
アデノウイルスおよびそれをコードしている核酸の新たな使用
【課題】アデノウイルスならびにそれをコードする核酸、ならびに組換え体発癌タンパク質の使用方法の提供。
【解決手段】ウイルスはその核内にYB−1を含まない細胞では複製を欠き、そして発癌遺伝子または発癌遺伝子産物、特に発癌遺伝子タンパク質をコードする核酸を有しており、この産物は少なくとも1つのウイルス遺伝子、好ましくはアデノウイルス遺伝子をトランス活性化する。上記遺伝子はE1B55kDa、E4orf6、E4orf3およびE3ADPを含む群から選ばれる。
【解決手段】ウイルスはその核内にYB−1を含まない細胞では複製を欠き、そして発癌遺伝子または発癌遺伝子産物、特に発癌遺伝子タンパク質をコードする核酸を有しており、この産物は少なくとも1つのウイルス遺伝子、好ましくはアデノウイルス遺伝子をトランス活性化する。上記遺伝子はE1B55kDa、E4orf6、E4orf3およびE3ADPを含む群から選ばれる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はアデノウイルスならびにそれをコードする核酸、ならびに組換え体発癌タンパク質の使用に関する。
【背景技術】
【0002】
腫瘍の治療には、現在多くの治療コンセプトが用いられている。手術に加え、化学療法および放射線治療がよく実施されている。しかしながらこれ技術全てに、無視できない副作用が伴う。複製特異的殺腫瘍ウイルスの使用が、腫瘍治療に新たなプラットホームを提供している。この方法では、腫瘍内選択的な上記ウイルスの複製が起こり、その結果ウイルスの複製、感染腫瘍細胞の溶解、および近接腫瘍細胞へのウイルスの放散が起こる。ウイルスの複製能力が腫瘍細胞に限定されることから、正常組織にはウイルスの複製は広がらず、従って溶解も起きない。
【0003】
現在、殺腫瘍を狙った複数のウイルス系が臨床試験にかけられている。かかるアデノウイルスの一例が、臨床第I相および第II相試験で良い結果を上げているD11520(Onyx−015)である(Khuri, Fら、Nature Medicine 6、879〜885、2000年)。Onyx−015は、E1B−55kDa遺伝子を完全に欠失したアデノウイルスである。このアデノウイルスが完全にE1B55kDaタンパク質を欠いていることは、p53欠失アデノウイルスベクターと一緒であれば複製すること、従って細胞を溶解することができるが(Kim, Dら、Proc.Am.Soc.Clin.Oncol.17、391a、1998年)、正常細胞を損傷することはないという発見に基づいている。より具体的には、E1B−55kDa遺伝子産物は、p53の阻害、ウイルスmRNAの輸送、および宿主細胞のタンパク質合成のスイッチングオフに関係している。p53の阻害は、p53とアデノウイルスがコードするE1B−55kDaタンパク質から成る複合体、および/またはE1B−55kDaとE4orf6.より成る複合体の形成を通じて起こる。TP53がコードするp53は、複合体制御メカニズムにとって開始点であり(Zambetti, G.Pら、FASEB J.7、855〜865、1993年)、p53はとりわけてもアデノウイルスのようなウイルスの細胞内複製が効率的に阻害する。TP53は全てのヒト腫瘍の約50%で欠失または突然変異しており、そのために化学療法または放射線治療では−望まれる−アポトーシスが起こらず、結果として腫瘍治療に成功しないことが多々ある。
【0004】
殺腫瘍アデノウイルスの別のコンセプトは、E1Aタンパク質が特殊な欠失型として存在するか、または1もしくはそれ以上の突然変異を含み、それらがRb/E2Fおよび/またはp107/E2Fおよび/またはp130/E2Fの結合に影響しない場合には、かかるアデノウイルスは感染細胞をS期に誘導せず、そして機能性のRbタンパク質を持たない腫瘍細胞内では複製できるという発見に基づいている。さらにE1Aタンパク質はN末端を欠失することも、そしてE1Aタンパク質のアミノ酸位置1〜76の領域に1またはそれ以上の突然変異を含むこともでき、その結果それぞれp300に対するE1Aの結合を阻害し、そしてそれにより腫瘍細胞内での選択的複製を提供することができる。このようなアプローチは欧州特許第0 931 830号に例示されている。かかるウイルスの例としてはAdΔ24、d1922−947、E1Ad/01/07およびCB016がある(Howe, J.A.ら、Molecular Therapy 2、485〜495、2000年; Fueyo、J.ら、Oncogene 19、2〜12、2000年; Heise, C.ら、Nature Medicine 6, 11341139、2001年; Balague, Cら、J.Virol.75、7602〜7611、2001年)。かくして、これら当分野既知の殺腫瘍に関するアデノウイルス系はE1Aタンパク質内に明瞭な欠失を含んでいるが、その場合この種の欠失は、機能性Rbタンパク質および非活性型Rbタンパク質とE2Fとから成る複合体がそれぞれ、効率的なインビボ(in vivo)での複製を遮断すること、そしてRb−欠失/突然変異細胞でのみアデノウイルスのインビボ複製を提供すること前提として作られている。これら従来技術によるアデノウイルス系は、E1Aに基づき、初期E2プロモータ(E2初期プロモータ)およびフリーE2F(Dyson, N.Genes & Development、12、2245〜2262、1998年)を用いてインビボ複製を制御している。
【0005】
殺腫瘍アデノウイルス系の別の形態は、腫瘍細胞内での選択的複製を提供するウイルス発癌遺伝子E1Aを特異的に発現させる選択的プロモータの使用に基づいている(Rodriguez, R.ら、Cancer Res.57、2259〜2563、1997年)。
【0006】
上記の如く、各コンセプトの基礎を成す作用様式に適った細胞背景を選択することが、アデノウイルスに関する殺腫瘍ウイルスの各種コンセプトにとって重要である。換言すれば、現在知られている各種アデノウイルス系は、明瞭な分子生物学的前提条件が揃っている場合にのみ使用することができるだろう。このことが、かかる系の使用を一部患者群に制限している。
【0007】
腫瘍疾患の治療に於ける特殊な問題は、細胞増殖抑制薬に対する腫瘍細胞の耐性に関する研究で特によく研究されている多剤耐性(英語でmultidrug resistance(MDR))と呼ばれる耐性を患者が発症することである(Gottesman and Pastan、Annu.Rev.Biochem.62、385〜427、1993年)。この問題は、膜結合輸送タンパク質、いわゆるABCトランスポータに属するP−糖タンパク質の過剰発現に基づいている(Stein, U.ら、JBC 276、28562〜69、2001年、J.Wijnholds、Novartis Found Symp.243、69〜79、2002年)。Bargou, R.C.らおよびOda, Y.ら、(Bargou, R.C.ら、Nature Medicine 3、447〜450、1997年; Clin.Cancer Res.4、2273〜2277、1998年)は、ヒト転写因子YB−1の核内局在がP−糖タンパク質の発現活性化に関係していることを示した。別の研究は、YB−1がUV照射、細胞分裂抑制薬投与(Koike, K.ら、FEBS Lett、390〜394、1997年)および高温(Stein, Uら、JBC 276、28562〜69、2001年)といった様々なストレス状態により核内に輸送されることを確認している。別の研究は、YB−1の核内局在が更に別のABCトランスポータに影響することを確認している。このABCトランスポータはMRP(英語でmultidrug resistance-relate potein:多剤耐性関連タンパク質)と呼ばれており、いわゆる非定型非P−糖タンパク質依存多剤耐性の形成に関係している(Stein、U.ら、JBC 276、28562〜69、2001年)。
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明の基調をなす課題は、具体的には殺腫瘍活性薬剤を用いて生物、より具体的にはヒトおよび患者群の処置を可能にする技術的教示および、特に手段を提供することである。本発明の基調をなす別の課題は、細胞分裂抑制薬に耐性である腫瘍疾患を持つ患者、特に多剤耐性を持つ患者に殺腫瘍を起こすのに好適である手段を提供することである。
【0009】
本発明によれば、第1の局面の課題は薬物製造に適したウイルス、好ましくはアデノウイルスを使用することで解決されるが、この時ウイルスは核内にYB−1を持たない細胞内では複製ができず、そしてウイルスは発癌遺伝子または発癌遺伝子産物、好ましくは発癌遺伝子タンパク質をコードしており、それはYB−1核陽性細胞でウイルス遺伝子の少なくとも1つ、好ましくはアデノウイルス遺伝子をトランス活性化し、そして遺伝子はE1B55kDa、E4orf6、E4orf3およびE3ADPを含む群より選ばれる。
【0010】
第2の局面では、課題は核内にYB−1を有する細胞での複製についてウイルス、好ましくはアデノウイルスを使用することで解決されるが、この時ウイルスは核内にYB−1を持たない細胞では複製できず、そしてウイルスは発癌遺伝子または発癌遺伝子産物、特には発癌遺伝子タンパク質をコードしており、それは少なくとも1つのウイルス遺伝子、好ましくはアデノウイルス遺伝子をトランス活性化し、そのとき遺伝子はE1B55kDa、E4orf6、E4orf3およびE3ADPを含む群より選ばれる。
【0011】
発明による上記2つの使用の実施態様では、ウイルス、好ましくはアデノウイルスは核内にYB−1を有する細胞で複製する。
【0012】
発明による2つの使用の別の実施態様では、ウイルス発癌遺伝子タンパク質はE1Aであり、そして/または発癌遺伝子はE1Aをコードする遺伝子であり、そして/または発癌遺伝子タンパク質はE1Aである。
【0013】
好適実施態様では、ウイルス発癌遺伝子タンパク質E1Aは機能的Rb腫瘍抑制遺伝子産物に結合できる。
【0014】
別の実施態様では、ウイルス発癌遺伝子タンパク質E1Aは、機能的Rb腫瘍抑制遺伝子産物に結合できない。
【0015】
発明による2つの使用の別実施態様では、ウイルス発癌遺伝子タンパク質E1AはYB−1の核内局在化を誘導しない。
【0016】
発明による2つの使用の更に別の実施態様では、薬物はその細胞がRb−陽性またはRb−陰性のいずれかである患者向けである。
【0017】
好適実施態様では、細胞は上記薬剤により影響を受ける状態の形成に関係する細胞である。
【0018】
発明による2つの使用の別実施態様では、細胞は核内でRb−陰性であり、そしてYB−1陽性であり、好ましくは細胞周期とは独立に核内でYB−1陽性である。
【0019】
発明による2つの使用の更に別の実施態様では、薬剤は腫瘍の処置を目的とする。
【0020】
発明による2つの使用の更に別の実施態様では、細胞、特に腫瘍またはその一部を形成する細胞は、薬剤耐性であり、特に多剤耐性、好ましくは抗腫瘍剤に対し耐性であり、そしてより好ましくは細胞増殖抑制薬に対し耐性である。
【0021】
発明による2つの使用の好適実施態様では、細胞は膜結合輸送タンパク質であるP−糖タンパク質および/またはMRPを発現、好ましくは過剰発現している。
【0022】
発明による2つの使用の別実施態様では、細胞はp53−陽性またはp53−陰性である。
【0023】
発明による2つの使用の実施態様の一つでは、発癌遺伝子タンパク質は、野生型発癌遺伝子タンパク質E1Aと比較して、1または複数の突然変異または欠失を有しており、その場合欠失はCR3領域の欠失およびN−末端の欠失、およびC−末端の欠失を含む群より選択されるのが好ましい。これに関連し、E1A発癌遺伝子タンパク質はRbに結合できることが好ましい。
【0024】
発明による2つの使用の別実施態様では、発癌遺伝子タンパク質は、野生型発癌遺伝子タンパク質と比較した場合、1または複数の突然変異または欠失を有しており、その場合欠失はCR1領域および/またはCR2領域であることが好ましい。発癌遺伝子タンパク質E1AがRbに結合できないことは発明の範囲内である。
【0025】
発明による2つの使用の実施態様の一つでは、ウイルス発癌遺伝子タンパク質、特にE1Aは、組織特異的および/または腫瘍特異的プロモータの制御下にある。
【0026】
発明による2つの使用の別実施態様では、ウイルス、特にアデノウイルスはYB−1をコードしている。
【0027】
発明による2つの使用の更に別の実施態様では、YB−1は組織特異的および/または腫瘍特異的プロモータの制御下にある。
【0028】
発明による2つの使用の好適実施態様では、ウイルス、特にアデノウイルスはE4orf6、E4orf3、E1B55KおよびアデノウイルスE3ADPタンパク質を含む群から選ばれるタンパク質を少なくとも1つコードしている。
【0029】
発明による2つの使用の別実施態様では、細胞は核内にYB−1を有しており、特に腫瘍もしくはその一部を形成する細胞は核内にYB−1を有している。
【0030】
発明による2つの使用の更なる実施態様では、腫瘍は核内へのYB−1輸送の誘導により核内にYB−1を有している。
【0031】
発明による2つの使用の好適実施態様では、核内へのYB−1の輸送は放射線照射、細胞増殖抑制薬の投与、および高温から成る群より選ばれた少なくとも1つの手段を通じ開始される。
【0032】
発明による2つの使用の特に好適な実施態様では、上記手段は細胞、臓器または生体に適用される。
【0033】
発明による2つの使用の好適実施態様では、ウイルス、特にアデノウイルスは、AdΔ24、dl922−947、E1Ad/01/07、d1119/1131、CB016、dl520、および機能的Rb腫瘍抑制遺伝子産物と結合できるウイルス性E1A発癌遺伝子の発現を欠いているウイルスを含む群より選ばれる。
【0034】
第三の局面では、課題はウイルス、好ましくはアデノウイルスを医薬の製造に使用することで解決されるが、この場合ウイルスは、好ましくはアデノウイルス、その複製がE2−後期プロモータの活性化を介したYB−1による手段を通し、もしくは手段により、好ましくはE2−後期プロモータの活性化を主に通じて制御されるようにデザインされている。実施態様の一つでは、YB−1はトランスジェニックYB−1または細胞質、特に細胞質脱制御型YB−1のいずれかである。好ましくは、トランスジェニックYB−1は、ベクター、好ましくはアデノウイルスによって細胞内で発現されるYB−1を意味する。E2−後期プロモータは野生型アデノウイルスに存在するアデノウイルスE2−後期プロモータ、またはここでトランス遺伝子の発現と関連付け記載されるE2−後期プロモータであるのが好ましい。
【0035】
第四の局面では、課題はウイルスおよび特にはアデノウイルスを核内にYB−1を有する細胞内での複製に使用することで解決されるが、この場合ウイルス、とりわけアデノウイルスは、複製がE2−後期プロモータの活性化を通じて、好ましくはE2−後期プロモータの活性化を主に通じてYB−1により制御されるようにデザインされる。実施態様の一つでは、YB−1はトランスジェニックYB−1または細胞質、特には細胞質脱制御YB−1のいずれかである。本明細書で用いる場合のトランスジェニックYB−1は、ベクター、好ましくはアデノウイルスによって細胞内で発現されるYB−1であるのが好ましい。E2−後期プロモータは野生型アデノウイルスに存在するアデノウイルスE2−後期プロモータ、またはここでトランス遺伝子の発現の使用と関連付け記載されるE2−後期プロモータであるのが好ましい。
【0036】
本発明の第三および/または第四の局面の好適実施態様では、アデノウイルスは本明細書に開示された如くにデザインされ、特には本発明に従った使用を目的としてデザインされる。
【0037】
第五の局面では、課題は次の特徴を有するウイルス発癌遺伝子タンパク質、特に単離されたウイルス発癌遺伝子タンパク質により解決される:
a)YB−1核陽性細胞内で、E1B−55K、E3ADPおよびE4orf6およびE4orf3を含む群から選ばれる少なくとも1つのウイルス遺伝子がトランス活性化されること;および
b)核内、特にウイルス発癌遺伝子タンパク質が存在している細胞の核内でのYB−1の誘導を欠くこと。
【0038】
実施態様の一つでは、ウイルス発癌タンパク質はE1Aである。
【0039】
更なる実施態様では、ウイルス発癌遺伝子タンパク質は、野生型発癌遺伝子タンパク質と比較した場合に1または複数個の突然変異または欠失を有しており、この場合の欠失はCR3領域の欠失、N−末端の欠失およびC−末端の欠失を含む群より選ばれるのが好ましい。
【0040】
実施態様の一つでは、ウイルス発癌遺伝子タンパク質を介したYB−1誘導は、E4ofr6および/またはE1B55kDが有核細胞内に存在しない場合には起こらない。
【0041】
この場合にはウイルス発癌遺伝子タンパク質はRbに結合できると解釈する。
【0042】
別実施態様では、ウイルス発癌遺伝子タンパク質は1または複数の突然変異または欠失を含み、この場合欠失はE1A発癌遺伝子タンパク質のCR1領域および/またはCR2領域内にあることが好ましい。この場合には、ウイルス発癌遺伝子タンパク質はRbに結合できないと解釈する。
【0043】
第六の局面では、発明は、ウイルス、特に本発明に従って使用されるアデノウイルスをコードする核酸を含み、そしてヘルパーウイルスの核酸を1つ含むウイルス複製系、好ましくはアデノウイルス複製系の使用であって、上記ヘルパーウイルスの核酸がYB−1をコードする核酸を含んでいる複製系に関する。
【0044】
実施態様の一つでは、ウイルス核酸、特にはアデノウイルス核酸、および/またはヘルパーウイルスの核酸は、複製可能なベクターとして存在する。
【0045】
第七の局面では、発明はウイルス、特にはアデノウイルスをコードする核酸を、発明に従って使用する医薬の製造、特には腫瘍処置を目的とする医薬の製造への使用に関する。
【0046】
実施態様の一つでは、細胞、特には腫瘍またはその一部を形成する細胞は、薬物、好ましくは抗ガン剤、より好ましくは細胞増殖抑制薬に対し耐性、特には多剤耐性である。
【0047】
第八の局面では、発明はウイルス、特にはアデノウイルスをコードする核酸を、発明に従って使用する、核内にYB−1を有する細胞内での複製に関する使用であって、ウイルスが核内にYB−1を持たない細胞に於いては複製欠失であり、そしてウイルスがYB−1核陽性細胞に於いては少なくとも1つのウイルス遺伝子、好ましくはアデノウイルス遺伝子をトランス活性化する発癌遺伝子もしくは発癌遺伝子産物をコードし、その場合の遺伝子はE1B55kDa、E4orf6、E4orf3およびE3ADPを含む群から選ばれる使用に関する。
【0048】
第九の局面では、課題はウイルス、好ましくはアデノウイルスをコードする核酸を、発明に従い使用し医薬の製造に用いることで解決されるが、そのためにウイルスはその複製がE2−後期プロモータの活性化を通じて、好ましくはE2−後期プロモータの活性化を主に通じて、YB−1により制御されるようにデザインされている。実施態様の一つでは、YB−1はトランスジェニックYB−1または細胞質、特には細胞質脱制御YB−1のいずれかである。ここで用いるトランスジェニックYB−1とは、ベクター、好ましくはアデノウイルスベクターによって細胞内で発現されるYB−1であることが好ましい。E2−後期プロモータは、野生型アデノウイルス内に存在するアデノウイルスE2−後期プロモータ、またはトランス遺伝子の発現の使用と関連してここに記載されるE2−後期プロモータであることが好ましい。
【0049】
第十の局面では、課題はウイルス、特にはアデノウイルスをコードする核酸の、細胞内での複製に関する発明に従って用いることによる使用により解決されるが、そのためにウイルスはその複製がE2−後期プロモータの活性化を通じて、好ましくはE2−後期プロモータの活性化を主に通じて、YB−1により制御されるようにデザインされている。実施態様の一つでは、YB−1はトランスジェニックYB−1または細胞質、特には細胞質脱制御YB−1のいずれかである。ここで用いるトランスジェニックYB−1はベクター、好ましくはアデノウイルスによって細胞内で発現されるYB−1であることが好ましい。E2−後期プロモータは、野生型アデノウイルス内に存在するアデノウイルスE2−後期プロモータ、またはトランス遺伝子の発現の使用と関連してここに記載されるE2−後期プロモータであることが好ましい。
【0050】
第十一の局面では、課題は前記核酸の一つを含むベクターを、本発明の第一または第二の局面に従い使用することで解決される。
【0051】
第十二の局面では、発明は、発明に従い用いられるウイルス、特にはアデノウイルスと細胞を接触すべきものであるか、および/またはそれらを用いて処置すべきものであるかを決定することを目的とした、細胞、腫瘍組織または患者の細胞の特徴付けのためのYB−1と相互作用する薬物の使用に関する。
【0052】
実施態様の一つでは、薬物は抗体、抗カリン(anticaline)、アプタマー、アプタザイムおよびシュピーゲルマー(spiegelmer)を含む群から選ばれる。
【0053】
第十三の局面では、課題は本発明によるウイルス発癌遺伝子タンパク質またはそれをコードする核酸を、本発明の第一および第二局面に従って用いられるウイルス、特にはアデノウイルスの製造に使用することで解決される。
【0054】
実施態様の一つでは、ウイルスはトランス遺伝子をコードする核酸を含む。
【0055】
別の実施態様では、ウイルスはトランス遺伝子の翻訳産物および/または転写産物を含む。
【0056】
好適実施態様では、アデノウイルス複製系の核酸および/またはヘルパーウイルスの核酸は、トランス遺伝子またはトランス遺伝子をコードする核酸を含む。
【0057】
更に別の実施態様では、核酸はトランス遺伝子またはトランス遺伝子をコードする核酸を含む。
【0058】
別の実施態様では、トランス遺伝子はプロドラッグ遺伝子、サイトカイン、アポトーシス誘導遺伝子、腫瘍抑制遺伝子、金属プロテアーゼインヒビターに関する遺伝子、および血管新生インヒビターに関する遺伝子を含む群より選ばれる。
【0059】
実施態様の一つでは、トランス遺伝子はsiRNA、アプタマー、アンチセンス分子およびリボザイムに関する核酸を含む群より選ばれ、この場合のsiRNA、アプタマー、アンチセンス分子および/またはリボザイムはあるターゲットとなる標的分子を持っている。
【0060】
更なる実施態様では、標的分子は耐性関連因子、抗アポトーシス因子、発癌遺伝子、血管新生因子、DNA合成酵素、DNA修復酵素、成長因子およびそれらのレセプター、転写因子、金属プロテアーゼ、特にマトリックス金属プロテアーゼ、およびウロキナーゼ
型プラスミノーゲン活性化因子を含む群より選ばれる。実施態様の一つでは、耐性関連因子は、P−糖タンパク質、MRPおよびGSTを含む、およびそれらをコードする核酸を含む群より選ばれるのが好ましい。実施態様の一つでは、抗アポトーシス因子は、BCL2を含み、更にそれをコードする核酸を含む群より選ばれる。実施態様の一つでは、発癌遺伝子はRas、特に突然変異型Ras、RbおよびMycを含み、またそれらをコードする核酸を含む群より選ばれる。実施態様の一つでは、血管新生因子はVEGFおよびHMGタンパク質を含む、ならびにそれらをコードする核酸を含む群より選ばれる。実施態様の一つでは、DNA合成酵素はテロメラーゼを含む、ならびにそれをコードする核酸を含む群より選ばれる。実施態様の一つでは、DNA修復酵素はKu−80を含み、またそれをコードする核酸を含む群より選ばれる。実施態様の一つでは、成長因子はPDGF、EGFおよびM−CSFを含み、またそれらをコードする核酸を含む群より選ばれる。実施態様の一つでは、レセプターは特には成長因子に関するレセプターであり、この場合の成長因子はPDGF、EGFおよびM−CSFを含み、またそれらをコードする核酸を含む群より選ばれる。実施態様の一つでは、転写因子はYB−1を含み、またそれをコードする核酸も含む群より選ばれる。実施態様の一つでは、金属プロテアーゼはマトリックス金属プロテアーゼであることが好ましい。好適実施態様では、金属プロテアーゼはMMP−1およびMMP−2を含み、またそれらをコードする核酸を含む群より選ばれる。実施態様の一つでは、ウロキナーゼ型プラスミノーゲン活性化因子は、uPa−Rを含み、またそれをコードする核酸を含む群より選ばれる。
【0061】
さらに別の実施態様では、医薬は少なくとも1つの医薬活性化合物を追加して含む。
【0062】
好適実施態様では、医薬的に活性な化合物は、サイトカイン、金属プロテアーゼインヒビター、血管新生インヒビター、細胞増殖抑制薬および細胞周期インヒビターを含む群から選ばれる。
【0063】
本発明はYB−1核陽性腫瘍細胞でのE1A−修飾アデノウイルスのDNA複製がE2−後期プロモータの活性化に基づいているという驚くべき発見に拠っている。本明細書で使用する場合のE1A−修飾アデノウイルスは、(a)YB−1核陰性細胞では複製しないか、またはYB−1核陰性細胞に於いては対応する野生型細胞に比べて複製の低下、好ましくは大きな低下を示し、(b)少なくとも1つのウイルス遺伝子をトランス活性化し、その場合の遺伝子は特にはE1B−55kDa、E4orf6、E4orf3およびE3ADPを含む群から選ばれ、そして/または(c)アデノウイルスを通じて細胞内YB−1を核内に移動しない、アデノウイルスである。場合によっては、本発明で使用するアデノウイルスは、アデノウイルスがコードするE1Aタンパク質の結合がRbへのE2Fの結合を妨害してRbとE2Fより成るそれぞれの複合体を解離することができるという特徴を更に有してもよい。上記特徴a)からc)のうちの少なくとも1つ、または複数、好ましくは特徴a)からc)の全てを有するアデノウイルスは、核内にYB−1を持たない細胞では複製できない。
【0064】
実施態様の一つでは、ここで用いる場合、大きな複製の低下とは特に、野生型と比較したときに2倍、好ましくは5倍、より好ましくは10倍、そして最も好ましくは100倍低下した複製を意味する。好適実施態様では、かかる複製の比較は、同一もしくは類似の細胞株、同一もしくは類似の感染ウイルス力価(感染多重度、MOIもしくはプラーク形成単位、pfu)および/または同一もしくは類似の一般的実験条件を用いて実施される。ここで使用する場合、複製とは、特には粒子形成を意味する。別の実施態様では、複製の評価はウイルス核酸合成の程度とすることができる。ウイルス核酸合成の程度を決定する方法、ならびに粒子形成を決定する方法は当業者周知である。
【0065】
ここに記載の発見、方法、使用または核酸、タンパク質、複製系等は、アデノウイルスに必ずしも限定されない。原則的には、かかる系はこれと共に含まれる他のウイルスにも存在する。
【0066】
本発明によるウイルスを使用する場合、または本発明により本明細書に記載のウイルスを使用する場合には、先行技術によれば、10〜100pfu/細胞に対し1〜10pfu/細胞の感染率で野生型の複製に匹敵する複製が実現できる。
【0067】
本明細書で使用する場合、細胞質YB−1は細胞によりコードされているYB−1、好ましくは細胞によりさらに発現されるYB−1を意味するものとし、この場合YB−1は細胞内に、アデノウイルス、好ましくは本書記載のアデノウイルスおよび/もしくはヘルペスウイルスが該当する細胞に感染する前に存在することが好ましい。しかしながら、細胞質YB−1が細胞内に導入された、または例えばウイルス、特にはアデノウイルスの感染といった外的方法を用いることによりかかる細胞が産生したYB−1である場合も本発明の範囲内である。
【0068】
以下これに結びつけることを望むことなしに、本発明者はE2−初期プロモータ、即ち初期E2プロモータは、本発明に従いここに使用されるウイルス複製との関係に於いて、ヒト細胞質E2F転写因子を通じて始動することはないと想定している。複製の始動は細胞のRbの状態とは無関係であり、即ちこのことは、ここに開示するウイルスを用い感染させた腫瘍細胞、およびその後好ましく溶解する腫瘍細胞は機能的Rbタンパク質および非活性型Rbタンパク質の両方を含んでよいことを意味する。さらに、アデノウイルスの複製は、ここに開示するアデノウイルスを使用した場合、またはここに開示の条件では、機能的p53タンパク質を必要とせず、またその存在の影響も受けない。その限りに於いて、技術的教示はAdΔ24、d1922−947、E1Ad/01/07、CB016型の殺癌もしくは殺腫瘍性アデノウイルスの、または例えば欧州特許第EP0 931 830号に記載のアデノウイルスの、あるいは無傷の機能的Rbタンパク質はインビボでの効果的複製にとって障害であり、従ってRb−陰性およびRb−突然変異細胞でのみアデノウイルスはインビボ複製するという仮説の下にE1Aタンパク質内に1または複数の欠失を導入したアデノウイルスの使用を基礎と成す原理とは、それぞれ異なっている。これらの先行技術のアデノウイルス系は、E1Aに基づき、初期E2プロモータ(E2初期プロモータ)および「無E2F」を利用してアデノウイルスのインビボ複製を制御するものである。しかしながら、これら先行技術ウイルスは、本発明に従って用いること、即ち細胞周期と無関係に核内にYB−1を含む細胞での複製に用いることもできるだろう。
【0069】
前記欧州特許第EP0 931 830号に記載のウイルス、特にアデノウイルスは本発明に従い使用出来るだろう。より具体的には、前記特許に記載のウイルスは複製できず、そして機能的なRb腫瘍抑制遺伝子産物と結合可能なウイルス発癌タンパク質を発現していない。このアデノウイルスは具体的には、機能的腫瘍抑制遺伝子産物、特にはRBを結合できるウイルスE1A発癌タンパク質の発現を欠いているアデノウイルスであろう。ウイルスE1A発癌タンパク質は例えば、p105Rbタンパク質、p130およびp107タンパク質の結合に関係する、Ad5のアミノ酸位置30〜85、ヌクレオチド位置697〜790にあるCR1ドメイン、ならびにAd5内アミノ酸位置120から139、ヌクレオチド位置920〜967にあるCR2ドメイン内に不活性化突然変異を含むことができる。アデノウイルスはタイプ2dl312から、またはアデノウイルスはタイプ5NTdl1010から指定することもできる。
【0070】
最終的に複製は核内にYB−1を含む細胞、即ちYB−1核陽性である細胞において、医薬製造、特には腫瘍疾患の処置用の医薬の製造を目的として発明のアデノウイルスを使用する場合、ならびに核内にYB−1を有する細胞での複製のために発明に従いアデノウイルスを使用する場合に、細胞周期とは無関係に起こることが好ましい。かかるアデノウイルスは核内にYB−1を持たず、YB−1は実質的に細胞質内にのみ存在している細胞では複製しないか、またはそのレベルが極めて低くなることに特に注意すべきである。この限りにおいて、これらウイルスが上手く複製するにはYB−1が核内に存在することが必要である。これは例えば、以下詳しく概要を示すように、核内にYB−1を発現させるか、または核内にYB−1が存在するようにする手段を細胞にこうじることで実現できる。そのような手段としては、例えばアデノウイルス遺伝子に加えてYb−1をコードする遺伝情報、特にYB−1の発現に関する遺伝情報も含んでいる、本発明に従い使用するアデノウイルスを介したYB−1のコード化および発現を挙げることができる。細胞核内へのYB−1の輸送、誘導または発現を起こすその他手段としては、細胞およびかかる細胞を含む生体への細胞増殖抑制薬投与、放射線照射、高温等のストレス条件がある。
【0071】
本発明に関連して、特に腫瘍溶解に関連して使用するアデノウイルスは更に、それらが核内にYB−1を持たない、換言すればYB−1核陰性である細胞では複製しないという特徴を持つ。
【0072】
発明で使用されるアデノウイルスは更に、それらが本明細書で発癌遺伝子タンパク質も呼ばれるウイルス発癌タンパク質をコードすることも特徴としており、この場合上記発癌タンパク質はE1Aであることが好ましく、そして発癌遺伝子タンパク質はウイルスの複製および/またはウイルス感染細胞の細胞溶解に影響を及ぼすことができるウイルス遺伝子の少なくとも1つを活性化できる。複製への影響は、ウイルスが、そのウイルスの発癌遺伝子タンパク質が存在しない場合に比べ、発癌遺伝子タンパク質が存在する場合により良く複製するものであることが好ましい。このプロセスは本明細書ではトランス活性化、そしてこのトランス活性化がE1Aを通じて行われる場合には特にE1Aトランス活性化とも呼ばれる。用語「トランス活性化する」または「トランス活性化」とは、各ウイルス発癌タンパク質がウイルス発癌タンパク質をコードする遺伝子そのものとは異なる1または複数のその他遺伝子の発現および/または転写に影響を及ぼす、即ちそれ/それらの発現および/または翻訳を好ましく制御する、そして特にはそれ/それらを活性化するプロセスを表す。かかるウイルス遺伝子はそれぞれE1B55kDa、E4orf6、E4orf3およびE3ADPならびにこれら遺伝子および遺伝子産物のいずれかの組み合わせであることが好ましい。
【0073】
さらに、場合によっては、発明に従い用いるアデノウイルスの特徴は、腫瘍抑制因子Rbにそして腫瘍抑制因子Rbと結合することである。原則的には、本発明に従い使用するアデノウイルスがRbに結合すること、またはRbに結合しないことは共に本発明の範囲内である。アデノウイルスに関する両実施態様は、処理対象となる細胞のRbの状態とは無関係に用いることができる。
【0074】
Rbに結合できなくするために、例えば次のようなE1A発癌タンパク質の欠失が考えられる:CR1領域内の欠失(Ad5内アミノ酸位置30〜85)およびCR2領域の欠失(AD5内のアミノ酸位置120〜139)。この様な欠失では、CR3領域はそのまま維持され、他の初期ウイルス遺伝子に対するトランス活性化機能を発揮できる。
【0075】
これに対し、E1AにRbへの結合能を付与する場合については、E1A発癌タンパク質に対する次の欠失が原理的に可能である:CR3領域(アミノ酸位置140〜185)の欠失;N−末端(アミノ酸位置1〜29)の欠失;アミノ酸位置85〜119の欠失;およびC−末端(アミノ酸位置186〜289)の欠失。ここに引用した領域は、RbへのE2Fの結合を妨害しない。しかし、トランス活性化機能は維持されるが、野生型Ad5に比べると低い。
【0076】
先行技術既知であるかかるウイルスは一般には複製欠失として知られている。しかし、本発明の利点は、かかるウイルスが好適な背景、特に細胞の背景があれば複製できることが認識されていることである。この種の好適細胞の背景は、核内にYB−1が存在すること、好ましくは核内にYB−1が細胞周期とは無関係に存在することより生じるか、または提供される。細胞または細胞系という用語は、ここで使用する場合、細胞の断片または細胞溶解物の断片、ならびにインビトロ、インビボまたはインシトゥ(in situ)に存在する細胞を含む。この限りにおいて細胞系または細胞という用語は、細胞培養体、組織培養体、臓器培養体、または単離された、グループ状態にある、または一部分である組織、臓器または生体であるインビボおよびインシトゥのその他組織または臓器内に存在する細胞、あるいは好ましくは生きている生物体内にその様な形で存在している細胞も含む。生体は好ましくは脊椎動物の生体であり、より好ましくは哺乳動物のものである。生体がヒトの生体であることが特に好ましい。
【0077】
更に、ここに提供する技術的教示に基づいて、ここに記載のアデノウイルスの複製特徴と、YB−1核陽性である細胞での従来技術のアデノウイルスの特徴とを持つ新規ウイルスを作成することも、本発明の範囲内である。換言すれば、既知アデノウイルスを基にして、ここに明示した本発明での使用に必要な特徴を有する更なるウイルスをデザインすることができる。
【0078】
本発明に関しては、発明で用いられる各種アデノウイルスの修飾E1A発癌タンパク質は、例えばE1B55K、E4orf3、E4orf6、E3ADPといった初期ウイルス遺伝子をYB−1核陽性細胞内でトランス活性化できるものである。これに関しては、その他の点についてウイルスゲノムにそれ以上の変更がないことが望ましく、各アデノウイルスはその他の点については野生型のアデノウイルスまたはその誘導体と同様でよい。
【0079】
本発明の意味での発癌遺伝子タンパク質のトランス活性化をコードしているか、またはかかる発癌遺伝子タンパク質を含むここに記載のウイルスは、たとえばアデノウイルスAdΔ24、dl922−947、E1Ad/01/07、CB106および/または欧州特許第EP0 931 380号に記載のアデノウイルスを含み、それらはそれぞれE1B、E2、E3および/またはE4といった初期遺伝子をトランス活性化でき、そして野生型のアデノウイルス、特には野生型Ad5と同等である。これらの例ではトランス活性化にはE1Aタンパク質の特定の領域が関係している。各種血清型アデノウイルスに於いてE1Aタンパク質中には極めて保存的である領域が3カ所ある。アミノ酸位置41〜80のCR1領域、アミノ酸位置120〜139のCR2領域、およびアミノ酸位置140〜188のCR3領域である。トランス活性化機能は主にE1Aの中にCR3領域が存在することに基づいている。CR3のアミノ酸配列は上記アデノウイルスについては変更がない。その結果、核内または細胞質内のYB−1の存在とは無関係に、初期遺伝子E1B、E2、E3およびE4は活性化される。
【0080】
しかし組換え体アデノウイルスd1520では、CR3領域は削除されている。即ちdl520はCR3領域のアミノ酸配列を含まない、いわゆるE1A12Sタンパク質を発現する。その結果d1520は特にE2領域に対し極めて弱いトランス活性化機能しか発揮できず、その結果YB−1核陰性細胞では複製しない。YB−1核陽性細胞では、YB−1はE2領域をトランス活性化し、それによりdl520は有効に複製できる。これがここでの開示の目的のための、それぞれdl520と同様のシステムおよび5120をベースとするシステムを用いることの根拠である。既報の両アデノウイルス群、即ちデルタ24(本明細書ではAdΔ24とも称する)とdl520間のより重要な違いは、d1520に関しては、初期遺伝子E1B、E3およびE4がYB−1核陰性細胞に比べてYB−1核陽性細胞でより強くトランス活性化されるという事実である。これに対しデルタ24との間には全く、または極僅かな違いしかない。しかしdl520の、より具体的にはE1A12Sタンパク質のトランス活性化効果は、野生型アデノウイルスに比べ大きく低下している。しかしこのトランス活性化は、実施例10に示す如く、YB−1核陽性細胞内での効果的複製を可能にするには十分である。本明細書に記載のE1Aタンパク質およびそれをコードする核酸のデザイン、特にそのEIAタンパク質が野生型発癌タンパク質E1Aに対し1もしくは複数の欠失、および/または突然を有しているような本明細書内のデザインであって、その欠失が好ましくはCR3領域およびN−末端の欠失およびC−末端の欠失を含む群から選ばれる一つであり、具体的にはdl520またはAdΔ24、dl922−924、E1Ad/01/07、CB106および/または欧州特許第EP0 931 830号に記載のアデノウイルスと関連し記載されるE1Aタンパク質の好適なデザインの実施態様がウイルス、特にアデノウイルスの実施態様であり、その複製はE2−後期プロモータの活性化を通じて、好ましくは主にE2−後期プロモータの活性化を通じて制御される。アデノウイルスのこの形態の複製を可能にするE1Aタンパク質の別の実施態様は、本明細書の開示に基づき、当業者により作り出すことができる。
【0081】
新たに構築され、本明細書では誘導体とも称され、且つ本発明にしがたい使用できる別のアデノウイルスは、一般的にはE1欠失、E1/E3欠失、および/またはE4欠失を有しており、即ち対応するアデノウイルスは機能的に活性なE1および/またはE3、および/またはE4発現産物および各産物をそれぞれ作り出すことができず、換言すれば、これらアデノウイルスは機能的に不活性なE1、E3および/またはE4発現産物のみ産生できるが、この時のそのような機能的に不活性なE1、E3および/またはE4発現産物とは、その転写レベルおよび/または翻訳レベルにおいて、たとえ発現産物として全く存在していない、あるいは野生型のアデノウイルスに存在する機能の少なくとも一つを欠いている形で存在していもよい。野生型アデノウイルスの発現産物の機能は当業者周知であり、そして例えばRussell, W, C、Journal of Virology、81、2573〜2604、2000年に記載されている。Russell(上記)はまた、参照によりここに組み込まれるアデノウイルスおよびアデノウイルスベクターの構築の原理についても記載している。上記修飾型E1A発癌タンパク質、E1B−55K、E4orf6、および/またはE3ADP(アデノウイルス崩壊タンパク質(ADP))(Tollefson, Aら、J.Virology、70、2296〜2306、1996年)をこの種のベクター内で、個々に、または組み合わせて発現させることも、本発明の範囲内である。これに関連して、ここに開示した個々に名前を挙げた遺伝子ならびにトランス遺伝子は、E1および/またはE3および/またはE4領域内にクローニングでき、好適プロモータの助けをかり、または好適プロモータの制御下に個別に発現させることができる。基本的には、領域E1、E3およびE4はアデノウイルス核酸内のクローニング部位として同等の好適性を有している。取り分けても好適なプロモータは本明細書の中でE1A、特に修飾型E1Aの制御および発現それぞれに関連付けて開示されているプロモータである。
【0082】
最後に実施態様の一つでは、本発明に従い用いられるアデノウイルスは、E1Bに関する、特にE1B19kDaに関する欠失体である。
【0083】
本明細書で使用する場合、欠失という用語はE1Bが野生型固有の特性の全てを有しているわけではないが、その特性の少なくとも一つを欠いている状態を意味する。
【0084】
ここに開示した発明に使用するアデノウイルスは、基本的には幾つかの実施態様については当分野周知である。本発明に用いられるアデノウイルスは、特に野生型と比較しここに示す技術的教示に従って変更する場合には、組換え体アデノウイルスであることが好ましい。本発明にとって必須でないアデノウイルス核酸を欠失または突然変異させることは、当業者の技術的水準内である。かかる欠失は、例えばここにも記す様にE3およびE4をコードする核酸の一部にかかっても良い。E4の欠失は、かかる欠失がタンパク質E4orf6を超えていないこと、換言すれば発明に使用するアデノウイルスはE4orf6をコードすることが特に好ましい。好適実施態様では、これらアデノウイルス核酸は更にウイルスカプシドに封入してもよく、即ち感染粒子を形成してもよい。このことは本発明での核酸の使用にも当てはまる。一般に、アデノウイルス系は単一または複数の発現産物について欠失してもよいことを記しておく。これに関連して、この現象はかかる発現産物をコードする核酸が完全に突然変異しているか、もしくは欠失しているか、または実質的に発現産物がもはや生じない程度まで突然変異しているか欠失しているかに基づいているか、あるいは核酸レベル(プロモータの欠失;シス作用要素)または翻訳系および転写系(トランス作用要素)のそれぞれについて、発現を制御するか、もしくは野生型とは異なる様式で作用するプロモータもしくは転写因子を欠いていることに基づいていることに注意しなければならない。具体的には、後者の観点は背景となる細胞に依存している。
【0085】
本発明によるアデノウイルスでは、さらに周知の如く新規アデノウイルスもまた、その他アデノウイルスに関しここに記した程度に使用することができる。発明による新規アデノウイルスは、ここに示す技術的教示より生ずる。特に好ましい例は、例えば図16および17に示すウイルスXvir03およびXvir03/01、実施例11および12に更に例示したデザイン原理である。
【0086】
ベクターXvir03の場合は、CMVプロモータを、IRES配列により隔てられているE1B55KおよびE4orf6の核酸をコードするE1領域内にクローニングする。これら2遺伝子の導入およびそこより産生される遺伝子産物それぞれによって、野生型ウイルスのものと実質同じ複製効率を得るが、その場合複製の選択性は細胞、特に腫瘍細胞について維持されるため、YB−1核陽性細胞、より具体的にはYB−1が脱制御状態にある細胞で複製が起こる。YB−1が脱制御状態にある細胞は、好ましくは正常細胞または非腫瘍細胞に比べて、好ましくはコンパートメント独立的(compartment-independent)に、YB−1の発現が上昇している細胞である。
【0087】
ウイルスXvir03の更なる発展形は、好適実施態様において、その中に治療遺伝子またはトランス遺伝子が特異的プロモータ、特に腫瘍特異的または組織特異的プロモータ制御下にクローニングされているウイルスXvir03/01である。E4領域が機能的に非活性であり、好ましくは欠失しているウイルスもまた上記ウイルスの範囲内である。ここに記したトランス遺伝子はまたE4領域内にクローニングしてもよく、この場合クローニングをE3領域内へのトランス遺伝子のクローニングに追加し、またはその代替して行っても良い。
【0088】
この様な治療遺伝子は、プロドラッグ遺伝子、サイトカインの遺伝子、アポトーシス誘導遺伝子、腫瘍抑制遺伝子、金属プロテアーゼインヒビターおよび/または血管新生インヒビターの遺伝子であろう。更にsiRNA、アプタマー、アンチセンスおよびリボザイムを、癌関連標的分子に向けて発現させてもよい。好ましくは、単一または複数の標的分子が、耐性関連因子、抗アポトーシス因子、発癌遺伝子、血管新生因子、DNA合成酵素、DNA修復酵素、増殖因子とそれらのレセプター、転写因子、金属プロテアーゼ、特にマトリックス金属プロテアーゼ、およびウロキナーゼ型プラスミノーゲン活性化因子を含む群より選ばれる。その好適実施態様は既に本明細書内に開示した。
【0089】
好適実施態様にて使用可能であろうプロドラッグ遺伝子は、例えばシトシンデアミナーゼ、チミジンキナーゼ、カルボキシペプチダーゼ、ウラシルホスホリボシルトランスフェラーゼ;プリンヌクレオシドホスホリラーゼ(PNP)である;Kimら、Trends in Molecular Medicine、8巻、No.4(Suppl)、2002年; Wybranietz.W.Aら、Gene Therapy、8、1654〜1664、2001年; Niculescu-Duvazら、Curr.Opin.Mol.Therapy、1、480.486、1999年; Koyamaら、Cancer Gene Therapy、7、1015〜1022、2000年; Rogersら、Human Gene Therapy、7、2235〜2245、1996年; Lockettら、Clinical Cancer Res、3、2075〜2080、1997年; Vijayakrishnaら、J.Pharmacol.And Exp.Therpeutics、304、1280〜1284、2003年。
【0090】
好適実施態様にて使用できるであろうサイトカインは、例えばGM−CSF、TNF−アルファ、IL−12、IL−2、IL−6、CSF、インターフェロン−ガンマである;Gene Therapy、Advances in Pharmacology、40巻、編集者:J.Thomas Augsut、Academic Press; ZhangとDegroot、Endocrinolgy、144、1393〜1398、2003年; Descampsら、J.Mol.Med、74、183〜189、1996年; Majumdarら、Cancer Gene Therapy、7, 1086〜1099、2000年。
【0091】
好適実施態様にて使用できるであろうアポトーシス誘導遺伝子は、例えばデコリン(decorin): Tralhaoら、FASEB J、17、464〜466、2003年;網膜芽細胞種94:Zhangら、Cancer Res、63、760〜765、2003年; BaxとBad: Zhangら、Hum.Gene Ther、20、2051〜2064、2002年;アポプチン(apoptin):NotebornとPietersen、Adv.Exp.Med.Biol、465、153〜161、2000);ADP:Tothら、Cancer Gene Therapy、10、193〜200、2003年; bcl-xs: Sumantranら、Cancer Res、55、2507〜2512、1995年;E4orf4:BraithwaiteとRussell、Apoptosis、6、359〜370、2001年;FasL、Apo−1およびTrail:Boehringer Manheim、Guide to Apoptotic Pathways、Araiら、PNAC、94、13862〜13867、1997年、Bims; Yamaguchiら、Gene Therapy、10、375〜385、2003年; GNR163: Oncology News、17 Juni、2000年、がある。
【0092】
好適実施態様にて使用できるであろう腫瘍抑制遺伝子は、例えばE1A、p53、p16、p21、p27、MDA−7がある。Opalkaら、Cell Tissues Organs、172、126〜132、2002年、Jiら、Cancer Res、59、3333〜3339、1999、Suら、Oncogene、22, 1164〜1180、2003年。
【0093】
好適実施態様にて使用できるであろう血管新生インヒビターは、例えばエンドスタチン、アンギオスタチン:Hajitouら、FASEB J、16, 1802〜1804、2002年およびVEGFに対する抗体がある(Ferrara、N、Semin Oncol 2002 Dec; 29(6 Suppl 16): 10−4。
【0094】
好適実施態様にて使用できるであろう金属プロテアーゼインヒビターは、例えばTimp-3、Ahonenら、Mol Therpy、5、705〜715、2002年; PAI-1; Soffら、J.Clin.Invest、96、2593〜2600、1995年; Timp-1、Brandt K.Curr.Gene Therapy、2、255〜271、2002年がある。
【0095】
siRNA(short ingerfering(短干渉)RNA)は塩基相補正により相互にハイブリダイゼーションする、即ち実質的に塩基が対を成しており、好ましくは最長50ヌクレオチド、好ましくは18〜30ヌクレオチド、より好ましくは25ヌクレオチド未満、そして最も好ましくは21、22もしくは23ヌクレオチドの長さを持つ2つのRNA鎖、好ましくは分離した2つのRNA鎖より成り、この時それらの特徴は単鎖のsiRNAであり、特に一本の鎖、好ましくは第二の単鎖とハイブリダイゼーションしてそれと塩基対合する単鎖の伸展した鎖である。siRNAは、特異的にmRNAの分解を誘導または媒介する。この時必要な特異性は、siRNAの配列とその結合部位から提供される。分解対象となる標的配列は第一または第二のsiRNA形成鎖に対し実質的に相補的である。正確な作用様式は不明であるが、siRNAは細胞が発生中にある特的の遺伝子座を阻害し、ウイルスからそれを保護するための生物学的戦略であると想定される。siRNAを介したRNA干渉は、タンパク質の特異的抑制に、または遺伝子特異的二本鎖RNAを導入することによるタンパク質発現の完全なノックアウトにも用いられる。高等生物では、19〜23ヌクレオチド長のsiRNAは特に、非特異的な防御反応、いわゆるインターロイキン反応の活性化をもたらさないので特に好ましい。3‘末端に対称的な2−nt長のオーバーハングを有する21ヌクレオチドから成る二本鎖RNAを迅速にトランスフェクションすることにより、哺乳動物細胞にRNA干渉を伝達でき、リボザイムやアンチセンス分子といった他技術に比べて高い効率を得ている(Elbashir、S.Harborth J. Lendeckel W. Yalvcin、A. Weber K Tuschl T: Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells、Nature 2001年、411: 494〜498)。標的遺伝子の発現を阻害するのに必要なRNA分子は極めて小数であった。特にsiRNA分子の一過性の干渉現象、および特異的配送特性を有するsiRNAが外部からの投与に限定されないようにするために、先行技術では内因性のsiRNAを発現できるベクターを使用している。例えば、センスおよびアンチセンス両方向の、ベクター内に導入される例えば9ヌクレオチド長のスペーサー配列により隔てられた、19ヌクレオチド長の標的配列を含む64ヌクレオチド長のオリゴヌクレオチドが提供されている。得られた転写体は、例えば19塩基対のステム構造を持つヘアピン構造に折りたたまれた。ループは細胞内で迅速に分解し、その結果機能的なsiRNAが生成する(Brummelkampら、Science、296、550〜553、2002年)。
【0096】
pRbおよびE2Fそれぞれの活性は、リン酸化によって制御されている。低リン酸化型のpRbは主にG1期とM期に存在している。これに対し、高リン酸化型のpRbはS期とG2期に存在している。E2Fは、E2Fと低リン酸化型pRbから成る複合体から、pRbのリン酸化によって放出される。E2Fと低リン酸化型pRbより成る複合体からE2Fが放出されると、E2F依存遺伝子の転写が起こる。E1Aタンパク質は低リン酸化型pRbに結合するだけでなく、その時はE1Aタンパク質のCR2領域を介したE1AとpRbとの結合も普通に起こっている。さらに、それはCR1領域にも、より低い親和性で結合する(Ben-IsraelとKleiberger、Frontiers in Bioscience、7、1369〜1395、2002年; HeltとGalloway、Carcinogenesis、24、159〜169、2003年)。
【0097】
本発明に従って用いるアデノウイルスの実施態様に於いて、そのアデノウイルスの一部を成すYB−1をコードする核酸はまた核内へのYB−1の輸送を媒介する核酸配列を含んでもよい。発明による核酸、アデノウイルスおよびアデノウイルス系、ならびに例えばOnyx−015、AdΔ24、dl922−947、E1Ad/01/07、CB016、dl520および欧州特許第EP0 931 830号に記載のアデノウイルスといった当分野既知のアデノウイルスは、アデノウイルスおよびアデノウイルス系として、即ち対応する核酸として、上記の如くまたはこれに関連する発明の核酸と組み合わせて使用できる。核輸送を媒介するのに好適な核酸配列は当業者周知であり、例えば(Whittaker、G.Rら、Virology、246、1〜23、1998年; Friedberg、E.C、TIBS 17、347、1992年; Jans、D.Aら、Bioessays 2000年6月; 22(6): 532〜44; Yoneda、Y、J.Biocehm.(Tokyo)1997年5月; 121(5): 811〜7; Boulikas、T、Crit.Rev.Eukaryot.Gene Expr.1993; 3(3): 193〜227; Lyons RH、Mol.Cell Biol、7、2451〜2456、1987年)に記載されている。核輸送媒介核酸配列に関連して様々な原理を利用できる。この様な原理の一つは、YB−1をシグナルペプチドとの融合タンパク質の形で形成して核内に導入すること、およびそれにより本発明のアデノウイルスの複製が起こることである。
【0098】
発明に使用するアデノウイルスのデザインを実現するであろう別の原理は、好ましくは細胞質での合成から始まってYB−1を細胞核内に誘導するか、またはYB−1を細胞核内に移動して、そこでウイルス複製を促進するトランスポーター配列と一緒にYB−1を与えることができる原理である。特に効果的な核輸送媒介核酸配列の例はHIVのTAT配列であり、これはEfthymiadis、A、Briggs、LJ、Jans、DA、JBC 273、1623〜1628、1998年にその他好適核酸配列と共に記載されている配列である。発明に使用するアデノウイルスが核内輸送をコードするペプチドをコードしている核酸配列を含むことは、本発明の範囲内である。
【0099】
YB−1が完全長、特に野生型YB−1に対応する形態で存在することは、本発明の範囲内である。YB−1が、例えば短形または断頭形(truncated form)の様な誘導体として用いられること、または存在することは、本発明の範囲内である。本発明の範囲内で誘導体として用いられる、または存在するYB−1は、E2−後期プロモータに結合でき、その結果アデノウイルスE2領域の遺伝子発現を活性化できるYB−1である。この種の誘導体は特に、本明細書に開示のYB−1誘導体を含む。更なる誘導体は、アミノ酸配列のN−末端、C−末端または内部にある単独または複数個のアミノ酸を欠失することで作られるだろう。
【0100】
これまでに記したアデノウイルスがコードする各種の発現遺伝子および遺伝子産物に関しては、これらを組み合わせてコードすることも、また組み合わせて発現することもそれぞれ可能である。
【0101】
アデノウイルスおよびアデノウイルス系という用語は、本発明内では実質的に同一の意味を持つものとする。アデノウイルスは特にカプシドおよび核酸を含む完全なウイルス粒子を意味するものとする。アデノウイルス系という用語は特に、核酸が野生型に対し変更されている事実に焦点を当てるものである。この種の変更は、プロモータ、制御配列および/またはオープンリーディングフレームといったコーディング配列の欠失および/または付加および/または突然変異に起因する、アデノウイルスゲノムの構造変化を含むことが好ましい。さらに、用語アデノウイルス系は、例えば遺伝子治療に用いられるベクターと結びつけて使用するのが好ましい。
【0102】
前述のコメントは、アデノウイルスおよびアデノウイルス系それぞれの使用ならびにデザインを含め、コーディング核酸にも当てはまり、またその逆も言える。
【0103】
本発明に関しては、本発明に使用するアデノウイルスおよびそれをコードする核酸それぞれは、かかる複製をもたらす、または別の核酸配列と組み合わせての複製をもたらすようないずれかのアデノウイルス核酸である。本明細書に説明したように、複製に必要な配列および/または遺伝子産物はヘルパーウイルスを利用して提供することができる。コーディング核酸配列と称するものであれば、そしてかかる核酸配列が既知であるものであれば、使用した配列と同一のものだけでなく、それより派生する配列もまた発明の範囲内である。この派生する配列という用語は、本明細書に於いては特に、非派生配列の機能に対応する機能を示す核酸またはポリペプチドである、遺伝子産物を依然として生ずる配列を意味する。このことは、当業者既知である簡単な通常試験により決定できる。かかる派生核酸配列の例は、同一遺伝子産物、特には同一アミノ酸配列をコードしているものの、遺伝子コードの縮重による異なる塩基配列を有しているものである。
【0104】
好適実施態様に於いて、本発明のアデノウイルスおよび本発明のアデノウイルス複製系、そして本発明でのそれらの使用に関してはそれぞれ、アデノウイルス核酸は発癌遺伝子タンパク質、特にE1Aタンパク質の発現を欠失しており、このことはそれが12SのE1Aタンパク質もしくは13SのE1Aタンパク質をコードしていないか、またはそれが12SのE1Aタンパク質と13SE1Aタンパク質のどちらもコードしていないか、または本明細書に記載のように修飾されていることを意味しており、そしてアデノウイルス複製システムがさらにヘルパーウイルスの核酸を含み、このヘルパーウイルスの核酸が発癌遺伝子タンパク質、特にE1Aタンパク質をコードする核酸配列を含み、さらにヘルパーウイルスの核酸が次の特徴を有すると同時にアデノウイルスに対し同特性を付与すること、即ちYB−1核陰性細胞では複製しないが、YB−1核陽性細胞では細胞周期と無関係に細胞内で複製し、少なくとも1つのウイルス遺伝子、特にはE1B55kDa、E4orf6、E4orf3および/またはE3ADPをトランス活性化し、そして/または細胞質YB−1を核内に移動しないという特性を付与することを意味する。ここに記したトランス遺伝子をヘルパーウイルスが個別または一緒にコードしていること、および/またはそれから発現することは本発明の範囲内である。
【0105】
本発明のこの様なアデノウイルス複製系の実施態様の一つでは、アデノウイルス核酸および/またはヘルパーウイルスの核酸は、さらに複製可能であるベクターとしても存在する。
【0106】
本発明に用いるアデノウイルスをコードするコーディング核酸配列がベクター内、好ましくは発現ベクター内に存在すること、そしてこの発現ベクターを本発明に従って使用することは、本発明の範囲内である。
【0107】
別の局面では本発明はさらに少なくとも2つのベクターを含むベクター群に関し、この場合上記ベクター群がここに記したアデノウイルス複製系を共に含んでおり、そして上記ベクター群は本発明に従い用いられる。アデノウイルス複製系の各構成成分は、個々のベクター、好ましくは発現ベクター内に配置されると解釈される。
【0108】
最後に、本発明はアデノウイルスについてここに記した同一の目的に関する細胞の使用についての別の局面に関するものであり、その場合細胞は発明に使用するここに記したアデノウイルスをコードする1または複数の核酸、および/または本発明による各アデノウイルス複製系、および/または各ベクターおよび/もしくはベクター群を含む。
【0109】
前記アデノウイルス構築体ならびに特にそれらの核酸およびそれをコードする核酸もまた細胞内、好ましくは腫瘍細胞に一部分導入してもよく、その場合各種個別構成成分が存在していることにより、それらはあたかも各構成成分は単一核酸および単一または複数のアデノウイルスに由来するように一つになって作用するだろう。
【0110】
発明に使用するアデノウイルス、アデノウイルス系、またはその一部分をコードする核酸は、ベクターとして存在するだろう。好ましくは、それらはウイルスベクターとして存在する。アデノウイルス核酸を含む核酸の場合、ウイルス粒子がベクターであることが好ましい。しかし、上記核酸がプラスミドベクター内にある場合も発明の範囲内である。いずれの場合も、ベクターは挿入した核酸の増幅、即ち挿入核酸の複製および場合によっては発現、それらの制御をもたらすそれぞれの要素を含んでいる。好適ベクター、特に発現ベクター、および対応する要素は当業者に周知であり、例えばGrunhaus, A、Horwitz, M.S、1994年、Adenoviruses as cloning vectors.In Rice, C.編集、Seminars in Virology, London: Saunders Scientific Publicationsに記載されている。
【0111】
ベクター群に関する発明の局面は、核酸の各種要素が必ずしも1つのベクターに含まれている必要がないという前述の実施態様に対応するものである。この場合、ベクター群は少なくとも2種類のベクターを含む。それ以外のことは、ベクターに関連して既に述べた事柄がベクターおよびベクター群にそのまま当てはまる。
【0112】
発明に用いられるアデノウイルスは、ここに開示した各種核酸および遺伝子産物により特徴付けられ、そしてそれ以外に含まれる要素は野生型のアデノウイルスと同様であり、全てが当業者周知である(Shenk、T.: Adnoviridae: The virus and their replication.Fields Virology、第三版、編集、Fields, B.N.、Knipe, D.M.、Howley, P.M.ら、Lippincott-Raven Publishers、Philadelphia、1996年、67章)。
【0113】
アデノウイルスの複製は極めて複雑な過程であり、一般にはヒト転写因子E2Fを利用する。ウイルス感染に際しては、まず「初期遺伝子」E1、E2、E3およびE4が発現する。「後期遺伝子」群はウイルスの構造タンパク質の合成を担っている。初期および後期遺伝子両方の活性化に関しては、異なるE1AおよびE1Bタンパク質をコードしている2つの転写単位E1AとE2Aから成るE1領域が重要であるが、それはE2、E3およびE4がそれらにより誘導されるからである(Nevins、J.R.Cell 26、213〜220、1981年)。更にE1Aタンパク質は休止細胞にDNA合成を誘導してS期に入らせることができる(上記BoulangerとBlair、1991年)。さらにそれらはRbクラスの腫瘍抑制因子と相互作用する(Whyte, P.ら、Nature 334、124〜127、1988年)。そうすると、細胞質転写因子であるE2Fが放出される。E2F因子は次に細胞遺伝子ならびにウイルス遺伝子の対応するプロモータ領域に結合し(特にアデノウイルスE2初期プロモータ)、転写および翻訳を開始させるだろう(Nevins、J.R.Science 258、424〜429、1992年)。
【0114】
E2領域の遺伝子産物は3種類の必須タンパク質をコードしていることから、それらは複製開始とその遂行に特に必要である。E2タンパク質の転写は2種類のプロモータ、ここではE2−初期プロモータもしくは初期E2プロモータとも呼ばれる「E2−初期E2F−依存」プロモータ、および「E2−後期」プロモータにより制御されている(SwaminathanとThimmapaya、The Molecular Repertoire of Adnoviruses III: Current Topics in Microbiology and Immunology、199巻、177〜194、Springer Verlag 1995年)。さらに、E4領域の産物は、E1AおよびE1B−55kDaタンパク質と一緒になり、それぞれE2Fの活性およびp53の安定性にとって重要な役割を果たす。例えば、E4領域にコードされているE4orf6/7タンパク質がE2FおよびDP1から成るヘテロダイマーとの直接相互作用によってプロモータはより活性化される(SwaminathanおよびThimmapay、JBC 258、736〜746、1996年)。更にp53は、E1B−55kDaとE4orf6から成る複合体により不活性化され(Steegenga, W.T.ら、Oncogene 16、349〜357、1998年)、溶解性の感染周期が成功して終了する。さらにE1B−55kDaタンパク質は、E4orf6タンパク質と相互作用してウイルスRNAの核外への運び出しを促進することからも更に重要な機能を果たしているが、この場合細胞固有のRNAは核内に維持される(BridgeおよびKetner、Virology 174、345〜353、1990年)。更に重要な発見は、E1B−55kDa/E4orf6から成るタンパク質複合体がいわゆる「ウイルス封入体」内に局在していることである。このことから、これら構造体が複製および転写部位であることが推測される(OrnellesおよびShenk、J.Virology 65、424〜429、1991年)。
【0115】
複製にとって重要である、特にアデノウイルスの放出に関し重要である別の領域はE3領域である。E3領域は、より具体的にはインビトロでのアデノウイルス感染周期にとって必須ではない、即ち細胞培養に必須ではない比較的小型の各種タンパク質に関する遺伝情報を含んでいる。しかしそれらは特に免疫制御およびアポトーシス機能を有していることから、インビボでの急性期および/または潜伏感染期間中のウイルスの生存にとって重要な役割を果たしている(Marshall S.Horwitz、Virololgie、279、1〜8、2001年; Russell、上記)。約11.6kDaの大きさを持つタンパク質が細胞死を誘導することが示されている。このタンパク質は、その機能故にADP−英語でアデノウイルス致死タンパク質(Adenovirus death protein)−と呼ばれている(Tollefson、J.Virology、70、2296〜2306、1996年)。このタンパク質は主に感染周期の後期に形成される。さらに、このタンパク質の過剰発現は感染細胞をより溶解する(Doroninら、J.Virology、74、6147〜6155、2000年)。
【0116】
更に、本発明者はE1A欠失ウイルス、即ち具体的には12SのE1Aタンパク質を持たず、そして13SのE1Aタンパク質を発現しないウイルスが、より高いMOIで極めて効率的に複製できることを知った(Nevins J.R.Cell 26、213〜220、1981年)が、しかしこのことを臨床的に応用できなかった。この現象は文献の中では「E1A−様活性」と呼ばれている。さらにE1Aがコードする5種類のタンパク質の内2種類のタンパク質、即ち12Sおよび13Sタンパク質が、それぞれ他のアデノウイルス遺伝子の発現を制御および誘導することも分かった(Nevins, J.R.Cell 26、213〜220、1981年; Boulanger、P.およびBlair, E.; Biochem.J.275、281〜299、1991年)。これに関連して、トランス活性化機能が主に13Sタンパク質のCR3領域から提供されるものであることが示された(Wong HKおよびZiff EB.J Virol.、68、4910〜20、1994年)。13Sタンパク質のCR1および/またはCR2領域、および/またはCR3領域を特異的に欠失したアデノウイルスは、大部分が複製欠失であるが、それでも一部の細胞株ではウイルス遺伝子をトランス活性化し、それぞれ特にE2領域を促進する(Wong HKおよびZiff EB.J Virol.、68、4910〜20、1994年; Mymryk, J.S.およびBayley, S.T.、Virus Research 33、89〜97、1994年)。
【0117】
野生型アデノウイルスを細胞、一般的には腫瘍細胞に感染させると、E1A、E1B−55KおよびE4orf6が介在してYB−1が核内に誘導され、ウイルス封入体の中に核内のE1B−55Kと共に局在するようになり、それがインビトロおよびインビボの両方での細胞核内に於けるウイルスの効率的複製を可能にする。これに関連して、E4orf6がE1B−55Kに結合すること(Weigel, S.およびDobbelstein, M.J. Virology、74、764〜772、2000年; Keith N.Leppard、Seminars in Virology、8、301〜307、1998年)、およびそれがE1B−55Kの核内への運び込みと分布とを媒介し、それによりそれぞれ最適なウイルス産生とアデノウイルス複製が提供されることが初期に既に見出されていた。本発明の効率的なウイルス複製は、E1A、E1B−55KおよびYB−1間の相互作用によって、そして、E1B−55K/E4orf6とYB−1とから成る複合体によって、およびYB−1と核内のE1B−55Kが、いわゆるウイルス封入体内に同時に局在することによって、ならびに個々に記したウイルスのYB−1核陽性細胞での複製に使用することによって、およびYB−1核陽性細胞が関係する疾患の治療を目的とした医薬の製造に使用することによって可能となる。この様な細胞の背景により可能となった複製は、細胞溶解、ウイルスの放出、および近接細胞の感染および溶解をもたらし、結果として腫瘍細胞および腫瘍に感染した場合には、それぞれ腫瘍の最終的な溶解、即ち癌溶解が起こる。
【0118】
YB−1は逆位CAAT配列、いわゆるY−ボックスに結合する保存性の強い因子の群に属する。それらは転写ならびに翻訳両レベルにおいて通常の様式で活性化する(Wolffe, A.P.Trends in Cell Biology 8、318〜323、1998年)。活性化によりY−ボックス依存制御経路の数が増加することが見出されているが、増殖およびアポトーシス関連遺伝子の阻害時でも数が増加することが見出されている(Swamynathan、S.K.ら、FASEB J.12、515〜522、1998年)。従って、YB−1はp53と直接相互作用しており(Okamoto、T.ら、Oncogene 19、6194〜6202、2000年)、Fasの遺伝子発現(Lasham, A.ら、Gene 252、1〜13、2000年)、MDRおよびMRP遺伝子発現(Stein, U.ら、JBC 276、28562〜69、2001年; Bargou, R.C.ら、Nature Medicine 3、447〜450、1997年)ならびにトポイソメラーゼおよび金属プロテアーゼの活性化(Mertens.P.R.ら、JBC 272, 22905〜22912、1997年; Shibao, L.ら、Int.J.Cancer 83、732〜737、1999年)に於いて重要な役割をはたしている。従って、YB−1はmRNAの安定性の制御(Chen, C-Y.ら、Genes & Development 14、1236〜1248、2000年)および修復プロセス(Ohga, T.ら、Cancer Res.56、4224〜4228、1996年)に関係している。
【0119】
腫瘍細胞でYB−1が核内に局在することにより、特に12SのE1Aタンパク質及び13SのE1Aタンパク質の両方ともが発現形態で存在しないかそして用いられない例(Holm, P.S.ら、JBC 277、10427〜10434、2002年)、および多剤耐性(多重耐性)でのタンパク質YB−1の過剰発現の例ではE1A独立にウイルス複製が起こる。更に、例えばE4orf6およびE1B−55Kの様なアデノウイルスタンパク質はウイルス複製に対し有利な影響を及ぼし(Goodrum, F.D.およびOrnelles, D.A、J.Virology 73、7474〜7488、1999年)、この時機能的E1Aタンパク質は他のウイルスの遺伝子産物(E4orf6、E3ADPおよびE1B−55Kの様な)のスイッチングに関係していること(Nevins J.R.、Cell 26、213〜220、1981年)が知られている。しかしこの様なことは13SのE1Aが存在しない先行技術のE1A−マイナスアデノウイルスでは起こらない。YB−1が核内に存在する多剤耐性細胞でのYB−1核内局在は、E1A−マイナスウイルスに複製および粒子形成を提供する。しかしこの場合、ウイルス複製および粒子形成の効率は、野生型のAd5に比べると数倍低い。腫瘍細胞の核内に既に存在しているYB−1と、または外部因子(細胞増殖抑制薬または放射線、または高温を作用させる)により腫瘍細胞内に誘導した、即ち特に細胞周期とは無関係に核内への存在を促すか、またはベクターを使ってトランス遺伝子として導入したYB−1と、アデノウイルス遺伝子を始動するが、ウイルスの複製は行わない系、好ましくはアデノウイルス系とを組み合わせると、驚くべきことにYB−1を通じてシステムが極めて効率的にウイルスの複製と粒子を形成し、癌溶解を誘導した。とりわけても好適な細胞増殖抑制薬は次の群に属するものである:ダウノマイシンおよびアドリアマイシンといったアントラサイクリン類;シクロホスファミドのようなアルキル化剤;エトポシドの様なアルカロイド;ビンクリスチンおよびビンブラスチンの様なビン−アルカロイド;例えば5−フルオロウラシルおよびメトトレキセートの様な代謝拮抗剤;例えばシス−プラチンの様な白金誘導体;カンホテシンの様なトポイソメラーゼインヒビター;および例えばタキソールの様なタキサン類である。ここに開示した、YB−1核陽性細胞内でのみ複製できるアデノウイルス、特に組換え体アデノウイルスは、ウイルス遺伝子E1B−55K、E4orf6、E4orf3およびE4ADPをトランス活性化するそれらの能力は、野生型アデノウイルス、特に野生型Ad5の持つ同様のトランス活性化能力に比べ制限されている。本発明者は、今回驚くべきことにこれら制限された活性化能力が対応する遺伝子によって、特にYB−1の核内局在と組み合わせて発現されるE1B−55KおよびE4orf6によって補完されることを見出した。本明細書の実施例に示す様に、ウイルス複製および粒子形成は共にこの様な条件では野生型アデノウイルスの複製および粒子形成に匹敵するレベルまで増加する。
【0120】
ここに記したアデノウイルスと結びつけた医薬、またはここに記したアデノウイルスを本発明に従い使用し製造される医薬は一般的には全身投与されるものと解釈されるが、かかる医薬を局所に適用するか、または配送することも本発明の範囲である。かかる使用は、本発明の医薬を使用する診断および/または予防および/または治療を目的として、特にある状態、典型的には疾患の形成に関係する細胞に上記アデノウイルスで感染すること、および特にそれら細胞内で複製すること意図して、好ましくは随時に行われる。
【0121】
かかる医薬は、好ましくは腫瘍疾患の処置に用いられる。腫瘍疾患の中では、腫瘍疾患の基礎を成すメカニズム、特に病理学的メカニズムによってYB−1が既に核内に局在している腫瘍、または外的手段により核内にYB−1が生じている腫瘍が特に好ましく、この場合の手段はYB−1を核内に運搬する、またはYB−1を核内に誘導する、またはYB−1を核内で発現するのに好適なものである。本明細書で使用する場合、腫瘍または腫瘍疾患という用語は、悪性および良性腫瘍の両方、ならびに各疾患を含むものとする。医薬はさらに少なくとも1つの医薬的に活性な化合物を含むものとする。かかる追加の医薬的に活性な化合物の種類および量は、かかる医薬が用いられる適用に依存する。医薬を腫瘍疾患の治療および/または予防に用いる場合、具体的にはシスプラスチンおよびタキソール、ダウノブラスチン、ダウノルビシン、アドリアマイシン、および/もしくはミトキサントロンのような細胞増殖抑制薬、またはここに記したその他の細胞増殖抑制薬あるいは細胞増殖抑制薬の群を使用する。
【0122】
本発明の医薬は様々な製剤形状に存在でき、液体形状が好ましい。更に、医薬は安定化剤、緩衝剤、保存剤、および医薬製剤分野当業者に周知であるこの種の作用剤を含むだろう。
【0123】
本発明者は驚くべきことに、ここに記載のウイルスを発明に従い、細胞周期とは無関係に核内にYB−1を持つ腫瘍に対し極めて高い成功率で応用できることを見出した。一般にYB−1は細胞質内、特に核周囲の原形質内に存在する。細胞周期のS期の間は、YB−1は正常細胞および腫瘍細胞両方の細胞核内に見出すことができる。しかしそれは修飾型アデノウイルスを用いたウイルスによる癌溶解を誘導するには十分ではない。この様な不良な結果は、先行技術に記載のこの種の弱毒化アデノウイルスの効率が比較的低いことに起因している。換言すれば、ここに記載の弱毒化または修飾型ウイルスを投与する際に、ウイルス性の癌溶解に必要な分子生物学的条件を与えさえすれば、かかるアデノウイルス系を使用する、特に高い効率で使用することができる。ここに記載様に、発明に従って用いられるAdΔ24、dl922−947、E1Ad/01/07、CB016、dl520および欧州特許第EP0 931 830号に記載の組換え体アデノウイルスといったアデノウイルスの場合は、上記条件は腫瘍疾患の例ではYB−1が細胞周期とは無関係に核局在する細胞に与えられる。このタイプの核内局在は、腫瘍細胞そのものに起因するものでも、またはここに記した発明による手段または薬物により誘導してもよい。即ち本発明は、発明のウイルス、特に先行技術に既に記載されている弱毒化または修飾型アデノウイルスを用いて良好に処置できる新規腫瘍および腫瘍疾患群、さらには患者を画定する。
【0124】
その一部は周知であり、そして本発明により治療可能であるアデノウイルスを用いて、または本明細書に初めて記載されたアデノウイルス、具体的にはEqAタンパク質内に突然変異および欠失を有するが、Rb/E2fの結合を妨害せず、そしてYB−1核陰性細胞では複製しないアデノウイルス、または本明細書内に規定された大きな複製低下を示し、および/または欠失型発癌タンパク質、特にE1Aを有する、例えばウイルスAdΔ24、dl922−947、E1Ad/01/07、CB106および欧州特許第EP0 931 830号に記載のアデノウイルスを用いて、本発明により治療可能なさらなる患者群は、ある条件を適用または放出することによりYB−1が核内に移動する、または核内に誘導する、または核内に運ばれることが確実である患者である。この患者群に関連してこの種のアデノウイルスを使用することは、ウイルス複製の誘導がYB−1の核内局在化とそれに続くYB−1のE2−後期プロモータへの結合に基づくという所見を根拠としている。この所見に基づけば、AdΔ24、dl922−947、E1Ad/01/07、CB106および欧州特許第EP0 931 830号に記載のアデノウイルスといった本明細書に開示するアデノウイルスは、YB−1核陽性である細胞および/またはYB−1が本発明に規定されるように脱制御されている細胞で複製できるだろう。その限りに於いてこれらアデノウイルスは、本発明に従って、これら特性を持つ細胞を含む疾患および患者群の処置に、特にこれら細胞が処置対象となる各疾患の成立に関係している時に用いることができる。本明細書に発明に有用であると記したアデノウイルス、そして使用するこれらウイルス、特に本明細書で初めて記載されたアデノウイルスを用いて処置可能であるさらなる患者群は、YB−1核陽性および/または以下記載の処置の結果としてYB−1核陽性である患者であり、この場合かかる処置は好ましくは医学的処置であり、そして/または各ウイルスの投与を伴い実施される患者である。YB−1核陽性患者が、腫瘍を形成する複数の細胞が細胞周期とは無関係に核内にYB−1を有する患者である場合も、本発明の範囲内である。とりわけこれら治療は、ここに記載の細胞増殖抑制薬投与と共に、そして/または腫瘍治療の一環として実施される。更に、放射線、好ましくは腫瘍療に使用するものである放射線は、この処置群に属する。放射線とは、具体的に高エネルギー照射を伴う放射線を意味し、好ましくは放射性放射線、好ましくは腫瘍治療に用いられる放射性放射線を意味する。高温および高温の適用、好ましくは腫瘍治療に用いられる高温も別の処置手段である。特に好ましい高温の実施態様は局所的適用である。最後にホルモン処置、特に腫瘍治療に用いられるホルモン処置は別の処置手段である。かかるホルモン処置に関連しては、抗エストロゲンおよび/または抗アンドロゲンが用いられる。タモキシフェンの様な抗エストロゲン剤は特に乳ガンの治療に、そして例えばフルタミドまたは酢酸シプロテロンの様な抗アンドロゲン剤は、前立腺癌の治療に用いられる。
【0125】
腫瘍を形成する細胞の一部が元から、または核内への誘導および能動的導入後にYB−1を含んでいること、または本開示の意味において脱制御したYB−1を含む場合も本発明の範囲内である。好ましくは、腫瘍形成細胞の約5%以上、即ち6、7、8%等がYB−1核陽性細胞またはYB−1が脱制御状態にある細胞である。YB−1の核内への局在は、外側から加えられたストレス、および局所的に加えられたストレスにより誘導できる。この誘導は、例えば放射線、特にUV照射、細胞増殖抑制薬、取り分けても本明細書に開示されたもの、および高温を用いて行うことができる。高温に関連しては、それは極めて特異的な様式、より具体的には局所に極めて特異的な様式で実現できることが必須であり、その結果YB−1はより特異的に細胞核内に局在するようになり、それによりアデノウイルスの複製に必要な条件、即ち細胞および腫瘍溶解に必要な条件が、好ましく局所に限定された形で提供される(Stein U、Jurchott K、Walther W、Bergmann S、Schlag PM、Royer HD.J Biol Chem.2001年、276(30)、28562〜9; Hu Z, Jin S, Scotto KW.J Biol Chem.2000年1月 28; 275(4): 2979〜85; Ohga T, Uchiumi T, Makino Y, Koike K、Wada M、Kuwano M, Kohno K.J Biol Chem.1998年、273(11): 5997〜6000)。
【0126】
かくして本発明の医薬は、好ましくは各腫瘍細胞に於いて、前処理または併用処置を通じてYB−1の輸送に影響を及ぼす形で患者および患者群に投与することもできるし、または、意図されていてもよい。
【0127】
この技術的教示に基づけば、当業者はその技術的水準の範囲内で、E1Aに例えば欠失または点突然変異を含む好適な修飾を実行して、発明の使用と関係して用いられるアデノウイルスの各種実施態様を生成することができる。
【0128】
既に説明した如く、発明に従い使用するアデノウイルスは、核内にYB−1を有する細胞および細胞系それぞれの中で複製できる。本発明に従って使用するアデノウイルスが複製でき、そして腫瘍を溶解できるかどうかという問題にとって、Rb、即ち網膜細胞芽腫腫瘍抑制産物の有無に関する細胞状態は無関係である。さらに、発明に従い上記アデノウイルスを使用することに関連して感染細胞、感染対象となる細胞、または処置対象となる細胞のp53の状態を考慮する必要はないが、それはここに開示したアデノウイルス系をYB−1核陽性細胞、即ち細胞周期とは無関係に核内にYB−1を持つ細胞と結びつけて使用することにより、p53に関する状態ならびにRbに関する状態が本明細書に開示する技術的教示の性能に影響しなくなるからである。
【0129】
発癌遺伝子および発癌遺伝子タンパク質はそれぞれ、特にE1Aは所有の天然アデノウイルスプロモータの制御下、および/または腫瘍もしくは組織特異的プロモータにより制御できる。好適な非アデノウイルス性のプロモータは、サイトメガロウイルスプロモータ、RSV(ラウス肉腫ウイルス)プロモータ、アデノウイルスベースドプロモータVaIおよび非ウイルス性YB−1プロモータ(Makino Y.ら、Nucleic Acids Res.1996年、15、1873〜1878)を含む群より選ぶことができる。ここに開示した発明の各局面およびいずれかの局面に関連して使用できるさらなるプロモータには、テロメラーゼプロモータ、アルファ−フェトプロテイン(AFP)プロモータ、癌胚抗原プロモータ(CEA)(Cao, G., Kuriyama, S., Gao, J., Mitoro, A., Cui, L., Nakatani, T., Zhang, X., Kikukawa, M., Pan, X., Fukui, H., Qi, Z. Int.J.Cancer、78、242〜247、1998年)、L−プラスチンプロモータ(Chung, I., Schwartz, PE., Crystal, RC., Pizzorno, G, Leavitt, J., Deisseroth, AB. Cancer Gene Therapy、6、90〜106、1999年)、アルギニンバソプレッシンプロモータ(Coulson, JM, Staley, J., Woll, PJ. British J.Cancer、80、1935〜1944、1999年)、E2fプロモータ(Tsukadaら、Cancer Res.、62、3428〜3477)、ウロプラキンIIプロモータ(Zhangら、Cancer Res.、62、3743〜3750、2002年)およびPSAプロモータ(Hallenbeck PL、Chang, YN、Hay, C, Golightly, D., Stewart, D., Lin, J., Phipps, S., Chiang, YL. Human Gene Therapy、10、1721〜1733, 1999年)がある。更にドイツ特許出願番号DE101 50 984.7に記載のYB−1依存型E2−後期プロモータは、本発明で使用可能なプロモータである。
【0130】
テロメラーゼプロモータはヒト細胞において極めて重要であることが知られている。故にテロメラーゼ活性はテロメラーゼ逆転写酵素遺伝子(hTERT)の転写制御を通じて制御されているが、上記遺伝子は酵素の触媒サブユニットである。テロメラーゼは85%のヒト腫瘍細胞で発現している。これに対し、大部分の正常細胞では発現していない。例外は生殖細胞と胚組織である(Braunstein, I.ら、Cancer Research、61、5529〜5536、2001年; Majumdar, A.S.ら、Gene Therpy 8、568〜578、2001年)。hTERTプロモータに関するより詳細な研究から、開始コドンからの距離が283bpおよび82bpのプロモータの断片が、腫瘍細胞での特異的発現に十分機能することを示した(Braunstein I.ら、; Majunmdar ASら、上記)。故にこのプロモータおよび上記特異的断片はそれぞれ、遺伝子、特にトランス遺伝子、好ましくはここに開示するトランス遺伝子の一つを腫瘍細胞内でのみ特異的に発現させるのに好適である。このプロモータが、腫瘍細胞のみにおいて、発癌遺伝子、好ましくはE1A発癌遺伝子の発現を可能にする。また、好適実施態様では、かかるアデノウイルスベクター内でのトランス遺伝子、特にE4rof6、E1B55kD、ADPおよびYB−1を含む群から選ばれるトランス遺伝子の発現は、これらプロモータのいずれかの制御下にある。トランス活性化発現遺伝子タンパク質、特にE1Aタンパク質のオープンリーディングフレームがアデノウイルス系の一つ、または複数の遺伝子産物と共にフレーム内に存在するものも本発明の範囲内である。しかし、トランス活性化E1Aタンパク質のオープンリーディングフレームは、それから独立してもよい。
【0131】
ここに記載のアデノウイルスを発明に従い用いて溶解するのに好適である細胞の特徴は、実施態様の一つでは、それらは耐性であり、好ましくは多剤または多重耐性である。ここで言う耐性とは、ここに記した細胞増殖抑制薬に対する耐性を意味するのが好ましい。この多剤耐性は、該当する細胞を決定するためのマーカーとして利用でき、従ってかかる多剤耐性を示す腫瘍および患者群について利用できる膜結合型輸送タンパク質P−糖タンパク質の発現、好ましくは過剰発現と共に現れる。ここで使用する場合の耐性という用語は、古典的耐性とも呼ばれるP−糖タンパク質介在耐性と、MRP介して伝達される耐性もしくはその他非P−糖タンパク質が介在する耐性を含む非定型耐性の両方を含む。YB−1の発現と相関するさらなるマーカーは、トポイソメラーゼIIアルファである。その場合、患者が発明のアデノウイルスの使用により良好に処置できるか否かを決定するスクリーニングに於いて、トポイソメラーゼIIアルファの発現は核内のYB−1の決定に代わる方法または追加の方法として使用できる。P−糖タンパク質と同様の様式で基本的に用いることができるさらなるマーカーはMRPである。少なくとも結腸直腸癌細胞または結腸直腸癌患者について想定されるさらなるマーカーは、例えばShibao K.ら、(Shibao Kら、Int, Cancer, 83、732〜737、1999年)が記載したようにPCNA(増殖細胞核抗原)である(Hasan S.ら、Nature、15、387〜391、2001年)。最後に、MDR(多剤耐性)の発現は、少なくとも乳ガンおよび骨肉腫細胞については、上記の意味でのマーカーである(Oda Yら、Clin.Cancer Res.、4、2273〜2277、1998年)。本発明に従い用いることができる可能性のあるさらなるマーカーはp73である(Kamiya、M.、Nakazatp, Y.、J Neurooncology 59、143〜149(2002年); Stieweら、J.Biol.Chem.、278、14230〜14236、2003年)。
【0132】
従って本発明の特別な利点は、臨床的な意味に於いてもはや処置できないと考えられる患者、および先行技術の方法による腫瘍疾患のそれ以上の処置が不可能であり、合理的に成功が期待できない患者、特に細胞増殖抑制薬の使用がもはや合理的には不可能であり、腫瘍に影響を及ぼすまたは腫瘍を縮小させるという意味でもはや良好に治療が実施できない患者についても、ここに記載の様にして本発明によるアデノウイルスを用いることで治療できることである。ここでは、腫瘍という用語は一般に核内にもともとYB−1を含んでいる、またはここに記載した様な外的な開放手段によって核内に、好ましくは細胞周期とは無関係にYB−1を含むようになった腫瘍または癌疾患を意味する。
【0133】
更に、ここに記載のウイルスは、一般的な腫瘍の処置にも用いることができる。好ましくは、これら腫瘍は、乳ガン、卵巣癌、前立腺癌、骨肉腫、神経芽腫、メラノーマ、小細胞肺ガンおよび結腸直腸癌を含む群より選ばれる。更なる腫瘍はここに記した様な耐性の腫瘍であり、好ましくは多重耐性である腫瘍であり、特には上記の群の多重耐性の腫瘍である。
【0134】
発明は更に別の局面では、修飾型アデノウイルスの一つ、即ち例えばAdΔ24、dl922−947、E1Ad/01/07、CB016または欧州特許第EP0 931 830号に記載のウイルスの様な本発明に使用するアデノウイルスの一つを用いて処置できる患者をスクリーニングする方法であって;
−腫瘍組織のサンプルを検査するステップ、および
−YB−1が細胞周期とは無関係に核内に局在しているか決定するステップ、を含む方法に関する。
【0135】
前記マーカーの存在はYB−1に代わるものとして、またはYB−1に追加するものとして検出できる。
【0136】
腫瘍組織またはその一部がYB−1を核内に含む場合、特に細胞周期とは無関係に含む場合、ここに開示したアデノウイルスは本発明の実施に従い用いることができる。
【0137】
本発明の方法の実施態様では、腫瘍組織の検査はYB−1に対する抗体、YB−1に対するアプタマー、YB−1に対するシュピーゲルマー(spiegelmer)、ならびにYB−1に対する抗カリン(anticaline)を含む群より選ばれる作用物質を用いることで実施される。基本的には対応するマーカカーについては同一の手段を作り、それに従い用いることができる。抗体、特にモノクローナル抗体の製造方法は当業者周知である。YB−1またはマーカーを特異的に検出するさらなる手段は、標的構造、本例ではYB−1または上記マーカーに高い親和性で結合するペプチドである。そのようなペプチドを作製するためにファージディスプレーのような周知方法が知られている。典型的には、開始時に個々のペプチドが8〜20アミノ酸の長さを持ち、ライブラリーのサイズが102〜1018、好ましくは108〜1015種類のペプチドであるペプチドライブラリーを準備する。標的分子に結合するポリペプチドの特殊な形態は抗カリンと呼ばれ、例えばそれはドイツ特許出願番号DE197 42 706号に記載されている。
【0138】
YB−1またはここに開示した対応するマーカーとの特異的結合に関する、従って細胞周期と無関係な細胞核内のYB−1局在を検出するためのさらなる手段は、いわゆるアプタマー、即ち単鎖または二本鎖としてRNAまたはDNAの形で存在し、標的分子に特異的に結合するD−核酸である。アプタマーの作成は、例えば欧州特許第EP0 533 838号に記載されている。特殊な形状をしたアプタマーはアプタザイムと呼ばれ、これは例えばPiagenau, N.ら(2000年)、Angew.Chem.Int.Ed., 39、no.29、4369〜4373項に記載されている。それらがアプタマー部分とは別にリボザイム部分を含み、アプタマー部分と結合している標的分子との結合または放出によって触媒活性を獲得し、核酸基質を開裂してシグナルを生成するという意味において、アプタマーの特別な実施態様である。
【0139】
さらなる形状のアプタマーはいわゆるシュピーゲルマー、即ちL−核酸から作られている標的分子結合核酸である。かかるシュピーゲルマーの製造方法は、例えば国際特許出願第WO98/08856号に記載されている。
【0140】
腫瘍組織のサンプルは、穿刺または手術により得ることができる。YB−1が細胞周期とは無関係に核内に局在しているか調べることは、顕微鏡技術および/または免疫組織分析、好ましくは抗体または上記その他手段を使って頻繁に実施される。核内のYB−1を検出する、そして特にYB−1が細胞周期とは無関係に核内に局在しているか検出するためのさらなる手段は当業者に周知である。例えば、YB−1の局在は、スクリーニング時に染色組織切片で容易に検出できる。核内にYB−1が存在する頻度は、局在が細胞周期とは無関係であることを既に示している。細胞周期とは無関係な核内YB−1を検出するための更なる選択肢は、YB−1を染色し、YB−1が核内に局在しているか検出し、そして細胞のフェーズを決定することである。この方法およびYB−1の検出は、YB−1を検出するための上記手段を用いて実行してもよい。これら手段は、当業者周知の方法により検出される。前記のYB−1と特異的に結合し、分析対象サンプル内、特には細胞内のその他構造には結合しない作用物質を用いることにより、これら手段を好適に標識する方法を用いてそれらの局在を、そしてそれらの持つYB−1への特異的結合故にYB−1の局在をも検出そして確立することができる。前記手段を標識する方法は、当業者周知である。
【0141】
以下、本発明を新規特徴、実施態様および利点を示す図面およびサンプルを参照しながら更に説明する。
【図面の簡単な説明】
【0142】
【図1】本明細書でAdE1/E3−マイナスと呼ばれる、E1/E3欠失アデノウイルスであるアデノウイルベクター、野生型アデノウイルスおよびアデノウイルスdl520の構造デザイン。
【図2】p300、p107およびp105の結合に関する、E1Aタンパク質の結合ドメイン。
【図3】E1/E3マイナスAd5と呼ばれるE1/E3欠失アデノウイルスAd5およびアデノウイルスdl520感染後の、核内にYB−1を持たないU20S細胞。
【図4】E1/E3マイナスAd5と呼ばれるE1/E3欠失アデノウイルスAd5およびアデノウイルスdl520感染後の、核内にYB−1を有する257RDB細胞。
【図5】アデノウイルスdl1119/1131感染後の257RDB細胞およびU2OS細胞。
【図6】多剤耐性細胞および細胞株257RDB、181RDB、MCF−7Ad内にそれぞれYB−1が存在するが、U2OSおよびHeLa細胞の核内にはYB−1が存在しないことを確認したEMSA分析の結果。
【図7】野生型アデノウイルス、アデノウイルスdl520およびアデノウイルスdl1119/1131のE1Aタンパク質の構造デザイン。
【図8】追加発現したウイルスタンパク質存在時のアデノウイルスの複製効率を絶対量で示した棒グラフ。
【図9】追加発現したウイルスタンパク質存在時のアデノウイルスの複製効率を増加率で示した棒グラフ。
【図10】ダウノルビシンを加えていない、およびダウノルビシン40ng/mlを加えた場合の、10および30pfu/細胞のdl520、およびコントロール(K)をそれぞれ感染させクリスタルバイオレット染色した後のU2OS細胞増殖ウエルを示す図。
【図11】ダウノルビシンを加えていない、およびダウノルビシン40ng/mlを加えた場合の、10および30pfu/細胞のdl5210、およびコントロール(K)をそれぞれ感染させクリスタルバイオレット染色した後のHeLa細胞増殖ウエルを示す図。
【図12】PBSおよびdl520でそれぞれ処理後の、起源(RDB257およびHeLa)の異なる腫瘍の腫瘍容積の経時変化を示すグラフ。
【図13】それぞれPBSおよび5×108pfuのdl520で処理した後に、RDB257細胞由来の腫瘍を発生した屠殺マウスの図。
【図14】dl520感染後のRDB257細胞およびHeLa細胞の細胞抽出物(皮下増殖した腫瘍)のサザンブロット分析の結果。
【図15】YB−1核陽性腫瘍細胞(257RDBおよび181RDB)およびYB−1核陰性腫瘍細胞(HeLa、U2OS)でのdl520および野生型アデノウイルスの複製効率および粒子形成を示す棒グラフ。
【図16】野生型アデノウイルスおよびアデノウイルスベクターAdXvir03の構造デザインを示す図。
【図17】アデノウイルスベクターAdXvir03/01の構造デザインを示す図。
【図18A】Ad312(20pfu/細胞)、Xvir03(5pfu/細胞)およびコントロール(非感染)に感染させ、クリスタルバイオレット染色を行った181RDB細胞(図18A)および272RDB細胞(図18B)増殖ウエル。クリスタルバイオレット染色は感染5日後に実施した。
【図18B】Ad312(20pfu/細胞)、Xvir03(5pfu/細胞)およびコントロール(非感染)に感染させ、クリスタルバイオレット染色を行った181RDB細胞(図18A)および272RDB細胞(図18B)増殖ウエル。クリスタルバイオレット染色は感染5日後に実施した。
【実施例1】
【0143】
発明により使用されるアデノウイルスが含むE1A修飾のタイプ
図1はAdE1/E3マイナス、即ちE1/E3欠失アデノウイルスと野生型アデノウイルスおよびアデノウイルスdl520の構造デザインを示している。
【0144】
アデノウイルスAdE1/E3マイナスは機能的E1Aまたは機能的E1BもしくはE3をコードする領域を有しておらず、本実験では毒性に関するコントロールに用いられる。
【0145】
野生型E1A遺伝子は、E1A RNAの異なる位置でのスプライシングから生じる合計で5種類のタンパク質をコードしている。とりわけ2種類のタンパク質、即ち289アミノ酸タンパク質および243アミノ酸タンパク質が生成される。dl520はE1A遺伝子のCR3ストレッチ内に欠失を持つために13S遺伝子産物を欠いているため、289アミノ酸タンパク質をコードしていない。発明に用いられるアデノウイルスdl520は当業者からは12S−E1Aウイルスと呼ばれる。先行技術周知のアデノウイルスdl347(WongおよびZiff、J.Virol.、68、4910〜4920、1994年)はまた12S−E1Aでもあり、これは本発明に使用することができる。
【0146】
13S−E1AのmRNAによりコードされる289アミノ酸タンパク質の中には、各種アデノウイルスサブタイプに保存されている3つの領域がある。これらはCR1、CR2およびCR3と呼ばれる。CR1およびCR2は両E1Aタンパク質(E1A 12SおよびE1A 13S)、即ち289アミノ酸および243アミノ酸タンパク質に存在するが、CR3領域は上記タンパク質のうちの大きい方だけに存在する。
【0147】
CR3領域はウイルス遺伝子、特にE1B、E2、E3およびE4の活性化に必要である。小型、即ち243アミノ酸タンパク質のみを含むウイルスは、極弱くしかウイルス遺伝子をトランス活性化できず、核内にYB−1を持たない細胞でのアデノウイルス複製を促進しない。YB−1は腫瘍細胞でのみ核内に存在すること、そしてその中にのみ見出すことができることから、このベクターは腫瘍特異的複製の誘導に好適である。
【0148】
dl520はCR3を欠失していることから、このアデノウイルスは本明細書で輸送とも呼ばれる細胞質YB−1の細胞核内への移動を行うことができず、それ故にYB−1核陰性である細胞では複製できないことから、このウイルスは本発明で使用可能なウイルスであり、そしてこのウイルスは本発明に必要なトランス活性化に使用できる。
【実施例2】
【0149】
細胞のRb状態に依存したアデノウイルスの作用様態
図2はp300,p107およびp105の結合に関係するE1Aタンパク質の結合ドメインを示している。P300およびP107は、細胞質結合タンパク質である。腫瘍抑制タンパク質である網膜芽細胞腫タンパク質(pRb)の結合は、CR1およびCR2を通じて媒介される。研究からpRbおよびp107/p300は細胞転写因子E2Fと組み合わさり効率的に転写制御することが示されている。野生型E1Aタンパク質はE2FのRbへの結合を妨害する。即ち放出されたE2FはE2初期プロモータに結合して、アデノウイルスの複製を誘導する。
【0150】
E1A発癌タンパク質内の特定の欠失がRb−陰性細胞で優先的に複製することが可能であり、そして本発明に用いることができる以下に述べるような組換え体アデノウイルスベクターを生成することが従来知られていた。例えばアデノウイルスベクターdl922−947は、CR2領域(アミノ酸位置122〜129)内に欠失を持ち、ベクターCB016はCR1(アミノ酸位置27〜80)およびCR2領域(アミノ酸位置122〜129)に欠失がある。ベクターE1Adl01/07はCR2領域(アミノ酸位置111〜123)内に欠失を持つ。更に、N−末端(アミノ酸位置4〜25)に追加の欠失があることから、タンパク質p300への結合は認められない。アデノウイルスベクターAdΔ24はCR2領域(アミノ酸位置120〜127)に欠失を持っている。欧州特許第EP0 931 830号に記載のアデノウイルスベクターはCR1領域よびCR2領域に欠失を含んでいる。
【0151】
E2F/RPの結合メカニズムおよびE1Aを媒介するE2Fの放出は、本発明の基礎を成すメカニズムとは基本的に異なるものである。先行技術での予想とは異なり、ウイルス複製にとって必須ではないが、重要であるのはRbタンパク質からのE2Fの放出ではなく、ヒト転写因子であるYB−1が核内に局在することである。この転写因子は、正常細胞に於いては、細胞周期の大部分について細胞質内にのみに存在している。アデノウイルスが感染すると、特定の条件の下にYB−1は核内に導かれるか、例えば乳ガン、卵巣癌、前立腺癌、骨肉腫、神経芽腫、メラノーマ、小細胞肺癌および結腸直腸癌を含むが、もとよりこれらに限定されないある種の腫瘍疾患といったある種の細胞システムでは核内に既に存在している。
【実施例3】
【0152】
U2OS細胞の感染
100,000個のU2OS細胞をウエルに播いた。翌日細胞に図3に示すような各種アデノウイルスを感染させた。感染は無血清DMEM培地500μl中で、37℃で1時間行った。続いて感染培地を取り除き、完全培地(10%FCS/DMEM)2mlと交換した。3日後にクリスタルバイオレット染色で分析を行った。
【0153】
図3から分かるように核内にYB−1を持たないU2OS細胞は、本発明に使用できる2種類のアデノウイルス、即ちE1/E3マイナスと呼ばれるE1/E3欠失ウイルスおよびアデノウイルスdl520で感染させた後にクリスタルバイオレット染色を行ったところ、溶解を示していなかった。この場合、まず培地を取り除いた。続いて細胞にクリスタルバイオレット液(50%ETOH、3%ホルムアルデヒド、5%酢酸、1%クリスタルバイオレット)を重層して室温で5〜10分間インキュベーションした。続いて6ウエル型プレートを水でよく濯いでから室温で乾かした。これから、本発明で使用するウイルスを誘導し、感染細胞を溶解させるには、YB−1の存在が必要であるという、本発明の基礎を成す発見が確認された。
【実施例4】
【0154】
257RDB細胞の感染
100,000個の257RDB細胞をウエルに播いた。翌日細胞に図4に示すような各種アデノウイルスを感染させた。感染は無血清DMEM培地500μl中で、37℃で1時間行った。続いて感染培地を取り除き、完全培地(10%FCS/DMEM)2mlと交換した。3日後にクリスタルバイオレット染色で分析を行った。
【0155】
この実験の結果を図4に示す。E1/E3を欠失したE1/E3マイナスと呼ばれるアデノウイルスは、核内にYB−1を持つ257RDB細胞への感染については、低MOI(pfu/細胞)では溶解を示さなかった。これに対し実施例3で示した、YB−1核陰性細胞では複製せず、同時に本発明に従い発癌遺伝子タンパク質をトランス活性化するE1Aをコードしているdl520は、40pfu/細胞というMOI(感染多重度)で実質完全な溶解をおこし、10pfu/細胞のMOIでも顕著な溶解を起こした。このことから、dl520およびここに記したdl1119/1131またはAdXvirといった同様のウイルスが必要とするMOIは、E1欠失またはE1/E3欠失アデノウイルスに比べ約1桁(10倍)低く、臨床使用が可能であると結論される。
【0156】
図7に示すように、dl520のタンパク質E1AはそのCR3領域を欠失することを特徴とし、その結果本発明による使用に求められるトランス活性化およびYB−1核陽性細胞での複製を起こす。
【実施例5】
【0157】
dl1119/1131による257RDBおよびU2OS細胞の感染
図5に示すように、E1Aタンパク質のアミノ酸4〜138およびそれをコードする核酸を欠失しており、さらにアミノ酸218の後に停止コドンを含むために発現する断頭型(truncate)E1Aタンパク質が完全なE1Aタンパク質のCR3領域を含んでいるアデノウイルスdl1119/1131をYB−1核陰性U2OS細胞に感染した場合、20pfu/細胞のMOIでは溶解は起きなかった。陰性コントロールには非感染細胞層を用いた。
【0158】
これに対し核内にYB−1を含む、即ちYB−1核陽性である、257RDBの様な細胞系では、アデノウイルスdl1119/1131の影響を受けて、20pfu/細胞のMOIで細胞層は実質的に完全に溶解した。この限りにおいて本実施例は、図7に示す様に例えばCR3領域のみを含み、そしてCR1領域およびCR2領域を欠いている修飾型E1A発癌遺伝子タンパク質が、発明によるアデノウイルスの複製に必要なYB−1核陽性細胞でのトランス活性化を提供していることを示す別の証拠である。即ちアデノウイルスdl1119/1131は、本発明に使用可能である別のアデノウイルスである。CR3領域に関してdl1119/1131と同様にデザインされているが、それと異なりCR1領域および/またはCR2領域も含んでいるウイルスを使用できることも、本発明の範囲内である。
【実施例6】
【0159】
多剤耐性細胞での核内YB−1の検出
本実施例は核内YB−1が転写因子としてmdr1プロモータ(多剤耐性プロモータ)内にあるY−ボックス(CAAT配列)に結合しなければならないという考察に基づいている。これを検出するために、いわゆるEMSA分析(電気泳動移動度シフトアッセイ)を行った。これに関連して、核タンパク質を単離し、続いてタンパク質1〜10μgを短いDNA鎖(オリゴ)と共の37℃でインキュベーションした。核内YB−1を決定するために以下のオリゴヌクレオチドを使用した;U2O3と反対のmdr1プロモータ(位置−86〜−67):TGAGGCTGATTGGCTGGGCA(Xボックスには下線を付した)。
【0160】
その前にこのDNA断片の5’末端を32Pで放射線標識した。続いて未変性ポリアクリルアミドゲルを用いて分離した。タンパク質YB−1がオリゴヌクレオチド内の配列に結合すると、その結合は、結合したオリゴヌクレオチドに比べて非結合のオリゴヌクレオチドはゲル中をより早く移動することから検出できる(Holm, P.S.ら、JBC 277、10427〜10434、2002年; Bargou、R.C.ら、Nature Medicine 3、447〜450、1997年)。
【0161】
図6に示すように、EMSA分析よりYB−1が多剤耐性細胞257RDB、181RDBおよびMCF−7Ad細胞の核内に存在すること、そして細胞株U2OSおよびHeLa細胞には存在しないことが分かる。
【0162】
実施例4および5に示した結果は、アデノウイルスdl520およびdl1119/1131が257RDBの様なYB−1核陽性細胞では複製してその溶解を誘導するが、U205では複製、溶解を誘導しないことを確認した。このことは、本発明のアデノウイルスの使用に関する所見が確認された。さらに結果は、野生型のアデノウイルスと比べて、修飾型または欠失型E1A遺伝子産物を介したYB−1核陽性細胞でのウイルス遺伝子の弱いトランス活性化が、例えば多剤耐性細胞を含む核内にYB−1が存在する細胞での良好な複製および良好な細胞溶解をもたらすこと、そしてここに記したアデノウイルスがかかる腫瘍の溶解に利用できることを確認している。
【実施例7】
【0163】
E1−マイナスアデノウイルスの複製効率上昇
本実施例は初期ウイルス遺伝子E1B−55KおよびE4orf6をプラスミドpE4orf6によるトランスフェクションおよびE1/E3欠失アデノウイルスAd−55Kによる感染で置換えられることを示す。Ad−55KはE1B−55KをE1にクローニングし、CMVの制御下に置いたE1/E3欠失ウイルスである。この置換は、AdYB−1、即ちYB−1を発現するアデノウイルスがこれら初期遺伝子を発現していないという事実、および本発明者が核内にYB−1を含む複製系でのこれら初期遺伝子を置換することで複製効率および粒子形成能率をそれぞれ野生型アデノウイルスであるAd5型のそれと同程度まで上げることができることを認識していたという事実より必要であった。
【0164】
以下を実施した:
リポフェクタミンを用いた各105個のU2OS細胞へのプラスミドpE4orf6のトランスフェクション。プラスミドpEorf6はCMV制御下にある初期ウイルス遺伝子E4orf6をコードするDNA配列を保持している。プラスミドpE4orf6をトランスフェクションした24時間後に、細胞にYB−1発現E1/E3欠失アデノウイルスAdYB−1(50pfu/細胞)およびE1/E3欠失E1B−55KアデノウイルスAd−55K(50pfu/細胞)を感染させた。Ad−55KはCMV制御下にあるウイルス遺伝子E1B−55Kをトランス遺伝子として保持するE1/E3欠失ウイルスである。
【0165】
続いて感染5日後(=トランスフェクション後)に細胞を培地(2ml)から取り出した。凍結と融解(凍結/融解)を交互に3回行って、単離細胞からのウイルス粒子を放出させた。次にプラークアッセイを293細胞を使って実施し、生成した感染粒子を決定した(プラーク形成単位/ml(pfu/ml))。結果は図8および9に示す。図8は絶対量で表した場合のプラークアッセイの結果である。AdYB−1単独感染例と最も大きな差は、プラスミドpE4orf6を感染させた例および2種類のウイルスAdYB−1およびAd−55Kを同時感染させた例との間で観察された。図9は図8の結果を、AdYB−1の複製効率の倍率として複製効率の増加の形で表したものである。細胞にプラスミドpE4orf6を感染させた後AdYB−1およびE1B−55K(Ad−55K)を感染させると、最大25倍高いpfu/mlを示した。
【0166】
これらの結果より、E1B−55KおよびE4orf6による置換はE1/E3−欠失アデノウイルスAdYB−1感染後に形成されるウイルス数(pfu/ml)を最大25倍増加すると結論できる。プラーク形成単位(pfu)生成に及ぼすE1B−55KおよびE4orf6の追加効果は、これら2遺伝子産物単独の効果に比べて有意に高かった。
【0167】
EGFPを発現するプラスミドを使ったコントロール実験は、選択した実験方法では、プラスミドpE4orf6でトランスフェクションされたのは僅かに細胞の10%だけであることを明瞭に示した。E1B−55KおよびE4orf6の両方を発現する細胞で形成された粒子の数は、ヒトアデノウイルス5型(野生型)のそれに匹敵した。このことから、E4orf6およびE1B−55Kの発現は、YB−1の核内局在と組み合わさると、アデノウイルス、特にE1A−欠失アデノウイルスの複製および粒子形成を、野生型Ad5のそれに匹敵する程度まで誘導できるという、本発明の基礎を成す発見が確認された。
【実施例8】
【0168】
細胞増殖抑制薬投与によるYB−1核陰性細胞では複製しないアデノウイルスのYB−1核陽性細胞での複製増加
各種細胞増殖抑制薬の添加がヒト転写因子YB−1の核内局在化を誘導することは、先行技術において知られていた。本発明者が見出したように、核内に局在したYB−1は、アデノウイルスのE2−後期プロモータの活性化によりアデノウイルスの複製をコントロールする。両効果の組み合わせを利用すれば特異的な腫瘍溶解を得ることができる。
【0169】
癌溶解アッセイを実施するに当たっては、以下の手順に従った:200、000個の細胞(それぞれHeLaおよびU2OS)を6ウエル型プレートの核ウエル内に接種した。翌日ダウノルビシン40ng/ml(最終濃度)を加えた。3時間インキュベーションした後、それぞれの細胞をdl520で、10〜30pfu/細胞の割合で感染させた。続いて細胞を、細胞増殖抑制薬を含まない培地でインキュベーションした。3〜5日後、細胞をクリスタルバイオレットで染色した。
【0170】
図10および11から分かるように、ダウノルビシンの添加はYB−1の核内局在化を通じてdl520の複製を誘導した。即ちdl520は、細胞増殖抑制薬と組み合わさった状態で、ダウノルビシン単独例に比べてより大きな癌溶解作用を発揮した。
【実施例9】
【0171】
dl520によるインビボの癌溶解
本インビボ試験で使用したHeLa(YB−1核陰性)および257RDB(YB−1核陽性)細胞は、無菌細胞培養条件にて拡大した。細胞をマウス(系統CD1NuNu)に注射して皮下腫瘍を形成させる前に、細胞をトリプシン処理して集め、DMEM培地(10%FCS)に加え、細胞数を測定してからPBSで1回洗浄した。続いて細胞を遠心分離してPBSを除き、細胞を希望数する細胞数になるよう新鮮PBS中に分配した。本研究で皮下注射した細胞の数は両細胞系統とも5×106細胞であった。注射は動物の一方の側腹部に行い、より区別し易いようにHeLa細胞は右側に、そして257RDB細胞は左側に注射した。腫瘍の増殖を週2回調べ、ノギスを使って腫瘍の長さおよび幅を測定した。その結果を用い、次式より腫瘍の容積を計算した:
3/4π*a/2*(b/2)2 a=長さ、b=幅
【0172】
腫瘍の大きさが200〜520mm3に達した時点で、ウイルスおよび陰性コントロールであるPBSをそれぞれ腫瘍内に投与した。注射量は同一で、1回50μlであった。この作業を3連続日繰り返した。投与したウイルスの総量は5×108pfuであった。次に腫瘍の増殖を毎週2回の割合で記録し続け、その容積を計算した。試験最終日にマウスを屠殺して、腫瘍を取り出し詳しく分析した。
【0173】
結果は図12および13に示した。
【0174】
図12は、腫瘍容積の経時変化を処理の種類別に示したグラフである。腫瘍がRDB257により形成された例では、PBS注射時に腫瘍は約438mm3から1466mm3へと有意に増大した。本発明に従い用いられるベクターdl520を作用させると、腫瘍の成長は有意に低下した。平均腫瘍サイズ344mm3から総容積543mm3へと、腫瘍サイズは僅かに21%増加しただけであった。
【0175】
本実施例では、RDB257を基本とする腫瘍にPBSを投与したのと同様にPBSを投与したHeLa細胞より成る腫瘍をコントロールとして用いた。HeLa細胞を基本とする、dl520で処理された腫瘍は、まだ有意な腫瘍成長の増加を示し、311mm3から1954mm3に増加した。
【0176】
図13はRDB257を用いて増殖した腫瘍を持つ、屠殺したヌードマウスの像である。本発明のアデノウイルスdl520を作用させた後に、腫瘍の顕著な縮小が起こったことが明瞭に分かる。本例では、腫瘍容積の減少も見られた(ウイルスdl520投与1日後:515mm3;ウイルスdl520投与30日後:350mm3)。
【実施例10】
【0177】
腫瘍DNAのサザンブロット
実施例9に於いて、発生中の腫瘍の中央部から採取した腫瘍サンプルよりDNAを抽出した。抽出にはQiagen社のDneasy Tissueキットを用いた。DNAの単離は製造元の指示書に従い実施した。それによれば、DNAは細胞からアルカリ溶解により放出された。続いて単離したDNAはカラムを使って精製される。次に単離したDNAの濃度を260nmの光度測定より決定した。分析はDNAサンプル2μgを用いて行い、サンプルは制限酵素Kpn I、10単位で消化した。次にサンプルを0.8%のアガロースゲルを用いて電気泳動により分離した。続いてDNAをナイロン膜上にブロッティングした(Schleicher & Schuell社のシステムを用いて実施)。膜上にブロッティングされたDNAを特異的な1501bpのDNAプローブとハイブリダイゼーションさせた。上記1501bpのDNAプローブは、Ad5配列をコードするE2A内にある3369bpのKpn I断片と特異的に結合する。プローブはPCR(プライマー:5’−GTC GGA GAT CAG ATC CGC GT、5’−GAT CCT CGT CGT CTT CGC TT)により準備し、32Pを用いて射線標識した。次に膜を洗い、フィルムに感光した。
【0178】
腫瘍DNAのサザンブロットの結果は図14に示した。分析より、レーン3、4および5が示すように、dl520だけが耐性細胞RDB257でインビトロ複製することが確認された。レーン1は陽性コントロールのAd−5dを、レーン6、7、および8はdl520を感染したHeLa細胞由来のDNAを示す。HeLa細胞はYB−1核陽性ではないため、ウイルスdl520は複製せず、そのためE2A配列は検出できなかった。
【0179】
dl520に関するさらなる結果を図15に示した。プラークアッセイを利用して、dl520および野生型のアデノウイルスを感染させた後の粒子形成(pfu/ml)を調べた。各種YB−1核陽性(257RDBおよび181RDB)腫瘍細胞、ならびにYB−1核陰性腫瘍細胞にdl520および野生型アデノウイルスを感染させた。
【0180】
次の手順で実施した:
100、000〜200、000個の各細胞を6個のウエルを持つプレート(6ウエルプレート)、FCSを10%含むL15培地(耐性細胞)およびDMEM(非耐性細胞)に接種した。24時間後にdl520および野生型アデノウイルスを感染させた(10pfu/細胞)。感染3日後(インフェクション後)、凍結と融解を3回繰り返して細胞浮遊液よりウイルス粒子を放出させた。次に293細胞を使ってプラークアッセイを行い、形成した感染粒子(プラーク形成単位/ml(pfu/ml))を決定した。結果を図15に示す。プラークアッセイの結果は、dl520がYB−1核陽性細胞(257RDBおよび181RDB)では野生型アデノウイルスの場合と同様に複製することを示した。この限りにおいて、本発明に従ってここに記載のアデノウイルスを使用する場合には、野生型アデノウイルスと同様の複製効率が観察できた。
【実施例11】
【0181】
アデノウイルスベクターXvir03の構造デザイン
図16はアデノウイルスベクターXvir03の構造デザインを示す。アデノウイルスXvir03は、いわゆるE1/E3欠失アデノウイルスである。このことは、アデノウイルス複製に機能するE1A、E1BおよびE3タンパク質が作られないことを意味する。E1領域の欠失は342〜3528の範囲である;アミノ酸位置27865〜30995のE3領域の欠失。本明細書で使用する場合、用語「E1欠失ウイルス」とは、E1がもはや機能的に作用しないウイルスを意味する。これは、その他の核酸配列およびアミノ酸配列は未変性な状態で不活性化することにより達成できるが、しかしこのことは欠失したE1領域がコードするタンパク質が様々なサイズを持つことを意味することにもなる。E1AおよびE1Bタンパク質およびそれらをコードする核酸を欠くことにより、E4orf6の様なE4領域は弱くにしか発現されないか(野生型アデノウイルスの1〜5%)、または全く発現されない。ウイルス遺伝子E1B55kおよびE4orf6はXvir03内に導入された異種のCMVプロモータ(Clontech: Plasmid pShuttle)によりE1領域内で発現する。上記CMVプロモータに代わって、E1Aの発現に関連して明細書中に開示したその他プロモータを用いることができる。両遺伝子のオープンリーディングフレームは、いわゆるIRES配列(内部リボソーム進入部位)(Plletier, J.およびSonenberg, N.Nature、1988年、334、320〜325)により互いに連結している。このエレメント(Novagen:pCITE)は1種類のmRNAから2種類のタンパク質を発現させる。
【0182】
ベクターは次のように作製した:
プラスミドE1B55k−pShuttleを、M.Dobelstein(マールブルグ大学)のpCGNE1Bより、XbaIおよびBfrIを用いてE1B55kのオープンリーディングフレームをClontech社製のpShuttleベクター内にクローニングして作製した。続いてpShuttle内のE1B55kをApaIを使って直線化し、末端を平滑末端とし、NheIを使って切断した。
【0183】
第二ベクターであるpcDNA3.1(+)(Invitroge)の場合は、それぞれに続いて、Novagen社製のpCITE−4a(+)を鋳型に用いTAクローニング法によりEcoRV切断部位にPCR産物であるIRESエレメントをクローン化し、そしてE4orf6はプラスミドpCMV−E4orf6(M.Dobelstein、マールブルグ大学)よりBamHIを用いてクローニングした=IRES−E4orf6−pcDNA3.1(+)。pcDNA3.1(+)内のIRES−E4orf6はNotIを使って直線化し、端部を平滑末端化してから断片IRES−E4orf6をNheIで切断した。断片IRES−E4orf6をオープンベクターE1B55k−pShuttle(平滑端、NheI)と連結した。続いてこうしてできたカセットを、I−CeuIおよびPI−SceIを用いて、E1B55k−IRES−E4orf6−pShuttleより、CMVプロモータおよびウシ成長ホルモン(BGH)−ポリAと共にΔE1、ΔE3Adeno−X−プラスミド(Clontech)内にクローニングし、AdcmvE1B/IRES/E4orf6と名付けた。次にアデノウイルスをメーカー(Clontech)指示書に従い作製した。PacIを使って直線化した、発現エレメントCMV−E1B55k−IRES−E4orf6−BGHポリAを有するアデノプラスミドをHEK293細胞内にトランスフェクションし、トランスフェクション11日後に剥離細胞を培地と共に取り除いてから凍結乾燥を繰り返しアデノウイルスを放出させた。
【0184】
上記ベクターは原理的には本発明での使用に関して、本明細書に記載したその他ウイルスと同様に好適である。具体的には、上記ベクターはYB−1核陽性細胞およびYB−1が脱制御状態にある細胞、即ちYB−1が正常細胞および非腫瘍細胞に比べ過剰発現している細胞での複製および溶解始動について好適である。このベクターの使用は、特に本発明に従い用いられるものとして本明細書に記載されているその他アデノウイルス、ならびに本明細書に開示されている本発明のその他アデノウイルスに関連し開示されている疾患、患者群または患者集団を適用とする。
【実施例12】
【0185】
アデノウイルスベクターXvir03/01の構造デザイン
図17から分かるように、Xvir03/01はXvir03を更に発展させたものである。例えばここに記した遺伝子の様な治療目的の遺伝子やトランス遺伝子をE3領域内にクローニングできる。さらに、E4領域内に欠失を導入してXvir03の発現カセットに由来するE4orf6との間で相同的組換えが起こるのを回避した。これによってより大型のトランス遺伝子をこの構築体内にクローニングすることが可能になった。欠失型のE3領域にはカセット導入に適したSacI、NdeIおよびNheI制限部位があり、ここに例えば治療用トランス遺伝子をクローニングすることができる。
【0186】
E3領域内への治療用遺伝子クローニングおよびE4領域内への欠失作製を目的としたプラスミドの調製:
Clontech社製pAdenoX−Plasmidには3’ITR領域の後に野生型アデノウイルスには無いSfuI向けの制限部位がある。E3〜E4領域をSpeI(位置23644)およびSfuIを使ってpAdenoX(Clontech)から取り出し、pcDNA3.1(+)(Invitrogen)内にトランスフェクションした=pcDNA3.1−E3Δ27865−30995−E4。E4ORF6の大部分、即ち33241〜33875をPstIを用いて取り出した=pcDNA3.1−E3Δ27865−30995、E4Δ33241−33875。Xvir03を更に発展させるために、pcDNA3.1−E3Δ27865−30995、E4Δ33241−33875由来の欠失E3/E4領域をSfuIおよびSpeIを使ってプラスミドpAdenoX内にクローニングした=pAdnoXE3Δ27865−30995、E4Δ33241−33875。
【0187】
次にXvir03について記載した発現カセットを、I−CeuIおよびPI−SceIを使ってE1B55k−IRES−E4orf6−pShuttleより、CMVプロモータおよびウシ成長ホルモン(BGH)−ポリAと共にpAdenoXE3Δ27865−30995、E4Δ33241−33875内にクローニングし、AdcmvE1B/IRES/E4orf6−ΔE4と命名した。次にアデノウイルスをメーカー(Clontech)の指示書に従い作製した。
【0188】
上記ベクターは原理的にはここに記載した本発明に従い使用される他ウイルスと同様に有用である。特に上記ベクターはYB−1核陽性細胞ならびにYB−1が脱制御状態にある、即ちYB−1が正常細胞および非腫瘍細胞に比べ過剰発現している細胞での複製、およびその限りにおいて溶解の誘導に好適である。このベクターは、本発明に従い用いられる本明細書記載の他アデノウイルス、および本明細書のアデノウイルスに関し開示されている疾患、患者群および患者集団にも使用できる。
【実施例13】
【0189】
257RDBおよび181RDB細胞でのXvir03の癌溶解作用
100、000個の細胞(257RDBおよび181RDB)を6個のウエルを持つプレート(6ウエルプレート)の各ウエルに接種した。翌日細胞に、図18に示すようにAd312(20pfu/ml)およびXvir03(5pfu/ml)を感染させた。感染は無血清DMEM培地500μl中で、37℃、1時間実施した。次に感染培地を取り除き、完全培地(10%FCS/DMEM)2mlと交換した。5日後にクリスタルバイオレット染色による分析を行った。結果を図18Aおよび18Bに示す。
【0190】
図18Aおよび18Bから明らかな様に、核内にYB−1を持つ多剤耐性細胞では、Ad312およびXvir03を感染させた場合、細胞のクリスタルバイオレット染色より示されるようにXvir03感染例のみ崩壊を示した。この場合まず培地を取り除いた。続いて細胞をクリスタルバイオレット液(50%ETOH、3%ホルムアルデヒド、5%酢酸、1%クリスタルバイオレット)で覆い、室温で5〜10分間インキュベーションした。次に6ウエルプレートを水でよく濯いでから室温で乾かした。
【0191】
本発明者は、本発明の意味においてアデノウイルスをトランス活性化しないE1A欠失ウイルス(例えばAd312)がより高いMOIでは極めて効率的に複製できるが(Nevins J.R.、Cell 26、213〜220、1981年)、しかしこれらは臨床的には応用できないことを承知していた。この現象は文献では「E1A−様活性」と呼ばれている。ここで用いるアデノウイルスAd312はE1A欠失ウイルスである。使用した力価(20pfu/ml)はまだ臨床応用には高すぎるが、この力価ではE1B55kおよびE4orf6といった初期アデノウイルス遺伝子は発現しないか、または極少量しか発現しない(Nevins J.R.、Cell 26、213〜220、1981年)。本明細書に既に記した様に、これら遺伝子およびタンパク質はウイルス複製において重要な役割をはたしており、それぞれアデノウイルスXvir03により発現される(図16)図18Aおよび18Bから分かるように、遺伝子E1B55kおよびE4orf6の発現は、必要とする感染力価(pfu/mlで表される)の低下を伴いながら効率的にウイルス複製および細胞溶解を起こすだろう。このことから、本発明の基礎となる発見、即ちYB−1の核内局在と組み合わさったE4orf6およびE1B−55Kの発現(且つE1Aが存在しない)は、極めて効率的なアデノウイルスの複製を誘導できることが確認される。これに必要な力価は僅かに1〜5pfu/mlに過ぎず、臨床的応用が可能である。
【0192】
このことから、本発明の基礎となる発見、即ち本発明に従い用いられるウイルスを用いて感染細胞を溶解させるには、核内にYB−1が存在すること、特に細胞周期と無関係に存在することが必要であることが確認される。
【0193】
前記明細書に開示した発明の特徴、特許請求の範囲ならびに図面は、個々およびいずれかの組み合わせの両方に於いて、その発明の各種実施態様での実現にとって極めて重要であろう。
【技術分野】
【0001】
本発明はアデノウイルスならびにそれをコードする核酸、ならびに組換え体発癌タンパク質の使用に関する。
【背景技術】
【0002】
腫瘍の治療には、現在多くの治療コンセプトが用いられている。手術に加え、化学療法および放射線治療がよく実施されている。しかしながらこれ技術全てに、無視できない副作用が伴う。複製特異的殺腫瘍ウイルスの使用が、腫瘍治療に新たなプラットホームを提供している。この方法では、腫瘍内選択的な上記ウイルスの複製が起こり、その結果ウイルスの複製、感染腫瘍細胞の溶解、および近接腫瘍細胞へのウイルスの放散が起こる。ウイルスの複製能力が腫瘍細胞に限定されることから、正常組織にはウイルスの複製は広がらず、従って溶解も起きない。
【0003】
現在、殺腫瘍を狙った複数のウイルス系が臨床試験にかけられている。かかるアデノウイルスの一例が、臨床第I相および第II相試験で良い結果を上げているD11520(Onyx−015)である(Khuri, Fら、Nature Medicine 6、879〜885、2000年)。Onyx−015は、E1B−55kDa遺伝子を完全に欠失したアデノウイルスである。このアデノウイルスが完全にE1B55kDaタンパク質を欠いていることは、p53欠失アデノウイルスベクターと一緒であれば複製すること、従って細胞を溶解することができるが(Kim, Dら、Proc.Am.Soc.Clin.Oncol.17、391a、1998年)、正常細胞を損傷することはないという発見に基づいている。より具体的には、E1B−55kDa遺伝子産物は、p53の阻害、ウイルスmRNAの輸送、および宿主細胞のタンパク質合成のスイッチングオフに関係している。p53の阻害は、p53とアデノウイルスがコードするE1B−55kDaタンパク質から成る複合体、および/またはE1B−55kDaとE4orf6.より成る複合体の形成を通じて起こる。TP53がコードするp53は、複合体制御メカニズムにとって開始点であり(Zambetti, G.Pら、FASEB J.7、855〜865、1993年)、p53はとりわけてもアデノウイルスのようなウイルスの細胞内複製が効率的に阻害する。TP53は全てのヒト腫瘍の約50%で欠失または突然変異しており、そのために化学療法または放射線治療では−望まれる−アポトーシスが起こらず、結果として腫瘍治療に成功しないことが多々ある。
【0004】
殺腫瘍アデノウイルスの別のコンセプトは、E1Aタンパク質が特殊な欠失型として存在するか、または1もしくはそれ以上の突然変異を含み、それらがRb/E2Fおよび/またはp107/E2Fおよび/またはp130/E2Fの結合に影響しない場合には、かかるアデノウイルスは感染細胞をS期に誘導せず、そして機能性のRbタンパク質を持たない腫瘍細胞内では複製できるという発見に基づいている。さらにE1Aタンパク質はN末端を欠失することも、そしてE1Aタンパク質のアミノ酸位置1〜76の領域に1またはそれ以上の突然変異を含むこともでき、その結果それぞれp300に対するE1Aの結合を阻害し、そしてそれにより腫瘍細胞内での選択的複製を提供することができる。このようなアプローチは欧州特許第0 931 830号に例示されている。かかるウイルスの例としてはAdΔ24、d1922−947、E1Ad/01/07およびCB016がある(Howe, J.A.ら、Molecular Therapy 2、485〜495、2000年; Fueyo、J.ら、Oncogene 19、2〜12、2000年; Heise, C.ら、Nature Medicine 6, 11341139、2001年; Balague, Cら、J.Virol.75、7602〜7611、2001年)。かくして、これら当分野既知の殺腫瘍に関するアデノウイルス系はE1Aタンパク質内に明瞭な欠失を含んでいるが、その場合この種の欠失は、機能性Rbタンパク質および非活性型Rbタンパク質とE2Fとから成る複合体がそれぞれ、効率的なインビボ(in vivo)での複製を遮断すること、そしてRb−欠失/突然変異細胞でのみアデノウイルスのインビボ複製を提供すること前提として作られている。これら従来技術によるアデノウイルス系は、E1Aに基づき、初期E2プロモータ(E2初期プロモータ)およびフリーE2F(Dyson, N.Genes & Development、12、2245〜2262、1998年)を用いてインビボ複製を制御している。
【0005】
殺腫瘍アデノウイルス系の別の形態は、腫瘍細胞内での選択的複製を提供するウイルス発癌遺伝子E1Aを特異的に発現させる選択的プロモータの使用に基づいている(Rodriguez, R.ら、Cancer Res.57、2259〜2563、1997年)。
【0006】
上記の如く、各コンセプトの基礎を成す作用様式に適った細胞背景を選択することが、アデノウイルスに関する殺腫瘍ウイルスの各種コンセプトにとって重要である。換言すれば、現在知られている各種アデノウイルス系は、明瞭な分子生物学的前提条件が揃っている場合にのみ使用することができるだろう。このことが、かかる系の使用を一部患者群に制限している。
【0007】
腫瘍疾患の治療に於ける特殊な問題は、細胞増殖抑制薬に対する腫瘍細胞の耐性に関する研究で特によく研究されている多剤耐性(英語でmultidrug resistance(MDR))と呼ばれる耐性を患者が発症することである(Gottesman and Pastan、Annu.Rev.Biochem.62、385〜427、1993年)。この問題は、膜結合輸送タンパク質、いわゆるABCトランスポータに属するP−糖タンパク質の過剰発現に基づいている(Stein, U.ら、JBC 276、28562〜69、2001年、J.Wijnholds、Novartis Found Symp.243、69〜79、2002年)。Bargou, R.C.らおよびOda, Y.ら、(Bargou, R.C.ら、Nature Medicine 3、447〜450、1997年; Clin.Cancer Res.4、2273〜2277、1998年)は、ヒト転写因子YB−1の核内局在がP−糖タンパク質の発現活性化に関係していることを示した。別の研究は、YB−1がUV照射、細胞分裂抑制薬投与(Koike, K.ら、FEBS Lett、390〜394、1997年)および高温(Stein, Uら、JBC 276、28562〜69、2001年)といった様々なストレス状態により核内に輸送されることを確認している。別の研究は、YB−1の核内局在が更に別のABCトランスポータに影響することを確認している。このABCトランスポータはMRP(英語でmultidrug resistance-relate potein:多剤耐性関連タンパク質)と呼ばれており、いわゆる非定型非P−糖タンパク質依存多剤耐性の形成に関係している(Stein、U.ら、JBC 276、28562〜69、2001年)。
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明の基調をなす課題は、具体的には殺腫瘍活性薬剤を用いて生物、より具体的にはヒトおよび患者群の処置を可能にする技術的教示および、特に手段を提供することである。本発明の基調をなす別の課題は、細胞分裂抑制薬に耐性である腫瘍疾患を持つ患者、特に多剤耐性を持つ患者に殺腫瘍を起こすのに好適である手段を提供することである。
【0009】
本発明によれば、第1の局面の課題は薬物製造に適したウイルス、好ましくはアデノウイルスを使用することで解決されるが、この時ウイルスは核内にYB−1を持たない細胞内では複製ができず、そしてウイルスは発癌遺伝子または発癌遺伝子産物、好ましくは発癌遺伝子タンパク質をコードしており、それはYB−1核陽性細胞でウイルス遺伝子の少なくとも1つ、好ましくはアデノウイルス遺伝子をトランス活性化し、そして遺伝子はE1B55kDa、E4orf6、E4orf3およびE3ADPを含む群より選ばれる。
【0010】
第2の局面では、課題は核内にYB−1を有する細胞での複製についてウイルス、好ましくはアデノウイルスを使用することで解決されるが、この時ウイルスは核内にYB−1を持たない細胞では複製できず、そしてウイルスは発癌遺伝子または発癌遺伝子産物、特には発癌遺伝子タンパク質をコードしており、それは少なくとも1つのウイルス遺伝子、好ましくはアデノウイルス遺伝子をトランス活性化し、そのとき遺伝子はE1B55kDa、E4orf6、E4orf3およびE3ADPを含む群より選ばれる。
【0011】
発明による上記2つの使用の実施態様では、ウイルス、好ましくはアデノウイルスは核内にYB−1を有する細胞で複製する。
【0012】
発明による2つの使用の別の実施態様では、ウイルス発癌遺伝子タンパク質はE1Aであり、そして/または発癌遺伝子はE1Aをコードする遺伝子であり、そして/または発癌遺伝子タンパク質はE1Aである。
【0013】
好適実施態様では、ウイルス発癌遺伝子タンパク質E1Aは機能的Rb腫瘍抑制遺伝子産物に結合できる。
【0014】
別の実施態様では、ウイルス発癌遺伝子タンパク質E1Aは、機能的Rb腫瘍抑制遺伝子産物に結合できない。
【0015】
発明による2つの使用の別実施態様では、ウイルス発癌遺伝子タンパク質E1AはYB−1の核内局在化を誘導しない。
【0016】
発明による2つの使用の更に別の実施態様では、薬物はその細胞がRb−陽性またはRb−陰性のいずれかである患者向けである。
【0017】
好適実施態様では、細胞は上記薬剤により影響を受ける状態の形成に関係する細胞である。
【0018】
発明による2つの使用の別実施態様では、細胞は核内でRb−陰性であり、そしてYB−1陽性であり、好ましくは細胞周期とは独立に核内でYB−1陽性である。
【0019】
発明による2つの使用の更に別の実施態様では、薬剤は腫瘍の処置を目的とする。
【0020】
発明による2つの使用の更に別の実施態様では、細胞、特に腫瘍またはその一部を形成する細胞は、薬剤耐性であり、特に多剤耐性、好ましくは抗腫瘍剤に対し耐性であり、そしてより好ましくは細胞増殖抑制薬に対し耐性である。
【0021】
発明による2つの使用の好適実施態様では、細胞は膜結合輸送タンパク質であるP−糖タンパク質および/またはMRPを発現、好ましくは過剰発現している。
【0022】
発明による2つの使用の別実施態様では、細胞はp53−陽性またはp53−陰性である。
【0023】
発明による2つの使用の実施態様の一つでは、発癌遺伝子タンパク質は、野生型発癌遺伝子タンパク質E1Aと比較して、1または複数の突然変異または欠失を有しており、その場合欠失はCR3領域の欠失およびN−末端の欠失、およびC−末端の欠失を含む群より選択されるのが好ましい。これに関連し、E1A発癌遺伝子タンパク質はRbに結合できることが好ましい。
【0024】
発明による2つの使用の別実施態様では、発癌遺伝子タンパク質は、野生型発癌遺伝子タンパク質と比較した場合、1または複数の突然変異または欠失を有しており、その場合欠失はCR1領域および/またはCR2領域であることが好ましい。発癌遺伝子タンパク質E1AがRbに結合できないことは発明の範囲内である。
【0025】
発明による2つの使用の実施態様の一つでは、ウイルス発癌遺伝子タンパク質、特にE1Aは、組織特異的および/または腫瘍特異的プロモータの制御下にある。
【0026】
発明による2つの使用の別実施態様では、ウイルス、特にアデノウイルスはYB−1をコードしている。
【0027】
発明による2つの使用の更に別の実施態様では、YB−1は組織特異的および/または腫瘍特異的プロモータの制御下にある。
【0028】
発明による2つの使用の好適実施態様では、ウイルス、特にアデノウイルスはE4orf6、E4orf3、E1B55KおよびアデノウイルスE3ADPタンパク質を含む群から選ばれるタンパク質を少なくとも1つコードしている。
【0029】
発明による2つの使用の別実施態様では、細胞は核内にYB−1を有しており、特に腫瘍もしくはその一部を形成する細胞は核内にYB−1を有している。
【0030】
発明による2つの使用の更なる実施態様では、腫瘍は核内へのYB−1輸送の誘導により核内にYB−1を有している。
【0031】
発明による2つの使用の好適実施態様では、核内へのYB−1の輸送は放射線照射、細胞増殖抑制薬の投与、および高温から成る群より選ばれた少なくとも1つの手段を通じ開始される。
【0032】
発明による2つの使用の特に好適な実施態様では、上記手段は細胞、臓器または生体に適用される。
【0033】
発明による2つの使用の好適実施態様では、ウイルス、特にアデノウイルスは、AdΔ24、dl922−947、E1Ad/01/07、d1119/1131、CB016、dl520、および機能的Rb腫瘍抑制遺伝子産物と結合できるウイルス性E1A発癌遺伝子の発現を欠いているウイルスを含む群より選ばれる。
【0034】
第三の局面では、課題はウイルス、好ましくはアデノウイルスを医薬の製造に使用することで解決されるが、この場合ウイルスは、好ましくはアデノウイルス、その複製がE2−後期プロモータの活性化を介したYB−1による手段を通し、もしくは手段により、好ましくはE2−後期プロモータの活性化を主に通じて制御されるようにデザインされている。実施態様の一つでは、YB−1はトランスジェニックYB−1または細胞質、特に細胞質脱制御型YB−1のいずれかである。好ましくは、トランスジェニックYB−1は、ベクター、好ましくはアデノウイルスによって細胞内で発現されるYB−1を意味する。E2−後期プロモータは野生型アデノウイルスに存在するアデノウイルスE2−後期プロモータ、またはここでトランス遺伝子の発現と関連付け記載されるE2−後期プロモータであるのが好ましい。
【0035】
第四の局面では、課題はウイルスおよび特にはアデノウイルスを核内にYB−1を有する細胞内での複製に使用することで解決されるが、この場合ウイルス、とりわけアデノウイルスは、複製がE2−後期プロモータの活性化を通じて、好ましくはE2−後期プロモータの活性化を主に通じてYB−1により制御されるようにデザインされる。実施態様の一つでは、YB−1はトランスジェニックYB−1または細胞質、特には細胞質脱制御YB−1のいずれかである。本明細書で用いる場合のトランスジェニックYB−1は、ベクター、好ましくはアデノウイルスによって細胞内で発現されるYB−1であるのが好ましい。E2−後期プロモータは野生型アデノウイルスに存在するアデノウイルスE2−後期プロモータ、またはここでトランス遺伝子の発現の使用と関連付け記載されるE2−後期プロモータであるのが好ましい。
【0036】
本発明の第三および/または第四の局面の好適実施態様では、アデノウイルスは本明細書に開示された如くにデザインされ、特には本発明に従った使用を目的としてデザインされる。
【0037】
第五の局面では、課題は次の特徴を有するウイルス発癌遺伝子タンパク質、特に単離されたウイルス発癌遺伝子タンパク質により解決される:
a)YB−1核陽性細胞内で、E1B−55K、E3ADPおよびE4orf6およびE4orf3を含む群から選ばれる少なくとも1つのウイルス遺伝子がトランス活性化されること;および
b)核内、特にウイルス発癌遺伝子タンパク質が存在している細胞の核内でのYB−1の誘導を欠くこと。
【0038】
実施態様の一つでは、ウイルス発癌タンパク質はE1Aである。
【0039】
更なる実施態様では、ウイルス発癌遺伝子タンパク質は、野生型発癌遺伝子タンパク質と比較した場合に1または複数個の突然変異または欠失を有しており、この場合の欠失はCR3領域の欠失、N−末端の欠失およびC−末端の欠失を含む群より選ばれるのが好ましい。
【0040】
実施態様の一つでは、ウイルス発癌遺伝子タンパク質を介したYB−1誘導は、E4ofr6および/またはE1B55kDが有核細胞内に存在しない場合には起こらない。
【0041】
この場合にはウイルス発癌遺伝子タンパク質はRbに結合できると解釈する。
【0042】
別実施態様では、ウイルス発癌遺伝子タンパク質は1または複数の突然変異または欠失を含み、この場合欠失はE1A発癌遺伝子タンパク質のCR1領域および/またはCR2領域内にあることが好ましい。この場合には、ウイルス発癌遺伝子タンパク質はRbに結合できないと解釈する。
【0043】
第六の局面では、発明は、ウイルス、特に本発明に従って使用されるアデノウイルスをコードする核酸を含み、そしてヘルパーウイルスの核酸を1つ含むウイルス複製系、好ましくはアデノウイルス複製系の使用であって、上記ヘルパーウイルスの核酸がYB−1をコードする核酸を含んでいる複製系に関する。
【0044】
実施態様の一つでは、ウイルス核酸、特にはアデノウイルス核酸、および/またはヘルパーウイルスの核酸は、複製可能なベクターとして存在する。
【0045】
第七の局面では、発明はウイルス、特にはアデノウイルスをコードする核酸を、発明に従って使用する医薬の製造、特には腫瘍処置を目的とする医薬の製造への使用に関する。
【0046】
実施態様の一つでは、細胞、特には腫瘍またはその一部を形成する細胞は、薬物、好ましくは抗ガン剤、より好ましくは細胞増殖抑制薬に対し耐性、特には多剤耐性である。
【0047】
第八の局面では、発明はウイルス、特にはアデノウイルスをコードする核酸を、発明に従って使用する、核内にYB−1を有する細胞内での複製に関する使用であって、ウイルスが核内にYB−1を持たない細胞に於いては複製欠失であり、そしてウイルスがYB−1核陽性細胞に於いては少なくとも1つのウイルス遺伝子、好ましくはアデノウイルス遺伝子をトランス活性化する発癌遺伝子もしくは発癌遺伝子産物をコードし、その場合の遺伝子はE1B55kDa、E4orf6、E4orf3およびE3ADPを含む群から選ばれる使用に関する。
【0048】
第九の局面では、課題はウイルス、好ましくはアデノウイルスをコードする核酸を、発明に従い使用し医薬の製造に用いることで解決されるが、そのためにウイルスはその複製がE2−後期プロモータの活性化を通じて、好ましくはE2−後期プロモータの活性化を主に通じて、YB−1により制御されるようにデザインされている。実施態様の一つでは、YB−1はトランスジェニックYB−1または細胞質、特には細胞質脱制御YB−1のいずれかである。ここで用いるトランスジェニックYB−1とは、ベクター、好ましくはアデノウイルスベクターによって細胞内で発現されるYB−1であることが好ましい。E2−後期プロモータは、野生型アデノウイルス内に存在するアデノウイルスE2−後期プロモータ、またはトランス遺伝子の発現の使用と関連してここに記載されるE2−後期プロモータであることが好ましい。
【0049】
第十の局面では、課題はウイルス、特にはアデノウイルスをコードする核酸の、細胞内での複製に関する発明に従って用いることによる使用により解決されるが、そのためにウイルスはその複製がE2−後期プロモータの活性化を通じて、好ましくはE2−後期プロモータの活性化を主に通じて、YB−1により制御されるようにデザインされている。実施態様の一つでは、YB−1はトランスジェニックYB−1または細胞質、特には細胞質脱制御YB−1のいずれかである。ここで用いるトランスジェニックYB−1はベクター、好ましくはアデノウイルスによって細胞内で発現されるYB−1であることが好ましい。E2−後期プロモータは、野生型アデノウイルス内に存在するアデノウイルスE2−後期プロモータ、またはトランス遺伝子の発現の使用と関連してここに記載されるE2−後期プロモータであることが好ましい。
【0050】
第十一の局面では、課題は前記核酸の一つを含むベクターを、本発明の第一または第二の局面に従い使用することで解決される。
【0051】
第十二の局面では、発明は、発明に従い用いられるウイルス、特にはアデノウイルスと細胞を接触すべきものであるか、および/またはそれらを用いて処置すべきものであるかを決定することを目的とした、細胞、腫瘍組織または患者の細胞の特徴付けのためのYB−1と相互作用する薬物の使用に関する。
【0052】
実施態様の一つでは、薬物は抗体、抗カリン(anticaline)、アプタマー、アプタザイムおよびシュピーゲルマー(spiegelmer)を含む群から選ばれる。
【0053】
第十三の局面では、課題は本発明によるウイルス発癌遺伝子タンパク質またはそれをコードする核酸を、本発明の第一および第二局面に従って用いられるウイルス、特にはアデノウイルスの製造に使用することで解決される。
【0054】
実施態様の一つでは、ウイルスはトランス遺伝子をコードする核酸を含む。
【0055】
別の実施態様では、ウイルスはトランス遺伝子の翻訳産物および/または転写産物を含む。
【0056】
好適実施態様では、アデノウイルス複製系の核酸および/またはヘルパーウイルスの核酸は、トランス遺伝子またはトランス遺伝子をコードする核酸を含む。
【0057】
更に別の実施態様では、核酸はトランス遺伝子またはトランス遺伝子をコードする核酸を含む。
【0058】
別の実施態様では、トランス遺伝子はプロドラッグ遺伝子、サイトカイン、アポトーシス誘導遺伝子、腫瘍抑制遺伝子、金属プロテアーゼインヒビターに関する遺伝子、および血管新生インヒビターに関する遺伝子を含む群より選ばれる。
【0059】
実施態様の一つでは、トランス遺伝子はsiRNA、アプタマー、アンチセンス分子およびリボザイムに関する核酸を含む群より選ばれ、この場合のsiRNA、アプタマー、アンチセンス分子および/またはリボザイムはあるターゲットとなる標的分子を持っている。
【0060】
更なる実施態様では、標的分子は耐性関連因子、抗アポトーシス因子、発癌遺伝子、血管新生因子、DNA合成酵素、DNA修復酵素、成長因子およびそれらのレセプター、転写因子、金属プロテアーゼ、特にマトリックス金属プロテアーゼ、およびウロキナーゼ
型プラスミノーゲン活性化因子を含む群より選ばれる。実施態様の一つでは、耐性関連因子は、P−糖タンパク質、MRPおよびGSTを含む、およびそれらをコードする核酸を含む群より選ばれるのが好ましい。実施態様の一つでは、抗アポトーシス因子は、BCL2を含み、更にそれをコードする核酸を含む群より選ばれる。実施態様の一つでは、発癌遺伝子はRas、特に突然変異型Ras、RbおよびMycを含み、またそれらをコードする核酸を含む群より選ばれる。実施態様の一つでは、血管新生因子はVEGFおよびHMGタンパク質を含む、ならびにそれらをコードする核酸を含む群より選ばれる。実施態様の一つでは、DNA合成酵素はテロメラーゼを含む、ならびにそれをコードする核酸を含む群より選ばれる。実施態様の一つでは、DNA修復酵素はKu−80を含み、またそれをコードする核酸を含む群より選ばれる。実施態様の一つでは、成長因子はPDGF、EGFおよびM−CSFを含み、またそれらをコードする核酸を含む群より選ばれる。実施態様の一つでは、レセプターは特には成長因子に関するレセプターであり、この場合の成長因子はPDGF、EGFおよびM−CSFを含み、またそれらをコードする核酸を含む群より選ばれる。実施態様の一つでは、転写因子はYB−1を含み、またそれをコードする核酸も含む群より選ばれる。実施態様の一つでは、金属プロテアーゼはマトリックス金属プロテアーゼであることが好ましい。好適実施態様では、金属プロテアーゼはMMP−1およびMMP−2を含み、またそれらをコードする核酸を含む群より選ばれる。実施態様の一つでは、ウロキナーゼ型プラスミノーゲン活性化因子は、uPa−Rを含み、またそれをコードする核酸を含む群より選ばれる。
【0061】
さらに別の実施態様では、医薬は少なくとも1つの医薬活性化合物を追加して含む。
【0062】
好適実施態様では、医薬的に活性な化合物は、サイトカイン、金属プロテアーゼインヒビター、血管新生インヒビター、細胞増殖抑制薬および細胞周期インヒビターを含む群から選ばれる。
【0063】
本発明はYB−1核陽性腫瘍細胞でのE1A−修飾アデノウイルスのDNA複製がE2−後期プロモータの活性化に基づいているという驚くべき発見に拠っている。本明細書で使用する場合のE1A−修飾アデノウイルスは、(a)YB−1核陰性細胞では複製しないか、またはYB−1核陰性細胞に於いては対応する野生型細胞に比べて複製の低下、好ましくは大きな低下を示し、(b)少なくとも1つのウイルス遺伝子をトランス活性化し、その場合の遺伝子は特にはE1B−55kDa、E4orf6、E4orf3およびE3ADPを含む群から選ばれ、そして/または(c)アデノウイルスを通じて細胞内YB−1を核内に移動しない、アデノウイルスである。場合によっては、本発明で使用するアデノウイルスは、アデノウイルスがコードするE1Aタンパク質の結合がRbへのE2Fの結合を妨害してRbとE2Fより成るそれぞれの複合体を解離することができるという特徴を更に有してもよい。上記特徴a)からc)のうちの少なくとも1つ、または複数、好ましくは特徴a)からc)の全てを有するアデノウイルスは、核内にYB−1を持たない細胞では複製できない。
【0064】
実施態様の一つでは、ここで用いる場合、大きな複製の低下とは特に、野生型と比較したときに2倍、好ましくは5倍、より好ましくは10倍、そして最も好ましくは100倍低下した複製を意味する。好適実施態様では、かかる複製の比較は、同一もしくは類似の細胞株、同一もしくは類似の感染ウイルス力価(感染多重度、MOIもしくはプラーク形成単位、pfu)および/または同一もしくは類似の一般的実験条件を用いて実施される。ここで使用する場合、複製とは、特には粒子形成を意味する。別の実施態様では、複製の評価はウイルス核酸合成の程度とすることができる。ウイルス核酸合成の程度を決定する方法、ならびに粒子形成を決定する方法は当業者周知である。
【0065】
ここに記載の発見、方法、使用または核酸、タンパク質、複製系等は、アデノウイルスに必ずしも限定されない。原則的には、かかる系はこれと共に含まれる他のウイルスにも存在する。
【0066】
本発明によるウイルスを使用する場合、または本発明により本明細書に記載のウイルスを使用する場合には、先行技術によれば、10〜100pfu/細胞に対し1〜10pfu/細胞の感染率で野生型の複製に匹敵する複製が実現できる。
【0067】
本明細書で使用する場合、細胞質YB−1は細胞によりコードされているYB−1、好ましくは細胞によりさらに発現されるYB−1を意味するものとし、この場合YB−1は細胞内に、アデノウイルス、好ましくは本書記載のアデノウイルスおよび/もしくはヘルペスウイルスが該当する細胞に感染する前に存在することが好ましい。しかしながら、細胞質YB−1が細胞内に導入された、または例えばウイルス、特にはアデノウイルスの感染といった外的方法を用いることによりかかる細胞が産生したYB−1である場合も本発明の範囲内である。
【0068】
以下これに結びつけることを望むことなしに、本発明者はE2−初期プロモータ、即ち初期E2プロモータは、本発明に従いここに使用されるウイルス複製との関係に於いて、ヒト細胞質E2F転写因子を通じて始動することはないと想定している。複製の始動は細胞のRbの状態とは無関係であり、即ちこのことは、ここに開示するウイルスを用い感染させた腫瘍細胞、およびその後好ましく溶解する腫瘍細胞は機能的Rbタンパク質および非活性型Rbタンパク質の両方を含んでよいことを意味する。さらに、アデノウイルスの複製は、ここに開示するアデノウイルスを使用した場合、またはここに開示の条件では、機能的p53タンパク質を必要とせず、またその存在の影響も受けない。その限りに於いて、技術的教示はAdΔ24、d1922−947、E1Ad/01/07、CB016型の殺癌もしくは殺腫瘍性アデノウイルスの、または例えば欧州特許第EP0 931 830号に記載のアデノウイルスの、あるいは無傷の機能的Rbタンパク質はインビボでの効果的複製にとって障害であり、従ってRb−陰性およびRb−突然変異細胞でのみアデノウイルスはインビボ複製するという仮説の下にE1Aタンパク質内に1または複数の欠失を導入したアデノウイルスの使用を基礎と成す原理とは、それぞれ異なっている。これらの先行技術のアデノウイルス系は、E1Aに基づき、初期E2プロモータ(E2初期プロモータ)および「無E2F」を利用してアデノウイルスのインビボ複製を制御するものである。しかしながら、これら先行技術ウイルスは、本発明に従って用いること、即ち細胞周期と無関係に核内にYB−1を含む細胞での複製に用いることもできるだろう。
【0069】
前記欧州特許第EP0 931 830号に記載のウイルス、特にアデノウイルスは本発明に従い使用出来るだろう。より具体的には、前記特許に記載のウイルスは複製できず、そして機能的なRb腫瘍抑制遺伝子産物と結合可能なウイルス発癌タンパク質を発現していない。このアデノウイルスは具体的には、機能的腫瘍抑制遺伝子産物、特にはRBを結合できるウイルスE1A発癌タンパク質の発現を欠いているアデノウイルスであろう。ウイルスE1A発癌タンパク質は例えば、p105Rbタンパク質、p130およびp107タンパク質の結合に関係する、Ad5のアミノ酸位置30〜85、ヌクレオチド位置697〜790にあるCR1ドメイン、ならびにAd5内アミノ酸位置120から139、ヌクレオチド位置920〜967にあるCR2ドメイン内に不活性化突然変異を含むことができる。アデノウイルスはタイプ2dl312から、またはアデノウイルスはタイプ5NTdl1010から指定することもできる。
【0070】
最終的に複製は核内にYB−1を含む細胞、即ちYB−1核陽性である細胞において、医薬製造、特には腫瘍疾患の処置用の医薬の製造を目的として発明のアデノウイルスを使用する場合、ならびに核内にYB−1を有する細胞での複製のために発明に従いアデノウイルスを使用する場合に、細胞周期とは無関係に起こることが好ましい。かかるアデノウイルスは核内にYB−1を持たず、YB−1は実質的に細胞質内にのみ存在している細胞では複製しないか、またはそのレベルが極めて低くなることに特に注意すべきである。この限りにおいて、これらウイルスが上手く複製するにはYB−1が核内に存在することが必要である。これは例えば、以下詳しく概要を示すように、核内にYB−1を発現させるか、または核内にYB−1が存在するようにする手段を細胞にこうじることで実現できる。そのような手段としては、例えばアデノウイルス遺伝子に加えてYb−1をコードする遺伝情報、特にYB−1の発現に関する遺伝情報も含んでいる、本発明に従い使用するアデノウイルスを介したYB−1のコード化および発現を挙げることができる。細胞核内へのYB−1の輸送、誘導または発現を起こすその他手段としては、細胞およびかかる細胞を含む生体への細胞増殖抑制薬投与、放射線照射、高温等のストレス条件がある。
【0071】
本発明に関連して、特に腫瘍溶解に関連して使用するアデノウイルスは更に、それらが核内にYB−1を持たない、換言すればYB−1核陰性である細胞では複製しないという特徴を持つ。
【0072】
発明で使用されるアデノウイルスは更に、それらが本明細書で発癌遺伝子タンパク質も呼ばれるウイルス発癌タンパク質をコードすることも特徴としており、この場合上記発癌タンパク質はE1Aであることが好ましく、そして発癌遺伝子タンパク質はウイルスの複製および/またはウイルス感染細胞の細胞溶解に影響を及ぼすことができるウイルス遺伝子の少なくとも1つを活性化できる。複製への影響は、ウイルスが、そのウイルスの発癌遺伝子タンパク質が存在しない場合に比べ、発癌遺伝子タンパク質が存在する場合により良く複製するものであることが好ましい。このプロセスは本明細書ではトランス活性化、そしてこのトランス活性化がE1Aを通じて行われる場合には特にE1Aトランス活性化とも呼ばれる。用語「トランス活性化する」または「トランス活性化」とは、各ウイルス発癌タンパク質がウイルス発癌タンパク質をコードする遺伝子そのものとは異なる1または複数のその他遺伝子の発現および/または転写に影響を及ぼす、即ちそれ/それらの発現および/または翻訳を好ましく制御する、そして特にはそれ/それらを活性化するプロセスを表す。かかるウイルス遺伝子はそれぞれE1B55kDa、E4orf6、E4orf3およびE3ADPならびにこれら遺伝子および遺伝子産物のいずれかの組み合わせであることが好ましい。
【0073】
さらに、場合によっては、発明に従い用いるアデノウイルスの特徴は、腫瘍抑制因子Rbにそして腫瘍抑制因子Rbと結合することである。原則的には、本発明に従い使用するアデノウイルスがRbに結合すること、またはRbに結合しないことは共に本発明の範囲内である。アデノウイルスに関する両実施態様は、処理対象となる細胞のRbの状態とは無関係に用いることができる。
【0074】
Rbに結合できなくするために、例えば次のようなE1A発癌タンパク質の欠失が考えられる:CR1領域内の欠失(Ad5内アミノ酸位置30〜85)およびCR2領域の欠失(AD5内のアミノ酸位置120〜139)。この様な欠失では、CR3領域はそのまま維持され、他の初期ウイルス遺伝子に対するトランス活性化機能を発揮できる。
【0075】
これに対し、E1AにRbへの結合能を付与する場合については、E1A発癌タンパク質に対する次の欠失が原理的に可能である:CR3領域(アミノ酸位置140〜185)の欠失;N−末端(アミノ酸位置1〜29)の欠失;アミノ酸位置85〜119の欠失;およびC−末端(アミノ酸位置186〜289)の欠失。ここに引用した領域は、RbへのE2Fの結合を妨害しない。しかし、トランス活性化機能は維持されるが、野生型Ad5に比べると低い。
【0076】
先行技術既知であるかかるウイルスは一般には複製欠失として知られている。しかし、本発明の利点は、かかるウイルスが好適な背景、特に細胞の背景があれば複製できることが認識されていることである。この種の好適細胞の背景は、核内にYB−1が存在すること、好ましくは核内にYB−1が細胞周期とは無関係に存在することより生じるか、または提供される。細胞または細胞系という用語は、ここで使用する場合、細胞の断片または細胞溶解物の断片、ならびにインビトロ、インビボまたはインシトゥ(in situ)に存在する細胞を含む。この限りにおいて細胞系または細胞という用語は、細胞培養体、組織培養体、臓器培養体、または単離された、グループ状態にある、または一部分である組織、臓器または生体であるインビボおよびインシトゥのその他組織または臓器内に存在する細胞、あるいは好ましくは生きている生物体内にその様な形で存在している細胞も含む。生体は好ましくは脊椎動物の生体であり、より好ましくは哺乳動物のものである。生体がヒトの生体であることが特に好ましい。
【0077】
更に、ここに提供する技術的教示に基づいて、ここに記載のアデノウイルスの複製特徴と、YB−1核陽性である細胞での従来技術のアデノウイルスの特徴とを持つ新規ウイルスを作成することも、本発明の範囲内である。換言すれば、既知アデノウイルスを基にして、ここに明示した本発明での使用に必要な特徴を有する更なるウイルスをデザインすることができる。
【0078】
本発明に関しては、発明で用いられる各種アデノウイルスの修飾E1A発癌タンパク質は、例えばE1B55K、E4orf3、E4orf6、E3ADPといった初期ウイルス遺伝子をYB−1核陽性細胞内でトランス活性化できるものである。これに関しては、その他の点についてウイルスゲノムにそれ以上の変更がないことが望ましく、各アデノウイルスはその他の点については野生型のアデノウイルスまたはその誘導体と同様でよい。
【0079】
本発明の意味での発癌遺伝子タンパク質のトランス活性化をコードしているか、またはかかる発癌遺伝子タンパク質を含むここに記載のウイルスは、たとえばアデノウイルスAdΔ24、dl922−947、E1Ad/01/07、CB106および/または欧州特許第EP0 931 380号に記載のアデノウイルスを含み、それらはそれぞれE1B、E2、E3および/またはE4といった初期遺伝子をトランス活性化でき、そして野生型のアデノウイルス、特には野生型Ad5と同等である。これらの例ではトランス活性化にはE1Aタンパク質の特定の領域が関係している。各種血清型アデノウイルスに於いてE1Aタンパク質中には極めて保存的である領域が3カ所ある。アミノ酸位置41〜80のCR1領域、アミノ酸位置120〜139のCR2領域、およびアミノ酸位置140〜188のCR3領域である。トランス活性化機能は主にE1Aの中にCR3領域が存在することに基づいている。CR3のアミノ酸配列は上記アデノウイルスについては変更がない。その結果、核内または細胞質内のYB−1の存在とは無関係に、初期遺伝子E1B、E2、E3およびE4は活性化される。
【0080】
しかし組換え体アデノウイルスd1520では、CR3領域は削除されている。即ちdl520はCR3領域のアミノ酸配列を含まない、いわゆるE1A12Sタンパク質を発現する。その結果d1520は特にE2領域に対し極めて弱いトランス活性化機能しか発揮できず、その結果YB−1核陰性細胞では複製しない。YB−1核陽性細胞では、YB−1はE2領域をトランス活性化し、それによりdl520は有効に複製できる。これがここでの開示の目的のための、それぞれdl520と同様のシステムおよび5120をベースとするシステムを用いることの根拠である。既報の両アデノウイルス群、即ちデルタ24(本明細書ではAdΔ24とも称する)とdl520間のより重要な違いは、d1520に関しては、初期遺伝子E1B、E3およびE4がYB−1核陰性細胞に比べてYB−1核陽性細胞でより強くトランス活性化されるという事実である。これに対しデルタ24との間には全く、または極僅かな違いしかない。しかしdl520の、より具体的にはE1A12Sタンパク質のトランス活性化効果は、野生型アデノウイルスに比べ大きく低下している。しかしこのトランス活性化は、実施例10に示す如く、YB−1核陽性細胞内での効果的複製を可能にするには十分である。本明細書に記載のE1Aタンパク質およびそれをコードする核酸のデザイン、特にそのEIAタンパク質が野生型発癌タンパク質E1Aに対し1もしくは複数の欠失、および/または突然を有しているような本明細書内のデザインであって、その欠失が好ましくはCR3領域およびN−末端の欠失およびC−末端の欠失を含む群から選ばれる一つであり、具体的にはdl520またはAdΔ24、dl922−924、E1Ad/01/07、CB106および/または欧州特許第EP0 931 830号に記載のアデノウイルスと関連し記載されるE1Aタンパク質の好適なデザインの実施態様がウイルス、特にアデノウイルスの実施態様であり、その複製はE2−後期プロモータの活性化を通じて、好ましくは主にE2−後期プロモータの活性化を通じて制御される。アデノウイルスのこの形態の複製を可能にするE1Aタンパク質の別の実施態様は、本明細書の開示に基づき、当業者により作り出すことができる。
【0081】
新たに構築され、本明細書では誘導体とも称され、且つ本発明にしがたい使用できる別のアデノウイルスは、一般的にはE1欠失、E1/E3欠失、および/またはE4欠失を有しており、即ち対応するアデノウイルスは機能的に活性なE1および/またはE3、および/またはE4発現産物および各産物をそれぞれ作り出すことができず、換言すれば、これらアデノウイルスは機能的に不活性なE1、E3および/またはE4発現産物のみ産生できるが、この時のそのような機能的に不活性なE1、E3および/またはE4発現産物とは、その転写レベルおよび/または翻訳レベルにおいて、たとえ発現産物として全く存在していない、あるいは野生型のアデノウイルスに存在する機能の少なくとも一つを欠いている形で存在していもよい。野生型アデノウイルスの発現産物の機能は当業者周知であり、そして例えばRussell, W, C、Journal of Virology、81、2573〜2604、2000年に記載されている。Russell(上記)はまた、参照によりここに組み込まれるアデノウイルスおよびアデノウイルスベクターの構築の原理についても記載している。上記修飾型E1A発癌タンパク質、E1B−55K、E4orf6、および/またはE3ADP(アデノウイルス崩壊タンパク質(ADP))(Tollefson, Aら、J.Virology、70、2296〜2306、1996年)をこの種のベクター内で、個々に、または組み合わせて発現させることも、本発明の範囲内である。これに関連して、ここに開示した個々に名前を挙げた遺伝子ならびにトランス遺伝子は、E1および/またはE3および/またはE4領域内にクローニングでき、好適プロモータの助けをかり、または好適プロモータの制御下に個別に発現させることができる。基本的には、領域E1、E3およびE4はアデノウイルス核酸内のクローニング部位として同等の好適性を有している。取り分けても好適なプロモータは本明細書の中でE1A、特に修飾型E1Aの制御および発現それぞれに関連付けて開示されているプロモータである。
【0082】
最後に実施態様の一つでは、本発明に従い用いられるアデノウイルスは、E1Bに関する、特にE1B19kDaに関する欠失体である。
【0083】
本明細書で使用する場合、欠失という用語はE1Bが野生型固有の特性の全てを有しているわけではないが、その特性の少なくとも一つを欠いている状態を意味する。
【0084】
ここに開示した発明に使用するアデノウイルスは、基本的には幾つかの実施態様については当分野周知である。本発明に用いられるアデノウイルスは、特に野生型と比較しここに示す技術的教示に従って変更する場合には、組換え体アデノウイルスであることが好ましい。本発明にとって必須でないアデノウイルス核酸を欠失または突然変異させることは、当業者の技術的水準内である。かかる欠失は、例えばここにも記す様にE3およびE4をコードする核酸の一部にかかっても良い。E4の欠失は、かかる欠失がタンパク質E4orf6を超えていないこと、換言すれば発明に使用するアデノウイルスはE4orf6をコードすることが特に好ましい。好適実施態様では、これらアデノウイルス核酸は更にウイルスカプシドに封入してもよく、即ち感染粒子を形成してもよい。このことは本発明での核酸の使用にも当てはまる。一般に、アデノウイルス系は単一または複数の発現産物について欠失してもよいことを記しておく。これに関連して、この現象はかかる発現産物をコードする核酸が完全に突然変異しているか、もしくは欠失しているか、または実質的に発現産物がもはや生じない程度まで突然変異しているか欠失しているかに基づいているか、あるいは核酸レベル(プロモータの欠失;シス作用要素)または翻訳系および転写系(トランス作用要素)のそれぞれについて、発現を制御するか、もしくは野生型とは異なる様式で作用するプロモータもしくは転写因子を欠いていることに基づいていることに注意しなければならない。具体的には、後者の観点は背景となる細胞に依存している。
【0085】
本発明によるアデノウイルスでは、さらに周知の如く新規アデノウイルスもまた、その他アデノウイルスに関しここに記した程度に使用することができる。発明による新規アデノウイルスは、ここに示す技術的教示より生ずる。特に好ましい例は、例えば図16および17に示すウイルスXvir03およびXvir03/01、実施例11および12に更に例示したデザイン原理である。
【0086】
ベクターXvir03の場合は、CMVプロモータを、IRES配列により隔てられているE1B55KおよびE4orf6の核酸をコードするE1領域内にクローニングする。これら2遺伝子の導入およびそこより産生される遺伝子産物それぞれによって、野生型ウイルスのものと実質同じ複製効率を得るが、その場合複製の選択性は細胞、特に腫瘍細胞について維持されるため、YB−1核陽性細胞、より具体的にはYB−1が脱制御状態にある細胞で複製が起こる。YB−1が脱制御状態にある細胞は、好ましくは正常細胞または非腫瘍細胞に比べて、好ましくはコンパートメント独立的(compartment-independent)に、YB−1の発現が上昇している細胞である。
【0087】
ウイルスXvir03の更なる発展形は、好適実施態様において、その中に治療遺伝子またはトランス遺伝子が特異的プロモータ、特に腫瘍特異的または組織特異的プロモータ制御下にクローニングされているウイルスXvir03/01である。E4領域が機能的に非活性であり、好ましくは欠失しているウイルスもまた上記ウイルスの範囲内である。ここに記したトランス遺伝子はまたE4領域内にクローニングしてもよく、この場合クローニングをE3領域内へのトランス遺伝子のクローニングに追加し、またはその代替して行っても良い。
【0088】
この様な治療遺伝子は、プロドラッグ遺伝子、サイトカインの遺伝子、アポトーシス誘導遺伝子、腫瘍抑制遺伝子、金属プロテアーゼインヒビターおよび/または血管新生インヒビターの遺伝子であろう。更にsiRNA、アプタマー、アンチセンスおよびリボザイムを、癌関連標的分子に向けて発現させてもよい。好ましくは、単一または複数の標的分子が、耐性関連因子、抗アポトーシス因子、発癌遺伝子、血管新生因子、DNA合成酵素、DNA修復酵素、増殖因子とそれらのレセプター、転写因子、金属プロテアーゼ、特にマトリックス金属プロテアーゼ、およびウロキナーゼ型プラスミノーゲン活性化因子を含む群より選ばれる。その好適実施態様は既に本明細書内に開示した。
【0089】
好適実施態様にて使用可能であろうプロドラッグ遺伝子は、例えばシトシンデアミナーゼ、チミジンキナーゼ、カルボキシペプチダーゼ、ウラシルホスホリボシルトランスフェラーゼ;プリンヌクレオシドホスホリラーゼ(PNP)である;Kimら、Trends in Molecular Medicine、8巻、No.4(Suppl)、2002年; Wybranietz.W.Aら、Gene Therapy、8、1654〜1664、2001年; Niculescu-Duvazら、Curr.Opin.Mol.Therapy、1、480.486、1999年; Koyamaら、Cancer Gene Therapy、7、1015〜1022、2000年; Rogersら、Human Gene Therapy、7、2235〜2245、1996年; Lockettら、Clinical Cancer Res、3、2075〜2080、1997年; Vijayakrishnaら、J.Pharmacol.And Exp.Therpeutics、304、1280〜1284、2003年。
【0090】
好適実施態様にて使用できるであろうサイトカインは、例えばGM−CSF、TNF−アルファ、IL−12、IL−2、IL−6、CSF、インターフェロン−ガンマである;Gene Therapy、Advances in Pharmacology、40巻、編集者:J.Thomas Augsut、Academic Press; ZhangとDegroot、Endocrinolgy、144、1393〜1398、2003年; Descampsら、J.Mol.Med、74、183〜189、1996年; Majumdarら、Cancer Gene Therapy、7, 1086〜1099、2000年。
【0091】
好適実施態様にて使用できるであろうアポトーシス誘導遺伝子は、例えばデコリン(decorin): Tralhaoら、FASEB J、17、464〜466、2003年;網膜芽細胞種94:Zhangら、Cancer Res、63、760〜765、2003年; BaxとBad: Zhangら、Hum.Gene Ther、20、2051〜2064、2002年;アポプチン(apoptin):NotebornとPietersen、Adv.Exp.Med.Biol、465、153〜161、2000);ADP:Tothら、Cancer Gene Therapy、10、193〜200、2003年; bcl-xs: Sumantranら、Cancer Res、55、2507〜2512、1995年;E4orf4:BraithwaiteとRussell、Apoptosis、6、359〜370、2001年;FasL、Apo−1およびTrail:Boehringer Manheim、Guide to Apoptotic Pathways、Araiら、PNAC、94、13862〜13867、1997年、Bims; Yamaguchiら、Gene Therapy、10、375〜385、2003年; GNR163: Oncology News、17 Juni、2000年、がある。
【0092】
好適実施態様にて使用できるであろう腫瘍抑制遺伝子は、例えばE1A、p53、p16、p21、p27、MDA−7がある。Opalkaら、Cell Tissues Organs、172、126〜132、2002年、Jiら、Cancer Res、59、3333〜3339、1999、Suら、Oncogene、22, 1164〜1180、2003年。
【0093】
好適実施態様にて使用できるであろう血管新生インヒビターは、例えばエンドスタチン、アンギオスタチン:Hajitouら、FASEB J、16, 1802〜1804、2002年およびVEGFに対する抗体がある(Ferrara、N、Semin Oncol 2002 Dec; 29(6 Suppl 16): 10−4。
【0094】
好適実施態様にて使用できるであろう金属プロテアーゼインヒビターは、例えばTimp-3、Ahonenら、Mol Therpy、5、705〜715、2002年; PAI-1; Soffら、J.Clin.Invest、96、2593〜2600、1995年; Timp-1、Brandt K.Curr.Gene Therapy、2、255〜271、2002年がある。
【0095】
siRNA(short ingerfering(短干渉)RNA)は塩基相補正により相互にハイブリダイゼーションする、即ち実質的に塩基が対を成しており、好ましくは最長50ヌクレオチド、好ましくは18〜30ヌクレオチド、より好ましくは25ヌクレオチド未満、そして最も好ましくは21、22もしくは23ヌクレオチドの長さを持つ2つのRNA鎖、好ましくは分離した2つのRNA鎖より成り、この時それらの特徴は単鎖のsiRNAであり、特に一本の鎖、好ましくは第二の単鎖とハイブリダイゼーションしてそれと塩基対合する単鎖の伸展した鎖である。siRNAは、特異的にmRNAの分解を誘導または媒介する。この時必要な特異性は、siRNAの配列とその結合部位から提供される。分解対象となる標的配列は第一または第二のsiRNA形成鎖に対し実質的に相補的である。正確な作用様式は不明であるが、siRNAは細胞が発生中にある特的の遺伝子座を阻害し、ウイルスからそれを保護するための生物学的戦略であると想定される。siRNAを介したRNA干渉は、タンパク質の特異的抑制に、または遺伝子特異的二本鎖RNAを導入することによるタンパク質発現の完全なノックアウトにも用いられる。高等生物では、19〜23ヌクレオチド長のsiRNAは特に、非特異的な防御反応、いわゆるインターロイキン反応の活性化をもたらさないので特に好ましい。3‘末端に対称的な2−nt長のオーバーハングを有する21ヌクレオチドから成る二本鎖RNAを迅速にトランスフェクションすることにより、哺乳動物細胞にRNA干渉を伝達でき、リボザイムやアンチセンス分子といった他技術に比べて高い効率を得ている(Elbashir、S.Harborth J. Lendeckel W. Yalvcin、A. Weber K Tuschl T: Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells、Nature 2001年、411: 494〜498)。標的遺伝子の発現を阻害するのに必要なRNA分子は極めて小数であった。特にsiRNA分子の一過性の干渉現象、および特異的配送特性を有するsiRNAが外部からの投与に限定されないようにするために、先行技術では内因性のsiRNAを発現できるベクターを使用している。例えば、センスおよびアンチセンス両方向の、ベクター内に導入される例えば9ヌクレオチド長のスペーサー配列により隔てられた、19ヌクレオチド長の標的配列を含む64ヌクレオチド長のオリゴヌクレオチドが提供されている。得られた転写体は、例えば19塩基対のステム構造を持つヘアピン構造に折りたたまれた。ループは細胞内で迅速に分解し、その結果機能的なsiRNAが生成する(Brummelkampら、Science、296、550〜553、2002年)。
【0096】
pRbおよびE2Fそれぞれの活性は、リン酸化によって制御されている。低リン酸化型のpRbは主にG1期とM期に存在している。これに対し、高リン酸化型のpRbはS期とG2期に存在している。E2Fは、E2Fと低リン酸化型pRbから成る複合体から、pRbのリン酸化によって放出される。E2Fと低リン酸化型pRbより成る複合体からE2Fが放出されると、E2F依存遺伝子の転写が起こる。E1Aタンパク質は低リン酸化型pRbに結合するだけでなく、その時はE1Aタンパク質のCR2領域を介したE1AとpRbとの結合も普通に起こっている。さらに、それはCR1領域にも、より低い親和性で結合する(Ben-IsraelとKleiberger、Frontiers in Bioscience、7、1369〜1395、2002年; HeltとGalloway、Carcinogenesis、24、159〜169、2003年)。
【0097】
本発明に従って用いるアデノウイルスの実施態様に於いて、そのアデノウイルスの一部を成すYB−1をコードする核酸はまた核内へのYB−1の輸送を媒介する核酸配列を含んでもよい。発明による核酸、アデノウイルスおよびアデノウイルス系、ならびに例えばOnyx−015、AdΔ24、dl922−947、E1Ad/01/07、CB016、dl520および欧州特許第EP0 931 830号に記載のアデノウイルスといった当分野既知のアデノウイルスは、アデノウイルスおよびアデノウイルス系として、即ち対応する核酸として、上記の如くまたはこれに関連する発明の核酸と組み合わせて使用できる。核輸送を媒介するのに好適な核酸配列は当業者周知であり、例えば(Whittaker、G.Rら、Virology、246、1〜23、1998年; Friedberg、E.C、TIBS 17、347、1992年; Jans、D.Aら、Bioessays 2000年6月; 22(6): 532〜44; Yoneda、Y、J.Biocehm.(Tokyo)1997年5月; 121(5): 811〜7; Boulikas、T、Crit.Rev.Eukaryot.Gene Expr.1993; 3(3): 193〜227; Lyons RH、Mol.Cell Biol、7、2451〜2456、1987年)に記載されている。核輸送媒介核酸配列に関連して様々な原理を利用できる。この様な原理の一つは、YB−1をシグナルペプチドとの融合タンパク質の形で形成して核内に導入すること、およびそれにより本発明のアデノウイルスの複製が起こることである。
【0098】
発明に使用するアデノウイルスのデザインを実現するであろう別の原理は、好ましくは細胞質での合成から始まってYB−1を細胞核内に誘導するか、またはYB−1を細胞核内に移動して、そこでウイルス複製を促進するトランスポーター配列と一緒にYB−1を与えることができる原理である。特に効果的な核輸送媒介核酸配列の例はHIVのTAT配列であり、これはEfthymiadis、A、Briggs、LJ、Jans、DA、JBC 273、1623〜1628、1998年にその他好適核酸配列と共に記載されている配列である。発明に使用するアデノウイルスが核内輸送をコードするペプチドをコードしている核酸配列を含むことは、本発明の範囲内である。
【0099】
YB−1が完全長、特に野生型YB−1に対応する形態で存在することは、本発明の範囲内である。YB−1が、例えば短形または断頭形(truncated form)の様な誘導体として用いられること、または存在することは、本発明の範囲内である。本発明の範囲内で誘導体として用いられる、または存在するYB−1は、E2−後期プロモータに結合でき、その結果アデノウイルスE2領域の遺伝子発現を活性化できるYB−1である。この種の誘導体は特に、本明細書に開示のYB−1誘導体を含む。更なる誘導体は、アミノ酸配列のN−末端、C−末端または内部にある単独または複数個のアミノ酸を欠失することで作られるだろう。
【0100】
これまでに記したアデノウイルスがコードする各種の発現遺伝子および遺伝子産物に関しては、これらを組み合わせてコードすることも、また組み合わせて発現することもそれぞれ可能である。
【0101】
アデノウイルスおよびアデノウイルス系という用語は、本発明内では実質的に同一の意味を持つものとする。アデノウイルスは特にカプシドおよび核酸を含む完全なウイルス粒子を意味するものとする。アデノウイルス系という用語は特に、核酸が野生型に対し変更されている事実に焦点を当てるものである。この種の変更は、プロモータ、制御配列および/またはオープンリーディングフレームといったコーディング配列の欠失および/または付加および/または突然変異に起因する、アデノウイルスゲノムの構造変化を含むことが好ましい。さらに、用語アデノウイルス系は、例えば遺伝子治療に用いられるベクターと結びつけて使用するのが好ましい。
【0102】
前述のコメントは、アデノウイルスおよびアデノウイルス系それぞれの使用ならびにデザインを含め、コーディング核酸にも当てはまり、またその逆も言える。
【0103】
本発明に関しては、本発明に使用するアデノウイルスおよびそれをコードする核酸それぞれは、かかる複製をもたらす、または別の核酸配列と組み合わせての複製をもたらすようないずれかのアデノウイルス核酸である。本明細書に説明したように、複製に必要な配列および/または遺伝子産物はヘルパーウイルスを利用して提供することができる。コーディング核酸配列と称するものであれば、そしてかかる核酸配列が既知であるものであれば、使用した配列と同一のものだけでなく、それより派生する配列もまた発明の範囲内である。この派生する配列という用語は、本明細書に於いては特に、非派生配列の機能に対応する機能を示す核酸またはポリペプチドである、遺伝子産物を依然として生ずる配列を意味する。このことは、当業者既知である簡単な通常試験により決定できる。かかる派生核酸配列の例は、同一遺伝子産物、特には同一アミノ酸配列をコードしているものの、遺伝子コードの縮重による異なる塩基配列を有しているものである。
【0104】
好適実施態様に於いて、本発明のアデノウイルスおよび本発明のアデノウイルス複製系、そして本発明でのそれらの使用に関してはそれぞれ、アデノウイルス核酸は発癌遺伝子タンパク質、特にE1Aタンパク質の発現を欠失しており、このことはそれが12SのE1Aタンパク質もしくは13SのE1Aタンパク質をコードしていないか、またはそれが12SのE1Aタンパク質と13SE1Aタンパク質のどちらもコードしていないか、または本明細書に記載のように修飾されていることを意味しており、そしてアデノウイルス複製システムがさらにヘルパーウイルスの核酸を含み、このヘルパーウイルスの核酸が発癌遺伝子タンパク質、特にE1Aタンパク質をコードする核酸配列を含み、さらにヘルパーウイルスの核酸が次の特徴を有すると同時にアデノウイルスに対し同特性を付与すること、即ちYB−1核陰性細胞では複製しないが、YB−1核陽性細胞では細胞周期と無関係に細胞内で複製し、少なくとも1つのウイルス遺伝子、特にはE1B55kDa、E4orf6、E4orf3および/またはE3ADPをトランス活性化し、そして/または細胞質YB−1を核内に移動しないという特性を付与することを意味する。ここに記したトランス遺伝子をヘルパーウイルスが個別または一緒にコードしていること、および/またはそれから発現することは本発明の範囲内である。
【0105】
本発明のこの様なアデノウイルス複製系の実施態様の一つでは、アデノウイルス核酸および/またはヘルパーウイルスの核酸は、さらに複製可能であるベクターとしても存在する。
【0106】
本発明に用いるアデノウイルスをコードするコーディング核酸配列がベクター内、好ましくは発現ベクター内に存在すること、そしてこの発現ベクターを本発明に従って使用することは、本発明の範囲内である。
【0107】
別の局面では本発明はさらに少なくとも2つのベクターを含むベクター群に関し、この場合上記ベクター群がここに記したアデノウイルス複製系を共に含んでおり、そして上記ベクター群は本発明に従い用いられる。アデノウイルス複製系の各構成成分は、個々のベクター、好ましくは発現ベクター内に配置されると解釈される。
【0108】
最後に、本発明はアデノウイルスについてここに記した同一の目的に関する細胞の使用についての別の局面に関するものであり、その場合細胞は発明に使用するここに記したアデノウイルスをコードする1または複数の核酸、および/または本発明による各アデノウイルス複製系、および/または各ベクターおよび/もしくはベクター群を含む。
【0109】
前記アデノウイルス構築体ならびに特にそれらの核酸およびそれをコードする核酸もまた細胞内、好ましくは腫瘍細胞に一部分導入してもよく、その場合各種個別構成成分が存在していることにより、それらはあたかも各構成成分は単一核酸および単一または複数のアデノウイルスに由来するように一つになって作用するだろう。
【0110】
発明に使用するアデノウイルス、アデノウイルス系、またはその一部分をコードする核酸は、ベクターとして存在するだろう。好ましくは、それらはウイルスベクターとして存在する。アデノウイルス核酸を含む核酸の場合、ウイルス粒子がベクターであることが好ましい。しかし、上記核酸がプラスミドベクター内にある場合も発明の範囲内である。いずれの場合も、ベクターは挿入した核酸の増幅、即ち挿入核酸の複製および場合によっては発現、それらの制御をもたらすそれぞれの要素を含んでいる。好適ベクター、特に発現ベクター、および対応する要素は当業者に周知であり、例えばGrunhaus, A、Horwitz, M.S、1994年、Adenoviruses as cloning vectors.In Rice, C.編集、Seminars in Virology, London: Saunders Scientific Publicationsに記載されている。
【0111】
ベクター群に関する発明の局面は、核酸の各種要素が必ずしも1つのベクターに含まれている必要がないという前述の実施態様に対応するものである。この場合、ベクター群は少なくとも2種類のベクターを含む。それ以外のことは、ベクターに関連して既に述べた事柄がベクターおよびベクター群にそのまま当てはまる。
【0112】
発明に用いられるアデノウイルスは、ここに開示した各種核酸および遺伝子産物により特徴付けられ、そしてそれ以外に含まれる要素は野生型のアデノウイルスと同様であり、全てが当業者周知である(Shenk、T.: Adnoviridae: The virus and their replication.Fields Virology、第三版、編集、Fields, B.N.、Knipe, D.M.、Howley, P.M.ら、Lippincott-Raven Publishers、Philadelphia、1996年、67章)。
【0113】
アデノウイルスの複製は極めて複雑な過程であり、一般にはヒト転写因子E2Fを利用する。ウイルス感染に際しては、まず「初期遺伝子」E1、E2、E3およびE4が発現する。「後期遺伝子」群はウイルスの構造タンパク質の合成を担っている。初期および後期遺伝子両方の活性化に関しては、異なるE1AおよびE1Bタンパク質をコードしている2つの転写単位E1AとE2Aから成るE1領域が重要であるが、それはE2、E3およびE4がそれらにより誘導されるからである(Nevins、J.R.Cell 26、213〜220、1981年)。更にE1Aタンパク質は休止細胞にDNA合成を誘導してS期に入らせることができる(上記BoulangerとBlair、1991年)。さらにそれらはRbクラスの腫瘍抑制因子と相互作用する(Whyte, P.ら、Nature 334、124〜127、1988年)。そうすると、細胞質転写因子であるE2Fが放出される。E2F因子は次に細胞遺伝子ならびにウイルス遺伝子の対応するプロモータ領域に結合し(特にアデノウイルスE2初期プロモータ)、転写および翻訳を開始させるだろう(Nevins、J.R.Science 258、424〜429、1992年)。
【0114】
E2領域の遺伝子産物は3種類の必須タンパク質をコードしていることから、それらは複製開始とその遂行に特に必要である。E2タンパク質の転写は2種類のプロモータ、ここではE2−初期プロモータもしくは初期E2プロモータとも呼ばれる「E2−初期E2F−依存」プロモータ、および「E2−後期」プロモータにより制御されている(SwaminathanとThimmapaya、The Molecular Repertoire of Adnoviruses III: Current Topics in Microbiology and Immunology、199巻、177〜194、Springer Verlag 1995年)。さらに、E4領域の産物は、E1AおよびE1B−55kDaタンパク質と一緒になり、それぞれE2Fの活性およびp53の安定性にとって重要な役割を果たす。例えば、E4領域にコードされているE4orf6/7タンパク質がE2FおよびDP1から成るヘテロダイマーとの直接相互作用によってプロモータはより活性化される(SwaminathanおよびThimmapay、JBC 258、736〜746、1996年)。更にp53は、E1B−55kDaとE4orf6から成る複合体により不活性化され(Steegenga, W.T.ら、Oncogene 16、349〜357、1998年)、溶解性の感染周期が成功して終了する。さらにE1B−55kDaタンパク質は、E4orf6タンパク質と相互作用してウイルスRNAの核外への運び出しを促進することからも更に重要な機能を果たしているが、この場合細胞固有のRNAは核内に維持される(BridgeおよびKetner、Virology 174、345〜353、1990年)。更に重要な発見は、E1B−55kDa/E4orf6から成るタンパク質複合体がいわゆる「ウイルス封入体」内に局在していることである。このことから、これら構造体が複製および転写部位であることが推測される(OrnellesおよびShenk、J.Virology 65、424〜429、1991年)。
【0115】
複製にとって重要である、特にアデノウイルスの放出に関し重要である別の領域はE3領域である。E3領域は、より具体的にはインビトロでのアデノウイルス感染周期にとって必須ではない、即ち細胞培養に必須ではない比較的小型の各種タンパク質に関する遺伝情報を含んでいる。しかしそれらは特に免疫制御およびアポトーシス機能を有していることから、インビボでの急性期および/または潜伏感染期間中のウイルスの生存にとって重要な役割を果たしている(Marshall S.Horwitz、Virololgie、279、1〜8、2001年; Russell、上記)。約11.6kDaの大きさを持つタンパク質が細胞死を誘導することが示されている。このタンパク質は、その機能故にADP−英語でアデノウイルス致死タンパク質(Adenovirus death protein)−と呼ばれている(Tollefson、J.Virology、70、2296〜2306、1996年)。このタンパク質は主に感染周期の後期に形成される。さらに、このタンパク質の過剰発現は感染細胞をより溶解する(Doroninら、J.Virology、74、6147〜6155、2000年)。
【0116】
更に、本発明者はE1A欠失ウイルス、即ち具体的には12SのE1Aタンパク質を持たず、そして13SのE1Aタンパク質を発現しないウイルスが、より高いMOIで極めて効率的に複製できることを知った(Nevins J.R.Cell 26、213〜220、1981年)が、しかしこのことを臨床的に応用できなかった。この現象は文献の中では「E1A−様活性」と呼ばれている。さらにE1Aがコードする5種類のタンパク質の内2種類のタンパク質、即ち12Sおよび13Sタンパク質が、それぞれ他のアデノウイルス遺伝子の発現を制御および誘導することも分かった(Nevins, J.R.Cell 26、213〜220、1981年; Boulanger、P.およびBlair, E.; Biochem.J.275、281〜299、1991年)。これに関連して、トランス活性化機能が主に13Sタンパク質のCR3領域から提供されるものであることが示された(Wong HKおよびZiff EB.J Virol.、68、4910〜20、1994年)。13Sタンパク質のCR1および/またはCR2領域、および/またはCR3領域を特異的に欠失したアデノウイルスは、大部分が複製欠失であるが、それでも一部の細胞株ではウイルス遺伝子をトランス活性化し、それぞれ特にE2領域を促進する(Wong HKおよびZiff EB.J Virol.、68、4910〜20、1994年; Mymryk, J.S.およびBayley, S.T.、Virus Research 33、89〜97、1994年)。
【0117】
野生型アデノウイルスを細胞、一般的には腫瘍細胞に感染させると、E1A、E1B−55KおよびE4orf6が介在してYB−1が核内に誘導され、ウイルス封入体の中に核内のE1B−55Kと共に局在するようになり、それがインビトロおよびインビボの両方での細胞核内に於けるウイルスの効率的複製を可能にする。これに関連して、E4orf6がE1B−55Kに結合すること(Weigel, S.およびDobbelstein, M.J. Virology、74、764〜772、2000年; Keith N.Leppard、Seminars in Virology、8、301〜307、1998年)、およびそれがE1B−55Kの核内への運び込みと分布とを媒介し、それによりそれぞれ最適なウイルス産生とアデノウイルス複製が提供されることが初期に既に見出されていた。本発明の効率的なウイルス複製は、E1A、E1B−55KおよびYB−1間の相互作用によって、そして、E1B−55K/E4orf6とYB−1とから成る複合体によって、およびYB−1と核内のE1B−55Kが、いわゆるウイルス封入体内に同時に局在することによって、ならびに個々に記したウイルスのYB−1核陽性細胞での複製に使用することによって、およびYB−1核陽性細胞が関係する疾患の治療を目的とした医薬の製造に使用することによって可能となる。この様な細胞の背景により可能となった複製は、細胞溶解、ウイルスの放出、および近接細胞の感染および溶解をもたらし、結果として腫瘍細胞および腫瘍に感染した場合には、それぞれ腫瘍の最終的な溶解、即ち癌溶解が起こる。
【0118】
YB−1は逆位CAAT配列、いわゆるY−ボックスに結合する保存性の強い因子の群に属する。それらは転写ならびに翻訳両レベルにおいて通常の様式で活性化する(Wolffe, A.P.Trends in Cell Biology 8、318〜323、1998年)。活性化によりY−ボックス依存制御経路の数が増加することが見出されているが、増殖およびアポトーシス関連遺伝子の阻害時でも数が増加することが見出されている(Swamynathan、S.K.ら、FASEB J.12、515〜522、1998年)。従って、YB−1はp53と直接相互作用しており(Okamoto、T.ら、Oncogene 19、6194〜6202、2000年)、Fasの遺伝子発現(Lasham, A.ら、Gene 252、1〜13、2000年)、MDRおよびMRP遺伝子発現(Stein, U.ら、JBC 276、28562〜69、2001年; Bargou, R.C.ら、Nature Medicine 3、447〜450、1997年)ならびにトポイソメラーゼおよび金属プロテアーゼの活性化(Mertens.P.R.ら、JBC 272, 22905〜22912、1997年; Shibao, L.ら、Int.J.Cancer 83、732〜737、1999年)に於いて重要な役割をはたしている。従って、YB−1はmRNAの安定性の制御(Chen, C-Y.ら、Genes & Development 14、1236〜1248、2000年)および修復プロセス(Ohga, T.ら、Cancer Res.56、4224〜4228、1996年)に関係している。
【0119】
腫瘍細胞でYB−1が核内に局在することにより、特に12SのE1Aタンパク質及び13SのE1Aタンパク質の両方ともが発現形態で存在しないかそして用いられない例(Holm, P.S.ら、JBC 277、10427〜10434、2002年)、および多剤耐性(多重耐性)でのタンパク質YB−1の過剰発現の例ではE1A独立にウイルス複製が起こる。更に、例えばE4orf6およびE1B−55Kの様なアデノウイルスタンパク質はウイルス複製に対し有利な影響を及ぼし(Goodrum, F.D.およびOrnelles, D.A、J.Virology 73、7474〜7488、1999年)、この時機能的E1Aタンパク質は他のウイルスの遺伝子産物(E4orf6、E3ADPおよびE1B−55Kの様な)のスイッチングに関係していること(Nevins J.R.、Cell 26、213〜220、1981年)が知られている。しかしこの様なことは13SのE1Aが存在しない先行技術のE1A−マイナスアデノウイルスでは起こらない。YB−1が核内に存在する多剤耐性細胞でのYB−1核内局在は、E1A−マイナスウイルスに複製および粒子形成を提供する。しかしこの場合、ウイルス複製および粒子形成の効率は、野生型のAd5に比べると数倍低い。腫瘍細胞の核内に既に存在しているYB−1と、または外部因子(細胞増殖抑制薬または放射線、または高温を作用させる)により腫瘍細胞内に誘導した、即ち特に細胞周期とは無関係に核内への存在を促すか、またはベクターを使ってトランス遺伝子として導入したYB−1と、アデノウイルス遺伝子を始動するが、ウイルスの複製は行わない系、好ましくはアデノウイルス系とを組み合わせると、驚くべきことにYB−1を通じてシステムが極めて効率的にウイルスの複製と粒子を形成し、癌溶解を誘導した。とりわけても好適な細胞増殖抑制薬は次の群に属するものである:ダウノマイシンおよびアドリアマイシンといったアントラサイクリン類;シクロホスファミドのようなアルキル化剤;エトポシドの様なアルカロイド;ビンクリスチンおよびビンブラスチンの様なビン−アルカロイド;例えば5−フルオロウラシルおよびメトトレキセートの様な代謝拮抗剤;例えばシス−プラチンの様な白金誘導体;カンホテシンの様なトポイソメラーゼインヒビター;および例えばタキソールの様なタキサン類である。ここに開示した、YB−1核陽性細胞内でのみ複製できるアデノウイルス、特に組換え体アデノウイルスは、ウイルス遺伝子E1B−55K、E4orf6、E4orf3およびE4ADPをトランス活性化するそれらの能力は、野生型アデノウイルス、特に野生型Ad5の持つ同様のトランス活性化能力に比べ制限されている。本発明者は、今回驚くべきことにこれら制限された活性化能力が対応する遺伝子によって、特にYB−1の核内局在と組み合わせて発現されるE1B−55KおよびE4orf6によって補完されることを見出した。本明細書の実施例に示す様に、ウイルス複製および粒子形成は共にこの様な条件では野生型アデノウイルスの複製および粒子形成に匹敵するレベルまで増加する。
【0120】
ここに記したアデノウイルスと結びつけた医薬、またはここに記したアデノウイルスを本発明に従い使用し製造される医薬は一般的には全身投与されるものと解釈されるが、かかる医薬を局所に適用するか、または配送することも本発明の範囲である。かかる使用は、本発明の医薬を使用する診断および/または予防および/または治療を目的として、特にある状態、典型的には疾患の形成に関係する細胞に上記アデノウイルスで感染すること、および特にそれら細胞内で複製すること意図して、好ましくは随時に行われる。
【0121】
かかる医薬は、好ましくは腫瘍疾患の処置に用いられる。腫瘍疾患の中では、腫瘍疾患の基礎を成すメカニズム、特に病理学的メカニズムによってYB−1が既に核内に局在している腫瘍、または外的手段により核内にYB−1が生じている腫瘍が特に好ましく、この場合の手段はYB−1を核内に運搬する、またはYB−1を核内に誘導する、またはYB−1を核内で発現するのに好適なものである。本明細書で使用する場合、腫瘍または腫瘍疾患という用語は、悪性および良性腫瘍の両方、ならびに各疾患を含むものとする。医薬はさらに少なくとも1つの医薬的に活性な化合物を含むものとする。かかる追加の医薬的に活性な化合物の種類および量は、かかる医薬が用いられる適用に依存する。医薬を腫瘍疾患の治療および/または予防に用いる場合、具体的にはシスプラスチンおよびタキソール、ダウノブラスチン、ダウノルビシン、アドリアマイシン、および/もしくはミトキサントロンのような細胞増殖抑制薬、またはここに記したその他の細胞増殖抑制薬あるいは細胞増殖抑制薬の群を使用する。
【0122】
本発明の医薬は様々な製剤形状に存在でき、液体形状が好ましい。更に、医薬は安定化剤、緩衝剤、保存剤、および医薬製剤分野当業者に周知であるこの種の作用剤を含むだろう。
【0123】
本発明者は驚くべきことに、ここに記載のウイルスを発明に従い、細胞周期とは無関係に核内にYB−1を持つ腫瘍に対し極めて高い成功率で応用できることを見出した。一般にYB−1は細胞質内、特に核周囲の原形質内に存在する。細胞周期のS期の間は、YB−1は正常細胞および腫瘍細胞両方の細胞核内に見出すことができる。しかしそれは修飾型アデノウイルスを用いたウイルスによる癌溶解を誘導するには十分ではない。この様な不良な結果は、先行技術に記載のこの種の弱毒化アデノウイルスの効率が比較的低いことに起因している。換言すれば、ここに記載の弱毒化または修飾型ウイルスを投与する際に、ウイルス性の癌溶解に必要な分子生物学的条件を与えさえすれば、かかるアデノウイルス系を使用する、特に高い効率で使用することができる。ここに記載様に、発明に従って用いられるAdΔ24、dl922−947、E1Ad/01/07、CB016、dl520および欧州特許第EP0 931 830号に記載の組換え体アデノウイルスといったアデノウイルスの場合は、上記条件は腫瘍疾患の例ではYB−1が細胞周期とは無関係に核局在する細胞に与えられる。このタイプの核内局在は、腫瘍細胞そのものに起因するものでも、またはここに記した発明による手段または薬物により誘導してもよい。即ち本発明は、発明のウイルス、特に先行技術に既に記載されている弱毒化または修飾型アデノウイルスを用いて良好に処置できる新規腫瘍および腫瘍疾患群、さらには患者を画定する。
【0124】
その一部は周知であり、そして本発明により治療可能であるアデノウイルスを用いて、または本明細書に初めて記載されたアデノウイルス、具体的にはEqAタンパク質内に突然変異および欠失を有するが、Rb/E2fの結合を妨害せず、そしてYB−1核陰性細胞では複製しないアデノウイルス、または本明細書内に規定された大きな複製低下を示し、および/または欠失型発癌タンパク質、特にE1Aを有する、例えばウイルスAdΔ24、dl922−947、E1Ad/01/07、CB106および欧州特許第EP0 931 830号に記載のアデノウイルスを用いて、本発明により治療可能なさらなる患者群は、ある条件を適用または放出することによりYB−1が核内に移動する、または核内に誘導する、または核内に運ばれることが確実である患者である。この患者群に関連してこの種のアデノウイルスを使用することは、ウイルス複製の誘導がYB−1の核内局在化とそれに続くYB−1のE2−後期プロモータへの結合に基づくという所見を根拠としている。この所見に基づけば、AdΔ24、dl922−947、E1Ad/01/07、CB106および欧州特許第EP0 931 830号に記載のアデノウイルスといった本明細書に開示するアデノウイルスは、YB−1核陽性である細胞および/またはYB−1が本発明に規定されるように脱制御されている細胞で複製できるだろう。その限りに於いてこれらアデノウイルスは、本発明に従って、これら特性を持つ細胞を含む疾患および患者群の処置に、特にこれら細胞が処置対象となる各疾患の成立に関係している時に用いることができる。本明細書に発明に有用であると記したアデノウイルス、そして使用するこれらウイルス、特に本明細書で初めて記載されたアデノウイルスを用いて処置可能であるさらなる患者群は、YB−1核陽性および/または以下記載の処置の結果としてYB−1核陽性である患者であり、この場合かかる処置は好ましくは医学的処置であり、そして/または各ウイルスの投与を伴い実施される患者である。YB−1核陽性患者が、腫瘍を形成する複数の細胞が細胞周期とは無関係に核内にYB−1を有する患者である場合も、本発明の範囲内である。とりわけこれら治療は、ここに記載の細胞増殖抑制薬投与と共に、そして/または腫瘍治療の一環として実施される。更に、放射線、好ましくは腫瘍療に使用するものである放射線は、この処置群に属する。放射線とは、具体的に高エネルギー照射を伴う放射線を意味し、好ましくは放射性放射線、好ましくは腫瘍治療に用いられる放射性放射線を意味する。高温および高温の適用、好ましくは腫瘍治療に用いられる高温も別の処置手段である。特に好ましい高温の実施態様は局所的適用である。最後にホルモン処置、特に腫瘍治療に用いられるホルモン処置は別の処置手段である。かかるホルモン処置に関連しては、抗エストロゲンおよび/または抗アンドロゲンが用いられる。タモキシフェンの様な抗エストロゲン剤は特に乳ガンの治療に、そして例えばフルタミドまたは酢酸シプロテロンの様な抗アンドロゲン剤は、前立腺癌の治療に用いられる。
【0125】
腫瘍を形成する細胞の一部が元から、または核内への誘導および能動的導入後にYB−1を含んでいること、または本開示の意味において脱制御したYB−1を含む場合も本発明の範囲内である。好ましくは、腫瘍形成細胞の約5%以上、即ち6、7、8%等がYB−1核陽性細胞またはYB−1が脱制御状態にある細胞である。YB−1の核内への局在は、外側から加えられたストレス、および局所的に加えられたストレスにより誘導できる。この誘導は、例えば放射線、特にUV照射、細胞増殖抑制薬、取り分けても本明細書に開示されたもの、および高温を用いて行うことができる。高温に関連しては、それは極めて特異的な様式、より具体的には局所に極めて特異的な様式で実現できることが必須であり、その結果YB−1はより特異的に細胞核内に局在するようになり、それによりアデノウイルスの複製に必要な条件、即ち細胞および腫瘍溶解に必要な条件が、好ましく局所に限定された形で提供される(Stein U、Jurchott K、Walther W、Bergmann S、Schlag PM、Royer HD.J Biol Chem.2001年、276(30)、28562〜9; Hu Z, Jin S, Scotto KW.J Biol Chem.2000年1月 28; 275(4): 2979〜85; Ohga T, Uchiumi T, Makino Y, Koike K、Wada M、Kuwano M, Kohno K.J Biol Chem.1998年、273(11): 5997〜6000)。
【0126】
かくして本発明の医薬は、好ましくは各腫瘍細胞に於いて、前処理または併用処置を通じてYB−1の輸送に影響を及ぼす形で患者および患者群に投与することもできるし、または、意図されていてもよい。
【0127】
この技術的教示に基づけば、当業者はその技術的水準の範囲内で、E1Aに例えば欠失または点突然変異を含む好適な修飾を実行して、発明の使用と関係して用いられるアデノウイルスの各種実施態様を生成することができる。
【0128】
既に説明した如く、発明に従い使用するアデノウイルスは、核内にYB−1を有する細胞および細胞系それぞれの中で複製できる。本発明に従って使用するアデノウイルスが複製でき、そして腫瘍を溶解できるかどうかという問題にとって、Rb、即ち網膜細胞芽腫腫瘍抑制産物の有無に関する細胞状態は無関係である。さらに、発明に従い上記アデノウイルスを使用することに関連して感染細胞、感染対象となる細胞、または処置対象となる細胞のp53の状態を考慮する必要はないが、それはここに開示したアデノウイルス系をYB−1核陽性細胞、即ち細胞周期とは無関係に核内にYB−1を持つ細胞と結びつけて使用することにより、p53に関する状態ならびにRbに関する状態が本明細書に開示する技術的教示の性能に影響しなくなるからである。
【0129】
発癌遺伝子および発癌遺伝子タンパク質はそれぞれ、特にE1Aは所有の天然アデノウイルスプロモータの制御下、および/または腫瘍もしくは組織特異的プロモータにより制御できる。好適な非アデノウイルス性のプロモータは、サイトメガロウイルスプロモータ、RSV(ラウス肉腫ウイルス)プロモータ、アデノウイルスベースドプロモータVaIおよび非ウイルス性YB−1プロモータ(Makino Y.ら、Nucleic Acids Res.1996年、15、1873〜1878)を含む群より選ぶことができる。ここに開示した発明の各局面およびいずれかの局面に関連して使用できるさらなるプロモータには、テロメラーゼプロモータ、アルファ−フェトプロテイン(AFP)プロモータ、癌胚抗原プロモータ(CEA)(Cao, G., Kuriyama, S., Gao, J., Mitoro, A., Cui, L., Nakatani, T., Zhang, X., Kikukawa, M., Pan, X., Fukui, H., Qi, Z. Int.J.Cancer、78、242〜247、1998年)、L−プラスチンプロモータ(Chung, I., Schwartz, PE., Crystal, RC., Pizzorno, G, Leavitt, J., Deisseroth, AB. Cancer Gene Therapy、6、90〜106、1999年)、アルギニンバソプレッシンプロモータ(Coulson, JM, Staley, J., Woll, PJ. British J.Cancer、80、1935〜1944、1999年)、E2fプロモータ(Tsukadaら、Cancer Res.、62、3428〜3477)、ウロプラキンIIプロモータ(Zhangら、Cancer Res.、62、3743〜3750、2002年)およびPSAプロモータ(Hallenbeck PL、Chang, YN、Hay, C, Golightly, D., Stewart, D., Lin, J., Phipps, S., Chiang, YL. Human Gene Therapy、10、1721〜1733, 1999年)がある。更にドイツ特許出願番号DE101 50 984.7に記載のYB−1依存型E2−後期プロモータは、本発明で使用可能なプロモータである。
【0130】
テロメラーゼプロモータはヒト細胞において極めて重要であることが知られている。故にテロメラーゼ活性はテロメラーゼ逆転写酵素遺伝子(hTERT)の転写制御を通じて制御されているが、上記遺伝子は酵素の触媒サブユニットである。テロメラーゼは85%のヒト腫瘍細胞で発現している。これに対し、大部分の正常細胞では発現していない。例外は生殖細胞と胚組織である(Braunstein, I.ら、Cancer Research、61、5529〜5536、2001年; Majumdar, A.S.ら、Gene Therpy 8、568〜578、2001年)。hTERTプロモータに関するより詳細な研究から、開始コドンからの距離が283bpおよび82bpのプロモータの断片が、腫瘍細胞での特異的発現に十分機能することを示した(Braunstein I.ら、; Majunmdar ASら、上記)。故にこのプロモータおよび上記特異的断片はそれぞれ、遺伝子、特にトランス遺伝子、好ましくはここに開示するトランス遺伝子の一つを腫瘍細胞内でのみ特異的に発現させるのに好適である。このプロモータが、腫瘍細胞のみにおいて、発癌遺伝子、好ましくはE1A発癌遺伝子の発現を可能にする。また、好適実施態様では、かかるアデノウイルスベクター内でのトランス遺伝子、特にE4rof6、E1B55kD、ADPおよびYB−1を含む群から選ばれるトランス遺伝子の発現は、これらプロモータのいずれかの制御下にある。トランス活性化発現遺伝子タンパク質、特にE1Aタンパク質のオープンリーディングフレームがアデノウイルス系の一つ、または複数の遺伝子産物と共にフレーム内に存在するものも本発明の範囲内である。しかし、トランス活性化E1Aタンパク質のオープンリーディングフレームは、それから独立してもよい。
【0131】
ここに記載のアデノウイルスを発明に従い用いて溶解するのに好適である細胞の特徴は、実施態様の一つでは、それらは耐性であり、好ましくは多剤または多重耐性である。ここで言う耐性とは、ここに記した細胞増殖抑制薬に対する耐性を意味するのが好ましい。この多剤耐性は、該当する細胞を決定するためのマーカーとして利用でき、従ってかかる多剤耐性を示す腫瘍および患者群について利用できる膜結合型輸送タンパク質P−糖タンパク質の発現、好ましくは過剰発現と共に現れる。ここで使用する場合の耐性という用語は、古典的耐性とも呼ばれるP−糖タンパク質介在耐性と、MRP介して伝達される耐性もしくはその他非P−糖タンパク質が介在する耐性を含む非定型耐性の両方を含む。YB−1の発現と相関するさらなるマーカーは、トポイソメラーゼIIアルファである。その場合、患者が発明のアデノウイルスの使用により良好に処置できるか否かを決定するスクリーニングに於いて、トポイソメラーゼIIアルファの発現は核内のYB−1の決定に代わる方法または追加の方法として使用できる。P−糖タンパク質と同様の様式で基本的に用いることができるさらなるマーカーはMRPである。少なくとも結腸直腸癌細胞または結腸直腸癌患者について想定されるさらなるマーカーは、例えばShibao K.ら、(Shibao Kら、Int, Cancer, 83、732〜737、1999年)が記載したようにPCNA(増殖細胞核抗原)である(Hasan S.ら、Nature、15、387〜391、2001年)。最後に、MDR(多剤耐性)の発現は、少なくとも乳ガンおよび骨肉腫細胞については、上記の意味でのマーカーである(Oda Yら、Clin.Cancer Res.、4、2273〜2277、1998年)。本発明に従い用いることができる可能性のあるさらなるマーカーはp73である(Kamiya、M.、Nakazatp, Y.、J Neurooncology 59、143〜149(2002年); Stieweら、J.Biol.Chem.、278、14230〜14236、2003年)。
【0132】
従って本発明の特別な利点は、臨床的な意味に於いてもはや処置できないと考えられる患者、および先行技術の方法による腫瘍疾患のそれ以上の処置が不可能であり、合理的に成功が期待できない患者、特に細胞増殖抑制薬の使用がもはや合理的には不可能であり、腫瘍に影響を及ぼすまたは腫瘍を縮小させるという意味でもはや良好に治療が実施できない患者についても、ここに記載の様にして本発明によるアデノウイルスを用いることで治療できることである。ここでは、腫瘍という用語は一般に核内にもともとYB−1を含んでいる、またはここに記載した様な外的な開放手段によって核内に、好ましくは細胞周期とは無関係にYB−1を含むようになった腫瘍または癌疾患を意味する。
【0133】
更に、ここに記載のウイルスは、一般的な腫瘍の処置にも用いることができる。好ましくは、これら腫瘍は、乳ガン、卵巣癌、前立腺癌、骨肉腫、神経芽腫、メラノーマ、小細胞肺ガンおよび結腸直腸癌を含む群より選ばれる。更なる腫瘍はここに記した様な耐性の腫瘍であり、好ましくは多重耐性である腫瘍であり、特には上記の群の多重耐性の腫瘍である。
【0134】
発明は更に別の局面では、修飾型アデノウイルスの一つ、即ち例えばAdΔ24、dl922−947、E1Ad/01/07、CB016または欧州特許第EP0 931 830号に記載のウイルスの様な本発明に使用するアデノウイルスの一つを用いて処置できる患者をスクリーニングする方法であって;
−腫瘍組織のサンプルを検査するステップ、および
−YB−1が細胞周期とは無関係に核内に局在しているか決定するステップ、を含む方法に関する。
【0135】
前記マーカーの存在はYB−1に代わるものとして、またはYB−1に追加するものとして検出できる。
【0136】
腫瘍組織またはその一部がYB−1を核内に含む場合、特に細胞周期とは無関係に含む場合、ここに開示したアデノウイルスは本発明の実施に従い用いることができる。
【0137】
本発明の方法の実施態様では、腫瘍組織の検査はYB−1に対する抗体、YB−1に対するアプタマー、YB−1に対するシュピーゲルマー(spiegelmer)、ならびにYB−1に対する抗カリン(anticaline)を含む群より選ばれる作用物質を用いることで実施される。基本的には対応するマーカカーについては同一の手段を作り、それに従い用いることができる。抗体、特にモノクローナル抗体の製造方法は当業者周知である。YB−1またはマーカーを特異的に検出するさらなる手段は、標的構造、本例ではYB−1または上記マーカーに高い親和性で結合するペプチドである。そのようなペプチドを作製するためにファージディスプレーのような周知方法が知られている。典型的には、開始時に個々のペプチドが8〜20アミノ酸の長さを持ち、ライブラリーのサイズが102〜1018、好ましくは108〜1015種類のペプチドであるペプチドライブラリーを準備する。標的分子に結合するポリペプチドの特殊な形態は抗カリンと呼ばれ、例えばそれはドイツ特許出願番号DE197 42 706号に記載されている。
【0138】
YB−1またはここに開示した対応するマーカーとの特異的結合に関する、従って細胞周期と無関係な細胞核内のYB−1局在を検出するためのさらなる手段は、いわゆるアプタマー、即ち単鎖または二本鎖としてRNAまたはDNAの形で存在し、標的分子に特異的に結合するD−核酸である。アプタマーの作成は、例えば欧州特許第EP0 533 838号に記載されている。特殊な形状をしたアプタマーはアプタザイムと呼ばれ、これは例えばPiagenau, N.ら(2000年)、Angew.Chem.Int.Ed., 39、no.29、4369〜4373項に記載されている。それらがアプタマー部分とは別にリボザイム部分を含み、アプタマー部分と結合している標的分子との結合または放出によって触媒活性を獲得し、核酸基質を開裂してシグナルを生成するという意味において、アプタマーの特別な実施態様である。
【0139】
さらなる形状のアプタマーはいわゆるシュピーゲルマー、即ちL−核酸から作られている標的分子結合核酸である。かかるシュピーゲルマーの製造方法は、例えば国際特許出願第WO98/08856号に記載されている。
【0140】
腫瘍組織のサンプルは、穿刺または手術により得ることができる。YB−1が細胞周期とは無関係に核内に局在しているか調べることは、顕微鏡技術および/または免疫組織分析、好ましくは抗体または上記その他手段を使って頻繁に実施される。核内のYB−1を検出する、そして特にYB−1が細胞周期とは無関係に核内に局在しているか検出するためのさらなる手段は当業者に周知である。例えば、YB−1の局在は、スクリーニング時に染色組織切片で容易に検出できる。核内にYB−1が存在する頻度は、局在が細胞周期とは無関係であることを既に示している。細胞周期とは無関係な核内YB−1を検出するための更なる選択肢は、YB−1を染色し、YB−1が核内に局在しているか検出し、そして細胞のフェーズを決定することである。この方法およびYB−1の検出は、YB−1を検出するための上記手段を用いて実行してもよい。これら手段は、当業者周知の方法により検出される。前記のYB−1と特異的に結合し、分析対象サンプル内、特には細胞内のその他構造には結合しない作用物質を用いることにより、これら手段を好適に標識する方法を用いてそれらの局在を、そしてそれらの持つYB−1への特異的結合故にYB−1の局在をも検出そして確立することができる。前記手段を標識する方法は、当業者周知である。
【0141】
以下、本発明を新規特徴、実施態様および利点を示す図面およびサンプルを参照しながら更に説明する。
【図面の簡単な説明】
【0142】
【図1】本明細書でAdE1/E3−マイナスと呼ばれる、E1/E3欠失アデノウイルスであるアデノウイルベクター、野生型アデノウイルスおよびアデノウイルスdl520の構造デザイン。
【図2】p300、p107およびp105の結合に関する、E1Aタンパク質の結合ドメイン。
【図3】E1/E3マイナスAd5と呼ばれるE1/E3欠失アデノウイルスAd5およびアデノウイルスdl520感染後の、核内にYB−1を持たないU20S細胞。
【図4】E1/E3マイナスAd5と呼ばれるE1/E3欠失アデノウイルスAd5およびアデノウイルスdl520感染後の、核内にYB−1を有する257RDB細胞。
【図5】アデノウイルスdl1119/1131感染後の257RDB細胞およびU2OS細胞。
【図6】多剤耐性細胞および細胞株257RDB、181RDB、MCF−7Ad内にそれぞれYB−1が存在するが、U2OSおよびHeLa細胞の核内にはYB−1が存在しないことを確認したEMSA分析の結果。
【図7】野生型アデノウイルス、アデノウイルスdl520およびアデノウイルスdl1119/1131のE1Aタンパク質の構造デザイン。
【図8】追加発現したウイルスタンパク質存在時のアデノウイルスの複製効率を絶対量で示した棒グラフ。
【図9】追加発現したウイルスタンパク質存在時のアデノウイルスの複製効率を増加率で示した棒グラフ。
【図10】ダウノルビシンを加えていない、およびダウノルビシン40ng/mlを加えた場合の、10および30pfu/細胞のdl520、およびコントロール(K)をそれぞれ感染させクリスタルバイオレット染色した後のU2OS細胞増殖ウエルを示す図。
【図11】ダウノルビシンを加えていない、およびダウノルビシン40ng/mlを加えた場合の、10および30pfu/細胞のdl5210、およびコントロール(K)をそれぞれ感染させクリスタルバイオレット染色した後のHeLa細胞増殖ウエルを示す図。
【図12】PBSおよびdl520でそれぞれ処理後の、起源(RDB257およびHeLa)の異なる腫瘍の腫瘍容積の経時変化を示すグラフ。
【図13】それぞれPBSおよび5×108pfuのdl520で処理した後に、RDB257細胞由来の腫瘍を発生した屠殺マウスの図。
【図14】dl520感染後のRDB257細胞およびHeLa細胞の細胞抽出物(皮下増殖した腫瘍)のサザンブロット分析の結果。
【図15】YB−1核陽性腫瘍細胞(257RDBおよび181RDB)およびYB−1核陰性腫瘍細胞(HeLa、U2OS)でのdl520および野生型アデノウイルスの複製効率および粒子形成を示す棒グラフ。
【図16】野生型アデノウイルスおよびアデノウイルスベクターAdXvir03の構造デザインを示す図。
【図17】アデノウイルスベクターAdXvir03/01の構造デザインを示す図。
【図18A】Ad312(20pfu/細胞)、Xvir03(5pfu/細胞)およびコントロール(非感染)に感染させ、クリスタルバイオレット染色を行った181RDB細胞(図18A)および272RDB細胞(図18B)増殖ウエル。クリスタルバイオレット染色は感染5日後に実施した。
【図18B】Ad312(20pfu/細胞)、Xvir03(5pfu/細胞)およびコントロール(非感染)に感染させ、クリスタルバイオレット染色を行った181RDB細胞(図18A)および272RDB細胞(図18B)増殖ウエル。クリスタルバイオレット染色は感染5日後に実施した。
【実施例1】
【0143】
発明により使用されるアデノウイルスが含むE1A修飾のタイプ
図1はAdE1/E3マイナス、即ちE1/E3欠失アデノウイルスと野生型アデノウイルスおよびアデノウイルスdl520の構造デザインを示している。
【0144】
アデノウイルスAdE1/E3マイナスは機能的E1Aまたは機能的E1BもしくはE3をコードする領域を有しておらず、本実験では毒性に関するコントロールに用いられる。
【0145】
野生型E1A遺伝子は、E1A RNAの異なる位置でのスプライシングから生じる合計で5種類のタンパク質をコードしている。とりわけ2種類のタンパク質、即ち289アミノ酸タンパク質および243アミノ酸タンパク質が生成される。dl520はE1A遺伝子のCR3ストレッチ内に欠失を持つために13S遺伝子産物を欠いているため、289アミノ酸タンパク質をコードしていない。発明に用いられるアデノウイルスdl520は当業者からは12S−E1Aウイルスと呼ばれる。先行技術周知のアデノウイルスdl347(WongおよびZiff、J.Virol.、68、4910〜4920、1994年)はまた12S−E1Aでもあり、これは本発明に使用することができる。
【0146】
13S−E1AのmRNAによりコードされる289アミノ酸タンパク質の中には、各種アデノウイルスサブタイプに保存されている3つの領域がある。これらはCR1、CR2およびCR3と呼ばれる。CR1およびCR2は両E1Aタンパク質(E1A 12SおよびE1A 13S)、即ち289アミノ酸および243アミノ酸タンパク質に存在するが、CR3領域は上記タンパク質のうちの大きい方だけに存在する。
【0147】
CR3領域はウイルス遺伝子、特にE1B、E2、E3およびE4の活性化に必要である。小型、即ち243アミノ酸タンパク質のみを含むウイルスは、極弱くしかウイルス遺伝子をトランス活性化できず、核内にYB−1を持たない細胞でのアデノウイルス複製を促進しない。YB−1は腫瘍細胞でのみ核内に存在すること、そしてその中にのみ見出すことができることから、このベクターは腫瘍特異的複製の誘導に好適である。
【0148】
dl520はCR3を欠失していることから、このアデノウイルスは本明細書で輸送とも呼ばれる細胞質YB−1の細胞核内への移動を行うことができず、それ故にYB−1核陰性である細胞では複製できないことから、このウイルスは本発明で使用可能なウイルスであり、そしてこのウイルスは本発明に必要なトランス活性化に使用できる。
【実施例2】
【0149】
細胞のRb状態に依存したアデノウイルスの作用様態
図2はp300,p107およびp105の結合に関係するE1Aタンパク質の結合ドメインを示している。P300およびP107は、細胞質結合タンパク質である。腫瘍抑制タンパク質である網膜芽細胞腫タンパク質(pRb)の結合は、CR1およびCR2を通じて媒介される。研究からpRbおよびp107/p300は細胞転写因子E2Fと組み合わさり効率的に転写制御することが示されている。野生型E1Aタンパク質はE2FのRbへの結合を妨害する。即ち放出されたE2FはE2初期プロモータに結合して、アデノウイルスの複製を誘導する。
【0150】
E1A発癌タンパク質内の特定の欠失がRb−陰性細胞で優先的に複製することが可能であり、そして本発明に用いることができる以下に述べるような組換え体アデノウイルスベクターを生成することが従来知られていた。例えばアデノウイルスベクターdl922−947は、CR2領域(アミノ酸位置122〜129)内に欠失を持ち、ベクターCB016はCR1(アミノ酸位置27〜80)およびCR2領域(アミノ酸位置122〜129)に欠失がある。ベクターE1Adl01/07はCR2領域(アミノ酸位置111〜123)内に欠失を持つ。更に、N−末端(アミノ酸位置4〜25)に追加の欠失があることから、タンパク質p300への結合は認められない。アデノウイルスベクターAdΔ24はCR2領域(アミノ酸位置120〜127)に欠失を持っている。欧州特許第EP0 931 830号に記載のアデノウイルスベクターはCR1領域よびCR2領域に欠失を含んでいる。
【0151】
E2F/RPの結合メカニズムおよびE1Aを媒介するE2Fの放出は、本発明の基礎を成すメカニズムとは基本的に異なるものである。先行技術での予想とは異なり、ウイルス複製にとって必須ではないが、重要であるのはRbタンパク質からのE2Fの放出ではなく、ヒト転写因子であるYB−1が核内に局在することである。この転写因子は、正常細胞に於いては、細胞周期の大部分について細胞質内にのみに存在している。アデノウイルスが感染すると、特定の条件の下にYB−1は核内に導かれるか、例えば乳ガン、卵巣癌、前立腺癌、骨肉腫、神経芽腫、メラノーマ、小細胞肺癌および結腸直腸癌を含むが、もとよりこれらに限定されないある種の腫瘍疾患といったある種の細胞システムでは核内に既に存在している。
【実施例3】
【0152】
U2OS細胞の感染
100,000個のU2OS細胞をウエルに播いた。翌日細胞に図3に示すような各種アデノウイルスを感染させた。感染は無血清DMEM培地500μl中で、37℃で1時間行った。続いて感染培地を取り除き、完全培地(10%FCS/DMEM)2mlと交換した。3日後にクリスタルバイオレット染色で分析を行った。
【0153】
図3から分かるように核内にYB−1を持たないU2OS細胞は、本発明に使用できる2種類のアデノウイルス、即ちE1/E3マイナスと呼ばれるE1/E3欠失ウイルスおよびアデノウイルスdl520で感染させた後にクリスタルバイオレット染色を行ったところ、溶解を示していなかった。この場合、まず培地を取り除いた。続いて細胞にクリスタルバイオレット液(50%ETOH、3%ホルムアルデヒド、5%酢酸、1%クリスタルバイオレット)を重層して室温で5〜10分間インキュベーションした。続いて6ウエル型プレートを水でよく濯いでから室温で乾かした。これから、本発明で使用するウイルスを誘導し、感染細胞を溶解させるには、YB−1の存在が必要であるという、本発明の基礎を成す発見が確認された。
【実施例4】
【0154】
257RDB細胞の感染
100,000個の257RDB細胞をウエルに播いた。翌日細胞に図4に示すような各種アデノウイルスを感染させた。感染は無血清DMEM培地500μl中で、37℃で1時間行った。続いて感染培地を取り除き、完全培地(10%FCS/DMEM)2mlと交換した。3日後にクリスタルバイオレット染色で分析を行った。
【0155】
この実験の結果を図4に示す。E1/E3を欠失したE1/E3マイナスと呼ばれるアデノウイルスは、核内にYB−1を持つ257RDB細胞への感染については、低MOI(pfu/細胞)では溶解を示さなかった。これに対し実施例3で示した、YB−1核陰性細胞では複製せず、同時に本発明に従い発癌遺伝子タンパク質をトランス活性化するE1Aをコードしているdl520は、40pfu/細胞というMOI(感染多重度)で実質完全な溶解をおこし、10pfu/細胞のMOIでも顕著な溶解を起こした。このことから、dl520およびここに記したdl1119/1131またはAdXvirといった同様のウイルスが必要とするMOIは、E1欠失またはE1/E3欠失アデノウイルスに比べ約1桁(10倍)低く、臨床使用が可能であると結論される。
【0156】
図7に示すように、dl520のタンパク質E1AはそのCR3領域を欠失することを特徴とし、その結果本発明による使用に求められるトランス活性化およびYB−1核陽性細胞での複製を起こす。
【実施例5】
【0157】
dl1119/1131による257RDBおよびU2OS細胞の感染
図5に示すように、E1Aタンパク質のアミノ酸4〜138およびそれをコードする核酸を欠失しており、さらにアミノ酸218の後に停止コドンを含むために発現する断頭型(truncate)E1Aタンパク質が完全なE1Aタンパク質のCR3領域を含んでいるアデノウイルスdl1119/1131をYB−1核陰性U2OS細胞に感染した場合、20pfu/細胞のMOIでは溶解は起きなかった。陰性コントロールには非感染細胞層を用いた。
【0158】
これに対し核内にYB−1を含む、即ちYB−1核陽性である、257RDBの様な細胞系では、アデノウイルスdl1119/1131の影響を受けて、20pfu/細胞のMOIで細胞層は実質的に完全に溶解した。この限りにおいて本実施例は、図7に示す様に例えばCR3領域のみを含み、そしてCR1領域およびCR2領域を欠いている修飾型E1A発癌遺伝子タンパク質が、発明によるアデノウイルスの複製に必要なYB−1核陽性細胞でのトランス活性化を提供していることを示す別の証拠である。即ちアデノウイルスdl1119/1131は、本発明に使用可能である別のアデノウイルスである。CR3領域に関してdl1119/1131と同様にデザインされているが、それと異なりCR1領域および/またはCR2領域も含んでいるウイルスを使用できることも、本発明の範囲内である。
【実施例6】
【0159】
多剤耐性細胞での核内YB−1の検出
本実施例は核内YB−1が転写因子としてmdr1プロモータ(多剤耐性プロモータ)内にあるY−ボックス(CAAT配列)に結合しなければならないという考察に基づいている。これを検出するために、いわゆるEMSA分析(電気泳動移動度シフトアッセイ)を行った。これに関連して、核タンパク質を単離し、続いてタンパク質1〜10μgを短いDNA鎖(オリゴ)と共の37℃でインキュベーションした。核内YB−1を決定するために以下のオリゴヌクレオチドを使用した;U2O3と反対のmdr1プロモータ(位置−86〜−67):TGAGGCTGATTGGCTGGGCA(Xボックスには下線を付した)。
【0160】
その前にこのDNA断片の5’末端を32Pで放射線標識した。続いて未変性ポリアクリルアミドゲルを用いて分離した。タンパク質YB−1がオリゴヌクレオチド内の配列に結合すると、その結合は、結合したオリゴヌクレオチドに比べて非結合のオリゴヌクレオチドはゲル中をより早く移動することから検出できる(Holm, P.S.ら、JBC 277、10427〜10434、2002年; Bargou、R.C.ら、Nature Medicine 3、447〜450、1997年)。
【0161】
図6に示すように、EMSA分析よりYB−1が多剤耐性細胞257RDB、181RDBおよびMCF−7Ad細胞の核内に存在すること、そして細胞株U2OSおよびHeLa細胞には存在しないことが分かる。
【0162】
実施例4および5に示した結果は、アデノウイルスdl520およびdl1119/1131が257RDBの様なYB−1核陽性細胞では複製してその溶解を誘導するが、U205では複製、溶解を誘導しないことを確認した。このことは、本発明のアデノウイルスの使用に関する所見が確認された。さらに結果は、野生型のアデノウイルスと比べて、修飾型または欠失型E1A遺伝子産物を介したYB−1核陽性細胞でのウイルス遺伝子の弱いトランス活性化が、例えば多剤耐性細胞を含む核内にYB−1が存在する細胞での良好な複製および良好な細胞溶解をもたらすこと、そしてここに記したアデノウイルスがかかる腫瘍の溶解に利用できることを確認している。
【実施例7】
【0163】
E1−マイナスアデノウイルスの複製効率上昇
本実施例は初期ウイルス遺伝子E1B−55KおよびE4orf6をプラスミドpE4orf6によるトランスフェクションおよびE1/E3欠失アデノウイルスAd−55Kによる感染で置換えられることを示す。Ad−55KはE1B−55KをE1にクローニングし、CMVの制御下に置いたE1/E3欠失ウイルスである。この置換は、AdYB−1、即ちYB−1を発現するアデノウイルスがこれら初期遺伝子を発現していないという事実、および本発明者が核内にYB−1を含む複製系でのこれら初期遺伝子を置換することで複製効率および粒子形成能率をそれぞれ野生型アデノウイルスであるAd5型のそれと同程度まで上げることができることを認識していたという事実より必要であった。
【0164】
以下を実施した:
リポフェクタミンを用いた各105個のU2OS細胞へのプラスミドpE4orf6のトランスフェクション。プラスミドpEorf6はCMV制御下にある初期ウイルス遺伝子E4orf6をコードするDNA配列を保持している。プラスミドpE4orf6をトランスフェクションした24時間後に、細胞にYB−1発現E1/E3欠失アデノウイルスAdYB−1(50pfu/細胞)およびE1/E3欠失E1B−55KアデノウイルスAd−55K(50pfu/細胞)を感染させた。Ad−55KはCMV制御下にあるウイルス遺伝子E1B−55Kをトランス遺伝子として保持するE1/E3欠失ウイルスである。
【0165】
続いて感染5日後(=トランスフェクション後)に細胞を培地(2ml)から取り出した。凍結と融解(凍結/融解)を交互に3回行って、単離細胞からのウイルス粒子を放出させた。次にプラークアッセイを293細胞を使って実施し、生成した感染粒子を決定した(プラーク形成単位/ml(pfu/ml))。結果は図8および9に示す。図8は絶対量で表した場合のプラークアッセイの結果である。AdYB−1単独感染例と最も大きな差は、プラスミドpE4orf6を感染させた例および2種類のウイルスAdYB−1およびAd−55Kを同時感染させた例との間で観察された。図9は図8の結果を、AdYB−1の複製効率の倍率として複製効率の増加の形で表したものである。細胞にプラスミドpE4orf6を感染させた後AdYB−1およびE1B−55K(Ad−55K)を感染させると、最大25倍高いpfu/mlを示した。
【0166】
これらの結果より、E1B−55KおよびE4orf6による置換はE1/E3−欠失アデノウイルスAdYB−1感染後に形成されるウイルス数(pfu/ml)を最大25倍増加すると結論できる。プラーク形成単位(pfu)生成に及ぼすE1B−55KおよびE4orf6の追加効果は、これら2遺伝子産物単独の効果に比べて有意に高かった。
【0167】
EGFPを発現するプラスミドを使ったコントロール実験は、選択した実験方法では、プラスミドpE4orf6でトランスフェクションされたのは僅かに細胞の10%だけであることを明瞭に示した。E1B−55KおよびE4orf6の両方を発現する細胞で形成された粒子の数は、ヒトアデノウイルス5型(野生型)のそれに匹敵した。このことから、E4orf6およびE1B−55Kの発現は、YB−1の核内局在と組み合わさると、アデノウイルス、特にE1A−欠失アデノウイルスの複製および粒子形成を、野生型Ad5のそれに匹敵する程度まで誘導できるという、本発明の基礎を成す発見が確認された。
【実施例8】
【0168】
細胞増殖抑制薬投与によるYB−1核陰性細胞では複製しないアデノウイルスのYB−1核陽性細胞での複製増加
各種細胞増殖抑制薬の添加がヒト転写因子YB−1の核内局在化を誘導することは、先行技術において知られていた。本発明者が見出したように、核内に局在したYB−1は、アデノウイルスのE2−後期プロモータの活性化によりアデノウイルスの複製をコントロールする。両効果の組み合わせを利用すれば特異的な腫瘍溶解を得ることができる。
【0169】
癌溶解アッセイを実施するに当たっては、以下の手順に従った:200、000個の細胞(それぞれHeLaおよびU2OS)を6ウエル型プレートの核ウエル内に接種した。翌日ダウノルビシン40ng/ml(最終濃度)を加えた。3時間インキュベーションした後、それぞれの細胞をdl520で、10〜30pfu/細胞の割合で感染させた。続いて細胞を、細胞増殖抑制薬を含まない培地でインキュベーションした。3〜5日後、細胞をクリスタルバイオレットで染色した。
【0170】
図10および11から分かるように、ダウノルビシンの添加はYB−1の核内局在化を通じてdl520の複製を誘導した。即ちdl520は、細胞増殖抑制薬と組み合わさった状態で、ダウノルビシン単独例に比べてより大きな癌溶解作用を発揮した。
【実施例9】
【0171】
dl520によるインビボの癌溶解
本インビボ試験で使用したHeLa(YB−1核陰性)および257RDB(YB−1核陽性)細胞は、無菌細胞培養条件にて拡大した。細胞をマウス(系統CD1NuNu)に注射して皮下腫瘍を形成させる前に、細胞をトリプシン処理して集め、DMEM培地(10%FCS)に加え、細胞数を測定してからPBSで1回洗浄した。続いて細胞を遠心分離してPBSを除き、細胞を希望数する細胞数になるよう新鮮PBS中に分配した。本研究で皮下注射した細胞の数は両細胞系統とも5×106細胞であった。注射は動物の一方の側腹部に行い、より区別し易いようにHeLa細胞は右側に、そして257RDB細胞は左側に注射した。腫瘍の増殖を週2回調べ、ノギスを使って腫瘍の長さおよび幅を測定した。その結果を用い、次式より腫瘍の容積を計算した:
3/4π*a/2*(b/2)2 a=長さ、b=幅
【0172】
腫瘍の大きさが200〜520mm3に達した時点で、ウイルスおよび陰性コントロールであるPBSをそれぞれ腫瘍内に投与した。注射量は同一で、1回50μlであった。この作業を3連続日繰り返した。投与したウイルスの総量は5×108pfuであった。次に腫瘍の増殖を毎週2回の割合で記録し続け、その容積を計算した。試験最終日にマウスを屠殺して、腫瘍を取り出し詳しく分析した。
【0173】
結果は図12および13に示した。
【0174】
図12は、腫瘍容積の経時変化を処理の種類別に示したグラフである。腫瘍がRDB257により形成された例では、PBS注射時に腫瘍は約438mm3から1466mm3へと有意に増大した。本発明に従い用いられるベクターdl520を作用させると、腫瘍の成長は有意に低下した。平均腫瘍サイズ344mm3から総容積543mm3へと、腫瘍サイズは僅かに21%増加しただけであった。
【0175】
本実施例では、RDB257を基本とする腫瘍にPBSを投与したのと同様にPBSを投与したHeLa細胞より成る腫瘍をコントロールとして用いた。HeLa細胞を基本とする、dl520で処理された腫瘍は、まだ有意な腫瘍成長の増加を示し、311mm3から1954mm3に増加した。
【0176】
図13はRDB257を用いて増殖した腫瘍を持つ、屠殺したヌードマウスの像である。本発明のアデノウイルスdl520を作用させた後に、腫瘍の顕著な縮小が起こったことが明瞭に分かる。本例では、腫瘍容積の減少も見られた(ウイルスdl520投与1日後:515mm3;ウイルスdl520投与30日後:350mm3)。
【実施例10】
【0177】
腫瘍DNAのサザンブロット
実施例9に於いて、発生中の腫瘍の中央部から採取した腫瘍サンプルよりDNAを抽出した。抽出にはQiagen社のDneasy Tissueキットを用いた。DNAの単離は製造元の指示書に従い実施した。それによれば、DNAは細胞からアルカリ溶解により放出された。続いて単離したDNAはカラムを使って精製される。次に単離したDNAの濃度を260nmの光度測定より決定した。分析はDNAサンプル2μgを用いて行い、サンプルは制限酵素Kpn I、10単位で消化した。次にサンプルを0.8%のアガロースゲルを用いて電気泳動により分離した。続いてDNAをナイロン膜上にブロッティングした(Schleicher & Schuell社のシステムを用いて実施)。膜上にブロッティングされたDNAを特異的な1501bpのDNAプローブとハイブリダイゼーションさせた。上記1501bpのDNAプローブは、Ad5配列をコードするE2A内にある3369bpのKpn I断片と特異的に結合する。プローブはPCR(プライマー:5’−GTC GGA GAT CAG ATC CGC GT、5’−GAT CCT CGT CGT CTT CGC TT)により準備し、32Pを用いて射線標識した。次に膜を洗い、フィルムに感光した。
【0178】
腫瘍DNAのサザンブロットの結果は図14に示した。分析より、レーン3、4および5が示すように、dl520だけが耐性細胞RDB257でインビトロ複製することが確認された。レーン1は陽性コントロールのAd−5dを、レーン6、7、および8はdl520を感染したHeLa細胞由来のDNAを示す。HeLa細胞はYB−1核陽性ではないため、ウイルスdl520は複製せず、そのためE2A配列は検出できなかった。
【0179】
dl520に関するさらなる結果を図15に示した。プラークアッセイを利用して、dl520および野生型のアデノウイルスを感染させた後の粒子形成(pfu/ml)を調べた。各種YB−1核陽性(257RDBおよび181RDB)腫瘍細胞、ならびにYB−1核陰性腫瘍細胞にdl520および野生型アデノウイルスを感染させた。
【0180】
次の手順で実施した:
100、000〜200、000個の各細胞を6個のウエルを持つプレート(6ウエルプレート)、FCSを10%含むL15培地(耐性細胞)およびDMEM(非耐性細胞)に接種した。24時間後にdl520および野生型アデノウイルスを感染させた(10pfu/細胞)。感染3日後(インフェクション後)、凍結と融解を3回繰り返して細胞浮遊液よりウイルス粒子を放出させた。次に293細胞を使ってプラークアッセイを行い、形成した感染粒子(プラーク形成単位/ml(pfu/ml))を決定した。結果を図15に示す。プラークアッセイの結果は、dl520がYB−1核陽性細胞(257RDBおよび181RDB)では野生型アデノウイルスの場合と同様に複製することを示した。この限りにおいて、本発明に従ってここに記載のアデノウイルスを使用する場合には、野生型アデノウイルスと同様の複製効率が観察できた。
【実施例11】
【0181】
アデノウイルスベクターXvir03の構造デザイン
図16はアデノウイルスベクターXvir03の構造デザインを示す。アデノウイルスXvir03は、いわゆるE1/E3欠失アデノウイルスである。このことは、アデノウイルス複製に機能するE1A、E1BおよびE3タンパク質が作られないことを意味する。E1領域の欠失は342〜3528の範囲である;アミノ酸位置27865〜30995のE3領域の欠失。本明細書で使用する場合、用語「E1欠失ウイルス」とは、E1がもはや機能的に作用しないウイルスを意味する。これは、その他の核酸配列およびアミノ酸配列は未変性な状態で不活性化することにより達成できるが、しかしこのことは欠失したE1領域がコードするタンパク質が様々なサイズを持つことを意味することにもなる。E1AおよびE1Bタンパク質およびそれらをコードする核酸を欠くことにより、E4orf6の様なE4領域は弱くにしか発現されないか(野生型アデノウイルスの1〜5%)、または全く発現されない。ウイルス遺伝子E1B55kおよびE4orf6はXvir03内に導入された異種のCMVプロモータ(Clontech: Plasmid pShuttle)によりE1領域内で発現する。上記CMVプロモータに代わって、E1Aの発現に関連して明細書中に開示したその他プロモータを用いることができる。両遺伝子のオープンリーディングフレームは、いわゆるIRES配列(内部リボソーム進入部位)(Plletier, J.およびSonenberg, N.Nature、1988年、334、320〜325)により互いに連結している。このエレメント(Novagen:pCITE)は1種類のmRNAから2種類のタンパク質を発現させる。
【0182】
ベクターは次のように作製した:
プラスミドE1B55k−pShuttleを、M.Dobelstein(マールブルグ大学)のpCGNE1Bより、XbaIおよびBfrIを用いてE1B55kのオープンリーディングフレームをClontech社製のpShuttleベクター内にクローニングして作製した。続いてpShuttle内のE1B55kをApaIを使って直線化し、末端を平滑末端とし、NheIを使って切断した。
【0183】
第二ベクターであるpcDNA3.1(+)(Invitroge)の場合は、それぞれに続いて、Novagen社製のpCITE−4a(+)を鋳型に用いTAクローニング法によりEcoRV切断部位にPCR産物であるIRESエレメントをクローン化し、そしてE4orf6はプラスミドpCMV−E4orf6(M.Dobelstein、マールブルグ大学)よりBamHIを用いてクローニングした=IRES−E4orf6−pcDNA3.1(+)。pcDNA3.1(+)内のIRES−E4orf6はNotIを使って直線化し、端部を平滑末端化してから断片IRES−E4orf6をNheIで切断した。断片IRES−E4orf6をオープンベクターE1B55k−pShuttle(平滑端、NheI)と連結した。続いてこうしてできたカセットを、I−CeuIおよびPI−SceIを用いて、E1B55k−IRES−E4orf6−pShuttleより、CMVプロモータおよびウシ成長ホルモン(BGH)−ポリAと共にΔE1、ΔE3Adeno−X−プラスミド(Clontech)内にクローニングし、AdcmvE1B/IRES/E4orf6と名付けた。次にアデノウイルスをメーカー(Clontech)指示書に従い作製した。PacIを使って直線化した、発現エレメントCMV−E1B55k−IRES−E4orf6−BGHポリAを有するアデノプラスミドをHEK293細胞内にトランスフェクションし、トランスフェクション11日後に剥離細胞を培地と共に取り除いてから凍結乾燥を繰り返しアデノウイルスを放出させた。
【0184】
上記ベクターは原理的には本発明での使用に関して、本明細書に記載したその他ウイルスと同様に好適である。具体的には、上記ベクターはYB−1核陽性細胞およびYB−1が脱制御状態にある細胞、即ちYB−1が正常細胞および非腫瘍細胞に比べ過剰発現している細胞での複製および溶解始動について好適である。このベクターの使用は、特に本発明に従い用いられるものとして本明細書に記載されているその他アデノウイルス、ならびに本明細書に開示されている本発明のその他アデノウイルスに関連し開示されている疾患、患者群または患者集団を適用とする。
【実施例12】
【0185】
アデノウイルスベクターXvir03/01の構造デザイン
図17から分かるように、Xvir03/01はXvir03を更に発展させたものである。例えばここに記した遺伝子の様な治療目的の遺伝子やトランス遺伝子をE3領域内にクローニングできる。さらに、E4領域内に欠失を導入してXvir03の発現カセットに由来するE4orf6との間で相同的組換えが起こるのを回避した。これによってより大型のトランス遺伝子をこの構築体内にクローニングすることが可能になった。欠失型のE3領域にはカセット導入に適したSacI、NdeIおよびNheI制限部位があり、ここに例えば治療用トランス遺伝子をクローニングすることができる。
【0186】
E3領域内への治療用遺伝子クローニングおよびE4領域内への欠失作製を目的としたプラスミドの調製:
Clontech社製pAdenoX−Plasmidには3’ITR領域の後に野生型アデノウイルスには無いSfuI向けの制限部位がある。E3〜E4領域をSpeI(位置23644)およびSfuIを使ってpAdenoX(Clontech)から取り出し、pcDNA3.1(+)(Invitrogen)内にトランスフェクションした=pcDNA3.1−E3Δ27865−30995−E4。E4ORF6の大部分、即ち33241〜33875をPstIを用いて取り出した=pcDNA3.1−E3Δ27865−30995、E4Δ33241−33875。Xvir03を更に発展させるために、pcDNA3.1−E3Δ27865−30995、E4Δ33241−33875由来の欠失E3/E4領域をSfuIおよびSpeIを使ってプラスミドpAdenoX内にクローニングした=pAdnoXE3Δ27865−30995、E4Δ33241−33875。
【0187】
次にXvir03について記載した発現カセットを、I−CeuIおよびPI−SceIを使ってE1B55k−IRES−E4orf6−pShuttleより、CMVプロモータおよびウシ成長ホルモン(BGH)−ポリAと共にpAdenoXE3Δ27865−30995、E4Δ33241−33875内にクローニングし、AdcmvE1B/IRES/E4orf6−ΔE4と命名した。次にアデノウイルスをメーカー(Clontech)の指示書に従い作製した。
【0188】
上記ベクターは原理的にはここに記載した本発明に従い使用される他ウイルスと同様に有用である。特に上記ベクターはYB−1核陽性細胞ならびにYB−1が脱制御状態にある、即ちYB−1が正常細胞および非腫瘍細胞に比べ過剰発現している細胞での複製、およびその限りにおいて溶解の誘導に好適である。このベクターは、本発明に従い用いられる本明細書記載の他アデノウイルス、および本明細書のアデノウイルスに関し開示されている疾患、患者群および患者集団にも使用できる。
【実施例13】
【0189】
257RDBおよび181RDB細胞でのXvir03の癌溶解作用
100、000個の細胞(257RDBおよび181RDB)を6個のウエルを持つプレート(6ウエルプレート)の各ウエルに接種した。翌日細胞に、図18に示すようにAd312(20pfu/ml)およびXvir03(5pfu/ml)を感染させた。感染は無血清DMEM培地500μl中で、37℃、1時間実施した。次に感染培地を取り除き、完全培地(10%FCS/DMEM)2mlと交換した。5日後にクリスタルバイオレット染色による分析を行った。結果を図18Aおよび18Bに示す。
【0190】
図18Aおよび18Bから明らかな様に、核内にYB−1を持つ多剤耐性細胞では、Ad312およびXvir03を感染させた場合、細胞のクリスタルバイオレット染色より示されるようにXvir03感染例のみ崩壊を示した。この場合まず培地を取り除いた。続いて細胞をクリスタルバイオレット液(50%ETOH、3%ホルムアルデヒド、5%酢酸、1%クリスタルバイオレット)で覆い、室温で5〜10分間インキュベーションした。次に6ウエルプレートを水でよく濯いでから室温で乾かした。
【0191】
本発明者は、本発明の意味においてアデノウイルスをトランス活性化しないE1A欠失ウイルス(例えばAd312)がより高いMOIでは極めて効率的に複製できるが(Nevins J.R.、Cell 26、213〜220、1981年)、しかしこれらは臨床的には応用できないことを承知していた。この現象は文献では「E1A−様活性」と呼ばれている。ここで用いるアデノウイルスAd312はE1A欠失ウイルスである。使用した力価(20pfu/ml)はまだ臨床応用には高すぎるが、この力価ではE1B55kおよびE4orf6といった初期アデノウイルス遺伝子は発現しないか、または極少量しか発現しない(Nevins J.R.、Cell 26、213〜220、1981年)。本明細書に既に記した様に、これら遺伝子およびタンパク質はウイルス複製において重要な役割をはたしており、それぞれアデノウイルスXvir03により発現される(図16)図18Aおよび18Bから分かるように、遺伝子E1B55kおよびE4orf6の発現は、必要とする感染力価(pfu/mlで表される)の低下を伴いながら効率的にウイルス複製および細胞溶解を起こすだろう。このことから、本発明の基礎となる発見、即ちYB−1の核内局在と組み合わさったE4orf6およびE1B−55Kの発現(且つE1Aが存在しない)は、極めて効率的なアデノウイルスの複製を誘導できることが確認される。これに必要な力価は僅かに1〜5pfu/mlに過ぎず、臨床的応用が可能である。
【0192】
このことから、本発明の基礎となる発見、即ち本発明に従い用いられるウイルスを用いて感染細胞を溶解させるには、核内にYB−1が存在すること、特に細胞周期と無関係に存在することが必要であることが確認される。
【0193】
前記明細書に開示した発明の特徴、特許請求の範囲ならびに図面は、個々およびいずれかの組み合わせの両方に於いて、その発明の各種実施態様での実現にとって極めて重要であろう。
【特許請求の範囲】
【請求項1】
ウイルスが核内にYB−1がない細胞では複製できず、そのときウイルスが発癌遺伝子または発癌遺伝子産物、特に発癌遺伝子タンパク質をコードしており、それが少なくとも1つのウイルス遺伝子、好ましくはアデノウイルス遺伝子をトランス活性化し、その時の遺伝子がE1B55kDa、E4ort6、E4orf3およびE3ADPを含む群から選ばれることを特徴とする、医薬製造へのウイルス、好ましくはアデノウイルスの使用。
【請求項2】
ウイルスが核内にYB−1がない細胞では複製できず、そのときウイルスが発癌遺伝子または発癌遺伝子産物、特に発癌遺伝子タンパク質をコードしており、それが少なくとも1つのウイルス遺伝子、好ましくはアデノウイルス遺伝子をトランス活性化し、その時の遺伝子がE1B55kDa、E4ort6、E4orf3およびE3ADPを含む群から選ばれることを特徴とする、核内にYB−1を示す細胞内における複製のためのウイルス、好ましくはアデノウイルスの使用。
【請求項3】
ウイルス、好ましくはアデノウイルスが、核内にYB−1を有する細胞内で複製することを特徴とする、請求項1または2に記載の使用。
【請求項4】
ウイルス発癌遺伝子タンパク質がE1Aであり、そして/または発癌遺伝子がE1Aをコードする遺伝子であり、そして/または発癌遺伝子タンパク質がE1Aであることを特徴とする、請求項1〜3のいずれか1項に記載の使用。
【請求項5】
ウイルス発癌遺伝子タンパク質E1Aが、機能的Rb腫瘍抑制遺伝子産物を結合できることを特徴とする、請求項4に記載の使用。
【請求項6】
ウイルス発癌遺伝子タンパク質E1Aが、機能的Rb腫瘍抑制遺伝子産物を結合できないことを特徴とする、請求項4に記載の使用。
【請求項7】
ウイルス発癌タンパク質E1Aが、YB−1の核内局在化を誘導しないことを特徴とする、請求項4〜6のいずれか1項に記載の使用。
【請求項8】
医薬が、細胞がRb陽性であるか、またはRb陰性である患者向けであることを特徴とする、請求項1または3〜7のいずれか1項に記載の使用。
【請求項9】
細胞が、医薬により影響を受ける状態の形成に関係している細胞であることを特徴とする、請求項8に記載の使用。
【請求項10】
細胞がRb陰性であり、細胞核がYB−1陽性であり、好ましくは細胞周期とは無関係に核内がYB−1陽性であることを特徴とする、請求項1〜9のいずれか1項に記載の使用。
【請求項11】
医薬が腫瘍の処置向けであることを特徴とする、請求項1または3〜10のいずれか1項に記載の使用。
【請求項12】
細胞、好ましくは腫瘍またはその一部を形成する細胞が、医薬、好ましくは抗癌剤、より好ましくは細胞増殖抑制薬に対し耐性、好ましくは多重耐性であることを特徴とする、請求項11に記載の使用。
【請求項13】
細胞が、膜結合輸送タンパク質であるP糖タンパク質の発現、好ましくは過剰発現を示すことを特徴とする、請求項12に記載の使用。
【請求項14】
細胞が、p53陽性であるか、またはp53陰性であることを特徴とする、請求項1〜13のいずれか1項に記載の使用。
【請求項15】
発癌遺伝子タンパク質が、野生型発癌遺伝子タンパク質E1Aと比較して1または複数の突然変異もしくは欠失を示し、このときの欠失がCR3ストレッチの欠失およびN−末端の欠失およびC−末端の欠失を含む群より好ましくは選ばれることを特徴とする、請求項5または7〜14のいずれか1項に記載の使用。
【請求項16】
E1A発癌遺伝子タンパク質がRbに結合できることを特徴とする、請求項15に記載の使用。
【請求項17】
発癌遺伝子タンパク質が、野生型発癌遺伝子タンパク質と比較して1または複数の突然変異もしくは欠失を含み、このときの欠失が好ましくはCR1領域および/またはCR2領域内の欠失であることを特徴とする、請求項6〜14のいずれか1項に記載の使用。
【請求項18】
発癌遺伝子タンパク質E1AがRbに結合できないことを特徴とする、請求項17に記載の使用。
【請求項19】
ウイルス発ガン遺伝子タンパク質、好ましくはE1Aが、組織および/または腫瘍特異的プロモータの制御下にあることを特徴とする、請求項1〜18のいずれか1項に記載の使用。
【請求項20】
ウイルス、好ましくはアデノウイルスが、YB−1をコードしていることを特徴とする、請求項1〜19のいずれか1項に記載の使用。
【請求項21】
YB−1が、組織特異的および/または腫瘍特異的プロモータの制御下にあることを特徴とする、請求項20に記載の使用。
【請求項22】
ウイルス、好ましくはアデノウイルスが、少なくとも1つのタンパク質をコードしており、この場合のタンパク質がE4orf6、E4orf3、E1B55kおよびアデノウイルスE3ADPタンパク質を含む群より選ばれることを特徴とする、請求項1〜21のいずれか1項に記載の使用。
【請求項23】
細胞が核内にYB−1を含むこと、好ましくは腫瘍またはその一部を形成する細胞が核内にYB−1を有することを特徴とする、請求項1〜22のいずれか1項に記載の使用。
【請求項24】
核内へのYB−1輸送の誘導後に、腫瘍がその核内にYB−1を含むことを特徴とする、請求項1〜23のいずれか1項に記載の使用。
【請求項25】
YB−1の輸送が、放射線、細胞増殖抑制薬および高温を含む群より選ばれる少なくとも1つの手段により始動することを特徴とする。請求項24に記載の使用。
【請求項26】
手段が細胞、器官、または生体に適用されることを特徴とする、請求項25に記載の使用。
【請求項27】
ウイルス、好ましくはアデノウイルスが、AdΔ24、dl922−947、E1Ad/01/07、dl1119/1131、CB 016、dl520および機能的Rb腫瘍抑制遺伝子産物を結合できる発現したウイルス発癌遺伝子を欠くウイルスを含む群より選ばれることを特徴とする、請求項1〜26のいずれか1項に記載の使用。
【請求項28】
医薬の製造のためのウイルス、好ましくはアデノウイルスの使用であって、当該ウイルス、好ましくはアデノウイルスがE2−後期プロモータの活性化を介してYB−1により複製が制御され、好ましくは活性化が主にE2−後期プロモータの活性化により制御されるようデザインされているウイルスの使用。
【請求項29】
核内にYB−1を有する細胞内における複製のためのウイルス、好ましくはアデノウイルスの使用であって、当該ウイルス、好ましくはアデノウイルスが、E2−後期プロモータの活性化を介してYB−1により複製が制御され、好ましくは活性化が主にE2−後期プロモータの活性化により制御されるようデザインされているウイルスの使用。
【請求項30】
以下の特徴:
a) 少なくとも1つのウイルス遺伝子をトランス活性化し、そのときウイルス遺伝子がE1B−55k、E3ADPおよびE4orf6およびE4orf3を含む群より選ばれること、ならびに
b)核内、好ましくはその中にウイルス発癌遺伝子タンパク質が存在している細胞の核内へのYB−1誘導を欠いていること、
を含むことを特徴とするウイルス発癌遺伝子タンパク質、好ましくは単離されたウイルス発癌遺伝子タンパク質。
【請求項31】
ウイルス発癌遺伝子タンパク質がE1Aであることを特徴とする、請求項30に記載のウイルス発癌遺伝子タンパク質。
【請求項32】
ウイルス発癌遺伝子タンパク質が、野生型発癌遺伝子タンパク質と比較して1または複数の突然変異または欠失を含み、このときの欠失がCR3領域の欠失およびN−末端の欠失およびC−末端の欠失を含む群より好ましくは選ばれることを特徴とする、請求項30または31に記載のウイルス発癌遺伝子タンパク質。
【請求項33】
Rbに結合できることを特徴とする、請求項32に記載のウイルス発癌遺伝子タンパク質。
【請求項34】
ウイルス発癌遺伝子タンパク質が、1または複数の突然変異または欠失を含み、このとき欠失が好ましくはE1A発癌遺伝子タンパク質のCR1領域および/またはCR2領域内の欠失であることを特徴とする、請求項30または31に記載のウイルス発癌遺伝子タンパク質。
【請求項35】
ウイルス発癌遺伝子タンパク質がRbに結合できないことを特徴とする、請求項34に記載のウイルス発癌遺伝子タンパク質。
【請求項36】
請求項1〜29のいずれか1項に記載のウイルス、好ましくはアデノウイルスをコードする核酸を含み、そしてヘルパーウイルスの核酸を含むウイルス複製系、好ましくはアデノウイルス複製系の使用であって、このときヘルパーウイルスの核酸がYB−1をコードする核酸を含む使用。
【請求項37】
ウイルス核酸、好ましくはアデノウイルスの核酸および/またはヘルパーウイルスの核酸が複製可能なベクターとして存在していることを特徴とする、請求項36に記載のウイルス複製系、好ましくはアデノウイルス複製系の使用。
【請求項38】
医薬の製造、好ましくは腫瘍の処置を目的とする医薬の製造のための、ウイルス、好ましくはアデノウイルスをコードする核酸の請求項1〜29のいずれか1項に記載の使用。
【請求項39】
細胞、好ましくは腫瘍もしくはその一部を形成する細胞が、医薬的活性作用物質、好ましくは抗癌剤、より好ましくは細胞増殖抑制薬に対する耐性、好ましくは多重耐性を有することを特徴とする、請求項38に記載の使用。
【請求項40】
ウイルスが核内にYB−1を持たない細胞では複製せず、そしてウイルスが少なくとも1つのウイルス遺伝子、好ましくはアデノウイルス遺伝子をトランス活性化する発癌遺伝子または発癌遺伝子産物をコードしており、そのとき遺伝子がE1B55kDa、E4orf6、E4orf3およびE3ADPを含む群から選ばれることを特徴とする、核内にYB−1を有する細胞における複製のための請求項1〜29のいずれか1項に記載の、ウイルス、好ましくはアデノウイルスをコードする核酸の使用。
【請求項41】
複製がE2−後期プロモータの活性化を通して、好ましくはE2−後期プロモータの活性化を通して主に制御されるようにウイルスがデザインされている、医薬の製造のための、請求項1〜29のいずれか1項に記載のウイルス、好ましくはアデノウイルスをコードする核酸の使用。
【請求項42】
複製がE2−後期プロモータの活性化を通して、好ましくはE2−後期プロモータの活性化を通して主に制御されるようにウイルスがデザインされている、細胞内における複製のための、請求項1〜29のいずれか1項に記載のウイルス、好ましくはアデノウイルスをコードする核酸の使用。
【請求項43】
請求項1〜29のいずれか1項に記載の使用を目的とする、請求項36〜42のいずれか1項に記載の核酸を含むベクターの使用。
【請求項44】
細胞、腫瘍組織の細胞または患者を、請求項1〜29のいずれか1項に記載のウイルス、好ましくはアデノウイルスと接触させるべきか、および/またはそれらにより処置すべきか決定するために、細胞、腫瘍組織の細胞または患者を特徴付けすることを目的とする、YB−1と相互作用する化合物の使用。
【請求項45】
手段が、抗体、アンチカリン、アプタマー、アプタザイムおよびシュピーゲルマーを含む群から選ばれることを特徴とする、請求項44に記載の使用。
【請求項46】
請求項1〜29のいずれか1項に記載された使用に関連して用いられ得るウイルス、好ましくはアデノウイルスの製造を目的とした、請求項30〜35のいずれか1項に記載のウイルス発癌遺伝子タンパク質またはそれをコードする核酸の使用。
【請求項47】
ウイルスが、トランス遺伝子をコードする核酸を含んでいる、請求項1〜29のいずれか1項に記載のウイルス、好ましくはアデノウイルスの使用。
【請求項48】
ウイルスが、トランス遺伝子の翻訳および/または転写産物を含んでいる、請求項1〜29のいずれか1項に記載のウイルス、好ましくはアデノウイルスの使用。
【請求項49】
アデノウイルス複製系の核酸および/またはヘルパーウイルスの核酸が、トランス遺伝子またはトランス遺伝子をコードする核酸を含んでいる、請求項36または37に記載のアデノウイルス複製系の使用。
【請求項50】
核酸が、トランス遺伝子またはトランス遺伝子をコードする核酸を含んでいる、請求項38〜42のいずれか1項に記載の核酸の使用。
【請求項51】
トランス遺伝子が、プロドラッグ遺伝子、サイトカイン、アポトーシス誘導遺伝子、腫瘍抑制因子遺伝子、金属プロテアーゼインヒビターの遺伝子、および血管新生インヒビターの遺伝子を含む群から選ばれる、請求項47〜50のいずれか1項に記載の使用。
【請求項52】
トランス遺伝子が、siRNA、アプタマー、アンチセンス分子およびリボザイムに関する核酸を含む群より選ばれ、そしてsiRNA、アプタマー、アンチセンス分子および/またはリボザイムが標的分子を標的している、請求項47〜50のいずれか1項に記載の使用。
【請求項53】
標的分子が、耐性関連因子、抗アポトーシス因子、発癌遺伝子、血管新生因子、DNA合成酵素、DNA修復酵素、成長因子、成長因子レセプター、転写因子、金属プロテアーゼ、好ましくはマトリックス金属タンパク質キナーゼ、およびウロキナーゼ型プラスミノーゲン活性化因子を含む群より選ばれる、請求項52に記載の使用。
【請求項54】
医薬が医薬的に活性な化合物を更に含む、請求項1〜53のいずれか1項に記載の使用。
【請求項55】
医薬的に活性な化合物が、サイトカイン、金属プロテアーゼインヒビター、血管新生インヒビター、細胞増殖抑制薬および細胞周期インヒビターを含む群より選ばれる、請求項54に記載の使用。
【請求項1】
ウイルスが核内にYB−1がない細胞では複製できず、そのときウイルスが発癌遺伝子または発癌遺伝子産物、特に発癌遺伝子タンパク質をコードしており、それが少なくとも1つのウイルス遺伝子、好ましくはアデノウイルス遺伝子をトランス活性化し、その時の遺伝子がE1B55kDa、E4ort6、E4orf3およびE3ADPを含む群から選ばれることを特徴とする、医薬製造へのウイルス、好ましくはアデノウイルスの使用。
【請求項2】
ウイルスが核内にYB−1がない細胞では複製できず、そのときウイルスが発癌遺伝子または発癌遺伝子産物、特に発癌遺伝子タンパク質をコードしており、それが少なくとも1つのウイルス遺伝子、好ましくはアデノウイルス遺伝子をトランス活性化し、その時の遺伝子がE1B55kDa、E4ort6、E4orf3およびE3ADPを含む群から選ばれることを特徴とする、核内にYB−1を示す細胞内における複製のためのウイルス、好ましくはアデノウイルスの使用。
【請求項3】
ウイルス、好ましくはアデノウイルスが、核内にYB−1を有する細胞内で複製することを特徴とする、請求項1または2に記載の使用。
【請求項4】
ウイルス発癌遺伝子タンパク質がE1Aであり、そして/または発癌遺伝子がE1Aをコードする遺伝子であり、そして/または発癌遺伝子タンパク質がE1Aであることを特徴とする、請求項1〜3のいずれか1項に記載の使用。
【請求項5】
ウイルス発癌遺伝子タンパク質E1Aが、機能的Rb腫瘍抑制遺伝子産物を結合できることを特徴とする、請求項4に記載の使用。
【請求項6】
ウイルス発癌遺伝子タンパク質E1Aが、機能的Rb腫瘍抑制遺伝子産物を結合できないことを特徴とする、請求項4に記載の使用。
【請求項7】
ウイルス発癌タンパク質E1Aが、YB−1の核内局在化を誘導しないことを特徴とする、請求項4〜6のいずれか1項に記載の使用。
【請求項8】
医薬が、細胞がRb陽性であるか、またはRb陰性である患者向けであることを特徴とする、請求項1または3〜7のいずれか1項に記載の使用。
【請求項9】
細胞が、医薬により影響を受ける状態の形成に関係している細胞であることを特徴とする、請求項8に記載の使用。
【請求項10】
細胞がRb陰性であり、細胞核がYB−1陽性であり、好ましくは細胞周期とは無関係に核内がYB−1陽性であることを特徴とする、請求項1〜9のいずれか1項に記載の使用。
【請求項11】
医薬が腫瘍の処置向けであることを特徴とする、請求項1または3〜10のいずれか1項に記載の使用。
【請求項12】
細胞、好ましくは腫瘍またはその一部を形成する細胞が、医薬、好ましくは抗癌剤、より好ましくは細胞増殖抑制薬に対し耐性、好ましくは多重耐性であることを特徴とする、請求項11に記載の使用。
【請求項13】
細胞が、膜結合輸送タンパク質であるP糖タンパク質の発現、好ましくは過剰発現を示すことを特徴とする、請求項12に記載の使用。
【請求項14】
細胞が、p53陽性であるか、またはp53陰性であることを特徴とする、請求項1〜13のいずれか1項に記載の使用。
【請求項15】
発癌遺伝子タンパク質が、野生型発癌遺伝子タンパク質E1Aと比較して1または複数の突然変異もしくは欠失を示し、このときの欠失がCR3ストレッチの欠失およびN−末端の欠失およびC−末端の欠失を含む群より好ましくは選ばれることを特徴とする、請求項5または7〜14のいずれか1項に記載の使用。
【請求項16】
E1A発癌遺伝子タンパク質がRbに結合できることを特徴とする、請求項15に記載の使用。
【請求項17】
発癌遺伝子タンパク質が、野生型発癌遺伝子タンパク質と比較して1または複数の突然変異もしくは欠失を含み、このときの欠失が好ましくはCR1領域および/またはCR2領域内の欠失であることを特徴とする、請求項6〜14のいずれか1項に記載の使用。
【請求項18】
発癌遺伝子タンパク質E1AがRbに結合できないことを特徴とする、請求項17に記載の使用。
【請求項19】
ウイルス発ガン遺伝子タンパク質、好ましくはE1Aが、組織および/または腫瘍特異的プロモータの制御下にあることを特徴とする、請求項1〜18のいずれか1項に記載の使用。
【請求項20】
ウイルス、好ましくはアデノウイルスが、YB−1をコードしていることを特徴とする、請求項1〜19のいずれか1項に記載の使用。
【請求項21】
YB−1が、組織特異的および/または腫瘍特異的プロモータの制御下にあることを特徴とする、請求項20に記載の使用。
【請求項22】
ウイルス、好ましくはアデノウイルスが、少なくとも1つのタンパク質をコードしており、この場合のタンパク質がE4orf6、E4orf3、E1B55kおよびアデノウイルスE3ADPタンパク質を含む群より選ばれることを特徴とする、請求項1〜21のいずれか1項に記載の使用。
【請求項23】
細胞が核内にYB−1を含むこと、好ましくは腫瘍またはその一部を形成する細胞が核内にYB−1を有することを特徴とする、請求項1〜22のいずれか1項に記載の使用。
【請求項24】
核内へのYB−1輸送の誘導後に、腫瘍がその核内にYB−1を含むことを特徴とする、請求項1〜23のいずれか1項に記載の使用。
【請求項25】
YB−1の輸送が、放射線、細胞増殖抑制薬および高温を含む群より選ばれる少なくとも1つの手段により始動することを特徴とする。請求項24に記載の使用。
【請求項26】
手段が細胞、器官、または生体に適用されることを特徴とする、請求項25に記載の使用。
【請求項27】
ウイルス、好ましくはアデノウイルスが、AdΔ24、dl922−947、E1Ad/01/07、dl1119/1131、CB 016、dl520および機能的Rb腫瘍抑制遺伝子産物を結合できる発現したウイルス発癌遺伝子を欠くウイルスを含む群より選ばれることを特徴とする、請求項1〜26のいずれか1項に記載の使用。
【請求項28】
医薬の製造のためのウイルス、好ましくはアデノウイルスの使用であって、当該ウイルス、好ましくはアデノウイルスがE2−後期プロモータの活性化を介してYB−1により複製が制御され、好ましくは活性化が主にE2−後期プロモータの活性化により制御されるようデザインされているウイルスの使用。
【請求項29】
核内にYB−1を有する細胞内における複製のためのウイルス、好ましくはアデノウイルスの使用であって、当該ウイルス、好ましくはアデノウイルスが、E2−後期プロモータの活性化を介してYB−1により複製が制御され、好ましくは活性化が主にE2−後期プロモータの活性化により制御されるようデザインされているウイルスの使用。
【請求項30】
以下の特徴:
a) 少なくとも1つのウイルス遺伝子をトランス活性化し、そのときウイルス遺伝子がE1B−55k、E3ADPおよびE4orf6およびE4orf3を含む群より選ばれること、ならびに
b)核内、好ましくはその中にウイルス発癌遺伝子タンパク質が存在している細胞の核内へのYB−1誘導を欠いていること、
を含むことを特徴とするウイルス発癌遺伝子タンパク質、好ましくは単離されたウイルス発癌遺伝子タンパク質。
【請求項31】
ウイルス発癌遺伝子タンパク質がE1Aであることを特徴とする、請求項30に記載のウイルス発癌遺伝子タンパク質。
【請求項32】
ウイルス発癌遺伝子タンパク質が、野生型発癌遺伝子タンパク質と比較して1または複数の突然変異または欠失を含み、このときの欠失がCR3領域の欠失およびN−末端の欠失およびC−末端の欠失を含む群より好ましくは選ばれることを特徴とする、請求項30または31に記載のウイルス発癌遺伝子タンパク質。
【請求項33】
Rbに結合できることを特徴とする、請求項32に記載のウイルス発癌遺伝子タンパク質。
【請求項34】
ウイルス発癌遺伝子タンパク質が、1または複数の突然変異または欠失を含み、このとき欠失が好ましくはE1A発癌遺伝子タンパク質のCR1領域および/またはCR2領域内の欠失であることを特徴とする、請求項30または31に記載のウイルス発癌遺伝子タンパク質。
【請求項35】
ウイルス発癌遺伝子タンパク質がRbに結合できないことを特徴とする、請求項34に記載のウイルス発癌遺伝子タンパク質。
【請求項36】
請求項1〜29のいずれか1項に記載のウイルス、好ましくはアデノウイルスをコードする核酸を含み、そしてヘルパーウイルスの核酸を含むウイルス複製系、好ましくはアデノウイルス複製系の使用であって、このときヘルパーウイルスの核酸がYB−1をコードする核酸を含む使用。
【請求項37】
ウイルス核酸、好ましくはアデノウイルスの核酸および/またはヘルパーウイルスの核酸が複製可能なベクターとして存在していることを特徴とする、請求項36に記載のウイルス複製系、好ましくはアデノウイルス複製系の使用。
【請求項38】
医薬の製造、好ましくは腫瘍の処置を目的とする医薬の製造のための、ウイルス、好ましくはアデノウイルスをコードする核酸の請求項1〜29のいずれか1項に記載の使用。
【請求項39】
細胞、好ましくは腫瘍もしくはその一部を形成する細胞が、医薬的活性作用物質、好ましくは抗癌剤、より好ましくは細胞増殖抑制薬に対する耐性、好ましくは多重耐性を有することを特徴とする、請求項38に記載の使用。
【請求項40】
ウイルスが核内にYB−1を持たない細胞では複製せず、そしてウイルスが少なくとも1つのウイルス遺伝子、好ましくはアデノウイルス遺伝子をトランス活性化する発癌遺伝子または発癌遺伝子産物をコードしており、そのとき遺伝子がE1B55kDa、E4orf6、E4orf3およびE3ADPを含む群から選ばれることを特徴とする、核内にYB−1を有する細胞における複製のための請求項1〜29のいずれか1項に記載の、ウイルス、好ましくはアデノウイルスをコードする核酸の使用。
【請求項41】
複製がE2−後期プロモータの活性化を通して、好ましくはE2−後期プロモータの活性化を通して主に制御されるようにウイルスがデザインされている、医薬の製造のための、請求項1〜29のいずれか1項に記載のウイルス、好ましくはアデノウイルスをコードする核酸の使用。
【請求項42】
複製がE2−後期プロモータの活性化を通して、好ましくはE2−後期プロモータの活性化を通して主に制御されるようにウイルスがデザインされている、細胞内における複製のための、請求項1〜29のいずれか1項に記載のウイルス、好ましくはアデノウイルスをコードする核酸の使用。
【請求項43】
請求項1〜29のいずれか1項に記載の使用を目的とする、請求項36〜42のいずれか1項に記載の核酸を含むベクターの使用。
【請求項44】
細胞、腫瘍組織の細胞または患者を、請求項1〜29のいずれか1項に記載のウイルス、好ましくはアデノウイルスと接触させるべきか、および/またはそれらにより処置すべきか決定するために、細胞、腫瘍組織の細胞または患者を特徴付けすることを目的とする、YB−1と相互作用する化合物の使用。
【請求項45】
手段が、抗体、アンチカリン、アプタマー、アプタザイムおよびシュピーゲルマーを含む群から選ばれることを特徴とする、請求項44に記載の使用。
【請求項46】
請求項1〜29のいずれか1項に記載された使用に関連して用いられ得るウイルス、好ましくはアデノウイルスの製造を目的とした、請求項30〜35のいずれか1項に記載のウイルス発癌遺伝子タンパク質またはそれをコードする核酸の使用。
【請求項47】
ウイルスが、トランス遺伝子をコードする核酸を含んでいる、請求項1〜29のいずれか1項に記載のウイルス、好ましくはアデノウイルスの使用。
【請求項48】
ウイルスが、トランス遺伝子の翻訳および/または転写産物を含んでいる、請求項1〜29のいずれか1項に記載のウイルス、好ましくはアデノウイルスの使用。
【請求項49】
アデノウイルス複製系の核酸および/またはヘルパーウイルスの核酸が、トランス遺伝子またはトランス遺伝子をコードする核酸を含んでいる、請求項36または37に記載のアデノウイルス複製系の使用。
【請求項50】
核酸が、トランス遺伝子またはトランス遺伝子をコードする核酸を含んでいる、請求項38〜42のいずれか1項に記載の核酸の使用。
【請求項51】
トランス遺伝子が、プロドラッグ遺伝子、サイトカイン、アポトーシス誘導遺伝子、腫瘍抑制因子遺伝子、金属プロテアーゼインヒビターの遺伝子、および血管新生インヒビターの遺伝子を含む群から選ばれる、請求項47〜50のいずれか1項に記載の使用。
【請求項52】
トランス遺伝子が、siRNA、アプタマー、アンチセンス分子およびリボザイムに関する核酸を含む群より選ばれ、そしてsiRNA、アプタマー、アンチセンス分子および/またはリボザイムが標的分子を標的している、請求項47〜50のいずれか1項に記載の使用。
【請求項53】
標的分子が、耐性関連因子、抗アポトーシス因子、発癌遺伝子、血管新生因子、DNA合成酵素、DNA修復酵素、成長因子、成長因子レセプター、転写因子、金属プロテアーゼ、好ましくはマトリックス金属タンパク質キナーゼ、およびウロキナーゼ型プラスミノーゲン活性化因子を含む群より選ばれる、請求項52に記載の使用。
【請求項54】
医薬が医薬的に活性な化合物を更に含む、請求項1〜53のいずれか1項に記載の使用。
【請求項55】
医薬的に活性な化合物が、サイトカイン、金属プロテアーゼインヒビター、血管新生インヒビター、細胞増殖抑制薬および細胞周期インヒビターを含む群より選ばれる、請求項54に記載の使用。
【図1】


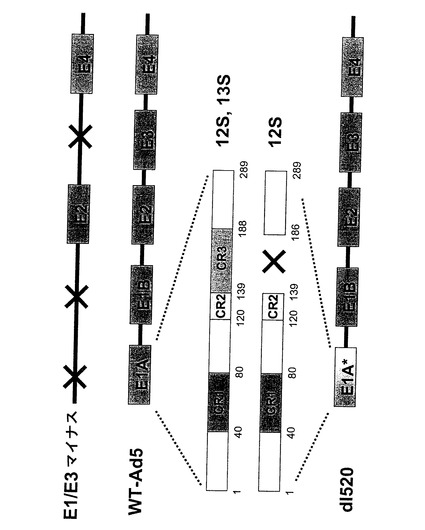
【図2】
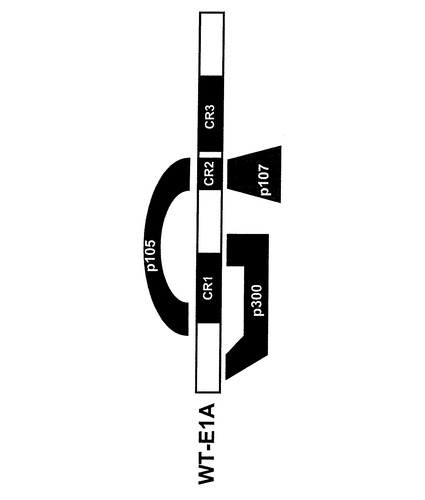
【図3】
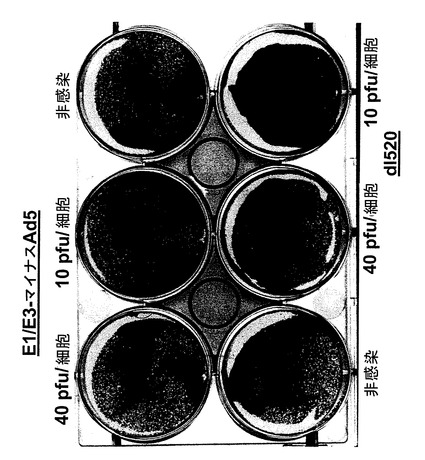
【図4】
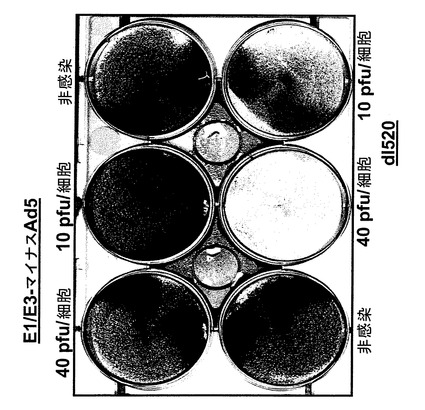
【図5】
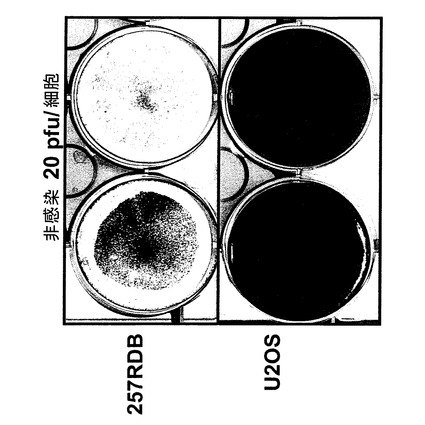
【図6】
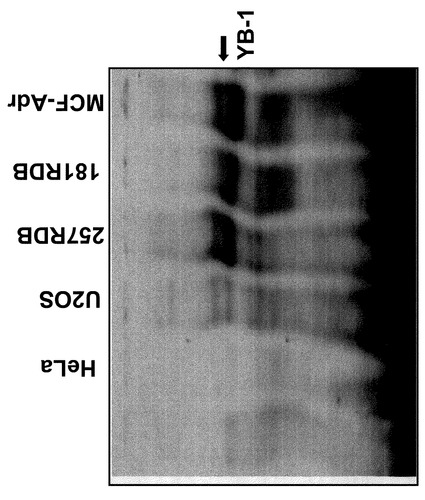
【図7】
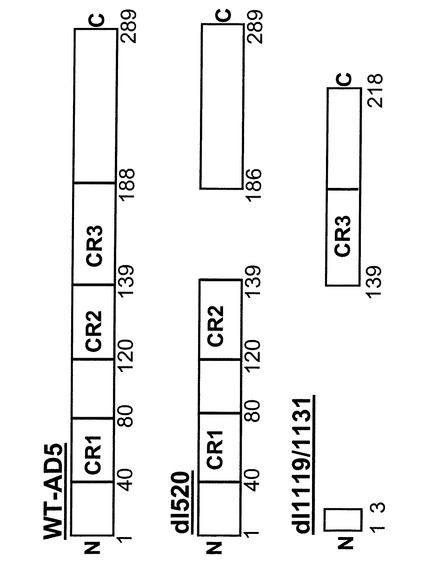
【図8】
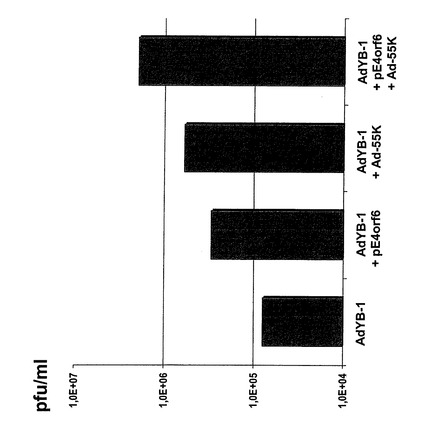
【図9】
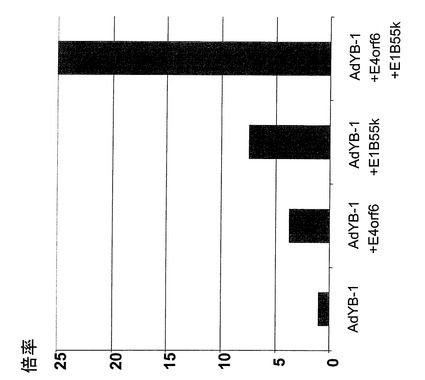
【図10】
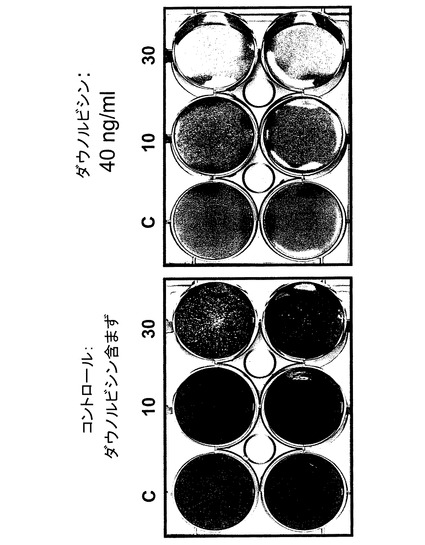
【図11】
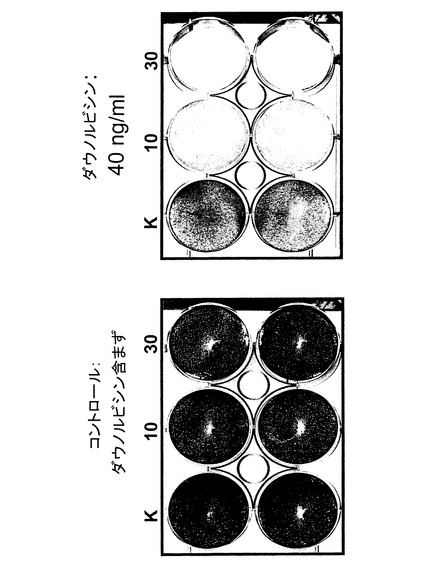
【図12】
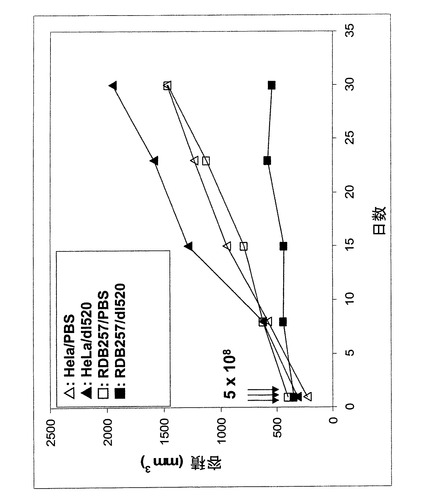
【図13】
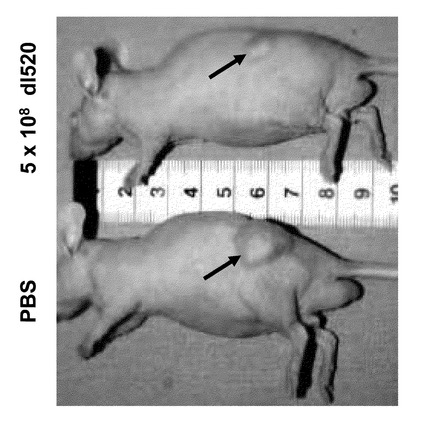
【図14】
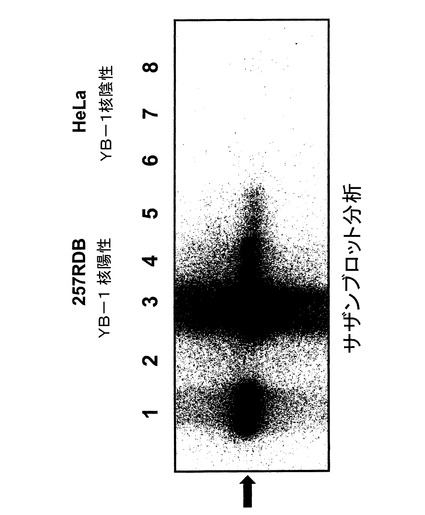
【図15】

【図16】
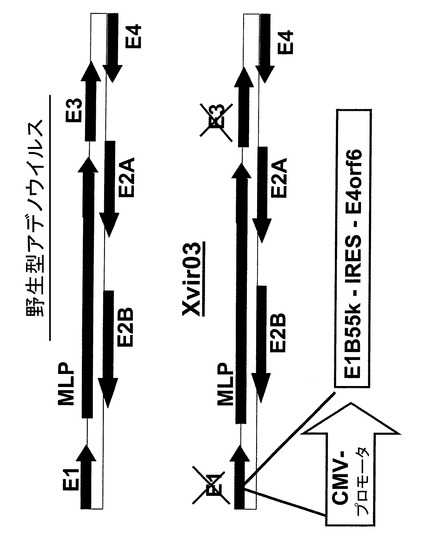
【図17】
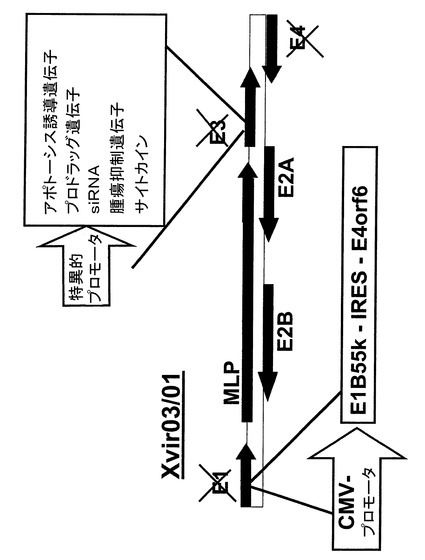
【図18A】

【図18B】
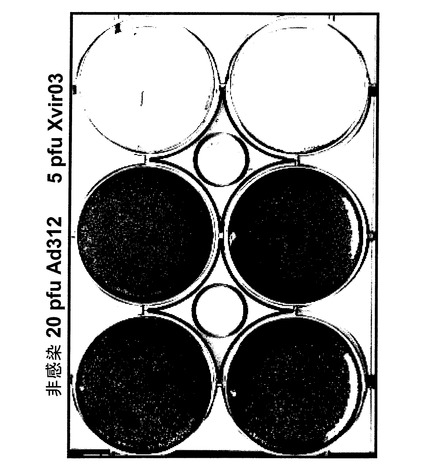
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18A】
【図18B】
【公開番号】特開2012−179058(P2012−179058A)
【公開日】平成24年9月20日(2012.9.20)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−116438(P2012−116438)
【出願日】平成24年5月22日(2012.5.22)
【分割の表示】特願2004−508113(P2004−508113)の分割
【原出願日】平成15年5月27日(2003.5.27)
【出願人】(501488619)
【氏名又は名称原語表記】HOLM,Per Sonne
【Fターム(参考)】
【公開日】平成24年9月20日(2012.9.20)
【国際特許分類】
【出願番号】特願2012−116438(P2012−116438)
【出願日】平成24年5月22日(2012.5.22)
【分割の表示】特願2004−508113(P2004−508113)の分割
【原出願日】平成15年5月27日(2003.5.27)
【出願人】(501488619)
【氏名又は名称原語表記】HOLM,Per Sonne
【Fターム(参考)】
[ Back to top ]