
アポトーシスから膵島細胞を保護するため、そしてその機能を保持するためのEIF−5A1のsiRNAの使用
本発明は、移植のためにドナー組織から分離された膵島の生存性、回収性、及び機能を改善する方法に関し、さらに具体的には、eIF−5A1 siRNAを使用して膵島の生存性及び機能性を向上することに関する。
【発明の詳細な説明】
【技術分野】
【0001】
ランゲルハンスの膵島は、膵臓内でインスリンを生産する細胞を含む多細胞実体である。平均的なヒトで、約百万の膵島を有し、そして膵臓内の細胞の合計の約2〜3%を占める。膵臓はランゲルハンスの膵島を含み、これは、インスリンを生成するβ細胞を取り囲んでいる。β細胞は、血中のグルコースレベルをモニターしており、そして精密に計測された量のインスリンを放出して、グルコースピークに均衡させる。I型及びII型糖尿病は、90%超のこれらのβ細胞が損傷を受けた場合に進行する。
【背景技術】
【0002】
1型糖尿病は、米国において2100万人の糖尿病患者の約10%が罹患するグルコース恒常性についての障害である。1型糖尿病は、膵島β細胞の実質的に完全な自己免疫破壊から生じ、生命維持のためのインスリン投与に患者を依存状態にする。1型糖尿病の発症機序は、免疫系の細胞と、膵島β細胞上に存在する抗原との間の複雑な相互作用に関わっている。この相互作用は、Th1細胞とマクロファージの活性化を導き、これは続いて他の因子のなかで特に、一酸化窒素産生を誘導するサイトカイン(IL1β、TNFα、及びIFNγ)であって、アポトーシス性及び壊死性の膵島β細胞死を引き起こすサイトカインを放出する。膵臓又は膵島移植に関するβ細胞「置換」戦略は、1型糖尿病を回復させる点で一定ではないが成功を収めたが、限られた数のソース(死体ドナー)を必要とし、そしてさらに、内在の自己免疫プロセスにより破壊される。
【0003】
結合マトリクス及び残りの外分泌組織からの膵島の分離又は単離は、研究室での実験及び移植目的の点で利点があり、そして有利である。膵島移植は、1型糖尿病の最も有望で、かつ生理的侵襲の少ない方法である。完全な膵臓組織ではなく膵島を移植することは、移植の容易さ、そして消化酵素の分泌に関わるドナー組織の膵臓外分泌機能が除去されるという確かな利点を有する。膵臓外分泌組織から膵島を遊離することは、膵島移植に影響する最初の及び重大な工程である。膵島単離における重要な目標は、十分な数の生存可能であり、機能を有しかつ強力な移植用の膵島を提供することである。
【0004】
「エドモントンプロトコル」は、健常な膵島を糖尿病患者に移植する。エドモントンプロトコルを用いた膵島移植が、Shapiro, Ryan, and Lakey, Clinical Islet Transplantation--State of the Art, Transplantation Proceedings, 33, pp. 3502-3503 (2001); Ryan et al., Clinical Outcomes and Insulin Secretion After Islet Transplantation With the Edmonton Protocol, Diabetes, Vol. 50, April 2001, pp. 710-719; and Ryan et al., Continued Insulin Reserve Provides Long-Term Glycemic Control, Diabetes, Vol. 51, July 2002, pp. 2148-2157に記載される。肝臓に配置されると、当該細胞は、血液供給を発達させ、そしてインスリンを生成する。エドモントン・プロトコルは、使用される方法に応じて7〜10の工程を含む。第一工程は、特定の酵素(リベラーゼ)を、ドナーの膵臓に導入することに関しており、当該酵素は膵臓組織を消化するが、膵島を消化しない。消化工程に続き、膵臓内の他の細胞から膵島を分離する幾つかの次なる工程が存在する。分離された膵島を、門脈として知られている肝臓の主な血管に移植する。肝臓は、損傷を受けた場合に、肝臓自体を再生することができ、新たな血管及び支持組織を構築できる。その結果、膵島を肝臓に移植した場合、新たな血管が形成して、膵島を支持すると信じられている。細胞が生成するインスリンは、周囲の血管を通して血流中に吸収され、そして体中に分布されて、血中のグルコースレベルを制御する。
【0005】
要するに、エドモントン・プロトコルの工程は、もろい三次元構造を有しかつ増殖及び生存には多量の酸素を必要とする膵島の生存性を損なわせる強制的プロセスを作り出す。当該プロセスの間に、膵島は、酸素デリバリーの非最適条件のため損傷を受けるか又は破壊されることがあり、所定のドナー膵臓から回収される健常な膵島の収率に影響する。さらに、ドナーの利用可能性により、膵島移植が極めて限定的なものになり;1人の患者においてインスリン独立性を得るためには、2つの膵臓が必要とされることが多い。
【0006】
ステロイドフリーの非糖尿病誘発性免疫抑制治療と併せた膵島移植は、1型糖尿病を患う患者を治療するために用いられてきた。しかしながら、このような治療は、脂質異常症及び高血圧の高いリスクを導くことがあり、そして長期間の研究によると、膵島生存性が損なわれることが示される。
【0007】
ヒトにおける膵島移植のエドモントンプロトコルにより、移植の1年後に、80%の患者が、インスリン独立性を達成するという短期間の顕著な成功が示された。しかしながら、5年目ではこの割合は、たった約10〜15%に低下する。移植片機能の進行的消失の理由は明らかではないが、進行性のサイトカイン媒介性炎症の結果として膵島が失われるようである。しかしながら、膵島移植は、1型糖尿病を患う患者の実行可能な治療選択のままであり、長期間のインスリン独立性が得られない患者であっても、血糖不安定性の低下、低血糖症の発症低下、及びインスリン量の低下などの他の重要な恩恵を受けることが特に結果により示されている。
【0008】
膵島移植後の初日は、移植された膵島について特に脆弱な期間である。最大で60%の移植された膵島が、移植後の初日に非自己抗原特異的アポトーシスを受ける。この初期の破壊は、糖尿病を回復するために十分な膵島が生存することを保証するために、2〜3個の死体ドナーを使用することを必要とすることの主な根本的理由となっている。炎症性サイトカイン、IL−1β、TNFα、及びIFNγの局所的生産は、移植された膵島の初期アポトーシスに関わっている。こうして、初期移植期間におけるサイトカインを制限する戦略は、十分な膵島を得るために必要となるドナーの数を限られたものにし、膵島生存性を高め、そして長期間機能する能力を延長する。本発明は、これらの戦略を提供する。
【発明の概要】
【0009】
本発明は、単離後の回収された膵島細胞の機能を保持する方法であって、膵島単離の前に、膵島細胞ドナーの膵島細胞にeIF−5A1 siRNAを投与することを含み、ここで当該eIF−5A1 siRNAは、膵島細胞においてeIF−5A1 の発現を抑制し、それにより膵島細胞におけるアポトーシスを抑制し、そして回収された膵島細胞の機能を保存する、前記方法を提供する。好ましい実施態様では、siRNAは、eIF−5A1 の以下のヌクレオチド配列を標的化する:
5'−AAAGGAATGACTTCCAGCTGA−3';
5'−AAGATCGTCGAGATGTCTACT−3';
5'−AAGGTCCATCTGGTTGGTATT−3';又は
5'−AAGCTGGACTCCTCCTACACA−3'
ある実施態様では、eIF−5A1 siRNAは、ヌクレオチド配列5'−AAAGGAAUGACUUCCAGCTGAdTdT−3'を含む。siRNAは、任意の許容される手段を介してドナーに投与されてもよい。ある実施態様では、siRNAは、腹腔内注射を介して膵島細胞ドナーへと投与される。
【0010】
本発明の別の実施態様は、膵島細胞がドナー回収プロセスの間にアポトーシスを引き起こすことを阻害する方法であって、膵島単離の前に膵島細胞ドナーへの腹腔内投与を介して膵島細胞ドナーへとeIF−5A1 siRNAを投与することを含み、ここで当該eIF−5A1 siRNAは、膵島細胞内におけるeIF−5A1 の発現を阻害し、それにより膵島細胞におけるアポトーシスを阻害する、前記方法を提供する。
【0011】
好ましい実施態様では、siRNAは、eIF−5A1 の以下のヌクレオチド配列:
5'−AAAGGAATGACTTCCAGCTGA−3';
5'−AAGATCGTCGAGATGTCTACT−3';
5'−AAGGTCCATCTGGTTGGTATT−3';又は
5'−AAGCTGGACTCCTCCTACACA−3'
を標的とする。ある実施態様では、eIF−5A1 siRNAは、ヌクレオチド配列:5'−AAAGGAAUGACUUCCAGCTGAdTdT−3'を含む。
【0012】
任意のsiRNA又はアンチセンスコンストラクトは、かかるコンストラクトが、膵島細胞においてeIF−5A1 の発現を阻害する限り、使用することができる。siRNAの投与は、任意の適切な経路であってもよい。代表的な投与方法は、膵島細胞ドナーの門脈を介したかん流、膵島細胞ドナーの門脈を介した流体力学的かん流、及び腹腔内注射を含む。
【0013】
本発明は、アポトーシスを阻害し、そして膵島細胞の機能を保存するための組成物であって、eIF−5A1 siRNAを含み、ここで当該siRNAが、eIF−5A1の発現を阻害し、それにより膵島細胞におけるアポトーシスを阻害する組成物も提供する。好ましいeIF−5A1 siRNAは上に論じられている。
【0014】
本発明は、膵島β細胞における標的タンパク質の発現を阻害する方法であって、β細胞回収前にβ細胞ドナーに対して、標的タンパク質をコードするmRNAに対して標的されたsiRNAコンストラクトを投与することを含み、ここで当該siRNAは、膵島β細胞中の標的タンパク質の発現を阻害する方法も提供する。
【0015】
本発明は、eIF−FAの発現を低下させることにより、誘導性一酸化窒素合成(iNOS)経路を介して、一酸化窒素(NO)の産生を低下させる方法を提供する。
【0016】
本発明は、iNOS経路を介して一酸化窒素の産生を低下させる医薬の製造のための、eIF−5A siRNAの使用をさらに提供する。
【図面の簡単な説明】
【0017】
【図1】図1は、門脈を介してeIF−5A siRNAをかん流した後における、β−アクチン、mAAT、及びeIF−5A1について行われたRT−PCRの結果を提供する。この図は、eIF−5A1 の発現が計測可能であり、そうして膵島に取り込まれたことを示す。
【図2】図2は、低速逆行性門脈かん流(slows regrograde portal vein perfusion)を示す。胆管(透明)及び門脈(赤)は、調製的ノット(preparatory knot)(暗色縫合)の準備ができている。針をノットの下に差し込み(矢印により方向を示す)、ノットの下を横断させ、そしてsiRNAを血管に放出して、これらは膵臓、脾臓、腸、及び遠位結腸の三分の一に到達する。
【図3】図3は、eIF−5A1 siRNAを膵島にかん流することにより、eIF−5A1 の発現の低下が誘導される(図には、eIF−5A1 のmRNAレベルの低下が示される)ことを示す。
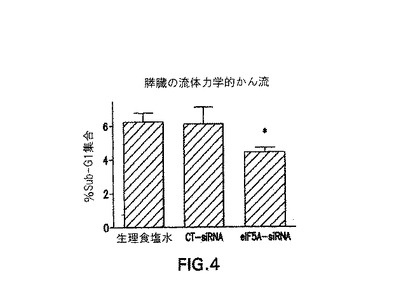
【図4】図4には、対照及び生理食塩水で処理された膵島に比べて、eIF−5A1 で処理された膵島細胞のアポトーシスを低下することが示される(ここで1群あたりn=2)。
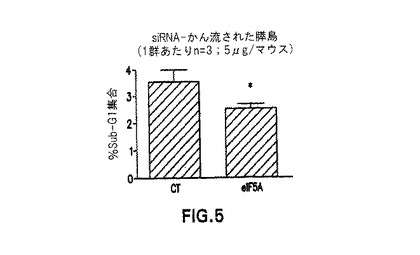
【図5】図5には、対照及び生理食塩水で処理された膵島に比較した場合に、eIF−5A1 siRNAで処理された膵島細胞のアポトーシスが低下することが示される(ここで1群辺りn=3)。
【図6】図6は、eIF−5A2に対して整列されたヒトeIF−5A1のヌクレオチド配列を提供する。
【図7】図7は、eIF−5A2に対して整列されたヒトeIF−5A1のアミノ酸配列を提供する。
【図8】図8は、代表的なアンチセンスオリゴヌクレオチドと供にヒトeIF−5A1のヌクレオチド配列を提供する。
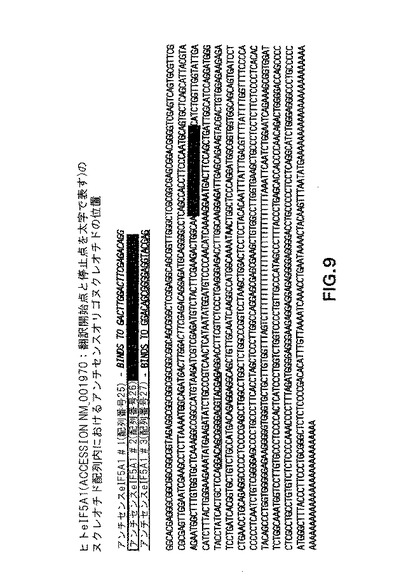
【図9】図9は、代表的なアンチセンスオリゴヌクレオチドと供にヒトeIF−5A1のヌクレオチド配列を提供する。
【図10A】図10A及び図10Bは、代表的なsiRNAと供にヒトeIF−5A1のヌクレオチド配列を提供する。
【図10B】図10A及び図10Bは、代表的なsiRNAと供にヒトeIF−5A1のヌクレオチド配列を提供する。
【図11】図11は、代表的なsiRNAと供にヒトeIF−5A1のヌクレオチド配列を提供する。
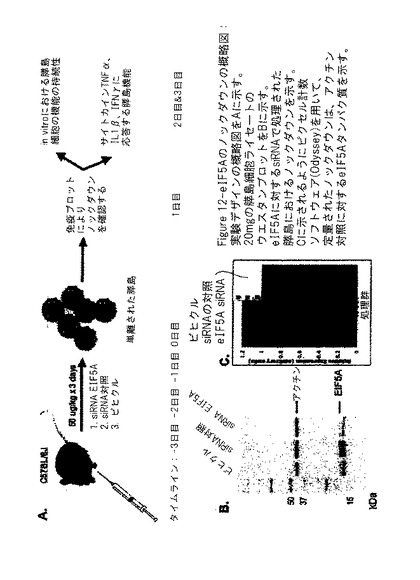
【図12】図12は、siRNA eIF−5A1が、膵島細胞において発現をノックダウン(減少)させることができるということを示す実験の模式図を示す。
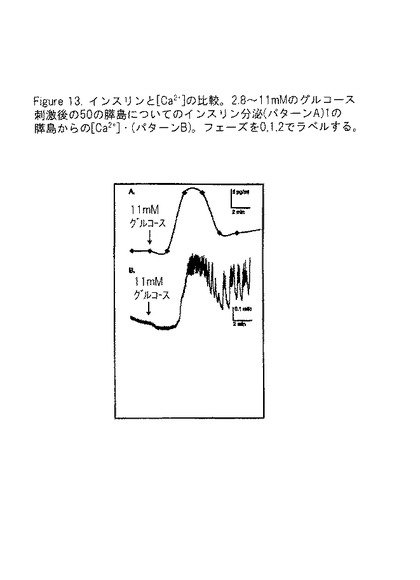
【図13】図13Aは、2.8から11mMのグルコース刺激の後の50個の膵島についてのインスリン分泌パターンを示し、そして図13Bは、2.8から11mMのグルコース刺激の後の単一膵島からの[Ca2+]iパターンを示す。この相は、0、1、2による。この図により、通常機能の膵島が、インスリン分泌の二相性のパターンでのグルコース刺激に応答することが示され、これは膵島細胞内カルシウム([Ca2+]i)における同様の変化に近似する。
【図14】図14は、カルシウム振動研究の結果を提供する。これらの図により、eIF−5A1 siRNAで処理され(そうしてeIF−5A1の発現が低下した)膵島細胞が、有意に強いグルコース応答及び全体の振動を示すということが示される。図14は、これらの処理された細胞が、長期間生存するばかりでなく、機能も保持するということを示す。
【図15】図15は、サイトカイン処理された染色済みの膵島細胞の異なる写真を示す。この写真により、対照又はビヒクル(生理食塩水のみ)で処理された膵島細胞よりも、eIF−5A1 siRNAで処理された膵島細胞の生存性が高く、そして死細胞が少ないということが示される。
【図16A】図16A及び16Bは、IP注射を介して膵島細胞ドナーに投与された場合にsiRNAが、膵島細胞に侵入することを4種の画像化技術を用いて示す。共焦点イメージングを用いて、単離膵島の3D再構成図を作成した。図16Aにより、0.9%の生理食塩水を1日1回、3日間腹腔内投与されたマウスから単離された膵島が、FITC自己蛍光を示すが、TRITCチャンネル(図ではCy3標識)において最低の蛍光しか示さないということが示される。
【図16B】図16A及び16Bは、IP注射を介して膵島細胞ドナーに投与された場合にsiRNAが、膵島細胞に侵入することを4種の画像化技術を用いて示す。共焦点イメージングを用いて、単離膵島の3D再構成図を作成した。図16Bにより、Cy−3標識された安定化siRNAを1日1回、3日間腹腔内投与されたC57Bl/6から単離された膵島が、Cy3標識の浸透を示すということが示される。
【図17A】図17(A)は、βTC3細胞が、2の異なるsiRNAコンストラクト(コンストラクトA及びB)、又は対照でトランスフェクトされ、そして指定のタンパク質について免疫ブロットに供された場合の結果を示す。
【図17B】図17(B)は、C57Bl/6マウスに同じsiRNAを1日1回、3日間腹腔内注射し、そして膵島を単離し、そしてコラゲナーゼ消化及び分画勾配遠心分離を介して精製した場合の結果を示す。膵島を2%SDS中で溶解し、そして免疫ブロットに供する。
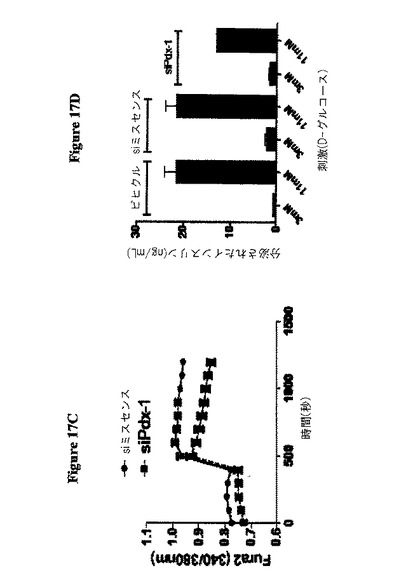
【図17C−D】図17(C)は、単離された膵島にFura2を30分間充填し、次に3mM D−グルコース中でイメージングした場合の結果を示す。膵島を11mM D−グルコースで300秒刺激し、そしてFura2比を、蛍光顕微鏡により連続してモニタリングした場合の結果を示す。図17(D)は、グルコース刺激されたインスリン分泌(GSIS)を、各処理群から得た50の膵島を用いて実行した結果を示す。
【図18A】図18A−Iは、eIF−5Aのノックダウンが、膵島機能及びex-vivoでの生存性を改善するということを示す。
【図18B】図18A−Iは、eIF−5Aのノックダウンが、膵島機能及びex-vivoでの生存性を改善するということを示す。
【図18C】図18A−Iは、eIF−5Aのノックダウンが、膵島機能及びex-vivoでの生存性を改善するということを示す。
【図18D−I】図18A−Iは、eIF−5Aのノックダウンが、膵島機能及びex-vivoでの生存性を改善するということを示す。
【図19A】図19A−Dは、サイトカイン誘導性のiNOSタンパク質の生産が、eIF−5A欠損細胞では見られないことを示す。
【図19B】図19A−Dは、サイトカイン誘導性のiNOSタンパク質の生産が、eIF−5A欠損細胞では見られないことを示す。
【図19C】図19A−Dは、サイトカイン誘導性のiNOSタンパク質の生産が、eIF−5A欠損細胞では見られないことを示す。
【図19D】図19A−Dは、サイトカイン誘導性のiNOSタンパク質の生産が、eIF−5A欠損細胞では見られないことを示す。
【発明を実施するための形態】
【0018】
真核生物翻訳開始因子5A(eIF−5A)は、ストレス誘導性の遺伝子の転写後調節において決定的な因子であることが明らかとなり、そして哺乳動物細胞においてサイトカイン媒介性のアポトーシスを促進することが明らかとなっている。eIF−5Aは、真核生物を通して極めて高度に保存されている小さな酸性タンパク質であり、そして固有のポリアミン由来のアミノ酸であるヒプシンを含むことが知られている唯一のタンパク質である。eIF−5Aは、mRNAプロセッシング、輸送、及び翻訳において役割を果たすことが提案されていた。しかしながら、どんなものであれ膵島における炎症カスケードにおけるその役割は、従前、特徴決定されていなかった。膵島は、グルコース及び脂質毒性、酸化ストレス、及び炎症性サイトカインに対して感受性が高い。膵島の細胞ストレスへの応答を媒介する点に関わる因子の役割を研究するため、本発明者は、低分子干渉RNA(siRNA)を用いてin vivoにおいて膵島の特定タンパク質を枯渇させるプロトコルを開発した。研究において、発明者らは、RNA干渉を用いてeIF−5Aタンパク質を膵島から枯渇させることにより、膵島におけるeIF−5Aの役割を決定することを試みた。
【0019】
しかしながら、おもに齧歯類膵島におけるRNA干渉研究における主要な試みでは、トランスフェクション効率が比較的低かった。従来の研究が、短いヘアピンRNAのウイルス媒介性のデリバリーを利用することを示した一方で、この技術は、適切なウイルスを作成するには時間がかかるものであり、そしてウイルス毒性を被るものであった。本発明者らは、安定化された低分子干渉RNA(siRNA)を繰り返し腹腔内注射することにより、マウス膵島から選択したタンパク質をin vivoで枯渇させる新たなプロトコルを発見した。このプロトコルは、膵島内へのsiRNAの顕著な浸透をもたらす。この技術は、よく特徴決定された膵臓転写因子Pdx1などの任意の所望の標的タンパク質を、膵島からうまく枯渇させるために使用することができる。この技術を用いて、発明者は、eIF−5Aタンパク質を膵島からうまく枯渇させ、そして誘導型一酸化窒素合成酵素(iNOS)をコードするmRNAの翻訳を促進することにより、eIF−5Aが、サイトカイン媒介性の膵島機能不全に寄与するということを示した。このデータは、iNOSのmRNAを含むmRNA(おそらく、サイトカイン媒介性のストレス応答に関与するmRNA)のサブセットの安定化及び/又は核輸送のために、eIF−5Aが膵島において必要であるというモデルを示唆する。
【0020】
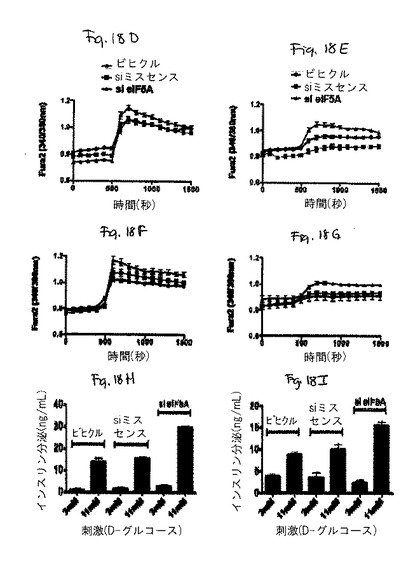
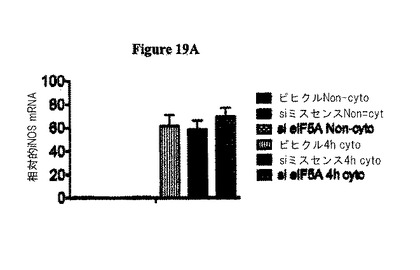
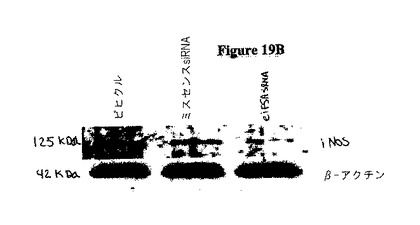
本発明者らは、in vivoでのsiRNA注射を使用して、サイトカイン媒介性ストレスにおける真核生物翻訳開始因子5A(eIF−5A)の役割を研究した。eIF−5Aは、ストレス誘導性遺伝子の翻訳において、決定的なレギュレーターとして他のシステムにおいて特徴決定されていた。eIF−5Aに対するsiRNAがマウスに注射された場合、単離された膵島においてeIF−5Aタンパク質レベルの50〜70%の低下が観察された。この低下に伴い、対照処理に比べて、グルコース刺激性のインスリン分泌及びCa2+可動化が30〜40%の増加すること観察された。単離された膵島を炎症性サイトカイン(IL1β、TNFα、IFNγ)のカクテルに暴露した場合に、膵島機能の相対的向上が、si−eIF−5Aで処理された動物において持続した。eIF−5Aノックダウンの後の膵島保護は、その生成物がグルコース応答性又はインスリン転写に関与する遺伝子(Slc2a2、Gck、Irs1、Nkx6−1、MafA、Pdx1、NeuroD1、及びSetd7)の発現変化を伴うことはない。図18Cを参照のこと。大事なことは、誘導性一酸化窒素合成酵素(iNOS)をコードするmRNAが、対照で処理された膵島及びsi−eIF−5A−処理された膵島の両方において、サイトカインに応答して、40倍超も上方制御されるが(図19Aを参照のこと)、iNOSタンパク質レベルが、si−eIF−5A処理された膵島において3〜5倍低いということである(図19Bを参照のこと)。これらのデータにより、si−eIF−5A処理された膵島における機能の向上が、二次的に一酸化窒素の生産を低下しうるということが示唆される。これにより、ストレスシグナルに応答した膵島β細胞において、eIF−5AがiNOS翻訳の制御を介して不可欠な役割を果たし、そして膵島細胞の生存性の保存並びに回収後及び移植後において機能する膵島細胞の能力の保存を狙った治療戦略について、eIF−5Aが実行可能な標的として役立ちうるという証拠を提供する。
【0021】
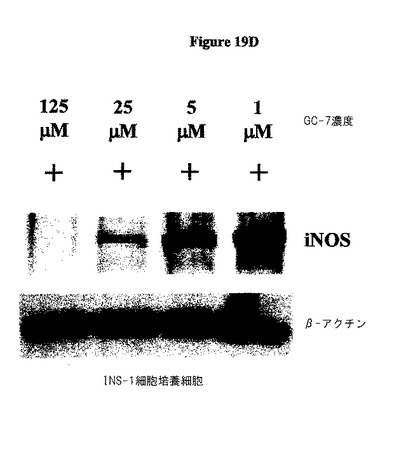
本発明者らは、(GC−7(DHSの阻害剤)を投与することによる)ヒプシン化の阻害を介したeIF−5A機能の遮断が、効果的にiNOSレベルを低下するということをさらに示した。図19Dを参照のこと。
【0022】
膵島へのsiRNAの取り込みが、逆行性の門脈閉塞を介した膵臓かん流により達成することができるということが以前に示されていた。Bradleyら、Transplantation Proceedings, 37, 233-236, 2005を参照のこと。簡潔に説明すると、Cy−3標識されたルシフェラーゼ(Luc)siRNA GL2 二本鎖を、リポフェクタミン2000でパッケージして、又はパッケージせずに用いて、そして尻尾静脈を通して(in vivo、マウス1匹あたり50μg)注射するか、又は逆行性門脈静脈閉塞(in situ, マウス1頭あたり2μg)により膵臓に直接注射した。膵臓を取得し、そしてIn situデリバリー後に24時間4℃で貯蔵するか、又はin vivoデリバリー後に4時間貯蔵し、そして膵島を単離し、そしてさらに試験前に16時間培養した。siRNAの分布を可視化するために、膵臓をインスリンについて染色し、そして蛍光顕微鏡の下で試験した。単離された膵島を直接蛍光顕微鏡で試験した。パッケージされていないsiRNAは、リポソームにパッケージされたsiRNAを用いて観察されるのと同程度に膵島に到達し、いわゆる「裸」のsiRNAデリバリーの報告に一致した。Lewis et al., Nat. Genet. 32: 107-108, Epub 2002 Jul 2029, 2002 and McCaffrey AP, et al., Nature 418: 38 - 39, 2002)。図1は、膵島細胞へのかん流が、膵島細胞への適切なデリバリーメカニズムを提供するということを示す。
【0023】
従って、本発明は、膵島細胞において標的タンパク質の発現を抑制する方法であって、標的タンパク質をコードするmRNAを標的とするsiRNAを、膵島細胞に投与することを含み、ここで当該siRNAが、膵島細胞において標的タンパク質の発現を阻害する、前記方法を提供する。任意の手段を介して、好ましくは膵島β細胞の単離の前に、膵島β細胞ドナーに腹腔内投与することを介して投与されてもよい。
【0024】
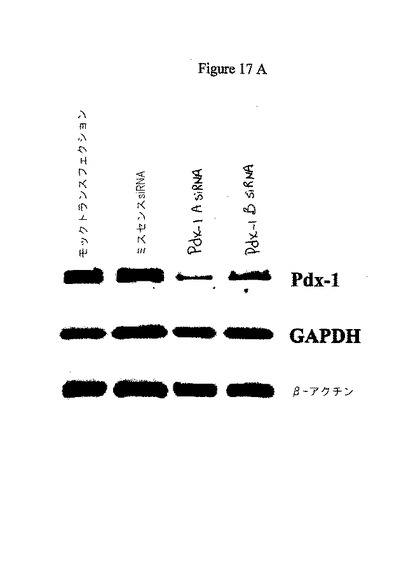
1の実施態様では、本発明者らは、安定化されたCy3−標識された二本鎖RNAをC57Bl/6マウスに三日間腹腔内注射することにより、共焦点顕微鏡で測定した場合に当該RNAが単離された膵島へと顕著に浸透することが明らかにされた。タンパク質ノックダウンについてこの技術を試験するために、Pdx1を標的とした安定化siRNAを用いた。Pdx1mRNAに対する2つの異なるsiRNAは、(免疫ブロット分析により評価した場合に)膵島においてPdx1タンパク質の50%及び90%ノックダウンをもたらした。図17A−Dを参照のこと。
【0025】
本発明は、膵島細胞においてeIF−5A1の発現を阻害する方法であって、eIF−5A1 siRNAを膵島細胞に投与することを含み、ここで当該eIF−5A1 siRNAが、膵島細胞においてeIF−5A1の発現を阻害する、方法を提供する。図16A及び図16Bは、IP注射を介した膵島細胞ドナーへの投与が、膵島細胞に対する適切なデリバリーメカニズムを提供するということを示した。図16Bは、安定化されたCy−3標識二本差RNAをC57Bl/6マウスに3日間の腹腔内注射することにより、共焦点顕微鏡で測定した場合に、当該RNAが単離膵島に顕著に浸透することを明らかにした。図12B及び18Bは、eIF−5A1 siRNAで処理された膵島細胞が、より少ないeIF−5A1 siRNAしか発現しないことを示す。
【0026】
eIF−5A1発現を抑制することにより、アポトーシスが阻害される。図4及び5は、単離前にeIF−5A1 siRNAで膵島細胞を処理することが、これらの細胞のアポトーシスを阻害するメカニズムを提供した(subG1期の細胞数の低下により示される)。したがって、本発明は、回収された膵島細胞において、アポトーシスを阻害する方法であって、eIF−5A1 siRNAを膵島β細胞ドナーに投与することを含み、ここで当該eIF−5A1 siRNAは、膵島細胞内におけるeIF−5A1の発現を阻害し、そしてここでeIF−5A1発現の抑制がアポトーシスを阻害する、方法を提供する。回収プロセスの前に、膵島細胞をeIF−5A1 siRNAで処理することが好ましい。
【0027】
本発明は、単離された後に回収された膵島細胞の機能を保存する方法であって、eIF−5A1 siRNAを膵島単離の前に膵島細胞ドナーの膵島細胞に投与することを含み、これが膵島細胞におけるeIF−5A1発現の抑制又は低下をもたらし、次に膵島細胞におけるアポトーシスを抑制し、さらにその機能を保存する方法をさらに提供する。膵島細胞の機能の保存は、膵島細胞が回収プロセスを生き残るということだけではなく、回収プロセス後においてその機能を維持する(細胞の機能は、長期間残存し、及び/又はそれらはeIF−5A1 siRNA処理されていない膵島細胞よりも優れたグルコース応答性を維持する)ということを意味する。
【0028】
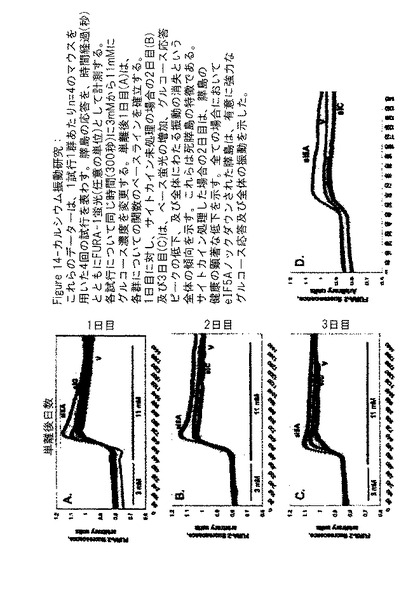
1の実施態様では、膵島機能及び全身の健康におけるeIF−5A1 ノックダウンの結果を調査するために、C57Bl/6雄マウスに、ビヒクル(0.9%生理食塩水)[群I]、対照siRNA(16.6mg/kg)[群II]、又はeIF−5A1 siRNA(16.6mg/kg[群III]のいずれかを腹腔内(IP)注射した。指定された処理の注射を、膵島回収(0日目)の前3日間(−3、−2、−1日目)毎日投与した。単離された膵島をコラゲナーゼ処理により精製し、そして試験の開始前16〜18時間静置した。回収後1日目に機能のベースラインを確立した後に、半分の膵島を、サイトカインカクテル(5ng/ml IL1β、10ng/ml TNF−α、100ng/ml INF−γ)に晒して、1型糖尿病において膵島が直面する条件を模倣した。実験設計の模式図及び膵島の配分を図12Aに示す。
【0029】
ウエスタンブロットにより、タンパク質ノックダウンについて膵島の各群から得たタンパク質ライセートを分析した。図12Bは、群IIIの処理を受けたマウス膵島におけるeIF−5A1の相対的ノックダウンを示し、これは図12Cにおいて定量化される。これらのデータにより、膵島内におけるeIF−5A1の発現のノックダウンをもたらすIP注射の効力が示される。
【0030】
回収された膵島における保存された機能の証拠は、カルシウム振動アッセイにより示される。本発明者らは、細胞内カルシウムが刺激及びストレスの指標であるということを決定した。ヒト及び齧歯類の両方において、通常通り機能する膵島は、図13Aに示されるようにインスリン分泌の2相性のパターンでグルコース刺激に応答する。一次近似として、この二相性パターンは、膵島の細胞内カルシウム[Ca2+]iにおける変化によく似ており、この変化は、第二相のプラトーに落ち着く第一相ピークからなる(図13B)。340及び380nm励起から放射される光の比として報告される[Ca2+]iの計測は、膵島機能のさらなる特徴を検出する優れた感度を提供する。グルコースに対する[Ca2+]i応答は、3つの相からなり(図13を参照のこと)、β細胞刺激と分泌との関係における「コンセンサスモデル」により記載されるプロセスを大まかに反映している。[Ca2+]iの初期の落ち込みは、グルコースがβ細胞に移動した後に、小胞体が[Ca2+]iを動員し、隔離するにつれ生じる(第0相)。次にグルコースは、解糖をとおして代謝され、そしてさらにトリカルボン酸(TCA)回路においてさらに代謝され、ATP生産をもたらす。ATP/ADPの十分な増加が生じて、KATP−イオンチャネルが閉じた場合、第一相インスリン分泌に関連するL型カルシウムチャネルが開くために、[Ca2+]iの急激な増加が生じる。この最初の第一相ピークの後に、[Ca2+]iは、プラトーに落ち着くが、プラトーは[Ca2+]i振動によりしばしば中断される。プラトーの高さは、グルコース濃度及び第二相インスリン分泌の割合に直接関連している。概して、イメージングに基づく[Ca2+]i計測は、個々の膵島のレベルで、動的なインスリン分泌の優れた一次近似となる。
【0031】
多くの因子は、グルコース刺激に対する通常の[Ca2+]i応答にマイナスの影響を与えることがある。グルコース代謝、ミトコンドリア及びATP産生、小胞体、イオンチャネルの機能、及び多種多様の他の問題の破綻は、グルコース刺激に関するカルシウム処理のダイナミクスに影響を与えることがある。[Ca2+]iの変化を頻繁にモニタリングすることにより、これらの活性態様における不具合は、静的インスリン分泌の標準計測により観察されるよりも頻繁に観察される。例えば、低いグルコースレベルにおけるベース[Ca2+]iの増加、及びグルコース刺激の間における[Ca2+]iの第0相の落ち込みの低下は、ER−ストレス又はおそらくイオンチャネル機能不全を指している可能性がある。このような不具合は、静的又は動的インスリン分泌を計測することによって容易に検出することはできないが、本明細書に記載される画像化技術は、これらの問題を指摘するために使用することができる。
【0032】
カルシウム振動及びグルコース刺激されたインスリン分泌研究においてサイトカインが存在する場合、及び存在しない場合の両方において、eIF−5A1 siRNA(16.6mg/kg)で処理された膵島[第3群(eIF−5A1ノックダウン膵島)]は、グルコースに対する高い応答性、並びに長期間の機能の維持(長期間の生存を示す)を示した。図14を参照のこと。従って、これらの膵島は、インスリンの高い生産を示すことが予期される(データー取得中)。生存/死亡染色した後に撮られた画像から得た元データーは、eIF−5A1 ノックダウンが膵島細胞の生存を延長するという概念を定性的に裏付ける(図15)。赤色膵島(死亡した膵島細胞の各にインターカレットするEtHD−1の結果)の数を、カルセイン−AMを蛍光カルセインへと能動的に切断する緑色膵島の数と比較することにより、膵島生存の定性的画像が確かめられる(図15)。
【0033】
本発明は、一酸化窒素の産生増加と関連する炎症性疾患に関与する細胞においてアポトーシスを阻害する方法をさらに提供する。例えば、リューマチ様関節炎(RA)などの炎症性疾患は、誘導性一酸化窒素合成(iNOS)経路の活性化のため、一酸化窒素(NO)の生産増加と関連する。動物モデルにおける研究により、NOが、関節炎症及び組織損傷の病原進行において通常の役割を果たすということが示唆される。なぜなら、関節炎の重篤度が、NOS阻害剤の投与により低下するからである。関節内に存在する幾つかの細胞(例えば滑膜線維芽細胞、内皮細胞、及び軟骨細胞)は、炎症性サイトカインにより誘導されて、インビトロでNOを産生することができる。さらに、局在化研究により、RAを煩う患者から得られた関節組織の滑膜表層細胞、軟骨細胞及び血管におけるiNOS発現が上方制御されることが示される。滑膜表層及び軟骨へのiNOSの局在化は、アポトーシスがRA、特に滑膜表層及び軟骨において特に増加することを示した他の研究の点でも関心が高い。リューマチ様関節におけるアポトーシスの原因となるメカニズムは、不明のままであるが、Fas抗原の発現が、RA滑膜細胞において増加し、そしてFas抗体が、インビトロにおける滑膜細胞のアポトーシス性の死亡を刺激することができるということが以前の研究者により示された。iNOS経路の活性化が、リューマチ性関節においてアポトーシスの刺激に関与するという仮説が検討された。なぜなら高レベルのNOが、インビトロにおいて多くの細胞型においてアポトーシスを刺激することが知られているからであり、そしてNOが、RAにおけるアポトーシスのメディエーターとして作用すると結論づけられた(R. J. van't Hof et al., Rheumatology 200, 39: 1004-1008を参照のこと)。
【0034】
このようなものとして、本発明はRAに関わる細胞(例えば、滑膜細胞及び軟骨細胞)においてアポトーシスを阻害する方法であって、eIF−5A1の発現を阻害し、それによりiNOS経路を介した一酸化窒素の産生を阻害又は低下させて、細胞のアポトーシスを阻害する方法を提供する。
【0035】
本発明は、さらに、細胞又は宿主におけるNO産生を阻害する方法であって、当該細胞又は宿主にeIF−5A1に対するsiRNAを投与することによる方法を提供する。siRNAは、eIF−5A1の発現を低下させ、そして次にiNOS経路の活性化を抑制することによりNOの産生を低下させる。
【0036】
本発明は、iNOS経路を介して一酸化窒素の産生を低下させる医薬の製造のための、eIF−5A siRNAの使用を提供する。
【0037】
eIF−5A1 の発現を抑制する任意のeIF−5A1 siRNAが使用されてもよい。「阻害」という語句は、対照細胞、つまりeIF−5A1 siRNAで処理されていない細胞において生じるレベルに比べて低下することを意味する。1の代表的、そして特に好ましいeIF−5A1 siRNAは、以下の配列:5'−AAAGGAAUGACUUCCAGCTGAdTdT−3'を含む。2005年11月28日に出願された同時係属米国出願11/293,391号(当該出願はその全てを本明細書に援用する)は、追加の代表的eIF−5A1 siRNAを提供し、そして他の細胞型においてeIF−5A1の発現を阻害するために使用され、そしてアポトーシスを阻害することが示された他のアンチセンスコンストラクトを提供する。当業者は、eIF−5A1 配列が与えられれば、他のeIF−5A1 siRNAを設計することができ、そして過度の実験を行うことなく、発現を阻害する能力について当該siRNAを容易に試験することができる。図6〜11は、eIF−5A1 の配列、代表的なeIF−5A1 siRNA及びアンチセンスコンストラクトを提供する。本発明の別の実施態様では、eIF−5A1 のアンチセンスコンストラクトを使用して、eIF−5A1 の発現を阻害し、そうして膵島細胞のアポトーシスを阻害し、並びにその機能を維持又は保存することもできる。
【0038】
好ましい実施態様では、eIF−5A1 siRNAは、eIF−5A1の以下のヌクレオチド配列:
5’−AAAGGAATGACTTCCAGCTGA−3';
5'−AAGATCGTCGAGATGTCTACT−3';
5'−AAGGTCCATCTGGTTGGTATT−3';及び
5'−AAGCTGGACTCCTCCTACACA−3'
を標的とする(図10を参照のこと)。特に好ましい実施態様では、siRNAは、eIF−5A1の以下の配列:5’−AAAGGAATGACTTCCAGCTGA−3'を標的とする。
【0039】
本発明は、ドナー回収プロセスの間に、膵島細胞がアポトーシスを引き起こすことを阻害するための方法を提供する。上に記載されるように、多くの膵島細胞は、回収された場合にアポトーシスを引き起こす。本発明者らは、回収する前に膵島細胞にeIF−5A1 siRNAを提供することが、アポトーシスに対する保護的利点を提供することを示した。eIF−5A1 siRNAは、膵島単離の前に、膵島細胞ドナーの膵島細胞に投与される。ドナー(及び膵島細胞)は、例えばヒト膵島細胞を含む任意の動物であっても良い。任意の投与方法が使用されてもよい。例えば、siRNAは、膵島細胞ドナーの門脈を通したかん流を介して投与されることもあるし、又は膵島細胞ドナーの門脈を介した流体力学的かん流を介して投与されることもある。実施例1を参照のこと。別の投与形態として腹腔内投与が挙げられる。実施例2、及び実施例3〜5を参照のこと。
【0040】
門脈を介したかん流は、胆管への挿管に類似しているが、針は反対の方向に向いている。門脈は、肝臓を引き出し、そしてマウスの左側へと内臓を移動させることにより露出する。調製的ノット(preperative knot)を門脈の周囲に作り、そして胆管を含める。血管に穿刺した後に、尖っていない針を膵臓にまで進め、そしてその周囲でノットを絞める。マウスモデルでは、1mlの生理食塩水又はsiRNA(5μg)をゆっくり放出し、針を取り出し、そしてノットを針の手前で閉じて、液体の漏出を妨げる。この時点で、マウスをひっくり返し、そして胆管を膵臓消化にアクセスさせた。膵臓にsiRNAを長時間適用してもよい。或いは、膵臓を取り出し、冷却状態を維持して、コラゲナーゼで処理することもできる。通常の膵島単離方法を行い、そして膵島(50)を16時間インキュベートしてもよい。
【0041】
本発明は、膵島細胞内でアポトーシスを阻害するための組成物であって、eIF−5A1 siRNAを含み、ここでsiRNAがeIF−5A1の発現を阻害し、それによりアポトーシスを阻害し、そして膵島細胞の機能を維持する。組成物は、他の又は追加のeIF−5A1 siRNAを上記のように含むこともある。好ましいsiRNAは、ヌクレオチド配列:5'−AAAGGAAUGACUUCCAGCTGAdTdT−3’を含む。
【実施例】
【0042】
実施例1:門脈かん流
マウス膵島は、eIF−5A1を発現する。全量RNAを、単離されたマウス膵島から抽出し、そしてRT−PCRを、β−アクチンとeIF−5Aについて行った(図1)。安静にして刺激を受けていない膵島は、eIF−5A1−mRNAの陽性レベルを示した。
【0043】
eIF−5A1−mRNAレベルは、eIF−5A1 siRNAデリバリー(門脈低速かん流)後に減少した。マウスに1mlのsiRNA(CT(対照)配列又はeIF−5A1、5mg)又は生理食塩水を、それぞれの群についてn=2で低速逆行性門脈かん流(図2))により導入した。膵管にコラゲナーゼを潅注することにより、膵臓を消化し、そして膵島を、Lewis et al., Proc. Natl. Acad. Sci. USA, 102: 12153-12158 Epub 12005 Aug. 12110, 2005に記載される通りに単離した。膵島(1マウスあたり50個)を16時間インキュベートした。全量RNAを抽出し、そしてβ−アクチン及びeIF−5A1についてRT−PCRを行った(図3)。eIF−5A1/β−アクチンのmRNAの比は、5.24(CT−siRNA)及び3.01(eIF−5A1 siRNA)であった。図3は、siRNAで処理された細胞においてeIF−5A1のmRNAレベルが低下したことを示す。この実験をn=3のマウスで繰り返し、そして膵島をRNA抽出のために3回インキュベートを行った;結果は、最初の観察と一致した。
【0044】
eIF−5A1 siRNAデリバリー(門脈流体力学的かん流)後におけるeIF−5A1mRNAレベルの低下及び膵島アポトーシス割合の低下
マウスに、1mlのsiRNA(CT又はeIF−5A1 5μg)又は生理食塩水を、流体力学的逆行性門脈かん流により導入した(1群あたりn=2)。当該かん流は5秒以内に完了した。膵管にコラゲナーゼを潅注することにより膵臓を消化し、そして膵島を単離した。膵島を16時間インキュベートし、次に分配した:1の群を、アポトーシスの評価のため、プロピジウムヨージドで染色し(1マウスあたり50の膵島)、そしてもう一方の群をRT−PCRで処理した(1マウスあたり25の膵島)。eIF−5A1/β−アクチンについてのmRNAのレベルは、CT−siRNA群において、eIF−5A1 siRNA群よりも高かった。アポトーシス率は、28.1%低下した(図4)。この実験を繰り返したところ(n=3)、アポトーシス率は、再び減少した(図5)。
【0045】
ビオチン化siRNAを用いた膵島かん流
ビオチン化siRNA(50μg)を、上に記載されるように膵島に還流した(低速かん流、n=1)。膵臓を染色用にホルマリン中で固定した。
【0046】
siRNA
siRNA分子は、Dharmacon, Lafayette, Co.により合成された。eIF−5A1 及び対照siRNAの配列はそれぞれ、5’CGGAAUGACUUCCAGCUGAdTdT 3’及び5'AGUCGACCUUCAGUAAGGCdTdT3'であった。
【0047】
RT−PCR
Qiagen RNeasyキットを用いて細胞から全量RNAを抽出した。eIF−5A1プライマーは次の通りである:
フォワード 5'−GAC AGT GGG GAG GTA CGA GA−3';
リバース 5'−GGG GTG AGG AAA ACC AAA AT−3’。
【0048】
プロピジウムヨージド(PI)アポトーシス染色
温和なトリプシン処理により膵島の単一細胞懸濁液を得た。PBSで細胞を洗浄し、そして0.3%サポニン、EDTA1mM、Rnase、1%アジド、1%FCS及び50μg/mlのPIをPBSに含むサポニン−PI混合液を加えた。細胞を完全にボルテックスし、そして暗所にて4℃でインキュベートした後にFACSによりsub−G1集合について分析した。
【0049】
実施例2:腹腔内注射
抗体及びsiRNA:
eIF−5A1 に対するマウスモノクローナル抗体を作成した。抗アクチンモノクローナル抗体(クローンC4、#69100)をMP Biomedicalsから購入した。ウエスタンブロットにおいて、Li−cor社から購入した近赤外フルオロフォア標識された二次抗体を用いた(IRDye800及びIRDye700)。eIF−5A1 特異的siRNAは、eIF−5A1の以下の配列:5'−AAAGGAATGACTTCCAGCTGA−3’に対するものである。コントロールsiRNAは転写配列:5'−AAAGTCGACCTTCAGTAAGGA−3'であり、そしていずれの既知のタンパク質についてもノックダウンを誘導しないことが前に示された。同時係属米国出願第11/725,470号及びPCT/US07/64424を参照のこと。これらの文献をその全てを本明細書に援用する。
【0050】
マウスへのsiRNAのIP(腹腔内)注射:
12匹の8〜10週齢のC57Bl/6雄マウスを、Charles River社から購入した。雌の発情サイクルに伴う代謝変化を避けるために、雄マウスを選択した。マウスをランダムに3つの処理群に割り当てた:第1群−ビヒクル処理(0.9%生理食塩水)、第2群−対照siRNA処理、第3群−eIF−5A1 siRNA処理。各マウスを、16.6mg/kg、又は(第一群の場合)同等の体積をIP注射を介して選択した処理に供した。膵島回収(0日目)の前に3日間(−3、−2、−1日目)毎日(午前11:00ころに)マウスに注射した。
【0051】
膵島単離とサイトカイン処理:
標準的単離技術及び動物実験委員会(IACUC)により承認されたプロトコルを用いてC57Bl/6マウスからの膵島の単離を行った。コラゲナーゼ消化まで膵臓を4.2mM炭酸水素ナトリウム及び1%BSA(Invitrogen)を添加したHBSS中に貯蔵した。分画遠心法により精製を行った。10%FBS及び5%ペニシリン/ストレプトマイシンを添加したフェノールレッドフリーDMEM溶液中に全ての膵島を維持した。培養液を37℃+5%CO2で5.5mMグルコース中においてインキュベートした。代謝試験(カルシウム振動)前に16〜18時間精製された膵島を回復させた。
【0052】
1日目の代謝試験の終わりに、生理的に関連のあるサイトカインの規定のカクテル(5mg/ml IL1β、10ng/mlのTNT−α、100ng/mlのINF−γ)を用いて、サイトカイン処理を適用した。サイトカイン処理された膵島を、次の試験まで24時間休ませ、そしてタンパク質分析を行った(2日目及び3日目)。
【0053】
免疫ブロットアッセイ:
(200mMのDTT及びベンゾナーゼを含むLaemmli緩衝液中で調製された)10μgの膵島細胞抽出物を、4〜20%のSDSポリアクリルアミドゲル(Invitrogen)で電気泳動することにより分離し、抗−eIF−5A1 マウスモノクローナル一次抗体(1:15000希釈)を用いて、免疫ブロット分析を行った。Odysseyシステム(Li−Cor Biosciences社)の近赤外技術を用いて画像化及び定量した。
【0054】
カルシウム振動アッセイ:
(上に記載されるように)コラゲナーゼ消化された8〜10週齢のC57Bl/6マウス膵臓から膵島を手で拾いあげた。Fura−2低グルコース溶液(3mM)中で20分間処理し、次に温度制御されたフローチャンバー中に配置した。低グルコース溶液(Furaを伴わない)を膵島上に流して、ベースカルシウム計測値(5分)を調べた。IP Lab4.0ソフトウェア(BD Biosciences)を用いてカルシウムの計測値を取得し、そして340〜380nmの放出光の比として報告する。ベースカルシウムレベルに達した後に、チャンバーを通して高グルコース(11mM)溶液を流して(15分間)、膵島の細胞内カルシウム応答を誘導する。カルシウム応答を、5秒間の間隔で、20分間の観察のあいだに測定した。試験を終える際、膵島を5%ブリーチ溶液中に配置する。
【0055】
実施例3:in vitro及びin vivoにおけるPdx−1に対するsiRNAの適用
βTC3細胞に、2種のPdx−1siRNAコンストラクト(コンストラクトA及びB)又は対照をトランスフェクトし、そしてタンパク質Pdx−1、GAPDH及びβ−アクチンについての免疫ブロットに供する。図17Aを参照のこと。C57Bl/6マウスに、同じsiRNAを1日1回3日間腹腔内注射し、そして膵島をコラゲナーゼ消化及び分画勾配遠心を介して単離及び精製した。膵島を2%SDS中に溶解し、そして免疫ブロットに供した。図17Bを参照のこと。30分間Fura2を単離膵島に充填し、次に3mM D−グルコース中で画像化した。〜300秒で11mM D−グルコースで膵島を刺激し、そしてFura2比を、連続して、蛍光顕微鏡によりモニタリングした。図17Cを参照のこと。グルコース刺激インスリン分泌(GSIS)を、各処理群から50の膵島を用いて行った。図17Dを参照のこと。
【0056】
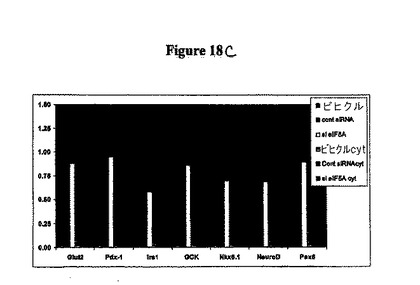
実施例4:eIF−5A1のノックダウンは、ex vivoでの膵島機能及び生存を改善する
eIF−5Aに対するsiRNAのin vitro投与が、Ex vivoでの膵島機能を改善することをこの実施例は示す。図18Aは、実験設計の模式図を提供する。膵島を処理後に単離し、そして抽出物を免疫ブロット分析に供した。図18Bを参照のこと。単離膵島をグルコース感知及びインスリン転写に必須である遺伝子についてRT−PCR分析に供した。遺伝子転写はサイトカイン処理(IFNg、IL−1b、TNFaと4時間)により抑制されるが、群間の差異は観察されなかった。図18Cを参照のこと。単離後24時間のsi−eIF−5A処理された膵島(図18Dを参照のこと)及び単離後48時間のsi−eIF−5A処理された膵島(図18Fを参照のこと)は、単離後24時間の対照処理された膵島(図18Dを参照のこと)及び単離後48時間の対照処理(図18Fを参照のこと)に比較して、グルコース刺激カルシウム(GSCa)応答の改善を示した。sieIF−5A処理された膵島は、単離後24時間のサイトカイン(図18Eを参照のこと)及び単離後48時間のサイトカイン(図18Gを参照のこと)の存在下で高いGSCaを示す。eIF−5A処理されたマウスから単離された膵島は、GSISにより評価された場合に、4時間のサイトカインインキュベーションがない場合(図18Hを参照のこと)とある場合(図18Hを参照のこと)の両方において、対照に比較して、グルコース刺激インスリン分泌(GSIS)の改善を示した。
【0057】
実施例5:eIF−5Aのノックダウンが、サイトカイン誘導性iNOS生産を無効にする
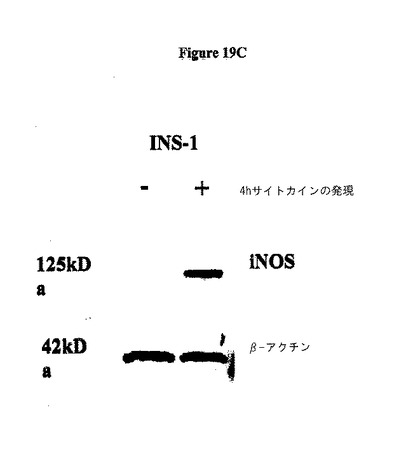
サイトカイン誘導性のiNOSタンパク質産生は、eIF−5A欠損細胞において生じない。サイトカインで処理された膵島を、iNOSmRNAについてRT−PCRに供した。全ての処理群は、サイトカイン暴露の際にiNOSmRNAの劇的な上方制御を示した。図19Aを参照のこと。サイトカイン処理された膵島を免疫ブロット分析に供した。注目すべきは、iNOSタンパク質は、サイトカイン暴露された際に対照膵島中において誘導されるが、si eIF−5A処理された膵島においては誘導されないという点である。図19Bを参照のこと。INS−1(832/13)β細胞を、サイトカインカクテルで4時間処理し、そして免疫ブロットに供した。細胞はiNOSタンパク質の同様の増加を示す。図19Cを参照のこと。INS−1細胞を、様々な濃度のGC−7で処理し、デオキシハイプシン合成酵素の阻害剤で一晩処理し、次にサイトカインの存在下で4時間インキュベートした。INS−1細胞は、iNOS生産及びCG−7濃度の逆相関を示し、活性eIF−5A産生を抑制が、iNOS翻訳を示唆する。図19Dを参照のこと。
【表1】

【表2】
【技術分野】
【0001】
ランゲルハンスの膵島は、膵臓内でインスリンを生産する細胞を含む多細胞実体である。平均的なヒトで、約百万の膵島を有し、そして膵臓内の細胞の合計の約2〜3%を占める。膵臓はランゲルハンスの膵島を含み、これは、インスリンを生成するβ細胞を取り囲んでいる。β細胞は、血中のグルコースレベルをモニターしており、そして精密に計測された量のインスリンを放出して、グルコースピークに均衡させる。I型及びII型糖尿病は、90%超のこれらのβ細胞が損傷を受けた場合に進行する。
【背景技術】
【0002】
1型糖尿病は、米国において2100万人の糖尿病患者の約10%が罹患するグルコース恒常性についての障害である。1型糖尿病は、膵島β細胞の実質的に完全な自己免疫破壊から生じ、生命維持のためのインスリン投与に患者を依存状態にする。1型糖尿病の発症機序は、免疫系の細胞と、膵島β細胞上に存在する抗原との間の複雑な相互作用に関わっている。この相互作用は、Th1細胞とマクロファージの活性化を導き、これは続いて他の因子のなかで特に、一酸化窒素産生を誘導するサイトカイン(IL1β、TNFα、及びIFNγ)であって、アポトーシス性及び壊死性の膵島β細胞死を引き起こすサイトカインを放出する。膵臓又は膵島移植に関するβ細胞「置換」戦略は、1型糖尿病を回復させる点で一定ではないが成功を収めたが、限られた数のソース(死体ドナー)を必要とし、そしてさらに、内在の自己免疫プロセスにより破壊される。
【0003】
結合マトリクス及び残りの外分泌組織からの膵島の分離又は単離は、研究室での実験及び移植目的の点で利点があり、そして有利である。膵島移植は、1型糖尿病の最も有望で、かつ生理的侵襲の少ない方法である。完全な膵臓組織ではなく膵島を移植することは、移植の容易さ、そして消化酵素の分泌に関わるドナー組織の膵臓外分泌機能が除去されるという確かな利点を有する。膵臓外分泌組織から膵島を遊離することは、膵島移植に影響する最初の及び重大な工程である。膵島単離における重要な目標は、十分な数の生存可能であり、機能を有しかつ強力な移植用の膵島を提供することである。
【0004】
「エドモントンプロトコル」は、健常な膵島を糖尿病患者に移植する。エドモントンプロトコルを用いた膵島移植が、Shapiro, Ryan, and Lakey, Clinical Islet Transplantation--State of the Art, Transplantation Proceedings, 33, pp. 3502-3503 (2001); Ryan et al., Clinical Outcomes and Insulin Secretion After Islet Transplantation With the Edmonton Protocol, Diabetes, Vol. 50, April 2001, pp. 710-719; and Ryan et al., Continued Insulin Reserve Provides Long-Term Glycemic Control, Diabetes, Vol. 51, July 2002, pp. 2148-2157に記載される。肝臓に配置されると、当該細胞は、血液供給を発達させ、そしてインスリンを生成する。エドモントン・プロトコルは、使用される方法に応じて7〜10の工程を含む。第一工程は、特定の酵素(リベラーゼ)を、ドナーの膵臓に導入することに関しており、当該酵素は膵臓組織を消化するが、膵島を消化しない。消化工程に続き、膵臓内の他の細胞から膵島を分離する幾つかの次なる工程が存在する。分離された膵島を、門脈として知られている肝臓の主な血管に移植する。肝臓は、損傷を受けた場合に、肝臓自体を再生することができ、新たな血管及び支持組織を構築できる。その結果、膵島を肝臓に移植した場合、新たな血管が形成して、膵島を支持すると信じられている。細胞が生成するインスリンは、周囲の血管を通して血流中に吸収され、そして体中に分布されて、血中のグルコースレベルを制御する。
【0005】
要するに、エドモントン・プロトコルの工程は、もろい三次元構造を有しかつ増殖及び生存には多量の酸素を必要とする膵島の生存性を損なわせる強制的プロセスを作り出す。当該プロセスの間に、膵島は、酸素デリバリーの非最適条件のため損傷を受けるか又は破壊されることがあり、所定のドナー膵臓から回収される健常な膵島の収率に影響する。さらに、ドナーの利用可能性により、膵島移植が極めて限定的なものになり;1人の患者においてインスリン独立性を得るためには、2つの膵臓が必要とされることが多い。
【0006】
ステロイドフリーの非糖尿病誘発性免疫抑制治療と併せた膵島移植は、1型糖尿病を患う患者を治療するために用いられてきた。しかしながら、このような治療は、脂質異常症及び高血圧の高いリスクを導くことがあり、そして長期間の研究によると、膵島生存性が損なわれることが示される。
【0007】
ヒトにおける膵島移植のエドモントンプロトコルにより、移植の1年後に、80%の患者が、インスリン独立性を達成するという短期間の顕著な成功が示された。しかしながら、5年目ではこの割合は、たった約10〜15%に低下する。移植片機能の進行的消失の理由は明らかではないが、進行性のサイトカイン媒介性炎症の結果として膵島が失われるようである。しかしながら、膵島移植は、1型糖尿病を患う患者の実行可能な治療選択のままであり、長期間のインスリン独立性が得られない患者であっても、血糖不安定性の低下、低血糖症の発症低下、及びインスリン量の低下などの他の重要な恩恵を受けることが特に結果により示されている。
【0008】
膵島移植後の初日は、移植された膵島について特に脆弱な期間である。最大で60%の移植された膵島が、移植後の初日に非自己抗原特異的アポトーシスを受ける。この初期の破壊は、糖尿病を回復するために十分な膵島が生存することを保証するために、2〜3個の死体ドナーを使用することを必要とすることの主な根本的理由となっている。炎症性サイトカイン、IL−1β、TNFα、及びIFNγの局所的生産は、移植された膵島の初期アポトーシスに関わっている。こうして、初期移植期間におけるサイトカインを制限する戦略は、十分な膵島を得るために必要となるドナーの数を限られたものにし、膵島生存性を高め、そして長期間機能する能力を延長する。本発明は、これらの戦略を提供する。
【発明の概要】
【0009】
本発明は、単離後の回収された膵島細胞の機能を保持する方法であって、膵島単離の前に、膵島細胞ドナーの膵島細胞にeIF−5A1 siRNAを投与することを含み、ここで当該eIF−5A1 siRNAは、膵島細胞においてeIF−5A1 の発現を抑制し、それにより膵島細胞におけるアポトーシスを抑制し、そして回収された膵島細胞の機能を保存する、前記方法を提供する。好ましい実施態様では、siRNAは、eIF−5A1 の以下のヌクレオチド配列を標的化する:
5'−AAAGGAATGACTTCCAGCTGA−3';
5'−AAGATCGTCGAGATGTCTACT−3';
5'−AAGGTCCATCTGGTTGGTATT−3';又は
5'−AAGCTGGACTCCTCCTACACA−3'
ある実施態様では、eIF−5A1 siRNAは、ヌクレオチド配列5'−AAAGGAAUGACUUCCAGCTGAdTdT−3'を含む。siRNAは、任意の許容される手段を介してドナーに投与されてもよい。ある実施態様では、siRNAは、腹腔内注射を介して膵島細胞ドナーへと投与される。
【0010】
本発明の別の実施態様は、膵島細胞がドナー回収プロセスの間にアポトーシスを引き起こすことを阻害する方法であって、膵島単離の前に膵島細胞ドナーへの腹腔内投与を介して膵島細胞ドナーへとeIF−5A1 siRNAを投与することを含み、ここで当該eIF−5A1 siRNAは、膵島細胞内におけるeIF−5A1 の発現を阻害し、それにより膵島細胞におけるアポトーシスを阻害する、前記方法を提供する。
【0011】
好ましい実施態様では、siRNAは、eIF−5A1 の以下のヌクレオチド配列:
5'−AAAGGAATGACTTCCAGCTGA−3';
5'−AAGATCGTCGAGATGTCTACT−3';
5'−AAGGTCCATCTGGTTGGTATT−3';又は
5'−AAGCTGGACTCCTCCTACACA−3'
を標的とする。ある実施態様では、eIF−5A1 siRNAは、ヌクレオチド配列:5'−AAAGGAAUGACUUCCAGCTGAdTdT−3'を含む。
【0012】
任意のsiRNA又はアンチセンスコンストラクトは、かかるコンストラクトが、膵島細胞においてeIF−5A1 の発現を阻害する限り、使用することができる。siRNAの投与は、任意の適切な経路であってもよい。代表的な投与方法は、膵島細胞ドナーの門脈を介したかん流、膵島細胞ドナーの門脈を介した流体力学的かん流、及び腹腔内注射を含む。
【0013】
本発明は、アポトーシスを阻害し、そして膵島細胞の機能を保存するための組成物であって、eIF−5A1 siRNAを含み、ここで当該siRNAが、eIF−5A1の発現を阻害し、それにより膵島細胞におけるアポトーシスを阻害する組成物も提供する。好ましいeIF−5A1 siRNAは上に論じられている。
【0014】
本発明は、膵島β細胞における標的タンパク質の発現を阻害する方法であって、β細胞回収前にβ細胞ドナーに対して、標的タンパク質をコードするmRNAに対して標的されたsiRNAコンストラクトを投与することを含み、ここで当該siRNAは、膵島β細胞中の標的タンパク質の発現を阻害する方法も提供する。
【0015】
本発明は、eIF−FAの発現を低下させることにより、誘導性一酸化窒素合成(iNOS)経路を介して、一酸化窒素(NO)の産生を低下させる方法を提供する。
【0016】
本発明は、iNOS経路を介して一酸化窒素の産生を低下させる医薬の製造のための、eIF−5A siRNAの使用をさらに提供する。
【図面の簡単な説明】
【0017】
【図1】図1は、門脈を介してeIF−5A siRNAをかん流した後における、β−アクチン、mAAT、及びeIF−5A1について行われたRT−PCRの結果を提供する。この図は、eIF−5A1 の発現が計測可能であり、そうして膵島に取り込まれたことを示す。
【図2】図2は、低速逆行性門脈かん流(slows regrograde portal vein perfusion)を示す。胆管(透明)及び門脈(赤)は、調製的ノット(preparatory knot)(暗色縫合)の準備ができている。針をノットの下に差し込み(矢印により方向を示す)、ノットの下を横断させ、そしてsiRNAを血管に放出して、これらは膵臓、脾臓、腸、及び遠位結腸の三分の一に到達する。
【図3】図3は、eIF−5A1 siRNAを膵島にかん流することにより、eIF−5A1 の発現の低下が誘導される(図には、eIF−5A1 のmRNAレベルの低下が示される)ことを示す。
【図4】図4には、対照及び生理食塩水で処理された膵島に比べて、eIF−5A1 で処理された膵島細胞のアポトーシスを低下することが示される(ここで1群あたりn=2)。
【図5】図5には、対照及び生理食塩水で処理された膵島に比較した場合に、eIF−5A1 siRNAで処理された膵島細胞のアポトーシスが低下することが示される(ここで1群辺りn=3)。
【図6】図6は、eIF−5A2に対して整列されたヒトeIF−5A1のヌクレオチド配列を提供する。
【図7】図7は、eIF−5A2に対して整列されたヒトeIF−5A1のアミノ酸配列を提供する。
【図8】図8は、代表的なアンチセンスオリゴヌクレオチドと供にヒトeIF−5A1のヌクレオチド配列を提供する。
【図9】図9は、代表的なアンチセンスオリゴヌクレオチドと供にヒトeIF−5A1のヌクレオチド配列を提供する。
【図10A】図10A及び図10Bは、代表的なsiRNAと供にヒトeIF−5A1のヌクレオチド配列を提供する。
【図10B】図10A及び図10Bは、代表的なsiRNAと供にヒトeIF−5A1のヌクレオチド配列を提供する。
【図11】図11は、代表的なsiRNAと供にヒトeIF−5A1のヌクレオチド配列を提供する。
【図12】図12は、siRNA eIF−5A1が、膵島細胞において発現をノックダウン(減少)させることができるということを示す実験の模式図を示す。
【図13】図13Aは、2.8から11mMのグルコース刺激の後の50個の膵島についてのインスリン分泌パターンを示し、そして図13Bは、2.8から11mMのグルコース刺激の後の単一膵島からの[Ca2+]iパターンを示す。この相は、0、1、2による。この図により、通常機能の膵島が、インスリン分泌の二相性のパターンでのグルコース刺激に応答することが示され、これは膵島細胞内カルシウム([Ca2+]i)における同様の変化に近似する。
【図14】図14は、カルシウム振動研究の結果を提供する。これらの図により、eIF−5A1 siRNAで処理され(そうしてeIF−5A1の発現が低下した)膵島細胞が、有意に強いグルコース応答及び全体の振動を示すということが示される。図14は、これらの処理された細胞が、長期間生存するばかりでなく、機能も保持するということを示す。
【図15】図15は、サイトカイン処理された染色済みの膵島細胞の異なる写真を示す。この写真により、対照又はビヒクル(生理食塩水のみ)で処理された膵島細胞よりも、eIF−5A1 siRNAで処理された膵島細胞の生存性が高く、そして死細胞が少ないということが示される。
【図16A】図16A及び16Bは、IP注射を介して膵島細胞ドナーに投与された場合にsiRNAが、膵島細胞に侵入することを4種の画像化技術を用いて示す。共焦点イメージングを用いて、単離膵島の3D再構成図を作成した。図16Aにより、0.9%の生理食塩水を1日1回、3日間腹腔内投与されたマウスから単離された膵島が、FITC自己蛍光を示すが、TRITCチャンネル(図ではCy3標識)において最低の蛍光しか示さないということが示される。
【図16B】図16A及び16Bは、IP注射を介して膵島細胞ドナーに投与された場合にsiRNAが、膵島細胞に侵入することを4種の画像化技術を用いて示す。共焦点イメージングを用いて、単離膵島の3D再構成図を作成した。図16Bにより、Cy−3標識された安定化siRNAを1日1回、3日間腹腔内投与されたC57Bl/6から単離された膵島が、Cy3標識の浸透を示すということが示される。
【図17A】図17(A)は、βTC3細胞が、2の異なるsiRNAコンストラクト(コンストラクトA及びB)、又は対照でトランスフェクトされ、そして指定のタンパク質について免疫ブロットに供された場合の結果を示す。
【図17B】図17(B)は、C57Bl/6マウスに同じsiRNAを1日1回、3日間腹腔内注射し、そして膵島を単離し、そしてコラゲナーゼ消化及び分画勾配遠心分離を介して精製した場合の結果を示す。膵島を2%SDS中で溶解し、そして免疫ブロットに供する。
【図17C−D】図17(C)は、単離された膵島にFura2を30分間充填し、次に3mM D−グルコース中でイメージングした場合の結果を示す。膵島を11mM D−グルコースで300秒刺激し、そしてFura2比を、蛍光顕微鏡により連続してモニタリングした場合の結果を示す。図17(D)は、グルコース刺激されたインスリン分泌(GSIS)を、各処理群から得た50の膵島を用いて実行した結果を示す。
【図18A】図18A−Iは、eIF−5Aのノックダウンが、膵島機能及びex-vivoでの生存性を改善するということを示す。
【図18B】図18A−Iは、eIF−5Aのノックダウンが、膵島機能及びex-vivoでの生存性を改善するということを示す。
【図18C】図18A−Iは、eIF−5Aのノックダウンが、膵島機能及びex-vivoでの生存性を改善するということを示す。
【図18D−I】図18A−Iは、eIF−5Aのノックダウンが、膵島機能及びex-vivoでの生存性を改善するということを示す。
【図19A】図19A−Dは、サイトカイン誘導性のiNOSタンパク質の生産が、eIF−5A欠損細胞では見られないことを示す。
【図19B】図19A−Dは、サイトカイン誘導性のiNOSタンパク質の生産が、eIF−5A欠損細胞では見られないことを示す。
【図19C】図19A−Dは、サイトカイン誘導性のiNOSタンパク質の生産が、eIF−5A欠損細胞では見られないことを示す。
【図19D】図19A−Dは、サイトカイン誘導性のiNOSタンパク質の生産が、eIF−5A欠損細胞では見られないことを示す。
【発明を実施するための形態】
【0018】
真核生物翻訳開始因子5A(eIF−5A)は、ストレス誘導性の遺伝子の転写後調節において決定的な因子であることが明らかとなり、そして哺乳動物細胞においてサイトカイン媒介性のアポトーシスを促進することが明らかとなっている。eIF−5Aは、真核生物を通して極めて高度に保存されている小さな酸性タンパク質であり、そして固有のポリアミン由来のアミノ酸であるヒプシンを含むことが知られている唯一のタンパク質である。eIF−5Aは、mRNAプロセッシング、輸送、及び翻訳において役割を果たすことが提案されていた。しかしながら、どんなものであれ膵島における炎症カスケードにおけるその役割は、従前、特徴決定されていなかった。膵島は、グルコース及び脂質毒性、酸化ストレス、及び炎症性サイトカインに対して感受性が高い。膵島の細胞ストレスへの応答を媒介する点に関わる因子の役割を研究するため、本発明者は、低分子干渉RNA(siRNA)を用いてin vivoにおいて膵島の特定タンパク質を枯渇させるプロトコルを開発した。研究において、発明者らは、RNA干渉を用いてeIF−5Aタンパク質を膵島から枯渇させることにより、膵島におけるeIF−5Aの役割を決定することを試みた。
【0019】
しかしながら、おもに齧歯類膵島におけるRNA干渉研究における主要な試みでは、トランスフェクション効率が比較的低かった。従来の研究が、短いヘアピンRNAのウイルス媒介性のデリバリーを利用することを示した一方で、この技術は、適切なウイルスを作成するには時間がかかるものであり、そしてウイルス毒性を被るものであった。本発明者らは、安定化された低分子干渉RNA(siRNA)を繰り返し腹腔内注射することにより、マウス膵島から選択したタンパク質をin vivoで枯渇させる新たなプロトコルを発見した。このプロトコルは、膵島内へのsiRNAの顕著な浸透をもたらす。この技術は、よく特徴決定された膵臓転写因子Pdx1などの任意の所望の標的タンパク質を、膵島からうまく枯渇させるために使用することができる。この技術を用いて、発明者は、eIF−5Aタンパク質を膵島からうまく枯渇させ、そして誘導型一酸化窒素合成酵素(iNOS)をコードするmRNAの翻訳を促進することにより、eIF−5Aが、サイトカイン媒介性の膵島機能不全に寄与するということを示した。このデータは、iNOSのmRNAを含むmRNA(おそらく、サイトカイン媒介性のストレス応答に関与するmRNA)のサブセットの安定化及び/又は核輸送のために、eIF−5Aが膵島において必要であるというモデルを示唆する。
【0020】
本発明者らは、in vivoでのsiRNA注射を使用して、サイトカイン媒介性ストレスにおける真核生物翻訳開始因子5A(eIF−5A)の役割を研究した。eIF−5Aは、ストレス誘導性遺伝子の翻訳において、決定的なレギュレーターとして他のシステムにおいて特徴決定されていた。eIF−5Aに対するsiRNAがマウスに注射された場合、単離された膵島においてeIF−5Aタンパク質レベルの50〜70%の低下が観察された。この低下に伴い、対照処理に比べて、グルコース刺激性のインスリン分泌及びCa2+可動化が30〜40%の増加すること観察された。単離された膵島を炎症性サイトカイン(IL1β、TNFα、IFNγ)のカクテルに暴露した場合に、膵島機能の相対的向上が、si−eIF−5Aで処理された動物において持続した。eIF−5Aノックダウンの後の膵島保護は、その生成物がグルコース応答性又はインスリン転写に関与する遺伝子(Slc2a2、Gck、Irs1、Nkx6−1、MafA、Pdx1、NeuroD1、及びSetd7)の発現変化を伴うことはない。図18Cを参照のこと。大事なことは、誘導性一酸化窒素合成酵素(iNOS)をコードするmRNAが、対照で処理された膵島及びsi−eIF−5A−処理された膵島の両方において、サイトカインに応答して、40倍超も上方制御されるが(図19Aを参照のこと)、iNOSタンパク質レベルが、si−eIF−5A処理された膵島において3〜5倍低いということである(図19Bを参照のこと)。これらのデータにより、si−eIF−5A処理された膵島における機能の向上が、二次的に一酸化窒素の生産を低下しうるということが示唆される。これにより、ストレスシグナルに応答した膵島β細胞において、eIF−5AがiNOS翻訳の制御を介して不可欠な役割を果たし、そして膵島細胞の生存性の保存並びに回収後及び移植後において機能する膵島細胞の能力の保存を狙った治療戦略について、eIF−5Aが実行可能な標的として役立ちうるという証拠を提供する。
【0021】
本発明者らは、(GC−7(DHSの阻害剤)を投与することによる)ヒプシン化の阻害を介したeIF−5A機能の遮断が、効果的にiNOSレベルを低下するということをさらに示した。図19Dを参照のこと。
【0022】
膵島へのsiRNAの取り込みが、逆行性の門脈閉塞を介した膵臓かん流により達成することができるということが以前に示されていた。Bradleyら、Transplantation Proceedings, 37, 233-236, 2005を参照のこと。簡潔に説明すると、Cy−3標識されたルシフェラーゼ(Luc)siRNA GL2 二本鎖を、リポフェクタミン2000でパッケージして、又はパッケージせずに用いて、そして尻尾静脈を通して(in vivo、マウス1匹あたり50μg)注射するか、又は逆行性門脈静脈閉塞(in situ, マウス1頭あたり2μg)により膵臓に直接注射した。膵臓を取得し、そしてIn situデリバリー後に24時間4℃で貯蔵するか、又はin vivoデリバリー後に4時間貯蔵し、そして膵島を単離し、そしてさらに試験前に16時間培養した。siRNAの分布を可視化するために、膵臓をインスリンについて染色し、そして蛍光顕微鏡の下で試験した。単離された膵島を直接蛍光顕微鏡で試験した。パッケージされていないsiRNAは、リポソームにパッケージされたsiRNAを用いて観察されるのと同程度に膵島に到達し、いわゆる「裸」のsiRNAデリバリーの報告に一致した。Lewis et al., Nat. Genet. 32: 107-108, Epub 2002 Jul 2029, 2002 and McCaffrey AP, et al., Nature 418: 38 - 39, 2002)。図1は、膵島細胞へのかん流が、膵島細胞への適切なデリバリーメカニズムを提供するということを示す。
【0023】
従って、本発明は、膵島細胞において標的タンパク質の発現を抑制する方法であって、標的タンパク質をコードするmRNAを標的とするsiRNAを、膵島細胞に投与することを含み、ここで当該siRNAが、膵島細胞において標的タンパク質の発現を阻害する、前記方法を提供する。任意の手段を介して、好ましくは膵島β細胞の単離の前に、膵島β細胞ドナーに腹腔内投与することを介して投与されてもよい。
【0024】
1の実施態様では、本発明者らは、安定化されたCy3−標識された二本鎖RNAをC57Bl/6マウスに三日間腹腔内注射することにより、共焦点顕微鏡で測定した場合に当該RNAが単離された膵島へと顕著に浸透することが明らかにされた。タンパク質ノックダウンについてこの技術を試験するために、Pdx1を標的とした安定化siRNAを用いた。Pdx1mRNAに対する2つの異なるsiRNAは、(免疫ブロット分析により評価した場合に)膵島においてPdx1タンパク質の50%及び90%ノックダウンをもたらした。図17A−Dを参照のこと。
【0025】
本発明は、膵島細胞においてeIF−5A1の発現を阻害する方法であって、eIF−5A1 siRNAを膵島細胞に投与することを含み、ここで当該eIF−5A1 siRNAが、膵島細胞においてeIF−5A1の発現を阻害する、方法を提供する。図16A及び図16Bは、IP注射を介した膵島細胞ドナーへの投与が、膵島細胞に対する適切なデリバリーメカニズムを提供するということを示した。図16Bは、安定化されたCy−3標識二本差RNAをC57Bl/6マウスに3日間の腹腔内注射することにより、共焦点顕微鏡で測定した場合に、当該RNAが単離膵島に顕著に浸透することを明らかにした。図12B及び18Bは、eIF−5A1 siRNAで処理された膵島細胞が、より少ないeIF−5A1 siRNAしか発現しないことを示す。
【0026】
eIF−5A1発現を抑制することにより、アポトーシスが阻害される。図4及び5は、単離前にeIF−5A1 siRNAで膵島細胞を処理することが、これらの細胞のアポトーシスを阻害するメカニズムを提供した(subG1期の細胞数の低下により示される)。したがって、本発明は、回収された膵島細胞において、アポトーシスを阻害する方法であって、eIF−5A1 siRNAを膵島β細胞ドナーに投与することを含み、ここで当該eIF−5A1 siRNAは、膵島細胞内におけるeIF−5A1の発現を阻害し、そしてここでeIF−5A1発現の抑制がアポトーシスを阻害する、方法を提供する。回収プロセスの前に、膵島細胞をeIF−5A1 siRNAで処理することが好ましい。
【0027】
本発明は、単離された後に回収された膵島細胞の機能を保存する方法であって、eIF−5A1 siRNAを膵島単離の前に膵島細胞ドナーの膵島細胞に投与することを含み、これが膵島細胞におけるeIF−5A1発現の抑制又は低下をもたらし、次に膵島細胞におけるアポトーシスを抑制し、さらにその機能を保存する方法をさらに提供する。膵島細胞の機能の保存は、膵島細胞が回収プロセスを生き残るということだけではなく、回収プロセス後においてその機能を維持する(細胞の機能は、長期間残存し、及び/又はそれらはeIF−5A1 siRNA処理されていない膵島細胞よりも優れたグルコース応答性を維持する)ということを意味する。
【0028】
1の実施態様では、膵島機能及び全身の健康におけるeIF−5A1 ノックダウンの結果を調査するために、C57Bl/6雄マウスに、ビヒクル(0.9%生理食塩水)[群I]、対照siRNA(16.6mg/kg)[群II]、又はeIF−5A1 siRNA(16.6mg/kg[群III]のいずれかを腹腔内(IP)注射した。指定された処理の注射を、膵島回収(0日目)の前3日間(−3、−2、−1日目)毎日投与した。単離された膵島をコラゲナーゼ処理により精製し、そして試験の開始前16〜18時間静置した。回収後1日目に機能のベースラインを確立した後に、半分の膵島を、サイトカインカクテル(5ng/ml IL1β、10ng/ml TNF−α、100ng/ml INF−γ)に晒して、1型糖尿病において膵島が直面する条件を模倣した。実験設計の模式図及び膵島の配分を図12Aに示す。
【0029】
ウエスタンブロットにより、タンパク質ノックダウンについて膵島の各群から得たタンパク質ライセートを分析した。図12Bは、群IIIの処理を受けたマウス膵島におけるeIF−5A1の相対的ノックダウンを示し、これは図12Cにおいて定量化される。これらのデータにより、膵島内におけるeIF−5A1の発現のノックダウンをもたらすIP注射の効力が示される。
【0030】
回収された膵島における保存された機能の証拠は、カルシウム振動アッセイにより示される。本発明者らは、細胞内カルシウムが刺激及びストレスの指標であるということを決定した。ヒト及び齧歯類の両方において、通常通り機能する膵島は、図13Aに示されるようにインスリン分泌の2相性のパターンでグルコース刺激に応答する。一次近似として、この二相性パターンは、膵島の細胞内カルシウム[Ca2+]iにおける変化によく似ており、この変化は、第二相のプラトーに落ち着く第一相ピークからなる(図13B)。340及び380nm励起から放射される光の比として報告される[Ca2+]iの計測は、膵島機能のさらなる特徴を検出する優れた感度を提供する。グルコースに対する[Ca2+]i応答は、3つの相からなり(図13を参照のこと)、β細胞刺激と分泌との関係における「コンセンサスモデル」により記載されるプロセスを大まかに反映している。[Ca2+]iの初期の落ち込みは、グルコースがβ細胞に移動した後に、小胞体が[Ca2+]iを動員し、隔離するにつれ生じる(第0相)。次にグルコースは、解糖をとおして代謝され、そしてさらにトリカルボン酸(TCA)回路においてさらに代謝され、ATP生産をもたらす。ATP/ADPの十分な増加が生じて、KATP−イオンチャネルが閉じた場合、第一相インスリン分泌に関連するL型カルシウムチャネルが開くために、[Ca2+]iの急激な増加が生じる。この最初の第一相ピークの後に、[Ca2+]iは、プラトーに落ち着くが、プラトーは[Ca2+]i振動によりしばしば中断される。プラトーの高さは、グルコース濃度及び第二相インスリン分泌の割合に直接関連している。概して、イメージングに基づく[Ca2+]i計測は、個々の膵島のレベルで、動的なインスリン分泌の優れた一次近似となる。
【0031】
多くの因子は、グルコース刺激に対する通常の[Ca2+]i応答にマイナスの影響を与えることがある。グルコース代謝、ミトコンドリア及びATP産生、小胞体、イオンチャネルの機能、及び多種多様の他の問題の破綻は、グルコース刺激に関するカルシウム処理のダイナミクスに影響を与えることがある。[Ca2+]iの変化を頻繁にモニタリングすることにより、これらの活性態様における不具合は、静的インスリン分泌の標準計測により観察されるよりも頻繁に観察される。例えば、低いグルコースレベルにおけるベース[Ca2+]iの増加、及びグルコース刺激の間における[Ca2+]iの第0相の落ち込みの低下は、ER−ストレス又はおそらくイオンチャネル機能不全を指している可能性がある。このような不具合は、静的又は動的インスリン分泌を計測することによって容易に検出することはできないが、本明細書に記載される画像化技術は、これらの問題を指摘するために使用することができる。
【0032】
カルシウム振動及びグルコース刺激されたインスリン分泌研究においてサイトカインが存在する場合、及び存在しない場合の両方において、eIF−5A1 siRNA(16.6mg/kg)で処理された膵島[第3群(eIF−5A1ノックダウン膵島)]は、グルコースに対する高い応答性、並びに長期間の機能の維持(長期間の生存を示す)を示した。図14を参照のこと。従って、これらの膵島は、インスリンの高い生産を示すことが予期される(データー取得中)。生存/死亡染色した後に撮られた画像から得た元データーは、eIF−5A1 ノックダウンが膵島細胞の生存を延長するという概念を定性的に裏付ける(図15)。赤色膵島(死亡した膵島細胞の各にインターカレットするEtHD−1の結果)の数を、カルセイン−AMを蛍光カルセインへと能動的に切断する緑色膵島の数と比較することにより、膵島生存の定性的画像が確かめられる(図15)。
【0033】
本発明は、一酸化窒素の産生増加と関連する炎症性疾患に関与する細胞においてアポトーシスを阻害する方法をさらに提供する。例えば、リューマチ様関節炎(RA)などの炎症性疾患は、誘導性一酸化窒素合成(iNOS)経路の活性化のため、一酸化窒素(NO)の生産増加と関連する。動物モデルにおける研究により、NOが、関節炎症及び組織損傷の病原進行において通常の役割を果たすということが示唆される。なぜなら、関節炎の重篤度が、NOS阻害剤の投与により低下するからである。関節内に存在する幾つかの細胞(例えば滑膜線維芽細胞、内皮細胞、及び軟骨細胞)は、炎症性サイトカインにより誘導されて、インビトロでNOを産生することができる。さらに、局在化研究により、RAを煩う患者から得られた関節組織の滑膜表層細胞、軟骨細胞及び血管におけるiNOS発現が上方制御されることが示される。滑膜表層及び軟骨へのiNOSの局在化は、アポトーシスがRA、特に滑膜表層及び軟骨において特に増加することを示した他の研究の点でも関心が高い。リューマチ様関節におけるアポトーシスの原因となるメカニズムは、不明のままであるが、Fas抗原の発現が、RA滑膜細胞において増加し、そしてFas抗体が、インビトロにおける滑膜細胞のアポトーシス性の死亡を刺激することができるということが以前の研究者により示された。iNOS経路の活性化が、リューマチ性関節においてアポトーシスの刺激に関与するという仮説が検討された。なぜなら高レベルのNOが、インビトロにおいて多くの細胞型においてアポトーシスを刺激することが知られているからであり、そしてNOが、RAにおけるアポトーシスのメディエーターとして作用すると結論づけられた(R. J. van't Hof et al., Rheumatology 200, 39: 1004-1008を参照のこと)。
【0034】
このようなものとして、本発明はRAに関わる細胞(例えば、滑膜細胞及び軟骨細胞)においてアポトーシスを阻害する方法であって、eIF−5A1の発現を阻害し、それによりiNOS経路を介した一酸化窒素の産生を阻害又は低下させて、細胞のアポトーシスを阻害する方法を提供する。
【0035】
本発明は、さらに、細胞又は宿主におけるNO産生を阻害する方法であって、当該細胞又は宿主にeIF−5A1に対するsiRNAを投与することによる方法を提供する。siRNAは、eIF−5A1の発現を低下させ、そして次にiNOS経路の活性化を抑制することによりNOの産生を低下させる。
【0036】
本発明は、iNOS経路を介して一酸化窒素の産生を低下させる医薬の製造のための、eIF−5A siRNAの使用を提供する。
【0037】
eIF−5A1 の発現を抑制する任意のeIF−5A1 siRNAが使用されてもよい。「阻害」という語句は、対照細胞、つまりeIF−5A1 siRNAで処理されていない細胞において生じるレベルに比べて低下することを意味する。1の代表的、そして特に好ましいeIF−5A1 siRNAは、以下の配列:5'−AAAGGAAUGACUUCCAGCTGAdTdT−3'を含む。2005年11月28日に出願された同時係属米国出願11/293,391号(当該出願はその全てを本明細書に援用する)は、追加の代表的eIF−5A1 siRNAを提供し、そして他の細胞型においてeIF−5A1の発現を阻害するために使用され、そしてアポトーシスを阻害することが示された他のアンチセンスコンストラクトを提供する。当業者は、eIF−5A1 配列が与えられれば、他のeIF−5A1 siRNAを設計することができ、そして過度の実験を行うことなく、発現を阻害する能力について当該siRNAを容易に試験することができる。図6〜11は、eIF−5A1 の配列、代表的なeIF−5A1 siRNA及びアンチセンスコンストラクトを提供する。本発明の別の実施態様では、eIF−5A1 のアンチセンスコンストラクトを使用して、eIF−5A1 の発現を阻害し、そうして膵島細胞のアポトーシスを阻害し、並びにその機能を維持又は保存することもできる。
【0038】
好ましい実施態様では、eIF−5A1 siRNAは、eIF−5A1の以下のヌクレオチド配列:
5’−AAAGGAATGACTTCCAGCTGA−3';
5'−AAGATCGTCGAGATGTCTACT−3';
5'−AAGGTCCATCTGGTTGGTATT−3';及び
5'−AAGCTGGACTCCTCCTACACA−3'
を標的とする(図10を参照のこと)。特に好ましい実施態様では、siRNAは、eIF−5A1の以下の配列:5’−AAAGGAATGACTTCCAGCTGA−3'を標的とする。
【0039】
本発明は、ドナー回収プロセスの間に、膵島細胞がアポトーシスを引き起こすことを阻害するための方法を提供する。上に記載されるように、多くの膵島細胞は、回収された場合にアポトーシスを引き起こす。本発明者らは、回収する前に膵島細胞にeIF−5A1 siRNAを提供することが、アポトーシスに対する保護的利点を提供することを示した。eIF−5A1 siRNAは、膵島単離の前に、膵島細胞ドナーの膵島細胞に投与される。ドナー(及び膵島細胞)は、例えばヒト膵島細胞を含む任意の動物であっても良い。任意の投与方法が使用されてもよい。例えば、siRNAは、膵島細胞ドナーの門脈を通したかん流を介して投与されることもあるし、又は膵島細胞ドナーの門脈を介した流体力学的かん流を介して投与されることもある。実施例1を参照のこと。別の投与形態として腹腔内投与が挙げられる。実施例2、及び実施例3〜5を参照のこと。
【0040】
門脈を介したかん流は、胆管への挿管に類似しているが、針は反対の方向に向いている。門脈は、肝臓を引き出し、そしてマウスの左側へと内臓を移動させることにより露出する。調製的ノット(preperative knot)を門脈の周囲に作り、そして胆管を含める。血管に穿刺した後に、尖っていない針を膵臓にまで進め、そしてその周囲でノットを絞める。マウスモデルでは、1mlの生理食塩水又はsiRNA(5μg)をゆっくり放出し、針を取り出し、そしてノットを針の手前で閉じて、液体の漏出を妨げる。この時点で、マウスをひっくり返し、そして胆管を膵臓消化にアクセスさせた。膵臓にsiRNAを長時間適用してもよい。或いは、膵臓を取り出し、冷却状態を維持して、コラゲナーゼで処理することもできる。通常の膵島単離方法を行い、そして膵島(50)を16時間インキュベートしてもよい。
【0041】
本発明は、膵島細胞内でアポトーシスを阻害するための組成物であって、eIF−5A1 siRNAを含み、ここでsiRNAがeIF−5A1の発現を阻害し、それによりアポトーシスを阻害し、そして膵島細胞の機能を維持する。組成物は、他の又は追加のeIF−5A1 siRNAを上記のように含むこともある。好ましいsiRNAは、ヌクレオチド配列:5'−AAAGGAAUGACUUCCAGCTGAdTdT−3’を含む。
【実施例】
【0042】
実施例1:門脈かん流
マウス膵島は、eIF−5A1を発現する。全量RNAを、単離されたマウス膵島から抽出し、そしてRT−PCRを、β−アクチンとeIF−5Aについて行った(図1)。安静にして刺激を受けていない膵島は、eIF−5A1−mRNAの陽性レベルを示した。
【0043】
eIF−5A1−mRNAレベルは、eIF−5A1 siRNAデリバリー(門脈低速かん流)後に減少した。マウスに1mlのsiRNA(CT(対照)配列又はeIF−5A1、5mg)又は生理食塩水を、それぞれの群についてn=2で低速逆行性門脈かん流(図2))により導入した。膵管にコラゲナーゼを潅注することにより、膵臓を消化し、そして膵島を、Lewis et al., Proc. Natl. Acad. Sci. USA, 102: 12153-12158 Epub 12005 Aug. 12110, 2005に記載される通りに単離した。膵島(1マウスあたり50個)を16時間インキュベートした。全量RNAを抽出し、そしてβ−アクチン及びeIF−5A1についてRT−PCRを行った(図3)。eIF−5A1/β−アクチンのmRNAの比は、5.24(CT−siRNA)及び3.01(eIF−5A1 siRNA)であった。図3は、siRNAで処理された細胞においてeIF−5A1のmRNAレベルが低下したことを示す。この実験をn=3のマウスで繰り返し、そして膵島をRNA抽出のために3回インキュベートを行った;結果は、最初の観察と一致した。
【0044】
eIF−5A1 siRNAデリバリー(門脈流体力学的かん流)後におけるeIF−5A1mRNAレベルの低下及び膵島アポトーシス割合の低下
マウスに、1mlのsiRNA(CT又はeIF−5A1 5μg)又は生理食塩水を、流体力学的逆行性門脈かん流により導入した(1群あたりn=2)。当該かん流は5秒以内に完了した。膵管にコラゲナーゼを潅注することにより膵臓を消化し、そして膵島を単離した。膵島を16時間インキュベートし、次に分配した:1の群を、アポトーシスの評価のため、プロピジウムヨージドで染色し(1マウスあたり50の膵島)、そしてもう一方の群をRT−PCRで処理した(1マウスあたり25の膵島)。eIF−5A1/β−アクチンについてのmRNAのレベルは、CT−siRNA群において、eIF−5A1 siRNA群よりも高かった。アポトーシス率は、28.1%低下した(図4)。この実験を繰り返したところ(n=3)、アポトーシス率は、再び減少した(図5)。
【0045】
ビオチン化siRNAを用いた膵島かん流
ビオチン化siRNA(50μg)を、上に記載されるように膵島に還流した(低速かん流、n=1)。膵臓を染色用にホルマリン中で固定した。
【0046】
siRNA
siRNA分子は、Dharmacon, Lafayette, Co.により合成された。eIF−5A1 及び対照siRNAの配列はそれぞれ、5’CGGAAUGACUUCCAGCUGAdTdT 3’及び5'AGUCGACCUUCAGUAAGGCdTdT3'であった。
【0047】
RT−PCR
Qiagen RNeasyキットを用いて細胞から全量RNAを抽出した。eIF−5A1プライマーは次の通りである:
フォワード 5'−GAC AGT GGG GAG GTA CGA GA−3';
リバース 5'−GGG GTG AGG AAA ACC AAA AT−3’。
【0048】
プロピジウムヨージド(PI)アポトーシス染色
温和なトリプシン処理により膵島の単一細胞懸濁液を得た。PBSで細胞を洗浄し、そして0.3%サポニン、EDTA1mM、Rnase、1%アジド、1%FCS及び50μg/mlのPIをPBSに含むサポニン−PI混合液を加えた。細胞を完全にボルテックスし、そして暗所にて4℃でインキュベートした後にFACSによりsub−G1集合について分析した。
【0049】
実施例2:腹腔内注射
抗体及びsiRNA:
eIF−5A1 に対するマウスモノクローナル抗体を作成した。抗アクチンモノクローナル抗体(クローンC4、#69100)をMP Biomedicalsから購入した。ウエスタンブロットにおいて、Li−cor社から購入した近赤外フルオロフォア標識された二次抗体を用いた(IRDye800及びIRDye700)。eIF−5A1 特異的siRNAは、eIF−5A1の以下の配列:5'−AAAGGAATGACTTCCAGCTGA−3’に対するものである。コントロールsiRNAは転写配列:5'−AAAGTCGACCTTCAGTAAGGA−3'であり、そしていずれの既知のタンパク質についてもノックダウンを誘導しないことが前に示された。同時係属米国出願第11/725,470号及びPCT/US07/64424を参照のこと。これらの文献をその全てを本明細書に援用する。
【0050】
マウスへのsiRNAのIP(腹腔内)注射:
12匹の8〜10週齢のC57Bl/6雄マウスを、Charles River社から購入した。雌の発情サイクルに伴う代謝変化を避けるために、雄マウスを選択した。マウスをランダムに3つの処理群に割り当てた:第1群−ビヒクル処理(0.9%生理食塩水)、第2群−対照siRNA処理、第3群−eIF−5A1 siRNA処理。各マウスを、16.6mg/kg、又は(第一群の場合)同等の体積をIP注射を介して選択した処理に供した。膵島回収(0日目)の前に3日間(−3、−2、−1日目)毎日(午前11:00ころに)マウスに注射した。
【0051】
膵島単離とサイトカイン処理:
標準的単離技術及び動物実験委員会(IACUC)により承認されたプロトコルを用いてC57Bl/6マウスからの膵島の単離を行った。コラゲナーゼ消化まで膵臓を4.2mM炭酸水素ナトリウム及び1%BSA(Invitrogen)を添加したHBSS中に貯蔵した。分画遠心法により精製を行った。10%FBS及び5%ペニシリン/ストレプトマイシンを添加したフェノールレッドフリーDMEM溶液中に全ての膵島を維持した。培養液を37℃+5%CO2で5.5mMグルコース中においてインキュベートした。代謝試験(カルシウム振動)前に16〜18時間精製された膵島を回復させた。
【0052】
1日目の代謝試験の終わりに、生理的に関連のあるサイトカインの規定のカクテル(5mg/ml IL1β、10ng/mlのTNT−α、100ng/mlのINF−γ)を用いて、サイトカイン処理を適用した。サイトカイン処理された膵島を、次の試験まで24時間休ませ、そしてタンパク質分析を行った(2日目及び3日目)。
【0053】
免疫ブロットアッセイ:
(200mMのDTT及びベンゾナーゼを含むLaemmli緩衝液中で調製された)10μgの膵島細胞抽出物を、4〜20%のSDSポリアクリルアミドゲル(Invitrogen)で電気泳動することにより分離し、抗−eIF−5A1 マウスモノクローナル一次抗体(1:15000希釈)を用いて、免疫ブロット分析を行った。Odysseyシステム(Li−Cor Biosciences社)の近赤外技術を用いて画像化及び定量した。
【0054】
カルシウム振動アッセイ:
(上に記載されるように)コラゲナーゼ消化された8〜10週齢のC57Bl/6マウス膵臓から膵島を手で拾いあげた。Fura−2低グルコース溶液(3mM)中で20分間処理し、次に温度制御されたフローチャンバー中に配置した。低グルコース溶液(Furaを伴わない)を膵島上に流して、ベースカルシウム計測値(5分)を調べた。IP Lab4.0ソフトウェア(BD Biosciences)を用いてカルシウムの計測値を取得し、そして340〜380nmの放出光の比として報告する。ベースカルシウムレベルに達した後に、チャンバーを通して高グルコース(11mM)溶液を流して(15分間)、膵島の細胞内カルシウム応答を誘導する。カルシウム応答を、5秒間の間隔で、20分間の観察のあいだに測定した。試験を終える際、膵島を5%ブリーチ溶液中に配置する。
【0055】
実施例3:in vitro及びin vivoにおけるPdx−1に対するsiRNAの適用
βTC3細胞に、2種のPdx−1siRNAコンストラクト(コンストラクトA及びB)又は対照をトランスフェクトし、そしてタンパク質Pdx−1、GAPDH及びβ−アクチンについての免疫ブロットに供する。図17Aを参照のこと。C57Bl/6マウスに、同じsiRNAを1日1回3日間腹腔内注射し、そして膵島をコラゲナーゼ消化及び分画勾配遠心を介して単離及び精製した。膵島を2%SDS中に溶解し、そして免疫ブロットに供した。図17Bを参照のこと。30分間Fura2を単離膵島に充填し、次に3mM D−グルコース中で画像化した。〜300秒で11mM D−グルコースで膵島を刺激し、そしてFura2比を、連続して、蛍光顕微鏡によりモニタリングした。図17Cを参照のこと。グルコース刺激インスリン分泌(GSIS)を、各処理群から50の膵島を用いて行った。図17Dを参照のこと。
【0056】
実施例4:eIF−5A1のノックダウンは、ex vivoでの膵島機能及び生存を改善する
eIF−5Aに対するsiRNAのin vitro投与が、Ex vivoでの膵島機能を改善することをこの実施例は示す。図18Aは、実験設計の模式図を提供する。膵島を処理後に単離し、そして抽出物を免疫ブロット分析に供した。図18Bを参照のこと。単離膵島をグルコース感知及びインスリン転写に必須である遺伝子についてRT−PCR分析に供した。遺伝子転写はサイトカイン処理(IFNg、IL−1b、TNFaと4時間)により抑制されるが、群間の差異は観察されなかった。図18Cを参照のこと。単離後24時間のsi−eIF−5A処理された膵島(図18Dを参照のこと)及び単離後48時間のsi−eIF−5A処理された膵島(図18Fを参照のこと)は、単離後24時間の対照処理された膵島(図18Dを参照のこと)及び単離後48時間の対照処理(図18Fを参照のこと)に比較して、グルコース刺激カルシウム(GSCa)応答の改善を示した。sieIF−5A処理された膵島は、単離後24時間のサイトカイン(図18Eを参照のこと)及び単離後48時間のサイトカイン(図18Gを参照のこと)の存在下で高いGSCaを示す。eIF−5A処理されたマウスから単離された膵島は、GSISにより評価された場合に、4時間のサイトカインインキュベーションがない場合(図18Hを参照のこと)とある場合(図18Hを参照のこと)の両方において、対照に比較して、グルコース刺激インスリン分泌(GSIS)の改善を示した。
【0057】
実施例5:eIF−5Aのノックダウンが、サイトカイン誘導性iNOS生産を無効にする
サイトカイン誘導性のiNOSタンパク質産生は、eIF−5A欠損細胞において生じない。サイトカインで処理された膵島を、iNOSmRNAについてRT−PCRに供した。全ての処理群は、サイトカイン暴露の際にiNOSmRNAの劇的な上方制御を示した。図19Aを参照のこと。サイトカイン処理された膵島を免疫ブロット分析に供した。注目すべきは、iNOSタンパク質は、サイトカイン暴露された際に対照膵島中において誘導されるが、si eIF−5A処理された膵島においては誘導されないという点である。図19Bを参照のこと。INS−1(832/13)β細胞を、サイトカインカクテルで4時間処理し、そして免疫ブロットに供した。細胞はiNOSタンパク質の同様の増加を示す。図19Cを参照のこと。INS−1細胞を、様々な濃度のGC−7で処理し、デオキシハイプシン合成酵素の阻害剤で一晩処理し、次にサイトカインの存在下で4時間インキュベートした。INS−1細胞は、iNOS生産及びCG−7濃度の逆相関を示し、活性eIF−5A産生を抑制が、iNOS翻訳を示唆する。図19Dを参照のこと。
【表1】
【表2】
【特許請求の範囲】
【請求項1】
単離後において回収された膵島細胞の機能を保存する方法であって、eIF−5A1 siRNAを、膵島の単離前に膵島細胞ドナーの膵島細胞に投与することを含み、ここで当該eIF−5A1 siRNAが、膵島細胞においてeIF−5A1の発現を抑制し、それにより膵島細胞におけるアポトーシスを阻害し、そして回収された膵島細胞の機能を保存する、前記方法。
【請求項2】
前記eIF−5A1 siRNAが、eIF−5A1の以下のヌクレオチド配列:
5’−AAAGGAATGACTTCCAGCTGA−3';
5'−AAGATCGTCGAGATGTCTACT−3';
5'−AAGGTCCATCTGGTTGGTATT−3';又は
5'−AAGCTGGACTCCTCCTACACA−3'
を標的とする、請求項1に記載の方法。
【請求項3】
前記eIF−5A1 siRNAが、以下のヌクレオチド配列:
5'−AAAGGAAUGACUUCCAGCTGAdTdT−3'
を含む、請求項1に記載の方法。
【請求項4】
前記siRNAが、膵島細胞ドナーへの腹腔内注射を介して投与される、請求項1に記載の方法。
【請求項5】
膵島細胞がドナー回収プロセスの間にアポトーシスを引き起こすことを阻害する方法であって、eIF−5A1 siRNAを、膵島単離前に膵島細胞ドナーへの腹腔内注射を介して膵島細胞ドナーにeIF−5A1 siRNAを投与することを含み、ここで当該eIF−5A1 siRNAは、膵島細胞におけるeIF−5A1 の発現を阻害し、それにより膵島細胞におけるアポトーシスを阻害する、前記方法。
【請求項6】
前記eIF−5A1 siRNAが、eIF−5A1 の以下のヌクレオチド配列:
5'−AAAGGAATGACTTCCAGCTGA−3';
5'−AAGATCGTCGAGATGTCTACT−3';
5'−AAGGTCCATCTGGTTGGTATT−3';又は
5'−AAGCTGGACTCCTCCTACACA−3'
を標的とする、請求項5に記載の方法。
【請求項7】
前記eIF−5A1 siRNAが、ヌクレオチド配列:5'−AAAGGAAUGACUUCCAGCTGAdTdT−3'を含む、請求項5に記載の方法。
【請求項1】
単離後において回収された膵島細胞の機能を保存する方法であって、eIF−5A1 siRNAを、膵島の単離前に膵島細胞ドナーの膵島細胞に投与することを含み、ここで当該eIF−5A1 siRNAが、膵島細胞においてeIF−5A1の発現を抑制し、それにより膵島細胞におけるアポトーシスを阻害し、そして回収された膵島細胞の機能を保存する、前記方法。
【請求項2】
前記eIF−5A1 siRNAが、eIF−5A1の以下のヌクレオチド配列:
5’−AAAGGAATGACTTCCAGCTGA−3';
5'−AAGATCGTCGAGATGTCTACT−3';
5'−AAGGTCCATCTGGTTGGTATT−3';又は
5'−AAGCTGGACTCCTCCTACACA−3'
を標的とする、請求項1に記載の方法。
【請求項3】
前記eIF−5A1 siRNAが、以下のヌクレオチド配列:
5'−AAAGGAAUGACUUCCAGCTGAdTdT−3'
を含む、請求項1に記載の方法。
【請求項4】
前記siRNAが、膵島細胞ドナーへの腹腔内注射を介して投与される、請求項1に記載の方法。
【請求項5】
膵島細胞がドナー回収プロセスの間にアポトーシスを引き起こすことを阻害する方法であって、eIF−5A1 siRNAを、膵島単離前に膵島細胞ドナーへの腹腔内注射を介して膵島細胞ドナーにeIF−5A1 siRNAを投与することを含み、ここで当該eIF−5A1 siRNAは、膵島細胞におけるeIF−5A1 の発現を阻害し、それにより膵島細胞におけるアポトーシスを阻害する、前記方法。
【請求項6】
前記eIF−5A1 siRNAが、eIF−5A1 の以下のヌクレオチド配列:
5'−AAAGGAATGACTTCCAGCTGA−3';
5'−AAGATCGTCGAGATGTCTACT−3';
5'−AAGGTCCATCTGGTTGGTATT−3';又は
5'−AAGCTGGACTCCTCCTACACA−3'
を標的とする、請求項5に記載の方法。
【請求項7】
前記eIF−5A1 siRNAが、ヌクレオチド配列:5'−AAAGGAAUGACUUCCAGCTGAdTdT−3'を含む、請求項5に記載の方法。
【図1】


【図2】

【図3】

【図4】
【図5】
【図6】

【図7】

【図8】

【図9】
【図10A】

【図10B】

【図11】

【図12】
【図13】
【図14】
【図15】

【図16A】

【図16B】
【図17A】
【図17B】

【図17C−D】
【図18A】

【図18B】

【図18C】
【図18D−I】
【図19A】
【図19B】
【図19C】
【図19D】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10A】
【図10B】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16A】
【図16B】
【図17A】
【図17B】
【図17C−D】
【図18A】
【図18B】
【図18C】
【図18D−I】
【図19A】
【図19B】
【図19C】
【図19D】
【公表番号】特表2010−536379(P2010−536379A)
【公表日】平成22年12月2日(2010.12.2)
【国際特許分類】
【出願番号】特願2010−521983(P2010−521983)
【出願日】平成20年8月20日(2008.8.20)
【国際出願番号】PCT/US2008/073642
【国際公開番号】WO2009/026317
【国際公開日】平成21年2月26日(2009.2.26)
【出願人】(502458132)セネスコ テクノロジーズ,インコーポレイティド (16)
【Fターム(参考)】
【公表日】平成22年12月2日(2010.12.2)
【国際特許分類】
【出願日】平成20年8月20日(2008.8.20)
【国際出願番号】PCT/US2008/073642
【国際公開番号】WO2009/026317
【国際公開日】平成21年2月26日(2009.2.26)
【出願人】(502458132)セネスコ テクノロジーズ,インコーポレイティド (16)
【Fターム(参考)】
[ Back to top ]