
アシル担体タンパク質に基づくタンパク質をラベルするための方法
広い多様性を有する様々なラベルによってアシル担体タンパク質(ACP)融合タンパク質をラベルするための方法を開示する。本方法は、ホロ-アシル担体タンパク質合成酵素(ACPS)またはその相同物を用いて、ラベルを補酵素A型基質からACP融合タンパク質へ転移することに基づく。本方法は、生体外および生体内の両方において、新しい物理的または化学的性質を融合タンパク質に導入する分子を融合タンパク質へ結合させることにより、融合タンパク質を検出し操作することを可能にする。そのようなラベルの例には、特に、分光学的プローブ即ちレポーター分子、親和性タグ、反応性ラジカルを生成する分子、架橋剤、タンパク質−タンパク質相互作用を媒介するリガンド、または融合タンパク質の固定化に適当な分子がある。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、基質から目的のタンパク質およびアシル担体タンパク質またはその断片を含む融合タンパク質へラベルを転移する方法、ならびに特にラベルされた融合タンパク質を検出しおよび/または操作することをさらに含む方法に関する。
【背景技術】
【0002】
複雑な生体系の理解の進歩は、その背後にある生体分子特にタンパク質の相互作用の特性決定に掛かっている。増大する数の生物におけるDNA配列決定により、その生物の読み取り枠(ORF)が明らかにされてきたが、その一方で、対応するタンパク質を生体内および生体外で特性決定できる可能性は限られている。この目的の実現を目指す戦略の大部分は、生体外での応用のための融合物の精製または生体内でのタンパク質の追跡を可能にする融合タンパク質を構築することに基礎を置いている。そのようなタグの例には、6×Hisタグ、グルタチオンS転移酵素、マルトース結合タンパク質、エピトープタグ、酵母ツーハイブリッドシステム、O6−アルキルグアニン-DNAアルキル転移酵素、分解ユビキチン、および緑色蛍光タンパク質(GFP)融合タンパク質が挙げられる。しかし、これらの方法はすべて様々な限界や不利益を有する。
【0003】
Gehring et al.(1997)および Lambalot and Walsh(1995)は、E.coliのホロ-アシル担体タンパク質合成酵素(ACPS)を用いて、アポ-アシル担体タンパク質(apo−ACP)の保存されているセリン残基に生体外で補助因子4’−ホスホパンテテイン(P−pant)を結合させて、ホロ-ACPを生成させる翻訳後修飾を触媒させることを記載している。P−pantの起源は補酵素Aである。Gehring et al.(1997)はさらにP−pant部分が修飾されているが依然としてACPSの基質として働くことができる補酵素Aの類似物質を用いて、修飾されたP−pantを補助因子とするホロ-ACPが得られることを示している。
【0004】
ACPSなどの単離されたホスホパンテテイニル転移酵素は国際特許出願WO 97/13845に記載されている。
【0005】
ラベルをO6−アルキルグアニン-DNAアルキル転移酵素(AGT)融合タンパク質に転移する方法、およびこの方法をAGT融合タンパク質の検出に用いることが国際特許出願WO 02/083937に記載されている。
【0006】
発明の開示
本発明は、目的のタンパク質を検出し、および/または操作する方法であって、目的のタンパク質およびアシル担体タンパク質(ACP)またはその断片を含む融合タンパク質を、ラベルされた補酵素A(CoA)型基質およびホロ-アシル担体タンパク質合成酵素(ACPS)もしくはその相同物と接触させること、その結果ACPSがラベルを融合タンパク質へ転移させ、また場合によっては、そのラベルを認識しおよび/または操作するために設計されたシステム中でそのラベルを用いて、得られたそのラベル付融合タンパク質を検出しおよび/またはさらに操作すること、を含む方法に関する。
【0007】
さらにまた、本発明は目的のタンパク質およびACPまたはその断片を含む融合タンパク質のそのような方法中での使用に関する。特に本発明の方法は、目的のタンパク質を精製もしくは固定化するため、または本発明の方法において目的のタンパク質をそれに結合されたラベルにより生体外もしくは生体内で連続的にモニターするために用いられる。
【0008】
本発明の融合タンパク質に組み込まれる目的のタンパク質は任意の種類のものでよく、それには二次構造、三次構造、もしくは四次構造を持つ、または持たない、任意の長さのタンパク質、ポリペプチドおよびペプチドが含まれる。
【0009】
本発明の方法で用いられる特別のラベルされた補酵素A(CoA)型基質は、CoA、または修飾されたCoAから、ラベル即ち検出可能なマーカーをさらに付加するための少なくとも1つの反応部位を有するリンカーを結合させることによって得ることができる。本発明はまた、そのような新規のラベルされた補酵素A型基質、それを製造する方法、そのような新規の補酵素A型基質の合成において有用な中間体、および本発明の方法中でのそれらの使用に関する。
【0010】
発明の詳細な説明
本発明は、目的のタンパク質を検出し、および/または操作する方法であって、目的のタンパク質およびアシル担体タンパク質(ACP)またはその断片を含む融合タンパク質を、ラベルされた補酵素A(CoA)型基質およびホロ-アシル担体タンパク質合成酵素(ACPS)もしくはその相同物と接触させること、その結果ACPSがラベルを融合タンパク質へ転移させ、また場合によっては、ラベルを認識しおよび/または操作するために設計されたシステム中でラベルを用いて、得られたラベル付融合タンパク質を検出し、および/またはさらに操作するようにすること、を含む方法に関する。
【0011】
ACPまたはACPドメインは、脂肪酸、ポリケチドの生合成および非リボソームペプチド合成において担体として作用する。ACPはホロ-アシル担体タンパク質合成酵素(ACPS)によって翻訳後修飾される。この酵素は補助因子4−ホスホパンテテイン(P−pant)を補酵素AからACPの保存されているセリン残基に転移する。ACPSはまたホスホパンテテイニル転移酵素とも呼ばれる。ACPSは、CoAのホスホパンテテイン部分のチオール基に対する修飾に関して比較的低い基質特異性を所持していることが示されている。これを活用して、ACP融合タンパク質をACPSおよびホスホパンテテイン部分を介してラベルを所持するCoA誘導体と共にインキュベートすることにより、特異的にラベルすることができる。ラベルが、このようにしてホスホパンテテイン部分と共にACPの保存されたセリン残基に転移される。ラベリングは融合タンパク質の性質とは独立である。
【0012】
ACPおよびACPSという語句は、ここでは任意の一対のタンパク質であって2つのうちの1つ(ACP)がCoA誘導体起源のホスホパンテテイン誘導体に対するアクセプターであり、他の1つ(ACPS)がホスホパンテテイン誘導体のACPへの転移を触媒するものを表す。
【0013】
ACPSおよびホスホパンテテイン転移酵素という語句は、E.coli由来のACPSと相同性のある多数のホスホパンテテイニル転移酵素が、脂肪酸合成に関与せず、むしろ自然産物の生合成に関与しているタンパク質を修飾するという事実にもかかわらず、ここでは交換できる語句であるとして用いる(Lambalot et al.,1996)。このようなホスホパンテテイニル転移酵素の例には:エンテロバクチン合成に関与するEntD;サーファクチン生合成に関与するSfpおよびPsf−1;グラミシジンS生合成に関与するGsp;リシン生合成に関与するLYS5;バシトラシン生合成に関与するBli;イチュリンA生合成に関与するLpa−14;および、ノシヘプチド生合成に関与するNshC、がある。本発明はE.coli由来のACPSと相同性のあるラベリング方式を持つ全てのACPSの使用を含む。
【0014】
ACPという用語は、Lambalot et al.(1996)によって定義されたように、E.coli由来のACPSまたはE.coli由来のACPSと相同性のある任意のACPSによって翻訳後にホスホパンテテイニル化されることとなる任意のタンパク質を表す。これには、脂肪酸合成に関与するタンパク質ばかりでなく、ポリケチド合成、非リボソームペプチド合成、アミノ酸合成、およびデプシペプチド合成に関与するタンパク質も含まれる。ACPSによる翻訳後修飾に加えて、これらのタンパク質は共通して種々の基質とアシルパンテテイニル・チオエステルを形成し、ホスホパンテテイニル部分がセリン残基に結合される。これらの自然の機能において、ACPは、多機能酵素(I型脂肪酸合成酵素のような)のドメイン、または別のタンパク質(II型脂肪酸合成酵素のような)のドメインであり得る。
【0015】
「検出する」とは、ラベルおよびそれに結合している目的のタンパク質を、ラベルを観測するために設計されたシステム中でラベルの性質に基づいて観察することを意味するものであり、また特別なラベルを認識し、ラベルの性質によって特別な環境でラベルを見出し、場合によっては、ラベルおよびそれに結合したタンパク質を定量化し、また場合によっては、ラベルの微細環境を決定することなどを含む。
【0016】
「操作する」とは、ラベルおよびそれに結合している目的のタンパク質を操作することを意味し、またラベルおよびそれに結合している目的のタンパク質を、ラベルを操作するために設計されたシステム中でラベルの性質に基づいて操作することを含み、ならびに化学的または生物学的環境から分離すること、別の化学的または生物学的環境へ導入すること、精製すること、即ち望ましくない副産物および不純物から分離すること、ラベルを固体の担体と反応させることによって固定化すること、ラベルおよび/または目的のタンパク質の性質を修飾するために化学的または生物学的試薬と接触させることなどを含む。
【0017】
ここで開示する方法は一般にある範囲の応用に適用でき、特異的におよび共有結合的に融合タンパク質を(1)ラベルされた融合タンパク質の環境の変化を感受し誘起することができるラベルによってラベルすることができ、(2)ラベルによって融合タンパク質に特異的に導入された物理的および/または化学的性質によって融合タンパク質を操作することを助けるラベルによってラベルすることができ、および/または(3)ラベルによって導入された性質を通じて融合タンパク質の精製を助けるラベルによってラベルすることができる。ここで開示する方法はACP融合タンパク質を生体外および生体内、例えば細胞中の両方でラベルするために用いることができる。
【0018】
さらにまた、本発明は目的のタンパク質とACPまたはその断片を含む融合タンパク質のそのような方法中での使用に関する。特に、本発明の方法は目的のタンパク質の精製と固定化に、または、目的のタンパク質を生体外および生体内において本発明の方法によりそれに結合されたラベルにより連続的にモニターすることに用いられる。
【0019】
一つの態様において、本発明は目的のタンパク質を生体外および生体内において連続的にモニターする方法を提供する。この方法では、目的のタンパク質とアシル担体タンパク質(ACP)またはその断片を含む融合タンパク質を、ラベルされた補酵素A(CoA)型基質およびホロ-アシル担体タンパク質合成酵素(ACPS)またはその相同物と接触させて、ACPSがラベルを融合タンパク質へ転移し、またラベルを、ラベルを認識するために設計されたシステム中で観察する。
【0020】
さらにある態様において、本発明は目的のタンパク質を生体外および生体内において操作する方法を提供する。そこでは目的のタンパク質およびアシル担体タンパク質(ACP)もしくはその断片を含む融合タンパク質を、ラベルされた補酵素A(CoA)型基質およびホロ-アシル担体タンパク質合成酵素(ACPS)もしくはその相同物と接触させて、その結果ACPSがラベルを融合タンパク質へ転移し、また融合タンパク質をラベルの物理的および/または化学的性質に基づいて操作する
【0021】
ある特別な態様においては、ラベルの物理的および/または化学的性質によってラベルされた融合タンパク質の効率的な精製が可能であり、したがって本発明は、本発明の方法の諸工程すなわち、ラベルの物理的および/または化学的性質を精製に利用し、その後融合タンパク質を切断して高純度の目的のタンパク質を供給する工程を遂行することによる、目的のタンパク質の精製方法を提供する。
【0022】
さらなる態様において、本発明は、目的のタンパク質およびアシル担体タンパク質(ACP)もしくはその断片を含む融合タンパク質を固体支持体に固定化する方法を提供する。この方法は融合タンパク質を、固体支持体に結合しているかまたは結合できるラベルされた補酵素A(CoA)型基質およびホロ-アシル担体タンパク質合成酵素(ACPS)もしくはその相同物と接触させることを含み、これにより、ホロ-アシル担体タンパク質合成酵素(ACPS)がラベルをACP融合タンパク質へ共有結合させるように転移し、ACP融合タンパク質はそれにより固体支持体に結合されるかまたはその後に結合され得る。ラベルが最初は固体支持体に結合されていない本発明の特別な態様において、本方法はさらに、ラベルされたACP融合タンパク質が固体支持体に固定化されるようにそれを固体支持体と接触させる工程を含んでもよい。これらの本発明の好ましい実施態様においてラベルは、転移されたときまたは次の反応において固体支持体に共有結合的に結合されてもよく、または特異的な結合対の一員であって、他の一員が固体支持体に共有結合的にもしくは任意の他の手段により結合しているか結合可能であってもよい。それは例えばビオチンおよびアビジンもしくはストレプトアビジンの特異的結合対を用いる場合である。
【0023】
さらなる態様において、本発明は、生体外はもちろん生体内においてもACP融合タンパク質をラベルする方法を提供する。生体内におけるACP融合タンパク質のラベリングという語句は、細胞外スペースに向いているACP融合タンパク質をラベルすることはもちろん、細胞内の全ての区画においてラベルすることを含む。もしACP融合タンパク質のラベリングが生体内で行なわれ、ACPに融合されたタンパク質が原形質膜タンパク質である場合には、融合タンパク質のACP部分は原形質膜の細胞質側または細胞外側のどちらに付着していてもよい。ラベリングを生体外で行なう場合は、融合タンパク質のラベリングは細胞抽出物中でまたはACP融合タンパク質を精製または濃縮した形で行うことができる。
【0024】
本発明は、所望のタンパク質へのラベルの特異的な付加を、その目的のタンパク質とアシル担体タンパク質(ACP)またはその断片との間で融合タンパク質を形成させることにより遂行できたという成果を基礎にしている。
【0025】
好ましい応用において、アシル担体タンパク質即ち「ACP」は、ホロ-アシル担体タンパク質合成酵素即ち「ACPS」によって、ラベルされた4’−ホスホパンテテインが適当な補酵素A(「CoA」)から融合タンパク質の一部を形成するACPもしくはその断片のセリン残基へ転移される方法で、修飾される性質を有する。好ましい実施態様において、ACPは、例えばRawling and Cronan, 1992およびその中の引用文献中に記載のE.coliのアシル担体タンパク質である。しかし、他のアシル担体タンパク質(ACP)、例えばGehring et al., 1997に記載されているStreptomyces種のACP、または適当にラベルされたCoA存在下で上に定義したACPSによって修飾される性質を有する任意のACPが知られており、本発明において使用できる。本発明においてACPはまた、野生型ACPの変異体であって1個以上のアミノ酸置換、欠失または付加によって野生型と異なり、しかしACPSにより触媒される反応において、ラベルされた4’−ホスホパンテテインに対するアクセプターとして作用する性質を依然として保持している変異体も含む。他のACP変異体を周知技術を用いて化学的に修飾してもよい。ACP変異体は周知のタンパク質工学技術を用いて、および/またはACPSによって触媒される反応におけるラベルされた4’−ホスホパンテテインの転移に対する新しいアクセプター配列を生成し選択するための分子進展を利用して生成することができる。ACP断片とは、ホスホパンテテイン誘導体と結合するセリン残基を含み、またそのようなホスホパンテテイン誘導体を受容する機能を残存させている断片である。
【0026】
好ましい実施態様において、ACPSは、例えばLambalot and Walsh,1995またはその中で引用された文献に記載されたE.coliのホロ-アシル担体タンパク質合成酵素である。しかし、他のホロ-アシル担体タンパク質合成酵素、例えばLambalot et al.,1996記載のBacillus subtilis由来のACPSまたは上で定義したACPを適当にラベルされたCoA存在下で修飾する性質を有するならば本発明において採用可能である任意の形のタンパク質なども知られている。これらのACPSはまたLambalot et al.,1996記載のように、ホスホパンテテイニル転移酵素としても知られており、また本発明はこの一般的な種別の酵素の使用を含む。本発明において、ホロ-アシル担体タンパク質合成酵素はまた野生型ACPSの変異体であって1個以上のアミノ酸置換、欠失または付加によって野生型と異なり、しかしラベルされた4’−ホスホパンテテインを特異的にACP融合タンパク質に転移する性質を依然として保持している変異体も含む。他のACPS変異体を周知技術を用いて化学的に修飾してもよい。ACPS変異体は周知のタンパク質工学技術を用いて、および/またはラベルされた4’−ホスホパンテテインを異なるアクセプター配列に転移するための新しい特異性を生成し選択する分子進展を利用して、生成することができる。
【0027】
ACP融合タンパク質のACPSによるラベリングのための、多くの考察を有利に織込む。最も大切なことは、ACP融合タンパク質のラベリングは、ラベルする前にACP融合タンパク質のACP部分がアポ型で存在することに依存していることである。もしACP融合タンパク質が、翻訳後修飾の基質としてこのACP融合タンパク質を受容する内在的ACPSを有する宿主において発現されているならば、ACP融合タンパク質は所望の修飾に対して少なくとも部分的に遮断される可能性がある。この望ましくないACP融合タンパク質の修飾を最小にするために種々の解決策が提案されている。第一に、宿主のACPSによって効率的に修飾されないACPを選択する、即ちACPが宿主の生化学と直交するように選択する。例えば、E.coli由来のACPは、ヒト細胞において融合タンパク質として発現されたときにヒトのACPSによって有意な程度には修飾されない(図3参照)。第二に、ACP融合タンパク質の過剰発現によってACP融合タンパク質の主にアポACPが形成されるようになる。
例えば、E.coliにおいてE.coli由来の6×His−ACPを過剰発現させることにより、これらの細胞内に内在性ACPSが存在するにもかかわらず、大部分は6×HisのアポACPが生成されることになる。
【0028】
ラベル反応のために、ACP融合タンパク質はまた、対応するACPSおよびCoA誘導体の両方と接触しなければならない。このことは生体内応用において、ACP融合タンパク質が細胞表面に表示されるか、またはACPSとCoA誘導体が微量注入などの技術を用いて目的の細胞内に導入されるかどちらかであることを意味する。
【0029】
本発明において、ACPとの融合タンパク質のタンパク質部分への言及は、二次構造、三次構造、四次構造を有するまたは有しないタンパク質、任意の長さのポリペプチドおよびペプチドが含まれることを意図したものであり、また好ましくは少なくとも12個のアミノ酸から2000個までのアミノ酸を、好ましくは50から1000個の間のアミノ酸から成る。本発明による目的のタンパク質は次の群から選択される。その群は、酵素、DNA結合タンパク質、転写調節タンパク質、膜タンパク質、核レセプタータンパク質、核局在信号タンパク質、タンパク質補助因子、抗体、膜ポンプタンパク質、膜チャネルタンパク質、膜担体タンパク質、運動タンパク質、シグナル伝達に含まれるタンパク質、核タンパク質、リボソームタンパク質、小単量体GTPアーゼ、ATP結合カセットタンパク質、細胞内構造タンパク質、特別な細胞内区画を標的とするタンパク質としての役割を担う配列を有するタンパク質、一般にラベルとしてまたは親和性タグとして用いられるタンパク質、および前述のタンパク質のドメインもしくはサブドメイン、から成る。ACP融合タンパク質は1つ以上の、例えば1つ、2つ、3つの、ACPのN−末端、C−末端、またはN−およびC−末端においてACPと融合した目的のタンパク質から成ればよい。
【0030】
より詳しく言えば、本発明による目的のタンパク質はつぎの群から選択される。
即ちその群は、
酵素、例えば、転移酵素(EC2)、より具体的にはメチル基以外のアルキルもしくはアリル基を転移する転移酵素(EC2.5)、特にグルタチオン転移酵素(EC2.5.1.18)、
あるいは、リンを含む基を転移する転移酵素であるキナーゼ(EC2.7)、例えば酵母のカゼインキナーゼ(EC2.7.1.37)またはチロシンタンパク質キナーゼ(EC2.7.1.112)のような、基質タンパク質のリン酸化標的部位としてセリンもしくはスレオニンを有するタンパク質キナーゼなど、特にアルコール基をアクセプターとするキナーゼ(EC2.7.1);
あるいは、例えば酸化還元酵素(EC1)、より具体的には過酸化物をアクセプターとする酸化還元酵素(EC1.11)、特にチトクロームCペルオキシダーゼ酵素(EC1.11.1.5);
あるいは、例えば加水分解酵素(EC3)、より具体的にはエステル結合に作用する加水分解酵素(EC3.1)、特にタンパク質リン酸モノエステル加水分解酵素などのリン酸モノエステル加水分解酵素(EC3.1.3);またはペプチダーゼもしくはプロテアーゼ(EC3.4)としても知られるペプチド結合を加水分解する加水分解酵素、特にカスパーゼ;
DNA結合タンパク質、より具体的にはmRNA合成を阻害するタンパク質因子である転写リプレッサータンパク質、特にE.coliにおいてmRNA合成を阻害するタンパク質因子、特にLexAタンパク質のDNA結合ドメイン;
転写調節タンパク質、より具体的には転写リプレッサータンパク質、特にトリプトファン/アスパラギン酸反復構造を含む転写リプレッサータンパク質;
膜タンパク質、例えば少なくとも1つの膜貫通へリックスを示す膜タンパク質、より具体的には小胞体(ER)膜由来の膜タンパク質、特にER膜貫通タンパク質Sec62などの特にERへのタンパク質移行活性を有する膜タンパク質;
あるいは、例えば7回膜貫通ヘリクッス(7−TM)タンパク質、より具体的には7−TMタンパク質がGタンパク質共役レセプター(GPCR)であるもの、特に哺乳類などの、例えばヒトのニューロキニン−1−レセプター(NK1)などの分子量約1kDaより上の高分子リガンドを結合する7−TMタンパク質;
あるいは、例えば細胞膜由来の膜貫通イオンチャネルタンパク質、特にリガンド作動性イオンチャネルタンパク質、より具体的には、セロトニンレセプター5−HT3などのセロトニン感受性のリガンド作動性イオンチャネルタンパク質;
あるいは、例えばイオンチャネルおよびGタンパク質共役レセプター以外の膜レセプター;
あるいは、例えば、特に酵母由来の、ペルオキシソームの膜タンパク質、タンパク質Pex15など;
核レセプタータンパク質、例えば転写因子ファミリー由来の核レセプタータンパク質、より具体的にはリガンド誘導可能な転写因子ファミリー由来の核レセプタータンパク質、特に例えばヒトエストロゲンレセプターhERなどのエストロゲンレセプターのようなステロイドファミリー由来の核レセプター;
サルウイルス40(SV40)由来の核局在信号などの核局在信号タンパク質;
例えば遺伝子構造中にユビキチン配列を含むタンパク質のようなタンパク質性補助因子;
小単量体GTPアーゼ、より具体的にはRasファミリーメンバーのような膜結合性小単量体GTPアーゼ;
例えば複数薬剤耐性タンパク質のようなATP結合カセット(ABC)タンパク質;
細胞内構造タンパク質、より具体的には細胞骨格タンパク質、より具体的にはヒト細胞質β−アクチン;
例えばゴルジ体、小胞体(ER)、ミトコンドリア、原形質膜、またはペルオキシソームへと、特別の細胞区画へタンパク質を指向させる役割の配列を有するタンパク質;
一般にラベルまたは親和性タグとして用いられるタンパク質、例えばUVもしくは可視光照射による励起によって蛍光信号を与える蛍光タンパク質、強化シアノ蛍光タンパク質(ECFP)として知られる蛍光タンパク質などの、特に緑色蛍光タンパク質(GFP)として知られるファミリー由来の蛍光タンパク質;
および上述タンパク質のドメインまたはサブドメイン、
である。
【0031】
さらに、本発明による目的のタンパク質は供給源に従って選択される。特に、目的のタンパク質には、ヒト、マウス、ラット、他の高等動物、真核生物種、バクテリア種、例えばサルモネラ、より具体的には、salmonella typhiもしくはsalmonella typhimurium、ミコバクテリア、より具体的には mycobacterium tuberculosis、ブドウ球菌、より具体的にはstaphylococcus aureusなどに存在するタンパク質、または、例えばヒト免疫不全ウイルス(HIV)、ヒトインフルエンザウイルス、肝炎ウイルス、またはコロナウイルスなどのウイルス源由来のタンパク質である。
【0032】
さらにまた、目的のタンパク質は、ガン、心臓血管病、精神障害、アルツハイマー、肥満、ウイルス感染、およびバクテリア感染などの、一定の病気におけるその役割に従って選択される。
【0033】
本発明の特定の実施態様において、融合タンパク質は、一方の側のE.coliのACPまたはそのようなACPのDNAの変異体およびACPのDNA配列のN−末端(N)もしくはC−末端(C)もしくはN−末端とC−末端側に結合された(上に挙げたような)目的のタンパク質をコードする配列とから生成され、本発明の融合タンパク質が導かれる。融合タンパク質はさらに適当なリンカー、例えば融合タンパク質中のACPと目的のタンパク質の間の、および/または2つの目的のタンパク質の間の適当な条件下で酵素切断を受けやすいリンカーを含む。そのようなリンカーの例には、DNA段階で適当な制限酵素によって切断されるリンカー、例えばBgl IIによって切断されるAGATCT、および/またはタンパク質段階で適当な酵素、例えばタバコエッチウイルスNla(TEV)プロテアーゼによって切断されるリンカーがある。
【0034】
融合タンパク質は原核生物宿主、好ましくはE.coli、または真核生物宿主、例えば酵母、真菌、昆虫、または哺乳動物細胞中で発現できる。
【0035】
本発明はまた、目的のタンパク質とACPもしくはその断片との新規な融合タンパク質に関する。
【0036】
本発明において、ラベルされたCoA型基質は、好ましくは次の一般式(I):
【化2】
または、次の一般式(II):
【化3】
を持つ、ラベルされた補酵素Aの誘導体である。式中、Rは補酵素Aとラベルを繋ぐリンカー基であり、また「ラベル」は、ここに記載した融合タンパク質の検出、精製および/または操作のために適当なラベル分子である。
【0037】
しかし、本発明は式(I)または(II)の基質に限られない。広い範囲の他の基質をACP融合タンパク質へラベルを転移するために使用できるからである。例えば、式(I)の化合物のプリン部の置換、糖部分またはパンテテイニル酸部分の修飾、が考えられる。
【0038】
式(I)または(II)の化合物中のリンカー基Rはラベルを補酵素Aに結合する柔軟なリンカーである。リンカー単位は構想した応用、即ちACPを含む融合タンパク質へのラベルの転移との関連で選ばれる。それらはまた適当な溶媒中で基質の溶解度を増加させる。用いたリンカーは実際の応用条件下で化学的に安定である。リンカーRはACPとの反応にもラベルの検出にも干渉せず、しかし補酵素A型基質がACPを含む融合タンパク質と反応した後のある時点で切断されるように構成することができる。
【0039】
リンカー基Rは1〜300個の炭素原子を有する直鎖または分岐鎖のアルキレン基であり、場合によっては、
(a)1個以上の炭素原子が酸素により置き換えられ、特に3個ごとの炭素原子が酸素に置き換えられ、例えば、1〜100個のエチレノキシ単位を有するポリエチレノキシ基を表し;
(b)1個以上の炭素原子が水素原子を持つ窒素原子に置き換えられ、また隣接する炭素原子がオキソにより置き換えられて、アミド官能基−NH−CO−を表し;
(c)1個以上の炭素原子が酸素に置き換えられ、また隣接する炭素原子がオキソで置き換えられ、エステル官能基−O−CO−を表し;
(d)隣接する2個の炭素原子間の結合が2重または3重結合であり、官能基−CH=CH−または−C≡C−を表し;
(e)1個以上の炭素原子がフェニレン、飽和または不飽和シクロアルキレン、飽和または不飽和ビシクロアルキレン、架橋ヘテロ芳香族または架橋飽和または不飽和ヘテロシクリル基に置き換えられており、
(f)隣接する2個の炭素原子がジスルフィド結合−S−S−により置き換えられており;
または、場合によっては置換基を含む、2個以上の、特に2個の、アルキレン基および/または上の(a)〜(f)で定義した修飾されたアルキレン基の組み合わせである。
【0040】
考慮した置換基は、例えば低級アルキル例えばメチル、低級アルコキシ例えばメトキシ、低級アシロキシ例えばアセトキシ、またはハロゲニル例えばクロロである。
【0041】
さらに考慮した置換基は、例えばα−アミノ酸がリンカーRに取り込まれた場合に得られる置換基であり、その場合炭素原子は(b)で定義したアミド官能基−NH−CO−で置き換えられる。そのようなアルキレン基の炭素鎖のリンカー部分において、Rは−(NH−CHR’−CO)n−基で置き換えられる。ここでnは1〜100の間であり、R’は種々のα−アミノ酸の残基を表す。
【0042】
さらなる置換基は光切断性リンカーRとなる置換基、例えばo−ニトロフェニル基である。特に、このo−ニトロフェニル置換基はアミド結合に隣接した炭素原子に、例えば−NH−CO−CH2−CH(o−ニトロフェニル)−NH−CO−基に存在する。
【0043】
上の(e)で定義した炭素原子と置き換えられるフェニレン基は、例えば1,2−、1,3−、または好ましくは1,4−フェニレンである。上の(e)で定義した炭素原子と置き換えられる飽和または不飽和シクロアルキレン基は、例えばシクロペンチレンまたはシクロヘキシレンまたはさらに、例えば1−または2−の位置で不飽和のシクロヘキシレンである。上の(e)で定義した炭素原子と置き換えられる飽和または不飽和ビシクロアルキレン基は、例えば、場合によっては、2−の位置で不飽和のまたは2−と5−の位置で二重に不飽和のビシクロ[2,2,1]ヘプチレンまたはビシクロ[2,2,2]オクチレンである。上の(e)で定義した炭素原子と置き換えられるヘテロ芳香族基は、例えばトリアゾリデン、好ましくは1,4−トリアゾリデン、またはイソオキサゾリデン、好ましくは3,5−イソオキサゾリデンである。上の(e)で定義した炭素原子と置き換えられる飽和または不飽和ヘテロシクリル基は、例えば2,5−テトラヒドロフランジイルまたは2,5−ジオギサンジイル、またはイソオキサゾリジネン、好ましくは3,5−イソオキサゾリジネンである。
【0044】
好ましいリンカー基Rは、例えば、場合によってはメチル、メトキシ、またはアセトキシ基で置換された1〜20個の炭素原子のアルキル鎖、または1〜20個のエチレノキシ基から成るポリエチレングリコール鎖などの、柔軟なリンカーである。
【0045】
基質のラベル部分は、当業者が融合タンパク質を応用しようと意図するその応用に応じて選択できる。ラベルの例には:
(1)蛍光団、発色団などの分光学的プローブ、磁気的プローブまたはコントラスト剤、あるいはまた電子顕微鏡技術で有用なプローブ;
(2)放射性ラベルされた分子;
(3)相手と特異的に結合できる特異的な結合対の一方の分子。そのような特異的結合対は周知であり、例えばアビジンまたはストレプトアビジンと結合できるビオチンである;
(4)他の生体分子と相互作用すると想定される分子;
(5)他の生体分子と相互作用すると想定される分子のライブラリー;
(6)例えばNadeau et al.,2002に記載された、当業者に周知の他の生体分子と架橋できる分子;
(7)H2O2とアスコルビン酸に曝露されるとヒドロキシラジカルを生成できる例えばHori et al.,2002に記載の繋留された金属キレートなどの分子;
(8)例えばJay et al.1999に記載のマラカイトグリーンなどの、光照射によって反応性ラジカルを生成できる分子;
(9)支持体がガラススライド、ミクロタイタープレートまたは熟練の当業者に周知の任意の一般ポリマーである、固形支持体に共有結合した分子;
(10)相補的な鎖と塩基対を形成できる核酸またはその誘導体;
(11)リピッドまたは膜に挿入される性質を有する他の疎水性分子;
(12)望ましい酵素学的、化学的、または物理学的性質を有する生体分子;
(13)上に挙げた性質の任意の組み合わせを有する分子;
が含まれる。
【0046】
ラベルされたCoA誘導体は、ホスホパンテテイニル部分がACP融合タンパク質へ転移することができ検出可能であるラベル、例えば蛍光団、発色団、磁気的プローブ、放射性ラベルされた分子、または任意の他の分光学的プローブなどのラベルを所持しており、そのラベルされたCoA誘導体を使用することによって、本発明を、生体外または細胞内もしくは細胞表面上(生体内)において、検出できるラベルをACP融合タンパク質へ特異的かつ共有結合的に結合させるために使用することが可能になる。これにより、生体内および生体外においてACP融合タンパク質の検出と特性決定が可能になる。生体内という語句は、細胞外スペースに向いているACP融合タンパク質のラベルはもちろん、細胞の全ての区画中でACP融合タンパク質をラベルすることを含む。この方法を、遺伝的に目的のタンパク質と融合させ、生きている細胞中でその研究をすることができるようにする緑色蛍光タンパク質(GFP)の応用と比較することができる。GFPとその突然変異体の欠点は、原則としてGFP中に存在する自然に存在する蛍光団の使用に限定されることである。細胞内(生体内)でのラベリングは、タンパク質の機能的な構造に小さな影響のみを与え、ACPのホスホパンテテイニル部分によりラベルされる機能を保持する一般的な固定手順によって細胞を固定した後にも利用できる。
【0047】
ホスホパンテテイニル部分がACPタンパク質に転移できるビオチンなどの親和性タグを有するようなラベル付CoA誘導体を使用することによって、本発明を親和性タグを融合タンパク質へ転移するために用いることが可能になり、融合タンパク質に親和性タグの結合相手が結合できるようになる。例として、ビオチンなどの親和性タグによってラベルされたCoA基質およびACPSを、ACP融合タンパク質を発現している細胞抽出液(バクテリアもしくは真核生物の)または精製されたACP融合タンパク質に加えることによって、親和性タグによる融合タンパク質の共有結合的修飾が可能になる。これによって次に、親和性タグとその結合相手との相互作用を利用して、例えばビオチンの場合は固定化されたアビジンもしくはストレプトアビジンとの相互作用によって、融合タンパク質の単離が可能になる。もしラベルがジスルフィド架橋などの切断可能な結合を有するリンカー基Rを介してACP融合タンパク質にリンクしているならば、またはもしリンカーが光切断性であれば、精製後にACP融合タンパク質から親和性タグを切り離すことができる。
【0048】
ACP融合タンパク質へ転移でき、また外部の刺激に曝露するとヒドロキシラジカルなどの反応性ラジカルを生成できるラベルをホスホパンテテイニル部分が所持するラベル付CoA誘導体の使用によって、目的のタンパク質およびその近くのタンパク質の立体構造の研究が可能になる。生成されたラジカルはACP融合タンパク質およびそのごく近傍のタンパク質を不活性化することができて、それのよってそれらのタンパク質の役割の研究が可能になる。そのようなラベルの例は、H2O2とアスコルビン酸に曝露することによりヒドロキシラジカルを生成するテザー金属キレート錯体、およびレーザー照射によってヒドロキシラジカルを生成するマラカイトグリーンなどの発色団である。ヒドロキシラジカルを生成するために発色団とレーザーの使用することは、例えばJay et al.,1998に記載されているように、発色団援用レーザー誘導不活性化(CALI)として周知である。CALIは、時間的に制御され空間的に解像された態様で特異的に細胞中の一定のタンパク質を特異的に不活性化するために使用する方法であって、また発色団とタンパク質が空間的に近傍にあることを基礎にしている。レーザー照射すると、発色団はヒドロキシラジカルを生成し、それは発色団から約100nm以内のそしてそれ以内のみの全てのタンパク質を不活性化する。現在までは、発色団は、目的のタンパク質に特異的な発色団でラベルされた抗体を微量注入することによって目的のタンパク質の空間的近傍にもたらされた。本発明において、マラカイトグリーンのような発色団でACP融合タンパク質をラベルし次にレーザー照射することによって、時間的に制御され空間的に解像された態様でACP融合タンパク質およびACP融合タンパク質と相互作用するタンパク質を不活性化することを可能にすることになる。この方法は生体内および生体外の両方で応用できる。
【0049】
類似の方法で、ACP融合タンパク質を繋留された金属キレートでラベルし、そしてACP融合タンパク質およびACP融合タンパク質と相互作用するタンパク質をH2O2およびアスコルビン酸に曝露することにより特異的に不活性化できる。この方法はACP融合タンパク質またはACP融合タンパク質のごく近傍のタンパク質の機能の研究に用いることができるばかりでなく、ACP融合タンパク質のごく近傍のタンパク質の同定にも用いることができる。ここで、ACP融合タンパク質のごく近傍のタンパク質を、特異的な抗体によりそのタンパク質の断片を検出することにより、高分解能二次元電気泳動ゲル上からそれらのタンパク質が消滅したことにより、または質量分析法もしくはN−末端分解によるタンパク質配列決定法などの分離および配列決定技術により切断されたタンパク質断片を同定することにより、同定できる。
【0050】
ホスホパンテテイニル部分がACP融合タンパク質に転移できるリガンドを有するラベル付CoA誘導体の使用により、本発明を、リガンドおよびタンパク質などのリガンドの結合相手を、特異的に融合タンパク質へ結合させるために用いることができることになる。もしリガンドがもう1つのタンパク質Yに結合し、タンパク質Yがラベルされた融合タンパク質と二量体を形成することが生物学的機能または測定可能な信号を生起させるならば、生物学的機能または測定可能な信号はラベルされたACP融合タンパク質の添加に依存する。もしACPが細胞表面に表示されているタンパク質に連結されているならば、リガンドとの相互作用が、リガンドによって修飾される他の分子(これにはリガンドによって修飾される、個々の生体分子または他の細胞、組織、および無傷の生物の一部としての生体分子、が含まれる)との接触を媒介する。
【0051】
ホスホパンテテイニル部分が担体の表面に共有結合的に結合している場合には、またはラベルが非共有結合的にそれ自身表面に結合している別の分子により結合され得る分子である場合には、ラベルされたCoA誘導体の使用によって、本発明を、固体支持体上にタンパク質配列(アレー)を構築するために使用することが可能になる。後者の手法の例は、ラベルがビオチンで、表面に結合している分子がストレプトアビジンまたはアビジンである。担体の可能な例はガラススライド、マイクロタイタープレートまたは任意の機能化ポリマーである。基質として用いられるCoA誘導体は担体上にラベルを介して固定化され、そして続くACPSが触媒するACP融合タンパク質の反応がラベルを融合タンパク質へ転移させ、担体上にそのような融合タンパク質を固定化させる。対応するCoA誘導体によって前処理された担体上に(様々な)ACP融合タンパク質をACPSと共に空間的に解像された態様でスポットすることによりタンパク質アレーの作製が可能になる。
【0052】
他のタンパク質に架橋することができる分子であるラベルにホスホパンテテイニル部分が共有結合的に結合している場合には、ラベルされたCoA誘導体の使用によって、本発明を、適当な環境における目的のタンパク質の相互作用を研究するために使用することが可能になる。そのような架橋リンカーの例は、マレイミド、活性エステルもしくはアジド、および熟練当業者に知られた他の分子、例えばNadeau et al.,2002に記載の分子などの官能基を含む分子である。そのようなラベルされたCoA誘導体をACPSの存在下で他のタンパク質と(生体内または生体外で)相互作用するACP融合タンパク質と接触させることによって、ACP融合タンパク質をそれが相互作用するタンパク質にラベルを介して共有結合的に架橋することができる。これによって、ACP融合タンパク質と相互作用するタンパク質の同定が可能になる。
【0053】
ホスホパンテテイニル部分がACPと膜レセプターから成るACP融合タンパク質へ転移されることが可能なリガンドを所持している場合の、ラベルされたCoA誘導体の使用は、ラベルされたレセプターの検出を可能にする。この細胞表面レセプターのラベリングはレセプターの内部移行の観察を可能にする。ある特別の応用はリガンド結合後の、例えば薬剤、新薬、または新薬候補によるレセプタータンパク質の内部移行の研究である。レセプターの内部移行は様々な方法により、例えば細胞膜から細胞内部へのラベルの移動を顕微鏡で検出することによって、またはラベルの蛍光の特徴が細胞内環境へ移動して変化することによって、検出できる。そのような変化はまた顕微鏡用具無しで検出することもできて、細胞外液に消光剤を加えることにより容易になる可能性がある。
【0054】
ホスホパンテテイニル部分が電子顕微鏡によって直接検出できるリガンド例えば電子密度の高いナノ粒子、を有するCoA誘導体の使用。ホスホパンテテイニル部分の電子顕微鏡による検出への別の応用は、例えばジアニシジンなどの芳香族アミンに光照射して酸化させ、高電子密度の沈殿を生じさせそれを次に電子顕微鏡で検出するためのエオシンなどの、光増感剤として使用されるラベルに基づいている。
【0055】
例として、本発明の実施態様を、添付した図を参照してここでより詳しく記載することにする。
【実施例】
【0056】
次の実施例および実験的手順は、当業者に本発明の完全な開示および実施方法の記載を提供するために記載するものであって、本発明の範囲の限定を意図するものではない。
【0057】
CoA−Btの合成
100μlのDMF中のビオチン-マレイミド(1mg、0.0022mmol)溶液に、90μl DMFおよび10μlの50mM Tris−HClpH7.5中の補酵素A二ナトリウム塩(1.79mg、0.0022mmol、1当量)溶液を加える。混合物を4時間室温で攪拌する。それを次にCH3CN/H2O 1:4で希釈し、小分けした500μlを調製用HPLCカラムに注入する:勾配(A=H2O 99%、CH3CN 1%、50mM NH4OAc/B=CH3CN)A/B95:5からA/B80:20までを2分間、A/B68:32までを7分間、A/B 20:80までを2分間行い、その後A/B 95:5に戻す。CoA−ビオチンの滞留時間は6.5分である。所望の生産物を含む分画を真空で濃縮し、DMSOに溶解し、そして純度を制御するために分析量を注入する。精製した分画を合わせる。CoA−ビオチンの濃度を260nmの吸収(ε(アデニン、260nm)=15,300[M-1cm-1])で決定する。CoA−Btの収量は0.895mg(33%)である。
ESI−MS(m/z)計算値1217.296[M(−1)]、測定値1217.2657[M(−1)]。
【0058】
【化4】
【0059】
CoA−Dgの合成
50μl DMF中のN−(ε−マレイミドカプロン酸)ヒドラジド2(1mg、0.0044mmol)溶液に、DMF/緩衝液混合液(70μl DMF/30μl 50mM Tris−Cl、pH7.5)中の補酵素A二ナトリウム塩(3.6mg、0.0044mmol、1当量)溶液を加える。反応を分析用HPLCによって追跡し(260nmで検出)反応の終了を証明する。続いて、50μl DMFおよび10μl Et3Nに溶解した3−アミノ−3−デオキシジゴキシゲニン ヘミスクシンアミド スクシンイミジル エステル(2.6mg、0.0044mmol、1当量)溶液を加え、反応混合液を4時間室温で攪拌する。反応物を調製用HPLCに掛け、望みの生成物を含む分画を真空で濃縮し、DMSOに溶解し、そして純度を分析用HPLCによって分析する。精製した生成物を含む分画を合わせる。
CoA−Dgの濃度をアデニンの吸光係数(ε(アデニン、260nm)=15,300[M-1cm-1])を使って決定する。CoA−Dgの収量は2.37mg(37%)である。
ESI−MS(m/z)計算値1462.3707[M(−1)]、測定値1462.481M(−1)]。
【0060】
【化5】
【0061】
CoA−Cy3の合成
100μl DMF中のCy3−マレイミド3(Pharmacia、1mg、0.00126mmol)溶液に、90μl DMFおよび10μl 50mM Tris−ClpH7.5中の補酵素A二ナトリウム塩(1.05mg、0.00126mmol、1当量)溶液を加える。混合液を4時間室温で攪拌する。それを次にCH3CN/H2O 1:4で希釈し、小分けした500μlを調製用HPLCカラムに注入する。勾配、A/B95:5からA/B90:10までを2分間、A/B 65:35までを15分間、A/B20:80までを2分間行い、その後A/B 95:5に戻す(AおよびBについては先の実施例参照)。CoA−Cy3の滞留時間は10分である。所望の生産物を含む分画を真空で濃縮し、DMSOに溶解し、そして純度を制御するために分析量を注入する。精製した分画を合併する。CoA−Cy3の濃度を549nmの吸収(ε(549nm)=150,000[M-1cm-1])で決定する。CoA−Cy3の収量は0.847mg(44%)である。
ESI−MS(m/z)計算値1519.370[M(-1)]、測定値1519.3071[M(-1)]。
【0062】
【化6】
【0063】
CoA−Cy5の合成
Cy5−マレイミド4(1mg、0.00122mmol)から出発するCoA−Cy5の合成をCoA−Cy3について記述したのと同様に行う。CoA−Cy5の濃度をCy5の吸光係数(ε(646nm)=250,000[M-1cm-1])を用いて決定する。CoA−Cy5の収量は0.828mg(44%)である。
ESI−MS(m/z)計算値1545.386[M(-1)]および772.189[M(-2)]、測定値771.6524[M(-2)]。
【0064】
【化7】
【0065】
6×His−ACPSのクローニング、発現、および精製
XL1−blue E.coliのACPS遺伝子を単一コロニーPCRによって増幅し、pET−15bプラスミド(Novagen)中にNdel制限部位(下線)付きの順方向プライマー5’−TCT GGT CAT ATG GCA ATA TTA GGT TTA GGC ACG G−3’およびXhol制限部位(下線)付きの逆方向プライマー5’−TCA AGT CTC GAG TTA ACT TTC AAT AAT TAC CGT GGC A−3’を用いてクローニングする。E.coliのACPSのN−末端に融合されたペプチドの配列(プラスミドpET−15b起源)は(1文字コードで)MGSSHHHHHHSSGLVPRGSHであり、これにACPSの最初のアミノ酸、メチオニン、が続く。この融合タンパク質を6×His−ACPSと呼ぶ。
【0066】
6×His−ACPSをコードするpET−15b(Novagen)に基づく発現ベクターを含むBL21 E.coli細胞の液体培養を吸光度OD600nm0.6となるまで成長させる。6×His−ACPSの発現を最終濃度1mMとなるようにIPTGを加えて誘導する。3.5時間220rpm、24℃でインキュベーションした後、培養物を10分間3000g 4℃で遠心する。沈殿を10mlの抽出緩衝液(150mM NaCl、5mMイミダゾール、50mM KH2PO4、pH8.0)に再懸濁し、PMSFとアプロチミンをそれぞれ最終濃度1mMおよび2μg/mlとなるように加える。リゾチームを1mg/mlとなるように加え、混合物を氷上で15分間インキュベーションしまた何度か反転させる。次に、それを10分間超音波で破壊する(95%出力、50%動作)。DNA分解酵素Iを最終濃度0.01mg/mlとなるよう加える。4℃で30分後、混合液を10分間18,000rpmで遠心する。
【0067】
タンパク質を精製するために、予め抽出緩衝液で3回洗浄しておいた350μlのNi−NTAを溶解物に加える。混合物を20分間氷上でインキュベーションし数度混ぜ合わせた。抽出混合物を次にプロピレンカラムに加え、流出に任せた。カラムを5×400μlのDNA溶出緩衝液(10mM Tris Cl、pH8.5)で洗い、次に2×5ml洗浄緩衝液(300mM NaCl、10mMイミダゾール、50mM KH2PO4、pH7.5)で洗浄する。タンパク質を流出させるために、溶出緩衝液(300mM NaCl、150mMイミダゾール、pH7.5)をカラムに加え、10分間インキュベーションし、次に流出液を集める。溶出は、150μl溶出緩衝液で段階的に続け、これ以上タンパク質がブラッドフォード(Bradford)分析で検出できなくなるまで行う。最後に合わせた溶出液を透析緩衝液(50mM HEPES、30%グリセロール、pH7.2)中で終夜透析し、残存する塩を除く。精製した6×His−ACPS(MW16.215kDa)を小分けにして−80℃で保存した。濃度をブラッドフォード分析で37.6μMと決定する。収量はBL21E.coli細胞培養1リットル当たり0.912mg。
【0068】
6×His−ACPのクローニング、発現、および精製
XL1−blue E.coliのACP遺伝子を、Ndel制限部位(下線)付きの順方向プライマー5’−GT CGG TAT CAT ATG AGC ACT ATC GAA GAA CG−3’、およびBamHI制限部位(下線)付きの逆方向プライマー5’−TCA TGC GGA TCC TTA CGC CTG GTG GCC GTT G−3’、を用いて、単一コロニーPCRによって増幅し、pET−15bプラスミド(Novagen)中にクローニングした。E.coliのACPのN−末端に融合されたペプチドの配列(6×His−ACPを生成する)は(1文字コードで)MGSSHHHHHHSSGLVPRGSHであり、これにACPの最初のアミノ酸、メチオニン、が続く。
【0069】
6×His−ACPをコードするpET−15b(Novagen)に基づく発現ベクターを含むBL21E.coli細胞の液体培養を吸光度OD600nm0.6となるまで成長させる。6×His−ACPの発現を最終濃度1mMとなるようにIPTGを加えて誘導する。3.5時間220rpm、37℃でインキュベーションした後、培養物を10分間3000g4℃で遠心する。6×His−ACPの精製のために沈殿を正確に上の実験で記述した通りに操作する。6×His−ACP(MW10.802kDa)の濃度をブラッドフォード分析で188μMと決定する。収量:BL21E.coli細胞培養1リットル当たり3.1mg。
【0070】
アポ-およびホロ-6×His−ACP間の比率を決定するために、精製した6×His−ACPをQ−Tof−Ultima(Micromass/Waters)を用いたESI−MS(pos.モード)により、場合によっては、Cap−LC(Waters)、Xterra RP−C4カラム(Waters,5μm、0.32×50mm;流速8μl/min)によるクロマトグラフィーに連結し、MaxEnt1−ソフトウエアによる逆畳み込みを行って、解析する。混合物のESI−MSが、調製品はホロ-およびアポ-ACPの混合物であることを証明した。最初のメチオニンを除く精製したアポ-6×His−ACPの質量は10.6720kDaであることが分かり(計算値10.6716kDa)、またホロ-6×His−ACPの質量は11.012kDa(計算値11.0119kDa)であることが分かる。LC−ESI−MSによって、ホロ-およびアポ-型に対応するピークの同定と積分が可能になる。ホロ-型の100分率は16%(滞留時間11.77分)、またアポ-型のそれは84%(滞留時間14.16分)である。
【0071】
6×His−ACP−haの発現および精製
E.coli ACPのN−末端に融合されたペプチドの配列は(1文字コードで)MGSSHHHHHHSSGLVPRGSHであり、これにACPの最初のアミノ酸、メチオニン、が続き、E.coli ACPのC−末端に融合されたペプチドの配列は(1文字コードで)TSRSYPYDVPDYARW(6×His−ACP−haを生成する)である。
【0072】
6×His−ACP−haをコードするpET−15bに基づく発現ベクターを含むBL21(DE3)E.coli細胞の液体培養を吸光度OD600nm0.6となるまで成長させる。6×His−ACP−haの発現を最終濃度1mMとなるようにIPTGを加えて誘導する。220rpm、24℃で3時間インキュベーションした後、培養物を3000g4℃で10分間遠心する。6×His−AcpSの精製に対して記述した通りに、6×His−ACPを精製する。6×His−ACP−ha(MW12.65kDa)の濃度をブラッドフォード分析で400μMと決定する。またタンパク質の総収量は振とうフラスコ培養1リットル当たり10mgである。
【0073】
CoA−Btおよび6×His−ACPSを用いた6×His−ACPの生体外ビオチン化
精製した6×His−ACP(1μM)を反応緩衝液(43μl、50mM Tris ClpH8.8)中で6×His−ACPS(0.2μM)およびMgCl2(10mM)と室温でインキュベートさせる。7.5μlを解析用に分取する。CoA−Btを最終濃度5μMとなるように加える。一定時間ごとに各7.5μlを分取する。分取したものを補酵素A(最終濃度1mM)で30秒間クエンチし、8.2μlSDS緩衝液2×を加える。試料を2分間95℃で加熱する。ビオチン化された6×His−ACPをストレプトアビジン-西洋わさびペルオキシダーゼ結合体(NEM)および化学発光ペルオキシダーゼ基質(Renaissance reagent plus, NEN)を用い、ウエスタン-ブロッティングにより検出する。ウエスタン-ブロットをイメージステーション(Kodak440)を用いて解析する。対照として、バックグラウンドをチェックし、また反応の特異性をチェックするために、各タンパク質1つのみを含む試料、および2つのタンパク質のただ1つおよびCoA−Btを含む試料も調製しビオチン化を上のように分析する。ビオチン化は3つの成分全ての存在に依存する。
【0074】
ゲルシフトアッセイによる6×His−ACPのビオチン化の定量化
精製した6×His−ACP(3μM)を室温でCoA−Bt(10μM)および精製した6×His−ACPS(5μM)と最終容積50μlの反応緩衝液(50mM Tris Cl、pH8.8、10mM MgCl2)中でインキュベートする。30分インキュベーション後、混合液を終夜TBS((10mM Tris Cl、150mM NaCl、pH7.9)に対して透析し、過剰のCoA−Btを除く。小分けした反応液を、最終濃度0.6μg/μlのストレプトアビジンと1時間インキュベートする。2%SDSのみを含む2×SDS試料緩衝液を試料に加え、そして試料を加熱せずにSDS−PAGEを直接行う。タンパク質をクーマシー染色で検出し、またビオチン化の程度を、ビオチン化されておらず、ストレプトアビジンとインキュベートしていない等量の6×His−ACPを含む試料とバンド強度を比較して推定する。
【0075】
CoAとCoA−ラベル間の競合の定量
精製された6×His−AcpS(0.4μM)を、室温で、CoA−BtまたはCoA−Dg(2μM)、種々の量のCoA(0.2、4、8、12、20、40、80μM)および精製6×His−ACP−ha(0.4μM)と20μl反応緩衝液(50mM Tris・Cl、pH7.5、10mM MgCl2)中でインキュベートする。25分後、各試料を20μlの2×SDS試料緩衝液を加えてクエンチし、95℃2分間加熱した。ラベルされた6×His−ACP−haを、ストレプトアビジン-西洋わさびペルオキシダーゼ結合体(1:12500希釈)または抗−ジゴキシゲニン抗体−西洋わさびペルオキシダーゼ結合体(1:500希釈)および化学発光ペルオキシダーゼ基質を用い、ウエスタン-ブロッティングにより検出する。各実験に対して、0μM CoAの信号強度を任意に1に設定する。
【0076】
酵母細胞表面上へのAga2−ACPのラベリング
酵母ゲノムDNA由来のAGA2配列を増幅し、PCR断片の両端に導入したEcoRIおよびSall制限部位を利用して、酵母発現ベクターpRS314のPcup1−プロモーター配列の後へ挿入する。ACP配列のPCR増幅によって導入したSallおよびAcc651制限部位を用いて、ACPをAGA2のうしろのフレームへクローニングする。ACPはHA−エピトープをコードする配列により延長される。Aga2pおよびACPを連結する配列は:FVDEMLYFQGMである。Aga2pの最後の残基とACPの最初の残基とに下線を付す。HA−エピトープを含むACPのC−末端配列は:QAYPYDVPDYAGである。ACPの最後の残基に下線を付す。PGAL1−プロモーター由来のAga1pとPCUP1−プロモーター由来のAga2−ACPとを発現する酵母系統EBY100(MATa ura3−52 trip1 leu2Δ1 his3Δ200 pep4::HIS3 can1 GAL pIU211:URA3)(Invitrogen, Carlsbad, CA)を、2%ガラクトースおよび0.1mM銅を含む選択培地10ml中で、細胞が1.4OD600単位のOD600になるまで生育したものを、2mlの水で洗い、0.2mlのラベリング緩衝液(50mM Tris Cl pH8.8、100mM NaCl、10mM MgCl2)に再懸濁する。CoA基質および6×His−ACPSをそれぞれ最終濃度10および1μM加える。室温で30分後、反応液を2mlPBSに希釈することによりラベリングを止める。細胞を2mlPBSで4回洗い、直接蛍光顕微鏡検査に付すか、または、20nM Qdot(登録商標)605ストレプトアビジン結合体(Milan Analytic AG, Switzerland)を含む0.2mlのQdot(登録商標)インキュベーション緩衝液中で20分間インキュベート後細胞を2mlPBSで4回洗浄して蛍光顕微鏡検査に付す。細胞を、63×油浸(開口数1.4)対物レンズを用いて、ツアイスAxiovert135蛍光顕微鏡(Carl Zeiss, Gottingen, Germany)で検査する。
【0077】
HEK293細胞上に表示されるACP−NK1融合タンパク質のラベリング
ACP−NK1の一時的発現のために、5−HT3−レセプターの信号配列(Sig5HT3)を短いDYVリンカーを介してACPのN−末端に融合し、NK1を短いTSリンカーを介してACPのC−末端に融合する。その結果の構成において、FLAGタグおよび6×HisタグもまたNK1のC−末端に結合させる。融合タンパク質に対応する遺伝子をpCEP4(Invitrogen)ベクターのNheIおよびBamHI部位に挿入する。HEK293細胞を2.2%ウシ胎児血清(GIBCO, BRL)を補充したDMEM/F12(Dulbeccoの修正Eagle培地; GIBCO BRL)中で生育させる。一時的トランスフェクションを記載(Nat Biotechnol 21,86-89 (2003))通りに行い、HEK293細胞を、ACP−NK1を発現するベクターと核を標的とするEGFP(EGFP−NLS3)を発現するベクターにより共トランスフェクションする。24時間後、細胞を10分間室温で、MgCl2(10mM)、6×His−AcpS(1μM)、およびCoA−Cy3、CoA−Cy5、またはCoA−Bt(各5μM)のいずれかを含むPBS緩衝液500μlとインキュベートする。細胞を次に過剰の基質を除くためにPBSで3回洗浄し、Cy3またはCy5でラベルする場合は直接にレーザー走査共焦点蛍光顕微鏡によって分析する。ビオチン化された細胞を、PBS中1μg/ml濃度のFluoroLink(登録商標)Cy5−ラベルしたストレプトアビジン(Amersham Biosciences)とインキュベートした後、PBSで3回洗浄する。レーザー走査共焦点蛍光顕微鏡像を、63×水浸(開口数1.2)対物レンズ付きのZeissLSM510顕微鏡(Carl Zeiss AG, Gottingen, Germany)の488nmアルゴン/クロプトンレーザー線、543nm HeNEレーザー線、または633nm HeNEレーザー線を用いて記録する。走査速度およびレーザー強度を蛍光プローブの光退色および細胞の損傷または形態学的変化を回避するように調整する。蛍光を、GFPに敏感なチャネルおよびそれぞれの色素に敏感なチャネルで解析する。試験した各細胞試料に対して、GFPチャネルにおいて明確な核ラベリングが観察され、また細胞膜領域において明確な膜ラベリングが、CoA−Cy3ラベル後、CoA−Cy5ラベル後、およびCoA−Btラベル後であって、かつそれに続くストレプトアビジン-Cy5染色後に観察される。膜染色はトランスフェクションされない細胞では観察されず、GFPは核内発現されないことを示しており、これはそれぞれCy3、Cy5、およびビオチンによるACP−NK1の特異的ラベリングを示している。一時的にHEK293細胞に発現されたACP−NK1レセプターをさらなる実験で、CoA−Cy5およびテトラメチルローダミンでラベルされたサブスタンスP(SP−rho)、即ちNK1の自然のリガンド、で共染色する。両物質はもっぱら膜領域を染色し、同一の染色領域となる。過剰のサブスタンスPによって1分以内にSP−rho染色が元に戻ることが、さらにラベリングの特異性とまたACP−NK1のリガンド結合機能性をも証明する。
【0078】
【表1】
【図面の簡単な説明】
【0079】
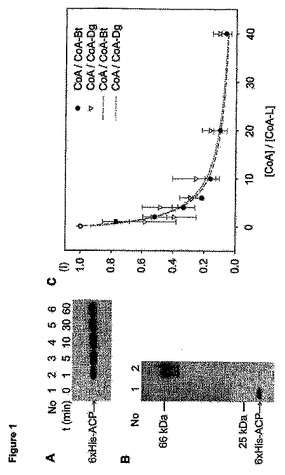
【図1】(A)6×His−ACP(1μM)、6×His−ACPS(0.2μM)およびCoA−Bt(5μM)の反応の解析:指定した時間(t)に一定量を反応混合液から分取し、ストレプトアビジン-ペルオキシダーゼ結合体を用いるウエスタンブロットにより6×His−ACPのビオチン化を精査する。(B)6×His−ACPのビオチン化の定量化:6×His−ACP(3μM)、6×His−ACPS(5μM)およびCoA−Bt(10μM)を30分間インキュベートし、透析し、分取した試料をストレプトアビジンとインキュベートしてSDS−PAGEに付す(第2列)。安定なビオチン-ストレプトアビジン複合体の形成がビオチン化タンパク質のゲルシフトをもたらす。ビオチン化の量を、第2列の6×His−ACPのバンド強度を、等濃度の6×His−ACPを含むがストレプトアビジンの無い試料(第1列)のバンド強度と比較して評価する。(C)CoAとCoA−Bt間のまたはCoAとCoA−Dg間の、ACPSの基質としての競合の定量:6×His−ACP(0.4μM)、6×His−ACPS(0.4μM)、CoA−BtまたはCoA−Dg(2μM)、および種々の濃度のCoA(0〜80μM)を30分間インキュベートし、ラベリングの程度をウエスタンブロットにより決定する。ウエスタンブロットの相対信号強度(I)を[CoA]/[CoA-L](L=ラベル)に対してプロットし、等式I=A/(1+BX)にフィットさせる。ここで、Xは[CoA]/[CoA-L]、Aは特定しない定数、Bは比(kcat/KM)CoA/(kcat/KM)CoA-Lである。これらの実験より、特異性定数の比が(kcat/KM)CoA/(kcat/KM)CoA-Btに対して0.48、(kcat/KM)CoA/(kcat/KM)CoA-Dgに対して0.35という結果となり、フリーのCoAと誘導体化されたCoAの間で重大な区別は無いことを明らかにする。
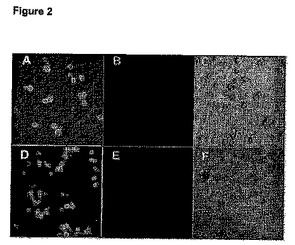
【図2】(A〜F)酵母の細胞表面上のACP融合タンパク質のラベリング。Cy3(A)またはストレプトアビジンでコートされた量子ドットにより追跡したビオチン(D)によってラベルされたAga2−ACPを発現した酵母細胞の蛍光顕微鏡写真。(B)は(A)と、(D)は(E)と同じであるが、但しAGA2−ACPを発現していない細胞である。(C)および(F)はそれぞれ(B)および(D)と同一の試料の透過顕微鏡写真である。これらの実験は、Aga2p−ACPを発現している酵母細胞のみがラベルされることを示す。
【技術分野】
【0001】
本発明は、基質から目的のタンパク質およびアシル担体タンパク質またはその断片を含む融合タンパク質へラベルを転移する方法、ならびに特にラベルされた融合タンパク質を検出しおよび/または操作することをさらに含む方法に関する。
【背景技術】
【0002】
複雑な生体系の理解の進歩は、その背後にある生体分子特にタンパク質の相互作用の特性決定に掛かっている。増大する数の生物におけるDNA配列決定により、その生物の読み取り枠(ORF)が明らかにされてきたが、その一方で、対応するタンパク質を生体内および生体外で特性決定できる可能性は限られている。この目的の実現を目指す戦略の大部分は、生体外での応用のための融合物の精製または生体内でのタンパク質の追跡を可能にする融合タンパク質を構築することに基礎を置いている。そのようなタグの例には、6×Hisタグ、グルタチオンS転移酵素、マルトース結合タンパク質、エピトープタグ、酵母ツーハイブリッドシステム、O6−アルキルグアニン-DNAアルキル転移酵素、分解ユビキチン、および緑色蛍光タンパク質(GFP)融合タンパク質が挙げられる。しかし、これらの方法はすべて様々な限界や不利益を有する。
【0003】
Gehring et al.(1997)および Lambalot and Walsh(1995)は、E.coliのホロ-アシル担体タンパク質合成酵素(ACPS)を用いて、アポ-アシル担体タンパク質(apo−ACP)の保存されているセリン残基に生体外で補助因子4’−ホスホパンテテイン(P−pant)を結合させて、ホロ-ACPを生成させる翻訳後修飾を触媒させることを記載している。P−pantの起源は補酵素Aである。Gehring et al.(1997)はさらにP−pant部分が修飾されているが依然としてACPSの基質として働くことができる補酵素Aの類似物質を用いて、修飾されたP−pantを補助因子とするホロ-ACPが得られることを示している。
【0004】
ACPSなどの単離されたホスホパンテテイニル転移酵素は国際特許出願WO 97/13845に記載されている。
【0005】
ラベルをO6−アルキルグアニン-DNAアルキル転移酵素(AGT)融合タンパク質に転移する方法、およびこの方法をAGT融合タンパク質の検出に用いることが国際特許出願WO 02/083937に記載されている。
【0006】
発明の開示
本発明は、目的のタンパク質を検出し、および/または操作する方法であって、目的のタンパク質およびアシル担体タンパク質(ACP)またはその断片を含む融合タンパク質を、ラベルされた補酵素A(CoA)型基質およびホロ-アシル担体タンパク質合成酵素(ACPS)もしくはその相同物と接触させること、その結果ACPSがラベルを融合タンパク質へ転移させ、また場合によっては、そのラベルを認識しおよび/または操作するために設計されたシステム中でそのラベルを用いて、得られたそのラベル付融合タンパク質を検出しおよび/またはさらに操作すること、を含む方法に関する。
【0007】
さらにまた、本発明は目的のタンパク質およびACPまたはその断片を含む融合タンパク質のそのような方法中での使用に関する。特に本発明の方法は、目的のタンパク質を精製もしくは固定化するため、または本発明の方法において目的のタンパク質をそれに結合されたラベルにより生体外もしくは生体内で連続的にモニターするために用いられる。
【0008】
本発明の融合タンパク質に組み込まれる目的のタンパク質は任意の種類のものでよく、それには二次構造、三次構造、もしくは四次構造を持つ、または持たない、任意の長さのタンパク質、ポリペプチドおよびペプチドが含まれる。
【0009】
本発明の方法で用いられる特別のラベルされた補酵素A(CoA)型基質は、CoA、または修飾されたCoAから、ラベル即ち検出可能なマーカーをさらに付加するための少なくとも1つの反応部位を有するリンカーを結合させることによって得ることができる。本発明はまた、そのような新規のラベルされた補酵素A型基質、それを製造する方法、そのような新規の補酵素A型基質の合成において有用な中間体、および本発明の方法中でのそれらの使用に関する。
【0010】
発明の詳細な説明
本発明は、目的のタンパク質を検出し、および/または操作する方法であって、目的のタンパク質およびアシル担体タンパク質(ACP)またはその断片を含む融合タンパク質を、ラベルされた補酵素A(CoA)型基質およびホロ-アシル担体タンパク質合成酵素(ACPS)もしくはその相同物と接触させること、その結果ACPSがラベルを融合タンパク質へ転移させ、また場合によっては、ラベルを認識しおよび/または操作するために設計されたシステム中でラベルを用いて、得られたラベル付融合タンパク質を検出し、および/またはさらに操作するようにすること、を含む方法に関する。
【0011】
ACPまたはACPドメインは、脂肪酸、ポリケチドの生合成および非リボソームペプチド合成において担体として作用する。ACPはホロ-アシル担体タンパク質合成酵素(ACPS)によって翻訳後修飾される。この酵素は補助因子4−ホスホパンテテイン(P−pant)を補酵素AからACPの保存されているセリン残基に転移する。ACPSはまたホスホパンテテイニル転移酵素とも呼ばれる。ACPSは、CoAのホスホパンテテイン部分のチオール基に対する修飾に関して比較的低い基質特異性を所持していることが示されている。これを活用して、ACP融合タンパク質をACPSおよびホスホパンテテイン部分を介してラベルを所持するCoA誘導体と共にインキュベートすることにより、特異的にラベルすることができる。ラベルが、このようにしてホスホパンテテイン部分と共にACPの保存されたセリン残基に転移される。ラベリングは融合タンパク質の性質とは独立である。
【0012】
ACPおよびACPSという語句は、ここでは任意の一対のタンパク質であって2つのうちの1つ(ACP)がCoA誘導体起源のホスホパンテテイン誘導体に対するアクセプターであり、他の1つ(ACPS)がホスホパンテテイン誘導体のACPへの転移を触媒するものを表す。
【0013】
ACPSおよびホスホパンテテイン転移酵素という語句は、E.coli由来のACPSと相同性のある多数のホスホパンテテイニル転移酵素が、脂肪酸合成に関与せず、むしろ自然産物の生合成に関与しているタンパク質を修飾するという事実にもかかわらず、ここでは交換できる語句であるとして用いる(Lambalot et al.,1996)。このようなホスホパンテテイニル転移酵素の例には:エンテロバクチン合成に関与するEntD;サーファクチン生合成に関与するSfpおよびPsf−1;グラミシジンS生合成に関与するGsp;リシン生合成に関与するLYS5;バシトラシン生合成に関与するBli;イチュリンA生合成に関与するLpa−14;および、ノシヘプチド生合成に関与するNshC、がある。本発明はE.coli由来のACPSと相同性のあるラベリング方式を持つ全てのACPSの使用を含む。
【0014】
ACPという用語は、Lambalot et al.(1996)によって定義されたように、E.coli由来のACPSまたはE.coli由来のACPSと相同性のある任意のACPSによって翻訳後にホスホパンテテイニル化されることとなる任意のタンパク質を表す。これには、脂肪酸合成に関与するタンパク質ばかりでなく、ポリケチド合成、非リボソームペプチド合成、アミノ酸合成、およびデプシペプチド合成に関与するタンパク質も含まれる。ACPSによる翻訳後修飾に加えて、これらのタンパク質は共通して種々の基質とアシルパンテテイニル・チオエステルを形成し、ホスホパンテテイニル部分がセリン残基に結合される。これらの自然の機能において、ACPは、多機能酵素(I型脂肪酸合成酵素のような)のドメイン、または別のタンパク質(II型脂肪酸合成酵素のような)のドメインであり得る。
【0015】
「検出する」とは、ラベルおよびそれに結合している目的のタンパク質を、ラベルを観測するために設計されたシステム中でラベルの性質に基づいて観察することを意味するものであり、また特別なラベルを認識し、ラベルの性質によって特別な環境でラベルを見出し、場合によっては、ラベルおよびそれに結合したタンパク質を定量化し、また場合によっては、ラベルの微細環境を決定することなどを含む。
【0016】
「操作する」とは、ラベルおよびそれに結合している目的のタンパク質を操作することを意味し、またラベルおよびそれに結合している目的のタンパク質を、ラベルを操作するために設計されたシステム中でラベルの性質に基づいて操作することを含み、ならびに化学的または生物学的環境から分離すること、別の化学的または生物学的環境へ導入すること、精製すること、即ち望ましくない副産物および不純物から分離すること、ラベルを固体の担体と反応させることによって固定化すること、ラベルおよび/または目的のタンパク質の性質を修飾するために化学的または生物学的試薬と接触させることなどを含む。
【0017】
ここで開示する方法は一般にある範囲の応用に適用でき、特異的におよび共有結合的に融合タンパク質を(1)ラベルされた融合タンパク質の環境の変化を感受し誘起することができるラベルによってラベルすることができ、(2)ラベルによって融合タンパク質に特異的に導入された物理的および/または化学的性質によって融合タンパク質を操作することを助けるラベルによってラベルすることができ、および/または(3)ラベルによって導入された性質を通じて融合タンパク質の精製を助けるラベルによってラベルすることができる。ここで開示する方法はACP融合タンパク質を生体外および生体内、例えば細胞中の両方でラベルするために用いることができる。
【0018】
さらにまた、本発明は目的のタンパク質とACPまたはその断片を含む融合タンパク質のそのような方法中での使用に関する。特に、本発明の方法は目的のタンパク質の精製と固定化に、または、目的のタンパク質を生体外および生体内において本発明の方法によりそれに結合されたラベルにより連続的にモニターすることに用いられる。
【0019】
一つの態様において、本発明は目的のタンパク質を生体外および生体内において連続的にモニターする方法を提供する。この方法では、目的のタンパク質とアシル担体タンパク質(ACP)またはその断片を含む融合タンパク質を、ラベルされた補酵素A(CoA)型基質およびホロ-アシル担体タンパク質合成酵素(ACPS)またはその相同物と接触させて、ACPSがラベルを融合タンパク質へ転移し、またラベルを、ラベルを認識するために設計されたシステム中で観察する。
【0020】
さらにある態様において、本発明は目的のタンパク質を生体外および生体内において操作する方法を提供する。そこでは目的のタンパク質およびアシル担体タンパク質(ACP)もしくはその断片を含む融合タンパク質を、ラベルされた補酵素A(CoA)型基質およびホロ-アシル担体タンパク質合成酵素(ACPS)もしくはその相同物と接触させて、その結果ACPSがラベルを融合タンパク質へ転移し、また融合タンパク質をラベルの物理的および/または化学的性質に基づいて操作する
【0021】
ある特別な態様においては、ラベルの物理的および/または化学的性質によってラベルされた融合タンパク質の効率的な精製が可能であり、したがって本発明は、本発明の方法の諸工程すなわち、ラベルの物理的および/または化学的性質を精製に利用し、その後融合タンパク質を切断して高純度の目的のタンパク質を供給する工程を遂行することによる、目的のタンパク質の精製方法を提供する。
【0022】
さらなる態様において、本発明は、目的のタンパク質およびアシル担体タンパク質(ACP)もしくはその断片を含む融合タンパク質を固体支持体に固定化する方法を提供する。この方法は融合タンパク質を、固体支持体に結合しているかまたは結合できるラベルされた補酵素A(CoA)型基質およびホロ-アシル担体タンパク質合成酵素(ACPS)もしくはその相同物と接触させることを含み、これにより、ホロ-アシル担体タンパク質合成酵素(ACPS)がラベルをACP融合タンパク質へ共有結合させるように転移し、ACP融合タンパク質はそれにより固体支持体に結合されるかまたはその後に結合され得る。ラベルが最初は固体支持体に結合されていない本発明の特別な態様において、本方法はさらに、ラベルされたACP融合タンパク質が固体支持体に固定化されるようにそれを固体支持体と接触させる工程を含んでもよい。これらの本発明の好ましい実施態様においてラベルは、転移されたときまたは次の反応において固体支持体に共有結合的に結合されてもよく、または特異的な結合対の一員であって、他の一員が固体支持体に共有結合的にもしくは任意の他の手段により結合しているか結合可能であってもよい。それは例えばビオチンおよびアビジンもしくはストレプトアビジンの特異的結合対を用いる場合である。
【0023】
さらなる態様において、本発明は、生体外はもちろん生体内においてもACP融合タンパク質をラベルする方法を提供する。生体内におけるACP融合タンパク質のラベリングという語句は、細胞外スペースに向いているACP融合タンパク質をラベルすることはもちろん、細胞内の全ての区画においてラベルすることを含む。もしACP融合タンパク質のラベリングが生体内で行なわれ、ACPに融合されたタンパク質が原形質膜タンパク質である場合には、融合タンパク質のACP部分は原形質膜の細胞質側または細胞外側のどちらに付着していてもよい。ラベリングを生体外で行なう場合は、融合タンパク質のラベリングは細胞抽出物中でまたはACP融合タンパク質を精製または濃縮した形で行うことができる。
【0024】
本発明は、所望のタンパク質へのラベルの特異的な付加を、その目的のタンパク質とアシル担体タンパク質(ACP)またはその断片との間で融合タンパク質を形成させることにより遂行できたという成果を基礎にしている。
【0025】
好ましい応用において、アシル担体タンパク質即ち「ACP」は、ホロ-アシル担体タンパク質合成酵素即ち「ACPS」によって、ラベルされた4’−ホスホパンテテインが適当な補酵素A(「CoA」)から融合タンパク質の一部を形成するACPもしくはその断片のセリン残基へ転移される方法で、修飾される性質を有する。好ましい実施態様において、ACPは、例えばRawling and Cronan, 1992およびその中の引用文献中に記載のE.coliのアシル担体タンパク質である。しかし、他のアシル担体タンパク質(ACP)、例えばGehring et al., 1997に記載されているStreptomyces種のACP、または適当にラベルされたCoA存在下で上に定義したACPSによって修飾される性質を有する任意のACPが知られており、本発明において使用できる。本発明においてACPはまた、野生型ACPの変異体であって1個以上のアミノ酸置換、欠失または付加によって野生型と異なり、しかしACPSにより触媒される反応において、ラベルされた4’−ホスホパンテテインに対するアクセプターとして作用する性質を依然として保持している変異体も含む。他のACP変異体を周知技術を用いて化学的に修飾してもよい。ACP変異体は周知のタンパク質工学技術を用いて、および/またはACPSによって触媒される反応におけるラベルされた4’−ホスホパンテテインの転移に対する新しいアクセプター配列を生成し選択するための分子進展を利用して生成することができる。ACP断片とは、ホスホパンテテイン誘導体と結合するセリン残基を含み、またそのようなホスホパンテテイン誘導体を受容する機能を残存させている断片である。
【0026】
好ましい実施態様において、ACPSは、例えばLambalot and Walsh,1995またはその中で引用された文献に記載されたE.coliのホロ-アシル担体タンパク質合成酵素である。しかし、他のホロ-アシル担体タンパク質合成酵素、例えばLambalot et al.,1996記載のBacillus subtilis由来のACPSまたは上で定義したACPを適当にラベルされたCoA存在下で修飾する性質を有するならば本発明において採用可能である任意の形のタンパク質なども知られている。これらのACPSはまたLambalot et al.,1996記載のように、ホスホパンテテイニル転移酵素としても知られており、また本発明はこの一般的な種別の酵素の使用を含む。本発明において、ホロ-アシル担体タンパク質合成酵素はまた野生型ACPSの変異体であって1個以上のアミノ酸置換、欠失または付加によって野生型と異なり、しかしラベルされた4’−ホスホパンテテインを特異的にACP融合タンパク質に転移する性質を依然として保持している変異体も含む。他のACPS変異体を周知技術を用いて化学的に修飾してもよい。ACPS変異体は周知のタンパク質工学技術を用いて、および/またはラベルされた4’−ホスホパンテテインを異なるアクセプター配列に転移するための新しい特異性を生成し選択する分子進展を利用して、生成することができる。
【0027】
ACP融合タンパク質のACPSによるラベリングのための、多くの考察を有利に織込む。最も大切なことは、ACP融合タンパク質のラベリングは、ラベルする前にACP融合タンパク質のACP部分がアポ型で存在することに依存していることである。もしACP融合タンパク質が、翻訳後修飾の基質としてこのACP融合タンパク質を受容する内在的ACPSを有する宿主において発現されているならば、ACP融合タンパク質は所望の修飾に対して少なくとも部分的に遮断される可能性がある。この望ましくないACP融合タンパク質の修飾を最小にするために種々の解決策が提案されている。第一に、宿主のACPSによって効率的に修飾されないACPを選択する、即ちACPが宿主の生化学と直交するように選択する。例えば、E.coli由来のACPは、ヒト細胞において融合タンパク質として発現されたときにヒトのACPSによって有意な程度には修飾されない(図3参照)。第二に、ACP融合タンパク質の過剰発現によってACP融合タンパク質の主にアポACPが形成されるようになる。
例えば、E.coliにおいてE.coli由来の6×His−ACPを過剰発現させることにより、これらの細胞内に内在性ACPSが存在するにもかかわらず、大部分は6×HisのアポACPが生成されることになる。
【0028】
ラベル反応のために、ACP融合タンパク質はまた、対応するACPSおよびCoA誘導体の両方と接触しなければならない。このことは生体内応用において、ACP融合タンパク質が細胞表面に表示されるか、またはACPSとCoA誘導体が微量注入などの技術を用いて目的の細胞内に導入されるかどちらかであることを意味する。
【0029】
本発明において、ACPとの融合タンパク質のタンパク質部分への言及は、二次構造、三次構造、四次構造を有するまたは有しないタンパク質、任意の長さのポリペプチドおよびペプチドが含まれることを意図したものであり、また好ましくは少なくとも12個のアミノ酸から2000個までのアミノ酸を、好ましくは50から1000個の間のアミノ酸から成る。本発明による目的のタンパク質は次の群から選択される。その群は、酵素、DNA結合タンパク質、転写調節タンパク質、膜タンパク質、核レセプタータンパク質、核局在信号タンパク質、タンパク質補助因子、抗体、膜ポンプタンパク質、膜チャネルタンパク質、膜担体タンパク質、運動タンパク質、シグナル伝達に含まれるタンパク質、核タンパク質、リボソームタンパク質、小単量体GTPアーゼ、ATP結合カセットタンパク質、細胞内構造タンパク質、特別な細胞内区画を標的とするタンパク質としての役割を担う配列を有するタンパク質、一般にラベルとしてまたは親和性タグとして用いられるタンパク質、および前述のタンパク質のドメインもしくはサブドメイン、から成る。ACP融合タンパク質は1つ以上の、例えば1つ、2つ、3つの、ACPのN−末端、C−末端、またはN−およびC−末端においてACPと融合した目的のタンパク質から成ればよい。
【0030】
より詳しく言えば、本発明による目的のタンパク質はつぎの群から選択される。
即ちその群は、
酵素、例えば、転移酵素(EC2)、より具体的にはメチル基以外のアルキルもしくはアリル基を転移する転移酵素(EC2.5)、特にグルタチオン転移酵素(EC2.5.1.18)、
あるいは、リンを含む基を転移する転移酵素であるキナーゼ(EC2.7)、例えば酵母のカゼインキナーゼ(EC2.7.1.37)またはチロシンタンパク質キナーゼ(EC2.7.1.112)のような、基質タンパク質のリン酸化標的部位としてセリンもしくはスレオニンを有するタンパク質キナーゼなど、特にアルコール基をアクセプターとするキナーゼ(EC2.7.1);
あるいは、例えば酸化還元酵素(EC1)、より具体的には過酸化物をアクセプターとする酸化還元酵素(EC1.11)、特にチトクロームCペルオキシダーゼ酵素(EC1.11.1.5);
あるいは、例えば加水分解酵素(EC3)、より具体的にはエステル結合に作用する加水分解酵素(EC3.1)、特にタンパク質リン酸モノエステル加水分解酵素などのリン酸モノエステル加水分解酵素(EC3.1.3);またはペプチダーゼもしくはプロテアーゼ(EC3.4)としても知られるペプチド結合を加水分解する加水分解酵素、特にカスパーゼ;
DNA結合タンパク質、より具体的にはmRNA合成を阻害するタンパク質因子である転写リプレッサータンパク質、特にE.coliにおいてmRNA合成を阻害するタンパク質因子、特にLexAタンパク質のDNA結合ドメイン;
転写調節タンパク質、より具体的には転写リプレッサータンパク質、特にトリプトファン/アスパラギン酸反復構造を含む転写リプレッサータンパク質;
膜タンパク質、例えば少なくとも1つの膜貫通へリックスを示す膜タンパク質、より具体的には小胞体(ER)膜由来の膜タンパク質、特にER膜貫通タンパク質Sec62などの特にERへのタンパク質移行活性を有する膜タンパク質;
あるいは、例えば7回膜貫通ヘリクッス(7−TM)タンパク質、より具体的には7−TMタンパク質がGタンパク質共役レセプター(GPCR)であるもの、特に哺乳類などの、例えばヒトのニューロキニン−1−レセプター(NK1)などの分子量約1kDaより上の高分子リガンドを結合する7−TMタンパク質;
あるいは、例えば細胞膜由来の膜貫通イオンチャネルタンパク質、特にリガンド作動性イオンチャネルタンパク質、より具体的には、セロトニンレセプター5−HT3などのセロトニン感受性のリガンド作動性イオンチャネルタンパク質;
あるいは、例えばイオンチャネルおよびGタンパク質共役レセプター以外の膜レセプター;
あるいは、例えば、特に酵母由来の、ペルオキシソームの膜タンパク質、タンパク質Pex15など;
核レセプタータンパク質、例えば転写因子ファミリー由来の核レセプタータンパク質、より具体的にはリガンド誘導可能な転写因子ファミリー由来の核レセプタータンパク質、特に例えばヒトエストロゲンレセプターhERなどのエストロゲンレセプターのようなステロイドファミリー由来の核レセプター;
サルウイルス40(SV40)由来の核局在信号などの核局在信号タンパク質;
例えば遺伝子構造中にユビキチン配列を含むタンパク質のようなタンパク質性補助因子;
小単量体GTPアーゼ、より具体的にはRasファミリーメンバーのような膜結合性小単量体GTPアーゼ;
例えば複数薬剤耐性タンパク質のようなATP結合カセット(ABC)タンパク質;
細胞内構造タンパク質、より具体的には細胞骨格タンパク質、より具体的にはヒト細胞質β−アクチン;
例えばゴルジ体、小胞体(ER)、ミトコンドリア、原形質膜、またはペルオキシソームへと、特別の細胞区画へタンパク質を指向させる役割の配列を有するタンパク質;
一般にラベルまたは親和性タグとして用いられるタンパク質、例えばUVもしくは可視光照射による励起によって蛍光信号を与える蛍光タンパク質、強化シアノ蛍光タンパク質(ECFP)として知られる蛍光タンパク質などの、特に緑色蛍光タンパク質(GFP)として知られるファミリー由来の蛍光タンパク質;
および上述タンパク質のドメインまたはサブドメイン、
である。
【0031】
さらに、本発明による目的のタンパク質は供給源に従って選択される。特に、目的のタンパク質には、ヒト、マウス、ラット、他の高等動物、真核生物種、バクテリア種、例えばサルモネラ、より具体的には、salmonella typhiもしくはsalmonella typhimurium、ミコバクテリア、より具体的には mycobacterium tuberculosis、ブドウ球菌、より具体的にはstaphylococcus aureusなどに存在するタンパク質、または、例えばヒト免疫不全ウイルス(HIV)、ヒトインフルエンザウイルス、肝炎ウイルス、またはコロナウイルスなどのウイルス源由来のタンパク質である。
【0032】
さらにまた、目的のタンパク質は、ガン、心臓血管病、精神障害、アルツハイマー、肥満、ウイルス感染、およびバクテリア感染などの、一定の病気におけるその役割に従って選択される。
【0033】
本発明の特定の実施態様において、融合タンパク質は、一方の側のE.coliのACPまたはそのようなACPのDNAの変異体およびACPのDNA配列のN−末端(N)もしくはC−末端(C)もしくはN−末端とC−末端側に結合された(上に挙げたような)目的のタンパク質をコードする配列とから生成され、本発明の融合タンパク質が導かれる。融合タンパク質はさらに適当なリンカー、例えば融合タンパク質中のACPと目的のタンパク質の間の、および/または2つの目的のタンパク質の間の適当な条件下で酵素切断を受けやすいリンカーを含む。そのようなリンカーの例には、DNA段階で適当な制限酵素によって切断されるリンカー、例えばBgl IIによって切断されるAGATCT、および/またはタンパク質段階で適当な酵素、例えばタバコエッチウイルスNla(TEV)プロテアーゼによって切断されるリンカーがある。
【0034】
融合タンパク質は原核生物宿主、好ましくはE.coli、または真核生物宿主、例えば酵母、真菌、昆虫、または哺乳動物細胞中で発現できる。
【0035】
本発明はまた、目的のタンパク質とACPもしくはその断片との新規な融合タンパク質に関する。
【0036】
本発明において、ラベルされたCoA型基質は、好ましくは次の一般式(I):
【化2】
または、次の一般式(II):
【化3】
を持つ、ラベルされた補酵素Aの誘導体である。式中、Rは補酵素Aとラベルを繋ぐリンカー基であり、また「ラベル」は、ここに記載した融合タンパク質の検出、精製および/または操作のために適当なラベル分子である。
【0037】
しかし、本発明は式(I)または(II)の基質に限られない。広い範囲の他の基質をACP融合タンパク質へラベルを転移するために使用できるからである。例えば、式(I)の化合物のプリン部の置換、糖部分またはパンテテイニル酸部分の修飾、が考えられる。
【0038】
式(I)または(II)の化合物中のリンカー基Rはラベルを補酵素Aに結合する柔軟なリンカーである。リンカー単位は構想した応用、即ちACPを含む融合タンパク質へのラベルの転移との関連で選ばれる。それらはまた適当な溶媒中で基質の溶解度を増加させる。用いたリンカーは実際の応用条件下で化学的に安定である。リンカーRはACPとの反応にもラベルの検出にも干渉せず、しかし補酵素A型基質がACPを含む融合タンパク質と反応した後のある時点で切断されるように構成することができる。
【0039】
リンカー基Rは1〜300個の炭素原子を有する直鎖または分岐鎖のアルキレン基であり、場合によっては、
(a)1個以上の炭素原子が酸素により置き換えられ、特に3個ごとの炭素原子が酸素に置き換えられ、例えば、1〜100個のエチレノキシ単位を有するポリエチレノキシ基を表し;
(b)1個以上の炭素原子が水素原子を持つ窒素原子に置き換えられ、また隣接する炭素原子がオキソにより置き換えられて、アミド官能基−NH−CO−を表し;
(c)1個以上の炭素原子が酸素に置き換えられ、また隣接する炭素原子がオキソで置き換えられ、エステル官能基−O−CO−を表し;
(d)隣接する2個の炭素原子間の結合が2重または3重結合であり、官能基−CH=CH−または−C≡C−を表し;
(e)1個以上の炭素原子がフェニレン、飽和または不飽和シクロアルキレン、飽和または不飽和ビシクロアルキレン、架橋ヘテロ芳香族または架橋飽和または不飽和ヘテロシクリル基に置き換えられており、
(f)隣接する2個の炭素原子がジスルフィド結合−S−S−により置き換えられており;
または、場合によっては置換基を含む、2個以上の、特に2個の、アルキレン基および/または上の(a)〜(f)で定義した修飾されたアルキレン基の組み合わせである。
【0040】
考慮した置換基は、例えば低級アルキル例えばメチル、低級アルコキシ例えばメトキシ、低級アシロキシ例えばアセトキシ、またはハロゲニル例えばクロロである。
【0041】
さらに考慮した置換基は、例えばα−アミノ酸がリンカーRに取り込まれた場合に得られる置換基であり、その場合炭素原子は(b)で定義したアミド官能基−NH−CO−で置き換えられる。そのようなアルキレン基の炭素鎖のリンカー部分において、Rは−(NH−CHR’−CO)n−基で置き換えられる。ここでnは1〜100の間であり、R’は種々のα−アミノ酸の残基を表す。
【0042】
さらなる置換基は光切断性リンカーRとなる置換基、例えばo−ニトロフェニル基である。特に、このo−ニトロフェニル置換基はアミド結合に隣接した炭素原子に、例えば−NH−CO−CH2−CH(o−ニトロフェニル)−NH−CO−基に存在する。
【0043】
上の(e)で定義した炭素原子と置き換えられるフェニレン基は、例えば1,2−、1,3−、または好ましくは1,4−フェニレンである。上の(e)で定義した炭素原子と置き換えられる飽和または不飽和シクロアルキレン基は、例えばシクロペンチレンまたはシクロヘキシレンまたはさらに、例えば1−または2−の位置で不飽和のシクロヘキシレンである。上の(e)で定義した炭素原子と置き換えられる飽和または不飽和ビシクロアルキレン基は、例えば、場合によっては、2−の位置で不飽和のまたは2−と5−の位置で二重に不飽和のビシクロ[2,2,1]ヘプチレンまたはビシクロ[2,2,2]オクチレンである。上の(e)で定義した炭素原子と置き換えられるヘテロ芳香族基は、例えばトリアゾリデン、好ましくは1,4−トリアゾリデン、またはイソオキサゾリデン、好ましくは3,5−イソオキサゾリデンである。上の(e)で定義した炭素原子と置き換えられる飽和または不飽和ヘテロシクリル基は、例えば2,5−テトラヒドロフランジイルまたは2,5−ジオギサンジイル、またはイソオキサゾリジネン、好ましくは3,5−イソオキサゾリジネンである。
【0044】
好ましいリンカー基Rは、例えば、場合によってはメチル、メトキシ、またはアセトキシ基で置換された1〜20個の炭素原子のアルキル鎖、または1〜20個のエチレノキシ基から成るポリエチレングリコール鎖などの、柔軟なリンカーである。
【0045】
基質のラベル部分は、当業者が融合タンパク質を応用しようと意図するその応用に応じて選択できる。ラベルの例には:
(1)蛍光団、発色団などの分光学的プローブ、磁気的プローブまたはコントラスト剤、あるいはまた電子顕微鏡技術で有用なプローブ;
(2)放射性ラベルされた分子;
(3)相手と特異的に結合できる特異的な結合対の一方の分子。そのような特異的結合対は周知であり、例えばアビジンまたはストレプトアビジンと結合できるビオチンである;
(4)他の生体分子と相互作用すると想定される分子;
(5)他の生体分子と相互作用すると想定される分子のライブラリー;
(6)例えばNadeau et al.,2002に記載された、当業者に周知の他の生体分子と架橋できる分子;
(7)H2O2とアスコルビン酸に曝露されるとヒドロキシラジカルを生成できる例えばHori et al.,2002に記載の繋留された金属キレートなどの分子;
(8)例えばJay et al.1999に記載のマラカイトグリーンなどの、光照射によって反応性ラジカルを生成できる分子;
(9)支持体がガラススライド、ミクロタイタープレートまたは熟練の当業者に周知の任意の一般ポリマーである、固形支持体に共有結合した分子;
(10)相補的な鎖と塩基対を形成できる核酸またはその誘導体;
(11)リピッドまたは膜に挿入される性質を有する他の疎水性分子;
(12)望ましい酵素学的、化学的、または物理学的性質を有する生体分子;
(13)上に挙げた性質の任意の組み合わせを有する分子;
が含まれる。
【0046】
ラベルされたCoA誘導体は、ホスホパンテテイニル部分がACP融合タンパク質へ転移することができ検出可能であるラベル、例えば蛍光団、発色団、磁気的プローブ、放射性ラベルされた分子、または任意の他の分光学的プローブなどのラベルを所持しており、そのラベルされたCoA誘導体を使用することによって、本発明を、生体外または細胞内もしくは細胞表面上(生体内)において、検出できるラベルをACP融合タンパク質へ特異的かつ共有結合的に結合させるために使用することが可能になる。これにより、生体内および生体外においてACP融合タンパク質の検出と特性決定が可能になる。生体内という語句は、細胞外スペースに向いているACP融合タンパク質のラベルはもちろん、細胞の全ての区画中でACP融合タンパク質をラベルすることを含む。この方法を、遺伝的に目的のタンパク質と融合させ、生きている細胞中でその研究をすることができるようにする緑色蛍光タンパク質(GFP)の応用と比較することができる。GFPとその突然変異体の欠点は、原則としてGFP中に存在する自然に存在する蛍光団の使用に限定されることである。細胞内(生体内)でのラベリングは、タンパク質の機能的な構造に小さな影響のみを与え、ACPのホスホパンテテイニル部分によりラベルされる機能を保持する一般的な固定手順によって細胞を固定した後にも利用できる。
【0047】
ホスホパンテテイニル部分がACPタンパク質に転移できるビオチンなどの親和性タグを有するようなラベル付CoA誘導体を使用することによって、本発明を親和性タグを融合タンパク質へ転移するために用いることが可能になり、融合タンパク質に親和性タグの結合相手が結合できるようになる。例として、ビオチンなどの親和性タグによってラベルされたCoA基質およびACPSを、ACP融合タンパク質を発現している細胞抽出液(バクテリアもしくは真核生物の)または精製されたACP融合タンパク質に加えることによって、親和性タグによる融合タンパク質の共有結合的修飾が可能になる。これによって次に、親和性タグとその結合相手との相互作用を利用して、例えばビオチンの場合は固定化されたアビジンもしくはストレプトアビジンとの相互作用によって、融合タンパク質の単離が可能になる。もしラベルがジスルフィド架橋などの切断可能な結合を有するリンカー基Rを介してACP融合タンパク質にリンクしているならば、またはもしリンカーが光切断性であれば、精製後にACP融合タンパク質から親和性タグを切り離すことができる。
【0048】
ACP融合タンパク質へ転移でき、また外部の刺激に曝露するとヒドロキシラジカルなどの反応性ラジカルを生成できるラベルをホスホパンテテイニル部分が所持するラベル付CoA誘導体の使用によって、目的のタンパク質およびその近くのタンパク質の立体構造の研究が可能になる。生成されたラジカルはACP融合タンパク質およびそのごく近傍のタンパク質を不活性化することができて、それのよってそれらのタンパク質の役割の研究が可能になる。そのようなラベルの例は、H2O2とアスコルビン酸に曝露することによりヒドロキシラジカルを生成するテザー金属キレート錯体、およびレーザー照射によってヒドロキシラジカルを生成するマラカイトグリーンなどの発色団である。ヒドロキシラジカルを生成するために発色団とレーザーの使用することは、例えばJay et al.,1998に記載されているように、発色団援用レーザー誘導不活性化(CALI)として周知である。CALIは、時間的に制御され空間的に解像された態様で特異的に細胞中の一定のタンパク質を特異的に不活性化するために使用する方法であって、また発色団とタンパク質が空間的に近傍にあることを基礎にしている。レーザー照射すると、発色団はヒドロキシラジカルを生成し、それは発色団から約100nm以内のそしてそれ以内のみの全てのタンパク質を不活性化する。現在までは、発色団は、目的のタンパク質に特異的な発色団でラベルされた抗体を微量注入することによって目的のタンパク質の空間的近傍にもたらされた。本発明において、マラカイトグリーンのような発色団でACP融合タンパク質をラベルし次にレーザー照射することによって、時間的に制御され空間的に解像された態様でACP融合タンパク質およびACP融合タンパク質と相互作用するタンパク質を不活性化することを可能にすることになる。この方法は生体内および生体外の両方で応用できる。
【0049】
類似の方法で、ACP融合タンパク質を繋留された金属キレートでラベルし、そしてACP融合タンパク質およびACP融合タンパク質と相互作用するタンパク質をH2O2およびアスコルビン酸に曝露することにより特異的に不活性化できる。この方法はACP融合タンパク質またはACP融合タンパク質のごく近傍のタンパク質の機能の研究に用いることができるばかりでなく、ACP融合タンパク質のごく近傍のタンパク質の同定にも用いることができる。ここで、ACP融合タンパク質のごく近傍のタンパク質を、特異的な抗体によりそのタンパク質の断片を検出することにより、高分解能二次元電気泳動ゲル上からそれらのタンパク質が消滅したことにより、または質量分析法もしくはN−末端分解によるタンパク質配列決定法などの分離および配列決定技術により切断されたタンパク質断片を同定することにより、同定できる。
【0050】
ホスホパンテテイニル部分がACP融合タンパク質に転移できるリガンドを有するラベル付CoA誘導体の使用により、本発明を、リガンドおよびタンパク質などのリガンドの結合相手を、特異的に融合タンパク質へ結合させるために用いることができることになる。もしリガンドがもう1つのタンパク質Yに結合し、タンパク質Yがラベルされた融合タンパク質と二量体を形成することが生物学的機能または測定可能な信号を生起させるならば、生物学的機能または測定可能な信号はラベルされたACP融合タンパク質の添加に依存する。もしACPが細胞表面に表示されているタンパク質に連結されているならば、リガンドとの相互作用が、リガンドによって修飾される他の分子(これにはリガンドによって修飾される、個々の生体分子または他の細胞、組織、および無傷の生物の一部としての生体分子、が含まれる)との接触を媒介する。
【0051】
ホスホパンテテイニル部分が担体の表面に共有結合的に結合している場合には、またはラベルが非共有結合的にそれ自身表面に結合している別の分子により結合され得る分子である場合には、ラベルされたCoA誘導体の使用によって、本発明を、固体支持体上にタンパク質配列(アレー)を構築するために使用することが可能になる。後者の手法の例は、ラベルがビオチンで、表面に結合している分子がストレプトアビジンまたはアビジンである。担体の可能な例はガラススライド、マイクロタイタープレートまたは任意の機能化ポリマーである。基質として用いられるCoA誘導体は担体上にラベルを介して固定化され、そして続くACPSが触媒するACP融合タンパク質の反応がラベルを融合タンパク質へ転移させ、担体上にそのような融合タンパク質を固定化させる。対応するCoA誘導体によって前処理された担体上に(様々な)ACP融合タンパク質をACPSと共に空間的に解像された態様でスポットすることによりタンパク質アレーの作製が可能になる。
【0052】
他のタンパク質に架橋することができる分子であるラベルにホスホパンテテイニル部分が共有結合的に結合している場合には、ラベルされたCoA誘導体の使用によって、本発明を、適当な環境における目的のタンパク質の相互作用を研究するために使用することが可能になる。そのような架橋リンカーの例は、マレイミド、活性エステルもしくはアジド、および熟練当業者に知られた他の分子、例えばNadeau et al.,2002に記載の分子などの官能基を含む分子である。そのようなラベルされたCoA誘導体をACPSの存在下で他のタンパク質と(生体内または生体外で)相互作用するACP融合タンパク質と接触させることによって、ACP融合タンパク質をそれが相互作用するタンパク質にラベルを介して共有結合的に架橋することができる。これによって、ACP融合タンパク質と相互作用するタンパク質の同定が可能になる。
【0053】
ホスホパンテテイニル部分がACPと膜レセプターから成るACP融合タンパク質へ転移されることが可能なリガンドを所持している場合の、ラベルされたCoA誘導体の使用は、ラベルされたレセプターの検出を可能にする。この細胞表面レセプターのラベリングはレセプターの内部移行の観察を可能にする。ある特別の応用はリガンド結合後の、例えば薬剤、新薬、または新薬候補によるレセプタータンパク質の内部移行の研究である。レセプターの内部移行は様々な方法により、例えば細胞膜から細胞内部へのラベルの移動を顕微鏡で検出することによって、またはラベルの蛍光の特徴が細胞内環境へ移動して変化することによって、検出できる。そのような変化はまた顕微鏡用具無しで検出することもできて、細胞外液に消光剤を加えることにより容易になる可能性がある。
【0054】
ホスホパンテテイニル部分が電子顕微鏡によって直接検出できるリガンド例えば電子密度の高いナノ粒子、を有するCoA誘導体の使用。ホスホパンテテイニル部分の電子顕微鏡による検出への別の応用は、例えばジアニシジンなどの芳香族アミンに光照射して酸化させ、高電子密度の沈殿を生じさせそれを次に電子顕微鏡で検出するためのエオシンなどの、光増感剤として使用されるラベルに基づいている。
【0055】
例として、本発明の実施態様を、添付した図を参照してここでより詳しく記載することにする。
【実施例】
【0056】
次の実施例および実験的手順は、当業者に本発明の完全な開示および実施方法の記載を提供するために記載するものであって、本発明の範囲の限定を意図するものではない。
【0057】
CoA−Btの合成
100μlのDMF中のビオチン-マレイミド(1mg、0.0022mmol)溶液に、90μl DMFおよび10μlの50mM Tris−HClpH7.5中の補酵素A二ナトリウム塩(1.79mg、0.0022mmol、1当量)溶液を加える。混合物を4時間室温で攪拌する。それを次にCH3CN/H2O 1:4で希釈し、小分けした500μlを調製用HPLCカラムに注入する:勾配(A=H2O 99%、CH3CN 1%、50mM NH4OAc/B=CH3CN)A/B95:5からA/B80:20までを2分間、A/B68:32までを7分間、A/B 20:80までを2分間行い、その後A/B 95:5に戻す。CoA−ビオチンの滞留時間は6.5分である。所望の生産物を含む分画を真空で濃縮し、DMSOに溶解し、そして純度を制御するために分析量を注入する。精製した分画を合わせる。CoA−ビオチンの濃度を260nmの吸収(ε(アデニン、260nm)=15,300[M-1cm-1])で決定する。CoA−Btの収量は0.895mg(33%)である。
ESI−MS(m/z)計算値1217.296[M(−1)]、測定値1217.2657[M(−1)]。
【0058】
【化4】
【0059】
CoA−Dgの合成
50μl DMF中のN−(ε−マレイミドカプロン酸)ヒドラジド2(1mg、0.0044mmol)溶液に、DMF/緩衝液混合液(70μl DMF/30μl 50mM Tris−Cl、pH7.5)中の補酵素A二ナトリウム塩(3.6mg、0.0044mmol、1当量)溶液を加える。反応を分析用HPLCによって追跡し(260nmで検出)反応の終了を証明する。続いて、50μl DMFおよび10μl Et3Nに溶解した3−アミノ−3−デオキシジゴキシゲニン ヘミスクシンアミド スクシンイミジル エステル(2.6mg、0.0044mmol、1当量)溶液を加え、反応混合液を4時間室温で攪拌する。反応物を調製用HPLCに掛け、望みの生成物を含む分画を真空で濃縮し、DMSOに溶解し、そして純度を分析用HPLCによって分析する。精製した生成物を含む分画を合わせる。
CoA−Dgの濃度をアデニンの吸光係数(ε(アデニン、260nm)=15,300[M-1cm-1])を使って決定する。CoA−Dgの収量は2.37mg(37%)である。
ESI−MS(m/z)計算値1462.3707[M(−1)]、測定値1462.481M(−1)]。
【0060】
【化5】
【0061】
CoA−Cy3の合成
100μl DMF中のCy3−マレイミド3(Pharmacia、1mg、0.00126mmol)溶液に、90μl DMFおよび10μl 50mM Tris−ClpH7.5中の補酵素A二ナトリウム塩(1.05mg、0.00126mmol、1当量)溶液を加える。混合液を4時間室温で攪拌する。それを次にCH3CN/H2O 1:4で希釈し、小分けした500μlを調製用HPLCカラムに注入する。勾配、A/B95:5からA/B90:10までを2分間、A/B 65:35までを15分間、A/B20:80までを2分間行い、その後A/B 95:5に戻す(AおよびBについては先の実施例参照)。CoA−Cy3の滞留時間は10分である。所望の生産物を含む分画を真空で濃縮し、DMSOに溶解し、そして純度を制御するために分析量を注入する。精製した分画を合併する。CoA−Cy3の濃度を549nmの吸収(ε(549nm)=150,000[M-1cm-1])で決定する。CoA−Cy3の収量は0.847mg(44%)である。
ESI−MS(m/z)計算値1519.370[M(-1)]、測定値1519.3071[M(-1)]。
【0062】
【化6】
【0063】
CoA−Cy5の合成
Cy5−マレイミド4(1mg、0.00122mmol)から出発するCoA−Cy5の合成をCoA−Cy3について記述したのと同様に行う。CoA−Cy5の濃度をCy5の吸光係数(ε(646nm)=250,000[M-1cm-1])を用いて決定する。CoA−Cy5の収量は0.828mg(44%)である。
ESI−MS(m/z)計算値1545.386[M(-1)]および772.189[M(-2)]、測定値771.6524[M(-2)]。
【0064】
【化7】
【0065】
6×His−ACPSのクローニング、発現、および精製
XL1−blue E.coliのACPS遺伝子を単一コロニーPCRによって増幅し、pET−15bプラスミド(Novagen)中にNdel制限部位(下線)付きの順方向プライマー5’−TCT GGT CAT ATG GCA ATA TTA GGT TTA GGC ACG G−3’およびXhol制限部位(下線)付きの逆方向プライマー5’−TCA AGT CTC GAG TTA ACT TTC AAT AAT TAC CGT GGC A−3’を用いてクローニングする。E.coliのACPSのN−末端に融合されたペプチドの配列(プラスミドpET−15b起源)は(1文字コードで)MGSSHHHHHHSSGLVPRGSHであり、これにACPSの最初のアミノ酸、メチオニン、が続く。この融合タンパク質を6×His−ACPSと呼ぶ。
【0066】
6×His−ACPSをコードするpET−15b(Novagen)に基づく発現ベクターを含むBL21 E.coli細胞の液体培養を吸光度OD600nm0.6となるまで成長させる。6×His−ACPSの発現を最終濃度1mMとなるようにIPTGを加えて誘導する。3.5時間220rpm、24℃でインキュベーションした後、培養物を10分間3000g 4℃で遠心する。沈殿を10mlの抽出緩衝液(150mM NaCl、5mMイミダゾール、50mM KH2PO4、pH8.0)に再懸濁し、PMSFとアプロチミンをそれぞれ最終濃度1mMおよび2μg/mlとなるように加える。リゾチームを1mg/mlとなるように加え、混合物を氷上で15分間インキュベーションしまた何度か反転させる。次に、それを10分間超音波で破壊する(95%出力、50%動作)。DNA分解酵素Iを最終濃度0.01mg/mlとなるよう加える。4℃で30分後、混合液を10分間18,000rpmで遠心する。
【0067】
タンパク質を精製するために、予め抽出緩衝液で3回洗浄しておいた350μlのNi−NTAを溶解物に加える。混合物を20分間氷上でインキュベーションし数度混ぜ合わせた。抽出混合物を次にプロピレンカラムに加え、流出に任せた。カラムを5×400μlのDNA溶出緩衝液(10mM Tris Cl、pH8.5)で洗い、次に2×5ml洗浄緩衝液(300mM NaCl、10mMイミダゾール、50mM KH2PO4、pH7.5)で洗浄する。タンパク質を流出させるために、溶出緩衝液(300mM NaCl、150mMイミダゾール、pH7.5)をカラムに加え、10分間インキュベーションし、次に流出液を集める。溶出は、150μl溶出緩衝液で段階的に続け、これ以上タンパク質がブラッドフォード(Bradford)分析で検出できなくなるまで行う。最後に合わせた溶出液を透析緩衝液(50mM HEPES、30%グリセロール、pH7.2)中で終夜透析し、残存する塩を除く。精製した6×His−ACPS(MW16.215kDa)を小分けにして−80℃で保存した。濃度をブラッドフォード分析で37.6μMと決定する。収量はBL21E.coli細胞培養1リットル当たり0.912mg。
【0068】
6×His−ACPのクローニング、発現、および精製
XL1−blue E.coliのACP遺伝子を、Ndel制限部位(下線)付きの順方向プライマー5’−GT CGG TAT CAT ATG AGC ACT ATC GAA GAA CG−3’、およびBamHI制限部位(下線)付きの逆方向プライマー5’−TCA TGC GGA TCC TTA CGC CTG GTG GCC GTT G−3’、を用いて、単一コロニーPCRによって増幅し、pET−15bプラスミド(Novagen)中にクローニングした。E.coliのACPのN−末端に融合されたペプチドの配列(6×His−ACPを生成する)は(1文字コードで)MGSSHHHHHHSSGLVPRGSHであり、これにACPの最初のアミノ酸、メチオニン、が続く。
【0069】
6×His−ACPをコードするpET−15b(Novagen)に基づく発現ベクターを含むBL21E.coli細胞の液体培養を吸光度OD600nm0.6となるまで成長させる。6×His−ACPの発現を最終濃度1mMとなるようにIPTGを加えて誘導する。3.5時間220rpm、37℃でインキュベーションした後、培養物を10分間3000g4℃で遠心する。6×His−ACPの精製のために沈殿を正確に上の実験で記述した通りに操作する。6×His−ACP(MW10.802kDa)の濃度をブラッドフォード分析で188μMと決定する。収量:BL21E.coli細胞培養1リットル当たり3.1mg。
【0070】
アポ-およびホロ-6×His−ACP間の比率を決定するために、精製した6×His−ACPをQ−Tof−Ultima(Micromass/Waters)を用いたESI−MS(pos.モード)により、場合によっては、Cap−LC(Waters)、Xterra RP−C4カラム(Waters,5μm、0.32×50mm;流速8μl/min)によるクロマトグラフィーに連結し、MaxEnt1−ソフトウエアによる逆畳み込みを行って、解析する。混合物のESI−MSが、調製品はホロ-およびアポ-ACPの混合物であることを証明した。最初のメチオニンを除く精製したアポ-6×His−ACPの質量は10.6720kDaであることが分かり(計算値10.6716kDa)、またホロ-6×His−ACPの質量は11.012kDa(計算値11.0119kDa)であることが分かる。LC−ESI−MSによって、ホロ-およびアポ-型に対応するピークの同定と積分が可能になる。ホロ-型の100分率は16%(滞留時間11.77分)、またアポ-型のそれは84%(滞留時間14.16分)である。
【0071】
6×His−ACP−haの発現および精製
E.coli ACPのN−末端に融合されたペプチドの配列は(1文字コードで)MGSSHHHHHHSSGLVPRGSHであり、これにACPの最初のアミノ酸、メチオニン、が続き、E.coli ACPのC−末端に融合されたペプチドの配列は(1文字コードで)TSRSYPYDVPDYARW(6×His−ACP−haを生成する)である。
【0072】
6×His−ACP−haをコードするpET−15bに基づく発現ベクターを含むBL21(DE3)E.coli細胞の液体培養を吸光度OD600nm0.6となるまで成長させる。6×His−ACP−haの発現を最終濃度1mMとなるようにIPTGを加えて誘導する。220rpm、24℃で3時間インキュベーションした後、培養物を3000g4℃で10分間遠心する。6×His−AcpSの精製に対して記述した通りに、6×His−ACPを精製する。6×His−ACP−ha(MW12.65kDa)の濃度をブラッドフォード分析で400μMと決定する。またタンパク質の総収量は振とうフラスコ培養1リットル当たり10mgである。
【0073】
CoA−Btおよび6×His−ACPSを用いた6×His−ACPの生体外ビオチン化
精製した6×His−ACP(1μM)を反応緩衝液(43μl、50mM Tris ClpH8.8)中で6×His−ACPS(0.2μM)およびMgCl2(10mM)と室温でインキュベートさせる。7.5μlを解析用に分取する。CoA−Btを最終濃度5μMとなるように加える。一定時間ごとに各7.5μlを分取する。分取したものを補酵素A(最終濃度1mM)で30秒間クエンチし、8.2μlSDS緩衝液2×を加える。試料を2分間95℃で加熱する。ビオチン化された6×His−ACPをストレプトアビジン-西洋わさびペルオキシダーゼ結合体(NEM)および化学発光ペルオキシダーゼ基質(Renaissance reagent plus, NEN)を用い、ウエスタン-ブロッティングにより検出する。ウエスタン-ブロットをイメージステーション(Kodak440)を用いて解析する。対照として、バックグラウンドをチェックし、また反応の特異性をチェックするために、各タンパク質1つのみを含む試料、および2つのタンパク質のただ1つおよびCoA−Btを含む試料も調製しビオチン化を上のように分析する。ビオチン化は3つの成分全ての存在に依存する。
【0074】
ゲルシフトアッセイによる6×His−ACPのビオチン化の定量化
精製した6×His−ACP(3μM)を室温でCoA−Bt(10μM)および精製した6×His−ACPS(5μM)と最終容積50μlの反応緩衝液(50mM Tris Cl、pH8.8、10mM MgCl2)中でインキュベートする。30分インキュベーション後、混合液を終夜TBS((10mM Tris Cl、150mM NaCl、pH7.9)に対して透析し、過剰のCoA−Btを除く。小分けした反応液を、最終濃度0.6μg/μlのストレプトアビジンと1時間インキュベートする。2%SDSのみを含む2×SDS試料緩衝液を試料に加え、そして試料を加熱せずにSDS−PAGEを直接行う。タンパク質をクーマシー染色で検出し、またビオチン化の程度を、ビオチン化されておらず、ストレプトアビジンとインキュベートしていない等量の6×His−ACPを含む試料とバンド強度を比較して推定する。
【0075】
CoAとCoA−ラベル間の競合の定量
精製された6×His−AcpS(0.4μM)を、室温で、CoA−BtまたはCoA−Dg(2μM)、種々の量のCoA(0.2、4、8、12、20、40、80μM)および精製6×His−ACP−ha(0.4μM)と20μl反応緩衝液(50mM Tris・Cl、pH7.5、10mM MgCl2)中でインキュベートする。25分後、各試料を20μlの2×SDS試料緩衝液を加えてクエンチし、95℃2分間加熱した。ラベルされた6×His−ACP−haを、ストレプトアビジン-西洋わさびペルオキシダーゼ結合体(1:12500希釈)または抗−ジゴキシゲニン抗体−西洋わさびペルオキシダーゼ結合体(1:500希釈)および化学発光ペルオキシダーゼ基質を用い、ウエスタン-ブロッティングにより検出する。各実験に対して、0μM CoAの信号強度を任意に1に設定する。
【0076】
酵母細胞表面上へのAga2−ACPのラベリング
酵母ゲノムDNA由来のAGA2配列を増幅し、PCR断片の両端に導入したEcoRIおよびSall制限部位を利用して、酵母発現ベクターpRS314のPcup1−プロモーター配列の後へ挿入する。ACP配列のPCR増幅によって導入したSallおよびAcc651制限部位を用いて、ACPをAGA2のうしろのフレームへクローニングする。ACPはHA−エピトープをコードする配列により延長される。Aga2pおよびACPを連結する配列は:FVDEMLYFQGMである。Aga2pの最後の残基とACPの最初の残基とに下線を付す。HA−エピトープを含むACPのC−末端配列は:QAYPYDVPDYAGである。ACPの最後の残基に下線を付す。PGAL1−プロモーター由来のAga1pとPCUP1−プロモーター由来のAga2−ACPとを発現する酵母系統EBY100(MATa ura3−52 trip1 leu2Δ1 his3Δ200 pep4::HIS3 can1 GAL pIU211:URA3)(Invitrogen, Carlsbad, CA)を、2%ガラクトースおよび0.1mM銅を含む選択培地10ml中で、細胞が1.4OD600単位のOD600になるまで生育したものを、2mlの水で洗い、0.2mlのラベリング緩衝液(50mM Tris Cl pH8.8、100mM NaCl、10mM MgCl2)に再懸濁する。CoA基質および6×His−ACPSをそれぞれ最終濃度10および1μM加える。室温で30分後、反応液を2mlPBSに希釈することによりラベリングを止める。細胞を2mlPBSで4回洗い、直接蛍光顕微鏡検査に付すか、または、20nM Qdot(登録商標)605ストレプトアビジン結合体(Milan Analytic AG, Switzerland)を含む0.2mlのQdot(登録商標)インキュベーション緩衝液中で20分間インキュベート後細胞を2mlPBSで4回洗浄して蛍光顕微鏡検査に付す。細胞を、63×油浸(開口数1.4)対物レンズを用いて、ツアイスAxiovert135蛍光顕微鏡(Carl Zeiss, Gottingen, Germany)で検査する。
【0077】
HEK293細胞上に表示されるACP−NK1融合タンパク質のラベリング
ACP−NK1の一時的発現のために、5−HT3−レセプターの信号配列(Sig5HT3)を短いDYVリンカーを介してACPのN−末端に融合し、NK1を短いTSリンカーを介してACPのC−末端に融合する。その結果の構成において、FLAGタグおよび6×HisタグもまたNK1のC−末端に結合させる。融合タンパク質に対応する遺伝子をpCEP4(Invitrogen)ベクターのNheIおよびBamHI部位に挿入する。HEK293細胞を2.2%ウシ胎児血清(GIBCO, BRL)を補充したDMEM/F12(Dulbeccoの修正Eagle培地; GIBCO BRL)中で生育させる。一時的トランスフェクションを記載(Nat Biotechnol 21,86-89 (2003))通りに行い、HEK293細胞を、ACP−NK1を発現するベクターと核を標的とするEGFP(EGFP−NLS3)を発現するベクターにより共トランスフェクションする。24時間後、細胞を10分間室温で、MgCl2(10mM)、6×His−AcpS(1μM)、およびCoA−Cy3、CoA−Cy5、またはCoA−Bt(各5μM)のいずれかを含むPBS緩衝液500μlとインキュベートする。細胞を次に過剰の基質を除くためにPBSで3回洗浄し、Cy3またはCy5でラベルする場合は直接にレーザー走査共焦点蛍光顕微鏡によって分析する。ビオチン化された細胞を、PBS中1μg/ml濃度のFluoroLink(登録商標)Cy5−ラベルしたストレプトアビジン(Amersham Biosciences)とインキュベートした後、PBSで3回洗浄する。レーザー走査共焦点蛍光顕微鏡像を、63×水浸(開口数1.2)対物レンズ付きのZeissLSM510顕微鏡(Carl Zeiss AG, Gottingen, Germany)の488nmアルゴン/クロプトンレーザー線、543nm HeNEレーザー線、または633nm HeNEレーザー線を用いて記録する。走査速度およびレーザー強度を蛍光プローブの光退色および細胞の損傷または形態学的変化を回避するように調整する。蛍光を、GFPに敏感なチャネルおよびそれぞれの色素に敏感なチャネルで解析する。試験した各細胞試料に対して、GFPチャネルにおいて明確な核ラベリングが観察され、また細胞膜領域において明確な膜ラベリングが、CoA−Cy3ラベル後、CoA−Cy5ラベル後、およびCoA−Btラベル後であって、かつそれに続くストレプトアビジン-Cy5染色後に観察される。膜染色はトランスフェクションされない細胞では観察されず、GFPは核内発現されないことを示しており、これはそれぞれCy3、Cy5、およびビオチンによるACP−NK1の特異的ラベリングを示している。一時的にHEK293細胞に発現されたACP−NK1レセプターをさらなる実験で、CoA−Cy5およびテトラメチルローダミンでラベルされたサブスタンスP(SP−rho)、即ちNK1の自然のリガンド、で共染色する。両物質はもっぱら膜領域を染色し、同一の染色領域となる。過剰のサブスタンスPによって1分以内にSP−rho染色が元に戻ることが、さらにラベリングの特異性とまたACP−NK1のリガンド結合機能性をも証明する。
【0078】
【表1】
【図面の簡単な説明】
【0079】
【図1】(A)6×His−ACP(1μM)、6×His−ACPS(0.2μM)およびCoA−Bt(5μM)の反応の解析:指定した時間(t)に一定量を反応混合液から分取し、ストレプトアビジン-ペルオキシダーゼ結合体を用いるウエスタンブロットにより6×His−ACPのビオチン化を精査する。(B)6×His−ACPのビオチン化の定量化:6×His−ACP(3μM)、6×His−ACPS(5μM)およびCoA−Bt(10μM)を30分間インキュベートし、透析し、分取した試料をストレプトアビジンとインキュベートしてSDS−PAGEに付す(第2列)。安定なビオチン-ストレプトアビジン複合体の形成がビオチン化タンパク質のゲルシフトをもたらす。ビオチン化の量を、第2列の6×His−ACPのバンド強度を、等濃度の6×His−ACPを含むがストレプトアビジンの無い試料(第1列)のバンド強度と比較して評価する。(C)CoAとCoA−Bt間のまたはCoAとCoA−Dg間の、ACPSの基質としての競合の定量:6×His−ACP(0.4μM)、6×His−ACPS(0.4μM)、CoA−BtまたはCoA−Dg(2μM)、および種々の濃度のCoA(0〜80μM)を30分間インキュベートし、ラベリングの程度をウエスタンブロットにより決定する。ウエスタンブロットの相対信号強度(I)を[CoA]/[CoA-L](L=ラベル)に対してプロットし、等式I=A/(1+BX)にフィットさせる。ここで、Xは[CoA]/[CoA-L]、Aは特定しない定数、Bは比(kcat/KM)CoA/(kcat/KM)CoA-Lである。これらの実験より、特異性定数の比が(kcat/KM)CoA/(kcat/KM)CoA-Btに対して0.48、(kcat/KM)CoA/(kcat/KM)CoA-Dgに対して0.35という結果となり、フリーのCoAと誘導体化されたCoAの間で重大な区別は無いことを明らかにする。
【図2】(A〜F)酵母の細胞表面上のACP融合タンパク質のラベリング。Cy3(A)またはストレプトアビジンでコートされた量子ドットにより追跡したビオチン(D)によってラベルされたAga2−ACPを発現した酵母細胞の蛍光顕微鏡写真。(B)は(A)と、(D)は(E)と同じであるが、但しAGA2−ACPを発現していない細胞である。(C)および(F)はそれぞれ(B)および(D)と同一の試料の透過顕微鏡写真である。これらの実験は、Aga2p−ACPを発現している酵母細胞のみがラベルされることを示す。
【特許請求の範囲】
【請求項1】
目的のタンパク質を検出し、および/または操作するための方法であって、目的のタンパク質およびアシル担体タンパク質(ACP)またはその断片を含む融合タンパク質を、ラベルされた補酵素A(CoA)型基質およびホロ-アシル担体タンパク質合成酵素(ACPS)もしくはその相同物と接触させ、その結果ACPSがラベルを融合タンパク質へ転移させること、ならびにそのラベルを検出しおよび/または操作するために設計されたシステムにおいてそのラベルを用いて、得られたそのラベルされた融合タンパク質を検出しおよび/またはさらに操作すること、を含む方法。
【請求項2】
得られたラベルされた融合タンパク質を、生体外もしくは生体内でラベルを連続的にモニターするために設計されたシステムにおいて連続的にモニターすることができる、請求項1記載の、目的のタンパク質を検出するための方法。
【請求項3】
請求項2記載の、生体外で目的のタンパク質を検出するための方法。
【請求項4】
請求項2記載の、生体内の細胞中の目的のタンパク質を検出するための方法。
【請求項5】
ラベルされた融合タンパク質を、ラベルを精製するために設計されたシステム中で精製し、およびさらなる工程において目的のタンパク質をラベルされた融合タンパク質から切断する、請求項1記載の、目的のタンパク質を操作するための方法。
【請求項6】
ラベルされた融合タンパク質を、ラベルを固定するために設計されたシステム中で固体支持体に固定する、請求項1記載の、目的のタンパク質を操作するための方法。
【請求項7】
ACPSがE.coli由来のACPSである、請求項1記載の、目的のタンパク質を操作するための方法。
【請求項8】
ACPS相同物が、EntD、Sfp、Psf−1、Gsp、LYS5、Bli、Lpa−14、およびノシヘプチド生合成に関与するNshCから成る群より選択される、請求項1記載の、目的のタンパク質を操作するための方法。本発明はラベリングスキームにおけるE.coli由来のACPSと相同的な、全てのACPSの使用を含む。
【請求項9】
ACPがE.coliのアシル担体タンパク質である、請求項1記載の、目的のタンパク質を操作するための方法。
【請求項10】
ACPがStreptomyces種由来である、請求項1記載の、目的のタンパク質を操作するための方法。
【請求項11】
ACPが野生型ACPの変異体であって、1個以上のアミノ酸置換、欠失または付加により野生型ACPと異なるが、ACPSにより触媒される反応において、ラベルされた4’−ホスホパンテテインに対するアクセプターとして働く性質を依然として保持している、請求項1記載の、目的のタンパク質を操作するための方法。
【請求項12】
ACPが、ホスホパンテテイン誘導体と結合するセリン残基を含むACP断片であって、かつそのようなホスホパンテテイン誘導体を受容する機能を保持している、請求項1記載の、目的のタンパク質を操作するための方法。
【請求項13】
目的のタンパク質が12〜5000個のアミノ酸から成ることを特徴とする、目的のタンパク質およびアシル担体タンパク質(ACP)またはその断片を含む融合タンパク質。
【請求項14】
目的のタンパク質が50〜1000個のアミノ酸から成ることを特徴とする、請求項13記載の融合タンパク質。
【請求項15】
目的のタンパク質が、酵素、DNA結合タンパク質、転写調節タンパク質、膜タンパク質、核レセプタータンパク質、核局在信号タンパク質、タンパク質補助因子、抗体、膜ポンプタンパク質、膜チャネルタンパク質、膜担体タンパク質、運動タンパク質、シグナル伝達に関与するタンパク質、核タンパク質、リボソームタンパク質、小単量体GTPアーゼ、ATP結合カセットタンパク質、細胞内構造タンパク質、タンパク質を特定の細胞内区画へ向かわせる原因である配列を有するタンパク質、一般にラベルとしてまたは親和性タグとして用いられるタンパク質、およびそのようなタンパク質のドメインもしくはサブドメインから成る群より選択されることを特徴とする、請求項13記載の融合タンパク質。
【請求項16】
目的のタンパク質が、ヒト、他の高等哺乳動物、真核生物種、バクテリア種、またはウイルス源に由来することを特徴とする、請求項13記載の融合タンパク質。
【請求項17】
目的のタンパク質が、サルモネラ、ミコバクテリア、ブドウ球菌、ヒト免疫不全ウイルス(HIV)、ヒトインフルエンザウイルス、肝炎ウイルス、またはコロナウイルスに由来することを特徴とする、請求項13記載の融合タンパク質。
【請求項18】
目的のタンパク質が、ガン、心臓血管病、精神障害、アルツハイマー、肥満、ウイルス感染、またはバクテリア感染において役割を果たすことを特徴とする、請求項13記載の融合タンパク質。
【請求項19】
適当な条件下において酵素切断を受けやすいリンカーを通じて連結されているACPおよび目的のタンパク質を含むことを特徴とする、請求項13記載の融合タンパク質。
【請求項20】
ACPおよびニューロキニン−1レセプターを含む請求項13記載の融合タンパク質。
【請求項21】
請求項1記載の方法における請求項13記載の融合タンパク質の使用。
【請求項22】
式(I)または(II):
【化1】
(式中、Rは補酵素Aとラベルを架橋するリンカー基であり;「ラベル」は、ラベルを検出し、および/または操作するために設計されたシステムにおいて融合タンパク質の検出および/または操作に適したラベル分子である)
のラベルされたCoA型基質。
【請求項23】
請求項22記載のラベルされたCoA型基質であって、Rは、1〜300個の炭素原子を有する直鎖または分岐鎖アルキレン基であり、場合によっては
(a)1個以上の炭素原子が酸素により置き換えられており;
(b)1個以上の炭素原子が水素原子を持つ窒素原子により置き換えられ、また隣接する炭素原子がオキソにより置き換えられて、アミド官能基−NH−CO−を表し;
(c)1個以上の炭素原子が酸素により置き換えられ、また隣接する炭素原子がオキソにより置き換えられて、エステル官能基−O−CO−を表し;
(d)2個の隣接する炭素原子間の結合が2重または3重結合であって、官能基−CH=CH−または−C≡C−を表し;
(e)1個以上の炭素原子がフェニレン、飽和または不飽和シクロアルキレン、飽和または不飽和ビシクロアルキレン、架橋ヘテロ芳香族または架橋飽和または不飽和ヘテロシクリル基により置き換えられており;
(f)2個の隣接する炭素原子がジスルフィド結合−S−S−により置き換えられており;または、場合によっては置換基を含む、2個以上の、特に2個の、アルキレン基および/または上の(a)〜(f)で定義された修飾されたアルキレン基の組み合わせである、請求項22記載のラベルされたCoA型基質。
【請求項24】
請求項22記載のラベルされたCoA型基質であって、「ラベル」が次:
(1)分光学的プローブ;
(2)放射性ラベルされた分子;
(3)相手と特異的に結合できる特異的な結合対の一方である分子。
(4)他の生体分子と相互作用すると想定される分子;
(5)他の生体分子と相互作用すると想定される分子のライブラリー;
(6)他の生体分子と架橋できる分子;
(7)H2O2とアスコルビン酸に曝露されるとヒドロキシルラジカルを生成できる分子;
(8)光照射により反応性ラジカルを生成できる分子;
(9)固形支持体に共有結合した分子;
(10)相補的な鎖と塩基対を形成できる核酸またはその誘導体;
(11)リピッドまたは膜に挿入される性質を有する他の疎水性分子;
(12)望ましい酵素学的、化学的、または物理的性質を有する生体分子;および
(13)(1)〜(12)の性質の任意の組み合わせを有する分子、
からなる群より選択される、請求項22記載のラベルされたCoA型基質。
【請求項25】
「ラベル」がビオチン、ジゴキシゲニン、Cy−3、およびCy−5から選択される、請求項22記載のラベルされたCoA型基質。
【請求項26】
請求項1記載の方法における請求項22記載のラベルされたCoA型基質の使用。
【請求項1】
目的のタンパク質を検出し、および/または操作するための方法であって、目的のタンパク質およびアシル担体タンパク質(ACP)またはその断片を含む融合タンパク質を、ラベルされた補酵素A(CoA)型基質およびホロ-アシル担体タンパク質合成酵素(ACPS)もしくはその相同物と接触させ、その結果ACPSがラベルを融合タンパク質へ転移させること、ならびにそのラベルを検出しおよび/または操作するために設計されたシステムにおいてそのラベルを用いて、得られたそのラベルされた融合タンパク質を検出しおよび/またはさらに操作すること、を含む方法。
【請求項2】
得られたラベルされた融合タンパク質を、生体外もしくは生体内でラベルを連続的にモニターするために設計されたシステムにおいて連続的にモニターすることができる、請求項1記載の、目的のタンパク質を検出するための方法。
【請求項3】
請求項2記載の、生体外で目的のタンパク質を検出するための方法。
【請求項4】
請求項2記載の、生体内の細胞中の目的のタンパク質を検出するための方法。
【請求項5】
ラベルされた融合タンパク質を、ラベルを精製するために設計されたシステム中で精製し、およびさらなる工程において目的のタンパク質をラベルされた融合タンパク質から切断する、請求項1記載の、目的のタンパク質を操作するための方法。
【請求項6】
ラベルされた融合タンパク質を、ラベルを固定するために設計されたシステム中で固体支持体に固定する、請求項1記載の、目的のタンパク質を操作するための方法。
【請求項7】
ACPSがE.coli由来のACPSである、請求項1記載の、目的のタンパク質を操作するための方法。
【請求項8】
ACPS相同物が、EntD、Sfp、Psf−1、Gsp、LYS5、Bli、Lpa−14、およびノシヘプチド生合成に関与するNshCから成る群より選択される、請求項1記載の、目的のタンパク質を操作するための方法。本発明はラベリングスキームにおけるE.coli由来のACPSと相同的な、全てのACPSの使用を含む。
【請求項9】
ACPがE.coliのアシル担体タンパク質である、請求項1記載の、目的のタンパク質を操作するための方法。
【請求項10】
ACPがStreptomyces種由来である、請求項1記載の、目的のタンパク質を操作するための方法。
【請求項11】
ACPが野生型ACPの変異体であって、1個以上のアミノ酸置換、欠失または付加により野生型ACPと異なるが、ACPSにより触媒される反応において、ラベルされた4’−ホスホパンテテインに対するアクセプターとして働く性質を依然として保持している、請求項1記載の、目的のタンパク質を操作するための方法。
【請求項12】
ACPが、ホスホパンテテイン誘導体と結合するセリン残基を含むACP断片であって、かつそのようなホスホパンテテイン誘導体を受容する機能を保持している、請求項1記載の、目的のタンパク質を操作するための方法。
【請求項13】
目的のタンパク質が12〜5000個のアミノ酸から成ることを特徴とする、目的のタンパク質およびアシル担体タンパク質(ACP)またはその断片を含む融合タンパク質。
【請求項14】
目的のタンパク質が50〜1000個のアミノ酸から成ることを特徴とする、請求項13記載の融合タンパク質。
【請求項15】
目的のタンパク質が、酵素、DNA結合タンパク質、転写調節タンパク質、膜タンパク質、核レセプタータンパク質、核局在信号タンパク質、タンパク質補助因子、抗体、膜ポンプタンパク質、膜チャネルタンパク質、膜担体タンパク質、運動タンパク質、シグナル伝達に関与するタンパク質、核タンパク質、リボソームタンパク質、小単量体GTPアーゼ、ATP結合カセットタンパク質、細胞内構造タンパク質、タンパク質を特定の細胞内区画へ向かわせる原因である配列を有するタンパク質、一般にラベルとしてまたは親和性タグとして用いられるタンパク質、およびそのようなタンパク質のドメインもしくはサブドメインから成る群より選択されることを特徴とする、請求項13記載の融合タンパク質。
【請求項16】
目的のタンパク質が、ヒト、他の高等哺乳動物、真核生物種、バクテリア種、またはウイルス源に由来することを特徴とする、請求項13記載の融合タンパク質。
【請求項17】
目的のタンパク質が、サルモネラ、ミコバクテリア、ブドウ球菌、ヒト免疫不全ウイルス(HIV)、ヒトインフルエンザウイルス、肝炎ウイルス、またはコロナウイルスに由来することを特徴とする、請求項13記載の融合タンパク質。
【請求項18】
目的のタンパク質が、ガン、心臓血管病、精神障害、アルツハイマー、肥満、ウイルス感染、またはバクテリア感染において役割を果たすことを特徴とする、請求項13記載の融合タンパク質。
【請求項19】
適当な条件下において酵素切断を受けやすいリンカーを通じて連結されているACPおよび目的のタンパク質を含むことを特徴とする、請求項13記載の融合タンパク質。
【請求項20】
ACPおよびニューロキニン−1レセプターを含む請求項13記載の融合タンパク質。
【請求項21】
請求項1記載の方法における請求項13記載の融合タンパク質の使用。
【請求項22】
式(I)または(II):
【化1】
(式中、Rは補酵素Aとラベルを架橋するリンカー基であり;「ラベル」は、ラベルを検出し、および/または操作するために設計されたシステムにおいて融合タンパク質の検出および/または操作に適したラベル分子である)
のラベルされたCoA型基質。
【請求項23】
請求項22記載のラベルされたCoA型基質であって、Rは、1〜300個の炭素原子を有する直鎖または分岐鎖アルキレン基であり、場合によっては
(a)1個以上の炭素原子が酸素により置き換えられており;
(b)1個以上の炭素原子が水素原子を持つ窒素原子により置き換えられ、また隣接する炭素原子がオキソにより置き換えられて、アミド官能基−NH−CO−を表し;
(c)1個以上の炭素原子が酸素により置き換えられ、また隣接する炭素原子がオキソにより置き換えられて、エステル官能基−O−CO−を表し;
(d)2個の隣接する炭素原子間の結合が2重または3重結合であって、官能基−CH=CH−または−C≡C−を表し;
(e)1個以上の炭素原子がフェニレン、飽和または不飽和シクロアルキレン、飽和または不飽和ビシクロアルキレン、架橋ヘテロ芳香族または架橋飽和または不飽和ヘテロシクリル基により置き換えられており;
(f)2個の隣接する炭素原子がジスルフィド結合−S−S−により置き換えられており;または、場合によっては置換基を含む、2個以上の、特に2個の、アルキレン基および/または上の(a)〜(f)で定義された修飾されたアルキレン基の組み合わせである、請求項22記載のラベルされたCoA型基質。
【請求項24】
請求項22記載のラベルされたCoA型基質であって、「ラベル」が次:
(1)分光学的プローブ;
(2)放射性ラベルされた分子;
(3)相手と特異的に結合できる特異的な結合対の一方である分子。
(4)他の生体分子と相互作用すると想定される分子;
(5)他の生体分子と相互作用すると想定される分子のライブラリー;
(6)他の生体分子と架橋できる分子;
(7)H2O2とアスコルビン酸に曝露されるとヒドロキシルラジカルを生成できる分子;
(8)光照射により反応性ラジカルを生成できる分子;
(9)固形支持体に共有結合した分子;
(10)相補的な鎖と塩基対を形成できる核酸またはその誘導体;
(11)リピッドまたは膜に挿入される性質を有する他の疎水性分子;
(12)望ましい酵素学的、化学的、または物理的性質を有する生体分子;および
(13)(1)〜(12)の性質の任意の組み合わせを有する分子、
からなる群より選択される、請求項22記載のラベルされたCoA型基質。
【請求項25】
「ラベル」がビオチン、ジゴキシゲニン、Cy−3、およびCy−5から選択される、請求項22記載のラベルされたCoA型基質。
【請求項26】
請求項1記載の方法における請求項22記載のラベルされたCoA型基質の使用。
【図1】

【図2】
【図2】
【公表番号】特表2007−509608(P2007−509608A)
【公表日】平成19年4月19日(2007.4.19)
【国際特許分類】
【出願番号】特願2006−530696(P2006−530696)
【出願日】平成16年5月19日(2004.5.19)
【国際出願番号】PCT/IB2004/001733
【国際公開番号】WO2004/104588
【国際公開日】平成16年12月2日(2004.12.2)
【出願人】(503183293)ウペエフエル・エコル・ポリテクニック・フェデラル・ドゥ・ローザンヌ (11)
【氏名又は名称原語表記】EPFL ECOLE POLYTECHNIQUEFEDERALE DE LAUSANNE
【Fターム(参考)】
【公表日】平成19年4月19日(2007.4.19)
【国際特許分類】
【出願日】平成16年5月19日(2004.5.19)
【国際出願番号】PCT/IB2004/001733
【国際公開番号】WO2004/104588
【国際公開日】平成16年12月2日(2004.12.2)
【出願人】(503183293)ウペエフエル・エコル・ポリテクニック・フェデラル・ドゥ・ローザンヌ (11)
【氏名又は名称原語表記】EPFL ECOLE POLYTECHNIQUEFEDERALE DE LAUSANNE
【Fターム(参考)】
[ Back to top ]