
アテローム性動脈硬化症を軽減するための、選択された乳酸菌の使用
アテローム性動脈硬化症及び他の心血管疾患を予防及び/又は治療するための、BSH活性を増加させ、その結果として血清LDLコレステロールを低下させ、同時に前炎症性サイトカインのTNF−αの濃度を減少させるその能力について選択された乳酸菌株、このような菌株の選択方法並びにこのような菌株を含む製品。

【発明の詳細な説明】
【技術分野】
【0001】
本明細書において、本発明は、アテローム性動脈硬化症及び他の心血管疾患を予防及び/又は治療するための、胆汁酸塩ヒドロラーゼ(BSH)活性を増加させ、その結果として血清LDLコレステロールを低下させ、同時に前炎症性サイトカインの腫瘍壊死因子(TNF−α)の濃度を減少させるその能力について選択された特定の乳酸菌株、このような菌株の選択方法並びにこのような菌株を含む製品を提供する。
【背景技術】
【0002】
プロバイオティクス
プロバイオティクスは、健康に有益な効果をもたらすことが示されている(Gorbach,S.L.2000.「プロバイオティクス及び胃腸の健康(Probiotics and gastrointestinal health.)」Am.J.Gastroenterol.95:S2〜S4)。多くのさまざまな活性がプロバイオティクスに帰するとされているが、これらの作用が達成される機序はよくわかっていない。これらの作用は、先天的及び後天的な免疫の増強(Gill,H.S.、K.J.Rutherfurd、J.Prasad、及びP.K Gopal.2000.「ラクトバチルス・ラムノサス(HN001)、ラクトバチルス・アシドフィルス(HN017)及びビフィドバクテリウム・ラクティス(HN019)による自然免疫及び後天性免疫の増強(Enhancement of natural and acquired immunity by Lactobacillus rhamnosus(HN001)、Lactobacillus acidophilus(HN017)and Bifidobacterium lactis(HN019).)」Br.J.Nutr.83:167〜176)、抗炎症性サイトカイン(IL−10)の産生の増強(Pessi,T.、Y.Sutas、M.Hurme及びE.Isolauri.2000.「ラクトバチルス・ラムノサスGGの経口投与後のアトピーの小児におけるインターロイキン−10の産生(Interleukin−10 generation in atopic children following oral Lactobacillus rhamnosus GG.)」Clin.Exp.Allergy 30:1804〜1808)、及び腸透過性の減少(Madsen,K.、A.Cornish、P.Soper、C.McKaigney、H.Jijon、C.Yachimec、J.Doyle、L.Jewell及びC.De Simone.2001.「プロバイオティック細菌は、マウス及びヒトの腸上皮細胞のバリア機能を増強させる(Probiotic bacteria enhance marine and human intestinal epithelial barrier function.)」Gastroenterology 121:580〜591)を含む。さまざまなラクトバチルス株は動物及びヒトの両方において、特によく研究されている。それらは、旅行者下痢(Marteau,P.R.、M.de Vrese、C.J.Cellier及びJ.Schrezenmeir.2001.「プロバイオティクスの使用による胃腸疾患からの保護(Protection from gastrointestinal diseases with the use of Probiotics.)」Am.J.Clin.Nutr.73:430S〜436S)、再発性クロストリジウム・ディフィシレ(Clostridium difficile)感染症(Gorbach,S.L.1987.「細菌性下痢及びその治療(Bacterial diarrhoea and its treatment.)」Lancet ii:1378〜1382)、ロタウィルス(Szajewska,H.、M.Kotowska、J.Z.Mrukowicz、M.Armanska及びW.Mikolajczyk.2001.「幼児の院内下痢症の予防における、ラクトバチルスGG(L.GG)の有効性(Efficacy of Lactobacillus GG(L.GG)in prevention of nosocomial diarrhea in infants.)」J.Pediatr.138:361〜365)及びヘリコバクター(Helicobacter)感染症(Mukai,T.、T.Asasaka、E.Sato、K.Mori、M.Matsumoto及びH Ohori.2002.「プロバイオティックなラクトバチルス・ロイテリによる、ヘリコバクター・ピロリの糖脂質受容体への結合の阻害(Inhibition of binding of Helicobacter pylori to the glycolipid receptors by probiotic Lactobacillus reuteni.)」FEMS Immunol.Med.Microbiol.32:105〜110)の予防及び治療に有効であり得る。マウスの腸から単離されたL.ロイテリ(L.Reuteri)は、IL−10トランスジェニック・ノックアウトマウスにおいて、大腸炎の発症を阻害した(Madsen,K.L.、J.S.Doyle、L.D.Jewell、M.M.Tavernini及びR.N.Fedorak.1999.「ラクトバチルス属種は、インターロイキン10遺伝子欠損マウスにおいて、大腸炎を予防する(Lactobacillus species prevents colitis in interleukin 10 gene−deficient mice.)」Gastroenterology 116:1107〜1114)。プロバイオティクス混合物を用いた臨床試験により、慢性回腸嚢炎において相当の改善が示された(Gionchetti,P.、F.Rizzello、A.Venturi、P.Brigidi、D.Matteuzzi、G.Bazzocchi、G.Poggioli、M.Miglioli及びM.Campieri.2000.「慢性回腸嚢炎を有する患者における維持療法としての経口細菌製剤療法:二重盲検の、プラセボを対照とした試験(Oral bacteriotherapy as maintenance treatment in patients with chronic pouchitis:a double−blind、placebo−controlled trial.)」Gastroenterology 119:305〜309)。
【0003】
免疫応答(Th−1/Th2/TR)
炎症は、サイトカインとして知られる細胞内シグナルタンパク質によって仲介され、これらはマクロファージ及び樹状細胞によって、上皮において、抗原性刺激に反応して産生される。上皮と抗原の間の接触において、上皮中の抗原提示細胞(樹状細胞を含む)が、ナイーブなマクロファージにシグナルを伝播し、ナイーブなマクロファージはその後いわゆるTh−1型応答で応答し、TNFα、IL−1、IL−6、IL−12を含む前炎症性サイトカインが、マクロファージによって産生される。これらのサイトカインは、ナチュラルキラー細胞、T細胞及び他の細胞を順番に刺激し、インターフェロンγ(IFNγ)が産生され、ナイーブなマクロファージは、Th−2型応答で抗原に応答することもあり得る。この応答は、IFNγによって抑制される。これらのTh−2型細胞は、IL−4、IL−5、IL−9及びIL−10などの抗炎症性サイトカインを産生する。
【0004】
IL−10は、IFNγの産生を阻害し、したがって免疫応答を抑制することが知られている。Th−1型及びTh−2型の細胞と、それらのそれぞれのサイトカインの産生とのバランスが、所与の抗原に対する炎症反応の程度を規定する。Th−2型細胞は、免疫系を介して免疫グロブリンの産生もまた刺激できる。TNFαの濃度が低減される場所である消化管における抗炎症活性は、上皮細胞の増強(腸壁内膜の完全性)、したがって消化管病原体及び毒素により起こる負の作用の減少に相関する。
【0005】
制御性T(TR)細胞は、免疫応答の不可欠な構成要素とみなされる。これらの細胞は、有害な免疫病理を最小にするために、まず防御的抗菌免疫を微調整すると思われる(Powrie F、Maloy KJ.2003.「レギュレーターの制御(Regulating the regulators)」、Science 299 1030〜1031)。TR細胞は、抗炎症性サイトカインのIL−10の濃度を増加させることが示された(Smits,H.H.、A.Engering、D.van der Kleij、E.C.de Jong、K.Schipper、T.M.van Capel、B.A.J.Zaat、M.Yazdanbakhsh、E.A.Wierenga、Y.van Kooyk及びL.Kapsenberg.2005.「選択的プロバイオティック細菌は、樹状細胞特異的細胞間接着分子3−捕捉非インテグリンを介して樹状細胞機能を調節することによって、インビトロでIL−10産生制御性T細胞を誘導する(Selective probiotic bacteria induce IL−10−producing regulatory T cells in vitro by modulating dendritic cell function through dendritic cell−specific intercellular adhesion molecule 3−grabbing nonintegrin.)」J Allergy Clin Immunol.115:1260〜1267)。TR細胞の発生及び活性化を調節する因子は、(自己免疫疾患を治療し、臓器の移植片の生存を増強するために)TR細胞の活性の方向か、又は(ワクチン接種及び腫瘍拒絶を強化するために)TR細胞の活性から離れる方向どちらかに平衡を移すことができるはずである(Walter J.Dobrogosz.「L.ロイテリを用いたヒトの健康の増強、プロバイオティック、イムノバイオティック及びイムノプロバイオティック(Enhancement of human health with L.reuteri、A Probiotic、Immunobiotic and Immunoprobiotic.)」NUTRAfoods.2005:4(2/3)15〜28)。
【0006】
プロバイオティクスの免疫調節作用
ラクトバチルス・ラムノサス(Lactobacillus rhamnosus)株GG(LGG)は、潜在的プロバイオティック剤であり、多くの研究により、腸管にコロニーを形成し、粘膜上皮及び免疫応答を調節するLGGの能力が実証されている。LGGは、単独定着ノトバイオートラットにおいて、腸細胞の増殖及び絨毛の大きさを増大させた(Banasaz,M.、E.Norin、R.Holma及びT.Midtvedt.2002.「ラクトバチルス・ラムノサスGGによる、単独定着ノトバイオートラットにおける腸細胞産生の増加(Increased enterocyte production in gnotobiotic rats mono−associated with Lactobacillus rhamnosus GG.)」Appl Environ Microbiol.68:3031〜3034)。LGGは、経口投与後に、エクスビボでマウスリンパ球応答の拡散もまた調節し(Kirjava(登録商標)inen,P.V.、H.S.ElNezami、S.J.Salminen、J.T.Ahokas及びP.F.Wright.1999.「マウスリンパ球増殖に関する、経口投与された、生存ラクトバチルス・ラムノサスGG及びプロピオニバクテリウム・フリューデンレイッヒイ亜種シャーマニィの作用(Effects of orally administered viable Lactobacillus rhamnosus GG and Propionibacterium freudenreichii subsp.sherinanii JS on mouse lymphocyte proliferation.)」Clin Diagn Lab Immunol 6:799〜802)、L.パラカセイ(L.paracasei)は、CD4+Tリンパ球の調節性サイトカインのプロフィールを改変する(Von der Weid T.、C.Bulliard及びE.J.Schiffrin.2001.「形質転換増殖因子β及びインターロイキン−10を産生する、低増殖能を有するCD4(+)T細胞の集団の乳酸菌による誘導(Induction by a lactic acid bacterium of a population of CD4(+)T cells with low proliferative capacity that produce transforming growth factor beta and interleukin−10.)」Clin Diagn Lab Immunol 8:695〜701)。適応免疫応答に加え、LGGは先天性免疫応答に対する影響を有する。LGGは核因子κB(NF−κB)、並びにヒトマクロファージにおけるシグナル伝達物質及び転写活性化因子(STAT)シグナル伝達経路を活性化し(Miettinen,M.、A.Lehtonen、I.Julkunen、及びS.Matikainen.2000.「乳酸桿菌及び連鎖球菌が、ヒトマクロファージにおいて、NF−κB及びSTATシグナル伝達経路を活性化する(Lactobacilli and Streptococci activate NF−kappa B and STAT signaling pathways in human macrophages.)」J Immunol 164:3733〜3740)、L.ラムノサス(L.rhamnosus)はマクロファージによるインターロイキン−12(IL−12)の産生を刺激する(Hessle,C.、L.A.Hanson及びA.E.Wold.1999.「ヒト胃腸粘膜由来の乳酸桿菌は、IL−12産生の強力な刺激因子である(Lactobacilli from human gastrointestinal mucosa are strong stimulators of IL−12 production.)」Clin Exp Immunol 116:276〜282)。LGGはまた小児においてIL−10のような免疫調節性サイトカインの産生も刺激し(Pessi,T.、Y.Sutas、M.Hurme及びE.Isolauri.2000.「ラクトバチルス・ラムノサスGGの経口投与後のアトピーの小児におけるインターロイキン−10の産生(Interleukin−l0 generation in atopic children following oral Lactobacillus rhamnosus GG.)」Clin Exp Allergy 30:1804〜1808)、インビボで前炎症性応答を制御できる。マクロファージ、樹状細胞及び好中球のような先天性免疫のエフェクター細胞は、大部分の炎症反応の一次駆動体である(Janeway,C.A.,Jr.及びR.Medzhitov.2002.「先天性免疫の認識(Innate immune recognition.)」Annu Rev Immunol 20:197〜216)。先天性免疫が、抗原に対する先天性応答及び後天性応答の両方の過程を、自己又は非自己として決定づけるという考えは、炎症の調節における先天性免疫の役割を強調している。
【0007】
米国特許出願公開第20020019043号は、サイトカインを産生するグラム陽性菌又はサイトカイン拮抗薬を産生するグラム陽性菌株を投与することによる、炎症性腸疾患の治療に関する。特定の実施形態において、サイトカイン又はサイトカイン拮抗薬は、IL−10、可溶性TNF−α受容体又は別のTNF−α拮抗薬、IL−12拮抗薬、インターフェロン−γ拮抗薬、IL−1拮抗薬などから選択される。特定の実施形態において、グラム陽性菌は、全般にサイトカイン、サイトカイン拮抗薬などを産生するように、遺伝子操作されている。
【0008】
L.ロイテリ(L.reuteri)の免疫調節作用
L.ロイテリ(L.reuteri)の免疫調節作用は、例えばプロバイオティック乳酸桿菌がそれらの免疫調節作用を、樹状細胞(DC)のTh1/Th2/Th3/Tr1/TR促進能力を調節することによって発揮することを示した、Christensenによって報告された(Christensen H.R.、H.Frokiaer及びJ.J.Pestka.2002.「マウスの樹状細胞において、乳酸桿菌はサイトカイン及び成熟表面マーカーの発現を特異的に調節する(Lactobacilli differentially modulate expression of cytokines and maturation surface markers in murine dendritic cells.)」J.Immunol.168:171〜178)。彼らは、マウスのDCが、L.ロイテリ(L.reuteri)株を含むさまざまなラクトバチルス株の共培養に曝された場合、それらは、サイトカインIL−6、IL−10、IL−12及びTNF−αの産生並びに表面マーカーのMHCクラスII及びCD86の上方制御が、濃度依存的方法で、特異的に調節されたことを示した。すべての乳酸桿菌が、DCの成熟の指標である、表面MHCクラスII及びCD86マーカーを上方制御した。これらの研究において特に注目に値するのは、L.ロイテリ(L.reuteri)(12246株)がIL−12の誘導因子としては弱いが、L.ジョンソニー(L.johnsonii)又はL.カゼイ(L.casei)と共培養した場合、後者の2種によって刺激された前炎症性サイトカインシグナルIL−12、IL−6及びTNF−αの産生を、特異的に阻害することであった。IL−10の産生は、これらの条件下でも変化しなかった。これらの発見により、「L.ロイテリ(L.reuteri)は、「危険なシグナル」を持たない抗原に対する耐性を支持し、同時にLPSのような危険なシグナルを介して認識される病原体に対する応答能力は損なわれないまま維持する、腸の樹状細胞発生の環境調節に寄与できる」という彼らの結論が導かれた。彼らはまた、いくつかのL.ロイテリ(L.reuteri)株が、抗炎症性IL−10を導入しながら、IL−12及びTNF−α(及びIL−6)の産生を下方制御するのに有効な、潜在的な標的の細かい治療であり得、したがって、前炎症腸サイトカイン環境の釣り合いをとる、別の治療的取り組みを提示すると結論付けた。
【0009】
Smitsは、これらの見解を拡大し、L.ロイテリ(L.reuteri)が、DCに制御性T(TR)細胞の産生を刺激させる能力を有することを示した。彼らは、インビトロで共培養した3種の異なるラクトバチルス属種及びヒト単球由来のDCを使用した。乳酸桿菌の2種、ヒトL.ロイテリ(L.reuteri)(ATCC 53609)株及びL.カゼイ(L.casei)は、これらのDCのTR細胞の発生を刺激するが、L.プランタラム(L.plantarur)株はしなかった。これらのTR細胞は、IL−10の産生濃度が増加することが示され、IL−10依存的にそばにいるT細胞の増殖を阻害できた(Smits,H.H.、A.Engering、D.van der Kleij、E.C.de Jong、K.Schipper、T.M.M.van Capel B.A.J.Zaat、M.Yazdanbakhsh、E.A.Wierenga、Y.van Kooyk及びL.Kapsenberg.2005.「選択的プロバイオティック細菌は、樹状細胞特異的細胞間接着分子3捕捉非インテグリンを介して、樹状細胞機能を調節することによって、インビトロでIL−10産生制御性T細胞を誘導する(Selective probiotic bacteria induce IL− 10−producing regulatory T cells in vitro by modulating dendritic cell function through dendritic cell−specific intercellular adhesion molecule 3−grabbing nonintegrin.)」J Allergy Clin Immunol.115:1260〜1267)。L.ロイテリ(L.reuteri)−DC相互作用に関するこれらの研究は、Horiによる、根底を覆す研究に関連するとみなされる(Hori S、T.Nomura及びS.Sakaguchi.2003.「転写因子Foxp3による、制御性T細胞発生の調節(Control of regulatory T cell development by the transcription factor Foxp3.)」Science.299:1057〜1061)並びにPasare及びMedzhitov(Pasare C.及びR.Medzhitov.2003.「樹状細胞によるCD4+Cd25+T細胞仲介抑制のToll経路依存性阻害(Toll pathway−dependent blockade of CD4+Cd25+ T cell−mediated suppression by dendritic cells.)」Science 299:1033〜1036)は、L.ロイテリ(L.reuteri)のイムノバイオティックな作用様式の1つに貴重な洞察を提供している。
【0010】
神経増殖因子(NGF)は、神経細胞に関するその活性に加えて、大腸炎のモデルを含むインビトロ及びインビボのいくつかの実験系において、有意な抗炎症作用を有する。Ma他(2004)は、L.ロイテリ(L.reuteri)の作用機序を、ヒト上皮細胞系において、サイトカイン及びNGFの合成並びにTNF−αに対するIL−8の応答を調査した。彼らは、L.ロイテリ(L.reuteri)は、強力な直接抗炎症活性を、ヒト上皮細胞において有し、それは摂取したプロバイオティクスの活性に関係すると思われると結論付けた。彼らはまた、L.ロイテリ(L.reuteri)は、独特の抗炎症性分子であるNGFを上方制御し、NF−κBの核への転移を阻害するとも結論付けた(Ma,D.、P.Forsythe及びJ.Bienenstock.2004.「生きたL.ロイテリは、腫瘍壊死因子αにより誘導されるインターロイキン8の発現の阻害作用に必須である(Live L.reuteri is essential for the inhibitory effect on tumor necrosis factor alpha−induced interleukin−8 expression.)」Infect.Immun.72:5308〜5314)。
【0011】
L.ロイテリ(L.reuteri)を含む、ラクトバチルス属種の多種多様な菌株は、プロバイオティック製剤に使用されている。ラクトバチルス・ロイテリ(Lactobacillus reuteri)は、動物の消化管の自然な常在菌の1つであり、ヒトを含む、健康な動物の腸において日常的に見出される。それは、抗菌作用を有することでも知られている。例えば、米国特許第5,439,678号、第5,458,875号、第5,534,253号、第5,837,238号及び第5,849,289号を参照されたい。L.ロイテリ(L.reuteri)細胞が、嫌気条件下で、グリセロールの存在下で増殖する場合、それらは、β−ヒドロキシ−プロピオンアルデヒド(3−HPA)として知られる抗菌物質を産生する。
【0012】
アテローム性動脈硬化症
動脈硬化性疾患及びその心血管結果は、米国その他における死亡及び罹患の主原因である。アテローム性動脈硬化症は、「粥(gruel)」又は「ベトベトしたもの(goo)」及び「硬化(hardening)」というギリシャ語に由来し、動脈の内壁にアテローム又は病巣が存在するとして定義される。プラークとしても知られる病巣は、脂肪性沈着物及び他の物質からなる。
【0013】
アテローム性動脈硬化症を特に危険にしているものは、重要な大動脈に対して特別な親和力を有することと思われる。プラークの詰まった病巣の一部が、動脈の内壁から破裂した場合、脂肪性材料が、心臓及び脳に直接供給している下流の小さい動脈に流れ込み、そこでそれらが詰まり、栄養及び酸素に富んだ血液がこれらの重要臓器に到達することを妨げる。完全な封鎖が起こった場合、心臓発作又は脳卒中が起こり得る(Little,W.C.、M.Constantinescu、R.J.Applegate、M.A.Kutcher、M.T.Burrows、F.R.Kahl及びW.P.Santamore.「冠動脈造影は、軽度から中程度の冠動脈疾患を有する患者のその後の心筋梗塞部位を予測できるか?(Can coronary angiography predict the site of a subsequent myocardial infarction in patients with mild−to−moderate coronary artery disease?)」Circulation.1988.78:1157〜66)。伝統的に、アテローム性動脈硬化症は、脂質代謝障害であると考えられてきた。アテローム性動脈硬化症に伴う危険因子は、LDLの血中濃度の高さ、ホモシステイン、高血圧、喫煙、肥満、糖尿病を含む。治療は、コレステロール濃度の調節、例えば特定の乳酸菌による胆汁塩の代謝の促進に焦点が当てられてきた。
【0014】
有効なプロバイオティクスとしての乳酸菌(LAB)を使用する可能性を評価する場合、多くの人が、胆汁酸の作用に耐えるLABのその能力を評価する必要があると考えている。胆汁酸は、肝臓においてコレステロールから合成され、胆嚢から十二指腸に、グリシン又はタウリンと抱合されて分泌される。それらの機能は、食物中の脂肪を乳化することである。ヒトにおける最も一般的な一次胆汁酸は、コール酸及びケノデオキシコール酸であり、これらは、肝臓におけるコレステロール代謝の主要な最終産物である。腸における細菌活性の結果として、これらの酸はその後脱抱合及び脱ヒドロキシル化などの化学的修飾を受け、そこでアミノ酸は抱合型から加水分解する(Cardona,M.E.、V.de Vanay、T.Midtvedt及びK.E.Nogin.「ノトバイオノートマウスにおけるプロバイオティクス。インビトロ及びインビボにおける、コレステロールのコプロスタノールへの転換及びインビトロにおける胆汁酸の脱抱合(Probiotics in gnotobiotic mice.Conversion of cholesterol to coprostanol in vitro and in vivo and bile acid deconjugation in vitro.)」Microb Ecol Health Dis.2000.12:219〜224;Dunne,C.、L.O’Mahony、L.Murphy、G.Thornton、D.Morrissey、S.O’Halloran、M.Feeney、S.Flynn、G.Fitzgerald、D.Daly、B.Kiely、G.C.O’Sullivan、F.Shanahan及びJ.K.Collins.「ヒト起源のプロバイオティック菌に関するインビトロの選択基準:インビボの発見との相関(In vitro selection criteria for probiotic bacteria of human origin:correlation with in vivo findings.)」Am J Clin Nutr.2001.73(suppl):386S〜392S)。いくつかの胃腸(GI)細菌、例えば、エンテロコッカス(Enterococcus)、ビフィドバクテリウム(Bifidobacterium)及びラクトバチルス(Lactobacillus)は、酵素の胆汁酸塩ヒドロラーゼ(BSH)を発現し、それは、抱合胆汁酸の加水分解を触媒し、遊離のグリシン又はタウリン及び非抱合型胆汁酸分子をもたらす(Tanaka,H.、K.Doesburg、T.Iwasaki及びI.Mierau.「胆汁塩ヒドロラーゼ活性のための乳酸菌のスクリーニング(Screening of lactic acid bacteria for bile salt hydrolase activity.)」J Dairy Sci.1999.82:2530〜2535;Bateup,J.M.、M.A.McConnell、H.F.Jenkinson及びG.W.Tannock.「胆汁塩ヒドロラーゼ活性、消化管のコロニー形成及びマウス宿主の増殖率に関する、ラクトバチルス株の比較(Comparison of Lactobacillus strains with respect to bile salt hydrolase activity、colonization of the gastrointestinal tract、and growth rate of the murine host.)」Appl Environ Microbiol.1995.61(3):1147〜1149;Tannock,G.W.、J.M.Bateup及びH.F.Jenkinson.「乳酸桿菌のインビトロの増殖に関するタウロコール酸ナトリウムの効果(Effect of sodium taurocholate on the in vitro growth of lactobacilli.)」Microb Ecol.1997.33:163〜167)。
【0015】
GI管において、BSHの発現がどのように細菌の機能に影響を与えるかについて、2つの主要な仮説がある。一方は、いくつかの細菌は胆汁塩を脱抱合し、アミノ酸のタウリンを電子受容体として使用するというもので、これに反して他方は、脱抱合型は、界面活性が低下して可溶性が低いので、酵素が胆汁塩の毒性を脱抱合によって減少させ、その結果細菌が保護されると述べている。抱合胆汁酸及び脱抱合胆汁酸の両方が、大腸菌(Escherichia coli)、クレブシエラ属種(Klebsiella sp)及びエンテロコッカス属種(Enterococcus sp)に対してインビトロで抗菌活性を示すことが見出されており、脱抱合型はより増殖阻害性である(Dunne,C.、L.O’Mahony、L.Murphy、G.Thornton、D.Morrissey、S.O’Halloran、M.Feeney、S.Flynn、G.Fitzgerald、C.Daly、B.Kiely、G.C.O’Sullivan、F.Shanahan及びJ.K.Collins.「ヒト起源のプロバイオティック菌に関するインビトロの選択基準:インビボの発見との相関(In vitro selection criteria for probiotic bacteria of human origin:correlation with in vivo findings.)」Am J Clin Nutr.2001.73(suppl):386S〜392S;Moser,S.A.及びD.C.Savage.「乳酸桿菌において、胆汁塩ヒドロラーゼ活性と抱合胆汁塩の毒性に対する耐性とは、関連のない特性である(Bile salt hydeolase activity and resistance to toxicity of conjugated bile salts are unrelated properties in lactobacilli.)」Appl Environ Microbiol.2001.67(8):3476〜3480)。
【0016】
発酵乳製品の強力なコレステロール低下作用は、コレステロールの胆汁酸との結合及びミセル形成の阻害によって説明できる。これらの製品中のプロバイオティック細菌がコレステロール低下作用を有し得る機序は、肝細胞によってコレステロールから作られた胆汁酸のコール酸及びデオキシコール酸を介する。これらは、グリシン及びタウリンと抱合し、小腸へ入り、そこで吸収され、肝臓へ向かう。再吸収の間に、抱合胆汁酸は腸においてミクロフローラに曝される。発酵食品中の細菌、例えば、乳酸桿菌及び連鎖球菌は、抱合胆汁酸を加水分解する。腸において高い胆汁塩ヒドロラーゼ活性を有するラクトバチルス(Lactobacillus)株は、胆汁の加水分解が促進できる可能性がある。このことは、コレステロールの転換率をより速くし、より多くの胆汁酸を産生する。インビボでコレステロールが低下するのは、脱抱合胆汁酸は、結腸において再吸収されないので、排泄物を介して胆汁酸が排出されるためである。このことは、新たな胆汁の合成を促進し、身体の胆汁のプールを一定に保つ(St−Onge M−P.、E.R.Farnworth及びP.J.H.Jones.2000.「発酵乳製品及び非発酵乳製品の摂取:コレステロールの濃度及び代謝に関する効果(Consumption of fermented and nonfermented dairy products:effects on cholesterol concentrations and metabolism.)」Am J Clin Nutr.71:674〜681)。
【0017】
胆汁酸の脱抱合は、血漿コレステロール濃度を低下させるであろう。しかし、これらの化合物は、大腸において、嫌気性細菌によって二次胆汁酸にさらに転換されることがあり、二次胆汁酸は、結腸癌の誘導因子候補として関係があるとされている。二次胆汁酸は、細胞系に対して有毒であり、それらは結腸粘膜において細胞毒性を発揮し、細胞増殖の増加をもたらすと考えられている。これらの過剰増殖性細胞は、変異原物質に対する感受性が増強されており、その結果結腸癌の危険性が増す(Hepner,G.、R.Fried、S.St.Jeor、L.Fusetti及びR.Morin.1979.「ヨーグルト及びミルクの高コレステロール作用(Hypercholesterolemic effect of yoghurt and milk.)」Am.J.Clin.Nutr.32:19〜24)。幸運なことに、乳酸菌は、脱抱合胆汁塩及び二次胆汁塩の可溶性を減少させると思われ、その結果、それらの生体利用率が低下する。De Boever他(2000)による研究は、L.ロイテリ(L.reuteri)が、細菌培養物中の胆汁塩の毒性を減少させたことを示した。さらに重要なことには、L.ロイテリ(L.reuteri)を加えることにより、赤血球の溶解に対するほぼ完全な耐性及びヒーラ細胞における胆汁塩の毒性の阻害がもたらされた(De Boever,P.、R.Wouters、L.Verschaeve、P.Berckmans、G.Schoeters及びW.Verstraete.「胆汁塩の細胞毒性に対する、胆汁塩ヒドロラーゼ活性ラクトバチルス・ロイテリの保護作用(Protective effect of the bile salt hydrolase−active Lactobacillus reuteri against bile salt cytotoxicity.)」Appl Microbiol Biotechnol.2000.53(6):709〜14)。
【0018】
免疫疾患であるアテローム性動脈硬化症
科学者は、アテローム性動脈硬化症の発症に関する新規なスキームを表しており、この病状が、前炎症性T細胞と、沈静しているものであるTRとのバランスの悪さによりもたらされ得ることを示唆している。このことは、第二回欧州血管ゲノム科学ネットワーク会議(Second European Vascular Genomics Network Conference)(EVGN Conference−ハンブルグ、2005年9月27〜30日)から現れた、興味をそそる科学的結果の1つである。これらの結果は、心疾患における炎症の役割に新しい洞察を提供し、血栓形成及び心臓発作をもたらす過程の新しい情報モデルの開発をもたらした。
【0019】
LDL粒子中の脂肪は内皮細胞を刺激し、単球及びTリンパ球を含むいくつかの細胞型の細胞浸潤を引き起こすので、アテローム性動脈硬化症は、内皮における脂肪線条の形成により始まる。単球は、内皮層と相互に作用し、内皮に硬く付着し、内皮細胞下の空間に遊走し、そこで単球はマクロファージに分化する。マクロファージは、サイトカインを含むさまざまな化学物質を放出する。増殖因子の産生が刺激され、これにより細胞増殖及び基質産生並びに基質の変性をもたらすメタロプロテイナーゼがもたらされる。したがって、マクロファージは、病巣の増殖に寄与し、不安定事象及び血栓事象に寄与し得る(Ross R.「アテローム性動脈硬化症−炎症性疾患(Athrosclerosis−An inflammatory disease.)」N Engl J Med.1999.340:115〜26)。Tリンパ球は、アテローム性動脈硬化症の全段階において存在することが示されている。それらの存在が、免疫応答との関連のさらなる証拠を提供する(Kol,A.及びP.Libby.1998.「感染性因子がアテローム性動脈硬化症に寄与し得る機序及びその臨床症状(The mechanisms by which infectious agents may contribute to atherosclerosis and its clinical manifestations.)」Trends Cardiovasc Med.8:191〜99;Andreotti,F.、F.Burzotta、A.Mazza、A.Manzoli、K.Robinson及びA.Maseri.1999.「ホモシステイン及び動脈閉塞性疾患:簡潔な総説(Homocysteine and arterial occlusive disease:a concise review.)」Cardiologia.44:341〜5)。
【0020】
炎症性物質産生の開始シグナルは、炎症シグナル伝達経路を活性化するいくつかの内在性分子を認識する、トール様受容体と呼ばれる受容体の関与に依存する(K.Edfeldt、J.Swedenborg、G.K.Hansson及びZ.Yan.2002.「ヒトアテローム性動脈硬化症の病巣におけるトール様受容体の発現:プラーク活性化のための可能な経路(Expression of Toll−Like Receptors in Human Atherosclerotic Lesions:A Possible Pathway for Plaque Activation)」Circulation.105:1158〜1161)。
【0021】
トール様受容体(TLR)は、微生物のモチーフを認識し、サイトカインの産生をもたらす一連の遺伝子を活性化する。伝統的に、TLRは、微生物感染のセンサーとみなされており、それらの役割は、炎症反応を誘導することである。しかし、TLRにより認識されるモチーフは、病原体に対して唯一ではなく、微生物のすべてのクラスによって共有される一般的モチーフであり、免疫系が、片利共生生物と病原菌とを、TLRを介してどのように識別するかは完全には理解されなかった。近年、TLRが、炎症反応の誘導におけるそれらの役割にもかかわらず、片利共生生物のミクロフローラを認識することによって腸内恒常性を維持する役割も担っているというデータが示された(Rakoff−Nahoum,S.、J.Paglino、F.Eslami−Varzaneh、S.Edberg及びR.Medzhitov.2004.「トール様受容体による片利共生生物のミクロフローラの認識は、腸内の恒常性にとって必要である。(Recognition of commensal microflora by toll−like receptors is required for intestinal homeostasis.)」Cell.23;118(2):229〜41)。
【0022】
炎症の血清マーカーが、心血管疾患の罹患及び死亡に関する独立した危険因子であることが確立されている。心血管疾患の終点にかかわる炎症性マーカーは、IL−6及びTNF−αなどの前炎症性サイトカイン、フィブリノゲン並びにC反応性タンパク質(CRP)を含む(Libby,P.、P.M.Ridker及びA.Maseri.2001.「炎症及びアテローム性動脈硬化症(Inflammation and atherosclerosis.)」Circulation.2002.105:1135〜1143;Ridker,P.M.「高感受性C反応性タンパク質:心血管疾患の一次予防における包括的リスク評価のための潜在的補助物質(High sensitivity C−reactive protein:potential adjunct for global risk assessment in the primary prevention of cardiovascular disease.)」Circulation.103:1813〜1818)。
【0023】
アテローム性動脈硬化症における肺炎クラミジア(C.pneumoaie)及びH.ピロリ(H.pylori)の役割
累積証拠により、アテローム性動脈硬化症が炎症性疾患であることが示唆されている。したがって、近年、感染性因子が心血管疾患の病因において役割を果たす可能性に多くの注目が集まっている。特定の感染性因子が、アテローム斑からそれらが単離されること、又は肺炎クラミジア(Chlamydia pneumoniae)、ヘリコバクター・ピロリ(Helicobacter pylori)、単純ヘルペスウィルス及びサイトメガロウィルスなどの生命体に関して、血清学的な陽性判定がでることに基づき、関与が示されている。
【0024】
プロスペクティブ研究は決定的な証拠の提供に欠けているが、肺炎クラミジア(C.pneumoniae)は、アテローム性動脈硬化症に最も強力にかかわっていると思われる。肺炎クラミジア(C.pneumoniae)は、死体解剖及び関節切除の標本から、並びに早期の病巣及び十分に発達した病巣の両方において単離されている。免疫学的細胞化学及び組織染色を用いた研究の場合、70〜100%の事例において、関連が見出されている。感染性因子がそれらの作用を発揮する可能な機序は、(i)内皮細胞、平滑筋細胞又はマクロファージに対する局所作用、或いは(ii)サイトカインの産生、単球の刺激及び凝集性高進の促進による全身作用を含み得る。
【0025】
コレステロール濃度の低下に関する従来の治療
血清コレステロール濃度の上昇は、アテローム性動脈硬化症及び冠動脈性心疾患に伴う危険因子であると長年にわたって認識されており、後者は西欧諸国における死亡の主原因である(Barr,D.P.、A.M.Russ及びH.A.Eder.1951.「ヒト血漿におけるタンパク質−脂質の関係 II.アテローム性動脈硬化症及び関連病態において(Protein−lipid relationship in human plasma.II.In atherosclerosis and related conditions.)」Am.J.Med.11:480〜493)。3−ヒドロキシメチルグルタリルコエンザイムA還元酵素阻害薬を含むコレステロールを低下させる多くの薬剤及び胆汁酸の正味の排出量を増加させる薬剤が、高コレステロール(HC)の個人を治療するために使用されてきた(Suckling,K.E.、G.M.Benson、B.Bond、A.Gee、A.Glen、C.Haynes及びB.Jackson.1991.「コレスチラミン治療を用いたハムスターにおける、コレステロールの低下及び胆汁酸の排出(Cholesterol lowering and bile acid excretion in the hamster with cholestyramine treatment.)」Atherosclerosis 89:183〜190)。
【0026】
しかし、これらの化合物の望ましくない副作用により、それらの治療的使用が懸念される。(Erkelens,D.W.、M.G.A.Baggen、J.J.Van Doormeal、M.Kettner、J.C.Koningsberger及びM.J.T.M.Mol.1988.「コレスチラミンと比較したシンバスタチンを用いた臨床経験(Clinical experience with simvastatin compared with cholestyramine.)」Drugs 39(Suppl.):87〜90)。
【0027】
コレステロール濃度を低下させる治療としての乳酸菌
これらの治療手段に加えて、プロバイオティック乳酸菌の摂取が、ヒトにおいて血清コレステロール濃度を低下させるより自然な方法であると思われる。いくつかの研究により、発酵乳製品の大量摂取(680から5000ml/d)の間に血清コレステロールの低下が報告されているが、これらの結果は、摂取のより現実的条件が推定できない(Mann,G.V.1977.「ヒトにおけるコレステロール血を低下させるヨーグルト中の因子(A factor in yogurt which lowers cholesterolemia in man.)」Atherosclerosis 26:335〜340;Mc Namara,D.J.、A.M.Lowell及びJ.E.Sabb.1989.「血中脂質が正常な男性における、血漿中脂質及びリポタンパク質の濃度に関する、ヨーグルト摂取の効果(Effect of yogurt intake on plasma lipid and lipoprotein levels in normolipidemic males.)」Atherosclerosis 79:167〜171)。
【0028】
Masseyは、成人男性において、ヨーグルトの摂取により、初めはコレステロールが10から12%有意に低下したが、2週間後、ヨーグルトの摂取を続けていたにも関わらず、濃度が対照値まで戻ったことを示した(Massey,L.1984.「ヒトの栄養摂取及び血清リポタンパク質に関する、ミルク及びヨーグルトの摂取を変えることによる効果(Effect of changing milk and yoghurt consumption on human nutrient intake and serum lipoprotein.)」J.Dairy Sci.67:255〜262)。類似の矛盾する結果が、ミルク及びその発酵製品を与えられた実験動物により得られた(Hepner,G.、R.S.T.Fried、S.Jeor、L.Fusetti及びR.Morin.1979.「ヨーグルト及びミルクのコレステロール低下作用(Hypocholesterolemic effect of yogurt and milk.)」Am.J.Clin.Nutr.32:19〜24;Rao,D.R.、C.B.Chawan及びS.R.Pulusani.1981.「ラットにおける、血漿コレステロール濃度及び肝臓コレステロール産生に関する、ミルク及びサーモフィルスミルクの影響(Influence of milk and thermophilus milk on plasma cholesterol levels and hepatic cholesterogenesis in rats.)」J.Food Sci.46:1339〜1341)。Raoは、ストレプトコッカス・サーモフィルス(Streptococcus thermophilus)により発酵させたミルクを与えられたラットのHC作用を報告した(Rao,D.R.、C.B.Chawan及びS.R.Pulusani.1981.「ラットにおける、血漿コレステロール濃度及び肝臓コレステロール産生に関する、ミルク及びサーモフィルスミルクの影響(Influence of milk and thermophilus milk on plasma cholesterol levels and hepatic cholesterogenesis in rats.)」J.Food Sci.46:1339〜1341)。Rodasは、ラクトバチルス・アシドフィルス(Lactobacillus acidophilus)を与えられたHCブタにおいて同様の作用を見出した(Rodas,B.Z.、S.E.Gilliland及びC.V.Maxwell.1996.「ラクトバチルス・アシドフィルスATCC 43121のコレステロール低下作用及び食餌によって誘導された高コレステロール血症のブタにおけるカルシウム(Hypocholesterolemic action of Lactobacillus acidophilus ATCC 43121 and calcium in swine with hypercholesterolemia induced by diet.)」J.Dairy Sci.79:2121〜2128)。
【0029】
以前に脂肪に富んだ食餌を与えられたマウスの血清中の、総コレステロール、トリグリセリド及び高比重リポタンパク質(HDL)と低比重リポタンパク質(LDL)との比に関する、L.ロイテリ(L.reuteri)CRL 1098の作用を調査する研究において、L.ロイテリ(L.reuteri)により、トリグリセリドが40%減少し、高比重リポタンパク質(HDL)と低比重リポタンパク質(LDL)との比は20%増加し、ナイーブなミクロフローラの脾臓及び肝臓への細菌転移はなかった(Taranto,M.P.、F.Sesma、A.P.Ruiz Holgado及びG.F.Valdez.1997.「胆汁塩ヒドロラーゼは、ラクトバチルス・ロイテリによるコレステロールの除去に関してカギとなる役割を果たす(Bile salts hydrolase plays a key role on cholesterol removal by Lactobacillus reuteri.)」Biotechnol.Left.9:245〜247)。これらのデータは、L.ロイテリ(L.reuteri)CRL 1098が、低細胞濃度でマウスのために有効なコレステロール低下補助物質であることを示唆している。しかし、本明細書中の本発明の開示とは異なり、コレステロールの低下は、BSH活性のみによるもので、BSH活性及び免疫調節作用との組み合わせによるものではなかった。
【0030】
コレステロール濃度を低下させる治療としての乳酸菌乳酸菌、免疫調節法
米国特許出願公開第20050169901号は、心血管障害を診断、予防及び治療するために、サイトカインの濃度又は活性を調節する方法に関する。このサイトカインの調節は、Th2からTh1へのサイトカインプロフィールの切り替えであり、切り替えが、Th1のサイトカインプロフィールから選択的に離れTNF−α産生の減少に向かう、本明細書の本発明とは異なる。この出願人らは、プロバイオティックとして、いくつかの異なる細菌属及び菌株について述べており、プロバイオティックが、TNF−α濃度の減少及び同時にBSH活性の増加に有効であるように選択された特定の乳酸菌である、本明細書の本発明とは異なる。
【0031】
Bukowskaは、高コレステロール血症の患者において、プロバイオティック細菌のラクトバチルス・プランタラム(Lactobacillus plantarum)299vの補給により、LDLコレステロール及びフィブリノゲンの濃度を有意に低下させることを示した(Bukowska H.、J.Pieczul−Mroz、M.Jastrzebska、K.Chelstowski及びM.Naruszewicz.1997.「中程度にコレステロールが上昇した対象における、ラクトバチルス・プランタラムを含む食餌を補給することに関するフィブリノゲン及びLDLコレステロールの減少(Decrease in fibrinogen and LDL−cholesterol levels upon supplementation of diet with Lactobacillus plantarum in subjects with moderately elevated cholesterol.)」Atherosclerosis.137:437〜8)。このことは、米国特許第6,214,336号にも記載されている。同じグループは、ラクトバチルス・プランタラム(Lactobacillus plantarum)を含む食餌の補給が、喫煙者の代謝障害の予防及び治療に寄与できることを示した。この好ましい作用は、食物繊維の細菌発酵によるプロピオン酸の産生に直接関連していると考えられる。彼らは、プロピオン酸が、おそらく、核転写因子Bを調節し、単球−マクロファージによる炎症性サイトカインの産生を低下させる、ペルオキシソーム増殖因子活性化受容体のイブプロフェンによる活性化に関係する、今に至るまで未知の機序を介して特異的抗炎症作用を発揮することを示唆する(M.Naruszewicz、M−L Johansson、D.Zapolska−Downar及びH.Bukowska、「喫煙者における心血管疾患の危険因子に関する、ラクトバチルス・プランタラム229vの作用(Effect of Lactobacillus plantarum 299v on cardiovascular disease risk factors in smokers.)」Am.J.Clinical Nutrition.2002.76:1249〜1255)。
【0032】
上記の本明細書の本発明とは異なり、参考文献は、BSH−活性の増加、同時にTNF−αの濃度の減少の両方を可能にする菌株を記載していない。
【0033】
前述のように、BSH活性の上昇は、血清コレステロール濃度を低下させ、その結果、アテローム性動脈硬化症の危険性を低下させることが長年にわたって周知であった。アテローム性動脈硬化症が炎症性疾患であることもまた、以前に実証されており、さまざまなサイトカインの調節が疾患を止めることが示唆されてきた。これらの発見を考慮に入れて、非病原性細菌株を、BSHの低下及び免疫調節性の両方に関して選択した。驚いたことに、BSH活性の増加を引き起こす菌株のいくつかは、前炎症性サイトカインのTNF−αの濃度を減少させることが同時に見出された、図1。本発明は、その結果として、例えば、L.ロイテリ(L.reuteri)ATCC−PTA4659、L.ロイテリ(L.reuteri)ATCC−6475又はL.コリニフォルミス(L.coryniformis)ATCC−PTA4660の、アテローム性動脈硬化症及び他の心血管疾患を予防及び/又は治療するための製品の製造のための使用並びに同じ方法で選択された他の菌株及びそれらの混合物に言及する。
【発明の概要】
【発明が解決しようとする課題】
【0034】
したがって、本発明の目的は、血清LDLコレステロールを低下させ、前炎症性サイトカインのTNF−αの濃度を減少させるその能力について選択された乳酸菌株を提供することである。他の目的及び有利性は、以下の開示及び添付の特許請求の範囲によりさらに完全に明らかになるであろう。
【課題を解決するための手段】
【0035】
本発明は、アテローム性動脈硬化症及び他の心血管疾患を予防及び/又は治療するための、BSH活性を増加させ、その結果として血清LDLコレステロールを低下させ、同時に前炎症性サイトカインのTNF−αの濃度を減少させるその能力について選択された特定の乳酸菌株、このような菌株の選択方法及びこのような菌株を含む製品を提供する。
【0036】
本発明の他の目的及び特徴は、以下の開示及び添付の特許請求の範囲によりさらに完全に明らかになるであろう。
【図面の簡単な説明】
【0037】
【図1】LPS活性化単球によるTNF−αの産生に対するラクトバチルス順化培地の作用を示す棒グラフである。菌株及び対照は24時間インキュベートした。
【発明を実施するための形態】
【0038】
本発明は、アテローム性動脈硬化症などにおいて、炎症を軽減し、BSH活性を増加させるその能力について選択された乳酸菌株を含む。このような菌株は、ラクトバチルス・ロイテリ(L.reuteri)ATCC−PTA4659を含み、これはブダペスト条約のもとに、American Type Culture Collection、10801 University Blvd、Manassas、VAに、2002年9月11日に寄託されている。ラクトバチルス・ロイテリ(L.reuteri)ATCC−PTA6475は、2004年12月21日に、ATCCに寄託された。これらの菌株の公衆への提供可能性に対するすべての制限は、特許が付与された場合に取り消し不能の条件で撤回されるであろう。これらの菌株に由来する全細胞又は成分、例えば、これらの菌株の無細胞培養物中に存在する、この能力を有する成分を含む、食品、栄養添加物、及び製剤などの製品、製剤又は医療用具は、当分野で公知のように、例えば、ラクトバチルス株のフリーズドライ培養物又はそれに由来する成分を含む硬ゼラチンカプセルとして製剤化することもできる。本明細書中で述べた菌株の混合物及び全細胞又はそれらの成分の混合物も本発明の範囲内である。
【0039】
実施例3において選択された菌株、例えば、L.ロイテリ(L.reuteri)ATCC PTA−6475を標準的ヨーグルトに加えた。L.ロイテリ(L.reuteri)ATCC PTA−6475株は、乳業において、ラクトバチルスを増殖するための標準的な方法を使用して増殖させ、凍結乾燥させた。この培養物を、その後、伝統的ヨーグルト培養物を使用して、あらかじめ発酵させたミルクに、10E+6 CFU/ヨーグルトのグラムの濃度で加え、アテローム性動脈硬化症の予防として人に使用した。ヨーグルト以外の他の摂取可能な支持材料は、例えば、ミルク、カード、発酵乳、ミルクベースの発酵製品、発酵シリアルベースの製品、ミルクベースの粉末である。
【0040】
適切なサイトカインを使用するモデル系を使用して、炎症を軽減又は増大させる因子を決定する。本明細書に提供された発明において、ヒト細胞ベースのアッセイを使用する。
【0041】
THP−1細胞は、白血病患者に由来するヒト単球細胞系であり、これらは、American Type Culture Collectionに保存されている(ATCC番号TIB202)。ヒトの宿主由来のこれらの細胞の起源により、それらは、ヒト胃腸免疫系とヒト共生細菌との相互作用の研究に、特に関係付けられる。
【0042】
本発明のデータは、特定の菌株、L.ロイテリ(L.reuteri)ATCC PTA−4659及びL.ロイテリ(L.reuteri)ATCC PTA−6475によるTNF−αの産生の強力な阻害並びにこの調節が、後期対数増殖期/定常増殖期の間に、これら2種の特定の菌株により増殖培地に放出される物質により仲介されることを示す。それどころか、L.ロイテリ(L.reuteri)の他の2種は、大腸菌毒に対する細胞の炎症反応を阻害できないだけでなく、それら自体が炎症反応を誘導した。
【0043】
胆汁塩を含む選択的de Man、Rogosa及びSharpe(MRS)固体培地を用いた直接プレート法を、胆汁塩ヒドロラーゼを排出する菌株をスクリーニングするために使用し、その基質に対する酵素特異的活性を確定した。BSH産生細菌の増殖は、加水分解及び培地の酸性化を起こす。加水分解は、コロニーの周りに沈殿した遊離の胆汁塩のハロー形成として起こる(Dashkevicz,M.P.及びS.D.Feighner.1989.「胆汁塩ヒドロラーゼ活性ラクトバチルス属種のための分化用培地の開発(Development of a differential medium for bile salt hydrolase−active Lactobacillus spp.)」Appl Environ Microbiol.55(1):11〜16)。
【0044】
本発明の特徴は、以下の実施例を参照することにより、より明確に理解されるであろうが、これらは本発明を限定するものではない。
【実施例】
【0045】
(実施例1)TNF−αの濃度を減少させる能力を有する菌株の評価
THP−1細胞を対照培地、又は選択されたL.ロイテリ(L.reuteri)株であるL.ロイテリ(L.reuteri)ATCC PTA−4659、L.ロイテリ(L.reuteri)ATCC PTA−4975、L.ロイテリ(L.reuteri)ATCC 55730及びL.ロイテリ(L.reuteri)株PTA−4965の増殖によって得られる順化培地(L−CM)のどちらかと共にインキュベートした。順化培地(L−CM)は、L.ロイテリ(L.reuteri)培養物それぞれの9時間培養物又は24時間培養物から得られる無細胞上清である。THP−1細胞は、3.5時間のインキュベーションの間に、対照培地又は大腸菌由来LPS(正常な炎症反応においてTNF−αの産生をもたらす)のどちらかを用いて刺激し、その後、細胞を除去し、上清を、ELISA技術を使用してTNF−αに関して分析した。
【0046】
材料:
THP−1単球白血病細胞系(ATCC、カタログ番号TIB202)
RPMI 1640培地(Gibco−Invitrogen)
ウシ胎児血清(Gibco−Invitrogen)
ペニシリン−ストレプトマイシン溶液(Sigma)
大腸菌、血清型O127:B8のリポ多糖類(Sigma、カタログ番号L3137)TNF−α/TNF−SFIIヒトDuoSet ELISA開発キット(R&D Systems、カタログ番号DY210)
ヒトIL−10 DuoSet、2ndジェネレーションキット(R&D Systems、カタログ番号DY217)
【0047】
方法:
THP−1単球細胞系を使用する。5%(v/v)のMRS培地及び5%(v/v)のラクトバチルス順化培地を適切なウェルに加える。ラクトバチルス順化培地は、MRS培地におけるラクトバチルス属種の24時間培養物の上清である。その後、順化培地を高速真空乾燥し、ペレットを等量の培地に再懸濁することによってpHを調整する。加湿チャンバーは、液体蒸発が最小になるように設計されているが、48時間のインキュベーション後、24ウェルプレート中の細胞懸濁液の容量は、約475μlまで減少する。
【0048】
100ng/mlの大腸菌、血清型O127:B8のリポ多糖類を適切なウェルに加え、これらを37℃、加湿、5%CO2のチャンバーにおいてインキュベートする。3.5時間のインキュベーション後、培養物を1.5mlの遠心分離管に回収し、1500RCFで5分間4℃において遠心分離機にかける。上清を回収する。
【0049】
サイトカインの発現をELISA(Quantikine TNF−α/TNF−SFII ヒトDuoSet)によって試験する。
【0050】
使用した培養培地は、10%のFBS、2%のペニシリン−ストレプトマイシンを含むRPMI1640であった。
【0051】
結果−実施例1
LPSをTHP−1細胞に、L−CMの不在下で加えることにより、3.5時間のインキュベート期間に130pg/mlのTNF−αの産生がもたらされた。これは、毒素に対するTHP−1細胞の予期された炎症反応である。L−CMの添加に対する対照として機能する、増殖培地(MRS)の添加により、132pg/mlのTNF−αの産生がもたらされ、したがって、MRSはLPSに対する反応に影響を与えなかった。L.ロイテリ(L.reuteri)ATCC PTA4659又はL.ロイテリ(L.reuteri)ATCC PTA6475由来の24時間L−CMの添加は、LPSにより刺激されたTNF−αの濃度を、それぞれわずか13及び11pg/mlに、劇的に低下させた。これは、LPSにより刺激されたTNF−αの産生が、それぞれ90及び93%阻害されたことを示す。
【0052】
反対に、L.ロイテリ(L.reuteri)ATCC55730及びL.ロイテリ(L.reuteri)株PTA−4965由来の24時間L−CMの存在下で、LPSは、LPSの不在下の濃度と比較して、TNF−αの有意な上昇をさらに誘導できた。LPSにより刺激されたTNF−αの産生は、L.ロイテリ(L.reuteri)ATCC55730及びL.ロイテリ(L.reuteri)株ATCC PTA−4965由来のL−CMの存在下にもかかわらず、それぞれ54%及び42%増加した(図1)。
【0053】
L.ロイテリ(L.reuteri)ATCC PTA4659又はL.ロイテリ(L.reuteri)ATCC PTA6475由来の9時間L−CMを用いて実施された同様の実験では、LPSにより刺激されたTNF−αの産生に関する阻害作用は、大幅に低いが、存在したことが実証された。したがって、L.ロイテリ(L.reuteri)株をより長くインキュベートし、後期対数増殖期/定常増殖期にL−CM収穫することにより、TNF−αの産生を阻害する効果が改善される。
【0054】
(実施例2)直接プレートアッセイ−細胞外BSH活性を用いた菌株の評価
ヒト乳酸菌の菌株を、酸素を制限した条件において、37℃で、MRSブロス(Acumedia Manufacturers、Inc.Baltimore、Maryland)中で一晩増殖させ、10%のグリセロール(BDH Laboratory Supplies、England)を含む乳酸桿菌を有する培地(LCM)に接種した。
【表1】
【0055】
培養物のストックを、さらなる使用のために−80℃で保存した。菌株は、BioGaia AB研究室並びにLund(スウェーデン)、Raleigh(NC、米国)及びLantbruksuniversitetet(農業大学)、Uppsala(スウェーデン)の収集株から入手した。
【0056】
細胞外BSH活性についてスクリーニングするために、菌株を、一晩培養物から、3mMの胆汁塩、GDCA Sigma、Steinheim、Germany)、TDCA(Sigma)、GCA(Sigma)及びTCA(Fluka,Sigma−Aldrich、Germany)を含むMRS−システイン(MRS−c)寒天(Acumedia)プレート上に、それぞれ画線した。プレートを37℃で48時間、嫌気的(AnaeroGen、Oxoid、UK)にインキュベートした。胆汁酸脱抱合の結果である沈殿を、視覚的に、その結果主観的に測定し、これをもとに、活性を、活性なし(−)、活性あり(+)とした。胆汁塩を加えていないMRS−c寒天プレートを増殖及び負の対照として使用した。
【0057】
(実施例3)BSH活性及び同時にTNF−αの濃度を減少させる能力を有する菌株の選択
【表2】
【0058】
上記の表のデータにより、L.ロイテリ(L.reuteri)のさまざまな菌株が、TNF−α及びBSHの産生に関してさまざまな作用を有し、L.ロイテリ(L.reuteri)ATCC PTA−4659株及びL.ロイテリ(L.reuteri)ATCC PTA−6475株が、アテローム性動脈硬化症への使用に特に適しているという驚くべき発見が確認された。
【0059】
(実施例4)順化培地の使用
実施例1における方法を使用して、TNF−αを有効に減少させる菌株に由来する順化培地、この実施例においては、L.ロイテリ(L.reuteri)ATCC PTA−4659に由来する培地を選択した。この培地を、菌株をde Man、Rogosa、Sharpe(MRS)(Difco、Sparks、MD)で増殖することによって、大規模に作った。乳酸菌の一晩培養物を、OD600が1.0になるように希釈し(およそ109細胞/mlを表す)、さらに1:10に希釈し、さらに24時間増殖した。細菌細胞を含まない順化培地を、8500rpmで10分間4℃において遠心分離機にかけることによって回収した。順化培地を細胞のペレットから分離し、その後0.22μmのろ過孔ユニット(Millipore、Bedford、Mass.)を介してろ過した。その後、順化培地を凍結乾燥し、標準的な方法を使用して、錠剤に製剤化した。この錠剤を、アテローム性動脈硬化症を有効に治療する薬剤として、ヒトに使用した。
【0060】
(実施例5)選択された抗炎症性ラクトバチルス・ロイテリ株の使用
実施例1及び2における方法を使用して、TNF−αを有効に減少させ、同時にBSH活性を増加させる菌株、この実施例においてはL.ロイテリ(L.reuteri)ATCC PTA−4659を選択した。L.ロイテリ(L.reuteri)株を、その後凍結乾燥し、標準的な方法を使用して、105〜109cfuの範囲でカプセルに製剤化した。このカプセルを、アテローム性動脈硬化症を有効に軽減する薬剤として、ヒトに使用した。
【0061】
(実施例6)アテローム性動脈硬化症において頸動脈プラークを減少させる、ラクトバチルス・ロイテリ
総計1059例の患者に、ベースライン及び1年の追跡時に効果的な超音波測定を実施した。ベースライン及び1年の追跡時において、同じ超音波画像系及びトランスデューサー(Acuson Xp10 128、ART改良型、7.5−MHzリニアアレイ・トランスデューサー搭載、開口サイズ38mm、SIEMENS)を使用した。B−モードの画像補正パラメーターを、固定値にプリセットし、いずれの調査の過程においても変更しない。仰臥位の対象を、頭をやや左に向けて、右の頸動脈を、いくつかの異なる超音波照射角度で、縦及び横の両方で、鎖骨のすぐ上から、分岐部に対して出来るだけ末梢側に離れてスキャンした。プラークを、血管壁の内腔への、隣接する内膜中膜厚(IMT)と比較して少なくとも50%の局所突起として定義する。各対象において、最大6個のプラークが、総頸動脈、分岐部及び内頸動脈の近く及び遠くの壁に、それぞれ記録された。それぞれのプラークに関して、トランスデューサーを用いて、血管壁に対して平行に、及び最大プラーク厚の点に対して出来るだけ垂直に、領域拡大選択を38mm×20mに設定して、さらなる画像を記録する。すべての記録は、Panasonic 7650ビデオプレーヤーで、Super VHSテープに実施した。
【0062】
ベースラインにおいて、1059例の男性にプラークが存在した(表1)。頸動脈プラーク面積は、いずれの年齢においても縮小した。ベースラインにおける平均総プラーク面積(SE)は、24.1mm2である。L.ロイテリ(L.reuteri)ATCC PTA−4659(108CFU)の1日容量を摂取後の、追跡期間において、すべての人が、総プラーク面積が縮小した。平均縮小は9.0mm2である。
【表3】
【0063】
本発明は特定の実施形態を参照して記載されているが、多くの変形、改善及び実施形態が可能であることが理解されると思われ、したがって、このような変形、改善及び実施形態もまた、本発明の精神及び範囲内であるとみなされる。
【0064】
【技術分野】
【0001】
本明細書において、本発明は、アテローム性動脈硬化症及び他の心血管疾患を予防及び/又は治療するための、胆汁酸塩ヒドロラーゼ(BSH)活性を増加させ、その結果として血清LDLコレステロールを低下させ、同時に前炎症性サイトカインの腫瘍壊死因子(TNF−α)の濃度を減少させるその能力について選択された特定の乳酸菌株、このような菌株の選択方法並びにこのような菌株を含む製品を提供する。
【背景技術】
【0002】
プロバイオティクス
プロバイオティクスは、健康に有益な効果をもたらすことが示されている(Gorbach,S.L.2000.「プロバイオティクス及び胃腸の健康(Probiotics and gastrointestinal health.)」Am.J.Gastroenterol.95:S2〜S4)。多くのさまざまな活性がプロバイオティクスに帰するとされているが、これらの作用が達成される機序はよくわかっていない。これらの作用は、先天的及び後天的な免疫の増強(Gill,H.S.、K.J.Rutherfurd、J.Prasad、及びP.K Gopal.2000.「ラクトバチルス・ラムノサス(HN001)、ラクトバチルス・アシドフィルス(HN017)及びビフィドバクテリウム・ラクティス(HN019)による自然免疫及び後天性免疫の増強(Enhancement of natural and acquired immunity by Lactobacillus rhamnosus(HN001)、Lactobacillus acidophilus(HN017)and Bifidobacterium lactis(HN019).)」Br.J.Nutr.83:167〜176)、抗炎症性サイトカイン(IL−10)の産生の増強(Pessi,T.、Y.Sutas、M.Hurme及びE.Isolauri.2000.「ラクトバチルス・ラムノサスGGの経口投与後のアトピーの小児におけるインターロイキン−10の産生(Interleukin−10 generation in atopic children following oral Lactobacillus rhamnosus GG.)」Clin.Exp.Allergy 30:1804〜1808)、及び腸透過性の減少(Madsen,K.、A.Cornish、P.Soper、C.McKaigney、H.Jijon、C.Yachimec、J.Doyle、L.Jewell及びC.De Simone.2001.「プロバイオティック細菌は、マウス及びヒトの腸上皮細胞のバリア機能を増強させる(Probiotic bacteria enhance marine and human intestinal epithelial barrier function.)」Gastroenterology 121:580〜591)を含む。さまざまなラクトバチルス株は動物及びヒトの両方において、特によく研究されている。それらは、旅行者下痢(Marteau,P.R.、M.de Vrese、C.J.Cellier及びJ.Schrezenmeir.2001.「プロバイオティクスの使用による胃腸疾患からの保護(Protection from gastrointestinal diseases with the use of Probiotics.)」Am.J.Clin.Nutr.73:430S〜436S)、再発性クロストリジウム・ディフィシレ(Clostridium difficile)感染症(Gorbach,S.L.1987.「細菌性下痢及びその治療(Bacterial diarrhoea and its treatment.)」Lancet ii:1378〜1382)、ロタウィルス(Szajewska,H.、M.Kotowska、J.Z.Mrukowicz、M.Armanska及びW.Mikolajczyk.2001.「幼児の院内下痢症の予防における、ラクトバチルスGG(L.GG)の有効性(Efficacy of Lactobacillus GG(L.GG)in prevention of nosocomial diarrhea in infants.)」J.Pediatr.138:361〜365)及びヘリコバクター(Helicobacter)感染症(Mukai,T.、T.Asasaka、E.Sato、K.Mori、M.Matsumoto及びH Ohori.2002.「プロバイオティックなラクトバチルス・ロイテリによる、ヘリコバクター・ピロリの糖脂質受容体への結合の阻害(Inhibition of binding of Helicobacter pylori to the glycolipid receptors by probiotic Lactobacillus reuteni.)」FEMS Immunol.Med.Microbiol.32:105〜110)の予防及び治療に有効であり得る。マウスの腸から単離されたL.ロイテリ(L.Reuteri)は、IL−10トランスジェニック・ノックアウトマウスにおいて、大腸炎の発症を阻害した(Madsen,K.L.、J.S.Doyle、L.D.Jewell、M.M.Tavernini及びR.N.Fedorak.1999.「ラクトバチルス属種は、インターロイキン10遺伝子欠損マウスにおいて、大腸炎を予防する(Lactobacillus species prevents colitis in interleukin 10 gene−deficient mice.)」Gastroenterology 116:1107〜1114)。プロバイオティクス混合物を用いた臨床試験により、慢性回腸嚢炎において相当の改善が示された(Gionchetti,P.、F.Rizzello、A.Venturi、P.Brigidi、D.Matteuzzi、G.Bazzocchi、G.Poggioli、M.Miglioli及びM.Campieri.2000.「慢性回腸嚢炎を有する患者における維持療法としての経口細菌製剤療法:二重盲検の、プラセボを対照とした試験(Oral bacteriotherapy as maintenance treatment in patients with chronic pouchitis:a double−blind、placebo−controlled trial.)」Gastroenterology 119:305〜309)。
【0003】
免疫応答(Th−1/Th2/TR)
炎症は、サイトカインとして知られる細胞内シグナルタンパク質によって仲介され、これらはマクロファージ及び樹状細胞によって、上皮において、抗原性刺激に反応して産生される。上皮と抗原の間の接触において、上皮中の抗原提示細胞(樹状細胞を含む)が、ナイーブなマクロファージにシグナルを伝播し、ナイーブなマクロファージはその後いわゆるTh−1型応答で応答し、TNFα、IL−1、IL−6、IL−12を含む前炎症性サイトカインが、マクロファージによって産生される。これらのサイトカインは、ナチュラルキラー細胞、T細胞及び他の細胞を順番に刺激し、インターフェロンγ(IFNγ)が産生され、ナイーブなマクロファージは、Th−2型応答で抗原に応答することもあり得る。この応答は、IFNγによって抑制される。これらのTh−2型細胞は、IL−4、IL−5、IL−9及びIL−10などの抗炎症性サイトカインを産生する。
【0004】
IL−10は、IFNγの産生を阻害し、したがって免疫応答を抑制することが知られている。Th−1型及びTh−2型の細胞と、それらのそれぞれのサイトカインの産生とのバランスが、所与の抗原に対する炎症反応の程度を規定する。Th−2型細胞は、免疫系を介して免疫グロブリンの産生もまた刺激できる。TNFαの濃度が低減される場所である消化管における抗炎症活性は、上皮細胞の増強(腸壁内膜の完全性)、したがって消化管病原体及び毒素により起こる負の作用の減少に相関する。
【0005】
制御性T(TR)細胞は、免疫応答の不可欠な構成要素とみなされる。これらの細胞は、有害な免疫病理を最小にするために、まず防御的抗菌免疫を微調整すると思われる(Powrie F、Maloy KJ.2003.「レギュレーターの制御(Regulating the regulators)」、Science 299 1030〜1031)。TR細胞は、抗炎症性サイトカインのIL−10の濃度を増加させることが示された(Smits,H.H.、A.Engering、D.van der Kleij、E.C.de Jong、K.Schipper、T.M.van Capel、B.A.J.Zaat、M.Yazdanbakhsh、E.A.Wierenga、Y.van Kooyk及びL.Kapsenberg.2005.「選択的プロバイオティック細菌は、樹状細胞特異的細胞間接着分子3−捕捉非インテグリンを介して樹状細胞機能を調節することによって、インビトロでIL−10産生制御性T細胞を誘導する(Selective probiotic bacteria induce IL−10−producing regulatory T cells in vitro by modulating dendritic cell function through dendritic cell−specific intercellular adhesion molecule 3−grabbing nonintegrin.)」J Allergy Clin Immunol.115:1260〜1267)。TR細胞の発生及び活性化を調節する因子は、(自己免疫疾患を治療し、臓器の移植片の生存を増強するために)TR細胞の活性の方向か、又は(ワクチン接種及び腫瘍拒絶を強化するために)TR細胞の活性から離れる方向どちらかに平衡を移すことができるはずである(Walter J.Dobrogosz.「L.ロイテリを用いたヒトの健康の増強、プロバイオティック、イムノバイオティック及びイムノプロバイオティック(Enhancement of human health with L.reuteri、A Probiotic、Immunobiotic and Immunoprobiotic.)」NUTRAfoods.2005:4(2/3)15〜28)。
【0006】
プロバイオティクスの免疫調節作用
ラクトバチルス・ラムノサス(Lactobacillus rhamnosus)株GG(LGG)は、潜在的プロバイオティック剤であり、多くの研究により、腸管にコロニーを形成し、粘膜上皮及び免疫応答を調節するLGGの能力が実証されている。LGGは、単独定着ノトバイオートラットにおいて、腸細胞の増殖及び絨毛の大きさを増大させた(Banasaz,M.、E.Norin、R.Holma及びT.Midtvedt.2002.「ラクトバチルス・ラムノサスGGによる、単独定着ノトバイオートラットにおける腸細胞産生の増加(Increased enterocyte production in gnotobiotic rats mono−associated with Lactobacillus rhamnosus GG.)」Appl Environ Microbiol.68:3031〜3034)。LGGは、経口投与後に、エクスビボでマウスリンパ球応答の拡散もまた調節し(Kirjava(登録商標)inen,P.V.、H.S.ElNezami、S.J.Salminen、J.T.Ahokas及びP.F.Wright.1999.「マウスリンパ球増殖に関する、経口投与された、生存ラクトバチルス・ラムノサスGG及びプロピオニバクテリウム・フリューデンレイッヒイ亜種シャーマニィの作用(Effects of orally administered viable Lactobacillus rhamnosus GG and Propionibacterium freudenreichii subsp.sherinanii JS on mouse lymphocyte proliferation.)」Clin Diagn Lab Immunol 6:799〜802)、L.パラカセイ(L.paracasei)は、CD4+Tリンパ球の調節性サイトカインのプロフィールを改変する(Von der Weid T.、C.Bulliard及びE.J.Schiffrin.2001.「形質転換増殖因子β及びインターロイキン−10を産生する、低増殖能を有するCD4(+)T細胞の集団の乳酸菌による誘導(Induction by a lactic acid bacterium of a population of CD4(+)T cells with low proliferative capacity that produce transforming growth factor beta and interleukin−10.)」Clin Diagn Lab Immunol 8:695〜701)。適応免疫応答に加え、LGGは先天性免疫応答に対する影響を有する。LGGは核因子κB(NF−κB)、並びにヒトマクロファージにおけるシグナル伝達物質及び転写活性化因子(STAT)シグナル伝達経路を活性化し(Miettinen,M.、A.Lehtonen、I.Julkunen、及びS.Matikainen.2000.「乳酸桿菌及び連鎖球菌が、ヒトマクロファージにおいて、NF−κB及びSTATシグナル伝達経路を活性化する(Lactobacilli and Streptococci activate NF−kappa B and STAT signaling pathways in human macrophages.)」J Immunol 164:3733〜3740)、L.ラムノサス(L.rhamnosus)はマクロファージによるインターロイキン−12(IL−12)の産生を刺激する(Hessle,C.、L.A.Hanson及びA.E.Wold.1999.「ヒト胃腸粘膜由来の乳酸桿菌は、IL−12産生の強力な刺激因子である(Lactobacilli from human gastrointestinal mucosa are strong stimulators of IL−12 production.)」Clin Exp Immunol 116:276〜282)。LGGはまた小児においてIL−10のような免疫調節性サイトカインの産生も刺激し(Pessi,T.、Y.Sutas、M.Hurme及びE.Isolauri.2000.「ラクトバチルス・ラムノサスGGの経口投与後のアトピーの小児におけるインターロイキン−10の産生(Interleukin−l0 generation in atopic children following oral Lactobacillus rhamnosus GG.)」Clin Exp Allergy 30:1804〜1808)、インビボで前炎症性応答を制御できる。マクロファージ、樹状細胞及び好中球のような先天性免疫のエフェクター細胞は、大部分の炎症反応の一次駆動体である(Janeway,C.A.,Jr.及びR.Medzhitov.2002.「先天性免疫の認識(Innate immune recognition.)」Annu Rev Immunol 20:197〜216)。先天性免疫が、抗原に対する先天性応答及び後天性応答の両方の過程を、自己又は非自己として決定づけるという考えは、炎症の調節における先天性免疫の役割を強調している。
【0007】
米国特許出願公開第20020019043号は、サイトカインを産生するグラム陽性菌又はサイトカイン拮抗薬を産生するグラム陽性菌株を投与することによる、炎症性腸疾患の治療に関する。特定の実施形態において、サイトカイン又はサイトカイン拮抗薬は、IL−10、可溶性TNF−α受容体又は別のTNF−α拮抗薬、IL−12拮抗薬、インターフェロン−γ拮抗薬、IL−1拮抗薬などから選択される。特定の実施形態において、グラム陽性菌は、全般にサイトカイン、サイトカイン拮抗薬などを産生するように、遺伝子操作されている。
【0008】
L.ロイテリ(L.reuteri)の免疫調節作用
L.ロイテリ(L.reuteri)の免疫調節作用は、例えばプロバイオティック乳酸桿菌がそれらの免疫調節作用を、樹状細胞(DC)のTh1/Th2/Th3/Tr1/TR促進能力を調節することによって発揮することを示した、Christensenによって報告された(Christensen H.R.、H.Frokiaer及びJ.J.Pestka.2002.「マウスの樹状細胞において、乳酸桿菌はサイトカイン及び成熟表面マーカーの発現を特異的に調節する(Lactobacilli differentially modulate expression of cytokines and maturation surface markers in murine dendritic cells.)」J.Immunol.168:171〜178)。彼らは、マウスのDCが、L.ロイテリ(L.reuteri)株を含むさまざまなラクトバチルス株の共培養に曝された場合、それらは、サイトカインIL−6、IL−10、IL−12及びTNF−αの産生並びに表面マーカーのMHCクラスII及びCD86の上方制御が、濃度依存的方法で、特異的に調節されたことを示した。すべての乳酸桿菌が、DCの成熟の指標である、表面MHCクラスII及びCD86マーカーを上方制御した。これらの研究において特に注目に値するのは、L.ロイテリ(L.reuteri)(12246株)がIL−12の誘導因子としては弱いが、L.ジョンソニー(L.johnsonii)又はL.カゼイ(L.casei)と共培養した場合、後者の2種によって刺激された前炎症性サイトカインシグナルIL−12、IL−6及びTNF−αの産生を、特異的に阻害することであった。IL−10の産生は、これらの条件下でも変化しなかった。これらの発見により、「L.ロイテリ(L.reuteri)は、「危険なシグナル」を持たない抗原に対する耐性を支持し、同時にLPSのような危険なシグナルを介して認識される病原体に対する応答能力は損なわれないまま維持する、腸の樹状細胞発生の環境調節に寄与できる」という彼らの結論が導かれた。彼らはまた、いくつかのL.ロイテリ(L.reuteri)株が、抗炎症性IL−10を導入しながら、IL−12及びTNF−α(及びIL−6)の産生を下方制御するのに有効な、潜在的な標的の細かい治療であり得、したがって、前炎症腸サイトカイン環境の釣り合いをとる、別の治療的取り組みを提示すると結論付けた。
【0009】
Smitsは、これらの見解を拡大し、L.ロイテリ(L.reuteri)が、DCに制御性T(TR)細胞の産生を刺激させる能力を有することを示した。彼らは、インビトロで共培養した3種の異なるラクトバチルス属種及びヒト単球由来のDCを使用した。乳酸桿菌の2種、ヒトL.ロイテリ(L.reuteri)(ATCC 53609)株及びL.カゼイ(L.casei)は、これらのDCのTR細胞の発生を刺激するが、L.プランタラム(L.plantarur)株はしなかった。これらのTR細胞は、IL−10の産生濃度が増加することが示され、IL−10依存的にそばにいるT細胞の増殖を阻害できた(Smits,H.H.、A.Engering、D.van der Kleij、E.C.de Jong、K.Schipper、T.M.M.van Capel B.A.J.Zaat、M.Yazdanbakhsh、E.A.Wierenga、Y.van Kooyk及びL.Kapsenberg.2005.「選択的プロバイオティック細菌は、樹状細胞特異的細胞間接着分子3捕捉非インテグリンを介して、樹状細胞機能を調節することによって、インビトロでIL−10産生制御性T細胞を誘導する(Selective probiotic bacteria induce IL− 10−producing regulatory T cells in vitro by modulating dendritic cell function through dendritic cell−specific intercellular adhesion molecule 3−grabbing nonintegrin.)」J Allergy Clin Immunol.115:1260〜1267)。L.ロイテリ(L.reuteri)−DC相互作用に関するこれらの研究は、Horiによる、根底を覆す研究に関連するとみなされる(Hori S、T.Nomura及びS.Sakaguchi.2003.「転写因子Foxp3による、制御性T細胞発生の調節(Control of regulatory T cell development by the transcription factor Foxp3.)」Science.299:1057〜1061)並びにPasare及びMedzhitov(Pasare C.及びR.Medzhitov.2003.「樹状細胞によるCD4+Cd25+T細胞仲介抑制のToll経路依存性阻害(Toll pathway−dependent blockade of CD4+Cd25+ T cell−mediated suppression by dendritic cells.)」Science 299:1033〜1036)は、L.ロイテリ(L.reuteri)のイムノバイオティックな作用様式の1つに貴重な洞察を提供している。
【0010】
神経増殖因子(NGF)は、神経細胞に関するその活性に加えて、大腸炎のモデルを含むインビトロ及びインビボのいくつかの実験系において、有意な抗炎症作用を有する。Ma他(2004)は、L.ロイテリ(L.reuteri)の作用機序を、ヒト上皮細胞系において、サイトカイン及びNGFの合成並びにTNF−αに対するIL−8の応答を調査した。彼らは、L.ロイテリ(L.reuteri)は、強力な直接抗炎症活性を、ヒト上皮細胞において有し、それは摂取したプロバイオティクスの活性に関係すると思われると結論付けた。彼らはまた、L.ロイテリ(L.reuteri)は、独特の抗炎症性分子であるNGFを上方制御し、NF−κBの核への転移を阻害するとも結論付けた(Ma,D.、P.Forsythe及びJ.Bienenstock.2004.「生きたL.ロイテリは、腫瘍壊死因子αにより誘導されるインターロイキン8の発現の阻害作用に必須である(Live L.reuteri is essential for the inhibitory effect on tumor necrosis factor alpha−induced interleukin−8 expression.)」Infect.Immun.72:5308〜5314)。
【0011】
L.ロイテリ(L.reuteri)を含む、ラクトバチルス属種の多種多様な菌株は、プロバイオティック製剤に使用されている。ラクトバチルス・ロイテリ(Lactobacillus reuteri)は、動物の消化管の自然な常在菌の1つであり、ヒトを含む、健康な動物の腸において日常的に見出される。それは、抗菌作用を有することでも知られている。例えば、米国特許第5,439,678号、第5,458,875号、第5,534,253号、第5,837,238号及び第5,849,289号を参照されたい。L.ロイテリ(L.reuteri)細胞が、嫌気条件下で、グリセロールの存在下で増殖する場合、それらは、β−ヒドロキシ−プロピオンアルデヒド(3−HPA)として知られる抗菌物質を産生する。
【0012】
アテローム性動脈硬化症
動脈硬化性疾患及びその心血管結果は、米国その他における死亡及び罹患の主原因である。アテローム性動脈硬化症は、「粥(gruel)」又は「ベトベトしたもの(goo)」及び「硬化(hardening)」というギリシャ語に由来し、動脈の内壁にアテローム又は病巣が存在するとして定義される。プラークとしても知られる病巣は、脂肪性沈着物及び他の物質からなる。
【0013】
アテローム性動脈硬化症を特に危険にしているものは、重要な大動脈に対して特別な親和力を有することと思われる。プラークの詰まった病巣の一部が、動脈の内壁から破裂した場合、脂肪性材料が、心臓及び脳に直接供給している下流の小さい動脈に流れ込み、そこでそれらが詰まり、栄養及び酸素に富んだ血液がこれらの重要臓器に到達することを妨げる。完全な封鎖が起こった場合、心臓発作又は脳卒中が起こり得る(Little,W.C.、M.Constantinescu、R.J.Applegate、M.A.Kutcher、M.T.Burrows、F.R.Kahl及びW.P.Santamore.「冠動脈造影は、軽度から中程度の冠動脈疾患を有する患者のその後の心筋梗塞部位を予測できるか?(Can coronary angiography predict the site of a subsequent myocardial infarction in patients with mild−to−moderate coronary artery disease?)」Circulation.1988.78:1157〜66)。伝統的に、アテローム性動脈硬化症は、脂質代謝障害であると考えられてきた。アテローム性動脈硬化症に伴う危険因子は、LDLの血中濃度の高さ、ホモシステイン、高血圧、喫煙、肥満、糖尿病を含む。治療は、コレステロール濃度の調節、例えば特定の乳酸菌による胆汁塩の代謝の促進に焦点が当てられてきた。
【0014】
有効なプロバイオティクスとしての乳酸菌(LAB)を使用する可能性を評価する場合、多くの人が、胆汁酸の作用に耐えるLABのその能力を評価する必要があると考えている。胆汁酸は、肝臓においてコレステロールから合成され、胆嚢から十二指腸に、グリシン又はタウリンと抱合されて分泌される。それらの機能は、食物中の脂肪を乳化することである。ヒトにおける最も一般的な一次胆汁酸は、コール酸及びケノデオキシコール酸であり、これらは、肝臓におけるコレステロール代謝の主要な最終産物である。腸における細菌活性の結果として、これらの酸はその後脱抱合及び脱ヒドロキシル化などの化学的修飾を受け、そこでアミノ酸は抱合型から加水分解する(Cardona,M.E.、V.de Vanay、T.Midtvedt及びK.E.Nogin.「ノトバイオノートマウスにおけるプロバイオティクス。インビトロ及びインビボにおける、コレステロールのコプロスタノールへの転換及びインビトロにおける胆汁酸の脱抱合(Probiotics in gnotobiotic mice.Conversion of cholesterol to coprostanol in vitro and in vivo and bile acid deconjugation in vitro.)」Microb Ecol Health Dis.2000.12:219〜224;Dunne,C.、L.O’Mahony、L.Murphy、G.Thornton、D.Morrissey、S.O’Halloran、M.Feeney、S.Flynn、G.Fitzgerald、D.Daly、B.Kiely、G.C.O’Sullivan、F.Shanahan及びJ.K.Collins.「ヒト起源のプロバイオティック菌に関するインビトロの選択基準:インビボの発見との相関(In vitro selection criteria for probiotic bacteria of human origin:correlation with in vivo findings.)」Am J Clin Nutr.2001.73(suppl):386S〜392S)。いくつかの胃腸(GI)細菌、例えば、エンテロコッカス(Enterococcus)、ビフィドバクテリウム(Bifidobacterium)及びラクトバチルス(Lactobacillus)は、酵素の胆汁酸塩ヒドロラーゼ(BSH)を発現し、それは、抱合胆汁酸の加水分解を触媒し、遊離のグリシン又はタウリン及び非抱合型胆汁酸分子をもたらす(Tanaka,H.、K.Doesburg、T.Iwasaki及びI.Mierau.「胆汁塩ヒドロラーゼ活性のための乳酸菌のスクリーニング(Screening of lactic acid bacteria for bile salt hydrolase activity.)」J Dairy Sci.1999.82:2530〜2535;Bateup,J.M.、M.A.McConnell、H.F.Jenkinson及びG.W.Tannock.「胆汁塩ヒドロラーゼ活性、消化管のコロニー形成及びマウス宿主の増殖率に関する、ラクトバチルス株の比較(Comparison of Lactobacillus strains with respect to bile salt hydrolase activity、colonization of the gastrointestinal tract、and growth rate of the murine host.)」Appl Environ Microbiol.1995.61(3):1147〜1149;Tannock,G.W.、J.M.Bateup及びH.F.Jenkinson.「乳酸桿菌のインビトロの増殖に関するタウロコール酸ナトリウムの効果(Effect of sodium taurocholate on the in vitro growth of lactobacilli.)」Microb Ecol.1997.33:163〜167)。
【0015】
GI管において、BSHの発現がどのように細菌の機能に影響を与えるかについて、2つの主要な仮説がある。一方は、いくつかの細菌は胆汁塩を脱抱合し、アミノ酸のタウリンを電子受容体として使用するというもので、これに反して他方は、脱抱合型は、界面活性が低下して可溶性が低いので、酵素が胆汁塩の毒性を脱抱合によって減少させ、その結果細菌が保護されると述べている。抱合胆汁酸及び脱抱合胆汁酸の両方が、大腸菌(Escherichia coli)、クレブシエラ属種(Klebsiella sp)及びエンテロコッカス属種(Enterococcus sp)に対してインビトロで抗菌活性を示すことが見出されており、脱抱合型はより増殖阻害性である(Dunne,C.、L.O’Mahony、L.Murphy、G.Thornton、D.Morrissey、S.O’Halloran、M.Feeney、S.Flynn、G.Fitzgerald、C.Daly、B.Kiely、G.C.O’Sullivan、F.Shanahan及びJ.K.Collins.「ヒト起源のプロバイオティック菌に関するインビトロの選択基準:インビボの発見との相関(In vitro selection criteria for probiotic bacteria of human origin:correlation with in vivo findings.)」Am J Clin Nutr.2001.73(suppl):386S〜392S;Moser,S.A.及びD.C.Savage.「乳酸桿菌において、胆汁塩ヒドロラーゼ活性と抱合胆汁塩の毒性に対する耐性とは、関連のない特性である(Bile salt hydeolase activity and resistance to toxicity of conjugated bile salts are unrelated properties in lactobacilli.)」Appl Environ Microbiol.2001.67(8):3476〜3480)。
【0016】
発酵乳製品の強力なコレステロール低下作用は、コレステロールの胆汁酸との結合及びミセル形成の阻害によって説明できる。これらの製品中のプロバイオティック細菌がコレステロール低下作用を有し得る機序は、肝細胞によってコレステロールから作られた胆汁酸のコール酸及びデオキシコール酸を介する。これらは、グリシン及びタウリンと抱合し、小腸へ入り、そこで吸収され、肝臓へ向かう。再吸収の間に、抱合胆汁酸は腸においてミクロフローラに曝される。発酵食品中の細菌、例えば、乳酸桿菌及び連鎖球菌は、抱合胆汁酸を加水分解する。腸において高い胆汁塩ヒドロラーゼ活性を有するラクトバチルス(Lactobacillus)株は、胆汁の加水分解が促進できる可能性がある。このことは、コレステロールの転換率をより速くし、より多くの胆汁酸を産生する。インビボでコレステロールが低下するのは、脱抱合胆汁酸は、結腸において再吸収されないので、排泄物を介して胆汁酸が排出されるためである。このことは、新たな胆汁の合成を促進し、身体の胆汁のプールを一定に保つ(St−Onge M−P.、E.R.Farnworth及びP.J.H.Jones.2000.「発酵乳製品及び非発酵乳製品の摂取:コレステロールの濃度及び代謝に関する効果(Consumption of fermented and nonfermented dairy products:effects on cholesterol concentrations and metabolism.)」Am J Clin Nutr.71:674〜681)。
【0017】
胆汁酸の脱抱合は、血漿コレステロール濃度を低下させるであろう。しかし、これらの化合物は、大腸において、嫌気性細菌によって二次胆汁酸にさらに転換されることがあり、二次胆汁酸は、結腸癌の誘導因子候補として関係があるとされている。二次胆汁酸は、細胞系に対して有毒であり、それらは結腸粘膜において細胞毒性を発揮し、細胞増殖の増加をもたらすと考えられている。これらの過剰増殖性細胞は、変異原物質に対する感受性が増強されており、その結果結腸癌の危険性が増す(Hepner,G.、R.Fried、S.St.Jeor、L.Fusetti及びR.Morin.1979.「ヨーグルト及びミルクの高コレステロール作用(Hypercholesterolemic effect of yoghurt and milk.)」Am.J.Clin.Nutr.32:19〜24)。幸運なことに、乳酸菌は、脱抱合胆汁塩及び二次胆汁塩の可溶性を減少させると思われ、その結果、それらの生体利用率が低下する。De Boever他(2000)による研究は、L.ロイテリ(L.reuteri)が、細菌培養物中の胆汁塩の毒性を減少させたことを示した。さらに重要なことには、L.ロイテリ(L.reuteri)を加えることにより、赤血球の溶解に対するほぼ完全な耐性及びヒーラ細胞における胆汁塩の毒性の阻害がもたらされた(De Boever,P.、R.Wouters、L.Verschaeve、P.Berckmans、G.Schoeters及びW.Verstraete.「胆汁塩の細胞毒性に対する、胆汁塩ヒドロラーゼ活性ラクトバチルス・ロイテリの保護作用(Protective effect of the bile salt hydrolase−active Lactobacillus reuteri against bile salt cytotoxicity.)」Appl Microbiol Biotechnol.2000.53(6):709〜14)。
【0018】
免疫疾患であるアテローム性動脈硬化症
科学者は、アテローム性動脈硬化症の発症に関する新規なスキームを表しており、この病状が、前炎症性T細胞と、沈静しているものであるTRとのバランスの悪さによりもたらされ得ることを示唆している。このことは、第二回欧州血管ゲノム科学ネットワーク会議(Second European Vascular Genomics Network Conference)(EVGN Conference−ハンブルグ、2005年9月27〜30日)から現れた、興味をそそる科学的結果の1つである。これらの結果は、心疾患における炎症の役割に新しい洞察を提供し、血栓形成及び心臓発作をもたらす過程の新しい情報モデルの開発をもたらした。
【0019】
LDL粒子中の脂肪は内皮細胞を刺激し、単球及びTリンパ球を含むいくつかの細胞型の細胞浸潤を引き起こすので、アテローム性動脈硬化症は、内皮における脂肪線条の形成により始まる。単球は、内皮層と相互に作用し、内皮に硬く付着し、内皮細胞下の空間に遊走し、そこで単球はマクロファージに分化する。マクロファージは、サイトカインを含むさまざまな化学物質を放出する。増殖因子の産生が刺激され、これにより細胞増殖及び基質産生並びに基質の変性をもたらすメタロプロテイナーゼがもたらされる。したがって、マクロファージは、病巣の増殖に寄与し、不安定事象及び血栓事象に寄与し得る(Ross R.「アテローム性動脈硬化症−炎症性疾患(Athrosclerosis−An inflammatory disease.)」N Engl J Med.1999.340:115〜26)。Tリンパ球は、アテローム性動脈硬化症の全段階において存在することが示されている。それらの存在が、免疫応答との関連のさらなる証拠を提供する(Kol,A.及びP.Libby.1998.「感染性因子がアテローム性動脈硬化症に寄与し得る機序及びその臨床症状(The mechanisms by which infectious agents may contribute to atherosclerosis and its clinical manifestations.)」Trends Cardiovasc Med.8:191〜99;Andreotti,F.、F.Burzotta、A.Mazza、A.Manzoli、K.Robinson及びA.Maseri.1999.「ホモシステイン及び動脈閉塞性疾患:簡潔な総説(Homocysteine and arterial occlusive disease:a concise review.)」Cardiologia.44:341〜5)。
【0020】
炎症性物質産生の開始シグナルは、炎症シグナル伝達経路を活性化するいくつかの内在性分子を認識する、トール様受容体と呼ばれる受容体の関与に依存する(K.Edfeldt、J.Swedenborg、G.K.Hansson及びZ.Yan.2002.「ヒトアテローム性動脈硬化症の病巣におけるトール様受容体の発現:プラーク活性化のための可能な経路(Expression of Toll−Like Receptors in Human Atherosclerotic Lesions:A Possible Pathway for Plaque Activation)」Circulation.105:1158〜1161)。
【0021】
トール様受容体(TLR)は、微生物のモチーフを認識し、サイトカインの産生をもたらす一連の遺伝子を活性化する。伝統的に、TLRは、微生物感染のセンサーとみなされており、それらの役割は、炎症反応を誘導することである。しかし、TLRにより認識されるモチーフは、病原体に対して唯一ではなく、微生物のすべてのクラスによって共有される一般的モチーフであり、免疫系が、片利共生生物と病原菌とを、TLRを介してどのように識別するかは完全には理解されなかった。近年、TLRが、炎症反応の誘導におけるそれらの役割にもかかわらず、片利共生生物のミクロフローラを認識することによって腸内恒常性を維持する役割も担っているというデータが示された(Rakoff−Nahoum,S.、J.Paglino、F.Eslami−Varzaneh、S.Edberg及びR.Medzhitov.2004.「トール様受容体による片利共生生物のミクロフローラの認識は、腸内の恒常性にとって必要である。(Recognition of commensal microflora by toll−like receptors is required for intestinal homeostasis.)」Cell.23;118(2):229〜41)。
【0022】
炎症の血清マーカーが、心血管疾患の罹患及び死亡に関する独立した危険因子であることが確立されている。心血管疾患の終点にかかわる炎症性マーカーは、IL−6及びTNF−αなどの前炎症性サイトカイン、フィブリノゲン並びにC反応性タンパク質(CRP)を含む(Libby,P.、P.M.Ridker及びA.Maseri.2001.「炎症及びアテローム性動脈硬化症(Inflammation and atherosclerosis.)」Circulation.2002.105:1135〜1143;Ridker,P.M.「高感受性C反応性タンパク質:心血管疾患の一次予防における包括的リスク評価のための潜在的補助物質(High sensitivity C−reactive protein:potential adjunct for global risk assessment in the primary prevention of cardiovascular disease.)」Circulation.103:1813〜1818)。
【0023】
アテローム性動脈硬化症における肺炎クラミジア(C.pneumoaie)及びH.ピロリ(H.pylori)の役割
累積証拠により、アテローム性動脈硬化症が炎症性疾患であることが示唆されている。したがって、近年、感染性因子が心血管疾患の病因において役割を果たす可能性に多くの注目が集まっている。特定の感染性因子が、アテローム斑からそれらが単離されること、又は肺炎クラミジア(Chlamydia pneumoniae)、ヘリコバクター・ピロリ(Helicobacter pylori)、単純ヘルペスウィルス及びサイトメガロウィルスなどの生命体に関して、血清学的な陽性判定がでることに基づき、関与が示されている。
【0024】
プロスペクティブ研究は決定的な証拠の提供に欠けているが、肺炎クラミジア(C.pneumoniae)は、アテローム性動脈硬化症に最も強力にかかわっていると思われる。肺炎クラミジア(C.pneumoniae)は、死体解剖及び関節切除の標本から、並びに早期の病巣及び十分に発達した病巣の両方において単離されている。免疫学的細胞化学及び組織染色を用いた研究の場合、70〜100%の事例において、関連が見出されている。感染性因子がそれらの作用を発揮する可能な機序は、(i)内皮細胞、平滑筋細胞又はマクロファージに対する局所作用、或いは(ii)サイトカインの産生、単球の刺激及び凝集性高進の促進による全身作用を含み得る。
【0025】
コレステロール濃度の低下に関する従来の治療
血清コレステロール濃度の上昇は、アテローム性動脈硬化症及び冠動脈性心疾患に伴う危険因子であると長年にわたって認識されており、後者は西欧諸国における死亡の主原因である(Barr,D.P.、A.M.Russ及びH.A.Eder.1951.「ヒト血漿におけるタンパク質−脂質の関係 II.アテローム性動脈硬化症及び関連病態において(Protein−lipid relationship in human plasma.II.In atherosclerosis and related conditions.)」Am.J.Med.11:480〜493)。3−ヒドロキシメチルグルタリルコエンザイムA還元酵素阻害薬を含むコレステロールを低下させる多くの薬剤及び胆汁酸の正味の排出量を増加させる薬剤が、高コレステロール(HC)の個人を治療するために使用されてきた(Suckling,K.E.、G.M.Benson、B.Bond、A.Gee、A.Glen、C.Haynes及びB.Jackson.1991.「コレスチラミン治療を用いたハムスターにおける、コレステロールの低下及び胆汁酸の排出(Cholesterol lowering and bile acid excretion in the hamster with cholestyramine treatment.)」Atherosclerosis 89:183〜190)。
【0026】
しかし、これらの化合物の望ましくない副作用により、それらの治療的使用が懸念される。(Erkelens,D.W.、M.G.A.Baggen、J.J.Van Doormeal、M.Kettner、J.C.Koningsberger及びM.J.T.M.Mol.1988.「コレスチラミンと比較したシンバスタチンを用いた臨床経験(Clinical experience with simvastatin compared with cholestyramine.)」Drugs 39(Suppl.):87〜90)。
【0027】
コレステロール濃度を低下させる治療としての乳酸菌
これらの治療手段に加えて、プロバイオティック乳酸菌の摂取が、ヒトにおいて血清コレステロール濃度を低下させるより自然な方法であると思われる。いくつかの研究により、発酵乳製品の大量摂取(680から5000ml/d)の間に血清コレステロールの低下が報告されているが、これらの結果は、摂取のより現実的条件が推定できない(Mann,G.V.1977.「ヒトにおけるコレステロール血を低下させるヨーグルト中の因子(A factor in yogurt which lowers cholesterolemia in man.)」Atherosclerosis 26:335〜340;Mc Namara,D.J.、A.M.Lowell及びJ.E.Sabb.1989.「血中脂質が正常な男性における、血漿中脂質及びリポタンパク質の濃度に関する、ヨーグルト摂取の効果(Effect of yogurt intake on plasma lipid and lipoprotein levels in normolipidemic males.)」Atherosclerosis 79:167〜171)。
【0028】
Masseyは、成人男性において、ヨーグルトの摂取により、初めはコレステロールが10から12%有意に低下したが、2週間後、ヨーグルトの摂取を続けていたにも関わらず、濃度が対照値まで戻ったことを示した(Massey,L.1984.「ヒトの栄養摂取及び血清リポタンパク質に関する、ミルク及びヨーグルトの摂取を変えることによる効果(Effect of changing milk and yoghurt consumption on human nutrient intake and serum lipoprotein.)」J.Dairy Sci.67:255〜262)。類似の矛盾する結果が、ミルク及びその発酵製品を与えられた実験動物により得られた(Hepner,G.、R.S.T.Fried、S.Jeor、L.Fusetti及びR.Morin.1979.「ヨーグルト及びミルクのコレステロール低下作用(Hypocholesterolemic effect of yogurt and milk.)」Am.J.Clin.Nutr.32:19〜24;Rao,D.R.、C.B.Chawan及びS.R.Pulusani.1981.「ラットにおける、血漿コレステロール濃度及び肝臓コレステロール産生に関する、ミルク及びサーモフィルスミルクの影響(Influence of milk and thermophilus milk on plasma cholesterol levels and hepatic cholesterogenesis in rats.)」J.Food Sci.46:1339〜1341)。Raoは、ストレプトコッカス・サーモフィルス(Streptococcus thermophilus)により発酵させたミルクを与えられたラットのHC作用を報告した(Rao,D.R.、C.B.Chawan及びS.R.Pulusani.1981.「ラットにおける、血漿コレステロール濃度及び肝臓コレステロール産生に関する、ミルク及びサーモフィルスミルクの影響(Influence of milk and thermophilus milk on plasma cholesterol levels and hepatic cholesterogenesis in rats.)」J.Food Sci.46:1339〜1341)。Rodasは、ラクトバチルス・アシドフィルス(Lactobacillus acidophilus)を与えられたHCブタにおいて同様の作用を見出した(Rodas,B.Z.、S.E.Gilliland及びC.V.Maxwell.1996.「ラクトバチルス・アシドフィルスATCC 43121のコレステロール低下作用及び食餌によって誘導された高コレステロール血症のブタにおけるカルシウム(Hypocholesterolemic action of Lactobacillus acidophilus ATCC 43121 and calcium in swine with hypercholesterolemia induced by diet.)」J.Dairy Sci.79:2121〜2128)。
【0029】
以前に脂肪に富んだ食餌を与えられたマウスの血清中の、総コレステロール、トリグリセリド及び高比重リポタンパク質(HDL)と低比重リポタンパク質(LDL)との比に関する、L.ロイテリ(L.reuteri)CRL 1098の作用を調査する研究において、L.ロイテリ(L.reuteri)により、トリグリセリドが40%減少し、高比重リポタンパク質(HDL)と低比重リポタンパク質(LDL)との比は20%増加し、ナイーブなミクロフローラの脾臓及び肝臓への細菌転移はなかった(Taranto,M.P.、F.Sesma、A.P.Ruiz Holgado及びG.F.Valdez.1997.「胆汁塩ヒドロラーゼは、ラクトバチルス・ロイテリによるコレステロールの除去に関してカギとなる役割を果たす(Bile salts hydrolase plays a key role on cholesterol removal by Lactobacillus reuteri.)」Biotechnol.Left.9:245〜247)。これらのデータは、L.ロイテリ(L.reuteri)CRL 1098が、低細胞濃度でマウスのために有効なコレステロール低下補助物質であることを示唆している。しかし、本明細書中の本発明の開示とは異なり、コレステロールの低下は、BSH活性のみによるもので、BSH活性及び免疫調節作用との組み合わせによるものではなかった。
【0030】
コレステロール濃度を低下させる治療としての乳酸菌乳酸菌、免疫調節法
米国特許出願公開第20050169901号は、心血管障害を診断、予防及び治療するために、サイトカインの濃度又は活性を調節する方法に関する。このサイトカインの調節は、Th2からTh1へのサイトカインプロフィールの切り替えであり、切り替えが、Th1のサイトカインプロフィールから選択的に離れTNF−α産生の減少に向かう、本明細書の本発明とは異なる。この出願人らは、プロバイオティックとして、いくつかの異なる細菌属及び菌株について述べており、プロバイオティックが、TNF−α濃度の減少及び同時にBSH活性の増加に有効であるように選択された特定の乳酸菌である、本明細書の本発明とは異なる。
【0031】
Bukowskaは、高コレステロール血症の患者において、プロバイオティック細菌のラクトバチルス・プランタラム(Lactobacillus plantarum)299vの補給により、LDLコレステロール及びフィブリノゲンの濃度を有意に低下させることを示した(Bukowska H.、J.Pieczul−Mroz、M.Jastrzebska、K.Chelstowski及びM.Naruszewicz.1997.「中程度にコレステロールが上昇した対象における、ラクトバチルス・プランタラムを含む食餌を補給することに関するフィブリノゲン及びLDLコレステロールの減少(Decrease in fibrinogen and LDL−cholesterol levels upon supplementation of diet with Lactobacillus plantarum in subjects with moderately elevated cholesterol.)」Atherosclerosis.137:437〜8)。このことは、米国特許第6,214,336号にも記載されている。同じグループは、ラクトバチルス・プランタラム(Lactobacillus plantarum)を含む食餌の補給が、喫煙者の代謝障害の予防及び治療に寄与できることを示した。この好ましい作用は、食物繊維の細菌発酵によるプロピオン酸の産生に直接関連していると考えられる。彼らは、プロピオン酸が、おそらく、核転写因子Bを調節し、単球−マクロファージによる炎症性サイトカインの産生を低下させる、ペルオキシソーム増殖因子活性化受容体のイブプロフェンによる活性化に関係する、今に至るまで未知の機序を介して特異的抗炎症作用を発揮することを示唆する(M.Naruszewicz、M−L Johansson、D.Zapolska−Downar及びH.Bukowska、「喫煙者における心血管疾患の危険因子に関する、ラクトバチルス・プランタラム229vの作用(Effect of Lactobacillus plantarum 299v on cardiovascular disease risk factors in smokers.)」Am.J.Clinical Nutrition.2002.76:1249〜1255)。
【0032】
上記の本明細書の本発明とは異なり、参考文献は、BSH−活性の増加、同時にTNF−αの濃度の減少の両方を可能にする菌株を記載していない。
【0033】
前述のように、BSH活性の上昇は、血清コレステロール濃度を低下させ、その結果、アテローム性動脈硬化症の危険性を低下させることが長年にわたって周知であった。アテローム性動脈硬化症が炎症性疾患であることもまた、以前に実証されており、さまざまなサイトカインの調節が疾患を止めることが示唆されてきた。これらの発見を考慮に入れて、非病原性細菌株を、BSHの低下及び免疫調節性の両方に関して選択した。驚いたことに、BSH活性の増加を引き起こす菌株のいくつかは、前炎症性サイトカインのTNF−αの濃度を減少させることが同時に見出された、図1。本発明は、その結果として、例えば、L.ロイテリ(L.reuteri)ATCC−PTA4659、L.ロイテリ(L.reuteri)ATCC−6475又はL.コリニフォルミス(L.coryniformis)ATCC−PTA4660の、アテローム性動脈硬化症及び他の心血管疾患を予防及び/又は治療するための製品の製造のための使用並びに同じ方法で選択された他の菌株及びそれらの混合物に言及する。
【発明の概要】
【発明が解決しようとする課題】
【0034】
したがって、本発明の目的は、血清LDLコレステロールを低下させ、前炎症性サイトカインのTNF−αの濃度を減少させるその能力について選択された乳酸菌株を提供することである。他の目的及び有利性は、以下の開示及び添付の特許請求の範囲によりさらに完全に明らかになるであろう。
【課題を解決するための手段】
【0035】
本発明は、アテローム性動脈硬化症及び他の心血管疾患を予防及び/又は治療するための、BSH活性を増加させ、その結果として血清LDLコレステロールを低下させ、同時に前炎症性サイトカインのTNF−αの濃度を減少させるその能力について選択された特定の乳酸菌株、このような菌株の選択方法及びこのような菌株を含む製品を提供する。
【0036】
本発明の他の目的及び特徴は、以下の開示及び添付の特許請求の範囲によりさらに完全に明らかになるであろう。
【図面の簡単な説明】
【0037】
【図1】LPS活性化単球によるTNF−αの産生に対するラクトバチルス順化培地の作用を示す棒グラフである。菌株及び対照は24時間インキュベートした。
【発明を実施するための形態】
【0038】
本発明は、アテローム性動脈硬化症などにおいて、炎症を軽減し、BSH活性を増加させるその能力について選択された乳酸菌株を含む。このような菌株は、ラクトバチルス・ロイテリ(L.reuteri)ATCC−PTA4659を含み、これはブダペスト条約のもとに、American Type Culture Collection、10801 University Blvd、Manassas、VAに、2002年9月11日に寄託されている。ラクトバチルス・ロイテリ(L.reuteri)ATCC−PTA6475は、2004年12月21日に、ATCCに寄託された。これらの菌株の公衆への提供可能性に対するすべての制限は、特許が付与された場合に取り消し不能の条件で撤回されるであろう。これらの菌株に由来する全細胞又は成分、例えば、これらの菌株の無細胞培養物中に存在する、この能力を有する成分を含む、食品、栄養添加物、及び製剤などの製品、製剤又は医療用具は、当分野で公知のように、例えば、ラクトバチルス株のフリーズドライ培養物又はそれに由来する成分を含む硬ゼラチンカプセルとして製剤化することもできる。本明細書中で述べた菌株の混合物及び全細胞又はそれらの成分の混合物も本発明の範囲内である。
【0039】
実施例3において選択された菌株、例えば、L.ロイテリ(L.reuteri)ATCC PTA−6475を標準的ヨーグルトに加えた。L.ロイテリ(L.reuteri)ATCC PTA−6475株は、乳業において、ラクトバチルスを増殖するための標準的な方法を使用して増殖させ、凍結乾燥させた。この培養物を、その後、伝統的ヨーグルト培養物を使用して、あらかじめ発酵させたミルクに、10E+6 CFU/ヨーグルトのグラムの濃度で加え、アテローム性動脈硬化症の予防として人に使用した。ヨーグルト以外の他の摂取可能な支持材料は、例えば、ミルク、カード、発酵乳、ミルクベースの発酵製品、発酵シリアルベースの製品、ミルクベースの粉末である。
【0040】
適切なサイトカインを使用するモデル系を使用して、炎症を軽減又は増大させる因子を決定する。本明細書に提供された発明において、ヒト細胞ベースのアッセイを使用する。
【0041】
THP−1細胞は、白血病患者に由来するヒト単球細胞系であり、これらは、American Type Culture Collectionに保存されている(ATCC番号TIB202)。ヒトの宿主由来のこれらの細胞の起源により、それらは、ヒト胃腸免疫系とヒト共生細菌との相互作用の研究に、特に関係付けられる。
【0042】
本発明のデータは、特定の菌株、L.ロイテリ(L.reuteri)ATCC PTA−4659及びL.ロイテリ(L.reuteri)ATCC PTA−6475によるTNF−αの産生の強力な阻害並びにこの調節が、後期対数増殖期/定常増殖期の間に、これら2種の特定の菌株により増殖培地に放出される物質により仲介されることを示す。それどころか、L.ロイテリ(L.reuteri)の他の2種は、大腸菌毒に対する細胞の炎症反応を阻害できないだけでなく、それら自体が炎症反応を誘導した。
【0043】
胆汁塩を含む選択的de Man、Rogosa及びSharpe(MRS)固体培地を用いた直接プレート法を、胆汁塩ヒドロラーゼを排出する菌株をスクリーニングするために使用し、その基質に対する酵素特異的活性を確定した。BSH産生細菌の増殖は、加水分解及び培地の酸性化を起こす。加水分解は、コロニーの周りに沈殿した遊離の胆汁塩のハロー形成として起こる(Dashkevicz,M.P.及びS.D.Feighner.1989.「胆汁塩ヒドロラーゼ活性ラクトバチルス属種のための分化用培地の開発(Development of a differential medium for bile salt hydrolase−active Lactobacillus spp.)」Appl Environ Microbiol.55(1):11〜16)。
【0044】
本発明の特徴は、以下の実施例を参照することにより、より明確に理解されるであろうが、これらは本発明を限定するものではない。
【実施例】
【0045】
(実施例1)TNF−αの濃度を減少させる能力を有する菌株の評価
THP−1細胞を対照培地、又は選択されたL.ロイテリ(L.reuteri)株であるL.ロイテリ(L.reuteri)ATCC PTA−4659、L.ロイテリ(L.reuteri)ATCC PTA−4975、L.ロイテリ(L.reuteri)ATCC 55730及びL.ロイテリ(L.reuteri)株PTA−4965の増殖によって得られる順化培地(L−CM)のどちらかと共にインキュベートした。順化培地(L−CM)は、L.ロイテリ(L.reuteri)培養物それぞれの9時間培養物又は24時間培養物から得られる無細胞上清である。THP−1細胞は、3.5時間のインキュベーションの間に、対照培地又は大腸菌由来LPS(正常な炎症反応においてTNF−αの産生をもたらす)のどちらかを用いて刺激し、その後、細胞を除去し、上清を、ELISA技術を使用してTNF−αに関して分析した。
【0046】
材料:
THP−1単球白血病細胞系(ATCC、カタログ番号TIB202)
RPMI 1640培地(Gibco−Invitrogen)
ウシ胎児血清(Gibco−Invitrogen)
ペニシリン−ストレプトマイシン溶液(Sigma)
大腸菌、血清型O127:B8のリポ多糖類(Sigma、カタログ番号L3137)TNF−α/TNF−SFIIヒトDuoSet ELISA開発キット(R&D Systems、カタログ番号DY210)
ヒトIL−10 DuoSet、2ndジェネレーションキット(R&D Systems、カタログ番号DY217)
【0047】
方法:
THP−1単球細胞系を使用する。5%(v/v)のMRS培地及び5%(v/v)のラクトバチルス順化培地を適切なウェルに加える。ラクトバチルス順化培地は、MRS培地におけるラクトバチルス属種の24時間培養物の上清である。その後、順化培地を高速真空乾燥し、ペレットを等量の培地に再懸濁することによってpHを調整する。加湿チャンバーは、液体蒸発が最小になるように設計されているが、48時間のインキュベーション後、24ウェルプレート中の細胞懸濁液の容量は、約475μlまで減少する。
【0048】
100ng/mlの大腸菌、血清型O127:B8のリポ多糖類を適切なウェルに加え、これらを37℃、加湿、5%CO2のチャンバーにおいてインキュベートする。3.5時間のインキュベーション後、培養物を1.5mlの遠心分離管に回収し、1500RCFで5分間4℃において遠心分離機にかける。上清を回収する。
【0049】
サイトカインの発現をELISA(Quantikine TNF−α/TNF−SFII ヒトDuoSet)によって試験する。
【0050】
使用した培養培地は、10%のFBS、2%のペニシリン−ストレプトマイシンを含むRPMI1640であった。
【0051】
結果−実施例1
LPSをTHP−1細胞に、L−CMの不在下で加えることにより、3.5時間のインキュベート期間に130pg/mlのTNF−αの産生がもたらされた。これは、毒素に対するTHP−1細胞の予期された炎症反応である。L−CMの添加に対する対照として機能する、増殖培地(MRS)の添加により、132pg/mlのTNF−αの産生がもたらされ、したがって、MRSはLPSに対する反応に影響を与えなかった。L.ロイテリ(L.reuteri)ATCC PTA4659又はL.ロイテリ(L.reuteri)ATCC PTA6475由来の24時間L−CMの添加は、LPSにより刺激されたTNF−αの濃度を、それぞれわずか13及び11pg/mlに、劇的に低下させた。これは、LPSにより刺激されたTNF−αの産生が、それぞれ90及び93%阻害されたことを示す。
【0052】
反対に、L.ロイテリ(L.reuteri)ATCC55730及びL.ロイテリ(L.reuteri)株PTA−4965由来の24時間L−CMの存在下で、LPSは、LPSの不在下の濃度と比較して、TNF−αの有意な上昇をさらに誘導できた。LPSにより刺激されたTNF−αの産生は、L.ロイテリ(L.reuteri)ATCC55730及びL.ロイテリ(L.reuteri)株ATCC PTA−4965由来のL−CMの存在下にもかかわらず、それぞれ54%及び42%増加した(図1)。
【0053】
L.ロイテリ(L.reuteri)ATCC PTA4659又はL.ロイテリ(L.reuteri)ATCC PTA6475由来の9時間L−CMを用いて実施された同様の実験では、LPSにより刺激されたTNF−αの産生に関する阻害作用は、大幅に低いが、存在したことが実証された。したがって、L.ロイテリ(L.reuteri)株をより長くインキュベートし、後期対数増殖期/定常増殖期にL−CM収穫することにより、TNF−αの産生を阻害する効果が改善される。
【0054】
(実施例2)直接プレートアッセイ−細胞外BSH活性を用いた菌株の評価
ヒト乳酸菌の菌株を、酸素を制限した条件において、37℃で、MRSブロス(Acumedia Manufacturers、Inc.Baltimore、Maryland)中で一晩増殖させ、10%のグリセロール(BDH Laboratory Supplies、England)を含む乳酸桿菌を有する培地(LCM)に接種した。
【表1】
【0055】
培養物のストックを、さらなる使用のために−80℃で保存した。菌株は、BioGaia AB研究室並びにLund(スウェーデン)、Raleigh(NC、米国)及びLantbruksuniversitetet(農業大学)、Uppsala(スウェーデン)の収集株から入手した。
【0056】
細胞外BSH活性についてスクリーニングするために、菌株を、一晩培養物から、3mMの胆汁塩、GDCA Sigma、Steinheim、Germany)、TDCA(Sigma)、GCA(Sigma)及びTCA(Fluka,Sigma−Aldrich、Germany)を含むMRS−システイン(MRS−c)寒天(Acumedia)プレート上に、それぞれ画線した。プレートを37℃で48時間、嫌気的(AnaeroGen、Oxoid、UK)にインキュベートした。胆汁酸脱抱合の結果である沈殿を、視覚的に、その結果主観的に測定し、これをもとに、活性を、活性なし(−)、活性あり(+)とした。胆汁塩を加えていないMRS−c寒天プレートを増殖及び負の対照として使用した。
【0057】
(実施例3)BSH活性及び同時にTNF−αの濃度を減少させる能力を有する菌株の選択
【表2】
【0058】
上記の表のデータにより、L.ロイテリ(L.reuteri)のさまざまな菌株が、TNF−α及びBSHの産生に関してさまざまな作用を有し、L.ロイテリ(L.reuteri)ATCC PTA−4659株及びL.ロイテリ(L.reuteri)ATCC PTA−6475株が、アテローム性動脈硬化症への使用に特に適しているという驚くべき発見が確認された。
【0059】
(実施例4)順化培地の使用
実施例1における方法を使用して、TNF−αを有効に減少させる菌株に由来する順化培地、この実施例においては、L.ロイテリ(L.reuteri)ATCC PTA−4659に由来する培地を選択した。この培地を、菌株をde Man、Rogosa、Sharpe(MRS)(Difco、Sparks、MD)で増殖することによって、大規模に作った。乳酸菌の一晩培養物を、OD600が1.0になるように希釈し(およそ109細胞/mlを表す)、さらに1:10に希釈し、さらに24時間増殖した。細菌細胞を含まない順化培地を、8500rpmで10分間4℃において遠心分離機にかけることによって回収した。順化培地を細胞のペレットから分離し、その後0.22μmのろ過孔ユニット(Millipore、Bedford、Mass.)を介してろ過した。その後、順化培地を凍結乾燥し、標準的な方法を使用して、錠剤に製剤化した。この錠剤を、アテローム性動脈硬化症を有効に治療する薬剤として、ヒトに使用した。
【0060】
(実施例5)選択された抗炎症性ラクトバチルス・ロイテリ株の使用
実施例1及び2における方法を使用して、TNF−αを有効に減少させ、同時にBSH活性を増加させる菌株、この実施例においてはL.ロイテリ(L.reuteri)ATCC PTA−4659を選択した。L.ロイテリ(L.reuteri)株を、その後凍結乾燥し、標準的な方法を使用して、105〜109cfuの範囲でカプセルに製剤化した。このカプセルを、アテローム性動脈硬化症を有効に軽減する薬剤として、ヒトに使用した。
【0061】
(実施例6)アテローム性動脈硬化症において頸動脈プラークを減少させる、ラクトバチルス・ロイテリ
総計1059例の患者に、ベースライン及び1年の追跡時に効果的な超音波測定を実施した。ベースライン及び1年の追跡時において、同じ超音波画像系及びトランスデューサー(Acuson Xp10 128、ART改良型、7.5−MHzリニアアレイ・トランスデューサー搭載、開口サイズ38mm、SIEMENS)を使用した。B−モードの画像補正パラメーターを、固定値にプリセットし、いずれの調査の過程においても変更しない。仰臥位の対象を、頭をやや左に向けて、右の頸動脈を、いくつかの異なる超音波照射角度で、縦及び横の両方で、鎖骨のすぐ上から、分岐部に対して出来るだけ末梢側に離れてスキャンした。プラークを、血管壁の内腔への、隣接する内膜中膜厚(IMT)と比較して少なくとも50%の局所突起として定義する。各対象において、最大6個のプラークが、総頸動脈、分岐部及び内頸動脈の近く及び遠くの壁に、それぞれ記録された。それぞれのプラークに関して、トランスデューサーを用いて、血管壁に対して平行に、及び最大プラーク厚の点に対して出来るだけ垂直に、領域拡大選択を38mm×20mに設定して、さらなる画像を記録する。すべての記録は、Panasonic 7650ビデオプレーヤーで、Super VHSテープに実施した。
【0062】
ベースラインにおいて、1059例の男性にプラークが存在した(表1)。頸動脈プラーク面積は、いずれの年齢においても縮小した。ベースラインにおける平均総プラーク面積(SE)は、24.1mm2である。L.ロイテリ(L.reuteri)ATCC PTA−4659(108CFU)の1日容量を摂取後の、追跡期間において、すべての人が、総プラーク面積が縮小した。平均縮小は9.0mm2である。
【表3】
【0063】
本発明は特定の実施形態を参照して記載されているが、多くの変形、改善及び実施形態が可能であることが理解されると思われ、したがって、このような変形、改善及び実施形態もまた、本発明の精神及び範囲内であるとみなされる。
【0064】
【特許請求の範囲】
【請求項1】
アテローム性動脈硬化症及び他の心血管疾患を予防及び/又は治療するための、BSH活性を増加させ、その結果として血清LDLコレステロールを低下させ、同時に前炎症性サイトカインのTNF−αの濃度を減少させる能力について選択されたラクトバチルス(Lactobacillus)株の生物学的に純粋な培養物。
【請求項2】
ラクトバチルス(Lactobacillus)株が、ラクトバチルス・ロイテリ(Lactobacillus reuteri)ATCC PTA−4659及びATCC PTA−6475並びにそれらの混合物からなる群から選択される、請求項1に記載の生物学的に純粋な培養物。
【請求項3】
アテローム性動脈硬化症における炎症の治療に有効な菌株を選択するための方法であって、ヒト由来のTHP−1単球細胞系を使用して、TNF−αの濃度を減少させるのに有効である菌株を同定するステップを含む方法。
【請求項4】
請求項1に記載のラクトバチルス(Lactobacillus)株の生物学的に純粋な培養物に由来する、アテローム性動脈硬化症に随伴する炎症を軽減する成分であって、前記菌株の増殖後の無細胞培養上清から得られ、TNF−α量を減少させる能力を有する成分。
【請求項5】
ラクトバチルス・ロイテリ(Lactobacillus reuteri)株ATCC PTA−4659又はATCC PTA−6475及びそれらの混合物の生物学的に純粋な培養物から単離された無細胞培養上清。
【請求項6】
摂取可能な支持体と、ラクトバチルス・ロイテリ(Lactobacillus reuteri)株ATCC PTA−4659及びATCC PTA−6475並びにそれらの混合物からなる群から選択されるラクトバチルス(Lactobacillus)株に由来する、アテローム性動脈硬化症に随伴する炎症を軽減する成分とを含む食品組成物。
【請求項7】
炎症を軽減する成分が、ラクトバチルス(Lactobacillus)株の生物学的に純粋な培養物の細胞を含む、請求項6に記載の食品組成物。
【請求項8】
ラクトバチルス(Lactobacillus)株が、ラクトバチルス・ロイテリ(Lactobacillus reuteri)ATCC PTA−4659及びATCC PTA−6475並びにそれらの混合物からなる群から選択される、請求項7に記載の食品組成物。
【請求項9】
医薬担体と、ラクトバチルス・ロイテリ(Lactobacillus reuteri)株ATCC PTA−4659及びATCC PTA−6475並びにそれらの混合物からなる群から選択されるラクトバチルス(Lactobacillus)株に由来する、アテローム性動脈硬化症に随伴する炎症を軽減する成分とを含む医薬組成物。
【請求項10】
前記成分が、ラクトバチルス(Lactobacillus)株の生物学的に純粋な培養物の細胞を含む、請求項10に記載の医薬組成物。
【請求項11】
ラクトバチルス(Lactobacillus)株が、ラクトバチルス・ロイテリ(Lactobacillus reuteri)ATCC PTA−4659及びATCC PTA−6475並びにそれらの混合物からなる群から選択される、請求項11に記載の医薬組成物。
【請求項12】
摂取可能な支持体と、ラクトバチルス・ロイテリ(Lactobacillus reuteri)株ATCC PTA−4659、ATCC PTA−6475及びそれらの混合物からなる群から選択されるラクトバチルス(Lactobacillus)株に由来する、アテローム性動脈硬化症に随伴する炎症を軽減する成分とを含む栄養補助剤。
【請求項13】
前記成分が、ラクトバチルス(Lactobacillus)株の生物学的に純粋な培養物の細胞を含む、請求項13に記載の栄養補助剤。
【請求項14】
食品組成物を調製するための方法であって、
a.請求項3に記載のラクトバチルス(Lactobacillus)株を選択するステップ、
b.前記菌株から抗炎症性成分を得るステップ、及び
c.前記成分を摂取可能な支持体に加えて、食品を提供するステップ
を含む方法。
【請求項15】
医薬組成物を調製するための方法であって、
a.請求項3に記載のラクトバチルス(Lactobacillus)株を選択するステップ、
b.前記菌株から抗炎症性成分を得るステップ、及び
c.前記成分を医薬担体に加えて、医薬組成物を提供するステップ
を含む方法。
【請求項16】
栄養補助剤を調製するための方法であって、
a.請求項3に記載のラクトバチルス(Lactobacillus)株を選択するステップ、
b.前記菌株から抗炎症性成分を得るステップ、及び
c.前記成分を摂取可能な支持体に加えて、栄養補助剤を提供するステップ
を含む方法。
【請求項17】
アテローム性動脈硬化症に随伴する炎症を治療又は予防するための薬剤であって、請求項1及び3に記載のラクトバチルス(Lactobacillus)株に由来する抗炎症性成分を含む薬剤。
【請求項18】
アテローム性動脈硬化症に随伴する炎症を治療又は予防するための方法であって、アテローム性動脈硬化症を軽減できることを特徴とする少なくとも1種のラクトバチルス(Lactobacillus)株を選択するステップ、及び前記少なくとも1種の菌株の細胞をヒトに投与するステップを含む方法。
【請求項19】
前記細胞が経口投与される、請求項18に記載の方法。
【請求項1】
アテローム性動脈硬化症及び他の心血管疾患を予防及び/又は治療するための、BSH活性を増加させ、その結果として血清LDLコレステロールを低下させ、同時に前炎症性サイトカインのTNF−αの濃度を減少させる能力について選択されたラクトバチルス(Lactobacillus)株の生物学的に純粋な培養物。
【請求項2】
ラクトバチルス(Lactobacillus)株が、ラクトバチルス・ロイテリ(Lactobacillus reuteri)ATCC PTA−4659及びATCC PTA−6475並びにそれらの混合物からなる群から選択される、請求項1に記載の生物学的に純粋な培養物。
【請求項3】
アテローム性動脈硬化症における炎症の治療に有効な菌株を選択するための方法であって、ヒト由来のTHP−1単球細胞系を使用して、TNF−αの濃度を減少させるのに有効である菌株を同定するステップを含む方法。
【請求項4】
請求項1に記載のラクトバチルス(Lactobacillus)株の生物学的に純粋な培養物に由来する、アテローム性動脈硬化症に随伴する炎症を軽減する成分であって、前記菌株の増殖後の無細胞培養上清から得られ、TNF−α量を減少させる能力を有する成分。
【請求項5】
ラクトバチルス・ロイテリ(Lactobacillus reuteri)株ATCC PTA−4659又はATCC PTA−6475及びそれらの混合物の生物学的に純粋な培養物から単離された無細胞培養上清。
【請求項6】
摂取可能な支持体と、ラクトバチルス・ロイテリ(Lactobacillus reuteri)株ATCC PTA−4659及びATCC PTA−6475並びにそれらの混合物からなる群から選択されるラクトバチルス(Lactobacillus)株に由来する、アテローム性動脈硬化症に随伴する炎症を軽減する成分とを含む食品組成物。
【請求項7】
炎症を軽減する成分が、ラクトバチルス(Lactobacillus)株の生物学的に純粋な培養物の細胞を含む、請求項6に記載の食品組成物。
【請求項8】
ラクトバチルス(Lactobacillus)株が、ラクトバチルス・ロイテリ(Lactobacillus reuteri)ATCC PTA−4659及びATCC PTA−6475並びにそれらの混合物からなる群から選択される、請求項7に記載の食品組成物。
【請求項9】
医薬担体と、ラクトバチルス・ロイテリ(Lactobacillus reuteri)株ATCC PTA−4659及びATCC PTA−6475並びにそれらの混合物からなる群から選択されるラクトバチルス(Lactobacillus)株に由来する、アテローム性動脈硬化症に随伴する炎症を軽減する成分とを含む医薬組成物。
【請求項10】
前記成分が、ラクトバチルス(Lactobacillus)株の生物学的に純粋な培養物の細胞を含む、請求項10に記載の医薬組成物。
【請求項11】
ラクトバチルス(Lactobacillus)株が、ラクトバチルス・ロイテリ(Lactobacillus reuteri)ATCC PTA−4659及びATCC PTA−6475並びにそれらの混合物からなる群から選択される、請求項11に記載の医薬組成物。
【請求項12】
摂取可能な支持体と、ラクトバチルス・ロイテリ(Lactobacillus reuteri)株ATCC PTA−4659、ATCC PTA−6475及びそれらの混合物からなる群から選択されるラクトバチルス(Lactobacillus)株に由来する、アテローム性動脈硬化症に随伴する炎症を軽減する成分とを含む栄養補助剤。
【請求項13】
前記成分が、ラクトバチルス(Lactobacillus)株の生物学的に純粋な培養物の細胞を含む、請求項13に記載の栄養補助剤。
【請求項14】
食品組成物を調製するための方法であって、
a.請求項3に記載のラクトバチルス(Lactobacillus)株を選択するステップ、
b.前記菌株から抗炎症性成分を得るステップ、及び
c.前記成分を摂取可能な支持体に加えて、食品を提供するステップ
を含む方法。
【請求項15】
医薬組成物を調製するための方法であって、
a.請求項3に記載のラクトバチルス(Lactobacillus)株を選択するステップ、
b.前記菌株から抗炎症性成分を得るステップ、及び
c.前記成分を医薬担体に加えて、医薬組成物を提供するステップ
を含む方法。
【請求項16】
栄養補助剤を調製するための方法であって、
a.請求項3に記載のラクトバチルス(Lactobacillus)株を選択するステップ、
b.前記菌株から抗炎症性成分を得るステップ、及び
c.前記成分を摂取可能な支持体に加えて、栄養補助剤を提供するステップ
を含む方法。
【請求項17】
アテローム性動脈硬化症に随伴する炎症を治療又は予防するための薬剤であって、請求項1及び3に記載のラクトバチルス(Lactobacillus)株に由来する抗炎症性成分を含む薬剤。
【請求項18】
アテローム性動脈硬化症に随伴する炎症を治療又は予防するための方法であって、アテローム性動脈硬化症を軽減できることを特徴とする少なくとも1種のラクトバチルス(Lactobacillus)株を選択するステップ、及び前記少なくとも1種の菌株の細胞をヒトに投与するステップを含む方法。
【請求項19】
前記細胞が経口投与される、請求項18に記載の方法。
【図1】

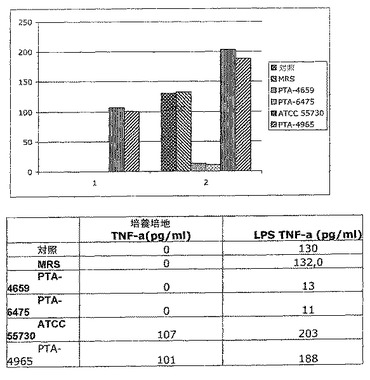
【公表番号】特表2010−523144(P2010−523144A)
【公表日】平成22年7月15日(2010.7.15)
【国際特許分類】
【出願番号】特願2010−502970(P2010−502970)
【出願日】平成20年3月5日(2008.3.5)
【国際出願番号】PCT/SE2008/050248
【国際公開番号】WO2008/127180
【国際公開日】平成20年10月23日(2008.10.23)
【出願人】(500155578)バイオガイア・エイビー (13)
【氏名又は名称原語表記】Biogaia AB
【Fターム(参考)】
【公表日】平成22年7月15日(2010.7.15)
【国際特許分類】
【出願日】平成20年3月5日(2008.3.5)
【国際出願番号】PCT/SE2008/050248
【国際公開番号】WO2008/127180
【国際公開日】平成20年10月23日(2008.10.23)
【出願人】(500155578)バイオガイア・エイビー (13)
【氏名又は名称原語表記】Biogaia AB
【Fターム(参考)】
[ Back to top ]