
アディポネクチン受容体及びそれをコードする遺伝子
ヒト及びマウスのアディポネクチン受容体を単離・同定し、アディポネクチン結合能を有する新規タンパク質を提供するとともに、該タンパク質を利用したアディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストのスクリーニング方法及びスクリーニング用キットを提供することを目的とし、この目的を達成するために、アディポネクチン結合能を有する新規タンパク質として、(a)配列番号2、4、6又は8に記載のアミノ酸配列からなるタンパク質、(b)配列番号2、4、6又は8に記載のアミノ酸配列において1又は複数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつアディポネクチン結合能を有するタンパク質を利用する。
【発明の詳細な説明】
【技術分野】
本発明は、アディポネクチン結合能を有する新規タンパク質、該タンパク質をコードする遺伝子、該遺伝子を含む組換えベクター、該組換えベクターを含む形質転換体、及び上記タンパク質に対する抗体又はその断片に関する。さらに、本発明は、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストのスクリーニング方法及びスクリーニング用キットに関する。
【背景技術】
肥満は、脂肪組織量の増加として定義され、糖尿病、高脂血症、冠動脈性心疾患等の心臓血管疾患及び代謝疾患の高リスクと関連する(Reaven,G.M.Diabetologia 38,3−13.(1995);Spiegelman,B.M.ら,Cell 87,377−389.(1996))。肥満及び2型糖尿病の顕著な特徴であるグルコース及び脂質の代謝異常は、筋肉、肝臓等のインスリン標的組織における脂質蓄積量の増加の原因となり、インスリン抵抗性を引き起こす(Ruderman,N.B.ら,Am.J.Physiol.276,E1−E18.(1999);Shulman,G.I.J.Clin.Invest.106,171−176.(2000))。脂肪組織は、エネルギー要求の変化に応答してトリグリセリド(TG)を蓄積するとともに遊離脂肪酸(FFA)/グリセロールを放出する部位としての役割を果たす(Spiegelman,B.M.,and Flier,J.S.Cell 87,377−389.(1996))。また、脂肪組織は、FFA(Shulman,G.I.J.Clin.Invest.106,171−176.(2000))、アディプシン(White,R.T.ら,J.Biol.Chem.267,9210−9213.(1992))、レプチン(Friedman,J.M.Nature 404,632−634.(2000))、プラスミノーゲンアクチベーターインヒビター−1(PAI−1)(Shimomura,I.ら,Nat.Med.2,800−803.(1996))、レジスチン(Steppan,C.M.ら,Nature 409,307−312.(2001))、腫瘍壊死因子α(TNF−α)(Hotamisligil,G.S.J.Intern.Med.245,621−625.(1999))等の「アディポカイン(adipokines)」(Matsuzawa,Y.ら,Ann.NY.Acad.Sci.892,146−154.(1999))と呼ばれる多数の生物活性物質を分泌する重要な内分泌器官として、様々なエネルギーホメオスタシスの調節に関与する。
アディポネクチン又はAcrp30(Hu,E.,Liang,P.ら,J.Biol.Chem.271,10697−10703.(1996)等)は、アディポサイト由来のホルモンであり、様々な生物学的機能を有している。肥満、2型糖尿病、冠動脈性心疾患は、アディポネクチンの血漿レベルの減少に関連しており、アディポネクチンはin vitroにおいて抗アテローム発生性を示すことが報告されている(Ouchi,N.ら,Circulation 103,1057−1063.(2001);Yokota,T.ら,Blood 96,1723−1732.(2000))。また、Acrp30の循環レベルの急増は、肝臓におけるグルコース産生を低下させることが報告されている(Berg,A.H.ら,Nat.Med.7,947−953.(2001);Combs,T.P.ら,J.Clin.Invest.108,1875−1881.(2001))。また、globular Acrp30は、筋肉における脂肪酸酸化を亢進させ、マウスにおける体重減少を引き起こすことが報告されている(Fruebis,J.ら,Proc.Natl.Acad.Sci.USA.98,2005−2010.(2001))。また、脂肪組織萎縮性マウス又は肥満マウスにおけるインスリン抵抗性が、globularドメインのみからなるアディポネクチン(globular adiponectin;gAd)で処理することにより、筋肉における脂肪酸酸化の亢進を介して改善したこと、及び全長アディポネクチンで処理することにより程度はgAdよりも低いが改善したことが報告されている(Yamauchi,T.ら,Nat.Med.7,941−946.(2001))。
さらに最近になって、骨格筋において、アディポネクチンがAMPキナーゼ(AMPK)を急性的に活性化し、その結果、脂肪酸酸化及びグルコース取り込みが亢進されることが報告されているとともに(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002))、アディポネクチンがPPARαを慢性的に活性化し、その結果、脂肪酸酸化が亢進されるが筋肉における組織TG含量は減少すること、そしてこれらの効果は全長アディポネクチンよりもgAdの方が大きいことが報告されている(Yamauchi,T.ら,J.Biol.Chem.278,2461−2468(2002))。また、興味深いことに、肝臓においては、全長アディポネクチンのみがAMPKを急性的に活性化し、その結果、糖新生に関連する分子を減少させるとともに脂肪酸酸化を亢進させる一方、全長アディポネクチンのみがAMPKを慢性的に活性化し、その結果、脂肪酸酸化を亢進させるとともに肝臓における組織TG量を減少させる。そして、これらの変化はいずれもin vivoにおけるインスリン感受性を増加させる(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002);Yamauchi,T.ら,J.Biol.Chem.278,2461−2468(2002))
これらのアディポネクチンの作用は、細胞表面の受容体によって媒介されると考えられるが、アディポネクチン受容体は未だ同定されておらず、骨格筋及び肝臓におけるアディポネクチン受容体が構造的及び機能的に異なるか否かも未だ不明である。なお、本発明者がアディポネクチン受容体をコードする遺伝子を同定した後、ホモロジー検索を行ったところ、ホモロジーを有する遺伝子として酵母YOL002cが発見された(Karpichev,I.V.ら,Journal of Biological Chemistry,277,19609−19617.(2002))。YOL002cは、脂肪酸酸化等の脂質の代謝経路において重要な役割を果たす7回膜貫通型タンパク質をコードする(Karpichev,I.V.ら,Journal of Biological Chemistry,277,19609−19617.(2002))。
【発明の開示】
本発明は、第一に、アディポネクチン結合能を有する新規タンパク質、該タンパク質をコードする遺伝子、該遺伝子を含む組換えベクター、該組換えベクターを含む形質転換体、及び上記タンパク質に対する抗体又はその断片を提供することを目的とする。
また、本発明は、第二に、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストのスクリーニング方法及びスクリーニング用キットを提供することを目的とする。
上記目的を達成するために、本発明は、以下のタンパク質、遺伝子、組換えベクター、形質転換体、抗体、並びにアディポネクチン受容体に対するアゴニスト又はアンタゴニストのスクリーニング方法及びスクリーニング用キットを提供する。
(1)以下の(a)又は(b)に示すタンパク質。
(a)配列番号2、4、6又は8に記載のアミノ酸配列からなるタンパク質
(b)配列番号2、4、6又は8に記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつアディポネクチン結合能を有するタンパク質
(2)前記(1)記載のタンパク質をコードする遺伝子。
(3)以下の(c)又は(d)に示すDNAを含む前記(2)記載の遺伝子。
(c)配列番号1、3、5又は7に記載の塩基配列からなるDNA
(d)配列番号1、3、5又は7に記載の塩基配列からなるDNAと相補的なDNAにストリンジェントな条件下でハイブリダイズし、かつアディポネクチン結合能を有するタンパク質をコードするDNA
(4)前記(2)又は(3)記載の遺伝子を含む組換えベクター。
(5)前記(4)記載の組換えベクターを含む形質転換体。
(6)前記(1)記載のタンパク質に反応し得る抗体又はその断片。
(7)試験物質と前記(1)記載のタンパク質とを接触させる工程を含む、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストのスクリーニング方法。
(8)前記(1)記載のタンパク質、前記(2)若しくは(3)記載の遺伝子、前記(4)記載の組換えベクター又は前記(5)記載の形質転換体を含む、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストのスクリーニング用キット。
【図面の簡単な説明】
図1(a)は、アディポネクチンのC2C12筋細胞への結合を示す図、図1(b)は、アディポネクチンの肝細胞への結合を示す図、図1(c)〜(e)は、FACS解析の結果を示す図、図1(f)は、AdipoR1遺伝子及びAdipoR2遺伝子の転写物の構造を示す模式図、図1(g)は、マウスの様々な組織に関するノーザンブロット解析の結果を示す図、図1(h)は、ヒトの様々な組織に関するノーザンブロット解析の結果を示す図である。
図2(a)は、AdipoR1及びAdipoR2の構造を示す模式図、図2(b)は、エピトープタグFLAGを有するAdipoR1及びAdipoR2をトランスフェクトした293T細胞の細胞溶解物を抗FLAG抗体で免疫染色した結果を示す図、図2(c)は、エピトープタグをN末端又はC末端に導入たAdipoR1又はAdipoR2をトランスフェクトした293T細胞におけるAdipoR1又はAdipoR2の細胞内局在を示す図、図2(d)は、AdipoR1及びAdipoR2の予測構造モデルを示す図である。
図3(a)は、AdipoR1又はAdipoR2をトランスフェクトした293T細胞への[125I]globular Adipo(gAd)の結合等温線(binding isotherm)を示す図、図3(b)は、当該293T細胞への[125I]全長Adipo(Ad)の結合等温線を示す図、図3(c)は、エピトープタグFLAG又はHAを有するAdipoR1又はAdipoR2をトランスフェクトした293T細胞の細胞溶解物を、抗FLAG抗体又は抗HA抗体を用いて免疫沈降した後、抗FLAG抗体又は抗HA抗体で免疫染色した結果を示す図、図3(d)は、マウスAdipoR1発現細胞又はBLT1発現細胞にglobular Adipo、全長Adipo、LTB4又はATPをチャレンジしたときの[Ca2+]iの変化を示す図、図3(e)は、ホルスコリンで処理した又は処理しないHEK−293細胞にcAMP又はcGMPを蓄積させた結果を示す図、図3(f)は、AdipoR1をトランスフェクトした293T細胞をglobular Adipo又は全長AdipoとインキュベートしたときのPPARαリガンド活性を示す図である。
図4(a)は、マウスAdipoR1 mRNA量を示す図、図4(b)はマウスAdipoR2 mRNA量を示す図、図4(c)及び(d)は、マウスAdipoR1又はマウスAdipoR2をトランスフェクトしたC2C12筋細胞に結合した[125I]globular Adipo(gAd)又は全長Adipo(Ad)の結合等温線を示す図、図4(e)は、マウスAdipoR1又はマウスAdipoR2をトランスフェクトしたC2C12筋細胞にLacZ又はDN−α2AMPKを含有するアデノウイルスを感染させ、globular Adipo又は全長Adipoで7時間処理したときのC2C12筋細胞内のPPARαリガンド活性を示す図、図4(f)は上記C2C12筋細胞内のin vitro脂肪酸酸化を示す図である。
図5(a)は、siRNA又はmockをトランスフェクトしたC2C12筋細胞内のマウスAdipoR1 mRNAの量を示す図、図5(b)は、上記C2C12筋細胞内のマウスAdipoR2 mRNAの量を示す図、図5(c)は、非標識globular Adipoの濃度を増加させることによって、二本鎖siRNAをトランスフェクトした細胞に結合する[125I]globular Adipoを置換した競合ラジオリガンド結合アッセイの結果を示す図、図5(d)は、非標識全長Adipoの濃度を増加させることによって、二本鎖siRNAをトランスフェクトした細胞に結合する[125I]全長Adipoを置換した競合ラジオリガンド結合アッセイの結果を示す図、図5(e)は、二本鎖siRNAをトランスフェクトしたC2C12筋細胞への[125I]globular Adipoの結合等温線を示す図、図5(f)は、二本鎖siRNAをトランスフェクトしたC2C12筋細胞への[125I]全長Adipoの結合等温線を示す図、図5(g)は、二本鎖siRNAをトランスフェクトしたC2C12筋細胞をglobular Adipo、全長Adipo又はWy−14,643と7時間インキュベートしたときのPPARαリガンド活性を示す図、図5(h)は、二本鎖siRNAをトランスフェクトしたC2C12筋細胞をglobular Adipo、全長Adipo又はWy−14,643と7時間インキュベートしたときのin vitro脂肪酸酸化を示す図、図5(i)は、二本鎖siRNAをトランスフェクトしたC2C12筋細胞をglobular Adipo、全長Adipo又はインスリンと7時間インキュベートしたときのグルコース取り込みを示す図である。
図6(a)は、gAdの肝細胞への特異的結合を示す図、図6(b)は、Adの肝細胞への特異的結合を示す図、図6(c)は、HAECにおけるヒトAdipoR1 mRNAの発現レベルを示す図、図6(d)は、HAECにおけるヒトAdipoR2 mRNAの発現レベルを示す図、図6(e)は、gAdのHAECへの特異的結合を示す図、図6(f)は、AdのHAECへの特異的結合を示す図である。
図7(a)は、二本鎖siRNAをトランスフェクトしたC2C12筋細胞への[125I]globular Adipoの結合等温線(binding isotherm)を示す図、図7(b)は、二本鎖siRNAをトランスフェクトしたC2C12筋細胞への[125I]全長Adipoの結合等温線(binding isotherm)を示す図、図7(c)は、図7(a)に示す結果に基づきスキャッチャードプロット解析を行った結果を示す図、図7(d)は、図7(b)に示す結果に基づきスキャッチャードプロット解析を行った結果を示す図である。
図8(a)は、AdipoR1をトランスフェクトした又はしていないC2C12細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、当該細胞の溶解物と抗リン酸化AMPK抗体とを反応させた結果を示す図、図8(b)は、AdipoR1をトランスフェクトした又はしていないC2C12細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、当該細胞の溶解物と抗リン酸化ACC抗体とを反応させた結果を示す図、図8(c)は、AdipoR1をトランスフェクトした又はしていないC2C12細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、当該細胞の溶解物と抗リン酸化p38 MAPK抗体とを反応させた結果を示す図、図8(d)は、AdipoR1をトランスフェクトした又はしていないC2C12細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、当該細胞の溶解物と抗リン酸化MAPK抗体とを反応させた結果を示す図、図8(e)は、AdipoR1をトランスフェクトした又はしていない肝細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、当該細胞の溶解物と抗リン酸化AMPK抗体とを反応させた結果を示す図、図8(f)は、AdipoR1をトランスフェクトした又はしていない肝細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、当該細胞の溶解物と抗リン酸化ACC抗体とを反応させた結果を示す図、図8(g)は、優性阻害型AMPキナーゼ(DN−AMPK)又はp38 MAPK特異的阻害剤SB203580の存在下におけるAdipoR1をトランスフェクトした又はしていないC2C12筋細胞の脂肪酸酸化を示す図、図8(h)は、優性阻害型AMPキナーゼ(DN−AMPK)又はp38 MAPK特異的阻害剤SB203580の存在下におけるAdipoR1をトランスフェクトした又はしていないC2C12筋細胞のグルコース取り込みを示す図である。
【発明を実施するための最良の形態】
本発明のタンパク質は、以下の(a)又は(b)に示すタンパク質である。
(a)配列番号2、4、6又は8に記載のアミノ酸配列からなるタンパク質(以下「タンパク質(a)」という。)
(b)配列番号2、4、6又は8に記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつアディポネクチン結合能を有するタンパク質(以下「タンパク質(b)」という。)
タンパク質(a)又は(b)は、アディポネクチン結合能を有するタンパク質である。「アディポネクチン結合能」とは、アディポネクチンと結合する能力、好ましくはアディポネクチンと特異的に結合する能力(すなわちアディポネクチン受容体が有するアディポネクチン結合能)を意味する。「アディポネクチン結合能」には、全長アディポネクチン又はglobularドメインのみからなるアディポネクチンのいずれに対する結合能も含まれる。アディポネクチン結合能を有するタンパク質には、全長アディポネクチンにのみ結合し得るタンパク質、globularドメインのみからなるアディポネクチンにのみ結合し得るタンパク質、全長アディポネクチン及びglobularドメインのみからなるアディポネクチンの両者に結合し得るタンパク質、全長アディポネクチン及びglobularドメインのみからなるアディポネクチンのいずれか一方に優先的に結合し得るタンパク質等が含まれる。
なお、「globularドメイン」とは、アディポネクチンのC末端側に約100アミノ酸強の長さで存在する糸状体類似の構造ドメインである。補体のClq等にもglobularドメインとホモロジーの高いドメインが存在する。アディポネクチンのglobularドメインは機能的には骨格筋等に強く作用して、脂肪酸を燃焼させ、脂肪が蓄積するのを抑制する作用を有しており、全長のアディポネクチンより数十分の一の低い濃度で同程度の強い活性を示す。
タンパク質(a)のうち、配列番号2又は4に記載のタンパク質はヒト由来のアディポネクチン受容体であり、配列番号6又は8に記載のタンパク質はマウス由来のアディポネクチン受容体である。また、配列番号2及び6記載のタンパク質は同一種類のアディポネクチン受容体(AdipoR1)であり、配列番号4及び8記載のタンパク質は、AdipoR1とは異なる種類のアディポネクチン受容体(AdipoR2)である。AdipoR1は、全長アディポネクチンよりもglobularドメインのみからなるアディポネクチンに対して比較的選択性を有する受容体であると考えられ、AdipoR2は、globularドメインのみからなるアディポネクチンよりも全長アディポネクチンに対して比較的選択性を有する受容体であると考えられる。
AdipoR1について、ヒト(配列番号2)及びマウス(配列番号6)間のアミノ酸レベルでの相同性は96.8%である。AdipoR2について、ヒト(配列番号4)及びマウス(配列番号8)間のアミノ酸レベルでの相同性は95.2%である。AdipoR1とAdipoR2の構造は類似しており、マウスにおけるAdipoR1(配列番号6)及びAdipoR2(配列番号8)の相同性は66.7%である。
生体において、AdipoR1はほとんどの組織で発現しており、特に骨格筋で高度に発現している一方、AdipoR2は肝臓で高度に発現している。また、AdipoR1及びAdipoR2は、ホモ又はヘテロマルチマーを形成していると考えられる。また、AdipoR1及びAdipoR2はともに、全長アディポネクチン及びglobularドメインのみからなるアディポネクチンと結合し、これらのアディポネクチンによるPPARα(Peroxisome proliferator−activated receptor α:ペルオキシソーム増殖因子活性化受容体α)リガンド活性及び脂肪酸酸化の亢進を媒介する。例えば、AdipoR1は、globularドメインのみからなるアディポネクチンによる筋細胞内のPPARαリガンド活性、脂肪酸酸化及びグルコース取り込みの増加を媒介すると考えられる。また、AdipoR2は、全長アディポネクチンによる肝細胞及び筋細胞内のPPARαリガンド活性及び脂肪酸酸化の増加を一部媒介すると考えられる。
AdipoR1及びAdipoR2はいずれも7個の膜貫通ドメインを有すると考えられる。配列番号2記載のアミノ酸配列(ヒトAdipoR1)のうち、136〜158番目、172〜194番目、207〜228番目、234〜255番目、267〜289番目、299〜321番目、336〜358番目のアミノ酸配列が膜貫通ドメインに相当すると考えられる。また、配列番号4記載のアミノ酸配列(ヒトAdipoR2)のうち、60〜82番目、96〜118番目、130〜152番目、158〜179番目、192〜214番目、222〜244番目、260〜282番目のアミノ酸配列が膜貫通ドメインに相当すると考えられる。また、配列番号6記載のアミノ酸配列(マウスAdipoR1)のうち、136〜158番目、172〜194番目、207〜228番目、234〜255番目、267〜289番目、299〜321番目、336〜358番目のアミノ酸配列が膜貫通ドメインに相当すると考えられる。また、配列番号8記載のアミノ酸配列(マウスAdipoR2)のうち、72〜94番目、108〜130番目、142〜164番目、170〜191番目、204〜226番目、234〜256番目、272〜294番目のアミノ酸配列が膜貫通ドメインに相当すると考えられる。
配列番号2、4、6又は8に記載のアミノ酸配列において欠失、置換又は付加されるアミノ酸の個数は、アディポネクチン結合能が保持され得る限り特に限定されるものではなく、その個数は1又は複数個、好ましくは1又は数個であり、その具体的な範囲は通常1〜100個、好ましくは1〜50個、さらに好ましくは1〜10個である。このとき、タンパク質(b)のアミノ酸配列は、タンパク質(a)のアミノ酸配列と通常60%以上、好ましくは80%以上、さらに好ましくは90%以上の相同性を有する。
配列番号2、4、6又は8に記載のアミノ酸配列において欠失、置換又は付加されるアミノ酸の位置は、アディポネクチン結合能が保持され得る限り特に限定されるものではない。
タンパク質(b)には、タンパク質(a)に対して人為的に欠失、置換、付加等の変異を導入したタンパク質の他、欠失、置換、付加等の変異が導入された状態で天然に存在するタンパク質や、それに対して人為的に欠失、置換、付加等の変異を導入したタンパク質も含まれる。欠失、置換、付加等の変異が導入された状態で天然に存在するタンパク質としては、例えば、ヒトを含む哺乳動物(例えば、ヒト、サル、ウシ、ヒツジ、ヤギ、ウマ、ブタ、ウサギ、イヌ、ネコ、マウス、ラット等)由来のタンパク質(これらの哺乳動物において多型によって生じ得るタンパク質を含む)が挙げられる。
タンパク質(a)及び(b)には、糖鎖が付加されたタンパク質及び糖鎖が付加されていないタンパク質のいずれもが含まれる。タンパク質に付加される糖鎖の種類、位置等は、タンパク質の製造の際に使用される宿主細胞の種類によって異なるが、糖鎖が付加されたタンパク質には、いずれの宿主細胞を用いて得られるタンパク質も含まれる。また、タンパク質(a)及び(b)には、その医薬的に許容される塩も含まれる。
タンパク質(a)又は(b)をコードする遺伝子は、例えば、ヒト、マウス等の哺乳動物の骨格筋、肝臓、心臓、マクロファージ、血管、脳、腎臓、肺、胎盤、脾臓、精巣、末梢血液、胸腺、腸管等の組織から抽出したmRNAを用いてcDNAライブラリーを作製し、配列番号1、3、5又は7に記載の塩基配列に基づいて合成したプローブを用いて、cDNAライブラリーから目的のDNAを含むクローンをスクリーニングすることにより得られる。以下、cDNAライブラリーの作製、及び目的のDNAを含むクローンのスクリーニングの各工程について説明する。
〔cDNAライブラリーの作製〕
cDNAライブラリーを作製する際には、例えば、ヒト、マウス等の哺乳動物の骨格筋、肝臓、心臓、マクロファージ、血管、脳、腎臓、肺、胎盤、脾臓、精巣、末梢血液、胸腺、腸管等の組織から全RNAを得た後、オリゴdT−セルロースやポリU−セファロース等を用いたアフィニティーカラム法、バッチ法等によりポリ(A+)RNA(mRNA)を得る。この際、ショ糖密度勾配遠心法等によりポリ(A+)RNA(mRNA)を分画してもよい。次いで、得られたmRNAを鋳型として、オリゴdTプライマー及び逆転写酵素を用いて一本鎖cDNAを合成した後、該一本鎖cDNAから二本鎖cDNAを合成する。このようにして得られた二本鎖cDNAを適当なクローニングベクターに組み込んで組換えベクターを作製し、該組換えベクターを用いて大腸菌等の宿主細胞を形質転換し、テトラサイクリン耐性、アンピシリン耐性を指標として形質転換体を選択することにより、cDNAのライブラリーが得られる。cDNAライブラリーを作製するためのクローニングベクターは、宿主細胞中で自立複製できるものであればよく、例えば、ファージベクター、プラスミドベクター等を使用できる。宿主細胞としては、例えば、大腸菌(Escherichia coli)等を使用できる。
大腸菌等の宿主細胞の形質転換は、塩化カルシウム、塩化マグネシウム又は塩化ルビジウムを共存させて調製したコンピテント細胞に、組換えベクターを加える方法等により行うことができる。なお、ベクターとしてプラスミドを用いる場合は、テトラサイクリン、アンピシリン等の薬剤耐性遺伝子を含有させておく。
cDNAライブラリーの作製にあたっては、市販のキット、例えば、SuperScript Plasmid System for cDNA Synthesis and Plasmid Cloning(Gibco BRL社製)、ZAP−cDNA Synthesis Kit(ストラタジーン社製)等を使用できる。
〔目的のDNAを含むクローンのスクリーニング〕
cDNAライブラリーから目的のDNAを含むクローンをスクリーニングする際には、配列番号1、3、5又は7に記載の塩基配列に基づいてプライマーを合成し、これを用いてポリメラーゼ連鎖反応(PCR)を行い、PCR増幅断片を得る。PCR増幅断片は、適当なプラスミドベクターを用いてサブクローニングしてもよい。PCRに使用するプライマーセットは特に限定されるものではなく、配列番号1、3、5又は7に記載の塩基配列に基づいて設計できる。
cDNAライブラリーに対して、PCR増幅断片をプローブとしてコロニーハイブリダイゼーション又はプラークハイブリダイゼーションを行うことにより、目的のDNAが得られる。プローブとしては、PCR増幅断片をアイソトープ(例えば、32P、35S)、ビオチン、ジゴキシゲニン、アルカリホスファターゼ等で標識したものを使用できる。目的のDNAを含むクローンは、抗体を用いたイムノスクリーニング等の発現スクリーニングによっても得られる。
取得されたDNAの塩基配列は、該DNA断片をそのまま、又は適当な制限酵素等で切断した後、常法によりベクターに組み込み、通常用いられる塩基配列解析方法、例えば、マキサム−ギルバートの化学修飾法、ジデオキシヌクレオチド鎖終結法を用いて決定できる。塩基配列解析の際には、通常、373A DNAシークエンサー(Perkin Elmer社製)等の塩基配列分析装置が用いられる。
タンパク質(a)又は(b)をコードする遺伝子は、タンパク質(a)又は(b)をコードするオープンリーディングフレームとその3’末端に位置する終止コドンとを含む。また、タンパク質(a)又は(b)をコードする遺伝子は、オープンリーディングフレームの5’末端及び/又は3’末端に非翻訳領域(UTR)を含むことができる。
タンパク質(a)をコードする遺伝子としては、例えば、配列番号1、3、5又は7に記載の塩基配列からなるDNAを含む遺伝子が挙げられる。ここで、配列番号1記載の塩基配列のうち1〜1125番目の塩基配列は、配列番号2記載のタンパク質をコードするオープンリーディングフレームであり、配列番号1記載の塩基配列のうち、翻訳開始コドンは1〜3番目の塩基配列に位置し、終止コドンは1126〜1128番目の塩基配列に位置する。また、配列番号3記載の塩基配列のうち1〜897番目の塩基配列は、配列番号4記載のタンパク質をコードするオープンリーディングフレームであり、配列番号3記載の塩基配列のうち、翻訳開始コドンは1〜3番目の塩基配列に位置し、終止コドンは898〜900番目の塩基配列に位置する。また、配列番号5記載の塩基配列のうち1〜1125番目の塩基配列は、配列番号6記載のタンパク質をコードするオープンリーディングフレームであり、配列番号5記載の塩基配列のうち、翻訳開始コドンは1〜3番目の塩基配列に位置し、終止コドンは1126〜1128番目の塩基配列に位置する。また、配列番号7記載の塩基配列のうち1〜933番目の塩基配列は、配列番号8記載のタンパク質をコードするオープンリーディングフレームであり、配列番号7記載の塩基配列のうち、翻訳開始コドンは1〜3番目の塩基配列に位置し、終止コドンは934〜936番目の塩基配列に位置する。
タンパク質(a)をコードする遺伝子の塩基配列は、タンパク質(a)をコードする限り特に限定されるものではなく、オープンリーディングフレームの塩基配列は、配列番号1、3、5又は7に記載の塩基配列に限定されるものではない。タンパク質(a)をコードする遺伝子は、その塩基配列に従って化学合成により得ることもできる。DNAの化学合成は、市販のDNA合成機、例えば、チオホスファイト法を利用したDNA合成機(島津製作所社製)、フォスフォアミダイト法を利用したDNA合成機(パーキン・エルマー社製)を用いて行うことができる。
タンパク質(b)をコードする遺伝子としては、例えば、配列番号1、3、5又は7に記載の塩基配列からなるDNAと相補的なDNAにストリンジェントな条件下でハイブリダイズし、かつアディポネクチン結合能を有するタンパク質をコードするDNAを含む遺伝子が挙げられる。
「ストリンジェントな条件」としては、例えば、42℃、2×SSC及び0.1%SDSの条件、好ましくは65℃、0.1×SSC及び0.1%SDSの条件が挙げられる。
配列番号1、3、5又は7に記載の塩基配列からなるDNAと相補的なDNAにストリンジェントな条件下でハイブリダイズするDNAの具体例としては、配列番号1、3、5又は7に記載の塩基配列からなるDNAと少なくとも60%以上、好ましくは80%以上、さらに好ましくは90%以上の相同性を有するDNAが挙げられる。
タンパク質(b)をコードする遺伝子は、例えば、配列番号1、3、5又は7に記載の塩基配列からなるDNAに、部位特異的変異誘発法等の公知の方法を用いて人為的に変異を導入することにより得られる。変異の導入は、例えば、変異導入用キット、例えば、Mutant−K(TAKARA社製)、Mutant−G(TAKARA社製)、TAKARA社のLA PCR in vitro Mutagenesisシリーズキットを用いて行うことができる。また、塩基配列が既に決定されているDNAについては化学合成によっても得られる。
タンパク質(a)又は(b)は、例えば、以下の工程に従って、それぞれのタンパク質をコードする遺伝子を宿主細胞中で発現させることにより製造できる。
〔組換えベクター及び形質転換体の作製〕
組換えベクターを作製する際には、目的とするタンパク質のコード領域を含む適当な長さのDNA断片を調製する。また、目的とするタンパク質のコード領域の塩基配列を、宿主細胞における発現に最適なコドンとなるように、塩基を置換したDNAを調製する。
このDNA断片を適当な発現ベクターのプロモーターの下流に挿入することにより組換えベクターを作製し、該組換えベクターを適当な宿主細胞に導入することにより、目的とするタンパク質を生産し得る形質転換体が得られる。上記DNA断片は、その機能が発揮されるようにベクターに組み込まれることが必要であり、ベクターは、プロモーターの他、エンハンサー等のシスエレメント、スプライシングシグナル、ポリA付加シグナル、選択マーカー(例えば、ジヒドロ葉酸還元酵素遺伝子、アンピシリン耐性遺伝子、ネオマイシン耐性遺伝子)、リボソーム結合配列(SD配列)等を含有できる。
発現ベクターとしては、宿主細胞において自立複製が可能なものであれば特に限定されず、例えば、プラスミドベクター、ファージベクター、ウイルスベクター等を使用できる。プラスミドベクターとしては、例えば、大腸菌由来のプラスミド(例えば、pRSET、pBR322、pBR325、pUC118、pUC119、pUC18、pUC19)、枯草菌由来のプラスミド(例えば、pUB110、pTP5)、酵母由来のプラスミド(例えば、YEp13、YEp24、YCp50)が挙げられ、ファージベクターとしては、例えば、λファージ(例えば、Charon4A、Charon21A、EMBL3、EMBL4、λgt10、λgt11、λZAP)が挙げられ、ウイルスベクターとしては、例えば、レトロウイルス、ワクシニアウイルス等の動物ウイルス、バキュロウイルス等の昆虫ウイルスが挙げられる。
宿主細胞としては、目的とする遺伝子を発現し得る限り、原核細胞、酵母、動物細胞、昆虫細胞、植物細胞等のいずれを使用してもよい。また、動物個体、植物個体、カイコ虫体等を使用してもよい。
細菌を宿主細胞とする場合、例えば、エッシェリヒア・コリ(Escherichia coli)等のエシェリヒア属、バチルス・ズブチリス(Bacillus subtilis)等のバチルス属、シュードモナス・プチダ(Pseudomonas putida)等のシュードモナス属、リゾビウム・メリロティ(Rhizobium meliloti)等のリゾビウム属に属する細菌を宿主細胞として使用できる。具体的には、Escherichia coli XL1−Blue、Escherichia coli XL2−Blue、Escherichia coli DH1、Escherichia coli K12、Escherichia coli JM109、Escherichia coli HB101等の大腸菌や、Bacillus subtilis MI 114、Bacillus subtilis 207−21等の枯草菌を宿主細胞として使用できる。この場合のプロモーターは、大腸菌等の細菌中で発現できるものであれば特に限定されず、例えば、trpプロモーター、lacプロモーター、PLプロモーター、PRプロモーター等の大腸菌やファージ等に由来するプロモーターを使用できる。また、tacプロモーター、lacT7プロモーター、letIプロモーターのように人為的に設計改変されたプロモーターも使用できる。
細菌への組換えベクターの導入方法としては、細菌にDNAを導入し得る方法であれば特に限定されず、例えば、カルシウムイオンを用いる方法、エレクトロポレーション法等を使用できる。
酵母を宿主細胞とする場合、サッカロミセス・セレビシエ(Saccharomycescerevisiae)、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)、ピヒア・パストリス(Pichia pastoris)等を宿主細胞として使用できる。この場合のプロモーターは、酵母中で発現できるものであれば特に限定されず、例えば、gal1プロモーター、gal10プロモーター、ヒートショックタンパク質プロモーター、MFα1プロモーター、PHO5プロモーター、PGKプロモーター、GAPプロモーター、ADHプロモーター、AOX1プロモーター等を使用できる。
酵母への組換えベクターの導入方法は、酵母にDNAを導入し得る方法であれば特に限定されず、例えば、エレクトロポレーション法、スフェロプラスト法、酢酸リチウム法等を使用できる。
動物細胞を宿主細胞とする場合、サル細胞COS−7、Vero、チャイニーズハムスター卵巣細胞(CHO細胞)、マウスL細胞、ラットGH3、ヒトFL細胞等を宿主細胞として使用できる。この場合のプロモーターは、動物細胞中で発現できるものであれば特に限定されず、例えば、SRαプロモーター、SV40プロモーター、LTR(Long Terminal Repeat)プロモーター、CMVプロモーター、ヒトサイトメガロウイルスの初期遺伝子プロモーター等を使用できる。
動物細胞への組換えベクターの導入方法は、動物細胞にDNAを導入し得る方法であれば特に限定されず、例えば、エレクトロポレーション法、リン酸カルシウム法、リポフェクション法等を使用できる。
昆虫細胞を宿主とする場合には、Spodoptera frugiperdaの卵巣細胞、Trichoplusia niの卵巣細胞、カイコ卵巣由来の培養細胞等を宿主細胞として使用できる。Spodoptera frugiperdaの卵巣細胞としてはSf9、Sf21等、Trichoplusia niの卵巣細胞としてはHigh 5、BTI−TN−5B1−4(インビトロジェン社製)等、カイコ卵巣由来の培養細胞としてはBombyx mori N4等が挙げられる。
昆虫細胞への組換えベクターの導入方法は、昆虫細胞にDNAを導入し得る限り特に限定されず、例えば、リン酸カルシウム法、リポフェクション法、エレクトロポレーション法等を使用できる。
〔形質転換体の培養〕
目的とするタンパク質をコードするDNAを組み込んだ組換えベクターを導入した形質転換体を通常の培養方法に従って培養する。形質転換体の培養は、宿主細胞の培養に用いられる通常の方法に従って行うことができる。
大腸菌や酵母等の微生物を宿主細胞として得られた形質転換体を培養する培地としては、該微生物が資化し得る炭素源、窒素源、無機塩類等を含有し、形質転換体の培養を効率的に行える培地であれば天然培地、合成培地のいずれを使用してもよい。
炭素源としては、グルコース、フラクトース、スクロース、デンプン等の炭水化物、酢酸、プロピオン酸等の有機酸、エタノール、プロパノール等のアルコール類を使用できる。窒素源としては、アンモニア、塩化アンモニウム、硫酸アンモニウム、酢酸アンモニウム、リン酸アンモニウム等の無機酸又は有機酸のアンモニウム塩、ペプトン、肉エキス、酵母エキス、コーンスチープリカー、カゼイン加水分解物等を使用できる。無機塩としては、リン酸第一カリウム、リン酸第二カリウム、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム、硫酸第一鉄、硫酸マンガン、硫酸銅、炭酸カルシウム等を使用できる。
大腸菌や酵母等の微生物を宿主細胞として得られた形質転換体の培養は、振盪培養又は通気攪拌培養等の好気的条件下で行う。培養温度は通常25〜37℃、培養時間は通常12〜48時間であり、培養期間中はpHを6〜8に保持する。pHの調整は、無機酸、有機酸、アルカリ溶液、尿素、炭酸カルシウム、アンモニア等を用いて行うことができる。また、培養の際、必要に応じてアンピシリン、テトラサイクリン等の抗生物質を培地に添加してもよい。
プロモーターとして誘導性のプロモーターを用いた発現ベクターで形質転換した微生物を培養するときには、必要に応じてインデューサーを培地に添加してもよい。例えば、lacプロモーターを用いた発現ベクターで形質転換した微生物を培養するときにはイソプロピル−β−D−チオガラクトピラノシド等を、trpプロモーターを用いた発現ベクターで形質転換した微生物を培養するときにはインドールアクリル酸等を培地に添加してもよい。
動物細胞を宿主細胞として得られた形質転換体を培養する培地としては、一般に使用されているRPMI1640培地、EagleのMEM培地、DMEM培地、Ham F12培地、Ham F12K培地又はこれら培地に牛胎児血清等を添加した培地等を使用できる。形質転換体の培養は、通常5%CO2存在下、37℃で3〜10日間行う。また、培養の際、必要に応じてカナマイシン、ペニシリン、ストレプトマイシン等の抗生物質を培地に添加してもよい。
昆虫細胞を宿主細胞として得られた形質転換体を培養する培地としては、一般に使用されているTNM−FH培地(ファーミンジェン社製)、Sf−900 II SFM培地(Gibco BRL社製)、ExCell400、ExCell405(JRHバイオサイエンシーズ社製)等を使用できる形質転換体の培養は、通常27℃で3〜10日間行う。また、培養の際、必要に応じてゲンタマイシン等の抗生物質を培地に添加してもよい。
目的とするタンパク質は、分泌タンパク質又は融合タンパク質として発現させることもできる。融合させるタンパク質としては、例えば、β−ガラクトシダーゼ、プロテインA、プロテインAのIgG結合領域、クロラムフェニコール・アセチルトランスフェラーゼ、ポリ(Arg)、ポリ(Glu)、プロテインG、マルトース結合タンパク質、グルタチオンS−トランスフェラーゼ、ポリヒスチジン鎖(His−tag)、Sペプチド、DNA結合タンパク質ドメイン、Tac抗原、チオレドキシン、グリーン・フルオレッセント・プロテイン等が挙げられる。
〔タンパク質の単離・精製〕
形質転換体の培養物より目的とするタンパク質を採取することにより、目的とするタンパク質が得られる。ここで、「培養物」には、培養上清、培養細胞、培養菌体、細胞又は菌体の破砕物のいずれもが含まれる。
目的とするタンパク質が形質転換体の細胞内に蓄積される場合には、培養物を遠心分離することにより、培養物中の細胞を集め、該細胞を洗浄した後に細胞を破砕して、目的とするタンパク質を抽出する。目的とするタンパク質が形質転換体の細胞外に分泌される場合には、培養上清をそのまま使用するか、遠心分離等により培養上清から細胞又は菌体を除去する。
こうして得られるタンパク質(a)又は(b)は、溶媒抽出法、硫安等による塩析法脱塩法、有機溶媒による沈殿法、ジエチルアミノエチル(DEAE)−セファロース、イオン交換クロマトグラフィー法、疎水性クロマトグラフィー法、ゲルろ過法、アフィニティークロマトグラフィー法等により精製できる。
タンパク質(a)又は(b)は、そのアミノ酸配列に基づいて、Fmoc法(フルオレニルメチルオキシカルボニル法)、tBoc法(t−ブチルオキシカルボニル法)等の化学合成法によっても製造できる。この際、市販のペプチド合成機を使用できる。
本発明の抗体又はその断片は、タンパク質(a)又は(b)に反応し得る抗体又はその断片である。ここで、「抗体」には、モノクローナル抗体及びポリクローナル抗体のいずれもが含まれ、「モノクローナル抗体及びポリクローナル抗体」には全てのクラスのモノクローナル抗体及びポリクローナル抗体が含まれる。また、「抗体」には、ウサギやマウス等の免疫動物にタンパク質(a)又は(b)のタンパク質を免疫して得られる抗血清、ヒト抗体、遺伝子組換えによって得られるヒト型化抗体も含まれる。また、「抗体の断片」には、Fab断片、F(ab)’2断片、単鎖抗体(scFv)等が含まれる。
本発明の抗体又はその断片は、タンパク質(a)又は(b)を免疫用抗原として利用することより作製できる。免疫用抗原としては、例えば、(i)タンパク質(a)又は(b)を発現している細胞又は組織の破砕物又はその精製物、(ii)遺伝子組換え技術を用いて、タンパク質(a)又は(b)をコードする遺伝子を大腸菌、昆虫細胞又は動物細胞等の宿主に導入して発現させた組換えタンパク質、(iii)化学合成したペプチド等を使用できる。
ポリクローナル抗体の作製にあたっては、免疫用抗原を用いて、ラット、マウス、モルモット、ウサギ、ヒツジ、ウマ、ウシ等の哺乳動物を免疫する。免疫動物は、抗体を容易に作製できることからマウスを利用することが好ましい。免疫の際には、抗体産生誘導する為に、フロイント完全アジュバント等の免疫助剤を用いてエマルジョン化した後、複数回の免疫することが好ましい。免疫助剤としては、フロイント完全アジュバント(FCA)の他、フロイント不完全アジュバント(FIA)、水酸化アルミニウムゲル等を利用できる。哺乳動物1匹当たりの抗原の投与量は、哺乳動物の種類に応じて適宜設定できるが、マウスの場合には通常50〜500μgである。投与部位は、例えば、静脈内、皮下、腹腔内等である。免疫の間隔は、通常、数日から数週間間隔、好ましくは4日〜3週間間隔で、合計2〜8回、好ましくは2〜5回免疫を行う。そして、最終免疫日から3〜10日後に、タンパク質(a)又は(b)に対する抗体力価を測定し、抗体力価が上昇した後に採血し、抗血清を得る。抗体力価の測定は、酵素免疫測定法(ELISA)、放射性免疫測定法(RIA)等により行うことができる。
抗血清から抗体の精製が必要とされる場合は、硫酸アンモニウムによる塩析、ゲルクロマトグラフィー、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー等の公知の方法を適宜選択して又はこれらを組み合わせて利用できる。
モノクローナル抗体の作製にあたっては、ポリクローナル抗体の場合と同様に免疫用抗原を用いて哺乳動物を免疫し、最終免疫日から2〜5日後に抗体産生細胞を採取する。抗体産生細胞としては、例えば、脾臓細胞、リンパ節細胞、胸腺細胞、末梢血細胞等が挙げられるが、脾臓細胞が一般的に利用される。
次いで、ハイブリドーマを得るために、抗体産生細胞とミエローマ細胞との細胞融合を行う。抗体産生細胞と融合させるミエローマ細胞としては、ヒト、マウス等の哺乳動物由来の細胞であって一般に入手可能な株化細胞を利用できる。利用する細胞株としては、薬剤選択性を有し、未融合の状態では選択培地(例えばHAT培地)で生存できず、抗体産生細胞と融合した状態でのみ生存できる性質を有するものが好ましい。ミエローマ細胞の具体例としては、P3X63−Ag.8.U1(P3U1)、P3/NSI/1−Ag4−1、Sp2/0−Ag14等のマウスミエローマ細胞株が挙げられる。
細胞融合は、血清を含まないDMEM、RPMI−1640培地等の動物細胞培養用培地中に、抗体産生細胞とミエローマ細胞とを所定の割合(例えば1:1〜1:10)で混合し、ポリエチレングリコール等の細胞融合促進剤の存在下で、又は電気パルス処理(例えばエレクトロポレーション)により融合反応を行う。
細胞融合処理後、選択培地を用いて培養し、目的とするハイブリドーマを選別する。次いで、増殖したハイブリドーマの培養上清中に、目的とする抗体が存在するか否かをスクリーニングする。ハイブリドーマのスクリーニングは、通常の方法に従えばよく、特に限定されるものではない。例えば、ハイブリドーマとして生育したウエルに含まれる培養上清の一部を採集し、酵素免疫測定法(ELISA)、放射性免疫測定法(RIA)等によってスクリーニングできる。
ハイブリドーマのクローニングは、例えば、限界希釈法、軟寒天法、フィブリンゲル法、蛍光励起セルソーター法等により行うことができ、最終的にモノクローナル抗体を産生するハイブリドーマを取得する。
取得したハイブリドーマからモノクローナル抗体を採取する方法としては、通常の細胞培養法等を利用することができる。細胞培養法においては、例えばハイブリドーマを10〜20%牛胎児血清含有RPMI−1640培地、MEM培地等の動物細胞培養培地中、通常の培養条件(例えば37℃,5%CO2濃度)で3〜10日間培養することにより、その培養上清からモノクローナル抗体を取得することができる。また、ハイブリドーマをマウス等の腹腔内に移植し、10〜14日後に腹水を採取し、当該腹水からモノクローナル抗体を取得することもできる。
モノクローナル抗体の精製が必要とされる場合は、硫酸アンモニウムによる塩析、ゲルクロマトグラフィー、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー等の公知の方法を適宜選択して又はこれらを組み合わせて利用できる。
モノクローナル抗体をヒトに投与する目的(抗体治療)で使用する場合には、免疫原性を低下させるため、ヒト抗体又はヒト型化抗体を使用することが好ましい。ヒト抗体又はヒト型化抗体は、例えば、免疫動物としてヒト抗体遺伝子を導入したマウス等を用いてハイブリドーマを作製することにより、また、ファージ上に抗体を提示したライブラリーを用いることにより取得できる。具体的には、ヒト抗体遺伝子のレパートリーを有するトランスジェニック動物に、抗原となるタンパク質、タンパク質発現細胞又はその溶解物を免疫して抗体産生細胞を取得し、これをミエローマ細胞と融合させたハイブリドーマを用いて目的のタンパク質に対するヒト抗体を取得できる(国際公開番号WO92−03918、WO93−2227、WO94−02602、WO96−33735及びWO96−34096参照)。また、複数の異なるヒトscFvをファージ上に提示させた抗体ライブラリーから、抗原となるタンパク質、タンパク質発現細胞又はその溶解物に結合する抗体を提示しているファージを選り分けることで、目的のタンパク質に結合するscFvを選択できる(Griffiths.等,EMBO J.12,725−734,1993)。
本発明のスクリーニング方法は、試験物質とタンパク質(a)又は(b)とを接触させる工程を含む。本発明のスクリーニング方法によれば、試験物質とタンパク質(a)又は(b)とを接触させ、試験物質がタンパク質(a)又は(b)に結合するか否かを判別することにより、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストをスクリーニングできる。スクリーニングされた物質は、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストとしての作用を実際に有するか否かを判別する工程にさらに供することが好ましい。
試験物質の種類は特に限定されるものではないが、例えば、高分子化合物、低分子化合物、細胞培養物、組織抽出物等が挙げられる。
スクリーニングの際には、タンパク質(a)又は(b)として、例えば、(i)タンパク質(a)又は(b)を発現している細胞若しくは組織又はそれらの処理物(ii)DNA組換え技術を用いて、タンパク質(a)又は(b)をコードするDNAを大腸菌、酵母、昆虫細胞、動物細胞等の宿主に導入して発現させた組換えタンパク質、(iii)化学合成したペプチド等を使用できる。タンパク質(a)又は(b)を発現している細胞又は組織は、タンパク質(a)又は(b)を内因性タンパク質として発現している細胞又は組織(例えば、筋細胞、骨格筋、心臓、マクロファージ、血管、脳、腎臓、肺、胎盤、脾臓、精巣、末梢血液、胸腺、腸管等)であってもよいし、外因性タンパク質として発現している細胞又は組織(例えば、タンパク質(a)又は(b)をコードするDNAを導入した大腸菌、酵母、昆虫細胞、動物細胞等)であってもよい。また、細胞又は組織の処理物は、細胞又は組織に、破砕、抽出、精製等の処理を施したものであり、細胞又は組織の処理物としては、例えば、細胞又は組織の細胞膜画分が挙げられる。
試験物質がアディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストとしての作用を有するか否かは、例えば、タンパク質(a)又は(d)に対する試験物質の結合量、試験物質とタンパク質(a)又は(b)との結合により生じる細胞応答の有無又は程度に基づいて判別できる。
タンパク質(a)又は(b)に対する試験物質の結合量は、例えば、標識した試験物質、試験物質に対する標識抗体等を使用して測定できる。標識としては、例えば、3H、14C、125I、35S、32P等の放射性同位体元素、蛍光色素等を使用できる。放射性同位体元素の放射活性は、例えば、液体シンチレーションカウンター、X線フィルム、イメージングプレート等を用いて測定でき、蛍光色素の蛍光強度は、例えば、CCDカメラ、蛍光スキャナー、分光蛍光光度計等を用いて測定できる。
試験物質とタンパク質(a)又は(b)との結合により生じる細胞応答としては、例えば、PPARαリガンド活性の亢進又は抑制、脂肪酸酸化の亢進又は抑制、グルコース取り込みの亢進又は抑制、細胞内pHの上昇又は低下、AMPキナーぜ活性の増加又は低下、AMPキナーゼのリン酸化の増加又は低下、p38 MAPキナーゼ活性の増加又は低下、p38 MAPキナーゼのリン酸化の増加又は低下、糖新生の促進又は抑制、脱共役蛋白の増加又は低下等が挙げられる。
タンパク質(a)又は(b)に対する試験物質の結合量の測定、及び細胞応答の有無又は程度の測定にあたっては、タンパク質(a)又は(b)の存在下及び不存在下における測定値を比較することが好ましい。
脂肪細胞は主要なインスリン感受性ホルモンとしてアディポネクチンを分泌し、脂肪細胞肥大はアディポネクチン分泌低下を惹起し、アディポネクチン分泌低下はインスリン抵抗性を惹起する。そして、インスリン抵抗性は、糖尿病、高脂血症、高血圧を惹起する。また、アディポネクチン分泌低下は動脈硬化を促進させる。したがって、アディポネクチン受容体に対するアゴニスト又はアンタゴニストとしてスクリーニングされた物質は、例えば、インスリン抵抗性改善薬、糖尿病予防・治療薬、高脂血症予防・治療薬、高血圧予防・治療薬、動脈硬化予防・治療薬、肥満予防・治療薬、抗炎症薬、骨粗鬆症予防・治療薬、抗癌剤等として使用できる。
これらの薬剤はスクリーニングされた物質のみから構成してもよいが、通常は、薬学的に許容され得る賦形剤その他任意の添加剤を用いて製剤化する。製剤化にあたっては、例えば、賦形剤、結合剤、崩壊剤、滑沢剤、安定剤、矯味矯臭剤、希釈剤、注射剤用溶剤等の添加剤を使用できる。投与経路としては、例えば、経口投与、非経口投与(例えば、皮下投与、筋肉内投与、腹腔内投与等)が挙げられ、投与剤形としては、例えば、噴霧剤、カプセル剤、錠剤、顆粒剤、シロップ剤、乳剤、座剤、注射剤、懸濁剤等が挙げられる。投与量及び投与回数は、目的とする作用効果、投与方法、治療期間、患者の年齢、体重、性別等により異なり、スクリーニングされた物質の種類に応じて適宜調節できる。
タンパク質(a)又は(b)、タンパク質(a)又は(b)をコードする遺伝子、該遺伝子を含有する組換えベクター又は該組換えベクターを含む形質転換体は、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストのスクリーニング用キットの構成要素として利用できる。これらは、タンパク質(a)又は(b)の供給源としてスクリーニング用キットに含まれる。
スクリーニング用キットは、タンパク質(a)又は(b)、タンパク質(a)又は(b)をコードする遺伝子、該遺伝子を含有する組換えベクター又は該組換えベクターを含む形質転換体を含む限り、いかなる形態であってもよく、各種試薬(例えば、緩衝液等)、測定機械器具、標識化合物、モデル動物、細胞株、細胞培養用培地等を含むことができる。
【実施例】
以下、実施例に関する本文及び図において、アディポネクチンを「Adipo」と、globularドメインのみからなるアディポネクチンを「globular Adipo」又は「gAd」と、全長アディポネクチンを「全長Adipo」又は「Ad」という場合がある。また、アディポネクチン受容体を「AdipoR」と、ヒト由来アディポネクチン受容体を「hAdipoR」と、マウス由来アディポネクチン受容体を「mAdipoR」という場合がある。また、「AdipoR1」及び「AdipoR2」はアミノ酸配列が異なるアディポネクチン受容体を表す。
1.実験方法
(1)レトロウイルスの作製及び感染
レトロウイルス含有上清を作製するために、リポフェクタミンPLUS(Life Technologies)を用いて、107個のPlat−E packaging細胞(Morita,S.ら,Gene Ther.7,1063−1066(2000))に、ヒト骨格筋cDNAライブラリー(Clontech)10μgを一過性にトランスフェクトした。24時間インキュベーションした後、上清(10mL)を回収した。Ba/F3細胞に、ポリブレン(hexadimethrine,Sigma)10μg/mLを添加した1/20倍希釈上清(推定m.o.i=0.3)を感染させた。6時間後、培養液を交換し、Ba/F3細胞を6日間培養して増殖させた後、冷凍保存又は細胞選別を行った。
(2)FACS解析及びcDNAの配列決定
FACS解析は、Stoecklinらの方法(Stoecklin,G.ら,EMBO J.21,4709−4718.(2000))に従って行った。細胞選別のために、ヒト骨格筋cDNAライブラリーをトランスフェクトしたBa/F3細胞1×107個からFACVantage(Becton Dickinson)を用いて細胞を濃縮した。細胞を回収し、増殖させた後、11日後にFACS解析に供した。選別された細胞をさらに増殖してFACS解析に供した。選別された細胞に組み込まれたcDNAの配列決定を行うために、レトロウイルスベクターに対する上流及び下流のプライマーを用いて、選別された細胞から抽出されたゲノムDNA 50ngを鋳型としてPCRを行い、得られたPCR増幅断片の配列決定を行った。
PCRは、Taqポリメラーゼ(Perkin−Elmer/Cetus)を用いて、94℃で1分間、56℃で2分間、72℃で3分間のサイクルを35サイクル行った。レトロウイルスベクターに対するプライマー(pLIBプライマー)は以下のとおりである。

PCR産物は、プライマーを除去した後、BigDye Terminater Kit(Applied Biosystems)を用いたdirect sequenceにより塩基配列を決定した。
(3)ノーザンブロット解析
ヒトの様々な組織(脳、心臓、骨格筋、大腸、胸腺、脾臓、腎臓、肝臓、小腸、胎盤、肺、末梢血白血球)のノーザンブロットフィルターI及びIIをClonetechより購入した。これらのフィルターを、4×SSC、5×Denhardt’s溶液、0.2%SDS、200mg/mLサケ精子DNA、及び50%ホルムアミドを含有するハイブリダイゼーション緩衝液中、42℃で24時間、[32P]dCTP標識したcDNAプローブ(ヒトAdipoR1 cDNA、マウスAdipoR1 cDNA、ヒト又はマウスAdipoR2 cDNAをそれぞれPstI/BstXI、BamHI/PstI、EcoRV/NotIを処理して得られた断片)とハイブリダイズさせた。0.1×SSC、0.1%SDS、65℃の条件でフィルターを洗浄した後、オートラジオグラフィーに供した。同様のノーザンブロット解析をマウスの様々な組織(脳、心臓、腎臓、肝臓、肺、骨格筋、脾臓、精巣)についても行った。
(4)哺乳動物細胞におけるタンパク質の発現及び性状決定
AdipoR1 cDNA又はAdipoR2 cDNAをpCXN2(Kinoshita,S.ら,Pharm.Res.15,1851−1856(1998))のEcoRV/NotI部位にライゲートすることにより、AdipoR1又はAdipoR2発現ベクターを構築した。HEK−293T(human embryonic kidney cells:ヒト胎児腎臓細胞)、HAEC(Normal Human Aortic Endothelial Cells:正常ヒト大動脈内皮細胞)及びC2C12筋細胞(マウス筋細胞株)をDMEM中で培養した。培養液には、10%ウシ胎児血清(FCS)を含有させた。DNAトランスフェクションは、いずれの細胞についても、リポフェクタミンPLUS(Gibco BRL)を用いたリポフェクションによって行った。
(5)C2C12筋細胞におけるRNA干渉(RNA interference)
Karpichevらの方法(Karpichev,I.V.ら,J.Biol.Chem.277,19609−19617(2002))に従い、二組のsiRNAを化学合成し、アニーリングさせ、リポフェクタミンPLUS(Life Technologies)を用いて、コンフルエントから2日後にウマ血清含有DMEMで4〜7日間培養して筋管細胞に分化させたC2C12筋細胞にトランスフェクトした。肝細胞又はHAECについても同様にsiRNAをトランスフェクトした。siRNAのトランスフェクション48時間後、細胞を溶解した。
無関係なコントロールsiRNA(siRNA unrelated)の塩基配列は次のとおりである。
マウスAdipoR1遺伝子のコーディング領域に対応するsiRNA(siRNA mAdipoR1)の塩基配列は次のとおりである。
マウスAdipoR2遺伝子のコーディング領域に対応するsiRNA(siRNA mAdipoR2)の塩基配列は次のとおりである。
ヒトAdipoR1遺伝子のコーディング領域に対応するsiRNA(siRNA hAdipoR1)の塩基配列は次のとおりである。
ヒトAdipoR2遺伝子のコーディング領域に対応するsiRNA(siRNA hAdipoR2)の塩基配列は次のとおりである。
(6)PPARα(Peroxisome proliferator−activated receptor α:ペルオキシソーム増殖因子活性化受容体α)リガンド活性の測定
大腸菌を用いて発現させたマウスglobular Adipo及び全長Adipoを、Yamauchiらの方法(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002))に従って精製した。Yamauchiらの方法(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002))に従って、分化したC2C12筋細胞又は単離した肝細胞を所定濃度のアディポネクチンで処理した。PPARαリガンド活性は、Yamauchiらの方法(Yamauchi,T.ら,J.Biol.Chem.278,2461−2468(2002))に従って、(UAS)×4−tk−LUCリポータープラスミド、GAL4−ratPPARα リガンド結合ドメイン発現プラスミド、及びβ−ガラクトシダーゼ発現プラスミド(内部コントロール)を用いて定量した。
(7)脂質及びグルコース代謝
Yamauchiらの方法(Yamauchi,T.ら,Nat.Med.7,941−946.(2001))に従って、細胞溶解物を用いて[1−14C]パルミチン酸からの[14C]CO2産生を測定した。また、Yamauchiらの方法(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002))に従って、グルコース取り込みを測定した。
(8)優性阻害型(dominant negative)AMPキナーゼ(AMPK)を用いた研究
α2 AMPK(45番目のリジン残基がアルギニン残基に置換された変異を含む)をコードするcDNAをDN−α2AMPKとして用いた(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002))。C2C12筋細胞に、コントロールMockベクター又はDN−α2AMPKを含有する等しい力価のアデノウイルスを感染させた。Yamauchiらの方法(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002))に従って、分化の誘導の5日後に、細胞を所定濃度のアディポネクチンで処理して、PPARαリガンド活性及び脂肪酸酸化を測定した。
(9)結合アッセイ
合成ヒト又はマウスAdipoのTyrを、Na125I(2000Ci/mmol,Amersham Pharmacia Biotech)の存在下、IODO−ビーズ(Pierce)により125Iで標識した。組換えglobular Adipo又は全長AdipoをNHS−LC−ビオチン(Pierce)を用いてビオチン化した。細胞を96wellプレート上に4.1×104個/wellの密度で播き、一晩培養した後、培養液を除去し、細胞を37℃で一晩、所定濃度の[125I]Adipo及び非標識競合物質を含有する結合アッセイ用緩衝液(HEPES緩衝化生理食塩水/0.1%ウシ血清アルブミン)とインキュベートした。その後、Yamauchiらの方法(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002);Yokomizo,T.ら,Nature 387,620−624(1997))に従って、細胞を氷冷リン酸緩衝化生理食塩水で3回洗浄し、0.1N NaOH/0.1%SDSで溶解し、細胞に結合した放射活性をγカウンターで測定した。
(10)蛍光顕微鏡による解析
AdipoR1又はAdipoR2の細胞内局在を、293T細胞を用いて、共焦点蛍光顕微鏡により評価した。細胞を1%パラホルムアルデヒドで固定した後、5倍希釈した透過緩衝液(permeabilization buffer)(Coulter)を用いて透過させて又は透過させずに、抗FLAG抗体(M2;30μg/mL)と22℃で1時間インキュベートした。次いで、AlexFluor 488を結合させた二次抗体10μg/mLとともにインキュベートした。次いで、ニコン顕微鏡及びクリプトン/アルゴンレーザー(488nm)を備えたレーザスキャンニング顕微鏡システムを用いて共焦点イメージングを行った。
(11)AdipoR1遺伝子及びAdipoR2遺伝子の転写物のリアルタイムPCRによる定量的解析
Heidらの方法(Heid,C.A.ら,Genome Res.6,986−994(1996))に従って、ヒトAdipoR1遺伝子、ヒトAdipoR2遺伝子、マウスAdipoR1遺伝子及びマウスAdipoR2遺伝子に関する4種類の転写物の定量をリアルタイムPCRによって行った。それぞれの転写物に関するプライマー及びプローブは以下のとおりである。PCR産物は、ABI PRISM7700 Sequence Detection System(Applied Biosystems)を用いて連続的に測定した。それぞれの転写物の相対量をアクチンの転写物量に対して標準化した。
[マウスAdipoR1遺伝子]
[マウスAdipoR2遺伝子]
[ヒトAdipoR1遺伝子]
[ヒトAdipoR2遺伝子]
(12)細胞内カルシウム濃度、cAMP量及びcGMP量の測定
細胞内Ca2+濃度は、Yokomizoらの方法(Yokomizo,T.ら,Nature 387,620−624(1997))に従って測定した。すなわち、細胞に、Hepes−Tyrode’s BSA緩衝液(25mM Hepes−NaOH(pH7.4),140mM NaCl,2.7mM KCl,1.0mM CaCl2,12mM NaHCO3,5.6mM D−グルコース,0.37mM NaH2PO4,0.49mM MgCl2,0.1%[wt/vol]脂肪酸不含BSA;Fraction V)に溶解した10μM Fura−2 AM(Dojin)を37℃で2時間接触させた。細胞を2回洗浄し、Hepes−Tyrode’s BSA緩衝液に106細胞/mLの濃度で懸濁させた。0.5mLの細胞懸濁液をCAF−100 system(Jasco)にアプライし、5μLのリガンドのエタノール溶液(LTB4用)又はPBS溶液(Adipo用)を加えた。340nm及び380nmの励起光によって発せられる500nmの蛍光の割合に基づいて細胞内Ca2+濃度を測定した。
また、cAMP量及びcGMP量は、Yokomizoらの方法(Yokomizo,T.ら,Nature 387,620−624(1997))に従い、アッセイキット(cAMP:Biotrak cAMP EIAシステム,cGMP:Biotrak cGMP EIAシステム)(Amersham Pharmacia Biotech)を用いて、製造業者のプロトコルに基づき測定した。
(13)AdipoR1及びAdipoR2の構造予測
Kyte−Doolitleのハイドロパシーインデックスを用いて、AdipoR1及びAdipoR2タンパク質のハイドロパシープロット(hydropathy plot)を行った。また、AdipoR1及びAdipoR2の構造モデルをSOSUIによって予測し、PRINTS(http://bioinf.man.ac.uk/dbbrowser/PRINTS/)によってコンセンサス配列を解析した。さらに、リン酸化部位をDNASIS Pro.によって解析した。さらに、AdipoR1/R2がその他のクラスのGPCRとホモロジーを有するか否かをhttp://cbrg.inf.ethz.ch/Server/AllAll.htmlに記載の方法により解析した。
(14)AMPキナーゼ(AMPK)、ACC、p38 MAPキナーゼ(p38 MAPK)及びMAPキナーゼ(MAPK)のリン酸化及びリン酸化量の測定
AMPK、ACC(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002))、p38 MAPK及びMAPK(Barger,P.M.ら,J.Biol.Chem.276,44495−44501(2001);Puigserver,P.ら,Mol.Cell 8,971−982(2001);Michael,L.F.ら,Proc.Natl.Acad.Sci.USA.98,3820−3825(2001))のリン酸化及びリン酸化量を、抗リン酸化AMPK抗体、抗リン酸化ACC抗体、抗リン酸化p38 MAPK抗体又は抗リン酸化MAPK抗体を利用したウェスタンブロッティング法により測定した。この際、AdipoR1をトランスフェクトした又はしていないC2C12細胞又は肝細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、各細胞の溶解物と各抗体とを反応させた。
2.結果
(1)AdipoR1及びAdipoR2の発現クローニング
筋肉において、globular Adipoは全長Adipoよりもインスリン抵抗性を改善させ、PPARα及び脂肪酸酸化を活性化させる(Fruebis,J.ら,Proc.Natl.Acad.Sci.U S A.98,2005−2010(2001);Yamauchi,T.ら,Nat.Med.7,941−946(2001);Yamauchi,T.ら,J.Biol.Chem.278,2461−2468(2002))。さらに、globular Adipoは全長AdipoよりもC2C12筋細胞に強く結合するとともに、肝細胞及び肝臓の膜よりも骨格筋の膜に強く結合する(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002))(図1a,b参照)。図1aは、globular Adipo又は全長AdipoとC2C12筋細胞との結合、図1bはglobular Adipo又は全長Adipoと肝細胞との結合を示す。細胞は、図示する濃度のビオチン化globular Adipo又は全長Adipoとともにインキュベートし、細胞表面に結合したビオチン化globular Adipo又は全長AdipoはELISAにより定量した。図中のそれぞれのバーは平均値±s.e.(n=3−5)を示し、「※」はP<0.05、「※※」はP<0.01を示す。
そこで、ヒト骨格筋のmRNA由来のcDNAを組み込んだレトロウイルスをBa/F3細胞に感染させて作製したライブラリーから、globular Adipoに対する結合能を有するタンパク質をスクリーニングすることにより、AdipoR1のcDNAの単離を試みた。
感染させたBa/F3細胞を回収し、ビオチン化したglobular Adipoとインキュベートした後、ストレプトアビジンを結合させたフィコエリトリン(phycoerythrin:PE)(赤色蛍光プローブ)で染色し、蛍光活性化セルソーター(fluorescence−activated cell sorting:FACS)に供した(図1c参照)。FACS解析の結果を図1c,d,eに示す。図1cは、感染後であるが、第一の選別前のBa/F3細胞を示し、図1dは、第三の選別前のBa/F3細胞を示し、図1eは、FITC(fluorescein isothiocyanate)を結合させたglobular Adipoとインキュベートした後であるが、第四の選別前のBa/F3細胞を示す。また、図1c,d,eにおいて、四角で囲まれた領域は、AdipoR1陽性細胞を示し、四角内の細胞は選別された細胞を示す。
選別されたglobular Adipo結合能を有する細胞を第二ラウンド目の選別に供した。再選別されたglobular Adipo結合能を有する細胞(図1d参照)を第三ラウンド目の選別に供し、再選別された細胞を直ちにFITC(fluorescein isothiocyanate)(緑色蛍光プローブ)を結合させたglobular Adipoとインキュベートした。赤色から緑色に変化した細胞がglobular Adipoに対する特異的結合部位を有する細胞であるから(図1e参照)、そのように変化した細胞のみを選別し、さらなる分析のために培養により増殖させた。これらの細胞から抽出したゲノムDNAを、ウイルスベクターに対するプライマーを用いたPCRに供し、配列決定した。
なお、上記のように細胞を二色(赤色及び緑色)で染色したのは、globular Adipoがその粘着性によって非特異的に結合した細胞を除去するためである。
配列決定されたヒトAdipoR1 cDNAの塩基配列を配列番号1に示し、それにコードされるタンパク質(ヒトAdipoR1)のアミノ酸配列を配列番号2に示す。また、C2C12筋細胞から同様の方法によって取得したマウスAdipoR1 cDNAの塩基配列を配列番号5に示し、それにコードされるタンパク質(マウスAdipoR1)のアミノ酸配列を配列番号6に示す。ヒトAdipoR1 cDNA(配列番号1)及びマウスAdipoR1 cDNA(配列番号5)は、375個のアミノ酸からなるタンパク質をコードする遺伝子であることが判明した(図1f参照)。ヒトAdipoR1及びマウスAdipoR1のアミノ酸配列の相同性は96.8%である(図1f参照)。図1fは、データベース(NIH−MGC Project and NCBI contig)におけるAdipoR1遺伝子及びAdipoR2遺伝子の転写物の模式的構造を示す。
これまでの知見(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002);Yamauchi,T.ら,J.Biol.Chem.278,2461−2468(2002))から、AdipoRには、globular Adipo又は全長Adipoに対する結合親和性が異なる2種類のタイプ、すなわち、骨格筋で発現するglobular Adipoに優先的に結合するタイプと、肝臓で発現する全長Adipoにのみ結合するタイプとが存在すると考えられる。
AdipoR1とホモロジーを有するタンパク質を調査したところ、ヒト及びマウスに関するデータベース(The Human Genome,http://www.ncbi.nlm.nih.gov/genome/guide/human/;Mouse Genome Resources,http://www.ncbi.nlm.nih.gov/genome/guide/mouse/)(Waterston,R.H.ら,Nature 420,520−562(2002);Okazaki,Y.ら,Nature 420,563−573(2002))において、AdipoR1 cDNAとは異なるオープンリーディングフレーム(ORF)を有する遺伝子が発見された。そこで、このcDNAをHepG2細胞(ヒト肝癌由来細胞株)のmRNAからクローニングし、配列決定し、このcDNAにコードされているタンパク質をAdipoR2と名付けた。配列決定されたヒトAdipoR2 cDNAの塩基配列を配列番号3に示し、それにコードされるタンパク質(ヒトAdipoR2)のアミノ酸配列を配列番号4に示す。また、C2C12筋細胞から同様の方法によって取得したマウスAdipoR2 cDNAの塩基配列を配列番号7に示し、それにコードされるタンパク質(マウスAdipoR2)のアミノ酸配列を配列番号8に示す。
ヒトAdipoR2及びマウスAdipoR2のアミノ酸配列の相同性は95.2%である。また、AdipoR1及びAdipoR2は構造的に高度に類似しており、マウスAdipoR1及びAdipoR2のアミノ酸配列の相同性は66.7%である。
SWISS−PROTにおいて、AdipoR1及びAdipoR2と相同性が高い哺乳動物のタンパク質は存在しなかったが、興味深いことに、AdipoR1及びAdipoR2をコードするcDNAは、酵母YOL002c(Karpichev,I.V.ら,J.Biol.Chem.277,19609−19617.(2002))とホモロジーを有していた。YOL002cは、脂肪酸酸化等の脂質の代謝経路において重要な役割を果たす7回膜貫通型タンパク質をコードすると報告されている(Karpichev,I.V.ら,J.Biol.Chem.277,19609−19617(2002))。種を問わず、7回膜貫通構造を有するAdipoR/YOL002cは、脂肪酸酸化等の脂肪代謝において重要な制御シグナルを媒介すると考えられる。
(2)AdipoR1及びAdipoR2の組織分布
マウスの様々な組織に関するノーザンブロット解析の結果を図1gに示し、ヒトの様々な組織に関するノーザンブロット解析の結果を図1hに示す。図1g中、レーン1は脳、レーン2は心臓、レーン3は腎臓、レーン4は肝臓、レーン5は肺、レーン6は骨格筋、レーン7は脾臓、レーン8は精巣の結果を示し、図1h中、レーン1は脳、レーン2は心臓、レーン3は骨格筋、レーン4は大腸、レーン5は胸腺、レーン6は脾臓、レーン7は腎臓、レーン8は肝臓、レーン9は小腸、レーン10は胎盤、レーン11は肺、レーン12は末梢血白血球の結果を示す。
ヒト及びマウスの様々な組織に関するノーザンブロット解析によって、上記データベースにおいて予測されたmRNAのサイズを有する2.0kbの一本の主要なバンドが同定され、AdipoR1がほとんどの組織で発現しており、骨格筋において高度に発現していることが明らかになった。また、上記データベースにおいて予測されたmRNAのサイズを有する4.0kbの一本の主要なバンドが同定され、AdipoR2が肝臓において高度に発現していることが明らかになった。
(3)AdipoR1及びAdipoR2の細胞内局在
マウスAdipoR1の推定アミノ酸配列(配列番号6)から、マウスAdipoR1は375個のアミノ酸からなるタンパク質であり、その分子量は42.4kDaと予測される。また、マウスAdipoR2の推定アミノ酸配列(配列番号8)から、マウスAdipoR2は311個のアミノ酸からなるタンパク質であり、その分子量は35.4kDaと予想される(図2a参照)。図2aは、ソフトウエアPRINTS(http://bioinf.man.ac.uk/dbbrowser/PRINTS/)によってAdipoRの配列をスキャンした結果であり、図中、下線領域は、AdipoR1及びAdipoR2の7個の膜貫通ドメインを示し、太字の下線部は、Gタンパク質結合受容体のメンバーに特徴的な保存モチーフを示す。また、図2aには、PKCリン酸化部位及びチロシンリン酸化部位も併せて示す。
AdipoR1及びAdipoR2の推定アミノ酸配列からAdipoR1及びAdipoR2は、7個の膜貫通ドメインを有するタンパク質であると予測される(図2a参照)。AdipoR1又はAdipoR2について、7個の膜貫通ドメインを有する公知の受容体(Waterston,R.H.ら,Nature 420,520−562(2002);Okazaki,Y.ら,Nature 420,563−573(2002);Wess,J.,FASEB.J.11,346−354(1997))とのアライメントを行ったが、Gタンパク質結合受容体(G−protein coupled receptor:GPCR)ファミリーに属するメンバーとのアミノ酸配列の相同性は低かった。AdipoR1及びAdipoR2は、Gタンパク質結合受容体ファミリーの特徴(例えば、保存アミノ酸、グリコシル化部位、Gタンパク質結合部位)(Wess,J.らFASEB.J.11,346−354(1997);Yokomizo,T.ら,Nature 387,620−624(1997);Scheer,A.ら,EMBO.J.15,3566−3578(1996))が欠落していた。Gタンパク質結合受容体ファミリーにおいて高度に保存されているアミノ酸のうち、AdipoR1及びAdipoR2の第一及び第二細胞外ループにおいて高度に保存されている2個のCys残基のうちの1つのみが存在していた。AdipoR1及びAdipoR2は、TM7の末端に存在する高度に保存されたAsn−Pro−Xaa2−Tyrモチーフを欠落していた。また、AdipoR1及びAdipoR2のTM3/細胞内ループ2トランジションに存在する高度に保存されたAsp−Arg−Tyrモチーフ(Wess,J.らFASEB.J.11,346−354(1997);Scheer,A.ら,EMBO.J.15,3566−3578(1996))も欠落していた。
エピトープタグFLAGで標識したヒト及びマウスのAdipoR1又はAdipoR2をHEK−293T細胞中で発現させ、抗FLAG抗体で免疫染色した。ヒト及びマウスAdipoR1及びAdipoR2は、予想された分子量を示した(図2b参照)。
マウスAdipoR1又はAdipoR2の細胞内局在及びトポロジーを調べるために、いずれかの末端にエピトープタグを有するAdipoR1又はAdipoR2のcDNAをHEK−293T細胞で発現させた(図2c参照)。図2c中、「intact」は細胞を透過させなかった場合、「permeabilized」は細胞を透過させた場合を表す。
エピトープタブをN末端に導入した場合には、細胞を透過させたときのみ、細胞表面に存在するAdipoR1及びAdipoR2を検出できた(図2c参照)。これに対して、エピトープタグをC末端に導入した場合には、細胞表面に存在するAdipoR1及びAdipoR2を検出できた(図2c参照)。これらの結果は、AdipoR1及びAdipoR2が7個の膜貫通ドメインを有する膜内在性タンパク質であり、そのN末端は膜内に存在し、そのC末端は膜外に存在することを示す(図2c,d参照)。これは、これまでに報告されている全てのGタンパク質結合受容体(Wess,J.らFASEB.J.11,346−354(1997);Yokomizo,T.ら,Nature 387,620−624(1997);Scheer,A.ら,EMBO.J.15,3566−3578(1996))と反対のトポロジーである。なお、図2dは、AdipoR1及びAdipoR2の予測構造モデルである。
(4)293T細胞におけるAdipoRの発現の効果
AdipoR1又はAdipoR2を細胞表面上に過剰発現させた293T細胞を用いて、globular Adipo又は全長Adipoにより亢進される結合活性及び細胞内シグナルを調べた。293T細胞におけるAdipoR1又はAdipoR2の発現は、globular Adipo及び全長Adipoの両者の結合を亢進させた(図3a,b参照)。図3aは、AdipoR1又はAdipoR2をトランスフェクトした293T細胞への[125I]globular Adipo(gAd)の結合等温線(binding isotherm)を示し、図3bは、当該293T細胞への[125I]全長Adipo(Ad)の結合等温線(binding isotherm)を示し、図中、□はMock、■はマウスAdipoR1、●はマウスAdipoR2に関する結果を示す。
次に、AdipoR1及びAdipoR2がマルチマー(multimer)を形成するかどうかを調べた。エピトープタグFLAGを有するAdipoR1及びエピトープタグHAを有するAdipoR1をともにHEK−293T細胞中で発現させた場合には、抗HA抗体を用いた免疫沈降において、エピトープタグFLAGを有するAdipoR1が検出された(図3c参照)。図3cは、エピトープタグFLAG又はHAを有するAdipoR1又はAdipoR2をトランスフェクトした293T細胞の細胞溶解物を、抗FLAG抗体(上側及び下側のパネル)を用いて免疫沈降(IP)した後、抗FLAG抗体(上側のパネル)又は抗HA抗体(下側のパネル)で免疫染色した結果であり、AdipoR1及びAdipoR2によるホモ−及びヘテロ−マルチマーの形成を示す。
さらに、エピトープタグHAを有するAdipoR1及びエピトープタグFLAGを有するAdipoR2をともにHEK−293T細胞中で発現させた場合には、抗HA抗体を用いた免疫沈降において、エピトープタグFLAGを有するAdipoR2が検出された(図3c参照)。これらの結果は、AdipoR1及びAdipoR2がホモ−及びヘテロ−マルチマーの両者を形成する可能性を示す。
GPCR又はLTB4受容体BLT1を発現させた細胞では、LTB4により細胞内カルシウムが増加し、AdipoR1を発現する細胞の発現レベルと類似する発現レベルを示したが(データは示さず)、AdipoR1を発現する細胞では、Adipoによる細胞内カルシウムの変化は観察されなかった(図3d参照)。図3dは、Fura−2/AMをロードしたマウスAdipoR1発現細胞又はBLT1発現細胞に、10μg/mL globular Adipo(gAd)、10μg/mL 全長Adipo(Ad)、1μM LTB4又は100μM ATPをチャレンジし、[Ca2+]iの変化を測定した結果である。
さらに、AdipoR1を発現する細胞において、AdipoR1はcAMP及びcGMPレベルに対して全く影響しないか、ほとんど影響を与えなかった(図3e参照)。図3eはホルスコリン(forskolin)で処理した又は処理しないHEK−293細胞にcAMP又はcGMPを蓄積させた結果であり、図中、「gAd0.01」、「gAd0.1」はそれぞれ0.01、0.1μg/mLのglobular Adipoを表し、「Ad1」、「Ad10」はそれぞれ1、10μg/mLの全長Adipoを表す。対照的に、AdipoR1の発現は、globular Adipo及び全長Adipoによる293T細胞内のPPARαリガンド活性の増加を増強した(図3f参照)。図3fは、AdipoR1をトランスフェクトした293T細胞を図示する濃度(μg/mL)のglobular Adipo又は全長AdipoとインキュベートしたときのPPARαリガンド活性を示し、図中、「gAd0.1」、「gAd0.5」、「gAd2.5」はそれぞれ0.1、0.5、2.5μg/mLのglobular Adipoを表し、「Ad1」、「Ad5」、「Ad25」はそれぞれ1、5、25μg/mLの全長Adipoを表す。また、図中、各バーは平均値±s.e.(n=3−5)を表し、「※」はP<0.05を表し、「※※」はP<0.01を表す。
(5)C2C12筋細胞におけるPPARα活性化及び脂肪酸酸化
C2C12筋細胞におけるAdipoR1の発現(図4a参照)は、globular Adipo及び全長Adipoの両方の結合を促進させ(図4c,d参照)、globular Adipo及び全長AdipoによるC2C12筋細胞内のPPARαリガンド活性(図4e参照)及び脂肪酸酸化(図4f参照)の増加と関連した。また、C2C12筋細胞におけるAdipoR2の発現(図4b参照)は、globular Adipo及び全長Adipoの両方の結合を促進させ(図4c,d参照)、globular Adipo及び全長AdipoによるC2C12筋細胞内の脂肪酸酸化の亢進(図4f)に関連した。図4aはマウスAdipoR1 mRNA量を示し、図4bはマウスAdipoR2mRNA量を示し、図中、□はMock、■はマウスAdipoR1、点線を引いた□はマウスAdipoR2に関する結果を示す。また、図4c及びdは、マウスAdipoR1又はマウスAdipoR2をトランスフェクトしたC2C12筋細胞に結合した[125I]globular Adipo(gAd)又は全長Adipo(Ad)の結合等温線(binding isotherm)を示し、図中、□はMock、■はマウスAdipoR1、●はマウスAdipoR2に関する結果を示す。また、図4eは、マウスAdipoR1又はマウスAdipoR2をトランスフェクトしたC2C12筋細胞に、LacZ又はDN−α2AMPKを含有するアデノウイルスを感染させ、図示する濃度(μg/mL)のglobular Adipo又は全長Adipoで7時間処理したときの、当該C2C12筋細胞内のPPARαリガンド活性を示し、図中、□はMock、■はマウスAdipoR1に関する結果を示す。また、図4fは、上記C2C12筋細胞内のin vitro脂肪酸酸化を示し、図中、□はMock、■はマウスAdipoR1、点線を引いた□はマウスAdipoR2に関する結果を示す。なお、各図中、各バーは平均値±s.e.(n=3−5)を表し、「※」はP<0.05を表し、「※※」はP<0.01を表す。
優性阻害型(dominant negative:DN)AMPキナーゼの発現は、globular Adipo及び全長Adipo誘導性であってAdipoR1発現依存的なPPARαリガンド活性の亢進に影響を与えなかった。これらの結果は、AdipoR1及びAdipoR2はともに、globular Adipo及び全長Adipoの結合を媒介し、globular Adipo及び全長AdipoによるPPARαリガンド活性及び脂肪酸酸化を亢進させることを示す。
(6)筋細胞内のAdipoRの結合及び作用に対するsiRNAの影響
筋細胞において、内因性のAdipoR1及びAdipoR2がAdipoの特異的結合及び代謝的作用を媒介しているか否かを調べるため、siRNAを用いてAdipoR1及びAdipoR2の発現を抑制した(図5a,b参照)。図5aは、siRNA又はmockをトランスフェクトしたC2C12筋細胞内のマウスAdipoR1 mRNAの量を示し、図5bは、当該C2C12筋細胞内のマウスAdipoR2 mRNAの量を示す。また、図5a,b中、レーン1はmock、レーン2は無関係なsiRNA、レーン3はマウスAdipoR1遺伝子に対するsiRNA、レーン4はマウスAdipoR2遺伝子に対するsiRNAを用いた場合の結果を示す。
C2C12筋細胞において、siRNAによりAdipoR1の発現を抑制すると(図5a参照)、globular Adipoの結合活性は全く観察されなくなり、全長Adipoの結合活性は一部低下した(図5c,d参照)。図5cは、非標識globular Adipoの濃度を増加させることによって、二本鎖siRNAをトランスフェクトした細胞に結合する[125I]globular Adipoを置換した競合ラジオリガンド結合アッセイの結果(n=4)であり、図5dは、非標識全長Adipoの濃度を増加させることによって、二本鎖siRNAをトランスフェクトした細胞に結合する[125I]全長Adipoを置換した競合ラジオリガンド結合アッセイの結果(n=4)である。
globular Adipo又は全長Adipoで7時間処理すると、C2C12細胞において、PPARαリガンド活性が増加し(図5g参照)、脂肪酸酸化が亢進し(図5h参照)、グルコース取り込みが亢進した(図5i参照)。図5gは、二本鎖siRNAをトランスフェクトしたC2C12筋細胞を、図示する濃度(μg/mL)のglobular Adipo又は全長Adipo、あるいは10−5M Wy−14,643(図中「Wy」と記載)と7時間インキュベートしたときのPPARαリガンド活性を示す。図5hは、二本鎖siRNAをトランスフェクトしたC2C12筋細胞を、図示する濃度(μg/mL)のglobular Adipo又は全長Adipo、あるいは10−5M Wy−14,643(図中「Wy」と記載)と7時間インキュベートしたときのin vitro脂肪酸酸化を示す。図5iは、二本鎖siRNAをトランスフェクトしたC2C12筋細胞を、図示する濃度(μg/mL)のglobular Adipo又は全長Adipo、あるいは10−7Mインスリン(図中「Ins」と記載)と7時間インキュベートしたときのグルコース取り込みを示す。図5g〜iにおいて、「gAd0.01」、「gAd0.03」、「gAd0.1」、「gAd0.5」、「gAd2.5」はそれぞれ0.01、0.03、0.1、0.5、2.5μg/mLのglobular Adipoを表し、「Ad0.1」、「Ad0.3」、「Ad1」、「Ad5」、「Ad25」はそれぞれ0.1、0.3、1、5、25μg/mLの全長Adipoを表す。図中、各バーは平均値±s.e.(n=3−5)を表し、「※」はP<0.05、「※※」はP<0.01を表す。また、図5g〜iにおいて、□は無関係なsiRNA、■はマウスAdipoR1に対するsiRNA、点線を引いた□はマウスAdipoR2に対するsiRNA、斜線を引いた□はマウスAdipoR1及びマウスAdipoR2に対するsiRNAを用いたときの結果を示す。
C2C12筋細胞において、siRNAによりAdipoR1の発現を抑制すると(図5a参照)、globular AdipoによるPPARαリガンド活性(図5g参照)、脂肪酸酸化(図5h参照)及びグルコース取り込み(図5i参照)の増加が減少した。これに対して、AdipoR1の発現を抑制しても、全長Adipoによるこれらの作用の有意な減少は観察されなかった。したがって、AdipoR1は、globular Adipoによる筋細胞内のPPARαリガンド活性、脂肪酸酸化及びグルコース取り込みの増加を媒介していると考えられる。
内因性のAdipoR2が、筋細胞におけるAdipoの特異的結合及び代謝的作用を媒介しているか否かを調べるため、siRNAを用いてAdipoR2の発現を抑制した(図5b参照)。C2C12筋細胞において、siRNAによってAdipoR2の発現を抑制すると(図5b参照)、globular Adipo及び全長Adipoの結合が一部減少した(図5e,f参照)。さらに、AdipoR2の発現抑制によって、全長AdipoによるPPARαリガンド活性(図5g参照)及び脂肪酸酸化(図5h参照)の亢進が部分的に減少した。したがって、AdipoR2は、全長Adipoによる筋細胞内のPARαリガンド活性、脂肪酸酸化の亢進を部分的に媒介していると考えられる。図5eは、二本鎖siRNAをトランスフェクトしたC2C12筋細胞への[125I]globular Adipoの結合等温線(binding isotherm)を示し、図5fは、二本鎖siRNAをトランスフェクトしたC2C12筋細胞への[125I]全長Adipoの結合等温線(binding isotherm)を示す。なお、図中、□は無関係なコントロールsiRNA、●はマウスAdipoR2に対するsiRNA、▲はマウスAdipoR1及びマウスAdipoR2に対するsiRNAを用いた場合の結果を示す。
AdipoR1は、globular Adipoに対して比較的選択性を有する受容体であり、AdipoR2は全長Adipoに対して比較的選択性を有する受容体であると考えられる。しかしながら、いずれのタイプのAdipoRの機能的発現を抑制しても、globular Adipo及び全長Adipoの両者に対する有意な影響が観察される。これらの結果は、AdipoR1及びAdipoR2は、ホモマルチマー及びヘテロマルチマーの両者を形成しているという結果(図3e参照)によって説明され得る。
興味深いことに、C2C12筋細胞において、AdipoR1及びAdipoR2の両者の発現をsiRNAにより同時に抑制すると、globular Adipo及び全長Adipoの両者の結合がほとんど観察されなくなるとともに(図5e,f参照)、globular Adipo及び全長AdipoによるPPARαリガンド活性(図5g参照)及び脂肪酸酸化(図5h参照)の増加がほとんど観察されなくなった。
(7)肝細胞内の結合及び作用に対するsiRNAの影響
肝細胞へのAdipoの結合を調べた。肝細胞は全長Adipoに対して特異的な結合を示した(図6b参照)。AdipoR1及びAdipoR2の発現によって、globular Adipo及び全長Adipoの肝細胞への結合が促進した(図6a,b参照)。逆に、肝細胞において、siRNAによりAdipoR2の発現を抑制すると、全長Adipoの結合が大きく減少した(図6b参照)。これらの結果から、AdipoR1は、globular Adipoに対して比較的選択性を有する受容体であり、AdipoR2は全長Adipoに対して比較的選択性を有する受容体であると考えられる。
正常ヒト大動脈内皮細胞(Normal Human Aortic Endothelial Cells:HAEC)へのAdipoの結合を調べた。AdipoR1及びAdipoR2の両者の発現をsiRNAにより同時に抑制すると、globular Adipoの結合は大幅に減少し(図6e参照)、全長Adipoの結合は一部減少した(図6f参照)。この結果は、AdipoR1及びAdipoRが、HAECにおいてもAdipoに対する受容体であることを示す。
(8)スキャッチャードプロット解析
図5e及びfに関する実験を再度より詳細に行って得られた結果を図7a及びbに示す。図7aは図5eに対応する図であり、二本鎖siRNAをトランスフェクトしたC2C12筋細胞への[125I]globular Adipoの結合等温線(binding isotherm)を示し、図7bは図5fに対応する図であり、二本鎖siRNAをトランスフェクトしたC2C12筋細胞への[125I]全長Adipoの結合等温線(binding isotherm)を示す。なお、図7a及びb中、□は無関係なコントロールsiRNA、■はマウスAdipoR1に対するsiRNA、●はマウスAdipoR2に対するsiRNA、▲はマウスAdipoR1及びマウスAdipoR2に対するsiRNAを用いた場合の結果を示す。
図7a及びbに示す結果に基づきスキャッチャードプロット解析を行った結果をそれぞれ図7c及びdに示す。
無関係なsiRNAをトランスフェクトしたC2C12筋細胞は、全長Adipoよりもglobular Adipoに強く結合した(図7a及びb参照)。
スキャッチャードプロット解析によって、globular Adipoに対する2種類の結合部位、すなわち高アフィニティー結合部位(Kd値:約0.06μg/mL,1.14nM gAd三量体等量)と中アフィニティー結合部位(Kd値:約0.80μg/mL,14.4nM gAd三量体等量)が存在する一方(図7c参照)、全長Adipoに対する2種類の結合部位、すなわち中アフィニティー結合部位(Kd値:約6.7μg/mL,49.1nM Ad六量体等量)と低アフィニティー結合部位(Kd値:約329.3μg/mL,2415nM Ad六量体等量)が存在することが明らかとなった(図7d参照)。
siRNAによりAdipoR1の発現を抑制すると、globular Adipoの結合はかなり減少したが(図7a参照)、全長Adipoの結合はわずかに減少しただけだった(図7b参照)。AdipoR1の発現を特異的に抑制すると、globular Adipoに対する高アフィニティー結合部位の活性は失われるが、globular Adipoに対する中アフィニティー結合部位の活性は影響されないことが、スキャッチャードプロット解析によって明らかとなった(図7c参照)。さらに、AdipoR1の発現を特異的に抑制すると、全長Adipoに対する中アフィニティー結合部位の活性は部分的に減少するが、全長Adipoに対する低アフィニティー結合部位の活性は失われることが、スキャッチャードプロット解析によって明らかとなった(図7d参照)。
AdipoR1とは対照的に、siRNAによりAdipoR2の発現を抑制すると、全長Adipoの結合はかなり減少したが(図7b参照)、globular Adipoの結合はわずかに減少した(図7a参照)。AdipoR2の発現を特異的に抑制すると、globular Adipoに対する高アフィニティー結合部位の活性は部分的に減少するが、globular Adipoに対する中アフィニティー結合部位の活性が失われることが、スキャッチャードプロット解析によって明らかとなった(図7c参照)。さらに、AdipoR2の発現を特異的に抑制すると、全長Adipoに対する中アフィニティー結合部位の活性は失われるが、全長Adipoに対する低アフィニティー結合部位の活性は影響されないことが、スキャッチャードプロット解析によって明らかとなった(図7d参照)。
(9)AMPK、ACC、p38 MAPK及びMAPKのリン酸化及びリン酸化量
図8aのパネルは、AdipoR1をトランスフェクトした(図中「AdipoR1」と記載)又はしていない(図中「Mock」と記載)C2C12細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、当該細胞の溶解物と抗リン酸化AMPK抗体とを反応させた結果を示し、図8bのパネルは、当該細胞の溶解物と抗リン酸化ACC抗体とを反応させた結果を示し、図8cのパネルは、当該細胞の溶解物と抗リン酸化p38 MAPK抗体とを反応させた結果を示し、図8dのパネルは、当該細胞の溶解物と抗リン酸化MAPK抗体とを反応させた結果を示す。図8eのパネルは、AdipoR1をトランスフェクトした(図中「AdipoR1」と記載)又はしていない(図中「Mock」と記載)肝細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、当該細胞の溶解物と抗リン酸化AMPK抗体とを反応させた結果を示し、図8fのパネルは、当該細胞の溶解物と抗リン酸化ACC抗体とを反応させた結果を示す。図8a〜fの各パネルの下側のグラフは、パネルの各位置におけるリン酸化量を示す。なお、図中、「pAMPK」はリン酸化AMPK、「pACC」はリン酸化ACC、「pp38 MAPK」はリン酸化p38 MAPK、「pp44MAPK」はリン酸化p44MAPK、「pp42 MAPK」はリン酸化p42 MAPKを表す。
AdipoR1をトランスフェクトしていないC2C12筋細胞において、globular Adipo及び全長Adipoの両者がAMPK、ACC及びp38 MAPKのリン酸化量を増加させたが、MAPK等の他のプロテインキナーゼのリン酸化量は増加させなかった(図8a〜d参照)。C2C12細胞におけるAdipoR1の発現は、globular AdipoによるAMPK、ACC及びp38 MAPKのリン酸化の亢進に関連していた(図8a〜d参照)。このことは、AdipoR1がglobular AdipoによるAMPK及びp38 MAPK活性化を媒介することを示唆する。
AdipoR1をトランスフェクトしていない肝細胞において、全長AdipoはAMPK活性化及びACCリン酸化を亢進させたが、globular Adipoは亢進させなった(図8e及びf参照)。肝細胞におけるAdipoR1の発現は、globular Adipo及び全長AdipoによるAMPK及びACCリン酸化の亢進に関連していた(図8e及びf参照)。このことは、AdipoR1がglobular Adipo及び全長AdipoによるAMPK及びACCリン酸化を媒介することができることを示唆する。
AdipoR1をトランスフェクトしていない(図8中「Mock」と記載)C2C12筋細胞において、globular Adipoにより亢進される脂肪酸酸化及びグルコース取り込みは、優性阻害型(dominant negative:DN)AMPK又はp38 MAPK特異的阻害剤 SB203580(Barger,P.M.ら,J.Biol.Chem.276,44495−44501(2001);Puigserver,P.ら,Mol.Cell 8,971−982(2001);Michael,L.F.ら,Proc.Natl.Acad.Sci.USA.98,3820−3825(2001))によって部分的に阻害された(図8g及びh参照)。C2C12筋細胞におけるAdipoR1の発現(図8中「mAdipoR1」と記載)は、globular Adipoによる脂肪酸酸化及びグルコース取り込みを亢進させたが、これもDN−AMPK又はSB203580によって部分的に阻害された(図8g及びh参照)。したがって、AdipoR1を介したglobular Adipoによる脂肪酸酸化及びグルコース取り込みの亢進は、C2C12筋細胞におけるAMPK及びp38 MAPKの両者の経路に関連していると考えられた。
以上の結果は、本実施例で取得されたAdipoR1 cDNA(配列番号1,5)及びAdipoR2 cDNA(配列番号3,7)が、それぞれ生物学的機能を有するAdipoR1(配列番号2,6)及びAdipoR2(配列番号4,8)をコードすることを示す。
【産業上の利用の可能性】
本発明によって、アディポネクチン結合能を有する新規タンパク質、該タンパク質をコードする遺伝子、該遺伝子を含有する組換えベクター、該組換えベクターを含む形質転換体及び上記タンパク質に対する抗体が提供される。また、本発明によって、上記タンパク質、遺伝子、組換えベクター又は形質転換体を利用した、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストのスクリーニング方法及びスクリーニング用キットが提供される。
【配列表】
【図1】
【技術分野】
本発明は、アディポネクチン結合能を有する新規タンパク質、該タンパク質をコードする遺伝子、該遺伝子を含む組換えベクター、該組換えベクターを含む形質転換体、及び上記タンパク質に対する抗体又はその断片に関する。さらに、本発明は、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストのスクリーニング方法及びスクリーニング用キットに関する。
【背景技術】
肥満は、脂肪組織量の増加として定義され、糖尿病、高脂血症、冠動脈性心疾患等の心臓血管疾患及び代謝疾患の高リスクと関連する(Reaven,G.M.Diabetologia 38,3−13.(1995);Spiegelman,B.M.ら,Cell 87,377−389.(1996))。肥満及び2型糖尿病の顕著な特徴であるグルコース及び脂質の代謝異常は、筋肉、肝臓等のインスリン標的組織における脂質蓄積量の増加の原因となり、インスリン抵抗性を引き起こす(Ruderman,N.B.ら,Am.J.Physiol.276,E1−E18.(1999);Shulman,G.I.J.Clin.Invest.106,171−176.(2000))。脂肪組織は、エネルギー要求の変化に応答してトリグリセリド(TG)を蓄積するとともに遊離脂肪酸(FFA)/グリセロールを放出する部位としての役割を果たす(Spiegelman,B.M.,and Flier,J.S.Cell 87,377−389.(1996))。また、脂肪組織は、FFA(Shulman,G.I.J.Clin.Invest.106,171−176.(2000))、アディプシン(White,R.T.ら,J.Biol.Chem.267,9210−9213.(1992))、レプチン(Friedman,J.M.Nature 404,632−634.(2000))、プラスミノーゲンアクチベーターインヒビター−1(PAI−1)(Shimomura,I.ら,Nat.Med.2,800−803.(1996))、レジスチン(Steppan,C.M.ら,Nature 409,307−312.(2001))、腫瘍壊死因子α(TNF−α)(Hotamisligil,G.S.J.Intern.Med.245,621−625.(1999))等の「アディポカイン(adipokines)」(Matsuzawa,Y.ら,Ann.NY.Acad.Sci.892,146−154.(1999))と呼ばれる多数の生物活性物質を分泌する重要な内分泌器官として、様々なエネルギーホメオスタシスの調節に関与する。
アディポネクチン又はAcrp30(Hu,E.,Liang,P.ら,J.Biol.Chem.271,10697−10703.(1996)等)は、アディポサイト由来のホルモンであり、様々な生物学的機能を有している。肥満、2型糖尿病、冠動脈性心疾患は、アディポネクチンの血漿レベルの減少に関連しており、アディポネクチンはin vitroにおいて抗アテローム発生性を示すことが報告されている(Ouchi,N.ら,Circulation 103,1057−1063.(2001);Yokota,T.ら,Blood 96,1723−1732.(2000))。また、Acrp30の循環レベルの急増は、肝臓におけるグルコース産生を低下させることが報告されている(Berg,A.H.ら,Nat.Med.7,947−953.(2001);Combs,T.P.ら,J.Clin.Invest.108,1875−1881.(2001))。また、globular Acrp30は、筋肉における脂肪酸酸化を亢進させ、マウスにおける体重減少を引き起こすことが報告されている(Fruebis,J.ら,Proc.Natl.Acad.Sci.USA.98,2005−2010.(2001))。また、脂肪組織萎縮性マウス又は肥満マウスにおけるインスリン抵抗性が、globularドメインのみからなるアディポネクチン(globular adiponectin;gAd)で処理することにより、筋肉における脂肪酸酸化の亢進を介して改善したこと、及び全長アディポネクチンで処理することにより程度はgAdよりも低いが改善したことが報告されている(Yamauchi,T.ら,Nat.Med.7,941−946.(2001))。
さらに最近になって、骨格筋において、アディポネクチンがAMPキナーゼ(AMPK)を急性的に活性化し、その結果、脂肪酸酸化及びグルコース取り込みが亢進されることが報告されているとともに(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002))、アディポネクチンがPPARαを慢性的に活性化し、その結果、脂肪酸酸化が亢進されるが筋肉における組織TG含量は減少すること、そしてこれらの効果は全長アディポネクチンよりもgAdの方が大きいことが報告されている(Yamauchi,T.ら,J.Biol.Chem.278,2461−2468(2002))。また、興味深いことに、肝臓においては、全長アディポネクチンのみがAMPKを急性的に活性化し、その結果、糖新生に関連する分子を減少させるとともに脂肪酸酸化を亢進させる一方、全長アディポネクチンのみがAMPKを慢性的に活性化し、その結果、脂肪酸酸化を亢進させるとともに肝臓における組織TG量を減少させる。そして、これらの変化はいずれもin vivoにおけるインスリン感受性を増加させる(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002);Yamauchi,T.ら,J.Biol.Chem.278,2461−2468(2002))
これらのアディポネクチンの作用は、細胞表面の受容体によって媒介されると考えられるが、アディポネクチン受容体は未だ同定されておらず、骨格筋及び肝臓におけるアディポネクチン受容体が構造的及び機能的に異なるか否かも未だ不明である。なお、本発明者がアディポネクチン受容体をコードする遺伝子を同定した後、ホモロジー検索を行ったところ、ホモロジーを有する遺伝子として酵母YOL002cが発見された(Karpichev,I.V.ら,Journal of Biological Chemistry,277,19609−19617.(2002))。YOL002cは、脂肪酸酸化等の脂質の代謝経路において重要な役割を果たす7回膜貫通型タンパク質をコードする(Karpichev,I.V.ら,Journal of Biological Chemistry,277,19609−19617.(2002))。
【発明の開示】
本発明は、第一に、アディポネクチン結合能を有する新規タンパク質、該タンパク質をコードする遺伝子、該遺伝子を含む組換えベクター、該組換えベクターを含む形質転換体、及び上記タンパク質に対する抗体又はその断片を提供することを目的とする。
また、本発明は、第二に、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストのスクリーニング方法及びスクリーニング用キットを提供することを目的とする。
上記目的を達成するために、本発明は、以下のタンパク質、遺伝子、組換えベクター、形質転換体、抗体、並びにアディポネクチン受容体に対するアゴニスト又はアンタゴニストのスクリーニング方法及びスクリーニング用キットを提供する。
(1)以下の(a)又は(b)に示すタンパク質。
(a)配列番号2、4、6又は8に記載のアミノ酸配列からなるタンパク質
(b)配列番号2、4、6又は8に記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつアディポネクチン結合能を有するタンパク質
(2)前記(1)記載のタンパク質をコードする遺伝子。
(3)以下の(c)又は(d)に示すDNAを含む前記(2)記載の遺伝子。
(c)配列番号1、3、5又は7に記載の塩基配列からなるDNA
(d)配列番号1、3、5又は7に記載の塩基配列からなるDNAと相補的なDNAにストリンジェントな条件下でハイブリダイズし、かつアディポネクチン結合能を有するタンパク質をコードするDNA
(4)前記(2)又は(3)記載の遺伝子を含む組換えベクター。
(5)前記(4)記載の組換えベクターを含む形質転換体。
(6)前記(1)記載のタンパク質に反応し得る抗体又はその断片。
(7)試験物質と前記(1)記載のタンパク質とを接触させる工程を含む、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストのスクリーニング方法。
(8)前記(1)記載のタンパク質、前記(2)若しくは(3)記載の遺伝子、前記(4)記載の組換えベクター又は前記(5)記載の形質転換体を含む、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストのスクリーニング用キット。
【図面の簡単な説明】
図1(a)は、アディポネクチンのC2C12筋細胞への結合を示す図、図1(b)は、アディポネクチンの肝細胞への結合を示す図、図1(c)〜(e)は、FACS解析の結果を示す図、図1(f)は、AdipoR1遺伝子及びAdipoR2遺伝子の転写物の構造を示す模式図、図1(g)は、マウスの様々な組織に関するノーザンブロット解析の結果を示す図、図1(h)は、ヒトの様々な組織に関するノーザンブロット解析の結果を示す図である。
図2(a)は、AdipoR1及びAdipoR2の構造を示す模式図、図2(b)は、エピトープタグFLAGを有するAdipoR1及びAdipoR2をトランスフェクトした293T細胞の細胞溶解物を抗FLAG抗体で免疫染色した結果を示す図、図2(c)は、エピトープタグをN末端又はC末端に導入たAdipoR1又はAdipoR2をトランスフェクトした293T細胞におけるAdipoR1又はAdipoR2の細胞内局在を示す図、図2(d)は、AdipoR1及びAdipoR2の予測構造モデルを示す図である。
図3(a)は、AdipoR1又はAdipoR2をトランスフェクトした293T細胞への[125I]globular Adipo(gAd)の結合等温線(binding isotherm)を示す図、図3(b)は、当該293T細胞への[125I]全長Adipo(Ad)の結合等温線を示す図、図3(c)は、エピトープタグFLAG又はHAを有するAdipoR1又はAdipoR2をトランスフェクトした293T細胞の細胞溶解物を、抗FLAG抗体又は抗HA抗体を用いて免疫沈降した後、抗FLAG抗体又は抗HA抗体で免疫染色した結果を示す図、図3(d)は、マウスAdipoR1発現細胞又はBLT1発現細胞にglobular Adipo、全長Adipo、LTB4又はATPをチャレンジしたときの[Ca2+]iの変化を示す図、図3(e)は、ホルスコリンで処理した又は処理しないHEK−293細胞にcAMP又はcGMPを蓄積させた結果を示す図、図3(f)は、AdipoR1をトランスフェクトした293T細胞をglobular Adipo又は全長AdipoとインキュベートしたときのPPARαリガンド活性を示す図である。
図4(a)は、マウスAdipoR1 mRNA量を示す図、図4(b)はマウスAdipoR2 mRNA量を示す図、図4(c)及び(d)は、マウスAdipoR1又はマウスAdipoR2をトランスフェクトしたC2C12筋細胞に結合した[125I]globular Adipo(gAd)又は全長Adipo(Ad)の結合等温線を示す図、図4(e)は、マウスAdipoR1又はマウスAdipoR2をトランスフェクトしたC2C12筋細胞にLacZ又はDN−α2AMPKを含有するアデノウイルスを感染させ、globular Adipo又は全長Adipoで7時間処理したときのC2C12筋細胞内のPPARαリガンド活性を示す図、図4(f)は上記C2C12筋細胞内のin vitro脂肪酸酸化を示す図である。
図5(a)は、siRNA又はmockをトランスフェクトしたC2C12筋細胞内のマウスAdipoR1 mRNAの量を示す図、図5(b)は、上記C2C12筋細胞内のマウスAdipoR2 mRNAの量を示す図、図5(c)は、非標識globular Adipoの濃度を増加させることによって、二本鎖siRNAをトランスフェクトした細胞に結合する[125I]globular Adipoを置換した競合ラジオリガンド結合アッセイの結果を示す図、図5(d)は、非標識全長Adipoの濃度を増加させることによって、二本鎖siRNAをトランスフェクトした細胞に結合する[125I]全長Adipoを置換した競合ラジオリガンド結合アッセイの結果を示す図、図5(e)は、二本鎖siRNAをトランスフェクトしたC2C12筋細胞への[125I]globular Adipoの結合等温線を示す図、図5(f)は、二本鎖siRNAをトランスフェクトしたC2C12筋細胞への[125I]全長Adipoの結合等温線を示す図、図5(g)は、二本鎖siRNAをトランスフェクトしたC2C12筋細胞をglobular Adipo、全長Adipo又はWy−14,643と7時間インキュベートしたときのPPARαリガンド活性を示す図、図5(h)は、二本鎖siRNAをトランスフェクトしたC2C12筋細胞をglobular Adipo、全長Adipo又はWy−14,643と7時間インキュベートしたときのin vitro脂肪酸酸化を示す図、図5(i)は、二本鎖siRNAをトランスフェクトしたC2C12筋細胞をglobular Adipo、全長Adipo又はインスリンと7時間インキュベートしたときのグルコース取り込みを示す図である。
図6(a)は、gAdの肝細胞への特異的結合を示す図、図6(b)は、Adの肝細胞への特異的結合を示す図、図6(c)は、HAECにおけるヒトAdipoR1 mRNAの発現レベルを示す図、図6(d)は、HAECにおけるヒトAdipoR2 mRNAの発現レベルを示す図、図6(e)は、gAdのHAECへの特異的結合を示す図、図6(f)は、AdのHAECへの特異的結合を示す図である。
図7(a)は、二本鎖siRNAをトランスフェクトしたC2C12筋細胞への[125I]globular Adipoの結合等温線(binding isotherm)を示す図、図7(b)は、二本鎖siRNAをトランスフェクトしたC2C12筋細胞への[125I]全長Adipoの結合等温線(binding isotherm)を示す図、図7(c)は、図7(a)に示す結果に基づきスキャッチャードプロット解析を行った結果を示す図、図7(d)は、図7(b)に示す結果に基づきスキャッチャードプロット解析を行った結果を示す図である。
図8(a)は、AdipoR1をトランスフェクトした又はしていないC2C12細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、当該細胞の溶解物と抗リン酸化AMPK抗体とを反応させた結果を示す図、図8(b)は、AdipoR1をトランスフェクトした又はしていないC2C12細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、当該細胞の溶解物と抗リン酸化ACC抗体とを反応させた結果を示す図、図8(c)は、AdipoR1をトランスフェクトした又はしていないC2C12細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、当該細胞の溶解物と抗リン酸化p38 MAPK抗体とを反応させた結果を示す図、図8(d)は、AdipoR1をトランスフェクトした又はしていないC2C12細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、当該細胞の溶解物と抗リン酸化MAPK抗体とを反応させた結果を示す図、図8(e)は、AdipoR1をトランスフェクトした又はしていない肝細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、当該細胞の溶解物と抗リン酸化AMPK抗体とを反応させた結果を示す図、図8(f)は、AdipoR1をトランスフェクトした又はしていない肝細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、当該細胞の溶解物と抗リン酸化ACC抗体とを反応させた結果を示す図、図8(g)は、優性阻害型AMPキナーゼ(DN−AMPK)又はp38 MAPK特異的阻害剤SB203580の存在下におけるAdipoR1をトランスフェクトした又はしていないC2C12筋細胞の脂肪酸酸化を示す図、図8(h)は、優性阻害型AMPキナーゼ(DN−AMPK)又はp38 MAPK特異的阻害剤SB203580の存在下におけるAdipoR1をトランスフェクトした又はしていないC2C12筋細胞のグルコース取り込みを示す図である。
【発明を実施するための最良の形態】
本発明のタンパク質は、以下の(a)又は(b)に示すタンパク質である。
(a)配列番号2、4、6又は8に記載のアミノ酸配列からなるタンパク質(以下「タンパク質(a)」という。)
(b)配列番号2、4、6又は8に記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつアディポネクチン結合能を有するタンパク質(以下「タンパク質(b)」という。)
タンパク質(a)又は(b)は、アディポネクチン結合能を有するタンパク質である。「アディポネクチン結合能」とは、アディポネクチンと結合する能力、好ましくはアディポネクチンと特異的に結合する能力(すなわちアディポネクチン受容体が有するアディポネクチン結合能)を意味する。「アディポネクチン結合能」には、全長アディポネクチン又はglobularドメインのみからなるアディポネクチンのいずれに対する結合能も含まれる。アディポネクチン結合能を有するタンパク質には、全長アディポネクチンにのみ結合し得るタンパク質、globularドメインのみからなるアディポネクチンにのみ結合し得るタンパク質、全長アディポネクチン及びglobularドメインのみからなるアディポネクチンの両者に結合し得るタンパク質、全長アディポネクチン及びglobularドメインのみからなるアディポネクチンのいずれか一方に優先的に結合し得るタンパク質等が含まれる。
なお、「globularドメイン」とは、アディポネクチンのC末端側に約100アミノ酸強の長さで存在する糸状体類似の構造ドメインである。補体のClq等にもglobularドメインとホモロジーの高いドメインが存在する。アディポネクチンのglobularドメインは機能的には骨格筋等に強く作用して、脂肪酸を燃焼させ、脂肪が蓄積するのを抑制する作用を有しており、全長のアディポネクチンより数十分の一の低い濃度で同程度の強い活性を示す。
タンパク質(a)のうち、配列番号2又は4に記載のタンパク質はヒト由来のアディポネクチン受容体であり、配列番号6又は8に記載のタンパク質はマウス由来のアディポネクチン受容体である。また、配列番号2及び6記載のタンパク質は同一種類のアディポネクチン受容体(AdipoR1)であり、配列番号4及び8記載のタンパク質は、AdipoR1とは異なる種類のアディポネクチン受容体(AdipoR2)である。AdipoR1は、全長アディポネクチンよりもglobularドメインのみからなるアディポネクチンに対して比較的選択性を有する受容体であると考えられ、AdipoR2は、globularドメインのみからなるアディポネクチンよりも全長アディポネクチンに対して比較的選択性を有する受容体であると考えられる。
AdipoR1について、ヒト(配列番号2)及びマウス(配列番号6)間のアミノ酸レベルでの相同性は96.8%である。AdipoR2について、ヒト(配列番号4)及びマウス(配列番号8)間のアミノ酸レベルでの相同性は95.2%である。AdipoR1とAdipoR2の構造は類似しており、マウスにおけるAdipoR1(配列番号6)及びAdipoR2(配列番号8)の相同性は66.7%である。
生体において、AdipoR1はほとんどの組織で発現しており、特に骨格筋で高度に発現している一方、AdipoR2は肝臓で高度に発現している。また、AdipoR1及びAdipoR2は、ホモ又はヘテロマルチマーを形成していると考えられる。また、AdipoR1及びAdipoR2はともに、全長アディポネクチン及びglobularドメインのみからなるアディポネクチンと結合し、これらのアディポネクチンによるPPARα(Peroxisome proliferator−activated receptor α:ペルオキシソーム増殖因子活性化受容体α)リガンド活性及び脂肪酸酸化の亢進を媒介する。例えば、AdipoR1は、globularドメインのみからなるアディポネクチンによる筋細胞内のPPARαリガンド活性、脂肪酸酸化及びグルコース取り込みの増加を媒介すると考えられる。また、AdipoR2は、全長アディポネクチンによる肝細胞及び筋細胞内のPPARαリガンド活性及び脂肪酸酸化の増加を一部媒介すると考えられる。
AdipoR1及びAdipoR2はいずれも7個の膜貫通ドメインを有すると考えられる。配列番号2記載のアミノ酸配列(ヒトAdipoR1)のうち、136〜158番目、172〜194番目、207〜228番目、234〜255番目、267〜289番目、299〜321番目、336〜358番目のアミノ酸配列が膜貫通ドメインに相当すると考えられる。また、配列番号4記載のアミノ酸配列(ヒトAdipoR2)のうち、60〜82番目、96〜118番目、130〜152番目、158〜179番目、192〜214番目、222〜244番目、260〜282番目のアミノ酸配列が膜貫通ドメインに相当すると考えられる。また、配列番号6記載のアミノ酸配列(マウスAdipoR1)のうち、136〜158番目、172〜194番目、207〜228番目、234〜255番目、267〜289番目、299〜321番目、336〜358番目のアミノ酸配列が膜貫通ドメインに相当すると考えられる。また、配列番号8記載のアミノ酸配列(マウスAdipoR2)のうち、72〜94番目、108〜130番目、142〜164番目、170〜191番目、204〜226番目、234〜256番目、272〜294番目のアミノ酸配列が膜貫通ドメインに相当すると考えられる。
配列番号2、4、6又は8に記載のアミノ酸配列において欠失、置換又は付加されるアミノ酸の個数は、アディポネクチン結合能が保持され得る限り特に限定されるものではなく、その個数は1又は複数個、好ましくは1又は数個であり、その具体的な範囲は通常1〜100個、好ましくは1〜50個、さらに好ましくは1〜10個である。このとき、タンパク質(b)のアミノ酸配列は、タンパク質(a)のアミノ酸配列と通常60%以上、好ましくは80%以上、さらに好ましくは90%以上の相同性を有する。
配列番号2、4、6又は8に記載のアミノ酸配列において欠失、置換又は付加されるアミノ酸の位置は、アディポネクチン結合能が保持され得る限り特に限定されるものではない。
タンパク質(b)には、タンパク質(a)に対して人為的に欠失、置換、付加等の変異を導入したタンパク質の他、欠失、置換、付加等の変異が導入された状態で天然に存在するタンパク質や、それに対して人為的に欠失、置換、付加等の変異を導入したタンパク質も含まれる。欠失、置換、付加等の変異が導入された状態で天然に存在するタンパク質としては、例えば、ヒトを含む哺乳動物(例えば、ヒト、サル、ウシ、ヒツジ、ヤギ、ウマ、ブタ、ウサギ、イヌ、ネコ、マウス、ラット等)由来のタンパク質(これらの哺乳動物において多型によって生じ得るタンパク質を含む)が挙げられる。
タンパク質(a)及び(b)には、糖鎖が付加されたタンパク質及び糖鎖が付加されていないタンパク質のいずれもが含まれる。タンパク質に付加される糖鎖の種類、位置等は、タンパク質の製造の際に使用される宿主細胞の種類によって異なるが、糖鎖が付加されたタンパク質には、いずれの宿主細胞を用いて得られるタンパク質も含まれる。また、タンパク質(a)及び(b)には、その医薬的に許容される塩も含まれる。
タンパク質(a)又は(b)をコードする遺伝子は、例えば、ヒト、マウス等の哺乳動物の骨格筋、肝臓、心臓、マクロファージ、血管、脳、腎臓、肺、胎盤、脾臓、精巣、末梢血液、胸腺、腸管等の組織から抽出したmRNAを用いてcDNAライブラリーを作製し、配列番号1、3、5又は7に記載の塩基配列に基づいて合成したプローブを用いて、cDNAライブラリーから目的のDNAを含むクローンをスクリーニングすることにより得られる。以下、cDNAライブラリーの作製、及び目的のDNAを含むクローンのスクリーニングの各工程について説明する。
〔cDNAライブラリーの作製〕
cDNAライブラリーを作製する際には、例えば、ヒト、マウス等の哺乳動物の骨格筋、肝臓、心臓、マクロファージ、血管、脳、腎臓、肺、胎盤、脾臓、精巣、末梢血液、胸腺、腸管等の組織から全RNAを得た後、オリゴdT−セルロースやポリU−セファロース等を用いたアフィニティーカラム法、バッチ法等によりポリ(A+)RNA(mRNA)を得る。この際、ショ糖密度勾配遠心法等によりポリ(A+)RNA(mRNA)を分画してもよい。次いで、得られたmRNAを鋳型として、オリゴdTプライマー及び逆転写酵素を用いて一本鎖cDNAを合成した後、該一本鎖cDNAから二本鎖cDNAを合成する。このようにして得られた二本鎖cDNAを適当なクローニングベクターに組み込んで組換えベクターを作製し、該組換えベクターを用いて大腸菌等の宿主細胞を形質転換し、テトラサイクリン耐性、アンピシリン耐性を指標として形質転換体を選択することにより、cDNAのライブラリーが得られる。cDNAライブラリーを作製するためのクローニングベクターは、宿主細胞中で自立複製できるものであればよく、例えば、ファージベクター、プラスミドベクター等を使用できる。宿主細胞としては、例えば、大腸菌(Escherichia coli)等を使用できる。
大腸菌等の宿主細胞の形質転換は、塩化カルシウム、塩化マグネシウム又は塩化ルビジウムを共存させて調製したコンピテント細胞に、組換えベクターを加える方法等により行うことができる。なお、ベクターとしてプラスミドを用いる場合は、テトラサイクリン、アンピシリン等の薬剤耐性遺伝子を含有させておく。
cDNAライブラリーの作製にあたっては、市販のキット、例えば、SuperScript Plasmid System for cDNA Synthesis and Plasmid Cloning(Gibco BRL社製)、ZAP−cDNA Synthesis Kit(ストラタジーン社製)等を使用できる。
〔目的のDNAを含むクローンのスクリーニング〕
cDNAライブラリーから目的のDNAを含むクローンをスクリーニングする際には、配列番号1、3、5又は7に記載の塩基配列に基づいてプライマーを合成し、これを用いてポリメラーゼ連鎖反応(PCR)を行い、PCR増幅断片を得る。PCR増幅断片は、適当なプラスミドベクターを用いてサブクローニングしてもよい。PCRに使用するプライマーセットは特に限定されるものではなく、配列番号1、3、5又は7に記載の塩基配列に基づいて設計できる。
cDNAライブラリーに対して、PCR増幅断片をプローブとしてコロニーハイブリダイゼーション又はプラークハイブリダイゼーションを行うことにより、目的のDNAが得られる。プローブとしては、PCR増幅断片をアイソトープ(例えば、32P、35S)、ビオチン、ジゴキシゲニン、アルカリホスファターゼ等で標識したものを使用できる。目的のDNAを含むクローンは、抗体を用いたイムノスクリーニング等の発現スクリーニングによっても得られる。
取得されたDNAの塩基配列は、該DNA断片をそのまま、又は適当な制限酵素等で切断した後、常法によりベクターに組み込み、通常用いられる塩基配列解析方法、例えば、マキサム−ギルバートの化学修飾法、ジデオキシヌクレオチド鎖終結法を用いて決定できる。塩基配列解析の際には、通常、373A DNAシークエンサー(Perkin Elmer社製)等の塩基配列分析装置が用いられる。
タンパク質(a)又は(b)をコードする遺伝子は、タンパク質(a)又は(b)をコードするオープンリーディングフレームとその3’末端に位置する終止コドンとを含む。また、タンパク質(a)又は(b)をコードする遺伝子は、オープンリーディングフレームの5’末端及び/又は3’末端に非翻訳領域(UTR)を含むことができる。
タンパク質(a)をコードする遺伝子としては、例えば、配列番号1、3、5又は7に記載の塩基配列からなるDNAを含む遺伝子が挙げられる。ここで、配列番号1記載の塩基配列のうち1〜1125番目の塩基配列は、配列番号2記載のタンパク質をコードするオープンリーディングフレームであり、配列番号1記載の塩基配列のうち、翻訳開始コドンは1〜3番目の塩基配列に位置し、終止コドンは1126〜1128番目の塩基配列に位置する。また、配列番号3記載の塩基配列のうち1〜897番目の塩基配列は、配列番号4記載のタンパク質をコードするオープンリーディングフレームであり、配列番号3記載の塩基配列のうち、翻訳開始コドンは1〜3番目の塩基配列に位置し、終止コドンは898〜900番目の塩基配列に位置する。また、配列番号5記載の塩基配列のうち1〜1125番目の塩基配列は、配列番号6記載のタンパク質をコードするオープンリーディングフレームであり、配列番号5記載の塩基配列のうち、翻訳開始コドンは1〜3番目の塩基配列に位置し、終止コドンは1126〜1128番目の塩基配列に位置する。また、配列番号7記載の塩基配列のうち1〜933番目の塩基配列は、配列番号8記載のタンパク質をコードするオープンリーディングフレームであり、配列番号7記載の塩基配列のうち、翻訳開始コドンは1〜3番目の塩基配列に位置し、終止コドンは934〜936番目の塩基配列に位置する。
タンパク質(a)をコードする遺伝子の塩基配列は、タンパク質(a)をコードする限り特に限定されるものではなく、オープンリーディングフレームの塩基配列は、配列番号1、3、5又は7に記載の塩基配列に限定されるものではない。タンパク質(a)をコードする遺伝子は、その塩基配列に従って化学合成により得ることもできる。DNAの化学合成は、市販のDNA合成機、例えば、チオホスファイト法を利用したDNA合成機(島津製作所社製)、フォスフォアミダイト法を利用したDNA合成機(パーキン・エルマー社製)を用いて行うことができる。
タンパク質(b)をコードする遺伝子としては、例えば、配列番号1、3、5又は7に記載の塩基配列からなるDNAと相補的なDNAにストリンジェントな条件下でハイブリダイズし、かつアディポネクチン結合能を有するタンパク質をコードするDNAを含む遺伝子が挙げられる。
「ストリンジェントな条件」としては、例えば、42℃、2×SSC及び0.1%SDSの条件、好ましくは65℃、0.1×SSC及び0.1%SDSの条件が挙げられる。
配列番号1、3、5又は7に記載の塩基配列からなるDNAと相補的なDNAにストリンジェントな条件下でハイブリダイズするDNAの具体例としては、配列番号1、3、5又は7に記載の塩基配列からなるDNAと少なくとも60%以上、好ましくは80%以上、さらに好ましくは90%以上の相同性を有するDNAが挙げられる。
タンパク質(b)をコードする遺伝子は、例えば、配列番号1、3、5又は7に記載の塩基配列からなるDNAに、部位特異的変異誘発法等の公知の方法を用いて人為的に変異を導入することにより得られる。変異の導入は、例えば、変異導入用キット、例えば、Mutant−K(TAKARA社製)、Mutant−G(TAKARA社製)、TAKARA社のLA PCR in vitro Mutagenesisシリーズキットを用いて行うことができる。また、塩基配列が既に決定されているDNAについては化学合成によっても得られる。
タンパク質(a)又は(b)は、例えば、以下の工程に従って、それぞれのタンパク質をコードする遺伝子を宿主細胞中で発現させることにより製造できる。
〔組換えベクター及び形質転換体の作製〕
組換えベクターを作製する際には、目的とするタンパク質のコード領域を含む適当な長さのDNA断片を調製する。また、目的とするタンパク質のコード領域の塩基配列を、宿主細胞における発現に最適なコドンとなるように、塩基を置換したDNAを調製する。
このDNA断片を適当な発現ベクターのプロモーターの下流に挿入することにより組換えベクターを作製し、該組換えベクターを適当な宿主細胞に導入することにより、目的とするタンパク質を生産し得る形質転換体が得られる。上記DNA断片は、その機能が発揮されるようにベクターに組み込まれることが必要であり、ベクターは、プロモーターの他、エンハンサー等のシスエレメント、スプライシングシグナル、ポリA付加シグナル、選択マーカー(例えば、ジヒドロ葉酸還元酵素遺伝子、アンピシリン耐性遺伝子、ネオマイシン耐性遺伝子)、リボソーム結合配列(SD配列)等を含有できる。
発現ベクターとしては、宿主細胞において自立複製が可能なものであれば特に限定されず、例えば、プラスミドベクター、ファージベクター、ウイルスベクター等を使用できる。プラスミドベクターとしては、例えば、大腸菌由来のプラスミド(例えば、pRSET、pBR322、pBR325、pUC118、pUC119、pUC18、pUC19)、枯草菌由来のプラスミド(例えば、pUB110、pTP5)、酵母由来のプラスミド(例えば、YEp13、YEp24、YCp50)が挙げられ、ファージベクターとしては、例えば、λファージ(例えば、Charon4A、Charon21A、EMBL3、EMBL4、λgt10、λgt11、λZAP)が挙げられ、ウイルスベクターとしては、例えば、レトロウイルス、ワクシニアウイルス等の動物ウイルス、バキュロウイルス等の昆虫ウイルスが挙げられる。
宿主細胞としては、目的とする遺伝子を発現し得る限り、原核細胞、酵母、動物細胞、昆虫細胞、植物細胞等のいずれを使用してもよい。また、動物個体、植物個体、カイコ虫体等を使用してもよい。
細菌を宿主細胞とする場合、例えば、エッシェリヒア・コリ(Escherichia coli)等のエシェリヒア属、バチルス・ズブチリス(Bacillus subtilis)等のバチルス属、シュードモナス・プチダ(Pseudomonas putida)等のシュードモナス属、リゾビウム・メリロティ(Rhizobium meliloti)等のリゾビウム属に属する細菌を宿主細胞として使用できる。具体的には、Escherichia coli XL1−Blue、Escherichia coli XL2−Blue、Escherichia coli DH1、Escherichia coli K12、Escherichia coli JM109、Escherichia coli HB101等の大腸菌や、Bacillus subtilis MI 114、Bacillus subtilis 207−21等の枯草菌を宿主細胞として使用できる。この場合のプロモーターは、大腸菌等の細菌中で発現できるものであれば特に限定されず、例えば、trpプロモーター、lacプロモーター、PLプロモーター、PRプロモーター等の大腸菌やファージ等に由来するプロモーターを使用できる。また、tacプロモーター、lacT7プロモーター、letIプロモーターのように人為的に設計改変されたプロモーターも使用できる。
細菌への組換えベクターの導入方法としては、細菌にDNAを導入し得る方法であれば特に限定されず、例えば、カルシウムイオンを用いる方法、エレクトロポレーション法等を使用できる。
酵母を宿主細胞とする場合、サッカロミセス・セレビシエ(Saccharomycescerevisiae)、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)、ピヒア・パストリス(Pichia pastoris)等を宿主細胞として使用できる。この場合のプロモーターは、酵母中で発現できるものであれば特に限定されず、例えば、gal1プロモーター、gal10プロモーター、ヒートショックタンパク質プロモーター、MFα1プロモーター、PHO5プロモーター、PGKプロモーター、GAPプロモーター、ADHプロモーター、AOX1プロモーター等を使用できる。
酵母への組換えベクターの導入方法は、酵母にDNAを導入し得る方法であれば特に限定されず、例えば、エレクトロポレーション法、スフェロプラスト法、酢酸リチウム法等を使用できる。
動物細胞を宿主細胞とする場合、サル細胞COS−7、Vero、チャイニーズハムスター卵巣細胞(CHO細胞)、マウスL細胞、ラットGH3、ヒトFL細胞等を宿主細胞として使用できる。この場合のプロモーターは、動物細胞中で発現できるものであれば特に限定されず、例えば、SRαプロモーター、SV40プロモーター、LTR(Long Terminal Repeat)プロモーター、CMVプロモーター、ヒトサイトメガロウイルスの初期遺伝子プロモーター等を使用できる。
動物細胞への組換えベクターの導入方法は、動物細胞にDNAを導入し得る方法であれば特に限定されず、例えば、エレクトロポレーション法、リン酸カルシウム法、リポフェクション法等を使用できる。
昆虫細胞を宿主とする場合には、Spodoptera frugiperdaの卵巣細胞、Trichoplusia niの卵巣細胞、カイコ卵巣由来の培養細胞等を宿主細胞として使用できる。Spodoptera frugiperdaの卵巣細胞としてはSf9、Sf21等、Trichoplusia niの卵巣細胞としてはHigh 5、BTI−TN−5B1−4(インビトロジェン社製)等、カイコ卵巣由来の培養細胞としてはBombyx mori N4等が挙げられる。
昆虫細胞への組換えベクターの導入方法は、昆虫細胞にDNAを導入し得る限り特に限定されず、例えば、リン酸カルシウム法、リポフェクション法、エレクトロポレーション法等を使用できる。
〔形質転換体の培養〕
目的とするタンパク質をコードするDNAを組み込んだ組換えベクターを導入した形質転換体を通常の培養方法に従って培養する。形質転換体の培養は、宿主細胞の培養に用いられる通常の方法に従って行うことができる。
大腸菌や酵母等の微生物を宿主細胞として得られた形質転換体を培養する培地としては、該微生物が資化し得る炭素源、窒素源、無機塩類等を含有し、形質転換体の培養を効率的に行える培地であれば天然培地、合成培地のいずれを使用してもよい。
炭素源としては、グルコース、フラクトース、スクロース、デンプン等の炭水化物、酢酸、プロピオン酸等の有機酸、エタノール、プロパノール等のアルコール類を使用できる。窒素源としては、アンモニア、塩化アンモニウム、硫酸アンモニウム、酢酸アンモニウム、リン酸アンモニウム等の無機酸又は有機酸のアンモニウム塩、ペプトン、肉エキス、酵母エキス、コーンスチープリカー、カゼイン加水分解物等を使用できる。無機塩としては、リン酸第一カリウム、リン酸第二カリウム、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム、硫酸第一鉄、硫酸マンガン、硫酸銅、炭酸カルシウム等を使用できる。
大腸菌や酵母等の微生物を宿主細胞として得られた形質転換体の培養は、振盪培養又は通気攪拌培養等の好気的条件下で行う。培養温度は通常25〜37℃、培養時間は通常12〜48時間であり、培養期間中はpHを6〜8に保持する。pHの調整は、無機酸、有機酸、アルカリ溶液、尿素、炭酸カルシウム、アンモニア等を用いて行うことができる。また、培養の際、必要に応じてアンピシリン、テトラサイクリン等の抗生物質を培地に添加してもよい。
プロモーターとして誘導性のプロモーターを用いた発現ベクターで形質転換した微生物を培養するときには、必要に応じてインデューサーを培地に添加してもよい。例えば、lacプロモーターを用いた発現ベクターで形質転換した微生物を培養するときにはイソプロピル−β−D−チオガラクトピラノシド等を、trpプロモーターを用いた発現ベクターで形質転換した微生物を培養するときにはインドールアクリル酸等を培地に添加してもよい。
動物細胞を宿主細胞として得られた形質転換体を培養する培地としては、一般に使用されているRPMI1640培地、EagleのMEM培地、DMEM培地、Ham F12培地、Ham F12K培地又はこれら培地に牛胎児血清等を添加した培地等を使用できる。形質転換体の培養は、通常5%CO2存在下、37℃で3〜10日間行う。また、培養の際、必要に応じてカナマイシン、ペニシリン、ストレプトマイシン等の抗生物質を培地に添加してもよい。
昆虫細胞を宿主細胞として得られた形質転換体を培養する培地としては、一般に使用されているTNM−FH培地(ファーミンジェン社製)、Sf−900 II SFM培地(Gibco BRL社製)、ExCell400、ExCell405(JRHバイオサイエンシーズ社製)等を使用できる形質転換体の培養は、通常27℃で3〜10日間行う。また、培養の際、必要に応じてゲンタマイシン等の抗生物質を培地に添加してもよい。
目的とするタンパク質は、分泌タンパク質又は融合タンパク質として発現させることもできる。融合させるタンパク質としては、例えば、β−ガラクトシダーゼ、プロテインA、プロテインAのIgG結合領域、クロラムフェニコール・アセチルトランスフェラーゼ、ポリ(Arg)、ポリ(Glu)、プロテインG、マルトース結合タンパク質、グルタチオンS−トランスフェラーゼ、ポリヒスチジン鎖(His−tag)、Sペプチド、DNA結合タンパク質ドメイン、Tac抗原、チオレドキシン、グリーン・フルオレッセント・プロテイン等が挙げられる。
〔タンパク質の単離・精製〕
形質転換体の培養物より目的とするタンパク質を採取することにより、目的とするタンパク質が得られる。ここで、「培養物」には、培養上清、培養細胞、培養菌体、細胞又は菌体の破砕物のいずれもが含まれる。
目的とするタンパク質が形質転換体の細胞内に蓄積される場合には、培養物を遠心分離することにより、培養物中の細胞を集め、該細胞を洗浄した後に細胞を破砕して、目的とするタンパク質を抽出する。目的とするタンパク質が形質転換体の細胞外に分泌される場合には、培養上清をそのまま使用するか、遠心分離等により培養上清から細胞又は菌体を除去する。
こうして得られるタンパク質(a)又は(b)は、溶媒抽出法、硫安等による塩析法脱塩法、有機溶媒による沈殿法、ジエチルアミノエチル(DEAE)−セファロース、イオン交換クロマトグラフィー法、疎水性クロマトグラフィー法、ゲルろ過法、アフィニティークロマトグラフィー法等により精製できる。
タンパク質(a)又は(b)は、そのアミノ酸配列に基づいて、Fmoc法(フルオレニルメチルオキシカルボニル法)、tBoc法(t−ブチルオキシカルボニル法)等の化学合成法によっても製造できる。この際、市販のペプチド合成機を使用できる。
本発明の抗体又はその断片は、タンパク質(a)又は(b)に反応し得る抗体又はその断片である。ここで、「抗体」には、モノクローナル抗体及びポリクローナル抗体のいずれもが含まれ、「モノクローナル抗体及びポリクローナル抗体」には全てのクラスのモノクローナル抗体及びポリクローナル抗体が含まれる。また、「抗体」には、ウサギやマウス等の免疫動物にタンパク質(a)又は(b)のタンパク質を免疫して得られる抗血清、ヒト抗体、遺伝子組換えによって得られるヒト型化抗体も含まれる。また、「抗体の断片」には、Fab断片、F(ab)’2断片、単鎖抗体(scFv)等が含まれる。
本発明の抗体又はその断片は、タンパク質(a)又は(b)を免疫用抗原として利用することより作製できる。免疫用抗原としては、例えば、(i)タンパク質(a)又は(b)を発現している細胞又は組織の破砕物又はその精製物、(ii)遺伝子組換え技術を用いて、タンパク質(a)又は(b)をコードする遺伝子を大腸菌、昆虫細胞又は動物細胞等の宿主に導入して発現させた組換えタンパク質、(iii)化学合成したペプチド等を使用できる。
ポリクローナル抗体の作製にあたっては、免疫用抗原を用いて、ラット、マウス、モルモット、ウサギ、ヒツジ、ウマ、ウシ等の哺乳動物を免疫する。免疫動物は、抗体を容易に作製できることからマウスを利用することが好ましい。免疫の際には、抗体産生誘導する為に、フロイント完全アジュバント等の免疫助剤を用いてエマルジョン化した後、複数回の免疫することが好ましい。免疫助剤としては、フロイント完全アジュバント(FCA)の他、フロイント不完全アジュバント(FIA)、水酸化アルミニウムゲル等を利用できる。哺乳動物1匹当たりの抗原の投与量は、哺乳動物の種類に応じて適宜設定できるが、マウスの場合には通常50〜500μgである。投与部位は、例えば、静脈内、皮下、腹腔内等である。免疫の間隔は、通常、数日から数週間間隔、好ましくは4日〜3週間間隔で、合計2〜8回、好ましくは2〜5回免疫を行う。そして、最終免疫日から3〜10日後に、タンパク質(a)又は(b)に対する抗体力価を測定し、抗体力価が上昇した後に採血し、抗血清を得る。抗体力価の測定は、酵素免疫測定法(ELISA)、放射性免疫測定法(RIA)等により行うことができる。
抗血清から抗体の精製が必要とされる場合は、硫酸アンモニウムによる塩析、ゲルクロマトグラフィー、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー等の公知の方法を適宜選択して又はこれらを組み合わせて利用できる。
モノクローナル抗体の作製にあたっては、ポリクローナル抗体の場合と同様に免疫用抗原を用いて哺乳動物を免疫し、最終免疫日から2〜5日後に抗体産生細胞を採取する。抗体産生細胞としては、例えば、脾臓細胞、リンパ節細胞、胸腺細胞、末梢血細胞等が挙げられるが、脾臓細胞が一般的に利用される。
次いで、ハイブリドーマを得るために、抗体産生細胞とミエローマ細胞との細胞融合を行う。抗体産生細胞と融合させるミエローマ細胞としては、ヒト、マウス等の哺乳動物由来の細胞であって一般に入手可能な株化細胞を利用できる。利用する細胞株としては、薬剤選択性を有し、未融合の状態では選択培地(例えばHAT培地)で生存できず、抗体産生細胞と融合した状態でのみ生存できる性質を有するものが好ましい。ミエローマ細胞の具体例としては、P3X63−Ag.8.U1(P3U1)、P3/NSI/1−Ag4−1、Sp2/0−Ag14等のマウスミエローマ細胞株が挙げられる。
細胞融合は、血清を含まないDMEM、RPMI−1640培地等の動物細胞培養用培地中に、抗体産生細胞とミエローマ細胞とを所定の割合(例えば1:1〜1:10)で混合し、ポリエチレングリコール等の細胞融合促進剤の存在下で、又は電気パルス処理(例えばエレクトロポレーション)により融合反応を行う。
細胞融合処理後、選択培地を用いて培養し、目的とするハイブリドーマを選別する。次いで、増殖したハイブリドーマの培養上清中に、目的とする抗体が存在するか否かをスクリーニングする。ハイブリドーマのスクリーニングは、通常の方法に従えばよく、特に限定されるものではない。例えば、ハイブリドーマとして生育したウエルに含まれる培養上清の一部を採集し、酵素免疫測定法(ELISA)、放射性免疫測定法(RIA)等によってスクリーニングできる。
ハイブリドーマのクローニングは、例えば、限界希釈法、軟寒天法、フィブリンゲル法、蛍光励起セルソーター法等により行うことができ、最終的にモノクローナル抗体を産生するハイブリドーマを取得する。
取得したハイブリドーマからモノクローナル抗体を採取する方法としては、通常の細胞培養法等を利用することができる。細胞培養法においては、例えばハイブリドーマを10〜20%牛胎児血清含有RPMI−1640培地、MEM培地等の動物細胞培養培地中、通常の培養条件(例えば37℃,5%CO2濃度)で3〜10日間培養することにより、その培養上清からモノクローナル抗体を取得することができる。また、ハイブリドーマをマウス等の腹腔内に移植し、10〜14日後に腹水を採取し、当該腹水からモノクローナル抗体を取得することもできる。
モノクローナル抗体の精製が必要とされる場合は、硫酸アンモニウムによる塩析、ゲルクロマトグラフィー、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー等の公知の方法を適宜選択して又はこれらを組み合わせて利用できる。
モノクローナル抗体をヒトに投与する目的(抗体治療)で使用する場合には、免疫原性を低下させるため、ヒト抗体又はヒト型化抗体を使用することが好ましい。ヒト抗体又はヒト型化抗体は、例えば、免疫動物としてヒト抗体遺伝子を導入したマウス等を用いてハイブリドーマを作製することにより、また、ファージ上に抗体を提示したライブラリーを用いることにより取得できる。具体的には、ヒト抗体遺伝子のレパートリーを有するトランスジェニック動物に、抗原となるタンパク質、タンパク質発現細胞又はその溶解物を免疫して抗体産生細胞を取得し、これをミエローマ細胞と融合させたハイブリドーマを用いて目的のタンパク質に対するヒト抗体を取得できる(国際公開番号WO92−03918、WO93−2227、WO94−02602、WO96−33735及びWO96−34096参照)。また、複数の異なるヒトscFvをファージ上に提示させた抗体ライブラリーから、抗原となるタンパク質、タンパク質発現細胞又はその溶解物に結合する抗体を提示しているファージを選り分けることで、目的のタンパク質に結合するscFvを選択できる(Griffiths.等,EMBO J.12,725−734,1993)。
本発明のスクリーニング方法は、試験物質とタンパク質(a)又は(b)とを接触させる工程を含む。本発明のスクリーニング方法によれば、試験物質とタンパク質(a)又は(b)とを接触させ、試験物質がタンパク質(a)又は(b)に結合するか否かを判別することにより、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストをスクリーニングできる。スクリーニングされた物質は、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストとしての作用を実際に有するか否かを判別する工程にさらに供することが好ましい。
試験物質の種類は特に限定されるものではないが、例えば、高分子化合物、低分子化合物、細胞培養物、組織抽出物等が挙げられる。
スクリーニングの際には、タンパク質(a)又は(b)として、例えば、(i)タンパク質(a)又は(b)を発現している細胞若しくは組織又はそれらの処理物(ii)DNA組換え技術を用いて、タンパク質(a)又は(b)をコードするDNAを大腸菌、酵母、昆虫細胞、動物細胞等の宿主に導入して発現させた組換えタンパク質、(iii)化学合成したペプチド等を使用できる。タンパク質(a)又は(b)を発現している細胞又は組織は、タンパク質(a)又は(b)を内因性タンパク質として発現している細胞又は組織(例えば、筋細胞、骨格筋、心臓、マクロファージ、血管、脳、腎臓、肺、胎盤、脾臓、精巣、末梢血液、胸腺、腸管等)であってもよいし、外因性タンパク質として発現している細胞又は組織(例えば、タンパク質(a)又は(b)をコードするDNAを導入した大腸菌、酵母、昆虫細胞、動物細胞等)であってもよい。また、細胞又は組織の処理物は、細胞又は組織に、破砕、抽出、精製等の処理を施したものであり、細胞又は組織の処理物としては、例えば、細胞又は組織の細胞膜画分が挙げられる。
試験物質がアディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストとしての作用を有するか否かは、例えば、タンパク質(a)又は(d)に対する試験物質の結合量、試験物質とタンパク質(a)又は(b)との結合により生じる細胞応答の有無又は程度に基づいて判別できる。
タンパク質(a)又は(b)に対する試験物質の結合量は、例えば、標識した試験物質、試験物質に対する標識抗体等を使用して測定できる。標識としては、例えば、3H、14C、125I、35S、32P等の放射性同位体元素、蛍光色素等を使用できる。放射性同位体元素の放射活性は、例えば、液体シンチレーションカウンター、X線フィルム、イメージングプレート等を用いて測定でき、蛍光色素の蛍光強度は、例えば、CCDカメラ、蛍光スキャナー、分光蛍光光度計等を用いて測定できる。
試験物質とタンパク質(a)又は(b)との結合により生じる細胞応答としては、例えば、PPARαリガンド活性の亢進又は抑制、脂肪酸酸化の亢進又は抑制、グルコース取り込みの亢進又は抑制、細胞内pHの上昇又は低下、AMPキナーぜ活性の増加又は低下、AMPキナーゼのリン酸化の増加又は低下、p38 MAPキナーゼ活性の増加又は低下、p38 MAPキナーゼのリン酸化の増加又は低下、糖新生の促進又は抑制、脱共役蛋白の増加又は低下等が挙げられる。
タンパク質(a)又は(b)に対する試験物質の結合量の測定、及び細胞応答の有無又は程度の測定にあたっては、タンパク質(a)又は(b)の存在下及び不存在下における測定値を比較することが好ましい。
脂肪細胞は主要なインスリン感受性ホルモンとしてアディポネクチンを分泌し、脂肪細胞肥大はアディポネクチン分泌低下を惹起し、アディポネクチン分泌低下はインスリン抵抗性を惹起する。そして、インスリン抵抗性は、糖尿病、高脂血症、高血圧を惹起する。また、アディポネクチン分泌低下は動脈硬化を促進させる。したがって、アディポネクチン受容体に対するアゴニスト又はアンタゴニストとしてスクリーニングされた物質は、例えば、インスリン抵抗性改善薬、糖尿病予防・治療薬、高脂血症予防・治療薬、高血圧予防・治療薬、動脈硬化予防・治療薬、肥満予防・治療薬、抗炎症薬、骨粗鬆症予防・治療薬、抗癌剤等として使用できる。
これらの薬剤はスクリーニングされた物質のみから構成してもよいが、通常は、薬学的に許容され得る賦形剤その他任意の添加剤を用いて製剤化する。製剤化にあたっては、例えば、賦形剤、結合剤、崩壊剤、滑沢剤、安定剤、矯味矯臭剤、希釈剤、注射剤用溶剤等の添加剤を使用できる。投与経路としては、例えば、経口投与、非経口投与(例えば、皮下投与、筋肉内投与、腹腔内投与等)が挙げられ、投与剤形としては、例えば、噴霧剤、カプセル剤、錠剤、顆粒剤、シロップ剤、乳剤、座剤、注射剤、懸濁剤等が挙げられる。投与量及び投与回数は、目的とする作用効果、投与方法、治療期間、患者の年齢、体重、性別等により異なり、スクリーニングされた物質の種類に応じて適宜調節できる。
タンパク質(a)又は(b)、タンパク質(a)又は(b)をコードする遺伝子、該遺伝子を含有する組換えベクター又は該組換えベクターを含む形質転換体は、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストのスクリーニング用キットの構成要素として利用できる。これらは、タンパク質(a)又は(b)の供給源としてスクリーニング用キットに含まれる。
スクリーニング用キットは、タンパク質(a)又は(b)、タンパク質(a)又は(b)をコードする遺伝子、該遺伝子を含有する組換えベクター又は該組換えベクターを含む形質転換体を含む限り、いかなる形態であってもよく、各種試薬(例えば、緩衝液等)、測定機械器具、標識化合物、モデル動物、細胞株、細胞培養用培地等を含むことができる。
【実施例】
以下、実施例に関する本文及び図において、アディポネクチンを「Adipo」と、globularドメインのみからなるアディポネクチンを「globular Adipo」又は「gAd」と、全長アディポネクチンを「全長Adipo」又は「Ad」という場合がある。また、アディポネクチン受容体を「AdipoR」と、ヒト由来アディポネクチン受容体を「hAdipoR」と、マウス由来アディポネクチン受容体を「mAdipoR」という場合がある。また、「AdipoR1」及び「AdipoR2」はアミノ酸配列が異なるアディポネクチン受容体を表す。
1.実験方法
(1)レトロウイルスの作製及び感染
レトロウイルス含有上清を作製するために、リポフェクタミンPLUS(Life Technologies)を用いて、107個のPlat−E packaging細胞(Morita,S.ら,Gene Ther.7,1063−1066(2000))に、ヒト骨格筋cDNAライブラリー(Clontech)10μgを一過性にトランスフェクトした。24時間インキュベーションした後、上清(10mL)を回収した。Ba/F3細胞に、ポリブレン(hexadimethrine,Sigma)10μg/mLを添加した1/20倍希釈上清(推定m.o.i=0.3)を感染させた。6時間後、培養液を交換し、Ba/F3細胞を6日間培養して増殖させた後、冷凍保存又は細胞選別を行った。
(2)FACS解析及びcDNAの配列決定
FACS解析は、Stoecklinらの方法(Stoecklin,G.ら,EMBO J.21,4709−4718.(2000))に従って行った。細胞選別のために、ヒト骨格筋cDNAライブラリーをトランスフェクトしたBa/F3細胞1×107個からFACVantage(Becton Dickinson)を用いて細胞を濃縮した。細胞を回収し、増殖させた後、11日後にFACS解析に供した。選別された細胞をさらに増殖してFACS解析に供した。選別された細胞に組み込まれたcDNAの配列決定を行うために、レトロウイルスベクターに対する上流及び下流のプライマーを用いて、選別された細胞から抽出されたゲノムDNA 50ngを鋳型としてPCRを行い、得られたPCR増幅断片の配列決定を行った。
PCRは、Taqポリメラーゼ(Perkin−Elmer/Cetus)を用いて、94℃で1分間、56℃で2分間、72℃で3分間のサイクルを35サイクル行った。レトロウイルスベクターに対するプライマー(pLIBプライマー)は以下のとおりである。
PCR産物は、プライマーを除去した後、BigDye Terminater Kit(Applied Biosystems)を用いたdirect sequenceにより塩基配列を決定した。
(3)ノーザンブロット解析
ヒトの様々な組織(脳、心臓、骨格筋、大腸、胸腺、脾臓、腎臓、肝臓、小腸、胎盤、肺、末梢血白血球)のノーザンブロットフィルターI及びIIをClonetechより購入した。これらのフィルターを、4×SSC、5×Denhardt’s溶液、0.2%SDS、200mg/mLサケ精子DNA、及び50%ホルムアミドを含有するハイブリダイゼーション緩衝液中、42℃で24時間、[32P]dCTP標識したcDNAプローブ(ヒトAdipoR1 cDNA、マウスAdipoR1 cDNA、ヒト又はマウスAdipoR2 cDNAをそれぞれPstI/BstXI、BamHI/PstI、EcoRV/NotIを処理して得られた断片)とハイブリダイズさせた。0.1×SSC、0.1%SDS、65℃の条件でフィルターを洗浄した後、オートラジオグラフィーに供した。同様のノーザンブロット解析をマウスの様々な組織(脳、心臓、腎臓、肝臓、肺、骨格筋、脾臓、精巣)についても行った。
(4)哺乳動物細胞におけるタンパク質の発現及び性状決定
AdipoR1 cDNA又はAdipoR2 cDNAをpCXN2(Kinoshita,S.ら,Pharm.Res.15,1851−1856(1998))のEcoRV/NotI部位にライゲートすることにより、AdipoR1又はAdipoR2発現ベクターを構築した。HEK−293T(human embryonic kidney cells:ヒト胎児腎臓細胞)、HAEC(Normal Human Aortic Endothelial Cells:正常ヒト大動脈内皮細胞)及びC2C12筋細胞(マウス筋細胞株)をDMEM中で培養した。培養液には、10%ウシ胎児血清(FCS)を含有させた。DNAトランスフェクションは、いずれの細胞についても、リポフェクタミンPLUS(Gibco BRL)を用いたリポフェクションによって行った。
(5)C2C12筋細胞におけるRNA干渉(RNA interference)
Karpichevらの方法(Karpichev,I.V.ら,J.Biol.Chem.277,19609−19617(2002))に従い、二組のsiRNAを化学合成し、アニーリングさせ、リポフェクタミンPLUS(Life Technologies)を用いて、コンフルエントから2日後にウマ血清含有DMEMで4〜7日間培養して筋管細胞に分化させたC2C12筋細胞にトランスフェクトした。肝細胞又はHAECについても同様にsiRNAをトランスフェクトした。siRNAのトランスフェクション48時間後、細胞を溶解した。
無関係なコントロールsiRNA(siRNA unrelated)の塩基配列は次のとおりである。
マウスAdipoR1遺伝子のコーディング領域に対応するsiRNA(siRNA mAdipoR1)の塩基配列は次のとおりである。
マウスAdipoR2遺伝子のコーディング領域に対応するsiRNA(siRNA mAdipoR2)の塩基配列は次のとおりである。
ヒトAdipoR1遺伝子のコーディング領域に対応するsiRNA(siRNA hAdipoR1)の塩基配列は次のとおりである。
ヒトAdipoR2遺伝子のコーディング領域に対応するsiRNA(siRNA hAdipoR2)の塩基配列は次のとおりである。
(6)PPARα(Peroxisome proliferator−activated receptor α:ペルオキシソーム増殖因子活性化受容体α)リガンド活性の測定
大腸菌を用いて発現させたマウスglobular Adipo及び全長Adipoを、Yamauchiらの方法(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002))に従って精製した。Yamauchiらの方法(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002))に従って、分化したC2C12筋細胞又は単離した肝細胞を所定濃度のアディポネクチンで処理した。PPARαリガンド活性は、Yamauchiらの方法(Yamauchi,T.ら,J.Biol.Chem.278,2461−2468(2002))に従って、(UAS)×4−tk−LUCリポータープラスミド、GAL4−ratPPARα リガンド結合ドメイン発現プラスミド、及びβ−ガラクトシダーゼ発現プラスミド(内部コントロール)を用いて定量した。
(7)脂質及びグルコース代謝
Yamauchiらの方法(Yamauchi,T.ら,Nat.Med.7,941−946.(2001))に従って、細胞溶解物を用いて[1−14C]パルミチン酸からの[14C]CO2産生を測定した。また、Yamauchiらの方法(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002))に従って、グルコース取り込みを測定した。
(8)優性阻害型(dominant negative)AMPキナーゼ(AMPK)を用いた研究
α2 AMPK(45番目のリジン残基がアルギニン残基に置換された変異を含む)をコードするcDNAをDN−α2AMPKとして用いた(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002))。C2C12筋細胞に、コントロールMockベクター又はDN−α2AMPKを含有する等しい力価のアデノウイルスを感染させた。Yamauchiらの方法(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002))に従って、分化の誘導の5日後に、細胞を所定濃度のアディポネクチンで処理して、PPARαリガンド活性及び脂肪酸酸化を測定した。
(9)結合アッセイ
合成ヒト又はマウスAdipoのTyrを、Na125I(2000Ci/mmol,Amersham Pharmacia Biotech)の存在下、IODO−ビーズ(Pierce)により125Iで標識した。組換えglobular Adipo又は全長AdipoをNHS−LC−ビオチン(Pierce)を用いてビオチン化した。細胞を96wellプレート上に4.1×104個/wellの密度で播き、一晩培養した後、培養液を除去し、細胞を37℃で一晩、所定濃度の[125I]Adipo及び非標識競合物質を含有する結合アッセイ用緩衝液(HEPES緩衝化生理食塩水/0.1%ウシ血清アルブミン)とインキュベートした。その後、Yamauchiらの方法(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002);Yokomizo,T.ら,Nature 387,620−624(1997))に従って、細胞を氷冷リン酸緩衝化生理食塩水で3回洗浄し、0.1N NaOH/0.1%SDSで溶解し、細胞に結合した放射活性をγカウンターで測定した。
(10)蛍光顕微鏡による解析
AdipoR1又はAdipoR2の細胞内局在を、293T細胞を用いて、共焦点蛍光顕微鏡により評価した。細胞を1%パラホルムアルデヒドで固定した後、5倍希釈した透過緩衝液(permeabilization buffer)(Coulter)を用いて透過させて又は透過させずに、抗FLAG抗体(M2;30μg/mL)と22℃で1時間インキュベートした。次いで、AlexFluor 488を結合させた二次抗体10μg/mLとともにインキュベートした。次いで、ニコン顕微鏡及びクリプトン/アルゴンレーザー(488nm)を備えたレーザスキャンニング顕微鏡システムを用いて共焦点イメージングを行った。
(11)AdipoR1遺伝子及びAdipoR2遺伝子の転写物のリアルタイムPCRによる定量的解析
Heidらの方法(Heid,C.A.ら,Genome Res.6,986−994(1996))に従って、ヒトAdipoR1遺伝子、ヒトAdipoR2遺伝子、マウスAdipoR1遺伝子及びマウスAdipoR2遺伝子に関する4種類の転写物の定量をリアルタイムPCRによって行った。それぞれの転写物に関するプライマー及びプローブは以下のとおりである。PCR産物は、ABI PRISM7700 Sequence Detection System(Applied Biosystems)を用いて連続的に測定した。それぞれの転写物の相対量をアクチンの転写物量に対して標準化した。
[マウスAdipoR1遺伝子]
[マウスAdipoR2遺伝子]
[ヒトAdipoR1遺伝子]
[ヒトAdipoR2遺伝子]
(12)細胞内カルシウム濃度、cAMP量及びcGMP量の測定
細胞内Ca2+濃度は、Yokomizoらの方法(Yokomizo,T.ら,Nature 387,620−624(1997))に従って測定した。すなわち、細胞に、Hepes−Tyrode’s BSA緩衝液(25mM Hepes−NaOH(pH7.4),140mM NaCl,2.7mM KCl,1.0mM CaCl2,12mM NaHCO3,5.6mM D−グルコース,0.37mM NaH2PO4,0.49mM MgCl2,0.1%[wt/vol]脂肪酸不含BSA;Fraction V)に溶解した10μM Fura−2 AM(Dojin)を37℃で2時間接触させた。細胞を2回洗浄し、Hepes−Tyrode’s BSA緩衝液に106細胞/mLの濃度で懸濁させた。0.5mLの細胞懸濁液をCAF−100 system(Jasco)にアプライし、5μLのリガンドのエタノール溶液(LTB4用)又はPBS溶液(Adipo用)を加えた。340nm及び380nmの励起光によって発せられる500nmの蛍光の割合に基づいて細胞内Ca2+濃度を測定した。
また、cAMP量及びcGMP量は、Yokomizoらの方法(Yokomizo,T.ら,Nature 387,620−624(1997))に従い、アッセイキット(cAMP:Biotrak cAMP EIAシステム,cGMP:Biotrak cGMP EIAシステム)(Amersham Pharmacia Biotech)を用いて、製造業者のプロトコルに基づき測定した。
(13)AdipoR1及びAdipoR2の構造予測
Kyte−Doolitleのハイドロパシーインデックスを用いて、AdipoR1及びAdipoR2タンパク質のハイドロパシープロット(hydropathy plot)を行った。また、AdipoR1及びAdipoR2の構造モデルをSOSUIによって予測し、PRINTS(http://bioinf.man.ac.uk/dbbrowser/PRINTS/)によってコンセンサス配列を解析した。さらに、リン酸化部位をDNASIS Pro.によって解析した。さらに、AdipoR1/R2がその他のクラスのGPCRとホモロジーを有するか否かをhttp://cbrg.inf.ethz.ch/Server/AllAll.htmlに記載の方法により解析した。
(14)AMPキナーゼ(AMPK)、ACC、p38 MAPキナーゼ(p38 MAPK)及びMAPキナーゼ(MAPK)のリン酸化及びリン酸化量の測定
AMPK、ACC(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002))、p38 MAPK及びMAPK(Barger,P.M.ら,J.Biol.Chem.276,44495−44501(2001);Puigserver,P.ら,Mol.Cell 8,971−982(2001);Michael,L.F.ら,Proc.Natl.Acad.Sci.USA.98,3820−3825(2001))のリン酸化及びリン酸化量を、抗リン酸化AMPK抗体、抗リン酸化ACC抗体、抗リン酸化p38 MAPK抗体又は抗リン酸化MAPK抗体を利用したウェスタンブロッティング法により測定した。この際、AdipoR1をトランスフェクトした又はしていないC2C12細胞又は肝細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、各細胞の溶解物と各抗体とを反応させた。
2.結果
(1)AdipoR1及びAdipoR2の発現クローニング
筋肉において、globular Adipoは全長Adipoよりもインスリン抵抗性を改善させ、PPARα及び脂肪酸酸化を活性化させる(Fruebis,J.ら,Proc.Natl.Acad.Sci.U S A.98,2005−2010(2001);Yamauchi,T.ら,Nat.Med.7,941−946(2001);Yamauchi,T.ら,J.Biol.Chem.278,2461−2468(2002))。さらに、globular Adipoは全長AdipoよりもC2C12筋細胞に強く結合するとともに、肝細胞及び肝臓の膜よりも骨格筋の膜に強く結合する(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002))(図1a,b参照)。図1aは、globular Adipo又は全長AdipoとC2C12筋細胞との結合、図1bはglobular Adipo又は全長Adipoと肝細胞との結合を示す。細胞は、図示する濃度のビオチン化globular Adipo又は全長Adipoとともにインキュベートし、細胞表面に結合したビオチン化globular Adipo又は全長AdipoはELISAにより定量した。図中のそれぞれのバーは平均値±s.e.(n=3−5)を示し、「※」はP<0.05、「※※」はP<0.01を示す。
そこで、ヒト骨格筋のmRNA由来のcDNAを組み込んだレトロウイルスをBa/F3細胞に感染させて作製したライブラリーから、globular Adipoに対する結合能を有するタンパク質をスクリーニングすることにより、AdipoR1のcDNAの単離を試みた。
感染させたBa/F3細胞を回収し、ビオチン化したglobular Adipoとインキュベートした後、ストレプトアビジンを結合させたフィコエリトリン(phycoerythrin:PE)(赤色蛍光プローブ)で染色し、蛍光活性化セルソーター(fluorescence−activated cell sorting:FACS)に供した(図1c参照)。FACS解析の結果を図1c,d,eに示す。図1cは、感染後であるが、第一の選別前のBa/F3細胞を示し、図1dは、第三の選別前のBa/F3細胞を示し、図1eは、FITC(fluorescein isothiocyanate)を結合させたglobular Adipoとインキュベートした後であるが、第四の選別前のBa/F3細胞を示す。また、図1c,d,eにおいて、四角で囲まれた領域は、AdipoR1陽性細胞を示し、四角内の細胞は選別された細胞を示す。
選別されたglobular Adipo結合能を有する細胞を第二ラウンド目の選別に供した。再選別されたglobular Adipo結合能を有する細胞(図1d参照)を第三ラウンド目の選別に供し、再選別された細胞を直ちにFITC(fluorescein isothiocyanate)(緑色蛍光プローブ)を結合させたglobular Adipoとインキュベートした。赤色から緑色に変化した細胞がglobular Adipoに対する特異的結合部位を有する細胞であるから(図1e参照)、そのように変化した細胞のみを選別し、さらなる分析のために培養により増殖させた。これらの細胞から抽出したゲノムDNAを、ウイルスベクターに対するプライマーを用いたPCRに供し、配列決定した。
なお、上記のように細胞を二色(赤色及び緑色)で染色したのは、globular Adipoがその粘着性によって非特異的に結合した細胞を除去するためである。
配列決定されたヒトAdipoR1 cDNAの塩基配列を配列番号1に示し、それにコードされるタンパク質(ヒトAdipoR1)のアミノ酸配列を配列番号2に示す。また、C2C12筋細胞から同様の方法によって取得したマウスAdipoR1 cDNAの塩基配列を配列番号5に示し、それにコードされるタンパク質(マウスAdipoR1)のアミノ酸配列を配列番号6に示す。ヒトAdipoR1 cDNA(配列番号1)及びマウスAdipoR1 cDNA(配列番号5)は、375個のアミノ酸からなるタンパク質をコードする遺伝子であることが判明した(図1f参照)。ヒトAdipoR1及びマウスAdipoR1のアミノ酸配列の相同性は96.8%である(図1f参照)。図1fは、データベース(NIH−MGC Project and NCBI contig)におけるAdipoR1遺伝子及びAdipoR2遺伝子の転写物の模式的構造を示す。
これまでの知見(Yamauchi,T.ら,Nat.Med.8,1288−1295.(2002);Yamauchi,T.ら,J.Biol.Chem.278,2461−2468(2002))から、AdipoRには、globular Adipo又は全長Adipoに対する結合親和性が異なる2種類のタイプ、すなわち、骨格筋で発現するglobular Adipoに優先的に結合するタイプと、肝臓で発現する全長Adipoにのみ結合するタイプとが存在すると考えられる。
AdipoR1とホモロジーを有するタンパク質を調査したところ、ヒト及びマウスに関するデータベース(The Human Genome,http://www.ncbi.nlm.nih.gov/genome/guide/human/;Mouse Genome Resources,http://www.ncbi.nlm.nih.gov/genome/guide/mouse/)(Waterston,R.H.ら,Nature 420,520−562(2002);Okazaki,Y.ら,Nature 420,563−573(2002))において、AdipoR1 cDNAとは異なるオープンリーディングフレーム(ORF)を有する遺伝子が発見された。そこで、このcDNAをHepG2細胞(ヒト肝癌由来細胞株)のmRNAからクローニングし、配列決定し、このcDNAにコードされているタンパク質をAdipoR2と名付けた。配列決定されたヒトAdipoR2 cDNAの塩基配列を配列番号3に示し、それにコードされるタンパク質(ヒトAdipoR2)のアミノ酸配列を配列番号4に示す。また、C2C12筋細胞から同様の方法によって取得したマウスAdipoR2 cDNAの塩基配列を配列番号7に示し、それにコードされるタンパク質(マウスAdipoR2)のアミノ酸配列を配列番号8に示す。
ヒトAdipoR2及びマウスAdipoR2のアミノ酸配列の相同性は95.2%である。また、AdipoR1及びAdipoR2は構造的に高度に類似しており、マウスAdipoR1及びAdipoR2のアミノ酸配列の相同性は66.7%である。
SWISS−PROTにおいて、AdipoR1及びAdipoR2と相同性が高い哺乳動物のタンパク質は存在しなかったが、興味深いことに、AdipoR1及びAdipoR2をコードするcDNAは、酵母YOL002c(Karpichev,I.V.ら,J.Biol.Chem.277,19609−19617.(2002))とホモロジーを有していた。YOL002cは、脂肪酸酸化等の脂質の代謝経路において重要な役割を果たす7回膜貫通型タンパク質をコードすると報告されている(Karpichev,I.V.ら,J.Biol.Chem.277,19609−19617(2002))。種を問わず、7回膜貫通構造を有するAdipoR/YOL002cは、脂肪酸酸化等の脂肪代謝において重要な制御シグナルを媒介すると考えられる。
(2)AdipoR1及びAdipoR2の組織分布
マウスの様々な組織に関するノーザンブロット解析の結果を図1gに示し、ヒトの様々な組織に関するノーザンブロット解析の結果を図1hに示す。図1g中、レーン1は脳、レーン2は心臓、レーン3は腎臓、レーン4は肝臓、レーン5は肺、レーン6は骨格筋、レーン7は脾臓、レーン8は精巣の結果を示し、図1h中、レーン1は脳、レーン2は心臓、レーン3は骨格筋、レーン4は大腸、レーン5は胸腺、レーン6は脾臓、レーン7は腎臓、レーン8は肝臓、レーン9は小腸、レーン10は胎盤、レーン11は肺、レーン12は末梢血白血球の結果を示す。
ヒト及びマウスの様々な組織に関するノーザンブロット解析によって、上記データベースにおいて予測されたmRNAのサイズを有する2.0kbの一本の主要なバンドが同定され、AdipoR1がほとんどの組織で発現しており、骨格筋において高度に発現していることが明らかになった。また、上記データベースにおいて予測されたmRNAのサイズを有する4.0kbの一本の主要なバンドが同定され、AdipoR2が肝臓において高度に発現していることが明らかになった。
(3)AdipoR1及びAdipoR2の細胞内局在
マウスAdipoR1の推定アミノ酸配列(配列番号6)から、マウスAdipoR1は375個のアミノ酸からなるタンパク質であり、その分子量は42.4kDaと予測される。また、マウスAdipoR2の推定アミノ酸配列(配列番号8)から、マウスAdipoR2は311個のアミノ酸からなるタンパク質であり、その分子量は35.4kDaと予想される(図2a参照)。図2aは、ソフトウエアPRINTS(http://bioinf.man.ac.uk/dbbrowser/PRINTS/)によってAdipoRの配列をスキャンした結果であり、図中、下線領域は、AdipoR1及びAdipoR2の7個の膜貫通ドメインを示し、太字の下線部は、Gタンパク質結合受容体のメンバーに特徴的な保存モチーフを示す。また、図2aには、PKCリン酸化部位及びチロシンリン酸化部位も併せて示す。
AdipoR1及びAdipoR2の推定アミノ酸配列からAdipoR1及びAdipoR2は、7個の膜貫通ドメインを有するタンパク質であると予測される(図2a参照)。AdipoR1又はAdipoR2について、7個の膜貫通ドメインを有する公知の受容体(Waterston,R.H.ら,Nature 420,520−562(2002);Okazaki,Y.ら,Nature 420,563−573(2002);Wess,J.,FASEB.J.11,346−354(1997))とのアライメントを行ったが、Gタンパク質結合受容体(G−protein coupled receptor:GPCR)ファミリーに属するメンバーとのアミノ酸配列の相同性は低かった。AdipoR1及びAdipoR2は、Gタンパク質結合受容体ファミリーの特徴(例えば、保存アミノ酸、グリコシル化部位、Gタンパク質結合部位)(Wess,J.らFASEB.J.11,346−354(1997);Yokomizo,T.ら,Nature 387,620−624(1997);Scheer,A.ら,EMBO.J.15,3566−3578(1996))が欠落していた。Gタンパク質結合受容体ファミリーにおいて高度に保存されているアミノ酸のうち、AdipoR1及びAdipoR2の第一及び第二細胞外ループにおいて高度に保存されている2個のCys残基のうちの1つのみが存在していた。AdipoR1及びAdipoR2は、TM7の末端に存在する高度に保存されたAsn−Pro−Xaa2−Tyrモチーフを欠落していた。また、AdipoR1及びAdipoR2のTM3/細胞内ループ2トランジションに存在する高度に保存されたAsp−Arg−Tyrモチーフ(Wess,J.らFASEB.J.11,346−354(1997);Scheer,A.ら,EMBO.J.15,3566−3578(1996))も欠落していた。
エピトープタグFLAGで標識したヒト及びマウスのAdipoR1又はAdipoR2をHEK−293T細胞中で発現させ、抗FLAG抗体で免疫染色した。ヒト及びマウスAdipoR1及びAdipoR2は、予想された分子量を示した(図2b参照)。
マウスAdipoR1又はAdipoR2の細胞内局在及びトポロジーを調べるために、いずれかの末端にエピトープタグを有するAdipoR1又はAdipoR2のcDNAをHEK−293T細胞で発現させた(図2c参照)。図2c中、「intact」は細胞を透過させなかった場合、「permeabilized」は細胞を透過させた場合を表す。
エピトープタブをN末端に導入した場合には、細胞を透過させたときのみ、細胞表面に存在するAdipoR1及びAdipoR2を検出できた(図2c参照)。これに対して、エピトープタグをC末端に導入した場合には、細胞表面に存在するAdipoR1及びAdipoR2を検出できた(図2c参照)。これらの結果は、AdipoR1及びAdipoR2が7個の膜貫通ドメインを有する膜内在性タンパク質であり、そのN末端は膜内に存在し、そのC末端は膜外に存在することを示す(図2c,d参照)。これは、これまでに報告されている全てのGタンパク質結合受容体(Wess,J.らFASEB.J.11,346−354(1997);Yokomizo,T.ら,Nature 387,620−624(1997);Scheer,A.ら,EMBO.J.15,3566−3578(1996))と反対のトポロジーである。なお、図2dは、AdipoR1及びAdipoR2の予測構造モデルである。
(4)293T細胞におけるAdipoRの発現の効果
AdipoR1又はAdipoR2を細胞表面上に過剰発現させた293T細胞を用いて、globular Adipo又は全長Adipoにより亢進される結合活性及び細胞内シグナルを調べた。293T細胞におけるAdipoR1又はAdipoR2の発現は、globular Adipo及び全長Adipoの両者の結合を亢進させた(図3a,b参照)。図3aは、AdipoR1又はAdipoR2をトランスフェクトした293T細胞への[125I]globular Adipo(gAd)の結合等温線(binding isotherm)を示し、図3bは、当該293T細胞への[125I]全長Adipo(Ad)の結合等温線(binding isotherm)を示し、図中、□はMock、■はマウスAdipoR1、●はマウスAdipoR2に関する結果を示す。
次に、AdipoR1及びAdipoR2がマルチマー(multimer)を形成するかどうかを調べた。エピトープタグFLAGを有するAdipoR1及びエピトープタグHAを有するAdipoR1をともにHEK−293T細胞中で発現させた場合には、抗HA抗体を用いた免疫沈降において、エピトープタグFLAGを有するAdipoR1が検出された(図3c参照)。図3cは、エピトープタグFLAG又はHAを有するAdipoR1又はAdipoR2をトランスフェクトした293T細胞の細胞溶解物を、抗FLAG抗体(上側及び下側のパネル)を用いて免疫沈降(IP)した後、抗FLAG抗体(上側のパネル)又は抗HA抗体(下側のパネル)で免疫染色した結果であり、AdipoR1及びAdipoR2によるホモ−及びヘテロ−マルチマーの形成を示す。
さらに、エピトープタグHAを有するAdipoR1及びエピトープタグFLAGを有するAdipoR2をともにHEK−293T細胞中で発現させた場合には、抗HA抗体を用いた免疫沈降において、エピトープタグFLAGを有するAdipoR2が検出された(図3c参照)。これらの結果は、AdipoR1及びAdipoR2がホモ−及びヘテロ−マルチマーの両者を形成する可能性を示す。
GPCR又はLTB4受容体BLT1を発現させた細胞では、LTB4により細胞内カルシウムが増加し、AdipoR1を発現する細胞の発現レベルと類似する発現レベルを示したが(データは示さず)、AdipoR1を発現する細胞では、Adipoによる細胞内カルシウムの変化は観察されなかった(図3d参照)。図3dは、Fura−2/AMをロードしたマウスAdipoR1発現細胞又はBLT1発現細胞に、10μg/mL globular Adipo(gAd)、10μg/mL 全長Adipo(Ad)、1μM LTB4又は100μM ATPをチャレンジし、[Ca2+]iの変化を測定した結果である。
さらに、AdipoR1を発現する細胞において、AdipoR1はcAMP及びcGMPレベルに対して全く影響しないか、ほとんど影響を与えなかった(図3e参照)。図3eはホルスコリン(forskolin)で処理した又は処理しないHEK−293細胞にcAMP又はcGMPを蓄積させた結果であり、図中、「gAd0.01」、「gAd0.1」はそれぞれ0.01、0.1μg/mLのglobular Adipoを表し、「Ad1」、「Ad10」はそれぞれ1、10μg/mLの全長Adipoを表す。対照的に、AdipoR1の発現は、globular Adipo及び全長Adipoによる293T細胞内のPPARαリガンド活性の増加を増強した(図3f参照)。図3fは、AdipoR1をトランスフェクトした293T細胞を図示する濃度(μg/mL)のglobular Adipo又は全長AdipoとインキュベートしたときのPPARαリガンド活性を示し、図中、「gAd0.1」、「gAd0.5」、「gAd2.5」はそれぞれ0.1、0.5、2.5μg/mLのglobular Adipoを表し、「Ad1」、「Ad5」、「Ad25」はそれぞれ1、5、25μg/mLの全長Adipoを表す。また、図中、各バーは平均値±s.e.(n=3−5)を表し、「※」はP<0.05を表し、「※※」はP<0.01を表す。
(5)C2C12筋細胞におけるPPARα活性化及び脂肪酸酸化
C2C12筋細胞におけるAdipoR1の発現(図4a参照)は、globular Adipo及び全長Adipoの両方の結合を促進させ(図4c,d参照)、globular Adipo及び全長AdipoによるC2C12筋細胞内のPPARαリガンド活性(図4e参照)及び脂肪酸酸化(図4f参照)の増加と関連した。また、C2C12筋細胞におけるAdipoR2の発現(図4b参照)は、globular Adipo及び全長Adipoの両方の結合を促進させ(図4c,d参照)、globular Adipo及び全長AdipoによるC2C12筋細胞内の脂肪酸酸化の亢進(図4f)に関連した。図4aはマウスAdipoR1 mRNA量を示し、図4bはマウスAdipoR2mRNA量を示し、図中、□はMock、■はマウスAdipoR1、点線を引いた□はマウスAdipoR2に関する結果を示す。また、図4c及びdは、マウスAdipoR1又はマウスAdipoR2をトランスフェクトしたC2C12筋細胞に結合した[125I]globular Adipo(gAd)又は全長Adipo(Ad)の結合等温線(binding isotherm)を示し、図中、□はMock、■はマウスAdipoR1、●はマウスAdipoR2に関する結果を示す。また、図4eは、マウスAdipoR1又はマウスAdipoR2をトランスフェクトしたC2C12筋細胞に、LacZ又はDN−α2AMPKを含有するアデノウイルスを感染させ、図示する濃度(μg/mL)のglobular Adipo又は全長Adipoで7時間処理したときの、当該C2C12筋細胞内のPPARαリガンド活性を示し、図中、□はMock、■はマウスAdipoR1に関する結果を示す。また、図4fは、上記C2C12筋細胞内のin vitro脂肪酸酸化を示し、図中、□はMock、■はマウスAdipoR1、点線を引いた□はマウスAdipoR2に関する結果を示す。なお、各図中、各バーは平均値±s.e.(n=3−5)を表し、「※」はP<0.05を表し、「※※」はP<0.01を表す。
優性阻害型(dominant negative:DN)AMPキナーゼの発現は、globular Adipo及び全長Adipo誘導性であってAdipoR1発現依存的なPPARαリガンド活性の亢進に影響を与えなかった。これらの結果は、AdipoR1及びAdipoR2はともに、globular Adipo及び全長Adipoの結合を媒介し、globular Adipo及び全長AdipoによるPPARαリガンド活性及び脂肪酸酸化を亢進させることを示す。
(6)筋細胞内のAdipoRの結合及び作用に対するsiRNAの影響
筋細胞において、内因性のAdipoR1及びAdipoR2がAdipoの特異的結合及び代謝的作用を媒介しているか否かを調べるため、siRNAを用いてAdipoR1及びAdipoR2の発現を抑制した(図5a,b参照)。図5aは、siRNA又はmockをトランスフェクトしたC2C12筋細胞内のマウスAdipoR1 mRNAの量を示し、図5bは、当該C2C12筋細胞内のマウスAdipoR2 mRNAの量を示す。また、図5a,b中、レーン1はmock、レーン2は無関係なsiRNA、レーン3はマウスAdipoR1遺伝子に対するsiRNA、レーン4はマウスAdipoR2遺伝子に対するsiRNAを用いた場合の結果を示す。
C2C12筋細胞において、siRNAによりAdipoR1の発現を抑制すると(図5a参照)、globular Adipoの結合活性は全く観察されなくなり、全長Adipoの結合活性は一部低下した(図5c,d参照)。図5cは、非標識globular Adipoの濃度を増加させることによって、二本鎖siRNAをトランスフェクトした細胞に結合する[125I]globular Adipoを置換した競合ラジオリガンド結合アッセイの結果(n=4)であり、図5dは、非標識全長Adipoの濃度を増加させることによって、二本鎖siRNAをトランスフェクトした細胞に結合する[125I]全長Adipoを置換した競合ラジオリガンド結合アッセイの結果(n=4)である。
globular Adipo又は全長Adipoで7時間処理すると、C2C12細胞において、PPARαリガンド活性が増加し(図5g参照)、脂肪酸酸化が亢進し(図5h参照)、グルコース取り込みが亢進した(図5i参照)。図5gは、二本鎖siRNAをトランスフェクトしたC2C12筋細胞を、図示する濃度(μg/mL)のglobular Adipo又は全長Adipo、あるいは10−5M Wy−14,643(図中「Wy」と記載)と7時間インキュベートしたときのPPARαリガンド活性を示す。図5hは、二本鎖siRNAをトランスフェクトしたC2C12筋細胞を、図示する濃度(μg/mL)のglobular Adipo又は全長Adipo、あるいは10−5M Wy−14,643(図中「Wy」と記載)と7時間インキュベートしたときのin vitro脂肪酸酸化を示す。図5iは、二本鎖siRNAをトランスフェクトしたC2C12筋細胞を、図示する濃度(μg/mL)のglobular Adipo又は全長Adipo、あるいは10−7Mインスリン(図中「Ins」と記載)と7時間インキュベートしたときのグルコース取り込みを示す。図5g〜iにおいて、「gAd0.01」、「gAd0.03」、「gAd0.1」、「gAd0.5」、「gAd2.5」はそれぞれ0.01、0.03、0.1、0.5、2.5μg/mLのglobular Adipoを表し、「Ad0.1」、「Ad0.3」、「Ad1」、「Ad5」、「Ad25」はそれぞれ0.1、0.3、1、5、25μg/mLの全長Adipoを表す。図中、各バーは平均値±s.e.(n=3−5)を表し、「※」はP<0.05、「※※」はP<0.01を表す。また、図5g〜iにおいて、□は無関係なsiRNA、■はマウスAdipoR1に対するsiRNA、点線を引いた□はマウスAdipoR2に対するsiRNA、斜線を引いた□はマウスAdipoR1及びマウスAdipoR2に対するsiRNAを用いたときの結果を示す。
C2C12筋細胞において、siRNAによりAdipoR1の発現を抑制すると(図5a参照)、globular AdipoによるPPARαリガンド活性(図5g参照)、脂肪酸酸化(図5h参照)及びグルコース取り込み(図5i参照)の増加が減少した。これに対して、AdipoR1の発現を抑制しても、全長Adipoによるこれらの作用の有意な減少は観察されなかった。したがって、AdipoR1は、globular Adipoによる筋細胞内のPPARαリガンド活性、脂肪酸酸化及びグルコース取り込みの増加を媒介していると考えられる。
内因性のAdipoR2が、筋細胞におけるAdipoの特異的結合及び代謝的作用を媒介しているか否かを調べるため、siRNAを用いてAdipoR2の発現を抑制した(図5b参照)。C2C12筋細胞において、siRNAによってAdipoR2の発現を抑制すると(図5b参照)、globular Adipo及び全長Adipoの結合が一部減少した(図5e,f参照)。さらに、AdipoR2の発現抑制によって、全長AdipoによるPPARαリガンド活性(図5g参照)及び脂肪酸酸化(図5h参照)の亢進が部分的に減少した。したがって、AdipoR2は、全長Adipoによる筋細胞内のPARαリガンド活性、脂肪酸酸化の亢進を部分的に媒介していると考えられる。図5eは、二本鎖siRNAをトランスフェクトしたC2C12筋細胞への[125I]globular Adipoの結合等温線(binding isotherm)を示し、図5fは、二本鎖siRNAをトランスフェクトしたC2C12筋細胞への[125I]全長Adipoの結合等温線(binding isotherm)を示す。なお、図中、□は無関係なコントロールsiRNA、●はマウスAdipoR2に対するsiRNA、▲はマウスAdipoR1及びマウスAdipoR2に対するsiRNAを用いた場合の結果を示す。
AdipoR1は、globular Adipoに対して比較的選択性を有する受容体であり、AdipoR2は全長Adipoに対して比較的選択性を有する受容体であると考えられる。しかしながら、いずれのタイプのAdipoRの機能的発現を抑制しても、globular Adipo及び全長Adipoの両者に対する有意な影響が観察される。これらの結果は、AdipoR1及びAdipoR2は、ホモマルチマー及びヘテロマルチマーの両者を形成しているという結果(図3e参照)によって説明され得る。
興味深いことに、C2C12筋細胞において、AdipoR1及びAdipoR2の両者の発現をsiRNAにより同時に抑制すると、globular Adipo及び全長Adipoの両者の結合がほとんど観察されなくなるとともに(図5e,f参照)、globular Adipo及び全長AdipoによるPPARαリガンド活性(図5g参照)及び脂肪酸酸化(図5h参照)の増加がほとんど観察されなくなった。
(7)肝細胞内の結合及び作用に対するsiRNAの影響
肝細胞へのAdipoの結合を調べた。肝細胞は全長Adipoに対して特異的な結合を示した(図6b参照)。AdipoR1及びAdipoR2の発現によって、globular Adipo及び全長Adipoの肝細胞への結合が促進した(図6a,b参照)。逆に、肝細胞において、siRNAによりAdipoR2の発現を抑制すると、全長Adipoの結合が大きく減少した(図6b参照)。これらの結果から、AdipoR1は、globular Adipoに対して比較的選択性を有する受容体であり、AdipoR2は全長Adipoに対して比較的選択性を有する受容体であると考えられる。
正常ヒト大動脈内皮細胞(Normal Human Aortic Endothelial Cells:HAEC)へのAdipoの結合を調べた。AdipoR1及びAdipoR2の両者の発現をsiRNAにより同時に抑制すると、globular Adipoの結合は大幅に減少し(図6e参照)、全長Adipoの結合は一部減少した(図6f参照)。この結果は、AdipoR1及びAdipoRが、HAECにおいてもAdipoに対する受容体であることを示す。
(8)スキャッチャードプロット解析
図5e及びfに関する実験を再度より詳細に行って得られた結果を図7a及びbに示す。図7aは図5eに対応する図であり、二本鎖siRNAをトランスフェクトしたC2C12筋細胞への[125I]globular Adipoの結合等温線(binding isotherm)を示し、図7bは図5fに対応する図であり、二本鎖siRNAをトランスフェクトしたC2C12筋細胞への[125I]全長Adipoの結合等温線(binding isotherm)を示す。なお、図7a及びb中、□は無関係なコントロールsiRNA、■はマウスAdipoR1に対するsiRNA、●はマウスAdipoR2に対するsiRNA、▲はマウスAdipoR1及びマウスAdipoR2に対するsiRNAを用いた場合の結果を示す。
図7a及びbに示す結果に基づきスキャッチャードプロット解析を行った結果をそれぞれ図7c及びdに示す。
無関係なsiRNAをトランスフェクトしたC2C12筋細胞は、全長Adipoよりもglobular Adipoに強く結合した(図7a及びb参照)。
スキャッチャードプロット解析によって、globular Adipoに対する2種類の結合部位、すなわち高アフィニティー結合部位(Kd値:約0.06μg/mL,1.14nM gAd三量体等量)と中アフィニティー結合部位(Kd値:約0.80μg/mL,14.4nM gAd三量体等量)が存在する一方(図7c参照)、全長Adipoに対する2種類の結合部位、すなわち中アフィニティー結合部位(Kd値:約6.7μg/mL,49.1nM Ad六量体等量)と低アフィニティー結合部位(Kd値:約329.3μg/mL,2415nM Ad六量体等量)が存在することが明らかとなった(図7d参照)。
siRNAによりAdipoR1の発現を抑制すると、globular Adipoの結合はかなり減少したが(図7a参照)、全長Adipoの結合はわずかに減少しただけだった(図7b参照)。AdipoR1の発現を特異的に抑制すると、globular Adipoに対する高アフィニティー結合部位の活性は失われるが、globular Adipoに対する中アフィニティー結合部位の活性は影響されないことが、スキャッチャードプロット解析によって明らかとなった(図7c参照)。さらに、AdipoR1の発現を特異的に抑制すると、全長Adipoに対する中アフィニティー結合部位の活性は部分的に減少するが、全長Adipoに対する低アフィニティー結合部位の活性は失われることが、スキャッチャードプロット解析によって明らかとなった(図7d参照)。
AdipoR1とは対照的に、siRNAによりAdipoR2の発現を抑制すると、全長Adipoの結合はかなり減少したが(図7b参照)、globular Adipoの結合はわずかに減少した(図7a参照)。AdipoR2の発現を特異的に抑制すると、globular Adipoに対する高アフィニティー結合部位の活性は部分的に減少するが、globular Adipoに対する中アフィニティー結合部位の活性が失われることが、スキャッチャードプロット解析によって明らかとなった(図7c参照)。さらに、AdipoR2の発現を特異的に抑制すると、全長Adipoに対する中アフィニティー結合部位の活性は失われるが、全長Adipoに対する低アフィニティー結合部位の活性は影響されないことが、スキャッチャードプロット解析によって明らかとなった(図7d参照)。
(9)AMPK、ACC、p38 MAPK及びMAPKのリン酸化及びリン酸化量
図8aのパネルは、AdipoR1をトランスフェクトした(図中「AdipoR1」と記載)又はしていない(図中「Mock」と記載)C2C12細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、当該細胞の溶解物と抗リン酸化AMPK抗体とを反応させた結果を示し、図8bのパネルは、当該細胞の溶解物と抗リン酸化ACC抗体とを反応させた結果を示し、図8cのパネルは、当該細胞の溶解物と抗リン酸化p38 MAPK抗体とを反応させた結果を示し、図8dのパネルは、当該細胞の溶解物と抗リン酸化MAPK抗体とを反応させた結果を示す。図8eのパネルは、AdipoR1をトランスフェクトした(図中「AdipoR1」と記載)又はしていない(図中「Mock」と記載)肝細胞を0.1μg/mL gAd又は1μg/mLとともに10分間インキュベートした後、当該細胞の溶解物と抗リン酸化AMPK抗体とを反応させた結果を示し、図8fのパネルは、当該細胞の溶解物と抗リン酸化ACC抗体とを反応させた結果を示す。図8a〜fの各パネルの下側のグラフは、パネルの各位置におけるリン酸化量を示す。なお、図中、「pAMPK」はリン酸化AMPK、「pACC」はリン酸化ACC、「pp38 MAPK」はリン酸化p38 MAPK、「pp44MAPK」はリン酸化p44MAPK、「pp42 MAPK」はリン酸化p42 MAPKを表す。
AdipoR1をトランスフェクトしていないC2C12筋細胞において、globular Adipo及び全長Adipoの両者がAMPK、ACC及びp38 MAPKのリン酸化量を増加させたが、MAPK等の他のプロテインキナーゼのリン酸化量は増加させなかった(図8a〜d参照)。C2C12細胞におけるAdipoR1の発現は、globular AdipoによるAMPK、ACC及びp38 MAPKのリン酸化の亢進に関連していた(図8a〜d参照)。このことは、AdipoR1がglobular AdipoによるAMPK及びp38 MAPK活性化を媒介することを示唆する。
AdipoR1をトランスフェクトしていない肝細胞において、全長AdipoはAMPK活性化及びACCリン酸化を亢進させたが、globular Adipoは亢進させなった(図8e及びf参照)。肝細胞におけるAdipoR1の発現は、globular Adipo及び全長AdipoによるAMPK及びACCリン酸化の亢進に関連していた(図8e及びf参照)。このことは、AdipoR1がglobular Adipo及び全長AdipoによるAMPK及びACCリン酸化を媒介することができることを示唆する。
AdipoR1をトランスフェクトしていない(図8中「Mock」と記載)C2C12筋細胞において、globular Adipoにより亢進される脂肪酸酸化及びグルコース取り込みは、優性阻害型(dominant negative:DN)AMPK又はp38 MAPK特異的阻害剤 SB203580(Barger,P.M.ら,J.Biol.Chem.276,44495−44501(2001);Puigserver,P.ら,Mol.Cell 8,971−982(2001);Michael,L.F.ら,Proc.Natl.Acad.Sci.USA.98,3820−3825(2001))によって部分的に阻害された(図8g及びh参照)。C2C12筋細胞におけるAdipoR1の発現(図8中「mAdipoR1」と記載)は、globular Adipoによる脂肪酸酸化及びグルコース取り込みを亢進させたが、これもDN−AMPK又はSB203580によって部分的に阻害された(図8g及びh参照)。したがって、AdipoR1を介したglobular Adipoによる脂肪酸酸化及びグルコース取り込みの亢進は、C2C12筋細胞におけるAMPK及びp38 MAPKの両者の経路に関連していると考えられた。
以上の結果は、本実施例で取得されたAdipoR1 cDNA(配列番号1,5)及びAdipoR2 cDNA(配列番号3,7)が、それぞれ生物学的機能を有するAdipoR1(配列番号2,6)及びAdipoR2(配列番号4,8)をコードすることを示す。
【産業上の利用の可能性】
本発明によって、アディポネクチン結合能を有する新規タンパク質、該タンパク質をコードする遺伝子、該遺伝子を含有する組換えベクター、該組換えベクターを含む形質転換体及び上記タンパク質に対する抗体が提供される。また、本発明によって、上記タンパク質、遺伝子、組換えベクター又は形質転換体を利用した、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストのスクリーニング方法及びスクリーニング用キットが提供される。
【配列表】
【図1】
【特許請求の範囲】
【請求項1】
以下の(a)又は(b)に示すタンパク質。
(a)配列番号2、4、6又は8に記載のアミノ酸配列からなるタンパク質
(b)配列番号2、4、6又は8に記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつアディポネクチン結合能を有するタンパク質
【請求項2】
請求項1記載のタンパク質をコードする遺伝子。
【請求項3】
以下の(c)又は(d)に示すDNAを含む請求項2記載の遺伝子。
(c)配列番号1、3、5又は7に記載の塩基配列からなるDNA
(d)配列番号1、3、5又は7に記載の塩基配列からなるDNAと相補的なDNAにストリンジェントな条件下でハイブリダイズし、かつアディポネクチン結合能を有するタンパク質をコードするDNA
【請求項4】
請求項2又は3記載の遺伝子を含む組換えベクター。
【請求項5】
請求項4記載の組換えベクターを含む形質転換体。
【請求項6】
請求項1記載のタンパク質に反応し得る抗体又はその断片。
【請求項7】
試験物質と請求項1記載のタンパク質とを接触させる工程を含む、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストのスクリーニング方法。
【請求項8】
請求項1記載のタンパク質、請求項2若しくは3記載の遺伝子、請求項4記載の組換えベクター又は請求項5記載の形質転換体を含む、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストのスクリーニング用キット。
【請求項1】
以下の(a)又は(b)に示すタンパク質。
(a)配列番号2、4、6又は8に記載のアミノ酸配列からなるタンパク質
(b)配列番号2、4、6又は8に記載のアミノ酸配列において、1又は複数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、かつアディポネクチン結合能を有するタンパク質
【請求項2】
請求項1記載のタンパク質をコードする遺伝子。
【請求項3】
以下の(c)又は(d)に示すDNAを含む請求項2記載の遺伝子。
(c)配列番号1、3、5又は7に記載の塩基配列からなるDNA
(d)配列番号1、3、5又は7に記載の塩基配列からなるDNAと相補的なDNAにストリンジェントな条件下でハイブリダイズし、かつアディポネクチン結合能を有するタンパク質をコードするDNA
【請求項4】
請求項2又は3記載の遺伝子を含む組換えベクター。
【請求項5】
請求項4記載の組換えベクターを含む形質転換体。
【請求項6】
請求項1記載のタンパク質に反応し得る抗体又はその断片。
【請求項7】
試験物質と請求項1記載のタンパク質とを接触させる工程を含む、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストのスクリーニング方法。
【請求項8】
請求項1記載のタンパク質、請求項2若しくは3記載の遺伝子、請求項4記載の組換えベクター又は請求項5記載の形質転換体を含む、アディポネクチン受容体に対するリガンド、アゴニスト又はアンタゴニストのスクリーニング用キット。
【図1】

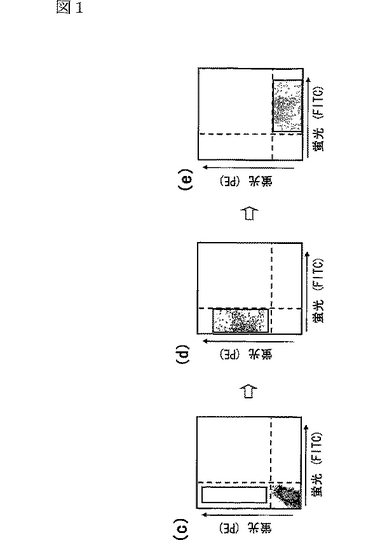
【図1】
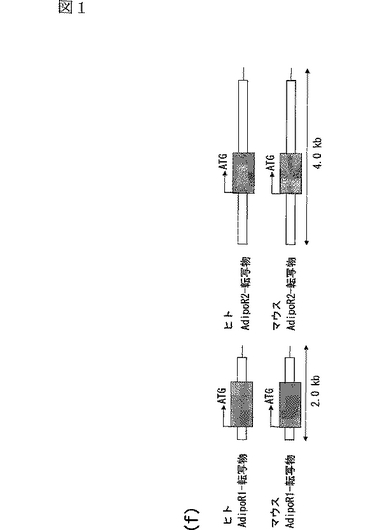
【図1】
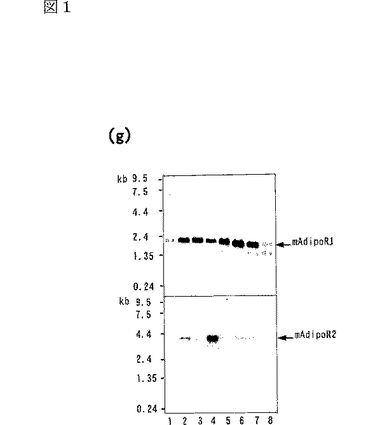
【図1】
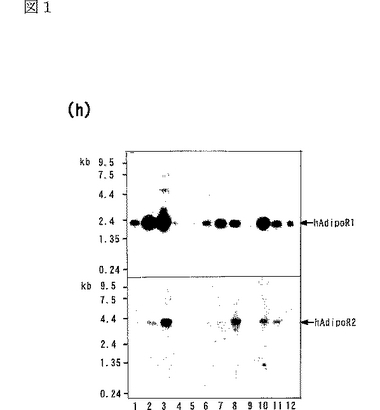
【図2】
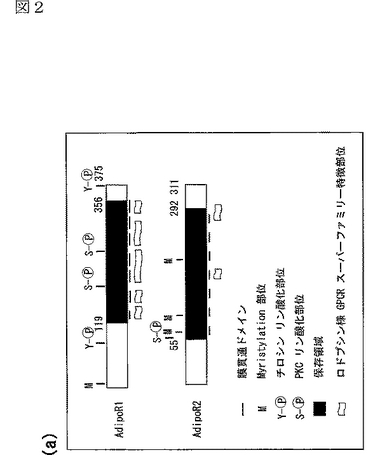
【図2】
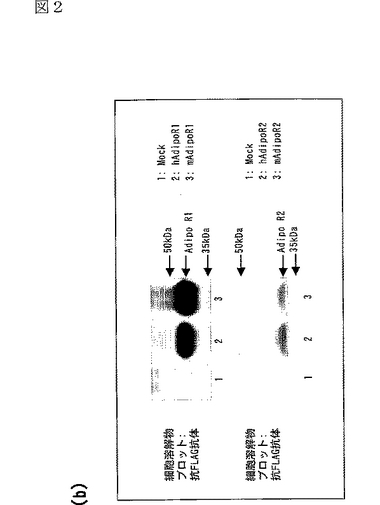
【図2】

【図2】
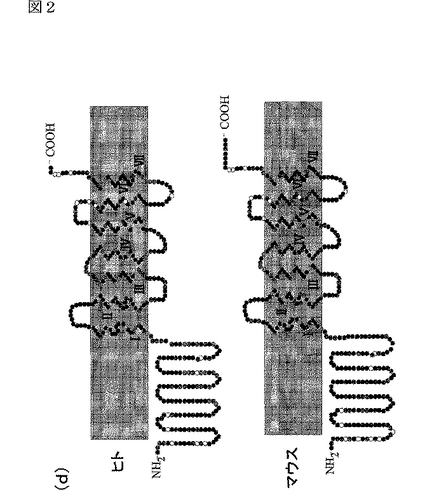
【図2】
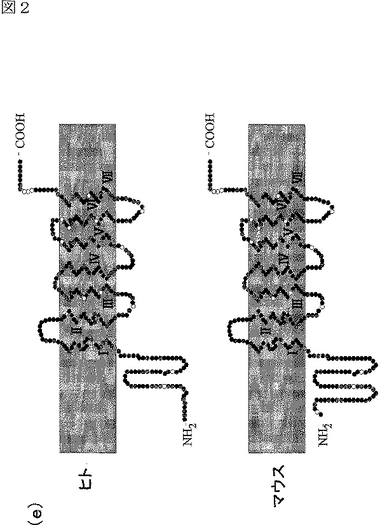
【図3】
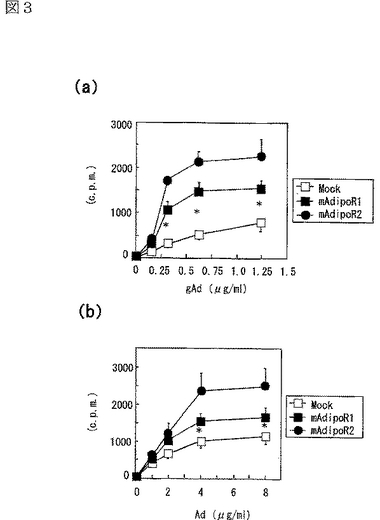
【図3】
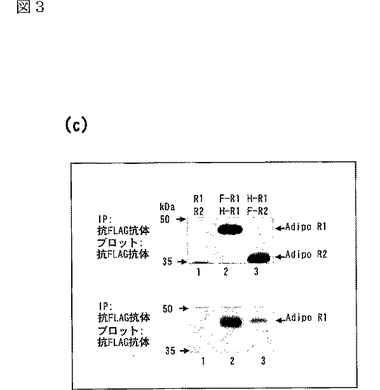
【図3】
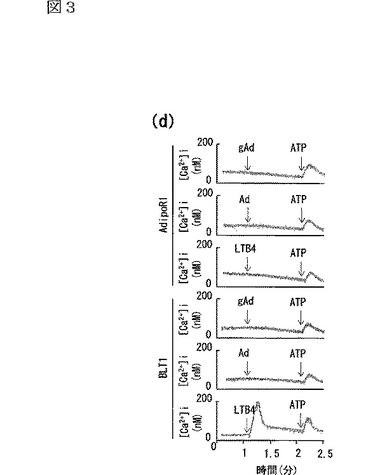
【図3】
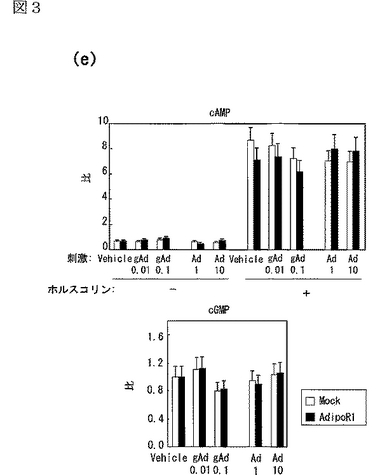
【図3】
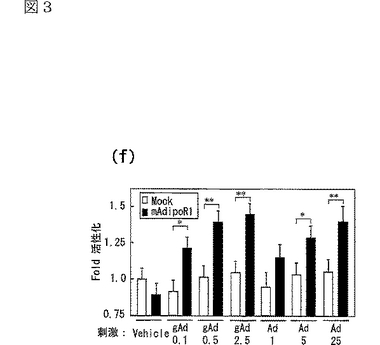
【図4】
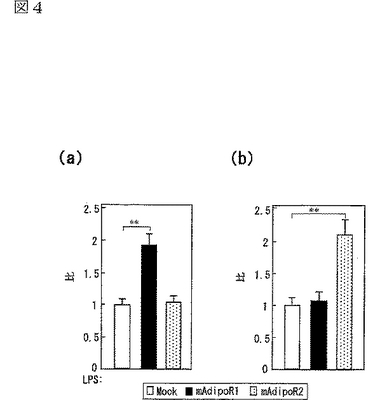
【図4】
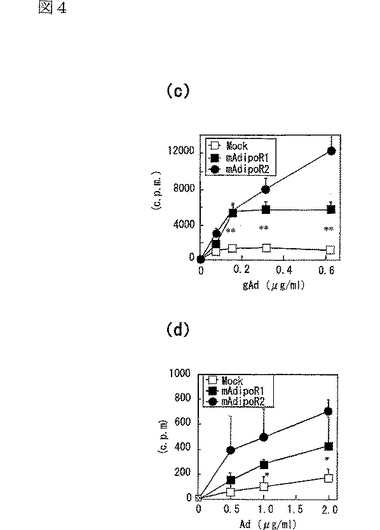
【図4】
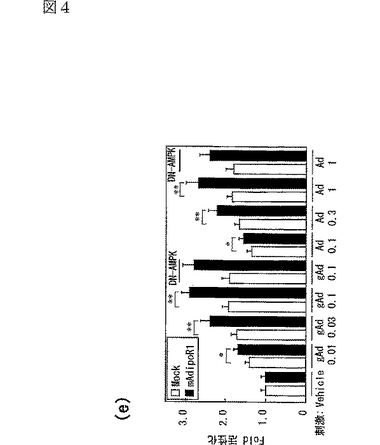
【図4】
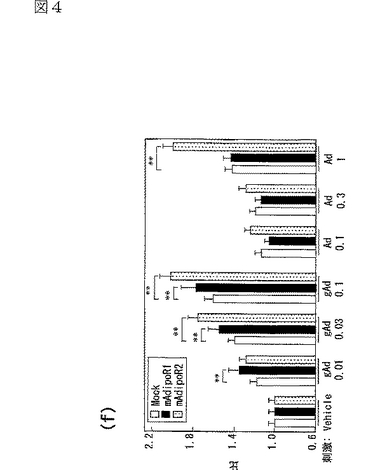
【図5】
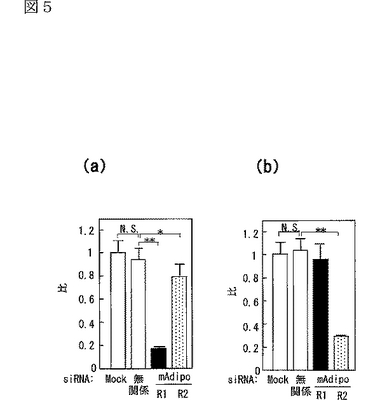
【図5】
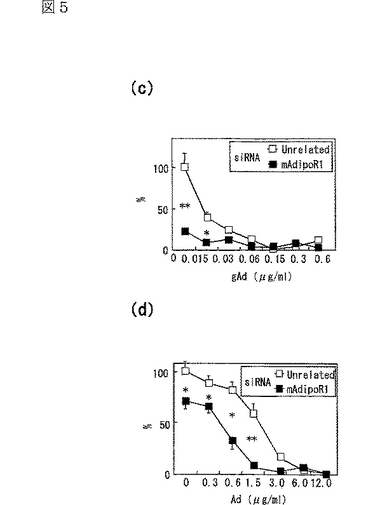
【図5】
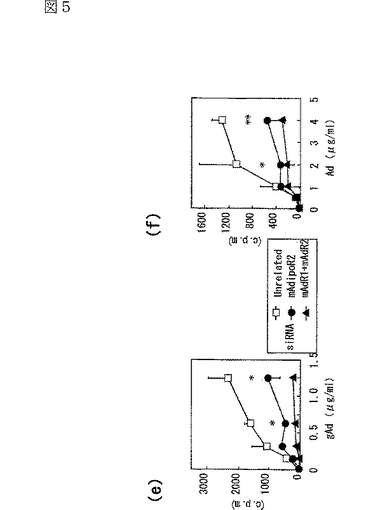
【図5】
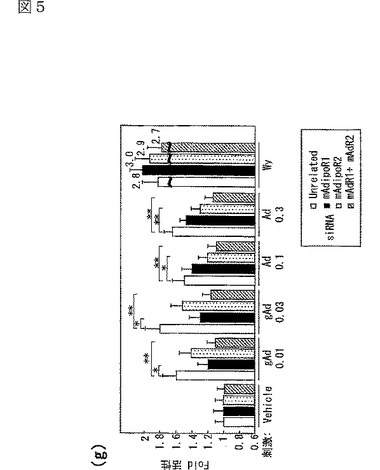
【図5】
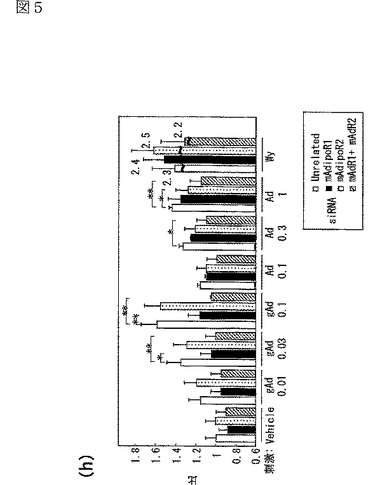
【図5】
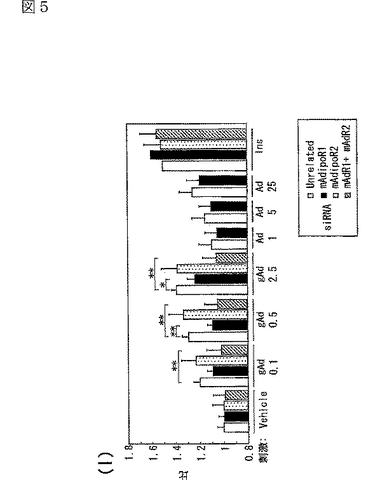
【図6】
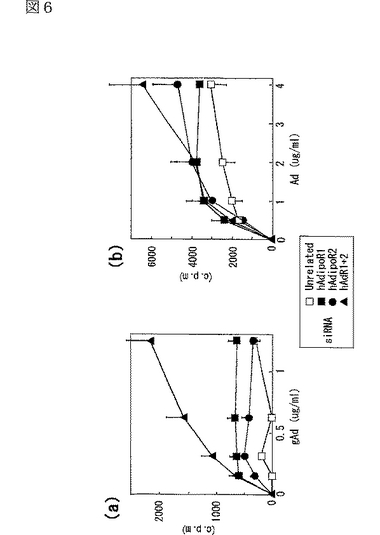
【図6】
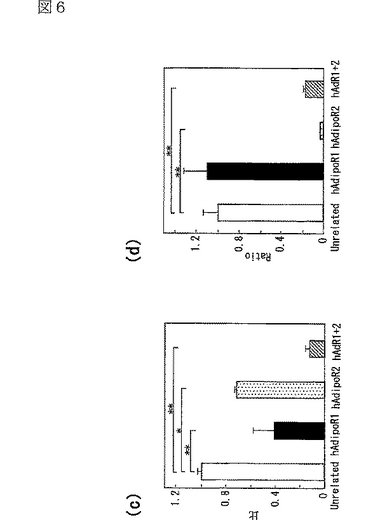
【図6】
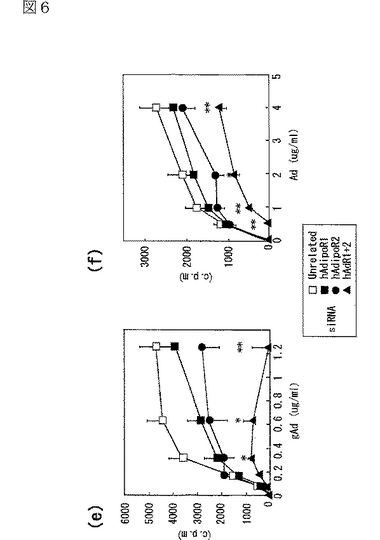
【図7】
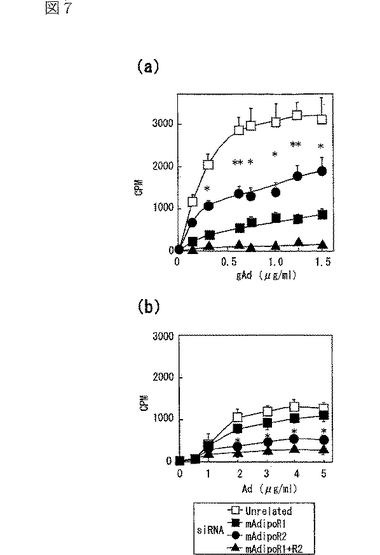
【図7】
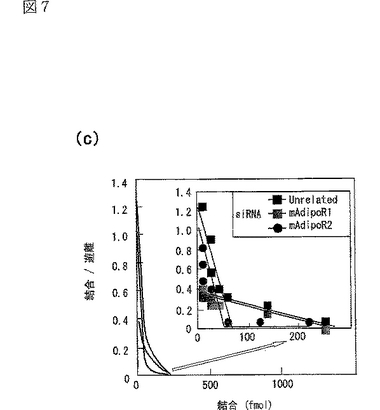
【図7】
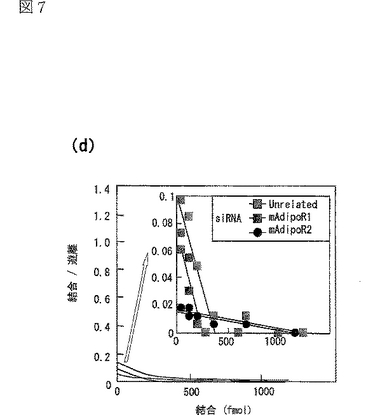
【図8】
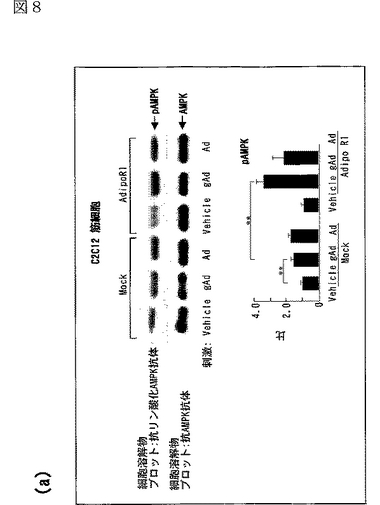
【図8】
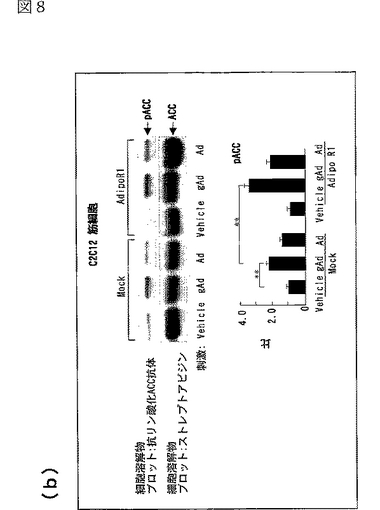
【図8】
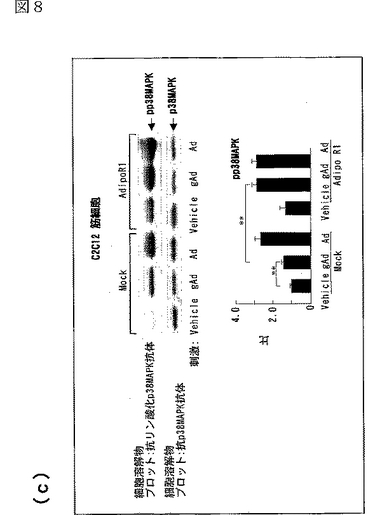
【図8】
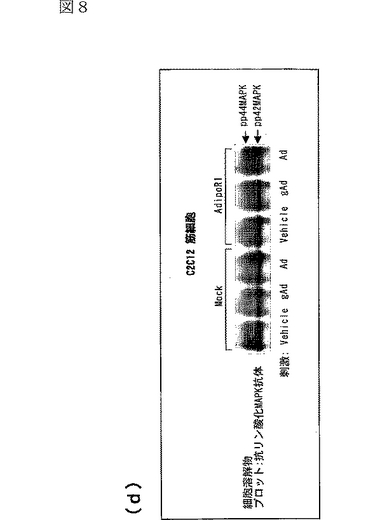
【図8】
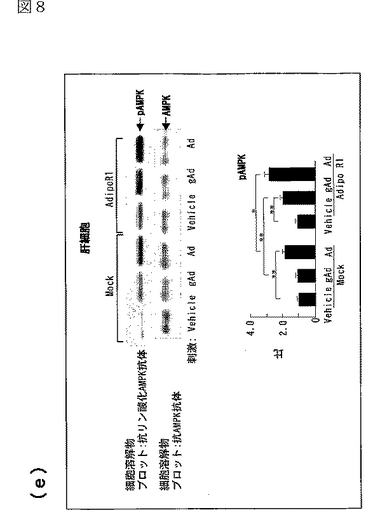
【図8】
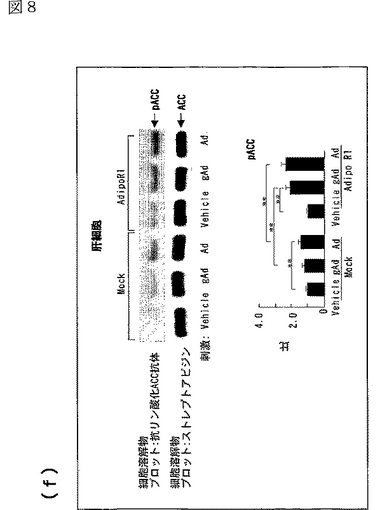
【図8】
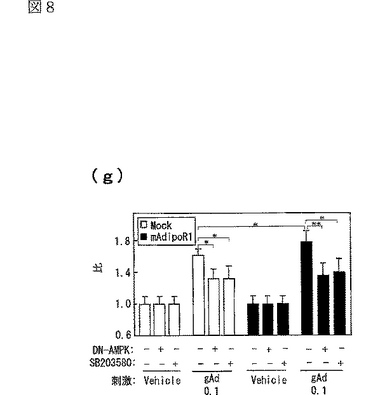
【図8】
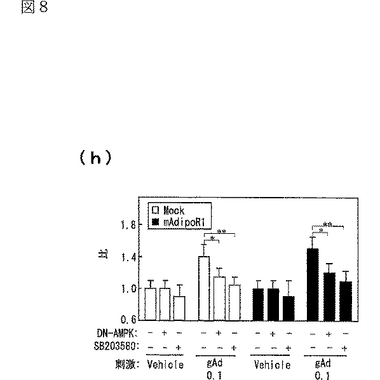
【図1】
【図1】
【図1】
【図2】
【図2】
【図2】
【図2】
【図2】
【図3】
【図3】
【図3】
【図3】
【図3】
【図4】
【図4】
【図4】
【図4】
【図5】
【図5】
【図5】
【図5】
【図5】
【図5】
【図6】
【図6】
【図6】
【図7】
【図7】
【図7】
【図8】
【図8】
【図8】
【図8】
【図8】
【図8】
【図8】
【図8】
【国際公開番号】WO2004/061108
【国際公開日】平成16年7月22日(2004.7.22)
【発行日】平成18年5月11日(2006.5.11)
【国際特許分類】
【出願番号】特願2004−564466(P2004−564466)
【国際出願番号】PCT/JP2003/007515
【国際出願日】平成15年6月12日(2003.6.12)
【出願人】(000003986)日産化学工業株式会社 (510)
【出願人】(899000024)株式会社東京大学TLO (50)
【Fターム(参考)】
【国際公開日】平成16年7月22日(2004.7.22)
【発行日】平成18年5月11日(2006.5.11)
【国際特許分類】
【国際出願番号】PCT/JP2003/007515
【国際出願日】平成15年6月12日(2003.6.12)
【出願人】(000003986)日産化学工業株式会社 (510)
【出願人】(899000024)株式会社東京大学TLO (50)
【Fターム(参考)】
[ Back to top ]