
アカゲザルHER2/neu、これをコードするヌクレオチド及びその使用
アカゲザルHER2/neuをコードするポリヌクレオチドを単離し、クローニングし、配列決定した。HER2/neuをコードする遺伝子は、一般に上皮由来のヒト癌の発症を伴う。本発明は、HER2/neuの異常発現が癌又はその発症を伴う場合に、HER2/neu腫瘍関連抗原によって発現されるタンパク質産物に対する免疫を惹起する又は増強するための組成物及び方法を提供する。本発明は、特に、rhHER2/neuを担持するアデノウイルスベクター構築物を提供し、癌を予防及び治療するためのワクチン及び医薬組成物におけるそれらの使用を開示する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、一般に癌の検出及び治療に関する。特に、本発明は、腫瘍関連ポリペプチドHER2/neuのアカゲザルホモログ、このタンパク質をコードする単離核酸分子、及びこのタンパク質をコードするDNAを含む組換えベクター及び宿主に関する。本発明はまた、アカゲザルHER2/neuを担持するアデノウイルスベクター構築物、及び癌を予防及び治療するためのワクチン及び医薬組成物におけるこれらの使用に関する。
【背景技術】
【0002】
癌は、典型的には、増殖因子及びそれらの受容体、癌遺伝子及び癌抑制遺伝子などの、細胞周期を維持する又は細胞増殖を制御することに寄与する遺伝子の調節不全を含む。これらの遺伝子の多くの産物は様々な腫瘍細胞の表面で発現され、従って、腫瘍関連抗原(TAA)と命名された。最近の証拠は、免疫応答を惹起することができる腫瘍関連抗原の存在を裏付けており、これらの分子はワクチン療法のための標的となっている。これらの遺伝子産物の多くは、より低いレベルではあるが、正常細胞でも発現されるので、腫瘍関連抗原を標的する多くの癌ワクチンは、免疫寛容の故に無効であることが証明された。
【0003】
HER2/neu癌原遺伝子(c−erbB−2とも呼ばれる)の産物は、上皮増殖因子受容体ファミリーの成員である膜貫通TAAである。HER2/neu遺伝子は、最初にラット神経膠芽細胞腫からクローン化され(Shihら、Nature 290:261−264(1981))、後にヒト細胞から単離されて、特性決定された(Coussensら、Science 230:1132−39(1985);Kingら、Science 229:974−76(1985))。現在まで、HER2/neuのサルホモログは得られていない。
【0004】
HER2/neuは、細胞増殖と分化に関与する4つの受容体から成る、受容体チロシンキナーゼのHERファミリーの成員としてさらに分類されてきた。HER受容体は、二量体として増殖因子リガンドに結合し、最終的に細胞増殖において重要な遺伝子の活性化を生じさせる細胞内シグナル伝達カスケードを開始させることにより、正常な細胞増殖を維持することに寄与する。幾つかのリガンドがHERファミリーの他の成員について特定されているが、HER2/neu受容体についての高親和性リガンドはまだ発見されていない(LohrischとPiccart,Semin.Oncol.28(6):補遺18:3−11(2001))。
【0005】
HER2/neu転写産物及びコードされる185kDタンパク質の低いレベルの発現が、皮膚及び乳房、及び胃腸管、生殖路及び尿路の組織を含む、様々な組織の健常成人上皮細胞において検出された(Pressら、Oncogene 5:963−962(1990))。より高いレベルのHER2/neu発現も、胚発生期の対応する胎児組織において検出された(Pressら、前出)。
【0006】
HER2/neuは、一般に乳癌、卵巣癌、子宮癌、結腸癌及び前立腺癌、及び肺の腺癌などの様々な悪性疾患において過剰発現される又は増幅される(DisisとCheever,Adv.Cancer Research 71:343−371(1997)において総説されている)。HER2/neuのこのような過剰発現は、癌患者についての予後不良及びより高い再発率と相関している(Slamonら、Science 244:707−712(1989))。
【0007】
HER2/neuの過剰発現に関連した悪性疾患に罹患している多くの癌患者は、以前にHER2/neu癌遺伝子のタンパク質産物に対して免疫応答を生じたことがあり、従ってHER2/neuは癌治療薬の開発のための免疫学的標的となっている。HER2/neuに対するこの免疫応答を利用した有効なワクチンは、この免疫を防御及び/又は予防的なレベルまで高め、自己寛容を克服するに違いない。
【0008】
HER2/neuは、種々の悪性疾患の免疫学的治療の開発のための標的として提案されてきた。種々の抗HERモノクローナル抗体が乳癌のための治療として検討されており、各々の抗体が様々なレベルの成功を明らかにしている(考察については、Yarden,Oncology 61(補遺2):1−13(2001)参照)。Amiciら(米国特許第6,127,344号)は、ヒトサイトメガロウイルスプロモーターに機能的に連結された完全長ヒトHER2/neu cDNAを含むベクターを投与することによってHER2/neuに対する免疫を誘導するための方法を開示している。CheeverとDisisは、HER2ペプチドでHER2/neuに関連する癌に対してヒトを免疫するための方法を開示している(米国特許第5,846,538号)。加えて、HER2/neuペプチドに基づくワクチンがげっ歯動物モデルにおいて試験されている(総説については、DisisとCheever,Advances in Cancer Research 71:343−371(1997)参照)。
【発明の開示】
【発明が解決しようとする課題】
【0009】
前述したHER2/neuクローンの特定にもかかわらず、有効であり且つ自己寛容によって妨げられない癌ワクチンの開発を可能にする、HER2/neuをコードするさらなる哺乳動物遺伝子を特定することは極めて望ましい。
【課題を解決するための手段】
【0010】
本発明は、配列番号2又は配列番号41に示す新規アカゲザルHER2/neuタンパク質(c−erbB−2とも呼ばれる、以下rhHER2/neuと称する)をコードするヌクレオチドの配列を含む単離又は精製核酸分子に関する。ここで開示するDNA分子は、選択宿主細胞にトランスフェクトしてもよく、前記組換え宿主細胞は、実質的なレベルの、発現される機能性rhHER2/neuタンパク質(配列番号2又は配列番号41)のためのソースを提供する。
【0011】
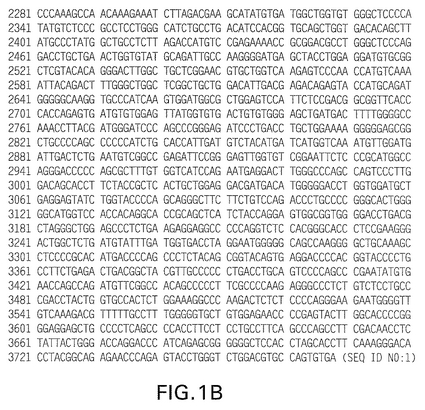
本発明はさらに、新規アカゲザルHER2/neuタンパク質を発現するmRNAをコードする単離核酸分子に関する。このDNA分子は、ここで配列番号1として開示するヌクレオチド配列を含む。本発明のこの部分の好ましい側面は図1に開示されており、図1は、新規rhHER2/neuタンパク質(配列番号2)をコードするDNA分子(配列番号1)を示す。
【0012】
本発明はまた、新規アカゲザルHER2/neuタンパク質を発現するmRNAをコードする単離核酸分子を提供する。このDNA分子は、ここで配列番号40として開示するヌクレオチド配列を含む。本発明のこの部分の好ましい側面は図5に開示されており、図5は、新規rhHER2/neuタンパク質(配列番号41)をコードするDNA分子(配列番号40)を示す。
【0013】
本発明はまた、本明細書全体を通じて開示する核酸分子を含む、原核及び真核生物の両方の、組換えベクター及び組換え宿主細胞に関する。
【0014】
本発明はさらに、(a)配列番号1又は配列番号40に示す核酸を含むベクターを適切な宿主細胞に導入すること;及び(b)前記宿主細胞を、前記アカゲザルHER2/neuタンパク質の発現を可能にする条件下で培養すること、を含む、組換え宿主細胞においてアカゲザルHER2/neuタンパク質を発現するための方法に関する。
【0015】
本発明の1つの好ましい側面は、図2に開示するアミノ酸配列(配列番号2)から成る、実質的に精製された形態のアカゲザルHER2/neuタンパク質である。
【0016】
本発明の1つの好ましい側面は、図6に開示するアミノ酸配列(配列番号41)から成る、実質的に精製された形態のアカゲザルHER2/neuタンパク質である。
【0017】
本発明のさらにもう1つの好ましい側面は、rhHER2/neuタンパク質を発現する、配列番号1又は配列番号40に示すヌクレオチド配列を含むDNA発現ベクターを含有する組換え宿主細胞から得られる、実質的に精製され、完全にプロセシングされた(タンパク質分解プロセシング、グリコシル化及び/又はリン酸化を含む)成熟rhHER2/neuタンパク質に関する。前記組換え宿主細胞は、哺乳動物細胞系などの真核生物宿主細胞であることが特に好ましい。
【0018】
本発明のもう1つの側面は、配列番号2又は配列番号41に示すアカゲザルHER2/neuタンパク質をコードするヌクレオチドの配列を含む単離核酸分子を含有するワクチンクチンベクターを哺乳動物に投与することを含む、癌を予防する又は治療する方法である。
【0019】
本発明はさらに、E1及びE3領域内の欠失及びE1領域内のインサートを有するアデノウイルスゲノムを含み、前記インサートは、(a)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド、及び(b)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む発現カセットを含有する、アデノウイルスワクチンベクターに関する。
【0020】
本発明はまた、プラスミド部分及び発現カセット部分を含有し、前記発現カセット部分は、(a)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及び(b)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含むワクチンプラスミドに関する。
【0021】
本発明のもう1つの側面は、(a)癌に羅患している哺乳動物に、i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド及びii)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む第一ベクターを導入すること;(b)あらかじめ定められた時間を経過させること;及び(c)前記哺乳動物に、i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド及びii)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む第二ベクターを導入すること、を含む、哺乳動物を癌から保護する又は癌に罹患している哺乳動物を治療する方法である。
【0022】
本明細書全体及び付属の特許請求の範囲において使用するとき、単数形態の「a」、「an」及び「the」は、文脈が明らかに異なる指示を命じない限り、複数の言及を包含する。
【0023】
本明細書全体及び付属の特許請求の範囲において使用するとき、以下の定義及び略語が適用される。
【0024】
「プロモーター」という用語は、RNAポリメラーゼが結合するDNA鎖上の認識部位を指す。プロモーターは、転写活性を開始させ、駆動するためにRNAポリメラーゼと開始複合体を形成する。前記複合体は、「エンハンサー」と称される活性化配列又は「サイレンサー」と称される抑制配列によって修飾され得る。
【0025】
「カセット」という用語は、発現されるべき核酸配列を含む本発明の配列を指す。前記カセットは、概念においてはカセットテープに類似し、各々のカセットはそれ自身の配列を有する。従って、カセットを交換することにより、ベクターは異なる配列を発現する。5’及び3’末端の制限部位の故に、カセットは容易に挿入する、除去する又はもう1つ別のカセットと置換することができる。
【0026】
「ベクター」という用語は、DNA断片を宿主生物又は宿主組織に導入することができる何らかの手段を指す。プラスミド、ウイルス(アデノウイルスを含む)、バクテリオファージ及びコスミドを含む様々な種類のベクターが存在する。
【0027】
アデノウイルスベクターに関して使用するとき「第一世代」という用語は、複製欠損である前記アデノウイルスベクターを表わす。第一世代アデノウイルスベクターは、典型的には欠失又は不活性化E1遺伝子領域を有し、好ましくは欠失又は不活性化E3遺伝子領域を有する。
【0028】
「pMRKAd5−rhHER2/neu」との名称は、E1及びE3領域の欠失したAd5アデノウイルスゲノムを含む、ここで開示するプラスミド構築物を指す。このプラスミドでは、E1領域は、イントロンAを伴わずにヒトCMVプロモーターの制御下にあり、続いてウシ成長ホルモンポリアデニル化シグナルが続く、E1と平行する方向のアカゲザルHER2/neuによって置換されている。
【0029】
「MRKAd5−rhHER2/neu」との名称は、制限及びPer.C6又はHEK293などのE1発現細胞系統へのトランスフェクションによるプラスミド配列の除去後にプラスミドpMRKAD5−rhHER2/neuから生成されるウイルスを指す。
【0030】
「pV1J−rhHER2/neu」との名称は、ヒトCMV最初期(IE)プロモーター及びイントロンA、完全長アカゲザルHER2/neu遺伝子、ウシ成長ホルモン由来のポリアデニル化及び転写終結配列、及び最小pUC骨格を含む、ここで開示するプラスミド構築物を指す。
【0031】
「rhHER2#1」という用語と交換可能に使用される、「第一アカゲザルHER2/neu DNA配列」という用語は、ここで実施例1において特定及び単離され、配列番号1に示されているアカゲザルHER2/neu配列を指す。この配列を翻訳して、配列番号2に示す「第一アカゲザルHER2/neuタンパク質」のアミノ酸配列を決定した。
【0032】
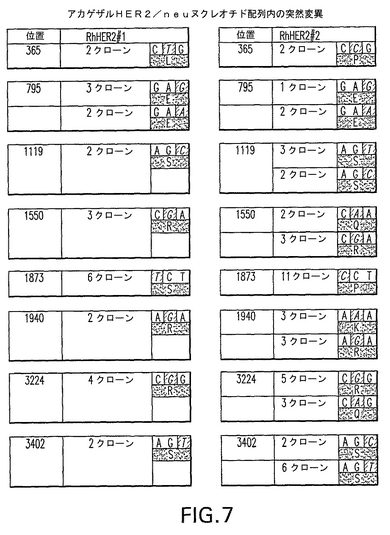
「rhHER2#2」という用語と交換可能に使用される、「第二アカゲザルHER2/neu DNA配列」という用語は、ここで実施例4において特定及び単離され、配列番号40に示されているアカゲザルHER2/neu配列を指す。このDNA分子は、実施例1で述べるDNA分子とは異なるアカゲザルから単離された。この配列を翻訳して、配列番号41に示す「第二アカゲザルHER2/neuタンパク質」のアミノ酸配列を演繹した。rhHER2#1とrhHER2#2ヌクレオチド及びアミノ酸配列の間の相違は図7において詳細する。
【0033】
「有効量」という用語は、免疫応答が生じるように、適切なレベルのポリペプチドを生産するのに十分なワクチン組成物を導入することを意味する。当業者は、このレベルが異なり得ることを認識する。
【0034】
「他の核酸を実質的に含まない」とは、他の核酸を少なくとも90%、好ましくは95%、より好ましくは99%、さらに一層好ましくは99.9%含まないことを意味する。交換可能に使用される、「他の核酸を実質的に含まない」、「実質的に精製された」、「単離核酸」又は「精製核酸」という用語はまた、他の細胞成分を除去して精製されたアカゲザルHER2/neuタンパク質についてのコード領域を含むDNA分子を指す。従って、他の核酸を実質的に含まないアカゲザルHER2/neuDNA試料は、その総核酸のパーセントとして、非アカゲザルHER2/neu核酸を10%、好ましくは5%、より好ましくは1%、さらに一層好ましくは0.1%しか含まない。所与のアカゲザルHER2/neuDNA試料が他の核酸を実質的に含まないかどうかは、例えば適切な染色法、例えば臭化エチジウム染色、と組み合わせたアガロースゲル電気泳動、などの核酸純度を評価する従来の手法によって、又は配列決定によって判定することができる。
【0035】
「他のタンパク質を実質的に含まない」又は「実質的に精製された」とは、他のタンパク質を少なくとも90%、好ましくは95%、より好ましくは99%、さらに一層好ましくは99.9%含まないことを意味する。従って、他のタンパク質を実質的に含まないアカゲザルHER2/neuタンパク質試料は、その総タンパク質のパーセントとして、非アカゲザルHER2/neuタンパク質を10%、好ましくは5%、より好ましくは1%、さらに一層好ましくは0.1%しか含まない。所与のアカゲザルHER2/neuタンパク質試料が他のタンパク質を実質的に含まないかどうかは、例えば適切な検出法、例えば銀染色又は免疫ブロット法、と組み合わせたドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動(SDS−PAGE)、などのタンパク質純度を評価する従来の手法によって判定することができる。
【0036】
交換可能に使用されるとき、「他のタンパク質を実質的に含まない」又は「実質的に精製された」又は「単離アカゲザルHER2/neuタンパク質」又は「精製アカゲザルHER2/neuタンパク質」という用語はまた、天然ソースから単離されたアカゲザルHER2/neuタンパク質を指す。「単離」又は「精製」という用語の使用は、アカゲザルHER2/neuタンパク質がその通常の細胞環境から取り出されていることを指示する。従って、単離アカゲザルHER2/neuタンパク質は、無細胞溶液中に存在し得る又はそれが天然に生じるのとは異なる細胞環境に置かれ得る。単離という用語は、単離アカゲザルHER2/neuタンパク質が存在する唯一のタンパク質であることを意味するのではなく、単離アカゲザルHER2/neuタンパク質が、rhHER2/neuタンパク質がインビボで天然に結合している他のタンパク質及び非アミノ酸物質(例えば核酸、脂質、炭水化物)を実質的に含まないことを意味する。従って、原核生物又は真核生物細胞において組換え発現され、天然では(すなわち介入処置なしでは)このrhHER2/neuタンパク質を発現しないこの宿主細胞から実質的に精製されたアカゲザルHER2/neuタンパク質は、言うまでもなく、ここで言及する状況下での「単離アカゲザルHER2/neuタンパク質」である。前述したように、単離又は精製rhHER2/neuタンパク質であるrhHER2/neuタンパク質試料は、実質的に他のタンパク質を含まず、及びその総タンパク質のパーセントとして、非アカゲザルHER2/neuタンパク質を10%、好ましくは5%、より好ましくは1%、さらに一層好ましくは0.1%しか含まない。
【0037】
「保存的アミノ酸置換」は、もう1つ別の、化学的に類似するアミノ酸残基による1個のアミノ酸残基の置換を指す。このような保存的置換の例は、疎水性残基(イソロイシン、ロイシン、バリン又はメチオニン)によるもう1つ別の疎水性残基の置換;1個の極性残基による同じ電荷のもう1つ別の極性残基の置換(例えばアルギニンによるリシンの置換;グルタミン酸によるアスパラギン酸の置換)である。
【0038】
「哺乳動物」という用語は、ヒトを含む何らかの哺乳動物を指す。
【0039】
「Ag」という略語は、抗原を指す。
【0040】
「Ab」及び「mAb」という略語は、それぞれ抗体及びモノクローナル抗体を指す。
【発明を実施するための最良の形態】
【0041】
HER2/neu腫瘍関連抗原をコードする遺伝子は、一般に上皮由来のヒト癌の発現を伴う。本発明は、HER2/neuの異常発現が癌又はその発症を伴う場合に、HER2/neu腫瘍関連抗原によって発現されるタンパク質産物に対する免疫を惹起する又は増強する組成物及び方法に関する。HER2/neuの異常発現と癌との随伴は、HER2/neuの異常発現は腫瘍の発症時に存在し、腫瘍が進行した後期には検出できないことがあるので、腫瘍発現の全ての時点でHER2/neuタンパク質が腫瘍組織において発現されることを必要としない。
【0042】
この目的ために、アカゲザルHER2/neuをコードするポリヌクレオチドを提供する。本発明の分子は、細胞媒介性免疫を通して上皮由来の癌に対する有効な免疫予防を提供するための組換えアデノウイルスワクチンにおいて使用し得る。インビボで脊椎動物に直接導入するとき、本発明のポリヌクレオチドは、霊長動物、イヌ及びヒトなどの哺乳動物を含む動物の体内でコードされるタンパク質の発現を誘導する。
【0043】
本発明は、配列番号2に示す新規rhHER2/neuタンパク質を発現するmRNAをコードするヌクレオチドの配列を含む単離核酸分子(ポリヌクレオチド)に関する。本発明はまた、配列番号41に示す新規rhHER2/neuタンパク質を発現するmRNAをコードするヌクレオチドの配列を含む単離核酸分子に関する。本発明の核酸分子は実質的に他の核酸を含まない。
【0044】
本発明の単離核酸分子は、一本鎖(コード又は非コード鎖)又は二本鎖であり得る、ゲノムDNA及び相補的DNA(cDNA)などのデオキシリボ核酸分子(DNA)、並びに合成一本鎖ポリヌクレオチドなどの合成DNAを包含し得る。本発明の単離核酸分子はまた、リボ核酸分子(RNA)を包含し得る。大部分のクローニングのために、DNAは好ましい核酸である。
【0045】
本発明の好ましいDNA分子は、図2に示す、配列番号2として表わされるアカゲザルHER2/neuタンパク質をコードする、図1に示す、ここで配列番号1として開示するヌクレオチド配列を含む。このrhHER2/neu核酸分子は、実施例1で詳細に述べるRT−PCRを通して特定された。配列番号1の795位の「R」の存在は、第一アカゲザルから単離されたクローンがその位置にA又はGのいずれかを含んでいたことを指示する。第一アカゲザルから単離された、795位に「A」を含む核酸分子をここでは配列番号42と称する。第一アカゲザルから単離された、795位に「G」を含む核酸分子をここでは配列番号43と称する。
【0046】
第二の好ましいDNA分子は、図6に示す、配列番号41として表わされるアカゲザルHER2/neuタンパク質をコードする、図5に示す、ここで配列番号40として開示するヌクレオチド配列を含む。前記単離cDNAクローン、関連ベクター、宿主、組換え細胞画分及び膜、及び発現された成熟形態のrhHER2/neuは、癌ワクチンの開発のために有用である。
【0047】
本発明はまた、新規rhHER2/neuタンパク質を発現するmRNAをコードする、配列番号1又は配列番号40の生物活性断片又は突然変異株を包含する。このような生物活性断片及び/又は突然変異株は、配列番号2及び配列番号41に示すrhHER2/neuタンパク質を含むがこれらに限定されない、rhHER2/neuタンパク質の薬理特性を少なくとも実質的に模倣するタンパク質又はタンパク質断片のいずれかをコードする。このようなポリヌクレオチドは、ヌクレオチド置換、欠失、付加、アミノ末端トランケーション及びカルボキシ末端トランケーションを含むが、必ずしもこれらに限定されない。本発明の突然変異株は、癌ワクチンの開発において有用であるように真核細胞において機能性rhHER2/neuタンパク質を発現するmRNA分子をコードする。
【0048】
本発明はまた、合成DNAのヌクレオチド配列が、配列番号1及び配列番号40のヌクレオチド配列とは有意に異なるが、まだ配列番号2又は配列番号41と同じrhHER2/neuタンパク質をコードする場合の、rhHER2/neuタンパク質をコードする合成DNAに関する。このような合成DNAは本発明の範囲内に包含されるものとする。
【0049】
従って、本発明は、同一タンパク質を発現する数多くのDNA分子を生じ得るコドンの重複性を開示する。本明細書のために、1またはそれ以上の置換されたコドンを担持する配列を縮重変異と定義する。また、発現されるタンパク質の最終物理特性を実質的に変化させない、DNA配列又は翻訳されたタンパク質のいずれかにおける突然変異も、本発明の範囲内に包含される。例えばバリンによるロイシン、アルギニンによるリシン又はアスパラギンによるグルタミンの置換は、ポリペプチドの機能性に変化を生じさせないと考えられる。
【0050】
ペプチドをコードするDNA配列は、天然で生じるペプチドのものとは異なる性質を有するペプチドをコードするように変化させ得ることが知られている。DNA配列を変化させる方法は、部位指定突然変異誘発を含むが、これに限定されない。変化する性質の例は、基質に対する酵素の親和性又はリガンドに対する受容体の親和性の変化を含むが、これらに限定されない。
【0051】
ストリンジェント条件下で配列番号1又は配列番号40にハイブリダイズするDNA配列は本発明に包含される。例として、限定ではなく、高ストリンジェンシーの条件を使用する手順は以下の通りである。DNAを含むフィルターのプレハイブリダイゼーションを、6XSSC、5Xデンハルト溶液及び100μg/ml変性サケ精子DNAから成る緩衝液中65℃で2時間から一晩実施する。フィルターを、100μg/ml変性サケ精子DNA及び5−20X106cpmの32P標識プローブを含むプレハイブリダイゼーション混合物中65℃で12から48時間ハイブリダイズさせる。2XSSC、0.1%SDSを含む溶液中37℃で1時間、フィルターの洗浄を行う。これに続いて0.1XSSC、0.1%SDS中50℃で45分間洗浄した後、オートラジオグラフィーに供する。高ストリンジェンシー条件を使用する他の手順は、5XSSC、5Xデンハルト溶液、50%ホルムアミド中42℃で12から48時間実施するハイブリダイゼーション工程又は0.2XSSPE、0.2%SDS中65℃で30から60分間実施する洗浄工程を含む。
【0052】
高ストリンジェンシーハイブリダイゼーションを実施するための前記手順で述べた試薬は、当技術分野において周知である。これらの試薬の組成物の詳細は、例えば本明細書中に参考として援用されているSambrookら、Molecular Cloning:A Laboratory Manual;Cold Spring Harbor Laboratory,Cold Spring Harbor,New York,1989に認められる。前記に加えて、使用し得る高ストリンジェンシーの他の条件は、当技術分野において周知である。
【0053】
本発明の1つの好ましい側面は、図2(配列番号2)に開示するアミノ酸の配列を含むアカゲザルHER2/neuタンパク質の実質的に精製された形態である。
【0054】
本発明のもう1つの好ましい側面は、図6(配列番号41)に開示するアミノ酸の配列を含むアカゲザルHER2/neuタンパク質の実質的に精製された形態である。
【0055】
本発明はまた、細胞外ドメイン及び細胞内ドメインなどの、rhHER2/neuの様々な機能性ドメイン、及びこれらの配列の少なくとも1つを含むハイブリッド分子に関する。
【0056】
本発明はまた、それらの変異が癌ワクチンの開発のために有用である診断、治療又は予防用途のタンパク質又はタンパク質断片を提供するような、アミノ酸置換、欠失、付加、アミノ末端トランケーション及びカルボキシ末端トランケーションを含む、必ずしもこれらに限定されない、配列番号2又は配列番号41に示すアミノ酸配列を含む、rhHER2/neuタンパク質の生物活性断片及び/又は突然変異株を包含する。
【0057】
本発明のアカゲザルHER2/neuタンパク質は、「成熟」タンパク質の形態であるか、又は融合タンパク質などのより大きなタンパク質の部分であり得る。分泌又はリーダー配列、プロ配列、多数のヒスチジン残基などの精製を助ける配列、又は組換え生産の間の安定性のための付加的な配列を含む、付加的アミノ酸配列を含むことがしばしば好都合である。
【0058】
本発明はまた、GFP(グリーン蛍光タンパク質)、MYCエピトープ、CST及びFcを含むがいかなる意味においてもこれらに限定されない、様々なマーカーに連結されたアカゲザルHER2/neuタンパク質の部分を発現する融合構築物を含むが、これらに限定されない、rhHER2/neu融合構築物に関する。このような融合構築物は、対象細胞系において発現させ、ここで開示するアカゲザルHER2/neuタンパク質の調節因子に関してスクリーニングするために使用し得る。
【0059】
本発明はさらに、本明細書全体を通じて開示する実質的に精製された核酸分子を含む組換えベクターに関する。これらのベクターはDNA又はRNAで構成され得る。クローニングの殆んどの目的のためには、DNAベクターが好ましい。典型的なベクターは、プラスミド、改変ウイルス、バクテリオファージ、コスミド、酵母人工染色体、及びrhHER2/neuタンパク質をコードすることができるエピソーム又は組込みDNAの他の形態を含む。個々の遺伝子導入又は他の用途のために適切なベクターを決定することは十分に当業者の理解範囲内である。
【0060】
rhHER2/neuタンパク質をコードするDNAを含む発現ベクターは、組換え宿主細胞におけるrhHER2/neuの発現のために使用し得る。発現ベクターは、クローニングベクター、改変クローニングベクター、特に設計されたプラスミド又はウイルスを含み得るが、これらに限定されない。また、所望に応じて細菌細胞において組換えrhHER2/neuを発現するために、様々な細菌発現ベクターを使用し得る。加えて、真菌細胞において組換えrhHER2/neuを発現するために、様々な真菌細胞発現ベクターを使用し得る。さらに、昆虫細胞において組換えタンパク質を発現するために、様々な昆虫細胞発現ベクターを使用し得る。
【0061】
本発明はまた、本発明の核酸分子を含むベクターで形質転換した又はトランスフェクトした宿主細胞に関する。組換え宿主細胞は、E.コリなどの細菌、酵母などの真菌細胞、ウシ、ブタ、サル及びげっ歯動物起源の細胞系統を含むがこれらに限定されない哺乳動物細胞、及びショウジョウバエ(Drosophia)及びカイコ由来の細胞系統を含むがこれらに限定されない昆虫細胞を含むが、これらに限定されない、原核又は真核細胞であり得る。このような組換え宿主細胞は、rhHER2/neu又は生物学的等価形態を生産するために適切な条件下で培養することができる。
【0062】
前述したように、rhHER2/neuタンパク質をコードするDNAを含む発現ベクターは、組換え宿主細胞におけるrhHER2/neuの発現のために使用し得る。従って、本発明のもう1つの側面は、(a)配列番号1又は配列番号40に示す核酸を含むベクターを適切な宿主細胞に導入すること;及び(b)前記宿主細胞を、前記アカゲザルHER2/neuタンパク質の発現を可能にする条件下で培養すること、を含む、組換え宿主細胞においてアカゲザルHER2/neuタンパク質を発現するための方法である。
【0063】
宿主細胞におけるrhHER2/neuの発現後、活性形態のrhHER2/neuタンパク質を提供するためにrhHER2/neuタンパク質を回収し得る。幾つかのrhHER2/neuタンパク質精製手順が使用可能であり、使用に適する。組換えrhHER2/neuタンパク質を、塩分画、イオン交換クロマトグラフィー、サイズ排除クロマトグラフィー、ヒドロキシアパタイト吸着クロマトグラフィー及び疎水性相互作用クロマトグラフィーの様々な組合せ又は個別適用によって細胞溶解産物及び抽出物から精製し得る。加えて、組換えrhHER2/neuタンパク質は、完全長rhHER2/neuタンパク質又はrhHER2/neuタンパク質のポリペプチド断片に特異的なモノクローナル又はポリクローナル抗体で作製した免疫アフィニティーカラムの使用によって他の細胞タンパク質から分離することができる。
【0064】
本発明の核酸は、ヒト細胞における前記タンパク質の効率的な発現を提供するように設計された配列を含む発現カセットに構築し得る。前記カセットは、好ましくは完全長rhHER2/neu遺伝子を、プロモーター及び終結配列などの、それに作動可能に連結された関連転写及び翻訳制御配列と共に含む。1つの好ましい実施態様では、前記プロモーターは、イントロンA配列を伴わないサイトメガロウイルスプロモーターであるが、当業者は、強力な免疫グロブリン又は他の真核生物遺伝子プロモーターなどの多くの他の公知のプロモーターのいずれもが使用し得ることを認識する。好ましい転写ターミネーターはウシ成長ホルモン終結因子であるが、他の公知の転写ターミネーターも使用し得る。CMV−BGHターミネーターの組合せは特に好ましい。
【0065】
本発明によれば、アカゲザルHER2/neu発現カセットをベクターに挿入する。前記ベクターは、好ましくはアデノウイルスベクターであるが、プロモーターに連結された線状DNA、又はアデノ関連ウイルス又は改変ワクシニアウイルスベクターなどの他のベクターも使用し得る。
【0066】
選択するベクターがアデノウイルスである場合は、いわゆる第一世代アデノウイルスベクターであることが好ましい。これらのアデノウイルスベクターは、非機能性E1遺伝子領域、好ましくは欠失アデノウイルスE1領域を有することを特徴とする。一部の実施態様では、アデノウイルスE1遺伝子が通常位置する部位に発現カセットを挿入する。加えて、これらのベクターは、場合により、非機能性又は欠失E3領域を有する。使用するアデノウイルスゲノムは、E1及びE3領域の両方を欠失している(△E1△E3)ことが望ましい。前記アデノウイルスを、293細胞又はPERC.6細胞などのウイルスE1遺伝子を発現する公知の細胞系統において、又は余分の(extra)タンパク質を発現するように一過性に又は安定に形質転換された293又はPERC.6細胞に由来する細胞系統において増殖させることができる。例えばテトラサイクリン調節可能プロモーター系などの制御された遺伝子発現を有する構築物を使用するとき、前記細胞系統は、調節系に関与する成分を発現し得る。このような細胞系統の一例はT−Rex−293であり、他は当技術分野において公知である。
【0067】
アデノウイルスベクターの操作上の便利さのために、アデノウイルスはシャトルプラスミド形態であり得る。本発明はまた、プラスミド部分及び、E1欠失及び場合によりE3欠失を有し、アカゲザルHER2/neuを含む発現カセットが挿入されたアデノウイルスゲノムを含有するアデノウイルス部分を含む、シャトルプラスミドベクターを対象とする。好ましい実施態様では、アデノウイルスベクターが容易に除去できるように、プラスミドのアデノウイルス部分に隣接する制限部位が存在する。前記シャトルプラスミドは、原核細胞又は真核細胞において複製し得る。
【0068】
本発明の好ましい実施態様では、発現カセットをpMRKAd5−HV0アデノウイルスプラスミドに挿入する(本明細書中に参照により組み込まれる、Eminiら、国際公開広報第WO02/22080号参照)。このプラスミドは、E1及びE3領域を欠失したAd5アデノウイルスゲノムを含む。pMRKAd5−HV0プラスミドの設計は、ウイルスパッケージングを最適化する上で重要であることが認められたエレメントを組み込むために5’シス作用性パッケージング領域をさらにE1遺伝子にまで延長することにより、先行アデノウイルスに比べて改善されており、高いウイルス増幅をもたらす。好都合には、この強化されたアデノウイルスベクターは、高い継代増殖後も遺伝的安定性を維持することができる。
【0069】
DNA構築物を作製し、精製するための分子生物学の標準手法は、本発明のアデノウイルス、シャトルプラスミド及びDNA免疫原の作製を可能にする。
【0070】
前述したベクターは、HER2/neu発現異常に関連する上皮由来の癌の発現を予防するため及び/又は既存の癌を治療するための免疫原性組成物及びワクチンにおいて使用し得る。このために、本発明の1つの側面は、配列番号2又は配列番号41に示すアカゲザルHER2/neuタンパク質をコードするヌクレオチドの配列を含む単離核酸分子を含有するワクチンベクターを哺乳動物に投与することを含む、癌を予防する又は治療する方法である。
【0071】
前述した方法によれば、前記ワクチンベクターは、哺乳動物における癌の治療又は予防のために投与し得る。本発明の1つの好ましい実施態様では、前記哺乳動物はヒトである。
【0072】
さらに、当業者は、前述した治療及び予防方法における使用のためにいかなる種類のベクターも選択し得る。好ましくは、前記ベクターはアデノウイルスベクター又はプラスミドベクターである。本発明の1つの好ましい実施態様では、前記ベクターは、アデノウイルスE1領域内の欠失、及び(a)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及び(b)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む発現カセットを含有するアデノウイルスE1領域内のインサートを有するアデノウイルスゲノムを含む、アデノウイルスベクターである。
【0073】
本発明はさらに、E1領域内の欠失、及び(a)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及び(b)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む発現カセットを含有するE1領域内のインサートを有するアデノウイルスゲノムを含む、アデノウイルスワクチンベクターに関する。
【0074】
本発明のこの側面の1つの好ましい実施態様では、前記アデノウイルスベクターはAd5ベクターである。
【0075】
もう1つの側面では、本発明は、プラスミド部分、及び(a)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及び(b)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む発現カセット部分を含有する、ワクチンプラスミドに関する。
【0076】
本発明の一部の実施態様では、高い免疫応答を誘導するために、ここで開示する組換えアデノウイルスワクチンを、プラスミドに基づくポリヌクレオチドワクチンと共に様々なプライム(初回抗原投与)/ブースト(追加抗原投与)の組合せにおいて使用する。この場合、前記の2つのベクターを「プライム&ブースト」レジメンにおいて投与する。例えば最初のタイプのベクターを投与し、次にあらかじめ定められた期間後、例えば1ヶ月後、2ヶ月後、6ヶ月後又は他の適切な間隔後に、第二のタイプのベクターを投与する。好ましくは、前記ベクターは同じポリヌクレオチド又はポリヌクレオチドの組合せをコードする発現カセットを担持する。プラスミドDNAも使用する実施態様では、前記ベクターは、哺乳動物又は昆虫細胞によって認識される1またはそれ以上のプロモーターを含むことが好ましい。1つの好ましい実施態様では、前記プラスミドは、ヒトCMVプロモーターなどの、但しこれに限定されない、強力なプロモーターを含む。アカゲザルHER2/neu遺伝子又は発現すべき他の遺伝子をこのようなプロモーターに連結する。このようなプラスミドの一例は、記述されている哺乳動物発現プラスミドV1Jnsである(本明細書中に参照により組み込まれる、J.Shiverら、DNA Vaccinesより、M.Liuら編集、N.Y.Acad.Sci.,N.Y.,772:198−208(1996))。
【0077】
前述したように、アデノウイルスベクターワクチン及びプラスミドワクチンは、免疫応答を誘導するために単一治療レジメンの一部として脊椎動物に投与し得る。このために、本発明は、(a)哺乳動物に、i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及びii)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む第一ベクターを導入すること;(b)あらかじめ定められた時間を経過させること;及び(c)前記哺乳動物に、i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及びii)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む第二ベクターを導入すること、を含む、哺乳動物を癌から保護する方法に関する。
【0078】
前述した保護の方法の1つの実施態様では、前記第一ベクターはプラスミドであり、及び前記第二ベクターはアデノウイルスベクターである。代替的実施態様では、前記第一ベクターはアデノウイルスベクターであり、及び前記第二ベクターはプラスミドである。
本発明はさらに、(a)上皮由来の癌に罹患している哺乳動物に、i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及びii)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む第一ベクターを導入すること;(b)あらかじめ定められた時間を経過させること;及び(c)前記哺乳動物に、i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及びii)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む第二ベクターを導入すること、を含む、上皮由来の癌に罹患している哺乳動物を治療する方法に関する。
【0079】
前述した治療方法の1つの実施態様では、前記第一ベクターはプラスミドであり、及び前記第二ベクターはアデノウイルスベクターである。代替的実施態様では、前記第一ベクターはアデノウイルスベクターであり、及び前記第二ベクターはプラスミドである。
【0080】
ワクチン受容者に導入すべき発現可能なDNA又は転写されたRNAの量は、部分的には、使用するプロモーターの強度及び発現される遺伝子産物の免疫原性に依存する。一般に、約1ngから100mg、好ましくは約10μgから300μgの免疫学的又は予防的有効用量のプラスミドワクチンベクターを筋組織に直接投与する。組換えアデノウイルスについての有効用量は、約106−1012粒子、好ましくは約107−1011粒子である。皮下注射、皮内導入、皮膚を通しての塗り(impression)、及び腹腔内、静脈内又は吸入送達などの他の投与方法も考慮される。また、ブースターワクチン接種を提供することも考慮される。本発明のワクチンの非経口導入と同時に又は導入後に、インターロイキン12タンパク質などのアジュバントによる静脈内、筋肉内、皮下などの非経口投与又は他の投与手段も好都合である。
【0081】
本発明のワクチンベクターは、裸であり得る、すなわち受容者の免疫系に影響を及ぼすいかなるタンパク質、アジュバント又は他の作用物質とも非結合であり得る。この場合、前記ワクチンベクターは、無菌食塩水又は無菌緩衝食塩水などの、但しこれらに限定されない、生理的に許容される溶液中に存在することが望ましい。また、アジュバント、サイトカイン、タンパク質又は他の担体などの免疫促進剤又は本発明の免疫原性組成物を投与することは好都合であると考えられる。従って、本発明は、本発明の組成物及び方法と組み合わせたこのような免疫促進剤の使用を包含する。免疫促進剤は、ここで使用するとき、基本的に外因性抗原に対する免疫応答(抗体及び/又は細胞媒介性)を高める又は増強する物質を指す。前記免疫促進剤は、DNA又はタンパク質の形態で投与することができる。GM−CSF、IFNα、破傷風毒素、IL12、B7.1、LFA−3及びICAM−1を含むがこれらに限定されない、様々な免疫促進剤のいずれかを、本発明のワクチン及び免疫原性組成物と共に使用し得る。前記免疫促進剤は当技術分野において周知である。カルシウムイオンなどの、但しこれに限定されない、DNAの細胞取り込みを助ける物質も使用し得る。これらの物質は、一般にトランスフェクション促進試薬及び医薬適合性の担体と称される。当業者は、個々の免疫促進剤又は医薬適合性の担体並びに適切な投与時間及び投与方法を決定することができる。
【0082】
様々な手順のいずれかを使用してrhHER2/neuをクローニングし得る。これらの方法は、(1)RACE PCRクローニング手法(Frohmanら、Proc.Natl.Acad.Sci.USA 85:8998−9002(1988))。5’及び/又は3’RACEは完全長cDNA配列を生成するために実施し得る。この戦略は、rhHER2/neu cDNAのPCR増幅のために遺伝子特異的オリゴヌクレオチドプライマーを使用することを含む。これらの遺伝子特異的プライマーは、多数の公的に使用し得る核酸及びタンパク質データベースを検索することによって特定されてきた、発現配列タグ(EST)ヌクレオチド配列の特定を通して設計される;(2)適切な発現ベクター系におけるrhHER2/neu含有cDNAライブラリーの構築後のrhHER2/neu cDNAの直接の機能的発現;(3)rhHER2/neuタンパク質のアミノ酸配列から設計した標識縮重オリゴヌクレオチドプローブで、バクテリオファージ又はプラスミドシャトルベクターにおいて構築したrhHER2/neu含有cDNAライブラリーをスクリーニングすること;(4)rhHER2/neuタンパク質をコードする部分的cDNAで、バクテリオファージ又はプラスミドシャトルベクターにおいて構築したrhHER2/neu含有cDNAライブラリーをスクリーニングすること。この部分的cDNAは、rhHER2/neuタンパク質に関連する他の増殖因子受容体についての公知のアミノ酸配列から縮重オリゴヌクレオチドプライマーを設計することを通して、rhHER2/neuDNA断片の特異的PCR増幅によって得られる;(5)哺乳動物rhHER2/neuタンパク質に相同性を有する部分的cDNA又はオリゴヌクレオチドで、バクテリオファージ又はプラスミドシャトルベクターにおいて構築したrhHER2/neu含有cDNAライブラリーをスクリーニングすること。この戦略も、前述したようにESTとして特定されたrhHER2/neu cDNAのPCR増幅のために遺伝子特異的オリゴヌクレオチドプライマーを使用することを含み得る;又は(6)rhHER2/neuをコードするヌクレオチド配列の完全長形態を単離するために、数多くの種類のcDNA及び/又はゲノムライブラリーの1つをスクリーニングするためのプローブとして使用するコード領域の部分を生成し、単離するために公知のRACE手法によって完全長cDNAを生成するか又はこれらの同じ公知のRACE手法によってコード領域の部分を生成するように、配列番号1又は配列番号40を鋳型として使用して5’及び3’遺伝子特異的オリゴヌクレオチドを設計すること、を含むが、これらに限定されない。
【0083】
他の種類のライブラリー、並びに他の細胞型又は他の種型から構築されたライブラリーが、rhHER2/neuをコードするDNA又はrhHER2/neuホモログを単離するために有用であり得ることは、当業者には容易に明白である。
【0084】
また、適切なcDNAライブラリーは、様々な上皮由来細胞などの、rhHER2/neu活性を有する細胞又は細胞系統から作製し得ることも、当業者には容易に明白である。rhHER2/neuをコードするcDNAを単離するためのcDNAライブラリーを作製するときに使用する細胞又は細胞系統の選択は、最初に、このような目的のために使用し得る何らかの公知のアッセイを用いて細胞に関連するrhHER2/neu活性を測定することによって実施し得る。
【0085】
cDNAライブラリーの作製は、当技術分野で周知の標準手法によって実施することができる。周知のcDNAライブラリー構築手法は、例えばSambrookら、Molecular Cloning:A Laboratory Manual;Cold Spring Harbor Laboratory,Cold Spring Harbor,New York,1989に認められる。相補的DNAライブラリーも、Clontech Laboratories,Inc.(Palo Alto,CA)及びStratagene(La Jolla,CA)を含むがこれらに限定されない、数多くの商業的供給元から入手し得る。
【0086】
本発明のDNA分子、RNA分子及び組換えタンパク質は、rhHER2/neuをスクリーニングし、そのレベルを測定するために使用し得る。前記組換えタンパク質、DNA分子及びRNA分子は、rhHER2/neuの検出及びタイピングに適するキットの作成に役立つ。このようなキットは、少なくとも1つの容器を厳重に閉じ込めて保持するのに適する区画化された担体を含む。前記担体は、rhHER2/neuを検出するのに適した組換えrhHER2/neu又は抗rhHER2/neu抗体などの試薬をさらに含む。前記担体はまた、標識抗原又は酵素基質等のような検出のための手段も含み得る。
【0087】
ここで言及する全ての公表文献は、本発明に関連して使用し得る方法及び材料を説明し、開示するために参照により組み込まれる。ここに含まれるいかなる内容も、本発明が先行発明によるこのような開示に先立つ権利がないことの是認と解釈されるべきではない。
【0088】
付属の図面を参照しながら本発明の好ましい実施態様を説明したが、本発明はそれらの厳密な実施態様に限定されないこと、及び付属の特許請求の範囲において定義される本発明の範囲及び精神から逸脱することなく当業者によって様々な変更及び修正が為され得ることは了解されねばならない。
【0089】
以下の実施例は本発明を例示するものであり、本発明を限定しない。
【実施例1】
【0090】
RT−PCRによるアカゲザルHER2/neu cDNAの単離
分子学的手順は、当技術分野において周知の標準手法(例えば本明細書中に参照により組み込まれる、Ausubelら、Short Protocols in Molecular Biology,F.M.,第2版、John Wiley & Sons(1992)及びSambrookら、Molecular Cloning,A Laboratory Manual、第2版、Cold Spring Harbor Laboratory Press(1989)参照)に従って実施した。
【0091】
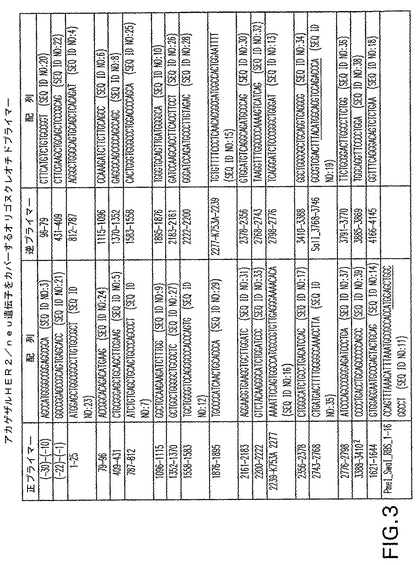
ヒト、ハムスター、イヌ及びラットからのHER2/neuヌクレオチド配列を、HER2/neu DNAの高度に保存された領域を特定するために整列させた。配列比較に基づき、HER2/neu遺伝子をカバーするオリゴヌクレオチドプライマーを、以下に述べる、逆転写酵素ポリメラーゼ連鎖反応(RT−PCR)によるアカゲザルHER2/neu cDNAの増幅のために設計した(図3参照)。
【0092】
2匹の異なるアカゲザル(macaca mulatta)からの結腸生検をDr.Willem Collignon(Biomedical Primate Research Centre(BPRC),Rijswijk,The Netherlands)より入手した。製造者の指示に従ってUltraSPec−II RNA単離システム(Biotecx,Houston,TX)を用いて各々の結腸生検からRNAを抽出し、精製した。アカゲザルHER2/neu遺伝子を単離するために、HER2/neu配列全体をカバーするRT−PCR増幅産物を、1匹のアカゲザルから単離した全RNAから生成した。
【0093】
逆転写工程を実施するために、製造者の指示に従ってSuperscript One−Step RT−PCR Amplification Kit for Long Templates(Life Technologies,Carlsbad,CA)を使用して全RNA試料を逆転写した。典型的には、RNA0.5−2.0μgを50μlの反応容量で逆転写酵素及び適切な緩衝液と混合した。試料を45℃で30分間インキュベートし、次に94℃で2分間インキュベートした。
【0094】
生じたcDNA鋳型を使用して、正プライマーと逆プライマーの種々の組合せを用いてPCR増幅を実施した(図3参照)。PCRはPerkin Elmer 2400サーモサイクラー(Perkin Elmer,Inc.,Wellesley,MA)において実施した。サイクリング条件は、94℃、15秒間の初期変性工程の35サイクル、次いでプライマーアニーリング工程及び伸長工程での終了から成った。プライマーアニーリング工程は、プライマー配列に依存して、50℃−51℃の範囲の温度で50秒間のインキュベーションから成った。伸長工程は、増幅産物の予想される長さに依存して、80秒間−100秒間の範囲の時間にわたる68℃でのインキュベーションから成った。前記35サイクルの後に72℃で7分間の広汎な延長工程(extensive elongation step)を実施した。
【0095】
QIAquick PCR Purification Kit(Qiagen,Hilden,Germany)を用いて増幅産物をゲル精製し、増幅に使用した同じプライマーで配列決定した。配列決定反応は、Cycle Sequencing Ready Reaction Kit(Applied Biosystems,Foster City,CA)を使用して、Big Dye Terminator化学を通して実施した。読取りはABI Prism 377 DNAシーケンサー(Applied Biosystem)を用いて実施した。
【0096】
HER2/neuコード領域全体を含む種々の増幅産物を配列決定することによって獲得したデータは、配列番号1としてここで開示する、アカゲザルHER2/neu配列(以下「第一アカゲザルHER2ヌクレオチド配列」又は「rhHER2#1」、図1参照)を特定した。配列番号1の795位の「R」の存在は、第一アカゲザルから単離されたクローンがその位置にA又はGのいずれかを含んでいたこと指示する。第一アカゲザルから単離された、795位に「A」を含む核酸分子をここでは配列番号42と称する。第一アカゲザルから単離された、795位に「G」を含む核酸分子をここでは配列番号43と称する。第一アカゲザルから単離されたDNAに基づき、配列番号1の795位の単一ヌクレオチド変化は、ここで配列番号2として開示する(図2参照)、生じたHER2/neuタンパク質のアミノ酸配列に影響を及ぼさなかった。
【0097】
前記RT−PCR反応から誘導された増幅産物を、ここではそれぞれBS又はCRと称する、ベクターBlueScript ks(+)又はpCRIIのいずれかにクローニングした。増幅産物の直接配列決定によって得たrhHER2/neuヌクレオチド配列を確認するために、生じたクローンを配列決定し、完全なアカゲザルHER2/neu cDNA配列を構築するために使用した。
【実施例2】
【0098】
完全なHER2/neu cDNA配列の構築
第一アカゲザルから得た一連のクローンをPCRによって構築し、集合させて、完全なアカゲザルHER2/neu cDNAを作製した。最初に、アカゲザルHER2/neu遺伝子の5’末端をカバーする4つのRT−PCR増幅産物を実施例1で述べたように生成した。前記クローンからのPCR増幅断片を、図4に示すように(反応1−4)、BS又はCRのいずれかにクローニングした。幾つかのクローンの配列分析は、クローニングされた配列がRT−PCR断片から得た配列と同じであることを確認した。
【0099】
前述した重複増幅産物を、以下の成分:前記断片の各々0.1−1.0pmol、Pfuポリメラーゼ(Stratagene,La Jolla,CA)及び適切な緩衝液、を組み合わせたPCR反応物100μl中で連結した。試料を、95℃、30秒間、次いで72℃、4分間から成る初期増幅サイクルに供した。
【0100】
生じた連結産物を、Pfuポリメラーゼ及び適切な緩衝液を用いて以下の熱プロフィール:95℃、30秒間、58℃、30秒間及び72℃、180秒間(30サイクル)により、PmeI_SwaI_RBS_1−16(配列番号11)及び1895−1876(配列番号10)プライマーで再び増幅した。このようにして得たPCR産物をゲル精製し、pCRIIベクターにクローニングして、クローン#1_CR_1−1895.7(−)(反時計回り方向)を作製した。幾つかのクローンの配列分析により、クローニングされた配列の同一性を確認した。
【0101】
第一アカゲザルからの結腸RNAを使用して、基本的に前述したように、RT−PCRによって付加的なPCR断片を生成した。このために使用したオリゴヌクレオチドプライマーは、1558−1583(配列番号12)及び2798−2776(配列番号13)であった(反応6、図4)。HER2/neu遺伝子の中央部分に位置するこの断片を、クローニングせずに、さらなる増幅のための鋳型として使用した。
【0102】
次に、bp1621−bp2277(図4、反応7、配列番号14、15)及びbp2239−bp2798(図4、反応8、配列番号16及び13)から、この領域をPCR増幅するためのプライマーを設計した。生じた産物をpCRIIベクターにクローニングした。タンパク質のチロシンキナーゼ活性を不活性化するために、KからAまでのアミノ酸579位を沈黙化するためのプライマーを設計した(配列番号15及び16)。幾つかのクローンの配列分析は、クローニングされた配列の同一性を確認した。
【0103】
第一アカゲザルからの結腸RNAを使用して、基本的に前述したように、RT−PCRによって付加的なPCR断片を生成した。このために使用したオリゴヌクレオチドプライマーは、2356−2378(配列番号17)及び4166−4145(配列番号18)であった(図4、反応9)。生じた産物をpCRIIベクターにクローニングした。幾つかのクローンの配列分析は、クローニングされた配列の同一性を確認した。
【0104】
以下のプライマーを使用して、前記の3つのクローンからDNA断片をPCR増幅した:1621−1644(配列番号14)及び2277−K−753−2248(配列番号15)、鋳型は#1_CR_1621−2277.2;2239−K756−2248(配列番号16)及び2798−2776(配列番号13)、鋳型は#1_CR_2239−2798.4;2356−2378(配列番号17)及びSalI_3768−3746(配列番号19)、鋳型は#1_CR_2356−4169.2。3つの産物を前述したようにPCRによって集合させた。生じた連結産物を以下のプライマー:1621−1644(配列番号14)及びSalI_3768−3746(配列番号19)を用いて増幅し、pCRIIベクターに時計周り方向にクローニングして、クローン#1_CR_1621−3768.8(以下クローン10.8)及び#1_CR_1621−3768.12(以下クローン10.12)を生成した(図4、反応10)。配列決定は、クローン10.12が3’領域内の3686位付近に突然変異を有することを明らかにした。同様に、クローン10.8は2666位付近に突然変異を有していた。クローン10.12からの508bp BstEII−SalI断片を、この領域内に突然変異を含まない、クローン10.8からの対応する領域で置き換えることにより、野生型配列を生成した。生じたクローンを#1_CR_1621−3768.128と命名した。
【0105】
クローン#1_CR_1621−3768.128(+)からのPmlI−XbaI断片を#1_CR_1_1895.7(−)のPmlI−SpeI部位にクローニングした。アカゲザルHER2/neuコード配列全体を含む、生じたプラスミドを#1_CR_1−3768(−)と命名した。配列分析により前記配列の同一性を確認した。
【実施例3】
【0106】
免疫原
遺伝子導入と免疫試験のために、PmeI及びSalIでの消化によってrhHER2/neuコード領域を#1_CR_1−3768(−)から切り出し、哺乳動物発現プラスミドpV1J_nsA(Montgomeryら、DNA Cell Biol.12(9):777−83(1993))のEcoRV及びSalI部位に挿入して、pV1J−rh−HER2/neuを生成した。
【0107】
アデノウイルスベクター構築のために、PmeI及びSalIでの消化によってrhHER2/neuコード領域を#1_CR_1−3768(−)から切り出し、ポリMRKAd5△E1シャトルプラスミドの対応する部位にクローニングして、pMRKAd5△E1−rhHER2/neuを生成した。シャトルベクターpMRKAd5△E1は、ヒトサイトメガロウイルス(HCMV)プロモーター(イントロンAなし)及びウシ成長ホルモンポリアデニル化シグナルを含む発現カセットと共に塩基対1−450及び塩基対3511−5798からのAd5配列を含む。このプラスミドを、大腸菌BJ5183細胞(Chartierら、J.Virol.70:4805−10(1996)を使用して、E1及びE3領域を含有する配列を除くすべてのAd5配列を含む、アデノウイルス骨格ベクターpMRKAd5HVOと組み換えた。
【0108】
ベクター生産のために、pMRKAd5−rhHER2/neuをPacIでの消化によって線状化し、Lipofectamine(Life Technologies,Rockville,MD)を用いてPerC.6細胞にトランスフェクトした。ウイルス力価を増幅するために5−6ウイルス継代を行い、大型ウイルス増幅を実施して1.3×1012物理的粒子(physical particles)(pp)の最終生産を得た。制限断片長多型(RFLP)分析によって示されたように、増幅したベクターから精製したウイルスゲノムにおいてゲノム再構成は検出できなかった。予想されたDNA断片は、平行して制限した、ウイルスゲノム内及び対照pMRKAd5−rhHER2/neuプラスミドの両方で認められた。
【実施例4】
【0109】
第二アカゲザルからのHER2/neuコードDNAの単離
第二アカゲザルからの結腸RNAを使用してRT−PCR産物の同様の配列決定分析を実施した(実施例1参照)。前記遺伝子全体を含む多くの異なる増幅産物を配列決定することによって獲得したデータは、ここで配列番号40として開示する、第二アカゲザルHER2/neu配列(以下「第二アカゲザルHER2ヌクレオチド配列」又は「rhHER2#2」、図5参照)を、ここで配列番号41として開示する演繹アミノ酸配列(以下「第二アカゲザルHER2アミノ酸配列」、実施例6参照)と共に特定した。
【0110】
実施例1におけるように、HER2/neuをカバーするプライマーを使用して第二アカゲザルからの全結腸RNAのRT−PCRによって増幅産物を生成した(図3参照)。生じた増幅産物をゲル精製し、配列決定した。さらに、これらの産物をBS又はCRベクターのいずれかにクローニングした。生じたクローンのDNA塩基配列決定分析により、増幅産物の直接配列決定によって得たrhHER2/neuヌクレオチド配列を確認した。
【0111】
rhHER2#1とrhHER2#2の間で8つの相違が検出された(詳細については、図7参照)。これらの突然変異のうち5つは、第一アカゲザルHER2アミノ酸配列(配列番号2)と比較したときタンパク質内にアミノ酸変化を導入する。注意すべきは、これらの突然変異のうち3つは、rhHER2#1タンパク質と比較したときrhHER2#2タンパク質内にアミノ酸変化を生じさせない。
【実施例5】
【0112】
rhHER2/neuによるアカゲザルの免疫
結腸直腸癌において発現されるヒト腫瘍抗原HER2/neuのアカゲザルホモログによるアカゲザル(Macaca mulatta)の免疫の効率を評価するため、Biomedical Primate Research Centre(BPRC),Rijswijk(The Netherlands)において免疫試験を実施した。試験は、アカゲザルHER2/neu抗原での免疫に対するB及びT細胞応答を評価するように設計した。
【0113】
最初の試験では、1つの群のサル(アカゲザル合計4匹;雄性2匹と雌性2匹)をヒト腫瘍抗原HER2/neuのアカゲザルホモログを発現するプラスミドDNAベクター又はアデノウイルスベクター(pV1J−rhHER2)で免疫した。初回免疫のために、0、4、8、12及び16週目にDNAの注射とそれに続く電気刺激により、筋肉内経路で(i.m.)プラスミドDNAによって動物をワクチン接種した。前記DNAは、pV1J−rhHER2プラスミドDNA5mgを含有する1ml溶液(2つの部位に分けて0.5ml/部位)として、体重2−5kgの動物に注射した。動物には麻酔下で(ケタミン/キシラジンの混合物)注射を行う。
【0114】
電気刺激のために、2トレーンの100矩形双極性パルス(各々1秒)を、3秒の全処置時間中1秒おきに送達した。パルス長は2ミリ秒/位相、パルス周波数及び振幅はそれぞれ100Hz及び100mA(定電流方式)であった。
【0115】
同じ群の動物を、アカゲザルHER2/neuを発現するアデノウイルス5(Ad5)の筋肉内注射によって追加免疫した。△E1−△E3、「第一世代」アデノウイルス(P2レベル)を使用した。1011ウイルス粒子(vp)の総量を24週目と28週目に注射した。
【0116】
36週目と40週目にアカゲザルHER2/neuを発現するアデノウイルス24(Ad24)の1011ウイルス粒子(vp)の筋肉内注射によってさらなる追加免疫を実施した。Ad5注射によって誘導される中和抗体はAd24感染に干渉しないので、Ad24を選択した。
【0117】
前記免疫プロトコールによって誘導されるHER2/neuに対する免疫応答を測定するため、総期間1年間にわたって4週間ごとに血液試料を採集した。HER2特異的細胞媒介性免疫応答をIFN−γ ELISPOTアッセイによって測定した。IFN−γ分泌性抗アカゲザルHER2 T細胞の数を、ペプチドのプールを使用してPBMCに関するELISPOTによって判定した。11残基が重複し、アカゲザルタンパク質配列全体をカバーする、各々15アミノ酸長の311ペプチドを、AからKまでのアルファベット文字で示した11のプール(N末端からC末端まで)と組み合わせた。PBMCを産生するIFN−γの頻度を、2つの異なる細胞濃度での2回の測定から導かれたスポットの平均値として算定した。数値は、スポット形成コロニー(SFC)/106全PBMCの数から、ペプチドの不在下で測定したバックグラウンド値(典型的には10SFC/106全脾細胞)を差し引いた値として表わした。算定結果は、IFN−γ ELISPOTによって測定したとき、4匹のサル全てが検出可能な細胞媒介性応答を示すことを示す。
【0118】
反応性を確認し、IFN−γを分泌するCD4+又はCD8+T細胞の頻度を測定する、IFN−γ+細胞内染色法(ICS)によってタイピングした。CD3+事象と小リンパ球で同時ゲーティングすることによってCD3+リンパ球を収集した。0.1%より高い数値を陽性とみなした。プールJについてサルRI504に関して0.19の陽性値(CD8+)を得た。検出可能な抗rhHER2抗体力価(>200)は検出されなかった。
【0119】
要約するに、アカゲザルHER2/neuを発現するプラスミドDNAの筋肉内注射は、アカゲザルにおいて寛容を打破し、rhHER2/neuに対する検出可能な細胞媒介性免疫応答を誘導する上で有効であった。
【実施例6】
【0120】
ヒト腫瘍抗原のアカゲザルホモログによるアカゲザルの免疫
全て結腸直腸癌において発現される、ヒト腫瘍抗原HER2/neu、Ep−CAM及びCEAのアカゲザルホモログによるアカゲザル(Macaca mulatta)の免疫の効率を評価するため、Biomedical Primate Research Centre(BPRC),Rijswijk(The Netherlands)において第二シリーズの免疫試験を実施した。プロトコールは、これらの腫瘍抗原の組合せに対するB及びT細胞応答を評価するように設計した。
【0121】
この試験では、4匹のアカゲザルから成る第二群(雄性2匹と雌性2匹)を、ヒト腫瘍抗原、Ep−CAM(pV1J−rhHER2)、CEA(pV1J−rhCEA)及びHER2/neu(pV1J−rhEpCAM)のアカゲザルホモログを発現する3つのプラスミドDNAベクターの混合物で免疫した。
【0122】
0、4、8、12及び16週目にプラスミドDNAの筋肉内注射とそれに続く電気刺激によって動物を初回免疫した。前記DNA注射は、体重2−5kgの動物に関して、プラスミドDNA6mg(3つのTAAの各々について2mg)を含有する1ml溶液(2つの部位に分けて0.5ml/部位)から成った。動物には麻酔下で(ケタミン/キシラジンの混合物)注射した。
【0123】
電気刺激のために、2トレーンの100矩形双極性パルス(各々1秒)を、3秒の全処置時間中1秒おきに送達した。パルス長は2ミリ秒/位相、パルス周波数及び振幅はそれぞれ100Hz及び100mA(定電流方式)であった。
【0124】
同じ群の動物を、アカゲザルHER2/neu(Ad5−rhHER2)、アカゲザルCEA(Ad5−rhCEA)及びアカゲザルEpCAM(Ad5−rhEpCAM)を発現する3つのAd5の混合物の注射によって追加免疫した。3×1011ウイルス粒子(vp)の総量を23週目と27週目に筋肉内注射した(3つのウイルスの各々について1×1011vp)。
【0125】
さらなる追加免疫を、アカゲザルHER2/neu(Ad24−rhHER2)、アカゲザルCEA(Ad24−rhCEA)及びアカゲザルEpCAM(Ad24−rhEpCAM)を発現する3つのAd24の混合物の注射によって実施した。3×1011ウイルス粒子(vp)の総量を36週目と40週目に筋肉内注射した(3つのウイルスの各々について1×1011vp)。
【0126】
前記免疫プロトコールを使用してHER2/neuに対する免疫応答を測定するため、総期間1年間にわたって4週間ごとに血液試料を採集した。細胞媒介性免疫応答をIFN−γ+ELISPOTによって測定し、体液性応答をELISAによって測定した。
【0127】
サルRI449及びRI519は、IFN−γ+ELISPOT分析によって測定したとき、検出可能なHER2特異的細胞媒介性応答を示した。同様の分析は、rhCEA及びrhEpCAMに対するいかなる応答も検出しなかった。検出可能な抗rhHER2抗体力価(>200)は検出されなかった。
【0128】
第三の試験では、4匹のアカゲザルを、0、2及び4週目にAd5誘導体の筋肉内注射によってAd5−rhHER2、Ad5−rhCEA及びAd5−rhEpCAMの混合物で免疫した。麻酔下で(ケタミン/キシラジンの混合物)、3×1011vp(3つのAd5ウイルスの各々について1011vp)を含有する1ml溶液(2つの部位に分けて0.5ml/部位)を体重2−5kgの動物に投与した。
【0129】
同じ群の動物を、24、26及び28週目にAd24−rhHER2、Ad24−rhCEA及びAd24−rhEpCAMの混合物(総量3×1011vp、3つのウイルスの各々について1011vp)の筋肉内注射によって追加免疫した。細胞媒介性応答をIFNγ ELISPOTアッセイによって測定した。サルRI514及びRI496に関して、反応性をIFN−γ+細胞内染色法(ICS)によってタイピングした。4匹のサルのうち3匹は検出可能な応答を示した。加えて、細胞媒介性応答が測定された3匹のサルにおいて200から500の範囲の抗rhHER2抗体力価が検出された。
【0130】
この免疫プロトコールはまた、寛容を打破し、抗rhCEA細胞媒介性及び体液性免疫応答を誘導する上でも有効であった。サルRI514は、IFN−γ ELISPOTアッセイによって測定可能な細胞媒介性応答を示した。細胞内染色法はこの応答を確認した。同じサルはまた、500から1000の範囲の抗rhCEA抗体力価を示した。これに対し、同様の分析はrhEpCAMに対するいかなる応答も検出しなかった。
【0131】
要するに、前記で論じた免疫プロトコールは、アカゲザルにおいてrhHER2/neu及びrhCEAに対する特異的免疫応答を誘導する上で有効であった。
【図面の簡単な説明】
【0132】
【図1A】図1は、配列番号1に示す、第一アカゲザルHER2/neu cDNAのヌクレオチド配列を示す(実施例1参照)。795位の「R」の存在は、「A」又は「G」のいずれかがその位置に存在することを指示する。
【図1B】図1は、配列番号1に示す、第一アカゲザルHER2/neu cDNAのヌクレオチド配列を示す(実施例1参照)。795位の「R」の存在は、「A」又は「G」のいずれかがその位置に存在することを指示する。
【図2】図2は、配列番号2に示す、アカゲザルHER2/neuタンパク質の予測アミノ酸配列を示す。示されているアミノ酸配列は、配列番号1として開示するヌクレオチド配列から演繹した。
【図3】図3は、RT−PCRによって一連のアカゲザルHER2/neu断片を作製するために使用した、HER2/neu遺伝子をカバーするオリゴヌクレオチドプライマーのヌクレオチド配列を開示する(実施例1参照)。「正」及び「逆」と表示した欄は、公表されているヒトHER2/neu配列(アクセッションM11730)に対するプライマーの位置を記載している。配列番号15及び16として開示するプライマーは、翻訳されたタンパク質のチロシンキナーゼ活性を不活性化するために、アミノ酸579位をKからAに突然変異させるために設計した。プライマー3388−3410及び3410−3388は、Ser(AGC)をコードする、それぞれ15位にC及び10位にGを含む配列を有する。これに対し、アカゲザルHER2/neu配列はその位置に、やはりSer(AGT)をコードする、Tを有する。これらのプライマーを配列決定のために使用したが、クローニングのためには使用しなかった。配列番号11及び19については、rhHER2/neuが始まる配列に下線を付している。
【図4】図4は、完全長アカゲザルHER2/neuクローンを構築するために使用したRT−PCRプライマーの配列を示す。最初の欄は、実施例2で述べる反応に対応する反応番号を列記している。各々の欄は、4番目の欄に列記するクローンを生成するために使用した正及び逆プライマーのセットを記載している。クローンの名称は、クローニングのために使用したベクター(pBluescript又はpCRIIに関して、それぞれBS又はCR)及び公表ヒトHER2/neu配列に対する配列の位置(数字で列記している)の両方を指示する。
【図5A】図5は、配列番号40に示す、第二アカゲザルHER2/neucDNAのヌクレオチド配列を示す(実施例4参照)。ヌクレオチド配列内の「R」の存在は、「A」又は「G」のいずれかがその位置に存在することを指示する。ヌクレオチド配列内の「Y」の存在は、「C」又は「T」のいずれかがその位置に存在することを指示する。第一アカゲザルHER2/neu(配列番号1)の対応する塩基とは異なるヌクレオチド塩基は、太字及び下線で表示している。
【図5B】図5は、配列番号40に示す、第二アカゲザルHER2/neucDNAのヌクレオチド配列を示す(実施例4参照)。ヌクレオチド配列内の「R」の存在は、「A」又は「G」のいずれかがその位置に存在することを指示する。ヌクレオチド配列内の「Y」の存在は、「C」又は「T」のいずれかがその位置に存在することを指示する。第一アカゲザルHER2/neu(配列番号1)の対応する塩基とは異なるヌクレオチド塩基は、太字及び下線で表示している。
【図6】図6は、配列番号41に示す、第二アカゲザルHER2/neuタンパク質の予測アミノ酸配列を示す(実施例4参照)。示されているアミノ酸配列は、配列番号40として開示するヌクレオチド配列から演繹した。517位の「X」は、「Q」(Gln)又は「R」(Arg)がその位置に存在し得ることを指示する。647位の「X」は、「K」(Lys)又は「R」(Arg)がその位置に存在し得ることを指示する。1075位の「X」は、「R」(Arg)又は「Q」(Gln)がその位置に存在し得ることを指示する。
【図7】図7は、第一アカゲザルHER2/neu DNA及びタンパク質配列(RhHER2#1、配列番号1及び2)と比較して、第二アカゲザルHER2/neuDNA及びタンパク質配列(RhHER2#2、配列番号40及び41)に存在する特定突然変異を詳細に示す。各々の表の最初の欄は、RhHER2#2とRhHER2#1の間で異なるヌクレオチドの位置を列記する。各々の表の2番目の欄は、単離して、それぞれRhHER2#1及びRhHER2#2の配列を決定するために使用したHER2/neu断片を担う特定クローンの番号を列記する。各々の表の3番目の欄は、相違が起こるコドンの配列を示しており、異なるヌクレオチドを強調している。コドンの下部は、灰色で強調した、生じるアミノ酸についての1文字アミノ酸記号である。
【配列表】
【技術分野】
【0001】
本発明は、一般に癌の検出及び治療に関する。特に、本発明は、腫瘍関連ポリペプチドHER2/neuのアカゲザルホモログ、このタンパク質をコードする単離核酸分子、及びこのタンパク質をコードするDNAを含む組換えベクター及び宿主に関する。本発明はまた、アカゲザルHER2/neuを担持するアデノウイルスベクター構築物、及び癌を予防及び治療するためのワクチン及び医薬組成物におけるこれらの使用に関する。
【背景技術】
【0002】
癌は、典型的には、増殖因子及びそれらの受容体、癌遺伝子及び癌抑制遺伝子などの、細胞周期を維持する又は細胞増殖を制御することに寄与する遺伝子の調節不全を含む。これらの遺伝子の多くの産物は様々な腫瘍細胞の表面で発現され、従って、腫瘍関連抗原(TAA)と命名された。最近の証拠は、免疫応答を惹起することができる腫瘍関連抗原の存在を裏付けており、これらの分子はワクチン療法のための標的となっている。これらの遺伝子産物の多くは、より低いレベルではあるが、正常細胞でも発現されるので、腫瘍関連抗原を標的する多くの癌ワクチンは、免疫寛容の故に無効であることが証明された。
【0003】
HER2/neu癌原遺伝子(c−erbB−2とも呼ばれる)の産物は、上皮増殖因子受容体ファミリーの成員である膜貫通TAAである。HER2/neu遺伝子は、最初にラット神経膠芽細胞腫からクローン化され(Shihら、Nature 290:261−264(1981))、後にヒト細胞から単離されて、特性決定された(Coussensら、Science 230:1132−39(1985);Kingら、Science 229:974−76(1985))。現在まで、HER2/neuのサルホモログは得られていない。
【0004】
HER2/neuは、細胞増殖と分化に関与する4つの受容体から成る、受容体チロシンキナーゼのHERファミリーの成員としてさらに分類されてきた。HER受容体は、二量体として増殖因子リガンドに結合し、最終的に細胞増殖において重要な遺伝子の活性化を生じさせる細胞内シグナル伝達カスケードを開始させることにより、正常な細胞増殖を維持することに寄与する。幾つかのリガンドがHERファミリーの他の成員について特定されているが、HER2/neu受容体についての高親和性リガンドはまだ発見されていない(LohrischとPiccart,Semin.Oncol.28(6):補遺18:3−11(2001))。
【0005】
HER2/neu転写産物及びコードされる185kDタンパク質の低いレベルの発現が、皮膚及び乳房、及び胃腸管、生殖路及び尿路の組織を含む、様々な組織の健常成人上皮細胞において検出された(Pressら、Oncogene 5:963−962(1990))。より高いレベルのHER2/neu発現も、胚発生期の対応する胎児組織において検出された(Pressら、前出)。
【0006】
HER2/neuは、一般に乳癌、卵巣癌、子宮癌、結腸癌及び前立腺癌、及び肺の腺癌などの様々な悪性疾患において過剰発現される又は増幅される(DisisとCheever,Adv.Cancer Research 71:343−371(1997)において総説されている)。HER2/neuのこのような過剰発現は、癌患者についての予後不良及びより高い再発率と相関している(Slamonら、Science 244:707−712(1989))。
【0007】
HER2/neuの過剰発現に関連した悪性疾患に罹患している多くの癌患者は、以前にHER2/neu癌遺伝子のタンパク質産物に対して免疫応答を生じたことがあり、従ってHER2/neuは癌治療薬の開発のための免疫学的標的となっている。HER2/neuに対するこの免疫応答を利用した有効なワクチンは、この免疫を防御及び/又は予防的なレベルまで高め、自己寛容を克服するに違いない。
【0008】
HER2/neuは、種々の悪性疾患の免疫学的治療の開発のための標的として提案されてきた。種々の抗HERモノクローナル抗体が乳癌のための治療として検討されており、各々の抗体が様々なレベルの成功を明らかにしている(考察については、Yarden,Oncology 61(補遺2):1−13(2001)参照)。Amiciら(米国特許第6,127,344号)は、ヒトサイトメガロウイルスプロモーターに機能的に連結された完全長ヒトHER2/neu cDNAを含むベクターを投与することによってHER2/neuに対する免疫を誘導するための方法を開示している。CheeverとDisisは、HER2ペプチドでHER2/neuに関連する癌に対してヒトを免疫するための方法を開示している(米国特許第5,846,538号)。加えて、HER2/neuペプチドに基づくワクチンがげっ歯動物モデルにおいて試験されている(総説については、DisisとCheever,Advances in Cancer Research 71:343−371(1997)参照)。
【発明の開示】
【発明が解決しようとする課題】
【0009】
前述したHER2/neuクローンの特定にもかかわらず、有効であり且つ自己寛容によって妨げられない癌ワクチンの開発を可能にする、HER2/neuをコードするさらなる哺乳動物遺伝子を特定することは極めて望ましい。
【課題を解決するための手段】
【0010】
本発明は、配列番号2又は配列番号41に示す新規アカゲザルHER2/neuタンパク質(c−erbB−2とも呼ばれる、以下rhHER2/neuと称する)をコードするヌクレオチドの配列を含む単離又は精製核酸分子に関する。ここで開示するDNA分子は、選択宿主細胞にトランスフェクトしてもよく、前記組換え宿主細胞は、実質的なレベルの、発現される機能性rhHER2/neuタンパク質(配列番号2又は配列番号41)のためのソースを提供する。
【0011】
本発明はさらに、新規アカゲザルHER2/neuタンパク質を発現するmRNAをコードする単離核酸分子に関する。このDNA分子は、ここで配列番号1として開示するヌクレオチド配列を含む。本発明のこの部分の好ましい側面は図1に開示されており、図1は、新規rhHER2/neuタンパク質(配列番号2)をコードするDNA分子(配列番号1)を示す。
【0012】
本発明はまた、新規アカゲザルHER2/neuタンパク質を発現するmRNAをコードする単離核酸分子を提供する。このDNA分子は、ここで配列番号40として開示するヌクレオチド配列を含む。本発明のこの部分の好ましい側面は図5に開示されており、図5は、新規rhHER2/neuタンパク質(配列番号41)をコードするDNA分子(配列番号40)を示す。
【0013】
本発明はまた、本明細書全体を通じて開示する核酸分子を含む、原核及び真核生物の両方の、組換えベクター及び組換え宿主細胞に関する。
【0014】
本発明はさらに、(a)配列番号1又は配列番号40に示す核酸を含むベクターを適切な宿主細胞に導入すること;及び(b)前記宿主細胞を、前記アカゲザルHER2/neuタンパク質の発現を可能にする条件下で培養すること、を含む、組換え宿主細胞においてアカゲザルHER2/neuタンパク質を発現するための方法に関する。
【0015】
本発明の1つの好ましい側面は、図2に開示するアミノ酸配列(配列番号2)から成る、実質的に精製された形態のアカゲザルHER2/neuタンパク質である。
【0016】
本発明の1つの好ましい側面は、図6に開示するアミノ酸配列(配列番号41)から成る、実質的に精製された形態のアカゲザルHER2/neuタンパク質である。
【0017】
本発明のさらにもう1つの好ましい側面は、rhHER2/neuタンパク質を発現する、配列番号1又は配列番号40に示すヌクレオチド配列を含むDNA発現ベクターを含有する組換え宿主細胞から得られる、実質的に精製され、完全にプロセシングされた(タンパク質分解プロセシング、グリコシル化及び/又はリン酸化を含む)成熟rhHER2/neuタンパク質に関する。前記組換え宿主細胞は、哺乳動物細胞系などの真核生物宿主細胞であることが特に好ましい。
【0018】
本発明のもう1つの側面は、配列番号2又は配列番号41に示すアカゲザルHER2/neuタンパク質をコードするヌクレオチドの配列を含む単離核酸分子を含有するワクチンクチンベクターを哺乳動物に投与することを含む、癌を予防する又は治療する方法である。
【0019】
本発明はさらに、E1及びE3領域内の欠失及びE1領域内のインサートを有するアデノウイルスゲノムを含み、前記インサートは、(a)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド、及び(b)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む発現カセットを含有する、アデノウイルスワクチンベクターに関する。
【0020】
本発明はまた、プラスミド部分及び発現カセット部分を含有し、前記発現カセット部分は、(a)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及び(b)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含むワクチンプラスミドに関する。
【0021】
本発明のもう1つの側面は、(a)癌に羅患している哺乳動物に、i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド及びii)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む第一ベクターを導入すること;(b)あらかじめ定められた時間を経過させること;及び(c)前記哺乳動物に、i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド及びii)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む第二ベクターを導入すること、を含む、哺乳動物を癌から保護する又は癌に罹患している哺乳動物を治療する方法である。
【0022】
本明細書全体及び付属の特許請求の範囲において使用するとき、単数形態の「a」、「an」及び「the」は、文脈が明らかに異なる指示を命じない限り、複数の言及を包含する。
【0023】
本明細書全体及び付属の特許請求の範囲において使用するとき、以下の定義及び略語が適用される。
【0024】
「プロモーター」という用語は、RNAポリメラーゼが結合するDNA鎖上の認識部位を指す。プロモーターは、転写活性を開始させ、駆動するためにRNAポリメラーゼと開始複合体を形成する。前記複合体は、「エンハンサー」と称される活性化配列又は「サイレンサー」と称される抑制配列によって修飾され得る。
【0025】
「カセット」という用語は、発現されるべき核酸配列を含む本発明の配列を指す。前記カセットは、概念においてはカセットテープに類似し、各々のカセットはそれ自身の配列を有する。従って、カセットを交換することにより、ベクターは異なる配列を発現する。5’及び3’末端の制限部位の故に、カセットは容易に挿入する、除去する又はもう1つ別のカセットと置換することができる。
【0026】
「ベクター」という用語は、DNA断片を宿主生物又は宿主組織に導入することができる何らかの手段を指す。プラスミド、ウイルス(アデノウイルスを含む)、バクテリオファージ及びコスミドを含む様々な種類のベクターが存在する。
【0027】
アデノウイルスベクターに関して使用するとき「第一世代」という用語は、複製欠損である前記アデノウイルスベクターを表わす。第一世代アデノウイルスベクターは、典型的には欠失又は不活性化E1遺伝子領域を有し、好ましくは欠失又は不活性化E3遺伝子領域を有する。
【0028】
「pMRKAd5−rhHER2/neu」との名称は、E1及びE3領域の欠失したAd5アデノウイルスゲノムを含む、ここで開示するプラスミド構築物を指す。このプラスミドでは、E1領域は、イントロンAを伴わずにヒトCMVプロモーターの制御下にあり、続いてウシ成長ホルモンポリアデニル化シグナルが続く、E1と平行する方向のアカゲザルHER2/neuによって置換されている。
【0029】
「MRKAd5−rhHER2/neu」との名称は、制限及びPer.C6又はHEK293などのE1発現細胞系統へのトランスフェクションによるプラスミド配列の除去後にプラスミドpMRKAD5−rhHER2/neuから生成されるウイルスを指す。
【0030】
「pV1J−rhHER2/neu」との名称は、ヒトCMV最初期(IE)プロモーター及びイントロンA、完全長アカゲザルHER2/neu遺伝子、ウシ成長ホルモン由来のポリアデニル化及び転写終結配列、及び最小pUC骨格を含む、ここで開示するプラスミド構築物を指す。
【0031】
「rhHER2#1」という用語と交換可能に使用される、「第一アカゲザルHER2/neu DNA配列」という用語は、ここで実施例1において特定及び単離され、配列番号1に示されているアカゲザルHER2/neu配列を指す。この配列を翻訳して、配列番号2に示す「第一アカゲザルHER2/neuタンパク質」のアミノ酸配列を決定した。
【0032】
「rhHER2#2」という用語と交換可能に使用される、「第二アカゲザルHER2/neu DNA配列」という用語は、ここで実施例4において特定及び単離され、配列番号40に示されているアカゲザルHER2/neu配列を指す。このDNA分子は、実施例1で述べるDNA分子とは異なるアカゲザルから単離された。この配列を翻訳して、配列番号41に示す「第二アカゲザルHER2/neuタンパク質」のアミノ酸配列を演繹した。rhHER2#1とrhHER2#2ヌクレオチド及びアミノ酸配列の間の相違は図7において詳細する。
【0033】
「有効量」という用語は、免疫応答が生じるように、適切なレベルのポリペプチドを生産するのに十分なワクチン組成物を導入することを意味する。当業者は、このレベルが異なり得ることを認識する。
【0034】
「他の核酸を実質的に含まない」とは、他の核酸を少なくとも90%、好ましくは95%、より好ましくは99%、さらに一層好ましくは99.9%含まないことを意味する。交換可能に使用される、「他の核酸を実質的に含まない」、「実質的に精製された」、「単離核酸」又は「精製核酸」という用語はまた、他の細胞成分を除去して精製されたアカゲザルHER2/neuタンパク質についてのコード領域を含むDNA分子を指す。従って、他の核酸を実質的に含まないアカゲザルHER2/neuDNA試料は、その総核酸のパーセントとして、非アカゲザルHER2/neu核酸を10%、好ましくは5%、より好ましくは1%、さらに一層好ましくは0.1%しか含まない。所与のアカゲザルHER2/neuDNA試料が他の核酸を実質的に含まないかどうかは、例えば適切な染色法、例えば臭化エチジウム染色、と組み合わせたアガロースゲル電気泳動、などの核酸純度を評価する従来の手法によって、又は配列決定によって判定することができる。
【0035】
「他のタンパク質を実質的に含まない」又は「実質的に精製された」とは、他のタンパク質を少なくとも90%、好ましくは95%、より好ましくは99%、さらに一層好ましくは99.9%含まないことを意味する。従って、他のタンパク質を実質的に含まないアカゲザルHER2/neuタンパク質試料は、その総タンパク質のパーセントとして、非アカゲザルHER2/neuタンパク質を10%、好ましくは5%、より好ましくは1%、さらに一層好ましくは0.1%しか含まない。所与のアカゲザルHER2/neuタンパク質試料が他のタンパク質を実質的に含まないかどうかは、例えば適切な検出法、例えば銀染色又は免疫ブロット法、と組み合わせたドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動(SDS−PAGE)、などのタンパク質純度を評価する従来の手法によって判定することができる。
【0036】
交換可能に使用されるとき、「他のタンパク質を実質的に含まない」又は「実質的に精製された」又は「単離アカゲザルHER2/neuタンパク質」又は「精製アカゲザルHER2/neuタンパク質」という用語はまた、天然ソースから単離されたアカゲザルHER2/neuタンパク質を指す。「単離」又は「精製」という用語の使用は、アカゲザルHER2/neuタンパク質がその通常の細胞環境から取り出されていることを指示する。従って、単離アカゲザルHER2/neuタンパク質は、無細胞溶液中に存在し得る又はそれが天然に生じるのとは異なる細胞環境に置かれ得る。単離という用語は、単離アカゲザルHER2/neuタンパク質が存在する唯一のタンパク質であることを意味するのではなく、単離アカゲザルHER2/neuタンパク質が、rhHER2/neuタンパク質がインビボで天然に結合している他のタンパク質及び非アミノ酸物質(例えば核酸、脂質、炭水化物)を実質的に含まないことを意味する。従って、原核生物又は真核生物細胞において組換え発現され、天然では(すなわち介入処置なしでは)このrhHER2/neuタンパク質を発現しないこの宿主細胞から実質的に精製されたアカゲザルHER2/neuタンパク質は、言うまでもなく、ここで言及する状況下での「単離アカゲザルHER2/neuタンパク質」である。前述したように、単離又は精製rhHER2/neuタンパク質であるrhHER2/neuタンパク質試料は、実質的に他のタンパク質を含まず、及びその総タンパク質のパーセントとして、非アカゲザルHER2/neuタンパク質を10%、好ましくは5%、より好ましくは1%、さらに一層好ましくは0.1%しか含まない。
【0037】
「保存的アミノ酸置換」は、もう1つ別の、化学的に類似するアミノ酸残基による1個のアミノ酸残基の置換を指す。このような保存的置換の例は、疎水性残基(イソロイシン、ロイシン、バリン又はメチオニン)によるもう1つ別の疎水性残基の置換;1個の極性残基による同じ電荷のもう1つ別の極性残基の置換(例えばアルギニンによるリシンの置換;グルタミン酸によるアスパラギン酸の置換)である。
【0038】
「哺乳動物」という用語は、ヒトを含む何らかの哺乳動物を指す。
【0039】
「Ag」という略語は、抗原を指す。
【0040】
「Ab」及び「mAb」という略語は、それぞれ抗体及びモノクローナル抗体を指す。
【発明を実施するための最良の形態】
【0041】
HER2/neu腫瘍関連抗原をコードする遺伝子は、一般に上皮由来のヒト癌の発現を伴う。本発明は、HER2/neuの異常発現が癌又はその発症を伴う場合に、HER2/neu腫瘍関連抗原によって発現されるタンパク質産物に対する免疫を惹起する又は増強する組成物及び方法に関する。HER2/neuの異常発現と癌との随伴は、HER2/neuの異常発現は腫瘍の発症時に存在し、腫瘍が進行した後期には検出できないことがあるので、腫瘍発現の全ての時点でHER2/neuタンパク質が腫瘍組織において発現されることを必要としない。
【0042】
この目的ために、アカゲザルHER2/neuをコードするポリヌクレオチドを提供する。本発明の分子は、細胞媒介性免疫を通して上皮由来の癌に対する有効な免疫予防を提供するための組換えアデノウイルスワクチンにおいて使用し得る。インビボで脊椎動物に直接導入するとき、本発明のポリヌクレオチドは、霊長動物、イヌ及びヒトなどの哺乳動物を含む動物の体内でコードされるタンパク質の発現を誘導する。
【0043】
本発明は、配列番号2に示す新規rhHER2/neuタンパク質を発現するmRNAをコードするヌクレオチドの配列を含む単離核酸分子(ポリヌクレオチド)に関する。本発明はまた、配列番号41に示す新規rhHER2/neuタンパク質を発現するmRNAをコードするヌクレオチドの配列を含む単離核酸分子に関する。本発明の核酸分子は実質的に他の核酸を含まない。
【0044】
本発明の単離核酸分子は、一本鎖(コード又は非コード鎖)又は二本鎖であり得る、ゲノムDNA及び相補的DNA(cDNA)などのデオキシリボ核酸分子(DNA)、並びに合成一本鎖ポリヌクレオチドなどの合成DNAを包含し得る。本発明の単離核酸分子はまた、リボ核酸分子(RNA)を包含し得る。大部分のクローニングのために、DNAは好ましい核酸である。
【0045】
本発明の好ましいDNA分子は、図2に示す、配列番号2として表わされるアカゲザルHER2/neuタンパク質をコードする、図1に示す、ここで配列番号1として開示するヌクレオチド配列を含む。このrhHER2/neu核酸分子は、実施例1で詳細に述べるRT−PCRを通して特定された。配列番号1の795位の「R」の存在は、第一アカゲザルから単離されたクローンがその位置にA又はGのいずれかを含んでいたことを指示する。第一アカゲザルから単離された、795位に「A」を含む核酸分子をここでは配列番号42と称する。第一アカゲザルから単離された、795位に「G」を含む核酸分子をここでは配列番号43と称する。
【0046】
第二の好ましいDNA分子は、図6に示す、配列番号41として表わされるアカゲザルHER2/neuタンパク質をコードする、図5に示す、ここで配列番号40として開示するヌクレオチド配列を含む。前記単離cDNAクローン、関連ベクター、宿主、組換え細胞画分及び膜、及び発現された成熟形態のrhHER2/neuは、癌ワクチンの開発のために有用である。
【0047】
本発明はまた、新規rhHER2/neuタンパク質を発現するmRNAをコードする、配列番号1又は配列番号40の生物活性断片又は突然変異株を包含する。このような生物活性断片及び/又は突然変異株は、配列番号2及び配列番号41に示すrhHER2/neuタンパク質を含むがこれらに限定されない、rhHER2/neuタンパク質の薬理特性を少なくとも実質的に模倣するタンパク質又はタンパク質断片のいずれかをコードする。このようなポリヌクレオチドは、ヌクレオチド置換、欠失、付加、アミノ末端トランケーション及びカルボキシ末端トランケーションを含むが、必ずしもこれらに限定されない。本発明の突然変異株は、癌ワクチンの開発において有用であるように真核細胞において機能性rhHER2/neuタンパク質を発現するmRNA分子をコードする。
【0048】
本発明はまた、合成DNAのヌクレオチド配列が、配列番号1及び配列番号40のヌクレオチド配列とは有意に異なるが、まだ配列番号2又は配列番号41と同じrhHER2/neuタンパク質をコードする場合の、rhHER2/neuタンパク質をコードする合成DNAに関する。このような合成DNAは本発明の範囲内に包含されるものとする。
【0049】
従って、本発明は、同一タンパク質を発現する数多くのDNA分子を生じ得るコドンの重複性を開示する。本明細書のために、1またはそれ以上の置換されたコドンを担持する配列を縮重変異と定義する。また、発現されるタンパク質の最終物理特性を実質的に変化させない、DNA配列又は翻訳されたタンパク質のいずれかにおける突然変異も、本発明の範囲内に包含される。例えばバリンによるロイシン、アルギニンによるリシン又はアスパラギンによるグルタミンの置換は、ポリペプチドの機能性に変化を生じさせないと考えられる。
【0050】
ペプチドをコードするDNA配列は、天然で生じるペプチドのものとは異なる性質を有するペプチドをコードするように変化させ得ることが知られている。DNA配列を変化させる方法は、部位指定突然変異誘発を含むが、これに限定されない。変化する性質の例は、基質に対する酵素の親和性又はリガンドに対する受容体の親和性の変化を含むが、これらに限定されない。
【0051】
ストリンジェント条件下で配列番号1又は配列番号40にハイブリダイズするDNA配列は本発明に包含される。例として、限定ではなく、高ストリンジェンシーの条件を使用する手順は以下の通りである。DNAを含むフィルターのプレハイブリダイゼーションを、6XSSC、5Xデンハルト溶液及び100μg/ml変性サケ精子DNAから成る緩衝液中65℃で2時間から一晩実施する。フィルターを、100μg/ml変性サケ精子DNA及び5−20X106cpmの32P標識プローブを含むプレハイブリダイゼーション混合物中65℃で12から48時間ハイブリダイズさせる。2XSSC、0.1%SDSを含む溶液中37℃で1時間、フィルターの洗浄を行う。これに続いて0.1XSSC、0.1%SDS中50℃で45分間洗浄した後、オートラジオグラフィーに供する。高ストリンジェンシー条件を使用する他の手順は、5XSSC、5Xデンハルト溶液、50%ホルムアミド中42℃で12から48時間実施するハイブリダイゼーション工程又は0.2XSSPE、0.2%SDS中65℃で30から60分間実施する洗浄工程を含む。
【0052】
高ストリンジェンシーハイブリダイゼーションを実施するための前記手順で述べた試薬は、当技術分野において周知である。これらの試薬の組成物の詳細は、例えば本明細書中に参考として援用されているSambrookら、Molecular Cloning:A Laboratory Manual;Cold Spring Harbor Laboratory,Cold Spring Harbor,New York,1989に認められる。前記に加えて、使用し得る高ストリンジェンシーの他の条件は、当技術分野において周知である。
【0053】
本発明の1つの好ましい側面は、図2(配列番号2)に開示するアミノ酸の配列を含むアカゲザルHER2/neuタンパク質の実質的に精製された形態である。
【0054】
本発明のもう1つの好ましい側面は、図6(配列番号41)に開示するアミノ酸の配列を含むアカゲザルHER2/neuタンパク質の実質的に精製された形態である。
【0055】
本発明はまた、細胞外ドメイン及び細胞内ドメインなどの、rhHER2/neuの様々な機能性ドメイン、及びこれらの配列の少なくとも1つを含むハイブリッド分子に関する。
【0056】
本発明はまた、それらの変異が癌ワクチンの開発のために有用である診断、治療又は予防用途のタンパク質又はタンパク質断片を提供するような、アミノ酸置換、欠失、付加、アミノ末端トランケーション及びカルボキシ末端トランケーションを含む、必ずしもこれらに限定されない、配列番号2又は配列番号41に示すアミノ酸配列を含む、rhHER2/neuタンパク質の生物活性断片及び/又は突然変異株を包含する。
【0057】
本発明のアカゲザルHER2/neuタンパク質は、「成熟」タンパク質の形態であるか、又は融合タンパク質などのより大きなタンパク質の部分であり得る。分泌又はリーダー配列、プロ配列、多数のヒスチジン残基などの精製を助ける配列、又は組換え生産の間の安定性のための付加的な配列を含む、付加的アミノ酸配列を含むことがしばしば好都合である。
【0058】
本発明はまた、GFP(グリーン蛍光タンパク質)、MYCエピトープ、CST及びFcを含むがいかなる意味においてもこれらに限定されない、様々なマーカーに連結されたアカゲザルHER2/neuタンパク質の部分を発現する融合構築物を含むが、これらに限定されない、rhHER2/neu融合構築物に関する。このような融合構築物は、対象細胞系において発現させ、ここで開示するアカゲザルHER2/neuタンパク質の調節因子に関してスクリーニングするために使用し得る。
【0059】
本発明はさらに、本明細書全体を通じて開示する実質的に精製された核酸分子を含む組換えベクターに関する。これらのベクターはDNA又はRNAで構成され得る。クローニングの殆んどの目的のためには、DNAベクターが好ましい。典型的なベクターは、プラスミド、改変ウイルス、バクテリオファージ、コスミド、酵母人工染色体、及びrhHER2/neuタンパク質をコードすることができるエピソーム又は組込みDNAの他の形態を含む。個々の遺伝子導入又は他の用途のために適切なベクターを決定することは十分に当業者の理解範囲内である。
【0060】
rhHER2/neuタンパク質をコードするDNAを含む発現ベクターは、組換え宿主細胞におけるrhHER2/neuの発現のために使用し得る。発現ベクターは、クローニングベクター、改変クローニングベクター、特に設計されたプラスミド又はウイルスを含み得るが、これらに限定されない。また、所望に応じて細菌細胞において組換えrhHER2/neuを発現するために、様々な細菌発現ベクターを使用し得る。加えて、真菌細胞において組換えrhHER2/neuを発現するために、様々な真菌細胞発現ベクターを使用し得る。さらに、昆虫細胞において組換えタンパク質を発現するために、様々な昆虫細胞発現ベクターを使用し得る。
【0061】
本発明はまた、本発明の核酸分子を含むベクターで形質転換した又はトランスフェクトした宿主細胞に関する。組換え宿主細胞は、E.コリなどの細菌、酵母などの真菌細胞、ウシ、ブタ、サル及びげっ歯動物起源の細胞系統を含むがこれらに限定されない哺乳動物細胞、及びショウジョウバエ(Drosophia)及びカイコ由来の細胞系統を含むがこれらに限定されない昆虫細胞を含むが、これらに限定されない、原核又は真核細胞であり得る。このような組換え宿主細胞は、rhHER2/neu又は生物学的等価形態を生産するために適切な条件下で培養することができる。
【0062】
前述したように、rhHER2/neuタンパク質をコードするDNAを含む発現ベクターは、組換え宿主細胞におけるrhHER2/neuの発現のために使用し得る。従って、本発明のもう1つの側面は、(a)配列番号1又は配列番号40に示す核酸を含むベクターを適切な宿主細胞に導入すること;及び(b)前記宿主細胞を、前記アカゲザルHER2/neuタンパク質の発現を可能にする条件下で培養すること、を含む、組換え宿主細胞においてアカゲザルHER2/neuタンパク質を発現するための方法である。
【0063】
宿主細胞におけるrhHER2/neuの発現後、活性形態のrhHER2/neuタンパク質を提供するためにrhHER2/neuタンパク質を回収し得る。幾つかのrhHER2/neuタンパク質精製手順が使用可能であり、使用に適する。組換えrhHER2/neuタンパク質を、塩分画、イオン交換クロマトグラフィー、サイズ排除クロマトグラフィー、ヒドロキシアパタイト吸着クロマトグラフィー及び疎水性相互作用クロマトグラフィーの様々な組合せ又は個別適用によって細胞溶解産物及び抽出物から精製し得る。加えて、組換えrhHER2/neuタンパク質は、完全長rhHER2/neuタンパク質又はrhHER2/neuタンパク質のポリペプチド断片に特異的なモノクローナル又はポリクローナル抗体で作製した免疫アフィニティーカラムの使用によって他の細胞タンパク質から分離することができる。
【0064】
本発明の核酸は、ヒト細胞における前記タンパク質の効率的な発現を提供するように設計された配列を含む発現カセットに構築し得る。前記カセットは、好ましくは完全長rhHER2/neu遺伝子を、プロモーター及び終結配列などの、それに作動可能に連結された関連転写及び翻訳制御配列と共に含む。1つの好ましい実施態様では、前記プロモーターは、イントロンA配列を伴わないサイトメガロウイルスプロモーターであるが、当業者は、強力な免疫グロブリン又は他の真核生物遺伝子プロモーターなどの多くの他の公知のプロモーターのいずれもが使用し得ることを認識する。好ましい転写ターミネーターはウシ成長ホルモン終結因子であるが、他の公知の転写ターミネーターも使用し得る。CMV−BGHターミネーターの組合せは特に好ましい。
【0065】
本発明によれば、アカゲザルHER2/neu発現カセットをベクターに挿入する。前記ベクターは、好ましくはアデノウイルスベクターであるが、プロモーターに連結された線状DNA、又はアデノ関連ウイルス又は改変ワクシニアウイルスベクターなどの他のベクターも使用し得る。
【0066】
選択するベクターがアデノウイルスである場合は、いわゆる第一世代アデノウイルスベクターであることが好ましい。これらのアデノウイルスベクターは、非機能性E1遺伝子領域、好ましくは欠失アデノウイルスE1領域を有することを特徴とする。一部の実施態様では、アデノウイルスE1遺伝子が通常位置する部位に発現カセットを挿入する。加えて、これらのベクターは、場合により、非機能性又は欠失E3領域を有する。使用するアデノウイルスゲノムは、E1及びE3領域の両方を欠失している(△E1△E3)ことが望ましい。前記アデノウイルスを、293細胞又はPERC.6細胞などのウイルスE1遺伝子を発現する公知の細胞系統において、又は余分の(extra)タンパク質を発現するように一過性に又は安定に形質転換された293又はPERC.6細胞に由来する細胞系統において増殖させることができる。例えばテトラサイクリン調節可能プロモーター系などの制御された遺伝子発現を有する構築物を使用するとき、前記細胞系統は、調節系に関与する成分を発現し得る。このような細胞系統の一例はT−Rex−293であり、他は当技術分野において公知である。
【0067】
アデノウイルスベクターの操作上の便利さのために、アデノウイルスはシャトルプラスミド形態であり得る。本発明はまた、プラスミド部分及び、E1欠失及び場合によりE3欠失を有し、アカゲザルHER2/neuを含む発現カセットが挿入されたアデノウイルスゲノムを含有するアデノウイルス部分を含む、シャトルプラスミドベクターを対象とする。好ましい実施態様では、アデノウイルスベクターが容易に除去できるように、プラスミドのアデノウイルス部分に隣接する制限部位が存在する。前記シャトルプラスミドは、原核細胞又は真核細胞において複製し得る。
【0068】
本発明の好ましい実施態様では、発現カセットをpMRKAd5−HV0アデノウイルスプラスミドに挿入する(本明細書中に参照により組み込まれる、Eminiら、国際公開広報第WO02/22080号参照)。このプラスミドは、E1及びE3領域を欠失したAd5アデノウイルスゲノムを含む。pMRKAd5−HV0プラスミドの設計は、ウイルスパッケージングを最適化する上で重要であることが認められたエレメントを組み込むために5’シス作用性パッケージング領域をさらにE1遺伝子にまで延長することにより、先行アデノウイルスに比べて改善されており、高いウイルス増幅をもたらす。好都合には、この強化されたアデノウイルスベクターは、高い継代増殖後も遺伝的安定性を維持することができる。
【0069】
DNA構築物を作製し、精製するための分子生物学の標準手法は、本発明のアデノウイルス、シャトルプラスミド及びDNA免疫原の作製を可能にする。
【0070】
前述したベクターは、HER2/neu発現異常に関連する上皮由来の癌の発現を予防するため及び/又は既存の癌を治療するための免疫原性組成物及びワクチンにおいて使用し得る。このために、本発明の1つの側面は、配列番号2又は配列番号41に示すアカゲザルHER2/neuタンパク質をコードするヌクレオチドの配列を含む単離核酸分子を含有するワクチンベクターを哺乳動物に投与することを含む、癌を予防する又は治療する方法である。
【0071】
前述した方法によれば、前記ワクチンベクターは、哺乳動物における癌の治療又は予防のために投与し得る。本発明の1つの好ましい実施態様では、前記哺乳動物はヒトである。
【0072】
さらに、当業者は、前述した治療及び予防方法における使用のためにいかなる種類のベクターも選択し得る。好ましくは、前記ベクターはアデノウイルスベクター又はプラスミドベクターである。本発明の1つの好ましい実施態様では、前記ベクターは、アデノウイルスE1領域内の欠失、及び(a)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及び(b)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む発現カセットを含有するアデノウイルスE1領域内のインサートを有するアデノウイルスゲノムを含む、アデノウイルスベクターである。
【0073】
本発明はさらに、E1領域内の欠失、及び(a)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及び(b)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む発現カセットを含有するE1領域内のインサートを有するアデノウイルスゲノムを含む、アデノウイルスワクチンベクターに関する。
【0074】
本発明のこの側面の1つの好ましい実施態様では、前記アデノウイルスベクターはAd5ベクターである。
【0075】
もう1つの側面では、本発明は、プラスミド部分、及び(a)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及び(b)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む発現カセット部分を含有する、ワクチンプラスミドに関する。
【0076】
本発明の一部の実施態様では、高い免疫応答を誘導するために、ここで開示する組換えアデノウイルスワクチンを、プラスミドに基づくポリヌクレオチドワクチンと共に様々なプライム(初回抗原投与)/ブースト(追加抗原投与)の組合せにおいて使用する。この場合、前記の2つのベクターを「プライム&ブースト」レジメンにおいて投与する。例えば最初のタイプのベクターを投与し、次にあらかじめ定められた期間後、例えば1ヶ月後、2ヶ月後、6ヶ月後又は他の適切な間隔後に、第二のタイプのベクターを投与する。好ましくは、前記ベクターは同じポリヌクレオチド又はポリヌクレオチドの組合せをコードする発現カセットを担持する。プラスミドDNAも使用する実施態様では、前記ベクターは、哺乳動物又は昆虫細胞によって認識される1またはそれ以上のプロモーターを含むことが好ましい。1つの好ましい実施態様では、前記プラスミドは、ヒトCMVプロモーターなどの、但しこれに限定されない、強力なプロモーターを含む。アカゲザルHER2/neu遺伝子又は発現すべき他の遺伝子をこのようなプロモーターに連結する。このようなプラスミドの一例は、記述されている哺乳動物発現プラスミドV1Jnsである(本明細書中に参照により組み込まれる、J.Shiverら、DNA Vaccinesより、M.Liuら編集、N.Y.Acad.Sci.,N.Y.,772:198−208(1996))。
【0077】
前述したように、アデノウイルスベクターワクチン及びプラスミドワクチンは、免疫応答を誘導するために単一治療レジメンの一部として脊椎動物に投与し得る。このために、本発明は、(a)哺乳動物に、i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及びii)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む第一ベクターを導入すること;(b)あらかじめ定められた時間を経過させること;及び(c)前記哺乳動物に、i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及びii)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む第二ベクターを導入すること、を含む、哺乳動物を癌から保護する方法に関する。
【0078】
前述した保護の方法の1つの実施態様では、前記第一ベクターはプラスミドであり、及び前記第二ベクターはアデノウイルスベクターである。代替的実施態様では、前記第一ベクターはアデノウイルスベクターであり、及び前記第二ベクターはプラスミドである。
本発明はさらに、(a)上皮由来の癌に罹患している哺乳動物に、i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及びii)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む第一ベクターを導入すること;(b)あらかじめ定められた時間を経過させること;及び(c)前記哺乳動物に、i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及びii)前記ポリヌクレオチドに作動可能に連結されたプロモーター、を含む第二ベクターを導入すること、を含む、上皮由来の癌に罹患している哺乳動物を治療する方法に関する。
【0079】
前述した治療方法の1つの実施態様では、前記第一ベクターはプラスミドであり、及び前記第二ベクターはアデノウイルスベクターである。代替的実施態様では、前記第一ベクターはアデノウイルスベクターであり、及び前記第二ベクターはプラスミドである。
【0080】
ワクチン受容者に導入すべき発現可能なDNA又は転写されたRNAの量は、部分的には、使用するプロモーターの強度及び発現される遺伝子産物の免疫原性に依存する。一般に、約1ngから100mg、好ましくは約10μgから300μgの免疫学的又は予防的有効用量のプラスミドワクチンベクターを筋組織に直接投与する。組換えアデノウイルスについての有効用量は、約106−1012粒子、好ましくは約107−1011粒子である。皮下注射、皮内導入、皮膚を通しての塗り(impression)、及び腹腔内、静脈内又は吸入送達などの他の投与方法も考慮される。また、ブースターワクチン接種を提供することも考慮される。本発明のワクチンの非経口導入と同時に又は導入後に、インターロイキン12タンパク質などのアジュバントによる静脈内、筋肉内、皮下などの非経口投与又は他の投与手段も好都合である。
【0081】
本発明のワクチンベクターは、裸であり得る、すなわち受容者の免疫系に影響を及ぼすいかなるタンパク質、アジュバント又は他の作用物質とも非結合であり得る。この場合、前記ワクチンベクターは、無菌食塩水又は無菌緩衝食塩水などの、但しこれらに限定されない、生理的に許容される溶液中に存在することが望ましい。また、アジュバント、サイトカイン、タンパク質又は他の担体などの免疫促進剤又は本発明の免疫原性組成物を投与することは好都合であると考えられる。従って、本発明は、本発明の組成物及び方法と組み合わせたこのような免疫促進剤の使用を包含する。免疫促進剤は、ここで使用するとき、基本的に外因性抗原に対する免疫応答(抗体及び/又は細胞媒介性)を高める又は増強する物質を指す。前記免疫促進剤は、DNA又はタンパク質の形態で投与することができる。GM−CSF、IFNα、破傷風毒素、IL12、B7.1、LFA−3及びICAM−1を含むがこれらに限定されない、様々な免疫促進剤のいずれかを、本発明のワクチン及び免疫原性組成物と共に使用し得る。前記免疫促進剤は当技術分野において周知である。カルシウムイオンなどの、但しこれに限定されない、DNAの細胞取り込みを助ける物質も使用し得る。これらの物質は、一般にトランスフェクション促進試薬及び医薬適合性の担体と称される。当業者は、個々の免疫促進剤又は医薬適合性の担体並びに適切な投与時間及び投与方法を決定することができる。
【0082】
様々な手順のいずれかを使用してrhHER2/neuをクローニングし得る。これらの方法は、(1)RACE PCRクローニング手法(Frohmanら、Proc.Natl.Acad.Sci.USA 85:8998−9002(1988))。5’及び/又は3’RACEは完全長cDNA配列を生成するために実施し得る。この戦略は、rhHER2/neu cDNAのPCR増幅のために遺伝子特異的オリゴヌクレオチドプライマーを使用することを含む。これらの遺伝子特異的プライマーは、多数の公的に使用し得る核酸及びタンパク質データベースを検索することによって特定されてきた、発現配列タグ(EST)ヌクレオチド配列の特定を通して設計される;(2)適切な発現ベクター系におけるrhHER2/neu含有cDNAライブラリーの構築後のrhHER2/neu cDNAの直接の機能的発現;(3)rhHER2/neuタンパク質のアミノ酸配列から設計した標識縮重オリゴヌクレオチドプローブで、バクテリオファージ又はプラスミドシャトルベクターにおいて構築したrhHER2/neu含有cDNAライブラリーをスクリーニングすること;(4)rhHER2/neuタンパク質をコードする部分的cDNAで、バクテリオファージ又はプラスミドシャトルベクターにおいて構築したrhHER2/neu含有cDNAライブラリーをスクリーニングすること。この部分的cDNAは、rhHER2/neuタンパク質に関連する他の増殖因子受容体についての公知のアミノ酸配列から縮重オリゴヌクレオチドプライマーを設計することを通して、rhHER2/neuDNA断片の特異的PCR増幅によって得られる;(5)哺乳動物rhHER2/neuタンパク質に相同性を有する部分的cDNA又はオリゴヌクレオチドで、バクテリオファージ又はプラスミドシャトルベクターにおいて構築したrhHER2/neu含有cDNAライブラリーをスクリーニングすること。この戦略も、前述したようにESTとして特定されたrhHER2/neu cDNAのPCR増幅のために遺伝子特異的オリゴヌクレオチドプライマーを使用することを含み得る;又は(6)rhHER2/neuをコードするヌクレオチド配列の完全長形態を単離するために、数多くの種類のcDNA及び/又はゲノムライブラリーの1つをスクリーニングするためのプローブとして使用するコード領域の部分を生成し、単離するために公知のRACE手法によって完全長cDNAを生成するか又はこれらの同じ公知のRACE手法によってコード領域の部分を生成するように、配列番号1又は配列番号40を鋳型として使用して5’及び3’遺伝子特異的オリゴヌクレオチドを設計すること、を含むが、これらに限定されない。
【0083】
他の種類のライブラリー、並びに他の細胞型又は他の種型から構築されたライブラリーが、rhHER2/neuをコードするDNA又はrhHER2/neuホモログを単離するために有用であり得ることは、当業者には容易に明白である。
【0084】
また、適切なcDNAライブラリーは、様々な上皮由来細胞などの、rhHER2/neu活性を有する細胞又は細胞系統から作製し得ることも、当業者には容易に明白である。rhHER2/neuをコードするcDNAを単離するためのcDNAライブラリーを作製するときに使用する細胞又は細胞系統の選択は、最初に、このような目的のために使用し得る何らかの公知のアッセイを用いて細胞に関連するrhHER2/neu活性を測定することによって実施し得る。
【0085】
cDNAライブラリーの作製は、当技術分野で周知の標準手法によって実施することができる。周知のcDNAライブラリー構築手法は、例えばSambrookら、Molecular Cloning:A Laboratory Manual;Cold Spring Harbor Laboratory,Cold Spring Harbor,New York,1989に認められる。相補的DNAライブラリーも、Clontech Laboratories,Inc.(Palo Alto,CA)及びStratagene(La Jolla,CA)を含むがこれらに限定されない、数多くの商業的供給元から入手し得る。
【0086】
本発明のDNA分子、RNA分子及び組換えタンパク質は、rhHER2/neuをスクリーニングし、そのレベルを測定するために使用し得る。前記組換えタンパク質、DNA分子及びRNA分子は、rhHER2/neuの検出及びタイピングに適するキットの作成に役立つ。このようなキットは、少なくとも1つの容器を厳重に閉じ込めて保持するのに適する区画化された担体を含む。前記担体は、rhHER2/neuを検出するのに適した組換えrhHER2/neu又は抗rhHER2/neu抗体などの試薬をさらに含む。前記担体はまた、標識抗原又は酵素基質等のような検出のための手段も含み得る。
【0087】
ここで言及する全ての公表文献は、本発明に関連して使用し得る方法及び材料を説明し、開示するために参照により組み込まれる。ここに含まれるいかなる内容も、本発明が先行発明によるこのような開示に先立つ権利がないことの是認と解釈されるべきではない。
【0088】
付属の図面を参照しながら本発明の好ましい実施態様を説明したが、本発明はそれらの厳密な実施態様に限定されないこと、及び付属の特許請求の範囲において定義される本発明の範囲及び精神から逸脱することなく当業者によって様々な変更及び修正が為され得ることは了解されねばならない。
【0089】
以下の実施例は本発明を例示するものであり、本発明を限定しない。
【実施例1】
【0090】
RT−PCRによるアカゲザルHER2/neu cDNAの単離
分子学的手順は、当技術分野において周知の標準手法(例えば本明細書中に参照により組み込まれる、Ausubelら、Short Protocols in Molecular Biology,F.M.,第2版、John Wiley & Sons(1992)及びSambrookら、Molecular Cloning,A Laboratory Manual、第2版、Cold Spring Harbor Laboratory Press(1989)参照)に従って実施した。
【0091】
ヒト、ハムスター、イヌ及びラットからのHER2/neuヌクレオチド配列を、HER2/neu DNAの高度に保存された領域を特定するために整列させた。配列比較に基づき、HER2/neu遺伝子をカバーするオリゴヌクレオチドプライマーを、以下に述べる、逆転写酵素ポリメラーゼ連鎖反応(RT−PCR)によるアカゲザルHER2/neu cDNAの増幅のために設計した(図3参照)。
【0092】
2匹の異なるアカゲザル(macaca mulatta)からの結腸生検をDr.Willem Collignon(Biomedical Primate Research Centre(BPRC),Rijswijk,The Netherlands)より入手した。製造者の指示に従ってUltraSPec−II RNA単離システム(Biotecx,Houston,TX)を用いて各々の結腸生検からRNAを抽出し、精製した。アカゲザルHER2/neu遺伝子を単離するために、HER2/neu配列全体をカバーするRT−PCR増幅産物を、1匹のアカゲザルから単離した全RNAから生成した。
【0093】
逆転写工程を実施するために、製造者の指示に従ってSuperscript One−Step RT−PCR Amplification Kit for Long Templates(Life Technologies,Carlsbad,CA)を使用して全RNA試料を逆転写した。典型的には、RNA0.5−2.0μgを50μlの反応容量で逆転写酵素及び適切な緩衝液と混合した。試料を45℃で30分間インキュベートし、次に94℃で2分間インキュベートした。
【0094】
生じたcDNA鋳型を使用して、正プライマーと逆プライマーの種々の組合せを用いてPCR増幅を実施した(図3参照)。PCRはPerkin Elmer 2400サーモサイクラー(Perkin Elmer,Inc.,Wellesley,MA)において実施した。サイクリング条件は、94℃、15秒間の初期変性工程の35サイクル、次いでプライマーアニーリング工程及び伸長工程での終了から成った。プライマーアニーリング工程は、プライマー配列に依存して、50℃−51℃の範囲の温度で50秒間のインキュベーションから成った。伸長工程は、増幅産物の予想される長さに依存して、80秒間−100秒間の範囲の時間にわたる68℃でのインキュベーションから成った。前記35サイクルの後に72℃で7分間の広汎な延長工程(extensive elongation step)を実施した。
【0095】
QIAquick PCR Purification Kit(Qiagen,Hilden,Germany)を用いて増幅産物をゲル精製し、増幅に使用した同じプライマーで配列決定した。配列決定反応は、Cycle Sequencing Ready Reaction Kit(Applied Biosystems,Foster City,CA)を使用して、Big Dye Terminator化学を通して実施した。読取りはABI Prism 377 DNAシーケンサー(Applied Biosystem)を用いて実施した。
【0096】
HER2/neuコード領域全体を含む種々の増幅産物を配列決定することによって獲得したデータは、配列番号1としてここで開示する、アカゲザルHER2/neu配列(以下「第一アカゲザルHER2ヌクレオチド配列」又は「rhHER2#1」、図1参照)を特定した。配列番号1の795位の「R」の存在は、第一アカゲザルから単離されたクローンがその位置にA又はGのいずれかを含んでいたこと指示する。第一アカゲザルから単離された、795位に「A」を含む核酸分子をここでは配列番号42と称する。第一アカゲザルから単離された、795位に「G」を含む核酸分子をここでは配列番号43と称する。第一アカゲザルから単離されたDNAに基づき、配列番号1の795位の単一ヌクレオチド変化は、ここで配列番号2として開示する(図2参照)、生じたHER2/neuタンパク質のアミノ酸配列に影響を及ぼさなかった。
【0097】
前記RT−PCR反応から誘導された増幅産物を、ここではそれぞれBS又はCRと称する、ベクターBlueScript ks(+)又はpCRIIのいずれかにクローニングした。増幅産物の直接配列決定によって得たrhHER2/neuヌクレオチド配列を確認するために、生じたクローンを配列決定し、完全なアカゲザルHER2/neu cDNA配列を構築するために使用した。
【実施例2】
【0098】
完全なHER2/neu cDNA配列の構築
第一アカゲザルから得た一連のクローンをPCRによって構築し、集合させて、完全なアカゲザルHER2/neu cDNAを作製した。最初に、アカゲザルHER2/neu遺伝子の5’末端をカバーする4つのRT−PCR増幅産物を実施例1で述べたように生成した。前記クローンからのPCR増幅断片を、図4に示すように(反応1−4)、BS又はCRのいずれかにクローニングした。幾つかのクローンの配列分析は、クローニングされた配列がRT−PCR断片から得た配列と同じであることを確認した。
【0099】
前述した重複増幅産物を、以下の成分:前記断片の各々0.1−1.0pmol、Pfuポリメラーゼ(Stratagene,La Jolla,CA)及び適切な緩衝液、を組み合わせたPCR反応物100μl中で連結した。試料を、95℃、30秒間、次いで72℃、4分間から成る初期増幅サイクルに供した。
【0100】
生じた連結産物を、Pfuポリメラーゼ及び適切な緩衝液を用いて以下の熱プロフィール:95℃、30秒間、58℃、30秒間及び72℃、180秒間(30サイクル)により、PmeI_SwaI_RBS_1−16(配列番号11)及び1895−1876(配列番号10)プライマーで再び増幅した。このようにして得たPCR産物をゲル精製し、pCRIIベクターにクローニングして、クローン#1_CR_1−1895.7(−)(反時計回り方向)を作製した。幾つかのクローンの配列分析により、クローニングされた配列の同一性を確認した。
【0101】
第一アカゲザルからの結腸RNAを使用して、基本的に前述したように、RT−PCRによって付加的なPCR断片を生成した。このために使用したオリゴヌクレオチドプライマーは、1558−1583(配列番号12)及び2798−2776(配列番号13)であった(反応6、図4)。HER2/neu遺伝子の中央部分に位置するこの断片を、クローニングせずに、さらなる増幅のための鋳型として使用した。
【0102】
次に、bp1621−bp2277(図4、反応7、配列番号14、15)及びbp2239−bp2798(図4、反応8、配列番号16及び13)から、この領域をPCR増幅するためのプライマーを設計した。生じた産物をpCRIIベクターにクローニングした。タンパク質のチロシンキナーゼ活性を不活性化するために、KからAまでのアミノ酸579位を沈黙化するためのプライマーを設計した(配列番号15及び16)。幾つかのクローンの配列分析は、クローニングされた配列の同一性を確認した。
【0103】
第一アカゲザルからの結腸RNAを使用して、基本的に前述したように、RT−PCRによって付加的なPCR断片を生成した。このために使用したオリゴヌクレオチドプライマーは、2356−2378(配列番号17)及び4166−4145(配列番号18)であった(図4、反応9)。生じた産物をpCRIIベクターにクローニングした。幾つかのクローンの配列分析は、クローニングされた配列の同一性を確認した。
【0104】
以下のプライマーを使用して、前記の3つのクローンからDNA断片をPCR増幅した:1621−1644(配列番号14)及び2277−K−753−2248(配列番号15)、鋳型は#1_CR_1621−2277.2;2239−K756−2248(配列番号16)及び2798−2776(配列番号13)、鋳型は#1_CR_2239−2798.4;2356−2378(配列番号17)及びSalI_3768−3746(配列番号19)、鋳型は#1_CR_2356−4169.2。3つの産物を前述したようにPCRによって集合させた。生じた連結産物を以下のプライマー:1621−1644(配列番号14)及びSalI_3768−3746(配列番号19)を用いて増幅し、pCRIIベクターに時計周り方向にクローニングして、クローン#1_CR_1621−3768.8(以下クローン10.8)及び#1_CR_1621−3768.12(以下クローン10.12)を生成した(図4、反応10)。配列決定は、クローン10.12が3’領域内の3686位付近に突然変異を有することを明らかにした。同様に、クローン10.8は2666位付近に突然変異を有していた。クローン10.12からの508bp BstEII−SalI断片を、この領域内に突然変異を含まない、クローン10.8からの対応する領域で置き換えることにより、野生型配列を生成した。生じたクローンを#1_CR_1621−3768.128と命名した。
【0105】
クローン#1_CR_1621−3768.128(+)からのPmlI−XbaI断片を#1_CR_1_1895.7(−)のPmlI−SpeI部位にクローニングした。アカゲザルHER2/neuコード配列全体を含む、生じたプラスミドを#1_CR_1−3768(−)と命名した。配列分析により前記配列の同一性を確認した。
【実施例3】
【0106】
免疫原
遺伝子導入と免疫試験のために、PmeI及びSalIでの消化によってrhHER2/neuコード領域を#1_CR_1−3768(−)から切り出し、哺乳動物発現プラスミドpV1J_nsA(Montgomeryら、DNA Cell Biol.12(9):777−83(1993))のEcoRV及びSalI部位に挿入して、pV1J−rh−HER2/neuを生成した。
【0107】
アデノウイルスベクター構築のために、PmeI及びSalIでの消化によってrhHER2/neuコード領域を#1_CR_1−3768(−)から切り出し、ポリMRKAd5△E1シャトルプラスミドの対応する部位にクローニングして、pMRKAd5△E1−rhHER2/neuを生成した。シャトルベクターpMRKAd5△E1は、ヒトサイトメガロウイルス(HCMV)プロモーター(イントロンAなし)及びウシ成長ホルモンポリアデニル化シグナルを含む発現カセットと共に塩基対1−450及び塩基対3511−5798からのAd5配列を含む。このプラスミドを、大腸菌BJ5183細胞(Chartierら、J.Virol.70:4805−10(1996)を使用して、E1及びE3領域を含有する配列を除くすべてのAd5配列を含む、アデノウイルス骨格ベクターpMRKAd5HVOと組み換えた。
【0108】
ベクター生産のために、pMRKAd5−rhHER2/neuをPacIでの消化によって線状化し、Lipofectamine(Life Technologies,Rockville,MD)を用いてPerC.6細胞にトランスフェクトした。ウイルス力価を増幅するために5−6ウイルス継代を行い、大型ウイルス増幅を実施して1.3×1012物理的粒子(physical particles)(pp)の最終生産を得た。制限断片長多型(RFLP)分析によって示されたように、増幅したベクターから精製したウイルスゲノムにおいてゲノム再構成は検出できなかった。予想されたDNA断片は、平行して制限した、ウイルスゲノム内及び対照pMRKAd5−rhHER2/neuプラスミドの両方で認められた。
【実施例4】
【0109】
第二アカゲザルからのHER2/neuコードDNAの単離
第二アカゲザルからの結腸RNAを使用してRT−PCR産物の同様の配列決定分析を実施した(実施例1参照)。前記遺伝子全体を含む多くの異なる増幅産物を配列決定することによって獲得したデータは、ここで配列番号40として開示する、第二アカゲザルHER2/neu配列(以下「第二アカゲザルHER2ヌクレオチド配列」又は「rhHER2#2」、図5参照)を、ここで配列番号41として開示する演繹アミノ酸配列(以下「第二アカゲザルHER2アミノ酸配列」、実施例6参照)と共に特定した。
【0110】
実施例1におけるように、HER2/neuをカバーするプライマーを使用して第二アカゲザルからの全結腸RNAのRT−PCRによって増幅産物を生成した(図3参照)。生じた増幅産物をゲル精製し、配列決定した。さらに、これらの産物をBS又はCRベクターのいずれかにクローニングした。生じたクローンのDNA塩基配列決定分析により、増幅産物の直接配列決定によって得たrhHER2/neuヌクレオチド配列を確認した。
【0111】
rhHER2#1とrhHER2#2の間で8つの相違が検出された(詳細については、図7参照)。これらの突然変異のうち5つは、第一アカゲザルHER2アミノ酸配列(配列番号2)と比較したときタンパク質内にアミノ酸変化を導入する。注意すべきは、これらの突然変異のうち3つは、rhHER2#1タンパク質と比較したときrhHER2#2タンパク質内にアミノ酸変化を生じさせない。
【実施例5】
【0112】
rhHER2/neuによるアカゲザルの免疫
結腸直腸癌において発現されるヒト腫瘍抗原HER2/neuのアカゲザルホモログによるアカゲザル(Macaca mulatta)の免疫の効率を評価するため、Biomedical Primate Research Centre(BPRC),Rijswijk(The Netherlands)において免疫試験を実施した。試験は、アカゲザルHER2/neu抗原での免疫に対するB及びT細胞応答を評価するように設計した。
【0113】
最初の試験では、1つの群のサル(アカゲザル合計4匹;雄性2匹と雌性2匹)をヒト腫瘍抗原HER2/neuのアカゲザルホモログを発現するプラスミドDNAベクター又はアデノウイルスベクター(pV1J−rhHER2)で免疫した。初回免疫のために、0、4、8、12及び16週目にDNAの注射とそれに続く電気刺激により、筋肉内経路で(i.m.)プラスミドDNAによって動物をワクチン接種した。前記DNAは、pV1J−rhHER2プラスミドDNA5mgを含有する1ml溶液(2つの部位に分けて0.5ml/部位)として、体重2−5kgの動物に注射した。動物には麻酔下で(ケタミン/キシラジンの混合物)注射を行う。
【0114】
電気刺激のために、2トレーンの100矩形双極性パルス(各々1秒)を、3秒の全処置時間中1秒おきに送達した。パルス長は2ミリ秒/位相、パルス周波数及び振幅はそれぞれ100Hz及び100mA(定電流方式)であった。
【0115】
同じ群の動物を、アカゲザルHER2/neuを発現するアデノウイルス5(Ad5)の筋肉内注射によって追加免疫した。△E1−△E3、「第一世代」アデノウイルス(P2レベル)を使用した。1011ウイルス粒子(vp)の総量を24週目と28週目に注射した。
【0116】
36週目と40週目にアカゲザルHER2/neuを発現するアデノウイルス24(Ad24)の1011ウイルス粒子(vp)の筋肉内注射によってさらなる追加免疫を実施した。Ad5注射によって誘導される中和抗体はAd24感染に干渉しないので、Ad24を選択した。
【0117】
前記免疫プロトコールによって誘導されるHER2/neuに対する免疫応答を測定するため、総期間1年間にわたって4週間ごとに血液試料を採集した。HER2特異的細胞媒介性免疫応答をIFN−γ ELISPOTアッセイによって測定した。IFN−γ分泌性抗アカゲザルHER2 T細胞の数を、ペプチドのプールを使用してPBMCに関するELISPOTによって判定した。11残基が重複し、アカゲザルタンパク質配列全体をカバーする、各々15アミノ酸長の311ペプチドを、AからKまでのアルファベット文字で示した11のプール(N末端からC末端まで)と組み合わせた。PBMCを産生するIFN−γの頻度を、2つの異なる細胞濃度での2回の測定から導かれたスポットの平均値として算定した。数値は、スポット形成コロニー(SFC)/106全PBMCの数から、ペプチドの不在下で測定したバックグラウンド値(典型的には10SFC/106全脾細胞)を差し引いた値として表わした。算定結果は、IFN−γ ELISPOTによって測定したとき、4匹のサル全てが検出可能な細胞媒介性応答を示すことを示す。
【0118】
反応性を確認し、IFN−γを分泌するCD4+又はCD8+T細胞の頻度を測定する、IFN−γ+細胞内染色法(ICS)によってタイピングした。CD3+事象と小リンパ球で同時ゲーティングすることによってCD3+リンパ球を収集した。0.1%より高い数値を陽性とみなした。プールJについてサルRI504に関して0.19の陽性値(CD8+)を得た。検出可能な抗rhHER2抗体力価(>200)は検出されなかった。
【0119】
要約するに、アカゲザルHER2/neuを発現するプラスミドDNAの筋肉内注射は、アカゲザルにおいて寛容を打破し、rhHER2/neuに対する検出可能な細胞媒介性免疫応答を誘導する上で有効であった。
【実施例6】
【0120】
ヒト腫瘍抗原のアカゲザルホモログによるアカゲザルの免疫
全て結腸直腸癌において発現される、ヒト腫瘍抗原HER2/neu、Ep−CAM及びCEAのアカゲザルホモログによるアカゲザル(Macaca mulatta)の免疫の効率を評価するため、Biomedical Primate Research Centre(BPRC),Rijswijk(The Netherlands)において第二シリーズの免疫試験を実施した。プロトコールは、これらの腫瘍抗原の組合せに対するB及びT細胞応答を評価するように設計した。
【0121】
この試験では、4匹のアカゲザルから成る第二群(雄性2匹と雌性2匹)を、ヒト腫瘍抗原、Ep−CAM(pV1J−rhHER2)、CEA(pV1J−rhCEA)及びHER2/neu(pV1J−rhEpCAM)のアカゲザルホモログを発現する3つのプラスミドDNAベクターの混合物で免疫した。
【0122】
0、4、8、12及び16週目にプラスミドDNAの筋肉内注射とそれに続く電気刺激によって動物を初回免疫した。前記DNA注射は、体重2−5kgの動物に関して、プラスミドDNA6mg(3つのTAAの各々について2mg)を含有する1ml溶液(2つの部位に分けて0.5ml/部位)から成った。動物には麻酔下で(ケタミン/キシラジンの混合物)注射した。
【0123】
電気刺激のために、2トレーンの100矩形双極性パルス(各々1秒)を、3秒の全処置時間中1秒おきに送達した。パルス長は2ミリ秒/位相、パルス周波数及び振幅はそれぞれ100Hz及び100mA(定電流方式)であった。
【0124】
同じ群の動物を、アカゲザルHER2/neu(Ad5−rhHER2)、アカゲザルCEA(Ad5−rhCEA)及びアカゲザルEpCAM(Ad5−rhEpCAM)を発現する3つのAd5の混合物の注射によって追加免疫した。3×1011ウイルス粒子(vp)の総量を23週目と27週目に筋肉内注射した(3つのウイルスの各々について1×1011vp)。
【0125】
さらなる追加免疫を、アカゲザルHER2/neu(Ad24−rhHER2)、アカゲザルCEA(Ad24−rhCEA)及びアカゲザルEpCAM(Ad24−rhEpCAM)を発現する3つのAd24の混合物の注射によって実施した。3×1011ウイルス粒子(vp)の総量を36週目と40週目に筋肉内注射した(3つのウイルスの各々について1×1011vp)。
【0126】
前記免疫プロトコールを使用してHER2/neuに対する免疫応答を測定するため、総期間1年間にわたって4週間ごとに血液試料を採集した。細胞媒介性免疫応答をIFN−γ+ELISPOTによって測定し、体液性応答をELISAによって測定した。
【0127】
サルRI449及びRI519は、IFN−γ+ELISPOT分析によって測定したとき、検出可能なHER2特異的細胞媒介性応答を示した。同様の分析は、rhCEA及びrhEpCAMに対するいかなる応答も検出しなかった。検出可能な抗rhHER2抗体力価(>200)は検出されなかった。
【0128】
第三の試験では、4匹のアカゲザルを、0、2及び4週目にAd5誘導体の筋肉内注射によってAd5−rhHER2、Ad5−rhCEA及びAd5−rhEpCAMの混合物で免疫した。麻酔下で(ケタミン/キシラジンの混合物)、3×1011vp(3つのAd5ウイルスの各々について1011vp)を含有する1ml溶液(2つの部位に分けて0.5ml/部位)を体重2−5kgの動物に投与した。
【0129】
同じ群の動物を、24、26及び28週目にAd24−rhHER2、Ad24−rhCEA及びAd24−rhEpCAMの混合物(総量3×1011vp、3つのウイルスの各々について1011vp)の筋肉内注射によって追加免疫した。細胞媒介性応答をIFNγ ELISPOTアッセイによって測定した。サルRI514及びRI496に関して、反応性をIFN−γ+細胞内染色法(ICS)によってタイピングした。4匹のサルのうち3匹は検出可能な応答を示した。加えて、細胞媒介性応答が測定された3匹のサルにおいて200から500の範囲の抗rhHER2抗体力価が検出された。
【0130】
この免疫プロトコールはまた、寛容を打破し、抗rhCEA細胞媒介性及び体液性免疫応答を誘導する上でも有効であった。サルRI514は、IFN−γ ELISPOTアッセイによって測定可能な細胞媒介性応答を示した。細胞内染色法はこの応答を確認した。同じサルはまた、500から1000の範囲の抗rhCEA抗体力価を示した。これに対し、同様の分析はrhEpCAMに対するいかなる応答も検出しなかった。
【0131】
要するに、前記で論じた免疫プロトコールは、アカゲザルにおいてrhHER2/neu及びrhCEAに対する特異的免疫応答を誘導する上で有効であった。
【図面の簡単な説明】
【0132】
【図1A】図1は、配列番号1に示す、第一アカゲザルHER2/neu cDNAのヌクレオチド配列を示す(実施例1参照)。795位の「R」の存在は、「A」又は「G」のいずれかがその位置に存在することを指示する。
【図1B】図1は、配列番号1に示す、第一アカゲザルHER2/neu cDNAのヌクレオチド配列を示す(実施例1参照)。795位の「R」の存在は、「A」又は「G」のいずれかがその位置に存在することを指示する。
【図2】図2は、配列番号2に示す、アカゲザルHER2/neuタンパク質の予測アミノ酸配列を示す。示されているアミノ酸配列は、配列番号1として開示するヌクレオチド配列から演繹した。
【図3】図3は、RT−PCRによって一連のアカゲザルHER2/neu断片を作製するために使用した、HER2/neu遺伝子をカバーするオリゴヌクレオチドプライマーのヌクレオチド配列を開示する(実施例1参照)。「正」及び「逆」と表示した欄は、公表されているヒトHER2/neu配列(アクセッションM11730)に対するプライマーの位置を記載している。配列番号15及び16として開示するプライマーは、翻訳されたタンパク質のチロシンキナーゼ活性を不活性化するために、アミノ酸579位をKからAに突然変異させるために設計した。プライマー3388−3410及び3410−3388は、Ser(AGC)をコードする、それぞれ15位にC及び10位にGを含む配列を有する。これに対し、アカゲザルHER2/neu配列はその位置に、やはりSer(AGT)をコードする、Tを有する。これらのプライマーを配列決定のために使用したが、クローニングのためには使用しなかった。配列番号11及び19については、rhHER2/neuが始まる配列に下線を付している。
【図4】図4は、完全長アカゲザルHER2/neuクローンを構築するために使用したRT−PCRプライマーの配列を示す。最初の欄は、実施例2で述べる反応に対応する反応番号を列記している。各々の欄は、4番目の欄に列記するクローンを生成するために使用した正及び逆プライマーのセットを記載している。クローンの名称は、クローニングのために使用したベクター(pBluescript又はpCRIIに関して、それぞれBS又はCR)及び公表ヒトHER2/neu配列に対する配列の位置(数字で列記している)の両方を指示する。
【図5A】図5は、配列番号40に示す、第二アカゲザルHER2/neucDNAのヌクレオチド配列を示す(実施例4参照)。ヌクレオチド配列内の「R」の存在は、「A」又は「G」のいずれかがその位置に存在することを指示する。ヌクレオチド配列内の「Y」の存在は、「C」又は「T」のいずれかがその位置に存在することを指示する。第一アカゲザルHER2/neu(配列番号1)の対応する塩基とは異なるヌクレオチド塩基は、太字及び下線で表示している。
【図5B】図5は、配列番号40に示す、第二アカゲザルHER2/neucDNAのヌクレオチド配列を示す(実施例4参照)。ヌクレオチド配列内の「R」の存在は、「A」又は「G」のいずれかがその位置に存在することを指示する。ヌクレオチド配列内の「Y」の存在は、「C」又は「T」のいずれかがその位置に存在することを指示する。第一アカゲザルHER2/neu(配列番号1)の対応する塩基とは異なるヌクレオチド塩基は、太字及び下線で表示している。
【図6】図6は、配列番号41に示す、第二アカゲザルHER2/neuタンパク質の予測アミノ酸配列を示す(実施例4参照)。示されているアミノ酸配列は、配列番号40として開示するヌクレオチド配列から演繹した。517位の「X」は、「Q」(Gln)又は「R」(Arg)がその位置に存在し得ることを指示する。647位の「X」は、「K」(Lys)又は「R」(Arg)がその位置に存在し得ることを指示する。1075位の「X」は、「R」(Arg)又は「Q」(Gln)がその位置に存在し得ることを指示する。
【図7】図7は、第一アカゲザルHER2/neu DNA及びタンパク質配列(RhHER2#1、配列番号1及び2)と比較して、第二アカゲザルHER2/neuDNA及びタンパク質配列(RhHER2#2、配列番号40及び41)に存在する特定突然変異を詳細に示す。各々の表の最初の欄は、RhHER2#2とRhHER2#1の間で異なるヌクレオチドの位置を列記する。各々の表の2番目の欄は、単離して、それぞれRhHER2#1及びRhHER2#2の配列を決定するために使用したHER2/neu断片を担う特定クローンの番号を列記する。各々の表の3番目の欄は、相違が起こるコドンの配列を示しており、異なるヌクレオチドを強調している。コドンの下部は、灰色で強調した、生じるアミノ酸についての1文字アミノ酸記号である。
【配列表】
【特許請求の範囲】
【請求項1】
配列番号2又は配列番号41に示すアカゲザルHER2/neuタンパク質をコードするヌクレオチドの配列を含む単離核酸分子。
【請求項2】
前記核酸がDNAである、請求項1に記載の単離核酸分子。
【請求項3】
前記ヌクレオチドの配列が、配列番号1、配列番号40、配列番号42又は配列番号43に示すヌクレオチドの配列を含む、請求項1に記載の単離核酸分子。
【請求項4】
請求項1に記載の核酸分子を含むベクター。
【請求項5】
請求項4に記載のベクターを含む宿主細胞。
【請求項6】
(a)請求項1に記載の核酸を含むベクターを適切な宿主細胞に導入すること;及び
(b)前記宿主細胞を、前記アカゲザルHER2/neuタンパク質の発現を可能にする条件下で培養すること
を含む、組換え宿主細胞においてアカゲザルHER2/neuタンパク質を発現するための方法。
【請求項7】
配列番号2又は配列番号41に示すアミノ酸の配列を含む、単離し、精製したアカゲザルHER2/neuポリペプチド。
【請求項8】
配列番号2又は配列番号41に示すアカゲザルHER2/neuタンパク質をコードするヌクレオチドの配列を含む単離核酸分子を含有するワクチンクチンベクターを哺乳動物に投与することを含む、癌を予防する又は治療する方法。
【請求項9】
前記哺乳動物がヒトである、請求項8に記載の方法。
【請求項10】
前記ベクターがアデノウイルスベクター又はプラスミドベクターである、請求項8に記載の方法。
【請求項11】
前記ベクターが、プラスミド部分及び発現カセットを含み、
前記発現カセットは、
(a)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及び
(b)前記ポリヌクレオチドに作動可能に連結されたプロモーター
を含む
プラスミドワクチンベクターである、請求項8に記載の方法。
【請求項12】
E1領域内の欠失及びE1領域内のインサートを有し、
前記インサートは、
(a)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及び
(b)前記ポリヌクレオチドに作動可能に連結されたプロモーター
を含む発現カセットを含有する
アデノウイルスゲノムを含む、アデノウイルスワクチンベクター。
【請求項13】
プラスミド部分及び発現カセット部分を含有し、
前記発現カセット部分は、
(a)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド、及び
(b)前記ポリヌクレオチドに作動可能に連結されたプロモーター
を含む、ワクチンプラスミド。
【請求項14】
(a)哺乳動物に、
(i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド、及び
(ii)前記ポリヌクレオチドに作動可能に連結されたプロモーター
を含む第一ベクターを導入すること;
(b)あらかじめ定められた時間を経過させること;及び
(c)前記哺乳動物に、
(i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド、及び
(ii)前記ポリヌクレオチドに作動可能に連結されたプロモーター
を含む第二ベクターを導入すること
を含む、哺乳動物を癌から保護する方法。
【請求項15】
前記第一ベクターがプラスミドであり、及び前記第二ベクターがアデノウイルスベクターである、請求項14に記載の方法。
【請求項16】
前記第一ベクターがアデノウイルスベクターであり、及び前記第二ベクターがプラスミドである、請求項14に記載の方法。
【請求項17】
(a)上皮由来の癌に罹患している哺乳動物に、
(i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及び
(ii)前記ポリヌクレオチドに作動可能に連結されたプロモーター
を含む第一ベクターを導入すること;
(b)あらかじめ定められた時間を経過させること;及び
(c)前記哺乳動物に、
(i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及び
(ii)前記ポリヌクレオチドに作動可能に連結されたプロモーター
を含む第二ベクターを導入すること
を含む、上皮由来の癌に罹患している哺乳動物を治療する方法。
【請求項18】
前記第一ベクターがプラスミドであり、及び前記第二ベクターがアデノウイルスベクターである、請求項17に記載の方法。
【請求項19】
前記第一ベクターがアデノウイルスベクターであり、及び前記第二ベクターがプラスミドである、請求項17に記載の方法。
【請求項20】
前記第一及び第二ベクターがアデノウイルスベクターである、請求項17に記載の方法。
【請求項1】
配列番号2又は配列番号41に示すアカゲザルHER2/neuタンパク質をコードするヌクレオチドの配列を含む単離核酸分子。
【請求項2】
前記核酸がDNAである、請求項1に記載の単離核酸分子。
【請求項3】
前記ヌクレオチドの配列が、配列番号1、配列番号40、配列番号42又は配列番号43に示すヌクレオチドの配列を含む、請求項1に記載の単離核酸分子。
【請求項4】
請求項1に記載の核酸分子を含むベクター。
【請求項5】
請求項4に記載のベクターを含む宿主細胞。
【請求項6】
(a)請求項1に記載の核酸を含むベクターを適切な宿主細胞に導入すること;及び
(b)前記宿主細胞を、前記アカゲザルHER2/neuタンパク質の発現を可能にする条件下で培養すること
を含む、組換え宿主細胞においてアカゲザルHER2/neuタンパク質を発現するための方法。
【請求項7】
配列番号2又は配列番号41に示すアミノ酸の配列を含む、単離し、精製したアカゲザルHER2/neuポリペプチド。
【請求項8】
配列番号2又は配列番号41に示すアカゲザルHER2/neuタンパク質をコードするヌクレオチドの配列を含む単離核酸分子を含有するワクチンクチンベクターを哺乳動物に投与することを含む、癌を予防する又は治療する方法。
【請求項9】
前記哺乳動物がヒトである、請求項8に記載の方法。
【請求項10】
前記ベクターがアデノウイルスベクター又はプラスミドベクターである、請求項8に記載の方法。
【請求項11】
前記ベクターが、プラスミド部分及び発現カセットを含み、
前記発現カセットは、
(a)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及び
(b)前記ポリヌクレオチドに作動可能に連結されたプロモーター
を含む
プラスミドワクチンベクターである、請求項8に記載の方法。
【請求項12】
E1領域内の欠失及びE1領域内のインサートを有し、
前記インサートは、
(a)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及び
(b)前記ポリヌクレオチドに作動可能に連結されたプロモーター
を含む発現カセットを含有する
アデノウイルスゲノムを含む、アデノウイルスワクチンベクター。
【請求項13】
プラスミド部分及び発現カセット部分を含有し、
前記発現カセット部分は、
(a)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド、及び
(b)前記ポリヌクレオチドに作動可能に連結されたプロモーター
を含む、ワクチンプラスミド。
【請求項14】
(a)哺乳動物に、
(i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド、及び
(ii)前記ポリヌクレオチドに作動可能に連結されたプロモーター
を含む第一ベクターを導入すること;
(b)あらかじめ定められた時間を経過させること;及び
(c)前記哺乳動物に、
(i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド、及び
(ii)前記ポリヌクレオチドに作動可能に連結されたプロモーター
を含む第二ベクターを導入すること
を含む、哺乳動物を癌から保護する方法。
【請求項15】
前記第一ベクターがプラスミドであり、及び前記第二ベクターがアデノウイルスベクターである、請求項14に記載の方法。
【請求項16】
前記第一ベクターがアデノウイルスベクターであり、及び前記第二ベクターがプラスミドである、請求項14に記載の方法。
【請求項17】
(a)上皮由来の癌に罹患している哺乳動物に、
(i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及び
(ii)前記ポリヌクレオチドに作動可能に連結されたプロモーター
を含む第一ベクターを導入すること;
(b)あらかじめ定められた時間を経過させること;及び
(c)前記哺乳動物に、
(i)アカゲザルHER2/neuタンパク質をコードするポリヌクレオチド;及び
(ii)前記ポリヌクレオチドに作動可能に連結されたプロモーター
を含む第二ベクターを導入すること
を含む、上皮由来の癌に罹患している哺乳動物を治療する方法。
【請求項18】
前記第一ベクターがプラスミドであり、及び前記第二ベクターがアデノウイルスベクターである、請求項17に記載の方法。
【請求項19】
前記第一ベクターがアデノウイルスベクターであり、及び前記第二ベクターがプラスミドである、請求項17に記載の方法。
【請求項20】
前記第一及び第二ベクターがアデノウイルスベクターである、請求項17に記載の方法。
【図1A】


【図1B】
【図2】

【図3】
【図4】

【図5A】

【図5B】

【図6】

【図7】
【図1B】
【図2】
【図3】
【図4】
【図5A】
【図5B】
【図6】
【図7】
【公表番号】特表2006−525787(P2006−525787A)
【公表日】平成18年11月16日(2006.11.16)
【国際特許分類】
【出願番号】特願2004−564235(P2004−564235)
【出願日】平成15年12月29日(2003.12.29)
【国際出願番号】PCT/EP2003/014997
【国際公開番号】WO2004/061105
【国際公開日】平成16年7月22日(2004.7.22)
【出願人】(501209427)イステイチユート・デイ・リチエルケ・デイ・ビオロジア・モレコラーレ・ピ・アンジエレツテイ・エツセ・ピー・アー (90)
【Fターム(参考)】
【公表日】平成18年11月16日(2006.11.16)
【国際特許分類】
【出願日】平成15年12月29日(2003.12.29)
【国際出願番号】PCT/EP2003/014997
【国際公開番号】WO2004/061105
【国際公開日】平成16年7月22日(2004.7.22)
【出願人】(501209427)イステイチユート・デイ・リチエルケ・デイ・ビオロジア・モレコラーレ・ピ・アンジエレツテイ・エツセ・ピー・アー (90)
【Fターム(参考)】
[ Back to top ]