
αグルコシドトランスポーター遺伝子及びその用途
【課題】マルトースおよびマルトトリオース資化能の高い醸造酵母、高濃度麦汁あるいはシロップ添加麦汁を用いた酒類の製造法などの提供。
【解決手段】αグルコシドトランスポーター遺伝子及びその用途、特に、マルトースおよびマルトトリオース資化能の高い醸造酵母、該酵母を用いて製造した酒類、その製造方法。さらに具体的には、醸造酵母のαグルコシドトランスポーターであるMPH2pをコードする遺伝子MPH2、特にビール酵母に特徴的なnonScMPH2遺伝子の発現量を高めることによって、マルトースおよびマルトトリオース資化能を向上させた酵母、当該酵母を用いた酒類の製造方法。
【解決手段】αグルコシドトランスポーター遺伝子及びその用途、特に、マルトースおよびマルトトリオース資化能の高い醸造酵母、該酵母を用いて製造した酒類、その製造方法。さらに具体的には、醸造酵母のαグルコシドトランスポーターであるMPH2pをコードする遺伝子MPH2、特にビール酵母に特徴的なnonScMPH2遺伝子の発現量を高めることによって、マルトースおよびマルトトリオース資化能を向上させた酵母、当該酵母を用いた酒類の製造方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、αグルコシドトランスポーター遺伝子及びその用途に関し、特に、マルトースおよびマルトトリオース発酵性に優れた醸造酵母、該酵母を用いて製造した酒類、その製造方法などに関する。さらに具体的には、本発明は、醸造酵母のαグルコシドトランスポーターであるMph2pをコードする遺伝子MPH2、特にビール酵母に特徴的なnonScMPH2遺伝子の発現量を高めることによって、マルトースおよびマルトトリオース資化能を向上させた酵母、当該酵母を用いた酒類の製造方法などに関する。
【背景技術】
【0002】
従来ビールの製造においては、エキス濃度11%前後の原麦汁を発酵させてアルコール濃度4.5〜5%前後のビールを得ている。ビールの生産性向上のために、世界中のビール会社で高濃度醸造法が採用されている。高濃度醸造法(highgravity brewing) とは、製造しようとするビールよりも高い原麦汁濃度のビールを与える麦汁を醗酵させた後、これを水で希釈して所定原麦汁濃度のビールとする方法である。その具体的方策としては、(1)通常発酵温度よりも高い温度を採用する、(2)麦汁への通気量を増加させる、(3)酵母の添加量を増加させる、およびそれらの組み合わせである。通常のビールを製造する場合には、これらの方法では原麦汁エキス濃度15%程度の高濃度醸造が限界であると言われている。
【0003】
原麦汁エキス濃度15%以上の高濃度醸造を行なった場合に生じる問題点として、まず一番に挙げられるのは、発酵中期から後期にかけての発酵速度の著しい低下である。麦汁中に含まれる糖質は、主にマルトース、マルトトリオース、グルコース、フラクトース、シュークロースである。酵母は、グルコース、フラクトース、シュークロースを先に資化し、その後にマルトース、マルトトリオースを資化する。従って、発酵中期から発酵後期にかけては、もろみ中の資化性糖はマルトース、マルトトリオースしか存在していない。しかもその存在比は、3:1と圧倒的にマルトースが多い。
【0004】
マルトースは、酵母のαグルコシドトランスポーターによって、細胞内に取り込まれた後、マルターゼによって2個のグルコースに加水分解され、エムブデン−マイヤーホッフ(Embden-Meyerhof)の経路を介して二酸化炭素とエタノールに転化する。この2つの酵素は、転写レベルで、マルトースによって誘導され、グルコースによって抑制される。
【0005】
麦汁中にはグルコースが、資化性糖の約17%含まれるため、酵母のマルトース及びマルトトリオース代謝系遺伝子は、発酵初期の段階でグルコースによる抑制を受け、マルトース及びマルトトリオースの資化が、大幅に遅れることになる。この現象は、高濃度醸造の際には、さらに顕著となり、マルトース及びマルトトリオース発酵が遅れるばかりでなく、発酵終了時に、残糖として多量のマルトース及びマルトトリオースが、資化されないまま残ってしまう。このような問題点によって、高濃度醸造、例えば通常の2倍の濃度の麦汁を醗酵させることは、従来の技術では不完全であった。
【0006】
特開平1−153082号公報(特許文献1)には、パン酵母によるドゥ発酵を改善するために、グルコースによる抑制を受けないアルコールデヒドロゲナーゼ遺伝子のプロモーターとマルターゼ遺伝子及びマルトーストランスポーター遺伝子とを含むプラスミドを導入したパン酵母を使用することが開示されている。一方、Saccharomyces cerevisiaeのマルトーストランスポーター遺伝子MAL6Tを醸造用酵母で高発現させ、高濃度醸造を実現させた例が報告されている(特開平6−245750号公報:特許文献2)。
【特許文献1】特開平1−153082号公報
【特許文献2】特開平6−245750号公報
【発明の開示】
【発明が解決しようとする課題】
【0007】
上述のとおり、発酵速度や製品の品質を損なうことなく、高濃度醸造を可能にする酵母が望まれていた。
【課題を解決するための手段】
【0008】
本発明者らは、上記課題を解決するため鋭意検討を重ねた結果、ビール酵母から既知のタンパク質より有利な効果を奏するαグルコシドトランスポーターをコードする遺伝子を同定・単離することに成功した。また、得られた遺伝子を酵母に導入し発現させた形質転換酵母を作製し、マルトース及びマルトトリオース資化が促進されることを確認して、本発明を完成した。
【0009】
すなわち本発明は、ビール酵母に特徴的に存在する新規なαグルコシドトランスポーター遺伝子、該遺伝子がコードするタンパク質、該遺伝子の発現が調節された形質転換酵母、該遺伝子の発現が調節された酵母を用いることによる酒類の製造方法などに関する。本発明は、具体的には、次に示すポリヌクレオチド、該ポリヌクレオチドを含有するベクター、該ベクターが導入された形質転換酵母、該形質転換酵母を用いる酒類の製造方法などを提供する。
【0010】
(1)以下の(a)〜(f)のいずれかに記載のポリヌクレオチド:
(a)配列番号:1の塩基配列からなるポリヌクレオチドを含有するポリヌクレオチド;
(b)配列番号:2のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(c)配列番号:2のアミノ酸配列において、1もしくは複数個のアミノ酸が欠失、置換、挿入および/または付加したアミノ酸配列からなり、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(d)配列番号:2のアミノ酸配列に対して60%以上の同一性を有するアミノ酸配列を有し、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(e)配列番号:1の塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;及び
(f)配列番号:2のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドの塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド。
【0011】
(2)以下の(g)〜(i)のいずれかである請求項1に記載のポリヌクレオチド:
(g)配列番号:2のアミノ酸配列又は配列番号:2のアミノ酸配列において、1〜10個のアミノ酸が欠失、置換、挿入および/または付加したアミノ酸配列からなり、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(h) 配列番号:2のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列を有し、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;及び
(i)配列番号:1の塩基配列からなるポリヌクレオチド、又は配列番号:1の塩基配列と相補的な塩基配列からなるポリヌクレオチドとハイストリンジェントな条件下でハイブリダイズし、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチドを含有するポリヌクレオチド。
(3)配列番号:1の塩基配列からなるポリヌクレオチドを含有する上記(1)に記載のポリヌクレオチド。
(4)配列番号:2のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有する上記(1)に記載のポリヌクレオチド。
(5)DNAである、請求項1〜4のいずれかに記載のポリヌクレオチド。
(6)上記(1)〜(5)のいずれかに記載のポリヌクレオチドにコードされるタンパク質。
【0012】
(7)上記(1)〜(5)のいずれかに記載のポリヌクレオチドを含有するベクター。
(7a)以下の(a)〜(c)の構成要素を含む発現カセットを含む上記(7)に記載のベクター:
(a)酵母細胞内で転写可能なプロモーター
(b)該プロモーターにセンス方向またはアンチセンス方向で結合した、上記(1)〜(5)のいずれかに記載のポリヌクレオチド;及び
(c)RNA分子の転写終結およびポリアデニル化に関し、酵母で機能するシグナル。
(8)上記(7)に記載のベクターが導入された形質転換酵母。
(9)上記(7)に記載のベクターを導入することによって、マルトースおよび/またはマルトトリオース資化能が増強された上記(8)に記載の醸造用酵母。
(10)上記(6)に記載のタンパク質の発現量を増加させることによってマルトースおよび/またはマルトトリオース資化能が向上した上記(9)に記載の醸造用酵母。
(11)上記(8)〜(10)のいずれかに記載の酵母を用いた酒類の製造方法。
(12)醸造する酒類が麦芽飲料である上記(11)に記載の酒類の製造方法。
(13)上記(11)または(12)に記載の方法で製造された酒類。
(14)配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の塩基配列に基づいて設計したプライマーまたはプローブを用いて、被検酵母のマルトースおよびマルトトリオース資化能について評価する方法。
(14a)上記(14)に記載の方法によって、マルトースおよびマルトトリオース資化能が高い酵母を選別する方法。
(14b)上記(14a)に記載の方法によって選別された酵母を用いて酒類(例えば、ビール)を製造する方法。
(15)被検酵母を培養し、配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の発現量を測定することによって、被検酵母のマルトースおよびマルトトリオース資化能を評価する方法。
(15a)上記(15)に記載の方法で、被検酵母を評価し、αグルコシドトランスポーター遺伝子の発現量が高い酵母を選別する、マルトースおよびマルトトリオース資化能の高い酵母を選別する方法。
(15b)上記(15a)に記載の方法によって選別された酵母を用いて酒類(例えば、ビール)を製造する方法。
(16)基準酵母および被検酵母を培養して配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の各酵母における発現量を測定し、基準酵母よりも該遺伝子が高発現している被検酵母を選択する、酵母の選択方法。
(17)
基準酵母および被検酵母を培養して各酵母における上記(6)に記載のタンパク質を定量し、基準酵母よりも該タンパク質量の多い被検酵母を選択する、酵母の選択方法。
【発明の効果】
【0013】
本発明の形質転換酵母を用いる酒類の製造法によれば、グルコースの存在下でもマルトース及びマルトトリオースの資化が抑制されず、この結果グルコースの消失を待たずにマルトース及びマルトトリオースが資化発酵されるため発酵速度が上昇し、高濃度麦汁によるビール醸造が可能となる。
【発明を実施するための最良の形態】
【0014】
本発明者らは、酵母のαグルコシドトランスポーターの活性を増大させることによって、さらに効率よくマルトース及びマルトトリオースを資化させることが可能であると考えた。このような着想に基づいて研究を重ね、特開2004-283169に開示の方法で解読したビール酵母ゲノム情報を基に、ビール酵母特有のαグルコシドトランスポーターをコードするnon-ScMPH2遺伝子を単離・同定した。この塩基配列を配列番号:1に示す。またこの遺伝子によりコードされるタンパク質のアミノ酸配列を配列番号:2に示す。
【0015】
1.本発明のポリヌクレオチド
まず、本発明は、(a)配列番号:1の塩基配列からなるポリヌクレオチドを含有するポリヌクレオチド(具体的には、DNA、以下、これらを単に「DNA」とも称する);及び(b)配列番号:2のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチドを提供する。本発明で対象とするDNAは、上記のビール酵母由来のαグルコシドトランスポーターをコードするDNAに限定されるものではなく、このタンパク質と機能的に同等なタンパク質をコードする他のDNAを含む。機能的に同等なタンパク質としては、例えば、(c)配列番号:2のアミノ酸配列において、1もしくは複数個のアミノ酸が欠失、置換、挿入および/または付加したアミノ酸配列からなり、かつαグルコシドトランスポーター活性を有するタンパク質が挙げられる。このようなタンパク質としては、配列番号:2のアミノ酸配列において、例えば、1〜100個、1〜90個、1〜80個、1〜70個、1〜60個、1〜50個、1〜40個、1〜39個、1〜38個、1〜37個、1〜36個、1〜35個、1〜34個、1〜33個、1〜32個、1〜31個、1〜30個、1〜29個、1〜28個、1〜27個、1〜26個、1〜25個、1〜24個、1〜23個、1〜22個、1〜21個、1〜20個、1〜19個、1〜18個、1〜17個、1〜16個、1〜15個、1〜14個、1〜13個、1〜12個、1〜11個、1〜10個、1〜9個、1〜8個、1〜7個、1〜6個(1〜数個)、1〜5個、1〜4個、1〜3個、1〜2個、1個のアミノ酸残基が欠失、置換、挿入および/または付加されたアミノ酸配列からなり、かつαグルコシドトランスポーター活性を有するタンパク質が挙げられる。上記アミノ酸残基の欠失、置換、挿入および/または付加の数は、一般的には小さい程好ましい。また、このようなタンパク質としては、(d)配列番号:2のアミノ酸配列と約60%以上、約70%以上、71%以上、72%以上、73%以上、74%以上、75%以上、76%以上、77%以上、78%以上、79%以上、80%以上、81%以上、82%以上、83%以上、84%以上、85%以上、86%以上、87%以上、88%以上、89%以上、90%以上、91%以上、92%以上、93%以上、94%以上、95%以上、96%以上、97%以上、98%以上、99%以上、99.1%以上99.2%以上99.3%以上99.4%以上99.5%以上99.6%以上99.7%以上99.8%以上99.9%以上の同一性を有するアミノ酸配列を有し、かつαグルコシドトランスポーター活性を有するタンパク質が挙げられる。上記相同性の数値は一般的に大きい程好ましい。なお、αグルコシドトランスポーター活性は、例えば特開平06-245750に記載の方法によって測定することができる。
【0016】
また、本発明は、(e)配列番号:1の塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;及び(f)配列番号:2のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドの塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチドも包含する。
【0017】
ここで、「ストリンジェントな条件下でハイブリダイズするポリヌクレオチド(DNA)」とは、配列番号:1の塩基配列と相補的な塩基配列からなるDNA又は配列番号:2のアミノ酸配列をコードするDNAの全部または一部をプローブとして、コロニーハイブリダイゼーション法、プラークハイブリダイゼーション法またはサザンハイブリダイゼーション法などを用いることにより得られるDNAをいう。ハイブリダイゼーションの方法としては、例えばMolecular Cloning 3rd Ed.、Current Protocols in Molecular Biology, John Wiley & Sons 1987-1997などに記載されている方法を利用することができる。
【0018】
本明細書でいう「ストリンジェントな条件」は、低ストリンジェントな条件、中ストリンジェントな条件及び高ストリンジェントな条件のいずれでもよい。「低ストリンジェントな条件」は、例えば、5×SSC、5×デンハルト溶液、0.5%SDS、50%ホルムアミド、32℃の条件である。また、「中ストリンジェントな条件」は、例えば、5×SSC、5×デンハルト溶液、0.5%SDS、50%ホルムアミド、42℃の条件である。「高ストリンジェントな条件」は、例えば、5×SSC、5×デンハルト溶液、0.5%SDS、50%ホルムアミド、50℃の条件である。これらの条件において、温度を上げるほど高い相同性を有するDNAが効率的に得られることが期待できる。ただし、ハイブリダイゼーションのストリンジェンシーに影響する要素としては温度、プローブ濃度、プローブの長さ、イオン強度、時間、塩濃度など複数の要素が考えられ、当業者であればこれら要素を適宜選択することで同様のストリンジェンシーを実現することが可能である。
【0019】
なお、ハイブリダイゼーションに市販のキットを用いる場合は、例えばAlkphos Direct Labelling Reagents(アマシャムファルマシア社製)を用いることができる。この場合は、キットに添付のプロトコルにしたがい、標識したプローブとのインキュベーションを一晩行った後、メンブレンを55℃の条件下で0.1% (w/v) SDSを含む1次洗浄バッファーで洗浄後、ハイブリダイズしたDNAを検出することができる。
【0020】
これ以外にハイブリダイズ可能なDNAとしては、FASTA、BLASTなどの相同性検索ソフトウェアにより、デフォルトのパラメータを用いて計算したときに、配列番号:2のアミノ酸配列をコードするDNAと約60%以上、約70%以上、71%以上、72%以上、73%以上、74%以上、75%以上、76%以上、77%以上、78%以上、79%以上、80%以上、81%以上、82%以上、83%以上、84%以上、85%以上、86%以上、87%以上、88%以上、89%以上、90%以上、91%以上、92%以上、93%以上、94%以上、95%以上、96%以上、97%以上、98%以上、99%以上、99.1%以上99.2%以上99.3%以上99.4%以上99.5%以上99.6%以上99.7%以上99.8%以上99.9%以上以上の同一性を有するDNAをあげることができる。
なお、アミノ酸配列や塩基配列の同一性は、カーリンおよびアルチュールによるアルゴリズムBLAST(proc. Natl. Acad. Sci. USA 872264-2268, 1990; proc Natl Acad Sci USA 90: 5873, 1993)を用いて決定できる。BLASTのアルゴリズムに基づいたBLASTNやBLASTXと呼ばれるプログラムが開発されている(Altschul SF, et al: J Mol Biol 215: 403, 1990)。BLASTNを用いて塩基配列を解析する場合は、パラメーターは、例えばscore=100、wordlength=12とする。また、BLASTXを用いてアミノ酸配列を解析する場合は、パラメーターは、例えばscore=50、wordlength=3とする。BLASTとGapped BLASTプログラムを用いる場合は、各プログラムのデフォルトパラメーターを用いる。
【0021】
2.本発明のタンパク質
本発明は、上記ポリヌクレオチド(a)〜(e)のいずれかにコードされるタンパク質も提供する。本発明の好ましいタンパク質は、配列番号:2のアミノ酸配列において、1もしくは複数個のアミノ酸が欠失、置換、挿入および/または付加したアミノ酸配列からなり、かつαグルコシドトランスポーター活性を有するタンパク質である。このようなタンパク質としては、配列番号:2のアミノ酸配列において、上記したような数のアミノ酸残基が欠失、置換、挿入および/または付加されたアミノ酸配列からなり、かつαグルコシドトランスポーター活性を有するタンパク質が挙げられる。また、このようなタンパク質としては、配列番号:2のアミノ酸配列と上記したような相同性を有するアミノ酸配列を有し、かつαグルコシドトランスポーター活性を有するタンパク質が挙げられる。このようなタンパク質は、「モレキュラークローニング第3版」、「カレント・プロトコールズ・イン・モレキュラー・バイオロジー」、“Nuc. Acids. Res., 10, 6487 (1982)”、“Proc. Natl. Acad. Sci. USA, 79, 6409 (1982)”、“Gene, 34, 315 (1985)”、“Nuc. Acids. Res., 13, 4431 (1985)”、“Proc. Natl. Acad. Sci. USA, 82, 488 (1985)”等に記載の部位特異的変異導入法を用いて、取得することができる。
【0022】
本発明のタンパク質のアミノ酸配列において1以上のアミノ酸残基が欠失、置換、挿入および/または付加されたとは、同一配列中の任意かつ1もしくは複数のアミノ酸配列中の位置において、1または複数のアミノ酸残基の欠失、置換、挿入及び/又は付加があることを意味し、欠失、置換、挿入及び付加のうち2種以上が同時に生じてもよい。
以下に、相互に置換可能なアミノ酸残基の例を示す。同一群に含まれるアミノ酸残基は相互に置換可能である。A群:ロイシン、イソロイシン、ノルロイシン、バリン、ノルバリン、アラニン、2-アミノブタン酸、メチオニン、o-メチルセリン、t-ブチルグリシン、t-ブチルアラニン、シクロヘキシルアラニン; B群:アスパラギン酸、グルタミン酸、イソアスパラギン酸、イソグルタミン酸、2-アミノアジピン酸、2-アミノスベリン酸; C群:アスパラギン、グルタミン; D群:リジン、アルギニン、オルニチン、2,4-ジアミノブタン酸、2,3-ジアミノプロピオン酸; E群:プロリン、3-ヒドロキシプロリン、4-ヒドロキシプロリン; F群:セリン、スレオニン、ホモセリン; G群:フェニルアラニン、チロシン。
【0023】
また、本発明のタンパク質は、Fmoc法(フルオレニルメチルオキシカルボニル法)、tBoc法(t-ブチルオキシカルボニル法)等の化学合成法のよっても製造することができる。また、アドバンスドケムテック社製、パーキンエルマー社製、ファルマシア社製、プロテインテクノロジーインストゥルメント社製、シンセセルーベガ社製、パーセプティブ社製、島津製作所等のペプチド合成機を利用して化学合成することもできる。
【0024】
3.本発明のベクター及びこれを導入した形質転換酵母
次に、本発明は、上記したポリヌクレオチドを含有するベクターを提供する。本発明のベクターは、上記(a)〜(i)のいずれかに記載のポリヌクレオチド(DNA)を含有するベクターに関する。また、本発明のベクターは、通常、(a)酵母細胞内で転写可能なプロモーター;(b)該プロモーターにセンス方向またはアンチセンス方向で結合した、上記(a)〜(i)のいずれかに記載のポリヌクレオチド(DNA);及び(c)RNA分子の転写終結およびポリアデニル化に関し、酵母で機能するシグナルを構成要素として含む発現カセットを含むように構成される。
【0025】
酵母に導入する際に用いるベクターとしては、多コピー型(YEp型)、単コピー型(YCp型)、染色体組み込み型(YIp型)のいずれもが利用可能である。例えば、YEp型ベクターとしてはYEp24 (J. R. Broach et al., Experimental Manipulation of Gene Expression, Academic Press, New York, 83, 1983) 、YCp型ベクターとしてはYCp50 (M. D. Rose et al., gene, 60, 237, 1987) 、YIp型ベクターとしてはYIp5 (K. Struhl et al., Proc. Natl. Acad. Sci. USP, 76, 1035, 1979) が知られており、容易に入手することができる。
酵母での遺伝子発現を調節するためのプロモーター/ターミネーターとしては、醸造用酵母中で機能するとともに、もろみ中のグルコース濃度に影響を受けなければ、任意の組み合わせでよい。例えばグリセルアルデヒド3リン酸デヒドロゲナーゼ遺伝子(TDH3)のプロモーター、3-ホスホグリセレートキナーゼ遺伝子(PGK1)のプロモーターなどが利用可能である。これらの遺伝子はすでにクローニングされており、例えばM. F. Tuite et al., EMBO J., 1, 603 (1982) に詳細に記載されており、既知の方法により容易に入手することができる。
【0026】
形質転換の際に用いる選択マーカーとしては、醸造用酵母の場合は栄養要求性マーカーが利用できないので、ジェネチシン耐性遺伝子(G418r)、銅耐性遺伝子(CUP1)(Marin et al., Proc. Natl. Acad. Sci. USA, 81, 337 1984)、セルレニン耐性遺伝子(fas2m, PDR4)(それぞれ猪腰淳嗣ら, 生化学, 64, 660, 1992; Hussain et et al., gene, 101, 149, 1991)などが利用可能である。上記のように構築されるベクターは、宿主酵母に導入される。宿主酵母としては、醸造用に使用可能な任意の酵母、例えばビール,ワイン、清酒等の醸造用酵母等が挙げられる。具体的には、サッカロマイセス(Saccharomyces)属等の酵母が挙げられるが、本発明においては、ビール酵母、例えばサッカロマイセス パストリアヌス(Saccharomyces pastorianus)W34/70等、サッカロマイセス カールスベルゲンシス(Saccharomyces carlsbergensis)NCYC453、NCYC456等、サッカロマイセス セレビシエ(Saccharomyces cerevisiae)NBRC1951、NBRC1952、NBRC1953、NBRC1954等が使用できる。さらにウイスキー酵母、例えばサッカロマイセス セレビシエNCYC90等、ワイン酵母、例えば協会ぶどう酒用1号、同3号、同4号等、清酒酵母、例えば協会酵母 清酒用7号、同9号等も用いることができるが、これに限定されない。本発明においては、ビール酵母、例えばサッカロマイセス パストリアヌスが好ましく用いられる。
【0027】
酵母の形質転換方法としては一般に用いられる公知の方法が利用できる。例えば、例えば、エレクトロポレーション法“Meth. Enzym., 194, p182 (1990)”、スフェロプラスト法“Proc. Natl. Acad. Sci. USA, 75 p1929(1978)”、酢酸リチウム法“J.Bacteriology, 153, p163(1983)”、Proc. Natl. Acad. Sci. USA, 75 p1929 (1978)、Methods in yeast genetics, 2000 Edition : A Cold Spring Harbor Laboratory Course Manualなどに記載の方法で実施可能であるが、これに限定されない。
より具体的には、宿主酵母を標準酵母栄養培地(例えばYEPD培地“Genetic Engineering. Vo1.1, Plenum Press, New York, 117(1979)”等)で、OD600nmの値が1〜6となるように培養する。この培養酵母を遠心分離して集め、洗浄し、濃度約1〜2Mのアルカリイオン金属イオン、好ましくはリチウムイオンで前処理する。この細胞を約30℃で、約60分間静置した後、導入するDNA(約1〜20μg)とともに約30℃で、約60分間静置する。ポリエチレングリコール、好ましくは約4,000ダルトンのポリエチレングリコールを、最終濃度が約20%〜50%となるように加える。約30℃で、約30分間静置した後、この細胞を約42℃で約5分間加熱処理する。好ましくは、この細胞懸濁液を標準酵母栄養培地で洗浄し、所定量の新鮮な標準酵母栄養培地に入れて、約30℃で約60分間静置する。その後、選択マーカーとして用いる抗生物質等を含む標準寒天培地上に植えつけ、形質転換体を取得する。 その他、一般的なクローニング技術に関しては、「モレキュラークローニング第3版」、“Methods in Yeast Genitics、A laboratory manual (Cold Spring Harbor Laboratory Press、Cold Spring Harbor, NY)”等を参照することができる。
【0028】
4.本発明の酒類の製法及びその製法によって得られる酒類
上述した本発明のベクターを製造対象となる酒類の醸造に適した酵母に導入し、その酵母を用いることによって高濃度麦汁を用いて短期間で酒類を製造することができる。対象となる酒類としては、これらに限定されないが、例えば、ビール、発泡酒などのビールテイストドリンクが挙げられる。
これらの酒類を製造する場合は、親株の代わりに本発明において得られた醸造酵母を用いる以外は公知の手法を利用することができる。したがって、原料、製造設備、製造管理等は従来法と全く同一でよく、発酵期間の短縮された酒類を製造するためのコストの増加はない。つまり、本発明によれば、既存の施設を用い、コストを増加させることなく製造することができる。
【0029】
5.本発明の酵母の評価方法
本発明は、配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の塩基配列に基づいて設計したプライマーまたはプローブを用いて、被検酵母のマルトースおよびマルトトリオース資化能について評価する方法に関する。プライマーまたはプローブを用いる評価方法の一般的手法は公知であり、例えば、WO01/040514号公報、特開平8−205900号公報などに記載されている。以下、この評価方法について簡単に説明する。
まず、被検酵母のゲノムを調製する。調製方法は、Hereford法や酢酸カリウム法など、公知の如何なる方法を用いることができる(例えば、Methods in Yeast Genetics, Cold Spring Harbor Laboratory Press, p130 (1990))。得られたゲノムを対象にして、αグルコシドトランスポーター遺伝子の塩基配列(好ましくは、ORF配列)に基づいて設計したプライマーまたはプローブを用いて、被検酵母のゲノムにその遺伝子あるいはその遺伝子に特異的な配列が存在するか否かを調べる。プライマーまたはプローブの設計は公知の手法を用いて行うことができる。
遺伝子または特異的な配列の検出は、公知の手法を用いて実施することができる。例えば、特異的配列の一部または全部を含むポリヌクレオチドまたはその塩基配列に対して相補的な塩基配列を含むポリヌクレオチドを一つのプライマーとして用い、もう一方のプライマーとしてこの配列よりも上流あるいは下流の配列の一部または全部を含むポリヌクレオチドまたはその塩基配列に対して相補的な塩基配列を含むポリヌクレオチドを用いて、PCR 法によって酵母の核酸を増幅し、増幅物の有無、増幅物の分子量の大きさなどを測定する。プライマーに使用するポリヌクレオチドの塩基数は、通常、10bp以上であり、15〜25bpであることが好ましい。
PCR 法の反応条件は、特に限定されないが、例えば、変性温度:90〜95℃、アニーリング温度:40〜60℃、伸長温度:60〜75℃、サイクル数:10回以上などの条件を用いることができる。得られる反応生成物はアガロースゲルなどを用いた電気泳動法等によって分離され、増幅産物の分子量を測定することができる。この方法により、増幅産物の分子量が特異部分のDNA 分子を含む大きさかどうかによって、その酵母のマルトースおよびマルトトリオース資化能について予測・評価する。また、増幅物の塩基配列を分析することによって、さらに上記性能についてより正確に予測・評価することが可能である。
【0030】
また、本発明においては、被検酵母を培養し、配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の発現量を測定することによって、被検酵母のマルトースおよびマルトトリオース資化能を評価することもできる。この場合は、被検酵母を培養し、αグルコシドトランスポーター遺伝子の産物であるmRNA又はたんぱく質を定量することによって可能である。mRNA又はたんぱく質の定量は、公知の手法を用いて行うことができる。例えば、mRNAの定量は例えばノーザンハイブリダイゼーションや定量的RT−PCRによって、たんぱく質の定量は例えばウエスタンブロッティングによって行うことができる(Current Protocols in Molecular Biology, John Wiley & Sons 1994-2003)。
また、基準酵母および被検酵母を培養して配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の各酵母における発現量を測定し、基準酵母よりも該遺伝子が高発現している被検酵母を選択する(即ち、複数の酵母を培養して配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の各酵母における発現量を測定し、該複数の酵母の中から遺伝子が高発現している酵母を選択する)ことによって酒類の醸造に好適な酵母を選択することができる。
【実施例】
【0031】
以下、実施例によって本発明の詳細を述べるが、本発明は以下の実施例に限定されるものではない。
【0032】
実施例1:新規αグルコシドトランスポーター(nonScMPH2)のクローニング
特開2004-283169に記載の比較データベースを用いて検索した結果、ビール酵母に特有の新規αグルコシドトランスポーター遺伝子、nonScMPH2を見出した(配列番号:1)。得られた塩基配列情報を基に、それぞれ全長遺伝子を増幅するためのプライマーnonScMPH2#F(配列番号:3)/nonScMPH2#R(配列番号:4)を設計し、ゲノム解読株サッカロマイセス パストリアヌス バイヘンステファン34/70株の染色体DNAを鋳型としたPCRによってnonScMPH2の全長遺伝子を含むDNA断片を取得した。
【0033】
上記のようにして得られたnonScMPH2遺伝子断片を、TAクローニングによってpCR2.1-TOPOベクター(インビトロジェン社製)に挿入した。nonScMPH2遺伝子の塩基配列をサンガーの方法 (F. Sanger, Science, 214, 1215, 1981) で分析し、塩基配列を確認した。
【0034】
実施例2:ビール試醸中のnonScMPH2遺伝子発現解析
ビール酵母サッカロマイセス パストリアヌスW34/70株を用いてビール試醸を行い、発酵中のビール酵母菌体から抽出したmRNAをビール酵母DNAマイクロアレイで検出した。
麦汁エキス濃度 12.69%
麦汁容量 70L
麦汁溶存酸素濃度 8.6ppm
発酵温度 15℃
酵母投入量 12.8×106cells/mL
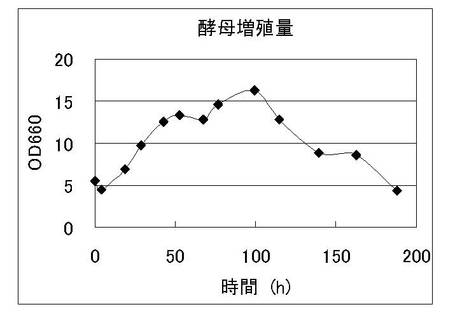
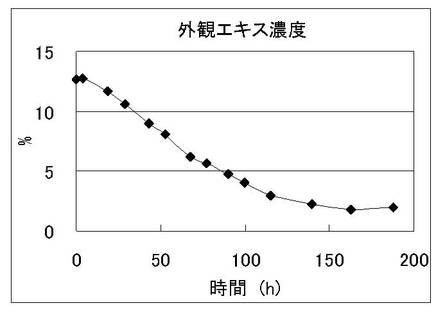
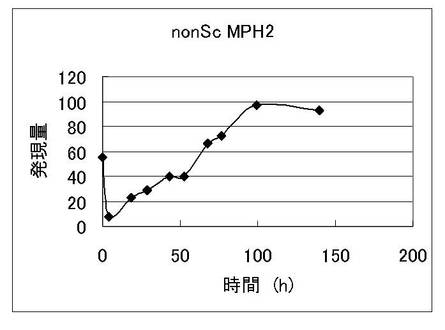
発酵液を経時的にサンプリングし、酵母増殖量(図1)、外観エキス濃度(図2)の経時変化を観察した。またこれと同時に酵母菌体をサンプリングし、調製したmRNAをビオチンラベルして、特開2004-283169に記載のビール酵母DNAマイクロアレイにハイブリダイズさせた。シグナルの検出はジーンチップオペレーティングシステム(GCOS;GeneChip Operating Software 1.0、アフィメトリクス社製)を用いて行った。nonScMPH2遺伝子の発現パターンを図3に示す。この結果より、通常のビール発酵においてnonScMPH2遺伝子が発現していることを確認した。
【0035】
実施例3:nonScMPH2遺伝子の構成的発現
実施例1に記載のnonScMPH2/pCR2.1-TOPOを制限酵素SacIおよびNotI消化し、タンパク質コード領域全長を含むDNA断片を調製する。この断片を制限酵素SacIおよびNotI処理したpYCGPYNotに連結させ、nonScMPH2構成的発現ベクターnonScMPH2/pYCGPYNotを構築する。pYCGPYNotはYCp型の酵母発現ベクターであり、導入された遺伝子はピルビン酸キナーゼ遺伝子PYK1のプロモーターによって構成的に発現される。酵母での選択マーカーとしてジェネチシン耐性遺伝子G418rを、また大腸菌での選択マーカーとしてアンピシリン耐性遺伝子Amprを含んでいる。
【0036】
上述の方法で作製した構成的発現ベクターを用い、特開平07-303475に記載された方法でサッカロマイセス パストリアヌス バイヘンステファン34/70株を形質転換する。ジェネチシン300mg/Lを含むYPD平板培地(1%酵母エキス、2%ポリペプトン、2%グルコース、2%寒天)で形質転換体を選択する。
【0037】
実施例4:ビール試験醸造におけるマルトース及びマルトトリオース資化量の測定
親株ならびに実施例3で得られたnonScMPH2高発現株を用いた発酵試験を以下の条件で行う。
麦汁エキス濃度 20%(12%麦汁に糖シロップを添加)
麦汁容量 1L
麦汁溶存酸素濃度 約 8ppm
発酵温度 15℃一定
酵母投入量 5g湿酵母菌体/L麦汁
発酵醪を経時的にサンプリングし、酵母増殖量(OD660)、エキス消費量、グルコース、マルトース、マルトトリオースの経時変化を調べる。
【産業上の利用可能性】
【0038】
本発明の酒類製造法によれば、酵母のマルトースおよびマルトトリオース資化能が増大するため、高濃度醸造においても短期間で酒類を製造することが可能となる。
【図面の簡単な説明】
【0039】
【図1】ビール試験醸造における酵母増殖量の経時変化を示す図である。横軸は発酵時間を、縦軸はOD660の値を示している。
【図2】ビール試験醸造におけるエキス消費量の経時変化を示す図である。横軸は発酵時間、縦軸は外観エキス濃度(w/w%)を示している。
【図3】ビール試験醸造中の酵母におけるnonScMPH2遺伝子の発現挙動を示す図である。横軸は発酵時間、縦軸は検出されたシグナル輝度を示している。
【配列表フリーテキスト】
【0040】
[配列番号:3] プライマー
[配列番号:4] プライマー
【技術分野】
【0001】
本発明は、αグルコシドトランスポーター遺伝子及びその用途に関し、特に、マルトースおよびマルトトリオース発酵性に優れた醸造酵母、該酵母を用いて製造した酒類、その製造方法などに関する。さらに具体的には、本発明は、醸造酵母のαグルコシドトランスポーターであるMph2pをコードする遺伝子MPH2、特にビール酵母に特徴的なnonScMPH2遺伝子の発現量を高めることによって、マルトースおよびマルトトリオース資化能を向上させた酵母、当該酵母を用いた酒類の製造方法などに関する。
【背景技術】
【0002】
従来ビールの製造においては、エキス濃度11%前後の原麦汁を発酵させてアルコール濃度4.5〜5%前後のビールを得ている。ビールの生産性向上のために、世界中のビール会社で高濃度醸造法が採用されている。高濃度醸造法(highgravity brewing) とは、製造しようとするビールよりも高い原麦汁濃度のビールを与える麦汁を醗酵させた後、これを水で希釈して所定原麦汁濃度のビールとする方法である。その具体的方策としては、(1)通常発酵温度よりも高い温度を採用する、(2)麦汁への通気量を増加させる、(3)酵母の添加量を増加させる、およびそれらの組み合わせである。通常のビールを製造する場合には、これらの方法では原麦汁エキス濃度15%程度の高濃度醸造が限界であると言われている。
【0003】
原麦汁エキス濃度15%以上の高濃度醸造を行なった場合に生じる問題点として、まず一番に挙げられるのは、発酵中期から後期にかけての発酵速度の著しい低下である。麦汁中に含まれる糖質は、主にマルトース、マルトトリオース、グルコース、フラクトース、シュークロースである。酵母は、グルコース、フラクトース、シュークロースを先に資化し、その後にマルトース、マルトトリオースを資化する。従って、発酵中期から発酵後期にかけては、もろみ中の資化性糖はマルトース、マルトトリオースしか存在していない。しかもその存在比は、3:1と圧倒的にマルトースが多い。
【0004】
マルトースは、酵母のαグルコシドトランスポーターによって、細胞内に取り込まれた後、マルターゼによって2個のグルコースに加水分解され、エムブデン−マイヤーホッフ(Embden-Meyerhof)の経路を介して二酸化炭素とエタノールに転化する。この2つの酵素は、転写レベルで、マルトースによって誘導され、グルコースによって抑制される。
【0005】
麦汁中にはグルコースが、資化性糖の約17%含まれるため、酵母のマルトース及びマルトトリオース代謝系遺伝子は、発酵初期の段階でグルコースによる抑制を受け、マルトース及びマルトトリオースの資化が、大幅に遅れることになる。この現象は、高濃度醸造の際には、さらに顕著となり、マルトース及びマルトトリオース発酵が遅れるばかりでなく、発酵終了時に、残糖として多量のマルトース及びマルトトリオースが、資化されないまま残ってしまう。このような問題点によって、高濃度醸造、例えば通常の2倍の濃度の麦汁を醗酵させることは、従来の技術では不完全であった。
【0006】
特開平1−153082号公報(特許文献1)には、パン酵母によるドゥ発酵を改善するために、グルコースによる抑制を受けないアルコールデヒドロゲナーゼ遺伝子のプロモーターとマルターゼ遺伝子及びマルトーストランスポーター遺伝子とを含むプラスミドを導入したパン酵母を使用することが開示されている。一方、Saccharomyces cerevisiaeのマルトーストランスポーター遺伝子MAL6Tを醸造用酵母で高発現させ、高濃度醸造を実現させた例が報告されている(特開平6−245750号公報:特許文献2)。
【特許文献1】特開平1−153082号公報
【特許文献2】特開平6−245750号公報
【発明の開示】
【発明が解決しようとする課題】
【0007】
上述のとおり、発酵速度や製品の品質を損なうことなく、高濃度醸造を可能にする酵母が望まれていた。
【課題を解決するための手段】
【0008】
本発明者らは、上記課題を解決するため鋭意検討を重ねた結果、ビール酵母から既知のタンパク質より有利な効果を奏するαグルコシドトランスポーターをコードする遺伝子を同定・単離することに成功した。また、得られた遺伝子を酵母に導入し発現させた形質転換酵母を作製し、マルトース及びマルトトリオース資化が促進されることを確認して、本発明を完成した。
【0009】
すなわち本発明は、ビール酵母に特徴的に存在する新規なαグルコシドトランスポーター遺伝子、該遺伝子がコードするタンパク質、該遺伝子の発現が調節された形質転換酵母、該遺伝子の発現が調節された酵母を用いることによる酒類の製造方法などに関する。本発明は、具体的には、次に示すポリヌクレオチド、該ポリヌクレオチドを含有するベクター、該ベクターが導入された形質転換酵母、該形質転換酵母を用いる酒類の製造方法などを提供する。
【0010】
(1)以下の(a)〜(f)のいずれかに記載のポリヌクレオチド:
(a)配列番号:1の塩基配列からなるポリヌクレオチドを含有するポリヌクレオチド;
(b)配列番号:2のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(c)配列番号:2のアミノ酸配列において、1もしくは複数個のアミノ酸が欠失、置換、挿入および/または付加したアミノ酸配列からなり、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(d)配列番号:2のアミノ酸配列に対して60%以上の同一性を有するアミノ酸配列を有し、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(e)配列番号:1の塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;及び
(f)配列番号:2のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドの塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド。
【0011】
(2)以下の(g)〜(i)のいずれかである請求項1に記載のポリヌクレオチド:
(g)配列番号:2のアミノ酸配列又は配列番号:2のアミノ酸配列において、1〜10個のアミノ酸が欠失、置換、挿入および/または付加したアミノ酸配列からなり、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(h) 配列番号:2のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列を有し、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;及び
(i)配列番号:1の塩基配列からなるポリヌクレオチド、又は配列番号:1の塩基配列と相補的な塩基配列からなるポリヌクレオチドとハイストリンジェントな条件下でハイブリダイズし、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチドを含有するポリヌクレオチド。
(3)配列番号:1の塩基配列からなるポリヌクレオチドを含有する上記(1)に記載のポリヌクレオチド。
(4)配列番号:2のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有する上記(1)に記載のポリヌクレオチド。
(5)DNAである、請求項1〜4のいずれかに記載のポリヌクレオチド。
(6)上記(1)〜(5)のいずれかに記載のポリヌクレオチドにコードされるタンパク質。
【0012】
(7)上記(1)〜(5)のいずれかに記載のポリヌクレオチドを含有するベクター。
(7a)以下の(a)〜(c)の構成要素を含む発現カセットを含む上記(7)に記載のベクター:
(a)酵母細胞内で転写可能なプロモーター
(b)該プロモーターにセンス方向またはアンチセンス方向で結合した、上記(1)〜(5)のいずれかに記載のポリヌクレオチド;及び
(c)RNA分子の転写終結およびポリアデニル化に関し、酵母で機能するシグナル。
(8)上記(7)に記載のベクターが導入された形質転換酵母。
(9)上記(7)に記載のベクターを導入することによって、マルトースおよび/またはマルトトリオース資化能が増強された上記(8)に記載の醸造用酵母。
(10)上記(6)に記載のタンパク質の発現量を増加させることによってマルトースおよび/またはマルトトリオース資化能が向上した上記(9)に記載の醸造用酵母。
(11)上記(8)〜(10)のいずれかに記載の酵母を用いた酒類の製造方法。
(12)醸造する酒類が麦芽飲料である上記(11)に記載の酒類の製造方法。
(13)上記(11)または(12)に記載の方法で製造された酒類。
(14)配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の塩基配列に基づいて設計したプライマーまたはプローブを用いて、被検酵母のマルトースおよびマルトトリオース資化能について評価する方法。
(14a)上記(14)に記載の方法によって、マルトースおよびマルトトリオース資化能が高い酵母を選別する方法。
(14b)上記(14a)に記載の方法によって選別された酵母を用いて酒類(例えば、ビール)を製造する方法。
(15)被検酵母を培養し、配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の発現量を測定することによって、被検酵母のマルトースおよびマルトトリオース資化能を評価する方法。
(15a)上記(15)に記載の方法で、被検酵母を評価し、αグルコシドトランスポーター遺伝子の発現量が高い酵母を選別する、マルトースおよびマルトトリオース資化能の高い酵母を選別する方法。
(15b)上記(15a)に記載の方法によって選別された酵母を用いて酒類(例えば、ビール)を製造する方法。
(16)基準酵母および被検酵母を培養して配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の各酵母における発現量を測定し、基準酵母よりも該遺伝子が高発現している被検酵母を選択する、酵母の選択方法。
(17)
基準酵母および被検酵母を培養して各酵母における上記(6)に記載のタンパク質を定量し、基準酵母よりも該タンパク質量の多い被検酵母を選択する、酵母の選択方法。
【発明の効果】
【0013】
本発明の形質転換酵母を用いる酒類の製造法によれば、グルコースの存在下でもマルトース及びマルトトリオースの資化が抑制されず、この結果グルコースの消失を待たずにマルトース及びマルトトリオースが資化発酵されるため発酵速度が上昇し、高濃度麦汁によるビール醸造が可能となる。
【発明を実施するための最良の形態】
【0014】
本発明者らは、酵母のαグルコシドトランスポーターの活性を増大させることによって、さらに効率よくマルトース及びマルトトリオースを資化させることが可能であると考えた。このような着想に基づいて研究を重ね、特開2004-283169に開示の方法で解読したビール酵母ゲノム情報を基に、ビール酵母特有のαグルコシドトランスポーターをコードするnon-ScMPH2遺伝子を単離・同定した。この塩基配列を配列番号:1に示す。またこの遺伝子によりコードされるタンパク質のアミノ酸配列を配列番号:2に示す。
【0015】
1.本発明のポリヌクレオチド
まず、本発明は、(a)配列番号:1の塩基配列からなるポリヌクレオチドを含有するポリヌクレオチド(具体的には、DNA、以下、これらを単に「DNA」とも称する);及び(b)配列番号:2のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチドを提供する。本発明で対象とするDNAは、上記のビール酵母由来のαグルコシドトランスポーターをコードするDNAに限定されるものではなく、このタンパク質と機能的に同等なタンパク質をコードする他のDNAを含む。機能的に同等なタンパク質としては、例えば、(c)配列番号:2のアミノ酸配列において、1もしくは複数個のアミノ酸が欠失、置換、挿入および/または付加したアミノ酸配列からなり、かつαグルコシドトランスポーター活性を有するタンパク質が挙げられる。このようなタンパク質としては、配列番号:2のアミノ酸配列において、例えば、1〜100個、1〜90個、1〜80個、1〜70個、1〜60個、1〜50個、1〜40個、1〜39個、1〜38個、1〜37個、1〜36個、1〜35個、1〜34個、1〜33個、1〜32個、1〜31個、1〜30個、1〜29個、1〜28個、1〜27個、1〜26個、1〜25個、1〜24個、1〜23個、1〜22個、1〜21個、1〜20個、1〜19個、1〜18個、1〜17個、1〜16個、1〜15個、1〜14個、1〜13個、1〜12個、1〜11個、1〜10個、1〜9個、1〜8個、1〜7個、1〜6個(1〜数個)、1〜5個、1〜4個、1〜3個、1〜2個、1個のアミノ酸残基が欠失、置換、挿入および/または付加されたアミノ酸配列からなり、かつαグルコシドトランスポーター活性を有するタンパク質が挙げられる。上記アミノ酸残基の欠失、置換、挿入および/または付加の数は、一般的には小さい程好ましい。また、このようなタンパク質としては、(d)配列番号:2のアミノ酸配列と約60%以上、約70%以上、71%以上、72%以上、73%以上、74%以上、75%以上、76%以上、77%以上、78%以上、79%以上、80%以上、81%以上、82%以上、83%以上、84%以上、85%以上、86%以上、87%以上、88%以上、89%以上、90%以上、91%以上、92%以上、93%以上、94%以上、95%以上、96%以上、97%以上、98%以上、99%以上、99.1%以上99.2%以上99.3%以上99.4%以上99.5%以上99.6%以上99.7%以上99.8%以上99.9%以上の同一性を有するアミノ酸配列を有し、かつαグルコシドトランスポーター活性を有するタンパク質が挙げられる。上記相同性の数値は一般的に大きい程好ましい。なお、αグルコシドトランスポーター活性は、例えば特開平06-245750に記載の方法によって測定することができる。
【0016】
また、本発明は、(e)配列番号:1の塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;及び(f)配列番号:2のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドの塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチドも包含する。
【0017】
ここで、「ストリンジェントな条件下でハイブリダイズするポリヌクレオチド(DNA)」とは、配列番号:1の塩基配列と相補的な塩基配列からなるDNA又は配列番号:2のアミノ酸配列をコードするDNAの全部または一部をプローブとして、コロニーハイブリダイゼーション法、プラークハイブリダイゼーション法またはサザンハイブリダイゼーション法などを用いることにより得られるDNAをいう。ハイブリダイゼーションの方法としては、例えばMolecular Cloning 3rd Ed.、Current Protocols in Molecular Biology, John Wiley & Sons 1987-1997などに記載されている方法を利用することができる。
【0018】
本明細書でいう「ストリンジェントな条件」は、低ストリンジェントな条件、中ストリンジェントな条件及び高ストリンジェントな条件のいずれでもよい。「低ストリンジェントな条件」は、例えば、5×SSC、5×デンハルト溶液、0.5%SDS、50%ホルムアミド、32℃の条件である。また、「中ストリンジェントな条件」は、例えば、5×SSC、5×デンハルト溶液、0.5%SDS、50%ホルムアミド、42℃の条件である。「高ストリンジェントな条件」は、例えば、5×SSC、5×デンハルト溶液、0.5%SDS、50%ホルムアミド、50℃の条件である。これらの条件において、温度を上げるほど高い相同性を有するDNAが効率的に得られることが期待できる。ただし、ハイブリダイゼーションのストリンジェンシーに影響する要素としては温度、プローブ濃度、プローブの長さ、イオン強度、時間、塩濃度など複数の要素が考えられ、当業者であればこれら要素を適宜選択することで同様のストリンジェンシーを実現することが可能である。
【0019】
なお、ハイブリダイゼーションに市販のキットを用いる場合は、例えばAlkphos Direct Labelling Reagents(アマシャムファルマシア社製)を用いることができる。この場合は、キットに添付のプロトコルにしたがい、標識したプローブとのインキュベーションを一晩行った後、メンブレンを55℃の条件下で0.1% (w/v) SDSを含む1次洗浄バッファーで洗浄後、ハイブリダイズしたDNAを検出することができる。
【0020】
これ以外にハイブリダイズ可能なDNAとしては、FASTA、BLASTなどの相同性検索ソフトウェアにより、デフォルトのパラメータを用いて計算したときに、配列番号:2のアミノ酸配列をコードするDNAと約60%以上、約70%以上、71%以上、72%以上、73%以上、74%以上、75%以上、76%以上、77%以上、78%以上、79%以上、80%以上、81%以上、82%以上、83%以上、84%以上、85%以上、86%以上、87%以上、88%以上、89%以上、90%以上、91%以上、92%以上、93%以上、94%以上、95%以上、96%以上、97%以上、98%以上、99%以上、99.1%以上99.2%以上99.3%以上99.4%以上99.5%以上99.6%以上99.7%以上99.8%以上99.9%以上以上の同一性を有するDNAをあげることができる。
なお、アミノ酸配列や塩基配列の同一性は、カーリンおよびアルチュールによるアルゴリズムBLAST(proc. Natl. Acad. Sci. USA 872264-2268, 1990; proc Natl Acad Sci USA 90: 5873, 1993)を用いて決定できる。BLASTのアルゴリズムに基づいたBLASTNやBLASTXと呼ばれるプログラムが開発されている(Altschul SF, et al: J Mol Biol 215: 403, 1990)。BLASTNを用いて塩基配列を解析する場合は、パラメーターは、例えばscore=100、wordlength=12とする。また、BLASTXを用いてアミノ酸配列を解析する場合は、パラメーターは、例えばscore=50、wordlength=3とする。BLASTとGapped BLASTプログラムを用いる場合は、各プログラムのデフォルトパラメーターを用いる。
【0021】
2.本発明のタンパク質
本発明は、上記ポリヌクレオチド(a)〜(e)のいずれかにコードされるタンパク質も提供する。本発明の好ましいタンパク質は、配列番号:2のアミノ酸配列において、1もしくは複数個のアミノ酸が欠失、置換、挿入および/または付加したアミノ酸配列からなり、かつαグルコシドトランスポーター活性を有するタンパク質である。このようなタンパク質としては、配列番号:2のアミノ酸配列において、上記したような数のアミノ酸残基が欠失、置換、挿入および/または付加されたアミノ酸配列からなり、かつαグルコシドトランスポーター活性を有するタンパク質が挙げられる。また、このようなタンパク質としては、配列番号:2のアミノ酸配列と上記したような相同性を有するアミノ酸配列を有し、かつαグルコシドトランスポーター活性を有するタンパク質が挙げられる。このようなタンパク質は、「モレキュラークローニング第3版」、「カレント・プロトコールズ・イン・モレキュラー・バイオロジー」、“Nuc. Acids. Res., 10, 6487 (1982)”、“Proc. Natl. Acad. Sci. USA, 79, 6409 (1982)”、“Gene, 34, 315 (1985)”、“Nuc. Acids. Res., 13, 4431 (1985)”、“Proc. Natl. Acad. Sci. USA, 82, 488 (1985)”等に記載の部位特異的変異導入法を用いて、取得することができる。
【0022】
本発明のタンパク質のアミノ酸配列において1以上のアミノ酸残基が欠失、置換、挿入および/または付加されたとは、同一配列中の任意かつ1もしくは複数のアミノ酸配列中の位置において、1または複数のアミノ酸残基の欠失、置換、挿入及び/又は付加があることを意味し、欠失、置換、挿入及び付加のうち2種以上が同時に生じてもよい。
以下に、相互に置換可能なアミノ酸残基の例を示す。同一群に含まれるアミノ酸残基は相互に置換可能である。A群:ロイシン、イソロイシン、ノルロイシン、バリン、ノルバリン、アラニン、2-アミノブタン酸、メチオニン、o-メチルセリン、t-ブチルグリシン、t-ブチルアラニン、シクロヘキシルアラニン; B群:アスパラギン酸、グルタミン酸、イソアスパラギン酸、イソグルタミン酸、2-アミノアジピン酸、2-アミノスベリン酸; C群:アスパラギン、グルタミン; D群:リジン、アルギニン、オルニチン、2,4-ジアミノブタン酸、2,3-ジアミノプロピオン酸; E群:プロリン、3-ヒドロキシプロリン、4-ヒドロキシプロリン; F群:セリン、スレオニン、ホモセリン; G群:フェニルアラニン、チロシン。
【0023】
また、本発明のタンパク質は、Fmoc法(フルオレニルメチルオキシカルボニル法)、tBoc法(t-ブチルオキシカルボニル法)等の化学合成法のよっても製造することができる。また、アドバンスドケムテック社製、パーキンエルマー社製、ファルマシア社製、プロテインテクノロジーインストゥルメント社製、シンセセルーベガ社製、パーセプティブ社製、島津製作所等のペプチド合成機を利用して化学合成することもできる。
【0024】
3.本発明のベクター及びこれを導入した形質転換酵母
次に、本発明は、上記したポリヌクレオチドを含有するベクターを提供する。本発明のベクターは、上記(a)〜(i)のいずれかに記載のポリヌクレオチド(DNA)を含有するベクターに関する。また、本発明のベクターは、通常、(a)酵母細胞内で転写可能なプロモーター;(b)該プロモーターにセンス方向またはアンチセンス方向で結合した、上記(a)〜(i)のいずれかに記載のポリヌクレオチド(DNA);及び(c)RNA分子の転写終結およびポリアデニル化に関し、酵母で機能するシグナルを構成要素として含む発現カセットを含むように構成される。
【0025】
酵母に導入する際に用いるベクターとしては、多コピー型(YEp型)、単コピー型(YCp型)、染色体組み込み型(YIp型)のいずれもが利用可能である。例えば、YEp型ベクターとしてはYEp24 (J. R. Broach et al., Experimental Manipulation of Gene Expression, Academic Press, New York, 83, 1983) 、YCp型ベクターとしてはYCp50 (M. D. Rose et al., gene, 60, 237, 1987) 、YIp型ベクターとしてはYIp5 (K. Struhl et al., Proc. Natl. Acad. Sci. USP, 76, 1035, 1979) が知られており、容易に入手することができる。
酵母での遺伝子発現を調節するためのプロモーター/ターミネーターとしては、醸造用酵母中で機能するとともに、もろみ中のグルコース濃度に影響を受けなければ、任意の組み合わせでよい。例えばグリセルアルデヒド3リン酸デヒドロゲナーゼ遺伝子(TDH3)のプロモーター、3-ホスホグリセレートキナーゼ遺伝子(PGK1)のプロモーターなどが利用可能である。これらの遺伝子はすでにクローニングされており、例えばM. F. Tuite et al., EMBO J., 1, 603 (1982) に詳細に記載されており、既知の方法により容易に入手することができる。
【0026】
形質転換の際に用いる選択マーカーとしては、醸造用酵母の場合は栄養要求性マーカーが利用できないので、ジェネチシン耐性遺伝子(G418r)、銅耐性遺伝子(CUP1)(Marin et al., Proc. Natl. Acad. Sci. USA, 81, 337 1984)、セルレニン耐性遺伝子(fas2m, PDR4)(それぞれ猪腰淳嗣ら, 生化学, 64, 660, 1992; Hussain et et al., gene, 101, 149, 1991)などが利用可能である。上記のように構築されるベクターは、宿主酵母に導入される。宿主酵母としては、醸造用に使用可能な任意の酵母、例えばビール,ワイン、清酒等の醸造用酵母等が挙げられる。具体的には、サッカロマイセス(Saccharomyces)属等の酵母が挙げられるが、本発明においては、ビール酵母、例えばサッカロマイセス パストリアヌス(Saccharomyces pastorianus)W34/70等、サッカロマイセス カールスベルゲンシス(Saccharomyces carlsbergensis)NCYC453、NCYC456等、サッカロマイセス セレビシエ(Saccharomyces cerevisiae)NBRC1951、NBRC1952、NBRC1953、NBRC1954等が使用できる。さらにウイスキー酵母、例えばサッカロマイセス セレビシエNCYC90等、ワイン酵母、例えば協会ぶどう酒用1号、同3号、同4号等、清酒酵母、例えば協会酵母 清酒用7号、同9号等も用いることができるが、これに限定されない。本発明においては、ビール酵母、例えばサッカロマイセス パストリアヌスが好ましく用いられる。
【0027】
酵母の形質転換方法としては一般に用いられる公知の方法が利用できる。例えば、例えば、エレクトロポレーション法“Meth. Enzym., 194, p182 (1990)”、スフェロプラスト法“Proc. Natl. Acad. Sci. USA, 75 p1929(1978)”、酢酸リチウム法“J.Bacteriology, 153, p163(1983)”、Proc. Natl. Acad. Sci. USA, 75 p1929 (1978)、Methods in yeast genetics, 2000 Edition : A Cold Spring Harbor Laboratory Course Manualなどに記載の方法で実施可能であるが、これに限定されない。
より具体的には、宿主酵母を標準酵母栄養培地(例えばYEPD培地“Genetic Engineering. Vo1.1, Plenum Press, New York, 117(1979)”等)で、OD600nmの値が1〜6となるように培養する。この培養酵母を遠心分離して集め、洗浄し、濃度約1〜2Mのアルカリイオン金属イオン、好ましくはリチウムイオンで前処理する。この細胞を約30℃で、約60分間静置した後、導入するDNA(約1〜20μg)とともに約30℃で、約60分間静置する。ポリエチレングリコール、好ましくは約4,000ダルトンのポリエチレングリコールを、最終濃度が約20%〜50%となるように加える。約30℃で、約30分間静置した後、この細胞を約42℃で約5分間加熱処理する。好ましくは、この細胞懸濁液を標準酵母栄養培地で洗浄し、所定量の新鮮な標準酵母栄養培地に入れて、約30℃で約60分間静置する。その後、選択マーカーとして用いる抗生物質等を含む標準寒天培地上に植えつけ、形質転換体を取得する。 その他、一般的なクローニング技術に関しては、「モレキュラークローニング第3版」、“Methods in Yeast Genitics、A laboratory manual (Cold Spring Harbor Laboratory Press、Cold Spring Harbor, NY)”等を参照することができる。
【0028】
4.本発明の酒類の製法及びその製法によって得られる酒類
上述した本発明のベクターを製造対象となる酒類の醸造に適した酵母に導入し、その酵母を用いることによって高濃度麦汁を用いて短期間で酒類を製造することができる。対象となる酒類としては、これらに限定されないが、例えば、ビール、発泡酒などのビールテイストドリンクが挙げられる。
これらの酒類を製造する場合は、親株の代わりに本発明において得られた醸造酵母を用いる以外は公知の手法を利用することができる。したがって、原料、製造設備、製造管理等は従来法と全く同一でよく、発酵期間の短縮された酒類を製造するためのコストの増加はない。つまり、本発明によれば、既存の施設を用い、コストを増加させることなく製造することができる。
【0029】
5.本発明の酵母の評価方法
本発明は、配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の塩基配列に基づいて設計したプライマーまたはプローブを用いて、被検酵母のマルトースおよびマルトトリオース資化能について評価する方法に関する。プライマーまたはプローブを用いる評価方法の一般的手法は公知であり、例えば、WO01/040514号公報、特開平8−205900号公報などに記載されている。以下、この評価方法について簡単に説明する。
まず、被検酵母のゲノムを調製する。調製方法は、Hereford法や酢酸カリウム法など、公知の如何なる方法を用いることができる(例えば、Methods in Yeast Genetics, Cold Spring Harbor Laboratory Press, p130 (1990))。得られたゲノムを対象にして、αグルコシドトランスポーター遺伝子の塩基配列(好ましくは、ORF配列)に基づいて設計したプライマーまたはプローブを用いて、被検酵母のゲノムにその遺伝子あるいはその遺伝子に特異的な配列が存在するか否かを調べる。プライマーまたはプローブの設計は公知の手法を用いて行うことができる。
遺伝子または特異的な配列の検出は、公知の手法を用いて実施することができる。例えば、特異的配列の一部または全部を含むポリヌクレオチドまたはその塩基配列に対して相補的な塩基配列を含むポリヌクレオチドを一つのプライマーとして用い、もう一方のプライマーとしてこの配列よりも上流あるいは下流の配列の一部または全部を含むポリヌクレオチドまたはその塩基配列に対して相補的な塩基配列を含むポリヌクレオチドを用いて、PCR 法によって酵母の核酸を増幅し、増幅物の有無、増幅物の分子量の大きさなどを測定する。プライマーに使用するポリヌクレオチドの塩基数は、通常、10bp以上であり、15〜25bpであることが好ましい。
PCR 法の反応条件は、特に限定されないが、例えば、変性温度:90〜95℃、アニーリング温度:40〜60℃、伸長温度:60〜75℃、サイクル数:10回以上などの条件を用いることができる。得られる反応生成物はアガロースゲルなどを用いた電気泳動法等によって分離され、増幅産物の分子量を測定することができる。この方法により、増幅産物の分子量が特異部分のDNA 分子を含む大きさかどうかによって、その酵母のマルトースおよびマルトトリオース資化能について予測・評価する。また、増幅物の塩基配列を分析することによって、さらに上記性能についてより正確に予測・評価することが可能である。
【0030】
また、本発明においては、被検酵母を培養し、配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の発現量を測定することによって、被検酵母のマルトースおよびマルトトリオース資化能を評価することもできる。この場合は、被検酵母を培養し、αグルコシドトランスポーター遺伝子の産物であるmRNA又はたんぱく質を定量することによって可能である。mRNA又はたんぱく質の定量は、公知の手法を用いて行うことができる。例えば、mRNAの定量は例えばノーザンハイブリダイゼーションや定量的RT−PCRによって、たんぱく質の定量は例えばウエスタンブロッティングによって行うことができる(Current Protocols in Molecular Biology, John Wiley & Sons 1994-2003)。
また、基準酵母および被検酵母を培養して配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の各酵母における発現量を測定し、基準酵母よりも該遺伝子が高発現している被検酵母を選択する(即ち、複数の酵母を培養して配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の各酵母における発現量を測定し、該複数の酵母の中から遺伝子が高発現している酵母を選択する)ことによって酒類の醸造に好適な酵母を選択することができる。
【実施例】
【0031】
以下、実施例によって本発明の詳細を述べるが、本発明は以下の実施例に限定されるものではない。
【0032】
実施例1:新規αグルコシドトランスポーター(nonScMPH2)のクローニング
特開2004-283169に記載の比較データベースを用いて検索した結果、ビール酵母に特有の新規αグルコシドトランスポーター遺伝子、nonScMPH2を見出した(配列番号:1)。得られた塩基配列情報を基に、それぞれ全長遺伝子を増幅するためのプライマーnonScMPH2#F(配列番号:3)/nonScMPH2#R(配列番号:4)を設計し、ゲノム解読株サッカロマイセス パストリアヌス バイヘンステファン34/70株の染色体DNAを鋳型としたPCRによってnonScMPH2の全長遺伝子を含むDNA断片を取得した。
【0033】
上記のようにして得られたnonScMPH2遺伝子断片を、TAクローニングによってpCR2.1-TOPOベクター(インビトロジェン社製)に挿入した。nonScMPH2遺伝子の塩基配列をサンガーの方法 (F. Sanger, Science, 214, 1215, 1981) で分析し、塩基配列を確認した。
【0034】
実施例2:ビール試醸中のnonScMPH2遺伝子発現解析
ビール酵母サッカロマイセス パストリアヌスW34/70株を用いてビール試醸を行い、発酵中のビール酵母菌体から抽出したmRNAをビール酵母DNAマイクロアレイで検出した。
麦汁エキス濃度 12.69%
麦汁容量 70L
麦汁溶存酸素濃度 8.6ppm
発酵温度 15℃
酵母投入量 12.8×106cells/mL
発酵液を経時的にサンプリングし、酵母増殖量(図1)、外観エキス濃度(図2)の経時変化を観察した。またこれと同時に酵母菌体をサンプリングし、調製したmRNAをビオチンラベルして、特開2004-283169に記載のビール酵母DNAマイクロアレイにハイブリダイズさせた。シグナルの検出はジーンチップオペレーティングシステム(GCOS;GeneChip Operating Software 1.0、アフィメトリクス社製)を用いて行った。nonScMPH2遺伝子の発現パターンを図3に示す。この結果より、通常のビール発酵においてnonScMPH2遺伝子が発現していることを確認した。
【0035】
実施例3:nonScMPH2遺伝子の構成的発現
実施例1に記載のnonScMPH2/pCR2.1-TOPOを制限酵素SacIおよびNotI消化し、タンパク質コード領域全長を含むDNA断片を調製する。この断片を制限酵素SacIおよびNotI処理したpYCGPYNotに連結させ、nonScMPH2構成的発現ベクターnonScMPH2/pYCGPYNotを構築する。pYCGPYNotはYCp型の酵母発現ベクターであり、導入された遺伝子はピルビン酸キナーゼ遺伝子PYK1のプロモーターによって構成的に発現される。酵母での選択マーカーとしてジェネチシン耐性遺伝子G418rを、また大腸菌での選択マーカーとしてアンピシリン耐性遺伝子Amprを含んでいる。
【0036】
上述の方法で作製した構成的発現ベクターを用い、特開平07-303475に記載された方法でサッカロマイセス パストリアヌス バイヘンステファン34/70株を形質転換する。ジェネチシン300mg/Lを含むYPD平板培地(1%酵母エキス、2%ポリペプトン、2%グルコース、2%寒天)で形質転換体を選択する。
【0037】
実施例4:ビール試験醸造におけるマルトース及びマルトトリオース資化量の測定
親株ならびに実施例3で得られたnonScMPH2高発現株を用いた発酵試験を以下の条件で行う。
麦汁エキス濃度 20%(12%麦汁に糖シロップを添加)
麦汁容量 1L
麦汁溶存酸素濃度 約 8ppm
発酵温度 15℃一定
酵母投入量 5g湿酵母菌体/L麦汁
発酵醪を経時的にサンプリングし、酵母増殖量(OD660)、エキス消費量、グルコース、マルトース、マルトトリオースの経時変化を調べる。
【産業上の利用可能性】
【0038】
本発明の酒類製造法によれば、酵母のマルトースおよびマルトトリオース資化能が増大するため、高濃度醸造においても短期間で酒類を製造することが可能となる。
【図面の簡単な説明】
【0039】
【図1】ビール試験醸造における酵母増殖量の経時変化を示す図である。横軸は発酵時間を、縦軸はOD660の値を示している。
【図2】ビール試験醸造におけるエキス消費量の経時変化を示す図である。横軸は発酵時間、縦軸は外観エキス濃度(w/w%)を示している。
【図3】ビール試験醸造中の酵母におけるnonScMPH2遺伝子の発現挙動を示す図である。横軸は発酵時間、縦軸は検出されたシグナル輝度を示している。
【配列表フリーテキスト】
【0040】
[配列番号:3] プライマー
[配列番号:4] プライマー
【特許請求の範囲】
【請求項1】
以下の(a)〜(f)のいずれかに記載のポリヌクレオチド:
(a)配列番号:1の塩基配列からなるポリヌクレオチドを含有するポリヌクレオチド;
(b)配列番号:2のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(c)配列番号:2のアミノ酸配列において、1もしくは複数個のアミノ酸が欠失、置換、挿入および/または付加したアミノ酸配列からなり、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(d) 配列番号:2のアミノ酸配列に対して60%以上の同一性を有するアミノ酸配列を有し、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(e)配列番号:1の塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;及び
(f)配列番号:2のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドの塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド。
【請求項2】
以下の(g)〜(i)のいずれかである請求項1に記載のポリヌクレオチド:
(g)配列番号:2のアミノ酸配列又は配列番号:2のアミノ酸配列において、1〜10個のアミノ酸が欠失、置換、挿入および/または付加したアミノ酸配列からなり、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(h) 配列番号:2のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列を有し、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;及び
(i)配列番号:1の塩基配列からなるポリヌクレオチド、又は配列番号:1の塩基配列と相補的な塩基配列からなるポリヌクレオチドとハイストリンジェントな条件下でハイブリダイズし、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチドを含有するポリヌクレオチド。
【請求項3】
配列番号:1の塩基配列からなるポリヌクレオチドを含有する請求項1に記載のポリヌクレオチド。
【請求項4】
配列番号:2のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有する請求項1に記載のポリヌクレオチド。
【請求項5】
DNAである、請求項1〜4のいずれかに記載のポリヌクレオチド。
【請求項6】
請求項1〜5のいずれかに記載のポリヌクレオチドにコードされるタンパク質。
【請求項7】
請求項1〜5のいずれかに記載のポリヌクレオチドを含有するベクター。
【請求項8】
請求項7に記載のベクターが導入された形質転換酵母。
【請求項9】
請求項7に記載のベクターを導入することによって、マルトースおよび/またはマルトトリオース資化能が向上した請求項8に記載の醸造用酵母。
【請求項10】
請求項6に記載のタンパク質の発現量を増加させることによって、マルトースおよび/またはマルトトリオース資化能が向上した請求項9に記載の醸造用酵母。
【請求項11】
請求項8から10のいずれかに記載の酵母を用いた酒類の製造方法。
【請求項12】
醸造する酒類が麦芽飲料である請求項11に記載の酒類の製造方法。
【請求項13】
請求項11または12に記載の方法で製造された酒類。
【請求項14】
配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の塩基配列に基づいて設計したプライマーまたはプローブを用いて、被検酵母のマルトースおよびマルトトリオース資化能について評価する方法。
【請求項15】
被検酵母を培養し、配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の発現量を測定することによって、被検酵母のマルトースおよびマルトトリオース資化能を評価する方法。
【請求項16】
基準酵母および被検酵母を培養して配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の各酵母における発現量を測定し、基準酵母よりも該遺伝子が高発現している被検酵母を選択する、酵母の選択方法。
【請求項17】
基準酵母および被検酵母を培養して各酵母における請求項6に記載のタンパク質を定量し、基準酵母よりも該タンパク質量の多い被検酵母を選択する、酵母の選択方法。
【請求項1】
以下の(a)〜(f)のいずれかに記載のポリヌクレオチド:
(a)配列番号:1の塩基配列からなるポリヌクレオチドを含有するポリヌクレオチド;
(b)配列番号:2のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(c)配列番号:2のアミノ酸配列において、1もしくは複数個のアミノ酸が欠失、置換、挿入および/または付加したアミノ酸配列からなり、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(d) 配列番号:2のアミノ酸配列に対して60%以上の同一性を有するアミノ酸配列を有し、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(e)配列番号:1の塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;及び
(f)配列番号:2のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドの塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド。
【請求項2】
以下の(g)〜(i)のいずれかである請求項1に記載のポリヌクレオチド:
(g)配列番号:2のアミノ酸配列又は配列番号:2のアミノ酸配列において、1〜10個のアミノ酸が欠失、置換、挿入および/または付加したアミノ酸配列からなり、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(h) 配列番号:2のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列を有し、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;及び
(i)配列番号:1の塩基配列からなるポリヌクレオチド、又は配列番号:1の塩基配列と相補的な塩基配列からなるポリヌクレオチドとハイストリンジェントな条件下でハイブリダイズし、かつαグルコシドトランスポーター活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチドを含有するポリヌクレオチド。
【請求項3】
配列番号:1の塩基配列からなるポリヌクレオチドを含有する請求項1に記載のポリヌクレオチド。
【請求項4】
配列番号:2のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有する請求項1に記載のポリヌクレオチド。
【請求項5】
DNAである、請求項1〜4のいずれかに記載のポリヌクレオチド。
【請求項6】
請求項1〜5のいずれかに記載のポリヌクレオチドにコードされるタンパク質。
【請求項7】
請求項1〜5のいずれかに記載のポリヌクレオチドを含有するベクター。
【請求項8】
請求項7に記載のベクターが導入された形質転換酵母。
【請求項9】
請求項7に記載のベクターを導入することによって、マルトースおよび/またはマルトトリオース資化能が向上した請求項8に記載の醸造用酵母。
【請求項10】
請求項6に記載のタンパク質の発現量を増加させることによって、マルトースおよび/またはマルトトリオース資化能が向上した請求項9に記載の醸造用酵母。
【請求項11】
請求項8から10のいずれかに記載の酵母を用いた酒類の製造方法。
【請求項12】
醸造する酒類が麦芽飲料である請求項11に記載の酒類の製造方法。
【請求項13】
請求項11または12に記載の方法で製造された酒類。
【請求項14】
配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の塩基配列に基づいて設計したプライマーまたはプローブを用いて、被検酵母のマルトースおよびマルトトリオース資化能について評価する方法。
【請求項15】
被検酵母を培養し、配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の発現量を測定することによって、被検酵母のマルトースおよびマルトトリオース資化能を評価する方法。
【請求項16】
基準酵母および被検酵母を培養して配列番号:1の塩基配列を有するαグルコシドトランスポーター遺伝子の各酵母における発現量を測定し、基準酵母よりも該遺伝子が高発現している被検酵母を選択する、酵母の選択方法。
【請求項17】
基準酵母および被検酵母を培養して各酵母における請求項6に記載のタンパク質を定量し、基準酵母よりも該タンパク質量の多い被検酵母を選択する、酵母の選択方法。
【図1】


【図2】
【図3】
【図2】
【図3】
【公開番号】特開2007−53921(P2007−53921A)
【公開日】平成19年3月8日(2007.3.8)
【国際特許分類】
【出願番号】特願2005−240349(P2005−240349)
【出願日】平成17年8月22日(2005.8.22)
【出願人】(000001904)サントリー株式会社 (319)
【Fターム(参考)】
【公開日】平成19年3月8日(2007.3.8)
【国際特許分類】
【出願日】平成17年8月22日(2005.8.22)
【出願人】(000001904)サントリー株式会社 (319)
【Fターム(参考)】
[ Back to top ]