
アセチルコエンザイムAトランスポータータンパク質をコードする核酸
【課題】単離された哺乳動物アセチルコエンザイムAトランスポーター(AT)タンパク質を提供する。
【解決手段】単離された哺乳動物アセチルコエンザイムAトランスポーター(AT)タンパク質、抗AT抗体、治療学的組成物、およびそれらをコードする核酸が提供される。本発明のATタンパク質を用いる、バイオアッセイおよび治療方法もまた、提供される。ATタンパク質は、ATタンパク質(好ましくはヒト)をまた特異的に認識する抗体によって特異的に認識される特定の配列を含むポリペプチド。単離されたATタンパク質は、天然のインビボ環境と通常関連する細胞性成分および/または夾雑物を含まない。
【解決手段】単離された哺乳動物アセチルコエンザイムAトランスポーター(AT)タンパク質、抗AT抗体、治療学的組成物、およびそれらをコードする核酸が提供される。本発明のATタンパク質を用いる、バイオアッセイおよび治療方法もまた、提供される。ATタンパク質は、ATタンパク質(好ましくはヒト)をまた特異的に認識する抗体によって特異的に認識される特定の配列を含むポリペプチド。単離されたATタンパク質は、天然のインビボ環境と通常関連する細胞性成分および/または夾雑物を含まない。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、国立研究所の国立ガン研究所によって与えられた、助成金番号R01 CA48737およびP01 CA 71932のもとで、政府の支援とともになされた。政府は本発明において一定の権利を有する。
発明の分野
本発明は、概して、分子生物学および酵素学に、ならびにより具体的には、膜を横切ってアセチルコエンザイムA(Ac-CoA)の輸送を調節するトランスポータータンパク質をコードする核酸に関する。
【背景技術】
【0002】
発明の背景
膜ガングリオシドにおける糖鎖の構造の多様性および複雑さは、いくつかの異なる種のシアル酸分子の出現によって、一部生じる。N-アセチルノイラミン酸およびN-グリコリルノイラミン酸は、脳中のガングリオシドにおいて、最も遍在するシアル酸であるが、それらのO-アセチル化形態はまた、LD1、B-系列ガングリオシド(GD3およびGT1bを含む)、ならびにC-系列(NeuAca2-8NeuAca2-8NeuAca2-3Gal-R)ガングリオシドにおいて、微量成分として見出される。いくつかの生物学的特性は、O-アセチル化によるシアル酸の修飾と関連すると仮定される。例えば、9-O-アセチル化ガングリオシドの発現は、神経細胞分化および移動と明らかに関連する(Stallcupら、ColdSpring Harbor Symp. Quant. Biol.、48:761-773(1983))。ラット脳におけるD1.1抗体によって検出される、9-O-アセチル化GD3は、生殖細胞領域に局在化されるが、有糸分裂後の細胞から消失することが見出された。これは、正常な成体脳および移動において、存在しない(Stallcupら、ColdSpring Harbor Symp. Quant. Biol.、48:761-773(1983))。ラット脳において、D1.1抗体によって検出される9-O-アセチル化GD3は、生殖細胞領域に局在化されるが、有糸分裂後の細胞から消失することが見出された。これは、正常な成体脳において、存在しない(Levineら、J.Neuros.、4:820-831(1984))。しかし、ハタオリマウス(weavermouse)において、成体脳における9-O-アセチル化ガングリオシドの持続的な発現が、小脳顆粒細胞移動における欠失と関連付けられた(Johnstoneら、J.Neurochem.、51:1655-1657(1988))。シアル酸残基におけるO-アセチル基の付着は、シアリダーゼのようなシアル酸代謝の酵素に対して、有意な効果を引き起こす。効果はまた、ウイルス結合、細胞接着、およびガングリオシドのシアル酸残基の免疫原性に対して見られる(概説のために、Varki、Glycobiology、2:25-40(1992)を参照のこと)。
【0003】
その重要性にもかかわらず、O-アセチル化機構は、分子レベルおよび遺伝子レベルで十分に理解されていない。Varkiのグループによってなされた一連の研究は、O-アセチル化ガングリオシドの生成は、単純なプロセスではなく、同じゴルジ体区分における、アクセプターガングリオシドGD3、アセチルCoA(Ac-CoA)トランスポーター、アセチルトランスフェラーゼの同時局在化を必要とすることを示す(Varki、Glycobiology、2:25-40(1992))。事実、インビトロアセチルトランスフェラーゼ活性の検出は、極めて困難であり、およびいったんインタクトな細胞膜調製物が、界面活性剤で処理されると、転移活性は、迅速に失われることが示された(VarkiおよびDiaz、J.Biol. Chem.、260:6600-6608(1985))。
【0004】
従って、O-アセチル化ガングリオシドの形成に関与する他のタンパク質因子を単離および特徴づけする、必要性が存在する。本発明は、この必要性を満たし、そして関連する利点もまた提供する。
【発明の開示】
【課題を解決するための手段】
【0005】
発明の要旨
本発明によれば、単離された哺乳動物アセチルコエンザイムAトランスポーター(AT)タンパク質が提供される。これらのATタンパク質、またはそのフラグメントは、抗AT抗体を生成するための免疫原として、またはこのようなタンパク質および/または抗体を含む治療学的組成物において、有用である。本発明のATタンパク質はまた、それに対するアゴニストおよびアンタゴニストを同定するためのバイオアッセイにおいて有用である。
【0006】
本発明によれば、新規のATタンパク質をコードする、単離された核酸がまた提供される。さらに、本発明の核酸を含むベクター、本発明の核酸にハイブリダイズするプローブ、本発明の核酸で形質転換された宿主細胞、本発明の核酸に対するアンチセンスオリゴヌクレオチド、および関連の組成物が、提供される。本明細書中に記載される核酸分子は、単離された組換えATタンパク質を容易に生成するために、当業者に公知の多様な組換え発現系に組込まれ得る。さらに、本発明の核酸分子は、所定のサンプル中のAT遺伝子またはmRNA転写物の存在および/または量についてアッセイするためのプローブとして有用である。本明細書中に記載される核酸分子、およびそのオリゴヌクレオチドフラグメントはまた、ATタンパク質をコードする核酸を増幅するためのPCR反応におけるプライマーおよび/または鋳型として有用である。また、本発明のタンパク質を発現するトランスジェニック非ヒト哺乳動物が提供される。
【0007】
本発明のATタンパク質と免疫反応性である抗体がまた、提供される。これらの抗体は、所定のサンプル(例えば、組識サンプル、ウエスタンブロットなど)に存在するATタンパク質のレベルを決定するための診断アッセイにおいて有用である。抗体はまた、粗細胞抽出物などからATタンパク質を精製するために使用され得る。さらに、これらの抗体は、インビボで、ATタンパク質の生物学的効果を調節するために治療学的に有用であると考えられる。
【0008】
種々の組識サンプルにおけるATタンパク質のレベルを決定するための方法および診断系がまた、提供される。これらの診断方法は、治療学的に有効な量の維持を容易にするために、治療学的に投与されるATタンパク質またはそのフラグメントのレベルをモニターするために使用され得る。これらの診断方法はまた、異常なレベルのまたは異常な構造のATタンパク質から生じる生理学的障害を診断するために使用され得る。
1.哺乳動物アセチルコエンザイムAトランスポーター(AT)タンパク質をコードする単離された核酸、そのスプライス改変体cDNA配列、またはそれらのフラグメント。
2.項目1に記載の核酸であって、前記哺乳動物ATタンパク質が、ヒト、ラット、マウス、ブタ、ヒツジ、イヌ、またはウシから選択される哺乳動物から単離される、核酸。
3.項目1に記載の核酸であって、該核酸が、以下:
(a)配列番号2に記載のアミノ酸配列をコードするDNA、または
(b)中程度のストリンジェントな条件下で(a)のDNAにハイブリダイズするDNAであって、ここで該DNAが、生物学的に活性なATをコードする、DNA、または
(c)上記の(a)もしくは(b)のいずれかに関するDNA縮重体であって、ここで該DNAが、生物学的に活性なATをコードする、DNA、または
(d)それらのスプライス改変体cDNA配列、
から選択される、核酸。
4.項目3に記載の核酸であって、該核酸が、配列番号1のヌクレオチド388〜2034のATコード部分に、高いストリンジェンシーの条件下でハイブリダイズする、核酸。
5.項目3に記載の核酸であって、該核酸のヌクレオチド配列が、配列番号1に記載のヌクレオチド388〜2034と、実質的に同じである、核酸。
6.項目3に記載の核酸であって、該核酸のヌクレオチド配列が、AT-1をコードし、および配列番号1に記載のヌクレオチド388〜2034と、同じである、核酸。
7.項目3に記載の核酸であって、該核酸がcDNAである、核酸。
8.項目3に記載の核酸を含む、ベクター。
9.項目3の核酸を含む、組換え細胞。
10.オリゴヌクレオチドであって、配列番号1に記載されるヌクレオチド配列の核酸の配列と、特異的にハイブリダイズし得る少なくとも14ヌクレオチドを含む、オリゴヌクレオチド。
11.項目10に記載のオリゴヌクレオチドであって、該オリゴヌクレオチドが、検出可能なマーカーで標識される、オリゴヌクレオチド。
12.アンチセンスオリゴヌクレオチドであって、項目3に記載の核酸によってコードされるmRNAに特異的に結合し得る、アンチセンスオリゴヌクレオチド。
13.AT cDNA配列の存在を検出するためのキットであって、項目11に記載の少なくとも1つのオリゴヌクレオチドを含む、キット。
14.膜を横切ってアセチルCoAを輸送し得ることによって特徴付けられる、単離されたアセチルコエンザイムAトランスポーター(AT)タンパク質。
15.項目14に記載のATタンパク質であって、該タンパク質のアミノ酸配列が、配列番号2に記載のタンパク質配列と実質的に同じ配列を含む、タンパク質。
16.項目15に記載のATタンパク質であって、配列番号2に記載のヒトAT-1タンパク質配列と同じアミノ酸を有する、タンパク質。
17.項目14に記載のATタンパク質であって、該タンパク質は、ヒトAT-1であるヌクレオチド配列によってコードされ、および項目1に記載のヌクレオチド配列と実質的に同じである、タンパク質。
18.項目17に記載のATタンパク質であって、該タンパク質が、ヒトATであり、および配列番号1に記載されるヌクレオチド配列によってコードされる、タンパク質。
19.項目14に記載のATタンパク質であって、該タンパク質が、配列番号1に記載されるヌクレオチド388〜2034と実質的に同じヌクレオチド配列を含むヌクレオチド配列によってコードされる、タンパク質。
20.ATタンパク質の発現のための方法であって、該方法が、該ATタンパク質の発現に適切な条件下で、項目9に記載の細胞を培養する工程を包含する、方法。
21.項目14に記載のATタンパク質との特異的反応性を有する、単離された抗ATタンパク質抗体。
22.項目21に記載の抗体であって、該抗体が、モノクローナル抗体である、抗体。
23.項目21に記載の抗体であって、該抗体が、ポリクローナル抗体である、抗体。
24.ヒトATタンパク質の発現を阻害するに有効な量の、項目12に記載のアンチセンスオリゴヌクレオチド、および細胞膜を通過し得る、受容可能な疎水性キャリアを含有する、組成物。
25.ATタンパク質をコードする外因性核酸を発現する、トランスジェニック非ヒト哺乳動物。
26.項目25に記載のトランスジェニック非ヒト哺乳動物であって、前記ATタンパク質をコードする核酸が、変異されており、およびそのように発現される該ATタンパク質が、ネイティブなATでない、トランスジェニック非ヒト哺乳動物。
27.項目25に記載のトランスジェニック非ヒト哺乳動物であって、該トランスジェニック非ヒト動物が、マウスである、トランスジェニック非ヒト哺乳動物。
28.哺乳動物ATタンパク質をコードする核酸を同定するための方法であって、該方法が、以下の工程:
項目10に記載のオリゴヌクレオチドと、核酸を含有するサンプルとを接触する工程であって、該接触する工程が、高いストリンジェンシーのハイブリダイゼーション条件下で達成される、工程、および
それにハイブリダイズする化合物を同定する工程、
を包含する、方法。
29.サンプル中のヒトATタンパク質の存在を検出するための方法であって、該方法が、項目21に記載の抗体と、試験サンプルとを接触する工程、抗体−AT複合体の存在を検出する工程、およびそれによって、該試験サンプル中のヒトATタンパク質の存在を検出する工程、を包含する、方法。
30.AT核酸の増幅のための1本鎖DNAプライマーであって、該プライマーが、配列番号1に記載の核酸配列に由来する核酸配列を含む、プライマー。
31.試験化合物が、項目14に記載のATタンパク質に対するアゴニストまたはアンタゴニストとして作用し得るか否かを評価するためのバイオアッセイであって、該バイオアッセイが、以下の工程:
(a)ATタンパク質またはその機能的に改変された形態を発現するDNAを含む細胞を培養する工程であって、
ここで該培養する工程が、ATタンパク質のAc-CoA輸送活性を調節する能力が決定されようとする、少なくとも1つの化合物の存在下で行われる、工程、およびその後
(b)Ac-CoAのレベルの増加または減少のいずれかについて、該細胞をモニターする工程、
を包含する、バイオアッセイ。
32.試験化合物が、項目14に記載のATタンパク質、または該ATタンパク質の機能的に改変された形態についてのアンタゴニストとして作用し得るか否かを評価するためのバイオアッセイであって、該バイオアッセイが、以下の工程:
(a)ATタンパク質またはその機能的に改変された形態を発現するDNAを含む細胞を培養する工程であって、
ここで該培養する工程が:
ATタンパク質のAc-CoA輸送活性を阻害する能力が決定されようとする、漸増濃度少なくとも1つの化合物、および
一定濃度のAc-CoA
の存在下で行われる、工程;ならびにその後、
(b)該細胞において、該化合物の濃度の関数として細胞内オルガネラに輸送されるAc-CoAのレベルを、モニターし、それによってAT輸送活性を阻害する該化合物の能力を示す工程、
を包含する、バイオアッセイ。
33.試験化合物が、項目14に記載のATタンパク質、または該ATタンパク質の機能的に改変された形態についてのアンタゴニストとして作用し得るか否かを評価するためのバイオアッセイであって、該バイオアッセイが、以下の工程:
(a)ATタンパク質またはその機能的に改変された形態を発現するDNAを含む細胞を培養する工程であって、
ここで該培養する工程が、
ATタンパク質のAc-CoA輸送活性を阻害する能力が決定されようとする、一定濃度の少なくとも1つの化合物、および
漸増濃度のAc-CoA
の存在下で行われる、工程;ならびにその後、
(b)該細胞において、該化合物の濃度の関数として細胞内オルガネラに輸送されるAc-CoAのレベルを、モニターし、それによってAT輸送活性を阻害する該化合物の能力を示す工程、
を包含する、バイオアッセイ。
34.試験化合物が、項目14に記載のATタンパク質、または該ATタンパク質の機能的に改変された形態についてのアゴニストとして作用し得るか否かを評価するためのバイオアッセイであって、該バイオアッセイが、以下の工程:
(a)ATタンパク質またはその機能的に改変された形態を発現するDNAを含む細胞を培養する工程であって、
ここで該培養する工程が、
ATタンパク質のAc-CoA輸送活性を増加する能力が決定されようとする、漸増濃度の少なくとも1つの化合物、および
一定濃度のAc-CoA
の存在下で行われる、工程;ならびにその後、
(b)該細胞において、該化合物の濃度の関数として細胞内オルガネラに輸送されるAc-CoAのレベルを、モニターし、それによってAT輸送活性を増加する該化合物の能力を示す工程、
を包含する、バイオアッセイ。
35.試験化合物が、項目14に記載のATタンパク質、または該ATタンパク質の機能的に改変された形態についてのアゴニストとして作用し得るか否かを評価するためのバイオアッセイであって、該バイオアッセイが、以下の工程:
(a)ATタンパク質またはその機能的に改変された形態を発現するDNAを含む細胞を培養する工程であって、
ここで該培養する工程が、
ATタンパク質のAc-CoA輸送活性を増加する能力が決定されようとする、一定濃度の少なくとも1つの化合物、および
漸増濃度のAc-CoA
の存在下で行われる、工程;およびその後、
(b)該細胞において、該化合物の濃度の関数として細胞内オルガネラに輸送されるAc-CoAのレベルを、モニターし、それによってAT輸送活性を増加する該化合物の能力を示す工程、
を包含する、バイオアッセイ。
36.ATタンパク質によって媒介されるAc-CoA輸送活性を調節するための方法であって、該方法が:
該ATタンパク質と、項目31によって同定されるアゴニストの調節するに有効な量とを、接触させる工程を、包含する、方法。
37.ATタンパク質によって媒介されるAc-CoA輸送活性を調節するための方法であって、該方法が:
該ATタンパク質と、項目31によって同定されるアンタゴニストの調節するに有効な量とを、接触させる工程を、包含する、方法。
38.異常な神経細胞分化、異常な神経細胞移動、神経芽腫、または黒色腫を処置する方法であって、該方法が、有効量の項目31によって同定される化合物を投与する工程を包含する、方法。
【発明を実施するための最良の形態】
【0009】
発明の詳細な説明
本発明によれば、単離された哺乳動物アセチルコエンザイムAトランスポーター(AT)タンパク質、ポリペプチド、および本発明の核酸によってコードされるそれらのフラグメントが提供される。本明細書中で使用されるように、句「AT」は、膜を横切ってアセチルコエンザイムA(Ac-CoA)を輸送し得る、哺乳動物ファミリー、好ましくはヒトの、単離されたおよび/または実質的に純粋なタンパク質をいう。本発明のATタンパク質はさらに、ガングリオシドGD3およびGT3のシアル酸残基のアセチル化を、間接的に促進する能力を有することによって、特徴付けられる。本発明のATタンパク質は、一次転写物の別のスプライシングによって生成されるmRNAによってコードされる、天然に存在するその対立遺伝子改変体を含み、そしてさらに、例えば、免疫原性などのような少なくとも1つの天然の生物学的活性を保持する、その活性なフラグメントを含む。
【0010】
本発明の別の実施態様において、本明細書中で言及されるATタンパク質は、ATタンパク質(好ましくはヒト)をまた特異的に認識する抗体によって特異的に認識されるそれらのポリペプチドであり、これは配列番号2に記載される配列を含む。本発明の単離されたATタンパク質は、天然のインビボ環境と通常関連する細胞性成分および/または夾雑物を含まない。
【0011】
本発明のタンパク質はさらに、少なくとも以下の細胞:心臓、脳、胎盤、肺、肝臓、骨格筋、腎臓、および膵臓において発現されることによって特徴付けられる。最も高いレベルの発現は、膵臓組識において検出された。3.3および4.3kbの大きさを有する、ATタンパク質の2つの主要なmRNA転写物は、試験した全ての組識において検出された(ノーザンブロットアッセイによって観察された)。2.7および3.4kbの大きさを有する2つのmRNA転写物は、AT-1でトランスフェクトされたHeLa細胞において強力に発現され、そして偽トランスフェクトされたHeLa細胞においてほとんど検出されなかった。従って、ATファミリーのタンパク質をコードするスプライス改変体cDNA転写物が、本発明によって明らかに意図される。
【0012】
AT-1(配列番号2)の予測されるアミノ酸配列は、神経伝達物質(概説については、HuchoおよびTsetlin、J.Neurochem.、66:1781-1792(1996)を参照のこと)、グルコース(Daviesら、Biochem.J.、266:799-808(1990))、アンモニア(Mariniら、EMBOJ.、12:3456-3463(1994))、およびヌクレオチド−糖類(Eckhardtら、Proc. Natl. Acad. Sci. USA、93:7572-7576(1996);Miuraら、J.Biochem.、120:236-241(1996);およびAbeijonら、Proc. Natl. Acad. Sci.USA、93:5963-5968(1996))についてのトランスポーターを含む多くのトランスポーターにおいて代表的に観察される、複数の膜貫通ドメインを有する。ほとんどのトランスポータータンパク質は、膜貫通α-ヘリックスを形成すると考えられる6〜12個の疎水性配列を含む。AT-1タンパク質の場合、少なくとも6個の膜貫通ドメインの存在が、Psort分析によって予測される。
【0013】
さらに、AT-1タンパク質は、転写因子の二量体化に関与することがもともと見出された(Landschulzら、Science,240:1759-1964(1988))、ロイシンジッパーモチーフを含む。非常に最近の研究は、CMP-NeuAc(Eckhardtら、Proc.Natl. Acad. Sci.USA、93:7572-7576(1996))、UDP-Gal(Miuraら、J.Biochem.、120:236-241(1996))、およびUDP-GlcNAc(Abeijonら、Proc.Natl.Acad.Sci.USA、93:5963-5968(1996))についてのトランスポーターのようなヌクレオチド−糖トランスポータータンパク質もまた、この構造を有することを示し、このことは、これらのタンパク質が、ゴルジ膜におけるホモ二量体として存在することを示唆する。近年、ラット肝臓ゴルジPAPSトランスポーター(Mandonら、Biochem.Biophy.Res.Commun.、225:932-938(1996))もまた、ホモ二量体であることが示されたが、ロイシンジッパー構造がこの特定のトランスポータータンパク質に存在するか否かは知られていない。従って、AT-1タンパク質は、複数の膜貫通ドメインを有するホモ二量体として機能し得ることが、本明細書中で意図される。しかし、AT-1タンパク質の分子量は、CMP-NeuAc、UDP-Ga1、およびUDP-GlcNAcについてのトランスポーターの分子量よりも実質的に大きく、このことは、AT-1タンパク質は、新規のATトランスポーターファミリーのメンバーを意味し得ることを示唆する。
【0014】
今までのところ同定された全てのヌクレオチド−糖トランスポーターは、ゴルジ体膜に局在化されることが見出されている(Miuraら、J. Biochem.、120:236-241(1996);Abeijonら、Proc. Natl.Acad.Sci.USA、93:5963-5968(1996);およびBhakdiら、Infect.Immun.、47:52-60(1985))。AT-1に対して特異的な抗体を用いる免疫組識学的研究は、AT-1タンパク質が、小胞体、ゴルジ、およびおそらくミトコンドリア膜を含む細胞質中に拡散して存在することを実証した。この観察は、アセチルトランスフェラーゼが、ゴルジ膜に富化されることが示されたので、ATタンパク質ファミリーは、アセチルトランスフェラーゼとして機能しないことを示す(Varki、Glycobiology、2:25-40(1992)およびVarkiおよびDiaz、J.Biol. Chem.、260:6600-6608(1985))。さらに、半インタクトな細胞を使用した、インビトロアッセイの結果(例えば、実施例VI、および図2を参照のこと)は、AT-1が、膜を横切ってAc-CoAを輸送するように機能するタンパク質ファミリーのメンバー(例えば、Ac-CoAトランスポーター)であることを示す。
【0015】
タンパク質および核酸データベース(GenBank、EMBLおよびSwiss-Prot)に対するホモロジー検索が、配列番号1に記載されるAT-1の核酸配列およびアミノ酸配列に相同な核酸配列およびアミノ酸配列について行われた。公知のタンパク質との有意な相同性は検出されなかったが、ホモロジー検索は、高い程度の相同性を有する2つの推定のタンパク質を同定した:35%のアミノ酸配列同一性を有する560アミノ酸を有する、S.cerevisiaeの推定のタンパク質[アクセス番号第Z36088(EMBL)および同第P38318(Swiss Prot)];ならびに47%のアミノ酸配列同一性を有する632アミノ酸を有する、Caenorhabditiselegans T26C5.3[EMBL、アクセス番号第Z50859]。従って、AT-1タンパク質は、普遍的であり、そして進化論的に保存されているようであり、そしてシアル酸の出現は、これらの2つの生物体において報告されていないので、ATタンパク質ファミリーは、シアル酸アセチルトランスフェラーゼとして機能しないようである。AT-1の転写物は、O-アセチル化ガングリオシドよりも広範囲に分配され、このことは、AT-1タンパク質以外の分子が、O-アセチル化GD3発現の調節を指図するという仮定を支持する。本明細書中に記載される結果は、AT-1タンパク質は、Ac-CoAトランスポーターとして機能すること、そしてこれは、ガングリオシドにおけるシアル酸のアセチル化プロセス以外のアセチル化プロセスにおいて、重要な役割を果たし得ることを、強力に示唆する。
【0016】
本明細書および請求の範囲における、DNA、RNA、ポリペプチド、またはタンパク質の修飾語としての、用語「単離された」および/または「精製された」の使用は、そのように設計されたDNA、RNA、ポリペプチド、またはタンパク質が、ヒトの手によってこのような形態において生成され、従ってそれらの天然のインビボ細胞環境から分離されることを意味する。このヒト介入の結果として、本発明の組換えのDNA、RNA、ポリペプチド、およびタンパク質は、それらが天然に存在するようなDNA、RNA、ポリペプチド、またはタンパク質ではないことが、本明細書中で記載する方法において有用である。
【0017】
本明細書中で使用されるように、「哺乳動物」は、本発明のATタンパク質が由来する多様な種(例えば、ヒト、ラット、マウス、ウサギ、サル、ヒヒ、ウシ、ブタ、ヒツジ、イヌ、ネコなど)をいう。
【0018】
現在好ましい本発明のATタンパク質は、配列番号2に記載のタンパク質配列と実質的に同じであるアミノ酸配列、ならびに生物学的に活性な、その改変された形態を含む。当業者は、上記配列の複数の残基が、得られるタンパク質種の生物学的活性を実質的に変化しないで、他の、化学的に、立体的に、および/または電子的に類似の残基と置換され得ることを認識する。さらに、本明細書中の配列番号2と実質的に同じ配列を含む、より大きなまたはより小さなポリペプチド配列(例えば、スプライス改変体、ATの活性なフラグメントなど)が、意図される。
【0019】
本明細書中で用いられるように、用語「実質的に同じアミノ酸配列」は、参照アミノ酸配列に関して少なくとも約70%の同一性を有し、および参照アミノ酸配列によって規定されるタンパク質の、比較可能な機能的なおよび生物学的な活性の特徴を保持するアミノ酸配列をいう。好ましくは、「実質的に同じアミノ酸配列」を有するタンパク質は、参照アミノ酸配列に関して少なくとも約80%、より好ましくは90%のアミノ酸同一性を有する;約95%を超えるアミノ酸配列同一性が、特に好ましい。しかし、スプライス改変体として生じる、記載されるレベル未満の配列同一性を含むか、または保存的なアミノ酸置換によって、もしくは縮重コドンの置換によって改変される、ポリペプチド(または、本明細書中で以前に言及された核酸)はまた、本発明の範囲内に包含される。本明細書中に開示される好ましいATタンパク質は、ヒトAT-1(配列番号2)である。
【0020】
用語「生物学的に活性な」または「機能的な」は、本発明のATタンパク質、またはそのポリペプチドフラグメントの修飾語として本明細書中で使用される場合、ATに起因される機能的な特徴の少なくとも1つを示すポリペプチドをいう。例えば、ATの1つの生物学的活性は、細胞膜、好ましくは細胞内オルガネラの膜(例えば、ゴルジ膜、小胞体膜、ミトコンドリア膜など)を横切ってアセチルコエンザイムA(Ac-CoA)を輸送する能力である。ATのさらに別の生物学的活性は、ガングリオシドGD3およびGT3のシアル酸残基のアセチル化を、間接的に促進する能力である。
【0021】
ATの別の生物学的活性は、ATに特異的に結合するポリクローナル抗体およびモノクローナル抗体の生成のための免疫原として作用する能力である。従って、ATをコードする本発明の核酸は、配列番号2に記載される配列(At-1)を含むATタンパク質(好ましくはヒト)をまた特異的に認識する抗体によって、特異的に認識されるポリペプチドをコードする。このような活性は、当業者に公知の任意の方法によってアッセイされ得る。例えば、ATcDNAによってコードされる試験ポリペプチドは、抗体を生成するために使用され得、次いで、抗体は、配列番号2に記載の配列を含むタンパク質に結合するそれらの能力についてアッセイされる。抗体が、実質的に同じ親和性で、試験ポリペプチド、および配列番号2に記載の配列を含むタンパク質に結合する場合、ポリペプチドは、必要とされる生物学的活性を保持する。
【0022】
本発明のATタンパク質は、当該分野において周知の多様な方法(例えば、本明細書中に記載される組換え発現系、沈殿、ゲル濾過、イオン交換クロマトグラフィー、逆相クロマトグラフィー、およびアフィニティークロマトグラフィーなど)によって単離され得る。他の周知の方法は、Deutscherら、Guideto Protein Purification: Methods in Enzymology 第182巻(Academic Press、(1990))(これは、本明細書中に参考として援用される)に記載される。あるいは、本発明の単離されたポリペプチドは、例えば、Sambrookら(前出、1989)に記載されるような、周知の組換え方法を使用して得られ得る。
【0023】
本発明のポリペプチドを調製するための手段の例は、当該分野において周知の方法を用いて、適切な宿主細胞(例えば、細菌細胞、酵母細胞、両生類細胞(すなわち、卵母細胞)、または哺乳動物細胞)においてATをコードする核酸を発現し、そして再び周知の方法を用いて、発現されたポリペプチドを回収することである。本発明のATポリペプチドは、本明細書中に記載されるような発現ベクターで形質転換された細胞から直接単離され得る。本発明のポリペプチド、生物学的に活性なフラグメント、およびそれらの機能的等価物はまた、化学合成によって生成され得る。例えば、合成ポリペプチドは、製造業者によって提供される化学製品を用いて、AppliedBiosystems、Inc. 430A型または431A型 自動ペプチド合成器(Foster City、CA)を使用して、生成され得る。
【0024】
また、用語ATによって含まれるものは、その活性なフラグメントまたはポリペプチドアナログである。用語「活性なフラグメント」は、この部分が、対応する全長タンパク質の特徴である活性を有することが提供される、全長ATタンパク質の部分であるペプチドフラグメントをいう。例えば、ATタンパク質の活性なフラグメント(例えば、細胞質ドメイン)は、Ac-CoAを結合するか、またはAc-CoAへの結合後にAc-CoAの膜輸送を媒介する能力のような活性を有し得る。免疫応答を誘発するATタンパク質の活性フラグメントの特徴は、抗AT抗体を得るために有用である。従って、本発明はまた、本発明のATタンパク質の活性なフラグメントを提供し、このフラグメントは、本明細書中に記載されるアッセイを使用して、同定され得る。
【0025】
用語「ポリペプチドアナログ」は、その1つ以上の残基が機能的に類似な残基で保存的に置換されている、本明細書中に具体的に示される配列に実質的に同一なアミノ酸残基配列を有し、そして本明細書中に記載されるようなATを模倣する能力を示す、任意のポリペプチドを含む。保存的置換の例としては、ある非極性(疎水性)残基(例えば、イソロイシン、バリン、ロイシン、もしくはメチオニン)での別の非極性残基の置換、ある極性(親水性)残基での別の極性残基の置換(例えば、アルギニンとリジンとの間、グルタミンとアスパラギンとの間、グリジンとセリンとの間)、ある塩基性残基(例えば、リジン、アルギニン、もしくはヒスチジン)での別の塩基性残基の置換、またはある酸性残基(例えば、アスパラギン酸もしくはグルタミン酸)での別の酸性残基の置換を含む。
【0026】
本明細書中で使用されるように、句「保存的置換」はまた、非誘導体化残基の代わりに化学的に誘導された残基の使用を含むが、ただしこのようなポリペプチドは、必要とされる結合活性を示す。句「化学的誘導体」は、官能側鎖基の反応によって化学的に誘導体化された1つ以上の残基を有する目的のポリペプチドをいう。このような誘導体化された分子は、例えば、遊離のアミノ基が、誘導体化され、アミン塩酸塩、p-トルエンスルホニル基、カルボベンズオキシ基、t-ブチルオキシカルボニル基、クロロアセチル基、またはホルミル基を形成するそれらの分子を含む。遊離のカルボキシル基は、誘導体化されて、塩、メチルおよびエチルエステル、または他のタイプのエステルもしくはヒドラジドを形成し得る。遊離のヒドロキシル基は、誘導体化されて、O-アシルまたはO-アルキル誘導体を形成し得る。ヒスチジンのイミダゾール窒素は、誘導体化されて、N-im-ベンジルヒスチジンを形成し得る。また、化学誘導体として含まれるものは、20個の標準アミノ酸の1つ以上の天然に存在するアミノ酸誘導体を含むそれらのペプチドである。例えば、4-ヒドロキシプロリンは、プロリンを置換し得;5-ヒドロキシリジンは、リジンを置換し得;3-メチルヒスチジンは、ヒスチジンを置換し得;ホモセリンは、セリンを置換し得;およびオルニチンは、リジンを置換し得る。本発明のポリペプチドはまた、必要とされる活性が維持される限り、配列が本明細書中に示されるポリペプチドの配列に比べて、残基の1つ以上の付加および/または欠失を有する任意のポリペプチドを含む。
【0027】
本発明はまた、受容可能なキャリア、ならびに任意の単離された、精製されたATポリペプチド、その活性なフラグメントもしくはポリペプチドアナログ、または精製された、成熟タンパク質およびその活性なフラグメントを、単独でまたは互いに組合わせて含む、組成物を提供する。これらのポリペプチドまたはタンパク質は、組換え的に得られ得るか、化学的に合成され得るか、または天然の供給源から精製され得る。本明細書中で使用されるように、用語「受容可能なキャリア」は、任意の標準的な薬学的キャリア(例えば、リン酸緩衝化生理食塩水溶液、水、および乳濁液(例えば、油/水、または水/油乳濁液)、ならびに種々のタイプの湿潤剤)を含む。
【0028】
本発明の別の実施態様によれば、本発明のAT(アセチル−コエンザイムA輸送体)タンパク質、およびそのフラグメントをコードする単離された核酸が提供される。本明細書中に記載される核酸分子は、このような核酸が当業者に公知の多様なタンパク質発現系に組込まれる場合、本発明のタンパク質を生成するために有用である。さらに、このような核酸分子またはそのフラグメントは、所定のサンプルにおけるAT遺伝子またはmRNA転写物の存在および/または量をアッセイするために、容易に検出可能な置換基で標識され得、そしてハイブリダイゼーションプローブとして使用され得る。本明細書中に記載される核酸分子、およびそのフラグメントはまた、本明細書中に記載される本発明のタンパク質をコードする遺伝子を増幅するためのPCR反応において、プライマーおよび/または鋳型として有用である。
【0029】
用語「核酸」(ポリヌクレオチドともいわれる)は、リボ核酸(RNA)またはデオキシリボ核酸(DNA)、プローブ、オリゴヌクレオチド、およびプライマーを含む。DNAは、相補的DNA(cDNA)またはゲノムDNA(例えば、ATタンパク質をコードする遺伝子)のいずれかであり得る。ATポリペプチドをコードする核酸を単離する1つの手段は、当該分野において周知の方法を使用して、天然のDNAプローブまたは人工的に設計されたDNAプローブで、哺乳動物ゲノムライブラリーをプローブすることである。AT遺伝子に由来するDNAプローブは、この目的のために特に有用である。ATポリペプチドをコードするDNAおよびcDNA分子は、以下により詳細に記載される方法によって、哺乳動物(例えば、ヒト、マウス、ラット、ウサギ、ブタなど)、もしくは他の動物供給源から相補的ゲノムDNA、cDNA、もしくはRNAを得るために、またはcDNAライブラリーもしくはゲノムライブラリーのスクリーニングによって関連のcDNAクローンもしくはゲノムクローンを単離するために使用され得る。核酸の例は、RNA、cDNA、またはATポリペプチドをコードする単離されたゲノムDNAである。このような核酸は、配列番号1(配列番号1の少なくともヌクレオチド388〜2034)に記載のものと実質的に同じヌクレオチド配列を有する核酸、またはそのスプライス改変体cDNA配列を含み得るが、これらに限定されない。
【0030】
本明細書中で使用されるように、本発明のレセプターをコードする特定のヌクレチド配列を記載するために使用される場合、句「スプライス改変体」または「選択的にスプライスされた」とは、周知の真核生物RNAスプライシングプロセスから生じるcDNA配列をいう。RNAスプライシングプロセスは、細胞質の成熟RNA分子を作製するための、真核生物一次RNA転写物からのイントロンの除去、およびエキソンの結合を含む。スプライス改変体ヌクレオチド配列を単離する方法は、当該分野において周知である。例えば、当業者は、本明細書中に記載されるようなcDNAライブラリーまたはゲノムライブラリーをスクリーニングするために、配列番号1のATをコードするcDNAに由来するヌクレオチドプローブを用い得る。
【0031】
本発明の1つの実施態様において、本明細書中に開示される本発明のATタンパク質をコードするcDNAは、配列番号1に記載されるような、実質的に同じヌクレオチド配列を含む。本発明の別の実施態様において、本発明のタンパク質をコードするcDNA分子は、配列番号1のヌクレオチド388〜2034と同じヌクレオチド配列を含む。
【0032】
本明細書中で用いられるように、用語「実質的に同じヌクレオチド配列」は、参照ポリヌクレオチドに、中程度にストリンジェントなハイブリダイゼーション条件下で参照ヌクレオチドにハイブリダイズするのに十分な同一性を有するDNAをいう。1つの実施態様において、参照ヌクレオチド配列と実質的に同じヌクレオチド配列を有するDNAは、配列番号2に記載されるアミノ酸配列と実質的に同じアミノ酸配列、または配列番号2を含むより大きなアミノ酸配列をコードする。別の実施態様において、参照ヌクレオチド配列と「実質的に同じヌクレオチド配列」を有するDNAは、参照ヌクレオチド配列に関して少なくとも60%の同一性を有する。参照ヌクレオチド配列に対して、少なくとも70%、より好ましくは少なくとも約90%、さらにより好ましくは少なくとも95%の同一性を有するDNAが、好ましい。
【0033】
本発明はまた、配列番号1に示される核酸とは異なるが、同じ表現型を有する核酸を含む。表現型が類似する核酸はまた、「機能的に等価な核酸」といわれる。本明細書中で使用されるように、句「機能的に等価な核酸」は、実質的に同じ様式において機能し、本明細書中で開示される核酸と同じタンパク質産物を生成する、僅かに重要なおよび重要でない配列の改変によって特徴付けられる核酸を包含する。特に、機能的に等価な核酸は、本明細書中に開示されるポリペプチドと同じであるポリペプチド、もしくは保存的アミノ酸改変を有するポリペプチドをコードするか、または配列番号2を含むより大きなポリペプチドをコードする。例えば、保存的改変は、別の非極性残基での非極性残基の置換、または同様に荷電された残基での荷電された残基の置換を含む。これらの改変は、タンパク質の三次構造を実質的に変化させない改変として、当業者によって認識される改変を含む。
【0034】
遺伝子コードの縮重によって、特定されたハイブリダイゼーション条件下で本発明の核酸に必ずしもハイブリダイズしない、ATポリペプチドをコードする核酸が、さらに提供される。本発明のATポリペプチドをコードする好ましい核酸は、配列番号2(すなわち、AT-1)に記載されるのと実質的に同じアミノ酸配列をコードするヌクレオチドから構成される。
【0035】
従って、本発明のATタンパク質をコードする例示的な核酸は、以下から選択され得る:
(a)配列番号2に記載されるアミノ酸配列をコードするDNA;
(b)中程度にストリンジェントな条件下で(a)のDNAにハイブリダイズするDNA、ここでこのDNAは、生物学的に活性なATをコードする;または
(c)上述の(a)または(b)のいずれかに関するDNA縮重体、ここでこのDNAは、生物学的に活性なATをコードする。
【0036】
ハイブリダイゼーションは、水素結合を介する、核酸の相補的な鎖(すなわち、センス:アンチセンス鎖、またはプローブ:標的−DNA)の互いへの結合をいい、染色体DNAにおいて天然に存在する結合に類似する。所定のプローブと標的−DNAとをハイブリダイズするために使用されるストリンジェンシーのレベルは、当業者によって容易に変更され得る。
【0037】
句「ストリンジェントなハイブリダイゼーション」とは、ポリ核酸配列ハイブリッドが安定である下での条件をいうために、本明細書中で用いられる。当業者に公知であるように、ハイブリッドの安定性は、ハイブリッドの融点(Tm)において反映される。一般に、ハイブリッドの安定性は、ナトリウムイオン濃度および温度の関数である。代表的に、ハイブリダイゼーション反応は、より低いストリンジェンシーの条件下で行われ、次いで変化するが、より高いストリンジェンシーの洗浄を行う。ハイブリダイゼーションのストリンジェンシーに対する参照は、このような洗浄条件に関する。
【0038】
本明細書中で使用されるように、句「中程度にストリンジェントなハイブリダイゼーション」は、標的DNAに対する約60%の同一性、好ましくは約75%の同一性、より好ましくは約85%の同一性を有する相補的核酸に、標的−DNAが結合することを許容する条件をいい;標的−DNAに対する約90%を超える同一性は、特に好ましい。好ましくは、中程度にストリンジェントな条件は、42℃で、50%ホルムアミド、5×デンハルト溶液、5×SSPE、0.2%SDSにおけるハイブリダイゼーション、次いで42℃で、0.5×SSPE、0.2% SDSにおける洗浄に等価である条件である。
【0039】
句「高いストリンジェンシーのハイブリダイゼーション」は、65℃で、0.018MNaClにおいて安定なハイブリッドを形成するそれらの核酸配列のみのハイブリダイゼーションを許容する条件をいう(すなわち、ハイブリッドが、65℃で、0.018MNaClにおいて安定でない場合、これは、本明細書中で意図されるように高いストリンジェンシー条件下で安定でない)。高いストリンジェンシー条件は、例えば、42℃で、50%ホルムアミド、5×デンハルト溶液、5×SSPE、0.2%SDSにおけるハイブリダイゼーション、次いで65℃で、0.1×SSPEおよび0.1% SDSにおける洗浄によって提供され得る。
【0040】
句「低いストリンジェンシーハイブリダイゼーション」は、37℃での10%ホルムアミド、5×デンハルト溶液、6×SSPE、0.2%SDSにおけるハイブリダイゼーション、次いで50℃での1×SSPE、0.2% SDSにおける洗浄に、等価な条件をいう。デンハルト溶液およびSSPE(例えば、Sambrookら、MolecularCloning、A Laboratory Manual、Cold Spring Harbor Laboratory Press、1989を参照のこと)は、他の適切なハイブリダイゼーション緩衝液と同様に、当業者に周知である。
【0041】
本明細書中で使用されるように、用語「縮重」は、参照核酸(例えば、配列番号1)から、少なくとも1ヌクレオチド異なるが、参照核酸と同じアミノ酸をコードするコドンをいう。例えば、3ヌクレオチド「UCU」、「UCC」、「UCA」、および「UCG」は、これらのコドンの4つは全て、アミノ酸のセリンをコードするので、互いに関して縮重である。
【0042】
本発明のポリペプチドをコードする好ましい核酸は、中程度にストリンジェント、好ましくは高いストリンジェンシーの条件下、配列番号1に記載の核酸配列の実質的な配列全体に、ハイブリダイズする。
【0043】
AT cDNAの任意の領域の部位特異的変異誘発は、変異体AT cDNAの生成に関して本明細書において意図される。例えば、TransformerMutagenesis Kit(Clontechから入手可能)は、AT cDNAに対して、様々なミスセンスおよび/またはノンセンス変異を構築するために使用され得るなど。
【0044】
本発明の核酸は、当該分野において周知の、様々な方法(例えば、配列番号1の種々の領域からオリゴヌクレオチドプライマーを使用して、PCR増幅を用いる、本明細書中に記載される方法など)によって、生成され得る。
【0045】
本発明のさらなる実施態様によれば、必要に応じて標識されるATコードcDNAまたはそのフラグメントが、関連の新規の哺乳動物ATタンパク質をコードするさらなる核酸配列について、ライブラリー(例えば、cDNA、ゲノムなど)をプローブするために用いられ得る。哺乳動物のcDNAライブラリーおよびゲノムライブラリー、好ましくはヒトライブラリーの構築は、当該分野において周知である。このようなcDNAライブラリーまたはゲノムライブラリーのスクリーニングは、約42℃未満の温度、約50%未満のホルムアミド濃度、および低い塩濃度への調整を含む、低いストリンジェンシー条件下で、最初に開始される。
【0046】
1つの実施態様において、プローブベースのスクリーニング条件は、約37℃の温度、約20%のホルムアミド濃度、および約5×標準生理食塩水クエン酸(SSC;3M塩化ナトリウム、0.3Mクエン酸ナトリウムを含む20×SSC、pH7.0)を含む。このような条件は、完全な相同性を必要とすることなく、プローブ配列と実質的な程度の類似性を有する配列の同定を許容する。句「実質的な類似性」は、少なくとも50%の相同性を共有する配列をいう。好ましくは、プローブと少なくとも約70%の相同性を有する配列の同定を許容するが、プローブと低い程度の相同性を有する配列に対して区別する、ハイブリダイゼーション条件が、選択される。結果として、配列番号1のヌクレオチド388〜2034と実質的に同じ(すなわち、類似の)ヌクレオチド配列を有する核酸が、得られる。
【0047】
本明細書中で使用されるように、プローブまたはプライマーとしてもまた本明細書中に言及される核酸「オリゴヌクレオチド」は、1本鎖DNAもしくはRNA、またはそのアナログであり、これは、配列番号1の任意に記載される任意の14個以上の連続的な塩基と同じである(またはこれに相補的である)少なくとも14個、好ましくは少なくとも20個、より好ましくは少なくとも50個の連続的な塩基を含むヌクレオチドの配列を有する。プローブを構築するために好ましい領域は、配列番号1の5’および/または3’コード領域を含む。さらに、本発明のATタンパク質の領域をコードする完全なcDNA、または配列番号1(AT)に対応する完全な配列は、プローブとして使用され得る。プローブは、本明細書中以後に記載されるように、当該分野において周知の方法によって標識され得、そして種々の診断キットにおいて使用され得る。
【0048】
本明細書中で使用されるように、種々の文法的形態における用語「標識」および「指示手段(indicatingmeans)」は、検出可能なシグナルの生成に、直接的または間接的のいずれかで関与する単一の原子および分子をいう。任意の標識または指示手段は、本発明の核酸プローブ、発現されるタンパク質、ポリペプチドフラグメント、または抗体分子に連結され得る。これらの原子または分子は、単独で、またはさらなる試薬と組合わせて使用され得る。このような標識は、臨床学的診断化学において、それ自体周知である。
【0049】
標識化手段は、変性を伴うことなく抗体または抗原に化学的に結合して、有用な免疫蛍光トレーサーである蛍光色素(染料)を形成する、蛍光標識化剤であり得る。免疫蛍光分析技術の記載は、DeLuca、「Immunofluorescence Analysis」、Antibody As a Tool、Marchalonisら編、JohnWiley & Sons,Ltd.、189〜231頁(1982)(これは、本明細書中に参考として援用される)において見出される。
【0050】
1つの実施態様において、指示基は、酵素(例えば、西洋ワサビペルオキシダーゼ(HRP)、グルコースオキシダーゼなど)である。別の実施態様において、放射活性エレメントは、標識化剤を用いられる。基質への標識の連結(すなわち、核酸プローブ、抗体、ポリペプチド、およびタンパク質の標識)は、当該分野において周知である。例えば、本発明の抗体は、培養培地において提供される放射標識されたアミノ酸の代謝取込みによって、標識され得る。例えば、Galfreら、Meth.Enzymol.、73:3-46(1981)を参照のこと。タンパク質結合または活性化された官能基によるカップリングの従来の手段は、特に適用可能である。例えば、Aurameasら、Scand.J.Immunol.、第8巻、補遺7:7-23(1978)、Rodwellら、Biotech.、3:889-894(1984)、および米国特許第4,493,795号を参照のこと。
【0051】
また、提供されるものは、mRNAの翻訳を妨げるように、ATポリペプチドをコードするmRNAの任意の部分と特異的に結合し得る配列を有するアンチセンスオリゴヌクレオチドである。アンチセンスオリゴヌクレオチドは、ATポリペプチドをコードするcDNAの配列の任意の部分と特異的に結合し得る配列を有し得る。本明細書中で使用されるように、句「特異的に結合する」は、相補的核酸配列を認識する、および相補的な塩基対間の水素結合の形成を介して、相補的核酸配列と二重らせんセグメントを形成する、核酸配列の能力を含む。アンチセンスオリゴヌクレオチドの例は、ヌクレオチドの化学アナログを含むアンチセンスオリゴヌクレオチドである。
【0052】
細胞膜を横切って通過し、そして翻訳を妨げるようにATポリペプチドをコードするmRNAと特異的に結合することによって、ATポリペプチドの発現を減少するに有効な、上記の、アンチセンスオリゴヌクレオチドの量、ならびに細胞膜を横切って通過し得る受容可能な疎水性キャリアを含む組成物がまた、本明細書において提供される。適切な疎水性キャリアは、例えば、米国特許第5,334,761号;同第4,889,953号;同第4,897,355号などにおいて記載される。細胞膜を横切って通過し得る受容可能な疎水性キャリアはまた、選択された細胞型に特異的なレセプターに結合し、そしてそれによって選択された細胞型の細胞によって取込まれる構造を含み得る。構造は、細胞型特異的レセプターに結合することが知られるタンパク質の部分であり得る。
【0053】
アンチセンスオリゴヌクレオチド組成物は、本発明のポリペプチドをコードするmRNAの翻訳を阻害するために有用である。合成オリゴヌクレオチド、または他のアンチセンス化学構造は、ATポリペプチドをコードするmRNAに結合し、そしてmRNAの翻訳を阻害するように設計され、および組識サンプルまたは被験体においてAT関連遺伝子の発現を阻害するための組成物として有用である。
【0054】
本発明の別の実施態様によれば、AT核酸配列の存在を検出するためのキットが提供され、少なくとも1つのオリゴヌクレオチド(例えば、本発明に従う、プローブまたはアンチセンスオリゴヌクレオチド)を含む。このようなキットは、AT遺伝子における変異、複製、欠失、再配置、または異数性を検出するために使用され得る。
【0055】
本発明は、ATポリペプチドをコードするmRNAの翻訳を阻害する、合成アンチセンスオリゴヌクレオチド組成物(本明細書中以後、SAOC)を用いることによって、これらのポリペプチドの発現のレベルを調節する手段を提供する。mRNAを認識し、そしてこれに選択的に結合するように設計された合成オリゴヌクレオチド、または他のアンチセンス化学構造は、ATコード鎖の部分または配列番号1に示されるヌクレオチド配列に相補的であるように構築される。SAOCは、注射によるもしくは直接的な腫瘍部位取込みによる、被験体への投与について、血流において安定であるように、または実験室の細胞培養条件において安定であるように、設計される。SAOCは、例えば、小さな、疎水性SAOC化学構造を設計することにより、SAOCを細胞膜を横切って通過し得るようにする、SAOCの物理的および化学的特性によって、またはSAOCを認識し、そしてこれを細胞へ輸送する、細胞における特異的な輸送系によって、細胞の細胞質に入るために細胞膜を横切って通過し得るように設計される。さらに、SAOCは、選択細胞集団内でのみSAOCを結合し、そしてこれを取込む、特異的な細胞取込み機構によって認識されるようにSAOCを標的化することによって、特定の選択された細胞集団のみへの投与について、設計され得る。
【0056】
例えば、SAOCは、上記のように、特定の細胞型においてのみ見出されるレセプターに結合するように設計され得る。SAOCはまた、配列番号1に示される配列内に含まれる配列に対応し得る、標的mRNA配列を認識し、これに選択的に結合するように設計される。SAOCは、標的mRNA配列に結合し、そして例えば、RNaseI消化によるmRNAの消化を誘導することによって、または翻訳調節因子またはリボソームの結合を妨げることによってmRNA標的配列の翻訳を阻害することによって、または、標的mRNAを分解するか、もしくは化学的に改変するかのいずれかの、他の化学構造(例えば、リボザイム配列または反応化学基)の包含によってのいずれかで、標的mRNA配列を不活化するように設計される。SAOCは、mRNA標的に対して指向される場合、このような特性が可能であることが示されている(Cohenら、TIPS、10:435(1989)およびWeintraub、Sci.American、January(1990)、40頁を参照のこと;これらの両方は、本明細書中に参考として援用される)。
【0057】
本発明のさらに別の実施態様によれば、適切な宿主細胞において、上記の核酸配列を発現することにより、本発明のATタンパク質を組換え産生するための方法が提供される。本明細書中に記載されるATタンパク質を生成するために適切である組換えDNA発現系は、当該分野において周知である。例えば、上記のヌクレオチド配列は、さらなる操作のためにベクターに組込まれ得る。本明細書中で使用されるように、ベクター(または、プラスミド)は、異種DNAの発現または複製のいずれかのために、細胞に異種DNAを導入するために使用される、不連続なエレメントをいう。
【0058】
適切な発現ベクターは、当該分野において周知であり、そして調節配列(例えば、DNAの発現を調節し得るプロモーター領域)に作動可能に連結されたこのようなDNAを発現し得るベクターを含む。従って、発現ベクターは、組換えDNAまたはRNA構築物(例えば、プラスミド、ファージ、組換えウイルス、もしくは適切な宿主細胞への取込みの際、挿入されたDNAの発現を生じる他のベクター)をいう。適切な発現ベクターは、当業者に周知であり、そして真核生物細胞および/または原核生物細胞において複製可能であるベクター、ならびにエピソームにとどまるベクターまたは宿主細胞ゲノムに組込まれるベクターを含む。さらに、ベクターは、ベクターが、多くのウイルスビリオン(例えば、レトロウイルス、ヘルペスウイルス、アデノウイルス)によってパッケージングされ、「ウイルスベクター」の形成を生じることを可能にする適切なパッケージングシグナルを含み得る。
【0059】
本明細書中に使用されるように、プロモーター領域は、DNAが作動可能に連結される、DNAの転写を制御するDNAのセグメントをいう。プロモーター領域は、RNAポリメラーゼ認識、結合、および転写開始に十分である特定の配列を含む。さらに、プロモーター領域は、RNAポリメラーゼのこの認識、結合、および転写開始活性を調節する配列を含む。これらの配列は、シス作用性であり得るか、またはトランス作用性因子に応答性であり得る。プロモーターは、調節の性質に依存して、構成的であり得るか、または調節され得る。本発明の実施における使用のために意図される例示的なプロモーターは、SV40初期プロモーター、サイトメガロウイルス(CMV)プロモーター、マウス乳房腫瘍ウイルス(MMTV)ステロイド誘導性プロモーター、モロニーマウス白血病ウイルス(MMLV)プロモーターなどを含む。
【0060】
本明細書中で使用されるように、用語「作動可能に連結される」は、DNAと、調節およびエフェクターヌクレオチド配列(例えば、プロモーター、エンハンサー、転写および翻訳の停止部位、ならびに他のシグナル配列)との機能的な関係をいう。例えば、プロモーターへのDNAの作動可能な連結は、DNAとプロモーターとの間の物理的および機能的な関係をいい、これによってこのようなDNAの転写が、DNAを特異的に認識し、結合し、そして転写するRNAポリメラーゼによって、プロモーターから開始される。
【0061】
本明細書中で使用されるように、発現は、ポリ核酸が、mRNAに転写され、そしてペプチド、ポリペプチド、またはタンパク質に翻訳されるプロセスをいう。ポリ核酸が、ゲノムDNAに由来する場合、発現は、適切な真核生物宿主細胞または生物が選択される場合、mRNAのスプライシングを含み得る。
【0062】
原核生物形質転換ベクターは、当該分野において周知であり、そしてpBlueskriptおよびファージλZAPベクター(Stratagene、La Jolla、CA)などを含む。他の適切なベクターおよびプロモーターは、1989年1月17日に発行された米国特許第4,798,885号(この開示は、その全体が本明細書中に参考として援用される)において詳細に開示される。
【0063】
E. coli細胞の形質転換についての他の適切なベクターは、pET発現ベクター(Novagen、米国特許第4,952,496号を参照のこと)(例えば、pET11a、これはT7プロモーター、T7ターミネーター、誘導性のE.coli lacオペレーター、およびlacリプレッサー遺伝子を含む;ならびにpET12a-c、これは、T7プロモーター、T7ターミネーター、およびE.coli ompT分泌シグナルを含む)を含む。別の適切なベクターは、pIN-IIIompA2(Duffaudら、Meth. in Enzymology、153:492-507、1987を参照のこと)であり、これは、lppプロモーター、lacUV5プロモーターオペレーター、ompA分泌シグナル、およびlacリプレッサー遺伝子を含む。
【0064】
例示的な、真核生物形質転換ベクターは、クローン化されたウシパピローマウイルスゲノム、マウスレトロウイルスのクローン化されたゲノム、および真核生物カセットを含み(例えば、pSV-2 gpt系(MulliganおよびBerg、Nature、277:108-114(1979))、Okayama-Bergクローニング系(Mol.Cell Biol.、2:161-170、(1982))、ならびにGenetics Institute(Science、228:810-815(1985))によって記載される発現クローニングベクターが利用可能であり、これは形質転換された真核生物細胞株において、目的のタンパク質の少なくともいくつかの発現の実質的な確実性を提供する。
【0065】
哺乳動物細胞のトランスフェクションのために、本発明のATをコードするDNAに連結され得る調節エレメントを含む、特に好ましベースベクターは、サイトメガロウイルス(CMV)プロモーターベースのベクター(例えば、pcDNA1(Invitrogen、SanDiego、CA))、MMTVプロモーターベースのベクター(例えば、pMAMNeo(Clontech、Palo Alto、CA)、およびpMSG(Pharmacia、Piscataway、NJ)、ならびにSV40プロモーターベースのベクター(例えば、pSVβ(Clontech、PaloAlto、CA)である。
【0066】
本発明の別の実施態様によれば、本発明の核酸分子(すなわち、DNAまたはmRNA)を含む「組換え細胞」が、提供される。適切な宿主細胞、好ましくは細菌細胞、およびより好ましくはE.coli細胞を形質転換する方法、ならびに異種タンパク質をコードする遺伝子を含むこの細胞を培養するために適用可能な方法は、当該分野において一般に知られている。例えば、Sambrookら、MolecularCloning:A Laboratory Manual(第2版)、Cold Spring Harbor Laboratory Press、ColdSpring Harbor、New York、USA(1989)を参照のこと。
【0067】
本発明の核酸を含む発現ベクターを、宿主細胞に導入(形質導入)して、形質導入された組換え細胞(すなわち、組換え異種核酸を含む細胞)を生成する例示的な方法は、当該分野において周知である(概説のために、Friedmann、Science、244:1275-1281(1989);Mulligan、Science、260:926-932(1993)を参照のこと、これらのそれぞれは、それらの全体が本明細書中に参考として援用される)。形質導入の例示的な方法は、例えば、ウイルスベクターを用いる感染(例えば、米国特許第4,405,712号および同第4,650,764号を参照のこと)、リン酸カルシウムトランスフェクション(米国特許第4,399,216号および同第4,634,665号)、デキストラン硫酸トランスフェクション、エレクトロポレーション、リポフェクション(例えば、米国特許第4,394,448号および同第4,619,794号を参照のこと)、サイトフェクション、粒子ビーズボンバードメントなどを含む。異種核酸は、必要に応じて、その染色体外(すなわち、エピソーム)での維持を許容する配列を含み得るか、または異種DNAは、宿主のゲノムへの取込みを引き起こされ得る(宿主における安定な維持を確実にするための代替の手段として)。
【0068】
本発明の実施において使用が意図される宿主生物として、異種タンパク質の組換え産生が行われた生物が挙げられる。このような宿主生物の例として、細菌(例えば、E. coli)、酵母(例えば、Saccharomyces cerevisiae、Candida tropicalis、Hansenulapolymorpha、および P. pastoris;例えば、米国特許第4,882,279号、同第4,837,184号、同第4,929,555号、および同第4,855,231号を参照のこと)、哺乳動物細胞(例えば、HEK293、CHO、およおびLtk−細胞)、昆虫細胞などが挙げられる。現在好ましい宿主生物は、細菌である。最も好ましい細菌は、E.coliである。
【0069】
1つの実施態様において、本発明のATタンパク質をコードする核酸が、哺乳動物細胞に、当該分野で周知の適切なウイルスベクター(例えば、レトロウイルスベクター、アデノウイルスベクターなど)を使用してインビボまたはインビトロのいずれかで送達され得る。さらに、本発明のATのインビボでの発現を制限するかまたは減少させることが所望される場合、本発明の核酸のアンチセンス鎖の導入が意図される。
【0070】
ウイルスに基づくシステムは、種々の細胞への比較的高レベルの異種核酸の導入が可能であるという利点を提供する。ATタンパク質をコードする本発明のAT核酸を哺乳動物細胞(例えば、血管組織区)に導入するための適切なウイルスベクターが、当該分野で周知である。これらのウイルスベクターとして、例えば、単純ヘルペスウイルスベクター(例えば、Gellerら、Science,241:1667-1669(1988))、ワクシニアウイルスベクター(例えば、Picciniら、Meth. in Enzymology, 153:545-563(1987);サイトメガロウイルスベクター(Mocarskiら、ViralVectors, Y. GluzmanおよびS.H. Hughes編、Cold Spring Harbor Laboratory, Cold SpringHarbor, N.Y., 1988, 78-84頁)、モロニーマウス白血病ウイルスベクター(Danosら、PNAS, USA, 85:6469(1980))、アデノウイルスベクター(例えば、Loganら、PNAS,USA, 81:3655-3659(1984);Jonesら、Cell, 17:683-689(1979);Berkner, Biotechniques, 6:616-626(1988);Cottenら、PNAS,USA 89,:6094-6098(1992);Grahamら、Meth. Mol. Biol., 7:109-127(1991))、アデノ随伴ウイルスベクター、レトロウイルスベクター(例えば、米国特許第4,405,712号および同第4,650,764号を参照のこと)などが挙げられる。特に好ましいウイルスベクターは、アデノウイルスベクターおよびレトロウイルスベクターである。
【0071】
例えば、本発明の1つの実施態様において、アデノウイルス−トランスフェリン/ポリリジン−DNA(TfAdpl−DNA)ベクター複合体(Wagnerら、PNAS, USA, 89:6099-6103(1992);Curielら、Hum.Gene Therapy, 3:147-154(1992);Gaoら、Hum. Gene Ther., 4:14-24(1993))が、異種AT核酸を哺乳動物細胞に導入するために使用される。本明細書中で記載される任意のプラスミド発現ベクターが、TfAdpl−DNA複合体中で使用され得る。
【0072】
本明細書中で使用される場合、「レトロウイルスベクター」とは、2つのレトロウイルスLTR間に存在する異種遺伝子をコードする発現カセットを有する、周知の遺伝子導入プラスミドをいう。レトロウイルスベクターは代表的には、レトロウイルスベクターまたはRNAが、適切なパッケージング細胞株中でウイルスビリオンへパッケージングされる、鋳型としてのレトロウイルスベクターを使用して転写されることを可能にする、適切なパッケージングシグナルを含む(例えば、米国特許第4,650,764号を参照のこと)。
【0073】
本明細書中での使用に適切なレトロウイルスベクターは、例えば、米国特許第5,252,479号、およびWIPO公開WO92/07573、WO 90/06997、WO 89/05345、WO 92/05266、およびWO 92/14829(本明細書中で参考として援用される)に記載されている。これらは、このようなレトロウイルスベクターを使用してヒト細胞に核酸を効率的に導入するための方法の記載を提供する。他のレトロウイルスベクターとして、例えば、マウス乳房腫瘍ウイルスベクター(例えば、Shacklefordら、PNAS,USA, 85:9655-9659(1988))などが挙げられる。
【0074】
本発明のなお別の実施態様に従って、本発明のATポリペプチドと特異的反応性を有する抗AT抗体が提供される。抗体の活性なフラグメントは、「抗体」の定義内に含まれる。本発明の抗体は、抗原としてATポリペプチド、タンパク質、またはその部分を使用して、当該分野で公知の方法によって産生され得る。例えば、ポリクローナル抗体およびモノクローナル抗体が、例えば、本明細書中で参考として援用される、HarlowおよびLane、Antibodies;ALaboratory Manual(Cold Spring Harbor Laboratory(1988))に記載されるような、当該分野で周知の方法によって産生され得る。本発明のATポリペプチドは、このような抗体を生成することにおいて、免疫原として使用され得る。あるいは、合成ペプチドが調製され(市販によって入手可能な合成機を使用して)、そして免疫原として使用され得る。アミノ酸配列は、それらが対応するポリペプチドの疎水性ドメインまたは親水性ドメインをコードするかどうかを決定するために、当該分野で周知の方法によって分析され得る。変化した抗体(例えば、キメラ、ヒト化、CDR-移植された、または二重機能性抗体)もまた、当該分野で周知の方法によって産生され得る。このような抗体はまた、例えば、Sambrookら、前出、ならびにHarlowおよびLane、前出、に記載されるような、ハイブリドーマ、化学合成、または組換え法によって産生され得る。抗ペプチドおよび抗融合タンパク質抗体の両方が、使用され得る。(例えば、Bahouthら、TrendsPharmacol. Sci. 12:338(1991);Ausubelら、Current Protocols in Molecular Biology、JohnWileyおよびSons, NY(1989)(本明細書中で参考として援用される)を参照のこと)。
【0075】
このように産生された抗体が、とりわけ、哺乳動物(好ましくは、ヒト、体のサンプル(例えば、組織または血液))中に存在するATタンパク質のレベルを検出するための診断方法およびシステムにおいて使用され得る。このような抗体はまた、本発明のATタンパク質のイムノアフィニティー精製またはアフィニティークロマトグラフィー精製のために使用され得る。さらに、kono方法は、細胞の表面または細胞内(例えば、核内)のいずれかのATポリペプチドの存在を検出するために、本明細書中で意図される。この方法は、ATポリペプチドに対する抗体の結合を可能にする条件下で、ATポリペプチドに特異的に結合する抗体と細胞とを接触させる工程、ATに結合した抗体の存在を検出する工程、およびそれによって細胞の表面または細胞内の本発明のポリペプチドの存在を検出する工程を包含する。このようなポリペプチドの検出に関して、抗体は、インビトロでの診断方法またはインビボでの画像診断方法のために使用され得る。
【0076】
サンプル中の標的ATポリペプチドのインビトロでの検出のために有用な免疫学的手順として、検出可能な抗体を使用するイムノアッセイが挙げられる。このようなイムノアッセイとして、例えば、当該分野で周知の、ELISA、Pandex微量蛍光定量アッセイ、凝集アッセイ、フローサイトメトリー、血清診断アッセイ、および免疫組織化学染色手順が挙げられる。抗体は、当該分野で周知の種々の手段によって検出可能にされ得る。例えば、検出可能なマーカーが、直接的または間接的に抗体に結合され得る。有用なマーカーとして、例えば、放射性ヌクレオチド、酵素、発蛍光団、色素原、および化学発光標識が挙げられる。
【0077】
本発明の抗AT抗体は、生存している動物、ヒト、または生物学的組織もしくはそれらから単離された液体中のATポリペプチドの活性を調節するために、本明細書中での使用が意図される。用語「調節」は、ATタンパク質の生物学的活性(例えば、ATのAc-CoA輸送活性)を増大(例えば、アゴニストを介して)または阻害(例えば、アンタゴニストを介して)する化合物の能力をいう。従って、キャリア、および、天然に存在するリガンドまたは他のAT結合タンパク質が、本発明のATポリペプチドに結合するのをブロックするために有効なATポリペプチドについて特異性を有する量の抗体を含有する組成物が、本明細書中で意図される。例えば、細胞の表面に存在し、そして配列番号2に示されるアミノ酸配列を含むATポリペプチドの細胞表面エピトープのアミノ酸配列と実質的に同じアミノ酸配列を有するATポリペプチド分子のエピトープを指向するモノクローナル抗体が、この目的のために有用であり得る。
【0078】
本発明はさらに、ATポリペプチドをコードする外因性の核酸を発現し得る、トランスジェニック非ヒト哺乳動物を提供する。本明細書中で使用される場合、句「外因性の核酸」は、宿主に対して天然ではないか、またはその天然の環境以外の宿主に存在する(例えば、遺伝子操作されたDNA構築物の一部として)核酸配列をいう。天然に存在するATのレベルに加えて、本発明のATタンパク質は、トランスジェニック哺乳動物中で、過剰発現され得るか、発現不足であり得るか、または不活性な変異形態で発現され得る(例えば、周知のノックアウトトランスジェニック)かのいずれかである。
【0079】
正常な活性を有さない(すなわち、天然のATを発現しない)ように変異されたATポリペプチドをコードする核酸を発現し得るトランスジェニック非ヒト哺乳動物もまた、提供される。本発明はまた、ATポリペプチドをコードするmRNAに相補的なアンチセンスmRNAに転写されるように配置された、ATポリペプチドをコードする核酸に相補的なアンチセンス核酸を含むゲノムを有する、トランスジェニック非ヒト哺乳動物を提供する。このアンチセンス核酸は、mRNAにハイブリダイズし、それによってその翻訳を減少させる。核酸は、誘導性プロモーターおよび/または組織特異的調節エレメントをさらに含み得、その結果、発現が誘導され得るか、または、特異的細胞型に制限され得る。核酸の例は、配列番号1に示されるコード配列と実質的に同じコード配列を有するDNAまたはcDNAである。非ヒトトランスジェニック哺乳動物の例は、トランスジェニックマウスである。
【0080】
ATポリペプチドの生理学的および作用的役割を説明する動物モデル系もまた、提供され、そしてトランスジェニック動物を作製することによって産生される。ここで、ATポリペプチドの発現は、種々の技術を使用して変更される。このような技術の例として、マイクロインジェクション、レトロウイルス感染、または当業者に周知の他の手段による、ATポリペプチドをコードする核酸の正常なまたは変異体バージョンの、トランスジェニック動物を産生するための適切な受精した胚への挿入が挙げられる。(例えば、Hoganら、Manipulatingthe Mouse Embryo:A Laboratory Manual(Cold Spring Harbor Laboratory,(1986)を参照のこと)。
【0081】
ATポリペプチドの発現の調節または構造を変化させるための、トランスジェニック動物中の天然の遺伝子座を有するAT遺伝子の変異体または正常なバージョンの相同組換えの使用もまた、本明細書中で意図される(Capecchiら、Science,244:1288(1989);Zimmerら、Nature, 338:150(1989)を参照のこと;これらは本明細書中で参考として援用される)。相同組換え技術は、当該分野で周知である。相同組換えによって、天然の(内因性の)遺伝子を、組換えまたは変異した遺伝子で置き換えて、天然の(内因性の)タンパク質を発現し得ないが、例えば、ATポリペプチドの変化した発現の結果である変異タンパク質を発現し得る動物を産生する。
【0082】
相同組換えとは対照的に、マイクロインジェクションは、宿主遺伝子を除去することなく宿主ゲノムに遺伝子を付加する。マイクロインジェクションは、内因性および外因性のATタンパク質の両方を発現し得るトランスジェニック動物を産生し得る。誘導性プロモーターは、トランスジーンの発現を調節する手段を提供するために、核酸のコード領域に連結され得る。組織特異的調節エレメントが、トランスジーンの組織特異的発現を可能にするために、コード領域に連結され得る。トランスジェニック動物モデル系は、特異的試薬(すなわち、タンパク質応答を活性化するかまたは阻害する、アゴニストおよびアンタゴニスト)の同定のための化合物のインビボでのスクリーニングに有用である。
【0083】
本発明の核酸、オリゴヌクレオチド(アンチセンスを含む)、それを含むベクター、形質転換された宿主細胞、ポリペプチド、およびそれらの組み合わせ、ならびに本発明の抗体が、化合物が、本発明のATポリペプチドに対する可能性のあるアゴニストまたはアンタゴニストとして作用するかどうかを決定するために、インビトロで化合物をスクリーニングするために使用され得る。これらのインビトロでのスクリーニングアッセイは、本発明のATポリペプチドの機能および活性に関する情報を提供する。これは、ポリペプチド、ペプチド、またはタンパク質の1つ以上の型と特異的に相互作用し得る化合物の同定および設計を導き得る。
【0084】
本発明のなお別の実施態様に従って、ATポリペプチドに結合する化合物(例えば、抗体、結合試薬など)を同定するための方法が提供される。例えば、本発明のATタンパク質は、競合結合アッセイにおいて使用され得る。このようなアッセイは、もしあれば、どの化合物が、ATタンパク質に結合し得るかを決定するために、多数の化合物を迅速なスクリーニングに適用し得る。続いて、より詳細なアッセイが、このような化合物が本発明のタンパク質の調節因子(例えば、アゴニストまたはアンタゴニスト)として作用するかどうかをさらに決定するために、結合することが見出されているこれらの化合物を用いて行われ得る。
【0085】
ATは、欠損している場合、異常な神経細胞の分化および/または移動に応答する遺伝子と考えられている。さらに、本発明の異常なレベル(例えば、より高いまたはより低いレベル)のATタンパク質、あるいは異常に作用するATタンパク質が、乳癌および神経芽細胞種および黒色腫のような外胚葉起源由来の癌を含むヒトの腫瘍に関連すると考えられている。
【0086】
従って、本発明の別の実施態様において、本発明のATポリペプチドの活性を調節する化合物を同定するためのバイオアッセイが提供される。この方法に従って、本発明のATポリペプチドに結合した膜は、試験化合物の存在下および非存在下でAc-CoAと接触される;ATタンパク質の活性は、試験化合物との接触に続いてモニターされ、そしてATタンパク質を有する膜を通過するAc-CoAの輸送を増大させるかまたは減少させるかのいずれかを引き起こすこれらの物質が、ATポリペプチドを調節するための機能的試薬として同定される。
【0087】
本発明の別の実施態様に従って、本発明のATポリペプチドを組換え的に発現する形質転換された宿主細胞(インタクトなまたは半インタクト(semi-intact)な細胞のいずれか)が、試験化合物と接触され、そして次いで、その調節効果が、試験化合物の存在下および非存在下でのAc-CoA(例えば、実施例VIに記載されるような放射性標識したAc-CoA)のATによって媒介される輸送を比較することによって、または試験細胞またはコントロール細胞(すなわち、ATポリペプチドを発現しない細胞)の応答を比較することによって、化合物の存在下に対して評価され得る。
【0088】
本明細書中で使用される場合、本発明のATポリペプチドの「活性を調節する」化合物またはシグナルは、ATポリペプチドの活性を変化させ、その結果、本発明のATポリペプチドの活性は、化合物またはシグナルの非存在下よりも、化合物またはシグナルの存在下で異なる化合物またはシグナルをいう。特に、このような化合物またはシグナルとして、アゴニストまたはアンタゴニストが挙げられる。アゴニストは、ATタンパク質の機能を活性化するかまたは増大させる化合物またはシグナルを含む。あるいは、アンタゴニストは、ATタンパク質の機能を阻害するか、そうでなければ低下して妨害する化合物またはシグナルを含む。代表的には、アンタゴニストの効果は、アゴニストによって誘導されるタンパク質活性のブロックとして観察される。アンタゴニストとして、競合および非競合アンタゴニストが挙げられる。競合アンタゴニスト(または競合ブロッカー)は、Ac-CoA輸送に特異的な部位と、またはその付近と相互作用する。非競合アンタゴニストまたはブロッカーは、本発明のATタンパク質のAc-CoA輸送領域とは異なる部位で相互作用することによってポリペプチドの機能を不活化する。
【0089】
当業者に理解されるように、AT活性を調節する化合物を同定するためのアッセイ方法は、一般に、コントロールとの比較を必要とする。「コントロール」の1つの型は、「コントロール」細胞または培養物が化合物に曝露されないという特徴を有する、化合物に曝された試験細胞または試験培養物と実質的に同じ処理をされた細胞または培養物である。例えば、「コントロール」細胞または培養物の型は、天然のタンパク質を発現しない「コントロール」細胞または培養物を除いて、トランスフェクトされた細胞と同一である細胞または培養物であり得る。従って、化合物に対するトランスフェクトされた細胞の応答が、同じ反応条件下で、同じ化合物に対する「コントロール」細胞または培養物の応答(またはその欠如)と比較される。
【0090】
従って、本発明の別の実施態様に従って、試験化合物が、ATタンパク質のアゴニストまたはアンタゴニストとして作用し得るかどうかを評価するためのバイオアッセイが提供される。ここで、上記のバイオアッセイは、以下を包含する:
(a)以下を含む細胞を培養する工程:
ATタンパク質またはその機能的に改変された形態を発現するDNA、
ここで、上記の培養工程は、ATタンパク質のAc-CoA輸送活性を調節するその能力が決定されていることが求められる少なくとも1つの化合物の存在下で行われ、そしてその後
(b)Ac-CoAのレベルにおける増大または低下のいずれかについて上記の細胞をモニターする工程。
【0091】
細胞内小器官への放射標識されたAc-CoAの輸送のような、Ac-CoAの取り込みを測定する当該分野で周知の方法が、ATタンパク質のアゴニストおよびアンタゴニストを同定するために本明細書中で記載されるバイオアッセイにおいて使用され得る。例えば、実施例VIで記載される方法は、哺乳動物宿主細胞中で発現される、組換えATタンパク質、またはその変異体および/もしくはそのアナログのAc-CoA輸送活性を評価するために使用され得る。
【0092】
本明細書中で使用される場合、「ATタンパク質のAc-CoA輸送活性を調節する能力」は、細胞内膜を通過するATタンパク質のAc-CoA輸送活性を誘導する(アゴニスト)か、または阻害する(アンタゴニスト)かのいずれかの能力を有する化合物をいう。
【0093】
本発明の別の実施態様において、試験化合物が本発明のATタンパク質のアンタゴニストとして作用し得るかどうか、または上記のATタンパク質の機能的に改変された形態であるかどうかを評価するためのバイオアッセイは、以下の工程を包含する:
(a)以下を含む細胞を培養する工程:
ATタンパク質またはその機能的に改変された形態を発現する、DNA,
ここで、上記の培養工程は、以下の存在下で行われる:
漸増濃度の少なくとも1つの、ATタンパク質のAc-CoA輸送能力を阻害するその能力を決定することを求められている化合物、および
固定された濃度のAc-CoA;
そして、その後の
(b)上記化合物の濃度の関数として細胞内小器官へ輸送されたAc-CoAのレベルを上記細胞中でモニターし、それによってAT輸送活性を阻害する上記化合物の能力を示す工程。
【0094】
上記のアンタゴニストバイオアッセイの工程(a)において、培養工程はまた、以下の存在下で行われる:
固定された濃度の少なくとも1つの、ATタンパク質のAc-CoA輸送活性を阻害するその能力を決定することが求められる化合物、
漸増濃度のAc-CoA。
【0095】
本明細書中で使用される場合、句「細胞内小器官」は、例えば、ゴルジ体、小胞体、ミトコンドリアなどをいう。本発明のバイオアッセイでの使用が意図される宿主細胞として、CV-1細胞、COS細胞、HeLa細胞などが挙げられる。本発明のバイオアッセイを行うための現在好ましい宿主細胞は、実施例VIに記載されるようなHeLa細胞である。
【0096】
本発明のなお別の実施態様において、ATタンパク質によって媒介されるAc-CoA輸送活性を調節するための方法もまた意図される。上記の方法は以下の工程を包含する:
上記のバイオアッセイによって同定されたアゴニストまたはアンタゴニストの有効な調節量とATタンパク質とを接触させる工程。
【0097】
異常な神経細胞の分化、異常な神経細胞の移動、神経芽細胞腫、または黒色腫を処置するための方法もまた、本明細書中で意図される。上記の方法は、本明細書中で記載される方法によって同定される有効量の化合物(アゴニストまたはアンタゴニスト)を投与する工程を包含する。このような化合物は、代表的には、生物学的に受容可能な組成物中で投与される。
【0098】
従って、本発明は、本明細書中で記載される治療方法を実施するために有用な治療用組成物を意図する。本発明の治療用組成物は、AT-1調節剤とともに生理学的に適合性のキャリア、または有効成分としてその中に溶解されたかまたは分散された、本明細書中で記載されるような坑AT-1抗体を含む。好ましい実施態様において、治療組成物は、治療目的で哺乳動物またはヒト患者に投与される場合、免疫原性ではない。
【0099】
本明細書中で使用される場合、用語「薬学的に受容可能な」、「生理学的に受容可能な」、およびその文法上のバリエーションは、それらが組成物、キャリア、希釈剤、および試薬をいう場合、互換的に使用され、そして吐き気、めまい、胃の不調などのような所望されない生理学的影響の産生を伴わずに哺乳動物に投与され得ることが示される。
【0100】
その中に溶解されたかまたは分散された有効成分を含有する薬理学的組成物の調製は、当該分野で周知である。代表的には、このような組成物は、液体の溶液または懸濁物のいずれかとして注射可能なものとして調製される;しかし、使用の前に液体中で、溶液または懸濁物に適切な固形の形態もまた、調製され得る。調製物はまた、乳濁物でもあり得る。
【0101】
有効成分は、薬学的に受容可能であり、そして本明細書中で記載される治療方法での使用に適切な量で有効成分と適合性である賦形剤と混合され得る。適切な賦形剤は、例えば、水、生理食塩水、デキストロース、グリセロール、エタノールなど、およびそれらの任意の2つ以上の組み合わせである。さらに、所望される場合は、組成物は、主要ではない量の補助物質(例えば、有効成分の有効性を増強する、湿潤剤、または乳化剤、pH緩衝剤など)を含み得る。
【0102】
本発明の治療用組成物として、本明細書中の成分の薬学的に受容可能な塩を含み得る。薬学的に受容可能な非毒性の塩として、例えば、以下のような無機酸を用いて形成される、酸添加塩(ポリペプチドの遊離のアミノ基を用いて形成される)が挙げられる:塩酸、臭化水素酸、過塩素酸、硝酸、チオシアン酸、硫酸、リン酸、酢酸、プロピオン酸、グリコール酸、乳酸、ピルビン酸、シュウ酸、マロン酸、コハク酸、マレイン酸、フマル酸、アントラニル酸、ケイ皮酸、ナフタレンスルホン酸、スルファニル酸など。
【0103】
遊離のカルボキシル基を用いて形成される塩はまた、例えば以下のような無機塩基から誘導され得る:水酸化ナトリウム、水酸化アンモニウム、水酸化カリウムなど;および例えば以下のような有機塩基から誘導される:モノ-、ジ-、およびトリ-アルキルアミンならびにトリ-アリールアミン(例えば、トリエチルアミン、ジイソプロピルアミン、メチルアミン、ジメチルアミンなど)、および必要に応じてエタノールアミン(例えば、エタノールアミン、ジエタノールアミンなど)で置換される。
【0104】
生理学的に寛容なキャリアは、当該分野で周知である。例示的な液体のキャリアは、有効成分および水に加えて他の物質を含まないか、または緩衝液(例えば、生理学的pHのリン酸ナトリウム、生理学的な生理食塩水、またはその両方(例えば、リン酸緩衝化生理食塩水))を含む、滅菌の水溶液である。なおさらに、水溶性のキャリアは、1つ以上の緩衝塩、および塩(例えば、塩化ナトリウムおよび塩化カリウム、デキストロース、ポリエチレングリコール、および他の溶質)を含み得る。
【0105】
液体の組成物はまた、水に加えて液体層を含み得、そして水を排除するために液体層を含み得る。例示的なさらなる液体層として、グリセリン、植物油(例えば、綿実油)、および水-油エマルジョンが挙げられる。
【0106】
本明細書中で記載されるように、「有効量」は、例えば、本発明のATタンパク質のAc-CoA輸送活性を調節するための、所望される治療効果を達成するように計算された予め決定された量である。必要とされる投与量は、特定の処置とともに、および所望される処置の期間とともに変化する;しかし、約10μg〜約1mg/kg体重/日の間の投与量が治療的処置に使用されることが予測される。本明細書中以下で議論されるような、デポー製剤または長期持続形態中でこのような化合物を投与することが、特に有用であり得る。治療有効量は、生理学的に受容可能な組成物中で投与される場合、代表的には、AT調節剤または本明細書中で同定される化合物の量である。そして約0.1μg/mlから約100μg/ml、好ましくは、約1.0μg/mlから約50μg/ml、より好ましくは、少なくとも約2μg/ml、および通常は5から10μg/mlの血漿濃度を達成するために十分である。治療用の本発明の坑AT抗体が、当業者の知識に従って、比例する適切な量で投与され得る。
【0107】
本明細書中で言及される全ての米国特許および全ての刊行物が、それらの全体を本明細書中で参考として援用するように引用される。本発明はここで、以下の限定的ではない実施例を参照して、さらに詳細に記載される。
【実施例】
【0108】
実施例
他に特に記載されない限りは、本発明は、例えば以下に記載されるような標準的な手順を使用して行った:Maniatisら、Molecular Cloning:A Laboratory Manual, Cold Spring HarborLaboratory Press、Cold Spring Harbor, New York, USA(1982);Sambrookら、MolecularCloning:A Laboratory Manual(第2版)、Cold Spring Harbor Laboratory Press, ColdSpring Harbor, New York, USA(1989);Davisら、 Basic Method in Molecular Biology,Elsevier Science Publishing, Inc., New York, USA(1986);またはMethods in Enzymology:Guideto Molecular Cloning Techniques 第152巻、S.L. BergerおよびA.R. Kimmerl編、Academic PressInc., San Diego, USA(1987)。
【0109】
実施例I
本明細書中で使用される抗体についての記載
C系列ポリシアロガングリオシドのNeuNAcα2-8NeuNAcα2-8NeuNAcα2-3Galβ1-R構造を有するGT3に特異的なモノクローナル抗体(mAb)M6703を、Hirabayashiら、1988,J. Biochem., 104:973-979(本明細書中で参考として援用される)に記載されるように調整した。GD3との反応が示されたmAb R24(Pukelら、1982,J. Exp. Med., 155:1133-1147、本明細書中で参考として援用される)を、アメリカンタイプカルチャーコレクションから入手した(ATCC#HB-8445)。9-O-アセチル化ジシアロガングリオシドを認識するmAb D1.1を、Levineら、1984, J. Neuroscience, 4:820-831(本明細書中で参考として援用される)に記載されるように確立し、そして9-O-アセチル化GT3と反応するmAB493D4は、S. Fujita博士(Mitsubishi Kasei Institute of Life Sciences)より提供された。AT-1タンパク質の合成のN末端の14アミノ酸ペプチド(配列番号2のアミノ酸1〜14)に対するポリクローナル抗体を、周知の方法を使用して調製した。
【0110】
安定にトランスフェクトされたレシピエント細胞の構築
この実施例は、ガングリオシドを産生するが、それらの9-O-Ac誘導体は産生しないトランスフェクトされた細胞株を得るための方法を提供する。この手順の目的は、ガングリオシドGD3およびGT3を発現するが、それらの9-O-アセチル化誘導体は発現しない細胞株を産生することである。9-O-アセチル化ガングリオシドの発現のために必要とされる新規の因子をクローニングするために、前駆体ガングリオシドを発現するが、対応する9-O-アセチル化ガングリオシドを発現しないレシピエント細胞を使用することが必要であった。研究した特異的ガングリオシドは、NeuAcα2-8NeuAcα2-3Galβ1-4Glcβ1-1’Cer(本明細書中以後、「GD3」)、9-O-Ac-GD3(末端のノイラミン酸の9位でアセチル化されている)、NeuAcα2-8NeuAcα2-8NeuAcα2-3Galβ1-4Glcβ1-1’Cer(本明細書中以後、「GT3」)、および9-O-Ac-GT3(末端のノイラミン酸の9位でアセチル化されている)であった。
【0111】
細胞株COS-1/GD3+(ガングリオシドGD3を安定に発現するが、9-O-Ac-GD3は発現しない)を、Nakayamaら(本明細書中で参考として援用される、J.Biol. Chem., 271:3684-3691(1996))によって以前に記載されているように調製した。そして、HeLa/GT3+(ガングリオシドGD3およびGT3を安定に発現するが、9-O-Ac-GD3または9-O-Ac-GT3は発現しない)を、以前に記載されているように(Nakayamaら、前出、1996)調製した。
【0112】
実施例II
AT-1をコードするcDNAの単離
哺乳動物発現ベクターに基づくcDNAライブラリーpcDNAI-SK-MEL-28(9-O-AD-GD3+(9-O-Ac-GD3+)を発現するヒト黒色腫SK-MEL-28細胞から単離したポリ(A)+RNAを使用して構築した)を、Invitrogen(San Diego, CA)より購入した(本明細書中で参考として援用されている、1996年1月16日に発行された米国特許第5,484,590号もまた参照のこと;本明細書中で参考として援用されている、BierhuzenおよびFukuda、Proc.Natl. Acad. Sci., U.S.A., 89:9326-9330(1992)を参照のこと)。
【0113】
約1.2×107個のCOS-1/GD3+細胞を、リポフェクトアミン(LifeTechnologies, Inc.)を使用して20μgのpcDNAI-SK-MEL-28でトランスフェクトした。60時間後、9-O-Ac-ガングリオシドを産生するトランスフェクトした細胞を、9-O-Ac-GD3に特異的に結合するモノクローナル抗体D1.1(本明細書中で参考として援用される、Levineら、J.Neuros., 4:820-831(1984))を使用して間接的な免疫蛍光によって染色した。蛍光標識した細胞を、FACStarセルソーター(BectonDickinson)を使用して蛍光活性化された細胞の分別を使用して単離した。
【0114】
9-O-Ac-GD3についてポジティブに染色されたCOS-1/GD3+細胞を回収し、そしてプラスミドDNAを、Hirt手順(本明細書中で参考として援用される、Hirt,J. Mol. Biol., 26:365-396(1967))を使用して単離した。プラスミドDNAを、アンピシリンおよびテトラサイクリンの存在下で、宿主細菌MC1061/P3に形質転換し、そして例えば、Sambrookら(MolecularCloning:A Laboratory Manual, Cold Spring Harbor, N.Y.(1989))に記載される周知の方法を使用して、細菌宿主から単離した。
【0115】
単離しプールしたプラスミドを、姉妹選択(sibling selection)(米国特許第5,484,590号を参照のこと)に供した。簡潔には、単離したプラスミドを、活性なプールが連続的に小さくなる、COS-1/GD3+細胞を使用する姉妹選択に供した。トランスフェクトした細胞を、モノクローナル抗体D1.1を使用して間接的な免疫蛍光顕微鏡法によってスクリーニングした。ポジティブに染色された細胞を回収し、そしてプラスミドDNAを抽出した。続く回の姉妹選択後、9-O-Ac-GD3の発現に必要とされる因子をコードする単一のプラスミドpcDNAI-AT-1を、単離した。この因子を、AT-1(配列番号1)と命名した。
【0116】
選択したクローニングした細胞に由来するプラスミドを単離し、そして挿入物のヌクレオチド配列を両方向で決定した。ヌクレオチド配列を、配列番号1(GenBank登録番号D88152)に示す。
【0117】
AT-1をコードするcDNAは、60,906Daの計算分子量を有する549アミノ酸のポリペプチド(配列番号2)をコードする1650塩基対を有するオープンリーディングフレームを含む。タンパク質および核酸のデータベース(GenBank,EMBLおよびSwiss-Prot)に対する相同性検索は、任意の既知のタンパク質とは有意な相同性を有さないことを示した。このことは、AT-1によってコードされるタンパク質が新規であることを示す。AT-1ポリペプチド配列のヒドロパシー分析(KyteおよびDoolittle(J.Mol. Biol., 157:105-132(1982))は、このポリペプチドがいくつかの可能性のある膜貫通ドメインを含むことを示した。(図1を参照のこと)。同様の分析を、プログラムPsort(Nakaiら、Genomics,14:897-911(1992)およびTmpred(HofmannおよびStoffel, Biol. Chem. Hoppe-Seyler, 347:166(1993))を使用して行った。これらの2つの分析は、AT-1ポリペプチドが、6から10個の膜貫通ドメインを有するIIIa型膜タンパク質構造を有することを示した。AT-1ポリペプチドは、アミノ酸残基144から165で膜貫通ドメイン中にロイシンジッパーモチーフを含む。ロイシンジッパーモチーフがしばしば、他のトランスポータータンパク質中に見出されることが注目されるはずである(例えば、Eckhardtら、1996、PNAS,USA, 93:7572-7576;Miuraら、1996、J. Biochem(Tokyo)、120:236-241;Abeijonら、1996,PNAS, USA, 93:5963-5968を参照のこと)。
【0118】
実施例III
ガングリオシドのO-アセチル化に対するAT-1の影響を特徴づける発現アッセイ
COS-1/GD3+細胞
COS-1/GD3+細胞を、pcDNAI(擬似トランスフェクション)またはpcDNAI-AT-1で一時的にトランスフェクトした。トランスフェクションの60時間後、トランスフェクトした細胞を固定し、そしてmAbR24(抗GD3)またはmAb D1.1(抗-9-O-アセチル化ジシアロガングリオシド)とともにインキュベーションすることによって、別々に免疫染色し、続いて、mAbR24についてはローダミン結合抗マウスIgG、またはmAb D1.1についてはフルオレセインイソチオシアネート結合抗マウスIgMとともにインキュベーションした。擬似トランスフェクタントとAT-1トランスフェクタントとの間のR24反応性において有意な差異は存在しなかったが、9-O-アセチル化GD3の発現は、AT-1トランスフェクタント中でmAbD1.1で強く検出された。弱い穿刺染色しか、擬似トランスフェクタントにおいて検出されなかった。これらの結果は、本発明のAT-1タンパク質がGD3の9-O-アセチル化を媒介する(影響を与える)ことを示す。
【0119】
HeLa/GT3+細胞
HeLa/GT3+細胞を、pcDNAI(コントロール)またはpcDNAI-AT-1のいずれかでトランスフェクトし、そして続いて、9-O-Ac-GT3に結合するmAb493D4で免疫染色した。HeLa/GT3+/AT-1でトランスフェクトされた細胞は、9-O-Ac-GT3についてポジティブに染色されたが、HeLa/GT3+/pcDNAIコントロール細胞は、検出可能な染色を示さなかった。HeLa/GT3+/AT-1細胞の染色を、アルカリ処理(0.1NのNaOHで15分間、室温)によって消した。これらの結果は、本発明のAT-1タンパク質がGT3の9-O-アセチル化を媒介する(影響を与える)ことを示す。
【0120】
安定にトランスフェクトされたHeLa/GT3+/AT-1細胞の確立
HeLa/GT3+細胞を、pcDNA-I-AT-1およびpSV2HyB(10:1)で同時トランスフェクトし(後者の発現は、ハイグロマイシンBに対する耐性を付与する)、次いで、ハイグロマイシンB耐性について選択した。トランスフェクトした細胞を、上記に議論したように、mAb493D4での免疫蛍光染色によって9-O-Ac-GT3の存在についてスクリーニングした。1つの安定にトランスフェクトしたクローンを単離し、そしてHeLa/GT3+/AT-1と命名した。
【0121】
全細胞内ガングリオシドの定量
AT-1でトランスフェクトした細胞がO-アセチル化ガングリオシドを合成することを確認するために、全ガングリオシドをHeLa/GT3+細胞から、ならびにそれらの一時的なおよび安定なAT-1トランスフェクタントから単離した。HeLa/GT3+細胞が単純なガングリオシド、GM3、GD3、GT3、および少量の推定のGQ3のみを含むことを見出し、これによって、これらを、薄層クロマトグラフィーを使用してAT-1でのトランスフェクション後のガングリオシド組成を試験するために理想的であるとした。
【0122】
分析的薄層クロマトグラフィーを、予めコーティングした薄層クロマトグラフィープレート(Kieselgel60, Merck)で行った。使用した溶媒系は、クロロホルム、メタノール、12mMのMgCl2を含有する水(5:4:1、容量比)であった。ガングリオシドをレゾルシノール/HCl試薬によって可視化した。5×105個のトランスフェクトした細胞からのガングリオシドの単離およびTLC-免疫染色を、Hirabayashiら、J.Biol. Chem. 104:973-979(1988)に記載されるように行った。細胞から精製されたガングリオシドを、プラスチックプレート(PolygramSil G, Nagel, Doren, Germany)上に載せ、そして上記と同じ条件下で展開させた。プレートを、mAb、続いてペルオキシダーゼ結合ヤギ抗マウスIgG抗体での免疫染色に供した。ペルオキシダーゼ活性を、4-クロロ-1-ナフトール/H2O2で可視化した。ガングリオシドを、アルカリ処理による脱アセチル化後に同時に分析した。
【0123】
結果は、O-Ac-GT3および推定のO-Ac-GQ3の発現が、AT-1で安定にトランスフェクトされた細胞中で強くポジティブであったことを示す。再度、ガングリオシドをアルカリ加水分解に供した場合には、ポジティブなスポットは観察されなかった。HeLa/GT3+およびHeLa/GT3+/AT-1細胞が類似の総ガングリオシド量を含むことに注目することが重要である。また、中性のガングリオシドおよび非アセチル化ガングリオシドの組成において目に見える差異は、薄層クロマトグラフィー分析を使用して検出されなかった。
【0124】
実施例IV
AT-1 mRNAのノーザンブロット分析
種々のヒト組織中のAT-1 mRNAの発現を、ノーザンブロット分析によって試験した。安定にトランスフェクトされたHeLa細胞から調製したポリ(A)+RNAのサンプルを、2.2Mのホルムアルデヒドを含有するアガロースゲル中で電気泳動し、そしてナイロンフィルター(HybondTM-N、Amersham)に移した。ヒトグリセルアルデヒド-3-リン酸デヒドロゲナーゼ(G3PDH)cDNAとのハイブリダイゼーションを、コントロール実験として行った。これらのブロットを、マルチプライムDNA標識システム(Amersham)による[α-32P]dCTPでの標識後に、AT-1のゲル精製したcDNA挿入物とハイブリダイズさせた。
【0125】
3.3および4.3kbの大きさを有するAT-1の2つの主要な転写物を、心臓、脳、胎盤、肺、肝臓、骨格筋、腎臓、およびすい臓を含む、試験した全ての組織中で検出した。興味深いことに、すい臓組織は、試験した組織の中でも最も強いシグナルを発現した。すい臓において、ランゲルハンス島は、mAb493D4およびmAb D1.1を使用して、O-アセチル化されたシアロ糖結合体を発現することが、免疫組織化学的染色によって示された。2.7および3.4kbの大きさを有するAT-1の2つの転写物は、AT-1でトランスフェクトされたHeLa細胞中で強く発現され、そして擬似トランスフェクトされたHeLa細胞中でかろうじて検出された。
【0126】
実施例V
AT-1タンパク質の免疫組織化学的局在
HeLa/GT3+/AT-1細胞中でのAT-1タンパク質の細胞内局在を研究するために、本発明者らは、合成ペプチド(AT-1のN末端に対応する配列番号2のアミノ酸残基1〜14)に対する特異的抗体を調製した。安定にトランスフェクトされた細胞HeLa/GT3+/AT-1およびHeLa/GT3+/pcDNAIを固定し、サポニンで透過性にし、そしてAT-1タンパク質に特異的な親和性精製したウサギ抗AT-1抗体とともにインキュベートした。続いて、フルオレセインイソチオシアネート結合抗ウサギIgGとともにインキュベートした。
【0127】
抗AT-1抗体を使用するHeLa/GT3+/AT-1細胞のウェスタンブロット分析によって、54kdの主要なポジティブバンドおよび58kdの弱い反応性スポットを明らかにした。親和性精製した抗AT-1ペプチドを使用する免疫蛍光分析は、このAT-1タンパク質が細胞質(小胞体、ゴルジ体、およびミトコンドリアを含む)に局在化されることを示した。この特異的染色は、予備免疫IgGを用いて、またはペプチド抗原の存在下では観察されなかった。
【0128】
実施例VI
半インタクトな細胞へのAc-CoAの取りこみ
培養された細胞からの小胞体またはゴルジ膜画分を調製することが困難であったので、細胞中のAc-CoAトランスポーター活性を、Bhakdiら(本明細書中で参考として援用されている、Infect. Immun., 47:52-60(1985));KuncanおよびSchlegel(本明細書中で参考として援用されている、J.Cell Biol., 67:160-173(1993));ならびに、(本明細書中で参考として援用されている、Kainら(J. Biol. Chem.,268:19640-19649(1993))の方法を使用して、半インタクトな、透過性にしたHeLa細胞を使用して試験した。ストレプトリジンO(本明細書中以後、「SLO」、GIBCOBRL)での処理によって、細胞内膜にいかなる損傷を与えることもなく、細胞の原形質膜に孔を生成する。これによって、原形質膜を通じてのAc-CoAの様な大きな分子の通過を可能にし、次いで、それらの細胞内の結果は以下のとおりであり得る。
【0129】
培養した細胞をトリプシン処理によって回収し、そしてPBSおよびカルシウム非含有マグネシウム非含有Hepes(HCMF:0.14MのNaCl−5mMのKCl−6mMのGlc−0.3mMのNa2HPO4−10mMのHepes、pH7.4)で連続して洗浄した。HCMF中で希釈した活性化したストレプトリジンO(SLO, GIBCO BRL)での28℃にて20分の処理後、細胞を洗浄し、そしてトランスポーター緩衝液(TB:25mMのHepes−KOH−75mMのKOAc−2.5mMのMgOAc−5mMのEGTA−1.8mMのCaCl2、pH7.2)中の[Ac-14C]Ac-CoAとともに、種々の時間で室温にてインキュベートした。反応を、氷冷TBの添加によってクエンチした。上清を遠心分離によって除去した後、ペレットをTBで3回洗浄し、1%のSDSで可溶化し、そして放射能を計数した。
【0130】
図2に示すように、半インタクトな細胞を[Ac-14C]Ac-CoA(35μM)とともにインキュベートした場合、放射能の取りこみの増大が、AT-1でトランスフェクトした細胞中で明らかに実証された。この効果は、擬似トランスフェクタントでは見られなかった。トランスポーターCoA(350μM)のインヒビターの添加の際に、放射能の取りこみは、コントロールの60%にまで減少した。これらの結果は、AT-1がAc-CoAトランスポーターをコードすることを強力に示唆する。
【0131】
本発明は、上記に提供される実施例を参照して記載されているが、種々の改変が、本発明の精神を逸脱することなく行われ得ることが理解されるはずである。従って、本発明は、請求の範囲によってのみ限定される。
(配列表)
【0132】
【表1】

【0133】
【表2】
【0134】
【表3】
【0135】
【表4】
【0136】
【表5】
【0137】
【表6】
【0138】
【表7】
【図面の簡単な説明】
【0139】
【図1】図1は、10のウインドウサイズを用いて、KyteおよびDoolitteの方法を使用して分析された、配列番号2(AT-1をコードする)の疎水性親水性プロットを示す。影付四角および黒く塗り四角は、それぞれ、TmpredおよびPsortプログラムによって予測される、膜貫通ドメインを示す。
【図2】図2Aは、実施例VIに記載されるような、[Ac-14C]Ac-CoAからの半インタクトな細胞への放射能の時間依存性の取込みを示す。白丸は、HeLa/GT3+/pcDNAIを、および黒丸は、HeLa/GT3+/AT-1を、示す。図2Bは、[Ac-14C]Ac-CoAからの半インタクトな細胞への放射能の時間依存性の取込みに対する、CoAの効果を示す。
【技術分野】
【0001】
本発明は、国立研究所の国立ガン研究所によって与えられた、助成金番号R01 CA48737およびP01 CA 71932のもとで、政府の支援とともになされた。政府は本発明において一定の権利を有する。
発明の分野
本発明は、概して、分子生物学および酵素学に、ならびにより具体的には、膜を横切ってアセチルコエンザイムA(Ac-CoA)の輸送を調節するトランスポータータンパク質をコードする核酸に関する。
【背景技術】
【0002】
発明の背景
膜ガングリオシドにおける糖鎖の構造の多様性および複雑さは、いくつかの異なる種のシアル酸分子の出現によって、一部生じる。N-アセチルノイラミン酸およびN-グリコリルノイラミン酸は、脳中のガングリオシドにおいて、最も遍在するシアル酸であるが、それらのO-アセチル化形態はまた、LD1、B-系列ガングリオシド(GD3およびGT1bを含む)、ならびにC-系列(NeuAca2-8NeuAca2-8NeuAca2-3Gal-R)ガングリオシドにおいて、微量成分として見出される。いくつかの生物学的特性は、O-アセチル化によるシアル酸の修飾と関連すると仮定される。例えば、9-O-アセチル化ガングリオシドの発現は、神経細胞分化および移動と明らかに関連する(Stallcupら、ColdSpring Harbor Symp. Quant. Biol.、48:761-773(1983))。ラット脳におけるD1.1抗体によって検出される、9-O-アセチル化GD3は、生殖細胞領域に局在化されるが、有糸分裂後の細胞から消失することが見出された。これは、正常な成体脳および移動において、存在しない(Stallcupら、ColdSpring Harbor Symp. Quant. Biol.、48:761-773(1983))。ラット脳において、D1.1抗体によって検出される9-O-アセチル化GD3は、生殖細胞領域に局在化されるが、有糸分裂後の細胞から消失することが見出された。これは、正常な成体脳において、存在しない(Levineら、J.Neuros.、4:820-831(1984))。しかし、ハタオリマウス(weavermouse)において、成体脳における9-O-アセチル化ガングリオシドの持続的な発現が、小脳顆粒細胞移動における欠失と関連付けられた(Johnstoneら、J.Neurochem.、51:1655-1657(1988))。シアル酸残基におけるO-アセチル基の付着は、シアリダーゼのようなシアル酸代謝の酵素に対して、有意な効果を引き起こす。効果はまた、ウイルス結合、細胞接着、およびガングリオシドのシアル酸残基の免疫原性に対して見られる(概説のために、Varki、Glycobiology、2:25-40(1992)を参照のこと)。
【0003】
その重要性にもかかわらず、O-アセチル化機構は、分子レベルおよび遺伝子レベルで十分に理解されていない。Varkiのグループによってなされた一連の研究は、O-アセチル化ガングリオシドの生成は、単純なプロセスではなく、同じゴルジ体区分における、アクセプターガングリオシドGD3、アセチルCoA(Ac-CoA)トランスポーター、アセチルトランスフェラーゼの同時局在化を必要とすることを示す(Varki、Glycobiology、2:25-40(1992))。事実、インビトロアセチルトランスフェラーゼ活性の検出は、極めて困難であり、およびいったんインタクトな細胞膜調製物が、界面活性剤で処理されると、転移活性は、迅速に失われることが示された(VarkiおよびDiaz、J.Biol. Chem.、260:6600-6608(1985))。
【0004】
従って、O-アセチル化ガングリオシドの形成に関与する他のタンパク質因子を単離および特徴づけする、必要性が存在する。本発明は、この必要性を満たし、そして関連する利点もまた提供する。
【発明の開示】
【課題を解決するための手段】
【0005】
発明の要旨
本発明によれば、単離された哺乳動物アセチルコエンザイムAトランスポーター(AT)タンパク質が提供される。これらのATタンパク質、またはそのフラグメントは、抗AT抗体を生成するための免疫原として、またはこのようなタンパク質および/または抗体を含む治療学的組成物において、有用である。本発明のATタンパク質はまた、それに対するアゴニストおよびアンタゴニストを同定するためのバイオアッセイにおいて有用である。
【0006】
本発明によれば、新規のATタンパク質をコードする、単離された核酸がまた提供される。さらに、本発明の核酸を含むベクター、本発明の核酸にハイブリダイズするプローブ、本発明の核酸で形質転換された宿主細胞、本発明の核酸に対するアンチセンスオリゴヌクレオチド、および関連の組成物が、提供される。本明細書中に記載される核酸分子は、単離された組換えATタンパク質を容易に生成するために、当業者に公知の多様な組換え発現系に組込まれ得る。さらに、本発明の核酸分子は、所定のサンプル中のAT遺伝子またはmRNA転写物の存在および/または量についてアッセイするためのプローブとして有用である。本明細書中に記載される核酸分子、およびそのオリゴヌクレオチドフラグメントはまた、ATタンパク質をコードする核酸を増幅するためのPCR反応におけるプライマーおよび/または鋳型として有用である。また、本発明のタンパク質を発現するトランスジェニック非ヒト哺乳動物が提供される。
【0007】
本発明のATタンパク質と免疫反応性である抗体がまた、提供される。これらの抗体は、所定のサンプル(例えば、組識サンプル、ウエスタンブロットなど)に存在するATタンパク質のレベルを決定するための診断アッセイにおいて有用である。抗体はまた、粗細胞抽出物などからATタンパク質を精製するために使用され得る。さらに、これらの抗体は、インビボで、ATタンパク質の生物学的効果を調節するために治療学的に有用であると考えられる。
【0008】
種々の組識サンプルにおけるATタンパク質のレベルを決定するための方法および診断系がまた、提供される。これらの診断方法は、治療学的に有効な量の維持を容易にするために、治療学的に投与されるATタンパク質またはそのフラグメントのレベルをモニターするために使用され得る。これらの診断方法はまた、異常なレベルのまたは異常な構造のATタンパク質から生じる生理学的障害を診断するために使用され得る。
1.哺乳動物アセチルコエンザイムAトランスポーター(AT)タンパク質をコードする単離された核酸、そのスプライス改変体cDNA配列、またはそれらのフラグメント。
2.項目1に記載の核酸であって、前記哺乳動物ATタンパク質が、ヒト、ラット、マウス、ブタ、ヒツジ、イヌ、またはウシから選択される哺乳動物から単離される、核酸。
3.項目1に記載の核酸であって、該核酸が、以下:
(a)配列番号2に記載のアミノ酸配列をコードするDNA、または
(b)中程度のストリンジェントな条件下で(a)のDNAにハイブリダイズするDNAであって、ここで該DNAが、生物学的に活性なATをコードする、DNA、または
(c)上記の(a)もしくは(b)のいずれかに関するDNA縮重体であって、ここで該DNAが、生物学的に活性なATをコードする、DNA、または
(d)それらのスプライス改変体cDNA配列、
から選択される、核酸。
4.項目3に記載の核酸であって、該核酸が、配列番号1のヌクレオチド388〜2034のATコード部分に、高いストリンジェンシーの条件下でハイブリダイズする、核酸。
5.項目3に記載の核酸であって、該核酸のヌクレオチド配列が、配列番号1に記載のヌクレオチド388〜2034と、実質的に同じである、核酸。
6.項目3に記載の核酸であって、該核酸のヌクレオチド配列が、AT-1をコードし、および配列番号1に記載のヌクレオチド388〜2034と、同じである、核酸。
7.項目3に記載の核酸であって、該核酸がcDNAである、核酸。
8.項目3に記載の核酸を含む、ベクター。
9.項目3の核酸を含む、組換え細胞。
10.オリゴヌクレオチドであって、配列番号1に記載されるヌクレオチド配列の核酸の配列と、特異的にハイブリダイズし得る少なくとも14ヌクレオチドを含む、オリゴヌクレオチド。
11.項目10に記載のオリゴヌクレオチドであって、該オリゴヌクレオチドが、検出可能なマーカーで標識される、オリゴヌクレオチド。
12.アンチセンスオリゴヌクレオチドであって、項目3に記載の核酸によってコードされるmRNAに特異的に結合し得る、アンチセンスオリゴヌクレオチド。
13.AT cDNA配列の存在を検出するためのキットであって、項目11に記載の少なくとも1つのオリゴヌクレオチドを含む、キット。
14.膜を横切ってアセチルCoAを輸送し得ることによって特徴付けられる、単離されたアセチルコエンザイムAトランスポーター(AT)タンパク質。
15.項目14に記載のATタンパク質であって、該タンパク質のアミノ酸配列が、配列番号2に記載のタンパク質配列と実質的に同じ配列を含む、タンパク質。
16.項目15に記載のATタンパク質であって、配列番号2に記載のヒトAT-1タンパク質配列と同じアミノ酸を有する、タンパク質。
17.項目14に記載のATタンパク質であって、該タンパク質は、ヒトAT-1であるヌクレオチド配列によってコードされ、および項目1に記載のヌクレオチド配列と実質的に同じである、タンパク質。
18.項目17に記載のATタンパク質であって、該タンパク質が、ヒトATであり、および配列番号1に記載されるヌクレオチド配列によってコードされる、タンパク質。
19.項目14に記載のATタンパク質であって、該タンパク質が、配列番号1に記載されるヌクレオチド388〜2034と実質的に同じヌクレオチド配列を含むヌクレオチド配列によってコードされる、タンパク質。
20.ATタンパク質の発現のための方法であって、該方法が、該ATタンパク質の発現に適切な条件下で、項目9に記載の細胞を培養する工程を包含する、方法。
21.項目14に記載のATタンパク質との特異的反応性を有する、単離された抗ATタンパク質抗体。
22.項目21に記載の抗体であって、該抗体が、モノクローナル抗体である、抗体。
23.項目21に記載の抗体であって、該抗体が、ポリクローナル抗体である、抗体。
24.ヒトATタンパク質の発現を阻害するに有効な量の、項目12に記載のアンチセンスオリゴヌクレオチド、および細胞膜を通過し得る、受容可能な疎水性キャリアを含有する、組成物。
25.ATタンパク質をコードする外因性核酸を発現する、トランスジェニック非ヒト哺乳動物。
26.項目25に記載のトランスジェニック非ヒト哺乳動物であって、前記ATタンパク質をコードする核酸が、変異されており、およびそのように発現される該ATタンパク質が、ネイティブなATでない、トランスジェニック非ヒト哺乳動物。
27.項目25に記載のトランスジェニック非ヒト哺乳動物であって、該トランスジェニック非ヒト動物が、マウスである、トランスジェニック非ヒト哺乳動物。
28.哺乳動物ATタンパク質をコードする核酸を同定するための方法であって、該方法が、以下の工程:
項目10に記載のオリゴヌクレオチドと、核酸を含有するサンプルとを接触する工程であって、該接触する工程が、高いストリンジェンシーのハイブリダイゼーション条件下で達成される、工程、および
それにハイブリダイズする化合物を同定する工程、
を包含する、方法。
29.サンプル中のヒトATタンパク質の存在を検出するための方法であって、該方法が、項目21に記載の抗体と、試験サンプルとを接触する工程、抗体−AT複合体の存在を検出する工程、およびそれによって、該試験サンプル中のヒトATタンパク質の存在を検出する工程、を包含する、方法。
30.AT核酸の増幅のための1本鎖DNAプライマーであって、該プライマーが、配列番号1に記載の核酸配列に由来する核酸配列を含む、プライマー。
31.試験化合物が、項目14に記載のATタンパク質に対するアゴニストまたはアンタゴニストとして作用し得るか否かを評価するためのバイオアッセイであって、該バイオアッセイが、以下の工程:
(a)ATタンパク質またはその機能的に改変された形態を発現するDNAを含む細胞を培養する工程であって、
ここで該培養する工程が、ATタンパク質のAc-CoA輸送活性を調節する能力が決定されようとする、少なくとも1つの化合物の存在下で行われる、工程、およびその後
(b)Ac-CoAのレベルの増加または減少のいずれかについて、該細胞をモニターする工程、
を包含する、バイオアッセイ。
32.試験化合物が、項目14に記載のATタンパク質、または該ATタンパク質の機能的に改変された形態についてのアンタゴニストとして作用し得るか否かを評価するためのバイオアッセイであって、該バイオアッセイが、以下の工程:
(a)ATタンパク質またはその機能的に改変された形態を発現するDNAを含む細胞を培養する工程であって、
ここで該培養する工程が:
ATタンパク質のAc-CoA輸送活性を阻害する能力が決定されようとする、漸増濃度少なくとも1つの化合物、および
一定濃度のAc-CoA
の存在下で行われる、工程;ならびにその後、
(b)該細胞において、該化合物の濃度の関数として細胞内オルガネラに輸送されるAc-CoAのレベルを、モニターし、それによってAT輸送活性を阻害する該化合物の能力を示す工程、
を包含する、バイオアッセイ。
33.試験化合物が、項目14に記載のATタンパク質、または該ATタンパク質の機能的に改変された形態についてのアンタゴニストとして作用し得るか否かを評価するためのバイオアッセイであって、該バイオアッセイが、以下の工程:
(a)ATタンパク質またはその機能的に改変された形態を発現するDNAを含む細胞を培養する工程であって、
ここで該培養する工程が、
ATタンパク質のAc-CoA輸送活性を阻害する能力が決定されようとする、一定濃度の少なくとも1つの化合物、および
漸増濃度のAc-CoA
の存在下で行われる、工程;ならびにその後、
(b)該細胞において、該化合物の濃度の関数として細胞内オルガネラに輸送されるAc-CoAのレベルを、モニターし、それによってAT輸送活性を阻害する該化合物の能力を示す工程、
を包含する、バイオアッセイ。
34.試験化合物が、項目14に記載のATタンパク質、または該ATタンパク質の機能的に改変された形態についてのアゴニストとして作用し得るか否かを評価するためのバイオアッセイであって、該バイオアッセイが、以下の工程:
(a)ATタンパク質またはその機能的に改変された形態を発現するDNAを含む細胞を培養する工程であって、
ここで該培養する工程が、
ATタンパク質のAc-CoA輸送活性を増加する能力が決定されようとする、漸増濃度の少なくとも1つの化合物、および
一定濃度のAc-CoA
の存在下で行われる、工程;ならびにその後、
(b)該細胞において、該化合物の濃度の関数として細胞内オルガネラに輸送されるAc-CoAのレベルを、モニターし、それによってAT輸送活性を増加する該化合物の能力を示す工程、
を包含する、バイオアッセイ。
35.試験化合物が、項目14に記載のATタンパク質、または該ATタンパク質の機能的に改変された形態についてのアゴニストとして作用し得るか否かを評価するためのバイオアッセイであって、該バイオアッセイが、以下の工程:
(a)ATタンパク質またはその機能的に改変された形態を発現するDNAを含む細胞を培養する工程であって、
ここで該培養する工程が、
ATタンパク質のAc-CoA輸送活性を増加する能力が決定されようとする、一定濃度の少なくとも1つの化合物、および
漸増濃度のAc-CoA
の存在下で行われる、工程;およびその後、
(b)該細胞において、該化合物の濃度の関数として細胞内オルガネラに輸送されるAc-CoAのレベルを、モニターし、それによってAT輸送活性を増加する該化合物の能力を示す工程、
を包含する、バイオアッセイ。
36.ATタンパク質によって媒介されるAc-CoA輸送活性を調節するための方法であって、該方法が:
該ATタンパク質と、項目31によって同定されるアゴニストの調節するに有効な量とを、接触させる工程を、包含する、方法。
37.ATタンパク質によって媒介されるAc-CoA輸送活性を調節するための方法であって、該方法が:
該ATタンパク質と、項目31によって同定されるアンタゴニストの調節するに有効な量とを、接触させる工程を、包含する、方法。
38.異常な神経細胞分化、異常な神経細胞移動、神経芽腫、または黒色腫を処置する方法であって、該方法が、有効量の項目31によって同定される化合物を投与する工程を包含する、方法。
【発明を実施するための最良の形態】
【0009】
発明の詳細な説明
本発明によれば、単離された哺乳動物アセチルコエンザイムAトランスポーター(AT)タンパク質、ポリペプチド、および本発明の核酸によってコードされるそれらのフラグメントが提供される。本明細書中で使用されるように、句「AT」は、膜を横切ってアセチルコエンザイムA(Ac-CoA)を輸送し得る、哺乳動物ファミリー、好ましくはヒトの、単離されたおよび/または実質的に純粋なタンパク質をいう。本発明のATタンパク質はさらに、ガングリオシドGD3およびGT3のシアル酸残基のアセチル化を、間接的に促進する能力を有することによって、特徴付けられる。本発明のATタンパク質は、一次転写物の別のスプライシングによって生成されるmRNAによってコードされる、天然に存在するその対立遺伝子改変体を含み、そしてさらに、例えば、免疫原性などのような少なくとも1つの天然の生物学的活性を保持する、その活性なフラグメントを含む。
【0010】
本発明の別の実施態様において、本明細書中で言及されるATタンパク質は、ATタンパク質(好ましくはヒト)をまた特異的に認識する抗体によって特異的に認識されるそれらのポリペプチドであり、これは配列番号2に記載される配列を含む。本発明の単離されたATタンパク質は、天然のインビボ環境と通常関連する細胞性成分および/または夾雑物を含まない。
【0011】
本発明のタンパク質はさらに、少なくとも以下の細胞:心臓、脳、胎盤、肺、肝臓、骨格筋、腎臓、および膵臓において発現されることによって特徴付けられる。最も高いレベルの発現は、膵臓組識において検出された。3.3および4.3kbの大きさを有する、ATタンパク質の2つの主要なmRNA転写物は、試験した全ての組識において検出された(ノーザンブロットアッセイによって観察された)。2.7および3.4kbの大きさを有する2つのmRNA転写物は、AT-1でトランスフェクトされたHeLa細胞において強力に発現され、そして偽トランスフェクトされたHeLa細胞においてほとんど検出されなかった。従って、ATファミリーのタンパク質をコードするスプライス改変体cDNA転写物が、本発明によって明らかに意図される。
【0012】
AT-1(配列番号2)の予測されるアミノ酸配列は、神経伝達物質(概説については、HuchoおよびTsetlin、J.Neurochem.、66:1781-1792(1996)を参照のこと)、グルコース(Daviesら、Biochem.J.、266:799-808(1990))、アンモニア(Mariniら、EMBOJ.、12:3456-3463(1994))、およびヌクレオチド−糖類(Eckhardtら、Proc. Natl. Acad. Sci. USA、93:7572-7576(1996);Miuraら、J.Biochem.、120:236-241(1996);およびAbeijonら、Proc. Natl. Acad. Sci.USA、93:5963-5968(1996))についてのトランスポーターを含む多くのトランスポーターにおいて代表的に観察される、複数の膜貫通ドメインを有する。ほとんどのトランスポータータンパク質は、膜貫通α-ヘリックスを形成すると考えられる6〜12個の疎水性配列を含む。AT-1タンパク質の場合、少なくとも6個の膜貫通ドメインの存在が、Psort分析によって予測される。
【0013】
さらに、AT-1タンパク質は、転写因子の二量体化に関与することがもともと見出された(Landschulzら、Science,240:1759-1964(1988))、ロイシンジッパーモチーフを含む。非常に最近の研究は、CMP-NeuAc(Eckhardtら、Proc.Natl. Acad. Sci.USA、93:7572-7576(1996))、UDP-Gal(Miuraら、J.Biochem.、120:236-241(1996))、およびUDP-GlcNAc(Abeijonら、Proc.Natl.Acad.Sci.USA、93:5963-5968(1996))についてのトランスポーターのようなヌクレオチド−糖トランスポータータンパク質もまた、この構造を有することを示し、このことは、これらのタンパク質が、ゴルジ膜におけるホモ二量体として存在することを示唆する。近年、ラット肝臓ゴルジPAPSトランスポーター(Mandonら、Biochem.Biophy.Res.Commun.、225:932-938(1996))もまた、ホモ二量体であることが示されたが、ロイシンジッパー構造がこの特定のトランスポータータンパク質に存在するか否かは知られていない。従って、AT-1タンパク質は、複数の膜貫通ドメインを有するホモ二量体として機能し得ることが、本明細書中で意図される。しかし、AT-1タンパク質の分子量は、CMP-NeuAc、UDP-Ga1、およびUDP-GlcNAcについてのトランスポーターの分子量よりも実質的に大きく、このことは、AT-1タンパク質は、新規のATトランスポーターファミリーのメンバーを意味し得ることを示唆する。
【0014】
今までのところ同定された全てのヌクレオチド−糖トランスポーターは、ゴルジ体膜に局在化されることが見出されている(Miuraら、J. Biochem.、120:236-241(1996);Abeijonら、Proc. Natl.Acad.Sci.USA、93:5963-5968(1996);およびBhakdiら、Infect.Immun.、47:52-60(1985))。AT-1に対して特異的な抗体を用いる免疫組識学的研究は、AT-1タンパク質が、小胞体、ゴルジ、およびおそらくミトコンドリア膜を含む細胞質中に拡散して存在することを実証した。この観察は、アセチルトランスフェラーゼが、ゴルジ膜に富化されることが示されたので、ATタンパク質ファミリーは、アセチルトランスフェラーゼとして機能しないことを示す(Varki、Glycobiology、2:25-40(1992)およびVarkiおよびDiaz、J.Biol. Chem.、260:6600-6608(1985))。さらに、半インタクトな細胞を使用した、インビトロアッセイの結果(例えば、実施例VI、および図2を参照のこと)は、AT-1が、膜を横切ってAc-CoAを輸送するように機能するタンパク質ファミリーのメンバー(例えば、Ac-CoAトランスポーター)であることを示す。
【0015】
タンパク質および核酸データベース(GenBank、EMBLおよびSwiss-Prot)に対するホモロジー検索が、配列番号1に記載されるAT-1の核酸配列およびアミノ酸配列に相同な核酸配列およびアミノ酸配列について行われた。公知のタンパク質との有意な相同性は検出されなかったが、ホモロジー検索は、高い程度の相同性を有する2つの推定のタンパク質を同定した:35%のアミノ酸配列同一性を有する560アミノ酸を有する、S.cerevisiaeの推定のタンパク質[アクセス番号第Z36088(EMBL)および同第P38318(Swiss Prot)];ならびに47%のアミノ酸配列同一性を有する632アミノ酸を有する、Caenorhabditiselegans T26C5.3[EMBL、アクセス番号第Z50859]。従って、AT-1タンパク質は、普遍的であり、そして進化論的に保存されているようであり、そしてシアル酸の出現は、これらの2つの生物体において報告されていないので、ATタンパク質ファミリーは、シアル酸アセチルトランスフェラーゼとして機能しないようである。AT-1の転写物は、O-アセチル化ガングリオシドよりも広範囲に分配され、このことは、AT-1タンパク質以外の分子が、O-アセチル化GD3発現の調節を指図するという仮定を支持する。本明細書中に記載される結果は、AT-1タンパク質は、Ac-CoAトランスポーターとして機能すること、そしてこれは、ガングリオシドにおけるシアル酸のアセチル化プロセス以外のアセチル化プロセスにおいて、重要な役割を果たし得ることを、強力に示唆する。
【0016】
本明細書および請求の範囲における、DNA、RNA、ポリペプチド、またはタンパク質の修飾語としての、用語「単離された」および/または「精製された」の使用は、そのように設計されたDNA、RNA、ポリペプチド、またはタンパク質が、ヒトの手によってこのような形態において生成され、従ってそれらの天然のインビボ細胞環境から分離されることを意味する。このヒト介入の結果として、本発明の組換えのDNA、RNA、ポリペプチド、およびタンパク質は、それらが天然に存在するようなDNA、RNA、ポリペプチド、またはタンパク質ではないことが、本明細書中で記載する方法において有用である。
【0017】
本明細書中で使用されるように、「哺乳動物」は、本発明のATタンパク質が由来する多様な種(例えば、ヒト、ラット、マウス、ウサギ、サル、ヒヒ、ウシ、ブタ、ヒツジ、イヌ、ネコなど)をいう。
【0018】
現在好ましい本発明のATタンパク質は、配列番号2に記載のタンパク質配列と実質的に同じであるアミノ酸配列、ならびに生物学的に活性な、その改変された形態を含む。当業者は、上記配列の複数の残基が、得られるタンパク質種の生物学的活性を実質的に変化しないで、他の、化学的に、立体的に、および/または電子的に類似の残基と置換され得ることを認識する。さらに、本明細書中の配列番号2と実質的に同じ配列を含む、より大きなまたはより小さなポリペプチド配列(例えば、スプライス改変体、ATの活性なフラグメントなど)が、意図される。
【0019】
本明細書中で用いられるように、用語「実質的に同じアミノ酸配列」は、参照アミノ酸配列に関して少なくとも約70%の同一性を有し、および参照アミノ酸配列によって規定されるタンパク質の、比較可能な機能的なおよび生物学的な活性の特徴を保持するアミノ酸配列をいう。好ましくは、「実質的に同じアミノ酸配列」を有するタンパク質は、参照アミノ酸配列に関して少なくとも約80%、より好ましくは90%のアミノ酸同一性を有する;約95%を超えるアミノ酸配列同一性が、特に好ましい。しかし、スプライス改変体として生じる、記載されるレベル未満の配列同一性を含むか、または保存的なアミノ酸置換によって、もしくは縮重コドンの置換によって改変される、ポリペプチド(または、本明細書中で以前に言及された核酸)はまた、本発明の範囲内に包含される。本明細書中に開示される好ましいATタンパク質は、ヒトAT-1(配列番号2)である。
【0020】
用語「生物学的に活性な」または「機能的な」は、本発明のATタンパク質、またはそのポリペプチドフラグメントの修飾語として本明細書中で使用される場合、ATに起因される機能的な特徴の少なくとも1つを示すポリペプチドをいう。例えば、ATの1つの生物学的活性は、細胞膜、好ましくは細胞内オルガネラの膜(例えば、ゴルジ膜、小胞体膜、ミトコンドリア膜など)を横切ってアセチルコエンザイムA(Ac-CoA)を輸送する能力である。ATのさらに別の生物学的活性は、ガングリオシドGD3およびGT3のシアル酸残基のアセチル化を、間接的に促進する能力である。
【0021】
ATの別の生物学的活性は、ATに特異的に結合するポリクローナル抗体およびモノクローナル抗体の生成のための免疫原として作用する能力である。従って、ATをコードする本発明の核酸は、配列番号2に記載される配列(At-1)を含むATタンパク質(好ましくはヒト)をまた特異的に認識する抗体によって、特異的に認識されるポリペプチドをコードする。このような活性は、当業者に公知の任意の方法によってアッセイされ得る。例えば、ATcDNAによってコードされる試験ポリペプチドは、抗体を生成するために使用され得、次いで、抗体は、配列番号2に記載の配列を含むタンパク質に結合するそれらの能力についてアッセイされる。抗体が、実質的に同じ親和性で、試験ポリペプチド、および配列番号2に記載の配列を含むタンパク質に結合する場合、ポリペプチドは、必要とされる生物学的活性を保持する。
【0022】
本発明のATタンパク質は、当該分野において周知の多様な方法(例えば、本明細書中に記載される組換え発現系、沈殿、ゲル濾過、イオン交換クロマトグラフィー、逆相クロマトグラフィー、およびアフィニティークロマトグラフィーなど)によって単離され得る。他の周知の方法は、Deutscherら、Guideto Protein Purification: Methods in Enzymology 第182巻(Academic Press、(1990))(これは、本明細書中に参考として援用される)に記載される。あるいは、本発明の単離されたポリペプチドは、例えば、Sambrookら(前出、1989)に記載されるような、周知の組換え方法を使用して得られ得る。
【0023】
本発明のポリペプチドを調製するための手段の例は、当該分野において周知の方法を用いて、適切な宿主細胞(例えば、細菌細胞、酵母細胞、両生類細胞(すなわち、卵母細胞)、または哺乳動物細胞)においてATをコードする核酸を発現し、そして再び周知の方法を用いて、発現されたポリペプチドを回収することである。本発明のATポリペプチドは、本明細書中に記載されるような発現ベクターで形質転換された細胞から直接単離され得る。本発明のポリペプチド、生物学的に活性なフラグメント、およびそれらの機能的等価物はまた、化学合成によって生成され得る。例えば、合成ポリペプチドは、製造業者によって提供される化学製品を用いて、AppliedBiosystems、Inc. 430A型または431A型 自動ペプチド合成器(Foster City、CA)を使用して、生成され得る。
【0024】
また、用語ATによって含まれるものは、その活性なフラグメントまたはポリペプチドアナログである。用語「活性なフラグメント」は、この部分が、対応する全長タンパク質の特徴である活性を有することが提供される、全長ATタンパク質の部分であるペプチドフラグメントをいう。例えば、ATタンパク質の活性なフラグメント(例えば、細胞質ドメイン)は、Ac-CoAを結合するか、またはAc-CoAへの結合後にAc-CoAの膜輸送を媒介する能力のような活性を有し得る。免疫応答を誘発するATタンパク質の活性フラグメントの特徴は、抗AT抗体を得るために有用である。従って、本発明はまた、本発明のATタンパク質の活性なフラグメントを提供し、このフラグメントは、本明細書中に記載されるアッセイを使用して、同定され得る。
【0025】
用語「ポリペプチドアナログ」は、その1つ以上の残基が機能的に類似な残基で保存的に置換されている、本明細書中に具体的に示される配列に実質的に同一なアミノ酸残基配列を有し、そして本明細書中に記載されるようなATを模倣する能力を示す、任意のポリペプチドを含む。保存的置換の例としては、ある非極性(疎水性)残基(例えば、イソロイシン、バリン、ロイシン、もしくはメチオニン)での別の非極性残基の置換、ある極性(親水性)残基での別の極性残基の置換(例えば、アルギニンとリジンとの間、グルタミンとアスパラギンとの間、グリジンとセリンとの間)、ある塩基性残基(例えば、リジン、アルギニン、もしくはヒスチジン)での別の塩基性残基の置換、またはある酸性残基(例えば、アスパラギン酸もしくはグルタミン酸)での別の酸性残基の置換を含む。
【0026】
本明細書中で使用されるように、句「保存的置換」はまた、非誘導体化残基の代わりに化学的に誘導された残基の使用を含むが、ただしこのようなポリペプチドは、必要とされる結合活性を示す。句「化学的誘導体」は、官能側鎖基の反応によって化学的に誘導体化された1つ以上の残基を有する目的のポリペプチドをいう。このような誘導体化された分子は、例えば、遊離のアミノ基が、誘導体化され、アミン塩酸塩、p-トルエンスルホニル基、カルボベンズオキシ基、t-ブチルオキシカルボニル基、クロロアセチル基、またはホルミル基を形成するそれらの分子を含む。遊離のカルボキシル基は、誘導体化されて、塩、メチルおよびエチルエステル、または他のタイプのエステルもしくはヒドラジドを形成し得る。遊離のヒドロキシル基は、誘導体化されて、O-アシルまたはO-アルキル誘導体を形成し得る。ヒスチジンのイミダゾール窒素は、誘導体化されて、N-im-ベンジルヒスチジンを形成し得る。また、化学誘導体として含まれるものは、20個の標準アミノ酸の1つ以上の天然に存在するアミノ酸誘導体を含むそれらのペプチドである。例えば、4-ヒドロキシプロリンは、プロリンを置換し得;5-ヒドロキシリジンは、リジンを置換し得;3-メチルヒスチジンは、ヒスチジンを置換し得;ホモセリンは、セリンを置換し得;およびオルニチンは、リジンを置換し得る。本発明のポリペプチドはまた、必要とされる活性が維持される限り、配列が本明細書中に示されるポリペプチドの配列に比べて、残基の1つ以上の付加および/または欠失を有する任意のポリペプチドを含む。
【0027】
本発明はまた、受容可能なキャリア、ならびに任意の単離された、精製されたATポリペプチド、その活性なフラグメントもしくはポリペプチドアナログ、または精製された、成熟タンパク質およびその活性なフラグメントを、単独でまたは互いに組合わせて含む、組成物を提供する。これらのポリペプチドまたはタンパク質は、組換え的に得られ得るか、化学的に合成され得るか、または天然の供給源から精製され得る。本明細書中で使用されるように、用語「受容可能なキャリア」は、任意の標準的な薬学的キャリア(例えば、リン酸緩衝化生理食塩水溶液、水、および乳濁液(例えば、油/水、または水/油乳濁液)、ならびに種々のタイプの湿潤剤)を含む。
【0028】
本発明の別の実施態様によれば、本発明のAT(アセチル−コエンザイムA輸送体)タンパク質、およびそのフラグメントをコードする単離された核酸が提供される。本明細書中に記載される核酸分子は、このような核酸が当業者に公知の多様なタンパク質発現系に組込まれる場合、本発明のタンパク質を生成するために有用である。さらに、このような核酸分子またはそのフラグメントは、所定のサンプルにおけるAT遺伝子またはmRNA転写物の存在および/または量をアッセイするために、容易に検出可能な置換基で標識され得、そしてハイブリダイゼーションプローブとして使用され得る。本明細書中に記載される核酸分子、およびそのフラグメントはまた、本明細書中に記載される本発明のタンパク質をコードする遺伝子を増幅するためのPCR反応において、プライマーおよび/または鋳型として有用である。
【0029】
用語「核酸」(ポリヌクレオチドともいわれる)は、リボ核酸(RNA)またはデオキシリボ核酸(DNA)、プローブ、オリゴヌクレオチド、およびプライマーを含む。DNAは、相補的DNA(cDNA)またはゲノムDNA(例えば、ATタンパク質をコードする遺伝子)のいずれかであり得る。ATポリペプチドをコードする核酸を単離する1つの手段は、当該分野において周知の方法を使用して、天然のDNAプローブまたは人工的に設計されたDNAプローブで、哺乳動物ゲノムライブラリーをプローブすることである。AT遺伝子に由来するDNAプローブは、この目的のために特に有用である。ATポリペプチドをコードするDNAおよびcDNA分子は、以下により詳細に記載される方法によって、哺乳動物(例えば、ヒト、マウス、ラット、ウサギ、ブタなど)、もしくは他の動物供給源から相補的ゲノムDNA、cDNA、もしくはRNAを得るために、またはcDNAライブラリーもしくはゲノムライブラリーのスクリーニングによって関連のcDNAクローンもしくはゲノムクローンを単離するために使用され得る。核酸の例は、RNA、cDNA、またはATポリペプチドをコードする単離されたゲノムDNAである。このような核酸は、配列番号1(配列番号1の少なくともヌクレオチド388〜2034)に記載のものと実質的に同じヌクレオチド配列を有する核酸、またはそのスプライス改変体cDNA配列を含み得るが、これらに限定されない。
【0030】
本明細書中で使用されるように、本発明のレセプターをコードする特定のヌクレチド配列を記載するために使用される場合、句「スプライス改変体」または「選択的にスプライスされた」とは、周知の真核生物RNAスプライシングプロセスから生じるcDNA配列をいう。RNAスプライシングプロセスは、細胞質の成熟RNA分子を作製するための、真核生物一次RNA転写物からのイントロンの除去、およびエキソンの結合を含む。スプライス改変体ヌクレオチド配列を単離する方法は、当該分野において周知である。例えば、当業者は、本明細書中に記載されるようなcDNAライブラリーまたはゲノムライブラリーをスクリーニングするために、配列番号1のATをコードするcDNAに由来するヌクレオチドプローブを用い得る。
【0031】
本発明の1つの実施態様において、本明細書中に開示される本発明のATタンパク質をコードするcDNAは、配列番号1に記載されるような、実質的に同じヌクレオチド配列を含む。本発明の別の実施態様において、本発明のタンパク質をコードするcDNA分子は、配列番号1のヌクレオチド388〜2034と同じヌクレオチド配列を含む。
【0032】
本明細書中で用いられるように、用語「実質的に同じヌクレオチド配列」は、参照ポリヌクレオチドに、中程度にストリンジェントなハイブリダイゼーション条件下で参照ヌクレオチドにハイブリダイズするのに十分な同一性を有するDNAをいう。1つの実施態様において、参照ヌクレオチド配列と実質的に同じヌクレオチド配列を有するDNAは、配列番号2に記載されるアミノ酸配列と実質的に同じアミノ酸配列、または配列番号2を含むより大きなアミノ酸配列をコードする。別の実施態様において、参照ヌクレオチド配列と「実質的に同じヌクレオチド配列」を有するDNAは、参照ヌクレオチド配列に関して少なくとも60%の同一性を有する。参照ヌクレオチド配列に対して、少なくとも70%、より好ましくは少なくとも約90%、さらにより好ましくは少なくとも95%の同一性を有するDNAが、好ましい。
【0033】
本発明はまた、配列番号1に示される核酸とは異なるが、同じ表現型を有する核酸を含む。表現型が類似する核酸はまた、「機能的に等価な核酸」といわれる。本明細書中で使用されるように、句「機能的に等価な核酸」は、実質的に同じ様式において機能し、本明細書中で開示される核酸と同じタンパク質産物を生成する、僅かに重要なおよび重要でない配列の改変によって特徴付けられる核酸を包含する。特に、機能的に等価な核酸は、本明細書中に開示されるポリペプチドと同じであるポリペプチド、もしくは保存的アミノ酸改変を有するポリペプチドをコードするか、または配列番号2を含むより大きなポリペプチドをコードする。例えば、保存的改変は、別の非極性残基での非極性残基の置換、または同様に荷電された残基での荷電された残基の置換を含む。これらの改変は、タンパク質の三次構造を実質的に変化させない改変として、当業者によって認識される改変を含む。
【0034】
遺伝子コードの縮重によって、特定されたハイブリダイゼーション条件下で本発明の核酸に必ずしもハイブリダイズしない、ATポリペプチドをコードする核酸が、さらに提供される。本発明のATポリペプチドをコードする好ましい核酸は、配列番号2(すなわち、AT-1)に記載されるのと実質的に同じアミノ酸配列をコードするヌクレオチドから構成される。
【0035】
従って、本発明のATタンパク質をコードする例示的な核酸は、以下から選択され得る:
(a)配列番号2に記載されるアミノ酸配列をコードするDNA;
(b)中程度にストリンジェントな条件下で(a)のDNAにハイブリダイズするDNA、ここでこのDNAは、生物学的に活性なATをコードする;または
(c)上述の(a)または(b)のいずれかに関するDNA縮重体、ここでこのDNAは、生物学的に活性なATをコードする。
【0036】
ハイブリダイゼーションは、水素結合を介する、核酸の相補的な鎖(すなわち、センス:アンチセンス鎖、またはプローブ:標的−DNA)の互いへの結合をいい、染色体DNAにおいて天然に存在する結合に類似する。所定のプローブと標的−DNAとをハイブリダイズするために使用されるストリンジェンシーのレベルは、当業者によって容易に変更され得る。
【0037】
句「ストリンジェントなハイブリダイゼーション」とは、ポリ核酸配列ハイブリッドが安定である下での条件をいうために、本明細書中で用いられる。当業者に公知であるように、ハイブリッドの安定性は、ハイブリッドの融点(Tm)において反映される。一般に、ハイブリッドの安定性は、ナトリウムイオン濃度および温度の関数である。代表的に、ハイブリダイゼーション反応は、より低いストリンジェンシーの条件下で行われ、次いで変化するが、より高いストリンジェンシーの洗浄を行う。ハイブリダイゼーションのストリンジェンシーに対する参照は、このような洗浄条件に関する。
【0038】
本明細書中で使用されるように、句「中程度にストリンジェントなハイブリダイゼーション」は、標的DNAに対する約60%の同一性、好ましくは約75%の同一性、より好ましくは約85%の同一性を有する相補的核酸に、標的−DNAが結合することを許容する条件をいい;標的−DNAに対する約90%を超える同一性は、特に好ましい。好ましくは、中程度にストリンジェントな条件は、42℃で、50%ホルムアミド、5×デンハルト溶液、5×SSPE、0.2%SDSにおけるハイブリダイゼーション、次いで42℃で、0.5×SSPE、0.2% SDSにおける洗浄に等価である条件である。
【0039】
句「高いストリンジェンシーのハイブリダイゼーション」は、65℃で、0.018MNaClにおいて安定なハイブリッドを形成するそれらの核酸配列のみのハイブリダイゼーションを許容する条件をいう(すなわち、ハイブリッドが、65℃で、0.018MNaClにおいて安定でない場合、これは、本明細書中で意図されるように高いストリンジェンシー条件下で安定でない)。高いストリンジェンシー条件は、例えば、42℃で、50%ホルムアミド、5×デンハルト溶液、5×SSPE、0.2%SDSにおけるハイブリダイゼーション、次いで65℃で、0.1×SSPEおよび0.1% SDSにおける洗浄によって提供され得る。
【0040】
句「低いストリンジェンシーハイブリダイゼーション」は、37℃での10%ホルムアミド、5×デンハルト溶液、6×SSPE、0.2%SDSにおけるハイブリダイゼーション、次いで50℃での1×SSPE、0.2% SDSにおける洗浄に、等価な条件をいう。デンハルト溶液およびSSPE(例えば、Sambrookら、MolecularCloning、A Laboratory Manual、Cold Spring Harbor Laboratory Press、1989を参照のこと)は、他の適切なハイブリダイゼーション緩衝液と同様に、当業者に周知である。
【0041】
本明細書中で使用されるように、用語「縮重」は、参照核酸(例えば、配列番号1)から、少なくとも1ヌクレオチド異なるが、参照核酸と同じアミノ酸をコードするコドンをいう。例えば、3ヌクレオチド「UCU」、「UCC」、「UCA」、および「UCG」は、これらのコドンの4つは全て、アミノ酸のセリンをコードするので、互いに関して縮重である。
【0042】
本発明のポリペプチドをコードする好ましい核酸は、中程度にストリンジェント、好ましくは高いストリンジェンシーの条件下、配列番号1に記載の核酸配列の実質的な配列全体に、ハイブリダイズする。
【0043】
AT cDNAの任意の領域の部位特異的変異誘発は、変異体AT cDNAの生成に関して本明細書において意図される。例えば、TransformerMutagenesis Kit(Clontechから入手可能)は、AT cDNAに対して、様々なミスセンスおよび/またはノンセンス変異を構築するために使用され得るなど。
【0044】
本発明の核酸は、当該分野において周知の、様々な方法(例えば、配列番号1の種々の領域からオリゴヌクレオチドプライマーを使用して、PCR増幅を用いる、本明細書中に記載される方法など)によって、生成され得る。
【0045】
本発明のさらなる実施態様によれば、必要に応じて標識されるATコードcDNAまたはそのフラグメントが、関連の新規の哺乳動物ATタンパク質をコードするさらなる核酸配列について、ライブラリー(例えば、cDNA、ゲノムなど)をプローブするために用いられ得る。哺乳動物のcDNAライブラリーおよびゲノムライブラリー、好ましくはヒトライブラリーの構築は、当該分野において周知である。このようなcDNAライブラリーまたはゲノムライブラリーのスクリーニングは、約42℃未満の温度、約50%未満のホルムアミド濃度、および低い塩濃度への調整を含む、低いストリンジェンシー条件下で、最初に開始される。
【0046】
1つの実施態様において、プローブベースのスクリーニング条件は、約37℃の温度、約20%のホルムアミド濃度、および約5×標準生理食塩水クエン酸(SSC;3M塩化ナトリウム、0.3Mクエン酸ナトリウムを含む20×SSC、pH7.0)を含む。このような条件は、完全な相同性を必要とすることなく、プローブ配列と実質的な程度の類似性を有する配列の同定を許容する。句「実質的な類似性」は、少なくとも50%の相同性を共有する配列をいう。好ましくは、プローブと少なくとも約70%の相同性を有する配列の同定を許容するが、プローブと低い程度の相同性を有する配列に対して区別する、ハイブリダイゼーション条件が、選択される。結果として、配列番号1のヌクレオチド388〜2034と実質的に同じ(すなわち、類似の)ヌクレオチド配列を有する核酸が、得られる。
【0047】
本明細書中で使用されるように、プローブまたはプライマーとしてもまた本明細書中に言及される核酸「オリゴヌクレオチド」は、1本鎖DNAもしくはRNA、またはそのアナログであり、これは、配列番号1の任意に記載される任意の14個以上の連続的な塩基と同じである(またはこれに相補的である)少なくとも14個、好ましくは少なくとも20個、より好ましくは少なくとも50個の連続的な塩基を含むヌクレオチドの配列を有する。プローブを構築するために好ましい領域は、配列番号1の5’および/または3’コード領域を含む。さらに、本発明のATタンパク質の領域をコードする完全なcDNA、または配列番号1(AT)に対応する完全な配列は、プローブとして使用され得る。プローブは、本明細書中以後に記載されるように、当該分野において周知の方法によって標識され得、そして種々の診断キットにおいて使用され得る。
【0048】
本明細書中で使用されるように、種々の文法的形態における用語「標識」および「指示手段(indicatingmeans)」は、検出可能なシグナルの生成に、直接的または間接的のいずれかで関与する単一の原子および分子をいう。任意の標識または指示手段は、本発明の核酸プローブ、発現されるタンパク質、ポリペプチドフラグメント、または抗体分子に連結され得る。これらの原子または分子は、単独で、またはさらなる試薬と組合わせて使用され得る。このような標識は、臨床学的診断化学において、それ自体周知である。
【0049】
標識化手段は、変性を伴うことなく抗体または抗原に化学的に結合して、有用な免疫蛍光トレーサーである蛍光色素(染料)を形成する、蛍光標識化剤であり得る。免疫蛍光分析技術の記載は、DeLuca、「Immunofluorescence Analysis」、Antibody As a Tool、Marchalonisら編、JohnWiley & Sons,Ltd.、189〜231頁(1982)(これは、本明細書中に参考として援用される)において見出される。
【0050】
1つの実施態様において、指示基は、酵素(例えば、西洋ワサビペルオキシダーゼ(HRP)、グルコースオキシダーゼなど)である。別の実施態様において、放射活性エレメントは、標識化剤を用いられる。基質への標識の連結(すなわち、核酸プローブ、抗体、ポリペプチド、およびタンパク質の標識)は、当該分野において周知である。例えば、本発明の抗体は、培養培地において提供される放射標識されたアミノ酸の代謝取込みによって、標識され得る。例えば、Galfreら、Meth.Enzymol.、73:3-46(1981)を参照のこと。タンパク質結合または活性化された官能基によるカップリングの従来の手段は、特に適用可能である。例えば、Aurameasら、Scand.J.Immunol.、第8巻、補遺7:7-23(1978)、Rodwellら、Biotech.、3:889-894(1984)、および米国特許第4,493,795号を参照のこと。
【0051】
また、提供されるものは、mRNAの翻訳を妨げるように、ATポリペプチドをコードするmRNAの任意の部分と特異的に結合し得る配列を有するアンチセンスオリゴヌクレオチドである。アンチセンスオリゴヌクレオチドは、ATポリペプチドをコードするcDNAの配列の任意の部分と特異的に結合し得る配列を有し得る。本明細書中で使用されるように、句「特異的に結合する」は、相補的核酸配列を認識する、および相補的な塩基対間の水素結合の形成を介して、相補的核酸配列と二重らせんセグメントを形成する、核酸配列の能力を含む。アンチセンスオリゴヌクレオチドの例は、ヌクレオチドの化学アナログを含むアンチセンスオリゴヌクレオチドである。
【0052】
細胞膜を横切って通過し、そして翻訳を妨げるようにATポリペプチドをコードするmRNAと特異的に結合することによって、ATポリペプチドの発現を減少するに有効な、上記の、アンチセンスオリゴヌクレオチドの量、ならびに細胞膜を横切って通過し得る受容可能な疎水性キャリアを含む組成物がまた、本明細書において提供される。適切な疎水性キャリアは、例えば、米国特許第5,334,761号;同第4,889,953号;同第4,897,355号などにおいて記載される。細胞膜を横切って通過し得る受容可能な疎水性キャリアはまた、選択された細胞型に特異的なレセプターに結合し、そしてそれによって選択された細胞型の細胞によって取込まれる構造を含み得る。構造は、細胞型特異的レセプターに結合することが知られるタンパク質の部分であり得る。
【0053】
アンチセンスオリゴヌクレオチド組成物は、本発明のポリペプチドをコードするmRNAの翻訳を阻害するために有用である。合成オリゴヌクレオチド、または他のアンチセンス化学構造は、ATポリペプチドをコードするmRNAに結合し、そしてmRNAの翻訳を阻害するように設計され、および組識サンプルまたは被験体においてAT関連遺伝子の発現を阻害するための組成物として有用である。
【0054】
本発明の別の実施態様によれば、AT核酸配列の存在を検出するためのキットが提供され、少なくとも1つのオリゴヌクレオチド(例えば、本発明に従う、プローブまたはアンチセンスオリゴヌクレオチド)を含む。このようなキットは、AT遺伝子における変異、複製、欠失、再配置、または異数性を検出するために使用され得る。
【0055】
本発明は、ATポリペプチドをコードするmRNAの翻訳を阻害する、合成アンチセンスオリゴヌクレオチド組成物(本明細書中以後、SAOC)を用いることによって、これらのポリペプチドの発現のレベルを調節する手段を提供する。mRNAを認識し、そしてこれに選択的に結合するように設計された合成オリゴヌクレオチド、または他のアンチセンス化学構造は、ATコード鎖の部分または配列番号1に示されるヌクレオチド配列に相補的であるように構築される。SAOCは、注射によるもしくは直接的な腫瘍部位取込みによる、被験体への投与について、血流において安定であるように、または実験室の細胞培養条件において安定であるように、設計される。SAOCは、例えば、小さな、疎水性SAOC化学構造を設計することにより、SAOCを細胞膜を横切って通過し得るようにする、SAOCの物理的および化学的特性によって、またはSAOCを認識し、そしてこれを細胞へ輸送する、細胞における特異的な輸送系によって、細胞の細胞質に入るために細胞膜を横切って通過し得るように設計される。さらに、SAOCは、選択細胞集団内でのみSAOCを結合し、そしてこれを取込む、特異的な細胞取込み機構によって認識されるようにSAOCを標的化することによって、特定の選択された細胞集団のみへの投与について、設計され得る。
【0056】
例えば、SAOCは、上記のように、特定の細胞型においてのみ見出されるレセプターに結合するように設計され得る。SAOCはまた、配列番号1に示される配列内に含まれる配列に対応し得る、標的mRNA配列を認識し、これに選択的に結合するように設計される。SAOCは、標的mRNA配列に結合し、そして例えば、RNaseI消化によるmRNAの消化を誘導することによって、または翻訳調節因子またはリボソームの結合を妨げることによってmRNA標的配列の翻訳を阻害することによって、または、標的mRNAを分解するか、もしくは化学的に改変するかのいずれかの、他の化学構造(例えば、リボザイム配列または反応化学基)の包含によってのいずれかで、標的mRNA配列を不活化するように設計される。SAOCは、mRNA標的に対して指向される場合、このような特性が可能であることが示されている(Cohenら、TIPS、10:435(1989)およびWeintraub、Sci.American、January(1990)、40頁を参照のこと;これらの両方は、本明細書中に参考として援用される)。
【0057】
本発明のさらに別の実施態様によれば、適切な宿主細胞において、上記の核酸配列を発現することにより、本発明のATタンパク質を組換え産生するための方法が提供される。本明細書中に記載されるATタンパク質を生成するために適切である組換えDNA発現系は、当該分野において周知である。例えば、上記のヌクレオチド配列は、さらなる操作のためにベクターに組込まれ得る。本明細書中で使用されるように、ベクター(または、プラスミド)は、異種DNAの発現または複製のいずれかのために、細胞に異種DNAを導入するために使用される、不連続なエレメントをいう。
【0058】
適切な発現ベクターは、当該分野において周知であり、そして調節配列(例えば、DNAの発現を調節し得るプロモーター領域)に作動可能に連結されたこのようなDNAを発現し得るベクターを含む。従って、発現ベクターは、組換えDNAまたはRNA構築物(例えば、プラスミド、ファージ、組換えウイルス、もしくは適切な宿主細胞への取込みの際、挿入されたDNAの発現を生じる他のベクター)をいう。適切な発現ベクターは、当業者に周知であり、そして真核生物細胞および/または原核生物細胞において複製可能であるベクター、ならびにエピソームにとどまるベクターまたは宿主細胞ゲノムに組込まれるベクターを含む。さらに、ベクターは、ベクターが、多くのウイルスビリオン(例えば、レトロウイルス、ヘルペスウイルス、アデノウイルス)によってパッケージングされ、「ウイルスベクター」の形成を生じることを可能にする適切なパッケージングシグナルを含み得る。
【0059】
本明細書中に使用されるように、プロモーター領域は、DNAが作動可能に連結される、DNAの転写を制御するDNAのセグメントをいう。プロモーター領域は、RNAポリメラーゼ認識、結合、および転写開始に十分である特定の配列を含む。さらに、プロモーター領域は、RNAポリメラーゼのこの認識、結合、および転写開始活性を調節する配列を含む。これらの配列は、シス作用性であり得るか、またはトランス作用性因子に応答性であり得る。プロモーターは、調節の性質に依存して、構成的であり得るか、または調節され得る。本発明の実施における使用のために意図される例示的なプロモーターは、SV40初期プロモーター、サイトメガロウイルス(CMV)プロモーター、マウス乳房腫瘍ウイルス(MMTV)ステロイド誘導性プロモーター、モロニーマウス白血病ウイルス(MMLV)プロモーターなどを含む。
【0060】
本明細書中で使用されるように、用語「作動可能に連結される」は、DNAと、調節およびエフェクターヌクレオチド配列(例えば、プロモーター、エンハンサー、転写および翻訳の停止部位、ならびに他のシグナル配列)との機能的な関係をいう。例えば、プロモーターへのDNAの作動可能な連結は、DNAとプロモーターとの間の物理的および機能的な関係をいい、これによってこのようなDNAの転写が、DNAを特異的に認識し、結合し、そして転写するRNAポリメラーゼによって、プロモーターから開始される。
【0061】
本明細書中で使用されるように、発現は、ポリ核酸が、mRNAに転写され、そしてペプチド、ポリペプチド、またはタンパク質に翻訳されるプロセスをいう。ポリ核酸が、ゲノムDNAに由来する場合、発現は、適切な真核生物宿主細胞または生物が選択される場合、mRNAのスプライシングを含み得る。
【0062】
原核生物形質転換ベクターは、当該分野において周知であり、そしてpBlueskriptおよびファージλZAPベクター(Stratagene、La Jolla、CA)などを含む。他の適切なベクターおよびプロモーターは、1989年1月17日に発行された米国特許第4,798,885号(この開示は、その全体が本明細書中に参考として援用される)において詳細に開示される。
【0063】
E. coli細胞の形質転換についての他の適切なベクターは、pET発現ベクター(Novagen、米国特許第4,952,496号を参照のこと)(例えば、pET11a、これはT7プロモーター、T7ターミネーター、誘導性のE.coli lacオペレーター、およびlacリプレッサー遺伝子を含む;ならびにpET12a-c、これは、T7プロモーター、T7ターミネーター、およびE.coli ompT分泌シグナルを含む)を含む。別の適切なベクターは、pIN-IIIompA2(Duffaudら、Meth. in Enzymology、153:492-507、1987を参照のこと)であり、これは、lppプロモーター、lacUV5プロモーターオペレーター、ompA分泌シグナル、およびlacリプレッサー遺伝子を含む。
【0064】
例示的な、真核生物形質転換ベクターは、クローン化されたウシパピローマウイルスゲノム、マウスレトロウイルスのクローン化されたゲノム、および真核生物カセットを含み(例えば、pSV-2 gpt系(MulliganおよびBerg、Nature、277:108-114(1979))、Okayama-Bergクローニング系(Mol.Cell Biol.、2:161-170、(1982))、ならびにGenetics Institute(Science、228:810-815(1985))によって記載される発現クローニングベクターが利用可能であり、これは形質転換された真核生物細胞株において、目的のタンパク質の少なくともいくつかの発現の実質的な確実性を提供する。
【0065】
哺乳動物細胞のトランスフェクションのために、本発明のATをコードするDNAに連結され得る調節エレメントを含む、特に好ましベースベクターは、サイトメガロウイルス(CMV)プロモーターベースのベクター(例えば、pcDNA1(Invitrogen、SanDiego、CA))、MMTVプロモーターベースのベクター(例えば、pMAMNeo(Clontech、Palo Alto、CA)、およびpMSG(Pharmacia、Piscataway、NJ)、ならびにSV40プロモーターベースのベクター(例えば、pSVβ(Clontech、PaloAlto、CA)である。
【0066】
本発明の別の実施態様によれば、本発明の核酸分子(すなわち、DNAまたはmRNA)を含む「組換え細胞」が、提供される。適切な宿主細胞、好ましくは細菌細胞、およびより好ましくはE.coli細胞を形質転換する方法、ならびに異種タンパク質をコードする遺伝子を含むこの細胞を培養するために適用可能な方法は、当該分野において一般に知られている。例えば、Sambrookら、MolecularCloning:A Laboratory Manual(第2版)、Cold Spring Harbor Laboratory Press、ColdSpring Harbor、New York、USA(1989)を参照のこと。
【0067】
本発明の核酸を含む発現ベクターを、宿主細胞に導入(形質導入)して、形質導入された組換え細胞(すなわち、組換え異種核酸を含む細胞)を生成する例示的な方法は、当該分野において周知である(概説のために、Friedmann、Science、244:1275-1281(1989);Mulligan、Science、260:926-932(1993)を参照のこと、これらのそれぞれは、それらの全体が本明細書中に参考として援用される)。形質導入の例示的な方法は、例えば、ウイルスベクターを用いる感染(例えば、米国特許第4,405,712号および同第4,650,764号を参照のこと)、リン酸カルシウムトランスフェクション(米国特許第4,399,216号および同第4,634,665号)、デキストラン硫酸トランスフェクション、エレクトロポレーション、リポフェクション(例えば、米国特許第4,394,448号および同第4,619,794号を参照のこと)、サイトフェクション、粒子ビーズボンバードメントなどを含む。異種核酸は、必要に応じて、その染色体外(すなわち、エピソーム)での維持を許容する配列を含み得るか、または異種DNAは、宿主のゲノムへの取込みを引き起こされ得る(宿主における安定な維持を確実にするための代替の手段として)。
【0068】
本発明の実施において使用が意図される宿主生物として、異種タンパク質の組換え産生が行われた生物が挙げられる。このような宿主生物の例として、細菌(例えば、E. coli)、酵母(例えば、Saccharomyces cerevisiae、Candida tropicalis、Hansenulapolymorpha、および P. pastoris;例えば、米国特許第4,882,279号、同第4,837,184号、同第4,929,555号、および同第4,855,231号を参照のこと)、哺乳動物細胞(例えば、HEK293、CHO、およおびLtk−細胞)、昆虫細胞などが挙げられる。現在好ましい宿主生物は、細菌である。最も好ましい細菌は、E.coliである。
【0069】
1つの実施態様において、本発明のATタンパク質をコードする核酸が、哺乳動物細胞に、当該分野で周知の適切なウイルスベクター(例えば、レトロウイルスベクター、アデノウイルスベクターなど)を使用してインビボまたはインビトロのいずれかで送達され得る。さらに、本発明のATのインビボでの発現を制限するかまたは減少させることが所望される場合、本発明の核酸のアンチセンス鎖の導入が意図される。
【0070】
ウイルスに基づくシステムは、種々の細胞への比較的高レベルの異種核酸の導入が可能であるという利点を提供する。ATタンパク質をコードする本発明のAT核酸を哺乳動物細胞(例えば、血管組織区)に導入するための適切なウイルスベクターが、当該分野で周知である。これらのウイルスベクターとして、例えば、単純ヘルペスウイルスベクター(例えば、Gellerら、Science,241:1667-1669(1988))、ワクシニアウイルスベクター(例えば、Picciniら、Meth. in Enzymology, 153:545-563(1987);サイトメガロウイルスベクター(Mocarskiら、ViralVectors, Y. GluzmanおよびS.H. Hughes編、Cold Spring Harbor Laboratory, Cold SpringHarbor, N.Y., 1988, 78-84頁)、モロニーマウス白血病ウイルスベクター(Danosら、PNAS, USA, 85:6469(1980))、アデノウイルスベクター(例えば、Loganら、PNAS,USA, 81:3655-3659(1984);Jonesら、Cell, 17:683-689(1979);Berkner, Biotechniques, 6:616-626(1988);Cottenら、PNAS,USA 89,:6094-6098(1992);Grahamら、Meth. Mol. Biol., 7:109-127(1991))、アデノ随伴ウイルスベクター、レトロウイルスベクター(例えば、米国特許第4,405,712号および同第4,650,764号を参照のこと)などが挙げられる。特に好ましいウイルスベクターは、アデノウイルスベクターおよびレトロウイルスベクターである。
【0071】
例えば、本発明の1つの実施態様において、アデノウイルス−トランスフェリン/ポリリジン−DNA(TfAdpl−DNA)ベクター複合体(Wagnerら、PNAS, USA, 89:6099-6103(1992);Curielら、Hum.Gene Therapy, 3:147-154(1992);Gaoら、Hum. Gene Ther., 4:14-24(1993))が、異種AT核酸を哺乳動物細胞に導入するために使用される。本明細書中で記載される任意のプラスミド発現ベクターが、TfAdpl−DNA複合体中で使用され得る。
【0072】
本明細書中で使用される場合、「レトロウイルスベクター」とは、2つのレトロウイルスLTR間に存在する異種遺伝子をコードする発現カセットを有する、周知の遺伝子導入プラスミドをいう。レトロウイルスベクターは代表的には、レトロウイルスベクターまたはRNAが、適切なパッケージング細胞株中でウイルスビリオンへパッケージングされる、鋳型としてのレトロウイルスベクターを使用して転写されることを可能にする、適切なパッケージングシグナルを含む(例えば、米国特許第4,650,764号を参照のこと)。
【0073】
本明細書中での使用に適切なレトロウイルスベクターは、例えば、米国特許第5,252,479号、およびWIPO公開WO92/07573、WO 90/06997、WO 89/05345、WO 92/05266、およびWO 92/14829(本明細書中で参考として援用される)に記載されている。これらは、このようなレトロウイルスベクターを使用してヒト細胞に核酸を効率的に導入するための方法の記載を提供する。他のレトロウイルスベクターとして、例えば、マウス乳房腫瘍ウイルスベクター(例えば、Shacklefordら、PNAS,USA, 85:9655-9659(1988))などが挙げられる。
【0074】
本発明のなお別の実施態様に従って、本発明のATポリペプチドと特異的反応性を有する抗AT抗体が提供される。抗体の活性なフラグメントは、「抗体」の定義内に含まれる。本発明の抗体は、抗原としてATポリペプチド、タンパク質、またはその部分を使用して、当該分野で公知の方法によって産生され得る。例えば、ポリクローナル抗体およびモノクローナル抗体が、例えば、本明細書中で参考として援用される、HarlowおよびLane、Antibodies;ALaboratory Manual(Cold Spring Harbor Laboratory(1988))に記載されるような、当該分野で周知の方法によって産生され得る。本発明のATポリペプチドは、このような抗体を生成することにおいて、免疫原として使用され得る。あるいは、合成ペプチドが調製され(市販によって入手可能な合成機を使用して)、そして免疫原として使用され得る。アミノ酸配列は、それらが対応するポリペプチドの疎水性ドメインまたは親水性ドメインをコードするかどうかを決定するために、当該分野で周知の方法によって分析され得る。変化した抗体(例えば、キメラ、ヒト化、CDR-移植された、または二重機能性抗体)もまた、当該分野で周知の方法によって産生され得る。このような抗体はまた、例えば、Sambrookら、前出、ならびにHarlowおよびLane、前出、に記載されるような、ハイブリドーマ、化学合成、または組換え法によって産生され得る。抗ペプチドおよび抗融合タンパク質抗体の両方が、使用され得る。(例えば、Bahouthら、TrendsPharmacol. Sci. 12:338(1991);Ausubelら、Current Protocols in Molecular Biology、JohnWileyおよびSons, NY(1989)(本明細書中で参考として援用される)を参照のこと)。
【0075】
このように産生された抗体が、とりわけ、哺乳動物(好ましくは、ヒト、体のサンプル(例えば、組織または血液))中に存在するATタンパク質のレベルを検出するための診断方法およびシステムにおいて使用され得る。このような抗体はまた、本発明のATタンパク質のイムノアフィニティー精製またはアフィニティークロマトグラフィー精製のために使用され得る。さらに、kono方法は、細胞の表面または細胞内(例えば、核内)のいずれかのATポリペプチドの存在を検出するために、本明細書中で意図される。この方法は、ATポリペプチドに対する抗体の結合を可能にする条件下で、ATポリペプチドに特異的に結合する抗体と細胞とを接触させる工程、ATに結合した抗体の存在を検出する工程、およびそれによって細胞の表面または細胞内の本発明のポリペプチドの存在を検出する工程を包含する。このようなポリペプチドの検出に関して、抗体は、インビトロでの診断方法またはインビボでの画像診断方法のために使用され得る。
【0076】
サンプル中の標的ATポリペプチドのインビトロでの検出のために有用な免疫学的手順として、検出可能な抗体を使用するイムノアッセイが挙げられる。このようなイムノアッセイとして、例えば、当該分野で周知の、ELISA、Pandex微量蛍光定量アッセイ、凝集アッセイ、フローサイトメトリー、血清診断アッセイ、および免疫組織化学染色手順が挙げられる。抗体は、当該分野で周知の種々の手段によって検出可能にされ得る。例えば、検出可能なマーカーが、直接的または間接的に抗体に結合され得る。有用なマーカーとして、例えば、放射性ヌクレオチド、酵素、発蛍光団、色素原、および化学発光標識が挙げられる。
【0077】
本発明の抗AT抗体は、生存している動物、ヒト、または生物学的組織もしくはそれらから単離された液体中のATポリペプチドの活性を調節するために、本明細書中での使用が意図される。用語「調節」は、ATタンパク質の生物学的活性(例えば、ATのAc-CoA輸送活性)を増大(例えば、アゴニストを介して)または阻害(例えば、アンタゴニストを介して)する化合物の能力をいう。従って、キャリア、および、天然に存在するリガンドまたは他のAT結合タンパク質が、本発明のATポリペプチドに結合するのをブロックするために有効なATポリペプチドについて特異性を有する量の抗体を含有する組成物が、本明細書中で意図される。例えば、細胞の表面に存在し、そして配列番号2に示されるアミノ酸配列を含むATポリペプチドの細胞表面エピトープのアミノ酸配列と実質的に同じアミノ酸配列を有するATポリペプチド分子のエピトープを指向するモノクローナル抗体が、この目的のために有用であり得る。
【0078】
本発明はさらに、ATポリペプチドをコードする外因性の核酸を発現し得る、トランスジェニック非ヒト哺乳動物を提供する。本明細書中で使用される場合、句「外因性の核酸」は、宿主に対して天然ではないか、またはその天然の環境以外の宿主に存在する(例えば、遺伝子操作されたDNA構築物の一部として)核酸配列をいう。天然に存在するATのレベルに加えて、本発明のATタンパク質は、トランスジェニック哺乳動物中で、過剰発現され得るか、発現不足であり得るか、または不活性な変異形態で発現され得る(例えば、周知のノックアウトトランスジェニック)かのいずれかである。
【0079】
正常な活性を有さない(すなわち、天然のATを発現しない)ように変異されたATポリペプチドをコードする核酸を発現し得るトランスジェニック非ヒト哺乳動物もまた、提供される。本発明はまた、ATポリペプチドをコードするmRNAに相補的なアンチセンスmRNAに転写されるように配置された、ATポリペプチドをコードする核酸に相補的なアンチセンス核酸を含むゲノムを有する、トランスジェニック非ヒト哺乳動物を提供する。このアンチセンス核酸は、mRNAにハイブリダイズし、それによってその翻訳を減少させる。核酸は、誘導性プロモーターおよび/または組織特異的調節エレメントをさらに含み得、その結果、発現が誘導され得るか、または、特異的細胞型に制限され得る。核酸の例は、配列番号1に示されるコード配列と実質的に同じコード配列を有するDNAまたはcDNAである。非ヒトトランスジェニック哺乳動物の例は、トランスジェニックマウスである。
【0080】
ATポリペプチドの生理学的および作用的役割を説明する動物モデル系もまた、提供され、そしてトランスジェニック動物を作製することによって産生される。ここで、ATポリペプチドの発現は、種々の技術を使用して変更される。このような技術の例として、マイクロインジェクション、レトロウイルス感染、または当業者に周知の他の手段による、ATポリペプチドをコードする核酸の正常なまたは変異体バージョンの、トランスジェニック動物を産生するための適切な受精した胚への挿入が挙げられる。(例えば、Hoganら、Manipulatingthe Mouse Embryo:A Laboratory Manual(Cold Spring Harbor Laboratory,(1986)を参照のこと)。
【0081】
ATポリペプチドの発現の調節または構造を変化させるための、トランスジェニック動物中の天然の遺伝子座を有するAT遺伝子の変異体または正常なバージョンの相同組換えの使用もまた、本明細書中で意図される(Capecchiら、Science,244:1288(1989);Zimmerら、Nature, 338:150(1989)を参照のこと;これらは本明細書中で参考として援用される)。相同組換え技術は、当該分野で周知である。相同組換えによって、天然の(内因性の)遺伝子を、組換えまたは変異した遺伝子で置き換えて、天然の(内因性の)タンパク質を発現し得ないが、例えば、ATポリペプチドの変化した発現の結果である変異タンパク質を発現し得る動物を産生する。
【0082】
相同組換えとは対照的に、マイクロインジェクションは、宿主遺伝子を除去することなく宿主ゲノムに遺伝子を付加する。マイクロインジェクションは、内因性および外因性のATタンパク質の両方を発現し得るトランスジェニック動物を産生し得る。誘導性プロモーターは、トランスジーンの発現を調節する手段を提供するために、核酸のコード領域に連結され得る。組織特異的調節エレメントが、トランスジーンの組織特異的発現を可能にするために、コード領域に連結され得る。トランスジェニック動物モデル系は、特異的試薬(すなわち、タンパク質応答を活性化するかまたは阻害する、アゴニストおよびアンタゴニスト)の同定のための化合物のインビボでのスクリーニングに有用である。
【0083】
本発明の核酸、オリゴヌクレオチド(アンチセンスを含む)、それを含むベクター、形質転換された宿主細胞、ポリペプチド、およびそれらの組み合わせ、ならびに本発明の抗体が、化合物が、本発明のATポリペプチドに対する可能性のあるアゴニストまたはアンタゴニストとして作用するかどうかを決定するために、インビトロで化合物をスクリーニングするために使用され得る。これらのインビトロでのスクリーニングアッセイは、本発明のATポリペプチドの機能および活性に関する情報を提供する。これは、ポリペプチド、ペプチド、またはタンパク質の1つ以上の型と特異的に相互作用し得る化合物の同定および設計を導き得る。
【0084】
本発明のなお別の実施態様に従って、ATポリペプチドに結合する化合物(例えば、抗体、結合試薬など)を同定するための方法が提供される。例えば、本発明のATタンパク質は、競合結合アッセイにおいて使用され得る。このようなアッセイは、もしあれば、どの化合物が、ATタンパク質に結合し得るかを決定するために、多数の化合物を迅速なスクリーニングに適用し得る。続いて、より詳細なアッセイが、このような化合物が本発明のタンパク質の調節因子(例えば、アゴニストまたはアンタゴニスト)として作用するかどうかをさらに決定するために、結合することが見出されているこれらの化合物を用いて行われ得る。
【0085】
ATは、欠損している場合、異常な神経細胞の分化および/または移動に応答する遺伝子と考えられている。さらに、本発明の異常なレベル(例えば、より高いまたはより低いレベル)のATタンパク質、あるいは異常に作用するATタンパク質が、乳癌および神経芽細胞種および黒色腫のような外胚葉起源由来の癌を含むヒトの腫瘍に関連すると考えられている。
【0086】
従って、本発明の別の実施態様において、本発明のATポリペプチドの活性を調節する化合物を同定するためのバイオアッセイが提供される。この方法に従って、本発明のATポリペプチドに結合した膜は、試験化合物の存在下および非存在下でAc-CoAと接触される;ATタンパク質の活性は、試験化合物との接触に続いてモニターされ、そしてATタンパク質を有する膜を通過するAc-CoAの輸送を増大させるかまたは減少させるかのいずれかを引き起こすこれらの物質が、ATポリペプチドを調節するための機能的試薬として同定される。
【0087】
本発明の別の実施態様に従って、本発明のATポリペプチドを組換え的に発現する形質転換された宿主細胞(インタクトなまたは半インタクト(semi-intact)な細胞のいずれか)が、試験化合物と接触され、そして次いで、その調節効果が、試験化合物の存在下および非存在下でのAc-CoA(例えば、実施例VIに記載されるような放射性標識したAc-CoA)のATによって媒介される輸送を比較することによって、または試験細胞またはコントロール細胞(すなわち、ATポリペプチドを発現しない細胞)の応答を比較することによって、化合物の存在下に対して評価され得る。
【0088】
本明細書中で使用される場合、本発明のATポリペプチドの「活性を調節する」化合物またはシグナルは、ATポリペプチドの活性を変化させ、その結果、本発明のATポリペプチドの活性は、化合物またはシグナルの非存在下よりも、化合物またはシグナルの存在下で異なる化合物またはシグナルをいう。特に、このような化合物またはシグナルとして、アゴニストまたはアンタゴニストが挙げられる。アゴニストは、ATタンパク質の機能を活性化するかまたは増大させる化合物またはシグナルを含む。あるいは、アンタゴニストは、ATタンパク質の機能を阻害するか、そうでなければ低下して妨害する化合物またはシグナルを含む。代表的には、アンタゴニストの効果は、アゴニストによって誘導されるタンパク質活性のブロックとして観察される。アンタゴニストとして、競合および非競合アンタゴニストが挙げられる。競合アンタゴニスト(または競合ブロッカー)は、Ac-CoA輸送に特異的な部位と、またはその付近と相互作用する。非競合アンタゴニストまたはブロッカーは、本発明のATタンパク質のAc-CoA輸送領域とは異なる部位で相互作用することによってポリペプチドの機能を不活化する。
【0089】
当業者に理解されるように、AT活性を調節する化合物を同定するためのアッセイ方法は、一般に、コントロールとの比較を必要とする。「コントロール」の1つの型は、「コントロール」細胞または培養物が化合物に曝露されないという特徴を有する、化合物に曝された試験細胞または試験培養物と実質的に同じ処理をされた細胞または培養物である。例えば、「コントロール」細胞または培養物の型は、天然のタンパク質を発現しない「コントロール」細胞または培養物を除いて、トランスフェクトされた細胞と同一である細胞または培養物であり得る。従って、化合物に対するトランスフェクトされた細胞の応答が、同じ反応条件下で、同じ化合物に対する「コントロール」細胞または培養物の応答(またはその欠如)と比較される。
【0090】
従って、本発明の別の実施態様に従って、試験化合物が、ATタンパク質のアゴニストまたはアンタゴニストとして作用し得るかどうかを評価するためのバイオアッセイが提供される。ここで、上記のバイオアッセイは、以下を包含する:
(a)以下を含む細胞を培養する工程:
ATタンパク質またはその機能的に改変された形態を発現するDNA、
ここで、上記の培養工程は、ATタンパク質のAc-CoA輸送活性を調節するその能力が決定されていることが求められる少なくとも1つの化合物の存在下で行われ、そしてその後
(b)Ac-CoAのレベルにおける増大または低下のいずれかについて上記の細胞をモニターする工程。
【0091】
細胞内小器官への放射標識されたAc-CoAの輸送のような、Ac-CoAの取り込みを測定する当該分野で周知の方法が、ATタンパク質のアゴニストおよびアンタゴニストを同定するために本明細書中で記載されるバイオアッセイにおいて使用され得る。例えば、実施例VIで記載される方法は、哺乳動物宿主細胞中で発現される、組換えATタンパク質、またはその変異体および/もしくはそのアナログのAc-CoA輸送活性を評価するために使用され得る。
【0092】
本明細書中で使用される場合、「ATタンパク質のAc-CoA輸送活性を調節する能力」は、細胞内膜を通過するATタンパク質のAc-CoA輸送活性を誘導する(アゴニスト)か、または阻害する(アンタゴニスト)かのいずれかの能力を有する化合物をいう。
【0093】
本発明の別の実施態様において、試験化合物が本発明のATタンパク質のアンタゴニストとして作用し得るかどうか、または上記のATタンパク質の機能的に改変された形態であるかどうかを評価するためのバイオアッセイは、以下の工程を包含する:
(a)以下を含む細胞を培養する工程:
ATタンパク質またはその機能的に改変された形態を発現する、DNA,
ここで、上記の培養工程は、以下の存在下で行われる:
漸増濃度の少なくとも1つの、ATタンパク質のAc-CoA輸送能力を阻害するその能力を決定することを求められている化合物、および
固定された濃度のAc-CoA;
そして、その後の
(b)上記化合物の濃度の関数として細胞内小器官へ輸送されたAc-CoAのレベルを上記細胞中でモニターし、それによってAT輸送活性を阻害する上記化合物の能力を示す工程。
【0094】
上記のアンタゴニストバイオアッセイの工程(a)において、培養工程はまた、以下の存在下で行われる:
固定された濃度の少なくとも1つの、ATタンパク質のAc-CoA輸送活性を阻害するその能力を決定することが求められる化合物、
漸増濃度のAc-CoA。
【0095】
本明細書中で使用される場合、句「細胞内小器官」は、例えば、ゴルジ体、小胞体、ミトコンドリアなどをいう。本発明のバイオアッセイでの使用が意図される宿主細胞として、CV-1細胞、COS細胞、HeLa細胞などが挙げられる。本発明のバイオアッセイを行うための現在好ましい宿主細胞は、実施例VIに記載されるようなHeLa細胞である。
【0096】
本発明のなお別の実施態様において、ATタンパク質によって媒介されるAc-CoA輸送活性を調節するための方法もまた意図される。上記の方法は以下の工程を包含する:
上記のバイオアッセイによって同定されたアゴニストまたはアンタゴニストの有効な調節量とATタンパク質とを接触させる工程。
【0097】
異常な神経細胞の分化、異常な神経細胞の移動、神経芽細胞腫、または黒色腫を処置するための方法もまた、本明細書中で意図される。上記の方法は、本明細書中で記載される方法によって同定される有効量の化合物(アゴニストまたはアンタゴニスト)を投与する工程を包含する。このような化合物は、代表的には、生物学的に受容可能な組成物中で投与される。
【0098】
従って、本発明は、本明細書中で記載される治療方法を実施するために有用な治療用組成物を意図する。本発明の治療用組成物は、AT-1調節剤とともに生理学的に適合性のキャリア、または有効成分としてその中に溶解されたかまたは分散された、本明細書中で記載されるような坑AT-1抗体を含む。好ましい実施態様において、治療組成物は、治療目的で哺乳動物またはヒト患者に投与される場合、免疫原性ではない。
【0099】
本明細書中で使用される場合、用語「薬学的に受容可能な」、「生理学的に受容可能な」、およびその文法上のバリエーションは、それらが組成物、キャリア、希釈剤、および試薬をいう場合、互換的に使用され、そして吐き気、めまい、胃の不調などのような所望されない生理学的影響の産生を伴わずに哺乳動物に投与され得ることが示される。
【0100】
その中に溶解されたかまたは分散された有効成分を含有する薬理学的組成物の調製は、当該分野で周知である。代表的には、このような組成物は、液体の溶液または懸濁物のいずれかとして注射可能なものとして調製される;しかし、使用の前に液体中で、溶液または懸濁物に適切な固形の形態もまた、調製され得る。調製物はまた、乳濁物でもあり得る。
【0101】
有効成分は、薬学的に受容可能であり、そして本明細書中で記載される治療方法での使用に適切な量で有効成分と適合性である賦形剤と混合され得る。適切な賦形剤は、例えば、水、生理食塩水、デキストロース、グリセロール、エタノールなど、およびそれらの任意の2つ以上の組み合わせである。さらに、所望される場合は、組成物は、主要ではない量の補助物質(例えば、有効成分の有効性を増強する、湿潤剤、または乳化剤、pH緩衝剤など)を含み得る。
【0102】
本発明の治療用組成物として、本明細書中の成分の薬学的に受容可能な塩を含み得る。薬学的に受容可能な非毒性の塩として、例えば、以下のような無機酸を用いて形成される、酸添加塩(ポリペプチドの遊離のアミノ基を用いて形成される)が挙げられる:塩酸、臭化水素酸、過塩素酸、硝酸、チオシアン酸、硫酸、リン酸、酢酸、プロピオン酸、グリコール酸、乳酸、ピルビン酸、シュウ酸、マロン酸、コハク酸、マレイン酸、フマル酸、アントラニル酸、ケイ皮酸、ナフタレンスルホン酸、スルファニル酸など。
【0103】
遊離のカルボキシル基を用いて形成される塩はまた、例えば以下のような無機塩基から誘導され得る:水酸化ナトリウム、水酸化アンモニウム、水酸化カリウムなど;および例えば以下のような有機塩基から誘導される:モノ-、ジ-、およびトリ-アルキルアミンならびにトリ-アリールアミン(例えば、トリエチルアミン、ジイソプロピルアミン、メチルアミン、ジメチルアミンなど)、および必要に応じてエタノールアミン(例えば、エタノールアミン、ジエタノールアミンなど)で置換される。
【0104】
生理学的に寛容なキャリアは、当該分野で周知である。例示的な液体のキャリアは、有効成分および水に加えて他の物質を含まないか、または緩衝液(例えば、生理学的pHのリン酸ナトリウム、生理学的な生理食塩水、またはその両方(例えば、リン酸緩衝化生理食塩水))を含む、滅菌の水溶液である。なおさらに、水溶性のキャリアは、1つ以上の緩衝塩、および塩(例えば、塩化ナトリウムおよび塩化カリウム、デキストロース、ポリエチレングリコール、および他の溶質)を含み得る。
【0105】
液体の組成物はまた、水に加えて液体層を含み得、そして水を排除するために液体層を含み得る。例示的なさらなる液体層として、グリセリン、植物油(例えば、綿実油)、および水-油エマルジョンが挙げられる。
【0106】
本明細書中で記載されるように、「有効量」は、例えば、本発明のATタンパク質のAc-CoA輸送活性を調節するための、所望される治療効果を達成するように計算された予め決定された量である。必要とされる投与量は、特定の処置とともに、および所望される処置の期間とともに変化する;しかし、約10μg〜約1mg/kg体重/日の間の投与量が治療的処置に使用されることが予測される。本明細書中以下で議論されるような、デポー製剤または長期持続形態中でこのような化合物を投与することが、特に有用であり得る。治療有効量は、生理学的に受容可能な組成物中で投与される場合、代表的には、AT調節剤または本明細書中で同定される化合物の量である。そして約0.1μg/mlから約100μg/ml、好ましくは、約1.0μg/mlから約50μg/ml、より好ましくは、少なくとも約2μg/ml、および通常は5から10μg/mlの血漿濃度を達成するために十分である。治療用の本発明の坑AT抗体が、当業者の知識に従って、比例する適切な量で投与され得る。
【0107】
本明細書中で言及される全ての米国特許および全ての刊行物が、それらの全体を本明細書中で参考として援用するように引用される。本発明はここで、以下の限定的ではない実施例を参照して、さらに詳細に記載される。
【実施例】
【0108】
実施例
他に特に記載されない限りは、本発明は、例えば以下に記載されるような標準的な手順を使用して行った:Maniatisら、Molecular Cloning:A Laboratory Manual, Cold Spring HarborLaboratory Press、Cold Spring Harbor, New York, USA(1982);Sambrookら、MolecularCloning:A Laboratory Manual(第2版)、Cold Spring Harbor Laboratory Press, ColdSpring Harbor, New York, USA(1989);Davisら、 Basic Method in Molecular Biology,Elsevier Science Publishing, Inc., New York, USA(1986);またはMethods in Enzymology:Guideto Molecular Cloning Techniques 第152巻、S.L. BergerおよびA.R. Kimmerl編、Academic PressInc., San Diego, USA(1987)。
【0109】
実施例I
本明細書中で使用される抗体についての記載
C系列ポリシアロガングリオシドのNeuNAcα2-8NeuNAcα2-8NeuNAcα2-3Galβ1-R構造を有するGT3に特異的なモノクローナル抗体(mAb)M6703を、Hirabayashiら、1988,J. Biochem., 104:973-979(本明細書中で参考として援用される)に記載されるように調整した。GD3との反応が示されたmAb R24(Pukelら、1982,J. Exp. Med., 155:1133-1147、本明細書中で参考として援用される)を、アメリカンタイプカルチャーコレクションから入手した(ATCC#HB-8445)。9-O-アセチル化ジシアロガングリオシドを認識するmAb D1.1を、Levineら、1984, J. Neuroscience, 4:820-831(本明細書中で参考として援用される)に記載されるように確立し、そして9-O-アセチル化GT3と反応するmAB493D4は、S. Fujita博士(Mitsubishi Kasei Institute of Life Sciences)より提供された。AT-1タンパク質の合成のN末端の14アミノ酸ペプチド(配列番号2のアミノ酸1〜14)に対するポリクローナル抗体を、周知の方法を使用して調製した。
【0110】
安定にトランスフェクトされたレシピエント細胞の構築
この実施例は、ガングリオシドを産生するが、それらの9-O-Ac誘導体は産生しないトランスフェクトされた細胞株を得るための方法を提供する。この手順の目的は、ガングリオシドGD3およびGT3を発現するが、それらの9-O-アセチル化誘導体は発現しない細胞株を産生することである。9-O-アセチル化ガングリオシドの発現のために必要とされる新規の因子をクローニングするために、前駆体ガングリオシドを発現するが、対応する9-O-アセチル化ガングリオシドを発現しないレシピエント細胞を使用することが必要であった。研究した特異的ガングリオシドは、NeuAcα2-8NeuAcα2-3Galβ1-4Glcβ1-1’Cer(本明細書中以後、「GD3」)、9-O-Ac-GD3(末端のノイラミン酸の9位でアセチル化されている)、NeuAcα2-8NeuAcα2-8NeuAcα2-3Galβ1-4Glcβ1-1’Cer(本明細書中以後、「GT3」)、および9-O-Ac-GT3(末端のノイラミン酸の9位でアセチル化されている)であった。
【0111】
細胞株COS-1/GD3+(ガングリオシドGD3を安定に発現するが、9-O-Ac-GD3は発現しない)を、Nakayamaら(本明細書中で参考として援用される、J.Biol. Chem., 271:3684-3691(1996))によって以前に記載されているように調製した。そして、HeLa/GT3+(ガングリオシドGD3およびGT3を安定に発現するが、9-O-Ac-GD3または9-O-Ac-GT3は発現しない)を、以前に記載されているように(Nakayamaら、前出、1996)調製した。
【0112】
実施例II
AT-1をコードするcDNAの単離
哺乳動物発現ベクターに基づくcDNAライブラリーpcDNAI-SK-MEL-28(9-O-AD-GD3+(9-O-Ac-GD3+)を発現するヒト黒色腫SK-MEL-28細胞から単離したポリ(A)+RNAを使用して構築した)を、Invitrogen(San Diego, CA)より購入した(本明細書中で参考として援用されている、1996年1月16日に発行された米国特許第5,484,590号もまた参照のこと;本明細書中で参考として援用されている、BierhuzenおよびFukuda、Proc.Natl. Acad. Sci., U.S.A., 89:9326-9330(1992)を参照のこと)。
【0113】
約1.2×107個のCOS-1/GD3+細胞を、リポフェクトアミン(LifeTechnologies, Inc.)を使用して20μgのpcDNAI-SK-MEL-28でトランスフェクトした。60時間後、9-O-Ac-ガングリオシドを産生するトランスフェクトした細胞を、9-O-Ac-GD3に特異的に結合するモノクローナル抗体D1.1(本明細書中で参考として援用される、Levineら、J.Neuros., 4:820-831(1984))を使用して間接的な免疫蛍光によって染色した。蛍光標識した細胞を、FACStarセルソーター(BectonDickinson)を使用して蛍光活性化された細胞の分別を使用して単離した。
【0114】
9-O-Ac-GD3についてポジティブに染色されたCOS-1/GD3+細胞を回収し、そしてプラスミドDNAを、Hirt手順(本明細書中で参考として援用される、Hirt,J. Mol. Biol., 26:365-396(1967))を使用して単離した。プラスミドDNAを、アンピシリンおよびテトラサイクリンの存在下で、宿主細菌MC1061/P3に形質転換し、そして例えば、Sambrookら(MolecularCloning:A Laboratory Manual, Cold Spring Harbor, N.Y.(1989))に記載される周知の方法を使用して、細菌宿主から単離した。
【0115】
単離しプールしたプラスミドを、姉妹選択(sibling selection)(米国特許第5,484,590号を参照のこと)に供した。簡潔には、単離したプラスミドを、活性なプールが連続的に小さくなる、COS-1/GD3+細胞を使用する姉妹選択に供した。トランスフェクトした細胞を、モノクローナル抗体D1.1を使用して間接的な免疫蛍光顕微鏡法によってスクリーニングした。ポジティブに染色された細胞を回収し、そしてプラスミドDNAを抽出した。続く回の姉妹選択後、9-O-Ac-GD3の発現に必要とされる因子をコードする単一のプラスミドpcDNAI-AT-1を、単離した。この因子を、AT-1(配列番号1)と命名した。
【0116】
選択したクローニングした細胞に由来するプラスミドを単離し、そして挿入物のヌクレオチド配列を両方向で決定した。ヌクレオチド配列を、配列番号1(GenBank登録番号D88152)に示す。
【0117】
AT-1をコードするcDNAは、60,906Daの計算分子量を有する549アミノ酸のポリペプチド(配列番号2)をコードする1650塩基対を有するオープンリーディングフレームを含む。タンパク質および核酸のデータベース(GenBank,EMBLおよびSwiss-Prot)に対する相同性検索は、任意の既知のタンパク質とは有意な相同性を有さないことを示した。このことは、AT-1によってコードされるタンパク質が新規であることを示す。AT-1ポリペプチド配列のヒドロパシー分析(KyteおよびDoolittle(J.Mol. Biol., 157:105-132(1982))は、このポリペプチドがいくつかの可能性のある膜貫通ドメインを含むことを示した。(図1を参照のこと)。同様の分析を、プログラムPsort(Nakaiら、Genomics,14:897-911(1992)およびTmpred(HofmannおよびStoffel, Biol. Chem. Hoppe-Seyler, 347:166(1993))を使用して行った。これらの2つの分析は、AT-1ポリペプチドが、6から10個の膜貫通ドメインを有するIIIa型膜タンパク質構造を有することを示した。AT-1ポリペプチドは、アミノ酸残基144から165で膜貫通ドメイン中にロイシンジッパーモチーフを含む。ロイシンジッパーモチーフがしばしば、他のトランスポータータンパク質中に見出されることが注目されるはずである(例えば、Eckhardtら、1996、PNAS,USA, 93:7572-7576;Miuraら、1996、J. Biochem(Tokyo)、120:236-241;Abeijonら、1996,PNAS, USA, 93:5963-5968を参照のこと)。
【0118】
実施例III
ガングリオシドのO-アセチル化に対するAT-1の影響を特徴づける発現アッセイ
COS-1/GD3+細胞
COS-1/GD3+細胞を、pcDNAI(擬似トランスフェクション)またはpcDNAI-AT-1で一時的にトランスフェクトした。トランスフェクションの60時間後、トランスフェクトした細胞を固定し、そしてmAbR24(抗GD3)またはmAb D1.1(抗-9-O-アセチル化ジシアロガングリオシド)とともにインキュベーションすることによって、別々に免疫染色し、続いて、mAbR24についてはローダミン結合抗マウスIgG、またはmAb D1.1についてはフルオレセインイソチオシアネート結合抗マウスIgMとともにインキュベーションした。擬似トランスフェクタントとAT-1トランスフェクタントとの間のR24反応性において有意な差異は存在しなかったが、9-O-アセチル化GD3の発現は、AT-1トランスフェクタント中でmAbD1.1で強く検出された。弱い穿刺染色しか、擬似トランスフェクタントにおいて検出されなかった。これらの結果は、本発明のAT-1タンパク質がGD3の9-O-アセチル化を媒介する(影響を与える)ことを示す。
【0119】
HeLa/GT3+細胞
HeLa/GT3+細胞を、pcDNAI(コントロール)またはpcDNAI-AT-1のいずれかでトランスフェクトし、そして続いて、9-O-Ac-GT3に結合するmAb493D4で免疫染色した。HeLa/GT3+/AT-1でトランスフェクトされた細胞は、9-O-Ac-GT3についてポジティブに染色されたが、HeLa/GT3+/pcDNAIコントロール細胞は、検出可能な染色を示さなかった。HeLa/GT3+/AT-1細胞の染色を、アルカリ処理(0.1NのNaOHで15分間、室温)によって消した。これらの結果は、本発明のAT-1タンパク質がGT3の9-O-アセチル化を媒介する(影響を与える)ことを示す。
【0120】
安定にトランスフェクトされたHeLa/GT3+/AT-1細胞の確立
HeLa/GT3+細胞を、pcDNA-I-AT-1およびpSV2HyB(10:1)で同時トランスフェクトし(後者の発現は、ハイグロマイシンBに対する耐性を付与する)、次いで、ハイグロマイシンB耐性について選択した。トランスフェクトした細胞を、上記に議論したように、mAb493D4での免疫蛍光染色によって9-O-Ac-GT3の存在についてスクリーニングした。1つの安定にトランスフェクトしたクローンを単離し、そしてHeLa/GT3+/AT-1と命名した。
【0121】
全細胞内ガングリオシドの定量
AT-1でトランスフェクトした細胞がO-アセチル化ガングリオシドを合成することを確認するために、全ガングリオシドをHeLa/GT3+細胞から、ならびにそれらの一時的なおよび安定なAT-1トランスフェクタントから単離した。HeLa/GT3+細胞が単純なガングリオシド、GM3、GD3、GT3、および少量の推定のGQ3のみを含むことを見出し、これによって、これらを、薄層クロマトグラフィーを使用してAT-1でのトランスフェクション後のガングリオシド組成を試験するために理想的であるとした。
【0122】
分析的薄層クロマトグラフィーを、予めコーティングした薄層クロマトグラフィープレート(Kieselgel60, Merck)で行った。使用した溶媒系は、クロロホルム、メタノール、12mMのMgCl2を含有する水(5:4:1、容量比)であった。ガングリオシドをレゾルシノール/HCl試薬によって可視化した。5×105個のトランスフェクトした細胞からのガングリオシドの単離およびTLC-免疫染色を、Hirabayashiら、J.Biol. Chem. 104:973-979(1988)に記載されるように行った。細胞から精製されたガングリオシドを、プラスチックプレート(PolygramSil G, Nagel, Doren, Germany)上に載せ、そして上記と同じ条件下で展開させた。プレートを、mAb、続いてペルオキシダーゼ結合ヤギ抗マウスIgG抗体での免疫染色に供した。ペルオキシダーゼ活性を、4-クロロ-1-ナフトール/H2O2で可視化した。ガングリオシドを、アルカリ処理による脱アセチル化後に同時に分析した。
【0123】
結果は、O-Ac-GT3および推定のO-Ac-GQ3の発現が、AT-1で安定にトランスフェクトされた細胞中で強くポジティブであったことを示す。再度、ガングリオシドをアルカリ加水分解に供した場合には、ポジティブなスポットは観察されなかった。HeLa/GT3+およびHeLa/GT3+/AT-1細胞が類似の総ガングリオシド量を含むことに注目することが重要である。また、中性のガングリオシドおよび非アセチル化ガングリオシドの組成において目に見える差異は、薄層クロマトグラフィー分析を使用して検出されなかった。
【0124】
実施例IV
AT-1 mRNAのノーザンブロット分析
種々のヒト組織中のAT-1 mRNAの発現を、ノーザンブロット分析によって試験した。安定にトランスフェクトされたHeLa細胞から調製したポリ(A)+RNAのサンプルを、2.2Mのホルムアルデヒドを含有するアガロースゲル中で電気泳動し、そしてナイロンフィルター(HybondTM-N、Amersham)に移した。ヒトグリセルアルデヒド-3-リン酸デヒドロゲナーゼ(G3PDH)cDNAとのハイブリダイゼーションを、コントロール実験として行った。これらのブロットを、マルチプライムDNA標識システム(Amersham)による[α-32P]dCTPでの標識後に、AT-1のゲル精製したcDNA挿入物とハイブリダイズさせた。
【0125】
3.3および4.3kbの大きさを有するAT-1の2つの主要な転写物を、心臓、脳、胎盤、肺、肝臓、骨格筋、腎臓、およびすい臓を含む、試験した全ての組織中で検出した。興味深いことに、すい臓組織は、試験した組織の中でも最も強いシグナルを発現した。すい臓において、ランゲルハンス島は、mAb493D4およびmAb D1.1を使用して、O-アセチル化されたシアロ糖結合体を発現することが、免疫組織化学的染色によって示された。2.7および3.4kbの大きさを有するAT-1の2つの転写物は、AT-1でトランスフェクトされたHeLa細胞中で強く発現され、そして擬似トランスフェクトされたHeLa細胞中でかろうじて検出された。
【0126】
実施例V
AT-1タンパク質の免疫組織化学的局在
HeLa/GT3+/AT-1細胞中でのAT-1タンパク質の細胞内局在を研究するために、本発明者らは、合成ペプチド(AT-1のN末端に対応する配列番号2のアミノ酸残基1〜14)に対する特異的抗体を調製した。安定にトランスフェクトされた細胞HeLa/GT3+/AT-1およびHeLa/GT3+/pcDNAIを固定し、サポニンで透過性にし、そしてAT-1タンパク質に特異的な親和性精製したウサギ抗AT-1抗体とともにインキュベートした。続いて、フルオレセインイソチオシアネート結合抗ウサギIgGとともにインキュベートした。
【0127】
抗AT-1抗体を使用するHeLa/GT3+/AT-1細胞のウェスタンブロット分析によって、54kdの主要なポジティブバンドおよび58kdの弱い反応性スポットを明らかにした。親和性精製した抗AT-1ペプチドを使用する免疫蛍光分析は、このAT-1タンパク質が細胞質(小胞体、ゴルジ体、およびミトコンドリアを含む)に局在化されることを示した。この特異的染色は、予備免疫IgGを用いて、またはペプチド抗原の存在下では観察されなかった。
【0128】
実施例VI
半インタクトな細胞へのAc-CoAの取りこみ
培養された細胞からの小胞体またはゴルジ膜画分を調製することが困難であったので、細胞中のAc-CoAトランスポーター活性を、Bhakdiら(本明細書中で参考として援用されている、Infect. Immun., 47:52-60(1985));KuncanおよびSchlegel(本明細書中で参考として援用されている、J.Cell Biol., 67:160-173(1993));ならびに、(本明細書中で参考として援用されている、Kainら(J. Biol. Chem.,268:19640-19649(1993))の方法を使用して、半インタクトな、透過性にしたHeLa細胞を使用して試験した。ストレプトリジンO(本明細書中以後、「SLO」、GIBCOBRL)での処理によって、細胞内膜にいかなる損傷を与えることもなく、細胞の原形質膜に孔を生成する。これによって、原形質膜を通じてのAc-CoAの様な大きな分子の通過を可能にし、次いで、それらの細胞内の結果は以下のとおりであり得る。
【0129】
培養した細胞をトリプシン処理によって回収し、そしてPBSおよびカルシウム非含有マグネシウム非含有Hepes(HCMF:0.14MのNaCl−5mMのKCl−6mMのGlc−0.3mMのNa2HPO4−10mMのHepes、pH7.4)で連続して洗浄した。HCMF中で希釈した活性化したストレプトリジンO(SLO, GIBCO BRL)での28℃にて20分の処理後、細胞を洗浄し、そしてトランスポーター緩衝液(TB:25mMのHepes−KOH−75mMのKOAc−2.5mMのMgOAc−5mMのEGTA−1.8mMのCaCl2、pH7.2)中の[Ac-14C]Ac-CoAとともに、種々の時間で室温にてインキュベートした。反応を、氷冷TBの添加によってクエンチした。上清を遠心分離によって除去した後、ペレットをTBで3回洗浄し、1%のSDSで可溶化し、そして放射能を計数した。
【0130】
図2に示すように、半インタクトな細胞を[Ac-14C]Ac-CoA(35μM)とともにインキュベートした場合、放射能の取りこみの増大が、AT-1でトランスフェクトした細胞中で明らかに実証された。この効果は、擬似トランスフェクタントでは見られなかった。トランスポーターCoA(350μM)のインヒビターの添加の際に、放射能の取りこみは、コントロールの60%にまで減少した。これらの結果は、AT-1がAc-CoAトランスポーターをコードすることを強力に示唆する。
【0131】
本発明は、上記に提供される実施例を参照して記載されているが、種々の改変が、本発明の精神を逸脱することなく行われ得ることが理解されるはずである。従って、本発明は、請求の範囲によってのみ限定される。
(配列表)
【0132】
【表1】
【0133】
【表2】
【0134】
【表3】
【0135】
【表4】
【0136】
【表5】
【0137】
【表6】
【0138】
【表7】
【図面の簡単な説明】
【0139】
【図1】図1は、10のウインドウサイズを用いて、KyteおよびDoolitteの方法を使用して分析された、配列番号2(AT-1をコードする)の疎水性親水性プロットを示す。影付四角および黒く塗り四角は、それぞれ、TmpredおよびPsortプログラムによって予測される、膜貫通ドメインを示す。
【図2】図2Aは、実施例VIに記載されるような、[Ac-14C]Ac-CoAからの半インタクトな細胞への放射能の時間依存性の取込みを示す。白丸は、HeLa/GT3+/pcDNAIを、および黒丸は、HeLa/GT3+/AT-1を、示す。図2Bは、[Ac-14C]Ac-CoAからの半インタクトな細胞への放射能の時間依存性の取込みに対する、CoAの効果を示す。
【特許請求の範囲】
【請求項1】
明細書に記載の発明。
【請求項1】
明細書に記載の発明。
【図1】

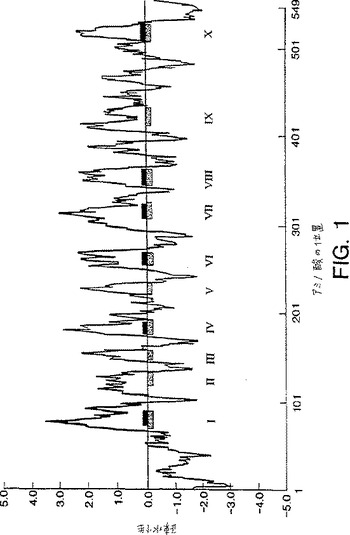
【図2】
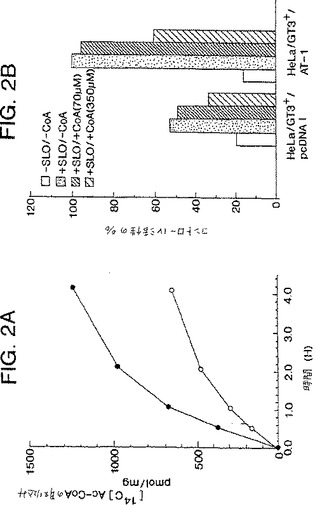
【図2】
【公開番号】特開2008−253257(P2008−253257A)
【公開日】平成20年10月23日(2008.10.23)
【国際特許分類】
【出願番号】特願2008−64996(P2008−64996)
【出願日】平成20年3月13日(2008.3.13)
【分割の表示】特願平10−532832の分割
【原出願日】平成9年1月31日(1997.1.31)
【出願人】(591180152)ザ バーナム インスティテュート (8)
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【Fターム(参考)】
【公開日】平成20年10月23日(2008.10.23)
【国際特許分類】
【出願日】平成20年3月13日(2008.3.13)
【分割の表示】特願平10−532832の分割
【原出願日】平成9年1月31日(1997.1.31)
【出願人】(591180152)ザ バーナム インスティテュート (8)
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【Fターム(参考)】
[ Back to top ]