
アセチル化アミノ糖及びアセチル化アミノ糖ヌクレオチド合成活性を有する耐熱性酵素
【課題】耐熱性で、かつアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する酵素を提供し、糖鎖合成の基質となるアセチル化アミノ糖ヌクレオチドを安定的に合成する。
【解決手段】
超好熱古細菌 Sulfolobus tokodaii からアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する酵素の候補遺伝子を見出し、該遺伝子を用いて遺伝子工学的手段により、耐熱性で、かつアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する酵素を製造するとともに、これを用いてアセチル化アミノ糖及び該アセチル化アミノ糖ヌクレオチドを安定的に合成する。
【解決手段】
超好熱古細菌 Sulfolobus tokodaii からアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する酵素の候補遺伝子を見出し、該遺伝子を用いて遺伝子工学的手段により、耐熱性で、かつアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する酵素を製造するとともに、これを用いてアセチル化アミノ糖及び該アセチル化アミノ糖ヌクレオチドを安定的に合成する。
【発明の詳細な説明】
【技術分野】
【0001】
本願発明は新規アミノ糖アセチル化活性及び該アセチル化アミノ糖の糖ヌクレオチド合成活性を有する耐熱性酵素、及び該酵素を用いて効率的にアセチル化アミノ糖及びその活性化体であるアセチル化アミノ糖ヌクレオチドを製造する方法に関する。
【背景技術】
【0002】
グルコサミン-1-リン酸(glucosamine-1-phosphate)にアセチル基を転移してN-アセチルグルコサミン-1-リン酸(N-acetylglucosamine-1-phosphate)を合成する活性は、UDP-N-アセチルグルコサミン(UDP-N-acetylglucosamine, UDP-GlcNAc)合成活性を有する酵素が共有する活性として、見出されている。このようなバイファンクショナル酵素としては、大腸菌(Escherichia coli)(非特許文献1、2参照)及びストレプトコッカス菌(Streptococcus pneumoniae) (非特許文献3参照)由来のGlmU (Glucosamine-1-phosphate uridyltransferase/Glucosamine-1-phosphate acetyltransferase)の詳しい性質や構造がすでに報告されている。GlmUは細胞表面の膜構造合成に必須なN-アセチルグルコサミン(N-acetylglucosamine)の活性体であるUDP-N-アセチルグルコサミンをフルクトース-6-リン酸(fructose-6-phosphate)から合成する代謝経路の最後の2段階の反応を触媒する二つの活性を併せ持つ酵素として見出されてきた。この酵素は、初めにフルクトース-6-リン酸から合成されてきたグルコサミン-1-リン酸にアセチルCoA(Acetyl-CoA)のアセチル基を転移することによってN-アセチルグルコサミン-1-リン酸を合成し、次にこの合成された糖とヌクレオシド三リン酸(UTP)を基質として、UDP-GlcNAcを生産する。その他にも、豚肝臓から同様の糖ヌクレオチドを合成する酵素が見出されているが、その多くが常温生物由来のため室温以上では極めて不安定で、活性は80℃程度の加熱処理により速やかに失活する。このため、使用時の滅菌等の処理が必要であったり、低温での注意深い保存が必要であった。
また、同様にN-アセチルガラクトサミン(N-Acetylgalactosamine)も膜構造の構築に於いて重要な糖であるが、この糖の合成経路に関しては、不明な点が多く、ガラクトサミン-1-リン酸(Galactosamine-1-phosphate)をアセチル化する酵素活性はこれまでに見出されていない。
【非特許文献1】Mengin-Lecreulx D and van Heijenoort J. " Copurification ofglucosamine-1-phosphate acetyltransferase and N-acetylglucosamine-1-phosphateuridyltransferase activities of Escherichia coli: characterization ofthe glmU gene product as a bifunctional enzyme catalyzing two subsequent stepsin the pathway for UDP-N-acetylglucosamine synthesis.“ (1994) J. Bacteriology,176, 5788-5795.
【非特許文献2】Brown K, Pompeo F, Dixon S, Mengin-Lecreulx D, Cambillau C and BourneY “Crystal structure of the bifunctional N-acetylglucosamine 1-phosphateuridyltransferase from Escherichia coli: a paradigm for the relatedpyrophosphorylase superfamily." (1999) EMBO J., 18, 4096-4107.
【非特許文献3】Sulzenbacher G, Gal L, Peneff C, Fassy F and Bourne Y. " Crystalstructure of Streptococcus pneumoniae N-acetylglucosamine-1-phosphateuridyltransferase bound to acetyl-coenzyme A reveals a novel active sitearchitecture." (2001) J. Biol. Chem., 276, 11844-11851.
【発明の開示】
【発明が解決しようとする課題】
【0003】
グルコサミン-1-リン酸及びガラクトサミン-1-リン酸をアセチル化する活性並びにそのアセチル化アミノ糖の活性化体であるアセチル化アミノ糖ヌクレオチドを合成する活性を有する耐熱性酵素が発見されれば、糖鎖合成の基質となる重要なアセチル化アミノ糖であるN-アセチルグルコサミン及びN-アセチルガラクトサミンについてのアセチル化アミノ糖ヌクレオチド体、すなわちヌクレオシド二リン酸結合体を安定に合成することが可能となる。これまで、糖鎖合成の際の基質として必要なこれらのアセチル化アミノ糖及びヌクレオシド三リン酸を基質として、結合反応を触媒出来る安定な酵素は存在しなかったので、渇望されていた。
【0004】
したがって、本発明の課題は、耐熱性を有し、かつグルコサミン-1-リン酸及びガラクトサミン-1-リン酸を基質として、それぞれのアセチル化アミノ糖-1-リン酸を合成し得る活性、並びにそれらのアセチル化アミノ糖-1-リン酸とヌクレオシド三リン酸を基質として、アセチル化アミノ糖ヌクレオチド(アセチル化アミノ糖−ヌクレオシド二リン酸結合体)を合成することが可能な新規酵素、さらに本酵素を用いたアセチル化アミノ糖及び該アセチル化アミノ糖ヌクレオチド合成法を提供することにある。
【課題を解決するための手段】
【0005】
本発明者は、以上のような課題を解決すべく、75 - 80℃で生育する超好熱古細菌 Sulfolobus tokodaii strain7に着目して、その遺伝子を大腸菌を使って発現させることによって酵素蛋白質を生産し、この酵素蛋白質が高温(80℃)で安定に存在するとともに目的とする活性を示すことを確認し、さらに、本酵素蛋白質を用いることにより目的とするアセチル化アミノ糖ヌクレオチド(アミノ糖−ヌクレオシド二リン酸結合体)を生産することもできることを見いだし、本発明を完成するに至ったものである。
【0006】
即ち、本発明は、以下の(1)〜(10)に係るものである。
(1)配列番号4に記載のアミノ酸配列を有するか、あるいは、配列番号4に記載のアミノ酸配列に一乃至数個のアミノ酸残基が欠失、置換、挿入又は付加されたアミノ酸配列を有し、かつアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有することを特徴とする、蛋白質。
(2)上記(1)に記載の蛋白質をコードするDNA。
(3)配列番号5に記載の塩基配列を有することを特徴とするDNA。
(4)配列番号5に記載のDNA とストリンジェントな条件下でハイブリダイズし、かつアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する蛋白質をコードすることを特徴とする、DNA。
(5)上記(2)〜(4)のいずれかに記載のDNAから選ばれるDNAがベクターに組み込まれていることを特徴とする組換え体DNA。
(6)上記(5)に記載の組換え体DNAが宿主細胞に導入されていることを特徴とする形質転換体。
(7)上記(6)に記載の形質転換体を培地に培養し、培養物からアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する蛋白質を採取することを特徴とする、アセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する蛋白質の製造方法。
(8)グルコサミン-1-リン酸及び/又はガラクトサミン-1-リン酸に、アセチルCoA存在下、上記(1)に記載の蛋白質を作用させることを特徴とする、N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸の製造方法。
(9)グルコサミン-1-リン酸及び/又はガラクトサミン-1-リン酸に、アセチルCoA存在下、上記(6)に記載の形質転換体の培養液あるいは培養物の処理物を作用させることを特徴とする、N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸の製造方法。
(10)グルコサミン-1-リン酸及び/又はガラクトサミン-1-リン酸に、アセチルCoA存在下、 上記(3)又は(4)に記載のDNAにコードされるタンパク質を作用させることを特徴とする、N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸の製造方法。
(11)N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸に、ヌクレオシド三リン酸の存在下、上記(1)に記載のタンパク質を作用させることを特徴とする、
N-アセチルグルコサミンおよび/又はN-アセチルガラクトサミンのヌクレオシド二リン酸結合体の製造方法。
(12)N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸に、ヌクレオシド三リン酸の存在下、上記(6)に記載の形質転換体の培養液あるいは培養物の処理物を作用させることを特徴とする、N-アセチルグルコサミンおよび/又はN-アセチルガラクトサミンのヌクレオシド二リン酸結合体の製造方法。
(13)N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸に、ヌクレオシド三リン酸の存在下、上記(2)〜(4)のいずれかに記載のDNAにコードされるタンパク質を作用させることを特徴とする、N-アセチルグルコサミンおよび/又はN-アセチルガラクトサミンのヌクレオシド二リン酸結合体の製造方法。
【発明の効果】
【0007】
本発明の酵素を使用すれば、N-アセチルグルコサミン-1-リン酸とN-アセチルガラクトサミン-1-リン酸の2種のアミノ糖アセチル化体が合成可能になると共に、さらに、UDP-GlcNAc、TDP-GlcNAc、及びUDP-GalNAcのようなアセチル化アミノ糖ヌクレオチドを合成することができる。このように本発明の酵素は、単一酵素でありながら広い合成活性を有し、かつ熱等に安定である。このような酵素特性は、既知の酵素にみられないものであり、新規なアセチル化アミノ糖ヌクレオチドの効率的な合成を可能とするものである。
一方、アセチル化アミノ糖ヌクレオチドは、糖タンパク質、糖脂質、多糖類の糖鎖合成にアセチル化アミノ糖供与体として機能するものであり、これらの糖鎖は、癌転移、器官発生あるいは細胞性免疫等に密接に関連するものとして近年注目されており、本発明は、これら糖鎖の人工的合成発展において、その貢献度は極めて大きい。
【発明を実施するための最良の形態】
【0008】
以下に、本願発明を具体的に説明する。
本発明の酵素は、好酸性好気性超好熱古細菌スルフォロバス、トーコーダイイ(Sulfolobus tokodaii)(JCM登録番号JCM10545)由来の酵素であり、該超好熱古細菌から本酵素活性を示すと推定した遺伝子領域を、PCR反応で増幅・抽出し、蛋白質発現プラスミドpET21bに挿入後、そのプラスミドにより形質転換した大腸菌を用いて得られたものである。生産された酵素は加熱処理およびカラムクロマトグラムで単離精製し、精製された酵素は、分子量が約44,000のタンパク質で、アセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を共に有する酵素である。本発明の酵素は、特に、グルコサミン-1-リン酸からN-アセチルグルコサミン-1-リン酸を合成する活性及びN-アセチルグルコサミン-1-リン酸からN-アセチルグルコサミンのヌクレオシド二リン酸結合体を合成する活性のほかに、ガラクトサミン-1-リン酸からN-アセチルガラクトサミン-1-リン酸を合成する活性及びN-アセチルガラクトサミン-1-リン酸からN-アセチルガラクトサミンのヌクレオシド二リン酸結合体を合成する活性を有する点に新規な特徴を有する。なお、これらヌクレオシド二リン酸結合体は、N-アセチルグルコサミン、N-アセチルガラクトサミンのそれぞれ1位にヌクレオシド二リン酸のリン酸基が結合しているものである。
【0009】
この酵素の半減期は、50mMトリス塩酸緩衝液(pH7.5)中で、80℃、40分以上であり、高い耐熱性を示した。
この酵素のアミノ酸配列およびその遺伝子DNA(ST0452)の塩基配列を、それぞれ配列表の配列番号4及び5に示す。
【0010】
本発明における酵素は、上記配列番号4に示されるアミノ酸配列を有するもののみに限定されず、該アミノ酸配列において、数個程度のアミノ酸残基が欠失、置換、挿入又は付加されたアミノ酸配列であっても、このアミノ酸配列を有する蛋白質が、上記酵素活性上の特徴に係るアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する限り、本発明に含まれる。また、本発明のこれら酵素遺伝子DNAについても、上記同配列番号5に示す塩基配列を有するもののみに限定されず、上記アミノ酸配列をコードするものを包含する。さらに上記配列番号5に示されるDNAにストリンジェントな条件下でハイブリダイズし、かつ目的とする上記酵素活性上の特徴に係るアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する蛋白質をコードするDNAも包含する。このストリンジェントな条件とは、ハイブリダイゼーション溶液1リットル中に52.59 g NaCl、26.46 g クエン酸ナトリウム、1 g フィコール(Type 400)、1 g ポリビニルピロリドン、1 g ウシ血清アルブミン、5 g SDS、1 g 断片化鮭精子DNA、500 ml ホルムアミドを含み、温度42℃で行う。その後の洗浄は、洗浄用溶液1リットル中に17.53 g NaCl、8.82 g クエン酸ナトリウム、5 g SDSを含み、温度68℃で行う条件である。
【0011】
本発明の酵素を得るには、通常の遺伝子工学的手法が適用でき、上記酵素遺伝子DNAを、例えば、pET21b、pHY481等の蛋白質発現プラスミドベクター等に挿入して組換えベクターを作製し、該組換えベクターを用いて宿主細胞を形質転換し、該形質転換体を培地で培養し、培養物、培養処理物あるいはこれら培養物から分離回収された形質転換体から、酵素を常法の蛋白質精製手段により精製し単離する。上記宿主細胞としては、大腸菌・枯草菌等が利用可能である。
【0012】
本発明においては、さらにこの酵素を用いて、アミノ糖-1-リン酸のアセチル化体を合成するが、この合成においては、アミノ糖-1-リン酸とアセチルCoAを含有する溶液に、該酵素を添加し、反応温度60℃〜95℃で反応させ、アセチル化アミノ糖-1-リン酸を得る。
アミノ糖-1-リン酸としては、例えば、グルコサミン-1-リン酸、ガラクトサミン-1-リン酸が挙げられる。
本発明においては、この酵素を用いて、アセチル化アミノ糖-1-リン酸の糖ヌクレオチド体を合成するが、この合成においては、アセチル化アミノ糖-1-リン酸とヌクレオシド三リン酸を含有する溶液に、該酵素を添加し、反応温度60℃〜95℃で反応させ、該アセチル化アミノ糖分子の糖ヌクレオチド体を得る。
アセチル化アミノ糖-1-リン酸としては、N-アセチルグルコサミン-1-リン酸、N-アセチルガラクトサミン-1-リン酸が挙げられ、ヌクレオシド三リン酸としては、N-アセチルグルコサミン-1-リン酸を基質とする場合にはTTP(チミジントリフォスフェート)及びUTP(ウリジントリフォスフェート)が挙げられ、N-アセチルガラクトサミン-1-リン酸を基質とする場合にはUTP(ウリジントリフォスフェート)が挙げられる。
【0013】
このアセチル化反応の式として、グルコサミン-1-リン酸とアセチルCoAからN-アセチルグルコサミン-1-リン酸を合成する場合について以下に示す。
【化1】

【0014】
このアセチル化アミノ糖ヌクレオチド合成反応の式として、N-アセチルグルコサミン-1-リン酸とUTPからUDP-N-アセチルグルコサミンを合成する場合について以下に示す。
【化2】
【0015】
また、この反応においては。上記精製した酵素のみならず、粗酵素であってもよい。例えば、宿主として枯草菌等分泌型の系を用いる場合には、培養液中に本酵素が生成蓄積され、大腸菌等の非分泌型の系を用いる場合には、菌体内に生成されるので、本酵素を含有する培養液あるいはその処理物、もしくは菌体破砕物等の培養処理物を用いて、アセチルグルコサミン-1-リン酸及び/又はアセチルガラクトサミン-1-リン酸、並びにアセチルグルコサミン及び/又はアセチルガラクトサミンのヌクレオシド二リン酸結合体を合成してもよい。
以下に、本発明の実施例を示すが、本発明実施例により限定されるものではない。
【実施例1】
【0016】
本バイファンクショナル酵素の製造
(1)菌の培養
好酸性好気性超好熱古細菌スルフォロバス、トーコーダイイ(JCM10545)は次の方法で培養した。
1.3gの(NH4)2SO4、0.28 gのKH2PO4、0.25 gのMgSO4・7H2O、0.07 gのCaCl2・2H2O、0.02 gのFeCl3・6H2O、1.8 mg のMnCl2・4H2O、4.5 mgのNa2B4O7・10H2O、0.22 mgのZnSO4・7H2O、0.05 mgのCuCl2・2H2O、0.03 mgのNa2MoO4・2H2O、0.03 mgのVOSO4・xH2O、0.01 mgのCoSO4・7H2O、1.0 gの酵母エキスを1Lの蒸留水に溶かし、この溶液のpHを3.5に10規定H2SO4溶液で調製した。加圧殺菌した後、JCM10545を植菌した。この培養液を80℃で1〜2日培養し、その後遠心分離し集菌した。
【0017】
(2)染色体DNAの調製
好酸性好気性超好熱古細菌スルフォロバス、トーコーダイイ(JCM10545)の染色体DNAは以下の方法により調製した。
培養終了後5000 rpm、10分間の遠心分離により菌体を集菌する。菌体を10 mM EDTA(pH 6.0)溶液で洗浄後、50 mM Tris/HCl-50 mM EDTA (pH 8.5)溶液を加えて細胞を溶解させる。さらに、0.5% Na-lauroylsarcosinate、1 mg/ml プロテアーゼKとなるように各々を加えた後、50℃で3時間保温する。フェノール処理を3回行った後、溶液を10 mM Tris-10 mM EDTA (pH 8.0)溶液に対して透析する。37℃で30分間のRNaseによるRNAの分解後、フェノールクロロフォルム溶液で処理した後、10 mM Tris-1 mM EDTA(pH 8.0)で透析を行う。
【0018】
(3)染色体DNAを含むショットガンライブラリークローンの作製
実施例2で得られた染色体DNAを超音波処理することにより断片化した後、アガロースゲル電気泳動により1kb及び2kb長のDNA断片を回収した。この断片をプラスミドベクターpUC118のHincII制限酵素部位に挿入したショットガンライブラリーを作製した。各ショットガンクローンの末端塩基配列を、ABI社製自動塩基配列読み取り装置377を用いて解読していった。各ショットガンクローンから得られた塩基配列を塩基配列自動連結ソフトSequencherを用いて連結編集し、本菌の全塩基配列を決定していった。
【0019】
(4)本バイファンクショナル酵素遺伝子の同定
上記手法で決定された好酸性好気性超好熱古細菌スルフォロバス、トーコーダイイのゲノム塩基配列の大型計算機による解析を行い、グルコサミン-1-リン酸アセチル化活性を有する可能性のあるタンパク質をコードする遺伝子(ST0452)を同定した。超好熱古細菌スルフォロバス、トーコーダイイのST0452遺伝子の開始コドンはATGで、401アミノ酸残基のタンパク質をコードする候補遺伝子として同定された。
【0020】
(5)発現プラスミドの構築
構造遺伝子領域の前後に制限酵素(NdeIとXhoI)サイトを構築する目的でDNAプライマーを合成し、PCRでその遺伝子の前後に制限酵素サイトを導入した。その際に合成されるタンパク質のC末端にヒスチジン残基をタグとして結合するように合成されるようにする場合とST0452遺伝子がコードするタンパク質のみを合成させるようにする場合とでプライマーの配列が異なる。
【0021】
Upper primer,
5'- ATAGCATATGAAGGCATTTATTCTTGCTGC -3'(配列番号1)
(下線部はNdeIサイトを示す)
Lower prime 1, ヒスチジン残基を結合させる場合
5'- TCAACTCGAGGACCTTGAAAAACTCACC-3'(配列番号2)
(下線部はXhoIサイトを示す)
Lower prime 2, ヒスチジン残基を結合させ無い場合
5'- TCAACTCGAGCTAGACCTTGAAAAACTCACC -3'(配列番号3)
(下線部はXhoIサイトを示す)
【0022】
Upper primerとLower primer1或いはLower primer2を組み合わせたPCR反応後、制限酵素(NdeIとXhoI)で完全分解(37℃で2時間)した後、その構造遺伝子領域断片を精製した。
制限酵素NdeIとXhoIで切断後精製したpET21b(Novagen社製)と上記の構造遺伝子(ST0452)領域断片とをT4リガーゼを用いて16℃、2時間反応させることによって連結した。連結したDNAの一部を大腸菌DH5αのコンピテントセルに導入し形質転換体のコロニーを得た。得られたコロニーからプラスミドをQIAprep Spin Miniprep Kit(QIAGEN社製)で精製し、塩基配列を確認して発現プラスミド、pET21b/ST0452-1及びpET21b/ST0452-2を得た。発現プラスミドpET21b/ST0452-1を用いるとST0452タンパク質はC末端にヒスチジンタグが付加された融合タンパク質として生産され、発現プラスミドpET21b/ST0452-2を用いるとC末端にヒスチジンタグが付加されないタンパク質として生産される。
【0023】
(6)組換え遺伝子の発現
大腸菌(E. coli BL21(DE3) CodonPlus RIL,、Novagen社製)のコンピテントセルを融解して、二本のファルコンチューブに各々0.1 mlづつ移す。その中に上記の2種の発現プラスミド10 ng分に相当する溶液を別々に加え氷中に30分間放置した後42℃でヒートショックを30秒間行い、そこにSOC培地0.9 mlを加え、37℃で1時間振とう培養する。その後、アンピシリンを含むLB寒天プレート上に適量まき、37℃で一晩培養し、形質転換体大腸菌 BL21(DE3) CodonPlus RIL/pET21b/ST0452-1及び形質転換体大腸菌 BL21(DE3) CodonPlus RIL/pET21b/ST0452-2を得た。
【0024】
当該形質転換体をアンピシリンを含むLB培地(2リットル)中で一晩37℃において培養した後、Isopropyl-b-D-thiogalactopyranoside(IPTG)を1 mMになるように加え、さらに30℃で5時間培養した。培養後遠心分離(6,000 rpm,20 分)により集菌を行った。
【0025】
(7)本バイファンクショナル酵素の精製
8リットル培養液から集菌した菌体に2倍量の40 mMトリス塩酸緩衝液(pH 8.0)、1錠のプロテアーゼ阻害剤(Complete EDTA-free, Roche社製)、0.5 mgのDNase RQ1(プロメガ社製)を加え懸濁液を得た。得られた懸濁液を超音波破砕し、75℃で10分保温した後、遠心分離(11,000 rpm、20分)により上清液を得た。この上清液を用いNi-カラム(Novagen, His・Bind metal chelation resin & His・Bind buffer kitを使用)による親和性クロマトグラムを行った。ここで得られた0.5 Mイミダゾール溶出画分(20 ml)を、再度75℃で10分加熱処理し、遠心分離(11,000 rpm、20分)により上清液を得た。次に、セントリプレップYM-50 (アミコン社)で2 mlまで濃縮し、これを20 mMトリス塩酸緩衝液(pH 8.0)で透析し、精製サンプルとした。
【実施例2】
【0026】
アセチル化アミノ糖の合成
(1)アミノ糖アセチル化反応(アセチルCoAからアミノ糖へのアセチル基の転移反応)
50 mM Tris緩衝液(pH7.5)、2 mM MgCl2、2 mM グルコサミン-1-リン酸又はガラクトサミン-1-リン酸、2 mM アセチルCoAからなる酵素反応液10 μlを2分間80℃でプレインキュベーションした後、その反応液中に実施例1で得られた精製酵素0.05 μgを加えた。この反応液を80℃で2分間保温することにより、反応を進行させた。40 μl の50 mM Tris緩衝液(pH 7.5)、6.4 M グアニジン塩酸を加えて反応を終了させた後に、50 μl の50 mM Tris緩衝液(pH7.5)、1 mM EDTA、0.5 mM 5,5’-dithio-bis(2-nitrobenzoic acid) [DTNB]を加えた。
【0027】
(2)アミノ糖アセチル化反応(アセチルCoAからアミノ糖へのアセチル基の転移反応)の測定
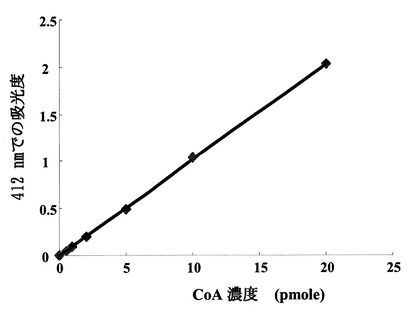
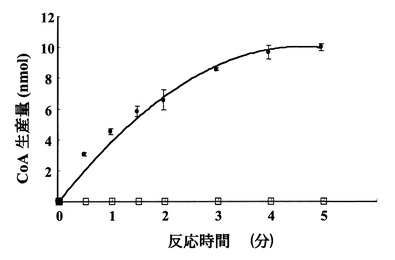
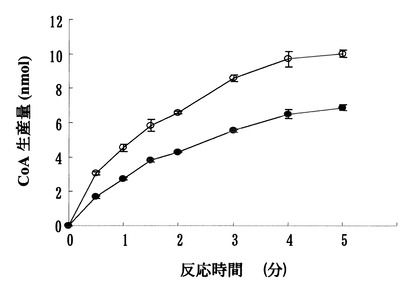
上記(1)の反応では、DTNBがCoAと反応し、4-ニトロチオフェノレートを生成するが、この分子は412 nmでの吸収の増大として検出できる。図1に示すように標準物質であるCoAを基質として加えた反応液では、CoAの量に比例した412 nmでの吸収の増大が見られた。標準CoAサンプル添加量を変化させたときの、吸光度と標準物質量は正確な比例関係にあり、この検量線を用いることにより反応生成物を定量出来る事が示された。さらに、図2に示すように、本バイファンクショナル酵素を添加後の時間に従った412 nmの吸光度の増幅が見られた事から、本酵素はグルコサミン-1-リン酸のアセチル化活性を有する事が確認された。
【実施例3】
【0028】
アセチル化アミノ糖ヌクレオチド(アセチル化アミノ糖−ヌクレオシド二リン酸結合体)の合成
(1)アセチル化アミノ糖ヌクレオチド(NDP-GlcNAc)合成反応(アセチル化アミノ糖とヌクレオチド結合反応)
50 mM Tris緩衝液(pH7.5)、2 mM MgCl2、2 mM N-アセチルグルコサミン-1-リン酸、1
mM TTPまたはUTPからなる酵素反応液10μlを2分間80℃でプレインキュベーションした後、その反応液中に実施例1で得られた精製酵素0.05 μgを加えた。この反応液を80℃で5分間保温することにより、反応を進行させた。5分後に100 μl の500 mM KH2PO4溶液を加える事により反応を停止させた。
【0029】
(2)アセチル化アミノ糖ヌクレオチド(NDP-GlcNAc)合成反応(アセチル化アミノ糖とヌクレオチドの結合反応)の測定
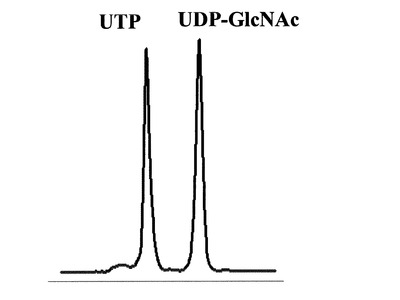
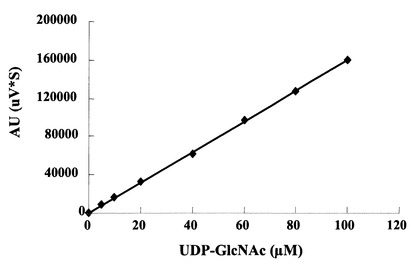
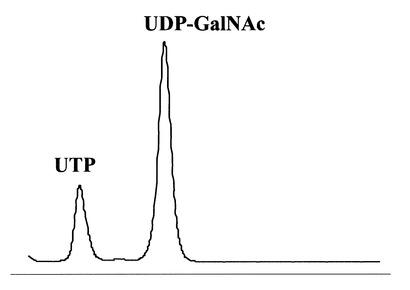
HPLCを用いて、反応生成物であるNDP-GlcNAcの量を、ヌクレオチド部分の紫外線の吸収を目安に測定した。図3に示すように標準物質であるUTP及びUDP-GlcNAcは、HPLCにおいて溶出位置が全く異なる。さらに、図4に示すように標準サンプル添加量を変化させた時の、ピークの面積と標準物質量は正確な比例関係にあり、この検量線を用いることにより反応生成物を定量出来る事が示された。
そこで、上記(1)で反応させたサンプルに関しても、HPLCで同様の解析を行った。
【0030】
(3)アセチル化アミノ糖ヌクレオチド(NDP-GalNAc)合成の逆反応(アセチル化アミノ糖ヌクレオチド分離反応)
50 mM Tris緩衝液(pH 7.5)、2 mM MgCl2、2 mM UDP-GalNAc、1 mM ピロリン酸からなる酵素反応液10 μlを2分間80℃でプレインキュベーションした後、その反応液中に実施例1で得られた精製酵素0.05 μgを加えた。この反応液を80℃で5分間保温することにより、反応を進行させた。5分後に100 μl の500 mM KH2PO4溶液を加える事により反応を停止させた。
【0031】
(4)アセチル化アミノ糖ヌクレオチド(NDP-GalNAc)合成の逆反応(アセチル化アミノ糖ヌクレオチドの分離反応)の測定
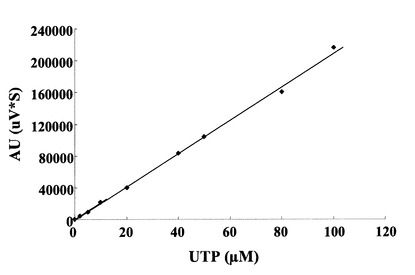
HPLCを用いて、反応生成物であるUTPの量を、ヌクレオチド部分の紫外線の吸収を目安に測定した。図5に示すように標準物質であるUTP及びUDP-GalNAcは、HPLCにおいて溶出位置が全く異なる。さらに、図6に示すように標準UTPサンプル添加量を変化させたときの、ピークの面積と標準物質量は正確な比例関係にあり、この検量線を用いることにより反応生成物を定量出来る事が示された。
そこで、上記(1)で反応させたサンプルに関しても、HPLCで同様の解析を行った。
【実施例4】
【0032】
酵素の性質
(1)タンパク質化学的性質
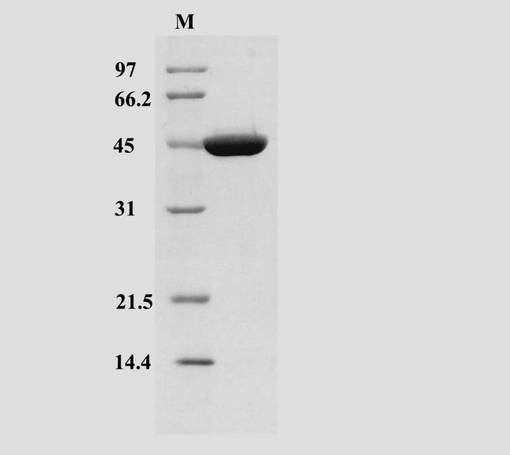
当該酵素は上記の精製プロセスで完全に精製され、SDS-PAGEで分子量約44 KDaの単一バンドを示した(図7)。当該酵素は401アミノ酸残基より構成され(配列番号4)、そのアミノ酸配列から予測される分子量は44,000 Daであった。
【0033】
(2)アミノ糖アセチル化合物合成活性(アミノ糖へのアセチルCoAのアセチル基の転移活性)
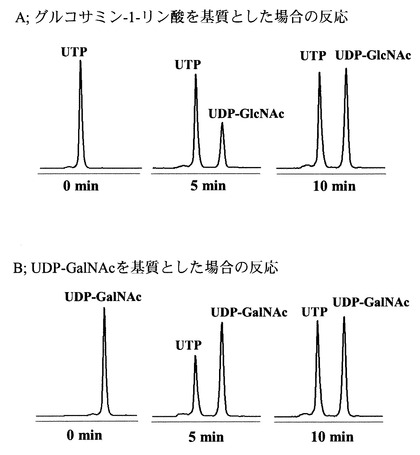
当該酵素は、図8に有るように80℃においては、グルコサミン-1-リン酸及びガラクトサミン-1-リン酸を基質としたどちらの場合にも高い活性を示した。
【0034】
(3)アミノ糖アセチル化反応におけるアミノ糖-1-リン酸基質の多様性
50 mM Tris緩衝液(pH 7.5)、2 mM MgCl2、2 mM アミノ糖-1-リン酸及び2 mM アセチルCoAからなる酵素反応液10μlを2分間80℃でプレインキュベーションした後、その反応液中に実施例1で得られた精製酵素0.05 μgを加えた。この反応液を80℃で2分間保温することにより、反応を進行させた。反応の進行は、実施例2の(2)に有るように412 nmの吸光度で測定した。結果を表1に示す。表1から明らかなように、本酵素はグルコサミン-1-リン酸以外にガラクトサミン-1-リン酸も基質として利用出来るが他の糖リン酸は利用出来ないことが示された。
【0035】
【表1】
【0036】
(4)アセチル化アミノ糖ヌクレオチド合成活性(アセチル化アミノ糖とヌクレオチドの結合活性)
当該酵素は、図9に有るように80℃においてはN-アセチルグルコサミン-1-リン酸及びUDP-GalNAcとピロリン酸を基質とした場合に、酵素活性を示した。
【0037】
(5)アセチル化アミノ糖ヌクレオチド合成活性に於けるアセチル化アミノ糖-1-リン酸基質の多様性
50 mM Tris緩衝液(pH 7.5)、2 mM MgCl2、1 mM UTP、2 mM N-アセチルグルコサミン-1-リン酸からなる酵素反応液10 μlを2分間80℃でプレインキュベーションした後、その反応液中に実施例1で得られた精製酵素0.05 μgを加えた。この酵素反応液を80℃で2分間保温することにより反応させた後、100 μl の500 mM KH2PO4溶液を加える事により反応を停止させた。反応の進行は、実施例3の(2)に有るようにHPLCで測定した。
50 mM Tris緩衝液(pH 7.5)、2 mM MgCl2、1 mM UTP、2 mM N-アセチルガラクトサミン-1-リン酸、1 mM ピロ燐酸からなる酵素反応液10 μlを2分間80℃でプレインキュベーションした後、その反応液中に実施例1で得られた精製酵素0.05 μgを加えた。この酵素反応液を80℃で2分間保温することにより反応させた後、100 μl の500 mM KH2PO4溶液を加える事により反応を停止させた。反応の進行は、実施例3の(4)に有るようにHPLCで測定した。
表2に示すように、本酵素はN-アセチルグルコサミン-1-リン酸以外にN-アセチルガラクトサミン-1-リン酸をアミノ糖ヌクレオチド合成活性の基質として利用出来ることが示された。
【0038】
【表2】
【0039】
(6)アセチル化アミノ糖ヌクレオチド合成活性に於けるヌクレオシド三リン酸基質の多様性
50 mM Tris緩衝液(pH 7.5)、2 mM MgCl2、1 mM NTP、及び2 mM N-アセチルグルコサミン-1-リン酸からなる酵素反応液10μl中に実施例1で得られた精製酵素0.05 μgを加えた。この酵素反応液を80℃で5分間保温することにより反応を進行させた後に、100 μl の500 mM KH2PO4溶液に加える事により反応を停止させた。反応の進行は、実施例3の(2)に有るようにHPLCで測定した。
または50 mM Tris緩衝液(pH 7.5)、2 mM MgCl2、1 mM NTP、2 mMガラクトサミン-1-リン酸、2 mMアセチルCoAからなる酵素反応液10μl中に実施例1で得られた精製酵素0.05 μgを加えた。この酵素反応液を80℃で5分間保温することにより反応を進行させた後に、100 μl の500 mM KH2PO4溶液に加える事により反応を停止させた。反応の進行は、実施例3の(2)に有るようにHPLCで測定した。
表3に示すように、本酵素はN-アセチルグルコサミン-1-リン酸を基質にする場合にはdTTPとUTPを、N-アセチルガラクトサミン-1-リン酸を基質とする場合にはUTPのみを基質として利用出来ることが示された。
【0040】
【表3】
【0041】
(7)熱安定性
50mM Tris緩衝液(pH7.5)、2 mM MgCl2、2 mM N-アセチルグルコサミン-1-リン酸、1
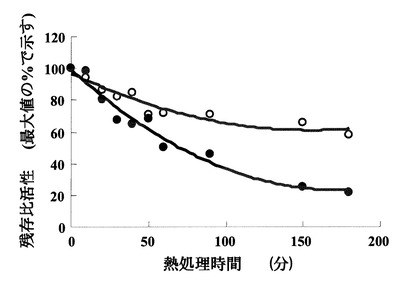
mM UTPからなる酵素反応液10 μl中に、あらかじめ80℃で5分10分20分30分40分60分90分120分180分間加熱した実施例1で得られた精製酵素0.05 μgを加えた。この酵素反応液を80℃で5分間保温することにより反応させた後に、100 μl の500 mM KH2PO4溶液に加える事により反応を停止させた。反応の進行は、実施例3の(2)に有るようにHPLCで測定した。その結果、図10に示すように、本酵素は80℃による180分間の加熱処理後でも、50%以上の活性を残すことから非常に安定で耐熱性が高いことが示された。
【図面の簡単な説明】
【0042】
【図1】DTNB試薬と標準CoAを反応させた場合の412nmでの吸収との関係を示す図である。
【図2】該酵素による時間に従った反応の進行(◆)及び酵素を加えない場合には反応が進行しないこと(◇)を示す図である。
【図3】HPLCによる、UTP、及びUDP-GlcNAc混合物の分離パターンを測定した図である。
【図4】HPLCを用いたUDP-GlcNAcの検量線を示す図である。
【図5】HPLCによる、UTP、及びUDP-GalNAc混合物の分離パターンを測定した図である。
【図6】HPLCを用いたUTPの検量線を示す図である。
【図7】精製されたST0452蛋白質のSDS-PAGEパターンを示す写真である。
【図8】ST0452酵素のグルコサミン-1-リン酸(○)及びガラクトサミン-1-リン酸(●)を基質として用いた場合の80℃におけるアセチル化活性量を示す図である。
【図9】ST0452酵素の80℃におけるアセチル化アミノ糖ヌクレオチド合成活性を示す図である。
【図10】ST0452蛋白質の80℃(○)及び95℃(●)での熱処理後における残存活性量を示す図である。
【技術分野】
【0001】
本願発明は新規アミノ糖アセチル化活性及び該アセチル化アミノ糖の糖ヌクレオチド合成活性を有する耐熱性酵素、及び該酵素を用いて効率的にアセチル化アミノ糖及びその活性化体であるアセチル化アミノ糖ヌクレオチドを製造する方法に関する。
【背景技術】
【0002】
グルコサミン-1-リン酸(glucosamine-1-phosphate)にアセチル基を転移してN-アセチルグルコサミン-1-リン酸(N-acetylglucosamine-1-phosphate)を合成する活性は、UDP-N-アセチルグルコサミン(UDP-N-acetylglucosamine, UDP-GlcNAc)合成活性を有する酵素が共有する活性として、見出されている。このようなバイファンクショナル酵素としては、大腸菌(Escherichia coli)(非特許文献1、2参照)及びストレプトコッカス菌(Streptococcus pneumoniae) (非特許文献3参照)由来のGlmU (Glucosamine-1-phosphate uridyltransferase/Glucosamine-1-phosphate acetyltransferase)の詳しい性質や構造がすでに報告されている。GlmUは細胞表面の膜構造合成に必須なN-アセチルグルコサミン(N-acetylglucosamine)の活性体であるUDP-N-アセチルグルコサミンをフルクトース-6-リン酸(fructose-6-phosphate)から合成する代謝経路の最後の2段階の反応を触媒する二つの活性を併せ持つ酵素として見出されてきた。この酵素は、初めにフルクトース-6-リン酸から合成されてきたグルコサミン-1-リン酸にアセチルCoA(Acetyl-CoA)のアセチル基を転移することによってN-アセチルグルコサミン-1-リン酸を合成し、次にこの合成された糖とヌクレオシド三リン酸(UTP)を基質として、UDP-GlcNAcを生産する。その他にも、豚肝臓から同様の糖ヌクレオチドを合成する酵素が見出されているが、その多くが常温生物由来のため室温以上では極めて不安定で、活性は80℃程度の加熱処理により速やかに失活する。このため、使用時の滅菌等の処理が必要であったり、低温での注意深い保存が必要であった。
また、同様にN-アセチルガラクトサミン(N-Acetylgalactosamine)も膜構造の構築に於いて重要な糖であるが、この糖の合成経路に関しては、不明な点が多く、ガラクトサミン-1-リン酸(Galactosamine-1-phosphate)をアセチル化する酵素活性はこれまでに見出されていない。
【非特許文献1】Mengin-Lecreulx D and van Heijenoort J. " Copurification ofglucosamine-1-phosphate acetyltransferase and N-acetylglucosamine-1-phosphateuridyltransferase activities of Escherichia coli: characterization ofthe glmU gene product as a bifunctional enzyme catalyzing two subsequent stepsin the pathway for UDP-N-acetylglucosamine synthesis.“ (1994) J. Bacteriology,176, 5788-5795.
【非特許文献2】Brown K, Pompeo F, Dixon S, Mengin-Lecreulx D, Cambillau C and BourneY “Crystal structure of the bifunctional N-acetylglucosamine 1-phosphateuridyltransferase from Escherichia coli: a paradigm for the relatedpyrophosphorylase superfamily." (1999) EMBO J., 18, 4096-4107.
【非特許文献3】Sulzenbacher G, Gal L, Peneff C, Fassy F and Bourne Y. " Crystalstructure of Streptococcus pneumoniae N-acetylglucosamine-1-phosphateuridyltransferase bound to acetyl-coenzyme A reveals a novel active sitearchitecture." (2001) J. Biol. Chem., 276, 11844-11851.
【発明の開示】
【発明が解決しようとする課題】
【0003】
グルコサミン-1-リン酸及びガラクトサミン-1-リン酸をアセチル化する活性並びにそのアセチル化アミノ糖の活性化体であるアセチル化アミノ糖ヌクレオチドを合成する活性を有する耐熱性酵素が発見されれば、糖鎖合成の基質となる重要なアセチル化アミノ糖であるN-アセチルグルコサミン及びN-アセチルガラクトサミンについてのアセチル化アミノ糖ヌクレオチド体、すなわちヌクレオシド二リン酸結合体を安定に合成することが可能となる。これまで、糖鎖合成の際の基質として必要なこれらのアセチル化アミノ糖及びヌクレオシド三リン酸を基質として、結合反応を触媒出来る安定な酵素は存在しなかったので、渇望されていた。
【0004】
したがって、本発明の課題は、耐熱性を有し、かつグルコサミン-1-リン酸及びガラクトサミン-1-リン酸を基質として、それぞれのアセチル化アミノ糖-1-リン酸を合成し得る活性、並びにそれらのアセチル化アミノ糖-1-リン酸とヌクレオシド三リン酸を基質として、アセチル化アミノ糖ヌクレオチド(アセチル化アミノ糖−ヌクレオシド二リン酸結合体)を合成することが可能な新規酵素、さらに本酵素を用いたアセチル化アミノ糖及び該アセチル化アミノ糖ヌクレオチド合成法を提供することにある。
【課題を解決するための手段】
【0005】
本発明者は、以上のような課題を解決すべく、75 - 80℃で生育する超好熱古細菌 Sulfolobus tokodaii strain7に着目して、その遺伝子を大腸菌を使って発現させることによって酵素蛋白質を生産し、この酵素蛋白質が高温(80℃)で安定に存在するとともに目的とする活性を示すことを確認し、さらに、本酵素蛋白質を用いることにより目的とするアセチル化アミノ糖ヌクレオチド(アミノ糖−ヌクレオシド二リン酸結合体)を生産することもできることを見いだし、本発明を完成するに至ったものである。
【0006】
即ち、本発明は、以下の(1)〜(10)に係るものである。
(1)配列番号4に記載のアミノ酸配列を有するか、あるいは、配列番号4に記載のアミノ酸配列に一乃至数個のアミノ酸残基が欠失、置換、挿入又は付加されたアミノ酸配列を有し、かつアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有することを特徴とする、蛋白質。
(2)上記(1)に記載の蛋白質をコードするDNA。
(3)配列番号5に記載の塩基配列を有することを特徴とするDNA。
(4)配列番号5に記載のDNA とストリンジェントな条件下でハイブリダイズし、かつアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する蛋白質をコードすることを特徴とする、DNA。
(5)上記(2)〜(4)のいずれかに記載のDNAから選ばれるDNAがベクターに組み込まれていることを特徴とする組換え体DNA。
(6)上記(5)に記載の組換え体DNAが宿主細胞に導入されていることを特徴とする形質転換体。
(7)上記(6)に記載の形質転換体を培地に培養し、培養物からアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する蛋白質を採取することを特徴とする、アセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する蛋白質の製造方法。
(8)グルコサミン-1-リン酸及び/又はガラクトサミン-1-リン酸に、アセチルCoA存在下、上記(1)に記載の蛋白質を作用させることを特徴とする、N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸の製造方法。
(9)グルコサミン-1-リン酸及び/又はガラクトサミン-1-リン酸に、アセチルCoA存在下、上記(6)に記載の形質転換体の培養液あるいは培養物の処理物を作用させることを特徴とする、N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸の製造方法。
(10)グルコサミン-1-リン酸及び/又はガラクトサミン-1-リン酸に、アセチルCoA存在下、 上記(3)又は(4)に記載のDNAにコードされるタンパク質を作用させることを特徴とする、N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸の製造方法。
(11)N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸に、ヌクレオシド三リン酸の存在下、上記(1)に記載のタンパク質を作用させることを特徴とする、
N-アセチルグルコサミンおよび/又はN-アセチルガラクトサミンのヌクレオシド二リン酸結合体の製造方法。
(12)N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸に、ヌクレオシド三リン酸の存在下、上記(6)に記載の形質転換体の培養液あるいは培養物の処理物を作用させることを特徴とする、N-アセチルグルコサミンおよび/又はN-アセチルガラクトサミンのヌクレオシド二リン酸結合体の製造方法。
(13)N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸に、ヌクレオシド三リン酸の存在下、上記(2)〜(4)のいずれかに記載のDNAにコードされるタンパク質を作用させることを特徴とする、N-アセチルグルコサミンおよび/又はN-アセチルガラクトサミンのヌクレオシド二リン酸結合体の製造方法。
【発明の効果】
【0007】
本発明の酵素を使用すれば、N-アセチルグルコサミン-1-リン酸とN-アセチルガラクトサミン-1-リン酸の2種のアミノ糖アセチル化体が合成可能になると共に、さらに、UDP-GlcNAc、TDP-GlcNAc、及びUDP-GalNAcのようなアセチル化アミノ糖ヌクレオチドを合成することができる。このように本発明の酵素は、単一酵素でありながら広い合成活性を有し、かつ熱等に安定である。このような酵素特性は、既知の酵素にみられないものであり、新規なアセチル化アミノ糖ヌクレオチドの効率的な合成を可能とするものである。
一方、アセチル化アミノ糖ヌクレオチドは、糖タンパク質、糖脂質、多糖類の糖鎖合成にアセチル化アミノ糖供与体として機能するものであり、これらの糖鎖は、癌転移、器官発生あるいは細胞性免疫等に密接に関連するものとして近年注目されており、本発明は、これら糖鎖の人工的合成発展において、その貢献度は極めて大きい。
【発明を実施するための最良の形態】
【0008】
以下に、本願発明を具体的に説明する。
本発明の酵素は、好酸性好気性超好熱古細菌スルフォロバス、トーコーダイイ(Sulfolobus tokodaii)(JCM登録番号JCM10545)由来の酵素であり、該超好熱古細菌から本酵素活性を示すと推定した遺伝子領域を、PCR反応で増幅・抽出し、蛋白質発現プラスミドpET21bに挿入後、そのプラスミドにより形質転換した大腸菌を用いて得られたものである。生産された酵素は加熱処理およびカラムクロマトグラムで単離精製し、精製された酵素は、分子量が約44,000のタンパク質で、アセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を共に有する酵素である。本発明の酵素は、特に、グルコサミン-1-リン酸からN-アセチルグルコサミン-1-リン酸を合成する活性及びN-アセチルグルコサミン-1-リン酸からN-アセチルグルコサミンのヌクレオシド二リン酸結合体を合成する活性のほかに、ガラクトサミン-1-リン酸からN-アセチルガラクトサミン-1-リン酸を合成する活性及びN-アセチルガラクトサミン-1-リン酸からN-アセチルガラクトサミンのヌクレオシド二リン酸結合体を合成する活性を有する点に新規な特徴を有する。なお、これらヌクレオシド二リン酸結合体は、N-アセチルグルコサミン、N-アセチルガラクトサミンのそれぞれ1位にヌクレオシド二リン酸のリン酸基が結合しているものである。
【0009】
この酵素の半減期は、50mMトリス塩酸緩衝液(pH7.5)中で、80℃、40分以上であり、高い耐熱性を示した。
この酵素のアミノ酸配列およびその遺伝子DNA(ST0452)の塩基配列を、それぞれ配列表の配列番号4及び5に示す。
【0010】
本発明における酵素は、上記配列番号4に示されるアミノ酸配列を有するもののみに限定されず、該アミノ酸配列において、数個程度のアミノ酸残基が欠失、置換、挿入又は付加されたアミノ酸配列であっても、このアミノ酸配列を有する蛋白質が、上記酵素活性上の特徴に係るアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する限り、本発明に含まれる。また、本発明のこれら酵素遺伝子DNAについても、上記同配列番号5に示す塩基配列を有するもののみに限定されず、上記アミノ酸配列をコードするものを包含する。さらに上記配列番号5に示されるDNAにストリンジェントな条件下でハイブリダイズし、かつ目的とする上記酵素活性上の特徴に係るアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する蛋白質をコードするDNAも包含する。このストリンジェントな条件とは、ハイブリダイゼーション溶液1リットル中に52.59 g NaCl、26.46 g クエン酸ナトリウム、1 g フィコール(Type 400)、1 g ポリビニルピロリドン、1 g ウシ血清アルブミン、5 g SDS、1 g 断片化鮭精子DNA、500 ml ホルムアミドを含み、温度42℃で行う。その後の洗浄は、洗浄用溶液1リットル中に17.53 g NaCl、8.82 g クエン酸ナトリウム、5 g SDSを含み、温度68℃で行う条件である。
【0011】
本発明の酵素を得るには、通常の遺伝子工学的手法が適用でき、上記酵素遺伝子DNAを、例えば、pET21b、pHY481等の蛋白質発現プラスミドベクター等に挿入して組換えベクターを作製し、該組換えベクターを用いて宿主細胞を形質転換し、該形質転換体を培地で培養し、培養物、培養処理物あるいはこれら培養物から分離回収された形質転換体から、酵素を常法の蛋白質精製手段により精製し単離する。上記宿主細胞としては、大腸菌・枯草菌等が利用可能である。
【0012】
本発明においては、さらにこの酵素を用いて、アミノ糖-1-リン酸のアセチル化体を合成するが、この合成においては、アミノ糖-1-リン酸とアセチルCoAを含有する溶液に、該酵素を添加し、反応温度60℃〜95℃で反応させ、アセチル化アミノ糖-1-リン酸を得る。
アミノ糖-1-リン酸としては、例えば、グルコサミン-1-リン酸、ガラクトサミン-1-リン酸が挙げられる。
本発明においては、この酵素を用いて、アセチル化アミノ糖-1-リン酸の糖ヌクレオチド体を合成するが、この合成においては、アセチル化アミノ糖-1-リン酸とヌクレオシド三リン酸を含有する溶液に、該酵素を添加し、反応温度60℃〜95℃で反応させ、該アセチル化アミノ糖分子の糖ヌクレオチド体を得る。
アセチル化アミノ糖-1-リン酸としては、N-アセチルグルコサミン-1-リン酸、N-アセチルガラクトサミン-1-リン酸が挙げられ、ヌクレオシド三リン酸としては、N-アセチルグルコサミン-1-リン酸を基質とする場合にはTTP(チミジントリフォスフェート)及びUTP(ウリジントリフォスフェート)が挙げられ、N-アセチルガラクトサミン-1-リン酸を基質とする場合にはUTP(ウリジントリフォスフェート)が挙げられる。
【0013】
このアセチル化反応の式として、グルコサミン-1-リン酸とアセチルCoAからN-アセチルグルコサミン-1-リン酸を合成する場合について以下に示す。
【化1】
【0014】
このアセチル化アミノ糖ヌクレオチド合成反応の式として、N-アセチルグルコサミン-1-リン酸とUTPからUDP-N-アセチルグルコサミンを合成する場合について以下に示す。
【化2】
【0015】
また、この反応においては。上記精製した酵素のみならず、粗酵素であってもよい。例えば、宿主として枯草菌等分泌型の系を用いる場合には、培養液中に本酵素が生成蓄積され、大腸菌等の非分泌型の系を用いる場合には、菌体内に生成されるので、本酵素を含有する培養液あるいはその処理物、もしくは菌体破砕物等の培養処理物を用いて、アセチルグルコサミン-1-リン酸及び/又はアセチルガラクトサミン-1-リン酸、並びにアセチルグルコサミン及び/又はアセチルガラクトサミンのヌクレオシド二リン酸結合体を合成してもよい。
以下に、本発明の実施例を示すが、本発明実施例により限定されるものではない。
【実施例1】
【0016】
本バイファンクショナル酵素の製造
(1)菌の培養
好酸性好気性超好熱古細菌スルフォロバス、トーコーダイイ(JCM10545)は次の方法で培養した。
1.3gの(NH4)2SO4、0.28 gのKH2PO4、0.25 gのMgSO4・7H2O、0.07 gのCaCl2・2H2O、0.02 gのFeCl3・6H2O、1.8 mg のMnCl2・4H2O、4.5 mgのNa2B4O7・10H2O、0.22 mgのZnSO4・7H2O、0.05 mgのCuCl2・2H2O、0.03 mgのNa2MoO4・2H2O、0.03 mgのVOSO4・xH2O、0.01 mgのCoSO4・7H2O、1.0 gの酵母エキスを1Lの蒸留水に溶かし、この溶液のpHを3.5に10規定H2SO4溶液で調製した。加圧殺菌した後、JCM10545を植菌した。この培養液を80℃で1〜2日培養し、その後遠心分離し集菌した。
【0017】
(2)染色体DNAの調製
好酸性好気性超好熱古細菌スルフォロバス、トーコーダイイ(JCM10545)の染色体DNAは以下の方法により調製した。
培養終了後5000 rpm、10分間の遠心分離により菌体を集菌する。菌体を10 mM EDTA(pH 6.0)溶液で洗浄後、50 mM Tris/HCl-50 mM EDTA (pH 8.5)溶液を加えて細胞を溶解させる。さらに、0.5% Na-lauroylsarcosinate、1 mg/ml プロテアーゼKとなるように各々を加えた後、50℃で3時間保温する。フェノール処理を3回行った後、溶液を10 mM Tris-10 mM EDTA (pH 8.0)溶液に対して透析する。37℃で30分間のRNaseによるRNAの分解後、フェノールクロロフォルム溶液で処理した後、10 mM Tris-1 mM EDTA(pH 8.0)で透析を行う。
【0018】
(3)染色体DNAを含むショットガンライブラリークローンの作製
実施例2で得られた染色体DNAを超音波処理することにより断片化した後、アガロースゲル電気泳動により1kb及び2kb長のDNA断片を回収した。この断片をプラスミドベクターpUC118のHincII制限酵素部位に挿入したショットガンライブラリーを作製した。各ショットガンクローンの末端塩基配列を、ABI社製自動塩基配列読み取り装置377を用いて解読していった。各ショットガンクローンから得られた塩基配列を塩基配列自動連結ソフトSequencherを用いて連結編集し、本菌の全塩基配列を決定していった。
【0019】
(4)本バイファンクショナル酵素遺伝子の同定
上記手法で決定された好酸性好気性超好熱古細菌スルフォロバス、トーコーダイイのゲノム塩基配列の大型計算機による解析を行い、グルコサミン-1-リン酸アセチル化活性を有する可能性のあるタンパク質をコードする遺伝子(ST0452)を同定した。超好熱古細菌スルフォロバス、トーコーダイイのST0452遺伝子の開始コドンはATGで、401アミノ酸残基のタンパク質をコードする候補遺伝子として同定された。
【0020】
(5)発現プラスミドの構築
構造遺伝子領域の前後に制限酵素(NdeIとXhoI)サイトを構築する目的でDNAプライマーを合成し、PCRでその遺伝子の前後に制限酵素サイトを導入した。その際に合成されるタンパク質のC末端にヒスチジン残基をタグとして結合するように合成されるようにする場合とST0452遺伝子がコードするタンパク質のみを合成させるようにする場合とでプライマーの配列が異なる。
【0021】
Upper primer,
5'- ATAGCATATGAAGGCATTTATTCTTGCTGC -3'(配列番号1)
(下線部はNdeIサイトを示す)
Lower prime 1, ヒスチジン残基を結合させる場合
5'- TCAACTCGAGGACCTTGAAAAACTCACC-3'(配列番号2)
(下線部はXhoIサイトを示す)
Lower prime 2, ヒスチジン残基を結合させ無い場合
5'- TCAACTCGAGCTAGACCTTGAAAAACTCACC -3'(配列番号3)
(下線部はXhoIサイトを示す)
【0022】
Upper primerとLower primer1或いはLower primer2を組み合わせたPCR反応後、制限酵素(NdeIとXhoI)で完全分解(37℃で2時間)した後、その構造遺伝子領域断片を精製した。
制限酵素NdeIとXhoIで切断後精製したpET21b(Novagen社製)と上記の構造遺伝子(ST0452)領域断片とをT4リガーゼを用いて16℃、2時間反応させることによって連結した。連結したDNAの一部を大腸菌DH5αのコンピテントセルに導入し形質転換体のコロニーを得た。得られたコロニーからプラスミドをQIAprep Spin Miniprep Kit(QIAGEN社製)で精製し、塩基配列を確認して発現プラスミド、pET21b/ST0452-1及びpET21b/ST0452-2を得た。発現プラスミドpET21b/ST0452-1を用いるとST0452タンパク質はC末端にヒスチジンタグが付加された融合タンパク質として生産され、発現プラスミドpET21b/ST0452-2を用いるとC末端にヒスチジンタグが付加されないタンパク質として生産される。
【0023】
(6)組換え遺伝子の発現
大腸菌(E. coli BL21(DE3) CodonPlus RIL,、Novagen社製)のコンピテントセルを融解して、二本のファルコンチューブに各々0.1 mlづつ移す。その中に上記の2種の発現プラスミド10 ng分に相当する溶液を別々に加え氷中に30分間放置した後42℃でヒートショックを30秒間行い、そこにSOC培地0.9 mlを加え、37℃で1時間振とう培養する。その後、アンピシリンを含むLB寒天プレート上に適量まき、37℃で一晩培養し、形質転換体大腸菌 BL21(DE3) CodonPlus RIL/pET21b/ST0452-1及び形質転換体大腸菌 BL21(DE3) CodonPlus RIL/pET21b/ST0452-2を得た。
【0024】
当該形質転換体をアンピシリンを含むLB培地(2リットル)中で一晩37℃において培養した後、Isopropyl-b-D-thiogalactopyranoside(IPTG)を1 mMになるように加え、さらに30℃で5時間培養した。培養後遠心分離(6,000 rpm,20 分)により集菌を行った。
【0025】
(7)本バイファンクショナル酵素の精製
8リットル培養液から集菌した菌体に2倍量の40 mMトリス塩酸緩衝液(pH 8.0)、1錠のプロテアーゼ阻害剤(Complete EDTA-free, Roche社製)、0.5 mgのDNase RQ1(プロメガ社製)を加え懸濁液を得た。得られた懸濁液を超音波破砕し、75℃で10分保温した後、遠心分離(11,000 rpm、20分)により上清液を得た。この上清液を用いNi-カラム(Novagen, His・Bind metal chelation resin & His・Bind buffer kitを使用)による親和性クロマトグラムを行った。ここで得られた0.5 Mイミダゾール溶出画分(20 ml)を、再度75℃で10分加熱処理し、遠心分離(11,000 rpm、20分)により上清液を得た。次に、セントリプレップYM-50 (アミコン社)で2 mlまで濃縮し、これを20 mMトリス塩酸緩衝液(pH 8.0)で透析し、精製サンプルとした。
【実施例2】
【0026】
アセチル化アミノ糖の合成
(1)アミノ糖アセチル化反応(アセチルCoAからアミノ糖へのアセチル基の転移反応)
50 mM Tris緩衝液(pH7.5)、2 mM MgCl2、2 mM グルコサミン-1-リン酸又はガラクトサミン-1-リン酸、2 mM アセチルCoAからなる酵素反応液10 μlを2分間80℃でプレインキュベーションした後、その反応液中に実施例1で得られた精製酵素0.05 μgを加えた。この反応液を80℃で2分間保温することにより、反応を進行させた。40 μl の50 mM Tris緩衝液(pH 7.5)、6.4 M グアニジン塩酸を加えて反応を終了させた後に、50 μl の50 mM Tris緩衝液(pH7.5)、1 mM EDTA、0.5 mM 5,5’-dithio-bis(2-nitrobenzoic acid) [DTNB]を加えた。
【0027】
(2)アミノ糖アセチル化反応(アセチルCoAからアミノ糖へのアセチル基の転移反応)の測定
上記(1)の反応では、DTNBがCoAと反応し、4-ニトロチオフェノレートを生成するが、この分子は412 nmでの吸収の増大として検出できる。図1に示すように標準物質であるCoAを基質として加えた反応液では、CoAの量に比例した412 nmでの吸収の増大が見られた。標準CoAサンプル添加量を変化させたときの、吸光度と標準物質量は正確な比例関係にあり、この検量線を用いることにより反応生成物を定量出来る事が示された。さらに、図2に示すように、本バイファンクショナル酵素を添加後の時間に従った412 nmの吸光度の増幅が見られた事から、本酵素はグルコサミン-1-リン酸のアセチル化活性を有する事が確認された。
【実施例3】
【0028】
アセチル化アミノ糖ヌクレオチド(アセチル化アミノ糖−ヌクレオシド二リン酸結合体)の合成
(1)アセチル化アミノ糖ヌクレオチド(NDP-GlcNAc)合成反応(アセチル化アミノ糖とヌクレオチド結合反応)
50 mM Tris緩衝液(pH7.5)、2 mM MgCl2、2 mM N-アセチルグルコサミン-1-リン酸、1
mM TTPまたはUTPからなる酵素反応液10μlを2分間80℃でプレインキュベーションした後、その反応液中に実施例1で得られた精製酵素0.05 μgを加えた。この反応液を80℃で5分間保温することにより、反応を進行させた。5分後に100 μl の500 mM KH2PO4溶液を加える事により反応を停止させた。
【0029】
(2)アセチル化アミノ糖ヌクレオチド(NDP-GlcNAc)合成反応(アセチル化アミノ糖とヌクレオチドの結合反応)の測定
HPLCを用いて、反応生成物であるNDP-GlcNAcの量を、ヌクレオチド部分の紫外線の吸収を目安に測定した。図3に示すように標準物質であるUTP及びUDP-GlcNAcは、HPLCにおいて溶出位置が全く異なる。さらに、図4に示すように標準サンプル添加量を変化させた時の、ピークの面積と標準物質量は正確な比例関係にあり、この検量線を用いることにより反応生成物を定量出来る事が示された。
そこで、上記(1)で反応させたサンプルに関しても、HPLCで同様の解析を行った。
【0030】
(3)アセチル化アミノ糖ヌクレオチド(NDP-GalNAc)合成の逆反応(アセチル化アミノ糖ヌクレオチド分離反応)
50 mM Tris緩衝液(pH 7.5)、2 mM MgCl2、2 mM UDP-GalNAc、1 mM ピロリン酸からなる酵素反応液10 μlを2分間80℃でプレインキュベーションした後、その反応液中に実施例1で得られた精製酵素0.05 μgを加えた。この反応液を80℃で5分間保温することにより、反応を進行させた。5分後に100 μl の500 mM KH2PO4溶液を加える事により反応を停止させた。
【0031】
(4)アセチル化アミノ糖ヌクレオチド(NDP-GalNAc)合成の逆反応(アセチル化アミノ糖ヌクレオチドの分離反応)の測定
HPLCを用いて、反応生成物であるUTPの量を、ヌクレオチド部分の紫外線の吸収を目安に測定した。図5に示すように標準物質であるUTP及びUDP-GalNAcは、HPLCにおいて溶出位置が全く異なる。さらに、図6に示すように標準UTPサンプル添加量を変化させたときの、ピークの面積と標準物質量は正確な比例関係にあり、この検量線を用いることにより反応生成物を定量出来る事が示された。
そこで、上記(1)で反応させたサンプルに関しても、HPLCで同様の解析を行った。
【実施例4】
【0032】
酵素の性質
(1)タンパク質化学的性質
当該酵素は上記の精製プロセスで完全に精製され、SDS-PAGEで分子量約44 KDaの単一バンドを示した(図7)。当該酵素は401アミノ酸残基より構成され(配列番号4)、そのアミノ酸配列から予測される分子量は44,000 Daであった。
【0033】
(2)アミノ糖アセチル化合物合成活性(アミノ糖へのアセチルCoAのアセチル基の転移活性)
当該酵素は、図8に有るように80℃においては、グルコサミン-1-リン酸及びガラクトサミン-1-リン酸を基質としたどちらの場合にも高い活性を示した。
【0034】
(3)アミノ糖アセチル化反応におけるアミノ糖-1-リン酸基質の多様性
50 mM Tris緩衝液(pH 7.5)、2 mM MgCl2、2 mM アミノ糖-1-リン酸及び2 mM アセチルCoAからなる酵素反応液10μlを2分間80℃でプレインキュベーションした後、その反応液中に実施例1で得られた精製酵素0.05 μgを加えた。この反応液を80℃で2分間保温することにより、反応を進行させた。反応の進行は、実施例2の(2)に有るように412 nmの吸光度で測定した。結果を表1に示す。表1から明らかなように、本酵素はグルコサミン-1-リン酸以外にガラクトサミン-1-リン酸も基質として利用出来るが他の糖リン酸は利用出来ないことが示された。
【0035】
【表1】
【0036】
(4)アセチル化アミノ糖ヌクレオチド合成活性(アセチル化アミノ糖とヌクレオチドの結合活性)
当該酵素は、図9に有るように80℃においてはN-アセチルグルコサミン-1-リン酸及びUDP-GalNAcとピロリン酸を基質とした場合に、酵素活性を示した。
【0037】
(5)アセチル化アミノ糖ヌクレオチド合成活性に於けるアセチル化アミノ糖-1-リン酸基質の多様性
50 mM Tris緩衝液(pH 7.5)、2 mM MgCl2、1 mM UTP、2 mM N-アセチルグルコサミン-1-リン酸からなる酵素反応液10 μlを2分間80℃でプレインキュベーションした後、その反応液中に実施例1で得られた精製酵素0.05 μgを加えた。この酵素反応液を80℃で2分間保温することにより反応させた後、100 μl の500 mM KH2PO4溶液を加える事により反応を停止させた。反応の進行は、実施例3の(2)に有るようにHPLCで測定した。
50 mM Tris緩衝液(pH 7.5)、2 mM MgCl2、1 mM UTP、2 mM N-アセチルガラクトサミン-1-リン酸、1 mM ピロ燐酸からなる酵素反応液10 μlを2分間80℃でプレインキュベーションした後、その反応液中に実施例1で得られた精製酵素0.05 μgを加えた。この酵素反応液を80℃で2分間保温することにより反応させた後、100 μl の500 mM KH2PO4溶液を加える事により反応を停止させた。反応の進行は、実施例3の(4)に有るようにHPLCで測定した。
表2に示すように、本酵素はN-アセチルグルコサミン-1-リン酸以外にN-アセチルガラクトサミン-1-リン酸をアミノ糖ヌクレオチド合成活性の基質として利用出来ることが示された。
【0038】
【表2】
【0039】
(6)アセチル化アミノ糖ヌクレオチド合成活性に於けるヌクレオシド三リン酸基質の多様性
50 mM Tris緩衝液(pH 7.5)、2 mM MgCl2、1 mM NTP、及び2 mM N-アセチルグルコサミン-1-リン酸からなる酵素反応液10μl中に実施例1で得られた精製酵素0.05 μgを加えた。この酵素反応液を80℃で5分間保温することにより反応を進行させた後に、100 μl の500 mM KH2PO4溶液に加える事により反応を停止させた。反応の進行は、実施例3の(2)に有るようにHPLCで測定した。
または50 mM Tris緩衝液(pH 7.5)、2 mM MgCl2、1 mM NTP、2 mMガラクトサミン-1-リン酸、2 mMアセチルCoAからなる酵素反応液10μl中に実施例1で得られた精製酵素0.05 μgを加えた。この酵素反応液を80℃で5分間保温することにより反応を進行させた後に、100 μl の500 mM KH2PO4溶液に加える事により反応を停止させた。反応の進行は、実施例3の(2)に有るようにHPLCで測定した。
表3に示すように、本酵素はN-アセチルグルコサミン-1-リン酸を基質にする場合にはdTTPとUTPを、N-アセチルガラクトサミン-1-リン酸を基質とする場合にはUTPのみを基質として利用出来ることが示された。
【0040】
【表3】
【0041】
(7)熱安定性
50mM Tris緩衝液(pH7.5)、2 mM MgCl2、2 mM N-アセチルグルコサミン-1-リン酸、1
mM UTPからなる酵素反応液10 μl中に、あらかじめ80℃で5分10分20分30分40分60分90分120分180分間加熱した実施例1で得られた精製酵素0.05 μgを加えた。この酵素反応液を80℃で5分間保温することにより反応させた後に、100 μl の500 mM KH2PO4溶液に加える事により反応を停止させた。反応の進行は、実施例3の(2)に有るようにHPLCで測定した。その結果、図10に示すように、本酵素は80℃による180分間の加熱処理後でも、50%以上の活性を残すことから非常に安定で耐熱性が高いことが示された。
【図面の簡単な説明】
【0042】
【図1】DTNB試薬と標準CoAを反応させた場合の412nmでの吸収との関係を示す図である。
【図2】該酵素による時間に従った反応の進行(◆)及び酵素を加えない場合には反応が進行しないこと(◇)を示す図である。
【図3】HPLCによる、UTP、及びUDP-GlcNAc混合物の分離パターンを測定した図である。
【図4】HPLCを用いたUDP-GlcNAcの検量線を示す図である。
【図5】HPLCによる、UTP、及びUDP-GalNAc混合物の分離パターンを測定した図である。
【図6】HPLCを用いたUTPの検量線を示す図である。
【図7】精製されたST0452蛋白質のSDS-PAGEパターンを示す写真である。
【図8】ST0452酵素のグルコサミン-1-リン酸(○)及びガラクトサミン-1-リン酸(●)を基質として用いた場合の80℃におけるアセチル化活性量を示す図である。
【図9】ST0452酵素の80℃におけるアセチル化アミノ糖ヌクレオチド合成活性を示す図である。
【図10】ST0452蛋白質の80℃(○)及び95℃(●)での熱処理後における残存活性量を示す図である。
【特許請求の範囲】
【請求項1】
配列番号4に記載のアミノ酸配列を有するか、あるいは、配列番号4に記載のアミノ酸配列に一乃至数個のアミノ酸残基が欠失、置換、挿入又は付加されたアミノ酸配列を有し、かつアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有することを特徴とする、蛋白質。
【請求項2】
請求項1記載の蛋白質をコードするDNA。
【請求項3】
配列番号5に記載の塩基配列を有することを特徴とするDNA。
【請求項4】
配列番号5に記載のDNA とストリンジェントな条件下でハイブリダイズし、かつアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する蛋白質をコードすることを特徴とするDNA。
【請求項5】
請求項2〜4のいずれかに記載のDNAから選ばれるDNAがベクターに組み込まれていることを特徴とする組換え体DNA。
【請求項6】
請求項5に記載の組換え体DNAが宿主細胞に導入されていることを特徴とする形質転換体。
【請求項7】
請求項6に記載の形質転換体を培地に培養し、培養物からアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する蛋白質を採取することを特徴とする、アセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する蛋白質の製造方法。
【請求項8】
グルコサミン-1-リン酸及び/又はガラクトサミン-1-リン酸に、アセチルCoA存在下、請求項1に記載の蛋白質を作用させることを特徴とする、N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸の製造方法。
【請求項9】
グルコサミン-1-リン酸及び/又はガラクトサミン-1-リン酸に、アセチルCoA存在下、請求項6に記載の形質転換体の培養液あるいは培養物の処理物を作用させることを特徴とする、N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸の製造方法。
【請求項10】
グルコサミン-1-リン酸及び/又はガラクトサミン-1-リン酸に、アセチルCoA存在下、 請求項3又は4に記載のDNAにコードされるタンパク質を作用させることを特徴とする、N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸の製造方法。
【請求項11】
N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸に、ヌクレオシド三リン酸の存在下、請求項1に記載のタンパク質を作用させることを特徴とする、N-アセチルグルコサミンおよび/又はN-アセチルガラクトサミンのヌクレオシド二リン酸結合体の製造方法。
【請求項12】
N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸に、ヌクレオシド三リン酸の存在下、請求項6に記載の形質転換体の培養液あるいは培養物の処理物を作用させることを特徴とする、N-アセチルグルコサミンおよび/又はN-アセチルガラクトサミンのヌクレオシド二リン酸結合体の製造方法。
【請求項13】
N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸に、ヌクレオシド三リン酸の存在下、請求項2〜4のいずれかに記載のDNAにコードされるタンパク質を作用させることを特徴とする、N-アセチルグルコサミンおよび/又はN-アセチルガラクトサミンのヌクレオシド二リン酸結合体の製造方法。
【請求項1】
配列番号4に記載のアミノ酸配列を有するか、あるいは、配列番号4に記載のアミノ酸配列に一乃至数個のアミノ酸残基が欠失、置換、挿入又は付加されたアミノ酸配列を有し、かつアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有することを特徴とする、蛋白質。
【請求項2】
請求項1記載の蛋白質をコードするDNA。
【請求項3】
配列番号5に記載の塩基配列を有することを特徴とするDNA。
【請求項4】
配列番号5に記載のDNA とストリンジェントな条件下でハイブリダイズし、かつアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する蛋白質をコードすることを特徴とするDNA。
【請求項5】
請求項2〜4のいずれかに記載のDNAから選ばれるDNAがベクターに組み込まれていることを特徴とする組換え体DNA。
【請求項6】
請求項5に記載の組換え体DNAが宿主細胞に導入されていることを特徴とする形質転換体。
【請求項7】
請求項6に記載の形質転換体を培地に培養し、培養物からアセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する蛋白質を採取することを特徴とする、アセチル化アミノ糖合成活性及びアセチル化アミノ糖ヌクレオチド合成活性を有する蛋白質の製造方法。
【請求項8】
グルコサミン-1-リン酸及び/又はガラクトサミン-1-リン酸に、アセチルCoA存在下、請求項1に記載の蛋白質を作用させることを特徴とする、N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸の製造方法。
【請求項9】
グルコサミン-1-リン酸及び/又はガラクトサミン-1-リン酸に、アセチルCoA存在下、請求項6に記載の形質転換体の培養液あるいは培養物の処理物を作用させることを特徴とする、N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸の製造方法。
【請求項10】
グルコサミン-1-リン酸及び/又はガラクトサミン-1-リン酸に、アセチルCoA存在下、 請求項3又は4に記載のDNAにコードされるタンパク質を作用させることを特徴とする、N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸の製造方法。
【請求項11】
N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸に、ヌクレオシド三リン酸の存在下、請求項1に記載のタンパク質を作用させることを特徴とする、N-アセチルグルコサミンおよび/又はN-アセチルガラクトサミンのヌクレオシド二リン酸結合体の製造方法。
【請求項12】
N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸に、ヌクレオシド三リン酸の存在下、請求項6に記載の形質転換体の培養液あるいは培養物の処理物を作用させることを特徴とする、N-アセチルグルコサミンおよび/又はN-アセチルガラクトサミンのヌクレオシド二リン酸結合体の製造方法。
【請求項13】
N-アセチルグルコサミン-1-リン酸及び/又はN-アセチルガラクトサミン-1-リン酸に、ヌクレオシド三リン酸の存在下、請求項2〜4のいずれかに記載のDNAにコードされるタンパク質を作用させることを特徴とする、N-アセチルグルコサミンおよび/又はN-アセチルガラクトサミンのヌクレオシド二リン酸結合体の製造方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図8】
【図9】
【図10】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図8】
【図9】
【図10】
【図7】
【公開番号】特開2008−148584(P2008−148584A)
【公開日】平成20年7月3日(2008.7.3)
【国際特許分類】
【出願番号】特願2006−337443(P2006−337443)
【出願日】平成18年12月14日(2006.12.14)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成16年度北海道大学委託研究「遺伝子情報解析に関する研究」委託研究、産業活力再生特別措置法第30条の適用を受ける特許出願
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成20年7月3日(2008.7.3)
【国際特許分類】
【出願日】平成18年12月14日(2006.12.14)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成16年度北海道大学委託研究「遺伝子情報解析に関する研究」委託研究、産業活力再生特別措置法第30条の適用を受ける特許出願
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]