
アナベナセンサリーロドプシンを利用したタンパク質発現法
【課題】発現量の正の制御(タンパク質量の増大)にも負の制御(タンパク質量の減少)にも利用できることに加え、発現量の時間的な制御も行え、しかも発現量の空間的な制御にも利用可能なタンパク質発現系及びその用途等を提供すること。
【解決手段】以下のステップ(1)及び(2)、即ち、(1)アナベナセンサリーロドプシン応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトと、遺伝子発現に必要な因子とを含み、且つその中でアナベナセンサリーロドプシンが発現している区画を用意するステップ;(2)前記アナベナセンサリーロドプシンを活性化可能な波長の光を前記区画内に照射するステップを含むタンパク質発現法が提供される。
【解決手段】以下のステップ(1)及び(2)、即ち、(1)アナベナセンサリーロドプシン応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトと、遺伝子発現に必要な因子とを含み、且つその中でアナベナセンサリーロドプシンが発現している区画を用意するステップ;(2)前記アナベナセンサリーロドプシンを活性化可能な波長の光を前記区画内に照射するステップを含むタンパク質発現法が提供される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はタンパク質発現法に関する。詳しくは、藍藻(アナベナ)由来のレチナールを発色団とする膜タンパク質(アナベナセンサリーロドプシン)を用い、光によりタンパク質発現量の増減をコントロールする方法及びシステムに関する。
【背景技術】
【0002】
細胞の生理機能発現に直接関与しているのは多くのタンパク質分子である。遺伝情報から正確に転写、翻訳されたタンパク質は、高次な機能構造をとり、それぞれの機能部位へと輸送されることで細胞は様々な生理機能を発現する。特定のタンパク質の量を人為的に制御すること(例えば遺伝子破壊体や過剰発現体を作製すること)は、タンパク質の機能を知る手段として或いは創薬の手段として極めて有用である。
【0003】
従来、タンパク質量を制御・操作する方法として、人為的に操作したプラスミドDNA(ベクターDNA)を用い、イソプロピル-β-チオガラクトピラノシド(IPTG)やアラビノースなどの化学物質によって、或いは温度変化により発現が調節されるプロモーター領域の下流に任意の目的タンパク質をプラスミド遺伝子上でつなげ、試薬や温度の変化を人為的に引き起こすことによってタンパク質量を増やすことが広く行われてきた(例えば非特許文献1を参照)。また、人工的に合成したRNAに相補的な配列を持つメッセンジャーRNA(mRNA)が細胞内で分解される特性を生かして標的タンパク質量を抑制する(減少させる)方法が広く一般に用いられてきた(RNAi法、例えば非特許文献2を参照)。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Artem Khlebnikov et al. (2000) J. Bacteriol. 182, 7029-2037.
【非特許文献2】RNAi: Mahmood-ur-Rahman, Ali I, Husnain T, Riazuddin S. (2008). “RNA interference: the story of gene silencing in plants and humans”. Biotechnol Adv. 26 (3): 202-209
【非特許文献3】Jensen PR,Westerhoff HV, Michelsen O (1993) The use of lac-type promoters in control analysis. Eur J Biochem 211:181-191
【非特許文献4】Lars H. et al. (1998) Curr. Microbiol. 36, 341-347
【非特許文献5】Simon Frantz, Nature Reviews Drug Discovery 5, 528-529 (July 2006)
【非特許文献6】Spudich JL, Lanyi JK. Curr Opin Cell Biol. 1996 Aug;8(4):452-7. Review.
【非特許文献7】Jung KH, Trivedi VD, Spudich JL. Mol Microbiol. 2003 Mar;47(6):1513-22.
【非特許文献8】Vogeley L, Sineshchekov OA, Trivedi VD, Sasaki J, Spudich JL, Luecke H. Science. 2004 Nov 19;306(5700):1390-3. Epub 2004 Sep 30.
【発明の概要】
【発明が解決しようとする課題】
【0005】
化学物質や温度による発現制御を行う発現系の場合、タンパク質量を増やすことが可能な一方、減らすことは困難である。また、化学物質や温度による刺激は細胞全体に作用するため発現部位の制御は困難であり、また、長時間の作用による細胞自体への影響(成長阻害など)もある(非特許文献3、4を参照)。RNAi法についても、タンパク質量を減らすことのみが可能であり、細胞への毒性や予期せぬ効果(例えば他のmRNAへの作用による他の生命現象への作用)もあり(非特許文献5を参照)、作用部位のコントロールも困難である。そこで本発明は、所望のタンパク質の発現量を制御可能なタンパク質発現系であって、発現量の正の制御(タンパク質量の増大)にも負の制御(タンパク質量の減少)にも利用できることに加え、発現量の時間的な制御も行え、しかも発現量の空間的な制御にも利用可能なタンパク質発現系及びその用途等を提供することを課題とする。
【課題を解決するための手段】
【0006】
細胞は、光を感じるために、光を認識(受容)出来るタンパク質を利用している。自然界には、可視部の光を感じるために、発色団と呼ばれる色素をタンパク質に取り込んだ、いわゆる光受容蛋白質が広く存在している。上記課題を解決すべく検討を進める中で本発明者らは、藍藻の一種であるアナベナの光受容蛋白質(アナベナセンサリーロドプシン、以下「ASR」と略称することがある)に着目した。ASRはビタミンAのアルデヒド型であるレチナールを発色団にもつタンパク質で、550 nm付近の光を吸収する。従来のレチナールを持つタンパク質群(ロドプシン)は、光駆動のイオンポンプもしくは光センサーの2つに大別されるが(非特許文献6)、従来のセンサー機能であるキナーゼ分子の調節には関わらない分子であると考えられている(非特許文献7)。証拠は無いものの、光依存的にタンパク質の増減をコントロールしうる転写翻訳反応に関わると推定されている(非特許文献8)。本発明者らは、ASRが光依存的にタンパク質の増減を制御できる可能性に注目し、光依存的にタンパク質発現制御が可能なタンパク質発現系の構築を試みた。具体的には、モデル生物であり且つタンパク質発現用の宿主として汎用性が高く、しかも光受容蛋白質を基本的には有しない大腸菌の中でASRを発現させるとともに、アナベナの各種色素タンパク質のプロモーター領域の下流にレポータータンパク質を連結させたプラスミドDNAを導入し、光照射によるレポータータンパク質の発現変化を調べた。その結果、フィコシアニン色素オペロンのプロモーター領域を利用した場合にはASRの励起波長の光の照射によってレポータータンパク質の発現量を増大できることが実証された。一方で、フィコエリスリン色素オペロンのプロモーター領域を利用すれば、同様の光照射によってレポータータンパク質の発現量を顕著に減少できることが明らかとなった。このように、ASR及びそれに応答性のプロモーター領域を利用することによって、タンパク質の発現量を制御できることが示された。特に、プロモーター領域の選択によって、発現量の増大のみならず、発現量の減少も可能である点は本発現系の汎用性及び優位性の高さを示す。本発現系によれば、任意のタンパク質の発現量を光依存的に制御可能であり、例えば、望む時間に(時間的な制御)、望む場所で(空間的な制御)特定のタンパク質を発現させることができる。従って、タンパク質生産手段やタンパク質の機能解明用ツールとしてはもちろんのこと、創薬(タンパク質の相互作用に基づくスクリーニング法への応用など)やタンパク質を利用した医療(生体の特定の部位へのタンパク質の送達・発現など)のためのツールとしても有用である。このように本発現系の利用価値は計り知れず、画期的なツールといえる。尚、本発現系は光による制御を行うことから、制御性に優れる(照射光源のオン/オフで制御可能)という利点に加え、発現の場として細胞等を用いた場合においては、当該細胞自体や周囲への影響が極めて少ないという利点がある。
【0007】
以上の成果及び考察に基づき、以下の発明を提供する。
[1]以下のステップ(1)及び(2)を含む、タンパク質発現法:
(1)アナベナセンサリーロドプシン応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトと、遺伝子発現に必要な因子とを含み、且つその中でアナベナセンサリーロドプシンが発現している区画を用意するステップ;及び、
(2)前記アナベナセンサリーロドプシンを活性化可能な波長の光を前記区画内に照射するステップ。
[2]アナベナセンサリーロドプシン遺伝子を含む発現コンストラクトから前記アナベナセンサリーロドプシンが供給される、[1]に記載のタンパク質発現法。
[3]前記区画が細胞からなる、[1]又は[2]に記載のタンパク質発現法。
[4]前記細胞が大腸菌、バシラス属細菌又はアナベナ属藍藻である、[3]に記載のタンパク質発現法。
[5]前記細胞のゲノム内にアナベナセンサリーロドプシン遺伝子が予め組み込まれており、該遺伝子が発現することによって前記アナベナセンサリーロドプシンが供給される、[3]又は[4]に記載のタンパク質発現法。
[6]前記アナベナセンサリーロドプシン遺伝子が恒常的に発現している、[5]に記載のタンパク質量発現法。
[7]前記区画が、その中に無細胞合成系が形成された区画である、[1]又は[2]に記載のタンパク質発現法。
[8]前記区画がリポソーム又はエマルジョンからなる、[1]、[2]又は[7]に記載のタンパク質発現法。
[9]前記アナベナセンサリーロドプシンが配列番号5の配列を含む、[1]〜[8]のいずれか一項に記載のタンパク質発現法。
[10]前記アナベナセンサリーロドプシンが、天然型のアミノ酸配列の一部を改変して得られた変異型であり、その光学特性が天然型と異なる、[1]〜[8]のいずれか一項に記載のタンパク質発現法。
[11]前記変異型が波長485nm〜575nmに吸収極大を有する、[10]に記載のタンパク質発現法。
[12]前記発現コンストラクトが、二以上の目的タンパク質遺伝子を発現可能な状態で含む、[1]〜[11]のいずれか一項に記載のタンパク質発現法。
[13]目的タンパク質遺伝子が異なる二以上の発現コンストラクトが前記発現コンストラクトとして前記区画内に内包されている、[1]〜[11]のいずれか一項に記載のタンパク質発現法。
[14]前記色素タンパク質プロモーターが、フィコエリスリン色素オペロン又はフィコシアニン色素オペロンのプロモーターである、[1]〜[13]のいずれか一項に記載のタンパク質発現法。
[15]前記色素タンパク質プロモーターが、アナベナ属由来のプロモーターである、[1]〜[14]のいずれか一項に記載のタンパク質発現法。
[16]前記色素タンパク質プロモーターが、配列番号1の配列又は配列番号2の配列を含む、[1]〜[13]のいずれか一項に記載のタンパク質発現法。
[17]遺伝子発現に必要な因子と、
アナベナセンサリーロドプシン、又は恒常的若しくは一過性に発現可能な状態のアナベナセンサリーロドプシン遺伝子と、
アナベナセンサリーロドプシン応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトと、
を含む区画からなる、タンパク質発現システム。
【図面の簡単な説明】
【0008】
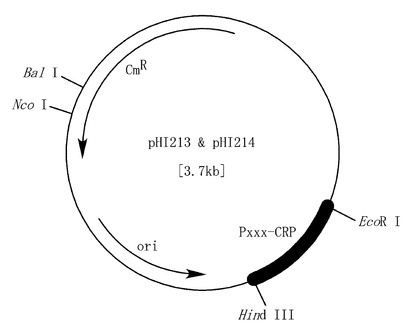
【図1】アナベナ推定プロモーター融合レポータータンパク質、cAMP受容体タンパク質(CRP)発現プラスミド。斜体は制限酵素を、CmRはクロラムフェニコール耐性遺伝子を表す。
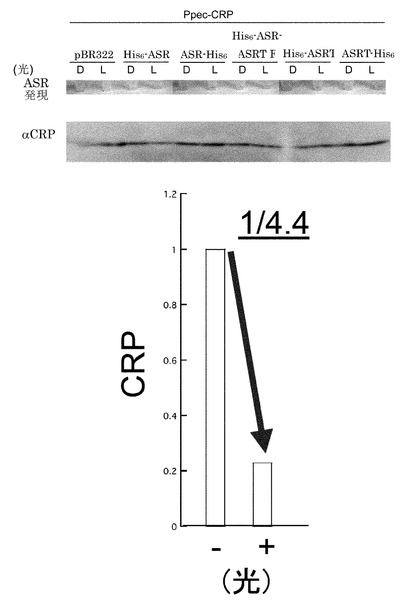
【図2】橙色光(600nm)照射による大腸菌内Ppec-CRP発現量の下方調節。Dは暗状態を、Lは光照射条件下での試料を表す。His6は、6個のHisが連続したいわゆるヒスチジンタグを意味し、ASRはアナベナセンサリーロドプシンを、ASRTは、アナベナセンサリーロドプシンの情報変換タンパク質を表す。His6-ASRは、タンパク質のN末端にヒスチジンタグを付加したものを、ASR-His6は、タンパク質のC末端にヒスチジンタグを付加したものを表す。pBR322は、プラスミドベクターのみのものを表す。ASRの発現は、ASRの持つ、赤紫色を、目で見て確認した(ASRの発現)。His6-ASRで、光照射条件下(L)で、CRPの量が減少している。複数回の実験により、その差を定量したものを下段に示した。
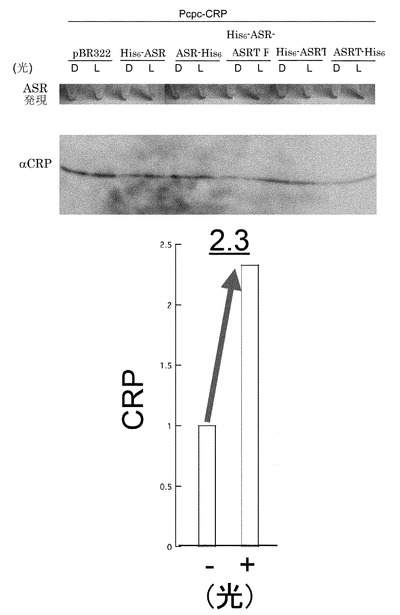
【図3】橙色光(600nm)照射による大腸菌内Pcpc-CRP発現量の上方調節。Dは暗状態を、Lは光照射条件下での試料を表す。His6は、6個のHisが連続したいわゆるヒスチジンタグを意味し、ASRはアナベナセンサリーロドプシンを、ASRTは、アナベナセンサリーロドプシンの情報変換タンパク質を表す。His6-ASRは、タンパク質のN末端にヒスチジンタグを付加したものを、ASR-His6は、タンパク質のC末端にヒスチジンタグを付加したものを表す。pBR322は、プラスミドベクターのみのものを表す。ASRの発現は、ASRの持つ、赤紫色を、目で見て確認した(ASRの発現)。His6-ASRで、光照射条件下(L)で、CRPの量が増加している。複数回の実験により、その差を定量したものを下段に示した。
【図4】色素タンパク質フィコエリスリン及びフィコシアニンの推定プロモーター領域含有500bp(配列番号1、配列番号2)を含む断片(Hind III-Kpn I)の配列(配列番号3、4)。大文字は制限酵素認識配列。
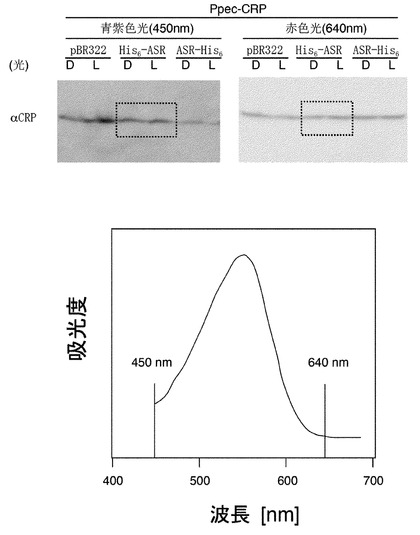
【図5】青紫色光450nmおよび赤色光640nm照射によるCRP量の増減変化。ASRが吸収することが出来ない光(下段:ASRの吸収特性(点線:野生型))では、実施例の4.で示した大腸菌内Ppec-CRPの顕著な発現量減少は起こらない。
【発明を実施するための形態】
【0009】
本発明の第1の局面は所望のタンパク質の発現量を制御可能なタンパク質発現法に関する。本発明のタンパク質発現法は、以下のステップ(1)及び(2)によって特徴付けられる。
(1)アナベナセンサリーロドプシン応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトと、遺伝子発現に必要な因子とを含み、且つその中でアナベナセンサリーロドプシンが発現している区画を用意するステップ
(2)前記アナベナセンサリーロドプシンを活性化可能な波長の光を前記区画内に照射するステップ
【0010】
ステップ(1)は、本発明の最大の特徴である「光によるタンパク質発現量の制御」が可能となるように、必要な材料や要素などを準備するステップである。当該ステップではその中に所定の発現コンストラクトと遺伝子発現用の因子を含み、且つその中でアナベナセンサリーロドプシン(ASR)が発現している区画を用意する。
【0011】
本発明では区画内に、第1の要素として、ASR応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトが含まれる。色素タンパク質プロモーターは、ASR応答性を示す限り特に限定されない。プロモーターの例を挙げると、フィコシアニン色素オペロンのプロモーター、フィコエリスリン色素オペロンのプロモーター、ASRオペロンのプロモーター、Kaiオペロンのプロモーター、フィコビリオペロンのプロモーターである。好ましくは、ASRによる良好な応答性を実現すべく、アナベナ属由来のプロモーターを採用する。ここで、色素タンパク質プロモーターの具体例を添付の配列表の配列番号1と配列番号2に示す。配列番号1は、アナベナ属フィコエリスリン色素オペロンの推定プロモーター領域を含む配列(オペロンの上流約500bp)であり、配列番号2はアナベナ属フィコシアニン色素オペロンの推定プロモーター領域を含む配列(オペロンの上流約500bp)である。ASRに対する応答性を保持する限り、これらの配列の一部(例えばオペロンの上流480bp、450bp、4200bp、390bp、360bp)を用いてもよい。応答性を保持するか否かの確認は、例えば、後述の実施例に示した発現系を利用して行えばよい。一方、上記配列に対して一部の塩基の置換、欠失、付加等の改変を施すことによって得られる配列を用いることも可能である。改変体が応答性を保持するか否かについても、例えば、後述の実施例に示した発現系を利用して容易に確認できる。
【0012】
発現コンストラクトは、ASR応答性の色素タンパク質プロモーターに加え、当該プロモーターの制御下にある目的タンパク質遺伝子を含む。「制御下にある」とは、「作動可能に連結している」と同義であり、通常、プロモーターの3'末端側に直接又は他の配列を介して目的タンパク質遺伝子が連結されることになる。
【0013】
「目的タンパク質遺伝子」とは、本発明の方法によって発現するタンパク質(即ち目的物)をコードする遺伝子である。目的や用途に応じて、任意の目的タンパク質遺伝子を採用することができる。目的タンパク質の例を挙げると、医薬や食品等の成分として有用なタンパク質(例えばインスリン、エリスロポエチン等のホルモン、インターフェロン、インスリン様成長因子、上皮成長因子(EGF)、線維芽細胞増殖因子(FGF)、インターロイキン類等の増殖因子・成長因子、抗体、酵素、)、試薬として有用なタンパク質(例えば各種酵素)、機能未知のタンパク質である。
【0014】
発現コンストラクトが、二以上の目的タンパク質遺伝子を発現可能な状態で含んでいてもよい。換言すれば、複数の目的タンパク質を発現可能な発現コンストラクトを採用してもよい。このようなコンストラクトは、例えば、複数のプロモーターやIRES(Internal Ribosome Entry Site)の使用によって構築できる。尚、異なる二以上の発現コンストラクトを併用することによって二以上の目的タンパク質が発現するように構成してもよい。この場合において、一部の発現コンストラクトのみ(例えば、二つの発現コンストラクトを使用するのであれば片方の発現コンストラクト)ASRの制御を受けるように構成してもよい。このように構成すれば、ASRによる発現制御と、他の手段による発現制御を併用した発現系となる。当該発現系は、複数のタンパク質の発現を個別に制御する目的に有用である。
【0015】
区画内には、第2の要素として、遺伝子発現に必要な因子が含まれる。「遺伝子発現に必要な因子」とは、遺伝子の発現、即ち遺伝子の転写及び翻訳が生じ、遺伝子の発現産物であるタンパク質が合成されるために必要な要素であり、RNAポリメラーゼ、転写基本因子、転写調節因子、転写伸長因子、アミノアシルtRNA合成酵素、各種アミノ酸、ATP、GTPなどのエネルギー源等が該当する。本発明では「遺伝子発現に必要な因子」が区画内に含まれることになるが、後述のように細胞を「区画」として用いた場合には、「遺伝子発現に必要な因子」として当該細胞に内在するものを用いることができる。即ち、大腸菌細胞のように、遺伝子発現に必要な一揃いの因子を備える場合、別途、「遺伝子発現に必要な因子」を供給する必要はない。但し、発現の安定化や効率化などを目的に一部又は全部の因子を補充することにしてもよい。また、一部の因子を欠損する細胞を採用した場合などは、当該因子を外部から補充すればよい。
【0016】
細胞ではなく、リポソーム等を用いて「区画」を形成した場合(詳細は後述する)には、通常、「遺伝子発現に必要な因子」として区画内に無細胞タンパク質合成系が構築される。本発明において無細胞合成系(無細胞転写/翻訳系)とは、生細胞を用いるのではく、生細胞由来の(或いは遺伝子工学的手法で得られた)リボソームや転写・翻訳因子などを用いて、鋳型である核酸からそれがコードするタンパク質をin vitroで合成することをいう。無細胞合成系では一般に、細胞破砕液を必要に応じて精製して得られる細胞抽出液が使用される。細胞抽出液には一般に、タンパク質合成に必要なリボソーム、開始因子などの各種因子、tRNAなどの各種酵素が含まれる。タンパク質の合成を行う際には、この細胞抽出液に各種アミノ酸、ATP、GTPなどのエネルギー源、クレアチンリン酸など、タンパク質の合成に必要なその他の物質を添加する。勿論、タンパク質合成の際に、別途用意したリボソームや各種因子、及び/又は各種酵素などを必要に応じて補充してもよい。
【0017】
タンパク質合成に必要な各分子(因子)を再構成した転写/翻訳系の開発も報告されている(Shimizu, Y. et al.: Nature Biotech., 19, 751-755, 2001)。この合成系では、バクテリアのタンパク質合成系を構成する3種類の開始因子、3種類の伸長因子、終結に関与する4種類の因子、各アミノ酸をtRNAに結合させる20種類のアミノアシルtRNA合成酵素、及びメチオニルtRNAホルミル転移酵素からなる31種類の因子の遺伝子を大腸菌ゲノムから増幅し、これらを用いてタンパク質合成系をin vitroで再構成している。このような再構成した合成系を利用することにしてもよい。
【0018】
無細胞タンパク質合成系には以下の利点がある。まず第1に、生細胞を維持する必要がないため操作性が良好で系の自由度も高い。したがって、目的のタンパク質の性質に応じて様々な修正や修飾を施した合成系を設計することが可能となる。次に、細胞系の合成では使用する細胞に毒性のあるタンパク質の合成は基本的にできないが、無細胞系ではそのような毒性のタンパク質であっても生産することができる。さらに、多種類のタンパク質を同時にかつ迅速に合成できることからハイスループット化が容易である。生産されるタンパク質の分離・精製が容易であるという利点も備え、これはハイスループット化に有利に働く。加えて、非天然型のアミノ酸を取り込ませるなどして非天然型タンパク質を合成することも可能であるという利点も併せ持つ。
【0019】
現在広く利用されている無細胞タンパク質合成系には以下のものがある。即ち、大腸菌S30抽出液の系(原核細胞の系)、コムギ胚芽抽出液の系(真核細胞の系)、及びウサギ網状赤血球可溶化物の系(真核細胞の系)である。これらの系はキットとしても市販されており、容易に利用することが可能である。
【0020】
歴史的には大腸菌S30抽出液の系の開発が最も古く、この系を利用して様々なタンパク質の合成が試みられてきた。大腸菌30S画分は、大腸菌の集菌、菌体破砕、精製の工程を経て調製される。大腸菌30S画分の調製及び、無細胞転写・翻訳共役反応はPrattらの方法(Pratt, J. M.: Chapter 7, in “Transcription and Translation: A practical approach”, ed. by B. D. Hames & S. J. Higgins, pp. 179-209, IRL Press, New York (1984))やEllmanらの方法(Ellman, J. et al.: Methods Enzymol., 202, 301-336(1991))を参考にして行うことができる。
【0021】
コムギ胚芽抽出液の系は、高品質の真核生物タンパク質を効率的に合成できるという利点を有し、大腸菌S30抽出液の系では合成が困難な真核生物のタンパク質を合成する際によく利用される。最近になって、種子胚乳成分を洗浄除去した胚芽から抽出液を調製することによって高効率かつ安定な合成系が構築されることが報告され注目を集めている(Madin, K. et al.: Proc. Natl. Acad. Sci. USA, 97: 559-564, 2000)。その後、高翻訳促進能を有するmRNA非翻訳配列、PCRを利用した多品目機能解析用のタンパク質合成法、専用高発現ベクターの構築などの技術開発が行われ(Sawasaki, T. et al.: Proc. Natl. Acad. Sci. USA, 99: 14652-14657, 2002)、様々な分野への応用が期待されている。
【0022】
コムギ胚芽抽出液は、コムギ胚芽をすり潰して遠心分離した後、上澄み液をゲルろ過で分離することによって得ることができる。翻訳反応については、Andersonらの方法(Anderson, C. W. et al.: Methods Enzymol., 101, 638-644(1983))を参考にできる。改良法についても報告されており、例えば河原崎らの方法(Kawarasaki, Y. et al.: Biotechnol. Prog., 16, 517-521(2000))やMadinらの方法(Madin, K. et al.: Proc. Natl. Acad. Sci. USA, 97: 559-564, 2000)等を参考にできる。その他、コムギ胚芽抽出液の系についてはWO 00/68412 A1、WO 01/27260 A1、WO 2002/024939 A1、WO 2005/063979 A1、特開平6-7134号公報、特開2002-529531号公報、特開2005-355513号公報、特開2006-042601号公報、特開2007-097438号公報、特開2008-029203号公報等が参考になる。
【0023】
ウサギ網状赤血球可溶化物の系はグロブリン生産に適する。ウサギ網状赤血球可溶化物は、ウサギにフェニルヒドラジンを数日間静脈注射して貧血状態とし、所定期間後(例えば第8日目)に採血し、その後溶血させた液から超遠心分離処理などを経て得られる。ウサギ網状赤血球可溶化物の調製法は、JacksonとHuntの方法(Jackson, R. J. and Hunt, T.: Methods Enzymol., 96, 50-74(1983))を参考にして行うことができる。
【0024】
尚、本発明の実施に際して利用できる無細胞合成系は上記のものに限られるものではなく、例えば大腸菌以外のバクテリアやコムギ以外の植物の抽出液、昆虫由来の抽出液、動物細胞由来の抽出液、又はゲノム情報を基に構築した系などを利用してもよい。
【0025】
本発明における区画内ではASRが発現している。ASRは光受容体として機能する。即ち、本発明では、ASRが光を受けて活性化し(詳細は後述のステップ(2)の欄で説明する)、発現コンストラクト中のプロモーターに作用する。
【0026】
区画内でのASRの発現は恒常的でも一過性でもよい。例えば、区画として細胞を利用し、当該細胞のゲノム内に予めASR遺伝子を組み込んでおくことによって、区画内でASRを発現させることができる。この態様ではステップ(1)として、例えば、宿主細胞のゲノムにASR(天然型又は変異型)を組み込むステップと、ASR応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトを当該宿主細胞に導入するステップとを行う。これら二つのステップは基本的には順不同であるが、操作の容易性及び効率の点から前者を先に行うことが好ましい。この態様において、ASR遺伝子を恒常的(構成的)プロモーターの制御下に配置しておけば、区画内でASRを恒常的に発現させることができる。ゲノム内にASR遺伝子を組み込むのではなく、恒常的プロモーターの制御下にあるASR遺伝子を含む発現コンストラクトを区画内に導入することによっても、区画内でのASRの恒常的発現を実現可能である。一方、組織特異的プロモーター、発育段階特異的プロモーター、誘導性プロモーター等を使用すれば、プロモーターの特性に応じた所望の発現(例えば、所望の時期に一過性に発現すること)が可能である。プロモーターの例を挙げると、lacプロモーター、tacプロモーター、trpプロモーター、T7プロモーター、α−アミラーゼプロモーター、β−アクチンプロモーター、L7プロモーター、CMVプロモーターである。
【0027】
宿主細胞のゲノム内に予めASR遺伝子を組み込んでおくのではなく、ASR遺伝子を含む発現コンストラクトの導入によってASRを発現させる場合にはステップ(1)として、例えば、宿主細胞内で機能するプロモーターの制御下にあるASR遺伝子を含む発現コンストラクトを宿主細胞に導入するステップと、ASR応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトを当該宿主細胞に導入するステップとを行う。これら二つのステップは基本的には順不同である。但し、前者のステップのみを行った宿主細胞を用意しておき、必要なときに後者のステップを行うという利用形態を採用すれば、任意のタンパク質の発現に利用できる汎用的な発現系を提供できることになる。
【0028】
光受容体として機能する限り、ASRは天然型でも変異型でもよい。天然型のASRのアミノ酸配列の一例を配列番号5に示す。また、当該アミノ酸配列をコードする遺伝子の配列を配列番号6に示す。天然型のASRは波長550nm付近に吸収極大を有する。天然型ASRの活性化には約500 nm〜約600 nmの波長の光を利用可能である。変異型(例えば波長485nm〜575nmに吸収極大を有する)を利用することにすれば、天然型の場合には利用不可能な又は活性化効率や細胞毒性などの点で利用し難い波長の光によってASRを活性化することが可能となる。これまで、ASRの類似タンパク質(いわゆる微生物型ロドプシン)において、様々な変異実験やレチナールの類似化合物を用いた発色団置換実験が行われ、様々な極大吸収波長を示すタンパク質が得られてきた。改変によって吸収極大が短波長側(例えば最大短波長化:498 nm)にシフトするように改変した変異体を使用することの利点は、例えば、汎用光源であるNd:YAGレーザー(532 nm)やアルゴンレーザー(488〜514 nm)の使用が可能なこと、及び有毒な紫外部の光により励起可能であり、細胞死が起こりながら実際の細胞や生体中でどのような事象が起きているのかを調べることも可能になることである。これとは逆に吸収極大が長波長側(例えば最大短波長化:575 nm)にシフトするように改変した変異体を使用することは、長波長光の組織透過性が高いことから、ヒトなど厚い組織(例えば皮膚など)に覆われている場合でもタンパク質を励起できる点などで有利である。これまでの研究によって、ロドプシンの吸収波長の決定に重要な部位(アミノ酸位置)が報告されている(例えば、Yokoyama S. Prog Retin Eye Res. 2000 Jul;19(4):385-419, 神取秀樹、加茂直樹 蛋白質核酸酵素 2002 Apr;47(5):620-5.を参照)。また、吸収波長のシフトに関する報告がいくつか存在する(例えばYokoyama S. Prog Retin Eye Res. 2000 Jul;19(4):385-419; 神取秀樹、加茂直樹 蛋白質核酸酵素 2002 Apr;47(5):620-5.; Shimono K, Hayashi T, Ikeura Y, Sudo Y, Iwamoto M, Kamo N. J Biol Chem. 2003 Jun 27;278(26):23882-9.を参照)。光学特性(特に吸収スペクトル)が天然型と異なる変異型を調製するためには、天然型ASRとASRの類似タンパク質との間で配列や立体構造などを比較し、天然型ASRの配列の中で吸収波長の決定に重要な部位を特定した上で、当該部位を標的として部位特異的変異誘発法(例えば、ストラタジーン社のクイックチェンジ(QuickChangeTM)を用いることができる)等による変異を加えればよい。
【0029】
本発明における用語「区画」は、タンパク質合成の場を与えるものであり、タンパク質合成及び本発明の作用・効果(即ち光によるタンパク質量の制御)の発揮に必要な因子ないし要素の保持及び維持を行う。典型的には、区画全体に亘ってその内部と外部を隔てる閉じた区画(閉鎖系)が用意される。但し、培養プレートのウェルに代表されるように、一部が開放した区画を用いることを妨げない。本発明の「区画」として例えば細胞が利用される。この態様では細胞をタンパク質合成の場として用いることになる。利用可能な細胞の例として、大腸菌などのグラム陰性菌、バシラス属細菌などのグラム陽性菌、藍藻(例えばアナベナ属)などの真正細菌全般、高度好塩菌などの古細菌、カビ、酵母や培養細胞などの真核生物など、が挙げられる。これらの中でも、本発明に好適なものとして、大腸菌、バシラス属細菌、藍藻が挙げられ、特に好適なものとして大腸菌が挙げられる。大腸菌は、汎用性が高い点、分裂速度が極めて速い点、遺伝子操作が容易である点、光感受性タンパク質がほとんど無い点等から、特に好適な細胞である。また、藍藻細胞(特にアナベナ属細胞)を採用した場合には、発現制御に利用する要素であるASRと細胞が同属又は近縁の関係になり、ASRの機能発揮に有利である。尚、人工的に構築した細胞(いわゆる人工細胞)を利用することにしてもよい。
【0030】
本発明の「区画」としてリポソーム、バイセル、エマルジョン、各種容器(培養プレート、培養皿、試験管、遠心管、遠心チューブ、マイクロテストチューブ)等を用いることもできる。エマルジョンとして例えばwater-in-oil(以下W/O)エマルジョンやW/O/W エマルジョン(double emulsion)が利用可能である。W/Oエマルジョンはin vitro compartmentalization(IVC)法に利用されている(Tawfik, D.S. & Griffiths, A.D.(1998)Nat. Biotechnol ., 7,652-656)。Tawfikらは、界面活性剤を加えた油相に水相を加え,撹拌することで形成されるW/O エマルジョンの水相を無細胞タンパク質合成反応の場としている。一方、Bernath らは、W/O/Wエマルジョンを利用したIVC法の開発にも成功している(Bernath, K., Hai, M., Mastrobattista, E., Griffiths, A.D., Magdassi, S., & Tawfik, D.S.(2004)Anal. Biochem., 325, 151-157)。尚、リポソームやエマルジョン等の人工材料を使用する場合、その修飾(糖修飾、抗体修飾、アルブミン修飾、ポリエチレングリコール(PEG)修飾など)が可能である。このような修飾は例えば、「区画」の安定化や最適化などの他、「区画」自体への機能の付加(例えば、生体の特定の部位へのタンパク質の送達・発現などに利用する場合における指向性の付加)などを目的として行われる。
【0031】
上で説明した通り、細胞ではなく、リポソーム等を用いて「区画」を形成する場合には、通常、無細胞タンパク質合成系を利用する。この態様においてはステップ(1)として、例えば、無細胞合成系を構成する因子等を区画(リポソーム等)内に用意するステップと、ASR用の発現コンストラクトと、ASR応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトを当該区画内に導入するステップとを行う。或いは、無細胞合成系を構成する因子等、ASR用の発現コンストラクト、ASR応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトを混合した後、リポソーム等を用いた区画化を行う。
【0032】
ステップ(1)に続くステップ(2)では、区画内で発現しているASRを活性化可能な波長の光を区画内に照射する。これによってASRが活性化し、区画内に存在する発現コンストラクト中のASR応答性プロモーターに作用する。その結果、プロモーターの特性に応じて、目的タンパク質遺伝子の転写が開始、促進、停止又は抑制される。例えば、フィコシアニン色素オペロンのプロモーターを用いた場合には、その制御下にある目的タンパク質遺伝子の転写が促進し、タンパク質の発現量が増大する(後述の実施例を参照)。また、フィコエリスリン色素オペロンのプロモーターを使用すれば、目的タンパク質の発現量が低減する(発現が抑制される)。このように本願発明によれば、目的タンパク質の発現量を光の照射という、簡便且つ容易でありしかも非侵襲的な手段によって制御可能である。
【0033】
照射する光の波長は、使用するASRに応じて決定すればよい。例えば波長550nm付近に吸収極大を有する天然型ASRを使用した場合は、約500 nm〜約600 nmの波長の光(具体的には例えば波長600nmの光)を照射すればよい。効率の点からは、ASRの吸収極大の波長の光を採用することが好ましい一方で、細胞毒性を回避するために、吸収極大よりも長波長側の光を照射するとよい。ASRを活性化可能な波長領域の光を含む限り、広スペクトル光、狭スペクトル光のいずれも利用可能である。また、光源についても特に制約はない。光源の例は冷陰極管、ハロゲンランプ、LED(発光ダイオード)、レーザーである。
【0034】
本発明の第2の局面は、本発明のタンパク質発現法に用いられるシステム(タンパク質発現システム)を提供する。本発明のタンパク質発現システムは、(1)遺伝子発現に必要な因子と、(2)アナベナセンサリーロドプシン、又は恒常的若しくは一過性に発現可能な状態のアナベナセンサリーロドプシン遺伝子と、(3)アナベナセンサリーロドプシン応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトと、を含む区画から構成される。本発明のタンパク質発現システムに用いる各要素については既述の通りであるので、対応する説明を援用することとし、ここでの重複した説明は省略する。本発明のタンパク質発現システムは、例えば適当な容器内に或いは容器を利用して(区画に容器を用いる場合)構築される。構築したタンパク質発現システムを生体に導入し、生体内で作動させることも可能である。このような利用形態は、タンパク質の機能解明やタンパク質を利用した医療(例えば生体の特定の部位へのタンパク質の送達・発現)の新たな手段を提供する。
以下、実施例(大腸菌を宿主として利用した発現実験)を用いて本発明を更に詳細に説明する。
【実施例】
【0035】
1.アナベナ推定プロモーター融合レポータータンパク質、cAMP receptor protein(CRP)発現プラスミドの作製(図1を参照)
Anabaena PCC7120株のゲノム(名古屋大学遺伝子実験施設の井原邦夫助教より分与)を鋳型として制限酵素Hind IIIおよびKpn I切断部位を含むプライマーDNAを用いてPCR法により増幅させた色素タンパク質(フィコエリスリンおよびフィコシアニン)コード領域上流の推定標的プロモーター領域(PpecおよびPcpc)を含有する増幅断片500bpを制限酵素(New England Biolabs、タカラバイオ株式会社)で切断した(Hind III-Kpn I)(図4)。切断断片をレポータータンパク質CRP発現プラスミドpHA7MK(アンピシリン耐性、鈴鹿医療科学大学の饗場弘二教授より分与、Aiba, H., Fujimoto, S. and Ozaki, N. (1982). “Molecular cloning and nucleotide sequencing of the gene for E. coli cAMP receptor protein”. Nucleic Acids Research 10: 1345-1361)のCRP開始コドン上流の制限酵素切断部位(Hind III-Kpn I)に挿入し、大腸菌DH5a株(Grant, S. G., Jessee, J., Bloom, F. R. and Hanahan, D. (1990). “Differential plasmid rescue from transgenic mouse DNAs into Escherichia coli methylation-restriction mutants”. Proc. Natl. Acad. Sci. USA 87: 4645-4649)に導入した。形質転換したDH5α株から一般的なアルカリ溶菌法によるミニプレップ法で作製したPpec-CRPおよびPcpc-CRP(以下まとめてPxxx-CRPと呼ぶ)発現プラスミドpHI103およびpHI104を回収した。pHI103およびpHI104のPxxx-CRPコード領域を含む制限酵素断片(Hind III-EcoR I)をpSU21(クロラムフェニコール耐性、Bartolome et al., 1991)のMCSの制限酵素部位(Hind III-EcoR I)に挿入し、大腸菌DH5α株に導入した。形質転換したDH5α株から一般的なアルカリ溶菌法によるミニプレップ法で作製したPxxx-CRP発現プラスミドpHI213およびpHI214(クロラムフェニコール耐性)を回収した。
【0036】
2.ASR発現大腸菌株のコンピテントセルの調製
His6-ASR発現プラスミドpKJ900(His6-ASR)、ASR-His6発現プラスミドpKJ900(ASR-His6)、His6-ASR-ASRT Fusion発現プラスミドpKJ900(His6-ASR-ASRT Fusion)、His6-ASR発現プラスミドpKJ900(His6-ASR)、ASRT-His6発現プラスミドpKJ900(ASRT-His6)(IPTG誘導型、アンピシリン耐性、以上のプラスミドは西江大学のKwang-Hwan Jung 准教授より分与、Jung, K. H., Trivedi, V. D. and Spudich, J. L. (2003). “Demonstration of a sensory rhodopsin in eubacteria”. Mol. Microbiol. 47: 1513-1522; Kawanabe, A., Furutani, Y., Yoon, S. R., Jung, K. H. and Kandori, H. (2008). “FTIR Study of the L Intermediate of Anabaena Sensory Rhodopsin: Structural Changes in the Cytoplasmic Region”. Biochemistry 47: 10033-10040)およびコントロールプラスミドpBR322(Bolivar, F., Rodriguez, R. L., Greene, P. J., Betlach, M. C., Heyneker, H. L., Boyer, H. W., Crosa, J. H. and Falkow, S. (1977). “Construction and characterization of new cloning vehicles. II. A multipurpose cloning system”. Gene 2(2): 95-113) をE. Coli TA341株(E. coli W3110 Dcrp、 鈴鹿医療科学大学の饗場弘二教授より分与、Sunohara, T., Jojima, K., Yamamoto, Y., Inada, T. and Aiba, H. (2004). “Nascent-peptide-mediated ribosome stalling at a stop codon induces mRNA cleavage resulting in nonstop mRNA that is recognized by tmRNA”. RNA 10: 378-386)のコンピテントセルに導入した。形質転換したASR発現プラスミド導入済みE. Coli TA341株を一般的な塩化カルシウム法で調製した。ここでHis6とは、アミノ酸の1種であるヒスチジンが6個連続した配列の事で、この配列に対する抗体(GE Healthcare)により、タンパク質をウェスタンブロッティングにより簡便に検出できること、Ni-NTA樹脂(GE Healthcare)などによりタンパク質を簡便に精製出来ることから付加したものである。
【0037】
3.ASRおよびPxxx-CRP発現大腸菌株の調製
1.で作製したPxxx-CRP発現プラスミドpHI213およびpHI214を2.で調製したASR発現プラスミド導入済みE. Coli TA341株のコンピテントセルに導入した。アンピシリン及びクロラムフェニコールを、それぞれ終濃度50μg/ml及び25μg/ml含む、通常の大腸菌用寒天培地(LB培地: 1% Bacto Tryptone、0.5% Yeast Extract、1% NaCl)にてセレクションし、目的菌株を得た。
【0038】
4.橙色光(600nm)照射による大腸菌内Ppec-CRP発現量の下方調節(図2)
3.で調製したASRおよびPpec-CRP発現大腸菌株を終濃度50μg/mlアンピシリンおよび25μg/mlクロラムフェニコールを含むLB培地で30℃、遮光状態で終夜、振盪培養した(一次培養)。一次培養した大腸菌を終濃度1mM IPTG, 10μM all-trans型レチナールを含むLB培地に終濃度50μg/mlアンピシリンおよび25μg/mlクロラムフェニコールを添加した溶液で25℃、8時間、二次培養した後、橙色光(600nm±10nmを透過するフィルター(Melles Griot)を通した白色光)を30分間照射しながら振盪速度を落として培養した。照射光源は、CL-250(ELMO)を用い、その際の光量を光量計により測定したところ、1.8mWであった。培養・橙色光(600nm)照射した菌を遠心(15,000rpm, 4℃, 4分)により集菌し、上清を除去した。沈澱に蒸留水、SDS-loading buffer(400 mM Tris-HCl buffer (pH 6.8)、6% Sodium dodecyl sulfate (SDS)、38.25% Glycerol、0.02% Bromophenol Blue)、2-メルカプトエタノールを添加後、100℃で4分間煮沸し、氷上で急冷してサンプルとした。調製したサンプルを一般的なSDS-PAGE(15%ゲル)、ウェスタンブロッティング(ウサギ抗CRP抗体(Ishizuka H., Hanamura A., Kunimura T. and Aiba H. (1993). “A lowered concentration of cAMP receptor protein caused by glucose is an important determinant for catabolite repression in Escherichia coli. Mol. Microbiol., 10: 341-350.)+ヤギ抗ウサギHRP抗体(Santa Cruz Biotechnology))を用いて解析し、橙色光(600nm)照射によるPpec-CRP量の変化を測定した。Ppec-CRP量は橙色光(600nm)照射により減少するという結果を得た。複数の実験による平均値は、1/4.4に低下するものであった。この結果は、光照射により人為的にタンパク質量を減少させる調節法の構築という点において、初の成果である。
【0039】
5.橙色光(600nm)照射による大腸菌内Pcpc-CRP発現量の上方調節(図3)
3.で調製したASRおよびPpec-CRP発現大腸菌株を終濃度50μg/mlアンピシリンおよび25μg/mlクロラムフェニコールを含むLB培地で30℃、遮光状態で終夜、振盪培養した(一次培養)。一次培養した大腸菌を終濃度1mM IPTG, 10μM all-trans 型レチナールを含むLB培地に終濃度50μg/mlアンピシリンおよび25μg/mlクロラムフェニコールを添加した溶液で25℃、8時間、二次培養した後、橙色光(600nm±10nmを透過するフィルター(Melles Griot)を通した白色光)を30分間照射しながら振盪速度を落として培養した。照射光源は、CL-250(ELMO)を用い、その際の光量を光量計により測定したところ、1.8mWであった。培養・橙色光(600nm)照射した菌を遠心(15,000rpm, 4℃, 4分)により集菌し、上清を除去した。沈澱に蒸留水、SDS-loading buffer(400 mM Tris-HCl buffer (pH 6.8)、6% Sodium dodecyl sulfate (SDS)、38.25% Glycerol、0.02% Bromophenol Blue)、2-メルカプトエタノールを添加後、100℃で4分間煮沸し、氷上で急冷してサンプルとした。調製したサンプルを一般的なSDS-PAGE(15%ゲル)、ウェスタンブロッティング(ウサギ抗CRP抗体(Ishizuka H., Hanamura A., Kunimura T. and Aiba H. (1993). “A lowered concentration of cAMP receptor protein caused by glucose is an important determinant for catabolite repression in Escherichia coli. Mol. Microbiol., 10: 341-350.)+ヤギ抗ウサギHRP抗体(Santa Cruz Biotechnology))を用いて解析し、橙色光(600nm)照射によるPcpc-CRP量の変化を測定した。Pcpc-CRP量は橙色光(600nm)照射により増加するという結果を得た。複数の実験による平均値は、2.3倍に増加するものであった。この結果は、光照射により人為的にタンパク質量を増加させる調節法の構築という点において、初の成果である。
【0040】
6.他の励起波長(450 nm, 640 nm)におけるCRP量の増減変化(図5)
3.で調製したASRおよびPpec-CRP発現大腸菌株を終濃度50 mg/ml アンピシリンおよび25 mg/ml クロラムフェニコールを含むLB培地で30℃、遮光状態で終夜、振盪培養した(一次培養)。一次培養した大腸菌を終濃度1 mM IPTG, 10 mM all-trans 型レチナールを含むLB培地に終濃度50 mg/ml アンピシリンおよび25 mg/ml クロラムフェニコールを添加した溶液で25℃、8時間、二次培養した後、青紫色光(450nm±8nm)もしくは赤色光(640nm±11nm)を透過するフィルター(Asahi Spectra)を通した白色光を30分間照射しながら振盪速度を落として培養した。照射光源は、MAX-302(Asahi Spectra)を用い、その際の光量を光量計により測定したところ、それぞれ1.2mW(青紫色光)および1.6mW(赤色光)であった。培養・光照射した菌を遠心(15,000rpm, 4℃, 4分)により集菌し、上清を除去した。沈澱に蒸留水、SDS-loading buffer(400 mM Tris-HCl buffer (pH 6.8)、6% Sodium dodecyl sulfate (SDS)、38.25% Glycerol、0.02% Bromophenol Blue)、2-メルカプトエタノールを添加後、100℃で4分間煮沸し、氷上で急冷してサンプルとした。調製したサンプルを一般的なSDS-PAGE(15%ゲル)、ウェスタンブロッティング(ウサギ抗CRP抗体(Ishizuka H., Hanamura A., Kunimura T. and Aiba H. (1993). “A lowered concentration of cAMP receptor protein caused by glucose is an important determinant for catabolite repression in Escherichia coli. Mol. Microbiol., 10: 341-350.)+ヤギ抗ウサギHRP抗体(Santa Cruz Biotechnology))を用いて解析し、青紫色光(450nm)もしくは赤色光(640nm)照射によるPpec-CRP量の変化を測定した。Ppec-CRP量は青紫色光(450nm)もしくは赤色光(640nm)照射では変化しないという結果を得た。この結果は、ASRが吸収しない波長の光では、上述の4.で示したタンパク質量の変化(減少)は起こらず、ASRが実際にこのような反応に寄与していることを示している。
【産業上の利用可能性】
【0041】
本発明のタンパク質発現法・発現システムによれば、任意のタンパク質の発現量を光で制御可能である。本発明はタンパク質生産手段或いはタンパク質発現制御手段として様々な用途で利用可能である。例えば、タンパク質の機能解明用ツールや創薬ツール、或いは医療用ツール等としての利用が想定される。
【0042】
この発明は、上記発明の実施の形態及び実施例の説明に何ら限定されるものではない。特許請求の範囲の記載を逸脱せず、当業者が容易に想到できる範囲で種々の変形態様もこの発明に含まれる。
本明細書の中で明示した論文、公開特許公報、及び特許公報などの内容は、その全ての内容を援用によって引用することとする。
【技術分野】
【0001】
本発明はタンパク質発現法に関する。詳しくは、藍藻(アナベナ)由来のレチナールを発色団とする膜タンパク質(アナベナセンサリーロドプシン)を用い、光によりタンパク質発現量の増減をコントロールする方法及びシステムに関する。
【背景技術】
【0002】
細胞の生理機能発現に直接関与しているのは多くのタンパク質分子である。遺伝情報から正確に転写、翻訳されたタンパク質は、高次な機能構造をとり、それぞれの機能部位へと輸送されることで細胞は様々な生理機能を発現する。特定のタンパク質の量を人為的に制御すること(例えば遺伝子破壊体や過剰発現体を作製すること)は、タンパク質の機能を知る手段として或いは創薬の手段として極めて有用である。
【0003】
従来、タンパク質量を制御・操作する方法として、人為的に操作したプラスミドDNA(ベクターDNA)を用い、イソプロピル-β-チオガラクトピラノシド(IPTG)やアラビノースなどの化学物質によって、或いは温度変化により発現が調節されるプロモーター領域の下流に任意の目的タンパク質をプラスミド遺伝子上でつなげ、試薬や温度の変化を人為的に引き起こすことによってタンパク質量を増やすことが広く行われてきた(例えば非特許文献1を参照)。また、人工的に合成したRNAに相補的な配列を持つメッセンジャーRNA(mRNA)が細胞内で分解される特性を生かして標的タンパク質量を抑制する(減少させる)方法が広く一般に用いられてきた(RNAi法、例えば非特許文献2を参照)。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Artem Khlebnikov et al. (2000) J. Bacteriol. 182, 7029-2037.
【非特許文献2】RNAi: Mahmood-ur-Rahman, Ali I, Husnain T, Riazuddin S. (2008). “RNA interference: the story of gene silencing in plants and humans”. Biotechnol Adv. 26 (3): 202-209
【非特許文献3】Jensen PR,Westerhoff HV, Michelsen O (1993) The use of lac-type promoters in control analysis. Eur J Biochem 211:181-191
【非特許文献4】Lars H. et al. (1998) Curr. Microbiol. 36, 341-347
【非特許文献5】Simon Frantz, Nature Reviews Drug Discovery 5, 528-529 (July 2006)
【非特許文献6】Spudich JL, Lanyi JK. Curr Opin Cell Biol. 1996 Aug;8(4):452-7. Review.
【非特許文献7】Jung KH, Trivedi VD, Spudich JL. Mol Microbiol. 2003 Mar;47(6):1513-22.
【非特許文献8】Vogeley L, Sineshchekov OA, Trivedi VD, Sasaki J, Spudich JL, Luecke H. Science. 2004 Nov 19;306(5700):1390-3. Epub 2004 Sep 30.
【発明の概要】
【発明が解決しようとする課題】
【0005】
化学物質や温度による発現制御を行う発現系の場合、タンパク質量を増やすことが可能な一方、減らすことは困難である。また、化学物質や温度による刺激は細胞全体に作用するため発現部位の制御は困難であり、また、長時間の作用による細胞自体への影響(成長阻害など)もある(非特許文献3、4を参照)。RNAi法についても、タンパク質量を減らすことのみが可能であり、細胞への毒性や予期せぬ効果(例えば他のmRNAへの作用による他の生命現象への作用)もあり(非特許文献5を参照)、作用部位のコントロールも困難である。そこで本発明は、所望のタンパク質の発現量を制御可能なタンパク質発現系であって、発現量の正の制御(タンパク質量の増大)にも負の制御(タンパク質量の減少)にも利用できることに加え、発現量の時間的な制御も行え、しかも発現量の空間的な制御にも利用可能なタンパク質発現系及びその用途等を提供することを課題とする。
【課題を解決するための手段】
【0006】
細胞は、光を感じるために、光を認識(受容)出来るタンパク質を利用している。自然界には、可視部の光を感じるために、発色団と呼ばれる色素をタンパク質に取り込んだ、いわゆる光受容蛋白質が広く存在している。上記課題を解決すべく検討を進める中で本発明者らは、藍藻の一種であるアナベナの光受容蛋白質(アナベナセンサリーロドプシン、以下「ASR」と略称することがある)に着目した。ASRはビタミンAのアルデヒド型であるレチナールを発色団にもつタンパク質で、550 nm付近の光を吸収する。従来のレチナールを持つタンパク質群(ロドプシン)は、光駆動のイオンポンプもしくは光センサーの2つに大別されるが(非特許文献6)、従来のセンサー機能であるキナーゼ分子の調節には関わらない分子であると考えられている(非特許文献7)。証拠は無いものの、光依存的にタンパク質の増減をコントロールしうる転写翻訳反応に関わると推定されている(非特許文献8)。本発明者らは、ASRが光依存的にタンパク質の増減を制御できる可能性に注目し、光依存的にタンパク質発現制御が可能なタンパク質発現系の構築を試みた。具体的には、モデル生物であり且つタンパク質発現用の宿主として汎用性が高く、しかも光受容蛋白質を基本的には有しない大腸菌の中でASRを発現させるとともに、アナベナの各種色素タンパク質のプロモーター領域の下流にレポータータンパク質を連結させたプラスミドDNAを導入し、光照射によるレポータータンパク質の発現変化を調べた。その結果、フィコシアニン色素オペロンのプロモーター領域を利用した場合にはASRの励起波長の光の照射によってレポータータンパク質の発現量を増大できることが実証された。一方で、フィコエリスリン色素オペロンのプロモーター領域を利用すれば、同様の光照射によってレポータータンパク質の発現量を顕著に減少できることが明らかとなった。このように、ASR及びそれに応答性のプロモーター領域を利用することによって、タンパク質の発現量を制御できることが示された。特に、プロモーター領域の選択によって、発現量の増大のみならず、発現量の減少も可能である点は本発現系の汎用性及び優位性の高さを示す。本発現系によれば、任意のタンパク質の発現量を光依存的に制御可能であり、例えば、望む時間に(時間的な制御)、望む場所で(空間的な制御)特定のタンパク質を発現させることができる。従って、タンパク質生産手段やタンパク質の機能解明用ツールとしてはもちろんのこと、創薬(タンパク質の相互作用に基づくスクリーニング法への応用など)やタンパク質を利用した医療(生体の特定の部位へのタンパク質の送達・発現など)のためのツールとしても有用である。このように本発現系の利用価値は計り知れず、画期的なツールといえる。尚、本発現系は光による制御を行うことから、制御性に優れる(照射光源のオン/オフで制御可能)という利点に加え、発現の場として細胞等を用いた場合においては、当該細胞自体や周囲への影響が極めて少ないという利点がある。
【0007】
以上の成果及び考察に基づき、以下の発明を提供する。
[1]以下のステップ(1)及び(2)を含む、タンパク質発現法:
(1)アナベナセンサリーロドプシン応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトと、遺伝子発現に必要な因子とを含み、且つその中でアナベナセンサリーロドプシンが発現している区画を用意するステップ;及び、
(2)前記アナベナセンサリーロドプシンを活性化可能な波長の光を前記区画内に照射するステップ。
[2]アナベナセンサリーロドプシン遺伝子を含む発現コンストラクトから前記アナベナセンサリーロドプシンが供給される、[1]に記載のタンパク質発現法。
[3]前記区画が細胞からなる、[1]又は[2]に記載のタンパク質発現法。
[4]前記細胞が大腸菌、バシラス属細菌又はアナベナ属藍藻である、[3]に記載のタンパク質発現法。
[5]前記細胞のゲノム内にアナベナセンサリーロドプシン遺伝子が予め組み込まれており、該遺伝子が発現することによって前記アナベナセンサリーロドプシンが供給される、[3]又は[4]に記載のタンパク質発現法。
[6]前記アナベナセンサリーロドプシン遺伝子が恒常的に発現している、[5]に記載のタンパク質量発現法。
[7]前記区画が、その中に無細胞合成系が形成された区画である、[1]又は[2]に記載のタンパク質発現法。
[8]前記区画がリポソーム又はエマルジョンからなる、[1]、[2]又は[7]に記載のタンパク質発現法。
[9]前記アナベナセンサリーロドプシンが配列番号5の配列を含む、[1]〜[8]のいずれか一項に記載のタンパク質発現法。
[10]前記アナベナセンサリーロドプシンが、天然型のアミノ酸配列の一部を改変して得られた変異型であり、その光学特性が天然型と異なる、[1]〜[8]のいずれか一項に記載のタンパク質発現法。
[11]前記変異型が波長485nm〜575nmに吸収極大を有する、[10]に記載のタンパク質発現法。
[12]前記発現コンストラクトが、二以上の目的タンパク質遺伝子を発現可能な状態で含む、[1]〜[11]のいずれか一項に記載のタンパク質発現法。
[13]目的タンパク質遺伝子が異なる二以上の発現コンストラクトが前記発現コンストラクトとして前記区画内に内包されている、[1]〜[11]のいずれか一項に記載のタンパク質発現法。
[14]前記色素タンパク質プロモーターが、フィコエリスリン色素オペロン又はフィコシアニン色素オペロンのプロモーターである、[1]〜[13]のいずれか一項に記載のタンパク質発現法。
[15]前記色素タンパク質プロモーターが、アナベナ属由来のプロモーターである、[1]〜[14]のいずれか一項に記載のタンパク質発現法。
[16]前記色素タンパク質プロモーターが、配列番号1の配列又は配列番号2の配列を含む、[1]〜[13]のいずれか一項に記載のタンパク質発現法。
[17]遺伝子発現に必要な因子と、
アナベナセンサリーロドプシン、又は恒常的若しくは一過性に発現可能な状態のアナベナセンサリーロドプシン遺伝子と、
アナベナセンサリーロドプシン応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトと、
を含む区画からなる、タンパク質発現システム。
【図面の簡単な説明】
【0008】
【図1】アナベナ推定プロモーター融合レポータータンパク質、cAMP受容体タンパク質(CRP)発現プラスミド。斜体は制限酵素を、CmRはクロラムフェニコール耐性遺伝子を表す。
【図2】橙色光(600nm)照射による大腸菌内Ppec-CRP発現量の下方調節。Dは暗状態を、Lは光照射条件下での試料を表す。His6は、6個のHisが連続したいわゆるヒスチジンタグを意味し、ASRはアナベナセンサリーロドプシンを、ASRTは、アナベナセンサリーロドプシンの情報変換タンパク質を表す。His6-ASRは、タンパク質のN末端にヒスチジンタグを付加したものを、ASR-His6は、タンパク質のC末端にヒスチジンタグを付加したものを表す。pBR322は、プラスミドベクターのみのものを表す。ASRの発現は、ASRの持つ、赤紫色を、目で見て確認した(ASRの発現)。His6-ASRで、光照射条件下(L)で、CRPの量が減少している。複数回の実験により、その差を定量したものを下段に示した。
【図3】橙色光(600nm)照射による大腸菌内Pcpc-CRP発現量の上方調節。Dは暗状態を、Lは光照射条件下での試料を表す。His6は、6個のHisが連続したいわゆるヒスチジンタグを意味し、ASRはアナベナセンサリーロドプシンを、ASRTは、アナベナセンサリーロドプシンの情報変換タンパク質を表す。His6-ASRは、タンパク質のN末端にヒスチジンタグを付加したものを、ASR-His6は、タンパク質のC末端にヒスチジンタグを付加したものを表す。pBR322は、プラスミドベクターのみのものを表す。ASRの発現は、ASRの持つ、赤紫色を、目で見て確認した(ASRの発現)。His6-ASRで、光照射条件下(L)で、CRPの量が増加している。複数回の実験により、その差を定量したものを下段に示した。
【図4】色素タンパク質フィコエリスリン及びフィコシアニンの推定プロモーター領域含有500bp(配列番号1、配列番号2)を含む断片(Hind III-Kpn I)の配列(配列番号3、4)。大文字は制限酵素認識配列。
【図5】青紫色光450nmおよび赤色光640nm照射によるCRP量の増減変化。ASRが吸収することが出来ない光(下段:ASRの吸収特性(点線:野生型))では、実施例の4.で示した大腸菌内Ppec-CRPの顕著な発現量減少は起こらない。
【発明を実施するための形態】
【0009】
本発明の第1の局面は所望のタンパク質の発現量を制御可能なタンパク質発現法に関する。本発明のタンパク質発現法は、以下のステップ(1)及び(2)によって特徴付けられる。
(1)アナベナセンサリーロドプシン応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトと、遺伝子発現に必要な因子とを含み、且つその中でアナベナセンサリーロドプシンが発現している区画を用意するステップ
(2)前記アナベナセンサリーロドプシンを活性化可能な波長の光を前記区画内に照射するステップ
【0010】
ステップ(1)は、本発明の最大の特徴である「光によるタンパク質発現量の制御」が可能となるように、必要な材料や要素などを準備するステップである。当該ステップではその中に所定の発現コンストラクトと遺伝子発現用の因子を含み、且つその中でアナベナセンサリーロドプシン(ASR)が発現している区画を用意する。
【0011】
本発明では区画内に、第1の要素として、ASR応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトが含まれる。色素タンパク質プロモーターは、ASR応答性を示す限り特に限定されない。プロモーターの例を挙げると、フィコシアニン色素オペロンのプロモーター、フィコエリスリン色素オペロンのプロモーター、ASRオペロンのプロモーター、Kaiオペロンのプロモーター、フィコビリオペロンのプロモーターである。好ましくは、ASRによる良好な応答性を実現すべく、アナベナ属由来のプロモーターを採用する。ここで、色素タンパク質プロモーターの具体例を添付の配列表の配列番号1と配列番号2に示す。配列番号1は、アナベナ属フィコエリスリン色素オペロンの推定プロモーター領域を含む配列(オペロンの上流約500bp)であり、配列番号2はアナベナ属フィコシアニン色素オペロンの推定プロモーター領域を含む配列(オペロンの上流約500bp)である。ASRに対する応答性を保持する限り、これらの配列の一部(例えばオペロンの上流480bp、450bp、4200bp、390bp、360bp)を用いてもよい。応答性を保持するか否かの確認は、例えば、後述の実施例に示した発現系を利用して行えばよい。一方、上記配列に対して一部の塩基の置換、欠失、付加等の改変を施すことによって得られる配列を用いることも可能である。改変体が応答性を保持するか否かについても、例えば、後述の実施例に示した発現系を利用して容易に確認できる。
【0012】
発現コンストラクトは、ASR応答性の色素タンパク質プロモーターに加え、当該プロモーターの制御下にある目的タンパク質遺伝子を含む。「制御下にある」とは、「作動可能に連結している」と同義であり、通常、プロモーターの3'末端側に直接又は他の配列を介して目的タンパク質遺伝子が連結されることになる。
【0013】
「目的タンパク質遺伝子」とは、本発明の方法によって発現するタンパク質(即ち目的物)をコードする遺伝子である。目的や用途に応じて、任意の目的タンパク質遺伝子を採用することができる。目的タンパク質の例を挙げると、医薬や食品等の成分として有用なタンパク質(例えばインスリン、エリスロポエチン等のホルモン、インターフェロン、インスリン様成長因子、上皮成長因子(EGF)、線維芽細胞増殖因子(FGF)、インターロイキン類等の増殖因子・成長因子、抗体、酵素、)、試薬として有用なタンパク質(例えば各種酵素)、機能未知のタンパク質である。
【0014】
発現コンストラクトが、二以上の目的タンパク質遺伝子を発現可能な状態で含んでいてもよい。換言すれば、複数の目的タンパク質を発現可能な発現コンストラクトを採用してもよい。このようなコンストラクトは、例えば、複数のプロモーターやIRES(Internal Ribosome Entry Site)の使用によって構築できる。尚、異なる二以上の発現コンストラクトを併用することによって二以上の目的タンパク質が発現するように構成してもよい。この場合において、一部の発現コンストラクトのみ(例えば、二つの発現コンストラクトを使用するのであれば片方の発現コンストラクト)ASRの制御を受けるように構成してもよい。このように構成すれば、ASRによる発現制御と、他の手段による発現制御を併用した発現系となる。当該発現系は、複数のタンパク質の発現を個別に制御する目的に有用である。
【0015】
区画内には、第2の要素として、遺伝子発現に必要な因子が含まれる。「遺伝子発現に必要な因子」とは、遺伝子の発現、即ち遺伝子の転写及び翻訳が生じ、遺伝子の発現産物であるタンパク質が合成されるために必要な要素であり、RNAポリメラーゼ、転写基本因子、転写調節因子、転写伸長因子、アミノアシルtRNA合成酵素、各種アミノ酸、ATP、GTPなどのエネルギー源等が該当する。本発明では「遺伝子発現に必要な因子」が区画内に含まれることになるが、後述のように細胞を「区画」として用いた場合には、「遺伝子発現に必要な因子」として当該細胞に内在するものを用いることができる。即ち、大腸菌細胞のように、遺伝子発現に必要な一揃いの因子を備える場合、別途、「遺伝子発現に必要な因子」を供給する必要はない。但し、発現の安定化や効率化などを目的に一部又は全部の因子を補充することにしてもよい。また、一部の因子を欠損する細胞を採用した場合などは、当該因子を外部から補充すればよい。
【0016】
細胞ではなく、リポソーム等を用いて「区画」を形成した場合(詳細は後述する)には、通常、「遺伝子発現に必要な因子」として区画内に無細胞タンパク質合成系が構築される。本発明において無細胞合成系(無細胞転写/翻訳系)とは、生細胞を用いるのではく、生細胞由来の(或いは遺伝子工学的手法で得られた)リボソームや転写・翻訳因子などを用いて、鋳型である核酸からそれがコードするタンパク質をin vitroで合成することをいう。無細胞合成系では一般に、細胞破砕液を必要に応じて精製して得られる細胞抽出液が使用される。細胞抽出液には一般に、タンパク質合成に必要なリボソーム、開始因子などの各種因子、tRNAなどの各種酵素が含まれる。タンパク質の合成を行う際には、この細胞抽出液に各種アミノ酸、ATP、GTPなどのエネルギー源、クレアチンリン酸など、タンパク質の合成に必要なその他の物質を添加する。勿論、タンパク質合成の際に、別途用意したリボソームや各種因子、及び/又は各種酵素などを必要に応じて補充してもよい。
【0017】
タンパク質合成に必要な各分子(因子)を再構成した転写/翻訳系の開発も報告されている(Shimizu, Y. et al.: Nature Biotech., 19, 751-755, 2001)。この合成系では、バクテリアのタンパク質合成系を構成する3種類の開始因子、3種類の伸長因子、終結に関与する4種類の因子、各アミノ酸をtRNAに結合させる20種類のアミノアシルtRNA合成酵素、及びメチオニルtRNAホルミル転移酵素からなる31種類の因子の遺伝子を大腸菌ゲノムから増幅し、これらを用いてタンパク質合成系をin vitroで再構成している。このような再構成した合成系を利用することにしてもよい。
【0018】
無細胞タンパク質合成系には以下の利点がある。まず第1に、生細胞を維持する必要がないため操作性が良好で系の自由度も高い。したがって、目的のタンパク質の性質に応じて様々な修正や修飾を施した合成系を設計することが可能となる。次に、細胞系の合成では使用する細胞に毒性のあるタンパク質の合成は基本的にできないが、無細胞系ではそのような毒性のタンパク質であっても生産することができる。さらに、多種類のタンパク質を同時にかつ迅速に合成できることからハイスループット化が容易である。生産されるタンパク質の分離・精製が容易であるという利点も備え、これはハイスループット化に有利に働く。加えて、非天然型のアミノ酸を取り込ませるなどして非天然型タンパク質を合成することも可能であるという利点も併せ持つ。
【0019】
現在広く利用されている無細胞タンパク質合成系には以下のものがある。即ち、大腸菌S30抽出液の系(原核細胞の系)、コムギ胚芽抽出液の系(真核細胞の系)、及びウサギ網状赤血球可溶化物の系(真核細胞の系)である。これらの系はキットとしても市販されており、容易に利用することが可能である。
【0020】
歴史的には大腸菌S30抽出液の系の開発が最も古く、この系を利用して様々なタンパク質の合成が試みられてきた。大腸菌30S画分は、大腸菌の集菌、菌体破砕、精製の工程を経て調製される。大腸菌30S画分の調製及び、無細胞転写・翻訳共役反応はPrattらの方法(Pratt, J. M.: Chapter 7, in “Transcription and Translation: A practical approach”, ed. by B. D. Hames & S. J. Higgins, pp. 179-209, IRL Press, New York (1984))やEllmanらの方法(Ellman, J. et al.: Methods Enzymol., 202, 301-336(1991))を参考にして行うことができる。
【0021】
コムギ胚芽抽出液の系は、高品質の真核生物タンパク質を効率的に合成できるという利点を有し、大腸菌S30抽出液の系では合成が困難な真核生物のタンパク質を合成する際によく利用される。最近になって、種子胚乳成分を洗浄除去した胚芽から抽出液を調製することによって高効率かつ安定な合成系が構築されることが報告され注目を集めている(Madin, K. et al.: Proc. Natl. Acad. Sci. USA, 97: 559-564, 2000)。その後、高翻訳促進能を有するmRNA非翻訳配列、PCRを利用した多品目機能解析用のタンパク質合成法、専用高発現ベクターの構築などの技術開発が行われ(Sawasaki, T. et al.: Proc. Natl. Acad. Sci. USA, 99: 14652-14657, 2002)、様々な分野への応用が期待されている。
【0022】
コムギ胚芽抽出液は、コムギ胚芽をすり潰して遠心分離した後、上澄み液をゲルろ過で分離することによって得ることができる。翻訳反応については、Andersonらの方法(Anderson, C. W. et al.: Methods Enzymol., 101, 638-644(1983))を参考にできる。改良法についても報告されており、例えば河原崎らの方法(Kawarasaki, Y. et al.: Biotechnol. Prog., 16, 517-521(2000))やMadinらの方法(Madin, K. et al.: Proc. Natl. Acad. Sci. USA, 97: 559-564, 2000)等を参考にできる。その他、コムギ胚芽抽出液の系についてはWO 00/68412 A1、WO 01/27260 A1、WO 2002/024939 A1、WO 2005/063979 A1、特開平6-7134号公報、特開2002-529531号公報、特開2005-355513号公報、特開2006-042601号公報、特開2007-097438号公報、特開2008-029203号公報等が参考になる。
【0023】
ウサギ網状赤血球可溶化物の系はグロブリン生産に適する。ウサギ網状赤血球可溶化物は、ウサギにフェニルヒドラジンを数日間静脈注射して貧血状態とし、所定期間後(例えば第8日目)に採血し、その後溶血させた液から超遠心分離処理などを経て得られる。ウサギ網状赤血球可溶化物の調製法は、JacksonとHuntの方法(Jackson, R. J. and Hunt, T.: Methods Enzymol., 96, 50-74(1983))を参考にして行うことができる。
【0024】
尚、本発明の実施に際して利用できる無細胞合成系は上記のものに限られるものではなく、例えば大腸菌以外のバクテリアやコムギ以外の植物の抽出液、昆虫由来の抽出液、動物細胞由来の抽出液、又はゲノム情報を基に構築した系などを利用してもよい。
【0025】
本発明における区画内ではASRが発現している。ASRは光受容体として機能する。即ち、本発明では、ASRが光を受けて活性化し(詳細は後述のステップ(2)の欄で説明する)、発現コンストラクト中のプロモーターに作用する。
【0026】
区画内でのASRの発現は恒常的でも一過性でもよい。例えば、区画として細胞を利用し、当該細胞のゲノム内に予めASR遺伝子を組み込んでおくことによって、区画内でASRを発現させることができる。この態様ではステップ(1)として、例えば、宿主細胞のゲノムにASR(天然型又は変異型)を組み込むステップと、ASR応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトを当該宿主細胞に導入するステップとを行う。これら二つのステップは基本的には順不同であるが、操作の容易性及び効率の点から前者を先に行うことが好ましい。この態様において、ASR遺伝子を恒常的(構成的)プロモーターの制御下に配置しておけば、区画内でASRを恒常的に発現させることができる。ゲノム内にASR遺伝子を組み込むのではなく、恒常的プロモーターの制御下にあるASR遺伝子を含む発現コンストラクトを区画内に導入することによっても、区画内でのASRの恒常的発現を実現可能である。一方、組織特異的プロモーター、発育段階特異的プロモーター、誘導性プロモーター等を使用すれば、プロモーターの特性に応じた所望の発現(例えば、所望の時期に一過性に発現すること)が可能である。プロモーターの例を挙げると、lacプロモーター、tacプロモーター、trpプロモーター、T7プロモーター、α−アミラーゼプロモーター、β−アクチンプロモーター、L7プロモーター、CMVプロモーターである。
【0027】
宿主細胞のゲノム内に予めASR遺伝子を組み込んでおくのではなく、ASR遺伝子を含む発現コンストラクトの導入によってASRを発現させる場合にはステップ(1)として、例えば、宿主細胞内で機能するプロモーターの制御下にあるASR遺伝子を含む発現コンストラクトを宿主細胞に導入するステップと、ASR応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトを当該宿主細胞に導入するステップとを行う。これら二つのステップは基本的には順不同である。但し、前者のステップのみを行った宿主細胞を用意しておき、必要なときに後者のステップを行うという利用形態を採用すれば、任意のタンパク質の発現に利用できる汎用的な発現系を提供できることになる。
【0028】
光受容体として機能する限り、ASRは天然型でも変異型でもよい。天然型のASRのアミノ酸配列の一例を配列番号5に示す。また、当該アミノ酸配列をコードする遺伝子の配列を配列番号6に示す。天然型のASRは波長550nm付近に吸収極大を有する。天然型ASRの活性化には約500 nm〜約600 nmの波長の光を利用可能である。変異型(例えば波長485nm〜575nmに吸収極大を有する)を利用することにすれば、天然型の場合には利用不可能な又は活性化効率や細胞毒性などの点で利用し難い波長の光によってASRを活性化することが可能となる。これまで、ASRの類似タンパク質(いわゆる微生物型ロドプシン)において、様々な変異実験やレチナールの類似化合物を用いた発色団置換実験が行われ、様々な極大吸収波長を示すタンパク質が得られてきた。改変によって吸収極大が短波長側(例えば最大短波長化:498 nm)にシフトするように改変した変異体を使用することの利点は、例えば、汎用光源であるNd:YAGレーザー(532 nm)やアルゴンレーザー(488〜514 nm)の使用が可能なこと、及び有毒な紫外部の光により励起可能であり、細胞死が起こりながら実際の細胞や生体中でどのような事象が起きているのかを調べることも可能になることである。これとは逆に吸収極大が長波長側(例えば最大短波長化:575 nm)にシフトするように改変した変異体を使用することは、長波長光の組織透過性が高いことから、ヒトなど厚い組織(例えば皮膚など)に覆われている場合でもタンパク質を励起できる点などで有利である。これまでの研究によって、ロドプシンの吸収波長の決定に重要な部位(アミノ酸位置)が報告されている(例えば、Yokoyama S. Prog Retin Eye Res. 2000 Jul;19(4):385-419, 神取秀樹、加茂直樹 蛋白質核酸酵素 2002 Apr;47(5):620-5.を参照)。また、吸収波長のシフトに関する報告がいくつか存在する(例えばYokoyama S. Prog Retin Eye Res. 2000 Jul;19(4):385-419; 神取秀樹、加茂直樹 蛋白質核酸酵素 2002 Apr;47(5):620-5.; Shimono K, Hayashi T, Ikeura Y, Sudo Y, Iwamoto M, Kamo N. J Biol Chem. 2003 Jun 27;278(26):23882-9.を参照)。光学特性(特に吸収スペクトル)が天然型と異なる変異型を調製するためには、天然型ASRとASRの類似タンパク質との間で配列や立体構造などを比較し、天然型ASRの配列の中で吸収波長の決定に重要な部位を特定した上で、当該部位を標的として部位特異的変異誘発法(例えば、ストラタジーン社のクイックチェンジ(QuickChangeTM)を用いることができる)等による変異を加えればよい。
【0029】
本発明における用語「区画」は、タンパク質合成の場を与えるものであり、タンパク質合成及び本発明の作用・効果(即ち光によるタンパク質量の制御)の発揮に必要な因子ないし要素の保持及び維持を行う。典型的には、区画全体に亘ってその内部と外部を隔てる閉じた区画(閉鎖系)が用意される。但し、培養プレートのウェルに代表されるように、一部が開放した区画を用いることを妨げない。本発明の「区画」として例えば細胞が利用される。この態様では細胞をタンパク質合成の場として用いることになる。利用可能な細胞の例として、大腸菌などのグラム陰性菌、バシラス属細菌などのグラム陽性菌、藍藻(例えばアナベナ属)などの真正細菌全般、高度好塩菌などの古細菌、カビ、酵母や培養細胞などの真核生物など、が挙げられる。これらの中でも、本発明に好適なものとして、大腸菌、バシラス属細菌、藍藻が挙げられ、特に好適なものとして大腸菌が挙げられる。大腸菌は、汎用性が高い点、分裂速度が極めて速い点、遺伝子操作が容易である点、光感受性タンパク質がほとんど無い点等から、特に好適な細胞である。また、藍藻細胞(特にアナベナ属細胞)を採用した場合には、発現制御に利用する要素であるASRと細胞が同属又は近縁の関係になり、ASRの機能発揮に有利である。尚、人工的に構築した細胞(いわゆる人工細胞)を利用することにしてもよい。
【0030】
本発明の「区画」としてリポソーム、バイセル、エマルジョン、各種容器(培養プレート、培養皿、試験管、遠心管、遠心チューブ、マイクロテストチューブ)等を用いることもできる。エマルジョンとして例えばwater-in-oil(以下W/O)エマルジョンやW/O/W エマルジョン(double emulsion)が利用可能である。W/Oエマルジョンはin vitro compartmentalization(IVC)法に利用されている(Tawfik, D.S. & Griffiths, A.D.(1998)Nat. Biotechnol ., 7,652-656)。Tawfikらは、界面活性剤を加えた油相に水相を加え,撹拌することで形成されるW/O エマルジョンの水相を無細胞タンパク質合成反応の場としている。一方、Bernath らは、W/O/Wエマルジョンを利用したIVC法の開発にも成功している(Bernath, K., Hai, M., Mastrobattista, E., Griffiths, A.D., Magdassi, S., & Tawfik, D.S.(2004)Anal. Biochem., 325, 151-157)。尚、リポソームやエマルジョン等の人工材料を使用する場合、その修飾(糖修飾、抗体修飾、アルブミン修飾、ポリエチレングリコール(PEG)修飾など)が可能である。このような修飾は例えば、「区画」の安定化や最適化などの他、「区画」自体への機能の付加(例えば、生体の特定の部位へのタンパク質の送達・発現などに利用する場合における指向性の付加)などを目的として行われる。
【0031】
上で説明した通り、細胞ではなく、リポソーム等を用いて「区画」を形成する場合には、通常、無細胞タンパク質合成系を利用する。この態様においてはステップ(1)として、例えば、無細胞合成系を構成する因子等を区画(リポソーム等)内に用意するステップと、ASR用の発現コンストラクトと、ASR応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトを当該区画内に導入するステップとを行う。或いは、無細胞合成系を構成する因子等、ASR用の発現コンストラクト、ASR応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトを混合した後、リポソーム等を用いた区画化を行う。
【0032】
ステップ(1)に続くステップ(2)では、区画内で発現しているASRを活性化可能な波長の光を区画内に照射する。これによってASRが活性化し、区画内に存在する発現コンストラクト中のASR応答性プロモーターに作用する。その結果、プロモーターの特性に応じて、目的タンパク質遺伝子の転写が開始、促進、停止又は抑制される。例えば、フィコシアニン色素オペロンのプロモーターを用いた場合には、その制御下にある目的タンパク質遺伝子の転写が促進し、タンパク質の発現量が増大する(後述の実施例を参照)。また、フィコエリスリン色素オペロンのプロモーターを使用すれば、目的タンパク質の発現量が低減する(発現が抑制される)。このように本願発明によれば、目的タンパク質の発現量を光の照射という、簡便且つ容易でありしかも非侵襲的な手段によって制御可能である。
【0033】
照射する光の波長は、使用するASRに応じて決定すればよい。例えば波長550nm付近に吸収極大を有する天然型ASRを使用した場合は、約500 nm〜約600 nmの波長の光(具体的には例えば波長600nmの光)を照射すればよい。効率の点からは、ASRの吸収極大の波長の光を採用することが好ましい一方で、細胞毒性を回避するために、吸収極大よりも長波長側の光を照射するとよい。ASRを活性化可能な波長領域の光を含む限り、広スペクトル光、狭スペクトル光のいずれも利用可能である。また、光源についても特に制約はない。光源の例は冷陰極管、ハロゲンランプ、LED(発光ダイオード)、レーザーである。
【0034】
本発明の第2の局面は、本発明のタンパク質発現法に用いられるシステム(タンパク質発現システム)を提供する。本発明のタンパク質発現システムは、(1)遺伝子発現に必要な因子と、(2)アナベナセンサリーロドプシン、又は恒常的若しくは一過性に発現可能な状態のアナベナセンサリーロドプシン遺伝子と、(3)アナベナセンサリーロドプシン応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトと、を含む区画から構成される。本発明のタンパク質発現システムに用いる各要素については既述の通りであるので、対応する説明を援用することとし、ここでの重複した説明は省略する。本発明のタンパク質発現システムは、例えば適当な容器内に或いは容器を利用して(区画に容器を用いる場合)構築される。構築したタンパク質発現システムを生体に導入し、生体内で作動させることも可能である。このような利用形態は、タンパク質の機能解明やタンパク質を利用した医療(例えば生体の特定の部位へのタンパク質の送達・発現)の新たな手段を提供する。
以下、実施例(大腸菌を宿主として利用した発現実験)を用いて本発明を更に詳細に説明する。
【実施例】
【0035】
1.アナベナ推定プロモーター融合レポータータンパク質、cAMP receptor protein(CRP)発現プラスミドの作製(図1を参照)
Anabaena PCC7120株のゲノム(名古屋大学遺伝子実験施設の井原邦夫助教より分与)を鋳型として制限酵素Hind IIIおよびKpn I切断部位を含むプライマーDNAを用いてPCR法により増幅させた色素タンパク質(フィコエリスリンおよびフィコシアニン)コード領域上流の推定標的プロモーター領域(PpecおよびPcpc)を含有する増幅断片500bpを制限酵素(New England Biolabs、タカラバイオ株式会社)で切断した(Hind III-Kpn I)(図4)。切断断片をレポータータンパク質CRP発現プラスミドpHA7MK(アンピシリン耐性、鈴鹿医療科学大学の饗場弘二教授より分与、Aiba, H., Fujimoto, S. and Ozaki, N. (1982). “Molecular cloning and nucleotide sequencing of the gene for E. coli cAMP receptor protein”. Nucleic Acids Research 10: 1345-1361)のCRP開始コドン上流の制限酵素切断部位(Hind III-Kpn I)に挿入し、大腸菌DH5a株(Grant, S. G., Jessee, J., Bloom, F. R. and Hanahan, D. (1990). “Differential plasmid rescue from transgenic mouse DNAs into Escherichia coli methylation-restriction mutants”. Proc. Natl. Acad. Sci. USA 87: 4645-4649)に導入した。形質転換したDH5α株から一般的なアルカリ溶菌法によるミニプレップ法で作製したPpec-CRPおよびPcpc-CRP(以下まとめてPxxx-CRPと呼ぶ)発現プラスミドpHI103およびpHI104を回収した。pHI103およびpHI104のPxxx-CRPコード領域を含む制限酵素断片(Hind III-EcoR I)をpSU21(クロラムフェニコール耐性、Bartolome et al., 1991)のMCSの制限酵素部位(Hind III-EcoR I)に挿入し、大腸菌DH5α株に導入した。形質転換したDH5α株から一般的なアルカリ溶菌法によるミニプレップ法で作製したPxxx-CRP発現プラスミドpHI213およびpHI214(クロラムフェニコール耐性)を回収した。
【0036】
2.ASR発現大腸菌株のコンピテントセルの調製
His6-ASR発現プラスミドpKJ900(His6-ASR)、ASR-His6発現プラスミドpKJ900(ASR-His6)、His6-ASR-ASRT Fusion発現プラスミドpKJ900(His6-ASR-ASRT Fusion)、His6-ASR発現プラスミドpKJ900(His6-ASR)、ASRT-His6発現プラスミドpKJ900(ASRT-His6)(IPTG誘導型、アンピシリン耐性、以上のプラスミドは西江大学のKwang-Hwan Jung 准教授より分与、Jung, K. H., Trivedi, V. D. and Spudich, J. L. (2003). “Demonstration of a sensory rhodopsin in eubacteria”. Mol. Microbiol. 47: 1513-1522; Kawanabe, A., Furutani, Y., Yoon, S. R., Jung, K. H. and Kandori, H. (2008). “FTIR Study of the L Intermediate of Anabaena Sensory Rhodopsin: Structural Changes in the Cytoplasmic Region”. Biochemistry 47: 10033-10040)およびコントロールプラスミドpBR322(Bolivar, F., Rodriguez, R. L., Greene, P. J., Betlach, M. C., Heyneker, H. L., Boyer, H. W., Crosa, J. H. and Falkow, S. (1977). “Construction and characterization of new cloning vehicles. II. A multipurpose cloning system”. Gene 2(2): 95-113) をE. Coli TA341株(E. coli W3110 Dcrp、 鈴鹿医療科学大学の饗場弘二教授より分与、Sunohara, T., Jojima, K., Yamamoto, Y., Inada, T. and Aiba, H. (2004). “Nascent-peptide-mediated ribosome stalling at a stop codon induces mRNA cleavage resulting in nonstop mRNA that is recognized by tmRNA”. RNA 10: 378-386)のコンピテントセルに導入した。形質転換したASR発現プラスミド導入済みE. Coli TA341株を一般的な塩化カルシウム法で調製した。ここでHis6とは、アミノ酸の1種であるヒスチジンが6個連続した配列の事で、この配列に対する抗体(GE Healthcare)により、タンパク質をウェスタンブロッティングにより簡便に検出できること、Ni-NTA樹脂(GE Healthcare)などによりタンパク質を簡便に精製出来ることから付加したものである。
【0037】
3.ASRおよびPxxx-CRP発現大腸菌株の調製
1.で作製したPxxx-CRP発現プラスミドpHI213およびpHI214を2.で調製したASR発現プラスミド導入済みE. Coli TA341株のコンピテントセルに導入した。アンピシリン及びクロラムフェニコールを、それぞれ終濃度50μg/ml及び25μg/ml含む、通常の大腸菌用寒天培地(LB培地: 1% Bacto Tryptone、0.5% Yeast Extract、1% NaCl)にてセレクションし、目的菌株を得た。
【0038】
4.橙色光(600nm)照射による大腸菌内Ppec-CRP発現量の下方調節(図2)
3.で調製したASRおよびPpec-CRP発現大腸菌株を終濃度50μg/mlアンピシリンおよび25μg/mlクロラムフェニコールを含むLB培地で30℃、遮光状態で終夜、振盪培養した(一次培養)。一次培養した大腸菌を終濃度1mM IPTG, 10μM all-trans型レチナールを含むLB培地に終濃度50μg/mlアンピシリンおよび25μg/mlクロラムフェニコールを添加した溶液で25℃、8時間、二次培養した後、橙色光(600nm±10nmを透過するフィルター(Melles Griot)を通した白色光)を30分間照射しながら振盪速度を落として培養した。照射光源は、CL-250(ELMO)を用い、その際の光量を光量計により測定したところ、1.8mWであった。培養・橙色光(600nm)照射した菌を遠心(15,000rpm, 4℃, 4分)により集菌し、上清を除去した。沈澱に蒸留水、SDS-loading buffer(400 mM Tris-HCl buffer (pH 6.8)、6% Sodium dodecyl sulfate (SDS)、38.25% Glycerol、0.02% Bromophenol Blue)、2-メルカプトエタノールを添加後、100℃で4分間煮沸し、氷上で急冷してサンプルとした。調製したサンプルを一般的なSDS-PAGE(15%ゲル)、ウェスタンブロッティング(ウサギ抗CRP抗体(Ishizuka H., Hanamura A., Kunimura T. and Aiba H. (1993). “A lowered concentration of cAMP receptor protein caused by glucose is an important determinant for catabolite repression in Escherichia coli. Mol. Microbiol., 10: 341-350.)+ヤギ抗ウサギHRP抗体(Santa Cruz Biotechnology))を用いて解析し、橙色光(600nm)照射によるPpec-CRP量の変化を測定した。Ppec-CRP量は橙色光(600nm)照射により減少するという結果を得た。複数の実験による平均値は、1/4.4に低下するものであった。この結果は、光照射により人為的にタンパク質量を減少させる調節法の構築という点において、初の成果である。
【0039】
5.橙色光(600nm)照射による大腸菌内Pcpc-CRP発現量の上方調節(図3)
3.で調製したASRおよびPpec-CRP発現大腸菌株を終濃度50μg/mlアンピシリンおよび25μg/mlクロラムフェニコールを含むLB培地で30℃、遮光状態で終夜、振盪培養した(一次培養)。一次培養した大腸菌を終濃度1mM IPTG, 10μM all-trans 型レチナールを含むLB培地に終濃度50μg/mlアンピシリンおよび25μg/mlクロラムフェニコールを添加した溶液で25℃、8時間、二次培養した後、橙色光(600nm±10nmを透過するフィルター(Melles Griot)を通した白色光)を30分間照射しながら振盪速度を落として培養した。照射光源は、CL-250(ELMO)を用い、その際の光量を光量計により測定したところ、1.8mWであった。培養・橙色光(600nm)照射した菌を遠心(15,000rpm, 4℃, 4分)により集菌し、上清を除去した。沈澱に蒸留水、SDS-loading buffer(400 mM Tris-HCl buffer (pH 6.8)、6% Sodium dodecyl sulfate (SDS)、38.25% Glycerol、0.02% Bromophenol Blue)、2-メルカプトエタノールを添加後、100℃で4分間煮沸し、氷上で急冷してサンプルとした。調製したサンプルを一般的なSDS-PAGE(15%ゲル)、ウェスタンブロッティング(ウサギ抗CRP抗体(Ishizuka H., Hanamura A., Kunimura T. and Aiba H. (1993). “A lowered concentration of cAMP receptor protein caused by glucose is an important determinant for catabolite repression in Escherichia coli. Mol. Microbiol., 10: 341-350.)+ヤギ抗ウサギHRP抗体(Santa Cruz Biotechnology))を用いて解析し、橙色光(600nm)照射によるPcpc-CRP量の変化を測定した。Pcpc-CRP量は橙色光(600nm)照射により増加するという結果を得た。複数の実験による平均値は、2.3倍に増加するものであった。この結果は、光照射により人為的にタンパク質量を増加させる調節法の構築という点において、初の成果である。
【0040】
6.他の励起波長(450 nm, 640 nm)におけるCRP量の増減変化(図5)
3.で調製したASRおよびPpec-CRP発現大腸菌株を終濃度50 mg/ml アンピシリンおよび25 mg/ml クロラムフェニコールを含むLB培地で30℃、遮光状態で終夜、振盪培養した(一次培養)。一次培養した大腸菌を終濃度1 mM IPTG, 10 mM all-trans 型レチナールを含むLB培地に終濃度50 mg/ml アンピシリンおよび25 mg/ml クロラムフェニコールを添加した溶液で25℃、8時間、二次培養した後、青紫色光(450nm±8nm)もしくは赤色光(640nm±11nm)を透過するフィルター(Asahi Spectra)を通した白色光を30分間照射しながら振盪速度を落として培養した。照射光源は、MAX-302(Asahi Spectra)を用い、その際の光量を光量計により測定したところ、それぞれ1.2mW(青紫色光)および1.6mW(赤色光)であった。培養・光照射した菌を遠心(15,000rpm, 4℃, 4分)により集菌し、上清を除去した。沈澱に蒸留水、SDS-loading buffer(400 mM Tris-HCl buffer (pH 6.8)、6% Sodium dodecyl sulfate (SDS)、38.25% Glycerol、0.02% Bromophenol Blue)、2-メルカプトエタノールを添加後、100℃で4分間煮沸し、氷上で急冷してサンプルとした。調製したサンプルを一般的なSDS-PAGE(15%ゲル)、ウェスタンブロッティング(ウサギ抗CRP抗体(Ishizuka H., Hanamura A., Kunimura T. and Aiba H. (1993). “A lowered concentration of cAMP receptor protein caused by glucose is an important determinant for catabolite repression in Escherichia coli. Mol. Microbiol., 10: 341-350.)+ヤギ抗ウサギHRP抗体(Santa Cruz Biotechnology))を用いて解析し、青紫色光(450nm)もしくは赤色光(640nm)照射によるPpec-CRP量の変化を測定した。Ppec-CRP量は青紫色光(450nm)もしくは赤色光(640nm)照射では変化しないという結果を得た。この結果は、ASRが吸収しない波長の光では、上述の4.で示したタンパク質量の変化(減少)は起こらず、ASRが実際にこのような反応に寄与していることを示している。
【産業上の利用可能性】
【0041】
本発明のタンパク質発現法・発現システムによれば、任意のタンパク質の発現量を光で制御可能である。本発明はタンパク質生産手段或いはタンパク質発現制御手段として様々な用途で利用可能である。例えば、タンパク質の機能解明用ツールや創薬ツール、或いは医療用ツール等としての利用が想定される。
【0042】
この発明は、上記発明の実施の形態及び実施例の説明に何ら限定されるものではない。特許請求の範囲の記載を逸脱せず、当業者が容易に想到できる範囲で種々の変形態様もこの発明に含まれる。
本明細書の中で明示した論文、公開特許公報、及び特許公報などの内容は、その全ての内容を援用によって引用することとする。
【特許請求の範囲】
【請求項1】
以下のステップ(1)及び(2)を含む、タンパク質発現法:
(1)アナベナセンサリーロドプシン応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトと、遺伝子発現に必要な因子とを含み、且つその中でアナベナセンサリーロドプシンが発現している区画を用意するステップ;及び、
(2)前記アナベナセンサリーロドプシンを活性化可能な波長の光を前記区画内に照射するステップ。
【請求項2】
アナベナセンサリーロドプシン遺伝子を含む発現コンストラクトから前記アナベナセンサリーロドプシンが供給される、請求項1に記載のタンパク質発現法。
【請求項3】
前記区画が細胞からなる、請求項1又は2に記載のタンパク質発現法。
【請求項4】
前記細胞が大腸菌、バシラス属細菌又はアナベナ属藍藻である、請求項3に記載のタンパク質発現法。
【請求項5】
前記細胞のゲノム内にアナベナセンサリーロドプシン遺伝子が予め組み込まれており、該遺伝子が発現することによって前記アナベナセンサリーロドプシンが供給される、請求項3又は4に記載のタンパク質発現法。
【請求項6】
前記アナベナセンサリーロドプシン遺伝子が恒常的に発現している、請求項5に記載のタンパク質量発現法。
【請求項7】
前記区画が、その中に無細胞合成系が形成された区画である、請求項1又は2に記載のタンパク質発現法。
【請求項8】
前記区画がリポソーム又はエマルジョンからなる、請求項1、2又は7に記載のタンパク質発現法。
【請求項9】
前記アナベナセンサリーロドプシンが配列番号5の配列を含む、請求項1〜8のいずれか一項に記載のタンパク質発現法。
【請求項10】
前記アナベナセンサリーロドプシンが、天然型のアミノ酸配列の一部を改変して得られた変異型であり、その光学特性が天然型と異なる、請求項1〜8のいずれか一項に記載のタンパク質発現法。
【請求項11】
前記変異型が波長485nm〜575nmに吸収極大を有する、請求項10に記載のタンパク質発現法。
【請求項12】
前記発現コンストラクトが、二以上の目的タンパク質遺伝子を発現可能な状態で含む、請求項1〜11のいずれか一項に記載のタンパク質発現法。
【請求項13】
目的タンパク質遺伝子が異なる二以上の発現コンストラクトが前記発現コンストラクトとして前記区画内に内包されている、請求項1〜11のいずれか一項に記載のタンパク質発現法。
【請求項14】
前記色素タンパク質プロモーターが、フィコエリスリン色素オペロン又はフィコシアニン色素オペロンのプロモーターである、請求項1〜13のいずれか一項に記載のタンパク質発現法。
【請求項15】
前記色素タンパク質プロモーターが、アナベナ属由来のプロモーターである、請求項1〜14のいずれか一項に記載のタンパク質発現法。
【請求項16】
前記色素タンパク質プロモーターが、配列番号1の配列又は配列番号2の配列を含む、請求項1〜13のいずれか一項に記載のタンパク質発現法。
【請求項17】
遺伝子発現に必要な因子と、
アナベナセンサリーロドプシン、又は恒常的若しくは一過性に発現可能な状態のアナベナセンサリーロドプシン遺伝子と、
アナベナセンサリーロドプシン応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトと、
を含む区画からなる、タンパク質発現システム。
【請求項1】
以下のステップ(1)及び(2)を含む、タンパク質発現法:
(1)アナベナセンサリーロドプシン応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトと、遺伝子発現に必要な因子とを含み、且つその中でアナベナセンサリーロドプシンが発現している区画を用意するステップ;及び、
(2)前記アナベナセンサリーロドプシンを活性化可能な波長の光を前記区画内に照射するステップ。
【請求項2】
アナベナセンサリーロドプシン遺伝子を含む発現コンストラクトから前記アナベナセンサリーロドプシンが供給される、請求項1に記載のタンパク質発現法。
【請求項3】
前記区画が細胞からなる、請求項1又は2に記載のタンパク質発現法。
【請求項4】
前記細胞が大腸菌、バシラス属細菌又はアナベナ属藍藻である、請求項3に記載のタンパク質発現法。
【請求項5】
前記細胞のゲノム内にアナベナセンサリーロドプシン遺伝子が予め組み込まれており、該遺伝子が発現することによって前記アナベナセンサリーロドプシンが供給される、請求項3又は4に記載のタンパク質発現法。
【請求項6】
前記アナベナセンサリーロドプシン遺伝子が恒常的に発現している、請求項5に記載のタンパク質量発現法。
【請求項7】
前記区画が、その中に無細胞合成系が形成された区画である、請求項1又は2に記載のタンパク質発現法。
【請求項8】
前記区画がリポソーム又はエマルジョンからなる、請求項1、2又は7に記載のタンパク質発現法。
【請求項9】
前記アナベナセンサリーロドプシンが配列番号5の配列を含む、請求項1〜8のいずれか一項に記載のタンパク質発現法。
【請求項10】
前記アナベナセンサリーロドプシンが、天然型のアミノ酸配列の一部を改変して得られた変異型であり、その光学特性が天然型と異なる、請求項1〜8のいずれか一項に記載のタンパク質発現法。
【請求項11】
前記変異型が波長485nm〜575nmに吸収極大を有する、請求項10に記載のタンパク質発現法。
【請求項12】
前記発現コンストラクトが、二以上の目的タンパク質遺伝子を発現可能な状態で含む、請求項1〜11のいずれか一項に記載のタンパク質発現法。
【請求項13】
目的タンパク質遺伝子が異なる二以上の発現コンストラクトが前記発現コンストラクトとして前記区画内に内包されている、請求項1〜11のいずれか一項に記載のタンパク質発現法。
【請求項14】
前記色素タンパク質プロモーターが、フィコエリスリン色素オペロン又はフィコシアニン色素オペロンのプロモーターである、請求項1〜13のいずれか一項に記載のタンパク質発現法。
【請求項15】
前記色素タンパク質プロモーターが、アナベナ属由来のプロモーターである、請求項1〜14のいずれか一項に記載のタンパク質発現法。
【請求項16】
前記色素タンパク質プロモーターが、配列番号1の配列又は配列番号2の配列を含む、請求項1〜13のいずれか一項に記載のタンパク質発現法。
【請求項17】
遺伝子発現に必要な因子と、
アナベナセンサリーロドプシン、又は恒常的若しくは一過性に発現可能な状態のアナベナセンサリーロドプシン遺伝子と、
アナベナセンサリーロドプシン応答性の色素タンパク質プロモーター及びその制御下にある目的タンパク質遺伝子を含む発現コンストラクトと、
を含む区画からなる、タンパク質発現システム。
【図1】


【図2】
【図3】
【図4】

【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2012−39885(P2012−39885A)
【公開日】平成24年3月1日(2012.3.1)
【国際特許分類】
【出願番号】特願2010−181053(P2010−181053)
【出願日】平成22年8月12日(2010.8.12)
【国等の委託研究の成果に係る記載事項】(出願人による申告)国等の委託研究の成果に係る特許出願(平成22年度、独立行政法人科学技術振興機構の戦略的創造研究推進事業個人型研究(さきがけ)における、研究領域「光の利用と物質材料・生命機能」の中で「光機能性・制御性蛋白質による細胞・個体操作」委託事業、産業技術力強化法19条の適用を受ける特許出願)
【出願人】(504139662)国立大学法人名古屋大学 (996)
【Fターム(参考)】
【公開日】平成24年3月1日(2012.3.1)
【国際特許分類】
【出願日】平成22年8月12日(2010.8.12)
【国等の委託研究の成果に係る記載事項】(出願人による申告)国等の委託研究の成果に係る特許出願(平成22年度、独立行政法人科学技術振興機構の戦略的創造研究推進事業個人型研究(さきがけ)における、研究領域「光の利用と物質材料・生命機能」の中で「光機能性・制御性蛋白質による細胞・個体操作」委託事業、産業技術力強化法19条の適用を受ける特許出願)
【出願人】(504139662)国立大学法人名古屋大学 (996)
【Fターム(参考)】
[ Back to top ]