
きのこを用いたバイオマスからの効率的エタノール生産システムの開発
【課題】 担子菌を用いた効率的なアルコール生産方法、アルコール飲料の生産方法、アルコール含有食品の生産方法、またはそれらに用いる種菌を明らかにする。
【解決手段】 シロアミタケ属の担子菌を用いて、炭素源からアルコールを生成する、アルコール生産方法を用いる。または、アラゲカワラタケ、カワラタケ、およびシロアミタケからなる群から選ばれる1種以上の担子菌を用いて、炭素源からアルコールを生成する、アルコール生産方法を用いる。
【解決手段】 シロアミタケ属の担子菌を用いて、炭素源からアルコールを生成する、アルコール生産方法を用いる。または、アラゲカワラタケ、カワラタケ、およびシロアミタケからなる群から選ばれる1種以上の担子菌を用いて、炭素源からアルコールを生成する、アルコール生産方法を用いる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、アルコールの生産方法、アルコール飲料の生産方法、アルコール含有食品の生産方法、またはそれらに用いる種菌に関する。
【背景技術】
【0002】
現在、世界のエネルギー消費は約9割が石油をはじめとする化石燃料に依存しており、エネルギー枯渇の窮地に立たされている。また、近年の原油価格の上昇に加え、炭酸ガスの増加や地球温暖化等の世界的な環境問題が注目されており、将来的に予想される化石燃料の枯渇および原油価格の高止まり等に先立ち、世界各国で化石燃料に代わる新たな再生可能エネルギーの開発が進められている。このような状況の下、バイオエタノールをはじめとするバイオ燃料の開発導入に係る動きがブラジル、米国、アジア各国を中心に各国で活発化するなど、バイオ燃料への期待が高まっている。しかしながら、これらの大半は、サトウキビやトウモロコシ等の食用や飼料作物を原料として生産されているため、現在、農家の転作等による余波が穀物市場の高騰を連鎖的に招いている。したがって、中長期的には、食用や飼料作物と競合しないバイオ燃料の実用化を目指して、廃木材や草本類等の未利用資源の活用が世界規模で重要となる。
【0003】
これに対して、木質系や草本系等のバイオマス資源から糖を生成し、その得られた糖から燃料として有用なエタノール等のアルコールを生成する技術が知られている。例えば、特許文献1および非特許文献1には、木質系バイオマス(または、リグノセルロース系植物材料)からバイオ燃料を生産する際の糖化処理に先駆けて、リグニン分解能を有する白色腐朽菌によりリグニンを分解する生物的処理(前処理)を行う方法について開示されている。
【0004】
また、菌類を用いて炭素源からアルコールを生産する研究もなされている。例えば、特許文献2〜4、非特許文献2〜6には、アガリクスタケ(Agaricus blazei)、ブナシメジ(Hypsizygus marmoreus)、マツタケ(Tricholoma matsutake)、ヒラタケ(Pleurotus ostreatus)、ミミナミハタケ(Lentinellus cochleatus)、およびエノキタケ(Flammulina velutipes)等の担子菌を用いて炭素源からアルコールを生産する方法が開示されている。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2008-6372号公報
【特許文献2】特開2001-286276号公報
【特許文献3】特開2004-298109号公報
【特許文献4】特開2006-223159号公報
【非特許文献】
【0006】
【非特許文献1】バイオエネルギー技術と応用展開、2003年10月31日発行、(株)シーエムシー出版、pp.165-171
【非特許文献2】Tokumitsu OKAMURA, Tomoko OGATA, Norie MINAMOTO, Tomomi TAKENO, Hiroko NODA, Shoko FUKUDA and Masahiro OHSUGI, "Characteristics of Wine Produced by Mushroom Fermentation", Bioscience, Biotechnology, and Biochemistry Vol. 65 (2001), No. 7 pp.1596-1600
【非特許文献3】Tokumitsu Okamura-Matsui, Tomomi Tomoda, Shoko Fukuda and Masahiro Ohsugi, "Discovery of alcohol dehydrogenase from mushrooms and application to alcoholic beverages", Journal of Molecular Catalysis B: Enzymatic、Volume 23, Issues 2-6, 1 September 2003, pp.133-144
【非特許文献4】化学工業日報、2007年10月22日発行、(株)化学工業日報社
【非特許文献5】'Use of whole crop sorghums as a raw material in consolidated bioprocessing bioethanol production using Flammulina velutipes.' Mizuno et al., Biosci Biotechnol Biochem. 2009 Jul;73(7):1671-3. Epub 2009 Jul 7.
【非特許文献6】'Properties of ethanol fermentation by Flammulina velutipes.' Mizuno et al., Biosci Biotechnol Biochem. 2009 Oct;73(10):2240-5. Epub 2009 Oct 7.
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、従来の木質系等のバイオマスからのアルコールの生産においては、バイオマスの主要成分であるセルロースやヘミセルロースを加水分解して糖を生成(糖化)する工程と、生成した糖を酵母や細菌により糖を発酵させることによりアルコールを生成する工程、の2工程が必要であった。したがって、バイオマスからのアルコール生産工程全体では、煩雑な作業が必要となり、コストおよび作業時間が嵩むという問題があった。
【0008】
また、上述した糖化する工程においては、硫酸による加水分解が必要となるため、処理コスト面および環境面で問題視されていた。また、生成したグルコース等の糖の過分解を抑制するために反応制御が必要であった。
【0009】
さらに、硫酸を用いる以外の糖化方法として、バイオマスに、(1)粉砕、蒸煮(蒸煮爆砕、蒸煮、熱水分解・加圧熱水処理等)、エネルギー線(電子線、γ線、マイクロウエーブ)の照射等の物理的方法で処理する物理的前処理、(2)酸(硫酸、亜硫酸、リン酸)、アルカリ(カセイソーダ、アンモニアなど)などを用いて化学的処理する化学的前処理、または、(3)白色腐朽菌等のリグニン分解菌を用いて行う生物的前処理、による前処理を行い、リグニン等の難分解性成分を分解した後に、セルラーゼのような糖化酵素を加えてセルロースを加水分解して糖にする酵素糖化方法がある。
【0010】
しかしながら、上述の(1)の前処理方法では、蒸煮のための高温・高圧装置や、電子線、γ線、マイクロウエーブ等のエネルギー線の照射装置などの高価な装置が必要なため、設備費が高くなり、さらに、前処理工程の管理を厳密に行う必要があった。また、上述の(2)の前処理方法においては、バイオマスの糖化または前処理に当たり、酸やアルカリ等の化学薬品を用いるため、それらの薬品に耐え得る設備を使用する必要があることから、設備費が高くなるという問題があった。そして、糖化または前処理に用いた酸やアルカリの中和処理とそれによって発生する大量の中和廃棄物(例えば、硫酸カルシウムなど)の処理、あるいは、前処理に用いた溶媒の除去を行う必要があるため、手間やコストが掛かり、しかも、環境面での問題があった。さらに、上述の(3)の前処理方法では、糖化酵素を用いるため、コストが掛かり、しかも、当該酵素による糖の過分解を抑制するために反応制御が必要であった。またこの方法は、他の前処理方法のような不都合は生じないが、酵素糖化工程およびその次工程である酵母等の糖の発酵工程のために、白色腐朽菌を除去する必要が生じ、手間が掛かるという問題があった。また、そもそも、白色腐朽菌は、アルコール生成を企図して導入されるものでなく、実際のアルコール生成は、酵母等の発酵により行われるので、上述の(3)の方法で用いられる白色腐朽菌のアルコール生成能については、特に考慮する必要はなかった。
【0011】
また、従来の担子菌を使用したアルコール生産方法では、キシロースのようなペントースに分類される糖をほとんどアルコールに発酵できなかった。そのため、バイオマスから得られるエタノール量も十分とはいえなかった。
【0012】
本発明は上記事情に鑑みてなされたものであり、特定の担子菌を用いた効率的なアルコール生産方法を提供することを目的とする。
【課題を解決するための手段】
【0013】
本発明によれば、シロアミタケ(Trametes)属の担子菌を用いて、炭素源からアルコールを生成する、アルコール生産方法が提供される。
【0014】
この生産方法は、後述する実施例で効率的にアルコールを生産するために利用できることが実証されている。そのため、この生産方法を用いれば、効率的にアルコールを生産できる。
【0015】
また本発明によれば、シロアミタケ属の担子菌を用いて、炭素源からアルコールを含有する液体を生成する、アルコール飲料の生産方法が提供される。
【0016】
ここで、後述する実施例でシロアミタケ属の担子菌を用いて、炭素源からアルコールを含有する液体を効率的に生成できることが実証されている。そのため、この生産方法を用いれば、効率的にアルコールを含有する液体を生産できる。
【0017】
また本発明によれば、シロアミタケ属の担子菌を用いて、炭素源からアルコールを含有する組成物を生成する、アルコール含有食品の生産方法が提供される。
【0018】
ここで、後述する実施例でシロアミタケ属の担子菌を用いて、炭素源からアルコールを含有する組成物を効率的に生成できることが実証されている。そのため、この生産方法を用いれば、効率的にアルコール含有食品を生産できる。
【0019】
また本発明によれば、シロアミタケ属の担子菌の菌糸と、菌糸を担持する担体と、を備えることを特徴とする、シロアミタケ属の担子菌を用いて炭素源からアルコールを生成するための種菌が提供される。
【0020】
この種菌は、後述する実施例で効率的にアルコールを生産できることが実証されている担子菌の菌糸を備えている。そのため、この種菌を用いれば、効率的にアルコールを生産できる。
【発明の効果】
【0021】
本発明によれば、アルコール生産効率および炭素源の資化性に優れた菌類により、バイオマス等から効率的なアルコール生産を実現することができる。
【図面の簡単な説明】
【0022】
【図1】図1(a)および図1(b)は、実施形態に係るシロアミタケ属の担子菌により炭素源からアルコールを生成する方法を説明するためのフローチャートである。
【図2】図2(a)、図2(b)および図2(c)は、実施形態に係るシロアミタケ属の担子菌を用いた種菌の構成を模式的に示した概念図である。
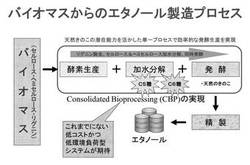
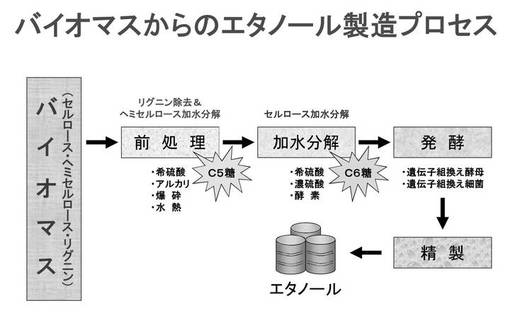
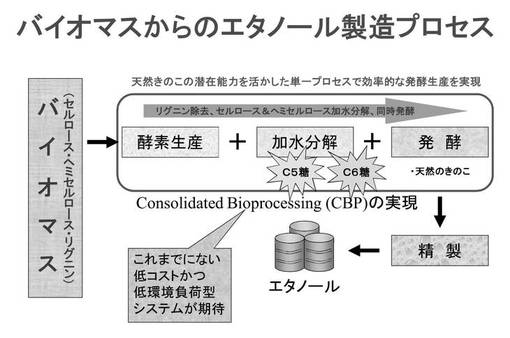
【図3】図3は、従来の典型的なバイオマスからのエタノール生産プロセスを説明するための概念図である。
【図4】図4は、実施形態に係るシロアミタケ属の担子菌を用いた、未利用バイオマス資源の再利用方法の一例を説明するための概念図である。
【図5】図5は、実施形態に係るシロアミタケ属の担子菌を用いた未利用バイオマス資源の燃料用アルコールとしての再利用のサイクルを説明するための概念図である。
【図6】図6は、シロアミタケ属の担子菌の培養および菌体回収の方法を説明するための実験プロトコルである。
【発明を実施するための最良の形態】
【0023】
本発明者らは、鋭意検討を重ねた結果、シロアミタケ属の担子菌が各種炭素源を糖化することにより糖を生成する工程、および糖を発酵することによりアルコールを生成する工程を単一で行うことができ、かつ高いアルコール生産性を有することを確認してバイオマスから直接アルコールが生産可能であることを見出し、本発明を完成するに至った。
【0024】
以下、本発明の実施の形態について詳細に説明する。なお、同様な内容については繰り返しの煩雑を避けるために、適宜説明を省略する。
【0025】
本発明の一実施形態は、シロアミタケ属の担子菌を用いて、炭素源からアルコールを生成する、アルコール生産方法である。この生産方法を適切に利用すれば、後述する実施例で実証されているように、効率的にアルコールを生産できる。上記シロアミタケ属の担子菌は特に限定されないが、例えばアラゲカワラタケ(Trametes hirsuta)、カワラタケ(Trametes versicolor)、シロアミタケ(Trametes suaveolens)、オオチリメンタケ(Trametes gibbosa)、ウサギタケ(Trametes trogiiBerkeley)、クジラタケ(Trametes orientalis)、ヤキフタケ(Trametes pubescens)、チリメンタケ(Trametes elegans )、Trametes lactinea を挙げることができる。この中でもアラゲカワラタケ、カワラタケ、およびシロアミタケは、フルクトース、マンノース、ガラクトース、セロビオース、マルトース、アラビノース、キシロース、スクロース、デンプン、CMC、小麦フスマ、稲わら、およびボールミル処理後の稲わらからアルコールを生産できることが後述する実施例で実証されている。このアラゲカワラタケ、カワラタケ、シロアミタケは、一般的にリグニン分解能を有し、世界的に分布している白色腐朽菌である。特に、アラゲカワラタケおよびカワラタケは、小麦フスマ、稲わら、およびボールミル処理後の稲わらのようなバイオマスからでも、高い濃度のアルコールを生産できることが後述する実施例で実証されている。
【0026】
また従来、担子菌を使用したアルコール生産方法では、キシロースのようなペントースに分類される糖をほとんどアルコールに発酵できなかった。そのため、バイオマスから得られるエタノール量も十分とはいえなかった。一方で、シロアミタケ属の担子菌はそのような糖からでもアルコールを生産できるため、より効率的にバイオマスからアルコールを生産できる。特に、アラゲカワラタケおよびカワラタケが、そのような糖を効率よくエタノールに変換できることが後述する実施例で実証されている。特にカワラタケで顕著であった。キシロースはヘミセルロースの一つであるキシランを構成するものとして植物に広く存在することが知られている。なお、キシロースを発酵できる酵母としてPichia stipitisが知られているが、エタノール耐性が低く、エタノール変換効率、グルコースの発酵効率も十分とはいえなかった。また、この酵母は糖化能を有しないため、バイオマスからエタノールを生産するには糖化工程を別に設けるか、遺伝子改変等の方法を行う必要があった。
【0027】
本実施形態では、アルコール生成にシロアミタケ属の担子菌を用いることを特徴の一つとする。一方で、以下に示すとおり、シロアミタケ属の担子菌のアルコール生成能についてはこれまで何ら報告されていなかった。例えば特許文献1および非特許文献1に記載されている担子菌は、上述した糖化工程における生物的前処理用として用いるものであって、直接的にアルコールを生成するために用いるものではない。したがって、特許文献1および非特許文献1では担子菌のアルコール生成能については考慮する必要がないので、当然のことながら、アルコール生成能については示唆すらされていない。また、特許文献2〜4、非特許文献2〜6においても、シロアミタケ属の担子菌を用いて炭素源からアルコールを生産することについては何ら記載されていない。
【0028】
即ち、シロアミタケ属の担子菌のアルコール生成能についてはこれまで何ら報告されておらず、本実施形態は、これまで存在しなかった優れたアルコール生産方法といえる。加えて、目、科、または属が異なればそれぞれの担子菌が持つ機能も異なるため、これらの文献から本実施形態のアルコール生産方法に想到することは困難である。
【0029】
また、本実施形態においては、シロアミタケ属の担子菌は、毒性はなく、遺伝子組換え体でなく、野生のものを用いることができる。したがって、本実施形態のアルコール生産方法では、安全性の高いアルコールの生産が実現可能となる。
【0030】
さらに、シロアミタケ属の担子菌は一般的に食用として汎用されていない。また、産業的に有効に利用されているという報告はなく、これまでほとんど利用されていなかった。そして、シロアミタケ属の担子菌は、成長が比較的早く、世界的に分布しており比較的入手しやすい。したがって、本実施形態に係るアルコール生産方法においては、用いるシロアミタケ属の担子菌は、少なくとも産業界と直接競合することは考え難く、資源として有用性および優位性に富むものである。
【0031】
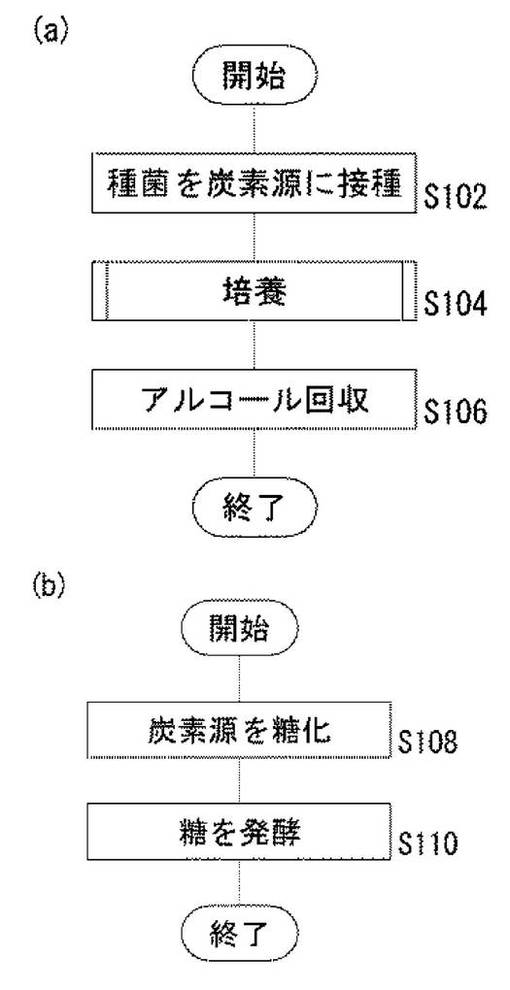
図1は、実施の形態に係るシロアミタケ属の担子菌により炭素源からアルコールを生成する方法を説明するためのフローチャートである。後述するようにシロアミタケ属の担子菌が、幅広い糖の資化性を有することを、本発明者らは見出している。
【0032】
具体的には、図1(a)に示すように、まず、シロアミタケ属の担子菌を木質系・草本系等のバイオマスまたは各種の糖などを含む炭素源に接種する(S102)。次いで、シロアミタケ属の担子菌の種菌を接種された炭素源を培養する(S104)。そして、シロアミタケ属の担子菌により炭素源から生成されたアルコールを含む液体を濾過などの手法により回収する(S106)。
【0033】
上述の炭素源が木質系等のバイオマスを含む場合には、上述の培養工程において、図1(b)に示すように、シロアミタケ属の担子菌により炭素源を糖化する(S108)。そして、糖化された炭素源をシロアミタケ属の担子菌により発酵してアルコールを生成する(S110)。
【0034】
なお、上述の発酵工程は、嫌気的条件であってもよく、好気的条件であってもよいが、微好気的条件または嫌気的条件が、アルコール生成効率の面から好ましい。
【0035】
また、上述の炭素源は、糖を含んでいてもよい。さらに、上述の炭素源は、グルコース、フルクトース、マンノース、ガラクトース、セロビオース、マルトース、アラビノース、キシロース、スクロース、およびラクトースからなる群より選ばれる一種以上の糖を含んでいてもよい。後述するように、シロアミタケ属の担子菌は、これらの糖に対する資化性を有していることを、本発明者らは見出している。さらに、上述の炭素源は、糖蜜、廃糖蜜、上白糖、黒糖、マルツエキスおよびデンプン等を含んでいてもよい。なお本実施形態に係るシロアミタケ属の担子菌は、複数の炭素源が混在した培地からアルコールを生成してもよい。培地中の個々の炭素源の質量%または容量%は、例えば1、2、3、5、10、20、50、80、または100%であってもよく、それらの値の範囲内またはそれらの値以上であってもよい。
【0036】
あるいは、上述の炭素源は、セルロース、木質系または草本系の材料(バイオマス等)を含んでいてもよい。セルロースとしては、綿リンター、木材パルプや溶解パルプ等から得られる植物系のセルロース、アセトバクター属(Acetobacter)等に属する微生物が生産するセルロース、再生セルロースおよび微結晶セルロース等が挙げられる。木質系の材料としては、建築廃材等の廃木材、製材残材、間伐材、林地残材などを含む木材、おがくず、古紙・廃紙等の紙類およびパルプ等を挙げることができる。草本系の材料としては、小麦フスマ等のフスマ、稲わら・麦わら等のわらおよびもみ殻などを含む農業残渣、バガス、刈り草および雑草類等を挙げることができる。なお、後述するようにシロアミタケ属の担子菌が、小麦フスマおよび稲わらを炭素源として直接的にアルコール生成能を有し、アルコール収率が高いことを本発明者らは見出している。また、後述する実施例でセルロース分解能を有していることが示唆されている。
【0037】
また上述の炭素源は生ゴミまたは廃棄飲料(以下、「生ゴミ等」と称することもある)を含んでいてもよい。上記生ゴミ等は、業務用または家庭用の生ゴミ等であってもよい。また、食堂、病院、学校、ホテル、食品工場、または農畜産場等から排出される生ゴミ等であってもよい。また、実験室や工場において調製した生ゴミ等であってもよい。また生ゴミは、廃棄食品または廃棄飲料を含んでいてもよい。なお、本明細書において廃棄食品は、廃棄された食品、食用としての商品価値がない食品、または消費期限後の食品を意味する。本明細書において廃棄飲料は、廃棄された飲料、飲用としての商品価値がない飲料、または消費期限後の飲料を意味する。廃棄食品は、例えば、廃棄野菜、廃棄果物、廃棄肉、廃棄魚、廃棄米飯、廃棄コーン、廃棄卵、または廃棄麺等を含んでいてもよい。廃棄飲料は、例えば、廃棄乳系飲料等を含んでいてもよい。また、生ゴミは破砕処理後の状態であってもよい。生ゴミの組成としては糖、リグニン、蛋白質、脂質、アミノ酸、有機酸、ビタミン類、またはミネラル等の無機物を含んでいてもよい。
【0038】
また上述の炭素源は、食品または乳系飲料を含んでいてもよい。食品は、加工食品または生鮮食品を含んでいてもよい。食品は、例えば野菜、果物、肉、魚、米飯、コーン、卵、または麺等を含んでいてもよい。乳系飲料は、例えば牛乳を含んでいてもよい。牛乳は、例えば明治おいしい牛乳(株式会社明治)等、市販の牛乳であってもよい。牛乳は、例えば成分無調整牛乳、または調整(例えば低脂肪、無脂肪、高脂肪)牛乳であってもよい。食品または乳系飲料は、消費期限内または消費期限後であってもよい。
【0039】
また、上述のシロアミタケ属の担子菌により生成されるアルコールは、エタノールを含んでいてもよい。後述するように、シロアミタケ属の担子菌は、炭素源からエタノールを生成することを、本発明者らは見出している。
【0040】
そして、上述のシロアミタケ属の担子菌により生成されるアルコールを用いて、アルコール飲料またはアルコール含有食品を生産してもよい。これらのアルコール飲料またはアルコール含有食品には、アルコール以外にも、シロアミタケ属の担子菌の生成する各種成分が含まれていてもよ
い。また、アルコール含有食品は、固体であってもよく、液体であってもよく、ゲル状体などであってもよい。
【0041】
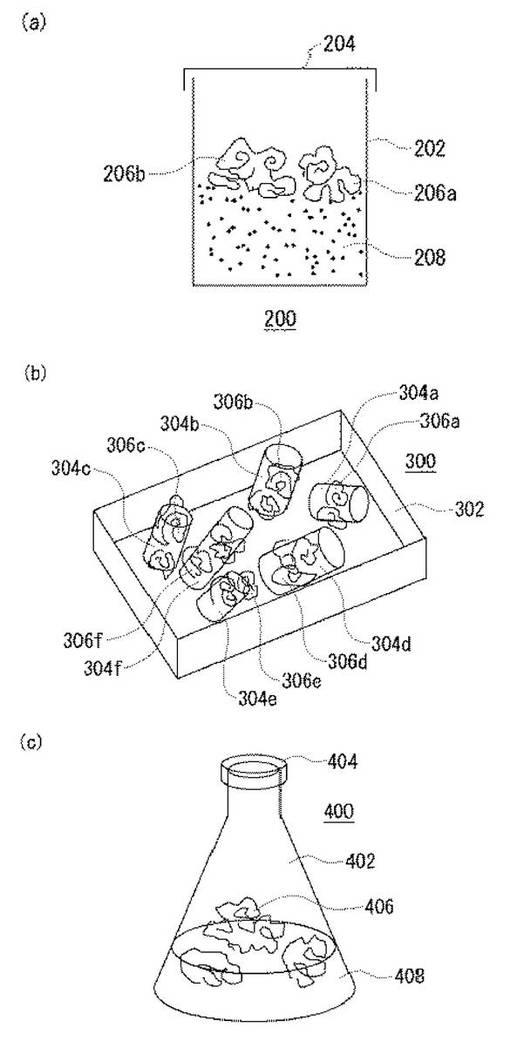
図2は、実施の形態に係るシロアミタケ属の担子菌を用いた種菌の構成を模式的に示した概念図である。種菌とは、研究対象の菌類を育て、増やす際の基になる培養物のことである。種菌は、アルコール生産用材料と言い換えることもできる。図2(a)は、おがくずを担体として用いたシロアミタケ属の担子菌の種菌である。この種菌200では、蓋204を備える容器202内に、おがくず208が敷き詰められている。このおがくず中にシロアミタケの菌糸206a、206bが担持されている。なお、シロアミタケ属の担子菌の菌糸は、図2(a)のように、きのこを形成している必要はない。
【0042】
図2(b)は、木材チップを担体として用いたシロアミタケ属の担子菌の種菌である。この種菌300では、容器302内に、コルク栓状の形状からなる木材チップ304a、304b、304c、304d、304e、304fが収納されている。これらの木材チップには、シロアミタケ属の担子菌の菌糸306a、306b、306c、306d、306e、306fが担持されている。なお、シロアミタケ属の担子菌の菌糸は、図2(b)のように、きのこを形成している必要はない。
【0043】
図2(c)は、液体培地を担体として用いたシロアミタケ属の担子菌の種菌である。この種菌400では、蓋404を備える容器402内に、各種の糖などの炭素源を含む液体培地408が収納されている。これらの液体培地には、シロアミタケ属の担子菌の菌糸406が担持されている。なお、シロアミタケ属の担子菌の菌糸は、図2(c)のように、きのこを形成している必要はない。
【0044】
上述の種菌は、菌類を用いて炭素源からアルコールを生成するための種菌であって、シロアミタケ属の担子菌の菌糸と、この菌糸を担持する担体とを備える。なお、この菌糸は、定常期の菌糸であってもよい。具体的には、この菌糸は、培養開始5〜7日経過後の菌糸であってもよい。
【0045】
以下、実施の形態に係るアルコール生産方法の作用効果についてさらに説明する。
シロアミタケ属の担子菌は、後述するように、カビや酵母と同等以上のアルコール生産能を有するので、優れたアルコール生産効率を実現することができる。
【0046】
また、シロアミタケ属の担子菌は、後述するように、グルコース、フルクトース、マンノース、ガラクトース、セロビオース、マルトース、アラビノース、キシロース、およびスクロースに対する資化性を有するので、担子菌類では困難であった幅広い種類の炭素源を用いてアルコール発酵を行うことができる。
【0047】
そのため、上述の実施の形態に係るアルコール生産方法によれば、アルコール生産効率および炭素源の資化性に優れるシロアミタケ属の担子菌を用いるため、アルコール生産方法の生産効率を向上し、炭素源の種類の幅を拡大することができる。
【0048】
図3は従来の典型的なバイオマスからのエタノール生産プロセスを説明するための概念図である。従来は、炭素源を糖化する工程と、糖を発酵する工程は別々のユニットによって行われていた。図4は、本実施形態に係るシロアミタケ属の担子菌を用いた、未利用バイオマス資源の再利用方法の一例を説明するための概念図である。この方法は、シロアミタケ属の担子菌が炭素源を糖化することにより糖を生成する工程と、シロアミタケ属の担子菌が糖を発酵することによりアルコールを生成する工程と、を含むため、従来の酸等による加水分解の工程を経ずに、糖化・発酵の両工程を行うことができる。このため、未利用バイオマス資源をシロアミタケ属の担子菌による糖化・発酵工程により効率よくエタノールに変換することができる。また、環境負荷、コストや手間の削減につながるのはいうまでもない。また糖化・発酵は同一容器で並行して、または同一容器で同時に行われてもよく、この場合、より効率的である。
【0049】
糖化と発酵を並行して行いアルコールを生産するプロセスは、一般的に統合バイオプロセス(consolidated bioprocessing :CBP)と呼ばれる。CBPは例えば[Lynd et al., Curr Opin Biotechnol. 2005 Oct;16(5):577-83.]、[van Zyl et al., Adv Biochem Eng Biotechnol. 2007;108:205-35.]、[Mizuno et al., Biosci Biotechnol Biochem. 2009 Jul;73(7):1671-3. Epub 2009 Jul 7.]などに記載されている。そして、本実施形態に係るシロアミタケ属の担子菌は、CBPによってアルコールを精製することができる。このCBPにおいては、リグニンやセルロース等を分解するために酵素や酸等を加えてもよいが、上記シロアミタケ属の担子菌を用いた場合には酵素や酸等を加えなくても、効率的にアルコールを生産可能である。なお、従来アルコール生産にはセルロースの分解が重要と考えられていたが、本実施形態においては、優れたリグニン分解能を有する上記シロアミタケ属の担子菌を用いたことが、従来予想しえない高いエタノール生産性を実現できた一因になっていると考えられる。リグニンは木質系または草本系の材料に含まれている。
【0050】
またこのCBPにおいては、本実施形態に係るシロアミタケ属の担子菌がリグニン、セルロース、またはヘミセルロースを分解し、さらに生成した単糖またはオリゴ糖を発酵することによってアルコールを生産してもよい。このとき、上記単糖はヘキソースまたはペントースであってもよく、グルコース、フルクトース、マンノース、ガラクトース、アラビノース、およびキシロースからなる群より選ばれる一種以上の糖であってもよい。また上記オリゴ糖はセロビオース、マルトース、およびスクロースからなる群より選ばれる一種以上の糖であってもよい。そしてこのようなCBPは、低コストで、低環境負荷である。なお、CBPと同様の原理は醸造酒の生産過程でも利用されており、その場合、並行複発酵とも呼ばれる。この並行複発酵も、糖化と発酵とが並行して行なわれる。
【0051】
ここで、木材や紙類などの木質系のバイオマスを酸糖化して得られる炭素源には、一般的にグルコースにくわえて、マンノース等が数%含まれる。酵母は、グルコースを好適に資化するが、マンノースに対する資化性は低い。一方、シロアミタケ属の担子菌は、酵母により資化することが困難なマンノースに対しても資化性を有するため、酵母による発酵後に残存するマンノースも資化することができる。そのため、木質系のバイオマスを糖酸化し、酵母およびシロアミタケ属の担子菌を組み合わせてアルコール発酵を行うことにより、資源のリサイクル効率およびアルコール生産効率を高めることができる。また、天然の酵母は、小麦フスマ等のバイオマスを直接的にエタノールに変換することはできない。これに対して、シロアミタケ属の担子菌は、後述するように、小麦フスマおよび稲わらから直接的にエタノールを生産することができる。したがって、実施の形態に係るアルコール生産方法においては、天然の酵母が利用することができないバイオマスからアルコールを直接的に生産することができる点でも、優位性および有用性に富むものである。
【0052】
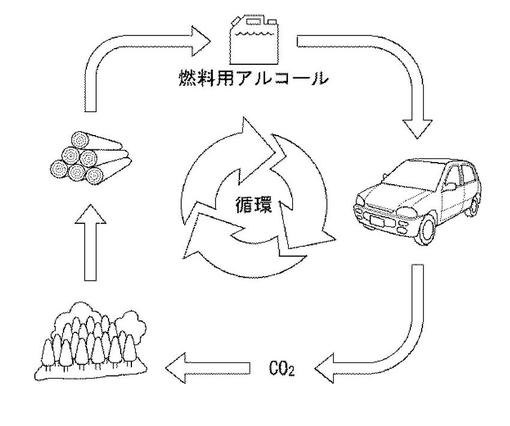
図5は、実施の形態に係るシロアミタケ属の担子菌を用いた、未利用バイオマス資源の燃料用アルコールとしての再利用のサイクルを説明するための概念図である。このサイクルは、未利用バイオマス資源をシロアミタケ属の担子菌による糖化・発酵工程により効率よくアルコールに変換することができるため、バイオマスと燃料用アルコールとの間で資源の循環システムを構築することができる。そのため、地球環境保全および産業の発展を両立しうるエネルギー供給システムを構築することができる。
【0053】
また、実施の形態に係るアルコール飲料の生産方法によれば、アルコール生産効率および炭素源の資化性に優れるシロアミタケ属の担子菌を用いるため、アルコール飲料の生産方法の生産効率を向上し、炭素源の種類の幅を拡大することができる。
【0054】
さらに、実施の形態に係るアルコール含有食品の生産方法によれば、アルコール生産効率および炭素源の資化性に優れるシロアミタケ属の担子菌を用いるため、アルコール含有食品の生産方法の生産効率を向上し、炭素源の種類の幅を拡大することができる。
【0055】
そして、実施の形態に係る種菌によれば、アルコール生産効率および炭素源の資化性に優れるシロアミタケ属の担子菌の菌糸を担体に担持させているため、菌類を用いて炭素源からアルコールを生成するための種菌として好適に用いることができる。
【0056】
以上、図面を参照して本発明の実施形態について述べたが、これらは本発明の例示であり、上記以外の様々な構成を採用することもできる。
【実施例】
【0057】
以下、本発明を実施例によりさらに説明するが、本発明はこれらに限定されるものではない。ただし、実施例中に表示される%は、特に断りのない限り質量パーセントを示す。
[使用菌株]
表1の菌株を準備した。表1の各菌株の28S rDNA塩基配列を解析し、GenBank/DDBJ/EMBL等の塩基配列データベースに基づく相同性検索を行ったところ、データベースに登録されている28S rDNA塩基配列とそれぞれ100%の相同率を示した。
【0058】
【表1】

【0059】
1)使用培地
使用培地としては、MYG培地もしくはT培地を用いた。下記に、各培地の基本的組成を示す。
MYG培地
グルコース* :0.4 %
Yeast extract :0.4 %
Malt extract :1.0 %
T培地
グルコース* :2.0 %
Yeast extract :1.0 %
KH2PO4 :1.0 %
(NH4)2SO4 :0.2 %
MgSO4・7H2O :0.05 %
*)グルコースについては、必要に応じて、他の炭素源に変更した。
【0060】
2)培養および菌体回収
図6は、カワラタケの培養方法を説明するための実験プロトコルである。まず、前培養ステップでは、カワラタケの平板培地から5mm四方の大きさの菌糸断片を切り取り、500ml容三角フラスコに入ったMYG培地50mlに加え、綿栓もしくはシリコ栓を用いて三角フラスコを好気状態にして、28℃で5〜7日間培養を行った。
【0061】
次いで、本培養ステップでは、500ml容三角フラスコにT培地を50ml加え、無菌的にろ過、洗浄したカワラタケの前培養菌糸を接種し、シリコンゴム栓を用いて三角フラスコを密栓状態にして、28℃で静置培養(微好気的件下での培養)を行った。
【0062】
続いて、培養液の回収ステップでは、経時的に次の手順で回収を行った。すなわち、培養液をサンプリングし、遠心分離後、0.22μmのフィルターでろ過した。培養濾液pHの測定、HPLC分析、および活性測定して、その後、冷凍保存した。表1に記載のカワラタケ以外の各菌株についても、カワラタケの場合と同様の手順で培養およびサンプルの回収を行なった。
【0063】
3)HPLC分析
表1に記載の各菌株の培養濾液をHPLC分析した。その分析条件は以下に示すとおりである。
(HPLC分析条件)
キャピラリーカラム :Shodex SUGAR SP0810またはShodex SUGAR KS−801
キャピラリーサイズ :8.0mmID×300mmL
流量 :0.6ml/min
カラム温度 :80℃
抽出液 :脱気蒸留水
試料 :10μl
【0064】
4)グルコース以外の炭素源を用いた培養
T培地の炭素源であるグルコースに代わり、同濃度のフルクトース、マンノース、ガラクトース、セロビオース、マルトース、アラビノース、キシロース、スクロース、デンプン(Starch)、CMC(カルボキシメチルセルロース)、小麦フスマ(Wheat bran)、稲わら(Rice straw)、ボールミル処理後の稲わら(BM-Rice straw)を用いて、その資化性とエタノール生産性を確かめた。手順は2)および3)と同様である。なお、ボールミル処理は遊星ボールミルGokin planet M2-3F(ナガオシステム社製)でジルコニア容器とジルコニアボールにより、トータル4時間(10分稼働、10分休止)行った。
【0065】
上記1)〜4)の結果を表2および表3に示す。各数値は3回実験を行った結果の平均値である。
【0066】
【表2】
【0067】
【表3】
【0068】
表2は、グルコースを基質としてアラゲカワラタケ(Trametes hirsuta)、カワラタケ(Trametes versicolor)を培養したときのエタノール生産率を、カビ(Rhizopus oryzae、Mucor corticolous)および酵母(Saccharomyces cerevisiae)と比較した表である。この表から、アラゲカワラタケやカワラタケがカビや酵母と比較して高いエタノール生産性を有していることがわかる。
【0069】
表3は、各種炭素源(20g/培地1L)を基質として、表1に記載の4種類の菌株(アラゲカワラタケ(Trametes hirsuta)、カワラタケ(Trametes versicolor)、シロアミタケ(Trametes suaveolens)、ヒラタケ(Pleurotus ostreatus)を5日間培養したときのエタノール生産量(g/培地1L)を表している。この表から、アラゲカワラタケ、カワラタケ、およびシロアミタケは、フルクトース、マンノース、ガラクトース、セロビオース、マルトース、アラビノース、キシロース、スクロース、デンプン(Starch)、CMC(カルボキシメチルセルロース)、小麦フスマ(Wheat bran)、稲わら(Rice straw)、およびボールミル処理後の稲わら(BM-Rice straw)を代謝することで、比較的高い濃度のエタノールを生産できることがわかる。このエタノール生産量は、特にグルコース、マンノース、セロビオース、デンプンで高い値を示した。
【0070】
また、カワラタケ、アラゲカワラタケはキシロース、小麦フスマ、稲わら、およびボールミル処理後の稲わらを基質としたときに、他の菌株に比べて顕著に高いエタノール生産量を示した。特に、通常のキノコを用いた場合、キシロースを炭素源にして2g/Lを超える量のエタノールを生産することは困難であるにも関わらず、カワラタケが7.2g/L、アラゲカワラタケが3.9g/Lの高い生産量を示したことは驚くべき結果であった。キシロースは種々のバイオマスに多く含まれる成分であるため、カワラタケやアラゲカワラタケを用いれば従来法よりも効率よくエタノールを種々のバイオマスから生産できると考えられる。なお、同様の実験を複数回行なったところ、カワラタケによるキシロースからのエタノール生産量は高いときで8.0g/Lを超えることもあった。また、アラゲカワラタケを用いた場合のキシロース、小麦フスマ、稲わら、およびボールミル処理後の稲わらからのエタノール生産量は、高いときでカワラタケの場合の9割程度の数値を示すこともあった。
【0071】
また、アラゲカワラタケ、カワラタケ、およびシロアミタケを用いた場合において、稲わらをボールミル処理しない場合(3.0、3.3、1.5)でも、ボールミル処理をした場合(3.4、3.7、1.7)に近い生産性が見られた。通常、ボールミルはセルロースの骨格を破壊することで糖化を促進させると考えられており、2倍くらいの効率でエタノール生産性を促進すると言われている。一方で本実施例においてアラゲカワラタケ、カワラタケ、およびシロアミタケを用いた場合には、これらの菌株がセルロースを分解したために、ボールミル処理の前後で生産性があまり変化しなかったと考えられる。従って、アラゲカワラタケ、カワラタケ、およびシロアミタケを用いたエタノール生産方法は、ボールミルのような特殊な破砕工程を行なわなくても効率的にエタノールを生産できる、優れた方法であるといえる。特に、ボールミルは大きなエネルギーを必要とする操作なので、省エネルギーの観点からも上記方法の意義は大きい。
【0072】
5)微好気培養、好気培養および嫌気培養
アラゲカワラタケ、カワラタケ、およびシロアミタケをグルコースを炭素源とするT培地で、微好気、好気、または嫌気培養を行い、エタノール生産性を調査した。微好気条件は上記2)と同様である。好気条件は上記2)の培養手順において、シリコンゴム栓に変えて、通気性を有するシリコ栓で蓋をして培養した。嫌気条件は上記2)の培養手順において、窒素ガスを充填してシリコンゴム栓で密栓状態にすることにより行った。その結果、アラゲカワラタケ、カワラタケ、およびシロアミタケは、微好気性および嫌気性条件下で培養したときに、同程度のエタノール生産量を示した。またアラゲカワラタケおよびカワラタケは、好気性条件下の場合には、微好気性条件下の7割程度のエタノール生産量を示した。
【0073】
6)バイオマスからの直接的なエタノール生産性の検討
カワラタケが、糖化処理を行うことなく、バイオマスから直接的に単一の工程によりエタノールを生産することができるか否かについて、またその生産性について、小麦フスマ培地および稲わら培地を炭素源として用いて検討を行った。
【0074】
小麦フスマは、製粉後に残った外皮と胚芽、胚乳の一部が混ざったもので、製粉時に30%ほど発生し、不溶性食物繊維としてヘミセルロース、セルロースおよびリグニンを含んでいる。一般的な小麦フスマの組成は、グルコース35.7%、キシロース17.9%、リグニン他46.4%である。小麦フスマは、その大半が家畜の飼料として利用されているのが現状であるが、国内の飼料の需要が伸び悩んでいることや、食品リサイクル法の実施に伴い、他の食品残渣の飼料化が行われることなどから、今後は小麦フスマの飼料としての需要が低下し、その価格が下落することも考えられる。そのため、飼料用以外での小麦フスマの有効利用が重要な課題となっていた(例えば、[樋口誠一、高橋学、山路明俊、「小麦由来機能性成分の新規利用技術の開発」、埼玉県産業技術総合センター研究報告 第5巻(2007)]参照)。
【0075】
また、稲わらは稲の茎を乾燥させたもので、不溶性食物繊維としてヘミセルロース、セルロースおよびリグニンを含んでいる。一般的な稲わらの組成は、グルコース29.4%、キシロース15.6%、リグニン他55%である。
【0076】
なお、小麦フスマ培地および稲わら培地は、上記1)のT培地において、グルコースを小麦フスマまたは稲わら培地に代えた以外は、基本的な組成は同様である。
【0077】
上記6)の結果、20gの小麦フスマから、培養4日後に5.2gのエタノール生産を確認した。これは、小麦フスマに含まれる発酵に利用可能なグルコースおよびキシロースの総量10.7gから、収率95%で変換されたと見積もられる。なお、上記収率とは小麦フスマに含まれるグルコースおよびキシロースが、全てエタノール生産に利用されたと仮定したときのエタノール量を100%として計算している。なお、グルコース100gからエタノールが生産されるときの反応は、C6H12O6(100g)→2C2H5OH(51.1g)+CO2(48.9g)で表すことができる。キシロース100gからエタノールが生産されるときの反応は、3C5H10O5(100g)→5C2H5OH(51.1g)+5CO2(48.9g)で表すことができる。仮に、1トンの小麦フスマがあった場合、535kg(発酵に利用可能な糖)×0.51×0.95(変換率)=259.2kg(328L)のエタノールが回収可能である。
【0078】
また、20gの稲わらから、培養3日後に3.3gのエタノール生産を確認した。これは、稲わらに含まれる発酵に利用可能なグルコースおよびキシロースの総量9.0gから、収率72%で変換されたと見積もられる。仮に、1トンの稲わらがあった場合、450kg(発酵に利用可能な糖)×0.51×0.72(変換率)=165.2kg(209L)のエタノールが回収可能である。
【0079】
また通常、硫酸等で前処理を行ない、従来の遺伝子組換え菌を用いて稲わらからエタノールを作った場合でも、収率は80%未満である。一方で、カワラタケを用いれば、硫酸等の化学処理をしなくても、従来の方法に近い72%という収率でエタノールが生産できることがわかった。
【0080】
さらに、カワラタケをアラゲカワラタケに変えて、同様の手順で小麦フスマまたは稲わらからの直接的なエタノール生産性を調査した。その結果、カワラタケを用いた場合に近い収率を示した。
【0081】
7)生ゴミからの直接的なエタノール生産性の検討
生ゴミを炭素源として、アラゲカワラタケ、カワラタケ、およびシロアミタケのエタノール生産能を検討した。生ゴミは、国立大学法人鳥取大学内(鳥取県鳥取市湖山町南4丁目101番地)の食堂において、生ゴミとして処分されたものをその日のうちに入手した。その後、生ゴミは4℃で保管した。この生ゴミには、野菜、果物、肉、魚、米飯、コーン等が含まれている。次に、生ゴミに水を加えて、生ゴミ培地を調製した(pHは未調整)。生ゴミ培地の調製は、生ゴミを食堂から入手後24時間以内に行った。この生ゴミ培地1L当たりの生ゴミは385gである。また、生ゴミの全糖量は約6%(23.1g)として見積もった。
【0082】
上記2)〜3)の手順において、T培地を生ゴミ培地に変えて、エタノール生産量を評価した。その結果を表4に示す。なお、表中の値はエタノール生産量(g/L)を示す。各数値は3回実験を行った平均値である。
【0083】
【表4】
【0084】
表4に示すとおり、カワラタケ、アラゲカワラタケ、およびシロアミタケを用いたことによって、別途糖化処理を行うことなく単一の工程で、生ゴミから高い濃度のエタノールを生産できた。具体的には、カワラタケの場合、385gの生ごみから培養5日後に8.4gのエタノール生産を確認した。これは、生ごみに含まれる全糖量23.1g(約6%含有として)から、エタノールが収率71%で変換されたと見積もられる。
【0085】
平均的な生ゴミの含水量は77±4%、全糖量は9±4%であることが知られている。従って、1トンの生ゴミがあったとすると、90kg(全糖量)×0.51(Glucoseを基準)×0.71(変換率)=33kg(41L)のエタノールが回収可能である。
【0086】
生ゴミは、日本では年間約2000万トンが排出され、そのうち約1700万トンがリサイクルされていない。人口30万〜40万人の都市の排出量としては、1日あたり約60トンと見積もられる。通常、ゴミ焼却費用は1トンあたり10,000円以上を要す。仮に、従来法で生ゴミからエタノール生産を試みたとしても、糖化と発酵を単一工程で行うことができず、コスト、エネルギー、作業時間が嵩むという問題がある。一方で、上記の通りカワラタケ、アラゲカワラタケ、またはシロアミタケを利用すれば、生ゴミからのエタノール生産において、糖化と発酵を単一工程で行えることがわかった。また、使用する生ゴミは、水と混ぜるだけでよく、pH調整等の特別な前処理を必要としなかった。従って、カワラタケ、アラゲカワラタケ、またはシロアミタケを利用した生ゴミからのエタノール生産方法は、これまでにない低コストかつ低環境負荷型のエタノール生産方法といえる。
【0087】
8)市販デンプン系食品からの直接的なエタノール生産性の検討
市販デンプン系食品を炭素源として、カワラタケのエタノール生産能を検討した。まず、市販のうどん(生麺)、中華そば(生麺)、または米飯に水を加えて、3種類の市販デンプン系食品培地を調製した(pHは未調整)。いずれも培地も、培地1L当たりの市販デンプン系食品含量は235gである。全糖量は、うどんの場合は約24%(57.1g)、中華そばの場合は約28%(66.4g)、米飯の場合は約37%(88.1g)として見積もった。なお、上記で使用した市販デンプン系食品は、いずれも消費期限から2日程度過ぎたものを使用した。
【0088】
上記2)〜3)の手順において、T培地を3種類の市販デンプン系食品培地に変えて、エタノール生産量を評価した。その結果を表5に示す。なお、表中の値はエタノール生産量(g/L)を示す。各数値は3回実験を行った平均値である。
【0089】
【表5】
【0090】
表5に示すとおり、カワラタケを用いたことによって、別途糖化処理を行うことなく単一の工程で、市販デンプン系食品から高い濃度のエタノールを生産できた。具体的には、238gのうどんから、培養21日後に28.8gのエタノール生産を確認した。また、238gの中華そばから、培養21日後に34.8gのエタノール生産を確認した。また、238gの米飯から、培養21日後に37.6gのエタノール生産を確認した。これらの結果はいずれも、含有する糖に対し、エタノールへの変換率が80%以上であったことを示している。以上の通り、カワラタケを用いたことによって、商品として価値のなくなった食品を無駄にせず、効率的にエタノールを生産することができた。また、使用する食品は、pH調整等の特別な前処理を必要としなかった。
【0091】
9)牛乳からの直接的なエタノール生産性の検討
牛乳を炭素源として、カワラタケのエタノール生産能を検討した。まず、市販の牛乳に水を容量比1:1で加えて、牛乳培地を調製した(pHは未調整)。この牛乳培地1L当たりのラクトースの量は23gである。上記牛乳は、消費期限から2日程度過ぎたものを使用した。
【0092】
上記2)〜3)の手順において、T培地を牛乳培地に変えて、エタノール生産量を評価した。その結果を表6に示す。なお、表中の値はエタノール生産量(g/L)を示す。各数値は2回実験を行った平均値である。括弧内の数値は培地中のラクトース残量(g/L)を示す。
【0093】
【表6】
【0094】
表6に示すとおり、カワラタケを用いたことによって、別途糖化処理を行うことなく単一の工程で、牛乳から高い濃度のエタノールを生産できた。具体的には、1Lの牛乳培地から、8日後に9.1gのエタノール生産を確認した。これは、牛乳に含まれるラクトース23gから、エタノールが収率70%以上で変換されたと見積もられる。以上の通り、カワラタケを用いたことによって、商品として価値のなくなった牛乳を無駄にせず、効率的にエタノールを生産することができた。また、使用する牛乳は、pH調整や乳脂肪分除去等の特別な前処理を必要としなかった。
【0095】
<結果の考察>
以上の結果を踏まえて考察する。本実施例のアルコール生産方法によれば、バイオマスを加水分解して糖を生成(糖化)する工程を行うことなく、単一の担子菌で、バイオマスから直接的に単一工程で効率的にエタノールを生産することができる。またこの生産方法は、従来法とは異なり、酸等による糖化処理を行わなくてもよいため、これに伴う環境面、設備面、作業面およびコスト面等での負荷が低減される。またこの生産方法によれば、ボールミル処理を行わなくても、ボールミル処理を行った場合に近いエタノール生産性が得られる。従って、ボールミル処理のような破砕工程にかかるエネルギーを削減できる。
【0096】
また、カワラタケおよびアラゲカワラタケを用いたアルコール生産方法によれば、キシロースを効率的に炭素源として利用できる。これにより、従来法よりも効率的にバイオマスからエタノールを生産することが可能になる。
【0097】
以上、本発明を実施例に基づいて説明した。この実施例はあくまで例示であり、種々の変形例が可能なこと、またそうした変形例も本発明の範囲にあることは当業者に理解されるところである。
【0098】
例えば、上述の実施例では、炭素源としてT培地、小麦フスマ培地、および稲わら培地等を含有する培地を用いているが、他の炭素源も同様に利用可能である。具体的には、木質系のバイオマスなどを炭素源として用いることもできる。
【産業上の利用可能性】
【0099】
以上のように、本発明で用いる担子菌は、アルコール生産効率および炭素源の資化性に優れるため、アルコール生産効率を向上し、アルコール生産方法、アルコール飲料の生産方法、アルコール含有食品の生産方法およびそれらに用いる種菌等として有用である。
【符号の説明】
【0100】
200 種菌
202 容器
204 蓋
206 菌糸
208 おがくず
300 種菌
302 容器
304 木材チップ
306 菌糸
400 種菌
402 容器
404 蓋
406 菌糸
408 液体培地
【技術分野】
【0001】
本発明は、アルコールの生産方法、アルコール飲料の生産方法、アルコール含有食品の生産方法、またはそれらに用いる種菌に関する。
【背景技術】
【0002】
現在、世界のエネルギー消費は約9割が石油をはじめとする化石燃料に依存しており、エネルギー枯渇の窮地に立たされている。また、近年の原油価格の上昇に加え、炭酸ガスの増加や地球温暖化等の世界的な環境問題が注目されており、将来的に予想される化石燃料の枯渇および原油価格の高止まり等に先立ち、世界各国で化石燃料に代わる新たな再生可能エネルギーの開発が進められている。このような状況の下、バイオエタノールをはじめとするバイオ燃料の開発導入に係る動きがブラジル、米国、アジア各国を中心に各国で活発化するなど、バイオ燃料への期待が高まっている。しかしながら、これらの大半は、サトウキビやトウモロコシ等の食用や飼料作物を原料として生産されているため、現在、農家の転作等による余波が穀物市場の高騰を連鎖的に招いている。したがって、中長期的には、食用や飼料作物と競合しないバイオ燃料の実用化を目指して、廃木材や草本類等の未利用資源の活用が世界規模で重要となる。
【0003】
これに対して、木質系や草本系等のバイオマス資源から糖を生成し、その得られた糖から燃料として有用なエタノール等のアルコールを生成する技術が知られている。例えば、特許文献1および非特許文献1には、木質系バイオマス(または、リグノセルロース系植物材料)からバイオ燃料を生産する際の糖化処理に先駆けて、リグニン分解能を有する白色腐朽菌によりリグニンを分解する生物的処理(前処理)を行う方法について開示されている。
【0004】
また、菌類を用いて炭素源からアルコールを生産する研究もなされている。例えば、特許文献2〜4、非特許文献2〜6には、アガリクスタケ(Agaricus blazei)、ブナシメジ(Hypsizygus marmoreus)、マツタケ(Tricholoma matsutake)、ヒラタケ(Pleurotus ostreatus)、ミミナミハタケ(Lentinellus cochleatus)、およびエノキタケ(Flammulina velutipes)等の担子菌を用いて炭素源からアルコールを生産する方法が開示されている。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2008-6372号公報
【特許文献2】特開2001-286276号公報
【特許文献3】特開2004-298109号公報
【特許文献4】特開2006-223159号公報
【非特許文献】
【0006】
【非特許文献1】バイオエネルギー技術と応用展開、2003年10月31日発行、(株)シーエムシー出版、pp.165-171
【非特許文献2】Tokumitsu OKAMURA, Tomoko OGATA, Norie MINAMOTO, Tomomi TAKENO, Hiroko NODA, Shoko FUKUDA and Masahiro OHSUGI, "Characteristics of Wine Produced by Mushroom Fermentation", Bioscience, Biotechnology, and Biochemistry Vol. 65 (2001), No. 7 pp.1596-1600
【非特許文献3】Tokumitsu Okamura-Matsui, Tomomi Tomoda, Shoko Fukuda and Masahiro Ohsugi, "Discovery of alcohol dehydrogenase from mushrooms and application to alcoholic beverages", Journal of Molecular Catalysis B: Enzymatic、Volume 23, Issues 2-6, 1 September 2003, pp.133-144
【非特許文献4】化学工業日報、2007年10月22日発行、(株)化学工業日報社
【非特許文献5】'Use of whole crop sorghums as a raw material in consolidated bioprocessing bioethanol production using Flammulina velutipes.' Mizuno et al., Biosci Biotechnol Biochem. 2009 Jul;73(7):1671-3. Epub 2009 Jul 7.
【非特許文献6】'Properties of ethanol fermentation by Flammulina velutipes.' Mizuno et al., Biosci Biotechnol Biochem. 2009 Oct;73(10):2240-5. Epub 2009 Oct 7.
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、従来の木質系等のバイオマスからのアルコールの生産においては、バイオマスの主要成分であるセルロースやヘミセルロースを加水分解して糖を生成(糖化)する工程と、生成した糖を酵母や細菌により糖を発酵させることによりアルコールを生成する工程、の2工程が必要であった。したがって、バイオマスからのアルコール生産工程全体では、煩雑な作業が必要となり、コストおよび作業時間が嵩むという問題があった。
【0008】
また、上述した糖化する工程においては、硫酸による加水分解が必要となるため、処理コスト面および環境面で問題視されていた。また、生成したグルコース等の糖の過分解を抑制するために反応制御が必要であった。
【0009】
さらに、硫酸を用いる以外の糖化方法として、バイオマスに、(1)粉砕、蒸煮(蒸煮爆砕、蒸煮、熱水分解・加圧熱水処理等)、エネルギー線(電子線、γ線、マイクロウエーブ)の照射等の物理的方法で処理する物理的前処理、(2)酸(硫酸、亜硫酸、リン酸)、アルカリ(カセイソーダ、アンモニアなど)などを用いて化学的処理する化学的前処理、または、(3)白色腐朽菌等のリグニン分解菌を用いて行う生物的前処理、による前処理を行い、リグニン等の難分解性成分を分解した後に、セルラーゼのような糖化酵素を加えてセルロースを加水分解して糖にする酵素糖化方法がある。
【0010】
しかしながら、上述の(1)の前処理方法では、蒸煮のための高温・高圧装置や、電子線、γ線、マイクロウエーブ等のエネルギー線の照射装置などの高価な装置が必要なため、設備費が高くなり、さらに、前処理工程の管理を厳密に行う必要があった。また、上述の(2)の前処理方法においては、バイオマスの糖化または前処理に当たり、酸やアルカリ等の化学薬品を用いるため、それらの薬品に耐え得る設備を使用する必要があることから、設備費が高くなるという問題があった。そして、糖化または前処理に用いた酸やアルカリの中和処理とそれによって発生する大量の中和廃棄物(例えば、硫酸カルシウムなど)の処理、あるいは、前処理に用いた溶媒の除去を行う必要があるため、手間やコストが掛かり、しかも、環境面での問題があった。さらに、上述の(3)の前処理方法では、糖化酵素を用いるため、コストが掛かり、しかも、当該酵素による糖の過分解を抑制するために反応制御が必要であった。またこの方法は、他の前処理方法のような不都合は生じないが、酵素糖化工程およびその次工程である酵母等の糖の発酵工程のために、白色腐朽菌を除去する必要が生じ、手間が掛かるという問題があった。また、そもそも、白色腐朽菌は、アルコール生成を企図して導入されるものでなく、実際のアルコール生成は、酵母等の発酵により行われるので、上述の(3)の方法で用いられる白色腐朽菌のアルコール生成能については、特に考慮する必要はなかった。
【0011】
また、従来の担子菌を使用したアルコール生産方法では、キシロースのようなペントースに分類される糖をほとんどアルコールに発酵できなかった。そのため、バイオマスから得られるエタノール量も十分とはいえなかった。
【0012】
本発明は上記事情に鑑みてなされたものであり、特定の担子菌を用いた効率的なアルコール生産方法を提供することを目的とする。
【課題を解決するための手段】
【0013】
本発明によれば、シロアミタケ(Trametes)属の担子菌を用いて、炭素源からアルコールを生成する、アルコール生産方法が提供される。
【0014】
この生産方法は、後述する実施例で効率的にアルコールを生産するために利用できることが実証されている。そのため、この生産方法を用いれば、効率的にアルコールを生産できる。
【0015】
また本発明によれば、シロアミタケ属の担子菌を用いて、炭素源からアルコールを含有する液体を生成する、アルコール飲料の生産方法が提供される。
【0016】
ここで、後述する実施例でシロアミタケ属の担子菌を用いて、炭素源からアルコールを含有する液体を効率的に生成できることが実証されている。そのため、この生産方法を用いれば、効率的にアルコールを含有する液体を生産できる。
【0017】
また本発明によれば、シロアミタケ属の担子菌を用いて、炭素源からアルコールを含有する組成物を生成する、アルコール含有食品の生産方法が提供される。
【0018】
ここで、後述する実施例でシロアミタケ属の担子菌を用いて、炭素源からアルコールを含有する組成物を効率的に生成できることが実証されている。そのため、この生産方法を用いれば、効率的にアルコール含有食品を生産できる。
【0019】
また本発明によれば、シロアミタケ属の担子菌の菌糸と、菌糸を担持する担体と、を備えることを特徴とする、シロアミタケ属の担子菌を用いて炭素源からアルコールを生成するための種菌が提供される。
【0020】
この種菌は、後述する実施例で効率的にアルコールを生産できることが実証されている担子菌の菌糸を備えている。そのため、この種菌を用いれば、効率的にアルコールを生産できる。
【発明の効果】
【0021】
本発明によれば、アルコール生産効率および炭素源の資化性に優れた菌類により、バイオマス等から効率的なアルコール生産を実現することができる。
【図面の簡単な説明】
【0022】
【図1】図1(a)および図1(b)は、実施形態に係るシロアミタケ属の担子菌により炭素源からアルコールを生成する方法を説明するためのフローチャートである。
【図2】図2(a)、図2(b)および図2(c)は、実施形態に係るシロアミタケ属の担子菌を用いた種菌の構成を模式的に示した概念図である。
【図3】図3は、従来の典型的なバイオマスからのエタノール生産プロセスを説明するための概念図である。
【図4】図4は、実施形態に係るシロアミタケ属の担子菌を用いた、未利用バイオマス資源の再利用方法の一例を説明するための概念図である。
【図5】図5は、実施形態に係るシロアミタケ属の担子菌を用いた未利用バイオマス資源の燃料用アルコールとしての再利用のサイクルを説明するための概念図である。
【図6】図6は、シロアミタケ属の担子菌の培養および菌体回収の方法を説明するための実験プロトコルである。
【発明を実施するための最良の形態】
【0023】
本発明者らは、鋭意検討を重ねた結果、シロアミタケ属の担子菌が各種炭素源を糖化することにより糖を生成する工程、および糖を発酵することによりアルコールを生成する工程を単一で行うことができ、かつ高いアルコール生産性を有することを確認してバイオマスから直接アルコールが生産可能であることを見出し、本発明を完成するに至った。
【0024】
以下、本発明の実施の形態について詳細に説明する。なお、同様な内容については繰り返しの煩雑を避けるために、適宜説明を省略する。
【0025】
本発明の一実施形態は、シロアミタケ属の担子菌を用いて、炭素源からアルコールを生成する、アルコール生産方法である。この生産方法を適切に利用すれば、後述する実施例で実証されているように、効率的にアルコールを生産できる。上記シロアミタケ属の担子菌は特に限定されないが、例えばアラゲカワラタケ(Trametes hirsuta)、カワラタケ(Trametes versicolor)、シロアミタケ(Trametes suaveolens)、オオチリメンタケ(Trametes gibbosa)、ウサギタケ(Trametes trogiiBerkeley)、クジラタケ(Trametes orientalis)、ヤキフタケ(Trametes pubescens)、チリメンタケ(Trametes elegans )、Trametes lactinea を挙げることができる。この中でもアラゲカワラタケ、カワラタケ、およびシロアミタケは、フルクトース、マンノース、ガラクトース、セロビオース、マルトース、アラビノース、キシロース、スクロース、デンプン、CMC、小麦フスマ、稲わら、およびボールミル処理後の稲わらからアルコールを生産できることが後述する実施例で実証されている。このアラゲカワラタケ、カワラタケ、シロアミタケは、一般的にリグニン分解能を有し、世界的に分布している白色腐朽菌である。特に、アラゲカワラタケおよびカワラタケは、小麦フスマ、稲わら、およびボールミル処理後の稲わらのようなバイオマスからでも、高い濃度のアルコールを生産できることが後述する実施例で実証されている。
【0026】
また従来、担子菌を使用したアルコール生産方法では、キシロースのようなペントースに分類される糖をほとんどアルコールに発酵できなかった。そのため、バイオマスから得られるエタノール量も十分とはいえなかった。一方で、シロアミタケ属の担子菌はそのような糖からでもアルコールを生産できるため、より効率的にバイオマスからアルコールを生産できる。特に、アラゲカワラタケおよびカワラタケが、そのような糖を効率よくエタノールに変換できることが後述する実施例で実証されている。特にカワラタケで顕著であった。キシロースはヘミセルロースの一つであるキシランを構成するものとして植物に広く存在することが知られている。なお、キシロースを発酵できる酵母としてPichia stipitisが知られているが、エタノール耐性が低く、エタノール変換効率、グルコースの発酵効率も十分とはいえなかった。また、この酵母は糖化能を有しないため、バイオマスからエタノールを生産するには糖化工程を別に設けるか、遺伝子改変等の方法を行う必要があった。
【0027】
本実施形態では、アルコール生成にシロアミタケ属の担子菌を用いることを特徴の一つとする。一方で、以下に示すとおり、シロアミタケ属の担子菌のアルコール生成能についてはこれまで何ら報告されていなかった。例えば特許文献1および非特許文献1に記載されている担子菌は、上述した糖化工程における生物的前処理用として用いるものであって、直接的にアルコールを生成するために用いるものではない。したがって、特許文献1および非特許文献1では担子菌のアルコール生成能については考慮する必要がないので、当然のことながら、アルコール生成能については示唆すらされていない。また、特許文献2〜4、非特許文献2〜6においても、シロアミタケ属の担子菌を用いて炭素源からアルコールを生産することについては何ら記載されていない。
【0028】
即ち、シロアミタケ属の担子菌のアルコール生成能についてはこれまで何ら報告されておらず、本実施形態は、これまで存在しなかった優れたアルコール生産方法といえる。加えて、目、科、または属が異なればそれぞれの担子菌が持つ機能も異なるため、これらの文献から本実施形態のアルコール生産方法に想到することは困難である。
【0029】
また、本実施形態においては、シロアミタケ属の担子菌は、毒性はなく、遺伝子組換え体でなく、野生のものを用いることができる。したがって、本実施形態のアルコール生産方法では、安全性の高いアルコールの生産が実現可能となる。
【0030】
さらに、シロアミタケ属の担子菌は一般的に食用として汎用されていない。また、産業的に有効に利用されているという報告はなく、これまでほとんど利用されていなかった。そして、シロアミタケ属の担子菌は、成長が比較的早く、世界的に分布しており比較的入手しやすい。したがって、本実施形態に係るアルコール生産方法においては、用いるシロアミタケ属の担子菌は、少なくとも産業界と直接競合することは考え難く、資源として有用性および優位性に富むものである。
【0031】
図1は、実施の形態に係るシロアミタケ属の担子菌により炭素源からアルコールを生成する方法を説明するためのフローチャートである。後述するようにシロアミタケ属の担子菌が、幅広い糖の資化性を有することを、本発明者らは見出している。
【0032】
具体的には、図1(a)に示すように、まず、シロアミタケ属の担子菌を木質系・草本系等のバイオマスまたは各種の糖などを含む炭素源に接種する(S102)。次いで、シロアミタケ属の担子菌の種菌を接種された炭素源を培養する(S104)。そして、シロアミタケ属の担子菌により炭素源から生成されたアルコールを含む液体を濾過などの手法により回収する(S106)。
【0033】
上述の炭素源が木質系等のバイオマスを含む場合には、上述の培養工程において、図1(b)に示すように、シロアミタケ属の担子菌により炭素源を糖化する(S108)。そして、糖化された炭素源をシロアミタケ属の担子菌により発酵してアルコールを生成する(S110)。
【0034】
なお、上述の発酵工程は、嫌気的条件であってもよく、好気的条件であってもよいが、微好気的条件または嫌気的条件が、アルコール生成効率の面から好ましい。
【0035】
また、上述の炭素源は、糖を含んでいてもよい。さらに、上述の炭素源は、グルコース、フルクトース、マンノース、ガラクトース、セロビオース、マルトース、アラビノース、キシロース、スクロース、およびラクトースからなる群より選ばれる一種以上の糖を含んでいてもよい。後述するように、シロアミタケ属の担子菌は、これらの糖に対する資化性を有していることを、本発明者らは見出している。さらに、上述の炭素源は、糖蜜、廃糖蜜、上白糖、黒糖、マルツエキスおよびデンプン等を含んでいてもよい。なお本実施形態に係るシロアミタケ属の担子菌は、複数の炭素源が混在した培地からアルコールを生成してもよい。培地中の個々の炭素源の質量%または容量%は、例えば1、2、3、5、10、20、50、80、または100%であってもよく、それらの値の範囲内またはそれらの値以上であってもよい。
【0036】
あるいは、上述の炭素源は、セルロース、木質系または草本系の材料(バイオマス等)を含んでいてもよい。セルロースとしては、綿リンター、木材パルプや溶解パルプ等から得られる植物系のセルロース、アセトバクター属(Acetobacter)等に属する微生物が生産するセルロース、再生セルロースおよび微結晶セルロース等が挙げられる。木質系の材料としては、建築廃材等の廃木材、製材残材、間伐材、林地残材などを含む木材、おがくず、古紙・廃紙等の紙類およびパルプ等を挙げることができる。草本系の材料としては、小麦フスマ等のフスマ、稲わら・麦わら等のわらおよびもみ殻などを含む農業残渣、バガス、刈り草および雑草類等を挙げることができる。なお、後述するようにシロアミタケ属の担子菌が、小麦フスマおよび稲わらを炭素源として直接的にアルコール生成能を有し、アルコール収率が高いことを本発明者らは見出している。また、後述する実施例でセルロース分解能を有していることが示唆されている。
【0037】
また上述の炭素源は生ゴミまたは廃棄飲料(以下、「生ゴミ等」と称することもある)を含んでいてもよい。上記生ゴミ等は、業務用または家庭用の生ゴミ等であってもよい。また、食堂、病院、学校、ホテル、食品工場、または農畜産場等から排出される生ゴミ等であってもよい。また、実験室や工場において調製した生ゴミ等であってもよい。また生ゴミは、廃棄食品または廃棄飲料を含んでいてもよい。なお、本明細書において廃棄食品は、廃棄された食品、食用としての商品価値がない食品、または消費期限後の食品を意味する。本明細書において廃棄飲料は、廃棄された飲料、飲用としての商品価値がない飲料、または消費期限後の飲料を意味する。廃棄食品は、例えば、廃棄野菜、廃棄果物、廃棄肉、廃棄魚、廃棄米飯、廃棄コーン、廃棄卵、または廃棄麺等を含んでいてもよい。廃棄飲料は、例えば、廃棄乳系飲料等を含んでいてもよい。また、生ゴミは破砕処理後の状態であってもよい。生ゴミの組成としては糖、リグニン、蛋白質、脂質、アミノ酸、有機酸、ビタミン類、またはミネラル等の無機物を含んでいてもよい。
【0038】
また上述の炭素源は、食品または乳系飲料を含んでいてもよい。食品は、加工食品または生鮮食品を含んでいてもよい。食品は、例えば野菜、果物、肉、魚、米飯、コーン、卵、または麺等を含んでいてもよい。乳系飲料は、例えば牛乳を含んでいてもよい。牛乳は、例えば明治おいしい牛乳(株式会社明治)等、市販の牛乳であってもよい。牛乳は、例えば成分無調整牛乳、または調整(例えば低脂肪、無脂肪、高脂肪)牛乳であってもよい。食品または乳系飲料は、消費期限内または消費期限後であってもよい。
【0039】
また、上述のシロアミタケ属の担子菌により生成されるアルコールは、エタノールを含んでいてもよい。後述するように、シロアミタケ属の担子菌は、炭素源からエタノールを生成することを、本発明者らは見出している。
【0040】
そして、上述のシロアミタケ属の担子菌により生成されるアルコールを用いて、アルコール飲料またはアルコール含有食品を生産してもよい。これらのアルコール飲料またはアルコール含有食品には、アルコール以外にも、シロアミタケ属の担子菌の生成する各種成分が含まれていてもよ
い。また、アルコール含有食品は、固体であってもよく、液体であってもよく、ゲル状体などであってもよい。
【0041】
図2は、実施の形態に係るシロアミタケ属の担子菌を用いた種菌の構成を模式的に示した概念図である。種菌とは、研究対象の菌類を育て、増やす際の基になる培養物のことである。種菌は、アルコール生産用材料と言い換えることもできる。図2(a)は、おがくずを担体として用いたシロアミタケ属の担子菌の種菌である。この種菌200では、蓋204を備える容器202内に、おがくず208が敷き詰められている。このおがくず中にシロアミタケの菌糸206a、206bが担持されている。なお、シロアミタケ属の担子菌の菌糸は、図2(a)のように、きのこを形成している必要はない。
【0042】
図2(b)は、木材チップを担体として用いたシロアミタケ属の担子菌の種菌である。この種菌300では、容器302内に、コルク栓状の形状からなる木材チップ304a、304b、304c、304d、304e、304fが収納されている。これらの木材チップには、シロアミタケ属の担子菌の菌糸306a、306b、306c、306d、306e、306fが担持されている。なお、シロアミタケ属の担子菌の菌糸は、図2(b)のように、きのこを形成している必要はない。
【0043】
図2(c)は、液体培地を担体として用いたシロアミタケ属の担子菌の種菌である。この種菌400では、蓋404を備える容器402内に、各種の糖などの炭素源を含む液体培地408が収納されている。これらの液体培地には、シロアミタケ属の担子菌の菌糸406が担持されている。なお、シロアミタケ属の担子菌の菌糸は、図2(c)のように、きのこを形成している必要はない。
【0044】
上述の種菌は、菌類を用いて炭素源からアルコールを生成するための種菌であって、シロアミタケ属の担子菌の菌糸と、この菌糸を担持する担体とを備える。なお、この菌糸は、定常期の菌糸であってもよい。具体的には、この菌糸は、培養開始5〜7日経過後の菌糸であってもよい。
【0045】
以下、実施の形態に係るアルコール生産方法の作用効果についてさらに説明する。
シロアミタケ属の担子菌は、後述するように、カビや酵母と同等以上のアルコール生産能を有するので、優れたアルコール生産効率を実現することができる。
【0046】
また、シロアミタケ属の担子菌は、後述するように、グルコース、フルクトース、マンノース、ガラクトース、セロビオース、マルトース、アラビノース、キシロース、およびスクロースに対する資化性を有するので、担子菌類では困難であった幅広い種類の炭素源を用いてアルコール発酵を行うことができる。
【0047】
そのため、上述の実施の形態に係るアルコール生産方法によれば、アルコール生産効率および炭素源の資化性に優れるシロアミタケ属の担子菌を用いるため、アルコール生産方法の生産効率を向上し、炭素源の種類の幅を拡大することができる。
【0048】
図3は従来の典型的なバイオマスからのエタノール生産プロセスを説明するための概念図である。従来は、炭素源を糖化する工程と、糖を発酵する工程は別々のユニットによって行われていた。図4は、本実施形態に係るシロアミタケ属の担子菌を用いた、未利用バイオマス資源の再利用方法の一例を説明するための概念図である。この方法は、シロアミタケ属の担子菌が炭素源を糖化することにより糖を生成する工程と、シロアミタケ属の担子菌が糖を発酵することによりアルコールを生成する工程と、を含むため、従来の酸等による加水分解の工程を経ずに、糖化・発酵の両工程を行うことができる。このため、未利用バイオマス資源をシロアミタケ属の担子菌による糖化・発酵工程により効率よくエタノールに変換することができる。また、環境負荷、コストや手間の削減につながるのはいうまでもない。また糖化・発酵は同一容器で並行して、または同一容器で同時に行われてもよく、この場合、より効率的である。
【0049】
糖化と発酵を並行して行いアルコールを生産するプロセスは、一般的に統合バイオプロセス(consolidated bioprocessing :CBP)と呼ばれる。CBPは例えば[Lynd et al., Curr Opin Biotechnol. 2005 Oct;16(5):577-83.]、[van Zyl et al., Adv Biochem Eng Biotechnol. 2007;108:205-35.]、[Mizuno et al., Biosci Biotechnol Biochem. 2009 Jul;73(7):1671-3. Epub 2009 Jul 7.]などに記載されている。そして、本実施形態に係るシロアミタケ属の担子菌は、CBPによってアルコールを精製することができる。このCBPにおいては、リグニンやセルロース等を分解するために酵素や酸等を加えてもよいが、上記シロアミタケ属の担子菌を用いた場合には酵素や酸等を加えなくても、効率的にアルコールを生産可能である。なお、従来アルコール生産にはセルロースの分解が重要と考えられていたが、本実施形態においては、優れたリグニン分解能を有する上記シロアミタケ属の担子菌を用いたことが、従来予想しえない高いエタノール生産性を実現できた一因になっていると考えられる。リグニンは木質系または草本系の材料に含まれている。
【0050】
またこのCBPにおいては、本実施形態に係るシロアミタケ属の担子菌がリグニン、セルロース、またはヘミセルロースを分解し、さらに生成した単糖またはオリゴ糖を発酵することによってアルコールを生産してもよい。このとき、上記単糖はヘキソースまたはペントースであってもよく、グルコース、フルクトース、マンノース、ガラクトース、アラビノース、およびキシロースからなる群より選ばれる一種以上の糖であってもよい。また上記オリゴ糖はセロビオース、マルトース、およびスクロースからなる群より選ばれる一種以上の糖であってもよい。そしてこのようなCBPは、低コストで、低環境負荷である。なお、CBPと同様の原理は醸造酒の生産過程でも利用されており、その場合、並行複発酵とも呼ばれる。この並行複発酵も、糖化と発酵とが並行して行なわれる。
【0051】
ここで、木材や紙類などの木質系のバイオマスを酸糖化して得られる炭素源には、一般的にグルコースにくわえて、マンノース等が数%含まれる。酵母は、グルコースを好適に資化するが、マンノースに対する資化性は低い。一方、シロアミタケ属の担子菌は、酵母により資化することが困難なマンノースに対しても資化性を有するため、酵母による発酵後に残存するマンノースも資化することができる。そのため、木質系のバイオマスを糖酸化し、酵母およびシロアミタケ属の担子菌を組み合わせてアルコール発酵を行うことにより、資源のリサイクル効率およびアルコール生産効率を高めることができる。また、天然の酵母は、小麦フスマ等のバイオマスを直接的にエタノールに変換することはできない。これに対して、シロアミタケ属の担子菌は、後述するように、小麦フスマおよび稲わらから直接的にエタノールを生産することができる。したがって、実施の形態に係るアルコール生産方法においては、天然の酵母が利用することができないバイオマスからアルコールを直接的に生産することができる点でも、優位性および有用性に富むものである。
【0052】
図5は、実施の形態に係るシロアミタケ属の担子菌を用いた、未利用バイオマス資源の燃料用アルコールとしての再利用のサイクルを説明するための概念図である。このサイクルは、未利用バイオマス資源をシロアミタケ属の担子菌による糖化・発酵工程により効率よくアルコールに変換することができるため、バイオマスと燃料用アルコールとの間で資源の循環システムを構築することができる。そのため、地球環境保全および産業の発展を両立しうるエネルギー供給システムを構築することができる。
【0053】
また、実施の形態に係るアルコール飲料の生産方法によれば、アルコール生産効率および炭素源の資化性に優れるシロアミタケ属の担子菌を用いるため、アルコール飲料の生産方法の生産効率を向上し、炭素源の種類の幅を拡大することができる。
【0054】
さらに、実施の形態に係るアルコール含有食品の生産方法によれば、アルコール生産効率および炭素源の資化性に優れるシロアミタケ属の担子菌を用いるため、アルコール含有食品の生産方法の生産効率を向上し、炭素源の種類の幅を拡大することができる。
【0055】
そして、実施の形態に係る種菌によれば、アルコール生産効率および炭素源の資化性に優れるシロアミタケ属の担子菌の菌糸を担体に担持させているため、菌類を用いて炭素源からアルコールを生成するための種菌として好適に用いることができる。
【0056】
以上、図面を参照して本発明の実施形態について述べたが、これらは本発明の例示であり、上記以外の様々な構成を採用することもできる。
【実施例】
【0057】
以下、本発明を実施例によりさらに説明するが、本発明はこれらに限定されるものではない。ただし、実施例中に表示される%は、特に断りのない限り質量パーセントを示す。
[使用菌株]
表1の菌株を準備した。表1の各菌株の28S rDNA塩基配列を解析し、GenBank/DDBJ/EMBL等の塩基配列データベースに基づく相同性検索を行ったところ、データベースに登録されている28S rDNA塩基配列とそれぞれ100%の相同率を示した。
【0058】
【表1】
【0059】
1)使用培地
使用培地としては、MYG培地もしくはT培地を用いた。下記に、各培地の基本的組成を示す。
MYG培地
グルコース* :0.4 %
Yeast extract :0.4 %
Malt extract :1.0 %
T培地
グルコース* :2.0 %
Yeast extract :1.0 %
KH2PO4 :1.0 %
(NH4)2SO4 :0.2 %
MgSO4・7H2O :0.05 %
*)グルコースについては、必要に応じて、他の炭素源に変更した。
【0060】
2)培養および菌体回収
図6は、カワラタケの培養方法を説明するための実験プロトコルである。まず、前培養ステップでは、カワラタケの平板培地から5mm四方の大きさの菌糸断片を切り取り、500ml容三角フラスコに入ったMYG培地50mlに加え、綿栓もしくはシリコ栓を用いて三角フラスコを好気状態にして、28℃で5〜7日間培養を行った。
【0061】
次いで、本培養ステップでは、500ml容三角フラスコにT培地を50ml加え、無菌的にろ過、洗浄したカワラタケの前培養菌糸を接種し、シリコンゴム栓を用いて三角フラスコを密栓状態にして、28℃で静置培養(微好気的件下での培養)を行った。
【0062】
続いて、培養液の回収ステップでは、経時的に次の手順で回収を行った。すなわち、培養液をサンプリングし、遠心分離後、0.22μmのフィルターでろ過した。培養濾液pHの測定、HPLC分析、および活性測定して、その後、冷凍保存した。表1に記載のカワラタケ以外の各菌株についても、カワラタケの場合と同様の手順で培養およびサンプルの回収を行なった。
【0063】
3)HPLC分析
表1に記載の各菌株の培養濾液をHPLC分析した。その分析条件は以下に示すとおりである。
(HPLC分析条件)
キャピラリーカラム :Shodex SUGAR SP0810またはShodex SUGAR KS−801
キャピラリーサイズ :8.0mmID×300mmL
流量 :0.6ml/min
カラム温度 :80℃
抽出液 :脱気蒸留水
試料 :10μl
【0064】
4)グルコース以外の炭素源を用いた培養
T培地の炭素源であるグルコースに代わり、同濃度のフルクトース、マンノース、ガラクトース、セロビオース、マルトース、アラビノース、キシロース、スクロース、デンプン(Starch)、CMC(カルボキシメチルセルロース)、小麦フスマ(Wheat bran)、稲わら(Rice straw)、ボールミル処理後の稲わら(BM-Rice straw)を用いて、その資化性とエタノール生産性を確かめた。手順は2)および3)と同様である。なお、ボールミル処理は遊星ボールミルGokin planet M2-3F(ナガオシステム社製)でジルコニア容器とジルコニアボールにより、トータル4時間(10分稼働、10分休止)行った。
【0065】
上記1)〜4)の結果を表2および表3に示す。各数値は3回実験を行った結果の平均値である。
【0066】
【表2】
【0067】
【表3】
【0068】
表2は、グルコースを基質としてアラゲカワラタケ(Trametes hirsuta)、カワラタケ(Trametes versicolor)を培養したときのエタノール生産率を、カビ(Rhizopus oryzae、Mucor corticolous)および酵母(Saccharomyces cerevisiae)と比較した表である。この表から、アラゲカワラタケやカワラタケがカビや酵母と比較して高いエタノール生産性を有していることがわかる。
【0069】
表3は、各種炭素源(20g/培地1L)を基質として、表1に記載の4種類の菌株(アラゲカワラタケ(Trametes hirsuta)、カワラタケ(Trametes versicolor)、シロアミタケ(Trametes suaveolens)、ヒラタケ(Pleurotus ostreatus)を5日間培養したときのエタノール生産量(g/培地1L)を表している。この表から、アラゲカワラタケ、カワラタケ、およびシロアミタケは、フルクトース、マンノース、ガラクトース、セロビオース、マルトース、アラビノース、キシロース、スクロース、デンプン(Starch)、CMC(カルボキシメチルセルロース)、小麦フスマ(Wheat bran)、稲わら(Rice straw)、およびボールミル処理後の稲わら(BM-Rice straw)を代謝することで、比較的高い濃度のエタノールを生産できることがわかる。このエタノール生産量は、特にグルコース、マンノース、セロビオース、デンプンで高い値を示した。
【0070】
また、カワラタケ、アラゲカワラタケはキシロース、小麦フスマ、稲わら、およびボールミル処理後の稲わらを基質としたときに、他の菌株に比べて顕著に高いエタノール生産量を示した。特に、通常のキノコを用いた場合、キシロースを炭素源にして2g/Lを超える量のエタノールを生産することは困難であるにも関わらず、カワラタケが7.2g/L、アラゲカワラタケが3.9g/Lの高い生産量を示したことは驚くべき結果であった。キシロースは種々のバイオマスに多く含まれる成分であるため、カワラタケやアラゲカワラタケを用いれば従来法よりも効率よくエタノールを種々のバイオマスから生産できると考えられる。なお、同様の実験を複数回行なったところ、カワラタケによるキシロースからのエタノール生産量は高いときで8.0g/Lを超えることもあった。また、アラゲカワラタケを用いた場合のキシロース、小麦フスマ、稲わら、およびボールミル処理後の稲わらからのエタノール生産量は、高いときでカワラタケの場合の9割程度の数値を示すこともあった。
【0071】
また、アラゲカワラタケ、カワラタケ、およびシロアミタケを用いた場合において、稲わらをボールミル処理しない場合(3.0、3.3、1.5)でも、ボールミル処理をした場合(3.4、3.7、1.7)に近い生産性が見られた。通常、ボールミルはセルロースの骨格を破壊することで糖化を促進させると考えられており、2倍くらいの効率でエタノール生産性を促進すると言われている。一方で本実施例においてアラゲカワラタケ、カワラタケ、およびシロアミタケを用いた場合には、これらの菌株がセルロースを分解したために、ボールミル処理の前後で生産性があまり変化しなかったと考えられる。従って、アラゲカワラタケ、カワラタケ、およびシロアミタケを用いたエタノール生産方法は、ボールミルのような特殊な破砕工程を行なわなくても効率的にエタノールを生産できる、優れた方法であるといえる。特に、ボールミルは大きなエネルギーを必要とする操作なので、省エネルギーの観点からも上記方法の意義は大きい。
【0072】
5)微好気培養、好気培養および嫌気培養
アラゲカワラタケ、カワラタケ、およびシロアミタケをグルコースを炭素源とするT培地で、微好気、好気、または嫌気培養を行い、エタノール生産性を調査した。微好気条件は上記2)と同様である。好気条件は上記2)の培養手順において、シリコンゴム栓に変えて、通気性を有するシリコ栓で蓋をして培養した。嫌気条件は上記2)の培養手順において、窒素ガスを充填してシリコンゴム栓で密栓状態にすることにより行った。その結果、アラゲカワラタケ、カワラタケ、およびシロアミタケは、微好気性および嫌気性条件下で培養したときに、同程度のエタノール生産量を示した。またアラゲカワラタケおよびカワラタケは、好気性条件下の場合には、微好気性条件下の7割程度のエタノール生産量を示した。
【0073】
6)バイオマスからの直接的なエタノール生産性の検討
カワラタケが、糖化処理を行うことなく、バイオマスから直接的に単一の工程によりエタノールを生産することができるか否かについて、またその生産性について、小麦フスマ培地および稲わら培地を炭素源として用いて検討を行った。
【0074】
小麦フスマは、製粉後に残った外皮と胚芽、胚乳の一部が混ざったもので、製粉時に30%ほど発生し、不溶性食物繊維としてヘミセルロース、セルロースおよびリグニンを含んでいる。一般的な小麦フスマの組成は、グルコース35.7%、キシロース17.9%、リグニン他46.4%である。小麦フスマは、その大半が家畜の飼料として利用されているのが現状であるが、国内の飼料の需要が伸び悩んでいることや、食品リサイクル法の実施に伴い、他の食品残渣の飼料化が行われることなどから、今後は小麦フスマの飼料としての需要が低下し、その価格が下落することも考えられる。そのため、飼料用以外での小麦フスマの有効利用が重要な課題となっていた(例えば、[樋口誠一、高橋学、山路明俊、「小麦由来機能性成分の新規利用技術の開発」、埼玉県産業技術総合センター研究報告 第5巻(2007)]参照)。
【0075】
また、稲わらは稲の茎を乾燥させたもので、不溶性食物繊維としてヘミセルロース、セルロースおよびリグニンを含んでいる。一般的な稲わらの組成は、グルコース29.4%、キシロース15.6%、リグニン他55%である。
【0076】
なお、小麦フスマ培地および稲わら培地は、上記1)のT培地において、グルコースを小麦フスマまたは稲わら培地に代えた以外は、基本的な組成は同様である。
【0077】
上記6)の結果、20gの小麦フスマから、培養4日後に5.2gのエタノール生産を確認した。これは、小麦フスマに含まれる発酵に利用可能なグルコースおよびキシロースの総量10.7gから、収率95%で変換されたと見積もられる。なお、上記収率とは小麦フスマに含まれるグルコースおよびキシロースが、全てエタノール生産に利用されたと仮定したときのエタノール量を100%として計算している。なお、グルコース100gからエタノールが生産されるときの反応は、C6H12O6(100g)→2C2H5OH(51.1g)+CO2(48.9g)で表すことができる。キシロース100gからエタノールが生産されるときの反応は、3C5H10O5(100g)→5C2H5OH(51.1g)+5CO2(48.9g)で表すことができる。仮に、1トンの小麦フスマがあった場合、535kg(発酵に利用可能な糖)×0.51×0.95(変換率)=259.2kg(328L)のエタノールが回収可能である。
【0078】
また、20gの稲わらから、培養3日後に3.3gのエタノール生産を確認した。これは、稲わらに含まれる発酵に利用可能なグルコースおよびキシロースの総量9.0gから、収率72%で変換されたと見積もられる。仮に、1トンの稲わらがあった場合、450kg(発酵に利用可能な糖)×0.51×0.72(変換率)=165.2kg(209L)のエタノールが回収可能である。
【0079】
また通常、硫酸等で前処理を行ない、従来の遺伝子組換え菌を用いて稲わらからエタノールを作った場合でも、収率は80%未満である。一方で、カワラタケを用いれば、硫酸等の化学処理をしなくても、従来の方法に近い72%という収率でエタノールが生産できることがわかった。
【0080】
さらに、カワラタケをアラゲカワラタケに変えて、同様の手順で小麦フスマまたは稲わらからの直接的なエタノール生産性を調査した。その結果、カワラタケを用いた場合に近い収率を示した。
【0081】
7)生ゴミからの直接的なエタノール生産性の検討
生ゴミを炭素源として、アラゲカワラタケ、カワラタケ、およびシロアミタケのエタノール生産能を検討した。生ゴミは、国立大学法人鳥取大学内(鳥取県鳥取市湖山町南4丁目101番地)の食堂において、生ゴミとして処分されたものをその日のうちに入手した。その後、生ゴミは4℃で保管した。この生ゴミには、野菜、果物、肉、魚、米飯、コーン等が含まれている。次に、生ゴミに水を加えて、生ゴミ培地を調製した(pHは未調整)。生ゴミ培地の調製は、生ゴミを食堂から入手後24時間以内に行った。この生ゴミ培地1L当たりの生ゴミは385gである。また、生ゴミの全糖量は約6%(23.1g)として見積もった。
【0082】
上記2)〜3)の手順において、T培地を生ゴミ培地に変えて、エタノール生産量を評価した。その結果を表4に示す。なお、表中の値はエタノール生産量(g/L)を示す。各数値は3回実験を行った平均値である。
【0083】
【表4】
【0084】
表4に示すとおり、カワラタケ、アラゲカワラタケ、およびシロアミタケを用いたことによって、別途糖化処理を行うことなく単一の工程で、生ゴミから高い濃度のエタノールを生産できた。具体的には、カワラタケの場合、385gの生ごみから培養5日後に8.4gのエタノール生産を確認した。これは、生ごみに含まれる全糖量23.1g(約6%含有として)から、エタノールが収率71%で変換されたと見積もられる。
【0085】
平均的な生ゴミの含水量は77±4%、全糖量は9±4%であることが知られている。従って、1トンの生ゴミがあったとすると、90kg(全糖量)×0.51(Glucoseを基準)×0.71(変換率)=33kg(41L)のエタノールが回収可能である。
【0086】
生ゴミは、日本では年間約2000万トンが排出され、そのうち約1700万トンがリサイクルされていない。人口30万〜40万人の都市の排出量としては、1日あたり約60トンと見積もられる。通常、ゴミ焼却費用は1トンあたり10,000円以上を要す。仮に、従来法で生ゴミからエタノール生産を試みたとしても、糖化と発酵を単一工程で行うことができず、コスト、エネルギー、作業時間が嵩むという問題がある。一方で、上記の通りカワラタケ、アラゲカワラタケ、またはシロアミタケを利用すれば、生ゴミからのエタノール生産において、糖化と発酵を単一工程で行えることがわかった。また、使用する生ゴミは、水と混ぜるだけでよく、pH調整等の特別な前処理を必要としなかった。従って、カワラタケ、アラゲカワラタケ、またはシロアミタケを利用した生ゴミからのエタノール生産方法は、これまでにない低コストかつ低環境負荷型のエタノール生産方法といえる。
【0087】
8)市販デンプン系食品からの直接的なエタノール生産性の検討
市販デンプン系食品を炭素源として、カワラタケのエタノール生産能を検討した。まず、市販のうどん(生麺)、中華そば(生麺)、または米飯に水を加えて、3種類の市販デンプン系食品培地を調製した(pHは未調整)。いずれも培地も、培地1L当たりの市販デンプン系食品含量は235gである。全糖量は、うどんの場合は約24%(57.1g)、中華そばの場合は約28%(66.4g)、米飯の場合は約37%(88.1g)として見積もった。なお、上記で使用した市販デンプン系食品は、いずれも消費期限から2日程度過ぎたものを使用した。
【0088】
上記2)〜3)の手順において、T培地を3種類の市販デンプン系食品培地に変えて、エタノール生産量を評価した。その結果を表5に示す。なお、表中の値はエタノール生産量(g/L)を示す。各数値は3回実験を行った平均値である。
【0089】
【表5】
【0090】
表5に示すとおり、カワラタケを用いたことによって、別途糖化処理を行うことなく単一の工程で、市販デンプン系食品から高い濃度のエタノールを生産できた。具体的には、238gのうどんから、培養21日後に28.8gのエタノール生産を確認した。また、238gの中華そばから、培養21日後に34.8gのエタノール生産を確認した。また、238gの米飯から、培養21日後に37.6gのエタノール生産を確認した。これらの結果はいずれも、含有する糖に対し、エタノールへの変換率が80%以上であったことを示している。以上の通り、カワラタケを用いたことによって、商品として価値のなくなった食品を無駄にせず、効率的にエタノールを生産することができた。また、使用する食品は、pH調整等の特別な前処理を必要としなかった。
【0091】
9)牛乳からの直接的なエタノール生産性の検討
牛乳を炭素源として、カワラタケのエタノール生産能を検討した。まず、市販の牛乳に水を容量比1:1で加えて、牛乳培地を調製した(pHは未調整)。この牛乳培地1L当たりのラクトースの量は23gである。上記牛乳は、消費期限から2日程度過ぎたものを使用した。
【0092】
上記2)〜3)の手順において、T培地を牛乳培地に変えて、エタノール生産量を評価した。その結果を表6に示す。なお、表中の値はエタノール生産量(g/L)を示す。各数値は2回実験を行った平均値である。括弧内の数値は培地中のラクトース残量(g/L)を示す。
【0093】
【表6】
【0094】
表6に示すとおり、カワラタケを用いたことによって、別途糖化処理を行うことなく単一の工程で、牛乳から高い濃度のエタノールを生産できた。具体的には、1Lの牛乳培地から、8日後に9.1gのエタノール生産を確認した。これは、牛乳に含まれるラクトース23gから、エタノールが収率70%以上で変換されたと見積もられる。以上の通り、カワラタケを用いたことによって、商品として価値のなくなった牛乳を無駄にせず、効率的にエタノールを生産することができた。また、使用する牛乳は、pH調整や乳脂肪分除去等の特別な前処理を必要としなかった。
【0095】
<結果の考察>
以上の結果を踏まえて考察する。本実施例のアルコール生産方法によれば、バイオマスを加水分解して糖を生成(糖化)する工程を行うことなく、単一の担子菌で、バイオマスから直接的に単一工程で効率的にエタノールを生産することができる。またこの生産方法は、従来法とは異なり、酸等による糖化処理を行わなくてもよいため、これに伴う環境面、設備面、作業面およびコスト面等での負荷が低減される。またこの生産方法によれば、ボールミル処理を行わなくても、ボールミル処理を行った場合に近いエタノール生産性が得られる。従って、ボールミル処理のような破砕工程にかかるエネルギーを削減できる。
【0096】
また、カワラタケおよびアラゲカワラタケを用いたアルコール生産方法によれば、キシロースを効率的に炭素源として利用できる。これにより、従来法よりも効率的にバイオマスからエタノールを生産することが可能になる。
【0097】
以上、本発明を実施例に基づいて説明した。この実施例はあくまで例示であり、種々の変形例が可能なこと、またそうした変形例も本発明の範囲にあることは当業者に理解されるところである。
【0098】
例えば、上述の実施例では、炭素源としてT培地、小麦フスマ培地、および稲わら培地等を含有する培地を用いているが、他の炭素源も同様に利用可能である。具体的には、木質系のバイオマスなどを炭素源として用いることもできる。
【産業上の利用可能性】
【0099】
以上のように、本発明で用いる担子菌は、アルコール生産効率および炭素源の資化性に優れるため、アルコール生産効率を向上し、アルコール生産方法、アルコール飲料の生産方法、アルコール含有食品の生産方法およびそれらに用いる種菌等として有用である。
【符号の説明】
【0100】
200 種菌
202 容器
204 蓋
206 菌糸
208 おがくず
300 種菌
302 容器
304 木材チップ
306 菌糸
400 種菌
402 容器
404 蓋
406 菌糸
408 液体培地
【特許請求の範囲】
【請求項1】
シロアミタケ(Trametes)属の担子菌を用いて、炭素源からアルコールを生成する、アルコール生産方法。
【請求項2】
前記担子菌が、アラゲカワラタケ(Trametes hirsuta)、カワラタケ(Trametes versicolor)、およびシロアミタケ(Trametes suaveolens)からなる群から選ばれる1種以上の担子菌を含む、請求項1に記載のアルコール生産方法。
【請求項3】
前記担子菌を用いて、前記炭素源を発酵させることによりアルコールを生成する、請求項1または2に記載のアルコール生産方法。
【請求項4】
前記炭素源が糖を含む、請求項1乃至3いずれかに記載のアルコール生産方法。
【請求項5】
前記炭素源が、フスマ、稲わら、セルロース、木材、おがくず、紙類、パルプ、わら、もみ殻、生ゴミ、食品、乳系飲料、廃棄乳系飲料、農業残渣、バガス、刈り草、および雑草類からなる群より選ばれる一種以上を含む、請求項1乃至4いずれかに記載のアルコール生産方法。
【請求項6】
前記炭素源が、生ゴミである、請求項1〜5いずれかに記載のアルコール生産方法。
【請求項7】
前記炭素源が、廃棄乳系飲料である、請求項1〜5いずれかに記載のアルコール生産方法。
【請求項8】
前記担子菌が前記炭素源を糖化することにより糖を生成する工程と、
前記担子菌が糖化により生成された前記糖を発酵することによりアルコールを生成する工程と、
を含む、請求項1乃至7いずれかに記載のアルコール生産方法。
【請求項9】
前記糖化と前記発酵が、同一容器で並行して行われる、請求項8に記載のアルコール生産方法。
【請求項10】
前記炭素源が、グルコース、フルクトース、マンノース、ガラクトース、セロビオース、マルトース、アラビノース、キシロース、およびスクロースからなる群より選ばれる一種以上の糖を含む、請求項1乃至7いずれかに記載のアルコール生産方法。
【請求項11】
統合バイオプロセスによってアルコールを生成する、請求項1乃至10いずれかに記載のアルコール生産方法。
【請求項12】
前記統合バイオプロセスが、前記担子菌を用いて前記炭素源に含まれるリグニンを分解する工程を含む、請求項11に記載のアルコール生産方法。
【請求項13】
前記統合バイオプロセスが、前記担子菌を用いて前記炭素源に含まれるセルロースおよびヘミセルロースを分解する工程を含む、請求項11または12に記載のアルコール生産方法。
【請求項14】
前記統合バイオプロセスは、
前記担子菌を用いてリグニン、セルロース、およびヘミセルロースを分解し、単糖またはオリゴ糖を生成する工程と、
前記担子菌を用いて前記単糖または前記オリゴ糖を発酵する工程と、
を含む、請求項11乃至請求項13に記載のアルコール生産方法。
【請求項15】
前記単糖がヘキソースまたはペントースを含み、
前記オリゴ糖がセロビオース、マルトース、およびスクロースからなる群より選ばれる一種以上の糖を含む、
請求項14に記載のアルコール生産方法。
【請求項16】
前記単糖がグルコース、フルクトース、マンノース、ガラクトース、アラビノース、およびキシロースからなる群より選ばれる一種以上の糖を含む、請求項15に記載のアルコール生産方法。
【請求項17】
前記担子菌がアラゲカワラタケまたはカワラタケである、請求項1乃至16いずれかに記載のアルコール生産方法。
【請求項18】
前記アラゲカワラタケまたはカワラタケを用いてペントースからアルコールを生成する、請求項17に記載のアルコール生産方法。
【請求項19】
前記ペントースがキシロースである、請求項18に記載のアルコール生産方法。
【請求項20】
前記アルコールが、エタノールを含む、請求項1から19に記載のアルコール生産方法。
【請求項21】
シロアミタケ属の担子菌を用いて、炭素源からアルコールを含有する液体を生成する、アルコール飲料の生産方法。
【請求項22】
シロアミタケ属の担子菌を用いて、炭素源からアルコールを含有する組成物を生成する、アルコール含有食品の生産方法。
【請求項23】
シロアミタケ属の担子菌の菌糸と、
菌糸を担持する担体と、
を備えることを特徴とする、シロアミタケ属の担子菌を用いて炭素源からアルコールを生成するための種菌。
【請求項1】
シロアミタケ(Trametes)属の担子菌を用いて、炭素源からアルコールを生成する、アルコール生産方法。
【請求項2】
前記担子菌が、アラゲカワラタケ(Trametes hirsuta)、カワラタケ(Trametes versicolor)、およびシロアミタケ(Trametes suaveolens)からなる群から選ばれる1種以上の担子菌を含む、請求項1に記載のアルコール生産方法。
【請求項3】
前記担子菌を用いて、前記炭素源を発酵させることによりアルコールを生成する、請求項1または2に記載のアルコール生産方法。
【請求項4】
前記炭素源が糖を含む、請求項1乃至3いずれかに記載のアルコール生産方法。
【請求項5】
前記炭素源が、フスマ、稲わら、セルロース、木材、おがくず、紙類、パルプ、わら、もみ殻、生ゴミ、食品、乳系飲料、廃棄乳系飲料、農業残渣、バガス、刈り草、および雑草類からなる群より選ばれる一種以上を含む、請求項1乃至4いずれかに記載のアルコール生産方法。
【請求項6】
前記炭素源が、生ゴミである、請求項1〜5いずれかに記載のアルコール生産方法。
【請求項7】
前記炭素源が、廃棄乳系飲料である、請求項1〜5いずれかに記載のアルコール生産方法。
【請求項8】
前記担子菌が前記炭素源を糖化することにより糖を生成する工程と、
前記担子菌が糖化により生成された前記糖を発酵することによりアルコールを生成する工程と、
を含む、請求項1乃至7いずれかに記載のアルコール生産方法。
【請求項9】
前記糖化と前記発酵が、同一容器で並行して行われる、請求項8に記載のアルコール生産方法。
【請求項10】
前記炭素源が、グルコース、フルクトース、マンノース、ガラクトース、セロビオース、マルトース、アラビノース、キシロース、およびスクロースからなる群より選ばれる一種以上の糖を含む、請求項1乃至7いずれかに記載のアルコール生産方法。
【請求項11】
統合バイオプロセスによってアルコールを生成する、請求項1乃至10いずれかに記載のアルコール生産方法。
【請求項12】
前記統合バイオプロセスが、前記担子菌を用いて前記炭素源に含まれるリグニンを分解する工程を含む、請求項11に記載のアルコール生産方法。
【請求項13】
前記統合バイオプロセスが、前記担子菌を用いて前記炭素源に含まれるセルロースおよびヘミセルロースを分解する工程を含む、請求項11または12に記載のアルコール生産方法。
【請求項14】
前記統合バイオプロセスは、
前記担子菌を用いてリグニン、セルロース、およびヘミセルロースを分解し、単糖またはオリゴ糖を生成する工程と、
前記担子菌を用いて前記単糖または前記オリゴ糖を発酵する工程と、
を含む、請求項11乃至請求項13に記載のアルコール生産方法。
【請求項15】
前記単糖がヘキソースまたはペントースを含み、
前記オリゴ糖がセロビオース、マルトース、およびスクロースからなる群より選ばれる一種以上の糖を含む、
請求項14に記載のアルコール生産方法。
【請求項16】
前記単糖がグルコース、フルクトース、マンノース、ガラクトース、アラビノース、およびキシロースからなる群より選ばれる一種以上の糖を含む、請求項15に記載のアルコール生産方法。
【請求項17】
前記担子菌がアラゲカワラタケまたはカワラタケである、請求項1乃至16いずれかに記載のアルコール生産方法。
【請求項18】
前記アラゲカワラタケまたはカワラタケを用いてペントースからアルコールを生成する、請求項17に記載のアルコール生産方法。
【請求項19】
前記ペントースがキシロースである、請求項18に記載のアルコール生産方法。
【請求項20】
前記アルコールが、エタノールを含む、請求項1から19に記載のアルコール生産方法。
【請求項21】
シロアミタケ属の担子菌を用いて、炭素源からアルコールを含有する液体を生成する、アルコール飲料の生産方法。
【請求項22】
シロアミタケ属の担子菌を用いて、炭素源からアルコールを含有する組成物を生成する、アルコール含有食品の生産方法。
【請求項23】
シロアミタケ属の担子菌の菌糸と、
菌糸を担持する担体と、
を備えることを特徴とする、シロアミタケ属の担子菌を用いて炭素源からアルコールを生成するための種菌。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】

【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2012−55307(P2012−55307A)
【公開日】平成24年3月22日(2012.3.22)
【国際特許分類】
【出願番号】特願2011−160196(P2011−160196)
【出願日】平成23年7月21日(2011.7.21)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 1.日本農芸化学会、社団法人日本農芸化学会、平成22年3月5日 2.スプリンガー、バイオテクノロジーレター、32巻7号 2010年3月18日
【出願人】(504150461)国立大学法人鳥取大学 (271)
【Fターム(参考)】
【公開日】平成24年3月22日(2012.3.22)
【国際特許分類】
【出願日】平成23年7月21日(2011.7.21)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 1.日本農芸化学会、社団法人日本農芸化学会、平成22年3月5日 2.スプリンガー、バイオテクノロジーレター、32巻7号 2010年3月18日
【出願人】(504150461)国立大学法人鳥取大学 (271)
【Fターム(参考)】
[ Back to top ]