
アデニル酸シクラーゼタンパク質またはその断片に挿入されたヒトパピローマウイルスエピトープを有する組換えタンパク質、およびその治療的使用
本発明は、1個または数個のHPV抗原の1個または数個のエピトープを有する1個または数個のポリペプチドを含む組換えタンパク質に関し、該ポリペプチドは、アデニル酸シクラーゼ(CyaA)タンパク質またはその断片の同じまたは異なる許容部位に挿入されており、該CyaA断片は、該アデニル酸シクラーゼが抗原提示細胞を標的化するという特性を保持している。本発明はまた、該組換えタンパク質をコードするポリヌクレオチドにも関する。組換えタンパク質またはポリヌクレオチドは、HPV感染またはその悪性作用に対する治療手段の設計ために使用できる。
【発明の詳細な説明】
【技術分野】
【0001】
本出願は、アデニル酸シクラーゼタンパク質またはその断片に挿入されたパピローマウイルスエピトープを有する組換えタンパク質に関する。
【0002】
したがって、本発明は、アデニル酸シクラーゼ(CyaA)タンパク質がタンパク質ベクターとして作用して、パピローマウイルス抗原、特にヒトパピローマウイルス抗原に由来するエピトープに対する免疫応答を誘起するような、組換えタンパク質に関する。
【0003】
本発明は、特に、エピトープを、真核細胞、好ましくは哺乳動物細胞、特にヒト細胞に送達するための、このようにして得られたタンパク質性ベクターの使用に関する。
【0004】
本発明はまた、本発明の組換えタンパク質をコードするポリヌクレオチドと、前記ポリヌクレオチドを含むベクター、並びに、前記ポリヌクレオチドまたはベクターを含む宿主細胞にも関する。
【0005】
本発明はまた、宿主におけるヒトパピローマウイルス感染の処置または予防のための、並びに、宿主、特に哺乳動物宿主における、ヒトパピローマウイルス感染から生じる悪性作用の処置または予防のための、上記の組換えタンパク質またはポリヌクレオチドの適用にも関する。特定の実施形態において、本発明は、免疫療法、特にパピローマウイルスの腫瘍特異的抗原に対する免疫療法に適した化合物の設計に有用な手段を提供する。
【0006】
数多くのヒトパピローマウイルス(HPV)タイプの中で、ハイリスクHPVと称されるものは、宿主において感染が持続した場合の上皮悪性疾患の発達に関連している(1)。世界中で2番目に最も広まっている婦人科癌である子宮頚部癌(1)では、大抵、HPV16およびHPV18 DNAが検出される(>99%)(2)。これらのウイルスの発癌能は、初期遺伝子、すなわち、E6およびE7遺伝子(その発現は、ウイルスの複製全周期において検出され、悪性形質転換の発症および存続に必要である)により発現される産物に起因する。しかし、これらのハイリスク発癌性HPVタイプによる肛門性器感染が高い頻度で起こることと、HPV関連悪性疾患を最終的に発症する個体の比率は低いこととは対照的であり、このことは、免疫応答によりハイリスクHPV感染が制御されていることを示唆する。いくつかの観察により、例えば、大半の悪性段階以前の病変の自発的減退(1)、CD4+T細胞およびマクロファージによる退行中の性器いぼの浸潤(1)、並びに、免疫抑制患者および免疫欠陥患者においてより多数の感染被験者が認められることなどにより(1)、この所説が強められた。さらに、HPV16−E6および/またはE7エピトープに対するCD4+およびCD8+T細胞応答が、HPV16関連悪性疾患と診断された患者の血液(4〜8)、並びに、健康個体の血液に(9、10)検出された。まとめると、これらの考察には、HPV16のE6および/またはE7タンパク質に標的化する免疫療法の開発に対する強い根拠が含まれていた。
【0007】
H−2Db HPV16−E749〜57拘束性エピトープに対する免疫応答を発生させることにより、C57BL/6マウスにおけるHPV16−E6および−E7陽性腫瘍原性細胞系の腫瘍増殖を予防するための、多くのワクチン戦略が開発されている。これらのワクチン接種アプローチは、プラスミドDNA、ウイルス性または細菌性ベクター、キメラウイルス様粒子、合成ペプチド、および組換えタンパク質を含んでいる(11)。残念なことに、該アプローチは、臨床的退行に関しては控えめに満足できる結果しかもたらさなかった(3)。したがって、細胞性応答を誘起するために、HPV−エピトープを免疫系に標的化させる新規ツールを評価することは依然として感心が高い。
【0008】
したがって、宿主において前記抗原に対する体液性および/または細胞性免疫応答の誘起が可能である条件において、HPV抗原のエピトープを標的細胞に送達するのに適した新規ベクターが必要である。本発明者らは、アデニル酸シクラーゼタンパク質が、このようなベクターを設計する上で興味深いことを見出した。Bordetella pertussisのアデニル酸シクラーゼタンパク質を使用して様々な観察がなされ、これにより、このタンパク質が、このような効率的なベクターの設計に適した基盤となる可能性があるという結論が得られた。
【0009】
Bordetella pertussisのアデニル酸シクラーゼ(CyaA)は、その触媒ドメインを、真核細胞のサイトゾルへと送達することができる(12)。したがって、CyaAの触媒部位に挿入されたCD4+およびCD8+T細胞エピトープは、抗原提示細胞(APC)の表面において、それぞれMHCクラスIIおよびI分子により処理され提示される(13)。さらに、CyaAは、近年、αMβ2インテグリン(CD11b/CD18)に特異的に結合し(14、第WO02/22169号)、これにより、これらのT細胞エピトープをCD11b+樹状細胞亜個体群に標的化することが実証された(15)。マウスを、適切なエピトープを有する組換えCyaAで免疫化すると、強力なCTL応答、致死的なウイルス攻撃に対する完全な防御、並びに効率的な予防的および治療的抗腫瘍免疫が誘導された(16、17)。アデニル酸シクラーゼ(CyaA)タンパク質は特徴づけられており、組換えDNA技術によるその調製に関して、特に第WO93/21324号または第WO02/22169号に開示されている。第WO02/22169号には、Cyaが包含する残基373〜1706の断片は、CD11b/CD18受容体との相互作用に実質的に必要とされる構造を含むことが記載されている。
【0010】
より具体的には、残基1166からアミノ酸残基1281の範囲のアミノ酸配列は、CD11b/CD18受容体との相互作用のための決定基を含み、より特定すると、残基1208から残基1243の範囲のアミノ酸配列は、毒素とCD11b/CD18の相互作用に重要であることが後記されている(第EP03291486.3および45)。
【0011】
本発明者らは、今回、HPV抗原のエピトープを、宿主(HPV感染およびその悪性形質転換に罹患している宿主を含む)の標的細胞、特に抗原提示細胞(APC)に送達できる、該エピトープを有する、すなわち含む、組換えCyaAタンパク質の作成のための条件を決定し評価した。
【0012】
したがって、本発明は、特に、1個または数個のHPV抗原の1個または数個のエピトープを有する1個または数個のポリペプチドを含む組換えタンパク質に関し、該ポリペプチドは、アデニル酸シクラーゼ(CyaA)タンパク質またはその断片の同じまたは異なる許容部位に挿入されており、該CyaA断片は、APC、特にCD11b/CD18細胞、例えば樹状細胞などのCyaAの標的細胞に標的化させるという、該アデニル酸シクラーゼタンパク質の特性を保持している。特定の実施形態において、この断片はまた、そこに挿入されたエピトープまたは該エピトープを含むポリペプチドの、標的細胞のサイトゾルへの転位を可能とするようなCyaAの特性を保持している。標的細胞のサイトゾルへのエピトープまたは該エピトープを含むポリペプチドの転位は、CyaA断片が、その触媒ドメインの転位を可能とするタンパク質ドメインを保持している場合に許容される。
【0013】
本発明の組換えタンパク質は、組換え技術を頼りにして調製できる。それはまた、合成、特に化学合成によっても得ることができる。したがって、「組換えタンパク質」なる語句は、キメラ形態のタンパク質を意味する。
【0014】
組換えタンパク質のCD11b/CD18細胞への標的化能は、特に、第EP03291486.3および(45)または第WO02/22169号に開示された方法にしたがってアッセイできる。さらに、組換えタンパク質の、該エピトープまたは該エピトープ含有ポリペプチドを標的細胞のサイトゾルへと転位する能力は、第WO02/22169号に記載の方法を適用することによりアッセイできる。
【0015】
特定の実施形態において、CyaA断片は、天然ではCyaA中で連続していない、CyaAの2個の異なる部分から構成されることができる。一例として、CyaAの触媒ドメイン、すなわち、CyaAのN末端部分の400アミノ酸残基、および、CD11b/CD18抗原提示細胞の標的化に必要とされるアミノ酸残基1208〜1243を含む断片を挙げることができる。
【0016】
先の定義において、「ポリペプチド」なる表現は、翻訳後修飾を受けたアミノ酸配列、特に、少なくとも6個のアミノ酸残基を有するアミノ酸配列、および、特に5〜500残基または約5〜約100、または約5〜約200、または約10〜約50残基、または約30もしくは約50〜200残基、または約100〜約210、または約100〜約200残基を有するアミノ酸配列を含む任意のアミノ酸配列(ただし、該アミノ酸配列は少なくとも1個のエピトープを含む)、すなわち、標的細胞、有利には宿主、特に哺乳動物宿主の標的細胞に送達した後に免疫応答を発生させることのできるアミノ酸配列を記載する。したがって、この定義によるポリペプチドは、エピトープ、さらには独特なエピトープに限定されても、または、数個の異なるもしくは同じエピトープを含んでいても、または、病原体の、すなわちヒトパピローマウイルスの全長抗原を包含していてもよい。本発明のエピトープは、体液性免疫応答および/または細胞性免疫応答、特にT細胞免疫応答に関与するアミノ酸配列を包含する。したがって、本発明の組換え分子のポリペプチド中のエピトープは、宿主においてAPC(抗原提示細胞)により処理されるもの、特に、クラスI MHC(主要組織適合性複合体)分子と会合して認識されるもの、例えば標的細胞がCD8+Tリンパ球であるエピトープ、またはクラスII MHC分子と会合して認識されるエピトープ、例えば標的細胞がCD4+Tリンパ球細胞であるものが挙げられる。
【0017】
特定の実施形態において、エピトープを有するポリペプチドは、様々な抗原に由来する、特に様々なHPV株の1タイプの抗原に由来する、または、様々なHPV株の数タイプの抗原に由来する、数個のエピトープを含む。したがって、HPV抗原に由来するポリペプチドは、多価、特に二価または三価であることができ、すなわち、数個の抗原に対する免疫応答が可能となる。
【0018】
本発明によれば、HPV抗原(これから1個または数個のエピトープを有するポリペプチドを設計できる)は、好ましくは、特にHPV感染後の悪性作用の発症および/または存続に関与するタンパク質に由来するものであり、いわゆる腫瘍抗原、すなわち、宿主において免疫応答を誘起し、宿主において抗体またはT細胞と特異的に反応できる、HPV感染に関連した腫瘍発達に関連した抗原を包含する。
【0019】
本発明に記載のエピトープを有するポリペプチドは、HPVの天然または成熟抗原から得られることができ、これには、抗原完全体を使用することにより、あるいは、全タンパク質ではなくむしろ、断片、特に抗原性断片、特に該抗原のエピトープを選択することにより、あるいは、特に、組換え分子中のCyaAタンパク質と会合させた場合に、宿主における免疫応答を誘導または誘起する能力を高めるために、該抗原またはその選択された抗原部分またはエピトープを改変することによることが含まれる。したがって、このようなエピトープの様々な可能な改変を説明するために、これらのポリペプチドは、抗原(これからエピトープが得られる)の天然または非天然フランキング配列によりフランキングされたエピトープを包含し、また、その免疫特性を高めるために化学的に改変されているエピトープまたはエピトープを含むアミノ酸配列を包含する。これらの改変は、CyaAタンパク質に関連して得られるポリペプチドの有効性を向上するために有益である可能性がある。
【0020】
いくつかの特定の改変が、本明細書で以後に例として開示されており、特に追加の陽性荷電のアミノ酸残基の挿入によるポリペプチド荷電の変化を包含する改変が含まれる。
【0021】
したがって、本発明のポリペプチドはまた、半合成または合成ポリペプチドも包含する。
【0022】
特定の実施形態によれば、HPV抗原に由来するポリペプチドは、各々または一緒になって、約5〜約500、または約5〜約100、または約5〜約200、例えば約10〜約50アミノ酸残基、または約30もしくは約50〜約200アミノ酸残基、または約100〜約210、または約100〜約200アミノ酸残基を含む。
【0023】
ポリペプチドは、特に、本発明の組換えタンパク質中で組換えられた場合に、抗原特異的応答を誘起できるように選択される。
【0024】
本発明の組換えタンパク質は、特に、分断された天然HPV抗原に存在するポリペプチドまたは数個のポリペプチドを含むように設計することができ、ここで、該分断は、該HPV抗原の酸性領域にける1個または数個のアミノ酸残基の欠失、および/または、アデニル酸シクラーゼの少なくとも2箇所の許容部位における該HPV抗原の少なくとも2個のポリペプチド断片の挿入からなる。
【0025】
この定義に包含される特定の分断は、アデニル酸シクラーゼの少なくとも2箇所の許容部位への、天然HPV抗原の少なくとも2個の断片の挿入により得られ、ここで、これらの少なくとも2個の断片は、天然抗原におけるその天然の位置と比べて逆であり、すなわち、天然抗原においてよりN末端側である断片は、CyaAタンパク質またはその断片に挿入されると、よりC末端側となり、その逆もある。
【0026】
アミノ末端断片およびカルボキシ末端断片の逆転は、E7Δ断片(すなわち、E7抗原の断片)で示されるように、特に癌免疫療法において、強力で長く続く防御免疫を誘導する際により効果的である可能性があることが観察されている。
【0027】
本発明によれば、アデニル酸シクラーゼ(CyaA)は、先に開示したように、全長タンパク質として、またはその断片として使用される。
【0028】
有利には、CyaAタンパク質またはその断片は、細胞、特に組換え細胞における、cyaA遺伝子およびcyaC遺伝子の両方の同時発現の結果である、タンパク質またはその断片である。実際に、標的細胞に対する侵襲性特性を有するために、CyaAは、cyaA遺伝子およびcyaC遺伝子の両方の発現により可能となる翻訳後修飾を受けている必要があることが示されている(第WO93/21324号)。
【0029】
本発明の特定の実施形態において、CyaAタンパク質の断片は、少なくとも約30個のアミノ酸残基を有する断片であり、約1300個まで、特に約500個までのアミノ酸残基、好ましくは約50〜約150個のアミノ酸残基を有することができ;該断片は、特定の実施形態において、CD11b/CD18標的細胞との相互作用のために、CyaAのアミノ酸残基1166〜1281、または、CyaAタンパク質のアミノ酸残基1208〜1243を含む。したがって、1個の具体的な断片は、天然タンパク質のC末端部分の全部または一部を包含し、この部分は、細胞膜および/またはCD11b/CD18受容体を標的化するための該タンパク質の結合に、および、細胞サイトゾルへのポリペプチド群に含まれるエピトープのその後の送達に関与している(12)。本発明に記載のCyaAタンパク質の1個の具体的な断片は、CyaAタンパク質のアミノ酸残基372〜1706を含む。別の具体的な断片は、アミノ酸残基225〜234が欠失しており、これにより1〜244および235〜1706を含むCyaA断片を与えるような、CyaAタンパク質に相当するものである。
【0030】
本発明の特定の実施形態において、アデニル酸シクラーゼタンパク質は、細菌性タンパク質である。好ましい実施形態において、CyaAタンパク質は、Bordetella種に由来している。
【0031】
本発明に記載の対象のBordetella種の中で、その1つが、Bordetella pertussisである。他の対象のBordetella株は、Bordetella parapertussisまたはBordetella bronchisepticaの株である。B.parapertussisのCyaAタンパク質の配列は、特にアクセッション番号NC002928.3(1740アミノ酸の配列として)およびParkhill J.ら(Nat.Genet.DOI、10(2003)に開示されており、B.bronchisepticaについては、Betsou Fら(Gene 1995、8月30日;162(1):165-6)に開示されている。
【0032】
Bordetella pertussisは、百日咳の原因物質であり、とりわけ、細菌の決定的な病原性因子であり、B.pertussis感染に対して防御性作用のある抗原の1つである、公知の百日咳毒素(PT)およびアデニル酸シクラーゼ毒素(CyaA)を含む数種の毒素を分泌している。
【0033】
Bordetella pertussisのアデニル酸シクラーゼタンパク質は、400アミノ酸残基のN末端触媒ドメインと、毒素が標的細胞膜に結合することとその後の細胞サイトゾルへの触媒部分の送達に関与している1306残基のC末端部分とを含む、1706残基の二機能性タンパク質として記載されている毒素である(12)。
【0034】
CyaAタンパク質は、不活性プロ毒素として合成され、これは、2個の内部リジン残基(リジン860および983)の翻訳後パルミトイル化により活性毒素へと変換される。この翻訳後修飾は、cyaA遺伝子と共に、アクセサリー遺伝子、すなわち、B.pertussis染色体上のcyaAの近くに位置するcyaCの発現が必要とされる。
【0035】
Bordetella pertussisのcyaAは、Glaser,P.ら、1988、Molecular Microbiology2(1)、19-30により、アミノ酸配列およびヌクレオチド配列として記載されている。したがって、B.pertussisのCyaAタンパク質のアミノ酸残基もしくは配列またはヌクレオチドもしくはヌクレオチド配列が本発明において引用される場合、その位置は、Glasserら、1988の該刊行物に開示された配列を参考にして示されている。
【0036】
本発明に記載の組換えタンパク質では、1個または数個のHPV抗原の1個または数個のエピトープを有するポリペプチドが、CyaAタンパク質の1個または数個の許容部位に挿入されている。
【0037】
本発明では、「許容部位」は、実質的にCyaAタンパク質の機能特性に影響を及ぼすことなく、特に実質的に細胞の標的化に、特にCyaAによるAPCの標的化に影響を及ぼすことなく(実質的にCD11b−CD18受容体への特異的結合に影響を及ぼすことなく、有利には、実質的に、標的細胞へのエピトープの転位の過程に関与するタンパク質のドメインに影響を及ぼすことなくを含む)、ポリペプチドを挿入できる、CyaAタンパク質の配列部位である。
【0038】
CyaA触媒ドメインの転位を可能とし、これによりこのような許容部位に挿入されたエピトープの転位を可能とする、Bordetella pertussisアデニル酸シクラーゼの許容部位には、残基137〜138(Val−Ala)、残基224〜225(Arg−Ala)、残基228〜229(Glu−Ala)、残基235〜236(Arg−Glu)、および残基317〜318(Ser−Ala)が挙げられるがこれに限定されない((44)Seboら、1995)。以下の追加の許容部位もまた、本発明の実施形態に含まれる:残基107〜108(Gly−His)、残基132〜133(Met−Ala)、残基232〜233(Gly−Leu)、335〜336(Gly−Gln)および336〜337(43)。
【0039】
他のBordetella種における対応する許容部位は、配列の比較および対応する残基の決定により定義できる。
【0040】
別の実施形態によれば、該ポリペプチドは、追加的にまたは代替的に、CyaAタンパク質またはその断片の一端および/または他の末端に挿入してもよい。
【0041】
本発明の目的のために使用するCyaAタンパク質の特定の断片は、1300個までのアミノ酸または約30〜約500個のアミノ酸残基、有利には約50〜約150個のアミノ酸残基を含むもの、特に、天然CyaAタンパク質のアミノ酸残基1166〜1281、有利には天然CyaAタンパク質の1208〜1243を包含するような断片である。
【0042】
したがって、本発明によれば、「ハイブリッドタンパク質」とも呼ばれるいわゆる組換えタンパク質をもたらすCyaAタンパク質へのポリペプチドの「挿入」は、特に、利用可能なDNA技術による遺伝子的挿入を包含する。または、「挿入」はまた、非遺伝子的挿入も包含され、これには、例えばCyaAもしくはその断片の一端において実施する共有結合、または非共有結合などの化学的挿入が含まれる。非遺伝子的挿入は、挿入するポリペプチドが合成または半合成である場合には特に関心が高い可能性がある。薬物をポリペプチドに結合させる方法は、当該技術分野において公知であり、例えば、N−ピリジルスルホニル活性化スルフヒドリルを使用するなどのジスルフィド連結が含まれる。
【0043】
特に、インビボで、Cyaの標的細胞へ、例えばACP、例えばCD11b/CD18細胞、特に該細胞のサイトゾルへと標的化させるために、化学的連結または遺伝子的挿入により、本発明のポリペプチドを特に含む分子を、CyaAに融合させることが可能である。実際に、所与のCD8+T細胞エピトープに相当する分子を、解毒したCyaAの触媒ドメインに、ジスルフィド結合または遺伝子的挿入により結合させると、遺伝子工学された分子は、インビボで特異的CTL応答を誘起することができ、これにより、該CD8+T細胞エピトープは、CD11b発現細胞のサイトゾルへと転位されたことが示される。
【0044】
特定の実施形態において、タンパク質性ベクターの製造に使用される組換えアデニル酸シクラーゼは、ジスルフィド結合により、該アデニル酸シクラーゼの触媒ドメイン内に位置する遺伝子的に挿入されたシステイン残基に化学的に結合されている本発明のポリペプチドを特に含む、システイン残基含有の1個以上の分子の挿入により特に改変されている、CyaAまたはその断片である。
【0045】
実際に、本発明のポリペプチドを特に含む複数の分子を、ジスルフィド結合により、触媒ドメイン内の様々な許容部位に位置する様々なシステイン残基に化学的に結合させることができる。
【0046】
宿主において免疫応答、特に細胞性免疫応答を誘起できる製品の設計に適した組換えタンパク質を提案するために、また特に、HPVに感染した宿主に観察される悪性作用に対する免疫応答を誘起できるような製品を設計するために、本発明者らは、高度に発癌性のHPV株、特にHPV16、HPV18、HPV31、HPV33、HPV35、HPV45、HPV52、またはHPV58から選択される株に由来する抗原に由来するエピトープを有するポリペプチドを得ることを提案した。
【0047】
これらの株の中で、HPV18およびHPV16が特に関心が高い。HPV16は、特に、哺乳動物宿主、特にヒトにおける子宮頚部癌の発症に関連しているために、HPVに感染した宿主の処置のための特定の標的である。
【0048】
これらのHPV株から出発して、本発明者らは、L1、L2、E1、E2、E4、およびE5タンパク質から選択される抗原に由来するエピトープを有するポリペプチドを得ることを提案する。
【0049】
代替的にまたは組み合わせて、本発明者らはまた、HPVのE6またはE7タンパク質に由来するエピトープを有する該ポリペプチドを得ることを提案する。
【0050】
本発明の特定の実施形態において、HPV16のE6もしくはE7タンパク質またはHPV18のE6もしくはE7タンパク質が、エピトープを有するポリペプチドの設計に使用される。
【0051】
HPV抗原に由来するポリペプチドの設計のために言及できる特定のHPVタンパク質は、HPVのE7タンパク質、特にHPV16またはHPV18のE7タンパク質である。本発明の実施形態によれば、該ポリペプチドは、様々なHPV株、特にHPV16およびHPV18の数個のE7タンパク質から得られる。例えば、該ポリペプチドは、HPV16およびHPV18の全長E7タンパク質、または、HPV16もしくはHPV18の各々のE7タンパク質の1個もしくは数個の断片(該断片の多量体、特に二量体を含む)から得られる。
【0052】
これらのHPVタンパク質およびそのアミノ酸およびヌクレオチド配列は、HPV16についてはSeedorf,Kら(ヒトパピローマウイルスタイプ16DNA配列、Virology、145:181-185、1985)、Cole S.T.、Danos O.(ヒトパピローマウイルスタイプ18ゲノムのヌクレオチド配列および比較解析。パピローマウイルスの系統発生並びにE6およびE7遺伝子産物の反復構造、J.Mol.Biol.193:599-606(1987))またはFernando GJら(子宮頚部癌に関連したヒトパピローマウイルスタイプ18(HPV18)のE7形質転換タンパク質のTヘルパーエピトープ、Virus Res.1995 Apr.36(1):1-13)に開示されている。
【0053】
E6およびE7タンパク質は、ウイルスの複製全周期を通じて特にHPV16またはHPV18により発現されている癌タンパク質であり、HPV株による感染後の宿主細胞の悪性形質転換の発症および存続に必要とされることが示されている。それ故、これらの両方の腫瘍特異的抗原が、養子CTL媒介免疫療法の可能性ある標的と考えられている。
【0054】
本発明の特定の実施形態によれば、該組換えタンパク質は、複数のポリペプチドを含み、その各々が、1個または数個のHPV抗原の1個または数個のエピトープを有する。
【0055】
例えば、このような複数のポリペプチドは、1個のHPV株、特にHPV16またはHPV18のE6およびE7タンパク質から得ることができる。別の例によれば、これらの複数のポリペプチドは、HPV16およびHPV18の両方に由来する、E6またはE7タンパク質から得られるエピトープを包含することができる。
【0056】
複数のポリペプチドはまた、対象のCyaAタンパク質の異なる許容部位に挿入された、1個のタンパク質、例えばE7またはE6タンパク質の断片群を有する様々なエピトープからなることができる。
【0057】
先の定義に記載の別の特定の組換えタンパク質は、エピトープを有する複数のポリペプチドが、HPV16のE7タンパク質の残基1〜29を含む断片、すなわち残基1〜29からなる断片、または残基42〜98を含む断片、すなわち残基42〜98からなる断片を包含するか、あるいは、複数のポリペプチドが、CyaAタンパク質の異なる許容部位に挿入された両方の断片を含むかまたはからなる、CyaA組換えタンパク質である。
【0058】
本発明に記載の別の組換えタンパク質は、複数のポリペプチドが、アミノ酸配列
【表2】

を有する断片を包含するようなタンパク質である。
【0059】
CyaAタンパク質の許容部位に挿入されたポリペプチドのアミノ酸残基の数は、全長抗原、特にHPVの全長E6またはE7タンパク質からなるポリペプチドを、CyaAタンパク質またはその断片に挿入できるようなものである。
【0060】
本発明の特定の実施形態によれば、組換えCyaAに含まれるポリペプチドは、CyaAのコドン224と235の間またはCyaAのコドン319と320の間に挿入された、E7タンパク質、特にHPV16のE7タンパク質である。
【0061】
別の実施形態において、本発明の組換えタンパク質は、複数のポリペプチド(そのいくつかは、1個または数個のHPVの1個または数個のエピトープを有するポリペプチドである)および他の病原体のエピトープを有する他のポリペプチドを含む。
【0062】
別の特定の実施形態において、本発明の組換えタンパク質はさらに、異なる病原性物質から派生した1個または数個のエピトープを含む。ChlamydiaまたはHIVレトロウイルス、またはHPV、HBV、HCV、アデノウイルスEBV、ヘルペスウイルス、HTLV.1ウイルスおよびCMVから派生したエピトープと、HPVから派生したエピトープとの会合は、特に関心が高い場合がある。
【0063】
本発明の別の特定の実施形態によれば、エピトープを有するポリペプチドは、例えば、配列内の負に荷電したアミノ酸残基の数を減少させるために、その天然アミノ酸配列と比べて改変されている。このような改変は、これらの負に荷電したアミノ酸残基のいくつかを除去することにより、または、いくつかの正に荷電したアミノ酸残基を、特にエピトープのフランキング残基として付加することによっても得ることができる。このようにより少ない数の負に荷電した残基を含むポリペプチドは、標的細胞のサイトゾルへのCyaAタンパク質の触媒ドメインの転位に好ましくあり得る。
【0064】
エピトープを有するポリペプチドはまた、CyaAに挿入される場合に折りたたまれないようにするように設計でき、これにより、標的細胞への組換えCyaAタンパク質の内部移行の効率が向上する。その含有アミノ酸の結果として折りたたみを受けたポリペプチド中のこのような折りたたみ解除は、例えば、システイン残基を除去または置き換えて、ポリペプチドの折りたたみに関与している可能性があるジスルフィド結合の形成を避けることにより達成することができる。いくつかの場合において、インビボでの再折りたたみを回避することのできる還元剤の存在下でそれらを調製することによりポリペプチドの折りたたみを防ぐことが可能である。
【0065】
特定の実施形態において、エピトープを有するポリペプチドは、潜在性エピトープであってもよい。
【0066】
本発明の特定の態様において、本発明者らは、実際に、(i)先に開示した定義に記載のアデニル酸シクラーゼ(CyaA)またはその断片と、(ii)1個または数個の抗原の1個または数個の抗原性断片を有するポリペプチドとを含む組換えタンパク質からなる、キメラタンパク質構造物は、組換え構造物における該抗原の潜在性エピトープを、それを提示する結果、免疫原性とすることができることが決定された。特に、特に治療目的(ワクチン接種目的を含む)のための、本発明に定義したCyaAまたはその断片と対象の抗原から得られたポリペプチドとを含む該キメラ構造物は、免疫原性とさせることができ、特に、宿主におけるT細胞応答、特にCTL応答を発生させることのできる、抗原の潜在性エピトープを含むことができる。
【0067】
したがって、本発明はまた、1個または数個の抗原の1個または数個のエピトープを有する1個または数個のポリペプチドを含む組換えタンパク質に関し、該ポリペプチドは、アデニル酸シクラーゼ(CyaA)またはその断片の同じまたは異なる許容部位に挿入されており、該CyaA断片は、抗原提示細胞を標的化するという該アデニル酸シクラーゼタンパク質の特性を保持しており、該エピトープの少なくとも1個は、亜優性潜在性T細胞エピトープ(subdominant cryptic T-cell epitope)であり、該組換えタンパク質は、該ポリペプチドに対する抗原特異的応答を誘起できる。
【0068】
特に、潜在性エピトープは、HPV抗原、特にHPV16および/またはHPV18抗原、特にE7抗原内に含まれる。
【0069】
このように定義した組換えタンパク質は、特に、HPV18E7タンパク質から得られたペプチド、すなわち、アミノ酸配列IDGVNHQHLを有するペプチドを含む。
【0070】
特定の実施形態によれば、潜在性エピトープは改変でき、例えば、最初の2箇所の位置において置換を有することができ、例えば配列ASGVNHQHLを有することができる。
【0071】
本発明は、特に、ペプチドIDGVNHQHLに関する。
【0072】
本発明はまた、この配列に置換を有するペプチド、特に1位および/または2位に置換を有するペプチド、特に配列ASGVNHQHLを有するペプチドに関する。
【0073】
本発明はまた、免疫原性特性を有する限り、特にT細胞を誘起できる限り、特にCTL応答を誘起できる限りにおいて、該ペプチドの変種を含む。
【0074】
有利には、本発明の組換えタンパク質を調製するために、CyaAタンパク質の酵素活性、すなわち、ATPからcAMPへと変換する能力は、不活性化されている。このような不活性化は、遺伝子不活性化の結果として得ることができる。一例として、遺伝子不活性化は、ジペプチドを、触媒部位の一部であるCyaAのアミノ酸配列の部位(例えば188と189の間)に導入する結果として得ることができる。このような不活性化CyaAタンパク質は、以下の実施例に実例を示す。
【0075】
本発明の組換えタンパク質は、有利には、細胞性免疫応答を誘起できる。該応答は、CTLおよびTh、特にTh1応答(CD4+T細胞応答および/またはCD8+T細胞応答を含む)を含む。
【0076】
組換えタンパク質がこの細胞性免疫応答を誘起できる能力は、特に、インビボにおいて腫瘍増殖を抑止するのにまたはさらには動物における腫瘍減退を可能とするのに十分であることが示された。
【0077】
本発明はまた、先に定義した組換えタンパク質をコードするポリヌクレオチドにも関する。
【0078】
本発明のポリヌクレオチドを発現ベクターに挿入して、本発明の組換えタンパク質の発現に適した組換え発現ベクターを提供できる。このような発現ベクターには、プラスミド、コスミド、ファージミド、ウイルスベクターが挙げられる。
【0079】
組換えベクターは、原核細胞、特に細菌における発現に適したものでも、または、真核細胞、特に哺乳動物細胞、有利にはヒト細胞における発現に適した発現ベクターでもよい。
【0080】
本発明は、特に、本発明に記載の組換えタンパク質をコードするプラスミドからなるベクターに関する、例えば:2004年3月18日にCNCM I−3191号でCNCM(パリ、フランス)に寄託されたpTRACE5−HPV16E7FULL(CyaAE5−HPV16E7FULLとも称される);2004年3月18日にCNCM I−3190号でCNCM(パリ、フランス)に寄託されたpTRACE5−HPV16E7Δ30〜42(CyaAE5−HPV16E7Δ30〜42とも称される)、または構造物pTRACE5−HPV16E749〜57。
【0081】
本発明はまた、本発明に記載のポリヌクレオチドまたはベクターで形質転換された、宿主細胞、特に原核細胞、または真核細胞、例えば哺乳動物細胞(ヒト細胞を含む)を含む。
【0082】
本発明は、特に、アクセッション番号CNCM I−3190号およびアクセッション番号CNCM I−3191号でCNCMに寄託された宿主細胞に関する。
【0083】
本発明はまた、活性物質として、先に定義した組換えタンパク質または先に定義したポリヌクレオチドまたは発現ベクターを含む、免疫原性組成物に関する。免疫原性組成物の該活性物質は、宿主への投与に適した生理的に許容されるビヒクル、賦形剤、担体、もしくは希釈剤と共に、またはその組合せと共に製剤化できる。
【0084】
免疫原性組成物は、有利には、哺乳動物宿主において、細胞性免疫応答、特にT細胞媒介免疫応答を誘導するように設計されている。好ましくは、細胞媒介細胞溶解免疫応答CTL、特にCD8+を誘導できる。
【0085】
本発明に記載の別の免疫原性組成物は、体液性免疫応答を誘導できるものである。
【0086】
本発明の免疫組成物が免疫応答を誘導する能力を高めるために、活性物質を、アジュバントおよび/または界面活性剤および/または免疫調節物質(例えばサイトカインまたはケモカイン)と合わせることは興味深い。
【0087】
アジュバントは、例えば、水層を含むエマルションの形態で一般に使用される、リポソーム、油層、例えばフロイントタイプのアジュバント、または、水不溶性無機塩、例えば水酸化アルミニウム、硫酸亜鉛、コロイド状水酸化鉄、リン酸カルシウム、または塩化カルシウムを含むことができる。
【0088】
本発明に記載の免疫原性組成物は、有利には、特に免疫療法のために、免疫応答を抗原刺激および/またはブーストすることにより、宿主の免疫応答を誘導するために使用されている。特に、本発明の免疫原性組成物は、宿主におけるHPV感染に起因する悪性形質転換の発症または存続の予防のために、あるいは、HPV感染、特にHPV−16またはHPV−18感染に起因する悪性形質転換に罹患する患者の処置のために興味深い。
【0089】
このような免疫治療組成物は、腫瘍誘発性状態をもたらす宿主における制御されていない細胞増殖の治療に、特に癌免疫療法に、特にHPV感染に関連した子宮頚部癌免疫療法において特に興味深い。それ故、それは、癌ウイルス感染に起因する悪性状態(腫瘍状態を含む)の処置に特に適した治療ワクチンの設計のための手段を提供する。
【0090】
本発明に使用する場合、「処置」または「治療処置」なる表現は、処置を受けている患者に有益な効果をもたらす、本出願で開示された化合物の効果を包含し、該効果は、細胞レベルまたは臨床レベルで観察され、処置の結果、患者の容態の改善もしくは寛解状態、または健康状態の回復が含まれる。処置する悪性状態が、制御されていない細胞増殖または腫瘍の発生もしくは持続である場合、有益な効果は、制御されていない増殖の安定化を含み得、あるいは、好ましくは、その予防、停止もしくは反転、または腫瘍の減退を含み得る。
【0091】
先に記載した悪性状態の処置を目的とした組成物は、有利には、約1〜約1000μgの組換えタンパク質、好ましくは約10〜約500μgの組換えタンパク質の量に対応する用量の活性物質を含むことができる。該組成物が活性物質として本発明の組換えタンパク質を含む場合、用量は、約0.05〜約10μgの組換えタンパク質、好ましくは約0.1〜約1μgのタンパク質を含むことができる。
【0092】
処置する状態に応じて、該組成物は、病変レベルで局所的に、1回または数回、例えば数日間の規則的な間隔で、例えば5〜10日間投与できる。また全身投与してもよい。
【0093】
本発明はまた、好ましくはヒト宿主における、先の定義に記載の組換えタンパク質、または、先に定義したポリヌクレオチド、または、該ポリヌクレオチドを含むベクター、および、適宜、免疫応答(細胞性免疫応答および/または体液性応答を含む)を誘起するための薬学的に許容されうるビヒクルを含む、ワクチン組成物、特に哺乳動物宿主、好ましくはヒトへの投与のために製剤化された組成物に関する。
【0094】
本発明はまた、HPV感染を予防または処置するための、本発明の組換えタンパク質またはポリヌクレオチドまたはベクターおよび薬学的に許容されうるビヒクルを含む、薬物組成物にも関する。
【0095】
別の実施形態によれば、薬物組成物は、宿主におけるHPV感染に起因する悪性形質転換の発症または存続の予防または処置のための、本発明に記載の組換えタンパク質またはポリヌクレオチドまたはベクターを、薬学的に許容されうるビヒクルと共に含む。
【0096】
癌免疫療法のための、組換えタンパク質またはポリヌクレオチドまたはベクターおよび薬学的に許容されうるビヒクルを含む薬物組成物。
【0097】
本発明はまた、宿主における免疫応答、すなわち、体液性および/または細胞性免疫応答を誘起するためのベクターとして使用するに適した、細菌タンパク質、特に細菌毒素(好ましくはその類毒素形態)またはその断片を含む組換えタンパク質(該タンパク質またはその断片は、1種類の癌ウイルスによる感染の処置のために、1個または数個の癌ウイルスの1個または数個の抗原の1個または数個のエピトープの挿入により改変されている)の患者における使用に関する。このような組換えタンパク質は、特に、悪性作用、特にこのような癌ウイルスによる感染により引き起こされる腫瘍の処置に提案されている。
【0098】
癌ウイルスの抗原のエピトープを運ぶためのベクターとして適した細菌タンパク質の例は、クレブシエラのOmpAまたは以下の毒素シガ毒素(そのβサブユニットを含む)(Haicher N.ら、J.Immunol.2000、165:3301-8)、炭素菌毒素(Goletz TJら、PNAS USA 1997、94:12059-64)、ジフテリア毒素(Stenmark H.ら、J.Cell.Biol.1991、113:1028-32)またはシュードモナス外毒素A(Donnelly JJ.ら、PNAS USA 1993、90:9530-4)である。細菌タンパク質への挿入のためのポリペプチドを調製するために抗原がエピトープを提供できる癌ウイルスには、HPV、HBV、HCV、アデノウイルスEBV、ヘルペスウイルス、HTLV.1ウイルス、およびCMVが挙げられる。
【0099】
CyaA組換えタンパク質について先に示した活性物質としての使用の記述は、他の細菌タンパク質および癌ウイルス抗原にも応用できる。
【0100】
本発明はまた、HPV感染の診断のための、または、このような感染を免疫モニタリングするためのキットに関し、これには、本発明に記載の組換えタンパク質、ポリヌクレオチド、または発現ベクターが含まれる。
【0101】
本発明はまた、患者におけるHPV感染の処置または予防のための、上記の本発明の組換えタンパク質、ポリヌクレオチド、またはベクターの使用に関する。
【0102】
本発明はまた、患者におけるHPV感染に起因する悪性形質転換の発症または存続に対する免疫療法のための、上記の本発明の組換えタンパク質、ポリヌクレオチド、またはベクターの使用に関する。
【0103】
本発明はまた、HPV感染をインビトロで診断する方法またはHPV感染を免疫モニタリングする方法に関し、該方法は、
哺乳動物、特にヒト患者から得られたT細胞を、本発明の組換えタンパク質にさらすこと、
T細胞の活性化の改変を検出すること
を含む。
【0104】
特定の実施形態において、該組換えタンパク質は、HPV感染の予防、または、HPV感染に罹患している宿主(このような感染に起因する腫瘍を有する宿主を含む)の処置のために使用できる。
【0105】
本発明はまた、キメラCyaA−ポリペプチドタンパク質に含まれるポリペプチドにおける未知または亜優性潜在性T細胞エピトープのスクリーニング方法であって、
ここで、CyaAは、先に開示されているアデニル酸シクラーゼまたはその断片であり、該方法は、
該キメラタンパク質を動物宿主に投与すること、
該宿主のT細胞応答、特にCTL応答を決定すること
を含む。
【0106】
本発明は、特に、本発明内で定義された組換えタンパク質に含まれるHPV抗原のポリペプチド中の未知または亜優性または潜在性T細胞エピトープ(特にCD8+T細胞エピトープ)をスクリーニングする方法に関し、該方法は、
該組換えタンパク質を動物宿主(非ヒト)に投与すること、
該宿主のT細胞応答、特にCTL応答を決定すること
を含む。
【0107】
本発明を特徴づけるさらなる特徴を、以下の実施例および図面に開示し説明する。
【0108】
実施例
実施例1
ここで、本発明者らは、HPV16のE7タンパク質の全長配列またはこのポリペプチドの亜断片(特に、H−2Db拘束性エピトープに対応するE7の残基49〜57およびHPV16−E7残基43〜98と1〜29を包含するペプチド)を含む、組換えCyaAを作成した。本発明者らは、C57BL/6マウスに注入した場合、これらのHPV16−E7組換えCyaAは、IFN−γの分泌により特徴づけられる特異的CTLおよびTh1応答を誘導できることを示した。さらに、治療的に試験した場合、これらの構造物は、TC−1細胞の皮下移植に対して100%以下の防御を提供できた。この研究は、初めて、インビボでの、ヒト腫瘍特異的抗原に対するCyaAにより媒介される抗腫瘍治療活性の実証を示した。
【0109】
材料および方法
マウスおよび細胞系
特定の病原体を含まない6〜10週令の雌C57BL/6マウスを、CER Janvier(Le Gesnet St-Isle、フランス)またはチャールズリバー(L'Arbresle、フランス)から得た。C57BL/6バックグラウンドへと育成したTAP1−/−(18)、MHCクラスII−/−(19)およびCD40−/−(20)もこの研究に使用した。動物を、水と食料が自由にとれ、病原体の存在しない条件下で、パスツール研究所動物施設で飼った。動物の関与する実験は、動物の世話の施設ガイドラインにしたがって実施した。HPV16E6およびE7タンパク質を発現しているT細胞(21)およびマウス胸腺EL4細胞(17)を、ATCCから得た。細胞を、10%の熱不活性化FCS、100U/mlのペニシリン、100μg/mlのストレプトマイシン、0.4mg/mlのジェネティシン(TC−細胞に対してのみ)、および5×105Mの2−メルカプトエタノール(ギブコBRL、Cergy-Pointoise、フランス)を補充したグルタマックスを含むRPMI1640中に維持した。
【0110】
ペプチド
HPV16−E7 H2−Db拘束性エピトープ(22)に対応する合成ペプチドE749〜57(RAHYNIVTF、一文字でアミノ酸をコード)並びにE749〜57CTLエピトープ(天然フランキング配列およびThエピトープ(太線)を含む)(23)に対応するE743〜77
【表3】
を、Neosystem(ストラスブール、フランス)から購入した。CpG ODN1826は、PROLIGO(パリ、フランス)から購入した。
【0111】
HPV16−E7エピトープを有する組換えB.pertussisアデニル酸シクラーゼの作成および精製
本文書で使用される組換えアデニル酸シクラーゼは、酵素的に不活性なCyaAをコードするプラスミドpTRACE5の誘導体(図1A)を使用することにより、E.coliにおいて発現させた(24)(25)。プラスミドpTRACE5は、酵素的に不活性で、それ故に細胞毒性であるB.pertussisCyaAの変種のための発現ベクターである。それはまた、CyaAの翻訳後のアシル化に必要とされるB.pertussisCyaCタンパク質も発現する。このプラスミドは、以前に記載されているpTRACGプラスミドの誘導体である(Gmiraら、2001、Res.Mic.152:889)。それは、cyaA DNA配列の5’部分内に位置するEcoRV部位へのヘキサヌクレオチドCTGCAGの挿入により得られた。これにより、触媒部位の不可欠な部位内のAsp188とIle189の間に、ジペプチドLeu−Glnが枠内に挿入された(Guermonprezら、2000、Meth.Enzymol.326:527)。
【0112】
プラスミドpTRACE5は、ColE1複製起点とアンピシリン耐性マーカーを有する。このプラスミドでは、cyaCおよび改変されたcyaA遺伝子が、λファージPrプロモーターの制御下で同じ転写単位に入れられている。pTRCAGプラスミドはまた、32℃より低い温度でλPrプロモーターにおける遺伝子転写を強力に抑制する熱感受性λリプレッサーcI857をコードしている。
【0113】
E.coli株のXL1−Blue(ストラタジーン、ラホーヤ、CA州)を、標準的なプロトコルにしたがって実施した全てのDNA操作において使用した(Maniatisら)。
【0114】
CyaA−E749〜57は、CyaAのコドン224と235の間に挿入された9アミノ酸長のポリペプチド配列(RAHYNIVTF)を含む。CyaA−E749〜57の発現プラスミドは、以下のように作成した。2つの合成オリゴヌクレオチド(MWG、Courtaboeuf、フランス)
【表4】
をアニーリングし、NheIおよびKpnIで消化したpTRACE5にライゲートした。CyaA−E7Fullは、HPV16−E7タンパク質の全配列、すなわち、酵素的に不活性なCyaAの同じ224位に挿入された98アミノ酸を含む。E7タンパク質をコードするDNA配列を、特異的プライマー
【表5】
を使用して、HPV16 DNA(先のSeedorf Kら)から増幅した。得られたPCR産物をNheIおよびKpnIで消化し、NheIおよびKpnIで切り出したpTRACE5にライゲートした。アニーリングしたオリゴヌクレオチド並びにHPV16−E7の全配列に存在するSspI部位により、挿入変異体の迅速な同定が可能となった。CyaA−E7Δ30〜42は、CyaAのコドン319と320の間に挿入されたHPV16−E7の最初の29アミノ酸残基、並びに、CyaAのコドン224と235の間に挿入されたHPV16−E7の残基43〜98を含む。CyaA−E7Δ30〜42のための発現プラスミドは、2ステップで作成した。HPV16−E7(のアミノ酸残基1〜29)をコードする第一のDNA断片を、標的DNAとして合成HPV16−E7遺伝子(E.coliにおける産生に最適化され、GTP Technologyにより設計、Labege、フランス)およびプライマー
【表6】
を使用してPCR増幅した。CyaAのコドン320〜372をコードする第二のDNA断片を、標的DNAとしてpTRACE5、および、プライマー
【表7】
を使用してPCR増幅した。これらの2個のDNA断片(一部重複する)を精製し、3回目のPCRでプライマーBTP5およびBTP8と合わせ、294bp長のDNA断片を増幅した。この断片を、AgeIおよびBstBIにより消化し、pTRACE5の対応する部位の間に挿入して、プラスミドpTRACE5−E1〜29を得た。その後、HPV16−E7のアミノ酸残基43〜98をコードするDNA断片を、標的DNAとしての合成HPV16−E7遺伝子、および、プライマー
【表8】
を使用してPCR増幅した。精製PCR断片をNheIおよびKpnIにより消化し、同じ制限酵素により消化したプラスミドpTRACE5−E71〜29にライゲートした。
【0115】
全ての組換えアデニル酸シクラーゼが、前記のように(26)、エシェリヒア・コリ株BLR(ノバゲン、WI州マジソン)において産生された。組換えタンパク質を、前記のように(26)、DEAE−セファロースおよびフェニル−セファロースクロマトグラフィーを含む2ステップ手順により封入体から均一に近くなるまで精製した(図1B)。フェニル−セファロースクロマトグラフィーに加えて、20mM Hepes−Na、pH7.5中の60%イソプロパノールを用いてのさらなる洗浄ステップを行ない、大半の混入LPSを排除した。LPS含量は、キットQCL−1000(Biowhittaker、Walkersville、MD州)を使用して決定した。精製組換えタンパク質を、SDS−ゲル分析により分析した。タンパク質濃度は、分子吸光係数142,000M−1cm−1を使用して280nmにおける吸光度から分光学的に決定した。
【0116】
組換えHPV16−E7タンパク質の作成および精製。
リクエストにより入手可能なHPV16−E7タンパク質(GTP technology)のDNA配列をコードするE.coli最適化cDNAを、pIVEX2.4bベクター(ロシュ・モレキュラー・バイオケミカルズ、Meylan、フランス)のNcoIとXhoI制限部位の間にサブクローニングした。その後、得られたプラスミドを、E.coli株BL21λDE3(ノバゲン)に形質転換した。0.5mMのイソプロピル−β−D−チオガラクトピラノシド(Euromedex、Souffelweyersheim、フランス)で誘導するとHis−Tag−HPV16−E7タンパク質が発現され、Ni−NTAアガロース(キアゲン、ヒルデン、ドイツ)を用いて製造業者の指示にしたがって精製した。LPS混入を除去するために、記載の通りに(27)イソプロパノールによる洗浄を使用した。
【0117】
イムノブロット
タンパク質をSDS−PAGEにより分離し、マウスモノクローナル抗HPV16E7抗体(Zymed、CA州サンフランシスコ)またはC57BL/6マウスにおいて調製したポリクローナル抗E.coli BLR血清でプローブした、ニトロセルロース膜(0.45μ、バイオラッド、Marnes la Coquette、フランス)にエレクトロトランスファーした。アルカリホスファターゼにコンジュゲートさせたヤギ抗マウス免疫グロブリン(Chemicon、Temecuka、CA)を用いて免疫複合体が検出され、5−ブロモ−4−クロロ−3−インドリルホスフェート/ニトロブルーテトラゾリウム(BCIP/NBT)(シグマ、MO州セントルイス)を用いて明確にした。
【0118】
マウス免疫化および腫瘍拒絶実験
動物を、PBS(ギブコBRL)に希釈した50μgの対照もしくはHPV16−E7組換えCyaAを1回静注するか、または、2回(各回10μg)皮内注射することにより免疫化した。皮内注射は、耳真皮に行なった(47)。インビトロでの解析のために、長期の応答の解析を除いて(このためにこの手順は、注射の3ヶ月後に行なった)、注射の7日後に、麻酔した動物(CO2)の脾摘除を行なった。腫瘍拒絶実験のために、マウスは、皮下に5×104個のTC−1細胞を投与され、腫瘍接種の1日後、5日後、または10日後にHPV16−E7組換えCyaAにより処置した。TC−1腫瘍増殖を、キャリパーを使用してモニタリングし、式V=(L×w2)/2(L:長さ、w:幅)を使用して立方ミリメートルで表現した(48)。
【0119】
インビトロでの細胞毒性アッセイ
免疫化マウスの脾臓細胞を、インビトロで、同系の照射し無処置の脾臓細胞の存在下で、完全培地(10%の熱で不活性化されたFCS、100U/mlのペニシリン、100μg/mlのストレプトマイシン、および5×10−5Mの2−メルカプトエタノールを補充したグルタマックスを含むRPMI1640)中で、5日間、1μg/mlのE749〜57またはE743〜77ペプチドで刺激した。これらのエフェクター細胞の細胞毒性活性を、TC−1細胞の5時間の51Cr放出アッセイで試験した。放射標識を以下のように行なった:7.5%CO2雰囲気中37℃で培養した指数関数的に増殖しているTC−1細胞を、素早くトリプシン処理し(トリプシン−EDTA、ギブコBRL)、100μCiの51Crと共に37℃で1時間インキュベートした。様々なE:T比を使用し、全てのアッセイを二重に行なった。各ウェルの上清に放出された放射能を測定した。特異的溶解率は、100×(実験的放出−自発的放出)/(最大放出−自発的放出)として計算した。最大放出は、10%トリトンX−405を標的細胞に加えることにより得、自発的放出は、完全培地のみの中でインキュベートした標的細胞を用いて得られた。最も高いE:T比で少なくとも20%の特異的溶解が観察された場合にマウスをレスポンダーと判断した。結果は、1群あたりのレスポンダーマウスの中間値±四分位範囲として表現した。
【0120】
分泌細胞のための、単一のIFN−γ産生細胞の酵素結合イムノスポットアッセイ
マルチスクリーンろ過プレート(96ウェル;ミリポア、Molshein、フランス)を、1mlあたり4μgのラット抗マウスγインターフェロン(IFN−γ)抗体(クローンR4−6A2;ファーミンゲン、CA州サンディエゴ)を用いて室温で一晩かけて被覆した。その後、プレートを洗浄し、完全培地で遮断した。免疫化マウスの脾臓細胞の連続2倍希釈液を、5×105個のγ照射された(2,500rad)同系支持細胞と共に加えた。細胞を、1μg/mlのE749〜57ペプチドと共にまたはその非存在下で36時間インキュベートした。十分に洗浄した後、プレートを、1mlあたり5μgのビオチニル化ラット抗マウスIFN−γ抗体(クローンXMG1.2;ファーミンゲン)と共にインキュベートし、その後、ストレプトアビジン−アルカリホスファターゼ(ファーミンゲン)と共にインキュベートすることにより明確にした。最後に、基質としてBCIP/NBTを使用してスポットを明確にした。IFN−γ産生細胞の数を、各ウェルにおけるスポット形成細胞(SFC)の数を計測することにより決定し(Bioreader、Karben、ドイツ)、結果を、脾臓1個あたりのSFCの全数として表現した(17)。
【0121】
酵素結合免疫吸着アッセイ(ELISA)
空のベクターCyaAE5により皮内免疫化したマウスから、30または90日後に採血し、個々のマウス血清を、ELISAにより抗体応答について試験した。マイクロプレート(Nunc、Roskilde、デンマーク)を、PBS中の空のベクターCyaAE5(3μg/ml)を用いて一晩かけて被覆した。PBS−tween20(0.1%)中で洗浄した後、希釈した血清を、ウェルに加え、37℃で1時間インキュベートした。PBS−tween20中で洗浄した後、プレートを、ヤギ抗マウスIgGペルオキシダーゼコンジュゲート(シグマ)と共に37℃で1時間インキュベートした。プレートを、o−フェニレンジアミンおよび過酸化水素(シグマ)を使用して展開した。硫酸を用いて反応を停止し、プレートを、ELISA読取機(Dynatech、Marnes la Coquette、フランス)を用いて492nmで解析した。結果は、希釈とA492をプロットした線形回帰分析により計算した抗体力価として表現した。力価は、1/100に希釈されたプール対照血清の吸光度の2倍をもたらす最も高い希釈度のlog10であると計算された。
【0122】
サイトカイン産生
免疫化マウスの脾臓細胞を、インビトロで、完全培地中で、10μg/mlのHisTag−HPV16−E7タンパク質または1μg/mlのE743〜77ペプチドを用いて72時間かけて刺激した。IFN−γおよびIL−5産生は、培養上清中で、前記のように(28)、サンドイッチ酵素結合免疫吸着アッセイ(ELISA)により決定された。全てのアッセイは、対応する組換えネズミサイトカイン(ファーミンゲン)を用いて標準化した。
【0123】
FACS(登録商標)解析
TC−1細胞は、特異的なFITC−コンジュゲートモノクローナル抗体(クローンKH95、ファーミンゲン、Le Pont de Claix、フランス)を使用するフローサイトメトリーによるMHCクラスI分子H−2Dbの発現レベルの解析についていずれかに記載されているように(29)処理した。
【0124】
統計的解析
様々な試料の量が少ないことを考慮して、ソフトウェアStatXact4(Cytel社、MA州ケンブリッジ)を使用するノンパラメトリック統計試験(30)を適用した。生存曲線を、Prismソフトウェア(GraphPadソフトウェア社、CA)を使用してプロットし、ソフトウェアのビルトインのlogランク検定を用いて比較した。データは、p<0.05で有意差があると判断した。
【0125】
結果
HPV16−E7エピトープを有する組換えアデノウイルスシクラーゼの作成および特徴づけ
CyaAのHPV16−E7特異的T細胞応答誘導能を試験するために、本発明者らは、3個の異なる組換え分子を作成した。CyaA−E749〜57は、酵素的に不活性な(したがって無毒性な)CyaAのコドン224と235の間に挿入された、以前に記載されたH−2Db拘束性CTLエピトープ(22)に対応する、9アミノ酸長のポリペプチド配列(RAHYNIVTF)を含む。CyaA−E7FUllは、酵素的に不活性なCyaAの同じ224位に挿入されたHPV16−E7タンパク質の全配列(98アミノ酸)を含む。CyaA−E7Δ30〜42は、酵素的に不活性なCyaAのコドン319と320の間に挿入されたHPV16−E7の最初の29アミノ酸残基、並びに、コドン224と235の間に挿入されたHPV16−E7の残基43〜98を含む。インビトロおよびインビボでのアッセイを可能とするために、CyaA構造物が産生され、精製されてほぼ均一とした(図1B)。LPS排除手順を精製プロトコル(26)に導入し、50μgあたり100単位未満の内毒素を含む組換えタンパク質を得た。CyaA−E7FullおよびCyaA−E7Δ30〜42におけるE7タンパク質の存在は、特異的なモノクローナル抗体(Zymed)を使用してウェスタンブロットにより確認された(図1C)。これに対し、H−2Db拘束性エピトープのみを含むCyaA−E749〜57は、抗HPV16−E7抗体により認識されなかった。3個の組換えCyaAの全体的な生化学的特性は、改変されておらず、これらの分子は、野生型アデニル酸シクラーゼと類似した溶血特性を示した(17)。
【0126】
HPV16−E7組換えCyaAでの免疫化は、E7特異的CTL応答を誘導した。
CyaAが、HPV16−E7エピトープに対するCTL応答を誘導できるかを試験するために、C57BL/6マウスに、50μgの様々なHPV16−E7組換えCyaAを1回静注することにより免疫化した。脾臓細胞を収集し、インビトロで、1μg/mlのE743〜77ペプチドで刺激した。TC−1細胞を溶解する能力を、5日後に、51Cr放出アッセイを使用して決定した。図2Aに示されているように、C57BL/6マウスにHPV16−E7組換えCyaAを1回静注して免疫化することにより、TC−1細胞に対する強力で特異的なCTL応答が誘導された。完全なHPV16−E7タンパク質(図2A、b)を含むCyaAまたはその欠失形態であるCyaA−E7Δ30〜42(図2A、c)で免疫化することにより、最小のH−2Db拘束性エピトープ(図2A、a)のみを含むCyaA−E749〜57により誘導されるものと比べてより高い最大CTL活性が得られたが、本発明者らは、本発明者らのデータから統計学的有意性を実証できなかった。ペプチドE749〜57をインビトロでの再刺激に使用した場合にも、類似の結果が得られた(データは示さず)。関連性のないエピトープ(OVA257〜264)を有する組換えCyaAでワクチン接種し、インビトロで、1μg/mlのE743〜77ペプチドで再刺激したマウスの脾臓細胞には、弱く非特異的なTC−1細胞溶解しか認められなかった(図2A、a)。インビボで、CyaAにより、OVA CD8+細胞エピトープ(SIINFEKL)をMHCクラスI分子に送達することは、TAP1機能に依存していることが以前に示されている(15)。本発明者らは、これがCyaA−E7Δ30〜42を使用しても当てはまるかどうかを試験した。図2A、dに示されているように、静注によりワクチン接種されたTAP1−/−マウスに由来するインビトロで刺激された脾臓細胞は、TC−1細胞を溶解できなかった。本発明者らはまた、インビボでのHPV16E7含有組換えCyaAによる、CTL応答の刺激における、CD4+T細胞による支持の必要性を試験した。以前の観察と一致して(15)、本発明者らは、CyaA−E7Δ30〜42を使用したMHCクラスII−/−マウスの静注ワクチン接種により、TC−1細胞に対する高いレベルの特異的CTL応答が誘導されることを観察した(図2A、e)。これに対し、本発明者らは、このモデルにおいて、CD40−/−マウスにおけるTC−1細胞に対するCTL応答のレベルが低いことを観察したので(図2A、f)、CD40シグナル伝達への幾分の依存性を観察した。
【0127】
生体外での組換えCyaAで免疫化したマウスにおけるHPV16−E7特異的脾臓細胞の発生頻度を推定するために、インビトロでのHPV16−E749〜57ペプチドでの刺激に応答してIFN−γを産生する細胞の数を、酵素結合イムノスポット(ELISPOT)により定量した。図2Bは、CyaE5−CysOVAで免疫化したマウスから得られたIFN−γを特異的に産生しているIFN−γ産生脾臓細胞の数は、CyaA−E749〜57で免疫化したものと比べて、僅かな差異しかなかったことを示す。これに対し、得られたIFN−γ産生脾臓細胞は、完全なHPV16−E7タンパク質またはその欠失形態を含むHPV16−E7組換えCyaAでワクチン接種したマウスにおいて、はるかにより多かった(p<0.05)。観察された応答は、エピトープ特異的であった。なぜなら、HPV16−E749〜57ペプチドにより刺激しない場合、これらのマウスの非常に僅かな脾臓細胞しか、IFN−γを産生しなかったからである(図2B)。これらの結果は、CyaAが、処理およびMHCクラスI経路へと提示するために、インビボにおいて、HPV16−E7タンパク質の免疫優性CD8+H−2Db拘束性T細胞エピトープを、免疫担当性細胞のサイトゾルへと送達でき、これにより強力なCTL応答を誘起できることを示す。以前の観察によれば(25、31)、本発明者らは、完全なHPV16−E7タンパク質またはその欠失形態および反転形態を有するCyaAもまた、強力なCTL応答を誘導できたので、CyaAは、大きなポリペプチド断片の挿入に耐えられることを確認した。本発明者らのデータはまた、これらの後者の分子は、マウスにおいて、有意により高い発生頻度のHPV16−E749〜57特異的応答を誘導したことを実証した。
【0128】
HPV16−E7組換えCyaAでの免疫化は、HPV16−E7特異的Th1応答を誘導した。
Th1応答は、細胞内病原体および腫瘍発達に対する防御において重要な役割を果たす(32、33)。それ故、本発明者らは、HPV16−E7組換えCyaAにより誘導されるT細胞応答のタイプを特徴づけた。C57BL/6マウスに、50μgの3種の異なるHPV16−E7組換えCyaAを1回静注することにより免疫化し、インビトロで脾臓細胞を10μg/mlの精製His−TagHPV16−E7タンパク質で刺激した後に、サイトカインの合成を決定した。図3に示されているように、完全なHPV16−E7タンパク質またはその欠失形態を有するCyaAでの免疫化により、高いレベルのIFN−γの産生および検出可能なレベルのIL−5が存在しないことにより特徴づけられる、Th1に似たプロファイルが得られた。この応答は特異的であった。なぜなら、CyaA−E7FUllまたはCyaA−E7Δ30〜42で免疫化した後に得られたIFN−γレベルは、CyaAE5−CysOVAでニセ免疫化したマウスで得られたものよりも有意に高かったからである(p<0.05)。しかし、これは、CyaA−E749〜57で免疫化したマウスの脾臓細胞には当てはまらなかった。1μg/mlのE743〜77ペプチドを用いて再刺激した場合に、類似の結果が得られた(図3A、挿入図)。
【0129】
合わせて考えると、これらの結果により、本発明者らの条件では、クラスII H−2b拘束性T細胞エピトープを含む完全HPV16−E7タンパク質を有するCyaAで得られたレベルは、クラス1H−2Db拘束性エピトープしか含まないCyaA−E749〜57で得られたレベルよりもはるかに高いので、CD4+T細胞は、IFN−γの分泌において重要な役割を果たしていることが示された。
【0130】
HPV16−E7組換えCyaAでの免疫化は、確立されたHPV16を発現している腫瘍の減退を誘導した。
強い免疫応答が得られたことを考慮して、その後、本発明者らは、インビボにおいて、HPV16−E6およびE7タンパク質H−2b腫瘍原性細胞系(TC−1細胞)(これはC57BL/6マウスの皮下に注射される)からなる前臨床モデルにおいて、HPV16−E7CyaAの治療活性を評価した。このモデルにおいては、腫瘍拒絶は、排他的に、E749〜57特異的CD8+T細胞により媒介されることが以前に示された(21、22、34、35)。したがって、5×104個のTC−1細胞を、C57BL/6マウスの右側腹に皮下注射し、50μgのCyaAE5−HPV16−E749〜57、−E7full、または−E7Δ30〜42を、1、5、または10日後にマウスに静注した。図4は、腫瘍移植の10日後に治療処置したマウスにおける腫瘍増殖を示す。特に、これらの条件において、100%の動物が、ワクチン接種を行なう時点までに、触診可能な腫瘍を発達させた。不必要な苦しみを避けるために、腫瘍サイズが1000mm3に到達した場合には動物を屠殺した。全ての非処置動物並びにニセCyaAE−cysOVAで処置したマウスは、最大49日間以内に、そのサイズ(>1000mm3)の腫瘍を発達させた。極めて対照的には、HPV16−E7組換えCyaAで処置した動物の大半が、実験期間中を通じて腫瘍がない状態を維持した(図4、C、D、E)。図5は、TC−1細胞を移植し、3つの異なる治療プロトコル(組換えCyaAを、TC−1移植の1、5、または10日後に注射した)を行なった、動物の生存プロットを示す。非処置またはニセ処置動物の生存時間の中間値は、31〜40日間であった。これに対し、HPV16−E7抗原を有するCyaAでワクチン接種したマウスの生存は、対照動物よりも有意に優れていた(p<0.05)。様々な構造物の防御活性の差異は確立できたが、統計学的有意性は確立できなかった。なぜなら、様々な試料のサイズが小さいからであった。CyaA−E749〜57および−E7Fullにより付与される腫瘍減退率が、認められる程には区別できなくても、CyaA−E7Δ30〜42は、腫瘍減退および増殖阻害の点において明らかに優れていた。なぜなら、3つ全ての治療スキームにおいて、防御率は、常に、90%よりも高かったからである。
【0131】
CyaA−E749〜57およびCyaA−E7Fullでワクチン接種した何匹かの動物は、実験の経過中、より後で腫瘍を発達させるようであった(図4(*)、データは示していない)。この現象は腫瘍エスケープ機序を反映する可能性があることを心に置き、本発明者らは、これらの動物から増殖中の腫瘍を外移植し、H−2Db発現について、FACS解析により、TC−1A1およびTC−1A2という名称のこれらの細胞系を解析した。図6に示されているように、その天然対応物に比べて、より後で増殖してくる腫瘍から外移植したTC−1A1およびA2細胞は、H−2Dbの発現を欠失しており、したがって、E749〜57特異的CD8+T細胞により検出されることができなくなっている可能性が高い。TC−1細胞は、インビボにおいて、選択のための抗生物質の負荷がない環境で増殖させたので、本発明者らはまた、TC−1A1およびA2細胞においてHPV16−E7の発現についてウェスタンブロットにより確認した。本発明者らは、TC−1およびTC−1A1およびA2細胞の間にこのタンパク質の発現の差異を全く見出すことができなかった(データは示さず)。
【0132】
合わせて考えると、これらの結果により、前臨床モデルにおいてHPV16を発現している腫瘍の減退を誘導するために適した治療ワクチンとしての、アデニル酸シクラーゼベクターの有効性が実証された。
【0133】
本発明者らは、臨床的に関心の高い別の注射経路を試験した。したがって、10μgのCyaA−E7Δ30〜42を、TC−1移植の10日後から始めて7日間間隔で2回皮内注射した。興味深いことに、全ての非処置およびニセ処置動物が腫瘍を発達させたが、本発明者らは、CyaA−E7Δ30〜42で処置した動物の全ての動物において腫瘍減退を観察した(図4B、a、b)。この治療的免疫化により、90日目においてCyaA−E7Δ30〜42処置マウスの100%の生存率が得られ、一方、非処置およびニセ処置動物の生存率の中間値は、それぞれ30日および32日であった(図4、c)。
【0134】
CyaA免疫化により誘導されるHPV16−E749〜57特異的CD8+T細胞の長期持続性
HPV16−E7組換えCyaAにより誘導される免疫応答の持続性を評価するために、3ヵ月後も治療実験にも関わらず生存しているマウスを屠殺し、その脾臓細胞を、インビトロで、1μg/mlのE743〜77ペプチドで5日間かけて刺激した。その後、TC−1細胞を溶解する能力を、51Cr放出アッセイにより決定した。図7に示されているように、HPV16−E743〜77ペプチドに対する特異的CTL応答が、依然として、3ヵ月前に免疫化した動物の脾臓細胞から実証された。30:1のエフェクター:標的の比での特異的溶解の最大パーセンテージに基づいて、免疫応答は、CyaA−E7Δ30〜42で処置した動物においてより強力であるようであるが、これらのデータから統計学的有意性は実証できなかった。このような長く続く免疫原性の生理的関連性を評価するために、残りの動物に、100日目に5×104個のTC−1細胞を用いて皮下に再攻撃した。このような条件下で、全ての未処置の年齢一致対照動物が腫瘍を発達させ、37.5日間という生存時間の中間値を示した(図8)。これに対し、HPV16−E7組換えCyaAで3ヶ月前に免疫化したマウスは、腫瘍発達から非常に有意に防御された。先に観察されているように、CyaAE−HPV16−E7Δ30〜42でワクチン接種した動物が、高いレベルの防御を示した。しかし、今回、最初のセットの治療実験で得られた結果の変動では、CyaA−E749〜57で処置した動物と、CyaA−E7FUllで処置した動物の間には顕著な防御の差異があり、後者の方が好ましいが、試料のサイズが少ないので、統計的有意性を明確に実証することはできなかった。これらの観察により、このモデルでは、完全なHPV16−E7タンパク質を有するCyaAにより提供されるT細胞の支持は、TC−1細胞に対する効果的で長期に持続する応答には重要であることが示唆された。
【0135】
合わせて考えると、本発明者らの結果により、HPV16−E7タンパク質のヘルパーエピトープ(DRAHYNIVTF)を有する50μgの組換えCyaAの1回の静注は、少なくとも6ヶ月の期間におよび、2回のTC−1腫瘍細胞攻撃に対する防御を付与できる、長期に持続するE749〜57特異的CD8+T細胞を誘導するのに十分であった。
【0136】
CyaA−E7Δ30〜42の治療効力は、CpG−ODN1826と共に投与されたペプチドの治療効力に匹敵した。
抗原送達システムとしてのCyaAの効力をより良く評価するために、本発明者らは、CyaA−E7Δ30〜42の治療効力を、CpG−ODN1826を補充したHPV16−E743〜77ペプチドの治療効力と比較した(37)。それ故、マウスに、5×104個のTC−1細胞を皮下注射し、10および17日後、皮内経路により、10μgのCyaA−E7Δ30〜42で、または、1μgのCpG−ODN1826と共に投与される10μgのHPV16−E743〜77ペプチドで、治療処置した。生存率は、これらの2つの群において類似していたが(図9)、CyaA−E7Δ30〜42で得られた結果は、CpG−ODN1826と混合されたHPV16−E743〜77ペプチドで得られた結果よりも僅かに良好であったが、統計学的に差異はなかった。特に、この結果は、モル基準で、CyaA−E7Δ30〜42に比べて50倍多いHPV16−E743〜77ペプチドを使用して得られた。単独で使用した場合、ペプチドHPV16−E743〜77は、TC−1腫瘍増殖に対して全く効果がなかった。
【0137】
CyaAベクターに対する事前免疫は、CyaA−E7Δ30〜42の治療効力に僅かにしか影響を及ぼさなかった。
臨床の場面では、効果的な細胞性免疫応答を得るために、病変を有する患者に、複数回のブースター投与をおそらく行なわなければならない。それ故、CyaAベクターに対する予備免疫が、腫瘍拒絶を引き起こす能力を損なわないことを実証することが不可欠であった。そうするためには、本発明者らは、5×104個のTC−1細胞を皮下注射する90または30日前に、7日間間隔で2回10μgの空ベクターCyaAE5でマウスを皮内免疫化した。7日間間隔の2回の10μgのCyaA−E7Δ30〜42の注射による治療処置を10日目に設定した。抗体応答の解析により、空ベクターで免疫化されたマウスは、TC−1注射時にCyaAに対して免疫があることが示された(図10A)。その後、本発明者らは、年齢の一致した未処置動物およびCyaA免疫動物において腫瘍拒絶を誘導する、CyaA−E7Δ30〜42処置の能力を比較した。CyaAに対するその免疫状態はどうであれ、CyaA−E7Δ30〜42で処置したマウスの大半が、実験を通じて腫瘍がないままであった(図10B)。30日目の免疫マウス群の中で1匹のみ、90日目の免疫マウス群において2匹のみが、腫瘍を発達させた(図10Bb、d、f)。これに対し、ニセ感染動物の100%が腫瘍を発達させ、屠殺された(図10B、a、c、e)。本発明者らは、抗CyaA抗体力価レベルと、TC−1腫瘍の発達の間の相関は全く観察しなかった(データは示さず)。さらに、CyaA−E8Δ30〜42処置マウスの生存曲線(図10B、b、d、f)は、統計学的に差異はなかった(p=0.324)。
【0138】
それ故、これらのデータにより、CyaAに対する既存免疫は、このベクターがその後に外来の所与の抗原に対する効果的な応答を誘導する能力に対して、殆ど作用を及ぼさないことが示された。
【0139】
考察
以前の研究により、Bordetella pertussisのアデニル酸シクラーゼは、インビボにおいて、CD4+およびCD8+T細胞エピトープを、樹状細胞のMHC−クラスIIおよびI提示経路に送達するための強力なツールであることが実証された。実験ネズミモデルにおいて、このシステムは、抗ウイルスおよび抗腫瘍防御をもたらす効果的なTh1およびCTL応答を引き起こすために使用されている(36)。HPV16関連の子宮頚部悪性疾患の処置のためのヒトにおけるCyaAの可能性ある適用の評価として、本発明者らは、このベクターが、インビボにおいて、HPV16のE7タンパク質のエピトープを効果的に送達することを実証し続けた。
【0140】
本発明者らは、HPV16の完全なE7タンパク質またはこのポリペプチドの亜断片(特に、残基49〜57に対応するH−2Db拘束性最小CTLエピトープを含む)を含む様々なHPV16組換えCyaAを作成した。本発明者らは、これらの異なる組換えタンパク質が、C57BL/6マウスに注射された時に、特異的かつ強力なCTL応答を抗原刺激できることを示した。本発明者らのデータにより、本発明者らはTAP1−/−マウスにおいてCyaA−E7Δ30〜42の注射によりCTLを刺激することができなかったので、CyaAによるCTLエピトープの送達には、完全に機能的なクラスI提示経路が必要であることが確認された(15)。CyaAにより媒介されるCTL刺激は、以前の結果と一致して、MHCクラスII−/−マウスで得られた効果的なCTL応答により示されるように、CD4+T細胞の存在とは独立していた(15)。ワクチンベクターとしてのこのCyaAの特徴は、CD4+T細胞数の変化を提示する免疫抑制患者または免疫欠陥患者のワクチン接種を考慮した場合に非常に重要である。しかし、CD40−/−マウスではCTL応答は低く、このことは、CTL刺激が、一部、CD40シグナル伝達に依存していることを示す。これらの観察により、HPV16−E7組換えCyaAは、プロフェッショナルAPCの直接刺激により、MHCクラスI拘束性CTLを直接的に誘起することが示唆された。それにも関わらず、CTL応答の最適な刺激を得るためには、CD40−CD40L相互作用が必要とされる。
【0141】
本発明者らは、HPV16−E7の最小H−2Db拘束性CTLエピトープの免疫原性を、おそらくとりわけ記載されたヘルパーエピトープDRAHYNIVTFを含む、完全もしくはΔ30〜42E7タンパク質の免疫原性と比較した。CTL刺激並びにCyaA−E7FUllおよびCyaAE−E7Δ30〜42により誘導されるHPV16−E7特異的脾臓細胞の発生頻度は、CTLエピトープE49〜57のみを有するCyaA−E749〜57により誘導されるものよりも優れていた。これらの観察により、CyaAによりCTLおよびThエピトープを同時に送達することにより、より強力なCTL応答がもたらされることが示された。これは、他のモデルにおいて、前臨床(37)および臨床レベル(38)で以前に公表されたデータと一致した。組換えCyaAによるHPV16−E7Thエピトープの送達は、さらに、インビトロで組換えHisTag−HPV16−E7タンパク質またはE743〜77ペプチドで再刺激したHPV16−E7特異的脾臓細胞により産生されるサイトカインの解析により証明された。実際に、本発明者らは、Thエピトープを含む組換えHPV16−E7CyaAでワクチン接種したマウスのみにおいて、IFN−γの特異的合成を観察した。本発明者らが、CyaA−E7FullおよびCyaA−E7Δ30〜42の1回静注により免疫化した後に観察した、IFN−γレベルが高いことおよびIL−5が全く分泌されないことにより特徴づけられる典型的なTh1プロファイルは、腫瘍免疫療法におけるこのベクターの関心の高さを強調する。
【0142】
これは、皮下で確立された腫瘍原性TC−1細胞に基づいた腫瘍拒絶モデルで試験した(21)。HPV16−E7組換えCyaAの免疫原性を実証する本発明者らのデータによれば、本発明者らは、これらの組換えタンパク質が、確立されたTC−1腫瘍の減退を誘導できることを観察した。CyaA−E7Δ30〜42は、90日間の期間におよび、生存の点で、CyaA−E749〜57およびCyaAE5−HPV16−E7Fullよりも優れていた。完全およびΔ30〜42E7タンパク質を有するCyaAは、CTL刺激能力、HPV16−E749〜57特異的脾臓細胞の発生頻度、およびIFNγ産生の点で、匹敵する結果を与えたので、本発明者らは、生存の点で、CyaA−E7Fullは、CyaA−E749〜57よりも優れていると期待した。より多くの数の試験動物が、この一見した矛盾を除外してくれるのに役立つ可能性が高い。しかし、CyaAの生化学に関する2つの側面をここで考察すべきである。第一に、CyaAの224〜235の領域の負に荷電したアミノ酸の存在は、真核細胞のサイトゾルへのCyaAの触媒ドメインの転位を阻害することが示された(39)。これに関して、酸性E7タンパク質(pKi=4.17)が、DCのサイトゾルへのCyaAのN末端ドメインの効率的な転位を妨害する可能性があった。CyaA−E7Δ30〜42は、DCへの送達に好ましいように、30〜42残基に位置する負に荷電したアミノ酸のストレッチ(DSSEEEDEIDGPA)を除去するように特別に設計され、さらに、2個の正に荷電したアミノ酸(KR)を、HPV16−E7の挿入されたN末端ドメインの各側に導入した。第二に、Gmiraら(25)は、CyaA内に挿入された異種タンパク質の折りたたみ解除は、組換えタンパク質が標的細胞へと内部移行するために必須であることを実証した。E7ポリペプチドの2個の異なる断片を、CyaA−E7Δ30〜42の2個の異なる許容部位へ挿入することにより、E7の再折りたたみが防がれ、したがって、標的細胞へのその転位は促進されるはずである。
【0143】
腫瘍拒絶実験の時間経過中に、何匹かのマウスは、確立されたTC−1腫瘍を以前に拒絶した後、より後になってHPV16陽性腫瘍を増殖させ始めた。FACS(登録商標)解析により、これらの腫瘍の細胞は、H−2Db分子を発現していなかったことが判明した。この観察により、より著しく腫瘍細胞を根絶し、エスケープ機序による腫瘍の再発を防ぐために、組換えCyaAワクチン接種の同様なブースター投与の可能性が考えられた。当該技術分野における他のチームのデータを心に置くと(37、40、41)、それは、注射された組換えHPV16−E7 CyaAの全量を100μg、すなわち0.56nmolに上げた、マウスにおけるCyaAワクチン接種のブースター投与に関連していた。この後者の観察を試験するための実験、並びに、CyaAの様々な投与法を試験するための実験を実施した。
【0144】
TC−1細胞で再攻撃した時、HPV16−E7組換えCyaAで免疫化した生存マウスは、選択的に防御された。これは、これらの動物の脾臓細胞中のHPV16−E749〜57CD8+T細胞の存在に関連していた。この観察は、図4で観察される遅い再発は、腫瘍エスケープ機序に起因し、E7タンパク質に対する免疫が徐々に衰えていくことに起因するものではないということが強まった。Thエピトープを含む組換えCyaAで免疫化したマウスのより良好な生存率により、同じ抗原に対するT細胞の支持が得られることにより、HPV16−E749〜57CD8+T細胞が効果的に想起されることが示された。これに関して、CD4+T細胞は、CD40Lを通じて、エフェクターCD8+T細胞に独特な分子的サインを刷り込む可能性があり、より向上した細胞機能を行なうための能力を付与する可能性があることが提唱された(42)。
【0145】
HPV関連新生物形成の処置のための新規免疫療法の試験のための確証されたモデルにおいて(21)、本発明者らは、CyaAが、確立腫瘍の減退を誘導し、並びに、長い期間におよぶ腫瘍原性攻撃に対する防御を与えるための、効率的なベクターであることを実証した。現在開発されている他のアプローチとは異なり(11)、CyaAを基礎とした免疫療法は、完全なタンパク質を挿入できるので、HLA拘束性エピトープを選択する必要がなく、ウイルスベクターおよび/または発癌の可能性のあるHPV DNA配列の使用が回避された。さらに、本発明者らは、CyaAの2個の異なる許容部位に挿入されたHPV−E7の2個の亜断片を含むCyaAを用いて最善の結果を得た。
【0146】
この後者の構造物が、文献に記載の全てのHPV16−E7 HLAクラスIおよびクラスIIエピトープを含むという事実により(8、46)、ワクチン適用にCyaA−E7Δ30〜42が選択されることが強まった。
【0147】
ここに提示したデータに基づいて、本発明者らは、HPV感染に関連した子宮頚部および肛門の異常形成を標的化した臨床試験において、HPV16−E7を含むCyaAの効力を試験する計画を立てた。
【0148】
実施例2
目的は、ヒトにおけるHPV16およびHPV18関連の悪性疾患を処置するための、HPV16およびHPV18 E7タンパク質の両方に標的化した二価治療ワクチンを製造することであった。それ故、CyaAE5−HPV16E7Δ30〜42およびCyaAE5−HPV18E7Δ32〜42と呼ばれるワクチン候補を、設計し、作成し、産生し、精製した。HHDマウスは、H−2Dbのα3膜貫通ドメインおよび細胞質ドメイン(D)に連結したHLA*0201のα1(H)およびα2(H)ドメインを含むHHD導入遺伝子を発現しているH−2D−/−β2m−/−ダブルノックアウトマウスであった(α1ドメインは、ヒトβ2−ミクログロブリンに連結している)。したがって、HHDマウスにより発現されるMHCクラスI分子のみが、改変されたHLA*0201分子であった(Pascolo et Lemonnier)。
【0149】
この実験の目標は、
1 組換えCyaAが、HPV16およびHPV18E7タンパク質のHLA−A2拘束性エピトープを送達できること、
2 1個のHPV組換えcyaAが他を上回る免疫優性の現象はないこと
を実証することであった。
【0150】
そうするために、HHDマウスに、50μgのCyaAE5−HPV16E7Δ30〜42(3匹のマウス)、CyaAE5−HPV18E7Δ32〜42(3匹のマウス)、またはCyaAE5−HPV16E7Δ30〜42+CyaAE5−HPV18E7Δ32〜42(5匹のマウス)を同じ(眼窩後の)注射(200μl中50μg)で静脈内にワクチン接種した。7日後、プールした脾臓細胞を、インビトロで、文献に記載またはSYFPEITHIソフトウェアから推定される、HPV16E7またはHPV18E7のHLA−A2ペプチドで再刺激した。
【0151】
CTL活性は、51Cr放出を使用して5日後にアッセイした。標的細胞は、異なる関連ペプチドをのせたまたはのせていない、HHD−EL4細胞であった。
【0152】
結果により、CyaAE5−HPV16E7Δ30〜42またはCyaAE5−HPV18E7Δ32〜42でのワクチン接種は、インビトロでの自己ペプチドによる再刺激後に、HPV16E7またはHPV18E7ペプチドをのせたEL4−HHD細胞の特異的溶解を誘導したことが示された。
【0153】
この結果によりまた、CyaAE5−HPV16E7Δ30〜42をCyaAE5−HPV18E7Δ32〜42と共に注射することにより、関連ペプチドに対する類似の応答が観察されたので、いずれのHPV組換えCyaAの免疫原性も損なわなかった。
【0154】
この結果により、CyaAE5−HPV16E7Δ30〜42およびCyaAE5−HPV18E7Δ32〜42は、インビボにおいて、それぞれのE7タンパク質のヒトHLA−A2拘束性エピトープに対する細胞毒性応答を誘導できることが実証された。
【0155】
マウス
特定の病原体を含まないHHDマウスを、パスツール研究所で育成した。11匹の6〜10週令の雄を、この実験に使用した。
【0156】
試薬および生物学的材料
試薬および緩衝液
RPMI1640培地−グルタマックス(インビトロジェン・ギブコ、参照番号:6187010)
エタノール70゜(Prolabo、参照番号:MC311631)
ペニシリン−ストレプトマイシン(インビトロジェン・ギブコ、参照番号:15140122)
ウシ血清アルブミン(FBS)(PERBIO、参照番号:CH30160.03)
βメルカプトエタノール(バイオラッド、参照番号:161−0710)
熱分解水
ブルートリパン(シグマ、T−8154)
51Cr
Triluxシンチラント(Wallac)
【0157】
ペプチド
5個の合成ペプチド(Neosystem、ストラスブール、フランス)を、51Cr放出アッセイの前の脾臓細胞のインビトロでの刺激に使用した:
HPV16−E7 HLA−A2拘束性エピトープ(1)に対応する、E711〜20(YMLDLQPETT、一文字でアミノ酸をコード、#253)、
HPV16−E7 HLA−A2拘束性エピトープ(1)に対応する、E782〜90(LLMGTLGIV、#258)、
HPV16−E7 HLA−A2拘束性エピトープ(1)に対応する、E786〜93(TLGIVCPI、#255)
SYFPEITHIソフトウェアにより予測されるHPV18−E7 HLA−A2拘束性エピトープに対応する、E77〜15(TLQDIVLHL、#251)、
HPV18−E7 HLA−A2拘束性エピトープ(2)に対応する、E786〜94(FQQLFLNTL、#257)。
【0158】
ペプチドは、無菌で無発熱原性の水(#253、#255、#251)または無発熱原性の水、0.1M NaHCO3;アセトニトリル(50/50)(#257および#258)で1mg/mlに希釈した。
【0159】
組換えアデニル酸シクラーゼ(CyaA)
2個のCyaAをこの実験において試験した:
CyaA−HPV16E7Δ30〜42
CyaA−HPV18E7Δ32〜42。
【0160】
これらのCyaAは、E.coli株BLRにおいて、酵素的に不活性なCyaA(CyaAE5)をコードするプラスミドpTRACE5の誘導体を使用することにより発現させた。プラスミドの作成、全ての組換えタンパク質の産生および精製は、HPV16−E7エピトープを有する組換えB.pertussisアデニル酸シクラーゼの作成および精製に関する実施例1において先に記載したように製造したが、エピトープは、本実験で使用したものであった。
【0161】
8M尿素中にそれぞれ1.22mg/mlおよび1.33mg/mlのCyaAを含むストック溶液(−20℃で保存)を解凍し、マウスに静注内投与する前に、PBS(ギブコBRL)で250μg/mlに希釈した。したがって、尿素の最終濃度は、それぞれ、1.6Mから1.5Mの間であった。
【0162】
細胞系
EL4−HHD細胞を、51CrCTLアッセイにおいて標的として使用した。これらの細胞を、完全培地中に維持した:10%熱不活性化FCS、100U/mlのペニシリン、100μg/mlのストレプトマイシン、および5×10−5Mのβ2−メルカプトエタノール(ギブコBRL、Cergy-Pontoise、フランス)を補充したグルタマックスを含むRPMI1640。
【0163】
方法
マウス免疫化
覚醒動物に、0.3mlのインスリンシリンジ(テルモ)を用いて、(200μl中50μgの)CyaAE5−HPV16E7Δ30〜42および/またはCyaAE5−HPV18E7Δ32〜42の1回の静脈内(眼窩後の)注射により免疫化した。
【0164】
実験計画
表1は、各群のマウスに投与するワクチン候補および処置を記載した。
【0165】
【表9】
【0166】
CTLアッセイ
免疫化により誘導される細胞性免疫応答をインビトロで評価するために、麻酔した動物(CO2)を、免疫化の7日後に脾摘除した。CTLアッセイの前に、各群の脾臓細胞をプールした。
【0167】
−CTLの刺激(1個の脾臓あたり2個のフラスコT25を準備)
−脾臓をサンプリングし、RPMI1640−グルタマックス1%Ab中でそれらを破粉した。
−デカント。
−1200rpmで10分間遠心分離。
−細胞を、2mlのRMPI1640−グルタマックス1%Ab−10%FCS−5.10−5M β−メルカプトエタノール(完全培地:CM)に再懸濁。
−各々のT25フラスコに、
10mlのCM
5×107個のエフェクター細胞
10μg/mlの最終濃度の関連ペプチド
を入れた。
フラスコを動かすことなく5日間インキュベート。
【0168】
−細胞毒性アッセイ
1)標的細胞:
−前日、標的細胞を3分の1または2分の1に希釈して、指数関数的な増殖期のそれらを収集。
−15mlのチューブに移す。
−1200rpmで10分間遠心分離。
−1mlのRPMI1640−1%Ab中に再懸濁。
−数を数えた。
−2個のチューブを準備:1個はペプチドを含み、1個はペプチドを含まない。
−150μl中に以下を再懸濁:
2個の中の1個のチューブにペプチド(50μモル)
3×106個の細胞につき100μCiの51Cr(50μl)。
150μlとするための培地。
−37℃の水浴中で1時間インキュベートし、15分毎に穏やかに振とう。
【0169】
2)エフェクター細胞
−約5mlの上清をパスツールピペットを用いて廃棄。
−ピペッティングにより細胞を完全に再懸濁。
−1200rpmで10分間遠心分離。
−1mlのCMに再懸濁。数を数えた。
−107個の細胞/mlに調整。
−次のエフェクター/標的比:200/1、100/1、50/1、25/1、12/1、6/1が得られるように希釈。
−100μl/U底マイクロタイタープレートのウェルで分配。
−標的細胞の調製を完成する間、37℃で7.5%CO2中でインキュベート。
【0170】
3)アッセイ:
−標的細胞を、10mlのRPMI1640−1%Abで洗浄。
−再度10mlのCMで洗浄。
−2mlのCMに再懸濁。
−数を数えた。
−105個の細胞/mlに調整。
−100μl/ウェルで分配。
−自発的放出のための6個のウェルを準備(CM中100μlに調整)。
−最大放出のための6個のウェルを準備(100μlの20%トリトンX−405を添加)
−37℃で7.5%CO2中で4〜5時間インキュベート。
【0171】
4)計測:
−2000rpmで5〜10分間遠心分離。
−100μlのTriluxシンチラントを、フロッピーP96マイクロタイタープレートに入れた。
−50μlの上清をサンプリングし、それをフロッピープレートに移した。
−プレートを、プラスチックフィルムテープで封をした。
−wallacカウンターで次の日に計測した。
【0172】
HPV16E7CTL特異的応答
CyaA−HPV16E7Δ30〜42により誘導されるCTL応答を図11に示した。
【0173】
HPV18E7CTL特異的応答
CyaA−HPV18E7Δ32〜42により誘導されるCTL応答を図12に示した。
【0174】
結論
CyaA−HPV16E7Δ30〜42およびCyaA−HPV18E7Δ32〜42の両方が、このキメラHHDモデルにおいて、それぞれのHLA−A2拘束性ペプチドに対して特異的なCTLを誘導できた。
【0175】
CyaA−HPV16E7Δ30〜42およびCyaA−HPV18E7Δ32〜42の同時注射により、各々の個々のCyaAがそれぞれの特異的HLA−A2拘束性ペプチドに対するCTLを誘導する能力は妨害されなかった。これにより、1つの構造物が他の構造物よりも上回る免疫優性を示す現象はなかったことが示された。これには、二価ワクチンの製造戦略に関する、重要な観察が含まれていた。
【0176】
実施例3
HPV18のE7タンパク質並びにHPV16およびHPV18のE7タンパク質を含む組換えアデニル酸シクラーゼの作成および免疫学的評価
肛門生殖管の癌は、女性における全ての癌の12%近くを占め、子宮頚部癌(CxCa)は世界中で二番目に最も頻繁に起こる婦人科の癌となっている。ヒトパピローマウイルス(HPV)感染がCxCaの原因である可能性があるという重要な観察は、その後、疫学的な研究により確認された。CxCaに関連する最も広まっているHPVのタイプは、HPV16およびHPV18である(それぞれ55%および12%の罹患率)(Clifford、2003)。より多くの人口を網羅するために、HPV16およびHPV18のE7を有する二価治療ワクチンを作成することが決定された。2つの可能性ある戦略が試験された:(i)第一の戦略は、一方はHPV16のE7を有する他方はHPV18のE7を有する、等モル量の2個の組換えCyaAを混合する;(ii)第二の戦略は、HPV16およびHPV18の両方のE7タンパク質を有する組換えCyaAを作成する。本リポートは、HPV18のE7または両方のウイルスのE7を有する組換えCyaAの作成を記載した。構造物の免疫原性は、CTLアッセイにおいて試験した。
【0177】
エピトープ同定
HPV18E7のアミノ酸配列を、H−2bエピトープの探索中に、SYFPEITHI(www.syfpeithi.de)にかけた。このソフトウェアは、アルゴリズムを使用して、画定されたMHC分子に結合する可能性があるエピトープを予測する。ソフトウェアは、25のスコアで、H−2Db分子により拘束される推定エピトープを選出した。通常、本発明者らは、そのスコアが22を超えた場合にエピトープであると判断した。それ故、推定ペプチドのIDGVNHQHLが、Neosystemにより合成された。
【0178】
組換えCyaAの作成
CyaA−HPV18E7、CyaA−HPV18E7Δ32〜42、CyaA−HPV16+18E7、CyaA−HPV16+18ΔE7を産生するためにプラスミドの作成に使用された戦略を示したスキームを、図13および14に示した。
【0179】
CTLアッセイ
この実験の目標は、インビボにおいて、完全なHPV18E7タンパク質またはその断片並びにHPV16およびHPV18E7タンパク質の両方を有する、組換えCyaAによるCTLの誘導を実証することであった。標的化されたH−2Dbエピトープは、コンピューター予測ソフトウェアから得られたHPV18E741〜49(IDGVNHQHL)およびHPV16E749〜57であった。C57BL/6マウスに、50μgのCyaA−CysOVA、CyaA−HPV18E7、CyaA−HPV18E7Δ32〜42、CyaA−HPV16+18E7、またはCyaA−HPV16+18ΔE7(各群において2匹)を静注によりワクチン接種した。7日後、プールした脾臓細胞を、インビトロで、ペプチドHPV18E741〜49(10μg/ml)、OVA257〜264(1μg/ml)またはHPV16E743〜77(1μg/ml)で再刺激した。CTL活性は、51Cr放出を使用して5日後にアッセイした。標的細胞は、HPV18E741〜49ペプチド(8nmol)をのせたまたはのせていないEL4細胞、あるいは、TC−1細胞であった。図中の説明文は:ワクチン接種/再刺激を示す。例:OVA/OVAは、CyaAE5−CysOVAでワクチン接種し、OVAペプチドで再刺激したマウスを意味する。
【0180】
これらの結果(図16)により、CyaA−HPV18E7Fullとは対照的に、CyaA−HPV18E7Δ32〜42は、インビボにおいて、処理およびMHCクラスI経路に提示するために、コンピューターで予測したHPV18−E7タンパク質のCD8+H−2Db拘束性T細胞エピトープを、免疫担当細胞のサイトゾルへと送達でき、これにより、強力なCTL応答を誘起できることが示された。CyaA−HPV18E7Δ32〜42が、HPV18E741〜49に対するCTL応答を刺激できることは奇妙である。なぜなら、この組換えCyaAは、ペプチドHPV18E741〜49の最初の2個のアミノ酸を欠失しているからである。しかし、HPV18E7断片43〜105の挿入部位においては、1個のアラニンおよび1個のセリンがあり、これにより、推定ペプチドは、ここで、IDGVNHQHLではなく、ASGVNHQHLであった。syfpeithiにかけると、このペプチドは、29のスコアで出て来た(天然のものは25)。それ故、本発明者らのクローニングにより、このペプチドの最初のアミノ酸を置換することにより、本発明者らは、潜在性から免疫原性へとすることができるようであった。これは、HPV18E7タンパク質のエピトープが、H−2Dbの関連で記載された始めてのケースであった。
【0181】
非特異的バックグラウンドレベルが高いにも関わらず、これらの結果によりまた、CyaA−HPV16+18E7FullおよびCyaA−HPV16+18E7Δは、インビボにおいて、処理およびMHCクラスI経路に提示するために、HPV16−E7タンパク質のH−2Db拘束性T細胞エピトープを、免疫担当細胞のサイトゾルへと送達でき、これにより、強力なCTL応答を誘起できることが示された。CyaA−HPV16+18E7Δは、CyaA−HPV16+18E7Fullと比べて高いその能力を示した。これらのデータによりまた、初めて、大きなポリペプチド断片(203までのアミノ酸)を有するCyaAは依然として免疫原性であることが示された。
【0182】
さらなる実験を実施し、図16に記載のことを確認した。各群の残りの2匹のマウスを、最初のものとは別々に処置した。実験設定は類似していた。結論は、CTLによるCyaAE5−HPV18E7Δ32〜42ワクチン接種マウスのEL4細胞の高い非特異的溶解レベルは、過剰のHPV18E741〜49ペプチドがフラスコに残っていることに起因することを除いて、図16のものと同一であった。
【0183】
またCyaA−HPV16+18ΔE7でワクチン接種したマウスの脾臓細胞ではTC−1細胞の溶解は認められたが、今回CyaA−HPV16+18E7でワクチン接種したマウスには認めれられなかった。この実験において、非特異的バックグラウンドは40%でピークに達した。それにも関わらず、図17から導き出せる結論は、図16から導いた結論を確認した、すなわち:
−CyaA−HPV18E7Δ32〜42は、インビボにおいて、処理およびMHCクラスI経路への提示のために、コンピューターで予測したHPV18−E7タンパク質のCD8+H−2Db拘束性T細胞エピトープを、免疫担当細胞のサイトゾルへと送達でき、これにより強力なCTL応答を誘起できる。これは、HPV18E7タンパク質のエピトープが、H−2Dbの関連で記載された最初のケースであった。
−大きなポリペプチド断片(203までのアミノ酸)を有する組換えCyaAは、依然として免疫原性であった。
【0184】
結論
HPV16およびHPV18のE7を有する二価治療ワクチンを作成することが決定された。2つの可能性ある戦略が考えられた:(i)第一の戦略は、一方はHPV16のE7を有する他方はHPV18のE7を有する、等モル量の2個の組換えCyaAを混合する;(ii)第二の戦略は、HPV16およびHPV18の両方のE7タンパク質を有する組換えCyaAを作成する。本結果は、HPV18のE7または両方のウイルスのE7を有する組換えCyaAの作成を記載した。構造物の免疫原性は、CTLアッセイにおいて試験した。
【0185】
本研究の主な結論は、考えられている両方の戦略が、実現可能であることである。なぜなら、HPV18E7の配列内に潜在性H−2Db拘束性エピトープが初めて判明したため、HPV18E7亜断片を有する組換えCyaAが機能的であるからである。さらに、HPV16および18ウイルスの両方のE7タンパク質(または亜断片)を有する組換えCyaAは、TC−1細胞により天然に提示されるH−2Db拘束性HPV16E7エピトープに対するCTL応答を刺激できるので、依然として最も興味深いことに免疫原性であった。
【0186】
【表10】
【図面の簡単な説明】
【0187】
【図1A】HPV16−E7組換えCyaAの作成および精製。(A)関連する制限酵素部位および挿入配列が示されている、pTRACE5のマップ図。(B)CyaAの酵素活性を阻害するジペプチドLQの挿入部位を示すCyaAの図解。HPV16−E7タンパク質挿入断片の位置も示されている。HPV16−E7 H−2b拘束性エピトープには下線を付している。
【図1B】HPV16−E7組換えCyaAの作成および精製。(C)HPV16−E7組換えCyaAのSDS−PAGE解析。5μgの精製タンパク質を、4〜15%SDSポリアクリルアミドゲル上で分離し、クーマシーブルーにより染色した。レーン1:野生型CyaA;レーン2:CyaA−E749〜57;レーン3:CyaA−E7FUll;レーン4:CyaA−E7Δ30〜42。(D)HPV16−E7組換えCyaAのウェスタンブロット解析。SDS−PAGE後、精製タンパク質を、ニトロセルロース膜にエレクトロトランスファーし、続いてこれをマウスモノクローナル抗HPV16−E7抗体でプローブした。レーン1、2:野生型CyaA(それぞれ2および0.4μg);レーン3、4、および5:それぞれ、CyaA−E749〜57、CyaA−E7FUllおよびCyaA−E7Δ30〜42、各々0.4μgのタンパク質。
【図2】組換えHPV16−E7CyaAによるT細胞応答の誘導。(A)C57BL/6(a、b、c)、TAP1−/−(d)、MHCクラスII−/−(e)、およびCD40−/−(f)マウスを、0日目に、50μgのCyaA−E749〜57(a)、CyaA−E7Full(b)、またはCyaA−E7Δ30〜42(c、d、eおよびf)の静注により免疫化した。数日後、動物を屠殺し、脾臓細胞を、インビトロで、1μg/mlのHPV16−E743〜77ペプチドを用いて、照射された同系脾臓細胞の存在下で5日間再刺激し、TC−1標的細胞(普通の四角)またはEL4(白抜きの四角)に対するエフェクターとして使用した。非関連エピトープOVA257〜264を有するCyaAE5−cysOVAで処置し、インビトロで1μg/mlのHPV16−E743〜77ペプチドを用いて、照射された同系脾臓細胞の存在下で5日間再刺激したマウスの脾臓細胞も表示する(a、普通の三角)。標的溶解は、51Cr放出により評価した。データは、特異的溶解値の中間パーセンテージ(n=動物の数を各グラフに示す)並びに四分位範囲を示す。(B)組換えHPV16−E7CyaAでの免疫化後のHPV16−E7特異的IFN−γ産生細胞の検出。C57BL/6マウスを、Aと同様に、CyaAE5−cysOVA(丸)、CyaA−E749〜57(四角)、CyaA−E7Full(三角)、またはCyaA−E7Δ30〜42(菱型)を用いて免疫化した。数日後、免疫化マウスから単離した脾臓細胞を、インビトロで、刺激せずに(すなわち、ペプチドなし、白抜きの記号)、または、1μg/mlのE749〜57ペプチド(普通の記号)を用いて刺激して、同系照射脾臓細胞の存在下で36時間培養した。データは、脾臓1個あたりのSFCの数として表現し、3回の独立した実験の個々のマウスの結果を、各群に対して示す。横棒は、各群の応答の中間値を示す。
【図3】組換えHPV16−E7CyaAは、HPV16E7特異的Th1応答を誘導する。(A)C57BL/6マウスを、未処置のままにするか(四角)、または、50μgのCyaAE5−cysOVA(丸)、CyaA−E749〜57(三角)、CyaA−E7Full(逆三角)、もしくはCyaA−E7Δ30〜42(菱型)を用いて静注により抗原刺激した。数日後、脾臓細胞を、インビトロで、10μg/mlのHis−Tag−HPV16−E7タンパク質で刺激し、上清を、IFN−γ含量について72時間後に試験した。4回の独立した実験の個々のマウスの結果を示し、2個の複製ウェルの上清に放出されたIFN−γの濃度として表現する。再刺激しなかった脾臓細胞で得られたバックグラウンドを差し引く。挿入図:1μg/mlのE743〜77ペプチドを用いたインビトロでの刺激。横棒は、各群の応答の中間値を示す。(B)上清をIL−5含有量について試験したこと以外は、(A)と同様。2回の独立した実験の個々のマウスの結果を示し、2個の複製ウェルの上清中に放出されたIL−5の濃度として表現する。
【図4A】組換えHPV16−E7CyaAを用いた治療的ワクチン接種は、確立された腫瘍を根絶する。(実験A)C57BL/6マウスを、0日目に、5×104個のTC−1腫瘍細胞を用いて移植した。10日目に、マウスを、CyaA−E749〜57(C)、CyaA−E7Full(D)、またはCyaA−E7Δ30〜42(E)の1回の静注により処置した。未処置のままのマウス(A)またはCyaAE5−cysOVAを注射したマウス(B)を対照として採用した。腫瘍サイズが1000mm3に達した時、または不必要な苦しみを避けるために動物の衛生状態により要求される場合にはいつでも(壊死腫瘍、20%を超える急速な体重減少)、マウスを屠殺した。後に進行性の腫瘍を発症したCyaA−E7Fullで処置した2匹のマウス(*)を、さらに調べるために屠殺した(図6参照)。
【図4B】組換えHPV16−E7CyaAを用いた治療的ワクチン接種は、確立された腫瘍を根絶する。(実験B)実験設定については(実験A)と同様。+10および+17日目に、10μgのCyaAE5−CysOVA(a、実線)または10μgのCyaA−E7Δ30〜42(b)を用いて耳真皮に治療的ワクチン接種を実施した。各曲線は、1匹の動物における腫瘍増殖を示す。2匹の未処置の動物が含まれていた(a、点線)。各々の四分の一区の右上には(a、b)、屠殺動物の数と、含まれる動物の全数が示されている。これらのマウスの生存曲線を示す(c)。未処置(白抜きの三角)、CyaAE5−CysOVAを用いたニセ処置(閉じた三角)、CyaA−E7Δ30〜42を用いた処置(丸)。
【図5】組換えHPV16−E7CyaAでの治療的ワクチン接種により、生存が延長される。治療的ワクチン接種は、図4に記載した通りに行なった。TC−1腫瘍細胞の注射時に、マウス(1群あたり5〜10匹)を、グラフに示されているように+1、+5、または+10日目にHPV16−E7組換えCyaAで免疫化した。マウスを未処置のまま(普通の四角、実線)、CyaAE5−cysOVAでニセ処置(白抜きの四角、点線)、またはCyaA−E749〜57(白抜きの三角)、CyaA−E7Full(白抜きの丸)、もしくはCyaA−E7Δ30〜42(白抜きの菱型)で処置した。+5日目の治療実験において、CyaA−E7FullおよびCyaA−E7Δ30〜42で処置した動物の生存曲線は、完全に重ね合わさることが特筆される。どの場合においても、組換えHPV16−E7CyaA処置動物の生存は、未処置またはニセ処置マウスと比べて有意に延びている(p<0.05)。
【図6】その後に増殖してきた腫瘍から外移植したTC−1腫瘍細胞は、H−2Db分子の発現を欠失している。腫瘍拒絶実験において、CyaA−E7Fullをワクチン接種した数匹の動物では、実験の時間経過中、後になって腫瘍が成長した(図4、*)。腫瘍を外移植するために、2匹の動物を屠殺した。これらの腫瘍細胞、TC−1A1およびA2並びに天然TC−1細胞を、H−2Db分子の発現レベルについてFACS(登録商標)により解析した(太線)。蛍光強度の中間値(MedFi)を示す。アイソタイプ対照で得られた結果を示す(グレーの陰影)。
【図7】組換えHPV16−E7CyaAで処置したマウスにおけるHPV16−E749〜57特異的CD8+T細胞の持続性。治療的実験セットにおいて、CyaA−E749〜57(A)、CyaA−E7Full(B)、またはCyaA−E7Δ30〜42(C)で免疫化し、TC−1移植にも関わらず生存しているC57BL/6マウスを屠殺し、脾臓細胞を、インビトロで、1μg/mlのHPV16−E743〜77ペプチドで、照射同系脾臓細胞の存在下で5日間再刺激した。標的溶解(TC−1、普通の四角;EL4、白抜きの四角)を、51Cr放出により評価した。データは、非特異的溶解の数値のパーセンテージの中間値(各群においてn=6)、並びに、四分位範囲を示す。エフェクターと標的の最大比において20%以上の特異的溶解により決定される対応する動物の数を、四分の一区の右上部分に示す。
【図8】組換えHPV16−E7CyaAにより誘導されるTC−1腫瘍増殖に対する長期防御。治療的実験セットにおいて、TC−1移植にも関わらず生存しているC57BL/6マウスに、100日目に、5×104個のTC−1細胞を再移植した。年齢の適合した未処置マウスを、対照として採用した(A)。CyaA−E749〜57、CyaA−E7Full、およびCyaA−E7Δ30〜42で最初に免疫化したマウスにおける腫瘍の増殖を示す(それぞれ、B、C、およびD)。腫瘍サイズが1000mm3に達した時、または動物の衛生状態により要求される場合にはいつでも、マウスを屠殺した。(E)未処置(白抜きの四角);もしくはCya−E749〜57(白抜きの三角);CyaA−E7Full(白抜きの丸);もしくは、CyaA−E7Δ30〜42(白抜きの菱型)で免疫化し、TC−1細胞を再移植した(移植日を0とする)動物の生存曲線。いずれの場合においても、組換えHPV16−E7CyaA処置動物の生存は、未処置またはニセ処置マウスに比べて有意に延びた(p<0.05)。
【図9】CyaA−E7Δ30〜42治療活性と、CpG ODN1826アジュバントの添加されたHPV16E743〜77の治療活性との比較。C57BL/6マウスに、0日目に、5×104個のTC−1腫瘍細胞を皮下移植した。+10および+17日目に、マウスに、10μgのHPV16−E743〜77(n=5、三角)、1μgのCpG−ODN1826(n=5、四角)、10μgのHPV16−E743〜77+1μgのCpG−ODN1826(n=5、菱型)、10μgのCyaA−CysOVA(n=3、逆三角)、または10μgのCyaA−E7Δ30〜42(n=7、丸)を1回皮内注射して処置した。腫瘍サイズが1000mm3を超えた時、または動物の衛生状態により要求される場合にはいつでも、マウスを屠殺した。
【図10】CyaA−E7Δ30〜42のTC−1腫瘍拒絶誘導能に対する、CyaAに対する予備免疫の効果の解析。(A)C57BL/6マウスは未処置のままとするか、または、90もしくは30日前に、7日間の間隔で10μgのCyaAE5を2回皮内注射することにより免疫化した。1日前に、動物の採血をし、血清を個々に抗CyaAE5 IgGの存在についてELISAにより評価した。結果は、希釈対A492をプロットした線形回帰分析により計算した個々の抗体力価として表現する。横棒は、各群の応答の中間値を示す。(B)未処置(a、b)、30日前にCyaAE5で免疫化(c、d)、および90日前にCyaAE5で免疫化した(e、f)動物に、0日目に、5×104個のTC−1腫瘍細胞を移植し、+10および+17日目に、10μgのCyaA−cysOVA(a、c、e)、または10μgのCyaA−E7Δ30〜42(b、d、f)を1回皮内注射することにより処置した。挿入図(b、d、f)は、0〜35日間の期間のクローズアップであり、全ての動物が、ワクチン接種をした時点で触診できる腫瘍を有していたことを示す。各曲線は、1匹の動物における腫瘍増殖を示す。腫瘍サイズが1000mm3を超えた時、または動物の衛生状態により要求される場合にはいつでも、マウスを屠殺した。各々の四分の一区の右上には、屠殺動物の数と、含まれる動物の全数が示されている。
【図11】HHDマウスにおけるCyaA−HPV16E7Δ30〜42によるCTL応答の誘導。EL4−HHD細胞へのペプチドの添加を、各グラフの上に示す。続く表には、カラムヘッダーVaccの下で注入されたCyaAのタイプ(HPV17E7またはHPV18E7に関する)、および、カラムヘッダーStimの下でインビトロでの再刺激に使用されたペプチドが示されている。見てとれるように、CyaA−HPV16E7Δ30〜42による免疫化後、本発明者らは、ペプチド#253のみに特異的なCTLを誘導することができた(右パネル)。インビトロでペプチド#255で再刺激した脾臓細胞は、ペプチド#253で被覆したEL4−HHD細胞に対して細胞毒性ではなかったため、このCTL活性は特異的であった(右パネル)。2個のその他のHLA−A2拘束性ペプチドに対するCTL特異的応答がないことは、異なる現象から生じている可能性がある:(i)ペプチド#253の免疫優性、(ii)EL4−HHD細胞のプロテアソームによりペプチド#255および#258が処理されないこと、(iii)ペプチド#258の溶解度が低く、このため本発明者らはアセトニトリル(50%)を使用しなければならず、これは細胞に対して毒性である可能性がある。最も興味深いことには、CyaA−HPV18E7Δ32〜42の同時注射は、CyaA−HPV16E7Δ30〜42のCTL誘導能を妨害しなかった(右パネル)。
【図12】HHDマウスにおけるCyaA−HPV18E7Δ32〜42によるCTL応答の誘導。EL4−HHD細胞へのペプチドの添加を、各グラフの上に示す。以下の表には、カラムヘッダーVaccの下で注入されたCyaAのタイプ(HPV16E7またはHPV18E7に関する)、および、カラムヘッダーStimの下でインビトロでの再刺激に使用されたペプチドが示されている。見てとれるように、CyaA−HPV16E7Δ30〜42による免疫化後、本発明者らは、ペプチド#251のみに特異的なCTLを誘導することができた(左パネル)。インビトロでペプチド#257で再刺激した脾臓細胞は、ペプチド#251で被覆したEL4−HHD細胞に対して細胞毒性ではなかったため、このCTL活性は特異的であった(左パネル)。HPV16E7 HLA−A2拘束性ペプチドと同様に、HPV18E7#257HLA−A2拘束性ペプチドに対するCTL特異的応答がないことは、異なる現象から生じている可能性がある:(i)ペプチド#251の免疫優性、(ii)EL4−HHD細胞のプロテアソームによりペプチド#257が処理されないこと、(iii)ペプチド#257の溶解度が低く、このため本発明者らはアセトニトリル(50%)を使用しなければならず、これは細胞に対して毒性である可能性がある。最も興味深いことには、CyaA−HPV16E7Δ30〜42の同時注射は、CyaA−HPV18E7Δ32〜42のCTL誘導能を妨害しなかった(左パネル)。
【図13】pTRACE5−E7HPV18の作成。
【図14A】pTRACE5−E7ΔHPV16+18の作成。 3つのプラスミド調製ステップを開示する。
【図14B】pTRACE5−E7ΔHPV16+18の作成。 3つのプラスミド調製ステップを開示する。
【図14C】pTRACE5−E7ΔHPV16+18の作成。 3つのプラスミド調製ステップを開示する。
【図15】CyaA−HPV18E7Full CyaA−HPV18E7Δ32〜42 CyaA−HPV16+18E7Full CyaA−HPV16+18E7Δをコードするポリヌクレオチドのマップ。
【図16】C57BL/6マウスにおけるHPV18E7を有する組換えCyaAによるCTLの誘導。
【図17】C57BL/6マウスにおけるHPV18E7を有する組換えCyaAによるCTLの誘導。寄託されている材料(I−3190およびI−3191)は、E.coli株BLRに含まれ、ルリアブロス(LB)培地中で増殖させることができ、100μg/mlのアンピシリンを含むLBに接種し、175rpmで振とうし照射しながら空気中で30℃でインキュベートできる。7〜10%DMSOを含むLB中での一晩の保存が可能である。
【技術分野】
【0001】
本出願は、アデニル酸シクラーゼタンパク質またはその断片に挿入されたパピローマウイルスエピトープを有する組換えタンパク質に関する。
【0002】
したがって、本発明は、アデニル酸シクラーゼ(CyaA)タンパク質がタンパク質ベクターとして作用して、パピローマウイルス抗原、特にヒトパピローマウイルス抗原に由来するエピトープに対する免疫応答を誘起するような、組換えタンパク質に関する。
【0003】
本発明は、特に、エピトープを、真核細胞、好ましくは哺乳動物細胞、特にヒト細胞に送達するための、このようにして得られたタンパク質性ベクターの使用に関する。
【0004】
本発明はまた、本発明の組換えタンパク質をコードするポリヌクレオチドと、前記ポリヌクレオチドを含むベクター、並びに、前記ポリヌクレオチドまたはベクターを含む宿主細胞にも関する。
【0005】
本発明はまた、宿主におけるヒトパピローマウイルス感染の処置または予防のための、並びに、宿主、特に哺乳動物宿主における、ヒトパピローマウイルス感染から生じる悪性作用の処置または予防のための、上記の組換えタンパク質またはポリヌクレオチドの適用にも関する。特定の実施形態において、本発明は、免疫療法、特にパピローマウイルスの腫瘍特異的抗原に対する免疫療法に適した化合物の設計に有用な手段を提供する。
【0006】
数多くのヒトパピローマウイルス(HPV)タイプの中で、ハイリスクHPVと称されるものは、宿主において感染が持続した場合の上皮悪性疾患の発達に関連している(1)。世界中で2番目に最も広まっている婦人科癌である子宮頚部癌(1)では、大抵、HPV16およびHPV18 DNAが検出される(>99%)(2)。これらのウイルスの発癌能は、初期遺伝子、すなわち、E6およびE7遺伝子(その発現は、ウイルスの複製全周期において検出され、悪性形質転換の発症および存続に必要である)により発現される産物に起因する。しかし、これらのハイリスク発癌性HPVタイプによる肛門性器感染が高い頻度で起こることと、HPV関連悪性疾患を最終的に発症する個体の比率は低いこととは対照的であり、このことは、免疫応答によりハイリスクHPV感染が制御されていることを示唆する。いくつかの観察により、例えば、大半の悪性段階以前の病変の自発的減退(1)、CD4+T細胞およびマクロファージによる退行中の性器いぼの浸潤(1)、並びに、免疫抑制患者および免疫欠陥患者においてより多数の感染被験者が認められることなどにより(1)、この所説が強められた。さらに、HPV16−E6および/またはE7エピトープに対するCD4+およびCD8+T細胞応答が、HPV16関連悪性疾患と診断された患者の血液(4〜8)、並びに、健康個体の血液に(9、10)検出された。まとめると、これらの考察には、HPV16のE6および/またはE7タンパク質に標的化する免疫療法の開発に対する強い根拠が含まれていた。
【0007】
H−2Db HPV16−E749〜57拘束性エピトープに対する免疫応答を発生させることにより、C57BL/6マウスにおけるHPV16−E6および−E7陽性腫瘍原性細胞系の腫瘍増殖を予防するための、多くのワクチン戦略が開発されている。これらのワクチン接種アプローチは、プラスミドDNA、ウイルス性または細菌性ベクター、キメラウイルス様粒子、合成ペプチド、および組換えタンパク質を含んでいる(11)。残念なことに、該アプローチは、臨床的退行に関しては控えめに満足できる結果しかもたらさなかった(3)。したがって、細胞性応答を誘起するために、HPV−エピトープを免疫系に標的化させる新規ツールを評価することは依然として感心が高い。
【0008】
したがって、宿主において前記抗原に対する体液性および/または細胞性免疫応答の誘起が可能である条件において、HPV抗原のエピトープを標的細胞に送達するのに適した新規ベクターが必要である。本発明者らは、アデニル酸シクラーゼタンパク質が、このようなベクターを設計する上で興味深いことを見出した。Bordetella pertussisのアデニル酸シクラーゼタンパク質を使用して様々な観察がなされ、これにより、このタンパク質が、このような効率的なベクターの設計に適した基盤となる可能性があるという結論が得られた。
【0009】
Bordetella pertussisのアデニル酸シクラーゼ(CyaA)は、その触媒ドメインを、真核細胞のサイトゾルへと送達することができる(12)。したがって、CyaAの触媒部位に挿入されたCD4+およびCD8+T細胞エピトープは、抗原提示細胞(APC)の表面において、それぞれMHCクラスIIおよびI分子により処理され提示される(13)。さらに、CyaAは、近年、αMβ2インテグリン(CD11b/CD18)に特異的に結合し(14、第WO02/22169号)、これにより、これらのT細胞エピトープをCD11b+樹状細胞亜個体群に標的化することが実証された(15)。マウスを、適切なエピトープを有する組換えCyaAで免疫化すると、強力なCTL応答、致死的なウイルス攻撃に対する完全な防御、並びに効率的な予防的および治療的抗腫瘍免疫が誘導された(16、17)。アデニル酸シクラーゼ(CyaA)タンパク質は特徴づけられており、組換えDNA技術によるその調製に関して、特に第WO93/21324号または第WO02/22169号に開示されている。第WO02/22169号には、Cyaが包含する残基373〜1706の断片は、CD11b/CD18受容体との相互作用に実質的に必要とされる構造を含むことが記載されている。
【0010】
より具体的には、残基1166からアミノ酸残基1281の範囲のアミノ酸配列は、CD11b/CD18受容体との相互作用のための決定基を含み、より特定すると、残基1208から残基1243の範囲のアミノ酸配列は、毒素とCD11b/CD18の相互作用に重要であることが後記されている(第EP03291486.3および45)。
【0011】
本発明者らは、今回、HPV抗原のエピトープを、宿主(HPV感染およびその悪性形質転換に罹患している宿主を含む)の標的細胞、特に抗原提示細胞(APC)に送達できる、該エピトープを有する、すなわち含む、組換えCyaAタンパク質の作成のための条件を決定し評価した。
【0012】
したがって、本発明は、特に、1個または数個のHPV抗原の1個または数個のエピトープを有する1個または数個のポリペプチドを含む組換えタンパク質に関し、該ポリペプチドは、アデニル酸シクラーゼ(CyaA)タンパク質またはその断片の同じまたは異なる許容部位に挿入されており、該CyaA断片は、APC、特にCD11b/CD18細胞、例えば樹状細胞などのCyaAの標的細胞に標的化させるという、該アデニル酸シクラーゼタンパク質の特性を保持している。特定の実施形態において、この断片はまた、そこに挿入されたエピトープまたは該エピトープを含むポリペプチドの、標的細胞のサイトゾルへの転位を可能とするようなCyaAの特性を保持している。標的細胞のサイトゾルへのエピトープまたは該エピトープを含むポリペプチドの転位は、CyaA断片が、その触媒ドメインの転位を可能とするタンパク質ドメインを保持している場合に許容される。
【0013】
本発明の組換えタンパク質は、組換え技術を頼りにして調製できる。それはまた、合成、特に化学合成によっても得ることができる。したがって、「組換えタンパク質」なる語句は、キメラ形態のタンパク質を意味する。
【0014】
組換えタンパク質のCD11b/CD18細胞への標的化能は、特に、第EP03291486.3および(45)または第WO02/22169号に開示された方法にしたがってアッセイできる。さらに、組換えタンパク質の、該エピトープまたは該エピトープ含有ポリペプチドを標的細胞のサイトゾルへと転位する能力は、第WO02/22169号に記載の方法を適用することによりアッセイできる。
【0015】
特定の実施形態において、CyaA断片は、天然ではCyaA中で連続していない、CyaAの2個の異なる部分から構成されることができる。一例として、CyaAの触媒ドメイン、すなわち、CyaAのN末端部分の400アミノ酸残基、および、CD11b/CD18抗原提示細胞の標的化に必要とされるアミノ酸残基1208〜1243を含む断片を挙げることができる。
【0016】
先の定義において、「ポリペプチド」なる表現は、翻訳後修飾を受けたアミノ酸配列、特に、少なくとも6個のアミノ酸残基を有するアミノ酸配列、および、特に5〜500残基または約5〜約100、または約5〜約200、または約10〜約50残基、または約30もしくは約50〜200残基、または約100〜約210、または約100〜約200残基を有するアミノ酸配列を含む任意のアミノ酸配列(ただし、該アミノ酸配列は少なくとも1個のエピトープを含む)、すなわち、標的細胞、有利には宿主、特に哺乳動物宿主の標的細胞に送達した後に免疫応答を発生させることのできるアミノ酸配列を記載する。したがって、この定義によるポリペプチドは、エピトープ、さらには独特なエピトープに限定されても、または、数個の異なるもしくは同じエピトープを含んでいても、または、病原体の、すなわちヒトパピローマウイルスの全長抗原を包含していてもよい。本発明のエピトープは、体液性免疫応答および/または細胞性免疫応答、特にT細胞免疫応答に関与するアミノ酸配列を包含する。したがって、本発明の組換え分子のポリペプチド中のエピトープは、宿主においてAPC(抗原提示細胞)により処理されるもの、特に、クラスI MHC(主要組織適合性複合体)分子と会合して認識されるもの、例えば標的細胞がCD8+Tリンパ球であるエピトープ、またはクラスII MHC分子と会合して認識されるエピトープ、例えば標的細胞がCD4+Tリンパ球細胞であるものが挙げられる。
【0017】
特定の実施形態において、エピトープを有するポリペプチドは、様々な抗原に由来する、特に様々なHPV株の1タイプの抗原に由来する、または、様々なHPV株の数タイプの抗原に由来する、数個のエピトープを含む。したがって、HPV抗原に由来するポリペプチドは、多価、特に二価または三価であることができ、すなわち、数個の抗原に対する免疫応答が可能となる。
【0018】
本発明によれば、HPV抗原(これから1個または数個のエピトープを有するポリペプチドを設計できる)は、好ましくは、特にHPV感染後の悪性作用の発症および/または存続に関与するタンパク質に由来するものであり、いわゆる腫瘍抗原、すなわち、宿主において免疫応答を誘起し、宿主において抗体またはT細胞と特異的に反応できる、HPV感染に関連した腫瘍発達に関連した抗原を包含する。
【0019】
本発明に記載のエピトープを有するポリペプチドは、HPVの天然または成熟抗原から得られることができ、これには、抗原完全体を使用することにより、あるいは、全タンパク質ではなくむしろ、断片、特に抗原性断片、特に該抗原のエピトープを選択することにより、あるいは、特に、組換え分子中のCyaAタンパク質と会合させた場合に、宿主における免疫応答を誘導または誘起する能力を高めるために、該抗原またはその選択された抗原部分またはエピトープを改変することによることが含まれる。したがって、このようなエピトープの様々な可能な改変を説明するために、これらのポリペプチドは、抗原(これからエピトープが得られる)の天然または非天然フランキング配列によりフランキングされたエピトープを包含し、また、その免疫特性を高めるために化学的に改変されているエピトープまたはエピトープを含むアミノ酸配列を包含する。これらの改変は、CyaAタンパク質に関連して得られるポリペプチドの有効性を向上するために有益である可能性がある。
【0020】
いくつかの特定の改変が、本明細書で以後に例として開示されており、特に追加の陽性荷電のアミノ酸残基の挿入によるポリペプチド荷電の変化を包含する改変が含まれる。
【0021】
したがって、本発明のポリペプチドはまた、半合成または合成ポリペプチドも包含する。
【0022】
特定の実施形態によれば、HPV抗原に由来するポリペプチドは、各々または一緒になって、約5〜約500、または約5〜約100、または約5〜約200、例えば約10〜約50アミノ酸残基、または約30もしくは約50〜約200アミノ酸残基、または約100〜約210、または約100〜約200アミノ酸残基を含む。
【0023】
ポリペプチドは、特に、本発明の組換えタンパク質中で組換えられた場合に、抗原特異的応答を誘起できるように選択される。
【0024】
本発明の組換えタンパク質は、特に、分断された天然HPV抗原に存在するポリペプチドまたは数個のポリペプチドを含むように設計することができ、ここで、該分断は、該HPV抗原の酸性領域にける1個または数個のアミノ酸残基の欠失、および/または、アデニル酸シクラーゼの少なくとも2箇所の許容部位における該HPV抗原の少なくとも2個のポリペプチド断片の挿入からなる。
【0025】
この定義に包含される特定の分断は、アデニル酸シクラーゼの少なくとも2箇所の許容部位への、天然HPV抗原の少なくとも2個の断片の挿入により得られ、ここで、これらの少なくとも2個の断片は、天然抗原におけるその天然の位置と比べて逆であり、すなわち、天然抗原においてよりN末端側である断片は、CyaAタンパク質またはその断片に挿入されると、よりC末端側となり、その逆もある。
【0026】
アミノ末端断片およびカルボキシ末端断片の逆転は、E7Δ断片(すなわち、E7抗原の断片)で示されるように、特に癌免疫療法において、強力で長く続く防御免疫を誘導する際により効果的である可能性があることが観察されている。
【0027】
本発明によれば、アデニル酸シクラーゼ(CyaA)は、先に開示したように、全長タンパク質として、またはその断片として使用される。
【0028】
有利には、CyaAタンパク質またはその断片は、細胞、特に組換え細胞における、cyaA遺伝子およびcyaC遺伝子の両方の同時発現の結果である、タンパク質またはその断片である。実際に、標的細胞に対する侵襲性特性を有するために、CyaAは、cyaA遺伝子およびcyaC遺伝子の両方の発現により可能となる翻訳後修飾を受けている必要があることが示されている(第WO93/21324号)。
【0029】
本発明の特定の実施形態において、CyaAタンパク質の断片は、少なくとも約30個のアミノ酸残基を有する断片であり、約1300個まで、特に約500個までのアミノ酸残基、好ましくは約50〜約150個のアミノ酸残基を有することができ;該断片は、特定の実施形態において、CD11b/CD18標的細胞との相互作用のために、CyaAのアミノ酸残基1166〜1281、または、CyaAタンパク質のアミノ酸残基1208〜1243を含む。したがって、1個の具体的な断片は、天然タンパク質のC末端部分の全部または一部を包含し、この部分は、細胞膜および/またはCD11b/CD18受容体を標的化するための該タンパク質の結合に、および、細胞サイトゾルへのポリペプチド群に含まれるエピトープのその後の送達に関与している(12)。本発明に記載のCyaAタンパク質の1個の具体的な断片は、CyaAタンパク質のアミノ酸残基372〜1706を含む。別の具体的な断片は、アミノ酸残基225〜234が欠失しており、これにより1〜244および235〜1706を含むCyaA断片を与えるような、CyaAタンパク質に相当するものである。
【0030】
本発明の特定の実施形態において、アデニル酸シクラーゼタンパク質は、細菌性タンパク質である。好ましい実施形態において、CyaAタンパク質は、Bordetella種に由来している。
【0031】
本発明に記載の対象のBordetella種の中で、その1つが、Bordetella pertussisである。他の対象のBordetella株は、Bordetella parapertussisまたはBordetella bronchisepticaの株である。B.parapertussisのCyaAタンパク質の配列は、特にアクセッション番号NC002928.3(1740アミノ酸の配列として)およびParkhill J.ら(Nat.Genet.DOI、10(2003)に開示されており、B.bronchisepticaについては、Betsou Fら(Gene 1995、8月30日;162(1):165-6)に開示されている。
【0032】
Bordetella pertussisは、百日咳の原因物質であり、とりわけ、細菌の決定的な病原性因子であり、B.pertussis感染に対して防御性作用のある抗原の1つである、公知の百日咳毒素(PT)およびアデニル酸シクラーゼ毒素(CyaA)を含む数種の毒素を分泌している。
【0033】
Bordetella pertussisのアデニル酸シクラーゼタンパク質は、400アミノ酸残基のN末端触媒ドメインと、毒素が標的細胞膜に結合することとその後の細胞サイトゾルへの触媒部分の送達に関与している1306残基のC末端部分とを含む、1706残基の二機能性タンパク質として記載されている毒素である(12)。
【0034】
CyaAタンパク質は、不活性プロ毒素として合成され、これは、2個の内部リジン残基(リジン860および983)の翻訳後パルミトイル化により活性毒素へと変換される。この翻訳後修飾は、cyaA遺伝子と共に、アクセサリー遺伝子、すなわち、B.pertussis染色体上のcyaAの近くに位置するcyaCの発現が必要とされる。
【0035】
Bordetella pertussisのcyaAは、Glaser,P.ら、1988、Molecular Microbiology2(1)、19-30により、アミノ酸配列およびヌクレオチド配列として記載されている。したがって、B.pertussisのCyaAタンパク質のアミノ酸残基もしくは配列またはヌクレオチドもしくはヌクレオチド配列が本発明において引用される場合、その位置は、Glasserら、1988の該刊行物に開示された配列を参考にして示されている。
【0036】
本発明に記載の組換えタンパク質では、1個または数個のHPV抗原の1個または数個のエピトープを有するポリペプチドが、CyaAタンパク質の1個または数個の許容部位に挿入されている。
【0037】
本発明では、「許容部位」は、実質的にCyaAタンパク質の機能特性に影響を及ぼすことなく、特に実質的に細胞の標的化に、特にCyaAによるAPCの標的化に影響を及ぼすことなく(実質的にCD11b−CD18受容体への特異的結合に影響を及ぼすことなく、有利には、実質的に、標的細胞へのエピトープの転位の過程に関与するタンパク質のドメインに影響を及ぼすことなくを含む)、ポリペプチドを挿入できる、CyaAタンパク質の配列部位である。
【0038】
CyaA触媒ドメインの転位を可能とし、これによりこのような許容部位に挿入されたエピトープの転位を可能とする、Bordetella pertussisアデニル酸シクラーゼの許容部位には、残基137〜138(Val−Ala)、残基224〜225(Arg−Ala)、残基228〜229(Glu−Ala)、残基235〜236(Arg−Glu)、および残基317〜318(Ser−Ala)が挙げられるがこれに限定されない((44)Seboら、1995)。以下の追加の許容部位もまた、本発明の実施形態に含まれる:残基107〜108(Gly−His)、残基132〜133(Met−Ala)、残基232〜233(Gly−Leu)、335〜336(Gly−Gln)および336〜337(43)。
【0039】
他のBordetella種における対応する許容部位は、配列の比較および対応する残基の決定により定義できる。
【0040】
別の実施形態によれば、該ポリペプチドは、追加的にまたは代替的に、CyaAタンパク質またはその断片の一端および/または他の末端に挿入してもよい。
【0041】
本発明の目的のために使用するCyaAタンパク質の特定の断片は、1300個までのアミノ酸または約30〜約500個のアミノ酸残基、有利には約50〜約150個のアミノ酸残基を含むもの、特に、天然CyaAタンパク質のアミノ酸残基1166〜1281、有利には天然CyaAタンパク質の1208〜1243を包含するような断片である。
【0042】
したがって、本発明によれば、「ハイブリッドタンパク質」とも呼ばれるいわゆる組換えタンパク質をもたらすCyaAタンパク質へのポリペプチドの「挿入」は、特に、利用可能なDNA技術による遺伝子的挿入を包含する。または、「挿入」はまた、非遺伝子的挿入も包含され、これには、例えばCyaAもしくはその断片の一端において実施する共有結合、または非共有結合などの化学的挿入が含まれる。非遺伝子的挿入は、挿入するポリペプチドが合成または半合成である場合には特に関心が高い可能性がある。薬物をポリペプチドに結合させる方法は、当該技術分野において公知であり、例えば、N−ピリジルスルホニル活性化スルフヒドリルを使用するなどのジスルフィド連結が含まれる。
【0043】
特に、インビボで、Cyaの標的細胞へ、例えばACP、例えばCD11b/CD18細胞、特に該細胞のサイトゾルへと標的化させるために、化学的連結または遺伝子的挿入により、本発明のポリペプチドを特に含む分子を、CyaAに融合させることが可能である。実際に、所与のCD8+T細胞エピトープに相当する分子を、解毒したCyaAの触媒ドメインに、ジスルフィド結合または遺伝子的挿入により結合させると、遺伝子工学された分子は、インビボで特異的CTL応答を誘起することができ、これにより、該CD8+T細胞エピトープは、CD11b発現細胞のサイトゾルへと転位されたことが示される。
【0044】
特定の実施形態において、タンパク質性ベクターの製造に使用される組換えアデニル酸シクラーゼは、ジスルフィド結合により、該アデニル酸シクラーゼの触媒ドメイン内に位置する遺伝子的に挿入されたシステイン残基に化学的に結合されている本発明のポリペプチドを特に含む、システイン残基含有の1個以上の分子の挿入により特に改変されている、CyaAまたはその断片である。
【0045】
実際に、本発明のポリペプチドを特に含む複数の分子を、ジスルフィド結合により、触媒ドメイン内の様々な許容部位に位置する様々なシステイン残基に化学的に結合させることができる。
【0046】
宿主において免疫応答、特に細胞性免疫応答を誘起できる製品の設計に適した組換えタンパク質を提案するために、また特に、HPVに感染した宿主に観察される悪性作用に対する免疫応答を誘起できるような製品を設計するために、本発明者らは、高度に発癌性のHPV株、特にHPV16、HPV18、HPV31、HPV33、HPV35、HPV45、HPV52、またはHPV58から選択される株に由来する抗原に由来するエピトープを有するポリペプチドを得ることを提案した。
【0047】
これらの株の中で、HPV18およびHPV16が特に関心が高い。HPV16は、特に、哺乳動物宿主、特にヒトにおける子宮頚部癌の発症に関連しているために、HPVに感染した宿主の処置のための特定の標的である。
【0048】
これらのHPV株から出発して、本発明者らは、L1、L2、E1、E2、E4、およびE5タンパク質から選択される抗原に由来するエピトープを有するポリペプチドを得ることを提案する。
【0049】
代替的にまたは組み合わせて、本発明者らはまた、HPVのE6またはE7タンパク質に由来するエピトープを有する該ポリペプチドを得ることを提案する。
【0050】
本発明の特定の実施形態において、HPV16のE6もしくはE7タンパク質またはHPV18のE6もしくはE7タンパク質が、エピトープを有するポリペプチドの設計に使用される。
【0051】
HPV抗原に由来するポリペプチドの設計のために言及できる特定のHPVタンパク質は、HPVのE7タンパク質、特にHPV16またはHPV18のE7タンパク質である。本発明の実施形態によれば、該ポリペプチドは、様々なHPV株、特にHPV16およびHPV18の数個のE7タンパク質から得られる。例えば、該ポリペプチドは、HPV16およびHPV18の全長E7タンパク質、または、HPV16もしくはHPV18の各々のE7タンパク質の1個もしくは数個の断片(該断片の多量体、特に二量体を含む)から得られる。
【0052】
これらのHPVタンパク質およびそのアミノ酸およびヌクレオチド配列は、HPV16についてはSeedorf,Kら(ヒトパピローマウイルスタイプ16DNA配列、Virology、145:181-185、1985)、Cole S.T.、Danos O.(ヒトパピローマウイルスタイプ18ゲノムのヌクレオチド配列および比較解析。パピローマウイルスの系統発生並びにE6およびE7遺伝子産物の反復構造、J.Mol.Biol.193:599-606(1987))またはFernando GJら(子宮頚部癌に関連したヒトパピローマウイルスタイプ18(HPV18)のE7形質転換タンパク質のTヘルパーエピトープ、Virus Res.1995 Apr.36(1):1-13)に開示されている。
【0053】
E6およびE7タンパク質は、ウイルスの複製全周期を通じて特にHPV16またはHPV18により発現されている癌タンパク質であり、HPV株による感染後の宿主細胞の悪性形質転換の発症および存続に必要とされることが示されている。それ故、これらの両方の腫瘍特異的抗原が、養子CTL媒介免疫療法の可能性ある標的と考えられている。
【0054】
本発明の特定の実施形態によれば、該組換えタンパク質は、複数のポリペプチドを含み、その各々が、1個または数個のHPV抗原の1個または数個のエピトープを有する。
【0055】
例えば、このような複数のポリペプチドは、1個のHPV株、特にHPV16またはHPV18のE6およびE7タンパク質から得ることができる。別の例によれば、これらの複数のポリペプチドは、HPV16およびHPV18の両方に由来する、E6またはE7タンパク質から得られるエピトープを包含することができる。
【0056】
複数のポリペプチドはまた、対象のCyaAタンパク質の異なる許容部位に挿入された、1個のタンパク質、例えばE7またはE6タンパク質の断片群を有する様々なエピトープからなることができる。
【0057】
先の定義に記載の別の特定の組換えタンパク質は、エピトープを有する複数のポリペプチドが、HPV16のE7タンパク質の残基1〜29を含む断片、すなわち残基1〜29からなる断片、または残基42〜98を含む断片、すなわち残基42〜98からなる断片を包含するか、あるいは、複数のポリペプチドが、CyaAタンパク質の異なる許容部位に挿入された両方の断片を含むかまたはからなる、CyaA組換えタンパク質である。
【0058】
本発明に記載の別の組換えタンパク質は、複数のポリペプチドが、アミノ酸配列
【表2】
を有する断片を包含するようなタンパク質である。
【0059】
CyaAタンパク質の許容部位に挿入されたポリペプチドのアミノ酸残基の数は、全長抗原、特にHPVの全長E6またはE7タンパク質からなるポリペプチドを、CyaAタンパク質またはその断片に挿入できるようなものである。
【0060】
本発明の特定の実施形態によれば、組換えCyaAに含まれるポリペプチドは、CyaAのコドン224と235の間またはCyaAのコドン319と320の間に挿入された、E7タンパク質、特にHPV16のE7タンパク質である。
【0061】
別の実施形態において、本発明の組換えタンパク質は、複数のポリペプチド(そのいくつかは、1個または数個のHPVの1個または数個のエピトープを有するポリペプチドである)および他の病原体のエピトープを有する他のポリペプチドを含む。
【0062】
別の特定の実施形態において、本発明の組換えタンパク質はさらに、異なる病原性物質から派生した1個または数個のエピトープを含む。ChlamydiaまたはHIVレトロウイルス、またはHPV、HBV、HCV、アデノウイルスEBV、ヘルペスウイルス、HTLV.1ウイルスおよびCMVから派生したエピトープと、HPVから派生したエピトープとの会合は、特に関心が高い場合がある。
【0063】
本発明の別の特定の実施形態によれば、エピトープを有するポリペプチドは、例えば、配列内の負に荷電したアミノ酸残基の数を減少させるために、その天然アミノ酸配列と比べて改変されている。このような改変は、これらの負に荷電したアミノ酸残基のいくつかを除去することにより、または、いくつかの正に荷電したアミノ酸残基を、特にエピトープのフランキング残基として付加することによっても得ることができる。このようにより少ない数の負に荷電した残基を含むポリペプチドは、標的細胞のサイトゾルへのCyaAタンパク質の触媒ドメインの転位に好ましくあり得る。
【0064】
エピトープを有するポリペプチドはまた、CyaAに挿入される場合に折りたたまれないようにするように設計でき、これにより、標的細胞への組換えCyaAタンパク質の内部移行の効率が向上する。その含有アミノ酸の結果として折りたたみを受けたポリペプチド中のこのような折りたたみ解除は、例えば、システイン残基を除去または置き換えて、ポリペプチドの折りたたみに関与している可能性があるジスルフィド結合の形成を避けることにより達成することができる。いくつかの場合において、インビボでの再折りたたみを回避することのできる還元剤の存在下でそれらを調製することによりポリペプチドの折りたたみを防ぐことが可能である。
【0065】
特定の実施形態において、エピトープを有するポリペプチドは、潜在性エピトープであってもよい。
【0066】
本発明の特定の態様において、本発明者らは、実際に、(i)先に開示した定義に記載のアデニル酸シクラーゼ(CyaA)またはその断片と、(ii)1個または数個の抗原の1個または数個の抗原性断片を有するポリペプチドとを含む組換えタンパク質からなる、キメラタンパク質構造物は、組換え構造物における該抗原の潜在性エピトープを、それを提示する結果、免疫原性とすることができることが決定された。特に、特に治療目的(ワクチン接種目的を含む)のための、本発明に定義したCyaAまたはその断片と対象の抗原から得られたポリペプチドとを含む該キメラ構造物は、免疫原性とさせることができ、特に、宿主におけるT細胞応答、特にCTL応答を発生させることのできる、抗原の潜在性エピトープを含むことができる。
【0067】
したがって、本発明はまた、1個または数個の抗原の1個または数個のエピトープを有する1個または数個のポリペプチドを含む組換えタンパク質に関し、該ポリペプチドは、アデニル酸シクラーゼ(CyaA)またはその断片の同じまたは異なる許容部位に挿入されており、該CyaA断片は、抗原提示細胞を標的化するという該アデニル酸シクラーゼタンパク質の特性を保持しており、該エピトープの少なくとも1個は、亜優性潜在性T細胞エピトープ(subdominant cryptic T-cell epitope)であり、該組換えタンパク質は、該ポリペプチドに対する抗原特異的応答を誘起できる。
【0068】
特に、潜在性エピトープは、HPV抗原、特にHPV16および/またはHPV18抗原、特にE7抗原内に含まれる。
【0069】
このように定義した組換えタンパク質は、特に、HPV18E7タンパク質から得られたペプチド、すなわち、アミノ酸配列IDGVNHQHLを有するペプチドを含む。
【0070】
特定の実施形態によれば、潜在性エピトープは改変でき、例えば、最初の2箇所の位置において置換を有することができ、例えば配列ASGVNHQHLを有することができる。
【0071】
本発明は、特に、ペプチドIDGVNHQHLに関する。
【0072】
本発明はまた、この配列に置換を有するペプチド、特に1位および/または2位に置換を有するペプチド、特に配列ASGVNHQHLを有するペプチドに関する。
【0073】
本発明はまた、免疫原性特性を有する限り、特にT細胞を誘起できる限り、特にCTL応答を誘起できる限りにおいて、該ペプチドの変種を含む。
【0074】
有利には、本発明の組換えタンパク質を調製するために、CyaAタンパク質の酵素活性、すなわち、ATPからcAMPへと変換する能力は、不活性化されている。このような不活性化は、遺伝子不活性化の結果として得ることができる。一例として、遺伝子不活性化は、ジペプチドを、触媒部位の一部であるCyaAのアミノ酸配列の部位(例えば188と189の間)に導入する結果として得ることができる。このような不活性化CyaAタンパク質は、以下の実施例に実例を示す。
【0075】
本発明の組換えタンパク質は、有利には、細胞性免疫応答を誘起できる。該応答は、CTLおよびTh、特にTh1応答(CD4+T細胞応答および/またはCD8+T細胞応答を含む)を含む。
【0076】
組換えタンパク質がこの細胞性免疫応答を誘起できる能力は、特に、インビボにおいて腫瘍増殖を抑止するのにまたはさらには動物における腫瘍減退を可能とするのに十分であることが示された。
【0077】
本発明はまた、先に定義した組換えタンパク質をコードするポリヌクレオチドにも関する。
【0078】
本発明のポリヌクレオチドを発現ベクターに挿入して、本発明の組換えタンパク質の発現に適した組換え発現ベクターを提供できる。このような発現ベクターには、プラスミド、コスミド、ファージミド、ウイルスベクターが挙げられる。
【0079】
組換えベクターは、原核細胞、特に細菌における発現に適したものでも、または、真核細胞、特に哺乳動物細胞、有利にはヒト細胞における発現に適した発現ベクターでもよい。
【0080】
本発明は、特に、本発明に記載の組換えタンパク質をコードするプラスミドからなるベクターに関する、例えば:2004年3月18日にCNCM I−3191号でCNCM(パリ、フランス)に寄託されたpTRACE5−HPV16E7FULL(CyaAE5−HPV16E7FULLとも称される);2004年3月18日にCNCM I−3190号でCNCM(パリ、フランス)に寄託されたpTRACE5−HPV16E7Δ30〜42(CyaAE5−HPV16E7Δ30〜42とも称される)、または構造物pTRACE5−HPV16E749〜57。
【0081】
本発明はまた、本発明に記載のポリヌクレオチドまたはベクターで形質転換された、宿主細胞、特に原核細胞、または真核細胞、例えば哺乳動物細胞(ヒト細胞を含む)を含む。
【0082】
本発明は、特に、アクセッション番号CNCM I−3190号およびアクセッション番号CNCM I−3191号でCNCMに寄託された宿主細胞に関する。
【0083】
本発明はまた、活性物質として、先に定義した組換えタンパク質または先に定義したポリヌクレオチドまたは発現ベクターを含む、免疫原性組成物に関する。免疫原性組成物の該活性物質は、宿主への投与に適した生理的に許容されるビヒクル、賦形剤、担体、もしくは希釈剤と共に、またはその組合せと共に製剤化できる。
【0084】
免疫原性組成物は、有利には、哺乳動物宿主において、細胞性免疫応答、特にT細胞媒介免疫応答を誘導するように設計されている。好ましくは、細胞媒介細胞溶解免疫応答CTL、特にCD8+を誘導できる。
【0085】
本発明に記載の別の免疫原性組成物は、体液性免疫応答を誘導できるものである。
【0086】
本発明の免疫組成物が免疫応答を誘導する能力を高めるために、活性物質を、アジュバントおよび/または界面活性剤および/または免疫調節物質(例えばサイトカインまたはケモカイン)と合わせることは興味深い。
【0087】
アジュバントは、例えば、水層を含むエマルションの形態で一般に使用される、リポソーム、油層、例えばフロイントタイプのアジュバント、または、水不溶性無機塩、例えば水酸化アルミニウム、硫酸亜鉛、コロイド状水酸化鉄、リン酸カルシウム、または塩化カルシウムを含むことができる。
【0088】
本発明に記載の免疫原性組成物は、有利には、特に免疫療法のために、免疫応答を抗原刺激および/またはブーストすることにより、宿主の免疫応答を誘導するために使用されている。特に、本発明の免疫原性組成物は、宿主におけるHPV感染に起因する悪性形質転換の発症または存続の予防のために、あるいは、HPV感染、特にHPV−16またはHPV−18感染に起因する悪性形質転換に罹患する患者の処置のために興味深い。
【0089】
このような免疫治療組成物は、腫瘍誘発性状態をもたらす宿主における制御されていない細胞増殖の治療に、特に癌免疫療法に、特にHPV感染に関連した子宮頚部癌免疫療法において特に興味深い。それ故、それは、癌ウイルス感染に起因する悪性状態(腫瘍状態を含む)の処置に特に適した治療ワクチンの設計のための手段を提供する。
【0090】
本発明に使用する場合、「処置」または「治療処置」なる表現は、処置を受けている患者に有益な効果をもたらす、本出願で開示された化合物の効果を包含し、該効果は、細胞レベルまたは臨床レベルで観察され、処置の結果、患者の容態の改善もしくは寛解状態、または健康状態の回復が含まれる。処置する悪性状態が、制御されていない細胞増殖または腫瘍の発生もしくは持続である場合、有益な効果は、制御されていない増殖の安定化を含み得、あるいは、好ましくは、その予防、停止もしくは反転、または腫瘍の減退を含み得る。
【0091】
先に記載した悪性状態の処置を目的とした組成物は、有利には、約1〜約1000μgの組換えタンパク質、好ましくは約10〜約500μgの組換えタンパク質の量に対応する用量の活性物質を含むことができる。該組成物が活性物質として本発明の組換えタンパク質を含む場合、用量は、約0.05〜約10μgの組換えタンパク質、好ましくは約0.1〜約1μgのタンパク質を含むことができる。
【0092】
処置する状態に応じて、該組成物は、病変レベルで局所的に、1回または数回、例えば数日間の規則的な間隔で、例えば5〜10日間投与できる。また全身投与してもよい。
【0093】
本発明はまた、好ましくはヒト宿主における、先の定義に記載の組換えタンパク質、または、先に定義したポリヌクレオチド、または、該ポリヌクレオチドを含むベクター、および、適宜、免疫応答(細胞性免疫応答および/または体液性応答を含む)を誘起するための薬学的に許容されうるビヒクルを含む、ワクチン組成物、特に哺乳動物宿主、好ましくはヒトへの投与のために製剤化された組成物に関する。
【0094】
本発明はまた、HPV感染を予防または処置するための、本発明の組換えタンパク質またはポリヌクレオチドまたはベクターおよび薬学的に許容されうるビヒクルを含む、薬物組成物にも関する。
【0095】
別の実施形態によれば、薬物組成物は、宿主におけるHPV感染に起因する悪性形質転換の発症または存続の予防または処置のための、本発明に記載の組換えタンパク質またはポリヌクレオチドまたはベクターを、薬学的に許容されうるビヒクルと共に含む。
【0096】
癌免疫療法のための、組換えタンパク質またはポリヌクレオチドまたはベクターおよび薬学的に許容されうるビヒクルを含む薬物組成物。
【0097】
本発明はまた、宿主における免疫応答、すなわち、体液性および/または細胞性免疫応答を誘起するためのベクターとして使用するに適した、細菌タンパク質、特に細菌毒素(好ましくはその類毒素形態)またはその断片を含む組換えタンパク質(該タンパク質またはその断片は、1種類の癌ウイルスによる感染の処置のために、1個または数個の癌ウイルスの1個または数個の抗原の1個または数個のエピトープの挿入により改変されている)の患者における使用に関する。このような組換えタンパク質は、特に、悪性作用、特にこのような癌ウイルスによる感染により引き起こされる腫瘍の処置に提案されている。
【0098】
癌ウイルスの抗原のエピトープを運ぶためのベクターとして適した細菌タンパク質の例は、クレブシエラのOmpAまたは以下の毒素シガ毒素(そのβサブユニットを含む)(Haicher N.ら、J.Immunol.2000、165:3301-8)、炭素菌毒素(Goletz TJら、PNAS USA 1997、94:12059-64)、ジフテリア毒素(Stenmark H.ら、J.Cell.Biol.1991、113:1028-32)またはシュードモナス外毒素A(Donnelly JJ.ら、PNAS USA 1993、90:9530-4)である。細菌タンパク質への挿入のためのポリペプチドを調製するために抗原がエピトープを提供できる癌ウイルスには、HPV、HBV、HCV、アデノウイルスEBV、ヘルペスウイルス、HTLV.1ウイルス、およびCMVが挙げられる。
【0099】
CyaA組換えタンパク質について先に示した活性物質としての使用の記述は、他の細菌タンパク質および癌ウイルス抗原にも応用できる。
【0100】
本発明はまた、HPV感染の診断のための、または、このような感染を免疫モニタリングするためのキットに関し、これには、本発明に記載の組換えタンパク質、ポリヌクレオチド、または発現ベクターが含まれる。
【0101】
本発明はまた、患者におけるHPV感染の処置または予防のための、上記の本発明の組換えタンパク質、ポリヌクレオチド、またはベクターの使用に関する。
【0102】
本発明はまた、患者におけるHPV感染に起因する悪性形質転換の発症または存続に対する免疫療法のための、上記の本発明の組換えタンパク質、ポリヌクレオチド、またはベクターの使用に関する。
【0103】
本発明はまた、HPV感染をインビトロで診断する方法またはHPV感染を免疫モニタリングする方法に関し、該方法は、
哺乳動物、特にヒト患者から得られたT細胞を、本発明の組換えタンパク質にさらすこと、
T細胞の活性化の改変を検出すること
を含む。
【0104】
特定の実施形態において、該組換えタンパク質は、HPV感染の予防、または、HPV感染に罹患している宿主(このような感染に起因する腫瘍を有する宿主を含む)の処置のために使用できる。
【0105】
本発明はまた、キメラCyaA−ポリペプチドタンパク質に含まれるポリペプチドにおける未知または亜優性潜在性T細胞エピトープのスクリーニング方法であって、
ここで、CyaAは、先に開示されているアデニル酸シクラーゼまたはその断片であり、該方法は、
該キメラタンパク質を動物宿主に投与すること、
該宿主のT細胞応答、特にCTL応答を決定すること
を含む。
【0106】
本発明は、特に、本発明内で定義された組換えタンパク質に含まれるHPV抗原のポリペプチド中の未知または亜優性または潜在性T細胞エピトープ(特にCD8+T細胞エピトープ)をスクリーニングする方法に関し、該方法は、
該組換えタンパク質を動物宿主(非ヒト)に投与すること、
該宿主のT細胞応答、特にCTL応答を決定すること
を含む。
【0107】
本発明を特徴づけるさらなる特徴を、以下の実施例および図面に開示し説明する。
【0108】
実施例
実施例1
ここで、本発明者らは、HPV16のE7タンパク質の全長配列またはこのポリペプチドの亜断片(特に、H−2Db拘束性エピトープに対応するE7の残基49〜57およびHPV16−E7残基43〜98と1〜29を包含するペプチド)を含む、組換えCyaAを作成した。本発明者らは、C57BL/6マウスに注入した場合、これらのHPV16−E7組換えCyaAは、IFN−γの分泌により特徴づけられる特異的CTLおよびTh1応答を誘導できることを示した。さらに、治療的に試験した場合、これらの構造物は、TC−1細胞の皮下移植に対して100%以下の防御を提供できた。この研究は、初めて、インビボでの、ヒト腫瘍特異的抗原に対するCyaAにより媒介される抗腫瘍治療活性の実証を示した。
【0109】
材料および方法
マウスおよび細胞系
特定の病原体を含まない6〜10週令の雌C57BL/6マウスを、CER Janvier(Le Gesnet St-Isle、フランス)またはチャールズリバー(L'Arbresle、フランス)から得た。C57BL/6バックグラウンドへと育成したTAP1−/−(18)、MHCクラスII−/−(19)およびCD40−/−(20)もこの研究に使用した。動物を、水と食料が自由にとれ、病原体の存在しない条件下で、パスツール研究所動物施設で飼った。動物の関与する実験は、動物の世話の施設ガイドラインにしたがって実施した。HPV16E6およびE7タンパク質を発現しているT細胞(21)およびマウス胸腺EL4細胞(17)を、ATCCから得た。細胞を、10%の熱不活性化FCS、100U/mlのペニシリン、100μg/mlのストレプトマイシン、0.4mg/mlのジェネティシン(TC−細胞に対してのみ)、および5×105Mの2−メルカプトエタノール(ギブコBRL、Cergy-Pointoise、フランス)を補充したグルタマックスを含むRPMI1640中に維持した。
【0110】
ペプチド
HPV16−E7 H2−Db拘束性エピトープ(22)に対応する合成ペプチドE749〜57(RAHYNIVTF、一文字でアミノ酸をコード)並びにE749〜57CTLエピトープ(天然フランキング配列およびThエピトープ(太線)を含む)(23)に対応するE743〜77
【表3】
を、Neosystem(ストラスブール、フランス)から購入した。CpG ODN1826は、PROLIGO(パリ、フランス)から購入した。
【0111】
HPV16−E7エピトープを有する組換えB.pertussisアデニル酸シクラーゼの作成および精製
本文書で使用される組換えアデニル酸シクラーゼは、酵素的に不活性なCyaAをコードするプラスミドpTRACE5の誘導体(図1A)を使用することにより、E.coliにおいて発現させた(24)(25)。プラスミドpTRACE5は、酵素的に不活性で、それ故に細胞毒性であるB.pertussisCyaAの変種のための発現ベクターである。それはまた、CyaAの翻訳後のアシル化に必要とされるB.pertussisCyaCタンパク質も発現する。このプラスミドは、以前に記載されているpTRACGプラスミドの誘導体である(Gmiraら、2001、Res.Mic.152:889)。それは、cyaA DNA配列の5’部分内に位置するEcoRV部位へのヘキサヌクレオチドCTGCAGの挿入により得られた。これにより、触媒部位の不可欠な部位内のAsp188とIle189の間に、ジペプチドLeu−Glnが枠内に挿入された(Guermonprezら、2000、Meth.Enzymol.326:527)。
【0112】
プラスミドpTRACE5は、ColE1複製起点とアンピシリン耐性マーカーを有する。このプラスミドでは、cyaCおよび改変されたcyaA遺伝子が、λファージPrプロモーターの制御下で同じ転写単位に入れられている。pTRCAGプラスミドはまた、32℃より低い温度でλPrプロモーターにおける遺伝子転写を強力に抑制する熱感受性λリプレッサーcI857をコードしている。
【0113】
E.coli株のXL1−Blue(ストラタジーン、ラホーヤ、CA州)を、標準的なプロトコルにしたがって実施した全てのDNA操作において使用した(Maniatisら)。
【0114】
CyaA−E749〜57は、CyaAのコドン224と235の間に挿入された9アミノ酸長のポリペプチド配列(RAHYNIVTF)を含む。CyaA−E749〜57の発現プラスミドは、以下のように作成した。2つの合成オリゴヌクレオチド(MWG、Courtaboeuf、フランス)
【表4】
をアニーリングし、NheIおよびKpnIで消化したpTRACE5にライゲートした。CyaA−E7Fullは、HPV16−E7タンパク質の全配列、すなわち、酵素的に不活性なCyaAの同じ224位に挿入された98アミノ酸を含む。E7タンパク質をコードするDNA配列を、特異的プライマー
【表5】
を使用して、HPV16 DNA(先のSeedorf Kら)から増幅した。得られたPCR産物をNheIおよびKpnIで消化し、NheIおよびKpnIで切り出したpTRACE5にライゲートした。アニーリングしたオリゴヌクレオチド並びにHPV16−E7の全配列に存在するSspI部位により、挿入変異体の迅速な同定が可能となった。CyaA−E7Δ30〜42は、CyaAのコドン319と320の間に挿入されたHPV16−E7の最初の29アミノ酸残基、並びに、CyaAのコドン224と235の間に挿入されたHPV16−E7の残基43〜98を含む。CyaA−E7Δ30〜42のための発現プラスミドは、2ステップで作成した。HPV16−E7(のアミノ酸残基1〜29)をコードする第一のDNA断片を、標的DNAとして合成HPV16−E7遺伝子(E.coliにおける産生に最適化され、GTP Technologyにより設計、Labege、フランス)およびプライマー
【表6】
を使用してPCR増幅した。CyaAのコドン320〜372をコードする第二のDNA断片を、標的DNAとしてpTRACE5、および、プライマー
【表7】
を使用してPCR増幅した。これらの2個のDNA断片(一部重複する)を精製し、3回目のPCRでプライマーBTP5およびBTP8と合わせ、294bp長のDNA断片を増幅した。この断片を、AgeIおよびBstBIにより消化し、pTRACE5の対応する部位の間に挿入して、プラスミドpTRACE5−E1〜29を得た。その後、HPV16−E7のアミノ酸残基43〜98をコードするDNA断片を、標的DNAとしての合成HPV16−E7遺伝子、および、プライマー
【表8】
を使用してPCR増幅した。精製PCR断片をNheIおよびKpnIにより消化し、同じ制限酵素により消化したプラスミドpTRACE5−E71〜29にライゲートした。
【0115】
全ての組換えアデニル酸シクラーゼが、前記のように(26)、エシェリヒア・コリ株BLR(ノバゲン、WI州マジソン)において産生された。組換えタンパク質を、前記のように(26)、DEAE−セファロースおよびフェニル−セファロースクロマトグラフィーを含む2ステップ手順により封入体から均一に近くなるまで精製した(図1B)。フェニル−セファロースクロマトグラフィーに加えて、20mM Hepes−Na、pH7.5中の60%イソプロパノールを用いてのさらなる洗浄ステップを行ない、大半の混入LPSを排除した。LPS含量は、キットQCL−1000(Biowhittaker、Walkersville、MD州)を使用して決定した。精製組換えタンパク質を、SDS−ゲル分析により分析した。タンパク質濃度は、分子吸光係数142,000M−1cm−1を使用して280nmにおける吸光度から分光学的に決定した。
【0116】
組換えHPV16−E7タンパク質の作成および精製。
リクエストにより入手可能なHPV16−E7タンパク質(GTP technology)のDNA配列をコードするE.coli最適化cDNAを、pIVEX2.4bベクター(ロシュ・モレキュラー・バイオケミカルズ、Meylan、フランス)のNcoIとXhoI制限部位の間にサブクローニングした。その後、得られたプラスミドを、E.coli株BL21λDE3(ノバゲン)に形質転換した。0.5mMのイソプロピル−β−D−チオガラクトピラノシド(Euromedex、Souffelweyersheim、フランス)で誘導するとHis−Tag−HPV16−E7タンパク質が発現され、Ni−NTAアガロース(キアゲン、ヒルデン、ドイツ)を用いて製造業者の指示にしたがって精製した。LPS混入を除去するために、記載の通りに(27)イソプロパノールによる洗浄を使用した。
【0117】
イムノブロット
タンパク質をSDS−PAGEにより分離し、マウスモノクローナル抗HPV16E7抗体(Zymed、CA州サンフランシスコ)またはC57BL/6マウスにおいて調製したポリクローナル抗E.coli BLR血清でプローブした、ニトロセルロース膜(0.45μ、バイオラッド、Marnes la Coquette、フランス)にエレクトロトランスファーした。アルカリホスファターゼにコンジュゲートさせたヤギ抗マウス免疫グロブリン(Chemicon、Temecuka、CA)を用いて免疫複合体が検出され、5−ブロモ−4−クロロ−3−インドリルホスフェート/ニトロブルーテトラゾリウム(BCIP/NBT)(シグマ、MO州セントルイス)を用いて明確にした。
【0118】
マウス免疫化および腫瘍拒絶実験
動物を、PBS(ギブコBRL)に希釈した50μgの対照もしくはHPV16−E7組換えCyaAを1回静注するか、または、2回(各回10μg)皮内注射することにより免疫化した。皮内注射は、耳真皮に行なった(47)。インビトロでの解析のために、長期の応答の解析を除いて(このためにこの手順は、注射の3ヶ月後に行なった)、注射の7日後に、麻酔した動物(CO2)の脾摘除を行なった。腫瘍拒絶実験のために、マウスは、皮下に5×104個のTC−1細胞を投与され、腫瘍接種の1日後、5日後、または10日後にHPV16−E7組換えCyaAにより処置した。TC−1腫瘍増殖を、キャリパーを使用してモニタリングし、式V=(L×w2)/2(L:長さ、w:幅)を使用して立方ミリメートルで表現した(48)。
【0119】
インビトロでの細胞毒性アッセイ
免疫化マウスの脾臓細胞を、インビトロで、同系の照射し無処置の脾臓細胞の存在下で、完全培地(10%の熱で不活性化されたFCS、100U/mlのペニシリン、100μg/mlのストレプトマイシン、および5×10−5Mの2−メルカプトエタノールを補充したグルタマックスを含むRPMI1640)中で、5日間、1μg/mlのE749〜57またはE743〜77ペプチドで刺激した。これらのエフェクター細胞の細胞毒性活性を、TC−1細胞の5時間の51Cr放出アッセイで試験した。放射標識を以下のように行なった:7.5%CO2雰囲気中37℃で培養した指数関数的に増殖しているTC−1細胞を、素早くトリプシン処理し(トリプシン−EDTA、ギブコBRL)、100μCiの51Crと共に37℃で1時間インキュベートした。様々なE:T比を使用し、全てのアッセイを二重に行なった。各ウェルの上清に放出された放射能を測定した。特異的溶解率は、100×(実験的放出−自発的放出)/(最大放出−自発的放出)として計算した。最大放出は、10%トリトンX−405を標的細胞に加えることにより得、自発的放出は、完全培地のみの中でインキュベートした標的細胞を用いて得られた。最も高いE:T比で少なくとも20%の特異的溶解が観察された場合にマウスをレスポンダーと判断した。結果は、1群あたりのレスポンダーマウスの中間値±四分位範囲として表現した。
【0120】
分泌細胞のための、単一のIFN−γ産生細胞の酵素結合イムノスポットアッセイ
マルチスクリーンろ過プレート(96ウェル;ミリポア、Molshein、フランス)を、1mlあたり4μgのラット抗マウスγインターフェロン(IFN−γ)抗体(クローンR4−6A2;ファーミンゲン、CA州サンディエゴ)を用いて室温で一晩かけて被覆した。その後、プレートを洗浄し、完全培地で遮断した。免疫化マウスの脾臓細胞の連続2倍希釈液を、5×105個のγ照射された(2,500rad)同系支持細胞と共に加えた。細胞を、1μg/mlのE749〜57ペプチドと共にまたはその非存在下で36時間インキュベートした。十分に洗浄した後、プレートを、1mlあたり5μgのビオチニル化ラット抗マウスIFN−γ抗体(クローンXMG1.2;ファーミンゲン)と共にインキュベートし、その後、ストレプトアビジン−アルカリホスファターゼ(ファーミンゲン)と共にインキュベートすることにより明確にした。最後に、基質としてBCIP/NBTを使用してスポットを明確にした。IFN−γ産生細胞の数を、各ウェルにおけるスポット形成細胞(SFC)の数を計測することにより決定し(Bioreader、Karben、ドイツ)、結果を、脾臓1個あたりのSFCの全数として表現した(17)。
【0121】
酵素結合免疫吸着アッセイ(ELISA)
空のベクターCyaAE5により皮内免疫化したマウスから、30または90日後に採血し、個々のマウス血清を、ELISAにより抗体応答について試験した。マイクロプレート(Nunc、Roskilde、デンマーク)を、PBS中の空のベクターCyaAE5(3μg/ml)を用いて一晩かけて被覆した。PBS−tween20(0.1%)中で洗浄した後、希釈した血清を、ウェルに加え、37℃で1時間インキュベートした。PBS−tween20中で洗浄した後、プレートを、ヤギ抗マウスIgGペルオキシダーゼコンジュゲート(シグマ)と共に37℃で1時間インキュベートした。プレートを、o−フェニレンジアミンおよび過酸化水素(シグマ)を使用して展開した。硫酸を用いて反応を停止し、プレートを、ELISA読取機(Dynatech、Marnes la Coquette、フランス)を用いて492nmで解析した。結果は、希釈とA492をプロットした線形回帰分析により計算した抗体力価として表現した。力価は、1/100に希釈されたプール対照血清の吸光度の2倍をもたらす最も高い希釈度のlog10であると計算された。
【0122】
サイトカイン産生
免疫化マウスの脾臓細胞を、インビトロで、完全培地中で、10μg/mlのHisTag−HPV16−E7タンパク質または1μg/mlのE743〜77ペプチドを用いて72時間かけて刺激した。IFN−γおよびIL−5産生は、培養上清中で、前記のように(28)、サンドイッチ酵素結合免疫吸着アッセイ(ELISA)により決定された。全てのアッセイは、対応する組換えネズミサイトカイン(ファーミンゲン)を用いて標準化した。
【0123】
FACS(登録商標)解析
TC−1細胞は、特異的なFITC−コンジュゲートモノクローナル抗体(クローンKH95、ファーミンゲン、Le Pont de Claix、フランス)を使用するフローサイトメトリーによるMHCクラスI分子H−2Dbの発現レベルの解析についていずれかに記載されているように(29)処理した。
【0124】
統計的解析
様々な試料の量が少ないことを考慮して、ソフトウェアStatXact4(Cytel社、MA州ケンブリッジ)を使用するノンパラメトリック統計試験(30)を適用した。生存曲線を、Prismソフトウェア(GraphPadソフトウェア社、CA)を使用してプロットし、ソフトウェアのビルトインのlogランク検定を用いて比較した。データは、p<0.05で有意差があると判断した。
【0125】
結果
HPV16−E7エピトープを有する組換えアデノウイルスシクラーゼの作成および特徴づけ
CyaAのHPV16−E7特異的T細胞応答誘導能を試験するために、本発明者らは、3個の異なる組換え分子を作成した。CyaA−E749〜57は、酵素的に不活性な(したがって無毒性な)CyaAのコドン224と235の間に挿入された、以前に記載されたH−2Db拘束性CTLエピトープ(22)に対応する、9アミノ酸長のポリペプチド配列(RAHYNIVTF)を含む。CyaA−E7FUllは、酵素的に不活性なCyaAの同じ224位に挿入されたHPV16−E7タンパク質の全配列(98アミノ酸)を含む。CyaA−E7Δ30〜42は、酵素的に不活性なCyaAのコドン319と320の間に挿入されたHPV16−E7の最初の29アミノ酸残基、並びに、コドン224と235の間に挿入されたHPV16−E7の残基43〜98を含む。インビトロおよびインビボでのアッセイを可能とするために、CyaA構造物が産生され、精製されてほぼ均一とした(図1B)。LPS排除手順を精製プロトコル(26)に導入し、50μgあたり100単位未満の内毒素を含む組換えタンパク質を得た。CyaA−E7FullおよびCyaA−E7Δ30〜42におけるE7タンパク質の存在は、特異的なモノクローナル抗体(Zymed)を使用してウェスタンブロットにより確認された(図1C)。これに対し、H−2Db拘束性エピトープのみを含むCyaA−E749〜57は、抗HPV16−E7抗体により認識されなかった。3個の組換えCyaAの全体的な生化学的特性は、改変されておらず、これらの分子は、野生型アデニル酸シクラーゼと類似した溶血特性を示した(17)。
【0126】
HPV16−E7組換えCyaAでの免疫化は、E7特異的CTL応答を誘導した。
CyaAが、HPV16−E7エピトープに対するCTL応答を誘導できるかを試験するために、C57BL/6マウスに、50μgの様々なHPV16−E7組換えCyaAを1回静注することにより免疫化した。脾臓細胞を収集し、インビトロで、1μg/mlのE743〜77ペプチドで刺激した。TC−1細胞を溶解する能力を、5日後に、51Cr放出アッセイを使用して決定した。図2Aに示されているように、C57BL/6マウスにHPV16−E7組換えCyaAを1回静注して免疫化することにより、TC−1細胞に対する強力で特異的なCTL応答が誘導された。完全なHPV16−E7タンパク質(図2A、b)を含むCyaAまたはその欠失形態であるCyaA−E7Δ30〜42(図2A、c)で免疫化することにより、最小のH−2Db拘束性エピトープ(図2A、a)のみを含むCyaA−E749〜57により誘導されるものと比べてより高い最大CTL活性が得られたが、本発明者らは、本発明者らのデータから統計学的有意性を実証できなかった。ペプチドE749〜57をインビトロでの再刺激に使用した場合にも、類似の結果が得られた(データは示さず)。関連性のないエピトープ(OVA257〜264)を有する組換えCyaAでワクチン接種し、インビトロで、1μg/mlのE743〜77ペプチドで再刺激したマウスの脾臓細胞には、弱く非特異的なTC−1細胞溶解しか認められなかった(図2A、a)。インビボで、CyaAにより、OVA CD8+細胞エピトープ(SIINFEKL)をMHCクラスI分子に送達することは、TAP1機能に依存していることが以前に示されている(15)。本発明者らは、これがCyaA−E7Δ30〜42を使用しても当てはまるかどうかを試験した。図2A、dに示されているように、静注によりワクチン接種されたTAP1−/−マウスに由来するインビトロで刺激された脾臓細胞は、TC−1細胞を溶解できなかった。本発明者らはまた、インビボでのHPV16E7含有組換えCyaAによる、CTL応答の刺激における、CD4+T細胞による支持の必要性を試験した。以前の観察と一致して(15)、本発明者らは、CyaA−E7Δ30〜42を使用したMHCクラスII−/−マウスの静注ワクチン接種により、TC−1細胞に対する高いレベルの特異的CTL応答が誘導されることを観察した(図2A、e)。これに対し、本発明者らは、このモデルにおいて、CD40−/−マウスにおけるTC−1細胞に対するCTL応答のレベルが低いことを観察したので(図2A、f)、CD40シグナル伝達への幾分の依存性を観察した。
【0127】
生体外での組換えCyaAで免疫化したマウスにおけるHPV16−E7特異的脾臓細胞の発生頻度を推定するために、インビトロでのHPV16−E749〜57ペプチドでの刺激に応答してIFN−γを産生する細胞の数を、酵素結合イムノスポット(ELISPOT)により定量した。図2Bは、CyaE5−CysOVAで免疫化したマウスから得られたIFN−γを特異的に産生しているIFN−γ産生脾臓細胞の数は、CyaA−E749〜57で免疫化したものと比べて、僅かな差異しかなかったことを示す。これに対し、得られたIFN−γ産生脾臓細胞は、完全なHPV16−E7タンパク質またはその欠失形態を含むHPV16−E7組換えCyaAでワクチン接種したマウスにおいて、はるかにより多かった(p<0.05)。観察された応答は、エピトープ特異的であった。なぜなら、HPV16−E749〜57ペプチドにより刺激しない場合、これらのマウスの非常に僅かな脾臓細胞しか、IFN−γを産生しなかったからである(図2B)。これらの結果は、CyaAが、処理およびMHCクラスI経路へと提示するために、インビボにおいて、HPV16−E7タンパク質の免疫優性CD8+H−2Db拘束性T細胞エピトープを、免疫担当性細胞のサイトゾルへと送達でき、これにより強力なCTL応答を誘起できることを示す。以前の観察によれば(25、31)、本発明者らは、完全なHPV16−E7タンパク質またはその欠失形態および反転形態を有するCyaAもまた、強力なCTL応答を誘導できたので、CyaAは、大きなポリペプチド断片の挿入に耐えられることを確認した。本発明者らのデータはまた、これらの後者の分子は、マウスにおいて、有意により高い発生頻度のHPV16−E749〜57特異的応答を誘導したことを実証した。
【0128】
HPV16−E7組換えCyaAでの免疫化は、HPV16−E7特異的Th1応答を誘導した。
Th1応答は、細胞内病原体および腫瘍発達に対する防御において重要な役割を果たす(32、33)。それ故、本発明者らは、HPV16−E7組換えCyaAにより誘導されるT細胞応答のタイプを特徴づけた。C57BL/6マウスに、50μgの3種の異なるHPV16−E7組換えCyaAを1回静注することにより免疫化し、インビトロで脾臓細胞を10μg/mlの精製His−TagHPV16−E7タンパク質で刺激した後に、サイトカインの合成を決定した。図3に示されているように、完全なHPV16−E7タンパク質またはその欠失形態を有するCyaAでの免疫化により、高いレベルのIFN−γの産生および検出可能なレベルのIL−5が存在しないことにより特徴づけられる、Th1に似たプロファイルが得られた。この応答は特異的であった。なぜなら、CyaA−E7FUllまたはCyaA−E7Δ30〜42で免疫化した後に得られたIFN−γレベルは、CyaAE5−CysOVAでニセ免疫化したマウスで得られたものよりも有意に高かったからである(p<0.05)。しかし、これは、CyaA−E749〜57で免疫化したマウスの脾臓細胞には当てはまらなかった。1μg/mlのE743〜77ペプチドを用いて再刺激した場合に、類似の結果が得られた(図3A、挿入図)。
【0129】
合わせて考えると、これらの結果により、本発明者らの条件では、クラスII H−2b拘束性T細胞エピトープを含む完全HPV16−E7タンパク質を有するCyaAで得られたレベルは、クラス1H−2Db拘束性エピトープしか含まないCyaA−E749〜57で得られたレベルよりもはるかに高いので、CD4+T細胞は、IFN−γの分泌において重要な役割を果たしていることが示された。
【0130】
HPV16−E7組換えCyaAでの免疫化は、確立されたHPV16を発現している腫瘍の減退を誘導した。
強い免疫応答が得られたことを考慮して、その後、本発明者らは、インビボにおいて、HPV16−E6およびE7タンパク質H−2b腫瘍原性細胞系(TC−1細胞)(これはC57BL/6マウスの皮下に注射される)からなる前臨床モデルにおいて、HPV16−E7CyaAの治療活性を評価した。このモデルにおいては、腫瘍拒絶は、排他的に、E749〜57特異的CD8+T細胞により媒介されることが以前に示された(21、22、34、35)。したがって、5×104個のTC−1細胞を、C57BL/6マウスの右側腹に皮下注射し、50μgのCyaAE5−HPV16−E749〜57、−E7full、または−E7Δ30〜42を、1、5、または10日後にマウスに静注した。図4は、腫瘍移植の10日後に治療処置したマウスにおける腫瘍増殖を示す。特に、これらの条件において、100%の動物が、ワクチン接種を行なう時点までに、触診可能な腫瘍を発達させた。不必要な苦しみを避けるために、腫瘍サイズが1000mm3に到達した場合には動物を屠殺した。全ての非処置動物並びにニセCyaAE−cysOVAで処置したマウスは、最大49日間以内に、そのサイズ(>1000mm3)の腫瘍を発達させた。極めて対照的には、HPV16−E7組換えCyaAで処置した動物の大半が、実験期間中を通じて腫瘍がない状態を維持した(図4、C、D、E)。図5は、TC−1細胞を移植し、3つの異なる治療プロトコル(組換えCyaAを、TC−1移植の1、5、または10日後に注射した)を行なった、動物の生存プロットを示す。非処置またはニセ処置動物の生存時間の中間値は、31〜40日間であった。これに対し、HPV16−E7抗原を有するCyaAでワクチン接種したマウスの生存は、対照動物よりも有意に優れていた(p<0.05)。様々な構造物の防御活性の差異は確立できたが、統計学的有意性は確立できなかった。なぜなら、様々な試料のサイズが小さいからであった。CyaA−E749〜57および−E7Fullにより付与される腫瘍減退率が、認められる程には区別できなくても、CyaA−E7Δ30〜42は、腫瘍減退および増殖阻害の点において明らかに優れていた。なぜなら、3つ全ての治療スキームにおいて、防御率は、常に、90%よりも高かったからである。
【0131】
CyaA−E749〜57およびCyaA−E7Fullでワクチン接種した何匹かの動物は、実験の経過中、より後で腫瘍を発達させるようであった(図4(*)、データは示していない)。この現象は腫瘍エスケープ機序を反映する可能性があることを心に置き、本発明者らは、これらの動物から増殖中の腫瘍を外移植し、H−2Db発現について、FACS解析により、TC−1A1およびTC−1A2という名称のこれらの細胞系を解析した。図6に示されているように、その天然対応物に比べて、より後で増殖してくる腫瘍から外移植したTC−1A1およびA2細胞は、H−2Dbの発現を欠失しており、したがって、E749〜57特異的CD8+T細胞により検出されることができなくなっている可能性が高い。TC−1細胞は、インビボにおいて、選択のための抗生物質の負荷がない環境で増殖させたので、本発明者らはまた、TC−1A1およびA2細胞においてHPV16−E7の発現についてウェスタンブロットにより確認した。本発明者らは、TC−1およびTC−1A1およびA2細胞の間にこのタンパク質の発現の差異を全く見出すことができなかった(データは示さず)。
【0132】
合わせて考えると、これらの結果により、前臨床モデルにおいてHPV16を発現している腫瘍の減退を誘導するために適した治療ワクチンとしての、アデニル酸シクラーゼベクターの有効性が実証された。
【0133】
本発明者らは、臨床的に関心の高い別の注射経路を試験した。したがって、10μgのCyaA−E7Δ30〜42を、TC−1移植の10日後から始めて7日間間隔で2回皮内注射した。興味深いことに、全ての非処置およびニセ処置動物が腫瘍を発達させたが、本発明者らは、CyaA−E7Δ30〜42で処置した動物の全ての動物において腫瘍減退を観察した(図4B、a、b)。この治療的免疫化により、90日目においてCyaA−E7Δ30〜42処置マウスの100%の生存率が得られ、一方、非処置およびニセ処置動物の生存率の中間値は、それぞれ30日および32日であった(図4、c)。
【0134】
CyaA免疫化により誘導されるHPV16−E749〜57特異的CD8+T細胞の長期持続性
HPV16−E7組換えCyaAにより誘導される免疫応答の持続性を評価するために、3ヵ月後も治療実験にも関わらず生存しているマウスを屠殺し、その脾臓細胞を、インビトロで、1μg/mlのE743〜77ペプチドで5日間かけて刺激した。その後、TC−1細胞を溶解する能力を、51Cr放出アッセイにより決定した。図7に示されているように、HPV16−E743〜77ペプチドに対する特異的CTL応答が、依然として、3ヵ月前に免疫化した動物の脾臓細胞から実証された。30:1のエフェクター:標的の比での特異的溶解の最大パーセンテージに基づいて、免疫応答は、CyaA−E7Δ30〜42で処置した動物においてより強力であるようであるが、これらのデータから統計学的有意性は実証できなかった。このような長く続く免疫原性の生理的関連性を評価するために、残りの動物に、100日目に5×104個のTC−1細胞を用いて皮下に再攻撃した。このような条件下で、全ての未処置の年齢一致対照動物が腫瘍を発達させ、37.5日間という生存時間の中間値を示した(図8)。これに対し、HPV16−E7組換えCyaAで3ヶ月前に免疫化したマウスは、腫瘍発達から非常に有意に防御された。先に観察されているように、CyaAE−HPV16−E7Δ30〜42でワクチン接種した動物が、高いレベルの防御を示した。しかし、今回、最初のセットの治療実験で得られた結果の変動では、CyaA−E749〜57で処置した動物と、CyaA−E7FUllで処置した動物の間には顕著な防御の差異があり、後者の方が好ましいが、試料のサイズが少ないので、統計的有意性を明確に実証することはできなかった。これらの観察により、このモデルでは、完全なHPV16−E7タンパク質を有するCyaAにより提供されるT細胞の支持は、TC−1細胞に対する効果的で長期に持続する応答には重要であることが示唆された。
【0135】
合わせて考えると、本発明者らの結果により、HPV16−E7タンパク質のヘルパーエピトープ(DRAHYNIVTF)を有する50μgの組換えCyaAの1回の静注は、少なくとも6ヶ月の期間におよび、2回のTC−1腫瘍細胞攻撃に対する防御を付与できる、長期に持続するE749〜57特異的CD8+T細胞を誘導するのに十分であった。
【0136】
CyaA−E7Δ30〜42の治療効力は、CpG−ODN1826と共に投与されたペプチドの治療効力に匹敵した。
抗原送達システムとしてのCyaAの効力をより良く評価するために、本発明者らは、CyaA−E7Δ30〜42の治療効力を、CpG−ODN1826を補充したHPV16−E743〜77ペプチドの治療効力と比較した(37)。それ故、マウスに、5×104個のTC−1細胞を皮下注射し、10および17日後、皮内経路により、10μgのCyaA−E7Δ30〜42で、または、1μgのCpG−ODN1826と共に投与される10μgのHPV16−E743〜77ペプチドで、治療処置した。生存率は、これらの2つの群において類似していたが(図9)、CyaA−E7Δ30〜42で得られた結果は、CpG−ODN1826と混合されたHPV16−E743〜77ペプチドで得られた結果よりも僅かに良好であったが、統計学的に差異はなかった。特に、この結果は、モル基準で、CyaA−E7Δ30〜42に比べて50倍多いHPV16−E743〜77ペプチドを使用して得られた。単独で使用した場合、ペプチドHPV16−E743〜77は、TC−1腫瘍増殖に対して全く効果がなかった。
【0137】
CyaAベクターに対する事前免疫は、CyaA−E7Δ30〜42の治療効力に僅かにしか影響を及ぼさなかった。
臨床の場面では、効果的な細胞性免疫応答を得るために、病変を有する患者に、複数回のブースター投与をおそらく行なわなければならない。それ故、CyaAベクターに対する予備免疫が、腫瘍拒絶を引き起こす能力を損なわないことを実証することが不可欠であった。そうするためには、本発明者らは、5×104個のTC−1細胞を皮下注射する90または30日前に、7日間間隔で2回10μgの空ベクターCyaAE5でマウスを皮内免疫化した。7日間間隔の2回の10μgのCyaA−E7Δ30〜42の注射による治療処置を10日目に設定した。抗体応答の解析により、空ベクターで免疫化されたマウスは、TC−1注射時にCyaAに対して免疫があることが示された(図10A)。その後、本発明者らは、年齢の一致した未処置動物およびCyaA免疫動物において腫瘍拒絶を誘導する、CyaA−E7Δ30〜42処置の能力を比較した。CyaAに対するその免疫状態はどうであれ、CyaA−E7Δ30〜42で処置したマウスの大半が、実験を通じて腫瘍がないままであった(図10B)。30日目の免疫マウス群の中で1匹のみ、90日目の免疫マウス群において2匹のみが、腫瘍を発達させた(図10Bb、d、f)。これに対し、ニセ感染動物の100%が腫瘍を発達させ、屠殺された(図10B、a、c、e)。本発明者らは、抗CyaA抗体力価レベルと、TC−1腫瘍の発達の間の相関は全く観察しなかった(データは示さず)。さらに、CyaA−E8Δ30〜42処置マウスの生存曲線(図10B、b、d、f)は、統計学的に差異はなかった(p=0.324)。
【0138】
それ故、これらのデータにより、CyaAに対する既存免疫は、このベクターがその後に外来の所与の抗原に対する効果的な応答を誘導する能力に対して、殆ど作用を及ぼさないことが示された。
【0139】
考察
以前の研究により、Bordetella pertussisのアデニル酸シクラーゼは、インビボにおいて、CD4+およびCD8+T細胞エピトープを、樹状細胞のMHC−クラスIIおよびI提示経路に送達するための強力なツールであることが実証された。実験ネズミモデルにおいて、このシステムは、抗ウイルスおよび抗腫瘍防御をもたらす効果的なTh1およびCTL応答を引き起こすために使用されている(36)。HPV16関連の子宮頚部悪性疾患の処置のためのヒトにおけるCyaAの可能性ある適用の評価として、本発明者らは、このベクターが、インビボにおいて、HPV16のE7タンパク質のエピトープを効果的に送達することを実証し続けた。
【0140】
本発明者らは、HPV16の完全なE7タンパク質またはこのポリペプチドの亜断片(特に、残基49〜57に対応するH−2Db拘束性最小CTLエピトープを含む)を含む様々なHPV16組換えCyaAを作成した。本発明者らは、これらの異なる組換えタンパク質が、C57BL/6マウスに注射された時に、特異的かつ強力なCTL応答を抗原刺激できることを示した。本発明者らのデータにより、本発明者らはTAP1−/−マウスにおいてCyaA−E7Δ30〜42の注射によりCTLを刺激することができなかったので、CyaAによるCTLエピトープの送達には、完全に機能的なクラスI提示経路が必要であることが確認された(15)。CyaAにより媒介されるCTL刺激は、以前の結果と一致して、MHCクラスII−/−マウスで得られた効果的なCTL応答により示されるように、CD4+T細胞の存在とは独立していた(15)。ワクチンベクターとしてのこのCyaAの特徴は、CD4+T細胞数の変化を提示する免疫抑制患者または免疫欠陥患者のワクチン接種を考慮した場合に非常に重要である。しかし、CD40−/−マウスではCTL応答は低く、このことは、CTL刺激が、一部、CD40シグナル伝達に依存していることを示す。これらの観察により、HPV16−E7組換えCyaAは、プロフェッショナルAPCの直接刺激により、MHCクラスI拘束性CTLを直接的に誘起することが示唆された。それにも関わらず、CTL応答の最適な刺激を得るためには、CD40−CD40L相互作用が必要とされる。
【0141】
本発明者らは、HPV16−E7の最小H−2Db拘束性CTLエピトープの免疫原性を、おそらくとりわけ記載されたヘルパーエピトープDRAHYNIVTFを含む、完全もしくはΔ30〜42E7タンパク質の免疫原性と比較した。CTL刺激並びにCyaA−E7FUllおよびCyaAE−E7Δ30〜42により誘導されるHPV16−E7特異的脾臓細胞の発生頻度は、CTLエピトープE49〜57のみを有するCyaA−E749〜57により誘導されるものよりも優れていた。これらの観察により、CyaAによりCTLおよびThエピトープを同時に送達することにより、より強力なCTL応答がもたらされることが示された。これは、他のモデルにおいて、前臨床(37)および臨床レベル(38)で以前に公表されたデータと一致した。組換えCyaAによるHPV16−E7Thエピトープの送達は、さらに、インビトロで組換えHisTag−HPV16−E7タンパク質またはE743〜77ペプチドで再刺激したHPV16−E7特異的脾臓細胞により産生されるサイトカインの解析により証明された。実際に、本発明者らは、Thエピトープを含む組換えHPV16−E7CyaAでワクチン接種したマウスのみにおいて、IFN−γの特異的合成を観察した。本発明者らが、CyaA−E7FullおよびCyaA−E7Δ30〜42の1回静注により免疫化した後に観察した、IFN−γレベルが高いことおよびIL−5が全く分泌されないことにより特徴づけられる典型的なTh1プロファイルは、腫瘍免疫療法におけるこのベクターの関心の高さを強調する。
【0142】
これは、皮下で確立された腫瘍原性TC−1細胞に基づいた腫瘍拒絶モデルで試験した(21)。HPV16−E7組換えCyaAの免疫原性を実証する本発明者らのデータによれば、本発明者らは、これらの組換えタンパク質が、確立されたTC−1腫瘍の減退を誘導できることを観察した。CyaA−E7Δ30〜42は、90日間の期間におよび、生存の点で、CyaA−E749〜57およびCyaAE5−HPV16−E7Fullよりも優れていた。完全およびΔ30〜42E7タンパク質を有するCyaAは、CTL刺激能力、HPV16−E749〜57特異的脾臓細胞の発生頻度、およびIFNγ産生の点で、匹敵する結果を与えたので、本発明者らは、生存の点で、CyaA−E7Fullは、CyaA−E749〜57よりも優れていると期待した。より多くの数の試験動物が、この一見した矛盾を除外してくれるのに役立つ可能性が高い。しかし、CyaAの生化学に関する2つの側面をここで考察すべきである。第一に、CyaAの224〜235の領域の負に荷電したアミノ酸の存在は、真核細胞のサイトゾルへのCyaAの触媒ドメインの転位を阻害することが示された(39)。これに関して、酸性E7タンパク質(pKi=4.17)が、DCのサイトゾルへのCyaAのN末端ドメインの効率的な転位を妨害する可能性があった。CyaA−E7Δ30〜42は、DCへの送達に好ましいように、30〜42残基に位置する負に荷電したアミノ酸のストレッチ(DSSEEEDEIDGPA)を除去するように特別に設計され、さらに、2個の正に荷電したアミノ酸(KR)を、HPV16−E7の挿入されたN末端ドメインの各側に導入した。第二に、Gmiraら(25)は、CyaA内に挿入された異種タンパク質の折りたたみ解除は、組換えタンパク質が標的細胞へと内部移行するために必須であることを実証した。E7ポリペプチドの2個の異なる断片を、CyaA−E7Δ30〜42の2個の異なる許容部位へ挿入することにより、E7の再折りたたみが防がれ、したがって、標的細胞へのその転位は促進されるはずである。
【0143】
腫瘍拒絶実験の時間経過中に、何匹かのマウスは、確立されたTC−1腫瘍を以前に拒絶した後、より後になってHPV16陽性腫瘍を増殖させ始めた。FACS(登録商標)解析により、これらの腫瘍の細胞は、H−2Db分子を発現していなかったことが判明した。この観察により、より著しく腫瘍細胞を根絶し、エスケープ機序による腫瘍の再発を防ぐために、組換えCyaAワクチン接種の同様なブースター投与の可能性が考えられた。当該技術分野における他のチームのデータを心に置くと(37、40、41)、それは、注射された組換えHPV16−E7 CyaAの全量を100μg、すなわち0.56nmolに上げた、マウスにおけるCyaAワクチン接種のブースター投与に関連していた。この後者の観察を試験するための実験、並びに、CyaAの様々な投与法を試験するための実験を実施した。
【0144】
TC−1細胞で再攻撃した時、HPV16−E7組換えCyaAで免疫化した生存マウスは、選択的に防御された。これは、これらの動物の脾臓細胞中のHPV16−E749〜57CD8+T細胞の存在に関連していた。この観察は、図4で観察される遅い再発は、腫瘍エスケープ機序に起因し、E7タンパク質に対する免疫が徐々に衰えていくことに起因するものではないということが強まった。Thエピトープを含む組換えCyaAで免疫化したマウスのより良好な生存率により、同じ抗原に対するT細胞の支持が得られることにより、HPV16−E749〜57CD8+T細胞が効果的に想起されることが示された。これに関して、CD4+T細胞は、CD40Lを通じて、エフェクターCD8+T細胞に独特な分子的サインを刷り込む可能性があり、より向上した細胞機能を行なうための能力を付与する可能性があることが提唱された(42)。
【0145】
HPV関連新生物形成の処置のための新規免疫療法の試験のための確証されたモデルにおいて(21)、本発明者らは、CyaAが、確立腫瘍の減退を誘導し、並びに、長い期間におよぶ腫瘍原性攻撃に対する防御を与えるための、効率的なベクターであることを実証した。現在開発されている他のアプローチとは異なり(11)、CyaAを基礎とした免疫療法は、完全なタンパク質を挿入できるので、HLA拘束性エピトープを選択する必要がなく、ウイルスベクターおよび/または発癌の可能性のあるHPV DNA配列の使用が回避された。さらに、本発明者らは、CyaAの2個の異なる許容部位に挿入されたHPV−E7の2個の亜断片を含むCyaAを用いて最善の結果を得た。
【0146】
この後者の構造物が、文献に記載の全てのHPV16−E7 HLAクラスIおよびクラスIIエピトープを含むという事実により(8、46)、ワクチン適用にCyaA−E7Δ30〜42が選択されることが強まった。
【0147】
ここに提示したデータに基づいて、本発明者らは、HPV感染に関連した子宮頚部および肛門の異常形成を標的化した臨床試験において、HPV16−E7を含むCyaAの効力を試験する計画を立てた。
【0148】
実施例2
目的は、ヒトにおけるHPV16およびHPV18関連の悪性疾患を処置するための、HPV16およびHPV18 E7タンパク質の両方に標的化した二価治療ワクチンを製造することであった。それ故、CyaAE5−HPV16E7Δ30〜42およびCyaAE5−HPV18E7Δ32〜42と呼ばれるワクチン候補を、設計し、作成し、産生し、精製した。HHDマウスは、H−2Dbのα3膜貫通ドメインおよび細胞質ドメイン(D)に連結したHLA*0201のα1(H)およびα2(H)ドメインを含むHHD導入遺伝子を発現しているH−2D−/−β2m−/−ダブルノックアウトマウスであった(α1ドメインは、ヒトβ2−ミクログロブリンに連結している)。したがって、HHDマウスにより発現されるMHCクラスI分子のみが、改変されたHLA*0201分子であった(Pascolo et Lemonnier)。
【0149】
この実験の目標は、
1 組換えCyaAが、HPV16およびHPV18E7タンパク質のHLA−A2拘束性エピトープを送達できること、
2 1個のHPV組換えcyaAが他を上回る免疫優性の現象はないこと
を実証することであった。
【0150】
そうするために、HHDマウスに、50μgのCyaAE5−HPV16E7Δ30〜42(3匹のマウス)、CyaAE5−HPV18E7Δ32〜42(3匹のマウス)、またはCyaAE5−HPV16E7Δ30〜42+CyaAE5−HPV18E7Δ32〜42(5匹のマウス)を同じ(眼窩後の)注射(200μl中50μg)で静脈内にワクチン接種した。7日後、プールした脾臓細胞を、インビトロで、文献に記載またはSYFPEITHIソフトウェアから推定される、HPV16E7またはHPV18E7のHLA−A2ペプチドで再刺激した。
【0151】
CTL活性は、51Cr放出を使用して5日後にアッセイした。標的細胞は、異なる関連ペプチドをのせたまたはのせていない、HHD−EL4細胞であった。
【0152】
結果により、CyaAE5−HPV16E7Δ30〜42またはCyaAE5−HPV18E7Δ32〜42でのワクチン接種は、インビトロでの自己ペプチドによる再刺激後に、HPV16E7またはHPV18E7ペプチドをのせたEL4−HHD細胞の特異的溶解を誘導したことが示された。
【0153】
この結果によりまた、CyaAE5−HPV16E7Δ30〜42をCyaAE5−HPV18E7Δ32〜42と共に注射することにより、関連ペプチドに対する類似の応答が観察されたので、いずれのHPV組換えCyaAの免疫原性も損なわなかった。
【0154】
この結果により、CyaAE5−HPV16E7Δ30〜42およびCyaAE5−HPV18E7Δ32〜42は、インビボにおいて、それぞれのE7タンパク質のヒトHLA−A2拘束性エピトープに対する細胞毒性応答を誘導できることが実証された。
【0155】
マウス
特定の病原体を含まないHHDマウスを、パスツール研究所で育成した。11匹の6〜10週令の雄を、この実験に使用した。
【0156】
試薬および生物学的材料
試薬および緩衝液
RPMI1640培地−グルタマックス(インビトロジェン・ギブコ、参照番号:6187010)
エタノール70゜(Prolabo、参照番号:MC311631)
ペニシリン−ストレプトマイシン(インビトロジェン・ギブコ、参照番号:15140122)
ウシ血清アルブミン(FBS)(PERBIO、参照番号:CH30160.03)
βメルカプトエタノール(バイオラッド、参照番号:161−0710)
熱分解水
ブルートリパン(シグマ、T−8154)
51Cr
Triluxシンチラント(Wallac)
【0157】
ペプチド
5個の合成ペプチド(Neosystem、ストラスブール、フランス)を、51Cr放出アッセイの前の脾臓細胞のインビトロでの刺激に使用した:
HPV16−E7 HLA−A2拘束性エピトープ(1)に対応する、E711〜20(YMLDLQPETT、一文字でアミノ酸をコード、#253)、
HPV16−E7 HLA−A2拘束性エピトープ(1)に対応する、E782〜90(LLMGTLGIV、#258)、
HPV16−E7 HLA−A2拘束性エピトープ(1)に対応する、E786〜93(TLGIVCPI、#255)
SYFPEITHIソフトウェアにより予測されるHPV18−E7 HLA−A2拘束性エピトープに対応する、E77〜15(TLQDIVLHL、#251)、
HPV18−E7 HLA−A2拘束性エピトープ(2)に対応する、E786〜94(FQQLFLNTL、#257)。
【0158】
ペプチドは、無菌で無発熱原性の水(#253、#255、#251)または無発熱原性の水、0.1M NaHCO3;アセトニトリル(50/50)(#257および#258)で1mg/mlに希釈した。
【0159】
組換えアデニル酸シクラーゼ(CyaA)
2個のCyaAをこの実験において試験した:
CyaA−HPV16E7Δ30〜42
CyaA−HPV18E7Δ32〜42。
【0160】
これらのCyaAは、E.coli株BLRにおいて、酵素的に不活性なCyaA(CyaAE5)をコードするプラスミドpTRACE5の誘導体を使用することにより発現させた。プラスミドの作成、全ての組換えタンパク質の産生および精製は、HPV16−E7エピトープを有する組換えB.pertussisアデニル酸シクラーゼの作成および精製に関する実施例1において先に記載したように製造したが、エピトープは、本実験で使用したものであった。
【0161】
8M尿素中にそれぞれ1.22mg/mlおよび1.33mg/mlのCyaAを含むストック溶液(−20℃で保存)を解凍し、マウスに静注内投与する前に、PBS(ギブコBRL)で250μg/mlに希釈した。したがって、尿素の最終濃度は、それぞれ、1.6Mから1.5Mの間であった。
【0162】
細胞系
EL4−HHD細胞を、51CrCTLアッセイにおいて標的として使用した。これらの細胞を、完全培地中に維持した:10%熱不活性化FCS、100U/mlのペニシリン、100μg/mlのストレプトマイシン、および5×10−5Mのβ2−メルカプトエタノール(ギブコBRL、Cergy-Pontoise、フランス)を補充したグルタマックスを含むRPMI1640。
【0163】
方法
マウス免疫化
覚醒動物に、0.3mlのインスリンシリンジ(テルモ)を用いて、(200μl中50μgの)CyaAE5−HPV16E7Δ30〜42および/またはCyaAE5−HPV18E7Δ32〜42の1回の静脈内(眼窩後の)注射により免疫化した。
【0164】
実験計画
表1は、各群のマウスに投与するワクチン候補および処置を記載した。
【0165】
【表9】
【0166】
CTLアッセイ
免疫化により誘導される細胞性免疫応答をインビトロで評価するために、麻酔した動物(CO2)を、免疫化の7日後に脾摘除した。CTLアッセイの前に、各群の脾臓細胞をプールした。
【0167】
−CTLの刺激(1個の脾臓あたり2個のフラスコT25を準備)
−脾臓をサンプリングし、RPMI1640−グルタマックス1%Ab中でそれらを破粉した。
−デカント。
−1200rpmで10分間遠心分離。
−細胞を、2mlのRMPI1640−グルタマックス1%Ab−10%FCS−5.10−5M β−メルカプトエタノール(完全培地:CM)に再懸濁。
−各々のT25フラスコに、
10mlのCM
5×107個のエフェクター細胞
10μg/mlの最終濃度の関連ペプチド
を入れた。
フラスコを動かすことなく5日間インキュベート。
【0168】
−細胞毒性アッセイ
1)標的細胞:
−前日、標的細胞を3分の1または2分の1に希釈して、指数関数的な増殖期のそれらを収集。
−15mlのチューブに移す。
−1200rpmで10分間遠心分離。
−1mlのRPMI1640−1%Ab中に再懸濁。
−数を数えた。
−2個のチューブを準備:1個はペプチドを含み、1個はペプチドを含まない。
−150μl中に以下を再懸濁:
2個の中の1個のチューブにペプチド(50μモル)
3×106個の細胞につき100μCiの51Cr(50μl)。
150μlとするための培地。
−37℃の水浴中で1時間インキュベートし、15分毎に穏やかに振とう。
【0169】
2)エフェクター細胞
−約5mlの上清をパスツールピペットを用いて廃棄。
−ピペッティングにより細胞を完全に再懸濁。
−1200rpmで10分間遠心分離。
−1mlのCMに再懸濁。数を数えた。
−107個の細胞/mlに調整。
−次のエフェクター/標的比:200/1、100/1、50/1、25/1、12/1、6/1が得られるように希釈。
−100μl/U底マイクロタイタープレートのウェルで分配。
−標的細胞の調製を完成する間、37℃で7.5%CO2中でインキュベート。
【0170】
3)アッセイ:
−標的細胞を、10mlのRPMI1640−1%Abで洗浄。
−再度10mlのCMで洗浄。
−2mlのCMに再懸濁。
−数を数えた。
−105個の細胞/mlに調整。
−100μl/ウェルで分配。
−自発的放出のための6個のウェルを準備(CM中100μlに調整)。
−最大放出のための6個のウェルを準備(100μlの20%トリトンX−405を添加)
−37℃で7.5%CO2中で4〜5時間インキュベート。
【0171】
4)計測:
−2000rpmで5〜10分間遠心分離。
−100μlのTriluxシンチラントを、フロッピーP96マイクロタイタープレートに入れた。
−50μlの上清をサンプリングし、それをフロッピープレートに移した。
−プレートを、プラスチックフィルムテープで封をした。
−wallacカウンターで次の日に計測した。
【0172】
HPV16E7CTL特異的応答
CyaA−HPV16E7Δ30〜42により誘導されるCTL応答を図11に示した。
【0173】
HPV18E7CTL特異的応答
CyaA−HPV18E7Δ32〜42により誘導されるCTL応答を図12に示した。
【0174】
結論
CyaA−HPV16E7Δ30〜42およびCyaA−HPV18E7Δ32〜42の両方が、このキメラHHDモデルにおいて、それぞれのHLA−A2拘束性ペプチドに対して特異的なCTLを誘導できた。
【0175】
CyaA−HPV16E7Δ30〜42およびCyaA−HPV18E7Δ32〜42の同時注射により、各々の個々のCyaAがそれぞれの特異的HLA−A2拘束性ペプチドに対するCTLを誘導する能力は妨害されなかった。これにより、1つの構造物が他の構造物よりも上回る免疫優性を示す現象はなかったことが示された。これには、二価ワクチンの製造戦略に関する、重要な観察が含まれていた。
【0176】
実施例3
HPV18のE7タンパク質並びにHPV16およびHPV18のE7タンパク質を含む組換えアデニル酸シクラーゼの作成および免疫学的評価
肛門生殖管の癌は、女性における全ての癌の12%近くを占め、子宮頚部癌(CxCa)は世界中で二番目に最も頻繁に起こる婦人科の癌となっている。ヒトパピローマウイルス(HPV)感染がCxCaの原因である可能性があるという重要な観察は、その後、疫学的な研究により確認された。CxCaに関連する最も広まっているHPVのタイプは、HPV16およびHPV18である(それぞれ55%および12%の罹患率)(Clifford、2003)。より多くの人口を網羅するために、HPV16およびHPV18のE7を有する二価治療ワクチンを作成することが決定された。2つの可能性ある戦略が試験された:(i)第一の戦略は、一方はHPV16のE7を有する他方はHPV18のE7を有する、等モル量の2個の組換えCyaAを混合する;(ii)第二の戦略は、HPV16およびHPV18の両方のE7タンパク質を有する組換えCyaAを作成する。本リポートは、HPV18のE7または両方のウイルスのE7を有する組換えCyaAの作成を記載した。構造物の免疫原性は、CTLアッセイにおいて試験した。
【0177】
エピトープ同定
HPV18E7のアミノ酸配列を、H−2bエピトープの探索中に、SYFPEITHI(www.syfpeithi.de)にかけた。このソフトウェアは、アルゴリズムを使用して、画定されたMHC分子に結合する可能性があるエピトープを予測する。ソフトウェアは、25のスコアで、H−2Db分子により拘束される推定エピトープを選出した。通常、本発明者らは、そのスコアが22を超えた場合にエピトープであると判断した。それ故、推定ペプチドのIDGVNHQHLが、Neosystemにより合成された。
【0178】
組換えCyaAの作成
CyaA−HPV18E7、CyaA−HPV18E7Δ32〜42、CyaA−HPV16+18E7、CyaA−HPV16+18ΔE7を産生するためにプラスミドの作成に使用された戦略を示したスキームを、図13および14に示した。
【0179】
CTLアッセイ
この実験の目標は、インビボにおいて、完全なHPV18E7タンパク質またはその断片並びにHPV16およびHPV18E7タンパク質の両方を有する、組換えCyaAによるCTLの誘導を実証することであった。標的化されたH−2Dbエピトープは、コンピューター予測ソフトウェアから得られたHPV18E741〜49(IDGVNHQHL)およびHPV16E749〜57であった。C57BL/6マウスに、50μgのCyaA−CysOVA、CyaA−HPV18E7、CyaA−HPV18E7Δ32〜42、CyaA−HPV16+18E7、またはCyaA−HPV16+18ΔE7(各群において2匹)を静注によりワクチン接種した。7日後、プールした脾臓細胞を、インビトロで、ペプチドHPV18E741〜49(10μg/ml)、OVA257〜264(1μg/ml)またはHPV16E743〜77(1μg/ml)で再刺激した。CTL活性は、51Cr放出を使用して5日後にアッセイした。標的細胞は、HPV18E741〜49ペプチド(8nmol)をのせたまたはのせていないEL4細胞、あるいは、TC−1細胞であった。図中の説明文は:ワクチン接種/再刺激を示す。例:OVA/OVAは、CyaAE5−CysOVAでワクチン接種し、OVAペプチドで再刺激したマウスを意味する。
【0180】
これらの結果(図16)により、CyaA−HPV18E7Fullとは対照的に、CyaA−HPV18E7Δ32〜42は、インビボにおいて、処理およびMHCクラスI経路に提示するために、コンピューターで予測したHPV18−E7タンパク質のCD8+H−2Db拘束性T細胞エピトープを、免疫担当細胞のサイトゾルへと送達でき、これにより、強力なCTL応答を誘起できることが示された。CyaA−HPV18E7Δ32〜42が、HPV18E741〜49に対するCTL応答を刺激できることは奇妙である。なぜなら、この組換えCyaAは、ペプチドHPV18E741〜49の最初の2個のアミノ酸を欠失しているからである。しかし、HPV18E7断片43〜105の挿入部位においては、1個のアラニンおよび1個のセリンがあり、これにより、推定ペプチドは、ここで、IDGVNHQHLではなく、ASGVNHQHLであった。syfpeithiにかけると、このペプチドは、29のスコアで出て来た(天然のものは25)。それ故、本発明者らのクローニングにより、このペプチドの最初のアミノ酸を置換することにより、本発明者らは、潜在性から免疫原性へとすることができるようであった。これは、HPV18E7タンパク質のエピトープが、H−2Dbの関連で記載された始めてのケースであった。
【0181】
非特異的バックグラウンドレベルが高いにも関わらず、これらの結果によりまた、CyaA−HPV16+18E7FullおよびCyaA−HPV16+18E7Δは、インビボにおいて、処理およびMHCクラスI経路に提示するために、HPV16−E7タンパク質のH−2Db拘束性T細胞エピトープを、免疫担当細胞のサイトゾルへと送達でき、これにより、強力なCTL応答を誘起できることが示された。CyaA−HPV16+18E7Δは、CyaA−HPV16+18E7Fullと比べて高いその能力を示した。これらのデータによりまた、初めて、大きなポリペプチド断片(203までのアミノ酸)を有するCyaAは依然として免疫原性であることが示された。
【0182】
さらなる実験を実施し、図16に記載のことを確認した。各群の残りの2匹のマウスを、最初のものとは別々に処置した。実験設定は類似していた。結論は、CTLによるCyaAE5−HPV18E7Δ32〜42ワクチン接種マウスのEL4細胞の高い非特異的溶解レベルは、過剰のHPV18E741〜49ペプチドがフラスコに残っていることに起因することを除いて、図16のものと同一であった。
【0183】
またCyaA−HPV16+18ΔE7でワクチン接種したマウスの脾臓細胞ではTC−1細胞の溶解は認められたが、今回CyaA−HPV16+18E7でワクチン接種したマウスには認めれられなかった。この実験において、非特異的バックグラウンドは40%でピークに達した。それにも関わらず、図17から導き出せる結論は、図16から導いた結論を確認した、すなわち:
−CyaA−HPV18E7Δ32〜42は、インビボにおいて、処理およびMHCクラスI経路への提示のために、コンピューターで予測したHPV18−E7タンパク質のCD8+H−2Db拘束性T細胞エピトープを、免疫担当細胞のサイトゾルへと送達でき、これにより強力なCTL応答を誘起できる。これは、HPV18E7タンパク質のエピトープが、H−2Dbの関連で記載された最初のケースであった。
−大きなポリペプチド断片(203までのアミノ酸)を有する組換えCyaAは、依然として免疫原性であった。
【0184】
結論
HPV16およびHPV18のE7を有する二価治療ワクチンを作成することが決定された。2つの可能性ある戦略が考えられた:(i)第一の戦略は、一方はHPV16のE7を有する他方はHPV18のE7を有する、等モル量の2個の組換えCyaAを混合する;(ii)第二の戦略は、HPV16およびHPV18の両方のE7タンパク質を有する組換えCyaAを作成する。本結果は、HPV18のE7または両方のウイルスのE7を有する組換えCyaAの作成を記載した。構造物の免疫原性は、CTLアッセイにおいて試験した。
【0185】
本研究の主な結論は、考えられている両方の戦略が、実現可能であることである。なぜなら、HPV18E7の配列内に潜在性H−2Db拘束性エピトープが初めて判明したため、HPV18E7亜断片を有する組換えCyaAが機能的であるからである。さらに、HPV16および18ウイルスの両方のE7タンパク質(または亜断片)を有する組換えCyaAは、TC−1細胞により天然に提示されるH−2Db拘束性HPV16E7エピトープに対するCTL応答を刺激できるので、依然として最も興味深いことに免疫原性であった。
【0186】
【表10】
【図面の簡単な説明】
【0187】
【図1A】HPV16−E7組換えCyaAの作成および精製。(A)関連する制限酵素部位および挿入配列が示されている、pTRACE5のマップ図。(B)CyaAの酵素活性を阻害するジペプチドLQの挿入部位を示すCyaAの図解。HPV16−E7タンパク質挿入断片の位置も示されている。HPV16−E7 H−2b拘束性エピトープには下線を付している。
【図1B】HPV16−E7組換えCyaAの作成および精製。(C)HPV16−E7組換えCyaAのSDS−PAGE解析。5μgの精製タンパク質を、4〜15%SDSポリアクリルアミドゲル上で分離し、クーマシーブルーにより染色した。レーン1:野生型CyaA;レーン2:CyaA−E749〜57;レーン3:CyaA−E7FUll;レーン4:CyaA−E7Δ30〜42。(D)HPV16−E7組換えCyaAのウェスタンブロット解析。SDS−PAGE後、精製タンパク質を、ニトロセルロース膜にエレクトロトランスファーし、続いてこれをマウスモノクローナル抗HPV16−E7抗体でプローブした。レーン1、2:野生型CyaA(それぞれ2および0.4μg);レーン3、4、および5:それぞれ、CyaA−E749〜57、CyaA−E7FUllおよびCyaA−E7Δ30〜42、各々0.4μgのタンパク質。
【図2】組換えHPV16−E7CyaAによるT細胞応答の誘導。(A)C57BL/6(a、b、c)、TAP1−/−(d)、MHCクラスII−/−(e)、およびCD40−/−(f)マウスを、0日目に、50μgのCyaA−E749〜57(a)、CyaA−E7Full(b)、またはCyaA−E7Δ30〜42(c、d、eおよびf)の静注により免疫化した。数日後、動物を屠殺し、脾臓細胞を、インビトロで、1μg/mlのHPV16−E743〜77ペプチドを用いて、照射された同系脾臓細胞の存在下で5日間再刺激し、TC−1標的細胞(普通の四角)またはEL4(白抜きの四角)に対するエフェクターとして使用した。非関連エピトープOVA257〜264を有するCyaAE5−cysOVAで処置し、インビトロで1μg/mlのHPV16−E743〜77ペプチドを用いて、照射された同系脾臓細胞の存在下で5日間再刺激したマウスの脾臓細胞も表示する(a、普通の三角)。標的溶解は、51Cr放出により評価した。データは、特異的溶解値の中間パーセンテージ(n=動物の数を各グラフに示す)並びに四分位範囲を示す。(B)組換えHPV16−E7CyaAでの免疫化後のHPV16−E7特異的IFN−γ産生細胞の検出。C57BL/6マウスを、Aと同様に、CyaAE5−cysOVA(丸)、CyaA−E749〜57(四角)、CyaA−E7Full(三角)、またはCyaA−E7Δ30〜42(菱型)を用いて免疫化した。数日後、免疫化マウスから単離した脾臓細胞を、インビトロで、刺激せずに(すなわち、ペプチドなし、白抜きの記号)、または、1μg/mlのE749〜57ペプチド(普通の記号)を用いて刺激して、同系照射脾臓細胞の存在下で36時間培養した。データは、脾臓1個あたりのSFCの数として表現し、3回の独立した実験の個々のマウスの結果を、各群に対して示す。横棒は、各群の応答の中間値を示す。
【図3】組換えHPV16−E7CyaAは、HPV16E7特異的Th1応答を誘導する。(A)C57BL/6マウスを、未処置のままにするか(四角)、または、50μgのCyaAE5−cysOVA(丸)、CyaA−E749〜57(三角)、CyaA−E7Full(逆三角)、もしくはCyaA−E7Δ30〜42(菱型)を用いて静注により抗原刺激した。数日後、脾臓細胞を、インビトロで、10μg/mlのHis−Tag−HPV16−E7タンパク質で刺激し、上清を、IFN−γ含量について72時間後に試験した。4回の独立した実験の個々のマウスの結果を示し、2個の複製ウェルの上清に放出されたIFN−γの濃度として表現する。再刺激しなかった脾臓細胞で得られたバックグラウンドを差し引く。挿入図:1μg/mlのE743〜77ペプチドを用いたインビトロでの刺激。横棒は、各群の応答の中間値を示す。(B)上清をIL−5含有量について試験したこと以外は、(A)と同様。2回の独立した実験の個々のマウスの結果を示し、2個の複製ウェルの上清中に放出されたIL−5の濃度として表現する。
【図4A】組換えHPV16−E7CyaAを用いた治療的ワクチン接種は、確立された腫瘍を根絶する。(実験A)C57BL/6マウスを、0日目に、5×104個のTC−1腫瘍細胞を用いて移植した。10日目に、マウスを、CyaA−E749〜57(C)、CyaA−E7Full(D)、またはCyaA−E7Δ30〜42(E)の1回の静注により処置した。未処置のままのマウス(A)またはCyaAE5−cysOVAを注射したマウス(B)を対照として採用した。腫瘍サイズが1000mm3に達した時、または不必要な苦しみを避けるために動物の衛生状態により要求される場合にはいつでも(壊死腫瘍、20%を超える急速な体重減少)、マウスを屠殺した。後に進行性の腫瘍を発症したCyaA−E7Fullで処置した2匹のマウス(*)を、さらに調べるために屠殺した(図6参照)。
【図4B】組換えHPV16−E7CyaAを用いた治療的ワクチン接種は、確立された腫瘍を根絶する。(実験B)実験設定については(実験A)と同様。+10および+17日目に、10μgのCyaAE5−CysOVA(a、実線)または10μgのCyaA−E7Δ30〜42(b)を用いて耳真皮に治療的ワクチン接種を実施した。各曲線は、1匹の動物における腫瘍増殖を示す。2匹の未処置の動物が含まれていた(a、点線)。各々の四分の一区の右上には(a、b)、屠殺動物の数と、含まれる動物の全数が示されている。これらのマウスの生存曲線を示す(c)。未処置(白抜きの三角)、CyaAE5−CysOVAを用いたニセ処置(閉じた三角)、CyaA−E7Δ30〜42を用いた処置(丸)。
【図5】組換えHPV16−E7CyaAでの治療的ワクチン接種により、生存が延長される。治療的ワクチン接種は、図4に記載した通りに行なった。TC−1腫瘍細胞の注射時に、マウス(1群あたり5〜10匹)を、グラフに示されているように+1、+5、または+10日目にHPV16−E7組換えCyaAで免疫化した。マウスを未処置のまま(普通の四角、実線)、CyaAE5−cysOVAでニセ処置(白抜きの四角、点線)、またはCyaA−E749〜57(白抜きの三角)、CyaA−E7Full(白抜きの丸)、もしくはCyaA−E7Δ30〜42(白抜きの菱型)で処置した。+5日目の治療実験において、CyaA−E7FullおよびCyaA−E7Δ30〜42で処置した動物の生存曲線は、完全に重ね合わさることが特筆される。どの場合においても、組換えHPV16−E7CyaA処置動物の生存は、未処置またはニセ処置マウスと比べて有意に延びている(p<0.05)。
【図6】その後に増殖してきた腫瘍から外移植したTC−1腫瘍細胞は、H−2Db分子の発現を欠失している。腫瘍拒絶実験において、CyaA−E7Fullをワクチン接種した数匹の動物では、実験の時間経過中、後になって腫瘍が成長した(図4、*)。腫瘍を外移植するために、2匹の動物を屠殺した。これらの腫瘍細胞、TC−1A1およびA2並びに天然TC−1細胞を、H−2Db分子の発現レベルについてFACS(登録商標)により解析した(太線)。蛍光強度の中間値(MedFi)を示す。アイソタイプ対照で得られた結果を示す(グレーの陰影)。
【図7】組換えHPV16−E7CyaAで処置したマウスにおけるHPV16−E749〜57特異的CD8+T細胞の持続性。治療的実験セットにおいて、CyaA−E749〜57(A)、CyaA−E7Full(B)、またはCyaA−E7Δ30〜42(C)で免疫化し、TC−1移植にも関わらず生存しているC57BL/6マウスを屠殺し、脾臓細胞を、インビトロで、1μg/mlのHPV16−E743〜77ペプチドで、照射同系脾臓細胞の存在下で5日間再刺激した。標的溶解(TC−1、普通の四角;EL4、白抜きの四角)を、51Cr放出により評価した。データは、非特異的溶解の数値のパーセンテージの中間値(各群においてn=6)、並びに、四分位範囲を示す。エフェクターと標的の最大比において20%以上の特異的溶解により決定される対応する動物の数を、四分の一区の右上部分に示す。
【図8】組換えHPV16−E7CyaAにより誘導されるTC−1腫瘍増殖に対する長期防御。治療的実験セットにおいて、TC−1移植にも関わらず生存しているC57BL/6マウスに、100日目に、5×104個のTC−1細胞を再移植した。年齢の適合した未処置マウスを、対照として採用した(A)。CyaA−E749〜57、CyaA−E7Full、およびCyaA−E7Δ30〜42で最初に免疫化したマウスにおける腫瘍の増殖を示す(それぞれ、B、C、およびD)。腫瘍サイズが1000mm3に達した時、または動物の衛生状態により要求される場合にはいつでも、マウスを屠殺した。(E)未処置(白抜きの四角);もしくはCya−E749〜57(白抜きの三角);CyaA−E7Full(白抜きの丸);もしくは、CyaA−E7Δ30〜42(白抜きの菱型)で免疫化し、TC−1細胞を再移植した(移植日を0とする)動物の生存曲線。いずれの場合においても、組換えHPV16−E7CyaA処置動物の生存は、未処置またはニセ処置マウスに比べて有意に延びた(p<0.05)。
【図9】CyaA−E7Δ30〜42治療活性と、CpG ODN1826アジュバントの添加されたHPV16E743〜77の治療活性との比較。C57BL/6マウスに、0日目に、5×104個のTC−1腫瘍細胞を皮下移植した。+10および+17日目に、マウスに、10μgのHPV16−E743〜77(n=5、三角)、1μgのCpG−ODN1826(n=5、四角)、10μgのHPV16−E743〜77+1μgのCpG−ODN1826(n=5、菱型)、10μgのCyaA−CysOVA(n=3、逆三角)、または10μgのCyaA−E7Δ30〜42(n=7、丸)を1回皮内注射して処置した。腫瘍サイズが1000mm3を超えた時、または動物の衛生状態により要求される場合にはいつでも、マウスを屠殺した。
【図10】CyaA−E7Δ30〜42のTC−1腫瘍拒絶誘導能に対する、CyaAに対する予備免疫の効果の解析。(A)C57BL/6マウスは未処置のままとするか、または、90もしくは30日前に、7日間の間隔で10μgのCyaAE5を2回皮内注射することにより免疫化した。1日前に、動物の採血をし、血清を個々に抗CyaAE5 IgGの存在についてELISAにより評価した。結果は、希釈対A492をプロットした線形回帰分析により計算した個々の抗体力価として表現する。横棒は、各群の応答の中間値を示す。(B)未処置(a、b)、30日前にCyaAE5で免疫化(c、d)、および90日前にCyaAE5で免疫化した(e、f)動物に、0日目に、5×104個のTC−1腫瘍細胞を移植し、+10および+17日目に、10μgのCyaA−cysOVA(a、c、e)、または10μgのCyaA−E7Δ30〜42(b、d、f)を1回皮内注射することにより処置した。挿入図(b、d、f)は、0〜35日間の期間のクローズアップであり、全ての動物が、ワクチン接種をした時点で触診できる腫瘍を有していたことを示す。各曲線は、1匹の動物における腫瘍増殖を示す。腫瘍サイズが1000mm3を超えた時、または動物の衛生状態により要求される場合にはいつでも、マウスを屠殺した。各々の四分の一区の右上には、屠殺動物の数と、含まれる動物の全数が示されている。
【図11】HHDマウスにおけるCyaA−HPV16E7Δ30〜42によるCTL応答の誘導。EL4−HHD細胞へのペプチドの添加を、各グラフの上に示す。続く表には、カラムヘッダーVaccの下で注入されたCyaAのタイプ(HPV17E7またはHPV18E7に関する)、および、カラムヘッダーStimの下でインビトロでの再刺激に使用されたペプチドが示されている。見てとれるように、CyaA−HPV16E7Δ30〜42による免疫化後、本発明者らは、ペプチド#253のみに特異的なCTLを誘導することができた(右パネル)。インビトロでペプチド#255で再刺激した脾臓細胞は、ペプチド#253で被覆したEL4−HHD細胞に対して細胞毒性ではなかったため、このCTL活性は特異的であった(右パネル)。2個のその他のHLA−A2拘束性ペプチドに対するCTL特異的応答がないことは、異なる現象から生じている可能性がある:(i)ペプチド#253の免疫優性、(ii)EL4−HHD細胞のプロテアソームによりペプチド#255および#258が処理されないこと、(iii)ペプチド#258の溶解度が低く、このため本発明者らはアセトニトリル(50%)を使用しなければならず、これは細胞に対して毒性である可能性がある。最も興味深いことには、CyaA−HPV18E7Δ32〜42の同時注射は、CyaA−HPV16E7Δ30〜42のCTL誘導能を妨害しなかった(右パネル)。
【図12】HHDマウスにおけるCyaA−HPV18E7Δ32〜42によるCTL応答の誘導。EL4−HHD細胞へのペプチドの添加を、各グラフの上に示す。以下の表には、カラムヘッダーVaccの下で注入されたCyaAのタイプ(HPV16E7またはHPV18E7に関する)、および、カラムヘッダーStimの下でインビトロでの再刺激に使用されたペプチドが示されている。見てとれるように、CyaA−HPV16E7Δ30〜42による免疫化後、本発明者らは、ペプチド#251のみに特異的なCTLを誘導することができた(左パネル)。インビトロでペプチド#257で再刺激した脾臓細胞は、ペプチド#251で被覆したEL4−HHD細胞に対して細胞毒性ではなかったため、このCTL活性は特異的であった(左パネル)。HPV16E7 HLA−A2拘束性ペプチドと同様に、HPV18E7#257HLA−A2拘束性ペプチドに対するCTL特異的応答がないことは、異なる現象から生じている可能性がある:(i)ペプチド#251の免疫優性、(ii)EL4−HHD細胞のプロテアソームによりペプチド#257が処理されないこと、(iii)ペプチド#257の溶解度が低く、このため本発明者らはアセトニトリル(50%)を使用しなければならず、これは細胞に対して毒性である可能性がある。最も興味深いことには、CyaA−HPV16E7Δ30〜42の同時注射は、CyaA−HPV18E7Δ32〜42のCTL誘導能を妨害しなかった(左パネル)。
【図13】pTRACE5−E7HPV18の作成。
【図14A】pTRACE5−E7ΔHPV16+18の作成。 3つのプラスミド調製ステップを開示する。
【図14B】pTRACE5−E7ΔHPV16+18の作成。 3つのプラスミド調製ステップを開示する。
【図14C】pTRACE5−E7ΔHPV16+18の作成。 3つのプラスミド調製ステップを開示する。
【図15】CyaA−HPV18E7Full CyaA−HPV18E7Δ32〜42 CyaA−HPV16+18E7Full CyaA−HPV16+18E7Δをコードするポリヌクレオチドのマップ。
【図16】C57BL/6マウスにおけるHPV18E7を有する組換えCyaAによるCTLの誘導。
【図17】C57BL/6マウスにおけるHPV18E7を有する組換えCyaAによるCTLの誘導。寄託されている材料(I−3190およびI−3191)は、E.coli株BLRに含まれ、ルリアブロス(LB)培地中で増殖させることができ、100μg/mlのアンピシリンを含むLBに接種し、175rpmで振とうし照射しながら空気中で30℃でインキュベートできる。7〜10%DMSOを含むLB中での一晩の保存が可能である。
【特許請求の範囲】
【請求項1】
1個または数個のHPV抗原の1個または数個のエピトープを有する1個または数個のポリペプチドを含む組換えタンパク質であって、前記ポリペプチドは、アデニル酸シクラーゼ(CyaA)タンパク質またはその断片の同じまたは異なる許容部位に挿入されており、前記CyaA断片は、抗原提示細胞を標的化するという前記アデニル酸シクラーゼタンパク質の特性を保持している、前記組換えタンパク質。
【請求項2】
CyaAタンパク質断片は、CD11b/CD18抗原提示細胞を標的化するという前記アデニル酸シクラーゼタンパク質の特性を保持している、請求項1に記載の組換えタンパク質。
【請求項3】
CyaAタンパク質断片は、標的化細胞のサイトゾルへの前記ポリペプチドのエピトープの転位または前記ポリペプチドの転位を可能とするというCyaAの特性を保持している、請求項1または2に記載の組換えタンパク質。
【請求項4】
CyaAタンパク質断片の同じまたは異なる許容部位に挿入されている1個または数個のHPV抗原の1個または数個のエピトープを有するポリペプチドを含む請求項1〜3のいずれかに記載の組換えタンパク質であって、前記断片は、約30〜約1300アミノ酸残基を含み、前記断片は、CyaAタンパク質のアミノ酸残基1208〜1243またはCyaAタンパク質のアミノ酸残基1166〜1281を包含する、前記組換えタンパク質。
【請求項5】
前記ポリペプチドおよび/または数個のポリペプチドが一緒になって、約5〜約500、または約5〜約200、または約10〜約50アミノ酸残基、または約30もしくは50〜200アミノ酸残基を含む、請求項1〜4のいずれか1項に記載の組換えタンパク質。
【請求項6】
前記ポリペプチドに対する抗原特異的応答を誘起することのできる、請求項5に記載の組換えタンパク質。
【請求項7】
1個または数個のエピトープを有する少なくとも1個のポリペプチドが、発癌性HPVに由来している、請求項1〜6のいずれか1項に記載の組換えタンパク質。
【請求項8】
1個または数個のエピトープを有するポリペプチドが、HPV16、HPV18、HPV31、HPV33、HPV35、HPV45、HPV52、またはHPV58に由来している、請求項1〜7のいずれか1項に記載の組換えタンパク質。
【請求項9】
1個または数個のエピトープを有するポリペプチドが、L1、L2、E1、E2、E4、およびE5タンパク質から選択されるHPV抗原に由来している、請求項1〜8のいずれか1項に記載の組換えタンパク質。
【請求項10】
1個または数個のエピトープを有するポリペプチドが、HPV16および/またはHPV18のE6またはE7タンパク質に由来している、請求項1〜9のいずれか1項に記載の組換えタンパク質。
【請求項11】
1個または数個のHPV抗原の1個または数個のエピトープを有する複数のポリペプチドを含む、請求項1〜10のいずれか1項に記載の組換えタンパク質。
【請求項12】
1個または数個のエピトープを有するポリペプチドが、HPV16のE7タンパク質に由来している、請求項9または11に記載の組換えタンパク質。
【請求項13】
1個または数個のエピトープを有するポリペプチドが、HPV18のE7タンパク質に由来している、請求項9または11に記載の組換えタンパク質。
【請求項14】
前記ポリペプチドが、全長E6またはE7タンパク質である、請求項9または11に記載の組換えタンパク質。
【請求項15】
CyaA配列またはその断片の異なる許容部位に挿入されている複数のポリペプチドを含む、請求項11〜14のいずれか1項に記載の組換えタンパク質。
【請求項16】
複数のポリペプチドが、HPV16のE7タンパク質の残基1〜29を含む断片もしくは残基42〜98を含む断片、または両方の断片を包含し、前記ポリペプチドは、CyaAタンパク質の異なる許容部位に挿入されている、請求項15に記載の組換えタンパク質。
【請求項17】
ポリペプチドが、アミノ酸配列
【表1】
を有する断片を包含する、請求項12に記載の組換えタンパク質。
【請求項18】
前記ポリペプチドまたはポリペプチド群が、分断された天然HPV抗原に存在し、前記分断は、前記HPV抗原の酸性領域における1個または数個のアミノ酸残基の欠失、および/または、アデニル酸シクラーゼの少なくとも2箇所の許容部位への前記HPV抗原の少なくとも2個のポリペプチド断片の挿入からなる、請求項1〜16のいずれか1項に記載の組換えタンパク質。
【請求項19】
1個または数個のHPV抗原の1個または数個のエピトープを有するポリペプチドが、得られる組換えタンパク質の免疫原性を高めるために改変されている、請求項1〜13または18のいずれか1項に記載の組換えタンパク質。
【請求項20】
CyaAタンパク質が、細菌性タンパク質である、請求項1〜19のいずれか1項に記載の組換えタンパク質。
【請求項21】
CyaAタンパク質が、Bordetella属に由来する、請求項1〜19のいずれか1項に記載の組換えタンパク質。
【請求項22】
CyaAタンパク質が、Bordetella pertussisに由来する、請求項1〜21のいずれか1項に記載の組換えタンパク質。
【請求項23】
CyaAタンパク質の酵素活性が不活性化されている、請求項1〜22のいずれか1項に記載の組換えタンパク質。
【請求項24】
CyaAタンパク質の酵素活性が遺伝子的に不活性化されている、請求項1〜22のいずれか1項に記載の組換えタンパク質。
【請求項25】
CyaAタンパク質の酵素活性が、シクラーゼ活性に関与するCyaAのアミノ酸配列部位にジペプチドが挿入されている結果、不活性化されている、請求項1〜22のいずれか1項に記載の組換えタンパク質。
【請求項26】
哺乳動物宿主において細胞性免疫応答を誘起できる、請求項1〜25のいずれか1項に記載の組換えタンパク質。
【請求項27】
哺乳動物宿主において体液性免疫応答を誘起できる、請求項1〜25のいずれか1項に記載の組換えタンパク質。
【請求項28】
CyaAタンパク質またはその断片の同じまたは異なる許容部位に挿入されている、様々なHPV抗原の1個または数個のエピトープを有する数個のポリペプチドを含む、請求項1〜27のいずれか1項に記載の組換えタンパク質。
【請求項29】
前記ポリペプチドが、それぞれHPV16およびHPV18に由来する全長E7ポリペプチドであり、前記ポリペプチドは、CyaAタンパク質の異なる許容部位に挿入されている、請求項28に記載の組換えタンパク質。
【請求項30】
pTRACE5−HPV16E7FULL(CNCM I−3191)またはpTRACE5−HPV16E7Δ30〜42(CNCM I−3190)から選択されるプラスミドに含まれる挿入物によりコードされる、請求項1〜9のいずれか1項に記載の組換えタンパク質。
【請求項31】
1個または数個の抗原の1個または数個のエピトープを有する1個または数個のポリペプチドを含む組換えタンパク質であって、前記ポリペプチドは、アデニル酸シクラーゼ(CyaA)タンパク質またはその断片の同じまたは異なる許容部位に挿入されており、前記CyaA断片は、抗原提示細胞を標的化するという前記アデニル酸シクラーゼタンパク質の特性を保持しており、前記エピトープの少なくとも1個は、亜優性潜在性T細胞エピトープであり、前記組換えタンパク質は、前記ポリペプチドに対する抗原特異的応答を誘起できる、前記組換えタンパク質。
【請求項32】
潜在性エピトープが、HPVのエピトープ、特にHPVのE7タンパク質のエピトープである、請求項31に記載の組換えタンパク質。
【請求項33】
潜在性エピトープは、HPV18 E7タンパク質のエピトープ、特に配列IDGVNHQHLを有するエピトープである、請求項32に記載の組換えタンパク質。
【請求項34】
請求項12〜33のいずれか1項に記載の組換えタンパク質をコードするポリヌクレオチド。
【請求項35】
前記ポリヌクレオチドが、ベクターpTRACE5−HPV16E7FULL(CNCM I−3191)またはpTRACE5−HPV16E7Δ30〜42(CNCM I−3190)に含まれる、請求項34に記載のポリヌクレオチド。
【請求項36】
請求項34に記載のポリヌクレオチドを含む、組換え発現ベクター。
【請求項37】
細菌で発現させるのに適した請求項36に記載の組換えベクター。
【請求項38】
真核細胞、特に哺乳動物細胞での発現に適した請求項36に記載の組換え発現ベクター。
【請求項39】
pTRACE5−HPV16E7FULL(CNCM I−3191)またはpTRACE5−HPV16E7Δ30〜42(CNCM I−3190)から選択される、請求項36に記載の組換えベクター。
【請求項40】
請求項34に記載のポリヌクレオチドまたは請求項36〜39のいずれか1項に記載の組換え発現ベクターを含む組換え宿主細胞。
【請求項41】
前記組換え宿主細胞が、I−3190またはI−3191号でCNCMに寄託されているE.coli細胞である、請求項40に記載の組換え宿主細胞。
【請求項42】
生理的に許容されるビヒクル、賦形剤、担体、および/または希釈剤と共に、請求項1〜33のいずれか1項に記載の組換えタンパク質または請求項34に記載のポリヌクレオチドまたは請求項36〜39のいずれか1項に記載のベクターを含む、免疫原性組成物。
【請求項43】
哺乳動物宿主において細胞性免疫応答を誘導する、請求項42に記載の免疫原性組成物。
【請求項44】
細胞媒介細胞溶解免疫応答を誘導する、請求項43に記載の免疫原性組成物。
【請求項45】
免疫応答が、体液性免疫応答を含む、請求項42〜44のいずれか1項に記載の免疫原性組成物。
【請求項46】
アジュバントおよび/または界面活性剤および/または免疫調節物質をさらに含む、請求項42〜45のいずれか1項に記載の免疫原性組成物。
【請求項47】
HPV感染を予防または処置するための、請求項42〜46に記載の免疫原性組成物。
【請求項48】
癌免疫療法のための請求項42〜46のいずれか1項に記載の免疫原性組成物。
【請求項49】
宿主におけるHPV感染に起因する悪性形質転換の発症または存続の予防または処置のための、請求項42〜46のいずれか1項に記載の免疫原性組成物。
【請求項50】
患者におけるHPV感染の処置または予防のための、請求項1〜30のいずれか1項に記載の組換えタンパク質または請求項34に記載のポリヌクレオチドまたは請求項36〜39のいずれか1項に記載のベクターの使用。
【請求項51】
患者におけるHPV感染に起因する悪性形質転換の発症または存続に対する免疫療法のための請求項50に記載の使用。
【請求項52】
宿主において細胞性および/または体液性免疫応答を誘起するのに適した、請求項1〜30のいずれか1項に記載の組換えタンパク質または請求項34に記載のポリヌクレオチドまたは請求項36〜39のいずれか1項に記載のベクターを含む、ワクチン組成物。
【請求項53】
HPV感染を予防または処置するための、請求項1〜33のいずれか1項に記載の組換えタンパク質または請求項34に記載のポリヌクレオチドまたは請求項36〜39のいずれか1項に記載のベクターを含む、薬物組成物。
【請求項54】
宿主におけるHPV感染に起因する悪性形質転換の発症または存続の予防または処置のための、請求項1〜33のいずれか1項に記載の組換えタンパク質または請求項34に記載のポリヌクレオチドまたは請求項36〜39のいずれか1項に記載のベクターを含む、薬物組成物。
【請求項55】
癌免疫療法のための、請求項1〜33のいずれか1項に記載の組換えタンパク質または請求項34に記載のポリヌクレオチドまたは請求項36〜39のいずれか1項に記載のベクターを含む、薬物組成物。
【請求項56】
請求項1〜33のいずれかに記載の組換えタンパク質または請求項34に記載のポリヌクレオチドまたは請求項36〜39のいずれかに記載のベクターを含む、HPV感染の診断キット、またはHPV感染を免疫モニタリングするためのキット。
【請求項57】
−哺乳動物、特にヒト患者から得られたT細胞を、請求項1〜33のいずれか1項に記載の組換えタンパク質にさらすこと、
−T細胞の活性化の改変を検出すること
を含む、HPV感染をインビトロで診断する方法または免疫モニタリングする方法。
【請求項58】
癌ウイルス感染の処置もしくは予防のための、または、このよう癌ウイルス感染に起因する悪性作用の処置もしくは予防のための、1個または数個の癌ウイルスの1個または数個の抗原の1個または数個のエピトープの挿入により改変される、細菌性タンパク質またはその断片を含む組換えタンパク質の使用。
【請求項59】
細菌性タンパク質は、毒素または類毒素である、請求項58に記載の組換えタンパク質の使用。
【請求項60】
キメラCyaA−ポリペプチドタンパク質に含まれるポリペプチド中の未知または亜優性潜在性T細胞エピトープのスクリーニング方法であって、
ここで、CyaAは、請求項1〜30のいずれかに開示されているアデニル酸シクラーゼまたはその断片であり、この方法は、
−前記キメラタンパク質を動物宿主に投与すること、
−前記宿主のT細胞応答、特にCTL応答を決定すること
を含む、前記スクリーニング方法。
【請求項61】
前記ポリペプチドが、HPV抗原、特にHPV16および/またはHPV18 E7タンパク質に由来する、請求項60に記載の方法。
【請求項1】
1個または数個のHPV抗原の1個または数個のエピトープを有する1個または数個のポリペプチドを含む組換えタンパク質であって、前記ポリペプチドは、アデニル酸シクラーゼ(CyaA)タンパク質またはその断片の同じまたは異なる許容部位に挿入されており、前記CyaA断片は、抗原提示細胞を標的化するという前記アデニル酸シクラーゼタンパク質の特性を保持している、前記組換えタンパク質。
【請求項2】
CyaAタンパク質断片は、CD11b/CD18抗原提示細胞を標的化するという前記アデニル酸シクラーゼタンパク質の特性を保持している、請求項1に記載の組換えタンパク質。
【請求項3】
CyaAタンパク質断片は、標的化細胞のサイトゾルへの前記ポリペプチドのエピトープの転位または前記ポリペプチドの転位を可能とするというCyaAの特性を保持している、請求項1または2に記載の組換えタンパク質。
【請求項4】
CyaAタンパク質断片の同じまたは異なる許容部位に挿入されている1個または数個のHPV抗原の1個または数個のエピトープを有するポリペプチドを含む請求項1〜3のいずれかに記載の組換えタンパク質であって、前記断片は、約30〜約1300アミノ酸残基を含み、前記断片は、CyaAタンパク質のアミノ酸残基1208〜1243またはCyaAタンパク質のアミノ酸残基1166〜1281を包含する、前記組換えタンパク質。
【請求項5】
前記ポリペプチドおよび/または数個のポリペプチドが一緒になって、約5〜約500、または約5〜約200、または約10〜約50アミノ酸残基、または約30もしくは50〜200アミノ酸残基を含む、請求項1〜4のいずれか1項に記載の組換えタンパク質。
【請求項6】
前記ポリペプチドに対する抗原特異的応答を誘起することのできる、請求項5に記載の組換えタンパク質。
【請求項7】
1個または数個のエピトープを有する少なくとも1個のポリペプチドが、発癌性HPVに由来している、請求項1〜6のいずれか1項に記載の組換えタンパク質。
【請求項8】
1個または数個のエピトープを有するポリペプチドが、HPV16、HPV18、HPV31、HPV33、HPV35、HPV45、HPV52、またはHPV58に由来している、請求項1〜7のいずれか1項に記載の組換えタンパク質。
【請求項9】
1個または数個のエピトープを有するポリペプチドが、L1、L2、E1、E2、E4、およびE5タンパク質から選択されるHPV抗原に由来している、請求項1〜8のいずれか1項に記載の組換えタンパク質。
【請求項10】
1個または数個のエピトープを有するポリペプチドが、HPV16および/またはHPV18のE6またはE7タンパク質に由来している、請求項1〜9のいずれか1項に記載の組換えタンパク質。
【請求項11】
1個または数個のHPV抗原の1個または数個のエピトープを有する複数のポリペプチドを含む、請求項1〜10のいずれか1項に記載の組換えタンパク質。
【請求項12】
1個または数個のエピトープを有するポリペプチドが、HPV16のE7タンパク質に由来している、請求項9または11に記載の組換えタンパク質。
【請求項13】
1個または数個のエピトープを有するポリペプチドが、HPV18のE7タンパク質に由来している、請求項9または11に記載の組換えタンパク質。
【請求項14】
前記ポリペプチドが、全長E6またはE7タンパク質である、請求項9または11に記載の組換えタンパク質。
【請求項15】
CyaA配列またはその断片の異なる許容部位に挿入されている複数のポリペプチドを含む、請求項11〜14のいずれか1項に記載の組換えタンパク質。
【請求項16】
複数のポリペプチドが、HPV16のE7タンパク質の残基1〜29を含む断片もしくは残基42〜98を含む断片、または両方の断片を包含し、前記ポリペプチドは、CyaAタンパク質の異なる許容部位に挿入されている、請求項15に記載の組換えタンパク質。
【請求項17】
ポリペプチドが、アミノ酸配列
【表1】
を有する断片を包含する、請求項12に記載の組換えタンパク質。
【請求項18】
前記ポリペプチドまたはポリペプチド群が、分断された天然HPV抗原に存在し、前記分断は、前記HPV抗原の酸性領域における1個または数個のアミノ酸残基の欠失、および/または、アデニル酸シクラーゼの少なくとも2箇所の許容部位への前記HPV抗原の少なくとも2個のポリペプチド断片の挿入からなる、請求項1〜16のいずれか1項に記載の組換えタンパク質。
【請求項19】
1個または数個のHPV抗原の1個または数個のエピトープを有するポリペプチドが、得られる組換えタンパク質の免疫原性を高めるために改変されている、請求項1〜13または18のいずれか1項に記載の組換えタンパク質。
【請求項20】
CyaAタンパク質が、細菌性タンパク質である、請求項1〜19のいずれか1項に記載の組換えタンパク質。
【請求項21】
CyaAタンパク質が、Bordetella属に由来する、請求項1〜19のいずれか1項に記載の組換えタンパク質。
【請求項22】
CyaAタンパク質が、Bordetella pertussisに由来する、請求項1〜21のいずれか1項に記載の組換えタンパク質。
【請求項23】
CyaAタンパク質の酵素活性が不活性化されている、請求項1〜22のいずれか1項に記載の組換えタンパク質。
【請求項24】
CyaAタンパク質の酵素活性が遺伝子的に不活性化されている、請求項1〜22のいずれか1項に記載の組換えタンパク質。
【請求項25】
CyaAタンパク質の酵素活性が、シクラーゼ活性に関与するCyaAのアミノ酸配列部位にジペプチドが挿入されている結果、不活性化されている、請求項1〜22のいずれか1項に記載の組換えタンパク質。
【請求項26】
哺乳動物宿主において細胞性免疫応答を誘起できる、請求項1〜25のいずれか1項に記載の組換えタンパク質。
【請求項27】
哺乳動物宿主において体液性免疫応答を誘起できる、請求項1〜25のいずれか1項に記載の組換えタンパク質。
【請求項28】
CyaAタンパク質またはその断片の同じまたは異なる許容部位に挿入されている、様々なHPV抗原の1個または数個のエピトープを有する数個のポリペプチドを含む、請求項1〜27のいずれか1項に記載の組換えタンパク質。
【請求項29】
前記ポリペプチドが、それぞれHPV16およびHPV18に由来する全長E7ポリペプチドであり、前記ポリペプチドは、CyaAタンパク質の異なる許容部位に挿入されている、請求項28に記載の組換えタンパク質。
【請求項30】
pTRACE5−HPV16E7FULL(CNCM I−3191)またはpTRACE5−HPV16E7Δ30〜42(CNCM I−3190)から選択されるプラスミドに含まれる挿入物によりコードされる、請求項1〜9のいずれか1項に記載の組換えタンパク質。
【請求項31】
1個または数個の抗原の1個または数個のエピトープを有する1個または数個のポリペプチドを含む組換えタンパク質であって、前記ポリペプチドは、アデニル酸シクラーゼ(CyaA)タンパク質またはその断片の同じまたは異なる許容部位に挿入されており、前記CyaA断片は、抗原提示細胞を標的化するという前記アデニル酸シクラーゼタンパク質の特性を保持しており、前記エピトープの少なくとも1個は、亜優性潜在性T細胞エピトープであり、前記組換えタンパク質は、前記ポリペプチドに対する抗原特異的応答を誘起できる、前記組換えタンパク質。
【請求項32】
潜在性エピトープが、HPVのエピトープ、特にHPVのE7タンパク質のエピトープである、請求項31に記載の組換えタンパク質。
【請求項33】
潜在性エピトープは、HPV18 E7タンパク質のエピトープ、特に配列IDGVNHQHLを有するエピトープである、請求項32に記載の組換えタンパク質。
【請求項34】
請求項12〜33のいずれか1項に記載の組換えタンパク質をコードするポリヌクレオチド。
【請求項35】
前記ポリヌクレオチドが、ベクターpTRACE5−HPV16E7FULL(CNCM I−3191)またはpTRACE5−HPV16E7Δ30〜42(CNCM I−3190)に含まれる、請求項34に記載のポリヌクレオチド。
【請求項36】
請求項34に記載のポリヌクレオチドを含む、組換え発現ベクター。
【請求項37】
細菌で発現させるのに適した請求項36に記載の組換えベクター。
【請求項38】
真核細胞、特に哺乳動物細胞での発現に適した請求項36に記載の組換え発現ベクター。
【請求項39】
pTRACE5−HPV16E7FULL(CNCM I−3191)またはpTRACE5−HPV16E7Δ30〜42(CNCM I−3190)から選択される、請求項36に記載の組換えベクター。
【請求項40】
請求項34に記載のポリヌクレオチドまたは請求項36〜39のいずれか1項に記載の組換え発現ベクターを含む組換え宿主細胞。
【請求項41】
前記組換え宿主細胞が、I−3190またはI−3191号でCNCMに寄託されているE.coli細胞である、請求項40に記載の組換え宿主細胞。
【請求項42】
生理的に許容されるビヒクル、賦形剤、担体、および/または希釈剤と共に、請求項1〜33のいずれか1項に記載の組換えタンパク質または請求項34に記載のポリヌクレオチドまたは請求項36〜39のいずれか1項に記載のベクターを含む、免疫原性組成物。
【請求項43】
哺乳動物宿主において細胞性免疫応答を誘導する、請求項42に記載の免疫原性組成物。
【請求項44】
細胞媒介細胞溶解免疫応答を誘導する、請求項43に記載の免疫原性組成物。
【請求項45】
免疫応答が、体液性免疫応答を含む、請求項42〜44のいずれか1項に記載の免疫原性組成物。
【請求項46】
アジュバントおよび/または界面活性剤および/または免疫調節物質をさらに含む、請求項42〜45のいずれか1項に記載の免疫原性組成物。
【請求項47】
HPV感染を予防または処置するための、請求項42〜46に記載の免疫原性組成物。
【請求項48】
癌免疫療法のための請求項42〜46のいずれか1項に記載の免疫原性組成物。
【請求項49】
宿主におけるHPV感染に起因する悪性形質転換の発症または存続の予防または処置のための、請求項42〜46のいずれか1項に記載の免疫原性組成物。
【請求項50】
患者におけるHPV感染の処置または予防のための、請求項1〜30のいずれか1項に記載の組換えタンパク質または請求項34に記載のポリヌクレオチドまたは請求項36〜39のいずれか1項に記載のベクターの使用。
【請求項51】
患者におけるHPV感染に起因する悪性形質転換の発症または存続に対する免疫療法のための請求項50に記載の使用。
【請求項52】
宿主において細胞性および/または体液性免疫応答を誘起するのに適した、請求項1〜30のいずれか1項に記載の組換えタンパク質または請求項34に記載のポリヌクレオチドまたは請求項36〜39のいずれか1項に記載のベクターを含む、ワクチン組成物。
【請求項53】
HPV感染を予防または処置するための、請求項1〜33のいずれか1項に記載の組換えタンパク質または請求項34に記載のポリヌクレオチドまたは請求項36〜39のいずれか1項に記載のベクターを含む、薬物組成物。
【請求項54】
宿主におけるHPV感染に起因する悪性形質転換の発症または存続の予防または処置のための、請求項1〜33のいずれか1項に記載の組換えタンパク質または請求項34に記載のポリヌクレオチドまたは請求項36〜39のいずれか1項に記載のベクターを含む、薬物組成物。
【請求項55】
癌免疫療法のための、請求項1〜33のいずれか1項に記載の組換えタンパク質または請求項34に記載のポリヌクレオチドまたは請求項36〜39のいずれか1項に記載のベクターを含む、薬物組成物。
【請求項56】
請求項1〜33のいずれかに記載の組換えタンパク質または請求項34に記載のポリヌクレオチドまたは請求項36〜39のいずれかに記載のベクターを含む、HPV感染の診断キット、またはHPV感染を免疫モニタリングするためのキット。
【請求項57】
−哺乳動物、特にヒト患者から得られたT細胞を、請求項1〜33のいずれか1項に記載の組換えタンパク質にさらすこと、
−T細胞の活性化の改変を検出すること
を含む、HPV感染をインビトロで診断する方法または免疫モニタリングする方法。
【請求項58】
癌ウイルス感染の処置もしくは予防のための、または、このよう癌ウイルス感染に起因する悪性作用の処置もしくは予防のための、1個または数個の癌ウイルスの1個または数個の抗原の1個または数個のエピトープの挿入により改変される、細菌性タンパク質またはその断片を含む組換えタンパク質の使用。
【請求項59】
細菌性タンパク質は、毒素または類毒素である、請求項58に記載の組換えタンパク質の使用。
【請求項60】
キメラCyaA−ポリペプチドタンパク質に含まれるポリペプチド中の未知または亜優性潜在性T細胞エピトープのスクリーニング方法であって、
ここで、CyaAは、請求項1〜30のいずれかに開示されているアデニル酸シクラーゼまたはその断片であり、この方法は、
−前記キメラタンパク質を動物宿主に投与すること、
−前記宿主のT細胞応答、特にCTL応答を決定すること
を含む、前記スクリーニング方法。
【請求項61】
前記ポリペプチドが、HPV抗原、特にHPV16および/またはHPV18 E7タンパク質に由来する、請求項60に記載の方法。
【図1A】

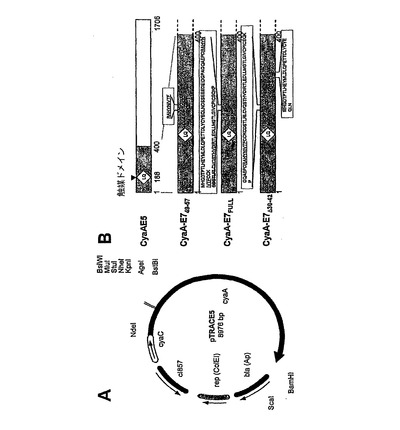
【図1(B)】
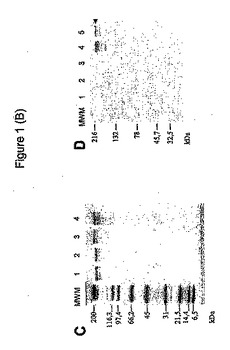
【図2】
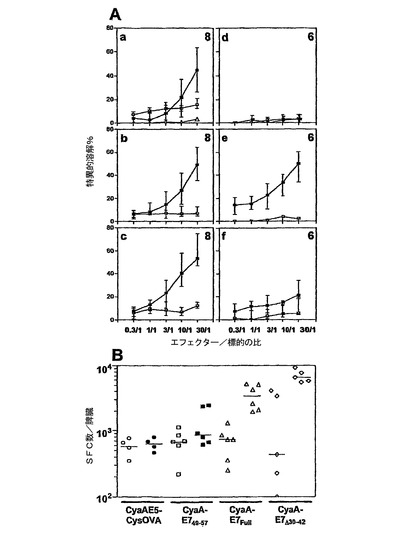
【図3】
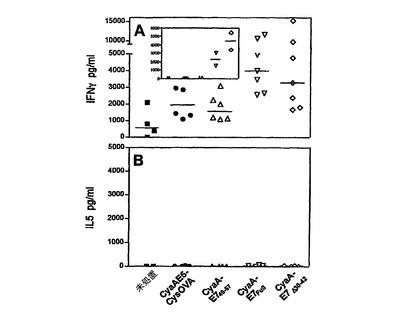
【図4A】
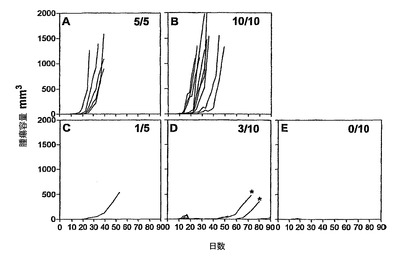
【図4B】
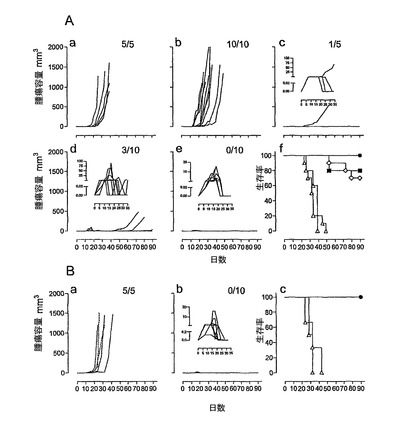
【図5】
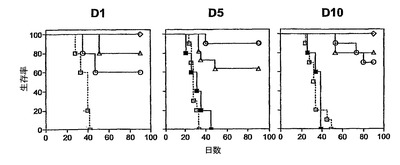
【図6】
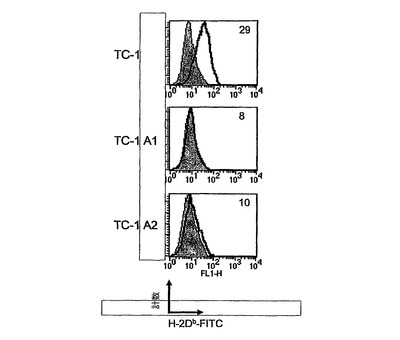
【図7】
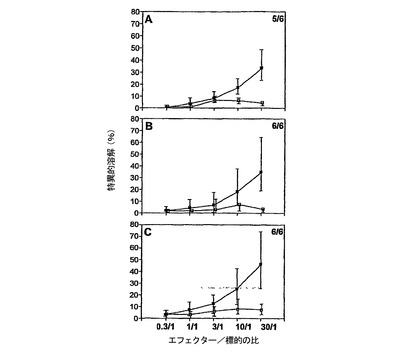
【図8】
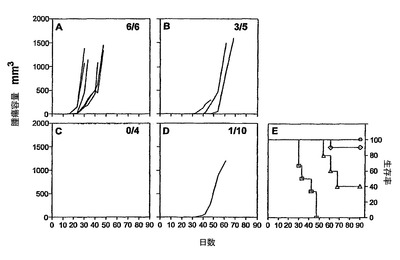
【図9】
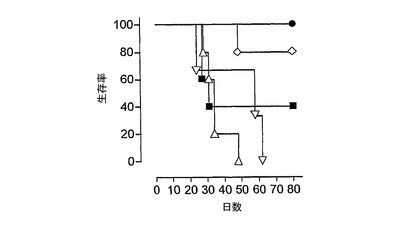
【図10】
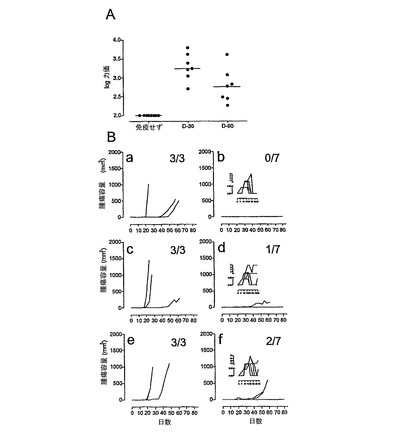
【図11】
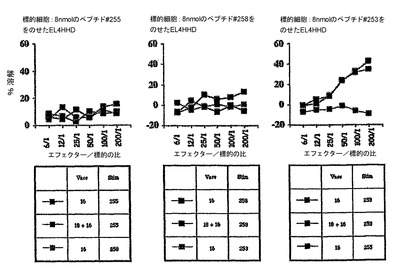
【図12】
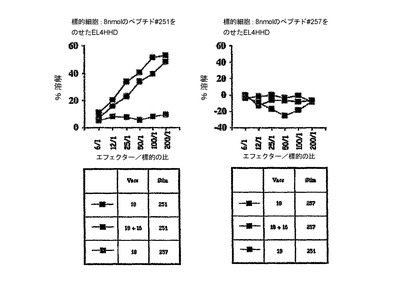
【図13】
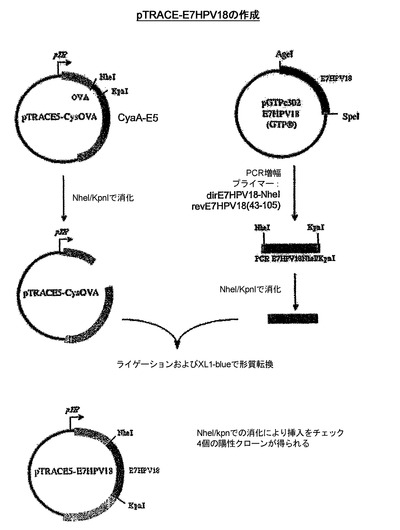
【図14A】
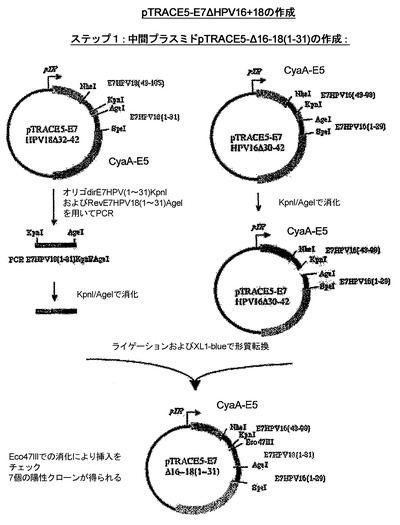
【図14B】
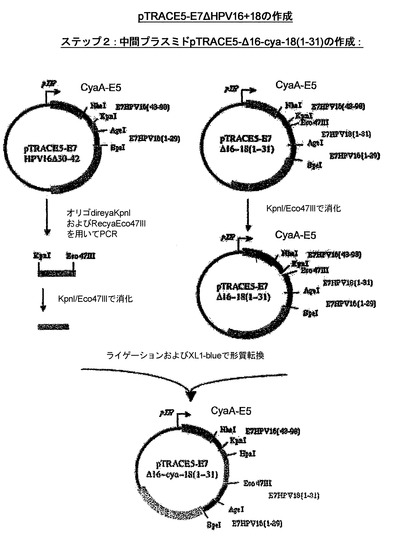
【図14C】
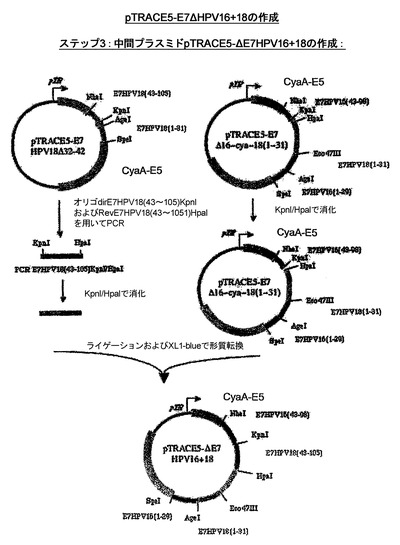
【図15】

【図16】
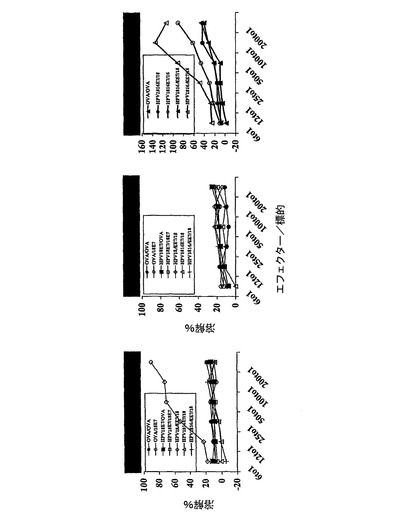
【図17】
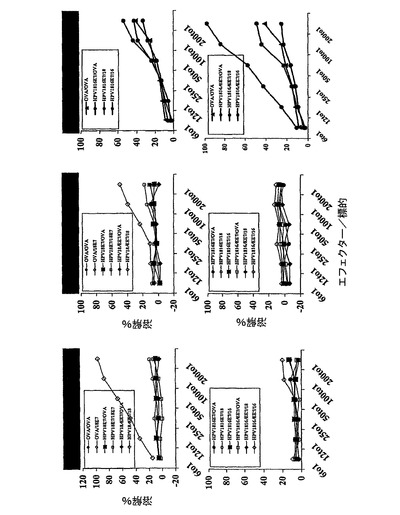
【図1(B)】
【図2】
【図3】
【図4A】
【図4B】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14A】
【図14B】
【図14C】
【図15】
【図16】
【図17】
【公表番号】特表2007−533307(P2007−533307A)
【公表日】平成19年11月22日(2007.11.22)
【国際特許分類】
【出願番号】特願2007−503308(P2007−503308)
【出願日】平成17年3月18日(2005.3.18)
【国際出願番号】PCT/EP2005/003452
【国際公開番号】WO2005/089792
【国際公開日】平成17年9月29日(2005.9.29)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.フロッピー
【出願人】(593218462)インスティチュート・パスツール (19)
【氏名又は名称原語表記】INSTITUT PASTEUR
【出願人】(506315871)
【氏名又は名称原語表記】BT PHARMA
【出願人】(591100596)アンスティチュ ナショナル ドゥ ラ サンテ エ ドゥ ラ ルシェルシュ メディカル (59)
【出願人】(595040744)サントル・ナショナル・ドゥ・ラ・ルシェルシュ・シャンティフィク (88)
【氏名又は名称原語表記】CENTRE NATIONAL DE LA RECHERCHE SCIENTIFIQUE
【Fターム(参考)】
【公表日】平成19年11月22日(2007.11.22)
【国際特許分類】
【出願日】平成17年3月18日(2005.3.18)
【国際出願番号】PCT/EP2005/003452
【国際公開番号】WO2005/089792
【国際公開日】平成17年9月29日(2005.9.29)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.フロッピー
【出願人】(593218462)インスティチュート・パスツール (19)
【氏名又は名称原語表記】INSTITUT PASTEUR
【出願人】(506315871)
【氏名又は名称原語表記】BT PHARMA
【出願人】(591100596)アンスティチュ ナショナル ドゥ ラ サンテ エ ドゥ ラ ルシェルシュ メディカル (59)
【出願人】(595040744)サントル・ナショナル・ドゥ・ラ・ルシェルシュ・シャンティフィク (88)
【氏名又は名称原語表記】CENTRE NATIONAL DE LA RECHERCHE SCIENTIFIQUE
【Fターム(参考)】
[ Back to top ]