
アカゲザル癌胎児性抗原、前記抗原をコードするヌクレオチド、及びその使用
アカゲザル癌胎児性抗原(rhCEA)をコードするDNAを単離し、クローニングし、配列決定した。CEAをコードする遺伝子は一般にヒト癌の発生に関連付けられる。本発明は異常CEA発現が癌又はその発生に関連付けられるCEA腫瘍関連抗原により発現される蛋白質産物に対する免疫を誘発又は強化するための組成物と方法を提供する。本発明は特にrhCEAをもつアデノウイルスベクター構築物を提供し、癌を予防及び治療するためのワクチン及び医薬組成物におけるその使用を開示する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は一般には癌の治療に関する。より詳細には、本発明は腫瘍関連ポリペプチド癌胎児性抗原のアカゲザルホモログ(本明細書ではrhCEAと言う)、この蛋白質をコードする単離核酸分子、並びにこの蛋白質をコードするDNAを含む組換えベクター及び宿主に関する。本発明はrhCEAをもつアデノウイルスベクター構築物と、癌の予防及び治療用ワクチン及び医薬組成物におけるその使用にも関する。
【背景技術】
【0002】
免疫グロブリンスーパーファミリー(IgSF)は細胞間接着等の種々の機能をもつ蛋白質をコードする多数の遺伝子から構成される。IgSF蛋白質は適正な分子間結合相互作用を維持するために重要な少なくとも1個のIg関連ドメインを含む。このような相互作用はIgSFメンバーの各種生体機能に必要であるため、多くのIgSF接着分子の破壊又は異常発現が多くのヒト疾患に関連付けられている。
【0003】
癌胎児性抗原(CEA)は細胞表面糖蛋白質から構成されるIgスーパーファミリーのサブファミリーに属する。CEAサブファミリーのメンバーはCEA関連細胞接着分子(CEACAM)として知られている。最近の科学文献では、CEA遺伝子はCEACAM5と改名されているが、蛋白質の呼称はCEAのままである。機能的には、CEACAMはホモタイプとヘテロタイプの両者の細胞間接着分子として作用することが示されている(Benchimolら,Cell 57:327−334(1989))。細胞接着に加え、CEAは細胞外マトリックスからの細胞の分離に起因する細胞死を抑制し、Bcl2やC−Myc等の所定のプロト癌遺伝子に関連する細胞形質転換に関与しているらしい(Berinstein,J.Clin Oncol.20(8):2197−2207(2002)参照)。
【0004】
胎児発生中と成人結腸粘膜ではCEAの正常発現が検出されている。CEA過剰発現は30年以上前にヒト結腸腫瘍で最初に検出され(Gold and Freedman,J.Exp.Med.121:439−462(1965))、その後、ほぼ全結腸直腸腫瘍で検出されている。更に、CEA過剰発現は高い百分率の膵臓腺癌、乳腺癌及び肺腺癌で検出可能である。これらの腫瘍種でCEA発現率が高いため、CEAはこれらの癌の管理と予後診断で広く臨床使用されている。
【0005】
CEA発現と転移増殖の相関により、CEAは結腸直腸癌治療の分子及び免疫介入のターゲットとしても認められている。CEAをターゲットとする治療アプローチの1つは抗CEA抗体の使用であり(Chesterら,Cancer Chemother.Pharmacol.46(Suppl):S8−S12(2000)参照)、別のアプローチはCEAワクチンを使用してCEAを発現する腫瘍を攻撃するように免疫系を活性化する方法である(詳細についてはBerinstein,前出参照)。
【発明の開示】
【発明が解決しようとする課題】
【0006】
ヒトCEAをコードする配列はクローニング及び特性決定されている(米国特許第5,274,087号;米国特許第5,571,710号;及び米国特許第5,843,761号。更にBeaucheminら,Mol.Cell.Biol.7:3221−3230(1987);Zimmermanら,Proc.Natl.Acad.Sci.USA 84:920−924(1987);Thompsonら Proc.Natl.Acad.Sci.USA 84(9):2965−69(1987)参照)。これらのCEA遺伝子が単離同定されているにも拘わらず、有効で自己寛容に妨害されない癌ワクチンの開発を実現できるようにCEAをコードする他の哺乳動物遺伝子を同定することが望ましい。
【課題を解決するための手段】
【0007】
本発明は配列番号2又は配列番号18に記載の新規アカゲザル癌胎児性抗原(以下、rhCEAと言う)をコードするヌクレオチド配列を含む単離又は精製核酸分子(ポリヌクレオチド)に関する。本明細書に開示するDNA分子は選択宿主細胞にトランスフェクトすることができ、組換え宿主細胞は発現される機能的rhCEA蛋白質(配列番号2及び配列番号18)の実質的レベルのソースを提供する。
【0008】
本発明は更に新規アカゲザルCEA蛋白質を発現するmRNAをコードする単離核酸分子に関し、このDNA分子は本明細書に配列番号1として開示するヌクレオチド配列を含む。アカゲザルCEAをコードするヌクレオチド配列を本明細書ではrhCEACAM5と言う。本発明のこの部分の1好適側面を図1Aに開示し、同図は新規rhCEA蛋白質(配列番号2)をコードするDNA分子(配列番号1)を示す。
【0009】
本発明の別の側面は新規アカゲザルCEA蛋白質(配列番号18)をコードする単離核酸分子であり、前記核酸分子は図1Bに示す配列番号5に記載のヌクレオチド配列を含む。
【0010】
本発明は更に本明細書全体に開示する核酸分子を含む原核及び真核両者の組換えベクター及び組換え宿主細胞に関する。
【0011】
本発明は更に組換え宿主細胞におけるアカゲザルCEA蛋白質の発現方法に関し、(a)配列番号1又は配列番号5に記載の核酸を含むベクターを適当な宿主細胞に導入する段階と;(b)前記アカゲザルCEA蛋白質の発現を可能にする条件下で宿主細胞を培養する段階を含む。
【0012】
本発明の1好適側面は図2A(配列番号2)に開示するアミノ酸配列から構成されるアカゲザルCEA蛋白質の実質精製形である。
【0013】
本発明の別の好適側面は図2B(配列番号18)に開示するアミノ酸配列から構成されるアカゲザルCEA蛋白質の実質精製形である。
【0014】
本発明の別の好適側面はrhCEA蛋白質を発現する配列番号1又は配列番号5に記載のヌクレオチド配列を含むDNA発現ベクターを含む組換え宿主細胞から得られ、実質精製され、完全にプロセシング(タンパク分解プロセシング、グリコシル化及び/又はリン酸化を含む)された成熟rhCEA蛋白質に関する。組換え宿主細胞は哺乳動物細胞株等の真核宿主細胞が特に好ましい。
【0015】
本発明の更に別の側面は単離核酸分子を含むワクチンベクターを哺乳動物に投与することを含む癌の予防又は治療方法であり、単離核酸分子は配列番号2又は配列番号18に記載のアカゲザル癌胎児性抗原(rhCEA)蛋白質をコードするヌクレオチド配列を含む。
【0016】
本発明は更に、E1領域に欠失とE1領域に挿入をもつアデノウイルスゲノムを含むアデノウイルスワクチンベクターに関し、挿入は(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含む。
【0017】
本発明は更に、プラスミド部分と発現カセット部分を含むワクチンプラスミドに関し、発現カセット部分は(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(b)ポリヌクレオチドに機能的に連結したプロモーターをを含む。
【0018】
本発明の別の側面は哺乳動物を癌から保護する方法又は癌の哺乳動物の治療方法であり、(a)i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第1のベクターを哺乳動物に導入する段階と;(b)所定時間経過させる段階と;(c)i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第2のベクターを哺乳動物に導入する段階を含む。
【0019】
本明細書と特許請求の範囲を通して使用する単数形は文脈からそうでないことが明白である場合を除き、複数も含む。
【0020】
本明細書と特許請求の範囲を通して以下の定義及び略語を適用する。
【0021】
「プロモーター」なる用語はRNAポリメラーゼが結合するDNA鎖上の認識部位を意味する。プロモーターはRNAポリメラーゼと開始複合体を形成し、転写活性を開始及び誘導する。複合体は「エンハンサー」と呼ばれる活性化配列又は「サイレンサー」と呼ばれる抑制配列により修飾することができる。
【0022】
「カセット」なる用語は発現させようとする核酸配列を含む本発明の配列を意味する。カセットはカセットテープと同様の概念であり、各カセットはそれ自体の配列をもつ。従って、カセットを交換することにより、ベクターは別の配列を発現する。5’及び3’末端の制限部位により、カセットは容易に挿入、除去又は別のカセットで置換することができる。
【0023】
「ベクター」なる用語はDNAフラグメントを宿主生物又は宿主組織に導入することができる手段を意味する。プラスミド、ウイルス(アデノウイルスを含む)、バクテリオファージ及びコスミド等の種々のベクターがある。
【0024】
アデノウイルスベクターに関して使用する「第1世代」なる用語はこのアデノウイルスベクターが複製欠損型であることを意味する。第1世代アデノウイルスベクターは一般に欠失又は不活化E1遺伝子領域をもち、欠失又は不活化E3遺伝子領域をもつことが好ましい。
【0025】
「pV1J−rhCEA」なる表記はイントロンAをもつヒトCMV極初期(IE)プロモーターと、完全長アカゲザルCEA遺伝子と、ウシ成長ホルモン由来ポリアデニル化及び転写終結配列と、最小pUCバックボーンを含む本明細書に開示するプラスミド構築物を意味する。
【0026】
「pMRK−Ad5−rhCEA」及び「MRK−rhCEA」なる表記はE1及びE3領域を欠失するAd5アデノウイルスゲノムを含む本明細書に開示する構築物を意味する。このプラスミドにおいて、E1領域はイントロンAをもたないヒトCMVプロモーターの制御下でE1と同じ向きに配置したアカゲザルCEA遺伝子と、その後に配置されたウシ成長ホルモンポリアデニル化シグナルで置換されている。
【0027】
「pBS−rhCEA」なる表記はpBluescriptII KS(+)プラスミドと完全長rhCEA遺伝子を含む本明細書に開示する構築物を意味する。
【0028】
「有効量」なる用語は免疫応答を生じるように十分なレベルのポリペプチドを生産するために十分なワクチン組成物を導入することを意味する。当業者に自明の通り、このレベルは可変である。
【0029】
「他の核酸を実質的に含まない」とは他の核酸を少なくとも90%、好ましくは95%、より好ましくは99%、更により好ましくは99.9%除去していることを意味する。「他の核酸を実質的に含まない」、「実質精製」、「単離核酸」又は「精製核酸」なる用語は交換可能に使用され、他の細胞成分を除去したアカゲザルCEA蛋白質のコーディング領域を含むDNA分子も意味する。従って、他の核酸を実質的に含まないアカゲザルCEA DNA調製物はその合計核酸の百分率として非アカゲザルCEA核酸の含有率が10%以下、好ましくは5%以下、より好ましくは1%以下、更により好ましくは0.1%以下である。所与アカゲザルCEA DNA調製物が他の核酸を実質的に含まないか否かは例えばアガロースゲル電気泳動と適当な染色法(例えば臭化エチジウム染色)の併用等の慣用核酸純度評価技術又は配列決定により判定することができる。
【0030】
「他の蛋白質を実質的に含まない」又は「実質精製」とは他の蛋白質を少なくとも90%、好ましくは95%、より好ましくは99%、更により好ましくは99.9%除去していることを意味する。従って、他の蛋白質を実質的に含まないアカゲザルCEA蛋白質調製物その合計蛋白質の百分率として非アカゲザルCEA蛋白質の含有率が10%以下、好ましくは5%以下、より好ましくは1%以下、更により好ましくは0.1%以下である。所与アカゲザルCEA蛋白質調製物が他の蛋白質を実質的に含まないか否かは例えばドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動(SDS−PAGE)と適当な検出法(例えば銀染色又はイムノブロッティング)の併用等の慣用蛋白質純度評価技術により判定することができる。
【0031】
「他の蛋白質を実質的に含まない」又は「実質精製」、又は「単離アカゲザルCEA蛋白質」又は「精製アカゲザルCEA蛋白質」なる用語は交換可能に使用され、天然ソースから単離されたアカゲザルCEA蛋白質も意味する。「単離」又は「精製」なる用語の使用はアカゲザルCEA蛋白質がその通常の細胞環境から取出されていることを示す。従って、単離アカゲザルCEA蛋白質は無細胞溶液中に存在していてもよいし、天然に存在するとは別の細胞環境に配置してもよい。単離なる用語は単離アカゲザルCEA蛋白質が唯一の存在する蛋白質であるという意味ではなく、単離rhCEA蛋白質が天然でrhCEA蛋白質とインビボ関連する他の蛋白質及び非アミノ酸物質(例えば核酸、脂質、炭水化物)を実質的に含まないという意味である。従って、原核又は真核細胞で組換え発現され、このrhCEA蛋白質を天然で(即ち介入なしには)発現しないこの宿主細胞から実質精製されたアカゲザルCEA蛋白質は当然のことながら本明細書に記載する任意状況下で「単離アカゲザルCEA蛋白質」である。上述のように、単離又は精製rhCEA蛋白質であるrhCEA蛋白質調製物は他の蛋白質を実質的に含まず、その合計蛋白質の百分率として非アカゲザルCEA蛋白質の含有率が10%以下、好ましくは5%以下、より好ましくは1%以下、更により好ましくは0.1%以下である。
【0032】
「保存アミノ酸置換」とはあるアミノ酸残基を別の化学的に類似するアミノ酸残基で置換することを意味する。このような保存置換の例は疎水性残基(イソロイシン、ロイシン、バリン、又はメチオニン)の相互置換、同一電荷の極性残基の相互置換(例えばリジン→アルギニン;アスパラギン酸→グルタミン酸)である。
【0033】
「rhCEA」とはアカゲザル癌胎児性抗原を意味する。
【0034】
「哺乳動物」なる用語はヒトを含む任意哺乳動物を意味する。
【0035】
「Ag」なる略語は抗原を意味する。
【0036】
「Ab」及び「mAb」なる略語は夫々抗体とモノクローナル抗体を意味する。
【0037】
「ORF」なる略語は遺伝子のオープンリーディングフレームを意味する。
【0038】
癌胎児性抗原(CEA)をコードする遺伝子は一般に腺癌発生に関連付けられる。本発明は異常CEA発現が癌又はその発生に関連付けられるCEA腫瘍関連抗原により発現される蛋白質産物に対する免疫を誘発又は強化するための組成物と方法に関する。異常CEA発現が腫瘍発症時に存在していても腫瘍進行後期に検出できない場合や、その逆の場合もあるので、異常CEA発現と癌の関連には、CEA蛋白質がその発生の全時点で腫瘍組織に発現される必要はない。
【0039】
この目的のために、アカゲザル癌胎児性抗原(rhCEA)をコードするポリヌクレオチドが提供される。本発明の分子は細胞性免疫により腺癌に対して有効な免疫予防を提供するために組換えアデノウイルス又はプラスミドワクチンで使用することができる。脊椎動物に直接インビボ導入する場合には、本発明のポリヌクレオチドはコードされる蛋白質の発現を霊長類、イヌ及びヒト等の哺乳動物を含む動物の体内に誘導する。
【0040】
本発明は配列番号2又は配列番号18に記載の新規rhCEA蛋白質を発現するmRNAをコードするヌクレオチド配列を含む単離核酸分子(ポリヌクレオチド)に関する。本発明の核酸分子は他の核酸を実質的に含まない。
【0041】
本発明の単離核酸分子としてはデオキシリボ核酸分子(DNA)が挙げられ、例えば1本鎖(コーディング鎖又は非コーディング鎖)でも2本鎖でもよいゲノムDNA及び相補的DNA(cDNA)や、合成1本鎖ポリヌクレオチド等の合成DNAが挙げられる。本発明の単離核酸分子はリボ核酸分子(RNA)でもよい。ほとんどのクローニング目的には、DNAが好適核酸である。
【0042】
本発明の1好適DNA分子は図2Aに示すアカゲザルCEA蛋白質(配列番号2)をコードする本明細書に配列番号1(図1A)として開示するヌクレオチド配列を含む。
【0043】
本発明の別の好適DNA分子は図2Bに配列番号18として示すアカゲザルCEA蛋白質をコードする本明細書に配列番号5(図1B)として開示するヌクレオチド配列(以下、「第2のrhCEA」DNA配列と言う)を含む。これらのrhCEA核酸分子は実施例2に詳述するようにRT−PCRにより同定した。第2のrhCEA DNA配列(配列番号5)は第1の配列と2ヌクレオチドが相違し、別のアカゲザルに由来する結腸組織からクローニングした。このDNA配列は第1のアカゲザルCEA蛋白質と2アミノ酸相違するアカゲザルCEA蛋白質をコードする。
【0044】
単離cDNAクローン、関連ベクター、宿主、組換えサブ細胞フラクション及び膜、並びに発現及び成熟形rhCEAは癌ワクチンの開発に有用である。
【0045】
本発明は新規rhCEA蛋白質を発現するmRNAをコードする配列番号1又は5の生体活性フラグメント又は突然変異体も含む。このような任意生体活性フラグメント及び/又は突然変異体はrhCEA蛋白質の医薬的性質と少なくとも実質的に類似する蛋白質又は蛋白質フラグメントをコードし、限定されないが、配列番号2又は配列番号18に記載のCEA蛋白質か挙げられる。このようなポリヌクレオチドとしては必ずしも限定されないが、ヌクレオチド置換、欠失、付加、アミノ末端短縮及びカルボキシ末端短縮が挙げられる。本発明の突然変異体は癌ワクチン開発に有用となるように真核細胞で機能的rhCEA蛋白質を発現するmRNA分子をコードする。
【0046】
本発明はrhCEA蛋白質をコードする合成DNAにも関し、合成DNAのヌクレオチド配列は配列番号1及び配列番号5のヌクレオチド配列と有意に相違するが、配列番号2又は配列番号18に記載のrhCEA蛋白質をコードする。このような合成DNAも本発明の範囲に含むものとする。
【0047】
従って、本発明は同一蛋白質を発現する多数のDNA分子を生じる可能性があるコドン冗長を開示する。本明細書の趣旨では、1個以上の置換コドンをもつ配列を縮重変異として定義する。発現される蛋白質の最終物性を実質的に変化させないDNA配列又は翻訳蛋白質の変異も本発明の範囲に含まれる。例えば、ロイシンをバリン、リジンをアルギニン、又はグルタミンをアスパラギンに置換してもポリペプチドの機能は変化し得ない。
【0048】
天然ペプチドの性質と異なる性質をもつペプチドをコードするようにペプチドをコードするDNA配列を改変できることが知られている。DNA配列の改変方法としては限定されないが、部位特異的突然変異誘発が挙げられる。改変される性質の例としては限定されないが、基質に対する酵素の親和性又はリガンドに対する受容体の親和性の改変が挙げられる。
【0049】
ストリンジェント条件下で配列番号1又は配列番号5とハイブリダイズするDNA配列も本発明に含まれる。例えば、限定されないが、高ストリンジェンシー条件を使用する方法は以下の通りである:6XSSC、5Xデンハルト溶液、及び変性サケ精子DNA100μg/mlから構成される緩衝液中でDNAを含むフィルターのプレハイプリタイゼーションを65℃で2時間〜一晩実施する。変性サケ精子DNA100μg/mlと32P標識プローブ5〜20X106cpmを含有するプレハイプリタイゼーション混合物中でフィルターを65℃で12〜48時間ハイブリダイズさせる。2XSSC、0.1%SDSを含有する溶液で37℃にて1時間フィルターの洗浄を行う。次に0.1XSSC、0.1%SDSで50℃にて45分間洗浄した後、オートラジオグラフィーにかける。高ストリンジェンシー条件を使用する他の方法としては、5XSSC、5Xデンハルト溶液、50%ホルムアミド中でハイブリダイゼーション段階を42℃で12〜48時間実施する方法や、0.2XSSPE、0.2%SDSで洗浄段階を65℃で30〜60分間実施する方法が挙げられる。
【0050】
高ストリンジェンシーハイブリダイゼーションを実施するための上記方法で記載した試薬は当分野で周知である。これらの試薬の組成の詳細は例えば参考資料として本明細書に組込むSambrookら,Molecular Cloning:A Laboratory Manual;Cold Spring Harbor Laboratory,Cold Spring Harbor,New York,1989に記載されている。上記以外に、使用可能な他の高ストリンジェンシー条件も当分野で周知である。
【0051】
本発明の1好適側面は図2Aに開示するアミノ酸配列(配列番号2)を含むアカゲザルCEA蛋白質の実質精製形である。
【0052】
本発明の別の好適側面は図2Bに開示するアミノ酸配列(配列番号18)を含むアカゲザルCEA蛋白質の実質精製形である。
【0053】
本発明はrhCEAの種々の機能的ドメインとこれらの配列の少なくとも1個を含むハイブリッド分子にも関する。CEA蛋白質はプロセシングされたリーダー配列をもつアミノ末端ドメインと疎水性カルボキシ末端ドメインを含む。CEAは更に3個のIg様内部ドメインを含む。CEAの細胞間接着機能にはN末端ドメインのサブドメインが必要であることがTaheriら(J.Biol.Chem.275(35):26935−26943(2000))により示されている。
【0054】
本発明は更に配列番号2又は配列番号18に記載のアミノ酸配列を含むrhCEA蛋白質の生体活性フラグメント及び/又は突然変異体を含み、必ずしも限定されないが、これらの突然変異が診断、治療又は予防用蛋白質又は蛋白質フラグメントを提供し、癌ワクチン開発に有用となるようなアミノ酸置換、欠失、付加、アミノ末端短縮及びカルボキシ末端短縮が挙げられる。
【0055】
本発明のアカゲザルCEA蛋白質は「成熟」蛋白質の形態でもよいし、融合蛋白質等の大型蛋白質の一部でもよい。分泌又はリーダー配列、プロ配列、精製を助長する配列(例えば多重ヒスチジン残基)、又は組換え生産中の安定性のための付加配列を含む付加アミノ酸配列を加えると有利なことが多い。
【0056】
本発明は更にrhCEA融合構築物に関し、限定されないが、種々のマーカー(限定されないが、GFP(緑色蛍光蛋白質)、MYCエピトープ、GST、及びFc)に連結したアカゲザルCEA蛋白質を発現する融合構築物が挙げられる。このような任意融合構築物を該当細胞株で発現させ、本明細書に開示するアカゲザルCEA蛋白質のモジュレーターをスクリーニングするために使用することができる。アカゲザルCEAに対する免疫応答を強化するように構築された融合構築物も含まれ、限定されないが、DOMやhsp70が挙げられる。
【0057】
本発明は更に本明細書を通して開示される実質精製核酸分子を含む組換えベクターにも関する。これらのベクターはDNA又はRNAから構成することができる。ほとんどのクローニング目的には、DNAベクターが好ましい。典型的ベクターとしてはプラスミド、改変ウイルス、バクテリオファージ、コスミド、酵母人工染色体、及びrhCEA蛋白質をコードすることができる他の形態のエピソーム又は組込みDNAが挙げられる。特定遺伝子導入又は他の用途に適したベクターを決定することは当業者が容易に実施可能である。
【0058】
組換え宿主でrhCEAを発現させるためにはrhCEA蛋白質をコードするDNAを含む発現ベクターを使用することができる。発現ベクターとしては限定されないが、クローニングベクター、改変クローニングベクター、特殊設計プラスミド又はウイルスが挙げられる。更に、所望により細菌細胞で組換えrhCEAを発現させるためには種々の細菌発現ベクターを使用することができる。更に、真菌細胞で組換えrhCEAを発現させるためには種々の真菌発現ベクターを使用することができる。更に、昆虫細胞で組換え蛋白質を発現させるためには種々の昆虫細胞発現ベクターを使用することができる。
【0059】
本発明は更に、本発明の核酸分子を含むベクターで形質転換又はトランスフェクトされた宿主細胞に関する。組換え宿主細胞は原核細胞でも真核細胞でもよく、限定されないが、大腸菌等の細菌、酵母等の真菌細胞、限定されないが、ウシ、ブタ、サル及び齧歯類に由来する細胞株等の哺乳動物細胞、並びに限定されないが、ショウジョウバエやカイコに由来する細胞株等の昆虫細胞が挙げられる。このような組換え宿主細胞はrhCEA又は生物学的に等価の形態を生産するのに適した条件下で培養することができる。
【0060】
上述のように、組換え宿主細胞でrhCEAを発現させるためにはrhCEA蛋白質をコードするDNAを含む発現ベクターを使用することができる。従って、本発明の別の側面は組換え宿主細胞におけるアカゲザルCEA蛋白質の発現方法であり、(a)配列番号1又は配列番号5に記載の核酸を含むベクターを適当な宿主細胞に導入する段階と;(b)前記アカゲザルCEA蛋白質の発現を可能にする条件下で宿主細胞を培養する段階を含む。
【0061】
宿主細胞でrhCEAの発現後、rhCEA蛋白質を回収し、活性形態のrhCEA蛋白質が得られる。数種のrhCEA蛋白質精製方法が利用可能であり、使用に適している。塩析分画、イオン交換クロマトグラフィー、サイズ排除クロマトグラフィー、ヒドロキシアパタイト吸着クロマトグラフィー及び疎水性相互作用クロマトグラフィーの各種組み合わせ又は個別適用により細胞溶解液及び抽出液から組換えrhCEA蛋白質を精製することができる。更に、全長rhCEA蛋白質又はrhCEA蛋白質のポリペプチドフラグメントに特異的なモノクローナル又はポリクローナル抗体から作製したイムノアフィニティーカラムの使用により他の細胞蛋白質から組換えrhCEA蛋白質を分離することもできる。
【0062】
本発明の核酸を融合し、ヒト細胞で蛋白質の効率的発現を可能にするように設計された配列を含む発現カセットとすることができる。カセットはプロモーターや終結配列等の関連転写及び翻訳調節配列を全長rhCEA遺伝子に機能的に連結することが好ましい。1好適態様では、プロモーターはイントロンA配列をもたないサイトメガロウイルスプロモーター(CMV)であるが、当業者に自明の通り、強力なイムノグロブリン又は他の真核遺伝子プロモーター等の多数の他の公知プロモーターも使用できる。1好適転写ターミネーターはウシ成長ホルモンターミネーターであるが、他の公知転写ターミネーターも使用できる。CMV−BGHターミネーターの組み合わせが特に好ましい。
【0063】
本発明によると、アカゲザルCEA発現カセットをベクターに挿入する。ベクターはアデノウイルスベクターが好ましいが、プロモーターに連結した直鎖DNAや、アデノ随伴ウイルス又は改変ワクチンウイルス、レトロウイルス又はレンチウイルスベクター等の他のベクターも使用できる。
【0064】
選択するベクターがアデノウイルスである場合には、ベクターは所謂第1世代アデノウイルスベクターであることが好ましい。これらのアデノウイルスベクターは非機能的E1遺伝子領域、好ましくは欠失アデノウイルスE1遺伝子領域をもつことを特徴とする。所定態様では、アデノウイルスE1遺伝子が正常に配置されている位置に発現カセットを挿入する。更に、これらのベクターは場合により非機能的又は欠失E3領域をもつ。使用するアデノウイルスゲノムはE1及びE3領域の両者を欠失していることが好ましい(ΔE1ΔE3)。アデノウイルスはウイルスE1遺伝子を発現する公知細胞株(例えば293細胞、又はERC.6細胞)又は別の蛋白質を発現するように一過的又もしくは安定的に軽質転換された293もしくPERC.6細胞に由来する細胞株で複製することができる。例えば、テトラサイクリン調節型プロモーターシステムのように制御下の遺伝子発現を可能にする構築物を使用する場合には、細胞株は調節システムに関与する成分を発現することができる。このような細胞株の1例はT−Rex−293であり、他の例も当分野で公知である。
【0065】
アデノウイルスベクターを操作し易くするために、アデノウイルスをシャトルプラスミド形とすることができる。本発明はプラスミド部分とアデノウイルス部分を含むシャトルプラスミドにも関し、アデノウイルス部分はE1を欠失し、場合によりE3も欠失しており、アカゲザルCEAを含む発現カセットを挿入したアデノウイルスゲノムを含む。好適態様では、アデノウイルスベクターを容易に除去できるようにプラスミドのアデノウイルス部分の両側に制限部位を配置する。シャトルプラスミドは原核細胞又は真核細胞で複製することができる。
【0066】
本発明の1好適態様では、発現カセットをpMRKAd5−HVOアデノウイルスプラスミドに挿入する(参照により本明細書に組込むEminiら,WO02/22080参照)。このプラスミドはE1領域とE3領域を欠失するAd5アデノウイルスゲノムを含む。pMRKAd5−HVOプラスミドの設計はウイルスパッケージングの最適化に重要であることが分かっているエレメントを組込むように5’シス作用性パッケージング領域を更にE1遺伝子まで延ばし、ウイルス増幅を強化することにより従来のアデノベクターを改善している。この強化型アデノウイルスベクターは高度の継代増殖後に遺伝的安定性を維持できるという利点がある。
【0067】
DNA構築物を作製及び精製するための標準分子生物学技術により、本発明のアデノウイルス、シャトルプラスミド及びDNA免疫原を作製することができる。
【0068】
上記ベクターは異常CEA発現に関連する腺癌発生の予防及び/又は既存癌の治療のための免疫原組成物及びワクチンで使用することができる。この点で、本発明の1側面は単離核酸分子を含むワクチンベクターを哺乳動物に投与することを含む癌の予防又は治療方法であり、単離核酸分子は配列番号2又は配列番号18に記載のアカゲザルCEA蛋白質をコードするヌクレオチド配列を含む。
【0069】
上記方法によると、任意哺乳動物の癌を治療又は予防するためにワクチンベクターを投与することができる。本発明の1好適態様では、哺乳動物はヒトである。
【0070】
更に、当業者は上記治療及び予防方法で使用するために任意の型のベクターを選択することができる。ベクターはアデノウイルスベクター又はプラスミドベクターが好ましい。本発明の1好適態様では、ベクターはアデノウイルスE1領域に欠失とアデノウイルスE1領域に挿入をもつアデノウイルスゲノムを含むアデノウイルスベクターであり、挿入は(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含む。
【0071】
本発明は更にE1領域に欠失とE1領域に挿入をもつアデノウイルスゲノムを含むアデノウイルスワクチンベクターに関し、挿入は(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含む。
【0072】
本発明のこの側面の1好適態様において、アデノウイルスベクターはAd5ベクターである。
【0073】
本発明の別の態様において、アデノウイルスベクターはAd6ベクターである。
【0074】
別の側面において、本発明はプラスミド部分と発現カセット部分を含むワクチンプラスミドに関し、発現カセット部分は(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(b)ポリヌクレオチドに機能的に連結したプロモーターを含む。
【0075】
本発明の所定態様において、本明細書に開示する組換えアデノウイルスワクチンは強化免疫応答を誘導するためにプラスミド型ポリヌクレオチドワクチンと各種プライム/ブーストの組み合わせで使用される。この場合、2種のベクターを「プライム及びブースト」レジメンで投与する。例えば、第1の型のベクターを投与した後に所定時間、例えば1カ月、2カ月、6カ月、又は他の適当な間隔後に第2の型のベクターを投与する。ベクターは同一ポリヌクレオチド又はポリヌクレオチド組み合わせをコードする発現カセットをもつことが好ましい。プラスミドDNAも使用する態様では、ベクターは哺乳動物又は昆虫細胞により認識される1種以上のプロモーターを含むことが好ましい。1好適態様では、プラスミドは限定されないが、CMVプロモーター等の強力プロモーターを含む。発現させるアカゲザルCEA遺伝子又は他の遺伝子をこのようなプロモーターと連結する。このようなプラスミドの1例は文献に記載されているような哺乳動物発現プラスミドV1Jnsである(参照により本明細書に組込むJ.Shiverら,DNA Vaccines,M.Liuら編,N.Y.Acad.Sci.,N.Y.,772:198−208(1996))。
【0076】
上述のように、アデノウイルスベクターワクチン及びプラスミドは免疫応答を誘導するために単独治療レジメンの一部として脊椎動物に投与することができる。この点で、本発明は哺乳動物を癌から保護する方法に関し、(a)i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第1のベクターを哺乳動物に導入する段階と;(b)所定時間経過させる段階と;(c)i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第2のベクターを哺乳動物に導入する段階を含む。
【0077】
上記保護方法の1態様において、第1のベクターはプラスミドであり、第2のベクターはアデノウイルスベクターである。代替態様において、第1のベクターはアデノウイルスベクターであり、第2のベクターはプラスミドである。
【0078】
本発明は更に腺癌の哺乳動物の治療方法に関し、(a)i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第1のベクターを哺乳動物に導入する段階と;(b)所定時間経過させる段階と;(c)i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第2のベクターを哺乳動物に導入する段階を含む。
【0079】
上記治療方法の1態様において、第1のベクターはプラスミドであり、第2のベクターはアデノウイルスベクターである。代替態様において、第1のベクターはアデノウイルスベクターであり、第2のベクターはプラスミドである。
【0080】
ワクチンレシピエントに導入される発現可能なDNA又は転写RNAの量は使用されるプロモーターの強度と発現される遺伝子産物の免疫原性に部分的に依存する。一般に、プラスミドワクチンベクター約1ng〜100mg、好ましくは約10μg〜300μgの免疫又は予防有効用量をマウス組織に直接投与する。組換えアデノウイルスの有効用量は粒子約106〜1012個、好ましくは約107〜1011個である。皮下注射、皮内導入、経皮圧入、及び他の投与方法(例えば腹腔内、静脈内、又は吸入送達)も考えられる。ブースターワクチン接種を行うことも考えられる。本発明のワクチンの非経口導入と同時又はその後にインターロイキン12蛋白質等のアジュバントを静脈内、筋肉内、皮下又は他の投与手段等で非経口投与しても有利である。
【0081】
本発明のワクチンベクターは裸のままでもよく、即ち蛋白質、アジュバント又はレシピエントの免疫系に影響を与える他の物質を伴っていなくてもよい。この場合に、ワクチンベクターを生理的に許容可能な溶液とすることが望ましく、限定されないが、滅菌食塩水又は滅菌緩衝食塩水が挙げられる。あるいは、アジュバント、サイトカイン、蛋白質、又は他のキャリヤー等の免疫刺激剤を本発明のワクチン又は免疫原組成物と共に投与すると有利である。従って、本発明は本発明の組成物及び方法とこのような免疫刺激剤の併用を含む。本明細書で使用する免疫刺激剤とは外来抗原に対する(抗体及び/又は細胞性)免疫応答を強化又は増強するほぼ任意物質を意味する。前記免疫刺激剤はDNA又は蛋白質の形態で投与することができる。種々の免疫刺激剤の任意のものを本発明のワクチン及び免疫原組成物と併用することができ、限定されないが、GM−CSF、IFNα、破傷風トキソイド、IL12、B7.1、LFA−3及びICAM−1が挙げられる。前記免疫刺激剤は当分野で周知である。DNAの細胞取込みを助長する物質も使用することができ、限定されないが、カルシウムイオンが挙げられる。これらの物質を一般にトランスフェクション助長剤及び医薬的に許容可能なキャリヤーと言う。当業者は特定の免疫刺激剤又は医薬的に許容可能なキャリヤーと許容可能な投与時間及び投与方法を決定することができよう。
【0082】
rhCEAをクローニングするためには種々の方法の任意のものを使用することができる。これらの方法としては限定されないが、以下の方法が挙げられる。(1)RACE PCRクローニング法(Frohmanら,Proc.Natl.Acad.Sci.USA 85:8998−9002(1988))。5’及び/又は3’RACEを実施して全長cDNA配列を作製することができる。このストラテジーはrhCEAcDNAのPCR増幅に遺伝子特異的オリゴヌクレオチドプライマーを使用する。これらの遺伝子特異的プライマーは多数の公共入手可能な核酸及び蛋白質データベースを検索することにより同定された発現配列タグ(EST)ヌクレオチド配列の同定により設計される;(2)適当な発現ベクターシステムでrhCEAを含むcDNAライブラリーの構築後にrhCEAcDNAの直接機能的発現;(3)バクテリオファージ又はプラスミドシャトルベクターで構築したrhCEAを含むcDNAライブラリーをrhCEA蛋白質のアミノ酸配列から設計した標識縮重オリゴヌクレオチドプローブでスクリーニング;(4)バクテリオファージ又はプラスミドシャトルベクターで構築したrhCEAを含むcDNAライブラリーを、rhCEA蛋白質をコードする部分cDNA蛋白質でスクリーニング。この部分cDNAはrhCEA蛋白質に関連する他の膜蛋白質について分かっているアミノ酸配列から縮重オリゴヌクレオチドプライマーを設計することによりrhCEA DNAフラグメントの特異的PCR増幅により得られる;(5)バクテリオファージ又はプラスミドシャトルベクターで構築したrhCEAを含むcDNAライブラリーを、哺乳動物rhCEA蛋白質と相同性をもつ部分cDNA又はオリゴヌクレオチドでスクリーニング。このストラテジーでは上記のようにESTとして同定されたrhCEAcDNAのPCR増幅に遺伝子特異的オリゴヌクレオチドプライマーを使用してもよい;あるいは(6)配列番号1を鋳型として5’及び3’遺伝子特異的オリゴヌクレオチドを設計し、公知RACE法により全長cDNAを作製できるようにするか、又はrhCEAをコードするヌクレオチド配列の全長形を単離するためにコーディング領域の一部を作製及び単離して多種のcDNA及び/又はゲノムライブラリーのうちの1個をスクリーニングするためのプローブとして使用するようにこれらの公知同一RACE法によりコーディング領域の一部を作製できるようにする。
【0083】
当業者に自明の通り、他の型のライブラリーや、他の細胞型又は種型から構築したライブラリーもrhCEAをコードするDNA又はrhCEAホモログを単離するのに有用である。他の型のライブラリーとしては限定されないが、他の細胞に由来するcDNAライブラリーが挙げられる。rhCEAをコードするcDNAを単離するためにcDNAライブラリーを構築するのに使用される細胞又は細胞株の選択はまずこのような目的に利用可能な任意公知アッセイを使用して細胞関連rhCEA活性を測定することにより実施することができる。
【0084】
cDNAライブラリーの構築は当業者に周知の標準技術により実施することができる。周知cDNAライブラリー構築技術は例えばSambrookら,Molecular Cloning:A Laboratory Manual;Cold Spring Harbor Laboratory,Cold Spring Harbor,New York,1989に記載されている。相補的DNAライブラリーは多数の商業ソースからも入手でき、限定されないが、Clontech Laboratories,Inc.(Palo Alto,CA)やStratagene(La Jolla,CA)が挙げられる。
【0085】
本発明のDNA分子、RNA分子、及び組換え蛋白質はrhCEAのレベルをスクリーニング及び測定するために使用することができる。組換え蛋白質、DNA分子、及びRNAはrhCEAの検出とタイピングに適したキットの構築に有用である。このようなキットは少なくとも1個の容器を緊密に拘束保持するのに適した区画化キャリヤーを含む。キャリヤーは更にrhCEAを検出するのに適した組換えrhCEA又は抗rhCEA抗体等の試薬を含む。キャリヤーは更に標識抗原又は酵素基質等の検出手段を加えることができる。
【0086】
本明細書に引用する全刊行物は本発明に関連して使用可能な方法及び材料を記載及び開示する目的で参照により組込む。本明細書の如何なる記載も本発明が先発明により前記開示以前の日付を享受する資格がないと認めるものであると解釈すべきではない。
【0087】
以上、添付図面を参照して本発明の好適態様を記載したが、当然のことながら本発明はこれらの厳密な態様に限定されず、当業者は特許請求の範囲に記載する本発明の範囲又は精神から逸脱せずに種々の変更及び変形を行うことができる。
【0088】
以下、実施例により本発明を例示するが、これらの実施例により発明を限定するものではない。
【実施例1】
【0089】
アカゲザルからのRNAの単離
分子操作は当分野で周知の標準操作に従って実施した(例えば、参照により本明細書に組込むAusubelら.Short Protocols in Molecular Biology,F.M.,第2版,John Wiley & Sons,(1992)及びSambrookら,Molecular Cloning,A Laboratory Manual,第2版,Cold Spring Harbor Laboratory Press(1989)参照)。
【0090】
アカゲザルCEAcDNAの単離用RNAを得るために、2頭の別個のアカゲザル(Macaca Mulatta)に由来する結腸試料を使用した。Biomedical Primate Research Center(BPRC,Rijswijk,オランダ)から凍結組織を入手した。アカゲザル結腸試料から全RNAを抽出するために、組織を機械的に粉砕し、Ultraspec RNA試薬(Biotecx Laboratories;Houston,TX)を製造業者の指示に従って添加した。精製RNAの完全性をホルムアルデヒド変性アガロースゲルにより確認した。試料を分画し、−80℃で保存した。
【実施例2】
【0091】
アカゲザルCEAcDNA増幅
ヒトCEAファミリーの全公知メンバーの5’及び3’非翻訳領域(UTR)に由来するヌクレオチド配列を整列させ、CEA DNAの高度保存領域を同定した(図3参照)。同定したCEA遺伝子ファミリー相同性に基づき、縮重オリゴヌクレオチドプライマーを設計し、アカゲザルCEAcDNAを以下に記載するように逆転写酵素ポリメラーゼ連鎖反応(RT−PCR)により増幅するようにPCR条件を最適化した。完全cDNAを増幅するために使用したプライマーは5’−RhCEA EcoRI 5’−CCGAATTCCGGACASAGCAGRCAGCAGRSACC−3’(配列番号3)及びCEA−8 RhXhoI 5’−CCGCTCGAGCGGCTGCTACATCAGAGCAACCCCAACC−3’(配列番号4)とした。SuperScript One−Step RT−PCR with Platinum Taqキット(インビトロgen;Carlsbad,CA)を使用して増幅を実施した。RNA1μg、両者プライマー200pmol、及び10% DMSO(終濃度)からなる反応容量100μlを使用した。
【0092】
逆転写段階を実施するために、2頭のアカゲザルの各々から単離した全RNA試料を45℃で30分間インキュベートした後、2分間94℃でインキュベートした。94℃15秒、52℃30秒、及び68℃2分20秒を1サイクルとし、得られた鋳型のPCR増幅を40サイクル実施した。
【0093】
CEACAM−5ホモログの予想寸法である約2100bpの増幅PCR産物を両者RNA試料から個々に獲得し、アガロースゲルから精製した。両者PCR産物の部分配列分析の結果、ヒトCEACAM−5との高い相同性が判明した。
【0094】
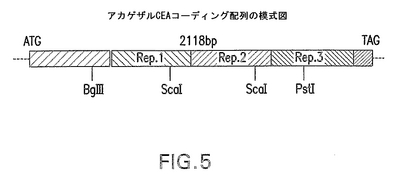
内部反復配列の高い相同性により、図5に示す制限部位を使用してDNAフラグメントを精製することにより完全遺伝子配列が得られた。各サルから得られたアカゲザルCEAヌクレオチド配列を本明細書の図1に開示し、配列番号1(以下、rhCEACAM−5と言う)及び配列番号5(以下、rhCEACAM−5#2と言う)に示す。CEAヌクレオチド配列分析の結果、705アミノ酸ポリペプチドをコードする2118ヌクレオチドのオープンリーディングフレーム(ORF)が判明した。2頭のアカゲザルから得られたrhCEAヌクレオチド配列を比較すると、2種の異なる蛋白質(図2A及び2B参照)をコードする2ヌクレオチド変異があることが判明した(図1A及び1B参照)。
【0095】
アカゲザルCEACAM−5ヌクレオチド配列(配列番号1)を更に公開ヒトCEACAM−5配列(配列番号6)に比較した処、ヌクレオチドレベルで88%の相同度が判明した(図6参照)。同様にアカゲザル(配列番号2)とヒト(配列番号7)のCEAポリペプチド配列を比較した処、アミノ酸レベルで78.9%の一致度が判明した(図7参照)。興味深いことに、アカゲザルCEAのカルボキシル末端にはヒトCEAに比較して3アミノ酸挿入が存在しており、恐らくグリコシルホスファチジルイノシトール(GPI)変異のシグナルに関係があると予想される。
【実施例3】
【0096】
λアカゲザルCEA特異的ライブラリーの作製とスクリーニング
RT−PCRにより得られた増幅rhCEA産物(実施例2参照)をEcoRI/XhoIで消化し、製造業者の指示に従ってZAP−CMV XRベクター(Stratagene;La Jolla,CA)にライゲートした。ライゲーション産物をGigapack III gold packaging extractと共にインキュベートし、得られたファージをXL−1 Blue MRF’細胞に感染させた。このCEA特異的一次ライブラリーを次に増幅し、〜1×106pfu/mlの力価を得た。ナイロンフィルターに転写することによりプラーク〜5×103個のスクリーニングを実施した。CEA分子の5’及び3’末端に対応する2種の異なるDNAプローブとフィルターをハイブリダイズさせた。二重陽性プラークをXL−1 Blue MRF’細胞中で切り出し、誘導された繊維状ファージをXL−OLR細胞中で増幅した。次にファージミドを増殖させ、制限消化により分析した。配列分析とGenbank比較の結果、ヒトCEACAM−5と最高の相同度であることが判明した。
【実施例4】
【0097】
プラスミド構築物とアデノウイルス作製
pCMV−script EXファージミドベクターからPstI/XhoIでRhCEAを切り出し、pBluescript II KSベクターに挿入し、pBS−RhCEAを得た。挿入部分を完全に配列決定した後、SmaI/XhoIフラグメントとしてpVIJnsAベクターにサブクローニングし、pVIJ−RhCEAを得た。同一フラグメントをpolyMRKベクターにサブクローニングすることによりアデノウイルス作製用シャトルプラスミドpMRK−RhCEAを得た。RhCEA及びE1フランキングAd5領域用発現カセットを含むpMRK−RhCEAからのPacI/StuIフラグメントをBJ5183大腸菌細胞でClaIにより直鎖化したpAd5又はpAd6に組換えた。得られたプラスミドをpAd5−RhCEA及びpAd6−RhCEAとした。両者プラスミドをPacIで切断し、アデノウイルスITRを遊離させ、PerC−6細胞にトランスフェクトした。連続継代によりウイルス増幅を実施した。標準CsCl精製プロトコールを使用してAd5−RhCEA及びAd6−RhCEAを精製し、A105緩衝液(5mM Tris pH8.0,1mM MgCl2,75mM NaCl,5%スクロース,0.005% Tween20)で十分に透析した。
【実施例5】
【0098】
インビトロ RhCEA発現及び検出
作製したベクターによるRhCEAの発現をウェスタンブロットとFACS分析により確認した。Lipofectamine 2000(Life Technologies;Carlsbad,CA)を使用してプラスミドをHeLa又はPerC.6細胞にトランスフェクトした。無血清培地で30分間37℃にてアデノウイルス感染を行った後に新鮮な培地を加えた。48時間インキュベーション後にヒトCEAに対するウサギポリクローナル血清(Fitzgerald,1500倍希釈液)を使用して全細胞溶解液をウェスタンブロットにより分析した。選択した全アカゲザルCEAクローンはHeLa細胞にトランスフェクトした場合に180〜200KDa蛋白質を発現した(図4参照)。
【0099】
FACS分析のために、細胞をトリプシンで剥離し、FACS緩衝液(PBS,1%FCS)に再懸濁した。250倍に希釈したウサギポリクローナル抗CEA抗体と共に30分間インキュベーション後に細胞を洗浄し、抗ウサギIgG−PEと共に30分間インキュベートし、最後にFACScalibur(Becton Dickinson,San Jose,CA)で分析した。
【実施例6】
【0100】
ペプチド
免疫した動物でアカゲザルCEAに対する細胞性免疫応答を分析するために、完全蛋白質に対応するように11アミノ酸がオーバーラップする15量体ペプチドを設計した。凍結乾燥アカゲザルCEAペプチドをBio−Synthesis,Inc.(Lewisville,TX)から購入し、DMSOに40mg/mlで再懸濁した。ペプチドをプールA(RhCEA−1〜RhCEA−34,34ペプチド);プールB(RhCEA−35〜RhCEA−79,45ペプチド);プールC(RhCEA−80〜RhCEA−124,48ペプチド);及びプールD(RhCEA−125〜RhCEA−173,53ペプチド)の4個のプールに分けた。終濃度はA=1.176mg/ml;プールB=0.888mg/ml;プールC=0.851mg/ml;プールD=0.769mg/mlとした。ペプチドとプールを−80℃で保存した。
【実施例7】
【0101】
rhCEA免疫によるマウスにおけるCEA特異的細胞性免疫応答の発生
CEA.Tgマウスはヒトに類似する組織分布でヒトCEAを自己抗原として発現するトランスジェニックマウスである。科学文献に広く実証されているように、これらのマウスは検出可能なCEA特異的血清抗体が存在せず、CEAに対するインビトロ脾臓T−cell応答を発生することができないことからCEA非応答性である。同種抗原をコードする異種遺伝子によるDNA免疫が同系メラノーマ細胞による腫瘍攻撃からマウスを保護することは多くの報告に示されている。異種DNAワクチン接種によりこのモデルで自己抗原に対する免疫応答を誘発できることを立証するために、CEA.アカゲザルCEA(異種)をコードするベクターでTgマウスを免疫した。
【0102】
C57BL/6マウス(H−2b)はCharles River(Lecco,イタリー)から購入した。CEA.tgマウス(H−2b)はHL Kaufman(Albert Einstein College of Medicine,New York)から入手し、標準条件下で飼育した。
【0103】
電気的遺伝子導入(EGT)のために、マウス大腿四頭筋にpVIJ−RhCEA50μgを外科的に接触させるか又は直接注入し、従来記載されているように電気刺激した(Rizzutoら.Proc.Natl.Acad.Sci.U.S.A.96(11):6417−22(1999))。アデノウイルス注入のために、Ad5−RhCEA 1×1010vpをマウス大腿四頭筋に注入した。
【0104】
マウスの大腿四頭筋にpVIJ−RhCEA50μgを注入し、注入直後に週1回ずつ4週間電気刺激した。C57BL/6マウスを対照として使用した。アカゲザルCEAに対する抗体がこれらのマウスに由来する血清中でウェスタンブロットにより検出され、体液性免疫応答が立証された。hCEAに対するマウスモノクローナルAbを陽性対照として使用し、免疫前血清とモック感染細胞抽出液を陰性対照として使用した(データは示さず)。重要な点として、ヒトCEA蛋白質に対する交差反応性抗体はアカゲザルCEA免疫群でしか測定することができず(図8)、平均力価は1:110であった。これらのデータによると、トランスジェニックマウスモデルでは異種DNAワクチン接種により寛容を解除することが可能である(抗CEA自己抗体として測定)。
【実施例8】
【0105】
抗体検出及び力価測定
眼窩後方採血により抗体力価測定用血清を採取した。ウェスタンブロット検出のために、Ad5−rhCEAを導入したHeLa細胞からの抽出液をSDS−pageゲルで泳動させ、ニトロセルロースフィルターに転写した。血清をプールし、4℃でのO/Nインキュベーション用に50倍に希釈した。抗マウスIgG−APコンジュゲート(Sigma,1:2500)を検出用に使用した。力価測定のために、コーティング用緩衝液(50mM NaHCO3 pH 9.4)で希釈したCEA(高純度CEA;Fitzgerald Industries International Inc.,Concord MA)100ng/ウェルでElisaプレート(Nunc maxisorp)をコーティングし、4℃でO/Nインキュベートした。次に5%BSAを添加したPBSでプレートを1時間37℃でブロックした。マウス血清をPBS5%BSAで希釈した(血清変換の評価には50倍希釈;力価値の評価には10倍〜31,250倍希釈)。免疫前血清をバックグラウンドとして使用した。希釈した血清を4℃でO/Nインキュベートした。PBS,1% BSA,0.05% tween 20で洗浄を行った。検出抗体(ヤギ抗マウスIgGペルオキシダーゼ,Sigma,St.Louis,MO)をPBS,5% BSAで2000倍に希釈し、シェーカーで2〜3時間室温にてインキュベートした。洗浄後、プレートをTMB基質(Pierce Biotechnology,Inc.,Rockford,IL)100μl/ウェルで展開した。1M H2SO4溶液25μl/ウェルで反応を停止し、プレートを450nm/620nmで読み取った。同一希釈倍率の自己免疫前血清の吸光度の少なくとも3倍の吸光度を生じる血清の限界希釈倍率として抗CEA血清力価を計算した。
【実施例9】
【0106】
IFN−γELISPOTアッセイ
96ウェルMAIPプレート(Millipore,Bedford,MA)に滅菌PBS中2.5μl/mlの精製ラット抗マウスIFN−γ(IgG1,クローンR4−6A2,Pharmingen,San Diego,CA)を100μl/ウェルの割合でコーティングした。滅菌PBSで洗浄後、プレートをR10培地200μl/ウェルで37℃にて少なくとも2時間ブロックした。
【0107】
脾細胞作製のために、屠殺したマウスから脾臓を滅菌摘出し、グリッドで細断した。0.1×PBS1mlを細胞ペレットに加え、15秒以内ボルテックスすることにより赤血球の浸透圧溶解を行った。次に2×PBS1mlを加え、PBS1×で容量を4mlにした。1200rpmで10分間室温にて遠心後、細胞ペレットをR10培地1mlに再懸濁し、生存細胞をカウントした。脾細胞を5×105個及び2×105個/ウェルの割合でプレーティングし、R10中各ペプチド1μg/mlを加え、20時間37℃のCO2インキュベーターでインキュベートした。コンカナバリンA(ConA)5μg/mlを各マウスで陽性内部対照として使用した。PBS,0.05% Tween 20で洗浄後、アッセイ緩衝液(PBS−5%FBS−0.005% Tween−20)で250倍に希釈したビオチン標識ラット抗マウスIFN−γ(Rat IgGl,クローンXMG1.2,Pharmingen,San Jose,CA)50μl/ウェルと共にプレートを4℃でO/Nインキュベートした。
【0108】
翌日、プレートを洗浄し、アッセイ緩衝液で2500倍に希釈したストレプトアビジン−APコンジュゲート(Pharmingen)と共に2時間室温でインキュベートした。十分に洗浄後、スポット展開が顕微鏡で観察されるまでNBT/B−CIP(Pierce Biotechnology)50μl/ウェルを加えることによりプレートを展開した。プレートを蒸留水で十分に洗浄することにより反応を停止した。プレートを完全に風乾し、自動ELISPOTリーダーを使用してスポットをカウントした。
【0109】
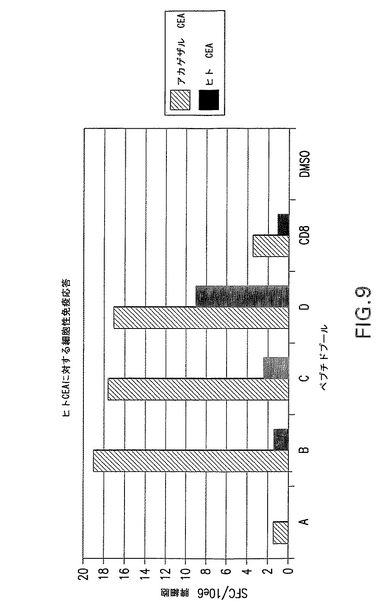
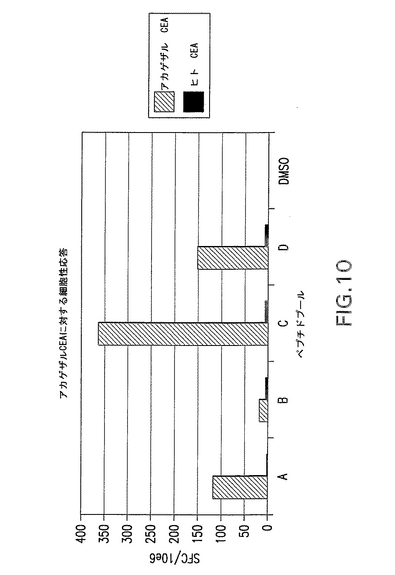
細胞性免疫応答のために、CEA.TgマウスにhCEA発現ベクター又はrhCEA発現ベクターをワクチン接種した。2群を分析し、第1群は最終DNA注入から21日後にELISPOTアッセイにより分析し、第2群はAd5−hCEA又はAd5−RhCEA 1×1010vpをブースター投与し、2週間後に分析した。その結果、ELISPOTにより測定した場合に4回のDNA注入後にhCEAに対する有意細胞性免疫応答は観察されなかった(図示せず)。他方、Ad5をブースター投与したマウスは顕著な応答増加を示し、CEAに対する免疫寛容の解除に一致した。この所見は、CEA自己抗原に有用な1つのワクチン接種プロトコールがEGTによるDNAの反復投与後にアデノウイルスブーストを行う方法(混合法)であることを示唆している。重要な点として、野生型マウスとトランスジェニックマウスのいずれにおいてもアカゲザルCEAの免疫はヒトCEAペプチドとの交差反応を生じ、逆の場合にも交差反応を生じた(データは示さず)。特に、ヒトCEAに対する免疫応答はrhCEAを免疫原として使用したトランスジェニックマウスのほうが著しく良好であった(図9参照)。これらの結果によると、アカゲザル(異種)遺伝子を使用してトランスジェニックマウスでCEAに対する良好な応答を得ることができる。アカゲザルCEAペプチドに対する応答を図10に示す。
【実施例10】
【0110】
rhCEAによるアカゲザルの免疫
結腸直腸腺癌で発現されるヒト腫瘍抗原CEAのアカゲザルホモログによるアカゲザル(macaca mulatta)の免疫効率を評価するために、Biomedical Primate Research Centre(BPRC,Rijswijk,オランダ)で免疫試験を実施した。この免疫試験はアカゲザルCEA抗原による免疫に対するB及びT細胞両者の応答を評価するように設計した。
【0111】
この試験(CV−1)では、アカゲザルCEACAM−5を発現するプラスミドDNAベクター及びアデノウイルスベクターで1群のサル(雄2頭と雌2頭)を免疫した。プライミングのために、rhCEAを発現するDNAを0、4、8、12、及び16週にDNA注入後に電気刺激により動物に筋肉内ワクチン接種した。DNA注入はプラスミドDNA5mgを含有する1ml溶液(2部位に分けて各部位0.5mlずつ)を体重2〜5キロの動物に注入した。麻酔下に動物に注入した(ケタミン/キシラジン混合物)。
【0112】
電気刺激については、方形バイポーラパルス100バルス2系列(各1秒)を1秒間隔で発生し、合計3秒間処理した。パルス長は2msec/位相とし、パルス周波数及び振幅は夫々100Hz及び100mA(定電流モード)とした。
【0113】
上記免疫プロトコールを使用してCEAに対する免疫応答を測定するために、血液試料を4週間おきに採取した。細胞性応答はIFNγ Elispotアッセイにより測定し、体液性応答はELISAアッセイにより測定した。16週で有意免疫応答が得られなかったので、rhCEAを発現するAd5を使用して更に2回注入した(24週及び28週)。Ad5注入後に夫々ペプチドプールC及びプールB+Cに対応する2頭のサル(RI137及びCO12)でrhCEAに対する測定可能な免疫応答が検出された。細胞性免疫応答は両者サルで35週目に低下し始めた。
【0114】
DNA注入後に体液性免疫応答を経時的に追跡した。3頭のサル(CO12,RI311及びRI002)は1:143〜1:2099の良好な抗CEA抗体力価を示し、最初の注入から12〜16週間後にピークに達した。
【0115】
これらのデータによると、rhCEAをコードする遺伝子ベクターは霊長類でこの腫瘍抗原に対する免疫寛容を解除することができた。免疫応答には細胞性(処理したサルの50%)と体液性(処理したサルの75%)のどちらの免疫も検出された。
【実施例11】
【0116】
ヒト腫瘍関連抗原のアカゲザルホモログによるアカゲザルの免疫
いずれも結腸直腸腺癌で発現されるヒト腫瘍抗原HER2/neu、Ep−CAM及びCEAのアカゲザルホモログによるアカゲザル(macaca mulatta)の免疫効率を評価するために、第2系列の免疫試験を実施した。プロトコールはこれらの腫瘍抗原の組み合わせに対するB及びT細胞両者の応答を評価するように設計した。
【0117】
この試験では、ヒト腫瘍抗原Ep−CAM(pV1J−rhEpCAM)、CEA(pV1J−rhCEA)、及びHER2/neu(pV1J−rhHER2)のアカゲザルホモログを発現する3種のプラスミドDNAベクターの混合物で第2群のアカゲザル4頭(雄2頭と雌2頭)を免疫した。
【0118】
0、4、8、12、及び16週にプラスミドDNAの筋肉内注射後に電気刺激により動物をプライミングした。DNA注入は体重2〜5キロの動物にプラスミドDNA6mgを含有する1ml溶液(2部位に分けて各部位0.5mlずつ)を注入した。麻酔下に動物に注入した(ケタミン/キシラジン混合物)。
【0119】
電気刺激については、方形バイポーラパルス100バルス2系列(各1秒)を1秒間隔で発生し、合計3秒間処理した。パルス長は2msec/位相とし、パルス周波数及び振幅は夫々100Hz及び100mA(定電流モード)とした。
【0120】
アカゲザルCEA(Ad5−rhCEA)、アカゲザルHER2/neu(Ad5−rhHER2)、及びアカゲザルEpCAM(Ad5−rhEpCAM)を発現する3種のAd5の混合物の注入により同一群の動物をブーストした。合計量3×10exp11ウイルス粒子(vp)を23週と27週に注入した(3種のウイルスを各々1×10exp11 vp)。
【0121】
上記免疫プロトコールを使用して3種の腫瘍抗原に対する免疫応答を測定するために、血液試料を4週間おきに採取した。細胞性免疫応答はIFN−γ+ELISPOTアッセイにより測定し、体液性応答はELISAにより測定した。
【0122】
サルRI449及びRI519はIFN−γ ELISPOT分析により測定した場合に検出可能なHER2特異的細胞性応答を示した。同様の分析でrhCEA及びrhEpCAMに対する有意応答は検出されなかった。
【0123】
第3の試験では、0、2及び4週にAd5誘導体の筋肉内注射によりAd5−rhHER2、Ad5−rhCEA及びAd5−rhEpCAMの混合物でアカゲザル4頭を免疫した。3×10exp11 vp(3種のAd5ウイルスを各々10exp11)を含有する1ml溶液(2部位に分けて各部位0.5mlずつ)を体重2〜5キロの動物に麻酔下(ケタミン/キシラジン混合物)に投与した。
【0124】
細胞性応答はIFNγ ELISPOTアッセイにより測定した。Her2/Neuでは、4頭のサルのうちの3頭が検出可能な応答を示した。rhCEAとrhEpCAMでは有意細胞性応答は測定されなかった。
【0125】
まとめると、上記免疫プロトコールはアカゲザルでrhHER2/neuに対する特異的免疫応答を誘導するのに有効であった。3種の異なる腫瘍抗原をもつベクターによる同時免疫がrhCEAのみを免疫原として使用した試験1に比較してrhCEAに対する免疫応答を誘導するのに有効でない理由は不明である。理論に拘束する意図はないが、rhHER2/Neuの発現と免疫ドミナントエピトープの存在がサブドミナントrhCEA特異的T細胞の生産と発現を制限したのではないかと考えられる。
【図面の簡単な説明】
【0126】
【図1A】配列番号1(図1A)及び配列番号5(図1B)に記載のアカゲザルCEAcDNA分子のヌクレオチド配列を示す。実施例2参照。
【図1B】配列番号1(図1A)及び配列番号5(図1B)に記載のアカゲザルCEAcDNA分子のヌクレオチド配列を示す。実施例2参照。
【図2A】配列番号2(図2A)に記載の第1のアカゲザルCEA蛋白質と配列番号18(図2B)に記載の第2のアカゲザルCEA蛋白質の予想アミノ酸配列を示す。第1及び第2のアカゲザルCEA蛋白質間の2アミノ酸変異を図2Bに太字下線で示す。
【図2B】配列番号2(図2A)に記載の第1のアカゲザルCEA蛋白質と配列番号18(図2B)に記載の第2のアカゲザルCEA蛋白質の予想アミノ酸配列を示す。第1及び第2のアカゲザルCEA蛋白質間の2アミノ酸変異を図2Bに太字下線で示す。
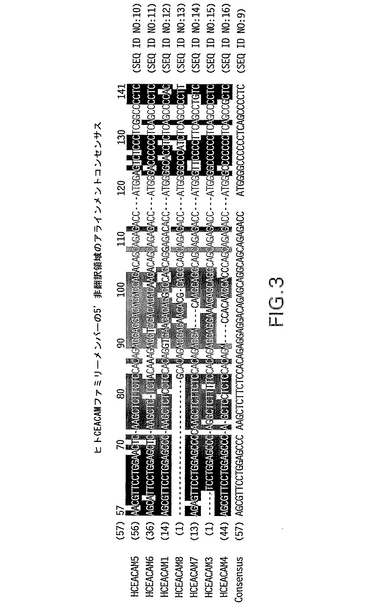
【図3】ヒトCEACAMファミリーメンバーの5’非翻訳領域のアラインメントを示す。指定配列を比較及び使用して実施例2に記載するように縮重プライマーを設計した。他のCEACAMファミリーメンバーの対応するヌクレオチドと同一のヌクレオチドを白抜き表示した。破線は配列のアラインメントを助長するためにスペースを加えたことを示す。GenBankに開示されているような各cDNA配列のヌクレオチド数を括弧内に示す。
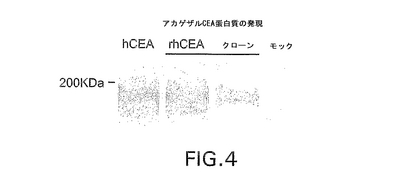
【図4】アカゲザルCEA蛋白質の発現を示す。λ−CEAライブラリーのスクリーニングにより得られたファージミドをHeLa細胞にトランスフェクトし、ヒトCEA蛋白質に対するウサギポリクローナル抗体を使用してウェスタンブロットを実施した。15個のクローンのうちの2個の発現を示す。
【図5】アカゲザルCEAコーディング領域の模式図を示す。内部反復配列を示し、遺伝子断片化用制限部位と配列を報告する。
【図6A】ヒト(配列番号6)及びアカゲザル(配列番号1)CEACAM−5ヌクレオチド配列のアラインメントを示す。2種のCEACAM−5配列間のヌクレオチド変異を太字で示す。
【図6B】ヒト(配列番号6)及びアカゲザル(配列番号1)CEACAM−5ヌクレオチド配列のアラインメントを示す。2種のCEACAM−5配列間のヌクレオチド変異を太字で示す。
【図6C】ヒト(配列番号6)及びアカゲザル(配列番号1)CEACAM−5ヌクレオチド配列のアラインメントを示す。2種のCEACAM−5配列間のヌクレオチド変異を太字で示す。
【図7】ヒト(配列番号7)及びアカゲザル(配列番号2)CEACAM−5オープンリーディングフレームのアラインメントを示す。2種のCEACAM−5配列間のアミノ酸変異を太字で示す。
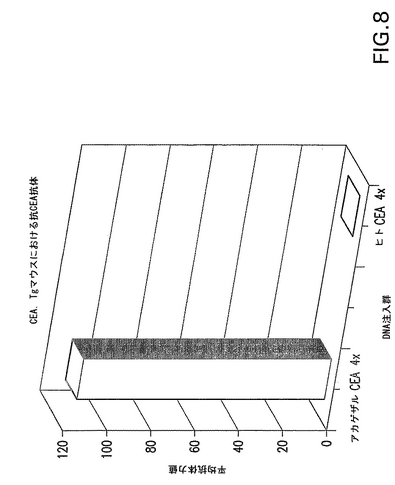
【図8】CEAトランスジェニックマウスにおけるヒトCEAに対する体液性応答を示す。アカゲザルCEAを免疫した群とヒトCEAを免疫した群の2つのマウス群の平均抗体力価を示す(実施例7)。
【図9】CEAトランスジェニックマウスにおけるヒトCEAに対する細胞性免疫応答を示す。hCEAを発現するベクター又はrhCEAを発現するベクターをCEAトランスジェニックマウスにワクチン接種した(実施例9)。
【図10】アカゲザル又はヒトCEAを免疫したCEAトランスジェニックマウスにおけるアカゲザルCEAペプチドに対する細胞性免疫応答を示す。
【配列表】

【技術分野】
【0001】
本発明は一般には癌の治療に関する。より詳細には、本発明は腫瘍関連ポリペプチド癌胎児性抗原のアカゲザルホモログ(本明細書ではrhCEAと言う)、この蛋白質をコードする単離核酸分子、並びにこの蛋白質をコードするDNAを含む組換えベクター及び宿主に関する。本発明はrhCEAをもつアデノウイルスベクター構築物と、癌の予防及び治療用ワクチン及び医薬組成物におけるその使用にも関する。
【背景技術】
【0002】
免疫グロブリンスーパーファミリー(IgSF)は細胞間接着等の種々の機能をもつ蛋白質をコードする多数の遺伝子から構成される。IgSF蛋白質は適正な分子間結合相互作用を維持するために重要な少なくとも1個のIg関連ドメインを含む。このような相互作用はIgSFメンバーの各種生体機能に必要であるため、多くのIgSF接着分子の破壊又は異常発現が多くのヒト疾患に関連付けられている。
【0003】
癌胎児性抗原(CEA)は細胞表面糖蛋白質から構成されるIgスーパーファミリーのサブファミリーに属する。CEAサブファミリーのメンバーはCEA関連細胞接着分子(CEACAM)として知られている。最近の科学文献では、CEA遺伝子はCEACAM5と改名されているが、蛋白質の呼称はCEAのままである。機能的には、CEACAMはホモタイプとヘテロタイプの両者の細胞間接着分子として作用することが示されている(Benchimolら,Cell 57:327−334(1989))。細胞接着に加え、CEAは細胞外マトリックスからの細胞の分離に起因する細胞死を抑制し、Bcl2やC−Myc等の所定のプロト癌遺伝子に関連する細胞形質転換に関与しているらしい(Berinstein,J.Clin Oncol.20(8):2197−2207(2002)参照)。
【0004】
胎児発生中と成人結腸粘膜ではCEAの正常発現が検出されている。CEA過剰発現は30年以上前にヒト結腸腫瘍で最初に検出され(Gold and Freedman,J.Exp.Med.121:439−462(1965))、その後、ほぼ全結腸直腸腫瘍で検出されている。更に、CEA過剰発現は高い百分率の膵臓腺癌、乳腺癌及び肺腺癌で検出可能である。これらの腫瘍種でCEA発現率が高いため、CEAはこれらの癌の管理と予後診断で広く臨床使用されている。
【0005】
CEA発現と転移増殖の相関により、CEAは結腸直腸癌治療の分子及び免疫介入のターゲットとしても認められている。CEAをターゲットとする治療アプローチの1つは抗CEA抗体の使用であり(Chesterら,Cancer Chemother.Pharmacol.46(Suppl):S8−S12(2000)参照)、別のアプローチはCEAワクチンを使用してCEAを発現する腫瘍を攻撃するように免疫系を活性化する方法である(詳細についてはBerinstein,前出参照)。
【発明の開示】
【発明が解決しようとする課題】
【0006】
ヒトCEAをコードする配列はクローニング及び特性決定されている(米国特許第5,274,087号;米国特許第5,571,710号;及び米国特許第5,843,761号。更にBeaucheminら,Mol.Cell.Biol.7:3221−3230(1987);Zimmermanら,Proc.Natl.Acad.Sci.USA 84:920−924(1987);Thompsonら Proc.Natl.Acad.Sci.USA 84(9):2965−69(1987)参照)。これらのCEA遺伝子が単離同定されているにも拘わらず、有効で自己寛容に妨害されない癌ワクチンの開発を実現できるようにCEAをコードする他の哺乳動物遺伝子を同定することが望ましい。
【課題を解決するための手段】
【0007】
本発明は配列番号2又は配列番号18に記載の新規アカゲザル癌胎児性抗原(以下、rhCEAと言う)をコードするヌクレオチド配列を含む単離又は精製核酸分子(ポリヌクレオチド)に関する。本明細書に開示するDNA分子は選択宿主細胞にトランスフェクトすることができ、組換え宿主細胞は発現される機能的rhCEA蛋白質(配列番号2及び配列番号18)の実質的レベルのソースを提供する。
【0008】
本発明は更に新規アカゲザルCEA蛋白質を発現するmRNAをコードする単離核酸分子に関し、このDNA分子は本明細書に配列番号1として開示するヌクレオチド配列を含む。アカゲザルCEAをコードするヌクレオチド配列を本明細書ではrhCEACAM5と言う。本発明のこの部分の1好適側面を図1Aに開示し、同図は新規rhCEA蛋白質(配列番号2)をコードするDNA分子(配列番号1)を示す。
【0009】
本発明の別の側面は新規アカゲザルCEA蛋白質(配列番号18)をコードする単離核酸分子であり、前記核酸分子は図1Bに示す配列番号5に記載のヌクレオチド配列を含む。
【0010】
本発明は更に本明細書全体に開示する核酸分子を含む原核及び真核両者の組換えベクター及び組換え宿主細胞に関する。
【0011】
本発明は更に組換え宿主細胞におけるアカゲザルCEA蛋白質の発現方法に関し、(a)配列番号1又は配列番号5に記載の核酸を含むベクターを適当な宿主細胞に導入する段階と;(b)前記アカゲザルCEA蛋白質の発現を可能にする条件下で宿主細胞を培養する段階を含む。
【0012】
本発明の1好適側面は図2A(配列番号2)に開示するアミノ酸配列から構成されるアカゲザルCEA蛋白質の実質精製形である。
【0013】
本発明の別の好適側面は図2B(配列番号18)に開示するアミノ酸配列から構成されるアカゲザルCEA蛋白質の実質精製形である。
【0014】
本発明の別の好適側面はrhCEA蛋白質を発現する配列番号1又は配列番号5に記載のヌクレオチド配列を含むDNA発現ベクターを含む組換え宿主細胞から得られ、実質精製され、完全にプロセシング(タンパク分解プロセシング、グリコシル化及び/又はリン酸化を含む)された成熟rhCEA蛋白質に関する。組換え宿主細胞は哺乳動物細胞株等の真核宿主細胞が特に好ましい。
【0015】
本発明の更に別の側面は単離核酸分子を含むワクチンベクターを哺乳動物に投与することを含む癌の予防又は治療方法であり、単離核酸分子は配列番号2又は配列番号18に記載のアカゲザル癌胎児性抗原(rhCEA)蛋白質をコードするヌクレオチド配列を含む。
【0016】
本発明は更に、E1領域に欠失とE1領域に挿入をもつアデノウイルスゲノムを含むアデノウイルスワクチンベクターに関し、挿入は(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含む。
【0017】
本発明は更に、プラスミド部分と発現カセット部分を含むワクチンプラスミドに関し、発現カセット部分は(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(b)ポリヌクレオチドに機能的に連結したプロモーターをを含む。
【0018】
本発明の別の側面は哺乳動物を癌から保護する方法又は癌の哺乳動物の治療方法であり、(a)i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第1のベクターを哺乳動物に導入する段階と;(b)所定時間経過させる段階と;(c)i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第2のベクターを哺乳動物に導入する段階を含む。
【0019】
本明細書と特許請求の範囲を通して使用する単数形は文脈からそうでないことが明白である場合を除き、複数も含む。
【0020】
本明細書と特許請求の範囲を通して以下の定義及び略語を適用する。
【0021】
「プロモーター」なる用語はRNAポリメラーゼが結合するDNA鎖上の認識部位を意味する。プロモーターはRNAポリメラーゼと開始複合体を形成し、転写活性を開始及び誘導する。複合体は「エンハンサー」と呼ばれる活性化配列又は「サイレンサー」と呼ばれる抑制配列により修飾することができる。
【0022】
「カセット」なる用語は発現させようとする核酸配列を含む本発明の配列を意味する。カセットはカセットテープと同様の概念であり、各カセットはそれ自体の配列をもつ。従って、カセットを交換することにより、ベクターは別の配列を発現する。5’及び3’末端の制限部位により、カセットは容易に挿入、除去又は別のカセットで置換することができる。
【0023】
「ベクター」なる用語はDNAフラグメントを宿主生物又は宿主組織に導入することができる手段を意味する。プラスミド、ウイルス(アデノウイルスを含む)、バクテリオファージ及びコスミド等の種々のベクターがある。
【0024】
アデノウイルスベクターに関して使用する「第1世代」なる用語はこのアデノウイルスベクターが複製欠損型であることを意味する。第1世代アデノウイルスベクターは一般に欠失又は不活化E1遺伝子領域をもち、欠失又は不活化E3遺伝子領域をもつことが好ましい。
【0025】
「pV1J−rhCEA」なる表記はイントロンAをもつヒトCMV極初期(IE)プロモーターと、完全長アカゲザルCEA遺伝子と、ウシ成長ホルモン由来ポリアデニル化及び転写終結配列と、最小pUCバックボーンを含む本明細書に開示するプラスミド構築物を意味する。
【0026】
「pMRK−Ad5−rhCEA」及び「MRK−rhCEA」なる表記はE1及びE3領域を欠失するAd5アデノウイルスゲノムを含む本明細書に開示する構築物を意味する。このプラスミドにおいて、E1領域はイントロンAをもたないヒトCMVプロモーターの制御下でE1と同じ向きに配置したアカゲザルCEA遺伝子と、その後に配置されたウシ成長ホルモンポリアデニル化シグナルで置換されている。
【0027】
「pBS−rhCEA」なる表記はpBluescriptII KS(+)プラスミドと完全長rhCEA遺伝子を含む本明細書に開示する構築物を意味する。
【0028】
「有効量」なる用語は免疫応答を生じるように十分なレベルのポリペプチドを生産するために十分なワクチン組成物を導入することを意味する。当業者に自明の通り、このレベルは可変である。
【0029】
「他の核酸を実質的に含まない」とは他の核酸を少なくとも90%、好ましくは95%、より好ましくは99%、更により好ましくは99.9%除去していることを意味する。「他の核酸を実質的に含まない」、「実質精製」、「単離核酸」又は「精製核酸」なる用語は交換可能に使用され、他の細胞成分を除去したアカゲザルCEA蛋白質のコーディング領域を含むDNA分子も意味する。従って、他の核酸を実質的に含まないアカゲザルCEA DNA調製物はその合計核酸の百分率として非アカゲザルCEA核酸の含有率が10%以下、好ましくは5%以下、より好ましくは1%以下、更により好ましくは0.1%以下である。所与アカゲザルCEA DNA調製物が他の核酸を実質的に含まないか否かは例えばアガロースゲル電気泳動と適当な染色法(例えば臭化エチジウム染色)の併用等の慣用核酸純度評価技術又は配列決定により判定することができる。
【0030】
「他の蛋白質を実質的に含まない」又は「実質精製」とは他の蛋白質を少なくとも90%、好ましくは95%、より好ましくは99%、更により好ましくは99.9%除去していることを意味する。従って、他の蛋白質を実質的に含まないアカゲザルCEA蛋白質調製物その合計蛋白質の百分率として非アカゲザルCEA蛋白質の含有率が10%以下、好ましくは5%以下、より好ましくは1%以下、更により好ましくは0.1%以下である。所与アカゲザルCEA蛋白質調製物が他の蛋白質を実質的に含まないか否かは例えばドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動(SDS−PAGE)と適当な検出法(例えば銀染色又はイムノブロッティング)の併用等の慣用蛋白質純度評価技術により判定することができる。
【0031】
「他の蛋白質を実質的に含まない」又は「実質精製」、又は「単離アカゲザルCEA蛋白質」又は「精製アカゲザルCEA蛋白質」なる用語は交換可能に使用され、天然ソースから単離されたアカゲザルCEA蛋白質も意味する。「単離」又は「精製」なる用語の使用はアカゲザルCEA蛋白質がその通常の細胞環境から取出されていることを示す。従って、単離アカゲザルCEA蛋白質は無細胞溶液中に存在していてもよいし、天然に存在するとは別の細胞環境に配置してもよい。単離なる用語は単離アカゲザルCEA蛋白質が唯一の存在する蛋白質であるという意味ではなく、単離rhCEA蛋白質が天然でrhCEA蛋白質とインビボ関連する他の蛋白質及び非アミノ酸物質(例えば核酸、脂質、炭水化物)を実質的に含まないという意味である。従って、原核又は真核細胞で組換え発現され、このrhCEA蛋白質を天然で(即ち介入なしには)発現しないこの宿主細胞から実質精製されたアカゲザルCEA蛋白質は当然のことながら本明細書に記載する任意状況下で「単離アカゲザルCEA蛋白質」である。上述のように、単離又は精製rhCEA蛋白質であるrhCEA蛋白質調製物は他の蛋白質を実質的に含まず、その合計蛋白質の百分率として非アカゲザルCEA蛋白質の含有率が10%以下、好ましくは5%以下、より好ましくは1%以下、更により好ましくは0.1%以下である。
【0032】
「保存アミノ酸置換」とはあるアミノ酸残基を別の化学的に類似するアミノ酸残基で置換することを意味する。このような保存置換の例は疎水性残基(イソロイシン、ロイシン、バリン、又はメチオニン)の相互置換、同一電荷の極性残基の相互置換(例えばリジン→アルギニン;アスパラギン酸→グルタミン酸)である。
【0033】
「rhCEA」とはアカゲザル癌胎児性抗原を意味する。
【0034】
「哺乳動物」なる用語はヒトを含む任意哺乳動物を意味する。
【0035】
「Ag」なる略語は抗原を意味する。
【0036】
「Ab」及び「mAb」なる略語は夫々抗体とモノクローナル抗体を意味する。
【0037】
「ORF」なる略語は遺伝子のオープンリーディングフレームを意味する。
【0038】
癌胎児性抗原(CEA)をコードする遺伝子は一般に腺癌発生に関連付けられる。本発明は異常CEA発現が癌又はその発生に関連付けられるCEA腫瘍関連抗原により発現される蛋白質産物に対する免疫を誘発又は強化するための組成物と方法に関する。異常CEA発現が腫瘍発症時に存在していても腫瘍進行後期に検出できない場合や、その逆の場合もあるので、異常CEA発現と癌の関連には、CEA蛋白質がその発生の全時点で腫瘍組織に発現される必要はない。
【0039】
この目的のために、アカゲザル癌胎児性抗原(rhCEA)をコードするポリヌクレオチドが提供される。本発明の分子は細胞性免疫により腺癌に対して有効な免疫予防を提供するために組換えアデノウイルス又はプラスミドワクチンで使用することができる。脊椎動物に直接インビボ導入する場合には、本発明のポリヌクレオチドはコードされる蛋白質の発現を霊長類、イヌ及びヒト等の哺乳動物を含む動物の体内に誘導する。
【0040】
本発明は配列番号2又は配列番号18に記載の新規rhCEA蛋白質を発現するmRNAをコードするヌクレオチド配列を含む単離核酸分子(ポリヌクレオチド)に関する。本発明の核酸分子は他の核酸を実質的に含まない。
【0041】
本発明の単離核酸分子としてはデオキシリボ核酸分子(DNA)が挙げられ、例えば1本鎖(コーディング鎖又は非コーディング鎖)でも2本鎖でもよいゲノムDNA及び相補的DNA(cDNA)や、合成1本鎖ポリヌクレオチド等の合成DNAが挙げられる。本発明の単離核酸分子はリボ核酸分子(RNA)でもよい。ほとんどのクローニング目的には、DNAが好適核酸である。
【0042】
本発明の1好適DNA分子は図2Aに示すアカゲザルCEA蛋白質(配列番号2)をコードする本明細書に配列番号1(図1A)として開示するヌクレオチド配列を含む。
【0043】
本発明の別の好適DNA分子は図2Bに配列番号18として示すアカゲザルCEA蛋白質をコードする本明細書に配列番号5(図1B)として開示するヌクレオチド配列(以下、「第2のrhCEA」DNA配列と言う)を含む。これらのrhCEA核酸分子は実施例2に詳述するようにRT−PCRにより同定した。第2のrhCEA DNA配列(配列番号5)は第1の配列と2ヌクレオチドが相違し、別のアカゲザルに由来する結腸組織からクローニングした。このDNA配列は第1のアカゲザルCEA蛋白質と2アミノ酸相違するアカゲザルCEA蛋白質をコードする。
【0044】
単離cDNAクローン、関連ベクター、宿主、組換えサブ細胞フラクション及び膜、並びに発現及び成熟形rhCEAは癌ワクチンの開発に有用である。
【0045】
本発明は新規rhCEA蛋白質を発現するmRNAをコードする配列番号1又は5の生体活性フラグメント又は突然変異体も含む。このような任意生体活性フラグメント及び/又は突然変異体はrhCEA蛋白質の医薬的性質と少なくとも実質的に類似する蛋白質又は蛋白質フラグメントをコードし、限定されないが、配列番号2又は配列番号18に記載のCEA蛋白質か挙げられる。このようなポリヌクレオチドとしては必ずしも限定されないが、ヌクレオチド置換、欠失、付加、アミノ末端短縮及びカルボキシ末端短縮が挙げられる。本発明の突然変異体は癌ワクチン開発に有用となるように真核細胞で機能的rhCEA蛋白質を発現するmRNA分子をコードする。
【0046】
本発明はrhCEA蛋白質をコードする合成DNAにも関し、合成DNAのヌクレオチド配列は配列番号1及び配列番号5のヌクレオチド配列と有意に相違するが、配列番号2又は配列番号18に記載のrhCEA蛋白質をコードする。このような合成DNAも本発明の範囲に含むものとする。
【0047】
従って、本発明は同一蛋白質を発現する多数のDNA分子を生じる可能性があるコドン冗長を開示する。本明細書の趣旨では、1個以上の置換コドンをもつ配列を縮重変異として定義する。発現される蛋白質の最終物性を実質的に変化させないDNA配列又は翻訳蛋白質の変異も本発明の範囲に含まれる。例えば、ロイシンをバリン、リジンをアルギニン、又はグルタミンをアスパラギンに置換してもポリペプチドの機能は変化し得ない。
【0048】
天然ペプチドの性質と異なる性質をもつペプチドをコードするようにペプチドをコードするDNA配列を改変できることが知られている。DNA配列の改変方法としては限定されないが、部位特異的突然変異誘発が挙げられる。改変される性質の例としては限定されないが、基質に対する酵素の親和性又はリガンドに対する受容体の親和性の改変が挙げられる。
【0049】
ストリンジェント条件下で配列番号1又は配列番号5とハイブリダイズするDNA配列も本発明に含まれる。例えば、限定されないが、高ストリンジェンシー条件を使用する方法は以下の通りである:6XSSC、5Xデンハルト溶液、及び変性サケ精子DNA100μg/mlから構成される緩衝液中でDNAを含むフィルターのプレハイプリタイゼーションを65℃で2時間〜一晩実施する。変性サケ精子DNA100μg/mlと32P標識プローブ5〜20X106cpmを含有するプレハイプリタイゼーション混合物中でフィルターを65℃で12〜48時間ハイブリダイズさせる。2XSSC、0.1%SDSを含有する溶液で37℃にて1時間フィルターの洗浄を行う。次に0.1XSSC、0.1%SDSで50℃にて45分間洗浄した後、オートラジオグラフィーにかける。高ストリンジェンシー条件を使用する他の方法としては、5XSSC、5Xデンハルト溶液、50%ホルムアミド中でハイブリダイゼーション段階を42℃で12〜48時間実施する方法や、0.2XSSPE、0.2%SDSで洗浄段階を65℃で30〜60分間実施する方法が挙げられる。
【0050】
高ストリンジェンシーハイブリダイゼーションを実施するための上記方法で記載した試薬は当分野で周知である。これらの試薬の組成の詳細は例えば参考資料として本明細書に組込むSambrookら,Molecular Cloning:A Laboratory Manual;Cold Spring Harbor Laboratory,Cold Spring Harbor,New York,1989に記載されている。上記以外に、使用可能な他の高ストリンジェンシー条件も当分野で周知である。
【0051】
本発明の1好適側面は図2Aに開示するアミノ酸配列(配列番号2)を含むアカゲザルCEA蛋白質の実質精製形である。
【0052】
本発明の別の好適側面は図2Bに開示するアミノ酸配列(配列番号18)を含むアカゲザルCEA蛋白質の実質精製形である。
【0053】
本発明はrhCEAの種々の機能的ドメインとこれらの配列の少なくとも1個を含むハイブリッド分子にも関する。CEA蛋白質はプロセシングされたリーダー配列をもつアミノ末端ドメインと疎水性カルボキシ末端ドメインを含む。CEAは更に3個のIg様内部ドメインを含む。CEAの細胞間接着機能にはN末端ドメインのサブドメインが必要であることがTaheriら(J.Biol.Chem.275(35):26935−26943(2000))により示されている。
【0054】
本発明は更に配列番号2又は配列番号18に記載のアミノ酸配列を含むrhCEA蛋白質の生体活性フラグメント及び/又は突然変異体を含み、必ずしも限定されないが、これらの突然変異が診断、治療又は予防用蛋白質又は蛋白質フラグメントを提供し、癌ワクチン開発に有用となるようなアミノ酸置換、欠失、付加、アミノ末端短縮及びカルボキシ末端短縮が挙げられる。
【0055】
本発明のアカゲザルCEA蛋白質は「成熟」蛋白質の形態でもよいし、融合蛋白質等の大型蛋白質の一部でもよい。分泌又はリーダー配列、プロ配列、精製を助長する配列(例えば多重ヒスチジン残基)、又は組換え生産中の安定性のための付加配列を含む付加アミノ酸配列を加えると有利なことが多い。
【0056】
本発明は更にrhCEA融合構築物に関し、限定されないが、種々のマーカー(限定されないが、GFP(緑色蛍光蛋白質)、MYCエピトープ、GST、及びFc)に連結したアカゲザルCEA蛋白質を発現する融合構築物が挙げられる。このような任意融合構築物を該当細胞株で発現させ、本明細書に開示するアカゲザルCEA蛋白質のモジュレーターをスクリーニングするために使用することができる。アカゲザルCEAに対する免疫応答を強化するように構築された融合構築物も含まれ、限定されないが、DOMやhsp70が挙げられる。
【0057】
本発明は更に本明細書を通して開示される実質精製核酸分子を含む組換えベクターにも関する。これらのベクターはDNA又はRNAから構成することができる。ほとんどのクローニング目的には、DNAベクターが好ましい。典型的ベクターとしてはプラスミド、改変ウイルス、バクテリオファージ、コスミド、酵母人工染色体、及びrhCEA蛋白質をコードすることができる他の形態のエピソーム又は組込みDNAが挙げられる。特定遺伝子導入又は他の用途に適したベクターを決定することは当業者が容易に実施可能である。
【0058】
組換え宿主でrhCEAを発現させるためにはrhCEA蛋白質をコードするDNAを含む発現ベクターを使用することができる。発現ベクターとしては限定されないが、クローニングベクター、改変クローニングベクター、特殊設計プラスミド又はウイルスが挙げられる。更に、所望により細菌細胞で組換えrhCEAを発現させるためには種々の細菌発現ベクターを使用することができる。更に、真菌細胞で組換えrhCEAを発現させるためには種々の真菌発現ベクターを使用することができる。更に、昆虫細胞で組換え蛋白質を発現させるためには種々の昆虫細胞発現ベクターを使用することができる。
【0059】
本発明は更に、本発明の核酸分子を含むベクターで形質転換又はトランスフェクトされた宿主細胞に関する。組換え宿主細胞は原核細胞でも真核細胞でもよく、限定されないが、大腸菌等の細菌、酵母等の真菌細胞、限定されないが、ウシ、ブタ、サル及び齧歯類に由来する細胞株等の哺乳動物細胞、並びに限定されないが、ショウジョウバエやカイコに由来する細胞株等の昆虫細胞が挙げられる。このような組換え宿主細胞はrhCEA又は生物学的に等価の形態を生産するのに適した条件下で培養することができる。
【0060】
上述のように、組換え宿主細胞でrhCEAを発現させるためにはrhCEA蛋白質をコードするDNAを含む発現ベクターを使用することができる。従って、本発明の別の側面は組換え宿主細胞におけるアカゲザルCEA蛋白質の発現方法であり、(a)配列番号1又は配列番号5に記載の核酸を含むベクターを適当な宿主細胞に導入する段階と;(b)前記アカゲザルCEA蛋白質の発現を可能にする条件下で宿主細胞を培養する段階を含む。
【0061】
宿主細胞でrhCEAの発現後、rhCEA蛋白質を回収し、活性形態のrhCEA蛋白質が得られる。数種のrhCEA蛋白質精製方法が利用可能であり、使用に適している。塩析分画、イオン交換クロマトグラフィー、サイズ排除クロマトグラフィー、ヒドロキシアパタイト吸着クロマトグラフィー及び疎水性相互作用クロマトグラフィーの各種組み合わせ又は個別適用により細胞溶解液及び抽出液から組換えrhCEA蛋白質を精製することができる。更に、全長rhCEA蛋白質又はrhCEA蛋白質のポリペプチドフラグメントに特異的なモノクローナル又はポリクローナル抗体から作製したイムノアフィニティーカラムの使用により他の細胞蛋白質から組換えrhCEA蛋白質を分離することもできる。
【0062】
本発明の核酸を融合し、ヒト細胞で蛋白質の効率的発現を可能にするように設計された配列を含む発現カセットとすることができる。カセットはプロモーターや終結配列等の関連転写及び翻訳調節配列を全長rhCEA遺伝子に機能的に連結することが好ましい。1好適態様では、プロモーターはイントロンA配列をもたないサイトメガロウイルスプロモーター(CMV)であるが、当業者に自明の通り、強力なイムノグロブリン又は他の真核遺伝子プロモーター等の多数の他の公知プロモーターも使用できる。1好適転写ターミネーターはウシ成長ホルモンターミネーターであるが、他の公知転写ターミネーターも使用できる。CMV−BGHターミネーターの組み合わせが特に好ましい。
【0063】
本発明によると、アカゲザルCEA発現カセットをベクターに挿入する。ベクターはアデノウイルスベクターが好ましいが、プロモーターに連結した直鎖DNAや、アデノ随伴ウイルス又は改変ワクチンウイルス、レトロウイルス又はレンチウイルスベクター等の他のベクターも使用できる。
【0064】
選択するベクターがアデノウイルスである場合には、ベクターは所謂第1世代アデノウイルスベクターであることが好ましい。これらのアデノウイルスベクターは非機能的E1遺伝子領域、好ましくは欠失アデノウイルスE1遺伝子領域をもつことを特徴とする。所定態様では、アデノウイルスE1遺伝子が正常に配置されている位置に発現カセットを挿入する。更に、これらのベクターは場合により非機能的又は欠失E3領域をもつ。使用するアデノウイルスゲノムはE1及びE3領域の両者を欠失していることが好ましい(ΔE1ΔE3)。アデノウイルスはウイルスE1遺伝子を発現する公知細胞株(例えば293細胞、又はERC.6細胞)又は別の蛋白質を発現するように一過的又もしくは安定的に軽質転換された293もしくPERC.6細胞に由来する細胞株で複製することができる。例えば、テトラサイクリン調節型プロモーターシステムのように制御下の遺伝子発現を可能にする構築物を使用する場合には、細胞株は調節システムに関与する成分を発現することができる。このような細胞株の1例はT−Rex−293であり、他の例も当分野で公知である。
【0065】
アデノウイルスベクターを操作し易くするために、アデノウイルスをシャトルプラスミド形とすることができる。本発明はプラスミド部分とアデノウイルス部分を含むシャトルプラスミドにも関し、アデノウイルス部分はE1を欠失し、場合によりE3も欠失しており、アカゲザルCEAを含む発現カセットを挿入したアデノウイルスゲノムを含む。好適態様では、アデノウイルスベクターを容易に除去できるようにプラスミドのアデノウイルス部分の両側に制限部位を配置する。シャトルプラスミドは原核細胞又は真核細胞で複製することができる。
【0066】
本発明の1好適態様では、発現カセットをpMRKAd5−HVOアデノウイルスプラスミドに挿入する(参照により本明細書に組込むEminiら,WO02/22080参照)。このプラスミドはE1領域とE3領域を欠失するAd5アデノウイルスゲノムを含む。pMRKAd5−HVOプラスミドの設計はウイルスパッケージングの最適化に重要であることが分かっているエレメントを組込むように5’シス作用性パッケージング領域を更にE1遺伝子まで延ばし、ウイルス増幅を強化することにより従来のアデノベクターを改善している。この強化型アデノウイルスベクターは高度の継代増殖後に遺伝的安定性を維持できるという利点がある。
【0067】
DNA構築物を作製及び精製するための標準分子生物学技術により、本発明のアデノウイルス、シャトルプラスミド及びDNA免疫原を作製することができる。
【0068】
上記ベクターは異常CEA発現に関連する腺癌発生の予防及び/又は既存癌の治療のための免疫原組成物及びワクチンで使用することができる。この点で、本発明の1側面は単離核酸分子を含むワクチンベクターを哺乳動物に投与することを含む癌の予防又は治療方法であり、単離核酸分子は配列番号2又は配列番号18に記載のアカゲザルCEA蛋白質をコードするヌクレオチド配列を含む。
【0069】
上記方法によると、任意哺乳動物の癌を治療又は予防するためにワクチンベクターを投与することができる。本発明の1好適態様では、哺乳動物はヒトである。
【0070】
更に、当業者は上記治療及び予防方法で使用するために任意の型のベクターを選択することができる。ベクターはアデノウイルスベクター又はプラスミドベクターが好ましい。本発明の1好適態様では、ベクターはアデノウイルスE1領域に欠失とアデノウイルスE1領域に挿入をもつアデノウイルスゲノムを含むアデノウイルスベクターであり、挿入は(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含む。
【0071】
本発明は更にE1領域に欠失とE1領域に挿入をもつアデノウイルスゲノムを含むアデノウイルスワクチンベクターに関し、挿入は(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含む。
【0072】
本発明のこの側面の1好適態様において、アデノウイルスベクターはAd5ベクターである。
【0073】
本発明の別の態様において、アデノウイルスベクターはAd6ベクターである。
【0074】
別の側面において、本発明はプラスミド部分と発現カセット部分を含むワクチンプラスミドに関し、発現カセット部分は(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(b)ポリヌクレオチドに機能的に連結したプロモーターを含む。
【0075】
本発明の所定態様において、本明細書に開示する組換えアデノウイルスワクチンは強化免疫応答を誘導するためにプラスミド型ポリヌクレオチドワクチンと各種プライム/ブーストの組み合わせで使用される。この場合、2種のベクターを「プライム及びブースト」レジメンで投与する。例えば、第1の型のベクターを投与した後に所定時間、例えば1カ月、2カ月、6カ月、又は他の適当な間隔後に第2の型のベクターを投与する。ベクターは同一ポリヌクレオチド又はポリヌクレオチド組み合わせをコードする発現カセットをもつことが好ましい。プラスミドDNAも使用する態様では、ベクターは哺乳動物又は昆虫細胞により認識される1種以上のプロモーターを含むことが好ましい。1好適態様では、プラスミドは限定されないが、CMVプロモーター等の強力プロモーターを含む。発現させるアカゲザルCEA遺伝子又は他の遺伝子をこのようなプロモーターと連結する。このようなプラスミドの1例は文献に記載されているような哺乳動物発現プラスミドV1Jnsである(参照により本明細書に組込むJ.Shiverら,DNA Vaccines,M.Liuら編,N.Y.Acad.Sci.,N.Y.,772:198−208(1996))。
【0076】
上述のように、アデノウイルスベクターワクチン及びプラスミドは免疫応答を誘導するために単独治療レジメンの一部として脊椎動物に投与することができる。この点で、本発明は哺乳動物を癌から保護する方法に関し、(a)i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第1のベクターを哺乳動物に導入する段階と;(b)所定時間経過させる段階と;(c)i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第2のベクターを哺乳動物に導入する段階を含む。
【0077】
上記保護方法の1態様において、第1のベクターはプラスミドであり、第2のベクターはアデノウイルスベクターである。代替態様において、第1のベクターはアデノウイルスベクターであり、第2のベクターはプラスミドである。
【0078】
本発明は更に腺癌の哺乳動物の治療方法に関し、(a)i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第1のベクターを哺乳動物に導入する段階と;(b)所定時間経過させる段階と;(c)i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第2のベクターを哺乳動物に導入する段階を含む。
【0079】
上記治療方法の1態様において、第1のベクターはプラスミドであり、第2のベクターはアデノウイルスベクターである。代替態様において、第1のベクターはアデノウイルスベクターであり、第2のベクターはプラスミドである。
【0080】
ワクチンレシピエントに導入される発現可能なDNA又は転写RNAの量は使用されるプロモーターの強度と発現される遺伝子産物の免疫原性に部分的に依存する。一般に、プラスミドワクチンベクター約1ng〜100mg、好ましくは約10μg〜300μgの免疫又は予防有効用量をマウス組織に直接投与する。組換えアデノウイルスの有効用量は粒子約106〜1012個、好ましくは約107〜1011個である。皮下注射、皮内導入、経皮圧入、及び他の投与方法(例えば腹腔内、静脈内、又は吸入送達)も考えられる。ブースターワクチン接種を行うことも考えられる。本発明のワクチンの非経口導入と同時又はその後にインターロイキン12蛋白質等のアジュバントを静脈内、筋肉内、皮下又は他の投与手段等で非経口投与しても有利である。
【0081】
本発明のワクチンベクターは裸のままでもよく、即ち蛋白質、アジュバント又はレシピエントの免疫系に影響を与える他の物質を伴っていなくてもよい。この場合に、ワクチンベクターを生理的に許容可能な溶液とすることが望ましく、限定されないが、滅菌食塩水又は滅菌緩衝食塩水が挙げられる。あるいは、アジュバント、サイトカイン、蛋白質、又は他のキャリヤー等の免疫刺激剤を本発明のワクチン又は免疫原組成物と共に投与すると有利である。従って、本発明は本発明の組成物及び方法とこのような免疫刺激剤の併用を含む。本明細書で使用する免疫刺激剤とは外来抗原に対する(抗体及び/又は細胞性)免疫応答を強化又は増強するほぼ任意物質を意味する。前記免疫刺激剤はDNA又は蛋白質の形態で投与することができる。種々の免疫刺激剤の任意のものを本発明のワクチン及び免疫原組成物と併用することができ、限定されないが、GM−CSF、IFNα、破傷風トキソイド、IL12、B7.1、LFA−3及びICAM−1が挙げられる。前記免疫刺激剤は当分野で周知である。DNAの細胞取込みを助長する物質も使用することができ、限定されないが、カルシウムイオンが挙げられる。これらの物質を一般にトランスフェクション助長剤及び医薬的に許容可能なキャリヤーと言う。当業者は特定の免疫刺激剤又は医薬的に許容可能なキャリヤーと許容可能な投与時間及び投与方法を決定することができよう。
【0082】
rhCEAをクローニングするためには種々の方法の任意のものを使用することができる。これらの方法としては限定されないが、以下の方法が挙げられる。(1)RACE PCRクローニング法(Frohmanら,Proc.Natl.Acad.Sci.USA 85:8998−9002(1988))。5’及び/又は3’RACEを実施して全長cDNA配列を作製することができる。このストラテジーはrhCEAcDNAのPCR増幅に遺伝子特異的オリゴヌクレオチドプライマーを使用する。これらの遺伝子特異的プライマーは多数の公共入手可能な核酸及び蛋白質データベースを検索することにより同定された発現配列タグ(EST)ヌクレオチド配列の同定により設計される;(2)適当な発現ベクターシステムでrhCEAを含むcDNAライブラリーの構築後にrhCEAcDNAの直接機能的発現;(3)バクテリオファージ又はプラスミドシャトルベクターで構築したrhCEAを含むcDNAライブラリーをrhCEA蛋白質のアミノ酸配列から設計した標識縮重オリゴヌクレオチドプローブでスクリーニング;(4)バクテリオファージ又はプラスミドシャトルベクターで構築したrhCEAを含むcDNAライブラリーを、rhCEA蛋白質をコードする部分cDNA蛋白質でスクリーニング。この部分cDNAはrhCEA蛋白質に関連する他の膜蛋白質について分かっているアミノ酸配列から縮重オリゴヌクレオチドプライマーを設計することによりrhCEA DNAフラグメントの特異的PCR増幅により得られる;(5)バクテリオファージ又はプラスミドシャトルベクターで構築したrhCEAを含むcDNAライブラリーを、哺乳動物rhCEA蛋白質と相同性をもつ部分cDNA又はオリゴヌクレオチドでスクリーニング。このストラテジーでは上記のようにESTとして同定されたrhCEAcDNAのPCR増幅に遺伝子特異的オリゴヌクレオチドプライマーを使用してもよい;あるいは(6)配列番号1を鋳型として5’及び3’遺伝子特異的オリゴヌクレオチドを設計し、公知RACE法により全長cDNAを作製できるようにするか、又はrhCEAをコードするヌクレオチド配列の全長形を単離するためにコーディング領域の一部を作製及び単離して多種のcDNA及び/又はゲノムライブラリーのうちの1個をスクリーニングするためのプローブとして使用するようにこれらの公知同一RACE法によりコーディング領域の一部を作製できるようにする。
【0083】
当業者に自明の通り、他の型のライブラリーや、他の細胞型又は種型から構築したライブラリーもrhCEAをコードするDNA又はrhCEAホモログを単離するのに有用である。他の型のライブラリーとしては限定されないが、他の細胞に由来するcDNAライブラリーが挙げられる。rhCEAをコードするcDNAを単離するためにcDNAライブラリーを構築するのに使用される細胞又は細胞株の選択はまずこのような目的に利用可能な任意公知アッセイを使用して細胞関連rhCEA活性を測定することにより実施することができる。
【0084】
cDNAライブラリーの構築は当業者に周知の標準技術により実施することができる。周知cDNAライブラリー構築技術は例えばSambrookら,Molecular Cloning:A Laboratory Manual;Cold Spring Harbor Laboratory,Cold Spring Harbor,New York,1989に記載されている。相補的DNAライブラリーは多数の商業ソースからも入手でき、限定されないが、Clontech Laboratories,Inc.(Palo Alto,CA)やStratagene(La Jolla,CA)が挙げられる。
【0085】
本発明のDNA分子、RNA分子、及び組換え蛋白質はrhCEAのレベルをスクリーニング及び測定するために使用することができる。組換え蛋白質、DNA分子、及びRNAはrhCEAの検出とタイピングに適したキットの構築に有用である。このようなキットは少なくとも1個の容器を緊密に拘束保持するのに適した区画化キャリヤーを含む。キャリヤーは更にrhCEAを検出するのに適した組換えrhCEA又は抗rhCEA抗体等の試薬を含む。キャリヤーは更に標識抗原又は酵素基質等の検出手段を加えることができる。
【0086】
本明細書に引用する全刊行物は本発明に関連して使用可能な方法及び材料を記載及び開示する目的で参照により組込む。本明細書の如何なる記載も本発明が先発明により前記開示以前の日付を享受する資格がないと認めるものであると解釈すべきではない。
【0087】
以上、添付図面を参照して本発明の好適態様を記載したが、当然のことながら本発明はこれらの厳密な態様に限定されず、当業者は特許請求の範囲に記載する本発明の範囲又は精神から逸脱せずに種々の変更及び変形を行うことができる。
【0088】
以下、実施例により本発明を例示するが、これらの実施例により発明を限定するものではない。
【実施例1】
【0089】
アカゲザルからのRNAの単離
分子操作は当分野で周知の標準操作に従って実施した(例えば、参照により本明細書に組込むAusubelら.Short Protocols in Molecular Biology,F.M.,第2版,John Wiley & Sons,(1992)及びSambrookら,Molecular Cloning,A Laboratory Manual,第2版,Cold Spring Harbor Laboratory Press(1989)参照)。
【0090】
アカゲザルCEAcDNAの単離用RNAを得るために、2頭の別個のアカゲザル(Macaca Mulatta)に由来する結腸試料を使用した。Biomedical Primate Research Center(BPRC,Rijswijk,オランダ)から凍結組織を入手した。アカゲザル結腸試料から全RNAを抽出するために、組織を機械的に粉砕し、Ultraspec RNA試薬(Biotecx Laboratories;Houston,TX)を製造業者の指示に従って添加した。精製RNAの完全性をホルムアルデヒド変性アガロースゲルにより確認した。試料を分画し、−80℃で保存した。
【実施例2】
【0091】
アカゲザルCEAcDNA増幅
ヒトCEAファミリーの全公知メンバーの5’及び3’非翻訳領域(UTR)に由来するヌクレオチド配列を整列させ、CEA DNAの高度保存領域を同定した(図3参照)。同定したCEA遺伝子ファミリー相同性に基づき、縮重オリゴヌクレオチドプライマーを設計し、アカゲザルCEAcDNAを以下に記載するように逆転写酵素ポリメラーゼ連鎖反応(RT−PCR)により増幅するようにPCR条件を最適化した。完全cDNAを増幅するために使用したプライマーは5’−RhCEA EcoRI 5’−CCGAATTCCGGACASAGCAGRCAGCAGRSACC−3’(配列番号3)及びCEA−8 RhXhoI 5’−CCGCTCGAGCGGCTGCTACATCAGAGCAACCCCAACC−3’(配列番号4)とした。SuperScript One−Step RT−PCR with Platinum Taqキット(インビトロgen;Carlsbad,CA)を使用して増幅を実施した。RNA1μg、両者プライマー200pmol、及び10% DMSO(終濃度)からなる反応容量100μlを使用した。
【0092】
逆転写段階を実施するために、2頭のアカゲザルの各々から単離した全RNA試料を45℃で30分間インキュベートした後、2分間94℃でインキュベートした。94℃15秒、52℃30秒、及び68℃2分20秒を1サイクルとし、得られた鋳型のPCR増幅を40サイクル実施した。
【0093】
CEACAM−5ホモログの予想寸法である約2100bpの増幅PCR産物を両者RNA試料から個々に獲得し、アガロースゲルから精製した。両者PCR産物の部分配列分析の結果、ヒトCEACAM−5との高い相同性が判明した。
【0094】
内部反復配列の高い相同性により、図5に示す制限部位を使用してDNAフラグメントを精製することにより完全遺伝子配列が得られた。各サルから得られたアカゲザルCEAヌクレオチド配列を本明細書の図1に開示し、配列番号1(以下、rhCEACAM−5と言う)及び配列番号5(以下、rhCEACAM−5#2と言う)に示す。CEAヌクレオチド配列分析の結果、705アミノ酸ポリペプチドをコードする2118ヌクレオチドのオープンリーディングフレーム(ORF)が判明した。2頭のアカゲザルから得られたrhCEAヌクレオチド配列を比較すると、2種の異なる蛋白質(図2A及び2B参照)をコードする2ヌクレオチド変異があることが判明した(図1A及び1B参照)。
【0095】
アカゲザルCEACAM−5ヌクレオチド配列(配列番号1)を更に公開ヒトCEACAM−5配列(配列番号6)に比較した処、ヌクレオチドレベルで88%の相同度が判明した(図6参照)。同様にアカゲザル(配列番号2)とヒト(配列番号7)のCEAポリペプチド配列を比較した処、アミノ酸レベルで78.9%の一致度が判明した(図7参照)。興味深いことに、アカゲザルCEAのカルボキシル末端にはヒトCEAに比較して3アミノ酸挿入が存在しており、恐らくグリコシルホスファチジルイノシトール(GPI)変異のシグナルに関係があると予想される。
【実施例3】
【0096】
λアカゲザルCEA特異的ライブラリーの作製とスクリーニング
RT−PCRにより得られた増幅rhCEA産物(実施例2参照)をEcoRI/XhoIで消化し、製造業者の指示に従ってZAP−CMV XRベクター(Stratagene;La Jolla,CA)にライゲートした。ライゲーション産物をGigapack III gold packaging extractと共にインキュベートし、得られたファージをXL−1 Blue MRF’細胞に感染させた。このCEA特異的一次ライブラリーを次に増幅し、〜1×106pfu/mlの力価を得た。ナイロンフィルターに転写することによりプラーク〜5×103個のスクリーニングを実施した。CEA分子の5’及び3’末端に対応する2種の異なるDNAプローブとフィルターをハイブリダイズさせた。二重陽性プラークをXL−1 Blue MRF’細胞中で切り出し、誘導された繊維状ファージをXL−OLR細胞中で増幅した。次にファージミドを増殖させ、制限消化により分析した。配列分析とGenbank比較の結果、ヒトCEACAM−5と最高の相同度であることが判明した。
【実施例4】
【0097】
プラスミド構築物とアデノウイルス作製
pCMV−script EXファージミドベクターからPstI/XhoIでRhCEAを切り出し、pBluescript II KSベクターに挿入し、pBS−RhCEAを得た。挿入部分を完全に配列決定した後、SmaI/XhoIフラグメントとしてpVIJnsAベクターにサブクローニングし、pVIJ−RhCEAを得た。同一フラグメントをpolyMRKベクターにサブクローニングすることによりアデノウイルス作製用シャトルプラスミドpMRK−RhCEAを得た。RhCEA及びE1フランキングAd5領域用発現カセットを含むpMRK−RhCEAからのPacI/StuIフラグメントをBJ5183大腸菌細胞でClaIにより直鎖化したpAd5又はpAd6に組換えた。得られたプラスミドをpAd5−RhCEA及びpAd6−RhCEAとした。両者プラスミドをPacIで切断し、アデノウイルスITRを遊離させ、PerC−6細胞にトランスフェクトした。連続継代によりウイルス増幅を実施した。標準CsCl精製プロトコールを使用してAd5−RhCEA及びAd6−RhCEAを精製し、A105緩衝液(5mM Tris pH8.0,1mM MgCl2,75mM NaCl,5%スクロース,0.005% Tween20)で十分に透析した。
【実施例5】
【0098】
インビトロ RhCEA発現及び検出
作製したベクターによるRhCEAの発現をウェスタンブロットとFACS分析により確認した。Lipofectamine 2000(Life Technologies;Carlsbad,CA)を使用してプラスミドをHeLa又はPerC.6細胞にトランスフェクトした。無血清培地で30分間37℃にてアデノウイルス感染を行った後に新鮮な培地を加えた。48時間インキュベーション後にヒトCEAに対するウサギポリクローナル血清(Fitzgerald,1500倍希釈液)を使用して全細胞溶解液をウェスタンブロットにより分析した。選択した全アカゲザルCEAクローンはHeLa細胞にトランスフェクトした場合に180〜200KDa蛋白質を発現した(図4参照)。
【0099】
FACS分析のために、細胞をトリプシンで剥離し、FACS緩衝液(PBS,1%FCS)に再懸濁した。250倍に希釈したウサギポリクローナル抗CEA抗体と共に30分間インキュベーション後に細胞を洗浄し、抗ウサギIgG−PEと共に30分間インキュベートし、最後にFACScalibur(Becton Dickinson,San Jose,CA)で分析した。
【実施例6】
【0100】
ペプチド
免疫した動物でアカゲザルCEAに対する細胞性免疫応答を分析するために、完全蛋白質に対応するように11アミノ酸がオーバーラップする15量体ペプチドを設計した。凍結乾燥アカゲザルCEAペプチドをBio−Synthesis,Inc.(Lewisville,TX)から購入し、DMSOに40mg/mlで再懸濁した。ペプチドをプールA(RhCEA−1〜RhCEA−34,34ペプチド);プールB(RhCEA−35〜RhCEA−79,45ペプチド);プールC(RhCEA−80〜RhCEA−124,48ペプチド);及びプールD(RhCEA−125〜RhCEA−173,53ペプチド)の4個のプールに分けた。終濃度はA=1.176mg/ml;プールB=0.888mg/ml;プールC=0.851mg/ml;プールD=0.769mg/mlとした。ペプチドとプールを−80℃で保存した。
【実施例7】
【0101】
rhCEA免疫によるマウスにおけるCEA特異的細胞性免疫応答の発生
CEA.Tgマウスはヒトに類似する組織分布でヒトCEAを自己抗原として発現するトランスジェニックマウスである。科学文献に広く実証されているように、これらのマウスは検出可能なCEA特異的血清抗体が存在せず、CEAに対するインビトロ脾臓T−cell応答を発生することができないことからCEA非応答性である。同種抗原をコードする異種遺伝子によるDNA免疫が同系メラノーマ細胞による腫瘍攻撃からマウスを保護することは多くの報告に示されている。異種DNAワクチン接種によりこのモデルで自己抗原に対する免疫応答を誘発できることを立証するために、CEA.アカゲザルCEA(異種)をコードするベクターでTgマウスを免疫した。
【0102】
C57BL/6マウス(H−2b)はCharles River(Lecco,イタリー)から購入した。CEA.tgマウス(H−2b)はHL Kaufman(Albert Einstein College of Medicine,New York)から入手し、標準条件下で飼育した。
【0103】
電気的遺伝子導入(EGT)のために、マウス大腿四頭筋にpVIJ−RhCEA50μgを外科的に接触させるか又は直接注入し、従来記載されているように電気刺激した(Rizzutoら.Proc.Natl.Acad.Sci.U.S.A.96(11):6417−22(1999))。アデノウイルス注入のために、Ad5−RhCEA 1×1010vpをマウス大腿四頭筋に注入した。
【0104】
マウスの大腿四頭筋にpVIJ−RhCEA50μgを注入し、注入直後に週1回ずつ4週間電気刺激した。C57BL/6マウスを対照として使用した。アカゲザルCEAに対する抗体がこれらのマウスに由来する血清中でウェスタンブロットにより検出され、体液性免疫応答が立証された。hCEAに対するマウスモノクローナルAbを陽性対照として使用し、免疫前血清とモック感染細胞抽出液を陰性対照として使用した(データは示さず)。重要な点として、ヒトCEA蛋白質に対する交差反応性抗体はアカゲザルCEA免疫群でしか測定することができず(図8)、平均力価は1:110であった。これらのデータによると、トランスジェニックマウスモデルでは異種DNAワクチン接種により寛容を解除することが可能である(抗CEA自己抗体として測定)。
【実施例8】
【0105】
抗体検出及び力価測定
眼窩後方採血により抗体力価測定用血清を採取した。ウェスタンブロット検出のために、Ad5−rhCEAを導入したHeLa細胞からの抽出液をSDS−pageゲルで泳動させ、ニトロセルロースフィルターに転写した。血清をプールし、4℃でのO/Nインキュベーション用に50倍に希釈した。抗マウスIgG−APコンジュゲート(Sigma,1:2500)を検出用に使用した。力価測定のために、コーティング用緩衝液(50mM NaHCO3 pH 9.4)で希釈したCEA(高純度CEA;Fitzgerald Industries International Inc.,Concord MA)100ng/ウェルでElisaプレート(Nunc maxisorp)をコーティングし、4℃でO/Nインキュベートした。次に5%BSAを添加したPBSでプレートを1時間37℃でブロックした。マウス血清をPBS5%BSAで希釈した(血清変換の評価には50倍希釈;力価値の評価には10倍〜31,250倍希釈)。免疫前血清をバックグラウンドとして使用した。希釈した血清を4℃でO/Nインキュベートした。PBS,1% BSA,0.05% tween 20で洗浄を行った。検出抗体(ヤギ抗マウスIgGペルオキシダーゼ,Sigma,St.Louis,MO)をPBS,5% BSAで2000倍に希釈し、シェーカーで2〜3時間室温にてインキュベートした。洗浄後、プレートをTMB基質(Pierce Biotechnology,Inc.,Rockford,IL)100μl/ウェルで展開した。1M H2SO4溶液25μl/ウェルで反応を停止し、プレートを450nm/620nmで読み取った。同一希釈倍率の自己免疫前血清の吸光度の少なくとも3倍の吸光度を生じる血清の限界希釈倍率として抗CEA血清力価を計算した。
【実施例9】
【0106】
IFN−γELISPOTアッセイ
96ウェルMAIPプレート(Millipore,Bedford,MA)に滅菌PBS中2.5μl/mlの精製ラット抗マウスIFN−γ(IgG1,クローンR4−6A2,Pharmingen,San Diego,CA)を100μl/ウェルの割合でコーティングした。滅菌PBSで洗浄後、プレートをR10培地200μl/ウェルで37℃にて少なくとも2時間ブロックした。
【0107】
脾細胞作製のために、屠殺したマウスから脾臓を滅菌摘出し、グリッドで細断した。0.1×PBS1mlを細胞ペレットに加え、15秒以内ボルテックスすることにより赤血球の浸透圧溶解を行った。次に2×PBS1mlを加え、PBS1×で容量を4mlにした。1200rpmで10分間室温にて遠心後、細胞ペレットをR10培地1mlに再懸濁し、生存細胞をカウントした。脾細胞を5×105個及び2×105個/ウェルの割合でプレーティングし、R10中各ペプチド1μg/mlを加え、20時間37℃のCO2インキュベーターでインキュベートした。コンカナバリンA(ConA)5μg/mlを各マウスで陽性内部対照として使用した。PBS,0.05% Tween 20で洗浄後、アッセイ緩衝液(PBS−5%FBS−0.005% Tween−20)で250倍に希釈したビオチン標識ラット抗マウスIFN−γ(Rat IgGl,クローンXMG1.2,Pharmingen,San Jose,CA)50μl/ウェルと共にプレートを4℃でO/Nインキュベートした。
【0108】
翌日、プレートを洗浄し、アッセイ緩衝液で2500倍に希釈したストレプトアビジン−APコンジュゲート(Pharmingen)と共に2時間室温でインキュベートした。十分に洗浄後、スポット展開が顕微鏡で観察されるまでNBT/B−CIP(Pierce Biotechnology)50μl/ウェルを加えることによりプレートを展開した。プレートを蒸留水で十分に洗浄することにより反応を停止した。プレートを完全に風乾し、自動ELISPOTリーダーを使用してスポットをカウントした。
【0109】
細胞性免疫応答のために、CEA.TgマウスにhCEA発現ベクター又はrhCEA発現ベクターをワクチン接種した。2群を分析し、第1群は最終DNA注入から21日後にELISPOTアッセイにより分析し、第2群はAd5−hCEA又はAd5−RhCEA 1×1010vpをブースター投与し、2週間後に分析した。その結果、ELISPOTにより測定した場合に4回のDNA注入後にhCEAに対する有意細胞性免疫応答は観察されなかった(図示せず)。他方、Ad5をブースター投与したマウスは顕著な応答増加を示し、CEAに対する免疫寛容の解除に一致した。この所見は、CEA自己抗原に有用な1つのワクチン接種プロトコールがEGTによるDNAの反復投与後にアデノウイルスブーストを行う方法(混合法)であることを示唆している。重要な点として、野生型マウスとトランスジェニックマウスのいずれにおいてもアカゲザルCEAの免疫はヒトCEAペプチドとの交差反応を生じ、逆の場合にも交差反応を生じた(データは示さず)。特に、ヒトCEAに対する免疫応答はrhCEAを免疫原として使用したトランスジェニックマウスのほうが著しく良好であった(図9参照)。これらの結果によると、アカゲザル(異種)遺伝子を使用してトランスジェニックマウスでCEAに対する良好な応答を得ることができる。アカゲザルCEAペプチドに対する応答を図10に示す。
【実施例10】
【0110】
rhCEAによるアカゲザルの免疫
結腸直腸腺癌で発現されるヒト腫瘍抗原CEAのアカゲザルホモログによるアカゲザル(macaca mulatta)の免疫効率を評価するために、Biomedical Primate Research Centre(BPRC,Rijswijk,オランダ)で免疫試験を実施した。この免疫試験はアカゲザルCEA抗原による免疫に対するB及びT細胞両者の応答を評価するように設計した。
【0111】
この試験(CV−1)では、アカゲザルCEACAM−5を発現するプラスミドDNAベクター及びアデノウイルスベクターで1群のサル(雄2頭と雌2頭)を免疫した。プライミングのために、rhCEAを発現するDNAを0、4、8、12、及び16週にDNA注入後に電気刺激により動物に筋肉内ワクチン接種した。DNA注入はプラスミドDNA5mgを含有する1ml溶液(2部位に分けて各部位0.5mlずつ)を体重2〜5キロの動物に注入した。麻酔下に動物に注入した(ケタミン/キシラジン混合物)。
【0112】
電気刺激については、方形バイポーラパルス100バルス2系列(各1秒)を1秒間隔で発生し、合計3秒間処理した。パルス長は2msec/位相とし、パルス周波数及び振幅は夫々100Hz及び100mA(定電流モード)とした。
【0113】
上記免疫プロトコールを使用してCEAに対する免疫応答を測定するために、血液試料を4週間おきに採取した。細胞性応答はIFNγ Elispotアッセイにより測定し、体液性応答はELISAアッセイにより測定した。16週で有意免疫応答が得られなかったので、rhCEAを発現するAd5を使用して更に2回注入した(24週及び28週)。Ad5注入後に夫々ペプチドプールC及びプールB+Cに対応する2頭のサル(RI137及びCO12)でrhCEAに対する測定可能な免疫応答が検出された。細胞性免疫応答は両者サルで35週目に低下し始めた。
【0114】
DNA注入後に体液性免疫応答を経時的に追跡した。3頭のサル(CO12,RI311及びRI002)は1:143〜1:2099の良好な抗CEA抗体力価を示し、最初の注入から12〜16週間後にピークに達した。
【0115】
これらのデータによると、rhCEAをコードする遺伝子ベクターは霊長類でこの腫瘍抗原に対する免疫寛容を解除することができた。免疫応答には細胞性(処理したサルの50%)と体液性(処理したサルの75%)のどちらの免疫も検出された。
【実施例11】
【0116】
ヒト腫瘍関連抗原のアカゲザルホモログによるアカゲザルの免疫
いずれも結腸直腸腺癌で発現されるヒト腫瘍抗原HER2/neu、Ep−CAM及びCEAのアカゲザルホモログによるアカゲザル(macaca mulatta)の免疫効率を評価するために、第2系列の免疫試験を実施した。プロトコールはこれらの腫瘍抗原の組み合わせに対するB及びT細胞両者の応答を評価するように設計した。
【0117】
この試験では、ヒト腫瘍抗原Ep−CAM(pV1J−rhEpCAM)、CEA(pV1J−rhCEA)、及びHER2/neu(pV1J−rhHER2)のアカゲザルホモログを発現する3種のプラスミドDNAベクターの混合物で第2群のアカゲザル4頭(雄2頭と雌2頭)を免疫した。
【0118】
0、4、8、12、及び16週にプラスミドDNAの筋肉内注射後に電気刺激により動物をプライミングした。DNA注入は体重2〜5キロの動物にプラスミドDNA6mgを含有する1ml溶液(2部位に分けて各部位0.5mlずつ)を注入した。麻酔下に動物に注入した(ケタミン/キシラジン混合物)。
【0119】
電気刺激については、方形バイポーラパルス100バルス2系列(各1秒)を1秒間隔で発生し、合計3秒間処理した。パルス長は2msec/位相とし、パルス周波数及び振幅は夫々100Hz及び100mA(定電流モード)とした。
【0120】
アカゲザルCEA(Ad5−rhCEA)、アカゲザルHER2/neu(Ad5−rhHER2)、及びアカゲザルEpCAM(Ad5−rhEpCAM)を発現する3種のAd5の混合物の注入により同一群の動物をブーストした。合計量3×10exp11ウイルス粒子(vp)を23週と27週に注入した(3種のウイルスを各々1×10exp11 vp)。
【0121】
上記免疫プロトコールを使用して3種の腫瘍抗原に対する免疫応答を測定するために、血液試料を4週間おきに採取した。細胞性免疫応答はIFN−γ+ELISPOTアッセイにより測定し、体液性応答はELISAにより測定した。
【0122】
サルRI449及びRI519はIFN−γ ELISPOT分析により測定した場合に検出可能なHER2特異的細胞性応答を示した。同様の分析でrhCEA及びrhEpCAMに対する有意応答は検出されなかった。
【0123】
第3の試験では、0、2及び4週にAd5誘導体の筋肉内注射によりAd5−rhHER2、Ad5−rhCEA及びAd5−rhEpCAMの混合物でアカゲザル4頭を免疫した。3×10exp11 vp(3種のAd5ウイルスを各々10exp11)を含有する1ml溶液(2部位に分けて各部位0.5mlずつ)を体重2〜5キロの動物に麻酔下(ケタミン/キシラジン混合物)に投与した。
【0124】
細胞性応答はIFNγ ELISPOTアッセイにより測定した。Her2/Neuでは、4頭のサルのうちの3頭が検出可能な応答を示した。rhCEAとrhEpCAMでは有意細胞性応答は測定されなかった。
【0125】
まとめると、上記免疫プロトコールはアカゲザルでrhHER2/neuに対する特異的免疫応答を誘導するのに有効であった。3種の異なる腫瘍抗原をもつベクターによる同時免疫がrhCEAのみを免疫原として使用した試験1に比較してrhCEAに対する免疫応答を誘導するのに有効でない理由は不明である。理論に拘束する意図はないが、rhHER2/Neuの発現と免疫ドミナントエピトープの存在がサブドミナントrhCEA特異的T細胞の生産と発現を制限したのではないかと考えられる。
【図面の簡単な説明】
【0126】
【図1A】配列番号1(図1A)及び配列番号5(図1B)に記載のアカゲザルCEAcDNA分子のヌクレオチド配列を示す。実施例2参照。
【図1B】配列番号1(図1A)及び配列番号5(図1B)に記載のアカゲザルCEAcDNA分子のヌクレオチド配列を示す。実施例2参照。
【図2A】配列番号2(図2A)に記載の第1のアカゲザルCEA蛋白質と配列番号18(図2B)に記載の第2のアカゲザルCEA蛋白質の予想アミノ酸配列を示す。第1及び第2のアカゲザルCEA蛋白質間の2アミノ酸変異を図2Bに太字下線で示す。
【図2B】配列番号2(図2A)に記載の第1のアカゲザルCEA蛋白質と配列番号18(図2B)に記載の第2のアカゲザルCEA蛋白質の予想アミノ酸配列を示す。第1及び第2のアカゲザルCEA蛋白質間の2アミノ酸変異を図2Bに太字下線で示す。
【図3】ヒトCEACAMファミリーメンバーの5’非翻訳領域のアラインメントを示す。指定配列を比較及び使用して実施例2に記載するように縮重プライマーを設計した。他のCEACAMファミリーメンバーの対応するヌクレオチドと同一のヌクレオチドを白抜き表示した。破線は配列のアラインメントを助長するためにスペースを加えたことを示す。GenBankに開示されているような各cDNA配列のヌクレオチド数を括弧内に示す。
【図4】アカゲザルCEA蛋白質の発現を示す。λ−CEAライブラリーのスクリーニングにより得られたファージミドをHeLa細胞にトランスフェクトし、ヒトCEA蛋白質に対するウサギポリクローナル抗体を使用してウェスタンブロットを実施した。15個のクローンのうちの2個の発現を示す。
【図5】アカゲザルCEAコーディング領域の模式図を示す。内部反復配列を示し、遺伝子断片化用制限部位と配列を報告する。
【図6A】ヒト(配列番号6)及びアカゲザル(配列番号1)CEACAM−5ヌクレオチド配列のアラインメントを示す。2種のCEACAM−5配列間のヌクレオチド変異を太字で示す。
【図6B】ヒト(配列番号6)及びアカゲザル(配列番号1)CEACAM−5ヌクレオチド配列のアラインメントを示す。2種のCEACAM−5配列間のヌクレオチド変異を太字で示す。
【図6C】ヒト(配列番号6)及びアカゲザル(配列番号1)CEACAM−5ヌクレオチド配列のアラインメントを示す。2種のCEACAM−5配列間のヌクレオチド変異を太字で示す。
【図7】ヒト(配列番号7)及びアカゲザル(配列番号2)CEACAM−5オープンリーディングフレームのアラインメントを示す。2種のCEACAM−5配列間のアミノ酸変異を太字で示す。
【図8】CEAトランスジェニックマウスにおけるヒトCEAに対する体液性応答を示す。アカゲザルCEAを免疫した群とヒトCEAを免疫した群の2つのマウス群の平均抗体力価を示す(実施例7)。
【図9】CEAトランスジェニックマウスにおけるヒトCEAに対する細胞性免疫応答を示す。hCEAを発現するベクター又はrhCEAを発現するベクターをCEAトランスジェニックマウスにワクチン接種した(実施例9)。
【図10】アカゲザル又はヒトCEAを免疫したCEAトランスジェニックマウスにおけるアカゲザルCEAペプチドに対する細胞性免疫応答を示す。
【配列表】
【特許請求の範囲】
【請求項1】
配列番号2に記載のアカゲザル癌胎児性抗原(rhCEA)蛋白質をコードするヌクレオチド配列を含む単離核酸分子。
【請求項2】
核酸がDNAである請求項1に記載の単離核酸分子。
【請求項3】
核酸がmRNAである請求項1に記載の単離核酸分子。
【請求項4】
核酸がcDNAである請求項1に記載の単離核酸分子。
【請求項5】
ヌクレオチド配列が配列番号1に記載のヌクレオチド配列を含む請求項1に記載の単離核酸分子。
【請求項6】
請求項1に記載の核酸分子を含むベクター。
【請求項7】
請求項6に記載のベクターを含む宿主細胞。
【請求項8】
組換え宿主細胞におけるアカゲザル癌胎児性抗原(CEA)蛋白質の発現方法であって、
(a)請求項1に記載の核酸を含むベクターを適当な宿主細胞に導入する段階と;
(b)前記アカゲザルCE蛋白質の発現を可能にする条件下で宿主細胞を培養する段階を含む前記方法。
【請求項9】
組換え宿主細胞におけるアカゲザル癌胎児性抗原(CEA)蛋白質の発現方法であって、
(a)請求項5に記載の核酸を含むベクターを適当な宿主細胞に導入する段階と;
(b)前記アカゲザルCEA蛋白質の発現を可能にする条件下で宿主細胞を培養する段階を含む前記方法。
【請求項10】
配列番号2に記載のアミノ酸配列を含む単離精製アカゲザルCEAポリペプチド。
【請求項11】
配列番号18に記載のアカゲザル癌胎児性抗原(rhCEA)蛋白質をコードするヌクレオチド配列を含む単離核酸分子。
【請求項12】
核酸がDNAである請求項11に記載の単離核酸分子。
【請求項13】
核酸がmRNAである請求項11に記載の単離核酸分。
【請求項14】
核酸がcDNAである請求項11に記載の単離核酸分。
【請求項15】
ヌクレオチド配列が配列番号5に記載のヌクレオチド配列を含む請求項11に記載の単離核酸分子。
【請求項16】
請求項11に記載の核酸分子を含むベクター。
【請求項17】
請求項16に記載のベクターを含む宿主細胞。
【請求項18】
組換え宿主細胞におけるアカゲザル癌胎児性抗原(CEA)蛋白質の発現方法であって、
(a)請求項11に記載の核酸を含むベクターを適当な宿主細胞に導入する段階と;
(b)前記アカゲザルCE蛋白質の発現を可能にする条件下で宿主細胞を培養する段階を含む前記方法。
【請求項19】
配列番号18に記載のアミノ酸配列を含む単離精製アカゲザルCEAポリペプチド。
【請求項20】
単離核酸分子を含むワクチンベクターを哺乳動物に投与することを含む癌の予防又は治療方法であって、単離核酸分子が配列番号2又は配列番号18に記載のアカゲザル癌胎児性抗原(rhCEA)蛋白質をコードするヌクレオチド配列を含む前記方法。
【請求項21】
哺乳動物がヒトである請求項20に記載の方法。
【請求項22】
ベクターがアデノウイルスベクター又はプラスミドベクターである請求項20に記載の方法。
【請求項23】
ベクターがアデノウイルスE1領域に欠失とアデノウイルスE1領域に挿入をもつアデノウイルスゲノムを含むアデノウイルスベクターであり、挿入が
(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;
(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含む請求項20に記載の方法。
【請求項24】
ベクターがプラスミド部分と、
(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;
(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含むプラスミドワクチンベクターである請求項20に記載の方法。
【請求項25】
E1領域に欠失とE1領域に挿入をもつアデノウイルスゲノムを含むアデノウイルスワクチンベクターであって、挿入が
(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;
(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含む前記アデノウイルスワクチンベクター。
【請求項26】
Ad5ベクターである請求項25に記載のアデノウイルスベクター。
【請求項27】
Ad6ベクターである請求項25に記載のアデノウイルスベクター。
【請求項28】
プラスミド部分と発現カセット部分を含むワクチンプラスミドであって、発現カセット部分が
(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;
(b)ポリヌクレオチドに機能的に連結したプロモーターを含む前記ワクチンプラスミド。
【請求項29】
哺乳動物を癌から保護する方法であって、
(a)(i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第1のベクターを哺乳動物に導入する段階と;
(b)所定時間経過させる段階と;
(c)(i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第2のベクターを哺乳動物に導入する段階を含む前記方法。
【請求項30】
第1のベクターがプラスミドであり、第2のベクターがアデノウイルスベクターである請求項29に記載の方法。
【請求項31】
第1のベクターがアデノウイルスベクターであり、第2のベクターがプラスミドである請求項29に記載の方法。
【請求項32】
結腸直腸癌の哺乳動物の治療方法であって、
(a)(i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第1のベクターを哺乳動物に導入する段階と;
(b)所定時間経過させる段階と;
(c)(i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第2のベクターを哺乳動物に導入する段階を含む前記方法。
【請求項33】
第1のベクターがプラスミドであり、第2のベクターがアデノウイルスベクターである請求項32に記載の方法。
【請求項34】
第1のベクターがアデノウイルスベクターであり、第2のベクターがプラスミドである請求項32に記載の方法。
【特許請求の範囲】
【請求項1】
配列番号2に記載のアカゲザル癌胎児性抗原(rhCEA)蛋白質をコードするヌクレオチド配列を含む単離核酸分子。
【請求項2】
核酸がDNAである請求項1に記載の単離核酸分子。
【請求項3】
核酸がmRNAである請求項1に記載の単離核酸分子。
【請求項4】
核酸がcDNAである請求項1に記載の単離核酸分子。
【請求項5】
ヌクレオチド配列が配列番号1に記載のヌクレオチド配列を含む請求項1に記載の単離核酸分子。
【請求項6】
請求項1に記載の核酸分子を含むベクター。
【請求項7】
請求項6に記載のベクターを含む宿主細胞。
【請求項8】
組換え宿主細胞におけるアカゲザル癌胎児性抗原(CEA)蛋白質の発現方法であって、
(a)請求項1に記載の核酸を含むベクターを適当な宿主細胞に導入する段階と;
(b)前記アカゲザルCE蛋白質の発現を可能にする条件下で宿主細胞を培養する段階を含む前記方法。
【請求項9】
組換え宿主細胞におけるアカゲザル癌胎児性抗原(CEA)蛋白質の発現方法であって、
(a)請求項5に記載の核酸を含むベクターを適当な宿主細胞に導入する段階と;
(b)前記アカゲザルCEA蛋白質の発現を可能にする条件下で宿主細胞を培養する段階を含む前記方法。
【請求項10】
配列番号2に記載のアミノ酸配列を含む単離精製アカゲザルCEAポリペプチド。
【請求項11】
配列番号8に記載のアカゲザル癌胎児性抗原(rhCEA)蛋白質をコードするヌクレオチド配列を含む単離核酸分子。
【請求項12】
核酸がDNAである請求項11に記載の単離核酸分子。
【請求項13】
核酸がmRNAである請求項11に記載の単離核酸分。
【請求項14】
核酸がcDNAである請求項11に記載の単離核酸分。
【請求項15】
ヌクレオチド配列が配列番号5に記載のヌクレオチド配列を含む請求項11に記載の単離核酸分子。
【請求項16】
請求項11に記載の核酸分子を含むベクター。
【請求項17】
請求項16に記載のベクターを含む宿主細胞。
【請求項18】
組換え宿主細胞におけるアカゲザル癌胎児性抗原(CEA)蛋白質の発現方法であって、
(a)請求項11に記載の核酸を含むベクターを適当な宿主細胞に導入する段階と;
(b)前記アカゲザルCE蛋白質の発現を可能にする条件下で宿主細胞を培養する段階を含む前記方法。
【請求項19】
配列番号8に記載のアミノ酸配列を含む単離精製アカゲザルCEAポリペプチド。
【請求項20】
単離核酸分子を含むワクチンベクターを哺乳動物に投与することを含む癌の予防又は治療方法であって、単離核酸分子が配列番号2又は配列番号8に記載のアカゲザル癌胎児性抗原(rhCEA)蛋白質をコードするヌクレオチド配列を含む前記方法。
【請求項21】
哺乳動物がヒトである請求項20に記載の方法。
【請求項22】
ベクターがアデノウイルスベクター又はプラスミドベクターである請求項20に記載の方法。
【請求項23】
ベクターがアデノウイルスE1領域に欠失とアデノウイルスE1領域に挿入をもつアデノウイルスゲノムを含むアデノウイルスベクターであり、挿入が
(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;
(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含む請求項20に記載の方法。
【請求項24】
ベクターがプラスミド部分と、
(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;
(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含むプラスミドワクチンベクターである請求項20に記載の方法。
【請求項25】
E1領域に欠失とE1領域に挿入をもつアデノウイルスゲノムを含むアデノウイルスワクチンベクターであって、挿入が
(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;
(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含む前記アデノウイルスワクチンベクター。
【請求項26】
Ad5ベクターである請求項25に記載のアデノウイルスベクター。
【請求項27】
Ad6ベクターである請求項25に記載のアデノウイルスベクター。
【請求項28】
プラスミド部分と発現カセット部分を含むワクチンプラスミドであって、発現カセット部分が
(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;
(b)ポリヌクレオチドに機能的に連結したプロモーターを含む前記ワクチンプラスミド。
【請求項29】
哺乳動物を癌から保護する方法であって、
(a)(i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第1のベクターを哺乳動物に導入する段階と;
(b)所定時間経過させる段階と;
(c)(i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第2のベクターを哺乳動物に導入する段階を含む前記方法。
【請求項30】
第1のベクターがプラスミドであり、第2のベクターがアデノウイルスベクターである請求項29に記載の方法。
【請求項31】
第1のベクターがアデノウイルスベクターであり、第2のベクターがプラスミドである請求項29に記載の方法。
【請求項32】
結腸直腸癌の哺乳動物の治療方法であって、
(a)(i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第1のベクターを哺乳動物に導入する段階と;
(b)所定時間経過させる段階と;
(c)(i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第2のベクターを哺乳動物に導入する段階を含む前記方法。
【請求項33】
第1のベクターがプラスミドであり、第2のベクターがアデノウイルスベクターである請求項32に記載の方法。
【請求項34】
第1のベクターがアデノウイルスベクターであり、第2のベクターがプラスミドである請求項32に記載の方法。
【請求項1】
配列番号2に記載のアカゲザル癌胎児性抗原(rhCEA)蛋白質をコードするヌクレオチド配列を含む単離核酸分子。
【請求項2】
核酸がDNAである請求項1に記載の単離核酸分子。
【請求項3】
核酸がmRNAである請求項1に記載の単離核酸分子。
【請求項4】
核酸がcDNAである請求項1に記載の単離核酸分子。
【請求項5】
ヌクレオチド配列が配列番号1に記載のヌクレオチド配列を含む請求項1に記載の単離核酸分子。
【請求項6】
請求項1に記載の核酸分子を含むベクター。
【請求項7】
請求項6に記載のベクターを含む宿主細胞。
【請求項8】
組換え宿主細胞におけるアカゲザル癌胎児性抗原(CEA)蛋白質の発現方法であって、
(a)請求項1に記載の核酸を含むベクターを適当な宿主細胞に導入する段階と;
(b)前記アカゲザルCE蛋白質の発現を可能にする条件下で宿主細胞を培養する段階を含む前記方法。
【請求項9】
組換え宿主細胞におけるアカゲザル癌胎児性抗原(CEA)蛋白質の発現方法であって、
(a)請求項5に記載の核酸を含むベクターを適当な宿主細胞に導入する段階と;
(b)前記アカゲザルCEA蛋白質の発現を可能にする条件下で宿主細胞を培養する段階を含む前記方法。
【請求項10】
配列番号2に記載のアミノ酸配列を含む単離精製アカゲザルCEAポリペプチド。
【請求項11】
配列番号18に記載のアカゲザル癌胎児性抗原(rhCEA)蛋白質をコードするヌクレオチド配列を含む単離核酸分子。
【請求項12】
核酸がDNAである請求項11に記載の単離核酸分子。
【請求項13】
核酸がmRNAである請求項11に記載の単離核酸分。
【請求項14】
核酸がcDNAである請求項11に記載の単離核酸分。
【請求項15】
ヌクレオチド配列が配列番号5に記載のヌクレオチド配列を含む請求項11に記載の単離核酸分子。
【請求項16】
請求項11に記載の核酸分子を含むベクター。
【請求項17】
請求項16に記載のベクターを含む宿主細胞。
【請求項18】
組換え宿主細胞におけるアカゲザル癌胎児性抗原(CEA)蛋白質の発現方法であって、
(a)請求項11に記載の核酸を含むベクターを適当な宿主細胞に導入する段階と;
(b)前記アカゲザルCE蛋白質の発現を可能にする条件下で宿主細胞を培養する段階を含む前記方法。
【請求項19】
配列番号18に記載のアミノ酸配列を含む単離精製アカゲザルCEAポリペプチド。
【請求項20】
単離核酸分子を含むワクチンベクターを哺乳動物に投与することを含む癌の予防又は治療方法であって、単離核酸分子が配列番号2又は配列番号18に記載のアカゲザル癌胎児性抗原(rhCEA)蛋白質をコードするヌクレオチド配列を含む前記方法。
【請求項21】
哺乳動物がヒトである請求項20に記載の方法。
【請求項22】
ベクターがアデノウイルスベクター又はプラスミドベクターである請求項20に記載の方法。
【請求項23】
ベクターがアデノウイルスE1領域に欠失とアデノウイルスE1領域に挿入をもつアデノウイルスゲノムを含むアデノウイルスベクターであり、挿入が
(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;
(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含む請求項20に記載の方法。
【請求項24】
ベクターがプラスミド部分と、
(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;
(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含むプラスミドワクチンベクターである請求項20に記載の方法。
【請求項25】
E1領域に欠失とE1領域に挿入をもつアデノウイルスゲノムを含むアデノウイルスワクチンベクターであって、挿入が
(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;
(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含む前記アデノウイルスワクチンベクター。
【請求項26】
Ad5ベクターである請求項25に記載のアデノウイルスベクター。
【請求項27】
Ad6ベクターである請求項25に記載のアデノウイルスベクター。
【請求項28】
プラスミド部分と発現カセット部分を含むワクチンプラスミドであって、発現カセット部分が
(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;
(b)ポリヌクレオチドに機能的に連結したプロモーターを含む前記ワクチンプラスミド。
【請求項29】
哺乳動物を癌から保護する方法であって、
(a)(i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第1のベクターを哺乳動物に導入する段階と;
(b)所定時間経過させる段階と;
(c)(i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第2のベクターを哺乳動物に導入する段階を含む前記方法。
【請求項30】
第1のベクターがプラスミドであり、第2のベクターがアデノウイルスベクターである請求項29に記載の方法。
【請求項31】
第1のベクターがアデノウイルスベクターであり、第2のベクターがプラスミドである請求項29に記載の方法。
【請求項32】
結腸直腸癌の哺乳動物の治療方法であって、
(a)(i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第1のベクターを哺乳動物に導入する段階と;
(b)所定時間経過させる段階と;
(c)(i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第2のベクターを哺乳動物に導入する段階を含む前記方法。
【請求項33】
第1のベクターがプラスミドであり、第2のベクターがアデノウイルスベクターである請求項32に記載の方法。
【請求項34】
第1のベクターがアデノウイルスベクターであり、第2のベクターがプラスミドである請求項32に記載の方法。
【特許請求の範囲】
【請求項1】
配列番号2に記載のアカゲザル癌胎児性抗原(rhCEA)蛋白質をコードするヌクレオチド配列を含む単離核酸分子。
【請求項2】
核酸がDNAである請求項1に記載の単離核酸分子。
【請求項3】
核酸がmRNAである請求項1に記載の単離核酸分子。
【請求項4】
核酸がcDNAである請求項1に記載の単離核酸分子。
【請求項5】
ヌクレオチド配列が配列番号1に記載のヌクレオチド配列を含む請求項1に記載の単離核酸分子。
【請求項6】
請求項1に記載の核酸分子を含むベクター。
【請求項7】
請求項6に記載のベクターを含む宿主細胞。
【請求項8】
組換え宿主細胞におけるアカゲザル癌胎児性抗原(CEA)蛋白質の発現方法であって、
(a)請求項1に記載の核酸を含むベクターを適当な宿主細胞に導入する段階と;
(b)前記アカゲザルCE蛋白質の発現を可能にする条件下で宿主細胞を培養する段階を含む前記方法。
【請求項9】
組換え宿主細胞におけるアカゲザル癌胎児性抗原(CEA)蛋白質の発現方法であって、
(a)請求項5に記載の核酸を含むベクターを適当な宿主細胞に導入する段階と;
(b)前記アカゲザルCEA蛋白質の発現を可能にする条件下で宿主細胞を培養する段階を含む前記方法。
【請求項10】
配列番号2に記載のアミノ酸配列を含む単離精製アカゲザルCEAポリペプチド。
【請求項11】
配列番号8に記載のアカゲザル癌胎児性抗原(rhCEA)蛋白質をコードするヌクレオチド配列を含む単離核酸分子。
【請求項12】
核酸がDNAである請求項11に記載の単離核酸分子。
【請求項13】
核酸がmRNAである請求項11に記載の単離核酸分。
【請求項14】
核酸がcDNAである請求項11に記載の単離核酸分。
【請求項15】
ヌクレオチド配列が配列番号5に記載のヌクレオチド配列を含む請求項11に記載の単離核酸分子。
【請求項16】
請求項11に記載の核酸分子を含むベクター。
【請求項17】
請求項16に記載のベクターを含む宿主細胞。
【請求項18】
組換え宿主細胞におけるアカゲザル癌胎児性抗原(CEA)蛋白質の発現方法であって、
(a)請求項11に記載の核酸を含むベクターを適当な宿主細胞に導入する段階と;
(b)前記アカゲザルCE蛋白質の発現を可能にする条件下で宿主細胞を培養する段階を含む前記方法。
【請求項19】
配列番号8に記載のアミノ酸配列を含む単離精製アカゲザルCEAポリペプチド。
【請求項20】
単離核酸分子を含むワクチンベクターを哺乳動物に投与することを含む癌の予防又は治療方法であって、単離核酸分子が配列番号2又は配列番号8に記載のアカゲザル癌胎児性抗原(rhCEA)蛋白質をコードするヌクレオチド配列を含む前記方法。
【請求項21】
哺乳動物がヒトである請求項20に記載の方法。
【請求項22】
ベクターがアデノウイルスベクター又はプラスミドベクターである請求項20に記載の方法。
【請求項23】
ベクターがアデノウイルスE1領域に欠失とアデノウイルスE1領域に挿入をもつアデノウイルスゲノムを含むアデノウイルスベクターであり、挿入が
(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;
(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含む請求項20に記載の方法。
【請求項24】
ベクターがプラスミド部分と、
(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;
(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含むプラスミドワクチンベクターである請求項20に記載の方法。
【請求項25】
E1領域に欠失とE1領域に挿入をもつアデノウイルスゲノムを含むアデノウイルスワクチンベクターであって、挿入が
(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;
(b)ポリヌクレオチドに機能的に連結したプロモーターを含む発現カセットを含む前記アデノウイルスワクチンベクター。
【請求項26】
Ad5ベクターである請求項25に記載のアデノウイルスベクター。
【請求項27】
Ad6ベクターである請求項25に記載のアデノウイルスベクター。
【請求項28】
プラスミド部分と発現カセット部分を含むワクチンプラスミドであって、発現カセット部分が
(a)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;
(b)ポリヌクレオチドに機能的に連結したプロモーターを含む前記ワクチンプラスミド。
【請求項29】
哺乳動物を癌から保護する方法であって、
(a)(i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第1のベクターを哺乳動物に導入する段階と;
(b)所定時間経過させる段階と;
(c)(i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第2のベクターを哺乳動物に導入する段階を含む前記方法。
【請求項30】
第1のベクターがプラスミドであり、第2のベクターがアデノウイルスベクターである請求項29に記載の方法。
【請求項31】
第1のベクターがアデノウイルスベクターであり、第2のベクターがプラスミドである請求項29に記載の方法。
【請求項32】
結腸直腸癌の哺乳動物の治療方法であって、
(a)(i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第1のベクターを哺乳動物に導入する段階と;
(b)所定時間経過させる段階と;
(c)(i)アカゲザルCEA蛋白質をコードするポリヌクレオチドと;(ii)ポリヌクレオチドに機能的に連結したプロモーターを含む第2のベクターを哺乳動物に導入する段階を含む前記方法。
【請求項33】
第1のベクターがプラスミドであり、第2のベクターがアデノウイルスベクターである請求項32に記載の方法。
【請求項34】
第1のベクターがアデノウイルスベクターであり、第2のベクターがプラスミドである請求項32に記載の方法。
【図1A】


【図1B】

【図2A】

【図2B】

【図3】
【図4】
【図5】
【図6A】

【図6B】

【図6C】

【図7】

【図8】
【図9】
【図10】
【図1B】
【図2A】
【図2B】
【図3】
【図4】
【図5】
【図6A】
【図6B】
【図6C】
【図7】
【図8】
【図9】
【図10】
【公表番号】特表2006−518202(P2006−518202A)
【公表日】平成18年8月10日(2006.8.10)
【国際特許分類】
【出願番号】特願2006−501784(P2006−501784)
【出願日】平成16年2月9日(2004.2.9)
【国際出願番号】PCT/EP2004/001181
【国際公開番号】WO2004/072287
【国際公開日】平成16年8月26日(2004.8.26)
【出願人】(501209427)イステイチユート・デイ・リチエルケ・デイ・ビオロジア・モレコラーレ・ピ・アンジエレツテイ・エツセ・ピー・アー (90)
【Fターム(参考)】
【公表日】平成18年8月10日(2006.8.10)
【国際特許分類】
【出願日】平成16年2月9日(2004.2.9)
【国際出願番号】PCT/EP2004/001181
【国際公開番号】WO2004/072287
【国際公開日】平成16年8月26日(2004.8.26)
【出願人】(501209427)イステイチユート・デイ・リチエルケ・デイ・ビオロジア・モレコラーレ・ピ・アンジエレツテイ・エツセ・ピー・アー (90)
【Fターム(参考)】
[ Back to top ]