
アジュバントワクチン製剤
本発明は、アジュバントワクチン製剤、特に鼻腔内送達のためのインフルエンザワクチンに関する。提供されるのは、(i)グラム陽性細菌から得られるペプチドグリカン微粒子と、(ii)少なくとも1つのインフルエンザウイルス抗原またはこの抗原性調製物とを含み、該抗原または抗原性調製物がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない、アジュバントインフルエンザワクチン製剤である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、アジュバントワクチン製剤、特に、例えば鼻腔内または筋肉内送達の際に粘膜性免疫応答を高めることができるインフルエンザワクチンに関する。
【背景技術】
【0002】
季節性インフルエンザは、依然として世界中で死亡率および罹患率の主な原因の一つである。年に1回のワクチン接種は、インフルエンザ感染を予防および制御する最も有効な戦略である。季節性インフルエンザワクチンは、来シーズンの流行予測株の予想に基づいて調製される。これらは、感染自体を予防しない非経口注射ワクチンであり、感染後の重症度および合併症を減少させる。非経口ワクチンは、血清中に中和IgG抗体を誘導することができるが、これらは、粘膜表面で作用する分泌IgA抗体を誘導することができない。対照的に、鼻腔内(i.n.)ワクチンは、全身性および粘膜性免疫応答の両方を誘導する可能性がある。粘膜表面の分泌IgA抗体は、インフルエンザウイルス感染の最初の標的である上皮細胞表面に病原体が結合する前に粘膜の表面で反応するため、感染を予防するのに極めて有効である。さらに、血清IgG抗体は、分泌IgA抗体よりも特異的に作用するため、連続変異ウイルス株(drifted viral strains)に対する有効性が低い。分泌IgA抗体は、インフルエンザウイルスのバリアント株に対する交差防御効果を有する。IgAの交差防御効果の正確な機構はいまだ不明であるが、この現象は感染を予防するのに大きな利点である。インフルエンザは、インフルエンザの2つの主な膜タンパク質、ヘマグルチニン(HA)およびノイラミ二ダーゼ(NA)の抗原特性を変化させる驚異的な能力を発揮する。これは、ヒト集団で確立された適応免疫応答から離れた連続選択により起こる。このウイルスの高い変異率のため、特定のワクチン製剤は通常、約1年間しか効かない。世界保健機関は、毎年ワクチンの内容物を調整して翌年に襲うウイルスの最も可能性の高い株を含有するようにしている。最近では、従来のワクチンは、3つの不活化インフルエンザウイルス(2つのA型株および1つのB型株)から成るワクチンである。この三価インフルエンザワクチンは、次回のインフルエンザシーズンに蔓延するとWHOにより予想されたインフルエンザ株に基づき、毎年再処方される。例えば、2007〜2008年シーズンの毎年更新される三価インフルエンザワクチンは、インフルエンザH3N2、H1N1、およびインフルエンザB型ウイルス由来のヘマグルチニン(HA)表面糖タンパク質成分から成る。
【0003】
ワクチンのi.n.送達の他の利点は、ワクチンの送達が、ワクチンの投与に訓練された医療従事者を必要とせず、このタイプのワクチンを針恐怖症の人に適したものにし、および針刺し事故の問題を回避する点である。さらに、粘膜免疫系は年少期に発達し、老化による影響を受けないことが報告されている(McElhaney JE. Vaccine 2005年7月8日;23 付録1:S10〜25頁; Szewczuk MRら Ann N Y Acad Sci 1983年6月30日;409:333〜44頁)。したがって、例えば鼻腔内インフルエンザ免疫化の付随する利点は、有効な免疫を全ての年齢層に潜在的に提供することができる点、および集団予防接種に使用することができる点である。経鼻または口腔咽頭経路を介し、および不活化インフルエンザ抗原を用いる、インフルエンザに対する免疫化のさまざまな概念は、皮下または筋肉内免疫の無針(needle-less)代替案として探究されている。無針アプローチを支持する実験データは動物モデルで生まれている。動物データにより支持されている、鼻腔内経路を介した免疫化のため不活化インフルエンザ抗原(化学的に不活化された全ウイルス粒子、もしくはスプリットウイルスなどのさらに加工されたウイルス成分、または精製表面抗原ヘマグルチニン(HA)および/もしくはノイラミ二ダーゼ(NA)など)を用いる概念は、不活化インフルエンザ抗原と併用してアジュバントもしくは免疫刺激剤のいずれかの使用を含み、または複数回のワクチン接種を必要とする。アジュバントは、これと混合された抗原の免疫原性を増強する任意の物質である。ヒトにおいて鼻腔内経路を介したインフルエンザに対する成功したワクチン接種は、(a)生(寒冷適応株)インフルエンザワクチン(FluMist(商標)、MedImmune Vaccines Inc社)、(b)大腸菌(E. coli)の易熱性毒素でアジュバントされたビロソーム(virosomal)インフルエンザワクチン(NasalFlu、Berna Biotech Ltd社)、または(c)多量の抗原および反復ワクチン接種の使用について報告されているのみである。生ワクチンは十分な免疫応答を誘導することができるが、生ウイルスであるという特異性は、安全性に対する別の懸念の原因となり、および上気道で必要とされるウイルス複製ラウンドによる副作用を誘導する可能性がある。また必要とされる貯蔵条件も、これらの製品の商業化を制限している。アジュバントとしての大腸菌HLTによる鼻腔内インフルエンザワクチンの使用と顔面神経麻痺(ベル麻痺)との強い関連は、市場からHLTアジュバントビロソームワクチンの撤退をもたらした。
【0004】
現在は、生弱毒化インフルエンザウイルスワクチン(LAIV)がi.n.投与用に市販されている。LAIVワクチンは、全身性および粘膜性免疫応答の両方を誘導することが示されている。しかし、LAIVワクチンは、2〜49歳の者についてのみFDAにより認可されており、ハイリスク集団(高齢者、子どもおよび慢性疾患患者)での使用については認可されていない(疾病対策予防センター。http://www.cdc.gov/flu/professionals/vaccination/pdf/targetpopchart.pdf ; Belshe RBら。Vaccine 2008年9月12日;26 付録4:D10〜6頁)。しかし、市販されたインフルエンザワクチンのほとんどは、集団全体にi.n.経路を介して安全に投与することができる不活化ワクチンである。これらのワクチンの欠点は、この経路を介して投与される場合、免疫原性が低いことを示した点である(Vaccine 2007年7月20日; 25 (29):5367〜73頁; Eylesら、BioDrugs 2000年1月;13(l):35〜59頁)。
【0005】
免疫原性を増加させるため、不活化インフルエンザワクチンは、i.n.経路を介して投与される場合、免疫応答を強化するアジュバントを必要とする。ウイルス様粒子(Matassov Dら。Viral Immunol 2007年9月;20(3):441〜52頁)、ISCOM (Sjolander Sら。Vaccine 2001年7月16日;19(28〜29):4072〜80頁)、脂質、核酸(Josephら。Vaccine 2006年5月1日;24(18):3990〜4006頁)および細菌成分(Haanら。Vaccine 2001年4月6日;19(20〜22):2898〜907頁; Planteら。Vaccine 2001年10月12日;20(1〜2):218〜25頁)のような幾つかのアジュバントが、i.n.免疫化用に現在開発中である。しかし、これらのアジュバント系の多くの開発が、安全性および規制上の懸念により阻まれている。例えばLT(大腸菌の易熱性毒素)のような強力な細菌アジュバントは、ヒトにおける重篤な副作用を示した(Mutschら。N Engl J Med 2004年2月26日;350(9):896〜903頁)。インフルエンザワクチンについて示唆されているように、アルミニウム塩アジュバントの封入は、製造中に余分な混合工程を必要とし、これにより製造全体が減速するだけでなく、これらの塩の封入はさまざまな問題と関連する。例えば、この不溶性は、吸収抗原が懸濁液から沈降することを意味し、そのためバルクワクチンから個々の用量を調製することは、余分な注意を必要とする。加えて、塩への抗原の結合は、最終ワクチンの品質管理を複雑にする。特に、インフルエンザワクチンの幾つかの力価試験は、非結合抗原を必要とするインビトロ免疫アッセイに基づいており、すなわちアジュバントへの吸着は、これらの試験が使用できないことを意味する。
【0006】
近年、免疫応答の表現型、すなわちTh1、Th2またはバランスの取れた応答(blanced response)に重点が置かれている。i.n.経路を介して投与されるサブユニットワクチンならびにキトサン、ISCOM、脂質、およびLTのような経鼻アジュバントの多くは、混合Th1/Th2型応答を誘導する。しかし、Th1応答は、1)より良好な感染防御をもたらす、および2) INF-γの分泌によるウイルス中和に役立つため、Th2または混合応答より優れていると考えられる。さらに、自然感染もまたTh1型の応答を誘導する。加えて、粘膜表面の分泌IgA抗体は感染予防に極めて有効であり、重要なことには、分泌IgA抗体はインフルエンザウイルスのバリアント株に対する交差防御効果を有し、粘膜免疫系は年少期に発達し、老化による影響を受けない。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】WO 02/101026
【特許文献2】US 6,896,887
【特許文献3】EP 1395648
【特許文献4】WO 2007/011216
【非特許文献】
【0008】
【非特許文献1】McElhaney JE. Vaccine 2005年7月8日;23 付録1:S10〜25頁
【非特許文献2】Szewczuk MRら Ann N Y Acad Sci 1983年6月30日;409:333〜44頁
【非特許文献3】疾病対策予防センター。http://www.cdc.gov/flu/professionals/vaccination/pdf/targetpopchart.pdf
【非特許文献4】Belshe RBら。Vaccine 2008年9月12日;26 付録4:D10〜6頁
【非特許文献5】Vaccine 2007年7月20日; 25 (29):5367〜73頁
【非特許文献6】Eylesら、BioDrugs 2000年1月;13(l):35〜59頁
【非特許文献7】Matassov Dら。Viral Immunol 2007年9月;20(3):441〜52頁
【非特許文献8】Sjolander Sら。Vaccine 2001年7月16日;19(28〜29):4072〜80頁
【非特許文献9】Josephら。Vaccine 2006年5月1日;24(18):3990〜4006頁
【非特許文献10】Haanら。Vaccine 2001年4月6日;19(20〜22):2898〜907頁
【非特許文献11】Planteら。Vaccine 2001年10月12日;20(1〜2):218〜25頁
【非特許文献12】Mutschら。N Engl J Med 2004年2月26日;350(9):896〜903頁
【非特許文献13】van Roosmalen MLら。Methods 2006年2月;38(2):144〜9頁
【非特許文献14】Audouy SAら。Vaccine 2007年3月22日;25(13):2497〜506頁
【非特許文献15】J Virol。1991年7月; 65(7): 3789〜3796頁
【非特許文献16】Amorij JPら。Vaccine 2007年12月21日;26(1):67〜76頁
【発明の概要】
【発明が解決しようとする課題】
【0009】
故に、ヒト用の強力、安全で、規制機関により容易に承認され得るアジュバントの必要性がある。好ましくは、分泌IgA抗体のような粘膜性免疫応答および/またはTh1型免疫に偏った応答を誘導することができるワクチンが望ましい。したがって、さらなるおよび改善されたアジュバントインフルエンザワクチン(パンデミック用およびパンデミック間期用の両方の)、好ましくは鼻腔内および/または筋肉内送達に適したワクチンを提供することが、本発明の目的である。さらなる目的は、簡便および費用効果の高い年に1回の再処方を可能にするインフルエンザワクチン調製の柔軟な方法を提供することである。
【課題を解決するための手段】
【0010】
上記の目標は、グラム陽性細菌から得られる不活化ペプチドグリカン粒子と共に抗原を同時処方することによって達成できることが見出された。該粒子は、鼻腔内投与されるサブユニットワクチンの免疫原性を増強するのに極めて有効であるだけでなく、分泌IgAを誘導し、バランスの取れた免疫応答からTh1偏向(Th1-skewed)免疫応答へと応答を調節する。鼻腔内送達は、サブユニットインフルエンザウイルス単独による従来の筋肉内免疫化と比較した場合、同程度の全身性免疫ならびにさらにはより優れた粘膜性および細胞媒介性免疫を誘導した。防御効果は、抗原と細菌粒子の単純な混合により達成することができた。
【0011】
したがって、本発明は、(i)グラム陽性細菌から得られるペプチドグリカン微粒子と、(ii)少なくとも1つの抗原またはこの抗原性調製物とを含み、該抗原または抗原性調製物がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない、アジュバントワクチン製剤に関する。いずれかの既知のまたはまだ発見されていない、例えばウイルス、細菌、寄生虫、真菌または酵母起源の防御抗原もしくはこの抗原性フラグメントが含まれ得る。
【0012】
1つの実施形態では、抗原は、B型肝炎表面抗原またはインフルエンザウイルス抗原などのウイルス抗原である。
【0013】
別の実施形態では、製剤は、細菌抗原、好ましくは少なくとも2つの細菌タンパク質性の抗原またはこの抗原性調製物を含み、抗原または抗原性調製物は、タンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない。2つ以上のタンパク質性抗原またはこの抗原性フラグメントのいずれかの既知のまたはまだ発見されていない防御的組み合わせ(肺炎連鎖球菌(Streptococcus pneumoniae)抗原PpmA、SlrA、IgA1プロテアーゼ、PspA、CbpA、PdBD等の組み合わせ、またはペスト菌(Yersinia pestis)抗原LcrV、F1、FliCの組み合わせ、またはネズミチフス菌(Salmonella typhimurium)、エルシニア・エンテロコリチカ(Yersinia enterocolitica)、赤痢菌(Shigella)のLcrV、IpaBおよびD、SipBおよびD、YopDなどのIII型分泌経路抗原の組み合わせ、または腸管毒素性大腸菌(enterotoxic Escherichia coli) (ETEC)のLTおよびST抗原の組み合わせ、または他の細菌タンパク質性抗原の組み合わせなど)が含まれ得る。1つの態様では、本発明は、PspA、CbpAおよび/またはPdBDを含む防御製剤を提供する。特に興味深いのは、細菌タンパク質性抗原がPspA、CbpAおよびPdBDである本発明の肺炎球菌三価ワクチン製剤である。グラム陽性細菌から得られるペプチドグリカン微粒子と混合すると、3抗原のこのカクテルは、肺炎連鎖球菌感染の鼻腔内チャレンジマウスモデルにおいて極めて良好な防御を与えることが見出された。驚くべきことに、混合後の防御活性は、タンパク質性のペプチドグリカン結合部分への融合を介して抗原が該粒子に結合される場合よりも高かった(本明細書の以下の実施例12および図16を参照のこと)。また、抗原PpmA、IgA1プロテアーゼ、PspA、CbpAおよびPsaAを含む五価製剤は、ペプチドグリカン粒子と混合された場合、結合される抗原と比べて相対的に低いことから、抗原の特定の組み合わせも防御活性に関連しているようである。実施例13および図17を参照のこと。
【0014】
さらなる実施形態では、製剤は、少なくとも1つの寄生虫抗原またはこの抗原性調製物を含み、抗原または抗原性調製物は、タンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない。熱帯熱マラリア原虫(Plasmodium falciparum)のスポロゾイト周囲表面抗原またはメロゾイト表面抗原などの、いずれかの既知のまたはまだ発見されていない防御寄生虫抗原またはこの抗原性フラグメントが含まれ得る。例示的な防御真菌抗原には、コクシジオイデス属(Coccidioides)亜種の抗原が含まれる。適切な酵母抗原は、カンジダ属(Candida)亜種の抗原である。同様に、提供されるのは、グラム陽性細菌から得られるペプチドグリカン微粒子、および少なくとも1つの多糖体抗原またはこの抗原性調製物を含み、該抗原または抗原性調製物がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない、アジュバントワクチン製剤である。肺炎連鎖球菌、インフルエンザ菌(Haemophilus influenza)、髄膜炎菌(Neisseria meningitides)、黄色ブドウ球菌(Staphylococcus aureus)莢膜多糖体または他の多糖体などの、いずれかの既知のまたはまだ発見されていない防御多糖体抗原またはこの抗原性フラグメントが含まれ得る。
【0015】
好ましい実施形態では、本発明は、(i)グラム陽性細菌から得られるペプチドグリカン微粒子と、(ii)少なくとも1つのインフルエンザウイルス抗原またはこの抗原性調製物とを含み、該抗原または抗原性調製物がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない、アジュバントインフルエンザワクチン製剤を提供する。別の実施形態では、本発明は、(i)グラム陽性細菌から得られるペプチドグリカン微粒子と、(ii)少なくとも1つのB型肝炎ウイルス抗原(例えばB型肝炎表面抗原(HBsAg)などのウイルスエンベロープタンパク質)、またはこの抗原性調製物とを含み、該抗原または抗原性調製物がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない、アジュバントB型肝炎ワクチン製剤を提供する。さらに別の実施形態では、本発明は、(i)グラム陽性細菌から得られるペプチドグリカン微粒子と、(ii)少なくとも1つの肺炎球菌抗原、好ましくはPdBD、より好ましくはPspA、CbpAおよびPdBDとを含み、該抗原がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない、アジュバント肺炎球菌ワクチン製剤を提供する。
【0016】
本発明によるワクチンにおいて使用するペプチドグリカン微粒子は、当技術分野で既知の方法により得ることができる。例えば、グラム陽性細菌の細胞壁材料を得る方法を開示するWO 02/101026およびUS 6,896,887を参照のこと。該方法は、細胞壁成分(前記細胞壁材料由来のタンパク質、(リポ)テイコ酸または炭水化物など)を除去することができる溶液により前記細胞壁材料を処理することを含み、前記細胞壁材料は球状ペプチドグリカン微粒子を本質的に含む。細胞壁材料は、グラム陽性細菌の大きさおよび形状を反映する球状ペプチドグリカン微粒子をもたらすため、機械的に破壊されなかった。粒子は生きておらず、インタクトな表面タンパク質および細胞内内容物が取り除かれている。しかし、厚いペプチドグリカン細胞壁は、インタクトなままであり、およびグラム陽性エンハンサー(Gram-positive enhancer) (GEM)粒子と呼ばれる、細菌の形をした大きさ約1μmのペプチドグリカン球(peptidoglycan sphere)を構成する構造的剛性を提供する。粘膜アジュバント開発の分野における重大な障害は、規制機関による承認を得るためにこの安全性を証明することである。本試験で使用された粒子は、ワクチン接種に関して評価される他のアジュバントおよび他の乳酸菌系と比較して安全に使用できる。粒子の製造中に、細菌は酸で処理され、これが遺伝物質の喪失をもたらす。遺伝物質の喪失は、細菌による粘膜層でのDNA脱落および感染の問題が回避されることから有益である。さらに、粒子は、乳製品の製造に使用される細菌から製造され、およびGRAS生物と見なされる。GEM粒子は、前臨床GLP毒性試験においてウサギで既に鼻腔内試験されており、有害事象は報告されなかった。したがって、GEM粒子は、ヒトでの粘膜使用に安全な候補アジュバントと見なすことができる。
【0017】
1つの実施形態では、ワクチン製剤は、食品用細菌、好ましくは乳酸菌から得られる微粒子を含む。好ましくは、微粒子は、非病原性の、非コロニー形成グラム陽性細菌である食品用細菌ラクトコッカス・ラクティス(Lactococcus lactis)から得られる。さらに、ラクトコッカス・ラクティスは規制機関によりヒト用に承認されており、GRAS (一般に安全と認められる(generally recognized as safe))生物と見なされている。1つの実施形態では、ペプチドグリカン粒子は、酸中のラクトコッカス・ラクティスを加熱し、この後リン酸緩衝液で洗浄して製造される(van Roosmalen MLら。Methods 2006年2月;38(2):144〜9頁)。
【0018】
粒子は、ペプチドグリカン結合ドメイン(AcmA細胞壁結合ドメインまたはこの相同体もしくは機能的誘導体のような)を含むタンパク質性物質と結合する改善された能力のため、マラリア原虫抗原および肺炎球菌抗原の粘膜ワクチン接種のための抗原担体として研究された。これらの研究は、GEM粒子に結合されGEM粒子上に提示された抗原が抗原単独よりも高い免疫応答を誘導することを示した。担体粒子における抗原の固定化および最適表面提示がアジュバント効果に重要であると、当技術分野では一般に考えられていた。
【0019】
本試験は驚くべきことに、抗原が粒子に結合された以前の試験とは対照的に、ペプチドグリカン粒子と抗原の単なる混合が抗原の免疫原性を著しく増強することを示す。しかも、特定のケースでは粒子への結合/固定化と比べて、粒子と混合するとより良好な防御活性を達成することができる。
【0020】
本明細書では表現「抗原または抗原性調製物がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない」は、本発明を従来技術と区別することが意図される。従来技術では、典型的にはAcmA細胞壁結合ドメインまたはこの相同体もしくは機能的誘導体の少なくとも1つの反復配列、しかし好ましくは2つまたは3つの反復配列を含む、「タンパク質アンカー」または「プロタン」(PA)とも呼ばれるタンパク質性物質に抗原を融合または結合して、抗原性部分がペプチドグリカン微粒子に結合される。例えば、EP 1395648は、AcmA型タンパク質アンカー融合を微生物の細胞壁材料に結合する方法を開示している。WO 2007/011216は、免疫原性担体に結合された少なくとも1つの二機能性ポリペプチドを含む抗原負荷免疫原性担体複合体に関し、前記二機能性ポリペプチドは、ペプチドグリカン結合ドメイン(PBD)を含み、PBDを通じてポリペプチドは前記担体に結合され、目的とする少なくとも1つの抗原が結合される抗原結合ドメイン(ABD)に融合される。
【0021】
好ましい実施形態では、抗原は、インフルエンザウイルス抗原である。本明細書で以下に示されるように、本発明によるGEMベースのi.n.インフルエンザワクチンは、Th1表現型に偏った応答を誘発することが驚くべきことに見出された。GEM粒子(ワクチンと単純に混合される)でアジュバントされた鼻腔内投与サブユニットワクチンは、防御の重要な相関であると考えられるHI価(>2log5.3)の防御レベルを誘導するのに、初回−ブーストワクチン接種戦略において使用され得ることが見出された。さらに、GEMベースのi.n.インフルエンザワクチンは、致死的チャレンジ後、完全に防御的となる。加えて、血清IgG結果は、GEM粒子がi.n.投与インフルエンザサブユニットワクチンの免疫原性を増強することを明らかに浮き彫りにする。十分な血清反応に加えて、GEMアジュバントi.n.ワクチンは強力な粘膜性免疫応答、すなわち呼吸粘膜におけるsIgAの分泌を誘発した。鼻粘膜における有意なレベルのsIgAの誘導は、GEM粒子が鼻粘膜で免疫増強物質として作用することを示す。鼻粘膜の免疫系は、鼻関連リンパ組織(NALT)から成る。NALTでは、抗原は、M細胞により取り込まれ、次いで抗原提示細胞に提示され、抗原提示細胞が今度は下部のBおよびT細胞に抗原フラグメントを提示する。この事象のカスケードは、インフルエンザウイルスに対する最初の先天性および適応免疫応答に必要とされる。本発明者らの結果は、GEM粒子と混合されたインフルエンザサブユニットワクチンによるi.n.免疫化は、ワクチンのみによるi.m.およびi.n.免疫化よりも鼻粘膜において高いsIgAレベルを誘導したことを示す。NALTにおけるsIgA抗体の誘導は、GEM粒子がインビトロ試験でTLR-2アゴニストとして作用することが知られていることから、GEM粒子に存在するペプチドグリカンのTLR-2 (Toll様受容体)との相互作用の結果であると考えられる。さらに、GEM粒子は、インビトロで樹状細胞およびマクロファージの成熟を活性化できることが知られている(Audouy SAら。Vaccine 2007年3月22日;25(13):2497〜506頁)。故に、TLR-2および樹状細胞の成熟の活性化はいずれも、より強力な粘膜性免疫応答に寄与した可能性がある。
【0022】
近年、免疫応答の表現型、すなわちTh1、Th2またはバランスの取れた応答に重点が置かれている。Th1応答は、1)より良好な感染防御をもたらす、および2) INF-γの分泌によるウイルス中和に役立つため、Th2または混合応答より優れていると考えられる。さらに、自然感染もまたTh1型の応答を誘導する。しかし、i.n.経路を介して投与されるサブユニットワクチンならびにキトサン、ISCOM、脂質およびLTのような経鼻アジュバントの多くは、混合Th1/Th2型応答を誘導する。対照的に、本発明によるi.n.インフルエンザワクチンは、Th1型に偏った応答を誘発した。故に、GEM粒子は、バランスの取れた応答からTh1偏向応答へと応答を調節する。さらに、本明細書に示されたワクチン製剤は、前処方されなければならない他のアジュバント系のほとんどと比べて、製造するのがはるかにより簡便である。これらの実験で使用された製剤は、GEM粒子を従来のサブユニットワクチンと混合して調製された。GEM粒子は、滅菌条件下で大量に製造することができ、および周囲温度で長時間貯蔵することができる。処方および投与の容易さは、i.n. GEMインフルエンザサブユニットワクチンを、パンデミックおよび流行状態におけるワクチン接種の有望な候補にする。
【図面の簡単な説明】
【0023】
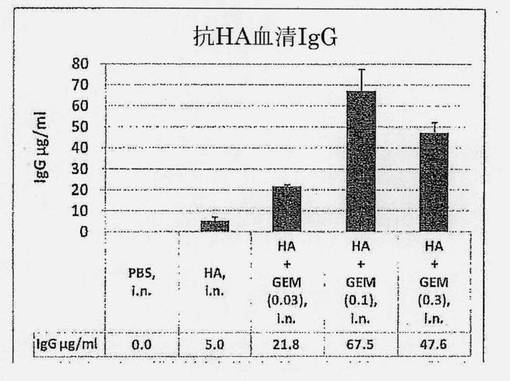
【図1】PBSまたはHA+種々の量のGEM粒子(乾燥重量で表された)で3回免疫化されたマウスにおける、μg/mlで表されるHA抗原(H1N1 A/Beijing)特異的血清総IgGを示す図である。誤差バーは標準誤差(SEM)を示す。
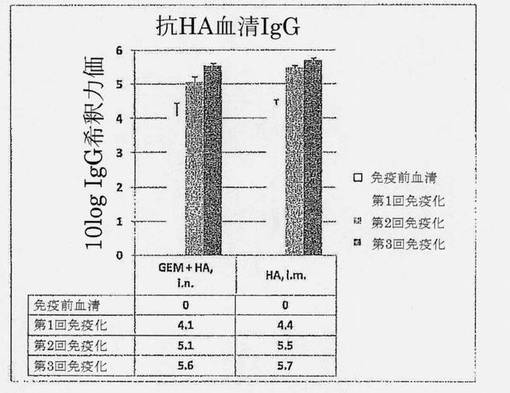
【図2】最初の免疫化後14、28および42日目(それぞれ、第1回、第2回および第3回免疫化)の種々のHA群、すなわちi.n. HA+GEMまたはi.m. HAにおける、HA抗原(H1N1 A/Beijing)特異的血清総IgG希釈力価の比較分析を示す図である。誤差バーはSEMを示す。
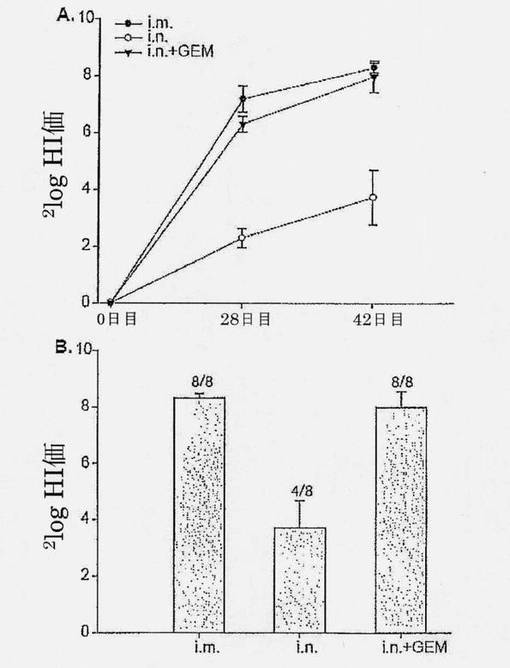
【図3】3回免疫化されたマウスの血清におけるHA抗原(H3N2 A/Wisconsin)特異的HI価を示す図である。A.最初の免疫化後0、28および42日目の種々のHA群、すなわちi.m.、i.n.およびi.n.+GEMにおけるHI価の比較分析。B.最初の免疫化後42日目の3つのHA群、すなわちi.m.、i.n.、i.n.+GEM間のHI価の比較分析。カラムの上の数字は1群当たりのレスポンダーの数を示す。誤差バーはSEMを示す。
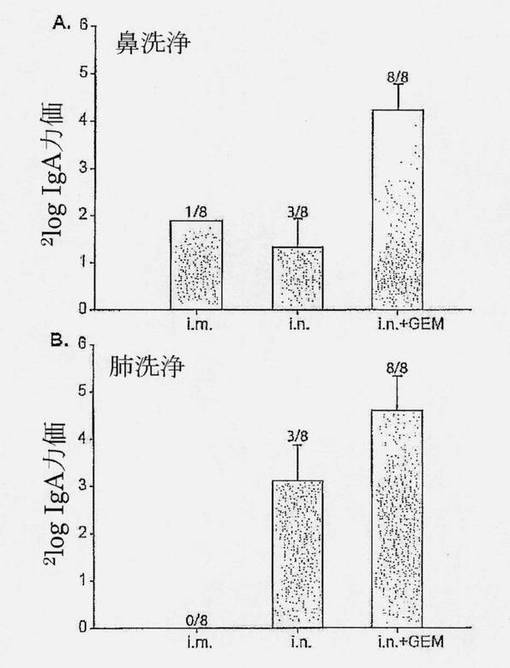
【図4】HA i.m.、i.n.またはi.n.+GEMで免疫化されたマウスの鼻洗浄(A)および肺洗浄(B)における、HA抗原(H3N2 A/Wisconsin)特異的sIgA力価を示す図である。カラムの上の数字は1群当たりのレスポンダーの数を示す。誤差バーはSEMを示す。
【図5】HA i.m.、i.n.またはi.n.+GEMで免疫化されたマウスの血清におけるHA抗原(H3N2 A/Wisconsin)特異的IgGサブタイプ力価を示す図である。IgG1 (A)、IgG2a (B)およびIgG2b (C)力価が決定された。アスタリスクは示された比較のP値<0.05を意味する。誤差バーはSEMを示す。
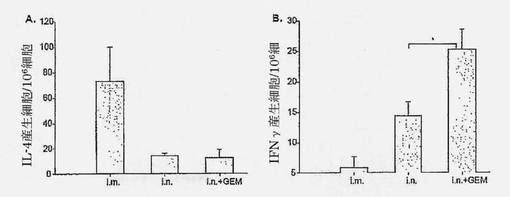
【図6】細胞媒介性免疫応答が、HA i.m.、i.n.またはi.n.+GEMで免疫化されたマウスにおけるサイトカイン放出プロフィール、すなわちIL-4 (A)、IFNy (B)を決定して決定されたことを示す図である。アスタリスクは示された比較のP値<0.05を意味する。誤差バーはSEMを示す。
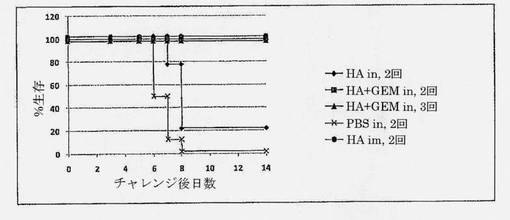
【図7】チャレンジ後の生存(%)を示す図である。動物は1用量当たり5μg HAでワクチン接種され、GEM含有ワクチンは1用量当たり0.3mg GEMを補充された。動物は最後のブースター免疫化の3週間後にチャレンジされ、14日間追跡された。5つのワクチン群間の比較分析。
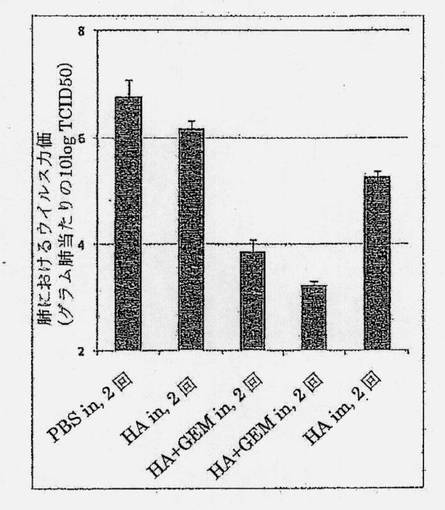
【図8】チャレンジ後の肺におけるウイルス力価(A/Puerto Rico/8/34 [PR8]、TCID50 [組織培養感染量]) (肺組織1グラム当たり)を示す図である。肺はチャレンジの4日後に分離された。5群間の比較分析。標準誤差(SEM)が誤差バーにより示されている。
【図9】経口HAまたは経口HA+GEMインフルエンザワクチンで免疫化されたマウスにおける、サブユニット抗原(A/Hiroshima [H3N2])特異的血清HI価を示す図である。本発明者らが1用量当たり20μg HAで3回免疫化したマウス。GEMワクチンは1用量当たり0.3mg GEMを含有した。*はp<0.05を示す。2Log5.3を超える力価が防御的である。標準誤差(SEM)が誤差バーにより示されている。
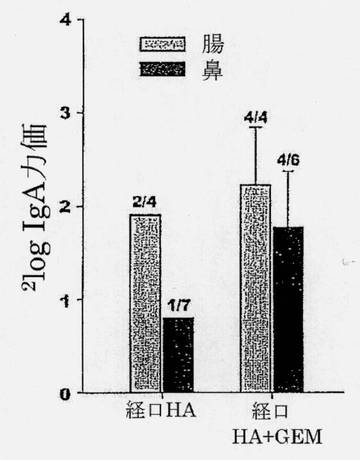
【図10】経口HAまたは経口HA+GEMインフルエンザワクチンで免疫化されたマウスの腸洗浄(グレーのバー)および鼻洗浄(黒のバー)における、サブユニット抗原(A/Hiroshima [H3N2])特異的sIgA力価を示す図である。カラムの上の数字は分析された動物数当たりのレスポンダーの数を示す。標準誤差(SEM)が誤差バーにより示されている。
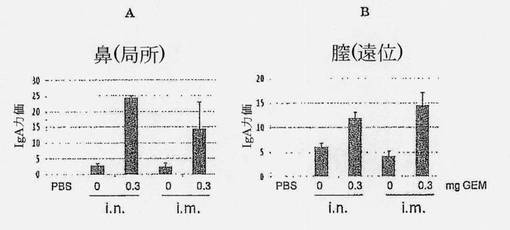
【図11】0.3mg GEM有りまたは無しの固定量のHA (5μg B/Shangdong/7/97)で、鼻腔内(パネルA)または筋肉内(パネルB)に3回(14日間隔)ワクチン接種されたメスマウスの鼻および膣洗浄における、HA特異的IgA力価を示す図である。洗浄試料は最後の免疫化の2週間後に採取された。標準誤差(SEM)が誤差バーにより示されている。
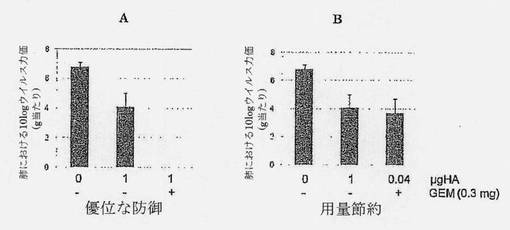
【図12】GEM無しのPBS (偽)、1μg HA (A/PuertoRico/8/34)またはGEMと共に処方された0.04μg HA (25分の1抗原)で2回ワクチン接種されたマウスの肺ウイルス力価を示す図である。最終用量の投与の2週間後、マウスはマウス適応A/PuertoRico/8/34でチャレンジされた。チャレンジの5日後、動物は屠殺され、肺が分離およびホモジナイズされ、ウイルス力価がMDCK細胞でのエンドポイント滴定により決定された。標準誤差(SEM)が誤差バーにより示されている。
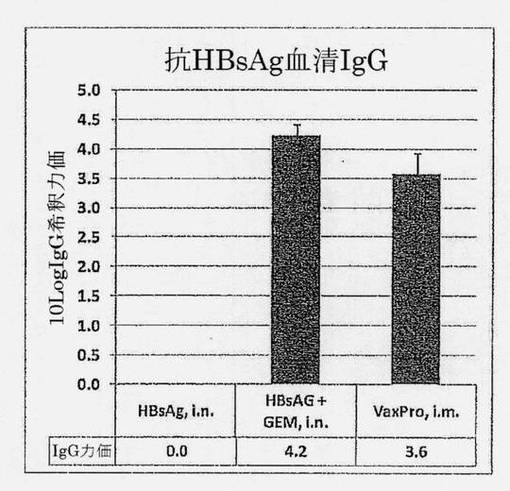
【図13】HBsAg単独(i.n.)、+ GEM (i.n.)またはVaxPro (i.m.)で3回免疫化されたC57BL6マウスの血清における、HBsAg抗原特異的IgG希釈力価を示す図である。誤差バーはSEMを示す。
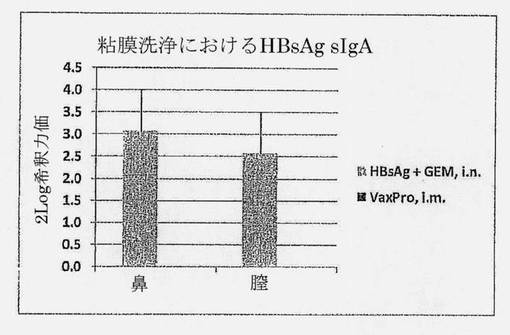
【図14】HBsAg+GEM (i.n.)またはVaxPro (i.m.)で3回免疫化されたC57BL6マウスの鼻および膣洗浄における、HBsAg抗原特異的sIgA力価を示す図である。誤差バーはSEMを示す。
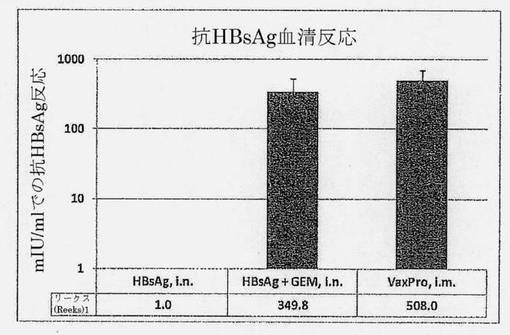
【図15】HBsAg単独(i.n.)、+GEM (i.n.)またはVaxPro (i.m.)で3回免疫化されたウィスターラットのmIU/mlとして測定されたHBsAg抗原特異的血清反応を示す図である。>10mIU/mlのレベルは防御的であると考えられる。誤差バーはSEMを示す。
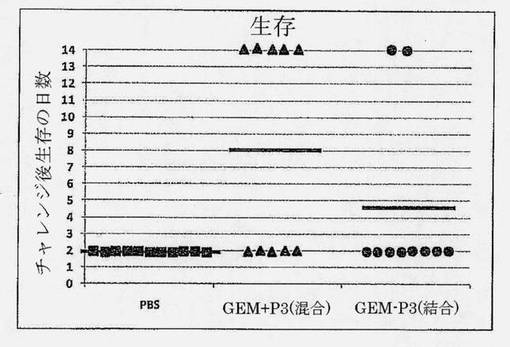
【図16】チャレンジ後数日における生存時間。全群の試験材料は鼻腔内(i.n.)適用された。マウスはPBS (偽免疫化)、GEMと混合された肺炎球菌P3タンパク質(PspA、CbpA、PdBD) (GEM+P3)、またはGEMに結合されたP3タンパク質(GEM-P3)で免疫化された。両方のワクチンは5μgの各抗原を含有した。各記号は動物1匹を表す。横線は平均を示す。
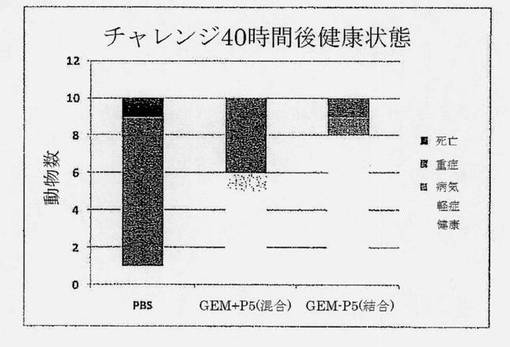
【図17】PBS (偽免疫化)、P5タンパク質と混合されたGEM(GEM+P5)、またはP5タンパク質が結合しているGEM (GEM-P5)で免疫化された、病原性肺炎連鎖球菌株D39 (血清型2)による鼻腔内チャレンジの40時間後のマウスの健康状態を示す図である。ワクチンは0.5μg IgA1prt、3μg PsaA、1.5μg CbpA、2μg PpmA、2μg PspAおよび0.3mg GEMを含有した。チャレンジの40時間後の健康状態は、ワクチンの防御能の尺度である。
【発明を実施するための形態】
【0024】
実施例1〜8に示されるように、本発明者らは、GEM粒子でアジュバントされたi.n.インフルエンザワクチンが、サブユニットインフルエンザワクチン単独によるi.m.免疫化と比べて、同程度の全身性免疫ならびにより優れた粘膜性および細胞媒介性免疫を誘導することを示す。特に、該ワクチンは、サブユニットインフルエンザワクチン単独によるi.m.免疫化と比べて、最初のブースター免疫化後にHI価により測定されるのと同程度の防御レベルの免疫を誘導する。重要なことには、該ワクチンは、上気道におけるインフルエンザ感染中の防御の最前線である、より高いsIgAレベルを誘導した。さらに、該ワクチンは、より優れた防御を提供すると考えられる偏向Th1型免疫応答を誘発した。加えて、これらの免疫応答は、GEMベースの鼻腔内インフルエンザワクチンで免疫化されたマウスの完全防御を提供することが示された。
【0025】
実施例7 (図9および10)は、経口投与インフルエンザワクチン組成物の有効性を示す。実施例8 (図11)は、筋肉内GEMベースのインフルエンザワクチンもまた、気道または他の粘膜層の粘膜内層において高いsIgAレベルを誘発するのに使用できることを示す。加えて、筋肉内経路は、通常の筋肉内ベンチマークワクチンの力価を著しく増加する、または大幅に抗原量を減少させる(抗原用量節約)ように、GEM粒子と混合されたインフルエンザワクチンに使用することができる(図12)。GEM粒子は、i.n.、i.m.または経口送達インフルエンザワクチンのための安全および強力なアジュバントと見なすことができる。
【0026】
他の適切なウイルス抗原には、呼吸器合胞体ウイルス(RSV)タンパク質、例えばRSV融合(F)および結合(G)糖タンパク質、またはキメラFGタンパク質などのこれらの関連部分もしくは組み合わせが含まれる(J Virol。1991年7月; 65(7): 3789〜3796頁)。RSV感染は、米国、欧州、オーストラリアおよび日本を含む、世界的に長年のおよび有害な問題となっている。RSV感染は、早産児、幼児、および高齢者において、および実際、弱まった免疫系を有する全ての個体に対して特に厄介である。1歳未満の子どもの約3人に2人、および1歳から4歳の間のほとんど全ての子どもが少なくとも1回はRSVに感染し、医学的注意を必要とすることなくほぼ回復していると推定される。しかし、5〜10%は、小児期後期の喘鳴および喘息様症状の素因と考えられる要因である、重篤な感染を長引かせている。ペプチドグリカン粒子との混合で使用される他の興味深い抗原には、ヒト免疫不全ウイルス(HIV)タンパク質、特に、gp120、gp140またはgp160のようなHIVエンベロープの表面で曝露される糖タンパク質が含まれる。Gp120は、侵入のための特異的細胞表面受容体を探し出すうえで極めて重要な役割を果たすことから、細胞へのウイルス侵入に不可欠である。
【0027】
本明細書で提供されるアジュバントインフルエンザワクチン製剤は、少なくとも1つのインフルエンザウイルス抗原またはこの抗原性調製物を含む。例えば、該ワクチン製剤は、インフルエンザタンパク質もしくはこのフラグメントおよび/またはインフルエンザタンパク質もしくはこのフラグメントを含む融合タンパク質(ペプチドグリカン結合ドメインに融合されないという条件で)を含む。本発明の異種タンパク質は、ヘマグルチニン抗原(HA)、ノイラミダーゼ抗原(NA)またはこれらの組み合わせを含む、目的とする任意のインフルエンザ抗原を含むことができる。好ましくは、インフルエンザ抗原は表面抗原であり、すなわちインフルエンザマトリックスタンパク質2 (M2e)の細胞外ドメインなどの構造抗原ではない。1つの実施形態では、インフルエンザ抗原は、M2e以外である。特定の態様では、インフルエンザワクチン製剤は、インフルエンザ抗原としてHAおよび/またはNAを含有する。さまざまな異なるインフルエンザHAおよびNAタンパク質(例えば、異なるサブタイプ、または株もしくは分離株由来の)のアミノ酸配列は、当技術分野で既知であり、GenBankなどの公開データベースにおいて入手可能である。好ましくは、ワクチン製剤は、少なくとも1つのHAサブタイプを含む。
【0028】
ワクチンで使用するインフルエンザウイルス株は、季節ごとに変化する。現在のパンデミック間期の時期では、ワクチンは典型的には、2つのインフルエンザA型株(H1N1およびH3N2)および1つのインフルエンザB型株を含み、三価ワクチンが典型である。本発明は、新たな「豚インフルエンザ」もしくは「メキシコインフルエンザ」H1などのパンデミック株、またはH2、H5、H7またはH9サブタイプ株(特にインフルエンザA型ウイルスの)などの他のパンデミック株(すなわちワクチンレシピエントおよび一般ヒト集団が免疫学的にナイーブである株)由来のウイルスも使用することができ、パンデミック株のインフルエンザワクチンは一価であってもよく、またはパンデミック株を補充された通常の三価ワクチンに基づいていてもよい。しかし、季節およびワクチンに含まれる抗原の性質に応じて、本発明は、1つまたは複数のHAサブタイプHI、H2、H3、H4、H5、H6、H7、H8、H9、H10、H11、H12、H13、H14、H15またはH16に対して防御することができる。例えば、1つの実施形態では、i.n.ワクチン製剤は、1インフルエンザ株当たり1から15μgの間のHAを含有する。
【0029】
特定の態様では、ワクチン製剤は、少なくとも2つのインフルエンザウイルス株、パンデミック発生に関連するまたはパンデミック発生に関連する可能性がある少なくとも1つの株由来のインフルエンザ抗原またはこの抗原性調製物を含む。
【0030】
ワクチン製剤に存在するペプチドグリカン粒子の量は、好ましくは、初回−ブーストワクチン接種戦略において防御レベルの赤血球凝集抑制(HI)価を誘導するのに十分である。例えば、本発明によるワクチン製剤は、インフルエンザウイルス抗原の1マイクログラム当たり0.001から1mg、好ましくは0.01から0.1mg微粒子(乾燥重量)を含んでもよい。ヒト用の例示的なi.n.ワクチン製剤は、以下を含む: 0.3〜2.5mg GEM (乾燥重量)、三価HA (卵、細胞、組換え) 3×1〜15μgまたは一価HA (パンデミック) 0.1〜15μg、0.05〜0.15M PBS pH6〜8。
【0031】
本発明の経鼻ワクチン組成物は、液体または粉末型組成物、特に、投与方法によりエアロゾル、滴剤、吸入器または吹送として処方することができ、粉末またはミクロスフェアが好ましい。点鼻剤用組成物は、防腐剤、粘性調節剤、浸透圧調節剤および緩衝液などの1つまたは複数の許容可能な賦形剤を含むことができる。しかし、本発明は、経鼻ワクチン製剤に限定されない。GEMの添加がi.m. HAワクチンに対し有効性を強化することが、驚くべきことに見出された。これは、用量節約戦略につながり得る。実施例8および9、図12を参照のこと。したがって、本発明はまた、ペプチドグリカン微粒子および(従来の)筋肉内HAワクチン調製物を含む組成物も提供する。
【0032】
さらに、GEMと混合されたインフルエンザHAは、経口投与した際に防御的な血清HI価をもたらした(実施例7、図9および10)。
【0033】
本発明のさらなる態様は、本明細書に開示されたワクチン製剤を含む容器に関する。1つの実施形態では、容器は、場合により使用説明書と共に提供される、エアロゾルまたは滴下送達システム(鼻腔内スプレー)の形態における装置などの鼻腔内調剤装置である。
【0034】
さらに、本発明は、方法が、本明細書上記に記載されたワクチン製剤を被験者に投与することを含む、被験者におけるインフルエンザ感染または疾患の予防方法を提供する。安全性のため、ワクチン製剤は、ヒトハイリスク集団での使用に特に適している。例えば、本発明はこれにより、高齢者、2歳までの子ども、または慢性疾患患者におけるインフルエンザ感染または疾患の予防のための簡便、安全な、信頼できる方法を提供する。予防方法は、ワクチン製剤の鼻腔内、経口または筋肉内送達、好ましくは鼻腔内送達を含んでもよい。調剤装置、例えばエアロゾルまたは滴下送達システムの形態における調剤装置を使用することが、極めて簡便である。
【0035】
ワクチンの投与量は、免疫応答を効果的に誘導することができる量として決定される。例えば、ヒトに対するワクチンの投与頻度は、1日1回から数回であり、投与量は1〜250μgおよび好ましくは2〜50μgである。
【0036】
本発明により調製されたワクチンは、子どもおよび大人の両方を治療するのに使用することができる。インフルエンザワクチンは、現在、6ヶ月齢から小児および成人予防接種での使用が推奨されている。故に、患者は1歳未満、1〜5歳、5〜15歳、15〜55歳、または少なくとも55歳であってもよい。ワクチンを受けるのに好ましい患者は、高齢者(例えば>50歳、>60歳、および好ましくは>65歳)、若齢者(例えば<5歳)、入院患者、医療従事者、兵役者および軍人、妊婦、慢性疾患患者、免疫不全患者、ワクチンを受ける7日前までに抗ウイルス化合物を服用した患者(例えばリン酸オセルタミビルなどのオセルタミビルまたはザナミビル化合物-以下を参照のこと)、卵アレルギーの人および/または国外旅行者である。しかし、理解されるように、ワクチンは、これらの群にのみ適しているわけではなく、集団においてより一般的に使用することができる。パンデミック株に関しては、全ての年齢群への投与が好ましい。
【0037】
同じく本発明の範囲内に包含されるのは、(a)グラム陽性細菌から得られるペプチドグリカン微粒子を提供する工程と、(b)少なくとも1つのインフルエンザ抗原またはこの抗原性調製物を提供する工程と、(c)微粒子と抗原を混合する工程とを含む、インフルエンザワクチン製剤を提供する方法である。(a)および(b)の両方の工程は、それ自体が当技術分野で知られている方法論を用いて実施することができる。工程(b)は、プロタンのようなペプチドグリカン結合ドメインへの抗原の融合または結合を必要としないことから、本発明の方法は、抗原がペプチドグリカン微粒子に結合するのに、抗原が最初に修飾されなければならない(例えばタンパク質性リンカー部分への融合により)従来技術方法よりもはるかに簡便および商業的に魅力的である。対照的に、本発明は、従来のサブユニットワクチンをそれ自体として用いて実践することができる。
【0038】
したがって、本発明はまた、インフルエンザワクチン製剤におけるアジュバントとしての、グラム陽性細菌から得られるペプチドグリカン微粒子の使用にも関し、前記製剤は、タンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されないインフルエンザウイルス抗原を含む。
【0039】
(実施例)
材料および方法
卵由来の株A/Wisconsin (H3N2)のインフルエンザ一価サブユニットワクチンおよび卵由来のA/Beijing (H1N1)スプリットウイルスワクチンを本試験で使用した。ワクチン中のヘマグルチニン(HA)の濃度は、単一放射免疫拡散アッセイを用いて決定した。
【0040】
ハンゼヌラ・ポリモルファ(Hansenula polymorpha)から単離した組換えHBsAg (ad/ay)を本試験で使用した。Sanofi Pasteur/MSD社からのHBVaxProをベンチマークHBsAgワクチン(40μg/ml)として使用した。GEM粒子は、以前に記載されたように製造した(Van Roosmalenら、Methods 2006年、2月;38(2):144〜9頁)。
【0041】
1.1免疫およびチャレンジ
動物実験は、オランダ動物防御法によって規定されたガイドラインに従って評価および承認された。Balb/c、C57BL6マウス(6〜8週齢)およびウィスターユニリーバ(Unilever)ラット(10週齢)はHarlan社、オランダから購入した。CD 1マウスはCharles River社、ドイツから購入した。マウスを各5〜10匹にグループ化した。ラット群は各4匹から成った。全てのマウス群は、5μgのHAによる0日目の初回ワクチン接種、ならびに14日目および28日目の2回のブースターワクチン接種、または5μgのHBsAgによる0日目、ならびに10日目および20日目の2回のブースターワクチン接種により免疫化した。鼻腔内マウス免疫化は、吸入麻酔下(イソフルラン/O2)、10μlのワクチンを両方の鼻孔に分けて行った。ラット群は、25μgのHBsAgによる0日目の初回ワクチン接種、ならびに10日目および20日目の2回のブースターワクチン接種により免疫化した。鼻腔内ラット免疫化は、30μlのワクチンによりマウスについてと似た方法で行った。筋肉内マウス群は、吸入麻酔下(イソフルラン/O2)、後部大腿筋に50μlワクチンを注入した。筋肉内ラット群は、200μlのワクチンを両方の後肢筋に分けて注入した。マウスおよびラットは、2回目のブースターワクチン接種の2週間後に屠殺した。動物を屠殺した後、Balb/cマウスの脾臓を回収し、この後5% FCS、1%ペニシリン/ストレプトマイシンおよび50μΜ β-メルカプトエタノールを含む補充IMDMグルタマックス培地で4℃で貯蔵した。マウスでの経口投与は、3回、すなわち0、14および28日目に行った。簡単には、0.3mg GEM粒子有りまたは無しの20μgサブユニットワクチンを、200μlの重炭酸ナトリウム溶液(3.2% w/v)で胃内投与した。経口投与は、ステンレス鋼経口ゾンデを用いて麻酔無しで実施した。
【0042】
チャレンジ実験では、インフルエンザHAワクチンで免疫化したマウスを、100プラーク形成単位(PFU)の株A/Puerto Rico/8/34 (高用量、1群当たり9匹)、または66 PFUの株A/Puerto Rico/8/34 (低用量、1群当たり4匹)により、最後のブースター免疫化の3週間後に鼻腔内チャレンジした(40μL)。
【0043】
チャレンジウイルスの鼻腔内投与は、O2/イソフルランの吸引による動物の軽麻酔下で行った。低用量を受け取った動物を、チャレンジ後4日目に屠殺し、インビトロ細胞ベースアッセイを用いて肺でのウイルス負荷の判定のため肺を分離した。手短かに言えば、ウイルス希釈と一緒にMDCK細胞をインキュベーター(37℃、5% CO2)で1時間インキュベートし、この後PBSで1回洗浄した。トリプシンを含有する新鮮培地(7,5μg/ml TPCKトリプシンを含む100μl培地)をウェルに添加した。細胞をインキュベーター(37℃、5% CO2)で72時間インキュベートし、この後上清を、50μl 1% (洗浄した)モルモット赤血球と一緒に丸底プレート(Costar社)に移した。混合物を室温で2時間インキュベートし、赤血球凝集を読み取った。赤血球凝集を依然として示す最大希釈の逆数が力価である。高用量を受け取った動物は、チャレンジ後14日目まで臨床徴候について追跡し、および許容できない苦しみのため動物が先に屠殺されない限り(人道的エンドポイント: 1日での10%体重減少、または嗜眠、乱れた毛並みおよび瀕死と組み合わせた複数日中の15%体重減少)、屠殺した。
【0044】
肺炎球菌免疫化のため、CD1マウスは、0.01mL (10μL)の用量を0、14および28日目に鼻腔内に受け取った。筋肉内群は、後肢の大腿筋(左、右および左を交互)に注入した0.04mL (40μL)の用量を0、14および28日目に受け取った。最終ブースター免疫化の3週間後、マウスを1×106 CFU肺炎連鎖球菌株TIGR4でチャレンジした。マウスを吸入麻酔(イソフルラン)により軽く麻酔している間に、肺炎球菌を50μL接種材料で鼻腔内に導入した。マウスは感染後に頻繁にモニターし、健康状態、体重および体温に基づく状態により採点した。血液中の細菌数は、チャレンジ後40時間で判定し、血液中に5.4×103 CFU/mL超を有するマウスと同様に、罹患したおよび屠殺する必要がある(人道的エンドポイント)マウスも屠殺した。残りのマウスは、罹患した場合または試験の終了時(チャレンジの14日後)に屠殺した。
【0045】
1.2血清採取および粘膜洗浄
血液試料は各ワクチン投与前に実験中3回取り出し、最終出血は最後のブースター投与の14日後の終了日に採血した。血清を5分間、1200×gで血液の遠心分離により得て、試料をこの後、さらなる分析まで-20℃で貯蔵した。
【0046】
鼻洗浄は、鼻咽頭を1ml PBS(プロテアーゼ阻害剤カクテルを補充した)で洗い流して得た。膣洗浄は、膣を100μl PBS(プロテアーゼ阻害剤カクテルを補充した)で洗い流して得た。100μlアリコートを取り出し、および黄色い200μlチップが取り付けられたピペットを用いて9回再導入した。洗浄液を清潔なバイアルに移し、-20℃で貯蔵した。腸洗浄は、シリコンチップを備えた1.2mm×38mmフレキシブルテフロン(登録商標)経口ゾンデを用いて、胃後方切開により十二指腸に挿管して実施した。洗浄前、空腸を結紮糸により回腸の前部で閉じた。次に、1mlシリンジを経口ゾンデに取り付け、洗浄は、1mlのPBSで十二指腸/空腸の洗い流しを繰り返して実施した。各試料採取後直ちに、洗浄(lavage)を10μl貯蔵溶液(プロテアーゼ阻害剤カクテルを補充した)と混合し、さらなる調製まで洗浄を氷上で維持した。洗浄試料を15分間、11,000×gで遠心分離し、上清を回収し、さらなる分析まで4℃で貯蔵した。
【0047】
1.3 ELISA
HA抗原に対する抗体反応を、血清IgG、IgG1、IgG2aおよびIgG2b希釈力価、粘膜分泌sIgAを決定するため、またはHA特異的IgGの量を決定するため、ELISAアッセイを用いて決定した。希釈力価については、プレートを200ngのHA/ウェルと共にインキュベートした。HAと共に一晩インキュベーション後、プレートを3%ウシ血清アルブミン(Sigma-Aldrich社、オランダ)でブロックした。次いでプレートを洗浄し、連続希釈における血清および粘膜試料と共に1.5時間、37℃でインキュベートした。次に、プレートを洗浄し、マウスIgG、IgG1、IgG2a、IgG2bおよびIgA (Southern Biotech社、バーミングハム、AL、USA)に対する西洋ワサビペルオキシダーゼ結合ヤギ抗体と共にインキュベートした。最後に、基質溶液(0.006% H2O2を含有する50mMリン酸緩衝液pH 5.6中0.02% 1,2-フェニレンジアミン-二塩酸塩)を添加し、プレートを室温で30分間、暗所でインキュベートした。反応を2M H2SO4の添加により停止し、490nmでの吸光度をベンチマークマイクロプレートリーダー(BioRad社、ハーキュリーズ、CA)により読み取った。報告された力価は、バックグラウンド補正後A490≧0.2に対応する、計算された試料希釈の逆数である。
【0048】
HA特異的血清IgGの量を決定するため、マイクロタイターウェルプレートを、200ng/100μl/ウェル H1N1 A/Beijingおよび較正曲線用の抗マウスIgGで被覆した。4℃で一晩インキュベーション後、プレートをコーティング緩衝液(0.5M炭酸−重炭酸塩緩衝液pH9.6〜9.8)で2回洗浄した。ブロッキングは、Protifar Plus (コーティング緩衝液中2.5%)により4℃で45分間実施した。コーティング緩衝液およびPBS/0.05%Tween20でプレートを4回洗浄後、血清および較正曲線をウェルに添加した。血清の連続希釈および較正曲線(マウスIgG1)を、4℃で1.5時間インキュベートした。この後、プレートをPBS/Tween20で3回洗浄した。西洋ワサビペルオキシダーゼ結合免疫グロブリン(ITK、Southern Biotech社)、PBS/Tween20中1:5000希釈、をウェルに添加し、4℃で1時間インキュベートした。プレートをPBS/Tween20で3回、および水で1回洗浄した後、プレートを基質溶液(0.006% H2O2を含有する50mMリン酸緩衝液pH 5.6中0.02% 1,2-フェニレンジアミン-二塩酸塩)を用いて30分間染色した。呈色反応を2M H2SO4により停止した。測定は493nmで実施した。
【0049】
HBsAg抗原に対する血清抗体反応を、IgG希釈力価を決定するためELISAアッセイを用いて決定した。この目的のため、PBS中2μg/ml HBsAgで被覆したELISAプレート、50μl/ウェルを添加し、37℃で1時間インキュベートした。プレートを洗浄緩衝液(PBS/0.1% tween20)により6×洗浄した。プレートをブロック緩衝液(PBS/1% BSA)、200μl/ウェルでブロックし、37℃で1時間インキュベートした。血清試料はブロック緩衝液で連続希釈し、50μl/ウェルを添加し、37℃で1時間インキュベートした。プレートを洗浄緩衝液により6×洗浄した。アルカリホスファターゼ(Southern Biotech社)に結合したヤギ抗ラットIgGを二次抗体(ブロック緩衝液中で1:3000希釈した)として使用し、50μl/ウェルを添加し、37℃で1時間インキュベートした。
【0050】
プレートを洗浄緩衝液(PBS/0.1% tween20)により6×洗浄した。基質緩衝液(10mMジエタノールアミン/0.5mM MgCl2 pH9.5)中p-ニトロフェニルリン酸二ナトリウム塩(Calbiochem社)を検出に使用し、測定は405nmで行った。力価は希釈力価として表し、免疫前基準の2倍のODを示す希釈として定義した。
【0051】
HBsAg特異的粘膜分泌sIgAを、IgG希釈力価を決定するためELISAアッセイを用いて決定した。この目的のため、ELISAプレートを前と同じように被覆し、洗浄し、ブロックした。粘膜洗浄はブロック緩衝液で連続希釈した。50μl/ウェルを添加し、37℃で1時間インキュベートした。プレートを洗浄緩衝液により6×洗浄した。ブロック緩衝液中で1:1000希釈した、西洋ワサビペルオキシダーゼ(Nordic Immunology社)に結合したヤギ抗ラットIgAを二次抗体として使用し、50μl/ウェルを添加し、37℃で1時間インキュベートした。プレートを洗浄緩衝液(PBS/0.1% tween20)により6×洗浄した。TMB (3,3',5,5'-テトラメチルベンジジン、Sigma社、ロット055K8208)を1ml DMSOに溶解し、9mlの0.05Mリン酸-クエン酸緩衝液、pH5.0を検出に使用した。2μlの30%過酸化水素を、使用の直前に基質緩衝溶液10mlごとに添加した。呈色反応を2M H2SO4により停止し、測定は450nmで行った。力価は希釈力価として表し、バックグラウンド(ブロッキング緩衝液と共にインキュベートしたHBsAg被覆)の3倍のODを示す希釈として定義した。
【0052】
1.4赤血球凝集抑制(HI)アッセイ
血清中のHI価は、以前に記載されているように決定した。簡単には、血清を56℃で30分間不活化した。非特異的赤血球凝集を減少させるため、25%カオリン懸濁液を不活化血清に添加した。1200×gでの遠心分離後、50μlの上清を96ウェル丸底プレート(Greiner社、Alphen a/d Rijn、オランダ)に2通りに移し、およびPBSで2倍に連続希釈した。次いで4赤血球凝集単位(HAU)のA/Wisconsinインフルエンザ不活化ウイルスを各ウェルに添加し、プレートを室温で40分間インキュベートした。最後に、50μlの1%モルモット赤血球細胞を各ウェルに添加し、室温で2時間インキュベートした。赤血球凝集を防止することができる最大希釈を、HI価として採点した。
【0053】
1.5 Abbott AxSYMシステムによるHBsAg特異的Ig力価決定
mIU/mlで表されるHBsAgに対する抗体の定量決定は、AxSYM AUSUBアッセイによりAbbott AxSYMシステムで行った。このアッセイは、固相としての微粒子上の組換えHBsAg (ad/ay)およびコンジュゲートとしての組換えHBsAgに結合したビオチンを用いる微粒子EIAである。次の工程では、アルカリホスファターゼ結合抗ビオチンを抗原サンドイッチに結合する。反応混合物を、微粒子が不可逆的に結合する不活性ガラス繊維マトリックスに移す。メチルウンベリフェリルリン酸を基質として使用し、最終産物メチルウンベリフェロンの蛍光を機器により読み取った。
【0054】
1.6 Elispot
Elispotアッセイは、前に記載されているように(Amorij JPら。Vaccine 2007年12月21日;26(1):67〜76頁)実施した。簡単には、96ウェルマイクロタイタープレート(Greiner社、Alphen a/d Rijn、オランダ)を、抗マウスインターフェロン-γ (IFN-γ)およびインターロイキン-4 (IL-4) (BD, Pharmingen社、エーレムボードヘム、ベルギー)と共に4℃で一晩インキュベートした。プレートをPBS/Tween (Sigma-Aldrich社、オランダ)で3回洗浄した後、プレートを37℃で1時間ブロック(PBS+4% BSA)し、脾臓細胞を、刺激ペプチドとしてのサブユニットワクチン有りまたは無しで濃度1×106細胞/ウェルでプレートに添加した。5% CO2、37℃で一晩インキュベーション後、細胞を冷水で溶解した。次に、プレートをPBS/Tweenで5回洗浄し、およびPBS+2% BSA中0.125μg/mlの濃度でビオチニル化抗マウスIFN-γおよびIL-4抗体(BD Pharmingen社)と共にインキュベートした。洗浄後、プレートを、ストレプトアビジンアルカリホスファターゼ(BD Pharmingen社)と共に37℃で1時間インキュベートした。最後に、PBS/Tweenで3回、およびPBSで2回洗浄した後、1mg/ml 5-ブロモ-4-クロロ-3-インドリルホスフェート、0.92% w/v 2-アミノ-2-メチル-1-プロパノール、0.08μl/ml TritonX-405、1M MgCl2および6mg/mlアガロースから成る基質溶液を用いて、スポットを発生させた。スポットは、Elispotリーダー(A. EL. VIS Elispotリーダー)を用いてカウントした。
【0055】
1.7統計分析
統計分析は、p< 0.05を最小有意水準としてStudentのt検定またはノンパラメトリックANOVA検定を用いて実施した。結果は、特に指示のない限り平均±標準誤差(SEM)で表す。
【0056】
(実施例1)鼻腔内HAワクチンにおけるGEMのアジュバント効果
鼻腔内HA (5μg H1N1 A/Beijing)に対する全身血清抗体反応の増強は、さまざまな量のGEM粒子(0、0.03、0.1および0.3mg乾燥重量)をHAに添加して鼻腔内マウスモデルで評価した。マウスは3回のワクチン投与を受け(各々2週間間隔および最後のブースター免疫化の2週間後)、血清試料が分析された。図1は、アジュバント無しのHAが鼻腔内投与経路を通じてほんの低レベルの全身IgG抗体(5.0μg/ml)を誘発することを示している。少量のGEM粒子(0.03mg)の添加は、ただでさえこのレベルを4倍に増加させる。最良の増強は、0.1mg GEM粒子の添加により、1ml当たり約67μg HA特異的IgGまで見出され、さらに多くのGEM粒子の添加によりさらに増加することはなかった。これらの結果は、GEM粒子をインフルエンザHAと混合することが、用量依存的方法で抗原特異的免疫応答を増強することを明らかに示している。
【0057】
(実施例2)筋肉内HAと比較したHAと混合した鼻腔内GEM
比較は、鼻腔内HA+GEMワクチンと、HAワクチンが投与される伝統的な方法、すなわち筋肉内経路を通じて投与される、アジュバント無しのHAの間で行った。マウスは、投与間が2週間間隔による、i.n. HA (5μg H1N1 A/Beijing)+GEM (0.15mg乾燥重量)またはi.m. HA (5μg)の3回の投与を受けた。鼻腔内および筋肉内ワクチンの免疫応答の大きさおよび動態を比較するため、HA特異的血清IgG力価を、各免疫の2週間後に採取した試料について決定した。図2は、i.n. HA+GEMワクチンの大きさおよび動態のいずれもがi.m. HAワクチンのものと類似することを明らかに示している。各投与後のi.n.ワクチンとi.m.ワクチンの反応の間に統計的有意差はない(各p値>0.05)。
【0058】
(実施例3)HAと混合した鼻腔内GEMは防御反応を誘発する
インフルエンザワクチンの防御能力は、HI価を測定して決定する。HI価は、i.n. HA (5μg H3N2 A/Wisconsin)、HA+GEM (0.3mg乾燥重量)、i.m. HAによる1回目および2回目のブースター免疫化後の全マウスについて決定した。
【0059】
図3は、従来のi.m.およびGEMアジュバントi.n.ワクチンがいずれも、1回目のブースター免疫化後に2log6を超える同程度のHI価に達したことを示している(p=0.2062)。これらの力価は、両方のケースで2log7から2log8の間の値まで増加し、2つの治療の間に有意差はない(p=0.7611)。サブユニットワクチン単独によるi.n.免疫化は、2回のブースター免疫化の後でさえ低いHI価を誘導した。さらに、動物の50%のみがi.n.サブユニットワクチンによる免疫後に応答したのに対し、全動物が2つの他のワクチン群で応答した。2log5.3を超えるHI価はヒトにおいて防御的であると考えられることから、これらの結果は、単回ブーストが、i.n. GEMアジュバントインフルエンザワクチンが防御免疫に達するのに十分であることを示している。サブユニットワクチン単独によるi.n.免疫化およびi.m.免疫化の両方と比べて、GEM粒子を含むサブユニットワクチンの製剤が強力な全身性免疫応答を誘導したことは、結果から明らかである。
【0060】
(実施例4)HAと混合した鼻腔内GEMの粘膜性免疫応答
i.n.免疫化は、気道、すなわちインフルエンザウイルスの侵入口において局所粘膜免疫を誘導する可能性があることが以前に報告されている。粘膜免疫の活性化は、下部のBおよびT細胞をプライミングし、粘膜部位でのsIgAの分泌をもたらす。それ故に、インフルエンザ特異的sIgA力価を、マウスの鼻および肺洗浄において決定した(図4)。
【0061】
i.m.免疫化は、マウスのほとんどで検出限界より下の鼻および肺洗浄においてsIgAレベルを誘発した(8マウス中1匹のみは鼻洗浄での応答を示した)。同様に、サブユニットワクチン単独によるi.n.免疫化は、肺および鼻洗浄において低いsIgA力価を示した(3/8レスポンダー)。対照的に、HA+GEMによるi.n.免疫化は、全マウスの鼻および肺洗浄において高いsIgA力価を誘導した。
【0062】
結論として、HA+GEMによるi.n.免疫化は、上および下気道の両方で強力な粘膜性免疫応答を誘導した。
【0063】
(実施例5)HAと混合した鼻腔内GEMの免疫応答の表現型
応答の表現型、すなわちTヘルパー1/Tヘルパー2比(Th1/Th2)を評価するため、IgGサブタイプ、IFN-γおよびIL-4応答を決定した。
【0064】
IgGサブタイププロファイリング(図5)は、サブユニットワクチン単独によるi.n.免疫化が低いIgG1、IgG2aおよびIgG2b応答を誘導することを示した。以前に報告されているように、サブユニットワクチンによるi.m.免疫化は高いIgG1応答を誘導したが、IgG2aおよびIgG2bをほとんど誘導しなかった。これは、Th2応答に偏った免疫応答を示している。i.m.免疫化と比較して、HA+GEMによるi.n.免疫化は、有意により高いIgG2a (p=0.042)およびIgG2b (p=0.030)およびより低いIgG1 (p=0.0135)応答を誘導した。これらの結果は、i.n. HA+GEMワクチンにより生じた抗体反応が、従来のi.m.ワクチンよりもTh1表現型に有意に偏向していることを示している。
【0065】
免疫応答の型(図6)を、免疫化マウスの抗原特異的IFN-γおよびIL-4産生脾臓細胞を決定してさらに評価した。サブユニットワクチンによるi.m.免疫化は、IFN-γ産生細胞よりも多数のIL-4産生細胞をもたらし、この場合もやはり優勢なTh2応答を示している。サブユニットワクチンによるi.n.免疫化は、より少数のIL-4産生細胞であるが、著しくより多数のIFN-γ産生細胞(図6)をもたらし、バランスの取れたTh1/Th2応答をもたらした。IFN-γ産生T細胞の増加は、HA+GEMによるi.n.免疫化後、一段と有意に(p=0.0373)より顕著であった。これは、バランスの取れたTh1/Th2から優勢なTh1応答への免疫応答の移行を示している。
【0066】
(実施例6)致死的チャレンジモデルにおけるHAと混合した鼻腔内GEMの防御
i.n. HA+GEMにより生じた免疫応答の防御能力を、致死的チャレンジモデルにおいて評価した。マウスを、PBS(偽免疫化)またはHA単独(2回)、HA+GEM (2回)もしくはHA+GEM (3回)でi.n.免疫化した。比較は、筋肉内に投与したHAベンチマークワクチンにより行った。この実験におけるHAは、株PR8 (H1N1)に由来した。用量は、1用量当たり5μg HAであり、GEMケースでは1用量当たり0.3mg GEMをワクチンに添加した。ワクチンは、2週間間隔で投与した。致死的チャレンジは、PR8の致死的用量による最後のブースター免疫化の3週間後に行った。チャレンジに対する防御を、群HA+GEM (i.n.2回、9/9生存者)、HA+GEM (i.n.3回、9/9生存者)およびHAベンチマーク対照(i.m.、9/9生存者)の動物について観察した[図7]。これらの群内の全動物は、チャレンジ後に臨床徴候(嗜眠、乱れた毛並みまたは背中が丸まった姿勢)を示さず、実験終了までの14日目まで生存した。防御は、体重減少の欠如と相関した(不図示)。
【0067】
対照的に、群HA i.n.およびPBS (偽免疫化、陰性対照)内のほとんどの動物は、それぞれ、3日目から、および4日目以降、重篤な体重減少を示し、重篤な臨床症状(体重<85%、嗜眠、乱れた毛並み、丸まった背中)のためチャレンジ後5日目から8日目に安楽死させた。
【0068】
チャレンジの4日後の肺におけるウイルス力価の決定は、HA+GEM (2または3回)によるi.n.ワクチン接種が、PBS陰性対照群と比べて、チャレンジの4日後の肺でウイルス力価の約1,000から10,000倍の減少をもたらすことを示した(図8)。チャレンジした際のウイルス力価の極めて限定された減少(4倍の減少)が、HAを単独でi.n.適用した場合に観察された。これは、防御を提供するのにGEMのアジュバント特性が必要とされることを示している。HA+GEM (2および3回)によるワクチン接種は、ベンチマーク陽性対照群(HA、i.m)と比べて、肺におけるウイルス力価の約20から100倍の改善をもたらした。ウイルス力価の減少は、ウイルスの排出の減少をもたらすことができ、集団防御(herd protection)を提供するうえで重要な要因であると考えられる。実施例4および5で示したようなi.n. HA+GEMワクチンにより生じた、気道の粘膜内層における局所IgAの存在および/またはより良好にバランスの取れたTh1/Th2型の免疫応答は、i.m.ベンチマークワクチンと比べて、観察された防御の優位性を説明することができよう。
【0069】
(実施例7)GEMと混合した経口HAは防御反応を誘発する
経口投与経路は、この簡便さのためワクチンにとって魅力的であるが、抗原が不活化または劣化されるため、有効性を欠く場合が多い。アジュバント無しのHAの経口投与は、防御的な血清HI応答および/または粘膜IgA応答を誘発するのに不十分であることが知られている。経口胃免疫化においてGEMをHAに添加する効果を、マウスモデルで分析した。H3N2 A/Hiroshimaサブユニット抗原HA (20μg/1回用量)を使用した。HA+GEMワクチンは、1用量当たりさらに0.3mg GEMを含有した。マウスを2週間間隔で3回免疫化し、最終免疫の2週間後の試料を分析した。免疫化の防御能力を比較するため、血清HI価を決定した。図9に示されているように、HA+GEMワクチンによる経口免疫は、GEM粒子無しの経口免疫よりも有意に高い(p<0.05) HI価を誘導した。経口HA+GEM群ではHI価は、防御のカットオフレベルの2log5.3を十分に超える、2log7超に達した。
【0070】
さらに、経口HA+GEMは、胃腸管においてかなりのレベルの粘膜IgAを惹起することができた(図10)。驚くべきことに、気道での強い局所IgA応答も動物のほとんどで誘発された。
【0071】
これらの結果は、GEMと混合した経口インフルエンザHAワクチンも、防御全身性免疫応答を誘発したこと、およびさらに気道を含む強力な粘膜応答を誘発することを示している。
【0072】
(実施例8)GEMと混合した筋肉内HAは粘膜表面での局所応答を誘発する
非経口ワクチンは通常、粘膜分泌IgAの産生を誘発しない。筋肉内免疫化マウスの粘膜試料の分析では、本発明者らは、HA+GEMを受けたマウスはが、鼻、肺および膣などの幾つかの粘膜組織で局所IgAを分泌することを驚くべきことに見出した。メスマウスに、固定量のHA (5μg B/Shangdong/7/97)、0.3mg GEM有りまたは無しにより鼻腔内または筋肉内に3回(間隔14日)ワクチン接種した。最後の免疫の2週間後、鼻および膣洗浄を実施し、IgA力価を特異的ELISAアッセイにより決定した。
【0073】
図11におけるデータは、HA+GEMの鼻腔内投与が局所IgA応答を効率的に誘導したことを示し、鼻洗浄におけるIgA力価として証明された。IgA力価は遠位でも誘導され、膣洗浄におけるIgA力価の出現として証明された。予想通り、HA単独の筋肉内投与は、関連局所IgA応答を誘導しない。驚くべきことに、HA+GEMの筋肉内投与は、鼻腔内投与後に達せられたものに迫る効率で鼻および膣の両方で関連IgA力価を誘導した。したがって、HA+GEMの筋肉内投与は、粘膜性免疫応答を誘導するのに使用することができる。
【0074】
(実施例9)GEMと混合したHAの筋肉内投与は十分な用量節約を支持する
筋肉内GEM+HAにより誘発された免疫応答が、インフルエンザHA抗原の用量節約を可能にするかどうかを決定するため、マウスを、PBS (偽治療)、GEM無しの1μg HA (A/PuertoRico/8/34)またはGEM (1用量当たり0.3mg)と共に処方した0.04μg HA (25倍少ない抗原)により2回ワクチン接種した。最終用量の投与の2週間後、マウスをマウス適応A/PuertoRico/8/34でチャレンジした。チャレンジの5日後、動物を屠殺し、肺を分離およびホモジナイズし、ウイルス力価をMDCK細胞でのエンドポイント滴定により決定した。
【0075】
図12、パネルAは、1μg HAによる動物の筋肉内ワクチン接種が、偽治療動物と比べて、1logを超える感染動物の肺におけるウイルス負荷の減少を提供することを示している。しかし、HA+GEMは、肺力価の完全な欠如により証明されるように、感染動物の肺におけるインフルエンザウイルスの複製に対し完全な防御を提供する。これらの結果は、ベンチマークi.m. HAと比べてi.m. HA+GEMワクチンの優位性を示している。
【0076】
ベンチマークi.m. HAについてと同じレベルの防御が、図12のパネルBに示されているように、わずか0.04μg HA (25倍少ない抗原)を含有するHA+GEM製剤で達成された。これは、大幅な抗原節約が、GEMを含む筋肉内インフルエンザワクチンを処方して達成できることを示している。
【0077】
(実施例10)鼻腔内GEMベースのB型肝炎ワクチンはマウスにおける強力な全身性IgGおよび局所IgA応答を誘発する
成体C57BL6マウスを、HBsAg抗原を含有するGEMベースのB型肝炎ワクチンで免疫化した。このケースでは、HBsAg [5μg]はGEM粒子[0.15mg乾燥重量]と混合した。GEM無しの等量のHBsAgも、比較のため使用した。ワクチンは、鼻腔内経路を通じて投与した。ミョウバンによるアジュバントである市販のHepBワクチンVaxProを、ベンチマークワクチンとして皮下投与した。血清IgGを、完全免疫化(3回用量、10日間隔により投与)後に測定した。図13は、鼻腔内ワクチンでのGEM粒子のアジュバント効果を明らかに示している。HBsAg特異的血清IgG応答は、HBsAg単独を鼻腔内投与した場合、測定不能であった。対照的に、HBsAg+GEMは、4.2の希釈力価を有する活発なHBsAg特異的血清IgG応答を誘発した。鼻腔内GEM-HBsAgワクチンは、皮下経路を通じて投与したベンチマークワクチンと似たHBsAg特異的IgGを誘発した(p=0.2290)。
【0078】
粘膜免疫の活性化は、粘膜部位でのsIgAの分泌をもたらす。この実験では、HBsAg特異的sIgAの局所分泌を、ワクチン接種部位(鼻)の洗浄および遠位粘膜部位(膣)での洗浄において測定した。
【0079】
図14は、sIgA応答がi.n. HBsAg+GEMワクチンを用いてのみ生じ、i.m. VaxProワクチンでは生じないことを明らかに示している。i.n. HBsAg+GEMワクチンは、膣の粘膜部位などの遠位粘膜部位でのsIgAの分泌でさえ生じる。
【0080】
(実施例11)ラットモデルにおける鼻腔内GEM-HBsAg B型肝炎ワクチンは防御レベルの血清抗体を誘発する
アジュバントB型肝炎ワクチンは、HBsAg抗原(25μg)をGEM (0.4mg)と混合して作製した。比較には、HBsAg抗原単独(25μg)、およびミョウバンと共に処方した同じ抗原を含有するベンチマークワクチン(VaxPro)。完全免疫は、10日間隔で投与した3回のワクチン投与から成った。最終血清を、最後のブースターの14日後に回収した。GEM-HBsAgおよびHBsAgは鼻腔内投与した。VaxProは、筋肉内経路を通じて投与した。B型肝炎ワクチンについては、防御の相関が知られている。血液血清の10mIU/mlより高い抗体レベルは防御的であると考えられ、防御に関する代理マーカーとして認められている。
【0081】
完全免疫化ラット(1群当たり4匹のウィスターラット)の血液血清を、1ml当たりのミリ国際単位(mIU/ml)で表されるHBsAg特異的抗体のレベルについて分析した。図15は、結果をまとめている。鼻腔内HBsAgは、反応を全く誘発しない。高いおよび防御レベルの抗体反応(mIU/ml>10)は、HBsAgをペプチドグリカン微粒子と共に処方した場合に、鼻腔内経路を通じて得られる。防御のレベルは、筋肉内経路を通じて投与したベンチマークワクチンVaxProと類似する(p=0.7715)。
【0082】
実施例10および11における結果は、抗原はGEM粒子に積極的に結合されないという事実にもかかわらず、強力な全身抗体反応および局所抗体反応が、鼻腔内GEMベースのB型肝炎HBsAgワクチンにおいて惹起されることを一貫して示している。
【0083】
(実施例12)三価肺炎球菌タンパク質ベースのGEMワクチンの防御能
比較は、タンパク質に混合または結合したGEMと共に処方した鼻腔内肺炎球菌タンパク質ベースのワクチン間で行った。3つの保存された肺炎球菌タンパク質(PspA、CbpA、PdBD)を、三価ワクチン、GEM+P3 (混合)およびGEM-P3 (結合)において使用した。マウスは、これらのワクチンまたは陰性対照(偽免疫化)としてのPBSにより、用量間が10日間隔で3回免疫化した。各GEMベースのワクチンは、1用量当たり5μgの各抗原および0.3mg GEMを含有した。最後のブースター免疫化の3週間後、マウスを、致死的用量肺炎連鎖球菌TIGR4 (血清型4)で鼻腔内チャレンジした。防御されないマウスは、チャレンジ後72時間以内に死亡した。マウスを、チャレンジ後14日間追跡した。マウスは、人道的エンドポイント(チャレンジの48時間後に1ml血液当たり>5.4×103コロニー形成単位(cfu)、体重<85%、嗜眠、乱れた毛並み、丸まった背中)に基づき、または試験終了時に安楽死させた。偽免疫化マウスはいずれも生存しなかった。GEM+P3 (混合)ワクチンで免疫化した群は、GEM-P3 (結合)ワクチンで免疫化した群(20%)よりも良好な生存(50%)を示すことが、驚くべきことに見出された(図16を参照のこと)。これらの結果は、P3タンパク質によるGEMワクチンは、これらのタンパク質をGEM粒子に混合した場合により効果的であることを明らかに示している。
【0084】
(実施例13)五価肺炎球菌タンパク質ベースのGEMワクチンの防御能
比較は、タンパク質に混合または結合したGEMと共に処方した鼻腔内肺炎球菌タンパク質ベースのワクチン間で行った。5つの保存された肺炎球菌タンパク質(PspA、PsaA、CbpA、PpmA、IgA1prt)を、五価ワクチン、GEM+P5 (混合)およびGEM-P5 (結合)において使用した。マウスは、これらのワクチンまたは陰性対照(偽免疫化)としてのPBSにより、用量間が10日間隔で3回免疫化した。各GEMベースのワクチンは、1用量当たり0.5μg IgA1prt、3μg PsaA、1.5μg CbpA、2μg PpmA、2μg PspAおよび0.3mg GEMを含有した。最後のブースター免疫化の3週間後、マウスを、致死的用量肺炎連鎖球菌D39 (血清型2)で鼻腔内チャレンジした。防御されないマウスは、チャレンジ後72時間以内に死亡した。マウスを、チャレンジの40時間後健康状態を、臨床症状(嗜眠、乱れた毛並み、丸まった背中)に基づき採点し、ワクチンの防御能力を測定するためのエンドポイントとして見なした。図17は、GEM-P5 (結合)ワクチンで免疫化した群では、10マウス中8匹が完全に健康なままであったのに対し、これがGEM+P5 (混合)ワクチン(5/10)についてはより少なく、陰性対照(1/10)については少ないことを示している。これらの結果は、P5タンパク質によるGEMワクチンが、これらのタンパク質をGEM粒子に結合した場合に、より効果的であることを明らかに示している。
【技術分野】
【0001】
本発明は、アジュバントワクチン製剤、特に、例えば鼻腔内または筋肉内送達の際に粘膜性免疫応答を高めることができるインフルエンザワクチンに関する。
【背景技術】
【0002】
季節性インフルエンザは、依然として世界中で死亡率および罹患率の主な原因の一つである。年に1回のワクチン接種は、インフルエンザ感染を予防および制御する最も有効な戦略である。季節性インフルエンザワクチンは、来シーズンの流行予測株の予想に基づいて調製される。これらは、感染自体を予防しない非経口注射ワクチンであり、感染後の重症度および合併症を減少させる。非経口ワクチンは、血清中に中和IgG抗体を誘導することができるが、これらは、粘膜表面で作用する分泌IgA抗体を誘導することができない。対照的に、鼻腔内(i.n.)ワクチンは、全身性および粘膜性免疫応答の両方を誘導する可能性がある。粘膜表面の分泌IgA抗体は、インフルエンザウイルス感染の最初の標的である上皮細胞表面に病原体が結合する前に粘膜の表面で反応するため、感染を予防するのに極めて有効である。さらに、血清IgG抗体は、分泌IgA抗体よりも特異的に作用するため、連続変異ウイルス株(drifted viral strains)に対する有効性が低い。分泌IgA抗体は、インフルエンザウイルスのバリアント株に対する交差防御効果を有する。IgAの交差防御効果の正確な機構はいまだ不明であるが、この現象は感染を予防するのに大きな利点である。インフルエンザは、インフルエンザの2つの主な膜タンパク質、ヘマグルチニン(HA)およびノイラミ二ダーゼ(NA)の抗原特性を変化させる驚異的な能力を発揮する。これは、ヒト集団で確立された適応免疫応答から離れた連続選択により起こる。このウイルスの高い変異率のため、特定のワクチン製剤は通常、約1年間しか効かない。世界保健機関は、毎年ワクチンの内容物を調整して翌年に襲うウイルスの最も可能性の高い株を含有するようにしている。最近では、従来のワクチンは、3つの不活化インフルエンザウイルス(2つのA型株および1つのB型株)から成るワクチンである。この三価インフルエンザワクチンは、次回のインフルエンザシーズンに蔓延するとWHOにより予想されたインフルエンザ株に基づき、毎年再処方される。例えば、2007〜2008年シーズンの毎年更新される三価インフルエンザワクチンは、インフルエンザH3N2、H1N1、およびインフルエンザB型ウイルス由来のヘマグルチニン(HA)表面糖タンパク質成分から成る。
【0003】
ワクチンのi.n.送達の他の利点は、ワクチンの送達が、ワクチンの投与に訓練された医療従事者を必要とせず、このタイプのワクチンを針恐怖症の人に適したものにし、および針刺し事故の問題を回避する点である。さらに、粘膜免疫系は年少期に発達し、老化による影響を受けないことが報告されている(McElhaney JE. Vaccine 2005年7月8日;23 付録1:S10〜25頁; Szewczuk MRら Ann N Y Acad Sci 1983年6月30日;409:333〜44頁)。したがって、例えば鼻腔内インフルエンザ免疫化の付随する利点は、有効な免疫を全ての年齢層に潜在的に提供することができる点、および集団予防接種に使用することができる点である。経鼻または口腔咽頭経路を介し、および不活化インフルエンザ抗原を用いる、インフルエンザに対する免疫化のさまざまな概念は、皮下または筋肉内免疫の無針(needle-less)代替案として探究されている。無針アプローチを支持する実験データは動物モデルで生まれている。動物データにより支持されている、鼻腔内経路を介した免疫化のため不活化インフルエンザ抗原(化学的に不活化された全ウイルス粒子、もしくはスプリットウイルスなどのさらに加工されたウイルス成分、または精製表面抗原ヘマグルチニン(HA)および/もしくはノイラミ二ダーゼ(NA)など)を用いる概念は、不活化インフルエンザ抗原と併用してアジュバントもしくは免疫刺激剤のいずれかの使用を含み、または複数回のワクチン接種を必要とする。アジュバントは、これと混合された抗原の免疫原性を増強する任意の物質である。ヒトにおいて鼻腔内経路を介したインフルエンザに対する成功したワクチン接種は、(a)生(寒冷適応株)インフルエンザワクチン(FluMist(商標)、MedImmune Vaccines Inc社)、(b)大腸菌(E. coli)の易熱性毒素でアジュバントされたビロソーム(virosomal)インフルエンザワクチン(NasalFlu、Berna Biotech Ltd社)、または(c)多量の抗原および反復ワクチン接種の使用について報告されているのみである。生ワクチンは十分な免疫応答を誘導することができるが、生ウイルスであるという特異性は、安全性に対する別の懸念の原因となり、および上気道で必要とされるウイルス複製ラウンドによる副作用を誘導する可能性がある。また必要とされる貯蔵条件も、これらの製品の商業化を制限している。アジュバントとしての大腸菌HLTによる鼻腔内インフルエンザワクチンの使用と顔面神経麻痺(ベル麻痺)との強い関連は、市場からHLTアジュバントビロソームワクチンの撤退をもたらした。
【0004】
現在は、生弱毒化インフルエンザウイルスワクチン(LAIV)がi.n.投与用に市販されている。LAIVワクチンは、全身性および粘膜性免疫応答の両方を誘導することが示されている。しかし、LAIVワクチンは、2〜49歳の者についてのみFDAにより認可されており、ハイリスク集団(高齢者、子どもおよび慢性疾患患者)での使用については認可されていない(疾病対策予防センター。http://www.cdc.gov/flu/professionals/vaccination/pdf/targetpopchart.pdf ; Belshe RBら。Vaccine 2008年9月12日;26 付録4:D10〜6頁)。しかし、市販されたインフルエンザワクチンのほとんどは、集団全体にi.n.経路を介して安全に投与することができる不活化ワクチンである。これらのワクチンの欠点は、この経路を介して投与される場合、免疫原性が低いことを示した点である(Vaccine 2007年7月20日; 25 (29):5367〜73頁; Eylesら、BioDrugs 2000年1月;13(l):35〜59頁)。
【0005】
免疫原性を増加させるため、不活化インフルエンザワクチンは、i.n.経路を介して投与される場合、免疫応答を強化するアジュバントを必要とする。ウイルス様粒子(Matassov Dら。Viral Immunol 2007年9月;20(3):441〜52頁)、ISCOM (Sjolander Sら。Vaccine 2001年7月16日;19(28〜29):4072〜80頁)、脂質、核酸(Josephら。Vaccine 2006年5月1日;24(18):3990〜4006頁)および細菌成分(Haanら。Vaccine 2001年4月6日;19(20〜22):2898〜907頁; Planteら。Vaccine 2001年10月12日;20(1〜2):218〜25頁)のような幾つかのアジュバントが、i.n.免疫化用に現在開発中である。しかし、これらのアジュバント系の多くの開発が、安全性および規制上の懸念により阻まれている。例えばLT(大腸菌の易熱性毒素)のような強力な細菌アジュバントは、ヒトにおける重篤な副作用を示した(Mutschら。N Engl J Med 2004年2月26日;350(9):896〜903頁)。インフルエンザワクチンについて示唆されているように、アルミニウム塩アジュバントの封入は、製造中に余分な混合工程を必要とし、これにより製造全体が減速するだけでなく、これらの塩の封入はさまざまな問題と関連する。例えば、この不溶性は、吸収抗原が懸濁液から沈降することを意味し、そのためバルクワクチンから個々の用量を調製することは、余分な注意を必要とする。加えて、塩への抗原の結合は、最終ワクチンの品質管理を複雑にする。特に、インフルエンザワクチンの幾つかの力価試験は、非結合抗原を必要とするインビトロ免疫アッセイに基づいており、すなわちアジュバントへの吸着は、これらの試験が使用できないことを意味する。
【0006】
近年、免疫応答の表現型、すなわちTh1、Th2またはバランスの取れた応答(blanced response)に重点が置かれている。i.n.経路を介して投与されるサブユニットワクチンならびにキトサン、ISCOM、脂質、およびLTのような経鼻アジュバントの多くは、混合Th1/Th2型応答を誘導する。しかし、Th1応答は、1)より良好な感染防御をもたらす、および2) INF-γの分泌によるウイルス中和に役立つため、Th2または混合応答より優れていると考えられる。さらに、自然感染もまたTh1型の応答を誘導する。加えて、粘膜表面の分泌IgA抗体は感染予防に極めて有効であり、重要なことには、分泌IgA抗体はインフルエンザウイルスのバリアント株に対する交差防御効果を有し、粘膜免疫系は年少期に発達し、老化による影響を受けない。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】WO 02/101026
【特許文献2】US 6,896,887
【特許文献3】EP 1395648
【特許文献4】WO 2007/011216
【非特許文献】
【0008】
【非特許文献1】McElhaney JE. Vaccine 2005年7月8日;23 付録1:S10〜25頁
【非特許文献2】Szewczuk MRら Ann N Y Acad Sci 1983年6月30日;409:333〜44頁
【非特許文献3】疾病対策予防センター。http://www.cdc.gov/flu/professionals/vaccination/pdf/targetpopchart.pdf
【非特許文献4】Belshe RBら。Vaccine 2008年9月12日;26 付録4:D10〜6頁
【非特許文献5】Vaccine 2007年7月20日; 25 (29):5367〜73頁
【非特許文献6】Eylesら、BioDrugs 2000年1月;13(l):35〜59頁
【非特許文献7】Matassov Dら。Viral Immunol 2007年9月;20(3):441〜52頁
【非特許文献8】Sjolander Sら。Vaccine 2001年7月16日;19(28〜29):4072〜80頁
【非特許文献9】Josephら。Vaccine 2006年5月1日;24(18):3990〜4006頁
【非特許文献10】Haanら。Vaccine 2001年4月6日;19(20〜22):2898〜907頁
【非特許文献11】Planteら。Vaccine 2001年10月12日;20(1〜2):218〜25頁
【非特許文献12】Mutschら。N Engl J Med 2004年2月26日;350(9):896〜903頁
【非特許文献13】van Roosmalen MLら。Methods 2006年2月;38(2):144〜9頁
【非特許文献14】Audouy SAら。Vaccine 2007年3月22日;25(13):2497〜506頁
【非特許文献15】J Virol。1991年7月; 65(7): 3789〜3796頁
【非特許文献16】Amorij JPら。Vaccine 2007年12月21日;26(1):67〜76頁
【発明の概要】
【発明が解決しようとする課題】
【0009】
故に、ヒト用の強力、安全で、規制機関により容易に承認され得るアジュバントの必要性がある。好ましくは、分泌IgA抗体のような粘膜性免疫応答および/またはTh1型免疫に偏った応答を誘導することができるワクチンが望ましい。したがって、さらなるおよび改善されたアジュバントインフルエンザワクチン(パンデミック用およびパンデミック間期用の両方の)、好ましくは鼻腔内および/または筋肉内送達に適したワクチンを提供することが、本発明の目的である。さらなる目的は、簡便および費用効果の高い年に1回の再処方を可能にするインフルエンザワクチン調製の柔軟な方法を提供することである。
【課題を解決するための手段】
【0010】
上記の目標は、グラム陽性細菌から得られる不活化ペプチドグリカン粒子と共に抗原を同時処方することによって達成できることが見出された。該粒子は、鼻腔内投与されるサブユニットワクチンの免疫原性を増強するのに極めて有効であるだけでなく、分泌IgAを誘導し、バランスの取れた免疫応答からTh1偏向(Th1-skewed)免疫応答へと応答を調節する。鼻腔内送達は、サブユニットインフルエンザウイルス単独による従来の筋肉内免疫化と比較した場合、同程度の全身性免疫ならびにさらにはより優れた粘膜性および細胞媒介性免疫を誘導した。防御効果は、抗原と細菌粒子の単純な混合により達成することができた。
【0011】
したがって、本発明は、(i)グラム陽性細菌から得られるペプチドグリカン微粒子と、(ii)少なくとも1つの抗原またはこの抗原性調製物とを含み、該抗原または抗原性調製物がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない、アジュバントワクチン製剤に関する。いずれかの既知のまたはまだ発見されていない、例えばウイルス、細菌、寄生虫、真菌または酵母起源の防御抗原もしくはこの抗原性フラグメントが含まれ得る。
【0012】
1つの実施形態では、抗原は、B型肝炎表面抗原またはインフルエンザウイルス抗原などのウイルス抗原である。
【0013】
別の実施形態では、製剤は、細菌抗原、好ましくは少なくとも2つの細菌タンパク質性の抗原またはこの抗原性調製物を含み、抗原または抗原性調製物は、タンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない。2つ以上のタンパク質性抗原またはこの抗原性フラグメントのいずれかの既知のまたはまだ発見されていない防御的組み合わせ(肺炎連鎖球菌(Streptococcus pneumoniae)抗原PpmA、SlrA、IgA1プロテアーゼ、PspA、CbpA、PdBD等の組み合わせ、またはペスト菌(Yersinia pestis)抗原LcrV、F1、FliCの組み合わせ、またはネズミチフス菌(Salmonella typhimurium)、エルシニア・エンテロコリチカ(Yersinia enterocolitica)、赤痢菌(Shigella)のLcrV、IpaBおよびD、SipBおよびD、YopDなどのIII型分泌経路抗原の組み合わせ、または腸管毒素性大腸菌(enterotoxic Escherichia coli) (ETEC)のLTおよびST抗原の組み合わせ、または他の細菌タンパク質性抗原の組み合わせなど)が含まれ得る。1つの態様では、本発明は、PspA、CbpAおよび/またはPdBDを含む防御製剤を提供する。特に興味深いのは、細菌タンパク質性抗原がPspA、CbpAおよびPdBDである本発明の肺炎球菌三価ワクチン製剤である。グラム陽性細菌から得られるペプチドグリカン微粒子と混合すると、3抗原のこのカクテルは、肺炎連鎖球菌感染の鼻腔内チャレンジマウスモデルにおいて極めて良好な防御を与えることが見出された。驚くべきことに、混合後の防御活性は、タンパク質性のペプチドグリカン結合部分への融合を介して抗原が該粒子に結合される場合よりも高かった(本明細書の以下の実施例12および図16を参照のこと)。また、抗原PpmA、IgA1プロテアーゼ、PspA、CbpAおよびPsaAを含む五価製剤は、ペプチドグリカン粒子と混合された場合、結合される抗原と比べて相対的に低いことから、抗原の特定の組み合わせも防御活性に関連しているようである。実施例13および図17を参照のこと。
【0014】
さらなる実施形態では、製剤は、少なくとも1つの寄生虫抗原またはこの抗原性調製物を含み、抗原または抗原性調製物は、タンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない。熱帯熱マラリア原虫(Plasmodium falciparum)のスポロゾイト周囲表面抗原またはメロゾイト表面抗原などの、いずれかの既知のまたはまだ発見されていない防御寄生虫抗原またはこの抗原性フラグメントが含まれ得る。例示的な防御真菌抗原には、コクシジオイデス属(Coccidioides)亜種の抗原が含まれる。適切な酵母抗原は、カンジダ属(Candida)亜種の抗原である。同様に、提供されるのは、グラム陽性細菌から得られるペプチドグリカン微粒子、および少なくとも1つの多糖体抗原またはこの抗原性調製物を含み、該抗原または抗原性調製物がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない、アジュバントワクチン製剤である。肺炎連鎖球菌、インフルエンザ菌(Haemophilus influenza)、髄膜炎菌(Neisseria meningitides)、黄色ブドウ球菌(Staphylococcus aureus)莢膜多糖体または他の多糖体などの、いずれかの既知のまたはまだ発見されていない防御多糖体抗原またはこの抗原性フラグメントが含まれ得る。
【0015】
好ましい実施形態では、本発明は、(i)グラム陽性細菌から得られるペプチドグリカン微粒子と、(ii)少なくとも1つのインフルエンザウイルス抗原またはこの抗原性調製物とを含み、該抗原または抗原性調製物がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない、アジュバントインフルエンザワクチン製剤を提供する。別の実施形態では、本発明は、(i)グラム陽性細菌から得られるペプチドグリカン微粒子と、(ii)少なくとも1つのB型肝炎ウイルス抗原(例えばB型肝炎表面抗原(HBsAg)などのウイルスエンベロープタンパク質)、またはこの抗原性調製物とを含み、該抗原または抗原性調製物がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない、アジュバントB型肝炎ワクチン製剤を提供する。さらに別の実施形態では、本発明は、(i)グラム陽性細菌から得られるペプチドグリカン微粒子と、(ii)少なくとも1つの肺炎球菌抗原、好ましくはPdBD、より好ましくはPspA、CbpAおよびPdBDとを含み、該抗原がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない、アジュバント肺炎球菌ワクチン製剤を提供する。
【0016】
本発明によるワクチンにおいて使用するペプチドグリカン微粒子は、当技術分野で既知の方法により得ることができる。例えば、グラム陽性細菌の細胞壁材料を得る方法を開示するWO 02/101026およびUS 6,896,887を参照のこと。該方法は、細胞壁成分(前記細胞壁材料由来のタンパク質、(リポ)テイコ酸または炭水化物など)を除去することができる溶液により前記細胞壁材料を処理することを含み、前記細胞壁材料は球状ペプチドグリカン微粒子を本質的に含む。細胞壁材料は、グラム陽性細菌の大きさおよび形状を反映する球状ペプチドグリカン微粒子をもたらすため、機械的に破壊されなかった。粒子は生きておらず、インタクトな表面タンパク質および細胞内内容物が取り除かれている。しかし、厚いペプチドグリカン細胞壁は、インタクトなままであり、およびグラム陽性エンハンサー(Gram-positive enhancer) (GEM)粒子と呼ばれる、細菌の形をした大きさ約1μmのペプチドグリカン球(peptidoglycan sphere)を構成する構造的剛性を提供する。粘膜アジュバント開発の分野における重大な障害は、規制機関による承認を得るためにこの安全性を証明することである。本試験で使用された粒子は、ワクチン接種に関して評価される他のアジュバントおよび他の乳酸菌系と比較して安全に使用できる。粒子の製造中に、細菌は酸で処理され、これが遺伝物質の喪失をもたらす。遺伝物質の喪失は、細菌による粘膜層でのDNA脱落および感染の問題が回避されることから有益である。さらに、粒子は、乳製品の製造に使用される細菌から製造され、およびGRAS生物と見なされる。GEM粒子は、前臨床GLP毒性試験においてウサギで既に鼻腔内試験されており、有害事象は報告されなかった。したがって、GEM粒子は、ヒトでの粘膜使用に安全な候補アジュバントと見なすことができる。
【0017】
1つの実施形態では、ワクチン製剤は、食品用細菌、好ましくは乳酸菌から得られる微粒子を含む。好ましくは、微粒子は、非病原性の、非コロニー形成グラム陽性細菌である食品用細菌ラクトコッカス・ラクティス(Lactococcus lactis)から得られる。さらに、ラクトコッカス・ラクティスは規制機関によりヒト用に承認されており、GRAS (一般に安全と認められる(generally recognized as safe))生物と見なされている。1つの実施形態では、ペプチドグリカン粒子は、酸中のラクトコッカス・ラクティスを加熱し、この後リン酸緩衝液で洗浄して製造される(van Roosmalen MLら。Methods 2006年2月;38(2):144〜9頁)。
【0018】
粒子は、ペプチドグリカン結合ドメイン(AcmA細胞壁結合ドメインまたはこの相同体もしくは機能的誘導体のような)を含むタンパク質性物質と結合する改善された能力のため、マラリア原虫抗原および肺炎球菌抗原の粘膜ワクチン接種のための抗原担体として研究された。これらの研究は、GEM粒子に結合されGEM粒子上に提示された抗原が抗原単独よりも高い免疫応答を誘導することを示した。担体粒子における抗原の固定化および最適表面提示がアジュバント効果に重要であると、当技術分野では一般に考えられていた。
【0019】
本試験は驚くべきことに、抗原が粒子に結合された以前の試験とは対照的に、ペプチドグリカン粒子と抗原の単なる混合が抗原の免疫原性を著しく増強することを示す。しかも、特定のケースでは粒子への結合/固定化と比べて、粒子と混合するとより良好な防御活性を達成することができる。
【0020】
本明細書では表現「抗原または抗原性調製物がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない」は、本発明を従来技術と区別することが意図される。従来技術では、典型的にはAcmA細胞壁結合ドメインまたはこの相同体もしくは機能的誘導体の少なくとも1つの反復配列、しかし好ましくは2つまたは3つの反復配列を含む、「タンパク質アンカー」または「プロタン」(PA)とも呼ばれるタンパク質性物質に抗原を融合または結合して、抗原性部分がペプチドグリカン微粒子に結合される。例えば、EP 1395648は、AcmA型タンパク質アンカー融合を微生物の細胞壁材料に結合する方法を開示している。WO 2007/011216は、免疫原性担体に結合された少なくとも1つの二機能性ポリペプチドを含む抗原負荷免疫原性担体複合体に関し、前記二機能性ポリペプチドは、ペプチドグリカン結合ドメイン(PBD)を含み、PBDを通じてポリペプチドは前記担体に結合され、目的とする少なくとも1つの抗原が結合される抗原結合ドメイン(ABD)に融合される。
【0021】
好ましい実施形態では、抗原は、インフルエンザウイルス抗原である。本明細書で以下に示されるように、本発明によるGEMベースのi.n.インフルエンザワクチンは、Th1表現型に偏った応答を誘発することが驚くべきことに見出された。GEM粒子(ワクチンと単純に混合される)でアジュバントされた鼻腔内投与サブユニットワクチンは、防御の重要な相関であると考えられるHI価(>2log5.3)の防御レベルを誘導するのに、初回−ブーストワクチン接種戦略において使用され得ることが見出された。さらに、GEMベースのi.n.インフルエンザワクチンは、致死的チャレンジ後、完全に防御的となる。加えて、血清IgG結果は、GEM粒子がi.n.投与インフルエンザサブユニットワクチンの免疫原性を増強することを明らかに浮き彫りにする。十分な血清反応に加えて、GEMアジュバントi.n.ワクチンは強力な粘膜性免疫応答、すなわち呼吸粘膜におけるsIgAの分泌を誘発した。鼻粘膜における有意なレベルのsIgAの誘導は、GEM粒子が鼻粘膜で免疫増強物質として作用することを示す。鼻粘膜の免疫系は、鼻関連リンパ組織(NALT)から成る。NALTでは、抗原は、M細胞により取り込まれ、次いで抗原提示細胞に提示され、抗原提示細胞が今度は下部のBおよびT細胞に抗原フラグメントを提示する。この事象のカスケードは、インフルエンザウイルスに対する最初の先天性および適応免疫応答に必要とされる。本発明者らの結果は、GEM粒子と混合されたインフルエンザサブユニットワクチンによるi.n.免疫化は、ワクチンのみによるi.m.およびi.n.免疫化よりも鼻粘膜において高いsIgAレベルを誘導したことを示す。NALTにおけるsIgA抗体の誘導は、GEM粒子がインビトロ試験でTLR-2アゴニストとして作用することが知られていることから、GEM粒子に存在するペプチドグリカンのTLR-2 (Toll様受容体)との相互作用の結果であると考えられる。さらに、GEM粒子は、インビトロで樹状細胞およびマクロファージの成熟を活性化できることが知られている(Audouy SAら。Vaccine 2007年3月22日;25(13):2497〜506頁)。故に、TLR-2および樹状細胞の成熟の活性化はいずれも、より強力な粘膜性免疫応答に寄与した可能性がある。
【0022】
近年、免疫応答の表現型、すなわちTh1、Th2またはバランスの取れた応答に重点が置かれている。Th1応答は、1)より良好な感染防御をもたらす、および2) INF-γの分泌によるウイルス中和に役立つため、Th2または混合応答より優れていると考えられる。さらに、自然感染もまたTh1型の応答を誘導する。しかし、i.n.経路を介して投与されるサブユニットワクチンならびにキトサン、ISCOM、脂質およびLTのような経鼻アジュバントの多くは、混合Th1/Th2型応答を誘導する。対照的に、本発明によるi.n.インフルエンザワクチンは、Th1型に偏った応答を誘発した。故に、GEM粒子は、バランスの取れた応答からTh1偏向応答へと応答を調節する。さらに、本明細書に示されたワクチン製剤は、前処方されなければならない他のアジュバント系のほとんどと比べて、製造するのがはるかにより簡便である。これらの実験で使用された製剤は、GEM粒子を従来のサブユニットワクチンと混合して調製された。GEM粒子は、滅菌条件下で大量に製造することができ、および周囲温度で長時間貯蔵することができる。処方および投与の容易さは、i.n. GEMインフルエンザサブユニットワクチンを、パンデミックおよび流行状態におけるワクチン接種の有望な候補にする。
【図面の簡単な説明】
【0023】
【図1】PBSまたはHA+種々の量のGEM粒子(乾燥重量で表された)で3回免疫化されたマウスにおける、μg/mlで表されるHA抗原(H1N1 A/Beijing)特異的血清総IgGを示す図である。誤差バーは標準誤差(SEM)を示す。
【図2】最初の免疫化後14、28および42日目(それぞれ、第1回、第2回および第3回免疫化)の種々のHA群、すなわちi.n. HA+GEMまたはi.m. HAにおける、HA抗原(H1N1 A/Beijing)特異的血清総IgG希釈力価の比較分析を示す図である。誤差バーはSEMを示す。
【図3】3回免疫化されたマウスの血清におけるHA抗原(H3N2 A/Wisconsin)特異的HI価を示す図である。A.最初の免疫化後0、28および42日目の種々のHA群、すなわちi.m.、i.n.およびi.n.+GEMにおけるHI価の比較分析。B.最初の免疫化後42日目の3つのHA群、すなわちi.m.、i.n.、i.n.+GEM間のHI価の比較分析。カラムの上の数字は1群当たりのレスポンダーの数を示す。誤差バーはSEMを示す。
【図4】HA i.m.、i.n.またはi.n.+GEMで免疫化されたマウスの鼻洗浄(A)および肺洗浄(B)における、HA抗原(H3N2 A/Wisconsin)特異的sIgA力価を示す図である。カラムの上の数字は1群当たりのレスポンダーの数を示す。誤差バーはSEMを示す。
【図5】HA i.m.、i.n.またはi.n.+GEMで免疫化されたマウスの血清におけるHA抗原(H3N2 A/Wisconsin)特異的IgGサブタイプ力価を示す図である。IgG1 (A)、IgG2a (B)およびIgG2b (C)力価が決定された。アスタリスクは示された比較のP値<0.05を意味する。誤差バーはSEMを示す。
【図6】細胞媒介性免疫応答が、HA i.m.、i.n.またはi.n.+GEMで免疫化されたマウスにおけるサイトカイン放出プロフィール、すなわちIL-4 (A)、IFNy (B)を決定して決定されたことを示す図である。アスタリスクは示された比較のP値<0.05を意味する。誤差バーはSEMを示す。
【図7】チャレンジ後の生存(%)を示す図である。動物は1用量当たり5μg HAでワクチン接種され、GEM含有ワクチンは1用量当たり0.3mg GEMを補充された。動物は最後のブースター免疫化の3週間後にチャレンジされ、14日間追跡された。5つのワクチン群間の比較分析。
【図8】チャレンジ後の肺におけるウイルス力価(A/Puerto Rico/8/34 [PR8]、TCID50 [組織培養感染量]) (肺組織1グラム当たり)を示す図である。肺はチャレンジの4日後に分離された。5群間の比較分析。標準誤差(SEM)が誤差バーにより示されている。
【図9】経口HAまたは経口HA+GEMインフルエンザワクチンで免疫化されたマウスにおける、サブユニット抗原(A/Hiroshima [H3N2])特異的血清HI価を示す図である。本発明者らが1用量当たり20μg HAで3回免疫化したマウス。GEMワクチンは1用量当たり0.3mg GEMを含有した。*はp<0.05を示す。2Log5.3を超える力価が防御的である。標準誤差(SEM)が誤差バーにより示されている。
【図10】経口HAまたは経口HA+GEMインフルエンザワクチンで免疫化されたマウスの腸洗浄(グレーのバー)および鼻洗浄(黒のバー)における、サブユニット抗原(A/Hiroshima [H3N2])特異的sIgA力価を示す図である。カラムの上の数字は分析された動物数当たりのレスポンダーの数を示す。標準誤差(SEM)が誤差バーにより示されている。
【図11】0.3mg GEM有りまたは無しの固定量のHA (5μg B/Shangdong/7/97)で、鼻腔内(パネルA)または筋肉内(パネルB)に3回(14日間隔)ワクチン接種されたメスマウスの鼻および膣洗浄における、HA特異的IgA力価を示す図である。洗浄試料は最後の免疫化の2週間後に採取された。標準誤差(SEM)が誤差バーにより示されている。
【図12】GEM無しのPBS (偽)、1μg HA (A/PuertoRico/8/34)またはGEMと共に処方された0.04μg HA (25分の1抗原)で2回ワクチン接種されたマウスの肺ウイルス力価を示す図である。最終用量の投与の2週間後、マウスはマウス適応A/PuertoRico/8/34でチャレンジされた。チャレンジの5日後、動物は屠殺され、肺が分離およびホモジナイズされ、ウイルス力価がMDCK細胞でのエンドポイント滴定により決定された。標準誤差(SEM)が誤差バーにより示されている。
【図13】HBsAg単独(i.n.)、+ GEM (i.n.)またはVaxPro (i.m.)で3回免疫化されたC57BL6マウスの血清における、HBsAg抗原特異的IgG希釈力価を示す図である。誤差バーはSEMを示す。
【図14】HBsAg+GEM (i.n.)またはVaxPro (i.m.)で3回免疫化されたC57BL6マウスの鼻および膣洗浄における、HBsAg抗原特異的sIgA力価を示す図である。誤差バーはSEMを示す。
【図15】HBsAg単独(i.n.)、+GEM (i.n.)またはVaxPro (i.m.)で3回免疫化されたウィスターラットのmIU/mlとして測定されたHBsAg抗原特異的血清反応を示す図である。>10mIU/mlのレベルは防御的であると考えられる。誤差バーはSEMを示す。
【図16】チャレンジ後数日における生存時間。全群の試験材料は鼻腔内(i.n.)適用された。マウスはPBS (偽免疫化)、GEMと混合された肺炎球菌P3タンパク質(PspA、CbpA、PdBD) (GEM+P3)、またはGEMに結合されたP3タンパク質(GEM-P3)で免疫化された。両方のワクチンは5μgの各抗原を含有した。各記号は動物1匹を表す。横線は平均を示す。
【図17】PBS (偽免疫化)、P5タンパク質と混合されたGEM(GEM+P5)、またはP5タンパク質が結合しているGEM (GEM-P5)で免疫化された、病原性肺炎連鎖球菌株D39 (血清型2)による鼻腔内チャレンジの40時間後のマウスの健康状態を示す図である。ワクチンは0.5μg IgA1prt、3μg PsaA、1.5μg CbpA、2μg PpmA、2μg PspAおよび0.3mg GEMを含有した。チャレンジの40時間後の健康状態は、ワクチンの防御能の尺度である。
【発明を実施するための形態】
【0024】
実施例1〜8に示されるように、本発明者らは、GEM粒子でアジュバントされたi.n.インフルエンザワクチンが、サブユニットインフルエンザワクチン単独によるi.m.免疫化と比べて、同程度の全身性免疫ならびにより優れた粘膜性および細胞媒介性免疫を誘導することを示す。特に、該ワクチンは、サブユニットインフルエンザワクチン単独によるi.m.免疫化と比べて、最初のブースター免疫化後にHI価により測定されるのと同程度の防御レベルの免疫を誘導する。重要なことには、該ワクチンは、上気道におけるインフルエンザ感染中の防御の最前線である、より高いsIgAレベルを誘導した。さらに、該ワクチンは、より優れた防御を提供すると考えられる偏向Th1型免疫応答を誘発した。加えて、これらの免疫応答は、GEMベースの鼻腔内インフルエンザワクチンで免疫化されたマウスの完全防御を提供することが示された。
【0025】
実施例7 (図9および10)は、経口投与インフルエンザワクチン組成物の有効性を示す。実施例8 (図11)は、筋肉内GEMベースのインフルエンザワクチンもまた、気道または他の粘膜層の粘膜内層において高いsIgAレベルを誘発するのに使用できることを示す。加えて、筋肉内経路は、通常の筋肉内ベンチマークワクチンの力価を著しく増加する、または大幅に抗原量を減少させる(抗原用量節約)ように、GEM粒子と混合されたインフルエンザワクチンに使用することができる(図12)。GEM粒子は、i.n.、i.m.または経口送達インフルエンザワクチンのための安全および強力なアジュバントと見なすことができる。
【0026】
他の適切なウイルス抗原には、呼吸器合胞体ウイルス(RSV)タンパク質、例えばRSV融合(F)および結合(G)糖タンパク質、またはキメラFGタンパク質などのこれらの関連部分もしくは組み合わせが含まれる(J Virol。1991年7月; 65(7): 3789〜3796頁)。RSV感染は、米国、欧州、オーストラリアおよび日本を含む、世界的に長年のおよび有害な問題となっている。RSV感染は、早産児、幼児、および高齢者において、および実際、弱まった免疫系を有する全ての個体に対して特に厄介である。1歳未満の子どもの約3人に2人、および1歳から4歳の間のほとんど全ての子どもが少なくとも1回はRSVに感染し、医学的注意を必要とすることなくほぼ回復していると推定される。しかし、5〜10%は、小児期後期の喘鳴および喘息様症状の素因と考えられる要因である、重篤な感染を長引かせている。ペプチドグリカン粒子との混合で使用される他の興味深い抗原には、ヒト免疫不全ウイルス(HIV)タンパク質、特に、gp120、gp140またはgp160のようなHIVエンベロープの表面で曝露される糖タンパク質が含まれる。Gp120は、侵入のための特異的細胞表面受容体を探し出すうえで極めて重要な役割を果たすことから、細胞へのウイルス侵入に不可欠である。
【0027】
本明細書で提供されるアジュバントインフルエンザワクチン製剤は、少なくとも1つのインフルエンザウイルス抗原またはこの抗原性調製物を含む。例えば、該ワクチン製剤は、インフルエンザタンパク質もしくはこのフラグメントおよび/またはインフルエンザタンパク質もしくはこのフラグメントを含む融合タンパク質(ペプチドグリカン結合ドメインに融合されないという条件で)を含む。本発明の異種タンパク質は、ヘマグルチニン抗原(HA)、ノイラミダーゼ抗原(NA)またはこれらの組み合わせを含む、目的とする任意のインフルエンザ抗原を含むことができる。好ましくは、インフルエンザ抗原は表面抗原であり、すなわちインフルエンザマトリックスタンパク質2 (M2e)の細胞外ドメインなどの構造抗原ではない。1つの実施形態では、インフルエンザ抗原は、M2e以外である。特定の態様では、インフルエンザワクチン製剤は、インフルエンザ抗原としてHAおよび/またはNAを含有する。さまざまな異なるインフルエンザHAおよびNAタンパク質(例えば、異なるサブタイプ、または株もしくは分離株由来の)のアミノ酸配列は、当技術分野で既知であり、GenBankなどの公開データベースにおいて入手可能である。好ましくは、ワクチン製剤は、少なくとも1つのHAサブタイプを含む。
【0028】
ワクチンで使用するインフルエンザウイルス株は、季節ごとに変化する。現在のパンデミック間期の時期では、ワクチンは典型的には、2つのインフルエンザA型株(H1N1およびH3N2)および1つのインフルエンザB型株を含み、三価ワクチンが典型である。本発明は、新たな「豚インフルエンザ」もしくは「メキシコインフルエンザ」H1などのパンデミック株、またはH2、H5、H7またはH9サブタイプ株(特にインフルエンザA型ウイルスの)などの他のパンデミック株(すなわちワクチンレシピエントおよび一般ヒト集団が免疫学的にナイーブである株)由来のウイルスも使用することができ、パンデミック株のインフルエンザワクチンは一価であってもよく、またはパンデミック株を補充された通常の三価ワクチンに基づいていてもよい。しかし、季節およびワクチンに含まれる抗原の性質に応じて、本発明は、1つまたは複数のHAサブタイプHI、H2、H3、H4、H5、H6、H7、H8、H9、H10、H11、H12、H13、H14、H15またはH16に対して防御することができる。例えば、1つの実施形態では、i.n.ワクチン製剤は、1インフルエンザ株当たり1から15μgの間のHAを含有する。
【0029】
特定の態様では、ワクチン製剤は、少なくとも2つのインフルエンザウイルス株、パンデミック発生に関連するまたはパンデミック発生に関連する可能性がある少なくとも1つの株由来のインフルエンザ抗原またはこの抗原性調製物を含む。
【0030】
ワクチン製剤に存在するペプチドグリカン粒子の量は、好ましくは、初回−ブーストワクチン接種戦略において防御レベルの赤血球凝集抑制(HI)価を誘導するのに十分である。例えば、本発明によるワクチン製剤は、インフルエンザウイルス抗原の1マイクログラム当たり0.001から1mg、好ましくは0.01から0.1mg微粒子(乾燥重量)を含んでもよい。ヒト用の例示的なi.n.ワクチン製剤は、以下を含む: 0.3〜2.5mg GEM (乾燥重量)、三価HA (卵、細胞、組換え) 3×1〜15μgまたは一価HA (パンデミック) 0.1〜15μg、0.05〜0.15M PBS pH6〜8。
【0031】
本発明の経鼻ワクチン組成物は、液体または粉末型組成物、特に、投与方法によりエアロゾル、滴剤、吸入器または吹送として処方することができ、粉末またはミクロスフェアが好ましい。点鼻剤用組成物は、防腐剤、粘性調節剤、浸透圧調節剤および緩衝液などの1つまたは複数の許容可能な賦形剤を含むことができる。しかし、本発明は、経鼻ワクチン製剤に限定されない。GEMの添加がi.m. HAワクチンに対し有効性を強化することが、驚くべきことに見出された。これは、用量節約戦略につながり得る。実施例8および9、図12を参照のこと。したがって、本発明はまた、ペプチドグリカン微粒子および(従来の)筋肉内HAワクチン調製物を含む組成物も提供する。
【0032】
さらに、GEMと混合されたインフルエンザHAは、経口投与した際に防御的な血清HI価をもたらした(実施例7、図9および10)。
【0033】
本発明のさらなる態様は、本明細書に開示されたワクチン製剤を含む容器に関する。1つの実施形態では、容器は、場合により使用説明書と共に提供される、エアロゾルまたは滴下送達システム(鼻腔内スプレー)の形態における装置などの鼻腔内調剤装置である。
【0034】
さらに、本発明は、方法が、本明細書上記に記載されたワクチン製剤を被験者に投与することを含む、被験者におけるインフルエンザ感染または疾患の予防方法を提供する。安全性のため、ワクチン製剤は、ヒトハイリスク集団での使用に特に適している。例えば、本発明はこれにより、高齢者、2歳までの子ども、または慢性疾患患者におけるインフルエンザ感染または疾患の予防のための簡便、安全な、信頼できる方法を提供する。予防方法は、ワクチン製剤の鼻腔内、経口または筋肉内送達、好ましくは鼻腔内送達を含んでもよい。調剤装置、例えばエアロゾルまたは滴下送達システムの形態における調剤装置を使用することが、極めて簡便である。
【0035】
ワクチンの投与量は、免疫応答を効果的に誘導することができる量として決定される。例えば、ヒトに対するワクチンの投与頻度は、1日1回から数回であり、投与量は1〜250μgおよび好ましくは2〜50μgである。
【0036】
本発明により調製されたワクチンは、子どもおよび大人の両方を治療するのに使用することができる。インフルエンザワクチンは、現在、6ヶ月齢から小児および成人予防接種での使用が推奨されている。故に、患者は1歳未満、1〜5歳、5〜15歳、15〜55歳、または少なくとも55歳であってもよい。ワクチンを受けるのに好ましい患者は、高齢者(例えば>50歳、>60歳、および好ましくは>65歳)、若齢者(例えば<5歳)、入院患者、医療従事者、兵役者および軍人、妊婦、慢性疾患患者、免疫不全患者、ワクチンを受ける7日前までに抗ウイルス化合物を服用した患者(例えばリン酸オセルタミビルなどのオセルタミビルまたはザナミビル化合物-以下を参照のこと)、卵アレルギーの人および/または国外旅行者である。しかし、理解されるように、ワクチンは、これらの群にのみ適しているわけではなく、集団においてより一般的に使用することができる。パンデミック株に関しては、全ての年齢群への投与が好ましい。
【0037】
同じく本発明の範囲内に包含されるのは、(a)グラム陽性細菌から得られるペプチドグリカン微粒子を提供する工程と、(b)少なくとも1つのインフルエンザ抗原またはこの抗原性調製物を提供する工程と、(c)微粒子と抗原を混合する工程とを含む、インフルエンザワクチン製剤を提供する方法である。(a)および(b)の両方の工程は、それ自体が当技術分野で知られている方法論を用いて実施することができる。工程(b)は、プロタンのようなペプチドグリカン結合ドメインへの抗原の融合または結合を必要としないことから、本発明の方法は、抗原がペプチドグリカン微粒子に結合するのに、抗原が最初に修飾されなければならない(例えばタンパク質性リンカー部分への融合により)従来技術方法よりもはるかに簡便および商業的に魅力的である。対照的に、本発明は、従来のサブユニットワクチンをそれ自体として用いて実践することができる。
【0038】
したがって、本発明はまた、インフルエンザワクチン製剤におけるアジュバントとしての、グラム陽性細菌から得られるペプチドグリカン微粒子の使用にも関し、前記製剤は、タンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されないインフルエンザウイルス抗原を含む。
【0039】
(実施例)
材料および方法
卵由来の株A/Wisconsin (H3N2)のインフルエンザ一価サブユニットワクチンおよび卵由来のA/Beijing (H1N1)スプリットウイルスワクチンを本試験で使用した。ワクチン中のヘマグルチニン(HA)の濃度は、単一放射免疫拡散アッセイを用いて決定した。
【0040】
ハンゼヌラ・ポリモルファ(Hansenula polymorpha)から単離した組換えHBsAg (ad/ay)を本試験で使用した。Sanofi Pasteur/MSD社からのHBVaxProをベンチマークHBsAgワクチン(40μg/ml)として使用した。GEM粒子は、以前に記載されたように製造した(Van Roosmalenら、Methods 2006年、2月;38(2):144〜9頁)。
【0041】
1.1免疫およびチャレンジ
動物実験は、オランダ動物防御法によって規定されたガイドラインに従って評価および承認された。Balb/c、C57BL6マウス(6〜8週齢)およびウィスターユニリーバ(Unilever)ラット(10週齢)はHarlan社、オランダから購入した。CD 1マウスはCharles River社、ドイツから購入した。マウスを各5〜10匹にグループ化した。ラット群は各4匹から成った。全てのマウス群は、5μgのHAによる0日目の初回ワクチン接種、ならびに14日目および28日目の2回のブースターワクチン接種、または5μgのHBsAgによる0日目、ならびに10日目および20日目の2回のブースターワクチン接種により免疫化した。鼻腔内マウス免疫化は、吸入麻酔下(イソフルラン/O2)、10μlのワクチンを両方の鼻孔に分けて行った。ラット群は、25μgのHBsAgによる0日目の初回ワクチン接種、ならびに10日目および20日目の2回のブースターワクチン接種により免疫化した。鼻腔内ラット免疫化は、30μlのワクチンによりマウスについてと似た方法で行った。筋肉内マウス群は、吸入麻酔下(イソフルラン/O2)、後部大腿筋に50μlワクチンを注入した。筋肉内ラット群は、200μlのワクチンを両方の後肢筋に分けて注入した。マウスおよびラットは、2回目のブースターワクチン接種の2週間後に屠殺した。動物を屠殺した後、Balb/cマウスの脾臓を回収し、この後5% FCS、1%ペニシリン/ストレプトマイシンおよび50μΜ β-メルカプトエタノールを含む補充IMDMグルタマックス培地で4℃で貯蔵した。マウスでの経口投与は、3回、すなわち0、14および28日目に行った。簡単には、0.3mg GEM粒子有りまたは無しの20μgサブユニットワクチンを、200μlの重炭酸ナトリウム溶液(3.2% w/v)で胃内投与した。経口投与は、ステンレス鋼経口ゾンデを用いて麻酔無しで実施した。
【0042】
チャレンジ実験では、インフルエンザHAワクチンで免疫化したマウスを、100プラーク形成単位(PFU)の株A/Puerto Rico/8/34 (高用量、1群当たり9匹)、または66 PFUの株A/Puerto Rico/8/34 (低用量、1群当たり4匹)により、最後のブースター免疫化の3週間後に鼻腔内チャレンジした(40μL)。
【0043】
チャレンジウイルスの鼻腔内投与は、O2/イソフルランの吸引による動物の軽麻酔下で行った。低用量を受け取った動物を、チャレンジ後4日目に屠殺し、インビトロ細胞ベースアッセイを用いて肺でのウイルス負荷の判定のため肺を分離した。手短かに言えば、ウイルス希釈と一緒にMDCK細胞をインキュベーター(37℃、5% CO2)で1時間インキュベートし、この後PBSで1回洗浄した。トリプシンを含有する新鮮培地(7,5μg/ml TPCKトリプシンを含む100μl培地)をウェルに添加した。細胞をインキュベーター(37℃、5% CO2)で72時間インキュベートし、この後上清を、50μl 1% (洗浄した)モルモット赤血球と一緒に丸底プレート(Costar社)に移した。混合物を室温で2時間インキュベートし、赤血球凝集を読み取った。赤血球凝集を依然として示す最大希釈の逆数が力価である。高用量を受け取った動物は、チャレンジ後14日目まで臨床徴候について追跡し、および許容できない苦しみのため動物が先に屠殺されない限り(人道的エンドポイント: 1日での10%体重減少、または嗜眠、乱れた毛並みおよび瀕死と組み合わせた複数日中の15%体重減少)、屠殺した。
【0044】
肺炎球菌免疫化のため、CD1マウスは、0.01mL (10μL)の用量を0、14および28日目に鼻腔内に受け取った。筋肉内群は、後肢の大腿筋(左、右および左を交互)に注入した0.04mL (40μL)の用量を0、14および28日目に受け取った。最終ブースター免疫化の3週間後、マウスを1×106 CFU肺炎連鎖球菌株TIGR4でチャレンジした。マウスを吸入麻酔(イソフルラン)により軽く麻酔している間に、肺炎球菌を50μL接種材料で鼻腔内に導入した。マウスは感染後に頻繁にモニターし、健康状態、体重および体温に基づく状態により採点した。血液中の細菌数は、チャレンジ後40時間で判定し、血液中に5.4×103 CFU/mL超を有するマウスと同様に、罹患したおよび屠殺する必要がある(人道的エンドポイント)マウスも屠殺した。残りのマウスは、罹患した場合または試験の終了時(チャレンジの14日後)に屠殺した。
【0045】
1.2血清採取および粘膜洗浄
血液試料は各ワクチン投与前に実験中3回取り出し、最終出血は最後のブースター投与の14日後の終了日に採血した。血清を5分間、1200×gで血液の遠心分離により得て、試料をこの後、さらなる分析まで-20℃で貯蔵した。
【0046】
鼻洗浄は、鼻咽頭を1ml PBS(プロテアーゼ阻害剤カクテルを補充した)で洗い流して得た。膣洗浄は、膣を100μl PBS(プロテアーゼ阻害剤カクテルを補充した)で洗い流して得た。100μlアリコートを取り出し、および黄色い200μlチップが取り付けられたピペットを用いて9回再導入した。洗浄液を清潔なバイアルに移し、-20℃で貯蔵した。腸洗浄は、シリコンチップを備えた1.2mm×38mmフレキシブルテフロン(登録商標)経口ゾンデを用いて、胃後方切開により十二指腸に挿管して実施した。洗浄前、空腸を結紮糸により回腸の前部で閉じた。次に、1mlシリンジを経口ゾンデに取り付け、洗浄は、1mlのPBSで十二指腸/空腸の洗い流しを繰り返して実施した。各試料採取後直ちに、洗浄(lavage)を10μl貯蔵溶液(プロテアーゼ阻害剤カクテルを補充した)と混合し、さらなる調製まで洗浄を氷上で維持した。洗浄試料を15分間、11,000×gで遠心分離し、上清を回収し、さらなる分析まで4℃で貯蔵した。
【0047】
1.3 ELISA
HA抗原に対する抗体反応を、血清IgG、IgG1、IgG2aおよびIgG2b希釈力価、粘膜分泌sIgAを決定するため、またはHA特異的IgGの量を決定するため、ELISAアッセイを用いて決定した。希釈力価については、プレートを200ngのHA/ウェルと共にインキュベートした。HAと共に一晩インキュベーション後、プレートを3%ウシ血清アルブミン(Sigma-Aldrich社、オランダ)でブロックした。次いでプレートを洗浄し、連続希釈における血清および粘膜試料と共に1.5時間、37℃でインキュベートした。次に、プレートを洗浄し、マウスIgG、IgG1、IgG2a、IgG2bおよびIgA (Southern Biotech社、バーミングハム、AL、USA)に対する西洋ワサビペルオキシダーゼ結合ヤギ抗体と共にインキュベートした。最後に、基質溶液(0.006% H2O2を含有する50mMリン酸緩衝液pH 5.6中0.02% 1,2-フェニレンジアミン-二塩酸塩)を添加し、プレートを室温で30分間、暗所でインキュベートした。反応を2M H2SO4の添加により停止し、490nmでの吸光度をベンチマークマイクロプレートリーダー(BioRad社、ハーキュリーズ、CA)により読み取った。報告された力価は、バックグラウンド補正後A490≧0.2に対応する、計算された試料希釈の逆数である。
【0048】
HA特異的血清IgGの量を決定するため、マイクロタイターウェルプレートを、200ng/100μl/ウェル H1N1 A/Beijingおよび較正曲線用の抗マウスIgGで被覆した。4℃で一晩インキュベーション後、プレートをコーティング緩衝液(0.5M炭酸−重炭酸塩緩衝液pH9.6〜9.8)で2回洗浄した。ブロッキングは、Protifar Plus (コーティング緩衝液中2.5%)により4℃で45分間実施した。コーティング緩衝液およびPBS/0.05%Tween20でプレートを4回洗浄後、血清および較正曲線をウェルに添加した。血清の連続希釈および較正曲線(マウスIgG1)を、4℃で1.5時間インキュベートした。この後、プレートをPBS/Tween20で3回洗浄した。西洋ワサビペルオキシダーゼ結合免疫グロブリン(ITK、Southern Biotech社)、PBS/Tween20中1:5000希釈、をウェルに添加し、4℃で1時間インキュベートした。プレートをPBS/Tween20で3回、および水で1回洗浄した後、プレートを基質溶液(0.006% H2O2を含有する50mMリン酸緩衝液pH 5.6中0.02% 1,2-フェニレンジアミン-二塩酸塩)を用いて30分間染色した。呈色反応を2M H2SO4により停止した。測定は493nmで実施した。
【0049】
HBsAg抗原に対する血清抗体反応を、IgG希釈力価を決定するためELISAアッセイを用いて決定した。この目的のため、PBS中2μg/ml HBsAgで被覆したELISAプレート、50μl/ウェルを添加し、37℃で1時間インキュベートした。プレートを洗浄緩衝液(PBS/0.1% tween20)により6×洗浄した。プレートをブロック緩衝液(PBS/1% BSA)、200μl/ウェルでブロックし、37℃で1時間インキュベートした。血清試料はブロック緩衝液で連続希釈し、50μl/ウェルを添加し、37℃で1時間インキュベートした。プレートを洗浄緩衝液により6×洗浄した。アルカリホスファターゼ(Southern Biotech社)に結合したヤギ抗ラットIgGを二次抗体(ブロック緩衝液中で1:3000希釈した)として使用し、50μl/ウェルを添加し、37℃で1時間インキュベートした。
【0050】
プレートを洗浄緩衝液(PBS/0.1% tween20)により6×洗浄した。基質緩衝液(10mMジエタノールアミン/0.5mM MgCl2 pH9.5)中p-ニトロフェニルリン酸二ナトリウム塩(Calbiochem社)を検出に使用し、測定は405nmで行った。力価は希釈力価として表し、免疫前基準の2倍のODを示す希釈として定義した。
【0051】
HBsAg特異的粘膜分泌sIgAを、IgG希釈力価を決定するためELISAアッセイを用いて決定した。この目的のため、ELISAプレートを前と同じように被覆し、洗浄し、ブロックした。粘膜洗浄はブロック緩衝液で連続希釈した。50μl/ウェルを添加し、37℃で1時間インキュベートした。プレートを洗浄緩衝液により6×洗浄した。ブロック緩衝液中で1:1000希釈した、西洋ワサビペルオキシダーゼ(Nordic Immunology社)に結合したヤギ抗ラットIgAを二次抗体として使用し、50μl/ウェルを添加し、37℃で1時間インキュベートした。プレートを洗浄緩衝液(PBS/0.1% tween20)により6×洗浄した。TMB (3,3',5,5'-テトラメチルベンジジン、Sigma社、ロット055K8208)を1ml DMSOに溶解し、9mlの0.05Mリン酸-クエン酸緩衝液、pH5.0を検出に使用した。2μlの30%過酸化水素を、使用の直前に基質緩衝溶液10mlごとに添加した。呈色反応を2M H2SO4により停止し、測定は450nmで行った。力価は希釈力価として表し、バックグラウンド(ブロッキング緩衝液と共にインキュベートしたHBsAg被覆)の3倍のODを示す希釈として定義した。
【0052】
1.4赤血球凝集抑制(HI)アッセイ
血清中のHI価は、以前に記載されているように決定した。簡単には、血清を56℃で30分間不活化した。非特異的赤血球凝集を減少させるため、25%カオリン懸濁液を不活化血清に添加した。1200×gでの遠心分離後、50μlの上清を96ウェル丸底プレート(Greiner社、Alphen a/d Rijn、オランダ)に2通りに移し、およびPBSで2倍に連続希釈した。次いで4赤血球凝集単位(HAU)のA/Wisconsinインフルエンザ不活化ウイルスを各ウェルに添加し、プレートを室温で40分間インキュベートした。最後に、50μlの1%モルモット赤血球細胞を各ウェルに添加し、室温で2時間インキュベートした。赤血球凝集を防止することができる最大希釈を、HI価として採点した。
【0053】
1.5 Abbott AxSYMシステムによるHBsAg特異的Ig力価決定
mIU/mlで表されるHBsAgに対する抗体の定量決定は、AxSYM AUSUBアッセイによりAbbott AxSYMシステムで行った。このアッセイは、固相としての微粒子上の組換えHBsAg (ad/ay)およびコンジュゲートとしての組換えHBsAgに結合したビオチンを用いる微粒子EIAである。次の工程では、アルカリホスファターゼ結合抗ビオチンを抗原サンドイッチに結合する。反応混合物を、微粒子が不可逆的に結合する不活性ガラス繊維マトリックスに移す。メチルウンベリフェリルリン酸を基質として使用し、最終産物メチルウンベリフェロンの蛍光を機器により読み取った。
【0054】
1.6 Elispot
Elispotアッセイは、前に記載されているように(Amorij JPら。Vaccine 2007年12月21日;26(1):67〜76頁)実施した。簡単には、96ウェルマイクロタイタープレート(Greiner社、Alphen a/d Rijn、オランダ)を、抗マウスインターフェロン-γ (IFN-γ)およびインターロイキン-4 (IL-4) (BD, Pharmingen社、エーレムボードヘム、ベルギー)と共に4℃で一晩インキュベートした。プレートをPBS/Tween (Sigma-Aldrich社、オランダ)で3回洗浄した後、プレートを37℃で1時間ブロック(PBS+4% BSA)し、脾臓細胞を、刺激ペプチドとしてのサブユニットワクチン有りまたは無しで濃度1×106細胞/ウェルでプレートに添加した。5% CO2、37℃で一晩インキュベーション後、細胞を冷水で溶解した。次に、プレートをPBS/Tweenで5回洗浄し、およびPBS+2% BSA中0.125μg/mlの濃度でビオチニル化抗マウスIFN-γおよびIL-4抗体(BD Pharmingen社)と共にインキュベートした。洗浄後、プレートを、ストレプトアビジンアルカリホスファターゼ(BD Pharmingen社)と共に37℃で1時間インキュベートした。最後に、PBS/Tweenで3回、およびPBSで2回洗浄した後、1mg/ml 5-ブロモ-4-クロロ-3-インドリルホスフェート、0.92% w/v 2-アミノ-2-メチル-1-プロパノール、0.08μl/ml TritonX-405、1M MgCl2および6mg/mlアガロースから成る基質溶液を用いて、スポットを発生させた。スポットは、Elispotリーダー(A. EL. VIS Elispotリーダー)を用いてカウントした。
【0055】
1.7統計分析
統計分析は、p< 0.05を最小有意水準としてStudentのt検定またはノンパラメトリックANOVA検定を用いて実施した。結果は、特に指示のない限り平均±標準誤差(SEM)で表す。
【0056】
(実施例1)鼻腔内HAワクチンにおけるGEMのアジュバント効果
鼻腔内HA (5μg H1N1 A/Beijing)に対する全身血清抗体反応の増強は、さまざまな量のGEM粒子(0、0.03、0.1および0.3mg乾燥重量)をHAに添加して鼻腔内マウスモデルで評価した。マウスは3回のワクチン投与を受け(各々2週間間隔および最後のブースター免疫化の2週間後)、血清試料が分析された。図1は、アジュバント無しのHAが鼻腔内投与経路を通じてほんの低レベルの全身IgG抗体(5.0μg/ml)を誘発することを示している。少量のGEM粒子(0.03mg)の添加は、ただでさえこのレベルを4倍に増加させる。最良の増強は、0.1mg GEM粒子の添加により、1ml当たり約67μg HA特異的IgGまで見出され、さらに多くのGEM粒子の添加によりさらに増加することはなかった。これらの結果は、GEM粒子をインフルエンザHAと混合することが、用量依存的方法で抗原特異的免疫応答を増強することを明らかに示している。
【0057】
(実施例2)筋肉内HAと比較したHAと混合した鼻腔内GEM
比較は、鼻腔内HA+GEMワクチンと、HAワクチンが投与される伝統的な方法、すなわち筋肉内経路を通じて投与される、アジュバント無しのHAの間で行った。マウスは、投与間が2週間間隔による、i.n. HA (5μg H1N1 A/Beijing)+GEM (0.15mg乾燥重量)またはi.m. HA (5μg)の3回の投与を受けた。鼻腔内および筋肉内ワクチンの免疫応答の大きさおよび動態を比較するため、HA特異的血清IgG力価を、各免疫の2週間後に採取した試料について決定した。図2は、i.n. HA+GEMワクチンの大きさおよび動態のいずれもがi.m. HAワクチンのものと類似することを明らかに示している。各投与後のi.n.ワクチンとi.m.ワクチンの反応の間に統計的有意差はない(各p値>0.05)。
【0058】
(実施例3)HAと混合した鼻腔内GEMは防御反応を誘発する
インフルエンザワクチンの防御能力は、HI価を測定して決定する。HI価は、i.n. HA (5μg H3N2 A/Wisconsin)、HA+GEM (0.3mg乾燥重量)、i.m. HAによる1回目および2回目のブースター免疫化後の全マウスについて決定した。
【0059】
図3は、従来のi.m.およびGEMアジュバントi.n.ワクチンがいずれも、1回目のブースター免疫化後に2log6を超える同程度のHI価に達したことを示している(p=0.2062)。これらの力価は、両方のケースで2log7から2log8の間の値まで増加し、2つの治療の間に有意差はない(p=0.7611)。サブユニットワクチン単独によるi.n.免疫化は、2回のブースター免疫化の後でさえ低いHI価を誘導した。さらに、動物の50%のみがi.n.サブユニットワクチンによる免疫後に応答したのに対し、全動物が2つの他のワクチン群で応答した。2log5.3を超えるHI価はヒトにおいて防御的であると考えられることから、これらの結果は、単回ブーストが、i.n. GEMアジュバントインフルエンザワクチンが防御免疫に達するのに十分であることを示している。サブユニットワクチン単独によるi.n.免疫化およびi.m.免疫化の両方と比べて、GEM粒子を含むサブユニットワクチンの製剤が強力な全身性免疫応答を誘導したことは、結果から明らかである。
【0060】
(実施例4)HAと混合した鼻腔内GEMの粘膜性免疫応答
i.n.免疫化は、気道、すなわちインフルエンザウイルスの侵入口において局所粘膜免疫を誘導する可能性があることが以前に報告されている。粘膜免疫の活性化は、下部のBおよびT細胞をプライミングし、粘膜部位でのsIgAの分泌をもたらす。それ故に、インフルエンザ特異的sIgA力価を、マウスの鼻および肺洗浄において決定した(図4)。
【0061】
i.m.免疫化は、マウスのほとんどで検出限界より下の鼻および肺洗浄においてsIgAレベルを誘発した(8マウス中1匹のみは鼻洗浄での応答を示した)。同様に、サブユニットワクチン単独によるi.n.免疫化は、肺および鼻洗浄において低いsIgA力価を示した(3/8レスポンダー)。対照的に、HA+GEMによるi.n.免疫化は、全マウスの鼻および肺洗浄において高いsIgA力価を誘導した。
【0062】
結論として、HA+GEMによるi.n.免疫化は、上および下気道の両方で強力な粘膜性免疫応答を誘導した。
【0063】
(実施例5)HAと混合した鼻腔内GEMの免疫応答の表現型
応答の表現型、すなわちTヘルパー1/Tヘルパー2比(Th1/Th2)を評価するため、IgGサブタイプ、IFN-γおよびIL-4応答を決定した。
【0064】
IgGサブタイププロファイリング(図5)は、サブユニットワクチン単独によるi.n.免疫化が低いIgG1、IgG2aおよびIgG2b応答を誘導することを示した。以前に報告されているように、サブユニットワクチンによるi.m.免疫化は高いIgG1応答を誘導したが、IgG2aおよびIgG2bをほとんど誘導しなかった。これは、Th2応答に偏った免疫応答を示している。i.m.免疫化と比較して、HA+GEMによるi.n.免疫化は、有意により高いIgG2a (p=0.042)およびIgG2b (p=0.030)およびより低いIgG1 (p=0.0135)応答を誘導した。これらの結果は、i.n. HA+GEMワクチンにより生じた抗体反応が、従来のi.m.ワクチンよりもTh1表現型に有意に偏向していることを示している。
【0065】
免疫応答の型(図6)を、免疫化マウスの抗原特異的IFN-γおよびIL-4産生脾臓細胞を決定してさらに評価した。サブユニットワクチンによるi.m.免疫化は、IFN-γ産生細胞よりも多数のIL-4産生細胞をもたらし、この場合もやはり優勢なTh2応答を示している。サブユニットワクチンによるi.n.免疫化は、より少数のIL-4産生細胞であるが、著しくより多数のIFN-γ産生細胞(図6)をもたらし、バランスの取れたTh1/Th2応答をもたらした。IFN-γ産生T細胞の増加は、HA+GEMによるi.n.免疫化後、一段と有意に(p=0.0373)より顕著であった。これは、バランスの取れたTh1/Th2から優勢なTh1応答への免疫応答の移行を示している。
【0066】
(実施例6)致死的チャレンジモデルにおけるHAと混合した鼻腔内GEMの防御
i.n. HA+GEMにより生じた免疫応答の防御能力を、致死的チャレンジモデルにおいて評価した。マウスを、PBS(偽免疫化)またはHA単独(2回)、HA+GEM (2回)もしくはHA+GEM (3回)でi.n.免疫化した。比較は、筋肉内に投与したHAベンチマークワクチンにより行った。この実験におけるHAは、株PR8 (H1N1)に由来した。用量は、1用量当たり5μg HAであり、GEMケースでは1用量当たり0.3mg GEMをワクチンに添加した。ワクチンは、2週間間隔で投与した。致死的チャレンジは、PR8の致死的用量による最後のブースター免疫化の3週間後に行った。チャレンジに対する防御を、群HA+GEM (i.n.2回、9/9生存者)、HA+GEM (i.n.3回、9/9生存者)およびHAベンチマーク対照(i.m.、9/9生存者)の動物について観察した[図7]。これらの群内の全動物は、チャレンジ後に臨床徴候(嗜眠、乱れた毛並みまたは背中が丸まった姿勢)を示さず、実験終了までの14日目まで生存した。防御は、体重減少の欠如と相関した(不図示)。
【0067】
対照的に、群HA i.n.およびPBS (偽免疫化、陰性対照)内のほとんどの動物は、それぞれ、3日目から、および4日目以降、重篤な体重減少を示し、重篤な臨床症状(体重<85%、嗜眠、乱れた毛並み、丸まった背中)のためチャレンジ後5日目から8日目に安楽死させた。
【0068】
チャレンジの4日後の肺におけるウイルス力価の決定は、HA+GEM (2または3回)によるi.n.ワクチン接種が、PBS陰性対照群と比べて、チャレンジの4日後の肺でウイルス力価の約1,000から10,000倍の減少をもたらすことを示した(図8)。チャレンジした際のウイルス力価の極めて限定された減少(4倍の減少)が、HAを単独でi.n.適用した場合に観察された。これは、防御を提供するのにGEMのアジュバント特性が必要とされることを示している。HA+GEM (2および3回)によるワクチン接種は、ベンチマーク陽性対照群(HA、i.m)と比べて、肺におけるウイルス力価の約20から100倍の改善をもたらした。ウイルス力価の減少は、ウイルスの排出の減少をもたらすことができ、集団防御(herd protection)を提供するうえで重要な要因であると考えられる。実施例4および5で示したようなi.n. HA+GEMワクチンにより生じた、気道の粘膜内層における局所IgAの存在および/またはより良好にバランスの取れたTh1/Th2型の免疫応答は、i.m.ベンチマークワクチンと比べて、観察された防御の優位性を説明することができよう。
【0069】
(実施例7)GEMと混合した経口HAは防御反応を誘発する
経口投与経路は、この簡便さのためワクチンにとって魅力的であるが、抗原が不活化または劣化されるため、有効性を欠く場合が多い。アジュバント無しのHAの経口投与は、防御的な血清HI応答および/または粘膜IgA応答を誘発するのに不十分であることが知られている。経口胃免疫化においてGEMをHAに添加する効果を、マウスモデルで分析した。H3N2 A/Hiroshimaサブユニット抗原HA (20μg/1回用量)を使用した。HA+GEMワクチンは、1用量当たりさらに0.3mg GEMを含有した。マウスを2週間間隔で3回免疫化し、最終免疫の2週間後の試料を分析した。免疫化の防御能力を比較するため、血清HI価を決定した。図9に示されているように、HA+GEMワクチンによる経口免疫は、GEM粒子無しの経口免疫よりも有意に高い(p<0.05) HI価を誘導した。経口HA+GEM群ではHI価は、防御のカットオフレベルの2log5.3を十分に超える、2log7超に達した。
【0070】
さらに、経口HA+GEMは、胃腸管においてかなりのレベルの粘膜IgAを惹起することができた(図10)。驚くべきことに、気道での強い局所IgA応答も動物のほとんどで誘発された。
【0071】
これらの結果は、GEMと混合した経口インフルエンザHAワクチンも、防御全身性免疫応答を誘発したこと、およびさらに気道を含む強力な粘膜応答を誘発することを示している。
【0072】
(実施例8)GEMと混合した筋肉内HAは粘膜表面での局所応答を誘発する
非経口ワクチンは通常、粘膜分泌IgAの産生を誘発しない。筋肉内免疫化マウスの粘膜試料の分析では、本発明者らは、HA+GEMを受けたマウスはが、鼻、肺および膣などの幾つかの粘膜組織で局所IgAを分泌することを驚くべきことに見出した。メスマウスに、固定量のHA (5μg B/Shangdong/7/97)、0.3mg GEM有りまたは無しにより鼻腔内または筋肉内に3回(間隔14日)ワクチン接種した。最後の免疫の2週間後、鼻および膣洗浄を実施し、IgA力価を特異的ELISAアッセイにより決定した。
【0073】
図11におけるデータは、HA+GEMの鼻腔内投与が局所IgA応答を効率的に誘導したことを示し、鼻洗浄におけるIgA力価として証明された。IgA力価は遠位でも誘導され、膣洗浄におけるIgA力価の出現として証明された。予想通り、HA単独の筋肉内投与は、関連局所IgA応答を誘導しない。驚くべきことに、HA+GEMの筋肉内投与は、鼻腔内投与後に達せられたものに迫る効率で鼻および膣の両方で関連IgA力価を誘導した。したがって、HA+GEMの筋肉内投与は、粘膜性免疫応答を誘導するのに使用することができる。
【0074】
(実施例9)GEMと混合したHAの筋肉内投与は十分な用量節約を支持する
筋肉内GEM+HAにより誘発された免疫応答が、インフルエンザHA抗原の用量節約を可能にするかどうかを決定するため、マウスを、PBS (偽治療)、GEM無しの1μg HA (A/PuertoRico/8/34)またはGEM (1用量当たり0.3mg)と共に処方した0.04μg HA (25倍少ない抗原)により2回ワクチン接種した。最終用量の投与の2週間後、マウスをマウス適応A/PuertoRico/8/34でチャレンジした。チャレンジの5日後、動物を屠殺し、肺を分離およびホモジナイズし、ウイルス力価をMDCK細胞でのエンドポイント滴定により決定した。
【0075】
図12、パネルAは、1μg HAによる動物の筋肉内ワクチン接種が、偽治療動物と比べて、1logを超える感染動物の肺におけるウイルス負荷の減少を提供することを示している。しかし、HA+GEMは、肺力価の完全な欠如により証明されるように、感染動物の肺におけるインフルエンザウイルスの複製に対し完全な防御を提供する。これらの結果は、ベンチマークi.m. HAと比べてi.m. HA+GEMワクチンの優位性を示している。
【0076】
ベンチマークi.m. HAについてと同じレベルの防御が、図12のパネルBに示されているように、わずか0.04μg HA (25倍少ない抗原)を含有するHA+GEM製剤で達成された。これは、大幅な抗原節約が、GEMを含む筋肉内インフルエンザワクチンを処方して達成できることを示している。
【0077】
(実施例10)鼻腔内GEMベースのB型肝炎ワクチンはマウスにおける強力な全身性IgGおよび局所IgA応答を誘発する
成体C57BL6マウスを、HBsAg抗原を含有するGEMベースのB型肝炎ワクチンで免疫化した。このケースでは、HBsAg [5μg]はGEM粒子[0.15mg乾燥重量]と混合した。GEM無しの等量のHBsAgも、比較のため使用した。ワクチンは、鼻腔内経路を通じて投与した。ミョウバンによるアジュバントである市販のHepBワクチンVaxProを、ベンチマークワクチンとして皮下投与した。血清IgGを、完全免疫化(3回用量、10日間隔により投与)後に測定した。図13は、鼻腔内ワクチンでのGEM粒子のアジュバント効果を明らかに示している。HBsAg特異的血清IgG応答は、HBsAg単独を鼻腔内投与した場合、測定不能であった。対照的に、HBsAg+GEMは、4.2の希釈力価を有する活発なHBsAg特異的血清IgG応答を誘発した。鼻腔内GEM-HBsAgワクチンは、皮下経路を通じて投与したベンチマークワクチンと似たHBsAg特異的IgGを誘発した(p=0.2290)。
【0078】
粘膜免疫の活性化は、粘膜部位でのsIgAの分泌をもたらす。この実験では、HBsAg特異的sIgAの局所分泌を、ワクチン接種部位(鼻)の洗浄および遠位粘膜部位(膣)での洗浄において測定した。
【0079】
図14は、sIgA応答がi.n. HBsAg+GEMワクチンを用いてのみ生じ、i.m. VaxProワクチンでは生じないことを明らかに示している。i.n. HBsAg+GEMワクチンは、膣の粘膜部位などの遠位粘膜部位でのsIgAの分泌でさえ生じる。
【0080】
(実施例11)ラットモデルにおける鼻腔内GEM-HBsAg B型肝炎ワクチンは防御レベルの血清抗体を誘発する
アジュバントB型肝炎ワクチンは、HBsAg抗原(25μg)をGEM (0.4mg)と混合して作製した。比較には、HBsAg抗原単独(25μg)、およびミョウバンと共に処方した同じ抗原を含有するベンチマークワクチン(VaxPro)。完全免疫は、10日間隔で投与した3回のワクチン投与から成った。最終血清を、最後のブースターの14日後に回収した。GEM-HBsAgおよびHBsAgは鼻腔内投与した。VaxProは、筋肉内経路を通じて投与した。B型肝炎ワクチンについては、防御の相関が知られている。血液血清の10mIU/mlより高い抗体レベルは防御的であると考えられ、防御に関する代理マーカーとして認められている。
【0081】
完全免疫化ラット(1群当たり4匹のウィスターラット)の血液血清を、1ml当たりのミリ国際単位(mIU/ml)で表されるHBsAg特異的抗体のレベルについて分析した。図15は、結果をまとめている。鼻腔内HBsAgは、反応を全く誘発しない。高いおよび防御レベルの抗体反応(mIU/ml>10)は、HBsAgをペプチドグリカン微粒子と共に処方した場合に、鼻腔内経路を通じて得られる。防御のレベルは、筋肉内経路を通じて投与したベンチマークワクチンVaxProと類似する(p=0.7715)。
【0082】
実施例10および11における結果は、抗原はGEM粒子に積極的に結合されないという事実にもかかわらず、強力な全身抗体反応および局所抗体反応が、鼻腔内GEMベースのB型肝炎HBsAgワクチンにおいて惹起されることを一貫して示している。
【0083】
(実施例12)三価肺炎球菌タンパク質ベースのGEMワクチンの防御能
比較は、タンパク質に混合または結合したGEMと共に処方した鼻腔内肺炎球菌タンパク質ベースのワクチン間で行った。3つの保存された肺炎球菌タンパク質(PspA、CbpA、PdBD)を、三価ワクチン、GEM+P3 (混合)およびGEM-P3 (結合)において使用した。マウスは、これらのワクチンまたは陰性対照(偽免疫化)としてのPBSにより、用量間が10日間隔で3回免疫化した。各GEMベースのワクチンは、1用量当たり5μgの各抗原および0.3mg GEMを含有した。最後のブースター免疫化の3週間後、マウスを、致死的用量肺炎連鎖球菌TIGR4 (血清型4)で鼻腔内チャレンジした。防御されないマウスは、チャレンジ後72時間以内に死亡した。マウスを、チャレンジ後14日間追跡した。マウスは、人道的エンドポイント(チャレンジの48時間後に1ml血液当たり>5.4×103コロニー形成単位(cfu)、体重<85%、嗜眠、乱れた毛並み、丸まった背中)に基づき、または試験終了時に安楽死させた。偽免疫化マウスはいずれも生存しなかった。GEM+P3 (混合)ワクチンで免疫化した群は、GEM-P3 (結合)ワクチンで免疫化した群(20%)よりも良好な生存(50%)を示すことが、驚くべきことに見出された(図16を参照のこと)。これらの結果は、P3タンパク質によるGEMワクチンは、これらのタンパク質をGEM粒子に混合した場合により効果的であることを明らかに示している。
【0084】
(実施例13)五価肺炎球菌タンパク質ベースのGEMワクチンの防御能
比較は、タンパク質に混合または結合したGEMと共に処方した鼻腔内肺炎球菌タンパク質ベースのワクチン間で行った。5つの保存された肺炎球菌タンパク質(PspA、PsaA、CbpA、PpmA、IgA1prt)を、五価ワクチン、GEM+P5 (混合)およびGEM-P5 (結合)において使用した。マウスは、これらのワクチンまたは陰性対照(偽免疫化)としてのPBSにより、用量間が10日間隔で3回免疫化した。各GEMベースのワクチンは、1用量当たり0.5μg IgA1prt、3μg PsaA、1.5μg CbpA、2μg PpmA、2μg PspAおよび0.3mg GEMを含有した。最後のブースター免疫化の3週間後、マウスを、致死的用量肺炎連鎖球菌D39 (血清型2)で鼻腔内チャレンジした。防御されないマウスは、チャレンジ後72時間以内に死亡した。マウスを、チャレンジの40時間後健康状態を、臨床症状(嗜眠、乱れた毛並み、丸まった背中)に基づき採点し、ワクチンの防御能力を測定するためのエンドポイントとして見なした。図17は、GEM-P5 (結合)ワクチンで免疫化した群では、10マウス中8匹が完全に健康なままであったのに対し、これがGEM+P5 (混合)ワクチン(5/10)についてはより少なく、陰性対照(1/10)については少ないことを示している。これらの結果は、P5タンパク質によるGEMワクチンが、これらのタンパク質をGEM粒子に結合した場合に、より効果的であることを明らかに示している。
【特許請求の範囲】
【請求項1】
(i)グラム陽性細菌から得られるペプチドグリカン微粒子と、(ii)少なくとも1つのインフルエンザウイルス抗原またはこの抗原性調製物とを含み、前記抗原または抗原性調製物がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない、アジュバントインフルエンザワクチン製剤。
【請求項2】
ヘマグルチニン抗原(HA)、ノイラミダーゼ抗原(NA)またはこれらの組み合わせを含む、請求項1に記載のワクチン製剤。
【請求項3】
少なくとも2つのインフルエンザウイルス株由来のインフルエンザ抗原またはこれらの抗原性調製物を含み、少なくとも1つの株がパンデミック発生に関連するもしくはパンデミック発生に関連する可能性がある、請求項1または2に記載のワクチン製剤。
【請求項4】
前記ワクチン製剤が、1インフルエンザ株当たり1から15μgの間のHAを含有する、請求項1から3のいずれか一項に記載のワクチン製剤。
【請求項5】
(i)グラム陽性細菌から得られるペプチドグリカン微粒子と、(ii)少なくとも1つの肺炎球菌抗原またはこの抗原性調製物とを含み、前記抗原または抗原性調製物がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない、アジュバント肺炎球菌ワクチン製剤。
【請求項6】
PspA、CbpAおよび/またはPdBDを含む、請求項5に記載のワクチン製剤。
【請求項7】
前記微粒子が食品用細菌、好ましくは乳酸菌、より好ましくはラクトコッカス・ラクティスから得られる、請求項1から6のいずれか一項に記載のワクチン製剤。
【請求項8】
抗原1マイクログラム当たり0.01から0.1ミリグラム微粒子(乾燥重量)を含む、請求項1から7のいずれか一項に記載のワクチン製剤。
【請求項9】
請求項1から8のいずれか一項に記載のワクチン製剤を含む、鼻腔内調剤装置。
【請求項10】
エアロゾルまたは滴下送達システムの形態での、請求項9に記載の調剤装置。
【請求項11】
インフルエンザワクチン製剤の製造におけるアジュバントとしてグラム陽性細菌から得られるペプチドグリカン微粒子の使用であって、前記製剤がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されないインフルエンザウイルス抗原を含む、使用。
【請求項12】
鼻腔内、筋肉内または経口ワクチン製剤における、請求項11に記載の使用。
【請求項13】
ワクチン製剤が、ヒトハイリスク集団において使用するために処方される、請求項11または12に記載の使用。
【請求項14】
ワクチンが、高齢者、2歳までの子どもおよび/または慢性疾患患者において使用するために処方される、請求項13に記載の使用。
【請求項15】
請求項1から4に記載のワクチンを被験者に投与することを含む、被験者におけるインフルエンザ感染または疾患の予防方法。
【請求項16】
ワクチン送達が鼻腔内または筋肉内である、請求項15に記載の方法。
【請求項17】
ワクチンが調剤装置により鼻腔内送達され、好ましくは調剤装置がエアロゾルまたは滴下送達システムの形態にある、請求項16に記載の方法。
【請求項1】
(i)グラム陽性細菌から得られるペプチドグリカン微粒子と、(ii)少なくとも1つのインフルエンザウイルス抗原またはこの抗原性調製物とを含み、前記抗原または抗原性調製物がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない、アジュバントインフルエンザワクチン製剤。
【請求項2】
ヘマグルチニン抗原(HA)、ノイラミダーゼ抗原(NA)またはこれらの組み合わせを含む、請求項1に記載のワクチン製剤。
【請求項3】
少なくとも2つのインフルエンザウイルス株由来のインフルエンザ抗原またはこれらの抗原性調製物を含み、少なくとも1つの株がパンデミック発生に関連するもしくはパンデミック発生に関連する可能性がある、請求項1または2に記載のワクチン製剤。
【請求項4】
前記ワクチン製剤が、1インフルエンザ株当たり1から15μgの間のHAを含有する、請求項1から3のいずれか一項に記載のワクチン製剤。
【請求項5】
(i)グラム陽性細菌から得られるペプチドグリカン微粒子と、(ii)少なくとも1つの肺炎球菌抗原またはこの抗原性調製物とを含み、前記抗原または抗原性調製物がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されない、アジュバント肺炎球菌ワクチン製剤。
【請求項6】
PspA、CbpAおよび/またはPdBDを含む、請求項5に記載のワクチン製剤。
【請求項7】
前記微粒子が食品用細菌、好ましくは乳酸菌、より好ましくはラクトコッカス・ラクティスから得られる、請求項1から6のいずれか一項に記載のワクチン製剤。
【請求項8】
抗原1マイクログラム当たり0.01から0.1ミリグラム微粒子(乾燥重量)を含む、請求項1から7のいずれか一項に記載のワクチン製剤。
【請求項9】
請求項1から8のいずれか一項に記載のワクチン製剤を含む、鼻腔内調剤装置。
【請求項10】
エアロゾルまたは滴下送達システムの形態での、請求項9に記載の調剤装置。
【請求項11】
インフルエンザワクチン製剤の製造におけるアジュバントとしてグラム陽性細菌から得られるペプチドグリカン微粒子の使用であって、前記製剤がタンパク質性のペプチドグリカン結合部分に融合または他の様式で共有結合されないインフルエンザウイルス抗原を含む、使用。
【請求項12】
鼻腔内、筋肉内または経口ワクチン製剤における、請求項11に記載の使用。
【請求項13】
ワクチン製剤が、ヒトハイリスク集団において使用するために処方される、請求項11または12に記載の使用。
【請求項14】
ワクチンが、高齢者、2歳までの子どもおよび/または慢性疾患患者において使用するために処方される、請求項13に記載の使用。
【請求項15】
請求項1から4に記載のワクチンを被験者に投与することを含む、被験者におけるインフルエンザ感染または疾患の予防方法。
【請求項16】
ワクチン送達が鼻腔内または筋肉内である、請求項15に記載の方法。
【請求項17】
ワクチンが調剤装置により鼻腔内送達され、好ましくは調剤装置がエアロゾルまたは滴下送達システムの形態にある、請求項16に記載の方法。
【図1】


【図2】
【図3】
【図4】
【図5】

【図6】
【図7】
【図8】
【図9】

【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【公表番号】特表2013−506659(P2013−506659A)
【公表日】平成25年2月28日(2013.2.28)
【国際特許分類】
【出願番号】特願2012−532031(P2012−532031)
【出願日】平成22年10月1日(2010.10.1)
【国際出願番号】PCT/NL2010/050639
【国際公開番号】WO2011/040811
【国際公開日】平成23年4月7日(2011.4.7)
【出願人】(510285171)
【Fターム(参考)】
【公表日】平成25年2月28日(2013.2.28)
【国際特許分類】
【出願日】平成22年10月1日(2010.10.1)
【国際出願番号】PCT/NL2010/050639
【国際公開番号】WO2011/040811
【国際公開日】平成23年4月7日(2011.4.7)
【出願人】(510285171)
【Fターム(参考)】
[ Back to top ]