR4配列を置換することもできる。mAb 15EVQ重鎖パラトープ残基と、続いて、HCDR3及びIGHJ1によりコードされたFR4配列を構成する約11〜13個の残基、例えば12個の残基を、IGHV5−51*01によりコードされたVh5フレームワークにグラフトすることができる。11〜13個の残基は、FR3領域(「CAR」)の終端とFR4領域(大部分のJH領域に対するWGQ)の初端の間に懸り、mAb 15EVQVh由来の4個の規定されたパラトープ残基を含む。他のVh5遺伝子由来の配列を使用することもでき、IGJH1の代わりに他のJh遺伝子のFR4配列を置換することもできる。別の例では、mAb 12QVQ/QSV軽鎖パラトープ残基を、IGJL2によりコードされたJ領域配列に直接連結したIGLV3−1*01によりコードされたVλ3フレームワークにグラフトすることができる。他のVλ3及びJλ遺伝子の配列を使用することもできる。LCDR3の長さは、約9〜11個の残基、例えば、10個の残基で維持される。これらの約9〜11個の残基は、FR3領域(大部分のVラムダ骨格に対する「YYC」)の終端とFR4領域(大部分のJL領域に対する「FGG」)の初端との間に懸り、mAb 12QVQ/QSV由来の3個の規定されたパラトープ残基を含む。mAb 12QVQ/QSV重鎖パラトープ残基と、続いてHCDR3及びIGJH1によりコードされたFR4配列を構成する約9〜11個の残基、例えば10個の残基を、IGHV6−1*01によりコードされたVh6フレームワークにグラフトすることができる。これらの約9〜11個の残基は、FR3領域(「CAR」)の終端とFR4領域(大部分のJH領域に対するWGQ)の初端との間に懸り、mAb 12QVQ/QSVVhからの4個の規定されたパラトープ残基を含む。IGHJ1の代わりに他のJh遺伝子のFR4配列を置換することもできる。TLR3への結合及び得られた抗体の生物活性を、標準的な方法を用いて評価することができる。mAb 15EVQ及びmAb 12QVQ/QSV軽鎖可変領域及び重鎖可変領域と、代表的なVκ1、Vh5、Vλ3、Vh6、Jκ、Jλ又はJh遺伝子とのアライメントを、図32〜35に示す。ラトープをグラフトした、作製された抗体を、バーニア残基又は親和性決定残基の置換により更に改変して、抗体の特性、例えば上述のように親和性を改善することができる。パラトープをグラフトした抗体がTLR3への結合を保持する限り、パラトープをグラフトした抗体中のフレームワークアミノ酸配列は、mAb 15EVQ又は12QVQ/QSVフレームワーク配列と70%、75%、80%、85%、90%、95%、96%、97%、98%、又は99%同一であってもよい。
【0120】
標準的な方法を用いて、パラトープ残基に追加して抗原結合部位由来の配列をグラフトすることもできる。例えば、完全なHCDR3又はLCDR3をグラフトしてもよい。
【0121】
本発明の別の態様は、単離された抗体又はそのフラグメントであって、特定の重鎖相補性決定領域(CDR)1、2及び3のアミノ酸配列、特定の軽鎖CDR1、2及び3のアミノ酸配列、特定の重鎖可変領域(VH)のアミノ酸配列、又は特定の軽鎖可変領域(VL)のアミノ酸配列を有するモノクローナル抗体とTLR3との結合について競合する、TLR3反応性を有することを特徴とする。本発明のモノクローナル抗体の例としては、配列番号216に示されるアミノ酸配列を有する重鎖可変領域及び配列番号41に示される軽鎖可変領域のアミノ酸配列を有する単離された抗体、並びに配列番号214に示されるアミノ酸配列を有する重鎖可変領域及び配列番号211に示される軽鎖可変領域のアミノ酸配列を有する抗体が挙げられる。
【0122】
TLR3との結合についての競合は公知の方法によってインビトロでアッセイすることができる。例えば、非標識抗体の存在下でのMSD Sulfo−Tag(商標)NHS−エステル標識抗体のTLR3への結合を、ELISAによって評価することができる。本発明の抗体の例としては、mAb 12、mAb 15及びmAb c1811がある(表3aを参照)。前述の抗TLR3抗体c1068及びその誘導体(国際公開第06/060513(A2)号)、TLR3.7(eBiosciences,カタログ番号14−9039)、並びにImgenex IMG−315A(Imgenex IMG−315A、ヒトTLR3のアミノ酸55〜70のVLTHNQLRRLPAANに対して生成されたもの)は、実施例5に示されるようにmAb 12、15又はc1811によるTLR3への結合と競合しない。
【0123】
本発明の別の態様は、TLR3反応性を有し、以下のa〜iの特性のうち少なくとも1つを有する、単離された抗体である;
a.Kd<10nMでヒトTLR3に結合する;
b.インビトロでのポリ(I:C)NF−κBレポーター遺伝子アッセイにおいて、1μg/mLで、ヒトTLR3の生物活性を50%超低減させる;
c.10μg/mLで、100ng/mL未満のポリ(I:C)で刺激されたBEAS−2B細胞からのIL−6又はCXCL5/IP−10の産生を60%超阻害する;
d.0.4μg/mLで、100ng/mL未満のポリ(I:C)で刺激されたBEAS−2B細胞からのIL−6又はCXCL5/IP−10の産生を50%超阻害する;
e.5μg/mLで、62.5ng/mLのポリ(I:C)で刺激されたNHBE細胞からのIL−6の産生を50%超阻害する;
f.1μg/mLで、62.5ng/mLのポリ(I:C)で刺激されたNHBE細胞からのIL−6の産生を50%超阻害する;
g.1μg/mLで、PBMC細胞による、ポリ(I:C)により誘導されたIFN−γ、IL−6又はIL−12の産生を20%超阻害する;
h.インビトロでのNF−κBレポーター遺伝子アッセイにおいて、IC50<10μg/mLで、カニクイザルのTLR3の生物活性を阻害する;
i.インビトロでのISREレポーター遺伝子アッセイにおいて、IC50<5μg/mLで、カニクイザルのTLR3の生物活性を阻害する。
【0124】
治療方法
本発明のTLR3アンタゴニスト、例えばTLR3抗体アンタゴニストは、免疫系を調節する目的で使用することができる。何らのかの特定の理論に束縛されることも望むものではないが、本発明のアンタゴニストは、TLR3へのリガンドの結合、TLR3の2量化、TLR3の細胞内移行、又はTLR3の輸送を防止又は低減することによって免疫系を調節し得るものと考えられる。本発明の方法を用いて任意の分類に属する動物患者を治療することができる。こうした動物の例としては、ヒト、齧歯類、犬、猫、及び家畜などの哺乳動物が挙げられる。例えば本発明の抗体は、TLR3活性の中和、炎症、炎症性及び代謝性疾患の治療に有用であるだけでなく、更にこうした治療のための薬剤の調製においても有用であり、その場合、薬剤は本明細書で定義する用量で投与されるように調製される。
【0125】
一般的に、本発明のTLR3抗体アンタゴニストの投与によって予防又は治療することが可能な炎症状態、感染にともなう状態、又は免疫介在性炎症性疾患には、サイトカイン又はケモカインによって介在されるもの、及びTLR3の活性化又はTLR3経路を介したシグナル伝達に全体的又は部分的に起因する状態が含まれる。こうした炎症性状態の例としては、敗血症にともなう状態、炎症性腸疾患、自己免疫性疾患、炎症性疾患、及び感染にともなう状態が挙げられる。更に、本発明のTLR3抗体アンタゴニストの投与によって癌、心血管及び代謝性の状態、神経病及び線維症の状態を予防又は治療することも可能であると考えられる。炎症は組織を冒し、全身性の症状となり得る。罹患する組織の例としては、気道、肺、消化管、小腸、大腸、結腸、直腸、心血管系、心組織、血管、関節、骨及び滑膜組織、軟骨、上皮、内皮、肝又は脂肪組織が挙げられる。全身性の炎症状態の例としては、サイトカインストームすなわち高サイトカイン血症、全身性炎症反応症候群(SIRS)、移植片対宿主病(GVHD)、急性呼吸窮迫症候群(ARDS)、重症急性呼吸器症候群(SARS)、劇症型抗リン脂質症候群、重篤なウイルス感染、インフルエンザ、肺炎、ショック、又は敗血症がある。
【0126】
炎症は、侵入する病原体を撃退するための生物による防御反応である。炎症は、多くの細胞性及び体液性のメディエーターが関与するカスケードイベントである。一方で炎症反応の抑制はホストを免疫的に無防備状態にし、炎症を放置した場合には慢性炎症性疾患(例えば、喘息、乾癬、関節炎、リウマチ様関節炎、多発性硬化症、炎症性腸疾患など)、敗血性ショック、及び多臓器不全などの重篤な合併症を引き起こし得る。重要な点として、これらの多様な疾患状態は、サイトカイン、ケモカイン、炎症性細胞、及びこれらの細胞によって分泌される他のメディエーターなどの共通の炎症性メディエーターを共有している。
【0127】
リガンドであるポリ(I:C)、2本鎖RNA又は内因性のmRNAによってTLR3が活性化されるとシグナル伝達経路が活性化され、これにより炎症性サイトカインの合成及び分泌、マクロファージ、顆粒球、好中球及び好酸球などの炎症性細胞の活性化及び動員、細胞死、並びに組織の破壊が引き起こされる。TLR3は、IL−6、IL−8、IL−12、TNF−α、MIP−1、CXCL5/IP−10及びRANTES、並びに免疫細胞の動員及び活性化に関わる他の炎症性サイトカイン及びケモカインの分泌を誘導することにより、自己免疫疾患及び他の炎症性疾患における組織の破壊に寄与する。TLR3のリガンドである内因性のmRNAは炎症時に壊死細胞から放出され、ポジティブフィードバックループを引き起こすことによってTLR3を活性化し、炎症及び更なる組織破壊を持続させる。TLR3抗体アンタゴニストのようなTLR3アンタゴニストはサイトカインの分泌を正常化し、炎症性細胞の動員を低減し、組織破壊及び細胞死を低減させると考えられる。したがって、TLR3アンタゴニストは炎症及び広範な炎症性状態を治療するうえで治療上の可能性を有するものである。
【0128】
炎症性状態の1つの例は、全身性炎症反応症候群(SIRS)、敗血性ショック又は多臓器不全症候群(MODS)が含まれ得る、敗血症にともなう状態である。ウイルス、細菌、真菌、又は寄生虫感染によって、及び壊死細胞によって放出される2本鎖RNAは敗血症の発症に寄与し得る。特定の理論によって束縛されることを望むものではないが、TLR3アンタゴニストによる治療は、敗血症にともなう炎症状態を有する患者における生存時間を延ばすか又は局所的な炎症性イベント(例えば肺の)が広がって全身症状となることを防止することにより、あるいは内在的な抗微生物活性を強化することにより、あるいは抗微生物剤と組み合わされる場合に相乗的な活性を示すことにより、あるいは病状に寄与する局所的炎症状態を抑制することにより、あるいは上記の任意の組み合わせにより、所定の治療効果を与え得るものと考えられる。こうした介入は、患者の生存を確実にするために必要とされる更なる治療(例えば基礎となる炎症の治療又はサイトカインレベルの低減)を可能とするうえで充分なものとなり得る。敗血症はD−ガラクトサミン及びポリ(I:C)を投与することによってマウスなどの動物でモデル化することができる。こうしたモデルにおいて、D−ガラクトサミンは、敗血症感作物質として機能する肝臓毒素であり、ポリ(I:C)は2本鎖RNAを模倣してTLR3を活性化させる敗血症誘導分子である。TLR3アンタゴニストによる治療は、敗血症のマウスモデルにおいてマウスの生存率を高め得るものであることから、TLR3アンタゴニストは敗血症の治療に有用であると考えられる。
【0129】
消化管の炎症は消化管の粘膜の炎症であり、急性及び慢性の炎症状態が含まれる。急性炎症は、短時間の発症及び好中球の浸潤又は流入によって一般的に特徴付けられる。慢性炎症は、比較的長期の発症及び単核球の浸潤又は流入によって一般的に特徴付けられる。粘膜層は、腸(小腸及び大腸を含む)、直腸、胃(胃部)壁、又は口腔の粘膜であってよい。慢性の消化管炎症状態の例としては、炎症性腸疾患(IBD)、外界からの刺激によって誘導される大腸炎(例えば、化学療法、放射線療法などの投与のような治療レジメンによって引き起こされるかあるいはこれにともなう(例えば副作用として)消化管炎症(例えば大腸炎))、感染性大腸炎、虚血性大腸炎、膠原性又はリンパ性大腸炎、壊死性腸炎、慢性肉芽腫症又はセリアック病などの状態における大腸炎、食品アレルギー、胃炎、感染性胃炎又は小腸結腸炎(例えばヘリコバクター・ピロリの感染による慢性活動性胃炎)、及び感染性病原体によって引き起こされる消化管炎症の他の形態がある。
【0130】
炎症性腸疾患(IBD)には、例えば潰瘍性大腸炎(UC)及びクローン病(CD)などの、一般に病因が不明の一群の慢性炎症性疾患が含まれる。臨床的及び実験的証拠は、IBDの病因が感受性遺伝子及び環境因子が関与する多因子的なものであることを示唆している。炎症性腸疾患では、組織の傷害は、腸内の微生物叢の、抗原に対する不適切又は過剰な免疫反応によってもたらされる。炎症性腸疾患の幾つかの動物モデルが存在している。最も広く用いられているモデルは、結腸に慢性炎症及び潰瘍形成を誘発する2,4,6−トリニトロベンゼンスルホン酸/エタノール(TNBS)誘発大腸炎モデル又はオキサゾロンモデルである(Neurathら、Intern.Rev.Immunol 19:51〜62、2000)。別のモデルでは、血性下痢、体重減少、結腸の短縮、及び好中球の浸潤をともなう粘膜の潰瘍形成として発現する急性大腸炎を誘発する硫酸デキストランナトリウム(DSS)を使用する。DSS誘発大腸炎は、リンパ組織過形成、陰窩傷害(focal crypt damage)、及び上皮潰瘍形成をともなう、基底膜内への炎症性細胞の浸潤によって組織学的に特徴付けられる(Hendricksonら、Clinical Microbiology Reviews 15:79〜94、2002)。別のモデルは、ナイーブなCD45RBhigh CD4 T細胞をRAG又はSCIDマウスに養子移入することを含む。このモデルでは、ドナーのナイーブT細胞がレシピエントの腸を攻撃することにより、ヒトの炎症性腸疾患に似た慢性腸炎症及び症状を引き起こす(Read及びPowrie、Curr.Protoc.Immunol.Chapter 15 unit 15.13、2001)。これらのモデルのいずれかにおいて本発明のアンタゴニストの投与を用いることによって、これらのアンタゴニストが、炎症性腸疾患などの腸内の炎症にともなう症状を改善し、疾患の経過を変化させる潜在的な有効性を評価することができる。IBDに対しては幾つかの治療選択肢が存在し、例えば抗TNFα抗体による治療法が、クローン病を治療する目的で10年前から用いられている(Van Asscheら、Eur.J.Pharmacol.Epub Oct 2009)。しかしながら、かなりの割合の患者が現在の治療法に応答しない(Hanauerら、Lancet 359:1541〜1549、2002;Hanauerら、Gastroenterology 130:323〜333、2006)ことから、難治性の患者集団を標的とした新たな治療法が求められている。
【0131】
炎症状態の別の一例は炎症性の肺状態である。炎症性の肺状態の例としては、ウイルス、細菌、真菌、寄生虫又はプリオンによる感染にともなうものなどの感染により誘発される肺状態、アレルゲンにより誘発される肺状態、アスベスト症、珪肺症、又はベリリウム肺症などの汚染物質により誘発される肺状態、胃吸引により誘発される肺状態、嚢胞性線維症などの遺伝的素因にともなう免疫調節不全炎症状態、及び人工呼吸器傷害などの物理的外傷により誘発される肺状態が挙げられる。これらの炎症状態には更に、喘息、気腫、気管支炎、慢性閉塞性肺疾患(COPD)、サルコイドーシス、ヒスチオサイトーシス、リンパ管筋腫症、急性肺傷害、急性呼吸窮迫症候群、慢性肺疾患、気管支肺異形成症、院外感染性肺炎、院内肺炎、人工呼吸器関連肺炎、敗血症、ウイルス性肺炎、インフルエンザ感染、パラインフルエンザ感染、ロタウイルス感染、ヒトメタニューモウイルス感染、呼吸系発疹ウイルス感染、及びアスペルギルス又は他の真菌感染も含まれる。感染にともなう炎症性疾患の例としては、重症肺炎、嚢胞性線維症、気管支炎、気道増悪、及び急性呼吸窮迫症候群(ARDS)を含むウイルス又は細菌性肺炎が挙げられる。こうした感染にともなう状態は、一次ウイルス感染及び二次ウイルス感染のように複数の感染をともなう場合がある。
【0132】
喘息は、気道過敏症(AHR)、気管支収縮、喘鳴、好酸球又は好中球性炎症、粘液分泌過多、上皮下線維症、及び高いIgEレベルによって特徴付けられる肺の炎症性疾患である。喘息の患者は、最も一般的には気道の微生物感染(例えばライノウイルス、インフルエンザウイルス、ヘモフィルスインフルエンザなど)により症状の悪化である「増悪」を経験する。喘息の発作は、環境因子(例えば、回虫、昆虫、動物(例えばネコ、イヌ、ウサギ、マウス、ラット、ハムスター、モルモット及び鳥)、真菌、空気汚染物質(例えば煙草の煙)、刺激性ガス、フューム、蒸気、エアゾール、化学物質、花粉、運動、又は冷気によって誘発される。喘息は別にして、肺を冒すいくつかの慢性炎症性疾患、例えば、慢性閉塞性肺疾患(COPD)、細菌性肺炎及び嚢胞性線維症は、気道への好中球浸潤により特徴付けられ(Lindenら、Eur.Respir.J.15:973〜977,2000;Rahmanら、Clin.Immunol.115:268〜276,2005)、COPD、アレルギー性鼻炎及び嚢胞性線維症などの疾患は、気道過敏症により特徴付けられる(Fahy及びO’Byrne、Am.J.Respir.Crit.Care Med.163:822〜823,2001)。喘息及び気道炎症の一般的に用いられる動物モデルとしては、オボアルブミンチャレンジモデル及びメタコリン感作モデル(Hesselら、Eur.J.Pharmacol.293:401〜412、1995)が挙げられる。培養したヒト気管支上皮細胞、気管支線維芽細胞又は気道平滑筋細胞からのサイトカイン及びケモカインの産生の阻害をインビトロモデルとして用いることもできる。本発明のアンタゴニストをこれらのモデルのいずれかに投与することによって、喘息、気道炎症、COPDなどの症状を改善し、経過を変化させるためのこれらのアンタゴニストの使用を評価することができる。
【0133】
本発明の方法によって予防又は治療し得る他の炎症状態及び神経障害は、自己免疫疾患によって引き起こされるものである。これらの状態及び神経障害としては、多発性硬化症、全身性エリテマトーデス、並びに、アルツハイマー病、パーキンソン病、ハンチントン病、双極性疾患、及び筋萎縮性側索硬化症(ALS)を含む神経変性及び中枢神経系(CNS)疾患、原発性胆汁性硬変、原発性硬化性胆管炎、非アルコール性脂肪性肝疾患/脂肪性肝炎、線維症、C型肝炎ウイルス(HCV)、及びB型肝炎ウイルス(HBV)などの肝臓疾患、糖尿病及びインスリン抵抗性、アテローム性動脈硬化症、脳出血、脳卒中、及び心筋梗塞などの心血管疾患、関節炎、関節リウマチ、乾癬性関節炎及び若年性関節リウマチ(JRA)、骨粗鬆症、変形性関節症、膵炎、線維症、脳炎、乾癬、巨細胞動脈炎、強直性脊椎炎、自己免疫性肝炎、ヒト免疫不全ウイルス(HIV)、炎症性皮膚状態、移植、癌、アレルギー、内分泌疾患、創傷治癒、他の自己免疫性疾患、気道過敏症、並びに、細胞、ウイルス又はプリオンによって介在される感染症又は疾患が挙げられる。
【0134】
変形性関節症、関節リウマチ、怪我による関節炎を有する関節などを含む関節炎は、本発明のアンタゴニストのような抗炎症性タンパク質の治療的使用によって利するところのある一般的な炎症状態である。例えば、関節リウマチ(RA)は全身を冒す全身性疾患であり、関節炎の最も一般的な形態の1つである。関節リウマチは組織の傷害を引き起こすため、TLR3リガンドが炎症部位に存在し得る。TLR3シグナル伝達の活性化によって、炎症を起こした関節の炎症及び更なる組織傷害が持続し得る。関節リウマチの幾つかの動物モデルが当該技術分野において知られている。例えばコラーゲン誘導性関節炎(CIA)モデルでは、ヒト関節リウマチとよく似た慢性炎症性関節炎をマウスに発症させる。本発明のTLR3アンタゴニストをCIAモデルマウスに投与することによって、疾患の症状を改善して疾患の経過を変化させるためのこれらのアンタゴニストの使用を評価することができる。
【0135】
真性糖尿病すなわち糖尿病は、複数の原因因子によって誘導され、高血糖によって特徴付けられる疾患プロセスのことを指す(LeRoithら(eds.)、Diabetes Mellitus,Lippincott−Raven Publishers,Philadelphia,Pa.U.S.A.1996、及び本明細書により引用されるすべての参考文献)。制御されない高血糖は、腎障害、神経障害、網膜障害、高血圧、脳血管疾患及び冠状動脈性心疾患などの微小血管及び大血管性疾患のリスクが増大するために死亡率が増加し、若年死をともなう。したがって、グルコースホメオスタシスの制御は糖尿病の治療にとって極めて重要なアプローチである。
【0136】
糖尿病を導く欠陥に基づき、糖尿病は、患者の膵線にインスリンを産生するβ細胞が存在しない場合に発症する1型糖尿病(インスリン依存性糖尿病、IDDM)と、β細胞のインスリン分泌不全及び/又はインスリンの作用に対する抵抗性を有する患者において発症する2型糖尿病(インスリン非依存性糖尿病、NIDDM)の2つの大きなグループに分類される。
【0137】
II型糖尿病は、絶対的ではなく、相対的なインスリン欠乏をともなうインスリン抵抗性によって特徴付けられる。インスリン抵抗性の患者では、身体はこの欠陥を補償するために異常に高量のインスリンを分泌する。インスリン抵抗性を補償し、血糖値を適正に制御するために不適切な量のインスリンが存在すると、境界型糖尿病の状態が現れる。多くの患者でインスリン分泌は更に低下し、血漿グルコース濃度が上昇し、糖尿病の臨床状態がもたらされる。肥満症にともなう炎症は、インスリン抵抗性、II型糖尿病、脂質代謝異常、及び心血管疾患の発症に深く関わっていることが示唆されている。肥満脂肪はマクロファージを動員及び維持し、TNF−α及びIL−6などの炎症性サイトカイン、遊離脂肪酸及びアディポカインを過剰に産生する場合があり、インスリンシグナル伝達を妨害してインスリン抵抗性を誘発し得る。マクロファージのTLR3活性化は、こうした脂肪の炎症誘発状態に寄与し得る。インスリン抵抗性の幾つかの動物モデルが知られている。例えば、食餌誘発肥満モデル(DIO)では、動物は体重増加をともなう高血糖症及びインスリン抵抗性を発症する。DIOモデルに本発明のTLR3アンタゴニストを投与することによって、2型糖尿病にともなう合併症を改善して疾患の経過を変化させるためのこれらのアンタゴニストの使用を評価することができる。
【0138】
癌の例としては、白血病、急性白血病、急性リンパ芽球性白血病(ALL)、B細胞又はT細胞のALL、急性骨髄性白血病(AmL)、慢性骨髄性白血病(CmL)、慢性リンパ球性白血病(CLL)、有毛細胞白血病、骨髄異形成症候群(MDS)、リンパ腫、ホジキン病、悪性リンパ腫、非ホジキンリンパ腫、バーキットリンパ腫、多発性骨髄腫、カポジ肉腫、直腸結腸癌、膵臓癌、腎細胞癌、乳癌、鼻咽頭癌、悪性組織球増殖症、悪性の腫瘍随伴症候群/高カルシウム血症、固形腫瘍、腺癌、扁平上皮細胞癌、肉腫、悪性黒色腫、特に転移性黒色腫、血管腫、転移性疾患、癌関連骨吸収、及び癌関連骨痛を含む(ただしこれらに限定されない)、細胞、組織、臓器、動物、又は患者における少なくとも1つの悪性疾患が挙げられる。
【0139】
心血管疾患の例としては、心臓性昏倒症候群、心筋梗塞、鬱血性心不全、卒中、虚血性卒中、出血、動脈硬化症、アテローム性硬化症、再狭窄、糖尿病性動脈硬化疾患、高血圧、動脈高血圧、腎血管性高血圧、失神、ショック、心血管系の梅毒、心不全、肺性心、原発性肺高血圧、心臓不整脈、心房異所性収縮、心房粗動、心房細動(持続性又は痙攣性)、潅流後症候群、心肺バイパス炎症反応、多源性(chaotic又はmultifocal)心房頻拍、規則的で狭いQRS頻拍、特異的不整脈、心室細動、ヒス束不整脈、房室ブロック、脚ブロック、心筋虚血性障害、冠動脈疾患、狭心症、心筋梗塞、心筋症、拡張型うっ血性心筋症、拘束型心筋症、心臓弁疾患、心内膜炎、心臓周囲疾患、心臓腫瘍、大動脈及び抹消動脈瘤、大動脈解離、大動脈の炎症、腹部大動脈及びその分枝の閉塞、抹消血管障害、閉塞性動脈障害、抹消性アテローム硬化疾患、閉塞性血栓血管炎、機能性抹消動脈障害、レイノー現象及び障害、先端チアノーゼ、先端紅痛症、静脈疾患、静脈血栓症、静脈瘤血管、動静脈瘤、リンパ水腫、脂肪性浮腫、不安定狭心症、再潅流傷害、ポンプ後症候群、及び虚血性再潅流傷害を含む(ただしこれらに限定されない)、細胞、組織、臓器、動物又は患者における心血管疾患が挙げられる。
【0140】
代表的な神経性疾患の例としては、神経変性疾患、多発性硬化症、偏頭痛、AIDS痴呆複合症、例えば多発性硬化症及び急性横断性脊髄炎などの髄鞘脱落性疾患;例えば皮質脊髄系の病変などの錐体外路及び小脳疾患;基底核の疾患又は小脳疾患;脳幹神経節の障害;例えばハンチントン舞踏病及び老年舞踏病などの運動亢進性運動障害;例えばCNSドーパミンレセプターを遮断する薬物によって誘導される運動障害などの薬物誘導性運動障害;例えばパーキンソン病などの運動低下性運動障害;進行性核上性麻痺;小脳の構造的病変;例えば脊髄性運動失調、フリードライヒ運動失調、小脳皮質変性、多発性系変性(Mencel、Dejerine−Thomas、Shi−Drager及びMachado−Joseph)などの脊髄小脳の変性;全身性疾患(Refsum’s疾患、無β−リポタンパク血症、麻痺、毛細管拡張症及びミトコンドリア性多系疾患);例えば多発性硬化症、急性横断性脊髄炎などの髄鞘脱落性コア疾患;及び神経性筋萎縮などの運動ユニットの障害(例えば筋委縮性測索硬化症、小児性脊髄性筋委縮及び若年性脊髄性筋委縮などの前角細胞変性);アルツハイマー病;中年のダウン症候群;びまん性レビー小体病;レビー小体型の老年性認知症;ウェルニッケ‐コルサコフ症候群;慢性アルコール中毒;クロイツフェルト・ヤコブ病;亜急性硬化性全脳炎、ハラーフォルデン・シュパッツ病;ボクサー痴呆を含む(ただしこれらに限定されない)細胞、組織、臓器、動物又は患者における神経疾患が挙げられる。
【0141】
線維症状態の例としては、肝臓線維症(アルコール性肝硬変、ウイルス性肝硬変、自己免疫性肝炎を含むがこれらに限定されない);肺線維症(強皮症、特発性肺線維症を含むがこれらに限定されない);腎臓線維症(強皮症、糖尿病性腎炎、糸球体腎炎、ループス腎炎を含むがこれらに限定されない);皮膚線維症(強皮症、肥厚性及びケロイド瘢痕、火傷を含むがこれらに限定されない);骨髄線維症;神経線維腫症;線維腫;腸線維症;及び外科手術による線維化付着などが挙げられる。こうした方法では、線維症は器官特異的な線維症又は全身性線維症であり得る。器官特異的な線維症は、肺線維症、肝線維症、腎線維症、心線維症、血管線維症、皮膚線維症、眼線維症、骨髄線維症、又はその他の線維症の少なくとも1つにともない得る。肺線維症は、特発性肺線維症、薬剤誘発性肺線維症、喘息、サルコイドーシス、又は慢性閉塞性肺疾患のうち少なくとも1つにともない得る。肝臓線維症は、肝硬変、住血吸虫症、又は胆管炎のうち少なくとも1つにともない得る。肝硬変は、アルコール性肝硬変、C型肝炎後肝硬変、原発性胆汁性肝硬変の中から選択され得る。胆管炎は硬化性胆管炎である。腎線維症は、糖尿病性腎障害又はループス糸球体腎炎にともない得る。心線維症は、心筋梗塞にともない得る。血管線維症は、血管形成術後動脈再狭窄、又はアテローム性動脈硬化にともない得る。皮膚線維症は、火傷瘢痕化、肥厚性瘢痕化、ケロイド、又は腎性線維性皮膚症にともない得る。眼線維症は、後眼窩線維症、白内障手術後又は増殖性硝子体網膜症にともない得る。骨髄線維症は、特発性骨髄線維症又は薬剤誘発性骨髄線維症にともない得る。他の線維症は、ペイロニー病、デュピュイトラン拘縮又は皮膚筋炎から選択され得る。全身性線維症は、全身性硬化症及び移植片体宿主病から選択され得る。
【0142】
投与/製薬学的組成物
TLR3活性の抑制が所望される病態の治療又は予防に効果的な剤の「治療上の有効量」は、標準的な研究技術により判定され得る。例えば、喘息、クローン病、潰瘍性大腸炎又は関節リウマチなどの炎症状態の治療又は予防において有効となる薬剤の用量は、本明細書に述べるモデルのような関連する動物モデルに薬剤を投与することによって決定することができる。
【0143】
更に、場合によりインビトロアッセイを用いて最適な用量範囲を特定することができる。特定の有効用量の選択は、当業者であれば幾つかの因子の考慮に基づいて(例えば臨床試験によって)決定することができる。こうした因子には、治療又は防止しようとする疾患、関与する症状、患者の体重、患者の免疫状態、及び当業者には周知の他の因子が含まれる。製剤に使用される正確な用量は、投与経路、及び疾患の重篤度にも依存し、医師の判断及び各患者の状況に基づいて決定されなければならない。有効用量は、インビトロ又は動物モデル試験系から導出される用量反応曲線から外挿することができる。
【0144】
本発明の方法では、TLR3アンタゴニストは単独で、又は少なくとも1種類の他の分子と組み合わせて投与することができる。こうした更なる分子は、他のTLR3アンタゴニスト分子か、又はTLR3受容体のシグナル伝達によって介在されない治療効果を有する分子であってよい。抗生剤、抗ウイルス剤、緩和剤、及びサイトカインのレベル又は活性を低減する他の化合物はこうした更なる分子の例である。
【0145】
本発明の薬剤の治療上の使用のための投与方法は、薬剤をホストに送達する任意の好適な経路でよい。これらの薬剤の製薬学的組成物は、例えば、皮内、筋肉内、腹腔内、静脈内、皮下、又は鼻腔内などの非経口投与に特に有用である。
【0146】
本発明の薬剤は、有効量の薬剤を製薬上許容され得る担体中の有効成分として含有する製薬学的組成物として調製することができる。「担体」なる用語は、活性化合物が一緒に投与される希釈剤、助剤、賦形剤、又は溶媒のことを指す。こうした医薬用溶媒は、落花生油、大豆油、鉱物油、ゴマ油などの、石油、動物、植物又は合成物由来の水及び油などの液体であってよい。例えば、0.4%生理食塩水及び0.3%グリシンを使用することができる。これらの溶液は滅菌され、粒子状物質を含まないものである。これらの溶液は、従来の公知の滅菌法(例えば濾過)によって滅菌することができる。組成物は、pH調整剤及び緩衝剤、安定化剤、増粘剤、潤滑剤、及び着色剤などの、適切な生理学的状態に必要とされるような、製薬上許容できる補助物質を含有し得る。そのような製剤処方中の本発明の剤の濃度は、幅広く、すなわち、約0.5重量%未満から通常は約1重量%、又は少なくとも約1重量%から、15又は20重量%まで変化させることができ、主に、選択された特定の投与様式による、必要とされる投与量、液体の容積及び粘度などに基づいて選択される。
【0147】
したがって、筋肉内注射用の本発明の製薬学的組成物は、1mLの滅菌緩衝水、及び約1ng〜約100mg、例えば約50ng〜約30mg、又はより好ましくは約5mg〜約25mgの本発明のTLR3アンタゴニストを含むように調製することができる。同様に、静脈内注射用の本発明の製薬学的組成物は、250mLの滅菌リンゲル溶液、及び約1mg〜約30mg、好ましくは5mg〜約25mgの本発明のアンタゴニストを含むように調製することができる。非経口投与可能な組成物を調製する実際の方法は周知のものであり、例えば、「Remington’s Pharmaceutical Science」、15th ed.、Mack Publishing Company,Easton,PAにより詳細に記載されている。
【0148】
本発明の抗体アンタゴニストは凍結乾燥して保存することができ、使用の前に適切な溶媒中で還元することができる。この方法は従来の免疫グロブリン及びタンパク質の作製において有効であることが示されており、当該技術分野では周知の凍結乾燥及び還元の技術を使用することができる。
【0149】
次に以下の具体的かつ非限定的な実施例を参照して本発明を説明する。
【実施例】
【0150】
(実施例1)
抗huTLR3アンタゴニストmAbの同定及び誘導
MorphoSys Human Combinatorial Antibody Library(HuCAL(登録商標))Goldファージディスプレイライブラリ(Morphosys AG,Martinsried,Germany)をヒト抗体フラグメントの供給源として使用し、C末端にポリヒスチジンタグを有するヒトTLR3(huTLR3)(配列番号4)のアミノ酸1〜703を発現させることによって生成した精製TLR3抗原に対してパニングさせ、固定化金属アフィニティークロマトグラフィーによって精製した。アミノ酸1〜703はhuTLR3の予想される細胞外ドメイン(ECD)と一致している。多様な抗体フラグメント群を固有のものとして同定、配列決定及び確認できるさまざまな方法において、TLR3タンパク質を提示することにより、huTLR3と特異的に結合したFabフラグメント(Fab)を選別した。手法の異なるパニングにより、62種類の候補物質(V領域配列が異なる)をhTLR3のECDに結合する固有の物質として同定した。
【0151】
huTLR3のECD結合物質として同定されたこれら62種類の候補物質を、抗炎症活性の特定に関するさまざまな細胞アッセイにより中和活性についてスクリーニングした。予備活性データ(下記実施例2を参照)を用いて、重鎖CDR2(HCDR2)及び軽鎖CDR3(LCDR3)のCDR成熟のための親候補物質として、62種類の候補物質からファミリー16〜19を形成する4種類の候補物質(Fab 16〜19)を選別した。これらの親候補物質の1つ(候補物質19)でHCDR2にN結合型グリコシル化部位が認められたため、この候補物質にSer→Ala(S→A)の変異を導入してこの部位を除去した。4種類の親候補物質をCDR成熟させた後、全部で15種類の子孫候補物質(候補物質1〜15)が同定され、これを下記実施例2で述べるように更なる特性評価に供した。19種類の候補物質のそれぞれに存在する軽鎖及び重鎖の可変領域のリストを上記表3に示した。ここで各候補物質は、Fabであるか完全長の抗体鎖としてクローニングされた(実施例3)ものであるかに応じてmAb 1〜19又はFab 1〜19と呼ぶものとする。発現ベクターの設計により、すべての候補物質について、可変領域の成熟したアミノ末端は重鎖ではQVEであり、軽鎖ではDIであった。これらの末端の好ましい配列は、それぞれの生殖細胞遺伝子内に存在し、候補配列と高い相同性を有する。ファミリー17及び18では、生殖細胞系の配列はVHでQVQ、VLでSYである。ファミリー19ではこの配列はVHでEVQ、VLでDIである。SY配列はλサブグループ3に固有であり、アミノ末端残基がS又はYのいずれかであることについての異種性が報告されている。したがって、有名なλサブグループ1由来のQSVコンセンサス末端は、ファミリー17及び18のVLのDIEに対するより好適な置換になるものと考えられる。これらの改変を、ファミリー18から候補物質9、10及び12に、ファミリー19から候補物質14及び15に導入した。このプロセスでは、これらの抗体のVH及びVL領域のコドンを最適化した。それぞれ、候補物質9、10及び11の軽鎖可変領域のN末端生殖細胞変異体のアミノ酸配列を配列番号209〜211に示し、候補物質9、10、12、14及び15の重鎖可変領域のN末端生殖細胞変異体のアミノ酸配列を配列番号212〜216に示す。本明細書では、これらの候補物質のN末端変異体を候補物質/mAb/Fab 9QVQ/QSV、10QVQ/QSV、12QVQ/QSV、14EVQ又は15EVQと呼ぶ。これらのN末端生殖細胞変異体をmAbとして発現させたところ、それぞれの親候補物質と比較した場合に、TLR3への結合についても、TLR3の生物活性を阻害する能力においても何ら影響を示さなかった(データは示されていない)。
【0152】
(実施例2)
インビトロでのTLR3アンタゴニスト活性の判定
上記に述べたCDRを成熟させた15種類の候補物質を潜在的なヒトの治療物質として選択し、結合範囲及び中和活性を調べた。4種類の親Fab(Fab 16〜19)及びCDR成熟させた15種類のFab(Fab 1〜15)又はこれらの非生殖細胞V領域変異体についての、活性のアッセイ及び結果を下記に述べる。
【0153】
NF−κB及びISREシグナル伝達カスケードの阻害
熱で不活化したFBSを添加したDMEM及びGlutaMax培地(Invitrogen,Carlsbad,CA)中で293T細胞を増殖させ、pNF−κB又はISREホタルルシフェラーゼレポータープラスミド30ng、pcDNA3.1ベクター13.5ng、phRL−TK 5ng、及びFLTLR3をコードするpCDNA(配列番号2)1.5ngをトランスフェクトした。phRL−TKプラスミドは、HSV−1チミジンキナーゼプロモーターによって制御されるウミシイタケルシフェラーゼ遺伝子(Promega,Madion,WI)を有している。TLR3抗体を30〜60分インキュベートした後、ポリ(I:C)(GE Healthcare,Piscataway,NJ)を加えた。各プレートを37℃で6時間又は24時間インキュベートした後、DualGloルシフェラーゼ試薬を加えて各プレートをFLUOstarプレートリーダーで読み取った。ホタルルシフェラーゼの相対発光量(RLU)をウミシイタケルシフェラーゼのRLUで割ることによって、正規化された値(ルシフェラーゼ比)を求めた。ホタルルシフェラーゼの産生を刺激するNF−κB又はISREシグナル伝達カスケードは、TLR3アゴニストであるポリ(I:C)(1μg/mL)による刺激前に、細胞を抗TLR3抗体(0.4、2.0及び10μg/mL)とインキュベートすることによって特異的に阻害された。NF−κBアッセイの結果を図1に示す。図中、5465をポジティブコントロール(抗ヒトTLR3 mAbを中和する)として、また抗ヒト組織因子mAb(859)をヒトIgG4アイソタイプコントロールとして用いて、ホタル/ウミシイタケ比の阻害率(%)を表す。0.4〜10μg/mLのmAb濃度で50%を上回る阻害率が得られた。c1068及びTLR3.7は、10μg/mLでTLR3の生物活性の約38%及び8%を阻害した。同様の結果がISREレポーター遺伝子アッセイで得られた(データは示されていない)。
【0154】
BEAS−2B細胞におけるサイトカイン放出
BEAS−2B細胞(SV−40で形質転換した正常なヒト気管支上皮細胞)を、I型コラーゲンをコーティングした培養皿に播種し、抗ヒトTLR3抗体の存在下又は非存在下でインキュベートした後、ポリ(I:C)を加えた。処理の24時間後に上澄み液を回収し、IL−6、IL−8、CCL−2/MCP−1、CCL5/RANTES及びCXCL10/IP−10の検出用にカスタムしたマルチプレックスビーズアッセイを用いて、サイトカイン及びケモカインレベルについてアッセイした。結果を、0.4、2.0及び10μg/mLのmAbで処理した後の、サイトカイン/ケモカインの個別の阻害率(%)として図2に示す。5465はポジティブコントロールであり、859はアイソタイプコントロールである。
【0155】
NHBE細胞におけるサイトカイン放出
サイトカインの放出を、正常なヒト気管支上皮(NHBE)細胞(Lonza,Walkersville,MD)においてもアッセイした。NHBE細胞を増殖させ、コラーゲンコーティングした培養皿に移して48時間インキュベートした後、培地を除去して0.2mLの新鮮な培地を補充した。次いで細胞を抗ヒトTLR3 mAbの存在下又は非存在下で60分間インキュベートした後、ポリ(I:C)を加えた。24時間後に上澄み液を回収し、−20℃で保存するか、又は直ちにIL−6のレベルについてアッセイした。結果を、投与量0.001〜50μg/mLでのmAb処理後の、IL−6分泌の阻害率(%)として図3にグラフで示す。5465はポジティブコントロールであり、859はアイソタイプコントロールである。大部分のmAbは1μg/mLでIL−6の産生の少なくとも50%を阻害し、5μg/mLで阻害率75%を達成した。
【0156】
PBMC細胞におけるサイトカイン放出
サイトカインの放出を、ヒト末梢血単核球細胞(PBMC)においてもアッセイした。ヒトドナーから全血をヘパリン回収試験管に回収し、これにFicoll−Paque Plus溶液を下側に層を形成するようにゆっくりと加えた。試験管を遠心してFicollの直ぐ上に白色の層を形成しているPBMCを回収して播種した。この後、PBMCを抗ヒトTLR3 mAbの存在下又は非存在下でインキュベートした後、25μg/mLのポリ(I:C)を加えた。24時間後に上澄み液を回収し、Luminex法によってサイトカインのレベルを調べた。5465をポジティブコントロールとし、hIgG4をアイソタイプコントロールとして用い、mAbの単回用量(0.4μg/mL)によるIFN−γ、IL−12及びIL−6の累積阻害率(%)として、結果を図4にグラフ化して示す。
【0157】
HASM細胞におけるサイトカイン放出
簡潔に言えば、500ng/mLのポリ(I:C)及び10ng/mLのTNF−αの相乗効果のある組み合わせを添加する前に、ヒト気道平滑筋(HASM)細胞を、抗ヒトTLR3 mAbsの存在又は非存在下でインキュベートした。24時間後に上澄み液を回収し、Luminex技術を用いて、サイトカインレベルを求めた。結果を、3段階の容量(0.4、2及び10μg/mL)でmAbを用いた場合のケモカインCCL5/RANTESのレベルとして図5にグラフで示す。5465はポジティブコントロールであり、hIgG4はアイソタイプコントロールである。
【0158】
huTLR3に結合することによりサイトカイン及びケモカインの放出を低減させる本発明の抗体の能力が、ヒト細胞におけるインビトロアッセイの結果によって確認された。
【0159】
(実施例3)
完全長の抗体コンストラクト
4種類の親Fab(候補物質番号16〜19)及び15種類の子孫Fab(候補物質番号1〜15)の重鎖をS229P Fc変異を有するヒトIgG4バックグラウンド上にクローニングした。候補物質9QVQ/QSV、10QVQ/QSV、12QVQ/QSV、14EVQ又は15EVQを、F235A/L236及びS229P Fc変異を有するヒトIgG4バックグラウンド上にクローニングした。
【0160】
成熟した完全長の重鎖アミノ酸配列を以下のように配列番号90〜102及び218〜220に示す。
【0161】
【表9】
【0162】
発現のために、これらの重鎖はMAWVWTLLFLMAAAQSIQA(配列番号103)のようなN末端のリーダー配列を有してもよい。リーダー配列を有する候補物質14EVQ及び15EVQの重鎖、及び成熟型(リーダー配列を有さない)をコードするヌクレオチド配列の例を、配列番号104及び105にそれぞれ示す。同様に、発現のために、本発明の抗体の軽鎖の配列は、MGVPTQVLGLLLLWLTDARC(配列番号106)のようなN末端のリーダー配列を有してもよい。リーダー配列を有するコドン最適化された候補物質15の軽鎖、及び成熟型(リーダー配列を有さない)をコードするヌクレオチド配列の例を、配列番号107及び108にそれぞれ示す。
【0163】
(実施例4)
抗TLR3 mAbの結合の特性評価
ヒトTLR3の細胞外ドメイン(ECD)に対するmAbの結合のEC50値をELISAによって求めた。ヒトTLR3のECDタンパク質をPBSで2μg/mLに希釈し、100μLの一定分量を96穴プレート(Corning Inc.,Acton,MA)の各ウェルに分配した。4℃で一晩インキュベートした後、PBSに0.05%のTween−20(Sigma−Aldrich)を加えた洗浄緩衝液によりプレートを3回洗浄した。ウェルを2%のI−Block(Applied Biosystems,Foster City,CA)及び0.05%のTween−20/PBSからなる200μLのブロッキング溶液によりブロッキングした。室温で2時間ブロッキングした後、プレートを3回洗浄し、続いてブロッキング緩衝液に希釈した抗TLR3 mAb候補物質1〜19を連続的に添加した。
【0164】
【表10】
【0165】
抗TLR3 mAbを室温で2時間インキュベートた後3回洗浄した。この後、ブロッキング緩衝液で1:4000に希釈したペルオキシダーゼ接合ヒツジ抗ヒトIgG(GE Health care,Piscataway,NJ)を加え、室温で1時間インキュベートした後、洗浄緩衝液で3回洗浄した。結合は、TMB−S(Fitzgerald Industries International,Inc.,Concord,MA)中で10〜15分間インキュベートすることによって検出した。反応を、2N H2SO4 25μLにより停止させ、及びSPECTRA Max分光光度計(Molecular Devices Corp.,Sunnyvale,CA)を用いて、吸光度を450nmで読み取り650nmでの値を差し引いた。EC50値をGraph Pad Prismプリズムソフトウェア(Graph Pad Software,Inc.,San Diego,CA)を使用して非線形回帰によって求めた。
【0166】
mAbを2.5μg/mL〜0.6pg/mLまで4倍ずつ希釈した100μLの連続希釈液とインキュベートすることにより、huTLR3に対する結合についてEC50値を求めた(表4)。抗ヒト組織因子mAb 859及びhu IgG4κをネガティブコントロールとして含めた。
【0167】
huTLR3 ECDに対する結合親和性もBiacore解析によって求めた。データ(示されていない)は、mAb 1〜19ではhuTLR3 ECDに対するKdが10-8M未満であることを示した。
【0168】
(実施例5)
競合的エピトープ結合
抗TLR3抗体競合グループ又は「エピトープビン」を決定するためにエピトープ結合実験を行った。
【0169】
競合的ELISAを行うため、実施例1に述べるようにして作製した5μL(20μg/mL)の精製ヒトTLR3 ECDタンパク質を、MSD High Bindプレート(Meso Scale Discovery,Gaithersburg,MD)の各ウェルに室温で2時間コーティングした。150μLの5% MSDブロッカーA緩衝液(Meso Scale Discovery)を各ウェルに加えて室温で2時間インキュベートした。プレートを0.1MのHEPES緩衝液(pH 7.4)で3回洗浄した後、標識した抗TLR3 mAbと異なる競合物質の混合液を加えた。標識した抗体(10nM)を、濃度を増加させた(1nM〜2μM)非標識抗TLR3抗体とインキュベートし、次いで25μLの量の混合液を、指定されたウェルに加えた。室温で穏やかに振盪しながら2時間インキュベートした後、プレートを0.1MのHEPES緩衝液(pH 7.4)で3回洗浄した。MSD Read緩衝液Tを蒸留水で希釈し(4倍)、150μL/ウェルの量で分配してSECTOR Imager 6000によって分析した。抗体を製造者(Meso Scale Discovery)の使用説明書に従ってMSD Sulfo−Tag(商標)NHS−エステルで標識した。
【0170】
以下の抗TLR3抗体を評価した。すなわち、MorphoSysヒト組み換え抗体ライブラリより入手したmAb 1〜19(表3に示すもの);c1068(国際公開第06/060513(A2)号に述べられるもの)、c1811(マウスTLR3タンパク質で免疫性を与えられたラットから作製されるハイブリドーマにより産生されるラット抗マウスTLR3 mAb)、TLR3.7(eBiosciences,San Diego,CA,カタログ番号14−9039)、及びIMG−315A(Imgenex,San Diego,CAより入手したヒトTLR3アミノ酸55〜70(VLNLTHNQLRRLPAAN)に対して生成したもの)。mAb 9、10、12、14及び15については、変異体9QVQ/QSV、10QVQ/QSV、12QVQ/QSV、14EVQ、又は15EVQをこの実験で使用した。
【0171】
競合アッセイに基づき、抗TLR3抗体を、次の5つの異なるビンに割り当てた。ビンA:mAb 1、2、13、14EVQ、15EVQ、16、19;ビンB:mAb 3、4、5、6、7、8、9QVQ/QSV、10QVQ/QSV、11、12QVQ/QSV、17、18;ビンC:抗体Imgenex IMG−315A;ビンD:抗体TLR3.7、c1068;及びビンE:抗体c1811。
【0172】
(実施例6)
エピトープマッピング
実施例5で述べた異なるエピトープビンからの代表的な抗体を更なるエピトープマッピングのために選択した。エピトープマッピングを、TLR3セグメント交換実験、変異誘発、H/D交換、及びインシリコタンパク質−タンパク質ドッキングを含む各種のアプローチを用いて行った(The Epitope Mapping Protocols,Methods in Molecular Biology,Volume 6,GlenE.Morrised.,1996)。
【0173】
TLR3セグメント交換TLR3ヒト−マウスキメラタンパク質を用いてTLR3上の全体の抗体結合ドメインの位置を特定した。ヒトTLR3タンパク質の細胞外ドメインを3つのセグメント(GenBank受諾番号NP_003256のヒトTLR3アミノ酸配列に基づくアミノ酸番号に従ってaa 1〜209、aa 210〜436、aa 437〜708)に分割した。ヒトTLR3のアミノ酸210〜436及び437〜708を対応するマウスのアミノ酸(マウスTLR3、GenBank受諾番号NP_569054、アミノ酸211〜437及び438〜709)と置き換えることによってMT5420キメラタンパク質を作製した。ヒトアミノ酸の位置437〜708をマウスTLR3のアミノ酸(マウスTLR3、GenBank受諾番号NP_569054、アミノ酸438〜709)で置き換えることによってMT6251キメラを作製した。すべてのコンストラクトを、標準的なクローニング手法を用いてpCEP4ベクター(Life Technologies,Carslbad,CA)で生成させた。各タンパク質をHEK293細胞内でV5−His6 C末端融合タンパク質として一過性に発現させ、実施例1で述べたようにして精製した。
【0174】
mAb c1068.mAb c1068は、ヒトTLR3 ECDに高親和性で結合したが、マウスTLR3には良好に結合しなかった。c1068は、MT5420及びMT6251の両方に結合する能力を喪失したことから、結合部位がWTヒトTLR3タンパク質のアミノ酸437〜708内に局在するということが示された。
【0175】
mAb 12QVQ/QSV:mAb 12QVQ/QSVは両方のキメラに結合したが、このことはmAb 12QVQ/QSVの結合部位が配列番号2に示される配列を有するヒトTLR3タンパク質のアミノ酸1〜209内に位置していることを示している。
【0176】
インシリコタンパク質−タンパク質ドッキングmAb 15EVQ(下記を参照)の結晶構造及び発表されているヒトTLR3構造(Bellら、J.Endotoxin Res.12:375〜378,2006)を、ドッキングの開始モデルとして使用するために、CHARMm(Brooksら、J.Computat.Chem.4:187〜217,1983)でエネルギー最小化した。タンパク質ドッキングは、ZDOCK 2.1(Chen及びWeng、Proteins 51:397(Accelrys,San Diego,CA)−408、2003)と同様のZDOCKpro 1.0により6°の角度グリッドで行った。ZDOCKアルゴリズム中のエネルギー項により、ヒトTLR3における既知のN結合グリコシル化部位のAsn残基(Asn 52、70、196、252、265、275、291、398、413、507及び636)(Sunら、J.Biol.Chem.281:11144〜11151,2006)を、抗体−抗原複合体インターフェースへの参加からブロックした。2000個の最初のポーズを出力してクラスター化し、ドッキングポーズを再精密化してRDOCKで再スコアリングを行った(Liら、Proteins 53:693〜707、2003)。最初のZDOCKスコアが最も高かった200個のポーズ及びRDOCKの上位200個のポーズを視覚的に調べた。
【0177】
Fab 15EVQの結晶化を20℃で蒸気拡散法によって行った(Benvenuti及びMangani、Nature Protocols 2:1633〜51、2007)。最初のスクリーニングを、96穴プレートでHydraロボットを使用してセットアップした。実験を、0.5μLのタンパク質溶液を0.5μLのリザーバ溶液に混合した液滴で構成した。液滴を90μLのリザーバ溶液に対して平衡化した。50mMのNaClを含む20mMのTris緩衝液(pH 7.4)中のFab溶液をAmicon Ultra−5kDaセルを使用して14.3mg/mLにまで濃縮した。スクリーニングは、Wizard I & II(Emerald BioSystems,Bainbridge Island,WA)及びインハウス結晶化スクリーンによって行った。Fab 12QVQ/QSVを類似の方法で結晶化させた。
【0178】
X線回折データを収集し、Osmic(商標)VariMax(商標)共焦光学系、Saturn944CCD検出器、及びX−stream(商標)2000低温冷却システムを備えたRigaku MicroMax(商標)−007HFs微小焦点X線発生装置(Rigaku,Woodlands,TX)を使用して処理した。回折強度を、1/2°の像当たり120秒の露光時間で270°(結晶回転)にわたって検出した。X線データをD*TREKプログラム(Rigaku)により処理した。構造を、Phaser又はCNXプログラム(Accelrys,SanDiego,CA)を使用して分子置換法によって決定した。すべてのデータを用いて、原子位置及び温度因子をREFMACによりFab 15EVQに対して15〜2.2Å及びFab 12QVQ/QSVに対して50〜1.9Åの分解能範囲で精密化した。3σのカットオフレベルを用いて、(Fo−Fc)電子密度ピークで水分子を追加した。すべての結晶学的計算を、プログラムのCCP4に適したプログラムにより行った(Collaborative Computational Project,Number 4.1994.The CCP4 suite:programs for protein crystallography.Acta Crystallogr.D50:760〜763)。COOTプログラム(Emsleyら、Acta Crystallogr.D60:2126〜2132、2004)を使用して、モデル調整を行った。
【0179】
解像されたmAb 15EVQの結晶構造によって、抗体結合部位が重鎖の多数の負に帯電した残基(D52、D55、E99、D106及びD109)によって特徴付けられることが示された。したがって、mAb 15EQVとTLR3との間の認識には正に帯電した残基が関与している可能性が高いと考えられた。実施したタンパク質−タンパク質ドッキングシミュレーションは、正に帯電した残基を複数含む、TLR3上の2個の大きなパッチが、抗体に対して優れた相補性を示すことを示唆するものであった。シミュレートしたTLR3−抗TLR3抗体複合体の界面内のTLR3上の残基は、R64、K182、K416、K467、Y468、R488、R489及びK493であった。
【0180】
変異誘発実験単一及び組み合わせの変異を、TLR3 ECDの表面残基の内の、上記でmAb 12及びmAb 15EVQのエピトープを含んでいることが特定された領域に導入し、この変異タンパク質を抗体結合性について試験した。
【0181】
ヒトTLR3のアミノ酸1〜703(ECD)をコードするヌクレオチド配列(配列番号4、GenBank受諾番号NP_003256)を標準的なプロトコールを用いてクローニングした。表5aに示されるオリゴヌクレオチドを用い、製造者のプロトコールに従ってStrategene Quickchange II XLキット(Stratagene,San Diego,CA)を使用して、すべての変異体を、部位特異的な変異の誘導によって作製した。変異はDNAの配列決定によって確認した。タンパク質はCMVプロモーターの制御下で、HEK293細胞内でC末端His−タグ融合タンパク質として発現させ、実施例1で述べたようにして精製した。
【0182】
結合アッセイヒトTLR3に対するmAb 12QVQ/QSV及びmAb 15EVQの結合活性及び生成された変異体をELISAによって評価した。プロセスを促進するため、mAb 15EVQの結合が予想される部位についての変異体を、mAb 12QVQ/QSVと共にC末端Hisタグを含むTLR3 ECD変異体と、HEK細胞にコトランスフェクトすることで同時発現させた後、金属アフィニティークロマトグラフィーにより精製した。回収された試料は、TLR3変異体とmAbとの複合体であった。このアプローチは、mAb 12QVQ/QSV及びmAb 15EVQの結合部位が互いに離れており、したがって一方の部位における点変異が他方の部位のエピトープに影響する可能性が低いことから実行可能であった。これらの複合体をELISA結合アッセイで使用した。PBSに加えた20μg/mL野生型TLR3 ECD又は変異タンパク質を各ウェル当たり5μLずつ、MSD HighBindプレート(Meso Scale Discovery,Gaithersburg,MD)にコーティングした。プレートを室温で60分間インキュベートし、ブロッキングした。
【0183】
【表11】
【0184】
相補的配列を有するアンチセンスオリゴヌクレオチドを使用し、MSDBlockerA緩衝液(Meso Scale Discovery,Gaithersburg,MD)で4℃で一晩変異誘発反応を行った翌日、プレートを洗浄し、MSD Sulfo−タグ標識したmAb 15EVQを500pM〜1pMの濃度で1.5時間添加した。洗浄後、MSD Read緩衝液Tを使用して標識抗体を検出し、SECTOR Imager 6000を使用してプレートを読み取った。ヒトTLR3及び変異体に対するmAb 12QVQ/QSVの結合活性を評価するため、mAb 15EVQと同時発現させ、検出抗体を標識mAb 12QVQ/QSVとした以外はmAb 15EVQについて述べたのと同様にして結合ELISAを行った。
【0185】
mAb 12QVQ/QSV:mAb 12QVQ/QSVの結合部位は、セグメント交換実験において決定されたようにヒトTLR3タンパク質のアミノ酸1〜209内に位置が特定された。以下のTLR3変異体について評価を行った:D116R、N196A、N140A、V144A、K145E、K147E、K163E、及びQ167A。野生型TLR3及びV144A変異体は、mAb 12QVQ/QSVに対して同等の結合性を示した(図6A)。抗体はTLR3 D116R変異体には結合せず、K145E変異体に対しては大幅に低い結合親和性を示した。したがって、TLR3上で近接して配置されている残基D116及びK145は、mAb 12QVQ/QSVに対する中心的なエピトープ部位と特定された(図7A)。
【0186】
mAb 12QVQ/QSV結合エピトープのこれら2つの重要な残基は、TLR3の外部ドメインのN末端セグメントの2本鎖RNA結合部位の面の近くに配置されている(Pirherら、Nature Struct.& Mol.Biol.,15:761〜763,2008)。完全なエピトープは、隣接領域に、実施した変異分析によっては明らかにされなかった他の残基を含むと考えられる。いずれの理論に束縛されることも望むものではないが、mAb 12QVQ/QSVがそのTLR3エピトープに結合すると、TLR外部ドメインへの2本鎖RNAの結合に直接的又は間接的に干渉することにより、受容体の2量化及び下流のシグナル伝達経路の活性化が妨害されるものと考えられる。
【0187】
mAb 15EVQ:以下のTLR3変異体について評価を行った。すなわち、R64E、K182E、K416E、Y465A、K467E、R488E、R489E、N517A、D536A、D536K、Q538A、H539A、H539E、N541A、E570R、K619A、K619E、2重変異体K467E/Y468A、3重変異体T472S/R473T/N474S、及び3重変異体R488E/R489E/K493E。野生型TLR3、R64E、K182E、K416E変異体及び3重変異体T472S/R473T/N474Sは、mAb 15EVQに対して同等の結合性を示した(図6B及び表5b)。抗体は、TLR3変異体K467E、R489E、K467E/Y468A及びR488E/R489E/K493Eには結合しなかった(図6B及び6C)。残りの変異体は中間的な結合性を示し、R488Eが最も高い作用を示した。これらの変異体のすべてがmAb 12QVQ/QSVに結合した。これらの結果は、残基K467及びR489がmAb 15EVQエピトープの重要な決定因子であることを示すものである。残基R488もエピトープに寄与した。これらの残基は、TLR3の同じ表面上に近接して配置されていた(図7A)。この結果はまた、いずれもK467、R488及びR489と同じ表面上に位置する残基Y465、Y468、N517、D536、Q538、H539、N541、E570、及びK619がエピトープに寄与することを示している。この結果は、mAb 15EVQを用いたH/D交換実験によって更に支持された。図7Aは、mAb 12QVQ/QSV及び15EVQ(黒)に対する結合エピトープ部位と、C1068 mAb(灰色)に対する結合エピトープ部位とがヒトTLR3の構造上で重なっていることを示している。mAb 15EVQに対するエピトープは、残基Y465、K467、Y468、R488、R489、N517、D536、Q538、H539、N541、E570、及びK619の範囲にわたっている。
【0188】
H/D交換実験H/D交換に対しては、抗体摂動の解析に使用した手順は、以前に述べたもの(Hamuroら、J.Biomol.Techniques 14:171〜182,2003;Hornら、Biochemistry 45:8488〜8498,2006)に類似し、若干の改変を加えたものである。組換えTLR3 ECD(C末端Hisタグを付加してSf9細胞で発現させて精製したもの)を重水素化水溶液中で所定の時間インキュベートして、交換可能な水素原子を重水素と交換した。重水素化TLR3 ECDを、固定されたmAb 15EVQを含むカラムで捕捉した後、水性緩衝液で洗浄した。逆交換した(back-exchanged)TLR3 ECDタンパク質をカラムから溶出し、重水素含有フラグメントの局在性をプロテアーゼ消化及び質量分析により調べた。参照対照として、TLR3 ECD試料を、抗体カラムで捕捉した後に重水素化水と接触させてから実験試料と同様にして洗浄及び溶出した点を除いて同様に処理した。抗体と結合する領域は比較的交換から保護される部位であり、そのため、基準TLR3 ECD試料と比較して高い割合の重水素を含むものと推測された。タンパク質の約80%を特定のペプチドにマッピングすることができた。mAb 15EVQによるTLR3 ECDのH/D交換の攪乱のマップを図7Bに示す。分かりやすさのために、TLR3の、mAb 15EVQに影響される部分の周囲のセグメントのみを示した。TLR3 ECDのアミノ末端及びカルボキシル末端に延びる、タンパク質の残りの部分は大きく影響されなかった。
【0189】
H/D交換実験により、配列番号2のペプチドセグメント465YNKYLQL471、514SNNNIANINDDmL526及び529LEKL532が、mAb 15EVQとの結合によってTLR3上の交換が特に変化する領域として特定された。その特性上、H/D交換は線形マッピング法であり、ペプチド内のどの残基が抗体結合によって最も影響されるかを定義することは通常できない。しかしながら、H/D交換と変異の結果が大きく重なっていることは、図7Aに示される表面がmAb 15の結合部位であることの更なる確証を与えるものである。この結合部位は、mAb c1068について過去に述べられているもの(国際公開第06/060513(A2)号)と同じ直線状アミノ酸配列領域内にあったが、まったく重ならない表面(図7A)上に位置することが判明し、これらの抗体間で交差競合が見られないことと一致する。
【0190】
mAb 15EVQ結合エピトープは、TLR3上のC末端セグメントにおいて二本鎖RNA結合部位に空間的に近位であった(Bellら、Proc.Natl.Acad.Sci.(USA)103:8792〜8797,2006;Ranjith−Kumarら、J.Biol.Chem,282:7668〜7678,2007;Liuら、Science 320:379〜381,2008)。いずれの特定の理論に束縛されることも望むものではないが、mAb 15EVQがそのTLR3エピトープと結合すると、リガンドである2本鎖RNA分子及び/又は2量体のもう一方との立体障害を生じ、リガンドの結合及びリガンドによって誘導される受容体の2量化が妨げられるものと考えられる。
【0191】
【表12】
【0192】
(実施例7)
熱安定性が向上した変異体の作製
結晶構造に基づいた機能改変(Structure−based engineering)を行って高い熱安定性を有する抗体の変異体を作製すると同時に、生物活性を維持しつつ免疫原性を最小に抑えようと試みた。
【0193】
mAb 15EVQを機能改変用に選択した。免疫原性を最小に抑えるため、構造的考慮に基づいて有益であることが予想される生殖細胞の変異のみを行った。mAb 15EVQのVL及びVHの配列(それぞれ配列番号41及び配列番号216)を、BLASTサーチを使用してヒト生殖細胞の遺伝子と並べた。同定された最近接の生殖細胞系列配列は、VH及びVLに対してそれぞれGenBank Acc.No.AAC09093及びX59318であった。生殖細胞のVH及びVLとmAb 15EVQのVH及びVL配列との間で以下の違いが特定された。すなわち、(VH)V34I、G35S、F50R、A61S、及びQ67H;(VL)G30S、L31S、及びA34N。特定された配列の違いをmAb 15EVQの結晶構造上にマッピングし、パッキング及び界面相互作用を変化させることが予測される残基を機能改変用に選択した。抗体の結晶構造(実施例6を参照)に基づいて、潜在的に構造を不安定にし得る残基を同定した。(1)小さな囲われた空洞がV34の近くのVHのコア中で同定された。この空洞は、Ileのようなわずかに大きな側鎖が収まるだけの充分な大きさを有していた。(2)VH CDR3のE99は、水素結合ネットワークに依らずVH/VL界面に埋め込まれていた。E99の負に帯電したカルボキシレート基は概ね疎水性環境にあり、周囲の残基と主としてファンデルワールス(vds)力による接触状態にあった。荷電基を埋め込むことは通常はエネルギー的に好ましくなく、したがって不安定化作用をもたらす。(3)VHのF50は、VH/VLの界面残基である。その芳香族側鎖は嵩張るため、ペアリングに負の影響を及ぼし得る。Fvの水素結合及びvdwパッキングネットワークを計算し、Pymolにより視覚的に調べた(www://_pymol_org)。VH及びVLドメイン内の埋め込まれた空洞はCaver(Petrekら、BMC Bioinformatics、7:316、2006)によって計算した。分子のグラフィック図をすべてPymolで作製した。QuickChange II XL部位特異的変異誘発キット(Stratagene,San Diego,CA)、Change−IT多重変異部位特異的変異誘発キット(USB Corporation,Cleveland,OH)、又はQuick Change II部位特異的変異誘発キット(Stratagene,San Diego,CA)を使用し、標準的なクローニング法によって、実施例3で述べたようにして作製したFabフラグメント又はIgG4完全ヒト抗体をコードする発現ベクターに変異を導入した。反応を、それぞれの製造者の推奨に従って行った。得られたクローンの配列を、確認のために配列決定し、得られた機能改変された変異体を、改変された重鎖又は軽鎖に従ってmAb s15−1〜15−10と命名した。mAb 15−10用の重鎖がmAb 15−6からのものであることを除いて、それぞれの変異体鎖(H又はL)を野生型mAb 15EVQL又はH鎖により発現させて、抗体を作製した。配列番号のリスト化:CDRに対して、mAb 15EVQ及びその機能改変された変異体に対する軽鎖及び重鎖並びに完全長重鎖及び軽鎖の可変領域を、表6に示す。表7は、それぞれの変異体を作製するためのプライマーを示す。
【0194】
【表13】
【0195】
TLR3に対するmAb 15−1〜15−9の結合性をELISA免疫アッセイによって評価した。ヒトTLR3 ECD(100μLの2μg/mL TLR3 ECD)を黒色のMaxisorbプレート(eBioscience)に4℃で一晩結合させた。プレートを洗浄し、ブロッキングし、希釈した抗体を各ウェル当たり50μLで二つ組複製で分注した。プレートを穏やかに振盪しながら室温で2時間インキュベートした。ルミネッセンスPOD基盤(Roche Applied Science,Mannheim,Germany,カタログ番号11 582 950 001)及びヤギ抗ヒトFc:HRP(Jackson ImmunoResearch,West Grove,PA,Cat.No.109−035−098)を用いて、結合を検出し、プレートをSpectraMaxプレートリーダー(Molecular Devices,Sunnyvale,CA)で読み取った。
【0196】
DSC実験を、MicroCalの自動VP−キャピラリーDSCシステム(MicroCal’s,LLC,Northampton,MA)で行って、基準細胞と試料細胞との間の温度差を継続的に測定して出力単位に較正した。試料は10℃〜95℃まで60℃/時の加熱速度で加熱した。前走査時間は15分とし、フィルタリング時間は10秒間とした。DSC実験で使用した濃度は約0.5mg/mLである。得られたサーモグラムの分析を、MicroCal Origin 7ソフトウェア(MicroCal,LLC)を用いて行った。
【0197】
【表14】
【0198】
作製した変異体の熱安定性(Tm)をDSCによって測定した(表8)。TLR3に対する抗体の変異体の結合性は親抗体の結合性と同等であった。
【0199】
【表15】
【0200】
(実施例8)
代替抗TLR3抗体の作製
実施例1で作製したヒト化抗体がマウスTLR3に対して充分な特異性もアンタゴニスト活性も有さなかったことから、本明細書ではmAb 5429と命名するキメラアンタゴニストのラット/マウス抗マウスTLR3抗体を作製して、さまざまなインビボモデルにおいてTLR3シグナル伝達を阻害する作用について評価を行った。この代替キメラmAb 5429及びその親ラット抗マウスTLR3抗体c1811はインビトロ及びインビボでマウスのTLR3シグナル伝達を阻害し、マウスのいくつかの疾患モデルにおいて発症機構を改善した。
【0201】
以下に考察するデータは、有害な炎症の誘発及び持続化におけるTLR3の役割を示唆するものであり、例えば高サイトカイン血症、喘息及び気道炎症、炎症性腸疾患及び関節リウマチ、ウイルス感染、及びII型糖尿病などの急性及び慢性の炎症状態におけるTLR3アンタゴニスト及びTLR3抗体アンタゴニストの治療的使用の根拠を与えるものである。
【0202】
代替mAb 5429の作製
通常の方法を用いて生成させた、遺伝子組み換え型マウスTLR3の外部ドメイン(配列番号162のアミノ酸1〜703、GenBank Acc.No.NP_569054)により、CDラットに免疫性を与えた。マウスTLR3に特異的な抗体力価を示す2匹のラット由来のリンパ球を、FO骨髄腫細胞に融合した。マウスTLR3に反応性のあるモノクローナル抗体のパネルを同定し、マウスルシフェラーゼレポーター及びマウス胎児線維芽細胞アッセイにおいて、インビトロアンタゴニスト活性について試験した。ハイブリドーマ株C1811Aを更なる研究のために選択した。ハイブリドーマによって分泌されるmAb c1811から、機能性可変領域の遺伝子を配列決定した。次いで、クローニングされた重鎖及び軽鎖可変領域遺伝子をプラスミド発現ベクターにそれぞれ挿入し、mAb 5429と表示されるキメララット/Balb C muIgG1/κ mAbを常法によって作製するためのコード配列を得た。抗体を実施例3で述べたようにして発現させた。mAb 5429の重鎖及び軽鎖可変領域のアミノ酸配列を配列番号164及び配列番号163にそれぞれ示し、重鎖及び軽鎖の完全長の配列を配列番号166及び配列番号165にそれぞれ示す。mAb c1811の重鎖及び軽鎖の完全長の配列を配列番号168及び配列番号167にそれぞれ示す。
【0203】
mAb 5429の特性評価
mAb 5429を、TLR3シグナル伝達に対する中和能力について一連のインビトロアッセイにおいて特性評価した。活性のアッセイ及び結果を以下に述べる。
【0204】
マウスルシフェラーゼレポーター遺伝子アッセイ
マウスTLR3のcDNA(配列番号161、GenBank受諾番号NM_126166)をマウス脾臓cDNA(BD Biosciences,Bedford,MA)からPCRによって増幅し、標準的な方法を用いてpCEP4ベクター(Life Technologies,Carslbad,CA)にクローニングした。200μLのHEK293T細胞を、完全DMEM中、4×104細胞/ウェルの濃度で96穴白色透明底プレートに播種し、30ngのpNF−κBホタルルシフェラーゼ(Stratagene,Carslbad,CA)又は30ngのpISREホタルルシフェラーゼ(BD Biosciences,Bedford,MA)、5ngのphRL−TKコントロールウミシイタケルシフェラーゼ(Promega Corp.,Madison,WI)レポータープラスミド、1.5ngの完全長マウスTLR3をコードするpCEP4、及び13.5ngの空のpcDNA3.1ベクター(Life Technologies,Carslbad,CA)を使用して、翌日Lipofectamine 2000(Invitrogen Corp.,Carslbad,CA)を用いて全DNA量を50ng/ウェルとし、トランスフェクションに使用した。トランスフェクションの24時間後、細胞を抗マウスTLR3抗体と新鮮な無血清DMEM中で37℃で30分〜1時間インキュベートした後、0.1又は1μg/μLのポリ(I:C)を加えた。24時間後にDual−Gloルシフェラーゼアッセイシステム(Promega,Madison,WI)を使用してプレートを回収した。相対的な光量単位を、FLUOstar OPTIMA多重検出リーダーを使用し、OPTIMALソフトウェア(BMG Labtech GmbH,Germany)により測定した。標準化された値(ルシフェラーゼ比)はホタルルシフェラーゼの相対光量単位(RLU)をウミシイタケルシフェラーゼのRLUで割ることによって得た。mAb 5429、並びにその親mAb c1811及びmAb 15(表3a)が、用量依存的にポリ(I:C)誘発NF−κB及びISRE活性化を低下させたことから(図8A及び8B)、TLR3の活性を中和するこれらの抗体の能力が実証された。ISREアッセイにおいて測定されたIC50は、mAb 5249、mAb 15及びmAb c1811についてそれぞれ0.5、22、及び0.7μg/mLであった。
【0205】
マウス胚性線維芽細胞(MEF)アッセイ
C57BL/6 MEF細胞をArtis Optimusより入手した(Opti−MEF(商標)C57BL/6−0001)。200μLのMEF培地(glutamax、10%熱失活FBS、1×NEAA、及び10μg/mLゲンタマイシンを加えたDMEM)中の細胞を、96穴平底プレート(BD Falcon)に20,000細胞/ウェルで播種した。インキュベーションはすべて、37℃/5%CO2で行った。播種の24時間後に、mAb 5429又はmAb c1811を各ウェルに加えた。プレートをmAbと1時間インキュベートした後、各ウェルにポリ(I:C)を1μg/mL加えた。24時間のインキュベーションの後、上澄み液を回収した。ビーズキット(Invitrogen Corp.,Carslbad,CA)を用い、サイトカインレベルを定量して、CXCL10/IP−10を製造者のプロトコールに従って検出した。結果をGraph Pad Prismソフトウェアを使用してグラフ化した。いずれの抗体もポリ(I:C)誘発CXCL10/IP−10レベルを用量依存的に低下させたことから、これらの抗体が、内因性のTLR3を中和してTLR3シグナル伝達を阻害する能力が実証された(図9)。
【0206】
フローサイトメトリー−表面染色
各群10匹ずつのC57BL/6及びTLR3ノックアウトマウス(TLR3 KO)(C57BL/6バックグラウンド;雌、8〜12週齢、Ace Animals,Inc.)に1mLの3%チオグリコレート培地(Sigma)を腹腔内投与し、96時間後にマウスを安楽死させ、各マウスから得た腹膜を10mLの滅菌PBSで洗浄した。チオグリコレートにより腹膜に誘導されたマクロファージをPBS中に再懸濁し、細胞の生存率をTrypan Blue染色によって評価した。細胞を遠心してペレットとし、250μLのFACS緩衝液(PBS−Ca2+−Mg2+、1%熱失活FBS、0.09%アジ化ナトリウム)に再懸濁し、濡れた氷上に保持した。CD16/32試薬(eBioscience)を10μg/106細胞に10分間使用してマクロファージ上のFc受容体をブロッキングした。細胞を100μL/ウェル中、106個細胞で分配して、表面染色を行った。Alexa−Fluor 647(Molecular Probe)−接合mAb c1811及びmAb 1679(TLR3特異性を有さないことからアイソタイプコントロールとして用いられるラット抗マウスTLR3抗体)を0.25μg/106個細胞で加え、氷上、暗所で30分間インキュベートした。細胞を洗浄し、250μLのFACS緩衝液に再懸濁した。生存率染色として7−AAD(BD Biosciences,Bedford,MA)を5μL/ウェルで30分を超えないように加えた後、FACS Caliburにより試料を取得して死細胞集団を検出した。Cell Quest Proソフトウェアを使用してFACS Caliburにより、試料を採取した。収集したデータを、FCS Expressを使用してヒストグラムを作成することによって解析した。
【0207】
C57BL/6及びTLR3KOマウスから得た、チオグリコレートで誘発されたマウス腹膜マクロファージへのmAb c1811の結合を、フローサイトメトリーにより評価して結合特異性を求めた。このキメラ抗体のマウスFc領域は、非特異的結合に寄与すると予期されたので、mAb 5429はこのアッセイで使用しなかった。mAb c1811は、TLR3KOマクロファージへの結合を示さず、C57BL/6腹膜マクロファージの細胞表面への結合を上昇させたことから、TLR3に対するmAbの特異性を示唆した。(図10)mAb 5429は、mAb c1811と同一の結合領域を有し、mAb c1811と同一の結合特異性を有すると推定される。
【0208】
(実施例9)
TLR3抗体アンタゴニストはTLR3介在型の全身性炎症を防御する
モデル
ポリ(I:C)誘発全身性サイトカイン/ケモカインモデルを、TLR3介在性全身性炎症のモデルとして用いた。このモデルでは、ポリ(I:C)(PIC)を腹腔内投与することで、TLR3によって一部介在される全身性のサイトカイン及びケモカイン応答を誘導した。
【0209】
雌C57BL/6マウス(8〜10週齢)又は雌TLR3ノックアウトマウス(C57BL/6バックグラウンド;8〜10週齢、Ace Animals、Inc.)に、mAb 5429を0.5mLのPBS中、10、20又は50mg/kgで、mAb c1811を、0.5mLのPBS中、2、10又は20mg/kgで、又は0.5mLのPBSを単独(溶媒コントロール)で皮下投与した。抗体投与の24時間後、マウスに、0.1mLのPBSに加えた50μgのポリ(I:C)(Amershamカタログ番号26−4732、ロット番号IH0156)を腹腔内投与した。ポリ(I:C)チャレンジの1及び4時間後に後眼窩より血液を採取した。全血より血清を調製してLuminexによりサイトカイン及びケモカインの濃度について解析した。
【0210】
結果
腹腔内投与されたポリ(I:C)は、TLR3ノックアウト動物においてケモカイン及びサイトカイン群の産生が大幅に低減されたことによって示されるように、TLR3によって一部介在される全身性のサイトカイン及びケモカイン応答を誘導した(表9A)。TLR3依存性のポリ(I:C)誘導型メディエーターは、ポリ(I:C)ポストチャレンジ1時間では、IL−6、KC、CCL2/MCP−1及びTNF−αであり、ポリ(I:C)ポストチャレンジ4時間では、IL−1α、CCL5/RANTES及びTNF−αであった。mAb c1811及びmAb 5429は、両方ともこれらのTLR3依存性のメディエーターのレベルを著しく低下させ、抗体がインビボでTLR3シグナル伝達を低下させる能力を実証した(表9B)。表9中の値は、6匹の動物/群±SEMのpg/mLでの平均のサイトカイン又はケモカイン濃度として示したものである。これらのデータは、TLR3の拮抗作用が、サイトカインストーム又は致死的ショックなどの状態における過剰なTLR3介在サイトカイン及びケモカインレベルを低減させるうえで有用となり得ることを示唆するものである。
【0211】
【表16】
【0212】
【表17】
【0213】
(実施例10)
TLR3抗体アンタゴニストは気道過敏症を軽減する。
モデル
気道過敏症をポリ(I:C)によって誘発させた。
【0214】
雌C57BL/6マウス(12週齢)又は雌TLR3ノックアウトマウス(C57BL/6バックグラウンド;12週齢、Ace Animals,Inc.)をイソフルランで麻酔し、50μLの滅菌PBS中のポリ(I:C)を数回投与量(10〜100μg)で鼻腔内投与した。各投与の間に24時間の休止時間を置いてポリ(I:C)(又はPBS)を3回マウスに投与した。最後のポリ(I:C)(又はPBS)の投与の24時間後に、全身プレチスモグラフィー(BUXCOシステム)を用いて肺機能及びメタコリンに対する気道過敏症を測定した。マウスを全身プレチスモグラフチャンバに入れ、少なくとも5分間順応させた。ベースラインの読み取りの後、投与量を増大させた霧状化したメタコリン(Sigma,St.Louis、MO)にマウスを曝露した。霧状化メタコリンを2分間投与した後、5分間のデータ収集時間、その後、10分間の休止時間を置いた後、メタコリンの用量を増大させた後続のチャレンジを行った。空気流の抵抗の増加をエンハンストポーズ(Enhanced Pause)(Penh)として測定し、5分間の記録時間にわたった平均のPenh値として表した(BUXCOシステム)。肺機能の測定の後、マウスを安楽死させ、肺にカニューレを挿管した。肺に1mLのPBSを注入し、流出液を回収することによって気管支肺胞洗浄(BAL)を行った。肺組織を切除して凍結した。BAL液を遠心(1200rpm、10分)し、細胞を含まない上澄み液を回収して、解析を行うまでの間−80℃で保存した。細胞ペレットを200μLのPBSに再懸濁して全細胞計数及び分画別の細胞計数を行った。製造者のプロトコール及びMultiplex Immunoassayキット(Millipore,Billercia,MA)に従って多重アッセイを行った。
【0215】
結果
過去の観察により、ポリ(I:C)の鼻腔内投与はマウスの肺機能にTLR3によって介在される障害を誘発し、全身プレチスモグラフィー(BUXCO)におけるベースラインのエンハンストポーズ(PenH)測定値が高くなり、霧状化されたメタコリンに対する応答性が高くなることが示されている(国際公開第06/060513(A2)号)。肺機能のこうした障害は、肺への好中球の動員、及び肺における炎症性サイトカイン/ケモカインのレベルの上昇をともなった。この実験では、肺機能におけるポリ(I:C)誘発障害におけるmAb 1811及びmAb 5429の作用を、ポリ(I:C)チャレンジに先立ってそれぞれの抗体を50mg/kgで皮下投与することによって評価した。
【0216】
TLR3によって介在される肺機能の障害は、ポリ(I:C)チャレンジに先立って動物をTLR3抗体アンタゴニストで処置することによって大幅に軽減された。抗TLR3抗体で処置された動物ではTLR3によって介在される、ベースラインPenH及びメタコリンに対する気道の感受性の増大を防止した(図11)。更に、抗TLR3抗体で処置した動物ではTLR3によって介在される、マウスの肺への好中球の動員及び気道におけるケモカインの生成を低減した。好中球の数(図12)及びCXCL10/IP−10のレベル(図13)は、回収した気管支肺胞洗浄液(BALF)から測定した。この実験を少なくとも3回繰り返して同様の結果を得た。図11、12、及び13に示されるデータは、1つの代表的な実験からのものである。各記号は1匹のマウスからのデータポイントを表し、横棒線は群の平均を示す。この実験によって、使用したモデルにおいて、全身投与されたTLR3抗体アンタゴニストが肺に達し、TLR3によって介在される肺機能の障害、気道内への好中球の浸潤、ケモカインの生成及び気道の炎症を軽減したことが示された。したがって、TLR3アンタゴニストは、喘息、アレルギー性鼻炎、慢性閉塞性肺疾患(COPD)、及び嚢胞性線維症などの、気道過敏症によって特徴付けられる呼吸器疾患の治療又は予防に有用であり得る。
【0217】
(実施例11)
TLR3抗体アンタゴニストは炎症性腸疾患を防御する。
モデル
DSS大腸炎モデルを炎症性腸疾患のモデルとして用いた。
【0218】
雌C57BL/6マウス(8週齢未満)又は雌TLR3ノックアウトマウス(C57BL/6バックグラウンド;8週齢未満で体重16.5g〜18g、Ace Animals,Inc)にγ線照射した餌を1日目から与えた。DSS(硫酸デキストラン)(MP Biomedicals,Aurora,OH,カタログ番号160110;35〜50kDa;18〜20%イオウ,ロット番号8247J)を、オートクレーブにかけた酸性化した飲料水中で5%の最終濃度まで希釈した。DSS水を5日間投与し、その後DSS水をただの水に置き換えた。マウスには実験全体を通じて自由に水を飲ませた。すべての水ボトルの重量を毎日測定して水の消費量を記録した。0、2、及び4日目にマウスに5mg/kg(0.1mL PBSに0.1mgを加えた)のmAb 5429、マウス抗TNF−α抗体、又はコントロールとしてのPBSを腹腔内投与した。マウスを実験の全体を通じて毎日監視し、0〜4日目及び7日目に体重を測定した。マウスを実験の2日目及び7日目に安楽死させた。腹腔を開き、上行結腸を盲腸と連結する位置で切断した。結腸を採取し、10%中性緩衝ホルマリン中で固定した。結腸をパラフィンに埋め込み、薄片にスライスし、H&E染色(Qualtek Molecular Labs,Santa Barbara,CA)した。下記に述べるように獣医病理学者によって結腸の組織病理学的評価を盲検により行った(PathoMetrix,San Jose,CA)。
【0219】
組織病理学的評価
大腸、結腸、及び直腸の2つの切片を評価し、以下の変化についてスコア化した。すなわち、(i)単一の細胞の壊死、(ii)上皮の潰瘍形成、(iii)上皮の脱落、(iv)陰窩腫瘍、(v)細胞増殖、(vi)陰窩細胞増殖、(vii)粘膜固有層における肉芽組織形成、(viii)粘膜下組織における肉芽組織、(ix)粘膜下炎症性細胞の浸潤物、好中球優位性、及び(x)粘膜下浮腫。
【0220】
重篤度の単一の全体的スコアを以下の基準に基づいて与えた。すなわち、
0−所見を認めず。
1−軽度、限局性、又は時折認められる。
2−軽度、多病巣である。
3−中度、頻繁だが限定された領域に認められる。
4−重度、提供された組織の多くの領域又は広範囲に認められる。
5−極めて重度、提供された組織の大部分に拡がっている。
【0221】
結果
過去の観察によれば、TLR3ノックアウト動物は、DSSの摂取によって誘発された炎症性腸疾患のモデルにおいて、野生型マウスと比較して大幅に軽減された組織病理学的所見を示すことが実証されており(国際公開第06/60513(A2)号)、このことはTLR3シグナル伝達がこのモデルにおける病因に一定の役割を有することを示唆するものである。壊死細胞から放出される共生生物のバクテリアRNA又は哺乳類RNAは、内因性リガンドとして作用して、TLR3シグナル伝達を刺激することができ(Karikoら、Immunity 23165〜231175 2005;Karikoら、J.Biol.Chem.279:12542〜12550 2004)、それゆえ、腸中の内因性リガンドによるTLR3刺激は、DSS大腸炎モデルにおける炎症を増進させ、及び持続させ得るということが報告された。
【0222】
複合組織病理学スコア(図14)によって評価されるように、DSSに曝露した動物における疾患重篤度は、抗TLR3抗体による処置によって改善された。図14は、疾患重篤度スコアについて平均、標準偏差及び95%信頼区間を横棒線として示している。抗TLR3抗体で処置した野生型のDSS曝露動物では、非処置の野生型動物と比較してスコアの有意な低下が認められた(p<0.05)。DSSに曝露したTLR3ノックアウト動物は、DSSによって誘発される変化から防御された。抗マウスTNF−α mAbを投与したDSS曝露動物では、DSSモデルにおいて組織病理学的所見の改善は認められなかった。したがって、DSSモデルは、抗TNF−αによる治療に応答しないヒト患者集団を対象とする治療法の評価において有用であり得、抗TLR3抗体を中和することは、抗TNF−αによる治療に応答しない炎症性腸疾患の患者にとって利益となる可能性を有すると考えられる。
【0223】
モデル
T細胞移植モデルを炎症性腸疾患のモデルとして用いた。このモデルでは、免疫応答能のあるマウスから得た制御性T細胞を含まないナイーブT細胞(腸粘膜の抗原提示細胞を攻撃する)の集団を移植することによって、SCIDマウスに腸の炎症を誘発した。
【0224】
ナイーブT細胞(CD4+CD45RBhighT細胞)をSCIDレシピエントに腹腔内注射することによって慢性大腸炎を誘発した。マウスに、PBS(500μL/マウス、腹腔内投与;溶媒コントロール)、mAb 5429(0.1mg/マウス、腹腔内投与)、又は抗TNF−α抗体(0.05mg/マウス、腹腔内投与;ポジティブコントロール)のいずれかを、T細胞の移植48時間後から開始して毎週2回、8週間の実験期間全体を通じて投与した。T細胞の移植から8週間後(又はマウスが初期の体重の15%以上を失った時点)に、動物を安楽死させて結腸を取り出した。結腸を固定し、パラフィンに埋め込んでH&E染色した。組織病理学的所見(細胞浸潤、陰窩腫瘍、上皮びらん、杯細胞喪失、及び腸壁の肥厚)を盲検的に定量的に評価した。
【0225】
結果
コントロール動物と比較した場合の組織病理学的スコアの合計の有意な低下(p<0.05)(図15A)によって評価されるように、疾患重篤度は、T細胞移植を受けた動物において抗TLR3抗体による処置に応じて改善された。スコアの合計には、陰窩腫瘍、潰瘍形成、好中球流入、杯細胞喪失、異常陰窩、粘膜固有層の炎症、及び経壁併発(transmural involvement)を評価した。陰窩腫瘍、潰瘍形成、及び好中球流入では有意な低下が認められた(いずれもp<0.05)(図15B)。抗TNFα抗体を、最適な効果を与えることが知られている用量でポジティブコントロールとして用いた。
【0226】
DSS及びT細胞移植モデルという2つの広く知られた炎症性腸疾患のモデルを用いた実験により、全身投与されたTLR3抗体アンタゴニストは腸粘膜に達し、2つの異なる病因機構によって誘発された消化管の炎症を軽減させることが実証された。したがって、TLR3アンタゴニストは、抗TNFα不応性の症例及び消化管の他の免疫介在性の病態を含む炎症性腸疾患の治療に有益であると考えられる。
【0227】
(実施例12)
TLR3抗体アンタゴニストはコラーゲン誘導性関節炎を防御する。
モデル
コラーゲン誘導性関節炎(CIA)モデルを関節リウマチのモデルとして用いた。
【0228】
雄B10RIIIマウス(6〜8週齢、Jackson Labs)を1群当たり15匹(関節炎群)又は1群当たり4匹(コントロール群)の群に分けた。関節炎群をイソフルランで麻酔し、II型コラーゲン(Elastin Products)、及び結核菌(M.tuberculosis)(Difco)を添加したフロイントの完全アジュバントを0及び15日目に投与した。12日目にII型コラーゲン誘導関節炎を発症したマウスを体重により治療群にランダム化し、mAb 5429(25mg/kg)、ネガティブコントロール抗体としてCVAM(マウスに既知の特異性を示さない組換えmAb)(5mg/kg)、又は抗TNF−α抗体(5mg/kg、ポジティブコントロール)を、12、17、及び22日目(d12、d17、d22)に皮下投与(SC)した。更に、マウスのコントロール群に溶媒(PBS)又はデキサメタゾン(0.5mg/kg、Dex、基準化合物)を12〜25日目まで毎日(QD)皮下投与(SC)した。動物を12〜26日まで毎日観察した。前肢及び後肢を臨床スコアシステム(下記に示す)によって評価した。実験26日目に動物を安楽死させ、組織病理学的所見を盲検的に評価した(下記にスコアシステムを述べる)。有効性評価は、動物の体重及び臨床的な関節炎スコアに基づいて行った。すべての動物が実験終了時まで生存した。
【0229】
前肢及び後肢の臨床的スコアの判定基準
0−正常
1−後肢又は前肢関節が冒される、すなわちわずかなびまん性紅斑及び腫脹を呈する。
2−後肢又は前肢関節が冒される、すなわち軽度のびまん性紅斑及び腫脹を呈する。
3−後肢又は前肢関節が冒される、すなわち中度のびまん性紅斑及び腫脹を呈する。
4−顕著なびまん性紅斑及び腫脹、すなわち4本の指関節が冒される。
5−足全体に重篤なびまん性紅斑及び重篤な腫脹を呈し、指を曲げられない。
【0230】
II型コラーゲン誘導関節炎を有するマウス関節の組織病理学的スコアリング方法
II型コラーゲン誘導関節炎の病変を有するマウスの肢又は足首関節をスコアリングする場合、変化の重篤度及び冒された個々の関節の数を考慮する。中手骨/中足骨/指又は足根骨/脛足根骨の多数の可能性のある関節のうち、1〜3個のみの足又は足首の関節が冒されている場合には、変化の重篤度に応じて下記のパラメータの1、2又は3の最大スコアが任意に与えられる。2個よりも多い関節が関与している場合には、最も重く冒された/大多数の関節に下記の基準を適用する。
【0231】
足スコアの臨床データは、1〜15日目についてAUCによって分析し、コントロールから阻害率(%)を計算した。
【0232】
炎症
0−正常
1−冒された関節の滑膜及び関節周囲組織に炎症性細胞が最小限に浸潤している。
2−軽度の浸潤を呈している。足の場合には、冒された関節に限定された軽度の浸潤を呈している。
3−中度の浸潤を呈している。足の場合には、冒された関節に限定された中度の浸潤を呈している。
4−大部分の領域が顕著な浮腫をともなう顕著な浸潤を呈している。
5−重篤な浮腫をともなう重篤なびまん性浸潤を呈している。
【0233】
パンヌス
0−正常
1−軟骨及び肋軟骨下骨にパンヌスが最小限に浸潤している。
2−冒された関節の硬組織の周辺帯の破壊をともなう浸潤を呈している。
3−冒された関節の中度の高組織破壊をともなう中度の浸潤を呈している。
4−大部分の関節で関節構造の顕著な破壊をともなう顕著な浸潤を呈している。
5−関節構造の全体的又はほぼ全体的な破壊をともなう重篤な浸潤がすべての関節を冒している。
【0234】
軟骨の傷害
0−正常
1−トルイジンブルー染色が最小限〜軽度に損なわれている。冒された関節に明らかな軟骨細胞の損失もコラーゲン破壊も認められない。
2−トルイジンブルー染色が軽度に損傷を受けている。冒された関節に限局性の軽度(表在性)の軟骨細胞の損失及び/又はコラーゲン破壊が認められる。
3−トルイジンブルー染色が中度に損傷を受けている。冒された関節に多病巣性の中度(深部又は中間層)の軟骨細胞の損失及び/又はコラーゲン破壊が認められる。
4−トルイジンブルー染色が顕著に損傷を受けている。大部分の関節に多病巣性の顕著な(深部〜深層)軟骨細胞の損失及び/又はコラーゲン破壊が認められる。
5−トルイジンブルー染色が重度に損傷を受けている。すべての関節に多病巣性の重度(深部〜タイドマーク)の軟骨細胞の損失及び/又はコラーゲン破壊が認められる。
【0235】
骨吸収
0−正常
1−小さい範囲の吸収が認められ、最小の程度に生じている。低倍率では容易に確認できず、冒された関節に破骨細胞をほとんど認めない。
2−より多くの範囲の吸収が認められ、軽度に生じている。低倍率では容易に確認できず、冒された関節により多くの破骨細胞が認められる。
3−皮質の厚さ全体の欠損をともなわない髄様骨梁(medullary trabecular)及び皮質骨の明らかな吸収が、中度に生じている。一部の髄様骨梁が損傷し、病変が低倍率で明瞭に確認できる。冒された関節により多くの破骨細胞が認められる。
4−残存する皮質表面の断面の歪みをしばしばともなう、皮質骨の厚さ全体の欠損が、顕著な程度に生じている。髄様骨の顕著な損失、多数の破骨細胞が認められる。大部分の関節が冒されている。
5−皮質骨の厚さ全体の欠損及びすべての関節の関節構造の破壊が認められる。重度。
【0236】
結果
デキサメタゾン(Dex)及び抗マウスTNF−α抗体をポジティブコントロールとして用い、PBSを溶媒コントロールとして用い、CVAMをネガティブコントロール抗体として用いた。すべての処置は、実験の12日目の関節疾患が進行する間に開始した。溶媒で処置した疾患コントロール動物では疾患発症率は実験22日目までに100%となった。溶媒又はCVAM抗体で処置したネガティブコントロール群では臨床スコアは最も高かった。Dex(d18〜d26についてp<0.05)、5mg/kgの抗TNF−α抗体(d18〜26についてp<0.05)、又は25mg/kgのmAb 5429(d18〜d23及びd25〜d26についてp<0.05)で処置した各群では有意に低い臨床スコアが認められた(図16)。曲線下面積(AUC)として表した臨床的関節炎スコアは、25mg/kgのmAb 5429(低下率43%)、5mg/kgの抗TNF−α抗体(52%)、又はDex(69%)による処置により、溶媒コントロールと比較して有意に低下した。図17は、各群についてAUCの平均及び標準偏差を示す。
【0237】
各処置の組織病理学的作用についても評価を行った。足の骨吸収は、25mg/kgのmAb 5429(低下率47%)による処置によって溶媒コントロールと比較して有意に低下した。5mg/kgの抗TNF−α抗体で処置したポジティブコントロールマウスでは、足の炎症(33%)、軟骨傷害(38%)、及び合計した足スコア(37%)は有意に低下した。Dexによる処置によって、足の組織病理学的パラメータのすべてが有意に低下した(合計スコアにおいて73%の低下率)。
【0238】
これらのデータは、TLR3抗体アンタゴニストが、CIAモデルにおいて臨床的及び組織病理学的疾患症状を改善することを示すものであり、関節リウマチの治療におけるTLR3アンタゴニストの使用を示唆するものである。
【0239】
(実施例14)
TLR3抗体アンタゴニストは急性致死性ウイルス感染を防御する。
モデル
インフルエンザAウイルスチャレンジモデルを急性致死性ウイルス感染のモデルとして使用した。
【0240】
1、4、8及び12日目に雌C57BL/6マウス(12週齢)又は雌TLR3ノックアウトマウス(C57BL/6バックグラウンド;12週齢、ACE Animals,Inc.、1群当たり15匹のマウス)に20mg/kgのmAb 5429、又はPBS単独を皮下投与した。0日目にイソフルランでマウスを麻酔し、インフルエンザA/PR/8/34ウイルス(ATCC、Rockland、MD、ロット番号218171)を25μLのPBSに加えたもの(105.55CEID50に相当)を鼻腔内投与した。動物を体重変化及び生存率について、14日間の期間にわたって1日2回観察した。臨床的スコアリングシステムを用いて疾患の進行のレベル及びインフルエンザAウイルス治療に対するわずかな改善を評価した。
【0241】
臨床スコア
0−正常。機敏で反応性があり、目に見える病気の兆候が認められない。
1−毛皮が逆立っている。歩行のわずかな低下が認められるか、又は認められない。
2−歩行時に背中が曲がっている。歩行をいやがる。呼吸困難である。
3−毛皮が逆立っている。呼吸困難である。運動失調。震えが生じている。
4−毛皮が逆立っている。そっと突いても歩行不可能。意識不明。触れると冷たく感じる。
5−死亡。
【0242】
結果
この実験では、生存率、毎日の臨床スコア、及び体重変化を評価した。mAb 5429(20mg/kg)を投与したインフルエンザA感染野生型マウス及びmAb 5429を投与しないインフルエンザA感染TLR3ノックアウトマウスのいずれも、インフルエンザウイルスを接種したC57BL/6マウスと比較して生存率が統計的に有意に増大した(それぞれp<0.001及びp<0.01)。このことは、TLR3の阻害又は欠損がインフルエンザによって誘発される死を予防し得ることを示すものである(図18)。臨床スコアは、20mg/kgのmAb 5429を投与した群、及びTLR3ノックアウト群において有意に低下した(図19)。マウスの体重をインフルエンザウイルス投与後14日間にわたって観察した。インフルエンザAウイルスを投与したC57BL/6マウスでは体重は安定的に減少した。しかしながら、20mg/kgのmAb 5429を投与したC57BL/6マウス及びTLR3ノックアウトマウスのいずれも、インフルエンザウイルスを接種した野生型C57BL/6マウスに対して有意に大きな体重を示した(図20)。これらの結果は、TLR3抗体アンタゴニストが急性致死性インフルエンザウイルス感染モデルにおける臨床症状及び死亡率を低下させることを示すものであり、TLR3アンタゴニストが急性の感染状態にあるヒトに防御を与えることを示唆するものである。
【0243】
(実施例15)
TLR3抗体アンタゴニストは高血糖を改善し、血症インスリンを低下させる。
モデル
食餌誘発型肥満(DIO)モデルを高血糖及びインスリン抵抗性、並びに肥満のモデルとして用いた。
【0244】
C57BL/6野生型動物(約3週齢、Jackson Labs)及びTLR3ノックアウト動物(C57BL/6バックグラウンド;約3週齢、Ace Animals,Inc.)に12〜16週間持続的に高脂肪の食餌を与えた。TLR3ノックアウト及び野生型C57BL/6マウスの両方に、通常の固形飼料又は60.9% kcalの脂肪及び20.8% kcalの炭水化物からなる高脂肪の食餌(Purina Test Dietカタログ番号58126)のいずれかを与えた。マウスには、水及び食餌を自由摂取させ、12時間/12時間の明暗サイクル中に維持した。各群の各マウスの体重を毎週測定した。mAb 5429を第1週に週2回、その後は全体で7週間にわたり、週1回腹腔内投与した。空腹時に後眼窩から採取した血清試料を、示した時点でのインスリンの測定に用いた。7週目に一晩絶食させた後、1.0mg/g(体重)のグルコースを腹腔内投与することによって耐糖試験を行った。更に、空腹時のインスリン及びグルコースレベルを測定した。
【0245】
以下の式を用いて、空腹時グルコースレベル及びインスリンレベル(12)に基づきHOMA−IRを式から求めた。すなわち、HOMA−IR=((空腹時グルコース(mmol/l)×空腹時インスリン(mU/l))/22.5(Wallaceら、Diabetes Care 27:1487〜1495、2004)。空腹時血中グルコース(血糖値)(BG)をグルコースオキシダーゼアッセイによって求めた。空腹時インスリンレベルをインスリンラット/マウスELISAキット(Crystal Chem、カタログ番号90060)を使用して求めた。
【0246】
結果
高脂肪の食餌を12〜16週間与えた後、野生型DIO動物は高血糖及び高インスリンとなった。mAb 5429による処置により、野生型DIO動物では耐糖能が改善したがTLR3ノックアウトDIO動物では改善が見られなかった。グルコースチャレンジの60、90、120及び180分後に、mAb 5429で処置した動物においてコントロール(PBSのみ)と比較して有意に低い血中グルコース濃度が観察された(図21A)。mAb 5429で処置した野生型DIO動物では、mAbを投与しない野生型DIOマウスと比較してAUCの約21%の低下が観察された。mAb 5429で処置した野生型DIO動物では空腹時インスリンレベルも低下した(図22)。TLR3ノックアウトDIO動物では、mAb 5429による処置により空腹時インスリンに改善は認められなかった。インスリン抵抗性指数(Homeostatic model assessment)(HOMA)分析は、mAb 5429で処置した野生型DIO動物においてインスリン感受性の改善を示したが、TLR3ノックアウトDIO動物では改善を示さなかった。HOMA−IR値は、野生型DIO、5mg/kgの野生型DIO(mAb 5429投与)、及び20mg/kgの野生型DIO(mAb 5429投与)動物で、それぞれ14.0±9.8、8.7±4.9、9.0±3.0であった。TLR3ノックアウトDIO動物では何の効果も認められなかった。
【0247】
この実験により、TLR3抗体アンタゴニストがDIOモデルにおいて体重の損失をともなわずにインスリン抵抗性を改善し、空腹時グルコースを低下させることが示されたが、このことは、TLR3アンタゴニストが高血糖症、インスリン抵抗性、及びII型糖尿病の治療に有用であり得ることを示唆するものである。
【0248】
(実施例16)
TLR3抗体アンタゴニストは細菌及びウイルス誘発炎症反応を防御する。
試薬
細菌性増悪したCOPD患者から単離された分類不能型ヘモフィルスインフルエンザ(Nontypeable Haemophilusインフルエンザ)(HNTHi)株35をDr.T.F.Murphy(Buffalo VA Medical Center,Buffalo,NY)より入手した。ヒトライノウイルス16を、アメリカ培養細胞系統保存機関(American Type Culture Collection)(ATCC)よりTCID(50)=2.8×107/mLのものを入手した。
【0249】
NTHi刺激アッセイ
NHBE細胞(Lonza,Wakersville,MD)をMicrotest 96穴組織培養プレート(BD Biosciences,Bedford,MA)に1×105/ウェルで播いた。寒天プレート上で16〜20時間増殖させたNTHiを増殖培地に約2×108cfu/mlで再懸濁し、100μg/mlのゲンタマイシンで30分間処理してから約2×107/ウェルでNHBEを含む96穴プレートに加えた。3時間後に上澄み液を除去し、抗体(0.08〜50μg/mLの最終濃度)を添加又は非添加の新鮮な増殖培地に交換した。更に24時間インキュベートした後、細胞上澄み液中のサイトカイン及びケモカインの存在を、Luminex 100IS多重蛍光分析器及びリーダーシステム(Luminex Corporation,Austin,TX)でサイトカイン25重ABビーズキット、ヒト(IL−1β、IL−1RA、IL−2、IL−2R、IL−4、IL−5、IL−6、IL−7、IL−8、IL−10、IL12p40p70、IL−13、IL−15、IL−17、TNF−α、IFN−α、IFN−γ、GM−CSF、MIP−1α、MIP−1β、IP−10、MIG、エオタキシン、RANTES及びMCP−1を含む)によって3重にアッセイした。
【0250】
ライノウイルス刺激アッセイ
NHBE細胞をMicrotest 96穴組織培養プレート(BD Biosciences,Bedford,MA)に1×105細胞/ウェルで播いた。翌日、抗体(0.08〜50μg/mLの最終濃度)をNHBE又はBEAS−2B細胞に加えて1時間インキュベートした後、10μL/ウェルのライノウイルスを加えた。更に24時間インキュベーションした後、細胞上澄み液中のサイトカイン及びケモカインの存在について上記に述べたようにluminexアッセイによってアッセイを行った。
【0251】
結果
mAb 15EVQが、NTHiにより誘導されたIP−10/CXCL10及びRANTES/CCL5の産生を用量依存的に阻害したのに対して、コントロール抗体であるヒトIgG4(Sigma,Louis,MO)はNTHiによる刺激に対して阻害作用を示さなかった(図23A)。mAb 15EVQは、ライノウイルスにより誘導されたCXCL10/IP−10及びCCL5/RANTESの産生も阻害した(図23B)。
【0252】
(実施例17)
TLR3抗体アンタゴニストは星状細胞における炎症反応を抑制する。
方法
2人のドナーから得た正常なヒト星状細胞(Lonza,Walkersville,MD)を24穴プレートに75,000細胞/ウェルで播き、一晩接着させた。翌日、星状細胞を200ng/mLのポリ(I:C)及び/又は10μg/mLのmAb 18で24時間処理した。サイトカインをLuminexによって測定した。
【0253】
結果
表10に示すように、ポリ(I:C)により誘導されたIL−6、IL−8、IL−12、IFN−α、IFN−γ、CXCL9/MIG、CCL3/MIP−1a、CCL4、CCL5/RANTES及びCXCL10/IP−10の産生はmAb 18によって阻害された。
【0254】
【表18】
【0255】
(実施例18)
TLR3抗体アンタゴニストは上皮細胞における炎症反応を抑制する。
方法
HUVEC細胞(Lonza,Walkersville,MD)をロンザ社の推奨する血清含有増殖培地中で培養した。細胞を無血清培地(Lonza,Walkersville,MD)に再懸濁し、96穴プレートに3×105細胞/mLで播き、37℃、5%CO2で24時間インキュベートした。ポリ(I:C)(GE Healthcare、Piscataway,NJ)を濃度を増加させながら(1.5〜100μg/mL)加え、更に24時間37℃でインキュベートした。サイトカイン阻害アッセイを行うため、mAb 15EVQを細胞に異なる濃度(0〜50μg/mL)で加えて30分間インキュベートし、その後、20μg/mLのポリ(I:C)を24時間加えた。細胞上澄み液を回収し、ヒトサイトカイン30重キット及びLuminex MAP技術(Invitrogen Corp.,Carslbad,CA)を用いてサイトカインレベルを測定した。sICAM−1の発現を測定するため、HUVEC細胞を20μg/mLのポリ(I:C)及び異なる濃度のmAb 15EVQ(0.8〜50μg/mL)で処理した。細胞上澄み液をELISA(R&D Systems)によりsICAM−1の発現について分析した。細胞の生存率をCellTiterGloキット(Promega,Madison,WI)により測定した。
【0256】
結果
HUVEC細胞は、ポリ(I:C)に応答して次のサイトカインを産生した:IL−1RA、IL−2、IL−2R、IL−6、IL−7、CXCL8/IL−8、IL−12(p40/p70)、IL−15、IL−17、TNF−α、IFN−α、IFN−γ、GM−CSF、CCL3/MIP−1α、CCL4/MIP−1β、CXCL10/IP−10、CCL5/RANTES、CCL2/MCP−1、VEGF、G−CSF、FGF−basic、及びHGF(表11)。mAb 15EVQは、ポリ(I:C)により誘導される全サイトカインのレベルを用量依存的に低下させた(表12)。ポリ(I:C)により誘導されるTNF−α、CCL2/MCP−1、CCL5/RANTES、及びCXCL10/IP−10の産生を低下させるmAb 15EVQの能力は、TLR3により介在される活性の阻害が、アテローム性動脈硬化症につながり得る白血球及びT細胞の浸潤に対する防御を与え得ることを示唆するものである。更に、mAb 15EVQによるVEGFの阻害は、各種の癌及び加齢性黄斑変性などの眼疾患における脈管形成を含む、VEGFによって介在される病態におけるTLR3の遮断の潜在的な効果を示唆するものである。
【0257】
TNF−α及びIFN−γは白血球の動員において機能し、かつ活性化された内皮上の接着分子の発現を増大させる(Doukasら、Am.J.Pathol.145:137〜47、1994;Poberら、Am.J.Pathol.133:426〜33、1988)。CCL2/MCP−1、CCL5/RANTES及びCXCL10/IP−10は、単球及びT細胞の動員に関与していることが示されており、アテローム性動脈硬化症の発症に寄与する(Lundgergら、Clin.Immunol.2009)。内皮細胞によるVEGFの生成は、各種の癌において脈管形成時の異常な組織増殖又は腫瘍との関連が示されている(Livengoodら、Cell.Immunol.249:55〜62、2007)。
【0258】
【表19】
【0259】
可溶性細胞内接着分子1(sICAM−1)はタンパク質切断によって生成される、内皮細胞活性化のマーカーである。ICAM−1は白血球の遊走及び活性化において重要な役割を有し、炎症時に内皮細胞及び上皮細胞において発現が上昇して、インテグリン分子LFA−1及びMac−1を介した白血球への接着を介在する。ポリ(I:C)は内皮細胞を活性化してsICAM−1の発現を上昇させたが、この発現上昇はmAb 15EVQによる処理によって低減した(図24A)。
【0260】
【表20】
【0261】
このことは、TLR3抗体アンタゴニストが白血球の輸送を阻害し、したがって炎症性細胞の流入によって引き起こされる組織障害を阻害し得ることを示す。
【0262】
生存率アッセイを行うため、上記で述べたようにHUVECを培養、播種し、ポリ(I:C)で刺激した。mAb 15EVQはポリ(I:C)により誘導されたHUVEC細胞の生存率の低下を用量依存的に回復させた(図24B)。
【0263】
内皮細胞の活性化の下方調節は、過剰な免疫細胞の浸潤を抑制し、炎症状態において増加するサイトカインによって引き起こされる組織障害を低減させ得る。炎症及び内皮細胞におけるサイトカイン及び接着分子の過剰発現は、アテローム性動脈硬化症及び高血圧の発症の主要な寄与因子である。これらのデータは、脈管炎及び内皮機能不全をともなう血管疾患などの血管の疾患におけるTLR3アンタゴニストの使用の潜在的な有用性を探求することの根拠を与えるものである。炎症及び過剰発現したサイトカインによって引き起こされる別の疾患として、免疫抑制患者及びHIV感染患者において一般的に見られ、カポジ肉腫ヘルペスウイルス(KSHV)によって引き起こされるカポジ肉腫(KS)がある。VEGF及びサイトカインの産生はKS細胞の生存に寄与する(Livengoodら、Cell Immunol.249:55〜62、2007)。TLR3アンタゴニストは、KS及び他の腫瘍にともなう血管新生のリスクを低減するうえで有用であるばかりでなく、細胞生存率の低下を防止し、内皮バリアの完全性を保護することによって、臓器不全及び敗血症などの重篤な炎症状態にともなう潜在的に重大な状態である血管漏出を防止するうえでも有用であり得る。TLR3拮抗作用は、また、フラビウイルス科(例えば、Dengue,yellow fever)、フロビウイルス科(Ebola,Marburg)、ブニヤウイルス科(例えば、Hantavirus,Nairovirus,Phlebovirus)、及びアレナウイルス科(例えば、Lujo,Lassa,Argentine,Bolivian,Venezuelan出血熱(Sihibamiyaら、Blood 113:714〜722,2009))の多数の属により引き起こされるウイルス出血熱などの内皮細胞病変を含むウイルス感染症においても有益でもあり得る。
【0264】
(実施例20)
TLR3抗体アンタゴニストのカニクイザル及びマウスTLR3との交差反応性
実施例2で述べたようにISREレポーター遺伝子アッセイを用いて、カニクイザル又はマウスTLR3に対する活性を評価した。カニクイザル(配列番号217)及びマウスTLR3 cDNA(配列番号161)を全血から増幅し、pCEP4ベクター(Clontech)の中にクローニングし、及び上述のように発現させた。mAb 15EVQは、ヒトTLR3 NF−κB及びISREアッセイにおけるそれぞれ0.44及び0.65μg/mLのIC50と比較して、cyno NF−κB及びISREアッセイにおいてそれぞれ4.18μg/mL及び1.74μg/mLのIC50を有していた。アイソタイプコントロール抗体はこれらのアッセイでは何の作用も示さなかった。
【0265】
(実施例21)
TLR3抗体アンタゴニストの治療用投与は、急性の致命的なウイルス感染症を防御する。
実施例14は、インフルエンザA感染症に対するTLR3抗体アンタゴニストによる予防的な治療(−1、4、8、及び12日目に投与)について述べている。この実施例は、TLR3抗体アンタゴニストの治療用投与(インフルエンザA感染後臨床症状の発症3日後)が生存率の向上において効果的であるということを実証している。
【0266】
モデル
動物へのmAb 5249の投与をインフルエンザAの感染後3日後に行ったこと、及び投与した動物が8週齢であったことを除いて、インフルエンザAウイルスチャレンジモデルを、実施例14に述べたように急性の致命的なウイルス感染症のモデルとして使用した。抗マウスIgG1アイソタイプコントロールmAbはBioLegendからのものであった。インフルエンザAの感染後3、7及び11日に動物に投与した。
【0267】
この実験では、生存率、毎日の臨床スコア、及び体重変化を評価した。mAb 5249を投与したC57Bl/6マウス及びTLR3 KOマウスは、抗マウスIgG1アイソタイプコントロールmAb及びインフルエンザウイルスを接種したC57BL/6マウスと比べて、両方とも生存率の統計的に有意な増加(p<0.028及びp<0.001)を示した(図25)。抗マウスIgG1アイソタイプコントロールmAb及びインフルエンザAを投与したC57BL/6マウスと比較した場合、mAb 5249を投与したC57BL/6マウス及びTLR3 KO動物において臨床的なスコアは低下し(図26)、体重は増加した(図27)。これらの結果は、TLR3抗体アンタゴニストが急性致死性インフルエンザウイルス感染モデルにおける臨床症状及び死亡率を低下させることを示すものであり、TLR3アンタゴニストが急性の感染状態にあるヒトに防御を与えることを示唆するものである。
【0268】
(実施例22)
X線結晶法によるTLR3抗体アンタゴニストのエピトープ及びパラトープ。
ヒトTLR3細胞外ドメインを、mAb 15EVQ、mAb 12QVQ/QSV及びmAb c1068のFabsと複合体の状態で結晶化させた。
【0269】
方法
タンパク質の発現及び精製。
TLR3 ECD(配列番号2のアミノ酸1〜703)の3つのFabの発現及び精製は上述の通りであった。
【0270】
TLR3 ECDの3つのFabの四次複合体の作製
1TLR3 ECD:1.1Fabのモル比に対応させて、4mgのヒトTLR3 ECDを2.4mgのそれぞれのFabと混合し、4℃で3.5時間インキュベートした。複合体をアニオン交換クロマトグラフ法により20mMのTris(pH 8.5)、10%のグリセロール(緩衝液A)で平衡化したMonoQ5/50GLカラム(GE Healthcare,Piscataway,NJ)で精製し、20mMのTris(pH 8.5)、10%のグリセロール、1MのNaCl(緩衝液B)により溶離した。1.74mL中のほぼ2.48mgの複合体を緩衝液Aにより10mLまで希釈し、カラム上に1mL/minで充填し、B濃度の0〜40%の直線的な勾配により40カラム容積にわたって溶離した。5回の連続的な精製操作を行った。ピーク1由来の区分をプールし、Amicon−15mLのUltra−30000 MWCO及びMicrocon 30000 MWCOにより20mMのTris(pH 8.5)/27mMのNaCl/10%グリセロール中に、14.49mg/mLまで濃縮した(吸光係数:A280(1mg/mL)=1.31)。
【0271】
結晶化
Corningプレート3550(Corning Inc.,Acton,MA)を用いて等容量のタンパク質及びリザーバ溶液を、シッティングドロップ法で分注するOryx4自動タンパク質結晶化ロボット(Douglas Instruments)を用いて、自動化結晶化スクリーニングを行った。初期のスクリーニングは、Hampton Crystal Screen HT(HR2−130,Hampton Research,Aliso Viejo,CA)により実施した。いくつかの条件から、小さい結晶を使用してシードを生成し、次いでこれをMicroseed−Matrix Screening(MMS)で使用した。小さい結晶を与える初期のスクリーニング条件に基づき、何回かの精密化を行った。MMSに使用するリザーバ条件は、精密化後に小さい結晶を与えた条件に基づくものであった:18〜28%のポリエチレングリコール(PEG)3350、1MのLiCl、pH4.5及び2.0〜2.9Mの(NH4)2SO4、5%のPEG400、pH4.5、及び探索したpH及び異なる添加物。成分を1タンパク質溶液:0.25シード原料:0.75リザーバ溶液の容量比で分注することにより、Oryx4自動タンパク質結晶化ロボット(Douglas Instruments)を用いて、MMS結晶化スクリーニングを行った。〜10Åの分解能で回折する結晶は、0.1Mの酢酸Na(pH 4.5)、2.9Mの(NH4)2SO4、5%メチル−ペンタン−ジオール(MPD)及び0.1Mの酢酸Na(pH 4.5)、26%のPEG3350、1MのLiClから成長させたものであった。
【0272】
結晶の分解能を改善するための試みとして、上記の条件を有するMMSを、Hampton Additive Screen HR2−428(Hampton Research,Aliso Viejo,CA)から選択された成分を容積比:1タンパク質溶液:0.125シード原料:0.2添加物溶液:0.675リザーバ溶液で用いる、添加物スクリーニングと組み合わせた。MMSと、0.1Mの酢酸Na(pH 4.5)、28%のPEG 3350、1MのLiCl、及び30mMのGly−Gly−Glyを含有する溶液からの添加物スクリーニングとの組み合わせを適用した後、Fabと複合体形成し、約5Åの分解能で回折する、TLR3 ECDのX線品質の結晶を得た。
【0273】
TLR3 ECD四次複合体のX線データ収集
X線データを収集するために、結晶(サイズ約1.0×0.5×0.1mm3)を、合成母液(0.1Mの酢酸Na(pH 4.5)、28% PEG 3350、1MのLiCl、16%グリセロール)中に数秒浸漬し、窒素流中100Kで瞬間凍結した。X線回折データを収集し、Osmic(商標)VariMax(商標)共焦光学系、Saturn 944 CCD検出器、及びX−stream(商標)2000低温冷却システムを備えたRigaku MicroMax(商標)−007HF微小焦点X線発生装置(Rigaku,Woodlands,TX)を使用して処理した。回折強度を、1/2度の像当たり1minの露出時間、5Åの最大分解能で250°(結晶回転)にわたって検出した。X線データを、プログラムD*TREK(Pflugrath,Acta Crystallographica Section D,55:1718〜1725,1999)により処理した。この結晶は、単位セルパラメーター:a=214.90Å、b=142.08Å、c=125.04Å、及びβ=103.17の単斜晶系空間群C2に属する。この非対称ユニットには1分子の複合体を含む。X線データ統計値を表13に示す。
【0274】
【表21】
【0275】
構造決定
TLR3 ECD−Fab 15EVQ−Fab 12QVQ/QSV−Fab c1068の結晶構造を、Phaser(Read,Acta Crystallogr.Crystallogr.D.Biol.Crystallogr.57:1373〜1382,2001)を用い分子置換により決定した。探索モデルはTLR3 ECD(すべてのグルカンを取り除いた、タンパク質データバンク(PDB)構造番号1ziw,Choeら、Science 309:581〜585,2005)であり、3つのFabの高分解能結晶構造を決定した(これらのFab構造に対する結晶データ及び精密化統計値の要約としては表13を参照のこと)。Fab 12QVQ/QSVのエルボー角度は、遊離形態におけるものから著しくはずれていることが判明した。エルボー角度を約5°間隔で調整することにより、一連のFab 12QVQ/QSVモデルを生成し、その1つが電子密度とよく一致することが判明した。構造精密化をPHENIX(Adamsら、J.Synchrotron Radiat.11:53〜55,2004)により行った。TLR3 ECDに対して構造を、Fab及び13個の剛体セグメント(精密化で使用される定義:30〜60、61〜108、109〜156、157〜206、207〜257、258〜307、308〜363、364〜415、416〜464、465〜514、515〜570、571〜618、619〜687)に対する剛体ドメイン(各V又はCドメイン)として、それぞれのFab剛体に対する1つのB因子及びTLR3 ECD全体に対する単一のB因子によって精密化した。
【0276】
Fab剛体のそれぞれに対して並進/回転/回転を伴った並進(TLS)精密化を導入し、配列番号2の残基330において、TLR3 ECDを2つのTLSセグメントに分割した。グリカン密度は15個のN−グリコシル化部位の10個に対して目視可能であった。次いで、ヒトTLR3細胞外ドメイン(Choeら、Science 309:581〜585,2005,PDB構造番号1ziw)の結晶構造からの炭水化物モデルを付加した。TLR3 ECD中の短い見えないセグメント(配列番号2の残基337〜342)に対する密度が剛体の精密化後に目視可能となった。それは、細胞外構造(2a0zBellら、Proc.Acad.Sci.(USA)102:10976〜10980,2005,PDB構造番号2a0z)からの対応するセグメントにより充填されていた。TLR3 ECDのC末端は、2a0zのそれに釣り合う追加の密度を含んでいた。次いで、このセグメント(配列番号2の647〜703)を2a0wの(残基647〜687)により置き換えた。このように、TLR3 ECDモデルは、TLR3構造体1ziwと2a0zとの間のハイブリッドであり、13個の剛体セグメント(アミノ酸範囲:30〜60、61〜108、109〜156、157〜206、207〜257、258〜307、308〜363、364〜415、416〜464、465〜514、515〜570、571〜618、619〜687)として精密化されたものであった。
【0277】
Fab 12QVQ/QSVのLCDR3は、遊離形態と異なる立体配置を見掛け上とっていた。マルチスタートシミュレーションアニーリングをPHENIXの標準的なパラメータにより行った。このLCDR3のモデルを電子密度マップで目視的に検査し、「最もよく合う」立体配置を元のモデル上に接合した。精密化のプロセスを、計算開始の前に脇に置いておいた反射の5%に対してRfreeによりモニターした。最終回では、それぞれの残基に対して1つのB因子を含めた。モデル検査、並びにFab及びタンパク質−タンパク質界面における側鎖のエルボー領域の手動による再組立てを、COOT(Emsleyら、Acta Crystallogr.D.Biol.Crystallogr.60:2126〜32,2004)を用いて行った。最終のRcryst及びRfreeは、5.0Å対する全15,792個の独立の反射のそれぞれについて26.8%及び30.0%であった。精密化統計値を表13及び14に示す。
【0278】
結果
TLR3 ECD−3つのFab四次複合体の分子構造
複合体の全体的な分子構造を図28に示す。非対称単位中に1つのTLR3 ECD及びそれぞれのFabの1分子が存在する。TLR3 ECDに対する構造モデルは、huTLR3(配列番号2)の30〜687個のすべての残基を含む。3つのFabに対しては、それぞれの非結合形からのすべての残基を、溶媒イオン及び水分子を除いて含めた。TLR3 ECD分子は、全体のトポロジー(1ziwに対して0.79Åのrms、613Cα’、及び2a0zに対して1.37Å、595Cα’)において以前に報告された構造と極めて類似している。このFab構造は、方法に記載のFab 12QVQ/QSVのLCDR3、並びにTLR3 ECD/Fab界面におけるエルボー領域及び一部の側鎖を除いて、それぞれの非結合形とすべて同一である。
【0279】
【表22】
【0280】
最高の分解能シェルに対する値は()中のものである。
a Rmerge=Σ|I−<I>|/ΣI、式中、Iは測定された反射の強度であり、<I>はこの反射のすべての測定値の平均強度である。
b Rcryst=Σ||Fobs|−|Fcalc||/Σ|Fobs|、式中、Fobs及びFcalcは測定及び計算された構造因子であり、Rfreeは精密化の前に反射のランダムに選択された5%の組に対して計算されたものである。
c ラマチャンドランプロットをMolProbity(Davis,I.W.ら、Nucleic Acids Res,32:W615−9,2004)により計算した。
【0281】
エピトープ及びパラトープ
TLR3 ECDと3つのFabとの間の結合に関与する残基を、図28Bに示す。Fab 12QVQ/QSVはTLR3 ECDのN末端近くで結合した。立体配置エピトープは、TLR3 LRR 3〜7からの残基(配列番号2のアミノ酸100〜221)から構成されるものであった。Fab 12QVQ/QSVの結合は、抗原及び抗体上でそれぞれほぼ928Å2及び896Å2を埋め込んだ。Fab 12QVQ/QSVに対しては、結晶構造は、次のTLR3(配列番号2)エピトープ残基:S115、D116、K117、A120、K139、N140、N141、V144、K145、T166、Q167、V168、S188、E189、D192、A195、及びA219を同定した。Fab 12QVQ/QSVに対しては、結晶構造は、次のパラトープ残基:軽鎖(配列番号211):G28、S29、Y30、Y31、E49、D50、Y90、D91及びD92、重鎖(配列番号214):N32、Q54、R56、S57、K58、Y60、Y104、P105、F106及びY107を同定した。
【0282】
Fab 15EVQ及びFab c1068は、C末端近くのLRR 15−23(配列番号2のアミノ酸406〜635)に懸る、非折りたたみ型エピトープを結合していた。(図28)。Fab 15EVQは、抗原及び抗体上でそれぞれ1080Å2及び1064Å2を埋め込み、Fab c1068は抗原及び抗体上でそれぞれ963Å2及び914Å2を埋め込んでいた。Fab 15EVQに対するエピトープは、配列番号2で示されるTLR3の残基K416、K418、L440、N441、E442、Y465、N466、K467、Y468、R488、R489、A491、K493、N515、N516、N517、H539、N541、S571、L595及びK619を覆う。Fab 15EVQに対しては、結晶構造は、次のパラトープ残基:軽鎖(配列番号41):Q27、Y32、N92、T93、L94及びS95、重鎖(配列番号216):W33、F50、D52、D55、Y57、N59、P62、E99、Y101、Y104及びD106を同定した。
【0283】
Fab c1068に対しては、結晶構造は、TLR3(配列番号2)上の次のエピトープ残基:E446、T448、Q450、R453、R473、N474、A477、L478、P480、S498、P499、Q503、P504、R507、D523、D524、E527、E530、及びK559を同定した。Fab c1068に対しては、結晶構造は、次のパラトープ残基、軽鎖:H30、N31、Y32、N50、E66、S67、G68(glyc)、重鎖:T30、T31、Y32、W33、H35、E50、N52、N54、N55、R57、N59、V99、M102、I103、及びT104を同定した。
【0284】
中和の機構及びTLR3機能との関連
mAb 15EVQ:mAb 15EVQエピトープは、TLR3残基N517、H539及びN541を含み、これらはC末端二本鎖RNA結合部位と折りたたみする。(Bellら、Proc.Natl.Acad.Sci.USA,103:8792〜7,2006).いかなる特定の理論からも束縛されることを望むものではないが、このように、mAb 15EVQは、リガンドに対して結合するTLR3に対して競合し、及びシグナル伝達単位の形成に必要とされる、リガンドにより誘起される受容体二量体化を防止すると考えられる。(Liuら、Science 320:379〜81,2008)。図29は、mAb 15EVQに対するこの直接の競合機構を図示する。抗体濃度に依存して、この機構は、ポリ(I:C)の全阻害又は二本鎖RNAにより誘起されるTLR3活性化を生じる。
【0285】
mAb 12QVQ/QSV及びmAb c1068:図30に示すように、これらの2つの抗体は二本鎖RNAリガンドとの直接的な衝突を持たない。したがって、これらがTLR3機能をmAb 15EVQのそれに類似の機構で中和することはありえない。Fabフラグメントもリガンドから離れて配向する(図30)。構造的に、mAb 12QVQ/QSV及びFab c1068は、両方とも機能を損なわずにシグナル伝達単位(SU)に結合することができる。立体的に、mAb分子の2つのFabフラグメントが、2つのTLR3分子を1つのSU中で同時に結合し、ひいては二本鎖RNA介在型TLR3二量体化を防止し得るということはありえない。いかなる特定の理論からも束縛されることを望むものではないが、TLR3へのmAb 12QVQ/QSV又はmAb c1068の結合が、抗体と隣接するシグナル伝達単位との間の立体的な衝突により、シグナル伝達単位のクラスター形成を防止すると考えられる。二本鎖RNAへのTLR3の結合は、二本鎖RNA:TLR3複合体により定義されるシグナル伝達単位に限定されない(Liuら、Science 320:379〜81,2008)。多数のSUのクラスター形成がシグナル伝達の増強を生じる得、又は効率的なTLR3シグナル伝達がこのクラスター形成を必要とし得る。mAb 12QVQ/QSV及びmAb c1068の配置が、クラスター形成をブロックし、TLR3活性の中和を生じることができる。それゆえ、抗体の最大中和効果は、抗体結合によるSUの分離の度合いに依存する。図30に図示するように、mAb 12QVQ/QSVは、mAb c1068よりも大きい分離を引き起こし、このことがmAb 12QVQ/QSVのより大きい効能に転換する可能性がある。このことは、mAb c1068及びmAb 15EVQが、飽和濃度においてそれぞれ約50%及び100%のTLR3中和を生じることができ、mAb 12QVQ/QSVが中間の活性を呈するという観察と一致する。このように、構造及びTLR3中和を組み合わせた研究は、二本鎖RNA:TLR3シグナル伝達単位がクラスターを形成して、効率的なシグナル伝達を達成する、TLR3シグナル伝達モデルを示唆する。mAb 12QVQ/QSV及びmAb c1068は、また、リガンド結合又は受容体二量体化を妨害せずにTLR3シグナル伝達を部分的に調節することができる抗体の類も定義する。
【特許請求の範囲】
【請求項1】
toll様受容体3(TLR3)の、配列番号2のアミノ酸残基;K416、K418、L440、N441、E442、Y465、N466、K467、Y468、R488、R489、A491、K493、N515、N516、N517、H539、N541、S571、L595、及びK619に結合する、単離された抗体又はそのフラグメント。
【請求項2】
toll様受容体3(TLR3)の、配列番号2のアミノ酸残基;S115、D116、K117、A120、K139、N140、N141、V144、K145、T166、Q167、V168、S188、E189、D192、A195、及びA219に結合する、単離された抗体又はそのフラグメント。
【請求項3】
重鎖可変領域及び軽鎖可変領域を含む抗体又はそのフラグメントであって、抗体が重鎖可変領域のChothia残基;W33、F50、D52、D54、Y56、N58、P61、E95、Y97、Y100及びD100b並びに軽鎖可変領域のChothia残基;Q27、Y32、N92、T93、L94及びS95により、配列番号2に示されるアミノ酸配列を有するTLR3に結合することを特徴とする、単離された抗体又はそのフラグメント。
【請求項4】
次のa〜iの特性のうち少なくとも1つを有する、請求項3に記載の単離された抗体;
a.Kd <10nMでヒトTLR3に結合する;
b.インビトロでのポリ(I:C)NF−κBレポーター遺伝子アッセイにおいて、1μg/mLで、ヒトTLR3の生物活性を50%超低減させる;
c.10μg/mLで、100ng/mL未満のポリ(I:C)で刺激されたBEAS−2B細胞からのIL−6又はCXCL10/IP−10の産生を60%超阻害する;
d.0.4μg/mLで、100ng/mL未満のポリ(I:C)で刺激されたBEAS−2B細胞からのIL−6又はCXCL10/IP−10の産生を50%超阻害する;
e.5μg/mLで、62.5ng/mLのポリ(I:C)で刺激されたNHBE細胞からのIL−6の産生を50%超阻害する;
f.1μg/mLで、62.5ng/mLのポリ(I:C)で刺激されたNHBE細胞からのIL−6の産生を50%超阻害する;
g.1μg/mLで、PBMCによる、ポリ(I:C)により誘導されたIFN−γ、IL−6又はIL−12の産生を20%超阻害する;
h.インビトロでのNF−kBレポーター遺伝子アッセイにおいて、IC50 <10μg/mLで、カニクイザルのTLR3の生物活性を阻害する;
i.インビトロでのISREレポーター遺伝子アッセイにおいて、IC50 <5μg/mLで、カニクイザルのTLR3の生物活性を阻害する。
【請求項5】
前記抗体が、アミノ酸12個分の長さの重鎖相補性決定領域(CDR)3(HCDR3)を含む、請求項3に記載の単離された抗体。
【請求項6】
前記抗体が、それぞれ配列番号82、86及び84で示されるアミノ酸配列と少なくとも90%同一のアミノ酸配列を有する重鎖相補性決定領域(CDR)1、2及び3(HCDR1、HCDR2、HCDR3)、並びにそれぞれ配列番号79、80及び87で示されるアミノ酸配列と少なくとも90%同一のアミノ酸配列を有する軽鎖相補性決定領域1、2及び3(LCDR1、LCDR2、LCDR3)を含む、請求項3に記載の単離された抗体。
【請求項7】
軽鎖可変領域カッパー1(Vκ1)フレームワーク(Vκ1)のアミノ酸配列と少なくとも90%同一の軽鎖フレームワーク及び重鎖可変領域Vh5フレームワーク(Vh5)のアミノ酸配列と少なくとも90%同一の重鎖フレームワークを含む、請求項3に記載の単離された抗体。
【請求項8】
前記Vk1フレームワークが、配列番号221で示されるアミノ酸配列を有するIGKV1−39*01によりコードされ、前記Vh5フレームワークが、配列番号222で示されるアミノ酸配列を有するIGHV5−51*01によりコードされる、請求項7に記載の単離された抗体。
【請求項9】
重鎖可変領域及び軽鎖可変領域を含む抗体又はそのフラグメントであって、抗体が重鎖可変領域のChothia残基;N31a、Q52、R52b、S53、K54、Y56、Y97、P98、F99及びY100、並びに軽鎖可変領域のChothia残基;G29、S30、Y31、Y32、E50、D51、Y91、D92及びD93により、配列番号2に示されるアミノ酸配列を有するTLR3に結合することを特徴とする、単離された抗体又はそのフラグメント。
【請求項10】
次のa〜gの特性のうち少なくとも1つを有する、請求項9に記載の単離された抗体;
a.Kd <10nMでヒトTLR3に結合する;
b.インビトロでのポリ(I:C)NF−κBレポーター遺伝子アッセイにおいて、1μg/mLで、ヒトTLR3の生物活性を50%超低減させる;
c.10μg/mLで、100ng/mL未満のポリ(I:C)で刺激されたBEAS−2B細胞からのIL−6又はCXCL10/IP−10の産生を60%超阻害する;
d.0.4μg/mLで、100ng/mL未満のポリ(I:C)で刺激されたBEAS−2B細胞からのIL−6又はCXCL10/IP−10の産生を50%超阻害する;
e.5μg/mLで、62.5ng/mLのポリ(I:C)で刺激されたNHBE細胞からのIL−6の産生を50%超阻害する;
f.1μg/mLで、62.5ng/mLのポリ(I:C)で刺激されたNHBE細胞からのIL−6の産生を50%超阻害する;
g.1μg/mLで、PBMCによる、ポリ(I:C)により誘導されたIFN−γ、IL−6又はIL−12の産生を20%超阻害する。
【請求項11】
前記抗体が、アミノ酸10個分の長さのHCDR3及びアミノ酸10個分の長さのLCDR3を含む、請求項9に記載の単離された抗体。
【請求項12】
前記抗体が、それぞれ配列番号70、77及び72で示されるアミノ酸配列と少なくとも90%同一のアミノ酸配列を有する、重鎖相補性決定領域(CDR)1、2及び3(HCDR1、HCDR2、HCDR3)、並びにそれぞれ配列番号67、68及び78で示されるアミノ酸配列と少なくとも90%同一のアミノ酸配列を有する軽鎖相補性決定領域1、2及び3(LCDR1、LCDR2、LCDR3)を含む、請求項9に記載の単離された抗体。
【請求項13】
軽鎖可変領域カッパー3(Vλ3)フレームワーク(Vλ3)のアミノ酸配列と少なくとも90%同一の軽鎖フレームワーク及び重鎖可変領域Vh6フレームワーク(Vh6)のアミノ酸配列と少なくとも90%同一の重鎖フレームワークを含む、請求項9に記載の単離された抗体。
【請求項14】
前記Vλ3フレームワークが、配列番号223で示されるアミノ酸配列を有するIGLV3−1*01によりコードされ、及びVh6フレームワークが、配列番号224で示されるアミノ酸配列を有するIGHV6−1*01によりコードされる、請求項13に記載の単離された抗体。
【請求項15】
前記抗体が、次のa〜eのいずれかである、請求項1又は9に記載の単離された抗体又はフラグメント;
a.完全なヒト型抗体;
b.ヒト適合性の抗体;
c.ポリエチレングリコール化抗体;
d.IgG4のイソタイプの抗体;
e.FcドメインがS229P、P235A又はL236A突然変異を含む抗体。
【請求項16】
請求項1又は9に記載の単離された抗体又はフラグメント及び製薬上許容され得る担体を含む、製薬学的組成物。
【請求項17】
請求項1又は9に記載の単離された抗体の治療上の有効量を、それらを必要とする患者に、炎症状態を処置又は予防する上で充分な時間にわたって投与することを含む、炎症状態の処置方法。
【請求項18】
前記炎症状態が、炎症性肺疾患、炎症性腸疾患、自己免疫疾患、全身性の炎症状態、又は関節リウマチである、請求項17に記載の方法。
【請求項19】
前記炎症性肺疾患が、喘息、慢性閉塞性肺疾患(COPD)、気道過敏症であるか、又は分類不能型のインフルエンザ菌により誘起される、請求項18に記載の方法。
【請求項20】
前記全身性の炎症状態が、サイトカインストームすなわち高サイトカイン血症、全身性炎症反応症候群(SIRS)、移植片対宿主病(GVHD)、急性呼吸窮迫症候群(ARDS)、重症急性呼吸器症候群(SARS)、劇症型抗リン脂質症候群、重篤なウイルス感染、インフルエンザ、肺炎、ショック、又は敗血症である、請求項18に記載の方法。
【請求項21】
前記炎症状態が消化管潰瘍をともなう、請求項17に記載の方法。
【請求項22】
前記消化管潰瘍が、炎症性大腸炎、虚血性大腸炎、コラーゲン形成大腸炎又はリンパ球性大腸炎又は壊死性腸炎をともなう、請求項21に記載の方法。
【請求項23】
治療上の有効量の請求項1又は9に記載の単離された抗体を、それらを必要とする患者に、II型糖尿病を処置する上で充分な時間にわたって投与することを含む、II型糖尿病、高血糖症又は高インスリン症の処置方法。
【請求項24】
治療上の有効量の請求項1又は9に記載の単離された抗体を、それらを必要とする患者に、ウイルス感染症を処置又は予防する上で充分な時間にわたって投与することを含む、ウイルス感染症の処置又は予防方法。
【請求項25】
前記ウイルス感染症がA型インフルエンザウイルス感染症である、請求項24に記載の方法。
【請求項1】
toll様受容体3(TLR3)の、配列番号2のアミノ酸残基;K416、K418、L440、N441、E442、Y465、N466、K467、Y468、R488、R489、A491、K493、N515、N516、N517、H539、N541、S571、L595、及びK619に結合する、単離された抗体又はそのフラグメント。
【請求項2】
toll様受容体3(TLR3)の、配列番号2のアミノ酸残基;S115、D116、K117、A120、K139、N140、N141、V144、K145、T166、Q167、V168、S188、E189、D192、A195、及びA219に結合する、単離された抗体又はそのフラグメント。
【請求項3】
重鎖可変領域及び軽鎖可変領域を含む抗体又はそのフラグメントであって、抗体が重鎖可変領域のChothia残基;W33、F50、D52、D54、Y56、N58、P61、E95、Y97、Y100及びD100b並びに軽鎖可変領域のChothia残基;Q27、Y32、N92、T93、L94及びS95により、配列番号2に示されるアミノ酸配列を有するTLR3に結合することを特徴とする、単離された抗体又はそのフラグメント。
【請求項4】
次のa〜iの特性のうち少なくとも1つを有する、請求項3に記載の単離された抗体;
a.Kd <10nMでヒトTLR3に結合する;
b.インビトロでのポリ(I:C)NF−κBレポーター遺伝子アッセイにおいて、1μg/mLで、ヒトTLR3の生物活性を50%超低減させる;
c.10μg/mLで、100ng/mL未満のポリ(I:C)で刺激されたBEAS−2B細胞からのIL−6又はCXCL10/IP−10の産生を60%超阻害する;
d.0.4μg/mLで、100ng/mL未満のポリ(I:C)で刺激されたBEAS−2B細胞からのIL−6又はCXCL10/IP−10の産生を50%超阻害する;
e.5μg/mLで、62.5ng/mLのポリ(I:C)で刺激されたNHBE細胞からのIL−6の産生を50%超阻害する;
f.1μg/mLで、62.5ng/mLのポリ(I:C)で刺激されたNHBE細胞からのIL−6の産生を50%超阻害する;
g.1μg/mLで、PBMCによる、ポリ(I:C)により誘導されたIFN−γ、IL−6又はIL−12の産生を20%超阻害する;
h.インビトロでのNF−kBレポーター遺伝子アッセイにおいて、IC50 <10μg/mLで、カニクイザルのTLR3の生物活性を阻害する;
i.インビトロでのISREレポーター遺伝子アッセイにおいて、IC50 <5μg/mLで、カニクイザルのTLR3の生物活性を阻害する。
【請求項5】
前記抗体が、アミノ酸12個分の長さの重鎖相補性決定領域(CDR)3(HCDR3)を含む、請求項3に記載の単離された抗体。
【請求項6】
前記抗体が、それぞれ配列番号82、86及び84で示されるアミノ酸配列と少なくとも90%同一のアミノ酸配列を有する重鎖相補性決定領域(CDR)1、2及び3(HCDR1、HCDR2、HCDR3)、並びにそれぞれ配列番号79、80及び87で示されるアミノ酸配列と少なくとも90%同一のアミノ酸配列を有する軽鎖相補性決定領域1、2及び3(LCDR1、LCDR2、LCDR3)を含む、請求項3に記載の単離された抗体。
【請求項7】
軽鎖可変領域カッパー1(Vκ1)フレームワーク(Vκ1)のアミノ酸配列と少なくとも90%同一の軽鎖フレームワーク及び重鎖可変領域Vh5フレームワーク(Vh5)のアミノ酸配列と少なくとも90%同一の重鎖フレームワークを含む、請求項3に記載の単離された抗体。
【請求項8】
前記Vk1フレームワークが、配列番号221で示されるアミノ酸配列を有するIGKV1−39*01によりコードされ、前記Vh5フレームワークが、配列番号222で示されるアミノ酸配列を有するIGHV5−51*01によりコードされる、請求項7に記載の単離された抗体。
【請求項9】
重鎖可変領域及び軽鎖可変領域を含む抗体又はそのフラグメントであって、抗体が重鎖可変領域のChothia残基;N31a、Q52、R52b、S53、K54、Y56、Y97、P98、F99及びY100、並びに軽鎖可変領域のChothia残基;G29、S30、Y31、Y32、E50、D51、Y91、D92及びD93により、配列番号2に示されるアミノ酸配列を有するTLR3に結合することを特徴とする、単離された抗体又はそのフラグメント。
【請求項10】
次のa〜gの特性のうち少なくとも1つを有する、請求項9に記載の単離された抗体;
a.Kd <10nMでヒトTLR3に結合する;
b.インビトロでのポリ(I:C)NF−κBレポーター遺伝子アッセイにおいて、1μg/mLで、ヒトTLR3の生物活性を50%超低減させる;
c.10μg/mLで、100ng/mL未満のポリ(I:C)で刺激されたBEAS−2B細胞からのIL−6又はCXCL10/IP−10の産生を60%超阻害する;
d.0.4μg/mLで、100ng/mL未満のポリ(I:C)で刺激されたBEAS−2B細胞からのIL−6又はCXCL10/IP−10の産生を50%超阻害する;
e.5μg/mLで、62.5ng/mLのポリ(I:C)で刺激されたNHBE細胞からのIL−6の産生を50%超阻害する;
f.1μg/mLで、62.5ng/mLのポリ(I:C)で刺激されたNHBE細胞からのIL−6の産生を50%超阻害する;
g.1μg/mLで、PBMCによる、ポリ(I:C)により誘導されたIFN−γ、IL−6又はIL−12の産生を20%超阻害する。
【請求項11】
前記抗体が、アミノ酸10個分の長さのHCDR3及びアミノ酸10個分の長さのLCDR3を含む、請求項9に記載の単離された抗体。
【請求項12】
前記抗体が、それぞれ配列番号70、77及び72で示されるアミノ酸配列と少なくとも90%同一のアミノ酸配列を有する、重鎖相補性決定領域(CDR)1、2及び3(HCDR1、HCDR2、HCDR3)、並びにそれぞれ配列番号67、68及び78で示されるアミノ酸配列と少なくとも90%同一のアミノ酸配列を有する軽鎖相補性決定領域1、2及び3(LCDR1、LCDR2、LCDR3)を含む、請求項9に記載の単離された抗体。
【請求項13】
軽鎖可変領域カッパー3(Vλ3)フレームワーク(Vλ3)のアミノ酸配列と少なくとも90%同一の軽鎖フレームワーク及び重鎖可変領域Vh6フレームワーク(Vh6)のアミノ酸配列と少なくとも90%同一の重鎖フレームワークを含む、請求項9に記載の単離された抗体。
【請求項14】
前記Vλ3フレームワークが、配列番号223で示されるアミノ酸配列を有するIGLV3−1*01によりコードされ、及びVh6フレームワークが、配列番号224で示されるアミノ酸配列を有するIGHV6−1*01によりコードされる、請求項13に記載の単離された抗体。
【請求項15】
前記抗体が、次のa〜eのいずれかである、請求項1又は9に記載の単離された抗体又はフラグメント;
a.完全なヒト型抗体;
b.ヒト適合性の抗体;
c.ポリエチレングリコール化抗体;
d.IgG4のイソタイプの抗体;
e.FcドメインがS229P、P235A又はL236A突然変異を含む抗体。
【請求項16】
請求項1又は9に記載の単離された抗体又はフラグメント及び製薬上許容され得る担体を含む、製薬学的組成物。
【請求項17】
請求項1又は9に記載の単離された抗体の治療上の有効量を、それらを必要とする患者に、炎症状態を処置又は予防する上で充分な時間にわたって投与することを含む、炎症状態の処置方法。
【請求項18】
前記炎症状態が、炎症性肺疾患、炎症性腸疾患、自己免疫疾患、全身性の炎症状態、又は関節リウマチである、請求項17に記載の方法。
【請求項19】
前記炎症性肺疾患が、喘息、慢性閉塞性肺疾患(COPD)、気道過敏症であるか、又は分類不能型のインフルエンザ菌により誘起される、請求項18に記載の方法。
【請求項20】
前記全身性の炎症状態が、サイトカインストームすなわち高サイトカイン血症、全身性炎症反応症候群(SIRS)、移植片対宿主病(GVHD)、急性呼吸窮迫症候群(ARDS)、重症急性呼吸器症候群(SARS)、劇症型抗リン脂質症候群、重篤なウイルス感染、インフルエンザ、肺炎、ショック、又は敗血症である、請求項18に記載の方法。
【請求項21】
前記炎症状態が消化管潰瘍をともなう、請求項17に記載の方法。
【請求項22】
前記消化管潰瘍が、炎症性大腸炎、虚血性大腸炎、コラーゲン形成大腸炎又はリンパ球性大腸炎又は壊死性腸炎をともなう、請求項21に記載の方法。
【請求項23】
治療上の有効量の請求項1又は9に記載の単離された抗体を、それらを必要とする患者に、II型糖尿病を処置する上で充分な時間にわたって投与することを含む、II型糖尿病、高血糖症又は高インスリン症の処置方法。
【請求項24】
治療上の有効量の請求項1又は9に記載の単離された抗体を、それらを必要とする患者に、ウイルス感染症を処置又は予防する上で充分な時間にわたって投与することを含む、ウイルス感染症の処置又は予防方法。
【請求項25】
前記ウイルス感染症がA型インフルエンザウイルス感染症である、請求項24に記載の方法。
【図1】

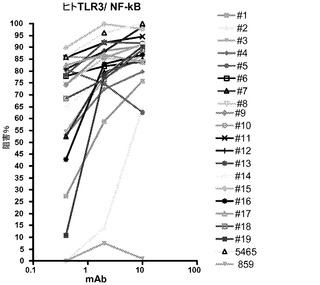
【図2A】
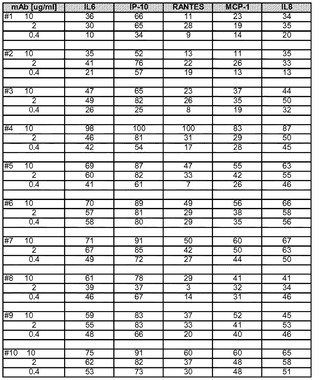
【図2B】
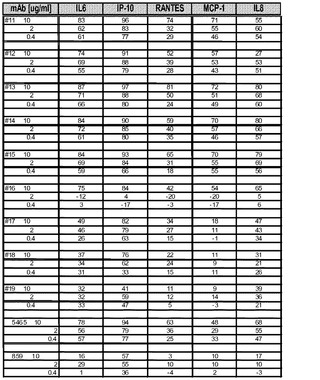
【図3A】
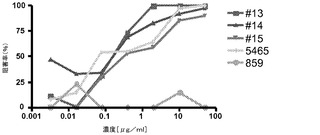
【図3B】
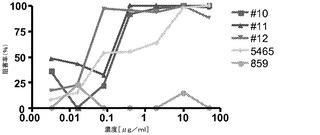
【図4】
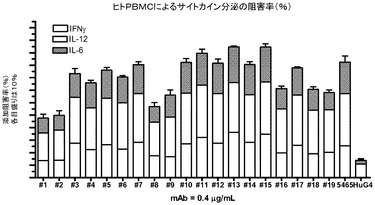
【図5A】
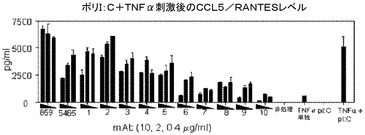
【図5B】
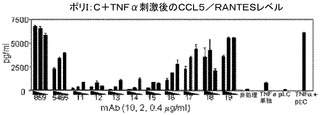
【図6A】
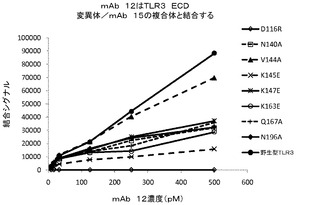
【図6B】
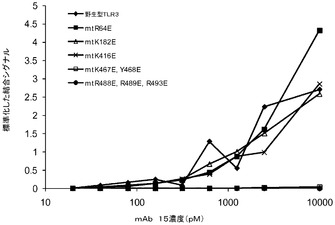
【図6C】
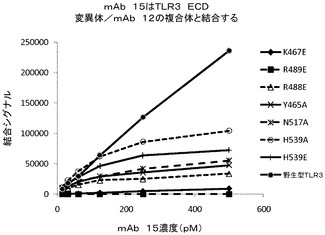
【図7A】
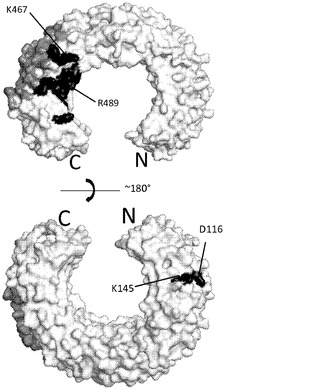
【図7B】
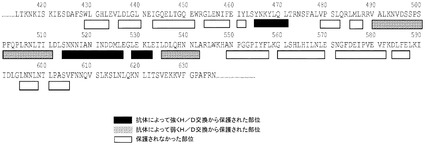
【図8A】
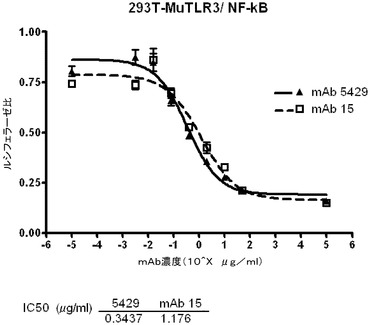
【図8B】
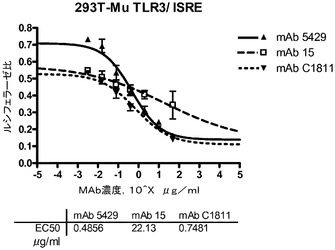
【図9】
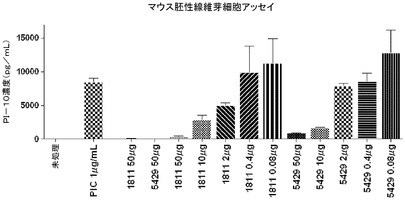
【図10】
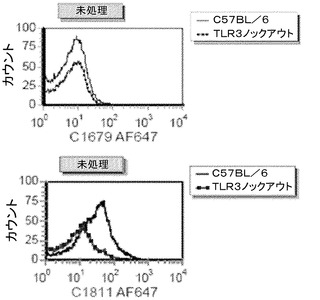
【図11】
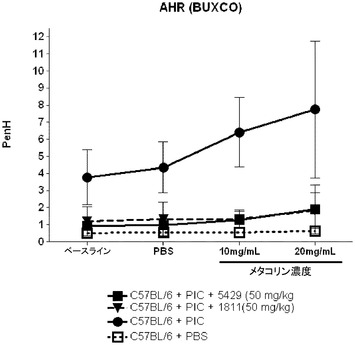
【図12】
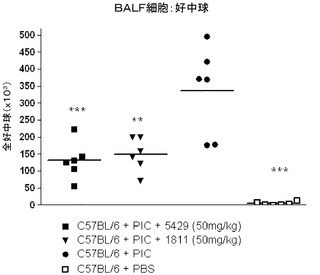
【図13】
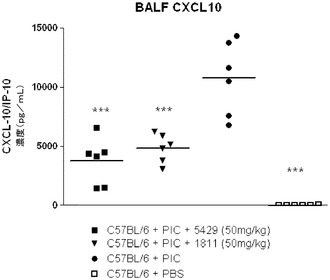
【図14】
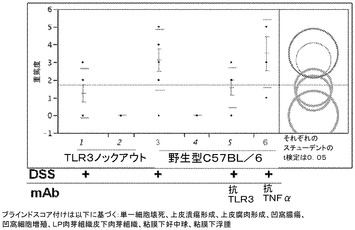
【図15】
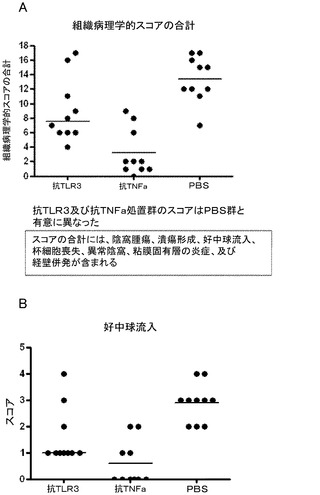
【図16】
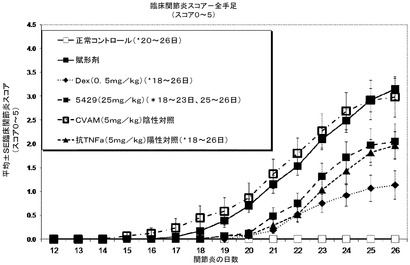
【図17】
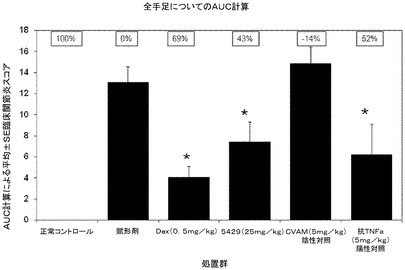
【図18】
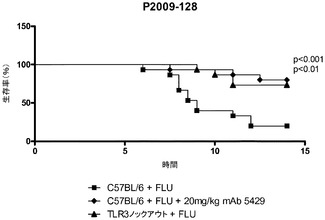
【図19】
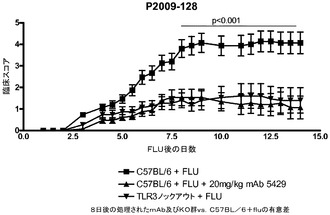
【図20】
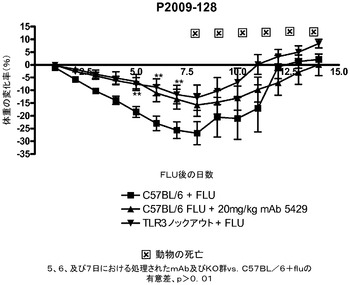
【図21】
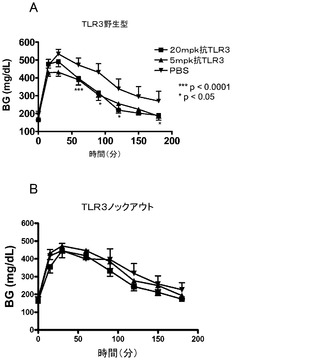
【図22】
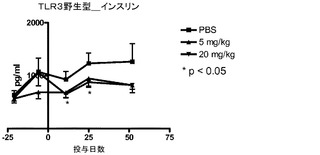
【図23】
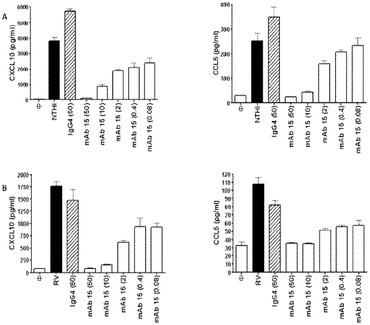
【図24】
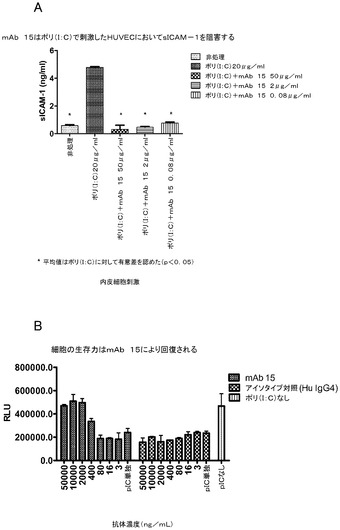
【図25】
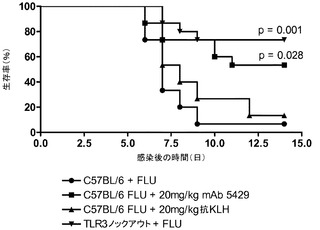
【図26】
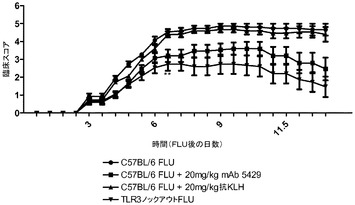
【図27】
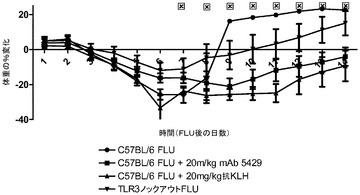
【図28】
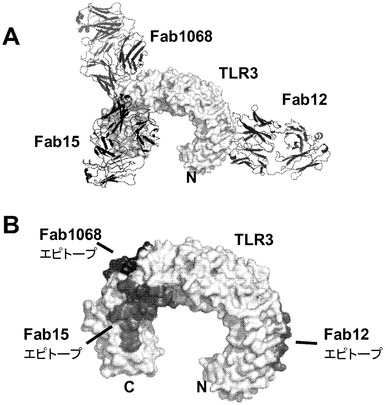
【図29】
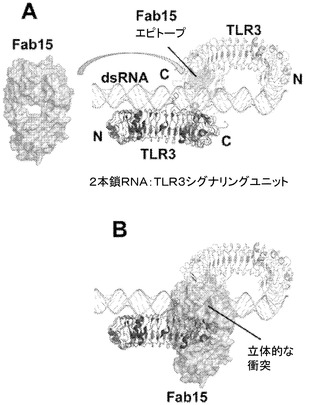
【図30】
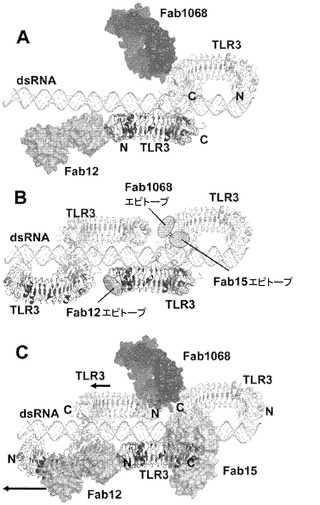
【図31】
【図32】

【図33】

【図34】
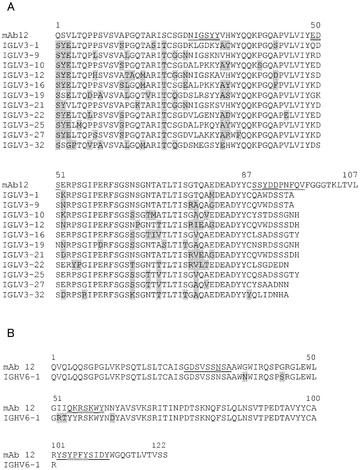
【図35】
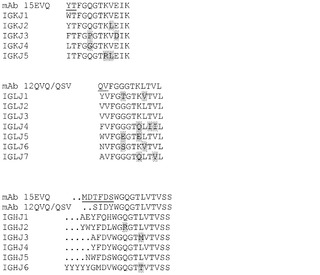
【図2A】
【図2B】
【図3A】
【図3B】
【図4】
【図5A】
【図5B】
【図6A】
【図6B】
【図6C】
【図7A】
【図7B】
【図8A】
【図8B】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【図34】
【図35】
【公表番号】特表2012−525425(P2012−525425A)
【公表日】平成24年10月22日(2012.10.22)
【国際特許分類】
【出願番号】特願2012−508728(P2012−508728)
【出願日】平成22年4月29日(2010.4.29)
【国際出願番号】PCT/US2010/032964
【国際公開番号】WO2010/127113
【国際公開日】平成22年11月4日(2010.11.4)
【出願人】(509087759)ヤンセン バイオテツク,インコーポレーテツド (77)
【Fターム(参考)】
【公表日】平成24年10月22日(2012.10.22)
【国際特許分類】
【出願日】平成22年4月29日(2010.4.29)
【国際出願番号】PCT/US2010/032964
【国際公開番号】WO2010/127113
【国際公開日】平成22年11月4日(2010.11.4)
【出願人】(509087759)ヤンセン バイオテツク,インコーポレーテツド (77)
【Fターム(参考)】
[ Back to top ]