
β−グルカナーゼ及びキシラナーゼの製造方法及び液体培地
【課題】キシランを含むセルロース資源の分解能力に優れたセルラーゼを低コストで製造すること。
【解決手段】(a)炭素源として加熱処理もアルカリ処理も行わない紙類由来のパルプ、及び(b)窒素源としてアンモニア態窒素またはアミノ態窒素を含む液体培地を用いて、トリコデルマ属に属する微生物を培養する工程を包含するβ−グルカナーゼ及びキシラナーゼの製造方法。
【解決手段】(a)炭素源として加熱処理もアルカリ処理も行わない紙類由来のパルプ、及び(b)窒素源としてアンモニア態窒素またはアミノ態窒素を含む液体培地を用いて、トリコデルマ属に属する微生物を培養する工程を包含するβ−グルカナーゼ及びキシラナーゼの製造方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はβ−グルカナーゼ及びキシラナーゼの製造方法及び液体培地に関する。
【背景技術】
【0002】
セルロース資源を有効利用するために、近年、セルロースを効率的に分解する方法が探索されている。自然界ではセルロースは主として微生物によって分解されており、細菌や糸状菌などの様々な微生物がセルロース分解酵素を生産することが知られている。
【0003】
これらの微生物は菌体外にセルロース分解酵素を分泌し、セルロースはその作用により、主に、セロオリゴ糖、セロビオースを経てグルコースへと分解される。セルロース分解酵素は、一般に、セルラーゼと呼ばれている。
【0004】
人工的にセルラーゼを製造しようとする場合、セルラーゼを分泌する微生物としては、トリコデルマ属が知られており、広く利用されている。そして、トリコデルマ属に属する微生物を炭素源及び窒素源などの栄養を含む培地を用いて培養し、セルラーゼを分泌させる方法も知られている。
【0005】
しかしながら、セルラーゼを製造するための従来の方法は、炭素源として使用できる材料に制限があり、高価な結晶セルロースであったり、仮に安価なセルロース資源であっても、加熱処理やアルカリ処理などの前処理を行う必要があり、比較的高いコストを要するものである。
【0006】
例えば、特許文献1は、古紙を硫酸第1鉄溶液中で蒸煮し、セルラーゼ生産菌を摂種可能なセルラーゼ生産用基質を開示している。また、特許文献2は、微粉砕したバガスを苛性アルカリで蒸煮し、次亜塩素酸塩溶液で処理し、セルラーゼ生産菌であるトリコデルマ・リーセイを摂種可能なセルラーゼ生産用基質の製造方法を開示している。
【0007】
また、これら従来の方法で得られるセルラーゼは主としてβ−グルカナーゼを含み、キシラナーゼ活性は低く、バガス及び稲わら等のような、キシランが含まれるセルロース資源の分解能力に劣っている。そのため、天然に存在する多様なセルロース資源の有効利用という目的のためには効果が低い。
【0008】
特許文献3は、固形構成成分の除去や非揮発成分の濃縮、濃縮物のオートクレーブ処理等の予備処理を行ったライムギの希薄アルコール蒸留廃液を用い、トリコデルマ属に属する微生物を培養してキシラナーゼを製造する方法を開示している。
【0009】
しかし、本技術で炭素源として用いられるライムギは入手が困難であり、更に複雑な前処理が必要であるために高いコストを要し、また、この方法ではβ−グルカナーゼの生成量が却って低下してしまう。
【0010】
尚、非特許文献1には、熱処理やアルカリ処理などの前処理を行わない紙類(新聞紙やオフィスペーパー)を用いたトリコデルマ・リーセイによる酵素産生の試験では、セルラーゼの生産性が低いことが示されている。
【0011】
加熱処理もアルカリ処理も行わない紙類をセルロース資源として用いβ−グルカナーゼ及びキシラナーゼを同時に高生産する成功例は今まで知られていない。
【先行技術文献】
【特許文献】
【0012】
【特許文献1】特開2003-137901
【特許文献2】特公平5-33984号公報
【特許文献3】特開平11-113568号公報
【非特許文献】
【0013】
【非特許文献1】Applied Biochemistry and Biotechnology pp. 237-245, Vol. 84-86, 2000
【発明の概要】
【発明が解決しようとする課題】
【0014】
本発明は上記従来の問題を解決するものであり、その目的とするところは、キシランを含むセルロース資源の分解能力に優れたセルラーゼを低コストで製造することにある。
【課題を解決するための手段】
【0015】
本発明は、(a)炭素源として加熱処理もアルカリ処理も行わない紙類由来のパルプ、及び(b)窒素源としてアンモニア態窒素またはアミノ態窒素を含む液体培地を用いて、トリコデルマ属に属する微生物を培養する工程を包含するβ−グルカナーゼ及びキシラナーゼの製造方法を提供する。
【0016】
ある一形態においては、前記パルプの前記液体培地中における初期濃度が2%W/V以上である。
【0017】
ある一形態においては、前記パルプの前記液体培地中における初期濃度が2〜7%W/Vである。
【0018】
ある一形態においては、前記アンモニア態窒素またはアミノ態窒素の前記液体培地中における初期濃度が50mM以上である。
【0019】
ある一形態においては、前記アンモニア態窒素またはアミノ態窒素の前記液体培地中における初期濃度が50〜660mMである。
【0020】
ある一形態においては、前記紙類が上質紙、更紙、コピー用紙、新聞紙及びダンボール紙からなる群から選択される少なくとも一種である。
【0021】
ある一形態においては、前記トリコデルマ属に属する微生物が、トリコデルマ・リーセイである。
【0022】
ある一形態においては、培養の過程において前記液体培地に対してパルプが追加される。
【0023】
また、本発明は、(a)炭素源として加熱処理もアルカリ処理も行わない紙類由来のパルプ、及び(b)窒素源としてアンモニア態窒素またはアミノ態窒素を含む液体培地であって、トリコデルマ属に属する微生物を培養するために用いられる液体培地を提供する。
【0024】
ある一形態においては、前記パルプは2%W/V以上含有される。
【0025】
ある一形態においては、前記アンモニア態窒素またはアミノ態窒素は50〜660mM含有される。
【0026】
また、本発明は、前記のいずれかに記載の方法により製造されたβ−グルカナーゼ及びキシラナーゼを提供する。
【0027】
また、本発明は、上記β−グルカナーゼ及びキシラナーゼを用いることを特徴とするセルロース資源の分解または糖化方法を提供する。
【発明の効果】
【0028】
本発明では、液体培地の炭素源として未処理の紙類を用いることができるので、低コスト、低エネルギーであり、環境にかかる負荷が少ない。また、セルラーゼとしてβ−グルカナーゼ及びキシラナーゼが同時に高生産されるため、バガスや稲わら等のキシランを含む天然セルロース資源の糖化に極めて有用である。特に、セルロース資源からエタノールを製造するバイオマスエタノール製造に有用である。
【図面の簡単な説明】
【0029】
【図1】コピー紙3%の培地における、硫酸アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図2】ダンボール紙3%の培地における、硫酸アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図3】新聞紙3%の培地における、硫酸アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図4】コピー紙3%の培地における、塩化アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図5】コピー紙3%の培地における、燐酸二アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図6】コピー紙3%の培地における、硝酸アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図7】コピー紙3%の培地における、アンモニアの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図8】コピー紙3%の培地における、尿素の濃度に対する培養上清液の酵素活性の変化を表すグラフである。
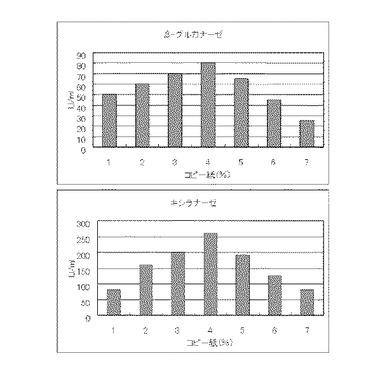
【図9】硫酸アンモニウム80mMの培地における、コピー紙の濃度に対する培養上清液の酵素活性の変化を表すグラフである。
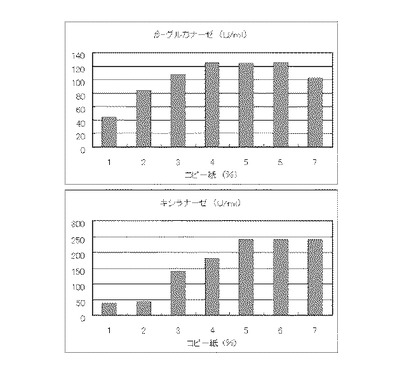
【図10】硫酸アンモニウム160mMの培地における、コピー紙の濃度に対する培養上清液の酵素活性の変化を表すグラフである。
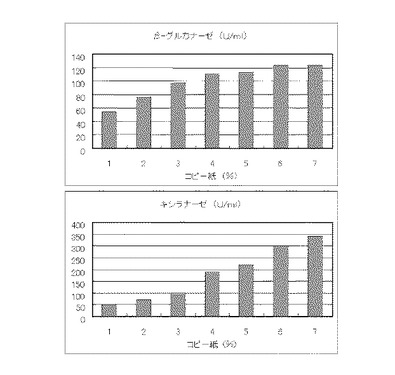
【図11】硫酸アンモニウム320mMの培地における、コピー紙の濃度に対する培養上清液の酵素活性の変化を表すグラフである。
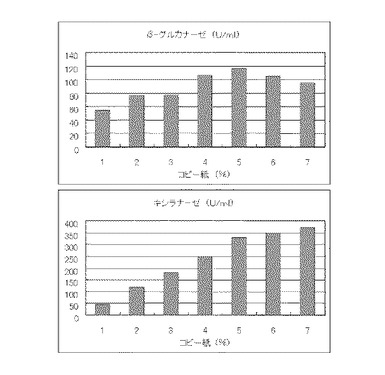
【図12】硫酸アンモニウム480mMの培地における、コピー紙の濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図13】アビセル1%の培地における、硫酸アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図14】硫酸アンモニウム160mMの培地における、アビセルの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図15】コピー紙1%の培地における、硫酸アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図16】コピー紙3%の培地における、硝酸カリウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図17】硫酸アンモニウム200mMの培地における、炭素源の種類に対する培養上清液の酵素活性の変化を表すグラフである。
【図18】使用済みコピー紙6%の培地における、硫酸アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図19】使用済みコピー紙6%及び硫酸アンモニウム45mMの培地における、pH調整用薬液の種類に対する培養上清液の酵素活性の変化を表すグラフである。
【図20】コピー紙3%の培地における、硫酸アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
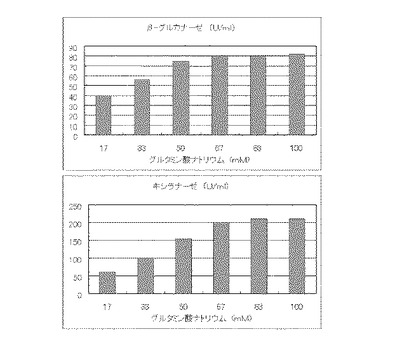
【図21】コピー紙3%の培地における、グルタミン酸ナトリウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
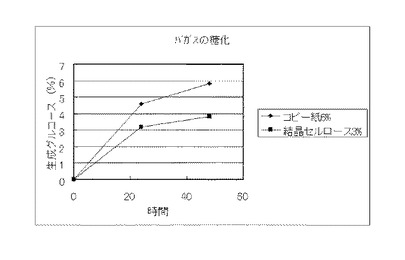
【図22】実施例13で得られたコピー紙6%の培地及び結晶セルロース3%の培地のそれぞれの上清液を用いてバガスを糖化した場合に、生成したグルコースの濃度を比較したグラフである。
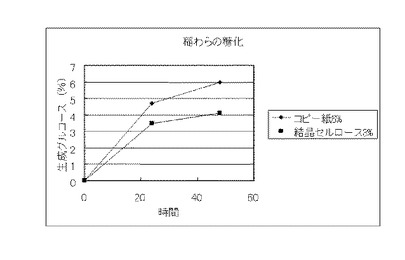
【図23】実施例13で得られたコピー紙6%の培地及び結晶セルロース3%の培地のそれぞれの上清液を用いて稲わらを糖化した場合に、生成したグルコースの濃度を比較したグラフである。
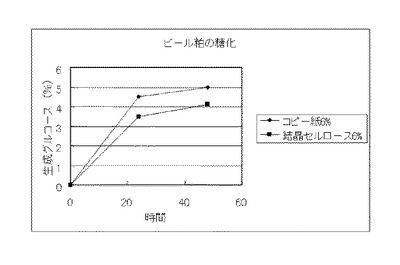
【図24】実施例13で得られたコピー紙6%の培地及び結晶セルロース3%の培地のそれぞれの上清液を用いてビール粕を糖化した場合に、生成したグルコースの濃度を比較したグラフである。
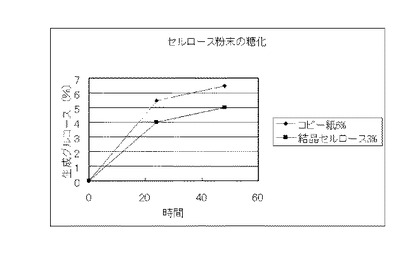
【図25】実施例13で得られたコピー紙6%の培地及び結晶セルロース3%の培地のそれぞれの上清液を用いてセルロース粉末を糖化した場合に、生成したグルコースの濃度を比較したグラフである。
【発明を実施するための形態】
【0030】
液体培地
本発明の液体培地はトリコデルマ属に属する微生物が生育する栄養を含む材料である。かかる液体培地は、以下の培地組成を水100mlに溶解及び懸濁した液体培地(一般に、マンデル培地と呼ばれる)を基に調整され、炭素源としてパルプ、及び窒素源としてアンモニア態窒素又はアミノ態窒素を含むものである。好ましい培地組成の一例を以下に示す。
【0031】
結晶セルロース(Fluka BioChemika製、商品名アビセルPH101):1g、(NH4)2SO4:0.14g、KH2PO4:1.5g、CaCl2・2H2O:0.03g、MgSO4・7H2O:0.03g、コーンスティープリカー:2mL、ツイーン80:0.1mL、微量元素液(H3BO4 6mg、(NH4)6Mo7O24・4H2O 26mg、FeCl3・6H2O 100mg、CuSO4・5H2O 40mg、MnCl2・4H2O 8mg、ZnSO4・7H2O 200mg液):0.1mL、水:100mLを含む(燐酸または水酸化ナトリウムでpH4.8に調整)
【0032】
パルプとは紙類の製造原料に用いられる繊維をいう。パルプの種類は化学パルプ及び古紙パルプのようなセルロース純度が高いものが好ましい。好ましいパルプは、紙類を分解、切断などして得られる紙類由来のパルプである。
【0033】
好ましい紙類の具体例としては、上質紙、更紙、コピー用紙、新聞紙及びダンボール紙などが挙げられる。紙類は好ましいパルプが含まれているものであればよく、印刷や筆記がなされたものや一般に古紙と呼ばれているものでもよい。例えば古書、雑誌及び使い古したノートの頁、チラシ、封筒、便箋、葉書、ティッシュ紙なども使用できる。
【0034】
パルプの液体培地中における濃度は2%W/V以上であることが好ましい。パルプの濃度が2%W/V未満であるとセルラーゼ、特にβ−グルカナーゼの生成量があまり増大しない場合がある。
【0035】
液体培地中のパルプの濃度は高ければ高いほどよい。すなわち、その上限は、液体培地の撹拌混合を行なうことができる限度の量である。なお、液体培地の攪拌混合を容易にするために、紙類はシュレッダーにより、裁断して用いることが好ましい。例えば、液体培地中のパルプの初期濃度の上限は、攪拌装置の性能に応じて20、15又は10%W/Vでありうる。一般的には、パルプの初期濃度の好ましい範囲は2〜7%W/V、好ましくは3〜5%W/Vである。
【0036】
アンモニア態窒素とはアンモニア又はアンモニア由来のアンモニウム塩に含まれている窒素をいう。また、アミノ態窒素とはアミン又はアミン由来のアミノ化合物に含まれている窒素をいう。アンモニア態窒素又はアミノ態窒素を含む化合物は、例えば、硫酸アンモニウム、硝酸アンモニウム、燐酸二アンモニウム、塩化アンモニウム、アンモニア水、尿素、アミノ酸およびその塩(例えば、グルタミン酸ナトリウム)である。
【0037】
これらのうち、窒素源として本発明の液体培地に用いるのに特に好ましい化合物は、硫酸アンモニウムである。その理由は、コストが低く入手が容易だからである。
【0038】
アンモニア態窒素又はアミノ態窒素の液体培地中における濃度は、アンモニウムのモル数として35〜660mMである。好ましくは、50〜580mMである。濃度が35mM未満であるとセルラーゼ、特にβ−グルカナーゼの生成量があまり増大しない場合がある。また、この初期濃度が660mMを超えると酵素の生産性が低下する。また、アンモニア態窒素又はアミノ態窒素の液体培地中における濃度は、液体培地中のパルプの濃度に応じて、増減させることが好ましく、例えば、パルプの濃度が3%W/Vである場合は、コスト等を考慮すると50mMが好ましい。
【0039】
β−グルカナーゼ及びキシラナーゼの製造方法
トリコデルマ属糸状菌はセルロースの糖化に必要なセルラーゼの生産菌として知られている。本発明に使用するトリコデルマ属に属する微生物はセルラーゼを生産するものであれば特に限定されない。好ましいトリコデルマ属に属する微生物はトリコデルマ・リーセイ又はトリコデルマ・ビリデである。特に好ましくは、トリコデルマ・リーセイである。
【0040】
糸状菌トリコデルマ・リーセイおよびトリコデルマ・ビリデの菌学的性質は、例えば、イー・ジー・シモンズ,アブストラクト・セカンド・インターナショナル・マイコロジカル・コングレス(E.G. Simmons, Abst. 2nd International Mycological Congress) 米国フロリダ州タンパ,1977年8月,618頁)に記載されている。
【0041】
液体培養には通常の通気撹拌培養装置が用いられ、上記液体培地を使用して、培養温度20〜33℃好ましくは、28〜30℃、培養pH4〜6で、4〜10日間培養する。培養の過程において液体培地に対してパルプを追加してもよい。培養の進行と共に培地中のパルプは分解されるため、炭素源を補うことによりセルラーゼの生成効率が向上する場合があるからである。パルプを追加する場合、追加の形態は連続式でも回分式でもよく、パルプの追加後も攪拌混合が可能であるように、追加の時期及び量を調節すればよい。
【0042】
パルプを追加する場合、アンモニア態窒素またはアミノ態窒素を必要に応じて、適宜追加してもよい。
【0043】
ついで、要すればこの培養液から遠心分離、濾過などの公知の方法によって菌体を除去して、トリコデルマ属糸状菌培養上清液が得られる。トリコデルマ属糸状菌培養液または培養上清液には目的とするセルラーゼ、すなわちβ−グルカナーゼ及びキシラナーゼが高濃度で含まれている。
【0044】
得られる培養液または培養上清液のβ−グルカナーゼ活性は30U/ml以上、好ましくは50U/ml以上、より好ましくは60U/ml以上、更に好ましくは70U/ml以上である。また、この培養液または培養上清液のキシラナーゼ活性は25U/ml以上、好ましくは30U/ml以上、より好ましくは40U/ml以上、更に好ましくは50U/ml以上である。培養液または培養上清液のβ−グルカナーゼ活性、キシラナーゼ活性のいずれかが上記下限より低下すると、天然に存在する多様なセルロース資源の有効利用という目的に対する効果が低くなる。
【0045】
尚、上記ヘミセルラーゼ活性は、oat spelts由来のキシランを基質とした酵素加水分解により生成した還元糖をDNSと反応させ、540nmの吸光度の増加で定量することができる。
【0046】
より具体的には1%キシラン基質溶液(シグマ社製 Xylan, from oat spelts を200nM酢酸緩衝液(pH4.5)に溶解)1.9mlに培養液または培養上清液0.1mlを加えて、40℃にて正確に10分間酵素反応を行なわせた後、DNS試薬(0.75%ジニトロサリチル酸、1.2%水酸化ナトリウム、22.5%酒石酸ナトリウムカリウム4水和物、0.3%乳糖1水和物を含む)4mlを加えてよく混合し、反応を停止する。反応停止液に含まれる還元糖量を定量するために、反応停止液を沸騰水浴中で15分間正確に加熱する。続いて、室温まで冷却した後、540nmの吸光度を測定することでキシロースに相当する還元糖量として定量する。1単位のヘミセルラーゼ活性は、40℃、10分間の反応条件下で、1分間に1μmolのキシロースに相当する還元糖を生成する酵素量として表す。
【0047】
セルロース資源の分解または糖化方法
本発明の方法により得られたβ−グルカナーゼ及びキシラナーゼは、セルロース資源を分解または糖化するのに有用である。ここでいうセルロース資源は、合成セルロースもしくは天然セルロース資源のどちらでも良い。合成セルロースとは、セルロース粉末として、流通しているものを表す。天然セルロース資源とは、バガス、稲わら、麦わら、ビール粕、木材などが挙げられる。本発明は、β-グルカナーゼおよびキシラナーゼを同時に高生産できるため、特に、バガス、稲わら、麦わら、ビール粕などの天然セルロース資源の糖化に優れている。
【0048】
セルロース資源の分解または糖化方法は、公知の方法を使用すればよく、特に制限されるものではないが、一例としては、基質としてセルロース資源を水性媒体中に懸濁させ、上記培養液または培養上清液を添加し、攪拌または振とうしながら、加温して糖化反応を行う方法が挙げられる。セルロース分解活性を示す上記培養液または培養上清液の代わりにその乾燥物、または乾燥物を水に分散もしくは溶解した液を用いてもよい。
【0049】
セルロース原料は、予め脱リグニンしておくことが好ましい。懸濁方法、攪拌方法、上記混合液の添加方法、添加順序、それらの濃度等の反応条件は、グルコースがより高収率で得られるよう適宜調整される。
【0050】
その際の、反応液のpH及び温度は、酵素が失活しない範囲内であればよく、一般的には、常圧で反応を行う場合、温度は30〜70℃、pHは3〜7の範囲でよい。また、この圧力、温度、pHについても、上記同様、グルコースがより高収率で得られるよう適宜調整されるものであるが、常圧で、酢酸またはリン酸緩衝液中で、温度50〜60℃、pH4〜6の範囲で行うことが好ましい。反応時間は一般に6〜147時間、好ましくは24〜72時間である。
【0051】
セルロースの糖化により、グルコースを含有する水溶液が得られる。得られた水溶液は、必要に応じて、脱色、脱塩、酵素除去等の精製処理を施すことができる。精製方法は、公知の方法であれば特に制限されないが、例えば、活性炭処理、イオン交換樹脂処理、クロマトグラフィー処理、精密ろ過、限外ろ過、逆浸透ろ過等の濾過処理、晶析処理等を使用してもよく、これらを単独で使用しても、2種以上を組み合わせてもよい。
【0052】
上記の方法で精製されたグルコースを主成分とする水溶液は、そのまま使用することができるが、必要に応じて、乾燥により固化させてもよい。乾燥方法は、公知の方法であれば特に制限されないが、例えば、噴霧乾燥、凍結乾燥、ドラム乾燥、薄膜乾燥、棚段乾燥、気流乾燥、真空乾燥等を使用してもよく、これらを単独で使用しても、2種以上を組み合わせてもよい。
【実施例】
【0053】
以下、本発明を実施例によってより具体的に説明するが、本発明はこれらの実施例に限定されるものではない。
【0054】
実施例1
トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させた。マンデル培地の炭素源である結晶セルロースをコピー用紙3%(3g/100ml)に置き換えて、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度がそれぞれ15mM、35mM、50mM、65mM、80mM、100mMまたは115mMになるように添加して燐酸及び水酸化ナトリウムでpH4.8に調整した100mlの液体培地を500ml容バッフル付三角フラスコに用意した。培養したトリコデルマ・リーセイの1白金耳をこの液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、上清液のβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。なお、コピー用紙はアルカリ処理や熱処理等の前処理をせず、シュレッダー(カール社デスクパーサーDS−4100)で2mmX7mmに裁断したのみで使用した。
【0055】
(酵素活性の測定)
前記で得られた培養液について酵素活性を測定した。
β-グルカナーゼ活性は、メガザイム社製のβ‐グルカナーゼ測定キットを用い、色素標識したβ-グルカンを基質とした酵素分解によって生じた染色断片を吸光度測定した。具体的には、アゾ大麦グルカン基質溶液0.1mlに培養液0.1mlを加えて、40℃にて正確に10分間酵素反応を行なわせた後、停止液〔4%酢酸ナトリウム、0.4%酢酸亜鉛、80%メチルセルソルブを含む(pH5)〕0.6mlを加えて5分放置し、反応を停止した。続いて遠心分離した後、上澄液を590nmの吸光度測定した。1単位のβ-グルカナーゼ活性は、40℃、10分間の反応条件下で、1分間に1μmolのグルコースに相当する還元糖を生成する酵素量として表した。
【0056】
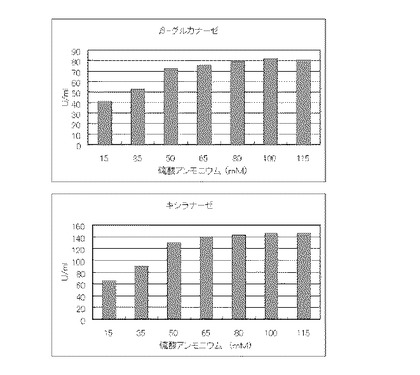
次に、キシラナーゼ活性は、oat spelts由来のキシランを基質とした酵素加水分解により生成した還元糖をDNSと反応させ、540nmの吸光度の増加で定量した。より具体的には1%キシラン基質溶液[シグマ社製Xylan, from oat speltsを200mM酢酸緩衝液(pH4.5)に溶解]1.9mlに培養液0.1mlを加えて、40℃にて正確に10分間酵素反応を行なわせた後、DNS試薬(0.75%ジニトロサリチル酸、1.2%水酸化ナトリウム、22.5%酒石酸ナトリウムカリウム4水和物、0.3%乳糖1水和物を含む)4mlを加えてよく混合し、反応を停止した。反応停止液に含まれる還元糖量を定量するために、反応停止液を沸騰水浴中で15分間正確に加熱した。続いて、室温まで冷却した後、540nmの吸光度を測定することでキシロースに相当する還元糖量として定量した。1単位のキシラナーゼ活性は、40℃、10分間の反応条件下で、1分間に1μmolのキシロースに相当する還元糖を生成する酵素量として表した。結果を図1に示す。
【0057】
実施例2
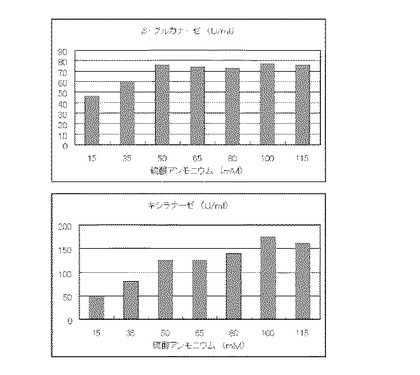
マンデル培地の炭素源である結晶セルロースをダンボール3%(3g/100ml)に置き換えて、実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図2に示す。
【0058】
実施例3
マンデル培地の炭素源である結晶セルロースを新聞紙3%(3g/100ml)に置き換えて、実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図3に示す。
【0059】
実施例4
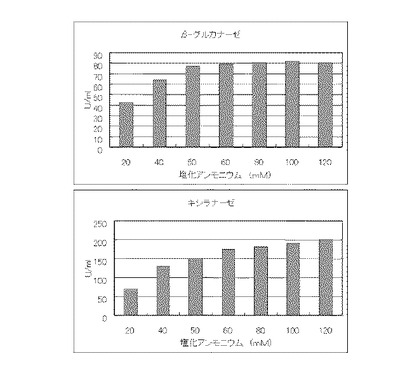
マンデル培地の炭素源である結晶セルロースをコピー紙3%(3g/100ml)に置き換えて、また窒素源である硫酸アンモニウムを塩化アンモニウムに置き換えてアンモニア態窒素のモル濃度がそれぞれ20mM、40mM、50mM、60mM、80mM、100mMまたは120mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図4に示す。
【0060】
実施例5
マンデル培地の炭素源である結晶セルロースをコピー紙3%(3g/100ml)に置き換えて、また窒素源である硫酸アンモニウムを燐酸二アンモニウムに置き換えてアンモニア態窒素のモル濃度がそれぞれ15mM、35mM、50mM、65mM、80mM、100mMまたは115mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図5に示す。
【0061】
実施例6
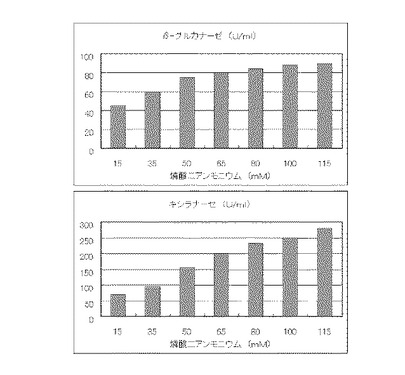
マンデル培地の炭素源である結晶セルロースをコピー紙3%(3g/100ml)に置き換えて、また窒素源である硫酸アンモニウムを硝酸アンモニウムに置き換えてアンモニア態窒素のモル濃度がそれぞれ12mM、24mM、36mM、48mM、60mM、72mMまたは84mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図6に示す。
【0062】
実施例7
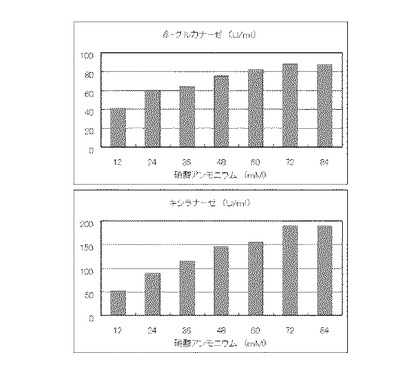
マンデル培地の炭素源である結晶セルロースをコピー紙3%(3g/100ml)に置き換えて、また窒素源である硫酸アンモニウムをアンモニア水に置き換えてモル濃度がそれぞれ15mM、30mM、45mM、65mM、75mM、90mMまたは105mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図7に示す。
【0063】
実施例8
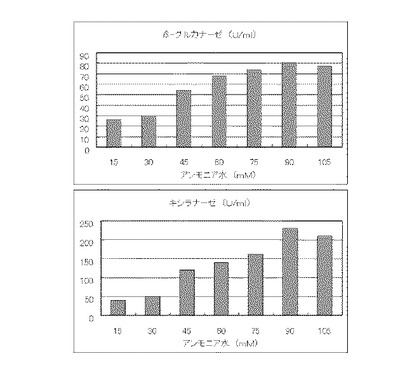
マンデル培地の炭素源である結晶セルロースをコピー紙3%(3g/100ml)に置き換えて、また窒素源である硫酸アンモニウムを尿素に置き換えてアミノ態窒素のモル濃度がそれぞれ17mM、33mM、50mM、67mM、83mMまたは100mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図8に示す。
【0064】
実施例9
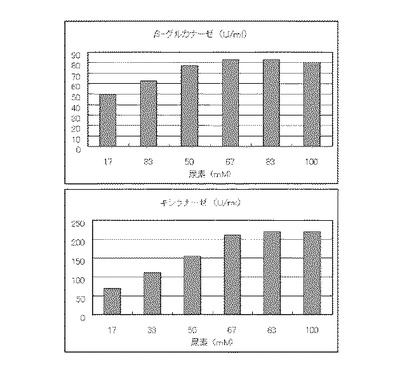
マンデル培地の炭素源である結晶セルロースをコピー紙に置き換えて濃度が1%、2%、3%、4%、5%、6%、7%になるように添加して、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度が80mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図9に示す。
【0065】
実施例10
マンデル培地の炭素源である結晶セルロースをコピー紙に置き換えて濃度が1%、2%、3%、4%、5%、6%、7%になるように添加して、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度が160mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図10に示す。
【0066】
実施例11
マンデル培地の炭素源である結晶セルロースをコピー紙に置き換えて濃度が1%、2%、3%、4%、5%、6%、7%になるように添加して、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度が320mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図11に示す。
【0067】
実施例12
マンデル培地の炭素源である結晶セルロースをコピー紙に置き換えて濃度が1%、2%、3%、4%、5%、6%、7%になるように添加して、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度が480mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図12に示す。
【0068】
参考例1
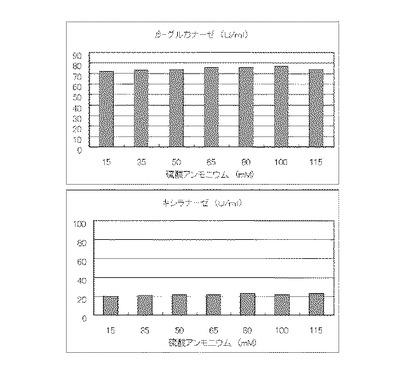
マンデル培地の炭素源である結晶セルロースの濃度を1%とし、窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度がそれぞれ15mM、35mM、50mM、65mM、80mM、100mMまたは115mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図13に示す。
【0069】
参考例2
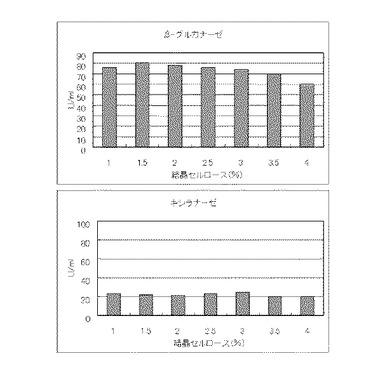
マンデル培地の炭素源である結晶セルロース濃度が1%、1.5%、2%、2.5%、3%、3.5%または4%になるように添加して、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度が160mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図14に示す。
【0070】
参考例3
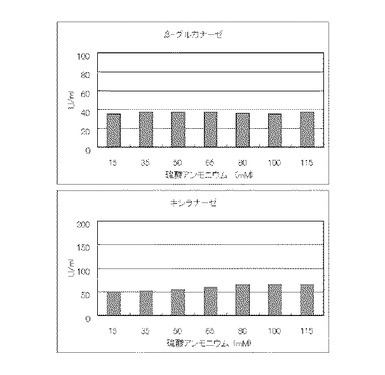
マンデル培地の炭素源である結晶セルロースをコピー紙1%(3g/100ml)に置き換えて添加して、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度がそれぞれ15mM、35mM、50mM、65mM、80mM、100mMまたは115mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図15に示す。
【0071】
参考例4
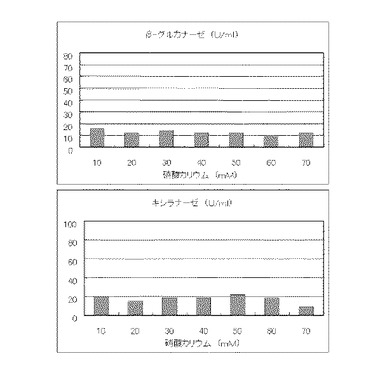
マンデル培地の炭素源である結晶セルロースをコピー紙3%(3g/100ml)に置き換えて添加して、また窒素源である硫酸アンモニウムを硝酸カリウムに置き換えてモル濃度が10mM、20mM、30mM、40mM、50mM、60mMまたは70mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図16に示す。
【0072】
実施例13
トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させた。その1白金耳を、マンデル培地100mlを含む500ml容バッフル付三角フラスコに接種して、28℃、180rpm、4日間振とう培養した。マンデル培地150mlを含む500ml容バッフル付三角フラスコに接種して、28℃、180rpm、2日間振とう培養して培養液を得た。マンデル培地の炭素源である結晶セルロースに置き換えて、コピー紙3%(30g/l)、6%(60g/l)又は結晶セルロース(アビセルPH101)3%を添加し、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度が200mMになるように添加し、ツイーン80のかわりにアデカノールLG-126(株式会社ADEKA製)6g/lを用いた培地3L及び本培養液を5L発酵槽(マルビシ製)に添加して、28℃で培養した。通気は1VVM、撹拌は450rpmで、またpHは2N水酸化ナトリウムおよび5倍希釈の燐酸によって培養期間中pH4.8に一定になるよう調整した。7日目に培養液を遠心分離し、その上清液のβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。その結果を図17に示す。
【0073】
なお、コピー紙は、シュレッダー(明光商会社製プリモ1400)で4mmX30mmに裁断して使用した。また、コピー紙6%(60g/l)は、一度に全量添加すると撹拌が不十分であったため、コピー紙と硫酸アンモニウムを培養1日目と3日目に半分ずつ分けて加えた。
【0074】
実施例14
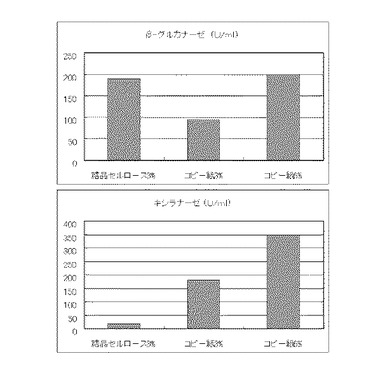
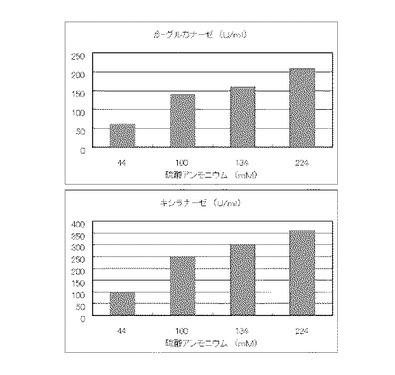
実施例13と同様の方法で5L発酵槽に、コピー紙6%(60g/l)を添加し、硫酸アンモニウムのアンモニア態窒素のモル濃度が44mM、100mM、134mMまたは224mMになるように添加し、酵素生産を調べた。その結果を図18に示す。
【0075】
実施例15
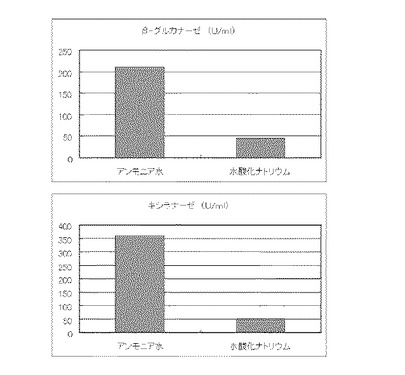
実施例13と同様の方法で5L発酵槽に、コピー紙6%(60g/l)と硫酸アンモニウムのアンモニア態窒素のモル濃度が45mMを添加し、培養期間中のpH調整用薬液を2N水酸化ナトリウムを使用した場合と、18%アンモニア水を使用した場合の酵素生産を比較した。このときのアンモニア水の培養期間中の投入量は123mMであった。その結果を図19に示す。
【0076】
実施例16
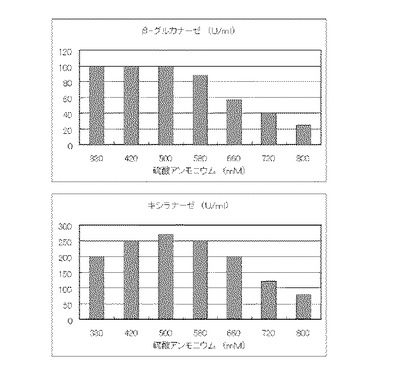
マンデル培地の炭素源である結晶セルロースをコピー紙3%(3g/100ml)に置き換えて、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度がそれぞれ330mM、420mM、500mM、580mM、660mM、720mMまたは800mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図20に示す。
【0077】
実施例17
マンデル培地の炭素源である結晶セルロースをコピー紙3%(3g/100ml)に置き換えて、また窒素源である硫酸アンモニウムをグルタミン酸ナトリウムに置き換えてアミノ態窒素のモル濃度がそれぞれ17mM、33mM、50mM、67mM、83mMまたは100mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図21に示す。
【0078】
実施例18
実施例13で得られたコピー紙6%の培地及び結晶セルロース3%の培地のそれぞれの上清液を用いて、セルロース資源の糖化試験を行った。糖化に供するセルロース資源は、セルロース粉末(日本製紙ケミカル製 商品名KCフロックW−50)、バガス、稲わらおよびビール粕を用いた。また、バガス、稲わらおよびビール粕は、それぞれ微粉砕し、0.3NのNaOHに懸濁して、120℃、15分間処理し、水で充分に洗浄後、乾燥し、脱リグニン処理を行い、糖化に供した。セルロース粉末は、そのまま糖化に供した。セルロース資源:0.8g、培養上清液:9ml、1M酢酸バッファー(pH4.8):0.2mlからなる液(セルロース資源8%液)を50℃、pH4.8、48時間、振とうさせて糖化し、生成したグルコースをグルコースCII-テストワコー(和光純薬工業)で測定した。結果を図22〜図25に示す。
【0079】
実施例及び参考例の結果から、炭素源として未処理の紙類を含み、窒素源としてアンモニア/アンモニウム塩を含む液体培地を用いて、トリコデルマ・リーセイを培養することにより、β−グルカナーゼ及びキシラナーゼが同時に高生産されることが示された。また、培地中の紙類の濃度を通常用いられる炭素源より上げるか、又は媒地中のアンモニア/アンモニウム塩濃度を特定範囲に調節することで、これらセルラーゼの生成量が顕著に増大することも示された。更に、得られた培養上清を用いて種々のセルロース資源を分解及び糖化できることも示された。
【産業上の利用可能性】
【0080】
セルロース資源、特にバガスや稲わら等の天然セルロース資源の糖化に極めて有用なβ−グルカナーゼ及びキシラナーゼを同時に高生産でき、セルロース資源からエタノールを製造するバイオマスエタノール製造に利用できる。
【技術分野】
【0001】
本発明はβ−グルカナーゼ及びキシラナーゼの製造方法及び液体培地に関する。
【背景技術】
【0002】
セルロース資源を有効利用するために、近年、セルロースを効率的に分解する方法が探索されている。自然界ではセルロースは主として微生物によって分解されており、細菌や糸状菌などの様々な微生物がセルロース分解酵素を生産することが知られている。
【0003】
これらの微生物は菌体外にセルロース分解酵素を分泌し、セルロースはその作用により、主に、セロオリゴ糖、セロビオースを経てグルコースへと分解される。セルロース分解酵素は、一般に、セルラーゼと呼ばれている。
【0004】
人工的にセルラーゼを製造しようとする場合、セルラーゼを分泌する微生物としては、トリコデルマ属が知られており、広く利用されている。そして、トリコデルマ属に属する微生物を炭素源及び窒素源などの栄養を含む培地を用いて培養し、セルラーゼを分泌させる方法も知られている。
【0005】
しかしながら、セルラーゼを製造するための従来の方法は、炭素源として使用できる材料に制限があり、高価な結晶セルロースであったり、仮に安価なセルロース資源であっても、加熱処理やアルカリ処理などの前処理を行う必要があり、比較的高いコストを要するものである。
【0006】
例えば、特許文献1は、古紙を硫酸第1鉄溶液中で蒸煮し、セルラーゼ生産菌を摂種可能なセルラーゼ生産用基質を開示している。また、特許文献2は、微粉砕したバガスを苛性アルカリで蒸煮し、次亜塩素酸塩溶液で処理し、セルラーゼ生産菌であるトリコデルマ・リーセイを摂種可能なセルラーゼ生産用基質の製造方法を開示している。
【0007】
また、これら従来の方法で得られるセルラーゼは主としてβ−グルカナーゼを含み、キシラナーゼ活性は低く、バガス及び稲わら等のような、キシランが含まれるセルロース資源の分解能力に劣っている。そのため、天然に存在する多様なセルロース資源の有効利用という目的のためには効果が低い。
【0008】
特許文献3は、固形構成成分の除去や非揮発成分の濃縮、濃縮物のオートクレーブ処理等の予備処理を行ったライムギの希薄アルコール蒸留廃液を用い、トリコデルマ属に属する微生物を培養してキシラナーゼを製造する方法を開示している。
【0009】
しかし、本技術で炭素源として用いられるライムギは入手が困難であり、更に複雑な前処理が必要であるために高いコストを要し、また、この方法ではβ−グルカナーゼの生成量が却って低下してしまう。
【0010】
尚、非特許文献1には、熱処理やアルカリ処理などの前処理を行わない紙類(新聞紙やオフィスペーパー)を用いたトリコデルマ・リーセイによる酵素産生の試験では、セルラーゼの生産性が低いことが示されている。
【0011】
加熱処理もアルカリ処理も行わない紙類をセルロース資源として用いβ−グルカナーゼ及びキシラナーゼを同時に高生産する成功例は今まで知られていない。
【先行技術文献】
【特許文献】
【0012】
【特許文献1】特開2003-137901
【特許文献2】特公平5-33984号公報
【特許文献3】特開平11-113568号公報
【非特許文献】
【0013】
【非特許文献1】Applied Biochemistry and Biotechnology pp. 237-245, Vol. 84-86, 2000
【発明の概要】
【発明が解決しようとする課題】
【0014】
本発明は上記従来の問題を解決するものであり、その目的とするところは、キシランを含むセルロース資源の分解能力に優れたセルラーゼを低コストで製造することにある。
【課題を解決するための手段】
【0015】
本発明は、(a)炭素源として加熱処理もアルカリ処理も行わない紙類由来のパルプ、及び(b)窒素源としてアンモニア態窒素またはアミノ態窒素を含む液体培地を用いて、トリコデルマ属に属する微生物を培養する工程を包含するβ−グルカナーゼ及びキシラナーゼの製造方法を提供する。
【0016】
ある一形態においては、前記パルプの前記液体培地中における初期濃度が2%W/V以上である。
【0017】
ある一形態においては、前記パルプの前記液体培地中における初期濃度が2〜7%W/Vである。
【0018】
ある一形態においては、前記アンモニア態窒素またはアミノ態窒素の前記液体培地中における初期濃度が50mM以上である。
【0019】
ある一形態においては、前記アンモニア態窒素またはアミノ態窒素の前記液体培地中における初期濃度が50〜660mMである。
【0020】
ある一形態においては、前記紙類が上質紙、更紙、コピー用紙、新聞紙及びダンボール紙からなる群から選択される少なくとも一種である。
【0021】
ある一形態においては、前記トリコデルマ属に属する微生物が、トリコデルマ・リーセイである。
【0022】
ある一形態においては、培養の過程において前記液体培地に対してパルプが追加される。
【0023】
また、本発明は、(a)炭素源として加熱処理もアルカリ処理も行わない紙類由来のパルプ、及び(b)窒素源としてアンモニア態窒素またはアミノ態窒素を含む液体培地であって、トリコデルマ属に属する微生物を培養するために用いられる液体培地を提供する。
【0024】
ある一形態においては、前記パルプは2%W/V以上含有される。
【0025】
ある一形態においては、前記アンモニア態窒素またはアミノ態窒素は50〜660mM含有される。
【0026】
また、本発明は、前記のいずれかに記載の方法により製造されたβ−グルカナーゼ及びキシラナーゼを提供する。
【0027】
また、本発明は、上記β−グルカナーゼ及びキシラナーゼを用いることを特徴とするセルロース資源の分解または糖化方法を提供する。
【発明の効果】
【0028】
本発明では、液体培地の炭素源として未処理の紙類を用いることができるので、低コスト、低エネルギーであり、環境にかかる負荷が少ない。また、セルラーゼとしてβ−グルカナーゼ及びキシラナーゼが同時に高生産されるため、バガスや稲わら等のキシランを含む天然セルロース資源の糖化に極めて有用である。特に、セルロース資源からエタノールを製造するバイオマスエタノール製造に有用である。
【図面の簡単な説明】
【0029】
【図1】コピー紙3%の培地における、硫酸アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図2】ダンボール紙3%の培地における、硫酸アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図3】新聞紙3%の培地における、硫酸アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図4】コピー紙3%の培地における、塩化アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図5】コピー紙3%の培地における、燐酸二アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図6】コピー紙3%の培地における、硝酸アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図7】コピー紙3%の培地における、アンモニアの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図8】コピー紙3%の培地における、尿素の濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図9】硫酸アンモニウム80mMの培地における、コピー紙の濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図10】硫酸アンモニウム160mMの培地における、コピー紙の濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図11】硫酸アンモニウム320mMの培地における、コピー紙の濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図12】硫酸アンモニウム480mMの培地における、コピー紙の濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図13】アビセル1%の培地における、硫酸アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図14】硫酸アンモニウム160mMの培地における、アビセルの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図15】コピー紙1%の培地における、硫酸アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図16】コピー紙3%の培地における、硝酸カリウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図17】硫酸アンモニウム200mMの培地における、炭素源の種類に対する培養上清液の酵素活性の変化を表すグラフである。
【図18】使用済みコピー紙6%の培地における、硫酸アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図19】使用済みコピー紙6%及び硫酸アンモニウム45mMの培地における、pH調整用薬液の種類に対する培養上清液の酵素活性の変化を表すグラフである。
【図20】コピー紙3%の培地における、硫酸アンモニウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図21】コピー紙3%の培地における、グルタミン酸ナトリウムの濃度に対する培養上清液の酵素活性の変化を表すグラフである。
【図22】実施例13で得られたコピー紙6%の培地及び結晶セルロース3%の培地のそれぞれの上清液を用いてバガスを糖化した場合に、生成したグルコースの濃度を比較したグラフである。
【図23】実施例13で得られたコピー紙6%の培地及び結晶セルロース3%の培地のそれぞれの上清液を用いて稲わらを糖化した場合に、生成したグルコースの濃度を比較したグラフである。
【図24】実施例13で得られたコピー紙6%の培地及び結晶セルロース3%の培地のそれぞれの上清液を用いてビール粕を糖化した場合に、生成したグルコースの濃度を比較したグラフである。
【図25】実施例13で得られたコピー紙6%の培地及び結晶セルロース3%の培地のそれぞれの上清液を用いてセルロース粉末を糖化した場合に、生成したグルコースの濃度を比較したグラフである。
【発明を実施するための形態】
【0030】
液体培地
本発明の液体培地はトリコデルマ属に属する微生物が生育する栄養を含む材料である。かかる液体培地は、以下の培地組成を水100mlに溶解及び懸濁した液体培地(一般に、マンデル培地と呼ばれる)を基に調整され、炭素源としてパルプ、及び窒素源としてアンモニア態窒素又はアミノ態窒素を含むものである。好ましい培地組成の一例を以下に示す。
【0031】
結晶セルロース(Fluka BioChemika製、商品名アビセルPH101):1g、(NH4)2SO4:0.14g、KH2PO4:1.5g、CaCl2・2H2O:0.03g、MgSO4・7H2O:0.03g、コーンスティープリカー:2mL、ツイーン80:0.1mL、微量元素液(H3BO4 6mg、(NH4)6Mo7O24・4H2O 26mg、FeCl3・6H2O 100mg、CuSO4・5H2O 40mg、MnCl2・4H2O 8mg、ZnSO4・7H2O 200mg液):0.1mL、水:100mLを含む(燐酸または水酸化ナトリウムでpH4.8に調整)
【0032】
パルプとは紙類の製造原料に用いられる繊維をいう。パルプの種類は化学パルプ及び古紙パルプのようなセルロース純度が高いものが好ましい。好ましいパルプは、紙類を分解、切断などして得られる紙類由来のパルプである。
【0033】
好ましい紙類の具体例としては、上質紙、更紙、コピー用紙、新聞紙及びダンボール紙などが挙げられる。紙類は好ましいパルプが含まれているものであればよく、印刷や筆記がなされたものや一般に古紙と呼ばれているものでもよい。例えば古書、雑誌及び使い古したノートの頁、チラシ、封筒、便箋、葉書、ティッシュ紙なども使用できる。
【0034】
パルプの液体培地中における濃度は2%W/V以上であることが好ましい。パルプの濃度が2%W/V未満であるとセルラーゼ、特にβ−グルカナーゼの生成量があまり増大しない場合がある。
【0035】
液体培地中のパルプの濃度は高ければ高いほどよい。すなわち、その上限は、液体培地の撹拌混合を行なうことができる限度の量である。なお、液体培地の攪拌混合を容易にするために、紙類はシュレッダーにより、裁断して用いることが好ましい。例えば、液体培地中のパルプの初期濃度の上限は、攪拌装置の性能に応じて20、15又は10%W/Vでありうる。一般的には、パルプの初期濃度の好ましい範囲は2〜7%W/V、好ましくは3〜5%W/Vである。
【0036】
アンモニア態窒素とはアンモニア又はアンモニア由来のアンモニウム塩に含まれている窒素をいう。また、アミノ態窒素とはアミン又はアミン由来のアミノ化合物に含まれている窒素をいう。アンモニア態窒素又はアミノ態窒素を含む化合物は、例えば、硫酸アンモニウム、硝酸アンモニウム、燐酸二アンモニウム、塩化アンモニウム、アンモニア水、尿素、アミノ酸およびその塩(例えば、グルタミン酸ナトリウム)である。
【0037】
これらのうち、窒素源として本発明の液体培地に用いるのに特に好ましい化合物は、硫酸アンモニウムである。その理由は、コストが低く入手が容易だからである。
【0038】
アンモニア態窒素又はアミノ態窒素の液体培地中における濃度は、アンモニウムのモル数として35〜660mMである。好ましくは、50〜580mMである。濃度が35mM未満であるとセルラーゼ、特にβ−グルカナーゼの生成量があまり増大しない場合がある。また、この初期濃度が660mMを超えると酵素の生産性が低下する。また、アンモニア態窒素又はアミノ態窒素の液体培地中における濃度は、液体培地中のパルプの濃度に応じて、増減させることが好ましく、例えば、パルプの濃度が3%W/Vである場合は、コスト等を考慮すると50mMが好ましい。
【0039】
β−グルカナーゼ及びキシラナーゼの製造方法
トリコデルマ属糸状菌はセルロースの糖化に必要なセルラーゼの生産菌として知られている。本発明に使用するトリコデルマ属に属する微生物はセルラーゼを生産するものであれば特に限定されない。好ましいトリコデルマ属に属する微生物はトリコデルマ・リーセイ又はトリコデルマ・ビリデである。特に好ましくは、トリコデルマ・リーセイである。
【0040】
糸状菌トリコデルマ・リーセイおよびトリコデルマ・ビリデの菌学的性質は、例えば、イー・ジー・シモンズ,アブストラクト・セカンド・インターナショナル・マイコロジカル・コングレス(E.G. Simmons, Abst. 2nd International Mycological Congress) 米国フロリダ州タンパ,1977年8月,618頁)に記載されている。
【0041】
液体培養には通常の通気撹拌培養装置が用いられ、上記液体培地を使用して、培養温度20〜33℃好ましくは、28〜30℃、培養pH4〜6で、4〜10日間培養する。培養の過程において液体培地に対してパルプを追加してもよい。培養の進行と共に培地中のパルプは分解されるため、炭素源を補うことによりセルラーゼの生成効率が向上する場合があるからである。パルプを追加する場合、追加の形態は連続式でも回分式でもよく、パルプの追加後も攪拌混合が可能であるように、追加の時期及び量を調節すればよい。
【0042】
パルプを追加する場合、アンモニア態窒素またはアミノ態窒素を必要に応じて、適宜追加してもよい。
【0043】
ついで、要すればこの培養液から遠心分離、濾過などの公知の方法によって菌体を除去して、トリコデルマ属糸状菌培養上清液が得られる。トリコデルマ属糸状菌培養液または培養上清液には目的とするセルラーゼ、すなわちβ−グルカナーゼ及びキシラナーゼが高濃度で含まれている。
【0044】
得られる培養液または培養上清液のβ−グルカナーゼ活性は30U/ml以上、好ましくは50U/ml以上、より好ましくは60U/ml以上、更に好ましくは70U/ml以上である。また、この培養液または培養上清液のキシラナーゼ活性は25U/ml以上、好ましくは30U/ml以上、より好ましくは40U/ml以上、更に好ましくは50U/ml以上である。培養液または培養上清液のβ−グルカナーゼ活性、キシラナーゼ活性のいずれかが上記下限より低下すると、天然に存在する多様なセルロース資源の有効利用という目的に対する効果が低くなる。
【0045】
尚、上記ヘミセルラーゼ活性は、oat spelts由来のキシランを基質とした酵素加水分解により生成した還元糖をDNSと反応させ、540nmの吸光度の増加で定量することができる。
【0046】
より具体的には1%キシラン基質溶液(シグマ社製 Xylan, from oat spelts を200nM酢酸緩衝液(pH4.5)に溶解)1.9mlに培養液または培養上清液0.1mlを加えて、40℃にて正確に10分間酵素反応を行なわせた後、DNS試薬(0.75%ジニトロサリチル酸、1.2%水酸化ナトリウム、22.5%酒石酸ナトリウムカリウム4水和物、0.3%乳糖1水和物を含む)4mlを加えてよく混合し、反応を停止する。反応停止液に含まれる還元糖量を定量するために、反応停止液を沸騰水浴中で15分間正確に加熱する。続いて、室温まで冷却した後、540nmの吸光度を測定することでキシロースに相当する還元糖量として定量する。1単位のヘミセルラーゼ活性は、40℃、10分間の反応条件下で、1分間に1μmolのキシロースに相当する還元糖を生成する酵素量として表す。
【0047】
セルロース資源の分解または糖化方法
本発明の方法により得られたβ−グルカナーゼ及びキシラナーゼは、セルロース資源を分解または糖化するのに有用である。ここでいうセルロース資源は、合成セルロースもしくは天然セルロース資源のどちらでも良い。合成セルロースとは、セルロース粉末として、流通しているものを表す。天然セルロース資源とは、バガス、稲わら、麦わら、ビール粕、木材などが挙げられる。本発明は、β-グルカナーゼおよびキシラナーゼを同時に高生産できるため、特に、バガス、稲わら、麦わら、ビール粕などの天然セルロース資源の糖化に優れている。
【0048】
セルロース資源の分解または糖化方法は、公知の方法を使用すればよく、特に制限されるものではないが、一例としては、基質としてセルロース資源を水性媒体中に懸濁させ、上記培養液または培養上清液を添加し、攪拌または振とうしながら、加温して糖化反応を行う方法が挙げられる。セルロース分解活性を示す上記培養液または培養上清液の代わりにその乾燥物、または乾燥物を水に分散もしくは溶解した液を用いてもよい。
【0049】
セルロース原料は、予め脱リグニンしておくことが好ましい。懸濁方法、攪拌方法、上記混合液の添加方法、添加順序、それらの濃度等の反応条件は、グルコースがより高収率で得られるよう適宜調整される。
【0050】
その際の、反応液のpH及び温度は、酵素が失活しない範囲内であればよく、一般的には、常圧で反応を行う場合、温度は30〜70℃、pHは3〜7の範囲でよい。また、この圧力、温度、pHについても、上記同様、グルコースがより高収率で得られるよう適宜調整されるものであるが、常圧で、酢酸またはリン酸緩衝液中で、温度50〜60℃、pH4〜6の範囲で行うことが好ましい。反応時間は一般に6〜147時間、好ましくは24〜72時間である。
【0051】
セルロースの糖化により、グルコースを含有する水溶液が得られる。得られた水溶液は、必要に応じて、脱色、脱塩、酵素除去等の精製処理を施すことができる。精製方法は、公知の方法であれば特に制限されないが、例えば、活性炭処理、イオン交換樹脂処理、クロマトグラフィー処理、精密ろ過、限外ろ過、逆浸透ろ過等の濾過処理、晶析処理等を使用してもよく、これらを単独で使用しても、2種以上を組み合わせてもよい。
【0052】
上記の方法で精製されたグルコースを主成分とする水溶液は、そのまま使用することができるが、必要に応じて、乾燥により固化させてもよい。乾燥方法は、公知の方法であれば特に制限されないが、例えば、噴霧乾燥、凍結乾燥、ドラム乾燥、薄膜乾燥、棚段乾燥、気流乾燥、真空乾燥等を使用してもよく、これらを単独で使用しても、2種以上を組み合わせてもよい。
【実施例】
【0053】
以下、本発明を実施例によってより具体的に説明するが、本発明はこれらの実施例に限定されるものではない。
【0054】
実施例1
トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させた。マンデル培地の炭素源である結晶セルロースをコピー用紙3%(3g/100ml)に置き換えて、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度がそれぞれ15mM、35mM、50mM、65mM、80mM、100mMまたは115mMになるように添加して燐酸及び水酸化ナトリウムでpH4.8に調整した100mlの液体培地を500ml容バッフル付三角フラスコに用意した。培養したトリコデルマ・リーセイの1白金耳をこの液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、上清液のβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。なお、コピー用紙はアルカリ処理や熱処理等の前処理をせず、シュレッダー(カール社デスクパーサーDS−4100)で2mmX7mmに裁断したのみで使用した。
【0055】
(酵素活性の測定)
前記で得られた培養液について酵素活性を測定した。
β-グルカナーゼ活性は、メガザイム社製のβ‐グルカナーゼ測定キットを用い、色素標識したβ-グルカンを基質とした酵素分解によって生じた染色断片を吸光度測定した。具体的には、アゾ大麦グルカン基質溶液0.1mlに培養液0.1mlを加えて、40℃にて正確に10分間酵素反応を行なわせた後、停止液〔4%酢酸ナトリウム、0.4%酢酸亜鉛、80%メチルセルソルブを含む(pH5)〕0.6mlを加えて5分放置し、反応を停止した。続いて遠心分離した後、上澄液を590nmの吸光度測定した。1単位のβ-グルカナーゼ活性は、40℃、10分間の反応条件下で、1分間に1μmolのグルコースに相当する還元糖を生成する酵素量として表した。
【0056】
次に、キシラナーゼ活性は、oat spelts由来のキシランを基質とした酵素加水分解により生成した還元糖をDNSと反応させ、540nmの吸光度の増加で定量した。より具体的には1%キシラン基質溶液[シグマ社製Xylan, from oat speltsを200mM酢酸緩衝液(pH4.5)に溶解]1.9mlに培養液0.1mlを加えて、40℃にて正確に10分間酵素反応を行なわせた後、DNS試薬(0.75%ジニトロサリチル酸、1.2%水酸化ナトリウム、22.5%酒石酸ナトリウムカリウム4水和物、0.3%乳糖1水和物を含む)4mlを加えてよく混合し、反応を停止した。反応停止液に含まれる還元糖量を定量するために、反応停止液を沸騰水浴中で15分間正確に加熱した。続いて、室温まで冷却した後、540nmの吸光度を測定することでキシロースに相当する還元糖量として定量した。1単位のキシラナーゼ活性は、40℃、10分間の反応条件下で、1分間に1μmolのキシロースに相当する還元糖を生成する酵素量として表した。結果を図1に示す。
【0057】
実施例2
マンデル培地の炭素源である結晶セルロースをダンボール3%(3g/100ml)に置き換えて、実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図2に示す。
【0058】
実施例3
マンデル培地の炭素源である結晶セルロースを新聞紙3%(3g/100ml)に置き換えて、実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図3に示す。
【0059】
実施例4
マンデル培地の炭素源である結晶セルロースをコピー紙3%(3g/100ml)に置き換えて、また窒素源である硫酸アンモニウムを塩化アンモニウムに置き換えてアンモニア態窒素のモル濃度がそれぞれ20mM、40mM、50mM、60mM、80mM、100mMまたは120mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図4に示す。
【0060】
実施例5
マンデル培地の炭素源である結晶セルロースをコピー紙3%(3g/100ml)に置き換えて、また窒素源である硫酸アンモニウムを燐酸二アンモニウムに置き換えてアンモニア態窒素のモル濃度がそれぞれ15mM、35mM、50mM、65mM、80mM、100mMまたは115mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図5に示す。
【0061】
実施例6
マンデル培地の炭素源である結晶セルロースをコピー紙3%(3g/100ml)に置き換えて、また窒素源である硫酸アンモニウムを硝酸アンモニウムに置き換えてアンモニア態窒素のモル濃度がそれぞれ12mM、24mM、36mM、48mM、60mM、72mMまたは84mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図6に示す。
【0062】
実施例7
マンデル培地の炭素源である結晶セルロースをコピー紙3%(3g/100ml)に置き換えて、また窒素源である硫酸アンモニウムをアンモニア水に置き換えてモル濃度がそれぞれ15mM、30mM、45mM、65mM、75mM、90mMまたは105mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図7に示す。
【0063】
実施例8
マンデル培地の炭素源である結晶セルロースをコピー紙3%(3g/100ml)に置き換えて、また窒素源である硫酸アンモニウムを尿素に置き換えてアミノ態窒素のモル濃度がそれぞれ17mM、33mM、50mM、67mM、83mMまたは100mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図8に示す。
【0064】
実施例9
マンデル培地の炭素源である結晶セルロースをコピー紙に置き換えて濃度が1%、2%、3%、4%、5%、6%、7%になるように添加して、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度が80mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図9に示す。
【0065】
実施例10
マンデル培地の炭素源である結晶セルロースをコピー紙に置き換えて濃度が1%、2%、3%、4%、5%、6%、7%になるように添加して、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度が160mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図10に示す。
【0066】
実施例11
マンデル培地の炭素源である結晶セルロースをコピー紙に置き換えて濃度が1%、2%、3%、4%、5%、6%、7%になるように添加して、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度が320mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図11に示す。
【0067】
実施例12
マンデル培地の炭素源である結晶セルロースをコピー紙に置き換えて濃度が1%、2%、3%、4%、5%、6%、7%になるように添加して、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度が480mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図12に示す。
【0068】
参考例1
マンデル培地の炭素源である結晶セルロースの濃度を1%とし、窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度がそれぞれ15mM、35mM、50mM、65mM、80mM、100mMまたは115mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図13に示す。
【0069】
参考例2
マンデル培地の炭素源である結晶セルロース濃度が1%、1.5%、2%、2.5%、3%、3.5%または4%になるように添加して、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度が160mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図14に示す。
【0070】
参考例3
マンデル培地の炭素源である結晶セルロースをコピー紙1%(3g/100ml)に置き換えて添加して、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度がそれぞれ15mM、35mM、50mM、65mM、80mM、100mMまたは115mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図15に示す。
【0071】
参考例4
マンデル培地の炭素源である結晶セルロースをコピー紙3%(3g/100ml)に置き換えて添加して、また窒素源である硫酸アンモニウムを硝酸カリウムに置き換えてモル濃度が10mM、20mM、30mM、40mM、50mM、60mMまたは70mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図16に示す。
【0072】
実施例13
トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させた。その1白金耳を、マンデル培地100mlを含む500ml容バッフル付三角フラスコに接種して、28℃、180rpm、4日間振とう培養した。マンデル培地150mlを含む500ml容バッフル付三角フラスコに接種して、28℃、180rpm、2日間振とう培養して培養液を得た。マンデル培地の炭素源である結晶セルロースに置き換えて、コピー紙3%(30g/l)、6%(60g/l)又は結晶セルロース(アビセルPH101)3%を添加し、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度が200mMになるように添加し、ツイーン80のかわりにアデカノールLG-126(株式会社ADEKA製)6g/lを用いた培地3L及び本培養液を5L発酵槽(マルビシ製)に添加して、28℃で培養した。通気は1VVM、撹拌は450rpmで、またpHは2N水酸化ナトリウムおよび5倍希釈の燐酸によって培養期間中pH4.8に一定になるよう調整した。7日目に培養液を遠心分離し、その上清液のβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。その結果を図17に示す。
【0073】
なお、コピー紙は、シュレッダー(明光商会社製プリモ1400)で4mmX30mmに裁断して使用した。また、コピー紙6%(60g/l)は、一度に全量添加すると撹拌が不十分であったため、コピー紙と硫酸アンモニウムを培養1日目と3日目に半分ずつ分けて加えた。
【0074】
実施例14
実施例13と同様の方法で5L発酵槽に、コピー紙6%(60g/l)を添加し、硫酸アンモニウムのアンモニア態窒素のモル濃度が44mM、100mM、134mMまたは224mMになるように添加し、酵素生産を調べた。その結果を図18に示す。
【0075】
実施例15
実施例13と同様の方法で5L発酵槽に、コピー紙6%(60g/l)と硫酸アンモニウムのアンモニア態窒素のモル濃度が45mMを添加し、培養期間中のpH調整用薬液を2N水酸化ナトリウムを使用した場合と、18%アンモニア水を使用した場合の酵素生産を比較した。このときのアンモニア水の培養期間中の投入量は123mMであった。その結果を図19に示す。
【0076】
実施例16
マンデル培地の炭素源である結晶セルロースをコピー紙3%(3g/100ml)に置き換えて、また窒素源である硫酸アンモニウムのアンモニア態窒素のモル濃度がそれぞれ330mM、420mM、500mM、580mM、660mM、720mMまたは800mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図20に示す。
【0077】
実施例17
マンデル培地の炭素源である結晶セルロースをコピー紙3%(3g/100ml)に置き換えて、また窒素源である硫酸アンモニウムをグルタミン酸ナトリウムに置き換えてアミノ態窒素のモル濃度がそれぞれ17mM、33mM、50mM、67mM、83mMまたは100mMになるように添加して実施例1と同様に液体培地を用意した。トリコデルマ・リーセイQM9414(NBRC 31329)をポテトデキストロース寒天培地上で28℃、7日間培養して胞子を充分形成させ、この1白金耳を液体培地に接種して、28℃、180rpm、7日間振とう培養した。7日目に培養液を遠心分離し、実施例1と同様にしてβ-グルカナーゼ活性およびキシラナーゼ活性を測定した。結果を図21に示す。
【0078】
実施例18
実施例13で得られたコピー紙6%の培地及び結晶セルロース3%の培地のそれぞれの上清液を用いて、セルロース資源の糖化試験を行った。糖化に供するセルロース資源は、セルロース粉末(日本製紙ケミカル製 商品名KCフロックW−50)、バガス、稲わらおよびビール粕を用いた。また、バガス、稲わらおよびビール粕は、それぞれ微粉砕し、0.3NのNaOHに懸濁して、120℃、15分間処理し、水で充分に洗浄後、乾燥し、脱リグニン処理を行い、糖化に供した。セルロース粉末は、そのまま糖化に供した。セルロース資源:0.8g、培養上清液:9ml、1M酢酸バッファー(pH4.8):0.2mlからなる液(セルロース資源8%液)を50℃、pH4.8、48時間、振とうさせて糖化し、生成したグルコースをグルコースCII-テストワコー(和光純薬工業)で測定した。結果を図22〜図25に示す。
【0079】
実施例及び参考例の結果から、炭素源として未処理の紙類を含み、窒素源としてアンモニア/アンモニウム塩を含む液体培地を用いて、トリコデルマ・リーセイを培養することにより、β−グルカナーゼ及びキシラナーゼが同時に高生産されることが示された。また、培地中の紙類の濃度を通常用いられる炭素源より上げるか、又は媒地中のアンモニア/アンモニウム塩濃度を特定範囲に調節することで、これらセルラーゼの生成量が顕著に増大することも示された。更に、得られた培養上清を用いて種々のセルロース資源を分解及び糖化できることも示された。
【産業上の利用可能性】
【0080】
セルロース資源、特にバガスや稲わら等の天然セルロース資源の糖化に極めて有用なβ−グルカナーゼ及びキシラナーゼを同時に高生産でき、セルロース資源からエタノールを製造するバイオマスエタノール製造に利用できる。
【特許請求の範囲】
【請求項1】
(a)炭素源として加熱処理もアルカリ処理も行わない紙類由来のパルプ、及び(b)窒素源としてアンモニア態窒素またはアミノ態窒素を含む液体培地を用いて、トリコデルマ属に属する微生物を培養する工程を包含するβ−グルカナーゼ及びキシラナーゼの製造方法。
【請求項2】
前記パルプの前記液体培地中における初期濃度が2%W/V以上である請求項1に記載のβ−グルカナーゼ及びキラナーゼの製造方法。
【請求項3】
前記パルプの前記液体培地中における初期濃度が2〜7%W/Vである請求項1又は2に記載のβ−グルカナーゼ及びキシラナーゼの製造方法。
【請求項4】
前記アンモニア態窒素またはアミノ態窒素の前記液体培地中における初期濃度が35〜660mM以上である請求項1〜3のいずれかに記載のβ−グルカナーゼ及びキシラナーゼの製造方法。
【請求項5】
前記アンモニア態窒素またはアミノ態窒素の前記液体培地中における初期濃度が50〜580mMである請求項1〜4のいずれかに記載のβ−グルカナーゼ及びキシラナーゼの製造方法。
【請求項6】
前記紙類が上質紙、更紙、コピー用紙、新聞紙及びダンボール紙からなる群から選択される少なくとも一種である請求項1〜5のいずれかに記載のβ−グルカナーゼ及びキシラナーゼの製造方法。
【請求項7】
前記トリコデルマ属に属する微生物が、トリコデルマ・リーセイである請求項1〜6のいずれかに記載のβ−グルカナーゼ及びキシラナーゼの製造方法。
【請求項8】
培養の過程において前記液体培地に対してパルプを追加する請求項1〜7に記載のβ−グルカナーゼ及びキシラナーゼの製造方法。
【請求項9】
(a)炭素源として加熱処理もアルカリ処理も行わない紙類由来のパルプ、及び(b)窒素源としてアンモニア態窒素またはアミノ態窒素を含む液体培地であって、トリコデルマ属に属する微生物を培養するために用いられる液体培地。
【請求項10】
前記パルプを2%W/V以上含有する請求項9に記載の液体培地。
【請求項11】
アンモニア態窒素またはアミノ態窒素を35〜660mM含有する請求項9又は10に記載の液体培地。
【請求項12】
請求項1〜8のいずれか1項に記載の方法により製造されたβ−グルカナーゼ及びキシラナーゼ。
【請求項13】
請求項12記載のβ−グルカナーゼ及びキシラナーゼを用いることを特徴とするセルロース資源の分解または糖化方法。
【請求項1】
(a)炭素源として加熱処理もアルカリ処理も行わない紙類由来のパルプ、及び(b)窒素源としてアンモニア態窒素またはアミノ態窒素を含む液体培地を用いて、トリコデルマ属に属する微生物を培養する工程を包含するβ−グルカナーゼ及びキシラナーゼの製造方法。
【請求項2】
前記パルプの前記液体培地中における初期濃度が2%W/V以上である請求項1に記載のβ−グルカナーゼ及びキラナーゼの製造方法。
【請求項3】
前記パルプの前記液体培地中における初期濃度が2〜7%W/Vである請求項1又は2に記載のβ−グルカナーゼ及びキシラナーゼの製造方法。
【請求項4】
前記アンモニア態窒素またはアミノ態窒素の前記液体培地中における初期濃度が35〜660mM以上である請求項1〜3のいずれかに記載のβ−グルカナーゼ及びキシラナーゼの製造方法。
【請求項5】
前記アンモニア態窒素またはアミノ態窒素の前記液体培地中における初期濃度が50〜580mMである請求項1〜4のいずれかに記載のβ−グルカナーゼ及びキシラナーゼの製造方法。
【請求項6】
前記紙類が上質紙、更紙、コピー用紙、新聞紙及びダンボール紙からなる群から選択される少なくとも一種である請求項1〜5のいずれかに記載のβ−グルカナーゼ及びキシラナーゼの製造方法。
【請求項7】
前記トリコデルマ属に属する微生物が、トリコデルマ・リーセイである請求項1〜6のいずれかに記載のβ−グルカナーゼ及びキシラナーゼの製造方法。
【請求項8】
培養の過程において前記液体培地に対してパルプを追加する請求項1〜7に記載のβ−グルカナーゼ及びキシラナーゼの製造方法。
【請求項9】
(a)炭素源として加熱処理もアルカリ処理も行わない紙類由来のパルプ、及び(b)窒素源としてアンモニア態窒素またはアミノ態窒素を含む液体培地であって、トリコデルマ属に属する微生物を培養するために用いられる液体培地。
【請求項10】
前記パルプを2%W/V以上含有する請求項9に記載の液体培地。
【請求項11】
アンモニア態窒素またはアミノ態窒素を35〜660mM含有する請求項9又は10に記載の液体培地。
【請求項12】
請求項1〜8のいずれか1項に記載の方法により製造されたβ−グルカナーゼ及びキシラナーゼ。
【請求項13】
請求項12記載のβ−グルカナーゼ及びキシラナーゼを用いることを特徴とするセルロース資源の分解または糖化方法。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【公開番号】特開2010−142221(P2010−142221A)
【公開日】平成22年7月1日(2010.7.1)
【国際特許分類】
【出願番号】特願2009−113765(P2009−113765)
【出願日】平成21年5月8日(2009.5.8)
【分割の表示】特願2009−500640(P2009−500640)の分割
【原出願日】平成20年12月17日(2008.12.17)
【出願人】(000000055)アサヒビール株式会社 (535)
【Fターム(参考)】
【公開日】平成22年7月1日(2010.7.1)
【国際特許分類】
【出願日】平成21年5月8日(2009.5.8)
【分割の表示】特願2009−500640(P2009−500640)の分割
【原出願日】平成20年12月17日(2008.12.17)
【出願人】(000000055)アサヒビール株式会社 (535)
【Fターム(参考)】
[ Back to top ]