
β−1,3−1,6−グルカンの定量方法
【課題】本発明は、完全に精製した酵素を用いる必要がなく、且つβ−1,3−1,6−グルカンを正確に定量できる方法を提供することを目的とする。また、本発明は、当該方法に用いることができるキットと、当該方法に対して有用な微生物を提供することも目的とする。
【解決手段】本発明に係るβ−1,3−1,6−グルカンの定量方法は、多糖類含有試料に特定のMitsuaria chitosanitabida種菌に由来する菌体外酵素液および特定のStreptomyces omiyaensis種菌に由来する菌体外酵素液を作用させる工程;および、生じたグルコースの量を測定する工程;を含むことを特徴とする。
【解決手段】本発明に係るβ−1,3−1,6−グルカンの定量方法は、多糖類含有試料に特定のMitsuaria chitosanitabida種菌に由来する菌体外酵素液および特定のStreptomyces omiyaensis種菌に由来する菌体外酵素液を作用させる工程;および、生じたグルコースの量を測定する工程;を含むことを特徴とする。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、β−1,3−1,6−グルカンを定量する方法、当該方法に用いることができるβ−1,3−1,6−グルカンの定量用キット、および当該方法に有用な菌体外酵素を産生する微生物に関するものである。
【背景技術】
【0002】
細胞は、グリコーゲンやデンプンなどエネルギーとなる貯蔵多糖のほか、構造多糖を産生する。構造多糖とは、細胞の表面を保護する細胞膜や細胞壁の構成成分である多糖であり、グルカン、マンナン、糖タンパク質、キチンなど様々な種類が確認されている。
【0003】
近年、かかる構造多糖の中でも、担子菌類の産生するものが優れた生理活性を有することから注目を集めている。
【0004】
担子菌類とは、担子胞子を生ずる高等菌類であり、松茸や舞茸、アガリクスと呼ばれる姫松茸などがある。この担子菌類が産生する構造多糖は、食物繊維としての効果のほか、抗腫瘍作用、抗菌作用、血糖・血清コレステロール降下作用などを示し、糖尿病などの生活習慣病やがんなどの予防に有効であるとの研究成果も報告されている。
【0005】
担子菌類が産生する構造多糖の中でも、β−1,3−グルコシド結合を主鎖に有しβ−1,6−グルコシド結合を側鎖に有するβ−1,3−1,6−グルカンは、とりわけ特徴的な生理活性を示す(非特許文献1)。
【0006】
しかし、構造多糖は優れた生理活性を有するものの、細胞壁などの中に存在するためにその抽出は非常に困難である。その上、たとえ抽出したとしてもその抽出物は水に対して不溶性または難溶性を示し、特有の色や不快臭を有し、品質が不安定であり非常に高価であるなどの問題を有する。
【0007】
それに対して、黒酵母といわれるAureobasidium pullulans種菌の中からは、ある条件下でβ−1,3−1,6−グルカンを菌体外へ放出するものが見出されている。しかも、β−1,3−1,6−グルカンを含む当該菌の培養液は、無味無臭で品質が安定しており、安価に製造できる。よって、当該菌から得られるβ−1,3−1,6−グルカンを機能性食品などとして利用する道が開かれつつある。
【0008】
しかし、β−1,3−1,6−グルカンを含む培養液などを食品の原料として利用する場合には、その含量を測定するなどして品質を評価する必要があるものの、正確な定量方法は未だ存在しない。
【0009】
例えば、財団法人日本食品分析センターが行っているβ−グルカンの定量方法は、アミラーゼやアミログルコシダーゼなどによる酵素処理の後、硫酸で全てのグルコシド結合を非特異的に加水分解し、生じたグルコースを定量するものである。しかしこの方法では、β−1,3−1,6−グルカンとそれ以外のβ−グルカンを区別することができない。さらに、グルコース以外の単糖を含むヘテログルカンなどに由来するグルコースまで分解され、測定されてしまう。従って当該方法は、β−1,3−1,6−グルカンの定量方法としては不適である。
【0010】
そのほか、一般的に使用されているβ−グルカンの定量キットは、連続したβ−1,3−グルコシド結合を認識して結合するタンパク質因子を利用するものであるため、試料中に含まれるβ−1,3−グルカンにおける側鎖の割合が多いほど、測定値の誤差は大きくなる。
【0011】
ところで、非特許文献2〜4のとおり、本発明者らが属する研究室では、土壌から単離したCeriporiopsis属微生物由来のβ−1,3−グルカナーゼとβ−1,6−グルカナーゼが、それぞれβ−1,3−グルコシド結合とβ−1,6−グルコシド結合に対して高い反応性を有することを見出している。よって、これら酵素を利用して試料中のβ−1,3−1,6−グルカン含量を正確に測定できると考えられる。
【非特許文献1】宮崎利夫,「多糖の構造と生理活性」朝倉書店(1990年)
【非特許文献2】野田洋彦,「高知大学大学院農学研究科農芸化学専攻醗酵及び醸造学研究室修士論文」(1994年)
【非特許文献3】中嶋英夫,「高知大学大学院農学研究科農芸化学専攻醗酵及び醸造学研究室修士論文」(1993年)
【非特許文献4】池上裕倫,「高知大学大学院農学研究科生物資源科学専攻応用微生物学研究室修士論文」(2004年)
【発明の開示】
【発明が解決しようとする課題】
【0012】
上述した様に、本発明者らが属する研究室では、反応性の高いβ−1,3−グルカナーゼとβ−1,6−グルカナーゼを産生するCeriporiopsis属微生物を見出しており、これら酵素を利用して試料中のβ−1,3−1,6−グルカン含量を正確に測定することが考えられる。
【0013】
しかし、酵素を完全に精製するには、各種クロマトグラフィを繰り返す必要があるなど手間やコストがかかるため、精製酵素を用いる定量方法は、工業的な実施のためには実用的であるとは到底いい難い。よって、菌体外に放出された酵素を完全に精製することなく定量に用いることが考えられる。ところが、上記Ceriporiopsis属微生物は非常に強いα−グルコシダーゼ活性を有するため、粗精製酵素中にはα−グルコシダーゼが混入するので、試料中にα−グルカンが存在しているとグルコースまで分解されてしまい、β−1,3−1,6−グルカン含量を正確に測定できなくなる。そのため、Ceriporiopsis属微生物由来の粗精製酵素を用いて試料中のβ−1,3−1,6−グルカンを定量するには、先ずα−グルコシダーゼなどによりα−グルカンを分解して除去してから、粗精製されたβ−1,3−グルカナーゼとβ−1,6−グルカナーゼでβ−グルカンを分解し、生じたグルコースを定量する必要がある。このような方法では、時間や手間がかかり非効率的である。
【0014】
そこで本発明では、完全に精製した酵素を用いる必要がなく、且つβ−1,3−1,6−グルカンを正確かつ効率的に定量できる方法を提供することを目的とする。また、本発明は、当該方法に用いることができるキットと、当該方法に対して有用な微生物を提供することも目的とする。
【課題を解決するための手段】
【0015】
本発明者らは、上記課題を解決すべく鋭意研究を重ねたところ、β−1,3−グルカナーゼの産生能を有し且つβ−1,3−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないMitsuaria chitosanitabida種菌と、β−1,6−グルカナーゼの産生能を有し且つβ−1,6−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないStreptomyces omiyaensis種菌に由来する菌体外酵素液を用いれば、試料中におけるα−グルカン、他のβ−グルカンおよびヘテログルカンの存在にかかわらず、β−1,3−1,6−グルカンを正確かつ効率的に定量できることを実証して、本発明を完成した。
【0016】
本発明に係るβ−1,3−1,6−グルカンの定量方法は、多糖類含有試料に、β−1,3−グルカナーゼの産生能を有し且つβ−1,3−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないMitsuaria chitosanitabida種菌に由来する菌体外酵素液、並びに、β−1,6−グルカナーゼの産生能を有し且つβ−1,6−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないStreptomyces omiyaensis種菌に由来する菌体外酵素液を作用させる工程;並びに、生じたグルコースの量を測定する工程;を含むことを特徴とする。
【0017】
上記本発明方法においては、上記M.chitosanitabida種菌としてM.chitosanitabida H1株(NITE AP−629)が好適であり、また、上記S.omiyaensis種菌としてS.omiyaensis SY26株(NITE AP−616)が好適である。上記菌株の優れた効果は、実験により実証されている。
【0018】
上記本発明方法においては、多糖類含有試料に、20℃以上、45℃以下で、上記M.chitosanitabida種菌に由来する菌体外酵素液と上記S.omiyaensis種菌に由来する菌体外酵素液とを同時に作用させることが好ましい。上記2種の菌体外酵素液は同時に多糖類含有試料へ作用させることが効率の面から好ましいが、それぞれの酵素液に含まれるβ−1,3−グルカナーゼとβ−1,6−グルカナーゼの至適温度は異なる。しかし20℃以上、45℃以下の温度範囲であれば、両酵素は良好にその作用効果を発揮することができ、多糖類含有試料に含まれるβ−1,3−1,6−グルカンを効率的に分解することが可能になる。
【0019】
本発明に係るβ−1,3−1,6−グルカンの定量用キットは、β−1,3−グルカナーゼの産生能を有し且つβ−1,3−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないM.chitosanitabida種菌に由来する菌体外酵素液またはその乾燥物;β−1,6−グルカナーゼの産生能を有し且つβ−1,6−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないS.omiyaensis種菌に由来する菌体外酵素液またはその乾燥物;並びにグルコース量測定用試薬;を含むことを特徴とする。
【0020】
また、本発明は、M.chitosanitabida H1株(NITE AP− 629)およびS.omiyaensis SY26株(NITE AP−616)に関する。
【発明の効果】
【0021】
従来、α−グルカンを含む多糖類含有試料中のβ−1,3−1,6−グルカンを正確に測定するには、先ずアミログルコシダーゼなどでα−グルカンを分解し除去してからβ−グルカン量を測定しなければならず、手間や時間がかかるものであった。また、従来方法では、他のβ−グルカンを含む試料中のβ−1,3−1,6−グルカンを正確に測定することは困難であった。しかし本発明方法によれば、完全に精製された酵素を用いなくとも、α−グルカンを事前に分解する必要も無く、β−1,3−1,6−グルカンを正確かつ効率的に定量することができる。また、本発明に係る定量用キットは、同様の効果を有する。さらに本発明に係る2種の菌株は、これら本発明に係る定量方法と定量用キットにとり非常に好適なものである。よって本発明は、その優れた生理活性が注目されているβ−1,3−1,6−グルカンを含む試料の品質を評価する指標になり得るものとして、産業上極めて有用である。
【発明を実施するための最良の形態】
【0022】
本発明に係るβ−1,3−1,6−グルカンの定量方法は、多糖類含有試料に、β−1,3−グルカナーゼの産生能を有し且つβ−1,3−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないM.chitosanitabida種菌に由来する菌体外酵素液、並びに、β−1,6−グルカナーゼの産生能を有し且つβ−1,6−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないS.omiyaensis種菌に由来する菌体外酵素液を作用させる工程;および、生じたグルコースの量を測定する工程;を含むことを特徴とする。
【0023】
本発明方法の測定対象である多糖類含有試料は、β−1,3−1,6−グルカンを含有するか或いは含有すると考えられるものであって、その含有量を測定すべきものであれば特にその種類などは制限されない。例えば、酵母の集菌体、培地、それらの乾燥物、抽出物、精製品、懸濁液、溶液;担子菌類自体や海藻類自体、それらの乾燥物、抽出物、粉砕物、生成物、懸濁液、溶液などを挙げることができる。
【0024】
本発明方法では、多糖類含有試料に、β−1,3−グルカナーゼの産生能を有し且つβ−1,3−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないM.chitosanitabida種菌に由来する菌体外酵素液、並びに、β−1,6−グルカナーゼの産生能を有し且つβ−1,6−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないS.omiyaensis種菌に由来する菌体外酵素液を作用させる。これら菌体外酵素液は、各β−グルカナーゼ活性を示す一方で、α−グルカンや他のβ−グルカンを分解しない。よって、これら菌体外酵素液を用いれば、酵素を精製しなくとも、多糖類含有試料中のβ−1,3−1,6−グルカンを特異的にグルコースまで分解することができる。
【0025】
なお、上記菌は、α−グルコシダーゼや、β−1,3−グルカナーゼおよびβ−1,6−グルカナーゼ以外のβ−グルカナーゼを全く産生しないものに限定されず、β−1,3−1,6−グルカンの正確な定量を阻害しない程度のこれら酵素活性を菌体外へ放出するものや、β−1,3−1,6−グルカナーゼから容易に分離除去できる程度のこれら酵素を産生して菌体外へ放出するものも含まれるものとする。具体的には、比活性が0.05unit/mg以下のα−グルコシダーゼ、またはβ−1,3−グルカナーゼおよびβ−1,6−グルカナーゼ以外のβ−グルカナーゼをその菌体外酵素液に放出する菌も含まれるものとする。但し、上記菌としては、これら酵素活性を実質的に示さないものが好適である。ここでいう実質的とは、菌体外酵素液におけるこれら酵素の活性が、下記実施例に記載の測定方法において、検出限界以下であることをいう。
【0026】
当該菌体外酵素液としては、上記M.chitosanitabida種菌および上記S.omiyaensis種菌の液体培養液から得た酵素液であって、グルコースやα−グルコシダーゼ、β−1,3−グルカナーゼおよびβ−1,6−グルカナーゼ以外のβ−グルカナーゼなどβ−1,3−1,6−グルカンの正確な定量を阻害する成分を含まないものをいう。即ち、上記菌の液体培養液に硫酸アンモニウムを加えて沈殿したタンパク質成分を分離したり、透析などにより、グルコースなどの低分子物質を除去することができる。また、上記菌の菌体外酵素液でもα−グルコシダーゼ活性等を示すことがあるが、その活性は極めて弱いので、硫酸アンモニウムの濃度を調整することにより、α−グルコシダーゼ等は容易に除去可能である。
【0027】
菌体外酵素液は、具体的には例えば以下のように調製することができる。先ず、上記菌を培養した液体培養液を遠心分離や濾過に付すことによって、菌体などの固体成分を除去する。得られた液体には、PMSF(フェニルメチルスルフォニルフルオリド)などのプロテアーゼ阻害剤を加えることが好ましい。次に、タンパク質成分を常法により分離する。例えば、得られた液体をある程度濃縮した上で緩衝液を加えた後、冷却しながら硫酸アンモニウムを加えることにより沈殿したタンパク質成分を分離すればよい。この際、予備実験などにより、β−1,3−グルカナーゼとβ−1,6−グルカナーゼが十分に沈殿し且つα−グルコシダーゼ等が沈殿しない硫酸アンモニウム濃度を事前に決定しておくと効率的である。得られたタンパク質成分からは、さらに透析などの簡便な方法により、所望のβ−グルカナーゼをある程度精製してもよい。但し、本発明で用いるM.chitosanitabida種菌とS.omiyaensis種菌は菌体外にα−グルコシダーゼ等を放出しないか或いはその放出量は極めて少ないので、クロマトグラフィなど工業的に適さない方法により酵素を精製する必要はない。
【0028】
上記菌体外酵素液は、保存などのためにいったん乾燥し、使用時に溶液としてもよい。但し、上記β−1,3−グルカナーゼとβ−1,6−グルカナーゼは、それぞれ70℃と50℃で失活する。よって、酵素液の濃縮や乾燥時においては、それぞれ温度を70℃、50℃以上に上げないことが好ましい。
【0029】
本発明においては、多糖類含有試料に上記M.chitosanitabida種菌由来の菌体外酵素液と上記S.omiyaensis種菌由来の菌体外酵素液を作用させるが、これら酵素液は同時に作用させてもよいし、逐次的に作用させてもよい。効率を考慮すれば、同時に作用させることが好ましい。
【0030】
上記菌体外酵素液におけるβ−1,3−グルカナーゼとβ−1,6−グルカナーゼの濃度は、反応混合液における多糖類の濃度などに応じて適宜調整すればよいが、例えば、反応混合液中に含まれる各β−グルカナーゼの活性として、β−1,3−1,6−グルカンであるラミナリンを基質に用いた場合で1.0unit/ml以上、5.0unit/ml以下程度、β−1,6−グルカンであるパスツランを基質に用いた場合で0.1unit/ml以上、3.0unit/ml以下程度とすることが好ましい。なお、本発明におけるunitとは、30℃で1時間に1μmolの還元糖を精製する酵素量の単位と定義する。
【0031】
上記菌体外酵素液のpHも適宜調整すればよいが、本発明者らによる実験ではpH6.0で良好な結果が得られているので、溶媒としてはpH5.0以上、7.5以下程度の緩衝液が好適である。
【0032】
上記酵素反応における反応温度も適宜調整すればよい。例えば、M.chitosanitabida H1株由来のβ−1,3−グルカナーゼの至適温度は約40℃であり、約30℃以上、約55℃以下であればその60%程度の活性を示す。また、S.omiyaensis SY26株由来のβ−1,6−グルカナーゼの至適温度は約30℃であり、約20℃以上、約45℃以下であればその60%程度の活性を示す。よって、各菌体外酵素液を逐次的に多糖類含有試料に作用させる場合には、各反応温度をそれぞれ約30℃以上、約55℃以下と、約20℃以上、約45℃以下に調整することが好ましい。また、上記M.chitosanitabida種菌由来の菌体外酵素液と上記S.omiyaensis種菌由来の菌体外酵素液を同時に多糖類含有試料に作用させる場合には、両グルカナーゼを有効に機能させるために、反応温度を20℃以上、45℃以下とすることが好ましい。より好ましくは、35℃以上、40℃以下で反応を行う。
【0033】
上記酵素反応の反応時間は、多糖類含有試料中に存在するβ−1,3−1,6−グルカンがグルコースまで十分に分解される時間とする。即ち、当該反応時間は、反応混合液に含まれるβ−1,3−1,6−グルカンの濃度、菌体外酵素液に含まれるβ−1,3−グルカナーゼとβ−1,6−グルカナーゼの力価、反応温度などにも依存するので、具体的には予備実験で反応混合液中へ数時間おきに数回菌体外酵素液を加えつつ生成されるグルコース量を経時的に測定し、酵素反応によりグルコースが新たに生成されないことが確認できなくなった時間とすればよい。通常は、β−1,3−1,6−グルカンを十分に分解するために一晩反応させることが好ましいが、菌体外酵素液量によっては2時間以上、10時間以下程度の短時間にて反応を終了させることも可能である。
【0034】
本発明で用いるM.chitosanitabida種菌としては、M.chitosanitabida H1株(以下、単に「H1株」という)が好適である。H1株は、下記の通り寄託機関に寄託されている。
(i) 受領機関の名称およびあて名
名称: 独立行政法人製品評価技術基盤機構 特許微生物寄託センター
あて名: 日本国 千葉県木更津市かずさ鎌足2−5−8
(ii) 受領日: 平成20年(2008年)8月12日
(iii) 受領番号: NITE AP−629
【0035】
H1株をWicherham寒天培地上で生育させた場合には、培養初期で白色のコロニーを形成し、成熟するにしたがって徐々に乳白色から黄色になる。また、光学顕微鏡を用いて観察した形態的特徴は、以下の通りである。即ち、幅0.7〜1.0μm、長さ1.0〜4.0μmのグラム陰性桿菌で、運動性とカタラーゼ活性を示す。以上の形態的特徴は、M.chitosanitabida種に属する細菌に当てはまる。H1株は好気性であり、30℃で良好に生育し、中性(pH6.8〜7.0)の培地で良好に増殖する。よって、H1株を培養する際の温度は、25℃以上、35℃以下程度が好適で、中性付近での培養が好適である。
【0036】
従来、β−1,3−グルカナーゼを菌体外へ放出する一方で、その他のβ−グルカナーゼやα−グルコシダーゼを実質的に放出しない菌は知られていなかった。よって、本発明に係るH1株は新規なものである。
【0037】
本発明で用いるS.omiyaensis種菌としては、S.omiyaensis SY26株(以下、単に「SY26株」という)が好適である。SY26株は、下記の通り寄託機関に寄託されている。
(i) 受領機関の名称およびあて名
名称: 独立行政法人製品評価技術基盤機構 特許微生物寄託センター
あて名: 日本国 千葉県木更津市かずさ鎌足2−5−8
(ii) 受領日: 平成20年(2008年)7月29日
(iii) 受領番号: NITE AP−616
【0038】
SY26株をWicherham寒天培地で生育させた場合には、培養初期で白色コロニーを形成し、成熟するにしたがって徐々にベージュから灰色のコロニーへと変化する。光学顕微鏡を用いて観察した形態的特徴は、以下の通りである。即ち、培地基質中において無色透明の基生菌糸が観察され、その直径は0.8〜1.5μmであり、菌糸は直線状で分岐しており、成熟するとその先端が切断されて胞子を形成する。胞子表面にはシワがなく、楕円形もしくは長楕円であり、その大きさは直径0.5〜1.0μm、長さは1μm〜4μmである。さらに、前記培地で生育した本菌では、胞子嚢、運動性の胞子、分節胞子は確認されず、メラニン生成は陰性である。以上の形態的特徴は、Streptomyces omiyaensis種に属する担子菌に当てはまる。
【0039】
また、SY26株は好気性であり、30℃で良好に生育し、中性(pH6.8〜pH7.0)の培地で良好に増殖した。よって、SY26株を培養する際の温度は、25℃以上、40℃以下程度が好適で、中性付近での培養が好適である。
【0040】
従来、β−1,6−グルカナーゼを菌体外へ放出する一方で、その他のβ−グルカナーゼやα−グルコシダーゼを実質的に放出しない菌は知られていなかった。よって、本発明に係るSY26株は新規なものである。
【0041】
上記菌体外酵素液による反応後、β−1,3−グルカナーゼとβ−1,6−グルカナーゼの作用により生じたグルコースの量を測定する。
【0042】
本発明に係るβ−1,3−グルカナーゼとβ−1,6−グルカナーゼは、それぞれβ−1,3−グルコシド結合とβ−1,6−グルコシド結合を特異的に切断する。よって、上記菌体外酵素液により、多糖類含有試料中に存在するβ−1,3−1,6−グルカンは選択的にグルコースまで分解されるので、測定されたグルコース量から、β−1,3−1,6−グルカンの量を把握することができる。
【0043】
具体的には、生成したグルコースの量は、例えばSomogyi−Nelson法(M.Somogyi,J.Biol.Chem.,195,p.19(1952)を参照)など遊離の還元糖を測定する方法により測定することが可能である。また、酵素法やHPLCなどでも測定可能である。酵素法としては、グルコース−6−リン酸デヒドロゲナーゼとヘキソキナーゼを組み合わせたG6PDH−HK法(F−キット,Roche社製)や、ムタロターゼとグルコースオキシダーゼを組み合わせたムタロターゼ・GOD法(グルコースCII−テストワコー,Wako社製)がある。また、HPLC分析用のカラムとしてはTSKgel Sugar AXG(東ソー社製)やSugar−PakTMI(Waters社製)など、グルコースを分析するのに適したカラムが多数存在するので、これを利用できる。
【0044】
本発明に係るβ−1,3−1,6−グルカンの定量用キットは、β−1,3−グルカナーゼの産生能を有し且つβ−1,3−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないM.chitosanitabida種菌に由来する菌体外酵素液またはその乾燥物;β−1,6−グルカナーゼの産生能を有し且つβ−1,6−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないS.omiyaensis種菌に由来する菌体外酵素液またはその乾燥物;並びに、
グルコース量測定用試薬;を含むことを特徴とする。
【0045】
上記M.chitosanitabida種菌に由来する菌体外酵素液と上記S.omiyaensis種菌に由来する菌体外酵素液は、本発明方法で用いるものと同様のものとすることができる。また、それらの乾燥物は、当該菌体外酵素液を乾燥したものであればよい。但し、上述したようにβ−1,3−グルカナーゼとβ−1,6−グルカナーゼは高温で失活するために、上記M.chitosanitabida種菌由来の菌体外酵素液は60℃以下で、上記S.omiyaensis種菌由来の菌体外酵素液は45℃以下で乾燥することが好ましい。
【0046】
グルコース量の測定用試薬としては、例えば、生じたグルコース量をSomogyi−Nelson法で測定する場合には、Somogyi試薬とNelson試薬、或いはこれらを調製するための化合物を挙げることができる。また酵素法を用いる場合には例えば、生じたグルコース量を酵素法で測定する場合にはグルコース−6−リン酸デヒドロゲナーゼとヘキソキナーゼを組み合わせたG6PDH−HK法(F−キット,Roche社製)や、ムタロターゼとグルコースオキシダーゼを組み合わせたムタロターゼ・GOD法(グルコースCII−テストワコー,Wako製)に類する試薬または、或いはこれらを調製するための化合物を挙げることができる。
【0047】
当該キットは、本発明に係るβ−1,3−1,6−グルカンの定量方法で用いることができる。
【実施例】
【0048】
以下、実施例を挙げて本発明をより具体的に説明するが、本発明はもとより下記実施例により制限を受けるものではなく、前・後記の趣旨に適合し得る範囲で適当に変更を加えて実施することも可能であり、それらはいずれも本発明の技術的範囲に含まれる。
【0049】
実施例1 β−1,3−グルカナーゼ産生菌の単離
高知大学農学部内から土壌試料を採取した。当該土壌試料(5g)に滅菌水(15mL)を加え、ボルテックスミキサーを用いて攪拌した。次いで、当該混合物を数分間静置し、上清(50μL)をマイクロピペッターで採取した。
【0050】
別途、カードラン(β−1,3−グルカン、2g)を水道水(200mL)に懸濁し、オートクレーブで121℃、20分間処理した。これをミキサーにかけ、生じたゲルを完全にすり潰した。これにポリペプトン(0.125g)と酵母エキス(0.125g)を溶解し、水道水を加えて500mLにメスアップした。さらに10gの寒天を加え、オートクレーブで121℃、20分間滅菌処理し、シャーレ上で固めることによって、カードラン平板培地を調製した。また、デンプン培地(0.4%デンプン、0.025%ポリペプトン、0.025%酵母エキス、2%寒天、pH7.0)も調製した。
【0051】
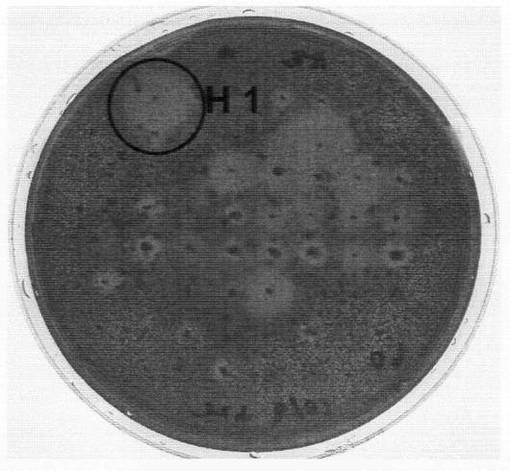
上記上清をカードラン平板培地に蒔き、スプレッダーで広げた後、30℃で1日間培養した。480の菌株を単離し、同平板培地に植菌してさらに30℃で2日間培養した。この培地を0.1%コンゴーレッド溶液で2〜3時間染色した後、流水中で2時間脱色してハロの形成を確認したところ、1菌株のみがハロを形成した。当該菌株をデンプン平板培地に塗布し、30℃で1日間培養した。この培地をヨウ素溶液で2〜3時間染色した後(中村道徳・貝沼圭二編,「生物化学実験法 19 澱粉・関連糖質実験法」学会出版センターを参照)、流水中で2時間脱色したが、ハロの形成は確認されなかった。カードラン平板培地で培養後のコンゴーレッド染色写真を図1に、デンプン培地で培養後のヨウ素染色写真を図2に示す。
【0052】
図1〜2のとおり、上記菌株はβ−1,3−グルカンの分解活性を示すのに対してデンプン(α−1,4−1,6−グルカン)に分解活性を示さないことから、β−1,3−グルカナーゼを産生する一方でα−グルコシダーゼは産生しないと考えられた。この菌株をH1株と仮に命名し、以後の実験に用いた。
【0053】
実施例2 H1株の顕微鏡観察
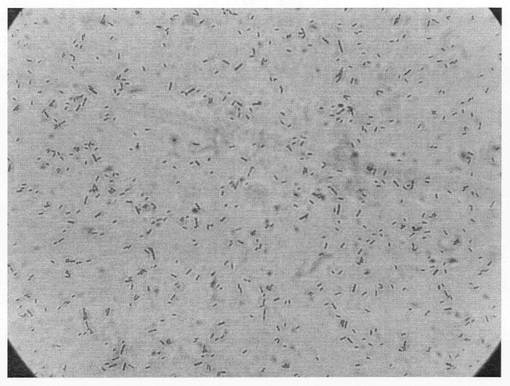
上記実施例1で得たH1株を顕微鏡(OLYMPUS社製、製品名「BH−2」)で1000倍に拡大し、観察した。拡大写真を図3に示す。
【0054】
図3のとおり、H1株は幅0.7〜1.0μm、長さ1.0〜4.0μmの桿菌であった。また、H1株はグラム陰性であり、カタラーゼ活性を示した。
【0055】
実施例3 H1株の16SrDNA塩基配列解析
(1) ポリメラーゼ連鎖反応
上記実施例1で得たH1株の属種を決定するために、16SrDNA塩基配列を解析した。具体的には、先ず10×ExTaq緩衝液(5μL)、dNTP、dTTP、dGTPおよびdCTPを各2.5mM含む混合液(8μL)、配列番号1〜2の各プライマーの5pmol/μL溶液(各1μL)を混合し、全量が49.5μLとなるように滅菌水に加えた。別途、MO BIO社製のUltra Clean Microbial DNA Isolation Kitを用いてH1株のゲノムDNA溶液を得た。当該ゲノムDNA溶液(1μL)か、或いは滅菌した爪楊枝によりH1株のコロニーから取得した少量の菌体を、上記溶液に加えて反応混合液とした。ポリメラーゼ連鎖反応には、DNAサーマルサイクラー(宝酒造社製)を使用した。反応条件は、以下のとおりである。
・ 熱変性 − 94℃,5分間
・ Ex Taq DNA ポリメラーゼ(宝酒造社製,2.5U)の添加
・ 熱変性 − 94℃,45秒間
・ プライマーのアニーリング − 55℃,1分間
・ DNA鎖の伸長反応 − 72℃,2分間
・ 2回目の熱変性以降のステップを35サイクル繰り返す
・ 熱処理 − 72℃,7分間
【0056】
(2) 16SrDNA断片の分離
上記ポリメラーゼ連鎖反応により増幅された16SrDNA断片を、アガロースゲル電気泳動により分離した。電気泳動に用いるアガロースゲルは、Sea Kem GTG アガロース(FMC)の1.5%溶液から調製した。泳動バッファーとしては、Howly緩衝液(40mM Tris,20mM酢酸ナトリウム,2mM EDTA,pH7.8)を用いた。分子量マーカーには、Hind IIIで処理したλDNAを使用した。
【0057】
電気泳動後、アガロースゲルを1μg/mLの臭化エチジウム溶液に10分間浸して染色することによりDNAを可視化し、紫外線を照射して写真撮影を行った。撮影後、目的の大きさと一致するバンドの部分を滅菌済みのかみそりで切り出し、メンブレンフィルター付きマイクロチューブ(宝酒造社製,SUPRECTM−01)に入れ、完全に凍結させた。37℃で5分間静置して融解した後、10,000rpmで10分間遠心分離し、200μLのTE緩衝液(0.1mM EDTA,10mM Tris−HCl,pH7.5)を加え、さらに10分間遠心分離した。2倍量の冷却エタノールと1/20量の3M酢酸ナトリウムを加え、−20℃で1時間静置した。その後、12,000rpmで10分間遠心分離し、エタノールを除去し、吸引乾燥することにより16SrDNA断片を含む混合物を得た。
【0058】
(3) 16SrDNA断片の塩基配列の決定
ポリメラーゼ連鎖反応により16SrDNA断片を増幅した上で、その塩基配列を解析した。具体的には、鋳型DNA(4μL)、5pmol/μLプライマー溶液(1μL)、BigDyeターミネーターver.3.1 5×シーケンシング緩衝液(1.5μL)、BigDyeターミネーターver.3.1(1μL)を混合し、滅菌水で全量を10μLにし、これを反応液とした。当該反応液に上記(2)で得た混合物を加え、下記反応条件でポリメラーゼ連鎖反応を行った。
・ 熱変性 − 96℃,1分間
・ 熱変性 − 96℃,10秒間
・ プライマーのアニーリング − 50℃,5秒間
・ DNA鎖の伸長反応 − 60℃,4分間
・ 2回目の熱変性以降のステップを25サイクル繰り返す
【0059】
次に、Sephadex G−50を用い、冷却した反応混合液から16SrDNA断片を精製した。得られた16SrDNA断片にHi−Diホルムアミド(10μL)を加え、ABI Prism 3100−Avant ジェネティックアナライザー(アプライドバイオシステム社製)により塩基配列を解析した。当該塩基配列を配列表の配列番号3に示す。
【0060】
得られた16SrDNA塩基配列は、M.chitosanitabida種菌の16SrDNA塩基配列と高い相同性を示すものであったことから、H1株はM.chitosanitabida種に属するものであると判断した。
【0061】
実施例4 H1株由来の粗酵素液の基質特異性
(1) H1株由来の粗酵素液の調製
試験管内で、7mLのWicherham培地(0.5%ペプトン,0.3%酵母エキス、0.3%麦芽エキス,1.0%グルコース,pH7.0)を作成した。当該培地に白金耳でH1株を植菌し、30℃で1日間振とう培養した。別途、生パン酵母培地(4.0%生パン酵母(協和発酵社製,ダイヤイースト))を調製し、当該培地に前培養したH1株を移し、30℃で2日間培養した。当該培養液を遠心分離(18,000rpm,20分間,4℃)し、沈殿が入らないよう上清を得た。この上清へ終濃度が0.1mMになるようにPhenylmethylsulfonyl fluoride(和光純薬社製,以下、「PMSF」と省略する)を加え、撹拌・氷冷しながら、乳鉢で粉砕した硫酸アンモニウムを飽和濃度の80%となるように加えた。撹拌・氷冷しながら一晩静置後、遠心分離(18,000rpm,20分間,4℃)し沈殿を得た。得られた沈殿を、0.1mM PMSFを含む少量の20mM酢酸ナトリウム緩衝液(pH6.0)に溶解した。これを透析膜(Viskase Companies,inc製)に入れ、2Lの同緩衝液を用いて透析することにより粗酵素液を得た。
【0062】
(2) 粗酵素液の活性測定
前項で得た粗酵素液と様々なグルカン基質を反応させ、生成した還元糖を検出することによりグルカナーゼ活性を測定した。具体的には、0.2M酢酸ナトリウム緩衝液(pH6.0)に、黒酵母β−グルカン(Aureobasidium sp.由来,株式会社ソフィより提供)、パスツラン(Calbiochem社製)、リケナン(シグマ社製)、ラミナリン(Eisenia araborea由来,半井化学薬品社製)、ラミナラン(Laminaria由来,東京化成工業社製)、カードラン(和光純薬社製)、セルロース(NIHONBASHI社製)、ルテオース(高知大農学部応用微生物学研究室より提供)、デンプン(和光純薬社製)、デキストラン(和光純薬社製)、プルラン(半井化学薬品社製)をそれぞれ10g/Lになるように溶解した。完全に溶解させるために、リケナンは熱湯中で煮沸し、また、パスツランはオートクレーブにより121℃で5分間高圧加熱した。なお、基質濃度はすべて0.5%に調整し、粗酵素溶液を加えてから30℃で1時間反応させた。反応後、5分間煮沸し、生成した還元糖量をSomogyi−Nelson法にて測定した(M.Somogyi,J.Biol.Chem.,195,p.19(1952)を参照)。
【0063】
詳しくは、KNaC4H4O6・4H2O(4.0g)、Na2HPO4・12H2O(7.1g)、CuSO4・5H2O(0.8g)、1M NaOH(10mL)およびNa2SO4・5H2O(18.0g)を蒸留水(100mL)に溶解し、37℃で24時間インキュベートした。次いで当該溶液を濾過し、得られた濾液をSomogyi試薬とした。また、(NH4)6Mo7O24・4H2O(5.0g)、H2SO4(4.2g)およびNa2HAsO4・7H2O(0.6g)を蒸留水(100mL)に溶解し、24時間インキュベートした。次いで当該溶液を濾過し、得られた濾液をNelson試薬とした。上記酵素反応溶液(1mL)にSomogyi試薬(1mL)を加え、湯鍋中にて正確に10分間反応させた後、即座に流水中で5分間水冷した。当該溶液にNelson試薬(1mL)と蒸留水(2mL)を加え、室温にて約10分間静置後、分光光度計(SHIMADZU社製,UV−1200)を用いて520nmにおける吸光度を測定した。なお、標準物質はグルコースを用いた。また、30℃で1時間に1μmolの還元糖を生成する酵素量を1unitと定義した。結果を表1に示す。
【0064】
【表1】

【0065】
上記結果のとおり、H1株から得られた粗酵素液は、β−1,3−グリコシド結合を含む黒酵母β−グルカン、ラミナリン、ラミナラン、リケナンおよびカードランに対して高い活性を示した。その一方で、β−1,6−グリコシド結合のみからなるパスツランとルテオースに対する活性は見られなかった。さらに、α−グルカンであるプルランに対する活性が観察されたものの、その比活性は低いものであった。
【0066】
以上の結果から、H1株由来の粗酵素液は、β−1,3−グリコシド結合を特異的に切断するβ−1,3−グルカナーゼを含む一方で、その他のβ−グルカナーゼを含まず、また、わずかにα−グルコシダーゼ活性を示すものの、デンプンとデキストランを分解できないことから、その活性は極めて低いことが明らかにされた。
【0067】
実施例5 H1株由来の菌体外酵素液の調製
(1) H1株由来の粗酵素液の調製
上記実施例4(1)と同様に、Mitsuaria chitosanitabida H1株をWicherham培地(70mL)で前培養した後、生パン酵母培地(1L)で本培養した。当該培養液を遠心分離(9,100rpm,20分間,4℃)し、沈殿が入らないようにして上清を得た。この上清に1/1,000量の0.1M PMSFと、1/5,000量の消泡剤を加え、エバポレーターで全量の約1/10量まで減圧濃縮した。次いで、終濃度が20mMになるようにリン酸カリウム緩衝液(pH7.4)を加え、撹拌・氷冷しながら、乳鉢で粉砕した硫酸アンモニウムを加えていき沈殿物を分離することにより、0〜20%画分、20〜40%画分、40〜60%画分、60〜80%画分を得た。各画分を20mM酢酸ナトリウム緩衝液(pH6.0)で透析後、β−1,3−グルカナーゼ活性を測定し、また、全タンパク質を定量した。詳しくは、E.araborea由来のラミナリンに酵素液を加え、30℃で1時間反応させた後、生じた還元糖量をSomogyi−Nelson法により定量することにより全活性と比活性を測定した。また、タンパク質はLowry法(Lowry,O.H.ら,J.Biol.Chem.,193,pp.265-275(1951)を参照)により定量した。結果を表2に示す。
【0068】
【表2】
【0069】
上記結果のとおり、全活性は60〜80%飽和画分で最も高く、比活性は20〜40%飽和画分で最も高かった。それに対して、0〜20%飽和画分は全活性と比活性が低かった。よって、以後の実験では20〜80%飽和画分を用いた。
【0070】
(2) H1株由来の菌体外酵素液の調製
0.1mM PMSFを含む20mM酢酸ナトリウム緩衝液(pH5.0)を使って、上記で得た粗酵素液の20〜80%飽和画分を透析した後、遠心分離(18,000rpm,20分間,4℃)したところ、ゲル状の沈殿が生じた。上記実施例4(2)と同様の方法により、E.araborea由来のラミナリンを基質として、生じたゲル状沈殿と上清のそれぞれのグルカナーゼ活性を測定したところ、ゲル状沈殿に非常に高い活性が見られた。当該ゲル状沈殿を20mM酢酸ナトリウム緩衝液(pH5.0)に懸濁してから遠心分離(18,000rpm,20分間,4℃)する操作を繰り返すことにより洗浄した。洗浄した粗酵素液の基質特異性に変化がないかを調べるために、上記実施例4(2)と同様の実験を再び行った。結果を表3に示す。
【0071】
【表3】
【0072】
表3に示す結果のとおり、得られた菌体外酵素液はβ−1,3−グリコシド結合を有する基質に対しては作用する一方で、α−1,4−グリコシド結合、α−1,6−グリコシド結合、β−1,4−グリコシド結合およびβ−1,6−グリコシド結合には作用しない。よって、当該H1株由来の菌体外酵素液は、β−1,3−グリコシド結合を特異的に切断する一方で、α−グルコシダーゼ活性やその他のβ−グルカナーゼ活性を有しないことが実証された。なお、当初はカラムクロマトグラフィによりさらに精製を進めてその酵素学的性質を調べる予定であったが、精製酵素液がゲル状であるためかかる精製は行うことができなかった。
【0073】
実施例6 H1株由来のβ−1,3−グルカナーゼの至適温度
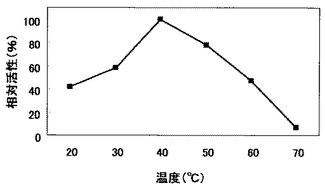
上記実施例5(2)で得た菌体外酵素液を用い、反応温度を20〜70℃に変更した以外は上記実施例5(1)と同様にして比活性を測定し、H1株由来のβ−1,3−グルカナーゼの至適温度を調べた。結果を図4に示す。
【0074】
図4のとおり、H1株由来のβ−1,3−グルカナーゼの至適温度は40℃であり、活性は50℃から徐々に下がり始め、70℃では活性はほとんど失われることが分かった。
【0075】
実施例7 H1株由来のβ−1,3−グルカナーゼの熱安定性
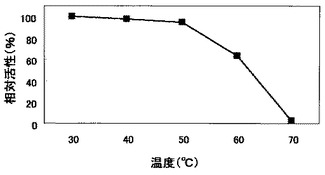
上記実施例5(2)で得た菌体外酵素液を30〜70℃で15分間処理した後に、氷上で急冷した。当該溶液を用い、上記実施例5(1)と同様にして比活性を測定し、H1株由来のβ−1,3−グルカナーゼの熱安定性を調べた。結果を図5に示す。
【0076】
図5のとおり、H1株由来のβ−1,3−グルカナーゼは50℃までは高い安定性を示すが、70℃では完全に失活することが分かった。
【0077】
実施例8 H1株由来のβ−1,3−グルカナーゼの反応生成物の同定
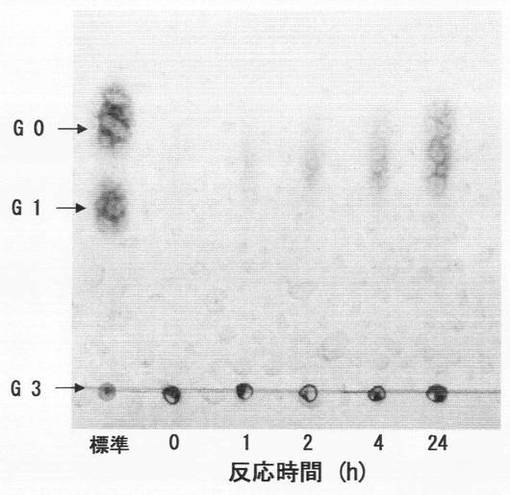
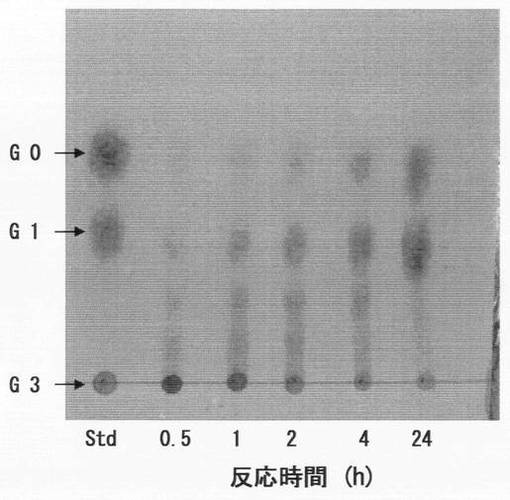
β−1,3−グルカンであるカードランに、上記実施例5(2)で得た菌体外酵素液を上記実施例4(2)と同様の反応条件で作用させ、反応混合液を経時的に分取し、反応生成物を薄層クロマトグラフィで分析した。具体的には、反応開始から0、1、2、4、24時間後に反応混合液を分取し、沸騰水で10分間加熱処理して酵素反応を止めた後、HPTLC Fertigplatten Kieselgel 60(Merck社製,10cm×10cm)を用いた薄層クロマトグラフィに供した。標準物質として1%グルコースを用い、展開溶媒にはn−ブタノール:エタノール:水=5:2:2の混合溶媒を用い、上昇法で行った。展開後、発色液として、ジフェニルアミン−アニリン−リン酸(2gジフェニルアミン,2mLアニリン,100mLアセトン,15mL 80%リン酸)をゲルプレート上に噴霧し、乾熱器中、100℃で10分間加熱し、出現したスポットを観察した。結果を図6に示す。図6中、「G3」はカードランを示し、「G1」はグルコース2分子がβ−1,6結合したゲンチオビオースであり、「G0」はグルコースである。
【0078】
図6のとおり、H1株由来のβ−1,3−グルカナーゼによる反応の最終生成物はグルコースであり、その生成量は経時的に増加した。よって、H1株由来のβ−1,3−グルカナーゼは、β−1,3−グルカンを構成単糖であるグルコースまで分解できるSタイプの酵素であることが確認された。
【0079】
実施例9 β−1,6−グルカナーゼ産生菌の単離
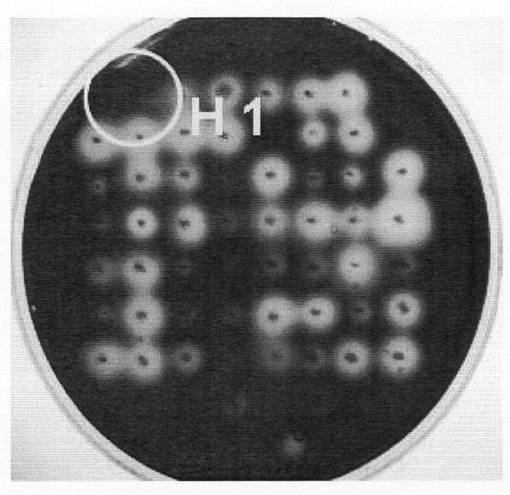
高知大学農学部内から土壌試料を採取した。当該土壌試料(5g)に滅菌水(15mL)を加え、ボルテックスミキサーを用いて攪拌した。次いで、当該混合物を数分間静置し、上清(50μL)をマイクロピペッターで採取した。当該上清をパスツラン平板培地(0.1%パスツラン(Calbiochem製,0.025%ポリペプトン、0.025%酵母エキス、2%寒天、pH7.0)に蒔き、スプレッダーで広げた後、30℃で2日間培養した。180の菌株を単離し、同平板培地に植菌してさらに30℃で4日間培養した。この培地を0.1%コンゴーレッド溶液で2〜3時間染色した後(Andres Solerら,Journal of microbiological methods,35,pp.245-251(1999)を参照)、流水中で2時間脱色した。さらに20mM酢酸ナトリウム緩衝液(pH5.5)に一晩浸すことでコンゴーレッドを変色させた後、ハロの形成を確認した。培地の写真を図7に示す。
【0080】
図7の結果のとおり、180の菌株中1菌株のコロニー周辺にハロらしきものが認められた。即ち、この菌株はβ−1,6−グルカンであるパスツランに対する分解活性を示すことから、β−1,6−グルカナーゼを産生していると考えられる。この菌株をSY26と命名し、以後の実験に用いた。
【0081】
実施例10 SY26株の顕微鏡観察
上記実施例9で得たSY26株を顕微鏡(OLYMPUS社製、製品名「BH−2」)で1000倍に拡大し、観察した。拡大写真を図8に示す。
【0082】
図8のとおり、SY26株は無色透明の基生菌糸を形成する。菌糸の直径は0.8〜1.5μmであり、直線状で分岐していた。また、当該菌糸は、成熟するとその先端が切断されて胞子を形成した。胞子表面にはシワがなく、楕円形もしくは長楕円であり、その大きさは直径0.5〜1.0μm、長さは1μm〜4μmであった。さらに、SY26株では、胞子嚢、運動性の胞子、分節胞子は確認されず、メラニン生成は陰性である。
【0083】
実施例11 SY26株の16SrDNA塩基配列解析
上記実施例3と同様にして、SY26株の16SrDNA塩基配列を解析した。当該塩基配列を配列表の配列番号4に示す。
【0084】
得られた16SrDNA塩基配列は、S.omiyaensis種菌の16SrDNA塩基配列と高い相同性を示すものであったことから、SY26株はS.omiyaensis種に属するものであると判断した。
【0085】
実施例12 SY26株由来の粗酵素液の基質特異性
本培養期間を6日間とした以外は上記実施例4(1)と同様にして、SY26株の培養液から粗酵素液を得た。さらに上記実施例4(2)と同様にして、当該粗酵素液の活性を測定した。結果を表4に示す。
【0086】
【表4】
【0087】
上記結果のとおり、SY26株から得られた粗酵素液は、β−1,6−グリコシド結合を含む黒酵母β−グルカン、ラミナリン、パスツランおよびルテオースに対して高い活性を示した。それに対して、β−1,6−グリコシド結合を含まず且つβ−1,3−グリコシド結合を含むリケナンとカードランに対する活性は見られなかった。さらに、α−グルコシダーゼ活性は見られなかった。
【0088】
以上の結果から、SY26株の粗酵素液は、β−1,6−グリコシド結合を特異的に切断できる一方で、α−グルコシダーゼ活性とその他のβ−グルカナーゼ活性を有しないことが明らかにされた。
【0089】
実施例13 SY26株由来の菌体外酵素液の調製
(1) SY26株由来の粗酵素液の調製
上記実施例4(1)と同様に、S.omiyaensis SY26株をWicherham培地(14mL)で前培養した後、生パン酵母培地(200mL)で本培養した。当該培養液から上記実施例5(1)と同様にして0〜20%画分、20〜40%画分、40〜60%画分、60〜80%画分を得た。また、基質としてラミナリンの代わりにパスツランを用いた以外は上記実施例5(1)と同様にして、β−1,6−グルカナーゼ活性を測定し、また、全タンパク質を定量した。結果を表5に示す。
【0090】
【表5】
【0091】
上記結果のとおり、全活性は20〜40%飽和画分で最も高く、比活性は40〜60%飽和画分で最も高かった。60〜80%飽和画分もわずかに活性を示したが、全活性と比活性の値が共に低かったため、以後の実験では20〜60%飽和画分を用いた。
【0092】
(2) β−1,6−グルカナーゼの精製
35mLのWicherham培地でSY26株を前培養した後に500mLの生パン酵母培地で本培養した以外は上記実施例13(1)と同様にして、培養液の20〜60%飽和画分を得た。
【0093】
0.1mM PMSFを含む20mM酢酸ナトリウム緩衝液(pH5.0)を使って、上記で得た粗酵素液の20〜60%飽和画分を透析した後、透析中に生じた沈殿物を遠心分離(18,000rpm,20分間,4℃)で取り除いたものを菌体外酵素液とし、上記実施例13(1)と同様に活性測定を行った。また、全タンパク質量をプロテインアッセイCBB溶液(ナカライテスク社製)により定量した。上記菌体外酵素液の全活性は40.9unit、全タンパク質量は3.24mg、比活性は3.89unit/mgであった。
【0094】
上記菌体外酵素液を、さらにゲル濾過クロマトグラフィにより精製した。具体的には、0.1mM PMSFを含む20mM酢酸ナトリウム緩衝液(pH5.0)で平衡化したSP−セファデックス C−50 カラム(ファルマシア社製,3.0×30cm)に上記粗酵素液を供した。非吸着タンパク質は同緩衝液で溶出させ、吸着タンパク質は0〜0.5Mの塩化ナトリウムを含む同緩衝液でリニアグラジエントにより溶出させ、5mL毎にフラクションを分取した。各フラクションの比活性を測定し、活性の高いフラクションを集めてアミコンメンブレンフィルターで6mLに濃縮した。0.1mM PMSFを含む20mMリン酸カリウム緩衝液(pH7.4)で透析後、透析中に生じた沈殿物を遠心分離(18,000rpm,20分間,4℃)で取り除いた。上記と同様にして活性測定とタンパク定量を行ったところ、全活性は29.8unit、全タンパク質量は2.23mg、比活性は13.3unit/mgであった。
【0095】
上記酵素液を、さらにイオン交換クロマトグラフィで精製した。詳しくは、0.1mM PMSFを含む20mMリン酸カリウム緩衝液(pH7.4)で平衡化したDEAE−トヨパール 650Mカラム(東ソー株式会社,3.0×30cm)に上記酵素液を供した。タンパク質を同緩衝液で溶出させ、5mL毎にフラクションを分取し、活性の高いフラクションを集めた。当該酵素液の全活性は18.3unit、全タンパク質量は0.31mg、比活性は58.2unit/mgであった。
【0096】
上記酵素液をCentriprepで約1mLまで濃縮し、さらにMicroconを用いて50μLまで濃縮した後、BioLogic Duo Flow(バイオラッド製)を用いたFPLCで精製した。具体的には、0.15Mの塩化ナトリウムを含む20mMリン酸カリウム緩衝液(pH7.4)で平衡化したSuperose 12 HR 10/30 カラムに濃縮した酵素液を供した。同緩衝液を通液し続けてタンパク質を溶出させて0.5mL毎に分取し、活性の高いフラクションを集めてこれを精製酵素液とした。上記と同様にして当該精製酵素液の活性測定とタンパク定量を行ったところ、全活性は3.65unit、全タンパク質量は0.01mg、比活性は386unit/mgであった。
【0097】
以上の各精製段階における全タンパク質量などのデータを、表6にまとめる。
【0098】
【表6】
【0099】
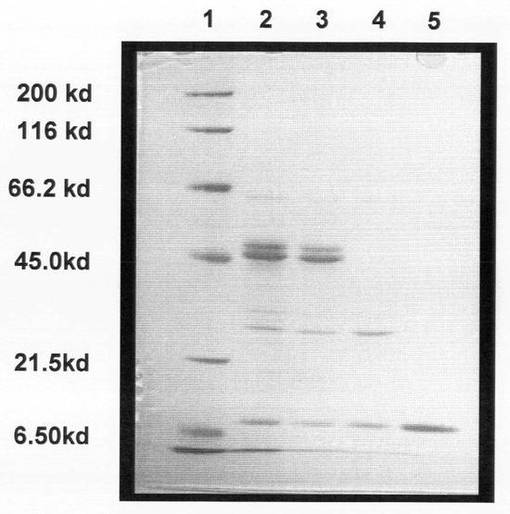
また、上記各精製段階の酵素液を、Laemmliの方法(U.K.Laemmli,Nature,277,pp.680-685(1970)を参照)に従って分析した。電流は、濃縮ゲル時は10mA、分離ゲル時は20mAとした。また、分子質量測定の際のマーカーとして、ミオシン(200kD)、β−ガラクトシダーゼ(116kD)、ウシ血清アルブミン(66kD)、卵白アルブミン(45kD)、炭酸脱水酵素(31kD)、トリプシン阻害剤(21.5kD)、リゾチーム(14.4kD)、アプロチニン(6.5kD)からなるタンパク質マーカー(ナカライテスク社製)を用いた。泳動終了後は、0.25%クマシーブリリアントブルーR−250を含むメタノール:酢酸:水=5:1:88の混合溶液にゲルを浸し、振とうしながら50℃で30分間染色した。その後、メタノール:酢酸:水=5:7:88の混合溶液に浸し、振とうしながら50℃で脱色した。脱色したゲルは、デジタルカメラを用いて画像を保存し、ゲル乾燥機で乾燥させて保存した。結果を図9に示す。
【0100】
図9のとおり、最終的に得られた精製酵素液中には、分子質量が10kDのタンパク質のみが含まれていることが分かる。
【0101】
実施例14 SY26株由来のβ−1,6−グルカナーゼの至適温度
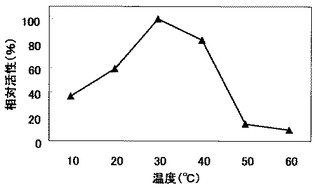
上記実施例13(2)で得たSuperose 12 HR 10/30 カラム溶出画分である精製酵素液を用い、反応温度を10〜60℃に変更した以外は上記実施例13(1)と同様にして比活性を測定し、SY26株由来のβ−1,6−グルカナーゼの至適温度を調べた。結果を図10に示す。
【0102】
図10のとおり、SY26株由来のβ−1,6−グルカナーゼの至適温度は30℃であり、40℃では30℃の場合の83%の活性を示したが、50℃以上になると活性は急激に失われることが分かった。
【0103】
実施例15 SY26株由来のβ−1,6−グルカナーゼの熱安定性
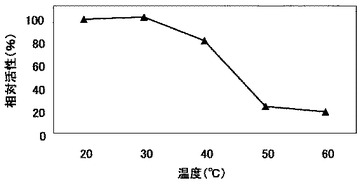
上記実施例13(2)で得たSuperose 12 HR 10/30 カラム溶出画分である精製酵素液を20〜60℃で15分間処理した後に、氷上で急冷した。当該溶液を用い、上記実施例13(1)と同様にして比活性を測定し、SY26株由来のβ−1,6−グルカナーゼの熱安定性を調べた。結果を図11に示す。
【0104】
図11のとおり、SY26株由来のβ−1,6−グルカナーゼは30℃までは高い安定性を示し、40℃では30℃の場合の約80%の活性を示したが、50℃では失活することが分かった。
【0105】
実施例16 SY26株由来のβ−1,6−グルカナーゼの反応生成物の同定
β−1,6−グルカンであるパスツランに、上記実施例13で精製したβ−1,6−グルカナーゼ含有溶液を上記実施例13(1)と同様の反応条件で作用させ、反応混合液を経時的に分取し、反応生成物を薄層クロマトグラフィで分析した。結果を図12に示す。図12中、「G3」はパスツランを示し、「G1」はグルコース2分子がβ−1,6結合したゲンチオビオースであり、「G0」はグルコースである。
【0106】
図12のとおり、SY26株由来のβ−1,6−グルカナーゼによる反応の最終生成物はグルコースであり、その生成量は経時的に増加した。よって、SY26株由来のβ−1,6−グルカナーゼは、β−1,6−グルカンを構成単糖であるグルコースまで分解できるSタイプの酵素であることが確認された。
【0107】
実施例17 β−1,3−1,6−グルカンの定量
天然由来のグルカン類としては、グルコースのみからなるホモグルカンであるα−グルカンとβ−グルカン、および他の糖と結合したヘテログルカンが存在する。従来方法は、これらグルカン類の内、α-グルカンを取り除いた残りの全てのグルカンを構成単糖まで分解してグルコースを定量するものであることから、有用なβ−1,3−1,6−グルカンのみを定量することはできなかった。
【0108】
一方、上記実験により、H1株とSY26株に由来する菌体外酵素液は、それぞれβ−1,3−グリコシド結合とβ−1,6−グリコシド結合を特異的に切断できることが実証されていることから、これら酵素液によりグルカンをグルコースまで完全に分解した後にグルコース量を測定すれば、事前にα−グルカンを分解除去しなくてもβ−1,3−1,6−グルカンを正確に定量できると期待できる。
【0109】
そこで、先ず、本発明に係る2種の菌体外酵素液を使って、天然由来のグルカン中のβ−1,3−1,6−グルカンを定量し、次に、当該グルカンからα−グルカンを除去した上で同様の操作を行い、両結果を比較することで、本発明方法の効果を評価した。
【0110】
(1) 多糖標品の調製
Aureobasidium pullulans AFO202株の培養液(ソフィ社より提供,3g)に蒸留水(27mL)を加えて10倍希釈培養液を作成した。これを遠心分離(18,000rpm,20分間,4℃)し、得られた上清(7mL)に2倍量のエタノールを加え、よく撹拌して多糖を沈殿させた。さらに、遠心分離(18,000rpm,20分間,4℃)後、上清を捨て、沈殿物をエタノール臭がしなくなるまで風乾した。風乾後、沈殿物を14mLの20mM酢酸ナトリウム緩衝液(pH4.5)に溶解し、これを多糖標品とした。
【0111】
フェノール硫酸法(Dubois,M.ら,Anal.Chem.,28,p.350(1956)を参照)を用い、上記多糖標品に含まれる全多糖量を測定した。具体的には、上記多糖標品(500μL)と蒸留水(500μL)とを混合し、さらに5%フェノール(1mL)を加えた。ここへ硫酸(5mL)を加え、室温で10分間冷却した。当該混合液を流水で20分間冷却した後、490nmにおける吸光度を測定することによって、多糖量を定量したところ、全多糖量は3.80g/Lであった。
【0112】
(2) 試料中にα−グルカンが含まれる場合
上記(1)で得た多糖標品(500μL)に、1M酢酸ナトリウム緩衝液(pH6.0,50μL)、H1株由来の菌体外酵素液(225μL)およびSY26株由来の菌体外酵素液(225μL)を混合し、40℃で6時間インキュベートした。なお、使用したH1株由来の菌体外酵素液の活性はラミナリン(E.araborea由来)に対して2.2unitであり、SY26株由来の菌体外酵素液の活性はパスツランに対して1.4unitと、上記多糖標品中のグルカンを完全に分解するに十分である。
【0113】
上記酵素反応後における反応混合液について、Somogyi−Nelson法により還元糖量を測定した。結果を表7に示す。
【0114】
(3) 試料中にα−グルカンが含まれない場合
アミログルコシダーゼ(Rhizopus sp由来,シグマ社製)を0.2M酢酸ナトリウム水溶液(pH4.5)に1μg/mLの濃度で溶解し、アミログルコシダーゼ溶液を得た。上記多糖標品(3.6mL)とアミログルコシダーゼ溶液(0.4mL)をファルコンチューブに入れ、55℃で一晩インキュベートした。その後、5分間煮沸し、酵素反応を終了させた。このアミログルコシダーゼ処理で生じた還元糖をSomogyi−Nelson法で定量し、α−グルカン量とした。この反応液(500μL)にH1株由来の菌体外酵素液(225μL)およびSY26株由来の菌体外酵素液(225μL)を加え、40℃で6時間反応させた。当該酵素反応後における反応混合液について、Somogyi−Nelson法により還元糖量を測定し、当該測定値から上記のα−グルカン量値を引いてβ−1,3−1,6−グルカン量とした。結果を表7に示す。
【0115】
【表7】
【0116】
α−グルカンを除去しない場合のβ−1,3−1,6−グルカン量は3.09g/Lであり、α−グルカンを除去した場合は3.18g/Lであって、この二つに大きな差はなかった。従って、本発明方法は、標品中にα−グルカンが存在したままでも正確にβ−1,3−1,6−グルカンを定量できることが分かった。
【0117】
実施例18 β−1,3−1,6−グルカンの定量
上記実施例17のとおり、本発明方法によれば標品中にα−グルカンが存在したままでも正確にβ−1,3−1,6−グルカンを定量できるが、反応溶液中のα−グルカン量がこれより多くなると、α−グルカンを除去しなければ正確にβ−1,3−1,6−グルカン量を正確に定量できない可能性もある。そこで、α−グルカンをより多く生産するA.pullulans 132株の培養液を試料とし、同様の実験を行った。
【0118】
(1) 多糖標品の調製
A.pullulans 132株(寺尾啓吾(高知大学農学部生物資源科学科),「応用微生物学研究室卒業論文」(2006)を参照)を白金耳で一掻き取り、5mLの米ヌカ含有液体培地(0.2%米ヌカ、0.2%アスコルビン酸、1%スクロース、pH5.2)に植菌し、25℃、125rpmで48時間振とうして前培養した。次いで、30mL容の三角フラスコに100mLの米ヌカ含有液体培地を作成し、ここへ上記前培養液(1mL)を加え、25℃、125rpmで72時間振とうして本培養した。なお、米ヌカ含有液体培地は、多糖の生産を促進する。
【0119】
A.pullulans AFO202株の培養液の代わりに上記本培養の培養液(3g)を用いた以外は上記実施例17(1)と同様にして、多糖標品を得た。当該多糖標品に含まれる多糖量を上記実施例17(1)と同様の方法で定量したところ、3.04g/Lであった。
【0120】
(2) β−1,3−1,6−グルカンの定量
上記多糖標品について、α−グルカンを除去しない場合と除去した場合において、β−1,3−1,6−グルカンの量を上記実施例17(2)〜(3)と同様に測定した。結果を表8に示す。
【0121】
【表8】
【0122】
上記結果のとおり、A.pullulans 132株の培養液に含まれるα−グルカンは、A.pullulans AFO202株の培養液に含まれるα−グルカンよりも多い。しかし、本発明方法により測定されるβ−1,3−1,6−グルカンの量は、α−グルカンを除去しない場合で0.92g/Lであり、α−グルカンを除去した場合は1.10g/Lであって、この二つに大きな差はなかった。
【0123】
上記実施例17〜18の結果から、H1株とSY26株にそれぞれ由来するβ−1,3−グルカナーゼおよびβ−1,6−グルカナーゼを用いた本発明方法によれば、試料中におけるα−グルカンの存在または非存在、そして混在するα−グルカンの存在量にかかわらず正確にβ−1,3−1,6−グルカンを定量できることが実証された。
【図面の簡単な説明】
【0124】
【図1】実施例1において、土壌試料から単離された菌株をカードラン平板培地で培養した後、コンゴーレッド染色した写真である。
【図2】実施例1において、土壌試料から単離された菌株をデンプン培地で培養した後、ヨウ素染色した写真である。
【図3】実施例1で得たH1株の顕微鏡写真である。
【図4】反応温度とH1株由来のβ−1,3−グルカナーゼの活性との関係を示すグラフである。
【図5】H1株由来のβ−1,3−グルカナーゼの活性と熱処理温度との関係を示すグラフである。
【図6】H1株由来のβ−1,3−グルカナーゼによるカードランの分解反応生成物の経時的変化を示す薄層クロマトグラフィの結果である。
【図7】実施例9において、土壌試料から単離された菌株をパスツラン平板培地で培養した後、コンゴーレッド染色した写真である。
【図8】実施例9で得たSY26株の顕微鏡写真である。
【図9】SY26株由来のβ−1,6−グルカナーゼの各精製段階のSDS−PAGEの結果である。図中、1は分子量マーカーレーンを示し、2は20〜60%飽和硫安沈殿レーンを示し、3はゲル濾過クロマトグラフィによる精製後のレーンを示し、4はイオン交換クロマトグラフィによる精製後のレーンを示し、5はFPLCによる精製後のレーンを示す。
【図10】反応温度とSY26株由来のβ−1,6−グルカナーゼの活性との関係を示すグラフである。
【図11】SY26株由来のβ−1,6−グルカナーゼの活性と熱処理温度との関係を示すグラフである。
【図12】SY26株由来のβ−1,6−グルカナーゼによるパスツランの分解反応生成物の経時的変化を示す薄層クロマトグラフィの結果である。
【技術分野】
【0001】
本発明は、β−1,3−1,6−グルカンを定量する方法、当該方法に用いることができるβ−1,3−1,6−グルカンの定量用キット、および当該方法に有用な菌体外酵素を産生する微生物に関するものである。
【背景技術】
【0002】
細胞は、グリコーゲンやデンプンなどエネルギーとなる貯蔵多糖のほか、構造多糖を産生する。構造多糖とは、細胞の表面を保護する細胞膜や細胞壁の構成成分である多糖であり、グルカン、マンナン、糖タンパク質、キチンなど様々な種類が確認されている。
【0003】
近年、かかる構造多糖の中でも、担子菌類の産生するものが優れた生理活性を有することから注目を集めている。
【0004】
担子菌類とは、担子胞子を生ずる高等菌類であり、松茸や舞茸、アガリクスと呼ばれる姫松茸などがある。この担子菌類が産生する構造多糖は、食物繊維としての効果のほか、抗腫瘍作用、抗菌作用、血糖・血清コレステロール降下作用などを示し、糖尿病などの生活習慣病やがんなどの予防に有効であるとの研究成果も報告されている。
【0005】
担子菌類が産生する構造多糖の中でも、β−1,3−グルコシド結合を主鎖に有しβ−1,6−グルコシド結合を側鎖に有するβ−1,3−1,6−グルカンは、とりわけ特徴的な生理活性を示す(非特許文献1)。
【0006】
しかし、構造多糖は優れた生理活性を有するものの、細胞壁などの中に存在するためにその抽出は非常に困難である。その上、たとえ抽出したとしてもその抽出物は水に対して不溶性または難溶性を示し、特有の色や不快臭を有し、品質が不安定であり非常に高価であるなどの問題を有する。
【0007】
それに対して、黒酵母といわれるAureobasidium pullulans種菌の中からは、ある条件下でβ−1,3−1,6−グルカンを菌体外へ放出するものが見出されている。しかも、β−1,3−1,6−グルカンを含む当該菌の培養液は、無味無臭で品質が安定しており、安価に製造できる。よって、当該菌から得られるβ−1,3−1,6−グルカンを機能性食品などとして利用する道が開かれつつある。
【0008】
しかし、β−1,3−1,6−グルカンを含む培養液などを食品の原料として利用する場合には、その含量を測定するなどして品質を評価する必要があるものの、正確な定量方法は未だ存在しない。
【0009】
例えば、財団法人日本食品分析センターが行っているβ−グルカンの定量方法は、アミラーゼやアミログルコシダーゼなどによる酵素処理の後、硫酸で全てのグルコシド結合を非特異的に加水分解し、生じたグルコースを定量するものである。しかしこの方法では、β−1,3−1,6−グルカンとそれ以外のβ−グルカンを区別することができない。さらに、グルコース以外の単糖を含むヘテログルカンなどに由来するグルコースまで分解され、測定されてしまう。従って当該方法は、β−1,3−1,6−グルカンの定量方法としては不適である。
【0010】
そのほか、一般的に使用されているβ−グルカンの定量キットは、連続したβ−1,3−グルコシド結合を認識して結合するタンパク質因子を利用するものであるため、試料中に含まれるβ−1,3−グルカンにおける側鎖の割合が多いほど、測定値の誤差は大きくなる。
【0011】
ところで、非特許文献2〜4のとおり、本発明者らが属する研究室では、土壌から単離したCeriporiopsis属微生物由来のβ−1,3−グルカナーゼとβ−1,6−グルカナーゼが、それぞれβ−1,3−グルコシド結合とβ−1,6−グルコシド結合に対して高い反応性を有することを見出している。よって、これら酵素を利用して試料中のβ−1,3−1,6−グルカン含量を正確に測定できると考えられる。
【非特許文献1】宮崎利夫,「多糖の構造と生理活性」朝倉書店(1990年)
【非特許文献2】野田洋彦,「高知大学大学院農学研究科農芸化学専攻醗酵及び醸造学研究室修士論文」(1994年)
【非特許文献3】中嶋英夫,「高知大学大学院農学研究科農芸化学専攻醗酵及び醸造学研究室修士論文」(1993年)
【非特許文献4】池上裕倫,「高知大学大学院農学研究科生物資源科学専攻応用微生物学研究室修士論文」(2004年)
【発明の開示】
【発明が解決しようとする課題】
【0012】
上述した様に、本発明者らが属する研究室では、反応性の高いβ−1,3−グルカナーゼとβ−1,6−グルカナーゼを産生するCeriporiopsis属微生物を見出しており、これら酵素を利用して試料中のβ−1,3−1,6−グルカン含量を正確に測定することが考えられる。
【0013】
しかし、酵素を完全に精製するには、各種クロマトグラフィを繰り返す必要があるなど手間やコストがかかるため、精製酵素を用いる定量方法は、工業的な実施のためには実用的であるとは到底いい難い。よって、菌体外に放出された酵素を完全に精製することなく定量に用いることが考えられる。ところが、上記Ceriporiopsis属微生物は非常に強いα−グルコシダーゼ活性を有するため、粗精製酵素中にはα−グルコシダーゼが混入するので、試料中にα−グルカンが存在しているとグルコースまで分解されてしまい、β−1,3−1,6−グルカン含量を正確に測定できなくなる。そのため、Ceriporiopsis属微生物由来の粗精製酵素を用いて試料中のβ−1,3−1,6−グルカンを定量するには、先ずα−グルコシダーゼなどによりα−グルカンを分解して除去してから、粗精製されたβ−1,3−グルカナーゼとβ−1,6−グルカナーゼでβ−グルカンを分解し、生じたグルコースを定量する必要がある。このような方法では、時間や手間がかかり非効率的である。
【0014】
そこで本発明では、完全に精製した酵素を用いる必要がなく、且つβ−1,3−1,6−グルカンを正確かつ効率的に定量できる方法を提供することを目的とする。また、本発明は、当該方法に用いることができるキットと、当該方法に対して有用な微生物を提供することも目的とする。
【課題を解決するための手段】
【0015】
本発明者らは、上記課題を解決すべく鋭意研究を重ねたところ、β−1,3−グルカナーゼの産生能を有し且つβ−1,3−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないMitsuaria chitosanitabida種菌と、β−1,6−グルカナーゼの産生能を有し且つβ−1,6−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないStreptomyces omiyaensis種菌に由来する菌体外酵素液を用いれば、試料中におけるα−グルカン、他のβ−グルカンおよびヘテログルカンの存在にかかわらず、β−1,3−1,6−グルカンを正確かつ効率的に定量できることを実証して、本発明を完成した。
【0016】
本発明に係るβ−1,3−1,6−グルカンの定量方法は、多糖類含有試料に、β−1,3−グルカナーゼの産生能を有し且つβ−1,3−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないMitsuaria chitosanitabida種菌に由来する菌体外酵素液、並びに、β−1,6−グルカナーゼの産生能を有し且つβ−1,6−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないStreptomyces omiyaensis種菌に由来する菌体外酵素液を作用させる工程;並びに、生じたグルコースの量を測定する工程;を含むことを特徴とする。
【0017】
上記本発明方法においては、上記M.chitosanitabida種菌としてM.chitosanitabida H1株(NITE AP−629)が好適であり、また、上記S.omiyaensis種菌としてS.omiyaensis SY26株(NITE AP−616)が好適である。上記菌株の優れた効果は、実験により実証されている。
【0018】
上記本発明方法においては、多糖類含有試料に、20℃以上、45℃以下で、上記M.chitosanitabida種菌に由来する菌体外酵素液と上記S.omiyaensis種菌に由来する菌体外酵素液とを同時に作用させることが好ましい。上記2種の菌体外酵素液は同時に多糖類含有試料へ作用させることが効率の面から好ましいが、それぞれの酵素液に含まれるβ−1,3−グルカナーゼとβ−1,6−グルカナーゼの至適温度は異なる。しかし20℃以上、45℃以下の温度範囲であれば、両酵素は良好にその作用効果を発揮することができ、多糖類含有試料に含まれるβ−1,3−1,6−グルカンを効率的に分解することが可能になる。
【0019】
本発明に係るβ−1,3−1,6−グルカンの定量用キットは、β−1,3−グルカナーゼの産生能を有し且つβ−1,3−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないM.chitosanitabida種菌に由来する菌体外酵素液またはその乾燥物;β−1,6−グルカナーゼの産生能を有し且つβ−1,6−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないS.omiyaensis種菌に由来する菌体外酵素液またはその乾燥物;並びにグルコース量測定用試薬;を含むことを特徴とする。
【0020】
また、本発明は、M.chitosanitabida H1株(NITE AP− 629)およびS.omiyaensis SY26株(NITE AP−616)に関する。
【発明の効果】
【0021】
従来、α−グルカンを含む多糖類含有試料中のβ−1,3−1,6−グルカンを正確に測定するには、先ずアミログルコシダーゼなどでα−グルカンを分解し除去してからβ−グルカン量を測定しなければならず、手間や時間がかかるものであった。また、従来方法では、他のβ−グルカンを含む試料中のβ−1,3−1,6−グルカンを正確に測定することは困難であった。しかし本発明方法によれば、完全に精製された酵素を用いなくとも、α−グルカンを事前に分解する必要も無く、β−1,3−1,6−グルカンを正確かつ効率的に定量することができる。また、本発明に係る定量用キットは、同様の効果を有する。さらに本発明に係る2種の菌株は、これら本発明に係る定量方法と定量用キットにとり非常に好適なものである。よって本発明は、その優れた生理活性が注目されているβ−1,3−1,6−グルカンを含む試料の品質を評価する指標になり得るものとして、産業上極めて有用である。
【発明を実施するための最良の形態】
【0022】
本発明に係るβ−1,3−1,6−グルカンの定量方法は、多糖類含有試料に、β−1,3−グルカナーゼの産生能を有し且つβ−1,3−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないM.chitosanitabida種菌に由来する菌体外酵素液、並びに、β−1,6−グルカナーゼの産生能を有し且つβ−1,6−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないS.omiyaensis種菌に由来する菌体外酵素液を作用させる工程;および、生じたグルコースの量を測定する工程;を含むことを特徴とする。
【0023】
本発明方法の測定対象である多糖類含有試料は、β−1,3−1,6−グルカンを含有するか或いは含有すると考えられるものであって、その含有量を測定すべきものであれば特にその種類などは制限されない。例えば、酵母の集菌体、培地、それらの乾燥物、抽出物、精製品、懸濁液、溶液;担子菌類自体や海藻類自体、それらの乾燥物、抽出物、粉砕物、生成物、懸濁液、溶液などを挙げることができる。
【0024】
本発明方法では、多糖類含有試料に、β−1,3−グルカナーゼの産生能を有し且つβ−1,3−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないM.chitosanitabida種菌に由来する菌体外酵素液、並びに、β−1,6−グルカナーゼの産生能を有し且つβ−1,6−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないS.omiyaensis種菌に由来する菌体外酵素液を作用させる。これら菌体外酵素液は、各β−グルカナーゼ活性を示す一方で、α−グルカンや他のβ−グルカンを分解しない。よって、これら菌体外酵素液を用いれば、酵素を精製しなくとも、多糖類含有試料中のβ−1,3−1,6−グルカンを特異的にグルコースまで分解することができる。
【0025】
なお、上記菌は、α−グルコシダーゼや、β−1,3−グルカナーゼおよびβ−1,6−グルカナーゼ以外のβ−グルカナーゼを全く産生しないものに限定されず、β−1,3−1,6−グルカンの正確な定量を阻害しない程度のこれら酵素活性を菌体外へ放出するものや、β−1,3−1,6−グルカナーゼから容易に分離除去できる程度のこれら酵素を産生して菌体外へ放出するものも含まれるものとする。具体的には、比活性が0.05unit/mg以下のα−グルコシダーゼ、またはβ−1,3−グルカナーゼおよびβ−1,6−グルカナーゼ以外のβ−グルカナーゼをその菌体外酵素液に放出する菌も含まれるものとする。但し、上記菌としては、これら酵素活性を実質的に示さないものが好適である。ここでいう実質的とは、菌体外酵素液におけるこれら酵素の活性が、下記実施例に記載の測定方法において、検出限界以下であることをいう。
【0026】
当該菌体外酵素液としては、上記M.chitosanitabida種菌および上記S.omiyaensis種菌の液体培養液から得た酵素液であって、グルコースやα−グルコシダーゼ、β−1,3−グルカナーゼおよびβ−1,6−グルカナーゼ以外のβ−グルカナーゼなどβ−1,3−1,6−グルカンの正確な定量を阻害する成分を含まないものをいう。即ち、上記菌の液体培養液に硫酸アンモニウムを加えて沈殿したタンパク質成分を分離したり、透析などにより、グルコースなどの低分子物質を除去することができる。また、上記菌の菌体外酵素液でもα−グルコシダーゼ活性等を示すことがあるが、その活性は極めて弱いので、硫酸アンモニウムの濃度を調整することにより、α−グルコシダーゼ等は容易に除去可能である。
【0027】
菌体外酵素液は、具体的には例えば以下のように調製することができる。先ず、上記菌を培養した液体培養液を遠心分離や濾過に付すことによって、菌体などの固体成分を除去する。得られた液体には、PMSF(フェニルメチルスルフォニルフルオリド)などのプロテアーゼ阻害剤を加えることが好ましい。次に、タンパク質成分を常法により分離する。例えば、得られた液体をある程度濃縮した上で緩衝液を加えた後、冷却しながら硫酸アンモニウムを加えることにより沈殿したタンパク質成分を分離すればよい。この際、予備実験などにより、β−1,3−グルカナーゼとβ−1,6−グルカナーゼが十分に沈殿し且つα−グルコシダーゼ等が沈殿しない硫酸アンモニウム濃度を事前に決定しておくと効率的である。得られたタンパク質成分からは、さらに透析などの簡便な方法により、所望のβ−グルカナーゼをある程度精製してもよい。但し、本発明で用いるM.chitosanitabida種菌とS.omiyaensis種菌は菌体外にα−グルコシダーゼ等を放出しないか或いはその放出量は極めて少ないので、クロマトグラフィなど工業的に適さない方法により酵素を精製する必要はない。
【0028】
上記菌体外酵素液は、保存などのためにいったん乾燥し、使用時に溶液としてもよい。但し、上記β−1,3−グルカナーゼとβ−1,6−グルカナーゼは、それぞれ70℃と50℃で失活する。よって、酵素液の濃縮や乾燥時においては、それぞれ温度を70℃、50℃以上に上げないことが好ましい。
【0029】
本発明においては、多糖類含有試料に上記M.chitosanitabida種菌由来の菌体外酵素液と上記S.omiyaensis種菌由来の菌体外酵素液を作用させるが、これら酵素液は同時に作用させてもよいし、逐次的に作用させてもよい。効率を考慮すれば、同時に作用させることが好ましい。
【0030】
上記菌体外酵素液におけるβ−1,3−グルカナーゼとβ−1,6−グルカナーゼの濃度は、反応混合液における多糖類の濃度などに応じて適宜調整すればよいが、例えば、反応混合液中に含まれる各β−グルカナーゼの活性として、β−1,3−1,6−グルカンであるラミナリンを基質に用いた場合で1.0unit/ml以上、5.0unit/ml以下程度、β−1,6−グルカンであるパスツランを基質に用いた場合で0.1unit/ml以上、3.0unit/ml以下程度とすることが好ましい。なお、本発明におけるunitとは、30℃で1時間に1μmolの還元糖を精製する酵素量の単位と定義する。
【0031】
上記菌体外酵素液のpHも適宜調整すればよいが、本発明者らによる実験ではpH6.0で良好な結果が得られているので、溶媒としてはpH5.0以上、7.5以下程度の緩衝液が好適である。
【0032】
上記酵素反応における反応温度も適宜調整すればよい。例えば、M.chitosanitabida H1株由来のβ−1,3−グルカナーゼの至適温度は約40℃であり、約30℃以上、約55℃以下であればその60%程度の活性を示す。また、S.omiyaensis SY26株由来のβ−1,6−グルカナーゼの至適温度は約30℃であり、約20℃以上、約45℃以下であればその60%程度の活性を示す。よって、各菌体外酵素液を逐次的に多糖類含有試料に作用させる場合には、各反応温度をそれぞれ約30℃以上、約55℃以下と、約20℃以上、約45℃以下に調整することが好ましい。また、上記M.chitosanitabida種菌由来の菌体外酵素液と上記S.omiyaensis種菌由来の菌体外酵素液を同時に多糖類含有試料に作用させる場合には、両グルカナーゼを有効に機能させるために、反応温度を20℃以上、45℃以下とすることが好ましい。より好ましくは、35℃以上、40℃以下で反応を行う。
【0033】
上記酵素反応の反応時間は、多糖類含有試料中に存在するβ−1,3−1,6−グルカンがグルコースまで十分に分解される時間とする。即ち、当該反応時間は、反応混合液に含まれるβ−1,3−1,6−グルカンの濃度、菌体外酵素液に含まれるβ−1,3−グルカナーゼとβ−1,6−グルカナーゼの力価、反応温度などにも依存するので、具体的には予備実験で反応混合液中へ数時間おきに数回菌体外酵素液を加えつつ生成されるグルコース量を経時的に測定し、酵素反応によりグルコースが新たに生成されないことが確認できなくなった時間とすればよい。通常は、β−1,3−1,6−グルカンを十分に分解するために一晩反応させることが好ましいが、菌体外酵素液量によっては2時間以上、10時間以下程度の短時間にて反応を終了させることも可能である。
【0034】
本発明で用いるM.chitosanitabida種菌としては、M.chitosanitabida H1株(以下、単に「H1株」という)が好適である。H1株は、下記の通り寄託機関に寄託されている。
(i) 受領機関の名称およびあて名
名称: 独立行政法人製品評価技術基盤機構 特許微生物寄託センター
あて名: 日本国 千葉県木更津市かずさ鎌足2−5−8
(ii) 受領日: 平成20年(2008年)8月12日
(iii) 受領番号: NITE AP−629
【0035】
H1株をWicherham寒天培地上で生育させた場合には、培養初期で白色のコロニーを形成し、成熟するにしたがって徐々に乳白色から黄色になる。また、光学顕微鏡を用いて観察した形態的特徴は、以下の通りである。即ち、幅0.7〜1.0μm、長さ1.0〜4.0μmのグラム陰性桿菌で、運動性とカタラーゼ活性を示す。以上の形態的特徴は、M.chitosanitabida種に属する細菌に当てはまる。H1株は好気性であり、30℃で良好に生育し、中性(pH6.8〜7.0)の培地で良好に増殖する。よって、H1株を培養する際の温度は、25℃以上、35℃以下程度が好適で、中性付近での培養が好適である。
【0036】
従来、β−1,3−グルカナーゼを菌体外へ放出する一方で、その他のβ−グルカナーゼやα−グルコシダーゼを実質的に放出しない菌は知られていなかった。よって、本発明に係るH1株は新規なものである。
【0037】
本発明で用いるS.omiyaensis種菌としては、S.omiyaensis SY26株(以下、単に「SY26株」という)が好適である。SY26株は、下記の通り寄託機関に寄託されている。
(i) 受領機関の名称およびあて名
名称: 独立行政法人製品評価技術基盤機構 特許微生物寄託センター
あて名: 日本国 千葉県木更津市かずさ鎌足2−5−8
(ii) 受領日: 平成20年(2008年)7月29日
(iii) 受領番号: NITE AP−616
【0038】
SY26株をWicherham寒天培地で生育させた場合には、培養初期で白色コロニーを形成し、成熟するにしたがって徐々にベージュから灰色のコロニーへと変化する。光学顕微鏡を用いて観察した形態的特徴は、以下の通りである。即ち、培地基質中において無色透明の基生菌糸が観察され、その直径は0.8〜1.5μmであり、菌糸は直線状で分岐しており、成熟するとその先端が切断されて胞子を形成する。胞子表面にはシワがなく、楕円形もしくは長楕円であり、その大きさは直径0.5〜1.0μm、長さは1μm〜4μmである。さらに、前記培地で生育した本菌では、胞子嚢、運動性の胞子、分節胞子は確認されず、メラニン生成は陰性である。以上の形態的特徴は、Streptomyces omiyaensis種に属する担子菌に当てはまる。
【0039】
また、SY26株は好気性であり、30℃で良好に生育し、中性(pH6.8〜pH7.0)の培地で良好に増殖した。よって、SY26株を培養する際の温度は、25℃以上、40℃以下程度が好適で、中性付近での培養が好適である。
【0040】
従来、β−1,6−グルカナーゼを菌体外へ放出する一方で、その他のβ−グルカナーゼやα−グルコシダーゼを実質的に放出しない菌は知られていなかった。よって、本発明に係るSY26株は新規なものである。
【0041】
上記菌体外酵素液による反応後、β−1,3−グルカナーゼとβ−1,6−グルカナーゼの作用により生じたグルコースの量を測定する。
【0042】
本発明に係るβ−1,3−グルカナーゼとβ−1,6−グルカナーゼは、それぞれβ−1,3−グルコシド結合とβ−1,6−グルコシド結合を特異的に切断する。よって、上記菌体外酵素液により、多糖類含有試料中に存在するβ−1,3−1,6−グルカンは選択的にグルコースまで分解されるので、測定されたグルコース量から、β−1,3−1,6−グルカンの量を把握することができる。
【0043】
具体的には、生成したグルコースの量は、例えばSomogyi−Nelson法(M.Somogyi,J.Biol.Chem.,195,p.19(1952)を参照)など遊離の還元糖を測定する方法により測定することが可能である。また、酵素法やHPLCなどでも測定可能である。酵素法としては、グルコース−6−リン酸デヒドロゲナーゼとヘキソキナーゼを組み合わせたG6PDH−HK法(F−キット,Roche社製)や、ムタロターゼとグルコースオキシダーゼを組み合わせたムタロターゼ・GOD法(グルコースCII−テストワコー,Wako社製)がある。また、HPLC分析用のカラムとしてはTSKgel Sugar AXG(東ソー社製)やSugar−PakTMI(Waters社製)など、グルコースを分析するのに適したカラムが多数存在するので、これを利用できる。
【0044】
本発明に係るβ−1,3−1,6−グルカンの定量用キットは、β−1,3−グルカナーゼの産生能を有し且つβ−1,3−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないM.chitosanitabida種菌に由来する菌体外酵素液またはその乾燥物;β−1,6−グルカナーゼの産生能を有し且つβ−1,6−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないS.omiyaensis種菌に由来する菌体外酵素液またはその乾燥物;並びに、
グルコース量測定用試薬;を含むことを特徴とする。
【0045】
上記M.chitosanitabida種菌に由来する菌体外酵素液と上記S.omiyaensis種菌に由来する菌体外酵素液は、本発明方法で用いるものと同様のものとすることができる。また、それらの乾燥物は、当該菌体外酵素液を乾燥したものであればよい。但し、上述したようにβ−1,3−グルカナーゼとβ−1,6−グルカナーゼは高温で失活するために、上記M.chitosanitabida種菌由来の菌体外酵素液は60℃以下で、上記S.omiyaensis種菌由来の菌体外酵素液は45℃以下で乾燥することが好ましい。
【0046】
グルコース量の測定用試薬としては、例えば、生じたグルコース量をSomogyi−Nelson法で測定する場合には、Somogyi試薬とNelson試薬、或いはこれらを調製するための化合物を挙げることができる。また酵素法を用いる場合には例えば、生じたグルコース量を酵素法で測定する場合にはグルコース−6−リン酸デヒドロゲナーゼとヘキソキナーゼを組み合わせたG6PDH−HK法(F−キット,Roche社製)や、ムタロターゼとグルコースオキシダーゼを組み合わせたムタロターゼ・GOD法(グルコースCII−テストワコー,Wako製)に類する試薬または、或いはこれらを調製するための化合物を挙げることができる。
【0047】
当該キットは、本発明に係るβ−1,3−1,6−グルカンの定量方法で用いることができる。
【実施例】
【0048】
以下、実施例を挙げて本発明をより具体的に説明するが、本発明はもとより下記実施例により制限を受けるものではなく、前・後記の趣旨に適合し得る範囲で適当に変更を加えて実施することも可能であり、それらはいずれも本発明の技術的範囲に含まれる。
【0049】
実施例1 β−1,3−グルカナーゼ産生菌の単離
高知大学農学部内から土壌試料を採取した。当該土壌試料(5g)に滅菌水(15mL)を加え、ボルテックスミキサーを用いて攪拌した。次いで、当該混合物を数分間静置し、上清(50μL)をマイクロピペッターで採取した。
【0050】
別途、カードラン(β−1,3−グルカン、2g)を水道水(200mL)に懸濁し、オートクレーブで121℃、20分間処理した。これをミキサーにかけ、生じたゲルを完全にすり潰した。これにポリペプトン(0.125g)と酵母エキス(0.125g)を溶解し、水道水を加えて500mLにメスアップした。さらに10gの寒天を加え、オートクレーブで121℃、20分間滅菌処理し、シャーレ上で固めることによって、カードラン平板培地を調製した。また、デンプン培地(0.4%デンプン、0.025%ポリペプトン、0.025%酵母エキス、2%寒天、pH7.0)も調製した。
【0051】
上記上清をカードラン平板培地に蒔き、スプレッダーで広げた後、30℃で1日間培養した。480の菌株を単離し、同平板培地に植菌してさらに30℃で2日間培養した。この培地を0.1%コンゴーレッド溶液で2〜3時間染色した後、流水中で2時間脱色してハロの形成を確認したところ、1菌株のみがハロを形成した。当該菌株をデンプン平板培地に塗布し、30℃で1日間培養した。この培地をヨウ素溶液で2〜3時間染色した後(中村道徳・貝沼圭二編,「生物化学実験法 19 澱粉・関連糖質実験法」学会出版センターを参照)、流水中で2時間脱色したが、ハロの形成は確認されなかった。カードラン平板培地で培養後のコンゴーレッド染色写真を図1に、デンプン培地で培養後のヨウ素染色写真を図2に示す。
【0052】
図1〜2のとおり、上記菌株はβ−1,3−グルカンの分解活性を示すのに対してデンプン(α−1,4−1,6−グルカン)に分解活性を示さないことから、β−1,3−グルカナーゼを産生する一方でα−グルコシダーゼは産生しないと考えられた。この菌株をH1株と仮に命名し、以後の実験に用いた。
【0053】
実施例2 H1株の顕微鏡観察
上記実施例1で得たH1株を顕微鏡(OLYMPUS社製、製品名「BH−2」)で1000倍に拡大し、観察した。拡大写真を図3に示す。
【0054】
図3のとおり、H1株は幅0.7〜1.0μm、長さ1.0〜4.0μmの桿菌であった。また、H1株はグラム陰性であり、カタラーゼ活性を示した。
【0055】
実施例3 H1株の16SrDNA塩基配列解析
(1) ポリメラーゼ連鎖反応
上記実施例1で得たH1株の属種を決定するために、16SrDNA塩基配列を解析した。具体的には、先ず10×ExTaq緩衝液(5μL)、dNTP、dTTP、dGTPおよびdCTPを各2.5mM含む混合液(8μL)、配列番号1〜2の各プライマーの5pmol/μL溶液(各1μL)を混合し、全量が49.5μLとなるように滅菌水に加えた。別途、MO BIO社製のUltra Clean Microbial DNA Isolation Kitを用いてH1株のゲノムDNA溶液を得た。当該ゲノムDNA溶液(1μL)か、或いは滅菌した爪楊枝によりH1株のコロニーから取得した少量の菌体を、上記溶液に加えて反応混合液とした。ポリメラーゼ連鎖反応には、DNAサーマルサイクラー(宝酒造社製)を使用した。反応条件は、以下のとおりである。
・ 熱変性 − 94℃,5分間
・ Ex Taq DNA ポリメラーゼ(宝酒造社製,2.5U)の添加
・ 熱変性 − 94℃,45秒間
・ プライマーのアニーリング − 55℃,1分間
・ DNA鎖の伸長反応 − 72℃,2分間
・ 2回目の熱変性以降のステップを35サイクル繰り返す
・ 熱処理 − 72℃,7分間
【0056】
(2) 16SrDNA断片の分離
上記ポリメラーゼ連鎖反応により増幅された16SrDNA断片を、アガロースゲル電気泳動により分離した。電気泳動に用いるアガロースゲルは、Sea Kem GTG アガロース(FMC)の1.5%溶液から調製した。泳動バッファーとしては、Howly緩衝液(40mM Tris,20mM酢酸ナトリウム,2mM EDTA,pH7.8)を用いた。分子量マーカーには、Hind IIIで処理したλDNAを使用した。
【0057】
電気泳動後、アガロースゲルを1μg/mLの臭化エチジウム溶液に10分間浸して染色することによりDNAを可視化し、紫外線を照射して写真撮影を行った。撮影後、目的の大きさと一致するバンドの部分を滅菌済みのかみそりで切り出し、メンブレンフィルター付きマイクロチューブ(宝酒造社製,SUPRECTM−01)に入れ、完全に凍結させた。37℃で5分間静置して融解した後、10,000rpmで10分間遠心分離し、200μLのTE緩衝液(0.1mM EDTA,10mM Tris−HCl,pH7.5)を加え、さらに10分間遠心分離した。2倍量の冷却エタノールと1/20量の3M酢酸ナトリウムを加え、−20℃で1時間静置した。その後、12,000rpmで10分間遠心分離し、エタノールを除去し、吸引乾燥することにより16SrDNA断片を含む混合物を得た。
【0058】
(3) 16SrDNA断片の塩基配列の決定
ポリメラーゼ連鎖反応により16SrDNA断片を増幅した上で、その塩基配列を解析した。具体的には、鋳型DNA(4μL)、5pmol/μLプライマー溶液(1μL)、BigDyeターミネーターver.3.1 5×シーケンシング緩衝液(1.5μL)、BigDyeターミネーターver.3.1(1μL)を混合し、滅菌水で全量を10μLにし、これを反応液とした。当該反応液に上記(2)で得た混合物を加え、下記反応条件でポリメラーゼ連鎖反応を行った。
・ 熱変性 − 96℃,1分間
・ 熱変性 − 96℃,10秒間
・ プライマーのアニーリング − 50℃,5秒間
・ DNA鎖の伸長反応 − 60℃,4分間
・ 2回目の熱変性以降のステップを25サイクル繰り返す
【0059】
次に、Sephadex G−50を用い、冷却した反応混合液から16SrDNA断片を精製した。得られた16SrDNA断片にHi−Diホルムアミド(10μL)を加え、ABI Prism 3100−Avant ジェネティックアナライザー(アプライドバイオシステム社製)により塩基配列を解析した。当該塩基配列を配列表の配列番号3に示す。
【0060】
得られた16SrDNA塩基配列は、M.chitosanitabida種菌の16SrDNA塩基配列と高い相同性を示すものであったことから、H1株はM.chitosanitabida種に属するものであると判断した。
【0061】
実施例4 H1株由来の粗酵素液の基質特異性
(1) H1株由来の粗酵素液の調製
試験管内で、7mLのWicherham培地(0.5%ペプトン,0.3%酵母エキス、0.3%麦芽エキス,1.0%グルコース,pH7.0)を作成した。当該培地に白金耳でH1株を植菌し、30℃で1日間振とう培養した。別途、生パン酵母培地(4.0%生パン酵母(協和発酵社製,ダイヤイースト))を調製し、当該培地に前培養したH1株を移し、30℃で2日間培養した。当該培養液を遠心分離(18,000rpm,20分間,4℃)し、沈殿が入らないよう上清を得た。この上清へ終濃度が0.1mMになるようにPhenylmethylsulfonyl fluoride(和光純薬社製,以下、「PMSF」と省略する)を加え、撹拌・氷冷しながら、乳鉢で粉砕した硫酸アンモニウムを飽和濃度の80%となるように加えた。撹拌・氷冷しながら一晩静置後、遠心分離(18,000rpm,20分間,4℃)し沈殿を得た。得られた沈殿を、0.1mM PMSFを含む少量の20mM酢酸ナトリウム緩衝液(pH6.0)に溶解した。これを透析膜(Viskase Companies,inc製)に入れ、2Lの同緩衝液を用いて透析することにより粗酵素液を得た。
【0062】
(2) 粗酵素液の活性測定
前項で得た粗酵素液と様々なグルカン基質を反応させ、生成した還元糖を検出することによりグルカナーゼ活性を測定した。具体的には、0.2M酢酸ナトリウム緩衝液(pH6.0)に、黒酵母β−グルカン(Aureobasidium sp.由来,株式会社ソフィより提供)、パスツラン(Calbiochem社製)、リケナン(シグマ社製)、ラミナリン(Eisenia araborea由来,半井化学薬品社製)、ラミナラン(Laminaria由来,東京化成工業社製)、カードラン(和光純薬社製)、セルロース(NIHONBASHI社製)、ルテオース(高知大農学部応用微生物学研究室より提供)、デンプン(和光純薬社製)、デキストラン(和光純薬社製)、プルラン(半井化学薬品社製)をそれぞれ10g/Lになるように溶解した。完全に溶解させるために、リケナンは熱湯中で煮沸し、また、パスツランはオートクレーブにより121℃で5分間高圧加熱した。なお、基質濃度はすべて0.5%に調整し、粗酵素溶液を加えてから30℃で1時間反応させた。反応後、5分間煮沸し、生成した還元糖量をSomogyi−Nelson法にて測定した(M.Somogyi,J.Biol.Chem.,195,p.19(1952)を参照)。
【0063】
詳しくは、KNaC4H4O6・4H2O(4.0g)、Na2HPO4・12H2O(7.1g)、CuSO4・5H2O(0.8g)、1M NaOH(10mL)およびNa2SO4・5H2O(18.0g)を蒸留水(100mL)に溶解し、37℃で24時間インキュベートした。次いで当該溶液を濾過し、得られた濾液をSomogyi試薬とした。また、(NH4)6Mo7O24・4H2O(5.0g)、H2SO4(4.2g)およびNa2HAsO4・7H2O(0.6g)を蒸留水(100mL)に溶解し、24時間インキュベートした。次いで当該溶液を濾過し、得られた濾液をNelson試薬とした。上記酵素反応溶液(1mL)にSomogyi試薬(1mL)を加え、湯鍋中にて正確に10分間反応させた後、即座に流水中で5分間水冷した。当該溶液にNelson試薬(1mL)と蒸留水(2mL)を加え、室温にて約10分間静置後、分光光度計(SHIMADZU社製,UV−1200)を用いて520nmにおける吸光度を測定した。なお、標準物質はグルコースを用いた。また、30℃で1時間に1μmolの還元糖を生成する酵素量を1unitと定義した。結果を表1に示す。
【0064】
【表1】
【0065】
上記結果のとおり、H1株から得られた粗酵素液は、β−1,3−グリコシド結合を含む黒酵母β−グルカン、ラミナリン、ラミナラン、リケナンおよびカードランに対して高い活性を示した。その一方で、β−1,6−グリコシド結合のみからなるパスツランとルテオースに対する活性は見られなかった。さらに、α−グルカンであるプルランに対する活性が観察されたものの、その比活性は低いものであった。
【0066】
以上の結果から、H1株由来の粗酵素液は、β−1,3−グリコシド結合を特異的に切断するβ−1,3−グルカナーゼを含む一方で、その他のβ−グルカナーゼを含まず、また、わずかにα−グルコシダーゼ活性を示すものの、デンプンとデキストランを分解できないことから、その活性は極めて低いことが明らかにされた。
【0067】
実施例5 H1株由来の菌体外酵素液の調製
(1) H1株由来の粗酵素液の調製
上記実施例4(1)と同様に、Mitsuaria chitosanitabida H1株をWicherham培地(70mL)で前培養した後、生パン酵母培地(1L)で本培養した。当該培養液を遠心分離(9,100rpm,20分間,4℃)し、沈殿が入らないようにして上清を得た。この上清に1/1,000量の0.1M PMSFと、1/5,000量の消泡剤を加え、エバポレーターで全量の約1/10量まで減圧濃縮した。次いで、終濃度が20mMになるようにリン酸カリウム緩衝液(pH7.4)を加え、撹拌・氷冷しながら、乳鉢で粉砕した硫酸アンモニウムを加えていき沈殿物を分離することにより、0〜20%画分、20〜40%画分、40〜60%画分、60〜80%画分を得た。各画分を20mM酢酸ナトリウム緩衝液(pH6.0)で透析後、β−1,3−グルカナーゼ活性を測定し、また、全タンパク質を定量した。詳しくは、E.araborea由来のラミナリンに酵素液を加え、30℃で1時間反応させた後、生じた還元糖量をSomogyi−Nelson法により定量することにより全活性と比活性を測定した。また、タンパク質はLowry法(Lowry,O.H.ら,J.Biol.Chem.,193,pp.265-275(1951)を参照)により定量した。結果を表2に示す。
【0068】
【表2】
【0069】
上記結果のとおり、全活性は60〜80%飽和画分で最も高く、比活性は20〜40%飽和画分で最も高かった。それに対して、0〜20%飽和画分は全活性と比活性が低かった。よって、以後の実験では20〜80%飽和画分を用いた。
【0070】
(2) H1株由来の菌体外酵素液の調製
0.1mM PMSFを含む20mM酢酸ナトリウム緩衝液(pH5.0)を使って、上記で得た粗酵素液の20〜80%飽和画分を透析した後、遠心分離(18,000rpm,20分間,4℃)したところ、ゲル状の沈殿が生じた。上記実施例4(2)と同様の方法により、E.araborea由来のラミナリンを基質として、生じたゲル状沈殿と上清のそれぞれのグルカナーゼ活性を測定したところ、ゲル状沈殿に非常に高い活性が見られた。当該ゲル状沈殿を20mM酢酸ナトリウム緩衝液(pH5.0)に懸濁してから遠心分離(18,000rpm,20分間,4℃)する操作を繰り返すことにより洗浄した。洗浄した粗酵素液の基質特異性に変化がないかを調べるために、上記実施例4(2)と同様の実験を再び行った。結果を表3に示す。
【0071】
【表3】
【0072】
表3に示す結果のとおり、得られた菌体外酵素液はβ−1,3−グリコシド結合を有する基質に対しては作用する一方で、α−1,4−グリコシド結合、α−1,6−グリコシド結合、β−1,4−グリコシド結合およびβ−1,6−グリコシド結合には作用しない。よって、当該H1株由来の菌体外酵素液は、β−1,3−グリコシド結合を特異的に切断する一方で、α−グルコシダーゼ活性やその他のβ−グルカナーゼ活性を有しないことが実証された。なお、当初はカラムクロマトグラフィによりさらに精製を進めてその酵素学的性質を調べる予定であったが、精製酵素液がゲル状であるためかかる精製は行うことができなかった。
【0073】
実施例6 H1株由来のβ−1,3−グルカナーゼの至適温度
上記実施例5(2)で得た菌体外酵素液を用い、反応温度を20〜70℃に変更した以外は上記実施例5(1)と同様にして比活性を測定し、H1株由来のβ−1,3−グルカナーゼの至適温度を調べた。結果を図4に示す。
【0074】
図4のとおり、H1株由来のβ−1,3−グルカナーゼの至適温度は40℃であり、活性は50℃から徐々に下がり始め、70℃では活性はほとんど失われることが分かった。
【0075】
実施例7 H1株由来のβ−1,3−グルカナーゼの熱安定性
上記実施例5(2)で得た菌体外酵素液を30〜70℃で15分間処理した後に、氷上で急冷した。当該溶液を用い、上記実施例5(1)と同様にして比活性を測定し、H1株由来のβ−1,3−グルカナーゼの熱安定性を調べた。結果を図5に示す。
【0076】
図5のとおり、H1株由来のβ−1,3−グルカナーゼは50℃までは高い安定性を示すが、70℃では完全に失活することが分かった。
【0077】
実施例8 H1株由来のβ−1,3−グルカナーゼの反応生成物の同定
β−1,3−グルカンであるカードランに、上記実施例5(2)で得た菌体外酵素液を上記実施例4(2)と同様の反応条件で作用させ、反応混合液を経時的に分取し、反応生成物を薄層クロマトグラフィで分析した。具体的には、反応開始から0、1、2、4、24時間後に反応混合液を分取し、沸騰水で10分間加熱処理して酵素反応を止めた後、HPTLC Fertigplatten Kieselgel 60(Merck社製,10cm×10cm)を用いた薄層クロマトグラフィに供した。標準物質として1%グルコースを用い、展開溶媒にはn−ブタノール:エタノール:水=5:2:2の混合溶媒を用い、上昇法で行った。展開後、発色液として、ジフェニルアミン−アニリン−リン酸(2gジフェニルアミン,2mLアニリン,100mLアセトン,15mL 80%リン酸)をゲルプレート上に噴霧し、乾熱器中、100℃で10分間加熱し、出現したスポットを観察した。結果を図6に示す。図6中、「G3」はカードランを示し、「G1」はグルコース2分子がβ−1,6結合したゲンチオビオースであり、「G0」はグルコースである。
【0078】
図6のとおり、H1株由来のβ−1,3−グルカナーゼによる反応の最終生成物はグルコースであり、その生成量は経時的に増加した。よって、H1株由来のβ−1,3−グルカナーゼは、β−1,3−グルカンを構成単糖であるグルコースまで分解できるSタイプの酵素であることが確認された。
【0079】
実施例9 β−1,6−グルカナーゼ産生菌の単離
高知大学農学部内から土壌試料を採取した。当該土壌試料(5g)に滅菌水(15mL)を加え、ボルテックスミキサーを用いて攪拌した。次いで、当該混合物を数分間静置し、上清(50μL)をマイクロピペッターで採取した。当該上清をパスツラン平板培地(0.1%パスツラン(Calbiochem製,0.025%ポリペプトン、0.025%酵母エキス、2%寒天、pH7.0)に蒔き、スプレッダーで広げた後、30℃で2日間培養した。180の菌株を単離し、同平板培地に植菌してさらに30℃で4日間培養した。この培地を0.1%コンゴーレッド溶液で2〜3時間染色した後(Andres Solerら,Journal of microbiological methods,35,pp.245-251(1999)を参照)、流水中で2時間脱色した。さらに20mM酢酸ナトリウム緩衝液(pH5.5)に一晩浸すことでコンゴーレッドを変色させた後、ハロの形成を確認した。培地の写真を図7に示す。
【0080】
図7の結果のとおり、180の菌株中1菌株のコロニー周辺にハロらしきものが認められた。即ち、この菌株はβ−1,6−グルカンであるパスツランに対する分解活性を示すことから、β−1,6−グルカナーゼを産生していると考えられる。この菌株をSY26と命名し、以後の実験に用いた。
【0081】
実施例10 SY26株の顕微鏡観察
上記実施例9で得たSY26株を顕微鏡(OLYMPUS社製、製品名「BH−2」)で1000倍に拡大し、観察した。拡大写真を図8に示す。
【0082】
図8のとおり、SY26株は無色透明の基生菌糸を形成する。菌糸の直径は0.8〜1.5μmであり、直線状で分岐していた。また、当該菌糸は、成熟するとその先端が切断されて胞子を形成した。胞子表面にはシワがなく、楕円形もしくは長楕円であり、その大きさは直径0.5〜1.0μm、長さは1μm〜4μmであった。さらに、SY26株では、胞子嚢、運動性の胞子、分節胞子は確認されず、メラニン生成は陰性である。
【0083】
実施例11 SY26株の16SrDNA塩基配列解析
上記実施例3と同様にして、SY26株の16SrDNA塩基配列を解析した。当該塩基配列を配列表の配列番号4に示す。
【0084】
得られた16SrDNA塩基配列は、S.omiyaensis種菌の16SrDNA塩基配列と高い相同性を示すものであったことから、SY26株はS.omiyaensis種に属するものであると判断した。
【0085】
実施例12 SY26株由来の粗酵素液の基質特異性
本培養期間を6日間とした以外は上記実施例4(1)と同様にして、SY26株の培養液から粗酵素液を得た。さらに上記実施例4(2)と同様にして、当該粗酵素液の活性を測定した。結果を表4に示す。
【0086】
【表4】
【0087】
上記結果のとおり、SY26株から得られた粗酵素液は、β−1,6−グリコシド結合を含む黒酵母β−グルカン、ラミナリン、パスツランおよびルテオースに対して高い活性を示した。それに対して、β−1,6−グリコシド結合を含まず且つβ−1,3−グリコシド結合を含むリケナンとカードランに対する活性は見られなかった。さらに、α−グルコシダーゼ活性は見られなかった。
【0088】
以上の結果から、SY26株の粗酵素液は、β−1,6−グリコシド結合を特異的に切断できる一方で、α−グルコシダーゼ活性とその他のβ−グルカナーゼ活性を有しないことが明らかにされた。
【0089】
実施例13 SY26株由来の菌体外酵素液の調製
(1) SY26株由来の粗酵素液の調製
上記実施例4(1)と同様に、S.omiyaensis SY26株をWicherham培地(14mL)で前培養した後、生パン酵母培地(200mL)で本培養した。当該培養液から上記実施例5(1)と同様にして0〜20%画分、20〜40%画分、40〜60%画分、60〜80%画分を得た。また、基質としてラミナリンの代わりにパスツランを用いた以外は上記実施例5(1)と同様にして、β−1,6−グルカナーゼ活性を測定し、また、全タンパク質を定量した。結果を表5に示す。
【0090】
【表5】
【0091】
上記結果のとおり、全活性は20〜40%飽和画分で最も高く、比活性は40〜60%飽和画分で最も高かった。60〜80%飽和画分もわずかに活性を示したが、全活性と比活性の値が共に低かったため、以後の実験では20〜60%飽和画分を用いた。
【0092】
(2) β−1,6−グルカナーゼの精製
35mLのWicherham培地でSY26株を前培養した後に500mLの生パン酵母培地で本培養した以外は上記実施例13(1)と同様にして、培養液の20〜60%飽和画分を得た。
【0093】
0.1mM PMSFを含む20mM酢酸ナトリウム緩衝液(pH5.0)を使って、上記で得た粗酵素液の20〜60%飽和画分を透析した後、透析中に生じた沈殿物を遠心分離(18,000rpm,20分間,4℃)で取り除いたものを菌体外酵素液とし、上記実施例13(1)と同様に活性測定を行った。また、全タンパク質量をプロテインアッセイCBB溶液(ナカライテスク社製)により定量した。上記菌体外酵素液の全活性は40.9unit、全タンパク質量は3.24mg、比活性は3.89unit/mgであった。
【0094】
上記菌体外酵素液を、さらにゲル濾過クロマトグラフィにより精製した。具体的には、0.1mM PMSFを含む20mM酢酸ナトリウム緩衝液(pH5.0)で平衡化したSP−セファデックス C−50 カラム(ファルマシア社製,3.0×30cm)に上記粗酵素液を供した。非吸着タンパク質は同緩衝液で溶出させ、吸着タンパク質は0〜0.5Mの塩化ナトリウムを含む同緩衝液でリニアグラジエントにより溶出させ、5mL毎にフラクションを分取した。各フラクションの比活性を測定し、活性の高いフラクションを集めてアミコンメンブレンフィルターで6mLに濃縮した。0.1mM PMSFを含む20mMリン酸カリウム緩衝液(pH7.4)で透析後、透析中に生じた沈殿物を遠心分離(18,000rpm,20分間,4℃)で取り除いた。上記と同様にして活性測定とタンパク定量を行ったところ、全活性は29.8unit、全タンパク質量は2.23mg、比活性は13.3unit/mgであった。
【0095】
上記酵素液を、さらにイオン交換クロマトグラフィで精製した。詳しくは、0.1mM PMSFを含む20mMリン酸カリウム緩衝液(pH7.4)で平衡化したDEAE−トヨパール 650Mカラム(東ソー株式会社,3.0×30cm)に上記酵素液を供した。タンパク質を同緩衝液で溶出させ、5mL毎にフラクションを分取し、活性の高いフラクションを集めた。当該酵素液の全活性は18.3unit、全タンパク質量は0.31mg、比活性は58.2unit/mgであった。
【0096】
上記酵素液をCentriprepで約1mLまで濃縮し、さらにMicroconを用いて50μLまで濃縮した後、BioLogic Duo Flow(バイオラッド製)を用いたFPLCで精製した。具体的には、0.15Mの塩化ナトリウムを含む20mMリン酸カリウム緩衝液(pH7.4)で平衡化したSuperose 12 HR 10/30 カラムに濃縮した酵素液を供した。同緩衝液を通液し続けてタンパク質を溶出させて0.5mL毎に分取し、活性の高いフラクションを集めてこれを精製酵素液とした。上記と同様にして当該精製酵素液の活性測定とタンパク定量を行ったところ、全活性は3.65unit、全タンパク質量は0.01mg、比活性は386unit/mgであった。
【0097】
以上の各精製段階における全タンパク質量などのデータを、表6にまとめる。
【0098】
【表6】
【0099】
また、上記各精製段階の酵素液を、Laemmliの方法(U.K.Laemmli,Nature,277,pp.680-685(1970)を参照)に従って分析した。電流は、濃縮ゲル時は10mA、分離ゲル時は20mAとした。また、分子質量測定の際のマーカーとして、ミオシン(200kD)、β−ガラクトシダーゼ(116kD)、ウシ血清アルブミン(66kD)、卵白アルブミン(45kD)、炭酸脱水酵素(31kD)、トリプシン阻害剤(21.5kD)、リゾチーム(14.4kD)、アプロチニン(6.5kD)からなるタンパク質マーカー(ナカライテスク社製)を用いた。泳動終了後は、0.25%クマシーブリリアントブルーR−250を含むメタノール:酢酸:水=5:1:88の混合溶液にゲルを浸し、振とうしながら50℃で30分間染色した。その後、メタノール:酢酸:水=5:7:88の混合溶液に浸し、振とうしながら50℃で脱色した。脱色したゲルは、デジタルカメラを用いて画像を保存し、ゲル乾燥機で乾燥させて保存した。結果を図9に示す。
【0100】
図9のとおり、最終的に得られた精製酵素液中には、分子質量が10kDのタンパク質のみが含まれていることが分かる。
【0101】
実施例14 SY26株由来のβ−1,6−グルカナーゼの至適温度
上記実施例13(2)で得たSuperose 12 HR 10/30 カラム溶出画分である精製酵素液を用い、反応温度を10〜60℃に変更した以外は上記実施例13(1)と同様にして比活性を測定し、SY26株由来のβ−1,6−グルカナーゼの至適温度を調べた。結果を図10に示す。
【0102】
図10のとおり、SY26株由来のβ−1,6−グルカナーゼの至適温度は30℃であり、40℃では30℃の場合の83%の活性を示したが、50℃以上になると活性は急激に失われることが分かった。
【0103】
実施例15 SY26株由来のβ−1,6−グルカナーゼの熱安定性
上記実施例13(2)で得たSuperose 12 HR 10/30 カラム溶出画分である精製酵素液を20〜60℃で15分間処理した後に、氷上で急冷した。当該溶液を用い、上記実施例13(1)と同様にして比活性を測定し、SY26株由来のβ−1,6−グルカナーゼの熱安定性を調べた。結果を図11に示す。
【0104】
図11のとおり、SY26株由来のβ−1,6−グルカナーゼは30℃までは高い安定性を示し、40℃では30℃の場合の約80%の活性を示したが、50℃では失活することが分かった。
【0105】
実施例16 SY26株由来のβ−1,6−グルカナーゼの反応生成物の同定
β−1,6−グルカンであるパスツランに、上記実施例13で精製したβ−1,6−グルカナーゼ含有溶液を上記実施例13(1)と同様の反応条件で作用させ、反応混合液を経時的に分取し、反応生成物を薄層クロマトグラフィで分析した。結果を図12に示す。図12中、「G3」はパスツランを示し、「G1」はグルコース2分子がβ−1,6結合したゲンチオビオースであり、「G0」はグルコースである。
【0106】
図12のとおり、SY26株由来のβ−1,6−グルカナーゼによる反応の最終生成物はグルコースであり、その生成量は経時的に増加した。よって、SY26株由来のβ−1,6−グルカナーゼは、β−1,6−グルカンを構成単糖であるグルコースまで分解できるSタイプの酵素であることが確認された。
【0107】
実施例17 β−1,3−1,6−グルカンの定量
天然由来のグルカン類としては、グルコースのみからなるホモグルカンであるα−グルカンとβ−グルカン、および他の糖と結合したヘテログルカンが存在する。従来方法は、これらグルカン類の内、α-グルカンを取り除いた残りの全てのグルカンを構成単糖まで分解してグルコースを定量するものであることから、有用なβ−1,3−1,6−グルカンのみを定量することはできなかった。
【0108】
一方、上記実験により、H1株とSY26株に由来する菌体外酵素液は、それぞれβ−1,3−グリコシド結合とβ−1,6−グリコシド結合を特異的に切断できることが実証されていることから、これら酵素液によりグルカンをグルコースまで完全に分解した後にグルコース量を測定すれば、事前にα−グルカンを分解除去しなくてもβ−1,3−1,6−グルカンを正確に定量できると期待できる。
【0109】
そこで、先ず、本発明に係る2種の菌体外酵素液を使って、天然由来のグルカン中のβ−1,3−1,6−グルカンを定量し、次に、当該グルカンからα−グルカンを除去した上で同様の操作を行い、両結果を比較することで、本発明方法の効果を評価した。
【0110】
(1) 多糖標品の調製
Aureobasidium pullulans AFO202株の培養液(ソフィ社より提供,3g)に蒸留水(27mL)を加えて10倍希釈培養液を作成した。これを遠心分離(18,000rpm,20分間,4℃)し、得られた上清(7mL)に2倍量のエタノールを加え、よく撹拌して多糖を沈殿させた。さらに、遠心分離(18,000rpm,20分間,4℃)後、上清を捨て、沈殿物をエタノール臭がしなくなるまで風乾した。風乾後、沈殿物を14mLの20mM酢酸ナトリウム緩衝液(pH4.5)に溶解し、これを多糖標品とした。
【0111】
フェノール硫酸法(Dubois,M.ら,Anal.Chem.,28,p.350(1956)を参照)を用い、上記多糖標品に含まれる全多糖量を測定した。具体的には、上記多糖標品(500μL)と蒸留水(500μL)とを混合し、さらに5%フェノール(1mL)を加えた。ここへ硫酸(5mL)を加え、室温で10分間冷却した。当該混合液を流水で20分間冷却した後、490nmにおける吸光度を測定することによって、多糖量を定量したところ、全多糖量は3.80g/Lであった。
【0112】
(2) 試料中にα−グルカンが含まれる場合
上記(1)で得た多糖標品(500μL)に、1M酢酸ナトリウム緩衝液(pH6.0,50μL)、H1株由来の菌体外酵素液(225μL)およびSY26株由来の菌体外酵素液(225μL)を混合し、40℃で6時間インキュベートした。なお、使用したH1株由来の菌体外酵素液の活性はラミナリン(E.araborea由来)に対して2.2unitであり、SY26株由来の菌体外酵素液の活性はパスツランに対して1.4unitと、上記多糖標品中のグルカンを完全に分解するに十分である。
【0113】
上記酵素反応後における反応混合液について、Somogyi−Nelson法により還元糖量を測定した。結果を表7に示す。
【0114】
(3) 試料中にα−グルカンが含まれない場合
アミログルコシダーゼ(Rhizopus sp由来,シグマ社製)を0.2M酢酸ナトリウム水溶液(pH4.5)に1μg/mLの濃度で溶解し、アミログルコシダーゼ溶液を得た。上記多糖標品(3.6mL)とアミログルコシダーゼ溶液(0.4mL)をファルコンチューブに入れ、55℃で一晩インキュベートした。その後、5分間煮沸し、酵素反応を終了させた。このアミログルコシダーゼ処理で生じた還元糖をSomogyi−Nelson法で定量し、α−グルカン量とした。この反応液(500μL)にH1株由来の菌体外酵素液(225μL)およびSY26株由来の菌体外酵素液(225μL)を加え、40℃で6時間反応させた。当該酵素反応後における反応混合液について、Somogyi−Nelson法により還元糖量を測定し、当該測定値から上記のα−グルカン量値を引いてβ−1,3−1,6−グルカン量とした。結果を表7に示す。
【0115】
【表7】
【0116】
α−グルカンを除去しない場合のβ−1,3−1,6−グルカン量は3.09g/Lであり、α−グルカンを除去した場合は3.18g/Lであって、この二つに大きな差はなかった。従って、本発明方法は、標品中にα−グルカンが存在したままでも正確にβ−1,3−1,6−グルカンを定量できることが分かった。
【0117】
実施例18 β−1,3−1,6−グルカンの定量
上記実施例17のとおり、本発明方法によれば標品中にα−グルカンが存在したままでも正確にβ−1,3−1,6−グルカンを定量できるが、反応溶液中のα−グルカン量がこれより多くなると、α−グルカンを除去しなければ正確にβ−1,3−1,6−グルカン量を正確に定量できない可能性もある。そこで、α−グルカンをより多く生産するA.pullulans 132株の培養液を試料とし、同様の実験を行った。
【0118】
(1) 多糖標品の調製
A.pullulans 132株(寺尾啓吾(高知大学農学部生物資源科学科),「応用微生物学研究室卒業論文」(2006)を参照)を白金耳で一掻き取り、5mLの米ヌカ含有液体培地(0.2%米ヌカ、0.2%アスコルビン酸、1%スクロース、pH5.2)に植菌し、25℃、125rpmで48時間振とうして前培養した。次いで、30mL容の三角フラスコに100mLの米ヌカ含有液体培地を作成し、ここへ上記前培養液(1mL)を加え、25℃、125rpmで72時間振とうして本培養した。なお、米ヌカ含有液体培地は、多糖の生産を促進する。
【0119】
A.pullulans AFO202株の培養液の代わりに上記本培養の培養液(3g)を用いた以外は上記実施例17(1)と同様にして、多糖標品を得た。当該多糖標品に含まれる多糖量を上記実施例17(1)と同様の方法で定量したところ、3.04g/Lであった。
【0120】
(2) β−1,3−1,6−グルカンの定量
上記多糖標品について、α−グルカンを除去しない場合と除去した場合において、β−1,3−1,6−グルカンの量を上記実施例17(2)〜(3)と同様に測定した。結果を表8に示す。
【0121】
【表8】
【0122】
上記結果のとおり、A.pullulans 132株の培養液に含まれるα−グルカンは、A.pullulans AFO202株の培養液に含まれるα−グルカンよりも多い。しかし、本発明方法により測定されるβ−1,3−1,6−グルカンの量は、α−グルカンを除去しない場合で0.92g/Lであり、α−グルカンを除去した場合は1.10g/Lであって、この二つに大きな差はなかった。
【0123】
上記実施例17〜18の結果から、H1株とSY26株にそれぞれ由来するβ−1,3−グルカナーゼおよびβ−1,6−グルカナーゼを用いた本発明方法によれば、試料中におけるα−グルカンの存在または非存在、そして混在するα−グルカンの存在量にかかわらず正確にβ−1,3−1,6−グルカンを定量できることが実証された。
【図面の簡単な説明】
【0124】
【図1】実施例1において、土壌試料から単離された菌株をカードラン平板培地で培養した後、コンゴーレッド染色した写真である。
【図2】実施例1において、土壌試料から単離された菌株をデンプン培地で培養した後、ヨウ素染色した写真である。
【図3】実施例1で得たH1株の顕微鏡写真である。
【図4】反応温度とH1株由来のβ−1,3−グルカナーゼの活性との関係を示すグラフである。
【図5】H1株由来のβ−1,3−グルカナーゼの活性と熱処理温度との関係を示すグラフである。
【図6】H1株由来のβ−1,3−グルカナーゼによるカードランの分解反応生成物の経時的変化を示す薄層クロマトグラフィの結果である。
【図7】実施例9において、土壌試料から単離された菌株をパスツラン平板培地で培養した後、コンゴーレッド染色した写真である。
【図8】実施例9で得たSY26株の顕微鏡写真である。
【図9】SY26株由来のβ−1,6−グルカナーゼの各精製段階のSDS−PAGEの結果である。図中、1は分子量マーカーレーンを示し、2は20〜60%飽和硫安沈殿レーンを示し、3はゲル濾過クロマトグラフィによる精製後のレーンを示し、4はイオン交換クロマトグラフィによる精製後のレーンを示し、5はFPLCによる精製後のレーンを示す。
【図10】反応温度とSY26株由来のβ−1,6−グルカナーゼの活性との関係を示すグラフである。
【図11】SY26株由来のβ−1,6−グルカナーゼの活性と熱処理温度との関係を示すグラフである。
【図12】SY26株由来のβ−1,6−グルカナーゼによるパスツランの分解反応生成物の経時的変化を示す薄層クロマトグラフィの結果である。
【特許請求の範囲】
【請求項1】
多糖類含有試料に、β−1,3−グルカナーゼの産生能を有し且つβ−1,3−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないMitsuaria chitosanitabida種菌に由来する菌体外酵素液、並びにβ−1,6−グルカナーゼの産生能を有し且つβ−1,6−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないStreptomyces omiyaensis種菌に由来する菌体外酵素液を作用させる工程;および
生じたグルコースの量を測定する工程;
を含むことを特徴とする、β−1,3−1,6−グルカンの定量方法。
【請求項2】
上記Mitsuaria chitosanitabida種菌として、Mitsuaria chitosanitabida H1株(NITE AP−629)を用いる請求項1に記載の定量方法。
【請求項3】
上記Streptomyces omiyaensis種菌として、Streptomyces omiyaensis SY26株(NITE AP−616)を用いる請求項1または2に記載の定量方法。
【請求項4】
多糖類含有試料に、20℃以上、45℃以下で、上記Mitsuaria chitosanitabida種菌に由来する菌体外酵素液と上記Streptomyces omiyaensis種菌に由来する菌体外酵素液とを同時に作用させる請求項1〜3の何れかに記載の定量方法。
【請求項5】
β−1,3−グルカナーゼの産生能を有し且つβ−1,3−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないMitsuaria chitosanitabida種菌に由来する菌体外酵素液またはその乾燥物;
β−1,6−グルカナーゼの産生能を有し且つβ−1,6−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないStreptomyces omiyaensis種菌に由来する菌体外酵素液またはその乾燥物;並びに
グルコース量測定用試薬;
を含むことを特徴とする、β−1,3−1,6−グルカンの定量用キット。
【請求項6】
Mitsuaria chitosanitabida H1株(NITE AP−629)。
【請求項7】
Streptomyces omiyaensis SY26株(NITE AP−616)。
【請求項1】
多糖類含有試料に、β−1,3−グルカナーゼの産生能を有し且つβ−1,3−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないMitsuaria chitosanitabida種菌に由来する菌体外酵素液、並びにβ−1,6−グルカナーゼの産生能を有し且つβ−1,6−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないStreptomyces omiyaensis種菌に由来する菌体外酵素液を作用させる工程;および
生じたグルコースの量を測定する工程;
を含むことを特徴とする、β−1,3−1,6−グルカンの定量方法。
【請求項2】
上記Mitsuaria chitosanitabida種菌として、Mitsuaria chitosanitabida H1株(NITE AP−629)を用いる請求項1に記載の定量方法。
【請求項3】
上記Streptomyces omiyaensis種菌として、Streptomyces omiyaensis SY26株(NITE AP−616)を用いる請求項1または2に記載の定量方法。
【請求項4】
多糖類含有試料に、20℃以上、45℃以下で、上記Mitsuaria chitosanitabida種菌に由来する菌体外酵素液と上記Streptomyces omiyaensis種菌に由来する菌体外酵素液とを同時に作用させる請求項1〜3の何れかに記載の定量方法。
【請求項5】
β−1,3−グルカナーゼの産生能を有し且つβ−1,3−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないMitsuaria chitosanitabida種菌に由来する菌体外酵素液またはその乾燥物;
β−1,6−グルカナーゼの産生能を有し且つβ−1,6−グルカナーゼ以外のβ−グルカナーゼ活性およびα−グルコシダーゼ活性を示さないStreptomyces omiyaensis種菌に由来する菌体外酵素液またはその乾燥物;並びに
グルコース量測定用試薬;
を含むことを特徴とする、β−1,3−1,6−グルカンの定量用キット。
【請求項6】
Mitsuaria chitosanitabida H1株(NITE AP−629)。
【請求項7】
Streptomyces omiyaensis SY26株(NITE AP−616)。
【図4】

【図5】
【図10】
【図11】
【図1】
【図2】
【図3】
【図6】
【図7】

【図8】

【図9】
【図12】
【図5】
【図10】
【図11】
【図1】
【図2】
【図3】
【図6】
【図7】
【図8】
【図9】
【図12】
【公開番号】特開2010−41957(P2010−41957A)
【公開日】平成22年2月25日(2010.2.25)
【国際特許分類】
【出願番号】特願2008−207967(P2008−207967)
【出願日】平成20年8月12日(2008.8.12)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 平成20年2月13日に高知大学大学院農学研究科発行の2007年度修士論文要旨において発表
【出願人】(594063474)株式会社ソフィ (3)
【出願人】(504174180)国立大学法人高知大学 (174)
【Fターム(参考)】
【公開日】平成22年2月25日(2010.2.25)
【国際特許分類】
【出願日】平成20年8月12日(2008.8.12)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 平成20年2月13日に高知大学大学院農学研究科発行の2007年度修士論文要旨において発表
【出願人】(594063474)株式会社ソフィ (3)
【出願人】(504174180)国立大学法人高知大学 (174)
【Fターム(参考)】
[ Back to top ]