
へテロマルチマー及び共通成分を有する多重特異性抗体の製造方法
【課題】所望するヘテロマルチマー多重特異性抗体の調製を提供する。
【解決手段】本発明は、二重特異性抗体、二重特異性免疫付着因子及び抗体-免疫付着因子キメラのようなヘテロマルチマーを製造する方法に関する。本発明はまた、その方法を用いて製造したヘテロマルチマーにも関する。一般に、該方法は抗体結合ドメインを有している各々のヘテロマー性ポリペプチドと結合した共通の軽鎖を有する多重特異性抗体を提供する。加えて本方法は、ヘテロマルチマー形成を促進する及びホモマルチマー形成を妨げるように第1のポリペプチドの界面と第2のポリペプチドの界面で特異的且つ相補的な相互作用を;及び/又は非天然存在ジスルフィド結合が該第1の及び第2のポリペプチド間に形成されるように、第1のポリペプチドの界面で遊離チオール含有残基を、及び第2のポリペプチドの界面中に相当する遊離チオール含有残基を、多重特異性抗体に導入することをさらに包含する。本方法は、望ましくないヘテロマルチマーとホモマルチマーに相対して望まれるヘテロマルチマーの増大した形成を与える。
【解決手段】本発明は、二重特異性抗体、二重特異性免疫付着因子及び抗体-免疫付着因子キメラのようなヘテロマルチマーを製造する方法に関する。本発明はまた、その方法を用いて製造したヘテロマルチマーにも関する。一般に、該方法は抗体結合ドメインを有している各々のヘテロマー性ポリペプチドと結合した共通の軽鎖を有する多重特異性抗体を提供する。加えて本方法は、ヘテロマルチマー形成を促進する及びホモマルチマー形成を妨げるように第1のポリペプチドの界面と第2のポリペプチドの界面で特異的且つ相補的な相互作用を;及び/又は非天然存在ジスルフィド結合が該第1の及び第2のポリペプチド間に形成されるように、第1のポリペプチドの界面で遊離チオール含有残基を、及び第2のポリペプチドの界面中に相当する遊離チオール含有残基を、多重特異性抗体に導入することをさらに包含する。本方法は、望ましくないヘテロマルチマーとホモマルチマーに相対して望まれるヘテロマルチマーの増大した形成を与える。
【発明の詳細な説明】
【技術分野】
【0001】
ヘテロマルチマー及び共通成分を有する多重特異性抗体の製造方法発明の分野本発明は、ヘテロマルチマー重鎖成分および共通する軽鎖成分を有する多重特異性抗体の作製方法に関する。このような多重特異性抗体として、二重特異性抗体、二重特異性免疫付着因子、ならびに本発明の方法を使用して作製される抗体−免疫付着因子キメラおよびヘテロマルチマーポリペプチドなどが挙げられる。
【背景技術】
【0002】
二重特異性抗体少なくとも2つの異なる抗原に対する結合特異性を有する二重特異性抗体(BsAb)は、インビトロおよびインビボでの免疫診断および治療に必要な標的化薬剤として、そして診断的な免疫アッセイに必要な標的化薬剤としての広範囲の診療適用における大きな可能性を有している。
【0003】
診断領域において、二重特異性抗体は、細胞表面分子の機能的特性を探る際に、そして細胞傷害性を媒介する様々なFcレセプターの能力を明らかにする際に非常に有用である(Fangerら、Crit.Rev.Immunol.12:101〜124(1992))。Nolanら、Biochem.Biophys.Acta.1040:1〜11(1990)は、BsAbに関する他の診断適用を記載する。特に、BsAbは、酵素免疫アッセイで使用される酵素を固定化するために構築することができる。これを達成するために、BsAbの一方のアーム(腕)は、酵素表面の特定のエピトープに結合するように設計することができ、その結果、結合による酵素阻害は生じない。BsAbのもう一方のアームは、所望の部位において高い酵素密度を確実にする固定化マトリックスに結合する。そのような診断的BsAbの例には、表面抗原を探し出すために使用された、Hammerlingら、J.Exp.Med.128:1461〜1473(1968)によって記載されたウサギ抗IgG/抗フェリチンBsAbが含まれる。
【0004】
ホースラディッシュペルオキシダーゼ(HRP)ならびにホルモンに対する結合特異性を有するBsAbもまた開発されている。BsAbに関する別の可能な免疫化学的適用には、二部位免疫アッセイにおけるその使用が含まれる。例えば、分析対象タンパク質の表面にある2つの異なるエピトープに結合する2つのBsAbか作製されている−一方のBsAbは複合体を不溶性マトリックスに結合させ、もう一方は指示酵素と結合する(Nolanら(上記)を参照)。
【0005】
二重特異性抗体はまた、ガンなどの様々な疾患のインビボ免疫診断またはインビトロ免疫診断を行うために使用することができる(Songsivilaiら、Clin.Exp.Immunol.79:315(1990))。BsAbのこのような診断的使用を容易にするために、BsAbの一方のアームは、腫瘍関連抗原と結合することができ、もう一方のアームは、放射性核種と強固に結合するキレート化剤などの検出可能なマーカーと結合することができる。このような方法を使用して、LeDoussalらは、結腸直腸ガンおよび甲状腺(thryoid)ガンの放射免疫検出に有用なBsAbを作製した。このBsAbは、ガン胎児抗原(CEA)と結合するアームを一方に有し、ジエチレントリアミン五酢酸(DPTA)と結合するアームをもう一方に有する。LeDoussalら、Int.J.Cancer Suppl.7:58〜62(1992)
およひLeDoussalら、J.Nucl.Med.34:1662〜1671(1993)を参照。Stickneyらは、放射免疫検出を使用してCEAを発現する結腸直腸ガンを検出するための方策を同様に記載する。この研究者らは、CEAならびにヒドロキシエチルチオ尿素−ベンジル−EDTA(EOTUBE)と結合するBsAbを記載する。Stickneyら、Cancer Res.51:6650〜6655(1991)を参照。
【0006】
二重特異性抗体はまた、標的(例えば、病原体または腫瘍細胞)と結合する一方のアーム、およびT細胞レセプターまたはFcγレセプターなどの細胞傷害性誘引因子分子と結合するもう一方のアームが提供されることによって、細胞傷害性の対象を変えることでヒトの治療に有用であり得る。従って、二重特異性抗体を使用して、患者の細胞性免疫防御機構を腫瘍細胞または感染性病原体に特異的に向けさせることができる。このような方策を使用して、FcγRIII(または、CD16)に結合する二重特異性抗体は、ナチュラルキラー(NK)細胞/大顆粒リンパ球(LGL)細胞によってインビトロでの腫瘍細胞殺傷を媒介することができ、インビボで腫瘍増殖を防止するのに効果的であることが明らかにされた。Segalら、Chem.Immunol.47:179(1989)およびSegalら、ガンの生物学的治療(Biologic Therapy ofCancer)2(4)、DeVitaら編、J.B.Lippincott、Philadelphia(1992)、1頁。同様に、FcγIIIと結合する一方のアームと、HER2レセプターに結合するもう一方のアームとを有する二重特異性抗体が、HER2抗原を過剰発現する卵巣腫瘍および乳腫瘍を治療するために開発された。(Hseih−Maら、Cancer Research52:6832〜6839(1992)およびWeinerら、Cancer Research53:94〜100(1993))。二重特異性抗体はまた、T細胞による殺傷を媒介することができる。通常、二重特異性抗体は、T細胞上のCD3複合体を腫瘍関連抗原に結合させる。抗p185HER2に結合した抗CD3からなる完全にヒト化されたF(ab')2BsAbを使用して、T細胞の標的化が、HER2レセプターを過剰発現する腫瘍細胞を殺傷するために行われた。Shalabyら、J.Exp.Med.175(1):217(1992)。二重特異性抗体は、いくつかの初期の臨床試験において調べられ、有望な結果が得られている。1つの臨床試験において、肺ガン、卵巣ガンまたは乳ガンの12名の患者が、抗CD3/抗腫瘍(MOC31)の二重特異性抗体で標的化された活性化Tリンパ球を注入することによって処置された。deLeijら、「二重特異性抗体および標的化細胞の細胞傷害性」、Romet−Lemonne、FangerおよびSegal編、Leinhart(1991)249頁。
【0007】
標的化された細胞において、腫瘍細胞の相当の局所的な溶解、穏和な炎症反応が誘導されたが、毒性の副作用または抗マウス抗体の応答は誘導されなかった。B細胞の悪性疾患の患者での抗CD3/抗CD19二重特異性抗体の非常に予備的な試験において、末梢の腫瘍細胞数の大きな減少もまた達成された。Clarkら、「二重特異性抗体および標的化細胞の細胞傷害性」、Romet−Lemonne、FangerおよびSegal編、Leinhart(1991)243頁。BsAbの治療的適用に関してはKroesenら、CancerImmunol.Immunother.37:400〜407(1993)、Kroesenら、Br.J.Cancer70:652〜661(1994)、およびWeinerら、J.Immunol.152:2385(1994)もまた参照。
【0008】
二重特異性抗体はまた、フィブリン溶解剤またはワクチンアジュバントとして使用することができる。さらに、このような抗体は、感染性疾患の処置において、(例えば、エフェタター細胞を、HIVウイルスまたはインフルエンザウイルスなどのウイルスに感染した細胞あるいはトキソプラズマゴンディイ(Toxoplasmagondii)などの原生動物に対して標的化するために)使用することができ、あるいはイムノトキシンを腫瘍細胞に送達するために、あるいは免疫複合体を細胞表面レセプターに標的化するために使用することができる(Fangerら(上記)を参照)。
【0009】
BsAbの使用は、BsAbを十分な量および純度で得ることが困難であるということによって著しく妨げられている。従来、二重特異性抗体は、ハイブリッド−ハイブリドーマ技術(MillsteinおよびCuello、Nature305:537〜539(1983))を使用して作製された。免疫グロブリンの重鎖および軽鎖は無作為に組み合わされるために、これらのハイブリドーマ(クアドローマ)は、10個の異なる抗体分子の混合物を産生する可能性があり、このうちの1個のみが、正しい二重特異性構造を有する(図1Aを参照)。この正しい分子の精製は、通常的にはアフィニティークロマトグラフィー工程によって行われているが、かなり面倒であり、生成物の収量は少ない。例えば、(Smith,W.ら(1992)Hybridoma4:87〜98;およびMassimo,Y.S.ら(1997)J.Immunol.Methods201:57〜66)を参照。従って、より大きな収量のBsAbを産生するための技術が開発されている。抗体フラグメントの化学的な連結を行うために、Brennanら、Science 229:81(1985)は、無傷の抗体をタンパク質分解的に切断して、F(ab')2フラグメントを得る方法を記載する。
【0010】
このようなフラグメントは、ジチオール複合化剤である亜ヒ酸ナトリウムの存在下で還元され、近傍のジチオールを安定化させ、分子間ジスルフィド結合が形成されないようにする。次いで、得られたFab’フラグメントは、チオニトロ安息香酸(TNB)誘導体に変換される。次いで、BsAbを形成するために、1つのFab’−TNB誘導体が、メルカプトエチルアミンを用いた還元によってFab’−チオールに再度変換され、もう1つのFab’−TNB誘導体の等モル量と混合されて、生成したBsAbは、酵素を選択的に固定化するための薬剤として使用することができる。
近年の進歩は、二重特異性抗体を得るために化学的に連結させることができるFab’−SHフラグメントを大腸菌から直接回収することを容易にした。Shalabyら、J.Exp.Med.175:217〜225(1992)は、p185HER2と結合する一方のアームおよびCD3と結合するもう一方のアームを有する完全にヒト化されたBsAbのF(ab')2分子の産生を記載する。BsAbを形成するために、それぞれのFab’フラグメントを大腸菌から別個に分泌させ、インビトロでの指向された化学的結合反応に供した。このようにして形成されたBsAbは、ヒト乳腫瘍標的に対するヒト細胞傷害性リンパ球の溶解活性を開始させるのと同様に、HER2レセプターを過剰発現する細胞および正常なヒトT細胞に結合することができた。Rodriguesら、Int.J.Cancers(Suppl.)7:45〜50(1992)もまた参照。
【0011】
BsAbフラクメントを作製し、組換え細胞の培養物から直接分離するための様々な技術もまた記載されている。例えば、二重特異性のF(ab')2ヘテロダイマーが、ロイシンジッパーを使用して産生されている(Kostelnyら、J.Immunol.148(5):1547〜1553(1992))。Fosタンパク質およびJunタンパク質に由来するロイシンジッパーペプチドが、遺伝子融合によって抗CD3抗体および抗インターロイキン−2レセプター(IL−2R)抗体のFab’部分に連結された。ホモダイマーの抗体をヒンジ部で還元してモノマーを形成させ、次いで、再酸化してヘテロダイマーの抗体を形成させた。このようなBsAbは、インビトロでHuT−102細胞を溶解するために、細胞傷害性T細胞を呼び寄せるのに非常に効果的であることが見出された。Hollingerら、PNAS(USA)90:6444〜6448(1993)により記載される「ジアボディ(diabody)」技術の出現は、BsAbフラダメントを作製するための代わりの機構をもたらした。このフラグメントは、短すぎて同じ鎖の表面にある2つのドメインの間での対形成ができないリンカーによって軽鎖の可変ドメイン(VL)に連結された重鎖の可変ドメイン(VH)を含む。従って、1つのフラグメントのVHドメインおよびVLドメインは、別のフラグメントの相補的なVHドメインおよびVLドメインと対形成することを強いられ、それによって2つの抗原結合部位が形成される。単鎖Fv(sFv)のダイマーの使用によるBsAbフラグメントを作製するための別の方策もまた報告されている。Gruberら、J.Immunol.152:5368(1994)を参照。この研究者らは、25アミノ酸残基のリンカーによって抗フルオレセイン抗体のVHドメインおよびVLドメインに連結されたT細胞レセプターに対する抗体のVHドメインおよびVLドメインを含む抗体を設計した。再生された分子は、フルオレセインおよびT細胞レセプターに結合し、フルオレセインがその表面に共有結合しているヒト腫瘍細胞を溶解させた。
【0012】
組換え細胞の培養物から直接回収することができる二重特異性抗体を作製するためのいくつかの技術が報告されているようである。しかし、完全長のBsAbは、その血清半減期がより長いと考えられることおよび可能なエフェクター機能のために、多くの診療的適用に関しては、BsAbフラグメントよりも好ましいとされ得る。
免疫付着因子(Immunoadhesins) 免疫付着因子(Ia)は、細胞表面レセプターまたはリガンド(「付着因子」(adhesin))などのタンパク質の結合ドメインを、免疫グロブリンの定常ドメインのエフェタター機能とともに併せ持つ抗体様分子である。免疫付着因子は、ヒト抗体の多くの貴重な化学的特性および生物学的特性を有し得る。免疫付着因子は、適当なヒト免疫グロブリンのヒンジ配列および定常ドメイン(Fc)配列に連結された所望の特異性を有するヒトのタンパク質配列から構築することができるので、目的とする結合特異性は、完全なヒト成分を使用して達成することができる。そのような免疫付着因子は、患者に対する免疫原性は最小であり、安全に長期間または繰り返し使用される。
【0013】
文献に報告されている免疫付着因子には下記が含まれる。T細胞レセプターの融合体(Gascoigneら、Proc.Natl.Acad.Sci.USA84:2936〜2940(1987));CD4の融合体(Caponら、Nature337:525〜531(1989);Trauneckerら、Nature 339:68〜70(1989);Zettmeisslら、DNACell Biol.USA 9:347〜353(1990);およびByrnら、Nature344:667〜670(1990));L−セレクチンまたはホーミングレセプターの融合体(Watsonら、J.Cell.Biol.110:2221〜2229(1990);およびWatsonら、Nature349:164〜167(1991));CD44の融合体(Aruffoら、Cell61:1303〜1313(1990));CD28およびB7の融合体(Linsleyら、J.Exp.Med.173:721〜730(1991));CTLA−4の融合体(Lisleyら、J.Exp.Med.174:561〜569(1991));CD22の融合体(Stamenkovicら、Cell66:1133〜1144(1991));TNFレセプターの融合体(Ashkenaziら、Proc.Natl.Acad.Sci.USA88:10535〜10539(1991);Lesslauerら、Eur.J.Immunol.27:2883〜2886(1991);およびPeppelら、J.Exp.Med.174:1483〜1489(1991));NPレセプターの融合体(Bennettら、J.Biol.Chem.266:23060〜23067(1991));インターフェロンγレセプターの融合体(Kurschnerら、J.Biol.Chem.267:9354〜9360(1992));4−1BBの融合体(Chalupnyら、PNAS(USA)89:10360〜10364(1992))およびIgEレセプターαの融合体(RidgwayおよびGorman、J.Cell.Biol.第115巻、抄録番号1448(1991))。
【0014】
治療的使用について記載されている免疫付着因子の例には、細胞表面CD4に対するHIVの結合を阻止するためのCD4−IgG免疫付着因子が含まれる。
【0015】
CD4−IgGを出産直前の妊娠女性に投与した第I相臨床試験から得られたデータにより、この免疫付着因子は、HIVの母−胎児間移動を防止することにおいて有用であり得ることが示唆されている。Ashkenaziら、Intern.Rev.Immunol.10:219〜227(1993)。腫瘍壊死因子(TNF)と結合する免疫付着因子もまた開発された。TNFは、敗血症性ショックの主要なメディエーターであることが明らかにされている前炎症性サイトカインである。敗血症性ショックのマウスモデルに基づいて、TNFレセプター免疫付着因子は、敗血症性ショックの処置において臨床的に使用するための候補物として有望であることが明らかにされた(Ashkenaziら、上記)。免疫付着因子は、治療以外にもまた使用される。例えば、L−セレクチンレセプター免疫付着因子は、末梢リンパ節の高内皮細静脈(HEV)の組織化学的染色を行うための試薬として使用された。この試薬はまた、L−セレクチンのリガンドを分離してその特徴づけを行うために使用された(Ashkenaziら、上記)。免疫付着因子構造の2つのアームが異なる特異性を有する場合、免疫付着因子は、二重特異性抗体に対する類推によって「二重特異性免疫付着因子」と呼ばれる。
【0016】
Dietschら、J.Immunol.Methods162:123(1993)は、接着分子(E−セレクチンおよびP−セレクチン)の細胞外ドメインを併せ持つそのような二重特異性免疫付着因子を記載する。結合性の研究により、そのようにして得られた二重特異性免疫グロブリン融合タンパク質は、それが誘導された単一特異性の免疫付着因子と比較して、骨髄様細胞株に対する結合能か高まったことが示された。
抗体−免疫付着因子のキメラ 抗体−免疫付着因子(Ab/Ta)キメラもまた文献に記載されている。このような分子は、免疫付着因子の結合領域を抗体の結合ドメインとともに併せ持つ。
【0017】
Bergら、PNAS(USA)88:4723〜4727(1991)は、マウスのCD4−IgGから誘導された二重特異性の抗体−免疫付着因子キメラを作製した。この研究者らは、2つのアームを有する四量体分子を構築した。一方のアームは、抗体軽鎖の定常ドメインと融合したCD4とともに、抗体重鎖の定常ドメインと融合したCD4からなった。もう一方のアームは、抗CD3抗体の完全な軽鎖とともに、抗CD3抗体の完全な重鎖からなった。CD4−IgGのアームによって、この二重特異性分子は、細胞傷害性T細胞の表面にあるCD3に結合する。細胞傷害性細胞およびHIV感染細胞を一緒にすると、HIV感染細胞が特異的に殺傷される。
Bergら(上記)は四量体構造の二重特異性分子を記載しているが、1個のCD4−IgG融合体のみを含有するトリマーのハイブリッド分子を作製することができる。Chamowら、J.Immunol.153:4268(1994)を参照。この構築物の第1のアームは、ヒト化された抗CD3κ軽鎖およびヒト化された抗CD3γ重鎖によって形成される。第2のアームは、IgGのFcドメインによるgp120の結合を担うCD4の細胞外ドメインの一部を併せ持つCD4−IgG免疫付着因子である。得られるAb/Iaキメラは、HIV感染細胞の殺傷を、純粋な細胞傷害性T細胞調製物、またはFcレセプターを産生する大顆粒状リンパ球エフェクター細胞をさらに含む全末梢血リンパ球(PBL)画分のいずれかを使用して媒介した。
【0018】
多重特異性抗体ヘテロマルチマーの製造においては、ホモマルチマーよりも、所望のヘテロマルチマーの産生を増大させることが望ましい。Fc含有のBsAbを得るために選択される現在の方法は、依然として、2つの抗体を同時に発現するハイブリッドハイブリドーマである(MilsteinおよびCuello、Nature 305:537〜540(1983))。
【0019】
ハイブリッドハイブリドーマにおいて、重(H)鎖は、典型的には、所望のヘテロダイマーと同様に、ホモダイマーを形成する。さらに、軽(L)鎖は、同系でない重鎖との誤った対形成を形成することが多い。従って、2つの抗体が同時に発現すると、重鎖および軽鎖の10個までの対形成が生じ得る(Suresh,M.R.ら、MethodsEnzymol.121:210〜228(1986))。これらの望ましくない鎖の対形成は、BsAbの産生を低下させ、そして重大で、ときには克服できない精製問題を必然的に強いる(Smithら(1992)上記、およびMassimoら(1997)上記)。
【0020】
抗体重鎖は、立体的に相補的な変異をマルチマー化ドメインのCH3ドメイン界面に導入することによるヘテロダイマー化(Ridgwayら、ProteinEng.9:617〜621(1996))、および本明細書中に記載されるファージディスプレーによる最適化を行うために以前に操作されている。改変されたCH3ドメインを含有する鎖によって、抗体/免疫付着因子ハイブリッド(Ab/Ia)の生成により判断されるように、約90%までのヘテロダイマーが生じる。ヘテロダイマー化した重鎖は、依然として、同系でない軽鎖との誤った対形成を形成し得るので、目的のBsAbの回収が妨げられる。
【0021】
発明の要旨本出願は、モノマー混合物から所望するヘテロマルチマー二重特異性抗体の形成を高めるのに役立つ方法を記載する。この方法は、共通する可変軽鎖を提供して二重特異性抗体のヘテロマーの各可変重鎖領域と相互作用させることによるヘテロオリゴマー化のために第1のポリペプチドと第2のポリペプチドとの間の界面を操作することによる。3つの可能なヘテロマルチマーおよびホモマルチマーが、第1のポリペプチドおよび第2のポリペプチドから形成され得る。そのようなポリペプチドのそれぞれは、同様に、第1の軽鎖および第2の軽鎖とそれぞれ会合する。これにより、鎖の対形成は合計で10通りが可能である(図1A)。所望のヘテロマルチマー形成を高める方法によって、その産生を、望ましくないヘテロマルチマーおよびホモマルチマーよりも大きく高めることができる。
【0022】
ヘテロマルチマー抗体の第1のポリペプチドと第2のポリペプチドとの間の好ましい界面は、抗体の定常ドメインのCH3ドメインの少なくとも一部を含む。
【0023】
界面で相互作用する第1および第2の各ポリペプチドのドメインは、マルチマー化ドメインと呼ばれる。マルチマー化ドメインは、好ましくは、特定の第1のポリペプチドと第2のポリペプチドとの問の相互作用を促進し、それによって所望のヘテロマルチマーの産生を増大させる(図1B)。相互作用は、空洞への隆起(protuberance-into-cavity)の相補的な領域の形成;天然に存在しないジスルフィド結合の形成;ロイシンジッパー;疎水性領域;および親水性領域によって接触面で促進され得る。「隆起」(protuberance)は、第1のポリペプチドの界面に由来する小さなアミノ酸側鎖を、より大きな側鎖(例えば、チロシンまたはトリプトファン)と置換することによって構築される。隆起と同一の大きさまたは類似する大きさの代替的な「空洞」(cavity)は、任意に、大きなアミノ酸側鎖を、より小さな側鎖(例えば、アラニンまたはトレオニン)と置換することによって第2のポリペプチドの界面に作製される。適切に配置され、そして適切な大きさを有する隆起または空洞が第1のポリペプチドまたは第2のポリペプチドのいずれかの界面に存在する場合、隣接する接触面において、それぞれ、対応する空洞または隆起の設計が必要とされるだけである。天然に存在しないジスルフィド結合は、第1のポリペプチドにおいて、天然に存在するアミノ酸を、システインなどの遊離チオールを含有する残基と置換して、遊離チオールが第2のポリペプチドの別の遊離チオール含有側鎖と相互作用し、そしてジスルフィド結合が第1のポリペプチドと第2のポリペプチドとの間で形成されるようにすることによって構築される(図1B)。
【0024】
大きな非免疫化ファージディスプレーライブラリー(Vaughan,T.J.ら(1996)Nature Biotechnology14:309〜314、これは参考としてその全体が本明細書中に組込まれる)に由来する単鎖Fvフラグメントは、V-遺伝子の使用を明らかにした。この場合、いくつかの生殖系列のV-遺伝子セグメントに由来するVH配列およびVL配列が優勢であり、ファミリーがレパートリーにおいて優勢であった。異なるパートナー鎖との組合せで特定の重鎖または軽鎖が見出されるレパートリーにおいて、鎖の入り交じった状態の例が認められた(Vaughan,T.J.ら(1996)上記)。
【非特許文献1】Fangerら、Crit.Rev.Immunol.12:101〜124(1992)
【発明の開示】
【発明が解決しようとする課題】
【0025】
所望するヘテロマルチマー多重特異性抗体の調製は、共通する軽鎖が多重特異性抗体の可変重鎖のそれぞれと対形成するように提供される場合に増強されることが本明細書中に開示される。共通する可変軽鎖の使用によって、抗原結合ドメインを形成するために正しく対形成しなければならないモノマーの数が減少する。これは、軽鎖の数を、(本発明が開示される前の二重特異性抗体または多重特異性抗体のそれぞれにおける)2つ以上の軽鎖から、(本発明の多重特異性抗体(図1Cを参照)における)1つの軽鎖に制限することによる。
【課題を解決するための手段】
【0026】
従って、本発明は、ヘテロマルチマー多重特異性抗体を調製する方法に関する。この抗体は、下記の1)および2)を含む:1)界面で接する第1のポリペプチドおよび第2のポリペプチド(および抗体の多重度に従ってさらなるポリペプチド)、ただし、第1のポリペプチドおよびさらなるポリペプチド(または、第1のポリペプチドおよび第2のポリペプチド)のそれぞれは、第1のポリペプチドと第2の(または、少なくとも1つのさらなる)ポリペプチドとの間の界面を形成するマルチマー化ドメインを含み、そしてこのマルチマー化ドメインは、第1のポリペプチドとさらなるポリペプチドとの間の安定な相互作用を促進する;および2)前記の第1のポリペプチドおよび少なくとも1つのさらなるポリペプチド(または、第2のポリペプチド)のそれぞれの結合ドメイン、ただし、各結合ドメインは可変重鎖および可変軽鎖を含み、第1のポリペプチドの可変軽鎖および第2のポリペプチドの可変軽鎖は共通するアミノ酸配列を有し、そしてそのような共通する配列は、前記ポリペプチドのそれぞれの元の軽鎖に対して、少なくとも80%、好ましくは少なくとも90%、より好ましくは少なくとも95%、最も好ましくは100%の配列同一性のアミノ酸配列同一性を有する。この方法は下記の工程を含む:(i)第1のポリペプチド、第2のポリペプチドおよび共通する軽鎖をコードする核酸を含む宿主細胞を培養し、この培養によって核酸を発現させる工程;および (ii)宿主細胞培養物から多重特異性抗体を回収する工程。
【0027】
本発明の関連する実施態様において、第1のポリペプチドをコードする核酸または第2のポリペプチドをコードする核酸、あるいはその両方は、その界面またはその一部をコードするように元の核酸から変更されている。
本発明の方法の別の実施態様において、第1のポリペプチドの界面は、第2のポリペプチドの界面の遊離チオール含有残基と相互作用するように配置され、そうすることによってジスルフィド結合が第1のポリペプチドと第2のポリペプチドとの間で形成される遊離チオール含有残基を含む。本発明により、第1のポリペプチドをコードする核酸は、遊離チオール含有残基をコードするように元の核酸から変更され、あるいは第2のポリペプチドをコードする核酸は、遊離チオール含有残基をコードするように元の核酸から変更され、あるいはその両方である。
【0028】
本発明の方法の別の実施態様において、第1のポリペプチドおよび少なくとも1つのさらなるポリペプチド(または、第2のポリペプチド)の両方をコードする核酸は、それぞれ、隆起および空洞をコードするように変更されている。第1のポリペプチドおよび第2のポリペプチドは、それぞれ、好ましくは、ヒトIgG1のCH3ドメインなどの抗体の定常ドメインを含む。
【0029】
別の態様において、本発明は、界面で接する第1のポリペプチドおよび第2のポリペプチドを含むヘテロマルチマー(二重特異性抗体、二重特異性免疫付着因子または抗体/免疫付着因子キメラなど)を提供する。第1のポリペプチドの界面は、少なくとも1つのさらなるポリペプチド(または、第2のポリペプチド)
【0030】
上のマルチマー化ドメインと相互作用するように配置されて、第1のポリペプチドと第2のポリペプチドとの界面を形成するマルチマー化ドメインを含む。本発明の好ましい実施態様において、マルチマー化ドメインは、特定の第1のポリペプチドと特定の第2のポリペプチドとの間の相互作用が促進されるように変更される。そのような変更には、隆起または空洞あるいはその両方の生成;天然に存在しないジスルフィド結合の生成;相補的な疎水性領域;および相補的な親水性領域の生成が含まれるが、これらに限定されない。ヘテロマルチマー多重特異性抗体は、薬学的に受容可能な担体をさらに含む組成物の形態で提供され得る。
【0031】
本発明はまた、前記のヘテロマルチマー多重特異性抗体をコードする核酸を含む宿主細胞に関する:この場合、第1のポリペプチドおよび少なくとも1つのさらなるポリペプチド(または、第2のポリペプチド)をコードするこの核酸は、1つのベクターまたは別個のベクターに存在する。宿主細胞は、ヘテロマルチマー多重特異性抗体を作製する方法で使用することができる。この方法は、核酸を発現するように宿主細胞を培養すること、およびその細胞培養物からヘテロマルチマー抗体を回収することを含む。
【0032】
さらなる態様において、本発明は、ヘテロマルチマー多重特異性抗体を調製する方法を提供し、この方法は下記の工程を含む:(a)少なくとも1つのさらなるポリペプチドの界面のアミノ酸と相互作用するように配置されているアミノ酸残基を第1のポリペプチドの界面に含む第1のポリペプチドをコードする第1の核酸を選択する工程。実施態様において、核酸は、相互作用するアミノ酸残基をコードするように元の核酸から変更されている。
【0033】
別の実施態様において、第1の核酸は、より大きな側鎖容量を有するアミノ酸をコードするように変更され、それによって、隆起か第1のポリペプチドに形成される;
(b)第2のポリペプチドをコードする第2の核酸を変更し、その結果、第2のポリペプチドの界面内のアミノ酸残基を、より小さな側鎖容量を有するアミノ酸残基と置換し、それによって、第2のポリペプチドに空洞を形成させる工程。
【0034】
この場合、前記の隆起は、この空洞と相互作用するように配置される;
(c)前記の第1の核酸および第2の核酸を宿主細胞に導入して、前記の第1の核酸および第2の核酸が発現されるように宿主細胞を培養する工程;および (d)細胞培養物から生成したヘテロマルチマー抗体を回収する工程。
【0035】
前記の抗体が組み込まれた多重特性抗体(二重特異性抗体など)を構築することもまた望ましいことであり得る。このような状況下では、元の軽鎖と対形成する場合、目的の第2の抗原に特異的に結合する重鎖を同定することは望ましい。
【0036】
Figiniらの方法(Figini,M.ら(1994)J.Mol.Biol.239:68〜78、これは参考としてその全体が本明細書中に組込まれる)を使用して、そのような重鎖を同定することができる。最初に、ファージライブラリーをグアニジン塩酸で処理して、元の軽鎖を解離させる。次に、ファージにディスプレーされた重鎖を、(透析などにより)変性剤を除くことによって目的の軽鎖と再構成させる。次いで、目的の第2の抗原に対して選別を行い、所望の重鎖を同定する。本発明はさらに、選択された軽鎖と対形成するように重鎖を選択するこの方法によって調製される多重特異性抗体、そのような抗体をコードする核酸、およびそのような核酸を含む宿主細胞を含む。
【0037】
本発明は、望ましくないヘテロマルチマーおよび/またはホモマルチマーなどの他の望ましくない最終産物よりもヘテロマルチマーの産生を増大させるための機構を提供する(図1A〜図1Cを参照)。組換え細胞の培養物から回収される所望のヘテロマルチマーの産生量は、好ましくは、副生成物の望ましくないヘテロダイマーまたはホモマルチマーと比較して、少なくとも80重量%を超え、好ましくは少なくとも90重量%を超える。
【発明を実施するための最良の形態】
【0038】
I.定義 一般に、下記の用語または表現は、本説明、実施例および請求項において使用される場合には下記の意味を有する。
「ヘテロマルチマー」,「ヘテロマルチマーポリペプチド」または「ヘテロマルチマー多重特異性抗体」は、少なくとも、第1のポリペプチドおよび第2のポリペプチドを含む分子である。この場合、第2のポリペプチドは、アミノ酸配列において、第1のポリペプチドと少なくとも1個のアミノ酸残基が異なる。ヘテロマルチマーは、少なくとも2つの異なるリガンドに対する複数の結合特異性または結合部位を有する。好ましくは、ヘテロマルチマーは、第1のポリペプチドおよび第2のポリペプチドによって形成される「ヘテロダイマー」を含むことができ、あるいはポリペプチドが、第1のポリペプチドおよび第2のポリペプチド以外に、さらに存在する場合には、より高次の三次構造を形成することができる。
【0039】
ヘテロマルチマーに関する例示的な構造には、ヘテロダイマー(例えば、Dietschら(上記)によって記載される二重特異性免疫付着因子)、ヘテロトリマー(例えば、Chamowら(上記)によって記載されるAb/Iaキメラ)、ヘテロ四量体(例えば二重特異性抗体)及び更なるオリゴマー構造が含まれる。
【0040】
本明細書中で使用されている「マルチマー化ドメイン」は、ヘテロマルチマーの各ポリペプチドの領域をいう。「マルチマー化ドメイン」は、ヘテロマルチマー複合体内のキメラ分子の安定な相互作用を促進する。マルチマー化ドメインは、好ましくは、特定の第1のポリペプチドと特定の第2のポリペプチドとの間の相互作用を促進し、それによって所望のヘテロマルチマーの形成が増強され、そして望ましくないヘテロマルチマーまたはホモマルチマーの形成の可能性が実質的に低下する。マルチマー化ドメインは、免疫グロブリン配列、ロイシンジッパー、疎水性領域、親水性領域、またはキメラなヘテロマルチマーのキメラ分子間での分子間ジスルフィド結合を形成する遊離チオールを介して相互作用することができる。遊離チオールは、ポリペプチド間のジスルフィド結合の形成を可能にする位置において、ポリペプチドの天然に存在する残基を、例えば、システインと置換することによって1つまたは複数の相互作用ポリペプチドの界面に導入することができる。マルチマー化ドメインは、免疫グロブリンの定常部を含むことができる。本発明において有用で可能なマルチマー化ドメインが、ハイブリッドの免疫グロブリンを記載する国際特許出願第PCT/US90/06849号(これは参考としてその全体が本明細書中に組込まれる)に開示されている。さらに、マルチマー化ドメインは、立体的な相互作用によって、安定な相互作用が促進されるだけでなく、モノマーの混合物に由来するホモダイマーよりもヘテロダイマーの形成がさらに促進されるように設計することができる。例えば、国際特許出願第PCT/US96/01598号(これは参考としてその全体が本明細書中に組込まれる)を参照。これは、ヘテロオリゴマー化のために第1のポリペプチドと第2のポリペプチドとの間の相互作用に関する「空洞への隆起」法を開示する。「隆起」は、第1のポリペプチドの界面に由来する小さなアミノ酸側鎖を、より大きな側鎖(例えば、チロシンまたはトリプトファン)と置換することによって構築される。隆起と同一の大きさまたは類似する大きさの代償的な「空洞」は、任意に、大きなアミノ酸側鎖を、より小さな側鎖(例えば、アラニンまたはトレオニン)と置換することによって第2のポリペプチドの界面に作製される。
【0041】
免疫グロブリン配列は、免疫グロブリンの定常ドメインであることが好ましいが、必ずしも、免疫グロブリンの定常ドメインである必要はない。本発明のキメラにおける免疫グロブリン部分は、IgG1、IgG2、IgG3またはIgG4のサブタイプ、IgA、IgE、IgDまたはIgMから得ることができるが、IgG1、IgG2、IgG3またはIgG4が好ましい。
【0042】
「遊離チオール含有化合物」は、本発明のポリペプチド界面のアミノ酸に組み込むことができるが、またはそのようなアミノ酸と反応することができる化合物であって、この化合物の遊離チオール部分は、本発明のさらなるポリペプチドの界面において遊離チオール部分と相互作用して、ジスルフィド結合が形成されるように配置されているそのような化合物を意味する。遊離チオール含有化合物は、好ましくは、システインである。
用語「標識されたエピトープ」は、本明細書で使用される場合、「標識ポリペプチド」に融合したキメラなヘテロ付着因子の全体またはそのフラグメントを含むキメラなポリペプチドを示す。標識ポリペプチドは、抗体が作製され得るエピトープを提供するのに十分な残基を有するが、キメラなヘテロ付着因子の活性を妨げないように十分に短い。標識ポリペプチドは、好ましくは、他にないほど非常に特徴的であり、その結果、それに対する抗体は、他のエピトープと実質的に交差反応しない。適切な標識ポリペプチドは、一般には、少なくとも6個のアミノ酸残基を有し、通常は、8個〜50個の間のアミノ酸残基(好ましくは、約9残基〜30残基の間)を有する。本発明の実施態様は、エピトープ標識に結合したキメラなヘテロ付着因子を含み、そのような標識を使用して、サンプル中の付着因子の検出またはサンプルからの付着因子の回収が行われる。
【0043】
本明細書中で使用されている「共通する軽鎖」または「軽鎖の共通するアミノ酸配列」は、本発明の多重特異性抗体における軽鎖のアミノ酸配列をいう。抗体パネルが、ファージディスプレーライブラリーの選別を行うことによって、少なくとも2つの異なる抗原に対して作製された。そのようなファージディスプレーライブラリーは、例えば、Vaughanら(1996)(上記)により記載されている(これは、ファージミドライブラリーを選択する方法を特に参照して、参考としてその全体が本明細書中に組込まれる)。軽鎖配列を、可変軽鎖アミノ酸配列に関して比較した。比較されたパネルに由来する有用な軽鎖は、少なくとも80%、好ましくは少なくとも90%、より好ましくは少なくとも95%、最も好ましくは100%の同一性のアミノ酸配列同一性を有する軽鎖である。共通する軽鎖配列は、比較された2つの軽鎖配列が近似するように考えられた配列である。比較される軽鎖が、アミノ酸レベルで100%の配列同一性である場合、共通する軽鎖は、軽鎖が多重特異性抗体の異なる結合ドメインにおいて機能するとして、選択されたライブラリーのクローンに由来する軽鎖と同一である。比較される軽鎖が上記と異なる場合、共通する軽鎖は、ライブラリーのクローンに由来する比較される軽鎖の1つまたはもう一方、あるいはその両方と異なり得る。共通する軽鎖が、ライブラリーのクローンの1つまたはもう一方と、あるいはその両方と異なる場合、異なる残基は、抗体軽鎖の抗原結合CDR残基の外側に存在することが好ましい。例えば、抗原結合CDR残基の位置は、配列定義(Kabatら(1991)上記)または構造定義(ChothiaおよびLesk(1987)J.Mol.Biol.196:901〜917)に従って決定することができる。
【0044】
本明細書中で用いている「アミノ酸配列同一性」は、1つの配列のアミノ酸が第2のアミノ酸配列のアミノ酸とどのくらい同じであるかの割合をいう。ポリペプチド鎖の間において100%の配列同一性は、鎖が同一であることを意味する。
【0045】
本明細書中で使用されている「ポリペプチド」は、一般に、約10個よりも多いアミノ酸を有するペプチドおよびタンパク質をいう。好ましくは、哺乳動物のポリペプチド(哺乳生物から最初に得られたポリペプチド)が使用され、より好ましくは、培地に直接分泌されるポリペプチドである。細菌のポリペプチドの例には、例えば、アルカリホスファターゼおよびβ−ラクタマーゼが含まれる。哺乳動物のポリペプチドの例には、レニン、成長ホルモンなどの分子が含まれ、下記が含まれる:ヒト成長ホルモン;ウシ成長ホルモン;成長ホルモン放出因子;
副甲状腺ホルモン;甲状腺刺激ホルモン;リポタンパク質;α−1抗トリプシン;インシュリンA鎖;インシュリンB鎖;プロインシュリン;卵胞刺激ホルモン;カルシトニン;黄体化ホルモン;グルカゴン;第VIIIC因子、第IX因子、組織因子およびフォンビルブランド因子などの凝固因子;プロテインCなどの抗凝固因子;心房性ナトリウム利尿因子;肺接触面活性物質;ウロキナーゼまたはヒトウリンまたは組織型プラスミノーゲン活性化因子(t−pA)などのプラスミノーゲン活性化因子;ボンベシン;トロンビン;造血成長因子;腫瘍壊死因子−αおよび腫瘍壊死因子−β;エンケファリナーゼ;RANTES(regulated on activation normallyT−cell expressed andsecreted);ヒトマクロファージ炎症タンパク質(MIPI−1−α);ヒト血清アルブミンなどの血清アルブミン;ミューラー阻害物質;レラキシンA鎖;レラキシンB鎖;プロレラキシン;マウス性腺刺激ホルモン関連ペプチド;β−ラクタマーゼなどの微生物タンパク質;DNase;インヒビン;アクチビン;血管内皮細胞増殖因子(VEGF);ホルモンまたは成長(増殖)因子のレセプター;インテグリン;プロテインAまたはプロテインD;リウマチ因子;骨由来神経栄養因子(BDNF)、ニューロトロフィン−3、ニューロトロフィン−4、ニューロトロフィン−5またはニューロトロフィン−6(NT−3、NT−4、NT−5またはNT−6)、あるいはNGF−βなどの神経増殖因子などの神経栄養因子;血小板由来増殖因子(PDGF);aFGFおよびbFGFなどの繊維芽細胞増殖因子;上皮増殖因子(EGF);TGF−β1、TGF−β2、TGF−β3、TGF−β4またはTGF−β5を含むTGF−αおよびTGF−βなどのトランスホーミング増殖因子(TGF);
インシュリン様増殖因子−Iおよびインシュリン様増殖因子−II(IGF−IおよびIGF−II);des(1−3)−IGF−I(脳IGF−I)、インシュリン様増殖因子結合タンパク質;CD−3、CD−4、CD−8およびCD−19などのCDタンパク質;エリスロポイエチン;骨誘導因子;イムノトキシン;骨形態形成タンパク質(BMP);インターフェロン−α、インターフェロン−βおよびインターフェロン−γなどのインターフェロン;コロニー刺激因子(CSF)類、例えば、M−CSF、GM−CSFおよびG−CSF;インターロイキン(IL)類、例えば、IL−1からIL−10;スーパーオキシドジスムターゼ;T細胞レセプター;表面膜タンパク質;崩壊促進因子;例えば、AIDSエンベロープの一部などのウイルス抗原;輸送タンパク質;ホーミングレセプター;アドレシン;調節タンパク質;抗体;および上記のポリペプチドのいずれかのフラグメント。
【0046】
「第1のポリペプチド」は、第2のポリペプチドと会合し得る任意のポリペプチドである。第1のポリペプチドおよび第2のポリペプチドは、「界面」(下記において定義)で接する。界面に加えて、第1のポリペプチドは、「結合ドメイン」(例えば、抗体の可変ドメイン、レセプター結合ドメイン、リガンド結合ドメインまたは酵素活性ドメイン)、またはCH2、CH1およびCLドメインを含む抗体の定常ドメイン(またはその一部)などのさらなるドメインを1つまたは複数含むことができる。通常、第1のポリペプチドは、抗体から誘導される少なくとも1つのドメインを含む。このようなドメインは、好都合なことに、抗体のCH3ドメインなどの定常ドメインであり、第1のポリペプチドの界面を形成することができる。例示的な第1のポリペプチドには下記が含まれる。抗体重鎖ポリペプチド、異種ポリペプチドの結合ドメインとともに抗体の定常ドメインを併せ持つキメラ(すなわち、免疫付着因子、下記の定義を参照)、レセプターポリペプチド(特に、別のレセプターポリペプチド、例えば、インターロイキン−8レセプター(IL−8R)およびインテグリンヘテロダイマー(例えば、LFA−IまたはGPIIIb/IIIa)とダイマーを形成するポリペプチド)、リガンドポリペプチド(例えば、神経増殖因子(NGF)、ニューロトロフィン−3(NT−3)および脳由来神経栄養因子(BDNF)−Arakawaら、J.Biol.Chem.269(45):27833〜27839(1994)およびRadziejewskiら、Biochem.32(48):1350(1993))、および抗体の可変ドメインポリペプチド(例えば、ジアボディ(diabody))。好ましい第1のポリペプチドは、免疫グロブリンの定常ドメインに融合した抗体の重鎖から選択され、この場合、定常ドメインは、本発明の第2のポリペプチドとの優先的な相互作用が促進されるように変更されている。
【0047】
「第2のポリペプチド」は、「界面」を介して第1のポリペプチドと会合し得る任意のポリペプチドである。界面に加えて、第2のポリペプチドは、「結合ドメイン」(例えば、抗体の可変ドメイン、レセプター結合ドメイン、リガンド結合ドメイン、または酵素活性ドメイン)、またはCH2ドメイン、CH1ドメインおよびCLドメインを含む抗体の定常ドメイン(または、その一部)などのさらなるドメインを含むことができる。通常、第2のポリペプチドは、抗体から誘導される少なくとも1つのドメインを含む。このようなドメインは、好都合なことに、抗体のCH3ドメインなどの定常領域であり、第2のポリペプチドの界面を形成することができる。例示的な第2のポリペプチドには下記が含まれる。抗体重鎖ポリペプチド、異種ポリペプチドの結合ドメインとともに抗体の定常ドメインを併せ持つキメラ(すなわち、免疫付着因子、下記の定義を参照)、レセプターポリペプチド (特に、別のレセプターポリペプチド、例えば、インターロイキン−8レセプター(IL−8R)およびインテグリンヘテロダイマー(例えば、LFA−IまたはGPIIIb/IIIa)とダイマーを形成するポリペプチド)、リガンドポリペプチド(例えば、神経増殖因子(NGF)、ニューロトロフィン−3(NT−3)および脳由来神経栄養因子(BDNF)−Arakawaら、J.Biol.Chem.269(45):27833〜27839(1994)およびRadziejewskiら、Biochem.32(48):1350(1993))、および抗体の可変ドメインポリペプチド(例えば、ジアボディ)。好ましい第2のポリペプチドは、免疫グロブリンの定常ドメインに融合した抗体の重鎖から選択されこの場合、この場合、定常ドメインは、本発明の第1のポリペプチドとの優先的な相互作用が促進されるように変更されている。
【0048】
「結合ドメイン」は、目的の分子(例えば、抗体、リガンド、レセプター、基質または阻害剤)との選択的な結合を担うポリペプチドの任意の領域を含む。例示的な結合ドメインには、抗体の可変ドメイン、レセプター結合ドメイン、リガンド結合ドメインおよび酵素活性ドメインが含まれる。好ましい実施態様において、結合ドメインは、免疫グロブリンの重鎖および軽鎖を含む。本発明の二重特異性抗体およびその作製方法により、二重特異性抗体の各結合ドメインに関する軽鎖は、共通する軽鎖であり、それによって重鎖および軽鎖の誤った対形成が存在する望ましくないヘテロマルチマーの形成が避けられる。
用語「抗体」は、それが本発明に関連する場合、目的の抗原のエピトープと結合するドメインを1つまたは複数含有するポリペプチドを意味するものとする。
【0049】
この場合、そのようなドメインは、抗体の可変部から誘導されるか、または抗体の可変部との配列同一性を有する。抗体の例には、全長の抗体、抗体フラグメント、単鎖分子、二重特異性分子または二機能性分子、ジアボディー、キメラ抗体(例えば、ヒト化抗体およびPRIMATIZEDTM抗体)および免疫付着因子が含まれる。「抗体フラグメント」には、Fv、Fv'、Fab、Fab’およびF(ab')2のフラグメントが含まれる。
【0050】
「ヒト化(された)」形態の非ヒト(例えば、齧歯類または霊長類)抗体は、非ヒト免疫グロブリンから誘導される最小の配列を含有する特異的でキメラな免疫グロブリン、免疫グロブリン鎖またはそのフラグメントである。その大部分において、ヒト化抗体は、受容者の相補性決定領域(CDR)に由来する残基が、所望の特異性、親和性および能力を有するマウス、ラット、ウサギまたは霊長類などの非ヒト種(ドナー抗体)のCDRに由来する残基によって置き換えられているヒト免疫グロブリン(受容体抗体)である。場合によっては、ヒト免疫グロブリンのFv枠組み構造領域(FR)残基は、対応する非ヒト残基によって置き換えられる。さらに、ヒト化抗体は、受容体抗体あるいは持ち込まれたCDR配列または枠組み構造配列のいずれにおいて見出されない残基を含むことができる。このような改変は、抗体の能力を改良して最大にするために行われる。一般に、ヒト化抗体は、少なくとも1つ、典型的には2つの可変ドメインの実質的に全てを含む。そのような可変ドメインにおいて、CDR領域の全て又は実質的に全ては、非ヒトの免疫グロブリンのCDR領域に対応し、そしてFR領域の全て又は実質的に全ては、ヒト免疫グロブリン配列のFR領域である。ヒト化抗体はまた、好ましくは、免疫グロブリンの定常部(Fc)、典型的には、ヒト免疫グロブリンの定常部(Fc)の少なくとも一部を含む。ヒト化抗体は、抗体の抗原結合領域が、目的の抗原でマカク(macaque)サルを免疫化することによって産生される抗体から誘導されるPRIMATIZEDTM抗体を含む。
【0051】
「多重特異性抗体」は、少なくとも2つの異なる抗原に対する結合特異性を有する分子である。そのような分子は、通常、2つの抗原と結合するだけである(すなわち、二重特異性抗体、BsAb)が、三重特異性抗体などのさらなる特異性を有する抗体も、本明細書中で使用される場合には本発明により含まれる。BsAbの例には、腫瘍細胞の抗原に対する一方のアームと、細胞傷害誘引因子分子に対するもう一方のアームとを有する下記のような二重特異性抗体が含まれる。例えば、抗FcγRI/抗CD15、抗p185HER2/FcγRIII(CD16)、抗CD3/抗悪性B細胞(1D10)、抗CD3/抗p185HER2、抗CD3/抗p97、抗CD3/抗腎細胞ガン、抗CD3/抗OVCAR−3、抗CD3/L−D1(抗結腸ガン)、抗CD3/抗メラニン細胞刺激ホルモンアナログ、抗EGFレセプター/抗CD3、抗CD3/抗CAMA1、抗CD3/抗CD19、抗CD3/MoV18、抗神経細胞付着因子分子(NCAM)/抗CD3、抗葉酸結合タンパク質(FBP)/抗CD3、抗汎カルシノーマ関連抗原(AMOC−31)/抗CD−3;腫瘍抗原に特異的に結合するアームと、毒素に結合するアームとを有するBsAb、例えば、抗サポリン/抗Id−1、抗CD22/抗サポリン、抗CD7/抗サポリン、抗CD38/抗サポリン、抗CEA/抗リシンA鎖、抗インターフェロン−α(INF−α)/抗ハイブリドーマイディオタイプ、抗CEA/抗ビンカアルカロイドなど;変換酵素によって活性化されるプロドラッグに対するBsAb、例えば、抗CD30/抗アルカリホスファターゼ(これは、マイトマイシンホスフェートプロドラッグのマイトマシンアルコールへの変換を触媒する)など;フィブリン溶解剤として使用することができるBsAb、例えば、抗フィブリン/抗組織プラスミノーゲン活性化因子(tPA)、抗フィブリン/抗ウロキナーゼ型プラスミノーゲン活性化因子(uPA)など;細胞表面レセプターに対して免疫複合体を標的化するためのBsAb、例えば、抗低密度リポタンパク質(LDL)/抗Fcレセプター(例えば、FcγRI、FcγRIIまたはFcγRIII)など;感染性疾患の治療において使用されるBsAb、例えは、抗CD3/抗単純ヘルペスウイルス (HSV)、抗T細胞レセプター:CD3複合体/抗インフルエンザ、抗FcγR/抗HIVなど;インビトロまたはインビボで腫瘍を検出するためのBsAb、例えば、抗CEA/抗EOTUBE、抗CEA/抗DPTA、抗p185HER2/抗ハプテン;ワクチンアジュバントとしてのBsAb(Fangerら(上記)を参照);および診断用具としてのBsAb、例えば、抗ウサギIgG/抗フェリチン、抗ホースラディッシュペルオキシダーゼ(HRP)/抗ホルモン、抗ソマトスタチン/抗サブスタンスP、抗HRP/抗FITC、抗CEA/抗β−ガラクトシダーゼ(Nolanら(上記)を参照)など。三重特異性抗体の例には、抗CD3/抗CD4/抗CD37、抗CD3/抗CD5/抗CD37および抗CD3/抗CD8/抗CD37が含まれる。
【0052】
本明細書中で使用されている用語「免疫付着因子」は、免疫グロブリンの定常ドメインのエフェクター機能とともに、異種タンパク質(「付着因子」、例えば、レセプター、リガンドまたは酵素)の「結合ドメイン」を併せ持つ抗体様の分子を示す。構造的には、免疫付着因子は、抗体の抗原認識部位および結合部位(抗原結合部位)以外である(すなわち、「異種である」)所望の結合特異性を有する付着因子のアミノ酸配列と、免疫グロブリンの定常ドメイン配列との融合体を含む。免疫付着因子における免疫グロブリンの定常ドメイン配列は、IgG1、IgG2、IgG3もしくはIgG4のサブタイプ、IgA、IgE、IgDまたはIgMなどの任意の免疫グロブリンから得ることができる。
【0053】
本明細書中で使用されている用語「リガンド結合ドメイン」は、定量的なリガンド結合能、および好ましくは対応する天然のレセプターの生物学的活性を少なくとも保持する任意の天然の細胞表面レセプターまたはその任意の領域もしくは誘導体をいう。特定の実施態様において、レセプターは、免疫グロブリンスーパーファミリーのメンバーと相同的な細胞外ドメインを有する細胞表面ポリペプチドに由来する。免疫グロブリンスーパーファミリーのメンバーではないが、それにもかかわらず、この定義によって具体的に含まれる他の代表的なレセプターは、サイトカインに対するレセプターであり、特にチロシンキナーゼ活性を有するレセプター(レセプターチロシンキナーゼ)であり、そしてヘマトポイエチンおよび神経増殖因子レセプタースーパーファミリーのメンバー、ならびに細胞付着因子分子、例えは、(E、LおよびP−)セレクチンである。
【0054】
用語「レセプター結合ドメイン」は、定量的なレセプター結合能、および好ましくは対応する天然のリガンドの生物学的活性を少なくとも保持するレセプターに対する任意の天然のリガンド(細胞付着因子分子を含む)またはその任意の領域もしくは誘導体を示すために使用される。この定義は、特に、具体的には、上記のレセプターに対するリガンドに由来する結合配列を含む。
【0055】
本明細書中で使用されている用語「多重特異性免疫付着因子」は、少なくとも2つの結合特異性を有する(すなわち、2つ以上の付着因子結合ドメインを併せ持つ)(上記に定義されている)免疫付着因子を示す。多重特異性免疫付着因子は、本質的には、国際特許公開第WO89/02922号(1989年4月6日公開)、欧州特許EP314,317(1989年5月3日公開)および米国特許第5,116,964号(1992年5月2日発行)に開示されているように、ヘテロダイマー、ヘテロトリマーまたはヘテロ四量体として組み立てることができる。好ましい多重特異性免疫付着因子は二重特異性である。二重特異性免疫付着因子の例には、CD4−TgG/INFレセプター−IgGおよびCD4−1gG/L−セレクチン−IgGが含まれる。後者の分子は、リンパ球ホーミングレセプター(LHR、L−セレクチン)のリンパ節結合機能と、CD4のHIV結合機能とを併せ持ち、HIV感染、関連する症状の予防または処置において診断薬としての適用の可能性が見出されている。
【0056】
「抗体−免疫付着因子キメラ(Ab/Iaキメラ)」は、(本出願において定義されている)少なくとも1つの免疫付着因子とともに、(上記に定義されている)抗体の少なくとも1つの結合ドメインを併せ持つ分子を含む。例示的なAb/Iaキメラは、Bergら(上記)およびChamowら(上記)によって記載されている二重特異性CD4−IgGキメラである。
【0057】
「界面」は、第2のポリペプチドの界面内において1つまたは複数の「接触」アミノ酸残基(または、アミノ酸でない他の基)と相互作用する第1のポリペプチドにおける「接触」アミノ酸残基(あるいは、炭水化物基、NADH、ピオチン、FADまたはヘム基などのアミノ酸でない他の基)を含む。好ましい界面は、可変ドメインまたは定常ドメイン(またはその領域)などの免疫グロブリンのドメインである。しかし、ヘテロマルチマーレセプターを形成するポリペプチド間の界面、またはNGF、NT−3およびBDNFなどの2つ以上のリガンドの間の界面も、この用語の範囲に含まれる。好ましい界面は、好ましくはIgG抗体から誘導され、最も好ましくはヒトIgG1抗体から誘導される免疫グロブリンのCH3ドメインを含む。
【0058】
「元の」アミノ酸残基は、元の残基よりも小さな側鎖容量または大きな側鎖容量を有し得る「移入」残基によって置換されるアミノ酸残基である。移入アミノ酸残基は、天然存在アミノ酸残基または非天然存在アミノ酸残基であり得るが、天然存在アミノ酸残基が好ましい。「天然存在」アミノ酸残基は、遺伝暗号によってコードされるアミノ酸残基、および国際特許出願第PCT/US96/01598号(これは参考としてその全体が本明細書中に取込まれる)の表1に列記されるアミノ酸残基である。「非天然存在」アミノ酸残基は、遺伝暗号によってコードされていないが、ポリペプチド鎖で隣のアミノ酸残基と共有結合することができる残基を意味する。非天然存在アミノ酸残基の例には、例えば、ノルロイシン、オルニチン、ノルバリン、ホモセリン、およびEllmanら、Meth.Enzym.202:301〜336(1991)に記載されるアミノ酸残基アナログなどの他のアミノ酸残基のアナログが挙げられる。天然に存在しないそのようなアミノ酸残基を作製するために、Norenら、Science244:182(1989)およびEllmanら(上記)の方法を使用することができる。簡単に記載すると、この方法は、非天然存在アミノ酸残基でサプレッサーtRNAを化学的に活性化し、その後、インビトロでRNAの転写および翻訳を行うことを含む。本発明の方法には、少なくとも1つの元のアミノ酸残基を置換することが含まれるが、2つ以上の元の残基を置換することができる。通常は、第1のポリペプチドまたは第2のポリペプチドの界面におけるすべての残基が、置換される元のアミノ酸残基を含むにすぎない。置換に好ましい残基は「埋もれている」。「埋もれている」は、残基が本質的には溶媒と接触できないことを意味する。移入残基は、好ましくは、酸化または誤ったジスルフィド結合の形成の可能性を避けるために、システインではない。
【0059】
「元の核酸」は、アミノ酸側鎖が、第1のポリペプチドと第2のポリペプチドとの間で界面で相互作用して、ポリペプチド間の安定な相互作用を促進させるアミノ酸がマルチマー化ドメイン内でコードされるように変更することができる、目的のポリペプチドをコードする核酸を意味する。そのような変更によって、空洞への隆起、天然に存在しないジスルフィド結合、ロイシンジッパー、疎水性相互作用および親水性相互作用のような安定な相互作用に限定されないが、そのような安定な相互作用を生じさせることができる。このような変更は、好ましくは、目的とする第1のポリペプチドと第2のポリペプチドとの間の特異的な相互作用を促進し、望ましくないヘテロマーの対形成またはホモマーの形成が生じる相互作用を効果的に除くように選択される。元の核酸または出発核酸は、天然に存在する核酸であり得るか、または以前に変更が行われた核酸(例えば、ヒト化抗体フラグメント)を含み得る。核酸を「変更する」は、元の核酸を、目的のアミノ酸残基をコードしているコドンの少なくとも1つの挿入、欠失または置換による遺伝子操作または変異処理を行うことを意味する。通常、元の残基をコードしているコドンは、移入残基をコードするコドンによって置換される。このようにDNAを遺伝子的に改変するための技術は、Mutagenesis:aPractical Approach、M.J.McPherson編、IRLPress、Oxford、UK(1991)に総説されており、例えば、部位特異的変異誘発法、カセット変異誘発法およびポリメラーゼ連鎖反応(PCR)変異誘発法を含む。
【0060】
隆起、空洞、または(ジスルフィド結合の形成に必要なシステイン残基などの)遊離チオールは、合成的手段によって、例えば、組換え技術、インビトロペプチド合成、前記の天然に存在しないアミノ酸残基を導入するためのそのような技術、ペプチドの酵素的または化学的な連結、あるいはこれらの技術のいくつかの組合せによって、第1のポリペプチドまたは第2のポリペプチドの界面に「導入する」ことができる。従って、「導入される」隆起、空洞、または遊離チオールは、「天然に存在しない」、すなわち、「非天然」である。これは、自然界または元のポリペプチド(例えば、ヒト化モノクローナル抗体)に存在しないことを意味する。
【0061】
隆起を形成するために移入アミノ酸残基は、比較的少数(例えば、約3個〜6個)の「回転異性体」を有することが好ましい。「回転異性体」は、アミノ酸側鎖のエネルギー的に有利な立体配座である。様々なアミノ酸残基の回転異性体の数は、PondersおよびRichards、J.Mol.Biol.193:775〜791(1987)に総説されている。
【0062】
「分離された」ヘテロマルチマーは、その天然の細胞培養環境の成分からの同定および分離および/または回収が行われたヘテロマルチマーを意味する。その天然環境の混入成分は、ヘテロマルチマーに関する診断使用または治療使用を妨害する物質であり、酵素、ホルモン、および他のタンパク質性または非タンパク質性の溶解物を含み得る。好ましい実施態様において、ヘテロマルチマーは、(1)ローリー法によって測定されるように95重量%を超えるように、最も好ましくは99重量%を超えるように、あるいは(2)スピニングカップ配列決定装置の使用によって、N末端または内部のアミノ酸配列の少なくとも15残基が得られるのに十分な程度に、あるいは(3)クマシーブルー染色、または好ましくは銀染色を使用して、還元条件下または非還元条件下でのSDS−PAGEにより均一になるまで精製される。
【0063】
本発明のヘテロマルチマーは、一般には、実質的に均一になるまで精製される。「実質的に均一」、「実質的に均一な形態」および「実質的な均一性」という表現は、生成物が、望ましくないポリペプチドの組合せ物に由来する副生成物(例えば、ホモマルチマー)を実質的に有していないことを示すために使用される。
【0064】
純度に関して表現される場合、実質的な均一性は、副生成物の量が、10%を超えないこと、好ましくは5%未満であること、より好ましくは1%未満であること、最も好ましくは0.5%未満であることを意味する。ただし、割合は重量比である。
【0065】
「制御配列」という表現は、特定の宿主生物において、機能的に連結されたコード配列を発現させるのに必要なDNA配列をいう。原核生物に適切な制御配列には、例えば、プロモーター、必要に応じてオペレーター配列、リボソーム結合部位、および可能であれば、未だあまりよく理解されていない他の配列が含まれる。真核生物細胞は、プロモーター、ポリアデニル化シグナルおよびエンハンサーを利用することが知られている。
【0066】
核酸は、核酸が別の核酸配列と機能的な関係におかれている場合に「機能的に連結されている」。例えば、プレ配列または分泌リーダーに関するDNAは、DNAが、ポリペプチドの分泌に参加しているプロタンパク質として発現される場合、ポリペプチドに関してDNAに機能的に連結されている;プロモーターまたはエンハンサーは、それが配列の転写に影響を及ぼす場合、コード配列に機能的に連結されている;あるいは、リボソーム結合部位は、それが翻訳を容易にするように配置されている場合、コード配列に機能的に連結されている。一般に、「機能的に連結されている」は、連結されているDNA配列が連続していることを、そして分泌リーダーの場合には、読み取り相で連続していることを意味する。
【0067】
しかし、エンハンサーは、連続している必要はない。連結は、都合の良い制限部位で結合させることによって達成される。そのような部位が存在しない場合、合成オリゴヌクレオチドアダプターまたはリンカーが、従来的な実施に従って使用される。
【0068】
II.ヘテロマルチマーの調製1.出発材料の調製 最初の工程として、第1のポリペプチドおよび第2のポリペプチド(およびヘテロマルチマーを形成する任意のさらなるポリペプチド)が選択される。通常、これらのポリペプチドをコードする核酸は分離しなければならず、その結果、核酸は、本明細書中で定義されているように、隆起または空洞またはその両方をコードするように変更することができる。しかし、変異は、合成的な手段を使用することによって、例えば、ペプチド合成を使用することによって導入することができる。同様に、移入残基が天然に存在しない残基である場合には、Norenら(上記)の方法を、そのような置換を有するポリペブチドを作製するために用いることができる。さらに、ヘテロマルチマーの一部は、細胞培養で組換え的に適切に作製され、そのような分子の他の部分は、上記のそのような技術によって作製される。
【0069】
抗体の分離および免疫付着因子の調製に関する技法が次に行われる。しかし、ヘテロマルチマーは、当技術分野で知られている技術を使用して、他のポリペプチドから形成され得るか、または他のポリペプチドを組み込むことができることが理解される。例えば、目的のポリペプチド(例えば、リガンド、レセプターまたは酵素)をコードする核酸は、ポリペプチドのmRNAを有し、そのmRNAを検出可能なレベルで発現していると考えられる組織から調製されたcDNAライブラリーから分離することができる。ライブラリーは、目的の遺伝子またはそれによってコードされるタンパク質を同定するために設計されたプローブ(例えば、抗体または約20塩基〜80塩基のオリゴヌクレオチドなど)を用いてスクリーニングされる。選択されたプローブによるcDNAライブラリーまたはゲノムライブラリーのスクリーニングは、Sambrookら、Molecular Cloning:A Laboratory Manual(NewYork:Cold Spring Harbor LaboratoryPress、1989)の第10章〜第12章に記載される標準的な手順を使用して行うことができる。
【0070】
(1)抗体の調製 抗体の産生に関する技術がいくつか記載されている。そのような技術には、モノクローナル抗体を作製するための従来のハイブリドーマ法、抗体(キメラ抗体、例えば、ヒト化抗体を含む)を作製するための組換え技術、トランスジェニック動物での抗体産生、および「完全なヒト」抗体を調製するために近年記載されたファージディスプレー技術が含まれる。これらの技術を下記に簡単に記載する。
【0071】
目的の抗原に対するポリクローナル抗体は、一般には、抗原およびアジュバントの皮下(sc)注射または腹腔内(ip)注射を多数回行うことによって動物に生起させることができる。抗原(または、標的アミノ酸配列を含有するフラグメント)を、免疫化される種において免疫原であるタンパク質(例えば、キーホルリンペットヘモシアニン、血清アルブミン、ウシサイログロブリン、またはダイズのトリプシン阻害剤)に、二官能性薬剤または誘導化剤を使用して結合させることは有用であり得る。そのような二官能性薬剤または誘導化剤は、例えば、マレイミドベンゾイルスルホスクシンイミドエステル(システイン残基を介するコンジュゲーション)、N−ヒドロキシスクシンイミド(リシン残基を介する)、グルタルアルデヒド、無水コハク酸、SOCl2、またはR1N=C=NR(ただし、RおよびR1は異なるアルキル基である)である。動物は、免疫原性のコンジュゲートまたは誘導体に対して、(ウサギまたはマウスに関して、それぞれ)1mgのコンジュゲートを3容量のフロイント完全アジュバントと一緒にして、その溶液を皮下に多数の部位に注射することによって免疫化される。1ヶ月後、動物は、コンジュゲートを含むフロイント完全アジュバントの最初の量の1/5〜1/10を用いて、多数の部位に皮下注射することによって追加免疫される。
【0072】
7日〜14日の後に、動物は採血され、血清を抗体力価についてアッセイする。
【0073】
動物は、力価がプラトーに達するまで追加免疫される。動物は、好ましくは、同じ抗原のコンジュゲートで追加免疫されるが、異なるタンパク質に結合させたコンジュゲートおよび/または異なる架橋剤によるコンジュゲートで追加免疫される。コンジュゲートはまた、タンパク質の融合体として、組換え細胞培養で作製することができる。同様に、ミョウバンなどの凝集化剤を使用して、免疫応答を増強することができる。
【0074】
モノクローナル抗体は、KohlerおよびMilstein、Nature256:495(1975)によって初めて記載されたハイブリドーマ法を使用して実質的に均一な抗体集団から得られるか、または、組換えDNA法 (Cabillyら、米国特許第4,816,567号)によって作製することができる。
【0075】
ハイブリドーマ法において、マウス、またはハムスターなどの他の適切な宿主動物を上記のように免疫化して、免疫化のために使用されたタンパク質に特異的に結合する抗体を産生するか、または産生し得るリンパ球が誘導される。あるいは、リンパ球をインビトロで免疫化することができる。次いで、ポリエチレングリコールなどの適切な融合化剤を使用してリンパ球をミエローマ細胞と融合して、ハイブリドーマ細胞を得る(Goding、MonoclonalAntibodies:Principles and Practice、59頁〜103頁(AcademicPress、1986))。このようにして調製されたハイブリドーマ細胞を、融合していない元のミエローマ細胞の生育または生存を阻害する物質を1つまたは複数含有することが好ましい適切な培養培地に播種して、増殖させる。例えば、元のミエローマ細胞が酵素ヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRTまたはHPRT)を欠失している場合、ハイブリドーマの培養培地は、典型的には、HGPRT欠損細胞の生育を妨げる物質であるヒポキサンチン、アミノプテリンおよびチミジンを含む(HAT培地)。好ましいミエローマ細胞は、効率よく融合し、選択された抗体産生細胞によって抗体の安定で高レベルの発現を維持し、HAT培地などの培地に対して感受性を有するミエローマ細胞である。このような細胞の中で、好ましいミエローマ細胞株は、ネズミのミエローマ細胞株であり、Salk Institute Cell Distribution Center(SanDiego、California、USA)から入手可能なMOPC−21およびMPC−11のマウス腫瘍から誘導されるミエローマ細胞株、ならびにAmerican Type Culture Collection(Rockville、Maryland、USA)から入手可能なSP−2細胞などである。ヒトのミエローマ細胞株およびマウス−ヒトのヘテロミエローマ細胞株もまた、ヒトのモノクローナル抗体の産生に関して記載されている(Kozbor、J.Immunol.、133:3001(1984);およびBrodeurら、Monoclonal Antibody Production Techniques andApplications、51頁〜63頁、Marcel Dekkar,Inc.、New York、1987)。ヒトモノクローナル抗体の産生技術に関しては、Boernerら、J.Immunol.、147(1)86:〜95(1991)および国際特許公開第WO91/17769号(1991年11月28日公開)もまた参照。ハイブリドーマ細胞が生育する培養培地は、目的の抗原に対するモノクローナル抗体の産生についてアッセイされる。
【0076】
好ましくは、ハイブリドーマ細胞によって産生されたモノクローナル抗体の結合特異性は、免疫沈降によって、あるいは放射免疫アッセイ(RIA)または酵素結合免疫吸着アッセイ(ELISA)などのインビトロ結合アッセイによって測定される。モノクローナル抗体の結合親和性は、例えば、MunsonおよびPollandのスキャッチャード分析(Anal.Biochem.107:220(1980))によって測定することができる。所望の特異性、親和性および/または活性の抗体を産生するハイブリドーマ細胞が同定された後に、クローンを限界稀釈法によってサブクローニングを行い、標準的な方法によって生育させることができる。Goding、Monoclonal Antibodies:Principles andPractice、59頁〜104頁(AcademicPress、1986)。この目的に適切な培養培地には、例えば、ダルベッコ改変イーグル培地またはRPMI−1640培地が含まれる。さらに、ハイブリドーマ細胞は、動物の体内において腹水腫瘍としてインビボで生育させることができる。サブクローンによって分泌されたモノクローナル抗体は、培養培地、腹水または血清から、例えば、プロテインA−セファロース、ヒドロキシルアパタイトクロマトグラフィー、ゲル電気泳動、透析またはアフィニティクロマトグラフィーなどの従来の免疫グロブリン精製手順によって適切に分離される。
【0077】
あるいは、免疫化したときに、内因性の免疫グロブリンを産生することなく、完全なレパートリーのヒト抗体を産生し得るトランスジェニック動物(例えば、マウス)の作製が現在可能である。例えば、キメラな生殖系列の変異マウスにおける抗体の重鎖連結領域(JH)遺伝子のホモ接合型欠失によって、内因性の抗体産生が完全に阻害されることが記載されている。ヒトの生殖系列の免疫グロブリン遺伝子列をそのような生殖系列の変異マウスに移すことによって、ヒト抗体が、抗原を投与したときに産生する。例えば、Jakobovitsら、Proc.Natl.Acad.Sci.USA、90:2551〜255(1993);Jakobovitsら、Nature362:255〜258(1993);Fishwild,D.M.ら(1996)Nat.Biotech14:845〜851;およびMendez,M.J.ら(1997)Nat.Genetics 15:146〜156を参照。
【0078】
さらなる実施態様において、抗体または抗体フラグメントは、McCaffertyら、Nature、348:522〜554(1990)に記載される技術を使用して作製された抗体ファージライブラリーから、目的の抗原を使用して、適切な抗体または抗体フラグメントに関する選択を行うことによって分離することができる。Clacksonら、Nature、352:624〜628(1991)およびMarksら、J.Mol.Biol.、222:581〜597(1991)は、それぞれ、ファージライブラリーを使用するマウス抗体およびヒト抗体の分離を記載する。その後の刊行物は、非常に大きなファージライブラリーを構築するための方法として、鎖シャッフリング(Markら、Bio/Technol.10:779〜783(1992))、ならびに結合感染およびインビボ組換えによる高い親和性(nM範囲)のヒト抗体の産生を記載する(Waterhousesら、Nuc.Acids.Res.21:2265〜2266(1993);Griffiths,A.D.ら(1994)EMBO J.13:3245〜3260;およびVaughanら(1996)上記)。従って、これらの技術は、本発明により含まれる「モノクローナル」抗体(特に、ヒト抗体)の分離に関する、従来のモノクローナル抗体ハイブリドーマ技術の実行可能な代替法である。
【0079】
本発明の抗体をコードするDNAは、従来の手順を使用して(例えば、ネズミ抗体の重鎖および軽鎖をコードする遺伝子に特異的に結合し得るオリゴヌクレオチドプローブを使用することによって)容易に分離され、そして配列決定される。本発明のハイブリドーマ細胞は、そのようなDNAの好ましい供給源として有用である。DNAが一旦分離されると、そのDNAは、発現ベクター内に配置することができ、次いで、サルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、またはそうでなければ免疫グロブリンタンパク質を産生しないミエローマ細胞などの宿主細胞にトランスフェクションされ、組換え宿主細胞においてモノクローナル抗体の合成物が得られる。DNAはまた、例えば、コード配列を、相同的なマウス配列の代わりに、ヒトの重鎖および軽鎖の定常ドメインに置換することによって改変することができる。Morrisonら、Proc.Nat.Acad.Sci.81:6851(1984)。そのようにして、本明細書中の抗抗原モノクローナル抗体の結合特異性を有する抗体が調製される。
【0080】
非ヒト抗体をヒト化するための方法は、当技術分野では十分に知られている。
【0081】
一般に、ヒト化抗体は、ヒトでない供給源からヒト化抗体に導入された1つまたは複数のアミノ酸残基を有する。ヒト化は、本質的には、Winterおよび共同研究者らの方法(Jonesら、Nature 321:522〜525(1986);Riechmannら、Nature332:323〜327(1988);Verhoeyenら、Science239:1534〜1536(1988))に従って、齧歯類のCDRまたはCDR配列をヒト抗体の対応配列に置換することによって行うことができる。従って、そのような「ヒト化」抗体は、キメラな抗体であり、(Cabilly、上記)。実質的には元のままでないヒトの可変ドメインは、ヒトでない種に由来する対応の配列によって置換されている。実際、典型的には、ヒト化抗体は、いくつかのCDR残基、および可能であれば、いくつかのFR残基が、齧歯類の抗体の類似する部位に由来する残基によって置換されているヒト抗体である。抗体は、抗原に対する高い親和性および他の有利な生物学的特性を保持してヒト化されることが重要である。このような目的を達成するために、好ましい方法に従って、ヒト化抗体が、元の配列およびヒト化配列の三次元モデルを使用して、元の配列および様々な考えられるヒト化配列を解析することによって調製される。三次元の免疫グロブリンモデルは、当業者には熟知されている。選択された候補の免疫グロブリン配列の可能性な三次元の立体配置構造を図示して表示するコンピュータープログラムを入手することができる。このような表示を検討することによって、候補の免疫グロブリン配列の機能発現における残基の可能な役割の分析、すなわち、候補の免疫グロブリンのその抗原に対する結合能に影響する残基の分析が可能になる。このように、FR残基は、コンセンサス配列および持ち込み配列からの選択および組合せを行うことができ、その結果、標的抗原に対する増大した親和性などの所望の抗体特性が達成される。さらなる詳細については、国際特許公開第WO92/22653号(1992年12月23日公開)を参照。
【0082】
(ii)免疫付着因子調製 免疫グロブリン(Ig)、およびその変異体が知られており、その多くが、組換え細胞培養によって調製されている。例えば、米国特許第4,745,055号;欧州特許第256,654号;Faulknerら、Nature 298:286 (1982);欧州特許第120,694号;欧州特許第125,023号;Morrison, J. Immun. 123:793 (1979);Koehlerら、Proc. Natl. Acad. Sci. USA 77:2197 (1980);Rasoら、Cancer Res.41:2073 (1981);Morrisonら, Ann. Rev.Immunol. 2:239 (1984);Morrison, Science229:1202 (1985);Morrisonら、Proc.Natl. Acad. Sci. USA 81:6851 (1984);欧州特許第255,694号;欧州特許第266,663号;および国際特許公開第88/03559号を参照。再編成された免疫グロブリン鎖も知られている。例えば、米国特許第4,444,878号;国際特許公開第88/03565号;および欧州特許第68,763号、ならびに、それらで引用されている参考文献を参照。
【0083】
付着因子結合ドメインの配列を、適当な免疫グロブリン定常ドメイン位配列に連結させて構築したキメラ(免疫付着因子)が当技術分野において知られている。
【0084】
文献で報告されている免疫付着因子には、T細胞レセプターとの融合タンパク質(Gascoigneら、Proc. Natl. Acad. Sci.USA 84:2936−2940 (1987));CD4との融合タンパク質(Caponら、Nature 337:525−531 (1989);Trauneckerら、Nature 339:68−70 (1989);Zettmeisslら、DNA Cell Biol.USA 9:347−353 (1990);およびBymら、Nature344:667−670(1990));L−セレクチン(ホーミンクルセプター)との融合タンパク質(Watsonら、J.CellBiol.110:2221−2229(1990);およびWatsonら、Nature349:164−167(1991));CD44との融合タンパク質(Aruffoら、Cell 61:1303−1313 (1990));CD28とB7との融合タンパク質(Linsleyら、J.Exp.Med.173:721−730 (1991));CTLA−4との融合タンパク質(Linsleyら、J.Exp.Med.174:561−569 (1991));CD22との融合タンパク質(Stamenkovicら、Cell66:1133−1144(1991));TNFレセプターとの融合タンパク質(Ashkenaziら、Proc.Natl.Acad.Sci.USA88:10535−10539 (1991);Lesslauerら,Eur.J.Immunol.27:2883−2886 (1991);およびPeppleら、J.Exp.Med.174:1483−1489 (1991));およびIgEレセプターαとの融合タンパク質(RidgwayとGovman、J.CellBiol.Vol.115,Abstract No.1448 (1991))などがある。
【0085】
もっとも単純で、もっとも素直な免疫付着因子設計は、付着因子の結合ドメイン(例えば、レセプターの細胞外ドメイン(ECD))を、免疫グロブリン重鎖のヒンジ領域とFc領域に結合させるものである。通常、本発明の免疫付着因子を調製するときには、付着因子の結合ドメインをコードする核酸がC末端になるように、免疫グロブリンの定常ドメイン配列のN末端をコードする核酸に融合させるが、N末端の融合も可能である。
【0086】
典型的には、このような融合配列において、コードされるキメラポリペプチドは、少なくとも、免疫グロブリン重鎖の定常ドメインのヒンジ、CH2、CH3の各ドメインの機能的活性を保持している。定常ドメインのFcドメインのC末端側で融合させたり、重鎖のCH1、または軽鎖の対応領域のすぐN末端側で融合させたりする。正確にどの位置で融合させるかは重要ではないが、特定の部位がよく知られていて、生物学的活性、分泌、またはIaの結合特性を最適なものにするときに選択される。
【0087】
好ましい実施態様において、免疫グロブリンG1(IgG1)のFcドメインのN末端に付着因子の配列を融合させる。重鎖の定常領域全体を付着因子の配列に融合させることも可能である。しかし、より好ましくは、化学的にIgGFcを画定するパパイン切断部位(すなわち、重鎖の定常領域の最初の残基を114とすると残基216にあたる)のすぐ上流にあるヒンジ領域から始まる配列、または、他の免疫グロブリンの同様の部位を融合に用いる。特に好ましい実施態様においては、付着因子のアミノ酸配列を、IgG1、IgG2、またはIgG3の重鎖の(a)ヒンジ領域、およびCH2とCH3からなる領域に、または、(b)CH1、ヒンジ、CH2、およびCH3ドメインに融合させる。融合させる正確な部位は重要ではなく、最適な部位を通常の実験によって決定することができる。
【0088】
双特異的免疫付着因子では、免疫付着因子はマルチマーとして集合し、ヘテロダイマーまたはヘテロ四量体として集合する。一般的には、これらの分子集合した免疫グロブリンは、既知のユニット構造をもっているはずである。基本的な4本鎖の構造単位は、IgG、IgD、およびIgEに見られる構造である。高分子量の免疫グロブリンでは、4本鎖の構造単位が繰り返される。すなわち、IgMは、一般的に、4本鎖の基本ユニットをジスルフィド結合によって結合させた五量体として存在する。IgAグロブリンと、場合によってはIgGグロブリンも、血清中ではマルチマーの形で存在することがある。マルチマーの場合、4本鎖ユニットは同じものであることもあれば、異なることもある。
【0089】
本明細書の範囲に含まれる集合免疫付着因子のさまざまな例を以下に概略的に示す:
(a)ACL-ACL;
(b)ACH-[ACH,ACL-ACH,ACL-VHCH,またはVLCL-ACH];
(c)ACL-ACH-[ACL-ACH,ACL-VHCH,VLCL-ACH,またはVLCL-VHCH];
(d)ACL-VHCH-[ACH,またはACL-VHCH,またはVLCL-ACH];
(e)VLCL-ACH-[ACL-VHCH,またはVLCL-ACH];および(f)[A-Y]n-[VLCL-VHCH]2;
ここで、Aは、それぞれ、同一か異なった付着因子のアミノ酸配列を表しており、 VLは、免疫グロブリン軽鎖の可変ドメインであり;
VHは、免疫グロブリン重鎖の可変ドメインであり;
CLは、免疫グロブリン軽鎖の定常ドメインであり;
CHは、免疫グロブリン重鎖の定常ドメインであり;
nは、1よりも大きい整数であり;
Yは、共有結合的な架橋剤の残基を示している。
【0090】
簡潔を期するために、上記の構造は、主要な特徴を示しているにすぎず、結合ドメイン(J)や、免疫グロブリンのその他のドメインは示されていない。さらに、ジスルフィド結合も示されていない。しかし、このようなドメインが結合活性にとって必要である場合には、それらが、免疫グロブリン分子の中で占める通常の位置に提示されるよう構成されている。
【0091】
または、キメラ重鎖を含む免疫グロブリンを得るために、免疫グロブリンの重鎖と軽鎖の配列の問に付着因子の配列を挿入することも可能である。この実施態様では、免疫グロブリンの各アームにおける、免疫グロブリン重鎖のヒンジとCH2ドメインの間か、またはCH2とCH3ドメインの間の3’末端に付着因子の配列を融合させる。同様の構築物が、Hoogenboomら、Mol.Immunol.28:1027−1037(1991)によって報告されている。
【0092】
免疫グロブリンの軽鎖が、付着因子−免疫グロブリン重鎖融合ポリペプチドに共有的に会合しているか、または、付着因子に直接融合するかして存在することもあろう。前者の場合、免疫グロブリン軽鎖をコードするDNAが、典型的には、付着因子−免疫グロブリン重鎖融合タンパク質をコードするDNAとともに発現する。分泌されるときに、ハイブリッドの重鎖と軽鎖が共有的に会合して、ジスルフィド会合で結ばれた免疫グロブリン重鎖−軽鎖の2組の組み合せを含む免疫グロブリン様構造物が提供される。このような構造物を調製するのに適した方法は、例えば、1989年3月28日公開の米国特許第4,816,567号に開示されている。
【0093】
好ましい実施態様において、本発明の免疫付着因子の構築に用いる免疫グロブリンの配列は、IgG免疫グロブリン重鎖の定常部位からとってくる。ヒトの免疫付着因子については、ヒトIgG1およびIgG3免疫グロブリンの配列が好ましい。IgG1を使用することの主な利点は、固定したプロテインA上で、IgG1免疫付着因子を効率的に精製できることにある。これに対して、IgG3の精製には、これよりもかなり使い勝手の悪い媒体であるプロテインGが必要となる。しかし、特定の免疫付着因子構築物のIgの融合相手を選ぶときには、免疫グロブリンのこの他の構造的性質および機能的性質も考慮すべきである。例えば、IgG3のヒンジは、より長く、可塑性もより高いため、IgG1に融合させると正しく折り畳まれて機能することができないような長い”付着因子”ドメインを受け入れることができる。もう一つ考慮すべきなのは、原子価である。IgG免疫付着因子は2価性のホモダイマーであるが、IgAおよびIgMなどのIgサブタイプは、それぞれ、基本的なIgホモダイマーユニットからなるダイマーと五量体を生じよう。インビボでの実用に供するために設計された免疫付着因子では、Fc領域によって特定される薬理動態的性質およびエフェクター機能も重要である。IgG1、IgG2およびIgG4はすべて、インビボでの半減期が21日で、補体系を活性化する相対的能力はそれぞれ異なる。IgG4は、補体をほとんど活性化せず、IgG2は、IgG1よりもかなり弱くしか補体活性を活性化しない。さらに、IgG1とは異なって、IgG2は、単核球または好中球上のFcレセプターには結合しない。補体の活性化に関しては、IgG3が最適であるが、インビボでの半減期が、他のIgGアイソタイプの約3分の1である。ヒトの治療薬として使用するために、免疫付着因子を設計するときに考慮すべき他の重要な点は、特定のアイソタイプがもつアロタイプの数である。一般的には、血清学的に区別されるアロタイプの数が少ないIgGのアイソタイプほど好ましい。例えば、IgG1は、血清学的に区別されるアロタイプ部位を4つしかもっておらず、これらのうち2つは(Gm1とGm2)はFc領域に位置しているが、このうちの1つの部位であるG1m1は、免疫原性がない。これに対して、IgG3には12の血清学的に区別されるアロタイプがあり、それらのすべてがFc領域中にあって、その内の3つの部位(G3m5、11および21)だけが免疫原性をもつ同一のアロタイプをもっている。したがって、γ1免疫付着因子よりも、γ3免疫付着因子方が免疫原としての可能性が高い。
【0094】
付着因子部分をコードするcDNA配列を、IgのcDNA配列にフレームを合わせて融合させることによって、もっとも好都合に免疫付着因子が構築される。しかし、ゲノムのIgフラグメントに融合させることもできる(例えば、Gascoigneら、上記;Aruffoら、Cell61:1303−1313(1990);およびStamenkovicら、Cell 66:1133−1144 (1991))参照)。後者のタイプの融合には、発現のためのIg調節配列が存在することが必要である。IgGの重鎖定常領域をコードするcDNAは、脾臓または末梢血のリンパ球由来のcDNAライブラリーからの公開された配列をもとに、ハイブリダイゼーションまたはポリメラーゼ連鎖反応(PCR)技術によって分離することができる。免疫付着因子の”付着因子”とIg部分をコードするcDNAを、選んだ宿主細胞の中で効率的に発現させるためのプラスミドベクターの中にひとつながりになるように挿入する。
【0095】
2.隆起および/または空洞の作出 隆起および/または空洞を形成するもととなる残基を選択するための最初の工程として、X線結晶解析やNMRなど、当技術分野において既知の技術を用いて、ヘテロマルチマーの三次元構造を得る。三次元構造に基づいて、当業者は、接触面の残基を同定することができる。
【0096】
好ましい接触面は、免疫グロブリン定常ドメインのCH3ドメインである。各種のIgGサブタイプの接触面残基と”埋もれた”残基が同定されていたため、IgG、IgA、IgD、IgE、およびIgMのCH3ドメインの接触面の残基で、取り入れる残基と置換するのに最適な残基などの残基が同定されている(例えば、PCT/US96/01598を参照、その全文が、参照してここに組み込まれる)。CH3接触面を改変するための基礎となるのは、X線結晶解析によって、ヒトIgG1重鎖の間に生じる、Fc領域での分子間結合は、CH3ドメイン間では広範なタンパク質/タンパク質間相互作用を含むのに対して、グリコシル化されたCH2ドメインでは、その糖部分によって結合するということが示されたことである。(Deisenhofer,Biochem.20:2361−2370 (1981))。さらに、哺乳動物細胞の中には、欠失によりCH2およびCH3ドメインが除去されないかぎり、2つの重鎖間では、抗体が発現されるときにジスルフィド結合が効率よく形成される(Kingら、Biochem.J.281:317 (1992))。したがって、重鎖の集合は、ジスルフィド結合を促進すると考えられ、その逆ではないと考えられる。これらの構造および機能に関するデータをまとめて考えると、抗体の重鎖の会合はCH3部位によってもたらされるという仮説に至る。さらに、CH3ドメイン間の接触面を改変して、異なった重鎖のヘテロマルチマーの形成を促進して、対応するホモマルチマーの集合を阻害することができるかもしれないという推測がなされた。
【0097】
本明細書で説明する実験によって、この方法を用いれば、ホモマルチマーよりもヘテロマルチマーの形成を促進することが可能なことが示された。したがって、目的のポリペプチドと抗体のCH3部位とを含むポリペプチド融合体を作出して、第1または第2のポリペプチドを形成させることができる。好ましいCH3ドメインは、ヒトIgG1のようなIgG抗体に由来するものである。
【0098】
隆起または空洞を形成するための候補となりうる接触面残基が同定されている。 ”埋もれた”残基を選んで置き換えることが好ましい。ある残基が埋込まれているか否かを判定するために、Leeら、J.Mol.Biol.55:379−400(1971)の表面接触プログラムを用いて、接触面にある残基と溶媒との接触可能性(SA)を計算することができる。そして、第1と第2のポリペプチドのそれぞれの残基について、一方のポリペプチドを除いた上で、SAを別々に計算することができる。そして、SA(ダイマー)−SA(モノマー)という等式を用いて、接触面のモノマー型とダイマー型の間の各残基のSAの差異を計算することができる。これによって、ダイマーを形成するときにSAが減少する残基のリストが示される。ダイマーの各残基のSAを、同じアミノ酸のトリペプチドGly−X−GlyのSAの理論値と比較する。ただし、X=目的のアミノ酸(Roseら、Science229:834−838(1985))。(a)モノマーに較べてダイマーでSA減少した残基、および(b)SAが、対応するトリペプチドにおけるSAの26%よりも少ない残基を接触面残基と考える。
【0099】
2つの分類を明らかにすることができる。すなわち、対応するトリペプチドに較べて10%よりも低いSAをもつ残基(すなわち、”埋もれた”残基)、および、対応するトリペプチドに較べて10%よりも高く25%よりは低いSAをもつ残基(すなわち、”一部埋もれた”残基)である(下記の表1参照)
【0100】
【表1】

【0101】
†残基番号は、IgG結晶構造にある通り(Deisenhofer,Biochemistry 20:2361-2370(1981))ポリペプチド鎖構造に対する残基置換の効果は、InsightTMプログラム(BiosymTechnologies)などの分子図形モデリングプログラムを用いて、調べることができる。このプログラムを用いて、第1のポリペプチドの接触面中に埋もれた残基で、側鎖容量の小さい残基を、例えば、より大きな側鎖容量をもつ残基(すなわち、隆起)に変えることができる。そして、第2のポリペプチドの接触面中の残基で、隆起に隣接する残基を調べて、空洞を形成するために適した残基を見つける。通常、この残基は側鎖容量が大きいので、側鎖容量の小さな残基と置き換える。一定の実施態様において、接触面の三次元構造を調べると、第1のポリペプチドの接触面上に適当な位置に置かれた、適当な大きさの隆起、または第2のポリペプチドの接触面上の空洞が明らかになるはずである。これらの例では、単一突然変異体、すなわち、合成によって導入された隆起または空洞をモデル化する必要があるだけである。
【0102】
第1および第2のポリペプチドがそれぞれCH3ドメインを含む場合に、置換することのできる元の残基を選択することに関しては、ヒトIgG1のCH3/CH3の接触面は、各表面から1090Å2埋もれた4つの逆平行β鎖上にある各ドメイン上の16の残基を含んでいる(Deisenhofer,上記、およびMiller、J.Mol.Biol.216:965(1990)。突然変異は、好ましくは、中央の逆平行β鎖上にある残基を標的とする。この目的は、作出した隆起が、相手方のCH3ドメインの相補的な空洞の中に入らずに、周囲の溶媒の中に突出して親和してしまう危険を最小限にするためである。
【0103】
分子モデリングによって、好ましい元/取り込み残基が一旦同定されたら、当技術分野において周知の技術を用いて、ポリペプチドの中にアミノ酸置換を導入する。通常は、ポリペプチドをコードするDNAは、突然変異誘発:実用的方法(Mutagenesis: a PracticalApproach)、上記、に記載されている技術を用いて遺伝子工学により改変する。
【0104】
オリゴヌクレオチドによる突然変異誘発が、第1または第2のポリペプチドをコードするDNAの置換変異体を調製するための好ましい方法である。この技術は、Adelmanら、DNA,2:183(1983)によって説明されているように、当技術分野において周知の技術である。簡単に述べると、所望の変異をコードしているオリゴヌクレオチドをDNA鋳型とハイブリダイズさせることによって、第1または第2のポリペプチドDNAを改変する。この鋳型DNAは、ヘテロマルチマーの未変更または天然のDNA配列を含む、一本鎖のプラスミドまたはバクテリオファージである。ハイブリダイゼーション後、DNAポリメラーゼを用いて、オリゴヌクレオチドプライマーを取り込んだ相補鎖で、ヘテロマルチマーDNAへの部位特異的な変化をコードする相補鎖である、鋳型に相補的な第2鎖の全長を合成する。
【0105】
Wellsら、Gene34:315(1985)が述べるところにしたがって、目的とするDNA領域を、相補的なオリゴヌクレオチドをアニーリングさせて作製した合成変異フラグメントで置換することによって、カセット突然変異誘発を行なうことができる。PCR突然変異も、第1または第2ポリペプチドDNAの変異配列を作製するのに適している。以下の考察においてはDNAを引用しているが、この技術は、RNAでも応用することができると解される。PCR技術とは、一般的には、次の手順をさす(Erlich, Science 252:1643−1650 (1991)、R. Higuchiが担当した章、p.61−70を参照)。
【0106】
隆起または空洞の突然変異以外にも、本発明は、ヘテロマルチマーのDNAの中に適当なヌクレオチド変異を導入するか、または所望のヘテロマルチマーポリペプチドを合成することによって調製することのできるヘテロマルチマーのアミノ酸配列の変異配列を含む。このような変異配列には、例えば、ヘテロマルチマーを形成する第1および第2のポリペプチドのアミノ酸配列中の残基の欠失、挿入、または置換が含まれる。最終的な構築物が望ましい抗原結合特性をもっているとすれば、最終的な構築物ができるように、欠失、挿入および置換を組み合わせる。グリコシル化部位の数と位置を変更したりして、アミノ酸を変えることによっても、ヘテロマルチマーの翻訳後のプロセッシングを変えることができる。
【0107】
突然変異を誘発するのに好ましい位置にある、ヘテロマルチマーポリペプチドの一定の残基または領域を同定するための有用な方法は、CunnighamとWells、Science 244:1081−1085(1989)によって述べられている“アラニンスキャニング突然変異誘発法”といわれるものである。ここで、標的残基の残基またはそのグループが同定され (例えば、Arg、Asp、His、およびGluなどの荷電性残基)、中性のアミノ酸または負に荷電したアミノ酸(もっとも好ましくは、アラニンまたはポリアラニン)で置き換えて、アミノ酸と、細胞の内外の周囲の水性環境との相互作用に影響を与える。そして、置換部位で、またはそれに対する、および/または別の変異残基を導入することによって、置換に対して機能的な感受性を示すドメインをさらに明確にすることができる。このように、アミノ酸配列変異を導入するための部位が予め決まっていても、突然変異そのものの性質を予め測ることはできない。
【0108】
通常、突然変異には、ヘテロマルチマーの非機能的な領域における保存的なアミノ酸置換が含まれる。突然変異の例を表2に示す。
【0109】
【表2】
【0110】
ヘテロマルチマーのポリペプチドの共有修飾も、本発明の範囲に含まれる。ヘテロマルチマーの共有修飾は、ヘテロマルチマー、またはそのフラグメントの標的アミノ酸残基を、N末端またはC末端の残基の選ばれた側鎖と反応することのできる有機的誘導化剤と反応させて、分子の中に導入することができる。ヘテロマルチマーポリペプチドの別のタイプの共有修飾で、本発明の範囲に含まれるものには、ポリペプチドの本来のグリコシル化パターンを改変することが含まれる。改変とは、もとのヘテロマルチマーの中に見られる糖部分を一つ以上欠失させるか、および/または、もとのヘテロマルチマーには存在しないグルコシル化部位を一つ以上付加することを意味する。ヘテロマルチマーポリペプチドへのグリコシル化部位の付加は、一つ以上のN結合型グリコシル化部位が含まれるよう、アミノ酸配列を改変することによって適宜行われる。一つ以上のセリンまたはトレオニン残基を、もとのヘテロマルチマー配列に付加または置換することによって改変を行なうこともできる(O結合型グリコシル化部位)。簡単にするために、好ましくは、DNAレベルでの変更によって、特に、所望のアミノ酸に翻訳されるコドンを作出するよう、ヘテロマルチマーポリペプチドをコードするDNAの予め決定された塩基突然変異を起こすことによって、ヘテロマルチマーのアミノ酸配列を改変する。ヘテロマルチマーポリペプチド上の糖部分の数を増加させる別の方法は、化学結合または酵素結合によってグリコシドをポリペプチドに結合させることである。これらの方法については、1987年9月11日発行の国際特許公開第87/05330号、およびAplinとWriston,CRCCrit.Rev.Biochem.pp.259−306(1981)に記載されている。ヘテロマルチマーに存在する糖部分の削除は、化学的または酵素的に行なうことができる。
【0111】
ヘテロマルチマーの別のタイプの共有修飾には、ヘテロマルチマーポリペプチドを、米国特許第4,640,835号;第4,496,689号;第4,301,144号;第4,670,417号;第4,791,192号;および第4,179,337号において提示されている方法で、例えば、ポリエチレングリコール、ポリプロピレングリコール、またはポリオキシアルキレンなど、さまざまな非タンパク質性ポリマーの一つに結合させることが含まれる。
【0112】
変異ヘテロマルチマーの特性を前もって予測することは困難であるため、回収された変異体をスクリーニングして、最適な変異体を選抜することが必要であると認められよう。
【0113】
3.共通の軽鎖をもつヘテロマルチマーの発現DNAを突然変異させて、本明細書で開示されているような共通の軽鎖を選抜した後、当技術分野において広く利用可能な組換え技術を用いて、この分子をコードするDNAを発現させる。しばしば、優れた発現系は、ヘテロマルチマーを適切にグルコシル化するための(例えば、グリコシル化されている抗体部位を含むヘテロマルチマーの場合など)、哺乳動物細胞の発現ベクターと宿主とを含んでいる。しかし、下で詳述するように、この分子は、原核生物の発現系で産生させることもできる。通常、一つのベクター、または別々のベクター上に存在する、第1ポリペプチド、第2ポリペプチド、共通の軽鎖ポリペプチド、および、ヘテロマルチマーを形成するのに必要な別のポリペプチドのすべてコードするDNAによっての宿主細胞を形質転換する。しかし、第1ポリペプチド、第2ポリペプチド、および共通の軽鎖ポリペプチド(ヘテロマルチマーの成分)を、別々の発現系で発現させ、発現したポリペプチドをインビトロで結合させることも可能である。
【0114】
ヘテロマルチマーと共通の軽鎖をコードするヌクレオチド(例えば、cDNAまたはゲノムDNA)を、さらなるクローニング(DNA増幅)または発現のために、複製可能なベクターの中に挿入する。多くのベクターを利用することができる。一般的に、ベクターの構成要素として、次のものを一つ以上含むが、これらに限定はされない。すなわち、シグナル配列、複製開始点、一つ以上のマーカー遺伝子、エンハンサー因子、プロモーター、および転写終結配列。
【0115】
ヘテロマルチマーの成分であるポリペプチドは、シグナル配列、または、それ以外で、成熟タンパク質またはポリペプチドのN末端に特異的な切断部位をもつポリペプチドに融合したポリペプチドとして作製することができる。一般的に、シグナル配列は、ベクターの成分であるか、または、ベクターの中に挿入されるDNAの一部であろう。好んで選ばれる異種由来のシグナル配列は、宿主細胞によって認識され、プロセッシングされる(すなわち、シグナルペプチダーゼによって開裂される)配列である。原核生物の宿主細胞では、シグナル配列を、例えば、アルカリホスファターゼ、ペニシリナーゼ、lpp、または熱安定性エンドトキシンIIの先導配列からなるグループから選択される、原核生物のシグナル配列で置き換えることができる。酵母での分泌を行なわせるためには、天然のシグナル配列を、例えば、酵母インベルターゼ、α因子の先導配列(サッカロマイセス(Saccharomyces)とクルイベロマイセス(Kluyveromyces)α因子の先導配列、後者については、1991年4月23日公開の米国特許第5,010,182号に記載されている)、または酸性ホスファターゼの先導配列、C.アルビカンス(C.albicans)のグルコアミラーゼの先導配列(1990年4月4日公開の欧州特許第362,179号)、または、1990年11月15日発行の国際特許公開第90/13646号に記載されているシグナルなどによって置換することができる。哺乳動物細胞で発現させるときには、本来のシグナル配列(例えば、抗体または付着因子のプレ配列で、通常、インビボで、これらの分子をヒト細胞から分泌させる配列)で十分であるが、哺乳動物の別のシグナル配列も適しているし、また、例えば、単純ヘルペスのgDシグナルのような、ウイルスの分泌を促す配列も適当であろう。このような前駆体領域に対するDNAを、読み枠がずれないように、ヘテロマルチマーを形成するポリペプチドをコードするDNAと連結させる。
【0116】
発現ベクターもクローニングベクターも、一つ以上の選ばれた宿主細胞の中でベクターの複製が可能になる塩基配列を含んでいる。一般的に、クローニングベクターにおいては、この配列は、宿主の染色体DNAとは独立に、ベクターが複製できるようにする配列であり、複製開始点または自律複製配列を含む配列である。このような配列は、さまざまな細菌、酵母、およびウイルスでよく知られている。プラスミドpBR322の複製開始点は、ほとんどのグラム陰性菌に適合し、2μプラスミドの開始点は酵母に適合し、また、さまざまなウイルスの開始点(SV40、ポリオーマ、アデノウイルス、VSV、またはBPV)は、哺乳動物細胞でのクローニングベクターとして有用である。一般的に、複製開始点という構成因子は、哺乳動物の発現ベクターには不要である(SV40の複製開始点は、初期プロモーターを含んでいるというだけの理由で一般的には用いられている)。
【0117】
発現ベクターとクローニングベクターは、選抜可能マーカーとも言われる選抜用遺伝子を持っていなければならない。典型的な選抜用遺伝子は、(a)例えば、アンピシリン、ネオマイシン、メトトレキセート、またはテトラサイクリンなどの抗生物質またはその他の毒素に対する抵抗性を付与するか、(b)栄養要求性欠損を相補するか、または(c)例えば、バシラス属(Bacilli)のD−アラニンラセマーゼをコードする遺伝子のように、複合培地からは摂取できない重要な栄養分を供給する。選抜計画の一例では、宿主細胞の増殖を停止させるための薬剤を利用する。異種由来の遺伝子によって形質転換するのに成功した細胞は、薬剤耐性を付与するタンパク質を産生し、それによって、選抜試験を生き残る。このような優性選抜の例では、ネオマイシン(Southernら、J.Molec.Appl.Genet.1:327(1982))、マイコフェノール酸(Mulliganら、Science209:1422(1980))、またはハイグロマイシン(Sugdenら、Mol.Cell.Biol.5:410−413(1985))という薬剤が用いられる。上記の3つの例は、それぞれ、G418もしくはネオマイシン(ゲネチシン(geneticin))、Xgpt(マイコフェノール酸)、またはハイグロマイシンという、適当な薬剤に対する抵抗性を持ち込むために、真核生物の遺伝子の調節下に置かれた細菌遺伝子が用いられている。
【0118】
哺乳動物にとって適当な選抜マーカーのもう一つの例は、DHFR、またはチミジンキナーゼのように、ヘテロマルチマーの塩基配列を取り込む能力のある細胞の同定を可能にする選抜マーカーである。マーカーを取り込んだために、形質転換体だけが、唯一、適応して生き残れるという選択圧の下に、哺乳動物細胞の形質転換体を置く。培地の中の選抜用薬剤の濃度を連続的に変化させ、それによって選抜用遺伝子とヘテロマルチマーをコードするDNAの両方の増幅がもたらされるような条件の下で形質転換体を培養することによって選択圧をかける。増幅されたDNAからは大量のヘテロマルチマーが合成される。この他に増幅可能な遺伝子の例には、メタロチオネイン−Iおよび−2、好ましくは、霊長類のメタロチオネイン遺伝子、アデノシンデアミナーゼ、オルニチンデカルボキシラーゼなどがある。
【0119】
例えば、まず、DHFRの競合的な拮抗剤であるメトトレキセート(Mtx)
を含む培養培地の中で、全部の形質転換細胞を培養して、DHFR選抜用遺伝子によって形質転換された細胞を同定する。野生型のDHFRを用いるときに、適当な宿主細胞は、UrlaubとChasin、Proc.Natl.Acad.Sci.USA77:4216(1980)による説明にしたがって、調製および増殖させたチャイニーズハムスター卵巣(CHO)細胞系のDHFR活性欠損株である。次に、形質転換された細胞を、濃度を上げたメトトレキセートに曝す。これによって、DHFR遺伝子のコピーが多数合成されるようになり、同時に、ヘテロマルチマーの成分をコードするDNAなど、発現ベクターを含むその他の遺伝子のコピーも多数合成されるようになる。この増幅技術は、例えば、ATCCCCL62、CHO−K1などの適当な宿主とともに用いることができ、Mtxに高い耐性をもつ変異DHFR遺伝子を用いるならば、内生的なDHFRがあったとしても構わない(欧州特許第117,060号)。
【0120】
または、ヘテロマルチマー、野生型DHFRタンパク質、およびアミノグリコシド3’−ホスホトランスフェラーゼ(APH)などの選抜マーカーをコードするDNA配列によって形質転換または同時形質転換された宿主細胞(特に、内生DHFRを含む野生型宿主)は、例えば、カナマイシン、ネオマイシン、またはG418などのアミノグリコシド系抗生物質選抜マーカーに対する選抜用薬剤を含む培地の中で細胞を増殖させることによって選抜することができる。米国特許第4,965,199号参照。
【0121】
酵母の中で用いるための適当な選抜用遺伝子は、酵母のプラスミドYRp7の中にあるtrp1遺伝子である。(Stinchcombら、Nature282:39(1979);Kingsmanら、Gene 7:141(1979);または、Tschemperら、Gene10:157(1980))。trp1遺伝子は、トリプトファンの中では増殖できない酵母の変異株、例えば、ATCC44076、またはPEP4−1に対する選抜マーカーを提供する(Jones、Genetics85:12(1977))。酵母宿主ゲノムにtrp1障害があることによって、トリプトファンなしで増殖させることによって、形質転換体を検出するための効果的な環境が提供される。同様に、Leu−2欠損酵母株(ATCC 20,622または38,626)は、Leu2遺伝子をもつ既知のプラスミドによって相補される。
【0122】
さらに、クルイベロマイセス属の酵母を形質転換するためには、1.6μm環状プラスミドpKD1由来のベクターを用いることができる。Bianchiら、Curr.Genet 12:185(1987)。さらに最近、組換えウシキモシンの大量生産のための発現系が報告された。Van denBerg、Bio/Technology8:135(1990)。クルイベロマイセス属の工業用菌株により成熟組換えヒト血清アルブミンを分泌させるための、安定したマルチコピー発現ベクターも開示されている(Fleerら、Bio/Technology 9:968−975(1991))。
【0123】
発現ベクターとクローニングベクターは、通常、宿主生物によって認識され、ヘテロマルチマーの核酸に機能的に結合しているプロモーターを含む。宿主に可能性のある、さまざまな細胞によって認識されるプロモーターが多数ある。制限酵素消化によって、もととなるDNAからプロモーターを除去し、分離したプロモーターをベクターに挿入にして、これらのプロモーターは、ヘテロマルチマーをコードするDNAに機能的に連結する。
原核生物宿主で使用するのに適したプロモーターには、β−ラクタマーゼおよびラクトースのプロモーター系(Changら、Nature275:615(1978);およびGoeddelら、Nature281:544(1979))、アルカリホスファターゼ、トリプトファン(trp)プロモーター系(Goeddelら、Nucleic AcidsRes.,8:4057(1980)、および欧州特許第36,776号)、および、tacプロモーターなどのハイブリッドプロモーター(deBoerら、Proc.Natl.Acad.Sci.USA80:21−25(1983))が含まれる。しかし、細菌のこれ以外の既知のプロモーターが適当である。それらの塩基配列は公開されているので、当業者は、必要な制限酵素部位をもたらすためのリンカーまたはアダプターを用いて、ヘテロマルチマーをコードするDNAに機能的にプロモーターを連結させることができる(Siebenlistら、Cell 20:269(1980))。また、細菌の系で使用するためのプロモーターは、ヘテロマルチマーをコードするDNAに機能的に連結したシャイン−ダルガーノ(S.D.)配列も含んでいる。
【0124】
プロモーターの配列は、真核生物で知られている。実質的にすべての真核生物遺伝子が、転写開始部位から約25から30塩基上流にあるATに富む領域を持っている。多くの遺伝子の転写開始点から上流70から80塩基のところに見つかっている別の配列とはCXCAAT領域である、ただし、Xを任意のヌクレオチドである。殆どの真核生物遺伝子の3’末端には、コーディング配列の3’末端にポリAテールを付加するためのシグナルらしいAATAAA配列がある。これらの配列はすべて、真核生物の発現ベクターの中に適当に挿入する。
【0125】
酵母宿主とともに使用するのに適したプロモーター配列の例には、3−ホスホグリセリン酸キナーゼのプロモーター(Hitzemanら、J.Biol.Chem.255:2073(1980))、または、エノラーゼ、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ、ヘキソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース−6−リン酸イソメラーゼ、3−ホスホグリン酸ムターゼ、ピルビン酸キナーゼ、トリオースリン酸イソメラーゼ、ホスホグルコースイソメラーゼ、およびグルコキナーゼなど、その他の解糖酵素のプロモーター(Hessら、J.Adv.Enzyme.Reg.7:149(1968);および、Holland,Biochemistry17:4900(1978))などである。
【0126】
この他の酵母プロモーターで、増殖条件によって転写を調節するという、さらに別の利点をもつ誘導的プロモーターは、アルコールデヒドロゲナーゼ2、イソシトクロムC、酸性ホスファターゼ、窒素代謝に関係する分解酵素、メタロチオネイン、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ、および、マルトースおよびガラクトース利用に関与する酵素のプロモーター領域である。酵母での発現に用いるのに適したベクターとプロモーターについては、Hitzemanら、欧州特許第73,657A号でさらに説明されている。また、酵母のエンハンサーも、酵母プロモーターとともに適宜用いられる。
哺乳動物の宿主細胞での、ベクターからのヘテロマルチマーの転写は、例えば、ポリオーマウイルス、鶏痘ウイルス(1989年7月5日発行の英国特許第2,211,504号)、アデノウイルス(アデノウイルス2など)、ウシポリオーマウイルス、トリ肉腫ウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルス、および、もっとも好ましくはシミアンウイルス40(SV40)などのウイルスのゲノムから得られたプロモーター、例えば、アクチンプロモーターまたは免疫グロブリンプロモーターなど、異種由来の哺乳動物のプロモーター、または、ヒートショックプロモーターによって調節される。
SV40ウイルスの前者および後者のプロモーターは、都合のよいことに、SV40ウイルスの複製開始点も含むSV40制限酵素フラグメントとして得られる。Fiersら、Nature 273:113 (1978);MulliganとBerg、Science 209:1422−1427(1980);Pavlakisら、Proc. Natl. Acad. Sci. USA 78:7398−7402 (1981)。ヒトサイトメガロウイルス最早期プロモーターは、都合よく、HindIIIE制限酵素フラグメントとして得られる。Greenawayら、Gene 18:355−360 (1982)。ウシポリオーマウイルスをベクターとして用いた、哺乳動物の宿主の中でDNAを発現させるための系が米国特許第4,419,446号に記載されている。
【0127】
この系を改変したものが、米国特許第4,601,978号に記載されている。
【0128】
また、免疫インターフェロンをコードするcDNAのサル細胞での発現については、Grayら、Nature295:503−508(1982);単純ヘルペスウイルス由来のチミジンキナーゼプロモーターの調節下での、ヒトβ−インターフェロンcDNAのマウス細胞における発現については、Reyesら、Nature 297:598−601 (1982);ヒトインターフェロンβ1遺伝子の培養マウス細胞とウサギ細胞における発現についてはCanaaniとBerg、Proc. Natl.Acad. Sci. USA 79:5166−5170 (1982);および、マウス肉腫ウイルスのLTR(長い末端反復配列)をプロモーターとして用いた、CV−1サル腎臓細胞、ニワトリ胚線維芽細胞、チャイニーズハムスター卵巣細胞、HeLa細胞、およひマウスNTH−3T3細胞における、細菌のCAT配列の発現についてはGormanら、Proc.Natl.Acad.Sci.USA 79:6777−6781(1982)も参照。
高等真核生物による、ヘテロマルチマー成分をコードするDNAの転写は、ベクターの中にエンハンサー配列を挿入することによって亢進することがよくある。エンハンサーは、方向や位置に比較的無関係であり、転写単位に対して5’側(Laiminsら、Proc.Natl.Acad.Sci.USA78:993(1981))、3’側(Luskyら、Mol.Cell.Bio.3:1108(1983))イントロンの内部(Banerjiら、Cell33:729(1983))、および、コーディング配列そのものの中(Osborneら、Mol.Cell.Bio.4:1293(1984))にも見つかっている。今では、哺乳動物の遺伝子(グロビン、エラスターゼ、アルブミン、α−フェトプロテイン、およびインシュリン)由来の多くのエンハンサー配列が知られている。しかし、典型的には、真核生物細胞のウイルスに由来するエンハンサーが使用される。例としては、複製開始点の後方側(100−270bp)にあるSV40エンハンサー、サイトメガロウイルス初期プロモーターのエンハンサー、複製開始点の後方側にあるポリオーマエンハンサー、およびアデノウイルスエンハンサーなどがある。また、真核生物のプロモーターを活性化するための促進因子については(Yanivら、Nature297:17−18(1982)を参照。エンハンサーは、ベクターの中のヘテロマルチマーをコードする配列の5’側か3’側の位置に挿入することができるが、好ましくは、プロモーターの5’側の部位に存在する。
【0129】
真核宿主細胞(酵母、菌類、昆虫、植物、動物、ヒト、またはその他の多細胞生物からの有核細胞)で用いられる発現ベクターは、転写の終結、およびmRNAの安定化に必要な配列も含んでいる。このような配列は、真核生物またはウイルスのDNAまたはcDNAの5’側、場合によっては3’側にある非翻訳領域から広く利用することができる。これらの領域には、ヘテロマルチマーをコードするmRNAの非翻訳部位のポリアデニル化フラグメントとして転写されるヌクレオチド分節が含まれる。
【0130】
上記で挙げた構成要素を一つ以上含む適当なベクターを構築するには、標準的な結紮技術を用いる。分離したプラスミドまたはDNAフラグメントは、必要とするプラスミドを作出するのに望ましい形になるように、開裂し、加工してから再連結する。
【0131】
構築したプラスミドの配列が正しいことを確認するための解析を行なうために、結紮混合液を用いて、大腸菌K12菌株294(ATCC31,446)を形質転換して、アンピシリン耐性またはテトラサイクリン耐性によって選抜された、形質転換に成功した株が適正なものである。形質転換からプラスミドを調製して、制限酵素消化によって解析し、および/または、Messingら、Nucleic AcidsRes.,9:309(1981)の方法、またはMaxamら、Methods in Enzymology65:499(1980)の方法によって配列決定を行なう。
【0132】
本発明の実施において特に有用なのは、ヘテロマルチマーをコードするDNAを哺乳動物細胞の中で一過性発現をもたらす発現ベクターである。一般的には、一過性発現には、宿主細胞が多くの発現ベクターのコピーをもち、次いで、発現ベクターによってコードされる所望のポリペプチドを大量に合成するように、宿主細胞の中で効率的に複製することのできる発現ベクターを使用することが含まれる。Sambrookら、上記、pp.16.17−16.22。適当な発現ベクターと宿主細胞を含む一過的発現系によって、クローニングされたDNAによってコードされているポリペプチドを便利に陽性のものを同定することと、所望の結合特性/親和性、または、本発明によって作出される、天然のジスルフィド結合をもたないヘテロマルチマーまたホモマルチマーに対して、所期のゲル泳動度をもつヘテロマルチマーを迅速にスクリーニングすることが可能になる。
【0133】
脊椎動物の組換え細胞培養でヘテロマルチマーを合成するように調整するのに適した、この他の方法、ベクター、および宿主細胞については、Gethingら、Nature 293:620−625(1981);Manteiら、Nature281:40−46(1979);欧州特許第117,060号;および欧州特許第117,058号に記載されている。哺乳動物の培養細胞でヘテロマルチマーを発現させるのに特に役立つプラスミドは、pRK5(欧州特許第307,247号)またはpSV16B(1991年6月13日発行のPCT国際特許公開第91/08291号)である。
【0134】
ヘテロマルチマーを発現させるために、どの宿主細胞系を選択するかは、主に、発現ベクターによって決まる。もう一つ考慮すべきなのは、必要とするタンパク質量である。ミリグラム単位の量を、一過的形質転換によって産生することができる。例えば、アデノウイルスEIA−によって形質転換された293ヒト胚腎臓細胞系を、ヘテロマルチマーの効率的な発現が可能になるよう、リン酸カルシウム法を修正した方法によって、pRK5を基本とするベクターで一過的に形質転換することができる。CDM8を基本とするベクターを用いて、DEAE−テキストラン法によってCOS細胞を形質転換することができる(Aruffoら、Cell 61:1303−1313(1990);および、Zettmeisslら、DNA CellBiol.(US)9:347−353(1990))。より大量のタンパク質が必要ならば、宿主細胞系を安定的に形質転換させた後、免疫付着因子を発現させることができる。例えは、pRK5を基本とするベクターを、ジヒドロ葉酸レダクターゼ(DHFR)をコードする別のベクターの存在下で、チャイニーズハムスター卵巣(CHO)細胞の中に導入して、G418に対する抵抗性を付与することができる。G418に抵抗性のクローンは、培養によって選抜することができる。これらのクローンを、DHFRのインヒビターであるメトトレキセートの量を増加させながら培養して、DHFRとヘテロマルチマーの配列をコードする遺伝子のコピー数が共に増幅されているクローンを選抜する。免疫付着因子が、N末端に疎水性の先導配列をもっていれば、形質転換された細胞によって加工されて分泌される可能性が高い。より複雑な構造をもつ免疫付着因子の発現には、特に適した宿主細胞が必要となろう。例えば、軽鎖またはJ鎖などの構成成分は、一定のミエローマまたはハイブリドーマの宿主細胞によって提供されよう(Gascoigneら、上記;およびMartinら、J.Virol.67:3561−3568(1993))。
【0135】
本明細書のベクターをクローニングし、発現させるのに適した、この他の宿主細胞は、上記のような原核生物、酵母、または、高等真核生物の細胞である。この目的に適した原核生物には、グラム陰性またはグラム陽性生物などの真性細菌で、例えば、腸内細菌である、例えば、大腸菌(E.coli)などのエシェリキア属(Escherichia)、エンテロバクター属(Enterobacter)、エルウィニア属(Erwinia)、クレブシアラ(Klebsiella)、プロテウス属(Proteus)、例えば、ネズミチフス菌(Salmonella typhimurium)などのサルモネラ属(Salmonella)、霊菌(Serratiamarcescans)などのセラティア属(Serratia)、およびシゲラ属(Shigella)、ならびに、枯草菌(B.subtilis)およびB.リケニフォルミス(B.licheniformis)(例えば、1989年4月12日公開のDD266,710で開示されているB.リケニフォルミス41P)などのバシラス属、緑膿菌(P.aeruginosa)などのシュードモナス属、およびストレプトマイセス属(Streptomyces)などである。好ましい大腸菌クローニング宿主の一つは大腸菌294(ATCC 31,446)であるが、その他、大腸菌B、大腸菌X1776(ATCC31,537)、および大腸菌W3110(ATCC27,325)などの菌株も適している。これらの例は、例示のためのものであり、制限のためのものではない。菌株W3110は、組換えDNA産物の発酵のための一般的な宿主菌株であるため、この菌株は、特に好ましい宿主、または宿主の親株である。好ましくは、宿主細胞は、最小量のタンパク質分解酵素を分泌するものでなければならない。例えば、W3110株を改変してタンパク質をコードする遺伝子に遺伝的変異を生じさせることができる。そのような宿主の例として、大腸菌W3110の27C7株などがある。27C7の完全な遺伝子型は、tonAΔptr3 phoAΔE15 Δ(argF−lac)169ompTΔdegP41kanrである。27C7株は、1991年10月30日に、ATCC番号55,244として、アメリカンタイプカルチャーコレクションに寄託された。または、1990年8月7日発行の米国特許第4,946,783号で開示されている、ペリプラズマプロテアーゼの変異体をもつ大腸菌の菌株を用いることもできる。または、例えば、PCR、または、別の核酸のポリメラーゼ反応などのクローニング法が適している。
【0136】
原核生物の他に、糸状菌類または酵母のような真核生物微生物が、ヘテロマルチマーをコードするベクターに適したクローニングおよび発現のための宿主である。サッカロマイセス・セレビシアエ(Saccharomyces cerevisiae)、すなわち一般的なパン酵母が、下等な真核生物宿主微生物の中で、もっとも一般的に用いられている。しかし、本明細書においては、分裂酵母(Schizosaccharomyces pombe)(BeachとNurse、Nature290:140(1981);1985年5月2日発行の欧州特許第139,383号);クルイベロマイセス属の宿主(米国特許第4,943,529号;Fleerら、上記)で、例えば、K.ラクティス(K.lactis)(MW98−8C,CBS683,CBS4574;Louvencourtら、J.Bacteriol.,737(1983))、K.フラジリス(K.fragiles)(ATCC 12,424)、K.ブルガリクス(K.bulgaricus) (ATCC 16,045)、K.ウィケラミイ(K.wicheramii)(ATCC24,178)、K.ワルティイ(K.waltii)(ATCC 56,500)、K.ドロソフィラルム(K.drosophilarum)(ATCC36,906;Van den Bergら、上記)、K.テルモトレランス(K.thermotolerans)、およびK.マルキサヌス(K.marxianus)など;ヤロウィア属(yarrowia)(欧州特許第402,226号);ピキア・パストリス(Pichiapastoris)(欧州特許第183,070号;Sreekrishnaら、J.BasicMicrobiol.28:265−278(1988));カンジダ属;トリコデルマ・レエシア(Trichodermareesia)(欧州特許第244,234号);ニューロスポラ・クラッサ(Neurosporacrassa)(Caseら、Proc.Natl.Acad.Sci.USA 76:5259−5263(1979));シュワニオマイセス・オクシデンタリス(Shwanniomycesoccidentalis)などのシュワニオマイセス属(1990年10月31日発行の欧州特許第394,538号);および、例えば、ニューロスポラ属、ペニシリウム属(Penicillium)、トリポタラジウム属(Tolypocladium)などの糸状菌(1991年1月10日発行の国際特許公開第91/00357号)、ならびに、アウペルギルス属(Aspergillus)宿主、例えば、A.ニドランス(A.nidulans) (Ballanceら、Biochem.Biophys.Res.Commun.112:284−289(1983);Tilburnら、Gene 26:205−221(1983);Yeltonら、Proc.Natl.Acad.Sci.USA81:1470−1474(1984)、およびA.ニガー(A. niger)(KellyとHynes,EMBOJ.4:475−479(1985))など、この他数多くの属、種、および株を用いることが可能であり、また有用でもある。
【0137】
グリコシル化されたヘテロマルチマーを発現させるのに適した宿主細胞は、多細胞生物由来のものである。そのような宿主細胞は、複雑なプロセッシングを行なうことができ、グリコシル化活性をもつ。原則として、脊椎動物由来でも、無脊椎動物由来でも、どのような高等真核生物の細胞培養を使うこともできる。無脊椎動物細胞の例としては、植物細胞、昆虫細胞などがある。多数のバキュロウイルス株とその変異株、および対応する昆虫宿主許容細胞として、スポドプテラ・フルギペルダ(Spodoptera frugiperda)(毛虫)、アエデス・アエギプチ(Aedesaegypti)(蚊)、アエデス・アルボピクタス(Aedes albopictus)(蚊)、ドロソフィラ・メラノガスター(Drosophilamelanogaster)(ショウジョウバエ)、およびボムビクス・モリ(Bombyxmori)などか確認されている。例えば、Luckowら、Bio/Technology6:47−55(1988);Setlowら編、遺伝子工学(GeneticEnginerring)第8巻より、Millerら、pp.277−279(プレナムパブリッシング社(PlenumPublishing)、1986年);および、Maedaら、Nature 315:592−594(1985)を参照。例えば、アウトグラファ・カリフォニカ(Autographacalifonica)NPVのL−1変異株、およびカイコNPV(Bombyx moriNPV)のBm−5株など、形質転換用のさまざまなウイルス株が一般に利用可能であり、そのようなウイルスは、本明細書において、本発明に記載されているウイルスとして、特に、スポドプテラ・フルギペルダ(spodoptera frugiperda)細胞の形質転換に用いることができる。
【0138】
ワタ、トウモロコシ、バレイショ、ダイズ、ペチュニア、トマト、およびタバコの植物培養細胞を宿主として利用することができる。典型的には、予め、ヘテロマルチマーDNAを持つように操作してあるアグロバクテリウム・ツメファシエンス(Agrobacteriumtumefaciens)菌の一定の菌株とインキュベートして、植物細胞を形質転換する。植物培養細胞をA.ツメファシエンスとインキュベートしている間に、ヘテロマルチマーをコードしているDNAが植物細胞宿主に移行して、それを形質転換し、適当な条件の下でヘテロマルチマーを発現させる。さらに、ノパリンシンターゼプロモーター、およびポリアデニル化シグナル配列のように、植物細胞と親和性のある調節配列とシグナル配列を利用することができる。Depickerら、J.Mol.Appl.
Gen.1:561(1982)。なお、T−DNA780遺伝子の上流領域から分離したDNA分節は、組換えDNAを含む植物組織において、植物で発現する遺伝子の転写レベルを活性化あるいは上昇させることができる。1989年6月21日発行の欧州特許第321,196号。
【0139】
好ましい宿主は脊椎動物の細胞であり、脊椎動物の細胞を培養して増殖させること(組織培養)は、近年では日常的な処理になっている(アカデミックプレス社 (Academic Press)、KruseとPatterson編、組織培養(TissueCulture)(1973))。有用な哺乳動物の宿主細胞系の例は、SV40で形質転換されたサル腎臓CV1系統(COS−7,ATCC CRL1651);ヒト胚腎臓系統(293、または、懸濁培養で増殖させるためにサブクローニングされた293細胞、Grahamら、J.Gen.Virol.36:59(1977));仔ハムスター腎臓細胞(BHK、ATCC CCL10);チャイニーズハムスター卵巣細胞/−DHFR(CHO、UrlaubとChasin、Proc.Natl.Acad.Sci.USA77:4216(1980));マウスセルトリ細胞(TM4、Mather,Biol.Repod.23:243−251(1980));サル腎臓細胞 (CV1 ATCC CCL 70);アフリカミドリザルの腎臓細胞(VERO−76、ATCCCRL−1587);ヒト子宮頚部ガン細胞(HELA、 ATCC CCL 2);イヌ腎臓細胞(MDCK,ATCC CCL34);バッファローラット肝細胞(BRL 3A,ATCCCRL 1442);ヒト肺細胞(W138,ATCC CCL 75);ヒト肝臓細胞(Hep G2,HB 8065);マウス乳ガン(MMT 060562,ATCC CCL 51);TRI細胞(Matherら、Annals N.Y.Acad.Sci.383:44−68(1982));MRC5細胞;FS4細胞;およびヒトヘパトーマ系(Hep G2)である。
【0140】
宿主細胞を本発明の上記発現ベクターまたはクローニングベクターによって形質転換して、プロモーターを誘導したり、形質転換体を選抜したり、または所望の配列をコードする遺伝子を増幅するのに適合するように修正した通常の培地で培養する。用いる宿主細胞によって、その細胞に適した標準的な技術を用いて形質転換を行なう。Sambrookら、上記の1.82節で説明されているように、一般的には、実質的な細胞壁という障壁を含む、原核生物またはその他の細胞に対して、塩化カルシウムを用いたカルシウム処理、またはエレクトロポレーションを用いる。Shawら、Gene23:315(1983)、および1989年6月29日発行の国際特許公開第89/05859号に記載されているようにして、アクロバクテリウム・ツメファシエンスによる感染を、一定の植物細胞を形質転換するために用いる。さらに、1991年1月10日発行の国際特許公開第91/00358号に記載されているようにして、超音波を用いて、植物を形質転換することができる。
【0141】
このような細胞壁をもたない哺乳動物細胞については、GrahamとvanderEb、Virology52:456−457(1978)のリン酸カルシウム法が好ましい。哺乳動物細胞宿主系の形質転換の一般的な態様を、1983年8月16日発行の米国特許第4,399,216号において、Axelが記述している。酵母への形質転換は、一般的には、VanSolingenら、J.Bact.130:946(1977)、およびHsiaoら、Proc.Natl.Acad.Sci.USA76:3829(1979)の方法にしたがって行われる。DNAを細胞の中に導入するために、核へのマイクロインジェクション、エレクトロポレーション、無処理の細胞とバクテリアのプロトプラストとの融合、または、例えば、ポリブレン、ポリオルニチンなどのポリカチオンのような別の方法を用いることもできる。哺乳動物細胞を形質転換するためのさまざまな技術については、Keownら、Methods in Enzymology(1989)、Keownら、Methods inEnzymology 185:527−537(1990)、およびMansourら、Nature 336:348−352(1988)を参照。
【0142】
本発明のヘテロマルチマーポリペプチドを産生するために用いた原核生物細胞を、Sambrookら、上記において、一般的に説明されているところにしたがって、適当な培地で培養する。
【0143】
本発明のヘテロマルチマーを産生するために用いた哺乳動物宿主細胞は、さまざまな培地出培養することができる。宿主細胞を培養するには、Ham’sF10(シグマ社(Sigma))、最小必須培地((MEM)、シグマ社)、RPMI−1640(シグマ社)、およびダルベッコの修正イーグル培地 ((DMEM)、シグマ社)などの市販の培地が適当である。さらに、それらの開示内容がすべて、参照してここに組み込まれる、HamとWallace,Meth.Enz.58:44(1979)、BarnesとSato、Anal.Biochem.102:255(1980)、米国特許第4,767,704号;第4,657,866号;第4,927,762号;または、第4,560,655号;国際特許公開第90/03430号;国際特許公開第87/00195号;米国特許第4,791,192号;および米国特許(U.S.Patent Re.)第30,985号;または、米国特許第5,122,469号に記載されている培地のいずれかを、宿主細胞用の培養培地として用いることができる。これらの培地はどれも、必要ならは、ホルモンおよび/またはその他の増殖因子(インシュリン、トランスフェリン、または上皮増殖因子など)、塩類(塩化ナトリウム、カルシウム、マグネシウム、およびリン酸)、緩衝液(HEPESなど)、ヌクレオシド(アデノシンおよびチミジンなど)、抗生物質(登録商標ゲンタマイシン薬剤)、微量要素(最終濃度がマイクロモルの単位で存在する無機化合物と定義される)、およびグルコースまたは同等のエネルギー源を添加することもできる。この他の添加物も、当業者に知られている適当な濃度で含むことができる。温度、pHなどの培養条件は、発現させるために選ばれた宿主細胞によって以前用いられた条件であり、当業者にとっては明らかである。
【0144】
一般的に、哺乳動物の細胞培養の生産性を最大にするための原理、プロトコール、および実際の技術については、IRLプレス社(IRLPress)、1991年、M.Butler編、哺乳動物細胞のバイオテクノロジー:実際的方法(Mammalian CellBiotechnology:aPractical Approach)に書かれている。
【0145】
本開示で言及されている宿主細胞は培養細胞だけでなく、宿主動物の中の細胞も含む。
4.ヘテロマルチマーの回収ヘテロマルチマーは、好ましくは、一般的に、分泌ポリペプチドとして、培養培地から回収するが、分泌シグナルなしに、直接産生されたときには、宿主細胞溶解物から回収する。ヘテロマルチマーに膜結合があるときには、適当な界面活性剤(例:Triton−X 100)を用いて膜から解離することができる。
【0146】
ヒトに由来しない組換え細胞の中でヘテロマルチマーが産生されたときには、ヒト由来のタンパク質やポリペプチドを全く含まない。しかし、組換え細胞のタンパク質やポリペプチドからヘテロマルチマーを精製して、ヘテロマルチマーについて実質的に均質な調製物を得る必要がある。最初の工程として、培養培地または細胞溶解物を普通に遠心分離して、微粒子化した細胞残滓を取り除く。
【0147】
抗体の定常部位をもつヘテロマルチマーは、ハイドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析、またはアフィニティークロマトグラフィーによって、都合のよいように精製することができるが、アフィニティークロマトグラフィーが好ましい精製技術である。ヘテロマルチマーがCH3部位を含むときは、登録商標ベーカーボンド ABX(Bakerbond ABX)樹脂(J.T.Baker,ニュージャージー州フィリップスバーグ (Phillipsburg))が、精製するために有用である。この他に、イオン交換カラムによる分画、エタノール沈殿、逆相HPLC、シリカによるクロマトグラフィー、ヘパリンセファロースによるクロマトグラフィー、陰イオンまたは陽イオン交換樹脂(ポリアスパラギン酸カラムなど)によるクロマトグラフィー、クロマトフォーカシング、SDS−PAGE、および硫安沈殿などのタンパク質精製技術も、回収するポリペプチドによっては利用可能である。親和性リガンドとしてのプロテインAの適合性は、キメラで用いられた免疫グロブリンFcドメインの種類およびアイソタイプによって異なる。プロテインAを用いて、ヒトγ1、γ2、またはγ4のH鎖に基づいて、免疫付着因子を精製することができる (Lindmarkら、J.Immunol.Meth.62:1−13(1983))。マウスのアイソタイプのすべてとヒトγ3(Gussら、EMBOJ.5:1567−1575(1986))には、プロテインGが推奨されている。親和性リガンドが結合する基質は、もっとも普通にはアガロースであるが、その他の基質を用いることもできる。調節された多孔性ガラスビーズ(controlledporeglass)またはポリ(スチレンジビニル)ベンジンのような物理的に安定した基質なら、アガロースを用いるよりも流速が早くなり、短い処理時間でできる。プロテインAまたはGアフィニティーカラムに免疫付着因子を結合させる条件は、Fc部位の特徴によって全面的に決定される。すなわち、その分子種とアイソタイプによって決まる。一般的には、適正なリガンドが選択されれば、直接、条件付けていない培養溶液からでも、効率的な結合が起こる。免疫付着因子の顕著な特徴の一つは、ヒトγ1分子で、同じFc型の抗体に較べて、プロテインAに対する結合能力が幾分低下することである。結合した免疫付着因子は、酸性pH(3.0かそれ以上)で、または、僅かにカオトロピックな塩を含む中性pHの緩衝液の中で効率的に溶出することできる。このアフィニティークロマトグラフィー工程によって、>95%純粋なヘテロマルチマー調製物を作製することができる。
【0148】
5.共通の軽鎖をもつヘテロマルチマーの多官能性抗体の使用ヘテロマルチマーについては、治療薬としての多くの応用が考えられる。例えば、細胞傷害性を切り換える(例:腫瘍細胞を殺す)ために、ワクチンのアジュバントして、血栓溶解剤を血餅まで輸送するために、標的部位(例えば、腫瘍)で、酵素的に活性化されたプロドラッグを転化するために、感染病を治療するために、免疫複合体を細胞表面へと向かわせるために、または、腫瘍細胞にイムノトキシンを輸送するためにヘテロマルチマーを用いることができる。例えば、クラスII主要組織適合遺伝子複合体であるモデル内皮抗原を標的とすることによって、抗体−リシンイムノトキシンによって、腫瘍の脈管構造を標的とすることができた(Burrows,F.J.とThorpe,P.E.(1993)Proc.Natl.Acad.Sci.USA 90:8996−9000)。
【0149】
抗−内皮イムノトキシンを別のイムノトキシンと組み合わせることによって、腫瘍細胞そのものに対する有意に高い効能が得られた(Burrows,F.J.とThorpe,P.E.(1993)上記)。最近、双特異的抗体を用いて、組織因子を腫瘍脈管構造に向かわせることに成功し、かなり高い抗腫瘍効果を生じる局所的な血栓症を引き起こした(Huang,Xら、(1997)Science275:547−550)。さらに、双特異的抗体を用いて、細胞傷害性T細胞に、インビトロで、標的乳ガン細胞およびB細胞リンパ腫細胞を殺させることに成功した(Zhu,Z.ら、(1996)Bio/Technology14:192−196;およびHolliger,P.ら、(1996)Protein. Engin.9:299−305)。
【0150】
望ましい精製度をもつヘテロマルチマーを、選択的に、生理学的に許容できる担体、賦形剤、または安定剤(レミントンの製薬科学(Remington’s Pharmaceutical Science)、第16版、Osol,A.編(1980))と混合して、凍結乾燥した固形または水溶液の形状で保存するために、ヘテロマルチマーの治療用処方剤を調製する。許容される担体、賦形剤、または安定剤は、用いられる投薬量と濃度では受容者にとって非毒性で、リン酸、クエン酸、およびその他の有機酸などを含む緩衝液;アスコルビン酸などの抗酸化剤;低分子量(約10残基未満)ポリペプチド;血清アルブミン、ゼラチン、または免疫グロブリンなどのタンパク質;ポリビニルピロリドンなどの疎水性ポリマー;グリシン、グルタミン、アスパラギン、アルギニン、またはリシンなどのアミノ酸;単糖類;二糖類;および、グルコース、マンノース、またはデキストリンなどの糖類;EDTAなどのキレート化剤;マンニトール、またはソルビトールなどの糖アルコール;ナトリウムなどの塩を形成する対イオン;および/または、トィーン(Tween)、プルロニクス(Pluronics)またはポリエチレングリコール(PEG)などの界面活性剤を含んでいる。
【0151】
ヘテロマルチマーは、例えば、コアセルベート化技術、または界面ポリマー化(例えば、ヒドロキシメチルセルロース、または、それぞれ、ゼラチン−マイクロカプセルおよびポリ−[メチルメトアシレート]マイクロカプセル)によって調製されたマイクロカプセルの中に取り込んだり、コロイド剤送達システム(例えば、リポソーム、アルブミンミクロスフィア、ミクロエマルジョン、ナノ粒子、およびナノカプセル)に取り込んだり、または、マクロエマルジョンの中に取り込んだりすることができる。このような技術については、レミントンの製薬科学(Remington’s PharmaceuticalScience)、上記で開示されている。
【0152】
インビボでの投与に用いるヘテロマルチマーは、滅菌されていなければならない。これは、凍結乾燥および再構成の前か後に、滅菌フィルター膜で濾過すれば簡単に行なえる。ヘテロマルチマーは、通常、凍結乾燥状態か水溶液の状態で保存する。
【0153】
治療用ヘテロマルチマー組成物は、一般的に、滅菌的に接触することのできる通路をもった容器、例えば、静脈注射用のバック、または皮下注射用の注射針によって貫通することのできる栓をもったバイアルの中に置かれる。
【0154】
ヘテロマルチマーの投与経路は、例えば、静脈、腹腔内、脳内、筋肉内、眼内、動脈内、または病巣内などの経路による注射または点滴のような既知の方法、または下記に示すような徐放性システムによる。ヘテロマルチマーは、点滴またはボーラス注射によって、連続的に投与する。
【0155】
徐放性調製物の適当な例は、タンパク質を含む固体疎水性ポリマーの半透過性基質で、例えば、フィルムまたはマイクロカプセルのような形のあるものになっている基質を含む。徐放性基質の例は、Langerら、J.Biomed.Mater.Res.15:167−277(1981)、およびLanger、Chem.Tech.12:98−105(1982)に記載されているポリエステル、ヒドロゲル(例えば、ポリ(2−ヒドロキシエチル−メトアクリル酸)、またはポリ(ビニルアルコール)、ポリアクチド(米国特許第3,773,919号、欧州特許第58,481号)、L−グルタミン酸とガンマエチル−L−グルタミン酸のコポリマー(Sidmanら、Biopolymers 22:547−556(1983))、非分解性エチレン−ビニル酢酸(Langerら、上記)(欧州特許第133,988号)、登録商標ルプロンデポット(LupronDepot)のような分解性の乳酸−グリコール酸コポリマー(乳酸−グリコール酸コポリマーと酢酸ロイプロリドからなる注射用ミクロスフィア)、およびポリ−D−(−)−3−ヒドロキシ酪酸(欧州特許第133,988号)などがある。
【0156】
エチレン−ビニル酢酸、および乳酸−グリコール酸などのコポリマーは、100日間にわたって分子を放出することが可能であるが、ある種のヒドロゲルでは、タンパク質を放出する期間が短くなる。カプセルに入ったタンパク質は、長時間体内に残留して、37℃で湿気に曝される結果、変性するか凝集して、生物学的活性を失うことになり、免疫原性にも変化が生じうる。関係するメカニズムによって、タンパク質を安定化させるための合理的な方策を考え出すことができる。例えば、凝集を起こすメカニズムが、チオ−ジスルフィド基の交換による分子内S−S結合形成であることが発見されていれば、スルフィドリル残基を修飾し、酸性溶液から凍結乾燥し、水分含有量を調節し、適当な添加剤を使用し、また、特異的なポリマー基質組成物を開発することによって、安定させることができる。
【0157】
また、徐放性ヘテロマルチマー組成物は、リポソームによって取り込まれたヘテロマルチマーでもよい。ヘテロマルチマーを含むリポソームは、それ自体は既知の方法:独国特許第3,218,121号;Epsteinら、Proc.Natl.Acad.Sci.USA 82:3688−3692(1985);Hwangら、Proc.Natl.Acad.Sci.USA77:4030−4034(1980);欧州特許第52,322号;欧州特許第36,676号;欧州特許第88,046号:欧州特許第143,949号;欧州特許第142,641号;日本国特許出願第83−118008号;米国特許第4,485,045号および第4,544,545号;ならびに欧州特許第102,324号によって調製される。通常、リポソームは、小さな(200−800オングストローム)均一ラメラ型で、脂質成分が約30mol.%コレステロールよりも高く、ヘテロマルチマー治療を最適なものにするために、選ばれた部位が調整されている。
【0158】
治療に用いるヘテロマルチマーの効果的な用量は、例えば、治療目的、投与経路、および患者の症状によって変わる。したがって、至適な治療効果を得るためには、治療者が用量を希釈したり、投与経路を変更することが必要となろう。典型的な毎日の投与量は、上記の要素によって、約1μg/kgから10mg/kgかそれよりも多い量になろう。典型的には、臨床医は、所期の効果を納めるまで、ヘテロマルチマーの投薬を続けることになろう。この治療の進行状況は、常法によって容易に観察される。
【0159】
本明細書に記述されているヘテロマルチマーは、酵素免疫測定法で用いることもできる。それを行なうためには、結合によって、酵素を阻害しないために、ヘテロマルチマーの一方のアームを、酵素上の特異的なエピトープに結合するように設計することができ、ヘテロマルチマーのもう一方のアームを、所望の部位で、酵素が確実に高濃度になるようにしながら、固定基質に結合するように設計することができる。このような診断用ヘテロマルチマーの例としては、IgG、およびフェリチンに対する特性を持ったヘテロマルチマー、および、ホースラディッシュパーオキシダーゼ(HRP)、ならびに、例えば、ホルモンに対する結合特異性をもつヘテロマルチマーなどがある。
【0160】
このヘテロマルチマーは、2部位免疫測定法に用いるよう設計することもできる。例えば、双特異的な2つのヘテロマルチマーを産生させて、解析するタンパク質上の2つの別々のエピトープに結合させるが、一方のヘテロマルチマーは、不溶性の基質と複合体になるように結合し、もう一方は、指示酵素に結合する。
【0161】
また、ヘテロマルチマーは、ガンなど、さまざまな病気のインビトロおよびインビボでの免疫診断に用いることもできる。この診断への使用を容易にするために、ヘテロマルチマーの一方のアームを腫瘍関連抗原に結合するように設計し、もう一方のアームを検出用マーカー(例えは、放射性核種に結合するキレート剤)に結合するように設計することができる。例えば、腫瘍関連抗原CEA、および二価性ハプテンに対して特異性をもつヘテロマルチマーは、結腸直腸ガンと甲状腺ガンを画像化するために用いることができる。この他にも、ヘテロマルチマーを、治療ではなく診断に使用できることは当業者にとって明らかであろう。
【0162】
診断に応用するためには、ヘテロマルチマーの少なくとも一方のアームが、典型的には、直接的または間接的に、検出可能な部分によって標識されている。例えば、検出可能な部分は、3H、14C、32P、35S、または125Iなどの放射性同位元素;フルオロセインイソチオシアナート、ローダミン、またはルシフェリンなどの蛍光化合物または化学発光化合物;または、アルカリホスファターゼ、β−ガラタトシダーゼ、またはホースラディッシュパーオキシダーゼ(HRP)などの酵素であろう。
【0163】
ヘテロマルチマーを検出可能な部分に別々に結合させるために、当技術分野において既知の方法を用いることができるが、そのような方法には、Hunterら、Nature 144:945(1962);Davidら、Biochemistry13:1014(1974);Painら、J.Immunol.Meth.40:219(1981);および、Nygen,J.Histochem.and Cytochem.30:407(1982)に記載されている方法などがある。
【0164】
本発明のヘテロマルチマーは、競合結合アッセイ法、直接および間接サンドイッチアッセイ法、および免疫沈殿法など、既知のアッセイ法に用いることができる。Zola、モノクローナル抗体:技術マニュアル(Monoclonal Antibodies:A Manual ofTechniques),pp.147−158(CRCプレス社(CRC Press,Inc.,1987)。
【0165】
競合結合アッセイ法は、限られた量のヘテロマルチマーへの結合をめぐって、標識された標準化合物が、解析対象の試験用サンプルと競合できることに基づいている。試験サンプル中の解析化合物の量は、ヘテロマルチマーに結合した標準化合物の量に反比例する。結合した標準化合物の量を容易に決定するために、ヘテロマルチマーは、一般的に、競合の前か後に不溶化して、ヘテロマルチマーに結合している標準化合物と解析化合物が、非結合のままの標準化合物と解析化合物とから都合良く分離できるようにする。
【0166】
このヘテロマルチマーは、それぞれが、検出すべきサンプルの異なった免疫原部分またはエピトープに結合することのできる2種類の分子を使用することを含むサンドイッチアッセイ法にとって、特に有用である。サンドイッチアッセイ法において、試験用サンプルの解析化合物は、固体担体上に固定されたヘテロマルチマーの一方のアームに結合していて、その後、ヘテロマルチマーのもう一方のアームが解析化合物に結合して、3つの部分からなる不溶性の複合体を形成する。例えば、米国特許第4,376,110号を参照。ヘテロマルチマー自体の第2のアームを、検出可能部分によって標識する(直接的サンドイッチアッセイ法)か、または、検出可能部分によって標識された抗免疫グロブリン抗体を用いて測定することができる(間接的サンドイッチアッセイ法)。例えば、サンドイッチアッセイ法の一つのタイプは、ELISA法であるが、この場合には、検出可能部分は酵素である。
【0167】
以下は、本発明を実施するための特定の実施態様の実施例である。実施例は、例示目的のだけに示されているのであって、いかなる意味でも、本発明の範囲を制限するためのものではない。
【0168】
本明細書において引用されている刊行物、特許、および特許出願は、上記のものも下記のものもすべてが参照してここに組み込まれる。
【実施例】
【0169】
計画がFc-含有BsAb(図1C)を製造するために提供される。この計画において、我々はそれがヘテロダイマー化であるがホモダイマー化されないように抗体重鎖のCH3を工作している。これは、論理的設計(Ridgwayら,上記(1996))及びここに記載した通りのファージディスプレー選択によって得られた立体的相補変異と共同してCH3ドメイン中に相互-鎖ジスルフィド結合を設けることによって達成された。両方の抗原結合特異性用の単軽鎖の使用は、軽鎖誤対合の問題を避ける(図1A- 1C)。同じ軽鎖を持った抗体は、非常に大きなヒトscFvライブラリーを選別することによって容易に分離された(Vaughan,T.J.ら, (1996)上記)。
【0170】
実施例1:
空洞への隆起(protuberance-into-cavity)ヘテロマルチマー免疫付着因子の生成 Chamowら,J.Imminol.153:4268(1994)により先に記載されたヒト化抗-CD3/CD4-IgGキメラは、回収できたヘテロマルチマーのパーセンテージを最大化するように構築した。空洞への隆起及び野生型CH3変異体は、ヒト化抗体-免疫付着因子キメラ(Ab/Ia)抗-CD3/CD4-IgGの形成を導くその能力を比較した。
【0171】
かくして変異体は、ヒト化抗-CD3抗体重鎖のCH3ドメインに及びミスマッチオリゴヌクレオチドを用いたサイト-誘導変異誘発(Kunkelら,Methods Enzymol.154:367(1987)とP.Carter,in Mutagenesis:a Practical Approach,M.J.McPherson,Ed.,IRL Press,Oxford,UK,pp.1-25(1991))によってCD4-IgG中で構築され、且つジデオキシヌクレオチド配列によって実証した (Sangerら,Proc.Natl.Acad.Sci.USA 74:5463(1977))。次の表3を参照
【0172】
【表3】
【0173】
残基T366は、パートナーCH3ドメイン上の残基Y407の水素結合間隔内にある。実際は、残基T366への主要な分子間接触は、残基Y407に対する及び逆の場合も同じである。一つの空洞への隆起ペアは、一つのCH3ドメイン中のT366Yとそのパートナードメイン中のY407Tの相互の変異を持ったこれらの残基を逆転することによって創製され、かくしてその界面で側鎖の容量が維持される。変異は、Kabat数付与システム(Kabatら,(1991)上記)を用いてその位置の次の野生型残基によって、そして一文字コードで残基の置換が定義される。複数変異は、コロンによって単一変異成分をリスト化することによって定義される。
【0174】
抗-CD3軽鎖(L)と重鎖(H)変異体(Shalabyら,J.Exp.Mcd.175:217(1992)とRodriges ら,Int.J.Cancer(上記)7:45(1992))をコードしているファージミドは、以前に記載された通り(Chamow ら,J.Immunol.153:4268(1994))、ファージミドをコードしているCD4-IgG変異体(Byrnら,Nature 344:667(1990))と一緒に、ヒト胎児性腎細胞,293S中に共移入(co-transfected)した。移入されたファージミドDNAの全量は固定した一方、異なるDNAの比率はAb/Iaキメラの生産を最大化するように変えた。投入DNAs(15μgトータル)Ia:重鎖:軽鎖の割合(質量で)は以下の通り:8:1:3;7:1:3;6:1:3;5:1:3;4:1:3;3:1:3;1:0:0。
【0175】
その生産物は、レーザーデンシトメトリースキャニングに続くSDS-PAGEによる分析の前にスタヒロコッカス蛋白質(ProSep A,BioProcessing Ltd,UK)を用いてアフィニティー精製した。重鎖DNAを超える過剰軽鎖は、制限化されるものからの軽鎖を回避するために用いた。生産物の同定は、アミノ末端配列決定に続きPVDF膜(Matsudaira,J.Biol.Chem.262:10035(1987))での電気ブロッティングによって証明した。
【0176】
重鎖及び野生型CH3を組み込んでいるIa用のそれと一緒に軽鎖用のファージミドの共移入は、予期した通り、Ab/Iaキメラ,IgGとIaホモダイマー生産物の混合物となる(Chamowら,J.Immunol.153:4268(1994))。抗体重鎖プラス軽鎖をコードしている投入DNAのより大きなフラクション又はホモダイマーに一致するより高いフラクションを回収した。Ia:H:Lの6:1:3の投入DNA比では、Iaホモダイマー(22.5%)とIgG(23.0%)の類似フラクションと共に54.5%のAb/Iaキメラを生じた。これらの比率は、分析方法によって導かれるバイアスを伴わず重鎖のランダムな組合せに続く各鎖の等モル発現から予期されたそれらと良く一致する:50%Ab/Iaキメラ、25% Iaホモダイマー及び25% IgG。
【0177】
野生型CH3を含有している鎖に対して、Ab/Iaキメラは、Y407T空洞とT366Y隆起変異をそれぞれ抗-CD3重鎖とCD4-IgGIaに含有せしめた共移入体から92%までの収率で回収された。抗体/免疫付着因子キメラの同様の収量は、もしこれらの相互変異が重鎖の隆起及びIa中の空洞により設けられたならば得られた。両方のケースにおいてモノマーは、隆起を含むが空洞のないその鎖で観測された。いずれか1つの理論に制限することなしに、T366Y隆起がY407T空洞よりもホモダイマー形成のためにより分裂されると確信する。Ab/Iaハイブリッドのフラクションは、隆起と空洞(Ab T366W,IaY407A)の両者のサイズを増加することによって有意に変更されることはない。第2の隆起と空洞ペア(Ab F405A,IaT394W)は、ホモダイマー化するIaT394W隆起変異体の予期しない傾向を補うための投入DNAの小さいフラクションを用いてAb/Iaの71%までを生じた。2つの独立した空洞への隆起変異体ペア(Ab T366Y:F405A、Ia T394W:Y407T)の組合せは、Ab T366Y,IaY407Tペア以上にはAb/Iaハイブリッドの生産を改善しない。
【0178】
T366YとY407T変異体ペアにより得られたAb/Iaキメラのフラクションは、試験した範囲にわたり、投入DNAsの比に実質的に非従属であった。さらに汚染している種は、mono S HR 5/5カラム(Pharmacia,Piscataway,NJ)上でイオン交換クロマトグラフィー(20mMトリス塩酸,pH8.0中0-300mM NaCl)によってAb/Iaキメラから容易に除去された。これは、AbとIaの相対的な発現レベルが一時的な発現系におけるよりも容易な操作性が劣る、安定な細胞系を用いた大量のAb/Iaキメラの製造のために良好であると予想する。
【0179】
同定した空洞への隆起変異体は、4又はそれ以下にまで下がった、可能性のある10の主要な種(Sureshら,Methods Enzymol.121:210(1990))から得られた生産物の混合物の複雑性を減じることによってFc-含有BsAbの潜在的な適用を増加することが予期される(図1A-1B)。T366YとY407Tが完全に保存され且つIgG1のCH3ドメイン界面で他の残基が高度に保存されることから、T366YとY407T変異体ペアは、他のヒトTgGアイソタイプ(IgG2,IgG3又はIgG4のような)のヘテロマルチマーを生産するために有用とされるであろう。
【0180】
実施例2:
ヘテロマルチマーの免疫付着因子中の非天然存在ジスルフィド結合の 生成 A. CH3相互鎖-ジスルフィド結合の設計 3つの基準を、パートナーCH3ドメイン間のジスルフィド結合を作るための残基のペアを同定するために使用した:i)Cα分離は、好ましくは自然ジスルフィド結合中に見出されるそれに類似している(5.0から6.8Å)(Srinivasan,N.等,Int.J.Peptides Protein Res.36:147-155(1990))。7.6Åまでの間隔は、主鎖移動を許すことおよび低い分解能の結晶構造での原子の位置を考慮することを許した(Deisenfofer,Biochemistry 20:2361-2370(1981))。ii)Cα原子は、2つのCH3ドメイン上の異なる残基上にあるだろう。iii)その残基はジスルフィド結合を与えるように位置付けられる(Srinivasan,N.ら,上記)。
【0181】
B. ジスルフィド結合のモデル化。ジスルフィド結合は、Insight II release 95.0(Biosym/MSI)を用いてhumAb4D5-Fvについて記載された通り(Rodriguesら,Cancer Res.55:63-70(1995))、ヒトIgG1Fc(Deisenhofer,上記)中にモデル化した。
【0182】
C. CH3変異体。変異は、以下の合成オリゴヌクレオチドを用いて、特定部位の突然変異誘発(Kunkelら,Methods Enzymol.154:367-382(1987))によってヒト化した抗-CD3重鎖又はCD4-IgGのCH3ドメイン中に導入した。
【0183】
【表4】
【0184】
変異は、置換アミノ酸の次の、アミノ酸残基と数(Kabatら,上記(1991)のEu数字付与スキーム)によって表した。多重変異は、コロンによって分けた単一の変異によって表される。変異体は、シーケナーゼversion 2.0(United States Biochemicals,Cleveland,OH)を用いるジデオキシヌクレオチド配列決定(Sangerら,上記(1977))によって確証した。
【0185】
D. ヘテロダイマー形成を増大する相互-鎖ジスルフィド。CH3ドメイン中に相互-鎖ジスルフィド結合を含んでいる6ペアの分子("ジスルフィド-CH3"変異体;v1-v6,表4)は、Ab/Iaハイブリッド、抗-CD3/CD4-IgG(Chamowら,上記(1994))の形成を導くその能力において親の分子と比較した。CD4-IgG及び抗-CD3重鎖変異体をコードしているプラスミドは、抗-CD3軽鎖をコードしている過剰のプラスミドと一緒に、293S細胞中に共移入した。ヘテロダイマーの収量は、Ia:H鎖:L鎖DNA比の範囲を持った移入により至適化した。Ab/Iaヘテロダイマー、IgG及びIaホモダイマー生産物は、ストレプトコッカス蛋白質Aアフィニティー-精製し、SDS-PAGEとレーザーデンシトメトリースキャニング(Ridgwayら,上記(1996))によって定量した。
【0186】
それぞれのジスルフィド-CH3ペアは、親の分子に類似した、3つの主要な種を生じる。しかしながら、ジスルフィド-CH3変異体からのAb/Iaヘテロマーは、CH3ドメイン中の相互-鎖ジスルフィドの形成に一致する、電気泳動の移動でシフトした。ジスルフィド結合形成の更なる証拠は、ヒンジにおいて相互-鎖ジスルフィドによって提供された。共有結合したAb/Iaハイブリッドは、ジスルフィドCH3変異体用の、しかしヒンジシステインがセリンに変異した野生型CH3ドメインを持った分子用ではないSDS-PAGEによって観測された。ジスルフィドCH3変異体を製造し、Y349C/S354'C,Y349C/E356'C,Y349C/E357'C,L351C/E354'C,T394C /E397'C,及びD399C/K392Cと定義した。唯一の変異体(D399C/K392'C)は、該変異体のSDS-PAGE分析による測定として野生型を越えるAb/Iaハイブリッドの収量を実質的に増加した(それぞれ76%対52%)。変異は、置換アミノ酸の次の、アミノ酸残基と数(Kabat ら,上記(1991)のEu数字付与スキーム)によって表した。CH3の第1及び第2のコピー中の変異は、それぞれ前後にスラッシュを付す。CH3の第2のコピー中の残基は、プライム符号(')によって表示される。この改善は、該変異体K392S/D399'Sが野生型に対して類似したAb/Ia収量とAb/Ia電気泳動移動性の両方を与えたことから、残基K392とD399の置換というよりむしろ、ジスルフィド結合形成を明らかに反映する。ホモダイマーは、ヘテロダイマーのCH3ドメイン中に相互-鎖ジスルフィド結合形成を優先して行うことを実証している、野生型Fcドメインの持つそれと同様に移動した。全てのジスルフィドCH3変異体は、293S細胞中の親分子とほぼ同じレベルで発現した。
【0187】
E. 空洞への隆起工学によって構築したジスルフィドは95%までヘテロダイマーの収率を増加する。最良のジスルフィドペアは、76%までヘテロダイマーのパーセンテージを増加し、空洞への隆起方法は87%までヘテロダイマーのパーセンテージを増加した(表4;Ridgwayら,(1996)上記も参照)。これら2つの方法は、ヘテロダイマー精製の確率を増加することの異なる原理による。従って、我々はヘテロダイマーにおける収率を更に改善することを意図して、その2つの方法を組み合わせた。L351C又はT394Cを含む、2つのモデル化したジスルフィドは、ジスルフィド結合ヘテロダイマー(L351C/S354'CとT394C/V397'C)と同じくジスルフィド結合ホモダイマーを潜在的に形成でき、かくしてそれの単一性を減じている。残る4つのジスルフィドペアは、ファージ選択したヘテロダイマー(変異体v9-v16)中に配置し、ヘテロダイマーの収率を分析した(表4)。ヘテロダイマーのほぼ95%の収率が得られた。再度、該ヘテロダイマーは、野生型及びv8変異体に匹敵する電気泳動移動シフトを示した。
【0188】
【表5】
【0189】
実施例3:
ヘテロマルチマー中のタンパク質-タンパク質相互作用を増大する相補の変異のための構造-誘導ファージディスプレー選択以下の方法は、マルチマー化ドメインを経て界面で相互作用するポリペプチト中の相補の変異の選択において有用である。その方法は、相補の空洞への隆起変異の選択にそれを適用するとして次に説明される。しかしながら、その実施例は制限するためのものではなく、その方法は非天然存在ジスルフィド結合、ロイシンジッパーモチーフ、疎水性相互作用、親水性相互作用などの形成に適当な変異の選択に同様に適用することができる。
【0190】
A. ファージディスプレー選択。ファージディスプレー方法は、安定なCH3ヘテロタイマーの選択用に発展し、図2中に概要が示される。その選択は、隆起変異体、T366W(Ridgwayら,上記(1996))、M13遺伝子IIIペプチドに融合したCH3の第2のコピーと共発現されるペプチドフラッグに融合した(gDペプチドフラッグ、例えばLasky,L.A.とDowbenko,D.J. (1984)DNA 3:23-29;及びBerman,P.W.ら,(1985)Science 227:1490-1492)を用いる。空洞変異体のライブラリーは、第1のCH3ドメイン上の隆起に最も近い残基の無作為化によってCH3のこの第2のコピー中に創製した。ファージディスプレー化安定CH3ヘテロダイマーは、次いで抗-フラッグAbを用いて補足した。
【0191】
CH3ファージディスプレーライブラリーの1.1x105の独立クローンは、PCRフラグメントによる自然CH3遺伝子のセグメントの置換によって構築した。
【0192】
そのフラグメントは、標準的な技術を用いて位置366、368と407で無作為化するために分解プライマーを用いてPCR増幅化により得られた。
【0193】
選択の2から5ラウンドの後、完全長クローンのフラクションは、一本鎖DNAのアガロースゲル電気泳動による判断として、それぞれ90%、60%、50%及び10%だった。完全長クローンを含むファージミドは、選択の5ラウンド後にゲル-精製した。二千の形質転換体がXL1-BLUETM細胞(Stratagene)を再形質転換後に得られた。
各クローンの平均>106コピーを、選択(panning)のラウンド当たりに使用した。かくして、該ライブラリーにおいて各クローンの多数のコピーが、たとえ幾つかの削除変異体が選択の間に生じたとしても、選択のために利用可能であると確信した。
【0194】
選択の7ラウンド後、得られたCH3変異体は、無作為化した残基でコンセンサスアミノ酸配列を研究した。実際に全てのクローンは、この位置でβ-ヒドロキシルの非常に強い優先性を示す残基 366でセリン又はトレオニンを有した。疎水性残基のための強い優先性は、バリンとアラニン優先化によって残基368と407で観測された。6の異なるアミノ酸の組み合わせは、11度回収された、三重変異体、T366S:L368A:Y407Vを含む、少なくとも2度回収された。先に定義されたヘテロダイマー、T366W/Y407'A(Ridgway,J.B.B.ら,(1996),上記)と同一の配列を有するこれらのファージ選択物はない。そのファージ選択物は、ドメイン界面残基の全側鎖容量において40-80Å3減少により判断されるように野生型CH3ホモダイマーよりも劣る緊密性で詰め込まれ得る。
【0195】
発現プラスミドpAK19(Carterら,1992)上にコードされたCH3変異体は、大腸菌33B6株中に導入し、培養槽中で高い細胞密度に増殖した大腸菌から創製した。DEAE-セファデックスFF、ABx及び Resource Sクロマトグラフィーによって精製したT366S:L368A:Y407V変異体は、SDS-PAGEの後に単一の主要なバンドを与えた。他のCH3変異体は、同様の精製により回収した。
【0196】
高解析度電気スプレー質量分析法によって測定した野生型CH3、T366S:L368A:Y407V、T366W及びY407A変異体の分子質量は、予期した通りであった。
【0197】
B. ファージ選択したヘテロダイマー安定性。CH3ヘテロダイマーの安定性は、酵素結合イムノソルベント検定法(ELISA)によって残るヘテロダイマーの希釈及び定量化に続き、グアニジン塩酸によって相当するファージを滴定することによって最初に評価した。CH3-ファージによるグアニジン塩酸分解アッセイは、選択物を直ちにスクリーンするための手段を提供する。
【0198】
ファージは、選択の7ラウンドの後の個別のクローンから及びコントロールベクター、pRA1からでも製造された。概略的には、XL1-BLUETM中のファージミドが、109pfu/ml M13K07の存在中で50μg/mlカルベニシリンと10μg/mlテトラサイクリンを含有する25ml LB培地に接種するために使用し、37℃で一晩インキュベーションした。その細胞は遠心分離(6000g、10分、4℃)によってペレット化した。ファージは、5mlの20%(w/v)PEG、2.5M NaClによって沈殿し遠心分離(12000g、10分、4℃)の後に上清から回収され、1mlのPBS中に再懸濁させた。PBS中0-6Mのグアニジン塩酸180μlが20μlファージ調製物に加えられ、5.0分間ほぼ〜25℃でインキュベーションした。各ファージサンプルのアリコート(20μl)は、水で十倍に希釈した。CH3のヘテロダイマーの存在は、5B6-被覆プレートを用い及び基質としてo-フェニレンジアミンを用いたホースラディッシュペルオキシダーゼに接合した抗-M13ポリクローナルAbによってファージを検出するELISAによって測定した。その反応は、2.5M H2SO450ml の添加によって抑止し、492nmで吸光度を測定した。その吸光度データは、その溶融の間にグアニジン塩酸濃度に対してプロットし、且つ Kaleidagraph 3.0.5(Synergy Software)を用いた非線形最小二乗法によって四つのパラメーターモデルに適用した。
【0199】
最も頻繁に回収されたヘテロダイマー、T366W/T366'S:L368'A:Y407'Vは、他のファージ-選択したヘテロダイマーと安定性において類似する。このファージ-選択したヘテロダイマーは、設計したヘテロダイマー、T366W/Y407'Aよりも有意により安定であるが、しかし野生型CH3よりは安定性に劣る。全てのCH3変異体、個別及び組み合わせの両方は、これら同じ分子が熱量測定法によって研究された条件下にサイズ排除クロマトグラフィーによってダイマーであることが見出された(1.75mg/ml、リン酸緩衝化生理食塩水(PBS)中)。唯一の例外は、CH3ダイマーよりも明らかに短い保持時間を有したT366S:L368A:Y407V変異体単独であった。
【0200】
T366Wの1:1混合物、隆起、及びT366S:L368A:Y407V、69.4℃で単独転移を持つ変異体溶融物は、サブユニット交換及び安定なヘテロダイマーの形成と密接に結びつく。一方、T366W隆起ホモダイマーは、T366W/T366'S:L368'A:Y407'V空洞への隆起ヘテロダイマー(ΔTm=−15.0℃)よりもより劣った安定性である。それ自身の上のT366S:L368A:Y407V空洞変異体は、加熱によって集合する傾向があり、なだらかな溶融転移を成さない。
【0201】
設計した空洞変異体、Y407Aは、T366W隆起変異体の不在及び存在でそれぞれ58.8℃及び65.4℃で溶融する。これは、T366W(ΔTm=11.0℃)又はY407A(ΔTm=6.6℃) ホモダイマーのいずれか一方よりもより大きな安定性を有するT366W/Y407'Aホモダイマーのサブユニット交換と形成と密接に結びつく。ファージ選択したヘテロダイマー、T366W/T366'S:L368'A:Y407'Vは、設計したヘテロダイマー、T366W/Y407'A,(ΔTm=4.0℃)よりも一層安定であるが、しかし、野生型CH3ホモダイマー(ΔTm=−11.0℃)よりも安定性が劣る。
【0202】
C. インビボでのファージ-選択した抗体免疫付着因子のマルチマー化。ファージ選択した及び設計したCH3変異体は、インビボでAb/Iaハイブリッド、抗-CD3/CD4-IgGの形成を導くその能力を比較した(Chamowら,(1994),上記。
【0203】
これは、CD4-IgGと一緒にヒト化した抗-CD3軽鎖(L)と重鎖の共発現により達成した。ヘテロダイマーとホモダイマーの形成は、SDS-PAGEとスキャニングレーザーデンシトメトリー(Ridgway,ら,(1996),上記)に続いてプロテインA精製によって評価した。Ab/Iaハイブリッドの比較できる収量が、抗-CD3重鎖が設計した隆起変異体を含んだ、T366WとIaがファージ-選択した変異体、T366S:L368A:Y407V、又は設計した空洞変異体、Y407Aのいずれかを含んだ共移入体から回収された(図3)。
【0204】
ファージ-選択した及び設計したCH3変異体は、次にホモダイマーを形成するためのその傾向において評価した。隆起変異体、T366Wは、相当する重鎖と軽鎖の共移入がIgGを越える過剰のHLモノマー(非-ジスルフィド結合IgGを含み得る)に至ることから、ホモダイマー化を明らかに非常に崩壊させた。一方、IgGしかしHLモノマーなしでは、野生型CH3ドメインを含む同じ抗体が観測された。空洞変異体、T366S:L368A:Y407Vは、相当するファージミドの遺伝子移入が幾つかのIaモノマーを持つ優性なIaダイマーの混合物に至ることから、多少ホモダイマー化を崩壊させる。空洞変異体、Y407Aは、Iaダイマーしかし相当するファージミドの移入後にIaモノマー無し、の存在による判断として、ホモダイマー化を最小限崩壊する。
【0205】
ここに記載したファージディスプレー方法は、安定したヘテロダイマーを形成するCH3変異体の援助での選択及び安定なホモダイマーを形成する変異体に対しての選択を与える。ホモダイマーに対するカウンター選択は、"遊離"CH3変異体が、利用可能なCH3変異体-遺伝子III融合タンパク質への結合のためにフラッグ化CH3ノブ変異体と競合するであろうことから生じる。遊離CH3変異体は、自然CH3変異体とM13遺伝子IIIの間のアンバー変異の結果として生じる。XL1-Blucのようなアンバー抑制宿主において、自然CH3遺伝子とM13遺伝子IIIの両方が創製されるだろう。
【0206】
グアニジン塩酸分解は、ファージ上のCH3ヘテロダイマー安定性を最初にスクリーニングするために有用なツールとして提供される。ファージは、5Mグアニジン塩酸にさらした後でも大腸菌への感染性を維持する(Figiniら,J.Mol.Biol.239:68-78(1994))。かくして、グアニジンは変異体選択の方法を増加するためにも有用となり得る。
【0207】
ファージディスプレーライブラリーの合理的な設計とスクリーニングは、ヘテロダイマー化を促進するためにホモダイマーのドメイン界面を再モデル化するための相補的なアプローチである。CH3ドメインのケースにおいて、設計した変異体は、ヘテロダイマー化を促進することを強化できるドメイン界面残基と同一視される。ファージディスプレーは、ヘテロダイマーを最も有効に形成する組み合わせのために固定した隆起の近くの3つの残基の置換をサーチするためにここに用いた。ファージ選択物は、ドメイン界面の更なる合理的な再設計を促進するために有効とされる一方、ここに記載したファージ選択方法は、タンパク質-タンパク質界面を再モデル化するためのその有用性を実証する。
【0208】
実施例4:
共通の軽鎖を有するヘテロマルチマー抗体又は抗体/免疫付着因子の生成と配置 以下の実施例は、本発明に従う同じ軽鎖を共有するヘテロマルチマー二重特異性抗体の調製とその標的抗原に結合する抗体の能力を実証する。
【0209】
A. 同じ軽鎖を共有する抗体の同定:抗体ライブラリーの比較は11の抗原に高まる。大きいヒト一本鎖Fv(scFV)抗体ライブラリー(Vaughanら(1996)上記)は、Ax1(ヒトレセプターチロシンキナーゼECD)、GCSF-R(ヒト顆粒球コロニー刺激因子レセプターECD)、IgE(ネズミIgE)、IgE-R(ヒトIgEレセプターα-鎖)、MPL(ヒトトロンボポイエチンレセプターチロシンキナーゼECD)、MusK(ヒト筋肉特異的レセプターチロシンキナーゼECD)、NpoR(ヒトオルファンレセプターNpoR ECD)、Rse(ヒトレセプターチロシンキナーゼ、Rse,ECD)、HER3(ヒトレセプターチロシンキナーゼHER3/c-erbB3 ECD)、Ob-R(ヒトレプチンレセプターECD)、及びVEGF(ヒト脈管内皮成長因子)、ECDは細胞外ドメインに関する、を含む11の抗原用の抗体特異性で選択した。各抗原に生じた抗体の集団からscFvフラグメントのためのヌクレオチド配列データは、相当するタンパク質配列から得るように翻訳した。VLは異列は、鎖の全てのペア様態組み合わせ間の同一性パーセンテージを計算するためにFengとDoolittle(1985,1987,1990)のアルゴリズムによりプログラム"アライン"を用いて比較した(Feng,D.F.とDoolittle,R.F. (1985)J.Mol.Evol.21:112-123;Feng,D.F.とDoolittle,R.F. (1987)J.Mol.Evol.25:351-360;及びFeng,D.F.とDoolittle,R.F.(1990)Method Enzymol.183:375-387)。
【0210】
各ペア様態軽鎖アミノ酸配列比較の配列同一パーセントの結果は、マトリックスフォーマット中に調整した(追記参照)。
【0211】
最もペア様態の比較品として、少なくとも1の共通軽鎖配列が見出された。表5は、117VLアミノ酸配列のアラインメントによって決定した同じ軽鎖を共有する(100%同一性)scFvの頻度を示しているVL鎖の比較である。例えば、そのエントリー4/9(HER3xOb-R、黒いボックスで表した)は、結合HER3が1又はそれ以上の抗-Ob-RクローンによってそれのVL配列を分配することが見出される一方、Ob-R結合9クローンは、1又はそれ以上の抗-HER3クローンによってそのVL配列を分配する、4クローンを表す。対角線上の参加物は、その集団中の1又はそれ以上のクローンによってVL配列を分ける集団内の抗体クローンの数を表す。例えば、MPLクローンの調査は、1又はそれ以上の他のMPLクローンによってそれのVL配列が分けられた5クローンを明らかにした。(IgE x Ax1)又は(NpoR x IgE-R)についてのような観測された共通の軽鎖の無いケースにおいて、少なくとも1の特異性と比較されるフラグメントの数は非常に少なかった(5以下)。見出された共通の軽鎖の数が与えられ、それは共通の軽鎖が、もし十分な数のクローンが比較されるなら、いずれかのVL比較物のために見出すことができることがありそうである。
【0212】
軽鎖のアミノ酸配列は、選択した共通の軽鎖に対しての配列相同性が98%と99%であった場合、異なったアミノ酸残基の位置によって決定された。図4は、Ax1(クローンAx1.78)、Rse(クローンRse.23,Rse.04,Rse.20,及びRse.15)、IgER(クローン IgER.MAT2C1G11)、Ob-R(クローンobr.4)、及びVEGF(クローンvegf.5)に特異性を持つ8つの異なる抗体のVL配列の比較である。配列の定義(Kabat,G.A.ら(1991)上記)又は構造の定義(ChothiaとLcsk,(1987)J.Mol.Biol.196:901-917) に従う抗原結合CDR残基の位置は、それぞれ下線と#によって示した。Ax1.78配列と異なる軽鎖残基は二重下線により示した。比較した9軽鎖の、6が同一である。Rse.04とobr.4の軽鎖(ほぼ99%配列相同性)は、抗原結合CDRsの外側の1残基で相違するRse.20の軽鎖(ほぼ98%配列相同性)は、抗原結合CDRsの外側の2残基で異なる。該アミノ酸残基の変化は、抗原結合性において僅かな又は影響なしを有し得る。かくしてこれらの軽鎖の配列類似性は、本発明の共通の軽鎖のためのその候補を作る。代替的に、期待される対になったscFv(例えばAx1.78)の軽鎖との98-99%相同性を有する軽鎖のように、本発明に従い、対になった軽鎖と置換でき且つ結合特異性を保持し得る。
B. 同じ軽鎖を共有する抗体の同定と軽鎖のそれを共有する二重特異性抗体の構築:抗-Ob-R/抗-HER3。
【0213】
ヒトレプチンレセプター(Ob-R)又はHER3/c-erbB3遺伝子生産物(HER3)の細胞外ドメインを結合したScFvフラグメントは、大きいヒトscFvファージライブラリーを用いた選別の3つのラウンドによって得られた(Vaughanら(1996),上記)。レプチンレセプター-IgGとHER3-IgG(1ml PBS中10μg)を、4℃で一晩、分離イムノチューブ(Nunc;Maxisorp)を被覆するために使用した。選別とファージ解放は、以下の修正を伴い、Vaugham(1996),上記により記載されたように実行した。1mg/mlの濃度で、ヒト化した抗体、 huMAb4D5-8(Carter,P.ら(1992)PNAS USA 89:4285-4289)又はヒト化した抗-IgE(Presta,L.ら(1993)J.Immunol.151:2623-2632)は、Fc-結合ファージを吸着するための各選別工程中に含めた。加えて、溶液の選別化(Hawkins,R.E.ら (1992)J.Mol.Biol.226:889-896)は、scFv結合レプチンレセプターを同定するためにも使用した。そのレプチンレセプターは、プロテインAセファロースクロマトグラフィーに続いて、工作したプロテアーゼ、ジェネナーゼ(Carter,P.ら (1989)Proteins:Structure,Function and Genetics 6:240-248)によるレプチンレセプター-IgGの位置特異的蛋白分解によってFcから分離した。そのレプチンレセプターはビオチン化し、それぞれ、選別の第1,第2,及び第3ラウンド用に100nM、25nM及び5nMの濃度で使用した。ファージ結合ビオチン化抗原は、ストレプタビジン-被覆パラ磁性ビーズ(Dynabeads,Dynal,Oslo,Norway)を用いて捕捉した。
【0214】
それぞれの選別のラウンド2及び3からのクローンは、ファージ及び相当する抗原及び制御免疫付着因子又は抗体をも用いるscFvELISAによってスクリーンした。抗原-陽性クローンの分割性は、プライマー、fdtetseqとPUC逆転(Vaughamら(1996),上記)を用いるscFv挿入のPCR-増幅及びBstNIによる消化(Marksら(1991)上記)によって分析した。BstNIフィンガープリント当たり1から5のクローンがPCR重リンクとmyc seq 10プライマー(Vaughamら(1996)上記)を用いたcycle-sequencedusing fluorescent dideoxy chain terminators(Applied Biosystems)した。サンプルは、Applied Biosystems Automated DNA Sequencerを用いて分析し且つ配列をSeqEdを用いて分析した。グアニジン塩酸抗体分解及びファージディスプレー選択と組み合わせた Figiniのインビトロでの鎖シャフティング方法が同じ軽鎖を有する抗体を選択する方法として有用であることにも注目される(Figini,M.ら (1994),上記、その全体が参照によりここに併合される)。
【0215】
上記の方法を用いて、11の異なる抗-HER3クローンと18抗-Ob-Rクローン(被覆した抗原を用いた選別からの11とビオチン化抗原による選別からの7)が得られた。そのクローンは、各結合ドメインと結合した軽鎖の配列を決定するための標準技術により配列決定した(図5)。その配列は、二重特異性抗体を構成するために使用した抗-Ob-Rクローン26と抗-HER3クローン18のVH及び共通のVL配列である(後記参照)。その残基は(Kabat,E.A.ら(1991)上記)に従い番号付けされる。配列の定義(Kabat,E.A.ら(1991)上記)又は構造の定義(ChothiaとLesk,(1987)J.Mol.Biol.(1987)196:901-917)は、それぞれ下線と上線によって示される。VH配列中の残基間の相同性は、*によって表示した。
【0216】
軽鎖の配列は、多重抗-Ob-Rクローンに対する多重抗-HER3クローンと比較した(図8及び表5)。11の抗-HER3クローン以外の4つが、1又はそれ以上の抗-Ob-Rレセプタークローンと同じVLを共有することが観測された。
【0217】
反対に、18の抗-Ob-Rクローン以外の9が抗-HER3クローンの一つとして同じVLを共有する(表5、黒い箱を参照)。
【0218】
【表6】
【0219】
抗-Ob-R/抗-HER3、共通の軽鎖を有する二重特異性抗体の構築は以下の通り実行した。相補の隆起と空洞、同じく第1及び第2のポリペプチド間の非天然存在ジスルフィド結合を有する変更したCH3第1及び第2のポリペプチドを、Fc-含有二重特異性抗体の構築において用いた。同じ軽鎖を、同じく各抗体からの重鎖を共有するクローンである、抗-Ob-Rクローン#26と抗-HER3クローン#18からのVLを、ここに記載した手順に従って二重特異性抗体を製造するために用いた。
この抗体は、非天然存在ジスルフィド結合を生成するために変更を欠くことによってのみそれと異なる二重特異性抗体に対応する明確な分子量において電気泳動移動性シフトを有した。非天然存在ジスルフィド結合を持った及び持たないヘテロダイマーの抗体変異体の8%SDS-PAGEゲルは、野生型ヘテロダイマーのほぼ230の見かけ上のMWから、一つの非天然ジスルフィド結合を有しているヘテロダイマーのほぼ200の見かけ上のMWまでの移動シフトを示した。そのMWシフトは非天然ジスルフィド結合を首尾良く形成するそれぞれの変異体のパーセントを与えるために十分であった。
二重特異性抗体のOb-R及びHER3の両方への特異的結合は、以下の方法のような標準ELISA法によって試験される。Ob-R結合は、Ob-R-Ig融合タンパク質として存在するOb-RによってELISAアッセイで実証される。該Ob-R-Ig融合タンパク質は、96-ウェルのミクロタイター板のウェル上に被覆され、そして二重特異性抗体が加えられる。
【0220】
そのウェルは、Ob-R-Igへの非特異的結合を取り除くために数回洗浄される。同じアッセイにおける第2の成分として、ビオチン化HER3-Ig融合タンパク質が加えられ、そしてビオチン化HER3-Ig融合タンパク質に結合するストレプタビジン-ホースラディッシュペルオキシダーゼ複合体によって測定される。結合は、過酸化水素及びTMBペルオキシダーゼ基質(Kirkegaard and Perry Laboratories,Gaithersburg,MD)の添加における着色の変化の生成により測定される。
【0221】
実際に記載した条件下、Ob-R-IgとHER3-Igの両方への二重特異性抗体の結合は、固定化Ob-R-Ig/二重特異性抗体/HER3-Igビオチン/検出可能に標識したストレプタビジンを含む複合体の良好な形成のためにミクロタイター表面上に固定化した検出可能な標識として観測されるであろう。Ob- R-Igと結合するが、HER3-Igとはしない抗体は、陰性の結果を提供するように、上記複合体を形成しない。同様に、HER3-Igと結合するが、Ob-R-Igとはしない抗体は、陰性の結果を提供するように、上記複合体を形成しない。一方、Ob-R-IgとHER3-Igの両方に結合することが予期される二重特異性抗体は、該アッセイにおいて陽性の結果を生じる複合体を形成し、共通の軽鎖を有する、HER3とOb-Rの両方に結合する、二重特異性抗体を表示する。
【0222】
抗-(Ob-R/HER3)二重特異性抗体の発現と精製は、以下の通り実行した。ヒト胎芽腎臓293S細胞は、抗-Ob-R重鎖、抗-HER3重鎖、又は上述した通りそれぞれの抗体に共通であったクローン26又は18からの軽鎖をそれぞれ独立にコードしている3つのプラスミドDNAによって移入した。それぞれの移入のために、軽鎖コード化DNAに対する重鎖コード化DNAの比率は、軽鎖が抗-Ob-R/抗-HER3二重特異性抗体の配列を制限しないであろうように1:3とした。両方の重鎖は、互いに関して1:1の比で移入した。12μgのトータルのプラスミドDNAは、次いでリン酸カルシウム沈降によって 293S細胞中に共移入(co-transfected)した(Gorman,C.,DNA Cloning,Vol.II,D.M.Glover,ed.,IRL Press,Oxford,p.143(1985))。その細胞は、タンパク質発現の増大を意図して増殖培地に加える前に洗浄した。Fc-含有タンパク質は、固定化したプロテインA(ProSep A,BioProcessing Ltd.,UK)を用いて細胞懸濁物から精製し、そしてPBS中に緩衝化-交換した。ヨードアセトアミドは、ジスルフィド結合の再シャフト化を防止するために50mMの最終濃度までタンパク質調製物に加えた。
【0223】
付加の実施例として、抗-(CD3/CD4)抗体/免疫付着物の発現と精製を以下の通り実行した。ヒト胎芽腎臓293S細胞は、3つのプラスミドDNAs、各プラスミドは抗-CD3軽鎖、抗-CD3 IgG1重鎖、又は抗-CD4 IgG1免疫付着因子を独立的にコードしている、によって移入した。それぞれの移入のために、重鎖コード化DNAに対する軽鎖コード化DNAの比率は、軽鎖が抗-CD3 IgGの整列のために制限されないであろうように、3:1とした。加えて、免疫付着因子が不十分に発現されるために、免疫付着因子コード化プラスミドの比率は、重鎖コード化プラスミドの過剰において加えた。試験したその比率は、免疫付着因子:重鎖:軽鎖が3:1:3から8:1:3間での範囲とした。トータルで10μgのプラスミドDNAは、移入前にPBSで細胞を洗浄し、次いでリン酸カルシウム沈降(Gorman,C.(1985),上記)により293S細胞中に共移入した。Fc-含有タンパク質は、固定化したプロテインA(ProSep A,BioProcessing Ltd.,UK)を用いて細胞懸濁物から精製し、そしてPBS中に緩衝化-交換した。ヨードアセトアミドは、ジスルフィド結合の再シャフト化を防止するために50mMの最終濃度までタンパク質調製物に加えた。
【0224】
上記それぞれの調製において、タンパク質サンプルは、8%ポリアクリルアミドゲル(Novex)で電気泳動し、Serva blueによる染色によって可視化した。ゲルは、マイナーな不純物の可視化及び定量化を与える努力において見せかけのバックグラウンドを無くすように脱- 染色化した。乾燥したゲルは、スキャニングデンシトメーター(GS-670,BioRad)によってスキャンし、そしてタンパク質製造物は Molecular Analystソフトウェアによって定量化した。
【0225】
CH3ドメイン中に導入した非天然 (工作した)ジスルフィド結合は、ヘテロダイマー形成を増大するためにここに開示されている。一つのペプチド、K392/D399'Cは、76%までのヘテロダイマー(表4、変異体v6)を精製することによってヘテロダイマー形成を増大した。さらに、相互-鎖ジスルフィド結合の存在が空洞への隆起技術と結び付けられた場合、ほぼ95%ヘテロダイマーが得られた(表4変異体v11,v12,及びv16)。かくして、二重特異性抗体の第1と第2のポリペプチド間のタンパク質/タンパク質相互作用を特異的に増加する本発明の方法は、望まれるヘテロマルチマーの収量を増加すると共に、望ましくないヘテロマルチマー又はホモマルチマーの形成を最小化する。
【0226】
加えて、電気泳動の移動性分析によって生産物ヘテロマルチマーを特徴付ける方法は、望ましくない生産物に対して、望まれるヘテロマルチマーの相対的な量の測定を与える。
【0227】
ここに記載したような共通の軽鎖の選択は、多重特異性抗体の可変の重鎖と軽鎖の間の誤対合の可能性を削除することによって望まれるヘテロマルチマーの収量を増加する。
【0228】
C. 同じ軽鎖を共有する抗体の同定とその軽鎖を共有している二重特異性抗体の構築:抗-Mpl/抗-HER3。
【0229】
本発明の別な二重特異性抗体の同定、構築及び発現がここに示される。この実施例のパートAとB中に記載した方法を、抗-Mpl/抗-HER3二重特異性抗体の製造用に利用した。
【0230】
この実施例のセクションA中に記載した方法(11の抗原を生じた抗体ライブラリーの比較)、上記、を用い、抗-HER3scFvのVHとVLアミノ酸配列を、ヒトトロンボポイエチンレセプター、c-Mplに結合する23 scFvで比較した。11の抗-HER3クローンの5つは、1又はそれ以上のMpl-結合クローンを持った同一のVLアミノ酸配列を共有する。反対に、23の抗-Mpl scFv以外の7つは、抗-HER3クローンの一つと同じVLを共有した(表5、上記、白い箱を参照)。一方、VHアミノ酸配列は、何れかの抗-Mplと抗-HER3クローンの間に40から90%の同一性レベルを持つ、より多く相違するものであった。
【0231】
その抗-Mpl scFv,12B5(Genbank accession number AF048775;配列番号:27)と抗HER3scFvクローンH6(Genbank accession number AF048774;配列番号:28)は、同一のVL配列及び実質的に異なるVH配列を利用する。これらのscFvフラグメントは、共有した軽鎖に対する有効なヘテロダイマー化ができる抗-Mpl/抗-HER3二重特異性IgG抗体を構築すること、同じく孔-中-ノブ変異体(ここに記載した)及びCH3ドメイン間に作られたジスルフィド結合の使用を通して用いた。同じL鎖を共有する抗体は、非-同系H鎖と対合するL鎖の問題を避けるために選択した。2つの天然存在ヒンジ領域ジスルフィド結合もまた存在した。その共通のL鎖は、変異体v11からのCH3変異を含んでいる2つのH鎖と共移入した。そのIgG生産物は、プロテインAアフィニティクロマトグラフィーによって精製し、且つ標準技法を用いてSDS-PAGEによって分析した。
【0232】
二重特異性IgG抗体(BsIgG)調製物は、IgG含有野生型CH3ドメインよりも、より大きい移動性を示す単一の主要バンドを生じる。電気泳動移動性におけるこの増加は、よりコンパクトなタンパク質種を形成するBsIgG中に作られたジスルフィド結合の形成と一致した。
【0233】
MplとHER3 ECD抗原の両方に結合する工作した抗-Mpl/抗-HER3BsIgG 抗体の能力は、以下の通りELISAを用いて評価した。全ての工程でPBS緩衝液を用い、96-ウェル板(Maxisorp,Nunc)の個々のウェルは、5μg/mlでHER3-IgG又はMpl-IgGで一晩被覆し、洗浄し次いで0.5%(w/v)BSAによって1時間ブロック化した。その最初の抗体は、変異体Y349C:T366S:L368A:Y407V/T366'W:S354'Cを含み、且つ変異したFc領域を持つ親の抗-Mpl又は抗 -HER3 IgGに一致する抗-Mplx抗-HER3BsIgGとした。最初の抗体(1μg/mL)は、ビオチン化HER3-IgG及びストレプタビジン-ホースラディッシュペルオキシダーゼ接合体 (Boehringer Mannheim)の1:5000希釈液と共に23℃で2時間、別個にインキュベーションし、次いで各ウェルに加え、23℃でさらに1時間インキュベーションした。ペルオキシダーゼ活性は、販売者(Kirkegaard and Perry Laboratories,Inc.,Gaithersburg,MD)によって指示された通りにTMB試薬によって検出した。
【0234】
予想した通り、抗-Mpl/抗-HER3 BsIgGは、個別にMplとHER3 ECD抗原のそれぞれに有効に且つ同時に、同じく両方の抗原に同時に結合した。これに対して、親の抗-Mplと親の抗-HER3 IgGは、同系抗原に一致するそれにのみ結合した(図6)。
【0235】
D. 工作したFc領域を含んでいる抗体は、有効な抗体-依存細胞介在細胞毒性の能力がある。
【0236】
本発明の実例化した二重特異性抗体の精製において用いた工作したFc領域(CH3変異体、上記)が有効な抗体-依存細胞介在細胞毒性の能力があることを実証するため、以下の実験を実行した。
【0237】
そのCH3変異体は、Lewis,G.D.ら(Lewis,G.D,ら(1993)Cancer Immunol.Immunother.37:255-263,その全部が参考により組込まれる)の方法を用いて示した通り、有能な抗体-依存細胞-介在細胞毒性(ADCC)をサポートする能力を維持する。概略的には、細胞毒性アッセイは、51Cr-標識したSK-BR-3及びHBL-100標的細胞(それぞれATCC受託番号HTB-30と45509)及びエフェクター細胞としてヒト末梢血液リンパ球によって実行した。しかしながら、Lewisらと異なり、そのリンパ球はIL-2で活性化されなかった。
【0238】
CH3変異体S354:T366W及びY349C:T366S:L368A:Y407Vは、Carterら(Carter,P.ら(1992)PNAS USA 89:4285-4289)により製造されたヒト化抗-HER2抗体、huMAb4D5-5のH鎖中に別個に導入した。再モデル化した及び野生型Fc領域を含む抗体は、HER2-過剰発現乳癌細胞系、SK-BR-3によりADCC中で類似の能力を有した(図7)。再モデル化した及び野生型抗体の両方は、通常の胸部上皮細胞系に対する低い活性と比較できることを示した。H鎖中の効果は、これらBsIgG'sが抗体-依存細胞-介在細胞毒性において機能するであろうことを予報する、結合ドメインの独立がある。
【0239】
本発明は、最も実践的、且つ好適な実施態様であるとして考慮される何れかにおいてここに示され且つ記載される。それは、しかしながら、発展は本発明の範囲内から作製できること、及び自明な修正がこの開示を熟読した当業者に生起することが認識される。ここに用いた全ての参考文献はそれの全体において参考によってここに組込まれる。
【図面の簡単な説明】
【0240】
【図1】図1A〜図1C。図1Aは、ホモマルチマー化よりもヘテロマルチマー化を増強させるための操作が行われない場合において、Fcを含有する二重特異性抗体の形成を示す略図である。図1Bは、所望のヘテロマルチマー化が、望ましくないヘテロマルチマー化およびホモマルチマー化よりも好ましいように、重(H)鎖が操作される場合に生じる対形成を示す略図である。図1Cは、同じ軽(L)鎖を有する抗体を選択して、同系でない重鎖と対形成する軽鎖の問題が回避される場合に生じる対形成を示す略図である。
【図2AB】図2Aは、ファージディスプレーベクターpRA2を使用するCH3ヘテロダイマーの選択反応図式を図示する。安定なCH3ヘテロダイマーをディスプレーするファージは、gDフラッグに対する抗体を使用して捕獲される。図2Bは、合成された遺伝子から発現されるCH3が、M13の遺伝子IIIタンパク質との融合タンパク質として、天然の遺伝子(Ellisonら、Nucleic Acids Res.10:4071〜4079(1982))から発現されるCH3の第2の複製物とともに同時に分泌されるジシストロン性オペロンを図示する。合成されたCH3遺伝子は、単純ヘルペスウイルスの糖タンパク質Dから誘導されるペプチド(gDフラッグ、Lasky,L.A.およびDowbenko,D.J.(1984)DNA3:23〜29;Berman,P.W.ら(1985)Science227:1490〜1492)、および部位特異的なプロテアーゼのケネナーゼ(Genenase)Jの開裂(G)部位(Carter,P.ら(1989)Proteins:Structure、Function and Genetics 6:240〜248)の上流に置かれる。
【図2C1】図2C1は、翻訳されるCH3遺伝子の残基に、KabatらのEu方式に従って番号を付けた図2Bのジシストロン性オペロンの核酸配列(配列番号2)である。「免疫学的に関心が持たれるタンパク質の配列」、第5版、第1巻、688〜696頁、NIH、Bethesda、MD(1991)。隆起の変異T366Wを示す。そのような残基は、天然のCH3遺伝子における無作為化のために標的化される(366、368および407)。
【図2C2】図2C2は、翻訳されるCH3遺伝子の残基に、KabatらのEu方式に従って番号を付けた図2Bのジシストロン性オペロンの核酸配列(配列番号2)である。「免疫学的に関心が持たれるタンパク質の配列」、第5版、第1巻、688〜696頁、NIH、Bethesda、MD(1991)。隆起の変異T366Wを示す。そのような残基は、天然のCH3遺伝子における無作為化のために標的化される(366、368および407)。
【図3】図3A〜図3C。図3Aおよび図3Bは、免疫付着因子(Ia)とともに抗体(Ab)の重鎖および軽鎖の共-移入から得られるプロテインA精製された生成物のSDS−PAGEをデンシトメトリー分析で走査した結果の棒グラフである。示したデータは、2つの独立した実験の平均である。x軸は、投入したDNAの質量比(Ia:H:L)を示し、y軸は、全生成物タンパク質に対する生成物マルチマーの各タイプの割合を示す。図3Cは、可能な生成物マルチマーを図示する。
【図4】図4は、Ax1、Rse、IgER、Ob−RおよびVEGFに対する特異性を有する8個の異なる抗体のVL配列の比較である。配列定義(Kabatら(1991)上記)または構造定義(Chothia,C.およびLesk,A.M.J.Mol.Biol.(1987)196:901〜917)に従った抗原結合CDR残基の位置を、それぞれ、下線および#によって示す。Ax1.78配列と異なる残基を二重下線によって示す。
【図5】図5は、選択された抗Ob−Rクローンおよび抗HER3クローンの重鎖および軽鎖の比較である。二重特異性抗体の構築に使用した抗Ob-Rクローン26および抗HER3クローン18のVH配列および共通するVL配列を示す。
【図6】図6。Mpl−IgGおよびHER3−IgGに対する同時結合を検出するためのサンドイッチELTSA。試験した抗体は、Y349C:T366S:L368A:Y407V/T366’W:S354’Cの変異を含有する抗Mpl×抗HER3のBsIgGであり、Fc領域を変異させた対応する元の抗Mplまたは抗HER3のIgGとともに使用した。
【図7】図7は、抗体依存性細胞媒介細胞傷害性(ADCC)の研究結果の棒グラフである。ADCCは、変異型(S354C:T366W/Y349’C:T366’S:L368’A:Y4O7’V)または野生型のFcあるいはアイソタイプが一致するコントロール抗体(E25、Presta,L.G.ら(1993)J.Immunol.151:2623〜2632)のいずれかを含有するhuMAb4D5−5(Carter,P.ら(1992)PNAS USA89:4285〜4289)によって媒介された。抗体(125ng/ml)を、ヒト末梢血単核エフェクター細胞およびSK−BR−3標的細胞とともに示した比でインキュベーションした。示したデータは、3連の測定値および3つの異なる実験の平均である。
【図8】図8は、HER3に対して生起された抗体の軽鎖とOb−Rに対して生起された抗体の軽鎖との間のアミノ酸配列同一性を示す行列である。配列同一性が100%である軽鎖を有する抗体を黒四角に示す。配列同一性が98%〜99%である軽鎖を有する抗体を白四角に示す。抗体クローンの同一性を行列の下部に示す。
【技術分野】
【0001】
ヘテロマルチマー及び共通成分を有する多重特異性抗体の製造方法発明の分野本発明は、ヘテロマルチマー重鎖成分および共通する軽鎖成分を有する多重特異性抗体の作製方法に関する。このような多重特異性抗体として、二重特異性抗体、二重特異性免疫付着因子、ならびに本発明の方法を使用して作製される抗体−免疫付着因子キメラおよびヘテロマルチマーポリペプチドなどが挙げられる。
【背景技術】
【0002】
二重特異性抗体少なくとも2つの異なる抗原に対する結合特異性を有する二重特異性抗体(BsAb)は、インビトロおよびインビボでの免疫診断および治療に必要な標的化薬剤として、そして診断的な免疫アッセイに必要な標的化薬剤としての広範囲の診療適用における大きな可能性を有している。
【0003】
診断領域において、二重特異性抗体は、細胞表面分子の機能的特性を探る際に、そして細胞傷害性を媒介する様々なFcレセプターの能力を明らかにする際に非常に有用である(Fangerら、Crit.Rev.Immunol.12:101〜124(1992))。Nolanら、Biochem.Biophys.Acta.1040:1〜11(1990)は、BsAbに関する他の診断適用を記載する。特に、BsAbは、酵素免疫アッセイで使用される酵素を固定化するために構築することができる。これを達成するために、BsAbの一方のアーム(腕)は、酵素表面の特定のエピトープに結合するように設計することができ、その結果、結合による酵素阻害は生じない。BsAbのもう一方のアームは、所望の部位において高い酵素密度を確実にする固定化マトリックスに結合する。そのような診断的BsAbの例には、表面抗原を探し出すために使用された、Hammerlingら、J.Exp.Med.128:1461〜1473(1968)によって記載されたウサギ抗IgG/抗フェリチンBsAbが含まれる。
【0004】
ホースラディッシュペルオキシダーゼ(HRP)ならびにホルモンに対する結合特異性を有するBsAbもまた開発されている。BsAbに関する別の可能な免疫化学的適用には、二部位免疫アッセイにおけるその使用が含まれる。例えば、分析対象タンパク質の表面にある2つの異なるエピトープに結合する2つのBsAbか作製されている−一方のBsAbは複合体を不溶性マトリックスに結合させ、もう一方は指示酵素と結合する(Nolanら(上記)を参照)。
【0005】
二重特異性抗体はまた、ガンなどの様々な疾患のインビボ免疫診断またはインビトロ免疫診断を行うために使用することができる(Songsivilaiら、Clin.Exp.Immunol.79:315(1990))。BsAbのこのような診断的使用を容易にするために、BsAbの一方のアームは、腫瘍関連抗原と結合することができ、もう一方のアームは、放射性核種と強固に結合するキレート化剤などの検出可能なマーカーと結合することができる。このような方法を使用して、LeDoussalらは、結腸直腸ガンおよび甲状腺(thryoid)ガンの放射免疫検出に有用なBsAbを作製した。このBsAbは、ガン胎児抗原(CEA)と結合するアームを一方に有し、ジエチレントリアミン五酢酸(DPTA)と結合するアームをもう一方に有する。LeDoussalら、Int.J.Cancer Suppl.7:58〜62(1992)
およひLeDoussalら、J.Nucl.Med.34:1662〜1671(1993)を参照。Stickneyらは、放射免疫検出を使用してCEAを発現する結腸直腸ガンを検出するための方策を同様に記載する。この研究者らは、CEAならびにヒドロキシエチルチオ尿素−ベンジル−EDTA(EOTUBE)と結合するBsAbを記載する。Stickneyら、Cancer Res.51:6650〜6655(1991)を参照。
【0006】
二重特異性抗体はまた、標的(例えば、病原体または腫瘍細胞)と結合する一方のアーム、およびT細胞レセプターまたはFcγレセプターなどの細胞傷害性誘引因子分子と結合するもう一方のアームが提供されることによって、細胞傷害性の対象を変えることでヒトの治療に有用であり得る。従って、二重特異性抗体を使用して、患者の細胞性免疫防御機構を腫瘍細胞または感染性病原体に特異的に向けさせることができる。このような方策を使用して、FcγRIII(または、CD16)に結合する二重特異性抗体は、ナチュラルキラー(NK)細胞/大顆粒リンパ球(LGL)細胞によってインビトロでの腫瘍細胞殺傷を媒介することができ、インビボで腫瘍増殖を防止するのに効果的であることが明らかにされた。Segalら、Chem.Immunol.47:179(1989)およびSegalら、ガンの生物学的治療(Biologic Therapy ofCancer)2(4)、DeVitaら編、J.B.Lippincott、Philadelphia(1992)、1頁。同様に、FcγIIIと結合する一方のアームと、HER2レセプターに結合するもう一方のアームとを有する二重特異性抗体が、HER2抗原を過剰発現する卵巣腫瘍および乳腫瘍を治療するために開発された。(Hseih−Maら、Cancer Research52:6832〜6839(1992)およびWeinerら、Cancer Research53:94〜100(1993))。二重特異性抗体はまた、T細胞による殺傷を媒介することができる。通常、二重特異性抗体は、T細胞上のCD3複合体を腫瘍関連抗原に結合させる。抗p185HER2に結合した抗CD3からなる完全にヒト化されたF(ab')2BsAbを使用して、T細胞の標的化が、HER2レセプターを過剰発現する腫瘍細胞を殺傷するために行われた。Shalabyら、J.Exp.Med.175(1):217(1992)。二重特異性抗体は、いくつかの初期の臨床試験において調べられ、有望な結果が得られている。1つの臨床試験において、肺ガン、卵巣ガンまたは乳ガンの12名の患者が、抗CD3/抗腫瘍(MOC31)の二重特異性抗体で標的化された活性化Tリンパ球を注入することによって処置された。deLeijら、「二重特異性抗体および標的化細胞の細胞傷害性」、Romet−Lemonne、FangerおよびSegal編、Leinhart(1991)249頁。
【0007】
標的化された細胞において、腫瘍細胞の相当の局所的な溶解、穏和な炎症反応が誘導されたが、毒性の副作用または抗マウス抗体の応答は誘導されなかった。B細胞の悪性疾患の患者での抗CD3/抗CD19二重特異性抗体の非常に予備的な試験において、末梢の腫瘍細胞数の大きな減少もまた達成された。Clarkら、「二重特異性抗体および標的化細胞の細胞傷害性」、Romet−Lemonne、FangerおよびSegal編、Leinhart(1991)243頁。BsAbの治療的適用に関してはKroesenら、CancerImmunol.Immunother.37:400〜407(1993)、Kroesenら、Br.J.Cancer70:652〜661(1994)、およびWeinerら、J.Immunol.152:2385(1994)もまた参照。
【0008】
二重特異性抗体はまた、フィブリン溶解剤またはワクチンアジュバントとして使用することができる。さらに、このような抗体は、感染性疾患の処置において、(例えば、エフェタター細胞を、HIVウイルスまたはインフルエンザウイルスなどのウイルスに感染した細胞あるいはトキソプラズマゴンディイ(Toxoplasmagondii)などの原生動物に対して標的化するために)使用することができ、あるいはイムノトキシンを腫瘍細胞に送達するために、あるいは免疫複合体を細胞表面レセプターに標的化するために使用することができる(Fangerら(上記)を参照)。
【0009】
BsAbの使用は、BsAbを十分な量および純度で得ることが困難であるということによって著しく妨げられている。従来、二重特異性抗体は、ハイブリッド−ハイブリドーマ技術(MillsteinおよびCuello、Nature305:537〜539(1983))を使用して作製された。免疫グロブリンの重鎖および軽鎖は無作為に組み合わされるために、これらのハイブリドーマ(クアドローマ)は、10個の異なる抗体分子の混合物を産生する可能性があり、このうちの1個のみが、正しい二重特異性構造を有する(図1Aを参照)。この正しい分子の精製は、通常的にはアフィニティークロマトグラフィー工程によって行われているが、かなり面倒であり、生成物の収量は少ない。例えば、(Smith,W.ら(1992)Hybridoma4:87〜98;およびMassimo,Y.S.ら(1997)J.Immunol.Methods201:57〜66)を参照。従って、より大きな収量のBsAbを産生するための技術が開発されている。抗体フラグメントの化学的な連結を行うために、Brennanら、Science 229:81(1985)は、無傷の抗体をタンパク質分解的に切断して、F(ab')2フラグメントを得る方法を記載する。
【0010】
このようなフラグメントは、ジチオール複合化剤である亜ヒ酸ナトリウムの存在下で還元され、近傍のジチオールを安定化させ、分子間ジスルフィド結合が形成されないようにする。次いで、得られたFab’フラグメントは、チオニトロ安息香酸(TNB)誘導体に変換される。次いで、BsAbを形成するために、1つのFab’−TNB誘導体が、メルカプトエチルアミンを用いた還元によってFab’−チオールに再度変換され、もう1つのFab’−TNB誘導体の等モル量と混合されて、生成したBsAbは、酵素を選択的に固定化するための薬剤として使用することができる。
近年の進歩は、二重特異性抗体を得るために化学的に連結させることができるFab’−SHフラグメントを大腸菌から直接回収することを容易にした。Shalabyら、J.Exp.Med.175:217〜225(1992)は、p185HER2と結合する一方のアームおよびCD3と結合するもう一方のアームを有する完全にヒト化されたBsAbのF(ab')2分子の産生を記載する。BsAbを形成するために、それぞれのFab’フラグメントを大腸菌から別個に分泌させ、インビトロでの指向された化学的結合反応に供した。このようにして形成されたBsAbは、ヒト乳腫瘍標的に対するヒト細胞傷害性リンパ球の溶解活性を開始させるのと同様に、HER2レセプターを過剰発現する細胞および正常なヒトT細胞に結合することができた。Rodriguesら、Int.J.Cancers(Suppl.)7:45〜50(1992)もまた参照。
【0011】
BsAbフラクメントを作製し、組換え細胞の培養物から直接分離するための様々な技術もまた記載されている。例えば、二重特異性のF(ab')2ヘテロダイマーが、ロイシンジッパーを使用して産生されている(Kostelnyら、J.Immunol.148(5):1547〜1553(1992))。Fosタンパク質およびJunタンパク質に由来するロイシンジッパーペプチドが、遺伝子融合によって抗CD3抗体および抗インターロイキン−2レセプター(IL−2R)抗体のFab’部分に連結された。ホモダイマーの抗体をヒンジ部で還元してモノマーを形成させ、次いで、再酸化してヘテロダイマーの抗体を形成させた。このようなBsAbは、インビトロでHuT−102細胞を溶解するために、細胞傷害性T細胞を呼び寄せるのに非常に効果的であることが見出された。Hollingerら、PNAS(USA)90:6444〜6448(1993)により記載される「ジアボディ(diabody)」技術の出現は、BsAbフラダメントを作製するための代わりの機構をもたらした。このフラグメントは、短すぎて同じ鎖の表面にある2つのドメインの間での対形成ができないリンカーによって軽鎖の可変ドメイン(VL)に連結された重鎖の可変ドメイン(VH)を含む。従って、1つのフラグメントのVHドメインおよびVLドメインは、別のフラグメントの相補的なVHドメインおよびVLドメインと対形成することを強いられ、それによって2つの抗原結合部位が形成される。単鎖Fv(sFv)のダイマーの使用によるBsAbフラグメントを作製するための別の方策もまた報告されている。Gruberら、J.Immunol.152:5368(1994)を参照。この研究者らは、25アミノ酸残基のリンカーによって抗フルオレセイン抗体のVHドメインおよびVLドメインに連結されたT細胞レセプターに対する抗体のVHドメインおよびVLドメインを含む抗体を設計した。再生された分子は、フルオレセインおよびT細胞レセプターに結合し、フルオレセインがその表面に共有結合しているヒト腫瘍細胞を溶解させた。
【0012】
組換え細胞の培養物から直接回収することができる二重特異性抗体を作製するためのいくつかの技術が報告されているようである。しかし、完全長のBsAbは、その血清半減期がより長いと考えられることおよび可能なエフェクター機能のために、多くの診療的適用に関しては、BsAbフラグメントよりも好ましいとされ得る。
免疫付着因子(Immunoadhesins) 免疫付着因子(Ia)は、細胞表面レセプターまたはリガンド(「付着因子」(adhesin))などのタンパク質の結合ドメインを、免疫グロブリンの定常ドメインのエフェタター機能とともに併せ持つ抗体様分子である。免疫付着因子は、ヒト抗体の多くの貴重な化学的特性および生物学的特性を有し得る。免疫付着因子は、適当なヒト免疫グロブリンのヒンジ配列および定常ドメイン(Fc)配列に連結された所望の特異性を有するヒトのタンパク質配列から構築することができるので、目的とする結合特異性は、完全なヒト成分を使用して達成することができる。そのような免疫付着因子は、患者に対する免疫原性は最小であり、安全に長期間または繰り返し使用される。
【0013】
文献に報告されている免疫付着因子には下記が含まれる。T細胞レセプターの融合体(Gascoigneら、Proc.Natl.Acad.Sci.USA84:2936〜2940(1987));CD4の融合体(Caponら、Nature337:525〜531(1989);Trauneckerら、Nature 339:68〜70(1989);Zettmeisslら、DNACell Biol.USA 9:347〜353(1990);およびByrnら、Nature344:667〜670(1990));L−セレクチンまたはホーミングレセプターの融合体(Watsonら、J.Cell.Biol.110:2221〜2229(1990);およびWatsonら、Nature349:164〜167(1991));CD44の融合体(Aruffoら、Cell61:1303〜1313(1990));CD28およびB7の融合体(Linsleyら、J.Exp.Med.173:721〜730(1991));CTLA−4の融合体(Lisleyら、J.Exp.Med.174:561〜569(1991));CD22の融合体(Stamenkovicら、Cell66:1133〜1144(1991));TNFレセプターの融合体(Ashkenaziら、Proc.Natl.Acad.Sci.USA88:10535〜10539(1991);Lesslauerら、Eur.J.Immunol.27:2883〜2886(1991);およびPeppelら、J.Exp.Med.174:1483〜1489(1991));NPレセプターの融合体(Bennettら、J.Biol.Chem.266:23060〜23067(1991));インターフェロンγレセプターの融合体(Kurschnerら、J.Biol.Chem.267:9354〜9360(1992));4−1BBの融合体(Chalupnyら、PNAS(USA)89:10360〜10364(1992))およびIgEレセプターαの融合体(RidgwayおよびGorman、J.Cell.Biol.第115巻、抄録番号1448(1991))。
【0014】
治療的使用について記載されている免疫付着因子の例には、細胞表面CD4に対するHIVの結合を阻止するためのCD4−IgG免疫付着因子が含まれる。
【0015】
CD4−IgGを出産直前の妊娠女性に投与した第I相臨床試験から得られたデータにより、この免疫付着因子は、HIVの母−胎児間移動を防止することにおいて有用であり得ることが示唆されている。Ashkenaziら、Intern.Rev.Immunol.10:219〜227(1993)。腫瘍壊死因子(TNF)と結合する免疫付着因子もまた開発された。TNFは、敗血症性ショックの主要なメディエーターであることが明らかにされている前炎症性サイトカインである。敗血症性ショックのマウスモデルに基づいて、TNFレセプター免疫付着因子は、敗血症性ショックの処置において臨床的に使用するための候補物として有望であることが明らかにされた(Ashkenaziら、上記)。免疫付着因子は、治療以外にもまた使用される。例えば、L−セレクチンレセプター免疫付着因子は、末梢リンパ節の高内皮細静脈(HEV)の組織化学的染色を行うための試薬として使用された。この試薬はまた、L−セレクチンのリガンドを分離してその特徴づけを行うために使用された(Ashkenaziら、上記)。免疫付着因子構造の2つのアームが異なる特異性を有する場合、免疫付着因子は、二重特異性抗体に対する類推によって「二重特異性免疫付着因子」と呼ばれる。
【0016】
Dietschら、J.Immunol.Methods162:123(1993)は、接着分子(E−セレクチンおよびP−セレクチン)の細胞外ドメインを併せ持つそのような二重特異性免疫付着因子を記載する。結合性の研究により、そのようにして得られた二重特異性免疫グロブリン融合タンパク質は、それが誘導された単一特異性の免疫付着因子と比較して、骨髄様細胞株に対する結合能か高まったことが示された。
抗体−免疫付着因子のキメラ 抗体−免疫付着因子(Ab/Ta)キメラもまた文献に記載されている。このような分子は、免疫付着因子の結合領域を抗体の結合ドメインとともに併せ持つ。
【0017】
Bergら、PNAS(USA)88:4723〜4727(1991)は、マウスのCD4−IgGから誘導された二重特異性の抗体−免疫付着因子キメラを作製した。この研究者らは、2つのアームを有する四量体分子を構築した。一方のアームは、抗体軽鎖の定常ドメインと融合したCD4とともに、抗体重鎖の定常ドメインと融合したCD4からなった。もう一方のアームは、抗CD3抗体の完全な軽鎖とともに、抗CD3抗体の完全な重鎖からなった。CD4−IgGのアームによって、この二重特異性分子は、細胞傷害性T細胞の表面にあるCD3に結合する。細胞傷害性細胞およびHIV感染細胞を一緒にすると、HIV感染細胞が特異的に殺傷される。
Bergら(上記)は四量体構造の二重特異性分子を記載しているが、1個のCD4−IgG融合体のみを含有するトリマーのハイブリッド分子を作製することができる。Chamowら、J.Immunol.153:4268(1994)を参照。この構築物の第1のアームは、ヒト化された抗CD3κ軽鎖およびヒト化された抗CD3γ重鎖によって形成される。第2のアームは、IgGのFcドメインによるgp120の結合を担うCD4の細胞外ドメインの一部を併せ持つCD4−IgG免疫付着因子である。得られるAb/Iaキメラは、HIV感染細胞の殺傷を、純粋な細胞傷害性T細胞調製物、またはFcレセプターを産生する大顆粒状リンパ球エフェクター細胞をさらに含む全末梢血リンパ球(PBL)画分のいずれかを使用して媒介した。
【0018】
多重特異性抗体ヘテロマルチマーの製造においては、ホモマルチマーよりも、所望のヘテロマルチマーの産生を増大させることが望ましい。Fc含有のBsAbを得るために選択される現在の方法は、依然として、2つの抗体を同時に発現するハイブリッドハイブリドーマである(MilsteinおよびCuello、Nature 305:537〜540(1983))。
【0019】
ハイブリッドハイブリドーマにおいて、重(H)鎖は、典型的には、所望のヘテロダイマーと同様に、ホモダイマーを形成する。さらに、軽(L)鎖は、同系でない重鎖との誤った対形成を形成することが多い。従って、2つの抗体が同時に発現すると、重鎖および軽鎖の10個までの対形成が生じ得る(Suresh,M.R.ら、MethodsEnzymol.121:210〜228(1986))。これらの望ましくない鎖の対形成は、BsAbの産生を低下させ、そして重大で、ときには克服できない精製問題を必然的に強いる(Smithら(1992)上記、およびMassimoら(1997)上記)。
【0020】
抗体重鎖は、立体的に相補的な変異をマルチマー化ドメインのCH3ドメイン界面に導入することによるヘテロダイマー化(Ridgwayら、ProteinEng.9:617〜621(1996))、および本明細書中に記載されるファージディスプレーによる最適化を行うために以前に操作されている。改変されたCH3ドメインを含有する鎖によって、抗体/免疫付着因子ハイブリッド(Ab/Ia)の生成により判断されるように、約90%までのヘテロダイマーが生じる。ヘテロダイマー化した重鎖は、依然として、同系でない軽鎖との誤った対形成を形成し得るので、目的のBsAbの回収が妨げられる。
【0021】
発明の要旨本出願は、モノマー混合物から所望するヘテロマルチマー二重特異性抗体の形成を高めるのに役立つ方法を記載する。この方法は、共通する可変軽鎖を提供して二重特異性抗体のヘテロマーの各可変重鎖領域と相互作用させることによるヘテロオリゴマー化のために第1のポリペプチドと第2のポリペプチドとの間の界面を操作することによる。3つの可能なヘテロマルチマーおよびホモマルチマーが、第1のポリペプチドおよび第2のポリペプチドから形成され得る。そのようなポリペプチドのそれぞれは、同様に、第1の軽鎖および第2の軽鎖とそれぞれ会合する。これにより、鎖の対形成は合計で10通りが可能である(図1A)。所望のヘテロマルチマー形成を高める方法によって、その産生を、望ましくないヘテロマルチマーおよびホモマルチマーよりも大きく高めることができる。
【0022】
ヘテロマルチマー抗体の第1のポリペプチドと第2のポリペプチドとの間の好ましい界面は、抗体の定常ドメインのCH3ドメインの少なくとも一部を含む。
【0023】
界面で相互作用する第1および第2の各ポリペプチドのドメインは、マルチマー化ドメインと呼ばれる。マルチマー化ドメインは、好ましくは、特定の第1のポリペプチドと第2のポリペプチドとの問の相互作用を促進し、それによって所望のヘテロマルチマーの産生を増大させる(図1B)。相互作用は、空洞への隆起(protuberance-into-cavity)の相補的な領域の形成;天然に存在しないジスルフィド結合の形成;ロイシンジッパー;疎水性領域;および親水性領域によって接触面で促進され得る。「隆起」(protuberance)は、第1のポリペプチドの界面に由来する小さなアミノ酸側鎖を、より大きな側鎖(例えば、チロシンまたはトリプトファン)と置換することによって構築される。隆起と同一の大きさまたは類似する大きさの代替的な「空洞」(cavity)は、任意に、大きなアミノ酸側鎖を、より小さな側鎖(例えば、アラニンまたはトレオニン)と置換することによって第2のポリペプチドの界面に作製される。適切に配置され、そして適切な大きさを有する隆起または空洞が第1のポリペプチドまたは第2のポリペプチドのいずれかの界面に存在する場合、隣接する接触面において、それぞれ、対応する空洞または隆起の設計が必要とされるだけである。天然に存在しないジスルフィド結合は、第1のポリペプチドにおいて、天然に存在するアミノ酸を、システインなどの遊離チオールを含有する残基と置換して、遊離チオールが第2のポリペプチドの別の遊離チオール含有側鎖と相互作用し、そしてジスルフィド結合が第1のポリペプチドと第2のポリペプチドとの間で形成されるようにすることによって構築される(図1B)。
【0024】
大きな非免疫化ファージディスプレーライブラリー(Vaughan,T.J.ら(1996)Nature Biotechnology14:309〜314、これは参考としてその全体が本明細書中に組込まれる)に由来する単鎖Fvフラグメントは、V-遺伝子の使用を明らかにした。この場合、いくつかの生殖系列のV-遺伝子セグメントに由来するVH配列およびVL配列が優勢であり、ファミリーがレパートリーにおいて優勢であった。異なるパートナー鎖との組合せで特定の重鎖または軽鎖が見出されるレパートリーにおいて、鎖の入り交じった状態の例が認められた(Vaughan,T.J.ら(1996)上記)。
【非特許文献1】Fangerら、Crit.Rev.Immunol.12:101〜124(1992)
【発明の開示】
【発明が解決しようとする課題】
【0025】
所望するヘテロマルチマー多重特異性抗体の調製は、共通する軽鎖が多重特異性抗体の可変重鎖のそれぞれと対形成するように提供される場合に増強されることが本明細書中に開示される。共通する可変軽鎖の使用によって、抗原結合ドメインを形成するために正しく対形成しなければならないモノマーの数が減少する。これは、軽鎖の数を、(本発明が開示される前の二重特異性抗体または多重特異性抗体のそれぞれにおける)2つ以上の軽鎖から、(本発明の多重特異性抗体(図1Cを参照)における)1つの軽鎖に制限することによる。
【課題を解決するための手段】
【0026】
従って、本発明は、ヘテロマルチマー多重特異性抗体を調製する方法に関する。この抗体は、下記の1)および2)を含む:1)界面で接する第1のポリペプチドおよび第2のポリペプチド(および抗体の多重度に従ってさらなるポリペプチド)、ただし、第1のポリペプチドおよびさらなるポリペプチド(または、第1のポリペプチドおよび第2のポリペプチド)のそれぞれは、第1のポリペプチドと第2の(または、少なくとも1つのさらなる)ポリペプチドとの間の界面を形成するマルチマー化ドメインを含み、そしてこのマルチマー化ドメインは、第1のポリペプチドとさらなるポリペプチドとの間の安定な相互作用を促進する;および2)前記の第1のポリペプチドおよび少なくとも1つのさらなるポリペプチド(または、第2のポリペプチド)のそれぞれの結合ドメイン、ただし、各結合ドメインは可変重鎖および可変軽鎖を含み、第1のポリペプチドの可変軽鎖および第2のポリペプチドの可変軽鎖は共通するアミノ酸配列を有し、そしてそのような共通する配列は、前記ポリペプチドのそれぞれの元の軽鎖に対して、少なくとも80%、好ましくは少なくとも90%、より好ましくは少なくとも95%、最も好ましくは100%の配列同一性のアミノ酸配列同一性を有する。この方法は下記の工程を含む:(i)第1のポリペプチド、第2のポリペプチドおよび共通する軽鎖をコードする核酸を含む宿主細胞を培養し、この培養によって核酸を発現させる工程;および (ii)宿主細胞培養物から多重特異性抗体を回収する工程。
【0027】
本発明の関連する実施態様において、第1のポリペプチドをコードする核酸または第2のポリペプチドをコードする核酸、あるいはその両方は、その界面またはその一部をコードするように元の核酸から変更されている。
本発明の方法の別の実施態様において、第1のポリペプチドの界面は、第2のポリペプチドの界面の遊離チオール含有残基と相互作用するように配置され、そうすることによってジスルフィド結合が第1のポリペプチドと第2のポリペプチドとの間で形成される遊離チオール含有残基を含む。本発明により、第1のポリペプチドをコードする核酸は、遊離チオール含有残基をコードするように元の核酸から変更され、あるいは第2のポリペプチドをコードする核酸は、遊離チオール含有残基をコードするように元の核酸から変更され、あるいはその両方である。
【0028】
本発明の方法の別の実施態様において、第1のポリペプチドおよび少なくとも1つのさらなるポリペプチド(または、第2のポリペプチド)の両方をコードする核酸は、それぞれ、隆起および空洞をコードするように変更されている。第1のポリペプチドおよび第2のポリペプチドは、それぞれ、好ましくは、ヒトIgG1のCH3ドメインなどの抗体の定常ドメインを含む。
【0029】
別の態様において、本発明は、界面で接する第1のポリペプチドおよび第2のポリペプチドを含むヘテロマルチマー(二重特異性抗体、二重特異性免疫付着因子または抗体/免疫付着因子キメラなど)を提供する。第1のポリペプチドの界面は、少なくとも1つのさらなるポリペプチド(または、第2のポリペプチド)
【0030】
上のマルチマー化ドメインと相互作用するように配置されて、第1のポリペプチドと第2のポリペプチドとの界面を形成するマルチマー化ドメインを含む。本発明の好ましい実施態様において、マルチマー化ドメインは、特定の第1のポリペプチドと特定の第2のポリペプチドとの間の相互作用が促進されるように変更される。そのような変更には、隆起または空洞あるいはその両方の生成;天然に存在しないジスルフィド結合の生成;相補的な疎水性領域;および相補的な親水性領域の生成が含まれるが、これらに限定されない。ヘテロマルチマー多重特異性抗体は、薬学的に受容可能な担体をさらに含む組成物の形態で提供され得る。
【0031】
本発明はまた、前記のヘテロマルチマー多重特異性抗体をコードする核酸を含む宿主細胞に関する:この場合、第1のポリペプチドおよび少なくとも1つのさらなるポリペプチド(または、第2のポリペプチド)をコードするこの核酸は、1つのベクターまたは別個のベクターに存在する。宿主細胞は、ヘテロマルチマー多重特異性抗体を作製する方法で使用することができる。この方法は、核酸を発現するように宿主細胞を培養すること、およびその細胞培養物からヘテロマルチマー抗体を回収することを含む。
【0032】
さらなる態様において、本発明は、ヘテロマルチマー多重特異性抗体を調製する方法を提供し、この方法は下記の工程を含む:(a)少なくとも1つのさらなるポリペプチドの界面のアミノ酸と相互作用するように配置されているアミノ酸残基を第1のポリペプチドの界面に含む第1のポリペプチドをコードする第1の核酸を選択する工程。実施態様において、核酸は、相互作用するアミノ酸残基をコードするように元の核酸から変更されている。
【0033】
別の実施態様において、第1の核酸は、より大きな側鎖容量を有するアミノ酸をコードするように変更され、それによって、隆起か第1のポリペプチドに形成される;
(b)第2のポリペプチドをコードする第2の核酸を変更し、その結果、第2のポリペプチドの界面内のアミノ酸残基を、より小さな側鎖容量を有するアミノ酸残基と置換し、それによって、第2のポリペプチドに空洞を形成させる工程。
【0034】
この場合、前記の隆起は、この空洞と相互作用するように配置される;
(c)前記の第1の核酸および第2の核酸を宿主細胞に導入して、前記の第1の核酸および第2の核酸が発現されるように宿主細胞を培養する工程;および (d)細胞培養物から生成したヘテロマルチマー抗体を回収する工程。
【0035】
前記の抗体が組み込まれた多重特性抗体(二重特異性抗体など)を構築することもまた望ましいことであり得る。このような状況下では、元の軽鎖と対形成する場合、目的の第2の抗原に特異的に結合する重鎖を同定することは望ましい。
【0036】
Figiniらの方法(Figini,M.ら(1994)J.Mol.Biol.239:68〜78、これは参考としてその全体が本明細書中に組込まれる)を使用して、そのような重鎖を同定することができる。最初に、ファージライブラリーをグアニジン塩酸で処理して、元の軽鎖を解離させる。次に、ファージにディスプレーされた重鎖を、(透析などにより)変性剤を除くことによって目的の軽鎖と再構成させる。次いで、目的の第2の抗原に対して選別を行い、所望の重鎖を同定する。本発明はさらに、選択された軽鎖と対形成するように重鎖を選択するこの方法によって調製される多重特異性抗体、そのような抗体をコードする核酸、およびそのような核酸を含む宿主細胞を含む。
【0037】
本発明は、望ましくないヘテロマルチマーおよび/またはホモマルチマーなどの他の望ましくない最終産物よりもヘテロマルチマーの産生を増大させるための機構を提供する(図1A〜図1Cを参照)。組換え細胞の培養物から回収される所望のヘテロマルチマーの産生量は、好ましくは、副生成物の望ましくないヘテロダイマーまたはホモマルチマーと比較して、少なくとも80重量%を超え、好ましくは少なくとも90重量%を超える。
【発明を実施するための最良の形態】
【0038】
I.定義 一般に、下記の用語または表現は、本説明、実施例および請求項において使用される場合には下記の意味を有する。
「ヘテロマルチマー」,「ヘテロマルチマーポリペプチド」または「ヘテロマルチマー多重特異性抗体」は、少なくとも、第1のポリペプチドおよび第2のポリペプチドを含む分子である。この場合、第2のポリペプチドは、アミノ酸配列において、第1のポリペプチドと少なくとも1個のアミノ酸残基が異なる。ヘテロマルチマーは、少なくとも2つの異なるリガンドに対する複数の結合特異性または結合部位を有する。好ましくは、ヘテロマルチマーは、第1のポリペプチドおよび第2のポリペプチドによって形成される「ヘテロダイマー」を含むことができ、あるいはポリペプチドが、第1のポリペプチドおよび第2のポリペプチド以外に、さらに存在する場合には、より高次の三次構造を形成することができる。
【0039】
ヘテロマルチマーに関する例示的な構造には、ヘテロダイマー(例えば、Dietschら(上記)によって記載される二重特異性免疫付着因子)、ヘテロトリマー(例えば、Chamowら(上記)によって記載されるAb/Iaキメラ)、ヘテロ四量体(例えば二重特異性抗体)及び更なるオリゴマー構造が含まれる。
【0040】
本明細書中で使用されている「マルチマー化ドメイン」は、ヘテロマルチマーの各ポリペプチドの領域をいう。「マルチマー化ドメイン」は、ヘテロマルチマー複合体内のキメラ分子の安定な相互作用を促進する。マルチマー化ドメインは、好ましくは、特定の第1のポリペプチドと特定の第2のポリペプチドとの間の相互作用を促進し、それによって所望のヘテロマルチマーの形成が増強され、そして望ましくないヘテロマルチマーまたはホモマルチマーの形成の可能性が実質的に低下する。マルチマー化ドメインは、免疫グロブリン配列、ロイシンジッパー、疎水性領域、親水性領域、またはキメラなヘテロマルチマーのキメラ分子間での分子間ジスルフィド結合を形成する遊離チオールを介して相互作用することができる。遊離チオールは、ポリペプチド間のジスルフィド結合の形成を可能にする位置において、ポリペプチドの天然に存在する残基を、例えば、システインと置換することによって1つまたは複数の相互作用ポリペプチドの界面に導入することができる。マルチマー化ドメインは、免疫グロブリンの定常部を含むことができる。本発明において有用で可能なマルチマー化ドメインが、ハイブリッドの免疫グロブリンを記載する国際特許出願第PCT/US90/06849号(これは参考としてその全体が本明細書中に組込まれる)に開示されている。さらに、マルチマー化ドメインは、立体的な相互作用によって、安定な相互作用が促進されるだけでなく、モノマーの混合物に由来するホモダイマーよりもヘテロダイマーの形成がさらに促進されるように設計することができる。例えば、国際特許出願第PCT/US96/01598号(これは参考としてその全体が本明細書中に組込まれる)を参照。これは、ヘテロオリゴマー化のために第1のポリペプチドと第2のポリペプチドとの間の相互作用に関する「空洞への隆起」法を開示する。「隆起」は、第1のポリペプチドの界面に由来する小さなアミノ酸側鎖を、より大きな側鎖(例えば、チロシンまたはトリプトファン)と置換することによって構築される。隆起と同一の大きさまたは類似する大きさの代償的な「空洞」は、任意に、大きなアミノ酸側鎖を、より小さな側鎖(例えば、アラニンまたはトレオニン)と置換することによって第2のポリペプチドの界面に作製される。
【0041】
免疫グロブリン配列は、免疫グロブリンの定常ドメインであることが好ましいが、必ずしも、免疫グロブリンの定常ドメインである必要はない。本発明のキメラにおける免疫グロブリン部分は、IgG1、IgG2、IgG3またはIgG4のサブタイプ、IgA、IgE、IgDまたはIgMから得ることができるが、IgG1、IgG2、IgG3またはIgG4が好ましい。
【0042】
「遊離チオール含有化合物」は、本発明のポリペプチド界面のアミノ酸に組み込むことができるが、またはそのようなアミノ酸と反応することができる化合物であって、この化合物の遊離チオール部分は、本発明のさらなるポリペプチドの界面において遊離チオール部分と相互作用して、ジスルフィド結合が形成されるように配置されているそのような化合物を意味する。遊離チオール含有化合物は、好ましくは、システインである。
用語「標識されたエピトープ」は、本明細書で使用される場合、「標識ポリペプチド」に融合したキメラなヘテロ付着因子の全体またはそのフラグメントを含むキメラなポリペプチドを示す。標識ポリペプチドは、抗体が作製され得るエピトープを提供するのに十分な残基を有するが、キメラなヘテロ付着因子の活性を妨げないように十分に短い。標識ポリペプチドは、好ましくは、他にないほど非常に特徴的であり、その結果、それに対する抗体は、他のエピトープと実質的に交差反応しない。適切な標識ポリペプチドは、一般には、少なくとも6個のアミノ酸残基を有し、通常は、8個〜50個の間のアミノ酸残基(好ましくは、約9残基〜30残基の間)を有する。本発明の実施態様は、エピトープ標識に結合したキメラなヘテロ付着因子を含み、そのような標識を使用して、サンプル中の付着因子の検出またはサンプルからの付着因子の回収が行われる。
【0043】
本明細書中で使用されている「共通する軽鎖」または「軽鎖の共通するアミノ酸配列」は、本発明の多重特異性抗体における軽鎖のアミノ酸配列をいう。抗体パネルが、ファージディスプレーライブラリーの選別を行うことによって、少なくとも2つの異なる抗原に対して作製された。そのようなファージディスプレーライブラリーは、例えば、Vaughanら(1996)(上記)により記載されている(これは、ファージミドライブラリーを選択する方法を特に参照して、参考としてその全体が本明細書中に組込まれる)。軽鎖配列を、可変軽鎖アミノ酸配列に関して比較した。比較されたパネルに由来する有用な軽鎖は、少なくとも80%、好ましくは少なくとも90%、より好ましくは少なくとも95%、最も好ましくは100%の同一性のアミノ酸配列同一性を有する軽鎖である。共通する軽鎖配列は、比較された2つの軽鎖配列が近似するように考えられた配列である。比較される軽鎖が、アミノ酸レベルで100%の配列同一性である場合、共通する軽鎖は、軽鎖が多重特異性抗体の異なる結合ドメインにおいて機能するとして、選択されたライブラリーのクローンに由来する軽鎖と同一である。比較される軽鎖が上記と異なる場合、共通する軽鎖は、ライブラリーのクローンに由来する比較される軽鎖の1つまたはもう一方、あるいはその両方と異なり得る。共通する軽鎖が、ライブラリーのクローンの1つまたはもう一方と、あるいはその両方と異なる場合、異なる残基は、抗体軽鎖の抗原結合CDR残基の外側に存在することが好ましい。例えば、抗原結合CDR残基の位置は、配列定義(Kabatら(1991)上記)または構造定義(ChothiaおよびLesk(1987)J.Mol.Biol.196:901〜917)に従って決定することができる。
【0044】
本明細書中で用いている「アミノ酸配列同一性」は、1つの配列のアミノ酸が第2のアミノ酸配列のアミノ酸とどのくらい同じであるかの割合をいう。ポリペプチド鎖の間において100%の配列同一性は、鎖が同一であることを意味する。
【0045】
本明細書中で使用されている「ポリペプチド」は、一般に、約10個よりも多いアミノ酸を有するペプチドおよびタンパク質をいう。好ましくは、哺乳動物のポリペプチド(哺乳生物から最初に得られたポリペプチド)が使用され、より好ましくは、培地に直接分泌されるポリペプチドである。細菌のポリペプチドの例には、例えば、アルカリホスファターゼおよびβ−ラクタマーゼが含まれる。哺乳動物のポリペプチドの例には、レニン、成長ホルモンなどの分子が含まれ、下記が含まれる:ヒト成長ホルモン;ウシ成長ホルモン;成長ホルモン放出因子;
副甲状腺ホルモン;甲状腺刺激ホルモン;リポタンパク質;α−1抗トリプシン;インシュリンA鎖;インシュリンB鎖;プロインシュリン;卵胞刺激ホルモン;カルシトニン;黄体化ホルモン;グルカゴン;第VIIIC因子、第IX因子、組織因子およびフォンビルブランド因子などの凝固因子;プロテインCなどの抗凝固因子;心房性ナトリウム利尿因子;肺接触面活性物質;ウロキナーゼまたはヒトウリンまたは組織型プラスミノーゲン活性化因子(t−pA)などのプラスミノーゲン活性化因子;ボンベシン;トロンビン;造血成長因子;腫瘍壊死因子−αおよび腫瘍壊死因子−β;エンケファリナーゼ;RANTES(regulated on activation normallyT−cell expressed andsecreted);ヒトマクロファージ炎症タンパク質(MIPI−1−α);ヒト血清アルブミンなどの血清アルブミン;ミューラー阻害物質;レラキシンA鎖;レラキシンB鎖;プロレラキシン;マウス性腺刺激ホルモン関連ペプチド;β−ラクタマーゼなどの微生物タンパク質;DNase;インヒビン;アクチビン;血管内皮細胞増殖因子(VEGF);ホルモンまたは成長(増殖)因子のレセプター;インテグリン;プロテインAまたはプロテインD;リウマチ因子;骨由来神経栄養因子(BDNF)、ニューロトロフィン−3、ニューロトロフィン−4、ニューロトロフィン−5またはニューロトロフィン−6(NT−3、NT−4、NT−5またはNT−6)、あるいはNGF−βなどの神経増殖因子などの神経栄養因子;血小板由来増殖因子(PDGF);aFGFおよびbFGFなどの繊維芽細胞増殖因子;上皮増殖因子(EGF);TGF−β1、TGF−β2、TGF−β3、TGF−β4またはTGF−β5を含むTGF−αおよびTGF−βなどのトランスホーミング増殖因子(TGF);
インシュリン様増殖因子−Iおよびインシュリン様増殖因子−II(IGF−IおよびIGF−II);des(1−3)−IGF−I(脳IGF−I)、インシュリン様増殖因子結合タンパク質;CD−3、CD−4、CD−8およびCD−19などのCDタンパク質;エリスロポイエチン;骨誘導因子;イムノトキシン;骨形態形成タンパク質(BMP);インターフェロン−α、インターフェロン−βおよびインターフェロン−γなどのインターフェロン;コロニー刺激因子(CSF)類、例えば、M−CSF、GM−CSFおよびG−CSF;インターロイキン(IL)類、例えば、IL−1からIL−10;スーパーオキシドジスムターゼ;T細胞レセプター;表面膜タンパク質;崩壊促進因子;例えば、AIDSエンベロープの一部などのウイルス抗原;輸送タンパク質;ホーミングレセプター;アドレシン;調節タンパク質;抗体;および上記のポリペプチドのいずれかのフラグメント。
【0046】
「第1のポリペプチド」は、第2のポリペプチドと会合し得る任意のポリペプチドである。第1のポリペプチドおよび第2のポリペプチドは、「界面」(下記において定義)で接する。界面に加えて、第1のポリペプチドは、「結合ドメイン」(例えば、抗体の可変ドメイン、レセプター結合ドメイン、リガンド結合ドメインまたは酵素活性ドメイン)、またはCH2、CH1およびCLドメインを含む抗体の定常ドメイン(またはその一部)などのさらなるドメインを1つまたは複数含むことができる。通常、第1のポリペプチドは、抗体から誘導される少なくとも1つのドメインを含む。このようなドメインは、好都合なことに、抗体のCH3ドメインなどの定常ドメインであり、第1のポリペプチドの界面を形成することができる。例示的な第1のポリペプチドには下記が含まれる。抗体重鎖ポリペプチド、異種ポリペプチドの結合ドメインとともに抗体の定常ドメインを併せ持つキメラ(すなわち、免疫付着因子、下記の定義を参照)、レセプターポリペプチド(特に、別のレセプターポリペプチド、例えば、インターロイキン−8レセプター(IL−8R)およびインテグリンヘテロダイマー(例えば、LFA−IまたはGPIIIb/IIIa)とダイマーを形成するポリペプチド)、リガンドポリペプチド(例えば、神経増殖因子(NGF)、ニューロトロフィン−3(NT−3)および脳由来神経栄養因子(BDNF)−Arakawaら、J.Biol.Chem.269(45):27833〜27839(1994)およびRadziejewskiら、Biochem.32(48):1350(1993))、および抗体の可変ドメインポリペプチド(例えば、ジアボディ(diabody))。好ましい第1のポリペプチドは、免疫グロブリンの定常ドメインに融合した抗体の重鎖から選択され、この場合、定常ドメインは、本発明の第2のポリペプチドとの優先的な相互作用が促進されるように変更されている。
【0047】
「第2のポリペプチド」は、「界面」を介して第1のポリペプチドと会合し得る任意のポリペプチドである。界面に加えて、第2のポリペプチドは、「結合ドメイン」(例えば、抗体の可変ドメイン、レセプター結合ドメイン、リガンド結合ドメイン、または酵素活性ドメイン)、またはCH2ドメイン、CH1ドメインおよびCLドメインを含む抗体の定常ドメイン(または、その一部)などのさらなるドメインを含むことができる。通常、第2のポリペプチドは、抗体から誘導される少なくとも1つのドメインを含む。このようなドメインは、好都合なことに、抗体のCH3ドメインなどの定常領域であり、第2のポリペプチドの界面を形成することができる。例示的な第2のポリペプチドには下記が含まれる。抗体重鎖ポリペプチド、異種ポリペプチドの結合ドメインとともに抗体の定常ドメインを併せ持つキメラ(すなわち、免疫付着因子、下記の定義を参照)、レセプターポリペプチド (特に、別のレセプターポリペプチド、例えば、インターロイキン−8レセプター(IL−8R)およびインテグリンヘテロダイマー(例えば、LFA−IまたはGPIIIb/IIIa)とダイマーを形成するポリペプチド)、リガンドポリペプチド(例えば、神経増殖因子(NGF)、ニューロトロフィン−3(NT−3)および脳由来神経栄養因子(BDNF)−Arakawaら、J.Biol.Chem.269(45):27833〜27839(1994)およびRadziejewskiら、Biochem.32(48):1350(1993))、および抗体の可変ドメインポリペプチド(例えば、ジアボディ)。好ましい第2のポリペプチドは、免疫グロブリンの定常ドメインに融合した抗体の重鎖から選択されこの場合、この場合、定常ドメインは、本発明の第1のポリペプチドとの優先的な相互作用が促進されるように変更されている。
【0048】
「結合ドメイン」は、目的の分子(例えば、抗体、リガンド、レセプター、基質または阻害剤)との選択的な結合を担うポリペプチドの任意の領域を含む。例示的な結合ドメインには、抗体の可変ドメイン、レセプター結合ドメイン、リガンド結合ドメインおよび酵素活性ドメインが含まれる。好ましい実施態様において、結合ドメインは、免疫グロブリンの重鎖および軽鎖を含む。本発明の二重特異性抗体およびその作製方法により、二重特異性抗体の各結合ドメインに関する軽鎖は、共通する軽鎖であり、それによって重鎖および軽鎖の誤った対形成が存在する望ましくないヘテロマルチマーの形成が避けられる。
用語「抗体」は、それが本発明に関連する場合、目的の抗原のエピトープと結合するドメインを1つまたは複数含有するポリペプチドを意味するものとする。
【0049】
この場合、そのようなドメインは、抗体の可変部から誘導されるか、または抗体の可変部との配列同一性を有する。抗体の例には、全長の抗体、抗体フラグメント、単鎖分子、二重特異性分子または二機能性分子、ジアボディー、キメラ抗体(例えば、ヒト化抗体およびPRIMATIZEDTM抗体)および免疫付着因子が含まれる。「抗体フラグメント」には、Fv、Fv'、Fab、Fab’およびF(ab')2のフラグメントが含まれる。
【0050】
「ヒト化(された)」形態の非ヒト(例えば、齧歯類または霊長類)抗体は、非ヒト免疫グロブリンから誘導される最小の配列を含有する特異的でキメラな免疫グロブリン、免疫グロブリン鎖またはそのフラグメントである。その大部分において、ヒト化抗体は、受容者の相補性決定領域(CDR)に由来する残基が、所望の特異性、親和性および能力を有するマウス、ラット、ウサギまたは霊長類などの非ヒト種(ドナー抗体)のCDRに由来する残基によって置き換えられているヒト免疫グロブリン(受容体抗体)である。場合によっては、ヒト免疫グロブリンのFv枠組み構造領域(FR)残基は、対応する非ヒト残基によって置き換えられる。さらに、ヒト化抗体は、受容体抗体あるいは持ち込まれたCDR配列または枠組み構造配列のいずれにおいて見出されない残基を含むことができる。このような改変は、抗体の能力を改良して最大にするために行われる。一般に、ヒト化抗体は、少なくとも1つ、典型的には2つの可変ドメインの実質的に全てを含む。そのような可変ドメインにおいて、CDR領域の全て又は実質的に全ては、非ヒトの免疫グロブリンのCDR領域に対応し、そしてFR領域の全て又は実質的に全ては、ヒト免疫グロブリン配列のFR領域である。ヒト化抗体はまた、好ましくは、免疫グロブリンの定常部(Fc)、典型的には、ヒト免疫グロブリンの定常部(Fc)の少なくとも一部を含む。ヒト化抗体は、抗体の抗原結合領域が、目的の抗原でマカク(macaque)サルを免疫化することによって産生される抗体から誘導されるPRIMATIZEDTM抗体を含む。
【0051】
「多重特異性抗体」は、少なくとも2つの異なる抗原に対する結合特異性を有する分子である。そのような分子は、通常、2つの抗原と結合するだけである(すなわち、二重特異性抗体、BsAb)が、三重特異性抗体などのさらなる特異性を有する抗体も、本明細書中で使用される場合には本発明により含まれる。BsAbの例には、腫瘍細胞の抗原に対する一方のアームと、細胞傷害誘引因子分子に対するもう一方のアームとを有する下記のような二重特異性抗体が含まれる。例えば、抗FcγRI/抗CD15、抗p185HER2/FcγRIII(CD16)、抗CD3/抗悪性B細胞(1D10)、抗CD3/抗p185HER2、抗CD3/抗p97、抗CD3/抗腎細胞ガン、抗CD3/抗OVCAR−3、抗CD3/L−D1(抗結腸ガン)、抗CD3/抗メラニン細胞刺激ホルモンアナログ、抗EGFレセプター/抗CD3、抗CD3/抗CAMA1、抗CD3/抗CD19、抗CD3/MoV18、抗神経細胞付着因子分子(NCAM)/抗CD3、抗葉酸結合タンパク質(FBP)/抗CD3、抗汎カルシノーマ関連抗原(AMOC−31)/抗CD−3;腫瘍抗原に特異的に結合するアームと、毒素に結合するアームとを有するBsAb、例えば、抗サポリン/抗Id−1、抗CD22/抗サポリン、抗CD7/抗サポリン、抗CD38/抗サポリン、抗CEA/抗リシンA鎖、抗インターフェロン−α(INF−α)/抗ハイブリドーマイディオタイプ、抗CEA/抗ビンカアルカロイドなど;変換酵素によって活性化されるプロドラッグに対するBsAb、例えば、抗CD30/抗アルカリホスファターゼ(これは、マイトマイシンホスフェートプロドラッグのマイトマシンアルコールへの変換を触媒する)など;フィブリン溶解剤として使用することができるBsAb、例えば、抗フィブリン/抗組織プラスミノーゲン活性化因子(tPA)、抗フィブリン/抗ウロキナーゼ型プラスミノーゲン活性化因子(uPA)など;細胞表面レセプターに対して免疫複合体を標的化するためのBsAb、例えば、抗低密度リポタンパク質(LDL)/抗Fcレセプター(例えば、FcγRI、FcγRIIまたはFcγRIII)など;感染性疾患の治療において使用されるBsAb、例えは、抗CD3/抗単純ヘルペスウイルス (HSV)、抗T細胞レセプター:CD3複合体/抗インフルエンザ、抗FcγR/抗HIVなど;インビトロまたはインビボで腫瘍を検出するためのBsAb、例えば、抗CEA/抗EOTUBE、抗CEA/抗DPTA、抗p185HER2/抗ハプテン;ワクチンアジュバントとしてのBsAb(Fangerら(上記)を参照);および診断用具としてのBsAb、例えば、抗ウサギIgG/抗フェリチン、抗ホースラディッシュペルオキシダーゼ(HRP)/抗ホルモン、抗ソマトスタチン/抗サブスタンスP、抗HRP/抗FITC、抗CEA/抗β−ガラクトシダーゼ(Nolanら(上記)を参照)など。三重特異性抗体の例には、抗CD3/抗CD4/抗CD37、抗CD3/抗CD5/抗CD37および抗CD3/抗CD8/抗CD37が含まれる。
【0052】
本明細書中で使用されている用語「免疫付着因子」は、免疫グロブリンの定常ドメインのエフェクター機能とともに、異種タンパク質(「付着因子」、例えば、レセプター、リガンドまたは酵素)の「結合ドメイン」を併せ持つ抗体様の分子を示す。構造的には、免疫付着因子は、抗体の抗原認識部位および結合部位(抗原結合部位)以外である(すなわち、「異種である」)所望の結合特異性を有する付着因子のアミノ酸配列と、免疫グロブリンの定常ドメイン配列との融合体を含む。免疫付着因子における免疫グロブリンの定常ドメイン配列は、IgG1、IgG2、IgG3もしくはIgG4のサブタイプ、IgA、IgE、IgDまたはIgMなどの任意の免疫グロブリンから得ることができる。
【0053】
本明細書中で使用されている用語「リガンド結合ドメイン」は、定量的なリガンド結合能、および好ましくは対応する天然のレセプターの生物学的活性を少なくとも保持する任意の天然の細胞表面レセプターまたはその任意の領域もしくは誘導体をいう。特定の実施態様において、レセプターは、免疫グロブリンスーパーファミリーのメンバーと相同的な細胞外ドメインを有する細胞表面ポリペプチドに由来する。免疫グロブリンスーパーファミリーのメンバーではないが、それにもかかわらず、この定義によって具体的に含まれる他の代表的なレセプターは、サイトカインに対するレセプターであり、特にチロシンキナーゼ活性を有するレセプター(レセプターチロシンキナーゼ)であり、そしてヘマトポイエチンおよび神経増殖因子レセプタースーパーファミリーのメンバー、ならびに細胞付着因子分子、例えは、(E、LおよびP−)セレクチンである。
【0054】
用語「レセプター結合ドメイン」は、定量的なレセプター結合能、および好ましくは対応する天然のリガンドの生物学的活性を少なくとも保持するレセプターに対する任意の天然のリガンド(細胞付着因子分子を含む)またはその任意の領域もしくは誘導体を示すために使用される。この定義は、特に、具体的には、上記のレセプターに対するリガンドに由来する結合配列を含む。
【0055】
本明細書中で使用されている用語「多重特異性免疫付着因子」は、少なくとも2つの結合特異性を有する(すなわち、2つ以上の付着因子結合ドメインを併せ持つ)(上記に定義されている)免疫付着因子を示す。多重特異性免疫付着因子は、本質的には、国際特許公開第WO89/02922号(1989年4月6日公開)、欧州特許EP314,317(1989年5月3日公開)および米国特許第5,116,964号(1992年5月2日発行)に開示されているように、ヘテロダイマー、ヘテロトリマーまたはヘテロ四量体として組み立てることができる。好ましい多重特異性免疫付着因子は二重特異性である。二重特異性免疫付着因子の例には、CD4−TgG/INFレセプター−IgGおよびCD4−1gG/L−セレクチン−IgGが含まれる。後者の分子は、リンパ球ホーミングレセプター(LHR、L−セレクチン)のリンパ節結合機能と、CD4のHIV結合機能とを併せ持ち、HIV感染、関連する症状の予防または処置において診断薬としての適用の可能性が見出されている。
【0056】
「抗体−免疫付着因子キメラ(Ab/Iaキメラ)」は、(本出願において定義されている)少なくとも1つの免疫付着因子とともに、(上記に定義されている)抗体の少なくとも1つの結合ドメインを併せ持つ分子を含む。例示的なAb/Iaキメラは、Bergら(上記)およびChamowら(上記)によって記載されている二重特異性CD4−IgGキメラである。
【0057】
「界面」は、第2のポリペプチドの界面内において1つまたは複数の「接触」アミノ酸残基(または、アミノ酸でない他の基)と相互作用する第1のポリペプチドにおける「接触」アミノ酸残基(あるいは、炭水化物基、NADH、ピオチン、FADまたはヘム基などのアミノ酸でない他の基)を含む。好ましい界面は、可変ドメインまたは定常ドメイン(またはその領域)などの免疫グロブリンのドメインである。しかし、ヘテロマルチマーレセプターを形成するポリペプチド間の界面、またはNGF、NT−3およびBDNFなどの2つ以上のリガンドの間の界面も、この用語の範囲に含まれる。好ましい界面は、好ましくはIgG抗体から誘導され、最も好ましくはヒトIgG1抗体から誘導される免疫グロブリンのCH3ドメインを含む。
【0058】
「元の」アミノ酸残基は、元の残基よりも小さな側鎖容量または大きな側鎖容量を有し得る「移入」残基によって置換されるアミノ酸残基である。移入アミノ酸残基は、天然存在アミノ酸残基または非天然存在アミノ酸残基であり得るが、天然存在アミノ酸残基が好ましい。「天然存在」アミノ酸残基は、遺伝暗号によってコードされるアミノ酸残基、および国際特許出願第PCT/US96/01598号(これは参考としてその全体が本明細書中に取込まれる)の表1に列記されるアミノ酸残基である。「非天然存在」アミノ酸残基は、遺伝暗号によってコードされていないが、ポリペプチド鎖で隣のアミノ酸残基と共有結合することができる残基を意味する。非天然存在アミノ酸残基の例には、例えば、ノルロイシン、オルニチン、ノルバリン、ホモセリン、およびEllmanら、Meth.Enzym.202:301〜336(1991)に記載されるアミノ酸残基アナログなどの他のアミノ酸残基のアナログが挙げられる。天然に存在しないそのようなアミノ酸残基を作製するために、Norenら、Science244:182(1989)およびEllmanら(上記)の方法を使用することができる。簡単に記載すると、この方法は、非天然存在アミノ酸残基でサプレッサーtRNAを化学的に活性化し、その後、インビトロでRNAの転写および翻訳を行うことを含む。本発明の方法には、少なくとも1つの元のアミノ酸残基を置換することが含まれるが、2つ以上の元の残基を置換することができる。通常は、第1のポリペプチドまたは第2のポリペプチドの界面におけるすべての残基が、置換される元のアミノ酸残基を含むにすぎない。置換に好ましい残基は「埋もれている」。「埋もれている」は、残基が本質的には溶媒と接触できないことを意味する。移入残基は、好ましくは、酸化または誤ったジスルフィド結合の形成の可能性を避けるために、システインではない。
【0059】
「元の核酸」は、アミノ酸側鎖が、第1のポリペプチドと第2のポリペプチドとの間で界面で相互作用して、ポリペプチド間の安定な相互作用を促進させるアミノ酸がマルチマー化ドメイン内でコードされるように変更することができる、目的のポリペプチドをコードする核酸を意味する。そのような変更によって、空洞への隆起、天然に存在しないジスルフィド結合、ロイシンジッパー、疎水性相互作用および親水性相互作用のような安定な相互作用に限定されないが、そのような安定な相互作用を生じさせることができる。このような変更は、好ましくは、目的とする第1のポリペプチドと第2のポリペプチドとの間の特異的な相互作用を促進し、望ましくないヘテロマーの対形成またはホモマーの形成が生じる相互作用を効果的に除くように選択される。元の核酸または出発核酸は、天然に存在する核酸であり得るか、または以前に変更が行われた核酸(例えば、ヒト化抗体フラグメント)を含み得る。核酸を「変更する」は、元の核酸を、目的のアミノ酸残基をコードしているコドンの少なくとも1つの挿入、欠失または置換による遺伝子操作または変異処理を行うことを意味する。通常、元の残基をコードしているコドンは、移入残基をコードするコドンによって置換される。このようにDNAを遺伝子的に改変するための技術は、Mutagenesis:aPractical Approach、M.J.McPherson編、IRLPress、Oxford、UK(1991)に総説されており、例えば、部位特異的変異誘発法、カセット変異誘発法およびポリメラーゼ連鎖反応(PCR)変異誘発法を含む。
【0060】
隆起、空洞、または(ジスルフィド結合の形成に必要なシステイン残基などの)遊離チオールは、合成的手段によって、例えば、組換え技術、インビトロペプチド合成、前記の天然に存在しないアミノ酸残基を導入するためのそのような技術、ペプチドの酵素的または化学的な連結、あるいはこれらの技術のいくつかの組合せによって、第1のポリペプチドまたは第2のポリペプチドの界面に「導入する」ことができる。従って、「導入される」隆起、空洞、または遊離チオールは、「天然に存在しない」、すなわち、「非天然」である。これは、自然界または元のポリペプチド(例えば、ヒト化モノクローナル抗体)に存在しないことを意味する。
【0061】
隆起を形成するために移入アミノ酸残基は、比較的少数(例えば、約3個〜6個)の「回転異性体」を有することが好ましい。「回転異性体」は、アミノ酸側鎖のエネルギー的に有利な立体配座である。様々なアミノ酸残基の回転異性体の数は、PondersおよびRichards、J.Mol.Biol.193:775〜791(1987)に総説されている。
【0062】
「分離された」ヘテロマルチマーは、その天然の細胞培養環境の成分からの同定および分離および/または回収が行われたヘテロマルチマーを意味する。その天然環境の混入成分は、ヘテロマルチマーに関する診断使用または治療使用を妨害する物質であり、酵素、ホルモン、および他のタンパク質性または非タンパク質性の溶解物を含み得る。好ましい実施態様において、ヘテロマルチマーは、(1)ローリー法によって測定されるように95重量%を超えるように、最も好ましくは99重量%を超えるように、あるいは(2)スピニングカップ配列決定装置の使用によって、N末端または内部のアミノ酸配列の少なくとも15残基が得られるのに十分な程度に、あるいは(3)クマシーブルー染色、または好ましくは銀染色を使用して、還元条件下または非還元条件下でのSDS−PAGEにより均一になるまで精製される。
【0063】
本発明のヘテロマルチマーは、一般には、実質的に均一になるまで精製される。「実質的に均一」、「実質的に均一な形態」および「実質的な均一性」という表現は、生成物が、望ましくないポリペプチドの組合せ物に由来する副生成物(例えば、ホモマルチマー)を実質的に有していないことを示すために使用される。
【0064】
純度に関して表現される場合、実質的な均一性は、副生成物の量が、10%を超えないこと、好ましくは5%未満であること、より好ましくは1%未満であること、最も好ましくは0.5%未満であることを意味する。ただし、割合は重量比である。
【0065】
「制御配列」という表現は、特定の宿主生物において、機能的に連結されたコード配列を発現させるのに必要なDNA配列をいう。原核生物に適切な制御配列には、例えば、プロモーター、必要に応じてオペレーター配列、リボソーム結合部位、および可能であれば、未だあまりよく理解されていない他の配列が含まれる。真核生物細胞は、プロモーター、ポリアデニル化シグナルおよびエンハンサーを利用することが知られている。
【0066】
核酸は、核酸が別の核酸配列と機能的な関係におかれている場合に「機能的に連結されている」。例えば、プレ配列または分泌リーダーに関するDNAは、DNAが、ポリペプチドの分泌に参加しているプロタンパク質として発現される場合、ポリペプチドに関してDNAに機能的に連結されている;プロモーターまたはエンハンサーは、それが配列の転写に影響を及ぼす場合、コード配列に機能的に連結されている;あるいは、リボソーム結合部位は、それが翻訳を容易にするように配置されている場合、コード配列に機能的に連結されている。一般に、「機能的に連結されている」は、連結されているDNA配列が連続していることを、そして分泌リーダーの場合には、読み取り相で連続していることを意味する。
【0067】
しかし、エンハンサーは、連続している必要はない。連結は、都合の良い制限部位で結合させることによって達成される。そのような部位が存在しない場合、合成オリゴヌクレオチドアダプターまたはリンカーが、従来的な実施に従って使用される。
【0068】
II.ヘテロマルチマーの調製1.出発材料の調製 最初の工程として、第1のポリペプチドおよび第2のポリペプチド(およびヘテロマルチマーを形成する任意のさらなるポリペプチド)が選択される。通常、これらのポリペプチドをコードする核酸は分離しなければならず、その結果、核酸は、本明細書中で定義されているように、隆起または空洞またはその両方をコードするように変更することができる。しかし、変異は、合成的な手段を使用することによって、例えば、ペプチド合成を使用することによって導入することができる。同様に、移入残基が天然に存在しない残基である場合には、Norenら(上記)の方法を、そのような置換を有するポリペブチドを作製するために用いることができる。さらに、ヘテロマルチマーの一部は、細胞培養で組換え的に適切に作製され、そのような分子の他の部分は、上記のそのような技術によって作製される。
【0069】
抗体の分離および免疫付着因子の調製に関する技法が次に行われる。しかし、ヘテロマルチマーは、当技術分野で知られている技術を使用して、他のポリペプチドから形成され得るか、または他のポリペプチドを組み込むことができることが理解される。例えば、目的のポリペプチド(例えば、リガンド、レセプターまたは酵素)をコードする核酸は、ポリペプチドのmRNAを有し、そのmRNAを検出可能なレベルで発現していると考えられる組織から調製されたcDNAライブラリーから分離することができる。ライブラリーは、目的の遺伝子またはそれによってコードされるタンパク質を同定するために設計されたプローブ(例えば、抗体または約20塩基〜80塩基のオリゴヌクレオチドなど)を用いてスクリーニングされる。選択されたプローブによるcDNAライブラリーまたはゲノムライブラリーのスクリーニングは、Sambrookら、Molecular Cloning:A Laboratory Manual(NewYork:Cold Spring Harbor LaboratoryPress、1989)の第10章〜第12章に記載される標準的な手順を使用して行うことができる。
【0070】
(1)抗体の調製 抗体の産生に関する技術がいくつか記載されている。そのような技術には、モノクローナル抗体を作製するための従来のハイブリドーマ法、抗体(キメラ抗体、例えば、ヒト化抗体を含む)を作製するための組換え技術、トランスジェニック動物での抗体産生、および「完全なヒト」抗体を調製するために近年記載されたファージディスプレー技術が含まれる。これらの技術を下記に簡単に記載する。
【0071】
目的の抗原に対するポリクローナル抗体は、一般には、抗原およびアジュバントの皮下(sc)注射または腹腔内(ip)注射を多数回行うことによって動物に生起させることができる。抗原(または、標的アミノ酸配列を含有するフラグメント)を、免疫化される種において免疫原であるタンパク質(例えば、キーホルリンペットヘモシアニン、血清アルブミン、ウシサイログロブリン、またはダイズのトリプシン阻害剤)に、二官能性薬剤または誘導化剤を使用して結合させることは有用であり得る。そのような二官能性薬剤または誘導化剤は、例えば、マレイミドベンゾイルスルホスクシンイミドエステル(システイン残基を介するコンジュゲーション)、N−ヒドロキシスクシンイミド(リシン残基を介する)、グルタルアルデヒド、無水コハク酸、SOCl2、またはR1N=C=NR(ただし、RおよびR1は異なるアルキル基である)である。動物は、免疫原性のコンジュゲートまたは誘導体に対して、(ウサギまたはマウスに関して、それぞれ)1mgのコンジュゲートを3容量のフロイント完全アジュバントと一緒にして、その溶液を皮下に多数の部位に注射することによって免疫化される。1ヶ月後、動物は、コンジュゲートを含むフロイント完全アジュバントの最初の量の1/5〜1/10を用いて、多数の部位に皮下注射することによって追加免疫される。
【0072】
7日〜14日の後に、動物は採血され、血清を抗体力価についてアッセイする。
【0073】
動物は、力価がプラトーに達するまで追加免疫される。動物は、好ましくは、同じ抗原のコンジュゲートで追加免疫されるが、異なるタンパク質に結合させたコンジュゲートおよび/または異なる架橋剤によるコンジュゲートで追加免疫される。コンジュゲートはまた、タンパク質の融合体として、組換え細胞培養で作製することができる。同様に、ミョウバンなどの凝集化剤を使用して、免疫応答を増強することができる。
【0074】
モノクローナル抗体は、KohlerおよびMilstein、Nature256:495(1975)によって初めて記載されたハイブリドーマ法を使用して実質的に均一な抗体集団から得られるか、または、組換えDNA法 (Cabillyら、米国特許第4,816,567号)によって作製することができる。
【0075】
ハイブリドーマ法において、マウス、またはハムスターなどの他の適切な宿主動物を上記のように免疫化して、免疫化のために使用されたタンパク質に特異的に結合する抗体を産生するか、または産生し得るリンパ球が誘導される。あるいは、リンパ球をインビトロで免疫化することができる。次いで、ポリエチレングリコールなどの適切な融合化剤を使用してリンパ球をミエローマ細胞と融合して、ハイブリドーマ細胞を得る(Goding、MonoclonalAntibodies:Principles and Practice、59頁〜103頁(AcademicPress、1986))。このようにして調製されたハイブリドーマ細胞を、融合していない元のミエローマ細胞の生育または生存を阻害する物質を1つまたは複数含有することが好ましい適切な培養培地に播種して、増殖させる。例えば、元のミエローマ細胞が酵素ヒポキサンチングアニンホスホリボシルトランスフェラーゼ(HGPRTまたはHPRT)を欠失している場合、ハイブリドーマの培養培地は、典型的には、HGPRT欠損細胞の生育を妨げる物質であるヒポキサンチン、アミノプテリンおよびチミジンを含む(HAT培地)。好ましいミエローマ細胞は、効率よく融合し、選択された抗体産生細胞によって抗体の安定で高レベルの発現を維持し、HAT培地などの培地に対して感受性を有するミエローマ細胞である。このような細胞の中で、好ましいミエローマ細胞株は、ネズミのミエローマ細胞株であり、Salk Institute Cell Distribution Center(SanDiego、California、USA)から入手可能なMOPC−21およびMPC−11のマウス腫瘍から誘導されるミエローマ細胞株、ならびにAmerican Type Culture Collection(Rockville、Maryland、USA)から入手可能なSP−2細胞などである。ヒトのミエローマ細胞株およびマウス−ヒトのヘテロミエローマ細胞株もまた、ヒトのモノクローナル抗体の産生に関して記載されている(Kozbor、J.Immunol.、133:3001(1984);およびBrodeurら、Monoclonal Antibody Production Techniques andApplications、51頁〜63頁、Marcel Dekkar,Inc.、New York、1987)。ヒトモノクローナル抗体の産生技術に関しては、Boernerら、J.Immunol.、147(1)86:〜95(1991)および国際特許公開第WO91/17769号(1991年11月28日公開)もまた参照。ハイブリドーマ細胞が生育する培養培地は、目的の抗原に対するモノクローナル抗体の産生についてアッセイされる。
【0076】
好ましくは、ハイブリドーマ細胞によって産生されたモノクローナル抗体の結合特異性は、免疫沈降によって、あるいは放射免疫アッセイ(RIA)または酵素結合免疫吸着アッセイ(ELISA)などのインビトロ結合アッセイによって測定される。モノクローナル抗体の結合親和性は、例えば、MunsonおよびPollandのスキャッチャード分析(Anal.Biochem.107:220(1980))によって測定することができる。所望の特異性、親和性および/または活性の抗体を産生するハイブリドーマ細胞が同定された後に、クローンを限界稀釈法によってサブクローニングを行い、標準的な方法によって生育させることができる。Goding、Monoclonal Antibodies:Principles andPractice、59頁〜104頁(AcademicPress、1986)。この目的に適切な培養培地には、例えば、ダルベッコ改変イーグル培地またはRPMI−1640培地が含まれる。さらに、ハイブリドーマ細胞は、動物の体内において腹水腫瘍としてインビボで生育させることができる。サブクローンによって分泌されたモノクローナル抗体は、培養培地、腹水または血清から、例えば、プロテインA−セファロース、ヒドロキシルアパタイトクロマトグラフィー、ゲル電気泳動、透析またはアフィニティクロマトグラフィーなどの従来の免疫グロブリン精製手順によって適切に分離される。
【0077】
あるいは、免疫化したときに、内因性の免疫グロブリンを産生することなく、完全なレパートリーのヒト抗体を産生し得るトランスジェニック動物(例えば、マウス)の作製が現在可能である。例えば、キメラな生殖系列の変異マウスにおける抗体の重鎖連結領域(JH)遺伝子のホモ接合型欠失によって、内因性の抗体産生が完全に阻害されることが記載されている。ヒトの生殖系列の免疫グロブリン遺伝子列をそのような生殖系列の変異マウスに移すことによって、ヒト抗体が、抗原を投与したときに産生する。例えば、Jakobovitsら、Proc.Natl.Acad.Sci.USA、90:2551〜255(1993);Jakobovitsら、Nature362:255〜258(1993);Fishwild,D.M.ら(1996)Nat.Biotech14:845〜851;およびMendez,M.J.ら(1997)Nat.Genetics 15:146〜156を参照。
【0078】
さらなる実施態様において、抗体または抗体フラグメントは、McCaffertyら、Nature、348:522〜554(1990)に記載される技術を使用して作製された抗体ファージライブラリーから、目的の抗原を使用して、適切な抗体または抗体フラグメントに関する選択を行うことによって分離することができる。Clacksonら、Nature、352:624〜628(1991)およびMarksら、J.Mol.Biol.、222:581〜597(1991)は、それぞれ、ファージライブラリーを使用するマウス抗体およびヒト抗体の分離を記載する。その後の刊行物は、非常に大きなファージライブラリーを構築するための方法として、鎖シャッフリング(Markら、Bio/Technol.10:779〜783(1992))、ならびに結合感染およびインビボ組換えによる高い親和性(nM範囲)のヒト抗体の産生を記載する(Waterhousesら、Nuc.Acids.Res.21:2265〜2266(1993);Griffiths,A.D.ら(1994)EMBO J.13:3245〜3260;およびVaughanら(1996)上記)。従って、これらの技術は、本発明により含まれる「モノクローナル」抗体(特に、ヒト抗体)の分離に関する、従来のモノクローナル抗体ハイブリドーマ技術の実行可能な代替法である。
【0079】
本発明の抗体をコードするDNAは、従来の手順を使用して(例えば、ネズミ抗体の重鎖および軽鎖をコードする遺伝子に特異的に結合し得るオリゴヌクレオチドプローブを使用することによって)容易に分離され、そして配列決定される。本発明のハイブリドーマ細胞は、そのようなDNAの好ましい供給源として有用である。DNAが一旦分離されると、そのDNAは、発現ベクター内に配置することができ、次いで、サルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、またはそうでなければ免疫グロブリンタンパク質を産生しないミエローマ細胞などの宿主細胞にトランスフェクションされ、組換え宿主細胞においてモノクローナル抗体の合成物が得られる。DNAはまた、例えば、コード配列を、相同的なマウス配列の代わりに、ヒトの重鎖および軽鎖の定常ドメインに置換することによって改変することができる。Morrisonら、Proc.Nat.Acad.Sci.81:6851(1984)。そのようにして、本明細書中の抗抗原モノクローナル抗体の結合特異性を有する抗体が調製される。
【0080】
非ヒト抗体をヒト化するための方法は、当技術分野では十分に知られている。
【0081】
一般に、ヒト化抗体は、ヒトでない供給源からヒト化抗体に導入された1つまたは複数のアミノ酸残基を有する。ヒト化は、本質的には、Winterおよび共同研究者らの方法(Jonesら、Nature 321:522〜525(1986);Riechmannら、Nature332:323〜327(1988);Verhoeyenら、Science239:1534〜1536(1988))に従って、齧歯類のCDRまたはCDR配列をヒト抗体の対応配列に置換することによって行うことができる。従って、そのような「ヒト化」抗体は、キメラな抗体であり、(Cabilly、上記)。実質的には元のままでないヒトの可変ドメインは、ヒトでない種に由来する対応の配列によって置換されている。実際、典型的には、ヒト化抗体は、いくつかのCDR残基、および可能であれば、いくつかのFR残基が、齧歯類の抗体の類似する部位に由来する残基によって置換されているヒト抗体である。抗体は、抗原に対する高い親和性および他の有利な生物学的特性を保持してヒト化されることが重要である。このような目的を達成するために、好ましい方法に従って、ヒト化抗体が、元の配列およびヒト化配列の三次元モデルを使用して、元の配列および様々な考えられるヒト化配列を解析することによって調製される。三次元の免疫グロブリンモデルは、当業者には熟知されている。選択された候補の免疫グロブリン配列の可能性な三次元の立体配置構造を図示して表示するコンピュータープログラムを入手することができる。このような表示を検討することによって、候補の免疫グロブリン配列の機能発現における残基の可能な役割の分析、すなわち、候補の免疫グロブリンのその抗原に対する結合能に影響する残基の分析が可能になる。このように、FR残基は、コンセンサス配列および持ち込み配列からの選択および組合せを行うことができ、その結果、標的抗原に対する増大した親和性などの所望の抗体特性が達成される。さらなる詳細については、国際特許公開第WO92/22653号(1992年12月23日公開)を参照。
【0082】
(ii)免疫付着因子調製 免疫グロブリン(Ig)、およびその変異体が知られており、その多くが、組換え細胞培養によって調製されている。例えば、米国特許第4,745,055号;欧州特許第256,654号;Faulknerら、Nature 298:286 (1982);欧州特許第120,694号;欧州特許第125,023号;Morrison, J. Immun. 123:793 (1979);Koehlerら、Proc. Natl. Acad. Sci. USA 77:2197 (1980);Rasoら、Cancer Res.41:2073 (1981);Morrisonら, Ann. Rev.Immunol. 2:239 (1984);Morrison, Science229:1202 (1985);Morrisonら、Proc.Natl. Acad. Sci. USA 81:6851 (1984);欧州特許第255,694号;欧州特許第266,663号;および国際特許公開第88/03559号を参照。再編成された免疫グロブリン鎖も知られている。例えば、米国特許第4,444,878号;国際特許公開第88/03565号;および欧州特許第68,763号、ならびに、それらで引用されている参考文献を参照。
【0083】
付着因子結合ドメインの配列を、適当な免疫グロブリン定常ドメイン位配列に連結させて構築したキメラ(免疫付着因子)が当技術分野において知られている。
【0084】
文献で報告されている免疫付着因子には、T細胞レセプターとの融合タンパク質(Gascoigneら、Proc. Natl. Acad. Sci.USA 84:2936−2940 (1987));CD4との融合タンパク質(Caponら、Nature 337:525−531 (1989);Trauneckerら、Nature 339:68−70 (1989);Zettmeisslら、DNA Cell Biol.USA 9:347−353 (1990);およびBymら、Nature344:667−670(1990));L−セレクチン(ホーミンクルセプター)との融合タンパク質(Watsonら、J.CellBiol.110:2221−2229(1990);およびWatsonら、Nature349:164−167(1991));CD44との融合タンパク質(Aruffoら、Cell 61:1303−1313 (1990));CD28とB7との融合タンパク質(Linsleyら、J.Exp.Med.173:721−730 (1991));CTLA−4との融合タンパク質(Linsleyら、J.Exp.Med.174:561−569 (1991));CD22との融合タンパク質(Stamenkovicら、Cell66:1133−1144(1991));TNFレセプターとの融合タンパク質(Ashkenaziら、Proc.Natl.Acad.Sci.USA88:10535−10539 (1991);Lesslauerら,Eur.J.Immunol.27:2883−2886 (1991);およびPeppleら、J.Exp.Med.174:1483−1489 (1991));およびIgEレセプターαとの融合タンパク質(RidgwayとGovman、J.CellBiol.Vol.115,Abstract No.1448 (1991))などがある。
【0085】
もっとも単純で、もっとも素直な免疫付着因子設計は、付着因子の結合ドメイン(例えば、レセプターの細胞外ドメイン(ECD))を、免疫グロブリン重鎖のヒンジ領域とFc領域に結合させるものである。通常、本発明の免疫付着因子を調製するときには、付着因子の結合ドメインをコードする核酸がC末端になるように、免疫グロブリンの定常ドメイン配列のN末端をコードする核酸に融合させるが、N末端の融合も可能である。
【0086】
典型的には、このような融合配列において、コードされるキメラポリペプチドは、少なくとも、免疫グロブリン重鎖の定常ドメインのヒンジ、CH2、CH3の各ドメインの機能的活性を保持している。定常ドメインのFcドメインのC末端側で融合させたり、重鎖のCH1、または軽鎖の対応領域のすぐN末端側で融合させたりする。正確にどの位置で融合させるかは重要ではないが、特定の部位がよく知られていて、生物学的活性、分泌、またはIaの結合特性を最適なものにするときに選択される。
【0087】
好ましい実施態様において、免疫グロブリンG1(IgG1)のFcドメインのN末端に付着因子の配列を融合させる。重鎖の定常領域全体を付着因子の配列に融合させることも可能である。しかし、より好ましくは、化学的にIgGFcを画定するパパイン切断部位(すなわち、重鎖の定常領域の最初の残基を114とすると残基216にあたる)のすぐ上流にあるヒンジ領域から始まる配列、または、他の免疫グロブリンの同様の部位を融合に用いる。特に好ましい実施態様においては、付着因子のアミノ酸配列を、IgG1、IgG2、またはIgG3の重鎖の(a)ヒンジ領域、およびCH2とCH3からなる領域に、または、(b)CH1、ヒンジ、CH2、およびCH3ドメインに融合させる。融合させる正確な部位は重要ではなく、最適な部位を通常の実験によって決定することができる。
【0088】
双特異的免疫付着因子では、免疫付着因子はマルチマーとして集合し、ヘテロダイマーまたはヘテロ四量体として集合する。一般的には、これらの分子集合した免疫グロブリンは、既知のユニット構造をもっているはずである。基本的な4本鎖の構造単位は、IgG、IgD、およびIgEに見られる構造である。高分子量の免疫グロブリンでは、4本鎖の構造単位が繰り返される。すなわち、IgMは、一般的に、4本鎖の基本ユニットをジスルフィド結合によって結合させた五量体として存在する。IgAグロブリンと、場合によってはIgGグロブリンも、血清中ではマルチマーの形で存在することがある。マルチマーの場合、4本鎖ユニットは同じものであることもあれば、異なることもある。
【0089】
本明細書の範囲に含まれる集合免疫付着因子のさまざまな例を以下に概略的に示す:
(a)ACL-ACL;
(b)ACH-[ACH,ACL-ACH,ACL-VHCH,またはVLCL-ACH];
(c)ACL-ACH-[ACL-ACH,ACL-VHCH,VLCL-ACH,またはVLCL-VHCH];
(d)ACL-VHCH-[ACH,またはACL-VHCH,またはVLCL-ACH];
(e)VLCL-ACH-[ACL-VHCH,またはVLCL-ACH];および(f)[A-Y]n-[VLCL-VHCH]2;
ここで、Aは、それぞれ、同一か異なった付着因子のアミノ酸配列を表しており、 VLは、免疫グロブリン軽鎖の可変ドメインであり;
VHは、免疫グロブリン重鎖の可変ドメインであり;
CLは、免疫グロブリン軽鎖の定常ドメインであり;
CHは、免疫グロブリン重鎖の定常ドメインであり;
nは、1よりも大きい整数であり;
Yは、共有結合的な架橋剤の残基を示している。
【0090】
簡潔を期するために、上記の構造は、主要な特徴を示しているにすぎず、結合ドメイン(J)や、免疫グロブリンのその他のドメインは示されていない。さらに、ジスルフィド結合も示されていない。しかし、このようなドメインが結合活性にとって必要である場合には、それらが、免疫グロブリン分子の中で占める通常の位置に提示されるよう構成されている。
【0091】
または、キメラ重鎖を含む免疫グロブリンを得るために、免疫グロブリンの重鎖と軽鎖の配列の問に付着因子の配列を挿入することも可能である。この実施態様では、免疫グロブリンの各アームにおける、免疫グロブリン重鎖のヒンジとCH2ドメインの間か、またはCH2とCH3ドメインの間の3’末端に付着因子の配列を融合させる。同様の構築物が、Hoogenboomら、Mol.Immunol.28:1027−1037(1991)によって報告されている。
【0092】
免疫グロブリンの軽鎖が、付着因子−免疫グロブリン重鎖融合ポリペプチドに共有的に会合しているか、または、付着因子に直接融合するかして存在することもあろう。前者の場合、免疫グロブリン軽鎖をコードするDNAが、典型的には、付着因子−免疫グロブリン重鎖融合タンパク質をコードするDNAとともに発現する。分泌されるときに、ハイブリッドの重鎖と軽鎖が共有的に会合して、ジスルフィド会合で結ばれた免疫グロブリン重鎖−軽鎖の2組の組み合せを含む免疫グロブリン様構造物が提供される。このような構造物を調製するのに適した方法は、例えば、1989年3月28日公開の米国特許第4,816,567号に開示されている。
【0093】
好ましい実施態様において、本発明の免疫付着因子の構築に用いる免疫グロブリンの配列は、IgG免疫グロブリン重鎖の定常部位からとってくる。ヒトの免疫付着因子については、ヒトIgG1およびIgG3免疫グロブリンの配列が好ましい。IgG1を使用することの主な利点は、固定したプロテインA上で、IgG1免疫付着因子を効率的に精製できることにある。これに対して、IgG3の精製には、これよりもかなり使い勝手の悪い媒体であるプロテインGが必要となる。しかし、特定の免疫付着因子構築物のIgの融合相手を選ぶときには、免疫グロブリンのこの他の構造的性質および機能的性質も考慮すべきである。例えば、IgG3のヒンジは、より長く、可塑性もより高いため、IgG1に融合させると正しく折り畳まれて機能することができないような長い”付着因子”ドメインを受け入れることができる。もう一つ考慮すべきなのは、原子価である。IgG免疫付着因子は2価性のホモダイマーであるが、IgAおよびIgMなどのIgサブタイプは、それぞれ、基本的なIgホモダイマーユニットからなるダイマーと五量体を生じよう。インビボでの実用に供するために設計された免疫付着因子では、Fc領域によって特定される薬理動態的性質およびエフェクター機能も重要である。IgG1、IgG2およびIgG4はすべて、インビボでの半減期が21日で、補体系を活性化する相対的能力はそれぞれ異なる。IgG4は、補体をほとんど活性化せず、IgG2は、IgG1よりもかなり弱くしか補体活性を活性化しない。さらに、IgG1とは異なって、IgG2は、単核球または好中球上のFcレセプターには結合しない。補体の活性化に関しては、IgG3が最適であるが、インビボでの半減期が、他のIgGアイソタイプの約3分の1である。ヒトの治療薬として使用するために、免疫付着因子を設計するときに考慮すべき他の重要な点は、特定のアイソタイプがもつアロタイプの数である。一般的には、血清学的に区別されるアロタイプの数が少ないIgGのアイソタイプほど好ましい。例えば、IgG1は、血清学的に区別されるアロタイプ部位を4つしかもっておらず、これらのうち2つは(Gm1とGm2)はFc領域に位置しているが、このうちの1つの部位であるG1m1は、免疫原性がない。これに対して、IgG3には12の血清学的に区別されるアロタイプがあり、それらのすべてがFc領域中にあって、その内の3つの部位(G3m5、11および21)だけが免疫原性をもつ同一のアロタイプをもっている。したがって、γ1免疫付着因子よりも、γ3免疫付着因子方が免疫原としての可能性が高い。
【0094】
付着因子部分をコードするcDNA配列を、IgのcDNA配列にフレームを合わせて融合させることによって、もっとも好都合に免疫付着因子が構築される。しかし、ゲノムのIgフラグメントに融合させることもできる(例えば、Gascoigneら、上記;Aruffoら、Cell61:1303−1313(1990);およびStamenkovicら、Cell 66:1133−1144 (1991))参照)。後者のタイプの融合には、発現のためのIg調節配列が存在することが必要である。IgGの重鎖定常領域をコードするcDNAは、脾臓または末梢血のリンパ球由来のcDNAライブラリーからの公開された配列をもとに、ハイブリダイゼーションまたはポリメラーゼ連鎖反応(PCR)技術によって分離することができる。免疫付着因子の”付着因子”とIg部分をコードするcDNAを、選んだ宿主細胞の中で効率的に発現させるためのプラスミドベクターの中にひとつながりになるように挿入する。
【0095】
2.隆起および/または空洞の作出 隆起および/または空洞を形成するもととなる残基を選択するための最初の工程として、X線結晶解析やNMRなど、当技術分野において既知の技術を用いて、ヘテロマルチマーの三次元構造を得る。三次元構造に基づいて、当業者は、接触面の残基を同定することができる。
【0096】
好ましい接触面は、免疫グロブリン定常ドメインのCH3ドメインである。各種のIgGサブタイプの接触面残基と”埋もれた”残基が同定されていたため、IgG、IgA、IgD、IgE、およびIgMのCH3ドメインの接触面の残基で、取り入れる残基と置換するのに最適な残基などの残基が同定されている(例えば、PCT/US96/01598を参照、その全文が、参照してここに組み込まれる)。CH3接触面を改変するための基礎となるのは、X線結晶解析によって、ヒトIgG1重鎖の間に生じる、Fc領域での分子間結合は、CH3ドメイン間では広範なタンパク質/タンパク質間相互作用を含むのに対して、グリコシル化されたCH2ドメインでは、その糖部分によって結合するということが示されたことである。(Deisenhofer,Biochem.20:2361−2370 (1981))。さらに、哺乳動物細胞の中には、欠失によりCH2およびCH3ドメインが除去されないかぎり、2つの重鎖間では、抗体が発現されるときにジスルフィド結合が効率よく形成される(Kingら、Biochem.J.281:317 (1992))。したがって、重鎖の集合は、ジスルフィド結合を促進すると考えられ、その逆ではないと考えられる。これらの構造および機能に関するデータをまとめて考えると、抗体の重鎖の会合はCH3部位によってもたらされるという仮説に至る。さらに、CH3ドメイン間の接触面を改変して、異なった重鎖のヘテロマルチマーの形成を促進して、対応するホモマルチマーの集合を阻害することができるかもしれないという推測がなされた。
【0097】
本明細書で説明する実験によって、この方法を用いれば、ホモマルチマーよりもヘテロマルチマーの形成を促進することが可能なことが示された。したがって、目的のポリペプチドと抗体のCH3部位とを含むポリペプチド融合体を作出して、第1または第2のポリペプチドを形成させることができる。好ましいCH3ドメインは、ヒトIgG1のようなIgG抗体に由来するものである。
【0098】
隆起または空洞を形成するための候補となりうる接触面残基が同定されている。 ”埋もれた”残基を選んで置き換えることが好ましい。ある残基が埋込まれているか否かを判定するために、Leeら、J.Mol.Biol.55:379−400(1971)の表面接触プログラムを用いて、接触面にある残基と溶媒との接触可能性(SA)を計算することができる。そして、第1と第2のポリペプチドのそれぞれの残基について、一方のポリペプチドを除いた上で、SAを別々に計算することができる。そして、SA(ダイマー)−SA(モノマー)という等式を用いて、接触面のモノマー型とダイマー型の間の各残基のSAの差異を計算することができる。これによって、ダイマーを形成するときにSAが減少する残基のリストが示される。ダイマーの各残基のSAを、同じアミノ酸のトリペプチドGly−X−GlyのSAの理論値と比較する。ただし、X=目的のアミノ酸(Roseら、Science229:834−838(1985))。(a)モノマーに較べてダイマーでSA減少した残基、および(b)SAが、対応するトリペプチドにおけるSAの26%よりも少ない残基を接触面残基と考える。
【0099】
2つの分類を明らかにすることができる。すなわち、対応するトリペプチドに較べて10%よりも低いSAをもつ残基(すなわち、”埋もれた”残基)、および、対応するトリペプチドに較べて10%よりも高く25%よりは低いSAをもつ残基(すなわち、”一部埋もれた”残基)である(下記の表1参照)
【0100】
【表1】
【0101】
†残基番号は、IgG結晶構造にある通り(Deisenhofer,Biochemistry 20:2361-2370(1981))ポリペプチド鎖構造に対する残基置換の効果は、InsightTMプログラム(BiosymTechnologies)などの分子図形モデリングプログラムを用いて、調べることができる。このプログラムを用いて、第1のポリペプチドの接触面中に埋もれた残基で、側鎖容量の小さい残基を、例えば、より大きな側鎖容量をもつ残基(すなわち、隆起)に変えることができる。そして、第2のポリペプチドの接触面中の残基で、隆起に隣接する残基を調べて、空洞を形成するために適した残基を見つける。通常、この残基は側鎖容量が大きいので、側鎖容量の小さな残基と置き換える。一定の実施態様において、接触面の三次元構造を調べると、第1のポリペプチドの接触面上に適当な位置に置かれた、適当な大きさの隆起、または第2のポリペプチドの接触面上の空洞が明らかになるはずである。これらの例では、単一突然変異体、すなわち、合成によって導入された隆起または空洞をモデル化する必要があるだけである。
【0102】
第1および第2のポリペプチドがそれぞれCH3ドメインを含む場合に、置換することのできる元の残基を選択することに関しては、ヒトIgG1のCH3/CH3の接触面は、各表面から1090Å2埋もれた4つの逆平行β鎖上にある各ドメイン上の16の残基を含んでいる(Deisenhofer,上記、およびMiller、J.Mol.Biol.216:965(1990)。突然変異は、好ましくは、中央の逆平行β鎖上にある残基を標的とする。この目的は、作出した隆起が、相手方のCH3ドメインの相補的な空洞の中に入らずに、周囲の溶媒の中に突出して親和してしまう危険を最小限にするためである。
【0103】
分子モデリングによって、好ましい元/取り込み残基が一旦同定されたら、当技術分野において周知の技術を用いて、ポリペプチドの中にアミノ酸置換を導入する。通常は、ポリペプチドをコードするDNAは、突然変異誘発:実用的方法(Mutagenesis: a PracticalApproach)、上記、に記載されている技術を用いて遺伝子工学により改変する。
【0104】
オリゴヌクレオチドによる突然変異誘発が、第1または第2のポリペプチドをコードするDNAの置換変異体を調製するための好ましい方法である。この技術は、Adelmanら、DNA,2:183(1983)によって説明されているように、当技術分野において周知の技術である。簡単に述べると、所望の変異をコードしているオリゴヌクレオチドをDNA鋳型とハイブリダイズさせることによって、第1または第2のポリペプチドDNAを改変する。この鋳型DNAは、ヘテロマルチマーの未変更または天然のDNA配列を含む、一本鎖のプラスミドまたはバクテリオファージである。ハイブリダイゼーション後、DNAポリメラーゼを用いて、オリゴヌクレオチドプライマーを取り込んだ相補鎖で、ヘテロマルチマーDNAへの部位特異的な変化をコードする相補鎖である、鋳型に相補的な第2鎖の全長を合成する。
【0105】
Wellsら、Gene34:315(1985)が述べるところにしたがって、目的とするDNA領域を、相補的なオリゴヌクレオチドをアニーリングさせて作製した合成変異フラグメントで置換することによって、カセット突然変異誘発を行なうことができる。PCR突然変異も、第1または第2ポリペプチドDNAの変異配列を作製するのに適している。以下の考察においてはDNAを引用しているが、この技術は、RNAでも応用することができると解される。PCR技術とは、一般的には、次の手順をさす(Erlich, Science 252:1643−1650 (1991)、R. Higuchiが担当した章、p.61−70を参照)。
【0106】
隆起または空洞の突然変異以外にも、本発明は、ヘテロマルチマーのDNAの中に適当なヌクレオチド変異を導入するか、または所望のヘテロマルチマーポリペプチドを合成することによって調製することのできるヘテロマルチマーのアミノ酸配列の変異配列を含む。このような変異配列には、例えば、ヘテロマルチマーを形成する第1および第2のポリペプチドのアミノ酸配列中の残基の欠失、挿入、または置換が含まれる。最終的な構築物が望ましい抗原結合特性をもっているとすれば、最終的な構築物ができるように、欠失、挿入および置換を組み合わせる。グリコシル化部位の数と位置を変更したりして、アミノ酸を変えることによっても、ヘテロマルチマーの翻訳後のプロセッシングを変えることができる。
【0107】
突然変異を誘発するのに好ましい位置にある、ヘテロマルチマーポリペプチドの一定の残基または領域を同定するための有用な方法は、CunnighamとWells、Science 244:1081−1085(1989)によって述べられている“アラニンスキャニング突然変異誘発法”といわれるものである。ここで、標的残基の残基またはそのグループが同定され (例えば、Arg、Asp、His、およびGluなどの荷電性残基)、中性のアミノ酸または負に荷電したアミノ酸(もっとも好ましくは、アラニンまたはポリアラニン)で置き換えて、アミノ酸と、細胞の内外の周囲の水性環境との相互作用に影響を与える。そして、置換部位で、またはそれに対する、および/または別の変異残基を導入することによって、置換に対して機能的な感受性を示すドメインをさらに明確にすることができる。このように、アミノ酸配列変異を導入するための部位が予め決まっていても、突然変異そのものの性質を予め測ることはできない。
【0108】
通常、突然変異には、ヘテロマルチマーの非機能的な領域における保存的なアミノ酸置換が含まれる。突然変異の例を表2に示す。
【0109】
【表2】
【0110】
ヘテロマルチマーのポリペプチドの共有修飾も、本発明の範囲に含まれる。ヘテロマルチマーの共有修飾は、ヘテロマルチマー、またはそのフラグメントの標的アミノ酸残基を、N末端またはC末端の残基の選ばれた側鎖と反応することのできる有機的誘導化剤と反応させて、分子の中に導入することができる。ヘテロマルチマーポリペプチドの別のタイプの共有修飾で、本発明の範囲に含まれるものには、ポリペプチドの本来のグリコシル化パターンを改変することが含まれる。改変とは、もとのヘテロマルチマーの中に見られる糖部分を一つ以上欠失させるか、および/または、もとのヘテロマルチマーには存在しないグルコシル化部位を一つ以上付加することを意味する。ヘテロマルチマーポリペプチドへのグリコシル化部位の付加は、一つ以上のN結合型グリコシル化部位が含まれるよう、アミノ酸配列を改変することによって適宜行われる。一つ以上のセリンまたはトレオニン残基を、もとのヘテロマルチマー配列に付加または置換することによって改変を行なうこともできる(O結合型グリコシル化部位)。簡単にするために、好ましくは、DNAレベルでの変更によって、特に、所望のアミノ酸に翻訳されるコドンを作出するよう、ヘテロマルチマーポリペプチドをコードするDNAの予め決定された塩基突然変異を起こすことによって、ヘテロマルチマーのアミノ酸配列を改変する。ヘテロマルチマーポリペプチド上の糖部分の数を増加させる別の方法は、化学結合または酵素結合によってグリコシドをポリペプチドに結合させることである。これらの方法については、1987年9月11日発行の国際特許公開第87/05330号、およびAplinとWriston,CRCCrit.Rev.Biochem.pp.259−306(1981)に記載されている。ヘテロマルチマーに存在する糖部分の削除は、化学的または酵素的に行なうことができる。
【0111】
ヘテロマルチマーの別のタイプの共有修飾には、ヘテロマルチマーポリペプチドを、米国特許第4,640,835号;第4,496,689号;第4,301,144号;第4,670,417号;第4,791,192号;および第4,179,337号において提示されている方法で、例えば、ポリエチレングリコール、ポリプロピレングリコール、またはポリオキシアルキレンなど、さまざまな非タンパク質性ポリマーの一つに結合させることが含まれる。
【0112】
変異ヘテロマルチマーの特性を前もって予測することは困難であるため、回収された変異体をスクリーニングして、最適な変異体を選抜することが必要であると認められよう。
【0113】
3.共通の軽鎖をもつヘテロマルチマーの発現DNAを突然変異させて、本明細書で開示されているような共通の軽鎖を選抜した後、当技術分野において広く利用可能な組換え技術を用いて、この分子をコードするDNAを発現させる。しばしば、優れた発現系は、ヘテロマルチマーを適切にグルコシル化するための(例えば、グリコシル化されている抗体部位を含むヘテロマルチマーの場合など)、哺乳動物細胞の発現ベクターと宿主とを含んでいる。しかし、下で詳述するように、この分子は、原核生物の発現系で産生させることもできる。通常、一つのベクター、または別々のベクター上に存在する、第1ポリペプチド、第2ポリペプチド、共通の軽鎖ポリペプチド、および、ヘテロマルチマーを形成するのに必要な別のポリペプチドのすべてコードするDNAによっての宿主細胞を形質転換する。しかし、第1ポリペプチド、第2ポリペプチド、および共通の軽鎖ポリペプチド(ヘテロマルチマーの成分)を、別々の発現系で発現させ、発現したポリペプチドをインビトロで結合させることも可能である。
【0114】
ヘテロマルチマーと共通の軽鎖をコードするヌクレオチド(例えば、cDNAまたはゲノムDNA)を、さらなるクローニング(DNA増幅)または発現のために、複製可能なベクターの中に挿入する。多くのベクターを利用することができる。一般的に、ベクターの構成要素として、次のものを一つ以上含むが、これらに限定はされない。すなわち、シグナル配列、複製開始点、一つ以上のマーカー遺伝子、エンハンサー因子、プロモーター、および転写終結配列。
【0115】
ヘテロマルチマーの成分であるポリペプチドは、シグナル配列、または、それ以外で、成熟タンパク質またはポリペプチドのN末端に特異的な切断部位をもつポリペプチドに融合したポリペプチドとして作製することができる。一般的に、シグナル配列は、ベクターの成分であるか、または、ベクターの中に挿入されるDNAの一部であろう。好んで選ばれる異種由来のシグナル配列は、宿主細胞によって認識され、プロセッシングされる(すなわち、シグナルペプチダーゼによって開裂される)配列である。原核生物の宿主細胞では、シグナル配列を、例えば、アルカリホスファターゼ、ペニシリナーゼ、lpp、または熱安定性エンドトキシンIIの先導配列からなるグループから選択される、原核生物のシグナル配列で置き換えることができる。酵母での分泌を行なわせるためには、天然のシグナル配列を、例えば、酵母インベルターゼ、α因子の先導配列(サッカロマイセス(Saccharomyces)とクルイベロマイセス(Kluyveromyces)α因子の先導配列、後者については、1991年4月23日公開の米国特許第5,010,182号に記載されている)、または酸性ホスファターゼの先導配列、C.アルビカンス(C.albicans)のグルコアミラーゼの先導配列(1990年4月4日公開の欧州特許第362,179号)、または、1990年11月15日発行の国際特許公開第90/13646号に記載されているシグナルなどによって置換することができる。哺乳動物細胞で発現させるときには、本来のシグナル配列(例えば、抗体または付着因子のプレ配列で、通常、インビボで、これらの分子をヒト細胞から分泌させる配列)で十分であるが、哺乳動物の別のシグナル配列も適しているし、また、例えば、単純ヘルペスのgDシグナルのような、ウイルスの分泌を促す配列も適当であろう。このような前駆体領域に対するDNAを、読み枠がずれないように、ヘテロマルチマーを形成するポリペプチドをコードするDNAと連結させる。
【0116】
発現ベクターもクローニングベクターも、一つ以上の選ばれた宿主細胞の中でベクターの複製が可能になる塩基配列を含んでいる。一般的に、クローニングベクターにおいては、この配列は、宿主の染色体DNAとは独立に、ベクターが複製できるようにする配列であり、複製開始点または自律複製配列を含む配列である。このような配列は、さまざまな細菌、酵母、およびウイルスでよく知られている。プラスミドpBR322の複製開始点は、ほとんどのグラム陰性菌に適合し、2μプラスミドの開始点は酵母に適合し、また、さまざまなウイルスの開始点(SV40、ポリオーマ、アデノウイルス、VSV、またはBPV)は、哺乳動物細胞でのクローニングベクターとして有用である。一般的に、複製開始点という構成因子は、哺乳動物の発現ベクターには不要である(SV40の複製開始点は、初期プロモーターを含んでいるというだけの理由で一般的には用いられている)。
【0117】
発現ベクターとクローニングベクターは、選抜可能マーカーとも言われる選抜用遺伝子を持っていなければならない。典型的な選抜用遺伝子は、(a)例えば、アンピシリン、ネオマイシン、メトトレキセート、またはテトラサイクリンなどの抗生物質またはその他の毒素に対する抵抗性を付与するか、(b)栄養要求性欠損を相補するか、または(c)例えば、バシラス属(Bacilli)のD−アラニンラセマーゼをコードする遺伝子のように、複合培地からは摂取できない重要な栄養分を供給する。選抜計画の一例では、宿主細胞の増殖を停止させるための薬剤を利用する。異種由来の遺伝子によって形質転換するのに成功した細胞は、薬剤耐性を付与するタンパク質を産生し、それによって、選抜試験を生き残る。このような優性選抜の例では、ネオマイシン(Southernら、J.Molec.Appl.Genet.1:327(1982))、マイコフェノール酸(Mulliganら、Science209:1422(1980))、またはハイグロマイシン(Sugdenら、Mol.Cell.Biol.5:410−413(1985))という薬剤が用いられる。上記の3つの例は、それぞれ、G418もしくはネオマイシン(ゲネチシン(geneticin))、Xgpt(マイコフェノール酸)、またはハイグロマイシンという、適当な薬剤に対する抵抗性を持ち込むために、真核生物の遺伝子の調節下に置かれた細菌遺伝子が用いられている。
【0118】
哺乳動物にとって適当な選抜マーカーのもう一つの例は、DHFR、またはチミジンキナーゼのように、ヘテロマルチマーの塩基配列を取り込む能力のある細胞の同定を可能にする選抜マーカーである。マーカーを取り込んだために、形質転換体だけが、唯一、適応して生き残れるという選択圧の下に、哺乳動物細胞の形質転換体を置く。培地の中の選抜用薬剤の濃度を連続的に変化させ、それによって選抜用遺伝子とヘテロマルチマーをコードするDNAの両方の増幅がもたらされるような条件の下で形質転換体を培養することによって選択圧をかける。増幅されたDNAからは大量のヘテロマルチマーが合成される。この他に増幅可能な遺伝子の例には、メタロチオネイン−Iおよび−2、好ましくは、霊長類のメタロチオネイン遺伝子、アデノシンデアミナーゼ、オルニチンデカルボキシラーゼなどがある。
【0119】
例えば、まず、DHFRの競合的な拮抗剤であるメトトレキセート(Mtx)
を含む培養培地の中で、全部の形質転換細胞を培養して、DHFR選抜用遺伝子によって形質転換された細胞を同定する。野生型のDHFRを用いるときに、適当な宿主細胞は、UrlaubとChasin、Proc.Natl.Acad.Sci.USA77:4216(1980)による説明にしたがって、調製および増殖させたチャイニーズハムスター卵巣(CHO)細胞系のDHFR活性欠損株である。次に、形質転換された細胞を、濃度を上げたメトトレキセートに曝す。これによって、DHFR遺伝子のコピーが多数合成されるようになり、同時に、ヘテロマルチマーの成分をコードするDNAなど、発現ベクターを含むその他の遺伝子のコピーも多数合成されるようになる。この増幅技術は、例えば、ATCCCCL62、CHO−K1などの適当な宿主とともに用いることができ、Mtxに高い耐性をもつ変異DHFR遺伝子を用いるならば、内生的なDHFRがあったとしても構わない(欧州特許第117,060号)。
【0120】
または、ヘテロマルチマー、野生型DHFRタンパク質、およびアミノグリコシド3’−ホスホトランスフェラーゼ(APH)などの選抜マーカーをコードするDNA配列によって形質転換または同時形質転換された宿主細胞(特に、内生DHFRを含む野生型宿主)は、例えば、カナマイシン、ネオマイシン、またはG418などのアミノグリコシド系抗生物質選抜マーカーに対する選抜用薬剤を含む培地の中で細胞を増殖させることによって選抜することができる。米国特許第4,965,199号参照。
【0121】
酵母の中で用いるための適当な選抜用遺伝子は、酵母のプラスミドYRp7の中にあるtrp1遺伝子である。(Stinchcombら、Nature282:39(1979);Kingsmanら、Gene 7:141(1979);または、Tschemperら、Gene10:157(1980))。trp1遺伝子は、トリプトファンの中では増殖できない酵母の変異株、例えば、ATCC44076、またはPEP4−1に対する選抜マーカーを提供する(Jones、Genetics85:12(1977))。酵母宿主ゲノムにtrp1障害があることによって、トリプトファンなしで増殖させることによって、形質転換体を検出するための効果的な環境が提供される。同様に、Leu−2欠損酵母株(ATCC 20,622または38,626)は、Leu2遺伝子をもつ既知のプラスミドによって相補される。
【0122】
さらに、クルイベロマイセス属の酵母を形質転換するためには、1.6μm環状プラスミドpKD1由来のベクターを用いることができる。Bianchiら、Curr.Genet 12:185(1987)。さらに最近、組換えウシキモシンの大量生産のための発現系が報告された。Van denBerg、Bio/Technology8:135(1990)。クルイベロマイセス属の工業用菌株により成熟組換えヒト血清アルブミンを分泌させるための、安定したマルチコピー発現ベクターも開示されている(Fleerら、Bio/Technology 9:968−975(1991))。
【0123】
発現ベクターとクローニングベクターは、通常、宿主生物によって認識され、ヘテロマルチマーの核酸に機能的に結合しているプロモーターを含む。宿主に可能性のある、さまざまな細胞によって認識されるプロモーターが多数ある。制限酵素消化によって、もととなるDNAからプロモーターを除去し、分離したプロモーターをベクターに挿入にして、これらのプロモーターは、ヘテロマルチマーをコードするDNAに機能的に連結する。
原核生物宿主で使用するのに適したプロモーターには、β−ラクタマーゼおよびラクトースのプロモーター系(Changら、Nature275:615(1978);およびGoeddelら、Nature281:544(1979))、アルカリホスファターゼ、トリプトファン(trp)プロモーター系(Goeddelら、Nucleic AcidsRes.,8:4057(1980)、および欧州特許第36,776号)、および、tacプロモーターなどのハイブリッドプロモーター(deBoerら、Proc.Natl.Acad.Sci.USA80:21−25(1983))が含まれる。しかし、細菌のこれ以外の既知のプロモーターが適当である。それらの塩基配列は公開されているので、当業者は、必要な制限酵素部位をもたらすためのリンカーまたはアダプターを用いて、ヘテロマルチマーをコードするDNAに機能的にプロモーターを連結させることができる(Siebenlistら、Cell 20:269(1980))。また、細菌の系で使用するためのプロモーターは、ヘテロマルチマーをコードするDNAに機能的に連結したシャイン−ダルガーノ(S.D.)配列も含んでいる。
【0124】
プロモーターの配列は、真核生物で知られている。実質的にすべての真核生物遺伝子が、転写開始部位から約25から30塩基上流にあるATに富む領域を持っている。多くの遺伝子の転写開始点から上流70から80塩基のところに見つかっている別の配列とはCXCAAT領域である、ただし、Xを任意のヌクレオチドである。殆どの真核生物遺伝子の3’末端には、コーディング配列の3’末端にポリAテールを付加するためのシグナルらしいAATAAA配列がある。これらの配列はすべて、真核生物の発現ベクターの中に適当に挿入する。
【0125】
酵母宿主とともに使用するのに適したプロモーター配列の例には、3−ホスホグリセリン酸キナーゼのプロモーター(Hitzemanら、J.Biol.Chem.255:2073(1980))、または、エノラーゼ、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ、ヘキソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース−6−リン酸イソメラーゼ、3−ホスホグリン酸ムターゼ、ピルビン酸キナーゼ、トリオースリン酸イソメラーゼ、ホスホグルコースイソメラーゼ、およびグルコキナーゼなど、その他の解糖酵素のプロモーター(Hessら、J.Adv.Enzyme.Reg.7:149(1968);および、Holland,Biochemistry17:4900(1978))などである。
【0126】
この他の酵母プロモーターで、増殖条件によって転写を調節するという、さらに別の利点をもつ誘導的プロモーターは、アルコールデヒドロゲナーゼ2、イソシトクロムC、酸性ホスファターゼ、窒素代謝に関係する分解酵素、メタロチオネイン、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ、および、マルトースおよびガラクトース利用に関与する酵素のプロモーター領域である。酵母での発現に用いるのに適したベクターとプロモーターについては、Hitzemanら、欧州特許第73,657A号でさらに説明されている。また、酵母のエンハンサーも、酵母プロモーターとともに適宜用いられる。
哺乳動物の宿主細胞での、ベクターからのヘテロマルチマーの転写は、例えば、ポリオーマウイルス、鶏痘ウイルス(1989年7月5日発行の英国特許第2,211,504号)、アデノウイルス(アデノウイルス2など)、ウシポリオーマウイルス、トリ肉腫ウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルス、および、もっとも好ましくはシミアンウイルス40(SV40)などのウイルスのゲノムから得られたプロモーター、例えば、アクチンプロモーターまたは免疫グロブリンプロモーターなど、異種由来の哺乳動物のプロモーター、または、ヒートショックプロモーターによって調節される。
SV40ウイルスの前者および後者のプロモーターは、都合のよいことに、SV40ウイルスの複製開始点も含むSV40制限酵素フラグメントとして得られる。Fiersら、Nature 273:113 (1978);MulliganとBerg、Science 209:1422−1427(1980);Pavlakisら、Proc. Natl. Acad. Sci. USA 78:7398−7402 (1981)。ヒトサイトメガロウイルス最早期プロモーターは、都合よく、HindIIIE制限酵素フラグメントとして得られる。Greenawayら、Gene 18:355−360 (1982)。ウシポリオーマウイルスをベクターとして用いた、哺乳動物の宿主の中でDNAを発現させるための系が米国特許第4,419,446号に記載されている。
【0127】
この系を改変したものが、米国特許第4,601,978号に記載されている。
【0128】
また、免疫インターフェロンをコードするcDNAのサル細胞での発現については、Grayら、Nature295:503−508(1982);単純ヘルペスウイルス由来のチミジンキナーゼプロモーターの調節下での、ヒトβ−インターフェロンcDNAのマウス細胞における発現については、Reyesら、Nature 297:598−601 (1982);ヒトインターフェロンβ1遺伝子の培養マウス細胞とウサギ細胞における発現についてはCanaaniとBerg、Proc. Natl.Acad. Sci. USA 79:5166−5170 (1982);および、マウス肉腫ウイルスのLTR(長い末端反復配列)をプロモーターとして用いた、CV−1サル腎臓細胞、ニワトリ胚線維芽細胞、チャイニーズハムスター卵巣細胞、HeLa細胞、およひマウスNTH−3T3細胞における、細菌のCAT配列の発現についてはGormanら、Proc.Natl.Acad.Sci.USA 79:6777−6781(1982)も参照。
高等真核生物による、ヘテロマルチマー成分をコードするDNAの転写は、ベクターの中にエンハンサー配列を挿入することによって亢進することがよくある。エンハンサーは、方向や位置に比較的無関係であり、転写単位に対して5’側(Laiminsら、Proc.Natl.Acad.Sci.USA78:993(1981))、3’側(Luskyら、Mol.Cell.Bio.3:1108(1983))イントロンの内部(Banerjiら、Cell33:729(1983))、および、コーディング配列そのものの中(Osborneら、Mol.Cell.Bio.4:1293(1984))にも見つかっている。今では、哺乳動物の遺伝子(グロビン、エラスターゼ、アルブミン、α−フェトプロテイン、およびインシュリン)由来の多くのエンハンサー配列が知られている。しかし、典型的には、真核生物細胞のウイルスに由来するエンハンサーが使用される。例としては、複製開始点の後方側(100−270bp)にあるSV40エンハンサー、サイトメガロウイルス初期プロモーターのエンハンサー、複製開始点の後方側にあるポリオーマエンハンサー、およびアデノウイルスエンハンサーなどがある。また、真核生物のプロモーターを活性化するための促進因子については(Yanivら、Nature297:17−18(1982)を参照。エンハンサーは、ベクターの中のヘテロマルチマーをコードする配列の5’側か3’側の位置に挿入することができるが、好ましくは、プロモーターの5’側の部位に存在する。
【0129】
真核宿主細胞(酵母、菌類、昆虫、植物、動物、ヒト、またはその他の多細胞生物からの有核細胞)で用いられる発現ベクターは、転写の終結、およびmRNAの安定化に必要な配列も含んでいる。このような配列は、真核生物またはウイルスのDNAまたはcDNAの5’側、場合によっては3’側にある非翻訳領域から広く利用することができる。これらの領域には、ヘテロマルチマーをコードするmRNAの非翻訳部位のポリアデニル化フラグメントとして転写されるヌクレオチド分節が含まれる。
【0130】
上記で挙げた構成要素を一つ以上含む適当なベクターを構築するには、標準的な結紮技術を用いる。分離したプラスミドまたはDNAフラグメントは、必要とするプラスミドを作出するのに望ましい形になるように、開裂し、加工してから再連結する。
【0131】
構築したプラスミドの配列が正しいことを確認するための解析を行なうために、結紮混合液を用いて、大腸菌K12菌株294(ATCC31,446)を形質転換して、アンピシリン耐性またはテトラサイクリン耐性によって選抜された、形質転換に成功した株が適正なものである。形質転換からプラスミドを調製して、制限酵素消化によって解析し、および/または、Messingら、Nucleic AcidsRes.,9:309(1981)の方法、またはMaxamら、Methods in Enzymology65:499(1980)の方法によって配列決定を行なう。
【0132】
本発明の実施において特に有用なのは、ヘテロマルチマーをコードするDNAを哺乳動物細胞の中で一過性発現をもたらす発現ベクターである。一般的には、一過性発現には、宿主細胞が多くの発現ベクターのコピーをもち、次いで、発現ベクターによってコードされる所望のポリペプチドを大量に合成するように、宿主細胞の中で効率的に複製することのできる発現ベクターを使用することが含まれる。Sambrookら、上記、pp.16.17−16.22。適当な発現ベクターと宿主細胞を含む一過的発現系によって、クローニングされたDNAによってコードされているポリペプチドを便利に陽性のものを同定することと、所望の結合特性/親和性、または、本発明によって作出される、天然のジスルフィド結合をもたないヘテロマルチマーまたホモマルチマーに対して、所期のゲル泳動度をもつヘテロマルチマーを迅速にスクリーニングすることが可能になる。
【0133】
脊椎動物の組換え細胞培養でヘテロマルチマーを合成するように調整するのに適した、この他の方法、ベクター、および宿主細胞については、Gethingら、Nature 293:620−625(1981);Manteiら、Nature281:40−46(1979);欧州特許第117,060号;および欧州特許第117,058号に記載されている。哺乳動物の培養細胞でヘテロマルチマーを発現させるのに特に役立つプラスミドは、pRK5(欧州特許第307,247号)またはpSV16B(1991年6月13日発行のPCT国際特許公開第91/08291号)である。
【0134】
ヘテロマルチマーを発現させるために、どの宿主細胞系を選択するかは、主に、発現ベクターによって決まる。もう一つ考慮すべきなのは、必要とするタンパク質量である。ミリグラム単位の量を、一過的形質転換によって産生することができる。例えば、アデノウイルスEIA−によって形質転換された293ヒト胚腎臓細胞系を、ヘテロマルチマーの効率的な発現が可能になるよう、リン酸カルシウム法を修正した方法によって、pRK5を基本とするベクターで一過的に形質転換することができる。CDM8を基本とするベクターを用いて、DEAE−テキストラン法によってCOS細胞を形質転換することができる(Aruffoら、Cell 61:1303−1313(1990);および、Zettmeisslら、DNA CellBiol.(US)9:347−353(1990))。より大量のタンパク質が必要ならば、宿主細胞系を安定的に形質転換させた後、免疫付着因子を発現させることができる。例えは、pRK5を基本とするベクターを、ジヒドロ葉酸レダクターゼ(DHFR)をコードする別のベクターの存在下で、チャイニーズハムスター卵巣(CHO)細胞の中に導入して、G418に対する抵抗性を付与することができる。G418に抵抗性のクローンは、培養によって選抜することができる。これらのクローンを、DHFRのインヒビターであるメトトレキセートの量を増加させながら培養して、DHFRとヘテロマルチマーの配列をコードする遺伝子のコピー数が共に増幅されているクローンを選抜する。免疫付着因子が、N末端に疎水性の先導配列をもっていれば、形質転換された細胞によって加工されて分泌される可能性が高い。より複雑な構造をもつ免疫付着因子の発現には、特に適した宿主細胞が必要となろう。例えば、軽鎖またはJ鎖などの構成成分は、一定のミエローマまたはハイブリドーマの宿主細胞によって提供されよう(Gascoigneら、上記;およびMartinら、J.Virol.67:3561−3568(1993))。
【0135】
本明細書のベクターをクローニングし、発現させるのに適した、この他の宿主細胞は、上記のような原核生物、酵母、または、高等真核生物の細胞である。この目的に適した原核生物には、グラム陰性またはグラム陽性生物などの真性細菌で、例えば、腸内細菌である、例えば、大腸菌(E.coli)などのエシェリキア属(Escherichia)、エンテロバクター属(Enterobacter)、エルウィニア属(Erwinia)、クレブシアラ(Klebsiella)、プロテウス属(Proteus)、例えば、ネズミチフス菌(Salmonella typhimurium)などのサルモネラ属(Salmonella)、霊菌(Serratiamarcescans)などのセラティア属(Serratia)、およびシゲラ属(Shigella)、ならびに、枯草菌(B.subtilis)およびB.リケニフォルミス(B.licheniformis)(例えば、1989年4月12日公開のDD266,710で開示されているB.リケニフォルミス41P)などのバシラス属、緑膿菌(P.aeruginosa)などのシュードモナス属、およびストレプトマイセス属(Streptomyces)などである。好ましい大腸菌クローニング宿主の一つは大腸菌294(ATCC 31,446)であるが、その他、大腸菌B、大腸菌X1776(ATCC31,537)、および大腸菌W3110(ATCC27,325)などの菌株も適している。これらの例は、例示のためのものであり、制限のためのものではない。菌株W3110は、組換えDNA産物の発酵のための一般的な宿主菌株であるため、この菌株は、特に好ましい宿主、または宿主の親株である。好ましくは、宿主細胞は、最小量のタンパク質分解酵素を分泌するものでなければならない。例えば、W3110株を改変してタンパク質をコードする遺伝子に遺伝的変異を生じさせることができる。そのような宿主の例として、大腸菌W3110の27C7株などがある。27C7の完全な遺伝子型は、tonAΔptr3 phoAΔE15 Δ(argF−lac)169ompTΔdegP41kanrである。27C7株は、1991年10月30日に、ATCC番号55,244として、アメリカンタイプカルチャーコレクションに寄託された。または、1990年8月7日発行の米国特許第4,946,783号で開示されている、ペリプラズマプロテアーゼの変異体をもつ大腸菌の菌株を用いることもできる。または、例えば、PCR、または、別の核酸のポリメラーゼ反応などのクローニング法が適している。
【0136】
原核生物の他に、糸状菌類または酵母のような真核生物微生物が、ヘテロマルチマーをコードするベクターに適したクローニングおよび発現のための宿主である。サッカロマイセス・セレビシアエ(Saccharomyces cerevisiae)、すなわち一般的なパン酵母が、下等な真核生物宿主微生物の中で、もっとも一般的に用いられている。しかし、本明細書においては、分裂酵母(Schizosaccharomyces pombe)(BeachとNurse、Nature290:140(1981);1985年5月2日発行の欧州特許第139,383号);クルイベロマイセス属の宿主(米国特許第4,943,529号;Fleerら、上記)で、例えば、K.ラクティス(K.lactis)(MW98−8C,CBS683,CBS4574;Louvencourtら、J.Bacteriol.,737(1983))、K.フラジリス(K.fragiles)(ATCC 12,424)、K.ブルガリクス(K.bulgaricus) (ATCC 16,045)、K.ウィケラミイ(K.wicheramii)(ATCC24,178)、K.ワルティイ(K.waltii)(ATCC 56,500)、K.ドロソフィラルム(K.drosophilarum)(ATCC36,906;Van den Bergら、上記)、K.テルモトレランス(K.thermotolerans)、およびK.マルキサヌス(K.marxianus)など;ヤロウィア属(yarrowia)(欧州特許第402,226号);ピキア・パストリス(Pichiapastoris)(欧州特許第183,070号;Sreekrishnaら、J.BasicMicrobiol.28:265−278(1988));カンジダ属;トリコデルマ・レエシア(Trichodermareesia)(欧州特許第244,234号);ニューロスポラ・クラッサ(Neurosporacrassa)(Caseら、Proc.Natl.Acad.Sci.USA 76:5259−5263(1979));シュワニオマイセス・オクシデンタリス(Shwanniomycesoccidentalis)などのシュワニオマイセス属(1990年10月31日発行の欧州特許第394,538号);および、例えば、ニューロスポラ属、ペニシリウム属(Penicillium)、トリポタラジウム属(Tolypocladium)などの糸状菌(1991年1月10日発行の国際特許公開第91/00357号)、ならびに、アウペルギルス属(Aspergillus)宿主、例えば、A.ニドランス(A.nidulans) (Ballanceら、Biochem.Biophys.Res.Commun.112:284−289(1983);Tilburnら、Gene 26:205−221(1983);Yeltonら、Proc.Natl.Acad.Sci.USA81:1470−1474(1984)、およびA.ニガー(A. niger)(KellyとHynes,EMBOJ.4:475−479(1985))など、この他数多くの属、種、および株を用いることが可能であり、また有用でもある。
【0137】
グリコシル化されたヘテロマルチマーを発現させるのに適した宿主細胞は、多細胞生物由来のものである。そのような宿主細胞は、複雑なプロセッシングを行なうことができ、グリコシル化活性をもつ。原則として、脊椎動物由来でも、無脊椎動物由来でも、どのような高等真核生物の細胞培養を使うこともできる。無脊椎動物細胞の例としては、植物細胞、昆虫細胞などがある。多数のバキュロウイルス株とその変異株、および対応する昆虫宿主許容細胞として、スポドプテラ・フルギペルダ(Spodoptera frugiperda)(毛虫)、アエデス・アエギプチ(Aedesaegypti)(蚊)、アエデス・アルボピクタス(Aedes albopictus)(蚊)、ドロソフィラ・メラノガスター(Drosophilamelanogaster)(ショウジョウバエ)、およびボムビクス・モリ(Bombyxmori)などか確認されている。例えば、Luckowら、Bio/Technology6:47−55(1988);Setlowら編、遺伝子工学(GeneticEnginerring)第8巻より、Millerら、pp.277−279(プレナムパブリッシング社(PlenumPublishing)、1986年);および、Maedaら、Nature 315:592−594(1985)を参照。例えば、アウトグラファ・カリフォニカ(Autographacalifonica)NPVのL−1変異株、およびカイコNPV(Bombyx moriNPV)のBm−5株など、形質転換用のさまざまなウイルス株が一般に利用可能であり、そのようなウイルスは、本明細書において、本発明に記載されているウイルスとして、特に、スポドプテラ・フルギペルダ(spodoptera frugiperda)細胞の形質転換に用いることができる。
【0138】
ワタ、トウモロコシ、バレイショ、ダイズ、ペチュニア、トマト、およびタバコの植物培養細胞を宿主として利用することができる。典型的には、予め、ヘテロマルチマーDNAを持つように操作してあるアグロバクテリウム・ツメファシエンス(Agrobacteriumtumefaciens)菌の一定の菌株とインキュベートして、植物細胞を形質転換する。植物培養細胞をA.ツメファシエンスとインキュベートしている間に、ヘテロマルチマーをコードしているDNAが植物細胞宿主に移行して、それを形質転換し、適当な条件の下でヘテロマルチマーを発現させる。さらに、ノパリンシンターゼプロモーター、およびポリアデニル化シグナル配列のように、植物細胞と親和性のある調節配列とシグナル配列を利用することができる。Depickerら、J.Mol.Appl.
Gen.1:561(1982)。なお、T−DNA780遺伝子の上流領域から分離したDNA分節は、組換えDNAを含む植物組織において、植物で発現する遺伝子の転写レベルを活性化あるいは上昇させることができる。1989年6月21日発行の欧州特許第321,196号。
【0139】
好ましい宿主は脊椎動物の細胞であり、脊椎動物の細胞を培養して増殖させること(組織培養)は、近年では日常的な処理になっている(アカデミックプレス社 (Academic Press)、KruseとPatterson編、組織培養(TissueCulture)(1973))。有用な哺乳動物の宿主細胞系の例は、SV40で形質転換されたサル腎臓CV1系統(COS−7,ATCC CRL1651);ヒト胚腎臓系統(293、または、懸濁培養で増殖させるためにサブクローニングされた293細胞、Grahamら、J.Gen.Virol.36:59(1977));仔ハムスター腎臓細胞(BHK、ATCC CCL10);チャイニーズハムスター卵巣細胞/−DHFR(CHO、UrlaubとChasin、Proc.Natl.Acad.Sci.USA77:4216(1980));マウスセルトリ細胞(TM4、Mather,Biol.Repod.23:243−251(1980));サル腎臓細胞 (CV1 ATCC CCL 70);アフリカミドリザルの腎臓細胞(VERO−76、ATCCCRL−1587);ヒト子宮頚部ガン細胞(HELA、 ATCC CCL 2);イヌ腎臓細胞(MDCK,ATCC CCL34);バッファローラット肝細胞(BRL 3A,ATCCCRL 1442);ヒト肺細胞(W138,ATCC CCL 75);ヒト肝臓細胞(Hep G2,HB 8065);マウス乳ガン(MMT 060562,ATCC CCL 51);TRI細胞(Matherら、Annals N.Y.Acad.Sci.383:44−68(1982));MRC5細胞;FS4細胞;およびヒトヘパトーマ系(Hep G2)である。
【0140】
宿主細胞を本発明の上記発現ベクターまたはクローニングベクターによって形質転換して、プロモーターを誘導したり、形質転換体を選抜したり、または所望の配列をコードする遺伝子を増幅するのに適合するように修正した通常の培地で培養する。用いる宿主細胞によって、その細胞に適した標準的な技術を用いて形質転換を行なう。Sambrookら、上記の1.82節で説明されているように、一般的には、実質的な細胞壁という障壁を含む、原核生物またはその他の細胞に対して、塩化カルシウムを用いたカルシウム処理、またはエレクトロポレーションを用いる。Shawら、Gene23:315(1983)、および1989年6月29日発行の国際特許公開第89/05859号に記載されているようにして、アクロバクテリウム・ツメファシエンスによる感染を、一定の植物細胞を形質転換するために用いる。さらに、1991年1月10日発行の国際特許公開第91/00358号に記載されているようにして、超音波を用いて、植物を形質転換することができる。
【0141】
このような細胞壁をもたない哺乳動物細胞については、GrahamとvanderEb、Virology52:456−457(1978)のリン酸カルシウム法が好ましい。哺乳動物細胞宿主系の形質転換の一般的な態様を、1983年8月16日発行の米国特許第4,399,216号において、Axelが記述している。酵母への形質転換は、一般的には、VanSolingenら、J.Bact.130:946(1977)、およびHsiaoら、Proc.Natl.Acad.Sci.USA76:3829(1979)の方法にしたがって行われる。DNAを細胞の中に導入するために、核へのマイクロインジェクション、エレクトロポレーション、無処理の細胞とバクテリアのプロトプラストとの融合、または、例えば、ポリブレン、ポリオルニチンなどのポリカチオンのような別の方法を用いることもできる。哺乳動物細胞を形質転換するためのさまざまな技術については、Keownら、Methods in Enzymology(1989)、Keownら、Methods inEnzymology 185:527−537(1990)、およびMansourら、Nature 336:348−352(1988)を参照。
【0142】
本発明のヘテロマルチマーポリペプチドを産生するために用いた原核生物細胞を、Sambrookら、上記において、一般的に説明されているところにしたがって、適当な培地で培養する。
【0143】
本発明のヘテロマルチマーを産生するために用いた哺乳動物宿主細胞は、さまざまな培地出培養することができる。宿主細胞を培養するには、Ham’sF10(シグマ社(Sigma))、最小必須培地((MEM)、シグマ社)、RPMI−1640(シグマ社)、およびダルベッコの修正イーグル培地 ((DMEM)、シグマ社)などの市販の培地が適当である。さらに、それらの開示内容がすべて、参照してここに組み込まれる、HamとWallace,Meth.Enz.58:44(1979)、BarnesとSato、Anal.Biochem.102:255(1980)、米国特許第4,767,704号;第4,657,866号;第4,927,762号;または、第4,560,655号;国際特許公開第90/03430号;国際特許公開第87/00195号;米国特許第4,791,192号;および米国特許(U.S.Patent Re.)第30,985号;または、米国特許第5,122,469号に記載されている培地のいずれかを、宿主細胞用の培養培地として用いることができる。これらの培地はどれも、必要ならは、ホルモンおよび/またはその他の増殖因子(インシュリン、トランスフェリン、または上皮増殖因子など)、塩類(塩化ナトリウム、カルシウム、マグネシウム、およびリン酸)、緩衝液(HEPESなど)、ヌクレオシド(アデノシンおよびチミジンなど)、抗生物質(登録商標ゲンタマイシン薬剤)、微量要素(最終濃度がマイクロモルの単位で存在する無機化合物と定義される)、およびグルコースまたは同等のエネルギー源を添加することもできる。この他の添加物も、当業者に知られている適当な濃度で含むことができる。温度、pHなどの培養条件は、発現させるために選ばれた宿主細胞によって以前用いられた条件であり、当業者にとっては明らかである。
【0144】
一般的に、哺乳動物の細胞培養の生産性を最大にするための原理、プロトコール、および実際の技術については、IRLプレス社(IRLPress)、1991年、M.Butler編、哺乳動物細胞のバイオテクノロジー:実際的方法(Mammalian CellBiotechnology:aPractical Approach)に書かれている。
【0145】
本開示で言及されている宿主細胞は培養細胞だけでなく、宿主動物の中の細胞も含む。
4.ヘテロマルチマーの回収ヘテロマルチマーは、好ましくは、一般的に、分泌ポリペプチドとして、培養培地から回収するが、分泌シグナルなしに、直接産生されたときには、宿主細胞溶解物から回収する。ヘテロマルチマーに膜結合があるときには、適当な界面活性剤(例:Triton−X 100)を用いて膜から解離することができる。
【0146】
ヒトに由来しない組換え細胞の中でヘテロマルチマーが産生されたときには、ヒト由来のタンパク質やポリペプチドを全く含まない。しかし、組換え細胞のタンパク質やポリペプチドからヘテロマルチマーを精製して、ヘテロマルチマーについて実質的に均質な調製物を得る必要がある。最初の工程として、培養培地または細胞溶解物を普通に遠心分離して、微粒子化した細胞残滓を取り除く。
【0147】
抗体の定常部位をもつヘテロマルチマーは、ハイドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析、またはアフィニティークロマトグラフィーによって、都合のよいように精製することができるが、アフィニティークロマトグラフィーが好ましい精製技術である。ヘテロマルチマーがCH3部位を含むときは、登録商標ベーカーボンド ABX(Bakerbond ABX)樹脂(J.T.Baker,ニュージャージー州フィリップスバーグ (Phillipsburg))が、精製するために有用である。この他に、イオン交換カラムによる分画、エタノール沈殿、逆相HPLC、シリカによるクロマトグラフィー、ヘパリンセファロースによるクロマトグラフィー、陰イオンまたは陽イオン交換樹脂(ポリアスパラギン酸カラムなど)によるクロマトグラフィー、クロマトフォーカシング、SDS−PAGE、および硫安沈殿などのタンパク質精製技術も、回収するポリペプチドによっては利用可能である。親和性リガンドとしてのプロテインAの適合性は、キメラで用いられた免疫グロブリンFcドメインの種類およびアイソタイプによって異なる。プロテインAを用いて、ヒトγ1、γ2、またはγ4のH鎖に基づいて、免疫付着因子を精製することができる (Lindmarkら、J.Immunol.Meth.62:1−13(1983))。マウスのアイソタイプのすべてとヒトγ3(Gussら、EMBOJ.5:1567−1575(1986))には、プロテインGが推奨されている。親和性リガンドが結合する基質は、もっとも普通にはアガロースであるが、その他の基質を用いることもできる。調節された多孔性ガラスビーズ(controlledporeglass)またはポリ(スチレンジビニル)ベンジンのような物理的に安定した基質なら、アガロースを用いるよりも流速が早くなり、短い処理時間でできる。プロテインAまたはGアフィニティーカラムに免疫付着因子を結合させる条件は、Fc部位の特徴によって全面的に決定される。すなわち、その分子種とアイソタイプによって決まる。一般的には、適正なリガンドが選択されれば、直接、条件付けていない培養溶液からでも、効率的な結合が起こる。免疫付着因子の顕著な特徴の一つは、ヒトγ1分子で、同じFc型の抗体に較べて、プロテインAに対する結合能力が幾分低下することである。結合した免疫付着因子は、酸性pH(3.0かそれ以上)で、または、僅かにカオトロピックな塩を含む中性pHの緩衝液の中で効率的に溶出することできる。このアフィニティークロマトグラフィー工程によって、>95%純粋なヘテロマルチマー調製物を作製することができる。
【0148】
5.共通の軽鎖をもつヘテロマルチマーの多官能性抗体の使用ヘテロマルチマーについては、治療薬としての多くの応用が考えられる。例えば、細胞傷害性を切り換える(例:腫瘍細胞を殺す)ために、ワクチンのアジュバントして、血栓溶解剤を血餅まで輸送するために、標的部位(例えば、腫瘍)で、酵素的に活性化されたプロドラッグを転化するために、感染病を治療するために、免疫複合体を細胞表面へと向かわせるために、または、腫瘍細胞にイムノトキシンを輸送するためにヘテロマルチマーを用いることができる。例えば、クラスII主要組織適合遺伝子複合体であるモデル内皮抗原を標的とすることによって、抗体−リシンイムノトキシンによって、腫瘍の脈管構造を標的とすることができた(Burrows,F.J.とThorpe,P.E.(1993)Proc.Natl.Acad.Sci.USA 90:8996−9000)。
【0149】
抗−内皮イムノトキシンを別のイムノトキシンと組み合わせることによって、腫瘍細胞そのものに対する有意に高い効能が得られた(Burrows,F.J.とThorpe,P.E.(1993)上記)。最近、双特異的抗体を用いて、組織因子を腫瘍脈管構造に向かわせることに成功し、かなり高い抗腫瘍効果を生じる局所的な血栓症を引き起こした(Huang,Xら、(1997)Science275:547−550)。さらに、双特異的抗体を用いて、細胞傷害性T細胞に、インビトロで、標的乳ガン細胞およびB細胞リンパ腫細胞を殺させることに成功した(Zhu,Z.ら、(1996)Bio/Technology14:192−196;およびHolliger,P.ら、(1996)Protein. Engin.9:299−305)。
【0150】
望ましい精製度をもつヘテロマルチマーを、選択的に、生理学的に許容できる担体、賦形剤、または安定剤(レミントンの製薬科学(Remington’s Pharmaceutical Science)、第16版、Osol,A.編(1980))と混合して、凍結乾燥した固形または水溶液の形状で保存するために、ヘテロマルチマーの治療用処方剤を調製する。許容される担体、賦形剤、または安定剤は、用いられる投薬量と濃度では受容者にとって非毒性で、リン酸、クエン酸、およびその他の有機酸などを含む緩衝液;アスコルビン酸などの抗酸化剤;低分子量(約10残基未満)ポリペプチド;血清アルブミン、ゼラチン、または免疫グロブリンなどのタンパク質;ポリビニルピロリドンなどの疎水性ポリマー;グリシン、グルタミン、アスパラギン、アルギニン、またはリシンなどのアミノ酸;単糖類;二糖類;および、グルコース、マンノース、またはデキストリンなどの糖類;EDTAなどのキレート化剤;マンニトール、またはソルビトールなどの糖アルコール;ナトリウムなどの塩を形成する対イオン;および/または、トィーン(Tween)、プルロニクス(Pluronics)またはポリエチレングリコール(PEG)などの界面活性剤を含んでいる。
【0151】
ヘテロマルチマーは、例えば、コアセルベート化技術、または界面ポリマー化(例えば、ヒドロキシメチルセルロース、または、それぞれ、ゼラチン−マイクロカプセルおよびポリ−[メチルメトアシレート]マイクロカプセル)によって調製されたマイクロカプセルの中に取り込んだり、コロイド剤送達システム(例えば、リポソーム、アルブミンミクロスフィア、ミクロエマルジョン、ナノ粒子、およびナノカプセル)に取り込んだり、または、マクロエマルジョンの中に取り込んだりすることができる。このような技術については、レミントンの製薬科学(Remington’s PharmaceuticalScience)、上記で開示されている。
【0152】
インビボでの投与に用いるヘテロマルチマーは、滅菌されていなければならない。これは、凍結乾燥および再構成の前か後に、滅菌フィルター膜で濾過すれば簡単に行なえる。ヘテロマルチマーは、通常、凍結乾燥状態か水溶液の状態で保存する。
【0153】
治療用ヘテロマルチマー組成物は、一般的に、滅菌的に接触することのできる通路をもった容器、例えば、静脈注射用のバック、または皮下注射用の注射針によって貫通することのできる栓をもったバイアルの中に置かれる。
【0154】
ヘテロマルチマーの投与経路は、例えば、静脈、腹腔内、脳内、筋肉内、眼内、動脈内、または病巣内などの経路による注射または点滴のような既知の方法、または下記に示すような徐放性システムによる。ヘテロマルチマーは、点滴またはボーラス注射によって、連続的に投与する。
【0155】
徐放性調製物の適当な例は、タンパク質を含む固体疎水性ポリマーの半透過性基質で、例えば、フィルムまたはマイクロカプセルのような形のあるものになっている基質を含む。徐放性基質の例は、Langerら、J.Biomed.Mater.Res.15:167−277(1981)、およびLanger、Chem.Tech.12:98−105(1982)に記載されているポリエステル、ヒドロゲル(例えば、ポリ(2−ヒドロキシエチル−メトアクリル酸)、またはポリ(ビニルアルコール)、ポリアクチド(米国特許第3,773,919号、欧州特許第58,481号)、L−グルタミン酸とガンマエチル−L−グルタミン酸のコポリマー(Sidmanら、Biopolymers 22:547−556(1983))、非分解性エチレン−ビニル酢酸(Langerら、上記)(欧州特許第133,988号)、登録商標ルプロンデポット(LupronDepot)のような分解性の乳酸−グリコール酸コポリマー(乳酸−グリコール酸コポリマーと酢酸ロイプロリドからなる注射用ミクロスフィア)、およびポリ−D−(−)−3−ヒドロキシ酪酸(欧州特許第133,988号)などがある。
【0156】
エチレン−ビニル酢酸、および乳酸−グリコール酸などのコポリマーは、100日間にわたって分子を放出することが可能であるが、ある種のヒドロゲルでは、タンパク質を放出する期間が短くなる。カプセルに入ったタンパク質は、長時間体内に残留して、37℃で湿気に曝される結果、変性するか凝集して、生物学的活性を失うことになり、免疫原性にも変化が生じうる。関係するメカニズムによって、タンパク質を安定化させるための合理的な方策を考え出すことができる。例えば、凝集を起こすメカニズムが、チオ−ジスルフィド基の交換による分子内S−S結合形成であることが発見されていれば、スルフィドリル残基を修飾し、酸性溶液から凍結乾燥し、水分含有量を調節し、適当な添加剤を使用し、また、特異的なポリマー基質組成物を開発することによって、安定させることができる。
【0157】
また、徐放性ヘテロマルチマー組成物は、リポソームによって取り込まれたヘテロマルチマーでもよい。ヘテロマルチマーを含むリポソームは、それ自体は既知の方法:独国特許第3,218,121号;Epsteinら、Proc.Natl.Acad.Sci.USA 82:3688−3692(1985);Hwangら、Proc.Natl.Acad.Sci.USA77:4030−4034(1980);欧州特許第52,322号;欧州特許第36,676号;欧州特許第88,046号:欧州特許第143,949号;欧州特許第142,641号;日本国特許出願第83−118008号;米国特許第4,485,045号および第4,544,545号;ならびに欧州特許第102,324号によって調製される。通常、リポソームは、小さな(200−800オングストローム)均一ラメラ型で、脂質成分が約30mol.%コレステロールよりも高く、ヘテロマルチマー治療を最適なものにするために、選ばれた部位が調整されている。
【0158】
治療に用いるヘテロマルチマーの効果的な用量は、例えば、治療目的、投与経路、および患者の症状によって変わる。したがって、至適な治療効果を得るためには、治療者が用量を希釈したり、投与経路を変更することが必要となろう。典型的な毎日の投与量は、上記の要素によって、約1μg/kgから10mg/kgかそれよりも多い量になろう。典型的には、臨床医は、所期の効果を納めるまで、ヘテロマルチマーの投薬を続けることになろう。この治療の進行状況は、常法によって容易に観察される。
【0159】
本明細書に記述されているヘテロマルチマーは、酵素免疫測定法で用いることもできる。それを行なうためには、結合によって、酵素を阻害しないために、ヘテロマルチマーの一方のアームを、酵素上の特異的なエピトープに結合するように設計することができ、ヘテロマルチマーのもう一方のアームを、所望の部位で、酵素が確実に高濃度になるようにしながら、固定基質に結合するように設計することができる。このような診断用ヘテロマルチマーの例としては、IgG、およびフェリチンに対する特性を持ったヘテロマルチマー、および、ホースラディッシュパーオキシダーゼ(HRP)、ならびに、例えば、ホルモンに対する結合特異性をもつヘテロマルチマーなどがある。
【0160】
このヘテロマルチマーは、2部位免疫測定法に用いるよう設計することもできる。例えば、双特異的な2つのヘテロマルチマーを産生させて、解析するタンパク質上の2つの別々のエピトープに結合させるが、一方のヘテロマルチマーは、不溶性の基質と複合体になるように結合し、もう一方は、指示酵素に結合する。
【0161】
また、ヘテロマルチマーは、ガンなど、さまざまな病気のインビトロおよびインビボでの免疫診断に用いることもできる。この診断への使用を容易にするために、ヘテロマルチマーの一方のアームを腫瘍関連抗原に結合するように設計し、もう一方のアームを検出用マーカー(例えは、放射性核種に結合するキレート剤)に結合するように設計することができる。例えば、腫瘍関連抗原CEA、および二価性ハプテンに対して特異性をもつヘテロマルチマーは、結腸直腸ガンと甲状腺ガンを画像化するために用いることができる。この他にも、ヘテロマルチマーを、治療ではなく診断に使用できることは当業者にとって明らかであろう。
【0162】
診断に応用するためには、ヘテロマルチマーの少なくとも一方のアームが、典型的には、直接的または間接的に、検出可能な部分によって標識されている。例えば、検出可能な部分は、3H、14C、32P、35S、または125Iなどの放射性同位元素;フルオロセインイソチオシアナート、ローダミン、またはルシフェリンなどの蛍光化合物または化学発光化合物;または、アルカリホスファターゼ、β−ガラタトシダーゼ、またはホースラディッシュパーオキシダーゼ(HRP)などの酵素であろう。
【0163】
ヘテロマルチマーを検出可能な部分に別々に結合させるために、当技術分野において既知の方法を用いることができるが、そのような方法には、Hunterら、Nature 144:945(1962);Davidら、Biochemistry13:1014(1974);Painら、J.Immunol.Meth.40:219(1981);および、Nygen,J.Histochem.and Cytochem.30:407(1982)に記載されている方法などがある。
【0164】
本発明のヘテロマルチマーは、競合結合アッセイ法、直接および間接サンドイッチアッセイ法、および免疫沈殿法など、既知のアッセイ法に用いることができる。Zola、モノクローナル抗体:技術マニュアル(Monoclonal Antibodies:A Manual ofTechniques),pp.147−158(CRCプレス社(CRC Press,Inc.,1987)。
【0165】
競合結合アッセイ法は、限られた量のヘテロマルチマーへの結合をめぐって、標識された標準化合物が、解析対象の試験用サンプルと競合できることに基づいている。試験サンプル中の解析化合物の量は、ヘテロマルチマーに結合した標準化合物の量に反比例する。結合した標準化合物の量を容易に決定するために、ヘテロマルチマーは、一般的に、競合の前か後に不溶化して、ヘテロマルチマーに結合している標準化合物と解析化合物が、非結合のままの標準化合物と解析化合物とから都合良く分離できるようにする。
【0166】
このヘテロマルチマーは、それぞれが、検出すべきサンプルの異なった免疫原部分またはエピトープに結合することのできる2種類の分子を使用することを含むサンドイッチアッセイ法にとって、特に有用である。サンドイッチアッセイ法において、試験用サンプルの解析化合物は、固体担体上に固定されたヘテロマルチマーの一方のアームに結合していて、その後、ヘテロマルチマーのもう一方のアームが解析化合物に結合して、3つの部分からなる不溶性の複合体を形成する。例えば、米国特許第4,376,110号を参照。ヘテロマルチマー自体の第2のアームを、検出可能部分によって標識する(直接的サンドイッチアッセイ法)か、または、検出可能部分によって標識された抗免疫グロブリン抗体を用いて測定することができる(間接的サンドイッチアッセイ法)。例えば、サンドイッチアッセイ法の一つのタイプは、ELISA法であるが、この場合には、検出可能部分は酵素である。
【0167】
以下は、本発明を実施するための特定の実施態様の実施例である。実施例は、例示目的のだけに示されているのであって、いかなる意味でも、本発明の範囲を制限するためのものではない。
【0168】
本明細書において引用されている刊行物、特許、および特許出願は、上記のものも下記のものもすべてが参照してここに組み込まれる。
【実施例】
【0169】
計画がFc-含有BsAb(図1C)を製造するために提供される。この計画において、我々はそれがヘテロダイマー化であるがホモダイマー化されないように抗体重鎖のCH3を工作している。これは、論理的設計(Ridgwayら,上記(1996))及びここに記載した通りのファージディスプレー選択によって得られた立体的相補変異と共同してCH3ドメイン中に相互-鎖ジスルフィド結合を設けることによって達成された。両方の抗原結合特異性用の単軽鎖の使用は、軽鎖誤対合の問題を避ける(図1A- 1C)。同じ軽鎖を持った抗体は、非常に大きなヒトscFvライブラリーを選別することによって容易に分離された(Vaughan,T.J.ら, (1996)上記)。
【0170】
実施例1:
空洞への隆起(protuberance-into-cavity)ヘテロマルチマー免疫付着因子の生成 Chamowら,J.Imminol.153:4268(1994)により先に記載されたヒト化抗-CD3/CD4-IgGキメラは、回収できたヘテロマルチマーのパーセンテージを最大化するように構築した。空洞への隆起及び野生型CH3変異体は、ヒト化抗体-免疫付着因子キメラ(Ab/Ia)抗-CD3/CD4-IgGの形成を導くその能力を比較した。
【0171】
かくして変異体は、ヒト化抗-CD3抗体重鎖のCH3ドメインに及びミスマッチオリゴヌクレオチドを用いたサイト-誘導変異誘発(Kunkelら,Methods Enzymol.154:367(1987)とP.Carter,in Mutagenesis:a Practical Approach,M.J.McPherson,Ed.,IRL Press,Oxford,UK,pp.1-25(1991))によってCD4-IgG中で構築され、且つジデオキシヌクレオチド配列によって実証した (Sangerら,Proc.Natl.Acad.Sci.USA 74:5463(1977))。次の表3を参照
【0172】
【表3】
【0173】
残基T366は、パートナーCH3ドメイン上の残基Y407の水素結合間隔内にある。実際は、残基T366への主要な分子間接触は、残基Y407に対する及び逆の場合も同じである。一つの空洞への隆起ペアは、一つのCH3ドメイン中のT366Yとそのパートナードメイン中のY407Tの相互の変異を持ったこれらの残基を逆転することによって創製され、かくしてその界面で側鎖の容量が維持される。変異は、Kabat数付与システム(Kabatら,(1991)上記)を用いてその位置の次の野生型残基によって、そして一文字コードで残基の置換が定義される。複数変異は、コロンによって単一変異成分をリスト化することによって定義される。
【0174】
抗-CD3軽鎖(L)と重鎖(H)変異体(Shalabyら,J.Exp.Mcd.175:217(1992)とRodriges ら,Int.J.Cancer(上記)7:45(1992))をコードしているファージミドは、以前に記載された通り(Chamow ら,J.Immunol.153:4268(1994))、ファージミドをコードしているCD4-IgG変異体(Byrnら,Nature 344:667(1990))と一緒に、ヒト胎児性腎細胞,293S中に共移入(co-transfected)した。移入されたファージミドDNAの全量は固定した一方、異なるDNAの比率はAb/Iaキメラの生産を最大化するように変えた。投入DNAs(15μgトータル)Ia:重鎖:軽鎖の割合(質量で)は以下の通り:8:1:3;7:1:3;6:1:3;5:1:3;4:1:3;3:1:3;1:0:0。
【0175】
その生産物は、レーザーデンシトメトリースキャニングに続くSDS-PAGEによる分析の前にスタヒロコッカス蛋白質(ProSep A,BioProcessing Ltd,UK)を用いてアフィニティー精製した。重鎖DNAを超える過剰軽鎖は、制限化されるものからの軽鎖を回避するために用いた。生産物の同定は、アミノ末端配列決定に続きPVDF膜(Matsudaira,J.Biol.Chem.262:10035(1987))での電気ブロッティングによって証明した。
【0176】
重鎖及び野生型CH3を組み込んでいるIa用のそれと一緒に軽鎖用のファージミドの共移入は、予期した通り、Ab/Iaキメラ,IgGとIaホモダイマー生産物の混合物となる(Chamowら,J.Immunol.153:4268(1994))。抗体重鎖プラス軽鎖をコードしている投入DNAのより大きなフラクション又はホモダイマーに一致するより高いフラクションを回収した。Ia:H:Lの6:1:3の投入DNA比では、Iaホモダイマー(22.5%)とIgG(23.0%)の類似フラクションと共に54.5%のAb/Iaキメラを生じた。これらの比率は、分析方法によって導かれるバイアスを伴わず重鎖のランダムな組合せに続く各鎖の等モル発現から予期されたそれらと良く一致する:50%Ab/Iaキメラ、25% Iaホモダイマー及び25% IgG。
【0177】
野生型CH3を含有している鎖に対して、Ab/Iaキメラは、Y407T空洞とT366Y隆起変異をそれぞれ抗-CD3重鎖とCD4-IgGIaに含有せしめた共移入体から92%までの収率で回収された。抗体/免疫付着因子キメラの同様の収量は、もしこれらの相互変異が重鎖の隆起及びIa中の空洞により設けられたならば得られた。両方のケースにおいてモノマーは、隆起を含むが空洞のないその鎖で観測された。いずれか1つの理論に制限することなしに、T366Y隆起がY407T空洞よりもホモダイマー形成のためにより分裂されると確信する。Ab/Iaハイブリッドのフラクションは、隆起と空洞(Ab T366W,IaY407A)の両者のサイズを増加することによって有意に変更されることはない。第2の隆起と空洞ペア(Ab F405A,IaT394W)は、ホモダイマー化するIaT394W隆起変異体の予期しない傾向を補うための投入DNAの小さいフラクションを用いてAb/Iaの71%までを生じた。2つの独立した空洞への隆起変異体ペア(Ab T366Y:F405A、Ia T394W:Y407T)の組合せは、Ab T366Y,IaY407Tペア以上にはAb/Iaハイブリッドの生産を改善しない。
【0178】
T366YとY407T変異体ペアにより得られたAb/Iaキメラのフラクションは、試験した範囲にわたり、投入DNAsの比に実質的に非従属であった。さらに汚染している種は、mono S HR 5/5カラム(Pharmacia,Piscataway,NJ)上でイオン交換クロマトグラフィー(20mMトリス塩酸,pH8.0中0-300mM NaCl)によってAb/Iaキメラから容易に除去された。これは、AbとIaの相対的な発現レベルが一時的な発現系におけるよりも容易な操作性が劣る、安定な細胞系を用いた大量のAb/Iaキメラの製造のために良好であると予想する。
【0179】
同定した空洞への隆起変異体は、4又はそれ以下にまで下がった、可能性のある10の主要な種(Sureshら,Methods Enzymol.121:210(1990))から得られた生産物の混合物の複雑性を減じることによってFc-含有BsAbの潜在的な適用を増加することが予期される(図1A-1B)。T366YとY407Tが完全に保存され且つIgG1のCH3ドメイン界面で他の残基が高度に保存されることから、T366YとY407T変異体ペアは、他のヒトTgGアイソタイプ(IgG2,IgG3又はIgG4のような)のヘテロマルチマーを生産するために有用とされるであろう。
【0180】
実施例2:
ヘテロマルチマーの免疫付着因子中の非天然存在ジスルフィド結合の 生成 A. CH3相互鎖-ジスルフィド結合の設計 3つの基準を、パートナーCH3ドメイン間のジスルフィド結合を作るための残基のペアを同定するために使用した:i)Cα分離は、好ましくは自然ジスルフィド結合中に見出されるそれに類似している(5.0から6.8Å)(Srinivasan,N.等,Int.J.Peptides Protein Res.36:147-155(1990))。7.6Åまでの間隔は、主鎖移動を許すことおよび低い分解能の結晶構造での原子の位置を考慮することを許した(Deisenfofer,Biochemistry 20:2361-2370(1981))。ii)Cα原子は、2つのCH3ドメイン上の異なる残基上にあるだろう。iii)その残基はジスルフィド結合を与えるように位置付けられる(Srinivasan,N.ら,上記)。
【0181】
B. ジスルフィド結合のモデル化。ジスルフィド結合は、Insight II release 95.0(Biosym/MSI)を用いてhumAb4D5-Fvについて記載された通り(Rodriguesら,Cancer Res.55:63-70(1995))、ヒトIgG1Fc(Deisenhofer,上記)中にモデル化した。
【0182】
C. CH3変異体。変異は、以下の合成オリゴヌクレオチドを用いて、特定部位の突然変異誘発(Kunkelら,Methods Enzymol.154:367-382(1987))によってヒト化した抗-CD3重鎖又はCD4-IgGのCH3ドメイン中に導入した。
【0183】
【表4】
【0184】
変異は、置換アミノ酸の次の、アミノ酸残基と数(Kabatら,上記(1991)のEu数字付与スキーム)によって表した。多重変異は、コロンによって分けた単一の変異によって表される。変異体は、シーケナーゼversion 2.0(United States Biochemicals,Cleveland,OH)を用いるジデオキシヌクレオチド配列決定(Sangerら,上記(1977))によって確証した。
【0185】
D. ヘテロダイマー形成を増大する相互-鎖ジスルフィド。CH3ドメイン中に相互-鎖ジスルフィド結合を含んでいる6ペアの分子("ジスルフィド-CH3"変異体;v1-v6,表4)は、Ab/Iaハイブリッド、抗-CD3/CD4-IgG(Chamowら,上記(1994))の形成を導くその能力において親の分子と比較した。CD4-IgG及び抗-CD3重鎖変異体をコードしているプラスミドは、抗-CD3軽鎖をコードしている過剰のプラスミドと一緒に、293S細胞中に共移入した。ヘテロダイマーの収量は、Ia:H鎖:L鎖DNA比の範囲を持った移入により至適化した。Ab/Iaヘテロダイマー、IgG及びIaホモダイマー生産物は、ストレプトコッカス蛋白質Aアフィニティー-精製し、SDS-PAGEとレーザーデンシトメトリースキャニング(Ridgwayら,上記(1996))によって定量した。
【0186】
それぞれのジスルフィド-CH3ペアは、親の分子に類似した、3つの主要な種を生じる。しかしながら、ジスルフィド-CH3変異体からのAb/Iaヘテロマーは、CH3ドメイン中の相互-鎖ジスルフィドの形成に一致する、電気泳動の移動でシフトした。ジスルフィド結合形成の更なる証拠は、ヒンジにおいて相互-鎖ジスルフィドによって提供された。共有結合したAb/Iaハイブリッドは、ジスルフィドCH3変異体用の、しかしヒンジシステインがセリンに変異した野生型CH3ドメインを持った分子用ではないSDS-PAGEによって観測された。ジスルフィドCH3変異体を製造し、Y349C/S354'C,Y349C/E356'C,Y349C/E357'C,L351C/E354'C,T394C /E397'C,及びD399C/K392Cと定義した。唯一の変異体(D399C/K392'C)は、該変異体のSDS-PAGE分析による測定として野生型を越えるAb/Iaハイブリッドの収量を実質的に増加した(それぞれ76%対52%)。変異は、置換アミノ酸の次の、アミノ酸残基と数(Kabat ら,上記(1991)のEu数字付与スキーム)によって表した。CH3の第1及び第2のコピー中の変異は、それぞれ前後にスラッシュを付す。CH3の第2のコピー中の残基は、プライム符号(')によって表示される。この改善は、該変異体K392S/D399'Sが野生型に対して類似したAb/Ia収量とAb/Ia電気泳動移動性の両方を与えたことから、残基K392とD399の置換というよりむしろ、ジスルフィド結合形成を明らかに反映する。ホモダイマーは、ヘテロダイマーのCH3ドメイン中に相互-鎖ジスルフィド結合形成を優先して行うことを実証している、野生型Fcドメインの持つそれと同様に移動した。全てのジスルフィドCH3変異体は、293S細胞中の親分子とほぼ同じレベルで発現した。
【0187】
E. 空洞への隆起工学によって構築したジスルフィドは95%までヘテロダイマーの収率を増加する。最良のジスルフィドペアは、76%までヘテロダイマーのパーセンテージを増加し、空洞への隆起方法は87%までヘテロダイマーのパーセンテージを増加した(表4;Ridgwayら,(1996)上記も参照)。これら2つの方法は、ヘテロダイマー精製の確率を増加することの異なる原理による。従って、我々はヘテロダイマーにおける収率を更に改善することを意図して、その2つの方法を組み合わせた。L351C又はT394Cを含む、2つのモデル化したジスルフィドは、ジスルフィド結合ヘテロダイマー(L351C/S354'CとT394C/V397'C)と同じくジスルフィド結合ホモダイマーを潜在的に形成でき、かくしてそれの単一性を減じている。残る4つのジスルフィドペアは、ファージ選択したヘテロダイマー(変異体v9-v16)中に配置し、ヘテロダイマーの収率を分析した(表4)。ヘテロダイマーのほぼ95%の収率が得られた。再度、該ヘテロダイマーは、野生型及びv8変異体に匹敵する電気泳動移動シフトを示した。
【0188】
【表5】
【0189】
実施例3:
ヘテロマルチマー中のタンパク質-タンパク質相互作用を増大する相補の変異のための構造-誘導ファージディスプレー選択以下の方法は、マルチマー化ドメインを経て界面で相互作用するポリペプチト中の相補の変異の選択において有用である。その方法は、相補の空洞への隆起変異の選択にそれを適用するとして次に説明される。しかしながら、その実施例は制限するためのものではなく、その方法は非天然存在ジスルフィド結合、ロイシンジッパーモチーフ、疎水性相互作用、親水性相互作用などの形成に適当な変異の選択に同様に適用することができる。
【0190】
A. ファージディスプレー選択。ファージディスプレー方法は、安定なCH3ヘテロタイマーの選択用に発展し、図2中に概要が示される。その選択は、隆起変異体、T366W(Ridgwayら,上記(1996))、M13遺伝子IIIペプチドに融合したCH3の第2のコピーと共発現されるペプチドフラッグに融合した(gDペプチドフラッグ、例えばLasky,L.A.とDowbenko,D.J. (1984)DNA 3:23-29;及びBerman,P.W.ら,(1985)Science 227:1490-1492)を用いる。空洞変異体のライブラリーは、第1のCH3ドメイン上の隆起に最も近い残基の無作為化によってCH3のこの第2のコピー中に創製した。ファージディスプレー化安定CH3ヘテロダイマーは、次いで抗-フラッグAbを用いて補足した。
【0191】
CH3ファージディスプレーライブラリーの1.1x105の独立クローンは、PCRフラグメントによる自然CH3遺伝子のセグメントの置換によって構築した。
【0192】
そのフラグメントは、標準的な技術を用いて位置366、368と407で無作為化するために分解プライマーを用いてPCR増幅化により得られた。
【0193】
選択の2から5ラウンドの後、完全長クローンのフラクションは、一本鎖DNAのアガロースゲル電気泳動による判断として、それぞれ90%、60%、50%及び10%だった。完全長クローンを含むファージミドは、選択の5ラウンド後にゲル-精製した。二千の形質転換体がXL1-BLUETM細胞(Stratagene)を再形質転換後に得られた。
各クローンの平均>106コピーを、選択(panning)のラウンド当たりに使用した。かくして、該ライブラリーにおいて各クローンの多数のコピーが、たとえ幾つかの削除変異体が選択の間に生じたとしても、選択のために利用可能であると確信した。
【0194】
選択の7ラウンド後、得られたCH3変異体は、無作為化した残基でコンセンサスアミノ酸配列を研究した。実際に全てのクローンは、この位置でβ-ヒドロキシルの非常に強い優先性を示す残基 366でセリン又はトレオニンを有した。疎水性残基のための強い優先性は、バリンとアラニン優先化によって残基368と407で観測された。6の異なるアミノ酸の組み合わせは、11度回収された、三重変異体、T366S:L368A:Y407Vを含む、少なくとも2度回収された。先に定義されたヘテロダイマー、T366W/Y407'A(Ridgway,J.B.B.ら,(1996),上記)と同一の配列を有するこれらのファージ選択物はない。そのファージ選択物は、ドメイン界面残基の全側鎖容量において40-80Å3減少により判断されるように野生型CH3ホモダイマーよりも劣る緊密性で詰め込まれ得る。
【0195】
発現プラスミドpAK19(Carterら,1992)上にコードされたCH3変異体は、大腸菌33B6株中に導入し、培養槽中で高い細胞密度に増殖した大腸菌から創製した。DEAE-セファデックスFF、ABx及び Resource Sクロマトグラフィーによって精製したT366S:L368A:Y407V変異体は、SDS-PAGEの後に単一の主要なバンドを与えた。他のCH3変異体は、同様の精製により回収した。
【0196】
高解析度電気スプレー質量分析法によって測定した野生型CH3、T366S:L368A:Y407V、T366W及びY407A変異体の分子質量は、予期した通りであった。
【0197】
B. ファージ選択したヘテロダイマー安定性。CH3ヘテロダイマーの安定性は、酵素結合イムノソルベント検定法(ELISA)によって残るヘテロダイマーの希釈及び定量化に続き、グアニジン塩酸によって相当するファージを滴定することによって最初に評価した。CH3-ファージによるグアニジン塩酸分解アッセイは、選択物を直ちにスクリーンするための手段を提供する。
【0198】
ファージは、選択の7ラウンドの後の個別のクローンから及びコントロールベクター、pRA1からでも製造された。概略的には、XL1-BLUETM中のファージミドが、109pfu/ml M13K07の存在中で50μg/mlカルベニシリンと10μg/mlテトラサイクリンを含有する25ml LB培地に接種するために使用し、37℃で一晩インキュベーションした。その細胞は遠心分離(6000g、10分、4℃)によってペレット化した。ファージは、5mlの20%(w/v)PEG、2.5M NaClによって沈殿し遠心分離(12000g、10分、4℃)の後に上清から回収され、1mlのPBS中に再懸濁させた。PBS中0-6Mのグアニジン塩酸180μlが20μlファージ調製物に加えられ、5.0分間ほぼ〜25℃でインキュベーションした。各ファージサンプルのアリコート(20μl)は、水で十倍に希釈した。CH3のヘテロダイマーの存在は、5B6-被覆プレートを用い及び基質としてo-フェニレンジアミンを用いたホースラディッシュペルオキシダーゼに接合した抗-M13ポリクローナルAbによってファージを検出するELISAによって測定した。その反応は、2.5M H2SO450ml の添加によって抑止し、492nmで吸光度を測定した。その吸光度データは、その溶融の間にグアニジン塩酸濃度に対してプロットし、且つ Kaleidagraph 3.0.5(Synergy Software)を用いた非線形最小二乗法によって四つのパラメーターモデルに適用した。
【0199】
最も頻繁に回収されたヘテロダイマー、T366W/T366'S:L368'A:Y407'Vは、他のファージ-選択したヘテロダイマーと安定性において類似する。このファージ-選択したヘテロダイマーは、設計したヘテロダイマー、T366W/Y407'Aよりも有意により安定であるが、しかし野生型CH3よりは安定性に劣る。全てのCH3変異体、個別及び組み合わせの両方は、これら同じ分子が熱量測定法によって研究された条件下にサイズ排除クロマトグラフィーによってダイマーであることが見出された(1.75mg/ml、リン酸緩衝化生理食塩水(PBS)中)。唯一の例外は、CH3ダイマーよりも明らかに短い保持時間を有したT366S:L368A:Y407V変異体単独であった。
【0200】
T366Wの1:1混合物、隆起、及びT366S:L368A:Y407V、69.4℃で単独転移を持つ変異体溶融物は、サブユニット交換及び安定なヘテロダイマーの形成と密接に結びつく。一方、T366W隆起ホモダイマーは、T366W/T366'S:L368'A:Y407'V空洞への隆起ヘテロダイマー(ΔTm=−15.0℃)よりもより劣った安定性である。それ自身の上のT366S:L368A:Y407V空洞変異体は、加熱によって集合する傾向があり、なだらかな溶融転移を成さない。
【0201】
設計した空洞変異体、Y407Aは、T366W隆起変異体の不在及び存在でそれぞれ58.8℃及び65.4℃で溶融する。これは、T366W(ΔTm=11.0℃)又はY407A(ΔTm=6.6℃) ホモダイマーのいずれか一方よりもより大きな安定性を有するT366W/Y407'Aホモダイマーのサブユニット交換と形成と密接に結びつく。ファージ選択したヘテロダイマー、T366W/T366'S:L368'A:Y407'Vは、設計したヘテロダイマー、T366W/Y407'A,(ΔTm=4.0℃)よりも一層安定であるが、しかし、野生型CH3ホモダイマー(ΔTm=−11.0℃)よりも安定性が劣る。
【0202】
C. インビボでのファージ-選択した抗体免疫付着因子のマルチマー化。ファージ選択した及び設計したCH3変異体は、インビボでAb/Iaハイブリッド、抗-CD3/CD4-IgGの形成を導くその能力を比較した(Chamowら,(1994),上記。
【0203】
これは、CD4-IgGと一緒にヒト化した抗-CD3軽鎖(L)と重鎖の共発現により達成した。ヘテロダイマーとホモダイマーの形成は、SDS-PAGEとスキャニングレーザーデンシトメトリー(Ridgway,ら,(1996),上記)に続いてプロテインA精製によって評価した。Ab/Iaハイブリッドの比較できる収量が、抗-CD3重鎖が設計した隆起変異体を含んだ、T366WとIaがファージ-選択した変異体、T366S:L368A:Y407V、又は設計した空洞変異体、Y407Aのいずれかを含んだ共移入体から回収された(図3)。
【0204】
ファージ-選択した及び設計したCH3変異体は、次にホモダイマーを形成するためのその傾向において評価した。隆起変異体、T366Wは、相当する重鎖と軽鎖の共移入がIgGを越える過剰のHLモノマー(非-ジスルフィド結合IgGを含み得る)に至ることから、ホモダイマー化を明らかに非常に崩壊させた。一方、IgGしかしHLモノマーなしでは、野生型CH3ドメインを含む同じ抗体が観測された。空洞変異体、T366S:L368A:Y407Vは、相当するファージミドの遺伝子移入が幾つかのIaモノマーを持つ優性なIaダイマーの混合物に至ることから、多少ホモダイマー化を崩壊させる。空洞変異体、Y407Aは、Iaダイマーしかし相当するファージミドの移入後にIaモノマー無し、の存在による判断として、ホモダイマー化を最小限崩壊する。
【0205】
ここに記載したファージディスプレー方法は、安定したヘテロダイマーを形成するCH3変異体の援助での選択及び安定なホモダイマーを形成する変異体に対しての選択を与える。ホモダイマーに対するカウンター選択は、"遊離"CH3変異体が、利用可能なCH3変異体-遺伝子III融合タンパク質への結合のためにフラッグ化CH3ノブ変異体と競合するであろうことから生じる。遊離CH3変異体は、自然CH3変異体とM13遺伝子IIIの間のアンバー変異の結果として生じる。XL1-Blucのようなアンバー抑制宿主において、自然CH3遺伝子とM13遺伝子IIIの両方が創製されるだろう。
【0206】
グアニジン塩酸分解は、ファージ上のCH3ヘテロダイマー安定性を最初にスクリーニングするために有用なツールとして提供される。ファージは、5Mグアニジン塩酸にさらした後でも大腸菌への感染性を維持する(Figiniら,J.Mol.Biol.239:68-78(1994))。かくして、グアニジンは変異体選択の方法を増加するためにも有用となり得る。
【0207】
ファージディスプレーライブラリーの合理的な設計とスクリーニングは、ヘテロダイマー化を促進するためにホモダイマーのドメイン界面を再モデル化するための相補的なアプローチである。CH3ドメインのケースにおいて、設計した変異体は、ヘテロダイマー化を促進することを強化できるドメイン界面残基と同一視される。ファージディスプレーは、ヘテロダイマーを最も有効に形成する組み合わせのために固定した隆起の近くの3つの残基の置換をサーチするためにここに用いた。ファージ選択物は、ドメイン界面の更なる合理的な再設計を促進するために有効とされる一方、ここに記載したファージ選択方法は、タンパク質-タンパク質界面を再モデル化するためのその有用性を実証する。
【0208】
実施例4:
共通の軽鎖を有するヘテロマルチマー抗体又は抗体/免疫付着因子の生成と配置 以下の実施例は、本発明に従う同じ軽鎖を共有するヘテロマルチマー二重特異性抗体の調製とその標的抗原に結合する抗体の能力を実証する。
【0209】
A. 同じ軽鎖を共有する抗体の同定:抗体ライブラリーの比較は11の抗原に高まる。大きいヒト一本鎖Fv(scFV)抗体ライブラリー(Vaughanら(1996)上記)は、Ax1(ヒトレセプターチロシンキナーゼECD)、GCSF-R(ヒト顆粒球コロニー刺激因子レセプターECD)、IgE(ネズミIgE)、IgE-R(ヒトIgEレセプターα-鎖)、MPL(ヒトトロンボポイエチンレセプターチロシンキナーゼECD)、MusK(ヒト筋肉特異的レセプターチロシンキナーゼECD)、NpoR(ヒトオルファンレセプターNpoR ECD)、Rse(ヒトレセプターチロシンキナーゼ、Rse,ECD)、HER3(ヒトレセプターチロシンキナーゼHER3/c-erbB3 ECD)、Ob-R(ヒトレプチンレセプターECD)、及びVEGF(ヒト脈管内皮成長因子)、ECDは細胞外ドメインに関する、を含む11の抗原用の抗体特異性で選択した。各抗原に生じた抗体の集団からscFvフラグメントのためのヌクレオチド配列データは、相当するタンパク質配列から得るように翻訳した。VLは異列は、鎖の全てのペア様態組み合わせ間の同一性パーセンテージを計算するためにFengとDoolittle(1985,1987,1990)のアルゴリズムによりプログラム"アライン"を用いて比較した(Feng,D.F.とDoolittle,R.F. (1985)J.Mol.Evol.21:112-123;Feng,D.F.とDoolittle,R.F. (1987)J.Mol.Evol.25:351-360;及びFeng,D.F.とDoolittle,R.F.(1990)Method Enzymol.183:375-387)。
【0210】
各ペア様態軽鎖アミノ酸配列比較の配列同一パーセントの結果は、マトリックスフォーマット中に調整した(追記参照)。
【0211】
最もペア様態の比較品として、少なくとも1の共通軽鎖配列が見出された。表5は、117VLアミノ酸配列のアラインメントによって決定した同じ軽鎖を共有する(100%同一性)scFvの頻度を示しているVL鎖の比較である。例えば、そのエントリー4/9(HER3xOb-R、黒いボックスで表した)は、結合HER3が1又はそれ以上の抗-Ob-RクローンによってそれのVL配列を分配することが見出される一方、Ob-R結合9クローンは、1又はそれ以上の抗-HER3クローンによってそのVL配列を分配する、4クローンを表す。対角線上の参加物は、その集団中の1又はそれ以上のクローンによってVL配列を分ける集団内の抗体クローンの数を表す。例えば、MPLクローンの調査は、1又はそれ以上の他のMPLクローンによってそれのVL配列が分けられた5クローンを明らかにした。(IgE x Ax1)又は(NpoR x IgE-R)についてのような観測された共通の軽鎖の無いケースにおいて、少なくとも1の特異性と比較されるフラグメントの数は非常に少なかった(5以下)。見出された共通の軽鎖の数が与えられ、それは共通の軽鎖が、もし十分な数のクローンが比較されるなら、いずれかのVL比較物のために見出すことができることがありそうである。
【0212】
軽鎖のアミノ酸配列は、選択した共通の軽鎖に対しての配列相同性が98%と99%であった場合、異なったアミノ酸残基の位置によって決定された。図4は、Ax1(クローンAx1.78)、Rse(クローンRse.23,Rse.04,Rse.20,及びRse.15)、IgER(クローン IgER.MAT2C1G11)、Ob-R(クローンobr.4)、及びVEGF(クローンvegf.5)に特異性を持つ8つの異なる抗体のVL配列の比較である。配列の定義(Kabat,G.A.ら(1991)上記)又は構造の定義(ChothiaとLcsk,(1987)J.Mol.Biol.196:901-917) に従う抗原結合CDR残基の位置は、それぞれ下線と#によって示した。Ax1.78配列と異なる軽鎖残基は二重下線により示した。比較した9軽鎖の、6が同一である。Rse.04とobr.4の軽鎖(ほぼ99%配列相同性)は、抗原結合CDRsの外側の1残基で相違するRse.20の軽鎖(ほぼ98%配列相同性)は、抗原結合CDRsの外側の2残基で異なる。該アミノ酸残基の変化は、抗原結合性において僅かな又は影響なしを有し得る。かくしてこれらの軽鎖の配列類似性は、本発明の共通の軽鎖のためのその候補を作る。代替的に、期待される対になったscFv(例えばAx1.78)の軽鎖との98-99%相同性を有する軽鎖のように、本発明に従い、対になった軽鎖と置換でき且つ結合特異性を保持し得る。
B. 同じ軽鎖を共有する抗体の同定と軽鎖のそれを共有する二重特異性抗体の構築:抗-Ob-R/抗-HER3。
【0213】
ヒトレプチンレセプター(Ob-R)又はHER3/c-erbB3遺伝子生産物(HER3)の細胞外ドメインを結合したScFvフラグメントは、大きいヒトscFvファージライブラリーを用いた選別の3つのラウンドによって得られた(Vaughanら(1996),上記)。レプチンレセプター-IgGとHER3-IgG(1ml PBS中10μg)を、4℃で一晩、分離イムノチューブ(Nunc;Maxisorp)を被覆するために使用した。選別とファージ解放は、以下の修正を伴い、Vaugham(1996),上記により記載されたように実行した。1mg/mlの濃度で、ヒト化した抗体、 huMAb4D5-8(Carter,P.ら(1992)PNAS USA 89:4285-4289)又はヒト化した抗-IgE(Presta,L.ら(1993)J.Immunol.151:2623-2632)は、Fc-結合ファージを吸着するための各選別工程中に含めた。加えて、溶液の選別化(Hawkins,R.E.ら (1992)J.Mol.Biol.226:889-896)は、scFv結合レプチンレセプターを同定するためにも使用した。そのレプチンレセプターは、プロテインAセファロースクロマトグラフィーに続いて、工作したプロテアーゼ、ジェネナーゼ(Carter,P.ら (1989)Proteins:Structure,Function and Genetics 6:240-248)によるレプチンレセプター-IgGの位置特異的蛋白分解によってFcから分離した。そのレプチンレセプターはビオチン化し、それぞれ、選別の第1,第2,及び第3ラウンド用に100nM、25nM及び5nMの濃度で使用した。ファージ結合ビオチン化抗原は、ストレプタビジン-被覆パラ磁性ビーズ(Dynabeads,Dynal,Oslo,Norway)を用いて捕捉した。
【0214】
それぞれの選別のラウンド2及び3からのクローンは、ファージ及び相当する抗原及び制御免疫付着因子又は抗体をも用いるscFvELISAによってスクリーンした。抗原-陽性クローンの分割性は、プライマー、fdtetseqとPUC逆転(Vaughamら(1996),上記)を用いるscFv挿入のPCR-増幅及びBstNIによる消化(Marksら(1991)上記)によって分析した。BstNIフィンガープリント当たり1から5のクローンがPCR重リンクとmyc seq 10プライマー(Vaughamら(1996)上記)を用いたcycle-sequencedusing fluorescent dideoxy chain terminators(Applied Biosystems)した。サンプルは、Applied Biosystems Automated DNA Sequencerを用いて分析し且つ配列をSeqEdを用いて分析した。グアニジン塩酸抗体分解及びファージディスプレー選択と組み合わせた Figiniのインビトロでの鎖シャフティング方法が同じ軽鎖を有する抗体を選択する方法として有用であることにも注目される(Figini,M.ら (1994),上記、その全体が参照によりここに併合される)。
【0215】
上記の方法を用いて、11の異なる抗-HER3クローンと18抗-Ob-Rクローン(被覆した抗原を用いた選別からの11とビオチン化抗原による選別からの7)が得られた。そのクローンは、各結合ドメインと結合した軽鎖の配列を決定するための標準技術により配列決定した(図5)。その配列は、二重特異性抗体を構成するために使用した抗-Ob-Rクローン26と抗-HER3クローン18のVH及び共通のVL配列である(後記参照)。その残基は(Kabat,E.A.ら(1991)上記)に従い番号付けされる。配列の定義(Kabat,E.A.ら(1991)上記)又は構造の定義(ChothiaとLesk,(1987)J.Mol.Biol.(1987)196:901-917)は、それぞれ下線と上線によって示される。VH配列中の残基間の相同性は、*によって表示した。
【0216】
軽鎖の配列は、多重抗-Ob-Rクローンに対する多重抗-HER3クローンと比較した(図8及び表5)。11の抗-HER3クローン以外の4つが、1又はそれ以上の抗-Ob-Rレセプタークローンと同じVLを共有することが観測された。
【0217】
反対に、18の抗-Ob-Rクローン以外の9が抗-HER3クローンの一つとして同じVLを共有する(表5、黒い箱を参照)。
【0218】
【表6】
【0219】
抗-Ob-R/抗-HER3、共通の軽鎖を有する二重特異性抗体の構築は以下の通り実行した。相補の隆起と空洞、同じく第1及び第2のポリペプチド間の非天然存在ジスルフィド結合を有する変更したCH3第1及び第2のポリペプチドを、Fc-含有二重特異性抗体の構築において用いた。同じ軽鎖を、同じく各抗体からの重鎖を共有するクローンである、抗-Ob-Rクローン#26と抗-HER3クローン#18からのVLを、ここに記載した手順に従って二重特異性抗体を製造するために用いた。
この抗体は、非天然存在ジスルフィド結合を生成するために変更を欠くことによってのみそれと異なる二重特異性抗体に対応する明確な分子量において電気泳動移動性シフトを有した。非天然存在ジスルフィド結合を持った及び持たないヘテロダイマーの抗体変異体の8%SDS-PAGEゲルは、野生型ヘテロダイマーのほぼ230の見かけ上のMWから、一つの非天然ジスルフィド結合を有しているヘテロダイマーのほぼ200の見かけ上のMWまでの移動シフトを示した。そのMWシフトは非天然ジスルフィド結合を首尾良く形成するそれぞれの変異体のパーセントを与えるために十分であった。
二重特異性抗体のOb-R及びHER3の両方への特異的結合は、以下の方法のような標準ELISA法によって試験される。Ob-R結合は、Ob-R-Ig融合タンパク質として存在するOb-RによってELISAアッセイで実証される。該Ob-R-Ig融合タンパク質は、96-ウェルのミクロタイター板のウェル上に被覆され、そして二重特異性抗体が加えられる。
【0220】
そのウェルは、Ob-R-Igへの非特異的結合を取り除くために数回洗浄される。同じアッセイにおける第2の成分として、ビオチン化HER3-Ig融合タンパク質が加えられ、そしてビオチン化HER3-Ig融合タンパク質に結合するストレプタビジン-ホースラディッシュペルオキシダーゼ複合体によって測定される。結合は、過酸化水素及びTMBペルオキシダーゼ基質(Kirkegaard and Perry Laboratories,Gaithersburg,MD)の添加における着色の変化の生成により測定される。
【0221】
実際に記載した条件下、Ob-R-IgとHER3-Igの両方への二重特異性抗体の結合は、固定化Ob-R-Ig/二重特異性抗体/HER3-Igビオチン/検出可能に標識したストレプタビジンを含む複合体の良好な形成のためにミクロタイター表面上に固定化した検出可能な標識として観測されるであろう。Ob- R-Igと結合するが、HER3-Igとはしない抗体は、陰性の結果を提供するように、上記複合体を形成しない。同様に、HER3-Igと結合するが、Ob-R-Igとはしない抗体は、陰性の結果を提供するように、上記複合体を形成しない。一方、Ob-R-IgとHER3-Igの両方に結合することが予期される二重特異性抗体は、該アッセイにおいて陽性の結果を生じる複合体を形成し、共通の軽鎖を有する、HER3とOb-Rの両方に結合する、二重特異性抗体を表示する。
【0222】
抗-(Ob-R/HER3)二重特異性抗体の発現と精製は、以下の通り実行した。ヒト胎芽腎臓293S細胞は、抗-Ob-R重鎖、抗-HER3重鎖、又は上述した通りそれぞれの抗体に共通であったクローン26又は18からの軽鎖をそれぞれ独立にコードしている3つのプラスミドDNAによって移入した。それぞれの移入のために、軽鎖コード化DNAに対する重鎖コード化DNAの比率は、軽鎖が抗-Ob-R/抗-HER3二重特異性抗体の配列を制限しないであろうように1:3とした。両方の重鎖は、互いに関して1:1の比で移入した。12μgのトータルのプラスミドDNAは、次いでリン酸カルシウム沈降によって 293S細胞中に共移入(co-transfected)した(Gorman,C.,DNA Cloning,Vol.II,D.M.Glover,ed.,IRL Press,Oxford,p.143(1985))。その細胞は、タンパク質発現の増大を意図して増殖培地に加える前に洗浄した。Fc-含有タンパク質は、固定化したプロテインA(ProSep A,BioProcessing Ltd.,UK)を用いて細胞懸濁物から精製し、そしてPBS中に緩衝化-交換した。ヨードアセトアミドは、ジスルフィド結合の再シャフト化を防止するために50mMの最終濃度までタンパク質調製物に加えた。
【0223】
付加の実施例として、抗-(CD3/CD4)抗体/免疫付着物の発現と精製を以下の通り実行した。ヒト胎芽腎臓293S細胞は、3つのプラスミドDNAs、各プラスミドは抗-CD3軽鎖、抗-CD3 IgG1重鎖、又は抗-CD4 IgG1免疫付着因子を独立的にコードしている、によって移入した。それぞれの移入のために、重鎖コード化DNAに対する軽鎖コード化DNAの比率は、軽鎖が抗-CD3 IgGの整列のために制限されないであろうように、3:1とした。加えて、免疫付着因子が不十分に発現されるために、免疫付着因子コード化プラスミドの比率は、重鎖コード化プラスミドの過剰において加えた。試験したその比率は、免疫付着因子:重鎖:軽鎖が3:1:3から8:1:3間での範囲とした。トータルで10μgのプラスミドDNAは、移入前にPBSで細胞を洗浄し、次いでリン酸カルシウム沈降(Gorman,C.(1985),上記)により293S細胞中に共移入した。Fc-含有タンパク質は、固定化したプロテインA(ProSep A,BioProcessing Ltd.,UK)を用いて細胞懸濁物から精製し、そしてPBS中に緩衝化-交換した。ヨードアセトアミドは、ジスルフィド結合の再シャフト化を防止するために50mMの最終濃度までタンパク質調製物に加えた。
【0224】
上記それぞれの調製において、タンパク質サンプルは、8%ポリアクリルアミドゲル(Novex)で電気泳動し、Serva blueによる染色によって可視化した。ゲルは、マイナーな不純物の可視化及び定量化を与える努力において見せかけのバックグラウンドを無くすように脱- 染色化した。乾燥したゲルは、スキャニングデンシトメーター(GS-670,BioRad)によってスキャンし、そしてタンパク質製造物は Molecular Analystソフトウェアによって定量化した。
【0225】
CH3ドメイン中に導入した非天然 (工作した)ジスルフィド結合は、ヘテロダイマー形成を増大するためにここに開示されている。一つのペプチド、K392/D399'Cは、76%までのヘテロダイマー(表4、変異体v6)を精製することによってヘテロダイマー形成を増大した。さらに、相互-鎖ジスルフィド結合の存在が空洞への隆起技術と結び付けられた場合、ほぼ95%ヘテロダイマーが得られた(表4変異体v11,v12,及びv16)。かくして、二重特異性抗体の第1と第2のポリペプチド間のタンパク質/タンパク質相互作用を特異的に増加する本発明の方法は、望まれるヘテロマルチマーの収量を増加すると共に、望ましくないヘテロマルチマー又はホモマルチマーの形成を最小化する。
【0226】
加えて、電気泳動の移動性分析によって生産物ヘテロマルチマーを特徴付ける方法は、望ましくない生産物に対して、望まれるヘテロマルチマーの相対的な量の測定を与える。
【0227】
ここに記載したような共通の軽鎖の選択は、多重特異性抗体の可変の重鎖と軽鎖の間の誤対合の可能性を削除することによって望まれるヘテロマルチマーの収量を増加する。
【0228】
C. 同じ軽鎖を共有する抗体の同定とその軽鎖を共有している二重特異性抗体の構築:抗-Mpl/抗-HER3。
【0229】
本発明の別な二重特異性抗体の同定、構築及び発現がここに示される。この実施例のパートAとB中に記載した方法を、抗-Mpl/抗-HER3二重特異性抗体の製造用に利用した。
【0230】
この実施例のセクションA中に記載した方法(11の抗原を生じた抗体ライブラリーの比較)、上記、を用い、抗-HER3scFvのVHとVLアミノ酸配列を、ヒトトロンボポイエチンレセプター、c-Mplに結合する23 scFvで比較した。11の抗-HER3クローンの5つは、1又はそれ以上のMpl-結合クローンを持った同一のVLアミノ酸配列を共有する。反対に、23の抗-Mpl scFv以外の7つは、抗-HER3クローンの一つと同じVLを共有した(表5、上記、白い箱を参照)。一方、VHアミノ酸配列は、何れかの抗-Mplと抗-HER3クローンの間に40から90%の同一性レベルを持つ、より多く相違するものであった。
【0231】
その抗-Mpl scFv,12B5(Genbank accession number AF048775;配列番号:27)と抗HER3scFvクローンH6(Genbank accession number AF048774;配列番号:28)は、同一のVL配列及び実質的に異なるVH配列を利用する。これらのscFvフラグメントは、共有した軽鎖に対する有効なヘテロダイマー化ができる抗-Mpl/抗-HER3二重特異性IgG抗体を構築すること、同じく孔-中-ノブ変異体(ここに記載した)及びCH3ドメイン間に作られたジスルフィド結合の使用を通して用いた。同じL鎖を共有する抗体は、非-同系H鎖と対合するL鎖の問題を避けるために選択した。2つの天然存在ヒンジ領域ジスルフィド結合もまた存在した。その共通のL鎖は、変異体v11からのCH3変異を含んでいる2つのH鎖と共移入した。そのIgG生産物は、プロテインAアフィニティクロマトグラフィーによって精製し、且つ標準技法を用いてSDS-PAGEによって分析した。
【0232】
二重特異性IgG抗体(BsIgG)調製物は、IgG含有野生型CH3ドメインよりも、より大きい移動性を示す単一の主要バンドを生じる。電気泳動移動性におけるこの増加は、よりコンパクトなタンパク質種を形成するBsIgG中に作られたジスルフィド結合の形成と一致した。
【0233】
MplとHER3 ECD抗原の両方に結合する工作した抗-Mpl/抗-HER3BsIgG 抗体の能力は、以下の通りELISAを用いて評価した。全ての工程でPBS緩衝液を用い、96-ウェル板(Maxisorp,Nunc)の個々のウェルは、5μg/mlでHER3-IgG又はMpl-IgGで一晩被覆し、洗浄し次いで0.5%(w/v)BSAによって1時間ブロック化した。その最初の抗体は、変異体Y349C:T366S:L368A:Y407V/T366'W:S354'Cを含み、且つ変異したFc領域を持つ親の抗-Mpl又は抗 -HER3 IgGに一致する抗-Mplx抗-HER3BsIgGとした。最初の抗体(1μg/mL)は、ビオチン化HER3-IgG及びストレプタビジン-ホースラディッシュペルオキシダーゼ接合体 (Boehringer Mannheim)の1:5000希釈液と共に23℃で2時間、別個にインキュベーションし、次いで各ウェルに加え、23℃でさらに1時間インキュベーションした。ペルオキシダーゼ活性は、販売者(Kirkegaard and Perry Laboratories,Inc.,Gaithersburg,MD)によって指示された通りにTMB試薬によって検出した。
【0234】
予想した通り、抗-Mpl/抗-HER3 BsIgGは、個別にMplとHER3 ECD抗原のそれぞれに有効に且つ同時に、同じく両方の抗原に同時に結合した。これに対して、親の抗-Mplと親の抗-HER3 IgGは、同系抗原に一致するそれにのみ結合した(図6)。
【0235】
D. 工作したFc領域を含んでいる抗体は、有効な抗体-依存細胞介在細胞毒性の能力がある。
【0236】
本発明の実例化した二重特異性抗体の精製において用いた工作したFc領域(CH3変異体、上記)が有効な抗体-依存細胞介在細胞毒性の能力があることを実証するため、以下の実験を実行した。
【0237】
そのCH3変異体は、Lewis,G.D.ら(Lewis,G.D,ら(1993)Cancer Immunol.Immunother.37:255-263,その全部が参考により組込まれる)の方法を用いて示した通り、有能な抗体-依存細胞-介在細胞毒性(ADCC)をサポートする能力を維持する。概略的には、細胞毒性アッセイは、51Cr-標識したSK-BR-3及びHBL-100標的細胞(それぞれATCC受託番号HTB-30と45509)及びエフェクター細胞としてヒト末梢血液リンパ球によって実行した。しかしながら、Lewisらと異なり、そのリンパ球はIL-2で活性化されなかった。
【0238】
CH3変異体S354:T366W及びY349C:T366S:L368A:Y407Vは、Carterら(Carter,P.ら(1992)PNAS USA 89:4285-4289)により製造されたヒト化抗-HER2抗体、huMAb4D5-5のH鎖中に別個に導入した。再モデル化した及び野生型Fc領域を含む抗体は、HER2-過剰発現乳癌細胞系、SK-BR-3によりADCC中で類似の能力を有した(図7)。再モデル化した及び野生型抗体の両方は、通常の胸部上皮細胞系に対する低い活性と比較できることを示した。H鎖中の効果は、これらBsIgG'sが抗体-依存細胞-介在細胞毒性において機能するであろうことを予報する、結合ドメインの独立がある。
【0239】
本発明は、最も実践的、且つ好適な実施態様であるとして考慮される何れかにおいてここに示され且つ記載される。それは、しかしながら、発展は本発明の範囲内から作製できること、及び自明な修正がこの開示を熟読した当業者に生起することが認識される。ここに用いた全ての参考文献はそれの全体において参考によってここに組込まれる。
【図面の簡単な説明】
【0240】
【図1】図1A〜図1C。図1Aは、ホモマルチマー化よりもヘテロマルチマー化を増強させるための操作が行われない場合において、Fcを含有する二重特異性抗体の形成を示す略図である。図1Bは、所望のヘテロマルチマー化が、望ましくないヘテロマルチマー化およびホモマルチマー化よりも好ましいように、重(H)鎖が操作される場合に生じる対形成を示す略図である。図1Cは、同じ軽(L)鎖を有する抗体を選択して、同系でない重鎖と対形成する軽鎖の問題が回避される場合に生じる対形成を示す略図である。
【図2AB】図2Aは、ファージディスプレーベクターpRA2を使用するCH3ヘテロダイマーの選択反応図式を図示する。安定なCH3ヘテロダイマーをディスプレーするファージは、gDフラッグに対する抗体を使用して捕獲される。図2Bは、合成された遺伝子から発現されるCH3が、M13の遺伝子IIIタンパク質との融合タンパク質として、天然の遺伝子(Ellisonら、Nucleic Acids Res.10:4071〜4079(1982))から発現されるCH3の第2の複製物とともに同時に分泌されるジシストロン性オペロンを図示する。合成されたCH3遺伝子は、単純ヘルペスウイルスの糖タンパク質Dから誘導されるペプチド(gDフラッグ、Lasky,L.A.およびDowbenko,D.J.(1984)DNA3:23〜29;Berman,P.W.ら(1985)Science227:1490〜1492)、および部位特異的なプロテアーゼのケネナーゼ(Genenase)Jの開裂(G)部位(Carter,P.ら(1989)Proteins:Structure、Function and Genetics 6:240〜248)の上流に置かれる。
【図2C1】図2C1は、翻訳されるCH3遺伝子の残基に、KabatらのEu方式に従って番号を付けた図2Bのジシストロン性オペロンの核酸配列(配列番号2)である。「免疫学的に関心が持たれるタンパク質の配列」、第5版、第1巻、688〜696頁、NIH、Bethesda、MD(1991)。隆起の変異T366Wを示す。そのような残基は、天然のCH3遺伝子における無作為化のために標的化される(366、368および407)。
【図2C2】図2C2は、翻訳されるCH3遺伝子の残基に、KabatらのEu方式に従って番号を付けた図2Bのジシストロン性オペロンの核酸配列(配列番号2)である。「免疫学的に関心が持たれるタンパク質の配列」、第5版、第1巻、688〜696頁、NIH、Bethesda、MD(1991)。隆起の変異T366Wを示す。そのような残基は、天然のCH3遺伝子における無作為化のために標的化される(366、368および407)。
【図3】図3A〜図3C。図3Aおよび図3Bは、免疫付着因子(Ia)とともに抗体(Ab)の重鎖および軽鎖の共-移入から得られるプロテインA精製された生成物のSDS−PAGEをデンシトメトリー分析で走査した結果の棒グラフである。示したデータは、2つの独立した実験の平均である。x軸は、投入したDNAの質量比(Ia:H:L)を示し、y軸は、全生成物タンパク質に対する生成物マルチマーの各タイプの割合を示す。図3Cは、可能な生成物マルチマーを図示する。
【図4】図4は、Ax1、Rse、IgER、Ob−RおよびVEGFに対する特異性を有する8個の異なる抗体のVL配列の比較である。配列定義(Kabatら(1991)上記)または構造定義(Chothia,C.およびLesk,A.M.J.Mol.Biol.(1987)196:901〜917)に従った抗原結合CDR残基の位置を、それぞれ、下線および#によって示す。Ax1.78配列と異なる残基を二重下線によって示す。
【図5】図5は、選択された抗Ob−Rクローンおよび抗HER3クローンの重鎖および軽鎖の比較である。二重特異性抗体の構築に使用した抗Ob-Rクローン26および抗HER3クローン18のVH配列および共通するVL配列を示す。
【図6】図6。Mpl−IgGおよびHER3−IgGに対する同時結合を検出するためのサンドイッチELTSA。試験した抗体は、Y349C:T366S:L368A:Y407V/T366’W:S354’Cの変異を含有する抗Mpl×抗HER3のBsIgGであり、Fc領域を変異させた対応する元の抗Mplまたは抗HER3のIgGとともに使用した。
【図7】図7は、抗体依存性細胞媒介細胞傷害性(ADCC)の研究結果の棒グラフである。ADCCは、変異型(S354C:T366W/Y349’C:T366’S:L368’A:Y4O7’V)または野生型のFcあるいはアイソタイプが一致するコントロール抗体(E25、Presta,L.G.ら(1993)J.Immunol.151:2623〜2632)のいずれかを含有するhuMAb4D5−5(Carter,P.ら(1992)PNAS USA89:4285〜4289)によって媒介された。抗体(125ng/ml)を、ヒト末梢血単核エフェクター細胞およびSK−BR−3標的細胞とともに示した比でインキュベーションした。示したデータは、3連の測定値および3つの異なる実験の平均である。
【図8】図8は、HER3に対して生起された抗体の軽鎖とOb−Rに対して生起された抗体の軽鎖との間のアミノ酸配列同一性を示す行列である。配列同一性が100%である軽鎖を有する抗体を黒四角に示す。配列同一性が98%〜99%である軽鎖を有する抗体を白四角に示す。抗体クローンの同一性を行列の下部に示す。
【特許請求の範囲】
【請求項1】
少なくとも2つの異なる結合ドメインを含む多重特異性抗体の製造方法であり、ここで、
第1の結合ドメインは第1の分子に結合し、且つ、第2の結合ドメインは前記第1の分子とは異なる第2の分子に結合し、
それぞれの結合ドメインは、(i)マルチマー化ドメインをさらに含むポリペプチドの一部である重鎖可変ドメイン及び(ii)軽鎖の可変ドメインを含み、
該第1の結合ドメインは、第1のポリペプチドの第1の重鎖可変ドメインを含み、該第2の結合ドメインは、第2のポリペプチドの第2の重鎖可変ドメインを含み、且つ該第1及び第2の重鎖可変ドメインは異なり、及び
該ポリペプチドは前記マルチマー化ドメインの相互作用によってマルチマー化され、
及びここで、
該多重特異性抗体は、第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに少なくとも80%のアミノ酸配列同一性を有し、相補性決定領域(CDR)の内側では同一であって、前記第1の結合ドメインが第1の軽鎖可変ドメインを含むか又は第2の軽鎖可変ドメインを含むかにかかわらず前記第1の結合ドメインは前記第1の分子に結合し、前記第2の結合ドメインが第2の軽鎖可変ドメインを含むか又は第1の軽鎖可変ドメインを含むかにかかわらず前記第2の結合ドメインは前記第2の分子に結合し、
工程:
(i)前記ポリペプチドと軽鎖をコードしている核酸を含む宿主細胞を培養すること、ここで該核酸が発現され且つ該ポリペプチドと軽鎖が生産され;及び
(ii)該宿主細胞培養物から多重特異性抗体を回収すること、
を含む方法。
【請求項2】
前記第1のポリペプチドのマルチマー化ドメインをコードしている核酸又は前記第2のポリペプチドのマルチマー化ドメインをコードしている核酸、又は両方が、コードされたアミノ酸配列を変更するような核酸の変更により提供される請求項1記載の方法。
【請求項3】
前記第1のポリペプチド、前記第2のポリペプチド、又は両方のマルチマー化ドメインをコードしている核酸が、前記第1及び/又は第2のポリペプチドのそれぞれの該マルチマー化ドメインが、前記第1及び/又は第2のポリペプチドの他方のマルチマー化ドメインのフリーのチオール含有残基とジスルフィド結合を形成するフリーのチオール含有残基を含むように変更される請求項2記載の方法。
【請求項4】
前記ポリペプチドのマルチマー化が、空洞への隆起相互作用を含み、該方法は工程(i)の前に:
より大きな側鎖容量を有する移入残基でアミノ酸残基を置換することによってコードされたポリペプチドのマルチマー化ドメイン中に隆起を生成するように核酸を置換することによって該第1のポリペプチドをコードしている核酸を用意すること、および
より小さな側鎖容量を有する移入残基でアミノ酸残基を置換することによってコードされたポリペプチドのマルチマー化ドメイン中に相補的な空洞を生成するように核酸を置換することによって該第2のポリペプチドをコードしている核酸を用意すること、をさらに含む請求項1記載の方法。
【請求項5】
ファージディスプレー選択によって、隆起を有するマルチマー化ドメインを含む第1のポリペプチド、空洞を有するマルチマー化ドメインを含む第2のポリペプチド、又は両方をコードする核酸を準備する請求項4記載の方法。
【請求項6】
より大きな側鎖容量を有する移入残基が、アルギニン(R)、フェニルアラニン(F)、チロシン(Y)、トリプトファン(W)、イソロイシン(I)及びロイシン(L)からなる群から選択される、請求項4記載の方法。
【請求項7】
より小さい側鎖容量を有する移入残基が、グリシン(G)、アラニン(A)、セリン(S)、トレオニン(T)及びバリン(V)からなる群から選択され、且つ該移入残基がシステイン(C)ではない、請求項4記載の方法。
【請求項8】
該第1及び第2のポリペプチドがそれぞれ抗体定常ドメインを含む請求項1記載の方法。
【請求項9】
該第1及び第2のポリペプチドがそれぞれ、CH3ドメインとIgG定常ドメインからなる群から選択される抗体定常ドメインを含む請求項8記載の方法。
【請求項10】
該多重特異性抗体が免疫付着因子である請求項1記載の方法。
【請求項11】
工程(i)に先行する工程をさらに含み、ここで前記核酸が宿主細胞内に導入される請求項1記載の方法。
【請求項12】
該多重特異性抗体が第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに少なくとも90%のアミノ酸配列同一性を有する請求項1記載の方法。
【請求項13】
該多重特異性抗体が第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに異なるが、互いに少なくとも95%のアミノ酸配列同一性を有する請求項12記載の方法。
【請求項14】
少なくとも2の異なる結合ドメインを含む多重特異性抗体であり、ここで
第1の結合ドメインは第1の分子に結合し、且つ、第2の結合ドメインは前記第1の分子とは異なる第2の分子に結合し、
それぞれの結合ドメインは、(i)マルチマー化ドメインをさらに含むポリペプチドの一部である重鎖可変ドメイン及び(ii)軽鎖の可変ドメインを含み、
該第1の結合ドメインは、第1のポリペプチドの第1の重鎖可変ドメインを含み、該第2の結合ドメインは、第2のポリペプチドの第2の重鎖可変ドメインを含み、且つ該第1及び第2の重鎖可変ドメインは異なり、及び
該ポリペプチドは前記マルチマー化ドメインの相互作用によってマルチマー化され、
及びここで、
前記多重特異性抗体は、第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに少なくとも80%のアミノ酸配列同一性を有し、相補性決定領域(CDR)の内側では同一であって、前記第1の結合ドメインが第1の軽鎖可変ドメインを含むか又は第2の軽鎖可変ドメインを含むかにかかわらず前記第1の結合ドメインは前記第1の分子に結合し、前記第2の結合ドメインが第2の軽鎖可変ドメインを含むか又は第1の軽鎖可変ドメインを含むかにかかわらず前記第2の結合ドメインは前記第2の分子に結合する、多重特異性抗体。
【請求項15】
該第1のポリペプチドのマルチマー化ドメイン又は該第2のポリペプチドのマルチマー化ドメイン、又は両方が、アミノ酸の変更により提供される請求項14記載の多重特異性抗体。
【請求項16】
前記第1又は第2のポリペプチドのそれぞれのマルチマー化ドメインが、前記第1又は第2のポリペプチドの他方のマルチマー化ドメインのフリーのチオール含有残基とジスルフィド結合を形成するフリーのチオール含有残基を含む請求項15記載の多重特異性抗体。
【請求項17】
前記ポリペプチドのマルチマー化が、空洞への隆起相互作用を含み且つ該第1のポリペプチドのマルチマー化ドメインが隆起を含み且つ第2のポリペプチドのマルチマー化ドメインが相補的な空洞を含む請求項15記載の多重特異性抗体。
【請求項18】
該隆起と空洞が、天然存在アミノ酸が該第1及び第2のポリペプチド中に移入される置換によって生成される請求項17記載の多重特異性抗体。
【請求項19】
該多重特異性抗体が第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに異なるが、互いに少なくとも90%のアミノ酸配列同一性を有する請求項14記載の多重特異性抗体。
【請求項20】
該多重特異性抗体が第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに異なるが、互いに少なくとも95%のアミノ酸配列同一性を有する請求項19記載の多重特異性抗体。
【請求項21】
請求項14記載の多重特異性抗体とキャリアとを含んでいる組成物。
【請求項22】
請求項14記載の多重特異性抗体をコードしている核酸を含む宿主細胞。
【請求項23】
宿主細胞が哺乳動物細胞である請求項22記載の宿主細胞。
【請求項24】
第1の分子に結合する第1の結合ドメインと、第1の分子とは異なる第2の分子に結合する第2の結合ドメインを含む多重特異性抗体の製造方法であり:
(a)第1の重鎖可変ドメインとマルチマー化ドメインを含む第1のポリペプチドをコードしている第1の核酸を選択すること、及び第2の重鎖可変ドメインとマルチマー化ドメインを含む第2のポリペプチドをコードしている第2の核酸を選択すること、ここで該第1及び第2の重鎖可変ドメインは異なっており、且つ前記第1及び第2のポリペプチドのそれぞれのマルチマー化ドメインは、前記第1及び第2のポリペプチドの他方のマルチマー化ドメインのアミノ酸残基と特異的に相互作用するアミノ酸残基を含み、それによって該第1及び第2のポリペプチド間に安定な相互作用を生成する;
(b)第1の軽鎖可変ドメイン-コード化核酸と第2の軽鎖可変ドメイン-コード化核酸を選択すること、ここで前記多重特異性抗体は、第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに少なくとも80%のアミノ酸配列同一性を有し、相補性決定領域(CDR)の内側では同一であって、前記第1の結合ドメインが第1の軽鎖可変ドメインを含むか又は第2の軽鎖可変ドメインを含むかにかかわらず前記第1の結合ドメインは前記第1の分子に結合し、前記第2の結合ドメインが第2の軽鎖可変ドメインを含むか又は第1の軽鎖可変ドメインを含むかにかかわらず前記第2の結合ドメインは前記第2の分子に結合する;
(c)前記ポリペプチドと軽鎖をコードしている核酸を宿主細胞中に導入すること、及び該細胞を、該核酸が発現され且つ該ポリペプチドと軽鎖が生産されるように培養すること;
(d)該細胞培養物から多重特異性抗体を回収すること、
を含む方法。
【請求項25】
該第1のポリペプチドをコードしている前記第1の核酸、該第2のポリペプチドをコードしている前記第2の核酸、又は両方が、コードされたアミノ酸配列を変更するように変えられた核酸から選択される請求項24記載の方法。
【請求項26】
該第1及び第2のポリペプチドが、空洞への隆起相互作用によって相互作用する請求項25記載の方法。
【請求項27】
該第1の核酸、該第2の核酸、又は両方が、そのコードされたアミノ酸配列中にフリーのチオール含有残基を移入するように変更される請求項25記載の方法。
【請求項28】
該第1及び第2のポリペプチドが抗体定常ドメインをそれぞれ含む請求項24記載の方法。
【請求項29】
抗体定常ドメインがCH3ドメインである請求項28記載の方法。
【請求項30】
抗体定常ドメインがヒトIgGからのものである請求項28記載の方法。
【請求項31】
該多重特異性抗体が第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに異なるが、互いに少なくとも90%のアミノ酸配列同一性を有する請求項24記載の方法。
【請求項32】
該多重特異性抗体が第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに異なるが、互いに少なくとも95%のアミノ酸配列同一性を有する請求項31記載の方法。
【請求項33】
ポリペプチドの混合物からヘテロマルチマー多重特異性抗体の形成を測定する方法であり、ここで該多重特異性抗体は少なくとも2の結合ドメインを含み、ここで
第1の結合ドメインは第1の分子に結合し、且つ、第2の結合ドメインは第2の分子に結合し、
それぞれの結合ドメインは、(i)マルチマー化ドメインをさらに含むポリペプチドの一部である重鎖可変ドメイン及び(ii)軽鎖の可変ドメインを含み、
該第1の結合ドメインは、第1のポリペプチドの第1の重鎖可変ドメインを含み、該第2の結合ドメインは、第2のポリペプチドの第2の重鎖可変ドメインを含み、且つ該第1及び第2の重鎖可変ドメインは異なり、及び
該ポリペプチドは前記マルチマー化ドメインの相互作用によってマルチマー化され、ここで前記第1又は第2のポリペプチドのぞれぞれのマルチマー化ドメインは、前記第1又は第2のポリペプチドの他方のマルチマー化ドメインのフリーのチオール含有残基とジスルフィド結合を形成するフリーのチオール含有残基を含み、
及びここで、
前記多重特異性抗体は、第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに少なくとも80%のアミノ酸配列同一性を有し、相補性決定領域(CDR)の内側では同一であって、前記第1の結合ドメインが第1の軽鎖可変ドメインを含むか又は第2の軽鎖可変ドメインを含むかにかかわらず前記第1の結合ドメインは前記第1の分子に結合し、前記第2の結合ドメインが第2の軽鎖可変ドメインを含むか又は第1の軽鎖可変ドメインを含むかにかかわらず前記第2の結合ドメインは前記第2の分子に結合し、
工程:
(a)それぞれの多重特異性抗体をゲルマトリックスにおいて移動させること;及び
(b)該第1及び第2ポリペプチド間に非天然存在ジスルフィド結合を有する多重特異性抗体に対応するバンドと、該第1及び第2のポリペプチド間に非天然存在ジスルフィド結合を欠いているヘテロマルチマーに対応する、遅く移動するバンドの相対量を測定すること、を含む方法。
【請求項34】
前記ポリペプチドのマルチマー化が、該マルチマー化ドメイン間の空洞への隆起相互作用によって促進される請求項33記載の方法。
【請求項35】
該多重特異性抗体が第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに異なるが、互いに少なくとも90%のアミノ酸配列同一性を有する請求項33記載の方法。
【請求項36】
該多重特異性抗体が第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに異なるが、互いに少なくとも95%のアミノ酸配列同一性を有する請求項35記載の方法。
【請求項37】
該多重特異性抗体が、抗-ヒトレプチンレセプターECD(Ob-R)/抗-ヒトレセプターチロシンキナーゼHER3及び抗-ヒトトロンボポイエチンレセプターチロシンキナーゼ(Mpl)/抗-ヒトレセプターチロシンキナーゼHER3からなる群から選択される請求項1記載の方法。
【請求項38】
抗-ヒトレプチンレセプターECD(Ob-R)/抗-ヒトレセプターチロシンキナーゼHER3及び抗-ヒトトロンボポイエチンレセプターチロシンキナーゼ(Mpl)/抗-ヒトレセプターチロシンキナーゼHER3からなる群から選択される請求項14記載の多重特異性抗体。
【請求項39】
請求項38記載の多重特異性抗体と担体を含む組成物。
【請求項40】
該多重特異性抗体が、抗-ヒトレプチンレセプターECD(Ob-R)/抗-ヒトレセプターチロシンキナーゼHER3及び抗-ヒトトロンボポイエチンレセプターチロシンキナーゼ(Mpl)/抗-ヒトレセプターチロシンキナーゼHER3からなる群から選択される請求項22記載の宿主細胞。
【請求項1】
少なくとも2つの異なる結合ドメインを含む多重特異性抗体の製造方法であり、ここで、
第1の結合ドメインは第1の分子に結合し、且つ、第2の結合ドメインは前記第1の分子とは異なる第2の分子に結合し、
それぞれの結合ドメインは、(i)マルチマー化ドメインをさらに含むポリペプチドの一部である重鎖可変ドメイン及び(ii)軽鎖の可変ドメインを含み、
該第1の結合ドメインは、第1のポリペプチドの第1の重鎖可変ドメインを含み、該第2の結合ドメインは、第2のポリペプチドの第2の重鎖可変ドメインを含み、且つ該第1及び第2の重鎖可変ドメインは異なり、及び
該ポリペプチドは前記マルチマー化ドメインの相互作用によってマルチマー化され、
及びここで、
該多重特異性抗体は、第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに少なくとも80%のアミノ酸配列同一性を有し、相補性決定領域(CDR)の内側では同一であって、前記第1の結合ドメインが第1の軽鎖可変ドメインを含むか又は第2の軽鎖可変ドメインを含むかにかかわらず前記第1の結合ドメインは前記第1の分子に結合し、前記第2の結合ドメインが第2の軽鎖可変ドメインを含むか又は第1の軽鎖可変ドメインを含むかにかかわらず前記第2の結合ドメインは前記第2の分子に結合し、
工程:
(i)前記ポリペプチドと軽鎖をコードしている核酸を含む宿主細胞を培養すること、ここで該核酸が発現され且つ該ポリペプチドと軽鎖が生産され;及び
(ii)該宿主細胞培養物から多重特異性抗体を回収すること、
を含む方法。
【請求項2】
前記第1のポリペプチドのマルチマー化ドメインをコードしている核酸又は前記第2のポリペプチドのマルチマー化ドメインをコードしている核酸、又は両方が、コードされたアミノ酸配列を変更するような核酸の変更により提供される請求項1記載の方法。
【請求項3】
前記第1のポリペプチド、前記第2のポリペプチド、又は両方のマルチマー化ドメインをコードしている核酸が、前記第1及び/又は第2のポリペプチドのそれぞれの該マルチマー化ドメインが、前記第1及び/又は第2のポリペプチドの他方のマルチマー化ドメインのフリーのチオール含有残基とジスルフィド結合を形成するフリーのチオール含有残基を含むように変更される請求項2記載の方法。
【請求項4】
前記ポリペプチドのマルチマー化が、空洞への隆起相互作用を含み、該方法は工程(i)の前に:
より大きな側鎖容量を有する移入残基でアミノ酸残基を置換することによってコードされたポリペプチドのマルチマー化ドメイン中に隆起を生成するように核酸を置換することによって該第1のポリペプチドをコードしている核酸を用意すること、および
より小さな側鎖容量を有する移入残基でアミノ酸残基を置換することによってコードされたポリペプチドのマルチマー化ドメイン中に相補的な空洞を生成するように核酸を置換することによって該第2のポリペプチドをコードしている核酸を用意すること、をさらに含む請求項1記載の方法。
【請求項5】
ファージディスプレー選択によって、隆起を有するマルチマー化ドメインを含む第1のポリペプチド、空洞を有するマルチマー化ドメインを含む第2のポリペプチド、又は両方をコードする核酸を準備する請求項4記載の方法。
【請求項6】
より大きな側鎖容量を有する移入残基が、アルギニン(R)、フェニルアラニン(F)、チロシン(Y)、トリプトファン(W)、イソロイシン(I)及びロイシン(L)からなる群から選択される、請求項4記載の方法。
【請求項7】
より小さい側鎖容量を有する移入残基が、グリシン(G)、アラニン(A)、セリン(S)、トレオニン(T)及びバリン(V)からなる群から選択され、且つ該移入残基がシステイン(C)ではない、請求項4記載の方法。
【請求項8】
該第1及び第2のポリペプチドがそれぞれ抗体定常ドメインを含む請求項1記載の方法。
【請求項9】
該第1及び第2のポリペプチドがそれぞれ、CH3ドメインとIgG定常ドメインからなる群から選択される抗体定常ドメインを含む請求項8記載の方法。
【請求項10】
該多重特異性抗体が免疫付着因子である請求項1記載の方法。
【請求項11】
工程(i)に先行する工程をさらに含み、ここで前記核酸が宿主細胞内に導入される請求項1記載の方法。
【請求項12】
該多重特異性抗体が第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに少なくとも90%のアミノ酸配列同一性を有する請求項1記載の方法。
【請求項13】
該多重特異性抗体が第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに異なるが、互いに少なくとも95%のアミノ酸配列同一性を有する請求項12記載の方法。
【請求項14】
少なくとも2の異なる結合ドメインを含む多重特異性抗体であり、ここで
第1の結合ドメインは第1の分子に結合し、且つ、第2の結合ドメインは前記第1の分子とは異なる第2の分子に結合し、
それぞれの結合ドメインは、(i)マルチマー化ドメインをさらに含むポリペプチドの一部である重鎖可変ドメイン及び(ii)軽鎖の可変ドメインを含み、
該第1の結合ドメインは、第1のポリペプチドの第1の重鎖可変ドメインを含み、該第2の結合ドメインは、第2のポリペプチドの第2の重鎖可変ドメインを含み、且つ該第1及び第2の重鎖可変ドメインは異なり、及び
該ポリペプチドは前記マルチマー化ドメインの相互作用によってマルチマー化され、
及びここで、
前記多重特異性抗体は、第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに少なくとも80%のアミノ酸配列同一性を有し、相補性決定領域(CDR)の内側では同一であって、前記第1の結合ドメインが第1の軽鎖可変ドメインを含むか又は第2の軽鎖可変ドメインを含むかにかかわらず前記第1の結合ドメインは前記第1の分子に結合し、前記第2の結合ドメインが第2の軽鎖可変ドメインを含むか又は第1の軽鎖可変ドメインを含むかにかかわらず前記第2の結合ドメインは前記第2の分子に結合する、多重特異性抗体。
【請求項15】
該第1のポリペプチドのマルチマー化ドメイン又は該第2のポリペプチドのマルチマー化ドメイン、又は両方が、アミノ酸の変更により提供される請求項14記載の多重特異性抗体。
【請求項16】
前記第1又は第2のポリペプチドのそれぞれのマルチマー化ドメインが、前記第1又は第2のポリペプチドの他方のマルチマー化ドメインのフリーのチオール含有残基とジスルフィド結合を形成するフリーのチオール含有残基を含む請求項15記載の多重特異性抗体。
【請求項17】
前記ポリペプチドのマルチマー化が、空洞への隆起相互作用を含み且つ該第1のポリペプチドのマルチマー化ドメインが隆起を含み且つ第2のポリペプチドのマルチマー化ドメインが相補的な空洞を含む請求項15記載の多重特異性抗体。
【請求項18】
該隆起と空洞が、天然存在アミノ酸が該第1及び第2のポリペプチド中に移入される置換によって生成される請求項17記載の多重特異性抗体。
【請求項19】
該多重特異性抗体が第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに異なるが、互いに少なくとも90%のアミノ酸配列同一性を有する請求項14記載の多重特異性抗体。
【請求項20】
該多重特異性抗体が第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに異なるが、互いに少なくとも95%のアミノ酸配列同一性を有する請求項19記載の多重特異性抗体。
【請求項21】
請求項14記載の多重特異性抗体とキャリアとを含んでいる組成物。
【請求項22】
請求項14記載の多重特異性抗体をコードしている核酸を含む宿主細胞。
【請求項23】
宿主細胞が哺乳動物細胞である請求項22記載の宿主細胞。
【請求項24】
第1の分子に結合する第1の結合ドメインと、第1の分子とは異なる第2の分子に結合する第2の結合ドメインを含む多重特異性抗体の製造方法であり:
(a)第1の重鎖可変ドメインとマルチマー化ドメインを含む第1のポリペプチドをコードしている第1の核酸を選択すること、及び第2の重鎖可変ドメインとマルチマー化ドメインを含む第2のポリペプチドをコードしている第2の核酸を選択すること、ここで該第1及び第2の重鎖可変ドメインは異なっており、且つ前記第1及び第2のポリペプチドのそれぞれのマルチマー化ドメインは、前記第1及び第2のポリペプチドの他方のマルチマー化ドメインのアミノ酸残基と特異的に相互作用するアミノ酸残基を含み、それによって該第1及び第2のポリペプチド間に安定な相互作用を生成する;
(b)第1の軽鎖可変ドメイン-コード化核酸と第2の軽鎖可変ドメイン-コード化核酸を選択すること、ここで前記多重特異性抗体は、第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに少なくとも80%のアミノ酸配列同一性を有し、相補性決定領域(CDR)の内側では同一であって、前記第1の結合ドメインが第1の軽鎖可変ドメインを含むか又は第2の軽鎖可変ドメインを含むかにかかわらず前記第1の結合ドメインは前記第1の分子に結合し、前記第2の結合ドメインが第2の軽鎖可変ドメインを含むか又は第1の軽鎖可変ドメインを含むかにかかわらず前記第2の結合ドメインは前記第2の分子に結合する;
(c)前記ポリペプチドと軽鎖をコードしている核酸を宿主細胞中に導入すること、及び該細胞を、該核酸が発現され且つ該ポリペプチドと軽鎖が生産されるように培養すること;
(d)該細胞培養物から多重特異性抗体を回収すること、
を含む方法。
【請求項25】
該第1のポリペプチドをコードしている前記第1の核酸、該第2のポリペプチドをコードしている前記第2の核酸、又は両方が、コードされたアミノ酸配列を変更するように変えられた核酸から選択される請求項24記載の方法。
【請求項26】
該第1及び第2のポリペプチドが、空洞への隆起相互作用によって相互作用する請求項25記載の方法。
【請求項27】
該第1の核酸、該第2の核酸、又は両方が、そのコードされたアミノ酸配列中にフリーのチオール含有残基を移入するように変更される請求項25記載の方法。
【請求項28】
該第1及び第2のポリペプチドが抗体定常ドメインをそれぞれ含む請求項24記載の方法。
【請求項29】
抗体定常ドメインがCH3ドメインである請求項28記載の方法。
【請求項30】
抗体定常ドメインがヒトIgGからのものである請求項28記載の方法。
【請求項31】
該多重特異性抗体が第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに異なるが、互いに少なくとも90%のアミノ酸配列同一性を有する請求項24記載の方法。
【請求項32】
該多重特異性抗体が第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに異なるが、互いに少なくとも95%のアミノ酸配列同一性を有する請求項31記載の方法。
【請求項33】
ポリペプチドの混合物からヘテロマルチマー多重特異性抗体の形成を測定する方法であり、ここで該多重特異性抗体は少なくとも2の結合ドメインを含み、ここで
第1の結合ドメインは第1の分子に結合し、且つ、第2の結合ドメインは第2の分子に結合し、
それぞれの結合ドメインは、(i)マルチマー化ドメインをさらに含むポリペプチドの一部である重鎖可変ドメイン及び(ii)軽鎖の可変ドメインを含み、
該第1の結合ドメインは、第1のポリペプチドの第1の重鎖可変ドメインを含み、該第2の結合ドメインは、第2のポリペプチドの第2の重鎖可変ドメインを含み、且つ該第1及び第2の重鎖可変ドメインは異なり、及び
該ポリペプチドは前記マルチマー化ドメインの相互作用によってマルチマー化され、ここで前記第1又は第2のポリペプチドのぞれぞれのマルチマー化ドメインは、前記第1又は第2のポリペプチドの他方のマルチマー化ドメインのフリーのチオール含有残基とジスルフィド結合を形成するフリーのチオール含有残基を含み、
及びここで、
前記多重特異性抗体は、第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに少なくとも80%のアミノ酸配列同一性を有し、相補性決定領域(CDR)の内側では同一であって、前記第1の結合ドメインが第1の軽鎖可変ドメインを含むか又は第2の軽鎖可変ドメインを含むかにかかわらず前記第1の結合ドメインは前記第1の分子に結合し、前記第2の結合ドメインが第2の軽鎖可変ドメインを含むか又は第1の軽鎖可変ドメインを含むかにかかわらず前記第2の結合ドメインは前記第2の分子に結合し、
工程:
(a)それぞれの多重特異性抗体をゲルマトリックスにおいて移動させること;及び
(b)該第1及び第2ポリペプチド間に非天然存在ジスルフィド結合を有する多重特異性抗体に対応するバンドと、該第1及び第2のポリペプチド間に非天然存在ジスルフィド結合を欠いているヘテロマルチマーに対応する、遅く移動するバンドの相対量を測定すること、を含む方法。
【請求項34】
前記ポリペプチドのマルチマー化が、該マルチマー化ドメイン間の空洞への隆起相互作用によって促進される請求項33記載の方法。
【請求項35】
該多重特異性抗体が第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに異なるが、互いに少なくとも90%のアミノ酸配列同一性を有する請求項33記載の方法。
【請求項36】
該多重特異性抗体が第1の軽鎖可変ドメイン及び第2の軽鎖可変ドメインを含み、第1の軽鎖可変ドメインと第2の軽鎖可変ドメインとは互いに異なるが、互いに少なくとも95%のアミノ酸配列同一性を有する請求項35記載の方法。
【請求項37】
該多重特異性抗体が、抗-ヒトレプチンレセプターECD(Ob-R)/抗-ヒトレセプターチロシンキナーゼHER3及び抗-ヒトトロンボポイエチンレセプターチロシンキナーゼ(Mpl)/抗-ヒトレセプターチロシンキナーゼHER3からなる群から選択される請求項1記載の方法。
【請求項38】
抗-ヒトレプチンレセプターECD(Ob-R)/抗-ヒトレセプターチロシンキナーゼHER3及び抗-ヒトトロンボポイエチンレセプターチロシンキナーゼ(Mpl)/抗-ヒトレセプターチロシンキナーゼHER3からなる群から選択される請求項14記載の多重特異性抗体。
【請求項39】
請求項38記載の多重特異性抗体と担体を含む組成物。
【請求項40】
該多重特異性抗体が、抗-ヒトレプチンレセプターECD(Ob-R)/抗-ヒトレセプターチロシンキナーゼHER3及び抗-ヒトトロンボポイエチンレセプターチロシンキナーゼ(Mpl)/抗-ヒトレセプターチロシンキナーゼHER3からなる群から選択される請求項22記載の宿主細胞。
【図1】

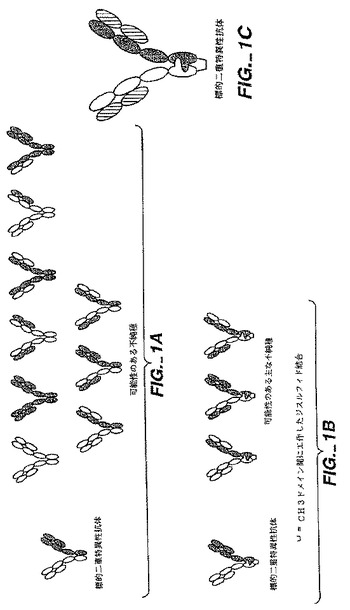
【図2AB】
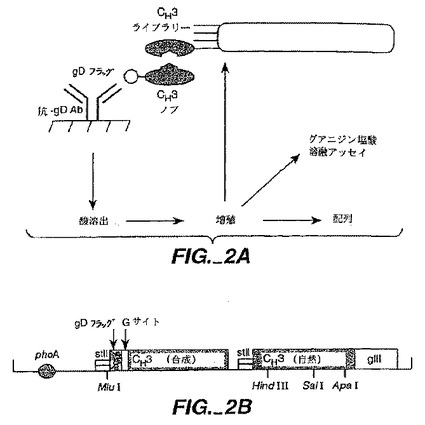
【図2C1】

【図2C2】

【図3】
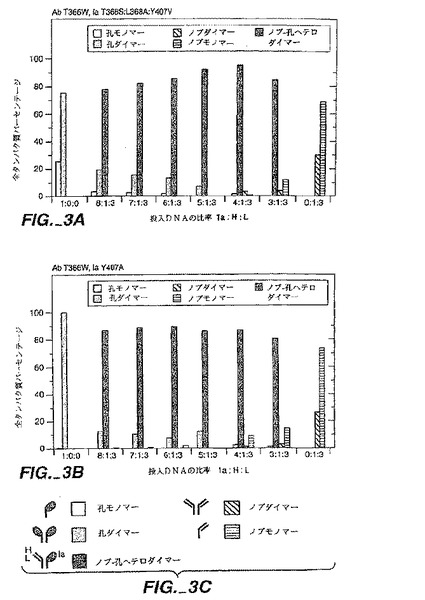
【図4】

【図5】

【図6】
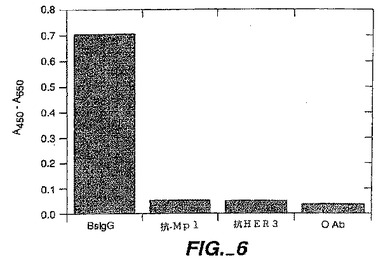
【図7】
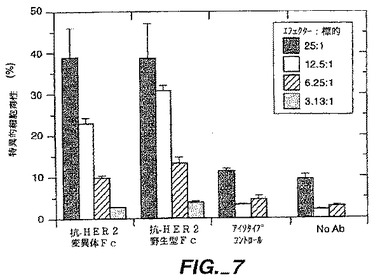
【図8】
【図2AB】
【図2C1】
【図2C2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2009−39122(P2009−39122A)
【公開日】平成21年2月26日(2009.2.26)
【国際特許分類】
【出願番号】特願2008−207339(P2008−207339)
【出願日】平成20年8月11日(2008.8.11)
【分割の表示】特願平10−548216の分割
【原出願日】平成10年4月30日(1998.4.30)
【出願人】(596168317)ジェネンテック・インコーポレーテッド (372)
【氏名又は名称原語表記】GENENTECH,INC.
【Fターム(参考)】
【公開日】平成21年2月26日(2009.2.26)
【国際特許分類】
【出願日】平成20年8月11日(2008.8.11)
【分割の表示】特願平10−548216の分割
【原出願日】平成10年4月30日(1998.4.30)
【出願人】(596168317)ジェネンテック・インコーポレーテッド (372)
【氏名又は名称原語表記】GENENTECH,INC.
【Fターム(参考)】
[ Back to top ]