
アデノウィルスE1A/E1B補完細胞株
本発明は、複製能のあるアデノウィルス(RCA)を実質的に含まないE1A/E1B欠損アデノウイルスの成長のためのアデノウィルスパッケージング細胞株を提供する。E1AおよびE1B遺伝子が、同一であっても異なっていてもよい非アデノウィルス異種プロモーターと作用可能に連結している安定的に組み込まれたE1AおよびE1B発現ベクターを含む。RCAを実質上含まないアデノウィルスであって、実質的な配列同一性をアデノウィルスE1AおよびE1Bプロモーターと共有するポリヌクレオチド配列を欠いたプロモーターと作用可能に連結しているアデノウィルスE1AおよびE1Bのコード配列を含む細胞株中で成長するアデノウィルス、を産生する方法も提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本願は2003年7月3日出願の米国特許出願第10/613,106号に対し優先権を主張するものであり、この内容は引用によりその全体が本明細書に組み入れられる。
【0002】
本発明は、組換えアデノウィルスベクター、例えば複製能のあるアデノウィルスベクター、腫瘍溶解性(oncolytic)アデノウィルスベクター、およびE1初期遺伝子領域の欠失を有する複製能欠陥アデノウィルスベクターの産生に役立つ新規なパッケージング細胞株に関する。
【背景技術】
【0003】
ベクターが介在する導入遺伝子の送達は、宿主中に欠けているかまたは不十分な量で存在するタンパク質または他の物質を補うことで遺伝病の治療に利用される。アデノウィルス(Ad)ベクターは、導入遺伝子送達にとってきわめて効率的な伝達体である。アデノウィルスベースの遺伝子導入ベクターは、細胞中への遺伝子導入にとってそれらを特に有用ならしめている複数の特徴を有しており、例えば、アデノウィルスの生態の特徴がよく調べられていること、アデノウィルスは周知のどのヒトの疾患とも関連性がないこと、アデノウィルスはそのDNAを宿主細胞中へ導入するのに効率的であること、このウィルスは広い宿主細胞分布域を有すること、および大量生産が達成されていること、である。少なくともE1領域が欠失し、注目の遺伝子によって置き換えられているヒトアデノウィルスベースのベクターは、遺伝子治療に広範囲に用いられてきた。遺伝子治療に現在用いられているアデノウィルスベクターは一般に複製する能力がなく、E1領域に欠失を有する。
【0004】
組換えアデノウィルスを潜在的に強力な遺伝子送達ベクターにならしめている特徴については広範囲に検討されてきた(Berkerの論文、Biotechniques 6: 616-629 (1988)ならびにKozarskyおよびWilsonの論文、Curr. Opin. Genet. Dev. 3: 499-503 (1993))。遺伝子の欠失を通じてであろうと、特定の組織に限定された複製を通じてであろうとアデノウィルスベクターの複製の制御は、アデノウィルスと関係するin vivo用途にとって特に重要である。
【0005】
複製性アデノウィルスは、in vivoでの選択的ターゲッティングおよび増幅を達成するように作り変えられてきた。条件付きで複製性且つ腫瘍溶解性のアデノウィルスは、癌の治療において大きな将来性を示している(Yu等の論文、Curr. Opin. Mol. Ther. 2002, Oct; 4 (5): 435-43、Bell等の論文、Curr. Gene Ther. 2002 May 2 (2): 243-54、Yoon等の論文、Curr. Cancer Drug Targets 2002 Aug; 1 (2): 85-107)。これら複製性アデノウィルスは、全身的に送達することができ、腫瘍細胞をターゲティングすることができ、また腫瘍特異的なやり方でそれらの細胞溶解効果を増幅することができ、それによって実質的な臨床的利益をもたらすことができる。Henderson他の米国特許第5,698,443号、Hallenbeck他の国際公開第WO96/17053号を参照されたい。このような系では細胞特異的転写調節エレメントがウィルス複製にとって不可欠の遺伝子の発現を制御しており、したがってウィルスの転写はそのエレメントが機能しうる細胞集団に限られる。例えば弱毒化した複製能のあるアデノウィルスは、そのウィルスがin vitroおよびin vivoで前立腺特異的抗原(PSA)発現細胞に対して選択的細胞毒性を示すアデノウィルス血清型5(Ad5)中のE1A転写単位の上流にPSAプロモーターおよびエンハンサー(PSE-TRE)を挿入することによって生成させてきた(Rodriguez等の論文、1997年Cancer Res. 57: 2559-2563)。
【0006】
腫瘍溶解性アデノウィルス、条件付きで複製性のアデノウィルス、および複製能欠陥アデノウィルスを含めた注目のアデノウィルスは、そのウィルスゲノムのE1初期遺伝子領域(遺伝地図単位1.30から9.24)中に遺伝子修飾部分を有するようにしばしば操作される。典型的な修飾には、E1初期遺伝子領域内の欠失および/またはE1Aプロモーターの置換え、導入遺伝子の導入などが含まれる。ヘルパーウィルスに依存しないアデノウィルスの産生は、ウィルス遺伝子産物を補完するパッケージング細胞株を必要とする可能性がある。
【0007】
研究および臨床試験用の組換えアデノウィルスベクターを産生するにはパッケージング細胞株は、アデノウィルスE1コード配列をトランスフェクションされる。この細胞株は、アデノウィルスDNA複製およびビリオン産生にとって直接または間接的に必要なトランスのE1AおよびE1B遺伝子産物を補充するのに十分なE1遺伝子産物を発現しなければならない。
【0008】
E1の補完は組換えアデノウィルスベクターの産生を可能にするが、宿主細胞中のトランスフェクションされたE1配列とそのアデノウィルスベクターの間の組換え事象が起こり、その結果、複製能のあるアデノウィルス(RCA)が生じる原因となる恐れがある。これは、大規模な産生およびそれに続く増殖の場合に特に問題であり、したがって治療に使用するためのアデノウィルス粒子ストックの調製において問題である。組換えと、組換えアデノウィルスベクター生産中のRCAの生成は、ウィルスストックを汚染するだけでなく、in vivo用途にこれらアデノウィルスベクターを使用することに関して問題がある。例えばShenk等の論文(1979)、Cold Springs Harb. Symp. Quant. Biol. 44 (1979) 367-375およびLochmullerの論文、Human Gene Therapy, 1994, 1485-1491に記載されているようにRCA生成の問題が知られてもうしばらくになる。
【0009】
入手可能なパッケージング細胞株は、一般にベクターからは欠失しているがウィルス複製には必要なアデノウィルス遺伝子を含有する。幾つかの事例では宿主細胞とアデノウィルスベクターの間の重複配列は完全には排除されない。例えばヒト胎児の腎臓から採った293細胞(Graham等の論文(1977)、J. general Virology 36: 59-74)は、アデノウィルスベクターを増殖するために広く用いられてきた。しかしながらこのアデノウィルスベクターゲノムと293細胞株の間の実質上の重複配列のせいで、複製能のあるアデノウィルス粒子の生成を引き起こす組換え事象が起こる。
【0010】
このベクターおよび細胞株に共通の配列の減少により、このパッケージング細胞株とベクターの間の組換え事象が原因する複製能のあるベクターの生成の可能性を低減させる改良がなされてきた(Fallaux等の論文(1998)、Human Gene Therapy 9: 1909-1917)。例えば米国特許第5,994,128号は、自然のE1Bプロモーター配列を保持すると同時にE1AとE1Bの両方またはその一方を補完する細胞株について記載している。PER.C6細胞株を用いて行った研究は、この細胞株とアデノウィルスベクターの間のたった1つの相同領域にもかかわらずRCAが生成し、細胞変性効果が細胞を用いた検定で観察されることを実証した(Kim等の論文(2001)、Exp. Mol. Med. 33 (3) 145-9)。分析するとこのRCA は、PER.C6細胞株を構築するために使用したプラスミド由来のPGKプロモーター−E1遺伝子を含有することが示された。残りの配列の重複のこの同じ問題は、293細胞に代わるものとして生成する他の細胞株についても当てはまる(例えばMassie他の米国特許第5,891,690号、Kovesdi他の国際公開第WO95/34671号、Kedan他の国際出願PCT/US95/15947号、Schiedner等の論文(2002)、Human Gene Therapy, 11: 2105-2116を参照されたい)。したがって細胞株とアデノウィルスベクターの間の望ましくない組換え事象の可能性が残っている。
【0011】
アデノウィルスベクター中の欠損部を補完するのに十分な、かつE1欠損アデノウィルスの成長を可能にするのに十分なE1AおよびE1Bコード配列を含むアデノウィルスパッケージング細胞株を提供する。このE1AおよびE1B配列は、天然または野生型アデノウィルスE1AおよびE1Bプロモーターと本質的な配列同一性を共有するポリヌクレオチド配列を欠いたプロモーターと作用可能に連結している。このようなパッケージング細胞株は、そのパッケージング細胞株ゲノムとアデノウィルスベクターの間の組換え事象の可能性が最小限のアデノウィルス粒子ストックを確実に産生する。本発明のパッケージング株を用いて産生されるウィルスストックは、意図する組換え遺伝子型を維持した最小限または検知できないレベルのRCAを特徴とする。
【0012】
本発明の1つの態様において、パッケージング細胞株は、E1AおよびE1B遺伝子が、同一であっても異なっていてもよい非アデノウィルス異種プロモーターと作用可能に連結している安定的に組み込まれたE1AおよびE1B発現ベクターを含む。
【0013】
本発明の別の態様において、RCAを実質上含まないアデノウィルスであって、実質的な配列同一性をアデノウィルスE1AおよびE1Bプロモーターと共有するポリヌクレオチド配列を欠いた細胞株中で成長するアデノウィルス、を産生する方法を提供する。
【0014】
定義:
別段の指示がない限り本明細書中で使用されるすべての技術的および科学的用語は、本発明の業界の技術者が使用するのと同じ意味を有する。当業界の定義および用語については、Sambrook等の著、Molecular Cloning: A Laboratory Manual (2nd Edition), Cold Spring Harbor Press, Plainview, N. Y., 1989およびAusubel FM等の著、Current Protocols in Molecular Biology, John Wiley & Sons, New York, N. Y. 1993は実務化向きである。本発明は、その記載されている特定の方法論、実験計画、および試薬には限定されず、したがってこれらは変えることができることを理解されたい。
【0015】
すべての特許、特許出願、刊行物(公開特許出願を含め)、および本明細書中で参照されるデータベース取得番号を含めた刊行物ならびに他の資料は、本発明の背景を明らかにするために、特にその実施に関して追加の細部を提供する場合に本明細書中で用いられる。これらすべての特許、特許出願、刊行物(公開特許出願を含め)、および本明細書中で参照されるデータベース取得番号を含めた刊行物ならびに他の資料は、それぞれが具体的かつ個々にその全体が参照により組み込まれて示されるのと同程度に参照により本明細書中に組み込まれる。
【0016】
「アデノウィルスパッケージング細胞」は、アデノウィルスゲノムまたは修飾ゲノムをパッケージしてウィルス粒子を産生することができる細胞である。それは欠けている遺伝子産物またはその同等物を提供することができる。したがってそのパッケージング細胞は、アデノウィルスゲノム中の欠失した遺伝子を補完する機能を提供することができ、またそれらアデノウィルスゲノムをアデノウィルス粒子中にパッケージすることができる。このような粒子の産生は、そのゲノムが複製され、かつ感染性ウィルスを組み立てるのに必要なそれらのタンパク質が産生されることを必要とする。この粒子はまた、そのウィルス粒子の成熟に必要なある種のタンパク質を必要とする可能性がある。このようなタンパク質は、そのベクターにより、またはそのパッケージング細胞により与えることができる。このパッケージング細胞株は、E1Aおよび/またはE1Bコード配列を含むようにアデノウィルス複製を許容する細胞株を遺伝子的に修飾することによって産生することができる。本発明のアデノウィルスパッケージング細胞株ではアデノウィルスE1AおよびE1Bコード配列は、天然のアデノウィルスE1AおよびE1Bプロモーターと実質的な配列同一性を共有するポリヌクレオチド配列を欠いたプロモーターと作用可能に連結している。
【0017】
「宿主細胞」には個別の細胞または細胞培養物が含まれ、これらは本発明のウィルスベクターのレシピエントとなることができ、又はレシピエントとなっていてもよい。宿主細胞には単一宿主細胞の子孫が含まれ、この子孫は自然、偶然、あるいは意図的な突然変異および/または変化のせいで元の親細胞と必ずしも完全に同一(形態または全DNA相補体において)でなくてもよい。宿主細胞には、本発明のアデノウィルスベクターをin vivoまたはin vitroにおいてトランスフェクションまたは感染させた細胞が含まれる。
【0018】
本明細書中で用いられる用語「アデノウィルス」および「アデノウィルス粒子」には、あらゆる群、亜群、および血清型を含めたヒトまたは動物に感染する任意のアデノウィルスを含めたアデノウィルスとして分類することができる、ありとあらゆるウィルスが含まれる。したがって本明細書中で用いられる「アデノウィルス」および「アデノウィルス粒子」は、ウィルス自体またはその誘導体を指し、すべての血清型と亜型ならびに天然に存在する品種および組換え品種の両方を包含する。一実施形態においてこのようなアデノウィルスはヒトの細胞に感染する。このようなアデノウィルスは野生型であってもよく、また当業界で知られているか、または本明細書中で開示した様々な方法で修飾されてもよい。このような修飾には、感染性ウィルスを作るためにその粒子中にパッケージングされるアデノウィルスゲノムに対する修飾が含まれる。このような修飾には、E1a、E1b、E2a、E2b、E3、またはE4コード領域の1または複数における欠失など、当業界で知られている欠失が挙げられる。本発明のアデノウィルスベクターの例には、限定しないが、DNA、アデノウィルスの外殻に包まれたDNA、別のウィルスまたはウィルス様品種(単純ヘルペスおよびAAVなど)中にパッケージングされたアデノウィルスDNA、リポソームに包まれたアデノウィルスDNA、ポリリジンと複合化させたアデノウィルスDNA、合成ポリカチオン分子と複合化させたアデノウィルスDNA、トランスフェリンと接合したアデノウィルスDNA、あるいはその抗原性を免疫的に「マスキング」するためにおよび/または半減期を増すためにPEGなどの化合物と複合化させたアデノウィルスDNA、あるいは非ウィルスタンパク質に接合したアデノウィルスDNA、が挙げられる。E1A/E1Bパッケージング株の生成に使用されるAAVベクターの例を図8および9に示す。
【0019】
本明細書中でアデノウィルスベクターに関して用いられる用語「複製能欠陥の」とは、ウィルスベクターがそのゲノムをそれ以上複製し、またパッケージすることができないことを意味する。例えば、ある被験者の細胞がrAAVビリオンに感染している場合、その異種遺伝子はその患者の細胞中で発現するが、その患者の細胞がAAV REPおよびCAP遺伝子と、アデノウィルスの補助機能遺伝子とが欠けているという事実により、そのrAAVは複製能欠陥があり、野生型AAVはその患者の細胞中で生成することができない。
【0020】
本明細書中で用いられる「パッケージングシステム」とは、組換えウィルスのパッケージングに関係するウィルスタンパク質をコードする遺伝子を含む一組のウィルス構築物を指す。一般にそのパッケージングシステムの構築物は、最後はパッケージング細胞中に取り込まれることになる。
【0021】
本明細書中でアデノウィルスベクターに関して用いられる用語「複製能のある」とは、そのウィルスベクターおよび粒子が或る型の細胞または組織中では優先的に複製するが、他の型ではずっと少ない程度かまたは全く複製しないことを意味する。本発明の1つの態様において、そのウィルスベクターおよび/または粒子は、充実性腫瘍および他の新生組織などの腫瘍細胞および/または異常増殖組織中で選択的に複製する。このようなウィルスは、「腫瘍溶解性ウィルス」または「腫瘍溶解性ベクター」と呼ぶことができ、また「細胞溶解性」または「細胞変性性」であり、標的細胞の「選択的細胞溶解」を行うと考えることができる。これらには、米国特許第5,677,178号、第5,698,443号、第5,871,726号、第5,801,029号、第5,998,205号、および第6,432,700号に開示されているウィルスが挙げられる。
【0022】
用語「ウィルス」、「ウィルス粒子」、「ベクター粒子」、「ウィルスベクター粒子」、および「ビリオン」は同義で用いられ、例えば本発明のウィルスベクターを感染性粒子の生成用の適切な細胞または細胞株に形質導入する場合に形成される感染性ウィルス粒子を意味するものと広範囲に理解されるべきである。本発明によるウィルス粒子は、核酸(例えばDNAまたはRNA)をin vitroまたはin vivoのいずれかで細胞中に伝達する目的に利用することができる。
【0023】
本明細書中で同義で用いられる用語「ポリヌクレオチド」および「核酸」は、任意の長さのポリマー形態のヌクレオチドであるリボヌクレオチドまたはデオキシリボヌクレオチドのいずれかを指す。これらの用語には、一本、二本、または三本鎖DNA、ゲノムDNA、cDNA、RNA、DNA−RNAハイブリッド、あるいはプリンおよびピリミジン塩基を含むポリマー、あるいは他の天然の化学的、生化学的に変性した塩基、非天然または誘導ヌクレオチド塩基を含むポリマーが含まれる。好ましくは本発明のベクターはDNAを含む。本明細書中で用いられる「DNA」には、塩基A、T、C、およびGだけでなく、それらの類似体またはこれら塩基の修飾形態、例えばメチル化ヌクレオチド、非荷電結合およびチオ酸塩などのヌクレオチド間修飾物、糖類似体の使用、ならびにポリアミドなどの変性および/または代替骨格構造のいずれかもまた含まれる。
【0024】
遺伝子または遺伝子フラグメント、エキソン、イントロン、mRNA、tRNA、rRNA、リボザイム、cDNA、組換えポリヌクレオチド、分枝ポリヌクレオチド、プラスミド、ベクター、任意の配列の単離DNA、任意の配列の単離RNA、核酸プローブ、およびプライマーは、ポリヌクレオチドの非限定的な例である。ポリヌクレオチドは、修飾ヌクレオチド、例えばメチル化ヌクレオチド、ヌクレオチド類似体、ウラシル、フルオロリボースおよびチオ酸塩などの他の糖類および結合基、ならびにヌクレオチド側鎖などを含むことができる。そのヌクレオチドの配列は非ヌクレオチド成分が介在していてもよい。ポリヌクレオチドは標識成分を接合させるなど、重合後さらに修飾されてもよい。この定義に含まれる他の種類の修飾は、キャップ、類似体による天然に存在するヌクレオチドのうちの1個または複数個の置換、およびタンパク質、金属イオン、標識成分、他のポリヌクレオチド、または固形支持体にそのポリヌクレオチドを付着させる手段の導入である。好ましくはこのポリヌクレオチドはDNAである。本明細書中で用いられる「DNA」には、塩基A、T、C、およびGが含まれるだけでなく、それらの類似体またはこれら塩基の修飾形態、例えばメチル化ヌクレオチド、非荷電結合およびチオ酸塩などのヌクレオチド間修飾物、糖類似体の使用、ならびにポリアミドなどの変性および/または代替骨格構造のいずれかもまた含まれる。
【0025】
核酸は、別の核酸配列と機能上の関係に置かれる場合、「作用可能に連結」している。例えばオペレーターまたはエンハンサーは、それがその配列の転写に影響する場合、コード配列と作用可能に連結している。通常、「作用可能に連結した」とは、それら結合しているDNA配列が隣接していることを意味する。しかしながらエンハンサーは隣接しなくてもよい。結合は、都合のよい制限部位においてライゲーションにより達成される。このような部位が存在しない場合、合成オリゴヌクレオチドアダプターまたはリンカーが従来のやり方に従って用いられる。
【0026】
用語「天然の」とは、野生型ウィルスまたは細胞のゲノム中に存在する遺伝子を指す。
【0027】
用語「天然に存在する」または「野生型」は、人間により人為的に生成されるものとはまったく異なるものとして自然界に見出すことができる物を記述するために用いられる。例えば、自然界の供給源から単離することができ、かつ実験室で人間によって故意に変性されなかった生物体(ウィルスを含めた)中に存在するタンパク質またはヌクレオチド配列は、天然に存在する。
【0028】
本明細書中で用いられる用語「プラスミド」は、染色体外でまたは宿主細胞の染色体(または染色体群)の一部として宿主細胞内で自己複製の能力があるDNA分子を指す。本明細書中の出発プラスミドは市販されており、公然と自由に利用可能であり、あるいは本明細書中に開示されているような利用可能なプラスミドからおよび/または出版物に記載されている手順に従って構築することができる。幾つかの事例では、当業者には明らかなように当業界で知られている他のプラスミドを本明細書中で記述したプラスミドと同義で用いることができる。
【0029】
本明細書中で用いられる用語「投与」または「導入」は、細胞中へのE1Aおよび/またはE1Bコード配列の安定組込みのための発現ベクターの送達を指す。ベクターはトランスフェクションによって細胞中に導入することができ、このトランスフェクションは一般に物理的手段(例えばリン酸カルシウムのトランスフェクション、電気穿孔、マイクロインジェクション、またはリポフェクション)、感染(一般には感染性病原体すなわちウィルスを経由する導入を指す)、または形質導入(一般には、ウィルスによる細胞の安定感染、またはウィルス病原体(例えばバクテリオファージ)を経由する或る微生物から別の微生物への遺伝物質の転移を意味する)による細胞中への異種DNAの挿入を意味する。前述のようにこのベクターは、プラスミド、ウィルス、または他の伝達体であることができる。
【0030】
本明細書中で核酸分子に関して用いられる用語「組換え体」は、組換えDNA技術を用いて結合されて子孫核酸分子になる核酸分子の複合体を指す。ウィルス、細胞、および生物体に関して本明細書中で用いられる用語「組換え」、「形質転換」、および「トタンスジェニック」は、異種核酸分子が導入された宿主のウィルス、細胞、または生物体を指す。この核酸分子はその宿主のゲノム中に安定的に組み込むことができるか、またはこの核酸分子は染色体外分子として存在することもできる。このような染色体外分子は、自己複製性であってもよい。これらの組換えウィルス、細胞、および生物体は、形質転換過程の最終産物だけでなく、それらの組換え子孫も包含すると考えられる。「非形質転換」、「非トタンスジェニック」、または「非組換え」宿主は、異種核酸分子を含有しない野生型のウィルス、細胞、または生物体を指す。
【0031】
「調節エレメント」は、ヌクレオチド配列の発現の制御と関係した配列である。調節エレメントには、プロモーター、エンハンサー、および終止コドンが含まれる。これらはまた、一般にそのヌクレオチド配列の適正な翻訳に必要な配列を包含する。
【0032】
用語「プロモーター」は、RNAポリメラーゼIIに対する結合部位を含有し、そのDNAの転写を開始するコード領域の上流に通常位置する非翻訳DNA配列を指す。このプロモーター領域は、遺伝子発現の調節遺伝子として働く他のエレメントもまた含むことができる。用語「最少プロモーター」は、不活性であるか、または上流の活性化エレメントの不在下において著しく低いプロモーター活性を有するプロモーターエレメント、特にTATAエレメントを指す。
【0033】
用語「エンハンサー」は本発明の意味の範囲内では、任意の遺伝エレメント、例えば或るコード配列と作用可能に連結した場合にプロモーター自体によってもたらされる転写活性化よりも大きい程度までそのプロモーターと作用可能に連結しているそのコード配列の転写を増大させる、すなわちそれがそのプロモーター由来の転写を増大させるヌクレオチド配列であることができる。
【0034】
語句「とハイブリダイズする」とは、その配列が複合混合物(例えば全細胞)のDNAまたはRNA中に存在する場合、ストリンジェントな条件下における特定のヌクレオチド配列のみとの分子の結合、二重らせん形成、またはハイブリダイゼーションを指す。「実質上結合する」とは、プローブ核酸と標的核酸の間の相補的ハイブリダイゼーションを指し、そのハイブリダイゼーション培地のストリンジェンシーを下げることによって適応することができる重要でない誤対合を容認して標的核酸配列の所望の検出を達成する。
【0035】
サザンおよびノーザンハイブリダイゼーションなどの核酸ハイブリダイゼーション実験の脈絡における「ストリンジェントなハイブリダイゼーション条件」および「ストリンジェントな洗浄条件」は配列に左右され、また様々な環境要因下で異なる。長い配列ほど高い温度でハイブリダイズする。核酸のハイブリダイゼーションに関する広範囲な手引きは、Tijssen著、(1993) Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probesの第1部第2章「Overview of principles of hybridization and the strategy of nucleic acid probe assays」Elsevier, New York中に見出される。一般に高ストリンジェントなハイブリダイゼーションおよび洗浄条件は、規定のイオン強度およびpHにおいてその特定の配列の融点(Tm)よりも約5℃から20℃(好ましくは5℃)低いように選択される。一般に高ストリンジェントな条件下ではプローブは、他の配列とではなく、その標的部分配列とハイブリダイズすることになる。
【0036】
用語「相補体」および「相補的」は、アンチパラレルヌクレオチド配列中の相補的塩基残基間で水素結合を形成するとき互いに組むことができるアンチパラレルヌクレオチド配列を含む2つのヌクレオチド配列を指す。
【0037】
用語「発現」は、細胞中の内因性遺伝子、導入遺伝子、またはコード領域の転写および/または翻訳を指す。
【0038】
本明細書中で用いられる「内部リボソーム侵入部位」または「IRES」は、ATGなどの開始コドンへのシストロン(タンパク質コード領域)の直接の内部リボソーム侵入を促進し、それによってキャップに影響されない遺伝子の翻訳を引き起こすエレメントを指す。例えばJackson R J、Howell M T、およびKaminski Aの論文(1990)、Trends Biochem Sci 15 (12): 447-83ならびにJackson R J、Howell M T、およびKaminski, Aの論文(1995)、RNA 1 (10): 985-1000を参照されたい。本発明は、開始コドンへのシストロンの直接の内部リボソーム侵入を促進することができる任意のエレメントを使用することを包含する。本明細書中で用いられる「IRESの翻訳制御下で」とは、翻訳がIRESと関連しており、キャップに影響されないやり方で進行することを意味する。本明細書中で用いられる用語「IRES」は、IRES配列の機能上の変異が開始コドンへのシストロンの直接の内部リボソーム侵入を促進することができる限り、その変異を包含する。
【0039】
本明細書中で言及される「自己プロセッシング切断部位」または「自己プロセッシング切断配列」は、翻訳時に自己プロセッシング切断部位を含むポリペプチドの急速な分子間(シス)切断を起こして不連続の成熟タンパク質またはポリペプチド産物の発現を引き起こすDNAまたはアミノ酸配列である。このような「自己プロセッシング切断部位」はまた、翻訳後のまたは翻訳に伴うプロセッシング切断部位、例えば2A部位、配列、またはドメインと呼ぶこともできる。2A部位、配列、またはドメインは、リボソームの活性を変えてエステル結合の加水分解を促進させ、それによって不連続な下流の翻訳産物の合成を進行させるやり方で翻訳複合体からポリペプチドを遊離することによって翻訳効果を実証する(Donnelly等の論文、J. Gen. Virol. 82: 1027-1041, 2001)。FMDV 2A領域による切断活性の発現にとって不可欠なアミノ酸残基を含む構築体が設計されている(Ryan等の論文(1991)、J. Gen. Virol. 72: 2727-2732およびFurler等の論文(2001)、Gene Therapy 8: 864-873)。2Aドメインはまた、ピコルナウィルス科のaphthovirideaおよびcardioviridaeから特徴が調べられた(Donnelly等の論文(1997)、J. Gen. Virol. 78: 13-21)。
【0040】
本明細書中で用いられる用語「E1A」は、2つの主要なRNAの発現生成物13Sおよび12Sを含むアデノウィルスE1A領域のすべての遺伝子産物を指す。これらは翻訳されてそれぞれ289(配列番号2)および243(配列番号3)のアミノ酸のポリペプチドになる。これら2種類のタンパク質はアミノ酸が46個異なり、Chow等の論文(1980)、Cold Spring Harb Symp Quant Biol. 44 Pt 1: 401-14およびChow等の論文(1979)、J. Mol. Biol. 134 (2): 265-303(これらは特に引用により本明細書に組み入れられる)に記載のようにそれらは12S mRNAからスプライシングされる。本発明の目的の場合、パッケージング細胞株はその289ポリペプチド、243ポリペプチド、または289および243ポリペプチドの両方を発現することができる。用語E1Aはまた、本明細書中では部分および変異E1Aコード配列に関して用いられる。
【0041】
本明細書中で用いられる用語「E1B」は、3つの主要なポリペプチドを含む19 kd(配列番号5)および55 kd(配列番号6)のアデノウィルスE1B領域のすべての遺伝子産物を指す。これらE1B 19 kdおよび55 kdタンパク質は、細胞の形質転換において重要である。本発明の目的では、パッケージング細胞株は19 kdポリペプチド、55 kdポリペプチド、または19 kdおよび55 kdポリペプチドの両方を発現することができる。用語「アデノウィルス許容」とは、そのアデノウィルスまたはアデノウィルスベクターが、その細胞環境中で全細胞内ウィルスのライフサイクルを終えることができることを意味する。これらの細胞は、初代細胞培養物、樹立細胞株等から得ることができる。霊長動物細胞、例えばヒトの細胞、サルの細胞などを含めた哺乳動物細胞が好ましい。様々な霊長動物細胞が好ましく、またこのようなヒトの胎児腎臓細胞がより好ましいが、ウィルスの複製を支持することができる任意の種類の細胞が本発明の実施において受け入れられる。用語E1Bはまた、本明細書中では部分および変異E1Bコード配列に関して用いられる。
【0042】
「複製」および「増殖」は同義で用いられ、アデノウィルスベクターを再生産または増殖する能力を指す。これらの用語は、当業界では十分に理解されている。本発明の目的の場合、複製はアデノウィルスタンパク質の産生と関係しており、一般にアデノウィルスの再生産を目的とする。複製は、ウィルスの収量検定、バースト検定、またはプラーク検定などの当業界で標準的なまた本明細書中で記述する検定法を用いて測定することができる。「複製」および「増殖」には、限定しないが、ウィルス遺伝子発現や、ウィルスのタンパク質、核酸、または他の成分の産生や、ウィルス構成要素をパッケージして完全なウィルスにすることや、細胞溶解を含めたウィルスの生産過程に直接または間接的に関係する任意の営みが含まれる。
【0043】
本発明の方法および組成物:
種々の方法および組成物を後述する。特定の方法を下記の考察において例示するが、複数の代替方法のいずれも適用可能であり、また本発明の実施において使用に適していることが分かる。また本発明のアデノウィルスベクターおよび方法の評価は、下記の診断および判定方法を含む当業界で標準的な手順を用いて行うことができる。
【0044】
本発明の実施においては別段の指示がない限り、当業者の範囲内にある細胞生態学、分子生物学(組換え技術を含めた)、微生物学、生化学、および免疫学の通常の技術を使用することになる。このような技術は、「Molecular Cloning: A Laboratory Manual」2nd edition (Sambrook等著、1989)、「Oligonucleotide Synthesis」(M. J. Gait編、1984)、「Animal Cell Culture」(R. I. Freshney編、1987)、「Methods in Enzymology」(Academic Press, Inc.)、「Handbook of Experimental Immunology」(D. M. WeirおよびC. C. Blackwell編)、「Gene Transfer Vectors for Mammalian Cells」(J. M. MillerおよびM. P. Calos編、1987)、「Current Protocols in Molecular Biology」(F. M. Ausubel等編、1987)、「PCR: The Polymerase Chain Reaction」(Mullis等編、1994)、および「Current Protocols in Immunology」(J. E. Coligan等編、1991) などの文献中で完全に説明されており、これらは特別にそれぞれ本明細書中に参照により組み込まれる。
【0045】
アデノウィルスに関係する技術に関しては、とりわけFelgnerおよびRingoldの論文(1989)、Nature 337: 387-388、BerkerおよびSharpの論文(1983)、Nucl. Acids Res. 11: 6003-6020、Grahamの論文(1984)、EMBO J. 3: 2917-2922、Bett等の論文(1993)、J. Virology 67: 5911-5921、Bett等の論文(1994)、Proc. Natl. Acad. Sci. USA 91: 8802-8806を参照されたい。
【0046】
アデノウィルスパッケージング株
E1AおよびE1B欠損アデノウィルスを補完および複製するのに十分であり、かつ野生型複製能のあるアデノウィルス(RCA)を生成させる可能性が最小限のアデノウィルスE1AおよびE1B配列を実現する、アデノウィルスパッケージング細胞株を提供する。本明細書中で用いられるRCAは、E1Aおよび/またはE1Bの発現のためにパッケージング細胞株による補完を必要としない複製能のあるアデノウィルスである。
【0047】
このパッケージング細胞株は、ヒトアデノウィルスE1AおよびE1Bタンパク質をコードする遺伝子配列を含む。この遺伝子配列は、天然配列またはそれらの変異型であることができる。本明細書中で用いられる実質的な配列同一性とは、その宿主細胞内の相同的組換えにとって十分な配列類似性レベルを指す。候補配列については、例えば注目の細胞において複製中に2つの配列に組換えがあるかないかを試験することによって経験的に組換えの有無を調べることができる。一般に配列は、連続した同一のポリヌクレオチド配列の約20個を超えるヌクレオチド、通例では連続した同一のポリヌクレオチド配列の約15個を超えるヌクレオチド、好ましくは連続した同一のポリヌクレオチド配列の約12個を超えるヌクレオチドが存在しない場合、実質的な配列同一性が欠けていることになる。
【0048】
参照配列は、通常はそのベクターが得られる元のアデノウィルス、例えばヒトアデノウィルス5、ヒトアデノウィルス2などであることになる。本発明のパッケージング細胞株中でE1AおよびE1Bの発現を駆動するプロモーターと、内因性のアデノウィルスE1AおよびE1Bプロモーターの間の実質的な配列同一性が欠けていることは、組換えおよびその結果としての複製能のあるアデノウィルス(RCA)の産生の可能性をできるだけ少なくするはずである。
【0049】
配列比較の場合、一般には一方の配列が試験配列をそれと比較する参照配列として働く。配列比較アルゴリズムを使用する場合、これら試験および参照配列をコンピュータにインプットし、必要ならば部分配列座標を指定し、配列アルゴリズムのプログラムパラメーターを指定する。次いで配列比較アルゴリズムが、指定したプログラムパラメーターに基づいて参照配列に対する試験配列の配列同一性のパーセントを計算する。
【0050】
比較のための配列の最適アライメントは、例えばSmithおよびWatermanの論文、Adv. Appl. Math. 2: 482 (1981) のローカルホモロジーアルゴリズムにより、あるいはNeedlemanおよびWunschの論文、J. Mol. Biol. 48: 443 (1970) の相同アライメントアルゴリズムにより、あるいはPearsonおよびLipmanの論文、Proc. Nat’l. Acad. Sci. USA 85: 2444 (1988)の類似性検索法により、あるいはこれらアルゴリズムのコンピュータ化されたインプリメンテーション(Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, Wis 中のGAP、BESTFIT、FASTA、およびTFASTA)により、あるいはNational Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/)を介して公けに利用できるソフトウェアを用いたBLASTアルゴリズム(Altschul等の論文、J. Mol. Biol. 215: 403-410 (1990))により、あるいは目視検査により行うことができる(全般的には後掲のAusubel等の論文を参照)。本発明の目的の場合、比較のための最適アライメントは、最も好ましくはSmithおよびWatermanの論文、Adv. Appl. Math. 2: 482 (1981) のローカルホモロジーアルゴリズムにより行われる。
【0051】
2つ以上の核酸またはタンパク質配列の脈絡における用語「同一の」、または「同一性」パーセントは、本明細書中で述べた配列比較アルゴリズムの一つ、例えばSmith−Watermanのアルゴリズムを用いて、または目視検査により測定した最大の対応関係について比較し、また位置合わせした場合に同一であるか、あるいは所定の割合の同一であるアミノ酸残基またはヌクレオチドを有する2つ以上の配列または部分配列を指す。
【0052】
多くのアデノウィルスE1AおよびE1Bタンパク質の配列、例えばヒトアデノウィルス11型、ヒトアデノウィルス41型、ヒトアデノウィルス12型、ヒトアデノウィルス5、ヒトアデノウィルス35、ヒトアデノウィルス41、ヒトアデノウィルス40、ヒトアデノウィルス4、ヒトアデノウィルス7、およびヒトアデノウィルス2由来の配列が知られており、また公けに利用できる。例えばE1Aポリペプチド配列、Genbank 取得番号AAN62486 (Ad11)、Q2AD5 (Ad5)、およびQ2AD2 (Ad2)、ならびにE1Bポリペプチド配列、Genbank 取得番号Q1AD25 (Ad5)およびQ1AD22 (Ad2) を参照されたい。E1AおよびE1Bコード配列の元はAd2由来のものが最も好ましい。Ad2を含む他のヒトおよび非ヒトアデノウィルス血清型もまた用いることができる。本発明のパッケージング株の調製に用いられるE1AおよびE1B配列は、E1AまたはE1Bのどちらかのプロモーター配列を含まない。通常このパッケージング株は、E1AおよびE1Bコード配列とは異なるアデノウィルス遺伝子配列を含まないはずである。
【0053】
E1AおよびE1Bコード配列の例は、GenBank取得番号M73260またはX02996のヌクレオチド560〜1545(本明細書中では配列番号1として示される)に対応するE1Aの2つのエキソン領域およびGenBank取得番号M73260またはX02996のヌクレオチド1682〜3825(本明細書中では配列番号4として示される)に対応する2つのE1Bコード配列を含む。多くのアデノウィルスゲノムの特徴が調べられ、使用のために入手できるので、本明細書中で提供されるアデノウィルス配列は好適な配列の単なる例に過ぎないことは当業者ならば理解するはずである。本発明の実施において用いられるE1AおよびE1Bコード配列の例を表1に示す。
【0054】
【表1】

【0055】
これらE1AおよびE1B配列は作用可能に非アデノウィルスプロモーターと結合している。このプロモーターは異種であってもよく、この場合、本明細書中では用語「異種」プロモーターはそのパッケージング細胞にとって天然でないプロモーター配列を意味するために用いられる。E1Aおよび/またはE1Bのコード配列を含むアデノウィルスベクターの複製の場合、そのパッケージング細胞株中のE1Aおよび/またはE1B用のプロモーターは、好ましくはアデノウィルスベクター中で作用可能にE1Aおよび/またはE1Bと結合したプロモーターとは異なる。
【0056】
注目のパッケージング細胞株では、アデノウィルスE1AおよびE1Bコード配列は、相同的組換えがあまり起こらないように、天然のアデノウィルスE1AおよびE1Bプロモーターと実質的な配列同一性を共有するポリヌクレオチド配列を欠いているプロモーターと作用可能に連結している。このようなパッケージング細胞株は、そのパッケージング細胞株と複製能欠損アデノウィルスベクターとの間の組換え事象がなく、それによってRCAの生成の可能性をできるだけ少なくするアデノウィルス粒子ストックを確実に生産する。
【0057】
本発明の一実施形態においてパッケージング細胞株は、安定的に組み込まれたE1AおよびE1B発現ベクターを含み、そのE1AおよびE1B遺伝子は非アデノウィルスプロモーターと作用可能に連結し、別個の発現ベクターを用いて導入されている。このプロモーターは、非アデノウィルス起源の強い構成性プロモーターであってもよい。1つの態様において、このE1Aと作用可能に連結したプロモーターは、E1Bと作用可能に連結したプロモーターと異なる。別の実施形態ではこのE1Aと作用可能に連結したプロモーターは、E1Bと作用可能に連結したプロモーターと同じものである。これらE1AおよびE1B遺伝子は、このようなプロモーターと全体的に調和して発現することができる。
【0058】
これらE1AおよびE1Bコード領域は、好ましくはパッケージング細胞株のゲノム中に安定的に組み込まれる。好ましい実施形態においてE1A組込み部位は、例えば別個の染色体上、同一の染色体の別個の領域上などでE1B組込み部位から物理的に引き離される。
【0059】
本発明の別の実施形態では、RCAを実質的に含まないアデノウィルスを生産する方法であって、当該アデノウィルスが、アデノウィルスE1AおよびE1Bプロモーターと実質的な配列同一性を共有するポリヌクレオチド配列を欠いた細胞株中で成長する方法、を提供する。
【0060】
本発明の細胞株中での複製にとって注目のアデノウィルスベクターは、複製に不可欠なアデノウィルス遺伝子、具体的にはE1AおよびE1B遺伝子の発現に欠陥がある。このようなベクターは、外因によりもたらされるウィルス遺伝子の不在下では増殖性感染に必要な十分なウィルス性タンパク質を産生することができない。E1AおよびE1Bの発現に欠陥のあるアデノウィルスベクターは、様々な遺伝的変化のせいで、例えばこれら遺伝子の一方または両方のコード配列の欠如、そのポリペプチドを作用不能にするコード配列中の突然変異、プロモーターまたはエンハンサー配列の変化などのせいで欠陥がある。
【0061】
本発明の幾つかの実施形態においてアデノウィルスベクターは、標的細胞型、例えば前立腺癌、肝臓癌などの標的腫瘍細胞中では複製応答能があるが、非標的細胞型中ではこのアデノウィルスはE1Aおよび/またはE1Bの発現に欠陥がある。例えばこのアデノウィルスベクターは、細胞型特異的、細胞状態特異的などである転写調節エレメントと作用可能に連結している複製にとって不可欠なアデノウィルス遺伝子を含むことができる。このようなベクターは、本明細書中で述べたような、例えばin vitroで多数のウィルス粒子を生成させるパッケージング細胞株中での成長から恩恵を受ける。
【0062】
E1AおよびE1Bを発現させるために用いられるプロモーター配列は、同一でも同一でなくてもよい。プロモーター配列が同一の場合、そのE1AおよびE1Bコード配列は全体的に協調して発現することができ、例えばこれら両方のコード配列が作用可能に単一プロモーターと結合し、IRESはそれら2つのコード配列間に存在する。
【0063】
本発明の一実施形態において一方または両方のプロモーターは調節可能なプロモーター、例えば金属類またはホルモン類(Brinster等の論文、Nature (1982), 296, 39-42)、あるいはホルモン類(Lee等の論文、P.N.A.S. USA (1988), 85, 1204-1208および(1981), 294, 228-232、Klock等の論文、Nature (1987), 329, 734-736、IsraelおよびKaufmanの論文、Nucleic Acids Res. (1989), 17, 2589-2604)などの物質で誘導できるプロモーターである。
【0064】
あるいはさらに別の実施形態においてこのプロモーターは構成性プロモーターである。これらプロモーターは、ポリオーマウィルス、鶏痘ウィルス、ウシ乳頭腫ウィルス、トリ肉腫ウィルス、サイトメガロウィルス、レトロウィルス、B型肝炎ウィルス、およびサルウィルス40(SV40)などのウィルスのゲノムから、あるいは異種哺乳動物プロモーター、例えばアクチンプロモーター、PGK(ホスホグリセリン酸キナーゼ)、または免疫グロブリンプロモーターから、あるいは熱ショックプロモーターからこのようなプロモーター宿主細胞株に適合するという条件で得ることができる。SV40ウィルスの初期および後期プロモーターは、好都合なことにまたSV40ウィルスの複製開始点をも含有するSV40制限フラグメントとして得られる。ヒトサイトメガロウィルスの即時型プロモーターは、便利にはHindIII E制限フラグメントとして得られる。
【0065】
例示的な1つの態様において、、MMLVのLTRは第一レトロウィルス発現ベクター中および第二レトロウィルス発現ベクター中のE1A遺伝子と作用可能に連結しており、このMMLVのLTRはE1B遺伝子と作用可能に連結しており、E1B遺伝子の発現を誘導するために用いられる。別の実施形態の例では、CAGプロモーターが第一発現ベクター中および第二発現ベクター中のE1A遺伝子のエキソン1およびエキソン2(配列番号1)と作用可能に連結しており、そのEF1−αプロモーターが19kおよび55k E1Bコード配列(配列番号4)の発現を誘導するために用いられる。
【0066】
高等真核生物による転写は多くの場合、エンハンサー配列をそのベクター中に挿入することによって増大する。エンハンサーはDNAのシス作用エレメントであり、一般に約10から300 bpであり、プロモーターに作用してその転写を増大させる。エンハンサーは相対的な配向にあり、位置と関係がなく、コード配列自体の内部だけでなくイントロン内部の転写単位の5′および3′側にあることが分かっている。多くのエンハンサーは、現在では哺乳動物遺伝子(グロブリン、エラスターゼ、アルブミン、α−フェトプロテイン、およびインスリン)から見分けられる。しかしながら一般には皆、真核細胞ウィルス由来のエンハンサーを使用するはずである。例には、複製起点の後期局面でのSV40エンハンサー、サイトメガロウィルスの初期プロモーターエンハンサー、複製起点の後期局面でのポリオーマエンハンサーなどが挙げられる。このエンハンサーは、そのコード配列の5′または3′位置でその発現ベクターにスプライシングすることができるが、好ましくはプロモーターの5′部位に位置する。
【0067】
(パッケージングアデノウィルス用細胞株の生成)
部位特異的DNA切断は、当業界で一般に理解されている条件および市販の制限酵素の製造者により指定される事項(例えばNew England Biolabs, Product Catalogを参照されたい)の下で、プラスミドまたは他のDNAを適切な制限酵素(または複数種の制限酵素)で処理することによって行われる。一般にプラスミドまたは他のDNA約1 μgは、緩衝液約20 μL中で1ユニットの酵素によって切断される。一般にはDNA基質の完全な消化を保証するために過剰の制限酵素が用いられる。変動を許容することができるが約37℃で約1時間から2時間のインキュベーション時間が実行可能である。それぞれインキュベーションの後、タンパク質をフェノール/クロロホルムで抽出することにより取り出し、続いてエーテル抽出し、エタノールで沈殿させることによって核酸を水性画分から回収することができる。望むなら標準的な技術を用いてポリアクリルアミドゲルまたはアガロースゲル電気泳動により切断フラグメントのサイズ選別を行うことができる。サイズ選別の全般的な説明は、Methods of Enzymology 65: 499-560 (1980)中に見出される(前掲のSambrookおよびRussellの著書もまた参照されたい)。
【0068】
制限切断フラグメントは、50 mMトリス(pH 7.6)、50 mM NaCl、6 mM MgCl2、6 mM DTT、および5〜10 μM dNTP中において20℃での約15から25分のインキュベーション時間を用いて4種類のデオキシヌクレオチド三リン酸(dNTP)の存在下で大腸菌DNAポリメラーゼI(クレノウ)の大きなフラグメントで処理することによって平滑末端化することができる。このクレノウフラグメントは5′粘着末端を埋めるが、4種類のdNTPが存在していても突出3’一本鎖に関しては取り込んだ後元に戻される。所望により、粘着末端の性質によって規定される制約内でそれらdNTPのうちのただ1種類を供給することにより、または選択したdNTPを用いて選択的修復を行うことができる。クレノウで処理した後、この混合物をフェノール/クロロホルムで抽出し、エタノールで沈殿させる。適切な条件下でSIヌクレアーゼまたはBal−31で処理すると任意の一本鎖部分の加水分解が起こる。
【0069】
ライゲーションは、次の標準的な条件および温度下、すなわち20 mMトリス−Cl(pH 7.5)、10 mM MgCl2、10 mM DTT、BSA 33 mg/mL、10 mM〜50 mM NaClと、40 μM ATP、0℃のT4 DNAリガーゼ0.01〜0.02(Weiss)ユニット(「粘着末端」ライゲーション用)か、または1 mM ATP、14℃のT4 DNAリガーゼ0.3〜0.6(Weiss)ユニット(「平滑末端」ライゲーション用)のいずれかとで体積15〜50 μLで行う。分子間「粘着末端」ライゲーションは一般に、全DNA濃度33〜100 μg/mL(5〜100 mMの合計の終濃度)で行われる。分子間「平滑末端」ライゲーション(一般に10〜30倍モル濃度過剰のリンカーを使用する)は、1 μMの合計の終濃度で行われる。
【0070】
E1AまたはE1Bを含むベクターは許容宿主細胞中に導入される。多くのこのようなベクター、例えばプラスミドベクター、ウィルスベクターなどが利用可能である。ベクターの構成成分には、限定しないが、複製起点、1種類または複数種類のマーカー遺伝子、エンハンサーエレメント、プロモーター、および転写末端配列のうちの1種類または複数種類を挙げることができる。
【0071】
E1A/E1Bコード配列の導入用の好ましい発現ベクターは、宿主細胞中に安定的に組み込むことができ、かつ高頻度で娘細胞中に保持されるものである。好ましい一実施形態においてこの発現ベクターはウィルス起源のものである。幾つかの組換えウィルスベクターが、本発明によるパッケージング細胞株を生産するために細胞中にE1A/E1Bコード配列を効果的に送達するのに有用であり、例えばレトロウィルスベクター、レンチウィルスベクター、アデノウィルス関連ベクター(AAV)、ヘルペスウィルスベクター、ポックスウィルスベクターなどが挙げられる。別の好ましい実施形態においてこの発現ベクターは、レトロウィルスから得られるプラスミドである。別の好ましい実施形態においてこの発現ベクターは、レンチウィルスから得られるプラスミドである。レトロウィルスと第二の非Adウィルス由来の配列を含むハイブリッドベクターもまた使用することができる。更に好ましい実施形態においてこの発現ベクターは、モロニーマウス白血病ウィルス(MMLV)から得られる、少なくとも7.5キロベースのクローニング能力を有するレトロウィルスベクターである。非ウィルス発現ベクターは、それらが宿主細胞ゲノム中への組込みを容易にする遺伝エレメントを含有する限り、代わりに使用することができる。
【0072】
E1Aおよび/またはE1Bポリペプチド産生用のコード配列を含む発現ベクターは、それらのゲノム中に安定的に組み込まれたE1Aおよび/またはE1Bコード配列を有する細胞を得るために、その特定のベクター/細胞株の組合せに適した方法を用いて大規模なアデノウィルスベクター生産に適した細胞株中に導入される。安定したトランスフェクションは、ゲノム中に安定的に組み込まれたトランスフェクトDNAを含有する娘細胞の集団からなる細胞株またはクローンの樹立によって実証される。
【0073】
細胞中へのウィルスまたは非ウィルスベクターの導入は、当業者により日常的に使用される標準的な技術を用いて行われる。本発明の発現ベクターはさらに、その細胞株に選択可能な形質を付与するものを含む非コードおよびコード配列を含有することができる。
【0074】
E1Aおよび/またはE1Bのコード配列を含む発現ベクターは、標準的なトランスフェクション法を用いて逐次的または同時に細胞中に導入する(前掲のSambrookの著書)か、あるいは最も好ましい実施形態では感染性ウィルス粒子にパッケージし、形質導入を介して細胞株中に導入することができる。容易に理解されるはずだが用語「導入した」は、その発現ベクターに適した形質導入およびトランスフェクションの方法を含む細胞中にDNA配列を送達するために使用される任意の方法(例えばDNAプラスミドに対する感染性粒子)を包含する。E1AおよびE1Bに対してそれぞれ別個の発現ベクターを用いてそれらを逐次的または同時に導入することにより、それぞれ別の場所においてパッケージング細胞ゲノム中でまとまるのでパッケージング細胞ゲノムとAdベクターの間の潜在的な組換え事象はさらに低減される。本発明はゲノム内で発現ベクターが空間的に離れているしている利点を提供し、RCAを生成させる、または組織特異的複製の喪失を生成させる恐れのある組換え事象の更なる低減を可能にする。
【0075】
本発明のパッケージング細胞株を用いて生産される複製能欠陥のあるおよび/または複製能のあるアデノウィルスベクターは、実質上RCAを含まない。実質上RCAを含まないとは、そのRCAの量が十分に低く、その結果、本発明のパッケージング細胞株を用いて生産されるアデノウィルスベクターのin vivo投与から何の毒性も生じないことを意味する。好ましくは、実質上RCAを含まないアデノウィルスベクター製剤は、患者線量当たりRCA粒子を約104個中約0個から約1個含有する。ここで一般的な患者線量はウィルス粒子約1012から約1013個である。しかしながら例として全ウィルス粒子1013個の患者線量は、組換えウィルス粒子を0から10、102、103、104、105、106、107、または5×107個含有することができ、またin vivo投与後に何の毒性も生じない限り実質上RCAを含まないとみなすことができる。
【0076】
細胞株
E1Aおよび/またはE1Bポリペプチド産生用のコード配列を含む発現ベクターを大規模なアデノウィルスベクターの生産に適した細胞株中に導入する。次いでこの細胞株を、プロモーターの誘発、形質転換細胞の選択、または所望の配列をコードする遺伝子の増幅に適するように変性した通常の栄養培地中で培養する。哺乳動物の宿主細胞は、様々な培地中で培養することができる。Ham’s F10(Sigma)、最少必須培地((MEM)、Sigma)、RPMI 1640(Sigma)、およびダルベッコ改変イーグル培地((DMEM)、Sigma)などの市販の培地が、これら宿主細胞を培養するのに適している。これら培地のいずれも必要に応じてホルモン類および/または他の成長因子(インスリン、トランスフェリン、または上皮増殖因子など)、塩類(塩化ナトリウム、カルシウム、マグネシウム、およびリン酸塩など)、緩衝液(HEPESなど)、ヌクレオシド類(アデノシンおよびチミジンなど)、抗生物質、微量元素、およびグルコースまたは同等のエネルギー源で補うことができる。任意の他の必要な補助剤もまた、当業者が知っているであろう適切な濃度で含まれることがある。温度、pHなどの培養条件は、発現のために選択される宿主細胞に関してさきに用いたものであり、当業者には明らかなはずである。本発明によるパッケージング細胞株の製造用に用いることができる宿主細胞の例には、限定しないが、A549、HeLa、MRC5、W138、CHO細胞、Vero細胞、ヒト胎児網膜細胞、またはそれら宿主細胞がアデノウィルスの成長を許容する限り任意の真核生物細胞が挙げられる。幾つかの好ましい宿主細胞株にはヒト腫瘍細胞株が挙げられる。好ましい実施形態においてこのパッケージング細胞株はPC−3細胞(ATCC番号CRL-1435)から得られる。これらPC−3細胞は転移性の前立腺細胞の腺癌から開始される。これらPC−3細胞は、無血清培地中での継代用に適応させることができるので臨床用Adベクターの大量生産には特に有利である。他の細胞型には、限定しないが、初代細胞培養物から得られる細胞、例えばヒト初代前立腺細胞、ヒト胎児網膜細胞、ヒト幹細胞が挙げられる。真核生物二倍体および異数体細胞株が本発明の範囲内に含まれる。
【0077】
好ましい細胞株は無血清培地に適応可能である。
【0078】
候補の細胞株は、当業界で周知の方法により、例えば感染していない細胞層あるいは1種または複数種のヘルパーウィルスに感染した細胞層をウィルス粒子と接触させ、続いてそれら細胞をインキュベートすることによりアデノウィルスの複製を支持する能力を試験することができる。ウィルスのプラークまたは細胞層中の無細胞領域の形成は、ある種のウィルス産物の発現によって引き起こされる細胞溶解の結果である。細胞溶解はウィルス複製を示唆している。
【0079】
アデノウィルス血清型
本発明によるパッケージング細胞株は、任意の既知のアデノウィルス血清型から得られる臨床用Adベクター、および2種類以上の血清型から得られる配列からなるキメラアデノウィルスの大量生産に役立つ。本発明は、すべてのアデノウィルス血清型由来のAdベクターの使用を意図している。アデノウィルス血清型1から47はAmerican Type Culture Collection(ATCC, Manassas, VA)から現在入手可能であり、また本発明は本発明のこのパッケージング細胞株がそのウィルスを複製する能力がある限り、任意の供給源から入手できる任意の他の血清型のアデノウィルスの生産に有用である。本発明によるパッケージング株を用いて生産することができるアデノウィルスは、ヒトまたは非ヒト起源のものであることができる。例えばアデノウィルスは、亜型A(例えば血清型12、18、31)、亜型B(例えば血清型3、7、11、14、16、21、34、35)、亜型C(例えば血清型1、2、5、6)、亜型D(例えば血清型8、9、10、13、15、17、19、20、22〜30、32、33、36〜39、42〜47)、亜型E(血清型4)、亜型F(血清型40、41)、または任意の他のアデノウィルス血清型のものであってもよい。
【0080】
本発明のパッケージング株を用いて生産される組換えアデノウィルスは、E1Aおよび/またはE1Bコード領域中に、欠損に加えて欠失または他の遺伝子変異を含むことができる。本発明に有用な組換えアデノウィルスは、任意選択で他の遺伝子変化、例えば導入遺伝子の封入などに耐えることができる。
【0081】
一実施形態においてこのパッケージング細胞株は、アデノウィルス複製およびアデノウィルス感染の両方を許容する。アデノウィルス血清型2および5の初代受容体は、Bergelson等(1997)(Science 275: 1320)によって同定され、CAR(コクサッキーウィルスおよびアデノウィルス受容体、GenBank取得番号HSU90716)と名付けられており、またRoelvink等(1998)によって亜型Bを除くすべてのアデノウィルス亜型に対する受容体であることが示されている。
【0082】
別の実施形態では本発明に基づくパッケージング株は、その粒子のキャプシドタンパク質、例えばその繊維状タンパク質のHIループまたはカルボキシル末端(C末端)中に、またはタンパク質IX中にリガンドまたは一本鎖抗体を含む修飾繊維状タンパク質などの中に含まれた標的リガンドを含むアデノウィルスベクターの生産に有用である。標的リガンドを含むアデノウィルスベクターは、例えば国際公開第WO00/67576号、第WO99/39734号、米国特許第6,683,170号、第6,555,368号、第5,922,315号、第5,543,328号、および第5,846,782号に記載されている。さらに別の実施形態において本発明に基づくパッケージング株は、米国特許出願第10/403,337号、国際公開第WO98/07877号、第WO01/92299号、米国特許第5,962,311号、第6,153,435号、第6,455,314号、第5,731,190号、第6,057,155号、第5,543,328号、第5,756,086号、第6,127,525号、第5,922,315号、およびWu等の論文(J. Virol. 2003 Jul 1; 77 (13): 7225-7235)中に例証されているものなど、繊維状タンパク質の他の突然変異を含むアデノウィルスベクターの生産に有用である。
【0083】
医薬組成物
本発明のパッケージング株を用いて生産されるアデノウィルスベクターは、例えば臨床用途に用いるために調合することができる。この溶離液を任意選択で濃縮し、また通常の方法により、例えば中空繊維濃縮器を用いてダイアフィルター濾過する。使用される最終製剤においてはそのウィルス試料を滅菌濾過することができる。例えばニトロセルロース膜フィルター、酢酸セルロース膜フィルター、PVDF(修飾ポリビニリデンフルオリド)膜フィルターなど、この目的に適した様々なフィルターが当業界で知られている。PVDF膜フィルター(例えばMillipore Millipakフィルター)が好ましい。
【0084】
滅菌濾過したウィルス懸濁液はin vitroまたはin vivoで使用するために調合される。これらの水性組成物は、医薬として許容される担体または水性媒体中に懸濁させた有効量のウィルスを含む。またこのような組成物は接種源と呼ぶこともできる。語句「医薬としてまたは薬理学的に許容できる」は、動物またはヒトに適宜投与した場合に有毒な、アレルギー性の、または他の厄介な反応を示さない分子の独立体および組成物を指す。本明細書中で用いられる「医薬として許容される担体」には、ありとあらゆる溶剤、分散媒体、剤皮、抗菌および抗カビ剤、等張化および吸収遅延剤などが含まれる。医薬として活性な物質に対してこのような媒体または薬品を使用することは当業界でよく知られている。任意の通常の媒体または薬品がその活性成分に適合しない場合を除いては治療用組成物中でそれを使用することが考えられる。補助的な活性成分もまた、この組成物に組み込むことができる。
【0085】
これら調合物には液状溶液または懸濁液として注射できる組成物が含まれ、また注射の前に液体状態の溶液または懸濁液にするのに適した固体形態もまた調製することができる。これらの製剤はまた乳化することもできる。このような目的の一般的な組成物は、医薬として許容される担体を含む。例えばこの組成物は、リン酸緩衝生理的食塩水1ミリリットル当たりヒト血清アルブミンを約100 mg含有することができる。他の医薬として許容される担体には塩類、防腐剤、緩衝剤などを含む水溶液、非毒性賦形剤が挙げられ、使用することができる。非水性溶媒の例は、プロピレングリコール、ポリエチレングリコール、植物油、およびオレイン酸エチルなどの注射可能な有機エステル類である。水性担体には、水、アルコール/水溶液、生理的食塩水、塩化ナトリウムなどの非経口伝達体、リンガーデキストロースなどが挙げられる。静脈内伝達体には流体と栄養の補充物が挙げられる。防腐剤には、抗菌物質、抗酸化剤、キレート化剤、および不活性ガスが挙げられる。医薬組成物中の様々な成分のpHおよび正確な濃度は、よく知られている要因に従って調整される。
【0086】
これら調合物は、所望の保管条件に対して最適化することができる。本発明の1つの態様において、特に臨床的使用のために処方されるウィルスの場合、それら試料は液体の形態で、好ましくは一般には約10℃未満、より一般には約5℃未満の冷温で保存される。このような条件の場合、保管にとって好ましい媒体は5%スクロース、1%グリシン、1 mM MgCl2、10 mMトリス、および少量の界面活性剤を含む。注目の界面活性剤の一つは非イオン界面活性剤、例えば約0.01%から約0.1%、好ましくは約0.05%の濃度のトゥイーン80、トゥイーン20などである。注目の他の界面活性剤には、例えば、濃度約5%から約10%、好ましくは約8%のLutrol F−68、Lutrol F−127などのポリエチレングリコールポリプロピレングリコールのポロキサマーブロックポリマーが挙げられる。
【0087】
例えば−20℃または−80℃で冷凍保存される試料の場合、好適な緩衝液は上記のとおりであるが、界面活性剤を含むことは安定性にとって一般にはあまり重要ではなく、省くことができる。約2%から約10%の濃度でグリセロールを含めることができる。
【0088】
本発明のこのウィルス粒子には、ヒトへの投与を含む治療養生法で用いられる古典的な医薬用製剤を挙げることができる。本発明による治療用組成物の投与は、その標的組織がそのルートを経て利用できる限り任意の普通のルートを経ることになる。これには、経口、経鼻、口内、直腸、膣、または局所的が含まれる。別法では投与は、通常位置の、皮内、皮下、筋内、腹膜内、または静脈内の注射によることになる。このような組成物は、生理学的に許容できる担体、緩衝液、または他の賦形剤を含む医薬として許容される組成物として普通は投与されることになる。腫瘍に対して適用する場合、直接の腫瘍内注射、切除後の腫瘍床への注射、局所(すなわちリンパ管)、または全身投与が考えられる。また疾患部位、例えば腫瘍または腫瘍部位へのカテーテルを介しての数時間または数日にわたる連続的灌流を行うことが望ましい場合もある。
【0089】
このアデノウィルスベクターの有効量を、限定しないが、生理的食塩水、適切な緩衝液、防腐剤、安定剤を含む医薬として許容される賦形剤中の組成物(なお同一組成物であってもなくてもよい)として患者に投与することができ、また制吐剤などの適切な薬剤と一緒に投与することもできる。有効量とは、臨床結果を含む有益なまたは望ましい結果をもたらすのに十分な量である。本発明の目的の場合、アデノウィルスベクターの有効量は、病状の進行を和らげ、改善し、安定化し、逆転し、速度を遅らせ、または先に延ばすのに十分な量である。一部の個体はこれらの治療に対して無反応性であり、その方法はこれら個体への投与を包含するものであることが分かる。与えられる量は、その個体の状態、病の程度、投与のルート、どれほどの用量が投与されるか、また所望の目標によって決まるはずである。
【0090】
治療薬の有効量は意図する目的、例えば(i)腫瘍細胞増殖の抑制、(ii)腫瘍細胞を排除または殺すこと、(iii)予防接種などに基づいて決められる。用語「単位用量」は、患者に使用するのに適した物理的に分離した単位を指し、各単位はその投与、すなわち適切なルートおよび治療養生法と共同して、上記で考察した所望の反応を生むように計算される所定量の治療用組成物を含有する。治療の回数および単位用量に応じた投与すべき量は、治療される患者、患者の状況、および希望する結果に左右される。
【0091】
或る特定の治療養生法の有効性の判定は、画像技術、血清腫瘍マーカーの分析、生検、および/または腫瘍に随伴する徴候の有無または改善の評価などの診断法を含めた当業界で知られている技術のいずれかにより求めることができる。或る所定の治療養生法は、有効性をできるだけ大きくするために適宜改変することができることは理解されるはずである。
【0092】
下記の実施例は、本発明をいかに構成し使用するかについての完全な開示および説明を当業者に提供するために述べられ、本発明者等がこの発明と考えるものの範囲を限定することを意図するものではなく、また下記の実験が行った実験の全体または唯一のものであることを意味することも意図しない。使用した数値(例えば量、温度など)に関して精度を保証する努力を行ったが、若干の実験誤差と偏差は斟酌されるべきである。別段の指示がない限り、部は重量部であり、分子量は重量平均分子量であり、温度は摂氏度であり、また圧力は大気圧またはほぼ大気圧である。
【0093】
本発明は、本発明者によって見出されまたは提案された特定の実施形態に関して記述されており、本発明の実施のための好ましい形態を含む。本開示内容に照らして本発明の意図する範囲から逸脱することなく例示の特定の実施形態について非常に多くの修正形態および変更形態を案出することができることは、当業者には理解されるはずである。例えばコドン重複性によりタンパク質配列に影響することなくその基礎をなすDNA中の変更を行うことができる。さらに生物学的な機能上の等価を考慮することにより種類または量的な生物学的活動に影響することなくタンパク質構造中の変更を行うことができる。あらゆるこのような修正形態は、添付の特許請求の範囲の範囲内に含まれるものである。
【実験】
【0094】
(実施例1)
E1A/E1Bパッケージング細胞株の構築:
MMLV発現ベクターの構築。モロニーマウス白血病ウィルス(MMLV)由来プラスミドをE1Aレトロウィルス発現ベクターおよびE1B レトロウィルス発現ベクターを構築するために使用した。具体的にはpRT43.2F3プラスミドが使用された。pRT43.2F3の構築については下記に要約し、完全には米国特許第5,686,279号に記載されている。
【0095】
pRT43.2F3。このレトロウィルスベクターは、レトロウィルスが産生されることになる細胞型中での効率的な転写を指示する修飾5′LTRを含有する。本発明のレトロウィルスベクターは、pZIPneoSVX(Cepko等の論文、Cell 37: 1053-1062 (1985))の新バージョンであるpZen(Johnson等の論文、EMBO Journal 8 (2): 441-448 (1989))を基にして作られ、発現されることになる遺伝子産物は、普通はその新カセット(前掲のCepko等の論文)が占める場所であるスプライス受容部の下流でクローニングされる。さらにMMLVのNar I部位までのウィルスのgag配列(ヌクレオチド1038)をパッケージングの改良のために加え(Armentano等の論文、J. Virol. 61: 11647-1650 (1987))、pZIPneoSVXのXho IからCla Iまでのフラグメントを欠失させた(前掲のCepko等の論文)。pIK1.1由来のEco RIからApa Iのポリリンカーをスプライス受容部の下流に挿入し、pIKプラスミド由来の挿入断片のレトロウィルス構築物中へのトランスフェクションを可能にした。得られたプラスミドはpRTD1.2と呼ばれ、5′および3′MMLV LTRの両方を含有する。このpZIPneoSVXの5′LTR U3領域をpIKMMSVのHindIII/Sac Iフラグメントから得られるMMSV U3で置き換えてpRTD4.2を生じさせた。
【0096】
pRTD2.2においてpZIPneoSVXの5′LTRのU3領域を、CMV即時型エンハンサー/プロモーターをコードするpIK1.1由来のHindIII/Sac Iフラグメントで置き換え、これをMMLVのヌクレオチド+1から+32(KpnI)と結合したHCMVプロモーターのヌクレオチド19(Sac I)から+1をコードするオリゴヌクレオチドによりMMLV R領域に融合させた(Schinnick等の論文、Nature 293: 543-548 (1980))。
【0097】
pRTD2.2の(750 bp)Sac IからBst EIIのフラグメントをLXSNの(736 bp)Sac IからBst EIIのフラグメントで置き換えることによってpRTD2.2SVGを構築した(MillerおよびRosmanの論文、BioTechniques 7: 980-990 (1989))。pRTD2.2の(1441 bp)Sac IからEco RIのフラグメントをLXSNの(1053 bp)Sac IからEco RIのフラグメントで置き換えることによってpRTD2.2SSAを構築した(前掲のMillerおよびRosmanの論文)。その5′末端へのApa I部位の付加反応によって付加されたpLXSN(GenBank取得番号M28248)のヌクレオチド2878〜2955をコードするオリゴヌクレオチドの合成によってpRTD2.2SVGE−を構築した。これを用いてポリリンカーのDNA配列3′および3′LTR中のNhe I部位の5′を含有するpRTD2.2 SVG のApa IからNhe Iのフラグメントを置き換えた。本発明のこれらレトロウィルスベクター構築物は、pBR322バックボーンを有し、かつpRTD2.2、pRTD4.2、pRTD2.2SVG、pRTD2.2SVGE−、およびpRTD2.2SSAを含む。
【0098】
SV40 T抗原を発現する細胞中でのプラスミドの複製を可能にするために、pRTD2.2の5′と3′LTRの間の配列を上記pIK1.1のSac IとEco RI部位の間でクローニングした。これはベクターpIKT2.2を形成するようにSV40の複製起点を含有した。pRTD2.2SVGのHCMVプロモーター中のSac I部位によってその5′末端で画定され、かつpIK1.1のSac IとEco RI部位の間のpRTD2.2SVGの3′LTRの750 bp下流に位置するEco RI部位によりその3′末端で画定されるフラグメントの挿入によってpIKT2.2SVGを構築した。pIKT2.2SVGF3の182塩基対のApa IからNhe Iのフラグメントを上記pRTD2.2SVGE−F3由来の80塩基対のApa IからNhe Iのフラグメントで置き換えることによってpIKT2.2SVGE−F3を構築した。
【0099】
3′LTRから約750塩基対下流に位置するEco RIからApa Iのポリリンカーを、Asc I認識部位を含有する合成オリゴヌクレオチドで置き換えることによってpIKT2.2SVGE−F3からpRT43.2F3を誘導した。さらにウィルスのgag配列の3′末端にあるNde I部位をオリゴヌクレオチド挿入によってXhoIに変換した。最初にpRT43.2F3中の3′LTRの除去と、PvuIIからXbaIまでの配列が欠失した3′LTRの挿入とによってpRT43.2F3からpRT43.3PGKF3を誘導した(MMLV、GenBank取得番号#J02255、ヌクレオチド番号7938〜8115)。さらにMMLVスプライス受容部領域をヒトホスホグリセリン酸キナーゼ遺伝子プロモーター(GenBank取得番号#M11958、ヌクレオチド2〜516)で置き換え、それをクローニングしてその5′末端にXho I部位とその3′末端にEco RI部位とを有するポリリンカーにした。
【0100】
E1A発現ベクター−rkat43.2E1A。rkat 43.2E1a(図1)は、レトロウィルスLTRの制御下でそのAd5 E1Aオープンリーディングフレームを発現するレトロウィルスベクターである。したがってアデノウィルスまたは哺乳動物の宿主細胞由来のプロモーターのどちらもAd E1Aの発現を方向づけるのには使用されない。rkat 43.2E1Aは、pRT43.2F3のCD4/ζコード配列(米国特許第5,686,279号およびRoberts等の論文、J. Immunology (1998) 161: 375-384)を、Ad5 E1Aオープンリーディングフレーム(Ad5ヌクレオチド548〜1575、GenBank取得X02992、配列番号1)をコードするDNA配列で置き換えることによって生じさせた。
【0101】
E1B発現ベクター−rkat 43.2E1B。rkat 43.2E1B′(図2)を pRT43.2F3から同様に生じさせた。このベクターは、レトロウィルスLTRの制御下でAd5 E1Bオープンリーディングフレーム(Ad5のヌクレオチド1682〜3825、GenBank取得X02996、配列番号4)を発現させ、アデノウィルスまたは哺乳動物の宿主細胞由来のプロモーターを含まない。rkat 43.2E1Aは、pRT43.2F3のCD4/ζコード配列(米国特許第5,686,279号および前掲のRoberts等の論文)を、Ad5 E1B mRNAをコードするcDNA配列で置き換えることによって生じさせた。
【0102】
一過性のレトロウィルス生産。E1AおよびE1B発現ベクターを含む感染性粒子を標準的な方法を用いて生産した。rkat 43.2E1Aまたはrkat 43.2E1BプラスミドにMCVecog/pおよび6.1CMVamphoenvを同時感染させることによって一過性のウィルス上澄みを調製した。次いで得られたウィルス上澄みMMLV−E1A (C5.03−.04と呼ぶ)およびMMLV−E1B(C5.05−.06と呼ぶ)を細胞の形質導入に使用した。
【0103】
A549細胞の形質導入。ナイーブA549細胞(ATCC番号CCL−185)をDMEM/High、10%ウシ胎児血清、1%グルタミン、および1%PenStrepを含む完全培地中で培養した。細胞にMMLV−E1AおよびMMLV E1Bウィルスをスピンオキュレーションにより同時感染させることによってアデノウィルスE1AおよびE1Bコード配列をA549細胞中に安定的に導入した。1.5×105個の細胞をE1A/E1Bウィルス上澄み1 mLおよびポリブレン8 μL/mL中に再度懸濁させた。次いでこの細胞およびウィルスの混合物を3400 rpm、34℃において4時間遠心分離した。最適なE1A/E1B比を確実にするために3つの異なるE1A/E1B比(E1A 25%/E1B 75%、E1A 50%/E1B 50%、E1A 75%/E1B 25%)をスピンオキュレーションに用いた。ウィルスの形質導入効率をモニターするための対照としてMMLV−グリーン蛍光タンパク質(GFP)ウィルスを含めた。スピンオキュレーションの後、これら3集団を完全培地で再度懸濁させ、6ウェルプレートに移し、5%インキュベーター中で37℃において8日間インキュベートした。
【0104】
E1A/E1B形質導入細胞の希釈クローニング。これら3集団を10 cm皿上で希釈クローニングした。培養状態で18日後、これら3集団の間にはっきりと違いが現れた。3集団由来のすべてのクローンを採取して96ウェルプレートに入れた。次いで同定した後に各プレートをトリプシン処理し、一方を機能的スクリーニング用に、また他方を機能クローンの修復用に使用することができるように2つの96ウェルプレートに分けた。細胞を5日間生育して増殖させた。
【0105】
E1補完に関する機能的スクリーニング。GFP導入遺伝子を担持するE1欠損組換えアデノウィルスを用いてアデノウィルス複製を支持する能力についてE1A/E1B形質導入クローンのそれぞれを試験した。候補クローンを含有する二倍にした96ウェルプレートのうちの一組にE1欠損Ad−GFPウィルスを100 μL/ウェル、M.O.I. 10で48時間感染させた。これらの細胞を3回の冷凍/解凍サイクルにかけてウィルス粒子を放出させた。これらのライゼートを用いてHuH7細胞に感染させた。感染3日後にHuH7細胞を採取し、FACSにより分析してGFPの発現を評価した。表2は、GFPの発現に関する複数のクローンの機能的特徴の結果を示す。この機能的特徴は、候補クローン由来のアデノGFP上澄みによるHuH7細胞の感染および最初のスクリーニングとしての増幅の評価に基づく。その上澄みがHuH7細胞中で高いGFP発現を示したクローンを、候補のE1A/E1B陽性パッケージングクローンとみなした。HuH7細胞を最大限度まで形質導入したこれらのクローンを、下記でさらに述べるように増殖させ、プラーク検定で力価測定した。
【0106】
【表2】
【0107】
E1A 50%/E1B 50%のE1A/E1Bウィルス比のスピンオキュレーション後に得られる細胞集団は、E1 Ad−GFPに感染させる前に生き残っている「健康な」クローン(形態によって決められる)の数が最も少なかったが、E1欠損ウィルスを補完することができるクローン産生の比率はより高かった。E1A 75%/E1B 25%の形質導入比から生まれるクローンは最もすぐれた全体的な成長特性を示したが、E1A 50%−E1B 50%クローンのE1欠損ウィルスで観察される同じ補完性をもたらさなかった。
【0108】
E1欠失および腫瘍溶解性アデノウィルスを補完する能力に関するクローンの評価。E1Aを欠失したGFP発現性の複製能欠損アデノウィルスに感染させることによりE1A/E1B補完的細胞を候補パッケージング細胞として評価した(E1A/E1B補完的細胞、親細胞、およびE1A/E1B陰性細胞を比較するウィルス収量検定を使用)。各クローンによるウィルス産生を比較することによってこれらのE1A/E1B補完的細胞をウィルス複製の支持能力に関して定量的に評価した。試験クローンにE1Aを欠失したGFP発現性の複製能欠損アデノウィルスをM.O.I. 5で4時間感染させ、新鮮な培養液を再度与え、72時間インキュベートした。細胞および培地を一緒に採取し、3回の冷凍/解凍にかけた。血清を含まない培地中で連続的な希釈を行い、293細胞上で検定した。それら細胞は、その24時間前に6ウェルプレート上に1ウェル当たり培養液4 mL当たり0.5×106個の細胞でプレーティングされた。これら試料を吸引し、各ウェルにアガロース(完全培地中0.8%アガロース)4 mLを覆いとして適用した。これらプレートを室温で放置して固化させ、プラークを展開させるためにインキュベートした。表3は、プラーク検定により得られた力価を示す。次いでその最もすぐれたクローンを、複製能のある(腫瘍溶解性)アデノウィルスベクターAFP−CG8900を用いた更なるプラーク検定で分析した(図2B)。これらの結果に基づいてそれぞれE1A/E1Bクローン51およびクローン139と呼ばれる2種類のクローンの特徴をさらに調べた。
【0109】
【表3】
【0110】
E1欠失アデノウィルスの産生ならびにE1クローン51および139の安定性。293細胞、PerC6細胞、ならびにE1A/E1Bクローン51および139から得られた細胞を成長させ、M.O.I.=75でE1欠失アデノGMCSFを感染させた。72時間後、細胞を採取した。ウィルスの粗ライゼートを3回の冷凍/解凍によって調製し、ウィルスの力価をHPLCにより求めた。感染後、293、PerC6、E1A/E1Bクローン51、およびクローン139細胞に週2回の継代を行った。Ad−GMの産生を継代1、10、および20世代(1、5、10週)で試験した。結果は、クローン51および139がPerC6および293細胞に匹敵する水準でE1欠損ウィルスを産生することができ、またクローン51およびクローン139細胞株によるウィルスの産生が安定していることを示した(図3)。
【0111】
E1A/E1B補完クローンから得られた細胞株を用いた腫瘍溶解性ウィルスの産生。A549細胞、293細胞、ならびにE1A/E1Bクローン51および139から得られた細胞を成長させ、4種類の複製能のある(腫瘍溶解性)アデノウィルス(CG8900、CG8840、OV945、およびOV1025)をM.O.I. 2で3〜4時間感染させた。72時間後、上澄みを採取し、これを用いて標準的なプラーク検定により293細胞に様々な希釈度で感染させた。感染を4時間進行するに任せ、次いでこの細胞をアガロース培地中で8〜11日間培養した。結果を表4に示す。CG8900、CG8840、OV945、およびOV1025は、E1AおよびE1Bを制御する細胞型特異的転写調節エレメントを含む複製能のあるアデノウィルスベクターである。
【0112】
【表4】
【0113】
初期および後期継代細胞中のE1A/E1B配列に関するクローン51および139のサザンブロット分析。ベクター内で一旦切断するBamH Iでクローン51、クローン139、および293細胞の初期および後期継代培養物を消化し、E1A(nts 827〜1340)またはE1B(nts 2805〜3329)特異的プローブを用いてサザンブロットにより分析した(図4AおよびB)。これらのブロットは、初期および後期の両継代細胞中へのE1AおよびE1B遺伝子の安定した組込みを示す。クローン139は、E1A遺伝子の2つのコピーとE1Bの1つのコピーを有するように見える。クローン51は、E1Aの4つのコピー(上側のバンドはダブレットである)とE1Bの2つのコピーを有する。星印は関連のあるバンドを示す。
【0114】
パッケージング細胞株中のゲノム組込みの特徴の記述。E1AおよびE1B遺伝子の同時形質導入に利用される別個の発現ベクターは、宿主細胞のゲノム中で異なる場所に組み込まれるべきであり、これは複製能欠損Adベクターと本発明のパッケージング細胞株の間の組換えの可能性をさらに低減させるように働く。クローン51と139、293、およびPerC6細胞由来のDNAを、E1A、E1B、および連続E1A−E1B配列に特効のあるプライマーを用いてPCRによりプローブした(図5)。プライマー1460.138.3/4(1460.138.3: 5′TGT GTC TAG AGA ATG CAA TAG 3′、1460.138.4: 5′GAT ATA TGT CGA CTG GCC TGG GGC GTT TAC AGC 3′)が、E1Aコード領域のc末端でAd5ゲノムのnts 1338から1542を増幅し、プライマー1460.138.5/6(1460.138.5: 5′GAC ATG CGT CGA CAT GGA GCG AAG AAA CCC ATC TG 3′、1460.138.6: 5′CCA TAG AAG CTT ACA CCG TGT AG 3′)が、nts 2019から2815(E1B 55kオープンリーディングフレームの大部分)を増幅する。すべての4つの細胞株から得られたサザンブロットの結果は、予想された個々のE1AおよびE1Bフラグメントが存在することを示した。E1Aのc末端からE1B 55kまでのAd5ゲノムが無傷であり、かつ293およびPerC6細胞株の場合と同様に連続的である場合には、1460.138.3/6はnts 1338から2815までの1477塩基対領域を増幅することになる。293およびPerC6細胞株由来のDNAのサザンブロットの結果は、1477塩基対領域が存在することを示した。一方、クローン51および139由来のDNAのサザンブロットの結果は、連続したE1A/E1Bゲノムがクローン中に存在しないことを裏付ける1477塩基対領域を示さなかった。
【0115】
クローン51および139のタンパク質分析。当業界で広く使用されている方法(例えばAntonおよびGrahamの論文、J. Virology, 69, 4600-4606, 1995、ならびに前掲のSambrookおよびRussellの著書)を用いてウェスタンブロット分析を行った。293、PerC6、A549、A549クローン51、およびA549クローン139細胞を10 cm組織培養皿にプレーティングした。72時間後、細胞を擦って上澄み中に入れ、ペレット化し、完全ミニプロテアーゼ阻害薬カクテル(Roche)を追加した溶解緩衝液(100 mM NaCl、20 mMトリス(pH 7.5)、10 mM EDTA、1%デオキシコール酸)中に再度懸濁させた。試料のタンパク質濃度をタンパク質検定キット(Bio-Rad)で推定した。E1Aの検出(図6A)の場合、各試料について合計タンパク質10 mgを4〜12%NuPage Novex Bis-Yris SDS-PAGEゲル(Invitrogen)上に載せ、NuPage MOPSランニング緩衝液中で分別した。画分をInvitrolon PVDF膜(Invitrogen)に移し、モノクローナルE1A一次抗体(Neomarkers)および西洋ワサビペルオキシダーゼ複合二次抗体によりプローブした。結合した抗体複合体を高感度化学発光(Amersham)により検出した。E1B 19KおよびE1B 55Kの検出は、下記の点を除いて上記と同様に行った(図6B)。各試料から得られた全タンパク質25 mgを分別した。使用した一次抗体は、E1B 19KおよびE1B 55K に対するモノクローナル抗体である(Oncogene Research Products)。これらの結果は、クローン51および139が本質的にPerC6と同等のE1A(38および46 kD)発現レベルであり、また293細胞について検出されるものよりも高いE1A発現レベルを有することを示した。これら2つのクローンにおけるE1B 19Kの産生もまた、293およびPerC6細胞で検出されるレベルと同等である。
【0116】
RCA検出検定。クローン51および139細胞を、従来からアデノウィルスの大量生産用に使用されている細胞株の293およびPerC6細胞と比較するためにRCAの検討を行った(Gao等の論文(2000)、Hum Gene Therapy 11: 213、Murakami等の論文(2002)、Hum Gene Thera.13: 909、Kim等の論文(2001)、Exp Mol Med 33: 145)。各細胞型を使用し、精製したE1A欠失Ad GM−CSFウィルスストックを20世代まで継代させた。クローン51、クローン139、および293細胞にM.O.I.=75でE1欠失GM−CSFウィルスを感染させた。72時間後、粗ライゼートを調製し、HPLCにより滴定し、これを同じ型のナイーブ細胞への第二ラウンドの感染(ナイーブ293細胞への293ライゼートなど)に用いた。各感染については同数の粒子を用いた。この感染サイクルは18回繰り返した。感染サイクル1、9、および18回においてアデノウィルスDNAをライゼートから精製し、E1A(nts 133〜696)領域に特異的なプライマーを用いてPCRにより増幅した(図7)。その予想されたPCR生成物は約0.56 Kbである。このRCA検定用に用いたPCRプライマーは、66.114.2: 5′−GTGGCGGAACACATGTAAGCG−3′および49.17.2: 5′−AGTTCGTGAAGGGTAGGTGGTTC−3′であった。E1A配列(RCAに起因)は、9サイクルまでの293細胞由来の継代中に検出された。クローン51および139から得られるアデノウィルスは、18サイクルを通してE1A配列に関して陰性であった。
【0117】
本発明は、例示した組換え発現ベクターおよび細胞株による範囲内に限定されるべきではなく、これらは本発明の一態様の実例として意図されている。上記の詳細な実施例および上述の実施形態は例示を意図しており、本発明の範囲を限定するものではないことを理解されたい。本発明の範囲内の他の態様、利点、および修正形態は、本発明が属する業界の技術者には明らかなはずである。
【図面の簡単な説明】
【0118】
【図1】アデノウィルスE1AおよびE1Bプロモーター配列を含まないMMLV−E1Aレトロウィルス発現カセットを示す図である。
【図2】アデノウィルスE1AおよびE1Bプロモーター配列を含まないMMLV−E1Bレトロウィルス発現カセットを示す図である。
【図3】293、PerC6、クローン51、およびクローン139細胞上でGM−CSFを発現する複製能欠陥アデノウィルスの産生を示す図である。
【図4A】クローン51および139から得られる初期および後期継代細胞のE1A(図4A)およびE1B(図4B)配列に関するサザンブロット分析の結果を示す図である(星印は関連のあるバンドを示す)。
【図4B】クローン51および139から得られる初期および後期継代細胞のE1A(図4A)およびE1B(図4B)配列に関するサザンブロット分析の結果を示す図である(星印は関連のあるバンドを示す)。
【図5】E1A(1460.138.3/4)、E1B(1460.138.5/6)、および連続E1A−E1B配列(1460.138.3/6)に特異的なPCRプラマーを用いてプローブしたクローン51および139、PerC6、293、およびA549細胞から得られるDNAのサザンブロット分析の結果を示す図である。
【図6A】E1A、E1B 19K、またはE1B 55kに向けられるモノクローナル抗体でプローブしたA549細胞、クローン51細胞、クローン139、PerC6細胞、および293細胞のライゼートを用いて行ったウェスタンブロット分析の結果を示す図である。
【図6B】E1A、E1B 19K、またはE1B 55kに向けられるモノクローナル抗体でプローブしたA549細胞、クローン51細胞、クローン139、PerC6細胞、および293細胞のライゼートを用いて行ったウェスタンブロット分析の結果を示す図である。
【図7】野生型アデノウィルス(wt Ad)、293細胞、クローン51、およびクローン139のライゼートから精製されたDNAをE1Aに特異的なプライマー(nts 133〜696)を用いてPCRにより増幅したRCA検出検定の結果を示す図である。
【図8】E1AおよびE1Bプロモーター配列を含まず、かつ5′から3′方向へ5′ITR、サイトメガロウィルスエンハンサー/ニワトリβアクチン/ウサギβグロブリンプロモーター(CAGプロモーター;Niwa等の論文(1991)、Gene 108 (2): 193-9)、E1Aのエキソン1およびエキソン2(配列番号1)、ウッドチャック転写後調節エレメント(WPRE)、子ウシ成長ホルモンポリA(BGHpA)配列、および3′ITRを含むAAV−E1A発現カセットを示す図である。
【図9】E1AおよびE1Bプロモーター配列を含まず、かつ5′から3′方向へ5′ITR、延長因子1−αプロモーター(EF1−α)のプロモーター(Kim等の論文(1990)、Gene 91 (2): 217-23およびGuo等の論文(1996)、Gene Ther. 3 (9): 802-10)とエンハンサー、19K E1Bコード配列と55K E1Bコード配列(配列番号4)、子ウシ成長ホルモンポリA(BGHpA)配列、および3′ITRを含むAAV−E1B発現カセットを示す図である。
【技術分野】
【0001】
本願は2003年7月3日出願の米国特許出願第10/613,106号に対し優先権を主張するものであり、この内容は引用によりその全体が本明細書に組み入れられる。
【0002】
本発明は、組換えアデノウィルスベクター、例えば複製能のあるアデノウィルスベクター、腫瘍溶解性(oncolytic)アデノウィルスベクター、およびE1初期遺伝子領域の欠失を有する複製能欠陥アデノウィルスベクターの産生に役立つ新規なパッケージング細胞株に関する。
【背景技術】
【0003】
ベクターが介在する導入遺伝子の送達は、宿主中に欠けているかまたは不十分な量で存在するタンパク質または他の物質を補うことで遺伝病の治療に利用される。アデノウィルス(Ad)ベクターは、導入遺伝子送達にとってきわめて効率的な伝達体である。アデノウィルスベースの遺伝子導入ベクターは、細胞中への遺伝子導入にとってそれらを特に有用ならしめている複数の特徴を有しており、例えば、アデノウィルスの生態の特徴がよく調べられていること、アデノウィルスは周知のどのヒトの疾患とも関連性がないこと、アデノウィルスはそのDNAを宿主細胞中へ導入するのに効率的であること、このウィルスは広い宿主細胞分布域を有すること、および大量生産が達成されていること、である。少なくともE1領域が欠失し、注目の遺伝子によって置き換えられているヒトアデノウィルスベースのベクターは、遺伝子治療に広範囲に用いられてきた。遺伝子治療に現在用いられているアデノウィルスベクターは一般に複製する能力がなく、E1領域に欠失を有する。
【0004】
組換えアデノウィルスを潜在的に強力な遺伝子送達ベクターにならしめている特徴については広範囲に検討されてきた(Berkerの論文、Biotechniques 6: 616-629 (1988)ならびにKozarskyおよびWilsonの論文、Curr. Opin. Genet. Dev. 3: 499-503 (1993))。遺伝子の欠失を通じてであろうと、特定の組織に限定された複製を通じてであろうとアデノウィルスベクターの複製の制御は、アデノウィルスと関係するin vivo用途にとって特に重要である。
【0005】
複製性アデノウィルスは、in vivoでの選択的ターゲッティングおよび増幅を達成するように作り変えられてきた。条件付きで複製性且つ腫瘍溶解性のアデノウィルスは、癌の治療において大きな将来性を示している(Yu等の論文、Curr. Opin. Mol. Ther. 2002, Oct; 4 (5): 435-43、Bell等の論文、Curr. Gene Ther. 2002 May 2 (2): 243-54、Yoon等の論文、Curr. Cancer Drug Targets 2002 Aug; 1 (2): 85-107)。これら複製性アデノウィルスは、全身的に送達することができ、腫瘍細胞をターゲティングすることができ、また腫瘍特異的なやり方でそれらの細胞溶解効果を増幅することができ、それによって実質的な臨床的利益をもたらすことができる。Henderson他の米国特許第5,698,443号、Hallenbeck他の国際公開第WO96/17053号を参照されたい。このような系では細胞特異的転写調節エレメントがウィルス複製にとって不可欠の遺伝子の発現を制御しており、したがってウィルスの転写はそのエレメントが機能しうる細胞集団に限られる。例えば弱毒化した複製能のあるアデノウィルスは、そのウィルスがin vitroおよびin vivoで前立腺特異的抗原(PSA)発現細胞に対して選択的細胞毒性を示すアデノウィルス血清型5(Ad5)中のE1A転写単位の上流にPSAプロモーターおよびエンハンサー(PSE-TRE)を挿入することによって生成させてきた(Rodriguez等の論文、1997年Cancer Res. 57: 2559-2563)。
【0006】
腫瘍溶解性アデノウィルス、条件付きで複製性のアデノウィルス、および複製能欠陥アデノウィルスを含めた注目のアデノウィルスは、そのウィルスゲノムのE1初期遺伝子領域(遺伝地図単位1.30から9.24)中に遺伝子修飾部分を有するようにしばしば操作される。典型的な修飾には、E1初期遺伝子領域内の欠失および/またはE1Aプロモーターの置換え、導入遺伝子の導入などが含まれる。ヘルパーウィルスに依存しないアデノウィルスの産生は、ウィルス遺伝子産物を補完するパッケージング細胞株を必要とする可能性がある。
【0007】
研究および臨床試験用の組換えアデノウィルスベクターを産生するにはパッケージング細胞株は、アデノウィルスE1コード配列をトランスフェクションされる。この細胞株は、アデノウィルスDNA複製およびビリオン産生にとって直接または間接的に必要なトランスのE1AおよびE1B遺伝子産物を補充するのに十分なE1遺伝子産物を発現しなければならない。
【0008】
E1の補完は組換えアデノウィルスベクターの産生を可能にするが、宿主細胞中のトランスフェクションされたE1配列とそのアデノウィルスベクターの間の組換え事象が起こり、その結果、複製能のあるアデノウィルス(RCA)が生じる原因となる恐れがある。これは、大規模な産生およびそれに続く増殖の場合に特に問題であり、したがって治療に使用するためのアデノウィルス粒子ストックの調製において問題である。組換えと、組換えアデノウィルスベクター生産中のRCAの生成は、ウィルスストックを汚染するだけでなく、in vivo用途にこれらアデノウィルスベクターを使用することに関して問題がある。例えばShenk等の論文(1979)、Cold Springs Harb. Symp. Quant. Biol. 44 (1979) 367-375およびLochmullerの論文、Human Gene Therapy, 1994, 1485-1491に記載されているようにRCA生成の問題が知られてもうしばらくになる。
【0009】
入手可能なパッケージング細胞株は、一般にベクターからは欠失しているがウィルス複製には必要なアデノウィルス遺伝子を含有する。幾つかの事例では宿主細胞とアデノウィルスベクターの間の重複配列は完全には排除されない。例えばヒト胎児の腎臓から採った293細胞(Graham等の論文(1977)、J. general Virology 36: 59-74)は、アデノウィルスベクターを増殖するために広く用いられてきた。しかしながらこのアデノウィルスベクターゲノムと293細胞株の間の実質上の重複配列のせいで、複製能のあるアデノウィルス粒子の生成を引き起こす組換え事象が起こる。
【0010】
このベクターおよび細胞株に共通の配列の減少により、このパッケージング細胞株とベクターの間の組換え事象が原因する複製能のあるベクターの生成の可能性を低減させる改良がなされてきた(Fallaux等の論文(1998)、Human Gene Therapy 9: 1909-1917)。例えば米国特許第5,994,128号は、自然のE1Bプロモーター配列を保持すると同時にE1AとE1Bの両方またはその一方を補完する細胞株について記載している。PER.C6細胞株を用いて行った研究は、この細胞株とアデノウィルスベクターの間のたった1つの相同領域にもかかわらずRCAが生成し、細胞変性効果が細胞を用いた検定で観察されることを実証した(Kim等の論文(2001)、Exp. Mol. Med. 33 (3) 145-9)。分析するとこのRCA は、PER.C6細胞株を構築するために使用したプラスミド由来のPGKプロモーター−E1遺伝子を含有することが示された。残りの配列の重複のこの同じ問題は、293細胞に代わるものとして生成する他の細胞株についても当てはまる(例えばMassie他の米国特許第5,891,690号、Kovesdi他の国際公開第WO95/34671号、Kedan他の国際出願PCT/US95/15947号、Schiedner等の論文(2002)、Human Gene Therapy, 11: 2105-2116を参照されたい)。したがって細胞株とアデノウィルスベクターの間の望ましくない組換え事象の可能性が残っている。
【0011】
アデノウィルスベクター中の欠損部を補完するのに十分な、かつE1欠損アデノウィルスの成長を可能にするのに十分なE1AおよびE1Bコード配列を含むアデノウィルスパッケージング細胞株を提供する。このE1AおよびE1B配列は、天然または野生型アデノウィルスE1AおよびE1Bプロモーターと本質的な配列同一性を共有するポリヌクレオチド配列を欠いたプロモーターと作用可能に連結している。このようなパッケージング細胞株は、そのパッケージング細胞株ゲノムとアデノウィルスベクターの間の組換え事象の可能性が最小限のアデノウィルス粒子ストックを確実に産生する。本発明のパッケージング株を用いて産生されるウィルスストックは、意図する組換え遺伝子型を維持した最小限または検知できないレベルのRCAを特徴とする。
【0012】
本発明の1つの態様において、パッケージング細胞株は、E1AおよびE1B遺伝子が、同一であっても異なっていてもよい非アデノウィルス異種プロモーターと作用可能に連結している安定的に組み込まれたE1AおよびE1B発現ベクターを含む。
【0013】
本発明の別の態様において、RCAを実質上含まないアデノウィルスであって、実質的な配列同一性をアデノウィルスE1AおよびE1Bプロモーターと共有するポリヌクレオチド配列を欠いた細胞株中で成長するアデノウィルス、を産生する方法を提供する。
【0014】
定義:
別段の指示がない限り本明細書中で使用されるすべての技術的および科学的用語は、本発明の業界の技術者が使用するのと同じ意味を有する。当業界の定義および用語については、Sambrook等の著、Molecular Cloning: A Laboratory Manual (2nd Edition), Cold Spring Harbor Press, Plainview, N. Y., 1989およびAusubel FM等の著、Current Protocols in Molecular Biology, John Wiley & Sons, New York, N. Y. 1993は実務化向きである。本発明は、その記載されている特定の方法論、実験計画、および試薬には限定されず、したがってこれらは変えることができることを理解されたい。
【0015】
すべての特許、特許出願、刊行物(公開特許出願を含め)、および本明細書中で参照されるデータベース取得番号を含めた刊行物ならびに他の資料は、本発明の背景を明らかにするために、特にその実施に関して追加の細部を提供する場合に本明細書中で用いられる。これらすべての特許、特許出願、刊行物(公開特許出願を含め)、および本明細書中で参照されるデータベース取得番号を含めた刊行物ならびに他の資料は、それぞれが具体的かつ個々にその全体が参照により組み込まれて示されるのと同程度に参照により本明細書中に組み込まれる。
【0016】
「アデノウィルスパッケージング細胞」は、アデノウィルスゲノムまたは修飾ゲノムをパッケージしてウィルス粒子を産生することができる細胞である。それは欠けている遺伝子産物またはその同等物を提供することができる。したがってそのパッケージング細胞は、アデノウィルスゲノム中の欠失した遺伝子を補完する機能を提供することができ、またそれらアデノウィルスゲノムをアデノウィルス粒子中にパッケージすることができる。このような粒子の産生は、そのゲノムが複製され、かつ感染性ウィルスを組み立てるのに必要なそれらのタンパク質が産生されることを必要とする。この粒子はまた、そのウィルス粒子の成熟に必要なある種のタンパク質を必要とする可能性がある。このようなタンパク質は、そのベクターにより、またはそのパッケージング細胞により与えることができる。このパッケージング細胞株は、E1Aおよび/またはE1Bコード配列を含むようにアデノウィルス複製を許容する細胞株を遺伝子的に修飾することによって産生することができる。本発明のアデノウィルスパッケージング細胞株ではアデノウィルスE1AおよびE1Bコード配列は、天然のアデノウィルスE1AおよびE1Bプロモーターと実質的な配列同一性を共有するポリヌクレオチド配列を欠いたプロモーターと作用可能に連結している。
【0017】
「宿主細胞」には個別の細胞または細胞培養物が含まれ、これらは本発明のウィルスベクターのレシピエントとなることができ、又はレシピエントとなっていてもよい。宿主細胞には単一宿主細胞の子孫が含まれ、この子孫は自然、偶然、あるいは意図的な突然変異および/または変化のせいで元の親細胞と必ずしも完全に同一(形態または全DNA相補体において)でなくてもよい。宿主細胞には、本発明のアデノウィルスベクターをin vivoまたはin vitroにおいてトランスフェクションまたは感染させた細胞が含まれる。
【0018】
本明細書中で用いられる用語「アデノウィルス」および「アデノウィルス粒子」には、あらゆる群、亜群、および血清型を含めたヒトまたは動物に感染する任意のアデノウィルスを含めたアデノウィルスとして分類することができる、ありとあらゆるウィルスが含まれる。したがって本明細書中で用いられる「アデノウィルス」および「アデノウィルス粒子」は、ウィルス自体またはその誘導体を指し、すべての血清型と亜型ならびに天然に存在する品種および組換え品種の両方を包含する。一実施形態においてこのようなアデノウィルスはヒトの細胞に感染する。このようなアデノウィルスは野生型であってもよく、また当業界で知られているか、または本明細書中で開示した様々な方法で修飾されてもよい。このような修飾には、感染性ウィルスを作るためにその粒子中にパッケージングされるアデノウィルスゲノムに対する修飾が含まれる。このような修飾には、E1a、E1b、E2a、E2b、E3、またはE4コード領域の1または複数における欠失など、当業界で知られている欠失が挙げられる。本発明のアデノウィルスベクターの例には、限定しないが、DNA、アデノウィルスの外殻に包まれたDNA、別のウィルスまたはウィルス様品種(単純ヘルペスおよびAAVなど)中にパッケージングされたアデノウィルスDNA、リポソームに包まれたアデノウィルスDNA、ポリリジンと複合化させたアデノウィルスDNA、合成ポリカチオン分子と複合化させたアデノウィルスDNA、トランスフェリンと接合したアデノウィルスDNA、あるいはその抗原性を免疫的に「マスキング」するためにおよび/または半減期を増すためにPEGなどの化合物と複合化させたアデノウィルスDNA、あるいは非ウィルスタンパク質に接合したアデノウィルスDNA、が挙げられる。E1A/E1Bパッケージング株の生成に使用されるAAVベクターの例を図8および9に示す。
【0019】
本明細書中でアデノウィルスベクターに関して用いられる用語「複製能欠陥の」とは、ウィルスベクターがそのゲノムをそれ以上複製し、またパッケージすることができないことを意味する。例えば、ある被験者の細胞がrAAVビリオンに感染している場合、その異種遺伝子はその患者の細胞中で発現するが、その患者の細胞がAAV REPおよびCAP遺伝子と、アデノウィルスの補助機能遺伝子とが欠けているという事実により、そのrAAVは複製能欠陥があり、野生型AAVはその患者の細胞中で生成することができない。
【0020】
本明細書中で用いられる「パッケージングシステム」とは、組換えウィルスのパッケージングに関係するウィルスタンパク質をコードする遺伝子を含む一組のウィルス構築物を指す。一般にそのパッケージングシステムの構築物は、最後はパッケージング細胞中に取り込まれることになる。
【0021】
本明細書中でアデノウィルスベクターに関して用いられる用語「複製能のある」とは、そのウィルスベクターおよび粒子が或る型の細胞または組織中では優先的に複製するが、他の型ではずっと少ない程度かまたは全く複製しないことを意味する。本発明の1つの態様において、そのウィルスベクターおよび/または粒子は、充実性腫瘍および他の新生組織などの腫瘍細胞および/または異常増殖組織中で選択的に複製する。このようなウィルスは、「腫瘍溶解性ウィルス」または「腫瘍溶解性ベクター」と呼ぶことができ、また「細胞溶解性」または「細胞変性性」であり、標的細胞の「選択的細胞溶解」を行うと考えることができる。これらには、米国特許第5,677,178号、第5,698,443号、第5,871,726号、第5,801,029号、第5,998,205号、および第6,432,700号に開示されているウィルスが挙げられる。
【0022】
用語「ウィルス」、「ウィルス粒子」、「ベクター粒子」、「ウィルスベクター粒子」、および「ビリオン」は同義で用いられ、例えば本発明のウィルスベクターを感染性粒子の生成用の適切な細胞または細胞株に形質導入する場合に形成される感染性ウィルス粒子を意味するものと広範囲に理解されるべきである。本発明によるウィルス粒子は、核酸(例えばDNAまたはRNA)をin vitroまたはin vivoのいずれかで細胞中に伝達する目的に利用することができる。
【0023】
本明細書中で同義で用いられる用語「ポリヌクレオチド」および「核酸」は、任意の長さのポリマー形態のヌクレオチドであるリボヌクレオチドまたはデオキシリボヌクレオチドのいずれかを指す。これらの用語には、一本、二本、または三本鎖DNA、ゲノムDNA、cDNA、RNA、DNA−RNAハイブリッド、あるいはプリンおよびピリミジン塩基を含むポリマー、あるいは他の天然の化学的、生化学的に変性した塩基、非天然または誘導ヌクレオチド塩基を含むポリマーが含まれる。好ましくは本発明のベクターはDNAを含む。本明細書中で用いられる「DNA」には、塩基A、T、C、およびGだけでなく、それらの類似体またはこれら塩基の修飾形態、例えばメチル化ヌクレオチド、非荷電結合およびチオ酸塩などのヌクレオチド間修飾物、糖類似体の使用、ならびにポリアミドなどの変性および/または代替骨格構造のいずれかもまた含まれる。
【0024】
遺伝子または遺伝子フラグメント、エキソン、イントロン、mRNA、tRNA、rRNA、リボザイム、cDNA、組換えポリヌクレオチド、分枝ポリヌクレオチド、プラスミド、ベクター、任意の配列の単離DNA、任意の配列の単離RNA、核酸プローブ、およびプライマーは、ポリヌクレオチドの非限定的な例である。ポリヌクレオチドは、修飾ヌクレオチド、例えばメチル化ヌクレオチド、ヌクレオチド類似体、ウラシル、フルオロリボースおよびチオ酸塩などの他の糖類および結合基、ならびにヌクレオチド側鎖などを含むことができる。そのヌクレオチドの配列は非ヌクレオチド成分が介在していてもよい。ポリヌクレオチドは標識成分を接合させるなど、重合後さらに修飾されてもよい。この定義に含まれる他の種類の修飾は、キャップ、類似体による天然に存在するヌクレオチドのうちの1個または複数個の置換、およびタンパク質、金属イオン、標識成分、他のポリヌクレオチド、または固形支持体にそのポリヌクレオチドを付着させる手段の導入である。好ましくはこのポリヌクレオチドはDNAである。本明細書中で用いられる「DNA」には、塩基A、T、C、およびGが含まれるだけでなく、それらの類似体またはこれら塩基の修飾形態、例えばメチル化ヌクレオチド、非荷電結合およびチオ酸塩などのヌクレオチド間修飾物、糖類似体の使用、ならびにポリアミドなどの変性および/または代替骨格構造のいずれかもまた含まれる。
【0025】
核酸は、別の核酸配列と機能上の関係に置かれる場合、「作用可能に連結」している。例えばオペレーターまたはエンハンサーは、それがその配列の転写に影響する場合、コード配列と作用可能に連結している。通常、「作用可能に連結した」とは、それら結合しているDNA配列が隣接していることを意味する。しかしながらエンハンサーは隣接しなくてもよい。結合は、都合のよい制限部位においてライゲーションにより達成される。このような部位が存在しない場合、合成オリゴヌクレオチドアダプターまたはリンカーが従来のやり方に従って用いられる。
【0026】
用語「天然の」とは、野生型ウィルスまたは細胞のゲノム中に存在する遺伝子を指す。
【0027】
用語「天然に存在する」または「野生型」は、人間により人為的に生成されるものとはまったく異なるものとして自然界に見出すことができる物を記述するために用いられる。例えば、自然界の供給源から単離することができ、かつ実験室で人間によって故意に変性されなかった生物体(ウィルスを含めた)中に存在するタンパク質またはヌクレオチド配列は、天然に存在する。
【0028】
本明細書中で用いられる用語「プラスミド」は、染色体外でまたは宿主細胞の染色体(または染色体群)の一部として宿主細胞内で自己複製の能力があるDNA分子を指す。本明細書中の出発プラスミドは市販されており、公然と自由に利用可能であり、あるいは本明細書中に開示されているような利用可能なプラスミドからおよび/または出版物に記載されている手順に従って構築することができる。幾つかの事例では、当業者には明らかなように当業界で知られている他のプラスミドを本明細書中で記述したプラスミドと同義で用いることができる。
【0029】
本明細書中で用いられる用語「投与」または「導入」は、細胞中へのE1Aおよび/またはE1Bコード配列の安定組込みのための発現ベクターの送達を指す。ベクターはトランスフェクションによって細胞中に導入することができ、このトランスフェクションは一般に物理的手段(例えばリン酸カルシウムのトランスフェクション、電気穿孔、マイクロインジェクション、またはリポフェクション)、感染(一般には感染性病原体すなわちウィルスを経由する導入を指す)、または形質導入(一般には、ウィルスによる細胞の安定感染、またはウィルス病原体(例えばバクテリオファージ)を経由する或る微生物から別の微生物への遺伝物質の転移を意味する)による細胞中への異種DNAの挿入を意味する。前述のようにこのベクターは、プラスミド、ウィルス、または他の伝達体であることができる。
【0030】
本明細書中で核酸分子に関して用いられる用語「組換え体」は、組換えDNA技術を用いて結合されて子孫核酸分子になる核酸分子の複合体を指す。ウィルス、細胞、および生物体に関して本明細書中で用いられる用語「組換え」、「形質転換」、および「トタンスジェニック」は、異種核酸分子が導入された宿主のウィルス、細胞、または生物体を指す。この核酸分子はその宿主のゲノム中に安定的に組み込むことができるか、またはこの核酸分子は染色体外分子として存在することもできる。このような染色体外分子は、自己複製性であってもよい。これらの組換えウィルス、細胞、および生物体は、形質転換過程の最終産物だけでなく、それらの組換え子孫も包含すると考えられる。「非形質転換」、「非トタンスジェニック」、または「非組換え」宿主は、異種核酸分子を含有しない野生型のウィルス、細胞、または生物体を指す。
【0031】
「調節エレメント」は、ヌクレオチド配列の発現の制御と関係した配列である。調節エレメントには、プロモーター、エンハンサー、および終止コドンが含まれる。これらはまた、一般にそのヌクレオチド配列の適正な翻訳に必要な配列を包含する。
【0032】
用語「プロモーター」は、RNAポリメラーゼIIに対する結合部位を含有し、そのDNAの転写を開始するコード領域の上流に通常位置する非翻訳DNA配列を指す。このプロモーター領域は、遺伝子発現の調節遺伝子として働く他のエレメントもまた含むことができる。用語「最少プロモーター」は、不活性であるか、または上流の活性化エレメントの不在下において著しく低いプロモーター活性を有するプロモーターエレメント、特にTATAエレメントを指す。
【0033】
用語「エンハンサー」は本発明の意味の範囲内では、任意の遺伝エレメント、例えば或るコード配列と作用可能に連結した場合にプロモーター自体によってもたらされる転写活性化よりも大きい程度までそのプロモーターと作用可能に連結しているそのコード配列の転写を増大させる、すなわちそれがそのプロモーター由来の転写を増大させるヌクレオチド配列であることができる。
【0034】
語句「とハイブリダイズする」とは、その配列が複合混合物(例えば全細胞)のDNAまたはRNA中に存在する場合、ストリンジェントな条件下における特定のヌクレオチド配列のみとの分子の結合、二重らせん形成、またはハイブリダイゼーションを指す。「実質上結合する」とは、プローブ核酸と標的核酸の間の相補的ハイブリダイゼーションを指し、そのハイブリダイゼーション培地のストリンジェンシーを下げることによって適応することができる重要でない誤対合を容認して標的核酸配列の所望の検出を達成する。
【0035】
サザンおよびノーザンハイブリダイゼーションなどの核酸ハイブリダイゼーション実験の脈絡における「ストリンジェントなハイブリダイゼーション条件」および「ストリンジェントな洗浄条件」は配列に左右され、また様々な環境要因下で異なる。長い配列ほど高い温度でハイブリダイズする。核酸のハイブリダイゼーションに関する広範囲な手引きは、Tijssen著、(1993) Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probesの第1部第2章「Overview of principles of hybridization and the strategy of nucleic acid probe assays」Elsevier, New York中に見出される。一般に高ストリンジェントなハイブリダイゼーションおよび洗浄条件は、規定のイオン強度およびpHにおいてその特定の配列の融点(Tm)よりも約5℃から20℃(好ましくは5℃)低いように選択される。一般に高ストリンジェントな条件下ではプローブは、他の配列とではなく、その標的部分配列とハイブリダイズすることになる。
【0036】
用語「相補体」および「相補的」は、アンチパラレルヌクレオチド配列中の相補的塩基残基間で水素結合を形成するとき互いに組むことができるアンチパラレルヌクレオチド配列を含む2つのヌクレオチド配列を指す。
【0037】
用語「発現」は、細胞中の内因性遺伝子、導入遺伝子、またはコード領域の転写および/または翻訳を指す。
【0038】
本明細書中で用いられる「内部リボソーム侵入部位」または「IRES」は、ATGなどの開始コドンへのシストロン(タンパク質コード領域)の直接の内部リボソーム侵入を促進し、それによってキャップに影響されない遺伝子の翻訳を引き起こすエレメントを指す。例えばJackson R J、Howell M T、およびKaminski Aの論文(1990)、Trends Biochem Sci 15 (12): 447-83ならびにJackson R J、Howell M T、およびKaminski, Aの論文(1995)、RNA 1 (10): 985-1000を参照されたい。本発明は、開始コドンへのシストロンの直接の内部リボソーム侵入を促進することができる任意のエレメントを使用することを包含する。本明細書中で用いられる「IRESの翻訳制御下で」とは、翻訳がIRESと関連しており、キャップに影響されないやり方で進行することを意味する。本明細書中で用いられる用語「IRES」は、IRES配列の機能上の変異が開始コドンへのシストロンの直接の内部リボソーム侵入を促進することができる限り、その変異を包含する。
【0039】
本明細書中で言及される「自己プロセッシング切断部位」または「自己プロセッシング切断配列」は、翻訳時に自己プロセッシング切断部位を含むポリペプチドの急速な分子間(シス)切断を起こして不連続の成熟タンパク質またはポリペプチド産物の発現を引き起こすDNAまたはアミノ酸配列である。このような「自己プロセッシング切断部位」はまた、翻訳後のまたは翻訳に伴うプロセッシング切断部位、例えば2A部位、配列、またはドメインと呼ぶこともできる。2A部位、配列、またはドメインは、リボソームの活性を変えてエステル結合の加水分解を促進させ、それによって不連続な下流の翻訳産物の合成を進行させるやり方で翻訳複合体からポリペプチドを遊離することによって翻訳効果を実証する(Donnelly等の論文、J. Gen. Virol. 82: 1027-1041, 2001)。FMDV 2A領域による切断活性の発現にとって不可欠なアミノ酸残基を含む構築体が設計されている(Ryan等の論文(1991)、J. Gen. Virol. 72: 2727-2732およびFurler等の論文(2001)、Gene Therapy 8: 864-873)。2Aドメインはまた、ピコルナウィルス科のaphthovirideaおよびcardioviridaeから特徴が調べられた(Donnelly等の論文(1997)、J. Gen. Virol. 78: 13-21)。
【0040】
本明細書中で用いられる用語「E1A」は、2つの主要なRNAの発現生成物13Sおよび12Sを含むアデノウィルスE1A領域のすべての遺伝子産物を指す。これらは翻訳されてそれぞれ289(配列番号2)および243(配列番号3)のアミノ酸のポリペプチドになる。これら2種類のタンパク質はアミノ酸が46個異なり、Chow等の論文(1980)、Cold Spring Harb Symp Quant Biol. 44 Pt 1: 401-14およびChow等の論文(1979)、J. Mol. Biol. 134 (2): 265-303(これらは特に引用により本明細書に組み入れられる)に記載のようにそれらは12S mRNAからスプライシングされる。本発明の目的の場合、パッケージング細胞株はその289ポリペプチド、243ポリペプチド、または289および243ポリペプチドの両方を発現することができる。用語E1Aはまた、本明細書中では部分および変異E1Aコード配列に関して用いられる。
【0041】
本明細書中で用いられる用語「E1B」は、3つの主要なポリペプチドを含む19 kd(配列番号5)および55 kd(配列番号6)のアデノウィルスE1B領域のすべての遺伝子産物を指す。これらE1B 19 kdおよび55 kdタンパク質は、細胞の形質転換において重要である。本発明の目的では、パッケージング細胞株は19 kdポリペプチド、55 kdポリペプチド、または19 kdおよび55 kdポリペプチドの両方を発現することができる。用語「アデノウィルス許容」とは、そのアデノウィルスまたはアデノウィルスベクターが、その細胞環境中で全細胞内ウィルスのライフサイクルを終えることができることを意味する。これらの細胞は、初代細胞培養物、樹立細胞株等から得ることができる。霊長動物細胞、例えばヒトの細胞、サルの細胞などを含めた哺乳動物細胞が好ましい。様々な霊長動物細胞が好ましく、またこのようなヒトの胎児腎臓細胞がより好ましいが、ウィルスの複製を支持することができる任意の種類の細胞が本発明の実施において受け入れられる。用語E1Bはまた、本明細書中では部分および変異E1Bコード配列に関して用いられる。
【0042】
「複製」および「増殖」は同義で用いられ、アデノウィルスベクターを再生産または増殖する能力を指す。これらの用語は、当業界では十分に理解されている。本発明の目的の場合、複製はアデノウィルスタンパク質の産生と関係しており、一般にアデノウィルスの再生産を目的とする。複製は、ウィルスの収量検定、バースト検定、またはプラーク検定などの当業界で標準的なまた本明細書中で記述する検定法を用いて測定することができる。「複製」および「増殖」には、限定しないが、ウィルス遺伝子発現や、ウィルスのタンパク質、核酸、または他の成分の産生や、ウィルス構成要素をパッケージして完全なウィルスにすることや、細胞溶解を含めたウィルスの生産過程に直接または間接的に関係する任意の営みが含まれる。
【0043】
本発明の方法および組成物:
種々の方法および組成物を後述する。特定の方法を下記の考察において例示するが、複数の代替方法のいずれも適用可能であり、また本発明の実施において使用に適していることが分かる。また本発明のアデノウィルスベクターおよび方法の評価は、下記の診断および判定方法を含む当業界で標準的な手順を用いて行うことができる。
【0044】
本発明の実施においては別段の指示がない限り、当業者の範囲内にある細胞生態学、分子生物学(組換え技術を含めた)、微生物学、生化学、および免疫学の通常の技術を使用することになる。このような技術は、「Molecular Cloning: A Laboratory Manual」2nd edition (Sambrook等著、1989)、「Oligonucleotide Synthesis」(M. J. Gait編、1984)、「Animal Cell Culture」(R. I. Freshney編、1987)、「Methods in Enzymology」(Academic Press, Inc.)、「Handbook of Experimental Immunology」(D. M. WeirおよびC. C. Blackwell編)、「Gene Transfer Vectors for Mammalian Cells」(J. M. MillerおよびM. P. Calos編、1987)、「Current Protocols in Molecular Biology」(F. M. Ausubel等編、1987)、「PCR: The Polymerase Chain Reaction」(Mullis等編、1994)、および「Current Protocols in Immunology」(J. E. Coligan等編、1991) などの文献中で完全に説明されており、これらは特別にそれぞれ本明細書中に参照により組み込まれる。
【0045】
アデノウィルスに関係する技術に関しては、とりわけFelgnerおよびRingoldの論文(1989)、Nature 337: 387-388、BerkerおよびSharpの論文(1983)、Nucl. Acids Res. 11: 6003-6020、Grahamの論文(1984)、EMBO J. 3: 2917-2922、Bett等の論文(1993)、J. Virology 67: 5911-5921、Bett等の論文(1994)、Proc. Natl. Acad. Sci. USA 91: 8802-8806を参照されたい。
【0046】
アデノウィルスパッケージング株
E1AおよびE1B欠損アデノウィルスを補完および複製するのに十分であり、かつ野生型複製能のあるアデノウィルス(RCA)を生成させる可能性が最小限のアデノウィルスE1AおよびE1B配列を実現する、アデノウィルスパッケージング細胞株を提供する。本明細書中で用いられるRCAは、E1Aおよび/またはE1Bの発現のためにパッケージング細胞株による補完を必要としない複製能のあるアデノウィルスである。
【0047】
このパッケージング細胞株は、ヒトアデノウィルスE1AおよびE1Bタンパク質をコードする遺伝子配列を含む。この遺伝子配列は、天然配列またはそれらの変異型であることができる。本明細書中で用いられる実質的な配列同一性とは、その宿主細胞内の相同的組換えにとって十分な配列類似性レベルを指す。候補配列については、例えば注目の細胞において複製中に2つの配列に組換えがあるかないかを試験することによって経験的に組換えの有無を調べることができる。一般に配列は、連続した同一のポリヌクレオチド配列の約20個を超えるヌクレオチド、通例では連続した同一のポリヌクレオチド配列の約15個を超えるヌクレオチド、好ましくは連続した同一のポリヌクレオチド配列の約12個を超えるヌクレオチドが存在しない場合、実質的な配列同一性が欠けていることになる。
【0048】
参照配列は、通常はそのベクターが得られる元のアデノウィルス、例えばヒトアデノウィルス5、ヒトアデノウィルス2などであることになる。本発明のパッケージング細胞株中でE1AおよびE1Bの発現を駆動するプロモーターと、内因性のアデノウィルスE1AおよびE1Bプロモーターの間の実質的な配列同一性が欠けていることは、組換えおよびその結果としての複製能のあるアデノウィルス(RCA)の産生の可能性をできるだけ少なくするはずである。
【0049】
配列比較の場合、一般には一方の配列が試験配列をそれと比較する参照配列として働く。配列比較アルゴリズムを使用する場合、これら試験および参照配列をコンピュータにインプットし、必要ならば部分配列座標を指定し、配列アルゴリズムのプログラムパラメーターを指定する。次いで配列比較アルゴリズムが、指定したプログラムパラメーターに基づいて参照配列に対する試験配列の配列同一性のパーセントを計算する。
【0050】
比較のための配列の最適アライメントは、例えばSmithおよびWatermanの論文、Adv. Appl. Math. 2: 482 (1981) のローカルホモロジーアルゴリズムにより、あるいはNeedlemanおよびWunschの論文、J. Mol. Biol. 48: 443 (1970) の相同アライメントアルゴリズムにより、あるいはPearsonおよびLipmanの論文、Proc. Nat’l. Acad. Sci. USA 85: 2444 (1988)の類似性検索法により、あるいはこれらアルゴリズムのコンピュータ化されたインプリメンテーション(Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, Wis 中のGAP、BESTFIT、FASTA、およびTFASTA)により、あるいはNational Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/)を介して公けに利用できるソフトウェアを用いたBLASTアルゴリズム(Altschul等の論文、J. Mol. Biol. 215: 403-410 (1990))により、あるいは目視検査により行うことができる(全般的には後掲のAusubel等の論文を参照)。本発明の目的の場合、比較のための最適アライメントは、最も好ましくはSmithおよびWatermanの論文、Adv. Appl. Math. 2: 482 (1981) のローカルホモロジーアルゴリズムにより行われる。
【0051】
2つ以上の核酸またはタンパク質配列の脈絡における用語「同一の」、または「同一性」パーセントは、本明細書中で述べた配列比較アルゴリズムの一つ、例えばSmith−Watermanのアルゴリズムを用いて、または目視検査により測定した最大の対応関係について比較し、また位置合わせした場合に同一であるか、あるいは所定の割合の同一であるアミノ酸残基またはヌクレオチドを有する2つ以上の配列または部分配列を指す。
【0052】
多くのアデノウィルスE1AおよびE1Bタンパク質の配列、例えばヒトアデノウィルス11型、ヒトアデノウィルス41型、ヒトアデノウィルス12型、ヒトアデノウィルス5、ヒトアデノウィルス35、ヒトアデノウィルス41、ヒトアデノウィルス40、ヒトアデノウィルス4、ヒトアデノウィルス7、およびヒトアデノウィルス2由来の配列が知られており、また公けに利用できる。例えばE1Aポリペプチド配列、Genbank 取得番号AAN62486 (Ad11)、Q2AD5 (Ad5)、およびQ2AD2 (Ad2)、ならびにE1Bポリペプチド配列、Genbank 取得番号Q1AD25 (Ad5)およびQ1AD22 (Ad2) を参照されたい。E1AおよびE1Bコード配列の元はAd2由来のものが最も好ましい。Ad2を含む他のヒトおよび非ヒトアデノウィルス血清型もまた用いることができる。本発明のパッケージング株の調製に用いられるE1AおよびE1B配列は、E1AまたはE1Bのどちらかのプロモーター配列を含まない。通常このパッケージング株は、E1AおよびE1Bコード配列とは異なるアデノウィルス遺伝子配列を含まないはずである。
【0053】
E1AおよびE1Bコード配列の例は、GenBank取得番号M73260またはX02996のヌクレオチド560〜1545(本明細書中では配列番号1として示される)に対応するE1Aの2つのエキソン領域およびGenBank取得番号M73260またはX02996のヌクレオチド1682〜3825(本明細書中では配列番号4として示される)に対応する2つのE1Bコード配列を含む。多くのアデノウィルスゲノムの特徴が調べられ、使用のために入手できるので、本明細書中で提供されるアデノウィルス配列は好適な配列の単なる例に過ぎないことは当業者ならば理解するはずである。本発明の実施において用いられるE1AおよびE1Bコード配列の例を表1に示す。
【0054】
【表1】
【0055】
これらE1AおよびE1B配列は作用可能に非アデノウィルスプロモーターと結合している。このプロモーターは異種であってもよく、この場合、本明細書中では用語「異種」プロモーターはそのパッケージング細胞にとって天然でないプロモーター配列を意味するために用いられる。E1Aおよび/またはE1Bのコード配列を含むアデノウィルスベクターの複製の場合、そのパッケージング細胞株中のE1Aおよび/またはE1B用のプロモーターは、好ましくはアデノウィルスベクター中で作用可能にE1Aおよび/またはE1Bと結合したプロモーターとは異なる。
【0056】
注目のパッケージング細胞株では、アデノウィルスE1AおよびE1Bコード配列は、相同的組換えがあまり起こらないように、天然のアデノウィルスE1AおよびE1Bプロモーターと実質的な配列同一性を共有するポリヌクレオチド配列を欠いているプロモーターと作用可能に連結している。このようなパッケージング細胞株は、そのパッケージング細胞株と複製能欠損アデノウィルスベクターとの間の組換え事象がなく、それによってRCAの生成の可能性をできるだけ少なくするアデノウィルス粒子ストックを確実に生産する。
【0057】
本発明の一実施形態においてパッケージング細胞株は、安定的に組み込まれたE1AおよびE1B発現ベクターを含み、そのE1AおよびE1B遺伝子は非アデノウィルスプロモーターと作用可能に連結し、別個の発現ベクターを用いて導入されている。このプロモーターは、非アデノウィルス起源の強い構成性プロモーターであってもよい。1つの態様において、このE1Aと作用可能に連結したプロモーターは、E1Bと作用可能に連結したプロモーターと異なる。別の実施形態ではこのE1Aと作用可能に連結したプロモーターは、E1Bと作用可能に連結したプロモーターと同じものである。これらE1AおよびE1B遺伝子は、このようなプロモーターと全体的に調和して発現することができる。
【0058】
これらE1AおよびE1Bコード領域は、好ましくはパッケージング細胞株のゲノム中に安定的に組み込まれる。好ましい実施形態においてE1A組込み部位は、例えば別個の染色体上、同一の染色体の別個の領域上などでE1B組込み部位から物理的に引き離される。
【0059】
本発明の別の実施形態では、RCAを実質的に含まないアデノウィルスを生産する方法であって、当該アデノウィルスが、アデノウィルスE1AおよびE1Bプロモーターと実質的な配列同一性を共有するポリヌクレオチド配列を欠いた細胞株中で成長する方法、を提供する。
【0060】
本発明の細胞株中での複製にとって注目のアデノウィルスベクターは、複製に不可欠なアデノウィルス遺伝子、具体的にはE1AおよびE1B遺伝子の発現に欠陥がある。このようなベクターは、外因によりもたらされるウィルス遺伝子の不在下では増殖性感染に必要な十分なウィルス性タンパク質を産生することができない。E1AおよびE1Bの発現に欠陥のあるアデノウィルスベクターは、様々な遺伝的変化のせいで、例えばこれら遺伝子の一方または両方のコード配列の欠如、そのポリペプチドを作用不能にするコード配列中の突然変異、プロモーターまたはエンハンサー配列の変化などのせいで欠陥がある。
【0061】
本発明の幾つかの実施形態においてアデノウィルスベクターは、標的細胞型、例えば前立腺癌、肝臓癌などの標的腫瘍細胞中では複製応答能があるが、非標的細胞型中ではこのアデノウィルスはE1Aおよび/またはE1Bの発現に欠陥がある。例えばこのアデノウィルスベクターは、細胞型特異的、細胞状態特異的などである転写調節エレメントと作用可能に連結している複製にとって不可欠なアデノウィルス遺伝子を含むことができる。このようなベクターは、本明細書中で述べたような、例えばin vitroで多数のウィルス粒子を生成させるパッケージング細胞株中での成長から恩恵を受ける。
【0062】
E1AおよびE1Bを発現させるために用いられるプロモーター配列は、同一でも同一でなくてもよい。プロモーター配列が同一の場合、そのE1AおよびE1Bコード配列は全体的に協調して発現することができ、例えばこれら両方のコード配列が作用可能に単一プロモーターと結合し、IRESはそれら2つのコード配列間に存在する。
【0063】
本発明の一実施形態において一方または両方のプロモーターは調節可能なプロモーター、例えば金属類またはホルモン類(Brinster等の論文、Nature (1982), 296, 39-42)、あるいはホルモン類(Lee等の論文、P.N.A.S. USA (1988), 85, 1204-1208および(1981), 294, 228-232、Klock等の論文、Nature (1987), 329, 734-736、IsraelおよびKaufmanの論文、Nucleic Acids Res. (1989), 17, 2589-2604)などの物質で誘導できるプロモーターである。
【0064】
あるいはさらに別の実施形態においてこのプロモーターは構成性プロモーターである。これらプロモーターは、ポリオーマウィルス、鶏痘ウィルス、ウシ乳頭腫ウィルス、トリ肉腫ウィルス、サイトメガロウィルス、レトロウィルス、B型肝炎ウィルス、およびサルウィルス40(SV40)などのウィルスのゲノムから、あるいは異種哺乳動物プロモーター、例えばアクチンプロモーター、PGK(ホスホグリセリン酸キナーゼ)、または免疫グロブリンプロモーターから、あるいは熱ショックプロモーターからこのようなプロモーター宿主細胞株に適合するという条件で得ることができる。SV40ウィルスの初期および後期プロモーターは、好都合なことにまたSV40ウィルスの複製開始点をも含有するSV40制限フラグメントとして得られる。ヒトサイトメガロウィルスの即時型プロモーターは、便利にはHindIII E制限フラグメントとして得られる。
【0065】
例示的な1つの態様において、、MMLVのLTRは第一レトロウィルス発現ベクター中および第二レトロウィルス発現ベクター中のE1A遺伝子と作用可能に連結しており、このMMLVのLTRはE1B遺伝子と作用可能に連結しており、E1B遺伝子の発現を誘導するために用いられる。別の実施形態の例では、CAGプロモーターが第一発現ベクター中および第二発現ベクター中のE1A遺伝子のエキソン1およびエキソン2(配列番号1)と作用可能に連結しており、そのEF1−αプロモーターが19kおよび55k E1Bコード配列(配列番号4)の発現を誘導するために用いられる。
【0066】
高等真核生物による転写は多くの場合、エンハンサー配列をそのベクター中に挿入することによって増大する。エンハンサーはDNAのシス作用エレメントであり、一般に約10から300 bpであり、プロモーターに作用してその転写を増大させる。エンハンサーは相対的な配向にあり、位置と関係がなく、コード配列自体の内部だけでなくイントロン内部の転写単位の5′および3′側にあることが分かっている。多くのエンハンサーは、現在では哺乳動物遺伝子(グロブリン、エラスターゼ、アルブミン、α−フェトプロテイン、およびインスリン)から見分けられる。しかしながら一般には皆、真核細胞ウィルス由来のエンハンサーを使用するはずである。例には、複製起点の後期局面でのSV40エンハンサー、サイトメガロウィルスの初期プロモーターエンハンサー、複製起点の後期局面でのポリオーマエンハンサーなどが挙げられる。このエンハンサーは、そのコード配列の5′または3′位置でその発現ベクターにスプライシングすることができるが、好ましくはプロモーターの5′部位に位置する。
【0067】
(パッケージングアデノウィルス用細胞株の生成)
部位特異的DNA切断は、当業界で一般に理解されている条件および市販の制限酵素の製造者により指定される事項(例えばNew England Biolabs, Product Catalogを参照されたい)の下で、プラスミドまたは他のDNAを適切な制限酵素(または複数種の制限酵素)で処理することによって行われる。一般にプラスミドまたは他のDNA約1 μgは、緩衝液約20 μL中で1ユニットの酵素によって切断される。一般にはDNA基質の完全な消化を保証するために過剰の制限酵素が用いられる。変動を許容することができるが約37℃で約1時間から2時間のインキュベーション時間が実行可能である。それぞれインキュベーションの後、タンパク質をフェノール/クロロホルムで抽出することにより取り出し、続いてエーテル抽出し、エタノールで沈殿させることによって核酸を水性画分から回収することができる。望むなら標準的な技術を用いてポリアクリルアミドゲルまたはアガロースゲル電気泳動により切断フラグメントのサイズ選別を行うことができる。サイズ選別の全般的な説明は、Methods of Enzymology 65: 499-560 (1980)中に見出される(前掲のSambrookおよびRussellの著書もまた参照されたい)。
【0068】
制限切断フラグメントは、50 mMトリス(pH 7.6)、50 mM NaCl、6 mM MgCl2、6 mM DTT、および5〜10 μM dNTP中において20℃での約15から25分のインキュベーション時間を用いて4種類のデオキシヌクレオチド三リン酸(dNTP)の存在下で大腸菌DNAポリメラーゼI(クレノウ)の大きなフラグメントで処理することによって平滑末端化することができる。このクレノウフラグメントは5′粘着末端を埋めるが、4種類のdNTPが存在していても突出3’一本鎖に関しては取り込んだ後元に戻される。所望により、粘着末端の性質によって規定される制約内でそれらdNTPのうちのただ1種類を供給することにより、または選択したdNTPを用いて選択的修復を行うことができる。クレノウで処理した後、この混合物をフェノール/クロロホルムで抽出し、エタノールで沈殿させる。適切な条件下でSIヌクレアーゼまたはBal−31で処理すると任意の一本鎖部分の加水分解が起こる。
【0069】
ライゲーションは、次の標準的な条件および温度下、すなわち20 mMトリス−Cl(pH 7.5)、10 mM MgCl2、10 mM DTT、BSA 33 mg/mL、10 mM〜50 mM NaClと、40 μM ATP、0℃のT4 DNAリガーゼ0.01〜0.02(Weiss)ユニット(「粘着末端」ライゲーション用)か、または1 mM ATP、14℃のT4 DNAリガーゼ0.3〜0.6(Weiss)ユニット(「平滑末端」ライゲーション用)のいずれかとで体積15〜50 μLで行う。分子間「粘着末端」ライゲーションは一般に、全DNA濃度33〜100 μg/mL(5〜100 mMの合計の終濃度)で行われる。分子間「平滑末端」ライゲーション(一般に10〜30倍モル濃度過剰のリンカーを使用する)は、1 μMの合計の終濃度で行われる。
【0070】
E1AまたはE1Bを含むベクターは許容宿主細胞中に導入される。多くのこのようなベクター、例えばプラスミドベクター、ウィルスベクターなどが利用可能である。ベクターの構成成分には、限定しないが、複製起点、1種類または複数種類のマーカー遺伝子、エンハンサーエレメント、プロモーター、および転写末端配列のうちの1種類または複数種類を挙げることができる。
【0071】
E1A/E1Bコード配列の導入用の好ましい発現ベクターは、宿主細胞中に安定的に組み込むことができ、かつ高頻度で娘細胞中に保持されるものである。好ましい一実施形態においてこの発現ベクターはウィルス起源のものである。幾つかの組換えウィルスベクターが、本発明によるパッケージング細胞株を生産するために細胞中にE1A/E1Bコード配列を効果的に送達するのに有用であり、例えばレトロウィルスベクター、レンチウィルスベクター、アデノウィルス関連ベクター(AAV)、ヘルペスウィルスベクター、ポックスウィルスベクターなどが挙げられる。別の好ましい実施形態においてこの発現ベクターは、レトロウィルスから得られるプラスミドである。別の好ましい実施形態においてこの発現ベクターは、レンチウィルスから得られるプラスミドである。レトロウィルスと第二の非Adウィルス由来の配列を含むハイブリッドベクターもまた使用することができる。更に好ましい実施形態においてこの発現ベクターは、モロニーマウス白血病ウィルス(MMLV)から得られる、少なくとも7.5キロベースのクローニング能力を有するレトロウィルスベクターである。非ウィルス発現ベクターは、それらが宿主細胞ゲノム中への組込みを容易にする遺伝エレメントを含有する限り、代わりに使用することができる。
【0072】
E1Aおよび/またはE1Bポリペプチド産生用のコード配列を含む発現ベクターは、それらのゲノム中に安定的に組み込まれたE1Aおよび/またはE1Bコード配列を有する細胞を得るために、その特定のベクター/細胞株の組合せに適した方法を用いて大規模なアデノウィルスベクター生産に適した細胞株中に導入される。安定したトランスフェクションは、ゲノム中に安定的に組み込まれたトランスフェクトDNAを含有する娘細胞の集団からなる細胞株またはクローンの樹立によって実証される。
【0073】
細胞中へのウィルスまたは非ウィルスベクターの導入は、当業者により日常的に使用される標準的な技術を用いて行われる。本発明の発現ベクターはさらに、その細胞株に選択可能な形質を付与するものを含む非コードおよびコード配列を含有することができる。
【0074】
E1Aおよび/またはE1Bのコード配列を含む発現ベクターは、標準的なトランスフェクション法を用いて逐次的または同時に細胞中に導入する(前掲のSambrookの著書)か、あるいは最も好ましい実施形態では感染性ウィルス粒子にパッケージし、形質導入を介して細胞株中に導入することができる。容易に理解されるはずだが用語「導入した」は、その発現ベクターに適した形質導入およびトランスフェクションの方法を含む細胞中にDNA配列を送達するために使用される任意の方法(例えばDNAプラスミドに対する感染性粒子)を包含する。E1AおよびE1Bに対してそれぞれ別個の発現ベクターを用いてそれらを逐次的または同時に導入することにより、それぞれ別の場所においてパッケージング細胞ゲノム中でまとまるのでパッケージング細胞ゲノムとAdベクターの間の潜在的な組換え事象はさらに低減される。本発明はゲノム内で発現ベクターが空間的に離れているしている利点を提供し、RCAを生成させる、または組織特異的複製の喪失を生成させる恐れのある組換え事象の更なる低減を可能にする。
【0075】
本発明のパッケージング細胞株を用いて生産される複製能欠陥のあるおよび/または複製能のあるアデノウィルスベクターは、実質上RCAを含まない。実質上RCAを含まないとは、そのRCAの量が十分に低く、その結果、本発明のパッケージング細胞株を用いて生産されるアデノウィルスベクターのin vivo投与から何の毒性も生じないことを意味する。好ましくは、実質上RCAを含まないアデノウィルスベクター製剤は、患者線量当たりRCA粒子を約104個中約0個から約1個含有する。ここで一般的な患者線量はウィルス粒子約1012から約1013個である。しかしながら例として全ウィルス粒子1013個の患者線量は、組換えウィルス粒子を0から10、102、103、104、105、106、107、または5×107個含有することができ、またin vivo投与後に何の毒性も生じない限り実質上RCAを含まないとみなすことができる。
【0076】
細胞株
E1Aおよび/またはE1Bポリペプチド産生用のコード配列を含む発現ベクターを大規模なアデノウィルスベクターの生産に適した細胞株中に導入する。次いでこの細胞株を、プロモーターの誘発、形質転換細胞の選択、または所望の配列をコードする遺伝子の増幅に適するように変性した通常の栄養培地中で培養する。哺乳動物の宿主細胞は、様々な培地中で培養することができる。Ham’s F10(Sigma)、最少必須培地((MEM)、Sigma)、RPMI 1640(Sigma)、およびダルベッコ改変イーグル培地((DMEM)、Sigma)などの市販の培地が、これら宿主細胞を培養するのに適している。これら培地のいずれも必要に応じてホルモン類および/または他の成長因子(インスリン、トランスフェリン、または上皮増殖因子など)、塩類(塩化ナトリウム、カルシウム、マグネシウム、およびリン酸塩など)、緩衝液(HEPESなど)、ヌクレオシド類(アデノシンおよびチミジンなど)、抗生物質、微量元素、およびグルコースまたは同等のエネルギー源で補うことができる。任意の他の必要な補助剤もまた、当業者が知っているであろう適切な濃度で含まれることがある。温度、pHなどの培養条件は、発現のために選択される宿主細胞に関してさきに用いたものであり、当業者には明らかなはずである。本発明によるパッケージング細胞株の製造用に用いることができる宿主細胞の例には、限定しないが、A549、HeLa、MRC5、W138、CHO細胞、Vero細胞、ヒト胎児網膜細胞、またはそれら宿主細胞がアデノウィルスの成長を許容する限り任意の真核生物細胞が挙げられる。幾つかの好ましい宿主細胞株にはヒト腫瘍細胞株が挙げられる。好ましい実施形態においてこのパッケージング細胞株はPC−3細胞(ATCC番号CRL-1435)から得られる。これらPC−3細胞は転移性の前立腺細胞の腺癌から開始される。これらPC−3細胞は、無血清培地中での継代用に適応させることができるので臨床用Adベクターの大量生産には特に有利である。他の細胞型には、限定しないが、初代細胞培養物から得られる細胞、例えばヒト初代前立腺細胞、ヒト胎児網膜細胞、ヒト幹細胞が挙げられる。真核生物二倍体および異数体細胞株が本発明の範囲内に含まれる。
【0077】
好ましい細胞株は無血清培地に適応可能である。
【0078】
候補の細胞株は、当業界で周知の方法により、例えば感染していない細胞層あるいは1種または複数種のヘルパーウィルスに感染した細胞層をウィルス粒子と接触させ、続いてそれら細胞をインキュベートすることによりアデノウィルスの複製を支持する能力を試験することができる。ウィルスのプラークまたは細胞層中の無細胞領域の形成は、ある種のウィルス産物の発現によって引き起こされる細胞溶解の結果である。細胞溶解はウィルス複製を示唆している。
【0079】
アデノウィルス血清型
本発明によるパッケージング細胞株は、任意の既知のアデノウィルス血清型から得られる臨床用Adベクター、および2種類以上の血清型から得られる配列からなるキメラアデノウィルスの大量生産に役立つ。本発明は、すべてのアデノウィルス血清型由来のAdベクターの使用を意図している。アデノウィルス血清型1から47はAmerican Type Culture Collection(ATCC, Manassas, VA)から現在入手可能であり、また本発明は本発明のこのパッケージング細胞株がそのウィルスを複製する能力がある限り、任意の供給源から入手できる任意の他の血清型のアデノウィルスの生産に有用である。本発明によるパッケージング株を用いて生産することができるアデノウィルスは、ヒトまたは非ヒト起源のものであることができる。例えばアデノウィルスは、亜型A(例えば血清型12、18、31)、亜型B(例えば血清型3、7、11、14、16、21、34、35)、亜型C(例えば血清型1、2、5、6)、亜型D(例えば血清型8、9、10、13、15、17、19、20、22〜30、32、33、36〜39、42〜47)、亜型E(血清型4)、亜型F(血清型40、41)、または任意の他のアデノウィルス血清型のものであってもよい。
【0080】
本発明のパッケージング株を用いて生産される組換えアデノウィルスは、E1Aおよび/またはE1Bコード領域中に、欠損に加えて欠失または他の遺伝子変異を含むことができる。本発明に有用な組換えアデノウィルスは、任意選択で他の遺伝子変化、例えば導入遺伝子の封入などに耐えることができる。
【0081】
一実施形態においてこのパッケージング細胞株は、アデノウィルス複製およびアデノウィルス感染の両方を許容する。アデノウィルス血清型2および5の初代受容体は、Bergelson等(1997)(Science 275: 1320)によって同定され、CAR(コクサッキーウィルスおよびアデノウィルス受容体、GenBank取得番号HSU90716)と名付けられており、またRoelvink等(1998)によって亜型Bを除くすべてのアデノウィルス亜型に対する受容体であることが示されている。
【0082】
別の実施形態では本発明に基づくパッケージング株は、その粒子のキャプシドタンパク質、例えばその繊維状タンパク質のHIループまたはカルボキシル末端(C末端)中に、またはタンパク質IX中にリガンドまたは一本鎖抗体を含む修飾繊維状タンパク質などの中に含まれた標的リガンドを含むアデノウィルスベクターの生産に有用である。標的リガンドを含むアデノウィルスベクターは、例えば国際公開第WO00/67576号、第WO99/39734号、米国特許第6,683,170号、第6,555,368号、第5,922,315号、第5,543,328号、および第5,846,782号に記載されている。さらに別の実施形態において本発明に基づくパッケージング株は、米国特許出願第10/403,337号、国際公開第WO98/07877号、第WO01/92299号、米国特許第5,962,311号、第6,153,435号、第6,455,314号、第5,731,190号、第6,057,155号、第5,543,328号、第5,756,086号、第6,127,525号、第5,922,315号、およびWu等の論文(J. Virol. 2003 Jul 1; 77 (13): 7225-7235)中に例証されているものなど、繊維状タンパク質の他の突然変異を含むアデノウィルスベクターの生産に有用である。
【0083】
医薬組成物
本発明のパッケージング株を用いて生産されるアデノウィルスベクターは、例えば臨床用途に用いるために調合することができる。この溶離液を任意選択で濃縮し、また通常の方法により、例えば中空繊維濃縮器を用いてダイアフィルター濾過する。使用される最終製剤においてはそのウィルス試料を滅菌濾過することができる。例えばニトロセルロース膜フィルター、酢酸セルロース膜フィルター、PVDF(修飾ポリビニリデンフルオリド)膜フィルターなど、この目的に適した様々なフィルターが当業界で知られている。PVDF膜フィルター(例えばMillipore Millipakフィルター)が好ましい。
【0084】
滅菌濾過したウィルス懸濁液はin vitroまたはin vivoで使用するために調合される。これらの水性組成物は、医薬として許容される担体または水性媒体中に懸濁させた有効量のウィルスを含む。またこのような組成物は接種源と呼ぶこともできる。語句「医薬としてまたは薬理学的に許容できる」は、動物またはヒトに適宜投与した場合に有毒な、アレルギー性の、または他の厄介な反応を示さない分子の独立体および組成物を指す。本明細書中で用いられる「医薬として許容される担体」には、ありとあらゆる溶剤、分散媒体、剤皮、抗菌および抗カビ剤、等張化および吸収遅延剤などが含まれる。医薬として活性な物質に対してこのような媒体または薬品を使用することは当業界でよく知られている。任意の通常の媒体または薬品がその活性成分に適合しない場合を除いては治療用組成物中でそれを使用することが考えられる。補助的な活性成分もまた、この組成物に組み込むことができる。
【0085】
これら調合物には液状溶液または懸濁液として注射できる組成物が含まれ、また注射の前に液体状態の溶液または懸濁液にするのに適した固体形態もまた調製することができる。これらの製剤はまた乳化することもできる。このような目的の一般的な組成物は、医薬として許容される担体を含む。例えばこの組成物は、リン酸緩衝生理的食塩水1ミリリットル当たりヒト血清アルブミンを約100 mg含有することができる。他の医薬として許容される担体には塩類、防腐剤、緩衝剤などを含む水溶液、非毒性賦形剤が挙げられ、使用することができる。非水性溶媒の例は、プロピレングリコール、ポリエチレングリコール、植物油、およびオレイン酸エチルなどの注射可能な有機エステル類である。水性担体には、水、アルコール/水溶液、生理的食塩水、塩化ナトリウムなどの非経口伝達体、リンガーデキストロースなどが挙げられる。静脈内伝達体には流体と栄養の補充物が挙げられる。防腐剤には、抗菌物質、抗酸化剤、キレート化剤、および不活性ガスが挙げられる。医薬組成物中の様々な成分のpHおよび正確な濃度は、よく知られている要因に従って調整される。
【0086】
これら調合物は、所望の保管条件に対して最適化することができる。本発明の1つの態様において、特に臨床的使用のために処方されるウィルスの場合、それら試料は液体の形態で、好ましくは一般には約10℃未満、より一般には約5℃未満の冷温で保存される。このような条件の場合、保管にとって好ましい媒体は5%スクロース、1%グリシン、1 mM MgCl2、10 mMトリス、および少量の界面活性剤を含む。注目の界面活性剤の一つは非イオン界面活性剤、例えば約0.01%から約0.1%、好ましくは約0.05%の濃度のトゥイーン80、トゥイーン20などである。注目の他の界面活性剤には、例えば、濃度約5%から約10%、好ましくは約8%のLutrol F−68、Lutrol F−127などのポリエチレングリコールポリプロピレングリコールのポロキサマーブロックポリマーが挙げられる。
【0087】
例えば−20℃または−80℃で冷凍保存される試料の場合、好適な緩衝液は上記のとおりであるが、界面活性剤を含むことは安定性にとって一般にはあまり重要ではなく、省くことができる。約2%から約10%の濃度でグリセロールを含めることができる。
【0088】
本発明のこのウィルス粒子には、ヒトへの投与を含む治療養生法で用いられる古典的な医薬用製剤を挙げることができる。本発明による治療用組成物の投与は、その標的組織がそのルートを経て利用できる限り任意の普通のルートを経ることになる。これには、経口、経鼻、口内、直腸、膣、または局所的が含まれる。別法では投与は、通常位置の、皮内、皮下、筋内、腹膜内、または静脈内の注射によることになる。このような組成物は、生理学的に許容できる担体、緩衝液、または他の賦形剤を含む医薬として許容される組成物として普通は投与されることになる。腫瘍に対して適用する場合、直接の腫瘍内注射、切除後の腫瘍床への注射、局所(すなわちリンパ管)、または全身投与が考えられる。また疾患部位、例えば腫瘍または腫瘍部位へのカテーテルを介しての数時間または数日にわたる連続的灌流を行うことが望ましい場合もある。
【0089】
このアデノウィルスベクターの有効量を、限定しないが、生理的食塩水、適切な緩衝液、防腐剤、安定剤を含む医薬として許容される賦形剤中の組成物(なお同一組成物であってもなくてもよい)として患者に投与することができ、また制吐剤などの適切な薬剤と一緒に投与することもできる。有効量とは、臨床結果を含む有益なまたは望ましい結果をもたらすのに十分な量である。本発明の目的の場合、アデノウィルスベクターの有効量は、病状の進行を和らげ、改善し、安定化し、逆転し、速度を遅らせ、または先に延ばすのに十分な量である。一部の個体はこれらの治療に対して無反応性であり、その方法はこれら個体への投与を包含するものであることが分かる。与えられる量は、その個体の状態、病の程度、投与のルート、どれほどの用量が投与されるか、また所望の目標によって決まるはずである。
【0090】
治療薬の有効量は意図する目的、例えば(i)腫瘍細胞増殖の抑制、(ii)腫瘍細胞を排除または殺すこと、(iii)予防接種などに基づいて決められる。用語「単位用量」は、患者に使用するのに適した物理的に分離した単位を指し、各単位はその投与、すなわち適切なルートおよび治療養生法と共同して、上記で考察した所望の反応を生むように計算される所定量の治療用組成物を含有する。治療の回数および単位用量に応じた投与すべき量は、治療される患者、患者の状況、および希望する結果に左右される。
【0091】
或る特定の治療養生法の有効性の判定は、画像技術、血清腫瘍マーカーの分析、生検、および/または腫瘍に随伴する徴候の有無または改善の評価などの診断法を含めた当業界で知られている技術のいずれかにより求めることができる。或る所定の治療養生法は、有効性をできるだけ大きくするために適宜改変することができることは理解されるはずである。
【0092】
下記の実施例は、本発明をいかに構成し使用するかについての完全な開示および説明を当業者に提供するために述べられ、本発明者等がこの発明と考えるものの範囲を限定することを意図するものではなく、また下記の実験が行った実験の全体または唯一のものであることを意味することも意図しない。使用した数値(例えば量、温度など)に関して精度を保証する努力を行ったが、若干の実験誤差と偏差は斟酌されるべきである。別段の指示がない限り、部は重量部であり、分子量は重量平均分子量であり、温度は摂氏度であり、また圧力は大気圧またはほぼ大気圧である。
【0093】
本発明は、本発明者によって見出されまたは提案された特定の実施形態に関して記述されており、本発明の実施のための好ましい形態を含む。本開示内容に照らして本発明の意図する範囲から逸脱することなく例示の特定の実施形態について非常に多くの修正形態および変更形態を案出することができることは、当業者には理解されるはずである。例えばコドン重複性によりタンパク質配列に影響することなくその基礎をなすDNA中の変更を行うことができる。さらに生物学的な機能上の等価を考慮することにより種類または量的な生物学的活動に影響することなくタンパク質構造中の変更を行うことができる。あらゆるこのような修正形態は、添付の特許請求の範囲の範囲内に含まれるものである。
【実験】
【0094】
(実施例1)
E1A/E1Bパッケージング細胞株の構築:
MMLV発現ベクターの構築。モロニーマウス白血病ウィルス(MMLV)由来プラスミドをE1Aレトロウィルス発現ベクターおよびE1B レトロウィルス発現ベクターを構築するために使用した。具体的にはpRT43.2F3プラスミドが使用された。pRT43.2F3の構築については下記に要約し、完全には米国特許第5,686,279号に記載されている。
【0095】
pRT43.2F3。このレトロウィルスベクターは、レトロウィルスが産生されることになる細胞型中での効率的な転写を指示する修飾5′LTRを含有する。本発明のレトロウィルスベクターは、pZIPneoSVX(Cepko等の論文、Cell 37: 1053-1062 (1985))の新バージョンであるpZen(Johnson等の論文、EMBO Journal 8 (2): 441-448 (1989))を基にして作られ、発現されることになる遺伝子産物は、普通はその新カセット(前掲のCepko等の論文)が占める場所であるスプライス受容部の下流でクローニングされる。さらにMMLVのNar I部位までのウィルスのgag配列(ヌクレオチド1038)をパッケージングの改良のために加え(Armentano等の論文、J. Virol. 61: 11647-1650 (1987))、pZIPneoSVXのXho IからCla Iまでのフラグメントを欠失させた(前掲のCepko等の論文)。pIK1.1由来のEco RIからApa Iのポリリンカーをスプライス受容部の下流に挿入し、pIKプラスミド由来の挿入断片のレトロウィルス構築物中へのトランスフェクションを可能にした。得られたプラスミドはpRTD1.2と呼ばれ、5′および3′MMLV LTRの両方を含有する。このpZIPneoSVXの5′LTR U3領域をpIKMMSVのHindIII/Sac Iフラグメントから得られるMMSV U3で置き換えてpRTD4.2を生じさせた。
【0096】
pRTD2.2においてpZIPneoSVXの5′LTRのU3領域を、CMV即時型エンハンサー/プロモーターをコードするpIK1.1由来のHindIII/Sac Iフラグメントで置き換え、これをMMLVのヌクレオチド+1から+32(KpnI)と結合したHCMVプロモーターのヌクレオチド19(Sac I)から+1をコードするオリゴヌクレオチドによりMMLV R領域に融合させた(Schinnick等の論文、Nature 293: 543-548 (1980))。
【0097】
pRTD2.2の(750 bp)Sac IからBst EIIのフラグメントをLXSNの(736 bp)Sac IからBst EIIのフラグメントで置き換えることによってpRTD2.2SVGを構築した(MillerおよびRosmanの論文、BioTechniques 7: 980-990 (1989))。pRTD2.2の(1441 bp)Sac IからEco RIのフラグメントをLXSNの(1053 bp)Sac IからEco RIのフラグメントで置き換えることによってpRTD2.2SSAを構築した(前掲のMillerおよびRosmanの論文)。その5′末端へのApa I部位の付加反応によって付加されたpLXSN(GenBank取得番号M28248)のヌクレオチド2878〜2955をコードするオリゴヌクレオチドの合成によってpRTD2.2SVGE−を構築した。これを用いてポリリンカーのDNA配列3′および3′LTR中のNhe I部位の5′を含有するpRTD2.2 SVG のApa IからNhe Iのフラグメントを置き換えた。本発明のこれらレトロウィルスベクター構築物は、pBR322バックボーンを有し、かつpRTD2.2、pRTD4.2、pRTD2.2SVG、pRTD2.2SVGE−、およびpRTD2.2SSAを含む。
【0098】
SV40 T抗原を発現する細胞中でのプラスミドの複製を可能にするために、pRTD2.2の5′と3′LTRの間の配列を上記pIK1.1のSac IとEco RI部位の間でクローニングした。これはベクターpIKT2.2を形成するようにSV40の複製起点を含有した。pRTD2.2SVGのHCMVプロモーター中のSac I部位によってその5′末端で画定され、かつpIK1.1のSac IとEco RI部位の間のpRTD2.2SVGの3′LTRの750 bp下流に位置するEco RI部位によりその3′末端で画定されるフラグメントの挿入によってpIKT2.2SVGを構築した。pIKT2.2SVGF3の182塩基対のApa IからNhe Iのフラグメントを上記pRTD2.2SVGE−F3由来の80塩基対のApa IからNhe Iのフラグメントで置き換えることによってpIKT2.2SVGE−F3を構築した。
【0099】
3′LTRから約750塩基対下流に位置するEco RIからApa Iのポリリンカーを、Asc I認識部位を含有する合成オリゴヌクレオチドで置き換えることによってpIKT2.2SVGE−F3からpRT43.2F3を誘導した。さらにウィルスのgag配列の3′末端にあるNde I部位をオリゴヌクレオチド挿入によってXhoIに変換した。最初にpRT43.2F3中の3′LTRの除去と、PvuIIからXbaIまでの配列が欠失した3′LTRの挿入とによってpRT43.2F3からpRT43.3PGKF3を誘導した(MMLV、GenBank取得番号#J02255、ヌクレオチド番号7938〜8115)。さらにMMLVスプライス受容部領域をヒトホスホグリセリン酸キナーゼ遺伝子プロモーター(GenBank取得番号#M11958、ヌクレオチド2〜516)で置き換え、それをクローニングしてその5′末端にXho I部位とその3′末端にEco RI部位とを有するポリリンカーにした。
【0100】
E1A発現ベクター−rkat43.2E1A。rkat 43.2E1a(図1)は、レトロウィルスLTRの制御下でそのAd5 E1Aオープンリーディングフレームを発現するレトロウィルスベクターである。したがってアデノウィルスまたは哺乳動物の宿主細胞由来のプロモーターのどちらもAd E1Aの発現を方向づけるのには使用されない。rkat 43.2E1Aは、pRT43.2F3のCD4/ζコード配列(米国特許第5,686,279号およびRoberts等の論文、J. Immunology (1998) 161: 375-384)を、Ad5 E1Aオープンリーディングフレーム(Ad5ヌクレオチド548〜1575、GenBank取得X02992、配列番号1)をコードするDNA配列で置き換えることによって生じさせた。
【0101】
E1B発現ベクター−rkat 43.2E1B。rkat 43.2E1B′(図2)を pRT43.2F3から同様に生じさせた。このベクターは、レトロウィルスLTRの制御下でAd5 E1Bオープンリーディングフレーム(Ad5のヌクレオチド1682〜3825、GenBank取得X02996、配列番号4)を発現させ、アデノウィルスまたは哺乳動物の宿主細胞由来のプロモーターを含まない。rkat 43.2E1Aは、pRT43.2F3のCD4/ζコード配列(米国特許第5,686,279号および前掲のRoberts等の論文)を、Ad5 E1B mRNAをコードするcDNA配列で置き換えることによって生じさせた。
【0102】
一過性のレトロウィルス生産。E1AおよびE1B発現ベクターを含む感染性粒子を標準的な方法を用いて生産した。rkat 43.2E1Aまたはrkat 43.2E1BプラスミドにMCVecog/pおよび6.1CMVamphoenvを同時感染させることによって一過性のウィルス上澄みを調製した。次いで得られたウィルス上澄みMMLV−E1A (C5.03−.04と呼ぶ)およびMMLV−E1B(C5.05−.06と呼ぶ)を細胞の形質導入に使用した。
【0103】
A549細胞の形質導入。ナイーブA549細胞(ATCC番号CCL−185)をDMEM/High、10%ウシ胎児血清、1%グルタミン、および1%PenStrepを含む完全培地中で培養した。細胞にMMLV−E1AおよびMMLV E1Bウィルスをスピンオキュレーションにより同時感染させることによってアデノウィルスE1AおよびE1Bコード配列をA549細胞中に安定的に導入した。1.5×105個の細胞をE1A/E1Bウィルス上澄み1 mLおよびポリブレン8 μL/mL中に再度懸濁させた。次いでこの細胞およびウィルスの混合物を3400 rpm、34℃において4時間遠心分離した。最適なE1A/E1B比を確実にするために3つの異なるE1A/E1B比(E1A 25%/E1B 75%、E1A 50%/E1B 50%、E1A 75%/E1B 25%)をスピンオキュレーションに用いた。ウィルスの形質導入効率をモニターするための対照としてMMLV−グリーン蛍光タンパク質(GFP)ウィルスを含めた。スピンオキュレーションの後、これら3集団を完全培地で再度懸濁させ、6ウェルプレートに移し、5%インキュベーター中で37℃において8日間インキュベートした。
【0104】
E1A/E1B形質導入細胞の希釈クローニング。これら3集団を10 cm皿上で希釈クローニングした。培養状態で18日後、これら3集団の間にはっきりと違いが現れた。3集団由来のすべてのクローンを採取して96ウェルプレートに入れた。次いで同定した後に各プレートをトリプシン処理し、一方を機能的スクリーニング用に、また他方を機能クローンの修復用に使用することができるように2つの96ウェルプレートに分けた。細胞を5日間生育して増殖させた。
【0105】
E1補完に関する機能的スクリーニング。GFP導入遺伝子を担持するE1欠損組換えアデノウィルスを用いてアデノウィルス複製を支持する能力についてE1A/E1B形質導入クローンのそれぞれを試験した。候補クローンを含有する二倍にした96ウェルプレートのうちの一組にE1欠損Ad−GFPウィルスを100 μL/ウェル、M.O.I. 10で48時間感染させた。これらの細胞を3回の冷凍/解凍サイクルにかけてウィルス粒子を放出させた。これらのライゼートを用いてHuH7細胞に感染させた。感染3日後にHuH7細胞を採取し、FACSにより分析してGFPの発現を評価した。表2は、GFPの発現に関する複数のクローンの機能的特徴の結果を示す。この機能的特徴は、候補クローン由来のアデノGFP上澄みによるHuH7細胞の感染および最初のスクリーニングとしての増幅の評価に基づく。その上澄みがHuH7細胞中で高いGFP発現を示したクローンを、候補のE1A/E1B陽性パッケージングクローンとみなした。HuH7細胞を最大限度まで形質導入したこれらのクローンを、下記でさらに述べるように増殖させ、プラーク検定で力価測定した。
【0106】
【表2】
【0107】
E1A 50%/E1B 50%のE1A/E1Bウィルス比のスピンオキュレーション後に得られる細胞集団は、E1 Ad−GFPに感染させる前に生き残っている「健康な」クローン(形態によって決められる)の数が最も少なかったが、E1欠損ウィルスを補完することができるクローン産生の比率はより高かった。E1A 75%/E1B 25%の形質導入比から生まれるクローンは最もすぐれた全体的な成長特性を示したが、E1A 50%−E1B 50%クローンのE1欠損ウィルスで観察される同じ補完性をもたらさなかった。
【0108】
E1欠失および腫瘍溶解性アデノウィルスを補完する能力に関するクローンの評価。E1Aを欠失したGFP発現性の複製能欠損アデノウィルスに感染させることによりE1A/E1B補完的細胞を候補パッケージング細胞として評価した(E1A/E1B補完的細胞、親細胞、およびE1A/E1B陰性細胞を比較するウィルス収量検定を使用)。各クローンによるウィルス産生を比較することによってこれらのE1A/E1B補完的細胞をウィルス複製の支持能力に関して定量的に評価した。試験クローンにE1Aを欠失したGFP発現性の複製能欠損アデノウィルスをM.O.I. 5で4時間感染させ、新鮮な培養液を再度与え、72時間インキュベートした。細胞および培地を一緒に採取し、3回の冷凍/解凍にかけた。血清を含まない培地中で連続的な希釈を行い、293細胞上で検定した。それら細胞は、その24時間前に6ウェルプレート上に1ウェル当たり培養液4 mL当たり0.5×106個の細胞でプレーティングされた。これら試料を吸引し、各ウェルにアガロース(完全培地中0.8%アガロース)4 mLを覆いとして適用した。これらプレートを室温で放置して固化させ、プラークを展開させるためにインキュベートした。表3は、プラーク検定により得られた力価を示す。次いでその最もすぐれたクローンを、複製能のある(腫瘍溶解性)アデノウィルスベクターAFP−CG8900を用いた更なるプラーク検定で分析した(図2B)。これらの結果に基づいてそれぞれE1A/E1Bクローン51およびクローン139と呼ばれる2種類のクローンの特徴をさらに調べた。
【0109】
【表3】
【0110】
E1欠失アデノウィルスの産生ならびにE1クローン51および139の安定性。293細胞、PerC6細胞、ならびにE1A/E1Bクローン51および139から得られた細胞を成長させ、M.O.I.=75でE1欠失アデノGMCSFを感染させた。72時間後、細胞を採取した。ウィルスの粗ライゼートを3回の冷凍/解凍によって調製し、ウィルスの力価をHPLCにより求めた。感染後、293、PerC6、E1A/E1Bクローン51、およびクローン139細胞に週2回の継代を行った。Ad−GMの産生を継代1、10、および20世代(1、5、10週)で試験した。結果は、クローン51および139がPerC6および293細胞に匹敵する水準でE1欠損ウィルスを産生することができ、またクローン51およびクローン139細胞株によるウィルスの産生が安定していることを示した(図3)。
【0111】
E1A/E1B補完クローンから得られた細胞株を用いた腫瘍溶解性ウィルスの産生。A549細胞、293細胞、ならびにE1A/E1Bクローン51および139から得られた細胞を成長させ、4種類の複製能のある(腫瘍溶解性)アデノウィルス(CG8900、CG8840、OV945、およびOV1025)をM.O.I. 2で3〜4時間感染させた。72時間後、上澄みを採取し、これを用いて標準的なプラーク検定により293細胞に様々な希釈度で感染させた。感染を4時間進行するに任せ、次いでこの細胞をアガロース培地中で8〜11日間培養した。結果を表4に示す。CG8900、CG8840、OV945、およびOV1025は、E1AおよびE1Bを制御する細胞型特異的転写調節エレメントを含む複製能のあるアデノウィルスベクターである。
【0112】
【表4】
【0113】
初期および後期継代細胞中のE1A/E1B配列に関するクローン51および139のサザンブロット分析。ベクター内で一旦切断するBamH Iでクローン51、クローン139、および293細胞の初期および後期継代培養物を消化し、E1A(nts 827〜1340)またはE1B(nts 2805〜3329)特異的プローブを用いてサザンブロットにより分析した(図4AおよびB)。これらのブロットは、初期および後期の両継代細胞中へのE1AおよびE1B遺伝子の安定した組込みを示す。クローン139は、E1A遺伝子の2つのコピーとE1Bの1つのコピーを有するように見える。クローン51は、E1Aの4つのコピー(上側のバンドはダブレットである)とE1Bの2つのコピーを有する。星印は関連のあるバンドを示す。
【0114】
パッケージング細胞株中のゲノム組込みの特徴の記述。E1AおよびE1B遺伝子の同時形質導入に利用される別個の発現ベクターは、宿主細胞のゲノム中で異なる場所に組み込まれるべきであり、これは複製能欠損Adベクターと本発明のパッケージング細胞株の間の組換えの可能性をさらに低減させるように働く。クローン51と139、293、およびPerC6細胞由来のDNAを、E1A、E1B、および連続E1A−E1B配列に特効のあるプライマーを用いてPCRによりプローブした(図5)。プライマー1460.138.3/4(1460.138.3: 5′TGT GTC TAG AGA ATG CAA TAG 3′、1460.138.4: 5′GAT ATA TGT CGA CTG GCC TGG GGC GTT TAC AGC 3′)が、E1Aコード領域のc末端でAd5ゲノムのnts 1338から1542を増幅し、プライマー1460.138.5/6(1460.138.5: 5′GAC ATG CGT CGA CAT GGA GCG AAG AAA CCC ATC TG 3′、1460.138.6: 5′CCA TAG AAG CTT ACA CCG TGT AG 3′)が、nts 2019から2815(E1B 55kオープンリーディングフレームの大部分)を増幅する。すべての4つの細胞株から得られたサザンブロットの結果は、予想された個々のE1AおよびE1Bフラグメントが存在することを示した。E1Aのc末端からE1B 55kまでのAd5ゲノムが無傷であり、かつ293およびPerC6細胞株の場合と同様に連続的である場合には、1460.138.3/6はnts 1338から2815までの1477塩基対領域を増幅することになる。293およびPerC6細胞株由来のDNAのサザンブロットの結果は、1477塩基対領域が存在することを示した。一方、クローン51および139由来のDNAのサザンブロットの結果は、連続したE1A/E1Bゲノムがクローン中に存在しないことを裏付ける1477塩基対領域を示さなかった。
【0115】
クローン51および139のタンパク質分析。当業界で広く使用されている方法(例えばAntonおよびGrahamの論文、J. Virology, 69, 4600-4606, 1995、ならびに前掲のSambrookおよびRussellの著書)を用いてウェスタンブロット分析を行った。293、PerC6、A549、A549クローン51、およびA549クローン139細胞を10 cm組織培養皿にプレーティングした。72時間後、細胞を擦って上澄み中に入れ、ペレット化し、完全ミニプロテアーゼ阻害薬カクテル(Roche)を追加した溶解緩衝液(100 mM NaCl、20 mMトリス(pH 7.5)、10 mM EDTA、1%デオキシコール酸)中に再度懸濁させた。試料のタンパク質濃度をタンパク質検定キット(Bio-Rad)で推定した。E1Aの検出(図6A)の場合、各試料について合計タンパク質10 mgを4〜12%NuPage Novex Bis-Yris SDS-PAGEゲル(Invitrogen)上に載せ、NuPage MOPSランニング緩衝液中で分別した。画分をInvitrolon PVDF膜(Invitrogen)に移し、モノクローナルE1A一次抗体(Neomarkers)および西洋ワサビペルオキシダーゼ複合二次抗体によりプローブした。結合した抗体複合体を高感度化学発光(Amersham)により検出した。E1B 19KおよびE1B 55Kの検出は、下記の点を除いて上記と同様に行った(図6B)。各試料から得られた全タンパク質25 mgを分別した。使用した一次抗体は、E1B 19KおよびE1B 55K に対するモノクローナル抗体である(Oncogene Research Products)。これらの結果は、クローン51および139が本質的にPerC6と同等のE1A(38および46 kD)発現レベルであり、また293細胞について検出されるものよりも高いE1A発現レベルを有することを示した。これら2つのクローンにおけるE1B 19Kの産生もまた、293およびPerC6細胞で検出されるレベルと同等である。
【0116】
RCA検出検定。クローン51および139細胞を、従来からアデノウィルスの大量生産用に使用されている細胞株の293およびPerC6細胞と比較するためにRCAの検討を行った(Gao等の論文(2000)、Hum Gene Therapy 11: 213、Murakami等の論文(2002)、Hum Gene Thera.13: 909、Kim等の論文(2001)、Exp Mol Med 33: 145)。各細胞型を使用し、精製したE1A欠失Ad GM−CSFウィルスストックを20世代まで継代させた。クローン51、クローン139、および293細胞にM.O.I.=75でE1欠失GM−CSFウィルスを感染させた。72時間後、粗ライゼートを調製し、HPLCにより滴定し、これを同じ型のナイーブ細胞への第二ラウンドの感染(ナイーブ293細胞への293ライゼートなど)に用いた。各感染については同数の粒子を用いた。この感染サイクルは18回繰り返した。感染サイクル1、9、および18回においてアデノウィルスDNAをライゼートから精製し、E1A(nts 133〜696)領域に特異的なプライマーを用いてPCRにより増幅した(図7)。その予想されたPCR生成物は約0.56 Kbである。このRCA検定用に用いたPCRプライマーは、66.114.2: 5′−GTGGCGGAACACATGTAAGCG−3′および49.17.2: 5′−AGTTCGTGAAGGGTAGGTGGTTC−3′であった。E1A配列(RCAに起因)は、9サイクルまでの293細胞由来の継代中に検出された。クローン51および139から得られるアデノウィルスは、18サイクルを通してE1A配列に関して陰性であった。
【0117】
本発明は、例示した組換え発現ベクターおよび細胞株による範囲内に限定されるべきではなく、これらは本発明の一態様の実例として意図されている。上記の詳細な実施例および上述の実施形態は例示を意図しており、本発明の範囲を限定するものではないことを理解されたい。本発明の範囲内の他の態様、利点、および修正形態は、本発明が属する業界の技術者には明らかなはずである。
【図面の簡単な説明】
【0118】
【図1】アデノウィルスE1AおよびE1Bプロモーター配列を含まないMMLV−E1Aレトロウィルス発現カセットを示す図である。
【図2】アデノウィルスE1AおよびE1Bプロモーター配列を含まないMMLV−E1Bレトロウィルス発現カセットを示す図である。
【図3】293、PerC6、クローン51、およびクローン139細胞上でGM−CSFを発現する複製能欠陥アデノウィルスの産生を示す図である。
【図4A】クローン51および139から得られる初期および後期継代細胞のE1A(図4A)およびE1B(図4B)配列に関するサザンブロット分析の結果を示す図である(星印は関連のあるバンドを示す)。
【図4B】クローン51および139から得られる初期および後期継代細胞のE1A(図4A)およびE1B(図4B)配列に関するサザンブロット分析の結果を示す図である(星印は関連のあるバンドを示す)。
【図5】E1A(1460.138.3/4)、E1B(1460.138.5/6)、および連続E1A−E1B配列(1460.138.3/6)に特異的なPCRプラマーを用いてプローブしたクローン51および139、PerC6、293、およびA549細胞から得られるDNAのサザンブロット分析の結果を示す図である。
【図6A】E1A、E1B 19K、またはE1B 55kに向けられるモノクローナル抗体でプローブしたA549細胞、クローン51細胞、クローン139、PerC6細胞、および293細胞のライゼートを用いて行ったウェスタンブロット分析の結果を示す図である。
【図6B】E1A、E1B 19K、またはE1B 55kに向けられるモノクローナル抗体でプローブしたA549細胞、クローン51細胞、クローン139、PerC6細胞、および293細胞のライゼートを用いて行ったウェスタンブロット分析の結果を示す図である。
【図7】野生型アデノウィルス(wt Ad)、293細胞、クローン51、およびクローン139のライゼートから精製されたDNAをE1Aに特異的なプライマー(nts 133〜696)を用いてPCRにより増幅したRCA検出検定の結果を示す図である。
【図8】E1AおよびE1Bプロモーター配列を含まず、かつ5′から3′方向へ5′ITR、サイトメガロウィルスエンハンサー/ニワトリβアクチン/ウサギβグロブリンプロモーター(CAGプロモーター;Niwa等の論文(1991)、Gene 108 (2): 193-9)、E1Aのエキソン1およびエキソン2(配列番号1)、ウッドチャック転写後調節エレメント(WPRE)、子ウシ成長ホルモンポリA(BGHpA)配列、および3′ITRを含むAAV−E1A発現カセットを示す図である。
【図9】E1AおよびE1Bプロモーター配列を含まず、かつ5′から3′方向へ5′ITR、延長因子1−αプロモーター(EF1−α)のプロモーター(Kim等の論文(1990)、Gene 91 (2): 217-23およびGuo等の論文(1996)、Gene Ther. 3 (9): 802-10)とエンハンサー、19K E1Bコード配列と55K E1Bコード配列(配列番号4)、子ウシ成長ホルモンポリA(BGHpA)配列、および3′ITRを含むAAV−E1B発現カセットを示す図である。
【特許請求の範囲】
【請求項1】
E1A/E1B欠損アデノウィルスベクターの複製を許容するアデノウィルスパッケージング細胞株であって、天然のアデノウィルスE1AまたはE1Bプロモーターとの実質的な配列同一性を欠くプロモーターと作用可能に連結したアデノウィルスE1Aコード配列およびアデノウィルスE1Bコード配列を含んで成る細胞株。
【請求項2】
前記アデノウィルスE1Aコード配列および前記アデノウィルスE1Bコード配列が安定的に前記細胞株に組み込まれている、請求項1に記載のアデノウィルスパッケージング細胞株。
【請求項3】
前記アデノウィルスE1Aコード配列および前記アデノウィルスE1Bコード配列が同一のプロモーターに作用可能に連結している、請求項2に記載のアデノウィルスパッケージング細胞株。
【請求項4】
前記アデノウィルスE1Aコード配列および前記アデノウィルスE1Bコード配列が同じプロモーターに作用可能に連結している、請求項2に記載のアデノウィルスパッケージング細胞株。
【請求項5】
前記アデノウィルスE1Aコード配列および前記アデノウィルスE1Bコード配列が異なるプロモーターに作用可能に連結している、請求項2に記載のアデノウィルスパッケージング細胞株。
【請求項6】
前記アデノウィルスE1Aコード配列および前記アデノウィルスE1Bコード配列が前記細胞株中の異なる部位に安定的に組み込まれている請求項5に記載のアデノウィルスパッケージング細胞株。
【請求項7】
前記細胞株がヒトの細胞株である、請求項6に記載のアデノウィルスパッケージング細胞株。
【請求項8】
前記細胞株が、アデノウィルス複製を許容するA549細胞、PC−3細胞、およびアデノウィルス産生を許容する一次細胞からなる群から選択される、請求項7に記載のアデノウィルスパッケージング細胞株。
【請求項9】
天然のアデノウィルスE1AまたはE1Bプロモーターとの実質的な配列同一性を欠く前記プロモーターが構成性プロモーターである、請求項1に記載のアデノウィルスパッケージング細胞株。
【請求項10】
天然のアデノウィルスE1AまたはE1Bプロモーターとの実質的な配列同一性を欠く前記プロモーターが調節可能なプロモーターである、請求項1に記載のアデノウィルスパッケージング細胞株。
【請求項11】
前記プロモーターがレトロウィルスプロモーターである、請求項9に記載のアデノウィルスパッケージング細胞株。
【請求項12】
前記アデノウィルスE1Aコード配列がアデノウィルス243遺伝子産物、289遺伝子産物、または243と289の両方の遺伝子産物をコードする、請求項1に記載のアデノウィルスパッケージング細胞株。
【請求項13】
前記アデノウィルスE1Aコード配列が配列番号1に示される配列を含む、請求項12に記載のアデノウィルスパッケージング細胞株。
【請求項14】
前記アデノウィルスE1Bコード配列がアデノウィルス19 Kd遺伝子産物、55 Kd遺伝子産物、または19と55 Kdの両方の遺伝子産物をコードする、請求項1に記載のアデノウィルスパッケージング細胞株。
【請求項15】
前記アデノウィルスE1Bコード配列が配列番号4に示される配列を含む、請求項14に記載のアデノウィルスパッケージング細胞株。
【請求項16】
アデノウィルスパッケージング細胞株であって、前記細胞株のゲノム中に安定的に組み込まれた第一発現ベクターおよび第二発現ベクターを含み、前記第一ベクターが非アデノウィルスプロモーターと作用可能に連結したアデノウィルスE1Aコード配列を含み、かつ前記第二ベクターが非アデノウィルスプロモーターと作用可能に連結したアデノウィルスE1Bコード配列を含む、細胞株。
【請求項17】
E1A/E1B欠損アデノウィルスベクターの複製を許容するアデノウィルスパッケージング細胞株の生産方法であって、
アデノウィルス複製を許容する細胞株中に、(i)天然のアデノウィルスE1AまたはE1Bプロモーターとの実質的な配列同一性を欠くプロモーターと作用可能に連結したアデノウィルスE1Aコード配列、及び(ii)天然のアデノウィルスE1AまたはE1Bプロモーターとの実質的な配列同一性を欠くプロモーターと作用可能に連結したアデノウィルスE1Bコード配列、を含む発現ベクターを導入するステップを含み、前記細胞株が複製能のあるアデノウィルス(RCA)を実質上含まない組換えアデノウィルスを産生することができる、方法。
【請求項18】
前記アデノウィルスE1Aコード配列および前記アデノウィルスE1Bコード配列が別々の発現ベクター上に存在する、請求項17に記載の方法。
【請求項19】
前記アデノウィルスE1Aコード配列および前記アデノウィルスE1Bコード配列が同じ発現ベクター上に存在する、請求項17に記載の方法。
【請求項20】
前記E1A発現ベクターがレトロウィルス発現ベクターである、請求項17に記載の方法。
【請求項21】
前記E1B発現ベクターがレトロウィルス発現ベクターである、請求項17に記載の方法。
【請求項22】
前記E1Aと前記E1Bの両方の発現ベクターがレトロウィルス発現ベクターである、請求項17に記載の方法。
【請求項23】
E1A/E1B欠損アデノウィルスを請求項1に記載のパッケージング細胞株中に導入するステップと、前記細胞株から複製能のあるアデノウィルス(RCA)を実質上含まないアデノウィルスの集団を回収するステップとを含む、E1A/E1B欠損アデノウィルスの生産方法。
【請求項1】
E1A/E1B欠損アデノウィルスベクターの複製を許容するアデノウィルスパッケージング細胞株であって、天然のアデノウィルスE1AまたはE1Bプロモーターとの実質的な配列同一性を欠くプロモーターと作用可能に連結したアデノウィルスE1Aコード配列およびアデノウィルスE1Bコード配列を含んで成る細胞株。
【請求項2】
前記アデノウィルスE1Aコード配列および前記アデノウィルスE1Bコード配列が安定的に前記細胞株に組み込まれている、請求項1に記載のアデノウィルスパッケージング細胞株。
【請求項3】
前記アデノウィルスE1Aコード配列および前記アデノウィルスE1Bコード配列が同一のプロモーターに作用可能に連結している、請求項2に記載のアデノウィルスパッケージング細胞株。
【請求項4】
前記アデノウィルスE1Aコード配列および前記アデノウィルスE1Bコード配列が同じプロモーターに作用可能に連結している、請求項2に記載のアデノウィルスパッケージング細胞株。
【請求項5】
前記アデノウィルスE1Aコード配列および前記アデノウィルスE1Bコード配列が異なるプロモーターに作用可能に連結している、請求項2に記載のアデノウィルスパッケージング細胞株。
【請求項6】
前記アデノウィルスE1Aコード配列および前記アデノウィルスE1Bコード配列が前記細胞株中の異なる部位に安定的に組み込まれている請求項5に記載のアデノウィルスパッケージング細胞株。
【請求項7】
前記細胞株がヒトの細胞株である、請求項6に記載のアデノウィルスパッケージング細胞株。
【請求項8】
前記細胞株が、アデノウィルス複製を許容するA549細胞、PC−3細胞、およびアデノウィルス産生を許容する一次細胞からなる群から選択される、請求項7に記載のアデノウィルスパッケージング細胞株。
【請求項9】
天然のアデノウィルスE1AまたはE1Bプロモーターとの実質的な配列同一性を欠く前記プロモーターが構成性プロモーターである、請求項1に記載のアデノウィルスパッケージング細胞株。
【請求項10】
天然のアデノウィルスE1AまたはE1Bプロモーターとの実質的な配列同一性を欠く前記プロモーターが調節可能なプロモーターである、請求項1に記載のアデノウィルスパッケージング細胞株。
【請求項11】
前記プロモーターがレトロウィルスプロモーターである、請求項9に記載のアデノウィルスパッケージング細胞株。
【請求項12】
前記アデノウィルスE1Aコード配列がアデノウィルス243遺伝子産物、289遺伝子産物、または243と289の両方の遺伝子産物をコードする、請求項1に記載のアデノウィルスパッケージング細胞株。
【請求項13】
前記アデノウィルスE1Aコード配列が配列番号1に示される配列を含む、請求項12に記載のアデノウィルスパッケージング細胞株。
【請求項14】
前記アデノウィルスE1Bコード配列がアデノウィルス19 Kd遺伝子産物、55 Kd遺伝子産物、または19と55 Kdの両方の遺伝子産物をコードする、請求項1に記載のアデノウィルスパッケージング細胞株。
【請求項15】
前記アデノウィルスE1Bコード配列が配列番号4に示される配列を含む、請求項14に記載のアデノウィルスパッケージング細胞株。
【請求項16】
アデノウィルスパッケージング細胞株であって、前記細胞株のゲノム中に安定的に組み込まれた第一発現ベクターおよび第二発現ベクターを含み、前記第一ベクターが非アデノウィルスプロモーターと作用可能に連結したアデノウィルスE1Aコード配列を含み、かつ前記第二ベクターが非アデノウィルスプロモーターと作用可能に連結したアデノウィルスE1Bコード配列を含む、細胞株。
【請求項17】
E1A/E1B欠損アデノウィルスベクターの複製を許容するアデノウィルスパッケージング細胞株の生産方法であって、
アデノウィルス複製を許容する細胞株中に、(i)天然のアデノウィルスE1AまたはE1Bプロモーターとの実質的な配列同一性を欠くプロモーターと作用可能に連結したアデノウィルスE1Aコード配列、及び(ii)天然のアデノウィルスE1AまたはE1Bプロモーターとの実質的な配列同一性を欠くプロモーターと作用可能に連結したアデノウィルスE1Bコード配列、を含む発現ベクターを導入するステップを含み、前記細胞株が複製能のあるアデノウィルス(RCA)を実質上含まない組換えアデノウィルスを産生することができる、方法。
【請求項18】
前記アデノウィルスE1Aコード配列および前記アデノウィルスE1Bコード配列が別々の発現ベクター上に存在する、請求項17に記載の方法。
【請求項19】
前記アデノウィルスE1Aコード配列および前記アデノウィルスE1Bコード配列が同じ発現ベクター上に存在する、請求項17に記載の方法。
【請求項20】
前記E1A発現ベクターがレトロウィルス発現ベクターである、請求項17に記載の方法。
【請求項21】
前記E1B発現ベクターがレトロウィルス発現ベクターである、請求項17に記載の方法。
【請求項22】
前記E1Aと前記E1Bの両方の発現ベクターがレトロウィルス発現ベクターである、請求項17に記載の方法。
【請求項23】
E1A/E1B欠損アデノウィルスを請求項1に記載のパッケージング細胞株中に導入するステップと、前記細胞株から複製能のあるアデノウィルス(RCA)を実質上含まないアデノウィルスの集団を回収するステップとを含む、E1A/E1B欠損アデノウィルスの生産方法。
【図1】

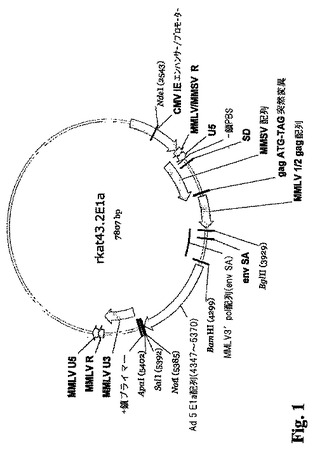
【図2】
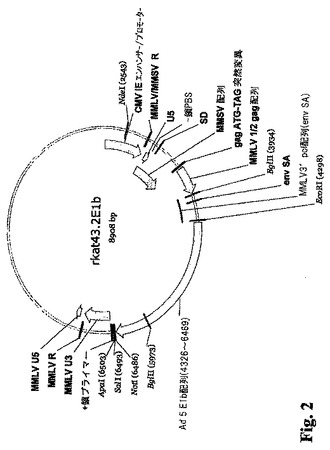
【図3】
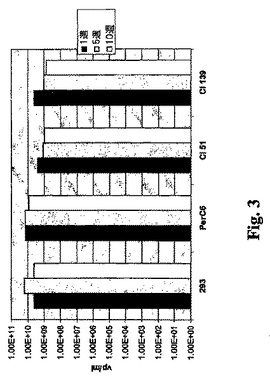
【図4A】

【図4B】
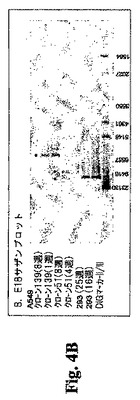
【図5】
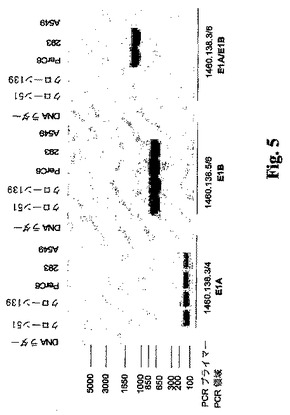
【図6A】
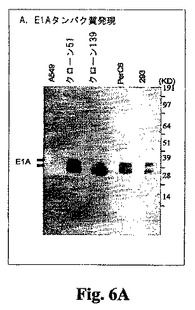
【図6B】
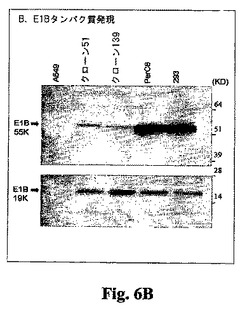
【図7】
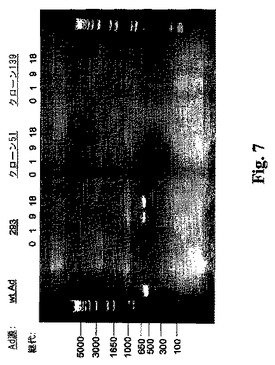
【図8】
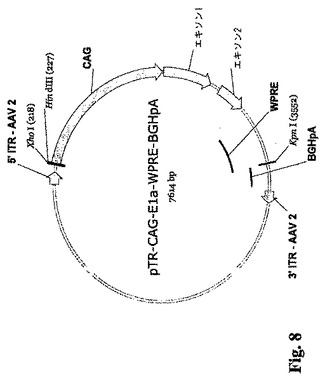
【図9】
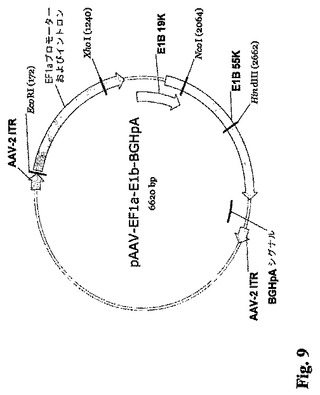
【図2】
【図3】
【図4A】
【図4B】
【図5】
【図6A】
【図6B】
【図7】
【図8】
【図9】
【公表番号】特表2007−527215(P2007−527215A)
【公表日】平成19年9月27日(2007.9.27)
【国際特許分類】
【出願番号】特願2006−518624(P2006−518624)
【出願日】平成16年5月28日(2004.5.28)
【国際出願番号】PCT/US2004/016867
【国際公開番号】WO2005/010146
【国際公開日】平成17年2月3日(2005.2.3)
【出願人】(597078983)セル ジェネシス インコーポレイテッド (21)
【Fターム(参考)】
【公表日】平成19年9月27日(2007.9.27)
【国際特許分類】
【出願日】平成16年5月28日(2004.5.28)
【国際出願番号】PCT/US2004/016867
【国際公開番号】WO2005/010146
【国際公開日】平成17年2月3日(2005.2.3)
【出願人】(597078983)セル ジェネシス インコーポレイテッド (21)
【Fターム(参考)】
[ Back to top ]