
β−ガラクトシダーゼ活性のないラクトバシラス・ブルガリカス突然変異株
【課題】β-ガラクトシダーゼをエンコードする遺伝子に欠失がなく、かつ有利な技術特性を有するエル・ブルガリカスの変異体を提供すること。
【解決手段】エル・ブルガリカスのβ-ガラクトシダーゼをエンコードする配列にナンセンス突然変異を有し、β-ガラクトシダーゼ活性を欠いている突然変異株であって、該変異株によって同化される少なくとも1つの糖を乳に補充した場合を除き、由来する野生型株より遅く乳中で成長し酸性化するエル・ブルガリカスの突然変異株(ただし、CNCMに番号I1968で1998年1月14日に寄託された株を除く)。
【解決手段】エル・ブルガリカスのβ-ガラクトシダーゼをエンコードする配列にナンセンス突然変異を有し、β-ガラクトシダーゼ活性を欠いている突然変異株であって、該変異株によって同化される少なくとも1つの糖を乳に補充した場合を除き、由来する野生型株より遅く乳中で成長し酸性化するエル・ブルガリカスの突然変異株(ただし、CNCMに番号I1968で1998年1月14日に寄託された株を除く)。
【発明の詳細な説明】
【技術分野】
【0001】
これらの株と発酵体は、グルコースを補足した乳から発酵乳製品を得るために用いることができる。
【0002】
本発明は、新規なブルガリカス変異株及び発酵乳製品を製造するためのその使用に関する。
【背景技術】
【0003】
ヨーグルトは、従来、ストレプトコッカス・テルモフィリス(Streptococcus thermophilus)とラクトバシラス・ブルガリカス(Lactobacillus bulgaricus)の組合わせで乳を発酵させて得られる。約40〜45℃の温度で発酵を行うあいだ、これらの細菌は、エネルギー基質として主にラクトースを用い、乳を凝固させる乳酸を生産する。pHが約4.8〜4.5の値に達すると、この発酵工程(「酸性化」とも呼ばれる)は、生成物を冷却することによって終結する。この生成物は、次いで残りの製造および包装処理のあいだ、そして消費されるまで冷やされつづける。
【0004】
しかし、冷却は、乳酸の発酵を完全には止めない。生成物が4℃で保持されいる場合でさえ、その酸性は、時間をかけて徐々に増すことが観察されている。
この現象は、後酸性化(postacidification)として公知であり、保存中の生成物の感覚受容の質(organoleptic quality)の低下の原因である。
後酸性化は、本質的に、細菌、主にエル・ブルガリカスが、制御された酸性化工程の終わりに生成物中に残っているラクトースを用いることから生じる。これを回避するために、ラクトースをほとんど又は全く発酵しないエル・ブルガリカス株の使用が提案されている。
【0005】
ラクトースの発酵に必須な酵素の1つは、ラクトースをグルコースとガラクトースに加水分解するβ-ガラクトシダーゼである。したがって、後酸性化しないエル・ブルガリカス株を得るために、この酵素活性が作用する人工的な突然変異体をつくるか、又は天然の突然変異体を選択することが提案されている。
【0006】
例えば、GENENCOR名義の欧州特許EP402450号は、β-ガラクトシダーゼ遺伝子の局在性突然変異によるエル・ブルガリカスの条件的突然変異体の製造を記載しており、ここで、40℃での発酵のあいだ活性なβ-ガラクトシダーゼは、発酵乳製品の保存条件に相当する温度又はpHでその活性を失う。
日本出願JP 90053437号は、ラクトース発酵能を完全に失っているエル・ブルガリカスの人工的な突然変異体の製造と所望のラクトース発酵能を有する天然の突然変異体の選択を記載している。しかし、これらの突然変異体は、培地にグルコースが補われると、ともにエス・テルモフィリスの存在下で正常に発育して酸性化することができる。これらの突然変異体の継代培養は、10継代培養後の乳欠乏グルコース中でもその酸性化特性を保持している。
【0007】
SOCIETE DES PRODUITS NESTLE名義の欧州特許 EP 0518 096号は、β-ガラクトシダーゼ遺伝子フラグメントの欠失を基準としてあらかじめ選択された後酸性化しにくいラクトバシラス・ブルガリカス突然変異体の、ヨーグルトを製造するための使用を提案している。これらの変異体のスクリーニングと特徴づけは、この欠失の有無が制限プロフィルで簡単に確認できるという事実のために容易である。また、この欠失は不可逆な突然変異であることが知られており、そのために、親株から安定的な突然変異株を容易に得ることができる。欧州特許EP 0518 096号は、このようにして選択された後酸性化が弱い突然変異体を2タイプ記載している。1つは、β-ガラクトシダーゼ遺伝子のみに作用する欠失を有する。それらはエス・テルモフィリスと組合わせて乳で培養すると、グルコースを加えていなくても、由来の野生型株に匹敵する成長と酸性化特性を示す。2つめは、β-ガラクトシダーゼ遺伝子の少なくとも1kb下流にわたって大きな欠失を有する。それらはエス・テルモフィリスと組合わすと、由来の野生型株よりもゆっくり成長し、あまり酸性化しない。グルコースの培養培地への添加は、その酸性化及び後酸性化特性にわずかに影響を及ぼすにすぎない。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】欧州特許EP402450号
【特許文献2】日本出願JP90053437号
【特許文献3】欧州特許EP0518096号
【発明の概要】
【発明が解決しようとする課題】
【0009】
β-ガラクトシダーゼが不活性な天然の突然変異体は、欠失変異体の場合よりも点突然変異体の場合に選別しにくく、かつ純粋な培養物として維持しにくい。これは、不活性タンパク質を産生する点突然変異の確立が低く、制限プロフィルを用いて点突然変異を局在化して特徴付けることが非常に難しく、かつ復帰割合が非常に高いためである。
【課題を解決するための手段】
【0010】
出願人は、ここで、β-ガラクトシダーゼをエンコードする遺伝子に欠失がなく、かつ有利な技術特性を有するエル・ブルガリカスの他の天然突然変異体を見出した。本発明に関して、ラクトースを同化できないナンセンス突然変異体は、野生型エル・ブルガリカスの培養物から単離される。乳での培養でエス・テルモフィリスと組合わせると、それは由来の野生型株よりもかなり遅く成長し、酸性化する。逆に、その成長とその酸性化は、乳にグルコースを補った際に、実質的に正常である。
【0011】
本発明の対象は、ラクトースオペロンのコード配列の1つ、特にβ-ガラクトシダーゼをエンコードする配列にナンセンスコドンを導入する突然変異を有することを特徴とする、β-ガラクトシダーゼ活性がないエル・ブルガリカスの突然変異株である。
【0012】
本発明によれば、エル・ブルガリカスの株は、パスツール研究所(25 rue du Docteur Roux, Paris)所有のCNCM (Collection Nationale de Cultures de Microorganisms [微生物培養物の国立コレクション])に、1998年1月14日、番号I1968でブダペスト条約にしたがって寄託された。
この株は、以下の形態学的かつ生化学的特徴を有する:
− 形態:グラム陽性微生物、固定化(immobile)、分離又は短鎖、無胞子、多形、薄状桿菌、
− 代謝:ホモ発酵、カタラーゼ(-)
− 糖の発酵:D-グルコース(+)、Dフルクトース(+)、D-マンノース(+)、エスクリン(esculine) (+)
【0013】
発明者らは、I1968突然変異体中のラクトースオペロンをシークエンスした。相当する配列を、番号SEQ ID No: 1で添付の配列リストに示す。翻訳産物(パーミアーゼ及びβ-ガラクトシダーゼ)の配列は、それぞれ番号SEQ ID No: 2 及びSEQ ID No: 3に示す。
この配列の解析によれば、2つの点突然変異が示されている:一方は、パーミアーゼ遺伝子中(配列SEQ ID No: 1の122位置)で、アミノ酸の変更を誘導し (Lys → Asn); 他方は、β-ガラクトシダーゼ遺伝子中(配列SEQ ID No: 1の4519位置)で、停止コドンを誘導する。その活性部位(464及び 531位置)は保存されているが、この突然変異体で産生されるβ-ガラクトシダーゼは不活性である。また、発明者らは、この突然変異が、グルコースを含有する培養培地での何代かの継代培養後に依然として安定であることを見出している。他方、グルコースのない培養培地で、このナンセンス突然変異は、約106の割合で非常に迅速に復帰する。
【0014】
本発明は、ラクトースを同化できず、かつI1968株から誘導される突然変異株も包含している。このような株は、例えばI1968株のラクトースオペロンに他の突然変異を部位特異的突然変異で誘導することによって、得ることができる。
【0015】
また、本発明の対象は、上記の本発明による少なくとも1つのエル・ブルガリカス株を含み、好ましくは少なくとも1つのエス・テルモフィリス株との組み合わせからなることを特徴とする、乳酸発酵体、特にヨーグルト発酵体である。
本発明による発酵体の製造には、ヨーグルトの製造に適したいずれのエス・テルモフィリス株も使用することができる。1以上のエス・テルモフィリス株は、最終製品に任意に付与されることが望まれる付加的な特徴を目的として選択することができる。
【0016】
一例として、本発明によるエル・ブルガリカス株と組合わせて使用できるエス・テルモフィリス株には、パスツール研究所(25 rue du Docteur Roux, Paris)所有のCNCM (Collection Nationale de Cultures de Microorganisms [微生物培養物の国立コレクション])に寄託された以下の株が挙げられる:
− 番号I1470で1994年8月25日に寄託された株、及び番号I-1620で1995年8月23日に寄託された株;これらの2株は、番号96/06924で欧州特許公報に記載されている;
− 番号I1520 及びI1521で1994年12月30日に寄託された株;これらの2株はPCT国際出願WO 96/20607号に記載されている;
− 番号I1630で1995年10月24日に寄託された株;この株の特徴は、PCT国際出願WO 96/01701号に記載されている。
【0017】
これらの株は、相互に、又は1以上の他の工業的なエス・テルモフィリス株と組合わせてもよい。
エス・テルモフィリス株は、従来のヨーグルト発酵と同じ方法及び同じ割合で、本発明によるエル・ブルガリカス株と組合わされる。本発明によるエル・ブルガリカス細菌群は、例えば細菌群全体の10〜90%、好ましくは20〜50%を占めていてもよい。
【0018】
また、本発明の対象は、本発明による少なくとも一つのエル・ブルガリカス株からなる発酵体を用いて、該株によって同化される少なくとも一つの糖の存在下で乳が発酵される工程からなることを特徴とする、発酵乳製品の製造法である。糖は詳しくはフルクトース、マンノースであってもよく、グルコースであることが好ましい。有利には、発酵乳製品はヨーグルトである。
本発明の方法は、酸性化工程を制御する主要な実施方法に関して、ヨーグルトの従来の製造法と同様である。特に、この酸性化は、20〜45℃、好ましくは30〜45℃の温度で「バッチワイズ」、つまり一つの発酵タンクを使用する単一工程で行われる。
【0019】
この制御された酸性化工程の期間は一般に約6〜24時間、好ましくは約6〜16時間である。それ故、期間は、ヨーグルトの従来の製造法の場合より長い(ヨーグルトでは44℃で3〜5時間)。詳しくは、本発明によるエル・ブルガリカス株は、エス・テルモフィリスと組合わせても、野生型株よりもかなり遅く成長し、酸性化する。
また、本発明によるエル・ブルガリカス株の成長速度と酸性化は、乳に加えられるグルコース量によって極めて著しく変わる。この性質は、発酵の最初に所望のグルコース量を単に加えることによって、その成長と酸性化を制御することができる。
【0020】
また、発明者らは、本発明によるエル・ブルガリカス株又は発酵体を用いる際に、pHが4.8〜4.5の範囲(これは従来法の場合では酸性化が停止するpH範囲に相当する)に達すると酸性化がかなり減速し、乳が発酵温度で維持されていたとしても、最小pHで安定化することを観察している。この最小pH値は、本質的に加えたグルコース量に依存している。
この特性により、ヨーグルトを製造する従来法で用いられた発酵を止めるための冷却段階を縮小、又は削除しさえすることができる。また、pHを測定して、発酵の停止に最適な時機を決定する必要がない。所定の発酵体と加えたグルコース量に対しては、過剰な酸性化の危険性なしに、最小pHに達するのに必要な時間の関数として算出される所定の期間の終わりに発酵を停止することができる。このため、発酵の終わりに最終pHの調和及び生成物のテキスチャーをより良く制御することができる。
【0021】
本発明の方法を実行するには、発酵前に乳に加えられるグルコース量は、望ましい酸性化の程度に応じて0.5〜10g/l、好ましくは0.5〜5g/lであることが有利である。
こうして得られる発酵生成物は、pHを低下させずに発酵温度に近い温度で数時間保存することができ、これにより、介在する冷蔵保存用装置を除き、発酵タンクの容量を増やすことができる。
【0022】
本発明の方法の実行により、長期保存中の発酵生成物における後酸性化を減じることができる。後酸性化の程度は、用いた発酵体組成物とグルコース量に応じて変化しうる。しかし、後酸性化は、従来の発酵体と方法で得られるヨーグルトの場合に見られるよりも常に明らかに低い。
例えば、発明者らにより行われた実験は、同じ保存条件(10℃で28日保存)下で、ΔpH(D0でのpHとD28でのpHの違い)が、本発明による発酵体を用いて得た生成物の場合には0.05〜0.4であるが、本発明によるエル・ブルガリカス株が野生型株で置換されている対照の発酵体の場合には常に0.7より高いことが示されている。
【0023】
この弱い後酸性化は、発酵株の良好な生存を伴う。保存の終わりに、本発明によって得られる発酵生成物中のエル・ブルガリカス群は、対照生成物中の群よりわずかに少ないにすぎない。
また、本発明の対象は、本発明の方法を実施して得られる発酵乳製品である。
これらの製品は、従来法を用いて得られる製品よりも長期間かつ高温で保存でき、保存のあいだ依然として安定した官能特性を有している。
【図面の簡単な説明】
【0024】
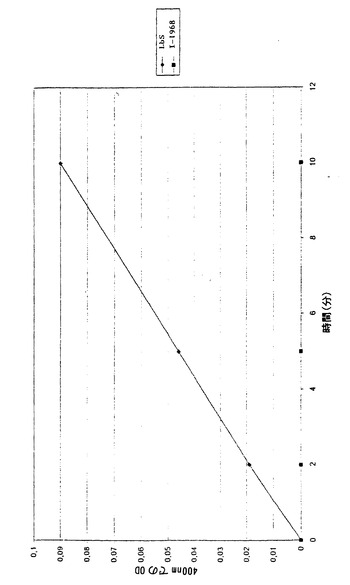
【図1】時間の関数として測定したLbS親株と本発明によるI-1968突然変異体のガラクトシダーゼ活性を示す。
【実施例】
【0025】
実施例1:本発明による突然変異体のβ-ガラクトシダーゼ活性に関する生化学的アッセイ
I-1968株のβ-ガラクトシダーゼ活性を、由来のエル・ブルガリカス野生株(以降、LbSとする)の活性と比較した。
細菌は、酸素定着剤(AnaerocultA, MERCK)の存在下、嫌気性ジャー(MERCK)中で、37℃でMRS寒天培地(MERCK)で一晩培養する。
【0026】
細菌10μlのループ(NUNC)を、滅菌水1mlに懸濁する。細菌は、ガラスのマイクロビーズ(直径0.5mm、BIOSPEC PRODUCTS)の存在下で1分当たり5000回転で20秒の激しい攪拌を2サイクルして溶解し、次いでクロロホルム0.15mlを加える。混合物は37℃で30分振盪し、4℃で滅菌水を用いて容量を2mlにする。次いで、β-ガラクトシダーゼ活性を測定する。細胞懸濁液0.2mlに、0.067M NaH2PO4緩衝液(pH 6.8) 1.2ml、t0でL-システイン(SIGMA) 0.05ml、O-ニトロフェニル-β-D-ガラクトピラノシド(SIGMA) 0.05mlを加える。0、2、5又は10分後に10% Na2CO3緩衝液1mlで酵素反応を停止し、反応培地を遠心分離後に、400nmでのODを上清について測定する。
【0027】
時間の関数として測定したLbS親株と本発明によるI-1968突然変異体のガラクトシダーゼ活性を、図1に示す。
これらの結果は、β-ガラクトシダーゼが、本発明による突然変異体において完全に不活性であることを示している。
【0028】
実施例2:エル・ブルガリカスI-1968突然変異株の安定性
I-1968突然変異株の安定性は、グルコースとラクトースの混合物又はラクトースのみのいずれかを炭素源として含有する培地で試験した。
グルコース含有MRS培地で得られたI-1968培養物は、酵母自己分解物(2g/l)を補足し、グルコース(20g/l)を補足しているか、もしくは補足していない殺菌乳で継代培養する。pHが5.2(乳の凝固)に達したら、各継代培養の試料を採取し、それについて、糖を発酵する細菌の能力ならびにβ-ガラクトシダーゼ活性の有無(X-galプレートアッセイ:白色コロニー=β-ガラクトシダーゼマイナス;青色コロニー=β-ガラクトシダーゼプラス)を分析した。
結果を表1に示す。
【0029】
【表1】

【0030】
これらの結果は、グルコースの存在下で、I-1968株がラクトースを使用できる株に復帰しないことを示している。逆に、唯一の炭素源としてラクトースを含有する培地では、I-1968株の本来の状態への急速な復帰が認められる。
【0031】
実施例3:エス・テルモフィリスと共生しているエル・ブルガリカスI-1968変異株の酸性化、後酸性化及び生存特性:一連のヨーグルトを製造する一方法(通気オーブン中での発酵)
本発明によるI-1968株をエス・テルモフィリスの種々の工業的な株と組合わせて、ヨーグルト発酵体を製造する(使用したエス・テルモフィリス株は、以降ST1、ST2及びST3とする)。
比較として、LbS親株を同株のエス・テルモフィリスと組合わせて、発酵体を製造する。
発酵体の製造には、以下の組成につき個別に1%で株を接種する。
【0032】
1リットルの組成:
スキムミルクパウダー 135g
酵母自己分解物 2g
蒸留水 920ml
グルコース (I-1968株のみ) 20g
水和: 10分
低温殺菌: 95℃で30分
次いで、乳を44℃に冷却して接種し、連鎖状球菌に対する85°D(ドルニック(Dornic)度)及び乳酸桿菌に対する80°Dの酸性が得られるまで、44℃で培養する。
【0033】
次に、培養物を冷却し、ストレプトコッカス・テルモフィリス80%とラクトバシラス・ブルガリカス20%からなる発酵体を得る。
こうして得られた発酵体を、以下の製品に接種するのに用いる。
1リットルの組成:
乳 99%
グルコース 0、1又は2g/l
水和: 10分
低温殺菌: 95℃で10分
次いで、乳を44℃に冷却し、1%で接種する。
各実験について、発酵体の組成と加えたグルコース量を、下記の表2に示す。
【0034】
【表2】
【0035】
接種後、乳を丸底フラスコに分配し、44℃の温度で培養する。培養のあいだ、酸性化プロフィルをモニターする。生成物は、冷蔵機で冷却(4℃で16時間)することによってpH4.6で凝固しない。
次いで、生成物を10分で保存試験に付す。この試験で、保存から1、14、21及び28日後にpHとドルニック酸性を測定する。
酸性化の結果(pH 4.6に達するまでの時間と24時間でのpH値)を以下の表3に示す。
【0036】
【表3】
【0037】
10℃での保存試験(pHとドルニック酸性の測定)と28日での生存試験(エス・テルモフィリス及びエル・ブルガリカスの群)の結果を、下記の表4に示す。
【0038】
【表4】
【0039】
これらの結果は、I-1968株を1以上のエス・テルモフィリス株と組合わせた共生を用いて製造したヨーグルトは、LbS親株との同じ共生に対して後酸性化をかなり減じるが、同時に、発酵の終わりに豊富な群を保持し、10℃で28日間良好に生存していることを示している。
攪拌ヨーグルト(stirred yogurt)を製造する場合には、酸性化を止め、44℃で少なくとも24時間約4.6〜4.5のpHを維持することによって、従来用いられているタンク中での冷却段階を縮小するか、または除くことができる。
【技術分野】
【0001】
これらの株と発酵体は、グルコースを補足した乳から発酵乳製品を得るために用いることができる。
【0002】
本発明は、新規なブルガリカス変異株及び発酵乳製品を製造するためのその使用に関する。
【背景技術】
【0003】
ヨーグルトは、従来、ストレプトコッカス・テルモフィリス(Streptococcus thermophilus)とラクトバシラス・ブルガリカス(Lactobacillus bulgaricus)の組合わせで乳を発酵させて得られる。約40〜45℃の温度で発酵を行うあいだ、これらの細菌は、エネルギー基質として主にラクトースを用い、乳を凝固させる乳酸を生産する。pHが約4.8〜4.5の値に達すると、この発酵工程(「酸性化」とも呼ばれる)は、生成物を冷却することによって終結する。この生成物は、次いで残りの製造および包装処理のあいだ、そして消費されるまで冷やされつづける。
【0004】
しかし、冷却は、乳酸の発酵を完全には止めない。生成物が4℃で保持されいる場合でさえ、その酸性は、時間をかけて徐々に増すことが観察されている。
この現象は、後酸性化(postacidification)として公知であり、保存中の生成物の感覚受容の質(organoleptic quality)の低下の原因である。
後酸性化は、本質的に、細菌、主にエル・ブルガリカスが、制御された酸性化工程の終わりに生成物中に残っているラクトースを用いることから生じる。これを回避するために、ラクトースをほとんど又は全く発酵しないエル・ブルガリカス株の使用が提案されている。
【0005】
ラクトースの発酵に必須な酵素の1つは、ラクトースをグルコースとガラクトースに加水分解するβ-ガラクトシダーゼである。したがって、後酸性化しないエル・ブルガリカス株を得るために、この酵素活性が作用する人工的な突然変異体をつくるか、又は天然の突然変異体を選択することが提案されている。
【0006】
例えば、GENENCOR名義の欧州特許EP402450号は、β-ガラクトシダーゼ遺伝子の局在性突然変異によるエル・ブルガリカスの条件的突然変異体の製造を記載しており、ここで、40℃での発酵のあいだ活性なβ-ガラクトシダーゼは、発酵乳製品の保存条件に相当する温度又はpHでその活性を失う。
日本出願JP 90053437号は、ラクトース発酵能を完全に失っているエル・ブルガリカスの人工的な突然変異体の製造と所望のラクトース発酵能を有する天然の突然変異体の選択を記載している。しかし、これらの突然変異体は、培地にグルコースが補われると、ともにエス・テルモフィリスの存在下で正常に発育して酸性化することができる。これらの突然変異体の継代培養は、10継代培養後の乳欠乏グルコース中でもその酸性化特性を保持している。
【0007】
SOCIETE DES PRODUITS NESTLE名義の欧州特許 EP 0518 096号は、β-ガラクトシダーゼ遺伝子フラグメントの欠失を基準としてあらかじめ選択された後酸性化しにくいラクトバシラス・ブルガリカス突然変異体の、ヨーグルトを製造するための使用を提案している。これらの変異体のスクリーニングと特徴づけは、この欠失の有無が制限プロフィルで簡単に確認できるという事実のために容易である。また、この欠失は不可逆な突然変異であることが知られており、そのために、親株から安定的な突然変異株を容易に得ることができる。欧州特許EP 0518 096号は、このようにして選択された後酸性化が弱い突然変異体を2タイプ記載している。1つは、β-ガラクトシダーゼ遺伝子のみに作用する欠失を有する。それらはエス・テルモフィリスと組合わせて乳で培養すると、グルコースを加えていなくても、由来の野生型株に匹敵する成長と酸性化特性を示す。2つめは、β-ガラクトシダーゼ遺伝子の少なくとも1kb下流にわたって大きな欠失を有する。それらはエス・テルモフィリスと組合わすと、由来の野生型株よりもゆっくり成長し、あまり酸性化しない。グルコースの培養培地への添加は、その酸性化及び後酸性化特性にわずかに影響を及ぼすにすぎない。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】欧州特許EP402450号
【特許文献2】日本出願JP90053437号
【特許文献3】欧州特許EP0518096号
【発明の概要】
【発明が解決しようとする課題】
【0009】
β-ガラクトシダーゼが不活性な天然の突然変異体は、欠失変異体の場合よりも点突然変異体の場合に選別しにくく、かつ純粋な培養物として維持しにくい。これは、不活性タンパク質を産生する点突然変異の確立が低く、制限プロフィルを用いて点突然変異を局在化して特徴付けることが非常に難しく、かつ復帰割合が非常に高いためである。
【課題を解決するための手段】
【0010】
出願人は、ここで、β-ガラクトシダーゼをエンコードする遺伝子に欠失がなく、かつ有利な技術特性を有するエル・ブルガリカスの他の天然突然変異体を見出した。本発明に関して、ラクトースを同化できないナンセンス突然変異体は、野生型エル・ブルガリカスの培養物から単離される。乳での培養でエス・テルモフィリスと組合わせると、それは由来の野生型株よりもかなり遅く成長し、酸性化する。逆に、その成長とその酸性化は、乳にグルコースを補った際に、実質的に正常である。
【0011】
本発明の対象は、ラクトースオペロンのコード配列の1つ、特にβ-ガラクトシダーゼをエンコードする配列にナンセンスコドンを導入する突然変異を有することを特徴とする、β-ガラクトシダーゼ活性がないエル・ブルガリカスの突然変異株である。
【0012】
本発明によれば、エル・ブルガリカスの株は、パスツール研究所(25 rue du Docteur Roux, Paris)所有のCNCM (Collection Nationale de Cultures de Microorganisms [微生物培養物の国立コレクション])に、1998年1月14日、番号I1968でブダペスト条約にしたがって寄託された。
この株は、以下の形態学的かつ生化学的特徴を有する:
− 形態:グラム陽性微生物、固定化(immobile)、分離又は短鎖、無胞子、多形、薄状桿菌、
− 代謝:ホモ発酵、カタラーゼ(-)
− 糖の発酵:D-グルコース(+)、Dフルクトース(+)、D-マンノース(+)、エスクリン(esculine) (+)
【0013】
発明者らは、I1968突然変異体中のラクトースオペロンをシークエンスした。相当する配列を、番号SEQ ID No: 1で添付の配列リストに示す。翻訳産物(パーミアーゼ及びβ-ガラクトシダーゼ)の配列は、それぞれ番号SEQ ID No: 2 及びSEQ ID No: 3に示す。
この配列の解析によれば、2つの点突然変異が示されている:一方は、パーミアーゼ遺伝子中(配列SEQ ID No: 1の122位置)で、アミノ酸の変更を誘導し (Lys → Asn); 他方は、β-ガラクトシダーゼ遺伝子中(配列SEQ ID No: 1の4519位置)で、停止コドンを誘導する。その活性部位(464及び 531位置)は保存されているが、この突然変異体で産生されるβ-ガラクトシダーゼは不活性である。また、発明者らは、この突然変異が、グルコースを含有する培養培地での何代かの継代培養後に依然として安定であることを見出している。他方、グルコースのない培養培地で、このナンセンス突然変異は、約106の割合で非常に迅速に復帰する。
【0014】
本発明は、ラクトースを同化できず、かつI1968株から誘導される突然変異株も包含している。このような株は、例えばI1968株のラクトースオペロンに他の突然変異を部位特異的突然変異で誘導することによって、得ることができる。
【0015】
また、本発明の対象は、上記の本発明による少なくとも1つのエル・ブルガリカス株を含み、好ましくは少なくとも1つのエス・テルモフィリス株との組み合わせからなることを特徴とする、乳酸発酵体、特にヨーグルト発酵体である。
本発明による発酵体の製造には、ヨーグルトの製造に適したいずれのエス・テルモフィリス株も使用することができる。1以上のエス・テルモフィリス株は、最終製品に任意に付与されることが望まれる付加的な特徴を目的として選択することができる。
【0016】
一例として、本発明によるエル・ブルガリカス株と組合わせて使用できるエス・テルモフィリス株には、パスツール研究所(25 rue du Docteur Roux, Paris)所有のCNCM (Collection Nationale de Cultures de Microorganisms [微生物培養物の国立コレクション])に寄託された以下の株が挙げられる:
− 番号I1470で1994年8月25日に寄託された株、及び番号I-1620で1995年8月23日に寄託された株;これらの2株は、番号96/06924で欧州特許公報に記載されている;
− 番号I1520 及びI1521で1994年12月30日に寄託された株;これらの2株はPCT国際出願WO 96/20607号に記載されている;
− 番号I1630で1995年10月24日に寄託された株;この株の特徴は、PCT国際出願WO 96/01701号に記載されている。
【0017】
これらの株は、相互に、又は1以上の他の工業的なエス・テルモフィリス株と組合わせてもよい。
エス・テルモフィリス株は、従来のヨーグルト発酵と同じ方法及び同じ割合で、本発明によるエル・ブルガリカス株と組合わされる。本発明によるエル・ブルガリカス細菌群は、例えば細菌群全体の10〜90%、好ましくは20〜50%を占めていてもよい。
【0018】
また、本発明の対象は、本発明による少なくとも一つのエル・ブルガリカス株からなる発酵体を用いて、該株によって同化される少なくとも一つの糖の存在下で乳が発酵される工程からなることを特徴とする、発酵乳製品の製造法である。糖は詳しくはフルクトース、マンノースであってもよく、グルコースであることが好ましい。有利には、発酵乳製品はヨーグルトである。
本発明の方法は、酸性化工程を制御する主要な実施方法に関して、ヨーグルトの従来の製造法と同様である。特に、この酸性化は、20〜45℃、好ましくは30〜45℃の温度で「バッチワイズ」、つまり一つの発酵タンクを使用する単一工程で行われる。
【0019】
この制御された酸性化工程の期間は一般に約6〜24時間、好ましくは約6〜16時間である。それ故、期間は、ヨーグルトの従来の製造法の場合より長い(ヨーグルトでは44℃で3〜5時間)。詳しくは、本発明によるエル・ブルガリカス株は、エス・テルモフィリスと組合わせても、野生型株よりもかなり遅く成長し、酸性化する。
また、本発明によるエル・ブルガリカス株の成長速度と酸性化は、乳に加えられるグルコース量によって極めて著しく変わる。この性質は、発酵の最初に所望のグルコース量を単に加えることによって、その成長と酸性化を制御することができる。
【0020】
また、発明者らは、本発明によるエル・ブルガリカス株又は発酵体を用いる際に、pHが4.8〜4.5の範囲(これは従来法の場合では酸性化が停止するpH範囲に相当する)に達すると酸性化がかなり減速し、乳が発酵温度で維持されていたとしても、最小pHで安定化することを観察している。この最小pH値は、本質的に加えたグルコース量に依存している。
この特性により、ヨーグルトを製造する従来法で用いられた発酵を止めるための冷却段階を縮小、又は削除しさえすることができる。また、pHを測定して、発酵の停止に最適な時機を決定する必要がない。所定の発酵体と加えたグルコース量に対しては、過剰な酸性化の危険性なしに、最小pHに達するのに必要な時間の関数として算出される所定の期間の終わりに発酵を停止することができる。このため、発酵の終わりに最終pHの調和及び生成物のテキスチャーをより良く制御することができる。
【0021】
本発明の方法を実行するには、発酵前に乳に加えられるグルコース量は、望ましい酸性化の程度に応じて0.5〜10g/l、好ましくは0.5〜5g/lであることが有利である。
こうして得られる発酵生成物は、pHを低下させずに発酵温度に近い温度で数時間保存することができ、これにより、介在する冷蔵保存用装置を除き、発酵タンクの容量を増やすことができる。
【0022】
本発明の方法の実行により、長期保存中の発酵生成物における後酸性化を減じることができる。後酸性化の程度は、用いた発酵体組成物とグルコース量に応じて変化しうる。しかし、後酸性化は、従来の発酵体と方法で得られるヨーグルトの場合に見られるよりも常に明らかに低い。
例えば、発明者らにより行われた実験は、同じ保存条件(10℃で28日保存)下で、ΔpH(D0でのpHとD28でのpHの違い)が、本発明による発酵体を用いて得た生成物の場合には0.05〜0.4であるが、本発明によるエル・ブルガリカス株が野生型株で置換されている対照の発酵体の場合には常に0.7より高いことが示されている。
【0023】
この弱い後酸性化は、発酵株の良好な生存を伴う。保存の終わりに、本発明によって得られる発酵生成物中のエル・ブルガリカス群は、対照生成物中の群よりわずかに少ないにすぎない。
また、本発明の対象は、本発明の方法を実施して得られる発酵乳製品である。
これらの製品は、従来法を用いて得られる製品よりも長期間かつ高温で保存でき、保存のあいだ依然として安定した官能特性を有している。
【図面の簡単な説明】
【0024】
【図1】時間の関数として測定したLbS親株と本発明によるI-1968突然変異体のガラクトシダーゼ活性を示す。
【実施例】
【0025】
実施例1:本発明による突然変異体のβ-ガラクトシダーゼ活性に関する生化学的アッセイ
I-1968株のβ-ガラクトシダーゼ活性を、由来のエル・ブルガリカス野生株(以降、LbSとする)の活性と比較した。
細菌は、酸素定着剤(AnaerocultA, MERCK)の存在下、嫌気性ジャー(MERCK)中で、37℃でMRS寒天培地(MERCK)で一晩培養する。
【0026】
細菌10μlのループ(NUNC)を、滅菌水1mlに懸濁する。細菌は、ガラスのマイクロビーズ(直径0.5mm、BIOSPEC PRODUCTS)の存在下で1分当たり5000回転で20秒の激しい攪拌を2サイクルして溶解し、次いでクロロホルム0.15mlを加える。混合物は37℃で30分振盪し、4℃で滅菌水を用いて容量を2mlにする。次いで、β-ガラクトシダーゼ活性を測定する。細胞懸濁液0.2mlに、0.067M NaH2PO4緩衝液(pH 6.8) 1.2ml、t0でL-システイン(SIGMA) 0.05ml、O-ニトロフェニル-β-D-ガラクトピラノシド(SIGMA) 0.05mlを加える。0、2、5又は10分後に10% Na2CO3緩衝液1mlで酵素反応を停止し、反応培地を遠心分離後に、400nmでのODを上清について測定する。
【0027】
時間の関数として測定したLbS親株と本発明によるI-1968突然変異体のガラクトシダーゼ活性を、図1に示す。
これらの結果は、β-ガラクトシダーゼが、本発明による突然変異体において完全に不活性であることを示している。
【0028】
実施例2:エル・ブルガリカスI-1968突然変異株の安定性
I-1968突然変異株の安定性は、グルコースとラクトースの混合物又はラクトースのみのいずれかを炭素源として含有する培地で試験した。
グルコース含有MRS培地で得られたI-1968培養物は、酵母自己分解物(2g/l)を補足し、グルコース(20g/l)を補足しているか、もしくは補足していない殺菌乳で継代培養する。pHが5.2(乳の凝固)に達したら、各継代培養の試料を採取し、それについて、糖を発酵する細菌の能力ならびにβ-ガラクトシダーゼ活性の有無(X-galプレートアッセイ:白色コロニー=β-ガラクトシダーゼマイナス;青色コロニー=β-ガラクトシダーゼプラス)を分析した。
結果を表1に示す。
【0029】
【表1】
【0030】
これらの結果は、グルコースの存在下で、I-1968株がラクトースを使用できる株に復帰しないことを示している。逆に、唯一の炭素源としてラクトースを含有する培地では、I-1968株の本来の状態への急速な復帰が認められる。
【0031】
実施例3:エス・テルモフィリスと共生しているエル・ブルガリカスI-1968変異株の酸性化、後酸性化及び生存特性:一連のヨーグルトを製造する一方法(通気オーブン中での発酵)
本発明によるI-1968株をエス・テルモフィリスの種々の工業的な株と組合わせて、ヨーグルト発酵体を製造する(使用したエス・テルモフィリス株は、以降ST1、ST2及びST3とする)。
比較として、LbS親株を同株のエス・テルモフィリスと組合わせて、発酵体を製造する。
発酵体の製造には、以下の組成につき個別に1%で株を接種する。
【0032】
1リットルの組成:
スキムミルクパウダー 135g
酵母自己分解物 2g
蒸留水 920ml
グルコース (I-1968株のみ) 20g
水和: 10分
低温殺菌: 95℃で30分
次いで、乳を44℃に冷却して接種し、連鎖状球菌に対する85°D(ドルニック(Dornic)度)及び乳酸桿菌に対する80°Dの酸性が得られるまで、44℃で培養する。
【0033】
次に、培養物を冷却し、ストレプトコッカス・テルモフィリス80%とラクトバシラス・ブルガリカス20%からなる発酵体を得る。
こうして得られた発酵体を、以下の製品に接種するのに用いる。
1リットルの組成:
乳 99%
グルコース 0、1又は2g/l
水和: 10分
低温殺菌: 95℃で10分
次いで、乳を44℃に冷却し、1%で接種する。
各実験について、発酵体の組成と加えたグルコース量を、下記の表2に示す。
【0034】
【表2】
【0035】
接種後、乳を丸底フラスコに分配し、44℃の温度で培養する。培養のあいだ、酸性化プロフィルをモニターする。生成物は、冷蔵機で冷却(4℃で16時間)することによってpH4.6で凝固しない。
次いで、生成物を10分で保存試験に付す。この試験で、保存から1、14、21及び28日後にpHとドルニック酸性を測定する。
酸性化の結果(pH 4.6に達するまでの時間と24時間でのpH値)を以下の表3に示す。
【0036】
【表3】
【0037】
10℃での保存試験(pHとドルニック酸性の測定)と28日での生存試験(エス・テルモフィリス及びエル・ブルガリカスの群)の結果を、下記の表4に示す。
【0038】
【表4】
【0039】
これらの結果は、I-1968株を1以上のエス・テルモフィリス株と組合わせた共生を用いて製造したヨーグルトは、LbS親株との同じ共生に対して後酸性化をかなり減じるが、同時に、発酵の終わりに豊富な群を保持し、10℃で28日間良好に生存していることを示している。
攪拌ヨーグルト(stirred yogurt)を製造する場合には、酸性化を止め、44℃で少なくとも24時間約4.6〜4.5のpHを維持することによって、従来用いられているタンク中での冷却段階を縮小するか、または除くことができる。
【特許請求の範囲】
【請求項1】
エル・ブルガリカスのβ-ガラクトシダーゼをエンコードする配列にナンセンス突然変異を有し、β-ガラクトシダーゼ活性を欠いている突然変異株であって、該変異株によって同化される少なくとも1つの糖を乳に補充した場合を除き、由来する野生型株より遅く乳中で成長し酸性化するエル・ブルガリカスの突然変異株(ただし、CNCMに番号I1968で1998年1月14日に寄託された株を除く)。
【請求項2】
同化される糖がグルコースである請求項1に記載のエル・ブルガリカスの突然変異株。
【請求項3】
該変異株によって同化される少なくとも1つの糖を乳に補充した場合を除き、由来する野生型株より少なくとも2倍遅く乳中で成長し酸性化する請求項1又は2に記載のエル・ブルガリカスの突然変異株。
【請求項4】
請求項1〜3のいずれか1つ記載の少なくとも1つのエル・ブルガリカス株を含んでなる乳酸発酵体。
【請求項5】
エル・ブルガリカス株が、少なくとも1つのエス・テルモフィリス株と組み合わされている請求項4に記載の乳酸発酵体。
【請求項6】
請求項4に記載の乳酸発酵体を用いて、前記変異株によって同化される少なくとも1つの糖の存在下で乳を発酵させる工程を含んでなる発酵乳製品の製造方法。
【請求項7】
同化される糖がグルコースである請求項6に記載の方法。
【請求項8】
発酵の抑制が乳製品を冷却しないで行われる請求項6又は7に記載の方法。
【請求項9】
請求項6〜8のいずれか1つに記載の方法を用いて得られる発酵乳製品。
【請求項10】
製品がヨーグルトである請求項9に記載の発酵乳製品。
【請求項1】
エル・ブルガリカスのβ-ガラクトシダーゼをエンコードする配列にナンセンス突然変異を有し、β-ガラクトシダーゼ活性を欠いている突然変異株であって、該変異株によって同化される少なくとも1つの糖を乳に補充した場合を除き、由来する野生型株より遅く乳中で成長し酸性化するエル・ブルガリカスの突然変異株(ただし、CNCMに番号I1968で1998年1月14日に寄託された株を除く)。
【請求項2】
同化される糖がグルコースである請求項1に記載のエル・ブルガリカスの突然変異株。
【請求項3】
該変異株によって同化される少なくとも1つの糖を乳に補充した場合を除き、由来する野生型株より少なくとも2倍遅く乳中で成長し酸性化する請求項1又は2に記載のエル・ブルガリカスの突然変異株。
【請求項4】
請求項1〜3のいずれか1つ記載の少なくとも1つのエル・ブルガリカス株を含んでなる乳酸発酵体。
【請求項5】
エル・ブルガリカス株が、少なくとも1つのエス・テルモフィリス株と組み合わされている請求項4に記載の乳酸発酵体。
【請求項6】
請求項4に記載の乳酸発酵体を用いて、前記変異株によって同化される少なくとも1つの糖の存在下で乳を発酵させる工程を含んでなる発酵乳製品の製造方法。
【請求項7】
同化される糖がグルコースである請求項6に記載の方法。
【請求項8】
発酵の抑制が乳製品を冷却しないで行われる請求項6又は7に記載の方法。
【請求項9】
請求項6〜8のいずれか1つに記載の方法を用いて得られる発酵乳製品。
【請求項10】
製品がヨーグルトである請求項9に記載の発酵乳製品。
【図1】

【公開番号】特開2010−166932(P2010−166932A)
【公開日】平成22年8月5日(2010.8.5)
【国際特許分類】
【出願番号】特願2010−109593(P2010−109593)
【出願日】平成22年5月11日(2010.5.11)
【分割の表示】特願2000−551011(P2000−551011)の分割
【原出願日】平成11年5月17日(1999.5.17)
【出願人】(500223925)
【氏名又は名称原語表記】COMPAGNIE GERVAIS DANONE
【Fターム(参考)】
【公開日】平成22年8月5日(2010.8.5)
【国際特許分類】
【出願日】平成22年5月11日(2010.5.11)
【分割の表示】特願2000−551011(P2000−551011)の分割
【原出願日】平成11年5月17日(1999.5.17)
【出願人】(500223925)
【氏名又は名称原語表記】COMPAGNIE GERVAIS DANONE
【Fターム(参考)】
[ Back to top ]